
UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE
CENTRO DE BIOCIÊNCIAS CURSO DE BIOMEDICINA
KAREN MYLENNA LUSTOSA PEREIRA
MODELO MURINO - Plasmodium berghei PARA ESTUDOS EM MALÁRIA EXPERIMENTAL
Natal 2016

Universidade Federal do Rio Grande do Norte - UFRN
Sistema de Bibliotecas - SISBI
Catalogação de Publicação na Fonte. UFRN - Biblioteca Setorial do Centro de Biociências - CB
Pereira, Karen Mylenna Lustosa.
Modelo murino - Plasmodium berghei para estudos em malária experimental / Karen Mylenna Lustosa Pereira. - Natal, 2016.
45 f.: il.
Monografia (Graduação) - Universidade Federal do Rio Grande do
Norte. Centro de Biociências. Curso de Biomedicina. Orientador: Prof. Dr. Valter Ferreira de Andrade Neto.
1. Malária - Monografia. 2. Plasmodium sp. - Monografia. 3. Modelo animal - Monografia. I. Andrade Neto, Valter Ferreira de.
II. Universidade Federal do Rio Grande do Norte. III. Título.
RN/UF/BSE-CB CDU 616.936

MODELO MURINO - Plasmodium berghei PARA ESTUDOS EM MALÁRIA
EXPERIMENTAL
por
Karen Mylenna Lustosa Pereira
Orientador: Prof. Dr. Valter Ferreira de Andrade Neto
Natal
2016
Monografia Apresentada à Coordenação do Curso de Biomedicina da Universidade Federal do Rio Grande do Norte, como Requisito Parcial à Obtenção do Título de Bacharel em Biomedicina.

UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE CENTRO DE BIOCIÊNCIAS
CURSO DE BIOMEDICINA
A Monografia MODELO MURINO - Plasmodium berghei PARA ESTUDOS EM MALÁRIA EXPERIMENTAL elaborada por Karen Mylenna Lustosa Pereira e aprovada por todos os membros da Banca examinadora foi aceita pelo Curso de Biomedicina e homologada pelos membros da banca, como requisito parcial à obtenção do título de BACHAREL EM BIOMEDICINA Natal, 13 de Dezembro de 2016
BANCA EXAMINADORA
_____________________________________________________ Valter Ferreira de Andrade Neto
Departamento de Microbiologia e Parasitologia - Centro de biociências UFRN
_____________________________________________________ Maria de Fátima de Souza
Departamento de Microbiologia e Parasitologia - Centro de biociências UFRN
____________________________________________________ Adriana Ferreira Uchoa
Departamento de Biologia Celular e Genética - Centro de biociências UFRN

AGRADECIMENTOS
Aos meus pais, por todo o esforço que fizeram para me manter nesta cidade, e
concluir a minha graduação.
Aos meus irmãos, principalmente a Karla, por aguentar quando eu começava a falar
sobre os assuntos das provas.
Ao professor Valter Ferreira de Andrade Neto, por ter me permitido participar de sua
família científica, o LABMAT, o que me permitiu aprender e evoluir com ela.
Agradeço também a todos os membros dessa família científica, os que ainda estão
presentes e os que já passaram.
Agradeço a todos os colegas de turma, por todas as experiências vividas nesses
quatro anos e meio.
Agradeço aos amigos e familiares, pela compreensão com minha ausência em
vários momentos importantes.

LISTA DE ABREVIATURAS
µL
A. albitarsis
A. aquasalis
A. bellator
A. cruzii
AGM
CID
EPO
Fd
GAF
H/mL
Hi
Hp
Ht
IFN-γ
IgG
IgM
IL1
IL6
IL8
IRA
LABMAT
LUMC
MC
MG
mg/dL
mmHg
NK
NO
OMS
PBS
P. berghei
Microlitro
Anopheles albitarsis
Anopheles aquasalis
Anopheles bellator
Anopheles cruzii
Anemia Grave na Malária
Coagulação Intravascular disseminada
Eritropoietina
Fator de diluição
Fator de ativação de gametócitos
Hemácias por mililitro
Hemácias infectadas contadas
Hemácias parasitadas
Hemácias totais contadas
Interferon gama
Imunoglobulina G
Imunoglobulina M
Interleucina 1
Interleucina 6
Interleucina 8
Insuficiência respiratória aguda
Laboratório de Biologia da Malária e Toxoplasmose
Leis Universitair Medisch Centrum
Malária cerebral
Malária grave
Miligramas por decilitro
Milímetros de mercúrio
Natural Killer
Oxido nítrico
Organização mundial da saúde
Tampão fosfato-salino
Plasmodium berghei

P. chabaudi
P. falciparum
P. knowlesi
P. malarie
P. ovale
P. vinckei
P. vivax
P. yoelii
SDRA
SIRS
TNF-α
TP
Plasmodium chabaudi
Plasmodium falciparum
Plasmodium knowlesi
Plasmodium malarie
Plasmodium ovale
Plasmodium vinckei
Plasmodium vivax
Plasmodium yoelii
Síndrome do Desconforto Respiratório Agudo
Síndrome de Resposta Infamatória Sistêmica
Fator de necrose tumoral alfa
Tempo de protrombina

vi
LISTA DE TABELAS
Tabela 1: Casos notificados de malária no Brasil no ano de 2015 (Fonte: DATASUS, 2016)..........................................................................................................................02
Tabela 2: Critérios para definir malária grave (Fonte: Monteiro, 2011).....................07
Tabela 3: Infecção por Plasmodium em diferentes linhagens de camundongos. (Fonte: Queiroz et al.; 2008).......................................................................................11
Tabela 4: cepas de Plasmodium berghei. (Fonte: LUMC, 2014)...............................14
Tabela 5: Média de parasitemia dos camundongos infectados.................................25

vii
LISTA DE FIGURAS
Figura 1: Ciclo biológico do Plasmodium sp. (Fonte: CDC, 2014)..........................04
Figura 2: Formas parasitárias das quatro espécies de Plasmodium murino mais utilizadas na pesquisa. Em A trofozoitos de Plasmodium P. chabaudi. Note a sincronicidade da infecção. Em B trofozoito de P. yoelii em reticulócitos. Em C formas parasitarias do P. vinckei. Notar a preferencia desse parasita por células maduras. Em D formas parasitárias do P. berghei (Fonte: Culleton, 2005)...............12
Figura 3: Formas parasitárias de P. berghei ANKA encontradas na circulação (Fonte: LUMC, 2014)..................................................................................................14 Figura 4: Ciclo de desenvolvimento do Plasmodium no mosquito: transformações do Plasmodium e tempo de duração de cada fase (Fonte: Ríos-Velásquez, 2014)..........................................................................................................................18 Figura 5: Câmara de Neubauer. As hemácias devem ser contadas no quadrante central, nos quatro quadrados menores da extremidade e no quadrado menor central.........................................................................................................................21
Figura 6: Formas trofozoitas do Plasmodium berghei ANKA (Fonte: Arquivo
pessoal).....................................................................................................................24
Figura 7: Trofozoitos e esquizontes de P. berghei ANKA (Fonte: Arquivo pessoal,
2016)..........................................................................................................................25
Figura 8: eritrócitos infectados com Plasmodium berghei ANKA. A seta aponta para
um eritrócito com trofozoito e apresentando protuberâncias em sua superfície....................................................................................................................26

viii
RESUMO
A malária é uma doença que apresenta um caráter infeccioso agudo, com picos febris, e possui como agente etiológico protozoários do gênero Plasmodium. Segundo dados da OMS, em 2015 ocorreram 214 milhões de casos em todo o mundo, que acarretaram 438.000 mortes. Dessas mortes, 90% ocorreram na África Subsaariana, e setenta e oito por cento dos óbitos são de crianças com menos de cinco anos de idade. O modelo animal passou a ser utilizado nos estudos de malária grave devido a dificuldades no acompanhamento de casos de malária cerebral humana, assim como a limitação no que concerne na analise dos processos patológicos. O Plasmodium berghei ANKA é utilizado em grande parte dos estudos que tratam da malária cerebral, buscando entender a cadeia complexa de interação parasito-hospedeiro, e as formas de contornar a gravidade gerada nas complicações da doença. O LABMAT trabalha com as cepas NK65 e ANKA do Plasmodium berghei, e as manutenções de infecção são feitas semanalmente utilizando camundongos Swiss. É observado por meio de esfregaços as formas parasitarias, assim como é determinado a parasitemia e a mortalidade dos animais infectados com as duas cepas. Esses animais podem vir a ser utilizados em posteriores estudos envolvendo a doença. Com esse entendimento, é possível chegar à produção de formas de combate ao parasito, seja por meio de desenvolvimento de fármacos eficazes no tratamento de indivíduos contaminados, seja no desenvolvimento de vacinas que podem, não somente gerar uma imunidade duradora, mas também interromper o ciclo biológico do parasito.
Palavras-chave: Malária, Plasmodium sp., Modelo animal

ix
ABSTRACT
Malaria is a disease that presents an acute infectious character, with febrile
peaks, and has as etiological agent protozoa of the genus Plasmodium. According to
WHO data, in 2015 there were 214 million cases worldwide, which resulted in
438,000 deaths. Of these deaths, 90% occurred in sub-Saharan Africa, and seventy-
eight percent of deaths are from children under five years of age. The animal model
has been used in the studies of severe malaria due to difficulties in the monitoring of
cases of human cerebral malaria, as well as the limitation in what concerns the
analysis of the pathological processes. Plasmodium berghei ANKA is used in large
part of the studies dealing with cerebral malaria, seeking to understand the complex
chain of host-parasite interaction, and ways to circumvent the severity generated in
the complications of the disease. The Laboratory of Malaria Biology and
Toxoplasmosis works with Plasmodium berghei strains NK65 and ANKA, and
infection maintenance is done weekly using Swiss mice. Parasite forms are observed
by means of smears, as well as the parasitemia and the mortality of the animals
infected with the two strains. These animals may be used in later studies involving
the disease. With this understanding, it is possible to produce ways to combat the
parasite, either through the development of effective drugs in the treatment of
infected individuals or in the development of vaccines that may not only generate
lasting immunity but also interrupt the cycle of the parasite.
Key words: Malaria, Plasmodium sp., Animal model

