Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DE JUIZ DE FORA
INSTITUTO DE CIÊNCIAS BIOLÓGICAS
DEPARATAMENTO DE PARASITOLOGIA,
MICROBIOLOGIA E IMUNOLOGIA
LEDA MARÍLIA FONSECA LUCINDA
MECANISMOS DE INDUÇÃO DA APOPTOSE POR
GLICOCORTICÓIDES E SEUS EFEITOS NO CÂNCER
JUIZ DE FORA
2009

LEDA MARÍLIA FONSECA LUCINDA
MECANISMOS DE INDUÇÃO DA APOPTOSE POR
GLICOCORTICÓIDES E SEUS EFEITOS NO CÂNCER
Monografia de Conclusão de Curso de
Especialização em Parasitologia e
Microbiologia e Imunologia do ICB da
UFJF, para obtenção do Título de
Especialista em Parasitologia,
Microbiologia e Imunologia.
Orientador: Prof. Dr Henrique C. Teixeira
JUIZ DE FORA
2009

Lucinda, Leda Marília Fonseca.
Mecanismos de indução da apoptose por glicocorticóides e seus efeitos no câncer / Leda Marília Fonseca Lucinda -- 2009.
44 f.
Trabalho de conclusão de curso (Especialização em Parasitologia, Microbiologia e Imunologia)-Universidade Federal de
Juiz de Fora, Juiz de Fora, 2009.
1. Apoptose. 2. Neoplasias. CDU 576.3

Leda Marília Fonseca Lucinda
MECANISMOS DE INDUÇÃO DA APOPTOSE POR GLICOCORTICÓIDES E SEUS EFEITOS NO CÂNCER
Monografia do Curso de Especialização em Parasitologia, Microbiologia e Imunologia do Instituto de Ciências Biológicas da Universidade Federal de Juiz de Fora, como parte dos requisitos para obtenção do título de Especialista. Aprovada em: 23/10 /2009 Banca Examinadora: Prof.Dr. Henrique Couto Teixeira Profa. Ms. – Michele Fernandes Rodrigues
Juiz de Fora 2009

Aos meus pais, Odaléa e José Jader, meu tio José Henrique e meu irmão
Garrone por terem sido meus mais fiéis amigos me apoiando e fortalecendo em
todos os momentos, inclusive os mais difíceis.
Ao meu marido Maycon, pela sua amizade, amor e extrema compreensão.

AGRADECIMENTOS
À Deus, que me deu forças, meios e coragem nos momentos mais difíceis,
para que eu sempre prosseguisse em minha vida sem pensar em desistir, e que me
iluminou durante toda essa caminhada.
Agradeço ao meu orientador professor Dr. Henrique Teixeira, por ter me dado
a grande oportunidade de ser sua aluna, pela sua paciência, dedicação
compreensão e competência na orientação deste trabalho.
À minha professora de língua inglesa Cassiana Monteiro Boya pelo auxílio na
tradução da língua inglesa e por seus ensinamentos.
Á todos os professores do Programa de Pós-Graduação em Parasitologia,
Microbiologia e Imunologia pelos ensinamentos importantes para a minha formação
acadêmica.
Aos meus grandes amigos da pós-graduação, Pollyanna, Matheus, Daniela e
Tatiane e aos demais colegas que caminharam junto comigo na busca de mais um
grande objetivo de nossas vidas.

RESUMO
A apoptose é um processo complexo que remove células danificadas ou
envelhecidas do corpo e ocorre em uma ampla variedade de organismos. A ativação
da apoptose é um importante mecanismo pelo qual drogas citotóxicas matam as
células tumorais. Defeitos ou problemas na sinalização da apoptose contribuem para
que as células tumorais se tornem resistentes. Esta revisão aborda o efeito do
tratamento com GCs na resistência de tumores sólidos ao tratamento convencional
(quimioterapia e radioterapia) e sua relação com a modulação da apoptose de
células tumorais. Os GCs são amplamente utilizados no tratamento de pacientes
com câncer devido a sua potente propriedade pró-apoptotica em células linfóides,
seu poder de redução de náuseas, e sua capacidade de diminuir a toxicidade do
tratamento quimioterápico. Entretanto, apesar dos GCs mediarem a proteção dos
tecidos normais, eles também podem proteger tumores sólidos da apoptose induzida
por agentes citotóxicos. Pouco se sabe sobre como os GCs afetam a resposta à
quimioterapia ou radioterapia em tumores sólidos não hematológicos. O aumento da
FOXO3A fosforilada em células cancerígenas pode explicar, em parte, a resistência
a apoptose nessas células. Foi descrito que células tumorais tratadas com diferentes
GCs, como a dexametasona, apresentaram alteraçóes morfológicas, como aumento
na expressão de CD38, CD44 e CD58 que podem implicar no aumento da
sobrevivência das células tumorais ao tratamento citotóxico. A dexametasona
também é responsável pela ativação dos receptores de GCs que aumentam a
expressão e a atividade de SGK-1 (proteína kinase1 induzida por glicocorticóide no
soro) e MKP-1 (MAPK fosfatase-1), que estão diretamente ligadas a redução da
apoptose. Portanto, é importante reconsiderar o amplo uso de GCs no tratamento de
pacientes com tumores sólidos, pois se por um lado podem apresentar um efeito
benéfico na redução da toxicidade de tecidos normais, podem, por outro lado,
aumentar a resistência a apoptose de células cancerígenas.
Palavras-Chave: Apopotose. Glicocorticóide. Câncer. Linfócitos.

ABSTRACT
Apoptosis is a complex process that removes injured or aging cells from the body
occurring in a wide variety of organisms. Activation of the apoptosis pathways is a
key mechanism by which cytotoxic drugs kill tumor cells. Defects or problems in
apoptosis signaling contribute to the resistance of tumors. This review explores the
effect of the treatment with glucocorticoids in the resistance of solid tumors to the
conventional treatment (chemotherapy and radiotherapy) as well as its relation with
the modulation of apoptosis in tumor cells. Glucocorticoids have been widely used in
the treatment of patients with cancer due to its potent pro-apoptotic property in
lymphoid cells, its reduction of nausea and capacity to diminish the toxicity of the
chemotherapy treatment. However, although glucocorticoids mediate the protection
of normal tissues, they also protect solid tumors from apoptosis induction by cytotoxic
agents. Little is known about how glucocorticoids affect the response to
chemotherapy or radiotherapy in solid, non-hematological tumors. Increased
phospho-FOXO3A level in cancer cells may explain in part the resistance to
apoptosis in these cells. It was described that tumor cells treated with different
glucocorticoids such as dexamethasone, showed morphological changes which
resulted in an increased expression of CD38, CD48 and CD58 prolonging the
survival of tumor cells during the cytotoxic therapy. The dexamethasone is also
responsible for the activation of the glucocorticoids receptors that increase the
expression and activity of SGK-1 (serum- and glucocorticoid-induced kinase-1) and
MKP-1 (MAPK phosphatase-1), which are directly connected to apoptosis reduction.
Thus, it is important to reconsider the widespread use of glucocorticoids in the
treatment of patients with solid tumors, as if on one hand they can present an
advantageous effect in the reduction of the toxicity in normal tissue, on the other
hand they can increase cancer cells resistance to apoptosis.
Key-Words: Apoptosis. Glucocorticoids. Cancer. Lynphocytes.

LISTA DE FIGURAS
Figura 1 - Duas maiores vias de apoptose em células de mamíferos........................17

LISTA DE ABREVIATURAS
AIF - “Apoptosis-inducing factor”/ fator indutor de apoptose
APAF1 - “Apoptotic protease activating factor 1”/ fator de ativação de proteases pró-
apoptóticos 1
ATP - Adenosina trifosfato
CED - “Cell death abnormal” / morte celular anormal
c-FLIPP - “FLICE-inhibitory protein”/ proteína inibidora de FLICE
DD- “Death domain”/ Domínio de morte
DED – “Death effector domain”/Efetor de domínio de morte
DISC - “Death-inducing signaling complex”/ complexo de sinalização indutor de
morte
DNA - Ácido desoxirribonucléico
FADD - “Fas-associated protein with death domain”/ domínio de morte associado ao
Fas
FasL - “Fas ligand”/Fas ligante
GCs - Glicocorticóides
IAPs - “ Inhibitor of apoptosis proteins”/ proteínas inibidoras da apoptose.
IқK - “Inhibitory kappa B kinase”/ inibidor de kappa B quinase
IL-2 - Interleucina -2
IL-6 - Interleucina - 6
IL-10 - Interleucina -10
mdm-2 - “murine double minute-2”/murina duplo minuto-2
MHC- “Major histocompability complex”/ Complexo de histocompatibilidade principal
MKP-1 - “MAPK phosphatase-1”/ MAPK fosfatase-1
NFk-B - “nuclear factor kappa B”/ fator de transcrição nuclear kappa B
SGK-1 - “Serum/ glucocorticoid regulated kinase 1”/ soro e quinase-1 dependente de
glicocorticóide
Smac/DIABLO - “Second mitochondria-derived activator of caspase/direct inhibitor of
apoptosis-binding protein with low pl” - segundo ativador mitocôndrial de caspase/
proteína de ligação-IAP direta com baixo pl.
TCR - “T cell receptor”/ receptor de células T

TGF-beta 1 - “Transforming growth factor beta 1”/ fator transformador de crescimento
beta 1
TNF - “Tumor necrosis factor”/ fator de necrose tumoral
TNFR - “Tumor necrosis factor receptor”/ receptor de fator de necrose tumoral
Treg – Células T regulatórias
UPR - “Unfolded Protein Response” / resposta as proteínas não dobradas

SUMÁRIO
1 INTRODUÇÃO....................................................................................................... 12
2 OBJETIVO ............................................................................................................. 15
3 REVISÃO DA LITERATURA ................................................................................ 16
3.1 APOPTOSE......................................................................................................... 16
3,1,1 Características morfológicas e bioquímicas......................................................19
3.1.2 Alteração da forma e estrutura celular..............................................................20
3.1.3 Controle genético da apoptose......................................................................... 21
3.1.4 Bcl-2................................................................................................................. 22
3.1.5 C-myc............................................................................................................... 23
3.1.6 p53.................................................................................................................... 24
3.1.7 Caspases como iniciadoras e executoras da apoptose................................... 24
3.1.8 Proteínas inibidoras da apoptose..................................................................... 25
3.2 APOPTOSE E CÂNCER..................................................................................... 26
3.3 GLICOCORTICÓIDES (GCs).............................................................................. 28
3.4 ATUAÇÃO DOS GLICOCORTICÓIDES (GCS) NA INDUÇÃO DA APOPTOSE
LINFÓCITOS E EM CÂNCERES.............................................................................. 29
4 DISCUSSÃO.......................................................................................................... 34
5 CONCLUSÃO ........................................................................................................ 36
6 REFERÊNCIAS...................................................................................................... 37

