
UNIVERSIDADE FEDERAL RURAL DA AMAZÔNIAINSTITUTO DA SAÚDE E PRODUÇÃO ANIMAL
PROGRAMA DE PÓS-GRADUAÇÃO EM SAÚDE E PRODUÇÃO ANIMAL NA AMAZÔNIA
ELLEN YASMIN EGUCHI MESQUITA
MORFOLOGIA DA PORÇÃO TUBULAR DO APARELHO DIGESTÓRIO DE PREGUIÇAS DE TRÊS DEDOS (Bradypus variegatus)
BELÉM2017

ELLEN YASMIN EGUCHI MESQUITA
MORFOLOGIA DA PORÇÃO TUBULAR DO APARELHO DIGESTÓRIO DE PREGUIÇAS DE TRÊS DEDOS (Bradypus variegatus).
Exame de qualificação apresentado para obtenção do grau de Doutor em Saúde e Produção Animal na Amazônia no. Programa de Pós-Graduação em saúde e produção animal na Amazônia. Universidade Federal Rural da Amazônia. Área de concentração: Saúde animal.
Orientador Prof. Dra. Érika Renata Branco.
BELÉM2017

ELLEN YASMIN EGUCHI MESQUITA
MORFOLOGIA DA PORÇÃO TUBULAR DO APARELHO DIGESTÓRIO DE PREGUIÇAS DE TRÊS DEDOS (Bradypus variegatus)
Exame de qualificação apresentado à Universidade Federal Rural da Amazônia, como parte das exigências do Curso de Doutorado em Saúde e Produção Animal na Amazônia, para
obtenção do título de Doutor.
Orientadora: Profa. Dra. Érika Renata Branco.
BANCA EXAMINADORA
___________________________________________________________Érika Renata Branco
Orientadora UNIVERSIDADE FEDERAL RURAL DA AMAZÔNIA - UFRA
____________________________________________________________Dra. Ana Rita Lima – 1º Examinador
UNIVERSIDADE FEDERAL RURAL DA AMAZÔNIA - UFRA
___________________________________________________________Dra.Elane Guerreiro Giese – 2º Examinador
UNIVERSIDADE FEDERAL RURAL DA AMAZÔNIA - UFRA
____________________________________________________________Dra. Fernanda Martins Hatano- 3º. Examinador
UNIVERSIDADE FEDERAL RURAL DA AMAZÔNIA - UFRA
___________________________________________________________Dra. Adriana Maciel de Castro Cardoso- 4º Examinador
UNIVERSIDADE FEDERAL RURAL DA AMAZÔNIA - UFRA

MORFOLOGIA DA PORÇÃO TUBULAR DO APARELHO DIGESTÓRIO DE PREGUIÇAS DE TRÊS DEDOS (Bradypus variegatus).
RESUMO: As adaptações para a dieta folívora arborícola de preguiças são evidentes logo na mastigação. Devido às especializações alimentares, as preguiças dispõem de uma gama de alterações morfofisiológicas no trato digestório e em consonância com o comportamento acrobático e a lateralidade realizada por esses Xenarthras, estes indivíduos representam um grupo diferenciado no que diz respeito a anatomia e fisiologia digestiva. Diante do exposto, objetivou-se investigar as características morfológicas do aparelho digestório da preguiça-comum, a fim de subsidiar o entendimento da digestão desses animais, bem como seu comportamento alimentar, e com isso auxiliar na preservação da espécie. Foram utilizados sete animais adultos, nos quais o esôfago, iniciou dorsal a laringe desviando a esquerda da traqueia, mantendo-se adjacente a mesma, ainda no antímero esquerdo, ao longo de toda a porção cervical. No tórax, percorreu o mediastino, cruzando o diafragma através do hiato esofágico, alcançando o estômago. Histologicamente, o esôfago seguiu um padrão peculiar que facilitou o entendimento do aparelho digestório do animal, sendo este com mucosa queratinizada e camada muscular estriada esquelética, apresentando musculatura lisa apenas na região transicional esôfago-estômago. Já a análise anatômica do estômago da preguiça permitiu caracterizar a divisão em sete compartimentos: saco cranial, saco central, fundo, saco caudal, divertículo e pré piloro I e II. O saco cranial caracterizou-se pela zona de inserção esofágica, sendo a primeira cavidade do estômago situada na região cranial, desviada a esquerda. Em seguida destacou-se o saco central posicionado na região mediana do corpo do animal, precedendo o saco caudal, que estava posicionado no antímero direito. O fundo era localizado na região crânio central do órgão de modo acima do divertículo, que sobrepunha os pré piloros I e II (inserção duodenal). Microscopicamente, os compartimentos revelaram dois padrões distintos: uma fração de tecido plano queratinizado estratificado baixo aglandular e uma fração glandular, de mucosa revestida por tecido epitelial glandular prismático baixo com formações de fossetas curtas e glândulas longas. Por fim concluímos até o presente momento que, a configuração única do tubo esofágico aliada a complexidade dos estômagos e comportamentos posturais pré e pós ingesta, a composição histológica e a taxa metabólica baixa inferem alguns dos comportamentos peculiares das preguiças da espécie Bradypus variegatus no que concerne também ao conhecimento do trânsito alimentar.
Palavras-chave: Esôfago, Estômago, Anatomia, Histologia, Xenarthra.

MORPHOLOGY OF TUBULAR PORTION OF DIGESTIVE TRACT OF THREE-TOED SLOTH (Bradypus variegatus)
ABSTRACT: The adaptations to the tree folivore diet of sloths are evident soon in chewing. Due to food specialties, sloths have a range of morphophysiological changes in the digestive tract and in agreement with the acrobatic behavior and laterality performed by these Xe-narthras, these individuals represent a differentiated group with respect to anatomy and diges-tive physiology. In view of the above, the objective was to investigate the morphological characteristics of the common sloth digestive apparatus, in order to support the understanding of the digestion of these animals, as well as their feeding behavior, and with that help in the preservation of the species. Seven adult animals were used, in which the esophagus started the larynx dorsal to the left of the trachea, remaining adjacent to it, still in the left antimer, along the entire cervical portion. In the thorax, it crossed the mediastinum, crossing the diaphragm through the esophageal hiatus, reaching the stomach. Histologically, the esophagus followed a peculiar pattern that facilitated the understanding of the animal's digestive apparatus, being this with keratinized mucosa and skeletal striated muscle layer, presenting smooth muscle only in the transitional esophagus-stomach region. The anatomical analysis of the stomach of laziness allowed us to characterize the division into seven compartments: cranial sac, central sac, fundus, caudal sac, diverticulum and pre pilorus I and II. The cranial sac was character-ized by the esophageal insertion zone, the first cavity of the stomach being located in the cra-nial region, deviated from the left. Next, the central bag positioned in the middle region of the animal's body was preceded by the caudal sac, which was positioned in the right antimer. The fundus was located in the central cranial region of the organ so above the diverticulum, which overlapped prepilorus I and II (duodenal insertion). Microscopically, the compartments re-vealed two distinct patterns: a flat agglutinated stratified keratinized flat tissue fraction and a glandular fraction of mucosa lined with low prismatic glandular epithelial tissue with short pits and long gland formations. Finally, we conclude that the unique configuration of the esophageal tube, combined with the complexity of the stomachs and posture behaviors before and after ingestion, the histological composition and the low metabolic rate infer some of the behaviors peculiar to the sloths of the Bradypus variegatus species, knowledge of food traffic.
Key words: Esophagus, Stomach, Anatomy, Histology, Xenarthra.

Sumário
1 INTRODUÇÃO..................................................................................................................................7
2 JUSTIFICATIVA................................................................................................................................9
3.1 OBJETIVO GERAL.......................................................................................................................10
3.2 OBJETIVOS ESPECÍFICOS.........................................................................................................10
4 CONTEXTUALIZAÇÃO.................................................................................................................11
4.1 –MORFOLOGIA DO APARELHO DIGESTÓRIO DE HERBÍVOROS......................................11
4.2 ALIMENTAÇÃO...........................................................................................................................12
REFERÊNCIAS...................................................................................................................................13
CAPITULO I: MORPHOLOGY OF THE ESOPHAGUS OF Bradypus variegatus VERSUS SWAL-LOWING MECHANISM (Submetido a Revista Ciência Animal Brasileira)......................................17
CAPITULO II – MORFOLOGIA x DINÂMICA DO ESTÔMAGO DE PREGUIÇA COMUM........28

7
1 INTRODUÇÃO
Uma das entraves que prejudicam a manutenção cativa de animais da Superordem
Xenarthra é o desconhecimento acerca da fisiologia do aparelho digestório, e para o auxílio no
entendimento, as diretrizes morfométricas são importantes no que diz respeito inclusive as
preferências alimentares (CARVALHO, 2014) .
A preguiça pode ser designada, zoologicamente, como um mamífero da classe
Eutheria, Superordem Xenarthra, e Família Bradypodidae. A ocorrência da espécie Bradypus
variegatus varia do norte de Honduras, passando pela sul da América Central. Na América, do
Sul varia desde a Colômbia até o oeste e sul da Venezuela, sul no Equador, leste do Peru e
Bolívia além de no Brasil (CHIARELLO, 2014).
A distribuição se sobrepõe com B. torquatus na parte central da Mata Atlântica
(HIRSCH e CHIARELLO, 2012). No Brasil, as espécies atualmente ocorrem em áreas
florestais da Amazônia, Mata Atlântica e possivelmente nas zonas de contato entre esses
biomas e Cerrado (CHIARELLO, 2014).
Conhecidos pela ecologia de movimentação lenta e baixa taxa metabólica, habitam
florestas e alimentam-se de folhas, ramos e brotos de várias plantas, frequentemente da
família Moraceae (CHIARELLO, 2008), Cecropiaceae e Clethraceae (URBANI & BOSQUE,
2007).
As preguiças do gênero Bradypus não se adaptam facilmente à vida em cativeiro,
pois são pouco resistentes. Quando removidas do seu habitat natural, sobrevivem por poucos
meses (CRANDALL, 1964). Preguiças de três dedos (Bradypus spp.) por serem considerados
folívoras muito estritas, se alimentam de uma gama limitada de espécies de árvores, o que as
torna mais difíceis de manter em cativeiro. Já as preguiças de dois dedos (Choloepus spp.)
costumam ter uma dieta mais ampla que inclui uma grande variedade de espécies de plantas e
são mais fáceis de manter em cativeiro (RAINES, 2005; VAUGHAN et al.., 2007).
A evolução e ecologia das preguiças são pontuadas por sua capacidade de sobreviver
na copa arbórea da região Neotropical e por supostamente apresentarem adaptações
fisiológicas e comportamentais para superar as limitações impostas por este ambiente, porém,
pouco se sabe sobre a capacidade da preguiça para adquirir nutrientes em folhas pobres
(BISCHOF et al., 2013) que compõem a maioria de sua dieta (HLADIK de 1978; SHIELS,
2006).
Foram desenvolvidos trabalhos mais recentes a respeito do conhecimento da
digestibilidade com outras espécies Xenarthras, como Myrmecophaga tridactyla (STAHL et

8
al.., 2012) e Choloepus didactylus (VENDL et al., 2015) para otimizar o manejo alimentar
desses animais, porém, ainda não há relatos no gênero Bradypus, possivelmente pelo
desconhecimento morfológico da espécie.
A incompreensão a respeito da fisiologia digestiva das preguiças do gênero
Bradypus dificulta o manejo em cativeiro desses animais, aumentando o número de
mortalidade por erros da nutrição. Este estudo baseado na morfologia visa além de contribuir
para pesquisas posteriores de digestibilidade aparente da espécie Bradypus variegatus,
fomentar o sucesso clínico quando os animais são criados na forma cativa ou semi-cativa.

