Embed Size (px)
Citation preview


Agradecimentos
Ao iniciar a apresentação do presente trabalho desejo expressar a minha sincera gratidão às
pessoas e instituições que colaboraram na sua concretização.
Ao meu orientador (de este e de outros trabalhos) Professor Doutor João Honrado, com quem
trabalho há já vários anos, por todo o seu apoio científico, sugestões e correcções, pela compreensão
e ajuda na transposição das fases mais complicadas.
À minha co-orientadora Doutora Maria José Vasconcelos, pela forma como me recebeu, por
me abrir as portas do mundo da detecção remota, pelo seu rigor científico, pelo seu interesse e
entusiasmo constantes.
Ao Doutor Luís Catarino coordenador do projecto POCI / BIA-BDE / 57965 / 2004 "Alterações
do coberto do solo e etapas de sucessão nas florestas do Cantanhez”, que me permitiu realizar este
trabalho, por me ter levado para a Guiné-Bissau, pelas suas sugestões, conselhos e partilha da sua
vasta informação sobre a Guiné-Bissau.
Ao Mestre Viriato Cassamá, autor de parte dos dados utilizados neste trabalho, por todo o seu
apoio e camaradagem quer em Portugal quer na Guiné.
A todos aqueles que tornaram a minha estadia na Guiné-Bissau tão extraordinária, e que, de
várias maneiras, contribuíram para o sucesso do trabalho de campo. Gostava de agradecer
particularmente ao Sr. Patrício Ribeiro, o português mais guineense que conheci, que me guiou à
chegada.
Ao IBAP (Instituto Biodiversidade Áreas Protegidas) e à AD (Acção para Desenvolvimento)
agradeço todo o apoio prestado na logística do trabalho de campo.
A todos aqueles que no IICT (Instituto Investigação Cientifica Tropical) e no ISA (Instituto
Superior de Agronomia) se interessaram e comentaram ou deram sugestões para o trabalho,
principalmente ao Doutor João Carreiras, pela ajuda com as misturas espectrais, e ao Engenheiro
Duarte Oom pela ajuda, sugestões e boa disposição.
A todos do CIBIO que se interessaram pelo trabalho, particularmente ao Doutor José Carlos
Brito pela sua disponibilidade e ajuda prestada no desenvolvimento da ENFA.
Ao Pedro Tarroso companheiro de mestrado pela sua ajuda e amizade.
Ao Doutor José Alberto Gonçalves pela ajuda com os SIG e na construção do MDT.
Aos meus pais e irmã pelo apoio e por vibrarem comigo na concretização desta quimera.
Por fim, um agradecimento especial à Catarina, pelo seu apoio e encorajamento constantes.

RESUMO
A compreensão dos aspectos estruturais e funcionais dos ecossistemas implica a realização
de estudos em distintas escalas espaciais, uma vez que alterações da paisagem como a desflorestação
ou urbanização podem modificar a natureza e os ritmos dos processos no seio dos ecossistemas. A
fragmentação da paisagem, por exemplo, pode alterar a taxas de recuperação dos ecossistemas, ao
condicionar a dispersão de espécies pioneiras. Devido a estes efeitos, numerosos estudos têm focado
as mudanças dos padrões espaciais que ocorrem em grandes extensões territoriais.
Com uma enorme capacidade de fornecer dados (comparáveis) a várias escalas e datas, a
detecção remota revela-se uma ferramenta poderosa para analisar as dinâmicas da paisagem. As
técnicas de detecção remota podem gerar, por exemplo, mapas de coberto do solo com distintas
resoluções espaciais e temporais, mapas estes que podem ser analisados com recurso a Sistemas de
Informação Geográfica e assim revelar qualitativa e quantitativamente as dinâmicas da paisagem.
No presente trabalho estudou-se a evolução da paisagem a partir de cartas de coberto do
solo obtidas por detecção remota, ao longo de 17 anos (1986-2003), na região do Cantanhez (Guiné
Bissau) (uma das regiões com maior biodiversidade da Africa Ocidental) bem como a influência dessa
dinâmica na adequabilidade de habitat para uma sub-espécie ameaçada de chimpanzé (Pan
troglodytes subsp. verus). Foram usadas métricas de paisagem para caracterizar a área de estudo em
quatro datas diferentes (1986, 1994, 2000 e 2003). Paralelamente, foi feita uma modelação da
adequabilidade de habitat para o chimpanzé, determinando espacialmente a área com maior
adequabilidade, e as classes de coberto de solo presentes na mesma. Finalmente, foi feita uma
análise da variação do habitat adequado para o chimpanzé ao longo do período temporal em estudo.
Os resultados revelaram uma área de máxima adequabilidade de habitat que compreende
12% do total da área de estudo e em que as classes de Floresta são maioritárias. Em relação à
evolução da paisagem, é notória a tendência para uma aglutinação de parcelas ao longo do tempo,
resultando, em termos gerais, numa progressiva simplificação da paisagem. De algum modo,
contrariando a tendência geral de estudos recentes da Ecologia da Paisagem, não foram obtidos
resultados que revelem um aumento da fragmentação da paisagem, mesmo nas classes florestais.
Ao contrário dos resultados de estudos anteriores relativos à África Ocidental, que indicam
sem excepção uma diminuição assinalável das áreas florestadas na região, os resultados da evolução
da paisagem para o Cantanhez revelam uma área com recuperação de áreas florestais e sem
evidências de aumento da fragmentação da paisagem. Os resultados da análise da paisagem indicam
que o habitat para chimpanzé na área de estudo não se encontra ameaçado, podendo mesmo a área
de habitat adequado ter aumentado entre 1986 e 2003.
- 2 -

ABSTRACT
A full understanding of structural and functional traits of ecosystems includes the
development of studies at multiple spatial scales, since landscape changes such as deforestation or
urbanization can modify the nature and rhythm of processes within those ecosystems. Fragmentation,
for example, can change the recovery capacity of ecosystems through its influence on dispersal of
pioneer species. Due to these effects, several studies have been focused on spatial changes over large
territories.
Due to its capacity of delivering (comparable) data over several spatial scales and from
distinct dates, remote sensing techniques have been shown to be a powerful tool in the analysis of
landscape dynamics. Remote sensing techniques can provide e.g. land cover maps with several spatial
and temporal scales which can be analysed in a GIS environment, allowing both qualitative and
quantitative analyses of landscape dynamics.
In this study, landscape changes were analysed over 17 years (1986-2003) from remote
sensing land cover maps for the Cantanhez region (Guinea Bissau), one of the richest areas of
Western Africa in terms of biodiversity. The influence of those changes in habitat suitability for a
threatened subspecies of chimpanzee (Pan troglodytes subsp. verus). Landscape metrics were used to
analyse the study area at four different dates (1986, 1994, 2000 and 2003). Also, a model for habitat
suitability was produced for chimpanzees, identifying the areas with higher suitability and the land
cover classes included in those areas. Finally, an analysis was made of the evolution of suitable
habitat for chimpanzees along the 17-year time period.
Results have revealed a forest-rich area of maximum habitat suitability that comprises 12% of
the total study area. Regarding landscape evolution, a patch coalescence trend has been noticed,
resulting in a general simplification of the landscape. Unlike most studies in Landscape Ecology, no
evidence was found for landscape fragmentation, even regarding forest classes.
Differently from previous studies in Western Africa, which have always reported a significant
decrease of forest areas, the results for landscape dynamics in Cantanhez indicate a recovery trend
for forest areas and do not show evidence of landscape fragmentation. The results of the landscape
analysis stress that the habitat for chimps is not threatened in the area, with suitable habitat probably
showing an increase between 1986 and 2003.
- 3 -

Índice
AGRADECIMENTOS...................................................................................................................................... - 1 - RESUMO........................................................................................................................................................... - 2 - ABSTRACT ....................................................................................................................................................... - 3 - 1. INTRODUÇÃO ........................................................................................................................................... - 5 - 2. MATERIAIS E MÉTODOS ....................................................................................................................... - 9 -
2.1 ÁREA DE ESTUDO ....................................................................................................................................... - 9 - 2.1.1 Localização ................................................................................................................................ - 9 - 2.1.2 Características biofísicas ...................................................................................................... - 9 - 2.1.3 Características socio-económicas mais significativas ............................................. - 11 -
2.2 BASES DE DADOS ..................................................................................................................................... - 12 - 2.2.1 Imagens do satélite Landsat ............................................................................................. - 12 - 2.2.2 Mapas de coberto do solo................................................................................................... - 13 - 2.2.3 Outras bases de dados ........................................................................................................ - 15 -
2.3 ANÁLISES NUMÉRICAS DA PAISAGEM ......................................................................................................... - 17 - 2.3.1 Introdução ............................................................................................................................... - 17 - 2.3.2 Métricas de análise da paisagem..................................................................................... - 18 - 2.3.3 Metodologia de análise ....................................................................................................... - 21 -
2.4 MODELO DE ADEQUABILIDADE DO HABITAT ............................................................................................... - 22 - 2.4.1 A técnica de modelação (ENFA) – breve introdução ................................................ - 22 - 2.4.2 O taxon alvo (Pan troglodytes) ........................................................................................ - 25 - 2.4.3 Recolha de dados e construção das variáveis a utilizar .......................................... - 26 - 2.4.4 Construção do modelo de adequabilidade do habitat para Pan troglodytes ... - 33 -
3. RESULTADOS .......................................................................................................................................... - 36 - 3.1 EVOLUÇÃO RECENTE DA PAISAGEM NA REGIÃO DO CANTANHEZ .................................................................. - 36 - 3.2 ADEQUABILIDADE DE HABITAT PARA PAN TROGLODYTES............................................................................ - 39 -
3.2.1 Análise global da adequabilidade de habitat na área de estudo.......................... - 39 - 3.2.2 Evolução da representação das classes florestais..................................................... - 44 -
4. DISCUSSÃO ............................................................................................................................................. - 50 - 4.1 ADEQUABILIDADE DE HABITAT PARA PAN TROGLODYTES NA REGIÃO DO CANTANHEZ .................................. - 50 - 4.2 EVOLUÇÃO DA PAISAGEM NA ÁREA DE ESTUDO (1986-2003) E SEU IMPACTO NA ADEQUABILIDADE DE HABITAT PARA PAN TROGLODYTES ................................................................................................................................ - 52 -
5. CONCLUSÕES E CONSIDERAÇÕES FINAIS................................................................................... - 55 - 5.1 PADRÕES DE EVOLUÇÃO DA PAISAGEM E ADEQUABILIDADE DE HABITAT PARA PAN TROGLODYTES ................ - 55 - 5.2 CRIAÇÃO DE UMA ÁREA PROTEGIDA NO CANTANHEZ .................................................................................. - 55 - 5.3 PRINCIPAIS CONTRIBUIÇÕES E PERSPECTIVAS FUTURAS ............................................................................. - 56 -
5. BIBLIOGRAFIA ....................................................................................................................................... - 58 -
- 4 -

1. INTRODUÇÃO
Uma das características mais impressionantes do mundo em que vivemos é a grande
diversidade de organismos que contem (Cox & Moore 2000). A biodiversidade é: the variability among
living organisms from all sources … and the ecological complexes of which they are part; this includes
diversity within species, between species, and of ecosystems (Convenção Internacional Diversidade
Biológica, 2003). A manutenção da biodiversidade tornou-se o principal objectivo do esforço da
conservação devido ao aumento da taxa de extinção, a mais alta de todos os períodos históricos
(Primack, 2002) e à degradação global dos ecossistemas (Myers et al., 2000; Spangenberg et al.,
2002; Beebee & Griffiths, 2005; Burney & Flannery, 2005).
A maioria das 25 regiões reconhecidas como tendo a maior biodiversidade no mundo
(compreendem 1,4% da superfície terrestre, 44% das plantas vasculares e 35% dos vertebrados)
estão situadas em países menos desenvolvidos (Myers et al., 2000), onde os recursos são escassos,
adicionando mais um constrangimento à conservação da biodiversidade. Com as dificuldades
causadas pela falta de dados e recursos muitas medidas de conservação são propostas recorrendo a
modelação predictiva (Pereira & Itami, 1991; Mladenoff et al., 1999; Brito et al., 1999; Root et al.,
2003; Kati et al., 2004). Segundo Guisan & Zimmermann (2000) a modelação predictiva é a análise e
quantificação das relações entre as espécies e o meio ambiente.
A Detecção Remota em combinação com os Sistemas de Informação Geográfica (SIG), podem
produzir dados ambientais com a mesma metodologia e com a resolução espacial adequada para
estudos em áreas remotas e/ou com falta de dados. As imagens provenientes de satélites são
actualmente o melhor método de produzir cartografia de vegetação e coberto do solo (Han et al.,
2004; Jansen & Di Gregorio, 2004). Este tipo de dados é largamente usado em modelos predictivos da
distribuição de espécies (Venier et al., 2004; Laurent et al., 2005), diversidade de espécies (Turner et
al., 2003; Virkkala et al., 2005), e adequabilidade de habitat para espécies (Hirzel et al., 2003; Brito et
al., 2002). Os custos da obtenção de dados com boa resolução espacial são muito menores usando
imagens de satélite em comparação com métodos de levantamento tradicionais, adicionalmente a
modelação da biodiversidade permite encontrar áreas com grande riqueza ou com perdas de
diversidade alarmantes com um esforço de amostragem menor em tempo e custos (Maes et al., 2003;
Lobo et al., 2004). A combinação das imagens de satélite com recurso a técnicas de modelação torna-
se assim muito útil em áreas remotas e menos desenvolvidas do globo (Rogers et al., 2002).
Existem dois tipos de modelos predictivos: modelos de distribuição potencial e modelos de
adequabilidade de habitat. Um modelo de distribuição potencial calcula a probabilidade de encontrar a
espécie numa determinada área enquanto as técnicas de modelação de adequabilidade de habitat
prevêem a adequabilidade potencial do habitat para a espécie (s) em estudo (Brotons et al., 2004).
Provavelmente a maior diferença entre estes tipos de modelos é que os primeiros requerem dados de
presença e ausência enquanto que os últimos utilizam só dados de presença (Hirzel et al., 2002).
- 5 -

As técnicas usadas mais frequentemente nos modelos de distribuição potencial são: a
regressão logística (Brito & Crespo, 2002; Balbontin 2005), modelos aditivos generalizados (Guisan
2002; Thuiller et al., 2004), análise de correspondência canónica (Jaberg & Guisan, 2001; Wang et al.,
2003), árvores de classificação (Franklin, 1998; Thuiller et al., 2003), análise discriminante (Boone et
al., 2000; Tatem et al., 2003) e redes neurais artificiais (Guégan et al., 1998; Foody & Cutler, 2003).
Na produção de mapas de adequabilidade de habitat utilizam-se, entre outras, a análise de
sobreposição (Brito et al., 1999; Alvares & Brito, 2006), Ecological-Niche Factor Analysis ou ENFA
(Hirzel et al., 2002), árvores de classificação (Andersen et al., 2000; Hansen et al., 2001) e Genetic
Algorithm for Rule-Set prediction ou GARP (Anserson et al., 2002). Todos estes métodos partilham um
conjunto de princípios similares (Guisan & Zimmermann 2000; Hirzel et al., 2002):
• A área de estudo é modelada como um mapa em formato raster composto de n
quadrados de igual tamanho.
• Dados eco-geográficos independentes descrevem quantitativamente algumas
características para cada célula (quadrado), como características topográficas (e.g.
altitude, declive), condições climáticas (e.g. temperatura, precipitação) ou características
da paisagem (e.g. coberto do solo, distância à Floresta mais próxima).
• Uma função dos dados eco-geográficos é calibrada para classificar as células como
presente/ausente ou adequado/não adequado para as espécies em estudo.
• O modelo só pode ser aplicado para uma determinada área geográfica sem erros
significativos.
O uso do solo e as alterações de coberto do mesmo são elementos fundamentais da direcção
das alterações globais (Vitousek 1992, Walker & SteVen 1996), apresentam também implicações
importantes em muitos temas da política internacional (Nunes & Auge 1999). As alterações do coberto
de solo são particularmente importantes nas terras baixas das regiões tropicais devido ao
alastramento de rápidas transformações na distribuição e características das florestas tropicais (Myers
1993, Houghton 1994, McGuffie et al., 1995). De facto a paisagem biofísica e características
antropogénicas estão frequentemente sujeitas a mudanças rápidas. Para melhor entender os
processos físicos e humanos em acção, é importante detectar e quantificar esses processos (Jensen &
Narumalani, 1992), devendo a análise ser baseada em períodos de tempo o mais longos possíveis
(Oldfield et al., 2000).
A investigação das alterações da cobertura da vegetação foca-se, por vezes, na descrição
geral da mesma, podendo usar dados de satélite como uma ferramenta qualitativa para estudar um
ecossistema específico (Carlson & Sanchez-Azofeifa, 1999). Rainis (2003) sugere que para além do
estudo da composição da paisagem também devem ser considerados o seu arranjo e distribuição para
a monitorização das mudanças. Durante várias décadas as medidas quantitativas dos padrões
espaciais (e.g. métricas de paisagem) foram usadas para ligar processos ecológicos e ambientais com
padrões encontrados na paisagem (Bunnell, 1997; Krummel et al., 1987; O’Neill et al., 1988; Prabhu
et al., 2001). As decisões tomadas sobre a gestão dos recursos requerem informação temporal e
espacial de modo a poderem ter efeitos duradouros sobre a paisagem. Consequentemente, o
- 6 -