1 INTRODUÇÃO
Os glicocorticóides (GCs) são uma classe de hormônios esteróides que
exercem um amplo papel na atividade antiinflamatória, imunossupressora e
antineoplásica e têm habilidade de induzir a apoptose em uma variedade de células,
incluindo linfócitos T e B (TUCKERMANN et al., 2005).
O efeito imunossupressor dos GCs se deve em parte a sua ação no bloqueio
da maturação de linfócito T maduros através da inibição da produção de IL-2 e da
inibição de expressão do receptor para interleucina-2 (IL-2). A morte dos timócitos
corticais após o tratamento com GCs é precedida por mudanças morfológicas
características da apoptose como condensação da cromatina nuclear, fragmentação
do DNA (ácido desoxirribonucléico) e outras (NIETO et al., 1991)
A apoptose é definida como uma série de eventos moleculares, envolvendo a
perda da integridade da membrana mitocondrial, condensação de cromatina,
fragmentação do DNA e geração de vesículas apoptóticas. Mediada por caspases,
esse processo culmina em um desarranjo orquestrado da célula e fagocitose da
célula morta (HENGARGTNER, 2000).
Para o desenvolvimento das células T, muitos dos rearranjos dos receptores
para antígenos (TCR - “T cell receptor”/receptor de células T) são inúteis, porque
estes receptores não podem se ligar aos alelos do MHC (“Major Histocompability
Complex”/ Complexo de Histocompatibilidade Principal) que estão presentes no
indivíduo, portanto a seleção positiva é um passo crucial que caracteriza a
diferenciação das células T restritas ao MHC, permitindo que somente células que
expressam TCR capazes de interagir com o complexo peptídeo-MHC próprio com
baixa afinidade continuem no processo de diferenciação, escapando da morte por
apoptose. Em seguida, as células T que interagem fortemente com o complexo
peptídio-MHC no timo também entram em apoptose, e somente células que
reconhecem com baixa avidez o complexo peptídeo-MHC são capazes de deixar o
timo, processo denominado seleção negativa, que direciona a resposta imune
especialmente contra antígenos microbianos estranhos (ABBAS, LICHTMAN e
POBER, 2003).
12

Os progenitores fracamente reativos que irão se tornam maduros, podem ser
encontrados nos órgãos linfóides secundários e participam da resposta imune a
antígenos estranhos (HOGQUIST et al., 2005). Entretanto, células T potencialmente
reativas podem escapar do processo de seleção negativa tornando inerente o perigo
de auto-imunidade. Felizmente, as células T potencialmente auto-reativas estão
sobre um forte controle na periferia (vasos e órgão periféricos). A característica da
tolerância central das células T é a deleção clonal; mecanismo que envolve a
apoptose dos progenitores de células T, que têm alta afinidade por antígenos
próprios, durante o processo de seleção positiva e negativa, no timo (PALMER,
2003). Desta forma, a apoptose é importante na regulação da resposta imune, uma
vez que na seleção das células T, apenas células que não reajam fortemente com
antígenos próprios podem completar o processo de maturação, e as células que
apresentarem alta afinidade a antígenos peptídicos próprios em associação com
moléculas do MHC irão sofrer apoptose e serão deletadas (ABBAS, LICHTMAN e
POBER, 2003).
Foi descrito que a diminuição da sinalização do GCs resulta em aumento da
sensibilidade da sinalização via TCR, causando a apoptose de timócitos que
normalmente seriam selecionados positivamente. Portanto os timócitos que são
selecionados positivamente nesta situação podem ter uma baixa avidez por
antígenos próprios/MHC, isto pode ser facilmente testável, uma vez que a alteração
na resposta ao GCs pode afetar a média e a variedade de TCR. Entretanto existem
evidência que altos níveis de GCs podem resgatar alguns timócitos com alta avidez
por antígenos próprios/MHC que normalmente deveriam ser deletados, ainda não se
tem certeza do mecanismo pelo qual estes timócitos sobrevivem (VACCHIO e
ASHWELL, 2000). Existem evidências de camundongos com autoimunidade
espontânea apresentarem elevados níveis de cortisol endógeno (WICH e
KROEMER,1993).
A apoptose de linfócitos induzida por GCs sintéticos é um importante
componente para protocolos terapêuticos usados no tratamento de várias formas de
linfomas e leucemias (WANG et al., 2006), e ainda a dexametasona é rotineiramente
dada as mulheres com câncer de mama antes que elas recebam quimioterapia
(REDONDO et al., 2007). Entretanto dados recentes demonstraram efeitos anti-
apoptóticos dos GCs nas células de carcinoma, prevenindo a citoxicidade induzida
por radiação e quimioterápicos que conduziriam a morte celular (RUTZ e HERR,
13

2004). Portanto os GCs conferem resistência a apoptose em pacientes com câncer
sólidos (não hematológicos), este efeito pode prejudicar a resposta ao tratamento
em pacientes com câncer.
Portanto o conhecimento dos mecanismos de indução de apoptose por GCs
deve ser investigado mais amplamente visto que sua utilização pode modular a
resposta de linfócitos T e interferir na indução da morte celular por apoptose em
pacientes em uso de radioterapia e quimioterápicos.
14

2 OBJETIVO
Este trabalho se propõe, através de uma revisão bibliográfica, avaliar os
mecanismos pelos quais os GCs interferem no tratamento com antineoplásicos,
considerando sua capacidade de induzir resistência a apoptose em alguns cânceres.
15

3 REVISÃO DA LITERATURA
3.1 APOPTOSE
A apoptose é uma forma de morte celular na qual uma seqüência de eventos
leva a eliminação de células sem a liberação de substâncias nocivas na área
circundante. É caracterizado pela formação de bolhas na membrana plasmática (os
“blebs”), pela condensação de cromatina, pelas alterações do citoplasma bem como
pela clivagem internucleossomal do DNA, ocorrida devido à ação de uma
endonuclease endógena (LEE et al., 1999). A apoptose desempenha um importante
papel na manutenção da homeostasia pela eliminação das células desnecessárias,
danificadas ou velhas (GALLUZI et al., 2007).
O termo “apoptose” vem do grego, onde o prefixo “apo” significa separação e
o sufixo “ptose” pode ser traduzido como queda, sendo associado com a queda das
folhas de uma árvore ou pétalas de flores. Esse termo é uma alusão à liberação de
pequenos corpos revestidos por membrana, resultantes da fragmentação da célula
apoptótica, denominados corpos apóptoticos (DUQUE-PARRA, 2005).
A ativação de morte celular programada pode ocorrer através de duas vias
principais. A via extrínseca, mediada pela ativação de receptores de morte celular
localizados na membrana citoplasmática. O FasL (“Fas ligand”/Fas ligante) e o TNF
(“tumor necrosis factor”/ fator de necrose tumoral) se ligam ao Fas e ao TNFR
(“tumor necrosis factor receptor”/ receptor de fator de necrose tumoral)
respectivamente, induzindo a formação do DISC (“death-inducing signaling
complex”/ complexo de sinalização indutor de morte), o DISC recruta a Caspase-8 e
ativa a cascata de ativação das procaspases (BUDIHARDJO et al., 1999). A via
intrínseca é ativada por estresse extracelular e/ou intracelular, como fatores de
privação de crescimento, hipóxia, dano ao DNA e indução de oncongene. Sinais que
são transferidos em resposta a este estresse convergem diretamente para a
mitocôndria, uma série de eventos bioquímicos é induzida e resulta na
permeabilização da membrana mitocondrial, onde há liberação de fatores
apoptogênicos como citorocromo c, AIF (“apoptosis-inducing factor”/ fator indutor de
16

apoptose), ATP (adenosina trifosfato) e proteínas de choque térmico. Ocorre a
formação do apoptossoma, uma proteína complexa que contém o citocromo c,
APAF1 (“apoptotic protease activating factor 1”/ fator de ativação de proteases pró-
apoptóticos 1) e caspase-9. Uma vez que a citocromo c é liberada, a cascata de
ativação da caspase é irreversível (GESKE e GERSCHENSON, 2001;
BUDIHARDJO et al., 1999). Portanto como resultado final de ambas as vias, ocorre
a ativação das caspases, proteases, que quebram proteínas celulares específicas e
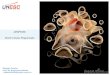
estão associadas à degradação do DNA (figura 1) (FISHER, 2001).
Figura 1: Duas maiores vias de apoptose em células de mamíferos adaptado de
HENGARTNER MO. The biochemistry of apoptosis. Nature, v.407, n.1, p. 685-687, 2000. A ligação do CD95 L ao CD95 induz a formação do complexo de sinalização indutor de morte
DISC, o complexo recruta via FADD, moléculas múltiplas de procaspases-8, induzindo a ativação da caspase-8, que pode ser bloqueada pelo recrutamento de c-FLIP. O mecanismo mitocondrial ocorre através da ativação de membros pró-apoptóticos da família Bcl-2. Proteínas pró-apoptóticas e anti-apoptóticas competem para regular a saída de citocromo c. se as pró-apoptóticas vencem, ocorre a
CD95L
Substrato apoptótico
Dano ao DNA
CD95
Citocromo C
Apoptossomo
Caspase-8
17