9
2 JUSTIFICATIVA
Literaturas clássicas relatam a dificuldade de manutenção cativa das preguiças do
gênero Bradypus em não se adaptarem facilmente pela baixa resistência, o que leva a uma
taxa de sobrevivência diminuída, sob alegação da dificuldade de mantença das exigências
tanto nutricionais, quanto ecológicas (CRANDALL, 1964; MONTGOMERY e SUNQUIST,
1975), além da escassa literatura sobre o manejo desse gênero.
O viés discutível em trabalhos a respeito da criação em cativeiro é a dificuldade de
manutenção comparada a preguiças de dois dedos (Choloepus spp.) que costumam ter uma
dieta mais ampla e são mais fáceis de cuidar na forma cativa (RAINES, 2005; VAUGHAN et
al., 2007).
A evolução e ecologia das preguiças do gênero Bradypus tem sido pesquisada em
algumas regiões da América do Sul (URBANI e BOSQUE, 2006; RÉUS e SOUZA, 2007) na
busca do entendimento da habilidade de adaptação fisiológica e comportamental desses
animais. Especula-se que para superar as limitações impostas por este estilo de vida, as
preguiças evoluíram várias adaptações anatômicas, fisiológicas e comportamentais
especializadas (CORK e FOLEY, 1991; GILMORE et al.., 2001; PAULI et al.., 2014), como
um estômago de três compartimentos (BAUCHOP, 1978; STEVENS e HUME, 1995).
Relatos antigos sobre cativeiro apontam que esses animais são muitas vezes
alimentados com dietas que não contêm folhas, mas arroz cozido, frutas e legumes (MERRIT,
1985), com digestibilidade mais elevada. O presuposto de que processos digestivos em
preguiças assemelham-se aos de outros mamíferos herbívoros ainda permanece sem
comprovação.
Os hábitos alimentares e a conformação corporal única dos espécimes dos Xenarthra
instigam a inferir, a partir da forma do tubo digestório os processos de digestão e absorção dos
nutrientes, e fornece subsídios para pesquisas aplicadas no campo da nutrição e das
enfermidades gastrointestinais (GAUDIN; MCDONALD, 2008; ENDO et al.., 2009).
As modificações mais extremas na forma de se alimentar afetam não só as estruturas
de mastigação e digestórias, mas também o comportamento, as taxas metabólicas e as funções
locomotoras (OWEN, 1857; MACALISTER, 1875; NAPLES, 1999).
Partindo então dessa dificuldade de estabelecer parâmetros de digestão e consequente
manejo nutricional, busca-se o entendimento da fisiologia digestiva por meio da morfologia
do aparelho digestório desses animais, além de subsidiar estudos de digestibilidade aparente e
dieta ideal estabelecida para preguiças comum em cativeiro.

10
3 OBJETIVOS
3.1 OBJETIVO GERAL
Avaliação morfofisiológica da porção tubular do aparelho digestório da preguiça de
três dedos (Bradypus variegatus) e sua implicação no manejo alimentar em animais criados de
forma cativa e semi-cativa.
3.2 OBJETIVOS ESPECÍFICOS
Descrição morfológica do esôfago dos animais para o entendimento do trânsito dos
alimentos;
Descrição morfológica do estômago dos animais, para o entendimento da digestão dos
alimentos;
Descrição dos intestinos dos animais, para entendimento da absorção dos nutrientes;
Análise em tempo real, por meio de exames de imagem do percurso do alimento e a
digestão dos espécimes estudados.

11
4 CONTEXTUALIZAÇÃO
4.1 –MORFOLOGIA DO APARELHO DIGESTÓRIO DE HERBÍVOROS
A morfologia dos animais claramente é afetada de acordo com os hábitos alimentares,
e as características da ingesta influem na forma de se alimentar, (SHIPLEY, 1999).
O aparelho digestório é responsável por digerir e degradar os alimentos em partes
menores, possibilitando gerar energia, crescimento e renovação celular. Os órgãos que
pertencem a esse aparelho são capazes de receber alimentos, fracioná-los química e
mecanicamente até seus componentes moleculares e então absorvê-los. Por fim, este aparelho
elimina resíduos excretados e que não foram absorvidos (KONIG E LIEBICH, 2016).
De forma geral, o herbívoros dependem de microrganismos simbióticos que podem
fermentar carboidratos estruturais presentes nas folhas, em subprodutos ricos em energia,
principalmente ácidos graxos voláteis (AGV's), que são usados como fonte de energia
primária (HUNGATE 1966, STEVENS 1988).
Nos primórdios dos relatos científicos sobre o processo de fermentação alimentar, as
abordagens alicerçavam-se em diferenças básicas em anatomia e fisiologia dos herbívoros
centrando-se numa dicotomia entre fermentadores de intestino anterior e posterior (JANIS,
1976; ALEXANDER, 1991). Mais recentemente, essa separação expandiu enfatizando as
diferenças entre os não ruminantes e os ruminantes fermentadores intestinais anteriores e a
flexibilidade no sistema de fermentação do intestino posterior (SCHWARM et al.., 2009;
CLAUSS et al.., 2010a).
Os animais herbívoros são classificados como ruminantes quando apresentam três
caracteristicas básicas: estômago poligastrico, fermentação bacteriana ativa para absorção de
nutrientes e regurgitação de ingesta (FRANDSON et al.., 2003). Apresentam o estômago
dividido em fração aglandular (rúmen, retículo e omaso) e fração glandular (abomaso)
contendo na composição do órgao os padrões cárdicos, fúndicos e pilóricos (EURELL e
FRAPPER, 2006; SAMUELSON, 2007).
Ruminantes não domésticos não necessariamente se encaixam no padrão proposto
anteriormente, como é o caso por exemplo dos Camelídeos, que exibem um padrão
morfológico diferente dos bovinos com o estômago apresentando dois ventrículos de
fermentação bacteriana diferentes do rúmen e retículo e um terceiro ventrículo semelhante ao

12
abomaso, dividido em três regiões (pseudocárdia, fundo e piloro) (WANG et al.., 2000;
ABDEL-MAGIED AND TAHA, 2003).
Hipopótamos pigmeus que também são considerados ruminantes apresentam o
estômago dividido em quatro cavidades, sendo as três primeiras aglandulares e a última
glandular, porém, com o padrão de apresentação totalmente diferente do ruminante clássico
(MCDONALD e HARTMAN, 1983).
Já no gênero Bradypus, alvo de nossa investigação, na década de 40 pesquisadores
destacavam um estômago grande, inicialmente dividido em quatro câmaras, representando
cerca de 20-30% do corpo do animal (BRITTON, 1941); nas três primeiras porções gástricas
ocorria a fermentação e na quarta, a ação de ácidos e enzimas (CORK & FOLEY, 1991).
Mais tarde, entre as décadas de, 1970, 1990 e 2000, pesquisadores interpretaram a
porção gástrica das preguiças de acordo com as adaptações anatomicas, fisiológicas e
comportamentais destes indivíduos e afirmaram que as preguiças apresentam um estômago
de três compartimentos com uma distenção intestinal com características ruminais
(BAUCHOP, 1978; CORK e FOLEY, 1991; STEVENS e HUME, 1995; GILMORE et al..,
2001; PAULI et al.., 2014).
4.2 ALIMENTAÇÃO
A alimentação das preguiças é a chave vital para a manutenção com sucesso destes
indivíduos criados em cativeiro, sendo necessário um esforço concentrado para se manter
espécies de plantas comestíveis, uma vez que a maioria da ingestão de água das preguiças
acontece pela ingesta das folhagens (NAGY e MONTGOMERY, 1980).
A dieta já descrita das preguiças em vida livre é bastante restrita, composta por
árvores e cipós com cerca de 30 espécies já relatadas, com cada indivíduo consumindo em
torno de 15 a 20 espécies/ano (CHIARELLO,1998; DRUMOND, 2010). No entanto, o
comportamento alimentar das preguiças, em geral, se dá de acordo com a preferencia, no que
diz respeito a questão nutricional e a seletividade. Pelo fato das folhas serem consideradas
alimentos energeticamente desfavoráveis, esses animais possuem uma clara preferência
manifestada por folhas jovens, que apresentam menores proporções de compostos de difícil
digestão como a celulose, taninos, alcaloides e lignina, e apresentam concentrações maiores
de açúcares, água e proteínas quando comparadas às folhas maduras (STRIER, 1992;
CHIARELLO, 1994; NOGUEIRA, 1996; CHIARELLO, 1998).

13
REFERÊNCIAS
ABDEL-MAGIED, EM.;TAHA, AAM. Morphological, morphometric and histochemical characterization of the gastric mucosa of the camel (Camelus dromedarius). Anat. Histol. Embryol., 32: 42-47, 2003.
ALEXANDER, R. McN. Optimization of gut structure and diet for higher vertebrate herbivores. Phil. Truns. R. Soc. (B): 249-255, 1991.
BAUCHOP, T. Digestion of leaves in vertebrate arboreal folivores. In The Ecology of Arboreal Folivores. Montgomery, G.G. (ed.). Washington, DC, USA: Smithsonian Institution Press, pp. 193–204, 1978.
BRITTON, S. W. Form and function in the sloth. Quart. Rev. Biol.,16(1):13-34, 1941.
CARVALHO, MM.;PIERI, NCG.; PEREIRA, KF.; et al.. Caracterização comparativa do intestino das espécies da Ordem Xenarthra. Pesq. Vet. Bras. vol.34 supl.1 Rio de Janeiro Dec, 2014.
CHIARELLO AG, MORAES-BARROS, N. Bradypus torquatus. The IUCN Red List of Threatened Species. Version 2014.2. www.iucnredlist.org.
CHIARELLO, A. G. Sloth ecology. An overview of field studies. Pp. 269–280 in The biology of the Xenarthra (S. F. Vizcaíno and W. J. Loughry, eds.). University Press of Florida, Gainesville, 2008.
CHIARELLO, A. G. Diet of the atlantic forest maned sloth Bradypus torquatus (Xenarthra: Bradypodidae). Journal of Zoology. London: v. 246, n. 1, p. 11-19, 1998.
CHIARELLO, A.G. Diet of the Brown Howler Monkey Alouatta fusca in a semi-deciduous Forest fragment of Southeastern Brazil. Primates, Tokyo, 35 (1): 25-34, 1994.
CLAUSS, M.; HUME, I.D.; HUMMEL, J. Evolutionary adaptations of ruminants and their potential relevance for modern production systems. Animal 4, 979-992, 2010a.
CORK, S.J.; FOLEY, W.J. Digestive and metabolic strategies of arboreal mammalian folivores in relation to chemical defenses in temperate and tropical forests. In: PALO, R.T.; ROBBINS, C.T. (Eds.) Plant defenses against mammalian herbivory. CRC Press, p.133-166, 1991.
CRANDALL, L. S. The management of wild mammals in captivity. Chicago: University of Chicago Press, p.187-190, 1964.
DRUMOND, GM., MACHADO, AB. M.; PAGLIA, AP. Livro Vermelho da Fauna Brasileira Ameaçada de Extinção, v II. Brasília. CIP Brasil, 908p, 2010.
ENDO, H.; KOMIYA, T; KAWADA, S.; et al.. Three-dimensional reconstruction of the xenarthrous process of the thoracic and lumber vertebrae in the giant anteater. Mammal Society of Japan, Tokyo, v. 34, n. 1, p. 1-6, 2009.

