Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DO RIO DE JANEIRO
FACULDADE DE MEDICINA
PROGRAMA DE PÓS-GRADUAÇÃO EM CLÍNICA MÉDICA
A VIA DE SINALIZAÇÃO HEDGEHOG REGULA A APOPTOSE E A
SECREÇÃO DE CITOCINAS NA LINHAGEM DE CÉLULAS EPITELIAIS DE
CARCINOMA DE CÓLON HUMANO HT-29
Dissertação de Mestrado apresentada ao
Programa de Pós-graduação em Clínica
Médica, Faculdade de Medicina da
Universidade Federal do Rio de Janeiro,
como parte dos requisitos necessários à
obtenção do título de Mestre em Medicina
(Clínica Médica)
Agnes Naomi Yoshimoto
Rio de Janeiro
Outubro 2011

ii
A VIA DE SINALIZAÇÃO HEDGEHOG REGULA A APOPTOSE E A SECREÇÃO DE
CITOCINAS NA LINHAGEM DE CÉLULAS EPITELIAIS DE CARCINOMA DE
CÓLON HUMANO HT-29
Agnes Naomi Yoshimoto
Dissertação de Mestrado apresentada ao
Programa de Pós-graduação em Clínica
Médica, Faculdade de Medicina da
Universidade Federal do Rio de Janeiro,
como parte dos requisitos necessários à
obtenção do título de Mestre em Medicina
(Clínica Médica)
Orientadores: Prof. Heitor Siffert Pereira de Souza
Prof a Morgana Teixeira Lima Castelo Branco
Aprovada em ____ de ______________ de ____.
___________________________________________________
Presidente da banca
Aprovado por
___________________________________________________
Prof:
___________________________________________________
Prof:
___________________________________________________
Prof:
Rio de Janeiro
Outubro 2011

iii
FICHA CATALOGRÁFICA
Yoshimoto, Agnes Naomi.
A via de sinalização hedgehog regula a apoptose e a secreção de citocinas na
linhagem de células epiteliais de carcinoma de cólon humano HT-29 / Agnes Naomi
Yoshimoto. Rio de Janeiro: UFRJ / Faculdade de Medicina, 2011.
xiv, 82f. ; 31 cm.
Orientadores: Heitor Siffert Pereira de Souza e Morgana Teixeira Lima
Castelo Branco.
Dissertação (Mestrado) – Universidade Federal do Rio de Janeiro,
Faculdade de Medicina, Programa de Pós-Graduação em Clínica Médica, 2011.
Referências bibliográficas: f.73 – 77.
1. Proteínas Hedgehog. 2. Linhagem de Célula. 3. Carcinoma. 4. Cólon. 5.
Neoplasias do Cólon. 6. Células Epiteliais. 7. Proliferação Celular. 8. Citocinas.
9. Apoptose. 10. Células HT-29. 11. Estudos Observacionais. 12. Clínica Médica
- Tese. I. Souza, Heitor Siffert Pereira de. II. Castelo Branco, Morgana Teixeira
Lima. III. Universidade Federal do Rio de Janeiro, Faculdade de Medicina, Pós-
Graduação em Clínica Médica. IV. Título.

iv
Dedico esta tese aos meus pais
Teruyoshi e Massako
que eu amo muito e que sempre me apoiaram
e ao meu marido Breno,
o amor da minha vida, que sempre esta ao meu lado.

v
AGRADECIMENTOS
Ao meu orientador Prof Heitor Siffert Pereira de Souza, que é um exemplo
como orientador, pesquisador e médico, que além de ter uma incrível paciência
comigo e todos os seus alunos, tem a humildade de explicar os temas mais difíceis
da forma mais simples e participar ativamente de todo o processo da tese,
proporcionando tranquilidade nos momentos primordiais.
À Profa Morgana Teixeira Lima Castelo Branco que é um exemplo de
organização e dedicação ao ensino, sempre participando e contribuindo para bons
resultados, também com sua presença tranquila, confiante e um sorriso amigo.
À Cesônia e ao Alyson que sempre socorrem todos os alunos da pós-
graduação mostrando amor, dedicação, paciência e acima de tudo boa vontade na
bancada, além de proporcionarem um bom ambiente de trabalho e comigo não foi
diferente.
Ao Claudio Bernadazi que me ajudou na técnica de PCR, sendo de
fundamental importância para o trabalho.
À Adriane, Carla e Claudio, que sempre se predispuseram a ajudar e a coletar
material quando necessário e trocaram idéias tanto relacionadas a tese quanto a
assuntos da vida.
Às alunas de iniciação científica que participaram da confecção da tese e dos
trabalhos em congressos como a Fernanda.
À família do meu marido que sempre me apoiou.
Ao meu marido que compreendeu minha ausência nas horas necessárias e
me apoiou ativamente.
Aos meus pais que são tudo para mim.

vi
Este trabalho foi realizado com recursos do CNPq (Conselho
Nacional de Desenvolvimento Científico e Tecnológico) e da
FAPERJ (Fundação de Amparo à Pesquisa do Estado do Rio de
Janeiro). Os resultados foram submetidos sob a forma de artigo
completo a revista Plos One, dia 15/09/2011, estando em análise.

vii
RESUMO
Introdução: A via hedgehog (Hh) está envolvida na embriogênese e em processos
fisiológicos incluindo sobrevivência e proliferação celular. Objetivos: células de
carcinoma de cólon humano HT-29 foram utilizadas para investigar a via de
sinalização Hh e as funções biológicas nas células epiteliais colônicas. Métodos:
Foram realizadas culturas de células HT-29 sob diferentes condições e exposição a
vários estímulos. A expressão dos componentes da via de sinalização Hh, proteínas
e genes relacionados, foram avaliados por imunofluorescência e por reação de
cadeia de polimerase (PCR) em tempo real. Viabilidade, proliferação celular e
apoptose foram avaliados, pelo método de 3-(4,5- Dimetiltiazol-2-yl)-2,5-
difeniltetrazolium bromida, tetrazol (MTT), por incorporação de Bromodeoxiuridina
(BrdU) e pela reação de Annexina -V/7-AAD, respectivamente. A produção de
quimiocinas no sobrenadante de culturas de células HT-29 foi avaliada por reação
imunoenzimática (ELISA). Resultados: Níveis de RNA mensageiro de Indian e
Sonic Hh e os fatores de transcrição Gli-1 e Gli-2 aumentaram após tratamento com
agonistas Hh e butirato, mas diminuiram após exposição a ciclopamina. A expressão
de proteínas morfogenéticas 4 e 7 (BMP4 e BMP7) diminui após o bloqueio da via
Hh com ciclopamina. A expressão da proteína Gli-1 aumenta após exposição com
agonistas e diminui após ciclopamina. A exposição a agonistas Hh promove a
redução da β-catenina e causa sua redistribuição subcelular. Níveis de interleucina 8
(IL-8) e proteína quimiotática de monócitos (MCP-1) diminuem após exposição a
agonistas Hh quando comparados a ciclopamina, lipopolissacarídeos (LPS),
inerferon (IFN-γ) ou fator de crescimento epitelial (EGF). A proliferação e a
viabilidade celular diminuem após o bloqueio da via Hh. Agonistas Hh revogaram a
apoptose induzida pelo anticorpo anti-CD95. Conclusão: A via Hh é um controlador
fundamental das células epiteliais colônicas, como demonstrado por seus efeitos
reduzindo sinais inflamatórios e antagonizando apoptose. A expressão diferenciada
de componentes da via Hh pode fundamentar anormalidades na reposta imune local,
na integridade da barreira epitelial, com potenciais implicações na homeostase para
o desenvolvimento de inflamação colônica e malignidade.

viii
ABSTRACT
Background & Aims: The Hedgehog (Hh) pathway is involved in
embryogenesis and physiologic processes including cell survival and proliferation.
Methods: The human colon carcinoma HT-29 cell line was used to investigate Hh
signaling and biological functions in colonic epithelial cells. HT-29 cells were cultured
under different conditions and exposed to various stimuli. The expression of Hh
pathway components, related proteins and genes were assessed by
immunofluorescence and protein chain reaction real-time (PCR). Viability, cell
proliferation and apoptosis were measured by the 3-(4,5- Dimethylthiazol-2-yl)-
2,5diphenyltetrazolium bromide, tetrazole (MTT) assay, Bromodeoxiuridine (BrdU)
uptake and Annexin-V/7-AAD staining, respectively. Chemokines production was
measured by enzyme-linked immunosorbent assay (ELISA) in culture supernatants.
Results Indian and Sonic Hh mRNA levels and the downstream transcription factors
Gli-1 and Gli-2 increased following treatment with Hh agonists and butyrate, but
decreased upon exposure to cyclopamine. Bone morphogen 4 and 7 (BMP4 and
BMP7) expression decreased after Hh blockade with cyclopamine. Gli-1 protein
expression increased after Hh agonists and decreased following cyclopamine.
Exposure to Hh agonists promoted β-catenin reduction and subcellular redistribution.
Levels of interleukin 8 (IL-8) and monocyte chemotactic protein (MCP-1) decreased
upon exposure to Hh agonists compared to cyclopamine, lipopolyssacharide (LPS),
interferon (IFN-γ) or epidermal growth factor (EGF). Cellular proliferation and cell
viability decreased following Hh blockade. Hh agonists abrogated the anti-CD95
induced apoptosis. Conclusions: Hh pathway is a key controller of colonic epithelial
cells, as demonstrated by its effect in dampening inflammatory signals and
antagonizing apoptosis. The differential expression of Hh components may underlie
abnormalities in the local immune response and in epithelial barrier integrity, with
potential homeostatic implications for the development of colonic inflammation and
malignancies.

ix
SUMÁRIO
Ficha catalográfica.......................................................................................................iii
Dedicatória...................................................................................................................iv
Agradecimentos............................................................................................................v
Recursos e aceitação..................................................................................................vi
Resumo.......................................................................................................................vii
Abstract......................................................................................................................viii
SUMÁRIO....................................................................................................................ix
Lista de figuras.............................................................................................................xi
Lista de anexo.............................................................................................................xii
Lista de siglas e abreviaturas.....................................................................................xiii
1 INTRODUÇÃO..........................................................................................................1
2 REVISÃO DE LITERATURA.....................................................................................4
2.1 A via de sinalização Hedghog................................................................................5
2.1.1.A via de sinalização Hh e o trato gastrointestinal................................................9
2.1.2 A via de sinalização Hh e o câncer gastrointestinal..........................................11
2.2 Linhagem de células HT-29..................................................................................12
3 OBJETIVOS............................................................................................................14
3.1 Objetivos gerais....................................................................................................15
3.2 Objetivos específicos............................................................................................15
4 MATERIAIS E MÉTODOS.......................................................................................16
4.1 Cultura de células.................................................................................................17
4.2 PCR em tempo real quantitativo (qRT-PCR)........................................................18
4.3 Reação de imunofluorescência e microscopia confocal.......................................19
4.4 Avaliação de citocinas..........................................................................................20
4.5 Avaliação de proliferação e viabilidade celular.....................................................21
4.6 Avaliação da apoptose celular..............................................................................23
4.7 Análise estatística.................................................................................................24
5 RESULTADOS........................................................................................................25
5.1 Expressão e modulação dos componentes da via hedghog na linhagem de
células HT-29.....................................................................................................26
5.2 Níveis subcelulares e distribuição de Gli1 e β-catenina nas células HT-
29.......................................................................................................................29

x
5.3 Efeito da via Hh na atividade inflamatória das células HT-
29.....................................................................................................................31
5.4 Efeito da via Hh na viabilidade e na proliferação das células HT-
29.....................................................................................................................33
5.5 Efeito da via Hh na apoptose mediada pelo anti-CD95 nas células HT-
29.....................................................................................................................35
6 DISCUSSÃO...........................................................................................................37
7. CONCLUSÃO.........................................................................................................42
8 REFERÊNCIA BIBLIOGRÁFICA.............................................................................44
9 anexo.......................................................................................................................53
9.1 anexo 1.................................................................................................................54

xi
LISTA DE FIGURAS
Figura 1 Esquema proposto da via de sinalização Hedgehog inibida por
Katoh&Katoh, 2006. ................................................................................................... 6
Figura 2 Esquema proposto da via de sinalização Hedgehog por Katoh&Katoh,
2006.. ......................................................................................................................... 8
Figura 3 Modulação gênica das células HT 29 após exposição a diferentes estímulos
................................................................................................................................. 28
Figura 4 Distribuição e níveis de Gli1 e β-catenina nas células HT-29 ..................... 30
Figura 5 Produção de citocinas pelas culturas de células HT-29 após exposição a
diferentes estímulos ................................................................................................. 32
Figura 6 Viabilidade e atividade proliferativa das células HT-29 após exposição a
diferentes estímulos ................................................................................................. 34
Figura 7 Relação entre apoptose e a atividade da via Hh nas células HT-29 ........... 36

xii
LISTA DE ANEXOS
Anexo 1 – Cópia do artigo enviado para publicação..................................................53

xiii
LISTA DE SIGLAS E ABREVIATURAS
Ac Anticorpo
APC Adenomatous polyposis coli
BMP4 Proteína morfogenética óssea 4
BMP7 Proteína morfogenética óssea 7
BrdU Bromodeoxiuridina
But Butirato
CKIα Caseina kinase I α
Cic Ciclopamina
DAPI 4’,6-diamidino-2-fenilindole
Dhh Dessert hedgehog
DMEM meio Dulbecco Eagle modificado
DMSO Dimetil sulfoxida
EGF Fator de crescimento epitelial
ELISA método imunoenzimático
FBS Soro bovino fetal
GAPDH Gliceraldeido 3 fosfato deidrogenase
GSK3β Glicogeneo sintase kinase 3β
HH Hedgehog
HHIP proteina de interação com Hedgehog
IFN Interferon
Ihh Indian hedgehog
IL Interleucina
LPS Lipopolissacarídeos
MCP1 proteína quimiotática de monócito 1
MTT 3-(4,5- Dimetiltiazol-2-yl)-2,5-difeniltetrazolium bromida, tetrazole
PBS solução salina tamponada com fosfato
PKA Proteína kinase A
Ptch Pacthed
Purm Purmorfamina
RT PCR transcriptase reversa – reação de cadeia de polimerase
Shh Sonic hedgehog
Smo Smoothened

xiv
STK36 Serina/treonina kinase
TGI Trato gastrointestinal
TMB Tetra metil benzidina
TNF Fator de necrose tumoral
VEGF Fator de crescimento vascular endotelial

1
INTRODUÇÃO

2
1. INTRODUÇÃO
O intestino possui uma função fisiológica essencial que é a manutenção da
barreira de proteção contra agentes que sejam potencialmente danosos ao nosso
organismo. A primeira linha de defesa nesta barreira é constituída por uma camada
única de células epiteliais que continuamente se renovam e interagem com o lúmen
e as células imunes da lâmina própria 1,2. O controle da renovação das células
epiteliais do cólon depende do equilíbrio dinâmico entre a proliferação celular e a
apoptose. O desequilíbrio desses mecanismos, como se sabe, pode resultar em
câncer3 e também em processos inflamatórios 4
A via de sinalização Hedgehog (Hh), de proteínas morfogenéticas, participam
de vários processos celulares fisiológicos, como embriogênese 5, formação de vários
órgãos6, reparo e homeostase de tecidos adultos7, regulação da transição epitélio-
mesenquimal8 e controle da sobrevivência e proliferação das células9.
A via canônica de Hh tem um papel no desenvolvimento do trato
gastrointestinal (TGI) normal, como já foi demonstrado em outros trabalhos, através
da regulação da diferenciação das vilosidades intestinais normais10 e das células
estroma-mesenquimais adjacentes11. No TGI normal de adutos, a indução da via Hh
aparentemente protege as células epiteliais diferenciadas da superfície vilosa,
regulando de forma negativa a sinalização da via canônica de Wnt nas células
basais das criptas intestinais12. Embora a ativação aberrante da via Hh tenha sido
demonstrada na oncogênese de tumores humanos de esôfago, estômago e
pâncreas 13,14, seu papel no câncer de cólon não está totalmente esclarecido.
A maioria dos tumores colorretais humanos esporádicos e hereditários se
originam da ativação da via de sinalização do Wnt através da mutação do genes
APC (adenomatous polyposis coli) ou β-catenina15. Há trabalhos que relatam que a
via de Hh atuaria de forma negativa ao efeito da via de Wnt na proliferação dos
colonócitos diferenciados12. Recentemente, surgiram algumas evidências do provável
envolvimento da via Hh na invasividade do câncer16, na carcinogênese no cólon17,18
e na disseminação metastática do câncer colorretal19. Estes achados se tornam
ainda mais relevantes diante da prevalência do câncer colorretal, sendo o segundo
câncer mais prevalente em países desenvolvidos e o terceiro em países em
desenvolvimento20. Apesar de avanços no diagnóstico e terapêutica, a mortalidade

3
em 5 anos continua elevada, em torno de 40%21. Entretanto, a participação da via
Hh na oncogênese de tumores de cólon necessita de melhor compreensão22,23.
Dessa forma, torna-se importante a investigação desta via no intuito de se encontrar
um alvo para tratamento ou marcadores biológicos, como por exemplo, proteínas
agonistas da via Hh (Shh e Ihh), proteínas que inibem a via Hh (HHIP1), proteínas
de receptores da via Hh (Ptch1) ou proteínas de transcrição da via HH (Gli1) que
pudessem apoiar o diagnóstico e a avaliação prognóstica.
As células HT-29, que são uma linhagem de células de carcinoma de cólon
humano, foram utilizadas através de várias condições experimentais para investigar
a via Hh e suas pontenciais funções biológicas (na proliferação e na apoptose
celular, assim como sua relação com outras vias de sinalização). Demonstramos a
presença e modulação de diversos componentes da via Hh e um efeito anti-
inflamatório e anti-apoptótico após a ativação da via Hh nas células HT-29.

4
REVISÃO DE LITERATURA

5
2. REVISÃO DE LITERATURA
2.1. A via de sinalização Hedghog
As vias de sinalização Hedgehog, BMP/TGF beta, WNT, FGF e Notch
participam da sinalização celular, fundamentais em uma variedade de processos
como embriogênese, homeostasia de tecidos adultos, reparo em processos
inflamatórios crônicos e carcinogênese. Estas vias se cruzam para constituir o
sistema de sinalização das células precursoras11,24,25.
As proteínas hedgehog (Hh) pertencem a uma família de moléculas de
sinalização intracelular26, originalmente identificadas em Drosophila27 com três
homólogos em vertebrados: Sonic Hedgehog (Shh), Indian Hedgehog (Ihh) e Desert
Hedghog (Dhh). Dentre estes, Shh tem sido a mais bem estudada. Os precursores
Hh sofrem modificação lipídica, gerando um domínio C-terminal e outro N-terminal.
Este último possuindo atividade sinalizadora28,29. Durante o processamento, a ação
da Hh acetiltransferase modifica a estrutura de Shh-N que sofre, ainda, palmitolação.
Esta, lhe confere a capacidade de permanecer associado à membrana e funcionar
como molécula de sinalização de curta distância, sendo capaz, ainda, de atuar na
interação entre células vizinhas30. Shh interage com um complexo receptor
composto de duas proteínas trans-membrana, patched (Ptch 1 e 2) e smoothened
(Smo). Ptch constitui a subunidade de ligação, enquanto que Smo, o receptor
transmembrana do tipo sete31 que é responsável pela transdução do sinal que, por
sua vez, é mediada pela família Gli de fatores de transcrição32,33. Na ausência da
sinalização por Hedgehog, a família de receptores Ptch inibem o transdutor de sinal
Smoothened34,35.
O Smo inativado leva a formação do complexo de degradação citoplasmático
Gli, no qual os membros da família Gli são fosforilados pelas caseina kinase I α
(CKIα), glicogêneo sintase kinase 3β (GSK3β) e proteína kinase A (PKA)36,37. As
moléculas de GLI fosforiladas são reconhecidas pela FBXW1/βTRCP1 e
FBXW11/βTRCP2 e encaminhadas para ubiquitinação. Após esse processo, o Gli é
parcialmente degradado para liberar sua porção N-terminal intacta que atua como
fator de transcrição repressor (Gli2 e 3)38,39,40 (Figura 1).

