Embed Size (px)
Citation preview

CENTRO UNIVERSITÁRIO DO MARANHÃO PRÓ-REITORIA DE PÓS-GRADUAÇÃO, PESQUISA E EXTENSÃO PROGRAMA DE PÓS-GRADUAÇÃO EM BIOLOGIA PARASITÁRIA
SÃO LUÍS
2012
VIVIANE MENEZES DE MENEZES
ADESÃO E FORMAÇÃO DE
BIOFILME POR ISOLADOS
CLÍNICOS DE Candida spp

ii
VIVIANE MENEZES DE MENEZES
ADESÃO E FORMAÇÃO DE BIOFILME POR ISOLADOS CLÍNICOS DE
Candida spp
Dissertação apresentada ao Programa de
Pós-Graduação em Biologia Parasitária
como requisito para a obtenção do título
de Mestre em Biologia Parasitária.
Orientador: Prof(a). Dr(a). Cristina de
Andrade Monteiro
Co-orientador: Prof(a). Dr(a.) Patricia
de Maria Silva Figueiredo
SÃO LUÍS
2012

iii
Menezes, Viviane Menezes
Adesão e Formação de Biofilme por isolados clínicos de Candida spp.
/ Viviane Menezes de Menezes. – São Luís, 2012.
102 f.
Orientador: Prof(a). Dr(a). Cristina de Andrade Monteiro
Dissertação (Mestrado em Biologia Parasitária) – Centro Universitário
do Maranhão, UNICEUMA, 2012.
1. Candida spp, fatores de virulência, adesão, biofilme, sítios
anatômicos.
CDU
1.1.1.1.1.1.1 CDU 616.24 – 002.5

iv
Viviane Menezes de Menezes
ADESÃO E FORMAÇÃO DE BIOFILME POR ISOLADOS CLÍNICOS DE
Candida spp
A Comissão julgadora da Defesa do Trabalho Final de Mestrado
em Biologia Parasitária, em sessão pública realizada no dia 09/04/2012,
considerou a candidata
( X ) APROVADA ( ) REPROVADA
_____________________________________________________ Profa. Dra. Rosimary de Jesus Gomes Turri
_____________________________________________________ Prof. Dr. Valério Monteiro Neto
_____________________________________________________ Profa. Dra. Patrícia de Maria Silva Figueiredo
_____________________________________________________ Profa. Dra. Cristina de Andrade Monteiro – orientadora

v
À Deus, por ser o meu Senhor, meu
Salvador, o Todo-Poderoso, o meu guia e que
me fortalece em todos os momentos.
Ao meu marido Neyderman Amorim,
meu grande companheiro e amigo. Seria
definitivamente impossível chegar até aqui
sem você. Agradeço pelos esforços e pela
compreensão em todo o desenvolvimento
deste trabalho, pela união e cuidados com
nossos filhos, nossa maior riqueza Mateus
Vitor, Marcos Levi e André Felipe, o meu
caçula, que chegou durante a finalização desta
etapa da minha vida. Amor é a palavra mais
simples e verdadeira que poderia dizer a
vocês.
À minha mãe, Valdecy Menezes, meu
pai, Antonio Carlos Menezes, que foram o
berço de toda minha criação e são os
responsáveis pelo meu caráter, meu irmão
Marcelo Menezes por seu auxílio nas horas
de necessidade.
À minha irmã Valéria Menezes por
sua ajuda inestimável, por todos os momentos
felizes e tristes, nos momentos de força e
fraqueza. Um muitíssimo obrigado é pouco
diante de tudo que passamos.

vi
AGRADECIMENTO ESPECIAL
À minha orientadora, Profa. Dra. Cristina de Andrade Monteiro, pelo
apoio, toda amizade e carinho, que me “adotou” durante este período e
contribuiu enormemente para minha formação critica e reflexiva diante da
ciência. Através de seu incentivo, ampliei meus conhecimentos, cresci
enquanto profissional e somei experiências jamais imagináveis antes de
nosso encontro, foi compreensiva e educadora. É impossível, apenas em
palavras, agradecer por todas as oportunidades, por todas as instruções,
pelo auxílio e paciência. A senhora é meu espelho como pesquisadora,
orientadora e educadora.

vii
AGRADECIMENTOS
À toda minha família, que sempre me apoiou na minhas decisões.
À minha co-orientadora, profa. Dra. Patrícia de Maria Silva Figueiredo,
pela cooperação, força, incentivo, contribuição científica e todos os
ensinamentos providos durante esta formação, inclusive as palavras
firmes que eram necessárias serem ditas.
Ao prof. Dr. Silvio Gomes Monteiro, quem colaborou significantemente
nas análises estatísticas e tão prontamente estava acessível a todos.
A todos do laboratório do Núcleo de Doenças Infecciosas e Parasitárias do
Centro Universitário do Maranhão pela grandiosa convivência, em especial
a Iven Neylla Farias Vale, que foi mais que uma colaboradora, foi
amiga, companheira, parceira em todos os momentos de pesquisa.
Aos meus colegas da segunda turma do Mestrado de Biologia Parasitária
Marcia Boor, Alícia, Roxana, Suzane, Inácio, Kênia, Ione e Adriana.
Aos diretores da Faculdade de Educação de Bacabal que investiram e
acreditaram no meu trabalho, deram apoio e oportunidade de crescimento
na carreira acadêmica e aos meus colegas docentes e técnicos pela
colaboração e palavras de incentivo em todo tempo.
Aos irmãos em Cristo da Igreja Batista Pioneira, em especial a minha
amiga Rosilene Assunção, que estiveram dando força e orando pelas
minhas vitórias.
A todos aqueles que contribuíram indiretamente durante mais esta etapa
de minha vida.

viii
“E, se algum de vós tem falta de
sabedoria, peça-a a Deus, que a
todos dá liberalmente”.
Tiago 5:1a

ix
RESUMO
A adesão e produção de biofilme têm sido apontadas como potenciais
fatores de virulência para desenvolvimento de candidíases e candidemias, pois tratam de mecanismos primários de infecção, principalmente em
pacientes imunocomprometidos. Estudos indicam que a presença de biofilme se constitui um mecanismo de resistência a ação dos antifúngicos
dificultando o tratamento dessas infecções. Esse estudo avaliou a
capacidade de adesão e formação de biofilme, em material inerte, in vitro, de 123 amostras de Candida isoladas de pacientes de diferentes sítios
anatômicos sendo 64 de C. albicans, 22 de C. tropicalis, 19 de C. parapsilosis e 18 de C. glabrata. As amostras foram cultivadas em meio
BHI incubadas a 37°C por 18h para adesão e, suplementadas com glicose, para formação de biofilme, a 37°C por 24h. Os ensaios de adesão foram
padronizados como negativo (-), menos de 01 célula por 70 campos lidos; fraca (+), apenas 01 a 10 células por 50 campos; moderada (++), acima
de 10 células por 30 campos ou forte (+++), acima de 25 células por 20 campos. A produção de biofilme foi determinada através da leitura de
absorbância em leitor de ELISA, classificadas como negativas, fraco (+1), moderado (+2), forte (+3) e muito forte (+4). Total de 93 (79%) dos
isolados foram aderentes sendo 42 (45%) com capacidade de adesão moderada ou forte. Os resultados de biofilmes mostraram 63 (51,2%) de
positividade sendo que 20 (91%) de C. tropicalis apresentaram biofilme
positivo, 55% destas no grupo +2 e +3, com capacidade significantemente maior que para as demais espécies (p < 0,0001). Os
isolados estudados também apresentaram padrões diferentes de agregação no material inerte sugerindo graus variáveis de patogenicidade
no gênero Candida. Verificou-se que amostras de C. não-albicans foram mais aderentes e mais produtoras de biofilme, sugerindo maior expressão
de fatores de virulência e, portanto maior incidência de infecções fúngicas por espécies de C. não albicans.
Palavras-chave: Candida spp, fatores de virulência, adesão, biofilme,
sítios anatômicos.

x
ABSTRACT
Adhesion and biofilm production have been identified as potential
virulence factors for developing candidiasis and candidemia, since they are primary mechanisms of infection, especially in immunocompromised
patients. Studies indicate that the presence of biofilm is a mechanism of action of antifungal resistance makes treating these infections. This study
evaluated the ability of adhesion and biofilm formation, in inert material,
in vitro, of 123 samples of Candida isolated from patients from different anatomical sites and 64 C. albicans, 22 C. tropicalis, 19 C. parapsilosis
and 18 C. glabrata. The samples were grown on BHI incubated at 37 ° C for 18 hours for membership, and supplemented with glucose for biofilm
formation at 37 ° C for 24h. The adhesion assays were standardized as negative (-), less than 01 cells per 70 fields read, weak (+), only 01 to 10
cells per 50 fields; moderate (++), above 10 cells per 30 fields or strong (+++), above 25 cells per 20 fields. The production of biofilm was
determined by absorbance reading in ELISA reader, classified as negative, weak (+1), moderate (+2), strong (+3) and very severe (+4). Total 93
(79%) isolates were adherent and 42 (45%) with ability to moderate or strong adhesion. The results of biofilms showed 63 (51.2%) positivity and
20 (91%) of C. tropicalis biofilms were positive, 55% of this group in +2 and +3, with a capacity significantly higher than for other species (p
<0.0001). All isolates also had different patterns of aggregation in the
inert material suggesting varying degrees of pathogenicity in the genus Candida. It was found that strains of non C. albicans were more adherent
and producing more biofilm, suggesting higher expression of virulence factors and therefore higher incidence of fungal infections by species of
non C. albicans.
Keywords: Candida spp, virulence factors, adhesion, biofilm, anatomical
sites.

xi
LISTA DE FIGURAS
FIGURA 1 Esquema das camadas da parede celular de leveduras
do gênero Candida. As manoproteínas são encontradas
em toda a parede celular. (CALDERONE, BRAUN, 1991)
05
FIGURA 2 Secreção e ancoramento na superfície celular de
adesinas fúngicas. (VERSTREPEN, KLIS, 2006)
08
FIGURA 3 Fases de formação do biofilme de Candida numa
superfície de dispositivo médico. 1 – Superfície do
cateter condicionado a adesão. 2 – Adesão primária
das leveduras. 3 – Formação das camadas basais das
microcolônias das leveduras e fixação à superfície. 4 –
Maturação do biofilme com leveduras, hifas, pseudo-
hifas envolvidos na matriz polimérica matriz polimérica
extracelular. Adaptado de Cardoso, 2004
14
FIGURA 4 Distribuição de frequência de isolados de diferentes
espécies de Candida em relação aos sítios de origem e
o nível de aderência.
27
FIGURA 5 Padrões de aderência em isolados de C. parapsilosis.
Isolado proveniente de sangue evidenciando um
aspecto difuso na aderência a lamínulas (a); Isolado
proveniente de urina evidenciando um aspecto
localizado de aderência a lamínulas (b); Isolado
proveniente de sangue evidenciando um aspecto
agregativo na aderência a lamínulas (c). Isolado
proveniente de urina evidenciando um aspecto pseudo-
hifal (d).
31
FIGURA 6 Estatistica descritiva e testes de Kruskal Wallis e
Student-Newman-Keuls para espécie em relação à
capacidade de formação de biofilme (Kruskal Wallis =
22.64, p < 0,0001; a,b,c Letras diferentes significa p <
36

xii
0,05 pelo teste de Student-Newman-Keuls)
FIGURA 7 Estatistica descritiva e testes de Kruskal Wallis e
Student-Newman-Keuls para sítio de origem em
relação à capacidade de formação de biofilme (Kruskal
Wallis = 2.33, p = 0.6746; a Letras iguais significa p ˃
0,05 pelo teste de Student-Newman-Keuls)
37
FIGURA 8 Estatistica descritiva e testes de Kruskal Wallis e
Student-Newman-Keuls para a capacidade de formação
de biofilme medida pela absorbância em relação à
espécie (Kruskal Wallis p < 0.001; a,b,c Letras diferentes
significa p < 0,05 pelo teste de Student-Newman-
Keuls)
39
FIGURA 9 Estatística descritiva e testes de Kruskal Wallis e
Student-Newman-Keuls para a capacidade de formação
de biofilme medida pela absorbância em relação à
espécie (Kruskal Wallis = 1.78, p = 0.7748)
40

xiii
LISTA DE TABELAS
TABELA 1 Manoproteínas (MP) de C. albicans. (Adaptado de
STURTEVAN, CALDERONE, 1997)
09
TABELA 2 Distribuição de frequência dos 123 isolados clínicos de
Candida obtidos de diferentes sítios de origem.
19
TABELA 3 Classificação do nível de capacidade de adesão dos
isolados de Candida.
25
TABELA 4 Níveis da capacidade de adesão dos isolados de
Candida em relação à espécie e ao sítio de origem
26
TABELA 5 Estatistica descritiva e teste de Kruskal Wallis da
adesão em relação à espécie.
28
TABELA 6 Estatistica descritiva e teste de Kruskal Wallis da
adesão em relação ao sítio.
29
TABELA 7 Distribuição do padrão de aderência das espécies de
Candida, em relação à espécie e ao sítio de origem.
32
TABELA 8 Capacidade de Formação de Biofilme dos isolados de
Candida, em relação à espécie e ao sítio de origem.
35

1
LISTA DE ABREVIATURAS E SIGLAS
ATCC American Type Culture Collection
ALS Agglutinin-like sequences
ABS Asorbância
BHI Infusão Cérebro Coração
CLSI Clinical and Laboratory Standards Institute
GPI Glicosil Fosfatidil Inositol
HWP 1 Hyphal wall protein
INT 1 Integrina 1
MP
NaCl
nm
PBS
SAP
DAS
Manoproteína
Cloreto de Sódio ou Solução salina
Nanômetro
Tampão fosfato salino
Proteases ácidas de aspartate
Sabouraud Dextrose Agar
UFC
µL
Unidades formadoras de colônias
Microlitros

2
SUMÁRIO
RESUMO ........................................................................................ viii
ABSTRACT ....................................................................................... ix
LISTA DE FIGURAS ........................................................................... x
LISTA DE TABELAS .......................................................................... xii
LISTA DE ABREVIATURAS E SIGLAS .................................................. xiii
1 INTRODUÇÃO ................................................................ 1
1.1 Infecções e Candida ..................................................... 1
1.2 Parede Celular .............................................................. 4
1.3 Virulência e capacidade de adesão ............................... 6
1.4 Formação de biofilme ................................................... 12
2 OBJETIVOS ................................................................... 17
2.1 Objetivo geral .............................................................. 17
2.2 Objetivos específicos ................................................... 17
3 METODOLOGIA............................................................. 18
3.1 Amostras ..................................................................... 18
3.2 Aderência em material inerte ....................................... 20
3.2.1 Preparação da suspensão de Candida para os ensaios de
adesão ..........................................................................
20
3.2.2 Aderência a lamínulas e análise microscópica ..................... 20
3.3 Formação de biofilme ................................................... 22
3.3.1 Preparação da suspensão de Candida para os ensaios de
biofilme .........................................................................
22
3.3.2 Análise da formação de biofilme ....................................... 22
3.4 Análise estatística ........................................................ 23
4 RESULTADOS ................................................................ 24
4.1 Capacidade de Adesão .................................................. 24
4.2 Formação de Biofilme ................................................... 33
5 DISCUSSÃO .................................................................. 41

3
6 CONCLUSÕES ............................................................... 50
REFERÊNCIAS ............................................................... 51
APENDICE A - Adherence ability and biofilm production by
clinical isolates of Candida species coming from different
sources .........................................................................
62
ANEXOS ....................................................................... 90

4
1 INTRODUÇÃO
1.1 Infecções e Candida
As espécies de leveduras de Candida existem como comensais no
organismo, estando presentes na microbiota humana desde o nascimento
sem causar infecção, coexistindo com o hospedeiro durante toda a sua
vida. Pode colonizar, principalmente, o trato gastrointestinal, cavidade
oral, vagina, aparelho respiratório, urinário, entre outras localizações
(COSTA, 2009a; CARDOSO, 2004).
A alteração destas leveduras comensais para um agente infeccioso
prejudicial ao organismo se dá devido principalmente aos fatores de
virulência do microorganismo e da reposta imunológica do hospedeiro,
tornando a Candida o fungo mais oportunista já descrito (RIBEIRO et al.,
2004). Infecções oportunistas em seres humanos frequentemente seguem
a supressão imune ou neutropenia (CARDOSO, 2004). De maneira
semelhante a todas as doenças infecciosas a adesão de um micróbio ao
tecido hospedeiro deve ocorrer antes que a infecção possa se estabelecer.
Infecções com espécies de Candida são invariavelmente oportunistas onde
uma falha na defesa do hospedeiro ocorre tais como perda de células
epiteliais de membranas e mucosas ou a redução no número de leucócitos
polimorfonucleares circulantes (KLOTZ, LIPKE, 2010).
Os fatores de risco que predispõe o aparecimento destes processos
infecciosos podem ser intrínsecos, quando associados ao próprio
hospedeiro, e extrínsecos quando incluem a ação terapêutica ou
medicamentosa como o uso de antibióticos de largo espectro, tratamentos
de quimioterapia de longa duração, a imunossupressão induzida do
sistema imunitário, o transplante de órgãos, os procedimentos cirúrgicos,
queimaduras, hemodiálise e outras situações de risco resultantes de
tratamentos hospitalares prolongados e intensivos (COLOMBO et al.,
2006; MEDRANO et al., 2006; MORGAN 2005; CHAKRAVARTHI et al.,
2010). Além de infecção pelo HIV, diabetes mellitus, desnutrição,

5
alcoolismo, uso de contraceptivos orais, tabagismo, estresse, pouca ou
falta de higiene são condições que favorecem o aparecimento ou a
recidiva da candidíase. As infecções fúngicas podem igualmente estar
associadas à utilização de dispositivos médicos como cateteres, próteses,
implantes entre outros (CHANDRA et al., 2001; YANG et al., 2003).
A partir da década de 80, a incidência de candidemias aumentou
substancialmente em hospitais terciários de todo o mundo
(WISPLINGHOFF et al., 2004), uma forte preocupação devido estas
infecções fúngicas invasivas serem difíceis de diagnosticar, prevenir e
tratar (COLOMBO, 2000; MEDRANO et al 2006; MORGAN, 2005; NUCCI,
2007; COLOMBO, 2002), e atualmente essas infecções têm emergido
como os maiores responsáveis pela morbidade e mortalidade nos
pacientes imunodeprimidos (PFALLER et al., 2000; PFALLER et al., 2008),
em pacientes com leucemia e transplantes de órgãos sólidos ou
receptores de células tronco (CHAKRAVARTHI, HALEAGRAHARA, 2011).
Cerca de 80% das infecções nosocomiais fúngicas estão associadas a
leveduras do gênero Candida, representando a maior causa de fungemia
com altas taxas de mortalidade (40%) (COLOMBO et al., 2006; COLOMBO
et al., 1999; MEDRANO et al., 2006). Candida albicans é o fungo mais
comumente encontrado em laboratórios microbiológicos hospitalares e é a
quarta causa mais comum em culturas de sangue positivas em pacientes
hospitalizados na América do Norte (EDMOND et al., 1999). Além disso,
20% ou mais de casos de candidemia são polimicrobianos envolvendo
uma ou mais bactérias e várias espécies de Candida (KLOTZ et al., 2007;
THORN et al., 2010) As infecções também levam a um aumento do tempo
de permanência hospitalar e maior custo de tratamento provavelmente
devido à resistência a drogas antifúngicas (TAMURA et al., 2007;
PFALLER, 1998). Esses fatos justificam a relevância do estudo deste
patógeno.
Candidemia é a mais frequente manifestação clínica de candidíase
invasiva e é uma causa significante de morbidade e mortalidade em
pacientes hospitalizados (CHAKRAVARTHI, HALEAGRAHARA, 2011). Nos

6
Estados Unidos as espécies de Candida, obtidas de infecções sanguíneas
adquiridas em hospitais, aparecem com uma frequência de 1,5 casos por
10.000 pacientes/dia (HAIJEH et al., 2004). Num estudo europeu a
frequência é levemente menor com 0,5 – 0,7 casos por 10.000
pacientes/dia (MARCHETTI et al., 2004).
Atualmente, o mais alto nível de incidência relatado de candidemia,
3,7 casos por 10.000 pacientes/dia veio de um estudo no Brasil feito por
Colombo et al. (2006).
No Brasil, Colombo et al. (2007) conduziram um estudo
epidemiológico reunindo dados sobre infecções de corrente sanguínea
documentados em quatro hospitais da cidade de São Paulo. Um total de
7038 episódios de bacteremias e fungemias ocorridos no período de um
ano foi avaliado. Candida spp. responderam por 4,3% do total das
infecções de corrente sanguínea. Frequência semelhante foi também
detectada no Hospital das Clínicas da Faculdade de Medicina de Botucatu
(HC/FMB), que de um total de 6417 episódios e amostras de culturas
positivas avaliadas, no período de janeiro de 1991 a dezembro de 1994,
Candida spp. foram isoladas em 222 (3,5%) culturas, as quais foram
oriundas principalmente das unidades de pediatria e berçário (SUGIZAKI
et al., 1998). Ruiz-Herrera et al. (2005) relataram que, nesse mesmo
hospital, as espécies mais frequentes foram C. albicans (38,7%) e C.
parapsilosis (30,7%).
Embora a C. albicans seja a principal espécie isolada de pacientes
com fungemia (PFALLER et al., 1998 a,b,c; SANDVEN et al., 1998;
KRCMÉRY JR V, KOVACICOVÁ G, 2000; CHAKRAVARTHI, HALEAGRAHARA,
2011), têm aumentado os relatos de infecções causadas por outras
espécies de Candida que não C. albicans (ARENDRUP et al., 2005;
COLOMBO et al., 2007). Em 1963, eram conhecidas apenas cinco espécies
de Candida causadoras de doenças humanas, C. albicans, C. parapsilosis,
C. tropicalis, C. stellatoidea e C. guilliermondii. Atualmente são conhecidas
cerca de 20 espécies de Candida implicadas em micoses superficiais ou
invasivas em humanos (DIGNANNI, SOLOMKIN, ANAISSIE, 2003).

7
As principais espécies de interesse clínico são: C. albicans, C.
parapsilosis, C. tropicalis, C. glabrata, C. krusei, C. guilliermondii e C.
lusitaniae. Entretanto, número progressivo de casos relacionados a
espécies emergentes de Candida tem sido descrito, envolvendo
isolamentos de C. dubliniensis, C. kefyr, C. rugosa, C. famata, C. utilis, C.
lipolytica, C. norvegensis, C. inconspicua entre outras (COLEMAN et al.,
1998). A frequência de outras espécies de Candida que não C. albicans
depende da população de pacientes estudada, da terapêutica utilizada, do
uso de antibióticos ou outras medidas adotadas (PFALLER, 1995; ABI-
SAID et al., 1997; NUCCI, COLOMBO, 2007; PFALLER et al., 2008).
Hinrichsen et al. (2009) avaliaram a distribuição das espécies de
Candida em dois hospitais terciários no Recife, nordeste do Brasil,
realizando um levantamento de todas as culturas positivas para Candida
processadas por um laboratório de Setembro de 2003 a Setembro de
2006. De um total de 1.279 amostras positivas para Candida, incluindo
culturas originadas de vários tipos de material biológico (sangue, urina,
traquéia, catéteres e outros), as espécies mais frequentes foram:
C.albicans (367), C. tropicalis (363), C. parapsilosis (147), C. glabrata
(81), C. krusei (30) e C. guillermondii (14). Os isolados foram obtidos de
746 pacientes hospitalizados. Um total de 221 hemoculturas positivas foi
detectado em 166 diferentes pacientes em ambos os hospitais, e 113
(68,1%) destes pacientes com hemoculturas positivas apresentavam
Candida em outros locais do corpo. Este estudo mostrou que outra espécie
de Candida, que não C. albicans, foi o principal agente isolado e prova a
importância da C. tropicalis em infecções fúngicas nosocomiais.
1.2 Parede Celular
A parede celular das leveduras não apenas tem a função de conferir
rigidez e proteção a célula, mas também está intimamente relacionada
com a mediação de interações entre o microorganismo e o ambiente

8
externo. Aproximadamente 80 a 90% da parede celular de Candida é
constituída por carboidratos havendo três constituintes que representam a
maioria dos polissarcarídeos de parede celular:
Polímeros de glucose com ramificações contendo ligações ß-
1,6 e ß-1,3;
Polímeros de N-acetil-D-glicosamina (GlcNac) ligados a
moléculas de quitina por ligações ß-1,4;
Polímeros de manose ligados covalentemente a
manoproteínas.
A parede celular contém ainda 6 a 25% de proteínas e 1 a 7% de
lipídeos. Os componentes estruturais são: os ß-glucanos e a quitina.
Polímeros de manose (manoproteínas) representam cerca de 40% dos
polissarcarídeos e são os principais constituintes da célula - grupo na qual
pertence as adesinas (CARDOSO, 2004; COSTA, 2009). A disposição das
camadas da parede celular de Candida está esquematizada na Figura 1.
FIGURA 1. Esquema das camadas da parede celular de leveduras do
gênero Candida. As manoproteínas são encontradas em
toda a parede celular. (CALDERONE, BRAUN, 1991)

9
1.3 Virulência e capacidade de adesão
A habilidade de C. albicans ser a espécie mais relacionada com os
processos de colonização e patogenicidade no homem é decorrente dos
seus fatores e propriedades de virulência como a acentuada capacidade de
dimorfismo e produção de exoenzimas facilitadoras da interação do fungo
às células do hospedeiro, propiciando a aderência, a formação de biofilme
e a penetração ao tecido afetado. Todas essas etapas são consideradas
relevantes na patogênese da candidíase (XU et al., 2000). Apesar de
haver estudos complexos destes fatores de virulência com relação à C.
albicans, os mesmos não são totalmente compreendidos em outras
espécies de Candida. Além disso, a variabilidade inter e intraespecífica
existente dentro do gênero Candida também são fatos que corroboram a
necessidade crescente e urgente de pesquisas relacionadas aos fatores de
virulência (YANG, 2003).
A capacidade que o microrganismo tem de aderir à superfície das
células do hospedeiro representa o primeiro estágio de patogênese e é
alcançada por uma combinação de interações específicas e não
específicas. Os mecanismos usados por espécies de Candida para aderir
são múltiplos e não têm sido definidos precisamente (GHANNOUM,
RADWAN, 1990; SAN MILLAN et al., 2000; VIDOTTO et al., 2003).
Doenças infecciosas como a candidíase são conhecidas por começar com a
fixação do patógeno a um alvo particular ao hospedeiro (CALDERONE,
FONZI, 2001). É claro, portanto que a inibição desta ligação constitui um
importante passo para controlar a infecção em um período precoce.
Adesão pode ocorrer a diferentes superfícies como às células e aos
polímeros inertes, e muitos fatores do hospedeiro podem alterar a
expressão destas adesinas ou sua ligação às células hospedeiras
influenciando na patogênese (SKERL et al., 1984). Alterações no
hospedeiro são geralmente necessárias para que microorganismos
comensais tornem-se patógenos oportunistas e desencadeiem a infecção
(YANG et al., 2003). A candidíase oral é uma das principais infecções que

10
acometem os pacientes em hospitais e os dispositivos prostéticos estão
entre as principais causas deste tipo de infecção, portanto a adesão de
espécies de Candida a materiais poliméricos é um dos principais objetivos
de estudo nestas leveduras (NIKAWA et al., 2003). É interessante
ressaltar que todos esses estudos são principalmente focados em
linhagens de Candida albicans, demonstrando a necessidade de estudos
acerca dos fatores primários de virulência em outras espécies de Candida.
O processo de adesão inclui fatores não biológicos, relacionado a
forças de van der Walls, interações hidrofóbicas e eletrostáticas; e fatores
biológicos que se referem a mecanismos moleculares específicos como a
expressão de adesinas, que são biomoléculas que promovem a aderência
(COTTER e KAVANAGH, 2000; CARDOSO, 2004).
Adesinas são proteínas importantes no processo de infecção, pois
são elas que fazem a ligação da Candida às células ou à superfície a ser
infectada (STURTEVANT, CALDERONE, 1997; CALDERONE, GOW, 2002).
Todas as adesinas fúngicas apresentam em comum três domínios
estruturais: o domínio C-terminal (com a âncora GPI - responsável pela
ligação da adesina a parede celular), a porção N-terminal que contém o
carboidrato ou peptídeo de ligação e o domínio central caracterizado por
uma região rica em aminoácidos serina e treonina codificados por uma
região de DNA extremamente conservada (VERSTREPEN, KLIS, 2006)
(Figura 2).

11
FIGURA 2. Secreção e ancoramento na superfície celular de adesinas
fúngicas (VERSTREPEN, KLIS, 2006).
Na parede das leveduras do gênero Candida inicia se o processo de
adesão, no qual há a integração do microorganismo com a superfície das
células hospedeiras. Para cada adesina há um receptor extracelular
específico que permite esta ligação. Uma série de proteínas tem sido
identificada como participante no reconhecimento de ligantes da célula
hospedeira. No entanto, a caracterização dos genes e a construção de
linhagens mutantes nulas para verificação das funções dos produtos
gênicos não foram totalmente realizadas (STURTEVAN, CALDERONE,
1997; GRUBB et al.,2008).
A tabela 1 a seguir relaciona as principais adesinas encontradas em
C. albicans e os ligantes de cada uma (STURTEVAN, CALDERONE, 1997).

