Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DO RIO DE JANEIRO CAMPUS UFRJ-MACAÉ Professor Aloísio Teixeira
Programa de Pós-graduação em Produtos Bioativos e Biociências
Envolvimento da proteína quinase B (AKT) na atividade de Hexoquinase do
carrapato Rhipicephalus microplus.
Luan Cordeiro Corrêa
Macaé – RJ
2017

II
Programa de Pós-graduação em Produtos Bioativos e Biociências
Envolvimento da proteína quinase B (AKT) na atividade de Hexoquinase do
carrapato Rhipicephalus microplus.
“Dissertação de Mestrado apresentada ao
Programa de Pós-gradução em Produtos
Bioativos e Biociências, Laboratório Integrado
de Ciências Morfofuncionais, Núcleo em
Ecologia e Desenvolvimento Socioambiental
de Macaé da Universidade Federal do Rio de
Janeiro, como parte das exigências para a
obtenção do título de Mestre em Ciências.”
Orientador: Prof. Dr. Rodrigo Nunes da Fonseca
Macaé – RJ
2017

III
Envolvimento da proteína quinase B (AKT) na atividade de Hexoquinase do
carrapato Rhipicephalus microplus.
LUAN CORDEIRO CORRÊA
“Dissertação de Mestrado apresentada ao
Programa de Pós-gradução em Produtos
Bioativos e Biociências, Laboratório Integrado
de Ciências Morfofuncionais, Núcleo em
Ecologia e Desenvolvimento Socioambiental
de Macaé da Universidade Federal do Rio de
Janeiro, como parte das exigências para a
obtenção do título de Mestre em Ciências.”
Aprovado em 11 de Agosto de 2017
Banca Examinadora:
Prof.(a) Dr(a). Cíntia Monteiro de Barros (UFRJ-Macaé)
Programa de Pós-graduação em Produtos Bioativos e Biociências
Universidade Federal do Rio de Janeiro – Campus UFRJ-Macaé Prof. Aluísio Teixeira
Prof. Dr. Petter Franco Entringer (UFRJ-Macaé)
Universidade Federal do Rio de Janeiro – Campus UFRJ-Macaé Prof. Aluísio Teixeira
___________________________________________________________________
Prof. Dr. Eldo Campos (UFRJ-Macaé)
Laboratório Integrado de Bioquímica - Hatisaburo Masuda
Universidade Federal do Rio de Janeiro – Campus UFRJ-Macaé Prof. Aluísio Teixeira

IV
___________________________________________________________________
Prof.(a) Dr.(a) Flavia Borges Mury – (UFRJ-Macaé)
Universidade Federal do Rio de Janeiro – Campus UFRJ-Macaé Prof. Aluísio Teixeira
(Suplente)
___________________________________________________________________
Prof. Dr. Jackson de Souza Menezes (UFRJ-Macaé)
Laboratório Integrado de Ciências Morfofuncionais (LICM)
Programa de Pós-graduação em Produtos Bioativos e Biociências
Universidade Federal do Rio de Janeiro – Campus UFRJ-Macaé Prof. Aluísio Teixeira
(Revisor)
___________________________________________________________________
Prof. Dr. Carlos Jorge Logullo de Oliveira (UFRJ-Macaé)
Laboratório de Química e Função de Proteínas e Peptídeos (LQFPP)
Universidade Estadual do Norte Fluminense Darcy Ribeiro (UENF)
(Co-orientador)
___________________________________________________________________
Prof. Dr. Rodrigo Nunes da Fonseca (UFRJ-Macaé)
Laboratório Integrado de Ciências Morfofuncionais (LICM)
Programa de Pós-graduação em Produtos Bioativos e Biociências
Universidade Federal do Rio de Janeiro – Campus UFRJ-Macaé Prof. Aluísio Teixeira
(Orientador)

V
Dedico este trabalho
A todos da minha família e amigos, por ficarem sempre ao meu lado me dando
forças e incentivos para continuar nessa jornada.

VI
AGRADECIMENTOS
Ao meu orientador Rodrigo Nunes, por ser tão amigo e compreensível;
Ao fantástico amigo e Co-orientador Carlos Logullo. “Ainda não esqueço os nossos
treinos de 10Km de corrida”;
Ao meu grande amigo Renato Martins pelos ensinamentos, brincadeiras, histórias,
bebedeiras, jogos e seus sábios conselhos, pois sem eles eu teria surtado;
Para a minha família LICM/UEA, pois o trabalho e todo esse esforço não aconteceria
sem o carinho de todos;
Ao Airton Luquêz, o meu herói, meu pai. Um grande homem, que sempre lutou ao
meu lado pelo meu sonho;
A minha mãe pelo amor, carinho, orações, palavras de conforto, abraços nos
momentos mais difíceis, sendo a mulher mais guerreira e batalhadora que já
conheci;
Ao meu irmão, por estar sempre na torcida por mim.
A mulher mais guerreira de todas, minha vó. Obrigado por tudo!
E a Mickaela, minha superamiga e namorada, obrigado por aguentar comigo tudo
que passamos até aqui.
A Camila por ter feito parte da minha formação e pela sua grande contribuição neste
trabalho.
Aos órgãos de fomento CAPES, CNPq, FAPERJ e INCT.

VII
RESUMO
O carrapato Rhipicephalus microplus acarreta diversos danos econômicos na
produção de leite e carne por ser o principal ectoparasito dos rebanhos de gado
bovino, transmitindo uma diversidade de patógenos. Os custos do controle químico
são elevados e representam riscos à saúde humana, contaminando a carne, o leite e
o meio ambiente. Para um controle mais eficiente sem prejuízos ao animal o
desenvolvimento de novos métodos torna-se necessário. A ampliação no
conhecimento do metabolismo energético deste carrapato, pode ajudar no
desenvolvimento de novas estratégias para o controle deste ectoparasita. Esse
trabalho tem como proposta estudar a possível regulação da proteína quinase B
(AKT) sobre a hexoquinase (HK) no carrapato R. microplus. Para tal, o silenciamento
gênico da AKT, utilizando a técnica de RNA dupla-fita, foi realizado em fêmeas
parcialmente ingurgitadas (paternógenas). As fêmeas de carrapato silenciadas para
esta enzima apresentaram uma maior taxa de ingurgitamento, e uma diminuição do
tamanho do ovário quando comparado ao controle. Visando utilizar um sistema in
vitro foi realizado o silenciamento gênico da AKT na linhagem celular embrionária
BME26. A atividade de HK aumentou significativamente na fração citoplasmática
quando realizamos o silenciamento. Tratamento com o inibidor específico de AKT
(10-DEBC) também levou ao aumento da atividade de HK na fração citoplasmática.
Análises de genes homólogos de HK obtidas a partir de buscas em transcriptomas
de carrapatos R. micropuls indicam a existência de duas isoformas de HK. Juntos
estes resultados indicam uma possível regulação da proteína quinase B (AKT) sobre
hexoquinase (HK), acrescentando à literatura importantes informações relativas à
fisiologia e ao metabolismo desse vetor de doenças.

VIII
ABSTRACT
The Rhipicephalus microplus tick causes several economic impacts in the
production of milk and meat, since it is the main ectoparasite of cattle herds, acting
as a vector of a diversity of pathogens. The high cost of chemical control pose risks
to human health, due to meat, milk and environmental contamination. For a more
efficient control with lower damage to the environment the development of new
methods becomes necessary. The amplification of our knowledge of the energetic
metabolism of this tick can help the development of news strategies in the control of
this ectoparasite. This study aims to study the regulation of protein kinase B (AKT) on
hexokinase (HK) in the R. microplus tick. To achieve this goal, AKT gene silencing
via RNAi was performed in partially engorged (parthenogenic) females. The tick
females silenced for this enzyme presented a higher engorgement rate, and a
decrease in the size of the ovary when compared to the control. In order to use an in
vitro system, the gene silencing of AKT was also performed in the BME26 embryonic
cell lineage. The activity of HK increased significantly in the cytoplasmic fraction
when we performed the silencing. Treatment with the specific inhibitor of AKT (10-
DEBC) also led to increased activity of HK in the cytoplasmic fraction. HK
homologous genes searches in R. microplus tick transcriptomes indicate the
existence of two HK isoforms. Altogether our preliminary results indicate a possible
regulation of the HK by the kinase B protein (AKT), Adding to the literature important
information regarding the physiology and metabolism of this disease vector.

IX
LISTA DE FIGURAS
Figura 1: O carrapato Rhipicephalus microplus parasitando seu hospedeiro (Bos
taurus), o boi. .............................................................................................................. 1
Figura 2: Representação do ciclo de vida do Carrapato R. microplus. ........................ 3
Figura 3: Utilização de acaricidas para o controle do carrapato Rhipicephalus
microplus. .................................................................................................................... 4
Figura 4: Estágios do desenvolvimento embrionário do carrapato Rhipicephalus
microplus. .................................................................................................................... 6
Figura 5: Imagem de microscopia de fluorescência em sobreposição com DIC
(contraste de interferência diferencial) das células BME26. ........................................ 8
Figura 6: Conservação evolutiva da via de sinalização por insulina em diversas
espécies. ................................................................................................................... 10
Figura 7: Esquema da reação catalisada pela Hexoquinase. ................................... 11
Figura 8: Esquema mostrando a ligação da hexoquinase na membrana externa da
mitocondria. ............................................................................................................... 12
Figura 9: Alinhamento da sequência de aminoácidos de HK de R. micróplus, I.
escapularis, D. melanogaster, H. sapiens, Mus musculus e T. castaneum. .............. 23
Figura 10: Análise filogenética da Hexoquinase de R. microplus, I. scapularis, D.
melanogaster, H. sapiens, Mus musculus e T. castaneum. ...................................... 24
Figura 11: Qualidade do fracionamento celular de embriões do carrapato R.
microplus. .................................................................................................................. 26
Figura 12: Atividade de Hexoquinase das frações citosólica e mitocondrial na
presença do inibidor 10-DEBC em embriões do carrapato R. microplus. ................. 27
Figura 13: Efeito do silenciamento gênico da proteína quinase B (AKT) no
desenvolvimento embrionário do carrapato Rhipicephalus microplus. ...................... 28
Figura 14: Qualidade do fracionamento na linhagem celular BME26. ....................... 29
Figura 15: Avaliação da atividade da Hexoquinase em células BME26 desafiadas
com inibidor 10-DEBC e silenciadas para AKT. ........................................................ 31

X
SUMÁRIO
1. INTRODUÇÃO .................................................................................................................................. 1
1.1 Carrapatos ............................................................................................................................... 1
1.2 O ciclo de vida do Rhipicephalus microplus ............................................................................ 2
1.3 Embriogênese do carrapato R. microplus ............................................................................... 5
1.4 Linhagem de células embrionárias BME26 ............................................................................. 7
1.5 Via de sinalização por insulina ................................................................................................ 9
1.6 Hexoquinase no metabolismo energético e sua regulação pela Akt .................................... 10
2 JUSTIFICATIVA ............................................................................................................................... 14
3 OBJETIVOS ..................................................................................................................................... 14
3.1 Objetivo geral ........................................................................................................................ 14
3.2 Objetivos específicos ............................................................................................................. 14
4 MATERIAIS E MÉTODOS ................................................................................................................ 15
4.1 Os embriões do carrapato Rhipicephalus microplus ............................................................. 15
4.2 Injeção do RNA de interferência em paternógenas .............................................................. 15
4.3 Alimentação Artificial ............................................................................................................ 15
4.4 Manutenção das células BME26 ........................................................................................... 16
4.5 Obtenção das frações citosólica e mitocondrial dos embriões e células embrionária do
carrapato R. microplus (BME26). ...................................................................................................... 16
4.6 Inibição da AKT por 10-DEBC de embriões e células embrionárias do carrapato R. microplus
(BME26) ............................................................................................................................................. 17
4.7 Atividade de Succinato Desidrogenase (SDH). ...................................................................... 17
4.8 Atividade da glicose-6-fosfato desidrogenase ...................................................................... 17
4.9 Atividade da Hexoquinase (HK) ............................................................................................. 18
4.10 Síntese de RNA de interferência (RNAi) para RmAKT ........................................................... 18
4.11 Silenciamento gênico nas células BME26 ............................................................................. 19
4.12 Ensaio de viabilidade celular ................................................................................................. 19
4.13 Análise da transcrição gênica de AKT e HK por qPCR ............................................................ 20

XI
4.14 Busca de sequencias similares a HK ...................................................................................... 20
4.15 Análise estatística .................................................................................................................. 21
5 RESULTADOS ................................................................................................................................. 22
5.1 Análise da homologia da sequência de aminoácidos da enzima Hexoquinase do carrapato R.
microplus. .......................................................................................................................................... 22
5.2 Análise filogenética da Hexoquinases de carrapato ............................................................. 24
5.3 Qualidade do fracionamento celular de embriões do carrapato Rhipicephalus microplus. . 25
5.4 Atividade de Hexoquinase das frações mitocondrial e citoplasmática de embriões do
carrapato R. microplus na presença do inibidor 10-DEBC. ............................................................... 26
5.5 Efeito do silenciamento gênico da proteína AKT no desenvolvimento do carrapato R.
microplus. .......................................................................................................................................... 27
5.6 Qualidade do fracionamento em linhagem celular BME26. ................................................. 29
5.7 Avaliação da atividade da Hexoquinase na linhagem celular BME26 desafiadas com inibidor
10-DEBC e silenciadas para AKT. ....................................................................................................... 30
6 DISCUSSÃO .................................................................................................................................... 33
7 CONCLUSÃO .................................................................................................................................. 38
8 REFERENCIAS BIBLIOGRÁFICAS ..................................................................................................... 39

XII
LISTA DE SIGLAS E ABREVIATURAS
ADP – Adenosina difosfato
AKT – Proteína quinase B
ANT – Translocador de nucleotídeo
ATP – Adenosina trifosfato
cDNA – Ácido desoxirribonucléico complementar
EDTA – Ácido etilenodiamino tetra – acético
H2O2 – Peróxido de hidrogênio
ISP – Via de sinalização por insulina
GLUTs – Transportadores de glicose
G6PDH – Glicose-6-fosfato desidrogenase
GSK3 – Glicogênio sintase quinase 3
HK – Hexoquinase
NAD – Nicotinamida adenina dinucleotídeo (forma oxidada)
NADH – Nicotinamida adenina dinucleotídeo (forma reduzida)
NADPH – Nicotinamida adenina dinucleotídeo fosfato (forma reduzida)
PBS – Solução fisiológica tamponada com Tris-fosfato
PEPCK - Fosfoenol Piruvatoquinase Carboxilase
PI3K – Fosfatidilinositol-4,5-bifosfato 3-quinase
PKB – Proteína quinase B
PKBα – Proteína quinase B alfa
RNA – Ácido ribonucleico
RNAi – Ácido ribonucleico por interferência
ROS – Espécie reativas de Oxigênio
Ser - Serina
Thr- Treonina
VDAC – Canal de ânion dependente de voltagem
µg – micrograma
µM – micromolar
µm – micrômetro

1
1. INTRODUÇÃO
1.1 Carrapatos
Os carrapatos são artrópodes ectoparasitas que se aderem à superfície do
corpo dos vertebrados terrestres (anfíbios, répteis, aves e mamíferos), e se
alimentam exclusivamente de sangue. Podem permanecer fixados à pele dos
hospedeiros por tempo prolongado, secretando substâncias que impedem a
coagulação sanguínea e diminuem a resposta inflamatória no local de fixação
(SONENSHINE, 1991).
As doenças transmitidas por esses artrópodes afetam primariamente os
animais (zoonoses), e os seres humanos são apenas eventualmente infectados,
geralmente quando entram em contato com ambientes rurais ou com animais
durante atividades de recreação ou de trabalho. Uma das principais espécies de
importância médico-veterinária no Brasil e nas Américas é o carrapato bovino
Rhipicephalus microplus (OIE, 2007; ANDREOTTI, 2010).
O carrapato R. microplus é um ectoparasita pertencente à família Ixodidae
(SONENSHINE et al., 2006), e é encontrado em quase todas as regiões do mundo,
com predominância em áreas tropicais e subtropicais, abrangendo regiões com
produção de gado de corte e leiteiro na América, África, Ásia e Austrália
(JOHNSTON et al., 1986), tendo como principal hospedeiro o bovino, embora seja
capaz de completar, eventualmente, o seu ciclo em búfalos, ovelhas, cavalos e
veados (SONENSHINE, 1991). O R. microplus acarreta diversos danos econômicos,
sendo considerado o principal ectoparasito dos rebanhos de gado bovino (HORN e
ARTECHE, 1985) (Figura 1).
Figura 1: O carrapato Rhipicephalus microplus parasitando seu hospedeiro (Bos taurus), o boi (Fonte: Embrapa.br, foto de Brito, Cristina).
Além da espoliação ao couro, causada por reações inflamatórias nos locais
de fixação do carrapato (SEIFERT et al., 1968), existe ainda uma grande perda na

