Embed Size (px)
Citation preview

Curso de Fisiologia 2007 Ciclo de Neurofisiologia Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida
120
FISIOLOGIA MUSCULAR
MUSCULATURA DO CORPO
Tendemos a pensar que movimento está sempre associado ao deslocamento do organismo, isto
é, a mudança dele de um lugar para o outro. Pensar assim é uma ilusão, pois vários movimentos estão sendo realizados enquanto estamos parados: os dedos coçando um ponto do corpo; o estômago misturando e propelindo o seu conteúdo; o coração ejetando e fazendo circular o sangue através do sistema circulatório, o diafragma e os músculos intercostais garantindo a respiração, etc.
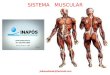
Os órgãos efetuadores de movimento do nosso corpo são formados de células musculares geradores de tensão mecânica e são de três tipos histológicos: fibras musculares estriadas esqueléticas e cardíacas e fibras musculares lisas.
Fibras musculares esqueléticas Fibras musculares cardíacas Fibras musculares lisas
Formam o tecido muscular esquelético, cujos músculos em sua maioria está associada a esqueletos; são responsáveis pela execução de atividades voluntárias e reflexas do organismo.
Formam o coração cujo órgão funciona como uma bomba ejetora cíclica, propulsionando o sangue através dos vasos.
Formam os demais órgãos viscerais do corpo (bexiga, trato gastrointestinal, útero, etc.). Como a musculatura cardíaca está apenas sob controle reflexo.
O músculo estriado esquelético está associado ao esqueleto e garante a execução de
movimentos e posturas do nosso corpo; faz exceção a musculatura que forma os esfíncteres (anais e vesicais) e a que está associada à expressão facial. A possibilidade de realizar os movimentos desejados torna-se possível graças ao controle voluntário que temos sobre a musculatura esquelética. A atividade das fibras musculares lisas e cardíacas não está sujeita ao domínio consciente. Distribuição e características da musculatura esquelética
Nos vertebrados, a musculatura esquelética corresponde aproximadamente a 40% do peso corporal e os demais tipos de tecido muscular (liso e cardíaco), 10%. Além de a musculatura esquelética expressar os nossos padrões de comportamento, desempenha outras funções como gerar calor (termorregulação) e, em condições de jejum prolongado, disponibilizar aminoácidos das proteínas musculares para o fígado (neoglicogênese). Os músculos podem movimentar partes do corpo porque cada extremidade de um músculo está presa a um osso por meio de um tendão que muitas vezes estende-se sobre uma articulação. Os músculos deslocam 2 ou mais ossos através de uma
articulação móvel, ou então, deslocam tecidos. A maioria das articulações possui dois grupos de músculos funcionalmente distintos: um que quando em atividade diminui o ângulo articular, e um outro de ação antagônica que aumenta o ângulo. Um movimento pode ser simples, envolvendo alguns grupos musculares ou muito complexos, envolvendo a ação coordenada de vários grupos. O grau de complexidade de um movimento está intimamente associado com o circuito nervoso que controla os diferentes grupos de músculos requeridos para expressar aquele comportamento.

Curso de Fisiologia 2007 Ciclo de Neurofisiologia Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida
121
Classificação funcional do músculo esquelético Os músculos são classificados funcionalmente como músculos flexores e extensores, abdutores e adutores, pronadores e supinadores. Do ponto de vista funcional, a maioria pode ser
classificada basicamente como músculos flexores que com a contração diminuem o ângulo articular e como extensores que aumentam. Nos animais quadrúpedes os músculos extensores dos membros anteriores e posteriores garantem que o corpo se mantenha em pé, sustentando o peso do corpo contra a gravidade. Por isso, os músculos extensores são chamados de músculos anti-gravitacionários. Nos seres humanos que são bípedes, os músculos extensores dos membros inferiores e do tronco desempenham este papel enquanto os membros anteriores são mantidos livres e independentes para realizarem outras tarefas como as de manipulação. Outra classificação que auxilia muito quando levamos em conta o controle que o SNC exerce sobre o movimento e a postura do corpo é a diferenciação entre músculos mediais (axiais e proximais) e laterais (distais). Os músculos da cintura pélvica e escapular que são chamados proximais participam do equilíbrio postural. Já os músculos distais dos membros e dos movimentos intrínsecos dos dedos, não participam primariamente da postura, mas de atividades relacionadas com a manipulação de objetos.
