Embed Size (px)
Citation preview

Universidade Federal do Piauí
Caracterização cariotípica de acessos de pimentas (Capsicum sp.)
Willame Rodrigues do Nascimento Sousa
Dissertação apresentada à Universidade Federal do Piauí como parte das exigências do Programa de Pós-Graduação em Genética e Melhoramento, para a obtenção do título de “Mestre”.
Teresina 2012

Willame Rodrigues do Nascimento Sousa Licenciado em Ciências Biológicas
Caracterização cariotípica de acessos de pimentas (Capsicum sp.)
Orientadora: Profa. Dra. ANA PAULA PERON
Co-orientadora:
Profa. Dra. ÂNGELA CELIS DE ALMEIDA LOPES
Dissertação apresentada à Universidade Federal do Piauí como parte das exigências do Programa de Pós-Graduação em Genética e Melhoramento, para a obtenção do título de “Mestre”.
Teresina 2012

FICHA CATALOGRÁFICA
Serviço de Processamento Técnico da Universidade Federal do Piauí
Biblioteca Comunitária Jornalista Carlos Castello Branco
S725c Sousa, Willame Rodrigues do Nascimento.
Caracterização cariotípica de acessos de pimentas (Capsicum sp.) [manuscrito] / Willame Rodrigues do Nascimento Sousa. – 2012.
45 f.
Cópia de computador (printout).
Dissertação (Mestrado) – Programa de Mestrado em Genética e Melhoramento.
“Orientadora: Profa. Dra Ana Paula Peron”

Caracterização cariotípica de acessos de pimentas (Capsicum sp.)
Willame Rodrigues do Nascimento Sousa
Licenciado em Ciências Biológicas
Aprovada em ___/___/2012
Comissão julgadora:
______________________________________________________
Prof. Dr. Reginaldo de Carvalho – UFRPE ______________________________________________________
Prof. Dr. Sérgio Emílio dos Santos Valente - CCN/UFPI
______________________________________________________ Profa. Dra. Ângela Celis de Almeida Lopes – CCN/UFPI
(Co-orientadora)
______________________________________________________ Profa. Dra. Ana Paula Peron - CSHN/UFPI
(Orientadora)

Aos meus queridos pais, Alípio e Nazaré, pelo exemplo de vida e por serem os melhores pais que eu poderia ter; aos meus irmãos: Socorro e Washington, por todo apoio, carinho, atenção e união; à minha querida companheira e esposa Sandra e aos filhos da minha vida: Ismael, Daniel e Davi.
Dedico.

AGRADECIMENTOS
Quero agradecer, em primeiro lugar, a DEUS, pela força e coragem durante toda
esta caminhada;
À Secretaria de Educação do Estado do Piauí, pela liberação no período da
execução do mestrado;
À Universidade Federal do Piauí e, aos professores do programa pelas sugestões e
ensinamentos e, à Coordenação do Programa de Pós-Graduação em Genética e
Melhoramento pela oportunidade de realização do curso;
À professora Drª Ana Paula Peron, pela dedicação, orientação, sugestões e críticas
que contribuíram não só para a realização deste trabalho, mas para meu
enriquecimento profissional e pessoal;
À Profª Drª Ângela Celis de Almeida Lopes, pela co-orientação neste estudo e pelo
fornecimento das sementes para a continuação deste trabalho;
À Profª Drª Francilene Leonel Campos, pela orientação na escolha do meu tema ao
me apresentar ao fabuloso mundo dos cromossomos, confiança, incentivo e apoio
em todas as oportunidades e pela valiosa amizade;
À Profª Drª Regina Lucia Ferreira Gomes, Coordenadora do Programa de
Capacitação a Docência (PROCAD/CAPES), pelo auxílio na estadia em Recife-PE;
Ao Prof. Dr. Reginaldo de Carvalho pela amizade, respeito e confiança construída ao
longo dos anos, e pela contribuição no meu crescimento profissional no curto
período em que estive sob a sua orientação no Laboratório de Genética Bioquímica
e Seqüenciamento de DNA (UFRPE). Meus sinceros agradecimentos;
Ao pesquisador e Prof. Dr. Francisco Rodrigues Freire Filho, pelo seu apoio,
conselho, incentivo e inspiração no amadurecimento dos meus conhecimentos,
conceitos que me levaram à execução e conclusão desse estudo, pela amizade que
se construiu além dos espaços da Universidade e por acreditar em mim;
Aos meus amigos do lado esquerdo do peito da 1ª turma de 2009 (a gente nunca
esquece) Verônica Brito, Mara Danielle, Isis Gomes, Claudia Oliveira, Michelly

Ferreira, Leane Fialho, Jeane Moura, Leonardo Castelo Branco e Sammya Vanessa,
pela amizade, coleguismo, paciência e incentivo em muitos momentos. Foi um
grande prazer em conhecê-los e tê-los como amigos;
Agradeço especialmente à minha grande amiga e irmã, Jeane Moura, que dispensa
comentários, pela confiança e amizade conquistada ao longo do curso;
Aos amigos da 2ª turma, Hendrie Ferreira Nunes, José Ribamar de Assunção Filho e
Éllida de Aguiar Silvestre, pessoas verdadeiras e de caráter irretocável. Minha
sincera admiração a vocês;
Aos amigos da 3ª Turma, Jaqueline Luz, Rosana Mendes, Camila Campêlo, João
Paulo Gomes, Kátia Carvalho, Kaline Gonzalez, Mariane de Moraes, pelo
companheirismo e boas risadas e pelas contribuições ao longo desta jornada;
Agradeço também à minha esposa, Sandra, que de forma especial e carinhosa me
deu força e coragem, me apoiando nos momentos de dificuldades, quero agradecer
também aos meus filhos, Ismael, Daniel e Davi, que embora não tivessem
conhecimento disto, mas iluminaram de maneira especial os meus pensamentos,
levando-me a buscar mais conhecimentos;
E não deixando de agradecer de forma afável e grandiosa aos meus pais, Alípio e
Nazaré, pelo companheirismo, carinho e pelo cuidado, possibilitando um caminhar
mais suave e menos tortuoso nesta estrada da vida, a quem eu rogo todas as noites
a minha existência;
Aos membros desta banca, desde já pela disposição em ler este trabalho e pelas
críticas que contribuirão nas publicações.

EPÍGRAFE
A glória não está em ficar em pé.
Esta em você se levantar a cada vez que você cair.
Quem espera que a vida
Seja feita de ilusão
Pode até ficar maluco
Ou morrer na solidão.
É preciso ter cuidado
Pra mais tarde não sofrer.
É preciso saber viver.
Toda pedra no caminho
Você pode retirar,
Numa flor que tem espinhos
Você pode se arranhar.
Se o bem e o mal existem
Você pode escolher.
É preciso saber viver.
(Roberto Carlos e Erasmo Carlos)

SUMÁRIO
RESUMO ................................................................................................................ VIII
ABSTRACT .............................................................................................................. IX
1. INTRODUÇÃO .................................................................................................... 10
2. REVISÃO DE LITERATURA ............................................................................... 11
2.1. Descrição botânica das espécies C. annuum, C. frutescens, C. chinense e C. baccatum ............................................................................................................ 11
2.2. Importância econômica das espécies C. annuum, C. frutescens, C. chinense e
C. baccatum ............................................................................................................. 15
2.3. Conservação e Diversidade genética das espécies C. annuum, C. frutescens,
C. chinense e C. baccatum ...................................................................................... 17
2.4. Citogenética de Capsicum sp. ........................................................................... 20
Referências .............................................................................................................. 24
3. CARACTERIZAÇÃO CARIOTÍPICA DE ACESSOS DE PIMENTAS
(Capsicum sp.)
Resumo .................................................................................................................... 30
Abstract .................................................................................................................... 31
3.1. Introdução ......................................................................................................... 32
3.2. Material e Métodos ............................................................................................ 34
3.2.1. Material genético ............................................................................................ 34
3.2.2. Avaliação citogenética .................................................................................... 34
3.2.3. Análise cariotípica .......................................................................................... 35
3.2.4. Análise dos dados .......................................................................................... 35
3.3. Resultados e Discussão .................................................................................... 36
3.4. Conclusões ........................................................................................................ 42
Referências .............................................................................................................. 43

viii
RESUMO
SOUSA, W. R. N. Caracterização cariotípica de acessos de pimentas (Capsicum sp.) 48p. Dissertação (Mestrado/Genética e Melhoramento) – UFPI, Teresina, 2012.
Capsicum é um gênero da família Solanaceae de significativa importância
econômica, nutricional, medicinal e ornamental em todo o mundo. De suas 25
espécies, quatro são extensivamente cultivadas no Brasil, que são: C. annuum, C.
frutescens, C. chinense e C. baccatum. No entanto, o conhecimento biológico sobre
estas plantas ainda é incipiente neste país, apesar de nos últimos anos ter ocorrido
um amplo empenho dos pesquisadores em estudos bioquímicos, moleculares e
citogenéticos com estas espécies. É fato que a análise do número e morfologia dos
cromossomos de espécies de importância econômica é de grande relevância, pois
geram subsídios importantes para programas de melhoramento genético. Assim,
este trabalho teve como objetivo caracterizar numérica e morfologicamente os
cromossomos de acessos das quatro espécies de Capsicum pertencentes à coleção
do Banco Ativo de Germoplasma de Capsicum da Universidade Federal do Piauí
(BGC-UFPI). Os cariótipos foram obtidos pelo método de esmagamento e coloração
com Giemsa de células dos meristemas das radículas dos acessos estudados. A
partir dos resultados verificou-se número cromossômico 2n = 2x = 24 para todas as
quatro espécies. Foi observado polimorfismo cromossômico para o acesso BGC 37,
da espécie C. frutescens, que apresentou 12 pares de cromossomos metacêntricos,
diferindo dos outros que apresentaram 11 pares de cromossomos metacêntricos e
um par de cromossomo sub-metacêntrico. Ainda foram visualizadas nos acessos
BGC 01 e BGC 37 constrições secundárias nos pares homólogos 01 e 12, e 6 e 11,
respectivamente. O índice de assimetria obtido mostrou que entre as espécies os
cariótipos analisados foram assimétricos, o que corrobora com a grande diversidade
genética descrita na literatura para as espécies estudadas.
Palavras-chave: Solanacea; citogenética; banco de germoplasma; morfometria.

viii
ABSTRACT
SOUSA, W. R. N. Characterization of karyotypic chili peppers (Capsicum sp.) 48p. Dissertation (Msc/Genetics and Breeding) – UFPI, Teresina, 2012.
Capsicum is a genus of the family Solanaceae of significant economic
importance, nutritional, medicinal and ornamental worldwide. Of its 25 species, four
are extensively cultivated in Brazil, which are: C. annuum, C. frutescens, C. chinense
and C. baccatum. However, the biological knowledge about these plants is still
incipient in this country, although in recent years have been a large commitment of
researchers in biochemical, molecular and cytogenetic with these species. It is a fact
that the analysis of the number and morphology of chromosomes of species of
economic importance is of great importance, because they generate important
information for breeding programs. This study aimed to characterize the morphology
and number of accesses chromosomes of four species of Capsicum from the
collection of the Active Germplasm Bank of Capsicum Federal University of Piauí
(UFPI-BGC). The karyotypes were obtained by the method of crushing and Giemsa
staining of cells of the radicle meristems of accessions. From the results it was
observed chromosome number 2n = 2x = 24 for all four species. Chromosomal
polymorphism was observed for access BGC 37, species C. frutescens, which
presented 12 pairs of metacentric chromosomes, differing from others who had 11
pairs of metacentric chromosomes and a pair of sub-metacentric chromosome. Even
were visualized in BGC 01 and BGC 37 this species secondary constrictions in
homologous pairs 01 and 12, and 6 and 11, respectively. The asymmetry index
obtained showed that among the species analyzed the karyotypes were asymmetric,
which confirms the great genetic diversity described in the literature for the species
studied.
Keywords: Solanacea; cytogenetics; genebank; morphometry.