6
Figura 1 Esquema proposto da via de sinalização Hedgehog inibida por (?) Katoh&Katoh, 2006. Quando a molécula Hedgehog está ausente, a família de receptores Ptch 1 e 2 inibem o transdutor de sinal Smo. O Smo inativado leva a formação do complexo de degradação citoplasmático Gli, no qual os membros da família Gli são fosforilados pelas *caseina kinase1α, glicogen sintase kinase 3β e proteina kinase A. Estes complexos são reconhecidos e levados à ubiquitinação e posterior degradação parcial do Gli para liberar sua porção N-terminal. Esta, atua como fator de transcrição repressor (Gli 2 e 3 ) nos genes alvos como GLI1, PTCH1, CCND2, FOXM1.
Quando o Hedgehog se liga a família de receptores Patched, liberam o sinal
de transdução Smo. Smo ativa serina/treonina kinase (STK36) para inibir a
degradação do complexo Gli e promover sua estabilização como molécula íntegra41.
A STK36 ativada também fosforila SUFU, inativando-o e promove o acúmulo nuclear
da molécula Gli íntegra42. SUFU foi identificada como principal inibidor da via de
sinalização Hh em mamíferos, inibindo o sinal da via e se ligando aos fatores de
transcrição Gli no citoplasma e no núcleo43. Gli1 apresenta apenas domínio de

7
ativação, enquanto que as proteínas Gli2 e Gli3 apresentam domínio de ativação e
repressão da via Hh. Parece que quando ativados, GLI2 e GLI 3 são transdutores
iniciais do sinal, necessários para induzir a expressão do Gli144,45. O Gli1 ativa a
transcrição da maioria dos genes alvo da via Hh. A ativação da via de sinalização
Hedgehog leva a ativação transcricional dependente do Gli de genes alvos, como
GLI1, PTCH1, CCND2, FOXL1 e JAG246,47 (Figura 2 ). Fox pertence a uma vasta
família de fatores de transcrição Winged-helix/Forkhead Box, que tem importante
papel na regulação de expressão de genes envolvidos na proliferação tecidual48. O
aumento da regulação do CCND2 e FOXM1 leva a proliferação de células alvo
através da progressão do ciclo celular38. A via de sinalização hedgehog é regulada
positivamente através do aumento do fator de transcrição Gli1, enquanto que o
aumento da regulação do Ptch1 estimula a via de forma negativa. As moléculas
CDON e BOC, que são proteínas transmembrana com domínio extracelular
imunoglobulina símile e fibronectina tipo III49, foram também relatados como co-
receptores da via Hh, que aumentariam sua atividade de sinalização.
A ciclopamina é um alcalóide esteroidal derivado da planta Veratrum
californicum, capaz de se ligar diretamente ao receptor Smo, resultando na inibição
da sinalização Hh (Figura 1). Esta molécula vem sendo utilizada em estudos para
modular a via50, apresentando citotoxicidade moderada, quando comparada com a
pequena molécula GANT61, que inibe GLI1 e GLI251.. Genes HHIP1 atuam como
inibidores da via Hh52,53. A proteína transmembrana HHIP (Hh interectaing protein)
liga-se aos ligantes Hh, receptores Ptch e atenua o sinal ao competir com as
proteínas Hh52,53,54.

8
Figura 2 Esquema proposto da via de sinalização Hedgehog por Katoh&Katoh, 2006. Hedgehog se liga a família de receptores Ptch 1 e 2 e libera o transdutor de sinal Smo, que estava reprimido pelo Ptch. Smo ativa a STK36 serina/treonina kinase para estabilizar os membros da família Gli (Gli1,2 e 3) para translocação nuclear. A via de sinalização Hh ativa os fatores de transcrição dependente de Gli que atuam nos genes alvos como GLI1, PTCH1, CCND2, FOXM1. A ciclopamina é um alcalóide capaz de se ligar ao receptor Smo e inibir a sinalização Hh, enquanto a purmorfamina se liga no receptor Smo e ativa a Via Hh.
A via de sinalização Shh tem sido considerada fundamental na regulação de
processos de desenvolvimento embrionário de vários órgãos e tecidos, como o trato
gastrointestinal26,55. Há estudos relacionando o processo inflamatório e o papel da
via Hh. A análise do ciclo celular de linfócitos T CD4, isolados do sangue e
estimulados com Shh, revelou que a adição exógena de Shh aumenta a proliferação
celular de células ativadas, via receptor de células T (TCR), ao passo que o bloqueio
com anticorpo anti-Shh diminui a proliferação56
. Evidências recentes revelam que a
reativação de Shh também tem sido observada em modelos de isquemia, nos quais

9
funcionaria como agente angiogênico. Embora não tenha sido observado efeito
direto sobre células endoteliais, a ativação da via Shh resultou em aumento do fluxo
sanguíneo, e da expressão de fatores tais como o fator de crescimento vascular
endotelial (VEGF)57. A reativação da via parece estar associada ao aparecimento de
vários tumores malignos58. De forma semelhante, a sinalização Hh indiretamente
induziria a transição epitélio-mesenquimal através do aumento dos múltiplos
reguladores Notch e TGF-β59. A compreensão da transição epitélio-mesenquimal é
importante, uma vez que ela está envolvida na embriogênese e na invasão e
metástase durante a carcinogênese60,61,62. Existem muitas controvérsias em relação
ao papel da via Hh em tumores do TGI baixo, e em sua participação nas células
epitlélias. Este, é um dos focos do nosso estudo.
2.1.1. A via de sinalização Hh e o trato gastrointestinal
A homeostasia do trato gastrointestinal de vertebrados requer interações
entre epitélio do endoderma e células mesenquimais derivadas do mesoderma. A
sinalização entre essas duas camadas teciduais é crucial para o desenvolvimento
adequado do intestino. Em estágios de desenvolvimento precoce, Shh e Ihh são
secretados pelo endoderma do intestino de mamíferos, indicando seu papel no
desenvolvimento63.
Defeitos na cascata de sinalização Hh foram relacionados a diversos defeitos
de nascimento, mutações de genes Shh e GLI3 da via demosntraram anormalidades
no TGI, como atresia de esôfago, má rotação intestinal e ânus imperfurado64. Análise
de camundongos com genes inativados Shh e Ihh no endoderma intestinal
demonstraram que a via Hh é essencial para o crescimento do mesênquima
adjacente ao epitélio no desenvolvimento intestinal e que as proteínas Hh atuam
como mitógenos parácrinos que promovem a expansão de progenitores
mesenquimais adjacentes, incluindo aqueles do compartimento de músculo liso65.
A via Hh está envolvida no desenvolvimento do TGI como documentado em
camundongos com fenótipo alterado com genes da via Hh inativados. Da mesma
forma má-formações no cólon foram observados em camundongos com os genes
SHH, IHH, GLI 2 e 3 inativados66. Em modelo animal de camundongos, in vitro e in
vivo também foi demonstrado que Ihh é expresso por colonócitos maduros e regula
sua diferenciação. A sinalização Hedgehog restringe a expressão de alvos da via

10
WNT/β catenina (que define o fenótipo das células progenitoras epiteliais colônicas),
na base das criptas colônicas10.
A expressão de componentes do sinal de transdução da via Hh e suas
proteínas também tem sido descrita em tecidos de adultos como estômago67,68 e
cólon10. No trato gastrointestinal SHH e PTCH1 são expressos nas células parietais
gástricas69 e foi visto que a proliferação de células parietais mediadas pelo SHH está
presente no mecanismo de reparo da mucosa gástrica durante a infecção crônica
pelo Helicobacter pylori, como visto no trabalho de Katoh e colaboradores (2005),
em um modelo de inflamação70. Van den Brink e colaboradores (2004) documentou
que IHH e RUNX3 são expressos em células diferenciadas na superfície do
estômago e intestino10. O estudo de Sasaki e colaboradores (2009) sugeriu a
presença de comunicação entre a via Hh e a transição epitélio-mesênquimal nas
células gástricas e câncer gástrico de tipo difuso71. Pouco se sabe sobre o
mecanismo que regula a transição do epitélio precursor do cólon em células
diferenciadas. O estudo de Van den Brink (2004)10 utilizou moléculas para avaliar a
diferenciação do epitélio colônico, villina, que é uma proteína do citoesqueleto que é
específica das microvilosidades, e a enzima anidrase carbônica IV da borda das
vilosidades. Os enterócitos de camundongos controles apresentaram localização
uniforme da villina na borda das microvilosidades e uma alta expressão da enzima
anidrase carbônica IV. Por outro lado, os enterócitos de camundongos tratados com
ciclopamina (inibidor da via Hh) apresentaram localização heterogênea da villina
com perda da expressão da villina na borda das microvilosidade e redistribuição da
villina no citoplasma, assim como não expressam mais a anidrase carbônica.
Os homológos em vertebrados BMP são coexpressados com genes Hh
durante o desenvolvimento. A sinalização BMP-RUNX3 induz a expressão do Hh na
superfície das células diferenciadas de estômago e intestino38. Existe a expressão
do BMP4 na base das criptas intestinais e no cólon de adultos o BMP2 é expresso
em colonócitos diferenciados. Foi demonstrado que a inibição do Hh, com
ciclopamina aumenta a expressão do BMP4, mas não altera a expressão do
BMP210.
A sinalização WNT tem um papel importante na manutenção do epitélio
intestinal38,72,73. A via WNT define o fenótipo das células epitélio colônico progenitor
e mutações no gene polipose adenomatose coli (APC) ativam a via WNT e causam

11
síndrome de polipose familar adenomatosa e a maioria dos cânceres esporádicos de
cólon15. A sinalização WNT é ativa na região progenitora ao redor das criptas
intestinais, enquanto que a via de sinalização hedgehog é presente na região
próxima da área diferenciada da superfície intestinal. Foi descrito que os colonócitos
terminalmente diferenciados produzem RNAm do gene IHH e a proteína Ihh10. A via
Hh inibiria a sinalização WNT e a proliferação das células intestinais e promoveria a
proliferação do epitélio de esôfago, estômago e pâncreas3.
Há relato que o receptor Ptch da via Hh é expresso no epitélio ao longo das
criptas intestinais e em células mesenquimais e que o Ihh poderia afetar diretamente
células alvo no epitélio e no mesênquima, possivelmente este processo esta
envolvido no processo de renovação, afetando as células precursoras66.
2.1.2. A via de sinalização Hh e o câncer gastrointestinal
Carcinogênese é um processo multifatorial, influenciado por predisposição
genética, fatores ambientais e idade74. O aumento transcricional de genes ligados às
vias carcinogênicas pode ocorrer durante processos inflamatórios para reparo de
lesão de tecidos, assim como a inativação epigenética de genes que regulam
negativamente essa via.
Sabe-se que a via Hh está constitutivamente ativada em alguns tipos de
câncer34. Mutações que levam a perda da função dos genes PTCH e SUFU e
mutações que aumentam a função do gene SMO tem se mostrado responsáveis
pela ativação independente das vias dos tumores: carcinoma basocelular,
meduloblastoma ou rabdomiossarcoma75. Também, foi observada amplificação do
gene GLI1 em gliomas malignos76. O gene HHIP que codifica a proteína que
interage com proteína Hh como inibidor da via está regulada negativamente em
tumores como gástrico, pancreático, e pulmonar 77
A via Hh é frequentemente ativada em câncer de esôfago, gástrico e
pancreático devido ao aumento transcricional dos ligantes da via Hh e ao
silenciamento epigenético do gene HHIP1. Contudo em alguns estudos a sinalização
Hh é raramente ativada no câncer coloretal, e em outros estudos a via Hh regularia
de forma negativa o sistema de sinalização WNT38.
Shh e IHH são expressos em todas as seis linhagens de câncer de esôfago
estudadas no trabalho de Berman e colaboradores (2003). Ptch e GLI1 são

12
expressos em 4 dos 6 tipos de câncer de esôfago. Shh, Ihh, PTCH1 e GLI1 são
expressos nos 6 tipos de linhagem de câncer gástrico e em 5 dos 6 tipos de
linhagem de câncer de pâncreas estudados. O possível mecanismo de interação da
via Hh inibindo a via Wnt vem sendo estudado, porém não está totalmente
esclarecido. Vários estudos tentam compreender o papel da via Hh no câncer de
cólon, mas os resultados são conflitantes. Mutações raras dos genes PTCH. e SMO
foram observados nesse tipo de tumor, mas elas parecem não afetar a atividade das
proteínas 78,79. O trabalho de Dourard e colaboradores (2006) sugeriu que a via Hh
pode ter um papel central na patogênese de câncer de cólon e a manutenção da via
Hh seria importante para a tumorigênese do câncer de cólon, mas a via não estaria
correlacionada ao tamanho e a localização do tumor. Foi demonstrado que o
aumento da expressão de RNAm do gene de SHH em células de adenocarcinoma
de cólon humano e na linhagem de células HT-29 aumentaram a expressão de
RNAm do gene GLI1 e RNAm do gene FOX1, que sabe-se promove proliferação
celular 22.
Há relato de trabalhos de que tumores poderiam expressar ligantes Hh,
ativando a via de sinalização nas células estromais vizinhas, demonstrando um
efeito parácrino na sinalização Hh, mas não descartando o papel da via em
subpopulações de células tumorais epiteliais, como células precursoras de câncer80
2.2. Linhagem de células HT-29
As células HT-29 são uma linhagem de células de carcinoma de cólon , que
começam a polarizar e formar microvilosidade apicais quando tratadas com butirato,
um ácido de cadeia curta81.Foi utilizada esta linhagem porque existe relato de que a
via de sinalização Hh regula a diferenciação das células epiteliais colônicas in vivo e
in vitro10.
Entretanto, há relatos contraditórios quanto à presença da via Hh nas células
HT-29. Sabe-se que quando as células HT-29 são tratadas com butirato, a proteína
Ihh é induzida e quando as células são tratadas com ciclopamina há uma redução
da estimulação que ocorre com a exposição ao butirato, ou seja, reduz os
marcadores que indicam a diferenciação do epitélio colônico como demonstrado com
a vilina10.De maneira controversa o trabalho de Chatel (2007) avaliou a expressão
gênica de SHH, IHH, PTCH, SMO, GLI1, GLI2, GLI3, SUFU and HHIP e as proteínas

13
Hh, Gli3 e Sufu em sete linhagens de células de câncer de cólon e em nenhuma
linhagem houve expressão de todos os genes envolvidos na ativação da via Hh,
além do tratamento da ciclopamina não modular a expressão os níveis dos genes
PTCH e GLI1, sugerindo que a via HH não estaria ativada naquelas linhagens de
células que incluem as células HT-2923. Como relatado anteriormente há dados
contraditórios nos estudos sendo considerado que as discrepâncias podem ocorrer
por modificações epigenéticas da expressão gênica. Também os autores do estudo
não puderam excluir que a via Hh é ativada em uma subpopulação de células
precursoras de câncer82,83.
Neste estudo, a investigação da expressão e modulação da via Hh na
linhagem de células HT-29 foi realizada através da identificação e quantificação dos
componentes da via Hh nas céulas HT-29 sem e com modulação de citocinas e
análogos e bloqueadores da via Hh; e foi avaliada a expressão de genes da via Hh e
outros genes de vias envolvidas por ela. O estudo da atividade biológica de Hh nas
células HT-29 foi realizada através da análise da expressão de citocinas pró-
inflamatórias e regulatórias, da mucosa intestinal tratada com adição ou bloqueio de
proteínas da via Hh, LPS, IFN-γ e EGF e da análise da proliferação e apoptose de
células epiteliais HT-29 tratadas com adição de análagos e bloqueio de proteínas da
via Hh.
Pretende-se esclarecer a participação da via Hh no câncer de cólon,
utilizando as células HT-29, que representam um modelo de estudo in vitro de
células epiteliais e cancerígenas diferenciadas, semelhante ao epitélio de superfície.

14
OBJETIVOS

15
3. OBJETIVOS
3.1. Objetivos gerais
Investigar a expressão e modulação da via Hh na linhagem de células
HT-29.
Investigar a atividade biológica de Hh nas células HT-29.
3.2. Objetivos específicos
Avaliar a expressão dos níveis de RNA mensageiro de moléculas da
via Hh após tratamentos específicos das células HT-29
Avaliar a expressão e localização da proteína Gli1 da via Hh e β-
catenina nas células HT-29 com tratamentos específicos
Dosar citocinas pró-inflamatórias nas células HT-29 após tratamentos
específicos
Avaliar a viabilidade e a proliferação das células HT-29 após
tratamentos específicos
Avaliar apoptose das células HT-29 após tratamentos específicos

16
MATERIAIS E MÉTODOS

17
4. MATERIAIS E MÉTODOS
4.1. Cultura de células
A linhagem de células HT-29 foi obtida do American Type Culture Collection
(ATCC, Rockville, MD, EUA) e mantida de acordo com as instruções da ATCC no
Banco de Células do Rio de Janeiro. Foram cultivadas no Laboratório de Imunologia
Celular em meio de cultivo Dulbecco’s modified Eagle’s medium (DMEM) (Invitrogen
Gibco, New York, NY, EUA), suplementado com 10% de soro fetal bovino inativado
(FBS) (Invitrogen Gibco, New York, NY, EUA), 4,5 g/L de glicose, 100 µg/ml
estreptomicina e 10000 U/ml de penicilina (todos da Sigma-Aldrich, St Louis, MO,
EUA) e incubadas por 24 h na estufa a 37oC com atmosfera úmida de 5% de CO2.
Após expansão e crescimento, as células HT-29 foram tripsinisadas com
tripsina a 0,3% (Gibco, New York, NY, EUA). Foram inativadas com solução salina,
centrifugadas e ressuspendidas em DMEM a 2,5 % de soro fetal bovino (FBS),
devido a sua interferência em concentrações mais altas na ação da ciclopamina.
Parte das células foram criopreservadas em criotubo com 106 células e
armazenadas no freezer a -800 C.
As células foram plaqueadas com 106 células por poço, com seus tratamentos
específicas para cada reação em placas de 24 poços para PCR real time, em placas
de 96 poços para ensaio de BrdU e os sobrenadante dos poços de culturas foram
armazenados a -200C para dosagem de citocinas. As células também foram
plaqueadas em lâminas para cultura de tecido (Lab Tek Nunc, Naperville, IL,EUA) e
para reação de imunofluorescência. As células HT-29 foram tratadas com moléculas
da via Hh: N-Shh recombinante humano da R&D Systems (Minneapolis, MN, EUA)
na concentração 500 ng/ml; ciclopamina (Sigma-Aldrich, St Louis, MO, EUA) na
concentração de 10 µM, sendo usado como diluente dimetil sulfoxida (DMSO)
(Merck, Darmstadt, Alemanha); purmorfamina, que é um agonista Hh químico (Alexis
Biochemicals, Plymounth, PA, EUA), diluído em DMSO na concentração de 2µM;
butirato de sódio foi acrescentado às culturas na concentração de 2.5µM, enquanto
que interferon gama (IFN-γ) e lipopolissacarídeo E coli O11:B4 (LPS) foram usados
na dose de 2 ng/ml (todos da Sigma-Aldrich,St. Louis, MO, EUA) e proteína

