Embed Size (px)
Citation preview

Desenvolvimento de Organismos Multicelulares 22Um animal ou planta inicia a sua vida como uma célula única – um óvulo fertilizado. Du-rante o desenvolvimento, esta célula divide-se repetidamente para produzir muitas células diferentes em um padrão final de complexidade e precisão espetaculares. Em última análise, o genoma determina o padrão, e o quebra-cabeça da biologia do desenvolvimento é enten-der como ele o faz.
O genoma normalmente é idêntico em todas as células; as células diferem não porque contenham informações genéticas diferentes, mas porque expressam conjuntos diferentes de genes. Esta expressão genética seletiva controla os quatro processos essenciais de cons-trução do embrião: (1) proliferação celular, produção de muitas células a partir de uma, (2) especialização celular, criação de células com diferentes características em diferentes posi-ções, (3) interações celulares, coordenação do comportamento de uma célula com o de suas vizinhas, e (4) movimentos celulares, rearranjo das células para formar tecidos e órgãos es-truturados (Figura 22-1).
Em um embrião em desenvolvimento, todos os processos estão acontecendo ao mes-mo tempo, em uma variedade caleidoscópica de maneiras diferentes, em partes distintas do organismo. Para entender as estratégias básicas do desenvolvimento, teremos que limitar o nosso foco. Em particular, precisamos entender o curso de eventos a partir do ponto de vista de uma célula individual e como o genoma atua nessa célula. Não há um oficial em coman-do mantendo-se fora do combate para direcionar as tropas; cada uma das milhões de células no embrião precisa tomar as suas próprias decisões, de acordo com a sua própria cópia de instruções genéticas e suas circunstâncias particulares.
A complexidade dos animais e das plantas depende de uma característica extraordiná-ria do sistema de controle genético. As células possuem uma memória: os genes que uma célula expressa e a maneira como ela se comporta dependem do seu passado e do seu am-biente presente. As células do corpo – as células musculares, os neurônios, as células da pele, as células do intestino, e assim por diante – mantêm as suas características especializadas não porque elas recebem continuamente as mesmas instruções do seu meio, mas porque elas retêm um registro dos sinais que as suas ancestrais receberam em um desenvolvimento embrionário inicial. Os mecanismos moleculares de memória celular foram introduzidos no Capítulo 7. Neste capítulo abordaremos as suas consequências.
MECANISMOS UNIVERSAIS DE DESENVOLVIMENTOANIMALExistem em torno de 10 milhões de espécies de animais, e eles são fantasticamente variados. Ninguém espera que o verme, a mosca, a águia e a lula gigante tenham sido gerados pelos mesmos mecanismos de desenvolvimento, assim como não se espera que os mesmos méto-dos tenham sido usados para fazer um sapato e um avião. Alguns princípios similares abstra-tos devem estar envolvidos, talvez, mas com certeza não as mesmas moléculas específicas.
Uma das revelações mais impressionantes dos últimos 10 ou 20 anos foi que as nossas suspeitas iniciais estavam erradas. De fato, muito da maquinaria básica de desenvolvimento é essencialmente a mesma, não somente em todos os vertebrados, mas também em todos os maiores filos de invertebrados. As moléculas reconhecidamente semelhantes e evoluti-vamente relacionadas definem nossos tipos celulares especializados, marcam as diferen-ças entre as regiões do corpo e auxiliam a criar o padrão corporal. As proteínas homólogas são, com frequência, funcionalmente intercambiáveis entre espécies muito diferentes. Uma proteína de camundongos produzida de maneira artificial em uma mosca pode, frequente-mente, realizar a mesma função da própria versão da proteína da mosca, e vice-versa, con-
Neste capítuloMECANISMOS 1305UNIVERSAIS DE DESENVOLVIMENTO ANIMAL
CAENORHABDITIS 1321ELEGANS: O DESENVOLVIMENTO A PARTIR DA PERSPECTIVA DE UMA CÉLULA INDIVIDUAL
DROSOPHILA E A 1328GENÉTICA MOLECULAR DA FORMAÇÃO DE PADRÕES: A GÊNESE DO PLANO CORPORAL
GENES SELETORES 1341HOMEÓTICOS E A FORMAÇÃO DE PADRÕES DO EIXO ÂNTEROPOSTERIOR
ORGANOGÊNESE E 1347A FORMAÇÃO DOS PADRÕES DOS ÓRGÃOS ACESSÓRIOS
MOVIMENTOS 1363CELULARES E A DETERMINAÇÃO DA FORMA DO CORPO DOS VERTEBRADOS
O CAMUNDONGO 1378
DESENVOLVIMENTO 1383NEURAL
DESENVOLVIMENTO 1398VEGETAL
Alberts_22.indd 1305Alberts_22.indd 1305 29.07.09 16:49:1229.07.09 16:49:12
1306 Alberts, Johnson, Lewis, Raff, Roberts & Walter
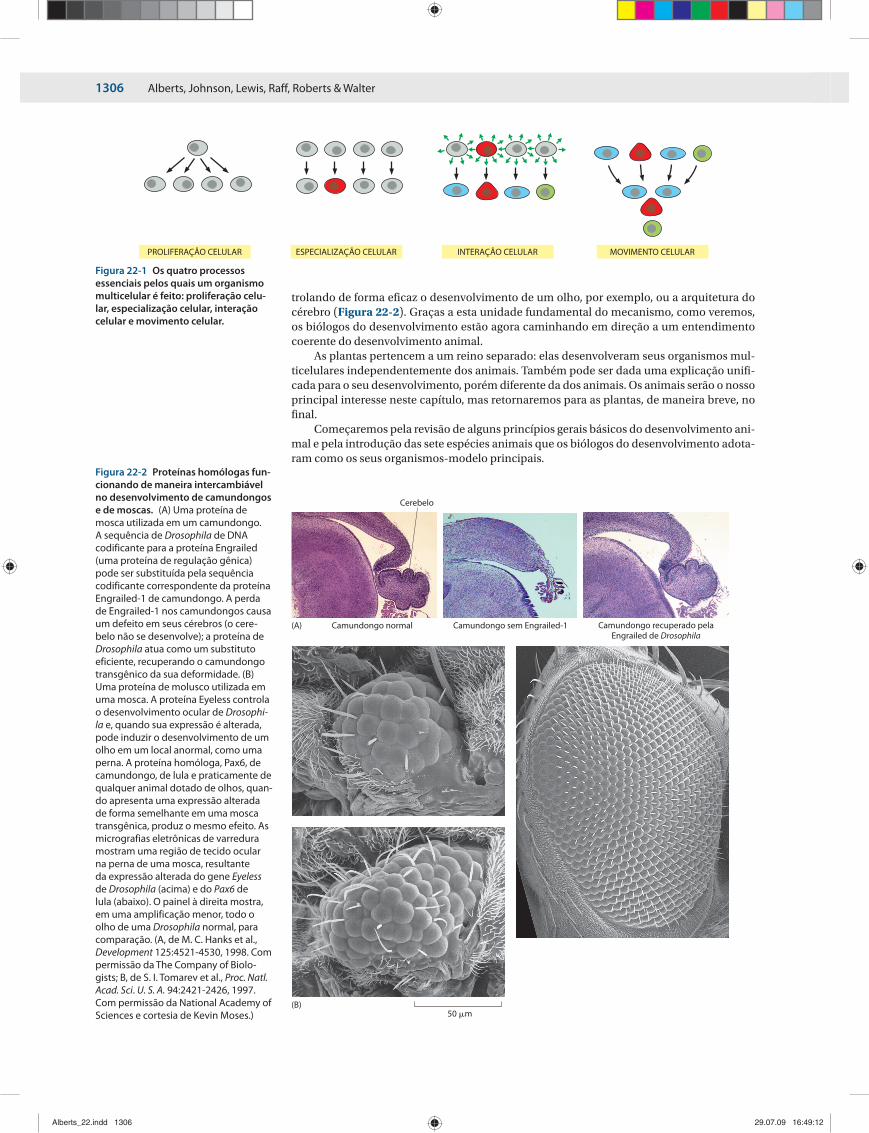
trolando de forma eficaz o desenvolvimento de um olho, por exemplo, ou a arquitetura do cérebro (Figura 22-2). Graças a esta unidade fundamental do mecanismo, como veremos, os biólogos do desenvolvimento estão agora caminhando em direção a um entendimento coerente do desenvolvimento animal.
As plantas pertencem a um reino separado: elas desenvolveram seus organismos mul-ticelulares independentemente dos animais. Também pode ser dada uma explicação unifi-cada para o seu desenvolvimento, porém diferente da dos animais. Os animais serão o nosso principal interesse neste capítulo, mas retornaremos para as plantas, de maneira breve, no final.
Começaremos pela revisão de alguns princípios gerais básicos do desenvolvimento ani-mal e pela introdução das sete espécies animais que os biólogos do desenvolvimento adota-ram como os seus organismos-modelo principais.
Figura 22-1 Os quatro processos essenciais pelos quais um organismo multicelular é feito: proliferação celu-lar, especialização celular, interação celular e movimento celular.
PROLIFERAÇÃO CELULAR ESPECIALIZAÇÃO CELULAR INTERAÇÃO CELULAR MOVIMENTO CELULAR
Figura 22-2 Proteínas homólogas fun-cionando de maneira intercambiável no desenvolvimento de camundongos e de moscas. (A) Uma proteína de mosca utilizada em um camundongo. A sequência de Drosophila de DNA codificante para a proteína Engrailed (uma proteína de regulação gênica) pode ser substituída pela sequência codificante correspondente da proteína Engrailed-1 de camundongo. A perda de Engrailed-1 nos camundongos causa um defeito em seus cérebros (o cere-belo não se desenvolve); a proteína de Drosophila atua como um substituto eficiente, recuperando o camundongo transgênico da sua deformidade. (B) Uma proteína de molusco utilizada em uma mosca. A proteína Eyeless controla o desenvolvimento ocular de Drosophi-la e, quando sua expressão é alterada, pode induzir o desenvolvimento de um olho em um local anormal, como uma perna. A proteína homóloga, Pax6, de camundongo, de lula e praticamente de qualquer animal dotado de olhos, quan-do apresenta uma expressão alterada de forma semelhante em uma mosca transgênica, produz o mesmo efeito. As micrografias eletrônicas de varredura mostram uma região de tecido ocular na perna de uma mosca, resultante da expressão alterada do gene Eyeless de Drosophila (acima) e do Pax6 de lula (abaixo). O painel à direita mostra, em uma amplificação menor, todo o olho de uma Drosophila normal, para comparação. (A, de M. C. Hanks et al., Development 125:4521-4530, 1998. Com permissão da The Company of Biolo-gists; B, de S. I. Tomarev et al., Proc. Natl. Acad. Sci. U. S. A. 94:2421-2426, 1997. Com permissão da National Academy of Sciences e cortesia de Kevin Moses.)
Cerebelo
Camundongo normal(A)
(B)
Camundongo sem Engrailed-1 Camundongo recuperado pelaEngrailed de Drosophila
50 �m
Alberts_22.indd 1306Alberts_22.indd 1306 29.07.09 16:49:1229.07.09 16:49:12
Biologia Molecular da Célula 1307
Os animais compartilham algumas características anatômicas básicasAs semelhanças entre as espécies animais em relação aos genes que controlam o desen-volvimento refletem a evolução dos animais a partir de um ancestral comum no qual esses genes já estavam presentes. Embora não saibamos como ele se parecia, o ancestral comum dos vermes, dos moluscos, dos insetos, dos vertebrados e de outros animais complexos ti-nha, necessariamente, muitos tipos celulares diferenciados que seriam reconhecidos por nós: células epidérmicas, por exemplo, formando uma camada externa protetora; células do intestino para absorver nutrientes da comida ingerida; células musculares para mover-se; neurônios e células sensoriais para controlar os movimentos. O corpo deve ter sido organi-zado com uma camada de pele cobrindo o exterior, uma boca para a alimentação e um tubo intestinal para reter e processar a comida – com músculos, nervos e outros tecidos arranja-dos no espaço entre a camada externa de pele e o tubo intestinal interno.
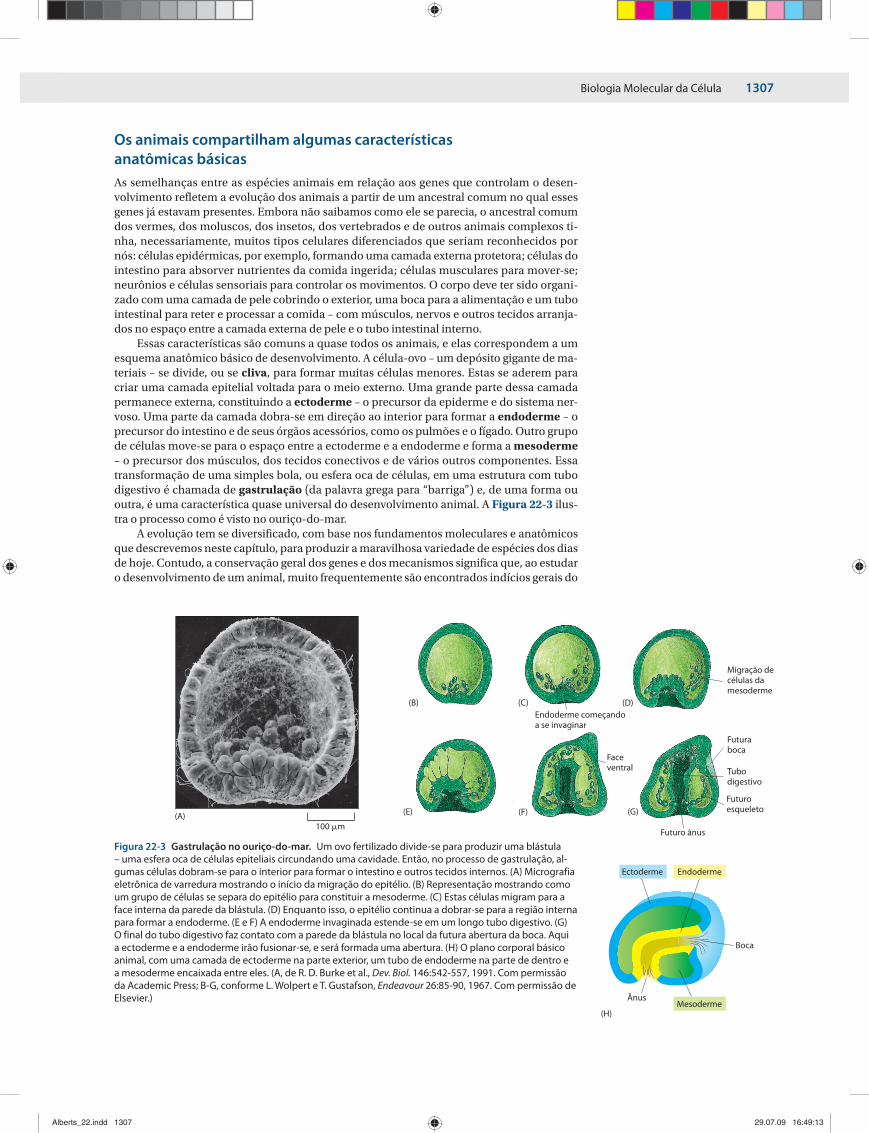
Essas características são comuns a quase todos os animais, e elas correspondem a um esquema anatômico básico de desenvolvimento. A célula-ovo – um depósito gigante de ma-teriais – se divide, ou se cliva, para formar muitas células menores. Estas se aderem para criar uma camada epitelial voltada para o meio externo. Uma grande parte dessa camada permanece externa, constituindo a ectoderme – o precursor da epiderme e do sistema ner-voso. Uma parte da camada dobra-se em direção ao interior para formar a endoderme – o precursor do intestino e de seus órgãos acessórios, como os pulmões e o fígado. Outro grupo de células move-se para o espaço entre a ectoderme e a endoderme e forma a mesoderme – o precursor dos músculos, dos tecidos conectivos e de vários outros componentes. Essa transformação de uma simples bola, ou esfera oca de células, em uma estrutura com tubo digestivo é chamada de gastrulação (da palavra grega para “barriga”) e, de uma forma ou outra, é uma característica quase universal do desenvolvimento animal. A Figura 22-3 ilus-tra o processo como é visto no ouriço-do-mar.
A evolução tem se diversificado, com base nos fundamentos moleculares e anatômicos que descrevemos neste capítulo, para produzir a maravilhosa variedade de espécies dos dias de hoje. Contudo, a conservação geral dos genes e dos mecanismos significa que, ao estudar o desenvolvimento de um animal, muito frequentemente são encontrados indícios gerais do
Figura 22-3 Gastrulação no ouriço-do-mar. Um ovo fertilizado divide-se para produzir uma blástula – uma esfera oca de células epiteliais circundando uma cavidade. Então, no processo de gastrulação, al-gumas células dobram-se para o interior para formar o intestino e outros tecidos internos. (A) Micrografia eletrônica de varredura mostrando o início da migração do epitélio. (B) Representação mostrando como um grupo de células se separa do epitélio para constituir a mesoderme. (C) Estas células migram para a face interna da parede da blástula. (D) Enquanto isso, o epitélio continua a dobrar-se para a região interna para formar a endoderme. (E e F) A endoderme invaginada estende-se em um longo tubo digestivo. (G) O final do tubo digestivo faz contato com a parede da blástula no local da futura abertura da boca. Aqui a ectoderme e a endoderme irão fusionar-se, e será formada uma abertura. (H) O plano corporal básico animal, com uma camada de ectoderme na parte exterior, um tubo de endoderme na parte de dentro e a mesoderme encaixada entre eles. (A, de R. D. Burke et al., Dev. Biol. 146:542-557, 1991. Com permissão da Academic Press; B-G, conforme L. Wolpert e T. Gustafson, Endeavour 26:85-90, 1967. Com permissão de Elsevier.)
(A)
(B) (D)(C)
(F) (G)(E)
(H)
100 �m
Endoderme começandoa se invaginar
Faceventral
Migração decélulas damesoderme
Futuraboca
Tubodigestivo
Futuroesqueleto
Futuro ânus
Ectoderme Endoderme
Boca
ÂnusMesoderme
Alberts_22.indd 1307Alberts_22.indd 1307 29.07.09 16:49:1329.07.09 16:49:13

1308 Alberts, Johnson, Lewis, Raff, Roberts & Walter
desenvolvimento de vários outros tipos de animais. Como resultado, os biólogos do desen-volvimento da atualidade, assim como os biólogos celulares, podem se dar ao luxo de estu-dar questões fundamentais na espécie que ofereça o caminho mais fácil para uma resposta.
Os animais multicelulares são ricos em proteínas que fazem a mediação das interações celulares e da regulação gênicaO sequenciamento de genomas revela a extensão das semelhanças moleculares entre as es-pécies. O verme nematoide Caenorhabditis elegans, a mosca Drosophila melanogaster e o vertebrado Homo sapiens são os primeiros três animais para os quais foi obtida a sequência completa do genoma. Na árvore familiar da evolução animal, eles estão muito distantes uns dos outros: a linhagem que leva aos vertebrados divergiu daquela que leva aos nematoides, aos insetos e aos moluscos mais de 600 milhões de anos atrás. Apesar disso, quando os 20 mil genes do C. elegans, os 14 mil genes da Drosophila e os 25 mil genes dos humanos são sistematicamente comparados uns com os outros, é observado que em torno de 50% dos genes de cada uma das espécies possuem homólogos claramente reconhecíveis em uma ou nas outras duas espécies. Em outras palavras, as versões reconhecíveis de pelo menos 50% de todos os genes humanos já estavam presentes no ancestral comum dos vermes, das moscas e dos humanos.
Obviamente, nem tudo é conservado: existem alguns genes com funções-chave no de-senvolvimento de vertebrados que não possuem homólogos no genoma de C. elegans ou de Drosophila, e vice-versa. Entretanto, o fato de existir uma grande proporção dos 50% dos genes que não possui homólogos identificáveis em outros filos não significa, simplesmente, que as suas funções são de menor importância. Embora esses genes não-conservados sejam transcritos e bem-representados em bibliotecas de DNA complementar (cDNA), os estudos de variabilidade de sequências de DNA e de aminoácidos dentro e entre as populações natu-rais indicam que esses genes podem, excepcionalmente, sofrer mutações sem comprometer seriamente a adaptabilidade; quando são inativados artificialmente, as consequências não são tão severas quanto seriam no caso dos genes que possuem homólogos em espécies que apresentam relações distantes. Uma vez que estes genes são livres para evoluir rapidamente, algumas dezenas de milhões de anos podem ser suficientes para destruir qualquer seme-lhança familiar, ou para permitir a sua perda do genoma.
Os genomas de diferentes classes de animais diferem também porque, como discutido no Capítulo 1, existem variações substanciais na extensão das duplicações gênicas: a quanti-dade de duplicações gênicas na evolução dos vertebrados tem sido particularmente grande; como resultado, um mamífero ou um peixe frequentemente possuem vários homólogos que correspondem a um único gene em um verme ou em uma mosca.
Apesar de tais diferenças, em uma primeira análise, podemos dizer que todos esses animais possuem um conjunto semelhante de proteínas a sua disposição para as suas funções-chave. Em outras palavras, eles constroem seus corpos usando, de maneira geral, o mesmo conjunto de partes moleculares.
Quais genes, então, são necessários para produzir um animal multicelular, além daque-les necessários para produzir uma única célula? A comparação dos genomas de animais com o de leveduras que se reproduzem por brotamento – um eucarioto unicelular – sugere que duas classes de proteínas são especialmente importantes para a organização multicelular. A primeira classe é a das moléculas transmembrana usadas para a adesão e a sinalização celular. Em torno de 2.000 genes de C. elegans codificam receptores de superfície celular, pro-teínas de adesão celular e canais iônicos que estão ausentes na levedura, ou presentes em número muito menor. A segunda classe é a das proteínas de regulação gênica: estas proteínas de ligação ao DNA são muito mais numerosas no genoma de C. elegans do que no de leve-dura. Por exemplo, a família básica hélice-alça-hélice possui 41 membros em C. elegans, 84 na Drosophila, 131 nos humanos e somente 7 nas leveduras, e outras famílias de regulado-res da expressão gênica também são dramaticamente superexpressas nos animais, quando comparadas a leveduras. Não é surpresa, portanto, que essas duas classes de proteínas sejam centrais para a biologia do desenvolvimento: como veremos, o desenvolvimento de animais multicelulares é dominado por interações célula-célula e pela expressão gênica diferencial.
Como discutido no Capítulo 7, microRNAs (miRNAs) também têm um papel significa-tivo no controle da expressão gênica durante o desenvolvimento, mas parecem ser de im-portância secundária quando comparados às proteínas. Dessa forma, um embrião mutante de peixe-zebra que não expresse a proteína Dicer, que é necessária à produção de miRNAs
Alberts_22.indd 1308Alberts_22.indd 1308 29.07.09 16:49:1329.07.09 16:49:13

Biologia Molecular da Célula 1309
funcionais, ainda iniciará o seu desenvolvimento quase normalmente, originando tipos ce-lulares especializados e uma organização do plano corporal mais ou menos correta, antes que as anomalias se tornem severas.
O DNA regulador define o programa de desenvolvimentoUm verme, uma mosca, um molusco e um mamífero compartilham muitos dos mesmos ti-pos celulares essenciais, e todos são dotados de uma boca, um intestino, um sistema nervoso e uma pele; contudo, além de umas poucas características básicas, eles parecem radicalmen-te diferentes em sua estrutura corporal. Se o genoma determina a estrutura do corpo e todos esses animais possuem esta coleção similar de genes, como podem ser tão diferentes?
As proteínas codificadas no genoma podem ser vistas como os componentes de um conjunto de ferramentas de construção. Muitas coisas podem ser construídas com este con-junto, assim como um conjunto de ferramentas de construção de crianças pode ser usado para fazer caminhões, casas, pontes, guindastes, e assim por diante, pela associação dos componentes em diferentes combinações. Alguns elementos, necessariamente, vão juntos – porcas com parafusos, rodas com pneus e eixos – mas a organização em grande escala do objeto final não é definida por essas estruturas. Ao contrário, ela é definida pelas instruções que acompanham os componentes e descrevem como eles devem ser montados.
Em grande parte, as instruções necessárias para produzir um animal multicelular es-tão contidas no DNA regulador não-codificante associado a cada gene. Como discutido no Capítulo 4, cada gene em um organismo multicelular está associado a milhares ou deze-nas de milhares de nucleotídeos de DNA não-codificante. Este DNA pode conter, dispersas nele, dúzias de elementos reguladores separados ou estimuladores – pequenos segmentos de DNA que servem como sítios de ligação para complexos específicos de proteínas de re-gulação gênica. Em termos gerais, como explicado no Capítulo 7, a presença de um dado módulo regulador desse tipo leva à expressão do gene sempre que o complexo de proteínas que reconhecem aquele segmento de DNA esteja apropriadamente montado na célula (em alguns casos, uma inibição ou um efeito mais complicado na expressão gênica é produzido em seu lugar). Se pudéssemos decifrar o conjunto completo de módulos reguladores asso-ciados a um gene, entenderíamos todas as condições moleculares diferentes sob as quais os produtos daquele gene devem ser produzidos. Este DNA regulador pode, assim, ser consi-derado como o definidor do programa sequencial de desenvolvimento: as regras passam de um estado para o próximo, enquanto as células proliferam e leem suas posições no embrião pela relação com as suas adjacências, ativando novos conjuntos de genes de acordo com as atividades das proteínas que elas correntemente contêm (Figura 22-4). Variações nas pró-prias proteínas, obviamente, também contribuem para as diferenças entre as espécies. No entanto, mesmo que o conjunto de proteínas codificado pelo genoma se mantenha com-pletamente inalterado, a variação no DNA regulador seria suficiente para originar tecidos e estruturas corporais radicalmente distintos.
Quando comparamos espécies animais com planos corporais semelhantes – diferentes vertebrados, como um peixe, um pássaro e um mamífero, por exemplo – observamos que os genes correspondentes normalmente possuem conjuntos semelhantes de módulos regula-dores: as sequências de DNA de muitos módulos individuais têm sido bem conservadas e são reconhecidas como homólogas nos diferentes animais. O mesmo é verdade se compa-ramos diferentes espécies de vermes nematoides ou diferentes espécies de insetos. Contu-do, quando comparamos regiões reguladoras de vertebrados com aquelas de vermes ou de
Figura 22-4 Como o DNA regulador define a sucessão de padrões de expressão gênica no desenvolvimen-to. Os genomas dos organismos A e B codificam o mesmo conjunto de proteí-nas, porém possuem DNAs reguladores diferentes. As duas células na figura co-meçam no mesmo estado, expressando as mesmas proteínas no estágio 1, mas passam para estados bem diferentes no estágio 2, devido ao arranjo distinto de módulos reguladores.
Estágio embrionário 2
Estágio embrionário 1
Gene 1 Gene 2 Gene 3
Gene 1 Gene 2 Gene 3
Módulos reguladores
TEMPO
Estágio embrionário 2
Estágio embrionário 1
Gene 1 Gene 2 Gene 3
Gene 1 Gene 2 Gene 3
TEMPO
Proteínareguladorado gene
CÉLULA NO ORGANISMO A CÉLULA NO ORGANISMO B RELACIONADO
Alberts_22.indd 1309Alberts_22.indd 1309 29.07.09 16:49:1329.07.09 16:49:13
1310 Alberts, Johnson, Lewis, Raff, Roberts & Walter
moscas, é difícil ver qualquer tipo de semelhança. As sequências codificantes de proteínas são indubitavelmente semelhantes, mas as sequências correspondentes de DNA regulador mostram-se muito diferentes. Este é o resultado esperado se diferentes planos de corpo são produzidos principalmente pela alteração do programa incorporado no DNA regulador, em-bora retendo a maior parte do mesmo conjunto de proteínas.
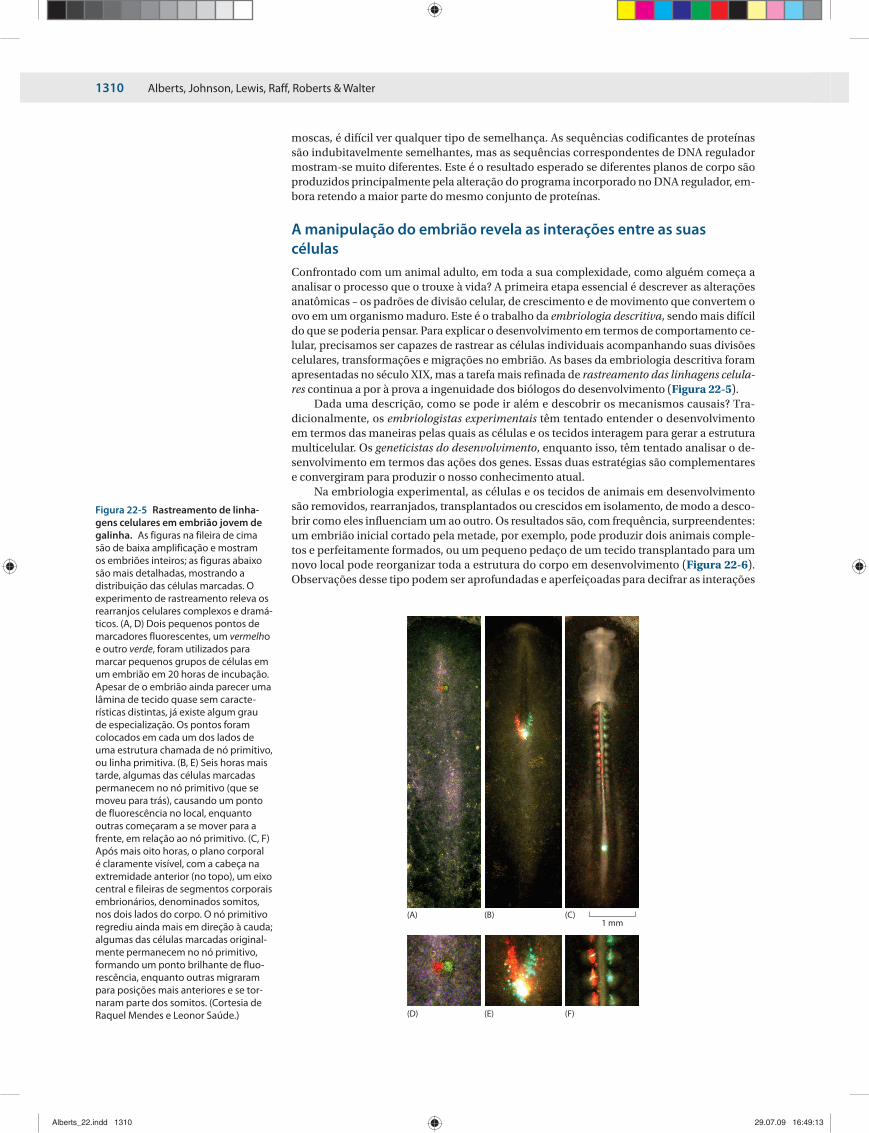
A manipulação do embrião revela as interações entre as suas célulasConfrontado com um animal adulto, em toda a sua complexidade, como alguém começa a analisar o processo que o trouxe à vida? A primeira etapa essencial é descrever as alterações anatômicas – os padrões de divisão celular, de crescimento e de movimento que convertem o ovo em um organismo maduro. Este é o trabalho da embriologia descritiva, sendo mais difícil do que se poderia pensar. Para explicar o desenvolvimento em termos de comportamento ce-lular, precisamos ser capazes de rastrear as células individuais acompanhando suas divisões celulares, transformações e migrações no embrião. As bases da embriologia descritiva foram apresentadas no século XIX, mas a tarefa mais refinada de rastreamento das linhagens celula-res continua a por à prova a ingenuidade dos biólogos do desenvolvimento (Figura 22-5).
Dada uma descrição, como se pode ir além e descobrir os mecanismos causais? Tra-dicionalmente, os embriologistas experimentais têm tentado entender o desenvolvimento em termos das maneiras pelas quais as células e os tecidos interagem para gerar a estrutura multicelular. Os geneticistas do desenvolvimento, enquanto isso, têm tentado analisar o de-senvolvimento em termos das ações dos genes. Essas duas estratégias são complementares e convergiram para produzir o nosso conhecimento atual.
Na embriologia experimental, as células e os tecidos de animais em desenvolvimento são removidos, rearranjados, transplantados ou crescidos em isolamento, de modo a desco-brir como eles influenciam um ao outro. Os resultados são, com frequência, surpreendentes: um embrião inicial cortado pela metade, por exemplo, pode produzir dois animais comple-tos e perfeitamente formados, ou um pequeno pedaço de um tecido transplantado para um novo local pode reorganizar toda a estrutura do corpo em desenvolvimento (Figura 22-6). Observações desse tipo podem ser aprofundadas e aperfeiçoadas para decifrar as interações
Figura 22-5 Rastreamento de linha-gens celulares em embrião jovem de galinha. As figuras na fileira de cima são de baixa amplificação e mostram os embriões inteiros; as figuras abaixo são mais detalhadas, mostrando a distribuição das células marcadas. O experimento de rastreamento releva os rearranjos celulares complexos e dramá-ticos. (A, D) Dois pequenos pontos de marcadores fluorescentes, um vermelho e outro verde, foram utilizados para marcar pequenos grupos de células em um embrião em 20 horas de incubação. Apesar de o embrião ainda parecer uma lâmina de tecido quase sem caracte-rísticas distintas, já existe algum grau de especialização. Os pontos foram colocados em cada um dos lados de uma estrutura chamada de nó primitivo, ou linha primitiva. (B, E) Seis horas mais tarde, algumas das células marcadas permanecem no nó primitivo (que se moveu para trás), causando um ponto de fluorescência no local, enquanto outras começaram a se mover para a frente, em relação ao nó primitivo. (C, F) Após mais oito horas, o plano corporal é claramente visível, com a cabeça na extremidade anterior (no topo), um eixo central e fileiras de segmentos corporais embrionários, denominados somitos, nos dois lados do corpo. O nó primitivo regrediu ainda mais em direção à cauda; algumas das células marcadas original-mente permanecem no nó primitivo, formando um ponto brilhante de fluo-rescência, enquanto outras migraram para posições mais anteriores e se tor-naram parte dos somitos. (Cortesia de Raquel Mendes e Leonor Saúde.)
1 mm(C)(B)(A)
(F)(E)(D)
Alberts_22.indd 1310Alberts_22.indd 1310 29.07.09 16:49:1329.07.09 16:49:13