10
1. INTRODUÇÃO
O gênero Capsicum, da família Solanaceae, apresenta importância agrícola
mundial e é largamente cultivada nas regiões temperadas e tropicais, não só devido
sua importância econômica como especiaria, mas também por seu valor nutricional,
ornamental e medicinal (POZZOBON et al., 2006).
Este gênero é composto por 31 espécies, sendo cinco classificadas como
domesticadas, que são: C. annuum L. (pimentão, pimenta doce), C. chinense Jaqc.
(pimenta-de-cheiro, pimenta-de-bode, murici), C. frutescens L. (pimenta malagueta)
e C. baccatum L. (dedo-de-moça, chapéu-de-frade), grandemente consumidas e
produzidas no Brasil, e a Capsicum pubescens L., não cultivada neste país
(LANNES et al., 2007). Destacam-se por terem boa adaptação às condições de
clima tropical e grande diversidade genética que é evidenciada, por exemplo, em
seus frutos e flores que apresentam variedades de tamanhos, formas e colorações
(HAVERROTH; NEGREIROS, 2011; ZENI; BOSIO, 2011).
O estudo morfoagronômico, citogenético, molecular e de divergências
genotípicas e fenotípicas de bancos de germoplasmas de Capsicum é de grande
significado para a agricultura por fornecer dados importantes para o entendimento
da organização e da estrutura das relações evolutivas entre as espécies, e também
por gerar informações importantes para os programas de melhoramento genético
destas hortícolas, como por exemplo, em esquemas de cruzamentos entre táxons
distintos, transferência de genes de interesse na seleção/criação de cultivares mais
produtivas, com melhores qualidades nutricionais e mais tolerantes a fatores bióticos
e abióticos (NOWACZYK et al., 2006; SUDRÉ et al., 2006; INCE et al., 2009).
Assim, este trabalho teve por objetivo analisar citogeneticamente acessos de
pimentas C. annuum, C. frutescens, C. chinense e C. baccatum provenientes do
Banco Ativo de Germoplasma de Capsicum (BGC) da Universidade Federal do
Piauí, no Campus Ministro Petrônio Portella, em Teresina-PI, por meio da contagem
cromossômica e da morfometria, com o intuito de detectar polimorfismos
cromossômicos numéricos e estruturais ampliando as informações citogenéticas
para este grupo de plantas, disponibilizando um maior número de dados quanto aos
recursos genéticos presentes em bancos de germoplasmas e auxiliar programas de
melhoramento genético voltados para este gênero.

11
2. REVISÃO DE LITERATURA
2.1 - Descrição botânica das espécies C. annuum, C. frutescens, C. chinense e
C. baccatum.
A taxonomia de Capsicum é considerada polêmica, devido, segundo Nuez et
al. (1996), à grande diversidade morfológica encontrada neste gênero, onde uma
combinação de caracteres diagnósticos é necessária para identificar e diferenciar as
espécies deste gênero (WALSH ; HOOT, 2001).
Ainda não há um consenso entre os taxonomistas quanto ao número de
espécies de Capsicum (BOSLAND, 1994; REIFSCHNEIDER, 2000; CARVALHO et
al., 2003). Entretanto, dentre as dezenas de espécies encontradas e descritas
Bosland (1994, 1996), Reifschneider (2000), Pickergill (1997) e Nuez et al. (1996)
relatam a existência de cinco espécies classificadas como domesticadas, que são:
C. annuum L., C. chinense Jacq., C. frutescens L., C. baccatum L. e C. pubescens
Ruiz & Pavon, a única das cinco espécies que não é cultivada no Brasil (RIBEIRO;
REIFSCHNEIDER, 2008).
As plantas deste gênero, em geral, são autógamas, o que facilita a sua
reprodução, embora a polinização cruzada também ocorra entre indivíduos dentro
da mesma espécie e entre espécies do gênero (CARVALHO; BIANCHETTI, 2008),
sendo facilitada por alterações morfológicas na flor, pela ação de insetos
polinizadores e por práticas de cultivo, como local, adensamento ou cultivo misto
(RIBEIRO; REIFSCHNEIDER, 2008).
Capsicum annuum é a espécie mais cultivada no Brasil, apesar de sua
domesticação ter ocorrido no México, e incluem as variedades mais populares do
gênero, como os pimentões e as pimentas doces (Figura 2.1). Segundo Carvalho e
Bianchetti (2008), estas plantas apresentam uma flor por nó, e na antese os
pedicelos podem ser eretos, pendentes ou inclinados. Geralmente, a corola é branca
sem manchas na base dos lobos das pétalas e as anteras são azuladas. Os cálices
dos frutos são pouco dentados e não possuem constrição anelar na junção do
pedicelo. Possuem várias cores e formas, com polpa firme, e sementes de cor clara.

12
Figura 2.1. Aspectos das folhagens, flores e frutos das pimentas Capsicum annuum.
Foto: Eliane Rodrigues Monteiro.
A C. frutescens inclui as pimentas malaguetas, extremamente picantes e
amplamente difundidas na América do Sul e Central, e em outras regiões tropicais e
subtropicais, como Ásia, África e Ilhas do Pacífico (YAMAMOTO; NAWATA, 2005).
(Figura 2.2). De acordo com Nuez et al. (1996) estas plantas tem sementes de cor
clara e flores solitárias em cada nó. Possuem pedicelos eretos na antese com as
flores deitadas. A corola é branca-esverdeada, sem manchas difusas na base das
pétalas e ligeiramente revoluta. O fruto tem bagas geralmente fusiformes e porte
ereto, e sua polpa é suave e extremamente fluída quando comparada às outras
espécies. No Brasil, o representante desta planta mais conhecido é a pimenta-
malagueta, grandemente apreciada em todas as regiões do país (NUEZ et al.,
1996).

13
Figura 2.2. Aspectos das folhagens, flores e frutos das pimentas Capsicum frutescens.
Foto: Eliane Rodrigues Monteiro.
Segundo Smith e Heiser (1957) a espécie apresenta a seguinte descrição
botânica: C. chinense Jacquin (syn. C. sinense), plantas arbustivas com 0,45 a 0,76
m de altura; hábito ereto, prostrado ou compacto; folhas e ramos essencialmente
glabros, pequena pubescência, folhas ovadas a ovado-lanceoladas de 10,5 cm,
largas, macias ou rugosas, de tonalidade verde claro a escuro; as flores aparecem
de 3 a 5 por nó, exceto em plantas depauperadas; o pedicelo é pendente, raramente
ereto, relativamente curto e grosso na antese; cálice sem dentes com forte
constrição na base e raramente sem constrição; corola verde-amarelada ou
raramente esbranquiçada, medindo de 0,5 a 1,0 cm de comprimento; anteras azuis,
púrpuras ou amareladas.
A principal característica morfológica que distingue C. chinense das demais
espécies cultivadas é a presença de uma constrição basal, localizada entre o cálice
e o pedicelo (NUEZ-VIÑALS et al., 2003; CARVALHO et al., 2003). Esta planta é
conhecida como pimenta-de-cheiro, pimenta-de-bode e murici e caracterizada por

14
sua forte pungência (REIFSCHENEIDER, 2000) (Figura 2.3). O nome dado a esta
espécie, “da China”, foi uma homenagem do holandês Kikolaus von Jacquinoist a
China, associando incorretamente o nome à sua origem, visto que nesta época já
tinha conhecimento de que estas plantas eram originárias das Américas (BOSLAND;
VOTAVA, 1999).
Figura 2.3. Aspectos das folhagens, flores e frutos das pimentas
Capsicum chinense. Foto: Eliane Rodrigues Monteiro.
Já a C. baccatum, representadas pelas pimentas dedo-de-moça e chapéu-de-
frade, são os tipos mais cultivados e consumidos no Brasil (Figura 2.4). A dedo-de-
moça é também conhecida como pimenta-vermelha, pimenta calabresa ou chifre-de-
veado. O seu cultivo, neste país, é realizado por pequenos, médios e grandes
produtores (CARVALHO et al., 2009).
Estas hortaliças são arbustivas, com cerca de 1m de altura. As flores se
apresentam em número de uma a duas. Na antese, os pedicelos são geralmente
eretos. A corola é branca e sempre apresenta um par de manchas amareladas ou
esverdeadas na base de cada lobo das pétalas. As anteras são amarelas.

15
Os cálices dos frutos maduros são evidentemente dentados e não possuem
constrição anelar na junção do pedicelo. Os frutos são de várias cores e formas,
geralmente pendentes, persistentes, com polpa firme, medem cerca de 1,0 a 1,5 cm
de diâmetro, de 8 a 10 cm de comprimento e possuem pungência suave e as
sementes são cor de palha. (CARVALHO et al., 2006; MOREIRA et al., 2006).
Figura 2.4 - Aspectos das folhagens, flores e frutos das pimentas Capsicum baccatum. Foto: Eliane Rodrigues Monteiro.
2.2- Importância econômica das espécies C. annuum, C. frutescens, C.
chinense e C. baccatum
As pimentas do gênero Capsicum são de grande importância econômica,
sendo largamente utilizadas na culinária tradicional de países latino-americanos,
como o Peru e o México (TOFANELLI et al., 2003). No Brasil, o cultivo de pimentas e
pimentões constitui uma importante atividade socioeconômica para o setor agrícola,
e ocorre em praticamente todas as regiões do país, sendo responsável por 13.000ha
de área cultivada anualmente, com produção aproximada de 296.000 toneladas de
frutos (REIFSCHNEIDER et al., 2000; HENZ, 2004; IBGE, 2006).
Estas hortaliças, com destaque para C. annuum, C. frutescens, C. chinense e
C. baccatum, são apreciadas na gastronomia mundial pelo realce picante que dão
aos alimentos salgados e saladas, no entanto, hoje em dia, também são utilizadas
em doces, geleias, bolos e sucos (HENZ; RIBEIRO, 2008). Assim, a sua importância
econômica tem se acentuado a cada dia e despertado o interesse de novos
produtores e consumidores em todo o mundo (KWON; KIM, 2009).
No Brasil o comércio destas quatro espécies de hortaliças é bastante
diversificado, indo desde o consumo interno in natura, na forma de condimentos,

16
temperos, flocos (calabresa), pó (páprica picante) e conservas de molhos líquidos,
até o segmento de exportação, em nível mundial, de páprica, pó de pimentão e
pimenta doce vermelha madura (HENZ, 2004; MOREIRA et al., 2006). No entanto, o
mercado de pimentas, de modo geral, neste país, é considerado secundário em
relação ao de outras hortaliças, apesar dos estados de Minas Gerais e São Paulo
serem grandes produtores e exportadores de pimentas e pimentões (HENZ;
RIBEIRO, 2008).
No Piauí, os dados sobre a produção agrícola de pimentas e pimentões são
muito imprecisos e irregulares já que são cultivadas por pequenos produtores em
agricultura familiar (IBGE, 2006). Apesar de grande parte das pimentas consumidas
neste estado serem importadas de outros estados, como Minas Gerais, São Paulo,
Ceará e Bahia, ocorrem um importante mercado paralelo das mesmas produzidas
por pequenos agricultores e comercializadas na forma in natura (HENZ; RIBEIRO,
2008). Assim, as pimentas processadas envolvem pequenas produções caseiras,
com a fabricação de molho artesanal, conservas, geleias, pimenta em pó, que são
vendidas para pequenas mercearias, padarias, feiras livre, lojas de produtos
agropecuários e supermercados. Outro aspecto importante
As pimentas C. annuum, C. frutescens, C. chinense e C. baccatum também
possuem grande importância nutricional por terem em sua constituição grandes
quantidades de proteínas, glicídios, lipídios, minerais, vitaminas, água e fibras que,
quando em proporções adequadas na dieta, ajudam na manutenção das funções
vitais do organismo (VAISHNAVA; WANG, 2003).
Capsicum é o único gênero da família Solanaceae que possui o alcalóide
capsaicina (8-metil-N-vanilil 1-6-nonamida), composto químico pungente sintetizado
e acumulado nos tecidos da placenta de seus frutos (BOSLAND; VOTAVA, 1999).
Este alcalóide possui extensa variedade de benefícios terapêuticos e é utilizado pela
população no combate ou amenização da síndrome de pós-mastectomia, urticária,
psoríase, neuropatia diabética, artrite, prurido vaginal, alergias, neuromas pós-
cirúrgicos (PALEVITCH; CRAKER, 1995; KNOTKOVA et al., 2008), regulação da
pressão arterial, metabolismo da gordura pela ação termogênica (LEUNG, 2008),
prevenção de câncer (SURH, 2003) e ação antibacteriana (TEWKSBURY et al.,
2008).
Ainda, segundo Leung (2008), a capsaicina em ação conjunta com o β-
caroteno e as vitaminas A e C atuam na diminuição do nível de gordura no sangue,