14
EURELL JA, FRAPPER BL, DELLMANN HD, editors. Textbook of veterinary histology. 6th ed. Oxford: Blackwell. p 173–177, 2006.
FRANDSON, RD.; WILKE, LW.; FAILS, DA. Anatomy and physiology of farm animal. 6.ed., Philadelphia: Williams and Wilkins, 2003, 501p
GAUDIN, TJ., MCDONALD HG. Morphology-based investigations of the phylogenetic relationships among extant and fossil xenarthrans; pp. 24–36 in S. F. Vizca´ıno and W. J. Loughry (eds.), The Biology of the Xenarthra. Florida University Press, Gainesville, Florida, 2008.
GILMORE, D. P., C. P. DA-COSTA, AND D. P. F. DUARTE. Sloth biology: an update on their physiological ecology, behavior and role as vectors of arthropods and arboviruses. Brazilian Journal of Medical and Biological Research 34:9–25, 2001.
HIRSCH, A., CHIARELLO AG. The endangered maned sloth Bradypus torquatus of the Brazilian Atlantic forest: a review and update of geographical distribution and habitat prefer-ences. Mammal Review 42: 35-54, 2011.
HUNGATE, R.E. The Rumen and its Microbes. Academic Press, N.Y, 1966.
JANIS, C. The evolutionary strategy of the equidae and the origins of rumen and caecal digestion. Evolution 30: 757 774, 1976.
KÖNIG, HORST ERICH; LIEBICH, HANS-GEORG. Anatomia dos animais domésticos: texto e atlas colorido. 6. ed. Porto Alegre: Artmed, 2016.
MACALISTER, 1875; MACALISTER, A. Report on the anatomy of insectivorous edentates. Proceedings of the Royal Irish Academy, Dublin, v. 25, p. 491-508, 1875.
MACDONALD, A.A. AND HARTMAN, W. Comparative and functional morphology of the stomach in the adult and newborn Pigmy Hippopotamus (Choeropsis liberiensis), Journal of Morphology 177; pp. 269-276, 1983.
MERRIT, D. A. The two-toed sloth, Choloepus hoffmanni Peters. In: G. G. Montgomery (ed.), The Evolution and Ecology of Armadillos, Sloths and Vermilinguas. Smithsonian Institution Press, Washington and London, pp. 333–341, 1985.
MONTGOMERY, GG.; SUNQUIST, ME. Impact of sloths on neotropical energy flow and nutrient cycling. In: GOLLEY, F.; MEDINA, E. (Ed.) Tropical ecological systems:trends in terrestrial and aquatic research. New York: Springer-Verlag, p.69-98,1975.
NAGY, KA.; MONTGOMERY, GG. Field metabolic rate, water flux, and food consumption in three-toed slots (Bradypus variegatus). Journal of Mammalogy, 61(3):465-472, 1980.
NAPLES, VL. Morphology, evolution and function of feeding in the giant anteater (Myrmecophaga tridactyla), Journal of Zoology, London, v. 249, p. 19- 41, 1999.
NOGUEIRA, CP. Ecologia e comportamento de femeas muriquis (Brachyteles arachnoids) em diferentes estagios reprodutivos [MA thesis]. [Guarulhos (Brazil)]: Universidade de

15
Guarulhos, 1996.
OWEN, R. On the Anatomy of the Great Anteater (Myrmecophaga jubata, Linn). Proceedings of The Zoological Society of London, London, v. 1857, p. 22-29, 1857.
PAULI, JN.; MENDOZA, JE.; STEFFAN, SA. et al.. A syndrome of mutualism reinforces the lifestyle of a sloth. Proceedings of the Royal Society B 281, 2013-3006, 2014.
RAINES, J. Captive Health and Husbandry of the Bradypodidae, Zoo Biology 24:557–568, 2005.
RÉUS, CL.; SOUZA, CM. Estrutura populacional de Bradypus tridactylus (Xenarthra, Bradypodidae) em fragmento florestal urbano no município de Manaus, Amazonas, Brasil Estud. Biol. 29(68/69):249-256, 2007.
SAMUELSON, DA. Textbook of veterinary histology. 1st Edn., Philadelphia, W. B. Saunders Co., PP: 323-335, 2007.
SCHWARM, A. et al.. More efficient mastication allows increasing intake without compro-mising digestibility or necessitating a larger gut: comparative feeding trials in banteng (Bos javanicus) and pygmy hippopotamus (Hexaprotodon liberiensis). Comp. Biochem. Physiol. A 152: 504512, 2009.
SHIPLEY, LA. Grazers and browsers: How digestive morphology affects diet selection. Pages 20- 27 IN K. L. Launchbaugh, J. C. Mosley, and K.D. Sanders, eds. Grazing behavior of livestock and wildlife. Idaho Forest, Wildlife, and Range Experiment Station Bulletin 70, 1999.
STAHL, M.; OSMANN, C.; ORTMANN, S. et al.. Energy intake for maintenance in a mammal with a low basal metabolism, the Giant anteater (Myrmecophaga tridactyla). Journal of Animal Physiology and Animal Nutrition 96, 818–824, 2012.
STEVENS, CE. Comparative Physiology of the Vertebrate Digestive System. Cambridge Univ. Press, N.Y. T, 1988.
STEVENS, E., HUME, ID. The mammalian gastrointestinal tract. In Comparative physiology of the vertebrate digestive system. New York, New York: Cambridge University Press, pp. 65-67, 1995.
STRIER, KB. Atelinae adaptations: behavioral strategies and ecological constraints. Am J Phys Anthropol 88:515–524, 1992.
URBANI, B., BOSQUE, C. Feeding ecology and postural behaviour of the three-toed sloth (Bradypus variegatus flaccidus) in northern Venezuela Mamm. biol. 72 (2007) 6 321–329, 2007.
VAUGHAN, C., RAMÍREZ, O., HERRERA, G. et al.. Spatial ecology and conservation of two sloth species in a cacao landscape in Limón, Costa Rica. Biodiversity and Conservation, vol. 16, p. 2293- 2310, 2007.

16
VENDL, C., FREI, S., DITTMANN, M. T., FURRER, S., OSMANN, C., ORTMANN, S., MUNN, A., KREUZER,M., CLAUSS, M. Digestive physiology, metabolism and methane productionof captive Linne’s two-toed sloths (Choloepus didactylus). Journal of Animal Physiology and Animal Nutrition, 2015.
WANG, JL; LAN, G; WANG, GX, et al.. (2000). Anatomical subdivisions of the stomach of the Bactrian camel (Camelus bactrianus). J. Morphol., 245: 161-176, 2000.

17
CAPITULO I: MORPHOLOGY OF THE ESOPHAGUS OF Bradypus variegatus VERSUS SWALLOWING MECHANISM (Submetido a Revista Ciência Animal Brasileira)
ABSTRACT: The adaptations for a folivore arboreal diet of sloths are evident right on chewing. Due to specialisation for grinding the fibrous foods, sloths have facial muscles reduced in number and complexity compared to the muscle of carnivores, and accordingly, the acrobatic behavior and laterality held by these Xenarthras make these individuals a differentiated group, including at the time of feeding. We aimed to investigate the morphological characteristics of the esophagus of the brown-throated sloth, in order to subsidize the understanding of the digestion of these animals, as well as its feeding behavior, in order to assist in the preservation of this species. We used seven adult animals, who lived in semi-captivity conditions and died by natural causes. After photo documentation of the esophagus topography, the organ was removed and samples of cervical, thoracic and abdominal parts. Topographically, the esophagus began dorsal to the larynx, and immediately diverted to the left of the trachea, adjacent to it, still on the left antimer throughout the entire cervical portion. In the thorax, has identified in the mediastinum, and then crossed the diaphragm through the esophageal hiatus, reaching the stomach. Histologically, the esophagus followed the same tissue pattern along the three parts (cervical, thoracic and abdominal) with minor modifications. The mucosa of the organ consisted by stratified keratinized squamous tissue, composed of a stratum corneum without stratification pattern, with formation of pleats supported only by keratin, succeeded by the lamina propria and submucosa of loose connective tissue without glands, absence of muscularis mucosae, striking muscular layer containing three layers (longitudinal, transversal and longitudinal), with a density of the muscles in the thoracic portion, and the entire organ composed of skeletal striated muscle with peripheral nucleus and multinucleated cells, adventitia by loose connective tissue, with the sympathetic nervous chain circumscribing the organ near the cardiac region.
Keywords: Anatomy, Digestive, Xenarthra, Sloths.
MORFOLOGIA DO ESÔFAGO DE PREGUIÇAS VERSUS MECANISMO DE
DEGLUTIÇÃO
As adaptações para a dieta folívora arborícola de preguiças são evidentes logo na mastigação. Devido às especializações para trituração dos alimentos fibrosos, as preguiças dispõem de músculos faciais reduzidos em número e complexidade em comparação com os músculos de carnívoros, e em consonância, o comportamento acrobático e a lateralidade realizada por esses Xenarthras fazem desses indivíduos um grupo diferenciado, inclusive no momento da alimentação. Diante do exposto, objetivou-se investigar as características morfológicas do esôfago da preguiça-comum, a fim de subsidiar o entendimento da digestão desses animais, bem como seu comportamento alimentar, e com isso auxiliar na preservação da espécie.Foram utilizados sete animais adultos que viviam em regime semi-cativo. Após fotodocumentação para registro da topografia, o órgão foi removido e amostras das porções cervical, torácica e abdominal foram fixadas para avalição pela microscopia de luz, seguindo protocolo de rotina histológica. Topograficamente, o esôfago iniciou dorsal a laringe e imediatamente desviou a esquerda da traqueia, mantendo-se adjacente a mesma, ainda no antímero esquerdo ao longo

18
de toda a porção cervical. No tórax, o esôfago foi identificado no mediastino e em seguida cruzou o diafragma através do hiato esofágico alcançando o estômago. Histologicamente, o esôfago seguiu o mesmo padrão tecidual ao longo das três porções (cervical, torácico e abdominal) com pequenas modificações. A mucosa do órgão constituía-se por tecido pavimentoso queratinizado estratificado, composto de um estrato córneo sem padrão de estratificação, com formações de pregas suportadas somente por queratina, sucedido pela lâmina própria e submucosa de tecido conjuntivo frouxo sem glândulas, ausência de muscular da mucosa, camada muscular marcante contendo três camadas (longitudinal, transversal e longitudinal), com adensamento da musculatura na porção torácica, e todo o órgão composto por musculatura estriada esquelética com núcleos periféricos, sendo as células multinucleadas, e adventícia por tecido conjuntivo.
Palavras-chave: Anatomia, digestiva, Xenarthra, preguiças
INTRODUCTION
To understand the diet of wild animals, is important to know the morphology of the
digestive tract. However, there is a literary shortage on the subject, which prevents knowing
the digestive processes and the adequate food for each species (PINHEIRO et al.. 2014). It is
well known though, that the digestive tube is very structurally variable in vertebrate animals,
even among species of the same genus (ZAMITH, 1952). Chewing, swallowing and food
transport of mammals also occurs differently from other vertebrates (ROMER; PARSONS,
1985), since the anatomical differences between species can reflect in the phylogenetic
adaptations of several diets (FOWLER, 1998).
In anatomy of the digestive tube, the first tubular portion covered by the food bolus is
the esophagus, which has the function to drive the food from the pharynx to the stomach
(SISSON, 1986) having three parts (cervical, thoracic and abdominal) and in some animals
already described, is composed by four layers: mucosa, submucosa, muscular and
adventitial/serosa (SAMUELSON, 2007).
The esophagus is geographically presented in herbivores as a relatively narrow tube
that begins dorsally to the cricoid cartilage of the larynx and accompanies the trachea along
the neck, leaning initially to the left and resuming the median dorsal position, after attaining
the thoracic region that, in this portion, features a deviation to the left of the aortic arch, and
reassumes the left antimer in the abdominal portion of stomach insertion. The food bolus
morphologically follows the tube architecture and, during this route, the food goes through
stages, until it reaches the stomach (DYCE, 2010).
Histologically, the esophagus of some animals, such as the swine for example, presents
stratified squamous epithelium, with the absence of keratin, and supported by a lamina propria

19
(SAMUELSON, 2007). However, in other species such as ruminants and equines, the
esophagus is keratinized, that is, with a layer of keratin on the surface of the epithelium,
probably related to the more fibrous diet performed by these animals (NASCIUTTI, 2016). In
addition to the epithelium and lamina propria, there is a delicate smooth muscle layer with
longitudinally oriented bundles, the muscularis mucosae, which delimits the mucosa of the
submucosa, which is adjacent to the dual smooth muscle layer and more externally follows
the adventitia, constituted of loose connective tissue (STISON and CALHOUN, 1982;
LANGER, 2003).
The primary functions of the digestive tube are the digestion and absorption of
essential nutrients to the metabolic processes of the animals, in addition to revealing general
eating habits, being useful for systematics. The digested nutrients are absorbed by the
epithelium and transferred to the circulation (HILDEBRAND and GOSLOW, 2006).
Sloths are among the few species of arboreal mammals with a strict folivore habit
(CORK; FOLEY, 1991), known for their slow movements and low metabolic rate, inhabiting
forests and feeding on leaves, branches and buds of several plants, often from the Moraceae
(CHIARELLO, 2008), Cecropiaceae and Clethraceae families (URBANI and BOSQUE,
2007), with leaves representing 94% of the items consumed (QUEIROZ, 1995).
The adaptations to the folivore arboreal diet of sloths are already evident in the
chewing process. Because of specializations for grinding fibrous foods, sloths have facial
muscles reduced in number and complexity when compared to the carnivorous muscles
(NAPLES, 1985) and accordingly, the acrobatic behavior and laterality performed by these
Xenarthras make these individuals a differentiated group, including at the time of feeding
(CONSENTINO, 2004; PINHEIRO, 2006).
In view of the above, our objective was to investigate the morphological
characteristics of the esophagus in brown-throated sloths, in order to subsidize the
understanding of the digestion of these animals, as well as their feeding behavior, and with
that help in the conservation of the species
MATERIAL AND METHODS
We used seven adult animals (4 females and 3 males), from the Bosque Rodrigues
Alves (SISBIO N ° 49221) (Belém - Pará - Brazil), that lived in semi-captivity conditions and
died by natural causes, not related to their digestive system. They were immediately frozen
and sent to the Laboratory of Morphological Animal Research (LaPMA) at the Federal Rural
University of Amazonia (UFRA-PA).

