Israelle Netto Freitas
Efeitos da suplementação com taurina sobre a obesidade e morfofunção pancreática
endócrina de camundongos machos alimentados com dieta hiperlipídica e em sua
prole de primeira geração
Macaé
2017
Universidade Federal do Rio de Janeiro Campus Macaé Professor Aloísio Teixeira
Programa de Pós-graduação em Produtos Bioativos e Biociências

Israelle Netto Freitas
Efeitos da suplementação com taurina sobre a obesidade e morfofunção pancreática
endócrina de camundongos machos alimentados com dieta hiperlipídica e em sua
prole de primeira geração
Dissertação apresentada ao Programa de Pós-graduação em Produtos Bioativos e Biociências da Universidade Federal do Rio de Janeiro como parte das exigências para obtenção do título de Mestre em Ciências. Orientadora: Prof.ª Drª Rosane Aparecida Ribeiro
Macaé
2017
Universidade Federal do Rio de Janeiro Campus Macaé Professor Aloísio Teixeira
Programa de Pós-graduação em Produtos Bioativos e Biociências

Efeitos da suplementação com taurina sobre a obesidade e morfofunção pancreática
endócrina de camundongos machos alimentados com dieta hiperlipídica e em sua
prole de primeira geração
Israelle Netto Freitas
Banca de defesa de dissertação apresentada ao Programa de Pós-graduação em Produtos Bioativos e Biociências da Universidade Federal do Rio de Janeiro como parte das exigências para obtenção do título de Mestre em Ciências. Orientadora: Prof.ª Drª. Rosane Aparecida Ribeiro
Apresentada em 30 de agosto de 2017
Comissão Examinadora:
____________________________________________
Prof. Drª. Cintia Monteiro de Barros
____________________________________________
Prof. Dra. Helene Nara Henriques Blanc
_____________________________________________
Prof. Drª. Kelse Tibau de Albuquerque
____________________________________________
Orientadora: Prof. Drª. Rosane Aparecida Ribeiro
Macaé
2017

Dedico esse trabalho a Deus, minha família, amigos e os professores que me auxiliaram nessa caminhada.

AGRADECIMENTOS
Em primeiro lugar, agradecer a Deus pela saúde e força para suportar os
momentos difíceis aos quais não me deixou desistir.
Aos meus pais, meus heróis e incentivadores por todo amor, carinho e por
acreditarem nos meus sonhos e fazerem dele, os próprios sonhos.
A Isabella, Isadora e Pedro pela amizade, cumplicidade, companheirismo e
amor.
Ao meu Nico, meu anjo, por toda fidelidade e alegria a qual me proporcionou
nesses lindos anos de convívio, saudade eterna.
A Liebe, por ser uma grande companheira nos meus piores momentos.
A minha família, em especial meus avós Antônio e Maria, por estarem sempre
dispostos a me ouvir e ajudar no que for necessário pra me fazer feliz.
Aos meus amigos pela paciência e ausência, e em especial Thiago, Valéria e
Renato pelo apoio, amizade, ajuda e ensinamentos durante todos esses anos.
A minha orientadora, Prof. Dra. Rosane Aparecida Ribeiro, por acreditar em
mim e por todo conhecimento proporcionado, confiança e paciência. O seu sucesso
profissional ultrapassa a falta de recursos, a minha eterna gratidão.
Aos meus colegas, em especial do Laboratório Integrado de Morfologia e
Laboratório Integrado de Bioquímica e os demais do NUPEM pelo convívio e ajuda
que foram fundamentais nessa caminhada.
Aos integrantes do Laboratório de Pâncreas Endócrino e Metabolismo da
UNICAMP e também à Prof. Dra. Maria Lucia Bonfleur da UNIOESTE- Cascavel, os
quais foram de grande importância no desenvolvimento das pesquisas.
À instituição UFRJ, pela oportunidade e todo aprendizado.
A FUNEMAC e FAPERJ, pela concessão da bolsa de estudos e por apoio nos
demais projetos do laboratório.
Aos animais minha gratidão, respeito e amor.
A todos que de alguma forma fizeram parte dessa trajetória, foram essenciais
para o meu crescimento pessoal e profissional.

Ninguém vence sozinho, nem no campo, nem na vida.
(Papa Francisco)
Os dias prósperos não vêm por acaso; nascem de muito trabalho e persistência.
(Henry Ford)

RESUMO
A condição metabólica paternal tem papel importante em predispor sua prole
a comorbidades como obesidade, intolerância à glicose e Diabetes mellitus. A prole
de fêmeas obesas suplementadas com o aminoácido L-taurina (Tau) não
desenvolveu obesidade, contudo tal relação ainda não foi evidenciada sobre o
impacto da obesidade paternal e a Tau sobre a indução da obesidade e
comorbidades na prole. Avaliamos a adiposidade, tolerância à glicose e a
morfofunção do pâncreas endócrino em camundongos machos tratados com dieta
hiperlipídica (HFD) e suplementados com Tau, bem como em sua
prole. Camundongos C57Bl/6 dos 30 aos 150 dias foram distribuídos nos grupos:
controle (CTL); CTL suplementado com 5% de Tau na água filtrada (CTAU); tratado
com dieta contendo 36% gordura (HFD); ou dieta HFD e Tau (HTAU). Aos 150 dias
foram acasalados com fêmeas CTL para obtenção da prole designada conforme o
tratamento dos pais. O consumo de dieta HFD aumentou o peso corporal, os
estoques de gordura abdominais, induziu intolerância à glicose e hipersecreção de
insulina. Pela primeira vez evidenciamos que o consumo de dieta HFD pelos pais foi
associado à redução na secreção de insulina em ilhotas pancreáticas de filhotes
HFD F1 quando incubadas com glicose. Ainda, o pâncreas desses filhotes teve
redução do número das células α, δ e do tamanho das células β. A suplementação
dada ao grupo paternal HTAU normalizou o número e o tamanho das células δ e β, e
aumentou o tamanho das células α no pâncreas da prole HTAU F1. O consumo de
HFD pelos pais promoveu ações deletérias sobre a secreção de insulina que estão
associadas a hipotrofia das células β e redução da proliferação das células α e δ.
Porém a suplementação com Tau atenuou as alterações funcionais no pâncreas
endócrino da prole macho F1.
Palavras-chave: aminoácido, epigenética, obesidade, pâncreas endócrino, secreção
de insulina.

ABSTRACT
The paternal metabolic condition plays an important role in predisposing their
offspring to comorbidities such as obesity, glucose intolerance and Diabetes mellitus.
Offspring of obese females supplemented with the amino acid L-taurine (Tau) did not
develop obesity, yet such a relationship has not yet been evidenced on and the
impact of paternal obesity and Tau on the induction of obesity and offspring
comorbidities. We evaluated the adiposity, glucose tolerance and morphofunction of
the endocrine pancreas in male mice treated with hyperlipidic diet (HFD) and
supplemented with Tau as well as in their offspring. C57Bl/6 mice from 30 to 150
days were divided into groups: control (CTL); CTL supplemented with 5% Tau in
filtered water (CTAU); Treated with diet containing 36% fat (HFD); Or HFD and Tau
(HTAU) diet. At 150 days, they were mated with CTL females to obtain offspring
designated according to the parents' treatment. The consumption of HFD diet
increased body weight, abdominal fat stores, induced glucose intolerance and insulin
hypersecretion. For the first time we showed that HFD dietary intake by parents was
associated with a reduction in insulin secretion in HFD F1 islets when incubated with
glucose. Moreover, the pancreas of these pups had a reduction in the number of α, δ
cells and the size of β cells. Supplementation in the HTAU paternal group normalized
the number and size of δ and β cells, and increased the size of the α cells in the
pancreas of HTAU offspring. The ingestion of HFD by parents promoted deleterious
actions on insulin secretion that are associated with β-cell hypotrophy and reduced
proliferation of α and δ cells. However, Tau given to parents which attenuated the
functional alterations in the endocrine pancreas of the male offspring F1.
Key words: amino acid, endocrine pancreas, epigenetics, insulin secretion and obesity.

LISTAS DE FIGURAS
Figura 1: Representação esquemática dos grupos experimentais. ........................... 33
Figura 2: Avaliação dos parâmetros de peso corporal e ingestão hídrica e de ração
nos camundongos dos grupos paternais. .................................................................. 40
Figura 3: Avaliação dos parâmetros da obesidade nos camundongos dos grupos
paternais.................................................................................................................... 41
Figura 4: Avaliação dos parâmetros de peso corporal e ingestão hídrica e de ração
nos camundongos dos grupos de prole macho F1. ................................................... 42
Figura 5: Avaliação dos parâmetros da obesidade na prole macho F1. .................... 43
Figura 6: Avaliação dos parâmetros de peso corporal e ingestão hídrica e de ração
nos camundongos dos grupos da prole fêmea F1. ................................................... 44
Figura 7: Avaliação dos parâmetros da obesidade na prole fêmea. .......................... 45
Figura 8: Avaliação da glicemia nos camundongos grupos
paternais.....................................................................................................................45
Figura 10: Avaliação da glicemia nos camundongos dos grupos da prole fêmea F1.
.................................................................................................................................. 50
Figura 11: Secreção estática de insulina estimulada por diferentes concentrações de
glicose em ilhotas pancreáticas isoladas de camundongos dos grupos paternais. ... 52
Figura 12: Secreção estática de insulina estimulada por diferentes concentrações de
glicose em ilhotas pancreáticas isoladas na prole macho F1. ................................... 53
Figura 13: Secreção estática de insulina estimulada por diferentes concentrações de
glicose em ilhotas pancreáticas isoladas na prole fêmea F1.................................... 54
Figura 14: Imuno-histoquimica para insulina, ou glucagon ou somatostatina no
pâncreas coletado da prole macho F1 ...................................................................... 56
Figura 15: Quantificação dos principais hormônios das ilhotas pancreáticas da prole
macho F1 .................................................................................................................. 57

LISTAS DE TABELAS
Tabela 1: Parâmetros bioquímicos plasmáticos no estado de jejum e alimentado
avaliados em camundongos dos grupos paternais ................................................... 47
Tabela 2: Parâmetros bioquímicos plasmáticos no estado de jejum e alimentado
avaliados na prole macho F1 .................................................................................... 49
Tabela 3: Parâmetros bioquímicos plasmáticos no estado de jejum e alimentado
avaliados na prole fêmea F1 ..................................................................................... 51
Tabela 4: Morfometria do pâncreas endócrino aos 90 dias de vida da prole macho F1
.................................................................................................................................. 58

LISTA DE QUADROS
Quadro 1: Composição das dietas............................................................................34

LISTAS DE ABREVIATURAS E SIGLAS
ABESO – Associação Brasileira para o Estudo da Obesidade e da Síndrome
Metabólica
Acetil-coA – Acetilcoenzima A
Akt – Proteína quinase B
AMPc – Adenosina Monofosfato Cíclica
ATP – Adenosina trifosfato
ATP/ADP – Adenosina trifosfato/ adenosina difosfato
AUC – Área abaixo da curva
Ca2+– Cálcio
[(Ca2+)i] – Cálcio intracelular
Cav – Canal de cálcio dependente da voltagem
CNA – Comprimento nasoanal
CHOL – Colesterol
CTL – Ração normoproteica
CTAU – Ração normoproteica e 5% de Tau
ACP – proteína carreadora de grupos acil
DAG – Diacilglicerol
DM – Diabetes mellitus
DM1 – Diabetes mellitus do tipo 1
DM2 – Diabetes mellitus do tipo 2
DNA – Ácido Desoxirribonucleico
EPM – Média erro padrão da média
GCK– Enzima glicoquinase
GHRH – Hormônio do crescimento
GIP – Peptídeo insulinotrópico dependente de glicose
GLP-1 – Peptídeo semelhante ao glucagon 1
GLUT 2– Transportador de glicose
GMPc – Guanosina-monofosfato cíclico
HAS – Hipertensão arterial sistêmica
H2O2 – Peróxido de hidrogênio
HFD- Dieta hiperlipídica

HTAU- Dieta hiperlipídica e 5% de Tau
IBGE – Instituto Brasileiro de Geografia e estatística
Icv – Intracerebroventricular
IDF- Federação internacional de diabetes
IGF-1 – Fator de crescimento semelhante à insulina tipo 1
IMC – Índice de massa corporal
Ip – Injeção intraperitoneal
ipGTT – Teste de tolerância à glicose intraperitoneal
ipITT– Teste de tolerância à insulina intraperitoneal
IR– Receptor de insulina
IRS – Substrato receptor de insulina
Irβ –Subunidade beta do receptor de insulina
JAK2/STAT3 – Proteína Janus quinase 2/transdutor de sinal e ativador de
transcrição 3
KATP – Canal de potássio dependente de ATP
KCAL– Quilocalorias
KCl – Cloreto de potássio
Il13rα2 – Gene receptor alfa 2 de interleucina 13
Malonil-coA – Malonil coenzima A
MTP – Proteína de transferência de triglicerídios microssomal
Na+ – Sódio
NADPH – Dinucleotídeo de adenina e nicotinamida fosfato
NOD – Camundongos não obesos diabéticos
OMS – Organização Mundial da Saúde
PAKT – Proteína quinase β fosforilada
PDX-1 – Pancreático e duodenal homeobox 1
PI – Fosfatidilinositol
PPARα- Receptor activado por proliferador de peroxissoma gama
RIA – Radioimunoensaio
RNAm – Ácido ribonucleico mensageiro
ROS – Espécies reativas de oxigênio
Tau – L-taurina , 2-aminoetanossulfônico
Toll- Receptores do tipo toll
TBS – Tampão tris-salina

TNFα – Fator de necrose tumoral

SUMÁRIO
1. INTRODUÇÃO .................................................................................................... 16 1.1 Obesidade ................................................................................................... 16
1.2 Modelos experimentais de obesidade ...................................................... 18
1.3 Secreção de insulina .................................................................................. 19
1.4 Obesidade e programação metabólica ..................................................... 22
1.5 L-taurina ...................................................................................................... 24
2. JUSTIFICATIVA .................................................................................................. 28 3. OBJETIVOS ........................................................................................................ 30
3.1 Objetivo geral ............................................................................................. 30
3.2 Objetivos específicos ................................................................................ 30
4. MATERIAIS E MÉTODOS .................................................................................. 31 4.1 Grupos experimentais ............................................................................... 31
4.2 Avaliação da ingestão de ração e de água e avaliação da obesidade ... 33
4.3 Teste intraperitoneal de tolerância à glicose (ipgtt) ................................ 34
4.4 Parâmetros bioquímicos plasmáticos ...................................................... 34
4.5 Isolamento de ilhotas pancreáticas e secreção estática de insulina .... 35
4.6 Morfologia do pâncreas ............................................................................. 36
4.7 Imuno-histoquímica para insulina, glucagon e somatostatina .............. 36
4.8 Análise estatística ...................................................................................... 37
5. RESULTADOS ................................................................................................... 39
5.1 Ingestão de ração e água e avaliação da obesidade nos grupos paternais e em sua respectiva prole .................................................................. 39
5.2 Avaliação da tolerância à glicose e dos parâmetros bioquímicos plasmáticos nos grupos paternais e em sua respectiva prole ........................ 45
5.3 Secreção estática de insulina nos grupos paternais e em sua respectiva prole.................................................................................................... 51
5.4 Morfologia e imuno-histoquímica no pâncreas da prole macho ........... 54
6. DISCUSSÃO ....................................................................................................... 59 6.1 Ações da suplementação com Tau no desenvolvimento da obesidade nos grupos paternais e na sua respectiva prole ............................................... 59
6.2 Ações da suplementação com Tau na homeostase glicêmica e nos parâmetros bioquímicos plasmáticos dos grupos paternais e de sua respectiva prole.................................................................................................... 64
6.3 Ações da suplementação com Tau na morfofunção pancreática endócrina tanto no grupo paternal quanto na sua prole .................................. 67
7. CONCLUSÕES ................................................................................................... 73 REFERÊNCIAS BIBLIOGRÁFICAS .......................................................................... 74 ANEXOS ................................................................................................................... 82

16
1. INTRODUÇÃO
1.1 Obesidade
A obesidade é uma doença de abrangência mundial, atualmente é uma
epidemia global (Organização Mundial de Saúde- OMS, 2016). É considerada
problema de saúde pública em diversos países do mundo, principalmente nas
sociedades ocidentais e industrializadas, atingindo diversas faixas etárias, em
ambos os sexos e níveis sociais (OMS, 2016), sendo caracterizada pelo grande
acúmulo de gordura corporal, resultante principalmente da combinação entre a
ingestão excessiva de alimentos e o sedentarismo (CHAVES et al., 2010; SOUZA et
al., 2010; TATEYA, KIM, TAMORI, 2013).
O excesso de peso pode ser medido pelo índice de massa corporal (IMC), o
qual é obtido entre a razão do peso e altura [peso (kg)/ altura2 (m2)], caso essa
relação dê valores acima ou igual a 25 e ≥ 30 kg/m2 indica que o indivíduo apresenta
sobrepeso ou obesidade, respectivamente (OMS, 2017), apesar das limitações
fornece uma medida útil para a população relativa ao excesso de peso ou
obesidade, uma vez que os valores de referências são os mesmos para ambos os
sexos e para adultos de todas as idades. No Brasil em 2015, cerca de 82 milhões de
pessoas adultas apresentaram o IMC 25 e ≥ 30 kg/m2. Em homens o sobrepeso ou
obesidade foram evidenciados em maior prevalência na faixa etária entre 25 e 29
anos perfazendo 50,4%. Contudo, as mulheres na faixa etária entre 35 e 44 anos
prevaleceram sobrepeso e obesidade equivalendo a 63,6% ultrapassando a
percentagem dos homens nessa mesma idade que foi 62,3% (Associação Brasileira
para o Estudo da Obesidade e da Síndrome Metabólica - ABESO, 2015). A literatura
sugere a hipótese de que a obesidade gera um ciclo vicioso, já que filhos de pais
obesos têm 80 a 90% de probabilidade de serem adultos obesos (JAMES et al.,
2001; SHANKAR et al., 2008).
Um fato ainda mais preocupante tem sido o aumento alarmante da obesidade
infantil. De acordo com o Instituto Brasileiro de Geografia e Estatística (IBGE) em
2015, 16,6% dos meninos, e 11,8% das meninas na faixa etária de 5 e 9 anos foram
considerados obesos. Na mesma faixa etária citada a prevalência de sobrepeso foi
de 34,8% e 32% em meninos e meninas, respectivamente (IBGE, 2015).
Mudanças nos hábitos e na rotina são os caminhos para evitar o aumento da
obesidade (OMS, 2016). Atualmente, as estimativas não são animadoras, de acordo
com o Ministério da Saúde, se o Brasil continuar nessa crescente estimativa de

17
sobrepeso e obesidade será o país mais obeso do mundo em 15 anos (VIGITEL,
2015). No mundo as estimativas também demonstram que pelo menos 2,8 milhões
de pessoas incluindo crianças e adultos morrem a cada ano, como resultado de
excesso de peso ou obesidade (OMS, 2014). A projeção é que, em 2025, cerca de
2,3 bilhões de adultos estejam com sobrepeso; e mais de 700 milhões, obesos. E o
número de crianças com sobrepeso e obesidade no mundo pode chegar a 75
milhões, caso nada seja feito para prevenir ou reverter esse quadro (OMS, 2016).
Na obesidade os fatores genéticos, metabólicos, endócrinos, neurais,
ambientais e comportamentais estão inter-relacionados para a sua manifestação e
manutenção (CHAVEY et al., 2014; SOMINSKY e SPENCER 2014; SOUBRYET et
al., 2015). Além de problemas sociais e psicológicos, a preocupação em torno do
crescimento da obesidade está relacionada a sua associação com diversas
condições mórbidas como; hipertensão arterial sistêmica (HAS), dislipidemias,
problemas ortopédicos, doenças cardiovasculares e diabetes mellitus (DM) do tipo 2
(DM2) (ENES et al., 2010, MALIK et al., 2013).
O DM é caracterizado por hiperglicemia, podendo ocorrer devido a defeitos
na secreção e/ou ação da insulina. Diversas condições podem levar ao DM, porém a
grande maioria dos casos está dividida em dois grupos, DM do tipo 1 (DM1),
resultante da destruição das células β que pode ocorrer por processo imunológico,
ou seja, pela formação de anticorpos pelo próprio organismo contra as células β
resultando na ausência da produção endógena de insulina (GLASTRAS et al.,
2016). Atualmente 90% dos pacientes diabéticos apresentam o DM2 (IDF,2017).
Este é caracterizado pela resistência à ação da insulina e disfunção ou falência das
células β (SPELLMAN, 2007; WONG et al., 2017).
Dados da OMS mostraram que o número de pessoas com DM no mundo
aumentou de 108 milhões em 1980, para 422 milhões em 2014, e estima que esse
número possa dobrar até 2030 (OMS, 2016). Anualmente mais de 1,5 milhões de
pessoas morre de DM, ocupando o quarto lugar no número de mortes no mundo,
ficando atrás das doenças cardiovasculares, do câncer e das doenças respiratórias
(OMS, 2016). De acordo com a Pesquisa Nacional de Saúde, realizada com parceria
entre IBGE e o Ministério da Saúde divulgada em 2015, foi mostrado que o DM
atingiu 6,2% da população adulta entre 18 e 74 anos, o que corresponde a 9 milhões
de brasileiros. As mulheres apresentaram maior proporção da doença do que os
homens, atingindo 7% o que corresponde a 5,4 milhões de mulheres e 5,4%,

18
correspondendo a 3,6 milhões de homens apresentando DM (IBGE, 2015; Ministério
da Saúde, 2015). Quanto maior a idade, maior a ocorrência dessa doença, por
exemplo, na faixa etária dos 65 a 74 anos, foi constatado que 19,9% dos brasileiros
eram diabéticos (IBGE, 2015).
A idade é um fator de risco maior para o desenvolvimento de DM2, porém, o
consumo de alimentos industrializados de alto valor calórico somados ao
sedentarismo, são atualmente determinantes para o aumento do número de
indivíduos com sobrepeso e obesidade diagnosticados com DM2 (ESPELAND et al.,
2014). Estima-se que entre 80 e 90% dos indivíduos acometidos por DM2 são
obesos, e o risco de desenvolver esta doença está diretamente associado ao
aumento do IMC (SARTORELLI, 2003; SCHIMIDT et al., 2009). O DM pode
acarretar em comorbidades tais como: insuficiência renal, doenças cardiovasculares,
acidente vascular cerebral, prejuízos visuais e neuropatia diabética (MORAIS et al.,
2009). Portanto, a associação obesidade e DM só tende a aumentar a
morbimortalidade da população.
1.2 Modelos experimentais de obesidade
Para melhor compreensão dos mecanismos fisiopatológicos envolvidos no
desenvolvimento da obesidade diversos modelos experimentais são utilizados.
Dentre os modelos estudados têm-se os que apresentam prejuízos genéticos como,
ratos Zucker fa/fa, camundongos ob/ob, que não expressam o gene da leptina;
modelos de obesidade por lesões eletrolíticas em regiões hipotalâmicas envolvidas
no controle da saciedade, como o núcleo ventromedial (SHIMIZUT et al., 2003). Mas
os mais utilizados para análises fisiopatológicas da obesidade são os modelos que
consomem dietas hipercalóricas, podendo ser a dieta de cafeteria ou dieta
hiperlipídica (HFD) devido à sua grande semelhança com a gênese e com as
respostas metabólicas decorrentes da obesidade em humanos (DESPRÉS et al.,
1994). A dieta HFD promove obesidade em roedores pelo alto consumo energético
(SHAFAT et al., 2009; KENNEDY et al., 2011). Conforme maior for o percentual de
gordura na dieta HFD, maior é o aumento do ganho de peso corporal devido à
quantidade de calorias fornecida pela dieta (DUARTE et al., 2006; WHITE et al.,
2013).
É descrito que na primeira semana de ingestão de dieta HFD os
camundongos apresentam elevação da glicemia (WINZELL et al., 2007). Ainda,
após oito semanas de ingestão da dieta HFD os camundongos desenvolvem

19
obesidade, resistência à insulina, intolerância à glicose, hiperleptinemia,
hiperinsulinemia, hipercolesterolemia, aumento de ácidos graxos circulantes e
hipersecreção de insulina (DUARTE et al., 2006; ESTADELA, 2004; MCLELLAN et
al., 2007). Essa hiperfunção acarreta em alterações plásticas do pâncreas (DUARTE
et al., 2006). Camundongos que ingeriram por três meses a dieta HFD apresentaram
aumento da massa das células β que foi principalmente relacionado a hiperplasia,
pois o tamanho das células β pancreáticas era significativamente maior (AHREN et
al., 2010). Outro estudo já demonstrou que a dieta HFD consumida durante seis
semanas levou a hiperplasia e hipertrofia das células β pancreáticas (RIBEIRO et al.,
2012). Sugere-se que esta resposta do pâncreas endócrino é uma tentativa de
normalizar o quadro de hiperglicemia e contrabalancear a resistência à insulina dos
tecidos periféricos, principalmente do músculo esquelético, tecido adiposo e fígado
(KAHN et al., 2006; PRENTKI e NOLAN, 2006, SOUZA-MELLO et al., 2014).
1.3 Secreção de insulina
O pâncreas é uma glândula mista, com função tanto endócrina quanto
exócrina. Em humanos, é um órgão achatado que mede aproximadamente 12,5 a 15
cm de comprimento, pesando aproximadamente 60 gramas, está situado na
curvatura do duodeno, retroperitoneal e transversalmente ao estômago e vai até o
baço (TORTORA e BRYAN et al., 2014). Nos roedores, entretanto o pâncreas
localiza-se difusamente aderido às alças intestinais de forma alongada estendendo-
se entre o duodeno e uma pequena parte encontra-se junto ao estômago e o baço
(COVELLI et al., 2006).
Aproximadamente 99% das células pancreáticas estão dispostas em
aglomerações chamadas ácinos. Estas células produzem enzimas digestivas que
fluem para o trato gastrointestinal pela rede de ductos e compõem a parte exócrina
do pâncreas. A porção endócrina representa 1 a 2% do peso total do órgão,
compreendendo 1 a 2 milhões de minúsculas aglomerações arredondadas de
células endócrinas que são denominadas de ilhotas pancreáticas ou ilhotas de
Langerhans e encontram-se distribuídas por todo o tecido (TORTORA e BRYAN et
al., 2014).
Histologicamente, a ilhota pancreática é revestida por uma delicada cápsula
de tecido conjuntivo, e seu parênquima é constituído por cordões das células
endócrinas entremeados por uma rica rede de capilares sanguíneos do tipo
fenestrado (ANDRALOJC et al., 2009).