18
recombinante do fator de crescimento epitelial humano (rhEGF), na concentraçao de
20 ng/ml, da Peprotech (Rocky Hill, NJ, EUA).
4.2. PCR em tempo real quantitativo (qRT-PCR)
Foram utilizadas amostras de 106 células HT-29 plaqueadas em 500µl de
DMEM completo por poço, e tratadas por 24 horas, a 37oC em estufa com atmosfera
úmida de CO2, com diluente (DMSO a 0.2%) (controle) sem butirato, com butirato,
com butirato mais ciclopamina, com ciclopamina, com cliclopamina mais
purmorfamina, com ciclopamina mais Shh, com Shh, com EGF, com INF-γ, com LPS
e com Purmorfamina.
Para a extração de RNAm foi utilizado o kit da Promega, Madison, WI, EUA,
utilizando o protocolo do fabricante. Foram adicionados às amostras de células HT-
29 solução de tampão de lise e tampão de diluição de RNA e a seguir foram
centrifugados por 10 min, a 14.000 rpm a 25oC. O sobrenadante foi extraído e
transferido para o microtubo próprio do kit e então misturado a etanol 95%,
novamente centrifugado por 1 min a 14000 rpm. Acrescentou-se solução de lavagem
SV RNA e centrifugou-se novamente por 1 min a 14.000 rpm. Misturou-se solução
de DNAse (yellow core +Mg Cl2+ DNAse) e incubou-se por 15 min a 25oC. Utilizou-
se a solução de paragem SV DNAse para interromper a reação e lavou-se o material
com solução de lavagem e água sem nuclease. Obteve-se 25 µl de solução de
RNAm.
Sintetizamos cDNA com o Kit da Applied biosystem. Associou-se aos 25 µl de
RNA m, 25 µl da solução com reverse transcription buffer, d NTP, RT random primer,
multiscribe tb reverse transcriptase e água sem Dnase. Incubou-se por 10 minutos a
250C e depois 120 minutos a 370C. O material foi estocado no freezer a -6oC.
Foi utilizado o aparelho de nanodrop 2000 UV-Vis espectofotômetro (Thermo
Scientific, Wiminton, DE, EUA) para quantificação de RNA e para avaliar a pureza
das amostras.
Foram analisados 2 µl de material de cDNA de cada amostra em duplicatas e
repetidas três vezes, através de placas customizadas de PCR da SABiosciences,
Frederick, MD, EUA com dez genes: homólogo do Indian hedgehog (Drosophila),
homólogo do Sonic hedghehog (Drosophila), GLI 1, GLI 2 , GLI 3, Homólogo do
Smoothened (Drosophila), homólogo Patched (Drosophila), HHIP (proteína de

19
interação hedgehog), WNT 1, BMP 4 (proteína morfogenética óssea 4), BMP7
(proteína morfogenética óssea 7), GAPDH (gliceraldeído 3 fosfato deidrogenase). O
RNAm do gene GAPDH foi escolhido para normalizar a quantificação dos outros
genes.
A quantificação dos níveis de RNAm foi realizada através da técnica de RT-
PCR (transcriptase reversa-reação de cadeia de polimerase), sendo utilizado ABI
Prism 7500 (Applied Biosystems, Foster City, CA, EUA) com o programa RT2 Tempo
RealTM SYBR Green/Rox PCR Master Mix (SABiosciences, Frederick, MD,EUA) que
realiza a quantificação relativa com o método da curva padrão relativa. Valores de
resultado positivos indicam regulação gênica positiva e valores negativos indicam
regulação gênica negativa.
4.3. Reação de imunofluorescência indireta e microscopia confocal
As células HT-29 foram plaqueadas em lâminas com poços para cultura (Lab
Tec, NUNC, Dinamarca), fixadas com paraformaldeído à 1%, permeabilizadas e
incubadas com o bloqueio, albumina do soro bovino a 2.5%, leite desnatado e soro
fetal bovino a 8%, por 2 horas em temperatura ambiente. Para retirar o excesso do
bloqueio as lâminas foram lavadas com PBS e Tween 20 a 0.05%. Em seguida
foram incubados por 1 hora em temperatura ambiente com os anticorpos primários
anti Gli1, anticorpo (Ac) de coelho policlonal na concentração de uso 5 mg/ml, 1:400
da Santa Cruz Biotechnology Inc, Santa Cruz, CA, EUA e anti β-catenina, anticorpo
policlonal de camundongo 1:500 da Millipore, Bedford, MA, EUA. As lâminas para
cultura de tecido foram lavadas com dois banhos de solução tampão PBS pH 7,4,
por 5 minutos cada, para remover o excesso de anticorpo primário não ligado.
Aplicou-se sobre os cortes o anticorpo (Ac) secundário conjugado AlexaR fluor 568
IgG conjugado a anti-coelho e AlexaR 633 conjugado a IgG anti-camundongo (todos
da Molecular Probes, Eugene, OR, EUA), tendo o cuidado de não permitir a
secagem dos cortes e proteger os mesmos da luz ambiente, deixando incubando
durante 30 minutos em temperatura ambiente. As amostras foram incubadas com
PBS sozinho ou com anticorpos secundários e serviram como controles de isotipos
negativos As lâminas foram montadas com meio de montagem Vecta shield
contendo 4’,6-diamidino-2-fenylindole (DAPI) (Vector Laboratories Inc, Burlingam ,
California, EUA). As lâminas de cultura de tecido foram armazenados em caixa

20
apropriada, ao abrigo da luz ambiente, estocadas no freezer -20°C até a
observação, que foi feito pelo microscópio confocal a laser da Leica TCS-SP5 AOBS
(Leica, Heidelberg, Alemanha) com analisador de imagem Leica.
4.4. Avaliação de citocinas
Placas com células HT-29 foram colocadas em cultura por 24 horas na
presença de DMSO, do peptídeo biologicamente ativo rShh amino-terminal,
purmorfamina, butirato, com e sem ciclopamina e IFN-γ, LPS e EGF. Os
sobrenadantes foram coletados e armazenados a -20oC para análise de citocinas,
pela técnica de ELISA (enzyme-linked immunosorbent assays). Utilizou-se kits para
as dosagens de interleucina 8 (IL-8) (BenderMedSystems, Viena, Áustria) e da
proteína quimiotátca de monócitos (MCP1) (e-Bioscience, San Diego, CA, EUA). A
quantidade de proteína total das monocamadas de células foi estimada pelo Kit de
proteína PierceR BCA (Thermo Scientific, Rockford, EUA) e usada para normalizar os
resultados.
Para a dosagem de IL-8, incubou-se 50 µl por poço de anticorpo de captura,
que é o anticorpo anti- IL8 humano na concentração de 5 µg/ml diluído 1:100. A
placa foi incubada durante a noite a temperatura ambiente, a seguir desprezou-se e
lavou-se os poços com solução de lavagem por 3 vezes. Adicionou-se 200 µl por
poço de solução bloqueadora e incubou-se por 1 hora a tempertura ambiente.
Desprezou-se e lavou-se os poços com solução de lavagem por 3 vezes. Diluiu-se
na placa o IL-8 humano recombinante em diluições seriadas de 2 vezes, de forma
que ficaram 50 µl de volume em cada poço. Adicionou-se 50µl de amostra
(sobrenadante de 24 horas) nos respectivos poços, conforme o desenho da placa.
Incubou-se a placa por 2 horas a temperatura ambiente. Desprezou-se e lavou-se
por 3 vezes. Adicionou-se 50µl por poço do anticorpo de detecção, que é o anticorpo
anti-IL8 conjugado a biotina na diluição de 1:1000. Incubou-se a noite a -40 C.
Lavou-se por 3 vezes novamente, adicionou-se 100µl de streptavidina HRP 1:10000
e incubou-se por 30 minutos. Lavou-se e adicionou-se 100 µl de tetra metil benzidina
(TMB) que reage com peroxidase conjugada em cada poço e incubou-se por 30
minutos. Parou-se a reação com solução de HCl 1N e fez-se a leitura da reação no
espectofotômetro na absorbância de 450 nM. Calculou-se a média dos valores de
absorbância para as amostras e criou-se uma curva standard.

21
Pela mesma técnica descrita anteriormente foram utilizadas como anticorpo
de captura o anticorpo purificado anti-MCP1 humano, o recombinate MCP 1 humano
(1 ug/ml), o anticorpo de detecção anti-MCP1 humano conjugado a biotina, a enzima
streptavidina HRP e o substrato TMB.
4.5. Avaliação de proliferação e viabilidade celular
Foi utilizado o Kit de reação de BrdU (Chemicon International, Temecula, CA,
EUA) para avaliar a proliferação celular. Para minimizar a morte celular espontânea,
as células HT-29 foram mantidas na concentração de 1×106 células/ml, sendo
diluídos a cada 2 a 3 dias em diluente fresco, para manter o crescimento celular
exponencial. Células HT-29 são lavadas com PBS e tripsinisadas, contadas e
plaqueadas na concentração de 2×105 células/mL, nas placas de 96 poços, com 100
µL por poço. Após aproximadamente 12 horas (overnight) as células foram tratadas
por 48 e 72 em três experimentos em triplicatas com tratamentos sem butirato e com
butirato, com meio sem BrdU, com BrdU, com DMSO, com ciclopamina, com
purmofamina, com rShh, com interferon gama, com lipopolissacarídeo e com EGF.
BrdU foi acrescentado aos poços com células HT-29, 4 horas antes do final
do período de leitura. Preparou-se a concentração estoque de BrdU 1:500, diluindo-
a em 500 vezes. Foram pipetados 20 µl do BrdU diluído nos poços apropriados.
Uma placa foi avaliada em 48 horas e outra placa em 72horas.
Para detectar o BrdU através do anticorpo monoclonal anti BrdU, foram
usados: solução de fixação de células para fixar e desnaturar o DNA, para incubar
as células a temperatura ambiente por 30 minutos. As placas foram centrifugadas
por 5 min a 1000 rpm, e foi realizado novamente o processo com solução de fixação.
Posteriormente as amostras foram lavadas com tampão de lavagem três vezes.
Acrescentou-se o anticorpo monoclonal de camundongo anti BrdU diluído 1:200 e
incubou-se por 1 hora em temperatura ambiente. Novamente utilizou-se solução de
lavagem por três vezes. Acrescentou-se então anticorpo secundário conjugado a
peroxidase (1:2000) e incubou-se por 30 minutos em temperatura ambiente. Lavou-
se novamente e adicionou-se o substrato TMB e incubou-se por 30 min em
temperatura ambiente no escuro. Os poços positivos na reação tiveram coloração
azulada. Aplicou-se a solução de interrupção e os poços positivos azuis tornaram-se

22
amarelados. Realizou-se a leitura da placa no espectofotômetro no comprimento de
onda de 450/550 nm.
Nesse método utiliza-se a bromodeoxiuridina (BrdU), que é um análogo da
timidina, que é incorporado na síntese das bandas de DNA nas células ativamente
em proliferação, sendo a sua incorporação na fase S do ciclo celular. Posteriormente
com a desnaturação parcial da dupla banda de DNA, o BrdU pode ser detectado
imunohistoquimicamente, permitindo avaliar a população de células que estão
sintetizando DNA.
Normalmente a determinação de proliferação celular é realizada pela
contagem de células viáveis após coloração com corante vital. Existem várias
técnicas, como a coloração com corante azul de trypan, que avalia a integridade da
membrana celular, mas não é um método sensível. O uso de timidina associado a
trítio é acurado, mas consome tempo e radiotividade. MTT amarelo (3-(4,5-
Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide, tetrazole) é reduzido em
formação roxa na mitocrôndria das células vivas. A absorbância dessa solução
colorida pode ser quantificada pela leitura do comprimento de onda de 500 a 600 nm
por um espectofotômetro. Essa reação de redução ocorre apenas quando as
enzimas mitocôndria redutases estão ativadas, e a conversão pode então estar
relacionada diretamente ao número de células viáveis. As enzimas das mitôcondrias
quebram o tetrazolium em cristais roxos insolúveis em soluções aquosas que podem
ser dissolvidas em isopropanol acidificado. Quando a quantidade de formação roxa
produzida pelas células tratadas por um agente é comparada a quantidade de
complexos roxos formados por um grupo de células controle não tratadas, a eficácia
do agente em causar a morte das células pode ser deduzida através de uma curva
dose-resposta. Soluções de MTT solubilizadas em culturas teciduais em fenol
vermelho tem cor amarela. Limitações do método: estado fisiológico das células,
variação da atividade mitocôndria desidrogenase. O método é útil na mensuração de
crescimento celular em reposta a mitôgenos.
Para avaliar a viabilidade celular foi utilizado o kit para MTT (Sigma-Aldrich, St
Louis, MO, USA). Foram feitos três experimentos utilizando-se placas de 96 poços
sendo as amostras de células HT-29 e seus tratamentos plaqueadas em triplicatas,
avaliadas em 24 e 48 horas. As amostras foram: controle, células mais ciclopamina,
células mais butirato, células com butirato mais ciclopamina, células mais

23
purmorfamina, células com purmorfamina mais ciclopamina, células com Shh,
células com Shh mais ciclopamina, células com INF-y, com EGF, com LPS, com
DMSO.
As células foram tripsinisadas e adicionadas 100 ul de células por poço no
primeiro dia, no segundo dia as células foram tratadas com os agentes em questão.
No terceiro dia adicionou-se 20 ul da solução MTT 5 mg/ml em cada poço. Incubou-
se por 3,5 horas a 37oC. Removeu-se o sobrenadante e adicionou-se 150 ul de
solvente e após 15 minutos fez se a leitura no espectofotômetro na absorbância de
590nm.
4.6. Avaliação da apoptose celular
Anexinas são uma família de proteínas de ligação de cálcio dependente de
fosfolipídios que preferencialmente se ligam a fosfatidilserina. Em condições
fisiológicas normais a fosfatidilserina é predominantemente localizada na
monocamada intra-celular. No início da apoptose a fosfatidilserina perde sua
distribuição assimétrica através da bicamada de fosfolipidios e é translocado para a
camada extracelular, marcando as células como alvos de fagocitose. Uma vez a
fosfatidilserina estando na camada externa da membrana ela pode ser detectada
pela annexina V fluorescente. A apoptose, que é uma forma de morte celular
programada, pode ocorrer em mamíferos através de duas vias principais: o receptor
CD95, também chamado de Fas, que é a chamada via da morte celular fisiológica e
através do p53 que é a via dita mitocondrial de destruição celular que envolve
principalmente as células com dano de DNA. O CD95 pertence a família dos
receptores de fator de necrose tumoral (TNF). Na via de apoptose mediada pelo Fas,
o primeiro evento que ocorre é o encontro do CD95 com seu ligante CD95L (FasL).,
que leva a formação de um complexo de sinalização indutor de morte.
As células HT-29 foram plaqueadas na concentração de 105 células /poço em
placas de 6 poços. Após a adesão das células nas 12 horas noturnas, as células
foram tratadas com diferentes reagentes, incluindo o anticorpo monoclonal
CD95/APO-1 (1:50) (Southern Biotech, Birmingham, AL, EUA), para induzir apoptose
(que recruta a molécula Fas - domínio proteico associado a morte celular e caspase
8 que induzem a sinalização de morte.), ou controle DMSO 0,2%, em triplicatas por
24 horas seguidas de lavagem com PBS, tripsinização e centrifugação. Para avaliar

24
a viabilidade celular no estágio inicial e tardio da apoptose, as células foram coradas
com o Kit da Annexina-V e kit 7-AAD (solução de coloração de viabilidade) ambos da
e-Biosciences, San Diego, CA, EUA. Rapidamente as células foram lavadas duas
vezes com PBS gelado e ressuspensas com reagente de ligação (1X10 6 células em
0.1ml) e 10µl de annexina conjugada a FITC V(fluorocromo) e 20 µl de 7-AAD foram
acrescentados. As células foram incubadas por 15 minutos no escuro, e adicionados
400µl de reagente de ligação e as células foram avaliadas em 1 hora por citometria
de fluxo. A aquisição foi realizada com FACSCalibur com CellQuest Pro software
(BD Biosciences, San Jose, CA) e analisado quantitativamente com o programa de
software Winlist (Verity Software House, Topsham, ME). Cada análise foi realizada
pelo menos por 30000 eventos.
As reações foram feitas em triplicatas, em grupos de controle com células
vivas, controle com células mortas, controle células mortas com anexina e anti-
CD95, controle com células mortas 7AAD e anti CD95, controle com célula viva
anexina mais 7AAD e controle célula morta com anexina mais 7AAD mais anti-CD95
e grupos com células mais butirato, células mais ciclopamina e butirato, células com
purmorfamina, células com ciclopamina mais purmorfamina, células com Shh,
células com ciclopamina e Shh, células com IFN-y, células com ciclopamina, células
com EGF, com LPS, com butirato mais anti-CD95, com Shh e anti-CD95,
purmorfamina com anti-CD95, com ciclopamina e anti-CD95 e controle.
4.7. Análise estatística
Os dados obtidos em cada etapa do projeto foram armazenados em planilhas
diferentes para posteriormente serem analisados com o auxílio de um programa de
análise estatística por computador SPSS para Windows (versão 10.1, SPSS Inc,
1989-1999, EUA).
Diferenças estatísticas entre os grupos experimentais foram avaliados com
teste de análise variada ANOVA no qual comparações múltiplas por pareamente
simultâneo foram realizadas pelo teste Dunnett T3. O nível de significância foi
estabelecido em p<0,05.

25
RESULTADOS

26
5. RESULTADOS
5.1. Expressão e modulação dos componentes da via hedgehog na linhagem
de células HT-29
Para investigar o papel da via de sinalização Hh em várias funções biológicas dos
enterócitos, usamos as células HT-29 que são uma linhagem de células de
carcinoma de cólon humano sob diferentes situações experimentais. Como existe
pouca informação sobre a via Hh e as células epiteliais intestinais, nós analisamos a
expressão e modulação dos componentes da via Hh nas células HT-29 por análise
de qRT-PCR. Analisamos a expressão gênica das células HT-29 sob o efeito de
estímulo direto e da inibição da via Hh e a exposição a produtos bacterianos
(lipopolissacarídeos, LPS), citocinas (interferon gama, IFN-γ) e fator de crescimento
epitelial (EGF). Foram avaliados genes envolvidos diretamente na via Hedgehog e
outros que possivelmente interagem com a via de sinalização (FIGURA 3A). Todos
os genes estudados foram demonstrados e ajustados em relação ao grupo controle
(normalizados arbritariamente a 1).
Indiam Hedgehog (Ihh) e Sonic Hedgheog (Shh) são ligantes da via Hh que
existem na forma intracellular e secretória e são responsáveis pela sinalização
parácrina da via Hh no intestino84
. Os níveis de RNAm do gene IHH e RNAm do
gene SHH aumentam significantemente após a exposição ao peptídio rShh ou ao
agonista da via, a purmorfamina (p<0,02) e reduzem após o tratamento das células
HT-29 com o antagonista da via Hh, a ciclopamina (p<0,02) (FIGURA 3 B).
Em relação aos fatores de transcrição Gli da via Hh, após tratamento das
células HT-29 com o peptídio rShh ou a purmorfamina observa-se um aumento
significativo dos níveis de RNAm dos genes GLI1 e GLI2 (p <0,02), que são os
principais ativadores dos genes alvos e indicadores da ativação da via Hh85. Níveis
de RNAm do gene GLI1 também aumentam após o tratamento das células HT-29
com butirato (ácido graxo de cadeia curta) (p<0,04), mas reduzem quando expostos
a ciclopamina (p<0,02) ou EGF (p<0,02). Níveis de RNAm do gene GLI3, que atua
como repressor da via Hh12
, não mudam significativamente, independente do
estímulo (FIGURA 3C).

