
UNIVERSIDADE FEDERAL DE SERGIPE
PROGRAMA DE PÓS-GRADUAÇÃO EM ECOLOGIA E CONSERVAÇÃO
VARIAÇÃO SAZONAL E LONGITUDINAL NA ECOLOGIA ALIMENTAR
DE CHIROPOTES SAGULATUS TRAIL, 1821 NA AMAZÔNIA
BRASILEIRA
Viviane Sodré Moura
Mestrado Acadêmico
São Cristóvão
Sergipe – Brasil
2016

Viviane Sodré Moura
VARIAÇÃO SAZONAL E LONGITUDINAL NA ECOLOGIA ALIMENTAR
DE CHIROPOTES SAGULATUS TRAIL, 1821 NA AMAZÔNIA
BRASILEIRA
Dissertação apresentada ao Programa de Pós-Graduação em Ecologia e Conservação da Universidade Federal de Sergipe, como requisito parcial para obtenção do título de Mestre em Ecologia. Orientador: Prof. Dr. Stephen Francis Ferrari. Co-Orientador: Prof. Dr. Fabiano Rodrigues de Melo.
SÃO CRISTÓVÃO
SERGIPE – BRASIL
2016

M929v
Moura, Viviane Sodré
Variação sazonal e longitudinal na ecologia alimentar de Chiropotes sagulatus Trail, 1821 na Amazônia brasileira / Viviane Sodré Moura ; orientador Stephen Francis Ferrari. – São Cristóvão, 2016.
63 f. : il.
Dissertação (mestrado em Ecologia e Conservação) – Universidade Federal de Sergipe, 2016.
1. Ecologia animal. 2. Dieta. 3. Animais - Comportamento. 4. Animais - Proteção. 5. Amazônia. I. Ferrari, Stephen Francis, orient. II. Título.
CDU 591.5(811.3)


DEDICATÓRIA
Dedico este trabalho a todos os cuxiús, especialmente aos do grupo Bacaba, do qual hoje
também faço parte. E à Liza Maria Veiga, que docemente abriu espaço e caminho para que eu
pudesse contribuir para o conhecimento a respeito dos cuxiús e para conhecer meu orientador,
Stephen Francis Ferrari. Por toda sua dedicação, entrega e gentileza em oferecer ajuda. E para
que tenha encontrado tudo o que lhe faltava nos braços da Mãe Divina. Gratidão.

AGRADECIMENTOS
Pai Celestial, Mãe, Amigo e Bem Amado Deus. Jesus Cristo, Baghavan Krishna,
Mahavatar Babaji, Lahiri Mahasaya, Swami Sri Yukteswar, Amado Gurudeva Paramahansa
Yogananda, gratidão porque todas as portas foram abertas e cuidadas pelo Infinito Amor que
transcende a matéria e flui de vocês diretamente a mim. Eu os reverencio.
A todos os que deixaram Deus cuidar de mim através de suas vidas, Gratidão. Cada
um que recebeu uma das minhas páginas amarelas tem um lugar especial na minha vida, e isto
extrapola o agradecimento pela ajuda acadêmica, pois são todos pedaços do meu coração
vivendo fora de mim.
Mãe e Pai, Irmã e João, gratidão porque vocês estavam aqui. Maria Eduarda, por ter
me tirado do estudo quando era necessário!
Thiago, o amor que foi escolhido para ser o companheiro da minha caminhada. A sua
família, tão querida que hoje também é minha. Gratidão por ser e por estar. Por aguardar com
amor e solidariedade durante as minhas ausências, e por cuidar com dedicação dos meus
afazeres na roça.
Steve, você disse sim e tudo isto foi possível. Gratidão por suas poucas palavras,
perfeitamente apropriadas para cada momento. Agora mesmo, durante os ajustes no texto e
interpretações a respeito dos dados que tenho em minha frente, Gratidão por estar presente,
quase que ao meu lado, orientando cada palavra e cada conceito. Escrever sob a sua
orientação bem humorada, paciente e muitíssimo competente, faz deste um trabalho mais
prazeroso.
Liza Maria Veiga, “Quem traz no nome esta marca, possui a estranha mania de ter fé
na vida, Maria, Maria”... Você continua sendo inspiração de competência, doçura. Gratidão
por ter sido chave para que portas fossem abertas, gratidão por ter sido porta, e fonte para
desfazer tantas dúvidas, quando eu ainda nem sabia o que significava direito uma pesquisa
comportamental. Como eu gostaria de te entregar pessoalmente minha página amarela...
Tati, por ter cuidado dos cuxiús após 2012, pelas conversas e confirmações a respeito
dos comportamentos. Por gentilmente doar as informações necessárias para este trabalho se
concretizar! Você me salva!!!!

Laboratório de Biologia da Conservação, Família Feliz, A Turma de Mestrado 2014.1,
Catingueiros do Arrocha, Amor... São os grupos com quem eu pude ser eu, com o meu
melhor e o meu pior. Por isso sim, Gratidão!!!!
Aos Deeksha Givers de Aracaju, especialmente Marizi Dória, lindas almas que me
acolheram com amor e luz, pela linda formatura e iniciação!!! Jay Bolo, gratidão porque esta
etapa se encerra com alegria, Sri Kalki Ki!!! Jay!!!!
Voltando ao tempo da Amazônia, Letícia, minha flor, eu faria tudo de novo para poder
não morrer de saudades de você! Pietro, Thiago, Branco, Tati, Cacá, Hugo, Vanessa, toda a
Confraria Vanilla etílica, foi muito melhor porque vocês estavam por perto. Seu João, meu
velhinho preferido, por ser suporte, alegria e amor pra vida inteira... Cleiton, Sarabá, Nego,
Colé, Edi, Eli, Careca, Serrão, gratidão por me ensinar a ler a Floresta Amazônica, desde as
primeiras letras, quando eu ainda era analfabeta. Bião (Fabiano Rodrigues de Melo) e Lelê
(Leandro Santana Moreira) pela parceria nessa pesquisa e oportunidade de trabalhar na
Amazônia e usar os dados coletados.
Ao PPEC da UFS, na pessoa da Juliana Cordeiro, que com belíssimo humor me
lembrou de cada compromisso. Aos professores do Programa, pela construção de minha
formação, gratidão!
Ao Keoma Coutinho, que sem me conhecer pessoalmente, foi gentil em ensinar a
análise do z binomial, esclarecendo até as minhas dúvidas mais primárias! Gratidão!
À Mari a e sua linda família, que durante grande parte da escrita, no tempo que a
Alegria ainda não tinha energia elétrica, cedeu gentilmente uma tomada para que eu ligasse o
computador. A caminhada diária de 1,5 km era recompensada pelo brilho no seu sorriso!
A CAPES pela concessão das bolsas, ao convênio FUNAPE UFG campus Jataí e a
MRN por ceder os dados coletados e apoio financeiro.
Pela chuva que caiu todas as vezes que era necessário desligar o computador na roça...
Gratidão.

Quando os olhos ainda não podem ver sozinhos, e a luz
que tudo permeia ainda não surgiu no horizonte, no resta
aguardar... É preciso estar pronto e atento para saber o
momento exato de dar a largada e seguir...
Para mais um dia de campo, ou para a vida.
(Vivi)

Resumo
Este estudo teve por objetivo avaliar variações sazonais e longitudinais no padrão de agrupamento e ecologia alimentar de um grupo de Chiropotes sagulatus, os cuxiús, na FLONA Saracá-Taquera, estado do Pará, Amazônia Brasileira. O grupo de estudo habituado, denominado Bacaba, foi monitorado entre Julho 2011 e Março 2012 (em todos os meses exceto Setembro) e Novembro 2014. Os dados comportamentais e de dieta foram coletados através do método de varredura instantânea, com duração de dois minutos e intervalo de 10 minutos, sendo coletadas amostras dos itens alimentares e realizada a contagem de indivíduos. O grupo Bacaba continha 67 indivíduos. Foram realizados 35 dias completos de observação e o orçamento de atividades foi Deslocamento (41,2%), Repouso (30,2%), Forrageamento (13,5%) e Alimentação (8,1%), Interações Sociais e Outros (6,9%). Foi realizado teste de Shapiro Wilk para a normalidade dos dados, o teste t de Student para os tamanhos de subgrupos e o teste z binomial para comparações de comportamento e dieta sazonal e longitudinal, além da similaridade de Jaccard para a composição taxonômica da dieta. Houve uma tendência para subgrupos maiores na estação chuvosa e no ano de 2011, mas a diferença não foi significativa. Deslocamento foi significativamente maior na estação chuvosa, quando os comportamentos de Alimentação e Forrageamento foram menores e as atividades de Repouso e Social foram significativamente maiores na estação seca. A dieta do grupo Bacaba apresentou alta ingestão de sementes para todo o período de estudo, e alta frequência de ingestão de sementes imaturas para as duas estações (59,9% estação seca e 47,7% estação chuvosa) com diferença significativa quando o item alimentar foi semente. Os itens partes florais e semente de fruto seco foram ingeridos apenas na estação seca. Todos os comportamentos apresentaram diferenças significativas entre anos, e foram mantidas as proporções dos itens alimentares na dieta mesmo com a baixa similaridade na composição taxonômica, mostrando a flexibilidade dos cuxiús para manter uma dieta de qualidade. As famílias mais importantes para a dieta do grupo foram Sapotaceae, Moraceae, Fabaceae, Chrysobalanaceae e Lecythidaceae, conhecidamente importantes para o gênero. Este estudo apresenta informações que reforçam os padrões comportamentais e de dieta para o gênero Chiropotes, corroborando a flexibilidade em sua ecologia alimentar e estratégia de forrageamento de maximização de energia.
Palavras chave: ecologia comportamental; dieta; conservação

Abstract
Seasonal and longitudinal variation in the feeding ecology of Chiropotes sagulatus Trail,
1821, in the Brazilian Amazon
The present study investigated the seasonal and longitudinal variation in the feeding ecology of a group of cuxiús, Chiropotes sagulatus, in the Saracá-Taquera National Forest, in the Brazilian Amazonian state of Pará. For the collection of quantitative behavioral data, the habituated study group, denominated the Bacaba group, was monitored between July 2011 and March 2012 (in all months except September) and November 2014. Data on the behavior and diet of the study group were collected on three to five days per month, based on instantaneous scan sampling, with a two-minute scan being conducted at 10-minute intervals, during which the number of individuals present was counted and samples of food items were collected. The Bacaba group contained 67 members. A total of 35 complete days of observation were conducted, and the general activity budget was Locomotion (41.2%), Rest (30.2%), Foraging (13.5%), and Feeding (8.1%), social interactions and others (6.9%). The normality of the data was evaluated using the Shapiro-Wilk test, variation in the size of subgroups was assessed using Student’s t, seasonal and longitudinal variation in the activity budget and composition of the diet was evaluated using binomial z scores, as well as Jaccard’s similarity index, for the comparison of the taxonomic composition of the diet. There was a tendency for the formation of larger subgroups during the rainy season of 2011, although the difference was not significant. Locomotion increased significantly during the rainy season, when Feeding and Foraging decreased, while Rest and Social Interactions increased significantly in the dry season. The diet of the Bacaba group was dominated by seeds throughout the study period, with high rates of ingestion of immature seeds being recorded during both the dry (59.9%) and rainy (47.7%) seasons, with a significant seasonal difference for seed feeding. Flower parts and the seeds of dry fruit were consumed only during the dry season. Significant differences were found between years in all behavior categories, although the composition of the diet was maintained, despite major modifications in its taxonomic composition, reflecting the capability of the cuxiús to obtain a high quality diet despite seasonal fluctuations in resource availability. The most important plant families in the study group’s diet were the Sapotaceae, Moraceae, Fabaceae, Chrysobalanaceae and Lecythidaceae, which are known to be important for Chiropotes. This study reconfirms the behavioral patterns and diet found in the genus Chiropotes, emphasizing the flexibility of its feeding ecology and energy-maximizing foraging strategy.
Key words: behavioral ecology; diet; conservation

SUMÁRIO Resumo .................................................................................................................................. xviii
Abstract .................................................................................................................................. xviii
LISTA DE FIGURAS ................................................................................................................ x
LISTA DE TABELAS .............................................................................................................. xi
1. Introdução.......................................................................................................................... 12
2. Objetivos ........................................................................................................................... 18
2.1 Geral ........................................................................................................................... 18
2.2 Específicos ................................................................................................................. 18
2.3 Hipóteses .................................................................................................................... 18
3. Material e Métodos............................................................................................................ 19
3.1 Área de estudo ........................................................................................................... 19
3.2 Definição do grupo de estudo .................................................................................... 22
4. Resultados ......................................................................................................................... 29
4.1 Esforço amostral ........................................................................................................ 29
4.2 Tamanho e composição do grupo .............................................................................. 29
4.3 Padrão de atividades .................................................................................................. 30
4.4 Dieta ........................................................................................................................... 32
4.5 Composição taxonômica da dieta .............................................................................. 35
5. Discussão ........................................................................................................................... 39
5.1 Tamanho e composição do grupo .............................................................................. 39
5.2 Padrão de atividades .................................................................................................. 41
5.3 Dieta ........................................................................................................................... 44
5.4 Composição da dieta .................................................................................................. 46
6. Conclusão .......................................................................................................................... 50
7. Referências Bibliográficas ................................................................................................ 51
8. APÊNDICES ..................................................................................................................... 58