Fontes de energia da contração muscular esquelética O suprimento energético imediato para a contração muscular é o ATP. Quando ele é hidrolisado, parte da energia liberada é utilizada para a geração de força mecânica e a outra parte é perdida na forma de calor. Como a disponibilidade de ATP é pequena nas fibras musculares, é necessária uma fonte adicional de energia como o fosfato de creatina que transfere o grupo fosfórico para o difosfato de adenosina e restaura rapidamente os níveis de ATP. Mas como a fonte de fosfato de creatina é também limitada, num exercício muscular mais prolongado o ATP passa a ser obtido pela oxidação completa de carboidratos e de ácidos graxos. A fonte de carboidrato muscular é armazenada na forma de glicogênio (cerca de 0,5 a 2% do peso muscular fresco) e contribui com cerca de 100 vezes mais do que a fonte de fosfato de creatina. Conforme as propriedades mecânicas das fibras musculares, será utilizado o metabolismo aeróbico ou anaeróbico como estudaremos mais tarde. Níveis de organização morfológica do músculo esquelético
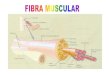
Devemos conhecer bem a organização morfológica da musculatura esquelética para compreendermos os mecanismos da contração muscular. O músculo esquelético é constituído de feixes paralelos de fibras musculares cujas células possuem diâmetro que variam de 10 a 80 µm e comprimentos de algumas a dezenas de mm. Cada fibra muscular é envolvida por uma membrana denominada sarcolema, constituída de uma membrana citoplasmática e uma camada fina de polissacarídeos e de fibrilas de colágeno. As fibras musculares afunilam-se em suas extremidades fundindo-se com os elementos fibrosos e tendinosos que se ligam aos ossos. Entre os feixes de fibras musculares estão os suprimentos sangüíneos, nervosos e tecidos adiposo.
Dentro da fibra muscular estão as miofibrilas (feixes cilíndricos de proteínas) dispostas paralelamente, apresentando padrões repetitivos de estrias transversais. Os pacotes de miofibrilas estão mergulhados no mioplasma intercalados por grânulos de glicogênio, enzimas glicolíticas e produtos intermediários, ATP, ADP, AMP, fosfato, fosfato de creatina, eletrólitos e outras estruturas subcelulares. O retículo endoplasmático das fibras musculares (retículo sarcoplasmático) armazena íons Ca++ e possui canais de Ca voltagem-dependentes. O retículo sarcoplasmático (RS) é uma estrutura volumosa (14% da fibra), formada de túbulos que se anastomosam e acompanham as miofibrilas; possuem extremidades que terminam em sacos (cisternas terminais). Neste ponto, são trespassados pelos túbulos transversais (túbulos T) que se originam do sarcolema formando uma estrutura tríade. Os túbulos T dispõem-se perpendicularmente ao RS e às miofibrilas, formando uma rede transversal e reticulada no interior da fibra muscular. Funcionalmente comunicam o meio extracelular transversalmente ao longo da fibra muscular.

Curso de Fisiologia 2007 Ciclo de Neurofisiologia Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida
122
Organização molecular do sarcômero
As miofibrilas possuem unidades repetitivas denominadas sarcômeros. O sarcômero constitui a unidade contrátil da fibra muscular e está limitada pelas linhas Z. No músculo de todos os vertebrados, o comprimento do sarcômero é semelhante sendo em torno de 3,5µm no estado de repouso e de 1,5µm quando contraído. Cada sarcômero apresenta uma organização molecular constituída de filamentos finos e filamentos grossos. A partir da linha Z, os filamentos finos estendem-se paralelamente para ambos os sentidos sendo intercalados pelos filamentos grossos, apenas na região central do sarcômero. Tal organização produz um padrão de bandas transversais que varia conforme o estado de contração muscular. Os filamentos grossos são constituídos de macromoléculas de miosina e os filamentos finos, de moléculas de actina, tropomiosina e troponina.