27
A via Hh é regulada por receptores transmembrana Ptch e Smo. Para a
ativação da via Hh, o Hh se liga ao Ptch liberando a repressão do Ptch sobre o Smo,
que se encontrava bloqueado86. Neste trabalho observamos que a expressão do
RNAm do gene PTCH1 aumenta com a exposição das células HT-29 ao rShh ou a
purmorfamina (p<0,02), mas os níveis reduzem significativamente após tratamento
com EGF (p<0,02). Os níveis de Smo não mudam significativamente nas células HT-
29 independente do tratamento (FIGURA 3D).
A expressão do WNT 1, componente da via de sinalização Wnt que
antagoniza a via Hh e que é expresso na maioria dos cânceres de cólon, diminui
significativamente após o tratamento das células HT-29 com o peptídio rShh, butirato
ou EGF (p<0,02), enquanto aumenta com a exposição ao IFN-γ (p<0,04).
A expressão do RNAm do gene HHIP, que é um gene alvo e um potente
inibidor da via Hh 77,87 também foi avaliado e foi visto que sua expressão reduz
significativamente após o tratamento das células HT-29 com butirato (p<0,04) ou
EGF (p<0,02) e aumenta após o tratamento com purmorfamina (p<0.02) (FIGURA
3E).
Proteínas morfogênicas ósseas são alvos da via Hh durante o
desenvolvimento88, e são conhecidas em contribuir na manutenção do nicho das
células precursoras intestinais89, sendo assim investigamos a possibilidade da
expressão epitelial de BMP nas células HT-29. O RNAm do gene BMP4 é conhecido
por estar presente nas células epiteliais intestinais e ser alvo da via Hh90 e da via
Wnt10 e BMP7 também é expresso no cólon, mas menos relacionado com o
epitélio91. Observamos que a expressão do BMP4 e do BMP7 diminuem
significativamente após o tratamento com IFN γ (p<0,04) ou EGF (p<0,02). A
exposição a ciclopamina sozinha também diminui BMP4 e BMP 7 (p<0,02) e seus
níveis são restaurados quando se adiciona o peptídio rShh, mas não com a
purmorfamina (p<0,05) (FIGURA 3F). Nenhum dos genes analizados foram
alterados após exposição das células HT-29 ao LPS.

28
Figura 3 Modulação gênica das células HT 29 após exposição a diferentes estímulos Mapa de calor representando a expressão gênica através de RT-qPCR. As células HT-29 foram tratadas com Shh (Sonic Hedgehog), Purm (Purmorfamina), But (butirato), LPS, IFN-γ, EGF, com ou sem adição de Cyc (ciclopamina), ou DMSO (diluente) por 24 horas (A). Histogramas de expressão gênica individual mudam nas HT-29: IHH e SHH (B); GLI1, GLI2 e GLI3 (C); SMO e PtcH (D); HHIP e Wnt (E); e BMP4 e BMP7 (F). Os gráficos usam as mesmas amostras usadas no A. Os valores representam as médias dos resultados de três experimentos independentes e foram normalizados pelo RNA mensageiro do gene GAPDH. Mudanças significantes em relação o grupo controle são destacadas.

29
5.2. Níveis subcelulares e distribuíção do Gli1 e β-catenina nas células HT-29
Para confirmar os achados encontrados, realizamos a imunofluorescência do
Gli1 com a análise de microscopia confocal. A proteína Gli1, fator de transcrição da
via Hh, encontrava-se presente em baixos níveis nas céulas HT-29, predominando
no citosol. Quando as células HT-29 eram expostas ao peptídio rShh, purmorfamina
(agonista Hh) ou butirato, os níveis de Gli1 aumentaram com uma relativa marcação
de distribuição subcelular para o núcleo. Um efeito oposto foi demonstrado com a
adição do antagonsita Hh, ciclopamina (FIGURA 4A). Como Gli1 é um dos
indicadores da ativação da via Hh, o aumento da expressão e a translocação nuclear
indicam que a via Hh está ativa e pode ser regulada nas células HT-29.
Considerando que a via Hh é conhecida por regular negativamente a via Wnt no
epitélio colônico10, estudamos possíveis interações entre a sinalização Wnt e a
expressão ou atividade da via Hh nas céulas HT-29. Através da microscopia
confocal, analisamos a expressão da β-catenina, um componente integral da via
Wnt, predominando no núcleo das céulas HT-29. A exposição dessas células ao
peptídio rShh durante 24 horas reduz a expressão da β-catenina e a restringe a
forma ligada a membrana (FIGURA 4B).
A análise densitométrica da densidade das colorações confirmaram que os
níveis de Gli1 foram significativamente maiores após exposição das células HT-29
tratadas com o peptídeo rShh, purmorfamina ou butirado, comparado aos diluentes
(*P<0,04) ou a ciclopamina (**P<0,02). A análise da coloração com β-catenina se
tornou significativamente menor após o tratamento da célula HT-29 com o peptideo
rShh (▲P<0,01) comparado com o diluente ou com a ciclopamina (FIGURA 4C).
Assim, concluímos que a sinalização Hh regula negativamente a sinalização Wnt nas
células HT-29 in vitro.

30
Figura 4 Distribuição e níveis de Gli1 e β-catenina nas células HT-29 Microscopia confocal de reações de imunofluorescência nas culturas de células HT-29 expostas a vários estímulos por 24 horas, demonstraram a distribuição nuclear e citoplasmática e níveis de Gli 1 (A) e β-catenina (B). Análises de densidade das colorações protêicas confirmaram que Gli 1 aumenta significativamente após tratamento com Shh (Sonic Hedgehog), Purm (purmorfamina) ou But (Butirato), comparado as células tratadas com DMSO (diluente)(*P<0,04), ou Ciclopamina (Cyc) (**P<0,02). A densidade da β-catenina reduz significativamente após o tratamento das células HT-29 com Shh (▲P<0,01) quando comparado ao DMSO ou cyc (C).O núcleo foi corado com DAPI (azul). Painel micrográfico foi representativo em 3-4 experimentos em cada condição (magnificação original X1000).

31
5.3. Efeitos da via Hh na atividade inflamatória das células HT-29
Para elucidar os efeitos funcionais da via Hh nas células HT-29, foram
analisadas a concentração das citocinas no sobrenadante das células HT-29 através
do método de ELISA. As células HT-29 foram colocadas em cultura na concentração
de 1x106/ml por poço e submetidas a diferentes estímulos como a exposição ao
peptídeo rShh, purmorfamina, butirato, LPS, IFN-γ, EGF, com ou sem adição de
ciclopamina ou DMSO (diluente- controle) por 24 horas.
Foram analisadas a interleucina 8 (IL-8) e a proteína quimiotática de
monócitos 1 (MCP1). Estes são mediadores inflamatórios, conhecidos por serem
produzidos por células epiteliais e atuarem principalmente como moléculas
quimioatrativas para leucócitos e monócitos92.
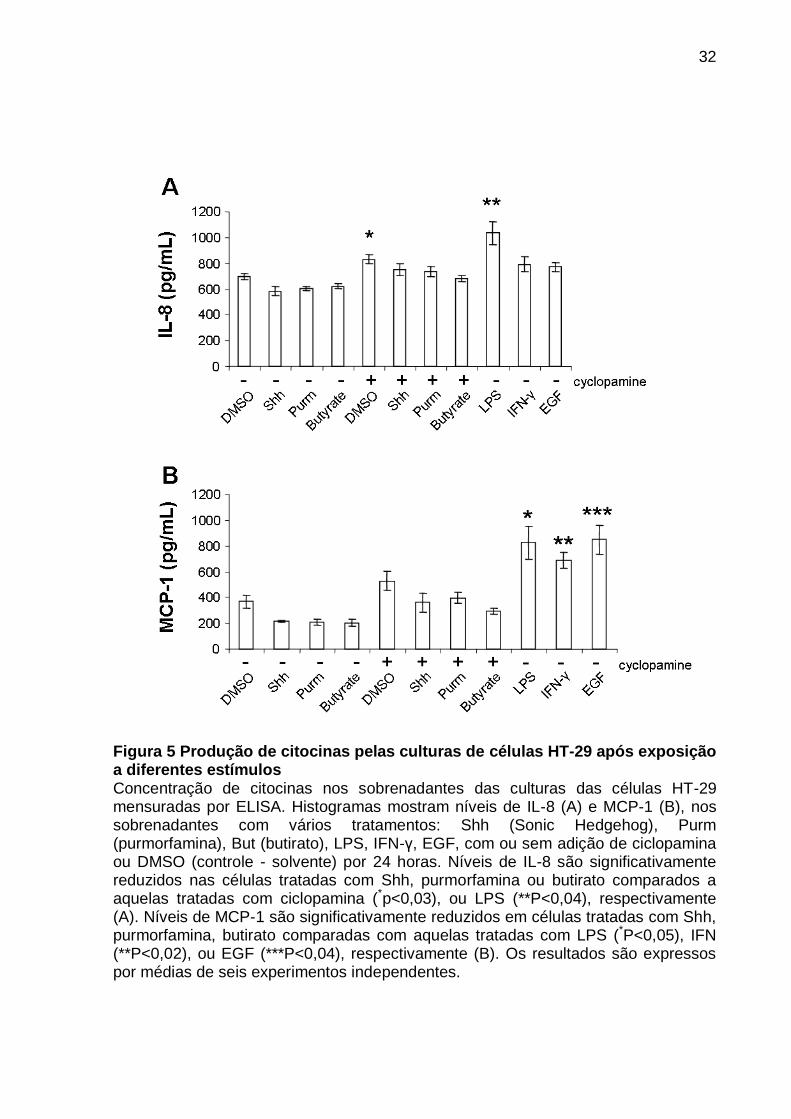
Os níveis de IL-8 foram significativamente reduzidos nas células HT-29
tratadas com peptídeo rShh, purmorfamina ou butirado comparado a aqueles
tratados com ciclopamina (*P<0,03) ou LPS (**P<0,04) respectivamente (FIGURA
5A). A produção de MCP1 diminui significativamente nas células tratadas com rShh,
purmorfamina ou butirato, quando comparado a aquelas expostas a LPS (*P<0,05),
INF-γ (**P<0,02) ou EGF (***P<0,04) respectivamente (FIGURA 5B).
As informações foram coletadas em 6 experimentos independentes.
32
Figura 5 Produção de citocinas pelas culturas de células HT-29 após exposição a diferentes estímulos Concentração de citocinas nos sobrenadantes das culturas das células HT-29 mensuradas por ELISA. Histogramas mostram níveis de IL-8 (A) e MCP-1 (B), nos sobrenadantes com vários tratamentos: Shh (Sonic Hedgehog), Purm (purmorfamina), But (butirato), LPS, IFN-γ, EGF, com ou sem adição de ciclopamina ou DMSO (controle - solvente) por 24 horas. Níveis de IL-8 são significativamente reduzidos nas células tratadas com Shh, purmorfamina ou butirato comparados a aquelas tratadas com ciclopamina (*p<0,03), ou LPS (**P<0,04), respectivamente (A). Níveis de MCP-1 são significativamente reduzidos em células tratadas com Shh, purmorfamina, butirato comparadas com aquelas tratadas com LPS (*P<0,05), IFN (**P<0,02), ou EGF (***P<0,04), respectivamente (B). Os resultados são expressos por médias de seis experimentos independentes.

33
5.4. Efeito da via Hh na viabilidade e na atividade proliferativa das células HT-
29
A viabilidade das células HT-29 foram analisadas em diferentes tempos e com
tratamentos diferentes, usando o método MTT. Embora não tenha sido observada
diferença nas células HT-29 tratadas nas primeiras 24 horas, a viabilidade celular
reduziu significativamente em 48 horas quando as células foram expostas a
ciclopamina (*P<0,04) (FIGURA 6A).
Com o propósito de analisar as mudanças da atividade proliferativa das
células HT-29 sob diferentes tratamentos, a incorporação celular do BrdU foi
analisada em 48 e 72 horas. Embora não tenha sido observada diferença
significativa entre os grupos tratados nas primeiras 48 horas, foram identificados
uma tendência para o aumento na incorporação do BrdU no grupo tratado com
ciclopamina. Contudo uma significativa redução na incorporação do BrdU foi
observada no grupo de células HT-29 expostas a ciclopamina de 48 para 72 horas
(**P<0,05). Após 72 horas, a incorporação do BrdU manteve-se nas células tratadas
com DMSO (diluente), enquanto que se tornou significativamente menor nos outros
grupos tratados (*P<0,05) (FIGURA 6B).
Todas as informações do grupo de MTT e de BrdU foram avaliadas com a
média de 3 experimentos independentes.

34
Figura 6 Viabilidade e atividade proliferativa das células HT-29 após exposição a diferentes estímulos A viabilidade das células HT-29 foi analisada em diferentes tempos, com diferentes tratamentos, usando o método MTT. Quando as células HT-29 foram expostas a ciclopamina, a viabilidade celular reduziu significativamente comparando 48 com 24 horas (*P<0,04) (A). Com o propósito de analisar as mudanças na atividade proliferativa nas células HT-29 sob diferentes tratamentos, a incorporação celular do BrdU foi avaliada em 48 e 72 horas. Após 72 horas, a incorporação do BrdU foi mantida nas células tratadas com o solvente (DMSO), enquanto ele significativamente reduz em todos os outros grupos de tratamento (*P<0,05). No grupo das células HT-29 expostas a ciclopamina, a incorporação de BrdU reduziu significativamente após 48 e 72 horas (**P<0,05) (B). Os resultados são expressos por médias de três experimentos independentes.

35
5.5. Efeito da via Hh na apoptose mediada pelo anti-CD95 nas células HT-29
Demonstração por citometria de fluxo da apoptose das células HT-29 após 24
horas expostas a diferentes estímulos, através da annexina V/7AA. Na figura 6,
observamos no quadrante inferior esquerdo células duplamente negativas, enquanto
que no quadrante inferior direito observamos células positivas para annexina V com
apoptose precoce e nos quadrantes superiores notamos células necróticas ou com
apoptose tardia. (FIGURA 7A)
Tratamento com peptideo rShh, purmorfamina ou butirato significativamente
anulam a apoptose induzida pelo anti-CD95 (anti Fas)(*P<0,01), que é parcialmente
restaurada pela adição de ciclopamina (FIGURA 7B).
Os dados foram analisados em 3 experimentos independentes.

36
Figura 7 Relação entre apoptose e a atividade da via Hh nas células HT-29 A demonstração da apoptose das células HT-29 por citometria de fluxo após 24 horas de exposição a diferentes estímulos foi avaliada pela reação de annexina-V/7-AAD. O quadrante inferior esquerdo indica células duplamente negativas, enquanto que o quadrante inferior direito indica células positivas para annexina-V (A). Tratamento com Shh, purmorfamina ou butirato significativamente impedem a apoptose induzida pelo anti-CD95 (*P<0,01), fato que é parcialmente restabelecido com a adição de ciclopamina (B). Os resultados são expressos por médias de três experimentos independentes.

37
DISCUSSÃO

38
6. DISCUSSÃO
Neste trabalho, demonstramos que a linhagem de células de carcinoma de
cólon em humanos, HT-29, expressam os componentes da via Hh, que podem ser
modulados por diferentes estímulos exógenos. Também demonstramos que o papel
biológico da via Hh nas células HT-29 envolve desde o controle da sobrevida das
células até a regulação de produção de citocinas. A via Hh tem sido demonstrada
em células e tecidos fetais, enquanto que no TGI de adutos, os componentes Hh são
frequentemente encontrados na camada de células epitelais sob certas condições
fisiológicas10,66. A via Hh parece também estar reativada em certos tipos de
cânceres13,93, nos quais a presença dos componentes Hh sugerem a participação da
via em mecanismos que regulam tanto a proliferação quanto a apoptose94. Assim
sendo, tivemos o interesse de investigar a via Hh e sua participação no controle da
homeostasia na sobrevida e funcão das células HT-29.
As células HT-29 tem sido utilizadas como modelo de estudo de células
epiteliais e também de vias de câncer e para o desenvolvimento de novos métodos
terapêuticos. Nesse trabalho analisamos os níveis básicos e o potencial de
modulação dos componentes da via Hh nas células HT-29 após a exposição de
diferentes estímulos exógenos. A via Hh tem ativamente participado na
carcinogênese e progressão dos tumores gastrointestinais12,13,22. Não obstante, uma
maior expressão das moléculas de sinalização Hh também foi descrita no câncer de
cólon19,95. No nosso trabalho evidenciamos a expressão dos componentes da via Hh
em células HT-29, em diferentes níveis, tanto na forma de RNAm, quanto de
proteína.
A expressão e modulação de genes da via Hh e outros genes indiretamente
associados a via Hh foram investigados por qRT PCR das células HT-29.
Demonstramos a presença do IHH, SHH, os principais indicadores da ativação da
via Hh, GLI1 e GLI2 e o PTCH1, e a habilidade de modular os níveis de RNAm
desses genes após o tratamento das células HT-29 com antagonista da via
(ciclopamina), ou após a específica estimulação com peptídeo rShh ou a
purmorfamina (agonista Hh). Semelhante ao estudo de Van den Brink e
colaboradores, (2004)10, observamos que RNAm do gene GLI1 aumenta após o
tratamento das células HT-29 com butirato, enquanto que os níveis diminuem após a

39
inibição com ciclopamina ou a exposição a EGF, que é fator de crescimento
conhecido pela sua importante participação na regeneração epitelial, ativação de
vias que aumentam a sobrevida nas células epiteliais intestinais96. É possível que a
via Hh esteja regulada negativamente ou seja ultrapassada por uma via alternativa
coordenada por sinais derivados do EGF. A exposição a produtos bacterianos ou
citocinas inflamatórias, apresenta impacto mínimo ou não causam influência na
modulação dos genes da via Hh nas células HT-29. Porém quando investigamos
proteínas morfogênicas de osso (BMP), observamos uma marcante regulação
negativa de BMP4 e BMP7 após a inibição da via Hh, mas os níveis são restaurados
com a adição do peptídio rShh às culturas de células HT-29. BMPs são morfógenos
da superfamília do TGF β, que demonstraram ter papel crítico na proliferação
epitelial e diferenciação terminal e maturação das células da linhagem secretória do
intestino89. Além disso, as BMPs são conhecidos alvos da via Hh durante o
desenvolvimento88, e as proteínas BMP4 e BMP7 foram identificados como
potenciais mediadores da interação epitélio-mesenquimal em resposta a sinalização
Hh no intestino de camundongos90. Assim, confirmamos aqui há expressão de BMPs
e a modulação pela sinalização Hh nas células HT-29 colônicas diferenciadas.
Demonstramos também que níveis de RNAm dos genes BMP4 e BMP7 reduzem
após a exposição de IFN-γ, provavelmente indicando uma regulação negativa em
resposta a sinais inflamatórios. A expressão WNT1, um antagonista da via Hh nas
células epiteliais, diminui acentuadamente após a exposição das células HT-29 ao
peptídio rShh e butirato, enquanto aumenta após a exposição ao IFN-γ. Os
componentes da via Wnt/β-catenina são conhecidos por estarem muito expressos na
maioria dos cânceres de cólon e nas células HT-29. Sua presença embasa o estado
hiperproliferativo, que parece estar neutralizado pela cascata da via de sinalização
Hh de forma parácrina e autócrina87. A resposta das células HT-29 ao IFNγ pode
indicar uma modulação epigenética da via Wnt por estímulo pró-inflamatório e pode
refletir uma conexão potencial entre inflamação e oncogênese nas células epiteliais
colônicas. Esses resultados, juntos, parecem revelar uma complexa interação de
múltiplos genes e vias de sinalização convergentes, envolvidos em processos
fisiológicos, como câncer e inflamação, que podem ser moduladas pela via Hh nas
células HT-29.