x
LISTA DE FIGURAS
Figura 1. Mapa de distribuição geográfica das cinco espécies do gênero Chiropotes. Adaptado de Silva-Júnior et al. (2013). .................................................................................................... 13
Figura 2. Mapa do Brasil destacando o estado do Pará (A) em vermelho. Localização da FLONA Saracá-Taquera no estado do Pará (B) e destaque do platô Bacaba em verde, área do presente estudo (C). Adaptado de Melo et al. (2010). .............................................................. 19
Figura 3. Histórico de temperatura (mínima e máxima °C) e precipitação (acumulado mm) para a região da FLONA Saracá-Taquera, PA entre 2001 e 2014. Fonte: CPTEC/INPE. ....... 20
Figura 4. Dados de pluviosidade (mm) e temperatura (°C) para a região da FLONA Saracá-Taquera, PA entre janeiro de 2011 e março de 2012. Fonte: CPTEC/INPE. ........................... 21
Figura 5. Vista do interior (A), borda (B) e dossel (C) da Vegetação no Platô Bacaba, FLONA Saracá-Taquera, PA. Fotos: Viviane Sodré. ............................................................................. 22
Figura 6. Mapa identificando o platô Bacaba e o sistema principal de trilhas utilizado para a pesquisa com o grupo de cuxiús. Fonte: Melo et al. 2012........................................................ 23
Figura 7. Demonstrativo de parte do tratamento dado às amostras botânicas, após a marcação das árvores de alimentação e coleta de material para posterior identificação. Na fotografia é possível observar a padronização no tratamento dos itens alimentares coletados, com detalhe da apresentação da abertura do fruto por parte dos Cuxiús, para a obtenção da parte da planta a ser ingerida. Foto Viviane Sodré. .......................................................................................... 27
Figura 8. Frequência relativa dos comportamentos apresentados pelo grupo de Chiropotes sagulatus (Bacaba) na FLONA Saracá-Taquera, PA entre julho/2011 e março/2012 (n=6926) (exceto set/2011)....................................................................................................................... 31
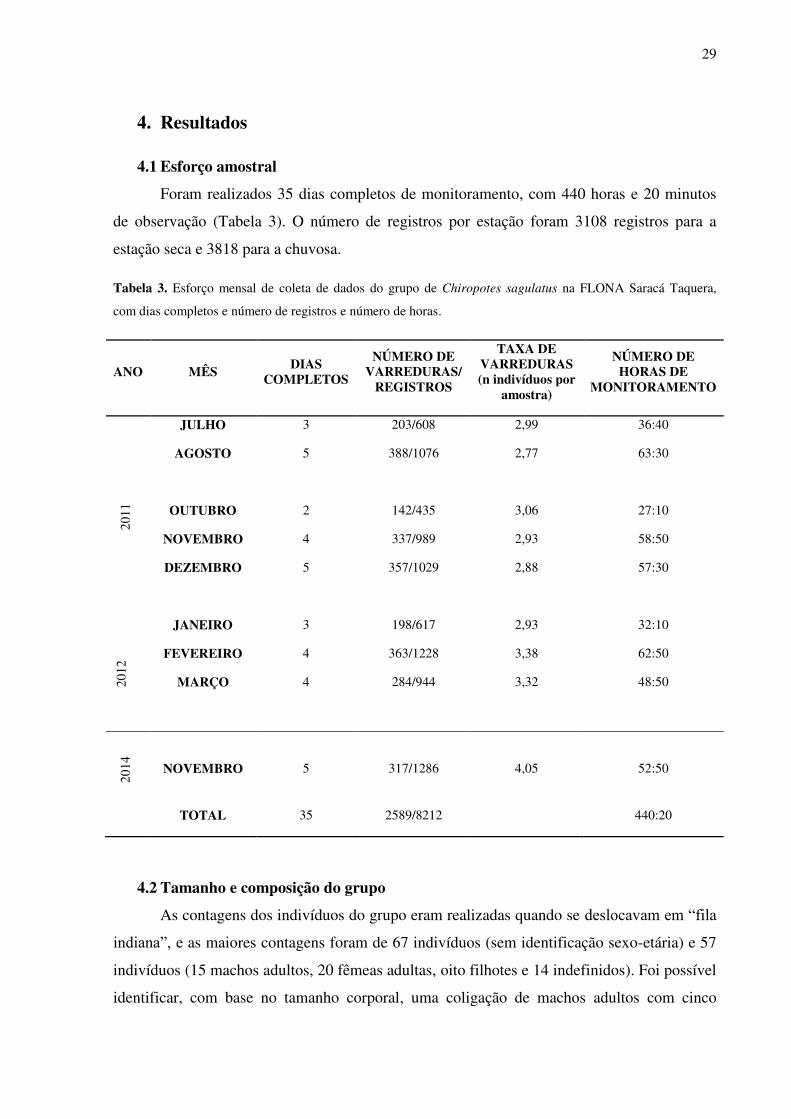
Figura 9. Frequência relativa dos itens alimentares explorados pelo grupo Bacaba na FLONA Saracá-Taquera, PA entre julho/2011 e março/2012 (n=900) (exceto set/2011). .................... 33
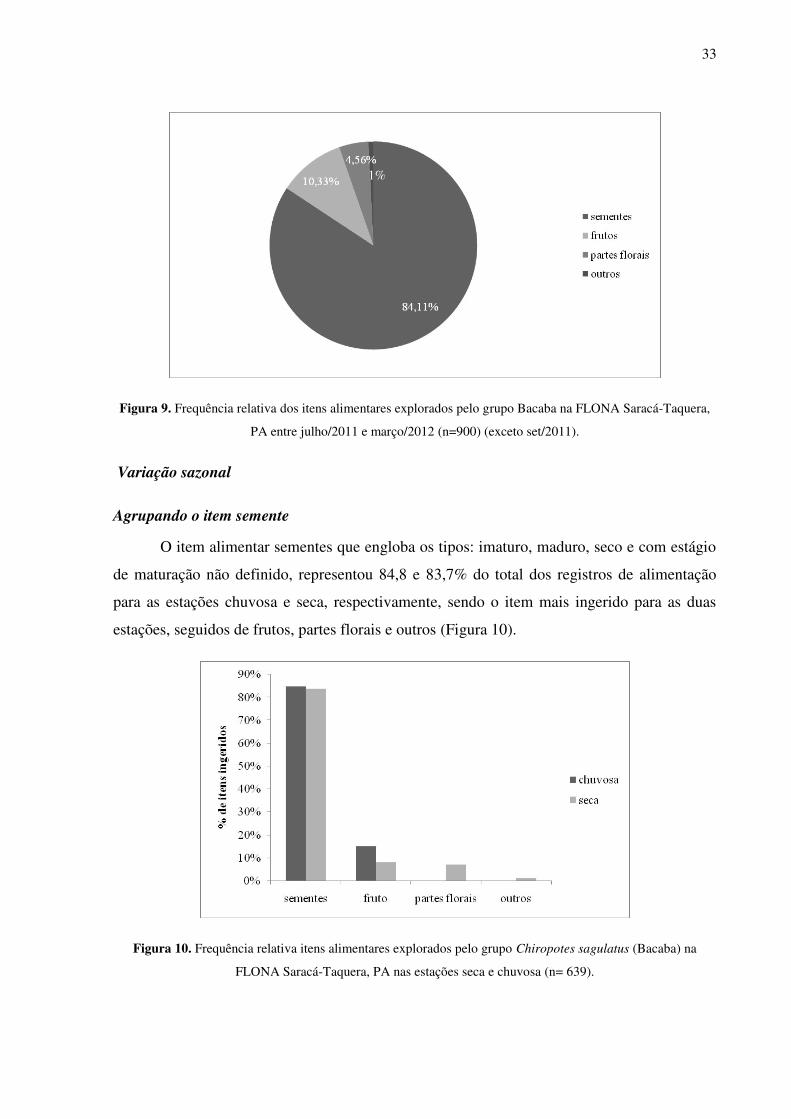
Figura 10. Frequência relativa itens alimentares explorados pelo grupo Chiropotes sagulatus (Bacaba) na FLONA Saracá-Taquera, PA nas estações seca e chuvosa (n= 639). .................. 33
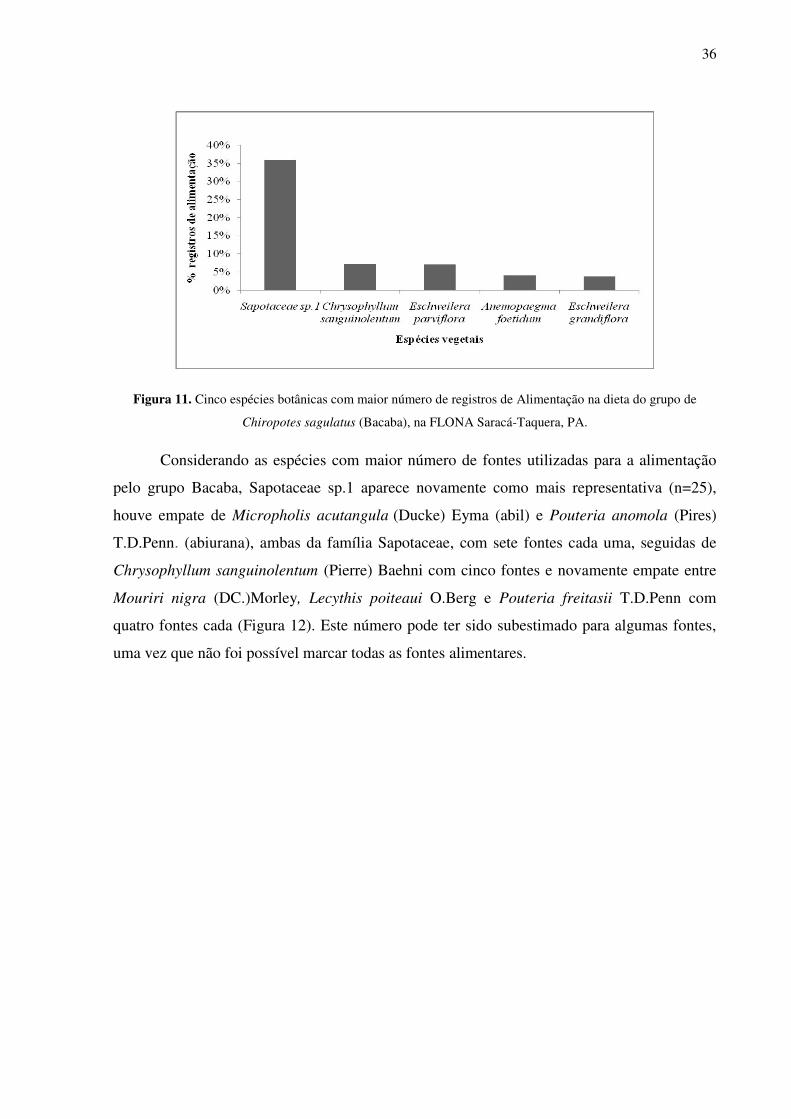
Figura 11. Cinco espécies botânicas com maior número de registros de Alimentação na dieta do grupo de Chiropotes sagulatus (Bacaba), na FLONA Saracá-Taquera, PA. ...................... 36
Figura 12. As cinco espécies botânicas com maior número de fontes exploradas para a alimentação do grupo de Chiropotes sagulatus (Bacaba), na FLONA Saracá-Taquera, PA. .. 37
Figura 13. Espécie da família Sapotaceae ainda não descrita consumida pelo grupo de Chiropotes sagulatus (Bacaba) de semente imatura a semente de fruto seco na FLONA Saracá-Taquera, PA. ................................................................................................................. 49

xi
LISTA DE TABELAS
Tabela 1. Etograma para coleta de dados comportamentais do grupo de Chiropotes sagulatus Bacaba. Adaptado de Veiga (2006). ......................................................................................... 25
Tabela 2. Classificação dos itens alimentares utilizados pelo grupo de Chiropotes sagulatus (Bacaba) na FLONA Saracá-Taquera, PA durante o período de estudo. ................................. 26
Tabela 3. Esforço mensal de coleta de dados do grupo de Chiropotes sagulatus na FLONA Saracá Taquera, com dias completos e número de registros e número de horas. .................... 29
Tabela 4. Orçamento de atividades do grupo de Chiropotes sagulatus (Bacaba) na FLONA Saracá-Taquera, PA. ................................................................................................................. 31
Tabela 6. Orçamento de atividades do grupo de Chiropotes sagulatus (Bacaba) na FLONA Saracá-Taquera, PA, para os meses de novembro/2011 e novembro/2014. ns=não significativo. ............................................................................................................................. 32
Tabela 5. Composição da dieta do grupo de Chiropotes sagulatus (Bacaba) na FLONA Saracá-Taquera, PA estações seca e chuvosa. z = escores do z binomial; *=não significativo. .................................................................................................................................................. 34
Tabela 7. Composição da dieta do grupo de Chiropotes sagulatus (Bacaba) na FLONA Saracá-Taquera, PA para os meses de novembro/2011 e novembro/2014. z = escores do z binomial; *=não significativo. .................................................................................................. 34
Tabela 8. Número de indivíduos por grupo em diferentes estudos com Chiropotes habitando floresta contínua. ...................................................................................................................... 39
Tabela 9. Resumo das contagens (média, min-max) de indivíduos de Chiropotes sagulatus por subgrupo para as estações seca e chuvosa e os meses de Novembro/2011 e Novembro/2014 na FLONA Saracá-Taquera, PA. ................................................................................................... 40
Tabela 10. Orçamento de atividades de Chiropotes em diferentes estudos. (Adaptado de Pinto, 2008). ........................................................................................................................................ 42
Tabela 11. Diferenças quanto ao número de famílias, gêneros e espécies vegetais consumidas por Chiropotes sagulatus em habitats de floresta contínua, em diferentes estudos. ** mais de um grupo por estudo. ................................................................................................................ 47

12
1. Introdução
As florestas tropicais são os ecossistemas mais ricos em biodiversidade do planeta
(Cuarón 2000). Ainda que ocupem originalmente somente 17 milhões de km², ou seja, menos
de 5% do planeta, estima-se que abriguem cerca de 50% de todas as espécies de seres vivos
existentes (Conservation Intenational 2007).
Dentre estas florestas, a Amazônia cobre cerca de sete milhões de km² e abrange o
maior trecho de floresta tropical existente (Haugaasen et al. 2005). Configura-se como um
mosaico de ecossistemas, e devido a sua grande extensão de floresta contínuas abriga uma das
maiores riquezas de flora e fauna do mundo (Capobianco et al. 2001; Conservation
International 2007). As chamadas florestas de “terra firme” são os ambientes que apresentam
os maiores níveis de diversidade local em táxons de plantas, vertebrados e invertebrados
(Haugaasen et al. 2005).
No começo do atual século, a região amazônica abrigava cerca de 75% (82) de todas
as espécies de primatas do Brasil (total de 110), apresentando a maior riqueza desse grupo em
todo o mundo (Rylands et al. 2000), sendo que novas espécies foram descritas ao longo dos
últimos anos (Rylands et al. 2011). A grande diversidade de primatas no bioma e suas
características específicas quanto aos padrões de atividade, uso de habitat, posicionamento no
estrato arbóreo, forrageamento e comportamento social (Bicca-Marques et al. 2006) os tornam
excelentes objetos de monitoramentos referentes às teorias ecológicas, sociais e
conservacionistas (Kappeler & Van Schaik 2002).
Pitheciidae é uma família de primatas neotropicais composta por duas subfamílias:
Callicebinae, formada pelo gênero mais basal Callicebus; e Pitheciinae, formada por gêneros
essencialmente amazônicos: Pithecia, Chiropotes e Cacajao (Silva-Júnior et al. 2013). Os
pitecíneos possuem características distintas dos demais macacos da infraordem Platyrrhini e
são conhecidos como “predadores de sementes”, sendo especializados morfologicamente para
a predação de sementes em menor grau em Pithecia e maior grau em Cacajao e Chiropotes
(van Roosmalen et al. 1988).
A revisão taxonômica mais recente para o gênero Chiropotes apresenta cinco espécies:
Chiropotes albinasus, C. chiropotes, C. sagulatus, C. satanas e C. utahickae (Silva-Júnior et
al. 2013) e esta será a classificação adotada aqui. Todos os cuxiús, primatas desse gênero são

13
parapátricos, com distribuições geográficas delimitadas por grandes rios (Figura 1), tais quais:
Amazonas, Madeira, Negro-Branco, Tocantins e Xingu. Os limites a oeste são bem definidos
pelo Solimões-Negro, Orinoco e Madeira-Jiparaná, mas os extremos sul e leste são menos
claros, estando relacionados com o ecótono Cerrado-Amazônia. A espécie Chiropotes
sagulatus Trail 1821, objeto desse estudo, ocorre no Brasil, Guiana Francesa, Guiana e
Suriname, e sua distribuição geográfica é definida por dois grandes rios: encontra-se ao norte
do baixo Amazonas, leste do rio Branco e no extremo oeste do sistema Negro-Branco
Figura 1. Mapa de distribuição geográfica das cinco espécies do gênero Chiropotes. Adaptado de Silva-Júnior et
al. (2013).
Chiropotes são primatas neotropicais de médio porte, com massa corporal variando de
2,5 a 3,6 kg quando adultos. Vivem em grandes grupos de estrutura social multi-macho multi-
fêmea que podem exceder 50 indivíduos (Gregory & Norconk 2013; Setz et al. 2013). O
comportamento de todas as espécies do gênero é marcado pelo hábito diurno e um longo
período de atividade diária (12h ou mais) sendo conhecidos por se deslocarem muito (com
distâncias diárias de até 8 km) e rapidamente, com um ritmo acelerado que influencia seu
padrão de atividades - passam mais tempo se deslocando do que se alimentando ou
descansando (Gregory & Norconk 2013; Setz et al. 2013). Os grandes grupos se dividem em
subgrupos ao longo do ano, os indivíduos escondem-se durante as atividades de repouso e
interações sociais (Veiga 2006) e, tendo preferência pelo dossel da floresta, raramente descem
ao chão (Barnett et al. 2012).