Biologia Molecular da Célula 1311
básicas célula-célula e as regras do comportamento celular. Os experimentos são mais fáceis de serem realizados em grandes embriões que sejam prontamente acessíveis para micro-cirurgias. Assim, as espécies mais usadas têm sido as aves – especialmente a galinha – e os anfíbios – particularmente a rã africana Xenopus laevis.
Os estudos de animais mutantes identificam os genes que controlam os processos do desenvolvimentoA genética do desenvolvimento inicia-se com o isolamento de animais mutantes cujo de-senvolvimento é anormal. Isso geralmente envolve uma sondagem genética, como descrito no Capítulo 8. Os animais parentais são tratados com um mutagênico químico ou com uma radiação ionizante para induzir mutações nas suas células germinativas, e grandes números da sua progênie são examinados. Os raros mutantes que mostram alguma anormalidade in-teressante no desenvolvimento – desenvolvimento alterado do olho, por exemplo – são se-lecionados para um estudo mais aprofundado. Dessa maneira, é possível descobrir os genes que são especificamente necessários ao desenvolvimento normal de qualquer característica escolhida. Pela clonagem e pelo sequenciamento de um gene encontrado dessa maneira, é possível identificar o seu produto proteico, investigar como ele funciona e começar uma análise do DNA regulador que controla a sua expressão.
A estratégia genética é mais fácil em pequenos animais com tempos de geração cur-tos que podem crescer em laboratório. O primeiro animal a ser estudado desse modo foi a mosca-das-frutas Drosophila melanogaster, a qual será estudada extensivamente a seguir. Contudo, a mesma estratégia tem sido bem sucedida no verme nematoide, Caenorhabditis elegans, no peixe-zebra, Danio rerio, e no camundongo, Mus musculus. Embora os humanos não sejam intencionalmente mutagenizados, eles são sondados para anormalidades em nú-meros enormes pelo sistema médico de saúde. Muitas mutações em humanos causam anor-malidades compatíveis com a vida, e as análises dos indivíduos afetados e das suas células fornecem indícios importantes sobre os processos do desenvolvimento.
Uma célula toma as decisões sobre o seu desenvolvimento muito antes de mostrar uma mudança visívelPor um simples olhar atento, ou com o auxílio de marcadores fluorescentes e outras técnicas de marcação celular, pode-se descobrir qual será o destino de determinada célula em um embrião, caso seja permitido a ele desenvolver-se normalmente. A célula pode ter como des-tino morrer, por exemplo, ou tornar-se um neurônio, ser parte de um órgão, como o pé, ou dar origem a uma progênie de células distribuídas por todo o corpo. Conhecer o destino ce-lular, nesse sentido, entretanto, é saber quase nada a respeito da característica intrínseca da célula. Em um extremo, a célula que é destinada a tornar-se, digamos, um neurônio pode já estar especializada de uma maneira que garanta que ela se tornará um neurônio, não impor-
Um embrião de 2 célulasdividido quase ao meiopor um grampo de cabelo
Transplante de um pequeno grupo de células
em um embrião hospedeiro
(A) (B) Figura 22-6 Alguns resultados surpre-endentes obtidos pela embriologia experimental. Em (A), um embrião anfíbio inicial é dividido praticamente em duas partes com um grampo de cabelo. Em (B), um embrião de anfíbio em um estágio um pouco mais tardio recebe um enxerto de um pequeno conjunto de células de outro embrião naquele estágio. As duas operações, bem-diferentes, induzem um único embrião a desenvolver-se em um par de gêmeos ligados (siameses). É também possível no experimento (A) dividir o embrião inicial em duas metades completamente separadas; dois girinos bem-formados inteiramente separados são então produzidos. (A, segundo H. Spemann, Embryonic Development and Induction. New Haven: Yale University Press, 1938; B, Segundo J. Holtfreter e V. Hamburger, in Analysis of Development [B.H. Willier, P. A. Weiss e V. Hamburger, eds.], p. 230-296. Philadelphia: Saun-ders, 1955.)
Alberts_22.indd 1311Alberts_22.indd 1311 29.07.09 16:49:1429.07.09 16:49:14

1312 Alberts, Johnson, Lewis, Raff, Roberts & Walter
tando o quanto o seu ambiente seja alterado; tal célula é considerada como determinada para o seu destino. No extremo oposto, a célula pode ser bioquimicamente idêntica a outras células fadadas a outros destinos, sendo a sua posição acidental a única diferença entre elas, o que expõe as células a influências futuras distintas.
O estado de determinação de uma célula pode ser testado transplantando-a para am-bientes alterados (Figura 22-7). Uma das conclusões-chave da embriologia experimental tem sido que, graças à memória celular, uma célula pode tornar-se determinada muito antes de mostrar algum sinal externo óbvio de diferenciação.
Entre os extremos de total determinação e completa indeterminação celular, há um amplo espectro de possibilidades. Uma célula pode, por exemplo, já estar levemente espe-cializada para o seu destino normal, com uma forte tendência para desenvolver-se naquela direção, mas ainda capaz de alterar-se e ter um destino diferente, se colocada em um am-biente suficientemente coercivo. (Alguns biólogos do desenvolvimento descreveriam esta célula como especificada ou comprometida, mas ainda não determinada.) Ou a célula pode estar determinada, digamos, como uma célula cerebral, mas ainda não determinada quanto a ser um componente neuronal ou glial do cérebro. E, frequentemente, parece que células adjacentes do mesmo tipo interagem e dependem de suporte mútuo para manter suas ca-racterísticas especializadas, de maneira que elas irão comportar-se como determinadas se mantidas juntas em um agrupamento, mas não determinadas se colocadas sozinhas e isola-das de suas companhias usuais.
As células relembram valores posicionais que refletem a sua localização no corpoEm muitos sistemas, muito antes de as células comprometerem-se com a diferenciação em um tipo celular específico, tornam-se regionalmente determinadas: ou seja, ativam e man-têm a expressão de genes que podem ser considerados como marcadores de posição ou de região do corpo. Esta característica posição-específica de uma célula é chamada de valor posicional e mostra seus efeitos na maneira como a célula se comporta em etapas subse-quentes da formação dos padrões.
O desenvolvimento da perna e da asa de galinha fornece um exemplo impressionante. Ambas, a perna e a asa do adulto, consistem em músculos, ossos, pele e assim por diante – quase exatamente a mesma gama de tecidos diferenciados. A diferença entre os dois mem-bros não reside nos tipos de tecidos, mas na maneira como estes tecidos estão arranjados no espaço. Como, então, essa diferença ocorre?
No embrião da galinha, a perna e a asa originam-se quase ao mesmo tempo, na forma de pequenos brotos no formato de língua que se projetam do flanco. As células nos dois pares de brotos dos membros parecem semelhantes e uniformemente indiferenciadas em um primeiro momento. Contudo, um simples experimento mostra que essa aparente seme-lhança é enganosa. Um pequeno bloco de tecido indiferenciado na base do broto da perna, da região que normalmente daria origem à coxa, pode ser cortado e enxertado na ponta do broto da asa. Surpreendentemente, o enxerto não dá origem à parte apropriada de ponta de asa, nem a um pedaço de tecido de coxa no local errado, mas a um dedo do pé (Figura 22-8). Esse experimento mostra que as células do broto da perna já estão previamente deter-
Figura 22-7 O teste-padrão para a de-terminação celular.
Doador
Transplante
Hospedeiro
Doador
Transplante
Hospedeiro
Após iniciar adiferenciação
Antes de iniciara diferenciação
DESTINO NORMAL NÃO-DETERMINADO DETERMINADO
Broto daperna
Brotoda asa
Asaresultante
Porção de tecido da mesoderme que formariaestruturas da coxa
Tecido presumivelmenteda coxa, enxertado naponta do broto da asa
Parte superior daasa e antebraço
Dedos do pé comgarras terminais
Figura 22-8 Provável tecido de coxa enxertado na ponta de um broto de asa de galinha, formando dedos do pé. (Segundo J.W. Saunders et al., Dev. Biol. 1:281-301, 1959. Com permissão da Academic Press.)
Alberts_22.indd 1312Alberts_22.indd 1312 29.07.09 16:49:1429.07.09 16:49:14
Biologia Molecular da Célula 1313
minadas como perna, mas ainda não irrevogavelmente comprometidas para vir a ser uma parte particular da perna: elas ainda podem responder a sinais no broto da asa, de maneira que formam estruturas apropriadas para a ponta do membro, em vez da base. O sistema de sinalização que controla as diferenças entre as partes do membro é, aparentemente, o mes-mo para a perna ou para a asa. A diferença entre os dois membros resulta da diferença nos estados internos das suas células no início do desenvolvimento dos membros.
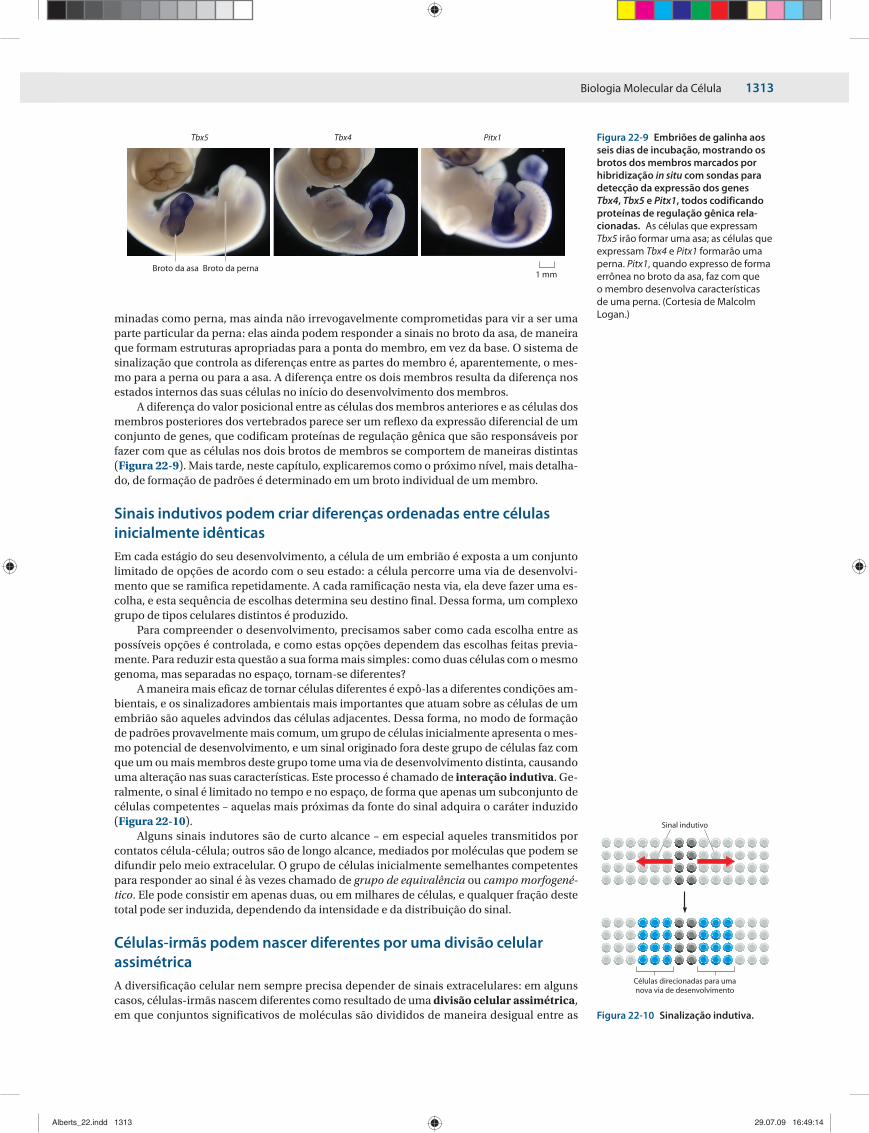
A diferença do valor posicional entre as células dos membros anteriores e as células dos membros posteriores dos vertebrados parece ser um reflexo da expressão diferencial de um conjunto de genes, que codificam proteínas de regulação gênica que são responsáveis por fazer com que as células nos dois brotos de membros se comportem de maneiras distintas (Figura 22-9). Mais tarde, neste capítulo, explicaremos como o próximo nível, mais detalha-do, de formação de padrões é determinado em um broto individual de um membro.
Sinais indutivos podem criar diferenças ordenadas entre células inicialmente idênticasEm cada estágio do seu desenvolvimento, a célula de um embrião é exposta a um conjunto limitado de opções de acordo com o seu estado: a célula percorre uma via de desenvolvi-mento que se ramifica repetidamente. A cada ramificação nesta via, ela deve fazer uma es-colha, e esta sequência de escolhas determina seu destino final. Dessa forma, um complexo grupo de tipos celulares distintos é produzido.
Para compreender o desenvolvimento, precisamos saber como cada escolha entre as possíveis opções é controlada, e como estas opções dependem das escolhas feitas previa-mente. Para reduzir esta questão a sua forma mais simples: como duas células com o mesmo genoma, mas separadas no espaço, tornam-se diferentes?
A maneira mais eficaz de tornar células diferentes é expô-las a diferentes condições am-bientais, e os sinalizadores ambientais mais importantes que atuam sobre as células de um embrião são aqueles advindos das células adjacentes. Dessa forma, no modo de formação de padrões provavelmente mais comum, um grupo de células inicialmente apresenta o mes-mo potencial de desenvolvimento, e um sinal originado fora deste grupo de células faz com que um ou mais membros deste grupo tome uma via de desenvolvimento distinta, causando uma alteração nas suas características. Este processo é chamado de interação indutiva. Ge-ralmente, o sinal é limitado no tempo e no espaço, de forma que apenas um subconjunto de células competentes – aquelas mais próximas da fonte do sinal adquira o caráter induzido (Figura 22-10).
Alguns sinais indutores são de curto alcance – em especial aqueles transmitidos por contatos célula-célula; outros são de longo alcance, mediados por moléculas que podem se difundir pelo meio extracelular. O grupo de células inicialmente semelhantes competentes para responder ao sinal é às vezes chamado de grupo de equivalência ou campo morfogené-tico. Ele pode consistir em apenas duas, ou em milhares de células, e qualquer fração deste total pode ser induzida, dependendo da intensidade e da distribuição do sinal.
Células-irmãs podem nascer diferentes por uma divisão celular assimétricaA diversificação celular nem sempre precisa depender de sinais extracelulares: em alguns casos, células-irmãs nascem diferentes como resultado de uma divisão celular assimétrica, em que conjuntos significativos de moléculas são divididos de maneira desigual entre as
Figura 22-9 Embriões de galinha aos seis dias de incubação, mostrando os brotos dos membros marcados por hibridização in situ com sondas para detecção da expressão dos genes Tbx4, Tbx5 e Pitx1, todos codificando proteínas de regulação gênica rela-cionadas. As células que expressam Tbx5 irão formar uma asa; as células que expressam Tbx4 e Pitx1 formarão uma perna. Pitx1, quando expresso de forma errônea no broto da asa, faz com que o membro desenvolva características de uma perna. (Cortesia de Malcolm Logan.)
Tbx5 Tbx4 Pitx1
Broto da asa Broto da perna1 mm
Sinal indutivo
Células direcionadas para umanova via de desenvolvimento
Figura 22-10 Sinalização indutiva.
Alberts_22.indd 1313Alberts_22.indd 1313 29.07.09 16:49:1429.07.09 16:49:14

1314 Alberts, Johnson, Lewis, Raff, Roberts & Walter
duas células no momento da divisão. Esta segregação assimétrica de moléculas (ou conjun-tos de moléculas) atua como determinante para um dos destinos celulares pela alteração di-reta ou indireta do padrão de expressão gênica na célula-filha que a contém (Figura 22-11).
As divisões assimétricas são particularmente comuns no início do desenvolvimento, quando o ovo fertilizado divide-se para originar células-filhas com destinos diferentes, mas elas também ocorrem em estágios mais tardios – na gênese das células nervosas, por exemplo.
A retroalimentação positiva pode originar assimetria onde não havia antesA sinalização indutiva e a divisão celular assimétrica representam duas estratégias distintas para a criação de diferenças entre as células. Ambas, no entanto, pressupõem uma assime-tria preexistente no sistema: a fonte do sinal indutivo deve estar localizada de forma que algumas células recebem o sinal forte e outras não; ou a célula-mãe já deve apresentar uma assimetria interna antes de se dividir. Muito frequentemente, o histórico do sistema assegura que alguma assimetria estará presente. Contudo, o que acontece se não estiver, ou se a assi-metria inicial for apenas sutil?
A resposta reside na retroalimentação positiva: pela retroalimentação positiva, um sis-tema que inicialmente era homogêneo e simétrico pode criar padrões espontaneamente, mesmo quando não houver um sinal externo organizado. E nos casos onde, como geralmen-te ocorre, o ambiente ou as condições iniciais imponham uma assimetria inicial fraca mas definitiva, a retroalimentação positiva provê os meios necessários para amplificar a assime-tria e criar um padrão de desenvolvimento.
Para ilustrar a ideia, considere um par de células adjacentes que iniciam em um estado similar e podem trocar sinais para influenciar o comportamento uma da outra (Figura 22-12). Quanto mais qualquer uma das células produzir o mesmo produto X, mais ela vai sinalizar para a célula vizinha que iniba sua produção de X. Este tipo de interação célula-célula é cha-mado de inibição lateral e origina um ciclo de retroalimentação positiva que tende a ampli-ficar qualquer diferença inicial entre as duas células. Esta diferença pode ser originada por condições impostas por algum fator externo anterior, ou simplesmente por flutuações aleató-rias espontâneas, ou “ruído” – uma característica inevitável do circuito do controle genético nas células, conforme discutido no Capítulo 7. Em qualquer um dos casos, a inibição lateral significa que, se a célula #1 sintetizar um pouco mais de X, ela fará com que a célula #2 sinte-tize menos; e como a célula #2 faz menos X, ela causa uma menor inibição na célula #1, o que permite que a quantidade de X na célula #1 aumente ainda mais; e assim sucessivamente, até que um estado de equilíbrio seja atingido, onde a célula #1 contém grandes quantidades de X e a célula #2 contém muito pouco.
Figura 22-11 Os dois modos de tornar células-irmãs diferentes.
1. Divisão assimétrica: as células-irmãs nascem diferentes
2. Divisão simétrica: as células-irmãs se tornam diferentes como resultado das influências que atuam sobre elas após o seu nascimento
Alberts_22.indd 1314Alberts_22.indd 1314 29.07.09 16:49:1529.07.09 16:49:15

Biologia Molecular da Célula 1315
Análises matemáticas mostram que este fenômeno depende da força do efeito da inibi-ção lateral: se ela for muito fraca, as flutuações irão desaparecer e não haverá efeito durador; mas se ela for forte e duradoura o suficiente, o efeito será autoamplificado de forma constante, rompendo a simetria inicial entre as duas células. A inibição lateral, frequentemente mediada pela troca de sinais nos pontos de contato célula-célula através da via de sinalização de Notch (como discutido no Capítulo 15), é um mecanismo comum de diversificação celular em teci-dos animais, fazendo com que células adjacentes se especializem de formas diferentes.
A retroalimentação positiva gera padrões, cria resultados tudo-ou-nada e provê memóriaProcessos similares de retroalimentação positiva podem operar sobre conjuntos maiores de células para originar diversos tipos de padrões espaciais. Por exemplo, uma substância A (um ativador de curto alcance) pode estimular sua própria produção nas células que a con-tenham e nas células adjacentes, enquanto pode também estimulá-las a produzir um sinal H (um inibidor de longo alcance) que se difunde amplamente e inibe a produção de A nas células localizadas a grandes distâncias. Se todas as células partirem de um estado inicial igual, mas um grupo de células ganhar certa vantagem pela produção um tanto maior de A do que o restante das células, a assimetria pode ser autoamplificada. A ativação de curto al-cance, combinada desta forma à inibição de longo alcance, pode colaborar para a formação de grupos de células que se tornem especializadas como centros sinalizadores localizados, em um tecido inicialmente homogêneo.
No polo oposto do espectro de magnitude, a retroalimentação positiva também pode ser o meio pelo qual as células individuais se tornam espontaneamente polarizadas e inter-namente assimétricas, por meio de sistemas de sinalização intracelular que tornam um sinal assimétrico inicial capaz de autoamplificação.
Por meio destas e de muitas outras variações sobre o tema da retroalimentação positiva, alguns princípios gerais se aplicam. Em cada um dos exemplos anteriores, a retroalimenta-ção positiva leva ao rompimento da simetria e a um fenômeno tudo-ou-nada. Se a retroali-mentação estiver abaixo de um certo limiar de força, as células se mantêm essencialmente no mesmo estado; se a retroalimentação estiver acima do limiar, elas se tornam muito dife-rentes. Acima deste limiar, o sistema tem estabilidade dupla ou é multiestável – ele se des-loca na direção de um ou outro resultado final, dentre os dois ou mais resultados possíveis altamente distintos, de acordo com qual das células (ou qual dos polos de uma única célula) ganhou a vantagem inicial.
A escolha entre resultados finais alternativos pode ser determinada por sinais externos que conferem a uma das células uma pequena vantagem inicial. Contudo, uma vez que a
Figura 22-12 Origem da assimetria pela retroalimentação positiva. Neste exemplo, duas células interagem, cada uma produzindo uma substância X que atua na outra célula, inibindo a produ-ção de X, um efeito conhecido como ini-bição lateral. Um aumento na produção de X em uma das células leva a uma re-troalimentação positiva que tende a au-mentar a produção de X nesta mesma célula, enquanto diminui a quantidade de X na célula vizinha. Isto pode originar uma instabilidade crescente, tornando as duas células radicalmente distintas. Por fim, o sistema se estabiliza em um dos dois estados opostos. A escolha fi-nal do estado representa uma forma de memória: uma pequena influência que inicialmente direcionou uma escolha não é mais necessária para manter o estado final estável.
X
X
XX
X XX
X
Uma flutuação transientecria uma pequena assimetria.
RETROALIMENTAÇÃO POSITIVAA assimetria é autoamplificada.
ESTABILIDADE DUPLAOs estados finais alternativos tudo-ou-nada representam uma memória estável.
XX
X XXX X
XX
X
XX
X
X XX
XXX
X
XX
XXX
XX
X XX
X
X XX X
XXX XXX X
Alberts_22.indd 1315Alberts_22.indd 1315 29.07.09 16:49:1529.07.09 16:49:15

1316 Alberts, Johnson, Lewis, Raff, Roberts & Walter
retroalimentação positiva tenha feito o seu trabalho, este sinal externo se torna irrelevante. O rompimento da simetria, uma vez estabelecido, é muito difícil de ser revertido: a retroali-mentação positiva faz com que o estado assimétrico escolhido seja autossustentado, mesmo quando o sinal inicial tenha desaparecido. Dessa forma, a retroalimentação positiva provê ao sistema uma memória dos sinais passados.
Todos estes efeitos da retroalimentação positiva – rompimento da simetria, resultados tudo-ou-nada, estabilidade dupla e memória – andam lado a lado e são encontrados repeti-das vezes no desenvolvimento dos organismos. Estes efeitos são fundamentais para a origem de padrões estáveis e fortemente delineados nas células em diferentes estados.
Um pequeno conjunto de vias de sinalização, utilizado repetidamente, controla o padrão de desenvolvimentoQuais, então, são as moléculas que atuam como sinais para coordenar a formação espacial de padrões em um embrião, seja para dar origem à assimetria de novo, ou para agir como indutores dos centros de sinalização estabelecidos para controlar a diversificação das células adjacentes? Em princípio, qualquer tipo de molécula extracelular poderia servir. Na prática, a maioria dos eventos indutivos conhecidos no desenvolvimento animal é governada por ape-nas uma família de proteínas de sinalização altamente conservadas, que são utilizadas repeti-damente em contextos diferentes. A descoberta deste vocabulário limitado que as células uti-lizam para comunicação durante o processo de desenvolvimento ocorreu nos últimos 10 a 20 anos como uma das grandes descobertas simplificadoras da biologia do desenvolvimento. Na Tabela 22-1, revisamos brevemente as seis principais famílias de proteínas de sinalização que atuam repetidamente como indutoras do desenvolvimento animal. Detalhes dos mecanismos intracelulares através dos quais estas moléculas atuam são encontrados no Capítulo 15.
O resultado final da maioria dos eventos de indução é uma alteração na transcrição do DNA na célula que responde ao sinal: alguns genes são ativados e outros são inibidos. Diferentes moléculas sinalizadoras ativam diferentes tipos de proteínas reguladoras de ge-nes. Além disso, o efeito de ativação de uma proteína reguladora de genes irá depender de quais outras proteínas reguladoras de genes também estiverem presentes em uma célula, uma vez que elas atuam em conjunto. Como resultado, diferentes tipos celulares em geral responderão de maneiras diferentes a um mesmo sinal, e células iguais frequentemente irão responder de maneiras diferentes a um mesmo sinal que seja iniciado em tempos distintos. A resposta dependerá de quais outras proteínas reguladoras de genes estiverem presentes antes da chegada do sinal – refletindo a memória celular dos sinais recebidos previamente, e de quais outros sinais a célula está recebendo no momento corrente.
Morfógenos são indutores de longo alcance que exercem efeitos graduadosMoléculas-sinal frequentemente parecem coordenar uma escolha simples tipo sim ou não: um efeito quando sua concentração é alta e outro quando sua concentração é baixa. A retro-
Tabela 22-1 Algumas proteínas-sinal são utilizadas repetidamente como indutoras do desenvolvimento animal
VIA DE SINALIZAÇÃO FAMÍLIA DE LIGANTES FAMÍLIA DE RECEPTORESINIBIDORES/MODULADORES EXTRACELULARES
Receptor tirosina- cinase (RTK) EGFFGF (Branchless)Efrinas
Receptores EGFReceptores FGF (Breathless)Receptores Eph
Argos
Superfamília TGF� TGF�BMP (Dpp)Nodal
Receptores TGF�Receptores BMP
chordin (Sog), noggin
Wnt Wnt (Wingless) Frizzled Dickkopf, CerberusHedgehog Hedgehog Patched, SmoothenedNotch Delta Notch Fringe
Apenas alguns exemplos representativos de cada classe de proteínas são listados – principalmente aqueles que são mencionados neste Capítulo. Nomes particulares para Drosophila são mostrados entre parênteses. Muitos dos componentes listados apresentam diversos homólogos distinguidos por números (FGF1, FGF2, etc.) ou por nomes compostos (Sonic hedgehog, Lunatic fringe). Outras vias de sinalização, incluindo as vias JAK/STAT, receptores nucleares de hormônios e receptores associados à proteína G, também desempenham um papel importante em alguns processos de desenvolvimento.
Alberts_22.indd 1316Alberts_22.indd 1316 29.07.09 16:49:1529.07.09 16:49:15
Biologia Molecular da Célula 1317
alimentação positiva faz com que as células respondam de forma tudo-ou-nada, de maneira que um resultado é obtido quando o sinal está abaixo de um dado valor crítico, e outro re-sultado quando está acima deste valor. Em muitos casos, no entanto, as respostas têm um ajuste mais fino: uma alta concentração pode, por exemplo, direcionar as células-alvo para uma via de desenvolvimento, uma concentração intermediária para uma outra via, e baixas concentrações podem induzir estas células a uma outra via possível. Um caso importante é aquele em que a molécula-sinal difunde-se a partir de um centro de sinalização localizado, criando um gradiente de concentração de sinal. As células a diferentes distâncias da fonte são direcionadas a comportarem-se em uma grande variedade de maneiras diferentes, de acordo com a concentração do sinal que elas recebem.
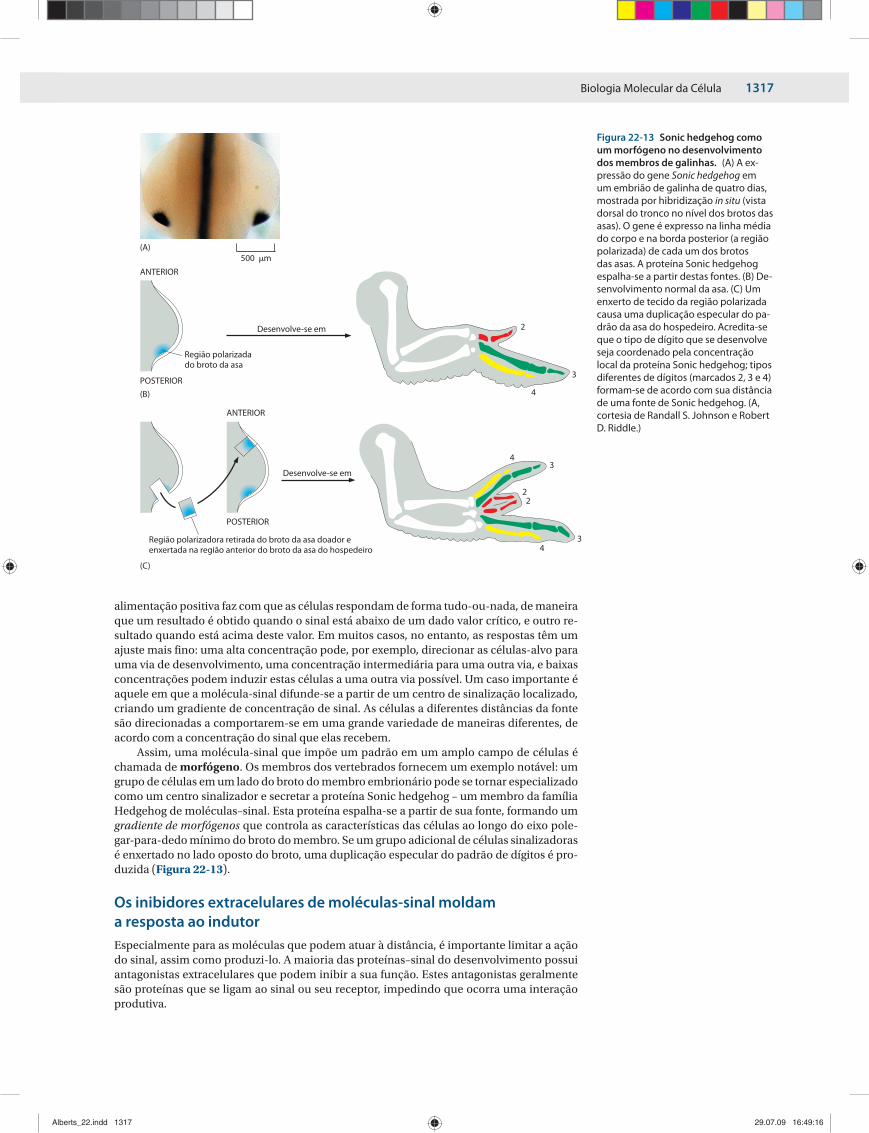
Assim, uma molécula-sinal que impõe um padrão em um amplo campo de células é chamada de morfógeno. Os membros dos vertebrados fornecem um exemplo notável: um grupo de células em um lado do broto do membro embrionário pode se tornar especializado como um centro sinalizador e secretar a proteína Sonic hedgehog – um membro da família Hedgehog de moléculas–sinal. Esta proteína espalha-se a partir de sua fonte, formando um gradiente de morfógenos que controla as características das células ao longo do eixo pole-gar-para-dedo mínimo do broto do membro. Se um grupo adicional de células sinalizadoras é enxertado no lado oposto do broto, uma duplicação especular do padrão de dígitos é pro-duzida (Figura 22-13).
Os inibidores extracelulares de moléculas-sinal moldam a resposta ao indutorEspecialmente para as moléculas que podem atuar à distância, é importante limitar a ação do sinal, assim como produzi-lo. A maioria das proteínas–sinal do desenvolvimento possui antagonistas extracelulares que podem inibir a sua função. Estes antagonis tas geralmente são proteínas que se ligam ao sinal ou seu receptor, impedindo que ocorra uma interação produtiva.
Figura 22-13 Sonic hedgehog como um morfógeno no desenvolvimento dos mem bros de galinhas. (A) A ex-pressão do gene Sonic hedgehog em um embrião de galinha de quatro dias, mostrada por hibridização in situ (vista dor sal do tronco no nível dos brotos das asas). O gene é expresso na linha média do corpo e na borda posterior (a região polarizada) de cada um dos brotos das asas. A proteína Sonic hed gehog espalha-se a partir destas fontes. (B) De-senvolvimento normal da asa. (C) Um enxerto de tecido da região polarizada causa uma dupli cação especular do pa-drão da asa do hospedei ro. Acredita-se que o tipo de dígito que se de senvolve seja coordenado pela concentração local da proteína Sonic hedgehog; tipos diferen tes de dígitos (marcados 2, 3 e 4) formam-se de acordo com sua distância de uma fonte de Sonic hedgehog. (A, cortesia de Randall S. Johnson e Robert D. Riddle.)
500 �m
ANTERIOR
POSTERIOR
ANTERIOR
POSTERIOR
(B)
(A)
(C)
Região polarizada do broto da asa
Região polarizadora retirada do broto da asa doador e enxertada na região anterior do broto da asa do hospedeiro
Desenvolve-se em
Desenvolve-se em
2
22
3
3
3
4
4
4
Alberts_22.indd 1317Alberts_22.indd 1317 29.07.09 16:49:1629.07.09 16:49:16
1318 Alberts, Johnson, Lewis, Raff, Roberts & Walter
Um número surpreendentemente grande de decisões no desenvolvimento é, na verda-de, re gulado por inibidores e não pela molécula-sinal primária. O sistema nervoso em um embrião de rã origina-se de um conjunto de células que é competente para formar tanto tecido neuronal quanto epiderme. Um tecido indutor libera a proteína chordin, a qual favo-rece a formação do tecido neuronal. A chordin não possui receptor próprio. Em vez disso, ela é um inibidor de proteínas–sinal da família BMP/TGF�, que induzem o desenvolvimento da epiderme e estão presentes por toda a região neuroepitelial onde os neurônios e a epiderme se formam. A indução do tecido neuronal é devida a um gradiente inibidor de um sinal an-tagonista (Figura 22-14).
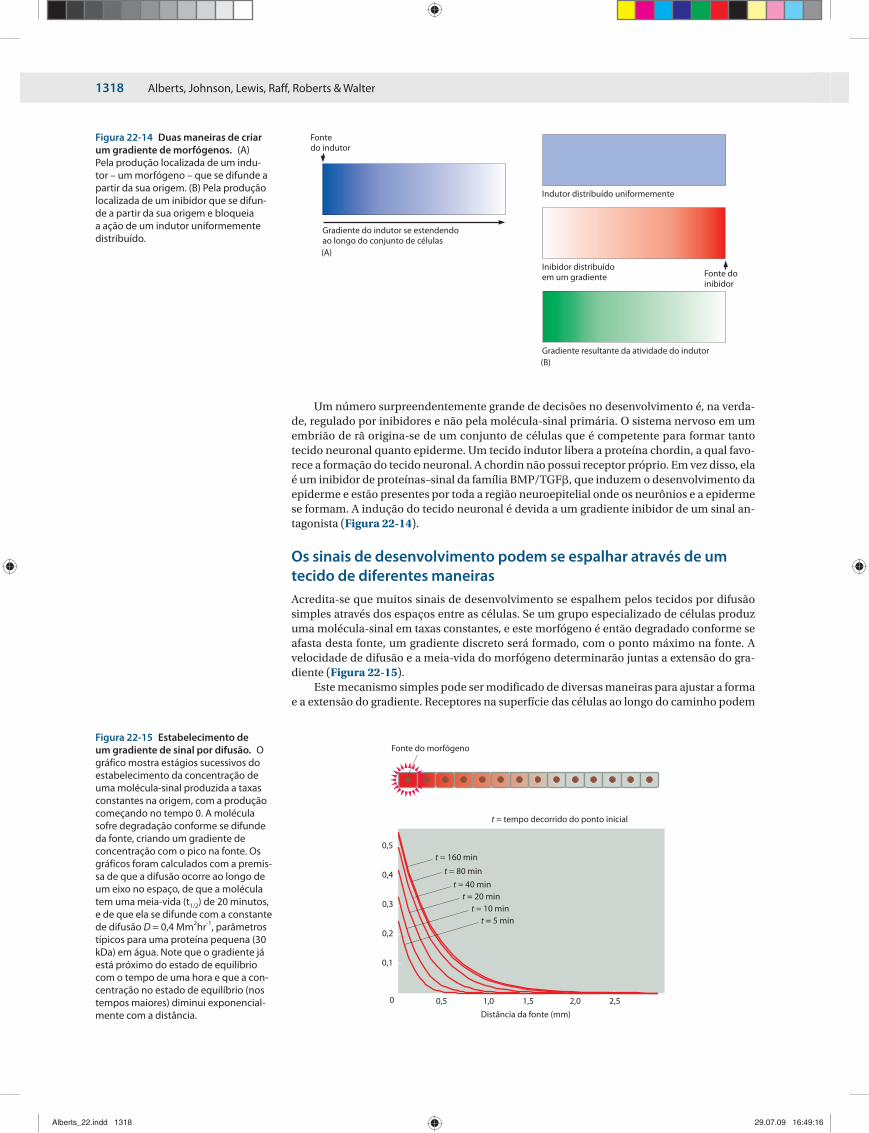
Os sinais de desenvolvimento podem se espalhar através de um tecido de diferentes maneirasAcredita-se que muitos sinais de desenvolvimento se espalhem pelos tecidos por difusão simples através dos espaços entre as células. Se um grupo especializado de células produz uma molécula-sinal em taxas constantes, e este morfógeno é então degradado conforme se afasta desta fonte, um gradiente discreto será formado, com o ponto máximo na fonte. A velocidade de difusão e a meia-vida do morfógeno determinarão juntas a extensão do gra-diente (Figura 22-15).
Este mecanismo simples pode ser modificado de diversas maneiras para ajustar a forma e a extensão do gradiente. Receptores na superfície das células ao longo do caminho podem
Figura 22-14 Duas maneiras de criar um gradiente de morfógenos. (A) Pela produção localizada de um indu-tor – um morfógeno – que se difunde a partir da sua origem. (B) Pela pro dução localizada de um inibidor que se difun-de a partir da sua origem e bloqueia a ação de um indutor uniformemente distribuído.
Indutor distribuído uniformemente
Inibidor distribuídoem um gradiente
Gradiente resultante da atividade do indutor(B)
(A)
Fonte doinibidor
Fontedo indutor
Gradiente do indutor se estendendoao longo do conjunto de células
Figura 22-15 Estabelecimento de um gradiente de sinal por difusão. O gráfico mostra estágios sucessivos do estabelecimento da concentração de uma molécula-sinal produzida a taxas constantes na origem, com a produção começando no tempo 0. A molécula sofre degradação conforme se difunde da fonte, criando um gradiente de concentração com o pico na fonte. Os gráficos foram calculados com a premis-sa de que a difusão ocorre ao longo de um eixo no espaço, de que a molécula tem uma meia-vida (t1/2) de 20 minutos, e de que ela se difunde com a constante de difusão D = 0,4 Mm2hr-1, parâmetros típicos para uma proteína pequena (30 kDa) em água. Note que o gradiente já está próximo do estado de equilíbrio com o tempo de uma hora e que a con-centração no estado de equilíbrio (nos tempos maiores) diminui exponencial-mente com a distância.
t = 160 min
t = 80 min
t = 40 mint = 20 mint = 10 min
t = 5 min
Distância da fonte (mm)
t = tempo decorrido do ponto inicial
Fonte do morfógeno
0,5 1,0 1,5 2,0 2,5
0,1
0,2
0,3
0,4
0,5
0
Alberts_22.indd 1318Alberts_22.indd 1318 29.07.09 16:49:1629.07.09 16:49:16