2
produção de leite e carne (SUTHERST, 1983) devido à perda de sangue do animal
hospedeiro. Os custos causados pelas perdas devidas a infestação por carrapatos
podem chegar a uma ordem de 3,24 bilhões de dólares por ano (GRISI et al., 2014)
seja na saúde humana, através dos resíduos de substâncias químicas, como os
acaricidas, na carne e no leite, ou ao meio ambiente, através da contaminação da
água e do solo (FERNANDES et al., 2006). Ainda, o R. microplus é um importante
vetor de doenças, como a tristeza parasitária bovina, causada por protozoários do
gênero Babesia e pela riquétsia do gênero Anaplasma (MCCOSKER, 1981; YOUNG
E MORZARIA, 1986). Além do mais, nenhum outro artrópode conhecido tem a
capacidade de ser vetor de tão ampla variedade de organismos patogênicos,
incluindo bactérias, vírus e fungos, que resultam frequentemente em infecções letais
(SONENSHINE et al., 2006).
1.2 O ciclo de vida do Rhipicephalus microplus
Existem duas etapas distintas no ciclo de vida do carrapato R. microplus,
uma fase parasitária em um único hospedeiro e uma fase não parasitária. A fase não
parasitária compreende os estágios em que a fêmea está completamente
ingurgitada (teleógina), ovo (embrião) e larva infestante. Cada fêmea de carrapato
pode colocar dois a três mil ovos que eclodem em duas a três semanas,
dependendo das condições de temperatura e umidade relativa do ar. A
embriogênese do R. microplus é completada após 21 dias e as larvas recém
eclodidas não tem poder infestante, pois precisam de um tempo para fortalecimento
da cutícula (quatro a seis dias). Após esse período, elas sobem nas hastes dos
capins, aguardando a passagem do hospedeiro para parasitá-lo (Figura 2).
A fase parasitária inicia-se quando a larva infestante se instala no
hospedeiro e dura em torno de 21 dias, apresentando três variações morfológicas:
larval, ninfa e adulto. A larva é bastante ativa, fixando-se na pele do bovino em
locais específicos. Uma vez fixada, a larva inicia sua alimentação e
consequentemente seu crescimento e desenvolvimento, atingindo o estágio de
metalarva. Após este estágio inicia-se uma nova fase no ciclo parasitário, a ninfa,
onde ocorre outras mudanças morfológicas. A diferenciação sexual ocorre em
seguida, em um período de inatividade chamado de metaninfa (Figura 2). Em torno
do 17º dia os machos já estão aptos à cópula podendo permanecer no bovino por

3
mais de 38 dias fecundando outras fêmeas (que já podem ser fecundadas passando
de metaninfa para neógina). Após um período de 3 dias ocorrem mudanças no
tegumento chegando ao estágio de partenógena (parcialmente ingurgitada), em
mais dois dias há um ingurgitamento completo onde a fêmea aumenta cerca de dez
vezes o seu tamanho em relação ao corpo do macho, ingerindo 300-500µl de
sangue (TATCHELL et al, 1972), chegando à fase teleógina (GONZALES et al.,
1974).
Figura 2: Representação do ciclo de vida do Carrapato R. microplus (modificado de GONZALES et al., 1974).
Atualmente são descritas diversas estratégias de controle do carrapato R.
microplus como o biológico, através de inimigos naturais, o rodizio destes animais no
manejo de pastagem, o uso de linhagens bovinas mais resistentes a ação
espoliativa deste carrapato e o controle imunológico através do uso de vacinas
contra o R. microplus por exemplo (PRUETT 1999). A indução de uma resposta
imunológica, em particular, tem sido testada por diferentes grupos de pesquisas há
três décadas (MCGOWAN et al., 1980; DA SILVA VAZ JR. et al., 1998;
WILLADSEN, 2006), fornecendo bases para futuras tentativas de utilização no
controle deste ectoparasita, mas a principal estratégia de controle é feita por meio de
acaricidas devido à eficiência e baixo custo de manejo (MEHLHORN et al., 2011). A
eficiência do método é comprovada, porém as populações de carrapatos estão cada
vez mais resistentes à aplicação desses compostos, e ainda comprometem a

4
qualidade da carne, do leite e da saúde humana com os resíduos químicos
provenientes de sua aplicação (WILLADSEN et al., 1996; WILLADSEN, 2006)
(Figura 3).
Figura 3: Utilização de acaricidas para o controle do carrapato Rhipicephalus microplus (Fonte: Embrapa.br, foto de La Falce, Marcos).
Com o avanço tecnológico e com o uso de ferramentas da biologia, como a
bioquímica e biologia molecular, é possível que possamos encontrar alternativas ao
uso de componentes químicos, como os acaricidas. Uma importante característica
dos carrapatos são as suas fases de resistência que lhes possibilitam uma maior
sobrevida no ambiente, garantindo o sucesso reprodutivo e o fechamento de seu
ciclo de vida. Carrapatos trioxenos (que realizam um ciclo de três hospedeiros), por
exemplo, tanto as larvas como as ninfas e adultos são estágios de resistência no
ambiente, uma vez que sua sobrevida é dependente das reservas energéticas
adquiridas do estágio anterior do ciclo de vida. O adulto é o estágio que por mais
tempo consegue sobreviver sem que encontre um hospedeiro, seguido pela ninfa, e
por último a larva, que apresenta a menor sobrevida em jejum. Os adultos de
Amblyomma spp, por exemplo, podem sobreviver em jejum sob condições naturais
por 12 a 24 meses, a ninfa por até 12 meses, e as larvas ao redor de 6 meses
(DIAMANT & STRICKLAND, 1965).
Outra importante fase de resistência desses artrópodes que acaba sendo
negligenciada é o estágio de ovo. Os ovos desses artrópodes consistem numa fase
de resistência, no sentido de que precisam lidar com recursos energéticos limitados,

5
depositados no ovo pela fêmea. E de fato, cada fêmea de carrapato pode colocar
dois a três mil ovos (SNOWBALL, 1957) que eclodem em duas a três semanas,
dependendo das condições de temperatura e umidade relativa do ar. Sendo assim, a
embriogênese do carrapato R. microplus pode ser uma fase do ciclo muito
importante na tentativa de encontrar possíveis alvos para o controle deste vetor.
1.3 Embriogênese do carrapato R. microplus
A embriogênese ocorre em um sistema fechado com os recursos
energéticos limitados (totalmente dependente do conteúdo do vitelo), ou seja, o
desenvolvimento do embrião ocorre na ausência do aporte de nutrientes externos, e
os nutrientes que favorecem seu desenvolvimento são de origem materna, onde a
fêmea deposita todo os nutrientes, incluindo lipídios, açucares e proteínas ao ovócito
antes da postura dos ovos, o que garante a diferenciação celular e o crescimento
(FAGOTTO 1990; SONG et al., 2006).
O desenvolvimento embrionário do R. microplus descrito Campos e seus
colaboradores (2006) demostraram etapas marcantes da embriogênese, como a
formação do blastoderma sincicial no quarto dia de desenvolvimento, formação do
blastoderma celular no sexto dia e a segmentação do embrião no sétimo dia.
Entretanto, dados recentes aos de Campos (2006), mostraram que a celularização
em embriões do carrapato R. microplus ocorre durante as primeiras 48 horas do
desenvolvimento (SANTOS et al., 2013). Além disso, neste mesmo trabalho, foram
observadas características tardias na embriogênese como por exemplo, formação
da cabeça, tórax, formação e retração do quarto par de patas (Figura 4). Esse
trabalho desenvolvido por Santos e colaboradores (2013) acabou, por tanto, por
caracterizar todos os estágios do desenvolvimento embrionário até o momento.
6
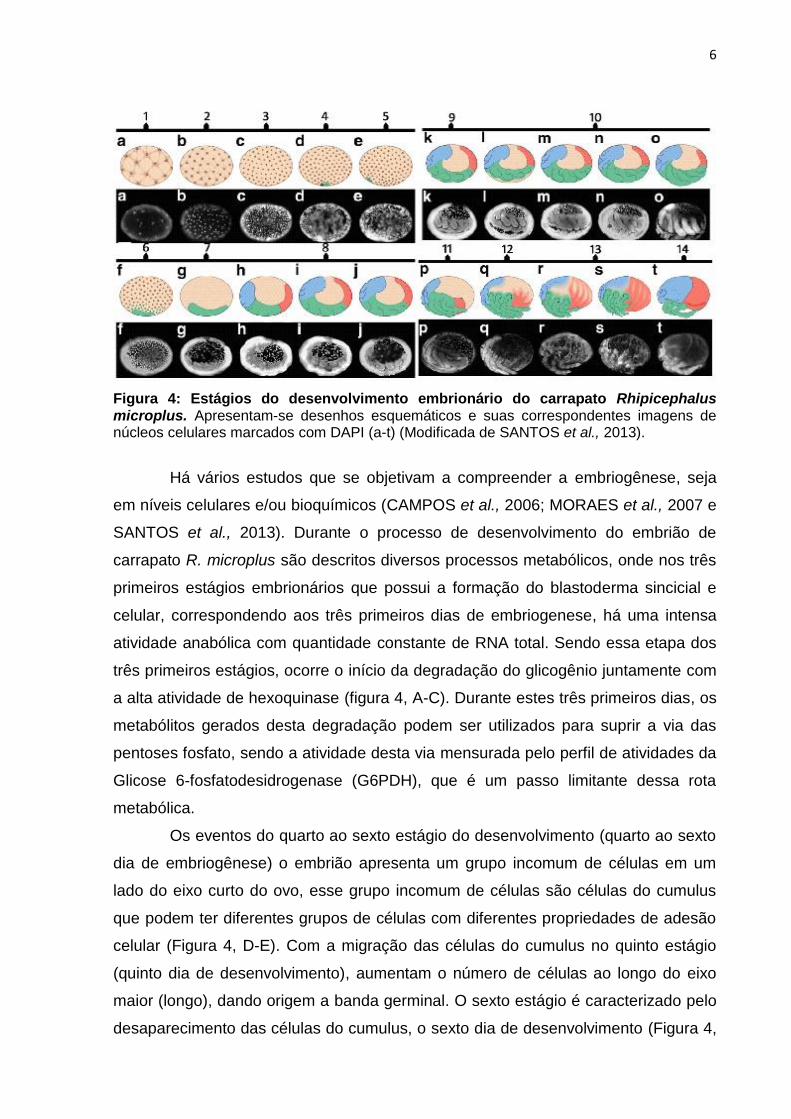
Figura 4: Estágios do desenvolvimento embrionário do carrapato Rhipicephalus microplus. Apresentam-se desenhos esquemáticos e suas correspondentes imagens de núcleos celulares marcados com DAPI (a-t) (Modificada de SANTOS et al., 2013).
Há vários estudos que se objetivam a compreender a embriogênese, seja
em níveis celulares e/ou bioquímicos (CAMPOS et al., 2006; MORAES et al., 2007 e
SANTOS et al., 2013). Durante o processo de desenvolvimento do embrião de
carrapato R. microplus são descritos diversos processos metabólicos, onde nos três
primeiros estágios embrionários que possui a formação do blastoderma sincicial e
celular, correspondendo aos três primeiros dias de embriogenese, há uma intensa
atividade anabólica com quantidade constante de RNA total. Sendo essa etapa dos
três primeiros estágios, ocorre o início da degradação do glicogênio juntamente com
a alta atividade de hexoquinase (figura 4, A-C). Durante estes três primeiros dias, os
metabólitos gerados desta degradação podem ser utilizados para suprir a via das
pentoses fosfato, sendo a atividade desta via mensurada pelo perfil de atividades da
Glicose 6-fosfatodesidrogenase (G6PDH), que é um passo limitante dessa rota
metabólica.
Os eventos do quarto ao sexto estágio do desenvolvimento (quarto ao sexto
dia de embriogênese) o embrião apresenta um grupo incomum de células em um
lado do eixo curto do ovo, esse grupo incomum de células são células do cumulus
que podem ter diferentes grupos de células com diferentes propriedades de adesão
celular (Figura 4, D-E). Com a migração das células do cumulus no quinto estágio
(quinto dia de desenvolvimento), aumentam o número de células ao longo do eixo
maior (longo), dando origem a banda germinal. O sexto estágio é caracterizado pelo
desaparecimento das células do cumulus, o sexto dia de desenvolvimento (Figura 4,

7
E-F), nesse momento ocorre a degradação de lipídios totais e a atividade de G6PDH
atinge o pico no quinto dia do desenvolvimento (Figura 4, E) devido a essa intensa
proliferação celular do embrião, fornecendo Ribose 5fosfato para síntese de
nucleotídeos (MORAES et al., 2007 e SANTOS et al., 2013).
Entre o sétimo e o nono estágio (sétimo ao décimo terceiro dia de
desenvolvimento) ocorrem a formação da cabeça, tórax e o quarto par de pata
(Figura 4, G-M). Até o sétimo dia de desenvolvimento (Figura 4, A-G) há diminuição
do conteúdo de glicogênio, seguido de aumento desse glicogênio até o décimo
segundo dia de desenvolvimento (Figura 4, G-I). Esse aumento de glicogênio até o
décimo segundo dia de desenvolvimento é correlacionado com o processo de
gliconeogênese, que é avaliada pelo conteúdo total de guanina, glicose e pela
enzima chave Fosfoenol Piruvatoquinase Carboxilase (PEPCK).
A PEPCK, juntamente com a glicose e guanina, mostrou subsequente
aumento até o décimo sétimo dia de desenvolvimento (correspondente ao décimo
segundo estágio), estágio no qual ocorre a regressão do quarto par de patas. A
partir do décimo segundo dia do desenvolvimento (Figura 4, I) ocorre o declínio do
glicogênio até o final da embriogênese, os produtos da degradação do glicogênio
podem estar “abastecendo” a síntese de novas proteínas, ou seja, a gliconeogênese
poderia estar utilizando aminoácidos para a resíntese de glicose até o final da
embriogênese (CAMPOS et al., 2006 e MORAES et al., 2007 e SANTOS et al.,
2013).
O estudo desse estágio de vida do carrapato R. microplus possibilita
entender como os embriões são capazes de coordenar todo o processo de obtenção
de energia, assim como o controle da mobilização e síntese das principais moléculas
de reservas.
1.4 Linhagem de células embrionárias BME26
Derivada de embriões de diferentes dias do carrapato R. microplus de uma
única fêmea ingurgitada, a linhagem celular BME26 é descrita desde agosto de 1981
(HOLMAN et al., 1980). O nível de diferenciação, assim como a origem tecidual
desta linhagem celular ainda é desconhecida, pois essa linhagem é proveniente de
fragmentos de tecido embrionário de diferentes dias da embriogênese.