12
TABELA 1. Manoproteínas (MP) de C. albicans (Adaptado de
STURTEVAN, CALDERONE, 1997).
Adesinas Ligantes
MP60 iC3b, C3d
MP60 Laminina, fibrinogênio, fibronectina
MP58 Fibrinogênio
MP70, 55, 42 C3H2O
MP130 iC3b
MP37/67 Laminina
Mudanças fenotípicas reversíveis podem ocorrer em linhagens de
Candida especialmente no que diz respeito às propriedades de adesão. Os
genes de adesão são ativados por diversos fatores, tais como: privação de
fontes de carbono e/ou nitrogênio no meio de cultura, mudanças de pH ou
alterações nos níveis de etanol (SAMPERMANS et al., 2005).
Als1p (FU et al., 1998) e Als3p (GAUR et al., 1999.), assim como
Als5p (HOYER et al., 1998a), são adesinas de C. albicans codificadas por
membros da grande família de genes ALS (Agglutinin-like sequences)
(HOYER, 2001) compostas por nove genes (HOYER et al., 1995, 1998;
GAUR , KLOTZ, 1999; HOYER, HECHT 2000, 2001).
A expressão heteróloga dos genes ALS em Saccharomyces
cerevisiae confere um fenótipo de aderência ao organismo, sugerindo que
proteínas Als funcionem na adesão às superfícies celulares (CALDERONE,
BRAUN, 1991; GAUR, KLOTZ, 1997; FU et al., 1998; HOYER et al., 2001).
Além disso, genes ALS são expressos diferencialmente sob uma
variedade de condições em C. albicans; de maneira similar ao que ocorre
com membros da família SAP (proteases ácidas de aspartato) (HOYER et

13
al., 1994; HOYER et al., 1998 ). Se essas famílias gênicas desempenham
uma função importante na patogênese de C. albicans, é possível que elas
também contribuam para a patogenicidade de outras espécies de Candida
clinicamente relevantes (HOYER et al., 2001).
Análises por Southern blot com sondas específicas de ALS tem
sugerido a presença de famílias gênicas ALS em C. dubliniensis e C.
tropicalis, sendo que três genes ALS já foram isolados para cada
organismo. Estes resultados preliminares também sugerem que há
número similar de genes ALS em C. albicans e C. dubliniensis (HOYER et
al., 2001).
Enquanto os genes ALS estão presentes em C. albicans, em
sequências de C. tropicalis estão em menor número e são pouco
relatados. Análises por Northern blot demonstraram que os mecanismos
que controlam a expressão de genes ALS em C. albicans e C.
dubluniensies são diferentes. Embora uma família ALS esteja presente em
cada organismo, análises filogenéticas desses genes em C. albicans, C.
dubliniensis e C. tropicalis indicam que existem diferenças entre as
espécies tanto na diversidade de sequências quanto no tamanho (HOYER
et al., 2001).
Outra proteína importante relacionada ao processo de adesão em
Candida é a integrina. A integrina 1 (Int1) é uma proteína de superfície de
C. albicans que apresenta alta similaridade com as integrinas de
vertebrados. A expressão do gene INT1 em Saccharomyces cerevisae foi
suficiente para torná-la aderente a células epiteliais humanas apesar de
normalmente essa espécie não apresentar tal característica. Além disso, a
mutação em INT1 em C. albicans suprime o desenvolvimento das hifas, a
adesão a células epiteliais e a virulência em camundongos (GALE et al.,
1998). O gene INT1 de C. albicans foi originalmente clonado devido a sua
similaridade com integrinas de leucócitos de vertebrados, adesinas que se
liga a proteínas da matriz extracelular e induzem mudanças morfológicas
em resposta a sinais extracelulares (YANG, 2003).

14
Outra importante adesina de C. albicans, a Hwp1 (Hyphal wall
protein), é encontrada na superfície dos tubos germinativos, e participa da
regulação de ligações a células epiteliais, por ser um substrato para
transglutaminases, está intimamente relacionada com a patogênese em
candidíase sistêmica (CHAFFIN et al., 1998).
Cepas que não apresentam o gene HWP1 não podem aderir de
forma eficiente em células epiteliais bucais e tem capacidade reduzida em
causar candidíase sistêmica em ratos (STAAB et al., 1999).
Muitos estudos têm descrito a aderência em C. albicans a vários
tipos celulares, mas pouco se conhece sobre a adesão em outras espécies
de Candida e o seu papel no reconhecimento da superfície celular do
hospedeiro (LIMA-NETO et al., 2009). Sugere-se que a capacidade de
adesão varia entre os isolados de diferentes espécies como sugere o
trabalho de Biasoli et al. (2002) onde foram apontadas diferenças entre C.
albicans, C. glabrata, C. krusei e C. lusitaniae.
A propriedade de aderência em C. glabrata parece ser diferente
daquela de C. albicans. Em C. glabrata a aderência às células epiteliais
humanas é mediada em grande parte por uma adesina única, Epa1p,
codificada pelo gene EPA1, que é uma lecitina dependente de Ca2+
(COMARCK et al., 1999). Epa1p é membro de uma grande classe de
glicosil fosfatidilinositol (GPI) ancoradas às proteínas da parede celular
(GPI-CWPs) encontradas em diversas espécies de fungos (KAPTEYN et al.,
1999; BRUNEAU et al., 2001). Estas proteoglicanas com ligações cruzadas
são caracterizadas pela presença de uma sequência de resíduos
hidrofóbicos, uma sequência C-terminal hidrofóbica sinalizada pela adição
de uma âncora GPI e, em muitos casos, a presença de uma serina C-
terminal com grande região rica em treonina de função desconhecida
(KAPTEYN et al., 1999).
O processo de adesão por ser o passo inicial e essencial para o
estabelecimento da doença (YANG, 2003), fator importantíssimo às
infecções por Candida, merece ser melhor estudado em outras espécies.

15
1.4 Formação de biofilme
Subsequente ao processo de adesão celular ocorre à formação de
biofilme, que está diretamente relacionado à morfologia e espécie
envolvida, seu potencial de virulência (SUZUKI, 2009), como a própria
superfície de contato e fatores ambientais (VIANI, 2007).
Biofilmes são complexos ecossistemas microbiológicos embebidos
em uma matriz de polímeros orgânicos, aderidos a uma superfície.
Donlan, Costerton (2002) fizeram revisão sobre biofilme e a matriz
extracelular, respectivamente, e foi definido que biofilme é a comunidade
séssil caracterizada por células que formam microcolônias aderentes a um
substrato, uma interface ou ainda a uma matriz exopolimérica de
substâncias extracelulares que conferem o fenótipo do biofilme; assim
permite uma proteção ambiental contra o estresse favorecendo o
desenvolvimento das infecções e uma resistência a agentes
antimicrobianos (BRANDA et al., 2005; DONLAN, 2001). A arquitetura do
biofilme fúngico é altamente heterogênea e diferencia-se do biofilme
bacteriano, mas poucos estudos descrevem sua composição e relevância
na patogênese das doenças, as quais tem grande importância clinica
devido ao aumento da prevalência das infecções fúngicas superficiais e
sistêmicas (CHANDRA et al.,2001).
A formação de biofilme em C. albicans é um processo gradual que se
inicia com a aderência a um substrato, seja ao próprio tecido do
hospedeiro ou ao dispositivo médico, resultando na formação de uma
camada basal confluente de células que se dividem e produzem hifas,
direcionadas para a região superior do biofilme (SOLL, 2008). Os agentes
microbianos presentes no biofilme diferem-se do comportamento na sua
forma planctônica tanto em relação ao crescimento quanto a resistência
aos antifúngicos (CARDOSO, 2004; SUZUKI, 2009). Estas células durante
o desenvolvimento do biofilme produzem uma matriz extracelular estável

16
de substâncias poliméricas (DOUGLAS, 2003; CHANDRA et al., 2001;
SOLL, 2008) que a composição da matriz inclui: carboidratos, proteínas,
fósforo, glicose e hexosaminas, mas a maior parte desse conteúdo ainda
não foi identificada (BAILLIE, DOUGLAS, 2000; BLANKENSHIP, MITCHELL,
2006).
A formação de biofilme está associada à capacidade de algumas
leveduras como C. parapsilosis colonizar o interior de cateteres e outros
dispositivos médicos, desse modo proporcionando um reservatório a partir
do qual o microorganismo pode entrar na corrente sanguínea em
indivíduos recebendo alimentação intravenosa (PFALLER et al., 1995).
Candida spp tem sido encontrada em diversos implantes médicos ou
dispositivos invasivos; C. albicans é o microorganismos comumente mais
isolado de cateter venoso central, urinário ou mesmo válvulas cardíacas
mecânicas, podendo ser proveniente de forma endógena ou exógena por
exposição aos profissionais de saúde ou infusantes contaminados
(DONLAN, 2001; HOTA, 2004).
Chandra et al. (2001) demonstraram que a formação do biofilme
possui três fases distintas de desenvolvimento, sendo a fase inicial, a fase
intermediária e a fase de maturação. A fase inicial ou adesão primária é a
aproximação da levedura à superfície de adesão e onde ocorrem os
processos físico-químicos, as interações biológicas ou especificas e não
biológicas ou não especificas citadas anteriormente. Na fase secundária há
o processo de formação das microcolônias, início da produção de
complexos extracelulares como as proteínas e os exopolissacarídeos e
fixação desta camada no substrato ou no tecido do hospedeiro. Na fase de
maturação há ainda a adição das hifas, pseudo-hifas e consolidação desta
matriz polimérica extracelular constituindo uma rede densa com canais de
água para a nutrição destas células. Já neste estágio final de maturação e
desenvolvimento do biofilme ocorre menor crescimento das leveduras, há
elevado crescimento das hifas e pseudo-hifas, com o envolvimento do
biofilme pela matriz polimérica extracelular, crescimento tridimensional e,
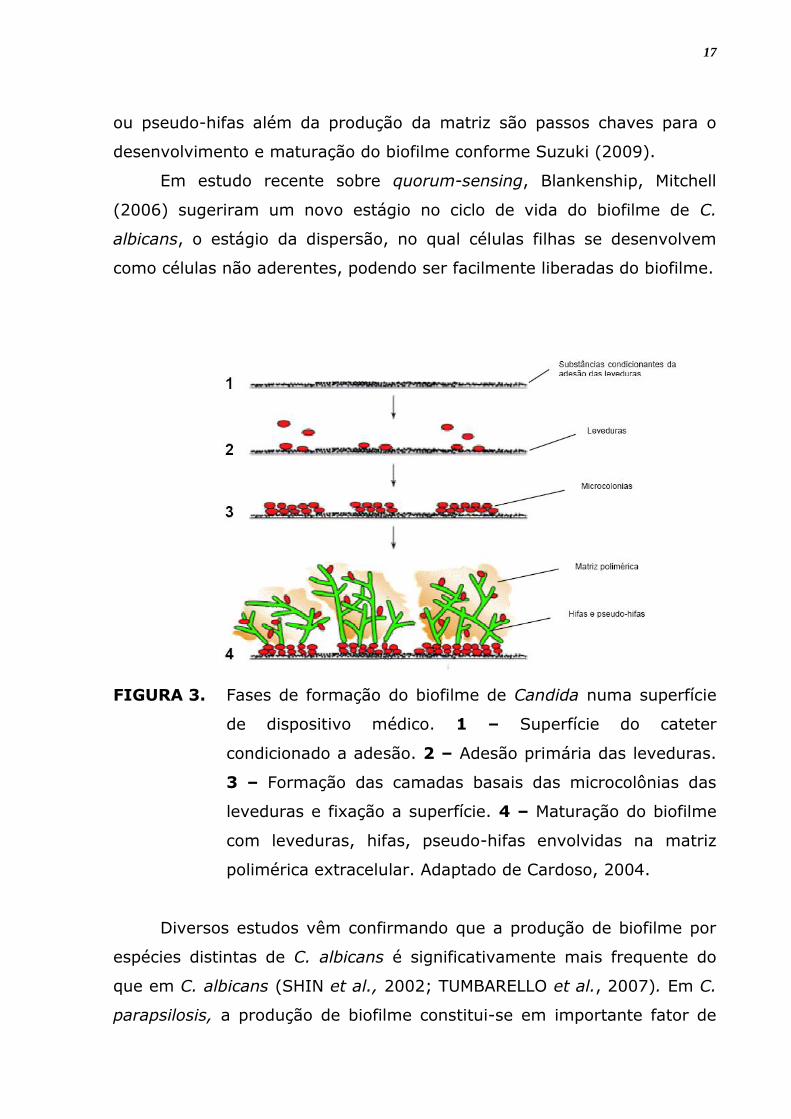
posterior, envelhecimento do biofilme (Figura 3). A diferenciação de hifas
17
ou pseudo-hifas além da produção da matriz são passos chaves para o
desenvolvimento e maturação do biofilme conforme Suzuki (2009).
Em estudo recente sobre quorum-sensing, Blankenship, Mitchell
(2006) sugeriram um novo estágio no ciclo de vida do biofilme de C.
albicans, o estágio da dispersão, no qual células filhas se desenvolvem
como células não aderentes, podendo ser facilmente liberadas do biofilme.
FIGURA 3. Fases de formação do biofilme de Candida numa superfície
de dispositivo médico. 1 – Superfície do cateter
condicionado a adesão. 2 – Adesão primária das leveduras.
3 – Formação das camadas basais das microcolônias das
leveduras e fixação a superfície. 4 – Maturação do biofilme
com leveduras, hifas, pseudo-hifas envolvidas na matriz
polimérica extracelular. Adaptado de Cardoso, 2004.
Diversos estudos vêm confirmando que a produção de biofilme por
espécies distintas de C. albicans é significativamente mais frequente do
que em C. albicans (SHIN et al., 2002; TUMBARELLO et al., 2007). Em C.
parapsilosis, a produção de biofilme constitui-se em importante fator de

18
virulência. C. parapsilosis é bem conhecida como causadora de fungemia e
candidíase invasiva associada à hiperalimentação parenteral, dispositivos
intravasculares e soluções oftálmicas contaminadas (PLOUFFE et al.,
1977; SOLOMON et al., 1984; O’DAY, HEAD, ROBINSON, 1987; WEMMS
et al., 1987; WEMMS, 1992). Vários fatores dão à C. parapsilosis uma
vantagem seletiva, incluindo a capacidade de proliferar-se em altas
concentrações de glicose e de aderência a materiais protéticos
(CRITCHLEY, DOUGLAS, 1985; WEMMS et al., 1987). Pfaller, Messer,
Hollis (1995), estudaram a produção de biofilme por amostras clínicas de
C. parapsilosis crescidas em meio de cultura contendo glicose e
verificaram que as isoladas de sangue e de cateter apresentaram maior
produção de biofilme do que amostras de outros sítios anatômicos.
A base molecular da formação e do desenvolvimento do biofilme
destes fungos ainda não está completamente compreendida, porém, já
está bem estabelecida que a interação da C. albicans com as células do
hospedeiro ou superfícies inertes resulta em alterações na expressão de
diferentes genes. Diferentes estudos têm descrito mudanças nos níveis de
expressão gênica durante o desenvolvimento do biofilme (MARCHAIS et
al., 2005, MURILLO et al., 2005).
Devido aos poucos estudos com relação à formação de biofilme que
tem sido considerado um fator potencial de virulência para outras espécies
de Candida (SHIN et al., 2002) e a composição do biofilme de leveduras
que parece ser única e às profundas implicações clínicas como resistência
intensificada a agentes microbianos e proteção contra defesas do
hospedeiro (RAMAGE et al., 2001) que são inerentes a capacidade de
formação de biofilmes são alguns dos fatos relevantes que justificam a
importância de estudos sobre esta propriedade. A propriedade de adesão
e formação de biofilmes bem como os demais fatores de virulência das
Candidas como dimorfismo, variabilidade fenotípica, produção de toxinas e
exoenzimas têm relevância para a instalação e estabilização da infecção
no organismo do hospedeiro, bem como para a sobrevivência e

19
acometimento de outros tecidos anteriormente não colonizados por este
patógeno (YANG et al., 2003).
A identificação de fatores de virulência únicos para uma
determinada espécie de Candida poderia proporcionar poderosos
esclarecimentos no processo patogênico com consequências diretas no
diagnóstico e terapia das infecções.

20
2 OBJETIVOS
2.1 Objetivo geral
Verificar a capacidade de aderência e formação de biofilme em
isolados clínicos de Candida provenientes de diferentes sítios anatômicos.
2.2 Objetivos específicos
• Determinar se há diferenças na capacidade de aderência
celular entre as espécies Candida;
• Comparar qualitativa e quantitativamente a capacidade de
formação de biofilme entre as espécies diferentes;
• Correlacionar sítio de origem com capacidade de adesão e
formação de biofilme.

21
3 METODOLOGIA
3.1 Amostras
Foram utilizados neste estudo um total de 123 isolados clínicos
representando as espécies de Candida tropicalis (22 amostras), C.
glabrata (18 amostras), C. parapsilosis (19 amostras) e C. albicans (64
amostras) provenientes de vários tipos de amostras clínicas como
secreções vaginais, urina, sangue, secreções traqueais e de ponta de
cateteres, todas obtidas de um laboratório particular da cidade de São
Luís – MA, gentilmente cedidas e depositadas na micoteca do Laboratório
de Micologia do Núcleo de Doenças Endêmicas e Parasitárias do
UNICEUMA (Tabela 2).
Todas as leveduras foram previamente identificadas pelo sistema
automatizado VITEK (bioMérieux).
Para comparação de dados, foi incluída nos experimentos a
linhagem padrão C. albicans ATCC18804 (American Type Culture
Collection, Rockville, Md).
As culturas estoque foram mantidas a -20ºC em caldo BHI (Brain
Heart Infusion - Acumedia Manufactures) que é um meio utilizado para
crescimento microbiológico. Após recuperação, as linhagens também
foram mantidas em meio líquido BHI, estocadas a 4ºC durante o período
experimental para proceder aos ensaios de adesão e formação de
biofilme.
22
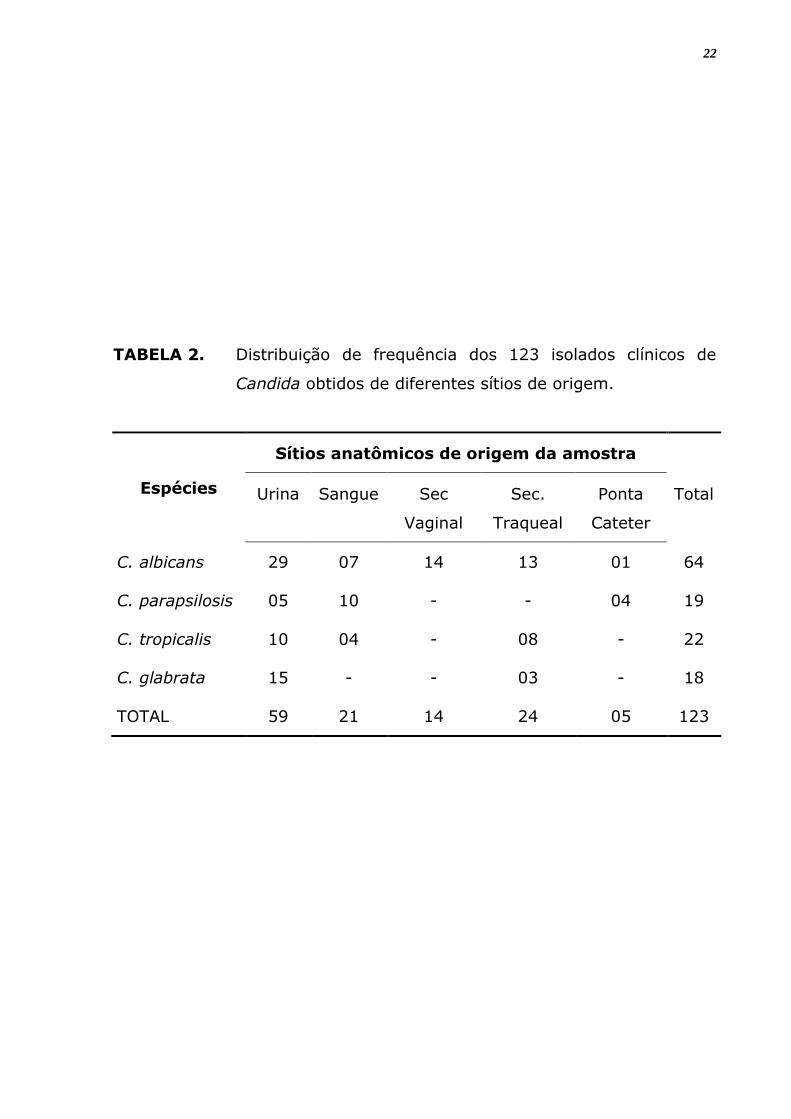
TABELA 2. Distribuição de frequência dos 123 isolados clínicos de
Candida obtidos de diferentes sítios de origem.
Espécies
Sítios anatômicos de origem da amostra
Urina Sangue Sec
Vaginal
Sec.
Traqueal
Ponta
Cateter
Total
C. albicans 29 07 14 13 01 64
C. parapsilosis 05 10 - - 04 19
C. tropicalis 10 04 - 08 - 22
C. glabrata 15 - - 03 - 18
TOTAL 59 21 14 24 05 123

23
3.2 Aderência em material inerte
3.2.1 Preparação da suspensão de Candida para os ensaios de adesão.
O inóculo das células de leveduras foi feito a partir de cultura
estoque, em meio agar Sabouraud Dextrose – SDA (Merck) que é
frequentemente utilizado para o crescimento de espécies de Candida, com
uma concentração de 30g/l. O meio SDA foi preparado e esterilizado a
121ºC durante 15 minutos antes de iniciarmos os ensaios. As culturas
celulares foram repicadas em placas de petri com meio sólido de SDA e
incubadas por 24 horas a 37ºC. Antes de cada ensaio de adesão, as
células foram submetidas a um processo de lavagem duas vezes com
solução tampão PBS (pH 7, 0.1 M) com agitação. As células foram
ressuspendidas em solução salina (NaCl 0.85 %) e os inóculos fúngicos
foram padronizados para aproximadamente 106 UFC/ml de acordo com a
turbidez do tubo a 0,5 da escala de McFarland (CLSI Clinical and
Laboratory Standards Institute – NCCLS, 2007).
3.2.2 Aderência a lamínulas e análise microscópica (Biasoli et al., 2002,
com adaptações).
Para a análise de adesão foi utilizado Biasoli et al.(2002) com
algumas modificações. Para os testes foram usados microplacas de
poliestireno de 24 poços (Zellkultur Testplate 24,Techno Plastic Products),
contendo em cada poço uma lamínula redonda estéril de vidro
(Glasscyto), adicionado 40ul do inóculo padronizado (106 UFC/ml) e 960 ul
de meio BHI incubados a 37°C por 18 horas. Para o controle do
experimento, somente lamínulas, sem inóculo, foram igualmente
processadas. Após incubação e remoção do meio foi procedida à lavagem
da microplaca com água destilada estéril para remoção das células não
aderentes e as lamínulas foram coradas com violeta cristal (1%) por 5
minutos. As lamínulas foram novamente lavadas duas vezes com água

24
estéril para remoção do excesso de corante e colocada em lâminas para
ser feita a leitura de microscopia de luz direta (NIKON Eclipse E100). A
determinação da aderência foi verificada pela contagem do número de
leveduras aderidas (Biasoli et al., 2002). A padronização da classificação
da capacidade de adesão das células em lamínulas foi realizada
considerando a contagem de células aderidas por campo(s). Para cada
lamínula do ensaio foi analisado um máximo de 70 campos e as células
aderidas foram contadas. O número de células obtido foi dado como
média de três experimentos realizados para cada isolado. Foi feita a
padronização e, posteriormente, analisada a capacidade de adesão celular
em material inerte sendo classificada como negativo, fraco (+), moderado
(++) ou forte (+++) (Tabela 3; VIANA et al., 2010, com adaptações).
Cada isolado foi analisado por dois observadores de forma independente.
A adesão celular de cada amostra foi padronizada como negativa se
menos que 01 (uma) célula estivesse aderida por campo, visualizados 70
(setenta) campos da lamínula; fraca se houve entre 01 a 10 (uma a dez)
células aderidas por campo em 50 (cinquenta) campos observados;
moderada se houve mais de 10 (dez) células por campo aderidas em 30
(trinta) campos e forte se foi igual ou maior que 25 (vinte e cinco) células
aderidas por campo em 20 (vinte) campos analisados. Não houve
discordância de resultados feita pelos dois observadores na visualização
por microscopia direta.
Verificou-se que as células de leveduras aderiram às lamínulas
seguindo padrões diferenciados de adesão, semelhante aos padrões de
aderência das enterobactérias Escherichia coli às células epiteliais
(CRAVIOTO et al., 1979). Desta forma, classificou- se o padrão da
capacidade de adesão das espécies de Candida em lamínulas em 04
(quatro) formas diferentes utilizando-se como referência a classificação de
E. coli feita por CRAVIOTO et al. (1979). Estes padrões serão
apresentados no item Resultados, visto ser a primeira vez que os mesmos
são definidos.

25
3.3 Formação de biofilme
3.3.1 Preparação da suspensão de Candida para os ensaios de biofilme.
O inóculo das células de leveduras foi feito a partir de cultura
estoque, em meio agar Sabouraud Dextrose – SDA (Merck) que é
frequentemente utilizado para o crescimento de espécies de Candida, com
uma concentração de 30g/l. O meio SDA foi preparado, suplementados
com 60g de glicose por litro para obtermos uma concentração final de 8%
de glicose no meio, esterilizado a 121ºC durante 15 minutos antes de
iniciarmos os ensaios. As culturas celulares foram repicadas em placas de
petri com meio sólido de SDA e incubadas por 18 horas a 37ºC. Antes de
cada ensaio para formação de biofilme, as células foram submetidas a um
processo de lavagem duas vezes com solução tampão PBS (pH 7, 0.1 M)
com agitação. As células foram ressuspendidas em solução salina (NaCl
0.85%) e os inóculos fúngicos foram padronizados para aproximadamente
106 UFC/ml de acordo com a turbidez do tubo a 0,5 da escala de
McFarland (CLSI Clinical and Laboratory Standards Institute – NCCLS,
2007).
3.3.2 Análise da formação de biofilme (SHIN et al., 2002, com
modificações).
Os testes de análise da formação de biofilme foi de acordo com
SHIN et al. (2002) feito algumas modificações. Foram utilizados em cada
experimento 20ul de suspensão do inóculo (106UFC/ml), estes colocados
em poços de microplacas de 96 poços acrescidos de 180 ul de BHI (Brain
Heart Infusion - Acumedia Manufactures) suplementado com 6% de
glicose. As microplacas foram incubadas a 37°C por 24 horas. Para
controle utilizou-se poços contendo meio BHI sem inóculo. Após a
incubação foi removido o líquido dos poços, a microplaca foi lavada três
vezes com água destilada estéril e então corada com violeta cristal (1%).

26
As microplacas foram novamente lavadas para remoção do excesso do
corante e por fim foram adicionados 200 uL de água destilada a cada poço
para leitura de absorbância . A formação de biofilme foi analisada pela
leitura das microplacas em uma leitora de microplacas medindo-se a
absorbância em comprimento de onda de 450 nm. Os valores de
absorbância foram dados como média de três (03) experimentos para
cada isolado e os dados foram expressos em porcentagem. A produção de
biofilme para cada isolado foi avaliado como negativo (% A < 5), fraco
(+1; 5 ≤ % A < 20), moderado (+2; 20 ≤ % A < 35), forte (+3; 35 ≤ %
A < 50) e muito forte (+4; % A ≥ 50). Cada isolado foi testado pelo
menos três vezes, para evitar qualquer discrepância nos valores de
absorbância obtidos de cada amostra.
3.4 Análise estatística
Os dados foram analisados através do programa estatístico Bioestat
5.0 (2007). A normalidade da variável numérica absorbância foi analisada
através do teste de Lilliefors. Como a distribuição não foi normal, ela foi
analisada através de teste não paramétrico. Juntamente com as variáveis
ordinais, a classificação do nível de biofilme e da adesão em relação à
espécie e ao sítio foram feitas através do teste não paramétrico de
Kruskal Wallis e a análise post-hoc foi feita pelo teste Student-Newman-
Keulls. O nível de significância (α) aplicado em todos os testes foi de 5%,
ou seja, considerou-se significativo quando p < 0,05.