17
possuem ação expectorante ajudando a descongestionar vias respiratórias (XU et
al., 2005), ação anti-inflamatória e antioxidante (TEWKSBURY et al., 2008). As
pimenteiras também são utilizadas como plantas ornamentais (Figura 2.5) em
função da sua folhagem variegada, do porte anão e dos frutos com diferentes
padrões de cores (MOREIRA et al., 2006).
Figura 2.5 - Pimentas ornamentais comercializadas em Teresina-PI, 2011.
Foto: Willame Rodrigues do Nascimento Sousa.
Apesar de sua importância, os dados estatísticos de produção e
comercialização de pimenta hortícola no Brasil são escassos em todas as fases do
seu sistema de produção e, a pouca informação disponível não reflete a realidade
econômica dessa hortaliça, visto que grande parte da produção é comercializada em
mercados regionais e locais que não fazem parte das estatísticas (DOMENICO et
al., 2010). A busca por maior produção e melhor qualidade de preços e custos
tem exigido dos produtores maior eficiência técnica e econômica na condução dos
sistemas de produção, utilização e conservação da diversidade genética destas
espécies.
2.3- Conservação e diversidade genética das espécies C. annuum, C.
frutescens, C. chinense e C. baccatum.
A origem das espécies C. annuum, C. frutescens, C. chinense e C. baccatum
suscita opiniões divergentes em todo mundo (INCE et al., 2009). Atualmente, a
informação mais aceita é que as espécies C. annuum e C. frutescens foram

18
domesticadas na América Central, e as espécies C. chinense, C. baccatum
juntamente com a C. pubescens foram domesticadas na América do Sul
(PICKERSGILL, 2007; KIM et al., 2011). Segundo relato de Crosby (2008), a maior
diversidade genética destas quatro espécies está na América do Sul e Central, onde
o México é o centro de diversidade primária de C. annuum, com centros secundários
no norte da América do Sul, Índia, região do Mediterrâneo e Sudeste Asiático.
O Brasil é um importante centro de diversidade para este gênero por abrigar
tanto espécies domesticadas, quanto semidomesticadas e silvestres (CARVALHO et
al., 2003; RÊGO et al., 2003). Porém, apesar destas espécies de pimentas terem
distribuição ampla, grande parte de sua variabilidade genética ainda não foi
explorada, o que as tornam de grande interesse para a construção de bancos de
germoplasma, para que se realizem estudos morfo-agronômicos, bioquímicos,
citogenéticos e moleculares com o intuito de se utilizar e conservar tais recursos
genéticos (SUDRÉ et al., 2006).
A caracterização e a avaliação de espécies silvestre, domesticada e semi-
domesticada de Capsicum são particularmente interessantes para curadores de
banco de genes, já que uma grande variabilidade, ainda não totalmente conhecido e
explorado, está disponível nestas espécies (SUDRÉ et al., 2006; INCE et al., 2009).
A ausência ou a discordância de informações sobre os acessos mantidos em
bancos de germoplasma motiva o baixo interesse e a importância dos melhoristas
em relação aos acessos conservados (BARBIERI, 2003). De acordo com Li et al.
(1998), os fatores que dificultam a utilização dos bancos de germoplasma são
vários, tais como a não disponibilidade dos acessos aos melhoristas,
desconhecimento sobre os acessos conservados e a ausência de informação dos
curadores sobre os programas realizados pelos melhoristas. O empenho nas
atividades de avaliação e caracterização dos acessos disponíveis em bancos de
germoplasma deve-se, a priori, sererm considerados entre as estratégias de
abordagem e manejo dos recursos genéticos no Brasil. Superada a escassez de
dados, o germoplasma terá mais proveito para os programas de melhoramento
genético (NEITZKE et al., 2010).
A caracterização e a avaliação dos acessos depositados colaboram na
apuração de dados do germoplasma disponível, localizando acessos duplicados,
estabelecimento de coleções nucleares e a identificação dos meios predominante de
reprodução dos acessos, bem como da ocorrência ou não de variabilidade intrínseca

19
em acessos individuais (VALLS, 2007). A caracterização morfológica é um método
que utiliza uma tabela descritiva, que trata de fornecer elementos mais detalhados
sobre o germoplasma conservado no banco, dispondo-o de uma forma mais eficaz
para o seu uso (RAMOS et al., 1999).
Assim, uma análise do germoplasma destas quatro espécies de Capsicum a
partir de análises morfoagronômicas, divergências genotípica e fenotípica e análise
de cromossomos é ideal para uma maior utilidade de seus genótipos em programas
de melhoramento genético, com o objetivo de desenvolver cultivares mais
produtivas, resistente a doenças (FERY; THIES, 1998; KIM et al., 2011) tolerantes
às adversidades das condições bióticas e abióticas e, com maior qualidade
nutricional na produção de β-caroteno, capsaicina e carotenoides em geral
(NOWACZYK et al., 2006; WALL et al., 2001).
Segundo Benko-Iseppon (2001), a caracterização citogenética de diferentes
acessos de bancos de germoplasma representa uma valorosa fonte de informações
para melhoristas e conservacionistas, permitindo um melhor gerenciamento tanto do
“pool” gênico como na escolha de recursos genéticos mais eficientes para os
programas de melhoramento genético.
A Universidade Federal do Piauí possui até o momento, em sua coleção de
germoplasma de Capsicum, cerca de 40 acessos, que representam espécies
domesticadas e semi-domesticadas, oriundas das regiões Nordeste e Sudeste do
Brasil.
Trabalhos de caracterização com a finalidade de identificação botânica e
estimação da divergência genética de 40 sub-amostras de Capsicum sp. por meio
de descritores morfoagronômicos, utilizando duas chaves morfológicas de
identificação e análise multivariada, foram realizados por Monteiro et al.(2008).
Dentre as 40 sub-amostras do banco ativo de germoplasma de Capsicum (BGC), da
Universidade Federal do Piauí, 38 foram identificadas como pertencente a quatro
espécies de Capsicum, com algumas subespécies.
Trajano (2009) avaliou o nível de resistência de dez subamostras de
Capsicum do BGC (UFPI) a sete isolados do fitopatógeno Pythium, onde as sub-
amostras C. annuum var. glabriusculum (BGC 11) foi classificada como altamente
resistente, e C. annuum var. glabriusculum (BGC 36) e C. chinense (BGC 23), como
resistentes, em relação a todas as espécies de Pythium testadas. As sub-amostras

20
C. frutescens (BGC 01), Capsicum sp (BGC 06) e C. chinense (BGC 48) foram
consideradas suscetíveis a todas as espécies de Pythium.
Trabalhos foram realizados com estes acessos tais como a identificação
botânica e divergência genética em pimentas Capsicum sp. por meio de descritores
morfoagronômicos, utilizando duas chaves morfológicas de identificação e análise
multivariada (MONTEIRO et al., 2008) e, o nível de resistência genética de dez
acessos de Capsicum sp., do BGC (UFPI) a sete isolados do fitopatógeno Pythium
sp. (TRAJANO, 2009).
2.4 – Citogenética de Capsicum sp.
A análise de cariótipos, abrangendo a avaliação de dados como número e
tamanho dos cromossomos, relação entre braços cromossômicos, presença de
constrição secundária, quantidade de heterocromatina, pode trazer informações
valiosas para comparar espécies ou examinar a variação entre indivíduos da mesma
espécie (MOSCONE et al., 1996; GUERRA et al., 1997; VENORA; PADULOSI,
1997).
O número cromossômico, assim como a morfometria, são características
amplamente utilizadas na citogenética (GUERRA, 2000) e citotaxonomia vegetal,
associadas com outras características citológicas, auxiliando na compreensão das
variações genéticas envolvidas na evolução de um táxon, como também na
delimitação taxonômica mais precisa das espécies (PEDROSA et al., 1999).
A utilização de parâmetros citogenético e morfométrico na sistemática vegetal
para melhor caracterização de espécies vegetais de interesse econômico, tem se
mostrado de grande importância no reconhecimento de raças, variedades e híbridos
(MIRZAIE-NODOUSHAN et al., 2006), bem como na compreensão das relações
filogenéticas dentro de pequenos táxons, como espécies e gêneros, e em níveis
superiores, como famílias e divisões (STEBBINS, 1971; GUERRA, 2000).
Os dados cromossômicos registrados para as pimentas e pimentões
destacam a presença universal de espécies diplóides com dois números básicos
cromossômicos, x = 12 e x = 13 (POZZOBON et al., 2006). O número de
cromossomos mais frequente encontrado no gênero é o 24 (2n=2x=24) com
cariótipos simétricos (Tabela 2.1), onde as espécies domesticadas C. annuum, C.
frutescens, C. chinense e C. baccatum pertencem a este grupo (MOSCONE, 1993,

21
1996, 2007). Mas, em outras espécies de pimentas como C. buforum, C.
campylopodium e C. cornutum, ocorrem cariótipos com 2n=2x=26, número
comumente encontrado em espécies selvagens da América do Sul (LANTERI;
PICKERGILL, 1993; POZZOBON et al., 2006; MOSCONE et al., 2007; TEODORO-
PARDO, 2007).
Moscone et al. (1996), analisaram os cariótipos de 15 acessos de Capsicum,
no qual foram utilizados um conjunto de técnicas de bandeamento por fluorocromo,
com o intuito de revelar as bandas das regiões de heterocromatina constitutivas, por
coloração com Giemsa, bandeamento-C e bandeamento por nitrato de prata para
localizar regiões organizadoras ativas do nucléolo (AgNORs). Os dados obtidos por
este pesquisadores contribuíram muito na assimilação de marcadores úteis para
identificações cromossômicas, permitindo uma análise cariosistemática deste
gênero, onde os padrões de bandeamento diferem entre os citótipos, espécies e
grupos, mostrando uma dinâmica de diferenciação cromossômica e divergência
evolutiva (MOSCONE et al., 1993, 1995, 1996).
Utilizando bandeamento com laranja de acridina em três acessos de C.
annuum, Almeida (2003), revelou uma banda fluorescente verde-amarelada,
Tabela 2.1. Números cromossômicos e fórmulas cariotípicas do gênero Capsicum.
Espécies 2n Fórmula cariotípica Referências Domesticadas C. annuum var.
annuum 24 12m; 10m+1sm+1a ALMEIDA (2003)
10m+1sm+1st MOSCONE et al. (1996) 11m+1a AARESTRUP (2001)
C. baccatum var. pendulum
24 11m+1st MOSCONE et al. (1996)
C. chinense 24 11m + 1st 11m+1sm; 10m+2sm
GUERRA ( 2001)
11m+1a; 11m+1sm 11m+ 1st
SOUZA (2008) MOSCONE et al. (1996)
C. frutescens 24 11m+1sm; 11m+1a SOUZA et al.(2011) MOSCONE et al. (1996)
11m+ 1st MOSCONE et al. (1996) MOSCONE et al. (1996)
Semidomesticadas C. annuum var. glabriusculum
24 11m+1sm MOSCONE et al. (1993, 2003)

22
subterminal no cromossomo 5, e duas bandas fortemente marcadas, flanqueando a
constrição secundária, no braço curto do cromossomo 11.
A obtenção de vários dados cromossômicos através da utilização das
técnicas convencionais e, em alguns casos a apresentação do cariótipo utilizando
corantes convencionais é insuficiente para uma caracterização intraespecífica e/ou
interespecífica de espécies, pelo fato da maioria das espécies apresentarem
cariótipos desfavoráveis, muitas vezes necessárias para a identificação individual
dos cromossomos. Embora haja uma preferência maior pelo uso de técnicas mais
sofisticadas de citogenética vegetal, o principal instrumento ou parâmetro
citogenético continua sendo a variabilidade cromossômica numérica. A análise da
variação cromossômica numérica de um táxon permite reconhecer o número básico
do grupo, o qual constitui o principal parâmetro citotaxonômico (GUERRA, 2000).
Souza et al. (2008) com o objetivo de caracterizar e analisar o cariótipo de
quatro genótipos de C. chinense, oriundas de regiões distintas do Brasil,
confirmaram o polimorfismo cromossômico de um acesso, no qual apresentaram
fórmula cariotípica 11m+1sm diferindo dos demais, que apresentaram 11m+1a. Do
mesmo modo, Guerra (2001) realizou contagens cromossômicas pela análise
citogenética por meio de método convencional em cromossomos mitóticos de 4
acessos de C. chinense. O comprimento dos cromossomos variou entre 2,15 a 4,52
µm, evidenciando em 3 acessos com 11 pares de cromossomos metacêntricos e
variação no último par entre um submetacêntrico ou subtelocêntrico, e um acesso
com 10 pares metacêntricos e dois submetacêntrico, configurando desta forma uma
assimetria cariotípica. Essas diferenças na morfologia ou no número de
cromossomos podem ocorrer em populações da mesma espécie ou em táxons
interespecíficos, e são denominados de citótipos ou raças cromossômicas
(MOSCONE et al., 2007).
O estudo morfológico dos cromossomos permite comparações entre
categorias taxonômicas detectando possíveis variações existentes, principalmente
no comprimento absoluto, na posição do centrômero, na presença de satélites e
constrições secundárias (MAYEDA, 1997). A constrição secundária é na maioria das
vezes notada em ao menos um dos cromossomos do conjunto haplóide da cada
espécie. Os cromossomos que contêm essa constrição são geralmente vistos,
durante a prófase, associados ao nucléolo desempenhando importantes
propriedades para o funcionamento celular (GUERRA, 1988).