40
Nas células HT-29, a expressão básica da proteína Gli1 foi baixa, mas
consistentemente aumentada tanto no citoplasma, quanto no núcleo, após a
exposição do peptídeo rShh, purmorfamina, e butirato, enquanto os níveis de Gli1
reduzem e desaparecem do núcleo após tratamento com ciclopamina. Em contraste
aos níveis altos constitutivos no carcinoma de estômago14 e mama97, a ativação Hh
pode ser modulada por estímulo exógeno nas células HT-29. Em paralelo,
demonstramos a expressão constitutiva baixa da β-catenina, membro da via Wnt nas
células HT-29. De forma adicional observamos um efeito oposto ao mesmo estímulo
exógeno na β-catenina comparado ao Gli1, em termos de níveis de proteína
subcelular e distribuição nas células HT-29. Já é bem estabelecido que a regulacão
dos componentes da via β-catenina/Wnt leva ao acúmulo nuclear de β-catenina e a
formação de tumor98. Contudo a ativação da β-catenina não é exclusivamente
relacionada às mutações dos componentes da via Wnt99, e também é induzida por
sinais pró-inflamatórios100. De acordo com um estudo prévio sugerindo a existência
de interações entre a via Wnt e a atividade Hh no epitélio colônico10, no nosso
estudo confirmamos a idéia de que a via Hh regula negativamente a via Wnt nas
células HT-29. Em particular, a exposição a ciclopamina resulta em acúmulo nuclear
de β-catenina, enquanto que a ativação de Hh leva a redução de β-catenina e a uma
predominante localização periférica citoplasmática. Esses achados parecem indicar
que a indução Hh nas células epiteliais na superfície vilosa intestinal provavelmente
exerce uma regulação negativa sobre a sinalização Wnt nas células basais das
criptas intestinais, protegendo as células terminalmente diferenciadas da ação
proliferativa Wnt38.
Para complementar, a superfície epitelial intestinal atua na absorção, na
barreira mecânica que separa o meio externo das células internas e essas células
estão envolvidas em processos inflamatórios, assim como são fontes de
citocinas101,102. Citocinas como se sabe são mediadores de inflamação, mas também
estão relacionados a atração quimiotática direta e migração de células metastáticas
para vasos sanguíneos103 e órgãos distantes104. No estudo, demonstramos que
níveis de IL-8, que é um potente quimiotático e agente ativador de neutrófilos da
subfamília de citocinas CXC92, diminuem após a ativação da via Hh, ao contrário do
bloqueio da via Hh ou a indução por LPS. De forma similar, MCP-1, citocina da
subfamília CC com atividade quimiotática para monócitos e outras células imunes105,

41
diminui após a ativação Hh, em oposição ao efeito do LPS, IFN-γ e EGF. Nossos
achados parecem corroborar a importância das células epiteliais colônicas como
elementos-chave na resposta a infecção e agressões 106 e a sinais das células
imunes da lâmina própria107. A secreção de citocinas resultante das células epiteliais
provavelmente constitui um sinal precoce nas reações imunes e inflamatórias e
podem representar a conexão entre a reposta imune inata e adaptativa, no intestino.
Esses dados sugerem que a modulação Hh na geração de citocinas por células
epiteliais podem afetar a gravidade e o potencial metastático do câncer colorretal.
O epitélio intestinal passa por um processo de renovação celular contínua,
envolvendo proliferação, diferenciação e morte celular das células precursoras do
epitélio intestinal, em um processo que requer modulação por um complexo
microambiente108. Neste contexto, cascatas de sinalização, incluindo as vias Wnt e
Hh que regulam a renovação de células precursoras normais, estão implicadas
também na carcinogênese6,9,87. As informações sobre a via Hh no câncer
colorretal13,18,93 e na linhagem de células cancerígenas são controversos10,16,23. Tem
sido proposto que estas discrepâncias podem ocorrer, em parte, por metodologias
distintas, análise de diferentes amostras patológicas e também por modificações
epigenéticas na expressão gênica. Neste estudo, identificamos uma tendência
precoce através da incorporação do BrdU, seguida por uma tardia redução
consistente na incorporação do BrdU após a inibição do Hh. Em um experimento
complementar, demonstramos a redução na viabilidade das células HT-29 também
após a inibição da via Hh com ciclopamina, semelhante a estudo prévio51. Para
entender o mecanismo de integração entre a via Hh e a desfecho das células HT-29,
estimulamos as células via sistema Fas/FasL para induzir a morte celular por
apoptose. Semelhante a outros estudos95 demonstramos que os agonistas Hh são
capazes de retardar a apoptose em linhagem de células de câncer de intestino.
Assim, em contraste a outro estudo22, propomos que a via Hh pode não ser
essencial para a proliferação de células de câncer de cólon muito diferenciadas
como as HT-29. Todavia, a viabilidade celular demonstrou ser dependente da via
Hh, corroborando o conceito que a via Hh ativa no epitélio colônico pode estar
envolvida na diferenciação das células e também na proteção contra apoptose.

42
CONCLUSÃO

43
7. CONCLUSÃO
Como relatado, existem estudos contraditórios quanto a participação da via
Hh nas células HT-29, então nosso estudo demonstrou que realmente a linhagem de
células HT-29 expressa o RNAm e a proteína Gli em níveis diferentes dos principais
componentes da via Hh e que estes podem ser modulados in vitro, como visto nas
reações de PCR e por imunofluorescência. Assim como genes (BMP) envolvidos na
proliferação e diferenciação epitelial e potenciais mediadores na transição epitélio-
mesenquimal são modulados por proteínas da via Hh e seus inibidores. Nossos
resultados parecem corroborar com os dados que sugerem que a indução da via Hh
nas células epiteliais na superfície vilosa intestinal exerce uma regulação negativa
sobre a sinalização Wnt das células basais das criptas intestinais. Concordamos que
a via Hh é uma via chave no controle da função das células epiteliais colônicas. A
investigação da atividade biológica da via Hh nas células HT-29 demonstrou seu
efeito em promover a redução dos sinais inflamatórios (células HT-29 moduladas
com agonistas da via Hh reduzem a produção de citocinas IL-8 e MCP-1) e
antagonizar a apoptose (visto nas reações Annexina Y/7-AAD anti CD95). A
atividade Hh nas células colônicas tem prováveis implicações homeostáticas no
desenvolvimento de inflamação colônica e malignidade, já que a expressão
diferencial dos componentes da via Hh podem causar anormalidades na reposta
imune local e na integridade da barreira epitelial. Esses resultados, juntos, ressaltam
a importância da ativação da via Hh como potencial alvo terapêutico em doença
inflamatória intestinal e câncer colorretal.
Propostas futuras: temos interesse de estudar melhor a transição epitélio-
mesenquimal, assim como estudos da via Hh em células de cólon humano normal e
com doença inflamatória intestinal para elucidar de forma mais detalhada o papel da
via Hh.

44
REFERÊNCIAS BIBLIOGRÁFICAS

45
8. REFERÊNCIAS BIBLIOGRÁFICAS
1. de Santa Barbara P, van den Brink GR, Roberts DJ. Development and differentiation of the intestinal epithelium. Cell Mol Life Sci 2003;60:1322-32.2.
2. Sansonetti PJ. War and peace at mucosal surfaces. Nat Rev Innumol 2004; 4:953-64
3. van den Brink GR, Hardwick JC. Hedgehog Wnteraction in colorectal cancer. Gut 2006;55:912-4.
4. Cho JH. The genetics and immunopathogenesis of inflammatory bowel disease. Nat Rev Immunol 2008;8:458-66.
5. Yu M, Gipp J, Yoon JW, Iannaccone P, Walterhouse D, Bushman W. Sonic hedgehog-responsive genes in the fetal prostate. J Biol Chem 2009;284:5620-9.
6. Kasper M, Regl G, Frischauf AM, Aberger F. GLI transcription factors: mediators of oncogenic Hedgehog signalling. Eur J Cancer 2006;42:437-45.
7. Lum L, Beachy PA. The Hedgehog response network: sensors, switches, and routers.Science 2004;304:1755-9.
8. Hooper JE, Scott MP. Communicating with Hedgehogs. Nat Rev Mol Cell Biol 2005;6:306-17.
9. Ruiz I Altaba A, Mas C, Stecca B. The Gli code: an information nexus regulating cell fate, stemness and cancer. Trends Cell Biol 2007;17:438-47
10. van den Brink GR, Bleuming SA, Hardwick JC, Schepman BL, Offerhaus GJ, Keller JJ, Nielsen C, Gaffield W, van Deventer SJ, Roberts DJ, Peppelenbosch MP. Indian Hedgehog is an antagonist of Wnt signaling in colonic epithelial cell differentiation. Nat Genet 2004;36:277-82
11. Alinger B, Kiesslich T, Datz C, Aberger F, Strasser F, Berr F, Dietze O, Kaserer K, Hauser-Kronberger C. Hedgehog signaling is involved in differentiation of normal colonic tissue rather than in tumor proliferation. Virchows Arch 2009;454:369-79.
12. Katoh Y, Katoh M. WNT antagonist, SFRP1, is Hedgehog signaling target. Int J Mol Med 2006;17:171-5. 14.
13. Berman DM, Karhadkar SS, Maitra A, Montes De Oca R, Gerstenblith MR, Briggs K, Parker AR, Shimada Y, Eshleman JR, Watkins DN, Beachy PA. Widespread requirement for Hedgehog ligand stimulation in growth of digestive tract tumours. Nature 2003;425:846-51.
14. Ohta M, Tateishi K, Kanai F, Watabe H, Kondo S, Guleng B, Tanaka Y, Asaoka Y, Jazag A, Imamura J, Ijichi H, Ikenoue T, Sata M, Miyagishi M, Taira K, Tada M, Kawabe T, Omata M. p53-Independent negative regulation of p21/cyclin-dependent

46
kinase-interacting protein 1 by the sonic hedgehog-glioma-associated oncogene 1 pathway in gastric carcinoma cells. Cancer Res 2005;65:10822-9.
15. Bienz M, Clevers H. Linking colorectal cancer to Wnt signaling. Cell 2000;103:311-20.
16. Varnat F, Duquet A, Malerba M, Zbinden M, Mas C, Gervaz P, Ruiz i Altaba A. Human colon cancer epithelial cells harbour active HEDGEHOG-GLI signalling that is essential for tumour growth, recurrence, metastasis and stem cell survival and expansion. EMBO Mol Med 2009;1:338-51.
17. Bian YH, Huang SH, Yang L, Ma XL, Xie JW, Zhang HW. Sonic hedgehog-Gli1 pathway in colorectal adenocarcinomas. World J Gastroenterol 2007;13:1659-65.
18. Yoshikawa K, Shimada M, Miyamoto H, Higashijima J, Miyatani T, Nishioka M, Kurita N, Iwata T, Uehara H. Sonic hedgehog relates to colorectal carcinogenesis. J Gastroenterol 2009;44:1113-7.
19. You S, Zhou J, Chen S, Zhou P, Lv J, Han X, Sun Y. PTCH1, a receptor of Hedgehog signaling pathway, is correlated with metastatic potential of colorectal cancer. Ups J Med Sci;115:169-75.
20. Pisani P, Bray F, Parkin DM. Estimates of the world-wide prevalence of câncer for 25 sites in the adult population. Int J Cancer. 2002;97:72-81).
21. Benson AB. Epidemiology, disease progressio, and economic burden of colorectal cancer. J Manag, care Pharm. 2007;13(6 SupplC):5-18
22. Douard R, Moutereau S, Pernet P, Chimingqi M, Allory Y, Manivet P, Conti M, Vaubourdolle M, Cugnenc PH, Loric S. Sonic Hedgehog-dependent proliferation in a series of patients with colorectal cancer. Surgery 2006;139:665-70.
23. Chatel G, Ganeff C, Boussif N, Delacroix L, Briquet A, Nolens G, Winkler R. Hedgehog signaling pathway is inactive in colorectal cancer cell lines. Int J Cancer 2007;121:2622-7.
24. Bailey J, Singh PK and Hollingsworth MA: Cancer metastasis facilitated by developmental pathways: Sonic hedgehog, Notch, and bone morphogenetic proteins. J Cell Biochem 2007; 102: 829-839
25. Katoh M: Networking of WNT, FGF, Notch, BMP, and Hedgehog signaling pathways during carcinogenesis.Stem Cell Rev 2007; 3: 30-38
26. Hammerschmidt M, Brook A, McMahon AP. The world according to hedgehog. Trends Genet 1997;13:14-21.
27. Nusslein-Volhard C, Wieschaus E. Mutations affecting segment number and polarity in Drosophila. Nature 1980;287:795-801.
28. Lee JJ, Ekker SC, von Kessler DP, Porter JA, Sun BI, Beachy PA. Autoproteolysis in hedgehog protein biogenesis. Science 1994;266:1528-1537

47
29. Chamoun Z, Mann RK, Nellen D, et al: Skinny hedgehog, an acyltransferase required for palmitoylation and activity of the hedgehog signal. Science 2001; 293: 2080-2084
30. Pepinsky RB, Zeng C, Wen D, Rayhorn P, Baker DP, Williams KP, Bixler SA, Ambrose CM, Garber EA, Miatkowski K, Taylor FR, Wang EA, Galdes A. Identification of a palmitic acid-modified form of human Sonic hedgehog. J Biol Chem 1998;273:14037-14045
31. Van den Heuvel M and Ingham PW: Smoothened encodes a receptor-like serpentine protein required for hedgehog signalling. Nature 1996; 382: 547-551
32. Murone M, Rosenthal A, de Sauvage FJ. Hedgehog signal transduction: from flies to vertebrates. Exp Cell Res 1999;253:25-33.
33. Theil T, Kaesler S, Grotewold L, Bose J, Ruther U. Gli genes and limb development. Cell Tissue Res 1999;296:75-83.
34. Pasca di Magliano M and Hebrok M: Hedgehog signalling in cancer formation and maintenance. Nat Rev Cancer 2003; 3: 903-911
35. Bijlsma MF, Spek CA and Peppelenbosch MP: Hedgehog: an unusual signal transducer. Bioessays 2004; 26: 387-394
36. Kinzler KW, Bigner SH, Bigner DD, et al: Identification of an amplified, highly expressed gene in a human glioma. Science 1987; 236: 70-73
37. Price MA: CKI, there's more than one: casein kinase I family members in Wnt and Hedgehog signaling. Genes Dev 2006; 20: 399-410
38. Katoh Y, Katoh M. Hedgehog signaling pathway and gastrointestinal stem cell signaling network (review). Int J Mol Med 2006;18:1019-23.
39. Jiang J and Struhl G: Regulation of the Hedgehog and Wingless signaling pathways by the F-box/WD40-repeat protein Slimb. Nature 1998; 391: 493-496
40. Koike J, Sagara N, Kirikoshi H, et al: Molecular cloning and characterization of the ßTRCP2 gene on chromosome 5q35.1.Biochem Biophys Res Commun 2000; 269: 103-109
41. Osterlund T, Everman DB, Betz RC, et al: The FU gene and its possible protein isoforms. BMC Genomics 2004; 5: 49
42. Taylor MD, Liu L, Raffel C, et al: Mutations in SUFU predispose to medulloblastoma. Nat Genet 2002; 31: 306-310
43. Svard J, Heby-Henricson K, Persson-Lek M, Rozell B, Lauth M, Bergstrom A, Ericson J, Toftgard R, Teglund S. Genetic elimination of Suppressor of fused reveals an essential repressor function in the mammalian Hedgehog signaling pathway. Dev Cell 2006;10:187–97

48
44. Park HL, Bai C, Platt KA, Matise MP, Beeghly A, Hui CC, Nakashima M, Joyner AL. Mouse Gli1 mutants are viable but have defects in SHH signaling in combination with a Gli2 mutation. Development 2000;127:1593-1605
45. Ikran MS, Neill GW, Regl G, Eichberger T, Frischauf AM, Aberger F, Quinn A, Philpott M. Gli2 is expressed in normal human epidermis and BCC and induces Gli1 expression by binding to its promoter. J Invest Dermatol 2004;122:1503-09
46. Yoon JW, Kita Y, Frank DJ, et al: Gene expression profiling leads to identification of GLI1-binding elements in target genes and a role for multiple downstream pathways in GLI1-induced cell transformation. J Biol Chem 2002; 277: 5548-5555
47. Hallikas O, Palin K, Sinjushina N, et al: Genome-wide prediction of mammalian enhancers based on analysis of transcription-factor binding affinity. Cell 2006; 124: 47-59
48. .Lehmann OJ, Sowden JC, Carlsson P, Fox’s in development and disease. Trends Genet 2003;19:338-11
49. Tenzen T, Allen BL, Cole F, et al: The cell surface membrane proteins Cdo and Boc are components and targets of the Hedgehog signaling pathway and feedback network in mice. Dev Cell 2006;10: 647-656
50. Taipale J, Chen JK, Cooper MK, Wang B, Mann RK, Milenkovic L,Scott MP, Beachy PA. Effects of oncogenic mutations in Smoothened and Patched can be reversed by cyclopamine.Nature 2000;406:1005–9.
51. Mazumdar T, DeVecchio J, Shi T, Jones J, Agyeman A, Houghton JA. Hedgehog signaling drives cellular survival in human colon carcinoma cells. Cancer Res 2011;71:1092-102
52. Chuang PT and McMahon AP: Vertebrate Hedgehog signaling modulated by induction of a Hedgehog-binding protein. Nature 1999; 397: 617-621
53. Katoh Y and Katoh M: Comparative genomics on HHIP family orthologs. Int J Mol Med 2006; 17: 391-395
54. Olsen CL, Hsu PP, Glienke J, Rubanyi GM, Brooks AR. Hedgehoginteracting protein is highly expressed in endothelial cells but downregulated during angiogenesis and in several human tumors. BMC Cancer 2004;4:43.
55. Fan H, Khavari PA. Sonic hedgehog opposes epithelial cell cycle arrest. J Cell Biol 1999;147:71-76.
56. Lowrey JA, Stewart GA, Lindey S, Hoyne GF, Dallman MJ, Howie SE, Lamb JR. Sonic hedgehog promotes cell cycle progression in activated peripheral CD4(+) T lymphocytes. J Immunol 2002;169:1869-1875.
57. Pola R, Ling LE, Silver M, Corbley MJ, Kearney M, Blake Pepinsky R, Shapiro R, Taylor FR, Baker DP, Asahara T, Isner JM. The morphogen Sonic hedgehog is an