A miosina corresponde a 55% da proteína miofibrilar e cada filamento grosso é formado por 200 a 400 dessas moléculas; é formada por duas cadeias: uma leve e outra pesada. Na cadeia pesada a cabeça possui dois sítios ativos: uma que possui alta afinidade pela actina e uma outra catalítica que hidrolisa o ATP. As moléculas de miosina formam um feixe em que as cabeças se destacam do eixo central e dispõem-se bem próximos aos filamentos de actina. Apesar da elevada afinidade das moléculas de miosina para com a actina, quando o músculo está em repouso (relaxado), a ligação entre elas está obstruída pelas moléculas de tropomiosina.
A actina constitui 20 a 25% da proteína miofibrilar e ocorre em duas formas (actina G e actina F) que, quando polimerizadas, formam cordões duplos helicoidais. Cada molécula de actina G liga-se com grande afinidade a um íon Ca++ e a uma molécula de ATP.

Curso de Fisiologia 2007 Ciclo de Neurofisiologia Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida
123
A tropomiosina é uma molécula formada por duas cadeias peptídicas separadas e que estão enroladas entre si.
A troponina é uma proteína globular com função reguladora e possui três subunidades: C que se liga a íons Ca++; I que é inibitória e T que se liga à tropomiosina. No músculo relaxado a tropomiosina está obstruindo o sítio de ligação da actina com a miosina. Antes de tratarmos do mecanismo de contração devemos incorporar outros conceitos.
Unidade motora
As fibras musculares esqueléticas só se contraem sob comando neural. Lembremos que a musculatura da cabeça e do pescoço é inervada por motoneurônios situados respectivamente no tronco encefálico e porções mais altas da medula cervical; todos os demais músculos do corpo são inervados por motoneurônios de origem medular.
Uma fibra muscular só recebe a inervação de um único motoneurônio, mas o motoneurônio pode inervar várias fibras musculares. Chamamos de unidade motora o motoneurônio e as fibras musculares por ele inervadas. O tamanho das unidades motoras varia: podemos encontrar unidades motoras cuja relação de inervação é baixa, isto é, um motoneurônio inervando 1700 fibras musculares ou unidades cuja relação é alta: um motoneurônio para algumas fibras musculares. As unidades que participam de movimentos finos possuem relação de inervação altas como nos músculos oculares e dos dedos (1 motoneurônio :10 fibras).
Os motoneurônios que inervam as fibras musculares são todos mielinizados e são do tipo Aα cujos axônios ao se aproximarem das fibras musculares ramificam-se e perdem a bainha de mielina. Os terminais se justapõe a uma depressão do sarcolema denominada placa motora. A região pós-sináptica da fibra muscular formam pregas juncionais e a região pré-sinaptica possui zonas ativas onde a acetilcolina (Ach), o NT exclusivo de todas as junções neuromusculares esqueléticas são secretadas. O mecanismo de neurotransmissâo é o mesmo já estudado na transmissão sináptica nervosa. Os receptores para a ACh são nicotínicos (ionotrópicos) e ao se ligarem a ACh abrem canais iônicos ocorrendo o influxo resultante de Na, ou seja, PEPS. Este potencial elétrico despolarizante é chamado de potencial de placa motora. A onda de despolarização propaga-se para fora da placa e abrem canais de Na e K voltagem dependentes distribuídos ao longo de todo o sarcolema. Assim são gerados PA na fibra muscular, cuja freqüência é proporcional a quantidade de ACh liberado.
Cada impulso nervoso libera cerca de 60 vesículas de Ach e cada vesícula contem 10.000 moléculas de Ach, quantidade suficiente para ativar os receptores pós-sinaticos e desencadear um potencial de placa completo. Entretanto, mesmo em repouso, pacotes de Ach são liberados espontaneamente, produzindo “microdespolarizaçôes” na membrana pós-sinaptica. A despolarização causada por um pacote de vesícula (quanta) é denominada potencial de placa em miniatura com amplitude de 0,5 mV , mas que não suficiente para causar o PA na fibra. Somente com a estimulação

Curso de Fisiologia 2007 Ciclo de Neurofisiologia Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida
124
nervosa e com a liberação de vários quanta de vesículas é que se torna possível a produção de um potencial de placa capaz de causar o PA. Miastenia grave é uma grave doença auto-imune em que o próprio organismo produz anticorpos contra os receptores nicotínicos. Como conseqüência, dificulta a transmissão sináptica causando fraqueza muscular (Veja discussão de caso na aula teórico-prática). Como ocorre a contração muscular esquelética? Visite o site http://www.blackwellscience.com/matthews/myosin.html
Quando o músculo não está sob estimulação nervosa, isto é, quando está relaxado a [Ca++] no mioplasma é insignificante em relação ao meio extracelular. Com a chegada do impulsos nervosos, a fibra muscular responde com PAs e a atividade elétrica propaga-se pelos túbulos T atingindo as cisternas do retículo sarcoplasmático. A função dos túbulos T é a de garantir a rápida propagação da onda de despolarização em direção às cisternas do RE. A despolarização dos túbulos T abre canais de Ca++ voltagem dependentes e, como conseqüência, o cálcio armazenado no interior do reticulo sarcoplasmatico flui passivamente para o mioplasma a favor do seu elevado gradiente de concentração.