14
De acordo com a Lista Vermelha da IUCN (International Union for the Conservation
of Nature) de espécies da fauna ameaçadas de extinção, quatro espécies do gênero Chiropotes
estão classificadas em algum grau: Criticamente em Perigo (C. satanas), Em Perigo (C.
albinasus e C. utahickae) e Menos Preocupante (C. chiropotes) (Veiga et al. 2008). Nesta
classificação, Chiropotes sagulatus ainda é reconhecida como subespécie de Chiropotes
chiropotes, podendo então considerá-la sob o status de “Menos preocupante”.
Chiropotes satanas e Chiropotes utahickae ainda apresentam os status de Em Perigo
e Vulnerável, respectivamente, sendo as principais ameaças a fragmentação e destruição de
habitats decorrentes de grandes empreendimentos de infra-estrutura e agropecuária,
especificamente as obras e explorações ocorridas em Tucuruí, Belo Monte, Grande Carajás, e
nas rodovias Transamazônica e Belém-Brasília (Machado et al. 2008). Embora a espécie
ainda não esteja tão impactada em termos gerais, algumas populações de Chiropotes
sagulatus têm ocorrência em regiões onde a extração de recursos minerais, como a bauxita,
depende da supressão da vegetação (Melo et al. 2010). Tal supressão acarreta a diminuição da
área disponível para os grupos, expondo as populações aos efeitos da fragmentação (Melo et
al. 2010).
A maior parte da dieta de Chiropotes é composta por frutos e partes de frutos, com
poucas variações entre espécies, e, diferente da maioria dos demais primatas frugívoros, eles
consomem grandes quantidades de sementes de frutos imaturos (Ayres 1989; Kinzey &
Norconk 1993; Port-Carvalho & Ferrari, 2004; Veiga 2006; Norconk et al. 2009; Boyle et al.
2012; Melo et al. 2012; Shaffer 2012; Shaffer 2013; Santos 2013). Primatas predadores de
sementes, como os cuxiús, usam duas estratégias para o forrageamento: (a) esclerocarpia,
definida como a extração de sementes usando dentição anterior (incisivos, caninos e/ou o
primeiro pré-molar na fileira de dentes) e as mãos, seguida pela mastigação de sementes pelos
molares (Kinzey & Norconk 1990) e (b) durofagia, a ingestão do fruto inteiro com
processamento mínimo pelas mãos, seguido de mastigação ou esmagando com dentição
posterior (Kay et al. 2013).
As adaptações para o “forrageamento esclerocarpico” em Chiropotes incluem fortes
músculos da mastigação e uma dentição anterior muito robusta, caracterizada por incisivos
procumbentes, caninos grandes e inclinados lateralmente, o que facilita a abertura de frutos
imaturos resistentes (Kinzey 1992; Kinzey & Norconk 1990). Essa capacidade de abrir frutos

15
muito duros e imaturos é comparável à de aves predadoras de sementes, como papagaios e
araras (Norconk et al. 1997).
A variação anual da composição da dieta de Chiropotes apresenta a utilização de
recursos como invertebrados, folhas, flores e cascas, além de um aumento no número de
espécies de sementes exploradas no período da seca (Ayres 1989; Kinzey & Norconk 1993;
Norconk 1996; Silva 2003; Norconk et al. 2009; Melo et al. 2012; Shaffer 2012), indicativo
de sua flexibilidade alimentar e possível preferência, quando na abundância de determinados
itens. O aumento no número de espécies e itens explorados durante o período da seca, que é
considerado o período de escassez de recursos para a maioria dos primatas frugívoros,
também é um padrão encontrado para primatas como Ateles belzebuth (Mourthé 2014)
Lagothrix cana (Peres 1994a), Saguinus spp. e Sapajus apella (Peres 1994b).
Em estudos com primatas, um importante aspecto a ser analisado são os fatores
ecológicos (bióticos e abióticos) que afetam sua abundância, história de vida, diversidade,
demografia e organização social (Rothman et al. 2012). Um fator que merece destaque por ser
comum tanto em regiões temperadas quanto nos trópicos é a variação sazonal do ambiente
(Hill 1997), ou seja, as modificações nos parâmetros ambientais no decorrer de um ano.
Todos os habitats tropicais são caracterizados pelas variações sazonais de precipitação e
temperatura (van Schaik & Brockman 2005) e estas podem apresentar padrões que excedem o
período anual, influenciando a maneira como as espécies se ajustam às mudanças ambientais
(Pereira 2006) ao longo do ano e entre anos distintos.
Adaptações comportamentais podem estar envolvidas na maneira como os primatas
lidam com a falta de recursos nos períodos de escassez (Di Fiore 2003), como expandir a
amplitude de sua dieta e incorporar recursos menos preferidos quando os recursos
preferenciais estão escassos (Chapman & Chapman 1990; Shaffer 2012). Dividir-se em
subgrupos ou alterar o tamanho da área de uso em função da distribuição espacial de alimento
e água e ainda se deslocar em distâncias menores do que em épocas de fartura são
características comuns aos membros da ordem Primates e ao gênero Chiropotes (Pianka 1994,
Veiga 2006, Boyle et al. 2009).
Desde os primeiros trabalhos com Chiropotes, como o de Ayres (1989) que agrupou
dados da dieta de Chiropotes albinasus nas estações seca e úmida para os anos de 1977 e
1978, já haviam sido encontrados efeitos da sazonalidade na dieta dos cuxiús, com predação

16
de sementes de um maior número de espécies na estação seca do que na chuvosa e inclusão de
outros itens alimentares na dieta. Outros trabalhos (Norconk 1996; Santos et al. 2013) com o
objetivo de confirmar se este padrão persiste de maneira longitudinal, ou seja, ao longo dos
anos, encontraram diferenças na ecologia alimentar em estações de anos distintos e, em alguns
deles, a variação foi relacionada de maneira significativa à mudanças nos padrões de
precipitação. Norconk (1996) concluiu que o período seco não pode ser considerado como um
período de escassez para cuxiús, já que o número de espécies de sementes exploradas durante
esta estação aumenta em relação ao período chuvoso. Já Pinto (2008) observou que a
estratégia de forrageamento de “maximização de energia” dos cuxiús os permite passar pelos
períodos de escassez aumentando o deslocamento diário, dividindo-se em subgrupos e
equilibrando a ingestão de nutrientes a partir do consumo de itens alimentares de composição
distinta, como sementes e polpa de frutos maduros.
Os cuxiús são conhecidos por ter extensas áreas de vida, que podem ultrapassar os
1000 ha (Setz et al. 2013), embora existam evidências de que as espécies do gênero
sobrevivam em áreas fragmentadas (Boyle et al. 2012) com menos de 3% do tamanho total
esperado (8 ha, Port-Carvalho & Ferrari 2004; 1-100 ha, Boyle & Smith 2010). Contudo,
Boyle & Smith (2010), afirmam que os cuxiús que vivem em pequenos fragmentos seguem
um padrão diferente dos que habitam florestas contínuas, sendo encontrados em pequenos
grupos, com maiores densidades, descansando com mais frequência, vocalizando e se
deslocando menos. Com relação à dieta, embora as espécies do gênero apresentem
flexibilidade na exploração de recursos alimentares, há baixa similaridade na dieta de grupos
que habitam os diferentes sítios tanto de áreas contínuas quanto fragmentadas (Boyle et al.
2015), o que justifica a necessidade de estudos a respeito de sua ecologia alimentar ao longo
da distribuição geográfica.
A flexibilidade do comportamento, dieta e padrão de agrupamento dos cuxiús (Boyle
2008; Boyle et al. 2009) podem refletir sua adaptação ao ambiente atual, permitindo sua
permanência temporariamente nesses locais. Como são predadores de sementes, a pressão
exercida sobre a comunidade vegetal em longo prazo pode, entre outros fatores, aumentar a
competição intra e interespecífica e levar ao declínio populacional. Assim, estudos como este,
que tenham como objetivo conhecer a ecologia alimentar dos cuxiús, são importantes para
definir os fatores limitantes para a sobrevivência destes primatas.

17
Chiropotes sagulatus é uma espécie que ainda apresenta status de conservação
favorável, mas se encontra sob pressão antrópica frente a demanda por recuros mineirais e
energéticos (Peetz 2001; Melo et al 2012). Conhecê-los no cenário atual permitirá o
aprofundamento a respeito de suas adaptações ao ambiente atual, alterações em sua ecologia,
e prever as ações necessárias para a manutenção de sua relação com o ambiente em que
vivem. A partir disso é que poderemos, no futuro, subsidiar ações que possam garantir a
conservação de seus habitats de maneira a favorecer a sua permanência nos mesmos.

18
2. Objetivos
2.1 Geral
O objetivo deste trabalho é avaliar a variação sazonal (ao longo das estações) e longitudinal
(entre dois anos) na ecologia alimentar de Chiropotes sagulatus em uma área de Floresta
Amazônica, norte do Brasil.
2.2 Específicos
Verificar se há variação sazonal e longitudinal no padrão de agrupamento do grupo de
estudo;
Avaliar a variação sazonal e longitudinal no padrão comportamental de C. sagulatus no
local de estudo;
Identificar a dieta do grupo estudado, verificando as possíveis variações sazonais e
longitudinais na exploração de itens alimentares;
Observar possíveis variações na composição taxonômica da dieta no padrão sazonal e
longitudinal.
2.3 Hipóteses
Para alcançar os objetivos propostos, foram testadas as seguintes hipóteses:
H1: O grupo apresenta padrão de agrupamento mais coeso na estação chuvosa do que na
estação seca e não há variação neste aspecto para o padrão longitudinal.
H2: O padrão comportamental do grupo de estudo varia entre estações e entre anos;
H3: A composição da dieta varia entre estações e entre anos diferentes;
H4: O número de espécies de sementes de frutos verdes presentes na dieta da estação seca é
maior do que na chuvosa. As espécies exploradas como fonte alimentar pelo grupo de estudo
são diferentes no padrão sazonal e longitudinal.

19
3. Material e Métodos
3.1 Área de estudo
FLONA Saracá-Taquera
A FLONA Saracá-Taquera está situada entre as coordenadas geográficas 01°20’ e
01°55’ S e 56°00’ e 57°15’ W, inserida nos municípios de Oriximiná, Faro e Terra Santa, no
Estado do Pará (Figura 2). É classificada como uma Unidade de Conservação de Uso
Sustentável e possui uma área de 429.600 ha (STCP 2001).
Figura 2. Mapa do Brasil destacando o estado do Pará (A) em vermelho. Localização da FLONA Saracá-
Taquera no estado do Pará (B) e destaque do platô Bacaba em verde, área do presente estudo (C). Adaptado de
Melo et al. (2010).
Clima e hidrografia
O clima da FLONA é do tipo “Af” segundo a classificação de Köppen, com uma
temperatura média de 26°C (STCP 2001; Moraes et al. 2005) e índice médio anual de
umidade relativa superior a 75%. Ocorrem dois períodos de precipitação, estação chuvosa de
dezembro a maio e estação seca de julho a outubro, e dois períodos de transição em novembro
e junho (STCP 2001).
A
B
C

20
Média histórica de clima
As médias climáticas históricas para a região da FLONA com precipitação e
temperatura do período de 2001 a 2014 são apresentadas na Figura 3, e foram utilizadas para a
definição das estações de 2011 e 2012. Como observado, de 2012 a 2014 houve uma
diminuição nos índices acumulados de precipitação, apresentando a maior queda da última
década.
Figura 3. Histórico de temperatura (mínima e máxima °C) e precipitação (acumulado mm) para a região da
FLONA Saracá-Taquera, PA entre 2001 e 2014. Fonte: CPTEC/INPE.
Definição de estações
Considerando as médias históricas, o clima do período de estudo seguiu o padrão
esperado para a área da FLONA, com precipitação acumulada para os 15 meses (jan a dez/11
e jan a março/12) de 3.334 mm, média mensal 222,29 mm e temperatura média anual 27,51°C
(Figura 4). As estações foram definidas como: chuvosa de dezembro a maio (1201,34 mm) e
seca de junho a novembro (590,03 mm). As médias mensais de temperatura variaram dentro
do esperado ao longo do ano.

21
Figura 4. Dados de pluviosidade (mm) e temperatura (°C) para a região da FLONA Saracá-Taquera, PA entre
janeiro de 2011 e março de 2012. Fonte: CPTEC/INPE.
Vegetação
A FLONA pertence à sub-região das Guianas (leste do Rio Negro e norte do rio
Amazonas) (STCP 2001) e está inserida no domínio da Floresta Amazônica, que é
caracterizada como a maior e mais rica formação florestal do planeta. Predominam as
formações vegetais Floresta Ombrófila Densa, Formações Pioneiras com Influência Fluvial e
Campinarana (Salomão et al. 2003). Apresenta duas tipologias vegetacionais principais: a
floresta de Terra Firme, ocorrendo no topo dos platôs, e a Floresta de Igapó, que ocorre nos
baixios (Salomão et al. 1997; Antonini & Oliveira 2007). A região de platôs é composta
basicamente por Floresta Ombrófila Densa Submontana de Platôs, onde o solo é latossolo
amarelo que constitui o material inerte predominante nas áreas de mineração da bauxita
(STCP 2001).
Segundo Salomão et al. (2002), a região do platô Bacaba, área alvo deste estudo é
classificada como Floresta Ombrófila Densa com Palmeiras (Figura 5), e trata-se de uma das
mais “belas facies” da Floresta Amazônica: rica em espécies e com alta biomassa (430 spp.
distribuídas em 51 famílias).

22
Figura 5. Vista do interior (A), borda (B) e dossel (C) da Vegetação no Platô Bacaba, FLONA Saracá-Taquera,
PA. Fotos: Viviane Sodré.
3.2 Definição do grupo de estudo
Fase preliminar
Esta pesquisa é parte do projeto “Monitoramento de Primatas na FLONA SARACÁ-
TAQUERA, PA”, um convênio da Universidade Federal de Goiás com a Mineração Rio do
Norte. A fase preliminar de mapeamento de trilhas para o censo populacional, definição de
grupos-alvo e habituação (que durou de 12 meses: agosto-2010 a julho-2011) foram
realizadas por Melo et al. (2010). O processo de habituação. Este estudo foi realizado no platô
Bacaba (Figura 6) com 215 ha de extensão, cuja região sofreu supressão integral da vegetação
do platô para a extração de bauxita entre os anos de 2010 e 2012. O platô está inserido na
“Zona de Mineração” estabelecida pelo plano de manejo da FLONA (STCP 2001).
A B C

23
Figura 6. Mapa identificando o platô Bacaba e o sistema principal de trilhas utilizado para a pesquisa com o
grupo de cuxiús. Fonte: Melo et al. 2012.
Escolha do grupo alvo
Na região do platô Bacaba havia dois grupos residentes de Chiropotes sagulatus, que
foram chamados de Leste e Oeste, de acordo com a face do platô mais utilizada. O grupo
Leste (atual Bacaba) foi escolhido para este estudo pelo maior número de encontros no
primeiro mês de habituação e por ocupar a porção da floresta onde não havia influência
antrópica, já que o grupo do outro lado do platô tem parte da área de vida ocupada por
ribeirinhos (para a moradia temporária e plantio de mandioca).
Coleta e análise de dados
Alguns contratempos como a dificuldade de encontrar o grupo (no mínimo três dias de
cada mês eram dedicados à procura), as chuvas intensas, a ausência dos cooperados
(mateiros), o avanço da supressão da vegetação e o comportamento dos primatas dificultaram
o cumprimento da meta de coletar cinco dias em cada mês. Os dados quantitativos foram
coletados durante 30 dias entre julho-2011 e março-2012 (exceto setembro-2011 quando não
houve coleta), e cinco dias em novembro-2014.
As análises gerais consideraram todos os dados (de julho-2011 a março-2012) para
melhor observação do padrão anual de atividades e dieta do grupo de estudo. As análises dos
padrões sazonais foram realizadas considerando os meses de dezembro/2011, janeiro,
fevereiro e março/2012 para estação chuvosa e julho, agosto, outubro e novembro/ 2012 para