liberação de uma série de moléculas do compartimento mitocondrial, principalmente citocromo c, que se associa ao APAF-1 e a procaspase-9 para formar o apoptossomo.
O mecanismo de receptor de morte e o mecanismo mitocondrial convergem para a ativação da caspase-3. A ativação da caspase-3 é antagonizada pelas IAPs, que são antagonizadas pela proteína Smac/DIABLO liberada pela mitocôndria. A ativação da caspase-3 leva a apoptose da célula e sua eliminação.
A sinalização e integração entre o receptor de morte e a via mitocondrial é feita através do Bid, um membro da família de moléculas pró-apoptóticas Bcl-2, a caspase-8 faz a mediação da clivagem da Bid, aumentando a atividade de morte celular, resultando na sua translocação para a mitocôndria, onde irá promover a liberação do citocromo c. Na maior parte das vezes, esta integração é mínima, e os dois mecanismos ocorrem independentemente um do outro.
As células que sofrem apoptose morrem de uma forma ordenada, sem
prejudicar as células vizinhas. A célula encolhe e condensa. O citoesqueleto sofre
colapso, o envelope nuclear é desmontado, e o DNA nuclear quebra-se em
fragmentos. A superfície celular é alterada, exibindo propriedades que levam a célula
moribunda a ser rapidamente fagocitada tanto pelas suas células vizinhas como por
fagócitos, em especial macrófagos, antes de ocorrer qualquer liberação do seu
conteúdo. Isso não somente evita danos conseqüentes da necrose celular, mas
permite que os componentes orgânicos da célula morta possam ser reciclados para
a célula que a ingeriu. A ocorrência da apoptose depende de uma família de
proteases que apresenta uma cisteína no sítio ativo e cliva suas proteínas alvo em
ácido aspártico específico. Sendo denominadas caspases. As caspases são
sintetizadas nas células na sua forma inativa, as quais são ativadas pela clivagem do
ácido aspártico por outras caspases. Uma vez ativada, as caspases clivam e então
ativam outras procaspases, resultando em uma cascata de amplificação proteolítica.
Algumas caspases ativadas clivam outras proteínas-chave na célula. Algumas
clivam a lâmina nuclear, por exemplo, causando a quebra irreversível da lâmina
nuclear, outras clivam uma proteína que normalmente mantém a enzima
degradadora do DNA na forma inativa, liberando a DNAse para digerir o DNA no
núcleo da célula. Assim, a célula desmancha-se rápida e ordenadamente e os
corpos apoptóticos são rapidamente fagocitados e digeridos por outras células
(BRASILEIRO-FILHO, 2004).
As procaspases são ativadas por proteínas adaptadoras que fazem com que
múltiplas cópias de procaspases específicas, chamadas iniciadoras, se unam e
formem um complexo. São dotadas de uma pequena atividade de protease e,
quando são forçadas a entrar no complexo, sua proximidade faz com que uma clive
a outra, desencadeando sua mútua ativação. Em outros casos a formação do
18

complexo causa uma mudança conformacional que ativa as procaspases. A caspase
no topo da cascata é ativada dentro de momentos, clivando a próxima procaspase
para estimular o sinal da morte, que se estende por toda a célula. A ativação da
procaspase pode ser desencadeada de fora da célula por uma ativação de
receptores da morte na superfície da célula. Quando as células estão danificadas,
podem também matar a si próprias desencadeando a agregação de procaspases e a
ativação de dentro da célula (ALBERTS et al., 2004).
Os corpos apoptóticos podem ser identificados ao microscópio óptico pela
coloração de hematoxilina e eosina, como massas redondas ovais com o citoplasma
intensamente eosinofílico e a cromatina condensada, e são comumente observados
dentro de fagolisossomos (GESKE e GERSCHENSON, 2001). Quantificação de
apoptose pela microscopia óptica é possível, entretanto a visualização das células
apoptóticas torna-se muito difícil pelo processo acontecer em poucas horas e as
células serem rapidamente fagocitadas (GAVRIELI, SHERMAN e BEM-SASSON,
1992). A microscopia eletrônica permite a análise estrutural de todos os estágios da
apoptose mas não é prático para análise de grandes áreas teciduais. Outros
métodos de detecção da apoptose vêm sendo amplamente utilizados, como a
técnica TUNEL para a identificação da fragmentação de DNA (KRAVTSOV, DANIEL
e KOURI,1999), as marcações imuno-histoquímicas para os diferentes tipos de
caspases e para fita simples de DNA (ssDNA), hibridização in situ, que é uma
técnica pela qual se identificam seqüências específicas de nucleotídeos em cortes
histológicos (LORO et al., 2003) e a citometria de fluxo através da marcação dupla
com Anexina V e iodeto de propídio (RODRIGUES et al, 2009).
3.1.1 Características morfológicas e bioquímicas
A morte celular programada ou apoptose pode ocorrer em células individuais
ou em grupos de células em áreas maiores e contínuas (como ocorre após lesões).
As células apoptóticas apresentam um padrão de alterações morfológicas muito
semelhante independente da situação investigada ou do agente utilizado para
induzi-la experimentalmente (KUMAR, ABBAS e FAUSTO, 2004). Porém a apoptose
não é o único tipo de morte celular, a necrose também é uma importante forma de
morte celular. A necrose resulta da ruptura da membrana celular e lise celular,
liberando moléculas que ativam a resposta inflamatória. A necrose é sempre
19

patológica, enquanto a apoptose é um processo fisiológico importante, mas que
pode ser ativado em situações patológicas (ALLEN, HUNTER e AGRAWAL,1997).
3.1.2 Alterações de forma e estrutura celular
Na apoptose as células se tornam mais arredondadas, perdem as junções de
adesão com as células vizinhas e com a matriz extracelular, podendo apresentar
bolhas ou vacúolos em seu citoplasma. No estágio mais avançado a célula se
condensa e se desintegra em corpos apoptóticos. A base para esta alteração
provavelmente envolve a reorganização de proteínas do citoesqueleto.
a) Alterações nucleares
Condensação da cromatina (picnose), ocorre marginação ao longo do
envoltório nuclear e mais tardiamente fragmentação do núcleo em várias partículas.
O DNA se fragmenta em fragmentos grandes de 50-300 kilobases, seguidos de
novas clivagens específicas entre nucleossomos por endonucleases dependentes
de Ca2+ e Mg2+, gerando fragmentos múltiplos de 180 a 200 pares de bases. Com a
desintegração da célula, estas partículas são empacotadas nos chamados corpos
apoptóticos, os quais são fagocitados por outras células (KUMAR, ABBAS e
FAUSTO, 2004).
b) Alterações das organelas
Maior parte das organelas se mantém morfologicamente intactas durante o
processo de apoptose. O retículo endoplasmático se torna dilatado, os ribossomos
são liberados do retículo endoplasmático rugoso se agregando uns aos outros. As
mitocôndrias inicialmente se mantêm intactas e posteriormente se condensam ou se
dilatam (KUMAR, ABBAS e FAUSTO, 2004).
Apesar da ausência de alterações estruturais significativas, atualmente sabe-
se que a maioria das organelas nas células apoptóticas sofrem alterações
bioquímicas sutis, particularmente proteólise parcial e permeabilização das
membranas. Ainda mais importante é que cada organela possui sensores que
detectam alterações específicas e ativam vias de transdução de sinais locais, estas
emitem sinais para comunicação entre organelas e finalmente podem ativar a
cascata apoptótica (FERRI e KROEMER, 2001).
O retículo endoplasmático participa da iniciação da apoptose através de pelo
menos dois mecanismos, a UPR (“Unfolded Protein Response” / resposta as
20

proteínas não dobradas) e pela sinalização de cálcio. Condições que comprometem
o dobramento protéico no retículo endoplasmático ativam a UPR, a qual pode corrigir
a falha no dobramento das proteínas ou ativar a apoptose. Perturbações nos níveis
de cálcio no retículo endoplasmático, assim como sua liberação rápida e intensa,
podem ser apoptogênicas (FERRI e KROEMER, 2001). O processo de apoptose no
retículo é regulado principalmente por proteínas da família Bcl-2 que serão descritas
mais à frente (FERRARI et al., 2002).
c) Alterações da membrana plasmática
A maioria das alterações da membrana é de natureza bioquímica e não
parecem envolver alterações morfológicas. Uma exceção é a perda de
microvilosidades, que parece ser decorrente da desfoforilização de proteínas que
normalmente estabilizam a ligação de microvilos da superfície celular às moléculas
de actina do citoesqueleto. Há ainda mudanças na fluidez da membrana devido a
modificações na síntese e degradação de lipídeos em conseqüência da ativação de
fosfolipases e esfingomielases (BRASILEIRO-FILHO, 2004).
A remoção dos restos celulares apoptóticos é feita por fagócitos profissionais,
e consiste no reconhecimento e subseqüente engolfamento destas estruturas. Para
que haja o reconhecimento pelo fagócito, as células apoptóticas expressam
diferentes moléculas na superfície sendo a principal o fosfolipídeo fosfatildilserina,
que reside normalmente no folheto interno da membrana citoplasmática e é
translocado para o folheto externo durante o processo de apoptose. Essa
translocação permite o reconhecimento destas células por macrófagos, resultando
na fagocitose sem que haja a liberação de componentes celulares pró-inflamatórios.
Assim as células apoptóticas são destruídas com um mínimo de comprometimento
do tecido adjacente e sem ativar os macrófagos a produzirem uma resposta
inflamatória (LAUBER et al., 2004).
3.1.3 Controle genético da apoptose
Estudos recentes têm contribuído significativamente para a compreensão dos
mecanismos moleculares controladores da morte celular por apoptose. Sua iniciação
e as inúmeras alterações celulares que fazem parte do processo de apoptose, bem
como sua inibição é dependente da atividade de genes determinados, embora os
mecanismos pelos quais estes genes atuam não estejam totalmente elucidados, já
21

se conhece parcialmente o mecanismo de controle genético da apoptose em
diferentes sistemas orgânicos e tecidos.
Os estudos envolvendo a participação de genes no controle de apoptose
iniciaram-se com nematódio Caenorhabditis elegans. A morte fisiológica nesses
organismos é controlada, principalmente, por 3 genes da família CED (“cell death
abnormal” / morte celular anormal): CED-3, CED-4 e CED-9, além de outras
proteínas (LIU e HENGARTNER, 1999). Nesses organismos o gene supressor da
apoptose CED-9 (homólogo ao gene humano Bcl-2) sempre está associado ao gene
CED-4 (homólogo à proteína APAF-1), o que impede a ativação de CED-3 (pró-
apoptótica). Quando a apoptose é iniciada, a proteína Bcl-1 (homóloga à proteína
humana Bax) se associa ao CED-9, liberando o CED-4 e levando a ativação do
CED-3. Em humanos, o processo é muito semelhante ao que ocorre com
Caenorhabditis elegans, ou seja, a proteína Bax se associa ao Bcl-2 induzindo a
liberação da APAF-1, ativando a caspase 9, induzindo a apoptose (HENGARTNER,
2000).
Os genes envolvidos no processo apoptótico podem agir inibindo ou
estimulando a apoptose, de acordo com a situação em que a célula ou tecido se
encontram, em decorrência disto eles são denominados anti-apoptóticos e pró-
apoptóticos respectivamente (LEE et al.,1999).
Entre os genes que apresentam ação anti-apoptótica está o Bcl-2, em
experimentos onde foram utilizados camundongos com expressão exarcebada
desse gene demonstraram um índice de morte celular diminuído no epitélio
seminífero (RODRÍGUEZ et al., 1997). Porém o gene Bcl-2 mesmo tendo essa
conhecida ação anti-apoptótica, sabe-se que ele não é capaz de inibir a apoptose
em quaisquer circunstâncias e parece não ter atividade em todos os tipos celulares
(COHEN et al., 1992). Em contrapartida, genes como c-myc, Bax, p53, e Fas fazem
parte dos genes pró-apoptóticos, que exercem importante função não só nas células
normais como também em células neoplásicas.
3.1.4 Bcl-2
Bcl-2 é uma família de proteínas de membrana mitocondrial, inicialmente
descrita em translocações cromossômicas de linfomas de células B foliculares
humanos (GESKE e GERSCHENSON, 2001). Os membros da família Bcl-2 são
22