mapeamento da paisagem ao longo do tempo, incorporando as relações espaciais e quantificando a
sua estrutura é componente crítica para um estudo de análise de alterações. Para melhor revelar as
dinâmicas da paisagem, o desafio da análise por detecção remota passa pela inferência de processos
ecologicamente significativos a partir dos instantâneos das imagens. Assim a integração da ecologia
da paisagem [estudo do como a estrutura e composição da paisagem afecta (os processos que
determinam) a abundância e distribuição dos organismos (Turner, 1989)], com a detecção remota e a
ciência de informação geográfica podem fornecer os meios para entender as dinâmicas da paisagem a
partir de dados temporalmente discretos com o auxílio das métricas de paisagem (Crews-Meyer,
2002).
A região em estudo (genericamente designada região do Cantanhez) situa-se no sul da Guiné-
Bissau, próxima da fronteira com a Republica da Guiné. A maior parte da área situa-se na última de
uma série de penínsulas que recortam o litoral do país. Delimitada no presente estudo pelas bacias
hidrográficas dos rios Cumbijã e Cacine, dois braços de mar que penetram no interior desta região e
lhe conferem um carácter recortado, repleto de enclaves. A navegabilidade desta zona torna-se difícil
devido aos numerosos bancos de areia, à grande variação das marés e à turbidez das águas, de uma
cor café com leite. O acesso à região por via terrestre faz-se utilizando uma pista, somente praticável
por veículos todo-o-terreno e camiões, tornando-se impraticável na estação das chuvas. Estas defesas
naturais explicam o carácter relativamente pouco perturbado da região do Cantanhez.
Podemos, ainda, encontrar nesta região as mais extensas e bem preservadas Florestas
Densas, localmente chamadas de Matos, da Guiné-Bissau. Alguns autores chegam mesmo a colocar a
hipótese de algumas destas florestas constituírem florestas primárias (Malaisse, 1996). O carácter
selvagem e a relativa extensão das suas florestas permitiu ao exército de libertação nacional controlar
a região de facto, bem antes do fim da guerra pela independência em 1974.
A região alberga uma comunidade de chimpanzés Pan troglodytes verus, uma das duas sub-
espécies de chimpanzé mais ameaçadas, ambas presentes na Africa Ocidental. Na realidade estas
sub-espécies encontram-se já extintas em 3 países: Benin, Togo e Burkina-Faso. Segundo Kormos,
R.& Boesch, C (2003), 75% dos chimpanzés da Africa Ocidental desapareceram nos últimos 30 anos.
Entre as principais ameaças que enfrentam, encontra-se a caça, captura de juvenis para servirem de
animais de estimação e sem dúvida a mais importante, a desflorestação e fragmentação do habitat.
Todas estas ameaças foram exacerbadas pela recente explosão demográfica da Africa Ocidental. Em
1988, os chimpanzés foram declarados extintos na Guiné-Bissau (Lee et al. 1988), estes relatos eram
claramente exagerados, já que dados mais recentes apontam para uma população entre 600 a 1000
indivíduos no país (Gippoliti & Dell’Omo 1996). A sua distribuição actual abrange o sudeste do país, na
região do Boé, entre o rio Corubal e a fronteira com a República da Guiné (Limoges 1989) e as
regiões de Quinara e Tombali no sudeste (Gippoliti & Dell’Omo 1996). Apesar de não ser objectivo
deste trabalho qualquer censo da população, foram georreferenciados na área de estudo (região
administrativa de Tombali) 178 pontos de ninhos de chimpanzé durante os trabalhos de campo.
Na Guiné-Bissau os chimpanzés (dari, na terminologia local) não são usados como alimento, por
serem considerados muito semelhantes aos humanos, “dari também é pecador" (humano), é uma
- 7 -

frase comum. Assim, a maior ameaça que enfrentam é a destruição e/ou fragmentação do habitat.
Actualmente a protecção dos chimpanzés e restante vida selvagem é da responsabilidade da Direcção
Geral das Florestas e Caça. O regulamento actual de caça (Decreto nº 21/1980) proíbe
completamente a caça nas reservas de caça. Parte da área de estudo e todo o Boé encontram-se
nesse estatuto (Chardonnet & Limoges, 1989). A Guiné-Bissau assinou a Convenção Internacional de
Comércio de Espécies Ameaçada (CITES), Convenção Diversidade Biológica e a Convenção de
Ramsar. As florestas do Cantanhez albergam outros primatas para além do chimpanzé como o
macaco colobo vermelho (Procolobus badius temminki), espécie que pode ocasionalmente fazer parte
da dieta do chimpanzé (Tutin et al., 2005).
Os rios (na realidade tratam-se de rias) da região albergam uma extraordinária
biodiversidade, com espécies emblemáticas como o manatim, o hipopótamo e várias espécies de
tartarugas. Esta conjugação de fauna e flora atribui à região de Cantanhez o estatuto de “hotspot” da
biodiversidade da Guiné-Bissau, e uma das mais ricas da Africa Ocidental. Este estatuto é referido por
vários autores, sendo considerada como Very Important Priority Area (Kormos & Boesch, 2003), para
o estudo e protecção dos chimpanzés. No entanto é importante referir que se trata de uma região
pouco explorada e com um grande potencial para futuros estudos. É assim fundamental preservar
esta área a curto, médio e longo prazo.
Neste contexto, o presente estudo teve como objectivo global a avaliação qualitativa e
quantitativa da paisagem (e sua evolução recente) na região do Cantanhez e a análise da sua
influência na sobrevivência e conservação do chimpanzé.
Constituiriam objectivos específicos do estudo:
• a caracterização quantitativa da paisagem actual nos seus aspectos composicionais e
estruturais;
• a modelação da adequabilidade do habitat para o chimpanzé no território;
• a análise quantitativa da evolução recente (1986-2003) da paisagem;
• a análise da influência da dinâmica temporal da paisagem na conservação do chimpanzé
na região do Cantanhez.
O presente estudo pretende, assim, contribuir para o conhecimento das dinâmicas de
evolução da paisagem nesta região da África Ocidental e para a preservação de um primata
ameaçado.
- 8 -

2. MATERIAIS E MÉTODOS
2.1 Área de estudo
2.1.1 Localização
A área de estudo localiza-se no extremo sudoeste da Guiné-Bissau (Figura 1). É delimitada
pelas bacias hidrográficas dos rios Cumbijã e Cacine e ocupa uma área terrestre de 2723 km2, a qual
inclui a península do Cantanhez, e os territórios situados a norte e a sul delimitados pelas linhas de
festo das duas bacias. Esta delimitação corresponde a uma fronteira natural para o estudo da
evolução da paisagem e do habitat do chimpanzé. Uma vez que as bacias hidrográficas constituem
unidades geomorfológicas às quais estão associados processos hidrológicos e de fluxos de matéria,
estas têm sido usadas como unidades de análise por vários autores em estudos semelhantes (FU et
al., 2005; ROTH et al., 1996).
No estudo realizado a utilização das bacias hidrográficas como unidades de paisagem em
análise apresenta algumas vantagens, pois: estas constituem uma delimitação natural (com fronteiras
determinadas pelo relevo) tornando-as vantajosas quando se pretende desenvolver uma análise da
paisagem (Turner et al., 2001), minimizando o corte artificial de parcelas; permitem a determinação
do balanço dos recursos i.e. nutrientes, carbono, sedimentos, este balanço reflecte as dinâmicas da
bacia e permite a determinação do efeito de diferentes usos do solo na bacia (Schreier & Brown,
2003); como se tratam de limites naturais e estáveis podem ser utilizados em estudos futuros.
2.1.2 Características biofísicas
A área de estudo ocupa terras baixas, com uma altitude máxima de 70m, um relevo com
alterações suaves e uma linha de costa muito recortada. A região do Cantanhez faz parte da grande
bacia sedimentar senegalo-mauritaniana meso-cenozóica que engloba sobretudo planícies costeiras.
Apresenta uma morfologia com colinas estreitas e alongadas, muito recortadas com encostas suaves
em vastas planícies aluviares (com origem no quaternário) de carácter lodoso ou arenoso, por onde
meandrizam as rias e seus múltiplos braços que as marés alagam na preia-mar (Henriques, 2000).
O clima da área de estudo é determinado pela migração sazonal da zona de convergência
intertropical (ZCIT), que origina duas grandes estações. A época seca ocorre de Dezembro a Maio,
mês em que a ZCIT atravessa a área em direcção norte, dando início à estação das chuvas que
termina em Novembro, mês em que a ZCIT atravessa de novo a área na sua migração para sul
(Catarino, 2004).
- 9 -

No contexto da Guiné-Bissau, trata-se da área com maior pluviosidade (Machado et al., 1972)
permitindo classificar o clima como húmido (tabela 1) (Fosberg et al., 1961). Estas condições
conferem a esta região a capacidade de albergar vegetação florestal de grande porte (Malaisse,
1996).
Figura 1: Localização da área de estudo no contexto da Africa Ocidental (direita) e da Guiné-Bissau (esquerda).
Segundo Anginot (1988), a região do Cantanhez apresenta quatro tipos principais de solos: a)
Ferralsolos: apresentam textura arenosa a areno-argilosa com fertilidade baixa e muito sensíveis a
degradação em caso de ausência de cobertura vegetal; b) Fluvisolos: solos de origem aluvial e
marinha, saturados de sal e pH elevado durante a maior parte do ano; contêm uma grande
percentagem de argila e apresentam uma fertilidade elevada; correspondem a áreas ocupada por
mangal e orizicultura inundada (bolanha); c) Gleisolos: solos hidromorfos temporários com pH ácido
são muitas vezes ocupados por horticultura e orizicultura; e d) Arenosolos: solos de formação aluvial,
de textura arenosa e intensivamente lixiviados, apresentam uma fertilidade muito baixa.
A área de estudo situa-se na zona mais florestada de um país que, segundo Amsallem (2001),
é o mais florestado da Africa Ocidental, com perto de 60% da sua extensão coberta por florestas. O
território da área de estudo contém as manchas de floresta densa mais setentrionais da África
Ocidental, tendo muitas das espécies aqui o seu limite setentrional de distribuição (Malaisse 1998;
Catarino 2004).
- 10 -

Tabela 1 – Características climáticas de Catió (adaptado de Catarino, 2004).
Cidade T (ºC) P (mm) EP (mm) Ia (%) Iu (%) Ih (%) Tipo de clima
Catió 26.7 2584 1619 45.2 102.9 75.8 B3A’w2a’
T: temperatura média anual; P: precipitação anual; EP: evapotranspiração potencial anual; Ia: índice de aridez =(D/EP) 100; Iu: índice de humidade =(S/EP) 100; Ih: índice hídrico =(Iu - 0.60 Ia) 100, sendo D: deficit anual de água e S: superavit anual de água. Símbolos dos tipos de clima: B3 clima húmido (60% <Ih≤80%); A’: clima megatérmico, com evapotranspiração potencial anual superior a 1140 mm; W2: índice de aridez superior a 33,3% (associado a índice hídrico positivo), revela grande défice de água no Inverno; a’: eficácia térmica no Verão inferior a 48% (Thornthwaite, 1948).
2.1.3 Características socio-económicas mais significativas
A área em estudo situa-se na região de Tombali, com a capital em Catió e uma densidade
populacional de 19 habitantes/km2 (a partir de uma estimativa da população, em 2004, de 91930
habitantes; Lourenço et al., 1994). Trata-se de uma densidade populacional relativamente baixa, em
comparação com a totalidade do país (Lourenço et al., 1994). A maior parte da população pertence a
uma das cinco principais etnias: Balanta, Fula, Nalu, Sosso e Tanda. As povoações, muitas vezes
constituídas por elementos de uma só etnia, surgem ao longo das vias de comunicação e são
tipicamente pequenas, albergando o maior aglomerado populacional (Iemberem) aproximadamente
2000 habitantes.
Na organização social mais típica (durante o século XX), as várias etnias apresentam uma
diferenciação de actividades e tarefas (Temudo, 1998). Os Balantas dominam a construção de diques
e portanto a produção de arroz de bolanha, os Fulas e Sossos dedicam-se mais à produção de arroz
de sequeiro e (as mulheres) de vários produtos hortícolas. Este sistema de produção (com
especialização em diferentes culturas) é depois complementado por um complicado, mas funcional,
sistema de trocas e empréstimos entre os habitantes da região (Temudo, 1998).
A base da alimentação é o arroz, de que as populações da área em estudo tradicionalmente
conseguem gerar excedentes, produzindo-o em dois sistemas:
- arroz de bolanha: cultivado em zonas baixas interiores em que a vegetação natural é a
lala (bolanha de água doce) ou das zonas estuarinas em que a vegetação natural é o
mangal (bolanha de água salgada); estes solos são relativamente férteis e a cultura
mantém-se por muitos anos no mesmo local; é este tipo de produção que permite gerar
alguns excedentes.
- arroz “pam-pam” ou de planalto: cultivado em sequeiro, no sistema dito itinerante, que
implica o arroteio de vegetação natural ou seminatural e eventualmente o derrube de
floresta.
A produção de caju, Anacardium occidentale aumentou significativamente nos últimos anos na
área de estudo e na generalidade do país convertendo-o no sexto produtor mundial. Este aumento,
com a consequente destruição da vegetação natural para o estabelecimento de pomares
- 11 -

monoespecíficos de caju, tem consequências ainda não totalmente avaliadas (Catarino, 2002). No
entanto, a área em estudo encontra-se entre as regiões da Guiné-Bissau menos afectadas por este
problema, apresentando uma maior cobertura de hortas tradicionais, com maior diversidade
específica, maior idade de implantação e maior biodiversidade (Catarino, 2002).
Em síntese, pode afirmar-se que a maior parte da população se dedica à prática da
agricultura, em que o uso de fertilizantes químicos ou orgânicos é praticamente inexistente, sendo
praticada em algumas culturas de sequeiro uma agricultura itinerante (“deslizante”). Este tipo de
exploração traz consequências para as áreas em que a vegetação natural se encontra em
recuperação, uma vez que os terrenos cultivados são utilizados por períodos de tempo relativamente
curtos, de dois a três anos, sendo depois deixados em pousio, procedendo-se então à queima ou
desmatação de terrenos que passaram por um período grande de pousio e entretanto já recuperaram
a fertilidade (Temudo, 1998). Por vezes, são também ocupadas novas áreas, que anteriormente
tinham cariz florestal. No entanto, a maior parte dos tipos de uso da floresta transformam-na ou
modificam-na, não a destruindo totalmente (Marsh et al., 1987). De facto, nos trópicos a maior parte
dos habitats florestais estão fragmentados e são usados como fonte de recursos (Pimm, 1986).
2.2 Bases de dados
2.2.1 Imagens do satélite Landsat
Foram utilizadas imagens de satélite para gerar os mapas de coberto do solo (Cassamá,
2006; tabela 2) e para gerar variáveis contínuas relacionadas com a estrutura das copas e densidade
da vegetação recorrendo a análise de misturas espectrais.
Em concreto, foram utilizadas imagens dos sensores Landsat TM e ETM+. As imagens
provenientes do satélite Landsat 5 (lançado em 1984) possuem um explorador de varrimento,
denominado TM (Thematic Mapper), com uma configuração especialmente ajustada para a produção
de cartografia temática, com um maior número de detectores por bandas em relação ao seu
antecessor (Landsat MMS). O satélite Landsat 7 (lançado em 1999) leva a bordo um novo sensor
ETM+ (Enhanced Thematic Mapper Plus), com melhorias nas características radiométricas e
geométricas. A resolução espacial (30m) e os comprimentos de onda das bandas utilizadas são iguais
nas imagens TM e ETM+ (Irons et al., 2006).
A superfície analisada tem uma área inferior à área de uma única cena Landsat, todavia situa-
se numa zona de confluência de trajectória do satélite, deste modo, foram usadas duas cenas Landsat
por ano estudado: as cenas Path 204/Row 052 e Path 203/Row 052 dos Landsat TM e ETM+. No total
o estudo incide em oito cenas, pois considera várias datas (1986, 1994, 2000 e 2003). Algumas destas
imagens (203/052 de 1986, 204/052 de 2000, 204/052 de 2003 e 203/052 de 2003) fazem parte de
um lote disponibilizado gratuitamente na Internet pela Global Land Cover Facility (GLCF) da
- 12 -

Universidade do Maryland, EUA (http:/glcf.umiacs.umd.edu/). As restantes imagens foram adquiridas
com recursos do Projecto POCI/BIA-BDE/57965/2004, Alterações do coberto do solo e etapas de
sucessão nas florestas do Cantanhez, coordenado pelo Doutor Luís Catarino.
Tabela 2 – Sensores, datas e cenas das imagens de satélite utilizadas.
Anos Sensores Cena TM/ETM
(203/052)
Cena TM/ETM
(204/052)
1986 TM 11/02/86 18/02/86
1994 TM 22/04/94 13/04/94
2000 ETM+ 30/04/00 20/03/00
2003 ETM+ 01/01/03 24/01/03
2.2.2 Mapas de coberto do solo
A produção dos mapas de coberto do solo foi anterior ao presente estudo no âmbito de uma
tese de mestrado anterior (Cassamá, 2006), realizada no contexto do projecto POCI/BIA-
BDE/57965/2004.
Para construção da legenda foi usado o Land Cover Classification System (LCCS) (Di Gregório
et al., 2000); sistema que se baseia nas características estruturais e funcionais da vegetação para a
definição de uma legenda hierárquica compatibilizável com os diversos sistemas de classificação do
coberto de solo adoptados em trabalhos semelhantes. De notar que não foi possível isolar alguns tipos
de coberto, como as hortas (pomares), que foram provavelmente classificadas como zonas florestais;
no entanto, tal eventualidade (actualmente em avaliação no IITC) não tem grande relevância para o
presente estudo, uma vez que estas hortas tradicionais são ecologicamente bastante semelhantes às
áreas florestais naturais. A partir das imagens Landsat referidas anteriormente e de um conjunto de
pontos recolhidos no terreno em 2005 (Cassamá, 2006), com o objectivo de reunir um conjunto de
pixeis representativos que descrevessem o comportamento espectral para cada classe de coberto de
solo; foi realizada uma classificação através da construção e aplicação do algoritmo de classificação
em árvore [desenvolvido por Breiman et al. (1984) e implementado no software CART; Steinberg &
Colla (1997)].
Para utilizar os dados presentes nos mapas de coberto de solo, foram efectuadas duas
operações recorrendo a um SIG (Sistema de Informação Geográfica): a) foram retirados todos os
pixeis que se encontravam fora da área de estudo definida pelas bacias hidrográficas dos rios Cumbijã
e Cacine; b) foram eliminadas todas as parcelas com menos de um hectare, sendo o espaço vazio
- 13 -

preenchido através da alocação destes pixeis a classes vizinhas seguindo uma regra de distância
euclidiana; este procedimento permite retirar da análise pixeis isolados e pequenas parcelas que
introduziriam uma considerável quantidade de ruído (Schowengerdt R., 2007). Estas operações foram
executadas para as quatro datas consideradas (1986, 1994, 2000 e 2003).
Os mapas resultantes incluem 10 classes (figura 2):
1. “Floresta densa”: Trata-se de uma formação natural com elevada densidade de árvores,
podendo atingir alturas superiores a 30 metros e com sobreposição praticamente total das
copas, o que origina um ambiente muito sombrio ao nível do solo em que o estrato
superior é dominante condicionando o desenvolvimento dos arbustos e herbáceas dos
outros estratos. Apresenta uma elevada cobertura total em que o somatório dos
diferentes estratos é superior a 100%, permitindo uma sobreposição de camadas de
órgãos fotossintéticos, o que reduz a quantidade de radiação solar que chega ao solo.
Esta configuração permite a existência de um microclima húmido propício ao
desenvolvimento das plantas após o fim da época das chuvas.
2. “Floresta aberta”: Formação vegetal natural ou seminatural, dominada por espécies
arbóreas, em que as copas não se sobrepõem. A densidade de árvores é inferior à da
Floresta densa e o seu porte atinge em geral entre 10 e 20 metros de altura. Em
consequência, os estratos de arbustos e de herbáceas apresentam algum
desenvolvimento. Na região do Cantanhez estas formações resultam normalmente de
degradação antropogénica, podendo ser consideradas como uma fase de regeneração da
Floresta densa. A composição florística das Florestas abertas alberga assim algumas
espécies em comum com a Floresta densa e outras de formações mais abertas.
3. “Savana arborizada”: Formação vegetal em que o estrato herbáceo tem uma cobertura
contínua e os estratos arbóreo e arbustivo apresentam uma cobertura esparsa. No
contexto da região em estudo representam formações secundárias resultantes da
degradação das florestas sendo a sucessão ecológica truncada pelas queimadas
frequentes. Na estação das chuvas a vegetação herbácea, pelas suas características, tem
um crescimento extremamente acelerado. No final desta estação, a vegetação herbácea
seca com a mesma rapidez do seu crescimento, tornando-se num combustível fácil.
4. “Savana herbácea”: Formação vegetal de carácter herbáceo, que ocupa áreas com
cotas altimétricas baixas com drenagem lenta e normalmente alagadas durante a época
das chuvas, acabando por secar na época seca. Localmente são designadas por lalas.
Trata-se de um ambiente hiper-sazonal, com contraste marcante entre a estação das
chuvas e a estação seca.
- 14 -