24
a estação seca. Novembro/2014 não foi incluso na análise geral e sazonal e a análise
longitudinal comparou novembro de 2011 com novembro de 2014.
Com o objetivo de estabelecer comparações com outros estudos realizados com
Chiropotes sp. (e.g. Veiga 2006; Pinto 2008), os dados foram obtidos através da metodologia
proposta por Altmann (1974) scan sampling, a partir da coleta de amostras por varredura
instantânea onde todos os animais no campo de visão do observador são registrados dentro do
período pré-determinado. Considerando a baixa visibilidade descrita em outros estudos com
cuxiús (Pinto et al. 2013) e a dificuldade de visualização também observada durante o
processo de habituação, as amostras (varreduras) tiveram duração de dois minutos e intervalo
de 10 minutos.
Para as amostragens do padrão de agrupamento, as contagens do número de indivíduos
no subgrupo foram realizadas através de observação direta em cada varredura e todas as vezes
que o grupo se subdividia ou se agrupava era realizada nova contagem. A análise da
normalidade de todos dados foi realizada através do teste de Shapiro Wilk através do
programa R 3.1.1. Como os dados eram paramétricos, o teste T com nível de significância de
0.05 foi utilizado para a comparação do número de indivíduos por subgrupo no padrão
sazonal e longitudinal, através do programa R 3.1.1. Além disso, o número de indivíduos por
amostra (taxa de avistamentos) foi calculado através do número total de registros dividido
pelo número total de varreduras do período (mês e estação, ano), a fim observar se houve
influência do número de indivíduos na varredura para os comportamentos e item ingerido.
Seguindo a formula:
Ta= nreg/nvarr
Onde: ta= taxa de avistamentos; nreg= número de registros para o período; nvarr=
número de varreduras para o período.
Para cada varredura foram marcados hora de início e o ponto (GPS) da localização
central do grupo e, para cada indivíduo durante a varredura foram anotadas: a classe sexo-
etária (cujas definições para o reconhecimento estão descritas no Apêndice A) e
comportamento (cujas classes foram definidas previamente no etograma (Tabela 1). Todas as
observações foram realizadas com auxílio de binóculo Canon 10x30is com estabilizador de
imagem, GPS Garmin 60csx e rangefinder Bushnell. Amostragem de todas as ocorrências foi

25
realizada sempre que houve ingestão de alimentos avulsos fora das varreduras, interações
sócio-sexuais (cópulas, agonismo, catação) e interações interespecíficas.
Tabela 1. Etograma para coleta de dados comportamentais do grupo de Chiropotes sagulatus Bacaba. Adaptado
de Veiga (2006).
COMPORTAMENTO CÓDIGO DESCRIÇÃO
DESLOCAMENTO DESL Qualquer atividade que envolva movimento (caminhar, andar, saltar).
REPOUSO REP Quando o indivíduo está parado e nenhuma outra atividade é observada.
FORRAGEAMENTO FOR Procura ou processamento de item alimentar.
ALIMENTAÇÃO AL Quando o indivíduo ingere o item alimentar ou água.
SOCIAL SOC Quando há interação entre dois ou mais indivíduos.
OUTROS OUT Atividades que não se encaixam nas demais categorias, como auto-catação.
Foi realizado o cálculo da freqüência relativa dos comportamentos para definir o
orçamento de atividades do grupo pesquisado, dividindo-se o somatório dos registros de todos
os comportamentos pelo número total de registros do comportamento em questão para o
respectivo período (anual, sazonal e longitudinal), seguindo a fórmula:
FR= Ntotal*100
nperíodo
onde: Ntotal= número total de registros de todos os comportamentos; nperíodo= número total
de registros do comportamento x para o período (anual, sazonal e longitudinal).
As comparações dos dados comportamentais no padrão sazonal e longitudinal foram
analisadas através da calculadora do teste z binomial no Programa Microsoft Excel
2007 com α = 0.01 para evitar erros do tipo I, seguindo Lima & Ferrari (2003). Este teste
foi também utilizado por Veiga (2006) e Rímoli et al. (2008). Os valores de p foram obtidos
através do Calculators for Statistical Table Entries http://vassarstats.net/tabs.html?#z.
Quando o comportamento registrado foi alimentação, as amostras dos itens ingeridos
foram coletadas para posterior identificação e classificados como sementes, folhas, frutos,
flores, brotos, cascas, dentre outros, a fim de qualificar a composição da dieta. Quando o item
era semente, estas foram divididas quanto ao estágio de maturação para refinar as análises da

26
dieta, já que os cuxiús se alimentam de uma vasta gama de sementes em diversos estágios,
conforme descrito na Tabela 2.
Tabela 2. Classificação dos itens alimentares utilizados pelo grupo de Chiropotes sagulatus (Bacaba) na
FLONA Saracá-Taquera, PA durante o período de estudo.
CATEGORIA ITENS INCLUÍDOS
SEMENTES Sementes sem identificação do estágio de maturação
SEFRV Sementes de frutos verdes (imaturas)
SEFRM Sementes de frutos maduros
SESECO Sementes cujo fruto já estava seco
FRUTOS Partes de frutos nos diversos estágios de maturação exceto sementes.
PARTES FLORAIS Qualquer parte de flores
OUTROS Folhas e casca. (Itens que apresentaram baixa frequência de ingestão)
Para avaliar a dieta do grupo de cuxiús pesquisado, foram calculadas as freqüências
relativas de consumo dos itens alimentares. As freqüências foram calculadas dividindo o
somatório de todos os registros de alimentação registrados dividido pelo número total de
registros do item alimentar em questão para o respectivo período (anual, sazonal e
longitudinal), seguindo a fórmula:
FR= Ntotal*100
nperíodo
onde: Ntotal= número total de registros de alimentação; nperíodo= número total de registros
do item alimentar x para o período (anual, sazonal e longitudinal).
As análises comparativas para o padrão de ingestão de itens alimentares sazonal e
longitudinal também foram realizadas pelo teste z binomial, seguindo o utilizado para os
dados comportamentais.
As árvores utilizadas como fontes alimentadas dentro e fora das varreduras foram
marcadas com plaqueta de metal contendo o número de identificação e fita zebrada, para
facilitar o encontro e identificação em dias posteriores ou estudos subsequentes. Não foi
possível coletar amostras de todas as fontes alimentares utilizadas pelo grupo Bacaba, porque

27
parte da equipe de campo envolvida não conseguiu cumprir essa meta e, neste caso, a
prioridade foi seguir o grupo.
As amostras dos itens alimentares foram acondicionadas em frascos plásticos com
solução de álcool 70% e devidamente fotografadas (Figura 7) com escala padronizada e
numeradas de acordo com o número da fonte. A identificação das plantas foi realizada por um
especialista vinculado ao Instituto Nacional de Pesquisas Amazônicas com base no material
coletado e revisita às fontes alimentares no campo. Após a identificação das espécies
utilizadas na dieta, estas foram divididas por mês, estação e ano coletado, para as posteriores
análises de composição taxonômica da dieta. Para avaliar a similaridade da dieta na análise
sazonal e longitudinal foi utilizado o Índice de Jaccard, através do programa Microsoft Excel
2007.
Figura 7. Demonstrativo de parte do tratamento dado às amostras botânicas, após a marcação das árvores de
alimentação e coleta de material para posterior identificação. Na fotografia é possível observar a padronização
no tratamento dos itens alimentares coletados, com detalhe da apresentação da abertura do fruto por parte dos
Cuxiús, para a obtenção da parte da planta a ser ingerida. Foto Viviane Sodré.
Como o número de registros dentro de cada varredura pode variar em função do
observador, do comportamento do grupo e da fonte alimentar, foi realizada uma abordagem
alternativa para a análise dos dados do orçamento de atividades e dieta, visando minimizar e
equilibrar as diferenças nos esforços amostrais que poderiam enviesar as análises do z
binomial. A média de indivíduos amostrados dentro de cada varredura variou entre os meses
(n=2,77 a 4,05), o que poderia sub ou superestimar os valores para determinados
comportamentos dentro do orçamento de atividades, principalmente os mais fáceis de
visualizar. Para ajustar estes valores, como análise alternativa, foi realizado o cálculo das
“proporções de comportamentos em cada varredura”, onde cada varredura tinha o peso 1 e as

28
categorias comportamentais tinham o valor representado de acordo com o número de registros
dentro da varredura, seguindo a fórmula:
P= 1/registros
Onde: P= proporções de comportamentos em cada varredura; 1= valor de cada varredura;
registros= número de registros dentro da varredura
Assim, se uma varredura foi composta pelo registro de 4 indivíduos, sendo um se
alimentando, dois se deslocando e um em repouso, na análise proporcional esta varredura teria
os valores 0,5 para deslocamento, 0,25 para alimentação e 0,25 para repouso.
Na composição da dieta, para diminuir a tendência de superestimar o uso de alguns
itens alimentares devido à visibilidade de grupos maiores em fontes maiores, foi realizada a
avaliação de “eventos” de alimentação. Quando vários indivíduos se alimentavam durante a
mesma varredura, dividiu-se o evento de valor 1 pelo número de indivíduos se alimentando
dentro da mesma varredura, e os itens alimentares tiveram o peso calculado de acordo com o
número de indivíduos ingerindo no mesmo evento, ao invés da contagem do número de
registros, seguindo a fórmula:
E= 1/nind, onde:
E= evento de alimentação; 1=valor da alimentação dentro da varredura; nind= número total de
indivíduos se alimentando na mesma varredura.
Assim, se em uma mesma varredura 6 indivíduos estavam se alimentando, este evento
apresentou valor 1. Se 3 indivíduos se alimentavam de semente de fruto verde, este item teve
peso 0,5, dois indivíduos se alimentavam de fruto maduro (0,33) e um se alimentava de casca
(0,17). Apesar de o ajuste ter apresentados valores absolutos mais refinados, não alterou as
representações dos comportamentos bem como dos itens alimentares ingeridos no valor total.
Como o cálculo das frequências relativas é mais utilizado nos estudos comportamentais,
optou-se por mantê-lo, a fim de melhorar a qualidade das comparações.
29
4. Resultados
4.1 Esforço amostral
Foram realizados 35 dias completos de monitoramento, com 440 horas e 20 minutos
de observação (Tabela 3). O número de registros por estação foram 3108 registros para a
estação seca e 3818 para a chuvosa.
Tabela 3. Esforço mensal de coleta de dados do grupo de Chiropotes sagulatus na FLONA Saracá Taquera,
com dias completos e número de registros e número de horas.
ANO MÊS DIAS
COMPLETOS
NÚMERO DE VARREDURAS/
REGISTROS
TAXA DE VARREDURAS (n indivíduos por
amostra)
NÚMERO DE HORAS DE
MONITORAMENTO
2011
JULHO 3 203/608 2,99 36:40
AGOSTO 5 388/1076 2,77 63:30
OUTUBRO 2 142/435 3,06 27:10
NOVEMBRO 4 337/989 2,93 58:50
DEZEMBRO 5 357/1029 2,88 57:30
2012
JANEIRO 3 198/617 2,93 32:10
FEVEREIRO 4 363/1228 3,38 62:50
MARÇO 4 284/944 3,32 48:50
2014
NOVEMBRO
5
317/1286
4,05
52:50
TOTAL 35 2589/8212 440:20
4.2 Tamanho e composição do grupo
As contagens dos indivíduos do grupo eram realizadas quando se deslocavam em “fila
indiana”, e as maiores contagens foram de 67 indivíduos (sem identificação sexo-etária) e 57
indivíduos (15 machos adultos, 20 fêmeas adultas, oito filhotes e 14 indefinidos). Foi possível
identificar, com base no tamanho corporal, uma coligação de machos adultos com cinco

30
indivíduos à qual foi dado o nome de “cinco machões”. Estes indivíduos eram vistos juntos
em todas as atividades, frequentemente se apresentavam como uma unidade distinta do
subgrupo e houve facilidade em observá-los porque eram proporcionalmente maiores do que
os demais machos adultos do grupo.
A média anual de indivíduos por subgrupo acompanhado foi de 23,46 indivíduos e o
padrão de agrupamento para as estações apresenta uma tendência para subgrupos maiores na
estação chuvosa do que na estação seca (n=25,6 ±11,9 e n=20,9 ±5,09 respectivamente), mas
a diferença não foi significativa (t = 1,4527, df = 20,859; p>0.05). Embora não tenha
apresentado diferença no tamanho de subgrupo entre as estações, a taxa de avistamentos por
varredura que na estação Chuvosa foi maior 3,11 ±0,42 do que na Seca 2,88 ±0,22.
Comparando o mês Novembro em 2011 e 2014, o número médio de indivíduos por
subgrupo foi maior no primeiro ano do que no segundo (n=20,25 ±6,65 e n=13 ±3,08
respectivamente), mas esta diferença também não foi significativa (t = 2,0137, df = 4,03;
p>0.05).
4.3 Padrão de atividades
A duração do período diário de atividades do grupo Bacaba variou entre 11h10m e 15h40m.
As primeiras vocalizações eram ouvidas cerca de 20 minutos antes de o sol nascer e o início
das atividades se deu entre 06h10m e 06h40m. O horário de se recolher para dormir variou
entre 18h20m e 19h10m.
Orçamento anual
Do total de 6926 registros, Deslocamento foi o comportamento observado com mais
frequência (41,2%; Figura 8).

31
Figura 8. Frequência relativa dos comportamentos apresentados pelo grupo de Chiropotes sagulatus (Bacaba)
na FLONA Saracá-Taquera, PA entre julho/2011 e março/2012 (n=6926) (exceto set/2011).
Variação sazonal
Na estação chuvosa, novamente a atividade de Deslocamento apresentou maior
frequência, com 50,2% dos registros, seguida de Repouso, Forrageamento e Alimentação. Já
na estação seca, Repouso foi um pouco maior que deslocamento e, juntos, somaram 62,4% do
total (Tabela 4). A diferença foi significativa para todos os comportamentos entre estações,
sendo Deslocamento maior na estação chuvosa, quando os comportamentos de Alimentação e
Forrageamento foram menores, e as atividades de Repouso e Social foram maiores na estação
seca.
Tabela 4. Orçamento de atividades do grupo de Chiropotes sagulatus (Bacaba) na FLONA Saracá-Taquera, PA.
Número de registros (% do total):
Categoria Estação seca Estação chuvosa Z P
DESLOCAMENTO 937 (30,2) 1919 (50,2) -12.950 <0.0001
REPOUSO 1000 (32,2) 1093 (28,6) 2.685 <0.01
ALIMENTAÇÃO 339 (10,9) 188 (4,9) 8.985 <0.0001
FORRAGEAMENTO 503 (16,2) 433 (11,3) 5.462 <0.0001
SOCIAL 262 (8,4) 162 (4,2) 7.010 <0.0001
OUTROS 66 (2,1) 24 (0,6) 5.431 <0.0001
TOTAL 3107 (100) 3819 (100)