27
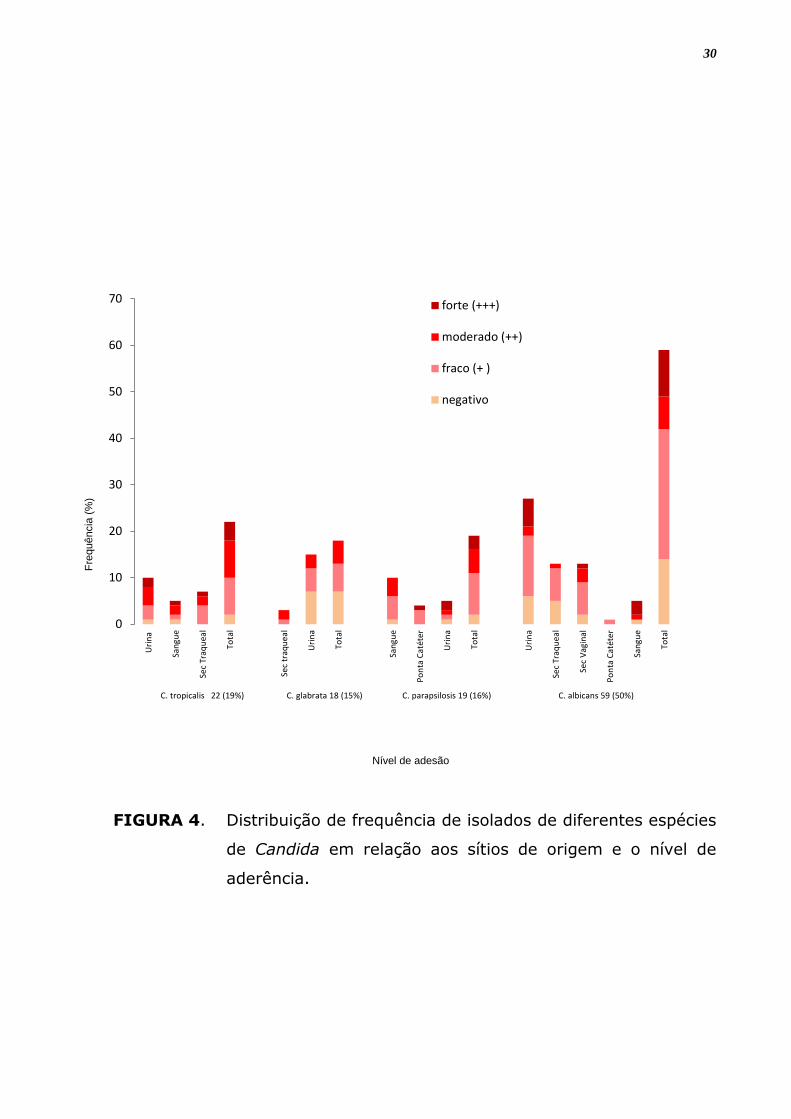
4 RESULTADOS
4.1 Capacidade de adesão
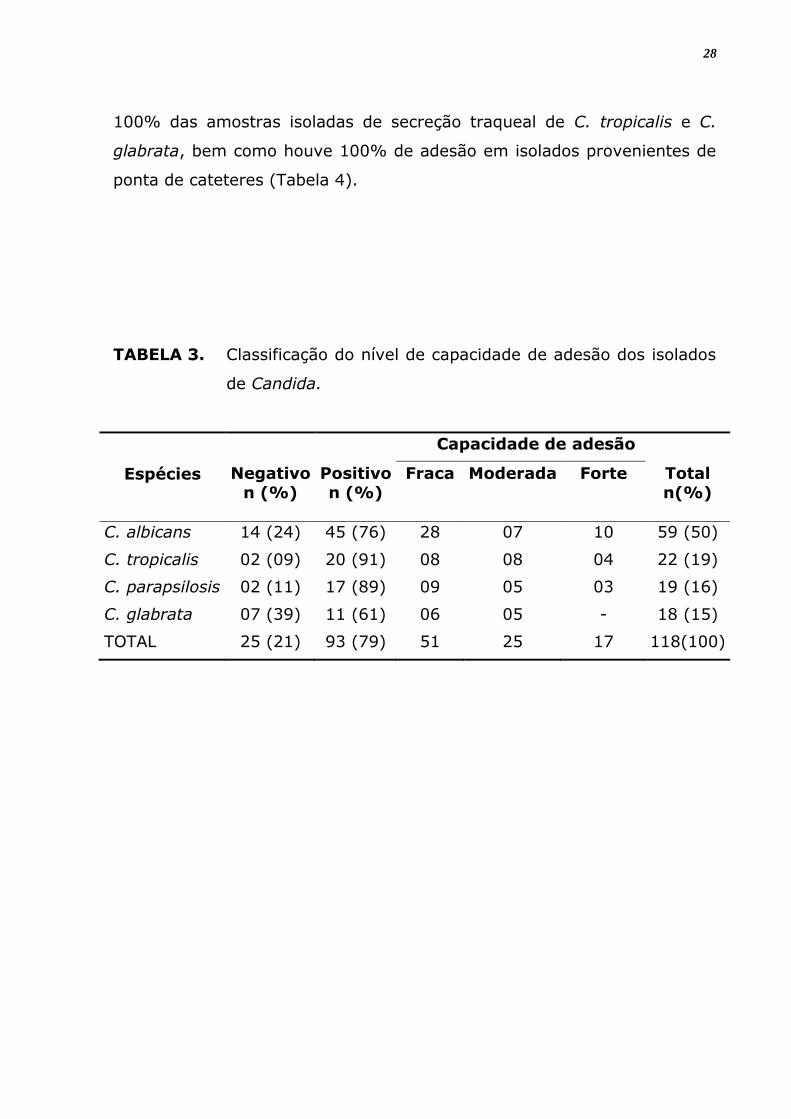
A tabela 3 mostra a classificação do nível de capacidade de adesão
dos isolados de Candida às lamínulas. De um total de 118 isolados
analisados para a capacidade de adesão 93 (79%) foram positivos para
esta propriedade. Dentre estes 81, 73% mostram uma capacidade de
aderência moderada ou fraca. É evidente que as 4 espécies diferentes de
Candida aderiram às lamínulas embora tenha-se observado diferenças no
nível de aderência entre elas. Dentre as espécies analisadas C. tropicalis
mostrou-se com maior capacidade de aderência a material inerte, onde
91% das linhagens foram positivas, seguida de C. parapsilosis com 89%
dos isolados apresentando aderência positiva, C. albicans com 76% e, por
último, C. glabrata com 61% de amostras que apresentaram aderência
em material inerte. Além da variação interespecífica verificou-se uma
variabilidade intraespecífica com relação á capacidade de adesão (Tabela
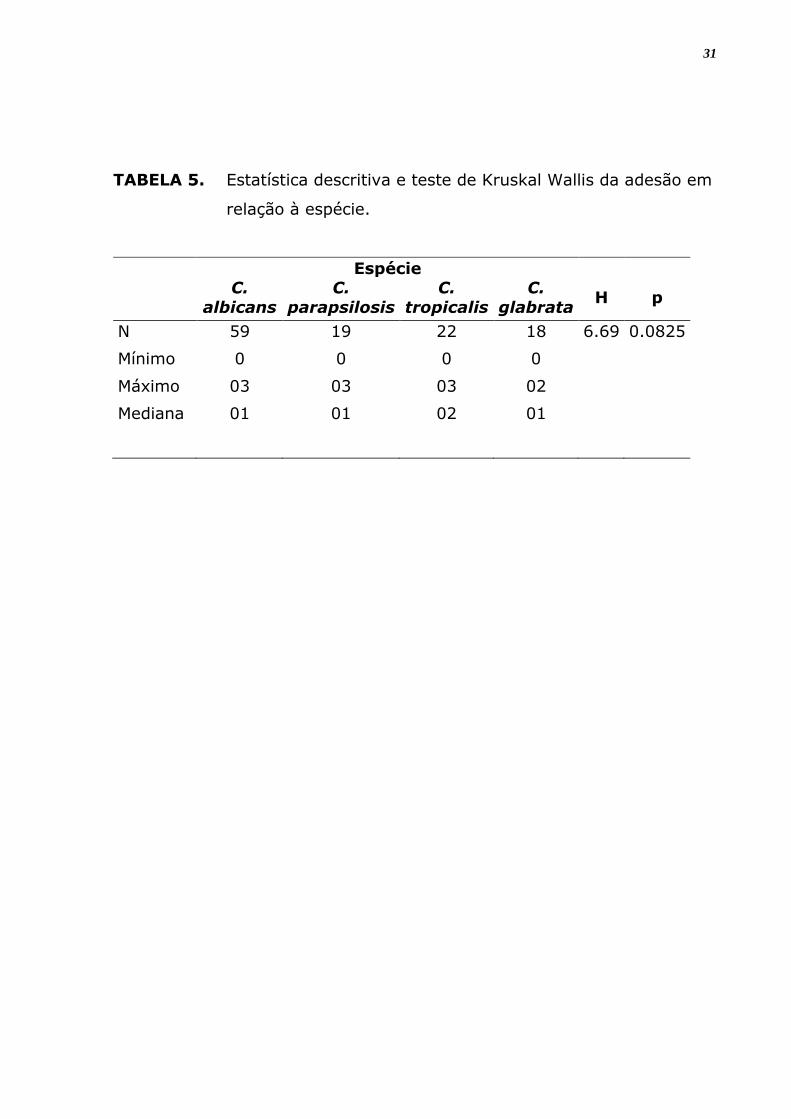
3). A análise estatística mostra que estas diferenças entre espécies e
isolados diferentes não são significantes (p > 0,05) (Figura 4).
A capacidade de adesão foi categorizada de acordo com o número
de células por campos lidos em fraca (+), apenas 01 a 10 células por 50
campos; moderada (++), acima de 10 células por 30 campos ou forte
(+++), acima de 25 células por 20 campos lidos. Foram poucos os
isolados (21%) que não apresentaram aderência nas lamínulas (Tabela 3;
Figura 4), sendo apenas 02 amostras das C. tropicalis e C. parapsilosis,
cada, seguido de 07 amostras de C. glabrata e 14 de C.albicans (Tabela
3). Independente do sítio de origem destas amostras houve diversidade
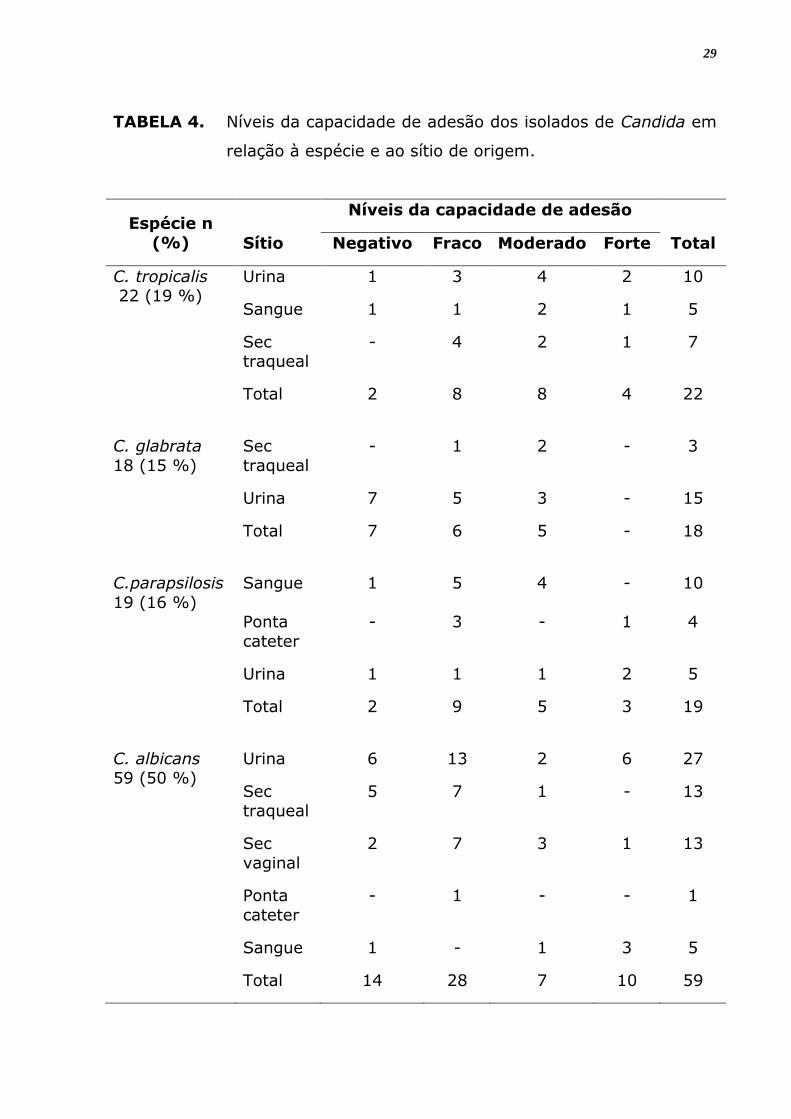
de capacidade de adesão para todas as espécies de Candida (Tabela 4).
Em relação ao número total de espécies analisadas estas diferenças não
foram significantes (Tabela 5), e entre o grau de positividade de adesão
em material inerte também não foi significante nem entre as espécies e
nem para os sítios de origem (Tabela 6). Notou-se que houve adesão em
28
100% das amostras isoladas de secreção traqueal de C. tropicalis e C.
glabrata, bem como houve 100% de adesão em isolados provenientes de
ponta de cateteres (Tabela 4).
TABELA 3. Classificação do nível de capacidade de adesão dos isolados
de Candida.
Espécies
Capacidade de adesão
Negativo n (%)
Positivo n (%)
Fraca
Moderada
Forte
Total n(%)
C. albicans 14 (24) 45 (76) 28 07 10 59 (50)
C. tropicalis 02 (09) 20 (91) 08 08 04 22 (19)
C. parapsilosis 02 (11) 17 (89) 09 05 03 19 (16)
C. glabrata 07 (39) 11 (61) 06 05 - 18 (15)
TOTAL 25 (21) 93 (79) 51 25 17 118(100)
29
TABELA 4. Níveis da capacidade de adesão dos isolados de Candida em
relação à espécie e ao sítio de origem.
Espécie n
(%)
Níveis da capacidade de adesão
Sítio Negativo Fraco Moderado Forte Total
C. tropicalis 22 (19 %)
Urina 1 3 4 2 10
Sangue 1 1 2 1 5
Sec traqueal
- 4 2 1 7
Total 2 8 8 4 22
C. glabrata
18 (15 %)
Sec
traqueal
- 1 2 - 3
Urina 7 5 3 - 15
Total 7 6 5 - 18
C.parapsilosis
19 (16 %)
Sangue 1 5 4 - 10
Ponta
cateter
- 3 - 1 4
Urina 1 1 1 2 5
Total 2 9 5 3 19
C. albicans 59 (50 %)
Urina 6 13 2 6 27
Sec traqueal
5 7 1 - 13
Sec
vaginal
2 7 3 1 13
Ponta
cateter
- 1 - - 1
Sangue 1 - 1 3 5
Total 14 28 7 10 59
30
0
10
20
30
40
50
60
70
Uri
na
San
gue
Sec
Traq
uea
l
Tota
l
Sec
traq
uea
l
Uri
na
Tota
l
San
gue
Po
nta
Cat
éter
Uri
na
Tota
l
Uri
na
Sec
Traq
uea
l
Sec
Vag
inal
Po
nta
Cat
éter
San
gue
Tota
l
C. tropicalis 22 (19%) C. glabrata 18 (15%) C. parapsilosis 19 (16%) C. albicans 59 (50%)
forte (+++)
moderado (++)
fraco (+ )
negativo
FIGURA 4. Distribuição de frequência de isolados de diferentes espécies
de Candida em relação aos sítios de origem e o nível de
aderência.
Fre
quência
(%
)
Nível de adesão
31
TABELA 5. Estatística descritiva e teste de Kruskal Wallis da adesão em
relação à espécie.
Espécie
C.
albicans
C.
parapsilosis
C.
tropicalis
C.
glabrata H p
N 59 19 22 18 6.69 0.0825
Mínimo 0 0 0 0
Máximo 03 03 03 02
Mediana 01 01 02 01

32
TABELA 6. Estatística descritiva e teste de Kruskal Wallis da adesão em
relação ao sítio.
Sítio
Sangue Urina Cateter
Sec
traqueal
Sec
vaginal H p
N 19 57 05 24 13 2.1 0.7177
Mínimo 0 0 01 0 0
Máximo 03 03 03 03 03
Mediana 02 01 01 01 01

33
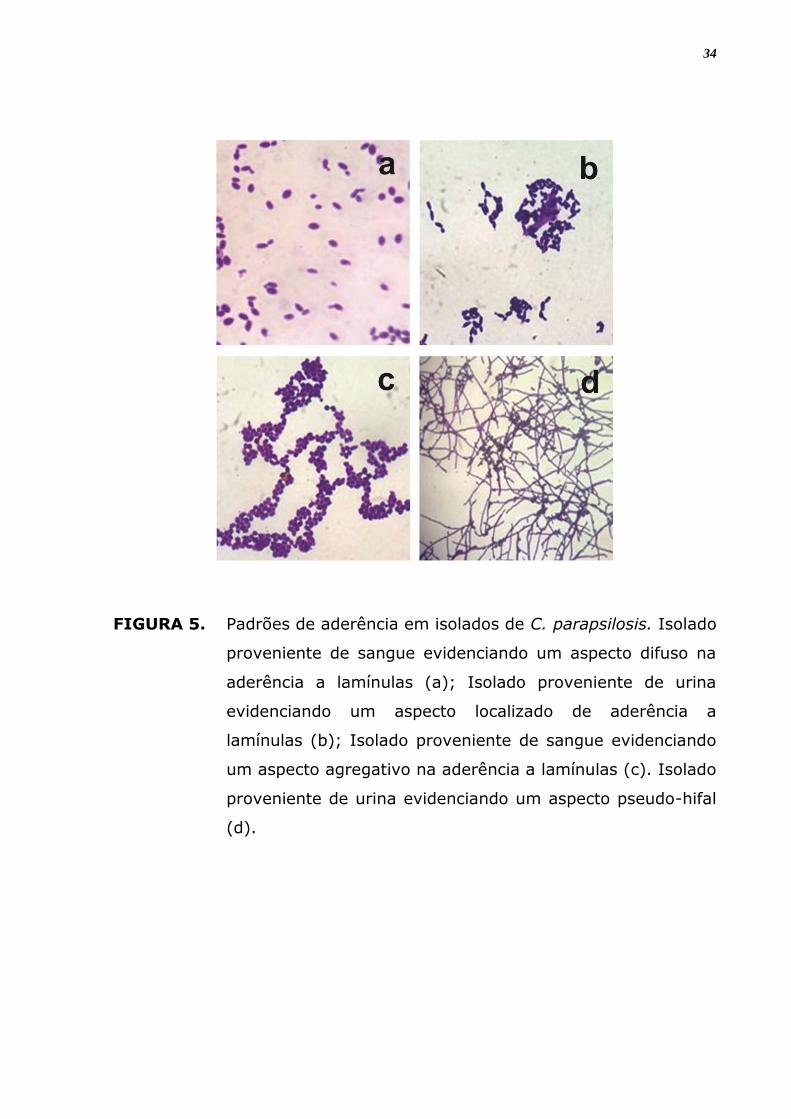
Foram verificados quatro padrões diferentes de organização celular
na adesão in vitro entre os isolados como demonstra a figura 5, foi
classificado como padrão difuso, localizado, agregativo e filamentoso ou
pseudo-hifal. No padrão difuso percebe-se que as células aderem por toda
a superfície da lamínula, espalhadas, sem formar agrupamentos celulares.
Na organização localizada, verifica-se formação de agrupamentos de
poucas células, bem definidos sobre a superfície; e no padrão agregativo,
o agrupamento celular é com um maior número de células aderidas umas
às outras formando microcolônias maiores, observa-se que há
empilhamentos das células leveduriformes em forma de cachos.
No quarto padrão de aderência há a formação filamentosa ou
pseudo-hifal por toda a superfície da lamínula.
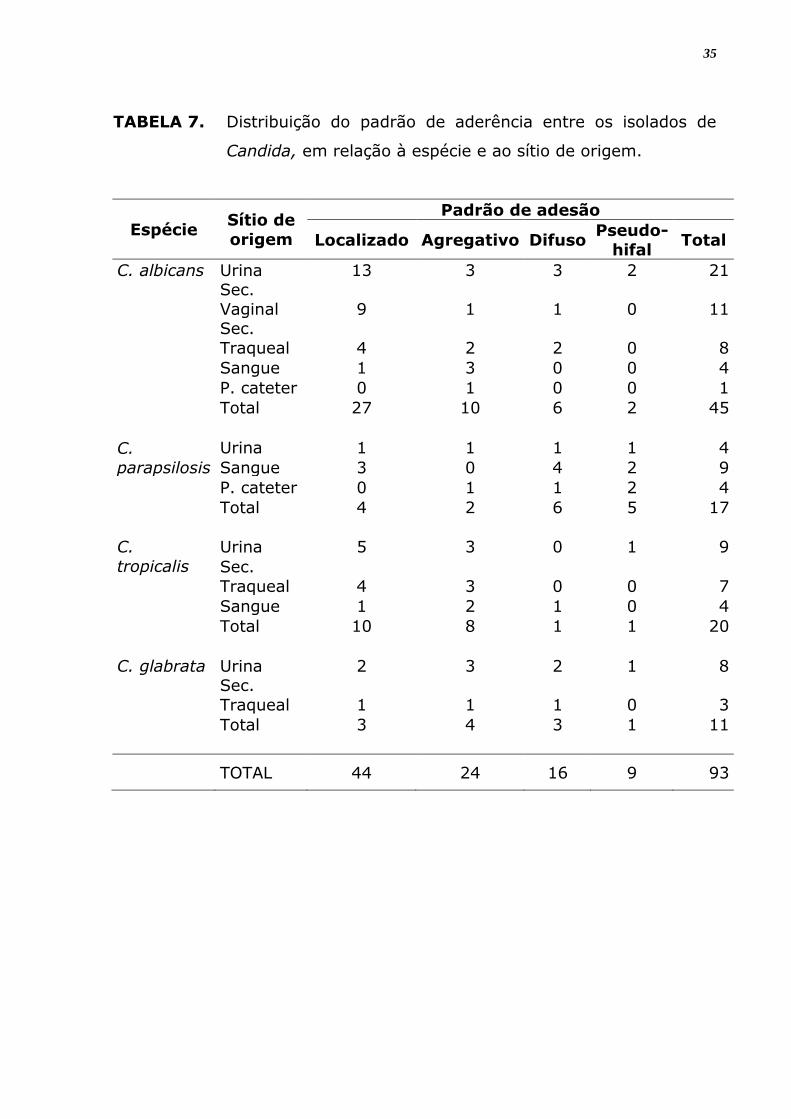
Dentre 93 isolados com capacidade de aderência, 44 apresentaram
padrão de aderência localizado (47%), seguido de 24 isolados que
demonstraram padrão agregativo (26%) e apenas 9 apresentaram padrão
pseudo-hifal (9,7%). Somente dois isolados de C. albicans apresentaram a
formação filamentosa ou pseudo-hifal. C. parapsilosis foi a espécie que
apresentou uma maior variabilidade de padrões entre os seus isolados, e
5 (29,4%) destes apresentaram o padrão de pseudo-hifas (Tabela 7). A
maioria dos isolados que mostrou padrão localizado ou agregativo é
proveniente de urina (Tabela 8).
34
FIGURA 5. Padrões de aderência em isolados de C. parapsilosis. Isolado
proveniente de sangue evidenciando um aspecto difuso na
aderência a lamínulas (a); Isolado proveniente de urina
evidenciando um aspecto localizado de aderência a
lamínulas (b); Isolado proveniente de sangue evidenciando
um aspecto agregativo na aderência a lamínulas (c). Isolado
proveniente de urina evidenciando um aspecto pseudo-hifal
(d).
35
TABELA 7. Distribuição do padrão de aderência entre os isolados de
Candida, em relação à espécie e ao sítio de origem.
Espécie Sítio de
origem
Padrão de adesão
Localizado Agregativo Difuso Pseudo-
hifal Total
C. albicans Urina 13 3 3 2 21
Sec.
Vaginal 9 1 1 0 11
Sec.
Traqueal 4 2 2 0 8
Sangue 1 3 0 0 4
P. cateter 0 1 0 0 1
Total 27 10 6 2 45
C.
parapsilosis
Urina 1 1 1 1 4
Sangue 3 0 4 2 9
P. cateter 0 1 1 2 4
Total 4 2 6 5 17
C.
tropicalis
Urina 5 3 0 1 9
Sec.
Traqueal 4 3 0 0 7
Sangue 1 2 1 0 4
Total 10 8 1 1 20
C. glabrata Urina 2 3 2 1 8
Sec.
Traqueal 1 1 1 0 3
Total 3 4 3 1 11
TOTAL 44 24 16 9 93

36
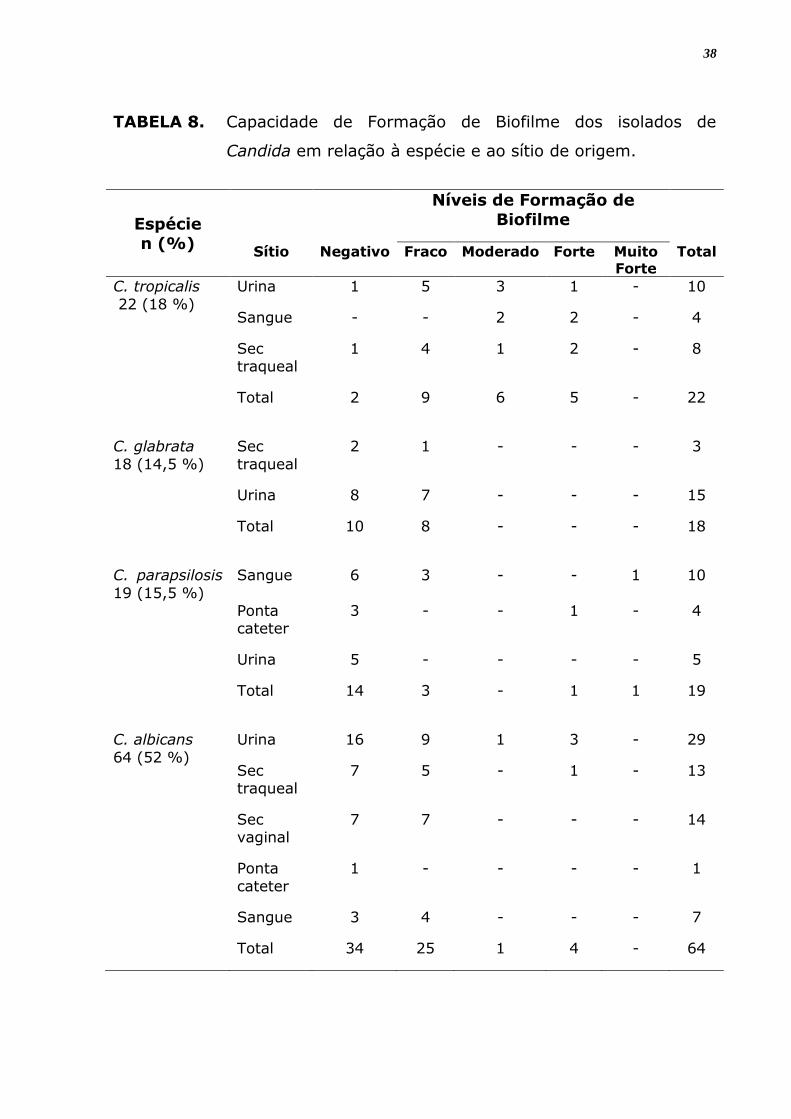
4.2 Formação de Biofilme
Os resultados de produção de biofilme para 123 isolados de espécies
de Candida obtidos de diferentes sítios estão na Tabela 9. Destes, um
total de 63 (51,2%) isolados analisados foram positivos para a formação
de biofilme em material inerte. Sendo que, 11 (17,5%) foram produtores
forte (3+) ou muito forte (4+), e 45 (71,4%) foram fracos produtores de
biofilme. Para os isolados de C. tropicalis, 20 amostras (91%) foram
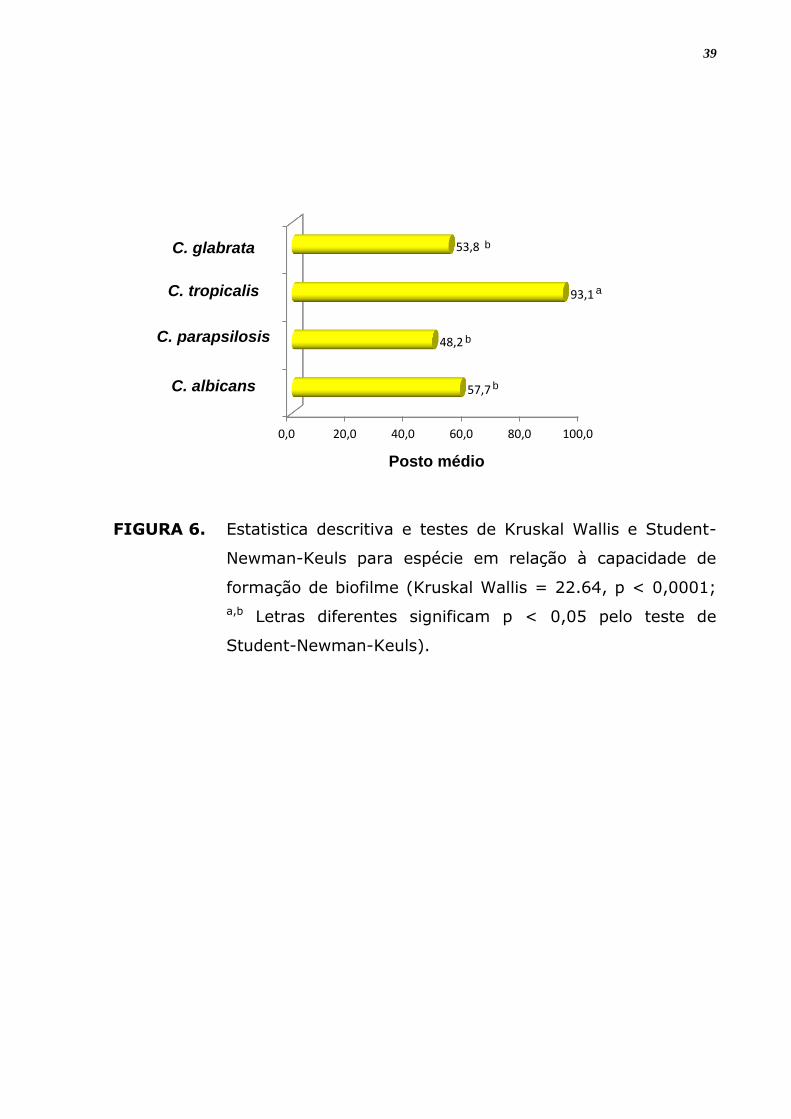
produtores de biofilme, sendo esta capacidade significantemente maior
que para as demais espécies (Figura 8; p < 0,0001). A capacidade de
formação de biofilme entre as demais espécies não foi estatisticamente
diferente (Figura 8).
Com relação à intensidade da capacidade de formação de biofilme, a
espécie com número maior de isolados fortemente produtores (3+) foi C.
tropicalis com 25% dos seus isolados, seguida de C. albicans com 13,3%.
C. glabrata mostrou 100% de fracos (1+) produtores de biofilme dentre
os isolados analisados.
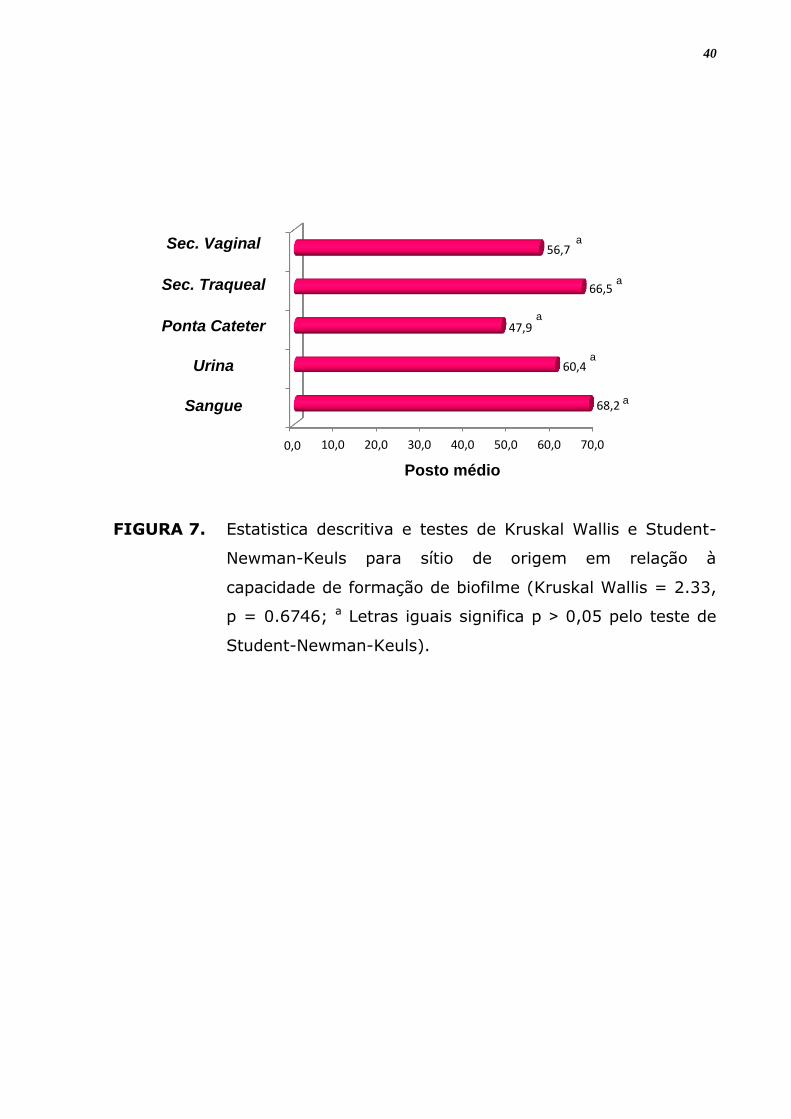
Considerando-se os isolados do mesmo sítio de origem, houve
produção de biofilme positiva para todos os sítios analisados só que com
expressão e intensidade variável de acordo como sítio e espécie avaliados.
Em relação a todos isolados com formação de biofilme positivo observou
maior positividade em isolados de urina (46,03%), seguidos dos
provenientes de secreção traqueal (22,22%) e de sangue (57,1%). Mas
analisando apenas amostras do mesmo sítio anatômico, percebeu-se que
a produção de biofilme foi mais frequente entre os isolados de secreção
traqueal (58,3%) seguidos das provenientes de sangue (57,1%) e urina
(49,2%), entretanto não houve diferença significante entre estes
resultados.
Dentre aquelas provenientes de secreção vaginal, 50% foram
produtoras, porém todas (100%) com capacidade fraca (1+); as do
sangue (57,1%) foram 41,7% formadoras moderada (2+) ou forte (3+)
ou muito forte (4+); já as de urina (49,2%) foram produtoras, sendo