5. “Mangal”: Formação vegetal de carácter lenhoso que ocupa zonas costeiras sob
influência das marés. É constituída por um reduzido número de espécies em que
dominam Avicennia germinans (L.) L., Rhizophora mangle L. e Rhizophora raemosa
G.Mey. Instala-se em substratos de vasas de declive suave. A riqueza biológica destes
ecossistemas costeiros faz com que sejam grandes "berçários" naturais, tanto para as
suas espécies vegetais características, como para peixes e outros animais. A vegetação
dos mangais permite a fixação das terras e as suas raízes funcionam como filtros na
retenção dos sedimentos, impedindo a erosão e ao mesmo tempo estabilizando a costa.
6. “Bolanha”: Áreas de cultivo de arroz alagado.
7. “Areias”: Depósitos sedimentares arenosos sem vegetação.
8. “Vasas”: Depósitos costeiros de sedimentos finos.
9. “Ardido”: Área queimada durante a estação seca até à data da colheita de cada imagem
de satélite (ver Anexo). Em geral, correspondem a antigas áreas de Floresta aberta ou
savana arborizada.
10. “Água”: Massas de água permanentes.
2.2.3 Outras bases de dados
A Altimetria usada no presente estudo provém da SRTM (Shuttle Radar Topography Mission)
que consiste num sistema de radar que sobrevoou a terra durante 11 dias (em Fevereiro de 2000) a
bordo do Space Shuttle Endeavour (Coltelli et al., 1996; Moreira et al., 1995; Werner, 2001), obtida a
partir do site do USGS (http://srtm.usgs.gov/). Esta informação foi posteriormente tratada em SIG de
modo a apresentar exactamente o mesmo número de linhas e colunas que as restantes fontes de
informação.
Foram recolhidos, durante o trabalho de campo de 2006, pontos (georreferenciados) dos
povoamentos (tabancas) e das estradas da maior parte da área de estudo. Estes pontos foram
transformados em mapas raster com software SIG de modo a apresentarem o mesmo tamanho e
características dos restantes mapas.
- 15 -

A
Figura 2: Aspectos da área de estudo: A) Pegada dede Floresta densa, D) Mangal, E) Mangal com FlorClorocebus aethiopicus em Floresta densa.
B
C
DF
EG
HPan troglodytes, B) Savana herbácea, C) Interior esta densa, F) Bolanha, G) Área de cultivo, H)
- 16 -

2.3 Análises numéricas da paisagem
2.3.1 Introdução
A avaliação das consequências das actividades humanas no ambiente pode ser expressa em
termos do uso do solo e de variações na estrutura da paisagem, que podem ser quantificadas através
de métricas descritivas dos padrões da paisagem (Turner et al., 2001). A Ecologia da Paisagem, por
comparação com os estudos ecológicos tradicionais, não assume que os sistemas em estudo são
homogéneos e tem em consideração o padrão espacial (Turner & Gardner, 1991). A quantificação dos
padrões da paisagem tem sido conduzida pela premissa de que estes estão ligados a processos
ecológicos, podendo estes ser previstos por aqueles padrões (Baskent & Jordan, 1995; Gustafson,
1998). Segundo King (2005), a paisagem é o “palco” onde se desenrola a dinâmica dos sistemas
ecológicos.
Reforçando a importância de descrever e quantificar o padrão espacial da paisagem, a
evolução e afirmação da Ecologia da Paisagem forneceu a matriz teórica para o desenvolvimento de
métricas de paisagem (O’Neill et al., 1995). Seguindo-se a um esforço inicial de O’Neill et al. (1988),
foram sendo propostos numerosos índices “candidatos”, quer por ecólogos da paisagem (Baker & Cai
1992), quer por processadores de imagens (Gonzales & Woods 1992). Na última década, diversos
autores reconheceram e afirmaram que o estudo e avaliação da estrutura da paisagem a diferentes
escalas espaciais, por aplicação de indicadores de paisagem, é provavelmente o método de maior
eficácia para a avaliação das condições ecológicas locais e regionais (e.g. Turner, 1989; O’Neill et al.,
1994). Actualmente, existem métricas baseadas na diversidade, na teoria da informação, na
geometria fractal e até na textura da imagem (Turner et al., 2001).
O estudo dos padrões e processos ecológicos à escala da paisagem tornou-se possível através
da disponibilidade de dados provenientes da detecção remota, os quais viabilizaram a análise da
evolução da estrutura da paisagem ao longo do tempo. Imagens de satélite podem ser classificadas
em classes de coberto do solo, proporcionando uma forma relativamente económica de estudar
grandes áreas (O’Neill et al., 1992). O desenvolvimento dos Sistemas de Informação Geográfica
(SIGs) forneceu os meios tecnológicos necessários à gestão de bases de dados espaciais relativas a
grandes áreas.
Actualmente, as capacidades técnicas da detecção remota associadas à tecnologia dos SIGs
fornecem a combinação ideal para medir e monitorizar o estado da paisagem, o que por sua vez
proporciona o contexto espacial para estudar as dinâmicas dos ecossistemas (O’Neill et al., 1997;
O’Neill 1999). Muitos dos indicadores propostos para avaliar a saúde dos ecossistemas, incluindo
indicadores para a produtividade e organização (Mageau et al., 1995), são passíveis de serem
monitorizados através de dados provenientes da detecção remota (Rapport et al., 1996; Jones et al.,
1997).
- 17 -

Existe actualmente uma grande variedade de métricas de paisagem e de programas
informáticos para o seu cálculo que são utilizados para quantificar diversos aspectos da
heterogeneidade espacial em mapas temáticos (O’Neill et al., 1988; Turner & Gardner, 1991;
McGarigal & Marks, 1995; Frohn, 1998). Este facto indica que a quantificação dessa heterogeneidade
permanece problemática, devido à complexidade do fenómeno e à insuficiente definição dos
componentes da paisagem (Li e Reynolds, 1994). Como resultado dos estudos efectuados para o
desenvolvimento de métodos para a quantificação da estrutura da paisagem, verificou-se que muitos
dos indicadores definidos apresentam uma elevada correlação entre si. Através de um estudo de
análise factorial, Riitters et al., (1995) analisaram 55 indicadores de paisagem para determinar a sua
independência estatística e concluíram que a informação contida nos 55 indicadores se resumia à
informação de apenas cinco dos indicadores testados (Dominância, Contágio, Dimensão Fractal, Razão
Média Perímetro/Área e Número de Classes). Assim, dada a complexidade do processo de selecção de
métricas, para o estudo da evolução da paisagem na região do Cantanhez utilizaram-se apenas
métricas de interpretação simples, com claras relações com a evolução e processos da paisagem e dos
ecossistemas.
Em síntese, no trabalho aqui apresentado, foram utilizados dois tipos de métricas:
• métricas de paisagem (globais, em que a paisagem é analisada como um todo);
• métricas de classe (em que são calculadas métricas para cada classe).
As métricas de parcela não foram utilizadas, uma vez que neste trabalho não se propunha
(devido à escala não ser adequada) um estudo detalhado de parcelas individuais na região do
Cantanhez. Nas tabelas 3 a 5, encontram-se listadas e sumariamente descritas (McGarigal et al.,
1995) as métricas seleccionadas para a análise da paisagem (e respectivas tendências evolutivas) na
região do Cantanhez.
2.3.2 Métricas de análise da paisagem
Para a análise da evolução da paisagem foram usadas as cartas de coberto do solo de 1986,
1994, 2000 e 2003 obtidas a partir das imagens de Landsat através de classificação supervisada (ver
2.2.2). A estas cartas foram retiradas as classes de coberto de solo em que não existe vegetação (a
água, as areias e as vasas). As classes retiradas da análise apenas sofrem os efeitos das marés, não
são influenciadas nem influenciam os processos ecológicos alvo da análise como a fragmentação e a
sua inclusão só acrescentaria ruído. De notar que o nos procedimentos o software utilizado retira
todas as parcelas que intersectam o limite da paisagem em estudo, acção que limita os erros gerados
pela alteração das parcelas produzidos pelo uso de uma “máscara” que limita a área de estudo.
- 18 -

Tabela 3: Métricas utilizadas no estudo da paisagem da região do Cantanhez.
Aspecto em análise Métrica de Classe Métrica de Paisagem
Nº de parcelas Nº de parcelas
Orla total Densidade de parcelas
Orla total
Áreas e Orlas
Índice da maior parcela
Área média da parcela
Média da dimensão fractal Forma Média Perímetro/Área
Média Perímetro/Área
Índice de Shannon
Índice de Simpson
Índice modificado de Simpson
Índice de equitabilidade de Shannon
Diversidade -
Índice de equitabilidade de Simpson
A forma e o tamanho das parcelas influenciam numerosos processos ecológicos. A forma da
parcela pode influenciar processos inter-parcelas como migrações e colonização de espécies arbóreas,
que se repercutem nas estratégias alimentares dos animais (Forman & Godron 1986). Muitos
processos ecológicos associados a padrões espaciais estudados pela ecologia da paisagem estão
relacionados com os efeitos de orla. A orla da floresta, por exemplo, afecta principalmente agentes
como o vento e a intensidade de luz que entra na parcela de floresta; estas modificações, em
combinação com alterações na dispersão de sementes e no herbívorismo, podem influenciar a
composição e estrutura da vegetação (Ranney et al. 1981).
A dimensão fractal reflecte a complexidade da forma ao longo de várias escalas espaciais (e
tamanhos das parcelas), contornando as limitações da razão simples entre perímetro e área, que é
afectada pelo tamanho médio das parcelas (i.e. para uma mesma forma geométrica, o aumento do
tamanho médio da parcela induz uma diminuição da razão perímetro/área). Na dimensão fractal um
valor próximo de 1 indica formas com perímetros muito simples (tais como quadrados) à medida que
forma da parcela apresenta uma maior complexidade os valores aumentam.
As medidas de diversidade têm sido usadas numa grande variedade de aplicações
ecológicas, tendo-se tornado muito populares na medição da diversidade específica de plantas e
animais. Estes índices de diversidade foram adoptados pela ecologia da paisagem para medir a
diversidade da composição da paisagem (O'Neill et al. 1988, Turner 1990). Os índices usados são
influenciados por dois factores, a riqueza e a equitabilidade das classes na paisagem. A riqueza refere-
se ao número de classes de parcelas presentes, a equitabilidade à distribuição relativa da área total
pelas diferentes classes de parcelas. A riqueza e a equitabilidade avaliam a composição e a estrutura
- 19 -

da diversidade, respectivamente. Os índices têm sensibilidades diferentes, podendo ponderar mais as
classes raras (índice de Shannon) ou as classes mais comuns (índice de Simpson).
Tabela 4: Breve descrição das métricas (indicadoras de áreas, orlas e formas) utilizadas para análise da paisagem da região do Cantanhez.
Nome Fórmula Unidades Comentários
Nº de parcelas
Adimensional Nº de parcelas presentes na paisagem
Densidade de
parcelas
Nº por 100
hectares
Nº de parcelas presentes em 100
hectares
Orla total
Metros Medida absoluta do total em metros
das orlas da paisagem ou de uma
classe
Área média da
parcela
Hectares
Média
Perímetro/Área
Adimensional Igual à média das razões entre os
perímetros das parcelas (m) e as suas
áreas (m2)
Média da
dimensão fractal
Adimensional Igual à média de 2 vezes o valor
logarítmico do perímetro da parcela
(m) dividido pelo valor logarítmico da
área da parcela (m2), o perímetro é
ajustado para corrigir o seu
enviezamento, provocado por se tratar
de um raster.
Tabela 5: Breve descrição das métricas (indicadoras de diversidade) utilizadas para análise da paisagem da região do Cantanhez.
Nome Fórmula Comentários
Índice de
Shannon
O índice de Shannon aumenta à medida que
aumenta o numero de diferentes classes de parcelas
e/ou a distribuição de áreas entre diferentes classes
de parcelas se torna mais equitativa. É mais sensível
a classes de parcelas mais raras, do que o índice de
Simpson.
Índice de
Simpson
O Índice de Simpson aproxima-se de 1 quanto maior
for a riqueza de classes de parcelas e/ou a
distribuição de áreas entre diferentes classes de
- 20 -

parcelas se torna mais equitativa. O valor representa
a probabilidade de 2 pixeis escolhidos ao acaso
serem de classes diferentes.
Índice
modificado de
Simpson
O Índice modificado de Simpson aumenta à medida
que aumenta a riqueza de classes de parcelas e/ou a
distribuição de áreas entre diferentes classes de
parcelas se torna mais equitativa.
Índice de
equitabilidade de
Shannon
Índice de equitabilidade de Shannon é igual a 1
quando as áreas das diferentes classes de parcelas
são iguais, à medida que as diferenças aumentam o
valor diminui.
Índice de
equitabilidade de
Simpson
Índice de equitabilidade de Simpson é igual a 1
quando as áreas das diferentes classes de parcelas
são iguais, à medida que as diferenças aumentam o
valor diminui.
Tabela 5 (cont.)
2.3.3 Metodologia de análise
Na realização do estudo das alterações da paisagem foram utilizados os mapas de coberto de
solo derivados das imagens Landsat (Figura 3). Estes mapas (em formato raster) contêm informação
do tipo de classe de coberto para cada célula, a partir da qual foram delimitadas as parcelas (áreas
continuas com o mesmo tipo de coberto de solo) utilizadas no cálculo das métricas de paisagem.
Cada uma das quatro cartas de coberto de solo, correspondendo às quatro datas em análise,
foi processada em SIG de modo a corresponder exactamente à mesma área contendo o mesmo
número de linhas e colunas, de modo a ser totalmente sobreponível. Este facto é importante uma vez
que se pretende analisar as alterações ocorridas ao longo do tempo, pelo que a componente espacial
tem de ser constante.
- 21 -

Figura 3: Imagem de pormenor de um mapa de coberto de solo; as diferentes cores no mapa correspondem à informação presente em cada célula sobre a classe de coberto do solo; cada célula (quadrado) tem 30 metros de lado. A informação contida em cada célula foi utilizada na delimitação das parcelas utilizadas no cálculo das métricas de análise da paisagem. No canto superior direito, imagem do mapa de coberto do solo para toda a área de estudo.
Para calcular as métricas, foi utilizado o programa FRAGSTATS (McGarigal & Marks 1995),
capaz de calcular várias estatísticas para as classes, calculando também métricas para a paisagem
total. Este programa permite escolher a configuração da vizinhança de cada célula, tendo sido usada a
regra das oito células (quatro ortogonais e quatro diagonais). Esta selecção permite que duas células
da mesma classe que se tocam diagonalmente possam ser consideradas da mesma parcela.
2.4 Modelo de adequabilidade do habitat
2.4.1 A técnica de modelação (ENFA) – breve introdução
No presente estudo, atendendo à dimensão da área a estudar, à dificuldade das deslocações
no terreno e ao tempo disponível para a recolha dos dados, não foi possível recolher informações
concretas sobre as ausências na distribuição do chimpanzé. A falta de informação acerca das áreas de
ausência confirmada da espécie alvo dificulta o uso de ferramentas clássicas de modelação ecológica,
como regressão logística ou árvores de classificação ou regressão (Guisan & Zimmermann, 2000).
Existe também o problema clássico de interpretar essas ausências, uma vez que uma “ausência” (não
- 22 -

houve observação) pode ter origem em pelo menos três causas: 1) a espécie está presente mas não
foi detectada, 2) o habitat é adequado, mas a espécie ainda/já não está presente, 3) o habitat, de
facto, não é adequado. Por estas razões, foi usada uma ferramenta de modelação que não requer
dados de ausência para identificar os factores ambientais que explicam a distribuição de Pan
troglodytes no contexto da área de estudo, assim como as áreas de maior adequabilidade de habitat.
A “Ecological Niche Factor Analysis” (ENFA, Hirzel et al., 2002) baseia-se na teoria do nicho
ecológico de Hutchinson (1957) e foi implementada recorrendo ao software Biomapper 3.1 (Hirzel et
al., 2004), seguindo os procedimentos delineados por Hirzel et al., (2002). Um dos objectivos desta
análise de factores (semelhante à análise de componentes principais) é transformar n variáveis
correlacionadas no mesmo número de factores não correlacionados. Estes factores explicam a mesma
quantidade da variância total, pelo que as análises subsequentes podem restringir-se a alguns
factores importantes (aqueles que explicam a maior parte da variância) sem perder muita informação.
A ENFA atinge este propósito calculando factores com significado ecológico.
Os dados necessários têm o formato de mapas raster, em que cada célula do mapa contém o
valor de uma variável, existindo um mapa para cada variável. Estes mapas das “variáveis
ecogeográficas” (EGVs) são quantitativos e contínuos. O mapa da distribuição conhecida da espécie
contém valores boleanos (0 ou 1), correspondendo ao valor 1 a presença da espécie em estudo na
célula. O valor 0 significa simplesmente a ausência de provas (de presença).
Um ponto no espaço multidimensional das variáveis pode representar cada célula. Admitindo
distribuições multinormais, a sua forma seria hiper-elipsóide. As células em que a espécie em estudo
existe constituem um subgrupo da distribuição global e definem um hiper-elipsóide mais pequeno
dentro do global. O primeiro factor da ENFA (marginalidade) é a linha que atravessa os centróides dos
dois elipsóides (Figuras 4 e 5). A marginalidade da espécie é a distância estandardizada entre estes
centróides (Hirzel et al., 2004).
Figura 4: Interpretação geométrica da ENFA. As células em análise da área de estudo são representadas num espaço tridimensional. O elipsóide maior (castanho) representa todas as células em análise, enquanto o elipsóide menor (roxo) representa o subconjunto de células em que a espécie estudada foi observada. A linha recta que atravessa os seus centróides é o factor marginalidade. De modo a extrair a variância associada com este factor, as coordenadas das células são projectadas num plano p perpendicular ao inicial, produzindo assim duas elipses. No caso do presente trabalho esta operação foi conduzida com 17 EGVs. (adaptado de Hirzel et al., 2004).
- 23 -

