Embed Size (px)
DESCRIPTION
Lotka volterra predação
Citation preview

Ecologia de Populações
Modelos de predador e presa Prof. Dr. Harold Gordon Fowler

Modelo genérico
),()(
),()(
yxehygdt
dy
yxhxfdt
dx
• f(x) crescimento da presa • g(y) mortalidade do predador • h(x,y) predação • e coeficiente da conversão de presas em biomassa do predador

Análise da estabilidade local
Jacobiana no equilibrio positivo
0
0*
*
*
by
axJ
detJ*>0 e trJ*=0 (centro)
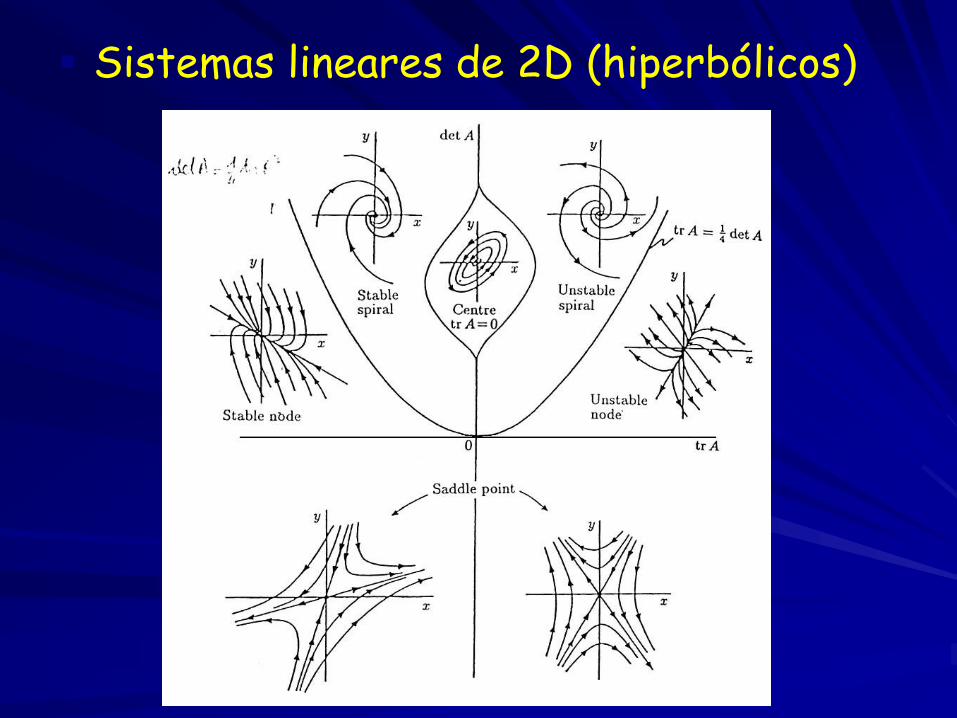
Sistemas lineares de 2D (hiperbólicos)

Análise da estabilidade local
Prova da existencia de trajetorias centrais (teorema da linearização)
Existência de um primeiro integral H(x,y) :
aybxyrxmyxH )ln()ln(),(

O Modelo de Lotka e Volterra com o crescimento logístico da presa
bxymydt
dy
axyK
xrx
dt
dx
1

Isoclinais nulos do modelo de Lotka-Volterra com o crescimento logístico
da presa

Modelo de Lotka-Volterra com o crescimento logístico da presa
bxymydt
dy
axyK
xrx
dt
dx
1
Pontos de equilíbrio : (0,0) (K,0) (x*,y*)