Sumário LISTA DE ABREVIATURAS ...................................................................................... iv
LISTA DE TABELAS ................................................................................................. vi
LISTA DE FIGURAS ................................................................................................. vii
RESUMO.................................................................................................................. viii
ABSTRACT ................................................................................................................ ix
1. INTRODUÇÃO ................................................................................................. 1
1.1 Malária ............................................................................................................................. 1
1.2 Biologia do Plasmodium e ciclo evolutivo ......................................................................... 2
1.3 Aspectos imunológicos .................................................................................................... 5
1.4 Malária grave ................................................................................................................... 6
1.4.1 Complicações clínicas ......................................................................................... 7
1.5 Modelo animal ..................................................................................................... 10
1.6 Introdução aos Plasmodium de roedores ....................................................................... 10
1.6.1 Plasmodium berghei ............................................................................................ 13
1.6.2Ultraestrutura, invasão celular de hospedeiros vertebrados e invertebrados ....... 16
2. OBJETIVOS ................................................................................................... 19
2.1 Objetivo geral ................................................................................................................. 19
2.2 Objetivos específicos ..................................................................................................... 19
3. METODOLOGIA ............................................................................................ 20
3.1 Infecção dos animais ..................................................................................................... 20
3.2 Preparação do inóculo ................................................................................................... 20
3.2.1 Determinação da porcentagem de hemácias parasitadas .................................... 20
3.2.2 Determinação da concentração de hemácias ...................................................... 21
3.2.3 Diluição do inóculo ............................................................................................... 21
3.3 Parasitemia dos animais ................................................................................................ 22
3.4 Congelamento de cepa .................................................................................................. 22
3.5 Descongelamento de cepa ............................................................................................ 23
4. RESULTADOS ............................................................................................... 24
4.1 Identificação de formas parasitárias ............................................................................... 24
4.1.1 Trofozoitos ........................................................................................................... 24
4.1.2 Esquizontes ......................................................................................................... 24
4.2 Crescimento parasitário ................................................................................................. 25
4.3 Mortalidade .................................................................................................................... 26
5. DISCUSSÃO .................................................................................................. 27

6. REFERÊNCIAS .............................................................................................. 29

1
1. INTRODUÇÃO 1.1 Malária
A malária é uma doença que apresenta um caráter infeccioso agudo, com
picos febris, e que é transmitida através da fêmea do Anopheles, no momento em
que o inseto infectado realiza o repasto sanguíneo. Apresenta-se como uma seria
situação problemática no âmbito da saúde publica, que afeta principalmente regiões
tropicais e subtropicais, com elevada taxa de infecção e mortalidade.
A malária possui como agente etiológico protozoários do gênero Plasmodium,
pertencentes ao filo Apicomplexa, família Plasmodiidae. O gênero se apresenta com
mais de 150 espécies que infectam diferentes hospedeiros vertebrados, sendo que
em geral as espécies são hospedeiro-específico (Singha; Daneshvar, 2013). As
espécies responsáveis por causar a infecção em humanos são o Plasmodium vivax,
Plasmodium falciparum, Plasmodium malarie, Plasmodium ovale e, mais
recentemente, o Plasmodium knowlesi passou a ser considerado a quinta espécie
causadora da malária em humanos, sendo que aparece como agente de infecções
principalmente no sudeste asiático. O P. vivax apresenta uma melhor distribuição se
comparado às outras espécies (Lacerda, 2007), enquanto que o P. falciparum
apresenta maior morbidade e mortalidade (Wright, 2010).
Segundo dados da OMS, em 2015 ocorreram 214 milhões de casos em todo
o mundo, que acarretaram 438.000 mortes. Dessas mortes, noventa por cento
sucederam na África Subsaariana, e setenta e oito por cento dos óbitos são de
crianças com menos de cinco anos de idade. Porém, entre os anos de 2000 e 2015
a taxa de mortalidade ocasionada pela malária diminuiu em sessenta por cento a
nível global, com o número de casos de infecção por malária caindo em trinta e sete
por cento.
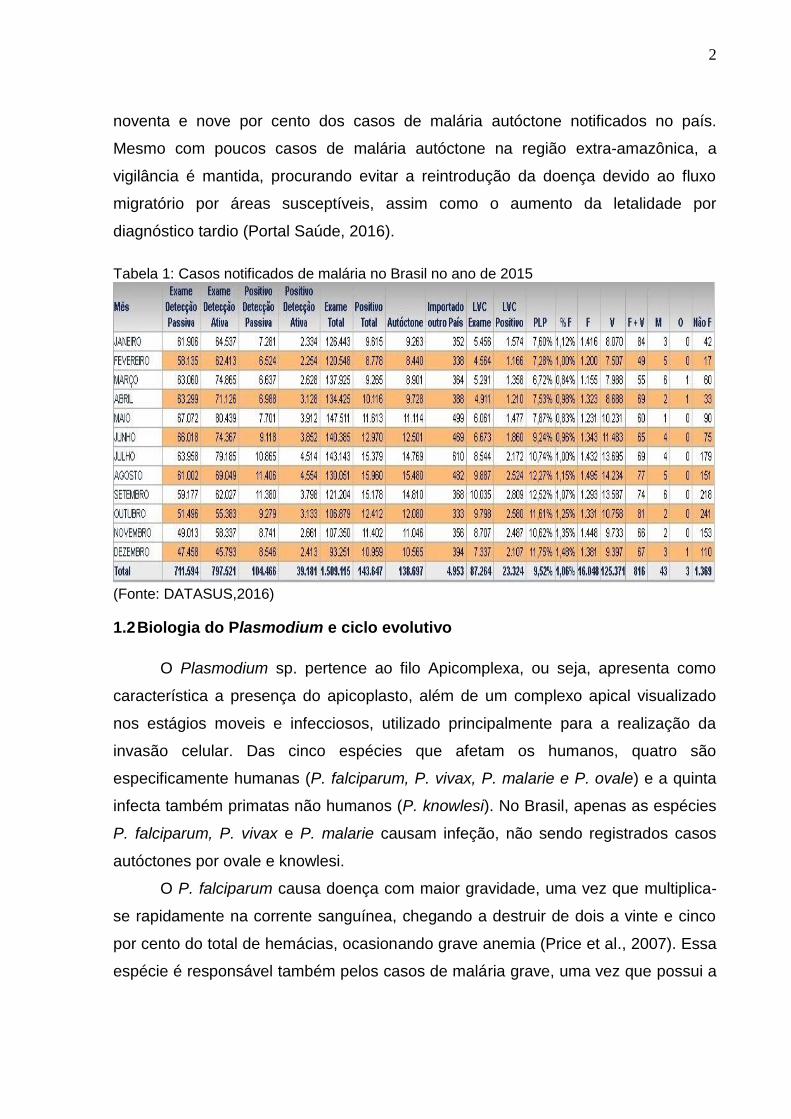
Já no Brasil, segundo dados do ministério da saúde, em 2015 foi registrado o
menor número de casos de malária dos últimos 35 anos, com a notificação de
143.647 casos em todo o território nacional, sendo que desses 138.697 foram de
casos autóctones e 4.953 importados de outros países (tabela 1). Já o número de
óbitos ficou em 26, uma redução de oitenta e nove por cento se comparado ao ano
de 2000. A área endêmica de transmissão da doença no Brasil compreende a região
da Amazônia legal, atingindo os estados de Acre, Amapá, Amazonas, Maranhão,
Mato Grosso, Pará, Rondônia, Roraima e Tocantins, sendo responsáveis por
2
noventa e nove por cento dos casos de malária autóctone notificados no país.
Mesmo com poucos casos de malária autóctone na região extra-amazônica, a
vigilância é mantida, procurando evitar a reintrodução da doença devido ao fluxo
migratório por áreas susceptíveis, assim como o aumento da letalidade por
diagnóstico tardio (Portal Saúde, 2016).
Tabela 1: Casos notificados de malária no Brasil no ano de 2015
(Fonte: DATASUS,2016)
1.2 Biologia do Plasmodium e ciclo evolutivo
O Plasmodium sp. pertence ao filo Apicomplexa, ou seja, apresenta como
característica a presença do apicoplasto, além de um complexo apical visualizado
nos estágios moveis e infecciosos, utilizado principalmente para a realização da
invasão celular. Das cinco espécies que afetam os humanos, quatro são
especificamente humanas (P. falciparum, P. vivax, P. malarie e P. ovale) e a quinta
infecta também primatas não humanos (P. knowlesi). No Brasil, apenas as espécies
P. falciparum, P. vivax e P. malarie causam infeção, não sendo registrados casos
autóctones por ovale e knowlesi.
O P. falciparum causa doença com maior gravidade, uma vez que multiplica-
se rapidamente na corrente sanguínea, chegando a destruir de dois a vinte e cinco
por cento do total de hemácias, ocasionando grave anemia (Price et al., 2007). Essa
espécie é responsável também pelos casos de malária grave, uma vez que possui a