A projecção das duas elipsóides num espaço hiperplano perpendicular ao factor marginalidade
(figura 4) garante que os factores subsequentes serão não correlacionados com o primeiro e diminui
uma dimensão ao espaço das variáveis (EGVs). Calculando o eixo que maximiza a razão variância
global/variância da espécie obtém-se o primeiro dos factores de especialização. Repetindo este
processo obtêm-se os factores seguintes, sendo que em cada repetição se retira uma dimensão ao
espaço até que a extracção dos n factores esteja completa. A quantidade de especialização registada
por cada factor vai diminuindo devido à extracção anterior. Todos os factores de especialização são
ortogonais, no sentido em que a distribuição do subgrupo de valores da espécie em qualquer factor
não está correlacionado com a sua distribuição nos outros.
Figura 5: A distribuição da espécie alvo em qualquer variável ecogeográficas (barras negras) pode diferir da distribuição de todo o conjunto de células da variável (barras cinzentas) em relação à sua média (mS ± mG), permitindo assim definir a marginalidade. Pode também diferir em relação ao desvio padrão (σS ± σG), permitindo definir a especialização. (adaptado de Hirzel et al., 2002)
Um número limitado dos primeiros factores contém geralmente a maior parte da informação.
O facto de serem poucos factores e independentes torna-os mais fáceis de usar do que as variáveis
originais. De notar que, seguindo este processo, os factores de nicho ecológico são muito
dependentes da dimensão da área analisada. Hirzel et al. (2002) desenvolveram a demonstração
matemática para este procedimento.
Em resumo o primeiro componente, chamado marginalidade, descreve a distância do “óptimo”
da espécie (usando os pontos dos ninhos) à média das variáveis ecogeográficas (EGVs) na área de
estudo, i.e. a direcção em que o nicho da espécie difere mais das condições disponíveis na área de
estudo (Hirzel et al., 2002). Um valor baixo na componente marginalidade (próximo de 0) indica que a
espécie tende a ocupar condições médias na área de estudo (generalista), enquanto um valor alto
(próximo de 1) indica uma tendência para viver em habitats mais raros na área de estudo. Os
coeficientes da matriz de resultados relacionados com o componente Marginalidade indicam a
correlação entre cada EGV e o componente. Quanto maior o valor absoluto do coeficiente, maior é o
contributo desse EGV para a marginalidade. Um valor positivo significa que a espécie “prefere” os
valores mais elevados desse EGV, enquanto um valor negativo significa que a espécie “prefere” os
valores baixos.
- 24 -

Os componentes subsequentes são chamados de componentes de Especialização e são
ordenados por ordem decrescente da quantidade de variância explicada. Eles descrevem o quanto
“especializada” é a espécie, em relação a variação dos habitats na área de estudo (Hirzel et al., 2002).
Assim, alguns dos primeiros componentes explicam a maior parte da informação total. Os valores para
a Especialização variam de 1 até ao infinito, tornando-se quase impossíveis de interpretar, por isso, é
mais fácil utilizar a Tolerância, que mede a “escolha” da espécie de entre os limites disponíveis de
EGVs. A Tolerância é definida como o inverso da especialização (1/S) e varia de 0 a 1, indicando se se
trata de uma espécie “especialista”, que ocupa condições muito particulares ou de uma espécie
“generalista”, que pode ocupar qualquer parte da área de estudo.
Os índices de adequabilidade de habitat, com uma escala de zero a cem, foram calculados
com o algoritmo das Medianas, este algoritmo assume que o melhor habitat está na mediana da
distribuição simétrica da espécie em cada factor.
2.4.2 O taxon alvo (Pan troglodytes)
O taxon alvo do estudo de adequabilidade do habitat é o chimpanzé (Pan troglodytes)
(Figura 6). Esta espécie inclui quatro sub-espécies (Figura 7): a ocidental P. t. ve us, as centrais P. t.
troglodytes e P.t. vellerosus, a oriental P. t. schweinfurthii (Napier & Napier 1967; Groves 2001)
r
A sub-espécie ocidental de chimpanzé (Pan troglodytes verus) tem uma distribuição que
abrange desde o Sudeste do Senegal até ao rio Níger (Butynski, 2003). Esta distribuição ampla (no
entanto, não contínua), conjugada com o facto de se tratar de um animal com elevado
desenvolvimento cultural, conduziu ao aparecimento de sub-culturas com influência em
comportamentos e hábitos assim como nas preferências ecológicas (Tutin et al., 2005). A diversidade
comportamental foi constatada durante o trabalho de campo, uma vez que os chimpanzés da região
de Cantanhez constroem quase sempre os seus ninhos em palmeiras (Elaeis guianensis Jacq.), um
comportamento pouco usual e diferente do dos seus “vizinhos” do Senegal e do centro e sul da
Republica da Guiné (Gippoliti, 1996). Este comportamento pode fazer parte das normas culturais do
grupo ou ser uma resposta às diferenças de habitat (Kormos et al., 2003). Assim, na área de estudo,
os chimpanzés constroem os seus ninhos preparando uma base de ramos sólidos e depois partindo ou
dobrando outros ramos e folhas mais delgados numa forma circular (Fruth & Hohmann 1996).
- 25 -

Figura 6: Fotografias de Pan troglodytes, obtidas pelo autor durante o trabalho de campo desenvolvido no âmbito do presente estudo.
Figura 7: Distribuição geográfica das sub-espécies de Pan troglodytes: 1. Pan troglodytes verus; 2. Pan troglodytes vellerosus; 3. Pan troglodytes troglodytes; 4. Pan troglodytes schweinfurthii (Keele et. al., 2006)
2.4.3 Recolha de dados e construção das variáveis a utilizar
Na construção do modelo de adequabilidade de habitat para Pan troglodytes verus, foram
utilizados dados recolhidos em trabalho de campo (presenças de ninhos e avistamentos) para
obtenção de uma amostra representativa da variável dependente – presença de chimpanzés, assim
como dados correspondentes às variáveis ambientais explicativas geradas a partir das diferentes
bases de dados disponíveis e acima descritas.
- 26 -

2.4.3.1 Dados de distribuição (variável dependente)
Os dados relativos à distribuição e ocupação de habitat dos chimpanzés foram recolhidos por
métodos quantitativos, não invasivos, nomeadamente através da recolha de pontos georreferenciados
(com recurso ao Global Positioning Sys em, GPS) de ninhos e avistamentos de chimpanzés (Figura 8),
durante a época seca (concretamente, entre 8 e 28 de Abril de 2006). Foram também usados pontos
recolhidos por C. Sousa e C. Casanova (projectos USFWS GA-0155 e POCI/Ant/57434/2004). Assim no
total foram utilizados 174 pontos de ninhos e avistamentos de chimpanzés.
t
Figura 8: Aspectos do trabalho de campo realizado na área de estudo (Abril 2006). Na fotografia da esquerda, recolha dos pontos com GPS. Na fotografia da direita pode observar-se um ninho de chimpanzé sobre Elaeis guineensis.
Foi usada, por motivos logísticos, uma combinação de levantamentos sistemáticos e
oportunistas, de maneira a optimizar o censo na área de estudo de uma espécie particularmente difícil
de observar/amostrar (Balcomb et al., 2000; Chapman & Lambert, 2000). Quando a amostragem
depende da estimação de indícios deixados pelos animais, neste caso ninhos, e não somente do
avistamento, são introduzidas fontes de erro. Por outro lado, com a metodologia utilizada é possível
aumentar enormemente o tamanho da amostra, melhorando a resolução estatística (White, 1994). O
impacto de eventuais fontes de erro na marcação de ninhos, como a idade do ninho, se é ou não
reutilizado, se apenas é usado durante o dia, etc. (Plumptre & Reynolds, 1996), não pôde ser avaliado
adequadamente. No entanto, no presente estudo, em que se pretende saber a adequabilidade de
habitat e não estimar densidades ou estrutura da população, estes tipos de erros foram considerados
a priori pouco relevantes.
- 27 -

2.4.3.2 Variáveis ambientais
A escolha das variáveis ambientais a utilizar teve por base a sua potencial importância
ecológica e a disponibilidade nas bases de dados existentes.
I) Fracções solo, sombra e vegetação
As análises de misturas espectrais têm como objectivo transformar os valores originais de
reflectância na superfície em proporções dos seus componentes, i. e., assume-se que a reflectância de
cada pixel de uma imagem Landsat é uma combinação linear das reflectâncias dos seus
componentes (Adams et al., 1995). Neste caso, e dado tratar-se de um território coberto
principalmente por vegetação, consideraram-se como componentes de um pixel a vegetação, o solo e
a sombra. Assim, a análise de misturas resulta em três imagens: a fracção vegetação, a fracção solo e
a fracção sombra, contendo cada pixel a proporção da componente a que se refere.
O uso desta técnica no presente trabalho tinha como objectivo inicial a produção de novos
algoritmos para a classificação das imagens Landsat. No entanto, os resultados mostraram que não
existia uma melhoria significativa entre o algoritmo gerado a partir das misturas espectrais e o
algoritmo já existente gerado a partir da classificação supervisada das imagens de satélite (Cassamá,
2006). Durante este processo de classificação foi notado que as imagens-fracção (sombra, solo e
vegetação) permitem destacar aspectos importantes do coberto vegetal, como a estrutura das copas
(fracção “sombra”) e a densidade da vegetação (fracções “solo” e “vegetação”). Como em estudos
anteriores (e.g. Carreiras, 2004), o modelo linear de misturas espectrais revelou-se eficaz no aumento
de interpretabilidade física dos resultados de classificação, relativamente ao que seria produzido
usando as regiões espectrais originais. Além disso, a utilização do modelo linear de misturas
espectrais permite a obtenção de valores quantitativos e contínuos de variáveis relevantes, as quais
foram utilizadas posteriormente no modelo de adequabilidade de habitat.
Para a computação do modelo linear de misturas espectrais foi utilizado o programa ENVI
3.6 (ITT, 2003), que possui um módulo com a implementação do respectivo algoritmo.
Resumidamente, foi construída uma biblioteca espectral em que constava o pixel mais próximo do
pixel puro de cada um dos três componentes (solo, vegetação e sombra). O programa utiliza essa
biblioteca para criar três imagens de valores contínuos (uma para cada componente) e uma imagem
de erro (RMS) (Figura 9).
O número de componentes (três: solo, vegetação e sombra) a utilizar na análise de
misturas espectrais tem de ser inferior ao número de bandas espectrais da imagem (sete, nas
imagens Landsat utilizadas) e todos as imagens fracção são utilizadas na produção dos raster finais
(Carreiras, 2004). O modelo linear de misturas espectrais pode ser utilizado com diferentes objectivos,
e.g.: estudos geológicos, oceanográficos, assim, de acordo com o fim pretendido, o tipo de
componentes utilizados varia. No caso em estudo, os componentes solo, sombra e vegetação foram
escolhidos por se adequarem a destacar aspectos importantes do coberto vegetal (Carreiras, 2004).
- 28 -

Figura 9: Imagens dos três componentes resultantes do modelo linear de misturas espectrais e do erro (root-mean-square). De cima para baixo e da direita para a esquerda: proporção de solo, proporção de sombra e proporção de vegetação obtidas pelo algoritmo do modelo (implementado no software ENVI).
- 29 -

II) Mapas produzidos a partir do coberto do solo
Uma das variáveis mais importantes a priori na análise de adequabilidade de habitat é o
tipo de coberto do solo. Assim, a relação entre a presença de Pan troglodytes e as classes de coberto
de solo poderá revelar tendências e preferências importantes na análise da adequabilidade de habitat
e na definição das áreas com maior importância na conservação desta espécie.
Os mapas originais do coberto de solo produzidos por Cassamá (2006) foram modificados,
para ser possível introduzi-los como variáveis, i.e. uma vez que se tratava de dados qualitativos, foi
necessário transformá-los em variáveis contínuas, de modo a poderem ser usados como EGVs. Assim,
com recurso ao programa ArcMap 9 (ESRI, 2004), foram calculadas separadamente, para cada pixel,
as distâncias euclidianas a cada uma das classes presentes no mapa de coberto (Figura 10),
originando 10 mapas com valores contínuos, em que cada célula contém o valor da distância à parcela
mais próxima da classe em análise.
Figura 10: Mapas originados a partir do mapa de coberto de solo de 2003 (Cassamá, 2006), representando a distância às parcelas das classes Floresta densa e Floresta aberta.
Os mapas originais de coberto de solo serviram também para calcular uma medida de
persistência para a classe “Floresta densa”, indicadora dos núcleos estáveis da floresta climácica do
território. A partir das datas disponíveis (1986, 1994, 2000 e 2003) foram seleccionadas as células que
ao longo destes 17 anos sempre pertenceram à classe Floresta densa, representando assim o co e
florestal que não foi alterado nas ultimas duas décadas.
r
Por fim, foi calculada uma medida da diversidade do coberto de solo, com recurso a uma
“moving window”, que indica para cada célula o valor da diversidade de classes de coberto numa área
- 30 -

quadrada de 2km e 5km de lado, de modo a se aproximarem da área média percorrida pelos
chimpanzés (Pontzer & Wrangham, 2006; Figura 11).
III) Altimetria e Diversidade morfológica do terreno
A variável “altimetria” está presente em praticamente todas as análises de modelação. No
presente estudo foi utilizado um modelo digital de terreno (MDT; Figura 12), produzido pela missão
SRTM (Coltelli et al., 1996; Moreira et al., 1995; Werner, 2001).
A distância percorrida por dia pelos chimpanzés apresenta grandes variações, dependendo
da região, tipos de habitat e disponibilidade de alimento (Pontzer & Wrangham, 2006). Essa distância
varia ainda com a composição do grupo, nomeadamente com o número total de indivíduos e com a
quantidade de juvenis. No entanto, estudos recentes apontam para um intervalo médio entre 1,5 e
3,5 km (Pontzer & Wrangham, 2006; Figura 11). Este intervalo foi utilizado para criar, a partir do
MDT, uma medida da diversidade morfológica do terreno utilizando duas “moving windows”, que
indicam para cada pixel o valor da diversidade de cotas altimétricas numa área de forma quadrada de
2km e 5km de lado (Figura 12).
Figura 11: Gráfico do movimento diário médio Vs. número de indivíduos do grupo (adaptado Pontzer & Wrangham, 2006).
- 31 -

Figura 12: Mapas da altimetria e da diversidade morfológica (usando uma “moving window” de 2km de lado) da área de estudo.
IV) Espacialização de métricas da paisagem
Através da utilização do software Patch analyst 3.0 (Rempel, 2006), disponível
gratuitamente no site do Centre fo Northern Forest Ecosystem Research, Lakehead University
(http://flash.lakeheadu.ca/~rrempel/patch/), foi possível construir e espacializar para a área de
estudo três variáveis contínuas: razão perímetro/área, dimensão fractal e índice de forma (Figura 13).
Trata-se de três métricas de paisagem que medem a complexidade da forma das parcelas:
r
1. O índice de forma é igual à soma do perímetro das parcelas dividida pela raiz quadrada da
área das parcelas e ajustada para quadrados padrão sendo dividida pelo número de parcelas;
assume o valor 1 quando a forma é o mais simples possível, ou seja um quadrado no caso de
se estar a analisar um raster ou um círculo no caso de se tratar de análise vectorial;
2. A dimensão fractal (ver 2.4.3), que também avalia a complexidade da forma da parcela,
aproxima-se de 1 para parcelas com perímetros simples, assumindo valores próximos de 2
quando a forma é mais complexa;
- 32 -