8
Esteves e colaboradores (2008) mostraram que essa linhagem celular foi
derivada da espécie Rhipicephalus microplus, através do sequenciamento parcial do
gene mitocondrial que codifica a proteína 16S, e a sequência desse gene é 100%
idêntica a sequência do gene do R. microplus.
Morfologicamente, culturas inicias pouco confluente apresentam formato de
fibroblastos (Figura 5 setas brancas), e à medida que a confluência aumenta as
células vão aderindo fortemente ao substrato adotando um formato arredondado
(Figura 5, seta vermelha) com um grande número de vesículas (Figura 5, C) (que
apresentam diferentes graus de acidificação), e também pode ser observado um
grande número de mitocôndrias (Figura 5, B) (ESTEVES et al., 2008).
Figura 5: Imagem de microscopia de fluorescência em sobreposição com DIC (contraste de interferência diferencial) das células BME26. Caracterização das células pela marcação de núcleos por DAPI (A), marcação de mitocôndrias por rodamina-123 (B) e a visualização das vesículas coradas com laranja de acridina (C). Setas brancas apontam formas de fibroblastos, e a seta vermelha mostra a forma arredondada das células. Barra com 20µm de escala (modificado de ESTEVES et al., 2008).
O isolamento e o estabelecimento da linhagem celular do carrapato pôde
proporcionar um modelo para diversos estudos na genômica funcional, na interação
com patógenos (Bactérias e vírus), transfecção, silenciamento por interferência de
RNA (RNAi) (MATTILA et al., 2006; BELL-SAKYI et al., 2007; KURTTI et al., 2008).
Recentemente nosso grupo demonstrou alterações metabólicas de células
embrionárias BME26 do carrapato R. microplus tratadas com diferentes
concentrações de glicose. Foi observada a ativação de uma gliconeogênese não
clássica quando as células BME26 foram supridas com elevada quantidade de
glicose, um programa metabólico similar ao observado para células de mamíferos
resistentes à insulina (MARTINS et al., 2015). Também foi demonstrado em outro
trabalho do grupo que as células BME26 responderam à insulina exógena
administrada no meio de cultura (ABREU et al., 2009). Esses trabalhos mostram que

9
não só o metabolismo de carboidratos possui uma regulação refinada como também
responde à via de sinalização de insulina. Dessa forma, o carrapato R. microplus
pode sofrer grandes adaptações metabólicas para garantir a sua sobrevivência e
desenvolvimento, mesmo em condições de estresse.
1.5 Via de sinalização por insulina
De modo geral o fluxo de glicose na célula é influenciado por fatores
exógenos, como a insulina, e a regulação desse fluxo dado pela cascata de
sinalização por insulina. Sendo assim, quando a via de sinalização por insulina é
iniciada ocorre uma cascata por fosforilação recrutando transportadores de glicose
(GLUTs) para membrana da célula, e pôr fim a internalização da glicose através
destes GLUTs. A insulina é um hormônio peptídico pertencente a superfamília de
peptídeos similares a insulina (insulin-like peptides). A resposta metabólica à insulina
é mediada primariamente pela PI3K (LE ROITH e ZICK, 2001), ativando assim uma
cascata de fosforilação, que é bastante conservada evolutivamente. Um desses
componentes da via de sinalização por insulina é a proteína quinase B (PKB,
também conhecida como AKT) (Figura 6). Em humanos estão descritas 3 isoformas
de Akt, ativadas mediante a sinalização por insulina, mas com papéis fisiológicos
não redundantes. A Akt1 (PKBα) é observada como necessária ao desenvolvimento
e crescimento normais, mas dispensável na manutenção da glicemia (CHO et al.,
2001a). Dentre essas funções tem-se mostrado uma possível participação da AKT
na montagem do complexo da Hexoquinase à membrana externa da mitocôndria.
(NATHAN MAJEWSKI et al., 2004). Todavia, esses estudos envolvendo a
hexoquinase mitocondrial estão sendo realizados em mamíferos, carecendo na
literatura de trabalhos em artrópodes.

10
Figura 6: Conservação evolutiva da via de sinalização por insulina em diversas espécies. A proteína quinase b (AKT) está marcada em cada espécie por retângulo vermelho (modificado de PORTE JR, et al., 2005).
1.6 Hexoquinase no metabolismo energético e sua regulação pela Akt
Nosso grupo tem avançado no estudo do metabolismo energético durante o
desenvolvimento embrionário do carrapato R. microplus, afim de compreender a
regulação do metabolismo energético, em particular o de glicose, na tentativa de
encontrar alvos que possam ser utilizados como controle (MORAES et al. 2007;
CAMPOS et al., 2006; ABREU et al., 2009). Os estudos mostram que o metabolismo
energético do embrião do carrapato é um processo bem mais dinâmico do que
anteriormente elucidado. O estudo desse estágio de vida do carrapato R. microplus
possibilita entender como os embriões são capazes de coordenar todo o processo
de obtenção de energia, assim como o controle da mobilização e síntese das
principais moléculas de reservas. A elevada demanda energética do embrião em
desenvolvimento é suprimida pelo catabolismo de biomoléculas, tais como

11
carboidratos e lipídeos, que também participarão de vias biossintéticas para garantir
o sucesso do desenvolvimento embrionário (THOMPSON e STEWART, 1997).
A hexoquinase é a primeira enzima envolvida na via glicolítica. No citosol
fosforila a molécula de glicose no sexto Carbono, aprisionando a molécula na célula,
formando glicose 6-fosfato (Figura 7) que pode ser utilizada pela glicólise para
sintetizar piruvato que será metabolizado no ciclo do ácido cítrico produzindo ATP.
Figura 7: Esquema da reação catalisada pela Hexoquinase. A glicose, uma vez fosforilada pela Hexoquinase, pode ser utilizada para a síntese de glicogênio, ou ser oxidada pela via glicolítica para produção de ATP ou pela via das pentoses fosfato para a produção de ribose e poder redutor na forma de NADPH.
Portanto, a inibição dessa enzima pode causar consequências graves ao
organismo, como a produção de espécie reativas de oxigênio e redução dos níveis
de ATP (FRAGA et al., 2013). Essa enzima é encontrada em vários tipos de
organismos, sendo altamente conservada (CARDENAS et al., 1998, FRAGA et al.,
2013). Sabe-se que a hexoquinase pode estar livre no citosol ou associada à
membrana externa da mitocôndria no R. microplus (DA-SILVA et al., 2004; FRAGA
et al., 2013). Em mamíferos a hexoquinase mitocondrial está envolvida na redução
da taxa de H2O2 sob condições hiperglicêmicas, atuando como um antioxidante
reduzindo a geração de ROS mitocondrial (SANTIAGO et al., 2008, MEYER et al.,
2006, DA-SILVA et al., 2004, KORSHUNOV et al., 1997; WEI et al., 1997). A
hexoquinase mitocondrial está associada a membrana externa da mitocôndria no
canal iônico de voltagem (VDAC) pelos quinze aminoácidos da parte N-terminal

12
hidrofóbica (ANJA SCHINDLER e EDAN FOLEY, 2013). Devido a essa propriedade
de associar com a membrana externa da mitocôndria, a hexoquinase atua ao lado
do translocador ADP-ATP (translocador de nucleotídeos de adenina - ANT), tendo
assim um fácil acesso ao ATP ofertado pela F1F0ATP-sintase formando ADP pela
reação catalisadora da hexoquinase (BEUTNER et al., 1996; BEUTNER et al.,1997;
BEUTNER et al., 1998). Este mecanismo de ciclagem do ADP explica o
envolvimento na redução da taxa H2O2 e redução na geração de ROS (Figura 8).
Figura 8: Esquema mostrando a ligação da hexoquinase na membrana externa da mitocondria (modificado de DA-SILVA et al., 2004).
Há diversos trabalhos que mostram que AKT é necessária para manter a
associação da Hexoquinase à membrada externa da mitocôndria, e essa associação
previne mudanças na permeabilidade da membrana externa da mitocôndria,
liberação do citocromo c em respostas aos estímulos apopitóticos (MAJEWSKI et al.,
2004; MIYAMOTO et al., 2008; DAVID et al., 2013). David e seus colaboradores
(2013) mostraram que a ligação da HK mitocondrial é regulada mediante
fosforilação, onde AKT fosforila no resíduo de treonina 473 da HK, conferindo
sobrevivência e resistência aos desafios citotóxicos. A AKT não só regula a ligação
da HK à mitocôndria de uma forma direta fosforilando a HK (ZUURBIER et al., 2005;
MIYAMOTO et al., 2008; SOUTHWORTH et al., 2007), essa regulação também pode
ser de uma forma indireta, sendo a regulação da AKT age de forma negativa sobre a

13
Glicogênio Sintase Quinase 3 (GSK3), onde a GSK3 é inibida por fosforilação em
Serina 21 ou Serina 9 por AKT (CROSS et al., 1995) permitindo a ligação da HK à
membrana externa da mitocôndria. Caso o contrário, com a GSK3 ativa, ocorre o
bloqueio da associação da HK à mitocôndria, pois a GSK3 fosforila o VDAC
impedindo a associação (PASTORINO et al., 2005; AMMAJI RAJALA, et al., 2013).
O nosso grupo mostrou que a enzima GSK3 tem um papel central na
regulação do metabolismo de carboidratos em embriões de carrapato R. microplus,
podendo ser mediada pela via PI3K/AKT sugerindo ser fortemente conservada de
uma forma que integra tanto o metabolismo de glicogênio quanto a sobrevivência
celular (MORAES et al., 2007; LOGULLO et al., 2009; ABREU et al., 2013;
MARTINS et al., 2015).
Em artrópodes a função da hexoquinase associada à mitocôndria não é bem
conhecida. Uma vez que a AKT é reguladora da hexoquinase mitocondrial em outros
modelos, uma interação similar da AKT sobre o complexo da hexoquinase da
membrana da mitocôndria pode ocorrer no carrapato Rhipicephalus microplus.

14
2 JUSTIFICATIVA
Afim de entender os processos celulares que conduzem a formação do
embrião do carrapato Rhipicephalus microplus, buscamos investigar o papel da AKT
sobre a Hexoquinase das frações citoplasmática e mitocondrial. Como consequencia
serão geradas modificações nas estruturas dos embriões acreditando que este
estudo contribuirá para o desenvolvimento de fármacos que visam controlar o
carrapato R. microplus.
3 OBJETIVOS
3.1 Objetivo geral
Estudar o envolvimento da AKT sobre a atividade da Hexoquinase nas
frações citoplasmática e mitocondrial, e sua relação com a morfogênese embrionária
do carrapato Rhipicephalus microplus.
3.2 Objetivos específicos
Analisar o grau de identidade e similaridade das sequências de
aminoácidos da HK do carrapato R. microplus com as sequências de aminoácidos
de HK de outros organismos através do alinhamento;
Observar as isoformas de HK do carrapato R. microplus através da
árvore filogenética utilizando as sequências de aminoácidos da HK do carrapato R.
microplus e de outros organismos para a construção da árvore filogenética;
Avaliar a atividade de Hexoquinase nos embriões e na linhagem celular
do carrapato Rhipicephalus microplus, com diferentes concentrações do inibidor
para AKT (10-DEBC) e diferentes concentrações de dupla-fita para AKT (dsAKT);
Observar os efeitos morfológicos a partir do silenciamento da AKT em
fêmeas parcialmente ingurgitadas e na linhagem de células BME26;
Caracterizar o fenótipo obtido pelo silenciamento através da avaliação
do ovário, oviposição, eclosão e desenvolvimento dos ovos das fêmeas silenciadas
para AKT;
Analisar a viabilidade celular por MTT das células desafiadas com
diferentes concentrações de dsAKT;

15
4 MATERIAIS E MÉTODOS
4.1 Os embriões do carrapato Rhipicephalus microplus
Os embriões foram obtidos a partir de fêmeas ingurgitadas enviadas pelo
professor Dr. Itabajara Vaz da Universidade Federal do Rio Grande do Sul. Os
Carrapatos que se desenvolvem por lá são colocados no dorso do gado, e estes
bovinos ficam isolados em estábulos. Assim que as fêmeas chegaram elas foram
mantidas em câmara climática (Visomes Plus) com temperatura e umidade
controlada, ou seja, a 28°C e 80% de umidade.
4.2 Injeção do RNA de interferência em paternógenas
Fêmeas no estágio de paternógenas foram removidas dos bovinos usadas
para a injeção de 4 μg (1μl) de dsRNA especifico para AKT na posição ventral com
seringa Hamilton (33 gauge needle). Carrapatos controle foram injetados com
tampão PBS (controle de injeção) e com dsRNA de um gene nao relacionado (β-
galactosidase) de E. coli. Após a injeção as paternóginas foram alimentadas
artificialmente e os parâmetros biológicos analisados como, o número de fêmeas
que ingurgitaram, o peso dos carrapatos, a ovoposição que ocorre em um período
de 10 dias, onde foram determinados por peso dos ovos postos, e a eclosão foi
verificada 40 dias após a ovoposição através dos pesos de larvas que eclodiram
(ARIANNE FABRES et al., 2010)
4.3 Alimentação Artificial
As fêmeas no estágio de paternógenas foram alimentadas com sangue de
um bovino não infestado com carrapato. O sangue foi coletado com sistema de
coleta a vácuo, com o anticoagulante heparina. As fêmeas foram fixadas em
bandejas com pesos aproximado entre 20 a 60 mg, e separadas em grupos controle,
controle negativo (injetadas com dsRNA de um gene não relacionado)
βgalactosidade de E. coli e injetadas com dsRNA especifico para AKT.
A cada três horas o capilar era trocado e as fêmeas permaneciam
alimentando-se por 28 horas na estufa, a 28°C, com 80% de umidade relativa. As
fêmeas paternóginas eram alimentadas com sangue previamente aquecido a 37°C
com ajuda de capilares posicionadas na peça bucal das fêmeas. Os ovos de cada
grupo foram coletados e mantidos nas mesmas condições.

16
4.4 Manutenção das células BME26
Neste trabalho o estabelecimento da manutenção das células BME26 teve
como base o trabalho publicado por Eliane Esteves em 2008. Essas células foram
cultivadas em garrafas de cultivo (25 cm2) com um volume de 5mL de meio completo
(L15b), sendo o meio trocado semanalmente. Antes das células serem utilizadas nos
experimentos elas foram ajustadas a 1 X 107 células em um volume final de 5mL de
meio completo, e para tal foi observada a densidade da cultura por câmara de
Newbauer e azul de Tripan (0,4% Sigma), deixando as células a 34 °C para
promover a duplicação e confluência da população de células. Assim que as células
tornaram confluentes nas garrafas de cultura elas foram utilizadas nos experimentos.
4.5 Obtenção das frações citosólica e mitocondrial dos embriões e
células embrionária do carrapato R. microplus (BME26).
O isolamento das frações foi realizado tanto para os ovos, quanto para as
células BME26 de acordo com a metodologia de Melissa et al., (2014) com
modificações. Os homogenatos dos ovos foram feitos em tampão de lise contendo, o
tampão de isolamento mitocondrial (Manitol 225mM, Sacarose 50mM, HEPES 5mM,
pH 7,3), Triton X-100 a 10% e Coquetel de inibidores de protease (Sigma). Os
embriões foram macerados com 1ml do tampão de lise em 114,28mg de ovos (com
uma mistura do primeiro ao sétimo dia de embriogênese), sendo este homogenato
centrifugado a 100 X g por 2 minutos, a 4° C, para retirar as cascas dos ovos. O
sobrenadante foi coletado para um novo tubo cônico de 1,5ml e o precipitado
descartado, o sobrenadante coletado foi passado em seringa de insulina por 10
vezes e novamente centrifugado a 300 X g por 6 minutos a 4° C. O sobrenadante foi
coletado e o precipitado descartado mais uma vez, o sobrenadante coletado foi
submetido a mais uma centrifugação a 1000 X g por 10 minutos a 4° C, onde se
precipita o núcleo, o precipitado foi descartado novamente e o sobrenadante mais
uma vez recolhido para um novo tubo cônico de 1,5ml e centrifugado a 12.000 X g
por 15 minutos a 4° C, etapa no qual o precipitado corresponde a fração mitocondrial
que foi posta em suspensão em 250µl de tampão de isolamento mitocondrial, e o
sobrenadante centrifugado em ultracentrífuga a 100.000 X g por 60 minutos a 4° C
para obtenção da fração citosólica.