49
indirect angiogenic agent upregulating two families of angiogenic growth factors. Nat Med 2001;7:706-711
58. Riobo NA, Lu K, Emerson CP Jr. Hedgehog signal transduction: signal integration and cross talk in development and cancer. Cell Cycle 2006;5:1612-1615.
59. YURIKO KATOH1 and MASARU KATOH Hedgehog signaling, epithelial-to-mesenchymal transition and miRNA (Review) Int Journal of Mol Med 2008; 22: 271-275
60. Thiery JP: Epithelial-mesenchymal transitions in tumour progression. Nature Rev Cancer 2002; 2: 442-454
61. Katoh M: Epithelial-mesenchymal transition in gastric cancer. Int J Oncol 2005 27: 1677-1683
62. Lee JM, Dedhar S, Kalluri R and Thompson EW: The epithelial-mesenchymal transition: new insights in signaling, development, and disease. J Cell Biol 2006; 172: 973-981
63. Sukegawa A, Narita T, Kameda T, Saitoh K, Nohno T, Iba H, Yasugi S, Fukuda K. Development. 2000; 127(9):1971-80
64. Nanni L, Ming JE, Bocian M et al (1999) The mutational spectrum of the sonic hedgehog gene in holoprosencephaly: SHH mutations cause a significant proportion of autosomal dominant holoprosencephaly. Hum Mol Genet 8:2479–2488.
65. Mao J, Kim BM, Rajurkar M, Shivdasani RA, McMahonAP. Hedgehog signaling controls mesenchymal growth in the developing mammalian digestive tract. Development. 2010; 137:1721-9)
66. Lees C, Howie S, Sartor RB, Satsangi J. The hedgehog signalling pathway in the gastrointestinal tract: implications for development, homeostasis, and disease. Gastroenterology 2005;129:1696-710
67. Dimmler A, Brabletz T, Hlubek F et al (2003) Transcription of sonic hedgehog, a potential factor for gastric morphogenesis and gastric mucosa maintenance, is up-regulated in acidic conditions.Lab Invest 83:1829–1837
68. Silberg DG, Kaestner KH (2001) Morphogenesis and maintenance of the gastric epithelium: a role for sonic hedgehog? Gastroenterology 121:485–487
69. Van den Brink GR, Hardwick JC, Tytgat GN, et al: Sonic hedgehog regulates gastric gland morphogenesis in man and mouse. Gastroenterology 2001; 121: 317-328
70. Katoh Y and Katoh M: Hedgehog signaling in gastric cancer.Cancer Biol Ther 2005; 4: 1050-1054
71. H Ohta1,4, K Aoyagi1, M Fukaya1, I Danjoh1, A Ohta1, N Isohata1, N Saeki1, H Taniguchi2, H Sakamoto1, T Shimoda3, T Tani4, T Yoshida1 and H Sasaki Cross

50
talk between hedgehog and epithelial–mesenchymal transition pathways in gastric pit cells and in diffuse-type gastric cancers British Journal of Cancer (2009) 100, 389 – 398
72. Katoh M: WNT and FGF gene clusters. Int J Oncol 2002; 21: 1269-1273
73. Gregorieff A, Pinto D, Begthel H, et al: Expression pattern of WNT signaling components in the adult intestine. Gastroenterology 2005; 129: 626-638
74. Katoh M: Bioinformatics for cancer management in the postgenome era. Technol Cancer Res Treat 2006; 5: 169-176
75. Evangelista M, Tian H, de Sauvage FJ. The hedgehog signaling pathway in cancer. Clin Cancer Res 2006;12:5924–8
76. Kinzler KW, Bigner SH, Bigner DD, et al: Identification of an amplified, highly expressed gene in a human glioma. Science 1987; 236: 70-73
77. Olsen CL, Hsu PP, Glienke J, et al: Hedgehog-interacting protein is highly expressed in endothelial cells, but down-regulated during angiogenesis and in several human tumors. BMC Cancer2004; 4: 43
78. Hu Z, Bonifas JM, Aragon G, Kopelovich L, Liang Y, Ohta S, Israel MA, Bickers DR, Aszterbaum M, Epstein EH, Jr. Evidence for lack of enhanced hedgehog target gene expression in common extracutaneous tumors. Cancer Res 2003;63:923–8
79. Zhu Y, James RM, Peter A, Lomas C, Cheung F, Harrison DJ, Bader SA. Functional Smoothened is required for expression of GLI3 in colorectal carcinoma cells. Cancer Lett 2004;207:205–14
80. Robert LY, Stephen EG, Suzie JS, Tracy T, Hua T, Cristina PA, Derek M, Ling Fu, Thomas J, Dara K, Michelle NP, Karen K, James CMJ, Lee LR, Frederic JS. A paracrine requirement for hedghog signalling in cancer. Nature. 2008; 455:406-10
81. Augeron, C. & Laboisse, C.L. Emergence of permanently differentiated cell clones in a human colonic cancer cell line in culture after treatment with sodium butyrate.Cancer Res1984;. 44, 3961–3969
82. Akiyoshi T, Nakamura M, Koga K, Nakashima H, Yao T, Tsuneyoshi M, Tanaka M, Katano M. Gli1, downregulated in colorectal cancers, inhibits proliferation of colon cancer cells involving Wnt signalling activation. Gut 2006;55:991–9
83. O’Brien CA, Pollett A, Gallinger S, Dick JE. A human colon cancer cell capable of initiating tumour growth in immunodeficient mice. Nature 2007;445:106–10.
84. Kolterud A, Grosse AS, Zacharias WJ, Walton KD, Kretovich KE, Madison BB, Waghray M, Ferris JE, Hu C, Merchant JL, Dlugosz AA, Kottmann AH, Gumucio DL. Paracrine Hedgehog signaling in stomach and intestine: new roles for hedgehog in gastrointestinal patterning. Gastroenterology 2009;137:618-28.

51
85. Bai J, Montell D. Eyes absent, a key repressor of polar cell fate during Drosophila oogenesis. Development 2002;129:5377-88
86. Kalderon D. Transducing the hedgehog signal. Cell 2000;103:371-4
87. Madison BB, Braunstein K, Kuizon E, Portman K, Qiao XT, Gumucio DL. Epithelial hedgehog signals pattern the intestinal crypt-villus axis. Development 2005;132:279-89.
88. Bitgood MJ, McMahon AP. Hedgehog and Bmp genes are coexpressed at many diverse sites of cell-cell interaction in the mouse embryo. Dev Biol 1995;172:126-38
89. Auclair BA, Benoit YD, Rivard N, Mishina Y, Perreault N. Bone morphogenetic protein signaling is essential for terminal differentiation of the intestinal secretory cell lineage. Gastroenterology 2007;133:887-96.
90. van Dop WA, Uhmann A, Wijgerde M, Sleddens-Linkels E, Heijmans J, Offerhaus GJ, van den Bergh Weerman MA, Boeckxstaens GE, Hommes DW, Hardwick JC, Hahn H, van den Brink GR. Depletion of the colonic epithelial precursor cell compartment upon conditional activation of the hedgehog pathway. Gastroenterology 2009;136:2195-203.
91. Kosinski C, Li VS, Chan AS, Zhang J, Ho C, Tsui WY, Chan TL, Mifflin RC, Powell DW, Yuen ST, Leung SY, Chen X. Gene expression patterns of human colon tops and basal crypts and BMP antagonists as intestinal stem cell niche factors. Proc Natl Acad Sci U S A 2007;104:15418-23.
92. Schall TJ, Bacon KB. Chemokines, leukocyte trafficking, and inflammation. Curr Opin Immunol 1994;6:865-73.
93. Oniscu A, James RM, Morris RG, Bader S, Malcomson RD, Harrison DJ. Expression of Sonic hedgehog pathway genes is altered in colonic neoplasia. J Pathol 2004;203:909-17.
94. Kobune M, Takimoto R, Murase K, Iyama S, Sato T, Kikuchi S, Kawano Y, Miyanishi K, Sato Y, Niitsu Y, Kato J. Drug resistance is dramatically restored by hedgehog inhibitors in CD34+ leukemic cells. Cancer Sci 2009;100:948-55.
95. Qualtrough D, Buda A, Gaffield W, Williams AC, Paraskeva C. Hedgehog signaling in colorectal tumour cells: induction of apoptosis with cyclopamine treatment. Int J Cancer 2004;110:831-7
96. Terzic J, Grivennikov S, Karin E, Karin M. Inflammation and colon cancer. Gastroenterology 2010;138:2101-14.
97. Kubo M, Nakamura M, Tasaki A, Yamanaka N, Nakashima H, Nomura M, Kuroki S, Katano M. Hedgehog signaling pathway is a new therapeutic target for patients with breast cancer. Cancer Res 2004;64:6071-4.

52
98. Kolligs FT, Bommer G, Goke B. Wnt/beta-catenin/tcf signaling: a critical pathway in gastrointestinal tumorigenesis. Digestion 2002;66:131-44.
99. Lu Z, Hunter T. Wnt-independent beta-catenin transactivation in tumor development. Cell Cycle 2004;3:571-3.
100. Oguma K, Oshima H, Aoki M, Uchio R, Naka K, Nakamura S, Hirao A, Saya H, Taketo MM, Oshima M. Activated macrophages promote Wnt signalling through tumour necrosis factor-alpha in gastric tumour cells. EMBO J 2008;27:1671-81.
101. Jung HC, Eckmann L, Yang SK, Panja A, Fierer J, Morzycka-Wroblewska E, Kagnoff MF. A distinct array of proinflammatory cytokines is expressed in human colon epithelial cells in response to bacterial invasion. J Clin Invest 1995;95:55-65.
102. Mazzucchelli L, Hauser C, Zgraggen K, Wagner HE, Hess MW, Laissue JA, Mueller C. Differential in situ expression of the genes encoding the chemokines MCP-1 and RANTES in human inflammatory bowel disease. J Pathol 1996;178:201-6.
103. Yu T, Wu Y, Helman JI, Wen Y, Wang C, Li L. CXCR4 promotes oral squamous cell carcinoma migration and invasion through inducing expression of MMP-9 and MMP-13 via the ERK signaling pathway. Mol Cancer Res 2011;9:161-72
104. Lu X, Kang Y. Chemokine (C-C motif) ligand 2 engages CCR2+ stromal cells of monocytic origin to promote breast cancer metastasis to lung and bone. J Biol Chem 2009;284:29087-96.
105. Rollins BJ. Chemokines. Blood 1997;90:909-28. 27 47.
106. Gewirtz AT, McCormick B, Neish AS, Petasis NA, Gronert K, Serhan CN, Madara JL. Pathogen-induced chemokine secretion from model intestinal epithelium is inhibited by lipoxin A4 analogs. J Clin Invest 1998;101:1860-9.
107. Kagnoff MF, Eckmann L. Epithelial cells as sensors for microbial infection. J Clin Invest 1997;100:6-10.
108. Walker RA, Martin CV. The aged breast. J Pathol 2007;211:232-40. 50.

53
ANEXOS

54
ANEXO 1
Title
Hedgehog Pathway Signaling Regulates Human Colon Carcinoma HT-29
Epithelial Cell Line Apoptosis and Cytokine Secretion.
Authors: 1,2Agnes N. Yoshimoto, 1Claudio Bernadazzi, 2Antonio José V.
Carneiro, 2Celeste C. S. Elia, 1Cesonia A. Martinusso, 3Grasiella M. Ventura,
4Morgana T. L. Castelo-Branco, 1,2Heitor S. P. de Souza
Affiliations: 1Laboratório Multidisciplinar de Pesquisa e 2Serviço de
Gastroenterologia, Departamento de Clínica Médica; 3Unidade de Microscopia
Confocal e 4Laboratório de Imunologia Celular, Instituto de Ciências Biomédicas;
Universidade Federal do Rio de Janeiro, Brazil.
Running title: Hedgehog pathway expression and function in HT-29 cells.
Keywords: Hedgehog pathway; HT-29 human colon carcinoma cell line;
chemokines; apoptosis; cell proliferation.
Corresponding author: Heitor S. P. de Souza, M.D., Laboratório
Multidisciplinar de Pesquisa (sub-solo), Hospital Universitário, Universidade Federal
do Rio de Janeiro. Rua Prof. Rodolpho Paulo Rocco 255, Ilha do Fundão, Rio de
Janeiro, RJ 21941-913, Brazil. Phone: 55 (21) 2562-2669, Fax: 55 (21) 2562-2669,
E-mail: [email protected]; [email protected]

55
Abstract
The Hedgehog (Hh) pathway is involved in embryogenesis and physiologic
processes including cell survival and proliferation. We used the human colon
carcinoma HT-29 cell line to investigate Hh signaling and biological functions in
colonic epithelial cells. HT-29 cells were cultured under different conditions and
exposed to various stimuli. The expression of Hh pathway components and related
genes and proteins were assessed by real-time PCR and immunofluorescence.
Viability, apoptosis and cell proliferation were measured by the MTT assay, Annexin-
V/7-AAD staining and BrdU uptake, respectively. Chemokines production was
measured by ELISA in culture supernatants. Indian and Sonic Hh mRNA levels and
the downstream transcription factors Gli-1 and Gli-2 increased following treatment
with Hh agonists and butyrate, but decreased upon exposure to cyclopamine. BMP4
and BMP7 expression decreased after Hh blockade with cyclopamine. Gli-1 protein
expression increased after Hh agonists and decreased following cyclopamine.
Exposure to Hh agonists promoted ß -catenin reduction and subcellular redistribution.
Levels of IL-8 and MCP-1 decreased upon exposure to Hh agonists compared
to cyclopamine, LPS, IFN-. or EGF. Cellular incorporation of BrdU and cell viability
decreased following Hh blockade. Hh agonists abrogated the anti-CD95 induced
apoptosis. Hh pathway is a key controller of colonic epithelial cells, as demonstrated
by its effect in dampening inflammatory signals and antagonizing apoptosis. The
differential expression of Hh components may underlie abnormalities in the local
immune response and in epithelial barrier integrity, with potential homeostatic
implications for the development of colonic inflammation and malignancies.

56
Introduction
An essential physiologic function of the intestine is the maintenance of a
protective barrier against potentially harmful gut luminal contents. The first line of
defense in this barrier is constituted by a single layer of epithelial cells that
continuously renew and interact with the lumen and the lamina propria immune cells
(1, 2). Control of cell renewal within the epithelial layer depends on a dynamic
equilibrium between cell proliferation and apoptosis, whereas imbalance between the
two mechanisms is known to result in cancer (3) and also in inflammatory conditions
(4).
The Hedgehog (Hh) pathway of morphogenic proteins is known to participate
in several physiologic cellular processes. Hh signaling has been associated with
embryogenesis (5), patterning of a variety of organs (6), adult tissue homeostasis
and repair (7), regulation of the epithelial-to-mesenchymal transition (8), and the
control of cell survival and proliferation (9). Activation of the canonical Hh signaling
pathway involves the binding of Hh ligands to the membrane receptor Patched
(PTCH1), resulting in the activation of the signaling cytoplasmic molecule
Smoothened (SMO). Once released from PTCH-mediated suppression, SMO
activates the Gli zinc finger transcription factors that ultimately regulate downstream
Hh target genes (6-8, 10).
The canonical Hh signaling cascade has also been shown to play a role in the
normal gastrointestinal development, where it regulates the differentiation of normal
intestinal villi (11), and the adjacent mesenchymal stromal cells (12). In the normal
adult gastrointestinal tract, induction of the Hh pathway appears to protect the
differentiated epithelial cells of the villous surface, counteracting the canonical Wnt
signaling in the basal cells of the crypt (13). Although aberrant activation of the Hh
pathway has been demonstrated in the oncogenesis of human esophageal, gastric,
and pancreatic cancers (14, 15), its role in colon cancer has not been sufficiently
clarified.
The majority of sporadic and hereditary human colorectal tumors are believed
to originate from constitutive activation of Wnt signaling by mutation of the APC or
ßcatenin genes (16), whereas the Hh pathway would oppose the proliferative effect
of Wnt pathway on differentiated colonocytes (13). Recently, there has been

57
evidence of the potential involvement of Hh signaling in cancer invasiveness (17), in
colonic carcinogenesis (18, 19), and in the metastatic dissemination of colorectal
cancer (20). Nevertheless, approaches to elucidate the role of the Hh pathway in
cancers are still limited, in particular in colon cancer where data have been
controversial (21, 22).
In order to investigate Hh signaling and its potential biological functions in
colonic epithelial cells, we used the epithelial HT-29 human colon carcinoma cell line
under various experimental conditions. We demonstrated the presence and
modulation of several Hh components and that Hh pathway activation has anti-
inflammatory and anti-apoptotic effects on HT-29 cells.

58
Materials and Methods
Cell cultures
Human colon adenocarcinoma cell line (HT-29) was obtained from the
American Type Culture Collection (ATCC, Rockville, MD, USA), and maintained
according to the ATCC’s instructions. Cells were grown as monolayers in Dulbecco’s
modified Eagle’s medium (DMEM) (Invitrogen Gibco, New York, NY, USA). Cells
were plated in 25 cm2 culture flasks with DMEM supplemented with heat inactivated
10% fetal bovine serum (FBS) (Invitrogen Gibco, New York, NY, USA), penicillin
(10,000 U/mL), streptomycin (100 µg /mL), 4.5 g/L glucose and L-glutamine, (all from
Sigma-Aldrich, St. Louis, MO, USA) at 37°C in a humidified atmosphere with 5%
CO2. After growth to confluence, cells were trypsinized and resuspended in DMEM
supplemented with 2.5% FBS. Part of the cells were frozen and maintained at -80oC.
Cells were then seeded at a density of 1×106 cells/mL and treated specifically for
each experiment in either 6-well, 24-well or 96-well microplates, as needed. Culture
supernatants were collected and maintained at -80oC for further chemokine
measurements. In parallel, cells were plated onto eight-chamber slides (Lab Tek
Nunc, Naperville, IL, USA) for immunofluorescence staining.
Reagents and treatments
Recombinant human N-Shh was purchased from R&D Systems (Minneapolis,
MN, USA), and used at 500 ng/mL; cyclopamine (Sigma-Aldrich, St. Louis, MO, USA)
was used as a selective chemical hedgehog inhibitor, and diluted in dimethyl
sulfoxide (DMSO) (control vehicle) (Merck, Darmstadt, Germany) at 10µM; chemical
hedgehog agonist purmorphamine (Alexis Biochemicals, Plymounth, PA, USA) was
diluted in DMSO at 2µM; sodium butyrate was added to cultures at 2.5 µM, while
interferon-gamma (IFN-γ.) and lipopolysaccharide (LPS) were used at 2 ng/mL (all
from Sigma-Aldrich, St. Louis, MO, USA); recombinant human epithelial growth factor
(rhEGF) was obtained from Peprotech (Rocky Hill, NJ, USA), and used at 20ng/mL.