23
Moscone et al. (1996, 2007) verificaram que o gênero possui diferentes
citótipos e divergem principalmente na fórmula cariotípica e no comprimento dos
cromossomos. Rohani et al. (2010) realizaram contagem cromossômica por meio de
método convencional em 10 genótipos de C. annuum L. e todos apresentaram um
par de cromossomos com satélite localizado na extremidade dos braços curtos dos
cromossomos 10, 11 e 12. Com o intuito de confirmar o número cromossômico e a
caracterização morfométrica dos cromossomos para estabelecer possíveis relações
de proximidade entre as espécies estudadas, verificaram que em ambas o número
diplóide é 2n=24. O maior comprimento total haplóide detectado foi de 51,65 µm,
enquanto a menor foi de 43,46 µm.
O cariótipo de uma espécie, em geral, é constante, e pode ser uma
característica útil na taxonomia. Contudo, a presença ou ausência de um ou mais
segmentos cromossômicos em indivíduos de uma população, conhecida como
polimorfismos, podem surgir e alterar o padrão cariotípico da espécie (SYBENGA,
1992).

24
Referências
AARESTRUP, J.R. Análise morfológica dos cromossomos de pimentão Capsicum annuum L. Dissertação (Mestrado em Genética e Melhoramento) Viçosa – MG, Universidade Federal de Viçosa – UFV, 143p, 2001. ALMEIDA, P.M. Caracterização morfológica e bandeamento com laranja de acridina dos cromossomos de pimentão (Capsicum annuum L.). Dissertação (Mestrado em Genética e Melhoramento) Viçosa – MG, Universidade Federal de Viçosa – UFV, 66p, 2003. BARBIERI, R.L. Conservação e uso de recursos genéticos vegetais. In: FREITAS, L.B.; BEREDl, F. (eds). Genética e evolução vegetal. Porto Alegre: UFRGS. p.403-413, 2003. BENKON-ISEPPON, A.M. Estudos moleculares e citogenéticos no Caupi e em espécies relacionadas: Avanços e perspectivas. EMBRAPA - Documentos 56, p.327-332, 2001. BOSLAND, P.W. New Mexico Climate Center. The Wonders of New Mexico Chile. New Mexico Journal of Science, v.34, 1994. BOSLAND, P.W. Capsicums: Innovative uses of an ancient crop. In: J. Janick (ed.), Progress in new crops. ASHS Press, Arlington, VA. p. 479-487, 1996. Disponível em: <http://www.hort.purdue.edu/newcrop/proceedings1996/v3-479.html>. Acesso em: 16 Ago. 2011. BOSLAND, P. W.; VOTAVA, E. J. Peppers: vegetable and spice Capsicums, New York: CABI Publishing, p.66-83, 1999. CARVALHO, S.I.C.; BIANCHETTI, L.B.; BUSTAMANTE, P.G.; SILVA, D.B. Catálogo de germoplasma de pimentas e pimentões (Capsicum spp.) da Embrapa Hortaliças. Brasília: Embrapa Hortaliças. 49p, 2003. CARVALHO, S.I.C.; BIANCHETTI, L.B.; RIBEIRO, C.S.C.; LOPES, C.A. Pimentas do Gênero Capsicum no Brasil. Brasília: Embrapa Hortaliças, (Embrapa Hortaliças. Documentos, 94). 27p, 2006. CARVALHO, S.I.C; BIANCHETTI, L.B. Botânica e recursos genéticos. In: RIBEIRO, C. S. C.; LOPES, C. A.; CARVALHO, S. I. C.; HENZ, G. P.;REIFSCHNEIDER, F. J. B. Pimentas Capsicum. Brasília: Embrapa Hortaliças. p.39–53, 2008. CARVALHO, S.I.C.; RIBEIRO, C.S.C.; HENZ, G.P.; REIFSCHNEIDER, F.J.B.‘BRS Mari’: Nova cultivar de pimenta dedo-de-moça para processamento. Horticultura Brasileira, v.27, p.571-573, 2009. CROSBY, K. M. Pepper. In: PROHENS, J.; NUEZ, F.; CARENAM, J. (eds) Vegetables II, 1st edn. Springer, New York, p.221–248, 2008.

25
DOMENICO, C.I.; LILLI, A.J.O.; MELO, A.M.T. Caracterização de componentes de produção de híbridos intra-específicos de pimenta-hortícola. In: CONGRESSO BRASILEIRO DE OLERICULTURA, 50. Anais, Guarapari: ABH. 2010. FERY, R. L.; THIES, J. A. PA-353, PA-398, and PA-426: Southern Root-Knot nematode-resistant Capsicum chinense Jacq. Germplasm lines. HortScience, v.33, n.4, p.760-761, 1998.
GUERRA, M. Introdução a citogenética geral. Rio de Janeiro: Ed. Guanabara Koogan,142p. 1988. GUERRA, M.; PEDROSA, A.; BARROS E SILVA, A.; CORNÉLIO, M.T.M.; SANTOS, K. G. B.; SOARES-FILHO, W.; et al. Chromosome number and secondary constriction variation in 51 accessions of a citrus germoplasm bank. Brazilian Journal of Genetics, v.20, p.489-496, 1997. GUERRA, M. Patterns of heterochromatin distribution in plant chromosomes. Genetics and Molecular Biology, Ribeirão Preto, v. 23, n. 4, p. 1029-1041, 2000. GUERRA, N.A. Estudios cromosómicos de cuatro selecciones de Capsicum chinense Jacq. Revista UDO Agrícola, v.1, n.1, p. 34-41, 2001. HAVERROTH, M.; NEGREIROS, P. R. M. Calendário agrícola, agrobiodiversidade e distribuição espacial de roçados Kulina (Madeja), Alto Rio Envira, Acre, Brasil. Sitientibus série Ciências Biológicas, v. 11, n. 2, p. 299-308, 2011.
HENZ, G. P. Perspectivas e potencialidades do mercado de pimentas. Anais do I Encontro Nacional do Agronegócio Pimentas (Capsicum spp.). I Mostra Nacional de Pimentas e Produtos Derivados. Anais. Embrapa Hortaliças, Brasília, 2004.
HENZ, G. P.; RIBEIRO, C. S. C. Mercado e comercialização. In: RIBEIRO, C. S. C.; LOPES, C. A.; CARVALHO, S. I. C.; HENZ, G. P.; REIFSCHNEIDER, F. J. B. Pimentas Capsicum. Brasília: Embrapa Hortaliças. p.15 – 24, 2008. INCE, A.G.; KARACA, M.; ONUS, A.N. Development and utilization of diagnostic DAMD-PCR markers for Capsicum accessions. Genetic Resources and Crop Evolution, v.56, p.211-221, 2009. INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA. CENSO AGROPECUÁRIO: Brasil, Grandes Regiões e Unidades da Federação. Segunda apuração. 2006. Disponível em:http://www.ibge.gov.br/Censos/Censoagropecuario 2006/SegundaApuração/censoagro20062apuracao.pdf. Acesso em: 7 fev. 2012. KIM, S. H.; YOON, J. B.; PARK, H. G. Inheritance of Anthracnose Resistance in a New Genetic Resource, Capsicum baccatum PI594137. Journal of Crop Science and Biotechnology, v.1, p.13 -16, 2011.
KNOTKOVA, H.; PAPPAGALLO, M.; SZALLASI, A. Capsaicin (TRPV1 Agonist) therapy for pain relief: farewell or revival? The Clinical Journal of Pain, v.24, p.142-154, 2008.

26
KWON, J. K.; KIM, B. D. Localization of 5S and 25S rRNA genes on somatic and meiotic chromosomes in Capsicum species of chili pepper. Molecules and Cells, OT, p.205-209, 2009.
LANNES, S.D.; FINGER, F.L.; SCHUELTER, A.R.; CASALI, V.W.D. Growth and quality of Brazilian accessions of Capsicum chinense fruits. Scientia Horticulturae, v.112, p.266-270, 2007. LANTERI, S.; PICKERSGILL, B. Chromosomal structural changes in Capsicum annuum L. and C. chinense JACQ. Euphytica, v.67, p.155-160, 1993. LEUNG, F.W. Capsaicin-sensitive intestinal mucosal afferent mechanism and body fat distribution. Life Sciences, v.83, p.1-5, 2008. LI, Y.; WANG J.; CAO, Y.; GAO, W.; FANG, J.; LOU, X. The use of genetic resources in crop improvement: Lessons from China. Genetic Resources and Crop Evolution, v.45, p.181-186, 1998. MAYEDA, L.Y. Estudo citogenético em dez táxons do gênero Passiflora L. (Passifloraceae). Dissertação (Mestrado em Genética e Melhoramento). Universidade de São Paulo: ESALQ. 89p. 1997. MIRZAIE-NODOUSHAN, H.; DEHGHANSHOAR, M.; MADDAH-AREFI, H.; ASADI- COROM, F. Karyotypic characteristics of several Bromus Species. Internation Journal of Agriculture and Biology, v.8, p.717–20, 2006. MONTEIRO, E.R.; BASTOS, E. M.; LOPES, A. C. A.; GOMES, R. L. F.; NUNES, J. A. R. Diversidade genética entre acessos de espécies cultivadas de pimentas. Ciência Rural, Santa Maria, v.40, n.2, p.288-283, fev. 2010. MOREIRA G.R.; CALIMAN, F.R.B.; SILVA, D.J.H.; RIBEIRO, C.S.C. Espécies e variedades de pimenta. Informe Agropecuário, v.27, p.16-29, 2006.
MOSCONE, E. A. Estudios cromossómicos en Capsicum (Solanaceae).In: Analisis cariotípica de C. parviflorum y C. annuum var. annuum. Kurtziana, v.22, n.2, p.9-18, 1993. MOSCONE, E.A.; LAMBROU, M.; HUNZIKER,A.T.; EHRENDORFER, F. Giemsa C-banded karyotypes in Capsicum (Solanaceae). Plant Systematics and Evolution, v.186, p.213–229, 1993. MOSCONE, E.A.; LOIDL, J.; EHRENDORFER,F.; HUNZIKER,A.T. Analysis of active nucleolus organizing regions in Capsicum (Solanaceae) by silver staining. American Journal of Botany. v.82, p.276–287, 1995. MOSCONE, E.A.; LAMBROU, M.; EHRENDORFER, F. Fluorescent chromosome banding in the cultivated species of Capsicum (Solanaceae). Plant Systematics and Evolution, v.202, p.37– 63, 1996.