20
Cada ilhota pancreática é constituída principalmente por quatro tipos
celulares: as células β que sintetizam e secretam insulina; as células α que são
responsáveis pela secreção do glucagon; as células δ que liberam somatostatina; e
as células PP que são responsáveis por secretar o polipeptídeo pancreático
(KANNO et al., 2002; ANDRALOJC et al., 2009; SUNDLER e HELLER, 2014). Com
relação à disposição ou ao arranjo das células endócrinas na ilhota, em humanos, as
células β, são as mais abundantes totalizando aproximadamente 48% a 59% do
número total de células, as células α em média representam 33% a 46% do total das
células da ilhota; as células δ, compõem em torno de 8% a 12% da massa celular; e
as células PP ocupam aproximadamente 5% da ilhota (CABRERA et al., 2006;
ABDULREDA et al., 2016). Nos roedores a organização das células endócrinas
difere das citadas acima em humanos. As células β ocupam a região central
perfazendo aproximadamente de 60-80% do volume total da ilhota, e as células α e
δ localizam-se na periferia da ilhota e representam de 15-20% e 10%,
respectivamente, do total de células. As células PP ocupam aproximadamente 1%, e
estão preferencialmente, dispostas na periferia da ilhota (ANDRALOJC et al., 2009;
ABDULREDA et al., 2016).
A secreção de insulina pelas células β pancreáticas é controlada
continuamente de acordo com as flutuações da concentração de nutrientes
circulantes, em especial, a glicose (HENQUIN et al., 2009). Os mecanismos
responsáveis pela secreção de insulina estimulada pela glicose iniciam-se com o
transporte deste açúcar para as células β, através do transportador de glicose
(GLUT)-2; a glicose é fosforilada à glicose-6-fosfato pela enzima glicoquinase (GCK)
e metabolizada gerando ATP. O resultado é o aumento da relação ATP/ADP
intracelular, que provoca o fechamento de canais de K+ sensíveis ao ATP (KATP),
reduzindo o efluxo de K+ e promovendo a despolarização da membrana, que resulta
na abertura de canais de Ca2+ sensíveis à voltagem e influxo deste cátion (PRENTKI
et al., 2013). A metabolização da glicose nas células β e a subsequente elevação da
concentração intracelular de Ca2+ ([Ca2+]i) podem ativar enzimas que produzirão
adenosina 3',5'-monofosfato cíclico (AMPC), diacilglicerol (DAG) e guanosina 3’,5’ -
monofosfato cíclico (GMPc) que contribuem para a amplificação do sinal iniciado
pela glicose (HENQUIN et al., 2009; TENGHOLM et al., 2009; PRENTKI et al.,
2013). Essa resposta potencializadora também pode ser desencadeada pela
liberação de neurotransmissores como acetilcolina e hormônios gastrointestinais

21
como GLP-1 (Peptídeo semelhante ao glucagon 1) e o peptídeo insulinotrópico
dependente de glicose (GIP) (HIRIART e AGUILAR-BRYAN, 2008; FUJITA et al.,
2016). Mas, ainda há neurotransmissores e hormônios que podem reduzir a
concentração de AMPC, como por exemplo, a noradrenalina e a somatostatina
(NESHER et al., 2002; MCCLENAGHAN, 2007; CALLAWAY et al., 2014).
Além da glicose, os ácidos graxos e aminoácidos também aumentam a
secreção de insulina (PESSIN et al., 2000; SHULMAN et al., 2009). Os aminoácidos
estimulam a secreção de insulina via geração de ATP derivado do seu metabolismo,
ou pela geração de compostos que ao serem degradados aumentam o fluxo de
intermediários para o ciclo de Krebs, como é o caso da L-glutamina (KOMATSU et
al., 2013). A transaminação da glutamina pela enzima glutaminase leva a geração
de amônio e glutamato. O glutamato por sua vez leva a síntese de outros
aminoácidos e de antioxidantes (TIRAPEGUI et al., 2009). Ainda, a glutamina pode
aumentar a síntese de proteínas de choque térmico em células β pancreáticas, fato
que regula a secreção de insulina (WU e MORRIS, 1998; BRENNAN et al., 2003;
CRUZAT, 2013). Outros aminoácidos também podem gerar intermediários para o
ciclo de Krebs, como é o caso da metabolização da L-alanina e L-arginina. Esses
aminoácidos aumentam a probabilidade da despolarização da célula β porque são
cotransportados com os íons Na+ (SENER et al., 2000). Estudos já evidenciaram
que esses aminoácidos ativam reguladores da síntese proteica e contribuem para
reconstrução de proteínas (SENER et al., 2000; NEWSHOLME et al., 2007). Na
obesidade a suplementação com L-arginina e L-alanina já mostrou ter ação na
redução significativa dos estoques de gordura abdominais e gordura corporal, além
de melhorar o controle da glicemia e liberação de adiponectina em indivíduos com
DM2 (LUCOTTI et al., 2006; ARAUJO et al., 2016). Segundo, ARAUJO e
colaboradores (2016) a suplementação com L-alanina foi eficaz para restaurar a
secreção de insulina em camundongos obesos.
Benefícios semelhantes já foram descritos em roedores suplementados com o
aminoácido L-taurina, ainda outro fato referenciado na literatura quanto à L-taurina é
o aumento da sensibilidade à insulina (CARNEIRO et al., 2009; RIBEIRO et al.,
2012; VETORAZZI, 2013). A L-taurina já mostrou controlar a homeostase da glicose
através da regulação da expressão de genes necessários para estimular a secreção
de insulina pelas células β, e assim melhora a sensibilidade periférica à insulina
(CARNEIRO et al., 2009). Foi visto também que a suplementação com L-taurina

22
dada a camundongos obesos restaura a secreção nesses roedores (RIBEIRO et al.,
2012).
Estudos também já mostraram que a suplementação com aminoácidos pode
ser uma forma eficaz para tratar ou prevenir a obesidade (RIBEIRO et al., 2012,
VETORAZZI, 2013; ARAUJO et al., 2016). Portanto a utilização de suplementos
nutricionais parece ser uma linha promissora para a prevenção e tratamento dessa
síndrome e de suas comorbidades.
1.4 Obesidade e programação metabólica
Atualmente, os estudos indicam um importante envolvimento biológico dos
pais na determinação do desenvolvimento da obesidade e programação metabólica
da prole, sendo que o meio nutricional ao qual os pais são expostos afeta o fenótipo
metabólico de seus descendentes (REUSENS et al., 2008; DUNN et al., 2009;
VICKERS, 2014). Conforme a hipótese de Barker as doenças evidenciadas na vida
adulta teriam origem fetal. A adaptação do feto ao meio interno maternal adverso
resultaria em mudanças permanentes na fisiologia e metabolismo do indivíduo
(HALES e BARKER, 2001). Esta hipótese é confirmada pelas evidências de que o
baixo peso ao nascer está associado com aumento do risco de desenvolvimento de
doenças como hipertensão, doença cardiovascular e DM2 (SARTORI et al., 1999;
LAL et al., 2003). A exposição a condições adversas como, a desnutrição, obesidade
e DM aumenta a ocorrência do desenvolvimento da obesidade e suas comorbidades
na vida adulta (LEVIN, 2008; METGES, 2009).
Estudos com roedores mostraram que o excesso de peso materno durante os
períodos de desenvolvimento da prole pode aumentar o risco de obesidade, nas
diferentes fases da vida (HOWIE, 2009). O consumo de dieta HFD materna por
camundongos a longo prazo, predispõe para elevações na pressão arterial e na
concentração de colesterol total em sua prole (ELAHI et al., 2009). Além disso, o
consumo de alimentos ricos em gordura apenas durante a gravidez e lactação já
predispõe a prole à resistência a insulina e esteatose hepática (GREGORIO et al.,
2010; ASHINO et al., 2012). Ainda, alterações no metabolismo da glicose induzidas
por ambiente intrauterino adverso podem se propagar não somente a primeira
geração mais para as próximas descendências (PINHEIRO et al., 2008).
O feto de ratas que foram alimentadas com dieta HFD durante a gestação não
demonstraram alteração do peso corporal ao nascimento, porém aos dez dias de
vida esses roedores apresentaram aumento do peso corporal, hiperleptinemia,

23
hiperinsulinemia, acompanhado de resistência à ação da leptina e insulina (TAYLOR
et al., 2005; CERF et al., 2008; GUPTA et al., 2009).
A dieta HFD consumida por fêmeas no período pré-acasalamento e período
gestacional, pode programar a fisiologia e o metabolismo da prole precocemente. Na
prole de ratos neonatos oriundos de mães alimentadas com dieta HFD evidenciou-
se hiperglicemia e prejuízos no desenvolvimento das células β (GNIULI et al., 2008).
Também já foi relatado alterações precoces no pâncreas da prole, incluindo um
remodelamento estrutural do pâncreas associadas ao aumento da massa de células
β, prejudicando o metabolismo da glicose, esses prejuízos podem levar a mudanças
adaptativas nas células β como hipertrofia, hiperplasia, aumento da síntese e
secreção de insulina na prole (GNIULI et al., 2008).
Em contrapartida, também já foi observado que em camundongos
provenientes de mães que comeram dieta HFD tiveram redução na massa de célula
β, e apresentaram adaptação compensatória nas células β remanescentes, levando
a hipertrofia e aumento da secreção de insulina nessas células β, o que está
relacionado à superestimulação ou efeitos diretos da hiperglicemia e hiperlipidemia
devido à exposição a um ambiente metabolicamente alterado durante seu
desenvolvimento (MASIELLO, 2006).
A prole de camundongos fêmea alimentadas com dieta HFD, durante a
gestação, apresentou redução da sensibilidade periférica à insulina e maior
crescimento linear, devido ao aumento da concentração plasmática do fator de
crescimento semelhante à insulina tipo 1 (IGF-1) associada ao aumento da
expressão gênica do hormônio de liberação do hormônio do crescimento (GHRH).
Este efeito foi decorrente da menor expressão gênica de repressores transcricionais
e redução da metilação da sequência codificadora do GHRH (DUNN et al., 2009).
Contudo, também já é estabelecido que não somente a condição maternal e
intrauterina pode levar à alterações importantes na condição fisiológica do novo
indivíduo (ZAMBRANO et al., 2006; NG et al., 2010). A influência paternal sobre o
desenvolvimento e aptidão da prole pode ser independente de qualquer interação
direta com descendência (CREAN e BONDURIANSKY, 2014). Recentemente,
considera-se que o meio interno ao qual os gametas são expostos durante sua
formação exerce influência no fenótipo da prole, sabe-se que tal programação ocorre
pela ativação de mecanismos epigenéticos, cujas alterações podem não ser visíveis

24
no fenótipo logo após o nascimento, mas que muitas vezes vêm a se manifestar na
forma de doenças na vida adulta (HOCHERA et al., 2016).
Os mecanismos epigenéticos que controlam essas alterações são
caracterizados por interferirem no padrão de metilação, fosforilação e acetilação do
DNA e das histonas, correlacionam-se com a conformação da cromatina, permitindo
que determinados genes sejam expressos ou silenciados, resultando na propagação
da alteração da atividade gênica de uma geração de células para a próxima
(HOWELL et al., 2009). Portanto o tipo de dieta consumida pelo pai exerce
implicações no desenvolvimento da adiposidade e prejuízo no metabolismo da
glicose na prole (MORRIS et al., 2014).
Em camundongos machos foi observado que o consumo da dieta HFD altera
a expressão gênica testicular (GHANAYEM et al., 2009). Já em humanos sabe-se
que o desenvolvimento da obesidade afeta os componentes do esperma, motilidade,
morfologia e aumenta o dano ao DNA dos espermatozoides (KASTURI et al., 2008).
Roedores que foram alimentados com HFD e acasalados com ratas
alimentadas com dieta CTL tiveram a prole fêmea com intolerância à glicose
associada a prejuízos na secreção de insulina, porém não houve diferença na
adiposidade (MORRIS et al., 2014).
A prole macho provenientes de pais que consumiram dieta HFD mostraram
redução do crescimento, apresentando menor peso corporal que a prole CTL, o que
foi relacionado com a menor concentração de IGF-1 nos pais obesos. Ainda, foram
observadas disfunção das células β no pâncreas endócrino da prole fêmea de pais
HFD, sendo as modificações acompanhadas por alteração na expressão de 77
genes relacionados ao transporte iônico, citoesqueleto, proliferação e apoptose nas
ilhotas pancreáticas (NG et al., 2010; YOUNGSON et al., 2011).
1.5 L-taurina
A L-taurina (Tau, 2-aminoetanossulfônico) é um aminoácido que está presente
em altas concentrações tanto no interior das células como no plasma de mamíferos
(HUXTABLE, 1992). Sua biossíntese ocorre principalmente nos hepatócitos e
adipócitos de humanos e roedores a partir dos aminoácidos metionina e cisteína
(TAPPAZ et al., 2004). A Tau também pode ser obtida através da ingestão de
alimentos, tais como leite e carne, principalmente de peixe e frutos do mar, e em
menor quantidade nos vegetais (TAPPAZ et al., 2004). É um aminoácido importante
em processos biológicos, tais como: desenvolvimento do sistema nervoso e retina,

25
modulação do Ca2+, estabilização de membranas, reprodução e imunidade
(ASSCHE et al., 2002). Evidências indicam que o mecanismo de ação da Tau possa
ocorrer pela combinação do aminoácido com vários tipos de canais iônicos,
transportadores e enzimas (HUXTABLE et al., 1992; SATOH e SPERELAKIS, 1998;
TRICARICO et al., 2000).
Além das várias funções já atribuídas à Tau, estudos têm mostrado que o
aminoácido possui um papel regulador sobre a secreção de insulina e homeostase
glicêmica. A sua concentração encontra-se reduzida no DM, como em ratos com
DM induzido por estreptozotocina (AERTS e VAN ASSCHE, 2001; COLIVICCHI et
al., 2004), e em roedores com resistência à insulina (ANURADHA e
BALAKRISHNAN, 1999; TSUBOYAMA-KASAOKA et al., 2006). Em pacientes
diabéticos as concentrações plasmáticas e plaquetárias de Tau apresentam
reduzidas, correlacionando-se com hiperglicemia e distúrbios na osmorregulação
celular (HANSEN et al., 2001). A deficiência de Tau também já foi relacionada com
algumas anormalidades, tais como a degeneração da retina, retardamento do
crescimento e disfunção cardíaca (HUXTABLE, 1992; AZUMA et al., 2012). Vários
estudos demonstraram que o tratamento com Tau é benéfico para a prevenção do
DM1 (ARANY et al., 2004), disfunção da célula β (KANUIK et al., 2007; OPRESCU
et al., 2007; XIAO et al., 2007) e DM2 (NAKAYA et al., 2000; NANDHINI et al., 2005;
TSUBOYAMA-KASAOKA et al., 2006).
No pâncreas, a Tau encontra-se acumulada cerca de cinco vezes mais nas
ilhotas pancreáticas em relação à porção exócrina do pâncreas, dentro da ilhota fica
estocado primariamente nas células α e δ do pâncreas endócrino, sugerindo que a
liberação de Tau possa ser necessária para que ocorra uma eficiente modulação do
processo de secreção de insulina em células β (BUSTAMANTE et al., 2001). O
aminoácido possui um papel regulador sobre a secreção de insulina e homeostase
glicêmica (CARNEIRO et al., 2009). Em situações de hiperglicemia a Tau aumenta a
secreção de insulina (KAPLAN et al., 2004) e a sensibilidade à insulina (NAKAYA et
al., 2000; TSUBOYAMA-KASAOKA et al., 2006). Em camundongos alimentados com
dieta HFD a suplementação com Tau preveniu a hiperfagia (CAMARGO et al.,
2013), o acúmulo de gordura (NARDELLI et al., 2011; BATISTA et al., 2013;
VETTORAZZI et al., 2014), aumentou a tolerância à glicose, melhorou a
sensibilidade à insulina e impediu a hipersecreção, hiperplasia e hipertrofia no
pâncreas endócrino de machos e fêmeas (RIBEIRO et al., 2009 e 2012). Em

26
roedores com resistência à insulina, a suplementação com Tau normalizou não
somente a concentração plasmática de insulina e glicose, mas também a
sensibilidade à insulina e hiperlipidemia (ANURADHA e BALAKRISHNAN, 1999;
NAKAYA et al., 2000; NANDHINI e ANURADHA, 2002).
Em ratos obesos e com resistência a insulina induzido pelo consumo de dieta
HFD por oito meses, a concentração plasmática de Tau também se encontrou
reduzida e a suplementação com Tau reduziu o ganho de peso corpóreo e de tecido
adiposo (TSUBOYAMA-KASAOKA et al., 2006). Dados clínicos mostraram que o
tratamento preventivo com Tau impediu a redução da secreção de insulina em
homens obesos e submetidos à infusão contínua com ácidos graxos por 48h (XIAO,
GIACCA e LEWIS, 2008). Foi observado também que indivíduos com sobrepeso
quando suplementados com Tau reduziram o peso corporal, e apresentaram menor
concentração de triglicerídeos comparados com o grupo não suplementado,
mostrando efeitos benéficos da Tau no metabolismo lipídico (ZHANG et al., 2004).
Contudo, não se sabe qual mecanismo que a Tau previne as alterações funcionais
no pâncreas endócrino, mas o tratamento in vivo com Tau mostrou que aumenta a
tolerância à glicose e sensibilidade à insulina além de aumentar a captação de cálcio
e expressão da subunidade β2 do canal de cálcio voltagem-dependente em ilhotas
pancreáticas (RIBEIRO et al., 2009), e este aumento da capacidade secretória da
célula β está relacionado com a regulação da expressão de genes e proteínas
importantes para o acoplamento estímulo/secreção da insulina, bem como, melhora
do manejo do pool extracelular e intracelular de íons Ca2+ (CARNEIRO et al., 2009;
RIBEIRO et al., 2010).Uma das ações mediadas pela Tau nas células β pancreáticas
é a de inibir a atividade dos canais de KATP resultando em aumento da [Ca2+]i, e
consequentemente da secreção de insulina (PARK et al., 2004).
Ainda no pâncreas, a Tau tem a capacidade de reduzir a taxa de apoptose
(EL IDRISSI, BOUKARROU e L'AMOREAUX, 2009; LIN et al., 2013) e atuar na
síntese de DNA, propiciando o desenvolvimento adequado do pâncreas endócrino
(BOUJENDAR et al., 2002). Experimentos in vitro e in vivo com ilhotas pancreáticas
de roedores suplementados com Tau apresentaram maior expressão da forma ativa
da proteína PDX-1 (essencial para síntese e expressão do GLUT-2 e da insulina),
modulando a homeostase da glicose e a sensibilidade à insulina, resultando na
regulação do número de genes envolvidos na função das células β pancreáticas,
ainda a Tau mostrou aumentar a fosforilação do IRβ no fígado e músculo

27
(CARNEIRO et al., 2009). Além disso, camundongos suplementados com Tau
apresentaram aumento da ativação da proteína quinase β (Akt) nos tecidos-alvo da
insulina (RIBEIRO et al., 2012). Essas ações da Tau podem estar relacionadas à
ativação de vias de sinalização relacionadas à sobrevivência e proliferação das
células β. Portanto, a Tau poderia manter a função pancreática endócrina
normalizada em filhotes de pais que ingerem dieta HFD por preservar a ativação de
vias reguladoras de diferenciação e sobrevivência celular como a Akt (BENES et al.,
1999; LAWRENCE et al., 2008).
É importante ressaltar que a Tau também mostrou efeitos sobre a regulação
da reprodução em machos, tendo envolvimento no controle da espermatogênese e
regulação da concentração plasmática de gonadotrofinas e testosterona (YANG et
al., 2006). A Tau já demonstrou regular os efeitos epigenéticos modulados pela
condição metabólica dos pais, e desta forma, impedir a transmissão desta para as
gerações posteriores. Neste sentido, CHERIF e colaboradores (1998) verificaram
que a secreção de insulina nas ilhotas pancreáticas apresentou-se reduzida em
fetos de ratas que foram submetidas à restrição proteica em resposta a nutrientes. E
que a suplementação com Tau durante a prenhez normaliza a secreção de insulina
e a proliferação celular, restaurando a síntese de DNA, a expressão de genes
envolvidos com o metabolismo e proliferação celular, reduzindo a apoptose e
normalizando a massa de células β nas ilhotas pancreáticas destes fetos e neonatos
de mães desnutridas (CHERIF et al., 1996 e 1998; MEREZAK et al., 2001 e 2004;
BOUJENDAR et al., 2002; KALBE et al., 2005; REUSENS et al., 2008). Em modelos
de restrição proteica que pré-dispõem ao DM2 quando administrado Tau in útero
observou-se a restauração da função como um todo da ilhota pancreática da prole
via regulação da expressão de fatores de crescimento, regulação da proliferação e
apoptose. Em relação ao DM1, a suplementação com Tau durante a gestação em
camundongos não obesos diabéticos (NOD) conferiu à prole destes animais, retardo
no aparecimento do DM, aumento da taxa de sobrevida e alteração na taxa de
proliferação e apoptose nas células (ARANY et al., 2004).

28
2. JUSTIFICATIVA
A obesidade é uma doença que pode trazer sérias consequências não
apenas para quem é obeso, mas também pode acarretar prejuízos à sua
descendência. O avanço do conhecimento acerca da obesidade tem sido
acompanhado de diferentes alternativas para o seu tratamento ou prevenção, dentre
essas alternativas existem as farmacológicas, suplementares ou até mesmo
cirúrgicas (JOHNSON et al., 2017). Mas pouco ainda se conhece sobre o aspecto
hereditário da obesidade e suas comorbidades, e como ela atua na propagação
dessas características para as próximas gerações.
Atualmente está bem estabelecido que além da condição maternal, a
condição paternal tem um papel importante em predispor sua descendência a
comorbidades como; intolerância a glicose, hiperglicemia, hiperlipidemia e prejuízos
funcionais no pâncreas endócrino (GUPTA et al., 2009; MORRIS et al., 2014). As
alternativas preventivas para obesidade devem contribuir não somente com a perda
de peso, mas prevenir ou evitar alterações morfofuncionais que são desencadeadas
por essa síndrome (SBEM, 2016). Evidências experimentais demonstram que a
obesidade desencadeia uma série de prejuízos que podem ser transmitidos para
gerações posteriores. Com isto, as buscas por estratégias terapêuticas para essas
alterações são importantes na prevenção e/ou tratamento da obesidade e suas
comorbidades, principalmente a sua propagação.
A suplementação com aminoácidos apresenta ações benéficas tanto para
quem o ingere, quanto na propagação para sua descendência. Dentre esses
aminoácidos temos a Tau, no qual já é descrito que a suplementação promove
aumento da secreção da insulina em resposta a nutrientes via melhora do influxo de
Ca2+ para as células β. A suplementação com Tau também mostrou prevenir
alterações morfológicas no pâncreas de camundongos HFD, impedindo a hipertrofia
e a hipersecreção induzidas pelo ambiente de resistência à insulina (RIBEIRO et al.,
2009; RIBEIRO et al., 2012). Contudo, ainda é pouco estudado o possível papel
preventivo da suplementação com a Tau ofertada aos pais na prevenção da
propagação de alterações para os seus filhotes. Porém, já foi demonstrado que em
ratos obesos a suplementação com Tau preveniu a reatividade vascular na sua prole
(LEÃO et al., 2015). Mas em relação ao aspecto metabólico e a secreção de
insulina nada ainda foi descrito acerca da influência paternal obesa sobre sua
descendência. Mas já se sabe dos prejuízos que obesidade acarreta na sua prole,

29
dentre eles foi evidenciado que ratos obesos acasalados com fêmeas alimentadas
com dieta CTL predispuseram a sua prole a intolerância à glicose (MORRIS et al.,
2014). Diante dos diversos efeitos benéficos apresentados, a Tau tornou-se alvo de
muitos estudos, visando esclarecer os seus mecanismos de ação e as suas
diferentes funções no organismo como agente protetor na propagação dos prejuízos
na secreção de insulina e na morfologia do pâncreas endócrino na prole de pais
obesos. Portanto, é necessário aumentar as investigações acerca do possível papel
preventivo da Tau sobre a morfofunção pancreática endócrina de descendentes de
camundongos machos obesos, pois não se sabe os efeitos da associação de dieta
HFD e Tau na prole, além disso, não se conhece qual é o papel regulador da Tau
nesses processos. Por isso nesse trabalho os possíveis ações do aminoácido na
prevenção da propagação de alterações da obesidade paternal para sua prole esta
sendo investigado.