Analise de estabilidade local
Jacobiano a equilíbrio positivo
0*
*
*
*
by
axK
rxJ
detJ*>0 e trJ*<0 (estável)
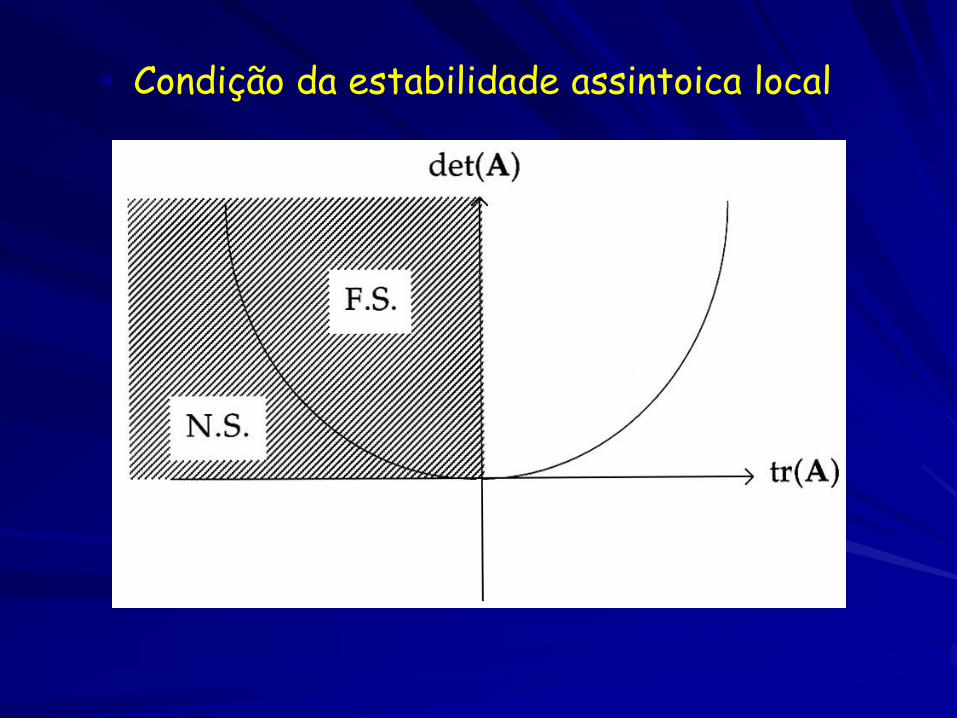
Condição da estabilidade assintoica local

Isoclinais de Lotka-Volterra

Campo de orientação do modelo de Lotka-Volterra

O modelo de Lotka-Volterra
Equação de crescimento da presa: dN = a N - b N P dt Equação do crescimento do predador: dN = c N P -- d P dt Onde N e P são as densidades da presa e predador
respectivamente. a e d são as taxas per capita de mudança na ausência do outro.
b e c são as taxas de mudança do predador e presa respectivamente que resultam da interação entre eles.
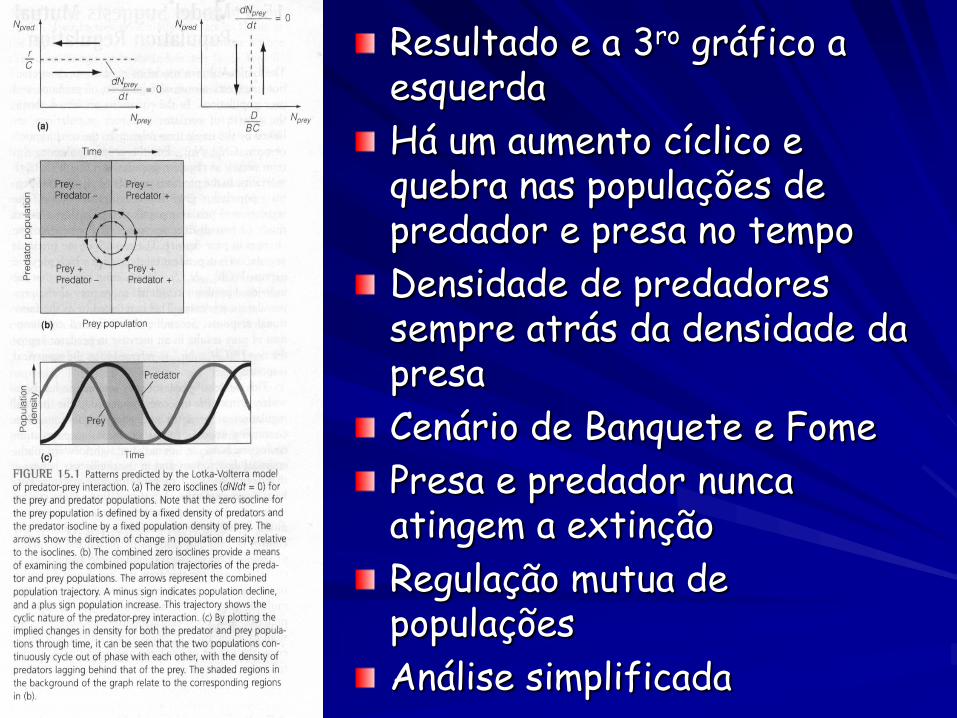
Resultado e a 3ro gráfico a esquerda
Há um aumento cíclico e quebra nas populações de predador e presa no tempo
Densidade de predadores sempre atrás da densidade da presa
Cenário de Banquete e Fome
Presa e predador nunca atingem a extinção
Regulação mutua de populações
Análise simplificada

Modelo sugere uma regulação mútua de populações
Resposta funcional: quanto mais presa, mais consumo ocorre
Resposta numérica: aumento do consumo resulta no aumento da reprodução do predador
Mas não é tão simples. Quais outros fatores podem estar envolvidos?
Exemplo. Cobertura vegetal

Prevê o comportamento cíclico - amplitude depende das condições iniciais
Isoclinal de Crescimento Zero Da presa
Isoclinal de crescimento Zero do predador

Coexistência de Predador e Presa
Sob quais condições existe uma coexistência estável de predador e presa?
O resultado é similar ao resultado com a competição entre duas espécies.