27
MOSCONE E.A.; BARANYI, M.; EBERT, I.; GREILHUBER, J.; EHRENDORFER,F.; HUNZIKER, A.T. Analysis of nuclear DNA content in Capsicum (Solanaceae) by flow cytometry and Feulgen densitometry. Annals of Botany, v.92, p.21–29, 2003. MOSCONE, E.A.; SCALDAFERRO, M.A.; GRABIELE, M.; CECCHINI, N.M.; SÁNCHEZ GARCÍA, Y.; JARRET, R.; DAVIÑA, J.R.; DUCASSE, D.A.; BARBOZA, G.E.; EHRENDORFER, F. The evolution of chili peppers (Capsicum – Solanaceae): a cytogenetic perspectives. Acta Horticulturae, v. 745, p.137-169, 2007. NEITZKE, R.S.; BARBIERI, R.L.; RODRIGUES, W.F.; CORREA, I.V.; CARVALHO, F.I.F de. Dissimilaridade genética entre acessos de pimenta com potencial ornamental. Hortic. Bras. vol.28, n.1, Brasília, Jan./Mar, 2010. NOWACZYK, P.; BANACH, M.; SIWICKA, J.; NOWACZYK, L. Capsaicinoid content of soft-flesh fruits of pepper Capsicum frutescens l. X C. annuum l. Interspecific hybrids – a short report. Polish Journal of Food and Nutrition Sciences, v.15/56, n.1, p.37–40, 2006. NUEZ, F.; GIL, R.; COSTA, J. Aspectos morfológicos y fisiológicos de la planta. In: El cultivo de pimentos, chiles y ajies. Ediciones Mundi – Prensa, Madri. p.60- 114, 1996. NUEZ-VIÑALS, F. N.; ORTEGA, R. G.; COSTA, J. C. El cultivo de pimientos, chiles y ajies. Madri: Mundi-Prensa, 607p. 2003. PALEVITCH, D.; CRAKER, L. E. “Nutritional and Medicinal Importance of Red Pepper (Capsicum spp.),” Journal of Herbs, Spices and Medicinal Plants, v.3, n.2, p.55-83, 1995. PEDROSA, A.; GITAÍ, J; SILVA, A.E.B.; FELIX, L.P.; GUERRA, M. Citogenética de angiospermas coletadas em Pernambuco. Acta Táxonotry Brasílica, São Paulo, n.13, p.49-60, 1999. PICKERSGILL, B. Genetic resources and breeding of Capsicum spp. Euphytica, v.96, p.129-133,1997. PICKERSGILL, B. Domestication of plants in the Americas: Insights from Mendelian and molecular genetics. Annals of Botany, v.100, p.92-940, 2007.
POZZOBON, M.T.; SCHIFINO-WITTMANN, M.T.E.; BIANCHETTI, L. B. Chromossome numbers in wild and semidomesticated Brazilian Capsicum L. (Solanaceae) species: do x=12 and x=13 represent two evolutionary lines?. Botanical Journal of the Linnean Society. v.151, p. 259-269, 2006. RAMOS, S.R.R.; QUEIRÓZ, M.A.; CASALI, V.W.D.; CRUZ, C.D. Recursos genéticos de Cucurbita moschata: caracterização morfológica de populações locais coletadas no Nordeste brasileiro. In: QUEIRÓZ, M.A.; GOEDERT, C.O.; RAMOS,S.R.R. (ed.). Recursos genéticos e melhoramento de plantas para o Nordeste brasileiro. (on line). Versão 1.0. Petrolina-PE: Embrapa Semi-Árido, Brasília-DF: Embrapa Recursos Genéticos e Biotecnologia, 1999.

28
RIBEIRO, C. S. C.; REIFSCHNEIDER, F. J. B. Genética e melhoramento. In: RIBEIRO, C. S. C.; LOPES, C. A.; CARVALHO, S. I. C.; HENZ, G. P.; REIFSCHNEIDER, F. J. B. Pimentas Capsicum. Brasília: Embrapa Hortaliças. p.55 – 69, 2008. RÊGO, E.R.; RÊGO, M.M.; CRUZ, C.D.; FINGER, F.L.; AMARAL, D.S.S.L. Genetic Diversity analysis of peppers: a comparison of discarding variables methods. Crop Breeding and Applied Biotechnology, Londrina, v.3, n.1, p.19-26, 2003. REIFSCHNEIDER, F.J.B. (Ed.). Capsicum: pimentas e pimentões no Brasil. Brasília: Embrapa Comunicação para Transferência de Tecnologia/Embrapa Hortaliças. 113p, 2000.
ROHANI, M.; MOHAMMADI, A.; KHOSROSHAHLI, M.; AHMADI, H.; DARANDEH, N. Karyotype analysis of several ecotypes of Capsicum annuum L. in Iran. Notulae Botanicae Horticultural and Agrobotanica Cluj, Cluj Napoca, v.38, n.1, p.177-180, 2010. SMITH, P.G.; HEISER, C.B. Taxonomy of Capsicum chinense Jacq. and the geographic distribution of the cultivated Capsicum species. Bulletim of the Torrey Botanical Club, v.84, n.6, p.413-420, 1957. SOUZA, S.A.M. Caracterização citogenética, química e molecular em acessos de Capsicum chinense Jacq. Dissertação (Mestrado em Genética e Melhoramento de Plantas) - Curso de Pós-graduação em Genética e Melhoramento de Plantas, Universidade Estadual do Norte Fluminense Darcy Ribeiro, Rio de Janeiro, RJ. 66p, 2008.
SOUZA, S.A.M.; MARTINS, K.C.; PEREIRA, T.N.S. Polimorfismo cromossômico em Capsicum chinense Jacq. Ciência Rural, v.41 n.10, p.1777-1783, 2011.
SUDRÉ, C.P.; CRUZ, C.D.; RODRIGUES, R.; RIVA, E.M.; AMARAL JÚNIOR, A.T.; SILVA, D.J.H.; PEREIRA, T.N.S. Variáveis multicategóricas na determinação da divergência genética entre acessos de pimenta e pimentão. Horticultura Brasileira, v.24, p.88-93, 2006. SURH, Y.J. Cancer chemoprevention with dietary phytochemicals. Nature Reviews Cancer, v.3, p.768-780, 2003. STEBBINS, G.L. Chromosomal evolution in higher plants. London: Edward Arnold. 216p, 1971.
SYBENGA, J. Cytogenetics in plant breeding, Berlin: Springer Verlag, 1992. 469p. TEODORO-PARDO, C.V.D. et al. Polimorfismo cromossômico en Capsicum annuum L.(Solanaceae) em recolectas de Puebla, Morelos y Quartenário, México. Agrociencia, v.41, p. 873-881, 2007. Disponível em: <http://www.colpos.mx/agrocien/agrociencia.htm>. Acesso em: 03 mar. 2012.

29
TEWKSBURY, J. J.; REAGAN, K.M.; MACHNICKI, N.J.; CARLO, T.A.; HAAK, D.C.; PENÃLOZA, A.L.; LEVEY, D.J. Evolutionary ecology of pungency in wild chilies. Proceedings of the National Academy of Sciences, v.105, p.11808–11811, 2008. TOFANELLI, M.B.D.; AMAYA-ROBLES, J.E.; RODRIGUES, J.D.; ONO, E.O. Ácido giberélico na produção de frutos partenocárpicos de pimenta. Horticultura Brasileira, Brasília, v.21, n.1, p.116-118, 2003. TRAJANO, H.M.R. Produção de pimenta (Capsicum spp.) e aspectos socioeconômicos das hortas comunitárias de Teresina, Piauí – Dissertação (Mestrado) – Desenvolvimento e Meio Ambiente da Universidade Federal do Piauí. 101p, 2009. VAISHNAVA, P.; WANG, D.H. Capsaicin sensitive-sensory nerves and blood pressure regulation. Current Medicinal Chemistry-Cardiovascular & Hematological Agents, v.1, p.177-188, 2003.
VALLS, J.F.M. Caracterização de recursos genéticos vegetais. In: NASS LL (Ed.) Recursos genéticos vegetais. Brasília: Embrapa Recursos Genéticos e Biotecnologia. p.281-305, 2007.
VENORA, G.; PADULOSI,S. Karyotypic analysis of wild taxa of V. unguiculata (L.) Walpers. Caryologia, v.50, p.125-138, 1997. WALL, M.M.; WADDELL, C.A.; BOSLAND, P. W. Variation in β-carotene and total carotenoid content in fruits of Capsicum. HortScience, v.36, n.4, p.746-749. 2001. WALSH, B.M.; HOOT, S.B. Phylogenetic relationships of Capsicum (Solanaceae) using dna sequences from two noncoding regions: the chloroplast atpb-rbcl spacer region and nuclear waxy introns. International Journal of Plant Sciences, v.162, n.6, p.1409–1418, 2001. XU, Q.; BARRIOS, C.A.; CUTRIGHT, T.; NEWBY, B.M. Assessment of antifouling effectiveness of two natural product anti-foulants by attachment study with freshwater bacteria. Environmental Science and Pollution Research, v.12, p.278-284, 2005. YAMAMOTO, S.; NAWATA, E. Capsicum frutescens L. in southeast and east Asia, and its dispersal routes into Japan. Economic Botany, v. 59, n. 1, p. 18-28, 2005. ZENI, A. L.; BOSIO, F. O uso de plantas medicinais em uma comunidade rural de Mata Atlântica – Nova Rússia, SC. Neotropical Biology and Conservation, v. 6, n.1, p.55-63, 2011.

30
3. CARACTERIZAÇÃO CARIOTÍPICA DE ACESSOS DE PIMENTAS (Capsicum
sp.)
RESUMO
Este trabalho teve por objetivo caracterizar numérica e morfologicamente os
cariótipos de acessos de Capsicum annuum, Capsicum chinense, Capsicum
frutescens e Capsicum baccatum pertencentes à coleção do Banco ativo de
Germoplasma de Capsicum da Universidade Federal do Piauí (BGC-UFPI), visto que
estas espécies têm grande importância econômica em todo mundo, e também pela
caracterização citogenética fornecer informações importantes para estudos de
taxonomia bem como para programas de melhoramento voltados para o gênero em
questão. Os cariótipos foram obtidos pelo método de esmagamento e coloração com
Giemsa de células dos meristemas das radículas dos acessos estudados. A partir
dos resultados verificou-se número cromossômico 2n = 2x = 24 para todas as quatro
espécies. Foi observado polimorfismo cromossômico para o acesso BGC 37 da
espécie C. frutescens, que apresentaram 12 pares de cromossomos metacêntricos,
diferindo dos outros que apresentaram 11 pares de cromossomos metacêntricos e 1
par de cromossomo sub-metacêntrico. Visualizou-se nos acessos BGC 01 e BGC 37
constrições secundárias nos homólogos 01 e 12, e 06 e 11, respectivamente. Os
cariótipos das espécies estudadas foram assimétricos entre si. Os resultados obtidos
neste estudo corroboram com a grande diversidade genética descrita na literatura
para este gênero.
PALAVRAS-CHAVES: pimenta; caracterização citogenética; banco de
germoplasma; morfometria.

31
Characterization of karyotypic chili peppers (Capsicum sp.)
ABSTRACT
The objective of this work was to numerically and morphologically characterize the
karyotypes of Capsicum annuum, Capsicum chinense, Capsicum frutescens and
Capsicum baccatum accessions in the collection of the active Capsicum sp.gene
bank of the Federal University of Piauí (BGC-UFPI), being that these species have
great economic importance throughout the world, and also because cytogenetic
characterization can provide important information for taxonomy studies as well as for
improvement programs focused on the genus in question. The karyotypes were
obtained from cells of the rootlet meristems of the studied accessions by the squash
method and staining with Giemsa Stain. From the results the chromosome number
2n = 2x = 24 was verified for all the four species. Chromosomal polymorphism was
observed for the accession BGC 37 of the C. frutescens species, that presented 12
pairs of metacentric chromosomes, differing from the others that presented 11 pairs
of metacentric chromosomes and 1 submetacentric chromosome pair. In the
accessions BGC 01 and BGC 37 secondary constrictions were seen in the 01 and
12, and 06 and 11 homologs, respectively. The karyotypes of the studied species
were asymmetrical among themselves. The results obtained in this study confirm the
high genetic diversity described in the literature for this genus.
Keywords: pepper; cytogenetics; genebank; morphometry.