30
3. OBJETIVOS
3.1 Objetivo geral:
Investigar os mecanismos da Tau que influenciam na propagação de
alterações morfofuncionais do pâncreas endócrino da prole de primeira geração de
camundongos machos que ingeriram dieta HFD.
3.2 Objetivos específicos:
Avaliar o desenvolvimento da obesidade em camundongos dos grupos
paternais alimentados com dieta HFD e suplementados ou não com Tau e em
sua primeira geração;
Verificar a tolerância à glicose nos grupos paternais e na sua prole fêmea e
macho;
Avaliar a secreção de insulina estimulada por glicose nos grupos paternais e
na sua prole macho e fêmea;
Avaliar a morfometria do pâncreas de camundongos machos provenientes de
pai obeso por dieta HFD suplementados ou não com Tau.

31
4. MATERIAIS E MÉTODOS
4.1 Grupos experimentais:
Camundongos machos C57Bl/6 foram mantidos no Biotério de
Experimentação de Roedores, localizado no Núcleo em Ecologia e Desenvolvimento
Socioambiental de Macaé (NUPEM), Polo Barreto, Campus UFRJ-Macaé sob
condições de luminosidade (8h-20h) e temperatura (21 ± 2o C) controladas. Todos os
procedimentos experimentais desse estudo foram aprovados pela Comissão de
Ética em Uso de Animais para a experimentação do Campus UFRJ-Macaé
(Certificado nº.: MAC031; Anexo 1). Dos 30 aos 150 dias de vida os camundongos
foram distribuídos nos grupos:
1) Controle (CTL): camundongos alimentados com ração CTL (PragSoluções
Serviços e Comércio Ltda., Jaú, SP, BRA) e água filtrada ad libitum;
2) CTL suplementado com Tau (CTAU): camundongos alimentados com ração
CTL e suplementados com 5%* (g/V) de Tau adicionada à água filtrada ad
libitum;
*Os camundongos foram suplementados com 5% de Tau adicionado a água filtrada
pois essa concentração de Tau foi utilizada em outros trabalhos e foi eficaz em
apresentar benefícios quando associada ao consumo de dieta HFD (RIBEIRO et al.,
2012).
3) Dieta Hiperlipídica (HFD): camundongos alimentados com ração contendo
36g% de gordura (PragSoluções Serviços e Comércio Ltda., Jaú, SP, BRA) e
água filtrada ad libitum;
4) HTAU: camundongos alimentados com dieta HFD e suplementados com 5%
de Tau adicionado à água filtrada ad libitum.
Aos 150 dias de vida os camundongos machos dos grupos anteriormente
relacionados foram submetidos ao acasalamento com fêmeas CTL de 150 dias para
obtenção da prole.
Os grupos de prole foram denominados conforme o tratamento dos pais em:
1) CTL-F1; 2) CTAU-F1; 3) HFD-F1 e 4) HTAU-F1 (representação dos grupos
experimentais na Fig. 1). Ao nascer, a prole de todos os grupos foi pesada e o
número de camundongos por ninhada foi registrado.
Os grupos de prole dos 30 aos 90 dias de vida foram alimentados com ração
padrão (Nuvital, Colombo, PR, Brasil) e água filtrada ad libitum. Por todo o período

32
experimental tanto o grupo paternal, quanto os grupos de prole tiveram livre acesso
à água e às respectivas dietas (composição detalhada abaixo no Quadro 1).
Obs: * Ração padrão e água ad libitum. Figura 1: Representação esquemática dos grupos experimentais. Grupos paternais CTL
(camundongos alimentados com ração CTL e água filtrada ad libitum), CTAU
(camundongos alimentados com ração CTL e suplementados com 5% (g/V) de Tau adicionada à água filtrada), HFD (camundongos alimentados com ração contendo 36g% de

33
gordura e água filtrada ad libitum) e HTAU (alimentados com a mesma ração do grupo HFD
e suplementados com 5% de Tau), aos 150 dias de vida foram acasalados com fêmeas CTL. Os grupos de prole foram designados de acordo com o tratamento dos pais: CTL-F1, CTAU-F1, HFD-F1 e HTAU-F1. Estes foram alimentados com ração padrão e água filtrada
ad libitum dos 30 aos 90 dias de vida.
Figura 1: Representação esquemática dos grupos experimentais.
Quadro 1: Composição das dietas
Ingredientes dieta AIN-93M CTL HFD
Caseína (84% de proteína)* 140 140 Amido 465,7 208,7 Dextrina 155 100 Sacarose 100 100 L-cistina 1,8 1,8 Fibra (microcelulose) 50 50 Óleo de soja 40 40 Banha - 312 Mistura de sais AIN93G** 35 35 Mistura de vitaminas AIN93G**
10 10
Cloridrato de colina 2,5 2,5
*Valores corrigidos em função do conteúdo de proteína na caseína.
** Composição detalhada dada por REEVES et al., 1993.
4.2 Avaliação da ingestão de ração e de água e avaliação da obesidade
Os camundongos dos grupos paternais, dos 30 aos 150 dias de vida, e dos
grupos de prole, dos 30 aos 90 dias, foram pesados uma vez por semana. O
consumo de ração foi medido semanalmente sendo a ração colocada menos a ração
restante dividida pelo número de camundongos por gaiola, e a ingestão de água foi
medida a cada dois dias e então, realizada a média do consumo hídrico diário.
Obteve-se a eficiência alimentar pela razão entre o total de ganho de peso corporal
(diferença entre o peso corporal final e o peso corporal inicial) dividido pelo consumo
total de ração durante todo o período experimental (DUIVENVOORDEN et al., 2005).
Ao final do período experimental, tanto nos camundongos dos grupos
paternais, quanto nos camundongos dos grupos de prole F1, foi feita as medidas
para avaliação da obesidade pelo peso corporal final e o comprimento nasoanal
(CNA) foram medidos para obtenção do índice de Lee [calculado por meio da raiz
cúbica do peso corporal (g) dividido pelo CNA (cm) x 1000] (BERNARDIS e
PATTERSON, 1968) que é o índice de massa corpórea utilizado em roedores. Em

34
seguida, os camundongos foram eutanasiados por decapitação. As gorduras
perigonadal e retroperitoneal foram dissecadas e pesadas para avaliação da
obesidade nos camundongos. Para análise dos dados da ingestão hídrica, de ração
e eficiência alimentar foi utilizado (n= 4) camundongos no grupo paternal por grupo
experimental, e na prole fêmea e macho F1 variou o (n= 4-16) camundongos por
grupo, o número de roedores da prole dependia do sucesso do nascimento no
mesmo período para realizar as medidas. Para os parâmetros da avaliação da
obesidade nos grupos paternais os registros esteve entre (n= 8-9) camundongos, e
para os grupos de prole macho e fêmea F1 (n= 9-16), valores similares aos citados
pela literatura que indica o número de amostras necessário para a adequada análise
estatística (NARDELLI et al., 2011).
4.3 Teste intraperitoneal de tolerância à glicose (ipgtt)
Aos 150 dias de vida dos camundongos dos grupos paternais, e aos 90 dias
dos camundongos dos grupos de prole fêmea e macho, foi realizado o ipGTT. Nos
roedores após 12h de jejum, o sangue foi coletado via caudal e então a glicemia
(tempo 0) medida utilizando-se um glicosímetro (Accu-Chek Advantage, Roche
Diagnostic, Suiça). Em seguida, os camundongos receberam uma injeção
intraperitoneal (ip) de glicose na concentração de 2g/kg de peso corporal. A glicemia
foi novamente verificada aos 15, 30, 60, 120 e 180 minutos após a administração da
glicose. Para adequada análise estatística desses dados obtidos no ipGTT foram
utilizados para o grupo paternal (n= 7-8) camundongos, e para os grupos da prole
fêmea F1 (n= 8-12) e para a prole macho F1 (n= 6-12) camundongos
correlacionando com outros estudos (RIBEIRO et al., 2009).
4.4 Parâmetros bioquímicos plasmáticos
Para a dosagem dos parâmetros bioquímicos plasmáticos, os experimentos
foram realizados aos 150 dias de vida dos grupos paternais e aos 90 dias de vida
dos grupos de prole. Na condição de alimentado, foram coletados 50 µL de sangue
via caudal e após dois dias foi realizada nova coleta de sangue, contudo após 12 h
de jejum dos mesmos camundongos. O sangue coletado foi centrifugado a 15204 x
g a 4°C, por 15 minutos, então o plasma separado foi armazenado a -20ºC e
posteriormente, direcionado à dosagem de insulina por radioimunoensaio (RIA)
(SCOTT et al., 1981), triglicerídeos, colesterol total usando kits colorimétricos

35
(Boehringer Mannhein®, Alemanha; Merck®, Alemanha; Laborclin, Pinhais, PR,
BRA) (NARDELLI et al., 2011). Para análise dos parâmetros bioquímicos foram
utilizados (n= 4) amostras por grupo experimental paternal e para os grupos de prole
variou (n= 5-19) na prole fêmea F1 e para a prole macho F1 foram usadas (n= 9-15)
amostras por grupo.
4.5 Isolamento de ilhotas pancreáticas e secreção estática de insulina
Após a eutanásia dos camundongos, tanto dos grupos paternais quanto dos
grupos de prole, foi realizada laparotomia para acesso ao ducto pancreático e sua
obstrução. Na porção distal do ducto colédoco foi introduzida uma agulha de insulina
(26 G 1/2) pela qual foi injetado no pâncreas 3 mL de solução de Hank’s (136 mM de
NaCl; 5,3 mM de KCl; 0,9 mM de MgSO4.7H2O; 0,3 mM de Na2HPO4.12H2O; 4,4
mM de KH2PO4; 1,2 mM de CaCl2.2H2O gaseificadas com 95% O2/5% CO2 , após
foi colocado 4,1 mM de NaHCO3 e em seguida acrescentado 1mM de glicose e
0,1% de albumina bovina (g/v), pH 7,4) contendo colagenase do tipo V (0,8 mg/mL),
para facilitar a retirada e iniciar a digestão do tecido exócrino. O pâncreas dissecado
foi transferido para um tubo de 15 mL e incubado a 37ºC por 18 minutos. As ilhotas
pancreáticas, completamente separadas do tecido acinar, foram coletadas uma a
uma, com auxílio de microscópio estereoscópico (Olympus Corporation model SZ2-
ILST,Tokyo,Japan). Grupos de 4 ilhotas pancreáticas de cada grupo experimental,
tanto paternal quanto da prole, foram transferidas para placas de cultura com 24
poços contendo 0,5 mL de solução de Kreb’s (115 mM de NaCl; 5 mM de KCl; 2,56
mM de CaCl2; 1 mM de MgCl2; 15 mM de HEPES suplementado com 5,6 mM de
glicose, gaseificado com 95% O2/5%, e após acrescentado 10 mM de NaHCO3 e
0,3% de albumina bovina (g/v), pH 7,4). A seguir as ilhotas pancreáticas foram
incubadas por 30 minutos em banho-maria, 37°C e continuamente gaseificadas com
95% O2/5% CO2. Após, a solução foi rapidamente removida e substituída por nova
solução de incubação contendo 2,8, 11,1 e 22,2 mM de glicose. Depois de 1h de
incubação, o sobrenadante foi removido, transferido para tubos de ensaio e
armazenado a –20°C. A dosagem de insulina foi posteriormente realizada por RIA,
utilizando-se anticorpo específico anti-insulina produzido em rato (doado gentilmente
por Dr. Leclerq-Meyer, Free University, Brussels, BEL), insulina de rato para traçar a
curva padrão (Crystal Chem. Inc., Downers Grove, IL, USA) e insulina recombinante
humana marcada com 125I (Amersham Biosciences, Little Chalfont, Bucks, UK). Para

36
separação da insulina livre da porção antígeno-anticorpo formada foi utilizado o
método do carvão ativado e dextran (SCOTT et al., 1981). Para dosagem de insulina
nos grupos paternais foi utilizado para cada condição de glicose avaliada o (n= 17-
40), e nos grupos da prole fêmea F1 (n= 24-47) e na prole macho F1 (n= 19-32)
amostras por condição de glicose em cada grupo experimental conforme a literatura
sugere (RIBEIRO et al., 2012).
4.6 Morfologia do pâncreas da prole macho F1
Após a eutanásia o pâncreas de cada camundongo da prole macho F1 foi
dissecado, pesado e então fixado em solução de Bouin (25% de formaldeído, 75%
de ácido pícrico saturado e 5% de ácido acético) por 16h, e depois lavado 1 vez com
cloreto de amônia e armazenado em solução de 70% de etanol. Para análise
morfométrica dos principais tipos celulares das ilhotas pancreáticas. Posteriormente,
o pâncreas foi desidratado em concentrações crescentes de etanol (80% de etanol
20min,90% de etanol 20min e 3 diferentes banhos de 15min cada com álcool 100%
e colocados em uma solução de álcool + xilol por 15min) diafanizado em 3 diferentes
soluções de xilol por 20min cada e posteriormente incluídos em 3 diferentes banhos
Paraplast® por 30min cada (Sigma-aldrich, St. Louis, MA, USA). Em sequência,
foram obtidas secções semi-seriadas de 5 μm, com espaçamento de 100 μm entre
os cortes, até esgotamento total do tecido.
4.7 Morfometria e imuno-histoquímica para insulina, glucagon e
somatostatina
Para realização da imuno-histoquímica, as secções seriadas das mesmas
porções do pâncreas foram selecionadas, desparafinizadas, hidratadas e, após
bloqueio da peroxidase endógena [tampão tris-salina (TBS) contendo 3% de H2O2] e
dos sítios inespecíficos (0,1% Tween 20 e 5% leite desnatado em TBS), foram
incubadas com anticorpo para insulina (1:100; Dako North America, Inc., CA, USA),
ou glucagon (1:50; Dako North America, Inc., CA, USA) ou somatostatina [(1:50,
Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA (sc-7819)], todos diluídos em
solução de TBS com 3% de leite, overnight à 4ºC. Após, as secções foram lavadas
com TBS e incubadas por 1h e 30 min com anticorpo secundário conjugado com
peroxidase [anti-cobaia, ou anti-coelho ou anti-cabra, respectivamente (1:1500;
Zymed Laboratories, Inc., San Francisco, CA, USA) diluidos em TBS com 1% de

37
leite]. A detecção da imunorreação foi realizada utilizando-se a solução 10% de DAB
(diaminobenzidina) e 0,2% de H2O2 em TBS (DAB; Sigma-aldrich, St. Louis, MA,
USA). Subsequentemente, as lâminas foram contra-coradas com solução de
hematoxilina Harris, e montadas com bálsamo sintético. Foram utilizadas 12 lâminas
por grupo de camundongos da prole macho F1 para cada tipo celular das ilhotas
pancreáticas (com mais de 5 células cada) avaliado, e então foram fotografadas nos
aumentos de 20x ou 40x em microscópio de luz Olympus BX51, acoplado a sistema
de captura de imagem (Olympus DP71, Olympus Optical do Brasil, São Paulo, SP,
BRA). A morfometria foi realizada utilizando-se o software Image J 1.50b
(http://imagej.nih.gov/ij/) calibrado com a régua conforme o aumento utilizado para
fazer as imagens, e os parâmetros avaliados foram: 1) área absoluta das ilhotas
pancreáticas, ou área absoluta das células β, ou α ou δ; 2) número de células β, ou
α ou δ; 3) massa (mg) de ilhotas pancreáticas e de células β, ou α ou δ [calculada
pelo total da área da ilhota ou das células β, ou α ou δ (µm2) dividido pela área da
secção do pâncreas (µm2), e após multiplicado pelo peso do pâncreas (mg)]; após
essas medidas estimamos a razão área de células β, ou α ou δ (µm2) dividido pelo
número das respectivas células (RIBEIRO et al., 2012). A quantificação das áreas foi
feita utilizando o recurso manual do Image J. O software foi calibrado com régua
micrométrica (1 mm) possibilitando a conversão dos valores obtidos em pixels para
um2. As medidas foram realizadas com a ferramenta de seleção a mão livre do
Image J, através da qual foi demarcada toda extensão da ilhota pancreática ou das
células que apresentaram marcação positiva para insulina, ou glucagon ou
somatostatina. Os números de células foram contados um a um. Para realização da
imuno-histoquímica foi utilizado (n= 3-4) pâncreas por grupo de prole macho F1, e os
mesmos pâncreas não foram utilizados para o experimento de secreção estática de
insulina.
4.8 Análise estatística
Os resultados foram expressos como média erro padrão da média (EPM).
As análises estatísticas foram realizadas utilizando-se o programa Prism 5.0
(GraphPad Software, USA). Todos os resultados foram submetidos a teste de
normalidade Shapiro-Wilk e então direcionados para testes paramétricos (ANOVA
de uma via seguido de pós-teste Newman Keuls) ou não paramétricos (Kruskal-

38
Wallis seguido de pós-teste Dunns). P < 0,05 foi adotado como critério de
significância.

39
5. RESULTADOS
5.1 Ingestão de ração e água e avaliação da obesidade nos grupos paternais
e em sua respectiva prole
Na figura 2A observa-se a evolução do peso corporal ao longo de doze
semanas em camundongos dos grupos paternais alimentados com dieta HFD
suplementados ou não com Tau. Camundongos HFD começaram a apresentar
maior peso corporal a partir da nona semana de ingestão de dieta HFD em relação
ao grupo CTL (P < 0,01; Fig. 2A). Porém, a partir da quinta semana, já tínhamos
observado que o grupo alimentado com dieta HFD apresentava maior peso corporal
em relação aos CTAU se mantendo até o final do tratamento (P < 0,01; Fig. 2A).
O ganho de peso corporal total dos camundongos dos grupos paternais foi
maior no grupo HFD comparado ao CTL (P < 0,05; Fig. 2B). A suplementação com
Tau não modificou esse parâmetro (Fig. 2B).
A ingestão hídrica do grupo paternal HFD não apresentou diferença em
relação ao grupo CTL (Fig. 2C), no entanto, observou-se redução do consumo de
água nos camundongos HFD e HTAU quando comparados ao grupo CTAU (P <
0,01; Fig. 2C). Não foram verificadas diferenças no consumo total em kcal de ração
ao longo do período experimental nos grupos paternais estudados (Fig. 2C). Ainda,
a eficiência alimentar foi similar no grupo HFD, quando comparado aos CTL (Fig.
2E). O tratamento com Tau não alterou a eficiência alimentar nos grupos estudados
(Fig. 2E).
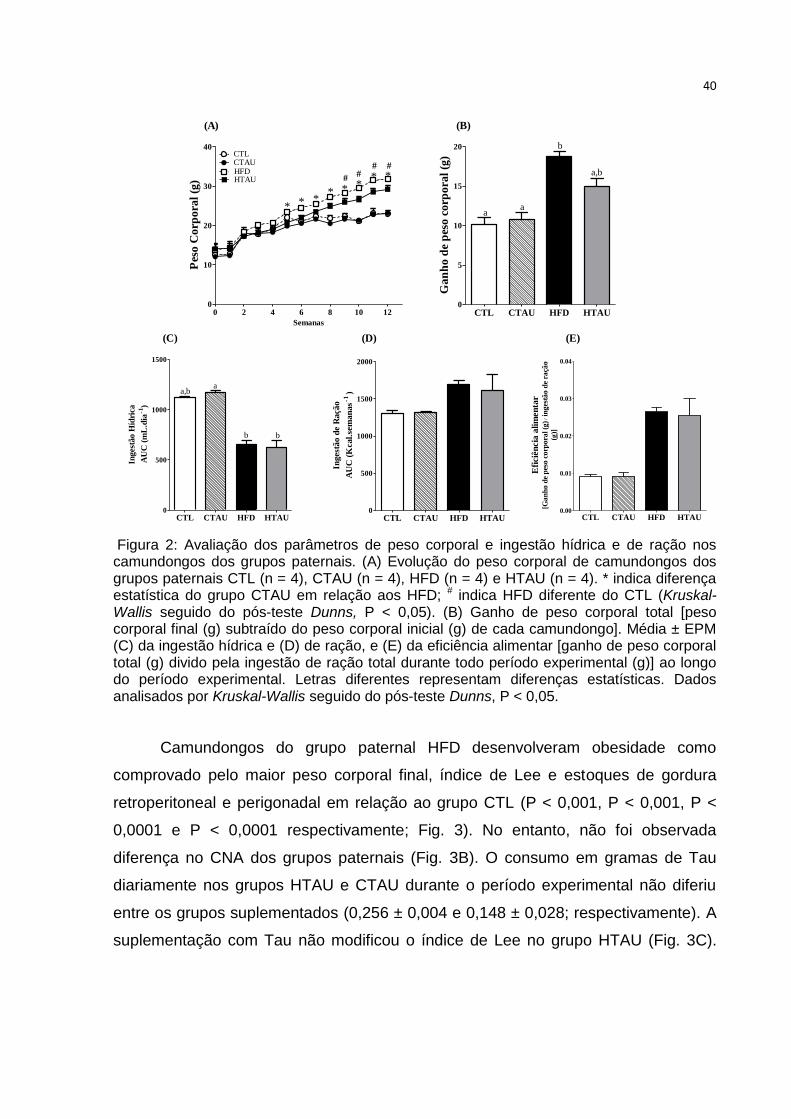
40
(A) (B)
0 2 4 6 8 10 120
10
20
30
40CTLCTAUHFDHTAU *
#
*
#
*
#
**
***
#
Semanas
Pes
o C
orp
ora
l (g
)
CTL CTAU HFD HTAU0
5
10
15
20
aa
b
a,b
Ga
nh
o d
e p
eso
co
rpo
ral
(g)
CTL CTAU HFD HTAU0
500
1000
1500
a,ba
b b
Inges
tão H
ídri
ca
AU
C (
mL
.dia
-1)
CTL CTAU HFD HTAU0
500
1000
1500
2000
Ingest
ão d
e R
ação
AU
C (
Kcal.
sem
an
as
-1)
CTL CTAU HFD HTAU0.00
0.01
0.02
0.03
0.04
Efi
ciê
ncia
ali
men
tar
[Gan
ho d
e p
eso c
orp
ora
l (g
)/ i
ng
estã
o d
e ra
ção
(g)]
(C) (D) (E)
Figura 2: Avaliação dos parâmetros de peso corporal e ingestão hídrica e de ração nos camundongos dos grupos paternais. (A) Evolução do peso corporal de camundongos dos grupos paternais CTL (n = 4), CTAU (n = 4), HFD (n = 4) e HTAU (n = 4). * indica diferença estatística do grupo CTAU em relação aos HFD; # indica HFD diferente do CTL (Kruskal-Wallis seguido do pós-teste Dunns, P < 0,05). (B) Ganho de peso corporal total [peso corporal final (g) subtraído do peso corporal inicial (g) de cada camundongo]. Média ± EPM (C) da ingestão hídrica e (D) de ração, e (E) da eficiência alimentar [ganho de peso corporal total (g) divido pela ingestão de ração total durante todo período experimental (g)] ao longo do período experimental. Letras diferentes representam diferenças estatísticas. Dados analisados por Kruskal-Wallis seguido do pós-teste Dunns, P < 0,05. Figura 2: Avaliação dos parâmetros de peso corporal e ingestão hídrica e de ração nos camundongos dos grupos paternais.
Camundongos do grupo paternal HFD desenvolveram obesidade como
comprovado pelo maior peso corporal final, índice de Lee e estoques de gordura
retroperitoneal e perigonadal em relação ao grupo CTL (P < 0,001, P < 0,001, P <
0,0001 e P < 0,0001 respectivamente; Fig. 3). No entanto, não foi observada
diferença no CNA dos grupos paternais (Fig. 3B). O consumo em gramas de Tau
diariamente nos grupos HTAU e CTAU durante o período experimental não diferiu
entre os grupos suplementados (0,256 ± 0,004 e 0,148 ± 0,028; respectivamente). A
suplementação com Tau não modificou o índice de Lee no grupo HTAU (Fig. 3C).