32
Variação longitudinal
A Tabela 5 apresenta os dados comportamentais do grupo Bacaba em novembro-2011
e novembro-2014. Foram encontradas diferenças significativas para Deslocamento, Repouso,
Forrageamento, Alimentação e Interação Social entre os dois anos. Os cuxiús do grupo
Bacaba dedicaram mais tempo às atividades de Repouso e Forrageamento em 2011 e ao
Deslocamento e Alimentação e Interação Social em 2014.
Tabela 5. Orçamento de atividades do grupo de Chiropotes sagulatus (Bacaba) na FLONA Saracá-Taquera, PA, para os meses de novembro/2011 e novembro/2014. ns=não significativo.
4.4 Dieta
Composição anual da dieta
A dieta do grupo Bacaba durante o período de estudos apresentou alta frequência de
ingestão de sementes, portanto “granívora”. Do total de 900 registros de alimentação
considerados, o item sementes, que compreende sementes em diversos estágios de maturação
apresentou maior frequência (84,11%), seguido de frutos (excluindo sementes), partes florais
e outros (Figura 9).
Número de registros (% do total):
Z
COMPORTAMENTOS Nov/11 Nov/14 p
DESLOCAMENTO 326 (33,0) 743 (57,8) -4.421 <0.0001
REPOUSO 325 (32,9) 113 (8,8) 6.276 <0.0001
FORRAGEAMENTO 191 (19,3) 6 (0,5) 7.997 <0.0001
ALIMENTAÇÃO 67 (6,8) 255 (19,8) 252.641 <0.0001
INTERAÇÃO SOCIAL 70 (7,1) 157 (12,2) -3.83303 <0.0001
OUTROS 9 (0,9) 12 (0,9) -0.05459 *0.4782
TOTAL 463 (100,0) 471 (100,0)
33
Figura 9. Frequência relativa dos itens alimentares explorados pelo grupo Bacaba na FLONA Saracá-Taquera,
PA entre julho/2011 e março/2012 (n=900) (exceto set/2011).
Variação sazonal
Agrupando o item semente
O item alimentar sementes que engloba os tipos: imaturo, maduro, seco e com estágio
de maturação não definido, representou 84,8 e 83,7% do total dos registros de alimentação
para as estações chuvosa e seca, respectivamente, sendo o item mais ingerido para as duas
estações, seguidos de frutos, partes florais e outros (Figura 10).
Figura 10. Frequência relativa itens alimentares explorados pelo grupo Chiropotes sagulatus (Bacaba) na
FLONA Saracá-Taquera, PA nas estações seca e chuvosa (n= 639).
1%

34
A composição sazonal da dieta (Tabela 6) apresentou alta frequência de ingestão de
sementes imaturas para as duas estações (59,9% estação seca e 47,7% estação chuvosa).
Houve diferença significativa para as estações quando o item alimentar foi semente: Semente
de Fruto Verde e Semente de Fruto Maduro foram mais frequentes na estação seca e semente
sem identificação do estágio de maturação, na estação chuvosa. Para os itens partes florais e
semente de fruto seco, a diferença foi absoluta, com ingestão apenas na estação seca.
Tabela 6. Composição da dieta do grupo de Chiropotes sagulatus (Bacaba) na FLONA Saracá-Taquera, PA estações seca e chuvosa. z = escores do z binomial; *=não significativo.
Número de registros (% of the total) coletados durante:
Categoria Estação seca Estação chuvosa Z p
PARTES FLORAIS 42 (7,2) 0
FRUTO 43 (7,5) 51 (15,2) 0.1724 *0.4315
SEFRV 347 (59,9) 157 (47,7) 4.0017 <0.0001
SEFRM 70 (12,3) 39 (11,2) 4.0640 <0.0001
SESECO 33 (5,8) 0 __ __
SE 36 (5,8) 82 (25,8) -3.1345 <0.001
OUTROS 9 (1,6) 0 __ __
TOTAL 580 (100) 329 (100)
Variação longitudinal
A dieta do grupo Bacaba em novembro de 2011 e 2014 apresentou alta frequência de
ingestão de Semente de Fruto Verde (61,7 e 66,5% respectivamente). Semente de fruto seco
apresentou novamente diferença absoluta, estando presente apenas em novembro de 2011.
Não houve diferenças significativas para os itens que estiveram presentes nos dois anos
observados (Tabela 7).
Tabela 7. Composição da dieta do grupo de Chiropotes sagulatus (Bacaba) na FLONA Saracá-Taquera, PA para
os meses de novembro/2011 e novembro/2014. z = escores do z binomial; *=não significativo.
Número de registros (% of the total) coletados durante:
Item alimentar Nov/11 Nov/14 Z p
SEFRV 66 (61,7) 169 (66,5) 0.031 *0.1625
SEFRM 10 (9,3) 56 (22,0) -1.895 *0.029

35
FRM 6 (5,6) 10 (3,9) 1.123 *0.1307
FLOR 3 (2,8) 4 (1,6) 1.068 *0.1425
SESECO 21 (19,6) 0 (0,0)
OUTROS 1 (0,9) 15 (5,9) *0.0399
TOTAL 107 (100) 254 (100)
4.5 Composição taxonômica da dieta
Foram marcadas 192 árvores alimentares, das quais 164 foram coletadas (58 dentro
das varreduras e 47 fora das varreduras) e identificadas em 105 espécies (Apêndice B),
pertencentes a 32 famílias botânicas e 70 gêneros. Dentre as 164 fontes alimentares
identificadas, as cinco famílias com maior número de fontes exploradas para Alimentação
pelo grupo Bacaba no período de estudo foram Sapotaceae (23,2%), Moraceae (9,1%),
Fabaceae (8,5%), Chrysobalanaceae (8,5%) e Lecythidaceae (8,5%). Os gêneros com maior
número de espécies presentes na dieta do grupo Bacaba foram Pouteria (n=8), Licania (n=7),
Eschweilera (n=5), Inga (n=4) e Micropholis (n=4).
Com relação às espécies, as seis que apresentaram maior número de registros de
alimentação representaram 61,2% do total de 528 registros (Figura 11). Sapotaceae sp.1, com
36,0% do total (n=190) é uma espécie ainda não identificada, que segundo o especialista é
uma espécie nova para a família. Chrysophyllum sanguinolentum (Pierre) Baehni (abil)
apresentou 7,2% do total de registros, seguido de Eschweilera parviflora (Aubl.) Miers (mata-
matá) 7,0%, Anemopaegma foetidum Bureau & K.Schum 4%, Eschweilera grandiflora
(Aubl.) Sandwith (mata-matá) 3,8% e Brosimum parinarioides Ducke (amapá) 3,2%.
36
Figura 11. Cinco espécies botânicas com maior número de registros de Alimentação na dieta do grupo de
Chiropotes sagulatus (Bacaba), na FLONA Saracá-Taquera, PA.
Considerando as espécies com maior número de fontes utilizadas para a alimentação
pelo grupo Bacaba, Sapotaceae sp.1 aparece novamente como mais representativa (n=25),
houve empate de Micropholis acutangula (Ducke) Eyma (abil) e Pouteria anomola (Pires)
T.D.Penn. (abiurana), ambas da família Sapotaceae, com sete fontes cada uma, seguidas de
Chrysophyllum sanguinolentum (Pierre) Baehni com cinco fontes e novamente empate entre
Mouriri nigra (DC.)Morley, Lecythis poiteaui O.Berg e Pouteria freitasii T.D.Penn com
quatro fontes cada (Figura 12). Este número pode ter sido subestimado para algumas fontes,
uma vez que não foi possível marcar todas as fontes alimentares.

37
Figura 12. As cinco espécies botânicas com maior número de fontes exploradas para a alimentação do grupo de
Chiropotes sagulatus (Bacaba), na FLONA Saracá-Taquera, PA.
Variação sazonal
Com relação ao número de espécies na dieta do grupo Bacaba, foram consumidas 37
espécies vegetais na estação seca e na estação chuvosa, apresentando baixa similaridade na
composição segundo o Índice de Jaccard, com apenas 14%. Foram exploradas 145 fontes
alimentares de 20 espécies de para o consumo de Semente de Fruto Verde na estação seca, e
120 fontes de 20 espécies na estação chuvosa. Mouriri nigra (D.C.) foi a espécie mais
representativa para a estação seca, com 32 fontes e Anemopaegma foetidum Bureau &
K.Schum com 29 fontes, para a estação chuvosa.
Variação longitudinal
A composição taxonômica variou entre novembro de 2011 e 2014. Em novembro do
primeiro ano, foram exploradas para a alimentação pelo grupo Bacaba nove espécies vegetais
pertencentes a seis gêneros, sendo as cinco espécies mais utilizadas Mouriri collocarpa Ducke
(32,8%), Sapotaceaae sp.1 (18,0%), Micropholis acutangula (Ducke) Eyma (14,8%), Goupia
glabra Aubl. (9,8%), Brosimum parinarioides Ducke (8,2%). Em 2014 apenas quatro gêneros
dos quais quatro espécies foram exploradas para a alimentação do grupo: Brosimum
guianense Aubl. Huber (53,3%), Micropholis acutangula (Ducke) Eyma (29,5%),
Oenocarpus distichus Mart. (9,5%) e Pouteria freitasii T.D.Penn. (7,6%). Apenas

38
Micropholis acutangula (Ducke) Eyma foi explorada pelo grupo nos dois anos de
amostragem e, segundo o Índice de Jaccard, houve baixa similaridade na composição da dieta
dos dois anos (8%).
Com relação às famílias, foram exploradas 5 famílias em 2011 e 4 famílias em 2014, e
Sapotaceae e Moraceae foram utilizadas nos dois anos. Pela ordem de importância de
exploração em 2011: Melastomataceae (41,0%) Sapotaceae (36,1%), Moraceae (11,5%),
Goupiaceae (9,8%) e Chrysobalanaceae (1,6%). Para 2014: Moraceae (53,3%), Sapotaceae
(37,1%) e Arecaceae (9,5%).

39
5. Discussão
5.1 Tamanho e composição do grupo
O grupo de primatas deste estudo apresentou o padrão de agrupamento do tipo fissão-
fusão, típico do gênero Chiropotes (Veiga 2006). A contagem do grupo Bacaba com 67
indivíduos é uma das maiores conhecidas para grupos de Chiropotes sagulatus habitando
floresta contínua. O número é bem próximo do maior número antes encontrado para a espécie
na Guyana por Shaffer (2013; Tabela 8), mas é metade do tamanho do grupo Bela Cruz,
também estudado na FLONA Saracá-Taquera cuja contagem (n=120) foi realizada
simultaneamente por dois pesquisadores (THG Alvim com. pess.).
Nosso estudo foi conduzido juntamente com a supressão da área de vida do grupo, e
com isso, houve o adensamento dos macacos na área em questão (Melo et al. 2012) que pode
ter refletido também na distribuição dos indivíduos dentro dos subgrupos. O platô Bela Cruz,
onde THG Alvim realizou a contagem e a área de estudo de Shaffer (2013) são regiões de
florestas contínuas que ainda não estavam sob pressão antrópica.
Apesar de estar dentro do encontrado por outros estudos para o gênero, o número
encontrado no presente estudo é subestimado, já que, raramente o grupo de cuxiús se
deslocava em “fila indiana” ou todos por uma única árvore, onde as contagens eram feitas
com mais acurácia. As chuvas com longo período de duração contribuíram para as
subestimativas na contagem de indivíduos no período Chuvoso deste estudo, e na seca, por
outro lado, o comportamento de “fissão-fusão” e deslocamento disperso, dificultou a
visibilidade do agrupamento e contagem dos indivíduos. A dificuldade em detectar os
indivíduos na floresta é comum aos estudos com pithecideos e devida ao comportamento
críptico, extensas áreas de vida e alta densidade de indivíduos nos grupos. Estas dificuldades
são consideradas os fatores para se conhecer tão pouco a respeito destes primatas (Pinto et al.
2013).
Tabela 8. Número de indivíduos por grupo em diferentes estudos com Chiropotes habitando floresta contínua.
Área de estudo Espécie N indivíduos Referência
FLONA Saracá-Taquera,PA,Brasil
Chiropotes sagulatus 67
Presente estudo
FLONA Saracá-Taquera,PA,Brasil
Chiropotes sagulatus 120
THG Alvim (com.pess.)

40
Rio Preto da Eva, AM, Brasil
Chiropotes sagulatus 30 Frazão (1992)
Upper Essequibo Conservation Concession
Guyana
Chiropotes sagulatus
65 Shaffer (2013)
O resultado do número de indivíduos por subgrupo na média anual (n=23) corroborou
as impressões obtidas em campo, de que o Grupo original se subdividia em três unidades
principais. As diferenças no número de indivíduos por subgrupo para as estações poderiam ter
sido confirmadas se a contagem total de indivíduos no subgrupo não fosse comprometida pelo
comportamento de “fissão-fusão” dos cuxiús (Veiga 2006). Apesar desses contratempos de
amostragem, há uma tendência para subgrupos maiores durante o período chuvoso, como
também foi observado por Veiga et al. 2006; Boyle 2008; Gregory & Norconk 2013) considerado
o período de abundância de frutos maduros nas florestas tropicais e período de abundância de
alimentos para os primatas frugívoros (Peres 1994b). Como já foi observado em outros
estudos (Ayres 1989, Norconk 1996, Veiga 2006) Chiropotes sagulatus complementa sua
dieta com polpa e arilo de frutos maduros durante a estação chuvosa, e a abundância deste
recurso pode ter influenciado na coesão do grupo nesta estação.
Nas observações longitudinais houve diferença entre o número médio de indivíduos
por subgrupo, com novembro de 2014 apresentando os números mais baixos, próximo ao
menor encontrado no primeiro ano (Tabela 9), mas como esperado, essa diferença não foi
significativa. Considerando que as amostras foram realizadas na mesma estação de anos
diferentes, era esperado que o padrão de agrupamento se mantivesse semelhante, sendo
influenciado pelas condições ambientais.
Tabela 9. Resumo das contagens (média, min-max) de indivíduos de Chiropotes sagulatus por subgrupo para as
estações seca e chuvosa e os meses de Novembro/2011 e Novembro/2014 na FLONA Saracá-Taquera, PA.
Estação; Ano Média ind. por subgrupo min-max ind. por
subgrupo
Seca 20,92±5,09 16-30
Chuvosa 25,68±11,92 10-57
Novembro/2011 20,25±6,65 12-27
Novembro/2014 13±3,08 8-15

41
5.2 Padrão de atividades
O padrão anual de atividades do grupo Bacaba apresentou o esperado para o gênero
Chiropotes, com maior porcentagem do tempo de atividade dedicado ao Deslocamento, e,
embora reforce o encontrado por Pinto (2008), Port-Carvalho & Ferrari (2004), Silva (2003),
Vieira (2005) e Veiga (2006), foi diferente do encontrado por Peetz (2001) para a mesma
espécie, cujo grupo dedicou maior parte do tempo à Alimentação. Embora haja um consenso a
respeito do padrão comportamento do gênero Chiropotes, com intenso deslocamento e
diferentes porcentagens alocadas aos comportamentos entre estações (Port-Carvalho & Ferrari
2004; Veiga 2006; Vieira 2005; Gregory & Norconk 2013 e Setz et al. 2013), algumas
modificações nos padrões comportamentais podem estar relacionadas mais a condições
ecológicas como o tamanho e qualidade do habitat do que com características das espécies.
Ambientes fragmentados, como os estudados por Peetz (2001), Silva (2003), Vieira
(2005) e Veiga (2006) podem oferecer menores áreas de vida, o que influenciaria na
diminuição do tempo de deslocamento, que por sua vez favoreceria o incremento de
comportamentos sociais e repouso. As espécies vegetais e sua distribuição nas florestas onde
os cuxiús habitam, associadas a determinadas características como os padrões de agrupamento
das espécies botânicas, influenciam diretamente na composição da dieta e, em sequência, o
tempo de deslocamento entre fontes, o tempo de manipulação dos itens alimentares e o
orçamento geral de atividades dos grupos.
As dificuldades na visibilidade dos cuxiús, dada a sua preferência pelo estrato superior
da floresta, intenso deslocamento, e comportamento críptico (Pinto et al. 2013) podem afetar a
taxa de registros por varredura, que podem desviar o padrão comportamental registrado.
Assim, uma abordagem proporcional dos dados, dando peso igual para as varreduras,
distribuído pelos diferentes comportamentos observados pode diminuir os vieses e possíveis
superestimativas para determinadas categorias de comportamentos mais simples de observar.
Neste estudo a abordagem alternativa não demonstrou diferenças importantes nas proporções
dos comportamentos, mas a observação proporcional dos dados foi importante para garantir o
refinamento e a fidelidade destes.
Além das variáveis inerentes aos grupos e seus ambientes, segundo Pinto (2008), a
grande variação entre os esforços amostrais e períodos de estudos com Chiropotes também
podem influenciar as diferenças encontradas para a porcentagem de tempo alocado às