3. A razão perímetro/área (ver 2.4.3) é o somatório das razões perímetro/área de todas as
parcelas a dividir pelo número de parcelas.
Figura 13: Espacialização de três métricas da paisagem (dimensão fractal, razão perímetro/área e índice de forma) na área de estudo.
2.4.4 Construção do modelo de adequabilidade do habitat para Pan
troglodytes
2.4.4.1 Pré-processamento dos dados
Os dados de distribuição (2.4.3.1), recolhidos com GPS durante o trabalho de campo, foram
transformados de informação pontual em mapas raster recorrendo ao programa IDRISI (Clark Labs,
2006). Este programa produz ficheiros ras er em formato aceite pelo programa de implementação da
ENFA, o BIOMAPPER. Estes mapas têm o mesmo número de células (com o mesmo tamanho,
30mx30m) dos mapas das variáveis ambientais (EGVs), estando a informação codificada como 1 para
as células com presença de ninhos e como 0 para as células sem presença de ninhos.
t
Os dados ambientais provenientes das bases acima referidas (2.4.2.2) foram trabalhados em
ambiente SIG, de modo a apresentarem o mesmo número de células e o mesmo tamanho de célula
(30mx30m), assim como o mesmo formato (IDRISI). Estes dados constituem as variáveis
ecogeográficas (EGVs) da ENFA e foram também verificados para discrepâncias, variabilidade, dados
não quantitativos e valores extremos. Por fim, procedeu-se a uma análise de colinearidade com todas
- 33 -

as EGVs e os dados de distribuição, de modo a não usar dados fortemente correlacionados (Figura
14).
Figura 14: Análise de colinearidade de todas as EGVs e dos dados de distribuição [UPGMA (Unweighted Pair Group Method with Arithmetic mean) executada no programa Biomapper]. A linha vermelha representa o limite máximo de correlação (0,85) entre as EGVs aceite no estudo realizado.
Os dados relativos as povoações e às hortas revelaram estar fortemente correlacionados com
os dados de distribuição, por isso foram retirados da análise; assim, as EGVs utilizadas na ENFA foram
as seguintes:
• a proporção de solo, sombra e vegetação provenientes da análise de misturas espectrais;
• as distâncias a: Floresta densa, Floresta aberta, savana arborizada, savana herbácea e
parcelas que sempre foram Floresta densa (1986-2003), provenientes dos mapas de
coberto de solo;
• a altimetria e a variação altimétrica provenientes do modelo digital do terreno;
• a razão perímetro/área, a dimensão fractal e o índice de forma provenientes da análise da
paisagem.
- 34 -

2.4.4.2 Validação cruzada
A precisão predictiva do mapa de adequabilidade de habitat gerado pelo ENFA foi avaliada por
um procedimento de validação-cruzada (Boyce et al., 2002). Os pontos de presença de ninhos foram
agrupados aleatoriamente em k subconjuntos do mesmo tamanho e mutuamente exclusivos. Cada
conjunto k-1 foi usado para gerar um modelo de adequabilidade de habitat e a porção, não usada no
modelo, utilizada para o validar como dados independentes. Este processo foi repetido k vezes,
deixando sempre de fora da construção do modelo um subconjunto de dados diferente. O processo
resultou em k mapas diferentes de adequabilidade de habitat, cuja comparação permite uma
avaliação do poder predictivo do modelo. Cada novo mapa foi reclassificado em i classes (no presente
estudo o número de partições usadas foi de cinco), em que cada classe ocupa uma proporção da área
de estudo (Ai) e compreende uma proporção dos pontos de validação (Ni), estes pontos de validação
são as observações que não entraram nos modelos do processo de validação-cruzada. A frequência
ajustada à área para cada classe foi calculada como Fi = Ni/Ai. O valor esperado para Fi seria 1 para
todas as classes se o modelo fosse completamente aleatório. No entanto, se o modelo for “robusto”, a
baixos valores de adequabilidade de habitat devem corresponder baixos valores de Fi (abaixo de 1) e
a altos valores de adequabilidade de habitat devem corresponder altos valores de Fi (acima de 1),
com um aumento monotónico entre ambos. A monotonicidade da curva foi medida aplicando o
“Spearman rank correlation” a Fi (Boyce et al., 2002; Hirzel et al., 2004).
O poder predictivo do modelo foi medido através do índice contínuo de Boyce (Boyce et al.,
2002), referido como o índice mais preciso na caracterização da capacidade predictiva de modelos que
só usam presenças (Hirzel et al., 2006).
- 35 -

3. RESULTADOS
3.1 Evolução recente da paisagem na região do Cantanhez
Tendo em vista a análise da evolução da paisagem na área de estudo, foi executada uma
análise numérica da paisagem global, i.e., incluindo todas as classes de coberto de solo elegíveis (ver
2.4.2), recorrendo ao programa informático FRAGSTATS. Foram assim calculadas métricas para aferir
atributos espaciais da paisagem relativos a densidade/orla, forma e diversidade das parcelas que
constituíam a paisagem da área de estudo nas quatro datas estudadas (1986, 1994, 2000 e 2003).
As métricas de paisagem relativas a áreas, orlas e respectivas densidades são
influenciadas pelo número e tamanho das parcelas e também pela quantidade de orlas criadas por
estas parcelas. A tabela 6 e a figura 15 sintetizam os valores obtidos para estas quatro métricas.
Tabela 6: Valores das métricas de paisagem relativas a área/densidade e orla.
Ano Nº de Parcelas Densidade de
parcelas* Orla total (km)
Área média da
parcela (hectares)
1986 128606 55,0 42938 1,8
1994 119091 52,0 44674 1,9
2000 119416 52,5 43731 1,9
2003 104318 46,7 41357 2,1
*(nº de parcelas por 100 hectares)
O número total de parcelas presentes na área de estudo apresenta um decréscimo no
horizonte temporal analisado, facto que implica (uma vez que a área total é sempre a mesma) uma
diminuição da densidade de parcelas. A partir destas duas métricas é possível inferir uma tendência
de aglutinação e coalescência das parcelas na paisagem. A observação da variação da orla total
confirma esta tendência, ou seja, a paisagem aparenta estar a tornar-se mais simples, com menor
densidade de parcelas e de orlas. O resultado desta tendência é o aumento gradual da área média da
parcela (ver Discussão).
A tabela 7 e a figura 16 sintetizam os valores obtidos para a dimensão fractal e a razão
perímetro/área, duas métricas que avaliam a forma média das parcelas. De notar que o significado
ecológico da forma das parcelas está relacionado com o efeito de orla, sendo as parcelas com forma
mais complexa caracterizadas por um efeito de orla mais intenso; a intensidade do efeito de orla
condiciona, por sua vez, a composição biológica das parcelas (Turner et al., 2001).
- 36 -
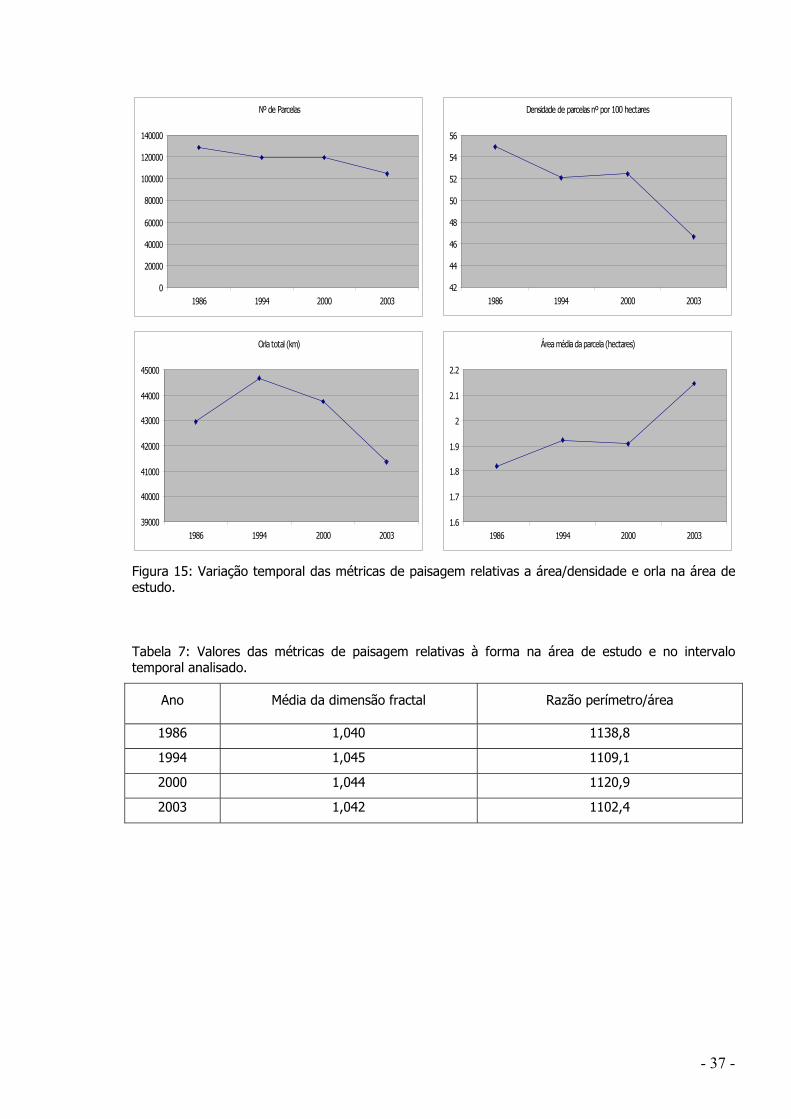
Nº de Parcelas
0
20000
40000
60000
80000
100000
120000
140000
1986 1994 2000 2003
Densidade de parcelas nº por 100 hectares
42
44
46
48
50
52
54
56
1986 1994 2000 2003
Orla total (km)
39000
40000
41000
42000
43000
44000
45000
1986 1994 2000 2003
Área média da parcela (hectares)
1.6
1.7
1.8
1.9
2
2.1
2.2
1986 1994 2000 2003
Figura 15: Variação temporal das métricas de paisagem relativas a área/densidade e orla na área de estudo.
Tabela 7: Valores das métricas de paisagem relativas à forma na área de estudo e no intervalo temporal analisado.
Ano Média da dimensão fractal Razão perímetro/área
1986 1,040 1138,8
1994 1,045 1109,1
2000 1,044 1120,9
2003 1,042 1102,4
- 37 -

média da dimensão fractal
1.0371.0381.039
1.041.0411.0421.043
1.0441.0451.046
1986 1994 2000 2003
média perímetro/área
1080
1090
1100
1110
1120
1130
1140
1150
1986 1994 2000 2003
Figura 16: Variação temporal das métricas de paisagem relativas à forma das parcelas na área de estudo.
As duas métricas analisadas medem a complexidade da forma da parcela. Ambas indiciam
uma tendência de simplificação da forma das parcelas ao longo do período de tempo analisado
(ver2.4.2), já identificada com as métricas de área e orla, e reforçando a constatação de uma
tendência para aglutinação das parcelas e simplificação da paisagem.
A tabela 8 e a figura 17 sintetizam os valores obtidos para cinco métricas de diversidade:
índices de diversidade de Shannon e de Simpson, índice modificado de Simpson e índices de
equitabilidade de Shannon e de Simpson.
Tabela 8: Valores das cinco métricas de diversidade utilizadas no estudo da área, no intervalo temporal analisado.
Ano Índice de
Shannon
Índice de
Simpson
Índice
modificado de
Simpson
Índice de
equitabilidade de
Shannon
Índice de
equitabilidade de
Simpson
1986 1,72 0,80 1,62 0,89 0,94
1994 1,80 0,82 1,72 0,92 0,96
2000 1,82 0,82 1,73 0,93 0,96
2003 1,68 0,80 1,57 0,86 0,92
Através da análise da tabela 8 e do gráfico da figura 17, é possível observar que não há
variações temporais significativas nos valores dos diversos índices. A paisagem da área de estudo
apresenta valores elevados nos índices de riqueza de classes. O índice de diversidade de Simpson
apresenta valores próximos (0,8) do máximo (1), indicando uma elevada riqueza (de classes de
coberto de solo). O índice de diversidade de Shannon também apresenta valores elevados, revelando
no entanto uma tendência para diminuir, como o número de classes é sempre o mesmo, esta
tendência indica que a paisagem está a ficar mais homogénea (ou menos rendilhada).
- 38 -

0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
1,8
2
1986 1994 2000 2003
Índice de Shannon
Índice de Simpson
Índice modificado deSimpsonÍndice de equitabilidade deShannonÍndice de equitabilidade deSimpson
Figura 17: Variação temporal das métricas de diversidade na área de estudo.
Como os índices de equitabilidade variam entre 0 e 1, os seus valores elevados (cerca de 0,9
para ambos os índices) na área de estudo indicam uma distribuição bastante equitativa das áreas
totais das diferentes classes (Ortega et al., 2004). Existe uma ligeira tendência para diminuição dos
valores destes índices entre 2000 e 2003 sugerindo que uma, ou mais classes, está (ão) a tornar-se
dominante (s) e/ou algumas classes apresentam menor representatividade.
3.2 Adequabilidade de habitat para Pan troglodytes
3.2.1 Análise global da adequabilidade de habitat na área de estudo
Os resultados da ENFA incluem eigenvalues (figura 18) e valores de componentes (ou
“Factor Scores”; tabela 9).
- 39 -

Figura 18: Eigenvectors das EGVs na ENFA realizada (os vectores 4, 5 e 6 correspondem as três primeiras EGVs da tabela 9).
Os resultados dos “factor scores” (tabela 9) indicam que (para a área em estudo) a espécie
Pan troglodytes ocupa habitats específicos, indicado pelo valor da Marginalidade 0,92 (ver 2.5.2.1). Os
resultados para o eixo 1, componente Marginalidade indicam a correlação entre cada EGV e o
componente. A tabela encontra-se organizada de forma decrescente em relação ao valor absoluto das
EGVs, quanto maior o valor absoluto do coeficiente, maior é o contributo desse EGV para a
marginalidade. Um valor positivo significa que a espécie “prefere” os valores mais elevados desse
EGV, enquanto um valor negativo significa que a espécie “prefere” os valores baixos. Assim é de
realçar a preferência da espécie: a) por áreas com variação topográfica, b) áreas de Floresta densa
com estabilidade nos últimos 17 anos, c) áreas de cariz florestal Floresta densa, Floresta aberta e
“savana arborizada”.
Os componentes subsequentes (Especialização) são ordenados por ordem decrescente da
quantidade de variância explicada (ver 2.5.2.1). No caso deste estudo, os três primeiros factores
explicam 96% da variância. Como os valores da Especialização não são interpretáveis (ver 2.5.2.1),
utiliza-se a Tolerância (inverso da especialização), cujos valores variam entre zero e um, podemos,
assim, constatar que o valor 0,21 indica que se trata de uma espécie especialista, ocupando condições
particulares da área de estudo.
- 40 -

Tabela 9: Valores dos componentes (“factor scores”) para as EGVs analisadas (ano 2003, excepto quando indicado).
EGVs Eixo 1 Eixo 2 Eixo 3
variação altimetria "moving window" 2km 0,46 -0,03 -0,10
distância a parcelas que sempre foram Floresta densa (1986-2003) -0,40 -0,39 -0,70
distância a Floresta densa -0,33 0,76 -0,15
distância a savana arborizada -0,30 0,06 0,26
distância a Floresta aberta -0,28 -0,49 0,59
variação das classes de coberto de solo "moving window" 5km 0,27 0,02 0,02
vegetação (Misturas espectrais) 0,24 -0,01 -0,02
razão perímetro/área (Patch analyst) 0,22 -0,01 -0,01
variação das classes de coberto de solo "moving window" 2km 0,19 -0,01 0,02
sombra (Misturas espectrais) -0,18 0,02 0,04
índice de forma (Patch analyst) -0,17 0,01 -0,01
distância a savana herbácea -0,16 -0,03 -0,05
variação das classes de coberto de solo "moving window" 1km 0,14 0,03 -0,01
distância a linha de costa -0,13 0,10 0,27
dimensão fractal (Patch analyst) -0,10 0,01 0,05
altimetria 0,09 0,07 -0,03
solo (Misturas espectrais) 0,02 -0,02 0,06
Marginalidade: 0,92
Especialização: 4,74
Tolerância (1/E): 0,21
A partir da matriz de resultados (tabela 9), foi produzido um mapa de adequabilidade de
habitat para Pan troglodytes na área de estudo (figura 19), usando o algoritmo das medianas. O
número de componentes incluídos na execução do mapa (3) foi decidido de acordo com a quantidade
de informação explicada pelo modelo (96%), i.e., a quantidade de variância explicada pela
Marginalidade e pela Especialização e a percentagem de Especialização explicada, ou seja, a
quantidade total de variância explicada pelo modelo. Seguindo os procedimentos delineados por Hirzel
(2002) e as recomendações incluídas no manual do programa Biomapper, o número de classes usadas
foi 3, sendo este número de classes, aquele que garantia uma melhor qualidade do modelo, qualidade
esta medida pelo índice contínuo de Boyce 0.91 +- 0.2 (Boyce et al., 2002). Por fim, o mapa de
adequabilidade de habitat foi reclassificado de acordo com as três classes de adequabilidade
consideradas (figura 19b).
- 41 -