41
CTL CTAU HFD HTAU0
10
20
30
40
50
a a b
bb
Pes
o C
orp
ora
l F
ina
l (g
)
CTL CTAU HFD HTAU0
100
200
300
400
a
b
aa,b
Índ
ice d
e L
ee
CTL CTAU HFD HTAU0
20
40
60
aa
bb
Go
rdu
ra p
erig
on
ad
al
(mg
/g p
eso
co
rpo
ral)
CTL CTAU HFD HTAU0
5
10
15
20
25
aa
b b
Gord
ura
ret
rop
erit
on
ial
(mg
/g p
eso c
orp
ora
l)
(A) (B)
CTL CTAU HFD HTAU0
5
10
15
CN
A (
cm)
(C)
(D) (E)
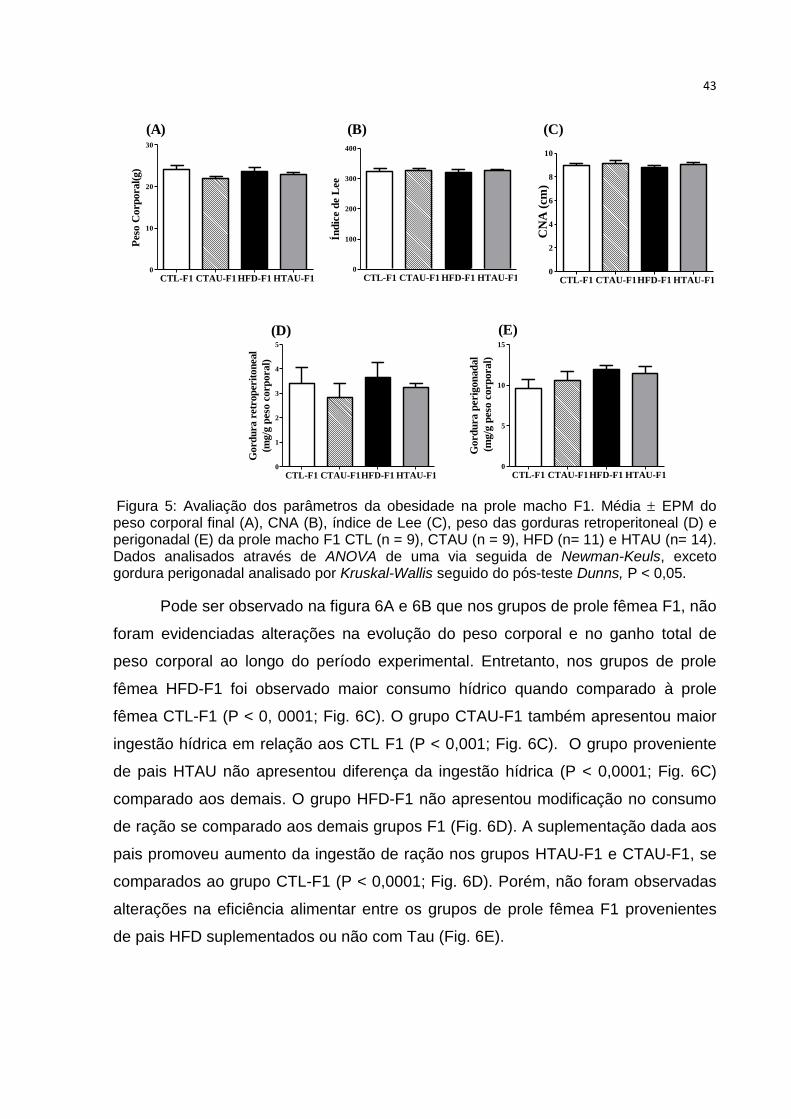
Figura 3: Avaliação dos parâmetros da obesidade nos camundongos dos grupos paternais.
(A) Peso corporal final, (B) CNA, (C) índice de Lee, (D) gordura retroperitoneal e (E) gordura
perigonadal de camundongos dos grupos paternais CTL (n = 9), CTAU (n = 8), HFD (n = 9)
e HTAU (n = 8). Letras diferentes indicam diferenças estatísticas. Dados de peso corporal e
gordura retroperitoneal analisados por ANOVA de uma via seguida de Newman-Keuls, e os
demais parâmetros analisados por Kruskal-Wallis seguido do pós-teste Dunns, P < 0,05.
Figura 3: Avaliação dos parâmetros da obesidade nos camundongos dos grupos paternais.
Os camundongos dos grupos paternais foram acasalados com fêmeas CTL
como já descrito em materiais e métodos (item 4.1). A prole obtida desses
cruzamentos não apresentou diferença no número de camundongos por ninhada
nos diferentes grupos experimentais (CTL 5,75 ± 0,45; CTAU 4,60 ± 0,60; HFD 6,33
± 0,42 e HTAU 5,20 ± 0,66). Ainda, não foram observadas alterações no peso médio
da ninhada dos grupos de prole F1 obtidos tanto de pai que ingeriu dieta HFD (2,76
± 0,10 g) ou que foi suplementado com Tau (HTAU 3,30 ± 0,30 e CTAU 3,10 ± 0,24
g), quando comparados à prole CTL (2,75 ± 0,14 g).
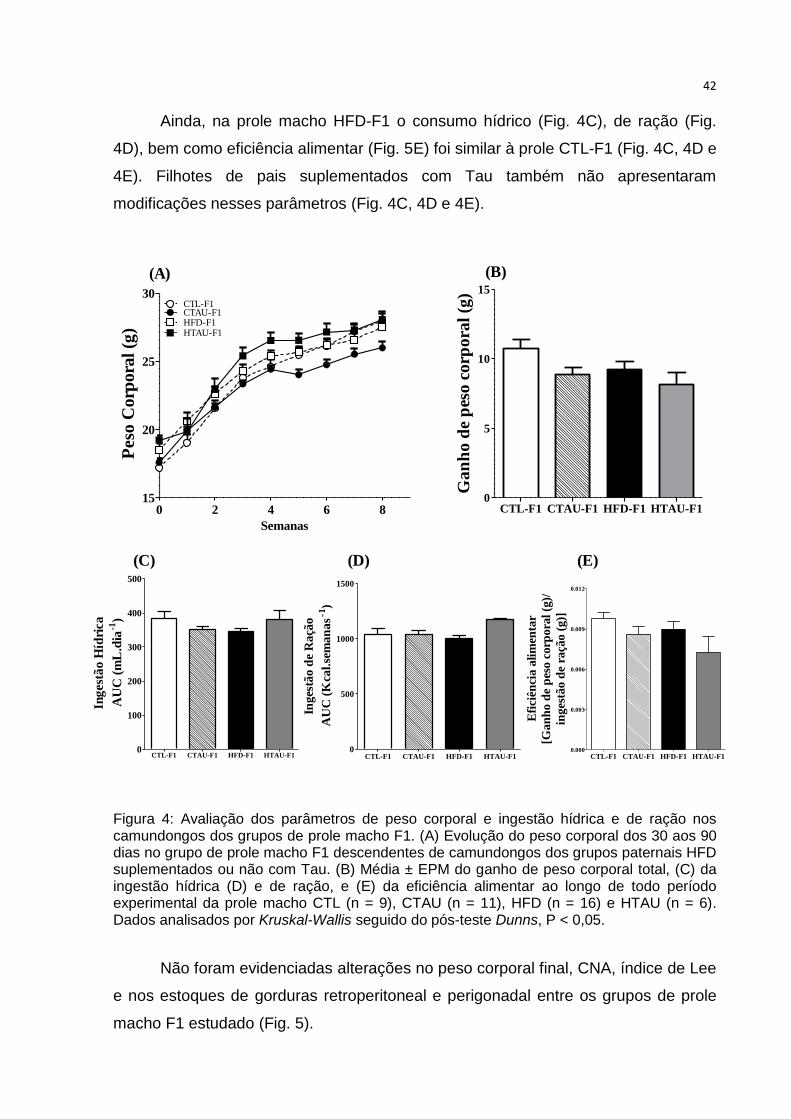
Na figura 4A e 4B não foram observadas diferenças na evolução ou ganho
total de peso corporal na prole macho F1 provenientes de pais HFD suplementados
ou não com Tau (Fig. 4A e 4B).
42
Ainda, na prole macho HFD-F1 o consumo hídrico (Fig. 4C), de ração (Fig.
4D), bem como eficiência alimentar (Fig. 5E) foi similar à prole CTL-F1 (Fig. 4C, 4D e
4E). Filhotes de pais suplementados com Tau também não apresentaram
modificações nesses parâmetros (Fig. 4C, 4D e 4E).
Figura 4: Avaliação dos parâmetros de peso corporal e ingestão hídrica e de ração nos camundongos dos grupos de prole macho F1. (A) Evolução do peso corporal dos 30 aos 90 dias no grupo de prole macho F1 descendentes de camundongos dos grupos paternais HFD suplementados ou não com Tau. (B) Média ± EPM do ganho de peso corporal total, (C) da ingestão hídrica (D) e de ração, e (E) da eficiência alimentar ao longo de todo período experimental da prole macho CTL (n = 9), CTAU (n = 11), HFD (n = 16) e HTAU (n = 6). Dados analisados por Kruskal-Wallis seguido do pós-teste Dunns, P < 0,05. Figura 4: Avaliação dos parâmetros de peso corporal e ingestão hídrica e de ração nos camundongos dos grupos de prole macho.
Não foram evidenciadas alterações no peso corporal final, CNA, índice de Lee
e nos estoques de gorduras retroperitoneal e perigonadal entre os grupos de prole
macho F1 estudado (Fig. 5).
0 2 4 6 815
20
25
30CTL-F1CTAU-F1HFD-F1HTAU-F1
Semanas
Peso
Corp
ora
l (g
)
CTL-F1 CTAU-F1 HFD-F1 HTAU-F10
5
10
15
Gan
ho d
e p
eso c
orp
ora
l (g
)
CTL-F1 CTAU-F1 HFD-F1 HTAU-F10
100
200
300
400
500
Ing
est
ão H
ídri
ca
AU
C (
mL
.dia
-1)
CTL-F1 CTAU-F1 HFD-F1 HTAU-F10
500
1000
1500
Ing
estã
o d
e R
açã
o
AU
C (
Kca
l.se
ma
na
s-1
)
CTL-F1 CTAU-F1 HFD-F1 HTAU-F10.000
0.003
0.006
0.009
0.012
Efi
ciên
cia a
lim
enta
r
[Gan
ho d
e p
eso c
orp
ora
l (g
)/
inges
tão d
e ra
ção (
g)]
(A) (B)
(C) (D) (E)
43
CTL-F1 CTAU-F1 HFD-F1 HTAU-F10
10
20
30
Pes
o C
orp
ora
l(g)
CTL-F1 CTAU-F1 HFD-F1 HTAU-F10
100
200
300
400
Índ
ice
de
Lee
CTL-F1 CTAU-F1HFD-F1 HTAU-F10
5
10
15
Gord
ura
per
igon
ad
al
(m
g/g
pes
o c
orp
ora
l)CTL-F1 CTAU-F1HFD-F1 HTAU-F1
0
1
2
3
4
5
Gord
ura
ret
rop
erit
on
eal
(mg/g
pes
o c
orp
oral)
(A) (B)
(D) (E)
CTL-F1 CTAU-F1HFD-F1 HTAU-F10
2
4
6
8
10
CN
A (
cm)
(C)
Figura 5: Avaliação dos parâmetros da obesidade na prole macho F1. Média EPM do peso corporal final (A), CNA (B), índice de Lee (C), peso das gorduras retroperitoneal (D) e perigonadal (E) da prole macho F1 CTL (n = 9), CTAU (n = 9), HFD (n= 11) e HTAU (n= 14). Dados analisados através de ANOVA de uma via seguida de Newman-Keuls, exceto gordura perigonadal analisado por Kruskal-Wallis seguido do pós-teste Dunns, P < 0,05. Figura 5: Avaliação dos parâmetros da obesidade na prole macho.
Pode ser observado na figura 6A e 6B que nos grupos de prole fêmea F1, não
foram evidenciadas alterações na evolução do peso corporal e no ganho total de
peso corporal ao longo do período experimental. Entretanto, nos grupos de prole
fêmea HFD-F1 foi observado maior consumo hídrico quando comparado à prole
fêmea CTL-F1 (P < 0, 0001; Fig. 6C). O grupo CTAU-F1 também apresentou maior
ingestão hídrica em relação aos CTL F1 (P < 0,001; Fig. 6C). O grupo proveniente
de pais HTAU não apresentou diferença da ingestão hídrica (P < 0,0001; Fig. 6C)
comparado aos demais. O grupo HFD-F1 não apresentou modificação no consumo
de ração se comparado aos demais grupos F1 (Fig. 6D). A suplementação dada aos
pais promoveu aumento da ingestão de ração nos grupos HTAU-F1 e CTAU-F1, se
comparados ao grupo CTL-F1 (P < 0,0001; Fig. 6D). Porém, não foram observadas
alterações na eficiência alimentar entre os grupos de prole fêmea F1 provenientes
de pais HFD suplementados ou não com Tau (Fig. 6E).

44
(A) (B)
0 2 4 6 812
14
16
18
20
22
24CTL-F1CTAU-F1HFD-F1HTAU-F1
Semanas
Pes
o C
orp
ora
l (g
)
CTL-F1 CTAU-F1 HFD-F1 HTAU-F10
2
4
6
8
Gan
ho d
e p
eso c
orp
ora
l (g
)
CTL-F1 CTAU-F1 HFD-F1 HTAU-F10
100
200
300
400
500
a
b ba,b
Ing
estã
o H
ídri
ca
AU
C (
mL
.dia
-1)
CTL-F1 CTAU-F1 HFD-F1 HTAU-F10
400
800
1200
aa,b
bb
Inges
tão d
e R
açã
o
AU
C (
Kca
l.se
man
as
-1)
CTL-F1 CTAU-F1 HFD-F1 HTAU-F10.000
0.005
0.010
0.015
0.020
0.025
Efi
ciên
cia
ali
men
tar
[Ga
nh
o d
e p
eso
co
rpo
ral
(g)/
ing
estã
o d
e ra
ção
(g
)]
(C) (D) (E)
Figura 6: Avaliação dos parâmetros de peso corporal e ingestão hídrica e de ração nos camundongos dos grupos de prole fêmea F1. (A) Evolução do peso corporal no grupo de prole fêmea F1 descendentes de camundongos dos grupos paternais HFD suplementados ou não com Tau. (B) Média ± EPM do ganho de peso corporal total, (C) da ingestão hídrica (D) e de ração, e (E) da eficiência alimentar ao longo de todo período experimental da prole macho CTL (n = 16), CTAU (n = 10), HFD (n = 6) e HTAU (n = 4). Dados da evolução do peso corporal analisados através de ANOVA de uma via seguida de Newman-Keuls, e os demais dados analisados por Kruskal-Wallis seguido do pós-teste Dunns, P < 0,05. Figura 6: Avaliação dos parâmetros de peso corporal e ingestão hídrica e de ração nos camundongos dos grupos de prole fêmea.
Nos grupos de prole fêmea F1 não foram observadas alterações no peso
corporal final, CNA, índice de Lee e nos estoques de gordura retroperitoneal e
perigonadal (Fig. 7).

45
CTL-F1 CTAU-F1 HFD-F1 HTAU-F10
10
20
30
Peso
Corp
ora
l(g)
CTL-F1 CTAU-F1 HFD-F1 HTAU-F10
100
200
300
400
Índ
ice d
e L
ee
CTL-F1 CTAU-F1 HFD-F1 HTAU-F10
5
10
15
Gord
ura
per
igon
ad
al
(m
g/g
pes
o c
orp
ora
l)
CTL-F1 CTAU-F1 HFD-F1 HTAU-F10
1
2
3
4
5
Gord
ura
ret
rop
erit
on
eal
(mg/g
pes
o c
orp
ora
l)
(A) (B)
(D) (E)
CTL-F1 CTAU-F1 HFD-F1 HTAU-F10
2
4
6
8
10
CN
A (
cm)
(C)
Figura 7: Avaliação dos parâmetros da obesidade na prole fêmea F1. Média EPM do peso corporal final (A), CNA (B), índice de Lee (C), peso das gorduras retroperitoneal (D) e perigonadal (E) aos 90 dias de vida da prole fêmea F1 CTL (n = 12), CTAU (n = 16), HFD (n= 11) e HTAU (n= 12). Dados analisados através de ANOVA de uma via seguida de Newman-Keuls, exceto gordura perigonadal analisado por Kruskal-Wallis seguido do pós-teste Dunns, P < 0,05. Figura 7: Avaliação dos parâmetros da obesidade na prole fêmea.
5.2 Avaliação da tolerância à glicose e dos parâmetros bioquímicos
plasmáticos nos grupos paternais e em sua respectiva prole
Os camundongos dos grupos paternais foram submetidos ao ipGTT. Na figura
8A é possível observar que após a administração ip de glicose, a concentração
plasmática do açúcar atingiu maior perfil nos grupos HFD e HTAU aos 30 minutos, e
nos grupos CTL e CTAU aos 15 minutos do ipGTT (Fig. 8A). Durante o teste,
camundongos HFD apresentaram maiores valores de glicose plasmática aos 30 e 60
minutos em relação ao grupo CTL (P < 0,001 e P < 0,05; Fig. 8A). Ainda,
camundongos HFD apresentaram maior total da glicemia durante o ipGTT, quando
comparado ao grupo CTL (P < 0,001; Fig. 8B). A suplementação com Tau não
modificou os valores da glicemia no grupo HTAU em relação aos demais grupos
(Fig. 8A). O total da glicemia expresso pela área abaixo da curva, não apresentou
diferença no grupo HTAU em relação aos demais (Fig. 8B).
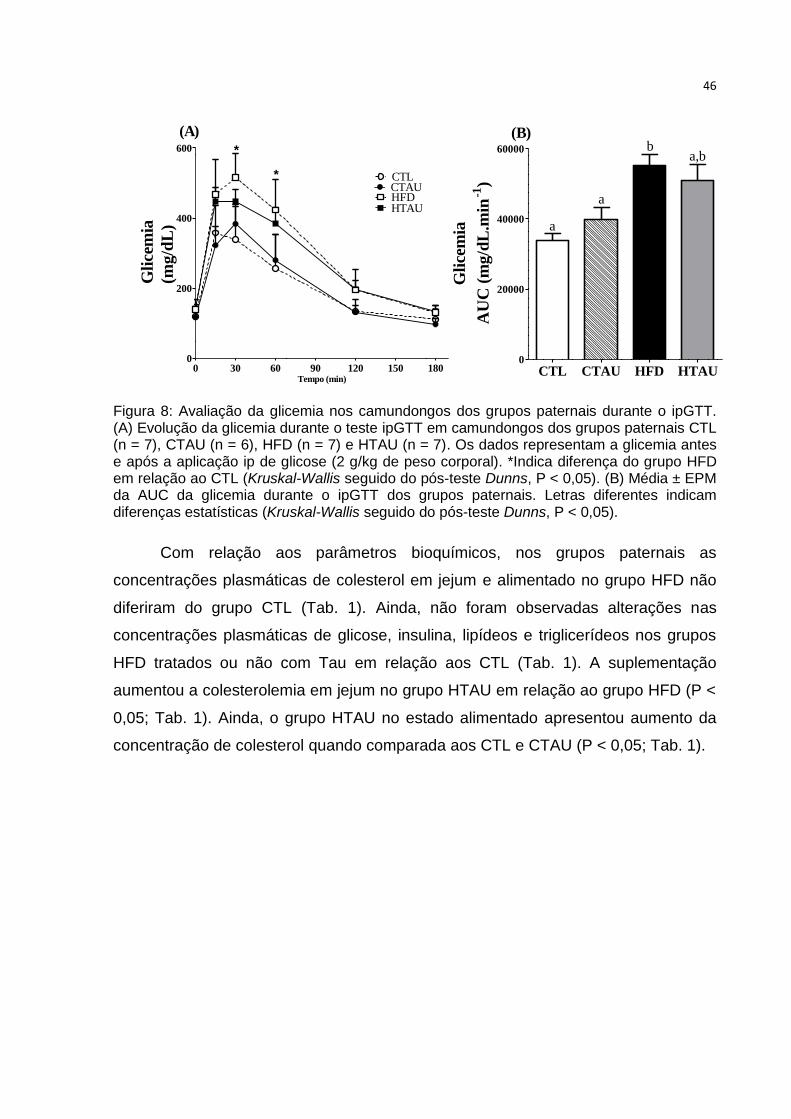
46
0 30 60 90 120 150 1800
200
400
600
CTAUHFDHTAU
CTL
Gli
cem
ia
(mg/d
L)
*
*
Tempo (min)
(A) (B)
CTL CTAU HFD HTAU0
20000
40000
60000
a
a
ba,b
Gli
cem
ia
AU
C (
mg/d
L.m
in-1
)
Figura 8: Avaliação da glicemia nos camundongos dos grupos paternais durante o ipGTT. (A) Evolução da glicemia durante o teste ipGTT em camundongos dos grupos paternais CTL (n = 7), CTAU (n = 6), HFD (n = 7) e HTAU (n = 7). Os dados representam a glicemia antes e após a aplicação ip de glicose (2 g/kg de peso corporal). *Indica diferença do grupo HFD em relação ao CTL (Kruskal-Wallis seguido do pós-teste Dunns, P < 0,05). (B) Média ± EPM da AUC da glicemia durante o ipGTT dos grupos paternais. Letras diferentes indicam diferenças estatísticas (Kruskal-Wallis seguido do pós-teste Dunns, P < 0,05).
Figura 8: Avaliação da glicemia nos camundongos dos grupos paternais durante o ipGTT.
Com relação aos parâmetros bioquímicos, nos grupos paternais as
concentrações plasmáticas de colesterol em jejum e alimentado no grupo HFD não
diferiram do grupo CTL (Tab. 1). Ainda, não foram observadas alterações nas
concentrações plasmáticas de glicose, insulina, lipídeos e triglicerídeos nos grupos
HFD tratados ou não com Tau em relação aos CTL (Tab. 1). A suplementação
aumentou a colesterolemia em jejum no grupo HTAU em relação ao grupo HFD (P <
0,05; Tab. 1). Ainda, o grupo HTAU no estado alimentado apresentou aumento da
concentração de colesterol quando comparada aos CTL e CTAU (P < 0,05; Tab. 1).