37
27,6% entre formadoras moderada (2+) ou forte (3+), com nenhuma
apresentando formação de biofilme muito forte. Os isolados de Candida
provenientes da ponta de cateter tiveram 80% das amostras não
formadoras de biofilme, com apenas 01 (20%) apresentando esta
capacidade, contudo muito forte (4+) (Tabela 9), entretanto não houve
diferença significante entre estes resultados (Figura 9; p = 0.6746).
38
TABELA 8. Capacidade de Formação de Biofilme dos isolados de
Candida em relação à espécie e ao sítio de origem.
Espécie
n (%)
Níveis de Formação de
Biofilme
Sítio Negativo Fraco Moderado Forte Muito Forte
Total
C. tropicalis
22 (18 %)
Urina 1 5 3 1 - 10
Sangue - - 2 2 - 4
Sec traqueal
1 4 1 2 - 8
Total 2 9 6 5 - 22
C. glabrata
18 (14,5 %)
Sec
traqueal
2 1 - - - 3
Urina 8 7 - - - 15
Total 10 8 - - - 18
C. parapsilosis
19 (15,5 %)
Sangue 6 3 - - 1 10
Ponta cateter
3 - - 1 - 4
Urina 5 - - - - 5
Total 14 3 - 1 1 19
C. albicans
64 (52 %)
Urina 16 9 1 3 - 29
Sec traqueal
7 5 - 1 - 13
Sec vaginal
7 7 - - - 14
Ponta
cateter
1 - - - - 1
Sangue 3 4 - - - 7
Total 34 25 1 4 - 64
39
0,0 20,0 40,0 60,0 80,0 100,0
C. Albicans
C. Parapsilosis
C. Tropicalis
C. Glabrata
57,7
48,2
93,1
53,8
Esp
éci
e
FIGURA 6. Estatistica descritiva e testes de Kruskal Wallis e Student-
Newman-Keuls para espécie em relação à capacidade de
formação de biofilme (Kruskal Wallis = 22.64, p < 0,0001;
a,b Letras diferentes significam p < 0,05 pelo teste de
Student-Newman-Keuls).
b
a
b
b
C. glabrata
C. tropicalis
C. parapsilosis
C. albicans
Posto médio
40
,0 10,0 20,0 30,0 40,0 50,0 60,0 70,0
Sangue
Urina
Ponta Cateter
Sec. Traqueal
Sec. Vaginal
68,2
60,4
47,9
66,5
56,7
Posto médio
Síti
o
FIGURA 7. Estatistica descritiva e testes de Kruskal Wallis e Student-
Newman-Keuls para sítio de origem em relação à
capacidade de formação de biofilme (Kruskal Wallis = 2.33,
p = 0.6746; a Letras iguais significa p ˃ 0,05 pelo teste de
Student-Newman-Keuls).
a
a
a
a
a
Posto médio
Sec. Vaginal
Sec. Traqueal
Ponta Cateter
Urina
Sangue
0,0

41
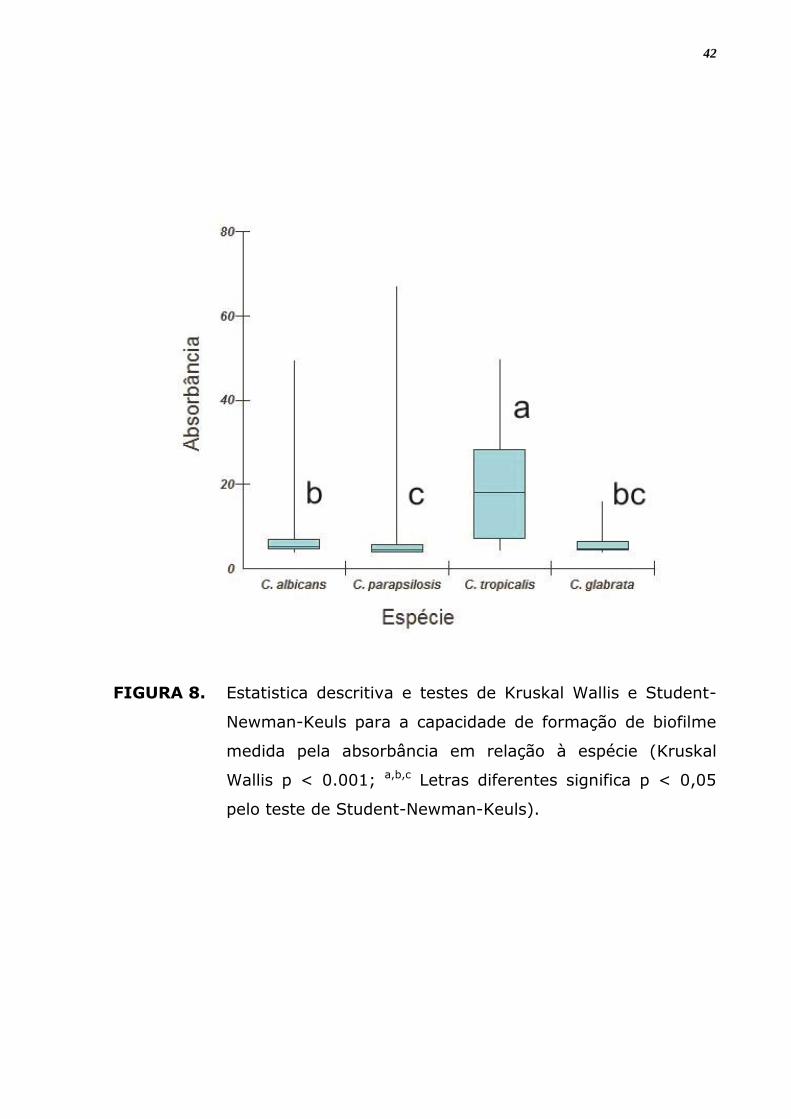
A análise dos dados quantitativos da formação de biofilme mostra
resultados diferentes e significantes para as espécies de Candida. Em
relação à espécie, C. tropicalis foi a maior produtora e C. parapsilosis, a
menor produtora (Figura 10; p < 0,05 pelo teste de Student-Newman-
Keuls).
Em relação ao sítio, a variação da capacidade de formação de
biofilme medida por valores de absorbância não foi significante entre as
diferentes origens (Figura 11; p = 0.7748).
42
FIGURA 8. Estatistica descritiva e testes de Kruskal Wallis e Student-
Newman-Keuls para a capacidade de formação de biofilme
medida pela absorbância em relação à espécie (Kruskal
Wallis p < 0.001; a,b,c Letras diferentes significa p < 0,05
pelo teste de Student-Newman-Keuls).
43
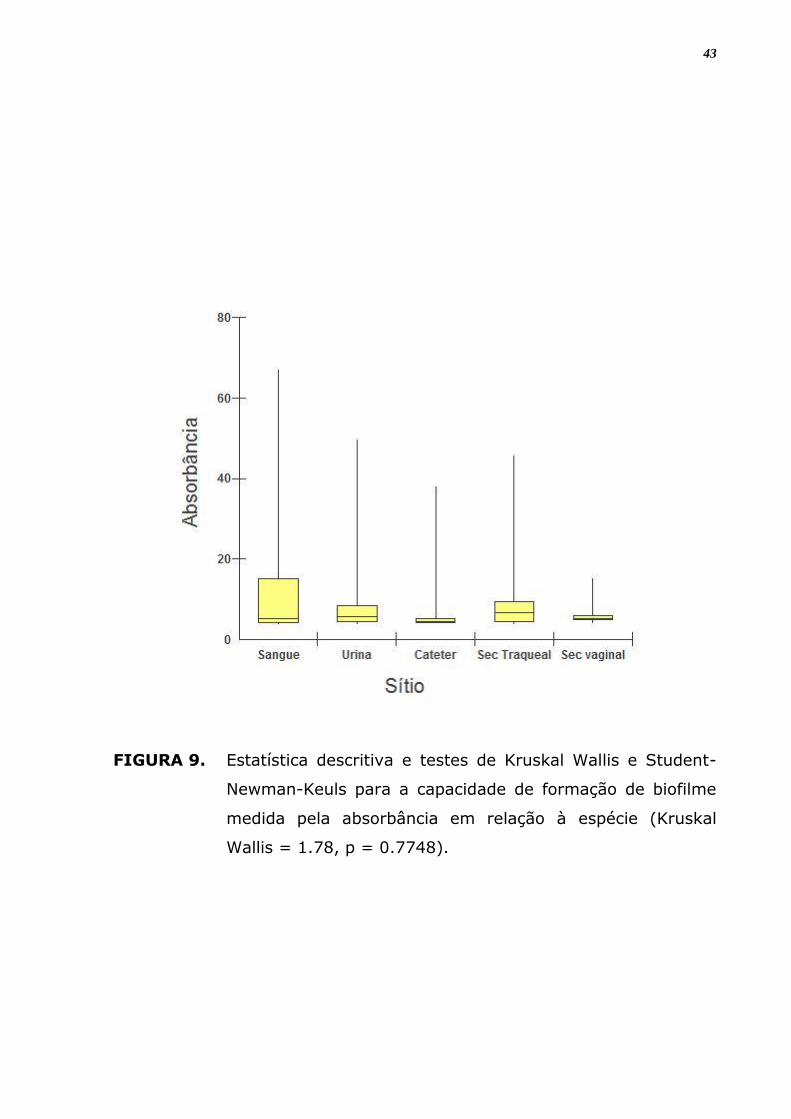
FIGURA 9. Estatística descritiva e testes de Kruskal Wallis e Student-
Newman-Keuls para a capacidade de formação de biofilme
medida pela absorbância em relação à espécie (Kruskal
Wallis = 1.78, p = 0.7748).

44
5 DISCUSSÃO
A aderência de Candida às superfícies mucosas de células
hospedeiras é um pré-requisito vital para o sucesso da colonização e
infecção. Esta ligação permite que os organismos evitem uma ação de
limpeza das secreções das mucosas ou seu deslizamento, e facilitem a
infecção (CHAFFIN et al., 1998, CANNON, CHAFFIN, 1999). Estudos
mostram que a capacidade de aderência nas diversas espécies de Candida
é diferente, isso explica que a colonização a superfícies mucosas são mais
frequentes em algumas espécies que em outras (JABRA, 2001;
REPENTIGNY, 2000) e a capacidade de aderir a materiais inertes pode ser
um indicativo de provável aderência a células vivas. Desta forma, o
estudo teve por objetivos analisar a capacidade de adesão e formação de
biofilme em isolados clínicos de Candida provenientes de sítios de origem
diferentes.
Nesta pesquisa, todas as espécies de Candida estudadas foram
capazes de adesão, o que está de acordo com outros estudos que relatam
a aderência de espécies de Candida a materiais inertes como silicone
(BUSSCHER et al., 1997; TAMURA et al., 2003; MORENO et al., 2009).
Entretanto, é importante notar que os isolados de outras espécies de
Candida que não C. albicans foram mais aderentes (81,4%) que os de C.
albicans (76,3%). C. tropicalis destacou-se entre as espécies não albicans
com 91% de amostras aderentes bem como destas 20% apresentaram
aderência forte (+++). Este resultado, em relação às demais espécies de
Candida que não C. albicans foi significante (p < 0,001). Da mesma
forma, Minage et al. (1985) observaram maior aderência nesta espécie de
Candida em material inerte, mas neste caso em polímeros de plásticos.
Moreno et al.(2009) verificaram a capacidade de aderência a material
inerte, neste caso em resina acrílica, para isolados de Candida, e dentre
as espécies analisadas, C. tropicalis apresentou maior capacidade de
adesão, seguida de C. dubliniensis e C. albicans. Este resultado é
relevante visto que em um estudo epidemiológico recente de 100 casos de

45
candidúria, no período entre 1999 e 2004, onde se considera a capacidade
de aderência a cateteres um fator significante e predisponente a este tipo
de infecção, em um hospital pediátrico no Brasil, 20 amostras de C.
tropicalis foram isoladas comparados às 56 de C. albicans, 11 de C.
glabrata e 4 de C. parapsilosis (SILVA et al., 2007). De fato, C. tropicalis
tem emergido como o segundo ou terceiro agente mais comum de
candidemia, principalmente em pacientes com câncer (WEINBERG et al.,
2005; NUCCI et al., 2007). Além disso, uma incidência elevada de C.
tropicalis como agente causal de infecções nosocomiais do trato urinário
tem sido relatado (RHO et al., 2004).
Em alguns estudos C. albicans tem sido a espécie com maior
capacidade de adesão a células do epitélio bucal que outras espécies de
Candida (BIASOLI et al., 2002; LYON, RESENDE, 2006; COSTA, 2009).
No estudo de Lyon e Resende (2006), por exemplo, a espécie com maior
capacidade de adesão foi C. albicans e, posteriormente, dentro das outras
espécies de Candida não que C. albicans foi C. tropicalis que apresentou
maior aderência, o que corrobora parcialmente com nossos achados.
Espécies de Candida que não C. albicans vem mostrando um
aumento no número de casos de fungemia em vários países (CHENG et
al., 2005; SHIN et al.,2002; BASSETTI et al., 2006; TAMURA et al, 2007);
provavelmente devido a maior expressão dos fatores de virulência, dentre
estes a capacidade de aderir e formar biofilme.
Dentre os isolados de C. tropicalis que foram aderentes positivos,
aqueles provenientes de secreção traqueal foram 100% aderentes, apesar
desta diferença não ter sido significante em relação à distribuição dos
sítios anatômicos. A alta capacidade de aderência apresentada por C.
tropicalis pode indicar maior virulência quando comparada com as demais
espécies, corroborando com Tamura et al. (2007) e Costa (2009), que
mostraram outras espécies de Candida que não C. albicans apresentando
maior propriedade de aderência do que a própria espécie de C. albicans,
sendo que no estudo de Tamura et al. (2007) os pesquisadores relataram
C. glabrata como a espécie de maior aderência e com capacidade de

46
formar biofilme, enquanto Costa (2009) mostra C. tropicalis
significativamente mais aderente que as demais espécies incluindo C.
albicans.
Importante ressaltar que em C. parapsilosis tem sido observado
fatores de virulência significantes como adesão a células e formação de
biofilme (BRANCHINI et al., 1994; NASCIMENTO, 2009) corroborando com
nossos achados dos ensaios de adesão em que C. parapsilosis foi a
segunda espécie com maior capacidade de adesão (89%), e 100% de
isolados de ponta de cateter foram aderentes. Embora C. parapsilosis seja
geralmente considerada uma das espécies de leveduras menos virulenta,
é uma das causas mais frequentes de candidemia associada à
hiperalimentação parenteral, dispositivos intravasculares e soluções
oftálmicas contaminadas (PLOUFFE et al., 1977; SOLOMON et al., 1984;
O’DAY, HEAD, ROBINSON, 1987; WEMMS et al., 1987; WEMMS, 1992).
Vários fatores dão à C. parapsilosis uma vantagem seletiva, incluindo a
capacidade de proliferar-se em altas concentrações de glicose e de
aderência a materiais protéticos (CRITCHLEY, DOUGLAS, 1985; WEMMS et
al., 1987). Surtos nosocomiais de C. parapsilosis também têm sido
descritos e sido atribuídos a transferência de levedura das mãos de
trabalhadores da saúde (BONASSOLI et al., 2005). Contudo em contraste
com os resultados de capacidade de adesão, C. parapsilosis foi a espécie
de Candida que menos expressou a propriedade de formação de biofilme
(26%), e os mesmos isolados provenientes da ponta de cateter foram
negativos para este fator de virulência (75%). Estes resultados
contrastam com os da literatura. Pfaller, Messer, Hollis (1995), estudaram
a produção de biofilme por amostras clínicas de C. parapsilosis crescidas
em meio de cultura contendo glicose e verificaram que as isoladas de
sangue e de cateter apresentaram maior produção de biofilme do que
amostras de outros sítios anatômicos. Nossos resultados mostram que
isolados de C. parapsilosis podem entrar na corrente sanguínea e causar
infecções mesmo sem formar biofilme nos dispositivos médicos já que
alguns destes isolados são provenientes de sangue.

47
Este estudo descreve, pela primeira vez, padrões diferenciados de
organização celular percebidos no ensaio de adesão mostrado pelas
espécies de Candida. Estes padrões foram categorizados de maneira
semelhante à classificação de linhagens diarréicas de E. coli. que mostram
padrões de aderência distintos quando aderidas às células HEp-2, HeLa ou
às lamínulas in vitro (CRAVIOTO et al., 1979; SCALETSKY et al., 1984).
Foram distinguidos quatro padrões de aderência: difuso, onde as
leveduras aderem por toda a superfície das lamínulas espaçadamente;
aderência localizada, que envolve grupos de leveduras, microcolônias, que
aderem a regiões localizadas nas lamínulas; aderência agregativa,
caracterizada por aglomerados de leveduras com uma aparência de tijolos
empilhados ou em cachos e o padrão de aderência filamentoso ou pseudo-
hifal, com o desenvolvimento de hifas ou pseudo-hifas por toda a
superfície da lamínula ou de maneira localizada. A maioria dos isolados
apresentou capacidade de aderência do tipo localizado (47%), seguido
daqueles que demonstraram padrão agregativo (26%). C. parapsilosis foi
a espécie que apresentou uma maior variabilidade de padrões entre os
seus isolados, sendo que destes 5 (29,4%) apresentaram o padrão de
pseudo-hifas. A maioria dos isolados que mostrou padrão localizado ou
agregativo é proveniente de urina. A grande quantidade de isolados
mostrando um padrão tipo localizado ou agregativo poderia ser explicado
devido à presença de adesinas na parede celular de Candida que seriam
responsáveis pela capacidade de aderência intercelular mostrada pelas
diferentes linhagens. Estes tipos de padrões de aderência também
facilitariam a formação de biofilme por estas espécies.
Em C. albicans as proteínas da família ALS medeiam vários modos
de adesão, por exemplo, adesão sem agregação celular com células
aderidas espaçadamente ou adesão seguida de agregação celular com
células formando grumos. O comportamento específico exibido por um
isolado depende do tipo de proteína expressa. Este fato, de certa forma,
corrobora com nossos achados onde evidenciamos escassez de um padrão
de aderência tipo difuso (17%) com a maioria das cepas evidenciando um

48
padrão tipo localizado ou agregativo (73%), onde ambos mostram
formação de colônias por união de várias células leveduriformes.
Regiões estruturais específicas das proteínas ALS em C. albicans
parecem estar envolvidas no fenômeno de agregação celular, como a
região de repetições em tandem e a região rica em resíduos de treonina
(KLOTZ, LIPKE, 2010). A região rica em treonina na Als5p que é composta
por 127 aminoácidos está envolvida na agregação (RAUCEO et al., 2006),
entretanto, Als7p e Als9p com 128 aminoácidos não proporcionam
agregação após adesão (HOYER 2001; KLOTZ et al., 2007). Da mesma
forma, a propriedade de agregação de várias proteínas Als de C. albicans
é proporcional ao número de repetições em tandem na molécula (RAUCEO
et al., 2006). Por exemplo, Alsp1 com 20 repetições induz a formação de
agregados maiores que aqueles induzidos por Als5p com somente 5
sequências repetidas (KLOTZ et al., 2007).
Somente dois isolados de C. albicans mostraram um padrão
filamentoso ou pseudo-hifal de aderência o que também foi observado
somente em 7 (7,5%) dos isolados das demais espécies de Candida que
não C. albicans. A capacidade de diferenciação entre as formas
leveduriformes para as formas filamentosas tem sido descrita em muitos
estudos e tem relação direta com a invasão tecidual do hospedeiro,
favorecendo a patogenicidade de Candida, permitindo a penetração mais
fácil ao tecido do hospedeiro (YANG, 2003), contudo não é essencial para
a patogênese (GOW, BROW, ODDS, 2002). Provavelmente a forma
filamentosa seja mais virulenta devido à maior extensão de contato entre
o microorganismo e a área a ser infectada, mas ambas as formas são
frequentemente encontradas tanto em estados comensais quanto em
processos infecciosos e mesmo as demais espécies de Candida que não C.
albicans que não formam hifas verdadeiras são potencialmente
patogênicas e estão envolvidas com a morbimortalidade por estes agentes
(ODDS, 2002; RIBEIRO, 2004).
A possibilidade de mudar o seu próprio fenótipo tem sido descrita
em alguns estudos em relação a C. albicans (YANG, 2003; HAYNES, 2001;

49
ÁLVARES et al., 2007; RIBEIRO et al., 2004) e pode modular alguns
fatores de virulência inclusive o dimorfismo celular e a aderência (SOLL,
2002). Entretanto, o conhecimento a respeito dos mecanismos que
possibilitam esta mudança fenotípica ainda precisa ser esclarecido
(COSTA, 2009b), provavelmente está associado à riqueza de nutrientes e
expressão de adesinas.
Investigação da presença e/ou expressão das Als ou de genes
responsáveis pela passagem para hifa nos isolados de Candida seriam
fundamentais para estabelecer correlações com os dados fenotípicos de
aderência obtidos nesta pesquisa.
Quanto à capacidade de formação de biofilmes o nosso estudo
mostra que C. tropicalis foi significantemente maior produtora de biofilme
dentre as espécies estudadas (p < 0,0001). A capacidade de formação de
biofilme entre as demais espécies não foi estatísticamente diferente. Estes
dados corroboram com os de Ferreira et al. (2006) que ao estudarem
fatores de virulência de C. tropicalis e C. albicans e compará-los,
consideraram a primeira espécie mais virulenta, pois apresentou
positividade maior nos testes realizados, incluindo a produção de biofilme
in vitro. Gasparetto et al. (2005) também encontraram maior e mais
significante produção de biofilme em outras espécies de Candida que não
C. albicans (64%), destes C. tropicalis destacou-se entre estas. Segundo
Shin et al. (2002) apesar da C. albicans ser patogênica e estabelecer
mecanismos para o estabelecimento de infecções sistêmicas, a sua
produção de biofilme foi menos frequentemente significante que as
demais espécies, independente do local da infecção, ou seja, do sítio
anatômico de origem e a C. tropicalis também foi a maior produtora de
biofilme seguida de C. parapsilosis, C. glabrata e C. albicans. Entretanto,
C. parapsilosis recebeu destaque quando foram analisados os isolados
sanguíneos, sendo estes isolados os maiores produtores de biofilme
dentre aqueles das demais espécies de Candida que não C. albicans,
diferentemente dos nossos achados onde todos os isolados de C. tropicalis
provenientes de sangue foram os que apresentaram maior formação de

50
biofilme (100%), seguidos pelos de C. albicans (57%) e, por último, os de
C. parapsilosis (40%). Não houve isolados sanguíneos de C. glabrata.
Não houve diferença significante em relação ao sítio de origem das
amostras e a capacidade de formação de biofilme, mas pudemos notar
que quase todas as amostras provenientes de ponta de cateter não foram
produtoras de biofilme, o que diverge da literatura que relata que
dispositivos médicos como cateteres são substratos preferenciais e
potenciais para a formação de biofilme (DONLAN, 2001; HOTA, 2004), e
para a espécie de C. parapsilosis onde os isolados urinários também foram
negativos para a formação de biofilme.
A suplementação do meio com glicose foi importante para os
achados em nosso experimento, dados que concordam com estudos
anteriores citados para a produção de biofilme, como os de Hawser,
Douglas (1994), Jain et al. (2007) e Uppluri et al. (2009) que já
observaram crescimento leveduriforme menor se o meio de cultivo não
fosse suplementado com este carboidrato, porém nossos resultados
divergem dos de Hawser, Douglas (1994) em relação que os mesmos
encontraram C. albicans como a mais patogênica em relação às outras
espécies de Candida. Estes achados são relevantes para pacientes
diabéticos que têm taxa elevada de glicose no sangue. De fato, infecções
fúngicas são mais comuns em pacientes diabéticos, particularmente
aquelas envolvendo espécies de Candida, ocasionalmente causando sérias
repercussões clínicas (NETO et al., 1995; OLIVEIRA et al., 2001).
Comparando nossos achados de aderência e formação de biofilme
percebe-se uma diferenciação na expressão destes em isolados de uma
mesma espécie. Tais resultados refletem diferenças fisiológicas inerentes
entre espécies e isolados e podem ter significância com relação ao
potencial patogênico. Somente isolados de C. tropicalis destacou-se tanto
quanto à capacidade de aderir–se ao material inerte bem como formar
biofilmes (91% em ambos os ensaios), os demais isolados das espécies
estudadas tiveram comportamentos diferentes nos testes realizados sendo
que aqueles de C. parapsilosis mostraram alta aderência (89%) em

51
contraste a baixa produção de biofilme (26%), da mesma forma os de C.
albicans e de C. glabrata apresentaram maior capacidade de aderir (76%
e 61 %, respectivamente) que em formar biofilmes (47% e 44%,
respectivamente) sugerindo que ambas as capacidades são etapas
distintas do processo infeccioso (RIBEIRO et al., 2004; ÁLVARES,
SVIDZINSKI, CONSOLARO, 2007) e que as espécies expressam diferentes
mecanismos para a formação de um biofilme maduro. Isto pode realmente
ser verdadeiro visto que Silva et al. (2009) estudando a caracterização de
biofilmes em outras espécies de Candida que não C. albicans em termos
de estrutura, composição da matriz e atividade metabólica revelou
diferenças significativas estruturais nos biofilmes no que diz respeito à
morfologia das células e à sua disposição espacial. A matriz dos biofilmes
de C. parapsilosis apresentou elevada quantidade de polissacarídeos e
baixa quantidade de proteínas. Contrariamente, as matrizes extraídas dos
biofilmes de C. tropicalis apresentaram reduzidas quantidades de
polissacarídeos e proteínas e; os de C. glabrata apresentaram elevada
quantidade dos dois componentes em estudo, mostrando que existem
diferenças intrínsecas em termos de atividade metabólica dos biofilmes.
Hansan et al. (2009) observaram que há diferença significante entre
produção de biofilme entre as espécies de Candida, sendo que usaram
dois métodos para a avaliação, evidenciando que a capacidade de
formação de biofilme in vitro é dependente da espécie da Candida. Seus
resultados divergiram dos nossos, pois na metodologia similar ao nosso
estudo, para estes autores, C. albicans foi a maior produtora de biofilme e
todos os isolados de C. glabrata apresentaram biofilme positivo, dados
não achados em nosso experimento.
Há poucos estudos que avaliam a produção de biofilme entre
espécies de Candida isoladamente, provenientes de diferentes sítios
anatômicos, sem agrupá-las como outras espécies que não C. albicans,
bem como correlacionar estes fatores de virulência das espécies de
Candida que tem relevância clínica em nossa região para podermos propor
medidas preventivas, diagnósticas e terapêuticas eficazes, e

52
consequentemente contribuir para redução da morbimortalidade causada
por espécies de Candida. Nossos achados demonstram que as espécies de
Candida testadas são capazes de aderir a superfícies inertes e formar
biofilmes e sugerem que estas propriedades estão fortemente ligadas a
patogenicidade destes agentes, mas que as mesmas são processos
distintos que precisam ser melhores compreendidos. Assim como, a
expressão destes fatores de virulência não está relacionada diretamente
com o sítio da infecção em si, porém com fatores específicos destes
microorganismos e, muito provavelmente, com a resposta do hospedeiro.
Estes achados são especialmente relevantes para todos os pacientes que
fazem uso de dispositivos médicos como cateteres, incluindo aqueles com
infecção do trato urinário, transplantados ou em unidades de tratamento
intensivo.
6 CONCLUSÕES
- A maioria das linhagens ensaiadas mostra capacidade de aderência
e formação de biofilme;
- Dentre a capacidade de aderir independente da espécie à maioria
apresentou fraca adesão, e na formação de biofilme, os isolados
foram principalmente fracos produtores de biofilme;
- C. tropicalis foi a espécie que mostrou maior capacidade de adesão
e formação de biofilme em relação às outras, enquanto que C.
glabrata foi aquela com menor expressão das propriedades de
adesão e C. parapsilosis menor expressão para a formação de
biofilme;
- Os sítios de origem das amostras não interferiram na expressão de
adesão e formação de biofilme entre as espécies de Candida;
- As amostras de Candida apresentaram padrões de aderência
distintos e classificados em um padrão difuso, localizado, agregativo
e filamentoso ou pseudo-hifal;

53
- O padrão de aderência mais frequente é o localizado e o menos
frequente é o pseudo- hifal;
- Somente dois isolados de C. albicans mostraram padrão de
aderência filamentoso ou pseudo-hifal e C. parapsilosis foi a espécie
que apresentou uma maior variabilidade de padrões entre os seus
isolados, e 05 (29,4%) destes apresentaram o padrão de pseudo-
hifas;
- A maioria dos isolados proveniente de ponta de cateter não foi
formador de biofilme.