Os íons Ca ligam-se co elevada afinidade aos sítios da troponina C, modificando a sua
organização espacial, tornando livre o sítio T que poderá se ligar à tropomiosina. Agora, o complexo formado pode desobstruir o sítio de ligação da actina com a miosina. A pronta formação do complexo actina-miosina forma uma ponte cruzada entre o filamento fino e o grosso.
Como a miosina tem um sítio catalítico para a hidrólise de ATP, esta união torna a energia química disponível para o dobramento da cabeça de miosina e os filamentos finos sofrem um ciclo de arraste para o centro, isto é, os filamentos finos deslizam-se sobre os grossos. Com a hidrólise do ATP, a molécula de miosina perde a afinidade pela actina, desliga-se e restabelecendo a sua posição original. Mas se há mais ATP e Ca++ disponíveis no mioplasma, o ciclo das pontes cruzadas se restabele e o deslizamento progride com o encurtamento cada vez maior do sarcômero. Assim, as fibras musculares, como um todo, encurtam-se. Cada ciclo pode mover o filamento fino cerca de uns 10nm e para cada molécula de troponina ativa, sete sítios fixadores de miosina são descobertos.

Curso de Fisiologia 2007 Ciclo de Neurofisiologia Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida
125
Denominamos acoplamento excitação-contração (ou acoplamento eletro-mecânico), ao processo em que a atividade elétrica da fibra muscular (PA) é transformada em atividade mecânica (encurtamento do sarcômero), ou seja, na conversão (transdução) de atividade elétrica em mecânica.
Relaxamento muscular
Terminada a contração o Ca++ deverá ser recaptado de volta para o reticulo sarcoplasmático ativamente por ATPases Ca/Mg dependentes. Como a afinidade dessa bomba pelo Ca++ é maior do que do íon pela troponina, o recolhimento ativo predomina. Enquanto não houver PA no sarcoplasma, a formação de novas pontes cruzadas não será possível e o músculo ficará no estado de relaxamento ou de repouso.
Rigor mortis Com a morte e, por conseqüência, com a falência sangüínea, o aporte de oxigênio e o controle nervoso deixam de chegar à musculatura. Mesmo assim, o músculo tem a disposição à via anaeróbica e contrairá formando complexos acto-miosinicos irreversíveis desde que haja cálcio disponível no mioplasma. Nesse estado, a musculatura atinge o rigor mortis, ou seja, estado de enrijecimento muscular generalizado. O resultado é um estado de contração máxima dos músculos sem relaxamento com o cadáver em estado de rigidez (temporário). O início do processo é bem variável, na dependência principalmente da reserva de glicogênio do músculo e a rigidez instalada ocorre simultaneamente em toda a musculatura. Durante a instalação da rigidez, ocorre uma queda brusca de pH (aumento de acido lático). Esse é problema muito sério quando se trata de carne para o consumo, pois poderá comprometer a sua qualidade. A sua instalação varia de 10 minutos a muitas horas após a morte e depende da temperatura. MECÂNICA DA CONTRAÇÃO MUSCULAR
O sistema contrátil depende da concentração de Ca++ disponível no
mioplasma, ou seja, da freqüência com que as fibras musculares da unidade motora estão sendo estimuladas. Para cada PA em uma unidade motora ocorre um abalo (espasmo muscular) ou uma breve contração seguida de relaxamento. O abalo começa uns 2ms após o inicio da despolarização e sua duração varia com tipo de fibras musculares: se são de contração rápida ou lenta. Cada abalo apresenta uma resposta mecânica do tipo tudo-ou-nada; se um segundo estimulo for aplicado antes que o relaxamento do primeiro abalo tenha começado, haverá uma superposição das respostas mecânicas que se denomina somação, resultando numa tensão maior.