3
capacidade de remodelar eritrócitos parasitados tornando-os mais agregativos, o
que pode vir a ocasionar tromboses e embolias (Maier et al., 2009).
A doença causada pelo P. vivax e pelo P. malarie são bastante semelhantes,
com um quadro de menor gravidade, apresentando anemia discreta, com alguns
casos de ruptura esplênica e síndrome nefrótica. A mortalidade aqui ocorre
raramente. Nos casos de infecção por P. vivax o tratamento é mais complicado que
na infecção por P. falciparum, uma vez que essa espécie possui a capacidade de
ficar adormecido no fígado sob a forma de hipnozoítos, que são formas
adormecidas, dificultando a eliminação. Porém é importante salientar que, segundo
Naing et al., (2014) o Plasmodium vivax está envolvido em casos de malária severa,
não sendo uma exclusividade somente do P. falciparum.
O gênero Anopheles é o principal transmissor da malária. Esses insetos
pertencem a ordem dos dípteros, família Culicidae. As espécies encontradas no
Brasil são o Anopheles darlingi, A. aquasalis, A. albitarsis, A. cruzii e A. bellator,
sendo que o A. darlingi é o principal vetor no Brasil, por causa de sua ampla
distribuição geográfica, antropofilia e a capacidade de ser infectado por espécies
diferentes. (FIOCRUZ, 2016).
O Plasmodium sp. apresenta ciclo de vida heteroxênico, ou seja, o ciclo
envolve um hospedeiro vertebrado (intermediário) e um invertebrado (definitivo), que
é um mosquito do gênero Anopheles (figura 1). A infecção no vertebrado se inicia
quando a fêmea infectada realiza o repasto sanguíneo e inocula esporozoítos no
hospedeiro. Esses esporozoítos uma vez na corrente sanguínea, seguem até os
hepatócitos, onde invadem essas células, amadurecem em esquizontes hepáticos,
tornando-se posteriormente merozoítos, quando rompem o hepatócito e são
liberados, envolvidos em membrana merossomal, na corrente sanguínea. P. vivax e
P. ovale possuem a capacidade de gerar hipnozoítos, permanecendo no fígado sob
a forma adormecida, e causar a doença semanas ou até anos depois da primeira
picada do mosquito. Uma vez livres do hepatócito, os merozoítos darão inicio a
multiplicação assexuada nos eritrócitos, ligando-se a superfície das hemácias,
passam para o interior, onde ficarão armazenados em vacúolos parasitóforos. Uma
vez nesses vacúolos podem seguir duas vias de desenvolvimento: diferenciar-se em
esquizontes e desenvolver novos merozoítos, que infectam novos eritrócitos, ou
produzir formar sexuais: gametócito masculino e feminino (Castro; Rodríguez, 2009).

4
A parte do ciclo que ocorre no fígado é denominada fase exoeritrocítica, enquanto
que a parte que ocorre nos eritrócitos é a fase eritrocítica. No processo de
diferenciação do merozoítos, os protozoários se diferenciam em trofozoitos e pelo
processo de esquizogonia se replicam até o rompimento das hemácias ocorrendo
assim os picos febris, sendo 24 horas para o P. knowlesi, 48 horas para P.
falciparum, P. vivax e P. ovale e 72 horas para P. malarie (Albrecht, 2008). O ciclo
no inseto ocorre quando a fêmea do mosquito ao realizar o repasto sanguíneo em
um hospedeiro infectado, ingere os gametócitos. Esses gametócitos, uma vez no
intestino dos insetos irão se diferenciar em gametas masculino e feminino, que uma
vez fecundados originam os zigotos. Os zigotos se transformam em oocinetos
móveis, que irão invadir o epitélio intestinal do mosquito até alcançar a lamina basal,
e posteriormente se converterão em esporozoítos, migrando para as glândulas
salivares do inseto, para iniciar um novo ciclo.
Figura 1: Ciclo biológico do Plasmodium sp.
(Fonte: CDC, 2013)

5
1.3 Aspectos imunológicos
Segundo Queiroz et al., (2008), o inicio da resposta imunológica se dá ainda
na fase exoeritrocítica, com a produção de citocinas pró-inflamatórias, como IFN-γ e
TNF-α, que agem como mediadores na eliminação do parasito através da via óxido
nítrico (NO) e da produção de interleucinas IL1, IL6 e IL8, assim como por meio das
células TCD8+ e TCD4+, que atuam através de ação citolítica ou por meio de
produção de citocinas pró-inflamatórias. Já na fase eritrocítica a resposta passa a
ser mediada por macrófagos e linfócitos TCD4+ auxiliares dos subtipos Th1 e Th2,
com produção de IFN-y, radicais de oxigênio e nitrogênio e produção de citocinas.
A exposição dos antígenos do parasito, no momento da ruptura dos
eritrócitos, é o que inicia as manifestações clássicas da malária, tais como febre
intermitente, náusea, cefaleia. Isso ocorre devido ação das citocinas inflamatórias
que geram danos teciduais e sistêmicos na tentativa de lutar contra a infecção. A
evolução da doença para a malária grave pode ocorrer em casos em que a doença é
negligenciada.
No momento em que ocorrem as primeiras esquizogônias sanguíneas
também da inicio a resposta humoral, devido aos antígenos parasitários lançados na
corrente sanguínea, antígenos estes que podem ser corpos residuais dos
esquizontes, glicoproteínas da superfície externa dos merozoítos, entre outros. A
resposta de fase aguda é direcionada para os diferentes estágios do
desenvolvimento do parasito, e as hemácias parasitadas apresentam em sua
membrana antígenos específicos do Plasmodium sp., o que auxilia no
reconhecimento pelo sistema imune do hospedeiro.
Um dos aspectos mais marcantes da infecção malárica é a
hipergamaglobulinemia, que se caracteriza por uma produção aumentada de IgG e
IgM (Mioto et al., 2012). A resposta imune protetora não é efetiva por causa da
diversidade e estrutura dos antígenos parasitários, o que acaba dificultando a
resposta imune. Além disso, os parasitos apresentam uma grande diversidade
antigênica, aumentando a capacidade de escape à resposta imune, adaptando-se
ao hospedeiro vertebrado.
A base da imunidade celular antimalárica é a hiperplasia do sistema
macrófagolinfóide. O principal dispositivo de defesa do organismo contra esses

6
parasitos é a fagocitose de muitos merozoítos e hemácias parasitadas, que ocorre
logo após cada esquizogonia (Mioto et al., 2012). O baço é o principal local no qual
ocorre essa limpeza, motivo pelo qual aumenta de volume (esplenomegalia), porém
no fígado e medula óssea pode ocorrer também essa limpeza, por meio do sistema
fagocítico mononuclear. As células auxiliares agem ativando linfócitos B e
aumentando resposta humoral, além de ativar macrófagos e células NK.
1.4 Malária grave
A gravidade da malária depende da relação entre hospedeiro (vulnerabilidade
e estado imunológico) e o Plasmodium sp. (espécie infectante e densidade
parasitária), (Gomes et al., 2011). Levando em consideração os aspectos do
hospedeiro, os idosos, as gestantes, crianças e os primoinfectados apresentam
maior chance de desenvolver as formas graves da doença. Ou seja, os fatores de
risco para o desenvolvimento da MG incluem idade superior a 65 anos, sexo
feminino (especialmente quando associado à gravidez), estado não imune,
condições médicas coexistentes, ausência de profilaxia antimalárica, atraso no
tratamento e o estado de gravidade da doença no momento da admissão (Trampuz
et al., 2003). A grande parte dos casos registrados de óbito por malária se deve a
infecção pelo P. falciparum, porém há registros de óbitos ocasionados pela infecção
por P. vivax.
De acordo com a OMS, os critérios utilizados para definir malária grave por P.
falciparum e por P. vivax são os mesmos, com diferença que na malária grave por P
vivax não é considerado o critério de hiperparasitemia e nem a densidade
parasitológica.
De acordo com alguns autores, a MG pode vir a ser definida como doença
multissistêmica, em que predomina resposta inflamatória a infecção, que gera uma
Síndrome de Resposta Inflamatória Sistêmica (SIRS), cuja manifestação clínica se
dá por meio de uma termo-desregulação, taquicardia, taquipineia, leucocitose em
que pode ou não ocorrer desvio a esquerda. Os achados clínicos de uma MG
incluem alguns aspectos, tais como alterações de consciência (em que pode ocorrer
o coma), prostração com fraqueza generalizada, dificuldade em alimentar-se,
respiração acidótica, choque, crises convulsivas múltiplas, icterícia, hemoglobinúria,
hemorragias espontâneas, edemas pulmonares, insuficiência renal. Além de