a
Figestde ma
b
ura 19: a) Adequabilidade de habitat para Pan troglodytes na área de udo. Os pontos indicam as ocorrências registadas durante os trabalhos campo. b) Mapa resultante da validação cruzada e reclassificação do pa de adequabilidade de habitat.
- 42 -
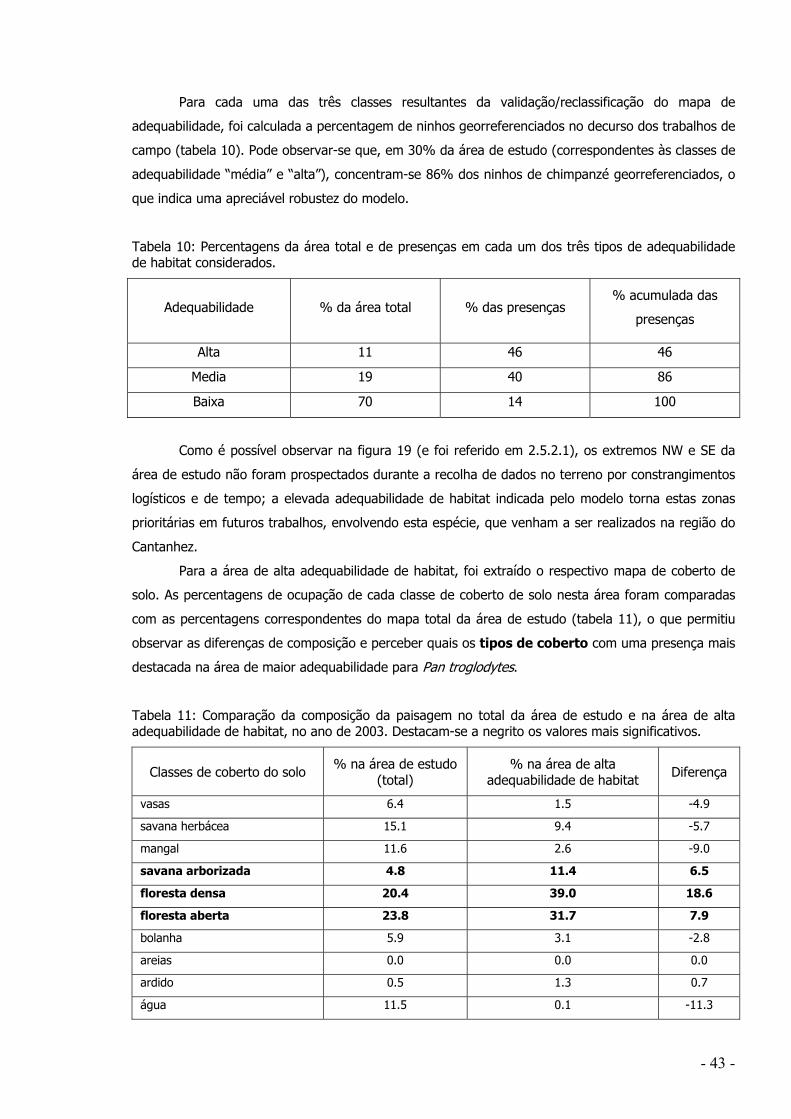
Para cada uma das três classes resultantes da validação/reclassificação do mapa de
adequabilidade, foi calculada a percentagem de ninhos georreferenciados no decurso dos trabalhos de
campo (tabela 10). Pode observar-se que, em 30% da área de estudo (correspondentes às classes de
adequabilidade “média” e “alta”), concentram-se 86% dos ninhos de chimpanzé georreferenciados, o
que indica uma apreciável robustez do modelo.
Tabela 10: Percentagens da área total e de presenças em cada um dos três tipos de adequabilidade de habitat considerados.
Adequabilidade % da área total % das presenças % acumulada das
presenças
Alta 11 46 46
Media 19 40 86
Baixa 70 14 100
Como é possível observar na figura 19 (e foi referido em 2.5.2.1), os extremos NW e SE da
área de estudo não foram prospectados durante a recolha de dados no terreno por constrangimentos
logísticos e de tempo; a elevada adequabilidade de habitat indicada pelo modelo torna estas zonas
prioritárias em futuros trabalhos, envolvendo esta espécie, que venham a ser realizados na região do
Cantanhez.
Para a área de alta adequabilidade de habitat, foi extraído o respectivo mapa de coberto de
solo. As percentagens de ocupação de cada classe de coberto de solo nesta área foram comparadas
com as percentagens correspondentes do mapa total da área de estudo (tabela 11), o que permitiu
observar as diferenças de composição e perceber quais os tipos de coberto com uma presença mais
destacada na área de maior adequabilidade para Pan troglodytes.
Tabela 11: Comparação da composição da paisagem no total da área de estudo e na área de alta adequabilidade de habitat, no ano de 2003. Destacam-se a negrito os valores mais significativos.
Classes de coberto do solo % na área de estudo (total)
% na área de alta adequabilidade de habitat Diferença
vasas 6.4 1.5 -4.9
savana herbácea 15.1 9.4 -5.7
mangal 11.6 2.6 -9.0
savana arborizada 4.8 11.4 6.5
floresta densa 20.4 39.0 18.6
floresta aberta 23.8 31.7 7.9
bolanha 5.9 3.1 -2.8
areias 0.0 0.0 0.0
ardido 0.5 1.3 0.7
água 11.5 0.1 -11.3
- 43 -

Os resultados da tabela 11 indicam que as classes de coberto relacionadas com as massas de
água (“mangal”, “água” e “vasas”) estão menos representadas na área de alta adequabilidade de
habitat do que no total da área de estudo, o que se compreende face aos hábitos terrestres de Pan
troglodytes. A classe “bolanha”, de maior influência antrópica (contudo, de igual modo relacionada
com a presença de massas de água), apresenta também uma menor representação, ainda que com
menor significado. Por outro lado, a classe savana herbácea, que ocorre principalmente em zonas
periodicamente alagadas (localmente conhecidas como “lalas”) manifesta uma redução significativa da
sua representação, em contraste com a savana arborizada.
Os tipos de coberto de solo com uma variação positiva significativa foram as classes com
características florestais, com destaque para a Floresta densa, que apresenta o maior diferencial
(+18,6%) entre a área de estudo e a área de maior adequabilidade de habitat para o chimpanzé. O
mesmo sucede, embora em menor escala coma Floresta aberta (+7,9%) e com a savana arborizada
(+6,5%). Esta variação positiva da representatividade das classes florestais é significativa, o que
justifica uma análise mais profunda da sua evolução recente.
3.2.2 Evolução da representação das classes florestais
Tendo em vista a análise do impacto da evolução recente da paisagem na região do
Cantanhez sobre a adequabilidade de habitat para Pan troglodytes, foi realizada uma nova análise
quantitativa (figura 20), desta vez focando a atenção, não na paisagem como um todo, mas em cada
uma das três classes de coberto com representação destacada na área de maior adequabilidade de
habitat para P. troglodytes (ver tabela 11). A análise da evolução da representação das três classes foi
executada comparando directamente os valores das métricas relativos à totalidade da área de estudo
e à área de maior adequabilidade de habitat.
Observa-se uma diminuição suave do número/densidade de parcelas, nas três classes na
área de maior adequabilidade, que indicia uma certa coalescência das parcelas. É observável uma
proporção constante das densidades de parcelas ao longo do tempo, apresentando a classe Floresta
aberta uma densidade de parcelas próxima do dobro da das outras duas classes, o que pode indicar
uma maior fragmentação deste tipo de coberto de solo. Quando a análise é executada para a
totalidade da área de estudo, as tendências de evolução das diferentes classes são diferentes. A
classe Floresta aberta apresenta uma diminuição suave da densidade de parcelas, a classe Floresta
densa apresenta um ligeiro aumento e a densidade de parcelas da classe “savana arborizada”
mantém-se praticamente inalterada.
Na análise de evolução da densidade de orlas, os valores apresentam uma certa
estabilidade, com uma ligeira tendência para a diminuição, excepção feita para a classe Floresta
aberta quando se analisa a totalidade da área de estudo. É de realçar a grande diferença entre os
valores obtidos para a totalidade da região e os valores para a área de maior adequabilidade de
habitat para Pan troglodytes. Apresentando esta última uma densidade muito menor de orlas,
- 44 -

provavelmente devido à hegemonia das classes florestais nesta área, que origina menos orlas do que
as outras classes de coberto de solo. Verifica-se também que o tamanho médio das parcelas florestais
é muito maior se incluirmos todas as classes de coberto de solo (ver figuras 15 e 20).
Quanto à área média das parcelas, a comparação entre os valores do total da área de
estudo e da área de maior adequabilidade de habitat revela valores muito díspares, o que se deverá
em parte, ao corte (artificial) das parcelas periféricas aquando da análise na área de maior
adequabilidade, resultando em uma diminuição importante do tamanho médio das parcelas. É, no
entanto, possível observar que em ambas as áreas há um aumento da área média das parcelas,
principalmente da classe Floresta densa, facto que corrobora a tendência para uma certa coalescência
das parcelas e mesmo para um aumento relativo das classes de coberto de cariz florestal. Observando
os gráficos da evolução da razão perímetro/área (indicadora da forma das parcelas), ressalta uma
propensão para a diminuição dos valores desta razão para as duas áreas analisadas, revelando-se
uma tendência para a simplificação da forma das parcelas. Essa tendência parece acentuar-se entre
2000 e 2003, sendo particularmente significativa para a classe Floresta Densa na área de maior
adequabilidade de habitat.
Nos últimos dois gráficos é possível observar a evolução da área ocupada por cada uma
das três classes, em relação ao total da área em análise. Assim, a classe Floresta densa exibe um
aumento (em ambos os gráficos) da área ocupada, a classe Floresta aberta apresenta um
comportamento mais irregular (no entanto, os valores de 2003 e 1986 são muito próximos), enquanto
a classe “savana arborizada” parece exibir uma ligeira propenção para a diminuição da área ocupada.
- 45 -
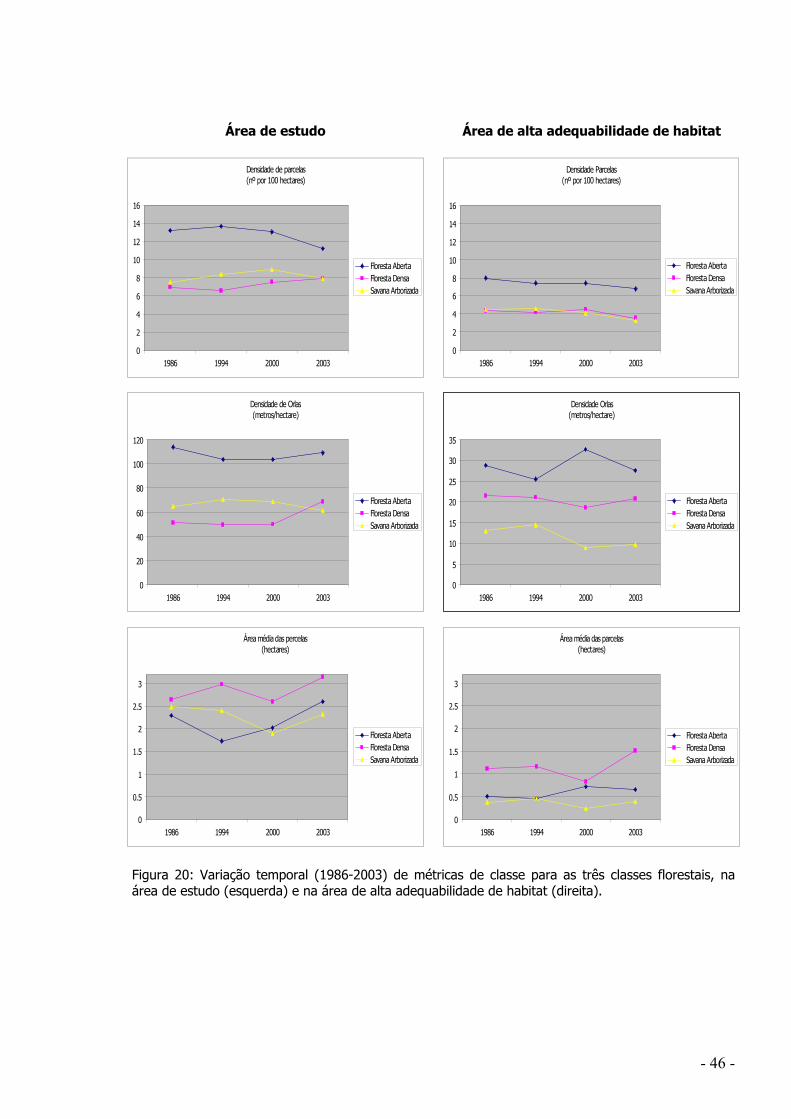
Área de estudo Área de alta adequabilidade de habitat
Densidade de parcelas(nº por 100 hectares)
0
2
4
6
8
10
12
14
16
1986 1994 2000 2003
Floresta Aberta Floresta DensaSavana Arborizada
Densidade Parcelas(nº por 100 hectares)
0
2
4
6
8
10
12
14
16
1986 1994 2000 2003
Floresta Aberta Floresta Densa Savana Arborizada
Densidade de Orlas(metros/hectare)
0
20
40
60
80
100
120
1986 1994 2000 2003
Floresta Aberta Floresta DensaSavana Arborizada
Densidade Orlas(metros/hectare)
0
5
10
15
20
25
30
35
1986 1994 2000 2003
Floresta Aberta Floresta Densa Savana Arborizada
Área média das percelas (hectares)
0
0.5
1
1.5
2
2.5
3
1986 1994 2000 2003
Floresta Aberta Floresta DensaSavana Arborizada
Área média das parcelas (hectares)
0
0.5
1
1.5
2
2.5
3
1986 1994 2000 2003
Floresta Aberta Floresta Densa Savana Arborizada
Figura 20: Variação temporal (1986-2003) de métricas de classe para as três classes florestais, na área de estudo (esquerda) e na área de alta adequabilidade de habitat (direita).
- 46 -

Média (perímetro/área)
1000
1050
1100
1150
1200
1250
1986 1994 2000 2003
Floresta Aberta Floresta DensaSavana Arborizada
Média (perímetro/área)
1000
1050
1100
1150
1200
1250
1986 1994 2000 2003
Floresta Aberta Floresta Densa Savana Arborizada
Percentagem de área ocupada pelas classes
0
5
10
15
20
25
30
35
40
45
1986 1994 2000 2003
Floresta AbertaFloresta DensaSavana Arborizada
Percentagem de área ocupada pelas classes
0
5
10
15
20
25
30
35
40
45
1986 1994 2000 2003
Floresta AbertaFloresta DensaSavana Arborizada
Figura 20 (cont.)
De notar que os valores correspondentes ao ano 2000 em algumas das métricas utilizadas
apresentam disparidades em relação aos valores das datas anteriores e de 2003. Embora não seja
objecto deste trabalho a análise das causas destas variações, julgamos que podem estar envolvidos
dois tipos de factores: a) diferenças nas características atmosféricas aquando da colheita da imagem
de 2000; b) perturbações causadas pelo aumento da população na área de estudo (Temudo &
Schiefer, 2003) na sequência do conflito armado que ocorreu no país em 1998-1999.
Das três classes de coberto de solo com cariz florestal analisadas, aquela que apresenta maior
percentagem de ocupação na área de maior adequabilidade e maior variação na comparação entre a
área de estudo e a área de maior adequabilidade de habitat é a classe Floresta densa (tabela 11).
Esta classe de coberto do solo pode considerar-se como a vegetação potencial para as condições
edafo-climáticas locais, apresentando grande diversidade específica de flora e fauna, assumindo um
enorme interesse como habitat e refúgio de inúmeras espécies. Trata-se das manchas de Floresta
densa semi-húmida mais setentrionais da Africa ocidental, podendo mesmo, alguns destes “matos”
(na terminologia local) constituir fragmentos de floresta primária (Malaisse, 1996).
Dada a importância desta classe para a ecologia global da região e para a espécie alvo do
estudo, foi executada uma análise temporal da evolução das suas orlas, ou seja, da identidade das
classes de coberto de solo com que a classe Floresta densa contacta (figura 21).
- 47 -

A partir da análise da figura 21, conclui-se que a classe Floresta densa contacta
maioritariamente com a classe Floresta aberta; é, no entanto, também possível observar uma
tendência geral para uma ligeira diminuição das orlas partilhadas por estas duas classes
(particularmente entre 1986 e 2000), podendo este facto resultar, pelo menos em parte, da evolução
de parcelas de Floresta aberta para Floresta densa. Esta observação é corroborada pelo aumento da
área média das parcelas de Floresta densa, verificado em toda área de estudo, mas mais notório na
área de maior adequabilidade de habitat (figura 20).
Para melhor discriminar a evolução das fronteiras, entre a classe Floresta densa e as outras
classes de coberto de solo, foi retirada do gráfico a classe Floresta aberta (figura 22). É assim possível
perceber que a percentagem de orlas partilhadas pelas classes “vasas”, “bolanha”, “ardido” e “água”
com a classe Floresta densa são escassas. As orlas partilhadas com as duas classes de savana sofrem
grandes alterações ao longo do tempo (no entanto, não aparentam ter uma tendência definida); de
notar que as alterações nos ecótonos Floresta densa, “savana” podem decorrer de erros de
classificação, uma vez que a maior dificuldade em descriminar classes ocorre precisamente entre a
Floresta aberta e a “savana arborizada” (Cassamá, 2006). A percentagem de orlas partilhadas com a
classe “mangal” está a aumentar, podendo indicar que o crescimento anteriormente registado da
classe Floresta densa também poderá estar a acontecer nas proximidades das massas de água (Rias).
Estes elementos sugerem uma tendência para o aumento da área de maior adequabilidade de
habitat para Pan troglodytes dentro da área de estudo ao longo do período estudado, com evidentes
implicações ao nível do estado de conservação da espécie.
0
10
20
30
40
50
60
70
80
90
1986 1994 2000 2003
vasas
savana arborizada
mangal
savana herbácea
floresta aberta
bolanha
ardido
água
Figura 21: Variação temporal da percentagem de orlas partilhadas entre a classe Floresta densa e as outras classes de coberto de solo, no total da área de estudo.
- 48 -

%
0
2
4
6
8
10
12
14
16
18
20
vasas savanaarborizada
mangal savanaherbácea
bolanha ardido água
1986
1994
2000
2003
Figura 22: Variação temporal da percentagem de orlas partilhadas entre a classe Floresta densa e as restantes classes de coberto de solo (com excepção da classe Floresta aberta).
- 49 -