17
As células BME26 também foram rompidas passando cerca de 10 vezes em
seringa de insulina, estas células homogeneizadas em tampão de lise com os
mesmos componentes utilizados para obtenção do homogeneizado de ovos. Os
próximos passos para obtenção das frações mitocondrial e citosólica das células são
os mesmos usados para obtenção destas frações em embriões a partir da
centrifugação a 300 X g por 6 minutos a 4° C.
4.6 Inibição da AKT por 10-DEBC de embriões e células embrionárias
do carrapato R. microplus (BME26)
No homogenato de embriões do primeiro ao sétimo dia do desenvolvimento
e no homogenato das células foram adicionado o inibidor da AKT 10-[4’-(N,N-
Diethylamino)butyl]-2-chlorophenoxazine hydrochloride (10-DEBC hydrochloride) nas
concentrações de 12µM, 24µM e 48µM e realizado o fracionamento celular, ou seja,
a separação de mitocôndria (fração mitocondrial) e citoplasma (fração
citoplasmática). A Atividade da Hexoquinase foi determinada nas respectivas frações
mitocondrial e citosólica obtidas do homogenato tratado de embriões e células.
4.7 Atividade de Succinato Desidrogenase (SDH).
Atividade de Succinato Desidrogenase foi determinada por adaptação do
método de Singer (1974) para avaliar a qualidade do fracionamento, utilizando 2,6
Diclorofenolindofenol (DCPIP) e Metasulfato de Fenazina (PMS) como aceitadores
artificiais de elétrons. A velocidade de redução do DCPIP foi monitorada em 600 nm.
O sistema de reação em volume final de 1 ml constituído de tampão fosfato de sódio
50 mM (pH 7,4), succinato de sódio 20 mM, EDTA 2 mM, Azida 10 mM, PMS 1mM e
20µl do homogenato (sendo padronizado por proteína). A reação aconteceu a 28 °C
em volume final de 1ml, e iniciou-se a leitura ao adicionar 180 µM de DCPIP (T0). Os
resultados são expressos em micromol de DCPIP reduzido.min-1.mg-1 de proteína,
utilizando o coeficiente de extinção molar Ɛ550 nm = 19.000 M-1.cm-1 para o DCPIP
reduzido.
4.8 Atividade da glicose-6-fosfato desidrogenase
A amostra (20ul de homogenato) foi adicionada em meio de reação
contendo Tris-HCl 50 mM pH 7,8, βNAD+ 6 mM e glicose 6 fosfato 100 mM. A

18
atividade a 30° C foi determinada espectrofotometricamente a 340 nm pela taxa de
formação de NAD(P)H de acordo com Worthington (1988).
4.9 Atividade da Hexoquinase (HK)
A amostra (20ul de homogenato) foi adicionada em meio de reação
contendo TRIS-HCl 20 mM pH 8. Em tubos de ensaio foram adicionados 385 uL de
meio de reação contendo MgCl2 6 mM, ATP 0,5 mM e β-NAD+ 0,5 mM, glicose 6-
fosfato desidrogenase 3 unidades/mL (Sigma-Aldrich Chemicals). A reação foi
disparada com glicose 2 mM. A leitura das amostras foi realizada em
espectrofotômetro a 340nm em intervalos de 1 minuto, durante 10 minutos de
reação. O ensaio foi feito à temperatura de 30°C. A atividade total de HK foi
determinada indiretamente pela produção de β-NADH, acoplada ao consumo de
glicose-6-fosfato pela glicose 6-fosfato desidrogenase. O método fundamenta-se na
detecção de β-NADH a 340 nm, que é proporcional à velocidade de formação de
glicose 6-fosfato no meio (GALINA et al., 1999). A atividade especifica foi obtida pela
razão entre a atividade total, proporcional à formação de β-NADH, pela quantidade
de proteínas (mg) na amostra.
4.10 Síntese de RNA de interferência (RNAi) para RmAKT
Para o silenciamento gênico da Rm-AKT tomamos como base o processo
descrito por Abreu e colaboradores (2013) e Fabres e colaboradores (2010). Como
controle negativo foi usada dsRNA para β-galactosidase de E. coli, gentilmente
doada pelo Professor Marcos H. Sorgine, do Instituto de Bioquímica Médica da
Universidade Federal do Rio de Janeiro. O tamanho do dsRNA sintetizado foi de 609
pares de bases (pb) para dsAKT.
O RNAi foi desenhado para o gene da AKT específico para o carrapato R.
microplus utilizando a plataforma de bioinformática primer 3. Foi selecionada uma
região apropriada do gene RmAKT utilizando uma sequência depositada em banco
de dados (acesso JX648548).
RNA total foi extraído de células usando o reagente Trizol (invitrogen) como
descrito pelo fabricante. O cDNA foi sintetizado a partir de 1 μg de RNA total. A
reação foi realizada na presença de RT Randon primer e transcriptase reversa
(High-capacity cDNA Reverse Transcripion Kit-Applied Biosystems), de acordo com

19
as instruções do fabricante. Depois da transcrição reversa, foram realizadas duas
etapas de reação de PCR utilizando primers para o gene AKT projetados como
descrito acima, sendo a primeira reação de PCR contendo um fragmento de 8
nucleotídeos (estratégia utilizada visando a técnica de hibridização in situ).
Iniciadores utilizados na primeira reação RmAKT-F: 5’-
ggccgccggGGCATTATCTACCGGGACCT-3’ e RmAKT-R: 5’-
cccgggccCTGCTGGAAGTACGGCTGTT-3’. Para a inserção da sequência do
promotor T7 para a síntese in vitro da dsRNA foram utilizados iniciadores chamados
de Universais, U-F: 5’-AGGGATCCTAATACGACTCACTATAGGGCCCGGGGC-3’ e
U-R: 5’-GAGAATTCTAATACGACTCACTATAGGGCCGCGG-3’. Os amplicons
gerados foram purificados do gel com o Kit de purificação (illustraTM GFXTM PCR
DNA and Gel Band Purification Kit) conforme o protocolo do fabricante. A síntese do
dsRNA foi realizada utilizando a enzima RNA polimerase T7 de acordo com as
especificações do fabricante (T7 RiboMAXTM Express RNAi System -Promega).
4.11 Silenciamento gênico nas células BME26
Células BME26 foram semeadas em placas de 24 poços (5×105
células/poço), acrescentando meio completo para um volume final de 500 μL. Após
de 24 horas de incubação a 34°C, o meio de cultura foi substituído com 200 μL de
meio fresco contendo de 4 e 6 μg de dsRNA por poço, e misturado por agitação
orbital suave. Após 24h com 200 μL de meio com a dsAKT, foram adicionados 300
μL de meio e as células foram incubadas novamente durante um período de 24
horas, para as análises de viabilidade celular e da expressão relativa dos alvos AKT
(para a confirmação do silenciamento) e da HK (LEONARDO ABREU et al., 2013).
4.12 Ensaio de viabilidade celular
Após os tratamentos com dsAKT nas células BME26, 50 μl de brometo de 3-
[4,5-dimetil-tiazol-2-il]-2,5-difeniltetrazólio (MTT) foi adicionado a cada poço. Depois
de 2 horas de incubação a 34°C, a solução foi completamente descartada e 1 ml de
álcool isopropílico com ácido clorídrico fumegante foi adicionado (0,15% de HCl)
para dissolver os cristais de formazan. A mistura foi transferida para tubos de 1.5 ml
e centrifugada a 6000 rpm durante 15 min.

20
Os sobrenadantes foram coletados em novos tubos e a absorbância foi
mesurada em 570 nm. Os valores de absorbância do tratamento controle foram
utilizados para a normalização (100% de viabilidade).
4.13 Análise da transcrição gênica de AKT e HK por qPCR
O RNA total foi extraído de células BME26 após as células serem
silenciadas com 4, 6 e 8 μg de dsAKT usando o reagente Trizol (Invitrogen) como
descrito pelo fabricante.
O cDNA foi sintetizado a partir de 1 μg de RNA total. A reação foi realizada
na presença de RT random primer e transcriptase reversa (High-capacity cDNA
Reverse Transcripion Kit – Applied Biosystems), de acordo com as instruções do
fabricante. Depois da reação da transcrição reversa, o produto de cada amostra foi
utilizada para a reação de PCR em tempo real usando Primers de genes específicos
com Kit Master SYBR Green (Roche). As análises de transcrição relativa foram
realizadas utilizando Step One Plus (Applied Biosystems, USA) e para as curvas de
calibração foram realizadas diluições seriadas do cDNA. Os valores de eficiência
para cada gene estudado foram superiores a 90% para o cálculo de transcrição
relativa. Os iniciadores utilizados, para RmAKT: 5’- GGCCAAAGCCATTCACCTTCA-
3’ Forward, 5’- CCTCCTCACTCGCCAACTTCTC-3’ Reverse (Tm= 58°C, amplicom
de 151 pares de base), e os iniciadores utilizados para RmHK:5’-
CATGGACAAAGAGCTTCAACTGCTC-3’ Forward, 5’-
GGAAAGCTCCCTTGACCAGGGTA-3’ Reverse (Tm=59°C, amplicon de 150 bp). Os
iniciadores específicos utilizados como gene housekeeping, o fator de alongamento
1A (Elf1A) (número de acesso EW679365) (Nijhof et al., 2009). A expressão relativa
foi determinada utilizando os valores de Ct de cada análise, em triplicata, na
expressão relativa no Software Tool-REST (PFAFFJ , 2001). Elf1A foi utilizado como
gene de referência em células silenciadas com β galactosidase, padronizando como
calibrador.
4.14 Busca de sequencias similares a HK
As sequencias da proteína HK1 de camundongo (número de acesso do
GeneBanK XP_021061143) foi utilizada em buscas em bancos de dados utilizando a
ferramenta BLAST (ALTSHULZ el al., 1990; ALTSHULZ et al., 1997). As sequências

21
de proteínas encontradas foram utilizadas para alinhamentos utilizando a ferramenta
CLUSTALW (LARKIN et al., 2007) e árvores filogenéticas utilizando a ferramenta
MEGA (EDGAR, 2004).
4.15 Análise estatística
Na avaliação da qualidade do fracionamento celular tanto em células quanto
em embriões, onde analisamos as atividades das enzimas SDH e G6PDH,
realizamos o teste não paramétrico (teste t). O número de amostras biológicas (n)
foram de 2 para as análises em embriões e de 3 para as análises em células.
Nas análises da atividade de HK em embriões e nas células BME26 que
foram inibidas com diferentes concentrações do inibidor para AKT (10-DEBC), e
silenciadas com diferentes concentrações de dupla-fita, foi utilizado o teste two way
anova e o Bonferroni como pós-teste. O número de amostras biológicas (n) foram 2.
Para as análises da viabilidade celular, os valores de absorbância do
tratamento controle foram utilizados para a normalização (100% de viabilidade). O
número de amostras biológicas (n) foram 3. Análise não paramétrica e pós teste de
Tukey foram usados para determinar diferenças estatísticas neste experimento.
Para as análises de expressão relativa por qRT-PCR foi usado o teste não
paramétrico (test t) com o objetivo de comparar as diferenças entre o grupo
considerado como controle e os grupos tratados. O número de amostras biológicas
(n) foi de 2.
Os dados são apresentados como média ± desvio padrão (DP). Valores de
p<0.05 foram considerados significativos. Todas as análises estatísticas foram
realizadas utilizando o programa Prism 5.0 (GraphPad Software, Inc., USA).

22
5 RESULTADOS
5.1 Análise da homologia da sequência de aminoácidos da enzima
Hexoquinase do carrapato R. microplus.

23
Figura 9: Alinhamento da sequência de aminoácidos de HK de R. micróplus, I. escapularis, D. melanogaster, H. sapiens, Mus musculus e T. castaneum. A primeira linha corresponde a sequência da HK presente em R. microplus. A marcação em preto e cinza correspondem ao grau de identidade e similaridade entre os aminoácidos. A caixa em azul representa os 15 aminoácidos da região N-terminal hidrofóbica. A caixa marrom representa o domínio regulatório, e a caixa verde o domínio catalítico.
A sequência de aminoácidos da HK de R. microplus (da primeira linha-R.
microplus_100.42) e dos outros organismos utilizados no alinhamento foram obtidas
do banco de dados GenBank/NCBI e FlyBase. Uma nova sequência de aminoácidos
de R. microplus da segunda linha (R.microplus_96.2) foi obtida a partir de um banco
de dados do transcriptoma de nosso grupo de pesquisa fornecido pelo Professor
Itabajara Vaz.
Tendo como base as características e funções das sequências de
aminoácidos das HKs de mamíferos (ANJA SCHINDLER e EDAN FOLEY., 2013), os
15 aminoácidos da parte N-terminal hidrofóbica das duas sequências de carrapato
não possuem um grau satisfatório de identidade e similaridade com as sequências
de mamíferos. Além do mais, podemos observar que existem duas regiões, uma N-
terminal e outra C-terminal que possuem alta similaridades entre si. Estas regiões

24
também possuem alta identidade e similaridade com as outras sequências de
aminoácidos de HK para os demais organismos (marcados em preto e cinza na
Figura 9). A atividade catalítica (caixa verde da Figura 9) está associada à região C-
terminal, enquanto a função regulatória está associada à região N-terminal, e cada
domínio pode se ligar a uma molécula de glicose e glicose 6-fosfato. De acordo com
o alinhamento esses domínios são bastante conservados entre os organismos. De
qualquer forma nossos resultados indicam a presença de duas isoformas codificadas
por dois genes distintos de HK em carrapato.
5.2 Análise filogenética da Hexoquinases de carrapato
Figura 10: Análise filogenética da Hexoquinase de R. microplus, I. scapularis, D. melanogaster, H. sapiens, Mus musculus e T. castaneum. As relações evolutivas foram inferidas utilizando o método de agrupamento de vizinhos (Neighbor-Joining). A figura mostra a árvore filogenética com o somatório das distâncias evolutivas dos galhos de 0.20. O alinhamento das sequências obtidas foi criado usando o Muscle no software 5.0 Mega (EDGAR, 2004).
A análise das relações filogenéticas das HKs utilizou as mesmas sequências
do alinhamento acima. A sequência de aminoácidos de HK de R. microplus
encontrada no banco de dados do NCBI (R. microplus 100.42 da Figura 10)