47
Tabela 1: Parâmetros bioquímicos plasmáticos no estado de jejum e alimentado avaliados em camundongos dos grupos paternais CTL, CTAU, HFD e HTAU.
Dados representam a média EPM [CTL (n = 4), CTAU (n = 4), HFD (n = 4), HTAU (n = 4)]. Letras diferentes indicam diferença estatística. Dados analisados através de Kruskal-Wallis seguido de pós-teste Dunns, P < 0,05. Tabela 1: Parâmetros bioquímicos plasmáticos no estado de jejum e alimentado avaliados em
camundongos aos 150 dias dos grupos paternais CTL, CTAU, HFD e HTAU.
Aos 90 dias de vida os grupos da prole macho F1 foram submetidos ao
ipGTT. Após a injeção de glicose, todos os grupos da prole macho F1 apresentaram
aumento da glicemia, atingindo os maiores valores aos 30 minutos e retornaram aos
valores basais aos 120 minutos depois da administração de glicose (Fig. 9A).
Durante o ipGTT os camundongos da prole macho HFD-F1 não apresentaram
alteração na tolerância à glicose quando comparados à prole CTL-F1 (Fig. 9A e 9B).
A suplementação com Tau dada aos pais HTAU não alterou esse parâmetro em sua
respectiva prole F1 (Fig. 9B).
CTL CTAU HFD HTAU
Glicemia (mg/dL)
Jejum 109 ±11 127 ± 8 140 ± 8 122 ± 10
Alimentado 154 ± 7 133 ± 6 165 ± 8 178 ± 12
Insulinemia (ng/mL)
Jejum 0,4 ± 0,1 0,5 ± 0,2 1,1 ± 0,5 0,3 ± 0,1
Alimentado
0,5 ± 0,1
1,4 ± 0,7 1,1 ± 0,3 0,8 ± 0,3
Triglicerídeos (mg/dL)
Jejum 53 ± 9 35 ± 3 36 ± 4 45 ± 2
Alimentado 75 ± 15 101 ±12 64 ± 9 93 ± 7
Colesterol (mg/dL)
Jejum 119 ± 5a, b 111 ± 11a, b 111 ± 4a 139 ± 5b
Alimentado 88 ± 6ª 72 ± 14ª 148 ±11a, b 223 ± 16b
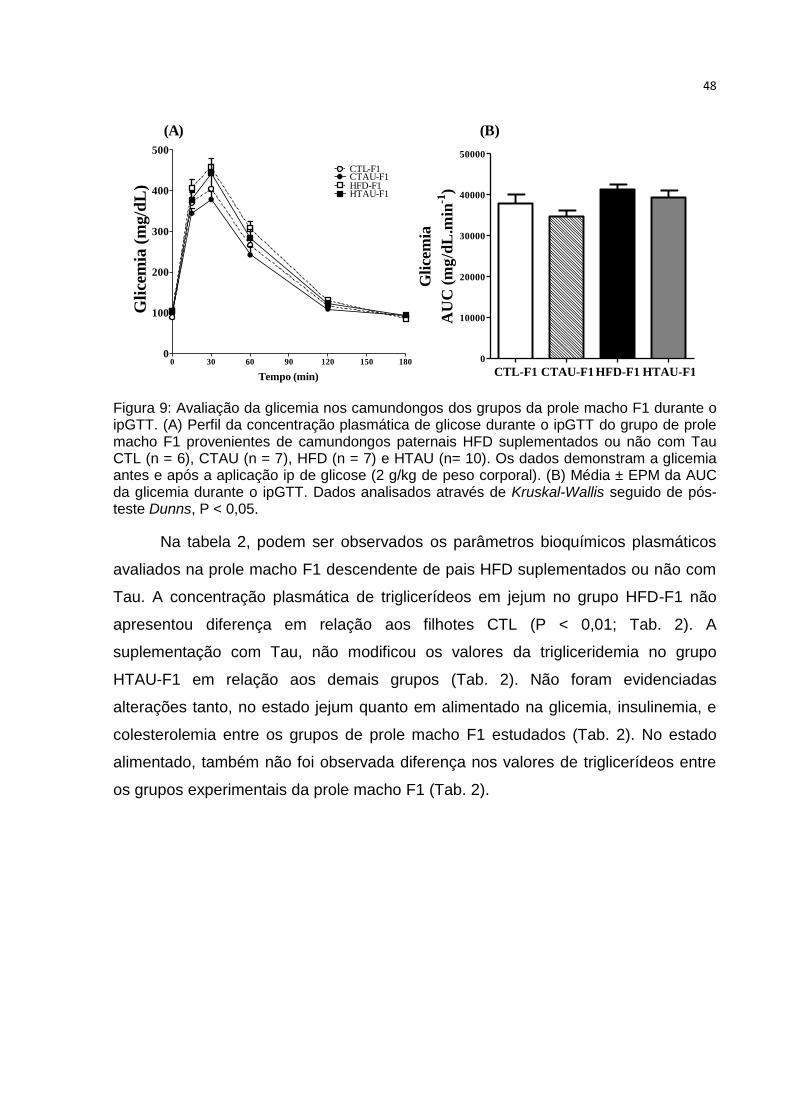
48
(A) (B)
0 30 60 90 120 150 1800
100
200
300
400
500
CTL-F1CTAU-F1HFD-F1HTAU-F1
Tempo (min)
Gli
cem
ia (
mg/d
L)
CTL-F1 CTAU-F1 HFD-F1 HTAU-F10
10000
20000
30000
40000
50000
Gli
cem
ia
AU
C (
mg
/dL
.min
-1)
Figura 9: Avaliação da glicemia nos camundongos dos grupos da prole macho F1 durante o ipGTT. (A) Perfil da concentração plasmática de glicose durante o ipGTT do grupo de prole macho F1 provenientes de camundongos paternais HFD suplementados ou não com Tau CTL (n = 6), CTAU (n = 7), HFD (n = 7) e HTAU (n= 10). Os dados demonstram a glicemia antes e após a aplicação ip de glicose (2 g/kg de peso corporal). (B) Média ± EPM da AUC da glicemia durante o ipGTT. Dados analisados através de Kruskal-Wallis seguido de pós-teste Dunns, P < 0,05. Figura 8: Avaliação da glicemia nos camundongos dos grupos da prole macho durante o ipGTT.
Na tabela 2, podem ser observados os parâmetros bioquímicos plasmáticos
avaliados na prole macho F1 descendente de pais HFD suplementados ou não com
Tau. A concentração plasmática de triglicerídeos em jejum no grupo HFD-F1 não
apresentou diferença em relação aos filhotes CTL (P < 0,01; Tab. 2). A
suplementação com Tau, não modificou os valores da trigliceridemia no grupo
HTAU-F1 em relação aos demais grupos (Tab. 2). Não foram evidenciadas
alterações tanto, no estado jejum quanto em alimentado na glicemia, insulinemia, e
colesterolemia entre os grupos de prole macho F1 estudados (Tab. 2). No estado
alimentado, também não foi observada diferença nos valores de triglicerídeos entre
os grupos experimentais da prole macho F1 (Tab. 2).

49
Tabela 2: Parâmetros bioquímicos plasmáticos no estado de jejum e alimentado avaliados na prole macho F1 descendentes de camundongos paternais HFD suplementados ou não com Tau.
Dados representam a média EPM [CTL (n = 9), CTAU (n = 9), HFD (n = 11), HTAU (n = 15)]. Letras diferentes indicam diferenças estatísticas. Dados analisados através de ANOVA de uma via seguida de Newman-Keuls, P < 0,05. Tabela 2: Parâmetros bioquímicos plasmáticos no estado de jejum e alimentado avaliados aos 90 dias de vida da prole macho descendentes de camundongos paternais HFD suplementados ou não com Tau.
Os camundongos da prole fêmea F1 com 90 dias de vida foram submetidos
ao ipGTT. Após a injeção de glicose todos os grupos da prole fêmea F1
apresentaram aumento da glicemia, atingindo os maiores valores aos 30 minutos e
retornando aos valores basais aos 120 minutos após administração do açúcar (Fig.
10A). O total da glicemia, expresso pela AUC durante o teste, não diferiu entre os
grupos de prole fêmea F1 provenientes de pais HFD suplementados ou não com
Tau (Fig. 10A e 10B).
CTL-F1 CTAU-F1 HFD-F1 HTAU-F1
Glicemia (mg/dL)
Jejum 106 ± 8 112 ± 8 111 ± 6 118 ± 6
Alimentado 156 ± 7 135 ± 4 151 ± 6 141 ± 5
Insulinemia (ng/mL)
Jejum 0,2 ± 0,03 0,2 ± 0,07 0,2 ± 0,06 0,2 ± 0,1
Alimentado 0,2 ± 0,07 0,5 ± 0,1 0,5 ± 0,1 0,4 ± 0,1
Triglicerídeos (mg/dL)
Jejum 83 ± 5a,b 67 ± 5a 90 ± 5b 77 ± 3a,b
Alimentado 84 ± 7 93 ± 5 77 ± 5 91 ± 7
Colesterol (mg/dL)
Jejum 78 ± 7 67 ± 9 79 ± 7 74 ± 7
Alimentado 88 ± 4 87 ± 10 95 ± 4 89 ± 8
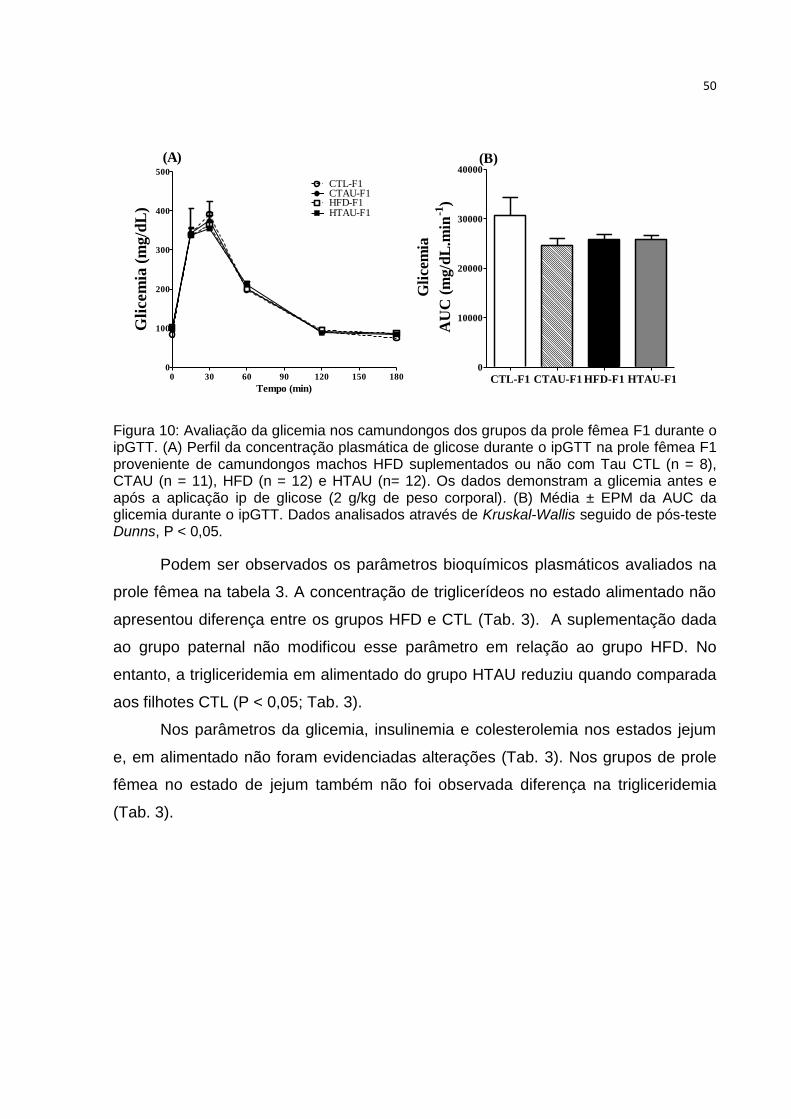
50
0 30 60 90 120 150 1800
100
200
300
400
500
CTL-F1CTAU-F1HFD-F1HTAU-F1
Tempo (min)
Gli
cem
ia (
mg/d
L)
CTL-F1 CTAU-F1 HFD-F1 HTAU-F10
10000
20000
30000
40000
Gli
cem
ia
AU
C (
mg/d
L.m
in-1
)
(A) (B)
Figura 10: Avaliação da glicemia nos camundongos dos grupos da prole fêmea F1 durante o ipGTT. (A) Perfil da concentração plasmática de glicose durante o ipGTT na prole fêmea F1 proveniente de camundongos machos HFD suplementados ou não com Tau CTL (n = 8), CTAU (n = 11), HFD (n = 12) e HTAU (n= 12). Os dados demonstram a glicemia antes e após a aplicação ip de glicose (2 g/kg de peso corporal). (B) Média ± EPM da AUC da glicemia durante o ipGTT. Dados analisados através de Kruskal-Wallis seguido de pós-teste Dunns, P < 0,05. Figura 9: Avaliação da glicemia nos camundongos dos grupos da prole fêmea durante o ipGTT.
Podem ser observados os parâmetros bioquímicos plasmáticos avaliados na
prole fêmea na tabela 3. A concentração de triglicerídeos no estado alimentado não
apresentou diferença entre os grupos HFD e CTL (Tab. 3). A suplementação dada
ao grupo paternal não modificou esse parâmetro em relação ao grupo HFD. No
entanto, a trigliceridemia em alimentado do grupo HTAU reduziu quando comparada
aos filhotes CTL (P < 0,05; Tab. 3).
Nos parâmetros da glicemia, insulinemia e colesterolemia nos estados jejum
e, em alimentado não foram evidenciadas alterações (Tab. 3). Nos grupos de prole
fêmea no estado de jejum também não foi observada diferença na trigliceridemia
(Tab. 3).
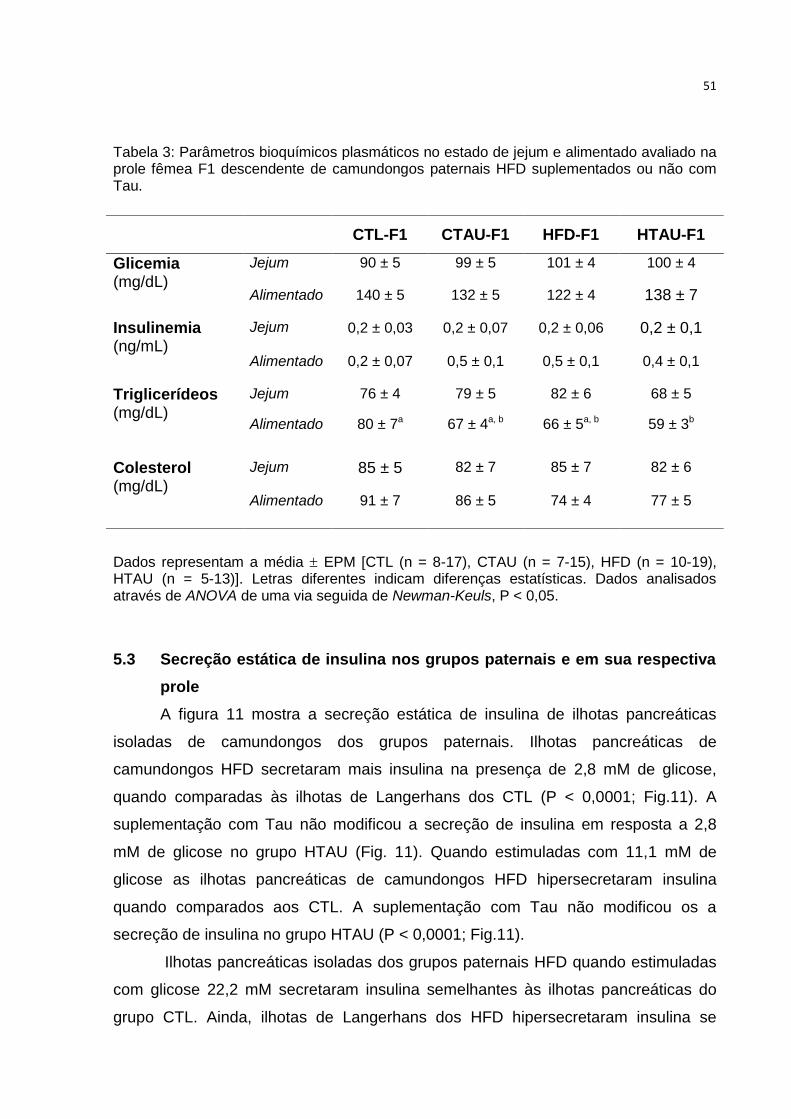
51
Tabela 3: Parâmetros bioquímicos plasmáticos no estado de jejum e alimentado avaliado na prole fêmea F1 descendente de camundongos paternais HFD suplementados ou não com Tau.
Dados representam a média EPM [CTL (n = 8-17), CTAU (n = 7-15), HFD (n = 10-19), HTAU (n = 5-13)]. Letras diferentes indicam diferenças estatísticas. Dados analisados através de ANOVA de uma via seguida de Newman-Keuls, P < 0,05. Tabela 3: Parâmetros bioquímicos plasmáticos no estado de jejum e alimentado avaliado aos 90 dias de vida da prole fêmea descendente de camundongos paternais HFD suplementados ou não com Tau.
5.3 Secreção estática de insulina nos grupos paternais e em sua respectiva
prole
A figura 11 mostra a secreção estática de insulina de ilhotas pancreáticas
isoladas de camundongos dos grupos paternais. Ilhotas pancreáticas de
camundongos HFD secretaram mais insulina na presença de 2,8 mM de glicose,
quando comparadas às ilhotas de Langerhans dos CTL (P < 0,0001; Fig.11). A
suplementação com Tau não modificou a secreção de insulina em resposta a 2,8
mM de glicose no grupo HTAU (Fig. 11). Quando estimuladas com 11,1 mM de
glicose as ilhotas pancreáticas de camundongos HFD hipersecretaram insulina
quando comparados aos CTL. A suplementação com Tau não modificou os a
secreção de insulina no grupo HTAU (P < 0,0001; Fig.11).
Ilhotas pancreáticas isoladas dos grupos paternais HFD quando estimuladas
com glicose 22,2 mM secretaram insulina semelhantes às ilhotas pancreáticas do
grupo CTL. Ainda, ilhotas de Langerhans dos HFD hipersecretaram insulina se
CTL-F1 CTAU-F1 HFD-F1 HTAU-F1
Glicemia (mg/dL)
Jejum 90 ± 5 99 ± 5 101 ± 4 100 ± 4
Alimentado 140 ± 5 132 ± 5 122 ± 4 138 ± 7
Insulinemia (ng/mL)
Jejum 0,2 ± 0,03 0,2 ± 0,07 0,2 ± 0,06 0,2 ± 0,1
Alimentado 0,2 ± 0,07 0,5 ± 0,1 0,5 ± 0,1 0,4 ± 0,1
Triglicerídeos (mg/dL)
Jejum 76 ± 4 79 ± 5 82 ± 6 68 ± 5
Alimentado 80 ± 7a 67 ± 4a, b 66 ± 5a, b 59 ± 3b
Colesterol (mg/dL)
Jejum 85 ± 5 82 ± 7 85 ± 7 82 ± 6
Alimentado 91 ± 7 86 ± 5 74 ± 4 77 ± 5
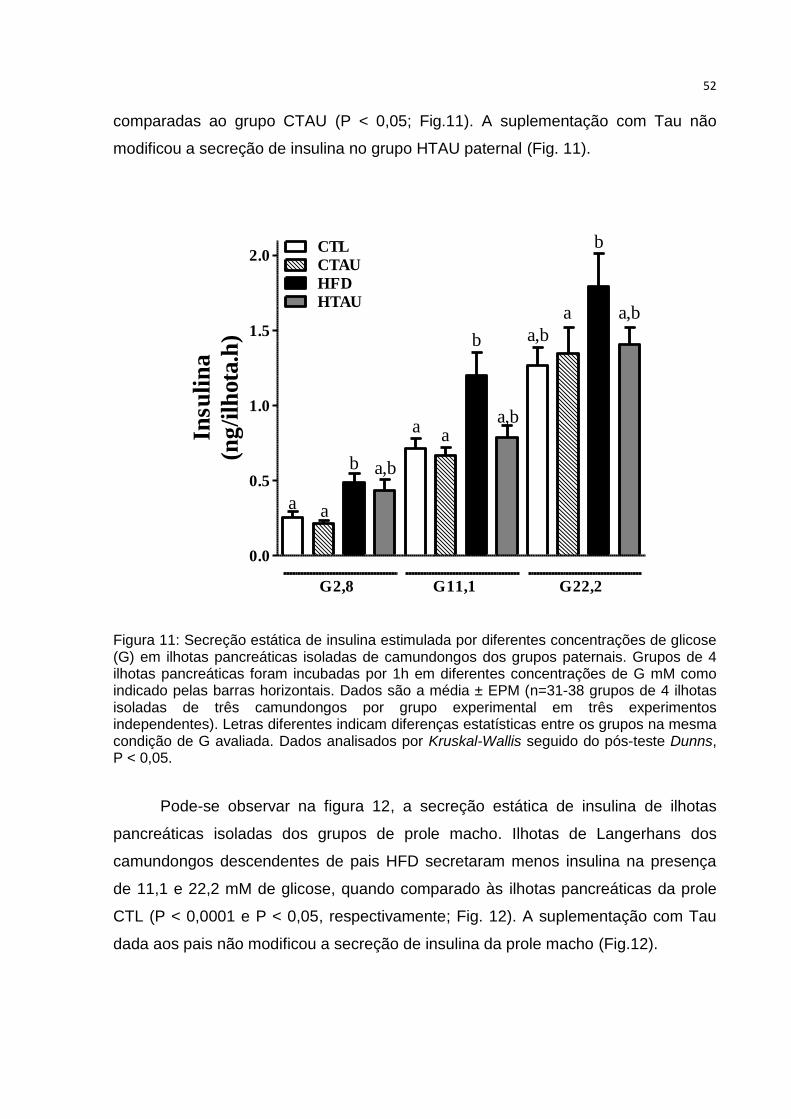
52
comparadas ao grupo CTAU (P < 0,05; Fig.11). A suplementação com Tau não
modificou a secreção de insulina no grupo HTAU paternal (Fig. 11).
0.0
0.5
1.0
1.5
2.0CTL
CTAU
HFD
HTAU
G2,8 G11,1 G22,2
a a
b a,b
a,b
a
b
a,b
aa
b
a,b
Insu
lin
a
(ng
/ilh
ota
.h)
Figura 11: Secreção estática de insulina estimulada por diferentes concentrações de glicose (G) em ilhotas pancreáticas isoladas de camundongos dos grupos paternais. Grupos de 4 ilhotas pancreáticas foram incubadas por 1h em diferentes concentrações de G mM como indicado pelas barras horizontais. Dados são a média ± EPM (n=31-38 grupos de 4 ilhotas isoladas de três camundongos por grupo experimental em três experimentos independentes). Letras diferentes indicam diferenças estatísticas entre os grupos na mesma condição de G avaliada. Dados analisados por Kruskal-Wallis seguido do pós-teste Dunns, P < 0,05. Figura 10: Secreção estática de insulina estimulada por diferentes concentrações de glicose (G) em ilhotas pancreáticas isoladas de camundongos dos grupos paternais.
Pode-se observar na figura 12, a secreção estática de insulina de ilhotas
pancreáticas isoladas dos grupos de prole macho. Ilhotas de Langerhans dos
camundongos descendentes de pais HFD secretaram menos insulina na presença
de 11,1 e 22,2 mM de glicose, quando comparado às ilhotas pancreáticas da prole
CTL (P < 0,0001 e P < 0,05, respectivamente; Fig. 12). A suplementação com Tau
dada aos pais não modificou a secreção de insulina da prole macho (Fig.12).
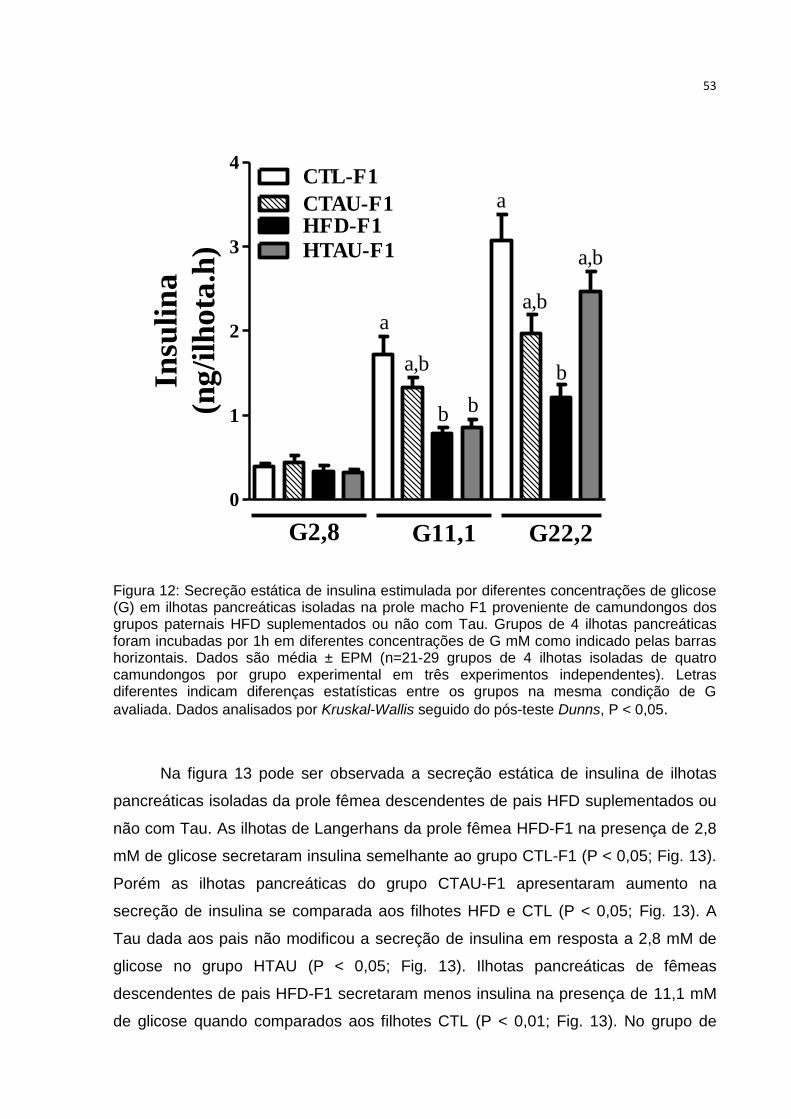
53
0
1
2
3
4CTL-F1
CTAU-F1
HTAU-F1
HFD-F1
G2,8 G11,1 G22,2
a
a,b
b b
a
a,b
b
a,b
Insu
lin
a
(ng
/ilh
ota
.h)
Figura 12: Secreção estática de insulina estimulada por diferentes concentrações de glicose (G) em ilhotas pancreáticas isoladas na prole macho F1 proveniente de camundongos dos grupos paternais HFD suplementados ou não com Tau. Grupos de 4 ilhotas pancreáticas foram incubadas por 1h em diferentes concentrações de G mM como indicado pelas barras horizontais. Dados são média ± EPM (n=21-29 grupos de 4 ilhotas isoladas de quatro camundongos por grupo experimental em três experimentos independentes). Letras diferentes indicam diferenças estatísticas entre os grupos na mesma condição de G
avaliada. Dados analisados por Kruskal-Wallis seguido do pós-teste Dunns, P < 0,05. Figura 11: Secreção estática de insulina estimulada por diferentes concentrações de glicose (G) em ilhotas pancreáticas isoladas na prole macho proveniente de camundongos dos grupos paternais HFD suplementados ou não com Tau.
Na figura 13 pode ser observada a secreção estática de insulina de ilhotas
pancreáticas isoladas da prole fêmea descendentes de pais HFD suplementados ou
não com Tau. As ilhotas de Langerhans da prole fêmea HFD-F1 na presença de 2,8
mM de glicose secretaram insulina semelhante ao grupo CTL-F1 (P < 0,05; Fig. 13).
Porém as ilhotas pancreáticas do grupo CTAU-F1 apresentaram aumento na
secreção de insulina se comparada aos filhotes HFD e CTL (P < 0,05; Fig. 13). A
Tau dada aos pais não modificou a secreção de insulina em resposta a 2,8 mM de
glicose no grupo HTAU (P < 0,05; Fig. 13). Ilhotas pancreáticas de fêmeas
descendentes de pais HFD-F1 secretaram menos insulina na presença de 11,1 mM
de glicose quando comparados aos filhotes CTL (P < 0,01; Fig. 13). No grupo de
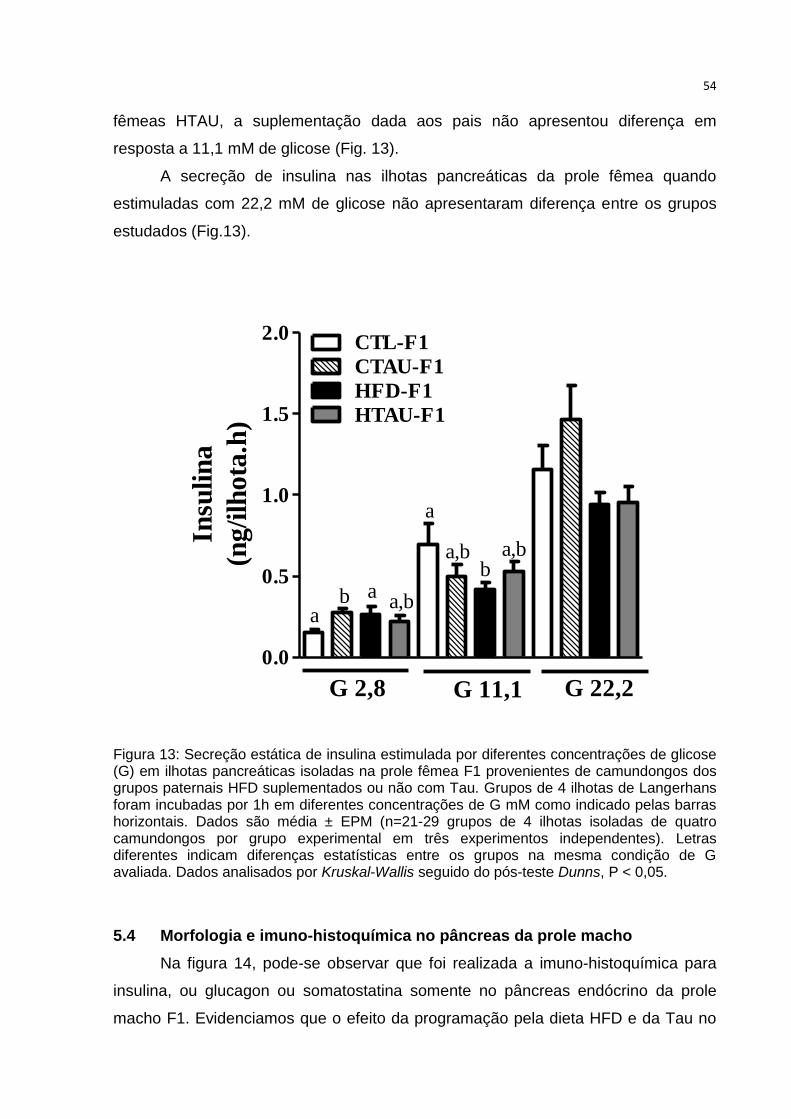
54
fêmeas HTAU, a suplementação dada aos pais não apresentou diferença em
resposta a 11,1 mM de glicose (Fig. 13).
A secreção de insulina nas ilhotas pancreáticas da prole fêmea quando
estimuladas com 22,2 mM de glicose não apresentaram diferença entre os grupos
estudados (Fig.13).
0.0
0.5
1.0
1.5
2.0 CTL-F1
CTAU-F1
HFD-F1
HTAU-F1
G 2,8 G 11,1 G 22,2
a
a,bb
a,b
ab a
a,b
Insu
lin
a
(ng/i
lhota
.h)
Figura 13: Secreção estática de insulina estimulada por diferentes concentrações de glicose (G) em ilhotas pancreáticas isoladas na prole fêmea F1 provenientes de camundongos dos grupos paternais HFD suplementados ou não com Tau. Grupos de 4 ilhotas de Langerhans foram incubadas por 1h em diferentes concentrações de G mM como indicado pelas barras horizontais. Dados são média ± EPM (n=21-29 grupos de 4 ilhotas isoladas de quatro camundongos por grupo experimental em três experimentos independentes). Letras diferentes indicam diferenças estatísticas entre os grupos na mesma condição de G avaliada. Dados analisados por Kruskal-Wallis seguido do pós-teste Dunns, P < 0,05. Figura 12: Secreção estática de insulina estimulada por diferentes concentrações de glicose (G) em ilhotas pancreáticas isoladas na prole fêmea provenientes de camundongos dos grupos paternais HFD suplementados ou não com Tau.
5.4 Morfologia e imuno-histoquímica no pâncreas da prole macho
Na figura 14, pode-se observar que foi realizada a imuno-histoquímica para
insulina, ou glucagon ou somatostatina somente no pâncreas endócrino da prole
macho F1. Evidenciamos que o efeito da programação pela dieta HFD e da Tau no