20
Initially the animals were thawed in running water for approximately six hours and
then, with the help of a basic dissecting material, we performed an incision in the menton
region, extending through the ventral midline of the neck, thorax and abdomen, until the pre-
umbilical region, separating the skin, musculature and the sternum, which was incised in the
costochondral joints, followed by exposure of the esophagus to outfall point, near the
stomach.
After photodocumentation to register the esophagus topography, the organ was
removed, and samples of the cervical, thoracic and abdominal portions were fixed in aqueous
solution of formaldehyde 10% for evaluation by light microscopy, following a histological
routine protocol of Hematoxylin-Eosin and Trichrome of Gomori (JUNQUEIRA and
JUNQUEIRA, 1983).
The nomenclatures adopted followed the standards of Nomina Anatomica
Veterinaria (International Committee on Veterinary Gross Anatomical Nomenclature, 2012).
RESULTS
Topographically, the esophagus began dorsally to the larynx, and immediately
diverted to the left side of the trachea, remaining adjacent to it, still in the left antimer along
the entire cervical portion. In the thorax, the esophagus was identified in the mediastinum, and
then crossed the diaphragm through the esophageal hiatus, reaching the stomach (Figure 1).

21
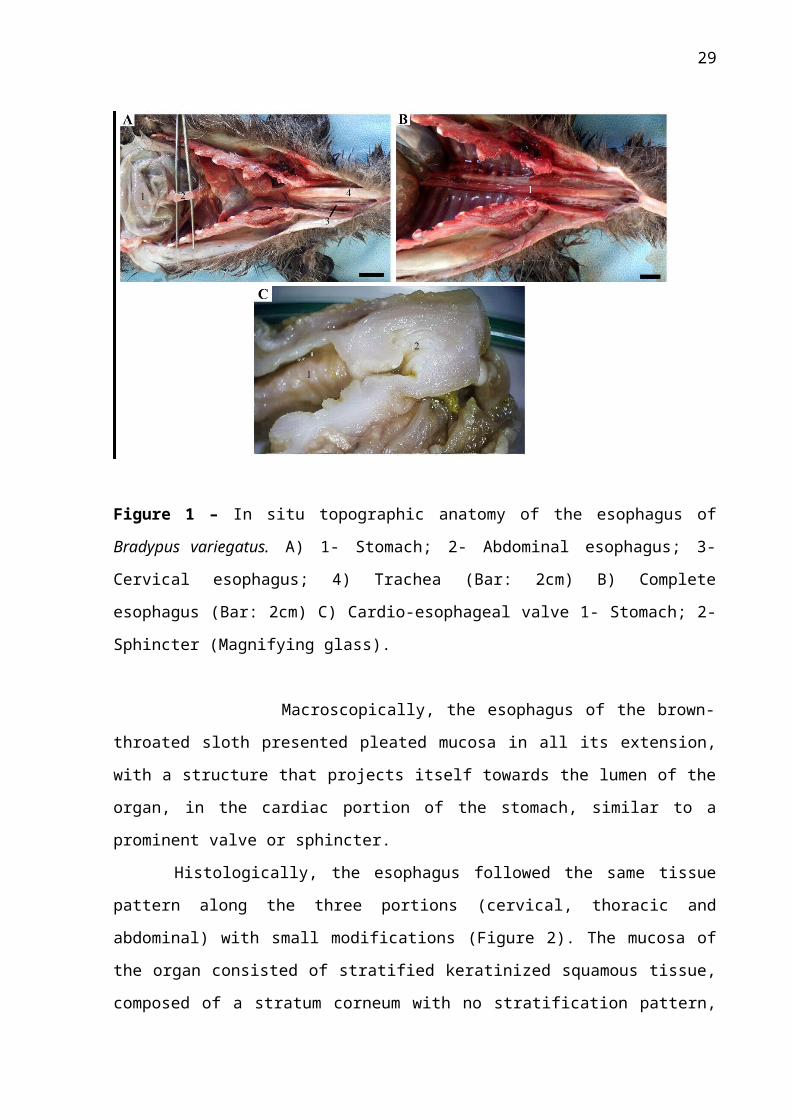
Figure 1 – In situ topographic anatomy of the esophagus of Bradypus variegatus. A) 1-
Stomach; 2- Abdominal esophagus; 3- Cervical esophagus; 4) Trachea (Bar: 2cm) B)
Complete esophagus (Bar: 2cm) C) Cardio-esophageal valve 1- Stomach; 2- Sphincter
(Magnifying glass).
Macroscopically, the esophagus of the brown-throated sloth presented pleated
mucosa in all its extension, with a structure that projects itself towards the lumen of the organ,
in the cardiac portion of the stomach, similar to a prominent valve or sphincter.
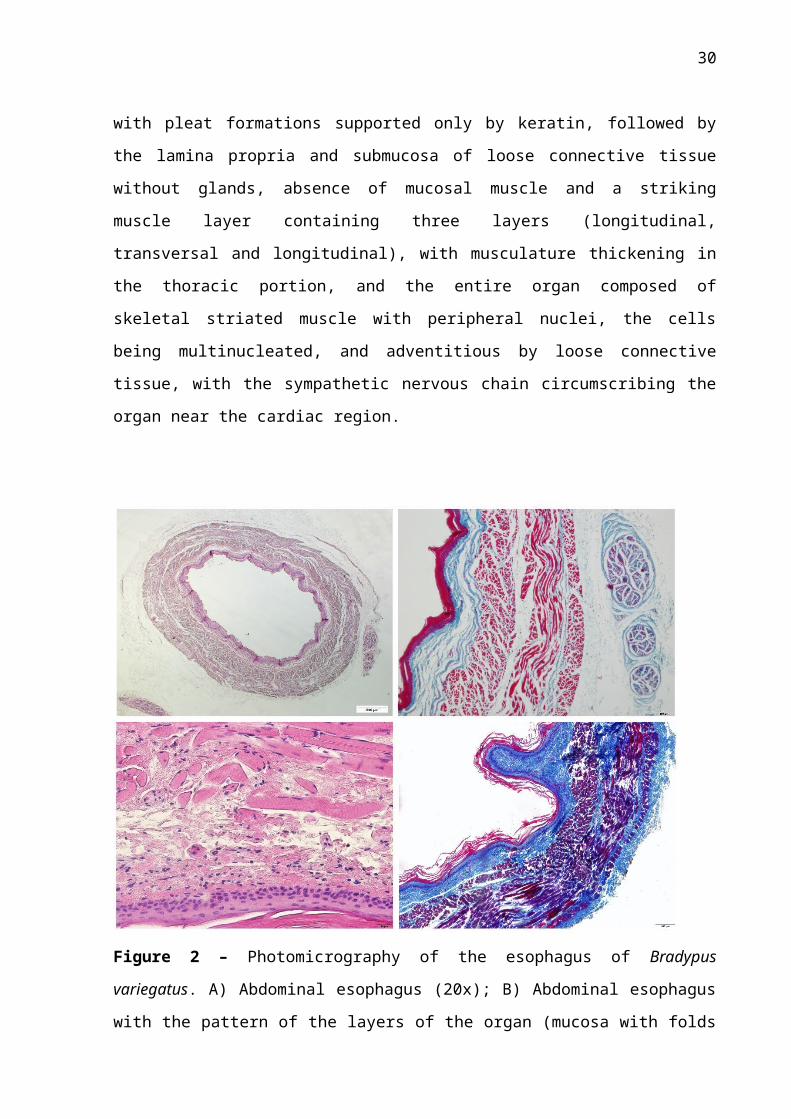
Histologically, the esophagus followed the same tissue pattern along the three
portions (cervical, thoracic and abdominal) with small modifications (Figure 2). The mucosa
of the organ consisted of stratified keratinized squamous tissue, composed of a stratum
corneum with no stratification pattern, with pleat formations supported only by keratin,
followed by the lamina propria and submucosa of loose connective tissue without glands,
absence of mucosal muscle and a striking muscle layer containing three layers (longitudinal,
transversal and longitudinal), with musculature thickening in the thoracic portion, and the
entire organ composed of skeletal striated muscle with peripheral nuclei, the cells being
multinucleated, and adventitious by loose connective tissue, with the sympathetic nervous
chain circumscribing the organ near the cardiac region.

22
Figure 2 – Photomicrography of the esophagus of Bradypus variegatus. A) Abdominal
esophagus (20x); B) Abdominal esophagus with the pattern of the layers of the organ (mucosa
with folds of keratin, muscular with the three layers, and serosa) and autonomic nervous
plexus; C) Muscle layer showing striations, characterizing the skeletal muscle tissue; D)
Crypts of epithelial tissue in the mucosa invading the light of the organ. (A: HE_20X
coloration; B HE_100X coloration; C: HE_400X coloration.) D) TG_100X coloration).
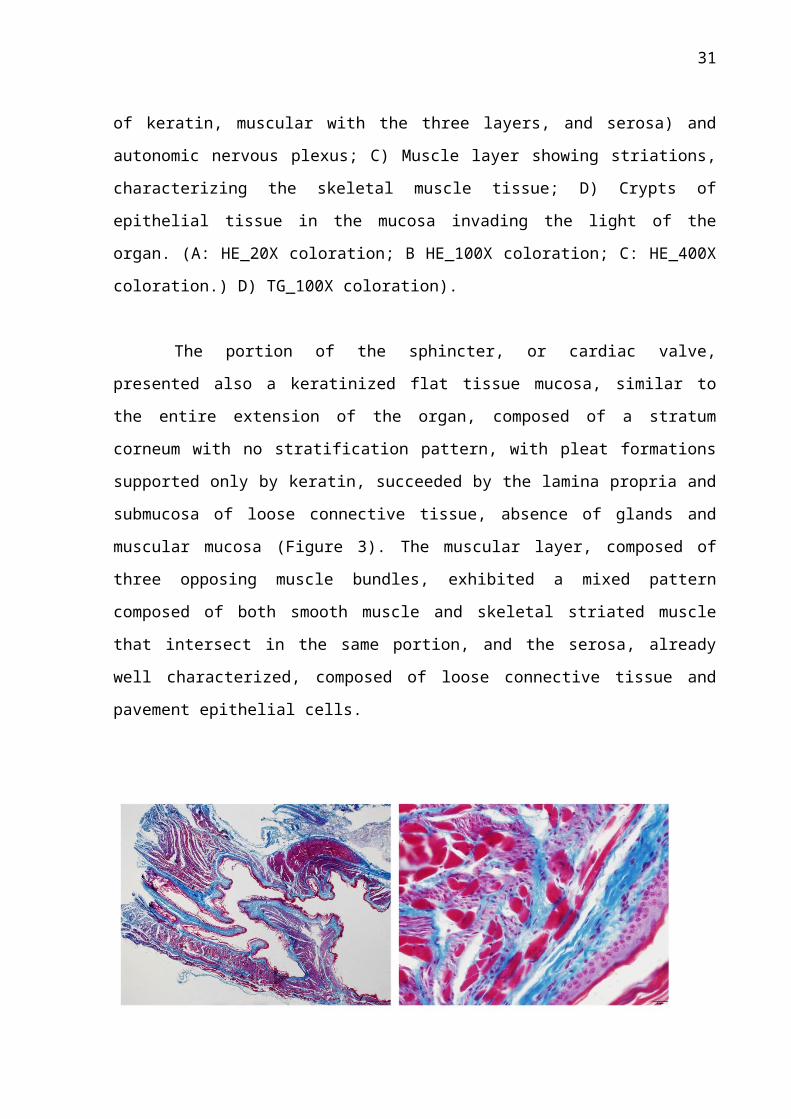
The portion of the sphincter, or cardiac valve, presented also a keratinized flat tissue
mucosa, similar to the entire extension of the organ, composed of a stratum corneum with no
stratification pattern, with pleat formations supported only by keratin, succeeded by the
lamina propria and submucosa of loose connective tissue, absence of glands and muscular
mucosa (Figure 3). The muscular layer, composed of three opposing muscle bundles,
exhibited a mixed pattern composed of both smooth muscle and skeletal striated muscle that
intersect in the same portion, and the serosa, already well characterized, composed of loose
connective tissue and pavement epithelial cells.