54
REFERÊNCIAS
ABI-SAID, D. et al. The epidemiology of hematogenous candidiasis caused
by different Candida species. Clinical Infectious Diseases, v. 24, p.1122–1128, 1997.
ALVARES, C. A.; SVIDZINSKI, T. I. E.; CONSOLARO, M. E. L. Candidíase vulvovaginal: fatores predisponentes do hospedeiro e virulência das
leveduras. Jornal Brasileiro de Patologia e Medicina Laboratorial, v. 43, n. 5, p. 319-327, out. 2007.
BAILLIE, G. S.; DOUGLAS L. J. Matrix polymers of Candida biofilms and
their possible role in biofilm resistance to antifungal agents. The Journal of Antimicrobial Chemotherapy, v.46, n.3, p.397-403, 2000.
BASSETTI, M. et al. Epidemiological trends in nosocomial candidemia in
intensive care. BMC Infectious Diseases., v. 6, n. 1, p.21, 2006.
BIASOLI, M. S.; TOSELLO, M. E.; MAGARÓ, H. M. Adherence of Candida strain isolated from the human gastrointestinal tract. Mycoses, v. 45, p.
465- 469, 2002.
BLANKENSHIP, J. R.; MITCHELL, A. P. How to build a biofilm: a fungal
perspective. Current Opinion in Microbiology, v.9, p.588–594, 2006.
BONASSOLI, L.; BERTOLI, M.; SVIDZINSKI, T. High frequency of Candida parapsilosis on the hands of healthy hosts. The Journal of Hospital
Infection, v. 59, n. 2, p.159–162, 2005.
BRANCHINI, M. L. et al. Genotypic variation and slime production among blood and catheter isolates of Candida parapsilosis. Journal of Clinical
Microbiology, v. 32, p. 452–456. 1994.
BRANDA, S. S. et al. Biofilms: the matrix revisited. Trends in Microbiology, v. 13, n.1, p. 20-26. jan. 2005.
BUSSCHER, H.; GEERTSEMA-DOORNBUSCH, G.; VAN DER MEI, H. Adhesion to silicone rubber of yeasts and bacteria isolated from voice
prostheses: influence of salivary conditioning films. Journal of Biomedical Materials Research, v. 34, n. 2, p. 201–209, 1997.
CALDERONE, R. A.; BRAUN, P. C. Adherence and Receptor Relationships
of Candida albicans. Microbiological and Molecular Biology Reviews, v. 55, n.1, p. 1-20, mar. 1991.

55
CALDERONE, R. A.; GOW, N. A. R. Host recognition by Candida species. In:, CALDERONE, R. A. (ed.). Candida and Candidiasis. Washington:
ASM Press, 2002, p. 67-86. CALDERONE, R. A.; FONZI, W. A. Virulence factors of Candida albicans.
Trends in Microbiology, v. 9, n.7, p. 327-335, jul. 2001.
CANNON, R.; CHAFFIN, W. Oral colonization by Candida albicans. Critical Reviews in Oral Biology and Medicine, v. 10, n. 3, p. 359–383, 1999.
CARDOSO, B. C. Efeito de antifúngicos em suspensões e biofilmes de Candida albicans e Candida dubliniensis. 2004. 85f. Dissertação
(Mestrado) Universidade do Minho Escola de Engenharia, Campos de Azurem, Guimarães, 2004.
CHAFFIN, W. L. et al. Cell Wall and Secreted Proteins of Candida albicans:
Identification, Function, and Expression. Microbiology and Molecular Biology Reviews, v. 62, n. 1, p. 130-180, mar. 1998.
CHAKRAVARTHI, S.; HALEAGRAHARA, N. A comprehensive review of the
occurrence and management of systemic Candidiasis as an opportunistic infection. Microbiology Journal, v. 1, p. 1-7, 2011.
CHAKRAVARTHI,S.; WEI, C.Z.; NAGARAJA, H.S. Increased susceptibility to
opportunistic renal candidiasis due to immunosuppression induced by
breast cancer cell lines. Science World Journal, v. 5, p.5-10, 2010.
CHANDRA J. et al. Biofilm Formation by the Fungal Pathogen Candida albicans: Development, Architecture, and Drug Resistance. Journal of
Bacteriology, v. 183, n. 18, p. 5385-5394, set. 2001.
CHENG M. F. et al. Risk factors for fatal candidemia caused by Candida albicans and non-albicans Candida species. BMC Infectious Diseases, v.
5, n. 22, p. 1-5, 2005.
CLSI. Clinical Laboratory Standards Institute: Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts; Approved Standard-
Second Edition M27-A2 National Committee for Clinical Laboratory Standards, Wayne, Pennsylvania, USA; 2007.
COLEMAN, D.C. et al. Importance of Candida species other than Candida albicans as opportunistic pathogens. Medical Mycology, Oxford, v.36,
p.156-165, 1998. Suppl 1.
COLOMBO, A. L. Epidemiology and treatment of hematogenous candidiasis: a Brazilian prospective. Brazilian Journal of Infectious
Diseases, v. 4, n. 3, p. 113-118, 2000.

56
COLOMBO, A. L. et al. Fluconazol susceptibility of Brazilian Candida Isolates Assessed by a Disk diffusion Method. Brazilian Journal of
Infectious Diseases, v. 6, n. 3, p. 118-123, 2002.
COLOMBO, A. L. et al. High rate of non-albicans candidemia in Brazilian tertiary care hospitals. Diagnostic Microbiology and Infectious
Disease, v. 34, p. 281–286, 1999.
COLOMBO, A. L. et al. Prospective observational study of candidemia in
São Paulo, Brazil: incidence rate, epidemiology and predictors of mortalily. Infection Control and Hospital Epidemiology, Thorofare, v. 28, n. 5,
p. 570-576, 2007.
COLOMBO, A.L.; NUCCI, M.; PARCK, B.J. Epidemiology of candidemia in Brazil: a nationwide sentinel surveillance of candidemia in eleven medical
centers. Journal of Clinical Microbiology, v. 44, n. 8, p. 2816-2823, 2006.
CORMACK, B.P.; GHORI, N.; FALKOW, S. An adhesin of the yeast
pathogen Candida glabrata mediating adherence to human epithelial cells. Science, v. 285, p. 578–582, 1999.
COSTA, C. R. Fatores de Virulência de isolados de Candida de
pacientes imunocomprometidos. Caracterização molecular de
Candida albicans suscetíveis e resistentes ao fluconazol. 2009. 86 f. Tese (Doutorado) – Universidade Federal do Goiás, Goiânia, 2009a.
COSTA, K. R. C. Aspectos fenotípicos e moleculares da adesão e
atividade enzimática de Candida sp isolada de pacientes com sinais clínicos de candidíase oral. 2009. 122 f. Tese (Doutorado) –
Faculdade de Ciências Farmacêuticas de Ribeirão Preto, Universidade de São Paulo, Riberão Preto, 2009b.
COTTER G., KAVANAGH K. Adherence mechanisms of Candida albicans.
British Journal of Biomedical Science, v. 57, p. 241-249, 2000.
CRAVIOTO, A. et al. An adhesive factor found in strains of Escherichia coli belonging to the traditional infantile enteropathogenic serotypes. Current
Microbiology, v. 3, n. 2, p.95–99, 1979.
CRITCHLEY I. A, DOUGLAS L. J. Differential adhesion of pathogenic
Candida species to epithelial and inert surfaces. FEMS Microbiology Letters, v.28, p.199-203, 1985.
DE REPENTIGNY, L. et al. Characterization of Binding of Candida albicans
to Small Intestinal Mucin and Its Role in Adherence to Mucosal Epithelial Cells. Infection and Immunity, v. 68, p. 3172–3179, jun. 2000.

57
DIGNANNI, M. C.; SOLOMKIN, J. S.; AMAISSIE, E. Candida. In: Anaissie E, McGinnis MR, Pfaller MA (eds). Medical Mycology, 1 ed. Filadélfia:
Churchill Livingstone, p. 195-239, 2003.
DONLAN, R. M. Biofilms and Device-Associated Infections. Emerging Infectious Diseases, v. 7, n. 2, p. 277-281, mar/abr. 2001.
DONLAN, R. M. COSTERTON, W. C. Biofilms: Survival Mechanisms of
Clinically Relevant Microorganisms. Clinical Microbiology Reviews, v.
15, n. 2, p. 167–193, abr. 2002.
DOUGLAS, L.J. Candida biofilms and their role in infection. Trends in Microbiology, v. 11, p. 30-36, 2003.
EDMOND, M. B. et al. Nosocomial bloodstream infections in United States
hospitals: a three-year analysis. Clinical Infectious Diseases, v. 29, p. 239-244, 1999.
FU, Y. et al. Expression of the Candida albicans gene ALS1 in
Saccharomyces cerevisiae induces adherence to endothelial and epithelial cells. Infection and Immunity., v. 66, p. 1783–1786, 1998.
GALE, C. A. Linkage of Adhesion, Filamentous Growth, and Virulence in
Candida albicans to a Single Gene, INT1. Science, v. 279, n. 5355, p.
1355-1358, fev. 1998.
GASPARETTO, A. et al. Produção de biofilme por leveduras isoladas de cavidade bucal de usuários de prótese dentária. Acta Scientiarum,
Health Sciences, v.27, n.1, p. 37-40, 2005.
GAUR, N. K., KLOTZ, S. A., HENDERSON, R. L. Overexpression of the Candida albicans ALA1 Gene in Saccharomyces cerevisiae Results in
Aggregation following Attachment of Yeast Cells to Extracellular Matrix Proteins, Adherence Properties Similar to Those of Candida albicans.
Infection and Immunity, v. 67, p. 6040-6047, 1999.
GAUR, N.K., KLOTZ, S.A. Expression, cloning, and characterization of a Candida albicans gene, ALA1, that confers adherence properties
upon Saccharomyces cerevisiae for extracellular matrix
proteins. Infection and Immunity, v. 65, p. 5289–5294, 1997.
GHANNOUM, M. A.; RADWAN, S. S. Candida adherence to epithelial cells. New York: CRC Press, 1990. 270p.
GOW, N. A.; BROW, A. J. P.; ODDS, F. O. Fungal morphogenesis and host
invasion. Current Opinion in Microbiology, v. 5, p. 366-371, jul. 2002.

58
GRUBB S. E. W. et al. Candida albicans-Endothelial Cell Interactions: a Key Step in the Pathogenesis of Systemic Candidiasis. Infection and
Immunity, v.76, n.10, p. 4370-4377, out. 2008.
HAJJEH, R.A.; SAFAIR, A.N.; HARRISON, L.H. Incidence of bloodstream infections due to Candida species and in vitro susceptibilities of isolates
collected from 1998 to 2000 in a population-based active surveillance program. Journal of Clinical Microbiology, v. 42, p. 1519-1527, 2004.
HANSAN, F. et al. Biofilm formation in clinical Candida isolates and its association with virulence. Microbes and Infection, v.11, p. 753-761,
maio. 2009.
HAWSER S.P., DOUGLAS L.J. Resistance of Candida albicans biofilms to antifungal agents in vitro. Antimicrobial Agents and Chemotherapy, v.
39, n. 9, p. 2128-2131, 1995.
HAYNES, K. Virulence in Candida species. Trends in Microbiology, Cambrigde, v. 9, n.12, p. 591-596, 2001.
HINRICHSEN, S. L. et al. Candida isolates in tertiary hospitals in
northeastern Brazil. Brazilian Journal of Microbiology, v. 40, n. 2, 2009.
HOTA, B. Contamination, Disinfection, and Cross-Colonization: are hospital surfaces reservoirs for nosocomial infection? Clinical Infectious
Diseases, v. 39, n.8, p. 1182-1189, 2004.
HOYER , L.L. et al. Characterization of Agglutinin-like Sequence Genes From Non-albicans Candida and Phylogenetic Analysis of the ALS Family.
Genetics, v. 157, n.4, p. 1555-1567, 2001.
HOYER, L. L. et al. A Candida albicans cyclic nucleotide phosphodiesterase: cloning and expression in Saccharomyces cerevisiae
and biochemical characterization of the recombinant enzyme. Microbiology, v. 140, p. 1533-1542, 1994.
HOYER, L. L. et al. Candida albicans ALS1: domains related to a
Saccharomyces cerevisiae sexual agglutinin separated by a repeating
motif. Molecular Microbiology, v. 15, n.1, p. 39-54, jan. 1995.
HOYER, L. L.; HECHT, J. E. The ALS5 gene of Candida albicans and analysis of the Als5p N-terminal domain. Yeast, v. 18, n.1, p. 49-60,
jan. 2001.
HOYER, L.L.; PAYNE, T.L.; HECHT, J.E. Identification of Candida albicans ALS2 and ALS4 and localization of Als proteins to the fungal cell surface.
Journal of Bacteriology, v. 180, p. 5334–5343, 1998.

59
JABRA-RIZK, M.; et al. Recovery of Candida dubliniensis and other yeasts from human immunodeficiency virus-associated periodontal lesions.
Journal of Clinical Microbiology, v. 39, n. 12, p. 4520, 2001.
JAIN, N. et al. Biofilm formation by and antifungal susceptibility of Candida isolates from urine. Applied and environmental microbiology, v. 73,
p. 1697, 2007.
KAPTEYN, J. C. et al. The contribuition of cell wall proteins to the
organization of the yeast cell wall. Biochimica et Biophysica Acta – general subjects, v. 146, n.2, p. 373-383, 1999.
KLOTZ, S. A.; LIPKE, P. N. The Perfect Adhesive. Current research,
technology and education topics in Applied Microbiology and Microbial Biotechnology. A. Mendez-Vilas (Ed). Formatex, v. 16, p.
838-844, 2010.
KLOTZ, S. A. et al. Candida albicans Als proteins mediate aggregation with bacteria and yeasts. Medical Microbiology, v. 45, p. 363-370, 2007.
KRCMERY, V. Jr; KOVACICOVÁ, G. Longitudinal 10-year prospective
survey of fungaemia in Slovak Republic: trends in etiology in 310 episodes. Slovak Fungaemia study group. Diagnostic microbiology and
infectious disease, v. 36, n. 1, p. 7-11, 2000.
LIMA-NETO, R. G. et al. Adherence of Candida albicans and Candida
parapsilosis to epithelial cells correlates with fungal cell surface carbohydrates. Mycoses, v. 54, n. 1, p. 23-29, set. 2009.
LION, J. P.; RESENDE, M.A. Evalution of adhesion to buccal epithelial cells
in Candida species obtained from denture wearers after exposure to fluconazole. Mycoses, Berlin, v. 50, p.21-24, 2006.
MARCHAIS, V. et al. DNA array analysis of Candida albicans gene
expression in response to adherence to polystyrene. FEMS Microbiology Letters, v.245, p.25-32, 2005.
MARCHETTI, O.; BILLE, J.; FLUKIGER, U. Epidemiology of candidemia in
Swiss tertiary care hospitals: secular trends; 1991-2000. Clinical
Infectious Diseases, v.38, p. 311-320, 2004.
MEDRANO, D. J. A.; BRILHANTE, R. S. N.; CORDEIRO, R. A. Candidemia in a Brazilian hospital: the importance of Candida parapsilosis. Revista do
Instituto de Medicina Tropical de São Paulo, v. 48, n. 1, p. 17-20, 2006.

60
MORENO, S. M. A. et al. Avaliação da adesão de Candida spp. em uma resina acrílica experimental. Revista de Odontologia da UNESP, v.38,
n. 89, out. 2009.
MORGAN, J. Global trends in candidemia: Review of reports from 1995–2005. Current Infectious Disease Reports, v. 7, n. 6, p. 429-439,
2005.
MURILLO L. A. et al. Genome-wide transcription profiling of the early
phase of biofilm formation by Candida albicans. Eukaryotic Cell, v.4, p.1562-1573, 2005.
NETO, M. M. et al. Infecção por fungos e Transplante Renal: Análise nos
primeiros 500 pacientes transplantados no Hospital das Clínicas da Faculdade de
Medicina de Ribeirão Preto – USP. Jornal Brasileiro de Nefrologia, v. 17, n. 3, p. 162-170, 1995.
NIKAWA H. The role of saliva and serum in Candida albicans biofilm
formation on denture acrylic surfaces. Microbial Ecology in Health and Disease, v.9, p. 35-48, 2003.
NUCCI, M.; COLOMBO, A. L. Candidemia due to Candida tropicalis:
clinical, epidemiologic, and microbiologic characteristics of 188 episodes
occurring in tertiary care hospitals. Diagnostic Microbiology and Infectious Disease., v. 58, n. 1, p. 77-82. maio. 2007.
O’DAY D. M., HEAD W. S., ROBINSON R. D. An outbreak of Candida
parapsilosis endophthalmitis: analysis of strains by enzyme profile and antifungal susceptibility. British Journal Ophthalmology, v.71, p.126-
129, 1987.
OLIVEIRA, R. D. R. de; MAFFEI, C. M. L.; MARTINEZ, R. Infecção urinária hospitalar por leveduras do gênero Candida. Revista da Associação
Médica Brasileira, São Paulo, v. 47, n. 3, p. 231-235, 2001.
PFALLER M. A. et al. Geographic and temporal trends in isolation and antifungal susceptibility of Candida parapsilosis: a global assessment from
the ARTEMIS DISK antifungal surveillance program, 2001 to 2005.
Journal of Clinical Microbiology, v.46, p.842–849, 2008.
PFALLER M. A. et al. International surveillance of bloodstream infections due to Candida species: frequency of occurrence and antifungal
susceptibilities of isolates collected in 1997 in the United States, Canada and South America for the SENTRY program. Journal of Clinical
Microbiology, v.36, p.1886-1889, 1998b.

61
PFALLER M. A. et al. National surveillance of nosocomial bloodstream infections due to species of Candida other than Candida albicans:
frequency of occurrence and antifungal susceptibility in the SCOPE program. Diagnostic Microbiology and Infectious Disease, v.39,
p.121-129, 1998a.
PFALLER M. A. et al. National surveillance of nosocomial bloodstream infections due to Candida albicans: frequency of occurrence and antifungal
susceptibility in the SCOPE program. Diagnostic Microbiology and
Infectious Disease, v.31, p.327-332, 1998c.
PFALLER M. A., MESSER S. A., HOLLIS R. J. Variations in DNA subtype, antifungal susceptibility, and slime production among clinical isolates of
Candida parapsilosis. Diagnostic Microbiology and Infectious Disease, v.21, p.9-14, 1995.
PFALLER M. A., MESSER S. A.; BOLMSTRO¨M, A. Evaluation of Etest for
Determining in vitro Susceptibility of Yeast Isolates to Amphotericin B. Diagnostic Microbiology and Infectious Disease, v.32, p.223-227,
1998.
PFALLER, M.A. et al. Bloodstream infections due to Candida species: SENTRY antimicrobial surveillance program in North America and Latin
America, 1997-1998. Antimicrobial Agents and Chemotherapy, v. 44,
n. 3, p. 747-51, 2000.
PLOUFFE J. F. et al. Nosocomial outbreak of Candida parapsilosis fungemia related to intravenous infusions. Archives of Internal Medcine, v.137,
p.1686-1689, 1977.
RAMAGE G. et al. Biofilm Formation by Candida dubliniensis. Journal of Clinical Microbiology, v. 39, n. 9, p. 3234-3240, set. 2001.
RAUCEO, J. M. et al. Threonine-rich repeats increase fibronectin binding in
the Candida albicans adhesin Als5p. Eukaryotic cell, v. 5, n. 10, p. 1664–1673, 2006.
RIBEIRO, M. A.; MIRANDAL, A. E.; PAULA, C. R. Prevalence and
exoenzyme secretion of Candida albicans isolates from oral and vagina
mucosas of HIV-infected women. Mycopathologia, v. 157, p. 255-261, 2004.
RUIZ - HERRERA J. et al. Molecular organization of the cell wall of
Candida albicans and its relation to pathogenicity. FEMS Yeast Research, v. 6, n. 1, p. 14-29, dez. 2005.

62
SAMPERMANS, S. et al. Flocculation onset in Saccharomyces cerevisiae: the role of nutrients. Journal of Applied Microbiology, v. 98, n. 2, p.
525-531, fev. 2005.
SAN MILLAN, R. et al. Effect of salivary secretory IgA on the adhesion of Candida albicans to polystyrene. Microbiology, v.146, n. 9, p. 2105-
2112, set. 2000.
SANDVEN, P. et al. Candidemia in Norway (1991 to 2003): results from a
nationwide study. Journal of Clinical Microbiology, v. 44, n. 6, p.1977-1981, 2006.
SCALETSKY, I.; SILVA, M.; TRABULSI, L.R. Distinctive patterns of
adherence of enteropathogenic Escherichia coli to HeLa cells. Infection and Immunity, v. 45, n. 2, p. 534, 1984.
SHIN J. H. et al. Biofilm production by isolates of Candida species
recovered from nonneutropenic patients: comparision of bloodstream isolates with isolates from other sources. Journal of Clinical
Microbiology, v.40, p.1244-1248, 2002.
SILVA, E.H. et al. Candiduria in a public hospital of São Paulo (1999-2004): characteristics of the yeast isolates. Revista do Instituto de
Medicina Tropical de São Paulo, v. 49, p. 349–353, 2007.
SILVA, S. et al. Biofilms of Non-Candida albicans Candida species:
quantification, structure and matrix composition. Medical Mycology, v. 47, p. 681-689. 2009.
SKERL, K.G.; CALDERONE, R. A. In vitro binding of Candida albicans yeast
cells to human fibronectin. Canadian Journal of Microbiology, Ottawa, v. 30, p. 221-227, 1984.
SOLL, D. R. Candida Biofilms: Is Adhesion Sexy? Current Biology, v. 26,
n.16, p. R717–20, ago. 2008.
SOLL, D. R. Candida commensalism and virulence: the evolution of phenotypic plasticity. Acta Tropica, v. 81, p. 101-110, 2002.
SOLOMON S. L. et al. An outbreak of Candida parapsilosis bloodstream infections in patients receiving parenteral nutrition. The Journal of
Infectious Diseases, v.149, p.98-102, 1984.
STAAB, J. F. et al. Adhesive and Mammalian Transglutaminase Substrate Properties of Candida albicans Hwp1. Science, v. 283, n. 5407, p. 1535-
1538, mar. 1999.

63
STURTEVANT, J. CALDERONE, R. Candida albicans adhesins: Biochemical aspects and virulence. Revista Iberoamericana de Micologia, v. 14, p.
90-97, 1997.
SUGIZAKI M. F. et al. Prevalence and in vitro antifungal susceptibility of Candida spp isolated from clinical specimens in São Paulo, Brazil. Revista
Iberoamericana de Micologia, v.15, p.16-18, 1998.
SUZUKI, L. C. Desenvolvimento de biofilme formado por Candida
albicans in vitro para estudo da terapia fotodinâmica. 2009. 49f. Dissertação (Mestrado). Instituto de Pesquisas Energéticas Nucleares.
Autarquia associada à Universidade de São Paulo. São Paulo, 2009.
TAMURA N. K. et al. Fatores de virulência de Candida spp isoladas de cateteres venosos e mãos de servidores hospitalares. Revista da
Sociedade Brasileira de Medicina Tropical, v. 40, n. 1, p. 91-93, jan/fev. 2007.
TAMURA N. K., GASPARETTO, A.; SVIDZINSKI, T. Evaluation of the
adherence of Candida species to urinary catheters. Mycopathologia, v. 156, n. 4, p. 269–272, 2003.
TUMBARELLO M. et al. Biofilm Production by Candida Species and
Inadequate Antifungal Therapy as Predictors of Mortality for Patients with
Candidemia. Journal of Clinical Microbiology, v.45, n.6, p. 1843-1850, 2007.
UPPULURI, P. et al. Characteristics of Candida albicans biofilms grown in a
synthetic urine medium. Journal of Clinical Microbiology, v. 47, n. 12, p. 4078, 2009.
VERSTREPEN, K. J.; KLIS, F. M. Flocculation, adhesion and biofilm
formation in yeasts. Molecular Microbiology, v. 60, n. 1, p. 5-15, abr. 2006.
VIANA, I. M. G. et al. Perfil epidemiológico de pacientes com baciloscopia
positiva para tuberculose pulmonar. Revista Brasileira de Clínica Médica, São Paulo, v. 8, n. 6, p. 505-508, nov/dez. 2010.
VIANI, P. R. C. Candida provenientes de Infecção Hospitalar isoladas de Pacientes internados em Hospital Infantil do Estado de
São Paulo e avaliadas por marcadores fenotípicos. 2007. 68f. Dissertação (Mestrado). Instituto de Ciências Biomédicas, Universidade de
São Paulo, 2007.

64
VIDOTTO, V. et al. Adherence of Candida albicans and candida dublniensis to buccal and vaginal cells. Revista Iberoamericana de Micologia., v.
20, p. 52-54, 2003.
WEINBERGER, M. et al. Characteristics of candidaemia with Candida albicans compared with non-albicans Candida species and predictors of
mortality. The Journal of Hospital Infection, v. 61, n. 2, p.146–154, 2005.
WEMMS J. J. Candida parapsilosis: epidemiology, pathogenicity, clinical manifestations, and antimicrobial susceptibility. Clinical Infectious
Diseases, v.14, p.756-766, 1992.
WEMMS J. J. et al. Candida parapsilosis fungemia associated with parenteral nutrition and contaminated blood pressure transducers.
Journal of Clinical Microbiology, v.25, p.1029-1032, 1987.
WISPLINGHOFF, H. et al. Nosocomial bloodstream infections in US hospitals: Analysis of 24,179 cases from a prospective nationwide
surveillance study. Clinical Infectious Diseases, v. 39, p. 309-317. 2004.
XU, J. et al. Clonal and spontaneous origins of fluconazole resistance in
Candida albicans. Journal of Clinical Microbiology, v. 38, p. 1214-
1220, 2000.
YANG, Y.L. Virulence factors of Candida species. Journal of Microbiology, Immunology, and Infection, v. 36, p. 223-228, 2003.