32
3.1 INTRODUÇÃO
As pimentas Capsicum sp, pertencentes a família Solanaceae, são
apreciadas por povos de diferentes culturas como especiarias ou hortaliças, e são
encontradas em regiões tropicais e temperadas de todo o mundo. Assim, possuem
importância significativa no mercado nacional e internacional de condimentos,
temperos e conservas, sendo cultivadas por pequenos, médios e grandes
produtores, e por sistemas integrados as grandes indústrias (FONSECA et al.,
2008). Também possuem expressiva representatividade na medicina fitoterápica,
principalmente em países da América do Sul e Latina, por terem altas concentrações
de vitaminas A e C, β-caroteno e capsaicinas, que em conjunto apresentam, entre
outros benefícios, ações antioxidante, antimicrobiana, anti-inflamatória e
hipocolesteremiante (COSTA et al., 2008, ZENI; BOSIO, 2011).
Em Capsicum encontram-se as espécies, classificadas como domesticadas:
C. annuum L. (pimentão, pimenta doce), C. chinense Jaqc. (pimenta-de-cheiro,
pimenta-de-bode, murici), C. frutescens L. (pimenta malagueta) e C. baccatum L.
(dedo-de-moça, chapéu-de-frade), que são grandemente consumidas e produzidas
no Brasil (LANNES et al., 2007). Destacam-se por terem ampla adaptação às
condições de clima tropical e grande diversidade biológica, sendo esta característica
evidenciada, por exemplo, em seus frutos que apresentam variedades de tamanhos,
formas e colorações (HAVERROTH; NEGREIROS, 2011, ZENI; BOSIO, 2011).
O Brasil é considerado um habitat natural destas pimentas, onde os principais
estados produtores são Minas Gerais, São Paulo, Rio de Janeiro, Ceará e Bahia
(IBGE, 2006). No entanto, o conhecimento sobre a diversidade genética destas
plantas, neste país, ainda é considerado incipiente, apesar de nas duas últimas
décadas ter ocorrido um grande empenho dos pesquisadores em estudos
bioquímicos, moleculares e citogenéticos com estas espécies (HAVERROTH;
NEGREIROS, 2011).
Estudos do número e morfologia de cromossomos de pimentas são de grande
relevância, pois geram dados importantes para a elucidação de fatos relacionados à
taxonomia do gênero Capsicum (SOUZA et al., 2011), contribuem para a
compreensão das variações genéticas envolvidas na evolução do mesmo
(MOSCONE et al., 2007), auxiliam na delimitação taxonômica mais precisa entre as
espécies cultivadas, semi-cultivadas e silvestres (PEREIRA et al., 2006,
CARVALHO; BIANCHETTI, 2008) e ampliam as perspectivas de conservação da

33
diversidade vegetal fornecendo informações que podem auxiliar programas de
melhoramentos genético deste gênero (SOUZA et al., 2011). Benko-Iseppon (2001)
cita que o estudo citogenético de diferentes acessos de bancos de germoplasmas de
Capsicum constitui-se em uma importante fonte de dados para melhoristas por
permitir um melhor gerenciamento do “pool gênico” e uma seleção mais eficiente dos
recursos genéticos de suas espécies.
Portanto, devido à relevância econômica e a ampla variabilidade genética
descrita na literatura para as espécies C. annuum, C. chinense, C. frutescens e C.
baccatum, bem como a necessidade de mais estudos sobre as mesmas, e
considerando a caracterização numérica e morfológica de cariótipos como uma
importante ferramenta para auxiliar estudos de taxonomia e programas de
melhoramento genético de espécies de interesse econômico, este estudo teve como
objetivo caracterizar e analisar o cariótipo de acessos provenientes da coleção do
Banco ativo de Germoplasma Capsicum da Universidade Federal do Piauí (BGC-
UFPI).

34
3.2 MATERIAL E MÉTODOS
3.2.1 Material Genético
Os acessos das espécies estudadas foram obtidos do Banco ativo de
Germoplasma de Capsicum da Universidade Federal do Piauí, localizado na cidade
de Teresina. Foram caracterizados citogeneticamente quatro acessos da espécie C.
annuum (BGC 34, BGC 36, BGC 39, BGC 59), dois acessos da espécie C. chinense
(BGC 07, BGC 49), dois acessos da espécie C. frutescens (BGC 01, BGC 37) e
quatro acessos da espécie C. baccatum (BGC 21, BGC 26, BGC 27, BGC 54).
Levou-se em consideração a distância geográfica entre as espécies, onde os
acessos BGC 39, BGC 36, BGC 01, BGC 37, BGC 21 e BGC 26 foram coletados em
Teresina (PI); os acessos BGC 34, BGC 54 e BGC 07 foram adquiridos em São
Raimundo Nonato (PI), Pedro II (PI) e Piripiri, respectivamente; o acesso BGC 49 foi
coletado na cidade de São Paulo (SP), e os acessos BGC 59 e BGC 27 foram
coletados em São Luís do Maranhão (MA) e em São Francisco (MA),
respectivamente.
Sementes destes acessos foram colocadas para germinar no laboratório de
Genética Bioquímica e Sequenciamento de DNA (LGBS) do Departamento de
Biologia da Universidade Federal Rural de Pernambuco (UFRPE), em janeiro de
2011, em placas de Petri revestidas com papel-filtro, diariamente umedecidos com
água destilada a temperatura ambiente, sob condições controlada de luminosidade,
durante cinco dias.
3.2.2 Avaliação citogenética
As análises citogenéticas foram realizadas no mesmo laboratório no primeiro
semestre de 2011. As radículas, com cerca de 0,5cm, dos diferentes acessos foram
coletadas e pré-tratadas com 8-hidroxiquinoleína (2mM) por 4 horas, a temperatura
ambiente. Em seguida foram lavadas em água destilada, fixadas em solução de
Carnoy – etanol absoluto e ácido acético glacial (3:1) – por 24 horas e conservadas
a -20ºC. Para o preparo das lâminas, as radículas fixadas foram lavadas em água
destilada e hidrolisadas em HCL 1N a temperatura ambiente, por 10 minutos.
Após a hidrólise, as radículas foram novamente lavadas em água destilada e,
em seguida, foram preparadas lâminas de cada acesso. Para o preparo de cada
lâmina destacou-se, com a ajuda de um bisturi, uma ponta de radícula que foi

35
macerada em uma gota de ácido acético glacial a 45%, e em seguida coberta com
lamínula. Posteriormente, as lamínulas foram removidas com nitrogênio, as lâminas
secas ao ar e coradas com Giemsa a 2%, por 10 a 15 minutos, e montadas com
Entellan (Merck®).
3.2.3 Análise cariotípica
As lâminas foram analisadas em microscópio Leica DM 2500, equipado com
microcâmera digital DC 345 FX acoplada a um microcomputador para captura direta
das imagens de interesse. As imagens foram obtidas utilizando-se o programa
Image-Pro Plus versão 5.1.
Cinco metáfases de cada acesso, as quais corresponderam as repetições,
foram utilizadas para a mensuração dos cromossomos, realizada com o auxílio do
Programa MicroMeasure 3.3 (REEVES; TEAR, 2011), para obtenção dos seguintes
valores médios: a) comprimento do braço longo: BL; b) comprimento do braço curto:
BC; c) comprimento total do cromossomo: Cti = BL + BC; d) comprimento total do
lote haploide: CTLH = ∑ Cti; e) comprimento relativo: r = (Cti/CTLH) x 100.
Com esses parâmetros realizou-se a identificação dos homólogos. Os
cromossomos foram classificados em metacêntrico, submetacêntrico, acrocêntrico e
telocêntrico, com base na razão entre braços (r) e no índice centromérico (ic),
proposto por Guerra (1986).
3.2.4 Análise dos dados
Os acessos foram comparados quanto aos parâmetros: comprimento total do
lote haplóide (CTLH) e comprimento total dos cromossomos entre acessos (Cti),
adotando o delineamento experimental inteiramente casualizado, com cinco
repetições. O modelo utilizado foi:
Yij = m + ti + e(ij), em que:
Yij: observação do acesso i, na repetição j = 1, 2, ..., 5
m: média geral
ti: efeito do acesso i, com i = 1, 2, ..., 12
e(ij): erro experimental associado à parcela que recebeu o tratamento i na repetição j;
Em seguida foi efetuada a análise de variância seguindo o teste de
comparação de médias de Tukey (1949) ao nível de 5% de significância. As análises
foram feitas por meio do software Genes (CRUZ, 2006).

36
3.3 RESULTADOS E DISCUSSÃO
A análise dos acessos de C. annuum, C. chinense, C. frutescens e C.
baccatum por meio da coloração com Giemsa permitiu boa descrição do número,
morfologia e tamanho cromossômico, tipo de núcleo interfásico e padrão de
condensação cromossômica. Os cromossomos, em sua grande maioria,
apresentaram-se sem a ocorrência de regiões diferenciadas ao longo de seu
comprimento, com exceção da região centromérica e da visualização, em acessos
de uma das espécies, de constrições secundárias.
Para os 12 acessos confirmaram-se metáfases 2n = 2x = 24 (Figura 3.1),
ploidia esta amplamente descrita na literatura para um grande número de espécies
de Capsicum (PICKERSGILL, 1997, GUERRA, 2001; POZZOBON; WITTMANN,
2006; MOSCONE et al., 2007; TEODORO-PARDO et al., 2007; SOUZA et al., 2011)
e muito encontrada na família Solanaceae (PICKERSGILL, 2007). Porém, em
algumas espécies silvestres, como C. buforum, C. capylopodium e C. cornutum
constatou-se ploidia 2n = 2x = 26 (POZZOBON; WITTMANN, 2006). Pozzobon e
Wittmann (2006) relatam que há duas linhas evolutivas distintas na diversificação
deste gênero, que mostram uma clara separação entre as espécies silvestres, com
número básico x = 13 e as domesticadas, com x = 12. Estes autores ainda relatam
que as x=13 são plantas mais antigas do qual se originou as x=12.
No entanto, Guerra (2001), utilizando a coloração com Giemsa, observou
fórmula cariotípica diferente às obtidas aqui em um estudo em que avaliou
polimorfismo em acessos de C. chinense, provenientes de diferentes estados da
Venezuela e verificou que muitos deles apresentaram fórmula 11M+ 1A. Já Souza et
al. (2011) observaram, por meio da citogenética convencional, fórmula 11M+1A para
acessos de C. chinense, provenientes de diferentes estados do Brasil. Sousa et al.
(2011) e Moscone et al. (1996), por bandeamento cromossômico, analisaram
padrões evolutivos em espécies de Capsicum e observaram para C. frutescens
fórmulas cariotípicas 11M+1SM e 11M+1A, respectivamente.
De acordo com Teodoro-Pardo et al. (2007), as diferentes fórmulas
cariotípicas para uma espécie podem ocorrer devido a variações genéticas entre as
populações, originadas das respostas dos genomas aos diferentes ambientes,
ampliando a variabilidade genética para o gênero.
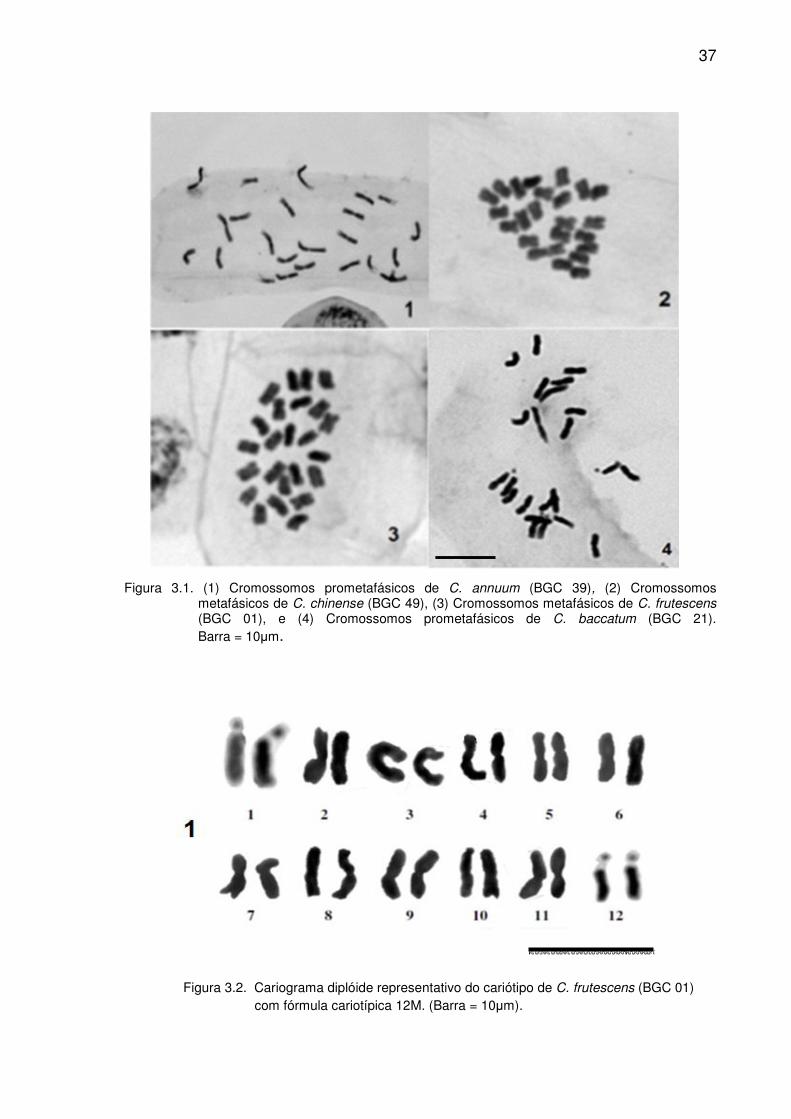
37
Figura 3.1. (1) Cromossomos prometafásicos de C. annuum (BGC 39), (2) Cromossomos
metafásicos de C. chinense (BGC 49), (3) Cromossomos metafásicos de C. frutescens (BGC 01), e (4) Cromossomos prometafásicos de C. baccatum (BGC 21). Barra = 10µm.
Figura 3.2. Cariograma diplóide representativo do cariótipo de C. frutescens (BGC 01) com fórmula cariotípica 12M. (Barra = 10µm).