Biologia Molecular da Célula 1319
capturar o morfógeno e promover a sua endocitose e degradação, diminuindo sua meia-vida efetiva. Ou ele pode se ligar a moléculas da matriz extracelular, reduzindo a sua taxa de difu-são efetiva. Em alguns casos, é como se o morfógeno fosse captado pelas células por endoci-tose e depois liberado novamente, apenas para ser captado e liberado por outras células, de forma que o sinal se espalha através de uma longa via intracelular.
Há ainda um outro mecanismo para a distribuição de sinal que depende de longos e finos filipódeos, ou citonemas, que se estendem por distâncias equivalentes a muitas vezes o diâmetro celular em alguns tecidos do epitélio. Uma célula pode enviar citonemas para fazer contato com outra célula distante, tanto para entregar quanto para receber um sinal desta célula. Dessa forma, por exemplo, uma célula pode realizar inibição lateral através da via Notch em um grande conjunto de células adjacentes.
Os programas que são intrínsecos a uma célula frequentemente definem o curso de tempo do seu desenvolvimentoSinais como os que acabamos de discutir desempenham um grande papel no controle do tempo dos eventos de desenvolvimento, mas seria errado imaginar que toda a mudança no desenvolvimento necessita de um sinal indutor para desencadeá-la. Muitos dos mecanis-mos que alteram características celulares são intrínsecos das células e não necessitam de sinais do ambiente celular: a célula progredirá no seu programa de desenvolvimento mesmo quando mantida em um ambiente constante. Existem muitos casos em que alguém pode-ria suspeitar de que algo deste tipo está ocorrendo no controle da duração do processo de desenvolvimento. Por exemplo, em um camundongo, as células progenitoras neuro nais no córtex cerebral continuam a dividir-se e a gerar neurônios por somente 11 ciclos celulares, e no macaco, por aproximadamente 28 ciclos, após os quais elas param. Diferentes tipos de neurônios são gerados em estágios distintos desse programa, sugerindo que, à medida que a célula progenitora envelhece, ela altera as especificações que fornece para as células da progênie em diferenciação.
No contexto de um embrião intacto, é difícil provar que tal curso de eventos é estrita-mente o resultado de um processo celular autônomo marcador de tempo, uma vez que o ambiente celular está se alterando. Os experimentos com células em cultura, entretanto, fornecem evidências claras. Por exemplo, as células progenitoras da glia isoladas do ner-vo óptico de um rato, sete dias após o nascimento, e cultivadas sob condições constantes em um meio apropriado irão manter a proliferação por um tempo estritamente limitado (cor respondente a um máximo de aproximadamente oito divisões celulares) e então irão dife renciar-se em oligodendrócitos (as células da glia que formam as bainhas de mielina ao redor dos axônios no cérebro), obedecendo a um padrão de tempo semelhante ao que elas teriam seguido se tivessem sido deixadas no seu lugar no embrião.
Os mecanismos moleculares responsáveis por estas alterações lentas nas condições internas da célula, realizadas no curso de dias, semanas, meses e mesmo anos, ainda não são conhecidos. Uma possibilidade é que eles reflitam mudanças progressivas no estado da cromatina (discutido no Capítulo 4).
Os mecanismos que controlam a escala de tempo de processos mais rápidos, apesar de ainda pouco conhecidos, não são um mistério. Mais adiante, discutiremos um exemplo – o oscilador de expressão gênica, conhecido como relógio de segmentação, que coordena a formação de somitos em embriões de vertebrados – os rudimentos das séries de vértebras, costelas e músculos associados.
Enquanto o embrião cresce, os padrões iniciais são estabelecidos em pequenos grupos de células e refinados por indução sequencialOs sinais que organizam o padrão espacial de um embrião em geral atuam sobre distâncias curtas e governam escolhas relati vamente simples. Um morfógeno, por exemplo, normal-mente atua sobre uma distância de menos de 1 mm – uma distância efetiva para difusão (ver Figura 22-15) – e direciona escolhas entre não mais do que uma porção de opções de desenvolvimento para as células nas quais ele atua. Contudo, os órgãos que eventualmente se desenvolvem são muito maiores e mais complexos do que isso.
A proliferação celular que se segue à especificação inicial é responsável pelo aumento em tamanho, enquanto que o refinamento do padrão inicial é explicado por uma série de induções locais que acrescentam níveis sucessivos de detalhes em um esboço inicialmen-
Alberts_22.indd 1319Alberts_22.indd 1319 29.07.09 16:49:1629.07.09 16:49:16

1320 Alberts, Johnson, Lewis, Raff, Roberts & Walter
te simples. Assim que dois tipos de células estão presentes, uma delas pode produzir um fator que induza um subconjunto de células vizinhas a se especializarem em uma ter ceira via. O terceiro tipo celular pode, por sua vez, sinalizar em resposta aos outros dois tipos celulares próximos, gerando um quarto e um quinto tipo celular, e assim por diante (Fi-gura 22-16).
Esta estratégia para a geração de um padrão progressivamente mais complicado é cha-mada de indução sequencial. É principalmente por meio de induções sequenciais que a estrutura do corpo de um animal em desenvolvimento, após ser primeiramente esboçada em miniatura, torna-se elaborada em detalhes progressivamente mais finos, enquanto o desenvolvi mento prossegue.
Nas seções que se seguem, focalizaremos em uma pequena seleção de organismos-mode-lo para ver como os princípios que citamos nesta primeira seção operam na prática. Começare-mos com o verme nematoide, Caenorhabditis elegans.
ResumoAs alterações óbvias no comportamento celular que observamos enquanto um organismo multice-lular desenvolve-se são os sinais exteriores de uma complexa computação molecular, dependente da me mória celular que está ocorrendo dentro das células enquanto elas recebem e processam os sinais de suas vizinhas e emitem sinais em resposta. O padrão final dos tipos celulares diferenciados é, dessa forma, o resultado de um programa mais oculto de especialização celular – um programa extensivamente utilizado na alteração dos padrões de expressão por proteínas de regulação gênica, dando a uma célula potencialidades diferentes das outras muito antes de a diferenciação terminal começar. Os biólogos do desenvolvimento procuram decifrar o programa oculto e relacioná-lo, por meio de experimentos genéticos e microcirúrgicos, aos sinais que as células trocam enquanto elas proliferam, interagem e movem-se.
Animais tão diferentes como vermes, moscas e humanos usam conjuntos semelhantes de pro-teínas para controlar o seu desenvolvimento, de maneira que o que descobrimos em um organismo frequentemente fornece informações sobre os outros. Um grupo de vias de sinalização célula-célula evolutivamente conservadas é usado repetitivamente, em dife rentes organismos e em tempos dis-tintos, para regular a criação de um padrão multicelu lar organizado. As diferenças no plano cor-poral parecem surgir em grande parte de dife renças no DNA regulador associado a cada gene. Este DNA desempenha uma função central na definição do programa sequencial de desenvolvimento, colocando genes em ação em tempos e em locais específicos, de acordo com o padrão de expressão gênica que estava presente em cada célula no estágio de desenvolvimento anterior.
As diferenças entre as células de um embrião surgem de várias maneiras. A retroalimentação positiva pode levar ao rompimento da simetria, criando uma diferença marcante e constante en-tre células inicialmente quase idênticas. Células-irmãs podem nascer diferentes como resultado de uma divisão celular assimétrica. Ou um grupo de células inicialmente semelhantes pode ser exposto a diferentes sinais indutivos de células localizadas fora do grupo; indutores de longo al-cance com efeitos gradativos, chamados de morfógenos, podem organizar padrões complexos. Por meio da memória celular, tais sinais temporários podem ter um efeito duradouro sobre o estado interno da célula, induzindo-a, por exemplo, a tornar-se determinada para um destino específico. Assim, as sequências de sinais simples atuando em tempos e em locais diferentes nas células em crescimento dão origem aos intricados e variados organismos multicelulares que povoam o mun-do ao nosso redor.
Figura 22-16 Formação de padrões por indução sequencial. Uma série de interações indutoras pode gerar muitos tipos celulares, ini ciando a partir de so-mente alguns.
B
A
A
C
B
A
D
C
E
B
C é induzidopelo sinal de B
atuando sobre A
D e E sãoinduzidos pelo
sinal de Catuando em A e B,respectivamente
Alberts_22.indd 1320Alberts_22.indd 1320 29.07.09 16:49:1729.07.09 16:49:17

Biologia Molecular da Célula 1321
CAENORHABDITIS ELEGANS: O DESENVOLVIMENTO A PARTIR DA PERSPECTIVA DE UMA CÉLULA INDIVIDUALO verme nematoide Caenorhabditis elegans é um organismo pequeno, relativamente sim-ples e precisamente estruturado. A anatomia de seu desenvolvimento tem sido descrita em extraordinário detalhe, e pode-se mapear a linhagem exata de cada célula no corpo. A se-quência genômica completa também é conhecida, e um grande número de fenótipos mu-tantes tem sido analisado para determinar funções gênicas. Se há algum animal mul ticelular cujo desenvolvimento deveríamos ser capazes de entender em termos de controle genético, é este.
Comparações de sequências de DNA indicam que, enquanto as linhagens que levam aos nematoides, aos insetos e aos vertebrados divergiram uma da outra ao redor da mesma época, a taxa de mudanças evolutivas na linhagem dos nematoides tem sido substancial-mente maior: os seus genes, a sua estrutura corporal e suas estratégias de desenvolvimento são mais divergentes dos nossos próprios do que dos da Drosophila. No entanto, no nível molecular, muitos dos seus mecanismos de desenvolvimento são similares em insetos e ver-tebrados, sendo coordenados por sistemas de genes homólogos. Se quisermos saber como um olho, um membro ou um coração se desenvolve, é preciso procurar estas respostas em outros locais: o C. elegans não possui estes órgãos. Contudo, em um nível mais fundamental, ele é bastante instrutivo: apresenta as questões gerais básicas do desenvolvimento animal de uma forma relati vamente simples e nos possibilita respondê-las em termos de função gênica e do comportamento das células individuais identificadas.
O Caenorhabditis elegans é anatomicamente simplesComo adulto, o C. elegans consiste em somente cerca de mil células somáticas e de 1.000 a 2.000 células germinativas (exatamente 959 núcleos celulares somáticos e aproxi madamente 2.000 células germinativas são encontrados em um sexo; exatamente 1.031 núcleos celulares somáticos e cerca de 1.000 células germinativas no outro) (Figura 22-17). A sua anatomia foi reconstruída, célula por célula, por microscopia eletrônica de seções seriadas. A estrutura do plano corporal do verme é simples: ele tem simetria aproximadamente bilateral, um corpo alongado composto dos mesmos tecidos básicos de outros animais (nervos, músculos, in-testino, pele), organizado com boca e cérebro na extremidade anterior e ânus na posterior. A parede externa do corpo é composta de duas camadas: a epiderme protetora, ou “pele”, e a camada muscular imediatamente abaixo. Um tubo de células endodermais forma o intesti-no. Um segundo tubo, localizado entre o intestino e a parede do corpo, constitui a gônada; a sua parede é composta de células somáticas, com as células germinativas dentro dela.
O verme C. elegans tem dois sexos – um hermafrodita e um macho. O hermafrodita pode ser visto simplesmente como uma fêmea que produz um número limitado de esperma: ela pode reproduzir-se tanto por autofecundação, usando o seu próprio esperma, como por fecundação cruzada após a transferência do esperma do macho pelo acasalamento. A au-tofecundação permite a um verme heterozigoto único produzir uma progênie homozigota. Esta é uma característica importante que auxilia a fazer do C. elegans um organismo excep-cionalmente conveniente para estudos genéticos.
Figura 22-17 Caenorhabditis ele-gans. É mostrada uma visão lateral de um adulto hermafrodita. (De J. E. Sulston e H. R. Horvitz, Dev. Biol. 56:110-156, 1977. Com permissão da Academic Press.)
1,2 mm
Faringe Oócitos
Intestino
Útero Vulva
Ovos Gônadas
Epiderme
MúsculosParede corporal
Ânus
ANTERIORDORSAL
VENTRAL
POSTERIOR
Alberts_22.indd 1321Alberts_22.indd 1321 29.07.09 16:49:1729.07.09 16:49:17
1322 Alberts, Johnson, Lewis, Raff, Roberts & Walter
Os destinos celulares no nematoide em desenvolvimento são quase perfeitamente previsíveisO C. elegans inicia a sua vida como uma única célula, o ovo fertilizado, o qual origina, por meio de repetidas divisões celulares, as 558 células que formam um pequeno verme dentro da casca do ovo. Após a eclosão, as divisões adicionais resultam no crescimento e na matu-ração sexual do verme, enquanto ele passa por quatro estágios larvais sucessivos, separados por mudas. Após a muda final para o estágio adulto, o verme hermafrodita inicia a produção de seus próprios ovos. A sequência inteira de desenvolvimento, de ovo a ovo, leva somente cerca de três dias.
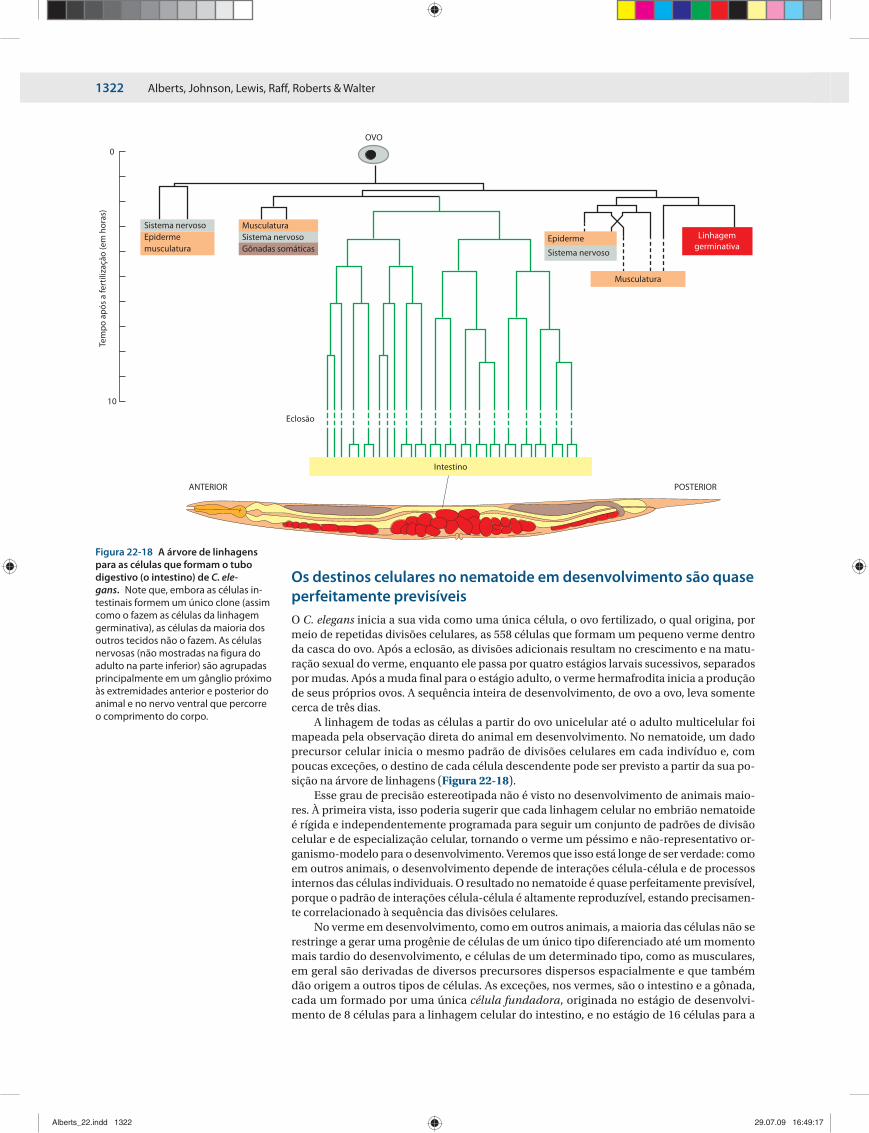
A linhagem de todas as células a partir do ovo unicelular até o adulto multicelular foi mapeada pela observação direta do animal em desenvolvimento. No nematoide, um dado precursor celular inicia o mesmo padrão de divisões celulares em cada indivíduo e, com poucas exceções, o destino de cada célula descendente pode ser previsto a partir da sua po-sição na árvore de linhagens (Figura 22-18).
Esse grau de precisão estereotipada não é visto no desenvolvimento de animais maio-res. À primeira vista, isso poderia sugerir que cada linhagem celular no embrião nematoi de é rígida e independentemente programada para seguir um conjunto de padrões de divi são celular e de especialização celular, tornando o verme um péssimo e não-representati vo or-ganismo-modelo para o desenvolvimento. Veremos que isso está longe de ser verda de: como em outros animais, o desenvolvimento depende de interações célula-célula e de processos internos das células individuais. O resultado no nematoide é quase perfeita mente previsível, porque o padrão de interações célula-célula é altamente reproduzível, estando precisamen-te correlacionado à sequência das divisões celulares.
No verme em desenvolvimento, como em outros animais, a maioria das células não se restringe a gerar uma progênie de células de um único tipo diferenciado até um momento mais tardio do desenvolvimento, e células de um determinado tipo, como as musculares, em geral são derivadas de diversos precursores dispersos espacialmente e que também dão origem a outros tipos de células. As exceções, nos vermes, são o intestino e a gônada, cada um formado por uma única célula fundadora, originada no estágio de desenvolvi-mento de 8 células para a linhagem celular do intestino, e no estágio de 16 células para a
Figura 22-18 A árvore de linhagens para as células que formam o tubo digestivo (o intestino) de C. ele-gans. Note que, embora as células in-testinais formem um único clone (as sim como o fazem as células da linhagem germi nativa), as células da maioria dos outros tecidos não o fazem. As células nervosas (não mos tradas na figura do adulto na parte inferior) são agrupadas principalmente em um gânglio próximo às extremi dades anterior e posterior do animal e no ner vo ventral que percorre o comprimento do corpo.
Intestino
ANTERIOR POSTERIOR
Sistema nervosoEpidermemusculatura
MusculaturaSistema nervosoGônadas somáticas
Epiderme
Sistema nervoso
Musculatura
Linhagemgerminativa
OVO
0
10
Tem
po
após
a fe
rtili
zaçã
o (e
m h
oras
)
Eclosão
Alberts_22.indd 1322Alberts_22.indd 1322 29.07.09 16:49:1729.07.09 16:49:17
Biologia Molecular da Célula 1323
linhagem de célula-ovo, ou linhagem germinativa. Contudo, em qualquer caso, a diver-sificação celular começa cedo, tão cedo quanto o ovo começa a se clivar: muito antes da diferenciação terminal, a célula começa a se encaminhar através de uma série de estágios intermediários de especialização, seguindo diferentes programas de acordo com sua loca-lização e suas interações com as células adjacentes. Como surgem estas diferenças iniciais entre as células?
Os produtos de genes de efeito materno organizam a divisão assimétrica do ovoO verme é semelhante à maioria dos animais na especificação inicial das células que irão eventualmente dar origem às células germinativas (ovos ou esperma). A linhagem germi-nativa dos vermes é produzida por uma série estrita de divisões celulares assimétricas do ovo fertilizado. A assimetria origina-se com um sinal do ambiente do ovo: o ponto de entrada do esperma define o futuro polo posterior do ovo alongado. As proteínas no ovo intera gem umas com as outras e organizam-se em relação a este ponto de maneira a criar uma assi-metria mais elaborada no interior da célula. As proteínas envolvidas são traduzidas prin-cipalmente a partir de produtos de mRNA acumulados dos genes da mãe. Como este RNA é produzido antes de o ovo ser posto, é somente o genótipo da mãe que determina o que acontece nos primeiros passos do desenvolvimento. Os genes que atuam desta maneira são chamados de genes de efeito materno.
Um subconjunto de genes de efeito materno é especificamente necessário para orga-nizar o padrão assimétrico do ovo nematoide. Estes são chamados de genes Par (defectivos em parti ção), e pelo menos seis foram identificados por rastreamento genético de mutantes em que o padrão tenha sido rompido. Os genes Par possuem homólogos em insetos e em vertebrados, onde desempenham papel fundamental na organização da polaridade da cé-lula, como discutido no Capítulo 19. De fato, uma das chaves para o entendimento atual dos mecanismos gerais envolvidos com a polaridade de células foi a descoberta destes genes por estudos em embriões de desenvolvimento inicial de C. elegans.
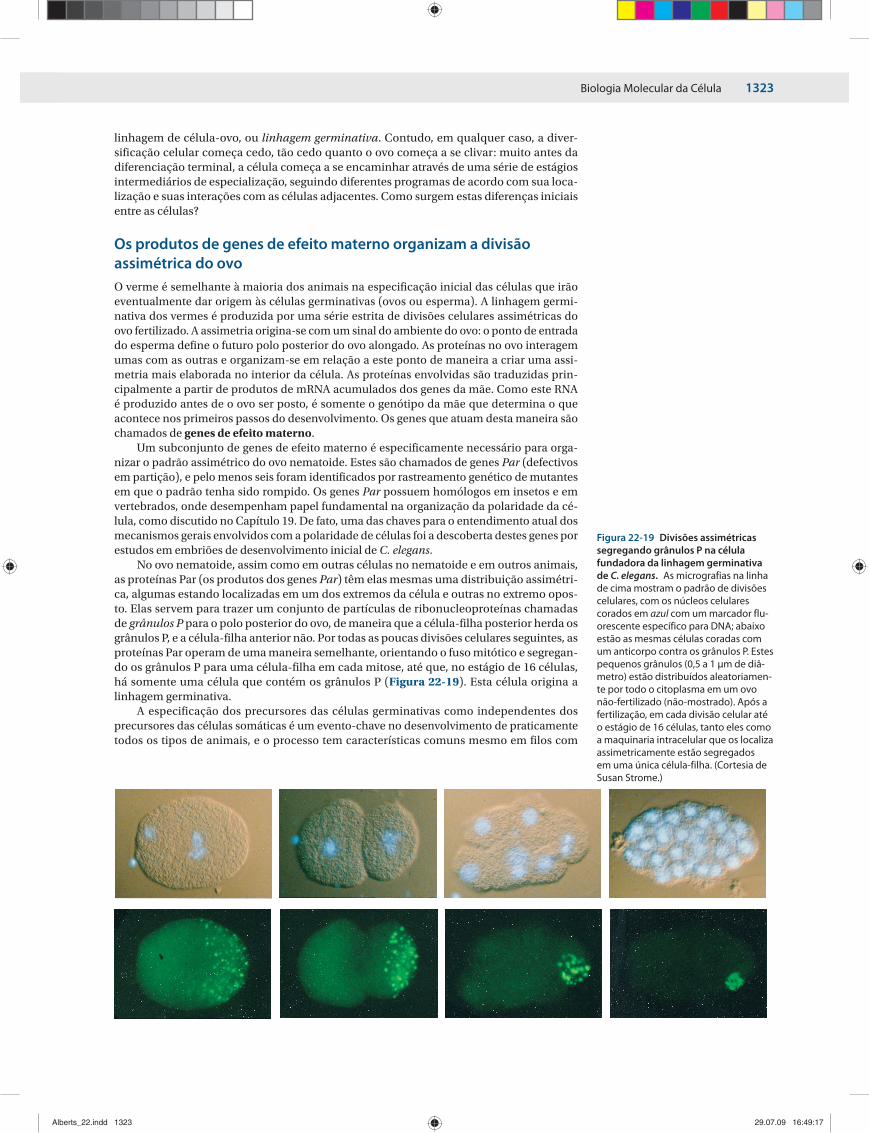
No ovo nematoide, assim como em outras células no nematoide e em outros animais, as proteínas Par (os produtos dos genes Par) têm elas mesmas uma distribuição assimétri-ca, algumas estando localizadas em um dos extremos da célula e outras no extremo opos-to. Elas servem para trazer um conjunto de partículas de ribonucleoproteínas chamadas de grânulos P para o polo posterior do ovo, de maneira que a célula-filha posterior herda os grânulos P, e a célula-filha anterior não. Por todas as poucas divisões celulares seguintes, as proteínas Par operam de uma maneira semelhante, orientando o fuso mitótico e segregan-do os grânulos P para uma célula-filha em cada mitose, até que, no estágio de 16 células, há somente uma célula que contém os grânulos P (Figura 22-19). Esta célula origina a linhagem germinativa.
A especificação dos precursores das células germinativas como independentes dos precur sores das células somáticas é um evento-chave no desenvolvimento de praticamente todos os tipos de animais, e o processo tem características comuns mesmo em filos com
Figura 22-19 Divisões assimétricas segregando grânulos P na célula fundadora da linhagem germinativa de C. elegans. As micrografias na linha de cima mostram o padrão de divisões celulares, com os núcleos celulares corados em azul com um marcador flu-orescente específico para DNA; abaixo estão as mesmas células coradas com um anticorpo contra os grânulos P. Estes pequenos grânulos (0,5 a 1 μm de diâ-metro) estão distribuídos aleatoriamen-te por todo o citoplasma em um ovo não-fertilizado (não-mostrado). Após a fertilização, em cada divisão celular até o estágio de 16 células, tanto eles como a maquinaria intracelular que os localiza assimetricamente estão segregados em uma única célula-filha. (Cortesia de Susan Strome.)
Alberts_22.indd 1323Alberts_22.indd 1323 29.07.09 16:49:1729.07.09 16:49:17