23
Figure 3 – Photomicrography of the cardio-esophageal valve of Bradypus variegatus. A)
General aspects in smaller increase with projection for the lumen of the organ B) Interspersed
mixed muscular pattern composed of smooth musculature and skeletal striated muscle (A:
HE_20X coloration; B HE_100X coloration).
DISCUSSION
The disposition of the brown-throated sloth’s esophagus, located exclusively in the
left antimer of the neck, presenting the rectilinear path and absence of flexure, differs from the
literature already described in both carnivores and domestic herbivores (GETTY et al.., 1986;
MURRAY, 1998; DYCE et al.., 2010). However, it resembles the one described by Dias
(2015) for the southern tamandua. This condition, together with the suspended feeding
behavior, with the pelvic limbs above and the head in a lower plane, supposedly clarifies the
need for a greater control of movement of the food bolus, since the presence of flexures,
usually described in domestic animals, would make this return difficult. This is not the case in
sloths, which in turn is justified by the presence of the cardio-esophageal valve that tends to
avoid the reflux process.
Moreover, this conformation without flexure and without deviation of the aortic arch
allows the acceleration of the metabolism of these animals at the moment of feeding since
there is a gravitational compression, as proposed by Duarte et al.. (2004), who affirm that the
posture of B. variegatus favors the increase of blood pressure, movements of systole and
diastole and, consequently, the cardiac rhythm of these animals.
The microscopy of the esophagus confirms that this organ has its own characteristics,
in which the mucosa reveals pleats disposed throughout the entire internal surface, different
from domestic animals in which pleats disappear in the abdominal portion (GENESER, 2003;

24
GARTNER e HIATT, 2007). In addition, we observed the absence of esophageal glands
which, together with the anatomical characteristics, confirms that the organ possibly presents
only mechanical functionality, as in some animals (MAGALHÃES, 2010).
This lack of lubrication based on the movement of anti gravitational deglutition
reinforced by the presence of the skeletal musculature, leads us to believe that there is some
control of the bolus of food, and the lubrication of this bolus must occur through salivation of
the animal.
The literature also points out that ruminants and dogs also have a muscular tunica
composed of skeletal muscles, but composed of two muscular bundles, just like the
morphological description of the esophagus of Hydrocherus hydrochaeris (the capybara),
which has shown that the muscular layer of the organ of these animals is also constituted by
two layers of skeletal striated muscle developed throughout the organ. This fact led the author
to attribute to this layer the importance of transporting remaining food particles after each
swallow, since motility in this organ is closely related to this layer (VELASQUEZ et al..,
2016).
According to our results, B. variegatus presents three muscular layers, which possibly
leads to an even more precise control of the esophageal movements of regurgitation when
compared to the ruminants that use this muscular domain for the alimentary return in the
process of rumination (BANKS, 1992).
At the end of the alimentary path until the stomach, B. variegatus present a cardiac
valve, presumably controlled by the autonomic nervous system in order to avoid reflux, since
the rectilinear anatomy would favor the event. In canines, the purpose of the cardiac valve is
already described, and consists of avoiding regurgitation since, as the alimentary bolus begins
the transit through the esophagus, there is a decrease on the pressure of the caudal esophageal
sphincter, relaxing it and allowing the passage of the food for the stomach. After the bolus has
passed, the sphincter contracts and creates a high pressure zone. The contraction avoids reflux
of gastric contents within the distal portion of the esophagus (HOFFER et al.., 1979; BANKS,
1992).
Possibly, B. variegatus follow a similar pattern of domestic animals in this aspect,
since the histology also shows the mixed composition of smooth and skeletal muscle tissue
and the connection of the autonomic nervous plexus. However, the cardiac portion differs
with respect to glands. In monogastric animals, the cardiac sphincter also has mucus-secreting
glands (mucosal branched tubular glands, whose secretion protects the esophagus wall from
gastric juice reflux) (GENESER, 2003; SAMUELSON, 2007; ROSS e PAWLINA, 2012). In

25
B. variegatus, these glands are absent, reiterating the organ's exclusive mechanical function
(FRAPE, 2004).
Finally, we conclude that the unique configuration of the esophageal tube together
with the complexity of pre and post ingestion postural behavior, the histological composition
and the low metabolic rate, infer in some of the peculiar behaviors of the sloths from the
Bradypus variegatus species, as well as in the knowledge of the food transit of these animals.
REFERENCES
CARRASCAL VELÁSQUEZ, JC.; ORTIZ BEDOYA, SA.; PETRO HERNÁNDEZ, VG. Caracterização microscópica das regiões esofágicas em Hydrochoerus hydrochaeris. Medicina Veterinaria Y Zootecnia, Mayo – agosto, 2016.
CHIARELLO, AG. Sloth ecology: an overview of field studies. In: The Biology of the Xenarthra, S. F. Vizcaíno e W. J. Loughry (eds.), University Press of Florida, Gainesville, 2008. p. 638– 671.CONSENTINO, LN. Aspectos do comportamento da preguiça-comum, Bradypus variegatus (Xenarthra, Bradipodidae) em uma área de semi-cativeiro no município de Valença- Rio de Janeiro. 38f. Dissertação (Mestrado em Biologia Animal) Universidade Federal Rural do Rio de Janeiro, Seropédica, 2004.
DIAS, GF.; SIQUEIRA, RAS.; FIRMINO, MO.; GUERRA, RR. Anatomia topográfica do sistema digestivo de tamanduá mirim (Tamadua tetradactyla, Linnaeus 1758) Anais de Foz do Iguaçu – PR, Brasil, 2015.
DUARTE, DPF; JAGUARIBE, AM.; PEDROSA, MAC.; CLEMENTINO, ACCR.; BARBOSA, AA.; SILVA, AFV.; GILMORE, DP.; DA COSTA, CP. Cardiovascular responses to lomomotor activity and feeding in unrestrained three-toed sloths, Bradypus variegatus. Brazilian Journal of Medical and Biological Research, Ribeirão Preto, 37, 10, 2004. p.1557-1561.
DYCE, KM.; SACK, WO.; WENSING, CJG. Tratado de anatomia veterinária. 3.ed. São Paulo: Saunders Elsevier, 2010. 813 p.
FONSECA, GAB.; HERMANN, G.; LEITE, YLR.; MITTERMEIER, RA.; RYLANDS, AB.; PATTON, JL. Lista anotada dos mamíferos do Brasil. Occasional Papers in Conservation Biology. Conservation International, Belo Horizonte, Brazil, 1996.
FRAPE, D. Equine nutrition and feeding. (3th ed.). Oxford: Blackwell Publishing, 2004.GARTNER, LP.; HIATT, JL. Tratado de Histologia em cores. 3.ed. Rio de Janeiro: Elsevier, 2007. pp. 373, 419, 421.
GENESER, F. Histologia: com bases moleculares. 3.ed. Buenos Aires: Médica Panamericana/ Rio de Janeiro: Guanabara Koogan, 2003. p. 125.
GETTY, R. Anatomia dos Animais Domésticos. Vol.2. 5ª ed. Guanabara Koogan, Rio de

26
Janeiro, 1986. p.1377-1391.
GOETSCH, E. The structure of the mammalian esophagus. The American journal of anatomy, Philadelphia, 10, 1910. p. 1-40.
JUNQUEIRA, LC.; JUNQUEIRA, LMMS. Técnicas básicas de citologia e histologia. São Paulo: Santos, 1983. 123p.
HAM, AW.; CORMACK, DH. Histologia. 8.ed. Rio de Janeiro: Guanabara Koogan, 1983. pp. 607-608, 623.
HOFER, RE., MACCOY, DM, QUICK, CB. Management of acquired achalasia in dogs. J Am Vet Med Assoc, v. 175, n. 8, 1979. p. 814-817, 1979.
LANGER, P. Lactation, weaning period, food quality, and digestive tract differentiations in eutheria. Evolution, Toronto, v. 57, n. 5, 2003. p. 1196-1215.
MURRAY, M.J. O esofago. In: REED, S.M. BAYLY, W.M., (ed). Medicina Interna Equine. Philadelphia, 1998. p. 524-529.
NASCIUTTI, LE.; NARCISO, MS.; LIMA, AVP.; GERLY, AC.; ORIÁ, RB. Histologia do Tubo Digestório. In: Sistema Digestório: Integração Básico-Clínica. São Paulo: Blucher, 2016. p. 273-314
PINHEIRO, SD. Comportamento alimentar da preguiça comum Bradypus variegatus (SCHINZ, 1825) (Xernarthra, Bradypodidae) no Parque Centenário de Barra Mansa – RJ. Universidade Federal de Juiz de Fora. Dissertação de Mestrado, 2006. 119p.
PINHEIRO, ACO.; LIMA, AR.; CARVALHO, AF.; PEREIRA, LC.; BRANCO, E. Aspectos morfológicos macro e microscópicos do estômago de tamanduá-mirim(Tamandua tetradactyla). Arq. Bras. Med. Vet. Zootec., v.66, n.4, 2014. p.1189-1196.
QUEIROZ, H.L. Preguiças e Guaribas, os mamíferos folívoros arborícolas do Mamirauá. v.2. Rio de Janeiro: CNPq e Sociedade Civil Mamirauá, 1995. 176p.
SAITO, CH.; BRASILEIRO, L. ALMEIDA, LE. TAVARES, MCH. Conflitos entre macacos-prego e visitantes no parque nacional de Brasília: Possíveis Soluções. Sociedade & Natureza. Uberlândia, 22 (3): 515,524, dezembro, 2010.
SAMUELSON, DA. Tratado de histologia veterinária. Trad. Newton da Cruz Rocha. Rio de Janeiro: Elsevier, 2007. p 87-189.
SCHUMMER, A.; NICKEL, R.; SACK, WO. The alimentary canal. In: SCHUMMER, A.; NICKEL, R.; SACK, W. O. The Viscera of Domestic Mammals, New York: Springer-Verlag , 1979. p. 99-202.
SISSON, S. Aparelho digestório. In: GETTY, R. Anatomia dos mamíferos domésticos. 5.ed. Rio de Janeiro: Guanabara, 7, 1986. p.101-107.
STINSON, AW.; CALHOUN, ML. Sistema digestivo. In: DELLMANN, H. D.; BROWN, E.