59
Indirect immunofluorescence staining and confocal laser microscopy
Cells were seeded onto eight-chamber slides, fixed, permeabilized, and
incubated for 2h at room temperature with 2.5% bovine serum albumin (BSA), 2.0%
skimmed milk, 8.0% fetal bovine serum (FBS) blocking buffer under shaking. Slides
were rinsed once with PBS and 0.05% Tween 20 and then incubated with
appropriately diluted primary antibodies in PBS. Cells were incubated with anti-Gli-1
rabbit antibody (Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA), and anti-ß-
catenin mouse antibody (Millipore, Bedford, MA, USA) for 1 h at room temperature.
After incubation, the slides were rinsed three times and incubated with Alexa® 488
conjugated anti-rabbit IgG, and Alexa® 633 conjugated anti-mouse IgG (all from
Molecular Probes, Eugene, OR, USA) for 30 min. at room temperature. Sections from
each sample were incubated with either PBS alone or secondary antibody and
served as negative isotype controls. Slides were air-dried, fixed for 5 min in a 1%
paraformaldehyde solution, and mounted in an antifading medium containing 4',6-
diamidino-2-phenylindole (DAPI) (Vector Labs, Inc., Burlingame, CA, USA).
Expression and localization of the proteins were observed with a Leica TCS-SP5
AOBS confocal laser scanning microscope (Leica, Heidelberg, Germany), for
capturing representative images of each sample.
RNA isolation and cDNA synthesis
The expression levels of selected genes were validated by qRT-PCR. Thus,
HT-29 cells were either untreated (vehicle control, 0.2% DMSO), or treated with
cyclopamine, purmorphamine, butyrate, LPS, EGF, or IFN-., for 24 hr at 37oC,
dissolved in DMSOcontaining medium. Total RNA isolation from cultured cells was
performed using SV Total RNA isolation systems (Promega, Madison, WI, USA),
following the manufacturer’s protocol. The Nanodrop 2000 UV-Vis
Spectrophotometer (Thermo Scientific, Wilmington, DE, USA) was used for
quantifying and determining the RNA purity of samples. Equal amounts of total RNA
were reverse transcribed using RT2 First Strand Kit (SABiosciences, Frederick, MD,
USA).

60
Quantitative Real-Time PCR (qRT-PCR)
To quantify the changes in mRNA levels, real-time RT-PCR was performed on
the ABI Prism 7500 (Applied Biosystems, Foster City, CA, USA) using RT2 Real
Time ™ SYBR Green/Rox PCR Master Mix (SABiosciences, Frederick, MD, USA).
For this purpose, we used a customized commercially available RT2 Profiler PCR
Array for detecting IHH, SHH, GLI-1, GLI-2, GLI-3, PTCH1, SMO, HHIP, WNT1,
BMP4, and BMP7 genes (SABiosciences, Frederick, MD, USA). Levels of mRNA
were normalized to the expression of glyceraldehydes phosphate dehydrogenase
(GAPDH). For data analysis the ..Ct method was used; determining the fold change
for all target genes in each sample with 95% confidence. Q-RT-PCR for each gene
was determined in duplicate, and each experiment was repeated at least three times.
A positive value indicates gene up-regulation and a negative value indicates gene
down-regulation. PCR cycles were performed according to the manufacturer's
instructions.
Chemokine Measurements
Culture supernatants were used for measuring the extra cellular concentration
of the chemokines IL-8 (Bender MedSystems, Vienna, Austria) and MCP-1 (e-
Bioscience, San Diego, CA, USA) by commercial sensitive enzyme-linked
immunosorbent assays (ELISA) method. The total protein content of cell monolayers
was estimated by the Pierce® BCA protein assay kit (Thermo Scientific, Rockford, IL,
USA), and used for normalizing the results. The minimum detectable concentration of
human IL-8 and MCP-1 was less than 5.0 ng/L.
Assessment of cell proliferation and viability
For the assessment of cell proliferation, we used the BrdU Cell Proliferation
Assay Kit (Chemicon International, Temecula, CA, USA). To minimize spontaneous
cell death, HT-29 cells were maintained at a concentration of 1×106 cells/mL by
diluting every 2-3 days in fresh medium to maintain exponential cell growth. HT-29
cells were rinsed with phosphate buffered saline (PBS), trypsinized, counted, and

61
seeded at a density of 2×105 cells/mL onto 96-well plates, with 100 µL per well. After
overnight attachment, cells were treated during 48 and 72h in triplicate with different
reagents, including rShh, cyclopamine, purmorphamine, butyrate, IFN-., EGF, or
LPS. BrdU was added to wells 4h prior to the plate reading at a wavelength of 450
nm.
For assessing cell viability, experiments with the same settings were
performed in parallel, and the labeling reagent 3-(4,5-dimethylthiazol-2-yl)-2,5-
diphenyltetrazolium bromide (MTT) (Sigma-Aldrich, St. Louis, MO, USA) was added
to wells, and the plates were read at a wavelength of 595 nm. The MTT test serves
as an indirect marker for proliferation and cell viability by measuring the mitochondrial
activity of cells.
Assessment of apoptosis
HT-29 cells were plated at a density of 105 cells/well in six-well plates. After
overnight attachment, cells were treated with different reagents, including the
antiCD95/APO-1 monoclonal antibody (Southern Biotech, Birmingham, AL, USA) to
induce apoptosis, or vehicle control (DMSO, 0.2%), in triplicate, for 24 hr, followed by
washing with PBS, trypsinization, and centrifugation. For the assessment of cell
viability and both early and late-stage apoptosis, cells were stained with the Annexin-
V kit and the 7-AAD Kit (both from e-Biosciences, San Diego, CA, USA). Briefly, cells
were washed twice in cold PBS, resuspended in binding buffer (1×106 cells in 0.1
ml), and 10 µl of FITCconjugated Annexin-V and 20µL of 7-AAD were added. Cells
were incubated for 15 min in the dark, an additional 400µL of binding buffer were
added, and the cells were analyzed within 1 hour by flow cytometry. Acquisition was
performed on a FACSCalibur using the CellQuest Pro software (BD Biosciences, San
Jose, CA, USA), and quantitatively analyzed using the Winlist software program
(Verity Software House, Topsham, ME, USA). Each analysis was performed on at
least 30,000 events.
Statistical analysis

62
Statistical analyses were performed using the statistical software SPSS for
Windows (Version 10.1, SPSS Inc., 1989-1999, USA). Statistical differences among
the experimental groups were evaluated with the one-way ANOVA test in which pair
wise multiple comparisons were carried out using the Dunnett’s T3 test. The level of
significance was set at p<0.05.
Results
Expression and modulation of Hh pathway components in HT-29 cells To
investigate the role of Hh signaling in various biological functions of colonic
enterocytes, we used the HT-29 human colon carcinoma cell line under different
experimental conditions. Because there is limited information on the Hh pathway in
the intestinal epithelial cells, we initially assessed the expressions and modulation of
Hh components in HT-29 cells by quantitative real-time PCR analyses. Thus, we
analyzed the effect of directly stimulating or inhibiting the Hh pathway, and the
exposure to bacterial products (LPS), cytokine (IFN-.), and growth-factor (EGF), in
the gene expression of HT29 cells. Several genes directly involved in the Hh pathway
and others which possibly interact with Hh signaling were profiled (Fig. 1A). All genes
studied were expressed as fold changes in relation to the control group (arbitrarily
normalized to 1).
Ihh and Shh constitute Hh ligands that exist in both intracellular and secreted
forms, and are responsible for the paracrine signaling of the Hh pathway in the
intestine (23). Levels of Ihh and Shh mRNA significantly increased after exposure to
rShh-peptide or to the Hh agonist purmorphamine (P < 0.02), and decreased after
treating HT-29 cells with the Hh antagonist cyclopamine (P < 0.02) (Fig. 1B).
In regard to the downstream Gli zinc-finger transcription factors of the Hh
pathway, treatment with rShh-peptide or purmorphamine significantly increased the
HT-29 levels of Gli-1 and Gli-2 mRNA (P < 0.02), the main activators of Hh target
genes and indicators of the Hh pathway activation (24). Levels of Gli-1 mRNA also
increased after treatment with the short chain fatty acid butyrate (P < 0.04), but
decreased upon exposure to cyclopamine (P < 0.02) or EGF (P < 0.02). Levels of Gli-
3, which acts mainly as a repressor of the Hh pathway (10) did not change
significantly, irrespective of the stimuli (Fig. 1C).

63
Hh signaling is regulated by transmembrane receptors Patched (Ptch) and
Smoothened (Smo). To activate the pathway, Hh binds Ptch releasing the repression
of Ptch over Smo (25). Here, we observed that the expression of Ptch1 increased
upon exposure to rShh-peptide or purmorphamine (P < 0.02), but levels were
significantly lower after EGF (P < 0.02). Levels of Smo did not change significantly in
HT-29 cells independently of the treatment (Fig. 1D).
The expression of Wnt1, component of the Wnt signaling pathway which
antagonizes the Hh pathway and is overexpressed in most colon cancers,
significantly decreased after rShh-peptide, butyrate, or EGF (P < 0.02), while it
increased upon exposure to IFN-. (P < 0.04). The expression of HHIP, a target gene
and a powerful inhibitor of the Hh pathway at the same time (26), was also assessed.
HHIP expression significantly decreased after treatment with butyrate (P < 0.04) or
EGF (P < 0.02), and increased after purmorphamine (P < 0.02) (Fig. 1E).
Because bone morphogenetic proteins are targets of Hh signaling during
development (27), and are also known to contribute to the maintenance of the
intestinal stem cell niche (28), we investigated the possible expression of epithelial
Bmps in HT-29 cells. We assessed the levels of mRNA for BMP4, known to be
present in intestinal epithelial cells and a target of both Hh (29) and Wnt (11)
pathways, and BMP7, also expressed in the colon but to a lesser extent within the
epithelium (30). We observed that the expression of BMP4 and BMP7 significantly
decreased after the treatment with IFN-. (P < 0.04) or EGF (P < 0.02). The exposure
to cyclopamine alone significantly downregulated BMP4 and BMP7 (P < 0.02) and
their levels were restored with the addition of rShh-peptide, but not with
purmorphamine (P < 0.05) (Fig. 1F). None of the genes analyzed were affected by
the exposure of HT-29 cells to LPS.
Subcellular levels and distribution of Gli-1 and ß-catenin in HT-29 cells
To further confirm these findings, we next performed immunocytochemistry for
Gli1, with confocal microscopy detection and analysis. Gli-1 protein, transcription
factor of the Hh pathway was present in low levels in HT-29 cells, predominantly in
the cytosol.

64
When HT-29 cells were exposed to rShh-peptide, purmorphamine (Hh
agonist), or butyrate, levels of Gli-1 increased with a marked relative subcellular
distribution into the nucleus. An opposing effect was shown with the addition of the
Hh antagonist cyclopamine (Fig. 2A). Because Gli-1 is one of the indicators of the Hh
pathway activation, enhanced expression and nuclear translocation indicate that Hh
pathway is active and can be regulated in HT-29 cells. Considering that Hh signaling
is known to negatively regulate Wnt signaling in the colonic epithelium (11), we were
interested in studying possible interactions between Wnt signaling and the Hh
expression or activity in HT-29 cells. Confocal microscopy detected a constitutive
expression of ß -catenin, an integral component of the Wnt pathway, predominantly
in the nucleus of HT-29 cells. Exposure of these cells to rShh-peptide during 24
hours reduced the expression of ßcatenin and restricted it to its membrane-bound
form (Fig. 2B). Densitometric analysis of staining densities confirmed that levels of
Gli-1 were significantly higher after exposure to rShh-peptide, purmorphamine, or
butyrate, compared to vehicle (*P < 0.04) or cyclopamine (**P< 0.02) treated cells. In
regard to ß-catenin staining, densities became significantly lower after exposure to
rShh-peptide (▲P < 0.01) compared to vehicle or cyclopamine treated cells (Fig. 2C).
Thus, we conclude that Hh signaling negatively regulates Wnt signaling in HT-29
colon cancer cells in vitro.
Effect of Hh pathway on the inflammatory activity of HT-29 cells
To elucidate the functional effects of Hh pathway on HT-29 cells, we first
examined the concentration of cytokines in culture supernatants of HT-29 cells
measured by ELISA. HT-29 cells were cultured at 1×106/mL per well, and subjected
to different stimuli such as the exposure to rShh-peptide, purmorphamine, butyrate,
LPS, IFN-., EGF, with or without the addition of cyclopamine or DMSO (vehicle
control) for 24 hours.
Interleukin-8 (IL-8) and monocyte chemoattractant protein-1 (MCP-1) are
inflammatory mediators known to be produced by epithelial cells and to act mainly as
chemoattractant molecules to leukocytes (31). Therefore, we analyzed the effect of
directly stimulating or inhibiting the Hh pathway, and the exposure to LPS, IFN-., and
EGF in cytokine production by HT-29 cells.

65
The levels of IL-8 were significantly lower in HT-29 cells treated with either
rShhpeptide, purmorphamine, or butyrate compared to those treated with
cyclopamine (*P < 0.03) or LPS (**P < 0.04), respectively (Fig. 3A). MCP-1
production decreased significantly in cells treated with rShh-peptide, purmorphamine,
or butyrate compared to the ones exposed to LPS (*P < 0.05), IFN-. (**P < 0.02) or
EGF (***P < 0.04), respectively (Fig. 3B). Data are expressed as the mean ± SEM of
6 independent experiments.
Effect of Hh pathway on survival and proliferative activity of HT-29 cells
HT-29 cell viability was analyzed at different time points with different
treatments, using the MTT assay. Although no difference was found among HT-29
cell treatments in the first 24 hours, cell viability decreased significantly at 48 hours
when cells were exposed to cyclopamine (*P<0.04) (Fig. 4A).
For the purpose of analyzing changes in the proliferative activity of HT-29 cells
upon different treatments, the cellular incorporation of BrdU was measured at 48 and
72 hours. Although no significant difference among the treatment groups was
detected in the first 48 hours, we identified a tendency for increase in BrdU
incorporation in the cyclopamine treated cells. However, a significant decrease in
BrdU incorporation was found within the group of cyclopamine exposed HT-29 cells,
from 48 to 72 hours (**P<0.05). After 72 hours, BrdU incorporation was maintained in
vehicle-treated cells (DMSO), while it became significantly lower in all other treatment
groups (*P<0.05) (Fig. 4B). All data in both the MTT and the BrdU assays are
expressed as the mean ± SEM of 3 independent experiments each.
Effect of Hh pathway on CD95-mediated apoptosis in HT-29 cells
Flow cytometric demonstration of HT-29 apoptosis after 24 hours of exposure
to different stimuli, as assessed by annexin-V/7-AAD. Left lower quadrant indicates
double negative cells, while right lower quadrant indicates early apoptotic annexin-V-
positive cells, and the upper quadrants indicate late apoptotic or necrotic cells (Fig.
5A). Treatment with rShh-peptide, purmorphamine or butyrate significantly abrogates
the anti-CD95 (anti-Fas) induced apoptosis (*P < 0.01), which is partially restored by

66
the addition of cyclopamine (Fig. 5B). Data are expressed as the mean ± SEM of 3
independent experiments.

67
Discussion
In this study, we show that the epithelial HT-29 human colon carcinoma cell
line expresses Hh pathway components which can be modulated by different
exogenous stimuli. In addition, we demonstrate that the biological role of the Hh
pathway in HT-29 cells ranges from the control of cell survival to the regulation of
cytokine production. The Hh pathway has been broadly detected in fetal tissues and
cells, whereas in the adult gastrointestinal tract Hh elements have been
predominantly found in the epithelial lining under physiological conditions (11, 32).
Nevertheless, the Hh pathway seems to be reactivated in several cancers (14, 33), in
which the presence of Hh components suggests the participation of this pathway in
mechanisms regulating either cell proliferation or apoptosis (34). Therefore, we were
interested in investigating whether Hh signaling would play a role in the homeostatic
control of HT-29 cells survival and function.
HT-29 cells constitute an epithelial colonic cancer cell line that has been
utilized as a model for studying epithelial cells, and also cancer pathways and for the
development of new therapeutic approaches. In this study, we analyzed the baseline
levels and the potential modulation of Hh pathway components in HT-29 cells upon
exposure to different exogenous stimuli. The Hh signaling pathway has been shown
to actively participate in the initiation and progression of gastrointestinal tumors (13,
14, 21). Nevertheless, overexpression of Hh signaling molecules has also been
reported in colon cancer (20, 35). In the present study, the expression of Hh
components in HT-29 colon cancer cell line was demonstrated both at the mRNA and
protein levels.
The expression and modulation of a selection of Hh pathway-related genes
and other indirectly associated genes were investigated by quantitative real-time
PCR in HT-29 cells. We demonstrated the presence of Ihh, Shh, the main indicators
of the Hh pathway activation Gli-1 and Gli-2, and of Ptch1, and the ability to modulate
mRNA levels of those genes following treatment of HT-29 cells with the Hh
antagonist cyclopamine or the specific stimulation with rShh-peptide or the Hh
agonist purmorphamine. Similar to another study, we also observed Gli-1 mRNA
upregulation after treatment with the short chain fatty acid butyrate (11), while levels

68
decreased after inhibition with cyclopamine or exposure to EGF. EGF is a growth
factor known to be important for epithelial regeneration, activating pro-survival
pathways in intestinal epithelial cells (36). It is possible that Hh pathway would be
downregulated or overridden by an alternative nonoverlapping pathway coordinated
by signals delivered by EGF. Of note, exposure to bacterial products or inflammatory
cytokine, had minimal or no impact on the Hh pathway gene modulation in HT-29
cells. However, when investigating epithelial bone morphogenetic proteins (Bmps) we
observed a marked downregulation of BMP4 and BMP7 following Hh inhibition, but
the levels were restored with the addition of rShh peptide to HT-29 cell cultures.
Bmps are morphogens of the transforming growth factor ß superfamily, which were
shown to play a critical role in epithelial proliferation and terminal differentiation and
maturation of cells from the secretory lineage of the intestine (28). Moreover, Bmps
are known targets of the Hh pathway during development (27), and BMP4 and BMP7
were identified as potential mediators of the mesenchymal-epithelial interaction in
response to Hh signaling within the mouse intestine (29). Therefore, here we further
confirm the expression of Bmps and the modulation by Hh signaling in differentiated
human colonic HT-29 cells. Furthermore, we showed that BMP4 and BMP7 mRNA
levels reduced after exposure to IFN-., probably indicating a downregulation in
response to inflammatory signals. The expression of Wnt1, an antagonist of the Hh
pathway in epithelial cells markedly decreased after exposure to rShh-peptide and
butyrate, while it increased upon exposure to IFN-.. Components of the Wnt/ß-
catenin pathway are known to be overexpressed in most colon cancers and in HT-29
cells their presence supports a deregulated hyperproliferative status, which appears
to be counteracted by the Hh signaling casacade in a paracrine and autocrine
fashion (26). Response of HT-29 cells to IFN-. may indicate epigenetic modulation of
the Wnt pathway by proinflammatory stimuli, and may reflect a potential link between
inflammation and oncogenesis in colonic epithelial cells. Taken together, these
results appear to unveil a complex interaction of multiple genes and converging
signaling pathways involved in physiologic processes, but also in cancer and in
inflammation, which can be modulated by the Hh cascade in HT-29 cells.
In HT-29 cells, the baseline expression of Gli1 protein was low but consistently
increased both in the cytoplasm and nucleus, following exposure to Shh peptide, the
Hh agonist purmorphamine, and the short chain fatty acid butyrate, while levels