1324 Alberts, Johnson, Lewis, Raff, Roberts & Walter
estrutu ras corporais muito diferentes. Dessa forma, na Drosophila, as partículas semelhantes aos grânulos P também são segregadas em uma extremidade do ovo e tornam-se incorpo-radas nas células precursoras da linhagem germinativa para a determinação do seu destino. Um fenômeno similar ocorre nos peixes e nas rãs. Nessas espécies, pode-se reconhe cer pelo menos algumas das mesmas proteínas no material que determina as células ger minativas, incluindo os homólogos de uma proteína de ligação ao RNA chamada de Vasa. Ainda é des-conhecido o modo como a Vasa e as suas proteínas associadas e moléculas de RNA atuam na definição da linhagem germinativa.
Os padrões progressivamente mais complexos são criados por interações célula-célulaO ovo do C. elegans, assim como de outros animais, é uma célula extraordinariamente gran-de, com espaço para a formação de padrões internos complexos. Além dos grânulos P, outros fatores são distribuídos em uma maneira ordenada ao longo do seu eixo ântero-posterior sob o controle das proteínas Par, que, assim, são alocadas para células diferentes enquanto o ovo passa por alguns dos primeiros ciclos de divisão celular. Essas divisões ocorrem sem cresci mento (uma vez que a alimentação não pode começar antes que a boca e o intestino tenham sido formados) e subdividem o ovo em células progressivamente menores. Muitos dos fatores que são localizados são proteínas de regulação gênica, as quais atuam direta-mente na célula que as herda para direcionar ou bloquear a expressão de genes específicos, adicionando diferen ças entre a célula e as suas vizinhas e comprometendo-a com um des-tino especializado.
Enquanto as primeiras poucas diferenças ao longo do eixo ântero-posterior do C. elegans são o resultado de divisões assimétricas, a formação de novos padrões, incluindo o padrão dos tipos celulares ao longo de outros eixos, depende de interações entre uma célula e outra. As linhagens celulares no embrião são tão reproduzíveis que as células individuais podem ser designadas com nomes e identificadas em cada animal (Figura 22-20); as células do estágio de quatro células, por exemplo, são chamadas de ABa e ABp (as duas células-irmãs anterio-res), e EMS e P2 (as duas células-irmãs posteriores). Como resultado das divisões assimétricas que acabamos de descrever, a célula P2 expressa uma proteína-sinal na sua superfície – Delta, uma proteína de nematoides homóloga ao ligante de Notch – enquanto as células ABa e ABp expressam o receptor transmembrana correspondente – um homólogo de Notch. A forma alongada da casca do ovo força essas células para um arranjo tal que a célula mais anterior, ABa, e a célula mais posterior, P2, não estão mais em contato uma com a outra. Assim, somen-te a célula ABp pode receber sinais da célula P2, tornando ABp diferente de ABa e definindo o futuro eixo dorso-ventral do verme (Figura 22-21).
Ao mesmo tempo, a P2 também expressa outra molécula-sinal, a proteína Wnt, a qual atua no receptor Wnt (uma proteína Frizzled) na membrana da célula EMS. Este sinal po-lariza a célula EMS em relação ao seu local de contato com P2, controlando a orientação do fuso mitótico. A célula EMS então se divide para originar duas células-filhas que se tornam comprometidas para destinos diferentes como resultado do sinal Wnt de P2. Uma filha, a célula MS, originará músculos e várias outras partes do corpo; a outra filha, a célula E, é a célula fundadora do intestino, comprometida em originar todas as células do intestino e de nenhum outro tecido (ver Figura 22-21).
Figura 22-20 O padrão de divisões celula res em um embrião jovem de C. elegans, in dicando os nomes e os destinos das célu las individuais. As célu-las que são irmãs são mostradas ligadas por uma linha preta curta. (Se gundo K. Kemphues, Cell 101:345-348, 2000. Com permissão de Elsevier.)
Ovo fertilizado
ANTERIOR POSTERIOR
P1AB(pele,neurônios,faringe eoutros)
P2
ABp
ABaEMS
E(intestino)
C(músculos,pele eneurônios)
D(músculos)
MS(músculos eoutras partesdo corpo)
P4(linhagemgerminativa)
Delta
Notch
Frizzled
Wnt
Aba
Aba
EMS
Futuracélula E
P2
P2
ABp
ABp
Futuracélula MS
Figura 22-21 Vias de sinalização celular controlando a designação de dife-rentes característi cas para as células em um embrião nematoide de quatro células. A célula P2 utiliza a via de sinalização de Notch para enviar um sinal indutor para a célula ABp, induzindo-a a adotar uma caracte rística especiali-zada. A célula ABa possui todo o aparato molecular para responder da mes-ma maneira ao mesmo sinal, mas ela não o faz, porque não está em contato com P2. Enquanto isso, um sinal Wnt da célula P2 induz a célula EMS a orientar o seu fuso mitótico e a gerar duas filhas que se tornam comprometidas com destinos diferentes, como resultado de suas exposições distintas à proteína Wnt – a célula MS e a célula E (a célula fundadora do intestino).
Alberts_22.indd 1324Alberts_22.indd 1324 29.07.09 16:49:1829.07.09 16:49:18

Biologia Molecular da Célula 1325
Tendo sido descrita a cadeia de causa e efeito no desenvolvimento inicial dos nema-toides, examinaremos agora alguns dos métodos que têm sido usados para decifrá-la.
A microcirurgia e a genética revelam a lógica do controle do desenvolvimento; a clonagem de genes e o seu sequenciamento revelam seus mecanismos molecularesPara descobrir os mecanismos causais, precisamos conhecer o potencial de desenvolvi-mento das células individuais no embrião. Em que momentos de suas vidas elas sofrem alterações internas decisivas que as determinam para um destino particular, e em que mo-mentos dependem dos sinais de outras células? No nematoide, a microcirurgia de micro-emissão laser pode ser usada para matar uma ou mais células vizinhas, e então observar diretamente como a célula comporta-se em circunstâncias alteradas. Alternativamente, as células de um embrião inicial podem ser empurradas e rearranjadas dentro da casca do ovo pelo uso de uma fina agulha. Por exemplo, as posições relativas de ABa e de ABp podem ser trocadas no estágio de desenvolvimento de quatro células. A célula ABa passa pelo que nor-malmente seria o programado para a célula ABp, e vice-versa, mostrando que as duas células inicialmente possuem o mesmo potencial de desenvolvimento e dependem dos sinais das suas vizinhas para torná-las diferentes. Uma terceira tática é remover a casca do ovo de um embrião inicial de C. elegans pela sua digestão com enzimas e, então, manipular as células em cultura. A existência de um sinal polarizante de P2 para EMS foi demonstrada dessa ma-neira.
Foram usadas sondagens genéticas para identificar genes envolvidos na interação das células P2-EMS. Procurou-se por linhagens mutantes de vermes nos quais nenhuma célu-la de intestino fosse induzida (chamados de mutantes Mom, porque possuem mais meso-derma, do inglês more mesoderm – o mesoderma sendo o destino de ambas as células-filhas de EMS, quando a indu ção falha). A clonagem e o sequenciamento dos genes Mom revela-ram que um codifica a proteína-sinal Wnt que é expressa na célula P2, e o outro codifica uma proteína Frizzled (um receptor Wnt) que é expressa na célula EMS. Uma segunda sondagem genética foi conduzida em busca de linhagens mutantes dos vermes com o fenótipo opos-to, nos quais células extras de intestino são induzidas (chamados de mutantes Pop, porque apresen tam uma faringe posterior, do inglês posterior pharynx). Um dos genes Pop (Pop1) codifica uma proteína de regulação gênica (um homólogo de LEF1/TCF) cuja atividade é diminuída pela sinalização de Wnt em C. elegans. Quando a atividade de Pop1 está ausente, ambas as filhas das células EMS comportam-se como se tivessem recebido o sinal Wnt de P2. Foram usados métodos genéticos semelhantes para identificar os genes cujos produtos medeiam a sinalização dependente de Notch de P2 para ABa.
Continuando neste caminho, é possível construir uma representação detalhada dos eventos decisivos no desenvolvimento do nematoide e da maquinaria geneticamente espe-cífica que os controlam.
As células alteram suas capacidades de resposta aos sinais do desenvolvimento ao longo do tempoA complexidade do corpo do nematoide adulto é alcançada por meio do uso repetido de uma série de mecanismos formadores de padrões, incluindo aqueles que acabamos de ver em ação no embrião inicial. Por exemplo, as divisões celulares com assimetria molecular são depen dentes das proteínas de regulação gênica Pop1, presentes durante todo o desen-volvimento de C. elegans, criando células-irmãs anteriores e posteriores com diferentes ca-racterísticas.
Como enfatizado anteriormente, enquanto os mesmos poucos tipos de sinais atuam repetidamente em tempos e locais distintos, os efeitos que eles provocam são diferentes por-que as células são programadas para responder diferentemente de acordo com a sua idade e o seu histórico. Vimos, por exemplo, que no estágio do desenvolvimento de quatro células, uma célula, ABp, altera seu potencial de desenvolvimento devido a um sinal recebido atra-vés da via de sinalização de Notch. No estágio de desenvolvimento de 12 células, as netas da célula ABp e as netas da célula ABa encontram outro sinal de Notch, desta vez oriundo de uma célula-neta EMS. A neta de ABa altera o seu estado interno em resposta a este sinal e inicia a formação da faringe. A neta de ABp não o faz – a exposição inicial ao sinal de Notch tornou-a não-responsiva. Portanto, em diferentes momentos na sua história, ambas as li-
Alberts_22.indd 1325Alberts_22.indd 1325 29.07.09 16:49:1829.07.09 16:49:18

1326 Alberts, Johnson, Lewis, Raff, Roberts & Walter
nhagens celulares ABa e ABp respondem a Notch, porém os resultados são diferentes. De alguma maneira, um sinal de Notch no estágio de 12 células induz a faringe, mas um sinal de Notch no estágio de quatro células possui outros efeitos – os quais incluem a prevenção da indução da faringe por este mesmo sinal em um estágio mais tardio. Este fenômeno, onde um mesmo mecanismo de sinalização tem efeitos distintos em estágios diferentes e em con-textos diferentes, é visto no desenvolvimento de todos os animais, e em todos eles a via de sinalização de Notch é utilizada desta forma, repetidamente.
Os genes heterocrônicos controlam o tempo no desenvolvimentoUma célula não precisa receber um sinal externo para alterar-se: um conjunto de molécu-las reguladoras dentro da célula pode provocar a produção de outro, e a célula pode então passar por uma série de diferentes estados de maneira autônoma. Esses estados diferem não somente em sua capacidade de resposta a sinais externos, mas também em outros aspec-tos da sua química interna, incluindo as proteínas que interrompem ou iniciam o ciclo de divisão celular. Dessa forma, os mecanismos internos da célula, junta mente com os sinais re cebidos no passado e no presente, ditam a sequência das alterações bio químicas na célula e o momento das suas divi sões celulares.
Os detalhes moleculares específicos dos me canismos que governam o programa tem-poral do desenvolvimento ainda são desconhecidos. Extraordinariamente, pouco se sabe, mesmo no embrião nematoide com o seu padrão rigidamen te previsível de divisões ce-lulares, a respeito de como a sequência das divisões celulares é con trolada. Entretanto, nos últimos estágios, quando a larva alimenta-se, cresce e sofre a muda para tornar-se um adulto, é possível identificar alguns dos genes que controlam os momentos dos eventos celulares. As mutações nesses genes causam fenótipos heterocrônicos: as células em uma larva de um estágio comportam-se como se pertencessem a uma larva de um estágio dife-rente, ou as células no adulto continuam dividindo-se como se pertencessem a uma larva (Figura 22-22).
Por meio de análises genéticas, pode-se determinar que os produtos dos genes hetero-crônicos agem em série, formando cascatas reguladoras. Curiosamente, dois genes no início das suas respectivas cascatas, chamados de Lin4 e Let7, não codificam proteínas, mas molé-culas de microRNAs – pequenas moléculas de RNA regulador não-traduzidas, com 21 ou 22 nucleotídeos. Estas atuam pela ligação a sequências complementares nas regiões não-codi-ficantes das molé culas de mRNA transcritas de outros genes heterocrônicos, inibindo, assim, sua tradução e promovendo a sua degradação, como discutido no Capítulo 7. O aumento dos níveis do RNA de Lin4 controla a progressão do comportamento celular do estágio 1 de larva para o comportamento celular do estágio 3 de larva; o aumento dos níveis do RNA de Let7 controla a progressão da larva tardia para o adulto. Na realidade, Lin4 e Let7 foram os
Figura 22-22 Mutações heterocrôni-cas no gene Lin-14 de C. elegans. São mostrados os efeitos em somente uma das muitas linhagens afetadas. A muta-ção com perda de função (recessiva) em Lin14 causa uma ocorrência prematura do padrão de divisão celular e as ca-racterísticas de diferenciação da larva tardia, de maneira que o animal alcança o seu estágio final de modo pre maturo e com um número anormalmente pe queno de células. A mutação com ganho de função (dominante) provoca o efeito oposto, induzindo as células a reiterarem os seus padrões de divi-são celular característicos do primeiro estágio larval, continuando por cinco ou seis ciclos de mudas e persistindo na produção de um tipo imaturo de cutí-cula. A cruz denota uma morte celular programada. As linhas verdes represen-tam as células que contêm a proteína Lin14 (que se liga ao DNA); as linhas vermelhas represen tam as células que não contêm essa proteína. No desenvol-vimento normal, o desaparecimen to de Lin14 é desencadeado pelo início da ali-mentação larval. (Segundo A. Ambros e H. R. Horvitz, Science 226:409-416, 1984, com permissão de AAAS; e P. Arasu, B. Wightman e G. Ruvkun, Growth Dev. Aging 5:1825-1833, 1991, com permis-são da Growth Publishing Co., Inc.)
Segundoestágiode larva
Quartoestágiode larva
Terceiroestágiode larva
Primeiroestágiode larva
TT T
Tiposelvagem
Mutante Lin14 comperda de função
Mutante Lin14 comganho de função
Tem
po
Alberts_22.indd 1326Alberts_22.indd 1326 29.07.09 16:49:1829.07.09 16:49:18

Biologia Molecular da Célula 1327
primeiros microRNAs a serem descritos nos animais: por meio de estudos da genética do de-senvolvimento em C. elegans foi descoberta a importância de toda essa classe de moléculas para a regulação gênica.
As moléculas de RNA que são idênticas ou quase idênticas ao RNA Let7 são encon tradas em muitas outras espécies, incluindo-se a Drosophila, o peixe-zebra e os humanos. Além disso, esses RNAs parecem atuar de maneira semelhante para regular o nível das suas mo-léculas de mRNA-alvo, e os próprios alvos são homólogos aos alvos do RNA Let7 do nema-toide. Em Drosophila, este sistema de moléculas parece estar envolvido na metamorfose da forma larval para mosca, mantendo conservado o seu papel de controlar o curso das transi-ções envolvidas no desenvolvimento.
As células não contam as divisões celulares para cronometrar seus programas internosUma vez que os passos da especialização celular têm de ser coordenados com as divi sões ce-lulares, frequentemente é sugerido que o ciclo de divisão celular deveria servir como um re-lógio no controle do ritmo dos outros eventos no desenvolvimento. Sob este ponto de vista, as alterações nos estados internos estariam condicionadas a sua passagem pelos ciclos de divi-são: a célula passaria para o próximo estado se sofresse mitose, por exemplo. Apesar de haver alguns casos onde as alterações no estado celular estão condicionadas aos eventos do ciclo celular, isto está longe de ser uma regra geral. As célu las dos embriões em desenvolvimento, sejam eles vermes, moscas ou vertebrados, normalmente mantêm seu cronograma-padrão de determinação e de diferenciação, mesmo quando o progresso pelo ciclo de divisão celular é bloqueado de maneira artificial. Existem neces sariamente algumas anormalidades, pelo menos porque uma célula única que não se dividiu não pode diferenciar-se de duas maneiras de uma só vez. Contudo, na maioria dos casos que têm sido estudados, parece claro que a célula altera o seu estado com o tempo, de forma mais ou menos independente da divisão celular, e que esta alteração de estado controla a decisão de dividir-se, assim como a decisão de quando e como se especializar.
Células selecionadas morrem por apoptose como parte do programa de desenvolvimentoO controle dos números celulares no desenvolvimento depende tanto da morte celular quanto da divisão celular. Um hermafrodita de C. elegans gera 1.030 núcleos somáticos celu-lares no curso do seu desenvolvimento, mas 131 das células morrem. Estas mortes celulares programadas ocorrem em um padrão absolutamente previsível. Em C. elegans, elas podem ser descritas em detalhes, pois é possível rastrear o destino de cada célula individual e ver quem morre, observando como cada vítima suicida sofre apoptose e é rapidamente engol-fada e digerida pelas células vizinhas (Figura 22-23). Em outros orga nismos, em que uma observação detalhada é mais difícil, tais mortes facilmente passam despercebidas; mas a morte celular por apoptose provavelmente seja o destino de uma fração substancial das cé-lulas produzidas pela maioria dos animais, desempenhando uma parte essencial na geração de um indivíduo com os tipos celulares certos, em números e locais certos, como discutido no Capítulo 18.
As sondagens genéticas em C. elegans têm sido cruciais na identificação dos genes que desencadeiam a apoptose e em salientar a sua importância no desenvolvimento. Desco-briu-se que três genes, chamados de Ced3, Ced4 e Egl1 (CED, morte celular anor mal, de cell death abnormal), são necessários para ocorrerem as 131 mortes celulares normais. Se esses genes são inativados por mutação, as células que normalmente são destinadas a morrer so-brevivem, diferenciando-se como tipos celulares reconhecidos, como neurônios. Por outro lado, a superex pressão ou a expressão em local errôneo dos mesmos genes causa a morte de
Figura 22-23 Morte celular apoptótica em C. elegans. A morte depende da expressão dos genes Ced3 e Ced4 na ausência da expressão de Ced9 – to-dos na própria célula que está mor rendo. O subsequente engolfamento e a remo ção dos restos dependem da expressão de ou tros genes nas células vizinhas.
A célula comete suicídio
A célula morta é englobada pelacélula vizinha
Os restos celularessão digeridos semdeixar resquícios
Alberts_22.indd 1327Alberts_22.indd 1327 29.07.09 16:49:1829.07.09 16:49:18

1328 Alberts, Johnson, Lewis, Raff, Roberts & Walter
muitas células que normalmente sobreviveriam. O mesmo efeito resulta de mutações que inati vam outro gene, Ced9, o qual normalmente reprime o programa de morte celular.
Todos esses genes codificam para componentes conservados da maquinaria de morte celular. Como descrito no Capítulo 18, o Ced3 codifica para um homólogo de caspases, en-quanto o Ced4, o Ced9 e o Egl1 são respectivamente homólogos de Apaf1, Bcl2 e Bad. Sem a compreensão obtida pela análise detalhada do desenvolvimento desse ver me nematoide transparente e geneticamente maleável, teria sido muito mais difícil desco brir esses genes e entender o processo de morte celular nos vertebrados.
ResumoO desenvolvimento do pequeno verme nematoide transparente e relativamente simples Caenorhab-ditis elegans é extraordinariamente reproduzível e tem sido descrito em deta lhes, pois uma célula em qualquer posição no corpo tem a mesma linhagem em todos os indivíduos, e essa linhagem é totalmente conhecida. Além disso, o seu genoma foi completa mente sequenciado. Assim, estraté-gias genéticas e técnicas microcirúrgicas podem ser combinadas para decifrar os mecanismos do desenvolvimento. Como em outros orga nismos, o desenvolvimento depende da ação recíproca de interações célula-célula e de processos celulares autônomos. O desenvolvimento inicia com uma divisão assimétrica do ovo fertilizado, dividindo-o em duas células menores contendo diferentes determinantes de destino celu lares. As filhas dessas células interagem através das vias de sinaliza-ção celulares de Notch e Wnt para criar uma série mais diversa de estados celulares. Enquanto isso, por divisões assimétricas adicionais, uma célula herda materiais do ovo que a determinam, em um estágio precoce, como progenitora da linhagem germinativa.
As sondagens genéticas identificam os conjuntos de genes responsáveis por estas e por etapas mais tardias do desenvolvimento, incluindo, por exemplo, os genes de morte celular que controlam a apoptose de um subconjunto específico de células como parte do programa normal de desenvolvi-mento. Os genes heterocrônicos que governam a duração dos eventos no desenvolvimento também foram identificados, embora, em geral, ainda tenhamos pouco entendimento sobre o controle tem-poral do desenvolvimento. Existem boas evidências, entretanto, de que o ritmo do desenvolvimento não é ajustado pela con tagem das divisões celulares.
DROSOPHILA E A GENÉTICA MOLECULAR DA FORMAÇÃO DE PADRÕES: A GÊNESE DO PLANO CORPORALA mosca Drosophila melanogaster (Figura 22-24), mais do que qualquer outro organismo, transformou o nosso conhecimento de como os genes governam a formação de padrões do corpo. A anatomia da Drosophila é mais complexa do que a de C. elegans, superando em
Figura 22-24 Drosophila melanogas-ter. Vista dorsal de uma mosca normal adulta. (A) Fotografia. (B) Desenho ilustrativo. (Fotografia cortesia de E. B. Lewis.)
(A)
(B)
Antena
Olho
Tórax
Cabeça
Haltere
Asa
Pata Abdome
Alberts_22.indd 1328Alberts_22.indd 1328 29.07.09 16:49:1929.07.09 16:49:19

Biologia Molecular da Célula 1329
cem vezes o seu número de células, e mostra paralelos mais óbvios com a nossa própria es-trutura corporal. Surpreendentemente, a mosca tem menos genes que o verme – cerca de 14 mil e 20 mil, respectivamente. Por outro lado, ela tem o dobro de DNA por gene (cerca de 10 mil nucleotídeos em média, se comparado com cerca de 5 mil do verme), a maior parte sen-do de DNA não-codificante. O conjunto de construção molecular tem um número menor de partes, mas as instruções de montagem – como especificado pelas sequências reguladoras no DNA não- codificante – parecem ser mais volumosas.
Décadas de estudo genético, culminando nas massivas sondagens genéticas, produzi-ram um catálogo dos genes de controle do desenvolvimento que definem o padrão espa-cial dos tipos celulares e das estruturas do corpo da mosca; e a biologia molecular tem nos fornecido ferramentas para observar esses genes em ação. Pela hibridização in situ, usan do sondas de DNA ou de RNA em embriões inteiros, ou pela coloração com anticorpos mar-cados para revelar a distribuição de proteínas específicas, pode-se observar diretamente como os estados internos das células são definidos pelos conjuntos de genes regu ladores que elas expressam em diferentes momentos do desenvolvimento. Além disso, pela análise de animais que são uma quimera de células mutantes e não–mutantes, pode-se descobrir como cada gene opera como parte de um sistema que espe cifica a organização do corpo.
A maioria dos genes que controlam o padrão do corpo da Drosophila tem contrapartes próximas em animais superiores, incluindo nós mesmos. De fato, muitos dos mecanismos básicos para a definição da estrutura do corpo e a formação dos padrões de órgãos e de teci-dos individuais são incrivelmente similares. Assim, surpreendentemente, a mosca for neceu a chave para o entendimento da genética molecular do nosso próprio desenvolvimento.
As moscas, assim como os vermes nematoides, são ideais para estudos genéticos: de baixo custo para criar, fáceis de mutar e rápidas em seu ciclo reprodutivo. Contudo, existe uma razão mais fundamental para elas serem tão importantes para os geneticistas do desen-volvimento. Como enfatizado anteriormente, como resultado das duplicações gênicas, os genomas dos vertebrados frequentemente contêm dois ou três genes homólogos que corres-pondem a um único gene na mosca. Uma mutação que destrua um desses genes muitas ve-zes não consegue revelar a função central do gene, pois os outros homólogos compartilham a função e mantêm-se ativos. Na mosca, com o seu conjunto gênico mais econômico, este fenômeno de redundância genética é menos prevalente. Assim, o fenômeno de uma única mutação na mosca, mais comumente, revela a função do gene mutante.
O corpo do inseto é construído como uma série de unidades segmentaresO cronograma de desenvolvimento da Drosophila, do ovo ao adulto, está resumido na Figu-ra 22-25. O período de desenvolvimento embrionário começa na fertilização e leva em torno de um dia, ao final do qual o embrião eclode da casca do ovo para tornar-se uma larva. A larva passa então por três estágios, ou ínstares, separados por mudas nas quais ela perde a sua camada antiga de cutícula e produz uma maior. No final do terceiro ínstar ela forma uma pupa. No interior da pupa ocorre um remodelamento radical do corpo – um pro cesso cha-mado de metamorfose. Eventualmente, cerca de nove dias após a fertilização, uma mosca adulta, ou imago, emerge.
A mosca consiste em uma cabeça, com boca, olhos e antenas, seguidas por três seg-mentos torácicos (numerados T1 a T3) e oito ou nove segmentos abdominais (numerados A1 a A9). Cada segmento, embora diferente dos outros, é produzido de acordo com um pla-no semelhante. O segmento T1, por exemplo, carrega um par de patas; T2 carrega um par de patas mais um par de asas, e T3 carrega um par de patas mais um par de halteres – pequenos balanceadores em forma de saliências arredondadas importantes para o voo, evoluídos a partir do segundo par de asas que os insetos mais primitivos possuem. A segmentação, qua-se repetitiva, desenvolve-se no embrião inicial durante as pri meiras horas após a fertilização (Figura 22-26), mas é mais óbvia na larva (Figura 22-27), na qual os segmentos parecem mais similares do que no adulto. No embrião, pode ser visto que os rudimentos da cabeça, ou pelo menos as futuras estruturas da boca do adulto, também são segmentares. Nas duas extremidades do animal, entretanto, existem estrutu ras terminais altamente especializadas que não são derivadas de segmentação.
Os limites entre os segmentos são tradicionalmente definidos por marcadores anatô-micos visíveis; contudo, na discussão dos padrões de expressão gênica, é conveniente de-senhar um con junto diferente de limites segmentares, definindo uma série de unidades
1 mm
Adulto
5 dias
1 dia
0 dias
FERTILIZAÇÃOOvo
DESENVOLVIMENTOEMBRIONÁRIO
ECLOSÃO
Larva
Três estágios larvais,separados por mudas
FORMAÇÃO DA PUPA
Pupa
METAMORFOSE
9 dias
Figura 22-25 Resumo do desenvol-vimento da Drosophila, do ovo até a mosca adulta.
Alberts_22.indd 1329Alberts_22.indd 1329 29.07.09 16:49:1929.07.09 16:49:19
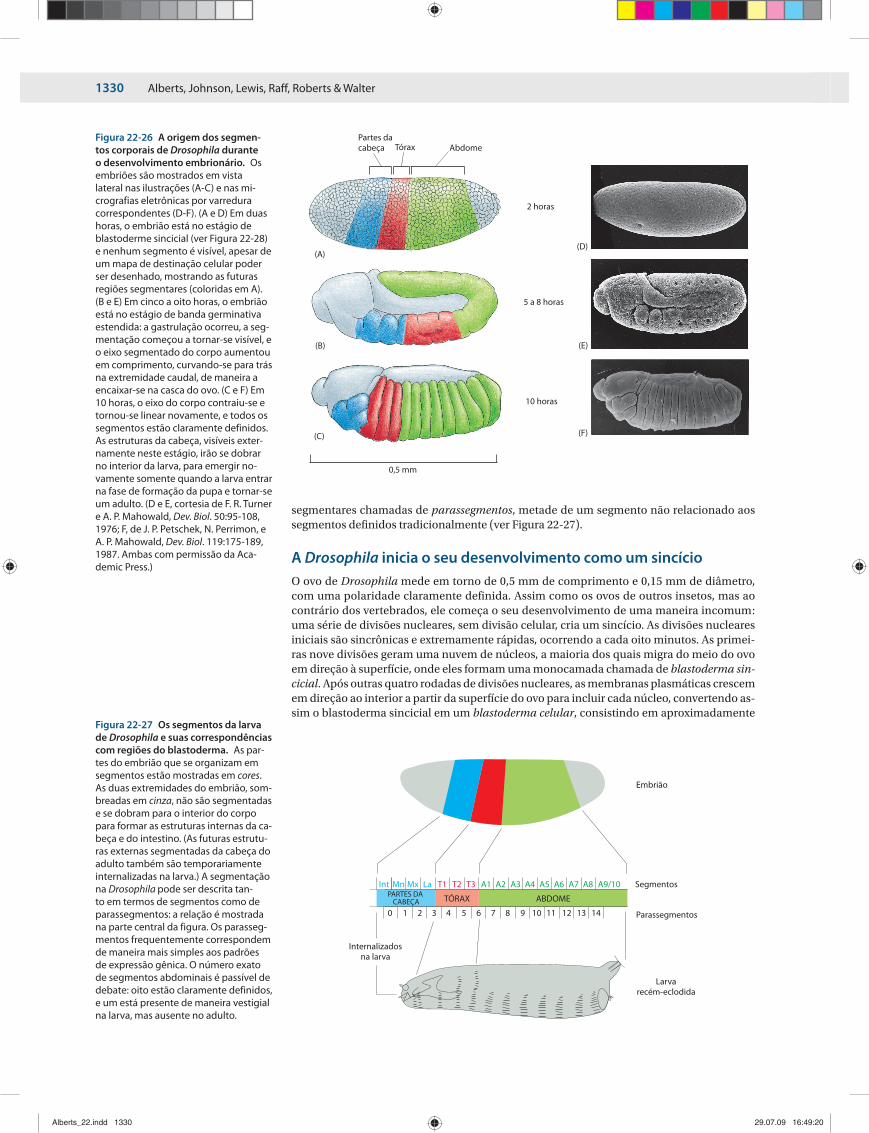
1330 Alberts, Johnson, Lewis, Raff, Roberts & Walter
segmentares cha madas de parassegmentos, metade de um segmento não relacionado aos segmentos definidos tradicionalmente (ver Figura 22-27).
A Drosophila inicia o seu desenvolvimento como um sincícioO ovo de Drosophila mede em torno de 0,5 mm de comprimento e 0,15 mm de diâmetro, com uma polaridade claramente definida. Assim como os ovos de outros insetos, mas ao contrário dos vertebrados, ele começa o seu desenvolvimento de uma maneira incomum: uma série de divisões nucleares, sem divisão celular, cria um sincício. As divisões nucleares iniciais são sincrônicas e extremamente rápidas, ocorrendo a cada oito minutos. As primei-ras nove divisões geram uma nuvem de núcleos, a maioria dos quais migra do meio do ovo em direção à superfície, onde eles formam uma monocamada chamada de blastoderma sin-cicial. Após outras quatro rodadas de divisões nucleares, as membranas plasmáticas crescem em direção ao interior a partir da superfície do ovo para incluir cada núcleo, convertendo as-sim o blastoderma sincicial em um blastoderma celu lar, consistindo em aproximadamente
Figura 22-26 A origem dos segmen-tos corporais de Drosophila durante o desenvolvimento embrionário. Os embriões são mostrados em vista lateral nas ilustrações (A-C) e nas mi-crografias eletrônicas por varredura correspondentes (D-F). (A e D) Em duas horas, o embrião está no estágio de blastoderme sincicial (ver Figura 22-28) e nenhum segmento é visível, apesar de um mapa de destinação celular poder ser desenhado, mostrando as futuras regiões segmentares (coloridas em A). (B e E) Em cinco a oito horas, o embrião está no estágio de banda germi nativa estendida: a gastrulação ocorreu, a seg-mentação começou a tornar-se visível, e o eixo seg mentado do corpo aumentou em comprimento, curvando-se para trás na extremidade caudal, de maneira a encaixar-se na casca do ovo. (C e F) Em 10 horas, o eixo do corpo contraiu-se e tor nou-se linear novamente, e todos os segmentos estão claramente definidos. As estruturas da ca beça, visíveis exter-namente neste estágio, irão se dobrar no interior da larva, para emergir no-vamente somente quando a larva entrar na fase de formação da pupa e tornar-se um adulto. (D e E, cortesia de F. R. Turner e A. P. Mahowald, Dev. Biol. 50:95-108, 1976; F, de J. P. Petschek, N. Perrimon, e A. P. Mahowald, Dev. Biol. 119:175-189, 1987. Ambas com permissão da Aca-demic Press.)
(D)
(E)
(F)
(A)
(B)
(C)
Partes dacabeça Tórax Abdome
0,5 mm
2 horas
5 a 8 horas
10 horas
Figura 22-27 Os segmentos da larva de Drosophila e suas correspondências com regiões do blastoderma. As par-tes do em brião que se organizam em segmentos estão mos tradas em cores. As duas extremidades do em brião, som-breadas em cinza, não são segmenta das e se dobram para o interior do corpo para formar as estruturas internas da ca-beça e do in testino. (As futuras estrutu-ras externas segmen tadas da cabeça do adulto também são tempo rariamente internalizadas na larva.) A segmentação na Drosophila pode ser descrita tan-to em termos de segmentos como de parasseg mentos: a relação é mostrada na parte central da figura. Os parasseg-mentos frequentemente correspondem de maneira mais simples aos pa drões de expressão gênica. O número exato de segmentos abdominais é passível de debate: oito estão claramente definidos, e um está presente de maneira vestigial na larva, mas ausente no adulto.
Mn Mx La T1 T2 T3 A1 A2 A3 A4 A5 A6 A7 A8 A9/10
1413121110987654321
Embrião
Parassegmentos
SegmentosInt
0
PARTES DA CABEÇA TÓRAX ABDOME
Larvarecém-eclodida
Internalizadosna larva
Alberts_22.indd 1330Alberts_22.indd 1330 29.07.09 16:49:2029.07.09 16:49:20
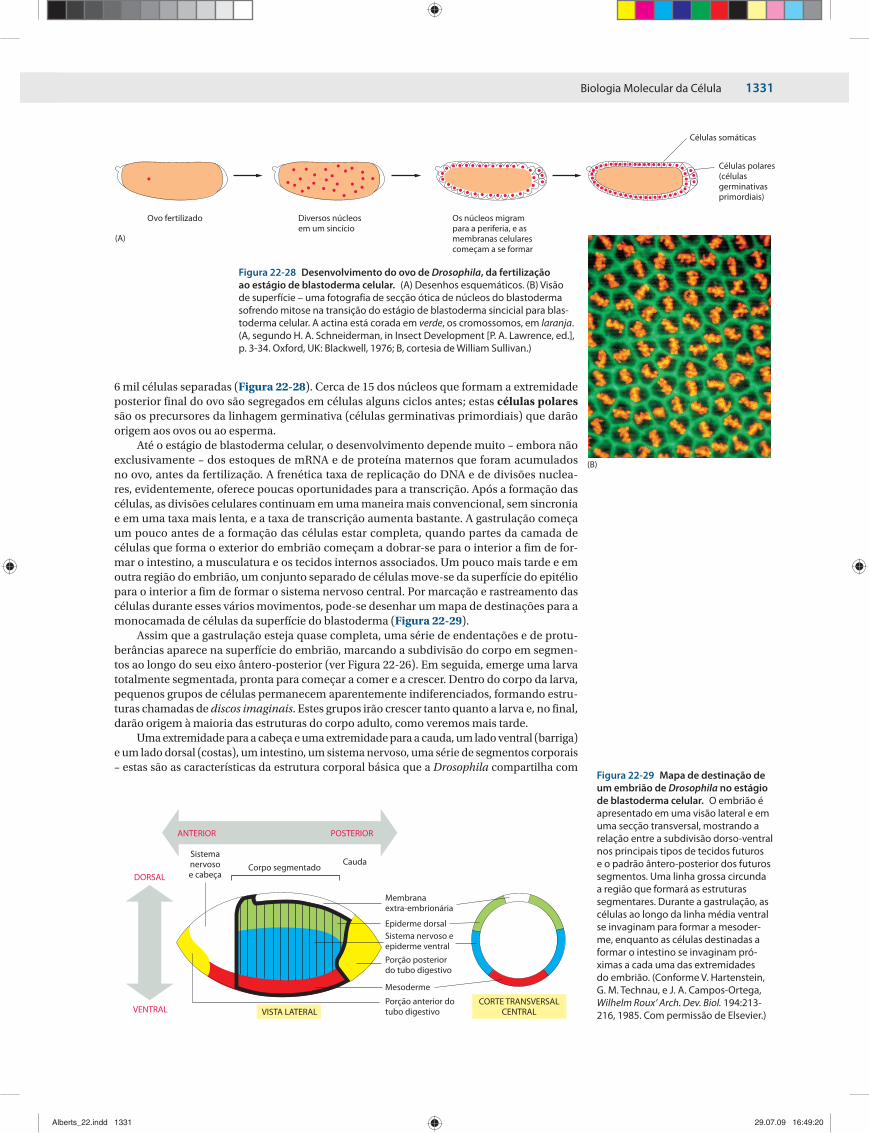
Biologia Molecular da Célula 1331
6 mil células separadas (Figura 22-28). Cerca de 15 dos núcleos que formam a extremidade posterior final do ovo são segregados em células alguns ciclos antes; estas células polares são os precursores da linhagem germinativa (células germinativas primordiais) que darão origem aos ovos ou ao esperma.
Até o estágio de blastoderma celular, o desenvolvimento depende muito – embora não exclusivamente – dos estoques de mRNA e de proteína maternos que foram acumulados no ovo, antes da fertilização. A frenética taxa de replicação do DNA e de divisões nuclea-res, evidentemente, oferece poucas oportunidades para a transcrição. Após a formação das células, as divisões celulares continuam em uma maneira mais convencional, sem sincronia e em uma taxa mais lenta, e a taxa de transcrição aumenta bastante. A gastrulação começa um pouco antes de a formação das células estar completa, quando partes da camada de células que forma o exterior do embrião começam a dobrar-se para o interior a fim de for-mar o intestino, a musculatura e os tecidos internos associados. Um pouco mais tarde e em outra região do embrião, um conjunto separado de células move-se da superfície do epitélio para o interior a fim de formar o sistema nervoso central. Por marcação e rastreamento das células durante esses vários movimentos, pode-se desenhar um mapa de destinações para a monocamada de células da superfície do blastoderma (Figura 22-29).
Assim que a gastrulação esteja quase completa, uma série de endentações e de protu-berâncias aparece na superfície do embrião, marcando a subdivisão do corpo em segmen-tos ao longo do seu eixo ântero-posterior (ver Figura 22-26). Em seguida, emerge uma larva totalmente segmentada, pronta para começar a comer e a crescer. Dentro do corpo da larva, pequenos grupos de células permanecem aparentemente indiferenciados, formando estru-turas chamadas de discos imaginais. Estes grupos irão crescer tanto quanto a larva e, no final, darão origem à maioria das estruturas do corpo adulto, como veremos mais tarde.
Uma extremidade para a cabeça e uma extremidade para a cauda, um lado ventral (barriga) e um lado dorsal (costas), um intestino, um sistema nervoso, uma série de seg mentos corporais – estas são as características da estrutura corporal básica que a Droso phila compartilha com
(A)
Ovo fertilizado Diversos núcleosem um sincício
Os núcleos migrampara a periferia, e asmembranas celularescomeçam a se formar
(B)
Células somáticas
Células polares(células germinativasprimordiais)
Figura 22-28 Desenvolvimento do ovo de Drosophila, da fertilização ao estágio de blastoderma celular. (A) Desenhos esquemá ticos. (B) Visão de superfície – uma fotografia de secção ótica de núcleos do blastoderma sofren do mitose na transição do estágio de blastoder ma sincicial para blas-toderma celular. A actina está corada em verde, os cromossomos, em la ranja. (A, segundo H. A. Schneiderman, in In sect Development [P. A. Lawrence, ed.], p. 3-34. Oxford, UK: Blackwell, 1976; B, cortesia de William Sullivan.)
ANTERIOR POSTERIOR
DORSAL
VENTRAL
Sistemanervosoe cabeça
Corpo segmentadoCauda
Membranaextra-embrionária
Epiderme dorsalSistema nervoso eepiderme ventral
Porção posteriordo tubo digestivo
Mesoderme
Porção anterior dotubo digestivoVISTA LATERAL
CORTE TRANSVERSALCENTRAL
Figura 22-29 Mapa de destinação de um embrião de Drosophila no estágio de blas toderma celular. O embrião é apresentado em uma visão lateral e em uma secção transversal, mostrando a relação entre a subdivisão dorso-ventral nos principais tipos de tecidos futuros e o padrão ântero-posterior dos futuros segmen tos. Uma linha grossa circunda a região que formará as estruturas segmentares. Durante a gas trulação, as células ao longo da linha média ven tral se invaginam para formar a mesoder-me, en quanto as células destinadas a formar o intestino se invaginam pró-ximas a cada uma das extremidades do embrião. (Conforme V. Hartens tein, G. M. Technau, e J. A. Campos-Ortega, Wilhelm Roux’ Arch. Dev. Biol. 194:213-216, 1985. Com permissão de Elsevier.)
Alberts_22.indd 1331Alberts_22.indd 1331 29.07.09 16:49:2029.07.09 16:49:20