7
achados laboratoriais que incluem hipoglicemia, acidose metabólica, anemia severa,
hiperparasitemia, hiperlactemia e hipovolemia (Tabela 2).
1.4.1 Complicações clínicas
Entre as complicações decorrentes da MG podem ser citados complicações
neurológicas (malária cerebral), anemia severa da malária, insuficiência respiratória
com acidose metabólica, quadro hiperglicêmico, insuficiência renal, hipotensão,
choque e distúrbios da coagulação.
A malária cerebral (MC) é a apresentação clínica mais comum e causa de
morte em adultos em áreas de baixa endemicidade (Trampuz et al., 2003). Os
sintomas iniciais da MC podem estar incluídos convulsão generalizada ou focal, com
sonolência e confusão mental. Os sintomas podem evoluir, levando o paciente a
apresentar um quadro de coma, que pode durar algumas horas ou até mesmo dias.
Acredita-se que uma das causas para o surgimento da MC e consequente
encefalopatia estão associados com sequestro eritrocítico de eritrócitos parasitados
em vênulas da circulação cerebral.
A Anemia Grave na Malária (AGM) ocorre devido a um conjunto de fatores,
em que a destruição de eritrócitos e a eritropoiese ineficaz exercem fundamental
papel na etiologia. Os eritrócitos são destruídos de modo direto pela infecção
parasitária, em decorrência da lise que ocorre quando os parasitos rompem a célula
após o ciclo de desenvolvimento. Porém não é só o processo de lise mediado pelo
parasito que acarreta toda a anemia. O sistema imune também exerce seu papel,
uma vez que os demais eritrócitos, mesmo não estando parasitados, sofrem
fagocitose e lise mediada pelo completo. Além disso, são observados contagem de
reticulócitos bem baixos, o que condiz com uma produção ineficaz de eritropoietina
(EPO) e em consequência produção insuficiente de células eritroides. Estudos de
Miller et al., (1994) e Casals-Pascual et al., (2006) demonstraram que há uma
associação entre os níveis elevados de TNF-α e IFN-γ, que são citocinas
inflamatórias, e a ineficaz produção de Epo. Estudos in vitro com cultura de células
humanas, mostraram que o TNF-α possui a capacidade de suprimir a proliferação de
células progenitoras da linhagem eritroide. Segunda a OMS o quadro de anemia
grave é confirmado por meio da dosagem de hemoglobina e do hematócrito. Em

8
crianças, a AGM apresenta hemoglobina <5 g/dL e um hematócrito < 15%. Já em
adultos, a hemoglobina se apresenta < 7 g/dL e hematócrito < 20% (OMS, 2004).
A insuficiência respiratória e a acidose metabólica ocorrem frequentemente na
malária grave e apresenta-se como um mau prognóstico da doença. O paciente
desenvolve a Síndrome do Desconforto Respiratório Agudo (SDRA), apresentando
sintomas como taquipneia, respiração profunda e ofegante, hipoxemia e por fim a
própria insuficiência respiratória (IRA), que leva ao paciente a necessidade de uso
de apoio ventilatório. Na MG, a IRA é definida como o aparecimento súbito de um
infiltrado pulmonar bilateral com desiquilíbrio da razão ventilação/perfusão < 300
mmHg e uma pressão da artéria pulmonar < 18 mmHg, sem evidências de
hipertensão arterial (Monteiro, 2011). A acidose metabólica severa tem sua origem
explicada na glicólise anaeróbica do hospedeiro e na interferência do sequestro
parasitário na microcirculação, além de produção aumentada de lactato,
hipovolemia, a insuficiência hepática e renal.
A hipoglicemia na MG apresenta-se com uma dosagem de glicose menor que
40 mg/dL. É bastante frequente, principalmente em crianças e grávidas. Tem como
sintomas dispneia, taquicardia, sudação profunda, coma, convulsões, alterações
posturais e ansiedade, sintomas estes que se confundem com a própria MG, o que
acaba gerando certa negligencia quanto ao diagnóstico. Segundo Weatherall et al.,
(2002) e Breman et al., (2010), crianças que apresentam concentrações normais de
insulina, porém desenvolvem quadro de hipoglicemia pode ter uma disfunção na
gliconeogênese hepática e o aumento do consumo de glicose nos tecidos periféricos
e pelos parasitos.
No quadro de insuficiência renal, a creatinina apresenta-se maior que 265
mmol, e o paciente desenvolve um quadro de oligúria ou anúria, podendo ser
necessário diálises. Apresenta-se comumente em adultos não imunes.
O quadro de colapso circulatório ocorre, segundo a OMS, quando a pressão
arterial sistólica encontra-se menor que 70 mmHG em adultos e 50 mmHg em
crianças. O quadro pode ocorrer de forma súbita, em conjunto com edema
pulmonar, acidose metabólica e hemorragia massiva.
O quadro de colapso circulatório ocorre, segundo a OMS, quando a pressão
arterial sistólica encontra-se menor que 70 mmHG em adultos e 50 mmHg em

9
crianças. O quadro pode ocorrer de forma súbita, em conjunto com edema
pulmonar, acidose metabólica e hemorragia massiva.
Tabela 2: Critérios para definir malária grave
(Fonte: Monteiro, 2011).
O quadro de colapso circulatório ocorre, segundo a OMS, quando a pressão
arterial sistólica encontra-se menor que 70 mmHG em adultos e 50 mmHg em
crianças. O quadro pode ocorrer de forma súbita, em conjunto com edema
pulmonar, acidose metabólica e hemorragia massiva.
Os distúrbios da coagulação constitui outro aparecimento presente nos casos
de MG. Segundo a OMS, a trombocitopenia é um achado que aparece de forma
constante nos casos de malária grave, porem não estão associados a episódios de
hemorragia. Os episódios hemorrágicos são decorrentes de distúrbios de
coagulação, tais como a Coagulação Intravascular Disseminada (CID). Na CID
aparece aumento dos produtos de degradação da fibrina (D-dímero), concentração

10
abaixo do normal de fibrinogênio, TP prolongado, associado com trombocitopenia.
Assim sendo, os indivíduos doentes apresentam hemorragias em mucosas, em
locais de punção e em casos graves podem apresentar necrose tecidual.
1.5 Modelo animal
O modelo animal passou a ser utilizado nos estudos de malária grave devido
a dificuldades no acompanhamento de casos de malária cerebral humana, assim
como a limitação no que concerne na analise dos processos patológicos. O modelo
mais utilizado é aquele que emprega camundongos infectados por diferentes
espécies de Plasmodium murinos adaptados (tabela 3), que acaba permitindo um
melhor estudo e elucidação de aspectos e características patogênicas da doença.
Porém é importante uma melhor analise dos dados obtidos do estudo com o
modelo animal, uma vez que apresentam algumas diferenças. Um exemplo seria
que no modelo murino o tipo predominante de célula sequestrado na microcirculação
cerebral é o leucócito, enquanto que na humana o tipo celular é o eritrócito (Queiroz
et al, 2008).
O Plasmodium berghei ANKA é utilizado em grande parte dos estudos que
tratam da malária cerebral, uma vez que apresenta divisão característica entre os
camundongos que são susceptíveis a infecção e aqueles que apresentam
resistência. Aqueles susceptíveis a infecção pode ocorrer o aparecimento de
angustia respiratória, anemia, nefrite e malária cerebral, achados semelhantes a
malária por P. falciparum. Já as espécies resistentes, não morrem pela MC, uma vez
que não a desenvolvem, morrendo em torno de vinte dias devido a grave anemia. A
infecção por P. berghei ANKA apresenta-se com um intenso processo inflamatório.
Outra espécie de Plasmodium utilizado no estudo de malária cerebral é o P. yoelii
17XL. A infecção por esse parasito se apresenta com um processo inflamatório
menos intenso que o P. berghei ANKA e sequestro de eritrócitos parasitados na
microcirculação. Já infecção por P. chabaudi pode cursar de maneira letal ou não,
dependendo da genética do hospedeiro.
1.6 Introdução aos Plasmodium de roedores
O isolamento da primeira cepa de Plasmodium de roedor foi feito por Vincke e
Lips, em 1948, na África central, mais precisamente no Zaire. O Plasmodium foi
encontrado em glândulas salivares da espécie silvestre de Anopheles dureni, e em