Coexistência de Predador e Presa
Nossa estratégia básica é – 1) escrever equações diferencias simples
que descrevem o crescimento das duas populações
– 2) definir o equilíbrio como o ponto onde as populações não mudam.
– 3) realizar uma análise do plano de fase usando os isoclinais das duas espécies.

Modelos de Predação
Nicholson e Bailey desenvolveram um modelo de hospedeiro – Parasitóide que renova as premissas:
- a mortalidade do predador é independente da densidade.
- a conversão de energia pelos predadores em nascimentos é retardada por uma geração.

Duas Equações Ligagas – O Modelo de Nicholson e Bailey :
Equação do crescimento do hospedeiro (presa): H t+1 = r H t e
(-a Pt)
Equação do crescimento do Parasitóide (predador): P t+1 = Pt [ 1 - e
(-a Pt) ] Onde H é o hospedeiro ou presa. P é o predador ou
parasitóide. t é o tempo, r é a taxa finita de aumento do hospedeiro. a é a taxa de parasitismo por cada parasitóide.

Duas Equações Ligagas – O Modelo de Nicholson e Bailey :
Equação de crescimento de hospedeiro (presa): H t+1 = H t e
(-a Pt)
Equação de crescimento de Parasitóide (predador): P t+1 = Pt [ 1 - e
(-a Pt) ] Se o número de hospedeiros retirado pelos parasitóides iguala a fração
ou presas que é o recrutamento, então a população parental fica igual. Se o parasitóide retira parte da população parental de hospedeiros então
a população de parasitóide diminua. Assim, a sobre-exploração por parasitóide/predador pode resultar em
oscilações maiores e a extinção possível de uma população ou outra

Modelo de Lotka-Volterra
Taxa de crescimento da poplação da presa: dH/dt = rH-pHP
H: tamanho da população da presa P: tamanho da população do predador p: proporção de encontros que terminam na matança (eficiencia de captura) r: taxa intrínseca de aumento da população de presa

Taxa de crescimento do predador: dP/dt = apHP-dP a: fração de energia da presa convertida em predadores novos p: proporção de encontros que resulta na matança (eficiencia de captura) d: taxa de mortalidade do predador
Modelo de Lotka-Volterra


O modelo de Lotka-Volterra pode gerar ciclos populacionais de predador e presa porque eles controlam reciprocamente o crescimento populacional do outro (efeito dinâmico) Porém, obtenção uma dinamica ciclica estável é rara com esse modelo simples
Modelo de Lotka-Volterra

Coexistência de Predador e Presa
Modelo da população de predador:
dPapHPdt
dP

Coexistência de Predador e Presa
Aqui, dP/dt é a taxa de crescimento do predador.
a = eficiência da produção do predador (proporçao da energia assimilada pelo predador que é convertida em biomassa nova do predador.
p= eficiência de ingestão do predador (proporção da presa disponível atualmente consumida).

Coexistência de Predador e Presa
H = densidade da presa.
d = taxa de mortalidade do predador. Na ausência da presa, a população do predador precisa cair a zero.

Coexistência de Predador e Presa
Para a população de presa:
pHPrHdt
dH

Coexistência de Predador e Presa
dH/dt = taxa de crescimento da população da presa.
H = densidade da presa.
p= eficiência de ingestão do predador (proporção da presa disponível atualmente consumida).
P é a densidade do predador.
r é a taxa de natalidade da presa.

Coexistência de Predador e Presa
Os nascimentos da presa sofrem diminuição pelas mortes (pHP).
Os encontros entre predadores e presas é o produto de seus números. Essa é a idéia da ‘movimentação browniana’.