38
Figura 3.3. Cariograma diplóide representativo do cariótipo de Capsicum frutescens
(BGC 37) com fórmula cariotípica 12M. (Barra = 10µm).
Estes mesmos autores relatam que o surgimento de polimorfismo a nível
cromossômico em indivíduos de uma mesma população pode alterar o padrão
cariotípico desses espécimes e originar raças cromossômicas distintas.
Para Moscone et al. (2007), as diferenças na morfologia, no tamanho e no
número cromossômico são comuns em populações da mesma espécie ou em táxon
interespecíficos, e são denominados citótipos ou raças cromossômicas.
Afirmam ainda que estas diferenças são frequentes no gênero Capsicum, o qual
possuem citótipos que diferem principalmente na fórmula cariotípica e tamanho dos
cromossomos. Ainda para os acessos BGC 01 e BGC 37, da espécie C. frutescens,
observou-se constrições secundárias nos pares homólogos 01 e 12, e 6 e 11,
respectivamente (Figuras 02 e 03). Moscone et al. (1996) relata que há constrições
secundárias em todas as espécies de Capsicum e que geralmente aparecem em
número de 1 a 4 em cada cariótipo.
Neste trabalho, o tamanho médio dos cromossomos, variou de 3,29 (BGC 49)
a 7,48µm (BGC 54) (Tabela 3.3). Este resultado mostra-se diferente aos dados
encontrados na literatura, como os relatados por Souza et al. (2011) que analisaram
quatro acessos de C. chinense de diferentes estados do Brasil e verificaram que o
tamanho médio dos cromossomos foi de 2,59 a 4,12µm, e os observados por
Teodoro-Pardo et al. (2007) que estudaram cariótipos de C. annuum de indivíduos

39
de três estados do México, e relataram valores variando de 1,6 a 8,4µm, 2,4 a 2,9
µm e 2,1 a 5,2 µm.
Para Wadt et al. (2004), a variação no tamanho médio dos cromossomos de
espécies de um mesmo gênero pode ser atribuída grandemente à diferenças nos
graus de condensação da cromatina dos genomas pesquisados durante as fases de
divisão celular. Neste trabalho, grande parte dos cariótipos estudados foi analisado
em prometáfase, onde os cromossomos ainda se mostram bem alongados (Figura
01 – prancha 1 e 4). Nesta fase os cromossomos estão se direcionando para a
condensação máxima da cromatina, que será bastante evidente na fase de
metáfase.
Tabela 3.1- Valores médios em micrômetros de ITC, IC, r, TF, e FC de quatro acessos da espécie C. annuum (BGC 34, BGC 36, BGC 39, BGC 59), dois acessos da espécie C. chinense (BGC 07, BGC 49), dois acessos da espécie C. frutescens (BGC 01, BGC 37) e quatro acessos da espécie C. baccatum (BGC 21, BGC 26, BGC 27, BGC 54).
ITC: intervalo do tamanho cromossômico, IC: índice centromérico, r: razão entre os braços do cromossomo , TF: índice de assimetria e FC: fórmula cariotípica.
Já em relação ao complemento cromossômico dos 12 acessos, observou-se
alta frequência de cromossomos metacêntricos, sendo esta morfologia comprovada
pelos valores médios de índices centroméricos (IC) e razão entre os braços dos
cromossomos (r) obtidos (Tabela 3.1) o que caracterizou cariótipos bastante
simétricos, com destaque ao acesso BGC 37, da espécie C. frutescens, que
Espécie/Acesso 2n ITC IC r TF(%) FC
C. annuum - BGC 34 24 6,92 - 3,64 45,34 1,22 45,61 11M+1SM
C. annuum - BGC 39 24 6,49 - 3,59 45,64 1,20 45,81 11M+1SM
C. annuum - BGC 36 24 7,05 - 3,69 45,53 1,21 45,77 11M+1SM
C. annuum - BGC 59 24 7,42 - 4,42 44,63 1,25 44,68 11M+1SM
C. chinense - BGC 07 24 6,60 - 4,56 45,80 1,19 45,97 11M+1SM
C. chinense - BGC 49 24 5,82 - 3,29 45,88 1,07 46,05 11M+1SM
C. frutescens - BGC 01 24 7,17 - 4,21 45,19 1,12 45,46 11M+1SM
C. frutescens - BGC 37 24 5,73 - 3,60 46,55 1,06 46,54 12M
C. baccatum - BGC 21 24 7,00 - 4,45 47,45 1,18 47,64 11M+1SM
C. baccatum - BGC 54 24 7,48 - 4,31 44,90 1,22 45,08 11M+1SM
C. baccatum - BGC 26 24 6,92 - 3,49 47,60 1,22 45,51 11M+1SM
C. baccatum - BGC 27 24 6,56 - 3,75 45,70 1,19 46,02 11M+1SM

40
apresentou somente cromossomos metacêntricos. Segundo Davide et al. (2007),
plantas com maior simetria cariotípica em relação as demais do mesmo gênero são
classificadas como mais antigas e as que possuem menor simetria classificam-se
como as mais recentes na evolução.
Aarestrup (2001) relata que apesar da maioria das espécies de Capsicum ser
2n=24 e apresentar grande similaridade quanto a morfologia dos cromossomos as
mesmas possuem grande variabilidade cariotípica intra e interespecífica. Isto pode
ser observado em relação aos resultados de índice de assimetria (TF%) entre os
cariótipos das espécies estudadas neste trabalho (Tabela 3.1) que variou de 44,68
(BGC 59) a 47,64% (BGC 21), valores estes menores que 50%, o que demonstra,
segundo Nardy et al. (2010) que os cariótipos são assimétricos entre si.
De acordo com Moraes et al. (2009), quanto maior o índice de assimetria
entre cariótipos de espécies de um mesmo gênero maior é a variabilidade genética
existente entre elas. Nas espécies estudadas aqui se pode tomar como exemplo de
variabilidade os diversos formatos e tamanho de frutos, onde em C. annuum o
formato do fruto é uma baga pequena com textura lisa (CARVALHO et al., 2003), em
C. chinense os frutos são alongados, medindo cerca de 1,0 a 1,5cm de diâmetro e
possuem textura lisa (NUEZ-VIÑALS, 2003), em C. frutescens os frutos são em
forma de bagas fusiformes lisas (BENTO et al., 2007), e em C. baccatum os frutos
se alteram em tamanho e forma, com 0,6cm de diâmetro, liso e arredondado, com
10cm de comprimento (CARVALHO et al. 2009).
De acordo com Carvalho e Bianchetti (2004), as flores também podem ser
utilizadas para se observar a variabilidade genética existente entre essas espécies
de pimentas, visto que existe uma ampla diversidade entre elas quando se considera
o número de flores por nó, posição da flor e do pedicelo, colorações da corola e da
antera, presença ou ausência de manchas nos lobos das pétalas e margem do
cálice.
Os acessos estudados foram agrupados pelo teste de Tukey, ao nível de 5%
(Tabela 3.2), e os resultados mostraram que não existem diferenças significativas
entre os acessos BGC 07, BGC 21, BGC 54, BGC 36, BGC 34 e BGC 27 em relação
aos parâmetros CTC e CTLH.
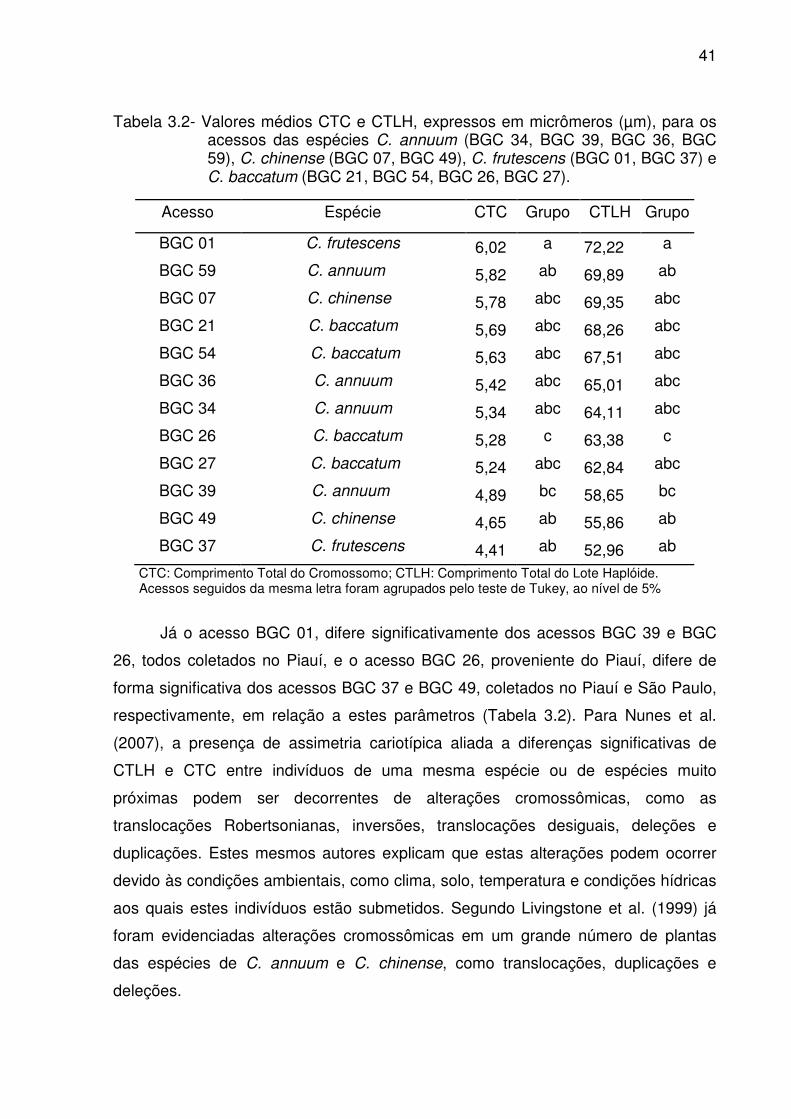
41
Tabela 3.2- Valores médios CTC e CTLH, expressos em micrômeros (µm), para os acessos das espécies C. annuum (BGC 34, BGC 39, BGC 36, BGC 59), C. chinense (BGC 07, BGC 49), C. frutescens (BGC 01, BGC 37) e C. baccatum (BGC 21, BGC 54, BGC 26, BGC 27).
Acesso Espécie CTC Grupo CTLH Grupo
BGC 01 C. frutescens 6,02 a 72,22 a
BGC 59 C. annuum 5,82 ab 69,89 ab
BGC 07 C. chinense 5,78 abc 69,35 abc
BGC 21 C. baccatum 5,69 abc 68,26 abc
BGC 54 C. baccatum 5,63 abc 67,51 abc
BGC 36 C. annuum 5,42 abc 65,01 abc
BGC 34 C. annuum 5,34 abc 64,11 abc
BGC 26 C. baccatum 5,28 c 63,38 c
BGC 27 C. baccatum 5,24 abc 62,84 abc
BGC 39 C. annuum 4,89 bc 58,65 bc
BGC 49 C. chinense 4,65 ab 55,86 ab
BGC 37 C. frutescens 4,41 ab 52,96 ab
CTC: Comprimento Total do Cromossomo; CTLH: Comprimento Total do Lote Haplóide. Acessos seguidos da mesma letra foram agrupados pelo teste de Tukey, ao nível de 5%
Já o acesso BGC 01, difere significativamente dos acessos BGC 39 e BGC
26, todos coletados no Piauí, e o acesso BGC 26, proveniente do Piauí, difere de
forma significativa dos acessos BGC 37 e BGC 49, coletados no Piauí e São Paulo,
respectivamente, em relação a estes parâmetros (Tabela 3.2). Para Nunes et al.
(2007), a presença de assimetria cariotípica aliada a diferenças significativas de
CTLH e CTC entre indivíduos de uma mesma espécie ou de espécies muito
próximas podem ser decorrentes de alterações cromossômicas, como as
translocações Robertsonianas, inversões, translocações desiguais, deleções e
duplicações. Estes mesmos autores explicam que estas alterações podem ocorrer
devido às condições ambientais, como clima, solo, temperatura e condições hídricas
aos quais estes indivíduos estão submetidos. Segundo Livingstone et al. (1999) já
foram evidenciadas alterações cromossômicas em um grande número de plantas
das espécies de C. annuum e C. chinense, como translocações, duplicações e
deleções.