25
encontra-se agrupada com a sequência de HK de outra espécie de carrapato, Ixodes
scapularis e das sequências de HK-2 de H. sapiens e Mus musculus.
Diferentemente da sequência encontrada no banco de dados do NCBI, a sequência
obtida do transcriptoma do grupo (R. microplus 96.2) não foi agrupada com as duas
sequências de carrapato. Tendo em vista que nossas análises preliminares não
identificaram duas HK em I. scapularis é possível que esta duplicação seja
específica de R. microplus. Nota-se que ocorreram duplicações independentes em
vertebrados, nos insetos como o T. castaneum e D. melanogaster, (Figura 10).
Assim, nossos resultados sugerem fortemente que mais de uma isoforma de HK
está presente em R. microplus que podem possuir funções diferentes.
5.3 Qualidade do fracionamento celular de embriões do carrapato
Rhipicephalus microplus.
As enzimas Succinato desidrogenase (SDH) e a Glicose 6 fosfato
desidrogenase (G6PDH) foram usadas como enzimas marcadoras para a qualidade
do fracionamento celular de embriões do carrapato R. microplus (descrito em
materiais e métodos) (Figura 11). SDH é uma flavoproteína intermembranar
encontrada na membrana interna da mitocôndria sendo uma proteína que faz parte
do complexo da cadeia respiratória e utilizada neste trabalho para determinar a
qualidade da fração citosólica. Atividade desta enzima é encontrada na fração
mitocondrial, porém não foi encontrada atividade na fração citosólica, demonstrando
assim, uma qualidade satisfatória do fracionamento e isolamento desta fração.
(Figura 11, A).
Dado em conjunto a atividade de SDH, a atividade G6PDH também foi
mensurada para a validação da qualidade do fracionamento. A G6PDH é uma
enzima reguladora da via das pentoses, convertendo glicose 6 fosfato em 6-
fosfogliconato, produzindo nesse processo NADPH. A G6PDH está presente no
citoplasma e neste trabalho é utilizada para determinar a pureza da fração
mitocondrial. A atividade desta enzima foi encontrada na fração citosólica e
consequentemente não foi detectada na fração mitocondrial dos embriões do
carrapato R. microplus, mostrando um fracionamento satisfatório e isolamento
mitocondrial. (Figura 11, B).
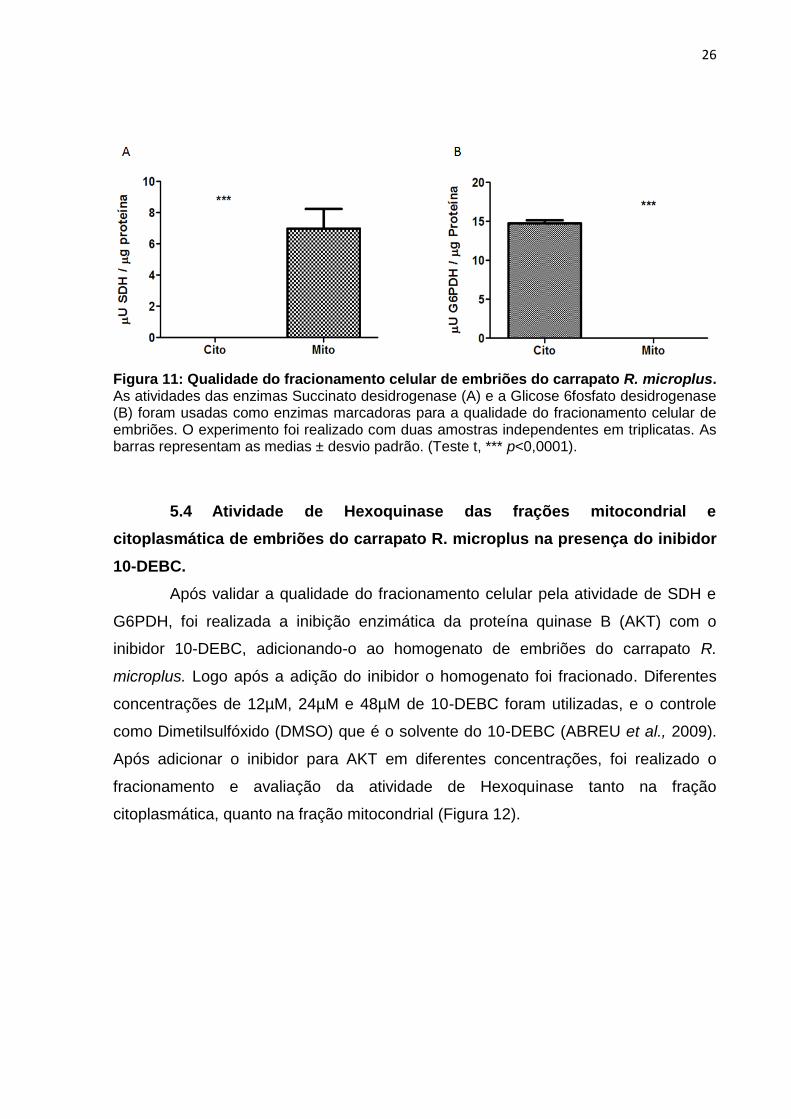
26
Figura 11: Qualidade do fracionamento celular de embriões do carrapato R. microplus. As atividades das enzimas Succinato desidrogenase (A) e a Glicose 6fosfato desidrogenase (B) foram usadas como enzimas marcadoras para a qualidade do fracionamento celular de embriões. O experimento foi realizado com duas amostras independentes em triplicatas. As barras representam as medias ± desvio padrão. (Teste t, *** p<0,0001).
5.4 Atividade de Hexoquinase das frações mitocondrial e
citoplasmática de embriões do carrapato R. microplus na presença do inibidor
10-DEBC.
Após validar a qualidade do fracionamento celular pela atividade de SDH e
G6PDH, foi realizada a inibição enzimática da proteína quinase B (AKT) com o
inibidor 10-DEBC, adicionando-o ao homogenato de embriões do carrapato R.
microplus. Logo após a adição do inibidor o homogenato foi fracionado. Diferentes
concentrações de 12µM, 24µM e 48µM de 10-DEBC foram utilizadas, e o controle
como Dimetilsulfóxido (DMSO) que é o solvente do 10-DEBC (ABREU et al., 2009).
Após adicionar o inibidor para AKT em diferentes concentrações, foi realizado o
fracionamento e avaliação da atividade de Hexoquinase tanto na fração
citoplasmática, quanto na fração mitocondrial (Figura 12).
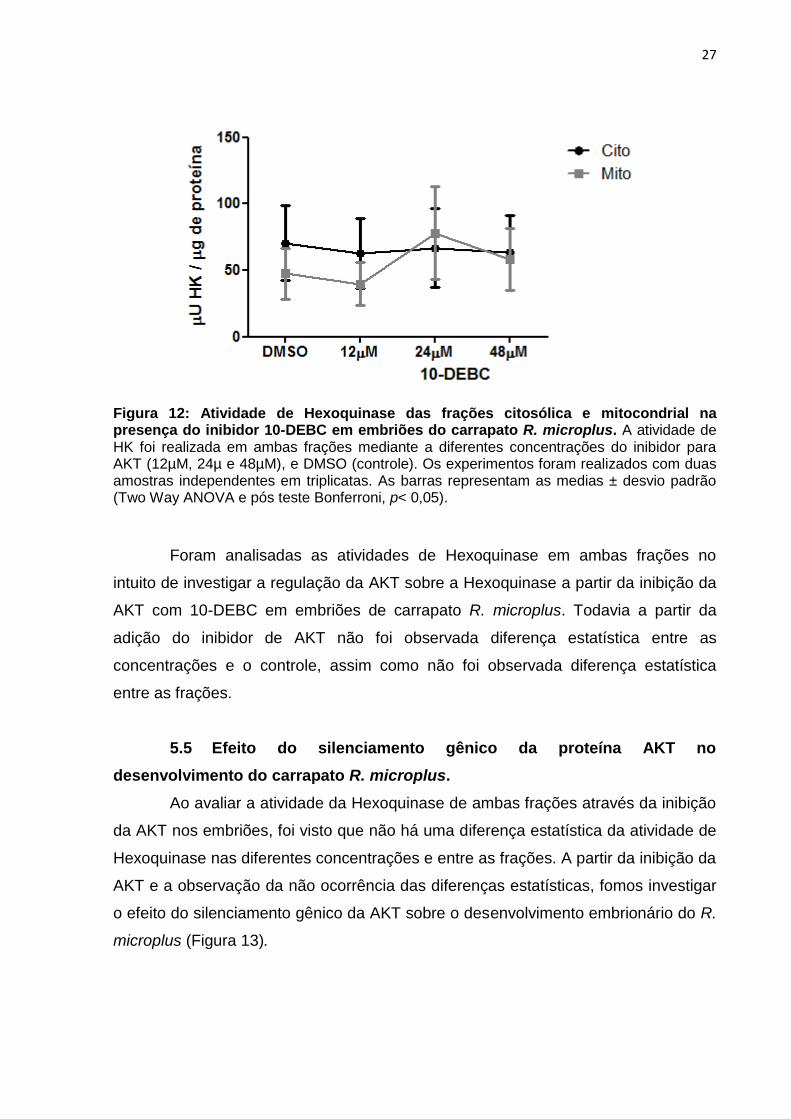
27
Figura 12: Atividade de Hexoquinase das frações citosólica e mitocondrial na presença do inibidor 10-DEBC em embriões do carrapato R. microplus. A atividade de HK foi realizada em ambas frações mediante a diferentes concentrações do inibidor para AKT (12µM, 24µ e 48µM), e DMSO (controle). Os experimentos foram realizados com duas amostras independentes em triplicatas. As barras representam as medias ± desvio padrão (Two Way ANOVA e pós teste Bonferroni, p< 0,05).
Foram analisadas as atividades de Hexoquinase em ambas frações no
intuito de investigar a regulação da AKT sobre a Hexoquinase a partir da inibição da
AKT com 10-DEBC em embriões de carrapato R. microplus. Todavia a partir da
adição do inibidor de AKT não foi observada diferença estatística entre as
concentrações e o controle, assim como não foi observada diferença estatística
entre as frações.
5.5 Efeito do silenciamento gênico da proteína AKT no
desenvolvimento do carrapato R. microplus.
Ao avaliar a atividade da Hexoquinase de ambas frações através da inibição
da AKT nos embriões, foi visto que não há uma diferença estatística da atividade de
Hexoquinase nas diferentes concentrações e entre as frações. A partir da inibição da
AKT e a observação da não ocorrência das diferenças estatísticas, fomos investigar
o efeito do silenciamento gênico da AKT sobre o desenvolvimento embrionário do R.
microplus (Figura 13).
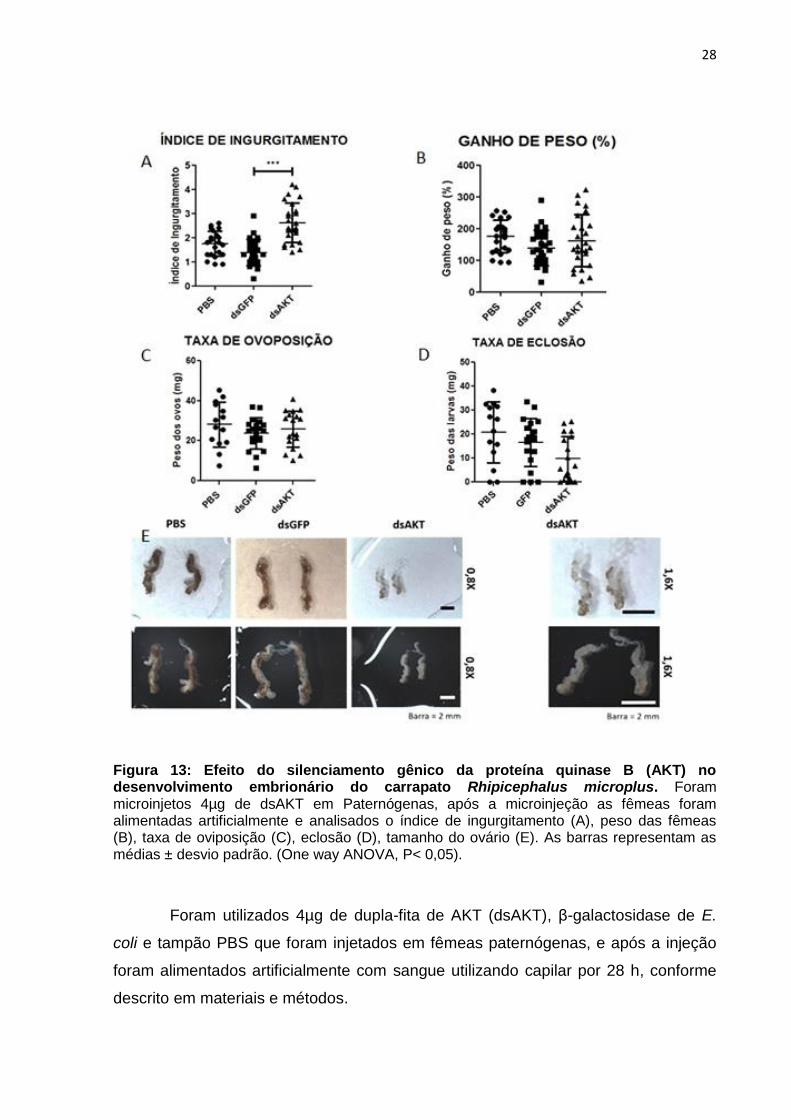
28
Figura 13: Efeito do silenciamento gênico da proteína quinase B (AKT) no desenvolvimento embrionário do carrapato Rhipicephalus microplus. Foram microinjetos 4µg de dsAKT em Paternógenas, após a microinjeção as fêmeas foram alimentadas artificialmente e analisados o índice de ingurgitamento (A), peso das fêmeas (B), taxa de oviposição (C), eclosão (D), tamanho do ovário (E). As barras representam as médias ± desvio padrão. (One way ANOVA, P< 0,05).
Foram utilizados 4µg de dupla-fita de AKT (dsAKT), β-galactosidase de E.
coli e tampão PBS que foram injetados em fêmeas paternógenas, e após a injeção
foram alimentados artificialmente com sangue utilizando capilar por 28 h, conforme
descrito em materiais e métodos.
29
Em relação ao ingurgitamento das fêmeas de carrapato após a alimentação
artificial, as fêmeas silenciadas para AKT apresentam um aumento na ingestão de
sangue se comparadas ao grupo controle (Figura 13, A). Enquanto a eclosão dos
ovos, ao peso dos carrapatos e a taxa de oviposição das fêmeas de carrapato após
a alimentação artificial não ocorreram diferenças significativa entre controle e o
grupo silenciado para AKT (Figura 13, B e C), indicando que a dsRNA para AKT não
interfere no ganho de peso, oviposição e nem na eclosão (Figura 13, D).
Além disso, as fêmeas silenciadas para AKT apresentaram uma redução no
tamanho do ovário quando comparado ao controle (Figura 13, E). Todavia, a
redução no tamanho do ovário em fêmeas silenciadas para AKT aparentemente não
interferiu na taxa de oviposição e eclosão quando comparados com o controle.
5.6 Qualidade do fracionamento em linhagem celular BME26.
Além dos estudos in vivo (como os ovos, por exemplo), o grupo vem
utilizando a linhagem celular BME26 para estudar alguns aspectos bioquímicos que
também estão envolvidos na embriogênese (CAMPOS et al., 2006; MORAES et al.,
2007; ABREU et al., 2009; ARIANNE FABRES et al., 2010; SANTOS et al., 2013).
Para estudarmos o envolvimento da AKT sobre a Hexoquinase na linhagem celular
BME26 também fracionamos as células utilizando os mesmos marcadores
bioquímicos usados em ovos para avaliarmos a qualidade do fracionamento,
descrito em materiais e métodos (Figura 14).
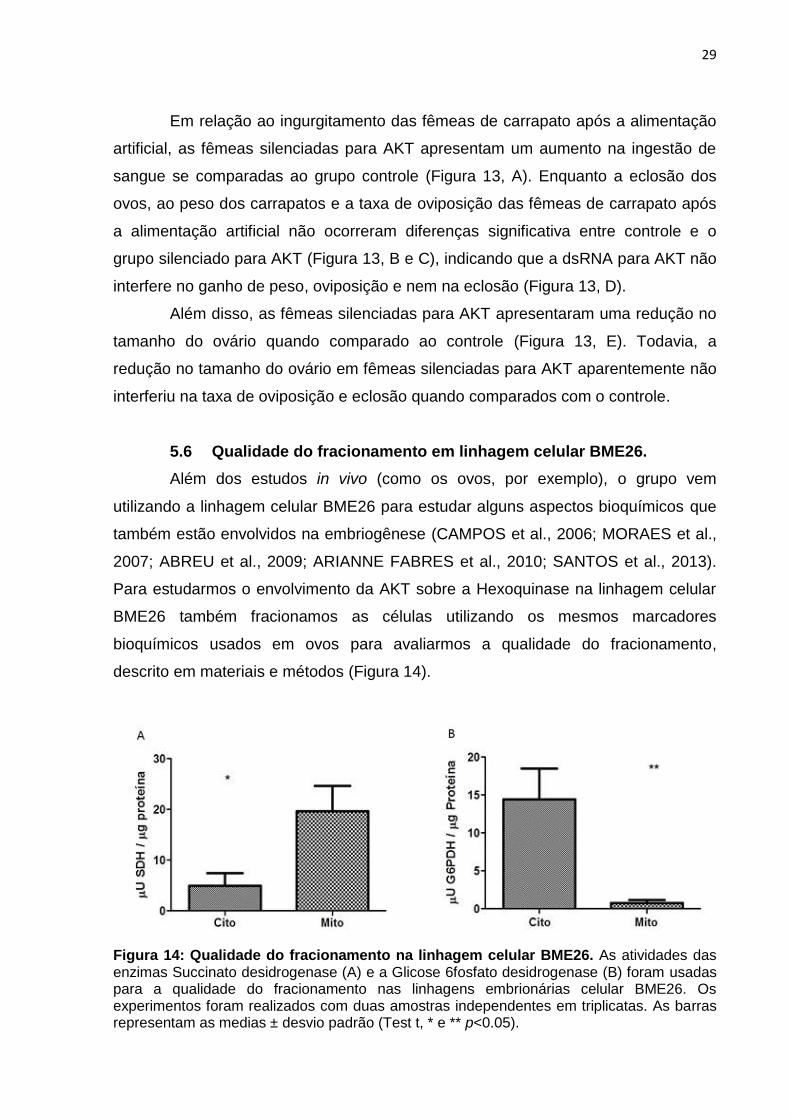
Figura 14: Qualidade do fracionamento na linhagem celular BME26. As atividades das enzimas Succinato desidrogenase (A) e a Glicose 6fosfato desidrogenase (B) foram usadas para a qualidade do fracionamento nas linhagens embrionárias celular BME26. Os experimentos foram realizados com duas amostras independentes em triplicatas. As barras representam as medias ± desvio padrão (Test t, * e ** p<0.05).