69
decreased and disappeared from the nucleus after treatment with cyclopamine. This
is in contrast with the high constitutive levels observed in gastric (15) and breast (37)
carcinomas, but still implies that Hh activation can be modulated by exogenous
stimuli in HT-29 cells. In parallel, we demonstrated a low constitutive expression of ß-
catenin, member of the Wnt pathway in HT-29 cells. Interestingly, we additionally
observed an opposing effect of the same exogenous stimuli on ß-catenin compared
to Gli-1, in terms of protein subcellular levels and distribution in HT-29 cells. It is well-
established that deregulation of the ßcatenin/Wnt pathway components leads to the
nuclear accumulation of ß-catenin and to tumor formation (38). However, ß -catenin
transactivation is not exclusively related to activating mutations of the Wnt pathway
components (39), and it was also shown to be induced by proinflammatory signals
(40). In accordance to a previous study suggesting the existence of interactions
between Wnt signaling and Hh activity in the colonic epithelium (11), here we confirm
the idea that Hh signaling negatively regulates the Wnt pathway in HT-29 cells. In
particular, exposure to cyclopamine resulted in the nuclear accumulation, while Hh
activation led to ß-catenin reduction and to a predominant peripheral cytoplasmic
localization. These findings seem to indicate that Hh induction in epithelial cells at the
villous surface would probably exert a negative feedback upon Wnt signaling in the
basal cells of the crypt, therefore protecting terminally differentiated cells from the
proliferative action of Wnt (41).
In addition to comprise a surface for absorption and a mechanical barrier
separating the external environment from the internal milieu, colonic epithelial cells
are also involved in inflammatory processes, and have been reported to be a source
of chemokines (42, 43). Chemokines are well-established mediators in inflammation,
but they also have been reported to direct chemotactic attraction of migrating or
invading malignant cells toward the blood vessels (44), and to control migration of
metastatic cells to distant organs (45). In this study, we demonstrated that the levels
of IL-8, a potent neutrophil chemotactic and activating agent of the CXC subfamily of
chemokines (31), decreased following Hh pathway activation, in contrast to Hh
blockade or to LPS induction. Similarly, MCP-1, of the CC subfamily with chemotactic
activity to monocytes and other immune cells (46), decreased following Hh activation,
in opposition to effects of LPS, IFN-., or EGF. Our findings appear to corroborate the
importance of colonic epithelial cells in the response to infection or injury (47), and to

70
signals from lamina propria immune cells (48). The resultant chemokine secretion
from epithelial cells probably constitutes an early signal in immune and inflammatory
reactions, and may represent a connection between innate and adaptive immune
responses within the gut. In conjunction, these data suggest that Hh modulation of
chemokine generation by epithelial cells might also affect the severity and the
metastatic potential of colorectal cancer.
The intestinal epithelium undergoes a continuous cellular turnover involving
intestinal epithelial stem cells proliferation, differentiation, and cell death, in a process
that requires modulation by a complex microenvironment (49). In this regard,
signaling cascades, including Wnt and Hh signaling pathways which regulate normal
stem cell renewal, are likely to be implicated in carcinogenesis as well (6, 9, 26).
Nevertheless, in the context of Hh signaling data on colorectal cancer (14, 19, 33),
and cancer cell lines (11, 17, 22) have been controversial. It has been proposed that
these discrepancies might be, at least in part, due to distinct methodologies, analysis
of unparalleled pathological samples, and also the consequences of epigenetic
modifications to gene expression. In this study, we identified an early tendency
toward BrdU incorporation, followed by a late consistent decrease in BrdU uptake
after Hh inhibition. In a complementary experiment, we demonstrated the reduction in
HT-29 cells viability also following Hh pathway inhibition with cyclopamine, in
accordance to a previous study (50). To further understand the mechanistic link
between Hh pathway and the fate of HT-29 cells, we stimulated cells via the
Fas/FasL system in order to mediate cell death by apoptosis. Similar to other studies
(35), we demonstrated that Hh agonists are able to abrogate apoptosis in a human
colon cancer cell line. Therefore, in contrast to another study (21), we propose that
Hh signaling may not be critical for the proliferation of colon cancer cells with high
differentiation status such as HT-29. Nevertheless, viability was shown to be
dependent on the Hh pathway, further supporting the concept that active Hh
signaling in the colonic epithelium might be involved in cell differentiation and also
the protection against apoptosis.
In conclusion, we show that the Hh pathway is a key controller of colonic
epithelial cell function, as demonstrated by its effect in promoting dampening of
inflammatory signals and antagonizing apoptosis. The Hh activity in colonic epithelial
cells has probable homeostatic implications for the development of colonic

71
inflammation and malignancies, since the differential expression of Hh components
may underlie abnormalities in the local immune response and in epithelial barrier
integrity. Taken together, these results highlight the importance of Hh activation as a
potential therapeutic target in inflammatory bowel disease and colorectal cancer.

72
Acknowledgements
The authors acknowledge the technical assistance of Alyson do Rosario Jr.
Grant support
This work was supported by grants from the Brazilian Research Council
(CNPq) and the FAPERJ (Fundação de Amparo à Pesquisa do Estado do Rio de
Janeiro).

73
References
1. de Santa Barbara P, van den Brink GR, Roberts DJ. Development and
differentiation of the intestinal epithelium. Cell Mol Life Sci 2003;60:1322-32.
2. Sansonetti PJ. War and peace at mucosal surfaces. Nat Rev Immunol
2004;4:953-64.
3. van den Brink GR, Hardwick JC. Hedgehog Wnteraction in colorectal
cancer. Gut 2006;55:912-4.
4. Cho JH. The genetics and immunopathogenesis of inflammatory bowel
disease. Nat Rev Immunol 2008;8:458-66.
5. Yu M, Gipp J, Yoon JW, Iannaccone P, Walterhouse D, Bushman W. Sonic
hedgehog-responsive genes in the fetal prostate. J Biol Chem 2009;284:5620-9.
6. Kasper M, Regl G, Frischauf AM, Aberger F. GLI transcription factors:
mediators of oncogenic Hedgehog signalling. Eur J Cancer 2006;42:437-45.
7. Lum L, Beachy PA. The Hedgehog response network: sensors, switches,
and routers. Science 2004;304:1755-9.
8. Hooper JE, Scott MP. Communicating with Hedgehogs. Nat Rev Mol Cell
Biol 2005;6:306-17.
9. Ruiz I Altaba A, Mas C, Stecca B. The Gli code: an information nexus
regulating cell fate, stemness and cancer. Trends Cell Biol 2007;17:438-47.
10. Katoh Y, Katoh M. Hedgehog target genes: mechanisms of carcinogenesis
induced by aberrant hedgehog signaling activation. Curr Mol Med 2009;9:873-86.
11. van den Brink GR, Bleuming SA, Hardwick JC, Schepman BL, Offerhaus
GJ, Keller JJ, Nielsen C, Gaffield W, van Deventer SJ, Roberts DJ, Peppelenbosch
MP. Indian Hedgehog is an antagonist of Wnt signaling in colonic epithelial cell
differentiation. Nat Genet 2004;36:277-82.
12. Alinger B, Kiesslich T, Datz C, Aberger F, Strasser F, Berr F, Dietze O,
Kaserer K, Hauser-Kronberger C. Hedgehog signaling is involved in differentiation of
normal colonic tissue rather than in tumor proliferation. Virchows Arch 2009;454:369-
79.
13. Katoh Y, Katoh M. WNT antagonist, SFRP1, is Hedgehog signaling target.
Int J Mol Med 2006;17:171-5.

74
14. Berman DM, Karhadkar SS, Maitra A, Montes De Oca R, Gerstenblith MR,
Briggs K, Parker AR, Shimada Y, Eshleman JR, Watkins DN, Beachy PA.
Widespread requirement for Hedgehog ligand stimulation in growth of digestive tract
tumours. Nature 2003;425:846-51.
15. Ohta M, Tateishi K, Kanai F, Watabe H, Kondo S, Guleng B, Tanaka Y,
Asaoka Y, Jazag A, Imamura J, Ijichi H, Ikenoue T, Sata M, Miyagishi M, Taira K,
Tada M, Kawabe T, Omata M. p53-Independent negative regulation of p21/cyclin-
dependent kinase-interacting protein 1 by the sonic hedgehog-glioma-associated
oncogene 1 pathway in gastric carcinoma cells. Cancer Res 2005;65:10822-9.
16. Bienz M, Clevers H. Linking colorectal cancer to Wnt signaling. Cell
2000;103:31120.
17. Varnat F, Duquet A, Malerba M, Zbinden M, Mas C, Gervaz P, Ruiz i
Altaba A. Human colon cancer epithelial cells harbour active HEDGEHOG-GLI
signalling that is essential for tumour growth, recurrence, metastasis and stem cell
survival and expansion. EMBO Mol Med 2009;1:338-51.
18. Bian YH, Huang SH, Yang L, Ma XL, Xie JW, Zhang HW. Sonic
hedgehog-Gli1 pathway in colorectal adenocarcinomas. World J Gastroenterol
2007;13:1659-65.
19. Yoshikawa K, Shimada M, Miyamoto H, Higashijima J, Miyatani T,
Nishioka M, Kurita N, Iwata T, Uehara H. Sonic hedgehog relates to colorectal
carcinogenesis. J Gastroenterol 2009;44:1113-7.
20. You S, Zhou J, Chen S, Zhou P, Lv J, Han X, Sun Y. PTCH1, a receptor of
Hedgehog signaling pathway, is correlated with metastatic potential of colorectal
cancer. Ups J Med Sci;115:169-75.
21. Douard R, Moutereau S, Pernet P, Chimingqi M, Allory Y, Manivet P, Conti
M, Vaubourdolle M, Cugnenc PH, Loric S. Sonic Hedgehog-dependent proliferation
in a series of patients with colorectal cancer. Surgery 2006;139:665-70.
22. Chatel G, Ganeff C, Boussif N, Delacroix L, Briquet A, Nolens G, Winkler
R. Hedgehog signaling pathway is inactive in colorectal cancer cell lines. Int J Cancer
2007;121:2622-7.
23. Kolterud A, Grosse AS, Zacharias WJ, Walton KD, Kretovich KE, Madison
BB, Waghray M, Ferris JE, Hu C, Merchant JL, Dlugosz AA, Kottmann AH, Gumucio

75
DL. Paracrine Hedgehog signaling in stomach and intestine: new roles for hedgehog
in gastrointestinal patterning. Gastroenterology 2009;137:618-28.
24. Bai J, Montell D. Eyes absent, a key repressor of polar cell fate during
Drosophila oogenesis. Development 2002;129:5377-88.
25. Kalderon D. Transducing the hedgehog signal. Cell 2000;103:371-4.
26. Madison BB, Braunstein K, Kuizon E, Portman K, Qiao XT, Gumucio DL.
Epithelial hedgehog signals pattern the intestinal crypt-villus axis. Development
2005;132:279-89.
27. Bitgood MJ, McMahon AP. Hedgehog and Bmp genes are coexpressed at
many diverse sites of cell-cell interaction in the mouse embryo. Dev Biol
1995;172:126-38.
28. Auclair BA, Benoit YD, Rivard N, Mishina Y, Perreault N. Bone
morphogenetic protein signaling is essential for terminal differentiation of the
intestinal secretory cell lineage. Gastroenterology 2007;133:887-96.
29. van Dop WA, Uhmann A, Wijgerde M, Sleddens-Linkels E, Heijmans J,
Offerhaus GJ, van den Bergh Weerman MA, Boeckxstaens GE, Hommes DW,
Hardwick JC, Hahn H, van den Brink GR. Depletion of the colonic epithelial precursor
cell compartment upon conditional activation of the hedgehog pathway.
Gastroenterology 2009;136:2195-203.
30. Kosinski C, Li VS, Chan AS, Zhang J, Ho C, Tsui WY, Chan TL, Mifflin RC,
Powell DW, Yuen ST, Leung SY, Chen X. Gene expression patterns of human colon
tops and basal crypts and BMP antagonists as intestinal stem cell niche factors. Proc
Natl Acad Sci U S A 2007;104:15418-23.
31. Schall TJ, Bacon KB. Chemokines, leukocyte trafficking, and inflammation.
Curr Opin Immunol 1994;6:865-73.
32. Lees C, Howie S, Sartor RB, Satsangi J. The hedgehog signalling pathway
in the gastrointestinal tract: implications for development, homeostasis, and disease.
Gastroenterology 2005;129:1696-710.
33. Oniscu A, James RM, Morris RG, Bader S, Malcomson RD, Harrison DJ.
Expression of Sonic hedgehog pathway genes is altered in colonic neoplasia. J
Pathol 2004;203:909-17.

76
34. Kobune M, Takimoto R, Murase K, Iyama S, Sato T, Kikuchi S, Kawano Y,
Miyanishi K, Sato Y, Niitsu Y, Kato J. Drug resistance is dramatically restored by
hedgehog inhibitors in CD34+ leukemic cells. Cancer Sci 2009;100:948-55.
35. Qualtrough D, Buda A, Gaffield W, Williams AC, Paraskeva C. Hedgehog
signalling in colorectal tumour cells: induction of apoptosis with cyclopamine
treatment. Int J Cancer 2004;110:831-7.
36. Terzic J, Grivennikov S, Karin E, Karin M. Inflammation and colon cancer.
Gastroenterology 2010;138:2101-14.
37. Kubo M, Nakamura M, Tasaki A, Yamanaka N, Nakashima H, Nomura M,
Kuroki S, Katano M. Hedgehog signaling pathway is a new therapeutic target for
patients with breast cancer. Cancer Res 2004;64:6071-4.
38. Kolligs FT, Bommer G, Goke B. Wnt/beta-catenin/tcf signaling: a critical
pathway in gastrointestinal tumorigenesis. Digestion 2002;66:131-44.
39. Lu Z, Hunter T. Wnt-independent beta-catenin transactivation in tumor
development. Cell Cycle 2004;3:571-3.
40. Oguma K, Oshima H, Aoki M, Uchio R, Naka K, Nakamura S, Hirao A,
Saya H, Taketo MM, Oshima M. Activated macrophages promote Wnt signalling
through tumour necrosis factor-alpha in gastric tumour cells. EMBO J 2008;27:1671-
81.
41. Katoh Y, Katoh M. Hedgehog signaling pathway and gastrointestinal stem
cell signaling network (review). Int J Mol Med 2006;18:1019-23.
42. Jung HC, Eckmann L, Yang SK, Panja A, Fierer J, Morzycka-Wroblewska
E, Kagnoff MF. A distinct array of proinflammatory cytokines is expressed in human
colon epithelial cells in response to bacterial invasion. J Clin Invest 1995;95:55-65.
43. Mazzucchelli L, Hauser C, Zgraggen K, Wagner HE, Hess MW, Laissue
JA, Mueller C. Differential in situ expression of the genes encoding the chemokines
MCP-1 and RANTES in human inflammatory bowel disease. J Pathol 1996;178:201-
6.
44. Yu T, Wu Y, Helman JI, Wen Y, Wang C, Li L. CXCR4 promotes oral
squamous cell carcinoma migration and invasion through inducing expression of
MMP-9 and MMP-13 via the ERK signaling pathway. Mol Cancer Res 2011;9:161-72.

77
45. Lu X, Kang Y. Chemokine (C-C motif) ligand 2 engages CCR2+ stromal
cells of monocytic origin to promote breast cancer metastasis to lung and bone. J Biol
Chem 2009;284:29087-96.
46. Rollins BJ. Chemokines. Blood 1997;90:909-28.
47. Gewirtz AT, McCormick B, Neish AS, Petasis NA, Gronert K, Serhan CN,
Madara JL. Pathogen-induced chemokine secretion from model intestinal epithelium
is inhibited by lipoxin A4 analogs. J Clin Invest 1998;101:1860-9.
48. Kagnoff MF, Eckmann L. Epithelial cells as sensors for microbial infection.
J Clin Invest 1997;100:6-10.
49. Walker RA, Martin CV. The aged breast. J Pathol 2007;211:232-40.
50. Mazumdar T, DeVecchio J, Shi T, Jones J, Agyeman A, Houghton JA.
Hedgehog signaling drives cellular survival in human colon carcinoma cells. Cancer
Res 2011;71:1092-102.

78
Figure Legends
Figure 1. Gene modulation in HT-29 cells upon exposure to different stimuli.
Heat map representation of gene expression determined by RT-qPCR. HT-29 cells
were treated with Shh (Sonic Hedgehog), Purm (purmorphamine), But (butyrate),
LPS, IFN-., EGF, with or without the addition of cyclopamine (Cyc), or DMSO
(vehicle) for 24 hours (A). Histograms of individual gene expression fold changes in
HT-29 cells: IHH and SHH (B); GLI1, GLI2 and GLI3 (C); SMO and PTCH1 (D); HHIP
and WNT1 (E); and BMP4 and BMP7 (F). The graphs use the same samples as in
(A). Values represent the means ± SEM of three independent experiments and are
normalized to the GAPDH RNA gene. Significant changes in relation to the control
group are highlighted.

79
Figure 2. Distribution and levels of Gli-1 and ß -catenin in HT-29 cells.
Confocal microscopy of cytospin preparations showing the relative nuclear and
cytoplasmic distribution and levels of Gli-1 (A) and ß-catenin (B) in HT-29 cells
exposed to different stimuli for 24 hours. Analysis of protein staining densities confirm
that Gli-1 significantly increases after treatment with Shh (Sonic Hedgehog), Purm
(purmorphamine), or But (butyrate), compared to cells treated with DMSO (vehicle)
(*P < 0.04), or Cyc (cyclopamine) (**P<0.02). Density of ß-catenin significantly
decreases upon treatment with Shh (.▲P<0.01) compared to either DMSO or Cyc
(C). Nuclei are stained with DAPI (blue). Micrograph panel is representative of 3-4
experiments for each condition (Original magnification ×1000).

80
Figure 3. Production of cytokines by HT-29 cell cultures upon exposure to
different stimuli. Concentration of cytokines in supernatants of 24 hour cultures of HT-
29 cells measured by ELISA. Histograms show the levels of IL-8 (A) and MCP-1 (B),
under various conditions: treatment with Shh (Sonic Hedgehog), Purm
(purmorphamine), Butyrate, LPS, IFN-., EGF, with or without the addition of
cyclopamine, or DMSO (vehicle control) for 24 hours. IL-8 levels are significantly
lower in cells treated with either Shh, purmorphamine, or butyrate compared to those
treated with cyclopamine (*P<0.03), or LPS (**P<0.04), respectively (A). MCP-1
levels are significantly lower in cells treated with either Shh, purmorphamine, or
butyrate compared to the ones treated with LPS (*P<0.05) IFN-. (**P<0.02), or EGF
(***P<0.04), respectively (B). Data are expressed as the mean ± SEM of 6
independent experiments.

81
Figure 4. Survival and proliferative activity of HT-29 cells upon exposure to
different stimuli. HT-29 cell viability was analyzed at different time points with different
treatments, using the MTT assay. When HT-29 cells were exposed to cyclopamine,
cell viability decreased significantly comparing 48 with 24 hours (*P<0.04) (A). For
the purpose of analyzing changes in the proliferative activity of HT-29 cells upon
different treatments, the cellular incorporation of BrdU was measured at 48 and 72
hours. After 72 hours, BrdU incorporation was maintained in vehicle-treated cells
(DMSO), while it significantly decreased in all other treatment groups (*P<0.05).
Within the group of cyclopamine exposed HT-29 cells, BrdU incorporation decreased
significantly from 48 to 72 hours (**P<0.05) (B). Data are expressed as the mean ±
SEM of 3 independent experiments.

82
Figure 5. Relationship between apoptosis and the activity of Hedgehog
pathway in HT-29 cells. Flow cytometric demonstration of HT-29 apoptosis after 24
hours of exposure to different stimuli, as assessed by annexin-V/7-AAD. Left lower
quadrant indicates double negative cells while right lower quadrant indicates
annexin-V-positive cells (A). Treatment with Shh, purmorphamine or butyrate
significantly abrogates the anti-CD95 induced apoptosis (*P<0.01), which is partially
restored by the addition of cyclopamine (B) Data are expressed as the mean ± SEM
of 3 independent experiments.





