42
principais atividades (Tabela 10). Neste caso, estas podem estar presentes até entre grupos da
mesma espécie.
Tabela 10. Orçamento de atividades de Chiropotes em diferentes estudos. (Adaptado de Pinto, 2008).
% comportamentos
Espécie n registros Alimentação Forrageio Deslocamento Repouso Social Referência
C. albinasus 22.373 23,8 0,5 36,2 27,5 8,8 Pinto
(2008)¹
C. satanas 419 19,8 - 58,5 13,8 7,9 Port-Carvalho & Ferrari (2004)²
C. satanas 7.715 25,8 2,9 35,4 26,4 8,5 Veiga
(2006) ³
C. satanas 8.033 30,0 4,2 26,1 23,2 15,2 Veiga
(2006)³
C. sagulatus 87.448 37,0 10,1 18,7 21,4 11,4 Peetz
(2001)*
C. utahickae 5.490 58,8 - 30,8 9,5 0,9 Santos (2002)³
C. utahickae 11.277 31,9 5,4 50,6 10,6 1,2 Vieira (2005)³
C. sagulatus 6.926 8,0 14,0 41,0 30,0 7,0 Presente estudo**
Local de Estudo: ¹FLONA Tapajós PA, Brasil; ²Imperatriz MA, Brasil; ³Tucuruí, PA, Brasil; * Lake Guri,
Venezuela; ** FLONA Saracá-Taquera, PA, Brasil.
No padrão sazonal, houve diferença significativa para todas as categorias
comportamentais do grupo Bacaba, corroborando a hipótese de que, durante a estação seca,
haveria aumento no tempo gasto com Alimentação e Repouso, favorecendo o comportamento
Social, e no período chuvoso aumento no tempo de Deslocamento entre fontes alimentares.
Veiga (2006) em seu estudo com Chiropotes satanas em Tucuruí também observou, para os
dois grupos do estudo, aumento nos comportamentos de Alimentação na estação seca e maior
tempo dedicado ao Deslocamento na estação chuvosa. Em estudos com Chiropotes sagulatus,
Peetz (2001) na Venezuela e Gregory (2011) no Suriname observaram que o maior tempo
dedicado à Alimentação foi também durante o período da seca. Apesar de Dezembro e Abril e
fevereiro e março, meses considerados secos no Suriname, serem os meses de chuva na
FLONA Saracá-Taquera, a espécie manteve o padrão ecológico influenciado pela

43
sazonalidade ambiental, ainda que as características florestais (fragmento florestal e floresta
contínua) fossem diferentes.
No padrão longitudinal, a porcentagem de registros de Deslocamento e Alimentação
aumentou enquanto repouso e forrageamento diminuíram de 2011 para 2014. O maior
Deslocamento, Repouso e Forrageamento do primeiro ano podem ter sido influenciados pela
composição da dieta, que será tratada posteriormente. Levando em consideração também as
mudanças ambientais no platô Bacaba, esta diferença comportamental entre os anos pode ser
um sinal da adaptação do grupo à mudança imediata do habitat, já que em 2014 a supressão
da vegetação que incluiu parte da área disponível para o grupo já havia sido encerrada, o que
poderia ser confirmado através das análises de utilização da área de vida. Assim, no primeiro
ano, o grupo Bacaba apresentou o comportamento mais semelhante a grupos vivendo em
áreas fragmentadas (Peetz 2001; Santos 2002) do que grupos habitando florestas contínuas,
embora a área (em hectares) que permaneceu disponível para o grupo seja maior do que os
fragmentos conhecidos que tem a presença de cuxiús.
Vieira (2005) comparando os dados de sua pesquisa com o mesmo grupo estudado por
Santos (2002) três anos antes, observou que o padrão comportamental dos cuxiús apresentou
diferenças longitudinais com maior alimentação no primeiro ano e maior deslocamento em
seu estudo de 2005, padrão contrário ao encontrado no presente estudo. Neste estudo as
diferenças orçamentárias foram atribuídas a alterações interanuais do ambiente, como período
de seca maior em um ano, que teria alterado o período de frutificação das espécies vegetais,
mas assim como no estudo de Vieira (2005), no presente estudo com o grupo Bacaba não foi
possível realizar avaliações a respeito da disponibilidade dos recursos no ambiente, não sendo
possível, portanto, afirmar que as variações orçamentárias tenham sido em função de padrões
fenológicos.
Outro aspecto a ser considerado é a diferença no tamanho de subgrupos e taxas de
avistamentos entre anos, que poderia subestimar os comportamentos crípticos ou superestimar
os comportamentos mais visíveis. Os dados apresentaram média de indivíduos por subgrupos
menores (n=13,0 contra n=20,25), mas com uma taxa de avistamentos maior em 2014 do que
em 2011 (n=4,05 contra n=2,93), e embora as diferenças não tenham sido significativas, estes
fatores, especialmente a taxa de avistamentos, podem ter influenciado nas diferenças entre
comportamentos.

44
5.3 Dieta
Assim como o padrão de atividades, a dieta dos primatas é fortemente influenciada por
flutuações na disponibilidade de recursos preferidos (Strier 2000). Dados do Inventário
Florestal realizado por Salomão et al. (2002) no platô Bacaba, apresentaram informações
sobre a área de estudo. Sapotaceae e Moraceae, que são famílias conhecidamente importantes
na dieta de Chiropotes, apresentaram maior riqueza de espécies (S=62 e S=32), seguidas de
Fabaceae (S=30). Arecaceae mostrou-se como a família mais abundante, seguida de
Sapotaceae, Burseraceae e Moraceae. Já com relação às espécies mais abundantes,
Oenocarpus bacaba Mart. (bacaba), que dá nome ao Platô foi a mais abundante, seguido de
Tetragastris panamensis (Engl.) Kuntze (breu branco) e Geissospermum sericeum Bth. &
Hook (acarirana). No estudo, Salomão et al. (2002) afirmaram que não há dominância de
quaisquer espécies, ou se existe é muito baixa, e que a Equabilidade é alta, com índices
próximos ao máximo esperado para o número de espécies amostradas.
Mesmo com alterações no padrão de ingestão de itens de acordo com as possíveis
mudanças na disponibilidade dos recursos no ambiente, a dieta dos primatas é geralmente
classificada levando em consideração os itens que eles consomem na maior parte do tempo
(Strier 2000). Neste caso, a dieta do grupo Bacaba pode ser classificada como “granívora”,
apresentando ingestão de altas quantidades de sementes ao longo de todo o período de estudo
(padrão anual, sazonal e longitudinal), similar ao encontrado por Ayres (1981), Kinzey &
Norconk (1993), Port-Carvalho & Ferrari (2004), Veiga & Ferrari (2006) e Santos (2013) para
o gênero. O consumo de sementes em diversos estágios de maturação, observado também
neste estudo, corrobora o esperado por sua classificação como “forrageador esclerocárpico”
(Kinzey 1992; Kinzey & Norconk 1990; Shaffer 2013) e é considerada uma estratégia adotada
pelos cuxiús para minimizar os efeitos da sazonalidade e escassez de recursos. Shaffer (2013)
ainda afirma que, mesmo sendo especialistas e com todo o aparato mecânico e morfológico
para a esclerocarpia, os cuxiús apresentam flexibilidade para enfrentar os períodos de escassez
de recursos como a estação seca através da inclusão de diferentes itens alimentares na dieta,
como partes reprodutivas de plantas e recursos animais, e que tal característica é comum a
muitos membros da ordem Primates.
Considerando o padrão sazonal, na estação seca houve alta taxa de ingestão de
sementes imaturas e também uma tendência ao aumento do tempo gasto com Alimentação,
mas não houve diferença no número de espécies exploradas para a extração de sementes

45
imaturas. A maior proporção de registros de ingestão de sementes imaturas nos meses de seca
também foi observada por outros estudos (Ayres 1989; Kinzey & Norconk 1993; Norconk
1996; Peetz 2001; Silva 2003; Norconk et al. 2009; Melo et al. 2012; Shaffer 2012; Santos
2013), reafirmando a proposição de Norconk (1996) de que o período da seca não deve ser
considerado período de escassez de recursos para Chiropotes.
Assim como no estudo de Santos (2002), não foram registrados itens animais na dieta
dos cuxiús do grupo Bacaba, mas a ausência desses itens na dieta pode ter sido devida à
dificuldade de observação, já que no período de habituação foi observado um evento de um
indivíduo adulto predando um invertebrado. Apesar disso, como nos demais estudos (Ayres
1989; Kinzey & Norconk 1993; Norconk 1996; Peetz 2001; Silva 2003; Norconk et al. 2009;
Melo et al. 2012; Shaffer 2012), a estação seca também foi marcada pela inclusão de itens
ausentes da dieta da estação chuvosa, como Semente de Fruto Seco, Partes Florais e Outros
(como casca, por exemplo), e embora a fenologia não tenha sido um parâmetro testado neste
estudo, podem ser chave para a sobrevivência do gênero às alterações na dinâmica de
frutificação das Florestas Tropicais.
Já na estação chuvosa o grupo apresentou o padrão esperado para os cuxiús,
corroborando a hipótese de que haveria maior ingestão de frutos, também encontrado por
Santos et al. (2013), e aumento também no número de registros de Deslocamento e menor
tempo dedicado às atividades de Forrageamento e Alimentação, corroborando a estratégia de
“maximização de energia” (Shaffer 2013; Strier 2000). Isso se deve ao fato de os frutos serem
recursos alimentares que tendem a estar mais dispersos no ambiente do que as folhas, mas que
também são mais ricos em calorias fáceis de digerir e reabastecer a energia gasta para
encontrá-los (Strier 2000). O maior Deslocamento dos cuxiús no período chuvoso pode estar
relacionado ao monitoramento constante das fontes alimentares para a obtenção dos recursos
no estágio de maturação favorável, onde a ingestão de compostos secundários de alguns frutos
também pode ser evitada. Este padrão comportamental dos macacos em florestas tropicais
está relacionado à distribuição e disponibilidade de recursos no ambiente, o que exige
adaptações para manter dietas de qualidade como a ingestão de outros itens alimentares não-
preferenciais, porém abundantes, conhecidos como alimentos “fallback” (Chapman et al.
2005).
O grupo Bacaba se alimentou de altas taxas de sementes nos dois anos e, da maneira
esperada, houve mais diferenças entre as estações ao longo do período de estudo do que entre

46
meses iguais de anos diferentes. Apesar de não ter havido diferenças nas proporções dos itens
alimentares da dieta dos dois anos, os cuxiús demonstraram uma capacidade de manter a
composição da dieta, no que diz respeito à proporção de itens, mas a partir da exploração
espécies diferentes nos dois anos. Isso reflete claramente a flexibilidade desses primatas para
enfrentar períodos de escassez de determinado recurso no ambiente, sendo favorecidos pela
capacidade que possuem em acessar diferentes itens alimentares.
As sementes de fruto seco ingeridas em 2011 eram pertencentes à Sapotaceae sp1, que
se mostrou como um importante recurso para os cuxiús, sendo explorada durante muitos
meses e em diversos estágios de maturação e contribuiu com quase 20% da dieta total de
novembro do primeiro ano, não sendo ingerida em 2014. No segundo ano, Sementes de
Frutos Maduros, que foram responsáveis por mais de 20% da dieta, eram pertencentes
especialmente a Brosimum guianense (Aubl.) Huber (pau-rainha), que também foi
responsável por mais da metade das fontes alimentares para 2014 e não foi utilizada para a
Alimentação em 2011.
Explorar as sementes de fruto seco pode ter elevado o tempo gasto com forrageamento
no primeiro ano, já que sementes, conforme observado por Peetz (2001) dependem de maior
tempo de manipulação para serem acessadas, diferente de polpa de Frutos Maduros, por
exemplo. E, considerando a abundância da família Arecaceae no platô Bacaba, e a utilização
das palmeiras Oenocarpus distichus Mart (bacaba-abacate) para a alimentação do grupo no
segundo ano, estas podem ter influenciado no maior o tempo gasto com Deslocamento entre
fontes, já que as palmeiras são árvores de pequeno porte, que suportam poucos indivíduos se
alimentando ao mesmo tempo.
5.4 Composição da dieta
Os cuxiús são conhecidos por se alimentar de um número grande de espécies vegetais,
o que é considerado por Peetz (2001) como uma estratégia adaptativa para evitar compostos
secundários desfavoráveis presentes nas espécies vegetais. Quando comparado com outros
estudos, o número o número de famílias, gêneros e espécies utilizadas como fontes
alimentares apresentado por este trabalho está dentro do encontrado pelos demais (Gregory
2011; Boyle et al. 2012; Melo 2012; Shaffer 2013) para a espécie C. sagulatus em floresta
contínua, conforme apresentado na Tabela 11. A tabela apresenta, em alguns casos,
compilação de dados de mais de um grupo por estudo.

47
Tabela 11. Diferenças quanto ao número de famílias, gêneros e espécies vegetais consumidas por Chiropotes
sagulatus em habitats de floresta contínua, em diferentes estudos. ** mais de um grupo por estudo.
DURAÇÃO DO ESTUDO
(MESES/ HORAS) N FAMÍLIAS N GÊNEROS N ESPÉCIES REFERÊNCIA
8/440 32 70 105 Presente estudo
17/535 47 115 244
Boyle et al. (2012)**
12/ 41 92 153
Melo 2012 (não publicados)**
12/540 32 112 Gregory (2011)
10/560 30 75 119 Shaffer (2013)
Com relação às famílias mais importantes para a dieta do grupo Bacaba, Sapotaceae,
Fabaceae e Lecythidaceae estiveram entre as cinco mais representativas. Todos os outros
estudos com Chiropotes apresentaram estas famílias entre as mais importantes para a
composição da dieta (e.g. Norconk 1996; Peetz 2001; Port-Carvalho & Ferrari 2004; Boyle
2008; Gregory 2011; Boyle et al. 2013; Shaffer 2013) na compilação de dados realizadas por
Boyle et al. (2015). Famílias como Lecythidaceae apresentam poucas adaptações químicas,
mas muitas adaptações mecânicas para evitar a predação de suas sementes, e a habilidade para
se alimentarem delas parece ser uma das características que define o nicho dos cuxiús
(Shaffer 2013). Em contrapartida, por ingerirem as sementes do fruto imaturo e as flores
destas famílias, a planta tem condição de alocar mais energia com os frutos restantes. Esta
adaptação tem levado autores a questionar se o limite geográfico destes macacos está limitado
pela extensão da viabilidade destas famílias (Norconk 2011; Ayres & Prance 2013; Shaffer
2013). No Platô Bacaba, Salomão et al. (2002) observaram que a família Sapotaceae
“sobressai-se entre quaisquer outras”, apresentando maior diversidade e dominância entre
todas as demais famílias e o número de espécies a coloca em segundo lugar no valor
percentual de importância para a região.
Os gêneros com maior número de espécies presentes na dieta do grupo Bacaba,
Pouteria, Licania, Eschweilera e Inga também estão entre os cinco mais importantes para
Chiropotes no trabalho de Boyle et al. (2015), definindo a preferência dos Chiropotes por
estes recursos alimentares quando disponíveis, mesmo quando diferenças locais na
configuração dos recursos. Com relação às espécies, Sapotaceae sp.1 e abil Chrysophyllum