fatores cruciais na regulação da apoptose. De acordo com critérios funcionais e
estruturais, são divididos em dois grupos, os membros antIAPsoptótico e os
membros pró-apoptóticos.
a)Grupos antipoptóticos: Bcl-2, Bcl-w, Bcl-xL.
b)Grupos pró-apoptóticos: Bad, Bak, Bax, Bid, Bim, Bcl-xS.
A proteína Bcl-2 bloqueia a liberação de citocromo C pela mitocôndria após
estímulo apoptogênico, impedindo a ativação de caspases. Mas o principal
mecanismo de ação destas moléculas é através da ligação e inibição das proteínas
pró-apoptóticas da família Bcl-2. Um dos mecanismos pelos quais estas proteínas
pró-apoptóticas controlam a apoptose é através da formação de poros na membrana
mitocondrial, permitindo a interação de várias proteínas envolvidas na regulação da
apoptose (REED et al., 1998).
Bax e Balk, originalmente localizadas no citoplasma, são translocadas para a
membrana mitocondrial externa após o início do processo apoptótico. Na membrana
elas sofrem alterações conformacionais que elevam a permeabilidade do poro
mitocondrial, levando a liberação de moléculas pró-apoptóticas, tais como citocromo
C, AIF e Smac/DIABLO (“second mitochondria-derived activator of caspase/direct
inhibitor of apoptosis-binding protein with low pl” - segundo ativador mitocôndrial de
caspase/ proteína de ligação-IAPS direta com baixo pl) (FAN et al., 2005).
As proteínas do grupo antIAPsoptótico podem se ligar seletivamente à
conformação ativa de Bax e prevenir sua inserção na membrana mitocondrial
externa, mantendo a permeabilidade normal do poro, e prevenindo assim, a
liberação dos fatores pró-apoptóticos (ANTONSSON e MARTINOU, 2000).
3.1.5 C-myc
O C-myc é um proto-oncongene que determina se a célula irá proliferar ou
entrar em processo de apoptose. Parece, que normalmente este gene programa a
célula para se desenvolver. No entanto, diante da ausência de fatores de
crescimento ou de um outro gene necessário para que este desenvolvimento ocorra,
a célula comete “suicídio”. Células que expressam esse gene e que, por algum
motivo, apresentam deficiência de algum outro fator estimulador de mitose,
interrompem seu desenvolvimento normal e entram em apoptose (COHEN et al.,
1992; FANIDI et al., 1992).
23

3.1.6 p53
O gene p53, um supressor de tumores que também regula a proliferação e a
morte celular, a expressão exarcebada da p53 pode tornar as células mais
susceptíveis a morte celular quando tratadas, por exemplo, com agentes
antineoplásicos (LANE, 1992; FANIDI et al., 1992).
Quando o DNA é danificado, a proteína p53 se acumula e sua função é parar
o ciclo celular na fase G1, dando mais tempo para o reparo da lesão antes que
ocorra a replicação do DNA. Porém, se este processo de reparo do DNA falhar, a
p53 ativa o processo de apoptose, prevenindo assim a replicação do DNA danificado
e o gradual acúmulo de lesões genéticas que poderia resultar em carcinogênese
(YU e ZHANG, 2005).
Depois que ocorre o reparo DNA, a p53 aumenta a transcrição da proteína
mdm-2 (“murine double minute-2”/murina duplo minuto-2) que age como inibidora da
p53. A mdm-2 se associa à p53, revertendo o bloqueio do ciclo celular promovendo
o avanço para a fase S (SHERR, 2000).
Quando os danos ao DNA não são passíveis de reparo, ocorre a ativação da
apoptose. Mutações no gene p53 resultam em um descontrole do ponto de
checagem de G1, possibilitando que células danificadas progridam para a fase S
sem reparar as lesões, ou entrar em apoptose (AGARWAL et al., 1998).
3.1.7 Caspases como iniciadores e executores da apo ptose
O reconhecimento de que os mecanismos de morte celular do nematódeo
Caenhorhbditis elegans são regulados da mesma maneira que ocorre na morte
celular da células humanas, foi extremamente importante para o entendimento da
apoptose (ELLIS et al., 1991)
O gene ced-3 é essencial para o desenvolvimento da morte celular em C.
elegans. O descobrimento que o ced-3 é uma cisteína protease, juntamente com a
clonagem e caracterização funcional do número dos homólogos ced-3 nos
mamíferos, teve um enorme impacto na ciência da apoptose. Estas proteínas, agora
chamadas de caspases, são reconhecidas como uma classe comum de proteínas
encontradas em diferentes organismos. É um consenso que em algumas situações
in vivo as células morrem por outros mecanismos além da ativação das caspases,
24

mas há dúvidas se as caspases são requeridas pelo menos nas mudanças
morfológicas mais proeminentes durante a apoptose (HACKER, 2000)
Estas importantes proteínas requeridas no processo de apoptose pertencem
à família da cisteínas proteases que têm a capacidade de reconhecer e clivar
substratos que possuam resíduos de aspartato (NICHOLSON e THORNBERRY,
1997). As caspases sinalizam para a apoptose e clivam estes substratos levando a
condensação e fragmentação nuclear, externalização de fosfolipídeos de membrana
que irão sinalizar para estas células serem fagocitadas por macrófagos
(NICHOLSON e THORNBERRY, 1997; BOATRIGHT e SALVASEN, 2003). São
conhecidas 14 caspases humanas, sendo que seis (caspases -3, -6, -7, -8, -9, -10)
participam da apoptose (BOATRIGHT e SALVASEN, 2003).
As enzimas podem interagir com receptores de membrana ou moléculas
adaptadoras que contenham domínios de morte, pois estes domínios também
existem nas caspases e a presença deles permite essa interação (NICHOLSON e
THORNBERRY, 1997). As caspases podem ser classificados de acordo com seu
pró-domínio e seu papel na apoptose. Caspases iniciadoras (Caspases -2, -8, -9, -10
e -12). possuem pró-domínios longos e estão envolvidas na iniciação da cascata
proteolítica. Caspases efetoras (Caspases-3 e -7). apresentam pró-domínios curtos
ou inexistentes, responsáveis pela clivagem de substratos (RUPNARAIN et al.,
2004). Entre os diversos substratos das caspases pode-se citar a mdm-2, uma
proteína que se liga à p53, mantendo-a no citoplasma. Ao ser clivada pelas
caspases, essa proteína libera a p53 que se transloca para o núcleo, ativando a
transcrição de genes pró-apoptóticos como o Bax (SCHULER, MAURER e
GOLDSTEIN, 2003).
3.1.8 Proteínas inibidoras da apoptose
As IAPs (“Inhibitor of apoptosis proteins”/ proteínas inibidoras da apoptose
são moléculas que exercem seu papel antiapoptótico através da capacidade de inibir
a atividade das caspases efetoras -3 e -7, da caspase iniciadora -9 e de modularem
o NF-ḳB (“nuclear factor kappa B”/ fator de transcrição nuclear kappa B)
(DEVERAUX e REED, 1999; SHI, 2002). Durante a apoptose, as IAPs são
removidas por uma proteína liberada da mitocôndria denominada Smac/DIABLO
(VERHAGEN et al., 2000; DU, FANG e WANG, 2000). Após o dano mitocôndrial
25

essa proteína é liberada para o citoplasma juntamente com o citocromo c, enquanto
o citocromo c liga-se a APAF-1 e ativa diretamente a caspase-9, Smac/DIABLO
remove as IAPs de sua ligação inibitória com as caspase (CHAI et al., 2000).
A c-FLIP (“FLICE-inhibitory protein”/ proteína inibidora de FLICE) também
inibe a apoptose através da sua ligação ao FADD (“Fas-associated protein with
death domain”/ domínio de morte associado ao Fas), que é uma proteína adaptadora
ligado ao Fas, prevenindo a ativação da caspase-8/Flice. Cinco diferentes membros
das IAPs já foram descritos: NAIP, XIAP, c-IAP-1, c-IAP-2 e suvivina (FERREIRA et
al., 2002).
3.2 APOPTOSE E CÂNCER
Embora o câncer exiba características muito heterogêneas, todos os tumores
malignos adquiriram a propriedade de crescer além dos limites impostos às células
normais. A expansão clonal de uma célula transformada depende de um descontrole
de sua capacidade proliferativa e de uma crescente incapacidade de morrer por
apoptose. Portanto, apesar de enorme variabilidade do câncer, evidências
demonstram que a resistência a apoptose é uma das características mais marcantes
da maioria dos tumores malignos (OKADA e MAK, 2004; GRIVICICH, REGNER e
ROCHA, 2007).
É de grande interesse científico e clínico a influência da apoptose na
sensibilidade de tumores ao tratamento contra o câncer, a maior razão para este
interesse é influenciada positivamente ou negativamente por uma variedade de
genes muitos deles sofrem mutações ou não estão funcionantes no câncer humano.
Entre os mais importantes estão o gene supressor p53 e os genes da família Bcl-2
(BROWN e WOUTERS, 1999). Algumas mutações permitem que célula resista ao
estímulo de morte, as células em um tumor em desenvolvimento precisam evitar a
morte celular que é induzida não somente pela expressão não regulada de
oncogenes, mas também pela limitação de fatores de crescimento, oxigênio ou
nutrientes. Somente superando estas ameaças as células cancerígenas podem
sobreviver e se tornarem invasivas e metastáticas. Os genes anti-apoptóticos Bcl-2 e
p53 têm demonstrado a importância das células cancerígenas de superar a
apoptose. A ativação da Bcl-2 pode levar a sobrevivência dos linfomas, uma vez que
26