65
APÊNDICE A - Adherence ability and biofilm production by clinical isolates of Candida species coming from different sources
(Full title)
Adherence ability and biofilm production by clinical isolates of Candida
species coming from different sources
(Short title)
Adherence ability and biofilm production by clinical isolates of Candida
species
Author(s): Viviane Menezes de Menezes - MENEZES, V.M1,2
Iven Neylla Farias Vale - VALE, I.N.F1
Valéria de Jesus Menezes de Menezes – MENEZES, V.J.M2
Silvio Gomes Monteiro - MONTEIRO, S.G1
Luís Henrique Bastos Gonçalves - GONÇALVES, L. H. B3
Patrícia Maria Silva Figueiredo - FIGUEIREDO, P.M.S.1
Cristina Andrade Monteiro - ANDRADE-MONTEIRO,C.1
Affiliation: 1- UNICEUMA - Centro Universitário do Maranhão, Pró-Reitoria
de Pós-Graduação, Pesquisa e Extensão, Rua Josué Montello,
n.1, Renascença II, CEP 65075120, São Luís, Maranhão,
Brasil. Telephone number: (98) 32144124
2- FEBAC - Faculdade de Educação de Bacabal, Coordenadoria
de Pós-Graduação, Pesquisa e Extensão, Rua 12 de Outubro,
n.377, BR 316, Centro, Bacabal, Maranhão, Brasil. Telephone
number: (99) 36213403
3- Florence - Instituto Florence de Ensino, Núcleo de Estudos,
Pesquisa e Extensão, Rua Rio Branco, n. 216, Centro, São
Luís, Maranhão, Brasil. Telephone number: (98) 3878 2120
Postal address:
UNICEUMA - Centro Universitário do Maranhão, Pró-Reitoria de
Pós-Graduação, Pesquisa e Extensão, Rua Josué Montello, n.1,
Renascença II, CEP 65075120, São Luís, Maranhão, Brasil.
Telephone: (98) 32144124 UniCeuma
E-mail: [email protected]
Key-words: Candida spp, virulence factors, adhesion, biofilm, anatomical
sites

66
Abstract
Adhesion and biofilm production have been identified as potential
virulence factors for developing candidiasis, since they are primary
mechanisms of infection, especially in immunocompromised patients. The
present study reports on an in vitro adherence assay and the ability of
biofilm formation of clinical isolates of Candida from many different
anatomical sites with the goal of comparing these virulence properties
among those isolates. Adherence ability and biofilm production were
quantified in 118 and 123 clinical strains of Candida, respectively. The
results showed that all isolates adhered to coverslips in a species- and
strain-dependent manner with C. glabrata showing the lowest and C.
tropicalis the highest levels of adhesion. Among 93 strains with the ability
to adhere, 44 presented a pattern of localized adherence (47.31%), 24
(25.8%) showed an aggregative pattern and only 9 had (9.7%) type
pseudo-hyphal. Of the 51.2% total biofilm-positive isolates, 71.4% and
17.5% were considered low and high biofilm producers, respectively.
Biofilm production by C. non C. albicans isolates was higher than that by
C. albicans, with C. tropicalis strains showing significantly the highest
biofilm production. Biofilm production was more frequent among secretion
tracheal isolates, however this variation was not significant among the
different sources.
INTRODUCTION
•
• Opportunistic infections in humans often follow immune suppression or neutropenia [1]. Like all infectious diseases adhesion of a microbe to
host tissue must occur before infection can become established. As with
many microorganisms, the ability of Candida albicans to adhere to surfaces is presumed to be an important survival trait [2,3]. C. albicans is
the most commonly encountered fungus in hospital microbiology laboratories and is the fourth most common cause of positive blood
cultures in hospitalized patients in North America [4]. Lastly, candidiasis is the most common opportunistic infection in HIV-infected individuals [5].
Although C. albicans is the main species isolated from patients with fungemia [6,7,8,9,10,11], recent reports indicate a trend toward an
increasing prevalence of infections caused by species of Candida other than C. albicans [12,13,14,15]. In Brazil the species more prevalent are C.
albicans, C. tropicalis, C. parapsilosis, C. glabrata, C. krusei and C. guillermondii [16,15] Candida albicans is well known in the clinical realm
for its ability to “stick” to damaged human tissue whether that be mucous membrane, bladder epithelium, heart valve or retinal vessel. In addition,
the capacity of tissue invasion and adhesion to artificial materials such as
probes, latex catheters, and several types of plastic material are

67
responsible for causing important infections [17]. It possesses cell surface proteins known as Agglutinin-like sequence (Als) proteins that are
necessary for continuous fungal residence in the dense microbial milieu on mucous membranes [18].
After adhesion, the fungus elaborates biofilm and aggregation can occur.
Biofilm is the preferred mode of social behavior and protects its members
against host defense mechanisms such as neutrophils. Biofilm formation
on medical devices has been described as an important source of infection
[19,20]. King et al. [21] demonstrated the prototypical C. albicans
adhesion phenotype of yeast cells covering buccal epithelial cells and
adhering to one another forming clumps of cells (aggregation). Adhesion
followed by aggregation has since been documented in numerous studies
involving a myriad of biological targets. When C. albicans encounters an
anchored protein, cell, or tissue, the fungus adheres and then aggregates.
The molecular explanation of this phenotype, i.e., adhesion and
aggregation, awaited the discovery of Als protein function [18].
Many previous studies have focused on C. albicans adherence and biofilms
[22,23,24,25] due to its well-recognized virulence. However, only a few
studies of adherence and biofilm generated by species other than C.
albicans have been reported [26,27,28,29].
Adhesion capacity probably varies among isolates of different species as
suggested by the work of Biasoli et al.[30] which pointed out differences
among C. albicans, C. glabrata, C. krusei and C. lusitaniae.
The analysis of the in vitro adherence of Candida species can contribute to
the understanding of the behavior of these organisms in a specific
infection process. Thus, the present study reports on an in vitro adherence
assay and the ability of biofilm formation of clinical isolates of C. albicans,
C. tropicalis, C. parapsilosis and C. glabrata from many different
anatomical sites with the goal of comparing these virulence properties
among those isolates.
MATERIALS AND METHODS
Strains and culture media
The strains of Candida spp. used in this study came from cultures of
hospitalized patients (both public and private hospitals in Sao Luis, State
of Maranhão, Brazil) suspected of being infected by microorganisms. The
isolates were obtained from 2007 to 2009 and belong to the collection of a
private laboratory and were kindly provided for this research. The
standard strain C. albicans ATCC 18804 (American Type Culture
Collection, Rockville, Md) was included in the experiments. The adherence

68
assay and biofilm formation was studied in clinical isolates of C. albicans
(64 strains), C. glabrata (18 strains), C. tropicalis (22 strains), C.
parapsilosis (19 strains). The isolates were previously identified using
CHROMagar Candida (CHROMagar, Paris, France) and by the automated
system VITEK (BioMeriux, Marcy-I’Etoile, France). The yeasts were
routinely grown in Sabouraud dextrose agar plates incubated at 37°C for
24h from stock cultures in BHI-glycerol and stored at 4°C during the
procedures.
Preparation of inoculums for the adherence assay and biofilm
formation
The inoculum of yeast cells was made from stock cultures and incubated
for 18 hours at 37°C in BHI (Brain Heart Infusion –Acumedia
Manufectures) or in liquid RPMI-1640 medium and standardized to
approximately 106 UFC/ml according to the 0.5 McFarland turbidity range
[31].
Adherence assay
For the adhesion assay, sterile round glass coverslips (Glasscyto) were
placed into 24 wells microtiter plates (TPP Zellkultur Testplatte 24F) and
40 µL of the standardized cell suspension (1 x 106 UFC/mL) plus 960 µL of
BHI medium were added to each well. For controls, glass coverslips were
similarly processed but in the absence of Candida cells. The microtiter
plates were incubated at 37°C for 18 h. After incubation, medium was
removed and the microtiter plates were washed once with ultrapure water
to remove non-adhered cells and the coverslips were stained with 1%
(v/v) of crystal violet for 5 minutes. The coverslips were then washed
twice with sterile water to remove the excess of staining and put onto
slides for visualization under light microscopy (NIKON Eclipse E100). The
standardization of the classification of the adhesion capacity of cells on
glass coverslips was performed considering the count of adhered cells by
fields [30, with adaptations]. For each slide a maximum of 70 fields were
analyzed and the adhered cells were counted. The number of adhered
cells obtained was registered as an average of three experiments realized
for each isolate. Adhesion ability in inert material for each isolate was
scored as negative, weak (+), moderate (++) or strong (+++) [32, with
adaptations]. Each isolate was analyzed by two observers independently.
The cellular adhesion was considered negative if less than 01 cell was
adhered per field, after analyzing 70 fields of the coverslip; weak if there

69
were between 01 and 10 adhered cells per field in 50 viewed fields;
moderate if there were more than 10 adhered cells per field in 30
observed fields and strong if there were an equal or more than 25 adhered
cells per field in 20 analyzed fields. There were no discrepant results
between the two observers in the direct microscopic visualization.
Determination of biofilm production
Biofilm production was determined by a method established by Shin et al.
[26] through spectrophotometric analysis. Broth Heart Infusion medium
was prepared according to the manufacturer’s instructions, and with
supplementation of 60 g of glucose per liter (final concentration, 80 g/liter
or 8%). Microorganisms were grown for 24 h at 37º C on Sabouraud
dextrose agar plates and suspensions of each strain of Candida species
were prepared on saline solution. We used C. albicans ATCC strain as
control in each experiment. After adjusting the turbidity of yeast
suspension each well of a 96-well microtitration plate (TPP Zellkultur
Testplatte 96F) was inoculated with aliquots of 20 µL of yeast cell
suspension and 180 µL of BHI. Plates were then incubated at 37ºC for 24
h without agitation. After this time the microtiter plate was washed three
times with distilled water and stained with crystal violet. After addition of
200 µL of distilled water in each well spectrophotometric readings were
performed at 450 nm with a microtiter plate reader (ANTHOS 2010). The
absorbance was measured as an average of three experiments for each
isolate and the data were expressed in percentage. Biofilm production for
each isolate was scored as either negative (% A < 5), weak (+1; 5 < % A
< 20), moderate (+2; 20 < % A < 35), strong (+3; 35 < % A < 50) and
very strong (+4; % A _> 50).
Statistical Analysis
The data were evaluated by the program BioEstat version 5.0 (2007). The
Lilliefors normality test was initially performed for the numeric variable
absorbance. Since this variable did not show normal distribution (p <
0.05), it was analyzed using the Kruskal-Wallis nonparametric variance
test. Along with the ordinal variables, the classification of biofilm and
adhesion levels in relation to species and to the anatomic site were made
through the nonparametric Kruskal-Wallis test and the analysis post-hoc
was done using the Student-Newman-Keulls test. The significance level
adopted in all tests was 5%; i.e., values were considered significant when
p < 0.05.

70
RESULTS
Adherence ability
Of a total of 118 analyzed isolates for the ability to adhere 93 (79%) were
positive for this property. Among these 81 (73%) showed a moderate or
weak ability of adherence (Table 1). Clearly, the four different species of
Candida adhered to coverslips although differences were observed in the
level of adhesion among them. Among the analyzed species, C. tropicalis
isolates showed greater capacity to adhere to inert material, with 91% of
the strains being positive, followed by C. parapsilosis with 89% of isolates
showing positive adherence, C. albicans with 76% and, finally, C. glabrata
with 61% of samples showing adherence on inert material. Besides the
interspecific variation we have noticed an intraspecific variability among
the isolates with respect to the capacity of adherence (Table 1). The
statistical analysis shows that these differences between species and
isolates are not significant (p> 0.05; Table 2).
The adhesion ability was categorized according to the number of cells read
per fields in weak (+), only 01 to 10 cells per 50 fields; moderate (++),
more than 10 cells per 30 fields or strong (+ + + ), above 25 cells per 20
fields read. Few isolates (21%) did not show any adherence on coverslips
(Table 1), being only 02 samples of C. tropicalis and C. parapsilosis, each
followed by 07 strains of C. glabrata and 14 of C. albicans (Table 1).
Regardless of site of origin of these samples there were differences in the
ability to adhere for all Candida species (Table 1). In relation to the total
number of species examined these differences were not significant (Table
2), and also, between the degree of positivity for adhesion on inert
material there has not been statistical significance either among the
species or the sites of origin (Table 3). However, it was noted that 100%
of C. tropicalis samples isolated from tracheal aspirates and 100% of C.
glabrata samples isolated from tip catheters were adherent (Table 1). Four different patterns of cellular organization of in vitro adhesion among
the isolates were verified, as shown in Figure 1. These patterns were
classified according to those proposed by Cravioto et al. [33] for
diahrreiogenic strains of E. coli. A diffuse adhesion (DA) pattern where the
yeast cells adhere over the entire surface of the glass coverslip, without
forming cells clumps. Localized adherence (LA) that involves clusters of
yeasts that adhere to localized regions of the coverslip. An aggregative
adhesion (AA) that is characterized by clumps of yeasts with a "stacked
brick" appearance that attached to the surface of the glass slide. There is

71
formation of filaments or pseudo-hyphae along the surface of coverslip in
a fourth pattern of adhesion (FA).
Among 93 strains with the ability to adhere, 44 presented a pattern of
localized adherence (47.31%), followed by 24 (25.8%) isolates that
showed aggregative pattern and only 9 (9.7%) had type pseudo-hyphal.
Only two isolates of C. albicans exhibited the filamentous or pseudo-
hyphal pattern. C. parapsilosis was the species that showed a greater
variability of adhesion patterns among their isolates and five of them
(29.4%) had the pseudo-hyphae pattern. Most isolates that showed the
localized or aggregative pattern came from urine (Table 4).
Biofilm formation
Table 5 presents the results for biofilm quantification for 123 Candida
isolates using spectrophotometric method. It was evident that all NCAC
species formed biofilms, although differences occurred depending on
species or strain. A total of 63 (51.2%) analyzed isolates were biofilm
positive. From these 11 (17.5%) were strong (3+) or very strong (4+)
biofilm formers, and 45 (71.4%) were weak formers. C. tropicalis isolates
were significantly higher producers of biofilm than those from others
species (91%, 20 of 22; p < 0.001 by the test of Student-Newman-Keuls;
Figure 2).
With regard to the intensity of the capacity of biofilm formation, the
species with the largest number of strong producer isolates (3 +) was C.
tropicalis (25% of its isolates), followed by C. albicans (13.3%). All C.
glabrata isolates that produced biofilm were weak (1 +) producers.
Biofilm production was detected in 50% of vaginal secretion isolates,
however all of them presented a weak (+1) ability. Among the strains
came from urine, 49.2% were biofilm producers, with 27.6% of them
showing moderate (2+) or strong (3+) production. Most Candida strains
coming from catheter tips were not biofilm producers (80%; Table 5).
Biofilm production was more frequent among isolates from tracheal
secretion (58.3%), followed by those from blood (57.1%), however there
was no significant difference among these results (Figure 3, p = 0.7748).
DISCUSSION
Infections of hospitalized patients with species others than C. albicans is
becoming increasingly prevalent [34, 35], especially in immune depressed
people. The increase in cases reported, is mainly caused by the increase
of the number of patients that are immunocompromised, the increase of

72
invasive surgery and antifungical therapy. Adherence of Candida to
mucosal surfaces of host cells is a vital prerequisite for successful
colonization and infection. Attachment enables the organisms to avoid
dislodgement by cleansing action of mucosal secretions and it facilitates
infection [reviewed by 36, 37]. Studies relate that the ability of adherence
shown by various Candida species is different among them. This fact could
explain why the colonization to mucosal surfaces are more common in
some species than in others [38,39] and ability to adhere to inert
materials may be indicative of a probable adherence to living cells. Thus,
the study aimed to analyze the adhesion and biofilm formation ability of
clinical isolates of Candida from different sites of origin onto glass
coverslips.
All Candida species were able to adhere to coverslips, which is in
agreement with other studies examining Candida species adherence to
inert materials like silicone rubber, for example [40,41,42]. However, it is
important to note that non-Candida albicans Candida (NCAC) isolates
adhered to a higher extent (81,4%) compared with those of C. albicans
(76,3%). C. tropicalis highlighted among NCAC species with 91% of
samples showing adhesion and of these 20% showed strong adhesion
(+++). These results, in relation to the others NCAC species was
significant (p < 0,001). Likewise, Minage et al.[43] observed greater
adhesion of these Candida species in inert material (plastic polymers).
Also, Moreno et al. [42]) verified the adherence ability of Candida isolates
to inert material (acrylic resin) and among the species analyzed, C.
tropicalis has shown the highest adhesion ability, followed by C.
dubliniensis and C. albicans. Our results are relevant because in a recent
epidemiological study of 100 candiduria cases (between 1999 and 2004),
in a paediatric hospital in Brazil, where the ability to adhere to catheters
was considered a significant and pre-predisposing factor to this type of
infection, 20 samples of C. tropicalis were isolated compared with 56 of C.
albicans, 11 of C. glabrata and 4 of C. parapsilosis [44)]. In fact, C.
tropicalis has emerged as the second or third most common agent of
candidemia, especially in patients with cancer [45,46]. In addition, a high
incidence of C. tropicalis as causative agent of nosocomial urinary tract
infections has been reported [47].
NCAC species has shown an increase in the number of cases of fungemia
in several countries [48,26,49,17] probably due to increased expression of
virulence factors, among them the ability to adhere and form biofilms.
Among the isolates of C. tropicalis that were positive for adherence, those
from tracheal aspirates were 100% adherent, although this difference was
not significant in relation to the distribution of anatomical sites. The high

73
capacity of adherence by C. tropicalis may indicate increased virulence of
this species when compared with others, corroborating data obtained by
Tamura et al. [17] and Costa [50] that NCAC species have higher
adhesion properties than C. albicans, being that in the study of Tamura et
al. [17] the researchers report C. glabrata as the species with the highest
ability to adhere and form biofilm, while Costa [50] shows isolates of C.
tropicalis significantly more adherent than those of other species including
C. albicans.
It is important to note that in C. parapsilosis cell adhesion and biofilm
formation have been observed as significant virulence factors [51,52],
corroborating our findings of the tests for adhesion where C. parapsilosis
was the second species with the highest adhesion ability (89%), and
100% of isolates from the catheter tip were adherent. Although C.
parapsilosis is generally regarded as one of the less virulent yeast species,
it is now a frequent cause of candidemia associated with parenteral
hyperalimentation and contaminated intravascular devices and ophthalmic
solutions [53,54,55,56,57]. Several factors give to C. parapsilosis a
selective advantage, including the ability to proliferate in high
concentrations of glucose and the adherence to prosthetic materials
[58,56]. Nosocomial outbreaks of C. parapsilosis have also been described
and have been attributed to transfer of yeast from the hands of healthcare
workers [59]. However in contrast with results obtained for adherence
capacity, C. parapsilosis was the species of Candida that least expressed
properties of biofilm formation (26%), and the same isolates from the
catheter tip were negative for this virulence factor (75%). These results
contrast with those of literature. Pfaller, Messer, Hollis [60] studied the
production of biofilms in clinical samples of C. parapsilosis grown in
culture mediums containing glucose and found that isolated from blood
and catheter, more biofilm was produced than samples from other
anatomical sites. Our results show that isolates of C. parapsilosis can
enter the bloodstream and cause infections even without forming a biofilm
on medical devices because some of these analyzed isolates were from
blood.
This study describes for the first time, different patterns of adherence to
glass coverslips shown by Candida species. These patterns were
categorized similarly to the classification of diarrheal strains of E. coli that
show distinct patterns of adhesion when adhered in vitro to HEp-2 and
HeLa cells, or to coverslips [33,61]. Four different patterns of cellular
organization of in vitro adhesion among the isolates were verified: a diffuse
adhesion (DA) pattern where the yeast cells adhere over the entire surface of
the glass coverslip, without forming cells clumps; localized adherence (LA)

74
that involves clusters of yeasts that adhere to localized regions of the
coverslip; an aggregative adhesion (AA) that is characterized by clumps of
yeasts with a "stacked brick" appearance that attached to the surface of the
glass slide and a filamentous or pseudo-hyphae (FA) pattern of adhesion
where there is formation of filaments or pseudo-hyphae along the surface
of coverslip. Most of the isolates showed ability to adhere from localized
type (47%), followed by those who have demonstrated aggregative
pattern (26%). C. parapsilosis was the species that showed a greater
variability of patterns among its isolates, and 5 (29.4%) of them showed
the pattern of pseudo-hyphae type. Most isolates that showed a localized
or aggregative pattern came from urine. The large number of isolates
showing a localized or aggregative pattern of adherence could be
explained by the presence of adhesins on the cell wall of Candida that
were responsible for intercellular adhesion ability displayed by different
strains. These types of adherence patterns would also facilitate biofilm
formation by these species.
In C. albicans ALS family of proteins mediate several ways of adhesion, for
example, cell adhesion without aggregation with sparsely adherent cells or
adhesion followed by cell-cell aggregation forming clumps. What specific
behavior is exhibited by an isolate depends on the type of protein
expressed. This fact, in a way, confirms our findings which show evidence
of a lack of diffuse pattern with most strains showing a localized or
aggregative pattern of adhesion, where both show the formation of
colonies by the union of several yeast cells.
Specific structural regions of ALS proteins in C. albicans appear to be
involved in the phenomenon of cell aggregation, as the region of tandem
repeats and the region rich in threonine residues [18]. The threonine-rich
region of Als5p which consists of 127 amino acids is involved in
aggregation [62], however, Als7p and Als9p with 128 amino acids do not
aggregate after adhesion [63,5]. In the same way, the aggregation
property of various C. albicans Als proteins is roughly proportional to the
number of tandem repeats in the molecule [62]. For example, Als1p with
20 tandem repeats forms aggregates larger than those found with Als5p
with only 5 tandem repeats [5].
Only two isolates of C. albicans showed a filamentous or pseudo-hifal
pattern of adherence which also was observed in only 7 (7.5%) isolates of
NCAC species. The ability to differentiate between the yeast form to
filamentous one has been described in many studies and is directly related
to the invasion of host tissue, favoring the pathogenicity of Candida,
allowing easier penetration of the host tissue [64] however it is not
essential for pathogenesis [65]. The filamentous form is probably more

75
virulent because of the greater length of contact between the
microorganism and the area to be infected, but both forms are often found
in both commensal state and in an infectious processes and even NCAC
species that do not form true hyphae are potentially pathogenic and are
involved in morbimortality by these agents [65,66].
The possibility of changing their own phenotype has been described in
some studies in relation to C. albicans [64,67,68,66] and can modulate
some virulence factors including dimorphism and cell adhesion [69].
However, knowledge about the mechanisms that enable this phenotypic
change remains to be elucidated [50], this change is probably associated
with a wealth (abundance) of nutrients and expression of adhesins.
Investigation of the presence and/or expression of Als or genes
responsible for the transition to hyphae in Candida isolates would be
critical to establish correlations with the phenotypic data obtained in this
study of adherence.
Regarding the ability to form biofilms the data obtained herein clearly
confirm reports from literature that different Candida species have
different abilities to produce biofilm in vitro and that NCAC species were
the highest biofilm producers [26,70]. The present study shows that C.
tropicalis was significantly the greater biofilm producer among the species
studied (p < 0.0001). The ability of biofilm formation among the others
species was not statistically different. These data also corroborate those
reported by Gasparetto et al. [71] that found greater and more significant
production of biofilms in NCAC species (64%), where the species C.
tropicalis stood out among these. According to Shin et al. [26] despite C.
albicans being pathogenic and capable of establishing mechanisms for the
occurrence of systemic infection, their biofilm production was significantly
less frequent than other species, independent of the anatomical site of
origin and C. tropicalis was also the largest producer of biofilm followed by
C. parapsilosis, C. glabrata and C. albicans. However, C. parapsilosis was
highlighted when their blood isolates were analyzed. These isolates were
the largest producers of biofilm in relation to those of other species of
Candida not C. albicans, unlike our findings where all isolates of C.
tropicalis from blood were those with the highest biofilm formation
(100%), followed by those of C. albicans (57%) and, finally, those of C.
parapsilosis (40%). There was no blood isolates of C. glabrata.
There was no significant difference in relation to the site of origin of the
samples and the ability of biofilm formation. However, almost all samples
from the catheter tip were not biofilm producers, a fact which differs from
the literature which says that medical devices like catheters are preferred
and potential substrates for biofilm formation [19,72].

76
Supplementation of the medium with glucose was important for the
findings in our experiment, this data agrees with previous studies for the
production of biofilm, such as those of Hawser, Douglas [73], Jain et al.
[27] and Uppluri et al. [74] that have already observed that the growth of
yeast is lower if the culture medium is not supplemented with this
carbohydrate. These findings are relevant for diabetic patients who have a
high rate of blood glucose. Indeed, fungal infections are more common in
diabetic patients, particularly those involving Candida species, occasionally
causing serious clinical consequences [75].
Comparing our findings of ability to adhere and biofilm formation one can
see a differential expression of those properties in isolates of the same
species. These results reflect inherent physiological differences among
species and isolates and may have significance with respect to pathogenic
potential. Only isolates of C. tropicalis stood out both in terms of ability to
adhere to inert material and to form biofilms (91% in both trials). The
other isolates of the species studied showed different behaviors in tests
such as those of C. parapsilosis that showed high adherence (89%) in
contrast to the low production of biofilm (26%), those of C. albicans and
C. glabrata that showed higher adhesion ability (76% and 61%
respectively) than to form biofilms (47% and 44% respectively)
suggesting that both capabilities are different stages of the infectious
process [(66),(68)] and that the species express different mechanisms for
the formation of a mature biofilm. This really can be true as Silva et al.
[76] studying the characterization of biofilms in NCAC species in terms of
structure, matrix composition and metabolic activity revealed significant
structural differences in biofilms with respect to the morphology of cells
and their spatial disposition. C. parapsilosis biofilm matrices have shown
high amounts of carbohydrate and relatively lower amounts of proteins.
Contrarily, C. tropicalis biofilms matrices were low in both carbohydrate
and protein content and those of C. glabrata have shown high amounts of
both components, showing that intrinsic differences should exist in terms
of metabolic activity of biofilms.
Hasan et al. [77], using two different methods, found that there is a
significant difference between the production of biofilms among species of
Candida, showing that the ability of biofilm formation in vitro is dependent
on the species of Candida. Their results differed from ours, because while
using a similar methodology, these authors found C. albicans to be the
largest producer of biofilm and all isolates of C. glabrata have shown
biofilm positive production, data not found in our experiments.
There are few studies that assess biofilm production among species of
NCAC from different anatomical sites without joining them as a unique

77
group. Also, there is no data about correlation among virulence factors of
Candida species that have clinical relevance in our region. This is
important because one can propose preventive, diagnostic and therapeutic
measures more effective and specific, and thus contribute to reduce
morbidity and mortality caused by Candida species. In summary, our
findings demonstrate that the tested Candida species were able to adhere
to inert surfaces and to form biofilms and suggest that these properties
are strongly linked to pathogenicity of these agents, but that they are
distinct processes that need to be better understood. They also shows that
the expression of these virulence factors are not directly related to the site
of infection itself, but with specific factors of these microorganisms and,
most likely, with the host response. These findings are especially relevant
to all patients who use medical devices like catheters, including those with
urinary tract infection, and who have received transplants or who are in
intensive care units.
Acknowledgements
The authors acknowledge FAPEMA (Fundação de Amparo à Pesquisa e ao
Desenvolvimento Científico e Tecnológico do Maranhão), for supporting
Viviane Menezes work through grant ATB-04191/10 Edital FAPEMA nº
026/2010 Bancada and Pró-Reitoria de Pós-Graduação, Pesquisa e
Extensão do Centro Universitário do Maranhão.
“None”
1. Meunier F, Aoun M, Bitar N. Candidemia in immunocompromised
patients. Clin Infect Dis 1992;14:120-125.
2. Calderone RA, Braun PC. Adherence and receptor relationships of
Candida albicans. Microbiol Mol Biol Rev 1991;55:1-20.
3. Klotz SA. Fungal adherence to the vascular compartment: a critical
step in the pathogenesis of disseminated candidiasis. Clin Infect Dis
1992;14:340-347.
4. Edmond MB, Wallace SE, McClish DK, et al. Nosocomial bloodstream
infections in United States hospitals: a three-year analysis. Clin Infect Dis
1999;29:239-244.
5. Klotz SA, Nguyen HC, Van Pham T, et al. Clinical features of
HIV/AIDS patients presenting to an inner city clinic in Ho Chi Minh City,
Vietnam. Int J STD AIDS 2007;18:482–485.