48
sanguinolentum (Pierre) Baehni, pertencentes à família Sapotaceae, apresentaram maior
número de registros e também maior número de fontes para a alimentação do grupo. Esta
presença da família Sapotaceae como importante fonte alimentar para os cuxiús no grupo
Bacaba em relação ao número de fontes, gênero e espécies demonstra que os cuxiús
provavelmente são favorecidos pela dominância desta na área de estudo. Apesar da inclusão
da família Bignoniaceae representada por Anemopaegma foetidum Bureau & K.Schum na
lista de seis espécies com maior número de registros de alimentação, Sapotaceae,
Lecythidaceae e Moraceae continuam presentes, como nas cinco famílias mais representativas
na dieta do grupo Bacaba, reforçando a importância destas também para este aspecto.
Além disso, as seis espécies com maior número de registros de Alimentação no nosso
trabalho apresentaram mais de 60% do total de registros, mostrando que apesar de sua
capacidade de explorar um grande número de espécies vegetais, algumas espécies são
preferencialmente exploradas quando disponíveis. No padrão sazonal, por exemplo,
Anemopaegma foetidum Bureau & K.Schum, Brosimum rubescens Taub., Chrysophyllum
sanguinolentum (Pierre) Baehni, Licania octandra (Hoffmanns. ex. Roem & Schult.) Kuntze,
Micropholis acutangula (Ducke) Eyma e Pouteria anomala (Pires) T.D.Penn. estiveram
presentes na estação chuvosa do período de estudo, e permaneceram durante a estação seca,
embora 16 outras espécies tenham sido incluídas para a dieta desta estação.
No padrão longitudinal, nossos dados apontaram que no primeiro ano
Melastomataceae foi a família mais representativa na dieta dos cuxiús do grupo Bacaba, com
mais de 40% do total de registros de alimentação, mas não foi explorada pelo grupo no
segundo ano. A espécie mais frequentemente utilizada para a alimentação em 2011 Mouriri
collocarpa Ducke também pertence à família, o que reforça a proposição de Santos et al.
(2013) de um padrão imprevisível de disponibilidade, em que os cuxiús presumivelmente
exploram os recursos mais abundantes em qualquer época, já que Melastomataceae não está
entre as 5 famílias mais representativas para a dieta nos estudos com Chiropotes. Além da
espécie comum à dieta, apenas o gênero Brosimum esteve presente nos registros alimentares
do grupo dos dois anos.
De Sapotaceae sp. 1 (Figura 13) foram exploradas pelos cuxiús as sementes imaturas,
o fruto em estágio maduro do qual extraíam a pouca polpa pressionando contra os lábios,
engolindo as sementes e a semente quando o fruto estava seco. O fato de ter sido explorada ao
longo dos meses e em diversos estágios de maturação pode colocar Sapotaceae sp.1 como um

49
recurso chave para a sua sobrevivência na FLONA Saracá-Taquera, já que várias áreas
disponíveis para os Chiropotes bem como para outras espécies da fauna e da flora tem sido
desmatados para a exploração vegetal. Assim estudos para a identificação das espécies, bem
como para verificar sua distribuição e de outras fontes alimentares na área de vida do grupo
são importantes e necessários para definir diretrizes e prioridades para a conservação desta
população.
Figura 13. Espécie da família Sapotaceae ainda não descrita consumida pelo grupo de Chiropotes sagulatus
(Bacaba) de semente imatura a semente de fruto seco na FLONA Saracá-Taquera, PA.

50
6. Conclusão
Este trabalho corrobora os padrões ecológicos já observados para o gênero Chiropotes,
que aparentam estar relacionadas à influência de características ambientais e não a
diferenças filogenéticas;
O padrão de agrupamento do grupo Bacaba é do tipo fissão-fusão e demonstrou
tendência para subgrupos maiores na estação chuvosa do que na seca, e também
maiores em 2011 do que em 2014;
Os parâmetros ecológicos de comportamento e dieta apresentaram variações sazonais
e longitudinais e refletiram a flexibilidade dos cuxiús enquanto maximizadores de
energia, com grande tempo gasto com Deslocamento e alta ingestão de sementes
imaturas;
As famílias botânicas mais importantes para a dieta do grupo Bacaba foram
Sapotaceae, Moraceae, Fabaceae, Chrysobalanaceae e Lecythidaecae, conhecidamente
importantes para o gênero Chiropotes em diferentes sítios, tanto em florestas
contínuas como em áreas fragmentadas;
Os cuxiús são primatas que desempenham um importante papel no ambiente, e o fato
de serem primatas granívoros levanta questões importantes a respeito de sua relação
com as espécies vegetais das quais se alimenta;
Direcionar estudos futuros quanto a este aspecto pode esclarecer um dos papeis
importantes representados pelos cuxiús na manutenção da diversidade botânica local,
em cada área de ocorrência das espécies;
A presente pesquisa oferece muitas indagações a serem respondidas nas próximas
análises e futuros trabalhos. Pesquisas futuras, e estudos de longo prazo com esta
população, que está sob a influência antrópica direta, favorecerão ainda mais o
conhecimento aprofundado a respeito da ecologia dos cuxiús e a definição de
protocolos confiáveis para sua conservação.

51
7. Referências Bibliográficas
Altmann J. 1974. Observational study of behavior: sampling methods. Behaviour 49: 227–267.
Antonini Y & Oliveira ML. 2007. Abelhas Euglossinae da Floresta Nacional de Saracá-Taquera em Porto Trombetas. In: Anais do VIII Congresso de Ecologia do Brasil, Caxambu - MG.
Ayres JM. 1989. Comparative feeding ecology of the Uakari and Bearded Saki, Cacajao and Chiropotes. Journal of Human Evolution 18: 697-716.
Ayres JM & Prance GT. 2013. On the distribution of Pitheciine monkeys and Lecythidaceae trees in Amazonia. In: Veiga LM, Barnett AA, Ferrari SF & Norconk MA (Org), Evolutionary Biology and Conservation of Titis, Sakis and Uacaris, Cambridge: Cambridge University Press, p. 127-139.
Barnett AA, Boyle SA, Norconk MA et al. 2012. Terrestrial activity in Pitheciins (Cacajao, Chiropotes, and Pithecia). American Journal of Primatology 74: 1106–1127.
Bicca-Marques JC, Silva VM & Gomes DF. 2006. Ordem Primates. In: Reis NR, Peracchi, AL, Pedro WA & Lima IP (Eds), Mamíferos do Brasil. Londrina: 437p.
Blom A, Cipottella C, Brunsting AMH & Prins HHT. 2004. Behavioral responses of gorillas to habituation in the Dzanga-Ndoki National Park, Central African Republic. International Journal of Primatology 25:179-196.
Boyle SA. The effects of forest fragmentation on primates in the Brazilian Amazon. 2008. 306f. Dissertação (Ph.D em Filosofia). Arizona State University, Tempe, AZ.
Boyle SA, Lourenço WA, da Silva LR, Smith AT. 2009. Travel and spatial patterns change when northern bearded saki monkeys (Chiropotes satanas chiropotes) inhabit forest fragments. International Journal of Primatology 30: 515–531.
Boyle SA, Smith AT. 2010. Behavioral modifications in northern bearded saki monkeys (Chiropotes satanas chiropotes) in forest fragments of central Amazonia. Primates. 51: 43–51.
Boyle SA, Zartman CE, Spironello WR, Smith AT. 2012. Implications of habitat fragmentation on the diet of bearded saki monkeys in central Amazonian Forest. Journal of Mammalogy 93(4): 959–976.
Boyle SA, Smith AT, Spironello WR & Zartman CE. 2013. The behavioral ecology of northern bearded sakis (Chiropotes satanas chiropotes) living in forest fragments of Central Brazilian Amazonia. In: Veiga LM, Barnett AA, Ferrari SF & Norconk MA (Org), Evolutionary Biology and Conservation of Titis, Sakis and Uacaris, Cambridge: Cambridge University Press, p. 255-261.

52
Boyle SA. 2015. The geography of choice: patterns in fruit consumption by the pitheciids Cacajao, Callicebus, Chiropotes, and Pithecia. American Journal of Primatology (artigo aceito para publicação).
Camargo CC, Porfírio S, Rylands AB, & Langguth A. 2008. Variação sazonal e longitudinal nos padrões de comportamento em uma população de Alouatta belzebul (Primates: Atelidae) do Nordeste brasileiro. In: Ferrari SF & Rímoli J (Eds), A Primatologia no Brasil 9, Aracaju: Biologia Geral e Experimental Universidade Federal de Sergipe, 201p.
Capobianco JPR, Veríssimo A, Moreira A, Sawyer D, Santos I & Pinto LP. 2001. Biodiversidade na Amazônia Brasileira: avaliação e ações prioritárias para a conservação, uso sustentável e repartição de benefícios. São Paulo: Instituto Socioambiental e Estação Liberdade. 540p.
Chapman CA & Chapman LJ. 1990. Dietary Variability in Primate Populations. Short Communication. PRIMATES 31(1): 121-128.
Chapman CA, Chapman LJ, Zanne AE, Poulsen JR & Clark CJ. 2005.A 12-year phenological record of Fruiting: Implications for frugivore populations and indicators of climate change. In: Dew JL and Boubli JP (Eds), Tropical Fruits and Frugivores: The Springer. Netherland, 5:75-92. Conservation International do Brasil. 2007. Corredor de biodiversidade do Amapá. São Paulo: Ipsis, 54p.
Cuarón AD. 2000. A Global Perspective on Habitat Disturbance and Tropical Rainforest Mammals. Conservation Biology 14 (6): 1574-1579.
Di Fiore A. 2003. Ranging behavior and foraging ecology of lowland woolly monkeys (Lagothrix lagotricha poeppigii) in Yasuní National Park, Ecuador. American Journal of Primatology 59: 47-66.
Frazão ER. Dieta e estratégia de forragear de Chiropotes satanas chiropotes (Cebidae: Primates) na Amazônia Central Brasileira. 1992. 116f. Dissertação (Mestrado em Ciências Biológicas, Ecologia) - Instituto Nacional de Pesquisas da Amazônia. Manaus, Amazonas.
Gregory TL. Socioecology of the guianan bearded saki, Chiropotes sagulatus. 2011. 177f. Tese (Doutorado em Filosofia) - Kent State University. Kent, Ohio.
Gregory TL & Norconk MA. 2013. Comparative socioecology of sympatric, free-ranging white-faced and bearded saki monkeys in Suriname: preliminary data. In: Veiga LM, Barnett AA, Ferrari SF & Norconk MA (Org), Evolutionary Biology and Conservation of Titis, Sakis and Uacaris, Cambridge: Cambridge University Press, p.285-294.
Gregory TL & Norconk MA . 2014. Bearded saki socioecology: affiliative male–male interactions in large, free-ranging primate groups in Suriname. Behaviour 493:533-531.

53
Haugaasen T, Peres, CA. 2005. Mammal assemblage structure in Amazon flooded and unflooded forests. Journal of Tropical Ecology 21(2): 133-145.
Hill DA. 1997. Seasonal variation in the feeding behavior and diet of japanese macaques (Macaca fuscata yakui) in lowland forest of Yakushima. American Journal of Primatology 43: 305–322.
Kappeler PM & Van Schaik CP. 2002. Evolution of primate social systems. International Journal of Primatology 23(4): 707-740.
Kay RF, Meldrum DJ & Takai M. 2013. Pitheciidae and other platyrrhine seed predators. In: Veiga LM, Barnett AA, Ferrari SF & Norconk MA (Org), Evolutionary Biology and Conservation of Titis, Sakis and Uacaris, Cambridge: Cambridge University Press, p.3-12.
Kinzey WG. 1992. Dietary and dental adaptations in the Pitheciinae. American Journal of Physical Anthropology 88:499-514.
Kinzey WG & Norconk MA. 1990. Hardness as a basis of fruit choice in two sympatric primates. American Journal of Physical Anthropology 81: 5-15.
Kinzey WG & Norconk MA. 1993. Phisical and chemical properties of fruit and seeds eaten by Pithecia and Chiropotes in Surinam and Venezuela. International Journal of Primatology 14(2): 207-227.
Lima EM & Ferrari SF. 2003. Diet of a free-ranging group of squirrel monkeys (Saimiri sciureus) in eastern Brazilian Amazonia. Folia Primatologica 74: 36-44.
Machado ABM, Drummond GM & Paglia AP. 2008. Livro vermelho da fauna brasileira ameaçada de extinção. Belo Horizonte, MG, Fundação Biodiversitas, 906p.
Melo FR, Moreira LS, Scarascia PO, Moura VS, Alvim THG & Silva LP. 2010. Monitoramento de duas espécies de primatas na Floresta Nacional de Saracá-Taquera, Pará. Relatório Técnico Anual. Jataí, UFG, 60p.
Melo FR, Moreira LS, Scarascia PO, Moura VS, Alvim THG & Silva LP. 2012. Monitoramento de duas espécies de primatas na Floresta Nacional de Saracá-Taquera, Pará. Relatório Técnico Anual. Jataí, UFG, 50p.
Moraes BC de, Costa JMN da, Costa ACL da, Costa MH. 2005. Variação espacial e temporal da precipitação no estado do Pará. Acta Amazônica 35(2): 207-214.
Mourthé IM da C. 2014. Response of frugivorous primates to changes in fruit supply.
Brazilian Journal of Biology 74 (3): 720-727.
Napier JR. 1976. Catalogue of Primates in the British Museum (Natural History), Part 1: Family Callitrichidae and Cebidae. London, British Museum (Natural History).

54
Norconk MA & Kinzey WG. 1994. Challenge of Neotropical frugivory: travel patterns of spider monkeys and bearded sakis. American Journal of Primatology 34: 171-183.
Norconk MA. 1996. Seasonal variation in the diets of white-faced and bearded sakis (Pithecia pithecia and Chiropotes satanas) in Guri Lake, Venezuela. In: Norconk MA, Rosenberger AL, Garber PA (Eds), Adaptative radiations of Neotropical Primates. New York: Plenum, p. 403-423.
Norconk MA, Wertis C, & Kinzey WG. 1997. Seed predation by monkeys and macaws in eastern Venezuela: preliminary findings. Primates 38(2): 177–184.
Norconk MA, Wright BW, Conklin-Brittain NL & Vinyard CJ. 2009. Mechanical and Nutritional Properties of Food as Factors in Platyrrhine Dietary Adaptations In: Garber PA, Estrada A, Bicca-Marques JC, Heymann EW & Strier KB (Eds), South American Primates, Developments in Primatology: Progress and Prospects. New York: Springer Science Business Media, p. 279-319.
Peetz, A. (2001). Ecology and social organization of the bearded saki Chiropotes satanas chiropotes (Primates: Pitheciinae) in Venezuela. Ecotropical Monographs, 1, 1–170.
Pereira LCM. Área de vida e padrões de deslocamento de Brachyteles arachnoides (e. Geoffroy, 1806) (Primates: Atelinae) em um fragmento florestal no município de Castro, estado do Paraná, Brasil. 2006. 120f. Dissertação (Mestrado em Engenharia Florestal) – Centro de Ciências Florestais e da Madeira. Universidade Federal do Paraná. Curitiba, Paraná.
Peres CA. 1994a. Diet and feeding ecology of Gray Wooly Monkey (Lagothrix lagotricha cana) in central Amazonia: comparisons with other Atelines. International Journal of Primatology 15(3): 333-372.
Peres CA. 1994b. Primate responses to phenological changes in an Amazonian Terra Firme Forest. Biotropica 26(1): 98-112.
Pianka ER. 1994. Evolutionary Ecology, Fifth Edition. New York: Harper and Row, 486p.
Pinto LP. Ecologia alimentar do cuxiú-de-nariz-vermelho Chiropotes albinasus Primates: Pitheciidae) na Floresta Nacional do Tapajós, Pará. 2008. 158f. Tese (Doutorado em Ecologia) – Instituto de Biologia. Universidade Estadual de Campinas. Campinas, São Paulo.
Pinto LP et al. 2013. Why we know so little: the challenges of fieldwork on the Pitheciids. In: Veiga LM, Barnett AA, Ferrari SF & Norconk MA (Org), Evolutionary Biology and Conservation of Titis, Sakis and Uacaris, Cambridge: Cambridge University Press, p. 145-150.
Port-Carvalho M, Ferrari SF. 2004. Occurrence and diet of the black bearded saki (Chiropotes satanas satanas) in the fragmented landscape of western Maranhão, Brazil. Neotropical Primates. 12(1): 17-21.