4. DISCUSSÃO
4.1 Adequabilidade de habitat para Pan troglodytes na região do
Cantanhez
Os resultados da ENFA indicam uma Marginalidade de 0,92, o que indica que, na área de
estudo o chimpanzé ocupa habitats específicos (i.e. não ocupa condições médias da área de
estudo), podendo, então afirmar-se que se trata de uma espécie especialista. Na análise dos
coeficientes da componente marginalidade (tabela 9), que indicam a correlação entre cada EGV e o
componente (2.4.1), é evidente a importância das classes de coberto de solo com cariz florestal, com
três das cinco EGVs que mais contribuem para a Marginalidade. Em todos os coeficientes destas EGVs
os valores negativos indicam que o chimpanzé prefere estar próximo das classes de coberto florestais.
Uma das EGVs que foi calculada através do uso dos quatro mapas de coberto de solo (1986, 1994,
2000 e 2003) e que pretende evidenciar as parcelas de Floresta densa, que não sofreram alterações
ao longo destes dezassete anos (2.4.3.2), apresenta uma contribuição para a Marginalidade ainda
maior. Facto este que poderá indicar uma preferência não só por parcelas de floresta, mas
particularmente por parcelas de Floresta densa com pouca perturbação. No entanto a EGV mais
relevante para este componente é a variação altimétrica (moving window de 2km). Esta variação
altimétrica pode estar correlacionada com a presença de Floresta e solos apropriados a Floresta. Na
figura 14 é possível observar uma forte correlação (embora não seja acima do limiar estipulado)
entre, as variáveis ligadas as classes florestais e a variação altimétrica.
O nível de conhecimento actual não permite uma indicação clara do porquê da
preponderância desta EGV, no entanto esta poderá dever-se a uma conjugação de factores entre os
quais: as áreas topograficamente mais complexas são menos atraentes para a agricultura, permitindo
uma maior estabilidade das classes de ocupação de cariz florestal.
Os componentes subsequentes ou componentes de Especialização estão ordenados por ordem
decrescente da quantidade de variância explicada e descrevem o quanto “especializada” é a espécie,
em relação a variação dos habitats na área de estudo (Hirzel et al., 2002). Como variam entre 1 até
ao infinito não são passíveis de interpretação. Deste modo, utiliza-se a Tolerância, que mede a
escolha da espécie de entre os limites disponíveis de EGVs e é definida como o inverso da
especialização (1/S). Neste caso, o valor de 0,21 indica-nos que a espécie escolhe condições
particulares de entre os limites disponíveis de EGVs na área de estudo, ou seja, está presente em
intervalos pequenos dentro da variação de cada EGV.
O mapa de adequabilidade de habitat produzido com a ENFA (Figura 19) foi reclassificado,
seguindo um procedimento (2.4.4.2) que permite a validação e produção de um novo mapa com três
classes de adequabilidade: baixa, média e alta (Figura 19), de modo a fornecer uma predição mais
honesta e relevante (Hirzel et al., 2006). Através da análise das percentagens das áreas ocupadas por
cada uma das três classes de adequabilidade e das percentagens de pontos de presenças, em cada
- 50 -

uma dessas classes (tabela 10), é possível afirmar que em 30% da área de estudo (817 km2),
correspondente às classes de adequabilidade média e alta, encontramos 86% dos pontos de
presença. Estes resultados sugerem duas observações: a) o modelo produzido pelo ENFA é capaz de
gerar resultados no mínimo congruentes; b) apesar de os resultados da Marginalidade e da Tolerância
nos indicarem que se trata de uma espécie relativamente especialista a área indicada como adequada
pelo mapa reclassificado é relevante e indica que dentro da área de estudo existem três áreas (NW,
central e SE) com possibilidade de comunicação entre elas e com boa adequabilidade para a espécie.
Estas três áreas correspondem às áreas com maior percentagem de ocupação por classes de coberto
de solo de cariz florestal.
Na comparação da composição da paisagem no total da área de estudo e na área de maior
adequabilidade de habitat em 2003 (tabela11), ressalta, mais uma vez, a correspondência entre as
classes de coberto florestais (que apresentam um acréscimo percentual de ocupação na área de maior
adequabilidade) e as indicadas como adequadas para o Chimpanzé. Esta relação liga a conservação
do chimpanzé à preservação das florestas do Cantanhez. Esta ligação é importante para a
conservação da biodiversidade, da área de estudo, que está incluída como Very Important Area
(Butynski, 2001), para o estudo e protecção dos chimpanzés (figura 23).
Figura 23: Mapa das áreas de conservação prioritárias para Chimpanzé (adaptado de Butynski, 2001).
A robustez do modelo produzido pela ENFA foi medida através do índice contínuo de Boyce
(Boyce et al., 2002) e apresenta valores de 0,91 +- 0,2, valores que indicam uma grande robustez
(Hirzel et al., 2006). Existem poucas publicações que usem dados de distribuição de chimpanzés. De
facto a falta de dados recolhidos in situ é tal que a espécie chegou a ser dada como extinta na Guiné-
Bissau na década de 90. No trabalho realizado usou-se pela primeira vez a ENFA de modo a aferir a
adequabilidade de habitat para Pan troglodytes, assim, não é possível comparar os resultados obtidos
com os de outros estudos. Por outro lado, a generalização dos resultados obtidos revela-se também
difícil, uma vez que a espécie alvo deste estudo apresenta variações culturais (Tutin et al., 2005),
ocorrendo comportamentos e respostas diferentes de população para população; deste modo, é
necessário todo o cuidado na extrapolação destes resultados para outros contextos.
- 51 -

4.2 Evolução da paisagem na área de estudo (1986-2003) e seu impacto
na adequabilidade de habitat para Pan troglodytes
A análise da evolução da paisagem foi executada com base em quatro mapas de coberto de
solo (1986, 1994, 2000, 2003). Nas análises iniciais foram utilizadas todas as classes de coberto de
solo elegíveis (ver 2.4.2).
Os resultados da tabela 6 e da Figura 15, correspondentes à análise das áreas e orlas das
parcelas, indicam uma tendência para uma simplificação e aglutinação da paisagem, entre 1986 e
2003. O número de parcelas total diminuiu o que, tal como se esperava, reduziu a densidade de
parcelas e aumentou a área média das parcelas. A quantidade de orla total também decresceu,
devendo-se este resultado, não só à diminuição do número de parcelas, mas também a uma
tendência para a simplificação das formas das parcelas (tabela 7 e Figura 20), o que vem reforçar os
resultados anteriores.
Os resultados obtidos indicam que na área de estudo e durante o período de tempo analisado,
não há indicações de fragmentação da paisagem, apresentando uma evolução no sentido de uma
menor complexidade da paisagem e da aglutinação de parcelas. Estes resultados podem ter
origem em várias causas diferentes, ou, provavelmente numa combinação de causas. Assim alguns
dos motivos poderão ser:
• O abandono de áreas previamente arroteadas, no decurso da guerra colonial (1963-1974).
Durante o trabalho de campo foram observadas zonas com 43 anos de regeneração (1963-
2006) cuja estrutura já era bastante similar à da Floresta densa embora com árvores de porte
menor e com maior quantidade de lianas. O abandono de algumas tabancas poderá ter
contribuído para o aumento da massa florestal. Autores da new ecology sugerem que a
população como elemento modelador da paisagem tem muitas vezes um papel positivo uma
vez que plantam algumas espécies nas proximidades dos povoamentos (com diferentes
objectivos como sombra, alimento ou como protecção dos fogos de savana) originando novas
“bolsas” de floresta (Fairhead & Leach, 1998).
• O relativo isolamento da região. A área de estudo tem vias de comunicação bastante
precárias o que dificulta o escoamento de todos os produtos e em particular de madeira, que
exige a utilização de veículos pesados. Apesar de existirem recentemente referências ao
derrube de árvores com utilização de motosseras próximo da área em estudo o que, a
confirmar-se, pode constituir uma ameaça a ter em conta, ainda há poucos meios mecânicos
e as zonas desmatadas não atingem uma grande dimensão.
• A existência de áreas designadas matos sag ados, que provavelmente não foram tocadas ao
longo de muitas dezenas de anos. Apesar de em geral serem zonas restritas e com pequena
superfície, a sua contribuição para a preservação das florestas é importante podendo servir de
reserva da biodiversidade e de origem para a recolonização de áreas degradadas.
r
- 52 -

Após o ENFA, a determinação da área de maior adequabilidade de habitat e da sua
comparação com a composição da paisagem na área de maior adequabilidade de solo (tabela 11),
emerge a importância das classes de coberto florestais, principalmente da classe Floresta densa que
aumenta a sua representatividade de 20 para 40%. De modo a melhor compreender o
comportamento destas relevantes classes florestais foi realizada uma nova análise da paisagem ao
nível da classe.
Os resultados obtidos na análise do comportamento de cada uma das classes de coberto
florestais ao longo do tempo (3.2.2) revelam uma tendência para a aglutinação de parcelas
(diminuição da densidade e aumento da área média das parcelas) e simplificação da forma
(diminuição da razão perímetro/área) das parcelas das classes florestais, particularmente da classe
Floresta densa. Estes resultados sugerem que embora haja evidência de que a desflorestação tem
sido drástica em muitos locais e em particular na Africa Ocidental (FAO, 1993), os dados por nós
obtidos para esta zona parecem revelar uma manutenção das florestas durante o período
estudado.
Para tentar perceber se existe uma tendência na evolução da relação entre a classe Floresta
densa e as restantes classes de coberto de solo foi analizada a evolução das orlas desta classe ao
longo do tempo. Os resultados mostram que ao longo da variação temporal não se verifica um padrão
definido na quantidade de orlas partilhadas com a classe Floresta densa, com a excepção do aumento
constante da quantidade de orlas partilhadas com a classe mangal, o que pode sugerir que pelo
menos parte do aumento de área da classe Floresta densa, se está a processar em áreas próximas de
massas de água (Rias).
Refira-se que, para obter a máxima robustez possível na análise da paisagem, utilizaram-se
imagens do satélite Landsat provenientes de sensores comparáveis (ref), i.e. Landsat TM e Landsat
ETM+, abdicando, em favor de uma maior fiabilidade e comparabilidade, de estender a análise por
um horizonte temporal maior. No interesse dessa mesma robustez todas as análises e operações
efectuadas (2.4.3), tiveram como objecto a totalidade das imagens com as mesmas premissas e
recorrendo ao mesmo sof ware. Os mapas de coberto de solo executados por Cassamá (2006), com
base nas mesmas imagens de satélite e utilizados neste estudo, apresentam resultados que indicam
que a classificação do coberto de solo tem uma alta fiabilidade (Cassamá, 2006) (Anexos). Por fim, é
de realçar a inexistência de contradições entre os resultados das várias métricas de paisagem
utilizadas e entre as tendências de evolução da paisagem detectadas neste trabalho em comparação
com o trabalho que lhe precedeu (Cassamá, 2006).
t
No entanto, deverão ser consideradas algumas limitações importantes à interpretação dos
resultados, nomeadamente a delicada separação das classes florestais e do complexo Floresta/Savana
em que a definição de floresta varia entre os diferentes estudos, o que provoca grandes discrepâncias
entre os resultados (Fairhead & Leach, 1998). Pode, também, existir alguma interferência
antropogénica nos resultados obtidos para o comportamento das classes de coberto de cariz florestal,
nomeadamente a possibilidade de em algumas plantações de caju (conhecidas localmente por hortas
de caju), feitas no fim dos anos 1980 e início dos anos 1990, a densificação do coberto vegetal do
- 53 -

solo, que está a ocorrer com o amadurecimento das plantas, pode fazer com que o sinal da vegetação
captado pelo satélite seja comparável ao da Floresta aberta e, em menor extensão, ao da Floresta
densa.
- 54 -

5. CONCLUSÕES E CONSIDERAÇÕES FINAIS
5.1 Padrões de evolução da paisagem e adequabilidade de habitat para
Pan troglodytes
Ao contrário dos resultados de estudos produzidos para a África Ocidental como um todo
(Aubréville, 1938; Gornitz & NASA, 1985; FAO, 1993; Pearce, 1997; Zheng et al., 1997), que indicam
sem excepção uma diminuição assinalável das áreas florestadas na região, os resultados da evolução
da paisagem para o Cantanhez revelam uma área com recuperação de áreas florestais e sem
evidências de aumento da fragmentação da paisagem, indicando que se trata de uma área ainda em
relativo bom estado de conservação.
Os resultados da análise da paisagem indicam que o habitat para chimpanzé na área de
estudo não se encontra muito ameaçado, podendo mesmo ter melhorado entre 1986 e 2003. Este
facto por si é importante mas torna-se ainda mais relevante se tivermos em conta o contexto regional
(Senegal e Republica da Guiné) e a sobrevivência da sub-espécie Pan troglodytes verus a médio
prazo. Dados recentes (Oates, 2006) apontam para uma diminuição dos efectivos populacionais de
chimpanzé na Africa Ocidental, que surge aliada a um isolamento das populações e consequente
perda de viabilidade. O Cantanhez pode assim emergir como uma área ecologicamente “saudável”,
capaz de manter populações viáveis e mesmo, seguindo as previsões da teoria de metapopulações e
metacomunidades (Gilpin & Hanski, 1991), servir como população de emigrantes (source) para as
áreas próximas onde a população de chimpanzés apresente sinais de declínio (sink), contribuindo para
a manutenção do fluxo génico e sobrevivência da espécie. De facto, se houver interesse por parte do
governo da Guiné-Bissau e da comunidade internacional, poderão vir a ser delineadas e
implementadas estratégias que permitam a movimentação da espécie numa escala transfronteiriça,
podendo o trabalho agora efectuado na análise da adequabilidade de habitat para Pan troglodytes
contribuir para a definição dessas estratégias.
5.2 Criação de uma área protegida no Cantanhez
A área de estudo apresenta muitos argumentos para constituir uma reserva de relevância
internacional de importantes valores naturais e um protótipo de desenvolvimento sustentável.
Contém as florestas sub-húmidas mais setentrionais de África (Malaisse, 1996), está incluída nos 9
sítios naturais importantes do ponto de vista da biodiversidade reconhecidos pelo World Conservation
Monitoring Centre (WCMC) do United Nations Environment Programme (UNEP). A zona costeira da
qual faz parte é uma das 200 eco-regiões mais importantes do mundo identificadas pela World
Wildlife Fundation (WWF, Projecto UICN, 2000) (Silva et al., 2001).
- 55 -

Os resultados do presente estudo indicam que a zona não apresenta uma degradação ou
fragmentação da paisagem preocupantes, corroborando resultados anteriores (Cassamá, 2006) que
indicam uma tendência para o aumento da área de floresta. A fauna presente inclui várias espécies
bandeira como: o chimpanzé (Pan troglodytes verus), hipopótamo (Hippopotamus amphibius),
manatim (Trichechus senegalenis), várias espécies de tartarugas marinhas (Silva et al., 2001). Por
todas estas características revela-se uma área com boas características para constituir uma área
protegida.
A constituição de uma área protegida na área de estudo contribuirá para a implementação de
várias acções com vista a promoção de práticas que permitam manter ou incrementar o bem-estar
das populações sem comprometer o estado dos ecossistemas e consequente preservação da
biodiversidade. Acções estas, que promovam a manutenção da Floresta Densa, ex-libris da região,
como por exemplo o incentivo [em detrimento das plantações de caju (Anacardium occidentale L.)],
das hortas tradicionais, sobretudo de cola (Cola nitida (Vent.) Schott E Endl.), um sistema de cultura
que parece bem adaptado às condições locais e permite manutenção da diversidade dos ecossistemas
florestais da região (Catarino, 2004).
A presença do chimpanzé conjugada com as práticas tradicionais, que o consideram muito
próximo dos humanos, não incluindo a espécie nos seus hábitos alimentares (Duvall et al. 2003), cria
um bom ambiente social para a conservação da espécie, que poderá ser encorajada com dividendos
económicos provenientes de um ecoturismo ainda incipiente, mas com um enorme potencial
ancorado nas manchas de Floresta Densa bem preservadas e na possibilidade de observação de
espécies como o chimpanzé e outra fauna emblemática. No contexto da Guiné-Bissau, esta região
poderá ainda ser uma das candidatas à implementação de projectos ao abrigo dos mecanismos de
desenvolvimento limpo, previstos no protocolo de Quioto (http://unfccc.int).
5.3 Principais contribuições e perspectivas futuras
O estudo realizado apresenta novos dados para a compreensão das dinâmicas recentes da
paisagem na região do Cantanhez. Corrobora os resultados que indicam aumento da área florestal, no
período estudado, obtidos por Cassamá (2006), sugerindo adicionalmente não haver indícios de
fragmentação da paisagem preocupantes. Apresenta também o primeiro mapa de adequabilidade de
habitat para chimpanzé e assim acrescenta mais dados que poderão ser usados na gestão e
conservação da espécie assim como na eventual criação de uma área protegida.
Na sequência deste estudo seria interessante aprofundar os resultados com novos projectos.
Um deles seria, certamente, complementar o estudo da evolução da paisagem com novos mapas de
coberto de solo derivados de imagens de satélite mais recentes e de datas intercalares (idealmente
1986, 1990, 1994, 1997, 2000, 2003 e 2006) para que a escala temporal tenha uma frequência de
datas constante (+- 3 a 4 anos), permitindo melhorar a resolução das modificações ocorridas.
- 56 -

Em relação à espécie estudada, os resultados deste trabalho poderão ser complementados
com mais trabalho de campo no levantamento de ninhos de chimpanzé usando os resultados do ENFA
como matriz destes levantamentos. Por exemplo, seria importante determinar se a espécie está
presente nas áreas de alta adequabilidade de habitat identificadas nos extremos NW e SE da área de
estudo que não apresentam pontos de presença (por dificuldades logísticas encontradas durante o
trabalho de campo). Ao realizar uma nova época de trabalho de campo com maior horizonte temporal
poderá ser utilizada uma grelha de amostragem que permita recolher sistematicamente pontos de
presença e ausência da espécie de forma a ser possível usar outros métodos de modelação da
distribuição da espécie (LMR, GLM, GAM) tendo em vista a comparação dos resultados obtidos com a
análise realizada neste trabalho (ENFA). Seria também interessante realizar estudos semelhantes em
outras áreas da Africa Ocidental e com outras sub-espécies de Pan troglodytes. A quantidade de
novos dados originada por semelhantes estudos permitiria um enorme avanço na compreensão das
apetências ecológicas da espécie, o que possibilitaria uma acção mais informada, tendo em vista a sua
conservação.
- 57 -

5. BIBLIOGRAFIA
Adams, J.B., Sabol, D.E., Kapos, V., Almeida Filho, R., Roberts, D.A., Smith, M.O., & Gillespie, A.R.
(1995). Classification of multispectral images based on fractions of endmembers: application to land-
cover change in the Brazilian Amazon. Remote Sensing of Environment, 52(2), 137-154.
Álvares, F. & Brito, J.C. (in press). Habitat requirements and potential areas of occurrence for Pine
Marten in North-western Portugal: conservation implications. In: Santos-Reis M., Birks, J., O´Doherty
E.O., & Proulx G. (Eds.), Martes in Carnivore Communities. Alpha Wildlife, Alberta.
Andersen, M.C., Watts, J.M., Yool, S. R., Fahnestock, P. B. (2000). Regression-tree modeling of desert
tortoise habitat in the central Mojave desert. Ecological Applications, 10 (3): 890-900.
Balbontin, J. (2005). Identifying suitable habitat for dispersal in Bonelli´s eagle: An important issue in
halting its decline in Europe. Biological Conservation. 126 (1): 74-83.
Balcomb SR., Chapman CA., Wrangham RW. (2000). Relationship between chimpanzee (Pan
troglodytes) density and large, fleshyfruit tree density. Am J Primatol 51: 197–204.
Beebee, T. G. C. & Griffiths, R. A. (2005). The amphibians decline crisis: A watershed for conservation
biology? Biological Conservation. 125 (3): 271-226.
Boone, J. D., McGwire, K. C., Otteson, E. W., DeBaca, R. S., Kuhn, E. A., Villard, P., Brussard, P. F. &
St. Jeor, S. C. (2000). Remote Sensing and Geographic Information Systems: Charting Sin Nombre
Virus Infections in Deer Mice. Emerging Infectious Diseases, 6 (3): 248-257.
Boyce, M.S., Vernier, P.R., Nielsen, S.E., Schmiegelow, F.K.A., (2002). Evaluating resource selection
functions. Ecological Modelling 157, 281–300.
Boyce, M.S., Vernier, P.R., Nielsen, S.E. & Schmiegelow, F.K.A. (2002). Evaluating resorce selection
functions. Ecological Modelling. 157 (2-3): 281-300.
Brito, J.C. & Crespo, E. G. (2002). Distributional analysis of two vipers (Vipera latastei and V. seoanei)
in a potential area of sympatry in the Northwestern Iberian Peninsula. In: Schuett , Hoggren, Douglas
& Greene (Eds.). Biology of the Vipers: 129-128. Eagle Mountain Publishing. LC.
Brito, J. C., Crespo, E. G., & Paulo, O. S. (1999). Modelling wildlife distributions: Logistic Multiple
Regression Vs. Overlap Analysis. Ecography. 22 (3): 251-260.
- 58 -