Coexistência de Predador e Presa
Em equilíbrio,
dPapHPdt
dP 0
pHPrHdt
dH 0

(c) 2001 by W. H. Freeman and
Company
Os sistemas de predador e presa podem ter mais de um estado estável
A presa é limitada pela disponibilidade de alimento e os efeitos do predador: – Algumas populações podem ter dois ou mais
pontos estáveis de equilíbrio, ou estados estáveis múltiplos:
Uma situação dessas aparece quando: – A presa exibe um padrão típico de dependência de
densidade (crescimento reduzido ao aproximar a capacidade de suporte)
– O predador exibe uma resposta funcional to Tipo III

(c) 2001 by W. H. Freeman and
Company
Três Equilíbrios O modelo de respostas de predador e presa a densidade de presa resulta em três estados estáveis de equilíbrio: – Um ponto estável A (densidade baixa da presa) onde:
Qualquer aumento da população de presa é compensado pelo aumento de eficiência da captura de presas pelo predador
– Um ponto não estável B (densidade intermédia da presa) onde:
O controle da presa muda da predação a limitação de recursos
– Um ponto estável C onde: A presa escapa de controle do predador e é regulada próxima a sua capacidade de suporte pela escassez de alimentos

(c) 2001 by W. H. Freeman and
Company
Implicações de Estados Estáveis Múltiplos
Os predadores podem controlar uma presa a um nível populacional baixo (ponto A no modelo), mas podem perder o potencial de regular a densidade da presa se essa aumenta acima do ponto B no modelo: – Um predador que controla uma praga agrícola pode
perder o controle dessa praga se o predador passa uma supressão por outros fatores temporalmente:
Uma vez que a população da praga excede o ponto B, aumentará a nível populacional no ponto C, independente da atividade do predador
Nesse ponto, a população de praga ficara elevada até outro fator reduz a população de praga embaixo do ponto B no modelo

Equilíbrio
pHPrH
dPapHP

Equilíbrio
Isoclinal do predador:
aP
dH

Equilíbrio
Isoclinal da presa:
p
rP

Equilíbrio
Como no caso de espécies competidorasl essas equações diferenciais não tem soluçõex explícitas. Precisamos graficar os isoclinais.
Isso produz o gráfico a seguir:


Equilíbrio
O comportamento desse sistema é intuitivo.
Produz o tipo de comportamento que pode ser presenciado no sistema de lobos e alces na Ilha de Royale.
O mesmo tipo de comportamento se apresenta no sistema de lince e lebre. Podemos fazer um gráfico:


Equilíbrio
Os tempos de retorno tem sentido.
Demora para que a população do predador alcançar a população da presa.
Os predadores não produzem predadores novos instantaneamente. Tampouco parem de reproduzir no mesmo instante.

Equilíbrio
Podemos tornar o modelo mais real.
Existe uma capacidade de suporte da população da presa, e provavelmente uma capacidade de suporte para o predador.
Também existe um efeito de Allee: alguma população mínima necessária para suster a população.


Equilíbrio
O sistema fica mais interessante. A localização exata do isoclinal do predador é importante.
Os resultados serão diferentes para sistemas nos quais o isoclinal do predador fica a esquerda ou direto do ‘corcundo’ do isoclinal da presa.


Equilíbrio Não Estável
No primeiro cenário, uma vez perturbado o sistema do ponto de equilíbrio, o sistema gira fora dos limites e resulta na extinção. Por que?

Equilíbrio Não Estável
Nesse caso, a corcunda fica ao direto do isoclinal do predador resultando num sistema não estável. Por que?
A população do predador é capaz de crescer ainda sob condições de densidade baixa da presa porque o predador é eficiente. Quando o predador fica menos eficiente, o isoclinal do predador muda ao direto


Equilíbrio Estável
Quando a ‘corcunda’ fica a esquerda, a região na qual a população do predador não cresce é maior.
As densidades muito elevadas da presa são necessárias para que o predador aumenta. Isso pode ser o resultado da cripse, ou o forrageio ineficiente do predador.

Modelo de Lotka-Volterra com o crescimento exponencial da
presa : coexistência

O modelo de Lotka-Volterra

O modelo de Lotka-Volterra

Modelo de Lotka-Volterra com o crescimento logístico da presa : r
extinção do predador

BifurcaçãoTranscritica
*xK
*xK (K,0) estável e (x*,y*) não estável e negativo
(K,0) e (x*,y*) são iguais
*xK (K,0) não estável e (x*,y*) estável e positivo

Perda de soluções periódicas
bxymydt
dy
axyK
xrx
dt
dx
1
x-y
0 0,3 0,6 0,9 1,2 1,5
x
0
1,6
3,2
4,8
6,4
8
y
x-y
0 0,3 0,6 0,9 1,2 1,5
x
0
4
8
12
16
20
y
coexistencia Extinção do predador

Resposta funcional Tipo I e II

O modelo de Holling
xD
bxymy
dt
dy
xD
axy
K
xrx
dt
dx
1

Existência de um ciclo de limites ( bifurcação super-crítica de Hopf)
22
22
yxyxdt
dy
yxxydt
dx
Coodenados Polares
1
2
dt
d
rrdt
dr