55
Rímoli J, Strier KB & Ferrari SF. 2008. Seasonal and longitudinal variation in the behavior of free-ranging black tufted capuchins Cebus nigritus (Goldfuss, 1809) in a fragment of Atlantic Forest in Southeastern Brazil. In: Ferrari SF & Rímoli J (Eds), A Primatologia no Brasil 9. Aracaju: Sociedade Brasileira de Primatologia, Biologia Geral e Experimental – UFS, p.130-146.
Rylands AB, Schneider H, Langguth A, Mittermeier RA, Groves CP & Rodriguez-Luna E. 2000. An assessment of the diversity of New World primates. Neotropical Primates 8(2): 61-93.
Rylands AB, Mittermeier RA, Silva-Júnior JS. 2011. Neotropical primates: taxonomy and recently described species and subspecies. International Zoo Yearbook 46: 11-24.
Rothman JM, Chapman CA, Van Soest PJ. 2012. Methods in Primate Nutritional Ecology: A User’s Guide. International Journal of Primatology 33: 542-566.
Salomão RP, Rosa NA, Ferraz J & Matos AH. 1997. Uso de parcelas permanentes em reflorestamentos de diversas idades para avaliação da recuperação de áreas mineradas, Porto Trombetas, Oriximiná, Pará. In: Simpósio Brasileiro de Recuperação de Áreas Degradadas, SINRAD. Ouro Preto, MG p.407-415.
Salomão R de P, Matos AH de, Corrêa M da S, Rosa N de A, Rosário C da S, Santos MR dos, Silva DF. 2002. Inventário Florestal em 205 hectares de Floresta Ombrófila Densa com Palmeiras, Platô Bacaba, da Floresta Nacional Saracá-Taquera/IBAMA, Porto Trombetas, Município de Oriximiná, Estado do Pará. Mineração Rio do Norte – MRN/Cooperativa de Tecnologia Organizacional – COOPERTEC. 148p.
Salomão RP, Matos AH, Rosa NA, Rosário CS & Santos MR. 2003. Inventário florestal em 407 hectares de floresta ombrófila densa, Platô Almeidas, da Floresta Nacional Saracá-Taqüera/IBAMA, Porto Trombetas, município de Oriximiná, estado do Pará. Belém: Mineração Rio do Norte, 267p.
Santos RR dos. Ecologia de cuxiús (Chiropotes satanas) na Amazônia Oriental: perspectivas para a conservação de populações fragmentadas. 2002. 64f. Tese (Mestrado em Zoologia) – Museu paraense Emílio Goeldi. Universidade Federal do Pará, Belém. Santos RR dos, Vieira TM & Ferrari SF. 2013. Feeding ecology of uta hick's bearded saki (Chiropotes utahickae) on a man-made island in southeastern brasilian amazonia: seasonal and longitudinal variation. In: Veiga LM, Barnett AA, Ferrari SF & Norconk MA (Org), Evolutionary Biology and Conservation of Titis, Sakis and Uacaris, Cambridge: Cambridge University Press, p. 250-254.
Setz EZF, Pinto LP, Bowler M, Barnett AA, Vié JC, Boubli JP & Norconk MA. 2013. Pitheciins: use of time and space. In: Veiga LM, Barnett AA, Ferrari SF & Norconk MA (Org), Evolutionary Biology and Conservation of Titis, Sakis and Uacaris, Cambridge: Cambridge University Press, p. 72-83.

56
Shaffer CA. Ranging behavior, group cohesiveness, and patch use in northern bearded sakis (Chiropotes sagulatus) in Guyana. 2012. 353f. Tese (Doutorado em Filosofia) - Department of Anthroplogy. Washington University, St. Louis, Missouri.
Shaffer CA. 2013. Feeding Ecology of Northern Bearded Sakis (Chiropotes sagulatus) in Guyana. American Journal of Primatology 75: 568–580.
Silva-Júnior JS & Figueiredo WMB. 2002. Revisão sistemática dos cuxiús, gênero Chiropotes Lesson, 1840 (Primates, Pitheciidae). In: Ferrari SF & Rímoli J (Eds), Livro de resumos do X Congresso Brasileiro de Primatologia. Aracaju: Sociedade Brasileira de Primatologia, Biologia Geral e Experimental – UFS, p. 21.
Silva-Júnior JS, Figueiredo WMB. & Ferrari SF 2013. Taxonomy and geographic distribution of the Pitheciidae. In: Veiga LM, Barnett AA, Ferrari SF & Norconk MA (Org), Evolutionary Biology and Conservation of Titis, Sakis and Uacaris, Cambridge: Cambridge University Press, p. 31-42.
Silva SSB. Comportamento alimentar do cuxiú-preto (Chiropotes satanas) na área de influência do reservatório da usina hidrelétrica de Tucuruí-Pará. 2003. 119f. Dissertação (Mestrado em Zoologia) – Museu Paraense Emilio Goeldi. Universidade Federal do Pará. Belém.
STCP Engenharia, Consultoria e Gerenciamento. 2001. Plano de manejo da floresta nacional de Saracá-Taqüera, Estado do Pará, Technical report. Curitiba, Brasil, s.n.
Strier KB. 2000. Primate behavioral ecology. Needham Heights: Allyn & Bacon. Needham, Massachusetts, 392p .
Tutin CEG & Fernandez F. 1991. Responses of wild chimpanzees and gorillas to the arrival of primatologists: behaviour observed during habituation. In: Chapman and Hall (Eds.) Primate Responses to Environmental Change. New York: Box, H.O., p. 187-197.
van Roosmalen MGM, Mittermeier RA, Fleagle JG. 1988. Diet of the northern bearded saki (Chiropotes satanas chiropotes): a neotropical seed predator. American Journal of Primatology 14(1): 11-35.
van Schaik CP & Brockman DK. 2005. Seasonality in primate ecology, reproduction and life history: an overview. In: Brockman DK & van Schaik CP (Eds.), Seasonality in Primates: Studies of Living and Extinct Human and Non-Human Primates. Cambridge: Cambridge University Press, p. 3-20.
Veiga LM. A ecologia e organização social do cuxiú-preto (Chiropotes satanas) no contexto de fragmentação de hábitat. 2006. 223f. Tese (Doutorado em Psicologia) - Centro de Filosofia e Ciências Humanas. Universidade Federal do Pará. Belém.
Veiga LM & Ferrari SF. 2006. Predation of arthropods by southern bearded sakis (Chiropotes satanas) in eastern Brazilian Amazonia. American Journal of Primatology, 68: 209–215.

57
Veiga, L.M., Pinto, L.P. & Ferrari, S.F. 2006. Fission-fusion sociality in bearded sakis (Chiropotes albinasus and Chiropotes satanas) in Brazilian Amazonia. Proc. XXIst Congresso Sociedade Internacional de Primatologia, Entebbe. 709p. Veiga LM, Silva-Júnior JS, Ferrari SF & Rylands AB. 2008. Chiropotes satanas. The IUCN Red List of Threatened Species. Version 2014.3. <www.iucnredlist.org>. Downloaded on 20 January 2015.
Vieira TM. Aspectos da ecologia do Cuxiú de Uta Hick, Chiropotes utahickae (Hershkovitz, 1985), com ênfase na exploração alimentar de espécies arbóreas da Ilha de Germoplasma Tucuruí – PA. 2005. 138f. Tese (Mestrado em Zoologia) - Museu Paraense Emílio Goeldi e Universidade Federal do Pará. Belém.

58
8. APÊNDICES
APÊNDICE A CLASSES SEXO-ETÁRIAS. Adaptado de Veiga (2006).
CLASSE SEXO ETÁRIA
CÓDIGO UTILIZADO
DEFINIÇÃO
FILHOTE
F
Indivíduos dependentes, carregados na região ventral e posteriormente dorsal da mãe. Apresentam a cauda preênsil nos
primeiros meses de vida. Esta classe tem duração de aproximadamente 12 meses .
JUVENIL
JUV
Indivíduos independentes, mas de porte físico menor do que os sub-adultos. Bulbos temporais, barba e órgãos genitais pouco
desenvolvidos, sendo difícil a identificação sexual.
FÊMEA SUB-ADULTA
FSA Indivíduo visivelmente menor do que a fêmea adulta. Os bulbos temporais, barba e pelagem da cauda são menos espessas.
Apresenta genitália de coloração rósea.
MACHO SUB-ADULTO
MSA Indivíduo visivelmente menor do que o adulto. Os bulbos temporais, barba e pelagem da cauda são menos espessas.
Apresenta bolsa escrotal distinta e de coloração rósea.
MACHO ADULTO MA Comprimento corporal de 400 a 480 mm. Apresentam barba e
bulbo temporal maiores do que das fêmeas e bolsa escrotal rosada, protuberante e bem desenvolvida.
FÊMEA ADULTA
FA Genitália bem desenvolvida. Bulbos temporais e barba são menos proeminente do que dos machos. Para o grupo Bacaba, apresentam pelagem dorsal mais amarelada do que os machos.
Durante o estro a região ano genital é avermelhada.
ADULTO A Comprimento corporal 380 a 480 mm.

59
APÊNDICE B. Lista das espécies vegetais utilizadas como fonte alimentar pelo grupo
Bacaba na FLONA Saracá-Taquera, PA durante o período de estudo.
FAMÍLIA ESPÉCIE NOME
POPULAR
ANACARDIACEAE Anacardium giganteumHanc. Ex Engl Cajuí
Astronium gracile Engl. Muiracatiara
ANNONACEAE Xylopia nitida Dunal Comida de
inhambu
APOCYNACEAE Couma guianensis Aublet. Sorva
Geisossopermum argentium Woodson Quinarana
Odontadenia verrucosa Willd. Ex Roem. & Schult.
Parahancornia fasciculata (Poir) Benoist pau-de-colher
Rhigospira quadrangularis (Mull. Arg.) Miers
ARALIACEAE Schefflera morototoni (Aubl.) Maguire, Steyerm. & Frodin Mandiocão
BIGNONIACEAE Anemopaegma foetidum Bureau & K.Schum. cipó-cururu
Nemora barcteosa cipó-de-cruz
CECROPIACEAE Pouruma minor Benoist Embaubarana
CHRYSOBALANACEAE
Couepia guianensis Aubl. Pajurá
Hirtella racemosa Lam.
Licania adolphoduckei Prance
Licania hypoleuca Benth. Caraipérana
Licania longistyla (Hook.f.) Fritsch uxi-de-cutia
Licania macrophylla Benth pajurá-pedra
Licania niloi Prance Caraipé
Licania octandra (Hoffmanns. ex Roem & Schult.) Kuntze Caraipé
CLUSIACEAE Caraipa sp.
Clusia amazônica Planch & Triana Apuí
Clusia insignis Apuí
Symphonia globulifera L. f. Ananim
COMBRETACEAE Buchenavia grandis Ducke tanimbuca
Nectandra cuspidata Nees & Mart.
DILLENIACEAE Pinzona coriácea Mart. & Zucc cipó-de-fogo
EUPHORBIACEAE Glycidendron amazonicum Ducke castanha-de-porco
Hevea guianensis Aubl. Siringueira
FABACEAE Abarema floribunda (Benth.) Barneby & J.W. Grimes Saboeira
Andira micrantha Ducke sucupira-amarela
Dimorphandra sp.
Enterolobium maximum Ducke Tamburil
Hymenaea courbaril L. Jatobá

60
Inga lateriflora Miq. inga-chichica
Inga longiflora Spruce Ingá
Inga paraensis Ducke Ingarana
Inga stipularis D.C. ingá-chichica
Macrolobium limbatum Benth.
Parkia ulei (harms) Kuhlm Faveira
Pseudopiptadenia suaveolens Miq. fava-timborana
Vatairea paraensis Ducke fava-margosa
GOUPIACEAE Goupia glabra Aubl. Cupiuba
HIPPOCRATEACEAE Tontelea congestiflora (A.C.Sm.)
HUMIRIACEAE Endopleura uchi (Huber) Cuatrec. uxi liso
Sacoglottis ceratocarpa Ducke Uxirana
Sacoglottis guianensis Benth. Achuá
Sacoglottis matogrosensis Malme Achuá
Vantanea macrocarpa Ducke Uxirana
ICACINACEAE Pleurisanthes emarginata Tiegh
Poaqueiba guianensis Aubl. Mari
IPPOCRATEACEAE Salacia impresifolia (Miers) A.C.Sm.
LAURACEAE Nectandra cuspidata Nees & Mart. Louro
Ocotea ceanotifolia Aubl. Louro
Ocotea spledens (Meisn.) Baill
LECYTIDACEAE Eschweilera atropetiolata S.A. Mori ripeiro-vermelho
Eschweilera grandiflora (Aubl.) Sandwith ripeiro-preto
Eschweilera parviflora (Aubl.) Miers castanha-de-
macaco
Eschweilera pedicellata (Rich.) S.A.Mori ripeiro-vermelho
Eschweilera rhododendrifolia (R. Kutnh) A.C.Sm. ripeiro-vermelho
Lecythis parvifructa S.A.Mori jarana
Lecythis poiteaui O.Berg Jarana
LOGANIACEAE Strychnos jobertiana Baillon
MALPIGHIACEAE Byrsonima crispa A. Juss Murucí
Hirea affinis
MELASTOMATACEAE Mouriri collocarpa Ducke Mamãozinho
Mouriri duckeana Morley Mamãozinho
Mouriri nigra (D.C.) Mamãozinho
MENISPERMACEAE Cissampeleos andromorpha D.C.
Scieadotenia sp.
MORACEAE Brosimum acutifolium ssp.interjectum (Huber) Ducke Mururé
Brosimum guianense (Aubl.) Ducke pau-rainha

61
Brosimum ovatifolium Ducke Garrote
Brosimum parinarioides Ducke Amapá
Brosimum rubescens Taub. pau-rainha
Brosimum rubescens rubescens Taub. pau-rainha
Ficus guianensis Desv. Ex Ham. Apuí
Ficus mathewsii Miq. mata-pau
Maquira sclerophylla (Ducke) C.C.Berg pau-tanino
MYRISTICACEAE Irianthera coriácea Ducke Cucuúba
Iryanthera ulei Warb. Cucuúba
Osteophloeum platyspermum (Spruce ex. A. DC.) viróla-vermelha
NYCTAGINACEAE Neea floribunda Poepp. & Endl. joão-mole
Neea madeirana Standl. joão-mole
OLACACEAE Minquartia guianensisAubl. Acariquara
OPILIACEAE Agonandra sylvatica Ducke
PASSIFLORACEAE Passiflora coccinea Aubl. Maracujá
SAPINDACEAE Matayba arborescens (Aubl.) Radlk Pitomba
Paullinia stipularis Benth. ex Radlk. Guaraná-bravo
Talisia cf. cucularis
SAPOTACEAE Chrysophylum sanguinolentum (Pierre) Baehni balata-brava
Chrysophylum ucuquirana-branca (Aubr’ev. & Pellegr.)
T.D.Penn.
Balata
Sapotaceae sp.1
Micropholis acutangula (Ducke) Eyma
Micropholis cassiquiarensis Abil-rosadinha
Micropholis mensalis (Baehni) Aubrév. abil-rosadinha
Micropholis venulosa (Mart. & Eichler) abil-rosadinha
Pouteria sp.
Pouteria anomala (Pires) T.D.Penn Abiurana
Pouteria cf. stipulifera
Pouteria freitasii T.D.Penn abiurana-vermelha
Pouteria manaosensis (Aubrév & Pellegr) T.D.Penn abil-peludo
Pouteria stipulifera T.D.Penn Rosadinha
Pouteria vernicosa T.D.Penn Abiurana
Pouteria virescens Baehni Abiurana
SIPARUNACEAE Siparuna glycicarapa (Ducke) Renner & Hausner Capitiú
Recommended

