11
1954 foi possível encontrar o parasita na circulação de roedores silvestres
Thamnomys surdaster, Praomys jacksoni e Leggada bela (Garnhan, 1966; Queiroz,
2007). O primeiro isolado foi, então, nomeado de Plasmodium berghei. A cepa NK65
foi isolada em 1964 e a cepa ANKA em 1966, ambas encontradas em Anopheles
dureni. Estudos tem mostrado a capacidade de Anopheles stephensi, Anopheles
gambiae e Aedes aegypti atuarem como vetores de P. berghei (Queiroz, 2007). A
partir de então, varias espécies e subespécies de Plasmodium de roedores foram
isoladas (Peters; Richards, 1984).
A primeira transmissão bem sucedida em laboratório, utilizando-se de
mosquitos Anopheles quadrimaculatus, foi feita em 1964 por Yoeli e colaboradores;
procedimento foi então modificado no ano de 1965 por Vanderberg e Yoeli, e em
1966 foi padronizado.
As espécies e subespécies de Plasmodium são divididas em dois grupos
principais: o grupo berghei, composto por Plasmodium berghei, P. yoelii e P. y.
nigeriensis, e o grupo vinckei contém P. vinckei e P. chabaudi.
Os estágios sanguíneos do grupo berghei apresentam predileção por
eritrócitos imaturos no sangue de roedores de laboratório, diferentemente daqueles
pertencentes ao grupo vinckei, que são caracterizados por não apresentarem tal
preferencia (Killick-Kendrick, 1978).
O P. chabaudi apresenta duas subespécies, o P. chabaudi chabaudi e P.
chabaudi adami. O parasito apresenta fácil repique em camundongos e ratos, tendo
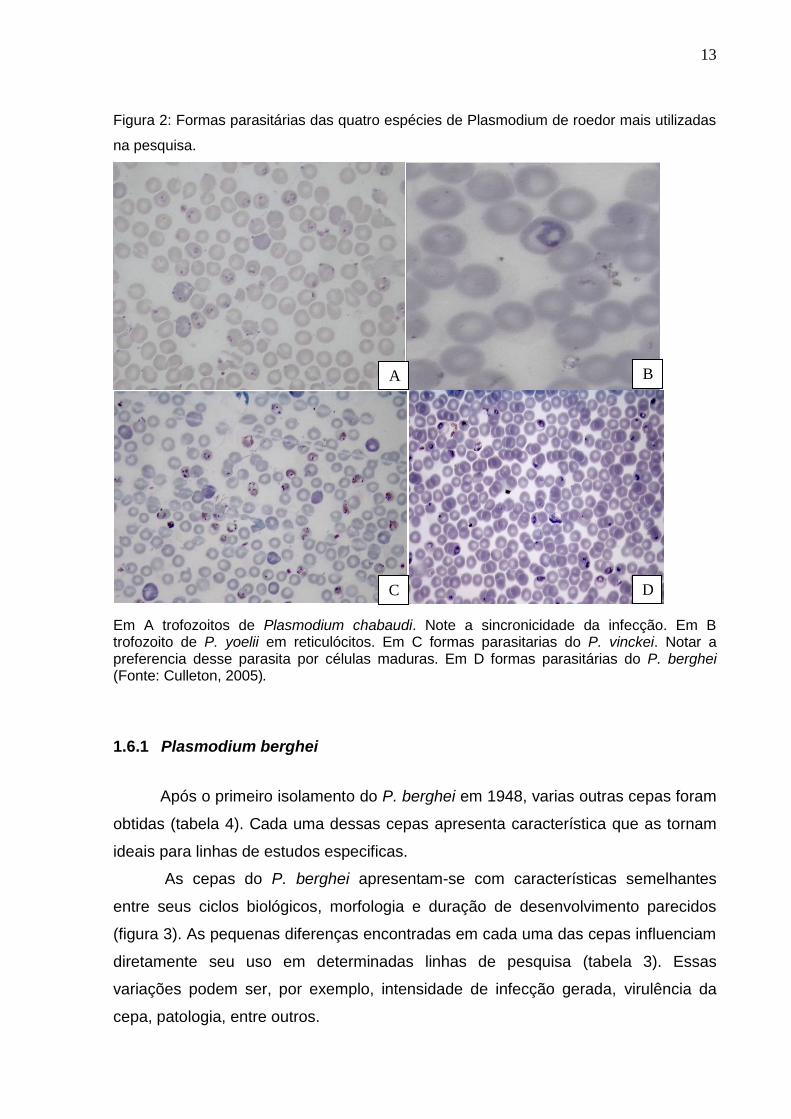
preferencia por parasitar eritrócitos maduros. Possui infecção síncrona, com
periocidade de 24 horas (figura 2A). O P. yoelii apresenta infecção assíncrona, com
periocidade de 22-25 horas, tendo preferencia por eritrócitos imaturos (figura 2B). O
P. vinckei foi isolado primeiramente em 1952, porem só teve seu reconhecimento
como espécie individual em 1975. Possui quatro subespécies reconhecidas, sendo
elas P. vinckei vinckei, P. vinckei petteri, P. vinckei lentum e P. vinckei brucechwatti.
Apresenta preferencia por eritrócitos maduros, com infecção síncrona (figura 2C).

12
Tabela 3: Infecção por Plasmodium em diferentes linhagens de camundongos.
(Fonte: Queiroz et al., 2008)
As particularidades de cada espécie de Plasmodium murino são o que os
torna ideais no estudo e sua relação com a malária humana. Como o P. chabaudi e
o seu uso principalmente no que concerne os mecanismos de resistência a
medicamentos e evasão do sistema imune. Uma das características dessa espécie é
a capacidade de gerar infecção crônica e não-letal. Já o P. berghei, como falado
anteriormente, é capaz de gerar uma infecção grave, por isso é o indicado em
estudos que envolve a malária grave e seus aspectos, como a malária cerebral, por
exemplo. O P. yoelii é mais utilizado em estudos sobre a biologia dos estágios extra
eritrocítico, tais como o estagio hepático e a ação do sistema imune nessas formas
(Otto et al., 2014).
13
Figura 2: Formas parasitárias das quatro espécies de Plasmodium de roedor mais utilizadas
na pesquisa.
Em A trofozoitos de Plasmodium chabaudi. Note a sincronicidade da infecção. Em B trofozoito de P. yoelii em reticulócitos. Em C formas parasitarias do P. vinckei. Notar a preferencia desse parasita por células maduras. Em D formas parasitárias do P. berghei (Fonte: Culleton, 2005).
1.6.1 Plasmodium berghei
Após o primeiro isolamento do P. berghei em 1948, varias outras cepas foram
obtidas (tabela 4). Cada uma dessas cepas apresenta característica que as tornam
ideais para linhas de estudos especificas.
As cepas do P. berghei apresentam-se com características semelhantes
entre seus ciclos biológicos, morfologia e duração de desenvolvimento parecidos
(figura 3). As pequenas diferenças encontradas em cada uma das cepas influenciam
diretamente seu uso em determinadas linhas de pesquisa (tabela 3). Essas
variações podem ser, por exemplo, intensidade de infecção gerada, virulência da
cepa, patologia, entre outros.
A B
C D

14
Figura 3: Formas parasitárias de P. berghei ANKA encontradas na circulação.
(Fonte: LUMC, 2014)
As cepas do P. berghei apresentam, em geral, preferência por parasitar
reticulócitos. Ainda que, foi visto que certas cepas apresentam uma maior predileção
por reticulócitos, como as cepas de NK65 em camundongo Swiss se comparadas as
cepas de ANKA (LUMC, 2014). Porém, quando a mudança é feita, e a comparação
ocorre em ratos Wistar, é observado que as cepas de P. berghei ANKA apresentam
maior predileção por reticulócitos que as cepas de NK65. É importante salientar que
o Plasmodium berghei também é capaz de infectar eritrócitos maduros.
O ciclo assexuado é em geral assincrônico, com os diferentes estágios
sanguíneos simultaneamente presentes no sangue no decorrer da infecção. Podem
ocorrer Infecções múltiplas de eritrócitos em que há poliparasitismo extensivo,
apresentando massa citoplasmática com aparência contínua, com aspecto de
esquizonte maduro (Queiroz, 2007).

15
A produção de merozoítos é variada de acordo com o estagio celular
parasitado. De uma forma geral, a infecção em eritrócitos maduros produz de 6 a 12
merozoítos enquanto que em reticulócitos, pode chegar a 18.
Os esquizontes eritrocíticos, principalmente de P. berghei ANKA costumam
desaparecer da circulação sanguínea sendo sequestrados no baço e pulmão,
podendo também ocorrer sequestro no tecido adiposo. Entre uma cepa e outra pode
ocorrer pequenas diferenças na quantidade de esquizontes circulantes. Por
exemplo, cepas de k173 em geral apresentam quantidades maiores de esquizontes
circulantes no sangue periférico que as cepas de ANKA.
O fator virulência da infecção depende em grande parte de fatores genéticos
do animal infectado. Uma infecção com P. berghei causa morte do animal em três
semanas em média, quando os animais podem morrer de complicações da doença,
como anemia grave. Porém, naqueles animais mais susceptíveis, a morte pode
ocorrer em torno de uma semana, devido complicações graves, como malária
cerebral. Outro fator que pode influenciar na infecção é dose de infecção.
camundongos infectados com uma concentração de 107, por exemplo, podem
apresentar evolução da doença mais rápida do que aqueles infectados com 105
células parasitadas. (ANDRADE-NETO, V. F., et al.).
A produção de gametócitos é influenciada por fatores ambientais. Algumas
estirpes aparentemente perdem a capacidade de produzir gametócitos. Tal fato é
visto em animais em que a passagem de cepa é feito por via sanguínea de um
animal para o outro, sem o estimulo do mosquito.
A duração da esporogonia no mosquito depende grandemente da
temperatura ambiental, e a invasão de glândulas salivares é diferente em cada um
dos hospedeiros invertebrados utilizados no laboratório, aja vista que foi observado
que ocorre uma invasão maior em A. stephensi e em A. quadrimaculatos. Já a
invasão de esporozoítos em células hepáticas varia de acordo com o roedor
utilizado, principalmente. Sendo que os roedores podem ser desde resistentes a
altamente susceptíveis a infecção hepática.