Burney, D.A. & Flannery, T.F. (2005). Fifty millennia of catastrophic extinctions after human contact.
Trends in Ecology & Evolution. 20 (7): 395-401.
Brotons, L., Thuiller, W., Araújo, M.B. & Hirzel (2004). Presence-absence versus presence-only
modelling methods for predicting bird habitat suitability. Ecography. 27: 437-448.
Clark Labs (2003) Idrisi Kilimanjaro. Clark Labs, Clark.
Chapman CA., Lambert JE., (2000). Habitat alteration and the conservation of African primates: case
study of Kibale National Park, Uganda. Am J P ima ol 50:169–186. r t
t
.
.
Chardonnet, P., & Limoges, B., (1989). Guinea Bissau. Antelopes: Global Survey and Regional Action
Plans, Par 3: West and Central Africa (ed. R. East).IUCN, Gland, Switzerland. pp. 35–37.
Chuvieco, E. S (2002). Teledeteción Ambiental, 1ª Edición, Ediciones Ariel Ciência Barcelona.
Coltelli, M., Fornaro, G., Franceschetti, G., Lanari, R., Migliaccio, M., Moreira, J. R., Papathanassiou, K.
P., Puglisi, G., Riccio, D., & Schwabisch, M. (1996). SIR-C/X-SAR multifrequency multipass
interferometry: A new tool for geological interpretation. Journal of Geophysical Research.
101(10):23127-23148.
Cox, C.B. & Moore, P.D. (2000). Biogeography. An evolutionary approach, 6th Edition. Blackwell
Publishing.
Crews-Meyer, K. A. (2002). Characterizing landscape dynamism using paneled-pattern metrics.
Photogrammetric Engineering and Remote Sensing, 68, 1031– 1040.
Di Gregorio, A. & Jansen, L. J. M., (2000). Land Cover Classification System, LCCS – classification
concepts and user manual. FAO, Roma.
ESRI (1996). ArcView 3 2. Environment Systems Research Institute, Inc.
ESRI (2004). ArcMap 9 0. Environmental Systems Research Institute Inc.
Fairhead James, Leach Melissa (1998) Reframing Deforestation: Global Analyses and Local Realities;
Studies in West Africa. Routledge, New York. 208pp.
FAO (1993). Summary of the Final Report of the Forest Resources Assessment 1990 for the Tropical
World. FAO, Rome.
- 59 -

Franklin, J. (1998). Predicting the distribution of shrub species in southern California from climate and
terrain-derived variables. Journal of Vegetation Science, 9 (5): 733-748.
Foddy, G.M. (2002) Status of land cover classification accuracy assessment. Remote Sensing o
Environment 80: 185-201.
f
, t
t
Foody, G. M. & Cutler, M. E. J. (2003). Tree biodiversity in protected and logged Bornean tropical rain
forests and its measurement by satellite Remote Sensing. Journal of Biogeography, 30 (7): 1053-
1056.
Forman, R. T. T., Godron M. (1986). Landscape Ecology. John Wiley and Sons. New York. 619 pp.
Fosberg, F.R., Garnier B.J. & Küchler A.W. (1961). Delimitation of the humid tropics. Geogr. Rev.
51(3): 333-347.
Fu, B. J., Zhao, W., Chen, l., Liu F. & Lü H. (2005). Eco-hydrological effects of landscape pattern
change. Landscape Ecology Engineering 1: 25-32.
Frohn, R. C. (1998). Remote Sensing for Landscape Ecology – New Metric Indicators for Monitoring,
Modeling and Assessment of Ecosys ems, Department of Geography, University of Cincinnati,
Cincinnati, Ohio.
Gilpin, M. & I. Hanski (1991). Metapopulation Dynamics: Empirical and Theoretical Investigations.
Academic Press, San Diego.
Gippoliti, S. & Dell’Omo, G. (1996). Primates of the Cantanhez Forest and the Cacine Basin, Guinea-
Bissau. Oryx, 30, 74–80.
Groves, C.P. (2001). Prima e Taxonomy. Smithsonian Institution Press, Washington D.C., USA.
Guégan, J. F., Lek, S. & Oberdorff, T. (1998). Energy availability and habitat heterogeneity predict
global riverine fish diversity. Nature, 391 (22): 382-348.
Guisan, A. (2002). Semi-quantitative response models for predicting the spatial distribution of plant
species. Predicting species occurrences: issues of accuracy and scale. Chapter 25:315-326. Island
Press, Covelo, California.
- 60 -

Guisan, A., Zimmermann, N.E. (2000). Predictive habitat distribution models in ecology. Ecological
Modelling 135, 147–186.
Han, K. S., Champeaux & Roujean J.L. (2004). A land cover classification product over France at 1 km
resolution using SPOT4/VEGETATION data. Remote Sensing of Environment. 92 (1): 52-66.
Hansen, M.J., Franklin, S. E., Woudsma, C.G., & Peterson, M. (2001). Caribou habitat mapping and
fragmentation analysis using Landsat Mss, TM, and GIS data in the North Columbia Mountains, British
Columbia, Canada. Remote Sensing of Environment, 77(1): 50-65.
Hirzel, A.H., Hausser, Chessel, D., Perrin, N. (2002). Ecological-niche factor analysis: how to compute
habitat-suitability maps without absence data?. Ecology 83, 2027–2036.
Hirzel, A.H., Hausser, J., Perrin, N. (2004). Biomapper 3.1. Division of Conservation Biology, University
of Bern. Available from: <http://www.unil.ch/biomapper>.
Houghton, R. A. (1994). The worldwide exten of land-use change. BioScience, 44, 305–313. t
I
t r t r
Irons, J. & Masek J. (2006). Requirements for a Landsat Data Continuity Mission, Photogrammetric
Engineering & Remote Sensing, vol. 72, no. 10, pp: 1102–1108.
Jaberg, C. & Guisan, A. (2001). Modeling the distribution of bats in relation to ladscape structure in a
temperate mountain environment. Journal of applied ecology, 38: 1169-1181.
Jansen, L. J. M. & Di Gregorio, A. (2004). Obtaining land-use information from a remotely sensed land
cover map: results from a case study in Lebanon. nternational Journal of Applied Earth Observation
and Geoinformation, 5 (2): 141-157.
Jensen, J. R. & Narumalani, S. (1992). Improved remote sensing and GIS reliability diagrams, image
genealogy diagrams, and thematic map legends to enhance communication. In e na ional A chives of
Photogrammetry and Remote Sensing, vol. 6(B6), 125– 132.
Kati, V., Devillers, P., Dufrene, M., Legakis, A., Vokou, D. & Lebrun, P. (2004). Hotspots,
complementarity or representativeness ? Designing optimal small-scale reserves for biodiversity
conservation. Biological Conservation. 120 (4): 471-480.
Keele (2006). Chimpanzee Reservoirs of Pandemic and Nonpandemic HIV-1. Scienceexpress, vol. 45,
231-324.
- 61 -

Kormos, R., Boesch, C (2003). Regional Action Plan for the Conservation of Chimpanzees in West
Africa. Center for Applied Biodiversity Science, Washington, DC.
Laurent, E. J., Shi, H., Gatziolis, D., LeBouton, J.P., Walters, M.B. & Liu, J. (2005). Using the spatial
and spectral precision of satellite imagery to predict wildlife occurrence patterns. Remote Sensing of
Environment, 97 (2): 249-262.
Lee, P.C., Thornback, J. & Bennett, E.L. (1988). Threatened Primates of Africa. The IUCN Red Data
Book. IUCN, Gland, Switzerland.
t t t t
t t
Limoges (1989). Resul ats de L’inven aire Faunis ique au Niveau National et Proposi ions de
Modification de la Loi sur la Chasse. DGFC/CECI/IUCN, Bissau.
Lobo, J. M., Jay-Robert, P. & Lumaret, J. P. (2004). Modelling the species richness distribution for
French Aphodiidae. Ecography. 27 (2):145-156.
Lourenço E. (1994). A las da língua por uguesa na história e no mundo. Imprensa Nacional Casa da
Moeda, Lisboa.
Machado J. S. S. (1972). Balanço hídrico da Província da Guiné. Boletim Cultural da Guiné Portuguesa
27(108): 753-779.
Maes, D., Gilbert, M., Titeux, N., Goffart, P. & Dennis, R. H. (2003). Prediction of butterfly diversity
hotspots in Belgium : a comparison of statistically focused and land use-focused models. Journal of
Biogeography. 30 (12): 1907-1920.
Marchesi P., Marchesi N., Fruth B. & Boesch C. (1995). Census and distribution of chimpanzees in
Cote d’Ivoire. Primates 36:591–607.
Marsh, C.W., Johns, A.D. & Ayres, J.M. (1987). Effects of habitat disturbance on rain forest primates.
Monographs in Primatology (Eds C.W. Marsh & R.A. Mittermeier). Alan R. Liss, pp. 83–107.
McGarigal, K., & Marks B. J. (1995). FRAGSTATS: spatial pattern analysis program for quantifying
landscape structure. USDA For. Serv. Gen. Tech. Rep. PNW-351.
McGarigal, K., Cushman S. A., Neel, M. C. & Ene E. (2002). FRAGSTATS: Spatial Pattern Analysis
Program for Categorical Maps. Computer software program produced by the authors at the University
of Massachusetts, Amherst.
- 62 -

McGuffie, K., Henderson-Sellers, A., Zhang, H., Durbidge, T. B., & Pitman, A. J. (1995). Global climate
sensitivity to tropical deforestation. Global and Planetary Change, 10, 97–128.
Mladenoff, D. J., Sickley, T. A. & Wydeven, A. P. (1999). Predicting gray wolf landscape
recolonization: Logistic regression models vs. new field data. Ecological Applications. 9 (1): 37-44.
Moreira, J., Schwabisch, M., Fornaro, G., Lanari, R., Bamler, R., Just, D., Steinbrecher, U., Breit, H.,
Eineder, M., Granceschetti, G., Geudtner. D., & Rinkel, H.( 1995). X-SAR interferometry: First results.
IEEE Transactions on Geoscience and Remote Sensing 33(4):950-956.
Myers, N., (1993). Tropical forests: the main deforestation fronts. Environmen al Conservation, 20, 9–
16.
t
Myers, N., Mittermeier, da Fonseca, G,B. & Kent, J. (2000). Biodiversity hotspots for conservation
priorities. Nature, 403 (6772):853-858.
Napier, J.R. & Napier, P.H. (1967). A Handbook of Living Primates. Academic Press, New York, USA.
Nunes, C., & Auge, J. I. (1999). Land-Use and Land-Cover Implementation Strategy. Stockholm: IGBP.
Oates J. F. (2006). Is the chimpanzee, Pan troglodytes, an endangered species? It depends on what
‘‘endangered’’ means. Primates 47: 102–112.
Oldfield, F., Dearing, J. A., Gaillard, M. J. & Bugmann, H. (2000). Ecosystem processes and human
dimensions - The scope and the future of HITE (Human Impacts on Terrestrial Ecosystems). Pages
Newsletter, 8, 21–23.
O’Neill, R.V., DeAngelis, D.L., Waide, J.B., Allen, T.F.H. (1986). A Hierarchical Concepte of
Ecosystems. Princeton University Press, Princeton, NJ.
O'Neill, R. V., Krummel, J. R., Gardner, R. H., Sugihara, G. B., Jackson, D. L., DeAngelis, B. T., Milne,
Turner M., Zygmunt, B., Christensen, Dale, V. H. & Graham, R. L. (1988). Indices of landscape
pattern. Landscape Ecology 1:153-162.
Ortega M., Elena-Roselló R., García Del Barrio J.M. (2004). Estimation of plant diversity at landscape
level. Environmental Monitoring and Assessment 95: 97-116.
- 63 -

Pereira, J. M. C. & Itami, R.M. (1991). GIS-based habitat modelling using logistic multiple regression:
a study of the Mt. Graham Red Squirrel. Photogrammetric Engineering & Remote Sensing. 57 (11):
1475-1486.
Pimm, S.L. (1986). Community stability and structure. Conservation Biology: the Science of Scarcity
and Diversity (Ed. M.E. Soule). Sinauer Associates, Sunderland, MA, pp. 309–329.
Primack, R.B. (2002). Essentials of Conservation Biology. Sinauer Associates, Ed.
Rogers, D. J., Randolph, S.E., Snow, R. W. & Hay, S. (2002). Satellite imagery in the study and
forecast of malaria. Nature. 415 (6872): 710-715.
Root, K. V., Akçakaya, R. & Ginzburg. L. (2003). A Multispecies Approach to Ecological Valuation and
Conservation. Conservation Biology. 17 (1): 196-206.
Roth, N. E., Allan, J. D. & Erickson D. L. (1996). Landscape influences on stream biotic integrity
assessed at multiple spatial scales. Lanscape Ecology 11(3): 141-156.
Schowengerdt Robert (2007). Remote Sensing 3rd. Academic Press. 515pp.
Silva, A., Cardoso, L. (2001). Um Tipo de Área protegida para Cantanhez. Gabinete de Planificação
Costeira, (UICN), Bissau.
Spangberg, J.H., Pfahl, S. & Deller, K. (2002). Towards indicators for institutional sustainability:
lessons from an analysis of Agenda 21. Ecological Indicators. 2 (1-2): 61-77.
Schreier, H. & Brown S. (2003). Building bridges between communities in watershed management.
Proceedings of the II Symposium on MMSEA, pp. 373-379. Kunming: Yunnan Science and Technology
Press.
Tatem, A. J., Baylis, M., Mellor, P. S., Purse, B. V., Capela, R., Pena, I. & Rogers, D. J. (2003).
Prediction of bluetongue vector distribution in Europe and north Africa using satellite imagery.
Veterinary Microbiology, 97 (1-2): 13-29.
Temudo, M. P. (1998). Inovação e mudança em sociedades rurais africanas. Gestão de recursos
naturais, saber local e insti uições de desenvolvimento. DissertaçãoDoutoramento ISA/UTL, Lisboa. t
Temudo, M. P. & Schiefer U. (2003) Disintegration and Resilience of Agrarian Societies in Africa.
Current Sociology, vol. 51(3/4): 393-416.
- 64 -

Thuiller, W., Araújo, M. B. & Lavorel, S. (2003). Generalized models vs. classification tree analysis:
Predicting spatial distributiona of plant species at different scales. Journal of Vegetation Science. 14
(5): 669-680.
Thuiller, W., Araújo, A. M., Brotons, L. & Lavorel, S. (2004). Effects of restricting environmental range
of dat to project current and future species distributions. Ecography. 27: 165-172.
Tutin, C., Stokes, E., Boesch, C., Morgan, D., Sanz, C., Reed, T., Blom, A., Walsh, P., Blake, S. &
Kormos, R. (2005). Regional Action Plan for the Conservation of Chimpanzees and Gorillas in Western
Equatorial A rica. Washington DC: Center for Applied Biodiversity Science at Conservation
International.
f
Turner, M. G., Ruscher, C. L. (1988). Changes in landscape patterns in Georgia, USA. Landscape
Ecology 1:241 – 258.
Turner, M. G. (1990). Spatial and temporal analysis of landscape patterns. Landscape Ecology 4:21-30
Turner, M.G., Gardner R. H. & O'Neill R. V. (2001). Landscape Ecology in Theory and Practice.
Springer-Verlag, New York, USA.
Venier, L. A., Pearce, J., Mckee, E., Mckenney, D. W. & Niemi, G. J. (2004). Climate and satellite-
derived land cover for predicting breeding bird distribution in the Great Lakes Basin. Journal of
Biogeography. 31 (2): 315-331.
Virkkala, R., Luoto, M, Heikkinen, R. & Leikola, N. (2005). Distribution patterns of boreal marshland
birds: modelling the relationships to land cover and climate. Journal of Biogeography. 32 (11): 1957-
1970.
Vitousek, P. M. (1992). Global environmental change: an introduction. Annual Review in Ecology and
Systematics, 23: 1–14.
Wang, H. G., Owen, R. D., Sánchez-Hernández, C. & De Lourdes Romero-Almaraz, M. (2003).
Ecological characterization of bat species distributions in Michoacán, México, using a geographic
information system. Global Ecology and Biogeography, 12 (1): 65-85.
Walker, B. H. & Steffen, W. L. (1996). GCTE science: objectives, structure and implementation. Global
Change and Terrestrial Ecosystems, Eds. Walker, B. & SteVen, W. Cambridge University Press.
- 65 -

Werner, M. (2001). Shuttle Radar Topography Mission (SRTM), Mission overview. Journal of
Telecommunication (Frequenz). 55: 75-79.
Werner, M. (2001). Status of the SRTM data processing: when will the world-wide 30 m DTM data be
available? Photogrammetic Week. 1:159-165.
White, L.J.T. (1994). Biomass of rainforest mammals in the Lope reserve, Gabon. J. Anim. Ecol. 63,
499-512.
Referências URL
http://srtm.usgs.gov/
www.umass.edu/landeco/research/fragstats/fragstats.html
http://www.unil.ch/biomapper
http://flash.lakeheadu.ca/~rrempel/patch/
http://www.wikipedia.org/
- 66 -

Anexos:
Tabela I: Percentagem de concordância (dados independentes) (Cassama, 2006)
Tabela II: Percentagem de concordância (dados de treino) (Cassama, 2006)

Figura I: Aspectos da biodiverdidade da área de estudo

Figura II: Fotografias de Pan troglodytes verus na área de estudo.





