42
3.4 Conclusões
Os resultados obtidos neste trabalho mostraram variabilidade entre as
espécies estudadas, corroborando aos dados da literatura científica, mesmo que
estas espécies apresentaram cariótipos bastante simétricos e com mesmo número
cromossômico. No entanto, estes acessos necessitam ser mais bem analisados
através de uma amostragem maior e de uma caracterização mais detalhada dos
cromossomos, por meio da distribuição de heterocromatina e da identificação de
sequências por hibridização in situ, com o intuito de verificar se as diferenças entre
as espécies caracterizam diferenciação intra e/ou interespecífica. Assim, também
será possível fazer a discriminação das espécies com a mesma fórmula cariotípica.
É importante ressaltar que atualmente no Estado do Piauí, os dados sobre a
produção agrícola de pimentas são imprecisos e irregulares, e a plantação desta
hortícola é feita apenas por pequenos produtores. A sua comercialização, na forma
in natura, envolve apenas pequenas produções caseiras ou artesanais de molhos,
conservas, geleias e pimenta em pó; e pequenas empresas que vendem sua
produção para feiras livres e pequenos supermercados. No entanto, este estado
possui fatores ambientais, como temperatura e solo, bastante propícios para a
produção em grande escala destas quatro espécies de Capsicum. Assim, tornam-se
importantes estudos, como os de citogenética, com o intuito de se conhecer melhor
a variabilidade presente nestas plantas e futuramente utilizar estas informações em
programas de melhoramento genético deste gênero direcionados ao Piauí.
Dessa forma, estes resultados, bem como os futuros trabalhos de citogenética
utilizando os acessos do BGC-UFPI, são de grande relevância para os programas
de melhoramento genético deste gênero no estado do Piauí, programas que, entre
outras características, visarão desenvolver cultivares mais resistentes a pragas e
doenças, com maior qualidade e produtividade, estimulando agricultores a produzir
estas hortaliças, gerar empregos com a mão de obra no plantio e, inicialmente,
atender as exigências do mercado deste estado.

43
Referências
AARESTRUP, J.R. Análise morfológica dos cromossomos de pimentão Capsicum annuum L. Dissertação (Mestrado em Genética e Melhoramento) Viçosa – MG, Universidade Federal de Viçosa – UFV, 143p, 2001. BENKON-ISEPPON, A.M. Estudos moleculares e citogenéticos no Caupi e em espécies relacionadas: Avanços e perspectivas. EMBRAPA - Documentos 56, p.327-332, 2001. BENTO, C. S.; SUDRÉ, C. P. RODRIGUES, R.; RIVA, E. M.; PEREIRA, M. G. Descritores qualitativos e multicategóricos na estimativa da variabilidade fenotípica entre acessos de pimentas. Scientia Agraria, v. 8, n. 2, 2007. CARVALHO, S.I.C.; BIANCHETTI, L.B.; BUSTAMANTE, P.G.; SILVA, D.B. Catálogo de germoplasma de pimentas e pimentões (Capsicum spp.) da Embrapa Hortaliças. Brasília: Embrapa Hortaliças. 49p, 2003. CARVALHO, S. I. C.; BIANCHETTI, L. B. Botânica. Embrapa hortaliças: versão eletrônica, 2004. Disponível em: <www.cnph.embrapa.br/sistprod/pimenta/botanica.htm>. Acesso em: 10 jun. 2011. CARVALHO, S.I.C.; BIANCHETTI, L.B. Botânica e recursos genéticos. In: RIBEIRO, C. S. C.; LOPES, C. A.; CARVALHO, S. I. C.; HENZ, G. P.; REIFSCHNEIDER, F. J. B. Pimentas Capsicum. Brasília: Embrapa Hortaliças. Cap. 5, p.39 – 53, 2008. CARVALHO, S.I.C; RIBEIRO, C.S.C; HENZ, G.P; REIFSCHNEIDER, F.J.B.‘BRS Mari’: nova cultivar de pimenta dedo-de-moça para processamento. Horticultura Brasileira. v.27, p.571-573, 2009. COSTA, L.V.; LOPES,T.G.; LOPES, R.; ALVES, S.R.M.; GAMA, A.S. Efeito das polinizações no vingamento de frutos de Capsicum chinense Jacq. Horticultura Brasileira. v. 26, p.1685-1689, 2008. CRUZ, C.D. Programa GENES: análise multivariada e simulação. Viçosa: UFV, 2006. 175p. DAVIDE, L. C.; TECHIO, V. H.; NENUES, J. D.; PEREIRA, A. V. Variação cromossômica numérica em Pennisetum. Ciência e Agrotecnologia, v. 31, n. 2, p. 398-405, 2007. FONSECA, R. M.; LOPES, R.; BARROS, W. S.; LOPES, M. T. G.; FERREIRA, F. M. Morphologic characterization and genetic diversity of Capsicum chinese Jacq. Accessions along the upper Rio Negro – Amazonas. Crop Breending and Applied Biotechnology, v. 81, p. 187-197, 2008.

44
GUERRA, M. Reviewing the chromosome nomenclature of Levan et al. Revista Brasileira de Genética, v.9, p.741-743, 1986. Disponível em: <http://web2.sbg.org.br/gmb/edicoesanteriores/v09n4/pdf/a16v09n4.pdf>. Acesso em: 10 jan. 2011. doi: 10.1590/S0100-84551997000400001. GUERRA, N.A. Estudios cromosómicos de cuatro selecciones de Capsicum chinense Jacq. Revista UDO Agrícola. v.1, n.1, p. 34-41, 2001. HAVERROTH, M.; NEGREIROS, P. R. M. Calendário agrícola, agrobiodiversidade e distribuição espacial de roçados Kulina (Madeja), Alto Rio Envira, Acre, Brasil. Sitientibus série Ciências Biológicas, v.11, n.2, p.299-308, 2011. INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA. CENSO AGROPECUÁRIO: Brasil, Grandes Regiões e Unidades da Federação. Segunda apuração. 2006. Disponível em:http://www.ibge.gov.br/Censos/Censoagropecuario 2006/SegundaApuração/censoagro20062apuracao.pdf. Acesso em: 7 fev. 2012. LANNES, S.D.; FINGER, F.L.; SCHUELTER, A.R.; CASALI, V.W.D. Growth and quality of Brazilian accessions of Capsicum chinense fruits. Scientia Horticulturae, v. 112, p.266-270, 2007. LIVINGSTONE, K. D.; LACKENEY, V. K.; BLAUTH, J. R.; WIJK, R. V.; JAHN, M. K. Genome mapping in Capsicum and the evolution of genome structure in the Solanaceae. Genetics, v. 152, p. 1183-1202, 1999. MORAES, B.N.G.; GROSS, M.C.; FELDBERG, E. Evolução cromossômica de Geophagus (Perciformes, Cichlidae) da Amazônia Central. Anais da XVIII Jornada de Iniciação Científica PIBIC CNPq/ FAPEAM/ INPA: p.270-272, 2009. MOSCONE, E. A. et al. The evolution of chili peppers (Capsicum – Solanaceae): a cytogenetic perspective. Acta Horticulturae, v.745, p.137-169, 2007. Disponível em: <http://www.wildchilli.eu/index.php/wild-chilli species/theevolution- of-chilli-peppers-new>. Acesso em: 5 mar. 2011. MOSCONE, E.A.; SCALDAFERRO, M.A.; GRABIELE, M.; CECCHINI, N.M.; SÁNCHEZ GARCÍA, Y.; JARRET, R.; DAVIÑA, J.R.; DUCASSE, D.A.; BARBOZA, G.E.; EHRENDORFER, F. The evolution of chili peppers (Capsicum – Solanaceae): a cytogenetic perspectives. Acta Horticulturae, v. 745, p.137-169, 2007. MOSCONE, E.A.; LAMBROU, M.; EHRENDORFER, F. Fluorescent chromosome banding in the cultivated species of Capsicum (Solanaceae). Plant Systematics and Evolution. v.202, p.37– 63. 1996. NARDY, M.; YUYAMA, L. N. A. A. R.; VANZELA, A. L. L. Chromossome banding patterns and localization of 5S and 45S rDNA sites in three shrub-tree species of Erythrina L. (Leguminosae: Papilionoideae) from Brazil. Revista Brasileira de Biociências, v. 8, n. 2, p. 149-152, 2010. NUNES, J.D. et al. Citogenética de Piper hipidinervum e Piper aduncum. Pesquisa Agropecuária Brasileira, Brasília, v. 42, n. 7, p. 1049-1052, July, 2007.

45
NUEZ-VIÑALS, F. N.; ORTEGA, R. G.; COSTA, J. C. El cultivo de pimientos, chiles y ajies. Madri: Mundi-Prensa, 607p. 2003. PEREIRA, L.P. et al. Número de cromossomos em populações de Achyrocline satureioides Lam. Ciência Rural, v.36, p.678-681, 2006. Disponível em: <http://www.scielo.br/pdf/ cr/v36n2/a52v36n2.pdf>. Acesso em: 2 abr. 2011. doi: 10.1590/S0103-84782006000200052. PICKERSGILL, B. Genetic resources and breeding of Capsicum spp. Euphytica v.96, 129-133,1997. PICKERSGILL, B. Domestication of plants in the Americas: insights from Mendelian and molecular genetics. Annals Botany, v.100, p. 925-940, 2007. POZZOBON, M. T.; WITTMANN, M. T. A meiotic study of the wild and semi-domesticated Brazilian species of genus Capsicum L. (Solanaceae). Cytologia, v.71, n. 3, p. 275-287, 2006. REEVES, A.; TEAR, J. MicroMensure for Windows, version 3.3. 2000. Disponível em: <http://www.colostate.edu/Depts/Biology/MicroMeasure>. Acesso em: 11 mar. 2011. SOUZA, S.A.M.; MARTINS, K.C.; PEREIRA, T.N.S. Polimorfismo cromossômico em Capsicum chinense Jacq. Ciência Rural v.41 n.10, p.1777-1783, 2011. TEODORO-PARDO, C.V.D. et al. Polimorfismo cromossômico en Capsicum annuum L.(Solanaceae) em recolectas de Puebla, Morelos y Quartenário, México. Agrociencia, v.41, p. 873-881, 2007. Disponível em: <http://www.colpos.mx/agrocien/agrociencia.htm>. Acesso em: 03 mar. 2012. TUKEY, J. W. One degree of freedon for non additivy. Biometrics, p. 232-242, 1949. WADT, L.H.O. de; EHRINGHAUS, C.; KAGEYAMA, P.Y. Genetic diversity of 'Pimenta Longa' genotypes (Piper spp., Piperaceae) of Embrapa Acre germplasm collection. Genetics and Molecular Biology, v.27, p.74-82, 2004. ZENI, A. L.; BOSIO, F. O uso de plantas medicinais em uma comunidade rural de Mata Atlântica – Nova Rússia, SC. Neotropical Biology and Conservation, v. 6, n. 1, p. 55-63, 2011. Disponível em: http://www.unisinos.br/index.php/neotropical/article/view/997.pdf. Doi: 10.4013/nbc.2011.61.07.





