a atividade oncogênica pode depender da atividade anti-apoptose do gene Bcl-2
(OKADA e MAK, 2004).
O gene de supressão tumoral p53 é importante na regulação da apoptose, a
deficiência da p53 acelera a tumorigênese (VOUSDEN e LU, 2002).
Além desses genes citados anteriormente, o gene Fas regula o sistema imune
através de sua função pró-apoptótica (NAGATA, 1999), a interrupção deste
mecanismo leva a linfoproliferação e cânceres hematopoiéticos (STRAUS et al.,
2001). Portanto, existem diferentes rotas para a inativação dos mecanismos de
sinalização pró-apoptótica no processo de tumorigênese.
A resistência a apoptose é um importante mecanismo no processo de
tumorigênese assim como no desenvolvimento de resistência de drogas anti-
tumorais. Muitas pesquisas sobre terapias para o câncer têm se focalizado em
desenvolver mecanismos para superar a resistência a apoptose pelas células
tumorais (RONINSON, BROUDE e CHANG, 2001). A análise da tumorigênese
revela que a capacidade de resistir à morte pode ser adquirida por diferentes
mecanismos e acontecer em vários momentos do desenvolvimento tumoral. Entre
estes, a resistência à morte por apoptose em células que escaparam do controle do
crescimento e da diferenciação normais exercidos por fatores solúveis ou por
contatos célula-célula ou célula-matriz extracelular até aquelas induzida por lesões
do DNA, por hipóxia ou por espécies reativas do oxigênio (ZÖRNIG et al., 2001).
A maioria dos tumores retêm a habilidade de sustentar seu crescimento
permanente ou sofrer tipos de morte celular não apoptóticas como necrose,
autofagia, senescência celular e outros (BROWN e WOUTERS, 1999). Esta
propriedade pode ser explorada para induzir a morte celular por mecanismos não
apoptóticos e confundir a sobrevivência dos clones resistentes às drogas. Importante
que a fração de células tumorais que respondem aos mecanismos de morte celular
não apoptóticos aumenta se a apoptose é inibida em resposta a agentes anti-
tumorais ou radiação. Portanto uma estratégia que deliberadamente induza a morte
celular não apoptótica pode ser tornar uma terapia padrão que pode contribuir
favoravelmente no desenlace do tratamentode tumores (RONINSON, BROUDE e
CHANG, 2001).
A elucidação de alguns mecanismos moleculares da apoptose abriram
perspectivas da modulação desses processos. As estratégias se baseiam em induzir
a morte nas células tumorais através do bloqueio de genes olligonucleotídeos
27

antisense e drogas convencionais, ou ainda a substituição de função desses genes
com o uso de moléculas recombinantes (NICHOLSON, 2000).
O oblimersen (G3139, Genasense, Genta, Berkeley, Heights, Nj) é um
oligonucleotídeo antisense anti Bcl-2 que está sendo testado em estudos clínicos da
fase III, tendo demonstrado alguns resultados positivos (KIM et al., 2006;
PAPADOPOULOS, 2006) Outro agente promissor é o ABT737, uma pequena
molécula que inibe as proteína Bcl-2, Bcl-XL, Bcl-w. Em estudo pré-clínico, o
ABT737 levou a completa regressão de tumor de pulmão de pequenas células
(OLTERSDORF et al., 2005).
3.3 GLICOCORTICÓIDES (GCs)
Os GCs são de grande valor no tratamento de desordens inflamatórias, como
reumatismo, artrite reumatóide, asma e dermatites, doenças auto-imunes, doença de
Chron. Suas propriedades antiinflamatórias e imunosupressoras têm feito deste
medicamento uma das drogas mais prescritas mundialmente, além de ser usado
como co-medicação em terapia contra o câncer. Os GCs possuem habilidade de
matar células linfóides o que levou a sua inclusão em todos protocolos de
quimioterapia para linfomas, sendo também utilizados como co-terapia em cânceres
sólidos. O GCs são eficientes para tratar os efeitos colaterais dos antineoplásicos
pois estimulam o apetite, reduzem os distúrbios eletrolíticos, previnem náusea,
êmese e reações tóxicas causadas pela citotoxicidade (RUTZ, 2004; MATTERN,
BUCHLER e HERR, 2007).
Os GCs são drogas com propriedades antiinflamatórias e imunossupressoras,
que induzem a apoptose de monócitos, macrófagos e linfócitos T que estão
envolvidos na resposta inflamatória e imunológica. A maioria dos seus efeitos são
mediadas pelos receptores de GCs. A sensibilidade aos GCs depende amplamente
de sua quantidade, ou da sua habilidade de se ligar ao seu receptor, causando a
modulação dos sítios genéticos para GCs. Condições clínicas de resistência tecido-
específica aos GCs ou de hipersensibilidade têm sido descritos em várias doenças
como, condições inflamatórias crônicas e auto-imunes e na síndrome metabólica
(LOGUI e FARIA, 2009).
28

Portanto a resposta clínica aos GCs varia muito em relação ao tipo de GCs,
dose e terapêutica, da doença e do tipo de célula alvo, e ainda depende da
sensibilidade de cada indivíduo aos GCs (LAMBERTS et al., 1996). Torna-se claro
que o reconhecimento das variações individuais e os aspectos da sensibilidade
órgão-tecido específico aos GCs são fundamentais para o sucesso de tratamentos
que envolvem o seu uso.
Foi demonstrado que drogas antiinflamatórias como os GCs são capazes de
suprimir a proliferação celular e induzir a apoptose em diferentes tipos de células,
afetando principalmente o ciclo celular e os fatores pró-apoptóticos (WEINSTEIN et
al., 1998; ZHANG et al., 2000; CORROYER et al., 2002). Na inflamação é importante
que os GCs possam induzir a apoptose nas células inflamatórias, enquanto
protegem as células dos tecidos circunjacentes. Entretanto, em pacientes com
câncer, especialmente aqueles com altos níveis de GCs, a habilidade dos GCs de
proteger as células cancerosas da apoptose se torna um fator prejudicial (MA et al.,
2008).
Além de influenciar negativamente no tratamento de alguns cânceres, os GCs
podem causar um impacto negativo sobre o tecido ósseo, uma vez que, diminuem a
formação óssea podendo levar a osteoporose. Um dos mecanismos responsáveis
pela indução de osteoporose pelos GCs seria a indução prematura de apoptose das
células osteoblásticas e de osteócitos (WEINSTEIN, 1998).
3.4 ATUAÇÃO DOS GLICOCORTICÓIDES (GCs) NA INDUÇÃO D E APOPTOSE
EM LINFÓCITOS E EM CÂNCERES
A maioria dos linfócitos no timo são sensíveis à indução de apoptose por GCs
in vivo. Entretanto uma pequena população de linfócitos tímicos maduros
permanecem não afetados mesmo com altas doses de GCs. Os receptores de GCs
nos linfócitos têm a mesma afinidade de ligação nos linfócitos sensíveis e
resistentes. A resistência aos GCs não é devido a alteração nestes receptores ou a
expressão de Bcl-2, mas sim a fatores tímicos exógenos e ou ao contato célula-
célula que provavelmente pode alterar a sinalização para o receptor de
glicocorticóide (OLDENBURG, EVANS-STORMS e CIDLOWSKI, 1997).
29

Os GCs têm efeitos profundos no tecidos linfóides, são capazes de alterar
muitas funções metabólicas celulares (transporte transmembrana de glicose,
aminoácidos, nucleosídeos, atividade RNA polimerase, produção de ATP) e
também ativam o processo de degradação do DNA causando a linfocitólise
(QUAGLINO e RONCHETTI, 2001). Aparentemente os GCs regulam uma
endonuclease que especificamente clivam o DNA do timócito. A indução de
apoptose pelos GCs não ocorre espontaneamente, a perda de viabilidade é
dependente da regulação genética, síntese de macromoléculas e atividade de
caspases. Os timócitos sofrem apoptose por outros mecanismos independentes de
caspase quando expostos aos GCs, diferentemente do que ocorre na morte celular
espontânea de timócitos, a morte destas células induzida por GCs parece ser
dependente de expressão gênica e das síntese de macromoléculas, uma vez que a
inibição de caspase não inibe a apoptose em timócitos expostos aos GCs (MANN.
HUGGES e CIDLOWSKI, 2000).
Tem sido descrito que os GCs induzem apoptose em linfócitos, enquanto
protegem algumas células cancerígenas da apoptose, por um mecanismo ainda
pouco conhecido (MA et al.,2008 ). Os GCs exercem um efeito dramático sobre
células de mamíferos. Esta classe de hormônios esteroidais tem efeito citotóxico
sobre certos tipos de células, como os linfócitos, células de mieolomas e células de
linfomas (FRANKFURT e ROSEN, 2004). Entretanto, recentemente, aumentaram as
evidências de que os GCs podem proteger uma variedade de células da apoptose,
como as células epiteliais das glândulas mamárias e células do folículo ovariano,
assim como células do câncer gástrico (MORAN et al., 2000). Os mecanismos pelos
quais os GCs afetam a apoptose ainda são pouco claros. Por exemplo, a atividade
mitocondrial dependente de proteosoma tem um importante papel regulador na
morte celular induzida pelo glicocorticóide dexametasona (MCLAUGHLIN et al.,
2003).
A inibição desta atividade afeta somente parcialmente a indução de apoptose
por GCs. Portanto, o mecanismo mitocondrial dependente de proteosoma não é o
único envolvido na apoptose induzida pelos GCs (MA et al.,2008).
As proteínas FOX são uma família de fatores de transcrição que
desempenham um importante papel na regulação da expressão de genes envolvidos
no crescimento celular, na proliferação, diferenciação e longevidade celular (GREER
e BRUNET, 2005).
30

O Foxp3 é um fator de transcrição expresso em células T regulatórias (Treg),
sendo necessário para o seu desenvolvimento e função (MARSON et al., 2007).
Além de ser expressa em células Treg, o Foxp3 é revelado também em células
tumorais. Sua localização nuclear desempenha um importante papel na formação de
complexos nucleares com fatores nucleares de ativação de células T, regulando,
dessa forma, a transcrição de vários genes como o da IL-10 (interleucina-10) que
conferem às Treg sua função supressora (KARANIKAS et al., 2008).
Corticoesteróides aumentam a expressão de Foxp3 assim como de IL-10 em células
T CD4. Porém o mesmo não foi observado em relação as células tumorais cultivadas
na presença de hidrocortisona. Neste estudo a expressão de Foxp3 foi a mesma na
presença e na ausência de hidrocortisona nas células tumorais. Esta diferença na
expressão de Foxp3 entre células tumorais e células TCD4 pode representar o
mecanismo pelo qual as células tumorais escapam da morte regulada pelos GCs
mediada pelos receptores de glicocorticóides, assim como ocorre nas células T
(RICARDI, CIFONE, MIGLIORATI, 1999).
A expressão de Foxp3 por células de carcinoma pancreático resultou na
inibição da proliferação e possivelmente na não ativação de células T CD4+ e CD25-
, podendo representar um mecanismo de escape do tumor (HINZ et al., 2007).
Nas células de mamíferos foram relatadas a presença da família de fatores de
transcrição forkhead incluindo FOXO1, FOXO3A e FOXO4. O FOXO3A,
especialmente regula genes que mediam sinais pró-apoptóticos que seguem
diferentes estímulos (GREER e BRUNET, 2005).
O FOXO3A não fosforilado é a forma ativa de FOXO3A, ele pode ser
translocado para o núcleo e induzir a transcrição de genes que codificam fatores pró-
apoptóticos e anti-proliferativos como FasL e devido a isso induzem a morte celular
através de mecanismos apoptóticos mediados pelo Fas (KOPS et al., 2002). Quando
o FOXO3A é fosforilado em múltiplos resíduos pelas proteínas quinases AKT e IқK
(“inhibitory kappa B kinase”/ inibidor de kappa B quinase), ela é excluída do núcleo,
o que pode proteger a célula da apoptose. A inativação do FOXO3A pela
fosforilização previne sua translocação para o núcleo reduzindo, então, a expressão
dos sítios genéticos pró-apoptóticos. Wu et al. (2006) observaram que a ativação
dos receptores de glicocorticóides é responsável pela fosforilização e inativação da
FOXO3A.
31