Equilíbrio estável

Na bifurcação
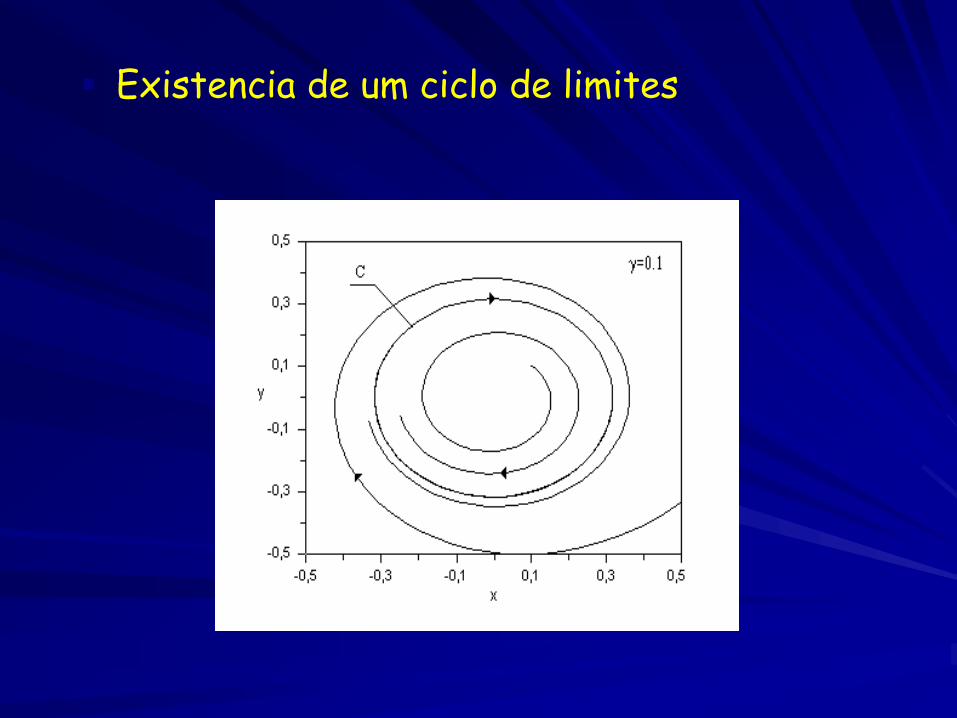
Existencia de um ciclo de limites

Bifurcação super-crítica de Hopf

A Teorema de Poincaré-Bendixson
Uma semi-orbita restrito no plano tende a : • um equilíbrio estável • um ciclo de limites • um gráfico de ciclo

Região de Armadilha

Região de Armadilha : Annulus

Exemplo de uma região de armadilha
xdt
dy
xxy
dt
dx
3
3
Modelo de Van der Pol (l>0)

O modelo de Holling
xD
bxymy
dt
dy
xD
axy
K
xrx
dt
dx
1

Isoclinais nulos do modelo de Holling

Caixa de Poincaré para o modelo de Holling

Modelo de Holling com ciclo de limites

Paradoxo de enriquecimento
Com o aumento de K : • Extinção do predador • Coexistencia do predador e presa (TC) • Equilibrio de predador e presa fica não estãvel (Hopf) • Ocorrencia de um ciclo de limite estável (variações grandes)

(c) 2001 by W. H. Freeman and
Company
Quais equilíbrios são prováveis?
Os modelos de predador e presa sugerem que: – A presa é mais provável estar em equilíbrio
relativamente alto ou relativamente baixo (ou tal vez ambos)
– Os equilíbrios a densidades intermédias da presa não são prováveis

Outros modelos de presa e predador
• Resposta funcional (Tipo III, dependente da razãot …) • Presa-predador-super-predador… • Níveis tróficos

Condições de estabilidade de Routh-Hurwitz
0... 1
1
2
2
1
1
n
n
n
nnn aaaa
00)(, ik
HRk
11aH
• Equações características
• Condições de estabilidade : M* l.a.s.
2
31
21 a
aaH
31
42
531
3
0
1
aa
aa
aaa
H

Condições de estabilidade de Routh-Hurwitz
032
2
1
3 aaa
011
trAaH
• Dimensão 2
• Dimensão 3
0det2 AtrA
0det3212
AaaaH
011 aH
03212 aaaH
033 aH

Exemplo de 3 níveis tróficos
dyzzzdt
dz
cyzbxymydt
dy
axyrxdt
dx
)1(

Modelo de competição inter-específica
2
1
21
2
2
22
2
1
2
12
1
1
11
1
1
1
K
xa
K
xxr
dt
dx
K
xa
K
xxr
dt
dx
Sistema transformado
buvvd
dv
avuud
du
1
1

O efeito de exploração sobre a Presa
( )H x hx
( )hx
H xc x
Analisa o modelo com duas funções não constantes de coleta na equalçao da
presa.
1.
2.