A aplicação de estímulos cada vez mais freqüentes produzirá contrações musculares sustentadas e uniformes denominada tétano. A força desenvolvida num tétano perfeito é de 4 vezes maior em relação a um abalo isolado. Durante a fase das somações registra-se o que se chama “fenômeno da escada”. Assim, uma maneira de aumentar a força de contração máxima de uma unidade motora é aumentando a freqüência de estimulação das unidades musculares.
Um músculo é constituído de várias unidades motoras. Assim, o desempenho mecânico do músculo como um todo pode aumentado recrutando-se a atividade de todas as unidades motoras. É por causa dessa organização morfo-funcional é podemos graduar precisamente a força muscular. Do contrário, se todas as fibras musculares de um músculo fossem inervadas pôr um único motoneurônio, o músculo sempre se contrairia de uma única vez. O recrutamento é realizado em uma determinada ordem onde as unidades motoras mais fracas (em termos de potencia mecânica) são recrutadas primeiro e depois as mais potentes.

Curso de Fisiologia 2007 Ciclo de Neurofisiologia Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida
126
Contrações isométricas e isotônicas A contração muscular é o estado de atividade mecânica. O músculo se comporta como um sistema de dois componentes no qual a parte contrátil (sarcômeros) está em série com o componente elástico (tendões e tecido conjuntivo). Quando os sarcômeros se encurtam a parte elástica é tracionada e se uma das extremidades do músculo for móvel, causará movimento e se ambas estão fixas, causará apenas tensão (ou estresse). Denominamos contrações isométricas quando a força está sendo exercida nos pontos de fixação muscular sem o encurtamento significativo das fibras musculares. Nesse tipo de contração, as pontes cruzadas se estabelecem aumentando a tensão ou a rigidez do músculo. O desenvolvimento deste tipo de força ocorre tipicamente nos músculos posturais ou quando um músculo atua contra uma força oposta. Apesar de não ocorrer trabalho externo, o músculo realizou o que
chamamos de trabalho interno. Quando as fibras musculares encurtam-se e uma carga externa é movida de um lugar para outro temos a realização de trabalho externo e dizemos que houve contração isotônica. Nesse caso, o músculo se encurtou gerando tensão mecânica constante (movimento).
No exemplo acima, quando o bíceps ergue a carga e realiza movimento “trabalha” isotonicamente, mas quando se tenta puxar o cordão que está fixo, não há movimento e o músculo trabalha isometricamente. Na prática, uma contração passa de isométrica a isotônica e vice-e-versa, o tempo todo. Relação entre a força máxima e o comprimento do músculo
A força máxima desenvolvida no tétano depende, entretanto, do comprimento inicial do músculo. O músculo possui elementos elásticos em série e em paralelo (todos os elementos situados entre as pontes cruzadas e as extremidades dos tendões). Quando um músculo é passivamente estirado ele tende a retornar ao seu comprimento de repouso. Trata-se da propriedade elástica do músculo que pode ser descrita numa curva de tensão passiva.
Quando um músculo executa uma contração isométrica, a tensão ativa gerada pelas pontes cruzadas é adicionada a tensão passiva. Na figura ao lado, a curva em tracejado é a curva de tensão ativa em função do comprimento e mostra que a faixa operacional do comprimento do músculo se encontra dentro de uma pequena porcentagem do
Contração isotônica Contração isométrica

Curso de Fisiologia 2007 Ciclo de Neurofisiologia Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida
127
comprimento em repouso. A tensão ativa é máxima quando próxima do comprimento em repouso; neste ponto a superposição dos filamentos finos e grossos é tal que todas as cabeças miosínicas podem formar pontes cruzadas. Se houver uma distensão passiva, o grau de superposição diminui e o número de pontes cruzadas diminui, reduzindo drasticamente a geração de força ativa. O mesmo se verifica quando o músculo tenta se contrair a partir de um grau de sobreposição já existente entre os filamentos. Em suma, os músculos esqueléticos estão adaptados a gerarem a sua força máxima a partir do comprimento de repouso e dentro de uma faixa muito estreita em torno desse comprimento. Essa faixa de desempenho ótimo é denominada faixa operacional real do músculo. Sistema de alavancas
Quando um músculo isolado é submetido a contrair-se suportando uma carga de 7 Kg, ele gerará uma força equivalente e puxará o objeto com um deslocamento 1cm (que corresponde ao encurtamento muscular). A associação do músculo com o esqueleto produz um sistema de alavancas que amplifica a força gerada pela contração muscular. Suponha que a mesma de carga de 7Kg seja movida pelo antebraço. Nesse caso o trabalho foi realizado pela contração do músculo bíceps e seus agonistas que fletiram o antebraço. O bíceps tem a inserção além da articulação móvel do cotovelo. Assim, a sua contração gera uma tensão mecânica deve ser multiplicada pela alavancagem do braço. Ainda que o músculo encurte apenas 1cm, a carga vai se mover muito mais, ou seja, 7cm e com bem menos força.