1332 Alberts, Johnson, Lewis, Raff, Roberts & Walter
muitos outros animais, incluindo os humanos. Começaremos nos so relato dos mecanismos de desenvolvimento da Drosophila considerando como esta estrutura corporal é produzida.
As sondagens genéticas definem os grupos de genes necessários aos aspectos específicos da formação dos padrões iniciaisPela realização de uma série de sondagens genéticas com base na mutagênese por satura ção (discutido no Capítulo 8), tem sido possível acumular uma coleção de mutantes de Drosophi-la que parecem incluir alterações em uma grande proporção dos genes que afetam o desen-volvimento. Mutações independentes no mesmo gene podem ser distinguidas de mutações em genes separados por um teste de complementação (ver Painel 8-1, p. 555), levando a um catálogo de genes classificados de acordo com os seus fenótipos mutantes. Nesse catálogo, um grupo de genes com fenótipos mutantes muito semelhantes frequentemente codificará um conjunto de proteínas que trabalham juntas para realizar uma função.
Algumas vezes as funções no desenvolvimento reveladas pelos fenótipos mutantes são aquelas que poderíamos esperar; outras vezes são uma surpresa. Uma sondagem genética em larga escala focando o desenvolvimento inicial da Drosophila revelou que os genes-chave formam um conjunto relativamente pequeno de classes funcionais definidas por seus fenó-tipos mutantes. Alguns – os genes de polaridade do ovo (Figura 22-30) – são necessários para
Figura 22-30 Domínios dos sistemas anterior, posterior e terminal dos ge-nes de polaridade do ovo. O diagrama superior mos tra os destinos das dife-rentes regiões do ovo/embrião inicial e indicam (em branco) as partes que fa-lham em desenvolver-se se os sistemas anterior, posterior ou terminal estão defectivos. A fileira do meio mostra esquematicamente o aparecimento de uma larva normal e de larvas mutantes que são defectivas em um gene do sis-tema anterior (p. ex., Bicoid), do sistema posterior (p. ex., Nanos) ou do sistema terminal (p. ex., Torso). A fileira de baixo dos desenhos mostra a aparência das larvas nas quais nenhum ou somente um dos três sistemas gênicos está funcional. As legendas abaixo de cada larva especificam quais sistemas estão in tactos (A P T para a larva normal, – P T para a larva onde o sistema anterior é defectivo, mas os sistemas posterior e terminal estão intactos, e assim por diante). A inativação de um sistema gê-nico particu lar causa a perda do conjun-to correspondente de estruturas do cor-po; as partes do corpo que se formam correspondem aos sistemas gênicos que permanecem funcionais. Note que as larvas com um defeito no sistema anterior ain da podem formar estruturas terminais na sua extremi dade anterior, mas estas são de um tipo normal mente encontrado na extremidade posterior do corpo e não na cabeça. (Ligeira-mente modificada de D. St. Johnston e C. Nuss lein-Volhard, Cell 68:201-219, 1992. Com permissão de Elsevier.)
Intestino e extremi-dade da cabeça
Partes da cabeça
Tórax
Abdome
Intestino e extre-midade posterior
ANTERIOR POSTERIOR TERMINAL
Normal Bicoid Nanos
A P T _ P T A _ T A P _
A ____ _ _ _ P T__
Torso
Alberts_22.indd 1332Alberts_22.indd 1332 29.07.09 16:49:2129.07.09 16:49:21

Biologia Molecular da Célula 1333
definir os eixos ântero-posterior e dorso-ventral do embrião e marcar as duas extremidades para destinos especiais por meio de mecanismos envolvendo interações entre o oócito e as células vizinhas no ovário. Outros, os genes gap, são necessários em regiões amplas e espe-cíficas ao longo do eixo ântero-posterior de um embrião inicial para permitir o seu desen-volvimento apropriado. Uma terceira categoria, os genes pair-rule, são necessários, mais sur-preendentemente, para o desenvolvimento de segmentos alternados do corpo. Uma quarta categoria, os genes de polaridade dos segmentos, é responsável pela organi zação do padrão ântero-posterior de cada segmento individual.
A descoberta desses quatro sistemas de genes, e a subsequente análise de suas funções (um trabalho ainda em andamento), foi uma famosa epopeia da genética do desenvolvi-mento. Ela teve um impacto revolucionário em toda a biologia do desenvolvimento ao apontar o caminho em direção a uma explicação sistemática e abrangente do controle genéti co do desenvolvimento embrionário. Nesta seção, resumiremos as conclusões re-lacionadas às fases iniciais do desenvolvimento da Drosophila, pois elas são específicas de insetos; abordaremos em maior extensão as partes do processo que ilustram princípios gerais.
As interações do oócito com seu ambiente definem os eixos do embrião: a função dosgenes de polaridade do ovoSurpreendentemente, as etapas mais iniciais do desenvolvimento animal estão entre as mais variáveis, mesmo dentro de um mesmo filo. Uma rã, uma galinha e um mamífero, por exem-plo, mesmo que desenvolvam-se de maneiras semelhantes mais tarde, produzem óvulos que diferem radicalmente em tamanho e em estrutura e começam o seu desenvolvimento com sequências diferentes de divisões celulares e eventos de especialização celular.
O estilo de desenvolvimento inicial que descrevemos para C. elegans é típico de mui-tas classes de animais. Por outro lado, o desenvolvimento inicial de Drosophila representa uma variação extrema. Os eixos principais do futuro corpo do inseto são definidos antes da fertilização por uma complexa troca de sinais entre o ovo não-fertilizado, ou oócito, e as células foliculares que o circundam no ovário (Figura 22-31). Então, na fase sincicial após a fertilização, uma quantidade excepcional de formação de padrões ocorre no conjunto de núcleos que se dividem rapidamente, antes da primeira divisão do ovo em células separa-das. Aqui, não há necessidade das formas comuns de comunicação célula-célula envol-vendo comunicação transmembrana; as regiões vizinhas do embrião inicial de Dro sophila podem comunicar-se por meio de proteínas de regulação e de moléculas de mRNA que se difundem ou que são ativamente transportadas através do citoplasma da célula gigante multinucleada.
Nos estágios anteriores à fertilização, o eixo ântero-posterior do futuro embrião se torna definido por três sistemas de moléculas que criam pontos de referência no oócito (Figura 22-32). Seguindo-se a fertilização, cada ponto de referência serve como um farol, fornecendo um sinal na forma de um gradiente de morfógenos, que organiza o processo de desenvolvimento na sua vizinhança. Dois desses sinais são gerados a partir de depósitos loca lizados de moléculas de mRNA específicas. A futura extremidade an terior do embrião contém uma grande concentração de mRNA para a proteína de regulação gênica chama-da de Bicoid; este mRNA é traduzido para produzir a proteína Bicoid, a qual se difunde
Figura 22-31 Um oócito de Drosophila no seu folículo. O oócito é derivado de uma célu la germinativa que se divide quatro vezes para originar uma família de 16 células que permane cem em comunicação umas com as outras atra vés de pontes citoplasmáticas (cin-za). Um mem bro do grupo da família torna-se o oócito, en quanto as outras células se tornam células auxi liares, que produzem muitos dos componen tes necessários pelo oócito e os transpor-tam para dentro dele através das pontes citoplasmáticas. As células foliculares que circundam parcialmente o oócito possuem uma ancestralidade separada. Como indicado, elas são as fontes dos sinais ter minal e ventral de polarização do ovo.
Célula auxiliar
Célula folicularOócito
Células folicularesfornecendo sinaisventraisCélulas foliculares
fornecendo sinaisterminais
Alberts_22.indd 1333Alberts_22.indd 1333 29.07.09 16:49:2129.07.09 16:49:21

1334 Alberts, Johnson, Lewis, Raff, Roberts & Walter
a partir de sua fonte para formar um gradiente de concentração com o seu máximo na extremidade anterior do ovo. A futura extremidade posterior do embrião contém uma alta concentração de mRNA para um regulador da tradu ção chamado de Nanos, o qual forma um gradiente posterior da mesma maneira. O terceiro sinal é gerado simetricamente em ambas as extre midades do ovo, pela ativação local de um receptor tirosina-cina se trans-membrana chamado de Torso. O receptor ativado exerce os seus efeitos em uma amplitu-de pequena, marcando os locais das estruturas terminais especializadas que irão formar as extremidades da cabeça e da cauda da futura larva e, também, definindo os rudimentos do futuro intestino. Os três conjuntos de genes responsáveis por esses determinantes lo-calizados são referidos como os conjuntos ante rior, posterior e terminal dos genes de polaridade do ovo.
Um quarto ponto de referência define o eixo dorso-ventral (ver Figura 22-32): uma pro-teína que é produzida pelas células foliculares abaixo da futura região ventral do em brião leva à ativação localizada de outro receptor transmembrana, chamado de Toll, na membra-na do oócito. Os genes necessários a essa função são chamados de genes dorso-ventrais de polaridade do ovo.
Todos os genes de polaridade do ovo, nessas quatro classes, são genes de efeito mater-no: é o genoma da mãe, não o genoma do zigoto, que é crítico. Assim, uma mosca cujos cro-mossomos são mutantes em ambas as cópias do gene Bicoid, mas que nasceu de uma mãe que possui uma cópia normal de Bicoid, desenvolve-se de maneira perfeitamente normal, sem nenhum defeito no padrão da cabeça. Entretanto, se aquela mosca-filha é uma fêmea, nenhum mRNA funcional de Bicoid pode ser depositado na porção anterior dos seus pró-prios ovos, e todos irão desenvolver-se em embriões sem cabeça, indepen dentemente do genótipo do pai.
Cada um dos quatro sinais de polaridade do ovo – fornecidos por Bicoid, Nanos, Torso e Toll – exerce seus efeitos pela regulação (direta ou indireta) da expressão de genes nos nú-cleos do blastoderma. O uso destas moléculas particulares para organizar o ovo não é uma característica geral do desenvolvimento inicial dos animais – na verdade, somente a Droso-phila e os insetos intimamente relacionados possuem um gene Bicoid. Toll foi aqui cooptado para a formação do padrão dorso-ventral; a sua função mais antiga e universal é a resposta imune inata, como discutido no Capítulo 24.
Apesar disso, o sistema de polaridade do ovo apresenta algumas características alta-mente conservadas. Por exemplo, a localização do mRNA de Nanos em uma extremidade do ovo está ligada e é dependente da localização dos determinantes da célula germinati va na-quele local, assim como em C. elegans. Mais adiante durante o desenvolvimento, enquanto o genoma do zigoto começa a atuar sob a influência do sistema de polaridade do ovo, mais semelhanças com outras espécies animais tornam-se aparentes. Utilizaremos o sistema dor-so-ventral para ilustrar este ponto.
Os genes de sinalização dorso-ventrais criam um gradiente de uma proteína nuclear de regulação gênicaA ativação localizada do receptor Toll no lado ventral do ovo controla a distribuição da Dorsal, uma proteína de regulação gênica que se encontra dentro do ovo. A proteína Dor-sal pertence à mesma família da proteína de regulação gênica NF�B de vertebrados (dis-cutida no Capítulo 15). A sua atividade regulada por Toll, assim como a de NF�B, depen de
Figura 22-32 A organização dos qua-tro sis temas de gradiente de polarida-de do ovo. Os receptores Toll e Torso são distribuídos por toda a membrana; o colorido nos diagramas à direita indica o local em que eles se tornam ati-vados pelos ligantes extracelulares.
SISTEMA DORSO-VENTRALSISTEMA TERMINALSISTEMA ANTERIORSISTEMA POSTERIOR
Receptores transmembrana (Toll)Receptores transmembrana (Torso)mRNA localizado (Bicoid ) mRNA localizado (Nanos )
Determinam• Ectoderme vs. mesoderme vs. endoderme• Estruturas terminais
Determinam• Células germinativas vs. células somáticas• Cabeça vs. cauda• Segmentos corporais
Alberts_22.indd 1334Alberts_22.indd 1334 29.07.09 16:49:2129.07.09 16:49:21
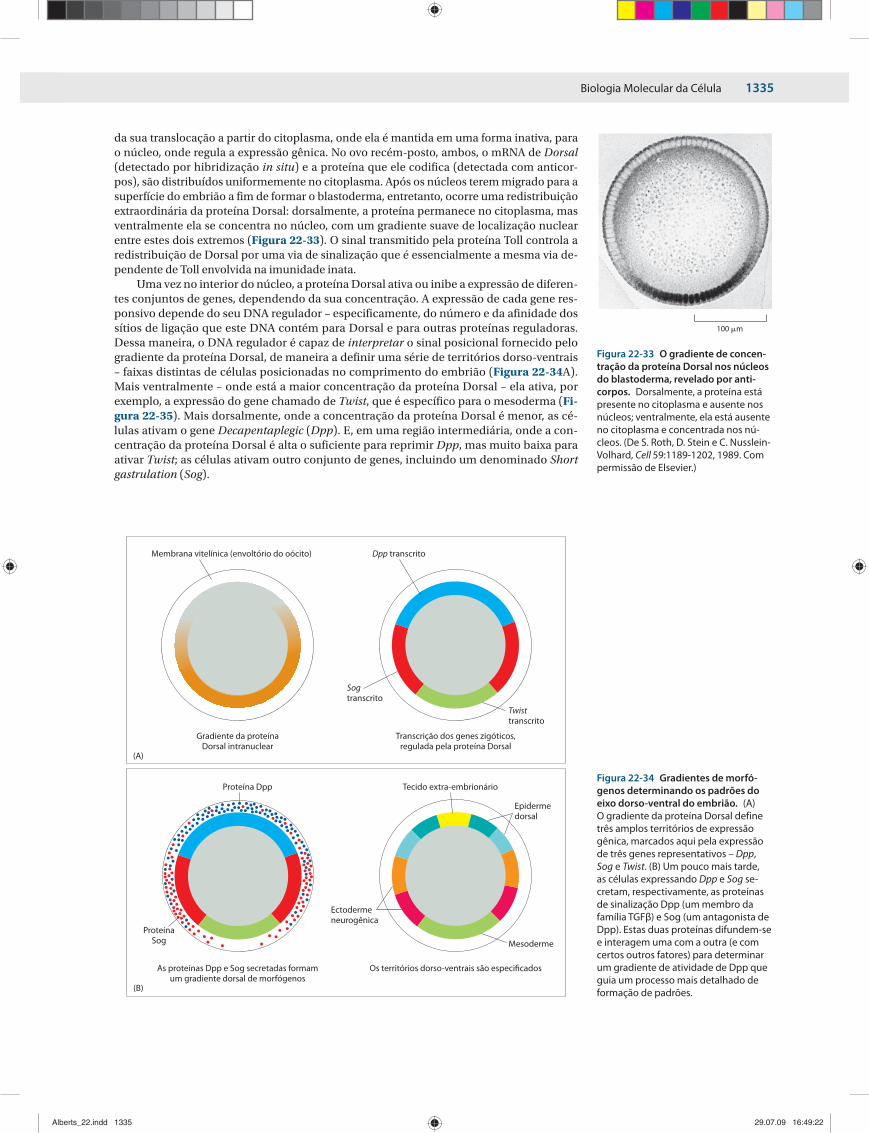
Biologia Molecular da Célula 1335
da sua translocação a partir do citoplasma, onde ela é mantida em uma forma inativa, para o núcleo, onde regula a expressão gênica. No ovo recém-posto, ambos, o mRNA de Dorsal (detectado por hibridização in situ) e a proteína que ele codifica (detectada com anticor-pos), são distribuídos uniformemente no citoplasma. Após os núcleos terem migra do para a superfície do embrião a fim de formar o blastoderma, entretanto, ocorre uma redistribuição extraordinária da proteína Dorsal: dorsalmente, a proteína permanece no citoplasma, mas ventralmente ela se concentra no núcleo, com um gradiente suave de localização nuclear entre estes dois extremos (Figura 22-33). O sinal transmitido pela proteína Toll controla a redistribuição de Dorsal por uma via de sinalização que é essen cialmente a mesma via de-pendente de Toll envolvida na imunidade inata.
Uma vez no interior do núcleo, a proteína Dorsal ativa ou inibe a expressão de diferen-tes conjuntos de genes, dependendo da sua concentração. A expressão de cada gene res-ponsivo depende do seu DNA regulador – especificamente, do número e da afinidade dos sítios de ligação que este DNA contém para Dorsal e para outras proteínas reguladoras. Dessa maneira, o DNA regulador é capaz de interpretar o sinal posicional fornecido pelo gradiente da proteína Dorsal, de maneira a definir uma série de territórios dorso-ventrais – faixas distintas de células posicionadas no comprimento do embrião (Figura 22-34A). Mais ventralmente – onde está a maior concentração da proteína Dorsal – ela ativa, por exemplo, a expressão do gene chamado de Twist, que é específico para o mesoderma (Fi-gura 22-35). Mais dorsalmente, onde a concentração da proteí na Dorsal é menor, as cé-lulas ativam o gene Decapentaplegic (Dpp). E, em uma região intermediária, onde a con-centração da proteína Dorsal é alta o suficiente para reprimir Dpp, mas muito baixa para ativar Twist; as células ativam outro conjunto de genes, incluindo um denominado Short gastrulation (Sog).
100 �m
Figura 22-33 O gradiente de concen-tração da proteína Dorsal nos núcleos do blasto derma, revelado por anti-corpos. Dorsalmen te, a proteína está presente no citoplasma e au sente nos núcleos; ventralmente, ela está ausen te no citoplasma e concentrada nos nú-cleos. (De S. Roth, D. Stein e C. Nusslein-Volhard, Cell 59:1189-1202, 1989. Com permissão de Elsevier.)
Figura 22-34 Gradientes de morfó-genos determinando os padrões do eixo dorso-ventral do embrião. (A) O gradiente da pro teína Dorsal define três amplos territórios de expressão gênica, marcados aqui pela expressão de três genes representativos – Dpp, Sog e Twist. (B) Um pouco mais tarde, as células expressan do Dpp e Sog se-cretam, respectivamente, as pro teínas de sinalização Dpp (um membro da famí lia TGFβ) e Sog (um antagonista de Dpp). Es tas duas proteínas difundem-se e interagem uma com a outra (e com certos outros fatores) para determinar um gradiente de atividade de Dpp que guia um processo mais detalhado de formação de padrões.
Membrana vitelínica (envoltório do oócito) Dpp transcrito
Sogtranscrito
Twisttranscrito
Gradiente da proteínaDorsal intranuclear
Transcrição dos genes zigóticos,regulada pela proteína Dorsal
(A)
As proteínas Dpp e Sog secretadas formamum gradiente dorsal de morfógenos
Os territórios dorso-ventrais são especificados
(B)
ProteínaSog
Proteína Dpp Tecido extra-embrionário
Epidermedorsal
Ectodermeneurogênica
Mesoderme
Alberts_22.indd 1335Alberts_22.indd 1335 29.07.09 16:49:2229.07.09 16:49:22

1336 Alberts, Johnson, Lewis, Raff, Roberts & Walter
Dpp e Sog produzem um gradiente de morfógenos secundário para refinar o padrão da parte dorsal do embriãoOs produtos dos genes regulados diretamente pela proteína Dorsal geram sinais locais que definem subdivisões mais finas do eixo dorso-ventral. Estes sinais atuam após a celulariza-ção e assumem a forma de moléculas de sinalização extracelular convencionais. Em particu-lar, Dpp codifica para a proteína secretada Dpp, a qual forma um gradiente de morfó genos na parte dorsal do embrião. O gene Sog, enquanto isso, codifica para outra proteína secreta-da que é produzida na ectoderme neurogênica e atua como um antagonista de Dpp. Os gra-dientes de difusão oposta dessas duas proteínas criam um gradiente acentuado de ativida de de Dpp. Os níveis mais altos de atividade de Dpp, em combinação com certos outros fato res, induzem o desenvolvimento do tecido mais dorsal de todos – a membrana extra-embrio-nária; os níveis intermediários induzem o desenvolvimento da ectoderme dorsal, e os níveis muito baixos permitem o desenvolvimento da ectoderme neurogênica (Figura 22-34B).
O eixo dorso-ventral dos insetos corresponde ao eixo ventro- dorsal dos vertebradosA Dpp é um membro da superfamília TGF� das moléculas de sinalização que também é importante nos vertebrados; a Sog é um homólogo da proteína chordin dos vertebrados. É surpreendente que um homólogo da Dpp, BMP4, e a chordin atuem juntos nos vertebra dos da mesma maneira que Dpp e Sog em Drosophila. Essas duas proteínas controlam o padrão dorso-ventral da ectoderme, com altos níveis de chordin definindo a região que é neuro-gênica, e altos níveis de atividade de BMP4 definindo a região que não é. Isto, combinado a outros paralelos moleculares, sugere que esta parte da estrutura corporal foi conservada entre os insetos e os vertebrados. Entretanto, o eixo está invertido, de manei ra que a parte dorsal na mosca corresponde à parte ventral no vertebrado (Figura 22-36). Em algum ponto da história evolutiva, parece que o ancestral de uma dessas classes de animais optou por viver a vida de cabeça para baixo.
Três classes de genes de segmentação refinam o padrão materno ântero-posterior e subdividem o embriãoApós os gradientes iniciais de Bicoid e de Nanos serem criados para definir o eixo ântero-pos-terior, os genes de segmentação refinam o padrão. As mutações em qualquer um dos genes de segmentação alteram o número de segmentos ou sua organização interna básica, sem afetar a polaridade global do embrião. Os genes de segmentação são expressos por subcon-juntos de células no embrião, de maneira que seus produtos são os primeiros com ponentes do genoma próprio do embrião, que não os do genoma materno, a contribuir para o desen-
Figura 22-35 A origem da mesoder-me a partir de células que expressam Twist. Os embriões foram fixados em estágios sucessivos, cortados transver-salmente e corados com um anticorpo contra a proteína Twist, uma proteína de regulação gênica da família bHLH. As células que expressam Twist movem-se para o interior do embrião para formar a mesoderme. (De M. Leptin, J. Casal, B. Grunewald e R. Reuter, Deve lopment Suppl. 23-31, 1992. Com permissão de The Company of Biologists.)
Figura 22-36 O plano corporal de verte brados como uma inversão dorso-ventral do plano corporal de in-setos. O mecanismo de determinação dos padrões dorso-ventrais em um em-brião vertebrado é discutido em mais detalhes posteriormente neste capítulo. Note a correspondência com re lação ao sistema circulatório, ao intestino e ao sistema nervoso. Em insetos, o sistema circulatório é representado por um coração tubular e um vaso sanguíneo dorsal principal, que bombeia sangue para os espaços dos tecidos por meio de um conjunto de aberturas e recebe o sangue de volta dos tecidos por meio de outro conjunto. Em contraste com os vertebrados, não há um sistema de vasos capilares para conter o sangue enquanto ele é percolado através dos tecidos. En tretanto, o desenvolvimento do coração depen de de genes homólo-gos nos vertebrados e nos insetos, re-forçando a relação entre os dois pla nos corporais. (Segundo E. L. Ferguson, Curr. Opin. Genet. Dev. 6:424-431, 1996. Com permissão de Elsevier.)
DORSAL
VENTRALINSETO VERTEBRADO
ÂnusSistema circulatório
Sistema circulatório Sistema nervoso central Intestino
BocaÂnus
BocaSistema nervoso central
Intestino
Alberts_22.indd 1336Alberts_22.indd 1336 29.07.09 16:49:2229.07.09 16:49:22