Ma et al. (2008) observaram que a redução da expressão de FOXO3A,
reduziu a apoptose de linfócitos pela metade, demonstrando como este fator de
transcrição é importante na indução de apoptose por GCs em células linfóides.
Várias linhagens de cânceres examinadas apresentaram aumento dos níveis de
fosfo-FOXO3A após tratamento com dexametasona. Estes dados revelam que a
dexametasona promove a fosforilização do FOXO3A, reduzindo a apoptose das
células linfóides. Em células cancerosas, a dexametasona não induz a apoptose,
como esperado, estas células resistentes a apoptose após o tratamento passam a
ter um alto poder de fosforilização da FOXO3A. A partir do exposto torna-se claro
que os mecanismos de apoptose são uma importante via pela qual as drogas
citotóxicas matam as células tumorais (DEBATIN, 2004). Em cânceres linfóides a
dexametasona induz uma resposta de apoptose rápida in vitro, que é antagonizada
pela interleucina-6 (IL-6) e outras citocinas.
Os benefícios da terapia com GCs em cânceres hematológicos são limitados
devido ao surgimento de resistência aos GCs principalmente na leucemia mielóide
aguda ou leucemia mielóide crônica (LOWENBERG, DOWNING, BURNETT, 1999)
Células de mieloma da linhagem RPMI8226 foram utilizados para testar a efetividade
citotóxica de diferentes GCs (dexametasona, prednisolona, metil-1prednisolona). As
células RPMI8226 têm alta expressão de CD38, que tem participação na adesão
celular. Foi observado redução na expressão de CD38 em células RPMI8226
resistentes a metil1-prednisolona e dexametasona (GENTY, DINE e DUFER, 2004).
A expressão de CD38 pode predizer o prognóstico e a progressão da doença
em pacientes com linfomas de células B (IBRAHIM et al., 2001). Além das alterações
na expressão de CD38 as células resistentes apresentaram aumento na expressão
de CD44 (GENTY, DINE e DUFER, 2004). O CD44 promove a adesão celular e
também pode aumentar a sobrevivência de células plasmáticas através da
aglomeração focal (MEDINA et al., 2002), um mecanismo que pode estar envolvido
no crescimento e alterações morfológicas observadas nas células RPMI8226. Ainda
ocorreu aumento da expressão de CD58 nas células resistentes que pode marcar os
estado de resistência aos GCs. O CD58 pode estimular as citocinas inflamatórias e
exercer uma importante influência sobre as interações inflamatórias que são alvos
do tratamento dos corticóides (KIRBY et al., 2000). Portanto a resistência aos GCs
nas células RPMI8226 é acompanhada por uma significante alteração fenotípica que
32

podem implicar no aumento da sobrevivência das células tumorais ao tratamento.
(GENTY, DINE e DUFER, 2004).
33

4 DISCUSSÃO
A partir desta revisão de literatura ficou evidente que a apoptose desempenha
um importante papel na manutenção e desenvolvimento dos seres vivos eliminando
células supérfluas ou defeituosas. A compreensão dos mecanismos apoptóticos
permitiu o desenvolvimento científico em vários aspectos, inclusive o
desenvolvimento de novas terapias contra o câncer. Medicamentos anti-neoplásicos
têm como estratégia a indução da morte das células tumorais e maior resposta ao
tratamento radioterápico (GRIVICICH, REGNER e ROCHA, 2007).
Os GCs são amplamente utilizados em combinação com quimioterápicos e
radioterapia em pacientes com tumores sólidos, pois os GCs reduzem náusea,
êmese, e protegem os tecidos contra os efeitos citótoxicos da terapia anti-neoplásica
(CHEN et al., 2009). Estes hormônios ainda são utilizados nos protocolos
terapêuticos para tratamento de várias formas de linfomas e leucemias. Agentes
quimioterápicos e irradiação-y induzem a sinalização apoptótica, que converge na
ativação das caspases iniciadoras (HERR et al., 2003), entretanto, já foi relatado que
os GCs apresentam efeitos anti-apoptóticos em células de tumores sólidos,
prevenindo a citotoxicidade de terapias convencionais (radioterapia, quimioterapia)
que induzem a morte celular (HERR e PIFTZENMAIER, 2006).
A inibição de moléculas chave para o receptor de morte em células de
carcinoma e da apoptose via mitocôndrial pela dexametasona, resulta no bloqueio
da atividade das caspases. A transferência direta de caspases restaurou a
sensibilidade a dexametasona em carcinomas in vitro e in vivo (HERR et al., 2003).
A dexametasona inibiu a apoptose induzida por 5-fluorouracil e cisplatina que
são agentes antineoplásicos e promoveu o crescimento de células de tumores
sólidos. Em contraste, nas células linfóides, induziu a apoptose e permitiu o efeito
citotóxico dos quimioterápicos (ZHANG et al., 2006). Herr et al. (2003) também
relataram que mesmo uma pequena exposição a dexametasona foi capaz de
reprimir a despolarização da membrana mitocondrial, inibindo a expressão e
atividade da caspase-9, levando a um decréscimo da atividade da caspase-3 efetora
34

em células de carcinomas expostas a cisplatina (agente antineoplásico) in vitro,
entretanto esta inibição não foi observada nas células linfóides
Ainda em relação ao tratamento combinado de GCs e quimioterápicos, Zhang
et al (2007) observou resistência ao tratamento com quimioterápico em 94% de
células de linhagem tumoral, 100% de células de transplante xenogênico de
carcinoma de próstata e 85% de células originadas de tumores sólidos ressecados.
A resistência pode ser devido à inibição de apoptose, promoção da viabilidade e
progressão celular. Apesar da resistência ter uma longa duração, ela se mostrou
reversível com a cessação da terapia com glicocorticóide.
A dexametasona ativa os receptores de GCs que aumentam a expressão e a
atividade de SGK-1 (“serum/glucocorticoid regulated kinase 1”/ soro e quinase-1
dependente de glicocorticóide) e MKP-1 (“MAPK phosphatase-1”/ MAPK fosfatase-
1). A super expressão de MKP-1 e SGK-1 está diretamente ligada a redução da
apoptose (WU et al., 2005).
Portanto, apesar de serem efetivos na indução de apoptose em linfócitos os
GCs podem proteger algumas células cancerígenas da apoptose por um mecanismo
ainda pouco conhecido. A resistência pode ser devido a alterações fenotípicas das
células tumorais (GENTY, DINE e DUFER, 2004). Chen et al (2009) relataram que
os GCs causam aumento da adesão da matriz extracelular que proporciona um
aumento na sinalização de integrina beta 1 e TGF-beta 1 (“transforming growth
factor beta 1”/ fator transformador de crescimento beta 1) que representam um
importante papel na indução de quimioresistência pelos GCs em células cancerígena
do ovário.
Além do efeito de quimioresistência dos GCs principalmente diminuindo a
apoptose de células tumorais eles são potentes imunossupressores. A
imunossupressão pode exacerbar o processo de metástase e acelerar o crescimento
tumoral. Juntos, o efeito imunossupressor e a indução de resistência a apoptose em
tumores sólidos, podem contribuir no aumento do potencial de metástase em
pacientes que recebem combinação de quimioterapia/radioterapia e uso de GCs
(ZHANG et al., 2006)
35

5 CONCLUSÃO
Em conclusão os GCs são amplamente usadas em altas doses na terapia de
leucemias e linfomas e são usados concomitantemente aos quimioterápicos. Porém,
estes medicamentos estão envolvidos na indução de resistência a apoptose na
maioria dos tumores sólidos submetidos ao tratamento com drogas anti-
cancerígenas e/ou radioterapia. Dessa forma, é importante se reconsiderar a ampla
utilização dos GCs no tratamento de pacientes com tumores sólidos.
36

6 REFERÊNCIAS
ABBAS, A.K.; LICHTMAN, A.H.; POBER, J.S. 2003. Imunologia Celular e Molecular. 4ª ed. Rio de Janeiro. Editora Revinter Rio de Janeiro , 125-130. AGARWAL, M.L.; TAYLOR, W.R.; CHERNOW, M.V.; CHERNOVA, O.B.; STARK, G.R. 1998. The p53 network. The Journal of Biology Chemestry, 273 :1-4. ALBERTS, B.; JOHNON, A.; LEWIS, J.; RAFF, M.; ROBERTS, K.; WALTER, P. 2004. Biologia molecular da célula. 4ª ed. Rio de Janeiro. Editora Artmed São Paulo , 1012-1014. ALLEN, R.T.; HUNTER, III W.J.; AGRAWAL, D.K. 1997. Morphological and biochemical characterization and analysis of apoptosis. Journal of Pharmacology and Toxicology Methods, 37 :215-228. ANTONSSON, B.; MARTINOU, J.C. 2000. The Bcl-2 protein family. Experimental Cell Research, 256 : 50-57. BOATRIGHT, K.M,; SALVESEN, G.S. 2003. Mechanism of caspase activation. Current Opinion in Cell Biology, 15 : 725-731. BRASILEIRO FILHO, G. 2004. Bogliolo- Patologia Geral, 3ª ed. Rio de Janeiro. Editora Guanabara Koogan , 367p. BROWN, J.M.; WOUTERS, B.G. 1999. Apoptosis, p53, and tumor cell sensitivity to anticancer agents. Cancer Research. 59 :1391-1399. BUDIHARDJO, I.; OLIVER, H.; LUTTER, M.; LUO, X.; WANG, X. 1999 Biochemical pathways of caspase activation during apoptosis. Annual Review of Cell and Developmental Biology, 15 : 269-290 CHAI, J.; DU, C.; WU, J.W.; KYRIN, S.; WANG, X.; SHI, Y. 2000. Structural and Biochemical basis of apoptotic activation by smac/DIABLO. Nature, 406 : 855-862. CHEN, Y.X.; WANG, Y.; FU, C.C.; DIAO, F.; SONG, L.N.; LI, Z.B.; YANG, R.; LU, J. 2009. Dexamethasone enhances cell resistance to chemotherapy through increasing adhesion to extracellular matrix in human ovarian cancer cells. Endocrine- Related Cancer IN PRESS. COHEN, G.M.; SUN, X.M.; SNOWDEN, R.T.; DINSDALE, D.; SKILLETER, D.N. 1992. Key morphologycal features of apoptosis may occur in the absence of internucleossomal DNA fragmentation. Biochemical Journal, 286 : 331-334.
37