O efeito de exploração sobre a Presa
Encontre os pontos de equilíbrio e determine a estabilidade local

O efeito de exploração sobre a Presa
Descobrir as bifurcações, órbitas periódicas e orbitas de conexão
Exemplo de uma Bifurcação de Hopf
Diagrama de Bifurcação da Equação Logística

Modelo: Esforço de Caça Constante
(1 )axy
x x x hxy x
bxy y d
y x
•A presa é caçada a uma taxa definida por uma função linear. •Existem dois pontos de equilíbrio no primeiro quadrante sob certos valores dos parametros. • um ponto representa a co-existencia das esppecies. •Esforço máximo de caça = 1

Bifurcações do Modelo
Bifurcação de Hopf
Bifurcação Transcritica

Conexão dos Orbitos

Modelo de Limitação da Caça
(1 )axy hx
x x xy x c x
bxy y d
y x
• A presa e caçada a uma taxa definida por uma função racional. •O modelo tem três equilíbrios que existem no primeiro quadrante sob certas condições. • Um desses pontos representa a coexistência das espécies

Bifurcações no Modelo de limitação da caça

Conclusões
A coexistência é possível com ambas políticas de exploração.
As bifurcações múltiplas e orbitas de conexão existem no equilíbrio de coexistência.
A caça máxima sustentável do modelo de esforço constante da caça

O modelo original prevê oscilações de predadores e presas observadas freqüentemente na natureza.
– Um aumento da taxa de natalidade da presa aumenta a densidade de equilíbrio do predador, mas não da presa
Essa previsão is borne out em experimentos simples com bactérias e bacteriófagos por Bohannan e Lenski.
Tem várias fraquezas, porém
– oscilações completamente neutras não são observadas na natureza – são artefatos da simplicidade do modelo
– modelo tem como premissa a eficiência de captura da presa é independente da densidade de presa – a presa não fica saciada
– Modelo tem como premissa nenhuma dependência de densidade da presa

(c) 2001 by W. H. Freeman and
Company
Modificação do Modelo de Lotka-Volterra para Predadores e
Presas Existem vários problemas com as equações de Lotka-Volterra: – A carência de qualquer força que tende
restaurar as populações ao equilíbrio conjunto:
Essa condição é conhecido como o equilíbrio neutro
– A carência da saciação do predador: Cada predador consume uma proporção constante da população da presa independente da densidade da presa

(c) 2001 by W. H. Freeman and
Company
Vários fatores reduzem as oscilações de predador e presa
Todas as condições a seguir tendem estabilizar os números de predadores e presas (no sentido de manter tamanhos populacionais de equilíbrio que não variam): – Ineficiência do predador
– Limitações dependentes de densidade do predador ou presa por fatores externos
– Presas alternativas para o predador
– Refúgios de predação em densidades baixas da presa
– Tempos de retorno curtos na resposta do predador a mudanças da abundancia da presa

(c) 2001 by W. H. Freeman and
Company
Influencias Destabilizantes
A presença de ciclos de predadores e presas indica influencias destabilizantes: – Tais influencias tipicamente retardam o tempo nas
interações entre predadores e presas: Período de desenvolvimento
Tempo necessário para a resposta numérica do predador
Tempo para induzir a resposta imune em indivíduos ou respostas induzidas nas plantas
– Quando as influencias destabilizantes pesam mais do que os estabilizantes, os ciclos populacionais podem acontecer

(c) 2001 by W. H. Freeman and
Company
Efeitos de Níveis Diferentes da Predação
Os predadores não eficientes não podem manter a presa a níveis baixos (presa limitada por recursos, principalmente).
O aumento da eficiência do predador adiciona um segundo ponto estável a densidade baixa da presa.
Um aumento adicional das respostas numéricas e funcionais do predador pode eliminar um ponto estável de densidade alta da presa
A predação intensa em todo nível pode forçar a extinção da presa

(c) 2001 by W. H. Freeman and
Company
Quando o predador força a presa a extinção?
É possível que o predador força a presa a extinção quando: – Os predadores e presas mantidos em
sistemas simples de laboratório
– O predador se mantêm em densidades altas pela disponibilidade de presas alternativas menos preferidas:
O controle biológico pode ser melhorado ao oferecer presas alternativas aos parasitóides e predadores

Como predadores (não prudentes) coexistir com presas naturalmente?
Natureza é complexa
Locais de esconderijo da presa
Meta-populações: populações podem “se esconder” de predadores
– Eventualmente os predadores encontram populações não exploradas e podem levar essas a extinção;
– Porém, as presas individuais dispersam continuamente e encontram locais onde não existem predadores.
– Exemplo e o universo de ácaros e laranjas de Huffaker e populações de Cactoblastis e Opuntia na Austrália
Escolhas alternativas de presas
– Podem estabilizar interações de predadores e presas por aliviar a pressão sobre cada espécie de presa
Coexistência de predadores e presas