27
M. Histologia veterinária. Rio de Janeiro: Guanabara Koogan, 1982. p. 164-211.
URBANI, B.; BOSQUE, C. Feeding ecology and postural behaviour of the three-toed sloth (Bradypus variegatus flaccidus) in northern Venezuela. Mammalian Biology, v 72, 2007. p. 321-329.
The authors declare no conflicts of interest.

28
CAPITULO II - MORFOLOGIA x DINÂMICA DO ESTÔMAGO DE PREGUIÇA COMUM (Bradypus variegatus)
RESUMO: Para superar as limitações impostas pelo estilo de vida, as preguiças, de modo geral, têm evoluído e criado adaptações anatômicas, fisiológicas e comportamentais. Este contexto envolve diretamente o aparelho digestório. Assim, objetivamos descrever morfologicamente o estômago da Bradypus variegatus para subsidiar uma melhor compreensão quanto a dieta e digestão desses animais, auxiliando na preservação desta espécie. Foram utilizados sete animais adultos que viviam em regime semi-cativo e identificados sete compartimentos: Saco cranial, saco central, saco caudal, fundo, divertículo, pré piloro I e pré piloro II. Topograficamente, o saco cranial caracterizou-se pela zona de inserção esofágica, sendo a primeira cavidade do estômago situada na região cranial, desviada a esquerda. Em seguida destacou-se o saco central posicionado na região mediana do corpo do animal, precedendo o saco caudal, que estava posicionado no antímero direito. O fundo era localizado na região crânio central do órgão de modo acima do divertículo, que sobrepunha os pré piloros I e II (inserção duodenal). Histologicamente, os compartimentos revelaram três padrões distintos: uma fração de tecido plano queratinizado estratificado baixo aglandular uma fração glandular, composta por glândulas da mucosa revestida por tecido epitelial glandular prismático baixo com formações de fossetas curtas e glândulas longas e uma porção glandular, composta de células principais e parietais (estômago químico). Por fim concluímos, até o presente momento, que a complexidade dos estômagos poligástricos, apresentando uma câmara fermentativa precedente a um estômago químico bem caracterizado, aliado a composição histológica torna esses indivíduos semelhantes aos ruminantes na morfologia gástrica, necessitando de estudos a respeito da fisiologia digestiva desses animais. Palavras-chave: Anatomia, Estômago, Histologia, Digestiva, Xenarthra.
INTRODUÇÃO
Na evolução do aparelho digestório, no que diz respeito aos hábitos alimentares,
tanto em onívoros, herbívoros e carnívoros, os órgãos dos animais sofreram várias adaptações
para atender as necessidades específicas (HILDEBRAND & GOSLOW 2006), e no caso de
alguns animais herbívoros destaca-se uma serosa peritoneal e uma rede vascular especializada
que constituem pré-estômagos, estômagos, cólon, mesentérios e várias adaptações da mucosa
(REZENDE et al., 2011).
Pelo hábito alimentar folívoro estrito, as preguiças do gênero Bradypus não se
adaptam facilmente à vida em cativeiro, tornando-as pouco resistentes. Assim, quando
removidas do seu habitat natural, sobrevivem por poucos meses (CRANDALL, 1964), uma

29
vez que se alimentam de uma gama limitada de espécies de árvores. As preguiças de dois
dedos (Choloepus spp.) costumam ter uma dieta mais ampla, pois inclui uma grande
variedade de espécies de plantas, sendo mais fáceis de manter em cativeiro (RAINES, 2005;
VAUGHAN et al., 2007).
Para superar as limitações impostas por este estilo de vida, as preguiças, de modo
geral, evoliuram e criaram adaptações anatômicas, fisiológicas e comportamentais (CORK E
FOLEY, 1991; GILMORE et al.., 2001; PAULI et al.., 2014), tais como um estômago
compartimentado com características ruminais (BAUCHOP, 1978; STEVENS E HUME,
1995).
Além disso, estudos anteriores apontaram que esses animais apresentam a menor
taxa de digestão quando comparado a outros mamíferos conhecidos (BRITTON, 1941;
FOLEY et al.., 1995), além de uma baixa taxa metabólica (MCNAB, 1978; NAGY e
MONTGOMERY, 1980), reduzida temperatura corporal e baixa densidade muscular
(MCNAB, 1978), o que pode explicar as necessidades energéticas reduzidas, permitindo a
esses animais subsistir em dietas pobres em calorias (DILL-MCFARLAND et al.., 2016),
porém, é necessário um estudo morfológico mais detalhado para que se possam fazer
inferências quanto a digestão desses animais.
Diante dos motivos supra expostos, e frente a importante atuação do médico
veterinário, não apenas na clínica e cirurgia, mas também no manejo nutricional de animais
selvagens, conhecer a morfologia do estômago da preguiça-comum, subsidiará melhor
compreensão quanto a dieta e digestão desses animais, auxiliando na preservação desta
espécie.
MATERIAL E MÉTODOS
Foram utilizados sete animais adultos (4 fêmeas e 3 machos), provenientes do
Bosque Rodrigues Alves Jardim Botânico da Amazônia (SISBIO N°49221) (Belém – Pará –
Brasil), que viviam em regime semi-cativo e foram a óbito por causas naturais, não correlatos
ao aparelho digestório, sendo imediatamente congelados e encaminhados ao Laboratório de
Pesquisa Morfológica Animal (LaPMA), da Universidade Federal Rural da Amazônia
(UFRA-PA).
Inicialmente os animais foram descongelados em água corrente, por período de
aproximadamente seis horas e em seguida, com auxílio de material básico de dissecação,
realizamos incisão na linha alba expondo a cavidade abdominal para visualização e

30
identificação do estômago de cada exemplar.
Tanto a porção final do esôfago, quanto a porção inicial do duodeno foram ligadas
com fio de algodão 0.0 e incisadas, para a extração do estômago, o qual foi cuidadosamente
lavado com solução fisiológica a fim de remover todo o conteúdo gástrico. Posteriormente,
parte do material foi destinado à macroscopia, sendo imerso em solução aquosa de
formaldeído a 10%, enquanto uma outra parte foi submetida a técnicas de microscopia de luz,
sendo previamente fixada em Davidson a fim de seguir o protocolo de rotina histológica. Para
as análises de microscopia eletrônica de varredura, foram utilizadas técnicas previamente
descritas por Murakami (1974), Watanabe e Yamada (1983), e Ohtani (1987).
As nomenclaturas adotadas seguiram os padrões do Nomina Anatomica Veterinaria
(International Committee on Veterinary Gross Anatomical Nomenclature, 2012).
Para o entendimento do trajeto alimentar, foram feitos exames de raio X de contraste
‘in vivo’ em um aparelho digital com contraste a base de sulfato de bário (BaSO4 -1ml/kg),
injetado no animal por via oral por meio de sonda esofágica e o exame sendo realizado em
intervalos de 10 minutos, por um período de 90 minutos. Foram adotados como critérios de
avaliação do exame o padrão das radiografias (60kv e 10mAs) em relação aos fatores de
exposição e o posicionamento adequado do animal na radiografia (VD). Também, a cobertura
da superfície mucosa promovida por uma fina camada de meio de contraste positivo, e a
distensão e transparência do estômago. O exame de ultrassom do órgão ‘ex situ’ transcorreu
utilizando um exemplar imerso em água para a obtenção da interface adequada.
Todos os procedimentos foram aprovados pelo comitê de ética da Universidade
Federal Rural da Amazônia sob n°013/2017 CEUA-UFRA.
RESULTADOS
Descrição macroscópica
A análise anatômica do estômago da preguiça permitiu verificar que este ocupa grande
parte da cavidade abdominal (FIGURA 1) sendo dividido em sete compartimentos: saco
cranial, saco central, fundo, saco caudal, divertículo, pré piloro I e II (Figura 2A).

31
FIGURA 1 – Fotomacrografia de corte longitudinal do tronco da Bradypus variegatus para visualização do
estômago in situ. A – Antímero esquerdo: 1- Coração, 2 – Estômago, 3 – Intestinos (barra 2cm); B – Antímero
esquerdo: 1 – saco cranial, 2 – saco central, 3- divertículo; C – Antímero direito: 1- saco cranial, 2 – saco central.
Barra de Escala em A, B e C: 2cm
Topograficamente, o saco cranial caracterizou-se pela zona de inserção esofágica,
sendo a primeira cavidade do estômago situada na região cranial, desviada a esquerda. Em
seguida destacou-se o saco central posicionado na região mediana do corpo do animal,
precedendo o saco caudal, que estava posicionado no antímero direito. O fundo era localizado
na região crânio central do órgão de modo contra-lateral ao divertículo, que sobrepunha os pré
piloros I e II (inserção duodenal) (Figura 2B).

32
FIGURA 2 – Estômago de Bradypus variegatus ‘in situ’ e ‘ex situ’. 1A: 1 - Saco cranial; 2 – Saco central; 3-
Saco caudal; 4 – Divertículo; 5 – Intestino (Barra 2cm). 1B: 1- Esôfago; 2- Saco cranial; 3 -Saco central; 4 –
Fundo; 5 – Divertículo; 6 – Saco caudal; 7 – Pré piloro I; 8 – Pré piloro II (Barra 2cm). 1C: 1 – Esôfago; 2 –
Inserção esofágica; 3- Fundo; 4 – Pré piloro II; 5 – Divertículo (Barra 2 cm).
Na abertura do órgão, verificou-se a face interna delimitada por válvulas que o
subdividiam em cavidades menores (Figura 2A). As áreas de saculação (saco cranial, central e
caudal) eram constituídas por uma mucosa pregueada, divididas por pilares que
circunscreviam cada compartimento, compondo a maior parte do estômago desses animais,
sendo o saco central delimitado de maneira mais proeminente, podendo ser visível com órgão
íntegro.
33
FIGURA 3 – Fotomacrografia interna do estômago de Bradypus variegatus. 1A: 1 - Saco cranial; 2 – Fundo; 3-
Divertículo; 4- Saco caudal; 5– Pré piloro I; 6 – Pré piloro II; 7 – Saco central (Barra 2cm). 1B – Pregas
diverticulares (bolsas) (Barra 1cm).
A região do fundo compôs a camada mais delgada do órgão, com ausência de pregas
no interior, diferenças macroscópicas de coloração e aspecto apresentando uma superfície lisa
e escurecida quando comparada com a região sacular, que compunha uma mucosa áspera e
clara, dividido por uma válvula densa.
34
Figure 4 – Ultrassonografia de estômago de preguiça Bradypus variegatus. Círculo: Válvula saco cranial –
fundo.
O divertículo revelou forma sacular projetada a partir do estômago, sendo bipartido e
mostrando na porção interna, diversas pregas tortuosas de aspecto labiríntico,
compartimentado por pequenas seções assimétricas (Figura 2B).
A porção final do estômago, já em direção aos intestinos era formada por duas partes
tubulares, sendo a primeira fração apresentando aspecto escurecido, delgado e liso
denominada pré piloro I, e a segunda fração subdividida por um esfíncter, revelando dois
padrões de mucosa e musculatura, com a primeira porção rugosa e parede fina, e a segunda
com a mucosa mais clara e musculatura densa com o lúmen estreito que precedeu o duodeno.
Descrição microscópica
Histologicamente, o saco cranial revelou, da porção interna para a mais externa,
mucosa de tecido plano queratinizado estratificado baixo, sendo o estrato córneo a região
mais desenvolvida e de padrão de estratificação ausente, sucedido pela região submucosa com
a discreta camada de tecido conjuntivo denso e frouxo, camada muscular marcante de padrão
aparente e fasciculado, e serosa espessada composta por tecido conjuntivo e revestida por
tecido epitelial plano, com a microscopia eletrônica confirmando os achados da microscopia
35
ótica (Figura 5).
Figure 5 – Fotomicrografia da porção queratinizada do estômago de preguiças Bradypus variegatus. A) Saco
cranial – mucosa cornificada, submucosa de tecido conjuntivo frouxo, muscular em feixes longitudinais B)
Padrão de distribuição muscular (feixes espessos) intervalados de tecido conjuntivo ( saco cranial); C) Superfície
mucosa recoberta de uma camada espessa de queratina (saco cranial) D) Saco central – mucosa com cristas de
queratina, submucosa de tecido conjuntivo acompanhando o padrão da mucosa, duas camadas musculares e
serosa padrão E) Lâmina muscular transversal com feixes recobertos por tecido conjuntivo (saco central); F)
Superfície sanfonada da mucosa do saco central com mudança de padrão em relação a porção anterior G) Saco
caudal – mucosa cornificada, submucosa padrão, dupla camada muscular (oblíquo e longitudinal) e serosa
padrão) Padrão muscular de dupla camada (transversal e longitudinal) do saco caudal H) Padrão de disposição de
queratina (saco caudal).
O saco central também exibiu uma mucosa com extrato córneo desenvolvido e sem
padrão de estratificação, porém, a camada muscular, apesar de padronização circular e
longitudinal, se mostrou menos pronunciada em comparação ao saco cranial, enquanto que na
região externa do saco, a camada serosa era semelhante ao supracitado.