55
pai foi similar sobre a secreção de insulina de ambas as proles então prosseguimos
avaliando o pâncreas da prole macho F1.
A análise morfométrica mostrou que o número de células β por ilhota foi
similar entre os diferentes pâncreas coletados dos grupos de prole macho estudados
(P < 0,01, Fig. 15A). Contudo, foi observado que o grupo de prole macho HFD-F1
apresentou redução do número de células α e δ por ilhota quando comparado à
prole CTL-F1 (P < 0,0001; Fig. 15B e 15C). A suplementação dada aos pais HTAU
não modificou o número de células β entre os grupos estudados de filhotes machos
(Fig. 15A). Porém, a suplementação reduziu o número de células α no grupo HTAU,
quando comparado ao CTL. Ainda, a Tau preveniu a redução do número de células
δ no pâncreas dos filhotes machos HTAU-F1 quando comparado ao grupo de prole
HFD (P < 0,0001; Fig. 15C). No entanto, os valores do número de células δ dos
filhotes HTAU-F1 foram similares aos do grupo de prole CTL-F1 (Fig. 15C).
Com relação ao tamanho das células endócrinas pancreáticas, o grupo HFD
apresentou células β menores em relação às CTL-F1 (P < 0,0001; Fig. 15D).
Contudo, o tamanho das células α e δ no grupo HFD não diferiram quando
comparadas ao dos filhotes CTL (Fig. 15E e 15F). A suplementação com Tau dada
ao grupo paternal impediu a redução do tamanho das células β na prole HTAU
quando comparada com os filhotes HFD (P < 0,0001; Fig. 15D), sendo similar ao
tamanho das células CTL (Fig. 15D). A Tau dada aos pais aumentou o tamanho das
células α em relação ao grupo de prole macho CTL-F1 (P < 0,01; Fig. 15E). Ainda, a
Tau dada aos pais reduziu o tamanho das células δ quando comparado aos filhotes
HFD (P < 0,01; Fig. 15F).
Na tabela 4, pode-se observar que as alterações celulares relatadas acima
não foram acompanhadas por modificações no peso do pâncreas, no número de
ilhotas pancreáticas por secção e o número de ilhotas pancreáticas analisadas entre
os grupos estudados (Tab. 4). No entanto, não foram observadas alterações na área
absoluta e massa das ilhotas pancreáticas, ou área de células β, ou α ou δ nos
grupos de filhotes machos estudados (Tab. 4).

56
Figura 14: Imuno-histoquímica para insulina, ou glucagon ou somatostatina no pâncreas coletado da prole macho F1 proveniente de pais HFD suplementados ou não suplementados com Tau. Insulina (A, D, G e J), glucagon (B, E, H e K) e somatostatina (C, F, I e L). (Barra de escala 100 μm). Figura 13: Imuno-histoquimica para insulina, ou glucagon ou somatostatina no pâncreas coletado da prole macho proveniente de pais HFD suplementados ou não com Tau.
INSULINA GLUCAGON SOMATOSTATINA
(A) (B) (C)
(F)
(I)
(L) (K)
(E)
(H) (G)
(D)
(J)

57
CTL-F1 CTAU-F1 HFD-F1 HTAU-F10
10
20
30
Nú
mero
de c
élu
las
CTL-F1 CTAU-F1 HFD-F1 HTAU-F10
5
10
15
20
25
a
b,c
c
a,b
Nú
mero
de c
élu
las
CTL-F1 CTAU-F1 HFD-F1 HTAU-F10
5
10
15
20
25
a
b b
a
Nú
mero
de c
élu
las
CTL-F1 CTAU-F1 HFD-F1 HTAU-F10
100
200
300
a
bb a
Áre
a d
e cé
lula
(
m2
)/n
º d
e cé
lula
CTL-F1 CTAU-F1 HFD-F1 HTAU-F10
100
200
300
400
500
a a,b
a,b
b
Áre
a d
e cé
lula
(
m2)/
nº
de
célu
la
CTL-F1 CTAU-F1 HFD-F1 HTAU-F10
50
100
150
200
250
a,b
a
b
a
Áre
a d
e cé
lula
(
m2)/
nº
de
célu
la
(A) (B) (C)
(F)(E)(D)
Figura 15: Quantificação dos principais tipos celulares das ilhotas pancreáticas da prole macho F1 provenientes de pais HFD suplementados ou não com Tau. Média ± EPM (n =3-4 pâncreas) (A e D) do número de células β e a razão da área de células β pelo número de células β, (B e E) do número de células α e razão da área de células α pelo número de células α, (C e F) do número de células δ e a razão da área de células δ pelo número de células δ. Dados analisados através de Kruskal-Wallis seguido do pós-teste Dunns, P <
0,05. Figura 14: Quantificação dos principais tipos celulares das ilhotas pancreáticas da prole macho proveniente de pais HFD suplementados ou não suplementados com Tau.
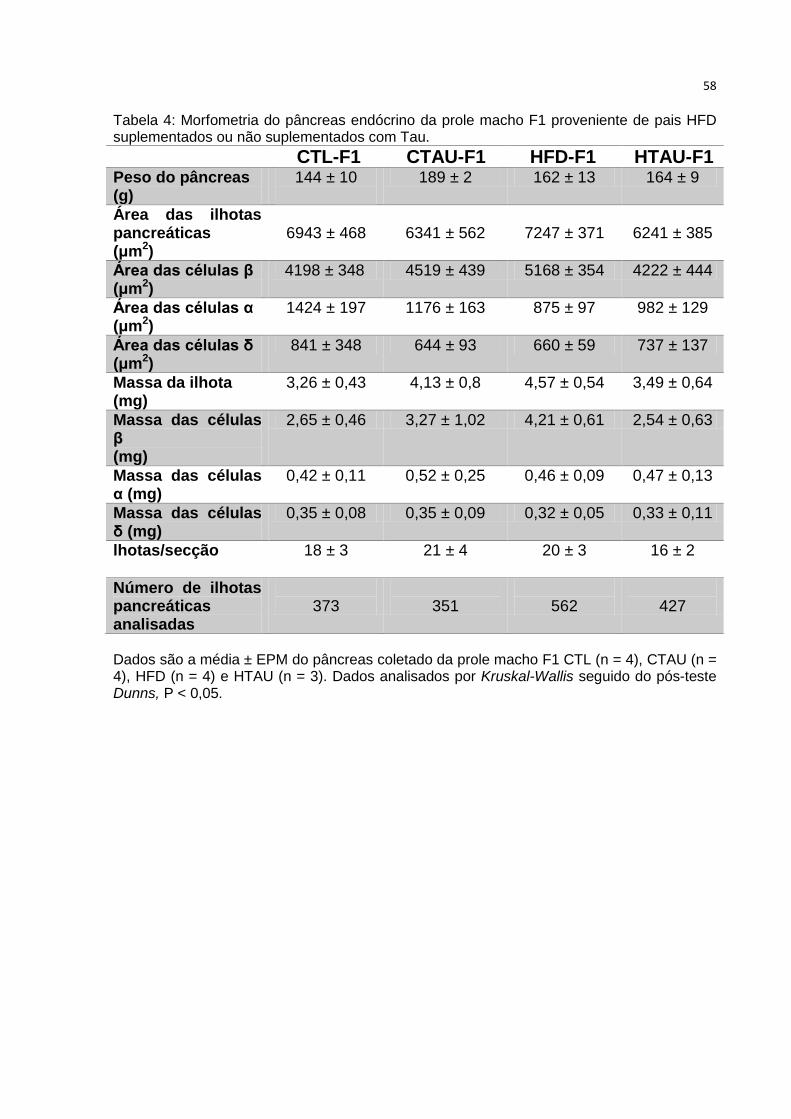
58
Tabela 4: Morfometria do pâncreas endócrino da prole macho F1 proveniente de pais HFD suplementados ou não suplementados com Tau.
CTL-F1 CTAU-F1 HFD-F1 HTAU-F1 Peso do pâncreas (g)
144 ± 10
189 ± 2 162 ± 13 164 ± 9
Área das ilhotas pancreáticas (μm2)
6943 ± 468
6341 ± 562
7247 ± 371
6241 ± 385
Área das células β (μm2)
4198 ± 348 4519 ± 439 5168 ± 354 4222 ± 444
Área das células α (μm2)
1424 ± 197 1176 ± 163 875 ± 97 982 ± 129
Área das células δ (μm2)
841 ± 348 644 ± 93 660 ± 59 737 ± 137
Massa da ilhota (mg)
3,26 ± 0,43 4,13 ± 0,8 4,57 ± 0,54 3,49 ± 0,64
Massa das células β (mg)
2,65 ± 0,46 3,27 ± 1,02 4,21 ± 0,61 2,54 ± 0,63
Massa das células α (mg)
0,42 ± 0,11 0,52 ± 0,25 0,46 ± 0,09 0,47 ± 0,13
Massa das células δ (mg)
0,35 ± 0,08 0,35 ± 0,09 0,32 ± 0,05 0,33 ± 0,11
lhotas/secção
18 ± 3 21 ± 4 20 ± 3 16 ± 2
Número de ilhotas pancreáticas analisadas
373
351
562
427
Dados são a média ± EPM do pâncreas coletado da prole macho F1 CTL (n = 4), CTAU (n = 4), HFD (n = 4) e HTAU (n = 3). Dados analisados por Kruskal-Wallis seguido do pós-teste Dunns, P < 0,05. Tabela 4: Morfometria do pâncreas endócrino da prole macho proveniente de pais HFD suplementados ou não suplementados com Tau.

59
6. DISCUSSÃO
Nesse estudo, observamos que o consumo de dieta HFD pelos pais foi eficaz
em induzir a obesidade, prejuízos na tolerância à glicose e hipersecreção de
insulina.
Pela primeira vez evidenciamos que a obesidade dos pais apesar de não
interferir na adiposidade e na tolerância a glicose em sua prole, promoveu prejuízos
na secreção de insulina. A prole fêmea e macho proveniente de pais HFD secretou
menos insulina em resposta a diferentes estímulos de glicose. Ainda, foi verificado
que os filhotes machos HFD apresentaram redução no número das células α e δ no
pâncreas, sem alterações no número de células β pancreáticas. Porém, os filhotes
machos HFD tiveram redução do tamanho das células β, mas essa alteração não foi
acompanhada de modificações no tamanho das células α e δ.
A suplementação com Tau não impediu a obesidade, a intolerância à glicose,
porém reduziu parcialmente a hipersecreção de insulina no grupo paternal HTAU.
Na prole proveniente de pais que consumiram dieta HFD e Tau, evidenciamos que a
Tau melhorou parcialmente a secreção de insulina, além disso, o número de células
δ e o tamanho das células β no pâncreas endócrino dos filhotes machos HTAU
foram similares ao da prole CTL. Ainda, o tamanho das células α na prole macho
HTAU foi maior. Sugerindo possível ação preventiva da Tau contra alterações
morfofuncionais no pâncreas endócrino que podem ser propagadas dos pais para
sua descendência.
Para melhor abordagem, os resultados dos grupos paternais e de suas proles
serão discutidos em tópicos.
6.1 Ações da suplementação com Tau no desenvolvimento da obesidade
nos grupos paternais e na sua respectiva prole
A dieta HFD tem sido amplamente utilizada na literatura para induzir a
obesidade em roedores, pois o consumo desse tipo de dieta retrata o atual cenário
mundial em que o excesso de lipídios presente na alimentação vem sendo
considerado um dos principais fatores que contribuem para a expansão da
obesidade na população (BRINGHENTI et al., 2016). Dessa forma a utilização da
dieta HFD pode contribuir para desvendar os mecanismos envolvidos na
fisiopatologia dessa doença e na propagação de prejuízos metabólicos para próxima

60
geração. Em nosso estudo, os camundongos machos C57Bl/6 foram alimentados
com dieta HFD por 16 semanas. A partir da nona semana de ingestão de dieta HFD,
os camundongos do grupo paternal começaram a apresentar aumento do peso
corporal e isso permaneceu até o final do tratamento. Ainda, de acordo com estudos
prévios (AHREN e PACINI, 2002; DE SOUZA et al., 2007; TSUBOYAMA-KASAOKA
et al., 2006; WINZELL et al., 2007; ARAUJO et al., 2007), a dieta HFD aumentou o
peso corporal final, o índice de Lee e os estoques de gordura. Esse tempo de
consumo da dieta HFD, foi usado em nosso estudo, por ter sido previamente
demonstrado pela literatura que além de induzir obesidade é tempo suficiente para
serem desencadeados prejuízos como a intolerância à glicose e a hiperfunção do
pâncreas endócrino (BOSE et al., 2008).
Apesar dos camundongos desenvolverem obesidade, nossos dados não
demonstraram modificações no consumo de ração e eficiência alimentar pela dieta
HFD. De acordo com SANTOS e VIANA (2007) camundongos HFD consomem
menor quantidade de ração devido ao maior aporte energético da dieta. Contudo,
como a ingestão de energia cronicamente excede o gasto energético, o resultado é
ganho de peso corporal e, consequentemente, obesidade (SANTOS e VIANA,
2007). Porém este dado ainda é discutido, DUARTE e colaboradores (2006),
mostraram que camundongos alimentados com dieta HFD apresentaram consumo
energético maior, mas sem modificação no consumo diário em gramas. Apesar dos
relatos contraditórios, tanto nos trabalhos que fazem correlação da dieta HFD com a
hiperfagia quanto nos que não correlacionam com essa condição, o consumo de
dieta HFD pode promover o aumento da adiposidade e apresentar maior propensão
de danos na tolerância à glicose (VELLOSO, 2009; ORNELLAS et al., 2015). Um
dos mecanismos pelos quais a dieta HFD contribui para esse prejuízo seria pelo
maior aporte de ácidos graxos saturados que podem mediar ações inflamatórias por
interagirem com os receptores do tipo Toll (TLR). Foi demonstrado que a interação
desses lipídeos com receptores TLR nas micróglias no hipotálamo estimula a
produção de citocinas pró-inflamatórias que promovem a destruição dos neurônios
responsáveis pelo controle do apetite e da termogênese (MILANSKI et al., 2009).
Os camundongos do grupo paternal HFD apresentaram redução do consumo
hídrico que pode estar relacionada ao aumento do consumo de lipídeos. Nesses
roedores a maior ativação da via lipogênica, pode contribuir para a produção pelo
metabolismo de água. A lipogênese ocorre principalmente quando a razão ATP/ADP

61
é alta, ou seja, o aumento de substratos para formação de ATP resulta em excesso
de acetil-CoA. Este é o precursor da biossíntese de ácido graxo, e a enzima acetil-
CoA carboxilase é uma das principais responsáveis por essa síntese, e age nesse
processo transferindo o grupo carboxila do bicarbonato para o acetil-CoA formando
assim, o malonil-CoA. A via lipogênica é composta por quatro etapas, na primeira
acontece à condensação do grupo malonil e acetil por ação da β-cetoacil-ACP
sintase formando acetoacetil-CoA e liberando CO2; em sequência há redução dos
grupos carbonila, onde o doador de elétron para acetoacetil-CoA é o NADPH,
formando assim o β-hidrobutiril-ACP; na terceira etapa ocorre a desidratação do β-
hidrobutiril-ACP pela β-hidroxiacil-ACP desidratase desencadeando a liberação de
água e formação de uma dupla ligação entre os carbonos (KHAN et al., 2016).
Ainda, na via lipogênica a última etapa tem a redução da dupla ligação, tendo
novamente o NADPH que é utilizado como doador de elétrons, o qual serve para
romper a dupla ligação da cadeia de ácidos graxos recém-formada para formar o acil
graxo. A cadeia acil-graxo cresce pela adição de grupos de dois átomos de carbono
doados pelo malonato e com perda de CO2. Sete ciclos de condensação e redução
produzem o grupo palmitoil saturado com 16 carbonos, liberando sete moléculas de
água (KHAN et al., 2016). Portanto, sugere-se que a ingestão de dieta HFD por
aumentar a lipogênese (IMAI et al., 2016), também promova maior produção
endógena de água reduzindo o consumo hídrico.
A suplementação com Tau em modelos experimentais de obesidade já
mostrou reduzir a síntese de lipídeos por diminuir a expressão da ácido graxo
sintase e acetil-CoA carboxilase no fígado (FUKUDA et al., 2011). A Tau demonstrou
também aumentar a expressão da carnitina palmitoil-transferase (CPT)-1α no fígado,
enzima envolvida com a primeira etapa do transporte dos ácidos graxos para a β-
oxidação (RUTHERFORD et al., 2010; CHANG et al., 2011; GENTILE et al., 2011;
VAVROVA et al., 2016). Segundo TSUBOYAMA-KASAOKA e colaboradores (2006),
em camundongos que não expressam leptina (ob/ob) ou que foram alimentados com
dieta HFD, à obesidade está associada à redução da concentração plasmática de
Tau. Em humanos com sobrepeso, ZHANG e colaboradores (2004) observaram que
a suplementação com o aminoácido reduz o peso corporal. Contudo, apesar dessas
diferentes observações da Tau contra a adiposidade, em nosso estudo, o
aminoácido não preveniu a obesidade no grupo paternal HTAU. Esses achados são
similares aos evidenciados em camundongos fêmeas Swiss alimentadas com HFD e