Biologia Molecular da Célula 1337
volvimento embrionário. Eles são chamados de genes de efeito zigótico, para distingui-los dos genes iniciais de efeito materno.
Os genes de segmentação distribuem-se em três grupos, de acordo com seus fenóti pos mutantes (Figura 22-37). É conveniente considerar estes três grupos como se suas ações ocorressem em sequência, apesar de, na realidade, suas funções se sobreporem. Primeiro, atuam em um conjunto de pelo menos seis genes gap, cujos produtos definem subdivisões não refina das do embrião. As mutações em um gene gap eliminam um ou mais grupos de segmentos adjacentes, e as mutações em genes gap distintos causam defeitos diferentes, po-rém par cialmente sobrepostos. Em um mutante Kruppel, por exemplo, a larva não apresenta oito segmentos, de T1 a A5.
Os próximos genes de segmentação a atuarem são um conjunto de oito genes pair- rule. As mutações nestes genes causam uma série de deleções afetando segmentos alter nados, deixando o embrião com somente a metade dos segmentos normais. Enquanto todos os mutantes pair-rule apresentam esta periodicidade de dois segmentos, eles diferem na po-sição precisa das deleções relativas aos limites segmentais ou parassegmentais. O mutante pair-rule Even-skipped (Eve), por exemplo, discutido no Capítulo 7, não apresenta todos os parassegmentos de números ímpares; o mutante pair-rule Fushi-tarazu (Ftz) não possui to-dos os parassegmentos de números pares, e o mutante pair-rule Hairy não apresenta uma série de regiões que são semelhantes em largura, mas que não têm relação com as unidades parassegmentais.
Finalmente, existem pelo menos 10 genes de polaridade segmentar. As mutações nesses genes produzem larvas com um número normal de segmentos, mas com uma par-te de cada segmento deletada e substituída por uma duplicação especular de todo ou de parte do segmento restante. Em mutantes Gooseberry, por exemplo, a metade posterior de cada segmento (ou seja, a metade anterior de cada parassegmento) é substituída por uma imagem aproximadamente especular de metade do segmento adjacente anterior (ver Figura 22-37).
Veremos mais tarde que, em paralelo com o processo de segmentação, um conjunto adicional de genes, os genes seletores homeóticos, serve para definir e preservar as dife renças entre um segmento e o próximo.
Os fenótipos dos vários mutantes de segmentação sugerem que os genes de segmenta-ção formam um sistema coordenado que subdivide o embrião progressivamente em domí-nios cada vez menores ao longo do eixo ântero-posterior, distinguido por padrões diferen-tes de expressão gênica. A genética molecular tem ajudado a revelar como este sistema fun-ciona.
A expressão localizada dos genes de segmentação é regulada por uma hierarquia de sinais posicionaisCerca de três quartos dos genes de segmentação, incluindo todos os genes gap e os genes pair-rule, codificam para proteínas de regulação gênica. As suas ações sobre outros genes
Figura 22-37 Exemplos de fenótipos de mutações afetando os três tipos de genes de segmentação. Em cada caso, as áreas som breadas em verde na larva normal (esquerda) es tão ausentes no mutante ou foram substituídas por du-plicações especulares das regiões não afe tadas. (Modificada de C. Nusslein-Volhard e E. Wies chaus, Nature 287:795-801, 1980. Com permissão de Macmillan Publishers Ltd.)
GENE GAP (Krüppel)GENE PAIR-RULE
(Even-skipped)GENE DE POLARIDADE
SEGMENTAR (Gooseberry)
Alberts_22.indd 1337Alberts_22.indd 1337 29.07.09 16:49:2229.07.09 16:49:22
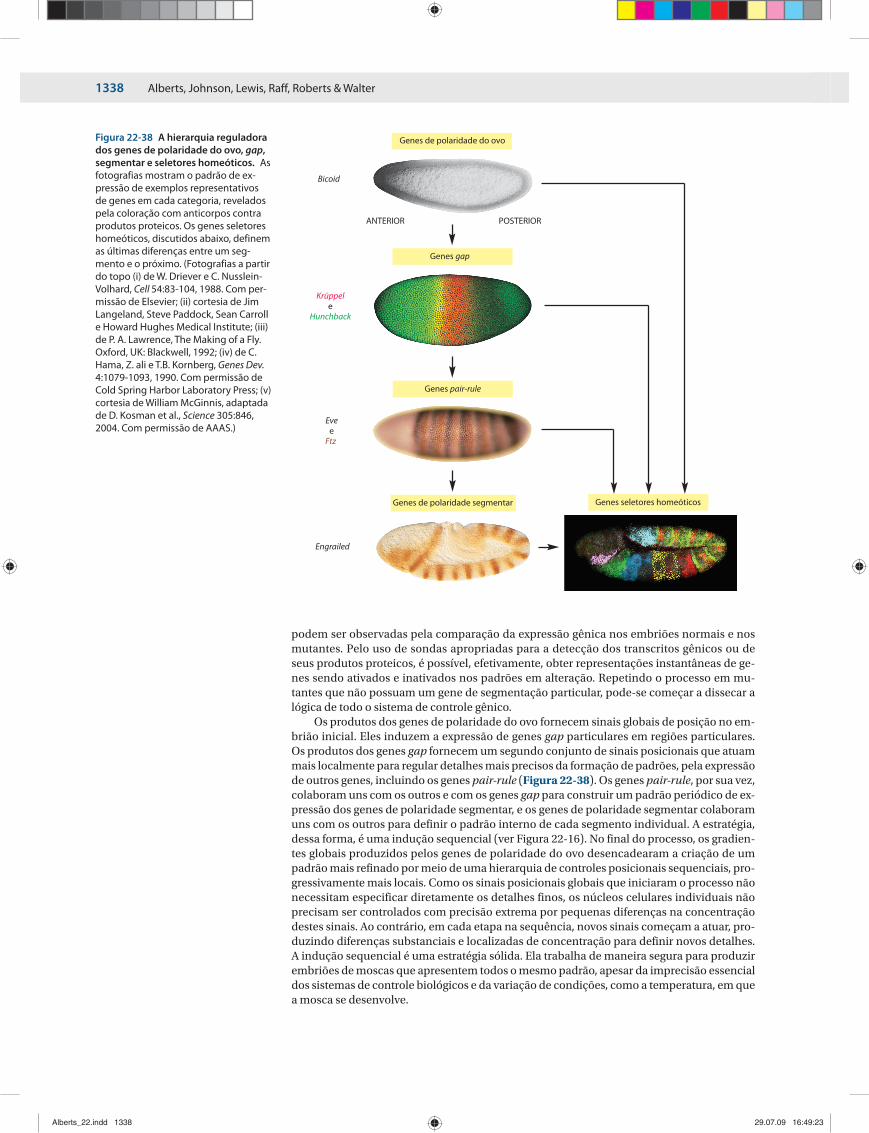
1338 Alberts, Johnson, Lewis, Raff, Roberts & Walter
podem ser observadas pela comparação da expressão gênica nos em briões normais e nos mutantes. Pelo uso de sondas apropriadas para a detecção dos trans critos gênicos ou de seus produtos proteicos, é possível, efetivamente, obter representa ções instantâneas de ge-nes sendo ativados e inativados nos padrões em alteração. Repe tindo o processo em mu-tantes que não possuam um gene de segmentação particular, pode-se começar a dissecar a lógica de todo o sistema de controle gênico.
Os produtos dos genes de polaridade do ovo fornecem sinais globais de posição no em-brião inicial. Eles induzem a expressão de genes gap particulares em regiões particula res. Os produtos dos genes gap fornecem um segundo conjunto de sinais posicionais que atuam mais localmente para regular detalhes mais precisos da formação de padrões, pela expressão de outros genes, incluindo os genes pair-rule (Figura 22-38). Os genes pair-rule, por sua vez, colaboram uns com os outros e com os genes gap para construir um padrão periódico de ex-pressão dos genes de polaridade segmentar, e os genes de polaridade segmentar colaboram uns com os outros para definir o padrão interno de cada segmen to individual. A estratégia, dessa forma, é uma indução sequencial (ver Figura 22-16). No final do processo, os gradien-tes globais produzidos pelos genes de polaridade do ovo desencadearam a criação de um padrão mais refinado por meio de uma hierarquia de controles posicionais sequenciais, pro-gressivamente mais locais. Como os sinais posici onais globais que iniciaram o processo não necessitam especificar diretamente os deta lhes finos, os núcleos celulares individuais não precisam ser controlados com precisão extrema por pequenas diferenças na concentração destes sinais. Ao contrário, em cada etapa na sequência, novos sinais começam a atuar, pro-duzindo diferenças substanciais e localiza das de concentração para definir novos detalhes. A indução sequencial é uma estratégia sólida. Ela trabalha de maneira segura para produzir embriões de moscas que apresentem todos o mesmo padrão, apesar da imprecisão essencial dos sistemas de controle biológicos e da variação de condições, como a temperatura, em que a mosca se desenvolve.
Figura 22-38 A hierarquia reguladora dos genes de polaridade do ovo, gap, seg mentar e seletores homeóticos. As fotografias mostram o padrão de ex-pressão de exemplos representativos de genes em cada ca tegoria, revelados pela coloração com anticor pos contra produtos proteicos. Os genes sele tores homeóticos, discutidos abaixo, definem as últimas diferenças entre um seg-mento e o pró ximo. (Fotografias a partir do topo (i) de W. Driever e C. Nuss lein-Volhard, Cell 54:83-104, 1988. Com per-missão de Elsevier; (ii) cortesia de Jim Langeland, Steve Paddock, Sean Carroll e Howard Hughes Medical Institute; (iii) de P. A. Lawrence, The Making of a Fly. Oxford, UK: Blackwell, 1992; (iv) de C. Hama, Z. ali e T.B. Kornberg, Genes Dev. 4:1079-1093, 1990. Com permissão de Cold Spring Harbor Laboratory Press; (v) cortesia de William McGinnis, adaptada de D. Kosman et al., Science 305:846, 2004. Com permissão de AAAS.)
Genes de polaridade do ovo
Bicoid
ANTERIOR
Genes gap
Krüppele
Hunchback
Genes pair-rule
Evee
Ftz
Genes de polaridade segmentar
Engrailed
Genes seletores homeóticos
POSTERIOR
Alberts_22.indd 1338Alberts_22.indd 1338 29.07.09 16:49:2329.07.09 16:49:23

Biologia Molecular da Célula 1339
A natureza modular do DNA regulador permite que os genes possuam múltiplas funções controladas independentementeO elaborado processo de formação de padrões recém-descrito depende de longos trechos de sequências de DNA não-codificante que controlam a expressão de cada um dos genes envolvidos. Essas regiões reguladoras ligam múltiplas cópias das proteínas de regu lação gê-nica produzidas pelos genes formadores de padrões expressos anteriormente. Assim como um mecanismo lógico de entrada e de saída, um gene individual é ativado e inativado de acordo com uma combinação particular de proteínas ligadas a suas regiões reguladoras em cada estágio do desenvolvimento. No Capítulo 7, descrevemos um gene de segmenta ção em particular – o gene pair-rule Even-skipped (Eve) – e discutimos como a decisão de transcrever o gene é feita de acordo com todas essas informações (ver Figura 7-55). Este exemplo pode ser mais elaborado para ilustrar alguns princípios importantes da formação dos padrões do desenvolvimento.
As faixas individuais da expressão de Eve dependem de módulos reguladores separa dos no DNA regulador de Eve. Assim, um módulo regulador é responsável pela expressão de Eve nas faixas 1 + 5, outro na faixa 2, outro nas faixas 3 + 7 e ainda outro nas faixas 4 + 6 (Figura 22-39). Cada módulo regulador define um conjunto dife rente de requisitos para a expressão gênica de acordo com a concentração dos produtos dos genes de polaridade do ovo e dos genes gap. Dessa forma, o DNA regulador de Eve serve para traduzir o padrão complexo e não-repetitivo das proteínas de polaridade do ovo e gap em um padrão periódico de expres-são de um gene pair-rule.
A organização modular do DNA regulador de Eve recém-descrito é típica da regula ção gênica de animais multicelulares e de plantas, e tem profundas implicações. Por meio da junção das sequências dos módulos que respondem a diferentes combinações de proteínas reguladoras, é possível gerar quase qualquer padrão de expressão gênica com base em qua-se qualquer outro. A presença dos módulos, além disso, permite ao DNA regulador definir padrões de expressão gênica que não são simplesmente complexos, mas cujas partes são independentemente ajustáveis. Uma alteração em um destes módulos reguladores pode al-terar uma parte do padrão de expressão, sem afetar o resto e sem necessitar de alterações nas proteínas reguladoras que levariam a repercussões na expres são de outros genes no geno-ma. Como descrito no Capítulo 7, é este DNA regulador que contém a chave para a organiza-ção complexa de plantas e de animais multicelulares, e suas propriedades tornam possível a adaptabilidade independente de cada parte da estru tura do corpo de um organismo no curso da evolução.
A maioria dos genes de segmentação também desempenha funções importantes em outros momentos e locais no desenvolvimento da Drosophila. O gene Eve, por exemplo, é expresso em subconjuntos de neurônios, em células precursoras musculares e em vários
Figura 22-39 Organização modular do DNA regulador do gene Eve. No experi mento mostrado, fragmentos clonados do DNA regulador foram ligados ao repórter LacZ (um gene bacteriano). Os embriões transgêni-cos contendo estas construções foram então coradas por hibridização in situ para revelar o padrão de expressão de LacZ (azul/preto) e con tracorado com um anticorpo anti-Eve (laranja) para mostrar as posições das faixas de expres são normais de Eve. Diferentes segmentos do DNA regulador de Eve (ocre) controlam a expressão gênica em regiões que cor respondem a diferentes partes do padrão de expressão normal de Eve. Dois segmentos em tandem con-trolam a expressão em um padrão que é a soma dos pa drões gerados por cada um deles individualmen te. Os módulos reguladores separados são res ponsáveis por diferentes momentos da expres são gênica, assim como diferentes localiza-ções: o painel mais à esquerda mostra a ação de um módulo que começa a agir mais tarde do que os outros ilustrados e controla a expressão em um subcon-junto de neurônios. (De M. Fujioka et al., Development 126:2527-538, 1999. Com permissão de The Com pany of Biologists.)
Codificação
Subconjunto de neurônios Faixas 4 e 6 Faixa 1 Faixa 5
Faixas 3 e 7
Faixa 2
3.000 pares de nucleotídeos
mRNA Eve
Precursoresmusculares
Faixas 1 e 5
Alberts_22.indd 1339Alberts_22.indd 1339 29.07.09 16:49:2429.07.09 16:49:24

1340 Alberts, Johnson, Lewis, Raff, Roberts & Walter
outros locais sob o controle de estimuladores adicionais (ver Figura 22-39). Pela adição de novos módulos a este DNA regulador, qualquer gene pode ser cooptado durante a evo-lução para novos propósitos em novos locais do corpo, sem detrimento de suas outras funções.
Os genes de polaridade do ovo, os genes gap e os genes pair-rule criam um padrão transiente que é relembrado por outros genesDentro das primeiras horas após a fertilização, os genes gap e os genes pair-rule são ativados. Seus produtos de mRNA aparecem primeiramente em padrões que somente se aproximam da situação final; então, em um curto período – por meio de uma série de ajustes interativos – a indefinida distribuição inicial dos produtos gênicos transforma-se em um sistema regular e nitidamente definido de faixas (Figura 22-40). Contudo, esse sistema por si só é instável e transiente. Enquanto o embrião avança pela gastru lação e além, o padrão regular segmentar dos produtos gênicos gap e pair-rule desintegram-se. As suas ações, entretanto, produziram um conjunto permanente de marcações – valores posicionais – nas células do blastoderma. Estas marcas posicionais estão gravadas na ativação persistente de certos genes de polarida-de segmentar e dos genes seletores homeóticos, os quais servem para manter a organização segmentar da larva e do adulto. O gene Engrailed de polaridade segmentar fornece um bom exemplo. Os seus transcritos de mRNA são vistos no blastoderma celular em uma série de 14 faixas, cada uma com apro ximadamente uma célula de largura, correspondendo às porções mais anteriores dos futu ros parassegmentos (Figura 22-41).
Os genes de polaridade segmentar são expressos em padrões que se repetem de um parassegmento para o próximo, e as suas faixas de expressão aparecem em uma relação fixa com as faixas de expressão dos genes pair-rule que os auxiliam na sua ativação. En-tretanto, a produção deste padrão em cada parassegmento depende de interações entre os próprios genes de polaridade segmentar. Essas interações ocorrem em etapas quando o blastoderma já se tornou totalmente dividido em células separadas, de maneira que a sinali zação célula-célula normal tem que entrar em ação. Um grande subconjunto de ge-nes de pola ridade segmentar codifica para componentes de duas vias de transdução de si-nais, a via Wnt e a via Hedgehog, incluindo as proteínas sinalizadoras secretadas Wingless
2,7 horas após a fertilização 3,5 horas após a fertilização
Figura 22-40 A formação das faixas de Ftz e de Eve no blastoderma de Drosophila. Ftz e Eve são ambos genes pair-rule. Os seus pa drões de expressão (mostrados em marrom para Ftz e em cinza para Eve) são inicialmente pouco definidos, mas rapidamente transfor-mam-se em fai xas nitidamente delinea-das. (De P. A. Lawrence, The Making of a Fly. Oxford, UK: Blackwell, 1992.)
Embrião de 5 horas100 �m
100 �mEmbrião de 10 horas
Adulto500 �m
Figura 22-41 O padrão de expressão de Engrailed, um gene de polaridade segmen tar. O padrão de Engrailed é mostrado em um embrião de cinco horas (no estágio de banda germinativa estendida), em um embrião de 10 horas e no adulto (cujas asas foram removidas nesta preparação). O padrão é revelado por um anticorpo (marrom) contra a proteína Engrailed (para os embriões de 5 e 10 horas) ou (para o adul to) pela construção de uma linhagem de Droso phila contendo as sequên-cias controladoras do gene Engrailed acopladas à sequência codificante do repórter LacZ, cujo produto é detectado his toquimicamente por meio de um produto azul da reação que ele catalisa. Note que o padrão de Engrailed, uma vez estabelecido, é preservado por toda a vida do animal. (De C. Hama, Z. Ali e T. B. Kornberg, Genes Dev. 4:1079-1093. Com permissão de Cold Spring Harbor Laboratory Press.)
Alberts_22.indd 1340Alberts_22.indd 1340 29.07.09 16:49:2429.07.09 16:49:24

Biologia Molecular da Célula 1341
(um membro da família Wnt) e Hedgehog. Estas são expressas em diferentes bandas de células que servem como centros de sinalização em cada parassegmento e atuam na ma-nutenção e no refino da expressão de outros genes de polaridade segmentar. Além disso, embora a sua expressão inicial seja determinada pelos genes pair-rule, as duas proteínas sinalizadoras regulam uma a expressão da outra em uma via mutuamente sustentada e continuam a auxiliar o desencadea mento da expressão de genes, como o Engrailed, preci-samente em seus locais corretos.
O padrão de expressão de Engrailed persistirá por toda a vida, muito depois de os si-nais que organizaram a sua produção terem desaparecido (ver Figura 22-41). Este exem plo ilustra não somente a subdivisão progressiva do embrião por meio de sinais cada vez mais precisamente localizados, mas também a transição entre os eventos de sinalização tempo-rários do desenvolvimento inicial e a posterior manutenção estável das informações do de-senvolvimento.
Além de regular os genes de polaridade segmentar, os produtos dos genes pair-rule co-laboram com os produtos dos genes gap para induzir a ativação precisamente localiza da de um conjunto adicional de marcações espaciais – os genes seletores homeóticos. São os genes seletores homeóticos que distinguem permanentemente um parassegmento de outro. Na próxima seção, examinaremos esses genes seletores em detalhe e discutiremos as suas funções na memória celular.
ResumoA mosca Drosophila tem sido o principal organismo, modelo para o estudo da genética do desen-volvimento animal. Assim como outros insetos, ela começa o seu desenvolvimen to com uma série de divisões nucleares, gerando um sincício, e grande parte da formação dos padrões iniciais ocorre nesta célula única gigante e multinucleada. O pa drão se origina das assimetrias no ovo, organiza-das tanto por depósitos localizados de mRNA dentro do ovo como por sinais das células folicula-res ao redor dele. As informa ções posicionais no embrião multinucleado são fornecidas por quatro gradientes intra celulares produzidos pelos produtos de quatro grupos de genes de efeito materno chama dos de genes de polaridade do ovo. Eles controlam quatro distinções fundamentais para a estrutura corporal dos animais: dorsal versus ventral, endoderme versus mesoderme e ectoderme, células germinativas versus células somáticas e cabeça versus parte posterior.
Os genes de polaridade do ovo operam pela produção de distribuições graduadas de proteí-nas de regulação gênica no ovo e no embrião inicial. Os gradientes ao longo do eixo ântero-pos-terior iniciam a expressão ordenada dos genes gap, dos genes pair-rule, dos genes de polari dade segmentar e dos genes seletores homeóticos. Estes, por meio de uma hierarquia de interações, tor-nam-se expressos em algumas regiões do embrião e não em outras, subdi vidindo progressivamen-te o blastoderma em uma série regular de unidades modulares repetidas chamadas de segmentos. Os padrões complexos de expressão gênica refletem a organização modular do DNA regulador, com estimuladores separados para cada gene individual responsável por partes separadas do seu padrão de expressão.
Os genes de polaridade segmentar começam a agir próximo ao final do processo de segmenta-ção, logo após o sincício ter se dividido em células separadas, e controlam a formação de padrões internos de cada segmento por sinalizações célula-célula por meio das vias de Wnt (Wingless) e Hedgehog. Isto leva a uma ativação localizada e persistente de genes como Engrailed, dando às células um registro da sua localização ântero-posterior dentro do segmento. Enquanto isso, um novo gradiente de si nalização célula-célula também é gerado ao longo do eixo dorso-ventral, com o membro da família TGF� Decapentaplegic (Dpp) e seu antagonista, Short gastrulation, atuando como morfógenos. Este gradiente auxilia a refinar a escolha das várias características das células em diferentes níveis dorso-ventrais. Sabe-se também que as proteínas homó logas controlam a for-mação de padrões do eixo ventro-dorsal nos vertebrados.
GENES SELETORES HOMEÓTICOS E A FORMAÇÃO DE PADRÕES DO EIXO ÂNTEROPOSTERIORÀ medida que o desenvolvimento progride, o corpo torna-se cada vez mais complexo. Em toda esta complexidade crescente existe, entretanto, uma característica simplificado-ra que coloca o entendimento de todo o processo de desenvolvimento a nosso alcance.
Alberts_22.indd 1341Alberts_22.indd 1341 29.07.09 16:49:2529.07.09 16:49:25

1342 Alberts, Johnson, Lewis, Raff, Roberts & Walter
Repetidamen te, em cada espécie e em cada nível de organização, as estruturas complexas são feitas pela repetição de alguns temas básicos, com variações. Assim, um número limita-do de tipos celulares básicos diferenciados, como as células musculares ou os fibroblastos, ressurge com varia ções individuais sutis em diferentes locais. Esses tipos celulares estão or-ganizados em uma variedade limitada de tipos de tecidos, como os músculos ou os tendões, os quais novamente estão repetidos com variações sutis em diferentes regiões do corpo. A partir de vários tecidos, órgãos como os dentes ou os dedos são produzidos – molares e inci-sivos, indicadores e pole gares e dedos dos pés – alguns tipos básicos de estrutura, repetidos com variações.
Em qualquer lugar em que encontremos este fenômeno de repetição modulada, pode-mos dividir o problema dos biólogos do desenvolvimento em duas questões: qual é o meca-nismo básico de construção comum a todos os objetos de uma dada classe, e como este me-canismo é modificado para originar as variações observadas? O embrião usa uma estratégia combinatória para gerar a sua complexidade, e podemos usar a estratégia com binatória para entendê-la.
Os segmentos do corpo do inseto fornecem um exemplo muito claro. Já havíamos es-boçado a maneira pela qual os rudimentos de um único segmento típico são construí dos. Precisamos agora considerar como um segmento é induzido a ser diferente de outro.
O código Hox especifica diferenças ântero-posterioresO primeiro vestígio de uma resposta genética para a questão de como cada segmento adquire a sua identidade individual surgiu há mais de 80 anos, com a descoberta da pri-meira de várias mutações na Drosophila que causavam perturbações bizarras na organi-zação da mosca adulta. No mutante Antennapedia, por exemplo, as pernas origi nam-se a partir da cabeça, no lugar das antenas (Figura 22-42), enquanto no mutante Bithorax, porções de um par de asas extras aparecem onde normalmente deveriam estar estruturas muito menores chamadas de halteres. Essas mutações transformam partes do corpo em estruturas apropriadas para outras posições e são chamadas de homeóticas. Um conjunto inteiro de genes seletores homeóticos determina o caráter ântero-posterior dos segmen-tos da mosca.
Os genes deste conjunto – oito deles na mosca – são relacionados uns com os outros como membros de uma família multigênica e pertencem a um ou a outro de dois agrupa-mentos gênicos conhecidos como complexo Bithorax e complexo Antennapedia. Os genes no complexo Bithorax controlam as diferenças entre os segmentos abdominais e torácicos do corpo, e aqueles do complexo Antennapedia controlam as diferenças entre os segmen-tos torácicos e os da cabeça. As comparações com outras espécies mostram que os mesmos genes estão presentes em essencialmente todos os animais, incluindo os humanos. Estas comparações também revelam que os complexos Antennapedia e Bithorax são as duas me-tades de uma única entidade, chamada de complexo Hox, que se tornou dividi da no curso da evolução da mosca e cujos membros operam de uma maneira coordenada para exercer o seu controle sobre o padrão cabeça-cauda do corpo.
Os genes seletores homeóticos codificam proteínas de ligação ao DNA que interagem com outras proteínas de regulação gênicaÀ primeira vista, cada gene seletor homeótico normalmente é expresso somente naquelas regiões que se desenvolvem de modo anormal quando o gene está mutado ou ausente. Os produtos desses genes podem, então, ser vistos como marcas de endereço molecular per-tencentes às células de cada parassegmento: eles são a encarnação física do valor posici-onal da célula. Se as marcas de endereçamento estiverem alteradas, o parassegmento com-porta-se como se estivesse localizado em outro lugar, e a deleção de todo o complexo resulta em uma larva cujos segmentos do corpo são todos semelhantes (Figura 22-43).
Um primeiro problema, portanto, é entender como os produtos dos genes seletores ho-meóticos atuam na maquinaria básica de formação de padrões segmentares para dar a cada segmento sua individualidade. Os produtos dos genes seletores homeóticos são pro teínas de regulação gênica, todas relacionadas umas às outras por possuírem um homeodomínio de ligação ao DNA altamente conservado (de 60 aminoácidos), discutido no Capítulo 7. O segmento correspondente na sequência de DNA é chamado de homeo box, do qual, pela abreviatura, o complexo Hox ganhou seu nome.
Figura 22-42 Mutação homeótica. A mosca mostrada aqui é um mutante Antennape dia. As suas antenas estão convertidas em estru turas de pernas por uma mutação na região re guladora do gene Antennapedia que o induz a ser expresso na cabeça. Compare com a mosca nor mal mostrada na Figura 21-24. (Cortesia de Mat thew Scott.)
Alberts_22.indd 1342Alberts_22.indd 1342 29.07.09 16:49:2529.07.09 16:49:25

Biologia Molecular da Célula 1343
Se os produtos dos genes seletores homeóticos são semelhantes nas suas regiões de li-gação ao DNA, como eles exercem efeitos diferentes de modo a fazer um parassegmento distinto do seguinte? A resposta parece estar, em grande parte, nas partes das proteínas que não se ligam diretamente ao DNA, mas interagem com outras proteínas nos comple xos de ligação ao DNA. Os diferentes parceiros nestes complexos atuam junto com as proteínas se-letoras homeóticas para determinar quais os sítios de ligação ao DNA serão reco nhecidos e se o efeito na transcrição naqueles sítios será de ativação ou repressão. Dessa maneira, os produtos dos genes seletores homeóticos combinam-se a outras proteínas de regulação gênica e modulam suas ações de maneira a dar a cada parassegmento os seus aspectos ca-racterísticos.
Os genes seletores homeóticos são expressos sequencialmente de acordo com a sua ordem no complexo HoxPara entender como o complexo Hox fornece valores posicionais às células, também preci-samos considerar como a expressão dos próprios genes Hox é regulada. As sequências codi-ficantes dos oito genes seletores homeóticos nos complexos Antennapedia e Bithorax es tão distribuídas em meio a uma quantidade muito maior – um total de aproximadamente 650 mil pares de nucleotídeos – de DNA regulador. Este DNA inclui sítios de ligação aos produtos dos genes de polaridade do ovo e dos genes de segmentação. O DNA regulador no complexo Hox atua como um intérprete dos múltiplos itens de informações posicionais fornecidos por todas estas proteínas de regulação gênica. Em resposta, é transcrito um conjunto parti cular de genes seletores homeóticos, apropriado para aquela localização.
No padrão de controle existe uma notável regularidade. A sequência na qual os genes estão ordenados ao longo do cromossomo, em ambos os complexos Antennapedia e Bitho-rax, corresponde quase exatamente à ordem na qual eles são expressos ao longo do eixo do corpo (Figura 22-44). Isso sugere que os genes são ativados em série por algum processo graduado – em duração ou em intensidade – ao longo do eixo do corpo e cuja ação se espa-lha gradualmente ao longo do cromossomo. De modo geral, o mais “pos terior” dos genes expressos em uma célula é o que domina, direcionando para uma diminuição da expressão dos genes “anteriores” previamente ativados e ditando a característi ca do segmento. Os me-canismos de regulação gênica subjacentes a esses fenômenos ain da não são compreendidos, mas as suas consequências são profundas. Veremos que a organização serial da expressão gênica no complexo Hox é uma característica fundamen tal que tem sido altamente conser-vada no curso da evolução.
Figura 22-43 O efeito da deleção da maio ria dos genes do complexo Bi-thorax. (A) Uma larva normal de Drosophila mostrada sob iluminação em campo escuro; (B) a larva mutan te com grande parte do complexo Bithorax deletada. No mutante, todos os parassegmentos posteriores a P5 têm a aparência de P5. (De G. Struhl, Nature 293:36-41, 1981. Com permissão de Macmillan Publishers Ltd.)
(A) (B)100 �m
Dfd
ScrDeformed
Sex combs reduced
Antennapedia P2
Ultrabithorax
Abdominal A
Abdominal B
Antp
Ubx
AbdA
AbdB
Cromossomo 3
ComplexoAntennapedia
ComplexoBithorax
Labial
Proboscipedia
Pb
Lab
Lab Dfd Scr Antp Ubx AbdA
AbdB
Figura 22-44 Os padrões de expressão comparados às localizações cromossô-micas dos genes do complexo Hox. O diagrama mostra a se quência dos genes em cada uma das duas subdivisões dos com plexos cromossômicos. Esta sequência corresponde, com poucas ex-ceções, à sequência espacial na qual os genes são expressos, conforme mostra-do na fotografia de um embrião no es-tágio de banda germinativa estendida, cerca de cinco horas após a fertilização. O embrião foi corado por hibridização in situ com diferentes sondas marcadas com cores distintas para detectar os produtos de mRNAs de diferentes ge-nes Hox. (Fotografia cortesia de William McGinnis, adaptada de D. Kosman et al., Science 305:846, 2004. Com permissão de AAAS.)
Alberts_22.indd 1343Alberts_22.indd 1343 29.07.09 16:49:2529.07.09 16:49:25

1344 Alberts, Johnson, Lewis, Raff, Roberts & Walter
Existem centenas de outros genes contendo homeobox no genoma da mosca – e de ou-tras espécies animais – mas a maioria está dispersa e não agrupada em complexos como o complexo Hox. Eles desempenham muitas funções diferentes na regulação gênica, mas uma proporção substancial apresenta funções afins às dos genes Hox: eles controlam as varia ções de um tema básico do desenvolvimento. As diferentes classes de neurônios, por exemplo, diferenciam-se frequentemente umas das outras pela expressão de genes específicos dessa grande superfamília.
O complexo Hox carrega um registro permanente das informações posicionaisO padrão espacial de expressão dos genes no complexo Hox é formado por sinais que atuam no início do desenvolvimento, mas as consequências são duradouras. Embora o padrão de expressão sofra ajustes complexos enquanto o desenvolvimento progride, o complexo Hox se comporta, em cada célula, como um registro permanente da posição ântero- posterior que a célula ocupou no embrião inicial. Assim, as células de cada segmento estão equipadas com uma memória de longa duração da sua localização ao longo do eixo ântero-posterior do corpo – em outras palavras, com um valor posicional ântero-posterior. Como veremos na próxima seção, esta memória impressa no complexo Hox controla a identi dade específica dos segmentos não somente nos segmentos larvais, mas também nas estruturas da mosca adulta, que são geradas em um estágio muito mais tardio em relação aos discos imaginais larvais e a outros abrigos de células imaginais precursoras na larva.
O mecanismo molecular da memória celular para esta informação posicional conta com dois tipos de informações reguladoras. Uma é originada dos próprios genes seletores home óticos: muitas das proteínas Hox autoativam a transcrição de seus próprios genes. Ou-tra informação crucial é originada em dois grandes conjuntos complementares de proteínas que controlam a estrutura da cromatina, chamados de grupo Polycomb e grupo Trithorax. Se estes reguladores estão defeituosos, o padrão de expressão dos genes seletores homeóti-cos é, no primeiro mo mento, formado de maneira correta, mas não é corretamente mantido conforme o embrião se desenvolve.
Os dois conjuntos de reguladores atuam de maneiras opostas. As proteínas do grupo Trithorax são necessárias para manter a transcrição dos genes Hox nas células em que a transcrição já foi ativada. Em contraste, as proteínas do grupo Polycomb formam comple-xos estáveis que se ligam à cromatina do complexo Hox e mantêm o estado reprimido nas células em que os genes Hox não foram ativados no seu momento crítico (Figura 22-45). A memó ria do desenvolvimento envolve modificações covalentes específicas nas histonas dos nucleossomos nas adjacências dos genes Hox, induzindo a uma mudança de estado da cromatina que pode ser perpetuada de uma geração de células para a próxima, conforme discutido nos Capítulos 4 e 7.
O eixo ântero-posterior é controlado pelos genes seletores Hox também nos vertebradosOs homólogos dos genes seletores homeóticos de Drosophila são encontrados em quase todas as espécies animais estudadas, dos cnidários (hidras) e nematoides até os moluscos e mamíferos. Notavelmente, esses genes frequentemente são agrupados em complexos semelhantes ao complexo Hox de insetos. No camun dongo, existem quatro desses com-plexos – chamados de complexos HoxA, HoxB, HoxC e HoxD – cada um em um cromos-somo diferente. Os genes indivi duais em cada complexo podem ser reconhecidos pe las suas sequências correspondentes de membros específicos do conjunto de genes de Dro-sophila. Na realidade, os genes Hox de mamíferos podem funcionar na Droso phila como substitutos parciais dos genes Hox correspondentes de Drosophila. Parece que cada um dos quatro complexos Hox de mamíferos é, grosseiramente falando, o equivalente a um complexo completo de inse tos (ou seja, o complexo Antennapedia mais o complexo Bitho-rax) (Figura 22-46).
A ordenação dos genes dentro de cada complexo Hox dos vertebrados é essencialmente a mesma do com plexo Hox de insetos, sugerindo que todos os quatro complexos dos verte-brados se originaram por dupli cações de um único complexo primordial e que pre servaram
Alberts_22.indd 1344Alberts_22.indd 1344 29.07.09 16:49:2729.07.09 16:49:27