30
A atividade da SDH (Figura 14, A) indica que foi possível o isolamento da
fração mitocondrial, com a atividade de SDH significativamente maior na fração
mitocondrial se comparada a fração citosólica. Em conjunto a atividade de SDH para
determinar a qualidade do fracionamento, a atividade de G6PDH (Figura 14, B)
também foi mensurada e é significativamente maior na fração citosólica se
comparado com a fração mitocondrial, mostrando uma qualidade satisfatória do
fracionamento celular e isolamento da fração mitocondrial. Estes resultados nos
permitem avaliar o padrão da atividade da HK em ambas frações sobre o
silenciamento e inibição da AKT das células embrionárias BME26.
5.7 Avaliação da atividade da Hexoquinase na linhagem celular
BME26 desafiadas com inibidor 10-DEBC e silenciadas para AKT.
Após validar a qualidade do fracionamento celular através da atividade das
enzimas SDH e G6PDH nas frações citoplasmáticas e mitocondriais das células
BME26, a atividade da HK em ambas frações foi avaliada diante a inibição e o
silenciamento da AKT afim de compreender a regulação da AKT sobre a HK (Figura
15).
Assim como no homogenato dos ovos do carrapato R. microplus, também
utilizamos o DMSO como controle e o inibidor 10-DEBC em diferentes
concentrações (12µM, 24µM e 48µM) para inibição da AKT na linhagem celular
BME26. Logo após a adição do DMSO ou do inibidor ao homogenato foi realizado o
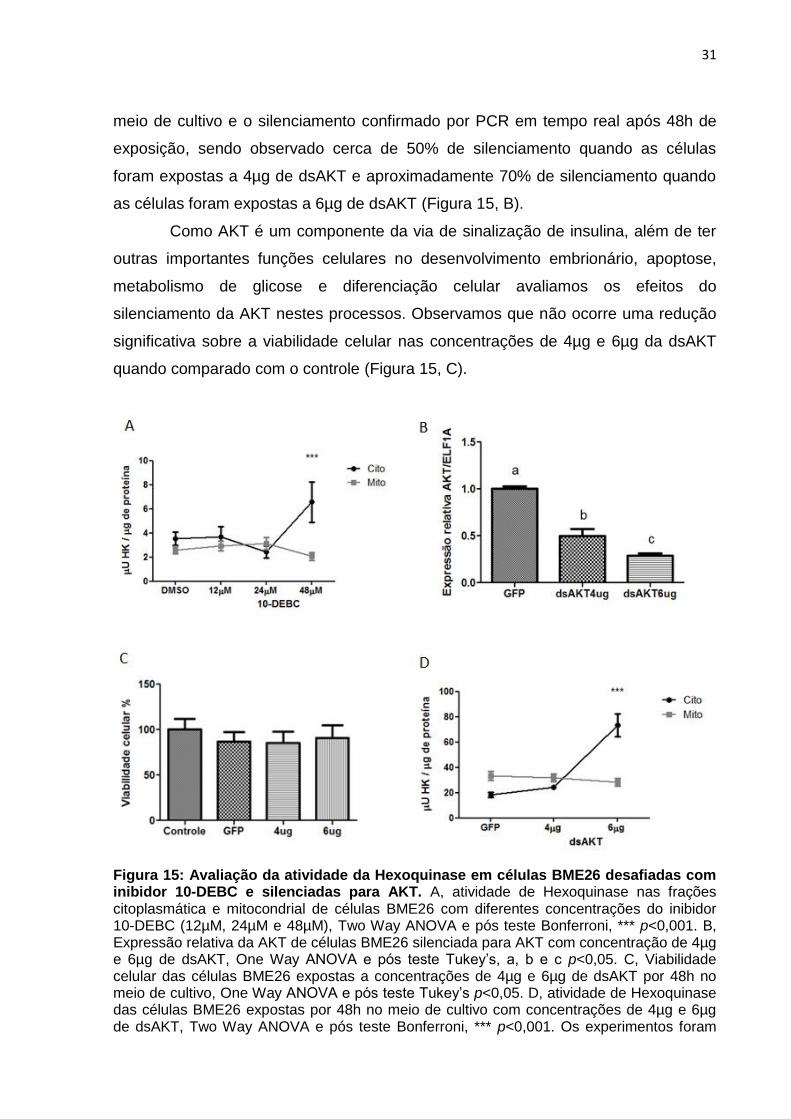
fracionamento (Figura 15, A). Em relação a atividade de HK das células BME26 com
concentrações de 12µM e 24µM do inibidor 10-DEBC não é observada uma
diferença estatística entre as frações, e nem entre as concentrações comparadas ao
controle. Entretanto, na concentração de 48µM do inibidor foi observado um
aumento significativo da atividade de HK na fração citoplasmática em relação a
fração mitocondrial se comparado as concentrações de 12µM, 24µM do inibidor 10-
DEBC e ao controle. Estes resultados sugerem que na utilização do inibidor na
concentração de 48µM possa ocorrer algum tipo de regulação da AKT sobre a HK.
Além da inibição da AKT em diferentes concentrações do inibidor 10-DEBC
em células BME26, também foi realizado o silenciamento gênico da AKT nas células
BME26 em duas concentrações de 4µg e 6µg de dupla fita para AKT (dsAKT), como
descrito em materiais e métodos. As células BME26 foram expostas a dsAKT no
31
meio de cultivo e o silenciamento confirmado por PCR em tempo real após 48h de
exposição, sendo observado cerca de 50% de silenciamento quando as células
foram expostas a 4µg de dsAKT e aproximadamente 70% de silenciamento quando
as células foram expostas a 6µg de dsAKT (Figura 15, B).
Como AKT é um componente da via de sinalização de insulina, além de ter
outras importantes funções celulares no desenvolvimento embrionário, apoptose,
metabolismo de glicose e diferenciação celular avaliamos os efeitos do
silenciamento da AKT nestes processos. Observamos que não ocorre uma redução
significativa sobre a viabilidade celular nas concentrações de 4µg e 6µg da dsAKT
quando comparado com o controle (Figura 15, C).
Figura 15: Avaliação da atividade da Hexoquinase em células BME26 desafiadas com inibidor 10-DEBC e silenciadas para AKT. A, atividade de Hexoquinase nas frações citoplasmática e mitocondrial de células BME26 com diferentes concentrações do inibidor 10-DEBC (12µM, 24µM e 48µM), Two Way ANOVA e pós teste Bonferroni, *** p<0,001. B, Expressão relativa da AKT de células BME26 silenciada para AKT com concentração de 4µg e 6µg de dsAKT, One Way ANOVA e pós teste Tukey’s, a, b e c p<0,05. C, Viabilidade celular das células BME26 expostas a concentrações de 4µg e 6µg de dsAKT por 48h no meio de cultivo, One Way ANOVA e pós teste Tukey’s p<0,05. D, atividade de Hexoquinase das células BME26 expostas por 48h no meio de cultivo com concentrações de 4µg e 6µg de dsAKT, Two Way ANOVA e pós teste Bonferroni, *** p<0,001. Os experimentos foram

32
realizados com duas amostras independentes em triplicatas. As barras representam as medias ± desvio padrão.
Por último ao observar que nenhuma das duas concentrações de dsAKT
afetou a viabilidade celular das células BME26, foi mensurada a atividade de HK em
ambas frações com o silenciamento de AKT nas concentrações de 4µg e 6µg de
dsAKT. Em células expostas a 4µg de dsAKT não houve diferença estatística na
atividade de HK quando comparado ao controle. Por outro lado, células expostas a
6µg de dsAKT demonstraram um aumento significativo na fração citoplasmática em
relação a fração mitocondrial quando comparado com o controle. Estes resultados
sugerem que possa ocorrer algum tipo de regulação da AKT sobre a HK quando as
células BME26 são expostas a 6µg de dsAKT. Este resultado é similar ao
previamente descrito na utilização do inibidor de AKT na concentração de 48µM.

33
6 DISCUSSÃO
São descritas diversas formas de controle do carrapato R. microplus na
tentativa de minimizar as perdas na área pecuária por esses ectoparasitas
hematófagos (MEHLHOM et al., 2011; SEIXAS et al., 2010; DE LA FUENTE et al.,
2007; WILLADSEN P. 1997). Dos mais diversos tipos de controle deste ectoparasita
os mais eficazes e os mais utilizados atualmente são as vacinas e os acaricidas. Os
acaricidas são grupos de produtos químicos que apresentam resultados imediatos,
porém contribuem para seleção de carrapatos mais resistentes (Figura 3). As
vacinas têm como estratégia um método imunológico, tornando gados mais
resistentes as ações espoliativas do carrapato.
Nosso grupo de pesquisa tem se esforçado em entender os processos
dinâmicos na utilização de nutrientes e a regulação metabólica em embriões de
carrapatos e mosquitos (MORAES et al., 2007; VITAL et al., 2010; ABREU et al.,
2009). Dentre as vias metabólicas estudadas, a gliconeogênese, a via de sinalização
de insulina e a síntese de glicogênio se destacam por sofrerem uma regulação
diferenciada (não-clássica) ao longo dos estágios embrionários. O metabolismo
dessas células embrionárias do carrapato (BME26) responde à insulina, ativando a
cascata de fosforilação PI3K/AKT, permitindo a internalização de glicose e
consequente aumento da atividade de Hexoquinase (HK) (ABREU et al., 2009 e
2012; MORAES et al., 2007). Essa relação entre via de sinalização de insulina (ISP)
e glicólise é bem estabelecida na literatura, entretanto nenhuma relação direta entre
AKT e HK foi investigada em embriões do R. microplus.
Vários estudos sugerem que a HK mitocondrial é regulada pela AKT,
mediando crescimento e sobrevivência celular em vertebrados (BRYSON et al.,
2002; GOTTLOB et al., 2001; MAJEWSKI et al., 2004; DA-SILVA et al., 2004). Foi
demonstrado que a ativação de AKT levava à inibição da dissociação da HK da
mitocôndria em células fibroblásticas de rato (GOTTLOB et al., 2001). A dissociação
da HK da membrana mitocondrial aumenta o potencial de membrana e reduz a
atividade do translocador ADP/ATP, gerando ROS e causando danos às células
(DA-SILVA et al., 2004).
No presente trabalho a atividade de HK foi investigada em frações
mitocondrial e citoplasmática na presença do inibidor da AKT (10-DEBC) afim de
determinar se haveriam diferenças cinéticas nos respectivos compartimentos em

34
embriões do carrapato R. microplus. Os dados mostram que existe atividade de HK
em frações mitocondriais em carrapatos R. microplus. Recentemente Fraga et al.
(2013) também mostrou atividade de HK em frações mitocondriais obtidas dos
estágios iniciais da embriogênese do carrapato R. microplus, sugerindo a existência
de uma isoforma mitocondrial dessa enzima. No entanto, a adição do inibidor de
AKT não afetou a atividade de HK em nenhuma das frações investigadas em
embriões do carrapato neste trabalho (Figura 12).
O inibidor de AKT também foi administrado em células embrionárias em
cultura (BME26) do carrapato R. microplus. Neste caso foi observada uma diferença
significativa da atividade de HK somente após a adição de 48uM do inibidor (Figura
15, A). Esta atividade foi mais elevada na fração citoplasmática quando comparado
à fração mitocondrial. Os resultados sugerem que as HKs presentes nas respectivas
frações sofrem uma regulação diferenciada na AKT. Em células HeLa já foram
demonstradas que a ação regulatória da AKT ocorre sobre a HK-I (ANJA
SCHINDLER e EDAN FOLEY, 2013). Entretanto, em outros trabalhos essa
regulação é demonstrada sobre a HK-II (NEARY et al., 2013; DAVID et al., 2013),
De fato, em humanos existem quatro isoformas de HK, podendo cada uma
ser regulada diferentemente. Dentre as quatro isoformas de mamíferos, as HK-I e
HK-II apresentam características como uma sequência N-terminal hidrofóbica que
confere o ancoramento à membrana externa da mitocôndria no canal iônico de
voltagem (VDAC). A HK-III vem sendo encontrada próximo ao núcleo, sendo o seu
papel desconhecido (WILSON et al., 2003; KATZEN e SCHIMKE et al., 1965). A HK-
II também foi localizada no núcleo quando a AKT foi inibida, mas o papel dessa
migração para o núcleo é ainda desconhecido (CATHERINE e PASTORINO, 2013).
Esses trabalhos reforçam que ainda não há um consenso se ambas as isoformas
são reguladas por AKT em células de mamífero, ou apenas uma das isoformas, ou
essas discrepâncias observadas são particularidades do câncer.
O fato é que essa enzima é altamente conservada e encontrada em vários
tipos de organismos (FRAGA et al., 2013; CARDENAS et al., 1998). Em alguns
artrópodes foi demonstrado a existência de mais de uma isoforma de HK. Através da
análise filogenética da hexoquinase neste trabalho, podemos notar que uma das
sequências de aminoácidos obtidas para HKs do R. microplus (depositada no Gene
Bank) possui maior proximidade da HK2 de H. sapiens e mus musculus, enquanto a

35
outra sequência (obtida do transciptoma do grupo) está mais distante
filogeneticamente da HK2, sugerindo que possam existir isoformas diferentes de
hexoquinase (Figura 10). Em Bombix mori identificaram quatro isoformas de HK,
expressas em diferentes tecidos (YANAGAWA, 1977); e em Tribolium castaneum
apenas duas isoformas dessa enzima (FRAGA et al., 2013); Drosophila
melanogaster apresentou três principais formas da Hexoquinase (Hex A, B e C)
presente em diferentes tecidos, com a Hex-A e Hex-B predominantemente
encontrada na forma adulta deste organismo (MADHAVAN et al., 1972; MOSER et
al., 1980; DAVID D. DUVERNELL e WALTER F. EANES 2000; JAYAKUMAR. et al.,
2001). Os nossos resultados sugerem que existe uma resposta diferenciada da
atividade de HK nas duas frações celulares estudadas (mitocondrial e
citoplasmática) quando a AKT foi inibida em células de carrapato (Figura 15, A).
A etapa seguinte foi avaliar a atividade de HK após o silenciamento gênico
da AKT em células BME26 para observar a regulação da AKT sobre HK.
Primeiramente foram utilizadas duas concentrações de dsAKT, 4µg e 6µg. Após a
validação do silenciamento por PCR em tempo real, observamos aproximadamente
50% de silenciamento de AKT com 4µg de dupla-fita, e aproximadamente 70% de
silenciamento quando as células foram mantidas com 6µg de dsAKT (Figura 15, B).
Trabalhos do próprio grupo (ABREU et al., 2013; Logullo et al., 2009) e trabalhos de
outros pesquisadores mostram que a AKT é uma enzima-chave para a manutenção
da sobrevivência celular, homeostase de glicose, crescimento e desenvolvimento do
indivíduo (CHO et al., 2001a; CHO et al., 2001b; CANTLEY, 2002; LIZCANO e
ALESSI, 2002; EASTON et al., 2005; TSCHOPP et al., 2005). Desta forma
analisamos a viabilidade celular por MTT a fim de investigar se uma dessas
concentrações de dsAKT afetaria a viabilidade celular, no entanto a viabilidade se
mantém nas concentrações de dupla-fita utilizadas quando comparadas ao controle
(Figura 15, C).
David J. Roberts et al. (2013) mostraram que células de cardiomiócitos de
ratos quando na presença de IGF-1 e de H2O2, a AKT é translocada para a
mitocôndria dessas células, conferindo proteção contra apoptose induzida por H2O2.
Outros trabalhos também mostram que AKT ativada inibe a liberação do citocromo c
e, consequentemente, a apoptose (MAJEWSKI et al., 2004; ROBEY e HAY, 2006).