78
6. Pfaller MA, Jones RN, Messer SA, Edmond MB, Wenzel RP. National
surveillance of nosocomial blood stream infection due to species of
Candida other than Candida albicans: frequency of occurrence and
antifungal susceptibility in the SCOPE Program. Diagn Microbiol Infect Dis
1998;30:121–129.
7. Pfaller MA, Jones RN, Messer SA, Edmond M, Wenzel RP. National
surveillance of nosocomial blood stream infection due to Candida albicans:
Frequency of Occurrence and Antifungal Susceptibility in the SCOPE
Program. Diagn Microbiol Infect Dis 1998;31:327–332.
8. Pfaller M, Jones R, Doern G, Sader H, Hollis R, Messer S, et al.
International surveillance of bloodstream infections due to Candida
species: frequency of occurrence and antifungal susceptibilities of isolates
collected in 1997 in the United States, Canada, and South America for the
SENTRY program. J Clin Microbiol 1998;36:1886-1889.
9. Sandven P, Bevanger L, Digranes A, et al. Candidemia in Norway
(1991 to 2003): results from a nationwide study. J Clin Microbiol
2006;44:1977-1981.
10. Krcmery Jr V, Kovacicová G, others. Longitudinal 10-year
prospective survey of fungaemia in Slovak Republic: trends in etiology in
310 episodes. Slovak Fungaemia study group. Diagn Microbiol Infect Dis
2000;36:7-11.
11. Chakravart S, Haleagraha N. A Comprehensive review of the
occurrence and management of systemic candidiasis as an opportunistic
infection. Microbiol J 2011;1:1–7.
12. Pfaller MA. Nosocomial candidiasis: emerging species, reservoirs,
and modes of transmission. Clin Infect Dis 1996;22:89-94.
13. Abi-Said D, Anaissie E, Uzun O, et al. The epidemiology of
hematogenous candidiasis caused by different Candida species. Clin Infect
Dis 1997;24:1122-1128.
14. Arendrup MC, Fuursted K, Gahrn-Hansen B, et al. Seminational
surveillance of fungemia in Denmark: notably high rates of fungemia and
numbers of isolates with reduced azole susceptibility. J Clin Microbiol
2005;43:4434-4440.
15. Colombo AL, Guimarães T, Silva LRBF, et al. Prospective
observational study of candidemia in Sao Paulo, Brazil: incidence rate,
epidemiology, and predictors of mortality. Infect Control Hosp Epidemiol

79
2007;28:570–576.
16. Hinrichsen SL, Falcão É, Vilella TAS, et al. Candida isolates in
tertiary hospitals in northeastern Brazil. Brazil J Microbiol 2009;40:325–
328.
17. Tamura NK, Negri MFN, Bonassoli LA, Svidzinski TIE. Virulence
factors for Candida spp recovered from intravascular catheters and
hospital workers’ hands(portuguese). Rev Soc Bras Med Trop.
2007;40:91–93.
18. Klotz SA, Lipke PN. The perfect adhesive. Proteins 2010;16:18.
19. Donlan RM, Costerton JW. Biofilms: survival mechanisms of clinically
relevant microorganisms. Clin Microbiol Rev 2002;15:167-193.
20. Douglas LJ. Candida biofilms and their role in infection. Trends
Microbiol 2003;11:30–36.
21. King RD, Lee JC, Morris AL. Adherence of Candida albicans and other
Candida species to mucosal epithelial cells. Infect Immun 1980;27:667-
674.
22. Baillie GS, Douglas LJ. Matrix polymers of Candida biofilms and their
possible role in biofilm resistance to antifungal agents. J Antimicrob
Chemother 2000;46:397–403.
22. Chandra J, Kuhn DM, Mukherjee PK, Hoyer LL, McCormick T,
Ghannoum MA. Biofilm formation by the fungal pathogen Candida
albicans: development, architecture, and drug resistance. J Bacteriol
2001;183:5385–5394.
23. Jin Y, Yip HK, Samaranayake YH, Yau JH, Samaranayake LP. Biofilm-
forming ability of Candida albicans is unlikely to contribute to high levels
of oral yeast carriage in cases of human immunodeficiency virus infection.
J Clin Microbiol 2003;41:2961-2967.
24. Ramage G, Martínez JP, López-Ribot JL. Candida biofilms on
implanted biomaterials: a clinically significant problem. FEMS Yeast Res
2006;6:979–986.
25. Shin JH, Kee SJ, Shin MG, et al. Biofilm Production by isolates of
Candida species recovered from nonneutropenic patients: comparison of
bloodstream isolates with isolates from other sources. J Clin Microbiol
2002;40:1244–1248.

80
27. ain N, Kohli R, Cook E, et al. Biofilm formation by and antifungal
susceptibility of Candida isolates from urine. Appl Environ Microbiol
2007;73:1693-1703.
27. Bizerra FC, Nakamura CV, Poersch C, et al. Characteristics of biofilm
formation by Candida tropicalis and antifungal resistance. FEMS Yeast Res
2008;8:442–450.
28. Lima-Neto RG, Beltrão EIC, Oliveira PC, Neves RP. Adherence of
Candida albicans and Candida parapsilosis to epithelial cells correlates
with fungal cell surface carbohydrates. Mycoses 2011;54:23-29.
30. Biasoli MS, Tosello ME, Magaró HM. Adherence of Candida strains
isolated from the human gastrointestinal tract. Mycoses. 2002;45:465–
469.
31. National Committee for Clinical Laboratory Standards (NCCLS).
Reference Methods for Broth Dilution Antifungal Susceptibility Testing of
Yeast; Approved Standard. Wayne, PA: NCCLS; 2002; 22:1-51. Document
M27-A2 .
32. Viana IMG, Sauaia N, Sauaia BA, Monteiro SG, Figueiredo PMS. Epidemiological profile of patients with positive sputum smears for
pulmonary tuberculosis (portuguese). Rev Soc Bras Clin Med 2010;8:505-
508.
33. Cravioto A, Gross RJ, Scotland SM, Rowe B. An adhesive factor
found in strains of Escherichia coli belonging to the traditional infantile
enteropathogenic serotypes. Curr Microbiol 1979; 3:95–99.
34. Enache-Angoulvant A, Hennequin C. Invasive Saccharomyces
infection: a comprehensive review. Clin Infect Dis 2005;41:1559-1568.
33. Muñoz P, Bouza E, Cuenca-Estrella M, et al. Saccharomyces
cerevisiae fungemia: an emerging infectious disease. Clin Infect Dis
2005;40:1625-1634.
34. Chaffin WL, Lopez-Ribot JL, Casanova M, Gozalbo D, Martinez JP.
Cell wall and secreted proteins of Candida albicans: identification,
function, and expression. Microbiol Mol Biol Rev 1998; 62:130-180.
37. Cannon R, Chaffin W. Oral colonization by Candida albicans. CROBM
1999;10:359–383.
36. Jabra-Rizk MA, Ferreira SMS, Sabet M, et al. Recovery of Candida
dubliniensis and other yeasts from human immunodeficiency virus-

81
associated periodontal lesions. J Clin Microbiol 2001;39:4520-4522.
37. De Repentigny L, Aumont F, Bernard K, Belhumeur P.
Characterization of binding of Candida albicans to small intestinal mucin
and its role in adherence to mucosal epithelial cells. Infect Immun 2000;
68:3172-3179.
38. Busscher H, Geertsema-Doornbusch G, Van der Mei H. Adhesion to
silicone rubber of yeasts and bacteria isolated from voice prostheses:
influence of salivary conditioning films. J Biomed Mater Res 1997;
34:201–209.
41. Tamura N, Gasparetto A, Svidzinski T. Evaluation of the adherence
of Candida species to urinary catheters. Mycopathologia 2003; 156:269–
272.
40. Moreno SMA, Vergani CE, Sanitá PV,et al. Adhesion of different
species of candida to an experimental hard chairside reline resin
(portuguese). Rev Odontol UNESP 2009; 38:89.
43. Minagi S, Miyake Y, Inagaki K, Tsuru H, Suginaka H. Hydrophobic
interaction in Candida albicans and Candida tropicalis adherence to
various denture base resin materials. Infect Immun 1985;47:11-14.
44. Silva EH, Ruiz LS, Matsumoto FE, et al. Candiduria in a public
hospital of São Paulo (1999-2004): characteristics of the yeast isolates.
Rev Inst Med Trop S Paulo 2007;49:349–353.
42. Weinberger M, Leibovici L, Perez S, et al. Characteristics of
candidaemia with Candida-albicans compared with non-albicans Candida
species and predictors of mortality. J Hosp Infect 2005;61:146–154.
43. Nucci M, Colombo AL. Candidemia due to Candida tropicalis: clinical,
epidemiologic, and microbiologic characteristics of 188 episodes occurring
in tertiary care hospitals. Diagn Microbiol Infect Dis 2007;58:77–82.
47. Rho J, Shin JH, Song JW, et al. Molecular investigation of two
consecutive nosocomial clusters of Candida tropicalis candiduria using
pulsed-field gel electrophoresis. J Microbiol 2004;42:80–86.
45. Cheng MF, Yang YL, Yao TJ, et al. Risk factors for fatal candidemia
caused by Candida albicans and non-albicans Candida species. BMC Infect
Dis 2005;5:22.
46. Bassetti M, Righi E, Costa A, et al. Epidemiological trends in
nosocomial candidemia in intensive care. BMC Infect Dis 2006;6:21.

82
50. Costa KRC. Aspectos fenotípicos e moleculares da adesão e
atividade enzimática de Candida sp isoladas de pacientes com sinais
clínicos de candidíase oral. Tese de Doutorado, Universidade de São
Paulo; 2009.
51. Branchini ML, Pfaller MA, Rhine-Chalberg J, Frempong T, Isenberg
HD. Genotypic variation and slime production among blood and catheter
isolates of Candida parapsilosis. J Clin Microbiol 1994; 32:452–456.
48. Nascimento ACMOB. Susceptibilidade antifúngica, produção de
biofilme e caracterização molecular do gene ALS3 em isolados de Candida
albicans e não albicans do Hospital das Clínicas. Dissertação de Mestrado,
UNESP, 2009.
49. Plouffe JF, Brown DG, Silva Jr J, et al. Nosocomial outbreak of
Candida parapsilosis fungemia related to intravenous infusions. Arch
Intern Med 1977; 137:1686-1689.
54. Solomon SL, Khabbaz RF, Parker RH, et al. An outbreak of Candida
parapsilosis bloodstream infections in patients receiving parenteral
nutrition. J Infect Dis 1984;149:98-102.
51. O’Day DM, Head WS, Robinson RD. An outbreak of Candida
parapsilosis endophthalmitis: analysis of strains by enzyme profile and
antifungal susceptibility. Br J Ophthalmol 1987;71:126–129.
52. Weems Jr JJ, Chamberland ME, Ward J, et al. Candida parapsilosis
fungemia associated with parenteral nutrition and contaminated blood
pressure transducers. J Clin Microbiol 1987; 25:1029–1032.
57. Weems JJ. Candida parapsilosis: epidemiology, pathogenicity,
clinical manifestations, and antimicrobial susceptibility. Clin Infect Dis
1992; 14:756-766.
54. Critchley IA, Douglas LJ. Differential adhesion of pathogenic Candida
species to epithelial and inert surfaces. FEMS Microbiol Lett 1985;28:199–
203.
55. Bonassoli LA, Bertoli M, Svidzinski TIE. High frequency of Candida
parapsilosis on the hands of healthy hosts. J Hosp Infect 2005;59:159–
162.
60. Pfaller MA, Messer SA, Hollis RJ. Variations in DNA subtype,
antifungal susceptibility, and slime production among clinical isolates of
Candida parapsilosis. Diagn Microbiol Infect Dis. 1995;21:9–14.

83
61. Scaletsky IC, Silva ML, Trabulsi LR. Distinctive patterns of adherence
of enteropathogenic Escherichia coli to HeLa cells. Infect Immun
1984;45:534-536.
58. Rauceo JM, De Armond R, Otoo H, et al. Threonine-rich repeats
increase fibronectin binding in the Candida albicans adhesin Als5p.
Eukaryot Cell 2006;5:1664–1673.
59. Hoyer LL. The ALS gene family of Candida albicans. Trends Microbiol
2001;9:176–180.
64. Yang YL. Virulence factors of Candida species. J Microbiol Immunol
Infect 2003;36:223–228.
61. Gow NAR, Brown AJP, Odds FC. Fungal morphogenesis and host
invasion. Curr Opin Microbiol 2002;5:366–371.
62. Ribeiro MA, Miranda AE, Gambale W, Rodrigues CP. Prevalence and
exoenzyme secretion by Candida albicans isolates from oral and vaginal
mucosas of HIV-infected women. Mycopathologia 2004 ;157:255–261.
67. Haynes K. Virulence in Candida species. Trends Microbiol
2001;9:591–596.
64. Álvares CA, Svidzinski TIE, Consolaro MEL. Vulvovaginal candidiasis:
susceptibility factors of the host and virulence of the yeasts (portuguese).
J Bras Patol Med Lab 2007;43:319–327.
65. Soll DR. Candida commensalism and virulence: the evolution of
phenotypic plasticity. Acta Trop 2002; 81:101–110.
70. Tumbarello M, Posteraro B, Trecarichi EM, et al. Biofilm production
by Candida species and inadequate antifungal therapy as predictors of
mortality for patients with candidemia. J Clin Microbiology 2007;45:1843-
1850.
71. Gasparetto A, Negri MFN, Paula CR, Svidzinski TIE. Biofilm
production by yeasts isolated from bucal cavity of prosthesis wearers. Acta
Sci Health Sci 2008; 27:37–40.
72. Weinstein RA, Hota B. Contamination, disinfection, and cross-
colonization: are hospital surfaces reservoirs for nosocomial infection? Clin
Infect Dis 2004;39:1182-1189.
73. Hawser SP, Douglas LJ. Resistance of Candida albicans biofilms to
antifungal agents in vitro. Antimicrob Agents Chemother 1995;39:2128-

84
2131.
74. Uppuluri P, Dinakaran H, Thomas DP, Chaturvedi AK, Lopez-Ribot JL.
Characteristics of Candida albicans biofilms grown in a synthetic urine
medium. J Clin Microbiol 2009;47:4078-4083.
75. Patterson JE, Andriole VT. Bacterial urinary tract infections in
diabetes. Infect Dis Clin North Am 1997;11:735-750.
76. Silva S, Henriques M, Martins A, et al. Biofilms of non-Candida
albicans Candida species: quantification, structure and matrix
composition. Med Mycol 2009;47:681–689.
77. Hasan F, Xess I, Wang X, Jain N, Fries BC. Biofilm formation in
clinical Candida isolates and its association with virulence. Microbes Infect
2009;11:753–761.
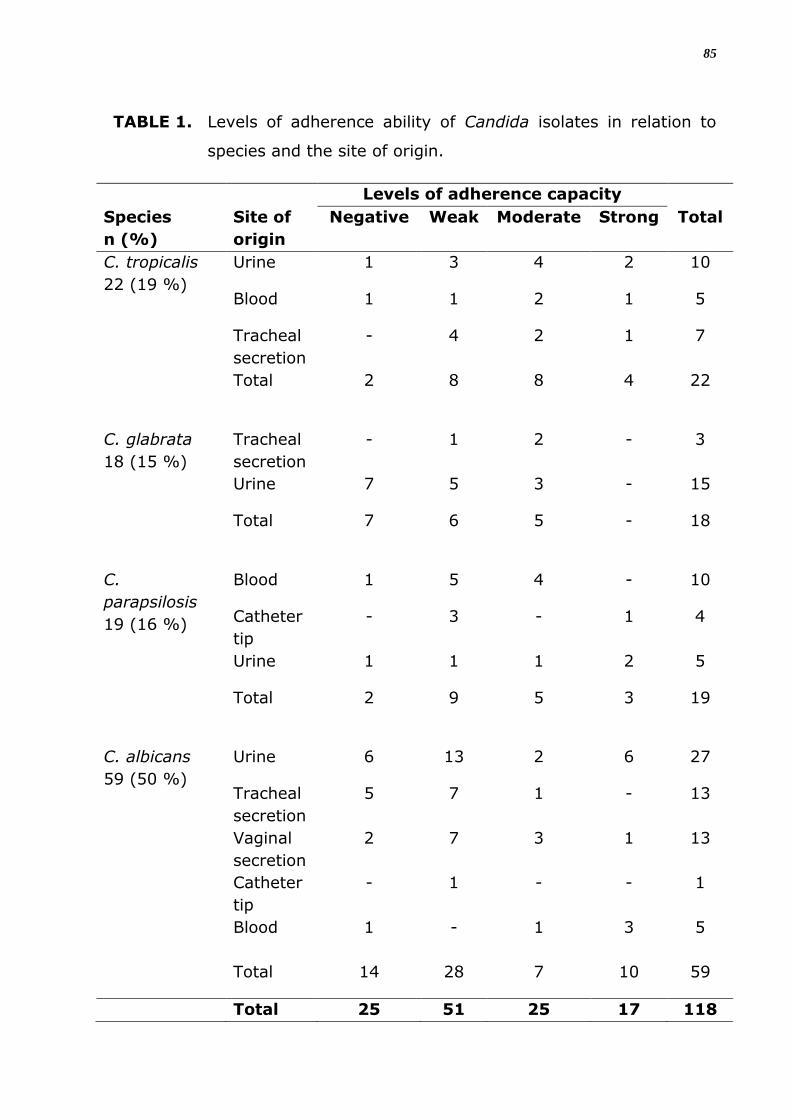
85
TABLE 1. Levels of adherence ability of Candida isolates in relation to
species and the site of origin.
Levels of adherence capacity
Species
n (%)
Site of
origin
Negative Weak Moderate Strong Total
C. tropicalis
22 (19 %)
Urine 1 3 4 2 10
Blood 1 1 2 1 5
Tracheal
secretion
- 4 2 1 7
Total 2 8 8 4 22
C. glabrata
18 (15 %)
Tracheal
secretion
- 1 2 - 3
Urine 7 5 3 - 15
Total 7 6 5 - 18
C.
parapsilosis
19 (16 %)
Blood 1 5 4 - 10
Catheter
tip
- 3 - 1 4
Urine 1 1 1 2 5
Total 2 9 5 3 19
C. albicans
59 (50 %)
Urine 6 13 2 6 27
Tracheal
secretion
5 7 1 - 13
Vaginal
secretion
2 7 3 1 13
Catheter
tip
- 1 - - 1
Blood 1 - 1 3 5
Total 14 28 7 10 59
Total 25 51 25 17 118
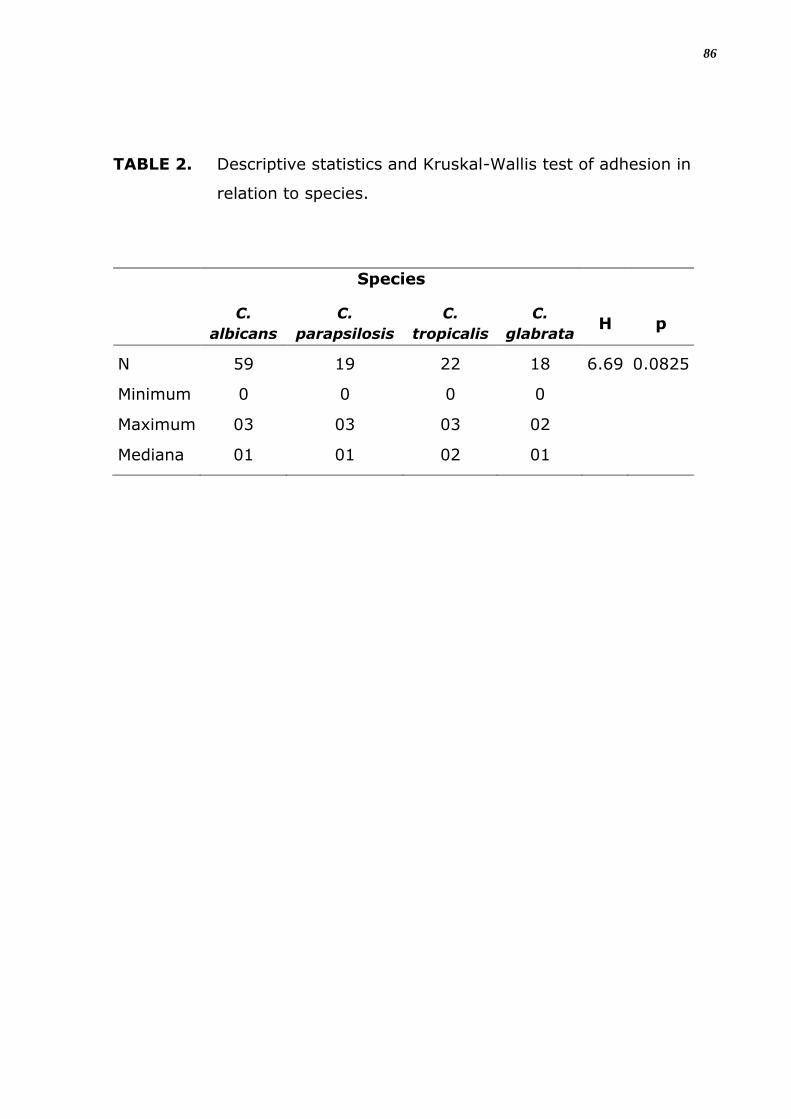
86
TABLE 2. Descriptive statistics and Kruskal-Wallis test of adhesion in
relation to species.
Species
C.
albicans
C.
parapsilosis
C.
tropicalis
C.
glabrata H p
N 59 19 22 18 6.69 0.0825
Minimum 0 0 0 0
Maximum 03 03 03 02
Mediana 01 01 02 01
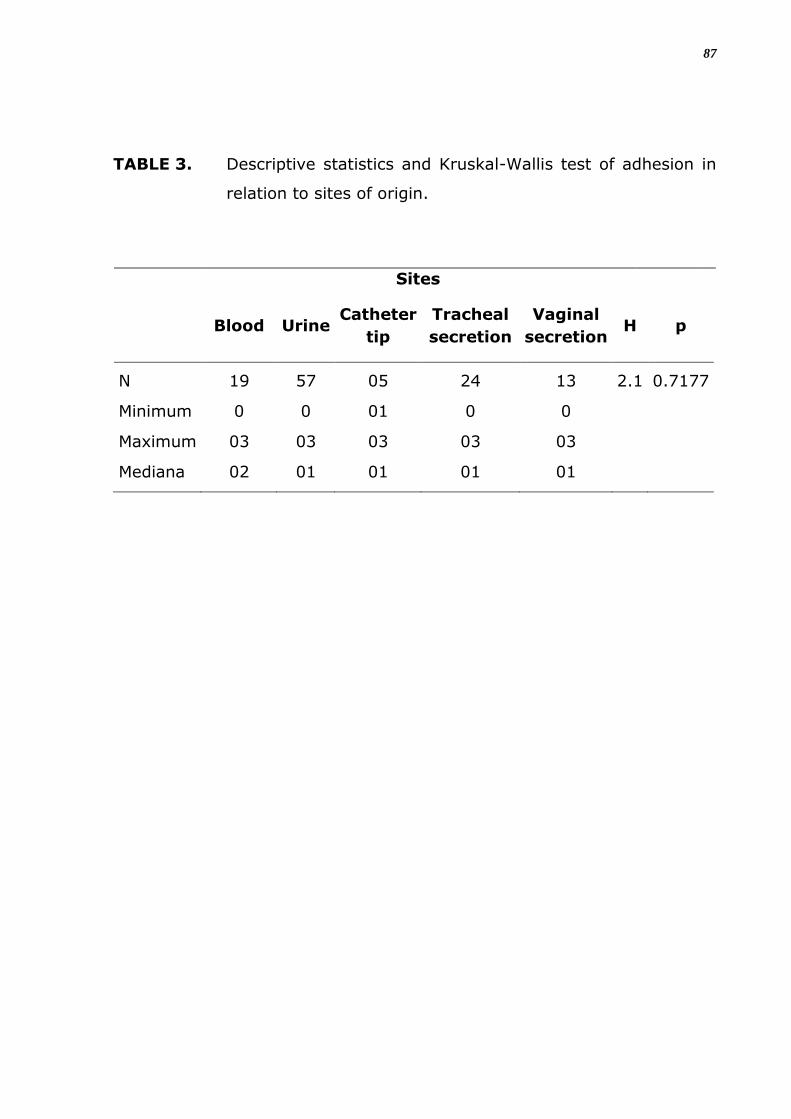
87
TABLE 3. Descriptive statistics and Kruskal-Wallis test of adhesion in
relation to sites of origin.
Sites
Blood Urine
Catheter
tip
Tracheal
secretion
Vaginal
secretion H p
N 19 57 05 24 13 2.1 0.7177
Minimum 0 0 01 0 0
Maximum 03 03 03 03 03
Mediana 02 01 01 01 01
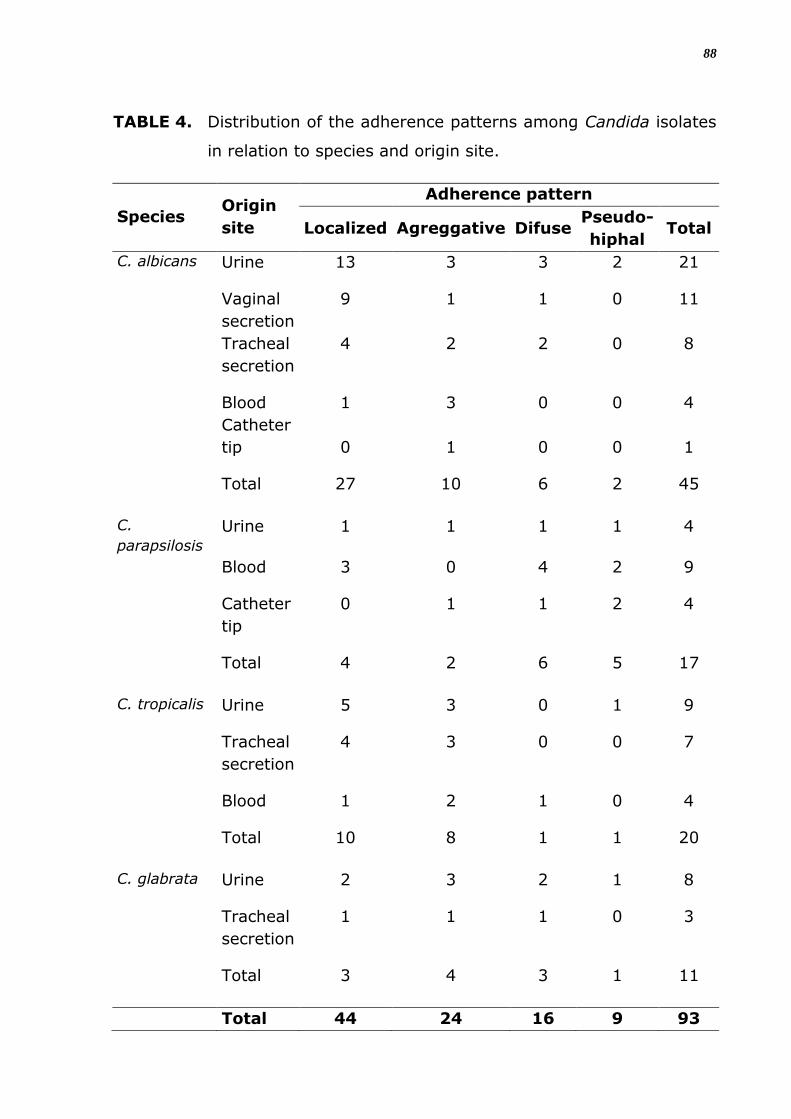
88
TABLE 4. Distribution of the adherence patterns among Candida isolates
in relation to species and origin site.
Species Origin
site
Adherence pattern
Localized Agreggative Difuse Pseudo-
hiphal Total
C. albicans Urine 13 3 3 2 21
Vaginal
secretion
9 1 1 0 11
Tracheal
secretion
4 2 2 0 8
Blood 1 3 0 0 4
Catheter
tip 0 1 0 0 1
Total 27 10 6 2 45
C.
parapsilosis
Urine 1 1 1 1 4
Blood 3 0 4 2 9
Catheter
tip
0 1 1 2 4
Total 4 2 6 5 17
C. tropicalis Urine 5 3 0 1 9
Tracheal
secretion
4 3 0 0 7
Blood 1 2 1 0 4
Total 10 8 1 1 20
C. glabrata Urine 2 3 2 1 8
Tracheal
secretion
1 1 1 0 3
Total 3 4 3 1 11
Total 44 24 16 9 93
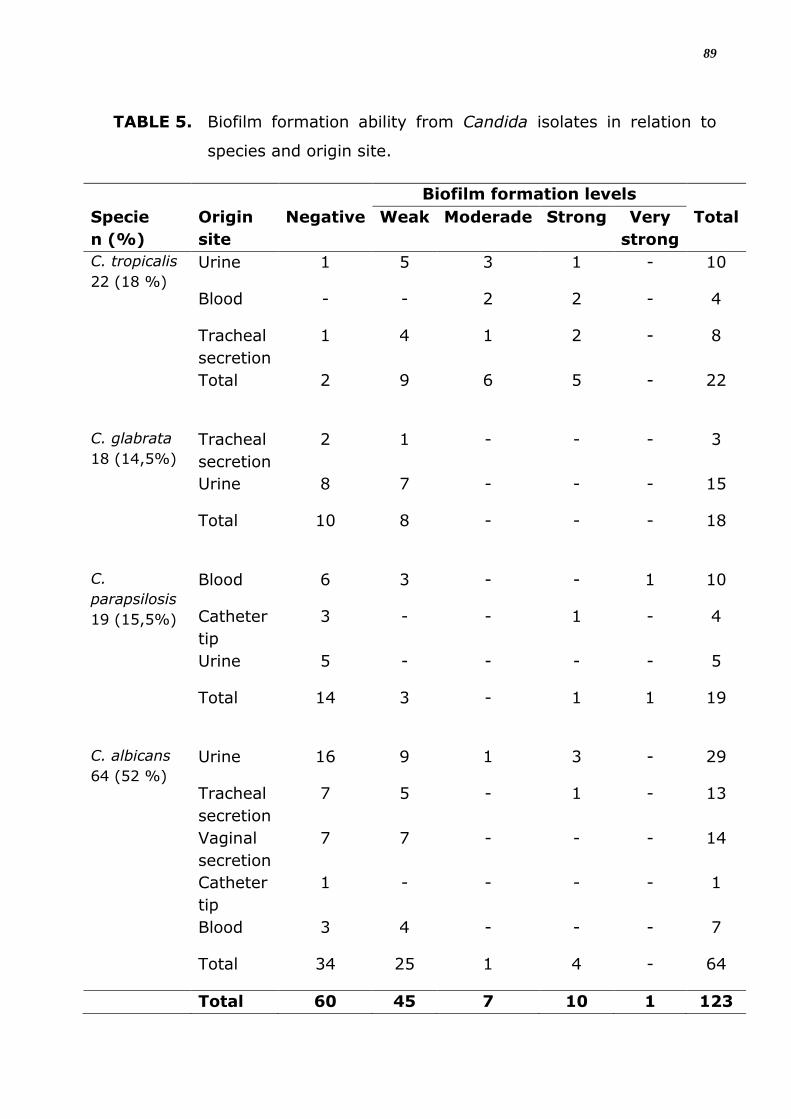
89
TABLE 5. Biofilm formation ability from Candida isolates in relation to
species and origin site.
Biofilm formation levels
Specie
n (%)
Origin
site
Negative Weak Moderade Strong Very
strong
Total
C. tropicalis
22 (18 %)
Urine 1 5 3 1 - 10
Blood - - 2 2 - 4
Tracheal
secretion
1 4 1 2 - 8
Total 2 9 6 5 - 22
C. glabrata
18 (14,5%)
Tracheal
secretion
2 1 - - - 3
Urine 8 7 - - - 15
Total 10 8 - - - 18
C.
parapsilosis
19 (15,5%)
Blood 6 3 - - 1 10
Catheter
tip
3 - - 1 - 4
Urine 5 - - - - 5
Total 14 3 - 1 1 19
C. albicans
64 (52 %)
Urine 16 9 1 3 - 29
Tracheal
secretion
7 5 - 1 - 13
Vaginal
secretion
7 7 - - - 14
Catheter
tip
1 - - - - 1
Blood 3 4 - - - 7
Total 34 25 1 4 - 64
Total 60 45 7 10 1 123
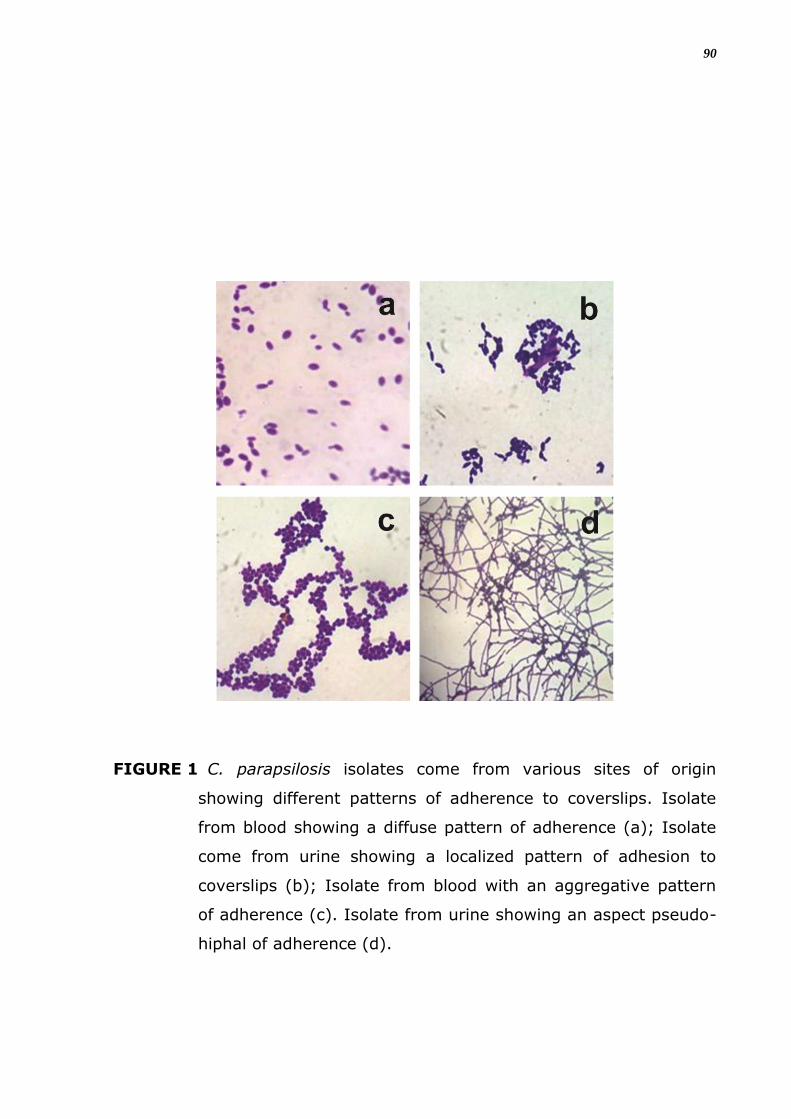
90
FIGURE 1 C. parapsilosis isolates come from various sites of origin
showing different patterns of adherence to coverslips. Isolate
from blood showing a diffuse pattern of adherence (a); Isolate
come from urine showing a localized pattern of adhesion to
coverslips (b); Isolate from blood with an aggregative pattern
of adherence (c). Isolate from urine showing an aspect pseudo-
hiphal of adherence (d).
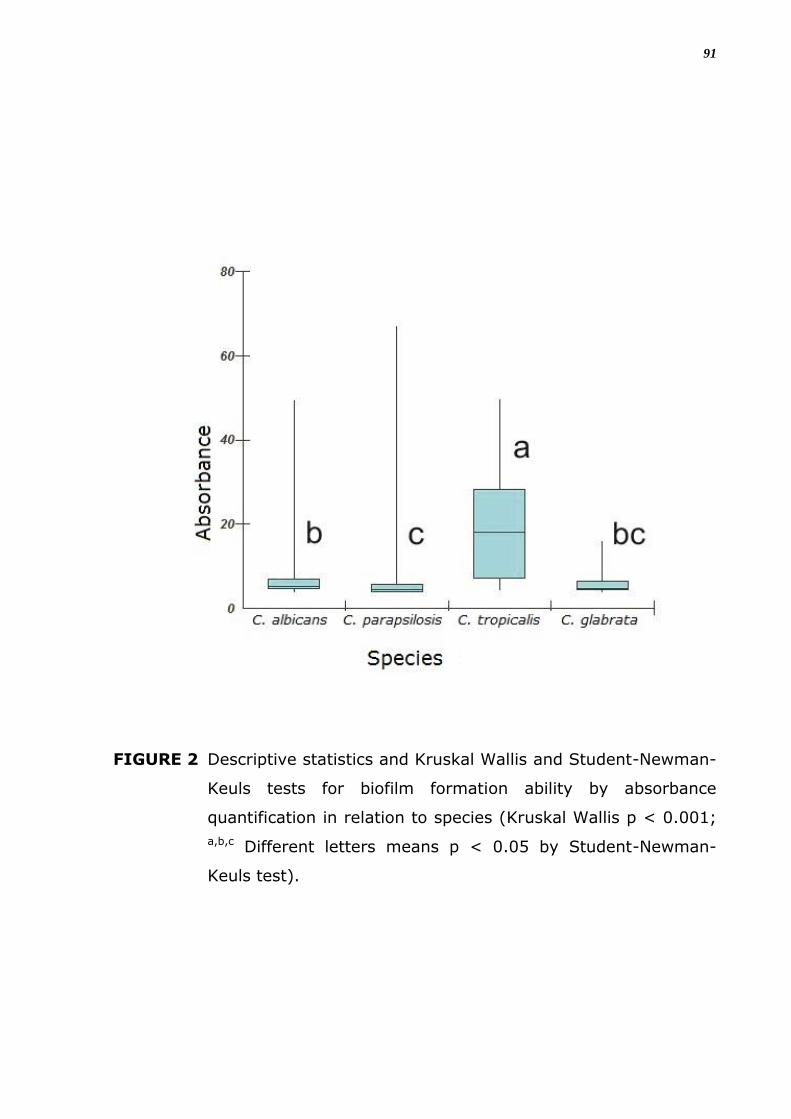
91
FIGURE 2 Descriptive statistics and Kruskal Wallis and Student-Newman-
Keuls tests for biofilm formation ability by absorbance
quantification in relation to species (Kruskal Wallis p < 0.001;
a,b,c Different letters means p < 0.05 by Student-Newman-
Keuls test).
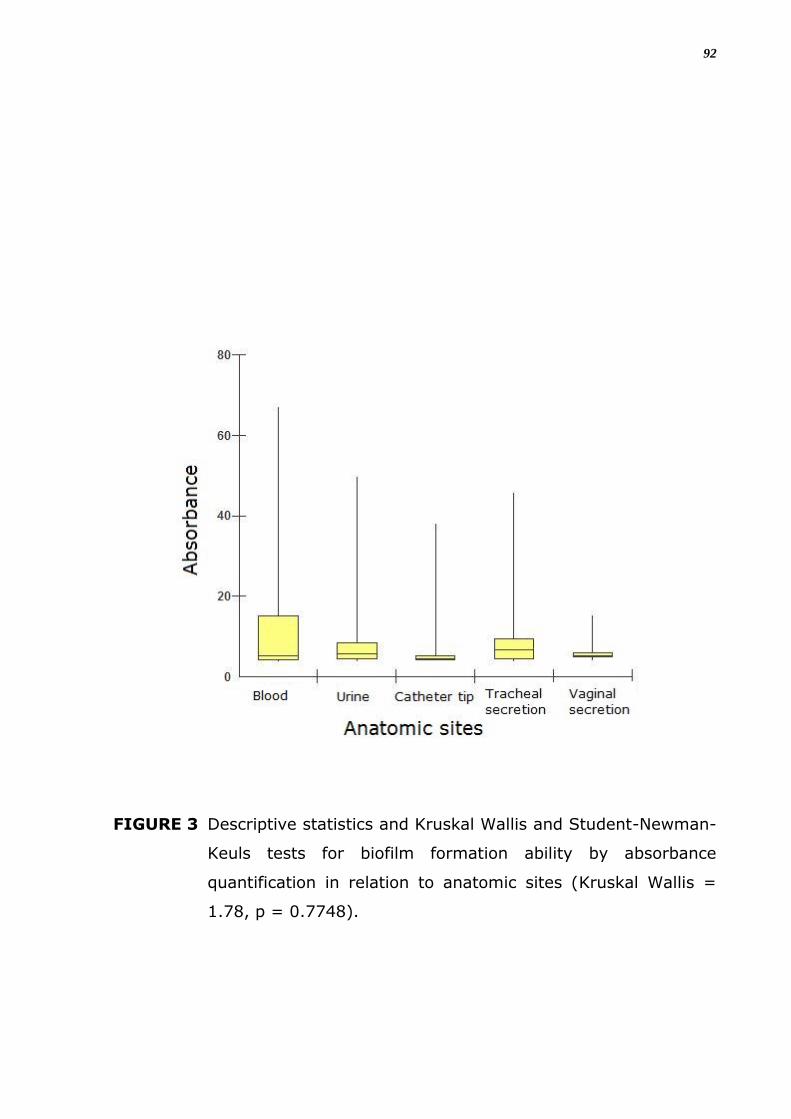
92
FIGURE 3 Descriptive statistics and Kruskal Wallis and Student-Newman-
Keuls tests for biofilm formation ability by absorbance
quantification in relation to anatomic sites (Kruskal Wallis =
1.78, p = 0.7748).