62
suplementadas com Tau que apresentaram melhora da homeostase da glicose sem
prevenção da obesidade (RIBEIRO et al., 2012).
Atualmente, sabe-se que a obesidade tanto maternal quanto paternal tem
forte influência sobre o desenvolvimento da obesidade na sua descendência. A
literatura tem muitos dados sobre os efeitos da obesidade materna e a programação
da adiposidade e prejuízos em sua prole. Porém os conhecimentos acerca da
influência da obesidade paterna sobre sua descendência ainda são escassos e
muitas vezes controversos. Foi demonstrado que a prole de primeira geração de
camundongos machos obesos por ingestão de dieta HFD apresentou elevado peso
corporal ao nascer (HIRAMATSU et al., 2016). Porém, já foi relatado que na idade
adulta, a prole de pais HFD não apresentou aumento do peso corporal
(HIRAMATSU et al., 2016). Ratas descendentes de roedores machos alimentados
com dieta HFD também não apresentaram alterações na adiposidade (NG et al.,
2010). Nossos resultados vão de encontro com esses últimos relatos, visto que a
prole macho e fêmea de pais HFD não apresentaram alteração no peso corporal ao
nascer e na adiposidade na vida adulta.
Os dados acerca da contribuição da obesidade paternal sobre as
modificações no padrão de ingestão alimentar e hídrica da prole também são
escassos. AKYOL e colaborados (2011) não evidenciaram diferença na ingestão de
ração e consumo de água na prole de ratos machos alimentados com dieta HFD.
NG e colaboradores (2010) também observaram que a ingestão paterna de dieta
HFD não alterou a eficiência energética na prole fêmea. O que é diferente do
observado na programação materna, na qual é frequente o aumento do consumo
energético em filhotes provenientes de fêmeas alimentadas com dieta HFD
(SAMUELSSON et al., 2008; NIVOIT et al., 2009). Em nosso estudo, em ambas as
proles não foram observadas diferenças no consumo de ração e eficiência alimentar
nos filhotes provenientes de pais HFD, porém na prole fêmea HFD foi observada
maior ingestão hídrica.
A literatura carece de dados acerca das alterações na ingestão hídrica em
filhotes de pais obesos. Porém, de acordo com achados em prenhas que passaram
por privação nutricional durante a gestação, o organismo do feto se adaptou a esse
ambiente intrauterino adverso. Ocorreu reprogramação na expressão dos genes
através de mecanismos epigenéticos que fazem com que o metabolismo da sua
prole se torne poupador e isso se mantém após o nascimento (CASTRO et al.,

63
2009). Portanto, é possível que tais modificações epigenéticas no metabolismo
possam modificar o consumo de água na prole, contudo, maiores estudos são
necessários para desvendar quais etapas do metabolismo em filhotes de pais
obesos por dieta HFD são modificados e promovem alterações no consumo hídrico.
Também não evidenciamos alterações no peso corporal e na adiposidade dos
filhotes provenientes de pais HTAU. Não há estudos prévios que abordem as ações
da Tau dada aos pais sobre a obesidade na prole. Contudo, existem informações
relatadas sobre os efeitos da suplementação materna com Tau sobre a prole. A
prole proveniente de camundongos CTL fêmeas suplementadas com Tau durante a
gestação apresentou prejuízos como maior mortalidade, hipoglicemia ao
nascimento, maior ganho de peso corporal, aumento dos estoques de gordura e
redução do peso do pâncreas (HULTMAN et al., 2007). Efeitos similares foram
observados na prole de ratas suplementadas com Tau durante a gestação
(BOUJENDAR et al., 2002; LI et., 2013). Portanto, a suplementação com Tau em
modelos experimentais CTL parece ser prejudicial para sua prole.
A suplementação com Tau tem ação direta sobre o hipotálamo. Segundo,
SOLON e colaboradores (2012) a administração intracerebroventricular (icv) de Tau
em ratos reduziu a expressão de peptídeos orexigênicos sem modificar a quantidade
de peptídeos anorexígenos, diminuindo a ingestão de alimentos. Este efeito foi
acompanhado por aumento da expressão da pAkt no hipotálamo. A Tau também
modificou a atividade hipotalâmica por ativar vias como da proteína Janus quinase 2
(JAK2)/STAT3 (Transdutor de sinal e ativador de transcrição 3) e o alvo da
rapamicina em mamíferos (mTOR)/proteína quinase dependente de AMP (AMPK)
(SOLON et al., 2012). Contraditoriamente a esses achados, em nosso estudo a prole
fêmea HTAU e CTAU apresentou aumento do consumo de ração, demonstrando
que alterações dependentes do gênero no consumo alimentar podem programar a
hiperfagia, o que merece mais estudos para compreender os mecanismos
epigenéticos que a Tau estaria modificando nesses roedores.

64
6.2 Ações da suplementação com Tau na homeostase glicêmica e nos
parâmetros bioquímicos plasmáticos dos grupos paternais e de sua
respectiva prole
Estudos demonstraram que o aumento da ingestão de gordura está associado
à redução da sensibilidade a insulina e ao aumento do risco de desenvolver DM
(CURI et al., 2002; KOWALSKI et al., 2017). O consumo de gorduras saturadas em
excesso pode modificar o funcionamento das células β pancreáticas reduzindo a
síntese e a secreção de insulina (LIN e SUN, 2010). O aumento da utilização de
ácidos graxos ao invés de glicose nos tecidos-alvo da insulina seria responsável
pela diminuição da oxidação de carboidratos e do transporte de glicose (WANG et
al., 2015). Isso ocorre porque a elevada β-oxidação com consequente aumento da
produção de acetil-CoA, resulta em inibição da piruvato desidrogenase e oxidação
do piruvato. Ao mesmo tempo, o aumento de citrato e ATP inibem a
fosfofrutoquinase e a glicólise, resultando em acúmulo da glicose-6-fosfato, a qual
inibe a atividade da hexoquinase, com redução no transporte e fosforilação da
glicose. Tais ações podem estar sendo desencadeadas nos camundongos do grupo
paternal HFD e contribuir para o prejuízo na homeostase da glicose nesses
roedores, visto que esse desvio no metabolismo desencadeia a intolerância à
glicose (SMERECZYŃSKI et al., 2016).
De acordo com CHEIK et al. (2006) os mecanismos que relacionam elevada
adiposidade à dislipidemias não estão completamente esclarecidos. Ainda, conforme
o tempo de ingestão da dieta HFD esses parâmetros diferem entre os estudos
(CHEIK et al., 2006; AKAMINE et al., 2010). Em nosso estudo os camundongos
paternais HFD não apresentaram modificações nas concentrações plasmáticas de
glicose, insulina, triglicerídeos e colesterol. ESTADELLA e colaboradores (2004)
também não observaram diferença na glicemia em roedores alimentados com dieta
HFD. Contudo, outros estudos registraram hiperglicemia no estado alimentado em
camundongos HFD (DUARTE et al., 2001; BERNARDES et al., 2004; CHEIK et al.,
2005). Em relação à insulina, os estudos também demonstraram que a
concentrações plasmáticas de insulina podem variar conforme o tempo de ingestão
da dieta HFD (CHEN e NYOMBA., 2003; ESTADELLA et al., 2004; CHEIK et al.,
2006; AKAMINE et al., 2010). Alguns autores registraram em ratos hiperinsulinemia
como efeito agudo da administração de dieta HFD, porém após dois meses desse

65
regime a insulinemia se normalizou (CHEN e NYOMBA., 2003; CHEIK et al., 2006).
Também foi visto hiperinsulinemia em ratas que consumiram a dieta HFD durante
quatro e seis meses (AKAMINE et al., 2010). Porém, a administração da dieta HFD
por período de um ano demonstrou reduzir as concentrações de insulina circulantes
(ESTADELLA et al., 2004).
As dislipidemias são desordens plasmáticas encontradas tanto em modelos
de obesidade experimental, quanto em humanos obesos (NARDELLI et al., 2011;
GÓMEZ-HERNÁNDEZ et al., 2016; CHOI et al., 2017). Porém, camundongos não
são modelos ideais para estudar dislipidemias, pois eles só expressam esse prejuízo
se houver algum tipo de modificação genética devido a elevada distribuição de
transportadores teciduais para ácidos graxos e colesterol, que promove a rápida
internalização desses compostos, bem como a utilização ou armazenamento
(CESARETTI et al., 2006; KIMURA et al., 2017, YANG et al., 2017). Possivelmente
por esse motivo em nosso estudo os camundongos dos grupos paternais HFD não
apresentaram diferença nas concentrações plasmáticas de lipídeos que foram
analisados.
Foi demonstrado que o aminoácido Tau regula a homeostase da glicose por
melhorar a secreção de insulina pelas células β e também aumentar a sensibilidade
periférica à insulina (CARNEIRO et al., 2009). A Tau aumenta as ações da insulina,
provavelmente, por interagir com o receptor de insulina (MATURO e KULAKOWSKI,
1988). Camundongos CTL que receberam injeção ip de 165 µM de Tau
apresentaram aumento da fosforilação do IRβ no fígado e músculo em valores
similares aos observados quando insulina foi administrada (CARNEIRO et al., 2009).
Mas, em nosso estudo a suplementação com Tau nos camundongos machos HTAU
não impediu a intolerância a glicose. De fato, as ações benéficas da Tau nem
sempre são acompanhadas por modificações na tolerância a glicose. NARDELLI e
colaboradores (2011) evidenciaram que ratos com obesidade hipotalâmica e
suplementados com Tau também demonstraram melhora no perfil lipídico
plasmático, porém sem modificação na intolerância à glicose.
A Tau pode aumentar a atividade da colesterol 7α-hidroxilase, enzima
limitante para síntese do ácido biliar, estimulando a conversão de colesterol para
ácido biliar, e essa ação demonstrou reduzir as concentrações plasmáticas e
hepáticas de colesterol em camundongos e ratos (MURAKAMI et al., 2002; CHEN et
al., 2016). Mas em nosso estudo os grupos suplementados com Tau apresentaram

66
aumento da colesterolemia. Esses dados se assemelham aos de SANTOS e
colaboradores (2015), que observaram em fêmeas ovariectomizadas alimentadas
com dieta HFD e suplementadas com Tau aumento nas concentrações plasmáticas
de colesterol, porém o mecanismo envolvido em tal efeito não foi relatado. Talvez a
suplementação prolongada com Tau desencadeie ações prejudiciais no metabolismo
do colesterol, porém é necessário mais estudo para investigar esse mecanismo.
Pouco se sabe sobre o papel da condição metabólica paternal influenciando
nas concentrações plasmáticas de insulina, glicose e triglicerídeos de sua prole (NG
et al., 2010; SOUBRY et al., 2013; FULLSTON et al., 2013). Estudo utilizando a
prole fêmea proveniente de ratos machos Sprague-Dawley que consumiram HFD,
não evidenciou alterações na adiposidade e concentrações plasmáticas de
triglicerídeos, insulina e glicose. SOUBRY, FULLSTON e colaboradores (2013)
também não observaram alterações nesses mesmos parâmetros em filhos de pais
obesos. Porém, ratas descendentes de pais HFD foram intolerantes à glicose devido
à redução na secreção de insulina (NG et al., 2010). Nossos dados se aproximam
dos previamente relatados, pois apesar de não evidenciarmos alterações na
tolerância à glicose, a prole de pais HFD apresentou redução da secreção de
insulina sem modificação nos parâmetros bioquímicos plasmáticos.
Em nosso trabalho também não evidenciamos alterações na tolerância à
glicose e nos parâmetros bioquímicos plasmáticos estudados dos grupos de proles
provenientes de pais suplementados com Tau. Porém, são escassos na literatura
estudos sobre os efeitos da suplementação com Tau dada aos pais sobre a
tolerância a glicose, e nas concentrações de insulina, glicose, colesterol e
triglicerídeos de sua prole. Contudo, foi observado que fetos provenientes de ratas
prenhas submetidas à restrição proteica, mas suplementadas com Tau tiveram
normalização da secreção de insulina e da proliferação das células β pancreáticas
(BOUJENDAR et al., 2002; KALBE et al., 2005; REUSENS et al., 2008). Esses
dados são indícios que o aminoácido Tau pode regular mecanismos epigenéticos
que previnem as ações deletérias propagadas dos pais para a sua prole.

67
6.3 Ações da suplementação com Tau na morfofunção pancreática
endócrina tanto no grupo paternal quanto na sua prole
Os prejuízos na ação da insulina estão associados ao acúmulo de lipídeos, à
inflamação tecidual, ativação de vias como do estresse de retículo endoplasmático,
entre outros, que comprometem a sensibilidade à insulina nos tecidos-alvo
(SAMUEL e SHULMAN, 2012). Em paralelo a resistência à insulina, alterações
morfológicas e funcionais acontecem nas ilhotas pancreáticas, resultando em
aumento na massa de células β e na secreção de insulina, o que ocasiona a
hiperinsulinemia (SEINO, SHIBASAKI e MINAMI, 2011).
A obesidade está fortemente associada à hipersecreção de insulina e
resistência à insulina. Alguns estudos já evidenciaram que o consumo da dieta HFD
por ratos e camundongos pode predispor à intolerância à glicose e hiperinsulinemia,
condições essas podendo estar associadas ao consumo de ácidos graxos oriundos
da dieta HFD (GREENBERG et al., 2006; BENOIT et al., 2013; ŠEFČÍKOVÁ et al.,
2016). Os mecanismos pelos quais os ácidos graxos plasmáticos podem promover
esses efeitos incluem o aumento da geração de metabolitos lipídicos, das espécies
reativas de oxigênio (ROS) e das citocinas pró-inflamatórias (BODEN et al., 2011).
A hiperinsulinemia compensatória devido à redução na ação do hormônio foi
comprovada em experimentos com ilhotas pancreáticas isoladas de roedores que
consumiram dieta HFD (MCLELLAN et al., 2007; GRAY et al., 2010). A hiperfunção
resulta em modificações morfológicas no pâncreas endócrino, na primeira semana
de ingestão da dieta HFD tanto por ratos, quanto por camundongos foi observado
aumento da proliferação e da massa das células β (ALONSO et al., 2007;
STAMATERIS et al., 2013). Após três meses de consumo da dieta HFD ainda foi
observado aumento da massa das células β, mas tal efeito foi relacionado à
hiperplasia das células insulares (AHRÉN et al., 2010). Há evidências de que a
proliferação da célula β declina com a idade, roedores apresentaram redução da
taxa de proliferação em dez vezes no quarto mês de vida quando comparada ao
primeiro mês de ingestão da dieta HFD (GOLSON et al., 2010; ROAT et al., 2014).
Em seis meses de consumo de dieta HFD, apesar de também ser evidenciado
aumento do tamanho das células β foram relatados danos estruturais relacionados à
menor expressão de genes envolvidos na resposta imune e na composição da
matriz extracelular (ROAT et al., 2014). Porém, quando o consumo da dieta HFD foi

68
de um ano foi evidenciada redução na apoptose das células β, o que possivelmente
pode contribuir para o aumento do volume (AHRÉN et al., 2010). Aos quatorzes
meses de administração da dieta HFD, foi obeservado ausência na capacidade de
proliferação da célula β em camundongos, condição que contribui para o declínio
funcional do pâncreas e predispõe ao DM (GOLSON et al., 2010; ROAT et al.,
2014). Portanto, o consumo da dieta HFD pode levar a prejuízos morfofuncionais
que são tempo dependentes para que ocorra declínio na função pancreática
endócrina no pâncreas endócrino e estabeleça o DM2.
Em nosso estudo, como previamente relatado por RIBEIRO et al. (2012) as
ilhotas pancreáticas de camundongos do grupo paternal HFD hipersecretaram
insulina em todas as concentrações estimulatórias de glicose estudadas. Contudo,
evidenciamos pela primeira vez que a ingestão de dieta HFD pelos pais promoveu
redução da secreção de insulina em sua prole. Apesar de poucos estudos
evidenciarem os efeitos da obesidade paternal sobre a fisiologia do pâncreas
endócrino de seus filhotes, estudo realizado por NG e colaboradores mostrou que o
consumo de dieta HFD pelos pais promoveu redução na secreção de insulina nos
filhotes fêmeas durante o ipGTT. Foi observado que a menor liberação da insulina
nesses filhotes está associada à redução da metilação do gene da subunidade alfa
2 do receptor para interleucina 13 (Il13rα2) que normalmente não é expresso nas
ilhotas de Langherans, este foi o primeiro achado de que há propagação
intergeracional de prejuízos metabólicos pelo consumo de dieta HFD pelos pais para
a sua prole (NG et al., 2010). O gene Il13rα2 quando expresso modula o
crescimento e invasão de várias linhas celulares de câncer de pâncreas e é
estimulado pela citocina pró-inflamatória TNF-α. Na obesidade é evidenciado que
essa citocina está aumentada em vários tecidos-alvo da insulina (GUIMARÃES et
al., 2007). Além disso, em roedores e humanos a concentração de leptina no plasma
está elevada devido ao aumento da adiposidade. Essa adipocitocina aumenta a
expressão do gene Il13rα2 no tecido adiposo (KAWAKITA et al., 2001; PINTO et al.,
2014) e em conjunto, a leptina e o Il13rα2, reduzem a expressão do gene da pró-
insulina e a secreção da insulina (MENDONÇA et al., 2015). Tanto a deficiência
quanto a resistência à leptina em ratos obesos estão acompanhadas de severa
resistência insulínica. Segundo GUIMARÃES et al. (2007), estudos realizados em
ratos evidenciaram que esse efeito parece ser mediado indiretamente pelo sistema
nervoso central. A leptina inibe a secreção de insulina por ativar canais de KATP o

69
que hiperpolariza a célula, além disso ativar a proteína quinase dependente de AMP,
e também promove a oxidação de triglicerídeos do tecido adiposo reduzindo o
acúmulo de gordura, condição que inibe a lipogênese e estimula a lipólise
(GUIMARÃES et al., 2007). Portanto, a condição metabólica do pai tem forte
propensão em propagar modificações epigenéticas que comprometem a função
pancreática endócrina da prole.
Nossos achados no pâncreas endócrino da prole HFD se assemelham
também aos mostrados na prole proveniente de fêmeas que foram submetidas à
restrição proteica durante a gestação e que apresentaram redução da secreção de
insulina (CHERIF et al.,1998 e 2001). Nesses filhotes foi observado menor massa de
células β devido à maior apoptose (BOUJENDAR et al., 2002). A origem molecular
dessas alterações, transmitidas da mãe para o feto, ainda não são totalmente
compreendidas, mas estudos recentes tem também apontado a epigenética como
uma possível explicação para esses fenômenos.
Com relação à suplementação com Tau, foi previamente demonstrado que
camundongos que ingeriram o aminoácido por 30 dias apresentaram aumento da
secreção de insulina em resposta à glicose (RIBEIRO et al., 2009). Mas, quando a
Tau foi administrada em fêmeas que consomem dieta HFD, foi evidenciado que o
aminoácido impediu a hiperfunção das células β (RIBEIRO et al., 2012). Evidências
da literatura mostraram, ainda, que o tratamento preventivo com Tau reestabelece a
secreção de insulina em homens obesos e submetidos à infusão contínua com
ácidos graxos por 48h (XIAO et al., 2008). Foi sugerido que o efeito preventivo da
Tau na disfunção da célula β, na presença de glicose ou ácidos graxos, é devido às
suas propriedades antioxidantes (KANIUK et al., 2007; OPRESCU et al., 2007; XIAO
et al., 2008). Como anteriormente citado, a Tau pode interagir com o IR, e em
camundongos obesos suplementados com Tau foi evidenciada maior relação
pAkt/Akt no fígado (RIBEIRO et al., 2012), sugerindo que a preservação da função
das células β induzida pela Tau também pode ser associada à melhora na
sensibilidade à insulina nos tecidos-alvo e possivelmente nas células β (ZAWALICH
et al., 2000 e 2002), pois a insulina autocrinamente aumenta a transcrição do seu
próprio gene (PERSAUD et al., 2008) e estimula a proliferação, mas inibe a
secreção do hormônio e a apoptose de células β pancreáticas (ELGHAZI et al.,
2006; LAWRENCE et al., 2008). Tal efeito pode estar associado com a ação
benéfica da Tau em prevenir a hipersecreção nas ilhotas pancreáticas de

70
camundongos alimentados com dieta HFD que foi relatado por RIBEIRO et al.,
(2012) e aqui observamos também tendência à normalização.
Ainda não se sabe os efeitos da obesidade paternal e a suplementação com
Tau sobre a condição metabólica da prole. Nosso estudo, portanto, pela primeira vez
demonstrou que a suplementação com Tau dada aos pais obesos HFD pode
atenuar a redução da secreção de insulina da prole HTAU. Previamente em estudos
com programação metabólica maternal foi observado que os fetos de ratas
submetidas à restrição proteica e suplementadas com Tau durante a gestação,
mostraram que o aminoácido normalizou a secreção de insulina e a proliferação
celular, reduzindo a apoptose e normalizando a massa de células β nas ilhotas
pancreáticas destes fetos e neonatos de mães desnutridas (MEREZAK et al., 2001 e
2004; BOUJENDAR et al., 2002; KALBE et al., 2005; REUSENS et al., 2008).
Em nosso estudo na prole, o consumo de dieta HFD pelos pais acarretou em
danos estruturais no pâncreas endócrino, promovendo redução no tamanho das
células β, e do número de células α e δ podendo estar associado a redução da
secreção de insulina na prole macho. Previamente, NG e colaboradores (2010)
evidenciaram que o pâncreas da prole proveniente de pais que comeram dieta HFD
apresentou redução da área das ilhotas pancreáticas e das células β, indicando que
esse regime nos pais prejudica a replicação das células β em seus descendentes.
Porém, esses pesquisadores também observaram aumento das ilhotas de
Langerhans pequenas na prole HFD, indicando uma resposta compensatória para
manter a massa de células β normal, e propuseram que essas células β são
suficientes para manter a normoglicemia e insulinemia em jejum, porém não foram
adequadas para a secreção de insulina estimulada por glicose (NG et al., 2010).
A literatura apresenta mais informações sobre o impacto da obesidade
maternal acarretando prejuízos na estrutura do pâncreas endócrino de sua prole.
Estudos evidenciaram que o pâncreas endócrino de filhotes machos provenientes de
mães que consumiram dieta HFD na gestação e lactação, apresentou maior área da
ilhota e hipertrofia das células β (GREGÓRIO et al., 2013). Segundo FETITA e
colaboradores (2006), o ambiente intrauterino rico em glicose induz hiperplasia nas
células β pancreáticas de camundongos neonatos, bem como promove o aumento
das concentrações de insulina no plasma. Também foram relatadas alterações
precoces no pâncreas da prole, incluindo remodelamento estrutural do pâncreas
associado ao aumento da massa de células β, prejudicando o metabolismo da

71
glicose, o que pode levar a mudanças adaptativas nas células β como hipertrofia,
hiperplasia, aumento da síntese e secreção de insulina na prole (GNIULI et al.,
2008). Portanto, com os nossos dados e as informações previamente relatadas
podemos observar que tanto o consumo de dieta HFD por mães quanto pelos pais
predispõe ao remodelamento estrutural do pâncreas endócrino da sua prole.
A suplementação com Tau mostrou evitar alterações ou restaurar a massa
pancreática endócrina na desnutrição (BOUJENDAR et al., 2002), obesidade
(RIBEIRO et al., 2012), DM1 (ARANY et al., 2004) e DM2 (LEE et al., 2011). Foi
evidenciado em camundongos NOD que a Tau reverteu a perda de massa de
células β por aumentar a proliferação celular (ARANY et al., 2004). O cultivo de
células de ratos com Tau promoveu aumento no número e tamanho das ilhotas
pancreáticas, por aumentar a área das células β, α e δ (L’AMOREAUX et al., 2010).
Outro estudo evidenciou que em camundongos ob/ob a suplementação com Tau
impede hipertrofia, o aumento da massa das ilhotas de Langerhans e das células β
no pâncreas, porém aumenta a massa das células δ e a sensibilidade da célula β à
somatostatina, essas descobertas sustentam a ideia de que a suplementação com
Tau modula o controle parácrino dos hormônios pancreáticos (SANTOS-SILVA et
al., 2015). RIBEIRO e colaboradores (2009) demonstraram que camundongos
suplementados com Tau apresentaram maior concentração plasmática e secreção
de glucagon. Como em nosso estudo observamos aumento do tamanho das células
α pancreáticas na prole HTAU, é possível que o aminoácido possa por efeitos
intergeracionais modificar também o funcionamento da célula α pancreática. Ainda, a
Tau parece exercer efeito parácrino aumentando o transporte, utilização e/ou
armazenamento de glicose e impedindo a hipersecreção de insulina. A literatura
sugere que o aumento da concentração de Tau no microambiente das ilhotas
pancreáticas, induzida pela suplementação, juntamente com a secreção de insulina
podem contribuir para a prevenção da hipertrofia e a hipersecreção das células β
que já foi relatada em camundongos fêmeas alimentadas com dieta HFD e
suplementadas com Tau (RIBEIRO et al., 2012). Em nosso estudo a suplementação
com Tau pelos pais normalizou o tamanho das células β e o número de células δ no
pâncreas da prole associando com a melhora na redução da secreção de insulina, o
que podemos sugerir que o aminoácido exerce ações intergeracionais, prevenindo
as alterações causadas pela obesidade paternal na sua prole.