Biologia Molecular da Célula 1345
sua organização básica. Mais surpreenden temente, quando os padrões de expressão dos ge-nes Hox são examinados no embrião de vertebrados por hibridização in situ, percebe-se que os membros de cada complexo são expressos em uma série cabeça -cauda ao longo do eixo do corpo, assim como em Droso phila (Figura 22-47). O padrão é mais claramente observa-do no tubo neural, mas também é visível em outros tecidos, especialmente na mesoder me. Com exceções menores, essa ordenação anatômica corresponde à ordenação cromos sômica dos genes em cada complexo, e os genes correspondentes nos quatro diferentes complexos Hox possuem domínios ântero-posteriores de expressão quase idênticos.
Os domínios de expressão gênica definem um sistema detalhado de correspondên-cias entre as regiões do corpo dos insetos e as regiões do corpo dos vertebrados (ver Figura 22-46). Os parassegmentos da mosca correspondem a uma série de segmentos semelhan-temente marcados na porção anterior do embrião de vertebrado. Estes segmentos estão mais claramente demarcados no cérebro posterior (ver Figuras 22-46 e 22-47), onde são chamados de rombômeros. Nos tecidos laterais ao cérebro posterior, a segmentação é vis-ta em uma série de arcos branquiais, proeminentes em todos os embriões de vertebrados – os precursores dos sistemas de brânquias em peixes e dos maxilares e das estruturas do pescoço em mamíferos; cada par de rombômeros no cérebro posterior corresponde a um arco branquial. No cérebro posterior, como na Drosophila, os limites dos domínios de expressão de muitos dos genes Hox estão alinhados com os limites dos segmentos ana-tômicos.
Os produtos dos genes Hox de mamíferos parecem especificar valores posicionais que controlam o padrão ântero-posterior de partes do cérebro posterior, do pescoço e do tron-co (assim como outras partes do corpo). Assim como em Drosophila, quando um gene Hox posterior é artificialmente expresso em uma região anterior, ele faz com que o tecido an-terior apresente características do tecido posterior. No entanto, a perda de um gene Hox posterior permite que o tecido posterior, onde ele normalmente seria expresso, adote uma característica anterior (Figura 22-48). As transformações observadas nos mutantes Hox de camundon gos frequentemente são incompletas, talvez devido à redundância entre os genes nos qua tro agrupamentos de genes Hox. Contudo, parece claro que a mosca e o ca-mundongo usam essencialmente a mesma maquinaria molecular para originar caracterís-ticas individuais para as regiões sucessivas ao longo de pelo menos uma parte do seu eixo ântero-posterior.
Figura 22-45 A ação dos genes do grupo Polycomb. (A) Fotografia de um embrião mu tante defeituoso para o gene Extra sex combs (Esc) e derivado de uma mãe que também não apre sentava este gene. O gene pertence ao grupo Polycomb. Essencialmente todos os seg-mentos foram transformados asseme-lhando-se ao seg mento abdominal mais posterior (compare com a Figura 22-43). No mutante, o padrão de ex pressão dos genes seletores homeóticos, que inicial-mente é bastante normal, torna-se tão ins tável que logo todos os genes estão ativados ao longo do eixo do corpo. (B) O padrão normal de ligação da proteína Polycomb aos cromosso mos gigantes da Drosophila, visualizado com um anticorpo contra Polycomb. A proteína está liga da ao complexo Antennapedia (ANT-C), ao com plexo Bithorax (BX-C) e a cerca de 60 outros locais. (A, de G. Struhl, Nature 293:36-41, 1981. Com permissão de Macmillan Publishers Ltd.; B, cortesia de B. Zink e R. Paro, Trends Genet. 6:416-421, 1990. Com permissão de Else vier.)
100 �m(A) (B)
ANT-C
BX-C
Alberts_22.indd 1345Alberts_22.indd 1345 29.07.09 16:49:2729.07.09 16:49:27
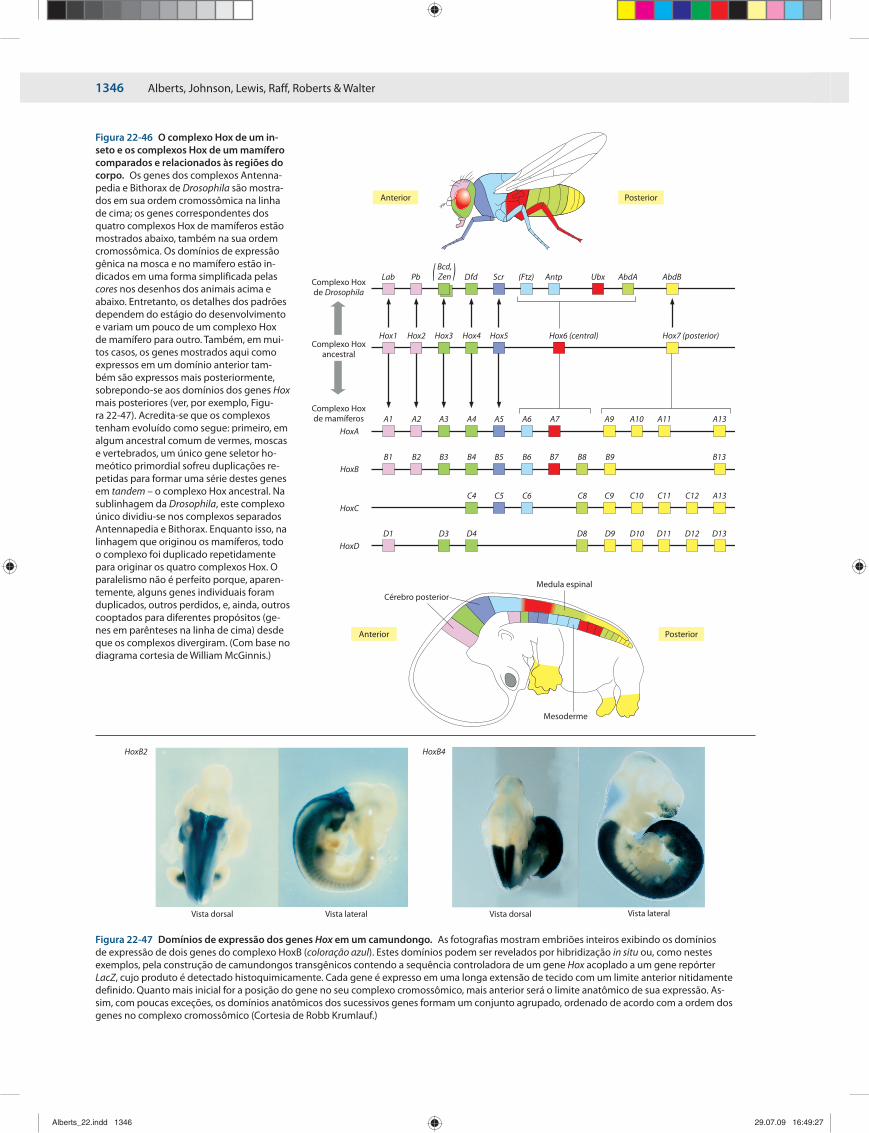
1346 Alberts, Johnson, Lewis, Raff, Roberts & Walter
Figura 22-46 O complexo Hox de um in-seto e os complexos Hox de um mamífe ro comparados e relacionados às regiões do corpo. Os genes dos complexos Antenna-pedia e Bithorax de Drosophila são mostra-dos em sua ordem cromossômica na linha de cima; os genes correspondentes dos quatro complexos Hox de mamíferos estão mostrados abaixo, tam bém na sua ordem cromossômica. Os domínios de expressão gênica na mosca e no mamífero estão in-dicados em uma forma simplificada pe las cores nos desenhos dos animais acima e abai xo. Entretanto, os detalhes dos padrões depen dem do estágio do desenvolvimento e variam um pouco de um complexo Hox de mamífero para outro. Também, em mui-tos casos, os genes mostrados aqui como expressos em um domí nio anterior tam-bém são expressos mais poste riormente, sobrepondo-se aos domínios dos genes Hox mais posteriores (ver, por exemplo, Figu-ra 22-47). Acredita-se que os complexos tenham evo luído como segue: primeiro, em algum ancestral comum de vermes, moscas e vertebrados, um único gene seletor ho-meótico primordial sofreu duplicações re-petidas para formar uma série destes genes em tandem – o complexo Hox an cestral. Na sublinhagem da Drosophila, este com plexo único dividiu-se nos complexos separa dos Antennapedia e Bithorax. Enquanto isso, na linhagem que originou os mamíferos, todo o com plexo foi duplicado repetidamente para originar os quatro complexos Hox. O paralelismo não é perfeito porque, aparen-temente, alguns genes in dividuais foram duplicados, outros perdidos, e, ainda, outros cooptados para diferentes propó sitos (ge-nes em parênteses na linha de cima) des de que os complexos divergiram. (Com base no diagrama cortesia de William McGinnis.)
HoxB2 HoxB4
Vista dorsal Vista dorsalVista lateral Vista lateral
Figura 22-47 Domínios de expressão dos genes Hox em um camundongo. As fotografias mostram embriões inteiros exibindo os domínios de expressão de dois genes do complexo HoxB (coloração azul). Estes domínios podem ser revelados por hibridização in situ ou, como nestes exemplos, pela construção de camundongos transgênicos contendo a sequência controladora de um gene Hox acoplado a um gene repórter LacZ, cujo produto é detectado histoquimicamente. Cada gene é expresso em uma longa extensão de tecido com um limite anterior nitidamente definido. Quanto mais inicial for a posição do gene no seu complexo cromossômico, mais anterior será o limite anatômico de sua expressão. As-sim, com poucas exceções, os domínios anatômicos dos sucessivos genes formam um conjunto agrupado, ordenado de acordo com a ordem dos genes no complexo cromossômico (Cortesia de Robb Krumlauf.)
Lab
Hox1 Hox2 Hox3 Hox4 Hox5 Hox6 (central) Hox7 (posterior)
Pb Dfd Scr (Ftz) Antp Ubx AbdA AbdB
A1 A2 A3 A4 A5 A7 A9 A10 A11 A13A6
B1 B2 B3 B4 B5 B7 B13B8 B9B6
C4 C5 C8 C9 C10 C11 C12 A13C6
D1 D3 D4 D13D8 D9 D10 D11 D12
Complexo Hoxde Drosophila
Complexo Hoxancestral
Complexo Hoxde mamíferos
HoxA
HoxB
HoxC
HoxD
Cérebro posterior
Medula espinal
Mesoderme
Anterior Posterior
Anterior Posterior
Bcd,Zen
Alberts_22.indd 1346Alberts_22.indd 1346 29.07.09 16:49:2729.07.09 16:49:27
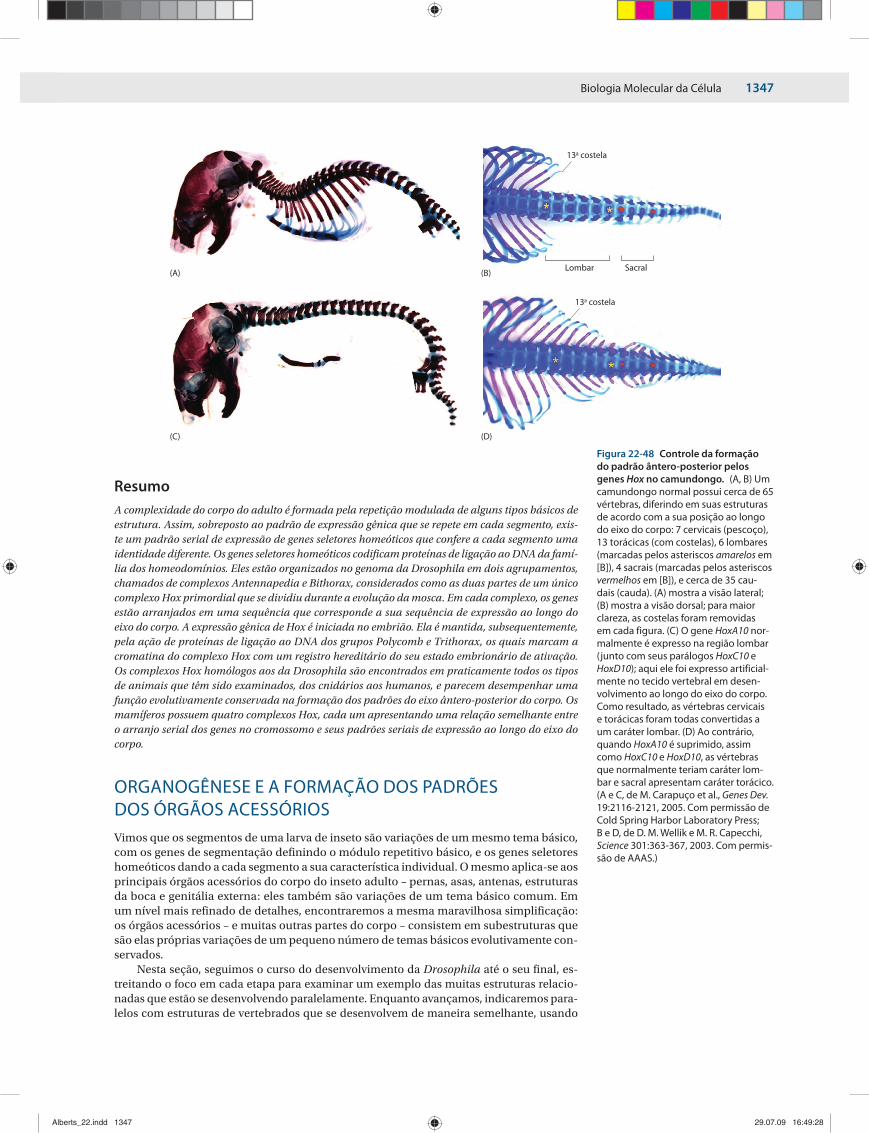
Biologia Molecular da Célula 1347
ResumoA complexidade do corpo do adulto é formada pela repetição modulada de alguns tipos básicos de estrutura. Assim, sobreposto ao padrão de expressão gênica que se repete em cada segmento, exis-te um padrão serial de expressão de genes seletores homeóticos que confere a cada segmento uma identidade diferente. Os genes seletores homeó ticos codificam proteínas de ligação ao DNA da famí-lia dos homeodomínios. Eles estão organizados no genoma da Drosophila em dois agrupamentos, chamados de complexos Antennapedia e Bithorax, considerados como as duas partes de um único complexo Hox primordial que se dividiu durante a evolução da mosca. Em cada complexo, os genes estão arranjados em uma sequência que corresponde a sua sequência de expressão ao longo do eixo do corpo. A expressão gênica de Hox é iniciada no embrião. Ela é mantida, subsequentemente, pela ação de proteínas de ligação ao DNA dos grupos Polycomb e Trithorax, os quais marcam a cromatina do complexo Hox com um registro hereditário do seu estado embrionário de ativação. Os complexos Hox homólogos aos da Drosophila são encon trados em praticamente todos os tipos de animais que têm sido examinados, dos cnidários aos humanos, e parecem desempenhar uma função evolutivamente conservada na formação dos padrões do eixo ântero-posterior do corpo. Os mamíferos possuem quatro complexos Hox, cada um apresentando uma relação semelhante entre o arranjo serial dos genes no cromossomo e seus padrões seriais de expressão ao longo do eixo do corpo.
ORGANOGÊNESE E A FORMAÇÃO DOS PADRÕES DOS ÓRGÃOS ACESSÓRIOSVimos que os segmentos de uma larva de inseto são variações de um mesmo tema básico, com os genes de segmentação definindo o módulo repetitivo básico, e os genes seletores homeóticos dando a cada segmento a sua característica individual. O mesmo aplica-se aos principais órgãos acessórios do corpo do inseto adulto – pernas, asas, antenas, estruturas da boca e genitália externa: eles também são variações de um tema básico comum. Em um nível mais refinado de detalhes, encontraremos a mesma maravilhosa simplificação: os ór gãos acessórios – e muitas outras partes do corpo – consistem em subestruturas que são elas próprias variações de um pequeno número de temas básicos evolutivamente con-servados.
Nesta seção, seguimos o curso do desenvolvimento da Drosophila até o seu final, es-treitando o foco em cada etapa para examinar um exemplo das muitas estruturas relacio-nadas que estão se desenvolvendo paralelamente. Enquanto avançamos, indicaremos para-lelos com estruturas de vertebrados que se desenvolvem de maneira semelhante, usando
Figura 22-48 Controle da formação do padrão ântero-posterior pelos genes Hox no camundongo. (A, B) Um camundongo normal possui cerca de 65 vértebras, diferindo em suas estruturas de acordo com a sua posição ao longo do eixo do corpo: 7 cervicais (pescoço), 13 torácicas (com costelas), 6 lombares (marcadas pelos asteriscos amarelos em [B]), 4 sacrais (marcadas pelos asteriscos vermelhos em [B]), e cerca de 35 cau-dais (cauda). (A) mostra a visão lateral; (B) mostra a visão dorsal; para maior clareza, as costelas foram removidas em cada figura. (C) O gene HoxA10 nor-malmente é expresso na região lombar (junto com seus parálogos HoxC10 e HoxD10); aqui ele foi expresso artificial-mente no tecido vertebral em desen-volvimento ao longo do eixo do corpo. Como resultado, as vértebras cervicais e torácicas foram todas convertidas a um caráter lombar. (D) Ao contrário, quando HoxA10 é suprimido, assim como HoxC10 e HoxD10, as vértebras que normalmente teriam caráter lom-bar e sacral apresentam caráter torácico. (A e C, de M. Carapuço et al., Genes Dev. 19:2116-2121, 2005. Com permissão de Cold Spring Harbor Laboratory Press; B e D, de D. M. Wellik e M. R. Capecchi, Science 301:363-367, 2003. Com permis-são de AAAS.)
(A) (B)
(C) (D)
13a costela
13a costela
Lombar Sacral
Alberts_22.indd 1347Alberts_22.indd 1347 29.07.09 16:49:2829.07.09 16:49:28

1348 Alberts, Johnson, Lewis, Raff, Roberts & Walter
não somente as mesmas estratégias gerais, mas muitos dos mesmos mecanismos molecu-lares específicos. Para evitar a interrupção da narrativa mais tarde, precisamos primeiro ex-plicar brevemente alguns métodos experimentais fundamentais, necessários para enfrentar um problema especial que surge quando tentamos descobrir como os genes controlam os es tágios mais tardios do desenvolvimento.
As mutações somáticas condicionais e induzidas tornam possível analisar funções gênicas tardias no desenvolvimentoComo anteriormente enfatizado, o mesmo gene pode ser usado repetidamente em muitas situações distintas – em diferentes regiões do corpo e em diferentes momentos. Com fre-quência, as mutações de perda de função alteram o desenvolvimento tão seriamente que o embrião ou a larva morre, privando-nos da oportunidade de observar como a mutação afetaria os pro cessos seguintes.
Uma maneira de contornar esse problema é estudar as mutações condicionais. Se ti-vermos, por exemplo, uma mutação sensível à temperatura no gene de interesse, podemos manter o animal durante o desenvolvimento inicial em uma temperatura baixa, em que o produto gênico funciona normalmente, e, então, desabilitar o produto gênico quando dese-jado, pela elevação da temperatura, a fim de descobrir as funções mais tardias.
Outros métodos envolvem a modificação do DNA em subconjuntos de células em está-gios mais tardios de desenvolvimento – um tipo de cirurgia genética de células individuais que permite que os grupos mutantes de células de um genótipo específico sejam gerados em um determinado momento do desenvolvimento. Este notável feito pode ser obtido por re-combinação somática induzida, e o organismo resultante é chamado de mosaico genético. Por meio dos mosaicos genéticos, podemos não apenas contornar o problema da letalidade quando a função de um gene é perturbada no organismo como um todo, mas também po-demos explorar as funções do gene na comunicação célula-célula, pela justaposição de cé-lulas mutantes e não-mutantes. Podemos testar, por exemplo, se a célula utiliza seu próprio produto gênico para a sinalização a células vizinhas, ou se ela recebe sinais destas outras células, ou nenhuma dessas alternativas. E, pela indução de alterações genéticas em mo-mentos diferentes, podemos determinar precisamente quando um gene atua para produzir um efeito particular.
Uma versão corrente desta técnica para indução de recombinação somática utiliza mos-cas transgênicas que tenham sido desenvolvidas para conter dois tipos de elementos gené-ticos derivados de leveduras: o gene FLP recombinase sítio-específico e a sequência-alvo FLP recombinase (FRT, FLP recombinase target). Tipicamente, o animal é homozigoto para uma inserção da sequência FRT próxima ao centrômero em um braço do cromossomo es-colhido, enquanto uma construção consistindo no gene Flp sob o controle de um promotor de choque térmi co é inserida em algum outro lugar do genoma. Se este embrião, ou larva transgênica, é submetido a um choque térmico (ou seja, exposto à alta temperatura por al-guns minutos), a expressão de Flp é induzida, e esta enzima catalisa permutações e recom-binações entre os cromossomos maternos e paternos no sítio FRT. Se o choque térmico é ajustado para ser suficientemente suave, este evento ocorrerá em somente uma ou poucas células, distribuídas ao acaso. Como explicado na Figura 22-49, se o animal também é hete-rozigoto para o gene de interesse em uma região cromossômica permutada, o processo pode resultar em um par de células-filhas que são homozigotas; uma recebendo duas cópias do alelo materno do gene, a outra recebendo duas cópias do alelo paterno. Cada uma dessas células-filhas irá normalmente crescer e dividir-se para dar origem a pequenas regiões clo-nais de progênie homozigota.
A ocorrência de permutação pode ser detectada se o animal é escolhido para ser tam-bém heterozigoto para uma mutação em um gene marcador que esteja no mesmo braço do cromossomo do gene de interesse e, dessa forma, sofre permutação juntamente a ele. As-sim, os clones de células mutantes homozigotas claramente identificados podem ser criados conforme a necessidade. Tanto FLP como FRT, ou os pares de elementos de recombinação análogos Cre e Lox, podem ser usados tam bém em outras configurações para ativar ou ina-tivar a expressão de um gene (ver Figura 5-79). Com estas técnicas, pode-se descobrir o que acontece, por exemplo, quando as células são induzidas a produzir uma molécula-sinal par-ticular em um local anormal, ou quando são privadas de um receptor particular.
Em vez de usar um promotor de choque térmico para direcionar a expressão da FLP re combinase, pode-se usar uma cópia da sequência reguladora de um gene no genoma
Alberts_22.indd 1348Alberts_22.indd 1348 29.07.09 16:49:2929.07.09 16:49:29

Biologia Molecular da Célula 1349
normal da mosca que seja expresso em algum momento e local interessantes. O evento de recombinação será então desencadeado, e as células mutantes serão criadas somente nos locais onde aquele gene normalmente é expresso. Uma variante dessa técnica usa empres-tada a maquinaria de regulação transcricional da levedura, em vez da maquinaria de re-combinação genética, para ativar ou inativar de maneira reversível um gene determinado da mosca, de acordo com o padrão normal de expressão de algum outro gene escolhido da mosca (Figura 22-50).
Assim, pela ativação ou inativação de funções gênicas em momentos e em locais espe-cíficos, os biólogos do desenvolvimento podem começar a decifrar o sistema de sinais espe-cificados geneticamente e as respostas que controlam a formação dos padrões de qualquer órgão do corpo.
As partes do corpo da mosca adulta desenvolvem-se a partir dos discos imaginaisAs estruturas externas da mosca adulta são formadas em grande parte de rudimentos cha-mados de discos imaginais – grupos de células que são deixadas de lado, aparentemente indiferenciadas, em cada segmento da larva. Os discos são bolsas de epitélio, de forma se-melhante a balões enrugados e achatados, e contíguas à epiderme (a camada superficial) da larva. Existem 19 discos, arranjados como nove pares em cada lado da larva mais um disco na linha média (Figura 22-51). Eles crescem e desenvolvem seu padrão interno enquanto a larva cresce, até que, finalmente, na metamorfose, viram pelo avesso (colocam a porção interna no lado exterior), estendem-se e diferenciam-se abertamente para formar a camada epidérmica do adulto. Os olhos e as antenas desenvolvem-se a partir de um par de discos, as asas e parte do tórax a partir de outro, o primeiro par de patas de outro, e assim por diante.
Figura 22-49 Criação de células mu-tantes pela recombinação somática induzida. Os diagramas seguem a destinação de um único par de cromos-somos homólogos, um do pai (som-breado), o outro da mãe (não-sombrea-do). Estes cromossomos possuem um elemento Frt (ver de) inserido próximo aos seus centrômeros e contêm um lócus para o gene de interesse – gene X – mais adiante ao longo do mesmo braço do cromossomo. O cromossomo paterno (neste exemplo) carrega o alelo selvagem do gene X (retângulo delimitado em vermelho), enquanto o cromossomo materno carrega um alelo mu tante recessivo (retângulo vermelho). A recom binação pela troca de DNA en-tre os cromosso mos materno e paterno, catalisada pela FLP recom binase, pode dar origem a um par de células–filhas, uma contendo duas cópias do tipo selvagem do gene X, a outra contendo duas cópias mutantes. Para auxiliar a identificação das células onde a re-combinação ocorreu, os cromossomos mater no e paterno podem ser escolhi-dos para portar diferentes marcadores genéticos (não-mostrado), capazes de gerar um produto visível, e posiciona-dos no cromossomo de maneira que a recombinação envolvendo o lócus mar-cador – resultando em uma alteração visível na aparên cia das células – possa ser considerada como um sinal seguro de que o gene X também sofreu recom-binação.
REPLICAÇÃOCROMOSSÔMICA
FLP CATALISA A PERMUTAÇÃOMITÓTICA E A RECOMBINAÇÃO
A CÉLULA SE DIVIDECélula homozigota parao gene mutante X
Célula homozigota parao gene normal X
A proliferação clonal gera regiões homozigotas na asa
UM SINAL INDUTORATIVA O PROMOTOR
DO GENE Flp
mRNA
Gene Flp
FLP recombinase
Célula da mosca heterozigota parao gene mutante X e homozigota
para Frt (alvo da FLP recombinase)Cromossomomaterno
Cromossomopaterno
Genemutante X
Frt
Alberts_22.indd 1349Alberts_22.indd 1349 29.07.09 16:49:2929.07.09 16:49:29

1350 Alberts, Johnson, Lewis, Raff, Roberts & Walter
Figura 22-50 A técnica Gal4/Uas para o controle da expressão gênica alterada em Drosophila. O método permite que se possa direcionar a expressão de um gene G escolhido nos locais e nos momentos onde um outro gene H de Drosophila normal-mente é expresso. (A) Um animal transgênico é criado com duas construções separadas inseridas no seu genoma. Um inserto consiste em uma sequência reguladora específica de leveduras, chamada de elemento Uas (de upstream activating sequence), acoplada a uma cópia da sequência codificante do gene G. O outro inserto contém a sequência codificante do gene de levedura Gal4, cujo produto é uma proteína de regulação gênica específica de levedura que se liga ao elemento Uas; este inserto Gal4 é colocado próximo à região reguladora do gene H e é controlado por ela. Onde quer que o gene H seja expresso normalmente, a proteína Gal4 também é produzida e induz a transcrição do gene G. (B) Embora se possa alcançar o mesmo resultado pela ligação de uma cópia da sequência reguladora H diretamente na sequência codificante G, a metodologia de Gal4/Uas possibilita uma estratégia que é mais eficiente em um plano maior. Duas “bibliotecas” separadas de moscas transgênicas são construídas; uma contendo insertos Gal4 controlados por uma variedade de sequências reguladoras de diferentes genes A, B, C, etc., a ou-tra contendo insertos Uas controlando uma variedade de sequências codificantes X, Y, Z, etc. Pelo cruzamento das moscas de uma biblioteca com moscas da outra, qualquer sequência codificante desejada pode ser funcionalmente acoplada a qualquer sequência reguladora desejada. Para gerar a biblioteca de moscas com inserções Gal4 em locais úteis, as moscas são primeira-mente produzidas com inserções Gal4 em localizações aleatórias do seu genoma. Estas moscas são então cruzadas com moscas contendo um elemento Uas ligado a um gene repórter com um produto facilmente detectável. A expressão do gene repórter re-vela se Gal4 foi inserido em um local que torna a sua expressão controlada por um estimulador interes sante; as moscas mostran-do padrões repórter interessantes são mantidas e estudadas. Esta técnica é chamada de técnica de armadilha de estimuladores, pois fornece uma maneira de caçar e caracterizar sequências reguladoras interessantes no genoma.
Gal4
Gal4
Gal4
gene XUas
gene YUas
gene ZUas
X
(B)(A) Qualquer combinação escolhida das sequências reguladoras (A, B, C, etc.)com as sequências codificantes (X, Y, Z, etc.)
Gene Gal4
Gene G
Proteína G
Sinais que ativam a expressãonormal do gene H
Sequênciareguladorado gene H
Expressãode Gal4
Expressãodo gene G
Proteína Gal4
ElementoUas
A
B
C
Lábio Clypeo-labrum
Pró-tóraxdorsal
Olho +antena
Pata Asa + tóraxdorsal
Haltere Genitais
Figura 22-51 Os discos imaginais na larva de Drosophila e as estruturas do adulto que eles originam. (Segundo J. W. Fris trom et al., in Problems in Biol-ogy: RNA in Deve lopment [E. W. Hanley, ed.], p. 382. Salt Lake City: University of Utah Press, 1969.)
Alberts_22.indd 1350Alberts_22.indd 1350 29.07.09 16:49:2929.07.09 16:49:29