Até aqui tivemos como premissa que as respostas de predadores a presas (e o oposto) são instantâneas
Efeito de tempos de retorno
Uma situação mais real inclua tempos de retorno (o tempo necessário para a presa consumida ser transforma em novos predadores, ou para os predadores morrer de fome)
A incorporação de tempos de retorno aos modelos geralmente tem efeitos destabiliziantes, resultando em oscilações maiores das populações de predador e presa
Harrison (1995) incorporou tempos de retorno a resposta numérica de Didinium consumindo presas de Paramecium Melhorou muito o ajuste dos modelos a flutuações reais de populações de predador e presa descritos por Luckinbill (1973)

Modelos de predação e experimentos simples de microcosmos prevêem:
Coexistência no equilíbrio estável, após ciclos de oscilações apagadas, ou dentro de ciclos de limites estáveis, ou instabilidade e falta de
coexistência, dependendo especialmente da biologia das espécies que interagem:
A resposta funcional dos predadores as presas (geralmente leva a instabilidade se não lineares)
Capacidade de suporte dos predadores e presas na ausência do outro (freqüentemente estabilizantes)
Refúgios para as presas (freqüentemente estabilizantes)
Especificidade do predador a presa (não estabilizante se a troca ocorre a uma densidade menor da presa, mas estabilizante se a troca ocorre em densidades maiores da presa)
E mais…


Dinâmica de um modelo dependente da razão de predadores e presas com diretrizes não constantes
de exploração

Modelos de Predadores e Presas
1925 e 1926: Lotka e Volterra independentemente propuseram um par de equações diferenciais que modelam a relação entre um predador solitário e uma presa solitária num ambiente:
x rx axy
y bxy cy
Variáveis e Parâmetros x – população da presa y – população do predador r – taxa intrínseca de aumento da presa a – coeficiente de predação b – taxa reprodutiva por 1 presa consumida c – taxa de mortalidade do predador

Modelo de dependência de razão de predadores e presas
(1 )axy
x x xy x
bxyy dy
y x
Parâmetros e variáveis x – população da presa y – população do predador a – taxa de captura da presa d – taxa de mortalidade do predador b – taxa de conversão do predador
Crescimento
Da presa predação
Mortalidade Do predador
Crescimento Do predador

O modelo de Lotka-Volterra
bxymydt
dy
axyrxdt
dx
• r taxa de crescimento da presa : lei de Malthus • m taxa de mortalidade do predador: mortalidade natural• lei de ação de massa • a e b coeficientes de predação : b=ea • e coeficiente de conversão de presa em biomassa do predador

( )H x hx
( )hx
H xc x
Comparar modelos com duas funções não constantes da exploração da presa.
1.
2.

Modelo de esforço constante de exploração
(1 )axy
x x x hxy x
bxy y d
y x
• A presa é explorada a uma taxa definida por uma função linear. •Existem dois equilíbrios no primeiro quadrante sob certos valores dos parâmetros. • Um dos pontos representa a coexistência da espécie. •Esforço máximo de exploração = 1
( )H x hx

O efeito de exploração sobre a Presa
(1 )axy
x x x hy x
bxyy dy
y x
( )H x hx
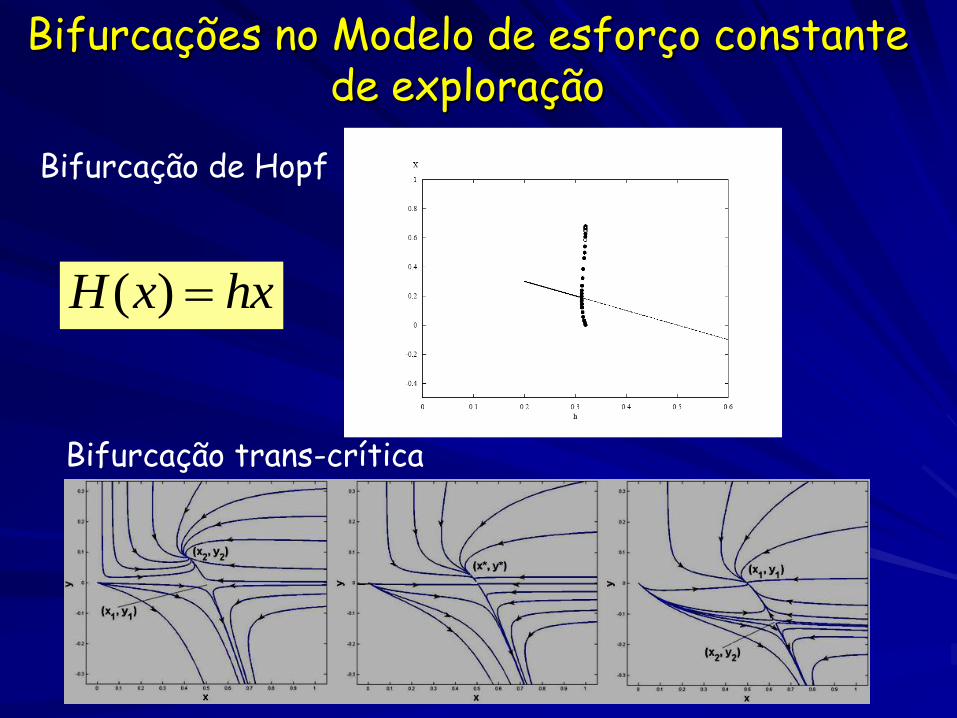
Bifurcações no Modelo de esforço constante de exploração
Bifurcação de Hopf
Bifurcação trans-crítica
( )H x hx