72
Foi evidenciado que a suplementação com Tau dada a prenhas com restrição
proteica foi eficaz em propagar modificações para seus descendentes, como
aumento da secreção de insulina das ilhotas pancreáticas do feto (CHERIF et al.,
1998). Ainda, quando fêmeas foram suplementadas com Tau impediu a redução da
massa de células β e também foi visto que a Tau melhora a vascularização das
ilhotas pancreáticas do feto (BOUJENDAR et al., 2002 e 2003). A suplementação
com Tau, tanto em fêmeas com restrição proteica quanto em machos alimentados
com dieta HFD parece ser eficaz em propagar efeitos benéficos na morfofunção
pancreática endócrina de seus descendentes.
Portanto, a obesidade pode estar associada a modificações programadas do
genoma, que regulam a atividade dos genes sem alteração da sequência primária
de DNA, sendo herdáveis ao longo das divisões celulares levando a alterações na
viabilidade dos espermatozóides, expressão genes como o Il13rα2 e a metilação do
DNA (GHANAYEM et al., 2010; FULLSTON et al., 2013). Na literatura foi mostrado
que as modificações epigenéticas em recém-nascidos filhos de pais obesos no
período da concepção e essas características podem propagar durante o
desenvolvimento e a gametogênese (SOUBRY et al., 2013). Consequentemente,
como o aminoácido Tau regula o estresse oxidativo, motilidade e sobrevivência dos
espermatozóides (YANG et al., 2010), sugere-se que o consumo de Tau pelo pais
pode estar prevenindo alterações epignéticas no pâncreas endócrino propagadas
pelos gametas como evidenciadas em nosso estudo. Porém mais estudos são
necessários para identificar quais genes podem estar modificados, e como a Tau
estaria prevenindo essas modificações propagadas a prole.

73
7. CONCLUSÕES
O consumo de dieta HFD pelos camundongos machos C57Bl/6 do grupo
paternal foi eficaz em desencadear a obesidade. Esses roedores apresentaram
aumento do peso corporal, dos estoques de gordura, e do índice de Lee.
Os camundongos HFD paternais foram intolerantes à glicose e
hipersecretaram insulina em resposta à glicose.
A suplementação com Tau não preveniu a obesidade, a intolerância à glicose,
e a hipersecreção de insulina no grupo paternal HTAU.
Nosso estudo demonstrou pela primeira vez que a ingestão de HFD pelo
grupo paternal reduziu a liberação da insulina frente à glicose em sua prole, sem
alterar a área e massa das células endócrinas, contudo reduziu o número das
células α, δ e o tamanho das células β por ilhota.
A suplementação com Tau dada aos pais foi eficaz normalizou o número de
células δ e o tamanho das células β nas ilhotas pancreáticas da prole HTAU.

74
REFERÊNCIAS BIBLIOGRÁFICAS
ABESO – Associação Brasileira para Estudo da Obesidade e Síndrome Metabólica. Disponível em: <http://www.abeso.org.br/atitude-saudavel/mapa-obesidade>. Acessado em 15 de fevereiro 2017.
AHREN B (2000). Autonomic regulation of islet hormone secretion implications for health and disease. Diabetologia 43(4):393–410. AHREN B, PACINI G (2002). Insufficient islet compensation to insulin resistance vs. reduced glucose effectiveness in glucose-intolerant mice. Am J Physiol Endocrinol Metab 283: E738–744. AHRÉN J, AHRÉN B & WIERUP N (2010). Increased β-cell volume in mice fed a high-fat diet: A dynamic study over 12 months, Islets, 2:6, 353-356 ANURADHA, C. V. and S. D. BALAKRISHNAN (1999). Taurine attenuates hypertension and improves insulin sensitivity in the fructose-fed rat, an animal model of insulin resistance. Can J Physiol Pharmacol,v.77, n.10, Oct, p.749-54. ARANY E, STRUTT B, ROMANUS P, REMACLE C, REUSENS B & HILL DJ (2004) Taurine supplement in early life altered islet morphology, decreased insulitis and delayed the onset of diabetes in non-obese diabetic mice. Diabetologia 47, 1831-1837. ASHINO NG, SAITO KN, SOUZA FD, NAKUTZ FS, ROMAN EA, VELLOSO LA, TORSONI AS, TORSONI MA (2012). Maternal high-fat feeding through pregnancy and lactation predisposes mouse offspring to molecular insulin resistance and fatty liver. J Nutr Biochem 23(4):341-8.
BATISTA TM, RIBEIRO RA, AMARAL AG, DE OLIVEIRA CA, BOSCHERO AC & CARNEIRO EM (2012). Taurine supplementation restores glucose and carbachol-induced insulin secretion in islets from low-protein diet rats: involvement of Ach-M3R, Synt 1 and SNAP-25 proteins. J Nutr Biochem 23, 306-312. BENOIT B, PLAISANCIÉ P, AWADA M, GÉLOEN A, ESTIENNE M, CAPEL F. High-fat diet action on adiposity, inflammation, and insulin sensitivity depends on the control low-fat diet. Nutr.Res. v.33, p.952-960, 2013. BERNARDIS, L.L., PATTERSON, B.D. (1968). Correlation between 'Lee index' and carcass fat content in weanling and adult female rats with hypothalamic lesions. The Journal of endocrinology 40, 527-528. BOUJENDAR S, REUSENS B, MEREZAK S, AHN MT, ARANY E, HILL D & REMACLE C (2002) Taurine supplementation to a low protein diet during foetal and early postnatal life restores a normal proliferation and apoptosis of rat pancreatic islets. Diabetologia 45, 856-866.

75
BOUJENDAR S, ARANY E, HILL D, REMACLE C, REUSENS B (2003) Taurine supplementation of a lowprotein diet fed to rat dams normalizes the vascularization of the fetal endocrine pancreas. J Nutr 133(9):2820–2825. BUSTAMANTE J, LOBO MV, ALONSO FJ, MUKALA NT, GINE E, SOLIS JM, TAMARIT RODRIGUEZ J & MARTIN DEL RIO R (2001) An osmotic-sensitive taurine pool is localized in rat pancreatic islet cells containing glucagon and somatostatin. Am J Physiol Endocrinol Metab 281, E1275-1285. CABRERA, O., BERMAN, D.M., KENYON, N.S., RICORDI, C., BERGGREN, P.O., CAICEDO, A., (2006). The unique cytoarchitecture of human pancreatic islets has implications for islet cell function. Proc Natl Acad Sci U S A 103, 2334-2339. CARNEIRO EM, LATORRACA MQ, ARAUJO E, BELTRA M, OLIVERAS MJ, NAVARRO M, BERNA G, BEDOYA FJ, VELLOSO LA, SORIA B & MARTIN F (2009) Taurine supplementation modulates glucose homeostasis and islet function. J Nutr Biochem 20, 503- 511. CHAVEY, A., AH KIOON, M. D., BAILBE, D., MOVASSAT, J., AND PORTHA, B. (2014). Maternal diabetes, programming of beta-cell disorders and intergenerational risk of type 2 diabetes. Diabetes Metab 40 (5): 323-30. CHEIK, NÁDIA CARLA; GUERRA, RICARDO LUÍS FERNANDES; VIANA, FABIANA PAVAN; ROSSI, ELIZEU ANTONIO; CARLOS, IRACILDA ZEPONNE; VENDRAMINI, REGINA; DUARTE, ANA CLÁUDIA GARCIA DE OLIVEIRA; DÂMASO, ANA RAIMUNDA. Efeito de Diferentes Frequências de Exercícios Físico na Dislipidemia e da Obesidade em Ratos Normo e Hipercolesterolêmicos. Revista Brasileira de Educação Física, 2006. CHERIF H, REUSENS B, AHN MT, HOET JJ & REMACLE C (1998) Effects of taurine on the insulin secretion of rat fetal islets from dams fed a low-protein diet. J Endocrinol 159, 341-348. CHERIF, H., B. REUSENS, S. DAHRI, C. REMACLE AND J. J. HOET(1996) Stimulatory effects of taurine on insulin secretion by fetal rat islets cultured in vitro. J Endocrinol, v.151, n.3, Dec, p.501-6. CNOP, M., N. WELSH, J. C. JONAS, A. JORNS, S. LENZEN E D. L. EIZIRIK (2005) Mechanisms of pancreatic beta-cell death in type 1 and type 2 diabetes: many differences, few similarities. Diabetes, v.54 Suppl 2, Dec, p.S97-107. DE SOUZA CT, ARAUJO EP, STOPPIGLIA LF, PAULI JR, ROPELLE E, ROCCO SA, MARIN RM, FRANCHINI KG, CARVALHEIRA JB, SAAD MJ, BOSCHERO AC, CARNEIRO EM, VELLOSO LA (2007). Inhibition of UCP2 expression reverses diet-induced diabetes mellitus by effects on both insulin secretion and action. FASEB J. 221:1153–1163. DUNN GA; BALE TL. (2009). Maternal high-fat diet promotes body length increases and insulin insensitivity in second-generation mice. Endocrinology 150: 4999-5009.

76
ENES, CARLA CRISTINA; SLATER, BETZABETH. Obesidade na adolescência e seus principais fatores determinados. Rev. bras. Epidemiol, v.13, n.1, p. 163-171, 2010.
ESPELAND MARK A., HENRY A. GLICK, ALAIN BERTONI, FREDERICK L. BRANCAT. Impact of an Intensive Lifestyle Intervention on Use and Cost of Medical Services Among Overweight and Obese Adults With Type 2 Diabetes: The Action for Health in Diabetes. Diabetes Care 2014 Sep; 37(9): 2548-2556. ESTADELLA D, OYAMA LM, DÂMASO AR, RIBEIRO EB, OLLER DO NASCIMENTO CM. Effect of palatable hyperlipidic diet on lipid metabolism of sedentary and exercised rats. Nutrition. 2004;20(2):218-24.
GHANAYEM BI, BAI R, KISSLING GE, TRAVLOS G, AND HOFFLER U. Biol Reprod 82: 96-104, 2009. GLASTRAS SJ, TSANG M, TEH R. Maternal Obesity Promotes Diabetic Nephropathy in Rodent Offspring. Scientific Reports. 2016;6:27769. GREENBERG AS, OBIN MS. Obesity and the role of adipose tissue in inflammation and metabolism. The American journal of clinical nutrition. 2006 Feb;83(2):461S-5S. PubMed PMID: 16470013. GNIULI D, CALCAGNO A, CARISTO ME (2008). Effects of high-fat diet exposure during fetal life on type 2 diabetes development in the progeny. J Lipid Res 49(09): 1936-1945. GUIMARAES, DE; SARDINHA, FLC; MIZURINI, DM, CARMO, MGT.Adipocitocinas: uma nova visão do tecido adiposo. Rev. Nutr. 2007, vol.20, n.5, pp.549-559. HENQUIN JC (2011).The dual control of insulin secretion by glucose involves triggering and amplifying pathways in beta cells. Diabetes Res Clin Pract 93 Suppl 1, S27-31. HIRIART M & AGUILAR-BRYAN L (2008) Channel regulation of glucose sensing in the pancreatic beta cell. Am J Physiol Endocrinol Metab 295, E1298- 1306. HOWELL, P. M. JR, LIU, S.; REN, S.; BEHLEN, C.; FODSTAD, O.; RIKER, A. I. (2009). Epigenetics in human melanoma. Cancer Control: Journal of the Moffitt Cancer Center, Tampa, v. 16, n. 6, p. 200- 218. HUXTABLE, R.J. (1992). Physiological actions of taurine. Physiological reviews 72, 101-163. Ito, T., Pastukh, V., Solodushko, V., Azuma, J., Schaffer, S.W., 2009. Effect of taurine on protein kinase C isoforms: role in taurine's actions? Advances in experimental medicine and biology 643, 3-11. INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA (IBGE) 2014 e 2015.

77
JAMES PT, LEACH R, KALAMARA E, SHAYEGHI M (2001). “The worldwide obesity epidemic”. Obes Res 9 (Suppl 4):S228-33. KAHN, S.E., HULL, R.L., UTZSCHNEIDER, K.M., (2006). Mechanisms linking obesity to insulin resistance and type 2 diabetes. Nature 444, 840-846. KALBE, L., A. LEUNDA, T. SPARRE, C. MEULEMANS, M. T. AHN, T. ORNTOFT, M. KRUHOFFER, B. REUSENS, J. NERUP E C. REMACLE (2005). Nutritional regulation of proteases involved in fetal rat insulin secretion and islet cell proliferation. Br J Nutr v.93, n.3, Mar, p.309-16. KANIUK, N. A., M. KIRALY, H. BATES, M. VRANIC, A. VOLCHUK AND J. H. BRUMELL (2007). Ubiquitinated-protein aggregates form in pancreatic beta cells during diabetes-induced oxidative stress and are regulated by autophagy. Diabetes v.56, n.4, Apr, p.930-9. KANNO, T., GOPEL, S.O., RORSMAN, P., WAKUI, M., (2002). Cellular function in multicellular system for hormone-secretion: electrophysiological aspect of studies on alpha-, beta- and delta-cells of the pancreatic islet. Neurosci Res 42, 79-90. L’AMOREAUX WJ, CUTTITTA C, SANTORA A, BLAIZE JF, TACHJADI J, EL IDRISSI A. Taurine regulates insulin release from pancreatic beta cell lines. Journal of Biomedical Science. 2010. LAWRENCE, M., C. SHAO, L. DUAN, K. MCGLYNN E M. H. COBB (2008). The protein kinases ERK1/2 and their roles in pancreatic beta cells. Acta Physiol (Oxf), v.192, n.1, Jan, p.11-7.
LI M, REYNOLDS CM, SLOBODA DM, GRAY C, VICKERS MH (2013) Effects of
Taurine Supplementation on Hepatic Markers of Inflammation and Lipid Metabolism in Mothers and Offspring in the Setting of Maternal Obesity. PLoS ONE 8(10): e76961. LUCAS, A. (1991). Programming by early nutrition in man. Ciba Found Symp 156:38-50. MALIK. VASANTI S., BARRY M. POPKIN, GEORGE A. BRAY, JEAN-PIERRE DESPRÉS, WALTER C. WILLETT and FRANK B. HU (2013). Sugar-Sweetened Beverages and Risk of Metabolic Syndrome and Type 2 Diabetes. American Diabetes Association 33(11): 2477-2483. MCLELLAN, KÁTIA CRISTINA PORTERO; BARBALHO, SANDRA MARIA; CATTALINI, MARINO and LERARIO, ANTONIO CARLOS (2007). Diabetes mellitus do tipo 2, síndrome metabólica e modificação no estilo de vida. Rev. Nutr. vol.20, n.5. MEIGS, J. B (2010). Epidemiology of Type 2 Diabetes and Cardiovascular Disease: Translation from Population to Prevention The Kelly West Award Lecture 2009. Diabetes care, 33(8), 1865-1871.

78
MEREZAK, S., A. A. HARDIKAR, C. S. YAJNIK, C. REMACLE E B. REUSENS (2001). Intrauterine low protein diet increases fetal beta-cell sensitivity to NO and IL-1 beta: the protective role of taurine. J Endocrinol,v.171, n.2, Nov, p.299-308. MEREZAK, S., B. REUSENS, A. RENARD, K. GOOSSE, L. KALBE, M. T. AHN, J. TAMARIT-RODRIGUEZ E C. REMACLE (2004). Effect of maternal low-protein diet and taurine on the vulnerability of adult Wistar rat islets to cytokines. Diabetologia, v.47, n.4, Apr, p.669-75. MINISTÉRIO DA SAÚDE (BR), Secretaria de Vigilância em Saúde, Departamento de Vigilância de Doenças e Agravos não Transmissíveis e promoção de Saúde. Vigitel Brasil 2015: vigilância de fatores de risco e proteção para doenças crônicas por inquérito telefônico. Brasília: Ministério da Saúde, 2015. NAKAYA Y, MINAMI A, HARADA N, SAKAMOTO S, NIWA Y & OHNAKA M (2000) Taurine improves insulin sensitivity in the Otsuka Long-Evans Tokushima Fatty rat, a model of spontaneous type 2 diabetes. Am J Clin Nutr 71, 54-58. NANDHINI, A. T., V. THIRUNAVUKKARASU, M. K. RAVICHANDRAN AND C. V. ANURADHA (2005). Effect of taurine on biomarkers of oxidative stress in tissues of fructose-fed insulin-resistant rats. Singapore Med J, v.46, n.2, Feb, p.82-7. NESHER, R., E. ANTEBY, M. YEDOVIZKY, N. WARWAR, N. KAISER E E. CERASI (2002). Beta-cell protein kinases and the dynamics of the insulin response to glucose. Diabetes, v.51 Suppl 1, p.S68-73. NG SF, LIN RC, LAYBUTT DR, BARRES R, OWENS JA, MORRIS MJ (2010). Chronic high-fat diet in fathers programs β-cell dysfunction in female rat offspring. Nature 467:963–966. OPRESCU, A. I., G. BIKOPOULOS, A. NAASSAN, E. M. ALLISTER, C. TANG, E. PARK, H. UCHINO, G. F. LEWIS,I. G. FANTUS, M. ROZAKIS-ADCOCK, M. B. WHEELER E A. GIACCA (2007). Free fatty acid-induced reduction in glucose-stimulated insulin secretion: evidence for a role of oxidative stress in vitro and in vivo. Diabetes, v.56, n.12, Dec, p.2927-37. PARK, E. J. , J. H. BAE, S. Y. KIM, J. G. LIM, W. K. BAEK, T. K. KWON, S. I. SUH, J. W. PARK, I. K. LEE, F. M. ASHCROFT AND D. K. SONG (2004). Inhibition of ATP-sensitive K+ channels by taurine through a benzamido-binding site on sulfonylurea receptor 1. Biochem Pharmacol, v.67, n.6, Mar 15, p.1089-96. PESSIN JE, SALTIEL AR (2000). Signaling pathways in insulin action: molecular targets of insulin resistance. J Clin Invest 106: 165-169. PRENTKI and NOLAN CJ (2006) Islet beta cell failure in type 2 diabetes. J Clin Invest 116, 1802-1812. REUSENS, B., T. SPARRE, L. KALBE, T. BOUCKENOOGHE, N. THEYS, M. KRUHOFFER, T. F. ORNTOFT, J. NERUP E C. REMACLE (2008). The intrauterine metabolic environment modulates the gene expression pattern in fetal rat islets:

79
prevention by maternal taurine supplementation. Diabetologia, v.51, n.5, May, p.836-45. RIBEIRO RA, BONFLEUR ML, AMARAL AG, VANZELA EC, ROCCO SA, BOSCHERO AC & CARNEIRO EM (2009). Taurine supplementation enhances nutrient-induced insulin secretion in pancreatic mice islets. Diabetes Metab Res Rev 25, 370- 379. RIBEIRO RA, VANZELA EC, OLIVEIRA CA, BONFLEUR ML, BOSCHERO AC, CARNEIRO EM (2010). Taurine supplementation: involvement of cholinergic/phospholipase C and protein kinase a pathways in potentiation of insulin secretion and Ca2+ handling in mouse pancreatic islets. The British journal of nutrition 104, 1148-1155. RIBEIRO RA, SANTOS-SILVA JC, VETTORAZZI JF, COTRIM BB, MOBIOLLI DD, BOSCHERO AC & CARNEIRO EM (2012). Taurine supplementation prevents morphophysiological alterations in high-fat diet mice pancreatic beta cells. Amino Acids 43(4):1791-801. SARTORELLI, DANIELA SAES AND FRANCO, LAÉRCIO JOEL. Tendências do diabetes mellitus no Brasil: o papel da transição nutricional. Cad. Saúde Pública [online]. 2003, vol.19, suppl.1, pp.S29-S36. SCHMIDT, M. I. et al. Doenças Crônicas não transmissíveis no Brasil: mortalidade, morbidade e fatores de risco. In: BRASIL. Ministério da Saúde. Departamento de Analise de Situação de Saúde. Secretaria de Vigilância em Saúde. Saúde Brasil 2009: Uma analise da situação de saúde e da Agenda Nacional e Internacional de Prioridades em Saúde. Brasília, 2010. SHAFAT, A.; MURRAY, B.; RUMSEY, D. Energy density in cafeteria diet induced hyperphagia in the rat. Appetite, v. 52, p. 34–38, 2009. SHANKAR K, ZHONG Y, KANG P (2011) Maternal obesity promotes a pro-inflammatory signature in rat uterus and blastocyst. Endocrinology 152, 4158-4170. SENER, A., L. C. BEST, A. P. YATES, M. M. KADIATA, E. OLIVARES, K. LOUCHAMI, H. JIJAKLI, L. LADRIERE E W. J. MALAISSE (2000). Stimulus-secretion coupling of arginine-induced insulin release: comparison between the cationic amino acid and its methyl ester. Endocrine, v.13, n.3, Dec, p.329-40. SHULMAN GI (2000) Cellular mechanisms of insulin resistance. J Clin Invest 106: 171- 176. SOLON CS, FRANCI D, IGNACIO-SOUZA LM, ROMANATTO T, ROMAN EA, ARRUDA AP (2012) Taurine enhances the anorexigenic effects of insulin in the hypothalamus of rats. Amino Acids 42:2403–2410. SOMINSKY, L., AND SPENCER, S. J. (2014) Eating behavior and stress: a pathway to obesity. Front Psychol 5, 434.

80
SOUBRY, A., MURPHY, S. K., WANG, F., HUANG, Z., VIDAL, A. C., FUEMMELER, B. F., KURTZBERG, J., MURTHA, A., JIRTLE, R. L., SCHILDKRAUT, J. M., AND HOYO, C. (2015). Newborns of obese parents have altered DNA methylation patterns at imprinted genes. Int J Obes (Lond) 39(4): 650–657. SOUZA, M. G., RIVERA, I. R., SILVA, M. A., CARVALHO, A. C. Relationship of obesity with high blood pressure in children and adolescents. Arquivos Brasileiros de Cardiologia, São Paulo, v. 94, n. 6, p. 714-9, 2010. STRAUB SG & SHARP GW (2002). Glucose-stimulated signaling pathways in biphasic insulin secretion. Diabetes Metab Res Rev 18, 451-463. TAPPAZ ML. (2004). Taurine biosynthetic enzymes and taurine transporter: molecular identification and regulations. Neurochem Res 29(1):83-96. TATEYA S, KIM F, TAMORI Y. (2013). Recent advances in obesity-induced inflammation and insulin resistance. Front Endocrinol 8; 4:93. TORTORA, Gerard, e BRYAN, Derrickson. Princípios de Anatomia e Fisiologia. 12ª edição. Rio de Janeiro: Editora Guanabara, 2014). TSUBOYAMA-KASAOKA N, SHOZAWA C, SANO K, KAMEI Y, KASAOKA S, HOSOKAWA Y & EZAKI O (2006). Taurine (2-aminoethanesulfonic acid) deficiency creates a vicious circle promoting obesity. Endocrinology 147, 3276-3284. VETTORAZZI JF, RIBEIRO RA, SANTOS-SILVA JC, BORCK PC, BATISTA TM, NARDELLI TR, BOSCHERO AC, CARNEIRO EM (2014). Taurine supplementation increases K channel protein content, improving Ca handling and insulin secretion in islets from malnourished mice fed on a high-fat diet. Amino acids 10.1007/s00726-014-1763-6.
VIGITEL MINISTÉRIO DA SAÚDE. Disponível em: http://portalsaude.saude.gov.br/images/pdf/2014/abril/30/Lancamento-Vigitel-28-04-ok.pdf 2014. Acessado em 15 de abril 2017.
WHITE CL, PURPERA MN, MORRISON CD. (2009). Maternal obesity is necessary for programming effect of high-fat diet on offspring. Am J Physiol Regul Integr Comp Physiol. May; 296(5):R1464-72. WHITEMAN, E. L., H. CHO E M. J. BIRNBAUM (2002). Role of Akt/protein kinase B in metabolism. Trends Endocrinol Metab, v.13, n.10, Dec, p.444-51. WONG ET, QIU B, SHI X, LIM J, BEZZI M, LOW D, ET AL.NUCKS is a positive transcriptional regulator of insulin signaling. Cell Rep. 2014:7:1876-86
WORLD HEALTH ORGANIZATION (WHO). Disponível em: http://www.who.int/mediacentre/factsheets/fs317/en/. Acessado em 27 de outubro 2016. Acessado em 19 de fevereiro 2017.

81
XIAO, C., A. GIACCA E G. F. LEWIS (2007). Oral taurine but not N-acetylcysteine ameliorates NEFA-induced impairment in insulin sensitivity and beta cell function in obese and overweight, non-diabetic men. Diabetologia, v.51, n.1, Jan, p.139-46. XU XF, and Du LZ. Chin Med J (Engl) 123: 2948-2954. YANG, SN.,BERGGREN (2006). The role of voltage-gated calcium channels in pancreatic beta-cell physiology and pathophysiology. Endocr Rev, v.27, n.6, Oct, p.621-76.

82
ANEXOS
Anexo 1 – Autorização da Comissão de Ética em Experimentação Animal da
Universidade Federal do Rio de Janeiro – Campus URJ – Macaé Prof. Aloísio
Teixeira em que consta protocolo com utilização de animais.
Recommended