16
Tabela 4: cepas de Plasmodium berghei.
(Fonte: LUMC, 2014)
1.6.2 Ultraestrutura e invasão celular de hospedeiros vertebrados e
invertebrados
A estrutura do P. berghei é bastante semelhante aos dos parasitas humanos.
Assim como os esporozoítos humanos, o P. berghei também apresenta a
capacidade de invadir hepatócitos. Resumidamente, a invasão se inicia por meio da
invaginação da membrana plasmática da célula hospedeira, formando um vacúolo
parasitóforo, que engloba o esporozoítos. Em um período que gira em torno de 47-
52 horas os esporozoítos evoluem até esquizonte maduro, podendo gerar 1500 a
8000 merozoítos. Após a ruptura da célula hepática os merozoítos são libertados na
corrente sanguínea passando a invadir glóbulos vermelhos. Estudos não
encontraram formas hipnozoítas de P. berghei no fígado.
Os merozoítos sanguíneos do P. berghei possuem semelhanças morfológica
com os merozoítos de Plasmodium humano, tendo conservadas características
relacionadas a invasão eritrocítica, tais como roptrias, micronemas, grânulos densos
e proteínas de superfície que desempenham importante papel na invasão da célula.
Após invadir o eritrócito, o merozoítos segue o seu desenvolvimento a
trofozoito, apresentando aumento no tamanho celular e citoplasmático. Assim como
na infecção humana, o trofozoito consome a hemoglobina, produzindo hemozoína,

17
que pode ser vista como grânulos pigmentares no citoplasma. O desenvolvimento
até atingir o estagio de trofozoito maduro leva cerca de 16 horas, e no final o
parasita duplica seu DNA, seguindo para a divisão celular, e entra no estagio de
esquizonte. Essa esquizogonia leva cerca de 6 a 8 horas, em que o parasito sofre
seguidas replicações de seu DNA, dividindo seu núcleo, por meio endomitótico, em
diversas vezes, originando uma célula sincicial com 8 a 24 núcleos. A duração total
do desenvolvimento sanguíneo gira em torno de 22 a 24 horas, e esquizontes em
eritrócitos maduros geralmente geram menos merozoítos que aqueles que se
apresentam em reticulócitos.
Já o período de desenvolvimento de gametócito de P. berghei leva em torno
de 36 horas. Há diferenças evidentes entre gametócitos de P. falciparum e P.
berghei, como a forma de banana para o primeiro e formas redondas a ovais no
segundo. De uma forma geral, a morfologia, duração de desenvolvimento e
produção de gametócitos são bastante semelhantes com as de outras espécies que
causam malária em humanos e em primatas não humanos.
O desenvolvimento do P. berghei em mosquitos é bastante estudado. Durante
o repasto sanguíneo, os mosquitos ingerem gametócitos masculinos e femininos, e,
uma vez dentro do intestino médio, esses gametócitos se transformam por meio de
gametogênese em microgametas, que são os gametas masculinos, e
macrogametas, gametas femininos. De uma forma geral, os macrogametas saem do
eritrócito e se transformam um único gameta esférico, enquanto os microgametas
sofrem uma serie de três ciclos de replicação do DNA, gerando oito microgametas,
processo esse conhecido por exflagelação (Ríos-Velásquez, 2014). Em pesquisas
foram relatados a presença de pelo menos três gatilhos iniciadores no processo de
diferenciação de gametócitos, que são: queda de temperatura do sangue em pelo
menos 5ºC em relação ao hospedeiro vertebrado, aumento de pH, que vai de 7,3 até
7,8-8,0 e a presença de fatores de ativação de gametócitos (GAF). O processo de
formação do gameta e morfologia são comparáveis nas espécies que infectam
roedores e humanos, além de que proteínas de superfície desses gametócitos são
conservadas nessas espécies (LUMC, 2014).
A fecundação dos macrogametas pelos microgametas e a formação do zigoto
ocorrem de forma rápida, geralmente de 2 a 3 horas após o repasto sanguíneo. Nas

18
seguintes horas, por volta de 10-30 horas, o zigoto se transforma em oocineto
móvel, migrando até as células do epitélio intestinal, e as atravessa, entrando em
contato com a lâmina basal, onde se transforma em oocisto. A formação desses
oocistos começa em torno de 24 horas após o repasto (figura 4). Os esporozoítos
são formados através de esporogonia e quando liberados circulam pela hemocele,
chegando até as glândulas salivares, onde as invade. Uma vez nas glândulas,
alcançam os ductos salivares do inseto, podendo ser inoculadas no hospedeiro
vertebrado no momento de um novo repasto sanguíneo (Ríos-Velásquez, 2014).
Figura 4: Ciclo de desenvolvimento do Plasmodium no mosquito: transformações do
Plasmodium e tempo de duração de cada fase
(Fonte: Ríos-Velásquez, 2014)

19
2 OBJETIVOS
2.1 Objetivo geral
Realizar a infecção in vivo de camundongos utilizando Plasmodium berghei.
2.2 Objetivos específicos
Descrever o Plasmodium berghei, espécie usada nas pesquisas in vivo com
malária no LABMAT
Estabelecer rotina de manutenções in vivo das cepas P. berghei NK65 e
ANKA
Identificar formas parasitárias em esfregaços sanguíneos
Observar e determinar curva de parasitemia
Determinar média de sobrevida dos roedores infectados.

20
3 METODOLOGIA
Os animais utilizados pelo LABMAT para o estudo in vivo da malária são
camundongos Swiss fêmeas, com idade em torno de 6-8 semanas, cedidos
principalmente pelo biotério central da UFRN. Foram mantidos no biotério da
Universidade Federal do Rio Grande do Norte, onde recebiam água e a alimentação
ad libitum. Seu uso foi aprovado pelo comitê de ética para uso de animais (CEUA -
UFRN nº 043/2010).
O laboratório trabalha com duas linhagens do Plasmodium berghei: cepas
NK65 e ANKA. O parasito é mantido por meio de passagens semanais, utilizando-se
de protocolo definido de infecção.
3.1 Infecção dos animais
A infeção para manutenção das cepas ocorre semanalmente, e para a
passagem são utilizados de três a cinco animais por caixa, para cada cepa. Os
animais são inoculados, via intraperitoneal, com uma concentração de 105 hemácias
parasitadas, por Plasmodium berghei NK65 e ANKA.
3.2 Preparação do inóculo
O inóculo que corresponde à concentração de 105 hemácias parasitadas
segue basicamente três etapas para a sua preparação, etapas essas que serão
descritas a seguir.
3.2.1 Determinação da porcentagem de hemácias parasitadas
Para a determinação da porcentagem foram utilizadas alíquota de sangue de
animais infectados obtido por meio de incisão na região terminal da cauda, e
posteriormente foi feito esfregaços sanguíneos, corados pelo método de Giemsa. A
parasitemia foi determinada utilizando o calculo:
%Hp = Hi * 100
Ht
onde Hp corresponde a porcentagem de hemácias parasitadas, Hi é o numero de
hemácias infectadas contadas e Ht é o total de hemácias.

21
3.2.2 Determinação da concentração de hemácias
Alíquotas do sangue analisado (de 3 gotas de sangue extraídas da cauda do
animal), foram diluídas em solução formada por um mL de PBS (tampão fosfato-
salino com pH em 7,4) e 100 µL de Citrato de sódio (3,8%, pH 7,22). As hemácias
foram contadas em câmara de Neubauer (figura 5), e a concentração foi
determinada utilizando-se o seguinte calculo:
H/mL = n*100*10*5*103,
onde H/ml corresponde ao numero de hemácias por mL de sangue, n é o numero de
hemácias contadas nos cinco quadrantes centrais da câmara, 100 é o inverso da
diluição, 10 é o inverso da profundidade da câmara, 5 corresponde a área contada e
103 conversão de mm3.
Figura 5: Câmara de Neubauer
As hemácias devem ser contadas no quadrante central, nos quatro quadrados menores da extremidade e no quadrado menor central.
3.2.3 Diluição do inóculo
Para determinar o numero de hemácias parasitadas, multiplica-se a
concentração de hemácias, pela porcentagem de parasitemia e divide-se por 100.
Cada animal deve receber uma concentração de 200 µL de solução com 105
hemácias parasitadas, por via intraperitoneal. Para tal é feito o calculo do fator de
diluição, que pode ser resumido da seguinte forma:

22
Fd = Hp*107
5*107
Com esse valor é calculado quanto de solução mãe (PBS/Citrato + alíquota de
sangue) deve ser utilizada para produzir uma nova solução com 107 células
infectadas. Para isso o volume final é importante. Por exemplo, para preparar uma
solução de 1 mL, deve-se pegar o valor final da solução e dividir pelo fator de
diluição, chegando a quantidade que seria necessário obter da solução 107.
sol107 = 1000
Fd
Partindo da solução inicial, segue-se uma sequencia de diluições seriadas até atingir
a concentração de 105 células parasitadas.
3.3 Parasitemia dos animais
A parasitemia é acompanhada por meio de esfregaços sanguíneos realizados
nos dias 3, 5, 7 e 9 pós-infecção.
3.4 Congelamento de Cepa
Também é feito no LABMAT o congelamento das cepas, para uso posterior. O
procedimento é simples.
Primeiro é determinado a parasitemia do animal infectado. Logo após, o
animal é anestesiado com solução de Ketamina e Xilazina numa proporção de 30%
e 70%. A profundidade da anestesia é avaliada utilizando a presença ou ausência de
resposta a um estimulo. Quando o animal não apresenta resposta a um estimulo
doloroso, é feito a crucificação e em seguida o sangue é obtido por meio de punção
cardíaca. A amostra sanguínea é coletada em seringa com citrato de sódio, e
colocada em criotubo contendo 0,5 mL de criopreservante glicerol. A amostra é
então, preservada em nitrogênio líquido, ou em ultrafreezer, até que venha a ser
utilizada no futuro.
Após a punção, os animais são eutanasiados por meio de deslocamento
cervical.