Orbitas de conexão
( )H x hx

Modelo de limitação a exploração
(1 )axy hx
x x xy x c x
bxy y d
y x
• A presa e caçada a uma taxa definida por uma função racional. •O modelo tem três equilíbrios que existem no primeiro quadrante sob certas condições. • Um desses pontos representa a coexistência das espécies
( )hx
H xc x

Bifurcações no Modelo de limitação a exploração
( )hx
H xc x

Resumo
Coexistência é possível sob os diretrizes de exploração.
Bifurcações múltiplas e orbitas de conexão existem no equilíbrio de coexistência.
A produção máxima sustentável calculada para o modelo de esforço constante de exploração

Images from Estes et al. 1998

Images from Estes et al. 1998

O modelo de Lotka-Volterra pode imitar os ciclos de predadores e presas
obeservados na natureza



Resultado e a 3ro gráfico a
esquerda
Há um aumento cíclico e
quebra nas populações de
predador e presa no tempo
Densidade de predadores
sempre atrás da densidade da
presa
Cenário de Banquete e Fome
Presa e predador nunca
atingem a extinção
Regulação mutua de
populações
Análise simplificada

Graphical analyses and stability of predator-prey systems Modifications of prey isocline (see lecture, text)
– Humped prey isocline
Why is it often hump-shaped? (Recall slope of logistic model)
Efeito de Allee em densidades baixas de presas
Stability depends on relative position of predator isocline
– Prey refuge from predator
Modifications of predator isocline
– Predator carrying capacity
– Predator interference (e.g., territoriality)
Factors that destabilize predator-prey interactions
– Time lags, predator efficiency
– Monophagous predator (inability to switch prey)

Estabilidade e Instabilidade Fatores que promovem a estabilidade
– Predadores não eficientes
– Dependência da densidade no predador ou na presa
– Predador troca a presa alternativa antes da extinção da presa
– Tempos de retorno pequenos em resposta a densidade da presa
Fatores que promovem a instabilidade
– Predadores muito eficientes
– Dependência da densidade inversa dos predadores ou presas
– Tempos de retorno longos em resposta a densidade do predador
– Ambientes simples, sem refúgios

Population Cycles in
Mathematical and Laboratory
Models Lotka Volterra assumes host population
grows exponentially, and population size is
limited by parasites, pathogens, and
predators:
dNh/dt = rhNh – pNhNp
rhNh = Exponential growth by host population.
– Opposed by:
P = rate of parasitism / predation.
Nh = Number of hosts.
Np = Number of parasites / predators.

Population Cycles in
Mathematical and Laboratory
Models Lotka Volterra assumes parasite/predator
growth rate is determined by rate of
conversion of food into offspring minus
mortality rate of parasitoid population:
dNp/dt = cpNhNp-dpNp
cpNhNp = Conversion rate of hosts into
offspring.
pNhNp = Rate at which exploiters destroy
hosts.
C = Conversion factor

O modelo de Lotka-Volterra imita os ciclos naturais do lince e da lebre

Seleção de um modelo Lotka-Volterra Clássico
(crescimento exponencial da presa, resposta funcional Tipo I do predador)
Não estável estruturalmente, mas interesse histórica
Lotka-Volterra (crescimento logístico da presa, resposta funcional Tipo I
do predador)
Não tem ciclos!
Rosenzweig-MacArthur (crescimento logístico da presa, resposta funcional Tipo II
do predador, saciação do predador)
Ciclos!





