Tipos de fibras musculares
�������������� ����������������������
� � � � � �� � � � � � ������ �������� ������ ��������� ������ � � ����������
���� � � �� � � � � � � �� �� �� �� � � � �� �
� � � ��� � � ���� � � � �� � �� � � �� � �� �� �� �� �� �� � � � ����
� ���� � �� ��� � � � �� � � ����� � �� �� �� �� �� � � � ����
! �" � � ��� �� � �! �� �� # � ��� � ��� � � � � � ��� �� � �� �� �� � � � ���� �
$ � � ��� � � �� � �� ��� ��� �� � ��� � � �� � % � � � �
% � � � ���� � �� �� �& � � �� �� � �& � � �� �� � �� �
� � �� �� � � �� � � �� �� ' ( �� ) � � � � � � �� � � � �� �
* � � � ��� � � �� �� ' ( �� ) �� � �� �� ���� �� �� �� � � � ����
+ ��' � �� �� ���� � �� �� ��� �� % , � � �
� � � �� �# � � � �- � �� � ! � � � � � ! � � � � � � � . � � � �
As fibras musculares diferem quanto às vias metabólicas preferenciais: há fibras musculares
“vermelhas” de contração lenta e fibras musculares “brancas” de contração rápida. Quando estamos de pé, alguns músculos estão tonicamente contraídos enquanto outros, intermitentemente. Há músculos que realizam movimentos rápidos e delicados e outros que são lentos e são mais grosseiros. Estas diferenças estão intimamente associadas com o padrão de resposta mecânica e do metabolismo celular. Cada motoneurônio inerva um conjunto de fibras musculares funcionalmente relacionadas. Assim, uma unidade motora vermelha possui contração lenta e é mecanicamente menos potente, ao contrário das unidades brancas que se são rápidas e mais potentes. Esta diferença está no fato de que as unidades vermelhas possuem menor relação de inervação e as fibras musculares são finas (menos miofilamentos contráteis) do que as unidades brancas. Em nosso cotidiano estamos geralmente recrutando unidades motoras vermelhas fracas e lentas, porém bastante resistentes à fadiga. Já as unidades brancas são recrutadas quando requeremos grande esforço muscular, rapidez e potência. Entretanto, há um tipo intermediário de fibras vermelhas que realizam contrações rápidas, de precisão e força moderada e de resistência. Os motoneurônios

Curso de Fisiologia 2007 Ciclo de Neurofisiologia Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida
128
destas respectivas fibras também variam em ordem crescente de tamanho. Esta organização tem 3 conseqüências importantes:
1) o recrutamento ordenado facilita a tarefa de modular a força de contração do músculo; 2) as unidades lentas e as intermediárias são as constantemente recrutadas e são sempre usadas. 3) Os motoneurônios maiores são recrutados nas emergências em que tarefas específicas de
grande esforço muscular são requeridas.
Exercício muscular
Quando realizamos exercício podemos optar por dois tipos de treinamento: para melhorar a
resistência ou a força. Para melhorar a resistência os exercícios recomendados são aqueles que recrutam fibras musculares aeróbicas e atividades de longa duração “trabalhando” os músculos com cargas leves. Assim, com o tempo a quantidade de mitocôndrias e a densidade capilar aumentarão.Com isso, a resistência ao fluxo sanguíneo será diminuída e a perfusão sanguínea melhorada, bem como, o condicionamento cárdio-respiratório. Para melhorar a força, os exercícios recomendados são os de alto impacto, ou seja, de curta duração com cargas pesadas e umas duas ou três vezes por semana. O resultado é o aumento de miofilamentos contráteis e hipertrofia dos músculos solicitados.