23
3.5 Descongelamento de Cepa
A amostra congelada é deixada em temperatura ambiente até atingir o
completo descongelamento. Em seguida é diluída em PBS em 1:2 e 1:4. Logo após
é administrado 200 µL de solução em cada animal.

24
4 RESULTADOS
4.1 Identificação de fases parasitárias
As formas parasitárias do Plasmodium berghei estão muito bem definidas na
literatura, tendo sua morfologia bastante conhecidas, assim como as características
apresentadas em cada momento da infecção.
4.1.1 Trofozoitos
De uma forma geral, as formas parasitárias das cepas NK65 e ANKA são
semelhantes.
Figura 6: Formas trofozoitas do Plasmodium berghei ANKA.
(fonte: Arquivo pessoal, 2016)
4.1.2 Esquizonte
Entre 16-18 horas pós-infecção é possível encontrar esquizontes na
circulação do animal. Geralmente esses esquizontes possuem em torno de 12 a 24
merozoítos. Os esquizontes de P. berghei ANKA sofrem sequestro e por esse
motivo são vistos bem menos na circulação.

25
Figura 7: Trofozoitos e esquizontes, P. berghei ANKA
(Fonte: Arquivo pessoal, 2016).
4.2 Crescimento parasitário
Os estudos de Vincke e Van de Bulcke em 1949 evidenciaram a capacidade
infectiva do Plasmodium berghei. Foi observado que a parasitemia aumenta
progressivamente, até a morte do animal, apresentando um platô.
A parasitemia dos animais infectados no LABMAT é acompanhada nos dias 3,
5, 7 e 9 pós-infecção, e a porcentagem média de hemácias parasitadas é anotada
(tabela 5).
Tabela 5: Média de parasitemia dos camundongos infectados
Dia pós-infecção Média de
parasitemia
0 dia 0
3º dia 5%
5º dia 10%
7º dia 15%
9º dia 20%
Nas cepas de P. berghei ANKA é observado também a alta capacidade
agregativa dos eritrócitos por meio da produção de knobs (figura 5). Esses knobs
são protuberâncias visíveis na superfície das hemácias parasitadas, que as torna
facilmente aderidas ao endotélio. Essa adesão pode ser vista na prática no momento
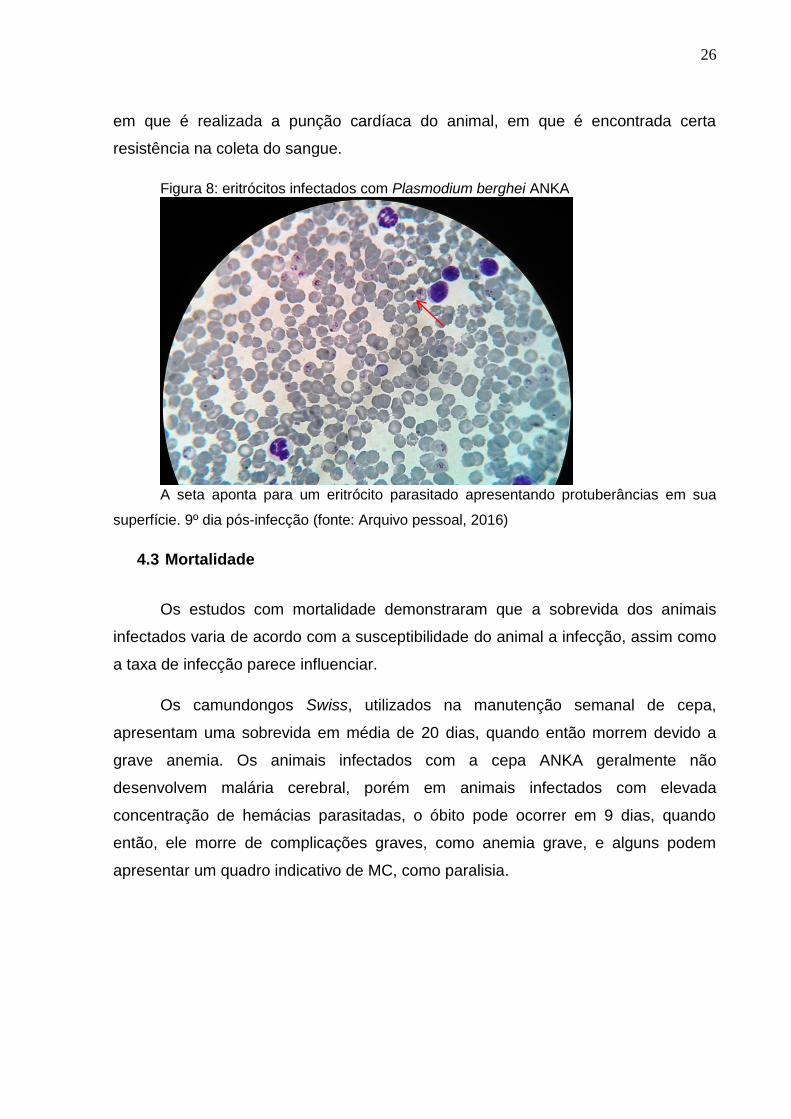
26
em que é realizada a punção cardíaca do animal, em que é encontrada certa
resistência na coleta do sangue.
Figura 8: eritrócitos infectados com Plasmodium berghei ANKA
A seta aponta para um eritrócito parasitado apresentando protuberâncias em sua
superfície. 9º dia pós-infecção (fonte: Arquivo pessoal, 2016)
4.3 Mortalidade
Os estudos com mortalidade demonstraram que a sobrevida dos animais
infectados varia de acordo com a susceptibilidade do animal a infecção, assim como
a taxa de infecção parece influenciar.
Os camundongos Swiss, utilizados na manutenção semanal de cepa,
apresentam uma sobrevida em média de 20 dias, quando então morrem devido a
grave anemia. Os animais infectados com a cepa ANKA geralmente não
desenvolvem malária cerebral, porém em animais infectados com elevada
concentração de hemácias parasitadas, o óbito pode ocorrer em 9 dias, quando
então, ele morre de complicações graves, como anemia grave, e alguns podem
apresentar um quadro indicativo de MC, como paralisia.

27
5 DISCUSSÃO
É sabido que a infecção por P. berghei apresenta um aspecto característico,
em que a parasitemia sanguínea tende a apresentar um aumento progressivo até se
chegar a um platô. Thurston, em 1950, demostrou que, independente da via de
infecção (intraperitoneal ou intravenosa) o curso da infecção segue o mesmo
caminho, seguindo o padrão da infecção até a morte do animal. Os estudos dele
demonstraram também que animais infectados com algo em torno de 15 milhões de
parasitas, por via intraperitoneal, e apresentavam parasitemia superior a 20% no
quinto dia pós-infecção evoluíam para a morte (Mercato; Coatney, 1951).
Em relação aos estudos desses autores (gráfico 1), a parasitemia medida em
nossos camundongos segue em concordância com os resultados encontrados. A
infecção segue um curso semelhante, com crescimento parasitário progressivo até o
alcance do platô, no qual a parasitemia se mantém estável. Os camundongos
infectados no LABMAT apresentam parasitemia semelhantes àquelas descritas na
literatura, com a morte dos animais ocorrendo por volta do oitavo dia pós-infecção
nos animais que desenvolviam a doença grave.
Gráfico 1: Percentual médio de parasitemia em Infecção por Plasmodium berghei em camundongos Swiss.
(Fonte: Mercado; Coatney, 1951).

28
Os estudos utilizando-se do modelo murino animal são importantes
ferramentas no auxilio a erradicação da malária no mundo. Tais estudos focam-se
desde o desenvolvimento de fármacos que visam o tratamento de pessoas
infectadas até o desenvolvimento de vacinas. Pesquisas com a malária cerebral têm
como objetivos entender o seu mecanismo e encontrar formas eficazes de
tratamento, seja por meio da utilização de um fármaco ou por meio de uma vacina.
Os estudos com vacinas, buscam formas de produzir imunidade duradora em
hospedeiros vertebrados ou produzir uma interrupção no ciclo do parasito,
interrompendo a transmissão seja para o mosquito, seja para o hospedeiro
vertebrado.
As pesquisas envolvendo os modelos murinos buscam entender a complexa
interação parasito-hospedeiro e as formas de contornar a gravidade gerada nas
complicações da doença. Com esse entendimento, é possível chegar a produção de
formas de combate ao parasito, seja por meio de desenvolvimento de fármacos
eficazes no tratamento de indivíduos contaminados, seja no desenvolvimento de
vacinas que podem, não somente gerar uma imunidade duradora, mas também
interromper o ciclo biológico do parasito. A meta da Organização Mundial da Saúde
é que em 2030, 90% dos casos de malária sejam reduzidas no mundo, e para isso,
os investimentos em pesquisas se fazem extremamente necessários.