CORROYER, S.; SCHITTNY, J.C.; DJONOV, V.; BURRI, P.H.; CLEMENT, A. 2002.Impairment of rat postnatal lung alveolar development by glucocorticoids: involvement of the p21CIP1 and p27KIP1 cyclin-dependent kinase inhibitors. Pediatric Research 51:169-76. DEBATIN, K.M. 2004. Apoptosis pathways in cancer and cancer therapy. Cancer Immunology Immunotherapy 53 :153-159. DEVERAUX, Q.L; REED, J.C. 1999. IAP family proteins--suppressors of apoptosis. Genes and Development, 13 :239-52 DU, C.; FANG, M.; LI, Y.; WANG, X. 2000. Smac, a mitochondrial protein that promotes cytochrome c-dependent caspase activation during apoptosis. Cell, 102:33-42. DUQUE-PARRA, J.E. Note on the origin and history of the term “apoptosis”. 2005. The Anatomical Record (Part B New Anat), 283B : 2-4. ELLIS, R.E; YUAB, J.Y; HORVITZ, H.A.R. 2001. Mechanism and function of cell death. Annual Review of Cell and Development Biology, 7 :663-698. FAN, T-J.; HAN, L.H.; CONG, R.S.; LIANG, J. 2005. Caspase family proteases and apoptosis. Acta Biochimica et Biophysica Sinica, 37 : 719-727. FANIDI, A.; HARRINGTON, E.A.; EVAN, G.I. 1992. Cooperative interaction between c-myc and bcl-2 proto oncogenes. Nature, 359 : 554-556. FERRARI, D.; PINTON, P.; SZABADKAI, G.; CHAMI, M.; CMAPANELLA, M.; POZZAN, T.; RIZZUTO, R. 2002. Endoplasmic reticulum, Bcl-2 and Ca2+ handling in apoptosis. Cell Calcium, 32 : 413-20. FERREIRA, C.G.; EPPING, M.; KRUYT, F.A.E; GIACCONE, G. 2002. Apoptosis: target of cancer therapy. Clinical Cancer Research, 8 : 20-34. FERRI, K.F.; KROEMER, G. 2001. Organelle-specific initiation of cell death pathways. Nature Cell Biology, 3 : 255-63. FISHER, D.E. 2001. Pathways of apoptosis and the modulation of cell death in cancer. Hematology/ Oncology Clinics of North america,15 :.931-956. FRANKFURT, O.; ROSEN, S.T. 2004. Mechanisms of glucocorticoid-induced apoptosis in hematologic malignancies. Current Opinion in Oncology , 16:553-563. GALLUZZI, L.; MAIURI, M.C.; VITALE, I.; ZISCHKA, H.; CASTEDO, M.; ZITVOGEL L; KROEMER, G. 2007. Cell death modalities: Classification and pathophysiological implications. Cell Death and Differentiation, 14 : 1237-1243. GAVRIELLI, Y.; SHERMAN, Y.; BEN-SASSON, S. 1992. Identification of programmed cell death in situ via specific labeling of nuclear DNA fragmentation. Journal of cell Biology, 119 : 493-502.
38

GENTY, V.; DINE, G.; DEAFER, J. 2004. Phenotypical alterations induced by glucocorticoids resistance in RPMI 8226 human myeloma cells. Leukemia Research, 28 : 307-313. GESKE, F.J.; GERSCHENSON, L.E. 2001.The bilogy of apoptosis. Human Pathology, 32 :1029-1038. GREER, E.L.; BRUNET, A. 2005. FOXO transcription factors at the interface between longevity and tumor suppression. Oncogene, 24 : 7410-7425. GRIVICICH, I.; REGNER, A.; ROCHA, A.B. 2007. Morte celular por apoptose. Revista Brasileira de Cancerologia, 55 : 335-343. HACKER, G. 2000. The morphology of apoptosis. Cell and Tissue Research, 301 : 5-17. HENGARTNER, M. O. 2000.The biochemistry of apoptosis. Nature, 407 : 770-776. HERR, I; PIFTZENMAIER, J. 2006. Glucocorticoid use in prostate câncer and other solid tumours: implications for effectiveness of cytotoxic treatment and metastases. Lancet Oncology, 7 :425-430. HERR, I.; UCUR, E,; HERZER, K.; OKOUOYO, S,; RIDDER, R.; KRAMMER, P.H.; DOEBERITZ, M.V.K.; DEBATIN, K.M. 2003. Glucocorticoid induces apoptosis resistance toward cancer therapy in carcinomas. Cancer Research, 63 : 3112-3120. HINZ, S.; PAGEROLS-RALUY, L.; OBERG, H.H.; AMMERPOHL, O.; GRÜSSEL, S.; SIPOS, B.; GRÜTZMANN, R.; PILARSKY, C.; UNGEFROREN, H.; SAEGER, H.D.; KLÖPPEL, G.; KABELITZ, D.; KALTHOFF, H. 2007. Foxp3 expression in pancreatic carcinoma cells as a novel mechanism of immune evasion in cancer. Cancer Research , 67:8344-8350. HOGGUIST, A.K.; BALDWIN, T.A.; JAMENSON, S.C. 2005. Central tolerance: learning self-control in the thymus. Nature, 5 : 772-781. IBRAHIM, S.; KEATING, M.; DO, K.A.; O'BRIEN, S.; HUH, Y.O.; JILANI, I.; LERNER, S.; KANTARJIAN, H.M.; ALBITAR, M. 2001. CD38 expression as an important prognostic factor in B-cell chronic lymphocytic leukemia. Blood, 98 :181-186. KARANIKAS, V.; SPELETAS, M.; ZAMANAKOU, M.; KALALA, F.; LOULES, G.; KERENIDI, T.; BARDA, A.K.; GOURGOULIANIS, K.I.; GERMENIS, A.E. 2008. Foxp3 expression in human cancer cells. Journal of Translation Medicine, 6 : 6-19. KIM, R.; EMI, M.; MATSUURA, K.; TANABE, K. 2006. Antisense and nonantisense effects of antisense Bcl-2 on multiple roles of Bcl-2 as a chemosensitizer in cancer therapy. Cancer Gene Therapy, 14 :1-11.
39

KIRBY, A.C.; CAHEN, P.; PORTER, S.R.; OLSEN, I. 2000. Soluble and cell-associated forms of the adhesion molecule LFA-3 (CD58) are differentially regulated by inflammatory cytokines. Cell Adhesion and communication, 7 :453-464. KOPS, G.J.; MEDEMA, R.H.; GLASSFORD, J.; ESSERS, M.A.; DIJKERS, P.F.; COFFER, P.J.; LAM, E.W.; BURGERING, B.M. 2002. Control of cell cycle exit and entry by protein kinase B-regulated forkhead transcription factors. Molecular and Cellular Biology 22 : 2025-2036. KRAVTSOV, V.D.; DANIEL, T.O.; KOURY, M.J. 1999. Comparative analysis of different methodological approaches to the in vitro study of drug induced apoptosis. American Journal of Pathology 155 :1327-1339. LAMBERTS, S.W.; HUIZENGA, A.T.; LANGE, P.; DE JONGE, F.H.; KOPER, J.W. 1996. Clinical aspects of glucocorticoid sensitivity. Steroids, 61 :157-160. LANE, D.P.1992. P53, Guardian of the genome. Nature, 358 :15-16. LAUBER, K., BLUMENTHAL, S.G.; WAIBEL, M.; WESSELBORG, S.2004. Clearance of apoptotic cells: getting rid of the corpses. Molecular Cell, 14 : 277-287.. LEE, J.; RICHBURG, J.H.; SHIPP, E.B.; MEISTRICH, M.L.; BOEKELHEIDE, K. 1999. The Fas system, a regulator of testicular germ cell apoptosis, is differentially up regulated in sertoli cells versus germ cell injury of the testis. Endocrinology, 140 : 852-858. LIU, Q.A.; HENGARTNER, M.O. 1999.The molecular mechanism of programmed cell death in C. elegans. Annals of the New York Academy Sciences, 887 : 92-104 LONGUI, C.A.; FARIA, C.D.C. 2009. Evaluation of glucocorticoid sensitivity and its potential clinical applicability. Hormone Research, 71 :305-309. LORO, L.L.; VINTERMYR, O. K.; JOHANNESSEN, A.C. 2003. Cell death regulation in oral squamous cell carcinoma: methodological considerations and clinical significance. Journal of Oral Pathology Medicine, 32 : 125-138. LOWENBERG, B,; DOWNING, J.R.; BURNETT, A. 1999. Acute myeloid leukaemia. New England Journal of Medicine, 34 :1051–1062. MA, J.; XIE, Y; SHI, Y.; QIN, W.; ZHAO, B., JIN, Y. 2008. Glucocorticoid-induced apoptosis requires FOXO3A activity. Biochemical and Biophysical Research communications, 377 : 894-898. MANN, C.L.; HUGHES, F.M. JR.; CIDLOWSKI, J.A. 2000. Delineation of the signaling pathways involved in glucocorticoid induced and spontaneous apoptosis of rat thymocytes. Endocrinology, 141 : 528-538. MARSON, A,; KRETSCHMER, K.; FRAMPTON, G.M.; JACOBSEN, E.S.; POLANSKY, J.K.; MACLSAAC, K.D.; LEVINE, S.S.; FRAENKEL, E.; BOEHMER,
40