Os atletas que realizarem exercícios predominantemente anaeróbicos (velocistas, etc) apresentam hipertrofia muscular e possuem massa muscular aumentada. Já os atletas maratonistas que investem em exercícios de resistência têm a capacidade de captação de oxigênio aumentada em até 20% e a uma baixa produção de ácido lático em relação ao indivíduo não treinado. No caso dos cultuadores físicos ocorrem duas alterações: aumento do diâmetro das fibras glicolíticas anaeróbicas e adição de colágenos e outros tecidos conjuntivos necessários para manter a tensão em resposta às cargas pesadas. Quando o regime dos exercícios é abandonado ocorre a atrofia por desuso. Já vimos que o ATP é a fonte de energia necessária para o músculo em atividade ou em repouso. O ATP origina-se de três fontes: metabolismo aeróbico ou anaeróbico e fosfato de creatina. Quando o músculo está em repouso, qualquer sobra de ATP é convertido em fosfato de creatina. Quando o músculo é submetido a um esforço moderado por longo período, o ATP provem do metabolismo aeróbico a partir da glicose (glicogênio muscular) ou o que é mais comum dos ácidos graxos. Já nos exercícios intenso de curta duração, o metabolismo anaeróbico e a degradação do fosfato de creatina fornecem energia por breve período de tempo (10 a 20s). Parte do acido lático formado é re-sintetizada em glicose no fígado e que poderá ser reutilizada pelos músculos anaeróbicos (Ciclo de Cori).
��� �
% , � � �
� � �� �
�� �� �� � � � ����
% , � � �
% , � � �
% , � ���
% , � ��� ��
% , � ��� ��
�� �� �� � � � �� � ��
������ � � ������� � ������� �� ������������
� �� ���� �� � � �� �# � � � �- � �� �
! � � � � �� � . � � � ��+ ��' � �� �� ����
� � �� ��) � � � ��� � �� �� � � �� � � �� �� ' ( ��
�� ����) �� � ��* � � � ��� � � �� �� ' ( ��
��� �� �� �+ �� . / # � � �� � �� �� � ���
� � �� ��� �� ��� � �� �� � � �� � � �� � � ' ( ��
������� �� ���) �� � ��� � �� � �� � ���� � � �
� ��� ��� �� �+ �� �� ��� � � ��� �
! � � � � � ��� � . � � � �� �% ���� � � �� � ��� �
� � �� � �) � � � � �+ �� � � �� � � � � �� � �
* �� �� �� � �0 � �� � � � �� ���� � �� �� � � ���� � ' ( �� �� �-�� � � �� ���� � �

Curso de Fisiologia 2007 Ciclo de Neurofisiologia Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida
129
Fadiga muscular A fadiga é uma condição temporária em que a força muscular não pode mais ser gerada pelo músculo por problemas metabólicos. A figura ao lado exemplifica tipos de fibras musculares com diferentes perfis de resistência: as RF fadigam-se muito rapidamente e as L são muito resistentes. Durante a fadiga, o músculo está em atividade, mas não consegue gerar força mecânica.
.
Desnervação
Quando os nervos motores são danificados, ocorrem fasciculações musculares (contrações
irregulares e pequenas) causadas pela liberação espontânea de Ach. Vários dias depois ocorre fibrilação (contrações assincrônicas das fibras musculares) causadas por um aumento de sensibilidade dos receptores nicotínicos. Cerca de 3 a 4 meses depois inicia-se o processo de atrofia muscular (diminuição do tamanho das fibras musculares). Após 1 a 2 anos as fibras musculares degeneradas são trocadas por tecido conjuntivo e tecido adiposo.
Se após a secção os motoneurônios forem estimulados adequadamente, pode ocorrer regeneração dos axônios e a recuperação motora. É interessante observar que os motoneurônios possuem fatores tróficos que regulam a atividade das fibras musculares que ele inerva: se fibras brancas tiverem os motoneurônios grandes trocados experimentalmente, por motoneurônios pequenos que inervam as fibras vermelhas, as suas propriedades metabólicas e mecânicas serão modificadas e vice-versa.