36
Ao observar que mesmo com 70% de silenciamento da AKT não ocorreu
uma diferença na viabilidade celular comparado ao controle, contudo nessa mesma
concentração de dupla-fita (6µg) foi possível observar um aumento significativo da
atividade de HK na fração citoplasmática se comparado à fração mitocondrial (Figura
15, D). Esses resultados de inibição e silenciamento de AKT sugerem que a
hexoquinase na fração citoplasmática pode sofrer uma regulação pela AKT, o que
não é observado na fração mitocondrial (Figura 15). Já foi visto em outros trabalhos
que quando a AKT está inativa, a GSK3 permanece ativa e fosforila o VDAC em Thr-
51 impedindo que HK se ligue ao VADC (CROSS et al., 1995; FENG et al., 2005;
PASTORINO et al., 2005). É provável que quando inibimos ou silenciamos a AKT, o
VADC é fosforilado, impedindo que a HK permaneça ancorada à membrana externa
da mitocôndria, e consequentemente seja liberada no citoplasma. Neary e Pastorino
(2013) relataram algo similar onde ocorre uma redistribuição da HK2 para o
citoplasma de células de câncer MDA-MB-231 quando AKT é inibida.
Anja Schindler e Edan Foley, 2013 e outros pesquisadores mostraram que a
hexoquinase ancora na membrana externa da mitocôndria utilizando 15
amonoácidos da região N-terminal hidrofóbica. Nossa análise por homologia das
sequências de aminoácidos das hexoquinases de carrapato com outros artrópodes e
mamíferos mostraram que os 15 aminoácidos da região N-terminal hidrofóbica das
sequências de carrapato não possuem um grau satisfatório de identidade e
similaridade com as sequencias de mamíferos, sugerindo que o acoplamento da HK
à membrana externa da mitocôndria em carrapatos R. microplus pode ocorrer em
outra região da proteína (Figura 9).
David J. Roberts et al. (2013) mostrou que a AKT pode fosforilar diretamente
a HK-II na Thr-473 levando ao ancoramento da mesma à membrana externa da
mitocôndria em mamíferos. Tendo em vista a possível regulação da AKT sobre a
hexoquinase, observamos através do alinhamento que o resíduo de aminoácido Thr-
473 (presente na HK-II de mamífero) não se encontra alinhado com as sequências
de hexoquinases do carrapato. Caso ocorra uma regulação direta da AKT sobre a
hexoquinase por fosforilação, esta regulação pode possivelmente ocorrer em outro
sítio de fosforilação (Figura 9).
Depois de observar esses eventos em cultura celular do carrapato R.
microplus, descrevendo o importante papel da AKT na regulação da atividade de

37
HK, e consequentemente na viabilidade celular, a dsAKT foi administrada em
fêmeas adultas partenógenas do carrapato R. microplus afim de determinar seus
efeitos na fisiologia desse organismo.
O silenciamento de AKT afetou o índice de ingurgitamento das fêmeas do
carrapato R. microplus. Após a administração da dupla-fita de RNAi para AKT as
fêmeas ingeriram significantemente mais sangue (Figura 13, A) que as fêmeas
controle, mas sem que isso afetasse o ganho de peso (Figura 13, B). Entretanto, o
efeito mais surpreendente do silenciamento de AKT foi observado no
desenvolvimento do ovário. Podemos notar que os ovários das fêmeas de carrapato
R. microplus tratadas com RNAi para AKT não desenvolveram (Figura 13, C),
diferentemente das fêmeas controle que obtiveram um desenvolvimento normal.
Esses resultados sugerem que a AKT tem um papel central na coordenação gênica
do metabolismo de glicose, propondo-a como um futuro alvo molecular para o
desenvolvimento de vacinas ou fármacos que visam controlar as populações desse
importante vetor de doenças.
Esses resultados em conjunto descrevem um panorama metabólico que
conduz a um melhor entendimento da modulação do metabolismo de glicose em
embriões desse importante vetor de doenças, além de contribuir para a identificação
de novos alvos para vacinas que integrem estratégias para o controle da espécie.
Atualmente métodos de controle alternativo interferem em fases preferenciais do
ciclo de vida do carrapato, como as ninfas, adultos e finalmente o embrião
(THOMPSON e STEWART, 1997). Embora esse último estádio seja negligenciado,
novos estudos na embriogênese de artrópodes, em particular focados no
metabolismo de carboidratos, têm conduzido a um melhor entendimento da dinâmica
de utilização de nutrientes desde a formação do ovócito até a mobilização de
reservas durante a formação do embrião (CAMPOS et al., 2006; VITAL et al., 2010;
MORAES et al., 2007). Esse estudo também contribui para preencher essa lacuna
na literatura e mostrar que a regulação do metabolismo de glicose na embriogênese
é processo mais dinâmico do que atualmente apreciado.

38
7 CONCLUSÃO
O alinhamento das sequencias de aminoácidos da Hexoquinase do
carrapato R. microplus com outros organismos mostrou domínios bastante
conservados.
A análise filogenética indica fortemente a presença de duas isoformas
codificadas por dois genes distintos de HK em carrapato.
Nas frações citoplasmática e mitocondrial de embriões desafiadas com
diferentes concentrações do inibidor para AKT (10-DEBC) não apresentaram
diferenças estatísticas das atividades de HK nas respectivas frações.
As fêmeas no estágio de paternógenas que foram silenciadas com 4µg de
dupla-fita para AKT ingeriram uma quantidade significativamente maior de sangue, e
apresentaram uma redução no tamanho do ovário se comparado ao controle.
Mesmo com a redução no tamanho do ovário não observamos diferenças
estatísticas na taxa de oviposição e nem na taxa de eclosão.
Ao utilizamos 48µM do inibidor para AKT (10-DEBC) nas células BME26
ocorre um aumento significativo da atividade de HK na fração citoplasmática, se
compararmos as concentrações de 12µM e 24µM do inibidor, e ao controle.
O silenciamento da AKT com a dupla-fita de dsAKT não afetaram a
viabilidade celular das células BME26.
Nas células silenciadas para AKT ocorre um aumento significativo da
atividade de HK na fração citoplasmática quando administramos 6µg de dupla-fita
para AKT no meio de cultivo.
Esses resultados, em conjunto, indicam uma possível regulação da AKT
sobre a HK, e sugere que essa regulação da AKT possa estar atuando em isoformas
diferes da HK em carrapatos R. microplus.

39
8 REFERENCIAS BIBLIOGRÁFICAS
ABREU, L.; FABRES, A.; ESTEVES, E.; MASUDA, A.; VAZ, I. S., JR.; DAFFRE, S.; LOGULLO, C. Exogenous insulin stimulates glycogen accumulation in Rhipicephalus (Boophilus) microplus embryo cell line BME26 via PI3K/AKT pathway. Comp. Biochem. Physiol. B, 153, 185–190. 2009. ALTSCHUL S. F., GISH W., MILLER W., MYERS E. W., LIPMAN D. J. Basic local alignment search tool. J. Mol. Biol. vol. 215 (pg. 403-410). 1990. ALTSCHUL S. F., MADDEN T. L., SCHÄFFER A. A., ZHANG J., ZHANG Z., MILLER W., LIPMAN D. J. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. Sep 1; 25(17): 3389–3402. 1997. ANDREOTTI, R. Situação atual da resistência do carrapato-do-boi Rhipicephalus (Boophilus) microplus aos acaricidas no Brasil [online], 1. ed. Campo Grande: Embrapa Gado de Corte, 2010. Disponível em: http://www.cnpgc.embrapa.br/publicacoes/doc/DOC180.pdf. Acesso em: 24 fev. 2016. ANJA SCHINDLER, EDAN FOLEY. Hexokinase 1 Blocks apoptotic signals at the mitochondria. Cellular Signalling. 25 2685-2692. 2013. ARIANNE FABRES, C.P. DE ANDRADE, M. GUIZZO, M.H.F. SORGINE, G.D.O. PAIVA-SILVA, A. MASUDA, I. D.S. VAZ JR., C. LOGULLO, Effect of GSK-3 activity, enzymatic inhibition and gene silencing by RNAi on tick oviposition and egg hatching, Parasitology 137 1537–1546. 2010. BELL-SAKYI,L., ZWEYGARTH,E., BLOUIN,E.F., GOULD,E.A., AND JONGEJAN,F. Tick cell lines: tools for tick and tick-borne disease research. Trends Parasitol. 23:450-457. 2007. BEUTNER G., RUCK A., RIEDE B., WELTE W. E BRDICZKA D. Complexes between kinases, mitochondrial porin and adenylate translocator in rat brain resemble the permeability transition pore. FEBS Lett. 396:2-3, 189-195. 1996. BEUTNER G., RUCK A., RIEDE B. E BRDICZKA D. Complexes between hexokinase, mitochondrial porin and adenylate translocator in brain: regulation of hexokinase, oxidative phosphorylation and permeability transition pore. Biochem.Soc.Trans. 25:1, 151-157. 1997. BEUTNER G., RUCK A., RIEDE B. E BRDICZKA D. Complexes between porin, hexokinase, mitochondrial creatine kinase and adenylate translocator display properties of the permeability transition pore. Implication for regulation of permeability transition by the kinases. Biochim. Biophys. Acta 1368:1, 7-18. 1998. BROGIOLO, W., STOCKER, H., IKEYA, T., RINTELEN, F., FERNANDEZ, R., AND HAFEN, E. An evolutionarily conserved function of the Drosophila insulin receptor and insulin-like peptides in growth control. Curr. Biol., 11:213-221. 2001

40
BROWN, M. R., CLARK, K. D., GULIA, M., ZHAO, Z., GARCZYNSKI, S. F., CRIM, J. W., Suderman, R. J., and Strand, M. R. An insulin-like peptide regulates egg maturation and metabolism in the mosquito Aedes aegypti. Proc. Natl. Acad.Sci. U.S.A, 105:5716-5721. 2008. CAMPOS E., MORAES J., FAÇANHA A.R., MOREIRA E., VALLE D., ABREU L., MANSO P.P., NASCIMENTO A., PELAJO-MACHADO M., LENZI H., MASUDA A., DA SILVA VAZ JR. I. & LOGULLO C. Kinetics of energy source utilization in Boophilus microplus (Canestrini, 1887) (Acari: Ixodidae) embryonic development. Veterinary Parasitology. 138(3-4): 49-357. 2006. CAMPOS, E., FAÇANHA, A., MORAES J., VAZ JR, I. S., AOI MASUDAB, CARLOS LOGULLO. A mitochondrial exopolyphosphatase activity modulated by phosphate demand in Rhipicephalus (Boophilus) microplus embryo. Insect Biochemistry and Molecular Biology 37 1103–1107. 2007. CANTLEY,L.C. (2002): The phosphoinositide 3-kinase pathway. Science, 296:1655-1657. CARDENAS, M. L., CORNISH-BOWDEN, A., URETA, T. Evolution and regulatory role of the hexokinases. Biochimica et biophysica acta. 1401: 242-64. 1998. CHO, H., THORVALDSEN,J.L., CHU,Q., FENG,F., AND BIRNBAUM,M.J. Akt1/PKBalpha is required for normal growth but dispensable for maintenance of glucose homeostasis in mice. J. Biol. Chem., 276:38349-38352. 2001a. CHO,H., MU,J., KIM,J.K., THORVALDSEN,J.L., CHU,Q., CRENSHAW,E.B., III, KAESTNER,K.H., Bartolomei,M.S., Shulman,G.I., and Birnbaum,M.J. Insulin resistance and a diabetes mellitus-like syndrome in mice lacking the protein kinase Akt2 (PKBbeta). Science, 292:1728-1731. 2001b. CROSS D.A., ALESSI, D.R., COHEN P., ANDJELKOVICH M., HEMMINGS B. A., Inhibition of glycogen synthase kinase-3 by insulin mediated by protein kinase B, Nature 378 785–789. 1995. DA-SILVA W. S., GOMEZ-PUYOU, A., DE GOMEZ-PUYOU M. T., MORENO-SANCHEZ, R., DE FELICE F. G., DE MEIS, L., et al. Mitochondrial bound hexokinase activity as a preventive antioxidant Defense - Steady-state ADP formation as a regulatory mechanism of membrane potential and reactive oxygen species generation in mitochondria. J Biol Chem. 279: 39846-55. 2004. DA SILVA VAZ JR. I., LOGULLO C., SORGINE M., VELLOSO F.F., ROSA DE LIMA M.F., GONZALES J. C., MASUDA H., OLIVEIRA P.L. & MASUDA A. Immunization of bovines with an aspartic proteinase precursor isolated from Boophilus microplus eggs. Veterinary Immunology and Immunopathology. 66(3-4): 331-341. 1998. DAVID D. DUVERNELL E WALTER F. EANES. Contrasting Molecular Population Genetics of Four Hexoquinase in Drosophila melanogaster, D. simulans and D. Yakuba. Genetics Society of America. 156: 1191-1201. 2000

41
DAVID J. ROBERTS, VALERIE P. TAN-SAH, JEFFERY M. SMITH, AND SHIGEKI MIYAMOTO AKT phosphorylates hk-ii at thr-473 and increases mitochondrial HK-ii association to protect cardiomyocytes; journal of biological chemistry volume 288 number 33 august 16, 2013. DE LA FUENTE J., ALMAZÁN C., CANALES M., PÉREZ DE LA LASTRA J.M., KOCAN K.M. & WILLADSEN P. A ten-year review of commercial vaccine performance for control of tick infestations on cattle. Animal Health Research Reviews/ Conference of Research Workers in Animal Disease. 8(1): 23-8. 2007. DE LA FUENTE J., ESTRADA-PEÑA A., VENZAL J.M., KOCAN K.M., SONENSHINE D.E. Overview: ticks as vectors of pathogens that cause disease in humans and animals. Front Biosci. 13(13): 6938-6946. http://dx.doi. org/10.2741/3200. PMid:18508706. 2008. DIAMANT, G.; STRICKLAND, R. K. Mannual of livestock ticks for animal disease eradication division personnel. Washington: USDA, Res. Serv., 141p. 1965. DICKSON, B. J. WIRING the brain with insulin. Science 300, 440–441. 2003. E. ESTEVES, F.A. LARA, D.M. LORENZINI, G.H.N. COSTA, A.H. FUKUZAWA, L.N. PRESSINOTTI, J.R.M.C. SILVA, J.A. FERRO, T.J. KURTTI, U.G. MUNDERLOH, S. DAFFRE, Cellular and molecular characterization of an embryonic cell line (BME26) from the tick Rhipicephalus (Boophilus) microplus, Insect Biochem. Mol. Biol. 38 568–580. 2008. EASTON,R.M., CHO,H., ROOVERS,K., SHINEMAN,D.W., MIZRAHI,M., FORMAN,M.S., LEE,V.M., SZABOLCS,M., DE,J.R., OLTERSDORF,T., LUDWIG,T., EFSTRATIADIS,A., AND BIRNBAUM,M.J. Role for Akt3/protein kinase Bgamma in attainment of normal brain size. Mol. Cell Biol., 25:1869-1878. 2005. EDGAR R.C. MUSCLE: a multiple sequence alignment method with reduced time and space complexity. BMC Bioinformatics. 5:113. 2004. FAGOTTO F. Yolk degradation in tick eggs: I. Occurrence of a cathepsin L-like acid proteinase in yolk spheres. Archives of Insect Biochemistry and Physiology. 14(4): 217-235. 1990 FENG, J., LUCCHINETTI, E., AHUJA, P., PASCH, T., PERRIARD, J.C., ZAUGG, M. Isoflurane postconditioning prevents opening of the mitochondrial permeability transition pore through inhibition of glycogen synthase kinase 3beta. Anesthesiology 103, 987–995. 2005. FERNANDES, E. K., COSTA, G. L., MORAES, A. M., ZAHNER, V., BITTENCOURT, V. R. Study on morphology, pathogenicity, and genetic variability of Beauveria bassiana isolates obtained from boophilus microplus tick. Parasitol. Res. 98, pp. 324-332. 2006. FISKE, C. F., SUBBAROW, Y., The colorimetric determination of phosphorus. J. Biol. Chem. 66, PP. 375-400. 1925.

42
FRAGA, A., MORAES, J., SILVA, J. R., COSTA, E. P., MENEZES, J., VAZ JR, I. S., CARLOS LOGULLO, DA FONSECA, R. N., E CAMPOS, E., Inorganic Polyphosphates Regulate Hexokinase Activity and Reactive Oxygen Species Generation in Mitochondria of Rhipicephalus (Boophilus) microplus Embryo. Int. J. Biol. Sci. Vol. 9, PP. 842-852. 2013. FRAGA, A., RIBEIRO, L., LOBATO, M., SANTOS V, SILVA J. R., REZENDE, G., et al. Glycogen and Glucose metabolism are essential for early embryonic development of the red flour beetle Tribolium castaneum. PloS One. 2013. GALINA, A., MARCELO, R., ALBUQUERQUE, M., PUYOU, A. G., MARIETTA T. GOMEZ PUYOU, M. T. G., AND DE MEIS, L. Different properties of the mitochondrial and cytosolic hexokinases in maize roots. Biochem. J. 309, 105-112 (Printed in Great Britain). 1995. GAROFALO, R. S., Genetic analysis of insulin signaling in Drosophila. Trends Endocrinol. Metab, 13:156-162. 2002. GEORGE, J. E., Present and future Technologies for tick control. Ann. N.Y. Acad. Sci. 916, PP. 583-588. 2000. GONZALES, J. C., SILVA, N.R., FRANCO, N. A vida livre do Boophilus microplus, Arq. Fac. Vet. UFRGS. RG., 3: 21-28. 1974. GOTTLOB, K., MAJEWSKI, N., KENNEDY, S., KANDEL, E., ROBEY, R.B., HAY, N., Inhibition of early apoptotic events by Akt/PKB is dependent on the first committed step of glycolysis and mitochondrial hexokinase. Genes Dev. 15, 1406–1418. 2001. GRISI L, LEITE RC, MARTINS JR, BARROS AT, ANDREOTTI R, CANÇADO PH, et al. Reassessment of the potential economic impact of cattle parasites in Brazil. Rev. Bras. Parasitol. Vet. 23(2): 150-156. http://dx.doi.org/10.1590/ S1984-29612014042. PMid:25054492. 2014. HIRO-AKI YNAGAWA. Tissue Distribution, Purifications, and Properties of Multiple Forms of Hexokinase in the Silkworm, Bombyx Mori. Insect Biocem. Vol. 8 pp. 293 to 305. 1977. HOLMAN, P.J., RONALD, N.C. A new tick cell line derived from Boophilus microplus. Res. Vet. Sci. 29, 383–387. 1980. HORN, S. C., ARTECHE, C.C.P., Situação parasitária da pecuária no Brasil. A hora Veterinária 23, PP. 12-32. 1985. HORN, S. C., Prováveis prejuízos causados pelos carrapatos. Ministério da Educação 2ª Ed. 1983. IBGE (Instituto Brasileiro de Geografia e Estatística). Disponível on line no endereço HTTP://www.ibge.gov.br/. 2015.