93
ANEXO A – Confirmação de Submissão do Artigo Científico
Preview
From: [email protected]
To:
[email protected], [email protected], [email protected], [email protected],
[email protected], [email protected],
[email protected], [email protected]
CC:
Subject: Manuscript has been submitted - TMMY-0092-2012
Body: @@date to be populated upon sending@@
Dear Menezes, Viviane Menezes de; Vale, Iven Neylla
Farias; Menezes, Valéria Jesus Menezes de; Monteiro, Silvio Gomes; Gonçalves, Luís Henrique Bastos;
Figueredo, Patricia Maria Silva; Monteiro, Cristina Andrade
You have been listed as a co-author on a manuscript
submitted to Medical Mycology. The manuscript title is "Adherence ability and biofilm production by clinical
isolates of Candida species coming from different sources". If you were not involved in the production of
this manuscript, please contact the editorial office at
[email protected]. Alternatively, if you are a co-author on this paper you do not have to take any further
action.
Thank you for your attention to this matter.
Sincerely,
Editor-in-Chief Medical Mycology
Date Sent: 23-Mar-2012

94
ANEXO B – Normas da Publicação da Medical Mycology
Official journal for The International Society for Human and
Animal Mycology (ISHAM) 2010 Impact Factor: 2.329
5-year Impact Factor: 2.284 Ranking: 5/22 (Mycology), 10/145 (Vet sciences)
© Thomson Reuters, Journal Citation Reports® 2011
Read More:
http://informahealthcare.com/page/Description?journalCode=mmy#Instructions
Instructions for Authors
The official publication of the International Society for Human and Animal Mycology (ISHAM), Medical Mycology, is an international journal, which
focuses on original and innovative studies of all aspects of medical, veterinary and environmental mycology. The topics include, but are not
limited to mycological, biochemical and molecular investigations of etiological agents of mycoses; aspects of pathogenesis, immunology, and
epidemiology of mycotic diseases; case reports of unusual medical or
veterinary fungal infections; laboratory approaches to the identification of fungal pathogens, antifungal therapy and prophylaxis; mode of action,
pharmacokinetics and assessments of new antifungal agents; investigations of the mycological aspects of the indoor environment, with
a focus on human and animal health. The aim of the journal is to present the best scientific reports from around the world and in so doing to
provide a comprehensive reference base for medical mycologists, microbiologists, clinicians, and environmental specialists.
Types of Papers Considered for Publication
1. Types of Papers
1.1.Original Papers. Manuscripts must have: (a) a Cover-Page which includes the full title and a short title, name(s) and affiliation(s) of all
author(s), full postal address, telephone and fax numbers, and email
address for the corresponding author and a list of up to five keywords; (b) a separate page for a Summary of no more than 200 words prepared
without sub headers; (c) text consisting of Introduction, Materials & Methods, Results, Discussion and References; (d) References must be
numbered sequentially in the order in which they first appear in the text, the numbers cited in the text within non-superscript, square brackets,
e.g., [1] and included in the same numerical order in the Reference section; (e) each Table should be submitted as a separate file (page) and
should have footnoted descriptions of all abbreviations contained in the

95
Table (see section below on Tables for more detailed information); (f) each Figure should be submitted as a separate file (page), with
appropriate figure legends allowing a reader to understand their contents without reference to the text (see section below on Figures for more
detailed information). Manuscripts MUST be in English, double-spaced, in no less than size 12 font, without line numbering and no more than 10
print pages (approximately 25 manuscript pages), including cover-page, abstract, text, references, and tables/figures.
1.2 Reviews. Authors must first electronically submit an outline of their proposed article for evaluation by the journal. The outline should be no
more than two, double-spaced pages in 12 font, in which the authors describe the objectives and contents of the report. The outline must be
submitted to the Reviews Editor, Wiley Schell: [email protected]. Once the proposal has been evaluated, the authors will be informed of the
results of the Review Editor’s initial consideration of their proposal. Reviews are NOT restricted by the formatting or length requirements of
Original Papers. NOTE - the journal does NOT accept uncommisioned reviews for publication.
1.3 Case Reports. Such articles must make a distinct, novel contribution
to the understanding of the etiologic agents, its clinical manifestations, and/or its treatment. They should NOT be based merely on the first
incidence of a known cosmopolitan or widely distributed etiologic agent in
the nation of the authors’ residence. This type of paper MUST include mycological information, e.g., the methods used to recover the etiologic
agent in culture, the procedures used in its identification, etc., as well as the clinical features of the patient. Reports describing new etiologic agents
should be substantiated by living cultures of the isolates deposited in an internationally recognized professional culture collection (i.e. one that has
full-time staff dedicated to receiving, preserving and shipping of cultures). In addition, it is recommended that discussions of unusual or rare fungal
pathogens be supported by the deposition of the organisms in recognized fungal collections. Alternatively, species identification may be supported
by the sequencing of recognized genes using appropriate primers and subsequent comparison of the obtained sequences with those in GenBank
or other recognized molecular databases. Nucleotide and/or amino acid sequence data should be deposited in Genbank or Swiss-Prot and the
accession numbers included in the Material and Methods section or in
figures of phylogenetic trees. Manuscripts MUST be in English and prepared as is an original article, EXCEPT that each should consist of a
Cover page, Abstract, Introduction, Case Report, Discussion and References. These papers should be no more than 10 print pages
(approximately 25 manuscript pages).
1.4 Short Communications. These manuscripts are to provide an opportunity for the presentation of preliminary or brief observations that
do not warrant an original paper. The manuscript should be prepared as is

96
an original paper, except they may be no more than 5 print pages (approximately 12 manuscript pages) including cover-page, abstract, text,
references, and tables/figures.
1.5 Letters to the Editor. Letters are to allow readers the opportunity to discuss issues related to previous published original articles, case reports
or short communications and should not be used for the presentation of the authors’ preliminary data from their own investigations. Letters should
be no more than 5 double-spaced manuscript pages (approximately 2
print pages) in not less than size 12 font, including references.
2. Submission of Manuscripts All submissions MUST be uploaded as Word files to the Medical Mycology
ScholarOne Manuscript site at mc.manuscriptcentral.com/tmmy. Users who have not previously submitted work through this site must create an
account from the link on the login page. Assistance with this and all other areas of the site is available in the User Guide, which is accessed via the
‘Get Help Now’ button at the top right corner of all ScholarOne Manuscript web pages.
Manuscript Structure
1. Preparation of Manuscripts
As indicated, manuscripts MUST be in English, double-spaced, in no less
than size 12 font, organized as appropriate for the manuscript type and have one (1) inch margins all around. Statistics and measurements
should be given in numerals when followed by a unit, e.g., mg, ml, etc. In contrast, numerals employed in other circumstance, e.g. two
patients, should be spelled out. All abbreviations, even those commonly used in medical mycology, e.g., Sabouraud glucose agar (SGA) must be
defined when first used in the abstract and text.
2. References In preparing your citations please note that software programs available
to format references DO NOT presently contain an option for Medical Mycology or for the use of the Vancouver system. Consequently, such
programs should be used cautiously and should be carefully reviewed by the authors to ensure that citations meet the style/format requirements of
the journal. References must be numbered sequentially in the order in
which they first appear in the text, the numbers cited in the text within square brackets, i.e., [1] and included in the same numerical order in the
Reference section. Citation numbers MUST not be indicated in superscript. Articles with six (6) or more authors are to be cited by the
names of the first three (3) authors, with the remaining authors included by et al. Journal names must be abbreviated, in italic font, without the use
of periods. The volume number must be in bold font, issue numbers should not be included and all page numbers must appear for each
citation (e.g. 105-107, NOT 105-7).

97
2.1. Journal article Author AB, Author CD, Author EF, et al. [if six or more] Title of paper. J
Title Abbrev 1995; 00: 000-000. If the title is in a language other than English, the authors should provide the English translation and the name
of the original language within a bracket following the citation.
2.2. Book chapter Author AB, Author CD, Author EF, et al. [if six or more] Chapter title. In:
Editor AB, Editor CD, eds. Book Title With Initial Uppercase Letters (as
indicated here and in italic font), 5th edn. Place: Publisher, 1995: 000-000.
2.3. Book
Author AB, Author CD (eds). Book Title With Initial Uppercase Letters (as indicated here and in italic font), 5th edn. Place: Publisher, 1995.
2.4. Thesis
Author AB. Paper title with lowercase initials to all words. PhD thesis, University, 1995.
2.5. Conference proceedings/Supplements
Author AB. Paper title. In: Editor AB, ed. Proceedings Title, place, date. Place: publisher, 1995: 000-000.
2.6. Meeting abstracts Abstracts of works presented at scientific meetings should be cited in
brackets within the text of the manuscripts rather than in the Reference sections. The citations should be formatted as: Author AB. Abstract title.
Name of meeting, its location, the date and abstract number.
2.7. Personal communications, Unpublished results etc References to personal communications, unpublished results and papers
submitted for publication (but not yet accepted) should only appear in brackets in the text, and in the following form: [A. B. Author, unpublished
results] or [C. D. Author, personal communication].
2.8. Journal article on the Internet Abood S. Quality improvement initiative in nursing homes: the ANA acts in
an advisory role. Am J Nurs [serial on the Internet]. 2002 Jun [cited 2002
Aug 12];102(6):[about3 p.]. Available from: <http://www. nursingworld.org/AJN/2002/june/Wawatch.htm>
2.9. Monograph on the Internet
Foley KM, Gelband H, editors. Improving palliative care for cancer [monograph on the Internet]. Washington: National Academy Press; 2001
[cited 2002 Jul 9]. Available from: <http://www.nap.edu/books/0309074029/html>

98
2.10. Homepage/Web site Cancer-Pain.org [homepage on the Internet]. New York: Association of
Cancer Online Resources, Inc.; c2000-01 [updated 2002 May 16; cited 2002 Jul 9]. Available from: http://www.cancer-pain.org
2.11. Part of a homepage/Web site
American Medical Association [homepage on the Internet]. Chicago: The Association; c1995-2002 [updated 2001 Aug 23; cited 2002 Aug 12]. AMA
Office of Group Practice Liaison; [about 2 screens]. Available from:
<http://www.ama-assn.org/ama/pub/category/1736. html>
2.12. Database on the Internet Open database:
Who’s Certified [database on the Internet]. Evanston (IL): The American Board of Medical Specialists. c2000-[cited 2001 Mar 8]. Available from:
<http://www.abms.org/newsearch.asp_/www.abms.org/newsearch.asp> Closed database:
Jablonski S. Online Multiple Congential Anomaly/Mental Retardation (MCA/MR) Syndromes [database on the Internet]. Bethesda (MD):
National Library of Medicine (US). c1999 [updated 2001 Nov 20; cited 2002 Aug 12]. Available from:
<http://www.nlm.nih.gov/mesh/jablonski/syndrome_title.html>
2.13. Part of a database on the Internet
MeSH Browser [database on the Internet]. Bethesda (MD): National Library of Medicine (US); 2002-[cited 2003 Jun 10]. Meta-analysis; unique
ID: D015201; [about 3 p.]. Available from: <http://www.nlm.nih.gov/mesh/MBrowser_/www.www.nlm.nih.gov/mesh
/MBrowser> Files updated weekly.
3. Tables Tables should present only essential data, must be numbered sequentially
in the order in which they first appear in the text, and all should be referred to in the text. Each table should be supplied as a separate file
and never imbedded within the body of the text. The table header should be brief and self explanatory and if it contains generic and species names,
the latter must be spelled out in full, e.g., Candida albicans, not C. albicans. Column headings should be brief and include units in
parentheses where applicable. Only horizontal rule lines should be used
and then only for headers and footers.
4. Figures/Images Images should be submitted as TIF, EPS, PDF or JPG (preferred) files.
Scanned images should be of a sufficient resolution, i.e., 300 dpi for halftones/color, 500 dpi for combination halftones and 1000–1200 dpi for
line art. Illustrations should be submitted separate from the manuscript text file, either as individual files or together in a single file. The journal
has a limited number of free colour pages within its annual page allowance

99
provided by the publisher. Authors should consult the editorial office at submission stage as they may be required to pay a fee to guarantee
colour reproduction in the print version of the journal. However, any figure submitted as a colour original may appear in colour within the
journal's on-line edition, without any charges being assessed to the authors. NO part of any image may be enhanced, removed, relocated, or
introduced after the capture of the original image unless the authors provide the editor with a clear and accurate description of the nature of
the image manipulation.
5. Nomenclature
Proposals of new fungal taxa MUST conform to the requirements of the current edition of the International Code of Botanical Nomenclature and be
supported, as indicated in the discussion of Case Report, by deposition of cultures and/or the unique nucleotide and/or amino acid sequences in
appropriate collections or databases, along with the new binomial. In addition, current binomials should appear in italics, must be spelled out in
full when first used in the abstract, text, figures/tables. Thereafter, a generic name may be abbreviated to the first initial only, e.g. Candida
albicans when first cited and C. albicans in the remainder of the abstract and text. Species names may NEVER be used without the generic full
name or abbreviation combined with the full name of the specie. For example, it must be Candida albicans or C. albicans, but never albicans, or
C. neoformans var. grubii but never var. grubii. The use of abbreviations
of generic and species names, e.g., (Ca) for C. albicans, in the text is NOT permitted.
6. Nucleotide and/or Amino Acid Sequence Data
Nucleotide and/or amino acid sequence data should be deposited and the GenBank/EMBL/DDBJ accession numbers should be included in the
Materials and Method Section of the manuscript. However, in a phylogenetic tree of the figure, the numbers should be shown along with
species names (e.g., Malassezia globosa AB099880) (No descriptions are needed in the Materials and Method Section in this case).
Required Statements
1. Humans in Research When reporting investigations involving human subjects, authors should
indicate in the manuscript if the procedures followed were in accordance
with the ethical standards of the responsible committee on human experimentation within their institutions and/or with the Helsinki
Declaration of 1975, as revised in 1983.
2. Animals in Research Manuscripts containing information related to the experimental use of
animals must clearly state that the studies complied with relevant professional and institutional animal welfare policies. Specifically, that
procedures involving animals conformed to the ILAR Guide for the Care

100
and Use of Laboratory Animals (1996 and later editions) of the Institute of Laboratory Animal Research, Commission on Life Sciences, National
Research Council (www.nap.edu/catalog/5140.html).
3. Safety Precautions Manuscripts describing the manipulation of cultures of fungal pathogens or
toxins included in the US Department of Health and Human Services and/or the US Department of Agriculture list of human or animal Select
Agents (available at www.selectagents.gov), or organisms that have been
classified as Risk Group III agents by their home country or by an international regulatory agency, MUST contain a discussion of the
biosafety procedures and laboratory containment facilities used and any additional precautions employed to ensure the safety of support
personnel.
4. Authorship Contributions Please note that you and your co-authors in submitting this manuscript
are confirming that all authors have (a) made a substantial contribution to conception and design, acquisition of data, and/or analysis and
interpretation of data, (b) participated in the drafting of the article or revising it critically for important intellectual content, (c) have read and
approved the final manuscript, AND (d) that the manuscript is not, either in whole or in part, currently under consideration by any other scientific
journal and has not been previously published in either hard copy or
electronic format. Submissions with ten (10) or more authors will only be considered if the corresponding authors provide, in their cover letters,
descriptions of each of the authors’ contributions. Manuscripts drafted or written, in whole or part, by individuals other than those indicated as
authors will NOT be considered for publication unless the contributions of these individuals are clearly and accurately presented in the
Acknowledgement section of the paper.
5. Acknowledgements The work of other individuals who assisted in the studies or in the
preparation of the submission must be specifically indicated in the Acknowledgment section of the paper.
6. Disclosures of Potential Conflicts of Interest
While there is no standard definition of Conflict of Interest, the United
States National Institutes of Health has published the following statement:
"It is the sole responsibility of authors to disclose any affiliation with any organization with a financial interest, direct or indirect, in the subject
matter or materials discussed in the manuscript (such as consultancies, employment, expert testimony, honoraria, speakers bureaus, retainers,
stock options or ownership) that may be perceived as potentially affecting the conduct or reporting of the work submitted. All sources of funding are
to be explicitly stated. If uncertain as to what might be considered a

101
potential conflict of interest, authors should err on the side of full disclosure" (i.e., when in doubt, provide full disclosure).
In addition, a conflict of interest may be perceived to exist in such
situations as when authors are affiliated with or employed by a private enterprise whose product or competitor’s product, e.g., drug, device,
diagnostic kit, is the focal point of the paper.
Information about potential conflict of interest should be clearly stated at
the point of submission. This may be made available to reviewers and may be published with the manuscript at the discretion of the Editors and
Publisher. The journal recommends that you and your co-authors include the sub header, “Conflict of Interest”, in the Acknowledgment section at
the end of your manuscript. If no such conflict exists, simply respond “None”. If you or your co-authors have received funding that meets the
criteria described in the paragraph above, the source(s) of the funds must be declared (NOTE – you are NOT required to disclose the amounts of
funding that you or your co-authors have received). Furthermore, authors must indicate their employment if describing their or other companies’
products.
The intent of this policy is not to prevent authors with these relationships from publishing work, but rather to adopt transparency such that readers
can make objective judgments on conclusions drawn.
Post Acceptance Information
1. Electronic Proofs When proofs are ready, corresponding authors will receive e-mail
notification with a password and Web address from which to download a PDF. Hard copies of proofs will NOT be mailed. To avoid delays in
publication, corrections to proofs must be returned, by fax or as an e-mail attachment, within 48 hours of receipt.
2. Offprints Corresponding authors will receive either fifty printed offprints or a PDF of
the final published version of the paper. Preferences should be stipulated when returning proofs.
3. Copyright It is a condition of publication that authors assign copyright or licence the
publication rights of their articles, including abstracts, to ISHAM. This
enables us to ensure full copyright protection and to disseminate the article, and the journal, to the widest possible readership in print and
electronic formats as appropriate. Authors may, of course, use the article elsewhere after publication without prior permission from Informa UK Ltd.,
provided that acknowledgement is given to the journal as the original source of publication, and that Informa Healthcare is notified so that
records show that its use was properly authorized. Authors retain a number of other rights under the Informa UK Ltd. rights policies
documents.

102
4. NIH Public Access Policy In consideration of the National Institutes of Health (NIH) and Wellcome
Trust Public Access Policies, Informa Healthcare acknowledges that the broad and open dissemination of NIH/Wellcome-funded research benefits
future scientific endeavour. Because we value the contribution our journals make to the body of scientific knowledge, our policies
accommodate authors who wish to submit to PubMed Central/UK PubMed Central.
1. Informa Healthcare will deposit funded work to PubMed Central/UK
PubMed Central for public access posting 12 months (NIH) or 6 months (Wellcome Trust) after final publication in electronic form.
2. 'Funded work' shall be defined as the final peer-reviewed manuscript, which was accepted for publication and that reflects any
author-agreed changes made in response to the peer review. 3. This embargo period begins upon final publication of the article in
print or electronic form (whichever is the sooner).
Read More: http://informahealthcare.com/page/Description?journalCode=mmy#Instru
ctions





