36
A do fundo apresentou mucosa revestida por tecido epitelial glandular prismático
baixo, com formações de fossetas curtas e glândulas longas e logo abaixo das glândulas
(produtoras de muco - padrão evidenciado na MEV), uma camada muscular da mucosa com
aspecto padrão. A submucosa apresentou-se irrigada e a camada muscular composta por duas
outras camadas mais delgadas (externa circular e interna longitudinal). Na região externa da
do fundo, a serosa também obedeceu ao padrão descrito anteriormente (Figura 5).
Figura 6 – Fotomicrografia da porção do fundo do estômago de preguiças Bradypus variegatus. A) Fundo –
mucosa com glândulas produtoras de muco e ausência de células oxínticas e pépticas; B) Transição saco
cranial/fundo – Válvula composta de tecido conjuntivo e tecido cornificado (queratina) (centro); a esquerda –
padrão glandular (fundo) e a direita, padrão cornificado (saco cranial); C) Transição saco cranial/fundo: abaixo –
padrão glandular (fundo) e acima, padrão cornificado (saco cranial). D) Fossetas e fovéolas do padrão glandular
da região do fundo.
Mais caudalmente, o divertículo evidenciou uma mucosa reticulada revestida por
tecido glandular prismático baixo de células da mucosa padrão, suportada por uma lâmina

37
basal e, muscular da mucosa com a camada muscular densa e serosa padrão (Figura 6).
Figura 7 – Fotomicrografia do divertículo do estômago de preguiças Bradypus variegatus. A) Divertículo em
menor aumento – padrão labiríntico do órgão com mucosas formando criptas para o lúmen, submucosa composta
de tecido conjuntivo acompanhando as criptas e dupla camada muscular circunscrevendo o órgão B) Mucosa
composta de criptas e cristas de células produtoras de muco e ausência de células oxínticas e pépticas, camada
muscular dupla (longitudinal interna e transversal externa) C) Cristas formadas de tecido conjuntivo com mucosa
para o lúmen do órgão D) Epitélio glandular da superfície diverticular.
Antecedendo a região pré pilórica e finalizando as saculações, o saco caudal
caracterizou-se pela mucosa composta por tecido epitelial plano estratificado queratinizado, e
suportado por uma lâmina própria, seguido por submucosa de tecido conjuntivo frouxo,
contendo inervação e vasos sanguíneos. A camada muscular era composta por tecido muscular
liso, disposto transversal e longitudinalmente e serosa constituída pelo mesmo padrão do
órgão.
Na região que precede os intestinos, o pré piloro I, era constituído por mucosa
glandular composta por tecido prismático baixo, composto de células parietais acidofílicas e
principais basofílicas em sua superfície, característica de estômago químico.
Por fim, o pré piloro II apresentava mucosa também aglandular, de tecido pavimentoso

38
plano intensamente queratinizado com cristas, a região muscular desenvolvida, em um único
feixe espesso e fasciculado, enquanto que a camada serosa revelou-se delicada, diferindo dos
compartimentos anteriores do órgão (Figura 7).
Figura 8 – Fotomicrografia da pré pilórica do estômago de preguiças Bradypus variegatus. A) pré piloro I –
Padrão tecidual com mucosa glandular (camadas mucosa, muscular da mucosa, submucosa, muscular e serosa);
B) Pré piloro - Mucosa composta de células oxínticas ou parietais (acidofílicas) nas criptas e pépticas ou
principais (basofílicas) na lamina basal; C) Transição pré piloro I/pré piloro II – padrão glandular químico
(direita – pré piloro I) e queratinizado musculo -fibroso (esquerda – pré piloro II) D) Pré piloro II – Mucosa

39
aglandular cornificada com submucosa espessa composta de tecido conjuntivo frouxo abundante e muscular
desenvolvida; E) Superfície glandular (pré piloro I); F) Válvula transição pré piloro I/pré piloro II; G) Superfície
cornificada com rosetas de queratina -pré piloro II (MEV_50X).
Exames de imagem e dinâmica gástrica
O trajeto após a injeção de contraste mostrou que o percurso alimentar, após o esôfago
foi na seguinte ordem: Saco cranial, saco central, fundo, divertículo, saco central, saco caudal
e pré piloros. O contraste seguiu direto para o saco cranial (área de inserção esofágica) no
tempo 0, avançou para o saco central. Em seguida, preencheu o fundo e posteriormente o
divertículo (10 min) e pré piloro I. Nos exames de 20 e 30 min, não houve preenchimento de
outras cavidades visíveis. Após 40 min, o saco caudal e pré piloro I começaram o processo de
pigmentação, não havendo grandes modificações até 80 min, sendo nítido apenas aos 90 min
a continuação do fluxo de contraste para as alças intestinais (Figura 4).
Figura 10 – Trânsito gástrico no raio X de contraste em preguiça comum (Bradypus
variegatus). A: 0 min; B: 10 min; C:20 min; D: 30 min; E: 40 min; F: 50 min; G: 60 min; H:
90 min; (1- Saco cranial; 2 – Fundo; 3- Divertículo; 4 – Saco central; 5 – Saco caudal; 6 Pré
piloro I; 7 – Pré piloro II).

40
DISCUSSÃO
A pluralidade da Superordem Xenarthra quanto a morfologia digestória é demonstrada
pela diversidade de conformações nas espécies já descritas e esses animais, mesmo que
pertencendo a mesma Superordem, apresentam características distintas, como observado nos
estudos de Pinheiro et al. (2014) em que descreveu a morfologia estomacal de Tamandua
tetradactyla, e de Menezes (2013) quanto a morfologia gástrica de Mymercophaga tridactyla,
sendo ambas as espécies com estômagos unicavitários em formato de “J”, equivalente a
diversas espécies domésticas. Rezende (2011) que descreveu o estômago e a vascularização
do trato digestório de B. tridactylus demonstrou um órgão de padrão multicavitário
semelhante ao do nosso estudo.
Essa diversidade pode dificultar o manejo cativo desses animais no que concerne a
nutrição, podendo diminuir a taxa de sucesso de reabilitação como retrata Diniz (2001), que
salienta a importância de se conhecer as características variáveis de anatomia, fisiologia,
comportamento, na tentativa de reduzir problemas clínicos, sendo a deficiência nutricional a
de maior incidência em preguiças (DINIZ, 1999).
Quanto a extensão do órgão, animais de hábitos folívoros tem, por natureza,
estômagos de grandes dimensões em relação aos outros órgãos do corpo, como evidenciado
por Britton (1941) o qual alega que este órgão abrange entre 20 a 30% da massa corporal em
preguiças da espécie Bradypus tridactylus, que foi ainda detalhada por outros autores
(GRASSÉ, 1955; LANGER,1979a, 1987; LANGER, 2002; GARCIA e LEAL; CLAUSS et
al.., 2005) e mais recentemente a Bradypus torquattus, descrita por Rezende (2011), que
tambem possui conformações semelhantes.
O aspecto macroscópico subsidia que cada porção apresenta uma mucosa
característica e consequentemente deve apresentar uma função específica, o que torna claro a
delimitação das câmaras, além de um sistema de válvulas que permite a individualização de
cada compartimento de acordo com curso alimentar, porém, a análise microscópica permite
inferir mais precisamente o número de cavidades, que nas Bradypus variegatus fica evidente
se tratarem de sete cavidades, contrariando o descrito por Cassano (2006), que descreveu o
estômago das preguiças do gênero Bradypus como grande e dividido em apenas quatro
câmaras, sendo as três primeiras para fermentação e a quarta para ação de ácidos e enzimas se
baseando apenas no aspecto macroscópico.

41
Inúmeras análises já foram documentadas quanto a anatomia do estômago de
preguiças do gênero Bradypus (GILMORE et al.., 2000; PACHECO et al.., 2007; REZENDE
et al.., 2011), porém, nenhuma evidencia exatamente o que ocorre no interior do órgão da
preguiça da espécie Bradypus variegatus, inclusive a correlação dos achados anatômicos com
a histologia. Kent (1954) analisando quanto a presença de glândulas descreveu três regiões:
cárdica, fúndica e pilórica, padrão este diferente do estômago de Bradypus variegatus, visto
que a disposição glandular não obedece ao padrão de animais monogástricos.
Já Bachop (1975) relatou que o estômago desses animais consiste em três câmaras
cardinais, divididas em pilares, compostas por um epitélio estratificado, uma camada direita
ou anterior composta por um saco cego que se estende até o divertículo, sendo composta por
glândulas e uma camada ventricular que vai até o piloro, no entanto, após analisarmos a
morfologia microscópica do estômago de B. variegatus evidenciamos sete compartimentos
valvulados e diferenciados não apenas microscopicamente, mas também quanto ao aspecto e
textura do conteúdo gástrico presente no momento do exame macroscópico, o que poderia
explicar esse número maior de compartimentos.
Mais recentemente, Mesquita (2015) em um estudo anatômico do estômago desses
animais, enumera seis compartimentos apenas macroscopicamente, unificando as porções
queratinizadas em duas partes (câmara cárdica e corpo do estômago), porém, o estudo
histológico sugere um padrão tanto de queratina quanto muscular diferente entre as camadas,
o que subsidia a existência de divisões e funcionalidades distintas.
As saculações gástricas (cranial, central e caudal) da B. variegatus seguem um padrão
de órgãos tubulares de animais herbívoros, que são responsáveis pelo trajeto do bolo
alimentar, com queratina evidente (PADYKULA, 1981; HAM e CORMACK, 1983). A
camada muscular densa permite uma movimentação intensa do conteúdo para facilitar a
degradação, visto que animais do gênero Bradypus são considerados primitivos, possuindo
apenas quatro pares de molar e uma mastigação deficiente (SANTANA et al., 2013). No
entanto, o esôfago da preguiça não possui dilatações, ocorrendo todo o processo nos
compartimentos estomacais (MESQUITA, 2018).
Os achados histológicos demonstram a similaridade das preguiças da espécie
Bradypus variegatus com animais poligástricos como os ruminantes que apresentam nas
saculações ruminais, uma mucosa estratificada fortemente queratinizada, recoberta por uma
camada muscular dividida por pilares favorecendo a contratilidade estomacal e a degradação
de folhas (DELLMANN E BROWN, 1982), possibilitando a proteção do epitélio, auxiliando
a exposição e metabolismo dos ácidos graxos, e subsidiando o local ideal para colonização