43
JOHNSTON, L. A., KEMP, D. H., PEARSON, R. D. Immunization of cattle against Boophilus microplus using extracts derived from adult female ticks: effects of induced immunity on tick populations. Int.J.Parasitol 16, pp. 27-34. 1986. KANDEL, E. S., AND HAY, N., The regulation and activities of the multifunctional serine/threonine kinase Akt/PKB. Exp.Cell Res., 253:210-229. 1999. KATZEN H. M. E SCHIMKE R. T. Multiple forms of hexokinase in the rat: tissue distribution, age dependency, and properties. Proc. Natl. Acad. Sci. U.S.A 54:4, 1218-1225. 1965. KORSHUNOV, S. S., SKULACHEV, V. P., STARKOV, A. A., High protonic potential actuates a mechanism of production of reactive oxygen species in mitochondria. Febs Lett.; 416: 15-8. 1997. KRIEGER, M. J., JAHAN, N., RIEHLE, M. A., CAO, C., AND BROWN,M.R. Molecular characterization of insulin-like peptide genes and their expression in the African malaria mosquito, Anopheles gambiae. Insect Mol. Biol., 13:305-315. 2004. KURTTI,T.J., MATTILA,J.T., HERRON,M.J., FELSHEIM,R.F., BALDRIDGE,G.D., BURKHARDT,N.Y., BLAZAR,B.R., HACKETT,P.B., MEYER,J.M., AND MUNDERLOH,U.G. Transgene expression and silencing in a tick cell line: A model system for functional tick genomics. Insect Biochem. Mol. Biol., 38:963-968. 2008. LARKIN M. A., BLACKSHIELDS G., BROWN N. P., CHENNA R., MCGETTIGAN P. A., MCWILLIAM H., VALENTIN F., WALLACE I. M., WILM A., LOPEZ R., THOMPSON J. D., GIBSON T. J., HIGGINS D. G. Clustal W and Clustal X version 2.0 Bioinformatics 23 (21): 2947-2948. 2007. LE ROITH, D. AND ZICK,Y. Recent advances in our understanding of insulin action and insulin resistance. Diabetes Care, 24:588-597. 2001. LEONARDO ARAUJO DE ABREU; CHRISTIANO CALIXTO; CAMILA FERNANDA WALTERO; BÁRBARA PITTA DELLA NOCE; NAFTALY WANG'OMBE GITHAKA; ADRIANA SEIXAS; LUÍS FERNANDO PARIZI; SATORU KONNAI; ITABAJARA DA SILVA VAZ JR; KAZUHIKO OHASHI; CARLOS LOGULLO. The conserved role of the AKT/GSK3 axis in cell survival and glycogen metabolism in Rhipicephalus (Boophilus) microplus embryo tick cell line BME26 Biochimica et Biophysica Acta 1830 2574–2582. 2013. LI, Z., NEUFELD, G. J., Isolation and characterization of mitochondrial F(1)-ATPase from crayfish (Orconectes virilis) gills. Comp. Biochem. Physiol. B: Biochem. Mol. Biol. 128, 325–338. 2001. LIZCANO,J.M. AND ALESSI,D.R. The insulin signalling pathway. Curr. Biol., 12:R236-R238. 2002. LOCKHEAD, P. A., COGHLAN, M., RICE, S. Q. J., SUTHERLAND, C. Inhibition of GSK-3 selectively reduces glucose-6-phosphatase and phosphoenolpyruvate carboxykinase gene expression. Diabetes 50, 937–946. 2001.

44
MADHAVAN, K., FOX, D.J., URSPRUNG, H., Developmental genetics of hexokinase isoenzymes in Drosophila melanogaster. Journal of Insect Physiology 18, 1523–1530. 1972. MAJEWSKI, N., NOGUEIRA, V., BHASKAR, P., COY, P.E., SKEEN, J.E., GOTTLOB, K., CHANDEL, N.S., THOMPSON, C.B., ROBEY, R.B., HAY, N., Hexokinase–mitochondria interaction mediated by Akt is required to inhibit apoptosis in the presence or absence of Bax and Bak. Mol. Cell 16, 819–830. 2004. MEHLHORN H., AL-RASHEID K.A., AL-QURAISHY S. & ABDEL-GHAFFAR F. Research and increase of expertise in arachno-entomology are urgently needed. Parasitology Research. 436(11): 2480-2487. 2011. MAPA (Ministério da Agricultura, pecuária e abastecimento). Disponivel on line no site http://www.agricultura.gov.br/noticias/producao-de-carne-no-brasil-aumenta-45-em-15-anos MATTILA, J.T., BALDRIDGE, G.D., FELSHEIM, R.E., BURKHARDT, N.Y., MUNDERLOH, U.G. AND KURTTI, T.J. RNAi-mediated silencing of a DsRed2- expressing Ixodes scapularis (Acari: Ixodidae) cell line. Society for In Vitro Biology Journal 42, 32-A. 2006. MCGOWAN, M. J., HOMER, J. T., O’DELL, G. V., MCNEW, R. W., BARKER, R. W., Performance of tick fed on rabbits inoculated with extracts derived from homogenized ticks Amblyomma maculatum Koch (Acarina: Ixodidae). J. Parasitol. 66, pp. 42-48. 1980. MEHLHORN H., AL-RASHEID K.A., AL-QURAISHY S. & ABDEL-GHAFFAR F. 2011. Research and increase of expertise in arachno-entomology are urgently needed. Parasitology Research. 436(11): 2480-2487. 2011. MELISSA K. MCCOY, ALICE KAGANOVICH, IAKOV N. RUDENKO, JINHUI DING AND MARK R. COOKSON. Hexokinase activity is required for recruitment of parkin to depolarized mitochondria. Human Molecular Genetics, Vol. 23, No. 1. 2014. MEYER, L. E., MACHADO, L. B., SANTIAGO, A. P., DA-SILVA, W. S., DE FELICE, F. G., Holub, O., et al. Mitochondrial creatine kinase activity prevents reactive oxygen species generation: antioxidant role of mitochondrial kinase-dependent ADP re-cycling activity. J Biol Chem; 281: 37361-71.doi:10.1074/jbc.M604123200. 2006. MICCOLI, L., OUDARD, S., SUREAU, F., POIRSON, F., DUTRILLAUX, B., POUPON,M.F., Intracellular pH governs the subcellular distribution of hexokinase in a glioma cell line. Biochem. J. 313 (Pt 3), 957–962. 1996. MICCOLI, L., BEURDELEY-THOMAS, A., DE, P.G., SUREAU, F., OUDARD, S., DUTRILLAUX, B., POUPON, M.F., Light-induced photoactivation of hypericin affects the energymetabolism of human glioma cells by inhibiting hexokinase bound to mitochondria. Cancer Res. 58, 5777–5786. 1998.

45
MIYAMOTO, S., MURPHY, A. N., BROWN, J. H. Akt mediates mitochondrial protection in cardiomyocytes through phosphorylation of mitochondrial hexokinase-II. Cell Death Differ. 15, 521–529. 2008. MORAES, J.; GALINA, A.; ALVARENGA, P.; REZENDE, G.; MASUDA, A.; VAZ, I.S., JR.; LOGULLO, C. Glucose metabolism during embryogenesis of the hard tick Boophilus microplus. Comp. Biochem. Physiol. A, 146, 528–533. 2007. MOSER, D., JOHNSON, L., LEE, C.-Y., Multiple forms of Drosophila hexokinase, purification, biochemical and immunological characterization. Journal of Biological Chemistry 255, 4673–4679. 1980. NATHAN MAJEWSKI, VERONIQUE NOGUEIRA, PRASHANTH BHASKAR, PLATINA E. COY, JENNIFER E. SKEEN, KATHRIN GOTTLOB, NAVDEEP S. CHANDEL, CRAIG B. THOMPSON, R.BROOKS ROBEY, NISSIM HAY. Hexokinase-mitochondria interaction mediated by Akt is required to inhibit apoptosis in the presence or absence of bax and bak. Molecular Cell, by Cell Press Vol. 16, 819–830, December 3, 2004. NEARY, C. I., AND PASTORINO, J. G., Akt Inhibition Promotes Hexokinase 2 Redistribution and Glucose Uptake in Cancer Cells. J. Cell. Physiol. 228: 1943–1948. 2013. NIJHOF, A.M., BALK, J.A., POSTIGO, M., JONGEJAN, F. Selection of reference genes for quantitative RT-PCR studies in Rhipicephalus (Boophilus) microplus and Rhipicephalus appendiculatus ticks and determination of the expression profile of Bm86. BMC Mol. Biol., 10:112. doi: 10.1186/1471-2199-10-112, 2009. OIE. Rhipicephalus (Boophilus) microplus – Southern Cattle Tick. 3 p., 2007. Disponível em: http://www.cfsph.iastate.edu/Factsheets/pdfs/boophilus_ microplus.pdf. Acesso em: 25 de fev. 2016. P. CYRIL JAYAKUMAR; YOGESH S. SHOUCHE; MILIND S. PATOLE. 2001. Cloning of two hexokinase isoenzyme sequences from Drosophila melanogaster. Insect Biochemistry and Molecular Biology 31 1165-1171. 2001. PASTORINO J. G., HOEK J. B. E SHULGA N. Activation of glycogen synthase kinase 3beta disrupts the binding of hexokinase II to mitochondria by phosphorylating voltage-dependent anion channel and potentiates chemotherapy-induced cytotoxicity. Cancer Res. 65:22, 10545-10554. 2005. PETER RJ, VAN DEN BOSSCHE P, PENZHORN BL, SHARP B. Tick, fly, and mosquito Control-Lessons from the past, solutions for the future. Vet Parasitol; 132(3-4): 205-215. 2005. PFAFFL M. W. A new mathematical model for relative quantification in realtime RT-PCR, Nucl. Acids Res. 2001. PRUETT J.H. Immunological control of arthropod ectoparasites - a review. International Journal for Parasitology. 29(1): 25-32.). 1999.

46
PORTAL BRASIL. Disponivel no site http://www.brasil.gov.br/economia-e-emprego/2015/10/rebanho-bovino-brasileiro-cresce-e-chega-a-212-3-milhoes-de-cabecas-de-gado RIEHLE, M. A., FAN, Y., CAO, C., AND BROWN,M.R. Molecular characterization of insulin-like peptides in the yellow fever mosquito, Aedes aegypti: expression, cellular localization, and phylogeny. Peptides, 27:2547-2560. 2006. ROBEY, R.B., HAY, N., Mitochondrial hexokinases: guardians of themitochondria. Cell Cycle 4, 654–658. 2005. ROBEY, R.B., HAY, N., Mitochondrial hexokinases, novel mediators of the antiapoptotic effects of growth factors and Akt. Oncogene 25, 4683–4696. 2006. SANTIAGO, A. P. S. A., CHAVES, E. A., OLIVEIRA, M. F., GALINA, A. Reactive oxygen species generation is modulated by mitochondrial kinases: Correlation with mitochondrial antioxidant peroxidases in rat tissues. Biochimie; 90: 1566-77. 2008. SEIFERT, G., SPRINGELL, P., TATCHELL, R. Radioactive studies on the feeding of larvae, nymphs and adults of the cattle tick Boophilus microplus (Canestrini), Parasitology, 58, pp 415–430, 1968. SEIXAS A., OLDIGES D.P., DA SILVA VAZ JR. I. & TERMIGNONI C. Endocrinologia e controle da vitelogênese em carrapatos. Acta Scientiae Veterinariae. 38(2):95-111. 2010. SIM, C., DENLINGER, D. L., Insulin signaling and FOXO regulate the overwintering diapause of the mosquito Culex pipiens. Proc Natl Acad Sci USA.; 105:6777–6781. PubMed: 18448677. 2008. SNOWBALL, G. J. Ecological observation on the cattle tick Boophilus microplus (Canestrini). Australian Journal of Agricultural Research. 8(4) 394-413, 1957. SONENSHINE, D. New York, Oxford University Press, Biology of ticks, Vol. 1.1991. SONENSHINE, D. E., KOCAN, K. M., DE LA, F. J. Tick control: further thoughts on a research agenda. Trends parasitol. 22, pp. 550-551. 2006. SONG J.L., WONG J.L. & WESSEL G.M. Oogenesis: Single cell development and differentation. DevelopmentalBiology. 300(1): 385-405. 2006. SOUTHWORTH, R., DAVEY, K.A., WARLEY, A., GARLICK, P.B. A reevaluation of the roles of hexokinase I and II in the heart. American Journal of Physiology - Heart and Circulatory Physiology. 292: 378–386. 2007. SUTHERST, R., MAYWALD, G., KERR, J., STEGEMEN, D. The effect of the cattle tick Boophilus microplus on the growth of Bos indicus x Bos Taurus steers. Aust. J. Agr. Res., 34, pp 317 – 327, 1983.

47
TATCHELL, R.J. CAENELL, R.; KEMP, D. H. Electrical studies on the feeding of the cattle tick, Boophilus microplus. Z. ParasitenK., 38: 32-44. 1972. TILGHMAN, S. M., et al. Hormonal regulation of phosphoenolpyruvate carboxykinase (GTP) in mammalian tissues. In Gluconeogenesis: Its Regulation in Mammalian Species (Hanson, R.W. and Mehlman, M.A., eds), pp. 47–91, John Wiley & Sons. 1976. THOMPSON, M. B., E STEWART, J. R., Embryonic metabolism and growth in lizards of the genus Eumeces. Comparative Biochemistry and Physiology, 118A: 647-654. 1997. TSCHOPP,O., YANG,Z.Z., BRODBECK,D., DUMMLER,B.A., HEMMINGS-MIESZCZAK,M., WATANABE,T., MICHAELIS,T., FRAHM,J., AND HEMMINGS,B.A. Essential role of protein kinase B gamma (PKB gamma/Akt3) in postnatal brain development but not in glucose homeostasis. Development, 132:2943-2954. 2005. WARBURG, O., On the origin of cancer cells. Scence; 123:309–314. 1956. WEI, Z., IBRAHIM, W., STCLAIR, D. K., CHOW, C. K. Increased mitochondrial generation of hydrogen peroxide in the skeletal muscle and liver of vitamin E-deficient mice. Faseb J; 11: 3380. 1997. WHARTON R. H. AND UTECH K. B. W. The relation between engorgement and dropping of Boophilus microplus (Canestrini) (Ixodidae) to the assessment of tick numbers on cattle. r. Aust. ent. Sot. 9, 171-18. 1970. WIKEL, S. K., BERGMAN, D., Tick-host immunology: significant advances and challenging opportunities. Parasitol. Today 13, pp. 383-389. 1997. WILLADSEN, P., Tick control: thoughts on a research agenda. Veterinary arasitology 138, 161–168. 2006 WILLADSEN P. Vaccines, genetics and chemicals in tick control: the Australian experience. Tropical Animal Health and Production. 29 (Suppl 4): 91-94. 1997. WILLADSEN, P., COBON, G., MCKENNA, R. V. Comparative vaccination of cattle against Boophilus microplus with recombinant antigen Bm86 alone or in combination with recombinant Bm91. Parasite Immunology 18, 241–246. 1996. WILSON J. E. Isozymes of mammalian hexokinase: structure, subcellular localization and metabolic function. J. Exp. Biol. 206: Pt 12, 2049-2057. 2003. ZUURBIER, C.J., EERBEEK, O., MEIJER, A.J. Ischemic preconditioning, insulin, and morphine all cause hexokinase redistribution. American Journal of Physiology – Heart and Circulatory Physiology. 289: 496-499. 2005.





