29
6 REFERÊNCIAS
BIOMEDCENTRAL. Clinical review: severe malaria. Disponível em: <http://ccforum.biomedcentral.com/articles/10.1186/cc2183>. Acesso em: 10 out. 2016.
BIREME. Dia mundial da malária 2016 enfoca a erradicação da doença. Disponível em: <http://www.paho.org/bireme/index.php?id=328%3adia-mundial-da-malaria-2016-enfoca-a-erradicacao-da-doenca&option=com_content>. Acesso em: 10 out. 2016.
BMC BIOLOGY. A comprehensive evaluation of rodent malaria parasite genomes and gene expression. Disponível em: <http://bmcbiol.biomedcentral.com/articles/10.1186/s12915-014-0086-0>. Acesso em: 15 out. 2016.
CASTRO, I.R.; RODRÍGUEZ, M.D.C.G. Análisis proteómico de Plasmodium, el agente causal de la malaria. Salud pública Méx, Cuernavaca, v. 51, supl. 3, p. 395-402, 2009.
CDC - DPX. Malaria. Disponível em: <https://www.cdc.gov/dpdx/malaria/>. Acesso em: 13 dez. 2016.
CULLETON, R. A. Pictorial Guide to Rodent Malaria Parasites. Edinburgh, 2005
DANTAS, G.R. Avaliação da Atividade antimalárica de extratos obtidos de algas marinhas no litoral do Rio Grande do Norte. 2012. p. 72. Dissertação para titulo de mestre, Universidade Federal do Rio Grande do Norte. Centro de biociências, Programa de Pós Graduação em Ciências Biológicas, 2012.
DELVES, MICHAEL et al. The Activities of Current Antimalarial Drugs on the Life Cycle Stages of Plasmodium: A Comparative Study with Human and Rodent Parasites. Ed. James G. Beeson. PLoS Medicine v. 9, p. 14, (2012).

30
FIOCRUZ. Malária. Disponível em: <http://www.fiocruz.br/bibmang/cgi/cgilua.exe/sys/start.htm?infoid=104&sid=106>. Acesso em: 08 out. 2016.
GOMES, A. P. et al. Malária grave por Plasmodium falciparum. Ver. Brasileira de Tererapia Intensiva [online]. vol.23, n.3, p. 358-369, 2011.
JHORAWAT, RAJESH, PANKAJ BENIWAL, e VINAY MALHOTRA. Plasmodium Vivax Induced Hemolytic Uremic Syndrome: An Uncommon Manifestation That Leads to a Grave Complication and Treated Successfully with Renal Transplantation. Tropical Parasitology. v. 5, p. 127–129, 2015.
Killick-Kendrick, R. Taxonomy, Zoogeography and Evolution. In:______; Petters, W. Rodent malaria. New York: Academic Press, 1978. p. 1-52
LACERDA, M. V. G. et al. The neglected Plasmodium vivax: are researchers from endemic areas really concerned about new treatment options?. Rev. Sociedade Brasileira de Medicina Tropical, [S.l.], v. 40, p. 489-490, 2007.
LUMC. Plasmodium berghei - model of malaria. Disponível em: <https://www.lumc.nl/org/parasitologie/research/malaria/berghei-model/>. Acesso em: 08 ago. 2016.
MAIER, A. G. Malaria parasite proteins that remodel the host erythrocyte. Nature reviews | Microbiology, v. 7, p. 341-354, 2009.
MALARIA JOURNAL. The relevance of non-human primate and rodent malaria models for humans. Disponível em: <http://malariajournal.biomedcentral.com/articles/10.1186/1475-2875-10-23>. Acesso em: 10 out. 2016.
MBENGUE, A; YAM, XY; BRAUN-BRENTON C. Human erythrocyte remodelling during Plasmodium falciparum malaria parasite growth and egress, British Journal of Haematology, 2012, v. 157, p. 171–179.

31
MERCADO, T.I; COATNEY, G.R. The course of the blood-induced plasmodium berghei infection in white mice, Federal Security Agency, Public Health Service, National Institutes of Health, Bethesda14,Marylan
MONTEIRO M.F.A.D. Efeito de factores do hospedeiro e parasitários na susceptibilidade à malária e gravidade da doença: estudo de Alguns Polimorfismos Eritrocitários e das Espécies de Plasmodium. 2011. p. 268 . Dissertação (Doutorado em medicina tropical) – Instituto de Higiene e Medicina Tropical, Universidade Nova de Lisboa, Lisboa, 2011.
NAING, CHO et al. Is Plasmodium Vivax Malaria a Severe Malaria?: A Systematic Review and Meta-Analysis. Ed. Hernando A. del Portillo. PLoS Neglected Tropical Diseases. V. 8, p. 11, (2014).
OTTO, THOMAS D.; et al. A Comprehensive Evaluation of Rodent Malaria Parasite Genomes and Gene Expression. BMC Biology, vol. 12, p. 86, 2014.
PETERS, W; RICHARDS, W. H.G.. “Antimalarial drugs i: Biological Background, Experimental Methods, and Drug Resistance”. New York: Springer-Verlag Berlin Heidelberg, 1 ed, p.491, 1984.
PORTAL DA SAÚDE. Situação epidemiológica - dados. Disponível em: <http://portalsaude.saude.gov.br/index.php/o-ministerio/principal/leia-mais-o-ministerio/662-secretaria-svs/vigilancia-de-a-a-z/malaria/11346-situacao-epidemiologica-dados>. Acesso em: 30 set. 2016.
PORTAL SAÚDE. Brasil registra menor numero de casos de malária nos últimos 35 anos. Disponível em: <http://portalsaude.saude.gov.br/index.php/cidadao/principal/agencia-saude/23368-brasil-registra-menor-numero-de-casos-de-malaria-nos-ultimos-35-anos>. Acesso em: 10 out. 2016.
PRICE, R. N.; et al. Vivax malaria: neglected and not benign. The American Journal of Tropical Medicine and Hygiene, p. 77-79, 2007.
QUEIROZ, N.L. Avaliação da resposta inflamatória cerebral em camundongos BALB/c e C57Bl/6 infectados por Plasmodium berghei cepa NK65. 2007. p. 93. Dissertação para título de mestre, Universidade Federal de Minas Gerais, Belo

32
Horizonte. Instituto de Ciências Biológicas, Programa de Pós-Graduação em Parasitologia, 2007.
______; TEIXEIRA M.M; TEIXEIRA A.L. Imunopatogênese da malária cerebral. Rev. Brasileira de Neurologia, vol. 44 (1), p. 13-19, 2008.
RAMIRO, R.S.; REECE, S.E; DARREN, J.O. Molecular Evolution and Phylogenetics of Rodent Malaria Parasites. BMC Evolutionary Biology v. 12, p. 219, 2012.
RÍOS-VELÁSQUEZ, C.M. Suscetibilidade e resposta imune de mosquitos Anopheles (Diptera: Culicidae) da Região Amazônica Brasileira quando infectados experimentalmente por Plasmodium vivax. 2014. p. 110. Tese para título de doutor em ciências, Biologia celular e molecular, Belo Horizonte, 2014.
ROCHA E SILVA L.F.; et al. In vivo antimalarial activity and mechanisms of action of 4-nerolidylcatechol derivatives. Antimicrob Agents Chemother. Vol. 59, e. 6, p. 3271–3280, 2015.
SHUTLER, D. et al. Rodent Malaria Parasites Plasmodium Chabaudi and P. Vinckei Do Not Increase Their Rates of Gametocytogenesis in Response to Mosquito Probing. Proceedings of the Royal Society B: Biological Sciences. p. 2397–2402, 2005.
SINGH, B.; DANESHVAR, C. Human Infections and Detection of Plasmodium knowlesi. Clinical Microbiology Reviews. v. 26, ed. 2, p. 165-184, 2013.
TAVARES, J. Atividade antimalárica in vivo e aspectos toxicológicos do composto 3,4 dihidroxibenzaldeído. Monografia (Graduação) – Universidade Federal do Rio Grande do Norte. Centro de Biociências, 52 f., 2015.
THE MALARIA UNIT. A pictorial guide to rodent malaria parasites. Disponível em:<http://www.tm.nagasakiu.ac.jp/malariaunit/culleton_lab/photos/pages/plasmodium_vinckei_files/rodentmalaria.pdf>. Acesso em: 10 out. 2016.
WANDERLEY, B.M.S. Atividade antiplasmódica e toxicológica de plantas medicinais usadas popularmente no Brasil: uma abordagem etnobotânica. 2012. 94 f.

33
Dissertação para titulo de mestre, Universidade Federal do Rio Grande do Norte. Centro de biociências, Programa de pós-graduação em Ciências biológicas. 2012.
WHO. Severe malaria. Tropical Medicine and International, published by John Wiley & Sons., v. 19 (Suppl. 1), p. 7–131, 2014.
WRIGHT, C. W. Recent developments in research on terrestrial plants used for the treatment of malaria. Nat Prod Rep, [S.l.], v. 27, p. 961-968, 2010.
Recommended

