H.V.; YOUNG, R.A. 2007. Foxp3 occupancy and regulation of key target genes during T-cell stimulation. Nature, 445 : 931-935. MATTERN, J.; BUCHLER, M.W.; HERR, I. 2007. Cell cycle arrest by glucocorticoids may protect normal tissue and solid tumors from cancer therapy. Cancer Biology and therapy, 6 : 1345-1354. MCLAUGHLIN, N.T.; GRIMM, L.; GOLDSBY, R.A.; OSBORNE, B.A, Glucocorticoid-induced apoptosis of thymocytes: requirement of proteasome-dependent mitochondrial activity, The Journal of Immunology, 170 : 2469–2478. MEDINA, F.; SEGUNDO, C.; CAMPOS-CARO, A.; GONZÁLEZ-GARCÍA, I.; BRIEVA JA. 2002. The heterogeneity shown by human plasma cells from tonsil, blood, and bone marrow reveals graded stages of increasing maturity, but local profiles of adhesion molecule expression. Blood, 99 :2154-2161. MORAN, T.J.; GRAY, S.; MIKOSZ, C.A.; CONZEN, S.D. 2000. The glucocorticoid receptor mediates survival signal in human mammary epithelial cells. Cancer Research, 60 :867-872. NAGATA, S. 1999. Fas ligand-induced apoptosis. Annual Review of Genetics, 33:29-55. NICHOLSON, D.W. 2000. From bench to clinic with apoptosis-based therapeutic agents. Nature, 407 : 810-816. NICHOLSON, D.W.; THORNBERRY, N.A. 1997. Caspases killer proteases. Trends in Biochemical Sciences, 22 : 299-306. NIETO, M.A.; GONZALEZ, F.; GAMBON, F.; DIAZ-ESPADA; LOPEZ-RIVAS A.1992. Apoptosis in human thymocytes after treatment with glucocorticoids. Clinical Experimental Immunology, 88 : 341-344. OKADA, H.; MAK, T.W. 2004. Pathways of apoptotic and non poptotic death in tumour cells. Nature Reviews Cancer, 4 : 592-603. OLDENBURG, N.B.; EVANS STORMS, R.B.; CIDLOWSKI, J.A. 1997. In vivo resistance to glucocorticoid induced apoptosis in rat thymocytes with normal steroid receptor function in vitro. Endocrinology, 138 : 810-818. OLTERSDORF, T.; ELMORE, S.W.; SHOEMAKER, A.R.; ARMSTRONG, R.C.; AUGERI, D.J.; BELLI, B.A.; BRUNCKO, M.; DECKWERTH, T.L.; DINGES, J.; HAJDUK, P.J.; JOSEPH, M.K.; KITADA, S.; KORSMEYER, S.J.; KUNZER, A.R.; LETAI, A.; LI, C.; MITTEN, M.J.; NETTESHEIM, D.G.; NG, S.; NIMMER, P.M.; O'CONNOR, J.M.; OLEKSIJEW, A.; PETROS, A.M.; REED, J.C.; SHEN, W.; TAHIR, S.K.; THOMPSON, C.B.; TOMASELLI, K.J.; WANG, B.; WENDT, M.D.; ZHANG, H.; FESIK, S.W.; ROSENBERG, S.H. 2005. An inhibitor of Bcl-2 family proteins induces regression of solid tumors. Nature, 435 :677-681.
41

PALMER, E. 2003.Negative selction-clearing out the bad apples from the T-cell repertoire. Nature Reviews Immunology, 3 : 383-391. PAPADOPOULOS, K. 2006. Targeting the Bcl-2 family in cancer therapy. Seminars Oncology, 33 :449-56. QUAGLINO, D.; RONCHETTI, I.P.2001. Cell death in the rat thymus: a mini review. Apoptosis, 6 : 389-401. REDONDO, M.; TÉLLEZ, T.; ROLDAN, M.J.; SERRANO, A.; GARCÍA-ARANDA M.; GLEAVE, M.E.; HORTAS, M.L.; MORELL, M. 2007. Anticlusterin treatment of breast cancer cells increases the sensitivities of chemotherapy and tamoxifen and counteracts the inhibitory action of dexamethasone on chemotherapy-induced cytotoxicity. Breast Cancer Research, 9 : R86 REED, J.C.; JURGENSMEIER, J.M.; MATSUYAMA, Y. 1998. Bcl-2 family proteins and mitochondria. Biochimica et Biophysica Acta, 281 : 127-137. RICCARDI, C.; CIFONE, M.G.; MIGLIORATI, G. 1999. Glucocorticoid hormoneinduced modulation of gene expression and regulation of Tcell death: role of GITR and GILZ, two dexamethasoneinduced genes. Cell Death and Differentiation , 6:1182-1189. RODRIGUES, M.F.; BARSANTE, M.M.; ALVES, C.C.S.; SOUZA, M.A.; FERREIRA, A.P.; MENDES, G.P.A.; TEIXEIRA, H.C. 2009. Apoptosis of macrophages during pulmonary Mycobacterium bovis infection: correlation with intracellular bacillary load and cytokine levels. Immunology, 128 : 691-699. RODRÍGUEZ, I.; ODY, C.; ARAKI; GARCIA, I.; VASSALE, P. 1997. An early and massive wave of germinal cell apoptosis is required for the development of functional spermatogenesis. The EMBO Journal, 16 : 2262-2270. RONINSON, I.B.; BROUDE, E.V.; CHANG, B.D. 2001. If not apoptosis, then what? Treatment-induced senescence and mitotic catastrophe in tumor cells. Drug Resistance Updates, 4 : 303-313. RUONARAIN, C.; DLAMINI, Z.; NAICKER, S.; BHOOLA, K. 2004. Colon cancer: genetics and apoptotic events. Journal of Biological Chemistry, 385 :449-464. RUTZ, H.P. 2002. Effects of corticosteroid use on treatment of solid tumours. Lancet, 360:1969-1970. 274-282. SCHULER, M.; MAURER, U.; GOLDSTEIN, J.C. 2003. p53 triggers apoptosis in oncongene-expressing fibroblast by the induction of Noxa and mitochondrial Ba translocation. Cell Death and Differentiation, 10 : 451-460. SHEER, C.J. 2000. The Pezcoller lecture: cancer cell cycles revisited. Cancer Research, 60 : 3689-3695.
42

SHY, Y. 2002. Apoptosome: the cellular engine for the activation of caspase-9. Structure, 10 : 285-288. STRAUS, S.E.; JAFFE, E.S.; PUCK; J.M.; DALE, J.K.; ELKON, K.B.; RÖSEN-WOLFF, A.; PETERS, A.M.; SNELLER, M.C.; HALLAHAN, C.W.; WANG, J.; FISCHER, R.E.; JACKSON, C.M.; LIN, A.Y.; BÄUMLER, C.; SIEGERT, E.; MARX, A.; VAISHNAW, A.K.; GRODZICKY, T.; FLEISHER, T.A.; LENARDO, M.J. 2001. The development of lymphomas in families with autoimmune lymphoproliferative syndrome with germline Fas mutations and defective lymphocyte apoptosis. Blood, 98:194-200. VACCHIO, M.S.; ASHWELL, J.D. Glucocorticoids and thymocyte development. Seminars in Immunology, 12 : 475-485. TUCKERMAN, J.P.; KLEIMAN, A.; MCPHERSON, K.G.; REICHARDT, H.M. 2005. Molecular mechanisms of glucocorticoids in the control of inflammation and lymphocyte apoptosis. Critical Reviews in Clinical Laboratory Sciences, 4 2: 71-104. VERHAGEN, A.M.; EKRET, P.G.; PAKUSCH, M.; SILKE, J.; CONNOLLY, L.M.; REID, G.E.; MORITZ, R.L.; SIMPSON, R.J.; VAUX, D.L. 2000. Identification of DIABLO, a mammalian protein that promotes apoptosis by binding to and antagonizing IAP proteins. Cell, 102 :43-53. VOUSDEN, K. H.; LU, X. 2002. Live or let die: the cell’s response to p53. Nature Review Cancer, 2 : 594–604. WANG, H.R.; GU, C.H.; ZHU; J.Y.; HAN, J.Y.; ZHONG, H.; CHEN, F.Y.; OUYANG, R. R. 2006. PNAS-2: a novel gene probably participating in leukemogenesis. Oncology, 71 : 423-9. WEINSTEIN, R.S.; JILKA, R.L.; PARTT, A.M., MANOLAGAS, S.C.1998. Inhibition of osteoblastogenesis and promotion of apoptosis of osteoblasts and osteocytes by glucocorticoids. Potential mechanisms of their deleterious effects on bone. The Journal of Clinical Investigation, 102 : 274-282. WICH, G.; KROEMER, G.1993. Immunoendocrine communication via the hypothalamo-pituitary-adrenal axis in autoimmune diseases. Endocrine Review, 14 : 539-563. WU, W.; ZOU, M.; BRICKLEY, D.R.; PEW, T.; CONZEN, S.D. 2006. Glucocorticoid receptor activation signals through forkhead transcription factor 3a in breast cancer cells. Molecular Endocrinology, 10 : 2304–2314. WU, W.; PEW, T.; ZOU, M.; PANG, D.; CONZEN, S.D. 2005. Glucocorticoid receptor-induced MAPK phosphatase-1 (MPK-1). Expression Inhibits Paclitaxel-associated MAPK activation and contributes to breast cancer cell survival. The Journal of Biological Chemestry, 280 : 4117-4124.
43

YU, J.; ZHANG, L. 2005. The transcriptional targets of p53 in apoptosis control. Biochemical and Biophysical Research Communications , 331: 851-858. ZHANG, C.; BECKERMANN, B.; KALLIFATIDIS, G.; LIU, Z.; RITTGEN, W.; EDLER, L.; BUCHLER, P.; DEBATIN, K.M.; BUCHLER, M.W.; FRIESS, H.; HERR, I. 2006. Corticosteroids induce chemotherapy resistance in the majority of tumour cells from bone, brain, breast, cervix, melanoma and neuroblastoma. International Journal of Oncology, 29 :1295-1301. ZHANG, G.; TU, C.; ZHANG, G.; ZHOU, G.; ZHENG, W. 2000. Indomethacin induces apoptosis and inhibits proliferation in chronic myeloid leukemia cells. Leukemia Research, 24:385-92. ZHANG, C.; WENGER, T.; MATTERN, J.; LLEA, S.; FREY, C.; GUTWEIN, P.; ALTEVOGT, P.; BODENMULLER, W.; GASSLER, N.; SCHNABEL, P.A.; DIENEMANN, H.; MARMÉ, A.; HOENFELNER, M.; HAFERKAMP, A.; PFITZENMAIER, J.; GRONE, H.J.; KOLB, A.; BUCHLER, M.W.; FRIESS, H.; RITTGEN, W.; EDLER, L.; DEBATIN, K.M.; KRAMMER, P.H.; RUTZ, H.P.; HERR, I. 2007. Clinical and mechanistic aspects of glucocorticoid-induced chemotherapy resistance in the majority of solid tumors. Cancer Biology and Therapy, 6 : 278-287. ZÖRNIG, M.; HUEBER, A.; BAUM, W.; EVAN, G. 2001. Apoptosis regulators and their role in tumorigenesis. Biochimica et Biophysica Acta, 1551 : F1-37.
44