42
bacteriana (SAMUELSON, 2007; DILL-MCFARLAND et al., 2016).
Essa similaridade, tanto anatômica quanto fisiologica com os herbívoros ruminantes,
foi reafirmado pelo aproveitamento de ácidos graxos de cadeia curta (AGCC) pela microbiota
bacteriana constituinte do estomago das preguicas, que também fazem a absorção fisiológica
desses ácidos a partir da fermentação nas câmaras estomacais, descritas por Dill-Mcfarland
(2016), a qual o mesmo raciocínio não se aplica a Koalas (BARKER et al.., 2013; ALFANO
et al.., 2015) e Alouatta caraya (AMATO et al.., 2013; AMATO et al.., 2014) que contam com
as colônias intestinais para o aproveitamento da biomassa das plantas ingeridas em nutrientes
absorvíveis.
O divertículo sugere também a presença de colonização bacteriana e a fermentação em
animais com ceco não fermentativo, o que provavelmente ocorre com o gênero Bradypus,
similar ao descrito em primatas das espécies Presbytis entellus e Presbytis cristatus, em que
são chamados de ‘Ruminant-like’ por se assemelharem com os ruminantes, porém, sem
ruminação efetiva (BAUCHOP e MARTUCCI, 1968). Essa similaridade foi descrita também
por Pacheco (2007), estudando microbiologia das porcões saculares e diverticular de
Bradypus tridactylus, observando que a carga bacteriana nessas porções se assemelha a
encontrada em ruminantes.
A existência de bactérias também sustenta a capacidade das preguiças de
‘desintoxicar’ plantas jovens consumidas, sendo essas bactérias responsáveis possivelmente
pela quebra dessas substâncias (KOHL et al.., 2014).
eOs pré piloros foram descritos morfologicamente nos animais da espécie Bradypus
torquarttus, divididos em parte glandular (pré piloro I) que se assemelha ao estômago
propriamente dito e parte musculo-fibrosa (pré piloro II) (REZENDE, 2011), com essa fração
glandular possivelmente responsável pela digestão enzimática, seguindo o padrão de outros
animais herbívoros descritos anteriormente, porém, estudos admitem que a funcionalidade
dele pode ser questionada quanto o percentual de participação digestiva pela distribuição de
lisozimas, sendo pouco abundante, o que difere dos ruminantes traçando um paralelo com a
região abomasal (PACHECO, 2007), entretanto se assemelha quanto a distribuição de células
quimicamente ativas produtoras de ácidos estomacais.
Ainda sobre a biodisponibilidade de nutrientes, aliadas a fermentação bacteriana e
comparativamente a bovinos no processo de ruminação, o processo pode ser análogo: A

43
preguiça, semelhante a ruminantes é herbívora, logo precisa da ação bacteriana para absorção
de nutrientes. Comparativamente, os estômagos sob forma de câmaras fermentativas com
bactérias ativas antes da ação química, torna esses indivíduos parentes muito próximos dos
ruminantes que biodisponibilizam os nutrientes através de fermentação de rúmen e retículo
com bactérias, sendo misturadas com o conteúdo ruminal, retorno a cavidade oral, mastigação
e posteriormente omaso e abomaso (digestão química) para absorção intestinal (KOLB,
1980), com os exames de raio X evidenciando os eventos equivalentes nesses animais.
CONCLUSÃO
Por fim concluímos, até o presente momento, que a complexidade dos estômagos
poligástricos, apresentando uma câmara fermentativa precedente a um estômago químico bem
caracterizado, aliado a composição histológica torna esses indivíduos semelhante aos
ruminantes na morfologia gástrica, necessitando de estudos a respeito da fisiologia digestiva
desses animais.
REFERÊNCIAS
ALFANO, N.; COURTIOL, A., VIELGRADER, H., TIMMS, P., ROCA, A.L., GREEN-WOOD, A.D. Variation in koala microbiomes within and between individuals: effect of body region and captivity status. Sci Rep 5, 2015.
AMATO, K.R., LEIGH, S.R., KENT, A., MACKIE, R.I., YEOMAN, C.J., STUMPF, R.M. et al.. The role of gut microbes in satisfying the nutritional demands of adult and juvenile wild, black howler monkeys (Alouatta pigra). American Journal of Physical Anthropology: n/a-n/a, 2014.
AMATO, K.R., YEOMAN, C.J., KENT, A., RIGHINI, N., CARBONERO, F., ESTRADA, A. et al.. Habitat degradation impacts black howler monkey (Alouatta pigra) gastrointestinal microbiomes. ISME J 7: 1344-1353, 2013.
BARKER, C.J., GILLETT, A., POLKINGHORNE, A., AND TIMMS, P. Investigation of the koala (Phascolarctos cinereus) hindgut microbiome via 16S pyrosequencing. Veterinary Mi-crobiology 167: 554-564, 2013.
BAUCHOP, P. Digestion of Leaves in Vertebrate Arboreal Folivores. In: MONTGOMERY G.G. (Eds). The Ecology of Arboreal Folivores. Smithsonian Institution Press, Washington, p.193-204, 1978.
BAUCHOP, T; MARTUCCI.R.W. Ruminant-Like Digestion of the Langur Monkey. Science,

44
v.161,pp.698-700,1968.
BRITTON, S.W. Form and function in the sloth. The Quarterly Review of Biology. v.16, p.190-207, 1941.
CASSANO, C.R. Ecologia e conservação da preguiça-de-coleira (Bradypus torquatus Illiger, 1811) no sul da Bahia. Dissertação de Mestrado, Universidade Estadual de Santa Cruz, 2006.
CORK, S.; FOLEY, W. Digestive and metabolic strategies of arboreal folivores in relation to chemical defences in temperate and tropical forests. In Plant Defenses Against Mammalian Herbivory. Palo, R., and Robbins, C. (eds). Boca Raton, FL: CRC Press, pp. 166- 175, 1991.
CRANDALL, LS. The management of wild mammals in captivity. Chicago: University of Chicago Press, 1964.
DILL-MCFARLAND, KA.; WEIMER, PJ.; PAULI, JN.; PEERY, MZ.; SUEN, G..Diet specialization selects for an unusual and simplified gut microbiota in two- and three-toed sloths. Environ Microbiol. May;18(5):1391-402, 2016.
DINIZ, LSM.. General medicine. In: Fowler M.E. (Ed.), Biology, Medicine and Surgery of South American Wild Animals. Iowa State University Press, p.402-422, 2001.
DINIZ, LSM.; OLIVEIRA PMA. Clinical problems of sloths (Bradypus sp and Choloepus sp) in captivity. J. Zoo Wildl. Med. 30:76-80, 1999.
FOLEY WJ, ENGELHARDT WV, CHARLES-DOMINIQUE P. The passage of digesta, par-ticle size, and in vitro fermentation rate in the three-toed sloth Bradypus tridactylus (Edentata: Bradypodidae). Journal of Zoology 236(4):681–696, 1995.
GILMORE, D., DA COSTA, C., AND DUARTE, D. Sloth biology: an update on the physio-logical ecology, behavior and role as vectors of arthropods and arboviruses. Braz J Med Biol Res 34: 9- 25, 2001.
GILMORE, D., DA COSTA, C., AND DUARTE, D. An update on the physiology of two- and three-toed sloths. Brazilian Journal of Medical and Biological Research 33: 129-146, 2000.
HAM, AW. CORMACK, DH. Histologia. 8ed. Rio de Janeiro: Guanabara Koogan. 543p, 1983.
HILDEBRAND, M. & G. GOSLOW. Celomas mesentéricos, p. 195-199. In: M. HILDEBRAND & G. GOSLOW (Eds). Análise da estrutura dos vertebrados. São Paulo, Atheneu, 637p, 2006.
KENT, G. C. Digestive system. In: Kent G. C. (Ed.). Comparative anatomy of the vertebrates. New York, McGraw-Hill Book Company, pp.305-329, 1954.
KOLB, E. Fisiologia veterinária. Rio de Janeiro: Guanabara Koogan. 612p, 1980.

45
KOHL, K.D., WEISS, R.B., COX, J., DALE, C., AND DENISE DEARING, M. Gut mi-crobes of mammalian herbivores facilitate intake of plant toxins. Ecology Letters 17: 1238-1246, 2014.
MACHADO, MRF. et al.. Morfologia do estômago do cervo-do-pantanal (Blastocerus dichotomus, Illiger 1815). Arquivo Brasileiro de Medicina Veterinária e Zootecnia, v. 67, n. 2, p. 424-432, 2015.
MCNAB BK. Energetics of arboreal folivores: physiological problems and ecological conse-quences of feeding on an ubiquitous food supply. In: Montgomery GG, ed. The ecology of ar-boreal folivores. Washington: Smithsonian University Press, 153–162, 1978.
MENEZES, LT. Morfologia do tubo digestório do tamanduá bandeira Myrmecophaga tridactyla (Pilosa: Myrmecophagidae) Dissertação de mestrado 79f - Universidade Federal de Uberlândia, 2013.
MESQUITA, EP.; ALBUQUERQUE, PV.; SANTOS, FC.; NASCIMENTO, JCS.; AMORIM, MJAAL.; MOURA, GJB. Estudo anatômico do estômago da Bradypus variegatus - Shinz, 1825 (Mammalia, Xenarthra) Ciênc. vet. tróp., Recife-PE, v.18 n 2, 2015.
MURAKAMI, T. A revised tannic-osmium method for noncoated scanning electron micro-scope specimens. Archivum Histologicum Japonicum, v. 36, p. 189-193, 1974.
NAGY KA, MONTGOMERY GG. Field metabolic rate, water flux, and food consumption in three-toed sloths (Bradypus variegatus). Journal of Mammalogy 61:465–472, 1980.
OHTANI, O. Three dimensional organization of the connective tissue fibers of the human pancreas: a scanning electron microscopy study of NaOH treated tissues. Archivum Histolog-icum Japonicum, v. 50, n. 5, p. 557-566, 1987.
PACHECO, M.A., CONCEPCIÓN, JL.; RANGEL, JDR.; RUIZ, MR.; MICHELANGELI, F.; DOMÍNGUEZ-BELLO, M.G., Stomach lysozymes of the three-toed sloth (Bradypus var-iegatus), an arboreal folivore from the Neotropics. Comparative Biochemistry and Physiol-ogy, Part A 147, p 808–819, 2007.
PADYKULA, HA. O trato digestivo. In: WEISS, L.; GREEP, R. O. Histologia. 4.ed. Rio de Janeiro: Guanabara Koogan, 1981. pp. 552-554.
PAULI, J. N.; MENDOZA, J. E.; STEFFAN, S. A.; CAREY, C. C.; WEIMER, P. J.; PEERY, M. Z. A syndrome of mutualism reinforces the lifestyle of a sloth. Proceedings of the Royal Society B 281, 2013-3006, 2014.
PINHEIRO, ACO.; LIMA, AR.; CARVALHO, AF.; PEREIRA, LC.; Branco, E.Aspectos morfológicos macro e microscópicos do estômago de tamanduá-mirim (Tamandua tetradactyla). Arq. Bras. Med. Vet. Zootec., v.66, n.4, p.1089-1096, 2014.
RAINES J. Captive health and husbandry of the Bradypodidae. Zoo Biology 24(6):557–568, 2005.
REZENDE; LC.; MONTEIRO; JM.; CARVALHO; P.; FERREIRA, JR.; MIGLINO,

46
MA. Morphology and Vascularization of the Gastric Compartments in Three-Toed Sloth (Bradypus torquatus Illiger, 1811). Int. J. Morphol. [online], vol.29, n.4, pp.1282-1290. 2011.
SANTANA, LNS.; BARBOSA, LVM.; TEIXEIRA, FB.; COSTA, AMP.; FERNANDES, LMP; LIMA, RR. Morphology of the Dentin Structure of Sloths Bradypus tridactylus: A Light and Scanning Electron Microscopy Investigation. Anatomia Histologia Embryologia, v 42p 410–414, 2013.
STEVENS, C.E., AND HUME, I.D. The mammalian gastrointestinal tract. In Comparative physiology of the vertebrate digestive system. New York, New York: Cambridge University Press, pp. 65-67, 1995.
VAUGHAN, C.; RAMIREZ, O.; HERRERA, G.; GURIES, R.. Spatial ecology and conserva-tion of two sloth species in a cacao landscape in Limon, Costa Rica. Biodiversity and Conser-vation :16, 2293– 2310, 2007.
WATANABE, I.; YAMADA, E. The fine structure of lamellated nerve endings found in the rat gingiva. Archivum Histologicum Japonicum, v. 46, n. 6, p. 173-182, 1983.
Recommended
















![AmazôNia [1]](https://img.document.onl/doc/110x75/55bdde59bb61eb3b668b4611/amazonia-1.jpg)