Biologia Molecular da Célula 1351
Os genes seletores homeóticos são essenciais para a memória da informação posicional nas células dos discos imaginaisAs células de um disco imaginal se parecem com qualquer outra, mas os experimentos de transplantes mostram que elas são de fato já determinadas regionalmente e não são equi-valentes. Se um disco imaginal é transplantado na posição de outro na larva, e a larva desen-volve-se até a metamorfose, o disco enxertado diferencia-se autonomamente em uma estru-tura apropriada para a sua posição original: um disco de asa origina estruturas da asa, um disco de halteres, estruturas de halteres, independentemente do seu novo local. Isso mostra que as células dos discos imaginais são controladas pela memória da sua posição original. Por um procedimento de transplante serial mais complexo, que deixa as células do disco imaginal proliferarem por um período mais extenso antes da diferenciação, pode-se mos trar que esta memória celular é estavelmente herdada (com raros lapsos) por um número inde-finidamente grande de gerações celulares.
Os genes seletores homeóticos são componentes essenciais do mecanismo de memó-ria. Se, em qualquer estágio do longo período que leva à diferenciação na metamorfose, ambas as cópias de um gene seletor homeótico forem eliminadas pela recombi nação so-mática induzida, a partir de um clone de células do disco imaginal que normalmente iriam expressar aquele gene, aquelas células irão diferenciar-se em estruturas incorretas, como se pertencessem a um segmento diferente do corpo. Essas e outras observações indicam que a memória de informação posicional de cada célula depende de uma atividade continuada dos genes seletores homeóticos. Essa memória, além disso, é expressa em uma forma celular autônoma – cada célula parecendo manter o seu estado individualmente, depen dendo da sua própria história e de seu próprio genoma.
Genes reguladores específicos definem as células que formarão um órgão acessórioPrecisamos agora examinar como um órgão acessório desenvolve o seu padrão interno. To-maremos a asa do inseto como exemplo.
O processo começa com os mecanismos iniciais de formação de padrões que já discu-timos. Os sistemas ântero-posterior e dorso-ventral de sinalizações no embrião inicial, em essência, delimitam uma grade ortogonal, no blastoderma, na forma de limites de expres-são gênica segmentares dorso-ventrais e ântero-posteriores, periodicamente espaçados. Em certos pontos da interseção desses limites, a combinação de genes expressos é tal que induz um agrupamento de células à via de formação dos discos imaginais.
Em termos moleculares, isso corresponde à ativação da expressão dos genes regulado-res definidores dos discos imaginais. Na maioria dos discos, o gene Distal-less é ativado. Ele codifica para uma proteína de regulação gênica que é essencial ao crescimento sustentado necessário para criar um órgão acessório alongado, como uma pata ou uma antena, com um eixo próximo-distal. Na ausência desse gene, tais órgãos acessórios não se formam, e quando ele é expresso artificialmente em locais anormais, podem ser produzidos órgãos acessórios em locais incorretos. O Distal-less é expresso em uma for ma semelhante nos membros em desenvolvimento e em outros órgãos acessórios da maio ria das espécies de invertebrados e de vertebrados que já foram examinadas (Figura 22-52). Para o disco do olho, outro gene, o
Figura 22-52 Expressão de Dis tal-less em patas em desenvolvimento e em órgãos acessórios relacionados de vá-rias espécies. (A) Uma larva de ouriço-do-mar. (B) Uma larva de mariposa. (A, de G. Panganiban et al., Proc. Natl. Acad. Sci. U. S. A. 94:5162– 5166, 1997. Com permissão da National Academy of Sciences; B, de G. Panganiban, L. Nagy e S.B. Carrol, Curr. Biol. 4:671-675, 1994. Com permissão de Elsevier.)
(B)(A)0,1 mm 0,1 mm
Alberts_22.indd 1351Alberts_22.indd 1351 29.07.09 16:49:2929.07.09 16:49:29
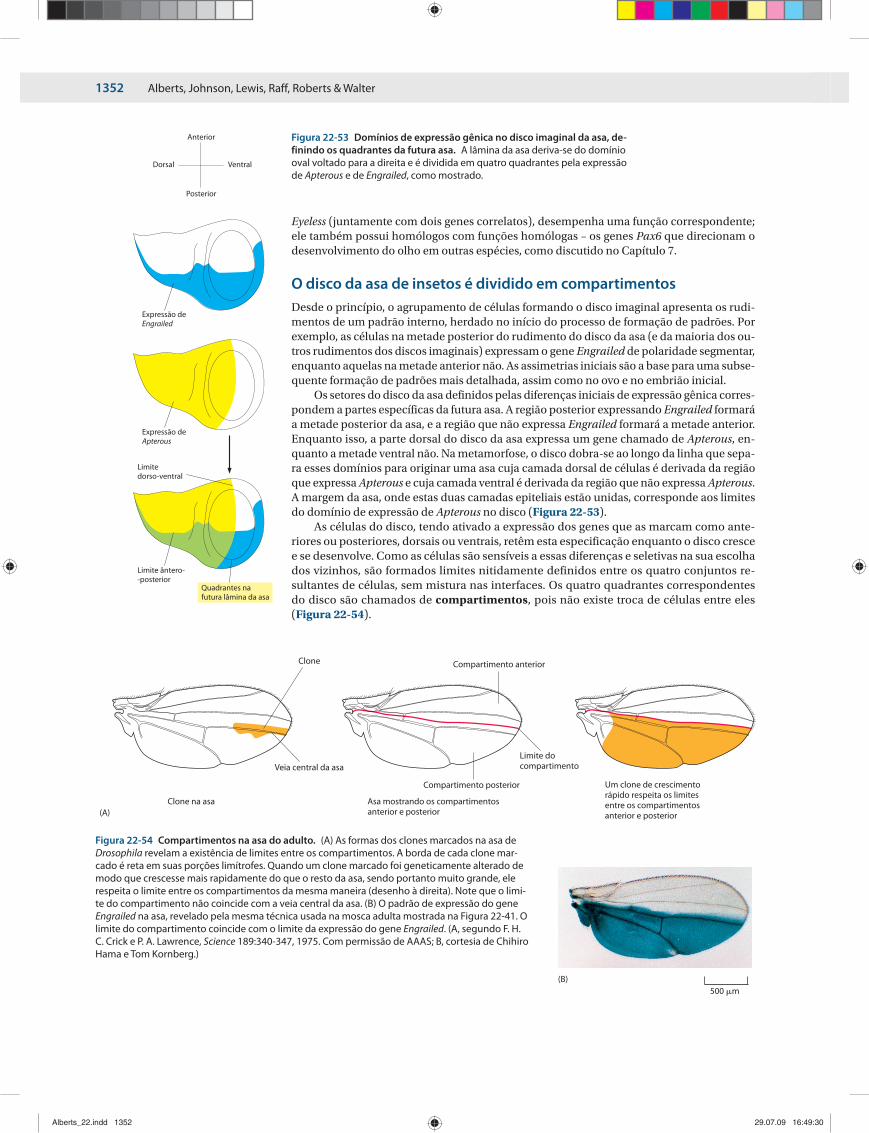
1352 Alberts, Johnson, Lewis, Raff, Roberts & Walter
Eyeless (juntamente com dois genes correlatos), desempenha uma função correspondente; ele também possui homólogos com funções homólogas – os genes Pax6 que direcionam o desenvolvimento do olho em ou tras espécies, como discutido no Capítulo 7.
O disco da asa de insetos é dividido em compartimentosDesde o princípio, o agrupamento de células formando o disco imaginal apresenta os rudi-mentos de um padrão interno, herdado no início do processo de formação de padrões. Por exem plo, as células na metade posterior do rudimento do disco da asa (e da maioria dos ou-tros rudimentos dos discos imaginais) expressam o gene Engrailed de polaridade segmentar, enquanto aquelas na metade anterior não. As assimetrias iniciais são a base para uma subse-quente formação de padrões mais detalhada, assim como no ovo e no embrião inicial.
Os setores do disco da asa definidos pelas diferenças iniciais de expressão gênica corres-pondem a partes específicas da futura asa. A região posterior expressando Engrailed formará a metade posterior da asa, e a região que não expressa Engrailed formará a meta de anterior. Enquanto isso, a parte dorsal do disco da asa expressa um gene chamado de Apterous, en-quanto a metade ventral não. Na metamorfose, o disco dobra-se ao longo da linha que sepa-ra esses domínios para originar uma asa cuja camada dorsal de células é derivada da região que expressa Apterous e cuja camada ventral é derivada da região que não expressa Apterous. A margem da asa, onde estas duas camadas epiteliais estão unidas, corresponde aos limites do domínio de expressão de Apterous no disco (Figura 22-53).
As células do disco, tendo ativado a expressão dos genes que as marcam como ante-riores ou posteriores, dorsais ou ventrais, retêm esta especificação enquanto o disco cres ce e se desenvolve. Como as células são sensíveis a essas diferenças e seletivas na sua escolha dos vizinhos, são formados limites nitidamente definidos entre os quatro conjun tos re-sultantes de células, sem mistura nas interfaces. Os quatro quadrantes corresponden tes do disco são chamados de compartimentos, pois não existe troca de células entre eles (Figura 22-54).
Figura 22-53 Domínios de expressão gênica no disco imaginal da asa, de-finindo os quadrantes da futura asa. A lâmina da asa deriva-se do domínio oval voltado para a direita e é dividida em quatro quadrantes pela expressão de Apterous e de Engrailed, como mostrado.
Expressão de Engrailed
Expressão deApterous
Limite ântero--posterior
Quadrantes nafutura lâmina da asa
Limite dorso-ventral
Anterior
Posterior
Dorsal Ventral
Clone
(A)
Compartimento anterior
Compartimento posterior
(B)
Clone na asa
Veia central da asa
Asa mostrando os compartimentosanterior e posterior
Limite docompartimento
Um clone de crescimentorápido respeita os limitesentre os compartimentosanterior e posterior
500 �m
Figura 22-54 Compartimentos na asa do adulto. (A) As formas dos clones marcados na asa de Drosophila revelam a existência de limites entre os compartimentos. A borda de cada clone mar-cado é reta em suas porções limítrofes. Quando um clone marcado foi geneticamente alterado de modo que crescesse mais rapidamente do que o resto da asa, sendo portanto muito grande, ele respeita o limite entre os compartimentos da mesma maneira (desenho à direita). Note que o limi-te do compartimento não coincide com a veia central da asa. (B) O padrão de expressão do gene Engrailed na asa, revelado pela mesma técnica usada na mosca adulta mos trada na Figura 22-41. O limite do compartimento coincide com o limite da expressão do gene Engrailed. (A, segundo F. H. C. Crick e P. A. Lawrence, Science 189:340-347, 1975. Com permissão de AAAS; B, cortesia de Chihiro Hama e Tom Kornberg.)
Alberts_22.indd 1352Alberts_22.indd 1352 29.07.09 16:49:3029.07.09 16:49:30

Biologia Molecular da Célula 1353
Quatro vias de sinalização da mesma família combinam-se para formar o padrão dos discos da asa: Wingless, Hedgehog, Dpp e NotchAo longo dos limites dos compartimentos – o limite ântero-posterior definido por Engrai led e o limite dorso-ventral definido por Apterous – células em diferentes estados con frontam-se umas com as outras e interagem para criar bandas estreitas de células espe cializadas. Estas células limítrofes produzem novos sinais para organizar o subsequente crescimento e a for-mação de padrões mais detalhados do órgão acessório.
As células no compartimento posterior da asa expressam a proteína de sinalização Hed-gehog, mas não respondem a ela. As células no compartimento anterior podem res ponder a Hedgehog. Uma vez que Hedgehog atua somente a uma curta distância, a via de recepção de sinais é ativada somente na estreita banda de células imediatamente anterior ao limite do compartimento, onde as células anteriores e posteriores estão justapostas. Estas células limítrofes respondem com a ativação da expressão de outra molécula sinalizadora, Dpp – a mesma proteína que encontramos previamente, na formação de padrões dorso-ventrais no embrião inicial (Figura 22-55). A Dpp atua no seu novo contexto em grande parte da mesma maneira que antes: ela difunde os seus efeitos para o exterior a partir das células limítrofes (por difusão, via citonemas, ou por meio de transferência célula-célula por exocitose e en-doxitose), produzindo um gradiente de morfógenos para controlar o subsequente padrão detalhado de crescimento e de expressão gênica.
Ocorrem eventos análogos no limite do compartimento dorso-ventral (ver Figura 22-55). Aqui, na futura margem da asa, uma comunicação de curta distância mediada pela via de Notch cria uma banda de células limítrofes que produzem outro morfógeno, a proteína Win-gless – o mesmo fator de sinalização, pertencendo à família Wnt, que atuou inicial mente na formação do padrão ântero-posterior de cada segmento embrionário. Os gradi entes de Dpp e de Wingless, juntamente com os outros sinais e assimetrias de expressão gênica que discutimos, combinam-se para conduzir a expressão de outros genes em localizações preci-samente definidas dentro de cada compartimento.
O tamanho de cada compartimento é regulado por interações entre as suas célulasUm dos mais misteriosos e pouco conhecidos aspectos do desenvolvimento animal é o con-trole do seu crescimento: por que cada parte do corpo cresce em um tamanho precisamen-
Figura 22-55 Sinais morfogenéticos cria dos nos limites dos comparti-mentos no disco imaginal da asa. (A) Criação da região de sinalização Dpp no limite do compartimento ântero--posterior por meio de uma interação mediada por Hedgehog entre as células anterio res e posteriores. De maneira análoga, uma inte ração mediada por Notch entre as células dor sais e ventrais cria uma região de sinalização Wingless (Wnt) ao longo do limite dorso-ven tral. (B) Os padrões de expressão observados de Dpp e de Wingless. Embora pareça claro que Dpp e Wingless atuem como morfógenos, ainda não está claro como eles se espalham a partir da sua fonte. Além disso, as célu las no disco imaginal são vistas emitindo longos citonemas, que podem lhes possibilitar a detecção de sinais a distância. Assim, a célula receptora pode enviar seus sensores para a fonte do sinal, em vez de o sinal mover-se para a célula receptora. (B, fo-tografias cortesia de Sean Carrol e Scott Weatherbee, de S. J. Day e P. A. Law-rence, Development 127:2977– 2987, 2000. Com permissão de The Company of Biologists.)
Dpp Wingless
(B)
Dpp
Compartimento anterior
(A)
Compartimentoposterior
A expressão de Engrailed defineo compartimento posterior
A Hedgehog no compartimentoposterior envia sinais decurto alcance para as célulasno compartimento anterior
Células anteriores no limiteentre os compartimentos expressam Dpp, um sinalizador de longo alcance
100 �m
Alberts_22.indd 1353Alberts_22.indd 1353 29.07.09 16:49:3029.07.09 16:49:30

1354 Alberts, Johnson, Lewis, Raff, Roberts & Walter
te definido? Esse problema é exemplificado de maneira marcante pelos discos imaginais de Drosophila. Por recombinação somática induzida, pode-se, por exemplo, criar um peque-no conjunto clonal de células que proliferam mais rapidamente do que as demais células no órgão em desenvolvimento. O clone pode crescer e ocupar quase a totalidade do com-partimento no qual ele está e, ainda assim, não ultrapassa o limite do comparti mento. Es-pantosamente, o seu crescimento rápido não apresenta quase nenhum efeito no tamanho final do compartimento, em sua forma ou mesmo nos detalhes do seu padrão interno (ver Figura 22-54). De alguma maneira, as células dentro do compartimento inte ragem umas com as outras determinando quando o seu crescimento deve parar, e cada com partimento comporta-se como uma unidade reguladora neste aspecto.
Uma primeira questão é se o tamanho do compartimento é regulado de maneira a con-ter um número determinado de células. Mutações nos componentes da maquinaria de con-trole do ciclo celular podem ser usadas para acelerar ou retardar a taxa de divisão celular, sem alterar a taxa de crescimento celular ou tecidual. Isso resulta em números anormais de pequenas células, ou o contrário, mas o tamanho – ou seja, a área – do com partimento prati-camente não é alterado. Assim, o mecanismo regulador parece depender de sinais que indi-cam a distância física entre uma parte do compartimento e a outra, e das respostas celulares que, de alguma forma, leem esses sinais de maneira a interromper o crescimento somente quando o espaçamento entre estas partes atingiu seu valor apropriado.
Este tipo de regulação do crescimento é demonstrado de forma notável na regeneração intercalar que ocorre quando partes separadas de um disco imaginal de Drosophila ou de uma pata em crescimento de barata são cirurgicamente enxertadas jun tas. Após o enxerto, as células na vizinhança da junção proliferam e completam as partes do padrão que deve-riam normalmente ficar entre elas, continuando o seu crescimento até que seja restaurado o espaçamento normal entre os pontos de referência (Figura 22-56). Os mecanismos que realizam isso permanecem um mistério, mas parece provável que sejam seme lhantes aos mecanismos que regulam o crescimento durante o desenvolvimento normal.
Qual mecanismo garante que cada pequeno pedaço do padrão dentro de um compar-timento cresça até o seu tamanho apropriado, apesar dos distúrbios locais na taxa de cres-cimento ou das condições iniciais? Os gradientes de morfógenos (de Dpp e Wingless, p. ex.) criam um padrão pela impo sição de características diferentes nas células em diferentes po-sições. Seria possível que as células em cada região possam, de alguma maneira, perceber o quão próximo está o espa çamento do padrão – o quão acentuado é o gradiente de alterações nas características da célula – e continuem o seu crescimento até que o tecido esteja espa-lhado até seu grau correto?
Essa ideia foi testada com a criação de clones de células do disco imaginal da asa em que os componentes subsequentes da via de sinalização Dpp estão expressos de maneira alterada, de forma a induzir um nível de ativação maior ou menor que o observado nas célu-las adjacentes. Do ponto de vista das células, as condições nos limites do clone mutante são equivalentes àquelas produzidas por um gradiente bastante acentuado de Dpp. O resultado é que as células nestas adjacências são estimuladas a se dividir em taxas aumentadas. Ao contrário, se o nível de sinalização de Dpp for tornado uniforme em uma região mediana do disco da asa em desenvolvimento, onde ele normalmente seria bastante acentuado, as divisões celulares são inibidas. Isso parece indicar que o gradiente do morfógeno realmente controla a taxa de proliferação. Contudo, se isso for verdadeiro, como as células percebem este gradiente?
A resposta não é conhecida, mas existem fortes evidências de que este mecanismo de-pende de sinais gerados nas junções célula-célula, onde células com diferentes graus de ati-vação da via do morfógeno fazem contato. Como discutido no Capítulo 19, mutações nos componentes juncionais, como as proteínas estruturais Discs-large (Dlg), ou no membro da
123456789
10
123456789
10
12389
10
123456789
10
Intercalação
Figura 22-56 Regeneração interca-lar. Quando porções não-correspon-dentes de uma pata em crescimento de barata são enxertadas jun tas, um novo tecido (verde) é intercalado (por prolife-ração celular) para preencher a lacuna entre os padrões das estruturas da pata, restaurando o seg mento da pata a seu tamanho e padrão normais.
Alberts_22.indd 1354Alberts_22.indd 1354 29.07.09 16:49:3029.07.09 16:49:30

Biologia Molecular da Célula 1355
superfamília das caderinas, Fat, podem levar a falhas dramáticas no controle do crescimen-to, permitindo que o disco da asa cresça muito além do seu tamanho normal apropriado. No caso da proteína Fat, um conjunto de outras moléculas, incluindo proteínas-cinase chama-das de Hippo e Warts, foi identificado como componente da via de sinalização que liga Fat na membrana celular, até o controle da expressão gênica, no núcleo. Os produtos dos genes--alvo incluem a ciclina E, reguladora do ciclo celular, e um inibidor da apoptose, assim como o microRNA Bantam, que parece ser parte essencial do mecanismo de controle do cresci-mento. Apesar destes fatos animadores, os mecanismos que controlam o tamanho de um órgão ainda são um mistério. Se pudermos descobrir como eles funcionam em Drosophila, poderemos ter alguma ideia de como ocorre o controle do tamanho dos órgãos em verte-brados, em nossa perplexidade acerca desta questão fundamental é ainda mais profunda. Para outros aspectos do desenvolvimento de órgãos, como discutiremos agora, as moscas e os vertebrados são inesperadamente similares em nível molecular, sugerindo que os seus mecanismos de controle do crescimento também possam ser similares.
Os padrões dos membros dos vertebrados são formados por mecanismos similaresOs membros dos vertebrados parecem muito diferentes dos membros dos insetos. A asa do inseto, por exemplo, consiste principalmente em duas camadas de epitélio de um pa drão elaborado, com pouco tecido entre elas. Em contraste, um membro de um vertebrado con-siste em um sistema elaboradamente padronizado de músculos, ossos e tecidos conectivos dentro de uma cobertura de epider me fina e estrutura de forma muito mais simples. Além disso, a evidência evolutiva sugere que o último ancestral comum entre os insetos e os verte-brados talvez não apresentasse pernas, nem braços, nem asas e nem nadadeiras, e que estes vários órgãos acessórios evoluíram independentemente no homem. E, ainda, quando exa-minamos os mecanismos moleculares que controlam o desenvolvimento dos membros dos vertebrados, encontramos um número surpreendente de similari dades com os membros dos insetos. Já mencionamos algumas dessas semelhanças, mas existem muitas outras: qua-se todas as moléculas que mencionamos na asa da mosca têm contrapartes nos membros dos vertebrados, embora estes sejam expressos em diferentes relações espaciais.
Os paralelismos têm sido mais cuidadosamente estudados no embrião de galinha. Como vimos anteriormente, cada perna ou asa de uma galinha origina-se de um broto de mem bro em forma de língua, consistindo em uma massa de células de tecido conectivo embrio nário, chamadas de células mesenquimais, encapsuladas em um envoltório de epitélio. Nessa es-trutura, encontra-se a expressão de homólogos de quase todos os genes que men cionamos em nossa explicação sobre a formação dos padrões de asa da Drosophila, in cluindo Distal-less, Wingless, Notch, Engrailed, Dpp e Hedgehog, desempenhando principalmente funções que parecem mais ou menos com as suas funções no disco imaginal da asa da Drosophila (Figura 22-57).
Os genes Hox, igualmente, fazem um aparecimento nos mem bros de insetos e de verte-brados. No órgão acessório dos insetos, os compartimentos anterior e posterior são distin-guidos pela expres são de diferentes genes do complexo Hox – um resultado do padrão de
ANTERIOR
POSTERIOR
DORSAL
VENTRALPROXIMAL
DISTAL
En1 (homólogo de Engrailed)
Wnt7a (homólogo de Wingless)
Lmx1 (homólogo de Apterous)
Bmp2 (homólogo de Dpp)
(B)(A)
500 �m
Crista apical ectodérmica,expressa Notch e secretaFGF4 e FGF8
Mesênquima posterior,secreta Sonic hedgehog
Figura 22-57 Moléculas que con-trolam a formação de padrões em um broto de membro de um verte-brado. (A) Um broto de asa de um embrião de galinha após quatro dias de incubação. A micrografia eletrônica de varredura mostra uma visão dorsal, com somi tos (os segmentos do tronco do embrião) visí veis à esquerda. Na margem distal do broto do membro, uma crista engrossada pode ser vista – a crista apical ectodérmica. (B) Padrões de ex pressão de proteínas-chave de sinalização e fa tores de regulação gê-nica no broto de membro de galinha. Os padrões estão representados es-quematicamente em dois planos imagi-nários de secção do broto do membro, um (ho rizontal) para mostrar o sistema dorso-ventral e o outro (vertical) para mostrar os sistemas ântero-posterior e próximo-distal. Sonic hed gehog, Bmp2 e Lmx1 são expressos no núcleo meso-dérmico do broto do membro; as outras moléculas no diagrama são expressas na sua co bertura epitelial. Quase todas as moléculas mos tradas possuem ho-mólogos que estão envolvi dos na for-mação dos padrões do disco da asa de Drosophila. (A, cortesia de Paul Martin.)
Alberts_22.indd 1355Alberts_22.indd 1355 29.07.09 16:49:3029.07.09 16:49:30
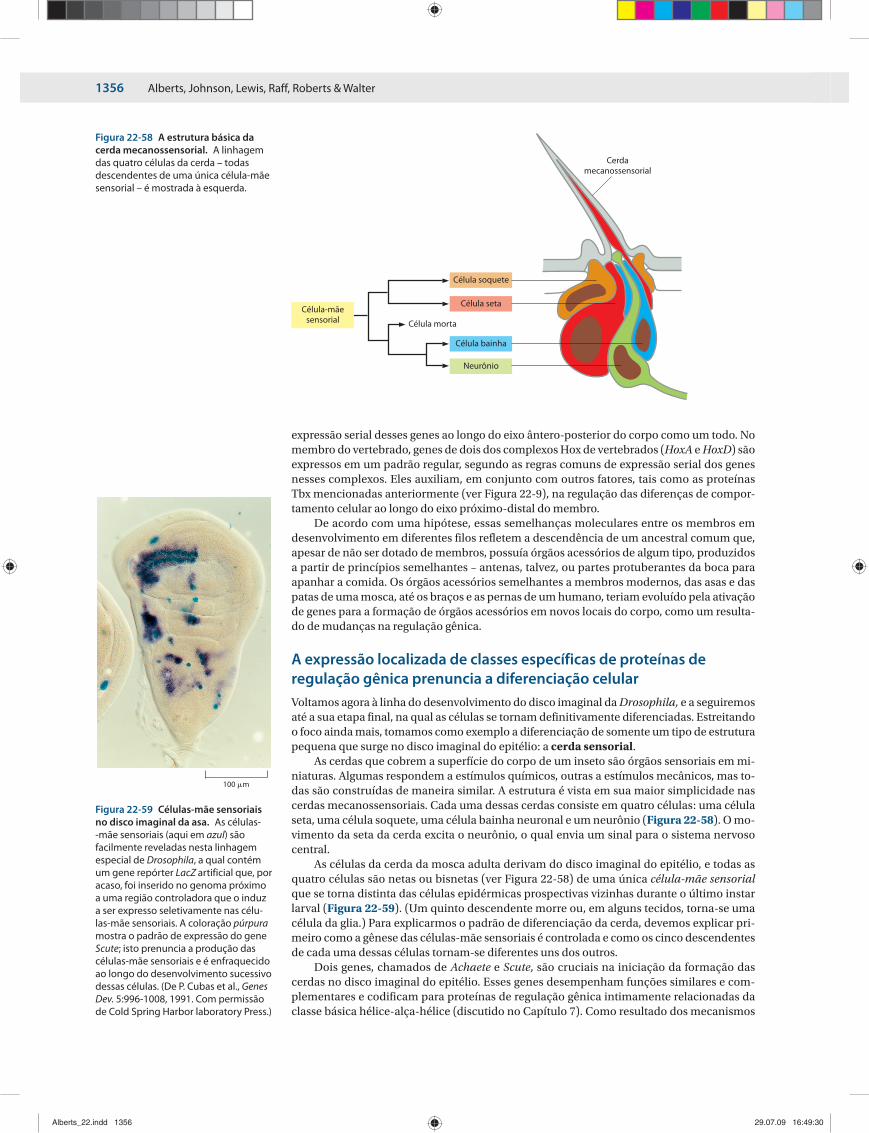
1356 Alberts, Johnson, Lewis, Raff, Roberts & Walter
expressão serial desses genes ao longo do eixo ântero-posterior do corpo como um todo. No membro do vertebrado, genes de dois dos complexos Hox de vertebrados (HoxA e HoxD) são ex pressos em um padrão regular, segundo as regras comuns de ex pressão serial dos genes nesses complexos. Eles auxiliam, em con junto com outros fatores, tais como as proteínas Tbx mencionadas anteriormente (ver Figura 22-9), na regulação das diferenças de compor-tamento celular ao longo do eixo próximo-distal do membro.
De acordo com uma hipótese, essas semelhanças moleculares entre os membros em desenvolvimento em diferentes filos refletem a descendência de um ancestral comum que, apesar de não ser dotado de membros, possuía órgãos acessórios de algum tipo, produ zidos a partir de princípios semelhantes – antenas, talvez, ou partes protuberantes da boca para apanhar a comida. Os órgãos acessórios semelhantes a membros modernos, das asas e das patas de uma mosca, até os braços e as pernas de um humano, teriam evoluído pela ativação de genes para a formação de órgãos acessórios em novos locais do corpo, como um resulta-do de mudanças na regulação gênica.
A expressão localizada de classes específicas de proteínas de regulação gênica prenuncia a diferenciação celularVoltamos agora à linha do desenvolvimento do disco imaginal da Drosophila, e a seguire mos até a sua etapa final, na qual as células se tornam definitivamente diferenciadas. Estreitando o foco ainda mais, tomamos como exemplo a diferenciação de somente um tipo de estrutura pequena que surge no disco imaginal do epitélio: a cerda sensorial.
As cerdas que cobrem a superfície do corpo de um inseto são órgãos sensoriais em mi-niaturas. Algumas respondem a estímulos químicos, outras a estímulos mecânicos, mas to-das são construídas de maneira similar. A estrutura é vista em sua maior simplicida de nas cerdas mecanossensoriais. Cada uma dessas cerdas consiste em quatro células: uma célula seta, uma célula soquete, uma célula bainha neuronal e um neurônio (Figura 22-58). O mo-vimento da seta da cerda excita o neurônio, o qual envia um sinal para o sistema nervoso central.
As células da cerda da mosca adulta derivam do disco imaginal do epitélio, e todas as quatro células são netas ou bisnetas (ver Figura 22-58) de uma única célula-mãe sensorial que se torna distinta das células epidérmicas prospectivas vizinhas durante o último instar larval (Figura 22-59). (Um quinto descendente morre ou, em alguns tecidos, torna-se uma célula da glia.) Para explicarmos o padrão de diferenciação da cerda, devemos explicar pri-meiro como a gênese das células-mãe sensoriais é controlada e como os cinco descendentes de cada uma dessas células tornam-se diferentes uns dos outros.
Dois genes, chamados de Achaete e Scute, são cruciais na iniciação da formação das cer das no disco imaginal do epitélio. Esses genes desempenham funções similares e com-plementares e codificam para proteínas de regulação gênica intimamente relacionadas da classe básica hélice-alça-hélice (discutido no Capítulo 7). Como resultado dos mecanismos
Célula soquete
Célula-mãesensorial
Célula seta
Célula bainha
Neurônio
Célula morta
Cerdamecanossensorial
Figura 22-58 A estrutura básica da cerda mecanossensorial. A linhagem das quatro cé lulas da cerda – todas descendentes de uma única célula-mãe sensorial – é mostrada à esquerda.
100 �m
Figura 22-59 Células-mãe sensoriais no disco imaginal da asa. As células--mãe sensoriais (aqui em azul) são facilmente reveladas nesta linhagem especial de Drosophila, a qual contém um gene repórter LacZ artificial que, por acaso, foi inserido no genoma próximo a uma região con troladora que o induz a ser expresso seletiva mente nas célu-las-mãe sensoriais. A coloração púrpura mostra o padrão de expressão do gene Scute; isto prenuncia a produção das células-mãe sen soriais e é enfraquecido ao longo do desen volvimento sucessivo dessas células. (De P. Cu bas et al., Genes Dev. 5:996-1008, 1991. Com permissão de Cold Spring Harbor laboratory Press.)
Alberts_22.indd 1356Alberts_22.indd 1356 29.07.09 16:49:3029.07.09 16:49:30

Biologia Molecular da Célula 1357
de for mação dos padrões do disco dos tipos que já discutimos, o Achaete e o Scute são ex-pressos no disco imaginal dentro das regiões em que as cerdas vão se formar. As mutações que eliminam a expressão desses genes em alguns dos locais comuns bloqueiam o desenvol-vimento das cerdas somente naqueles locais, e as mutações que causam a expressão em lo-cais adicionais anormais induzem as cerdas a desenvolverem-se lá. Contudo, a expressão de Achaete e de Scute é transitória, e somente algumas das células que inicialmente ex pressam os genes chegam a se tornar células-mãe sensoriais; as outras se tornam epiderme comum. O estado que é especificado pela expressão de Achaete e de Scute é chamado de pró-neural, e Achaete e Scute são chamados de genes pró-neurais. As células pró -neurais são induzidas a seguir a via neurossensorial da diferenciação, mas, como veremos, quais delas irão real-mente fazê-lo depende de interações competitivas entre elas.
A inibição lateral produz a distinção das células-mãe sensoriais nos agrupamentos pró-neuraisAs células que expressam genes pró-neurais ocorrem em grupos no disco imaginal epite-lial – um pequeno e isolado agrupamento de menos de 30 células para uma grande cerda isolada, e um amplo e contínuo conjunto de centenas ou de milhares de células para um campo de pequenas cerdas. No primeiro caso, somente um membro do agrupamento torna-se uma célula-mãe sensorial; no segundo caso, muitas células distribuídas por toda a região pró-neural o fazem. Em ambos os casos, cada célula-mãe sensorial é circunda da por células que inativam a expressão dos genes pró-neurais e são condenadas a diferenciarem-se como epiderme. Os experimentos com mosaicos genéticos mostram que isso se deve ao fato de que uma célula que se torna comprometida com a via de diferenciação da célula-mãe sensorial envia um sinal para as suas vizinhas não faze rem a mesma coisa: ela exerce uma inibição late-ral. Se uma célula que normalmente se tornaria uma célula-mãe sensorial for geneticamente incapacitada de fazê-lo, uma célula pró-neural adjacente, libertada da inibição lateral, irá se tornar uma célula-mãe sensorial em seu lugar.
A inibição lateral é mediada pela via de sinalização de Notch. Todas as células no agru-pamento inicialmente expressam o receptor transmembrana Notch e o seu ligante trans-membrana Delta. Em qualquer lugar que Delta ativar Notch, um sinal inibidor é enviado para a célula que expressa Notch; consequentemente, todas as células no agrupa mento ini-cialmente inibem umas às outras. Entretanto, acredita-se que o recebimento do sinal em uma dada célula diminua não somente a tendência daquela célula de se especializar como uma célula-mãe sensorial, mas também a sua habilidade de resposta por meio da liberação do sinal Delta. Isso cria uma situação competitiva, da qual uma única célula em cada peque-na região – a futura célula-mãe sensorial – emerge como ven cedora, remetendo um forte sinal inibidor para os seus vizinhos imediatos, mas não recebendo nenhum sinal deste tipo em troca (Figura 22-60). As consequências de uma falha desse mecanismo regulador estão mostradas na Figura 22-61.
A inibição lateral direciona a progênie da célula-mãe sensorial a diferentes destinações finaisO mesmo mecanismo de inibição lateral dependente de Notch opera repetidamente na formação das cerdas – não somente para forçar as vizinhas das células-mãe sensoriais a seguirem uma via diferente e se tornarem epidérmicas, mas também, mais tardiamente, para produzir filhas, netas e finalmente bisnetas da célula-mãe sensorial que expressem diferentes ge nes, de maneira a formar os distintos componentes da cerda. Em cada está-gio, a inibição lateral medeia uma interação competitiva que força células adjacentes a se compor tarem de maneiras contrastantes. Usando uma mutação Notch sensível à tempe-ratura, é possível desativar a sinalização Notch após a célula-mãe sensorial ter se diferen-ciado, mas antes de ela ter se dividido. A progênie então se distingue de maneira similar, originan do um agrupamento de neurônios no lugar dos quatro tipos distintos de células de uma cerda.
Como muitas outras competições, aquelas mediadas pela inibição lateral frequen-temente são fraudulentas: uma célula já inicia com uma vantagem que garante que ela será a vencedora. No desenvolvimento dos diferentes tipos celulares das cerdas sensoriais, uma tendência forte inicial é fornecida por uma assimetria em cada uma das divisões celulares da célula-mãe sensorial e de sua progênie. Uma proteína chamada de Numb (juntamente com
Alberts_22.indd 1357Alberts_22.indd 1357 29.07.09 16:49:3129.07.09 16:49:31
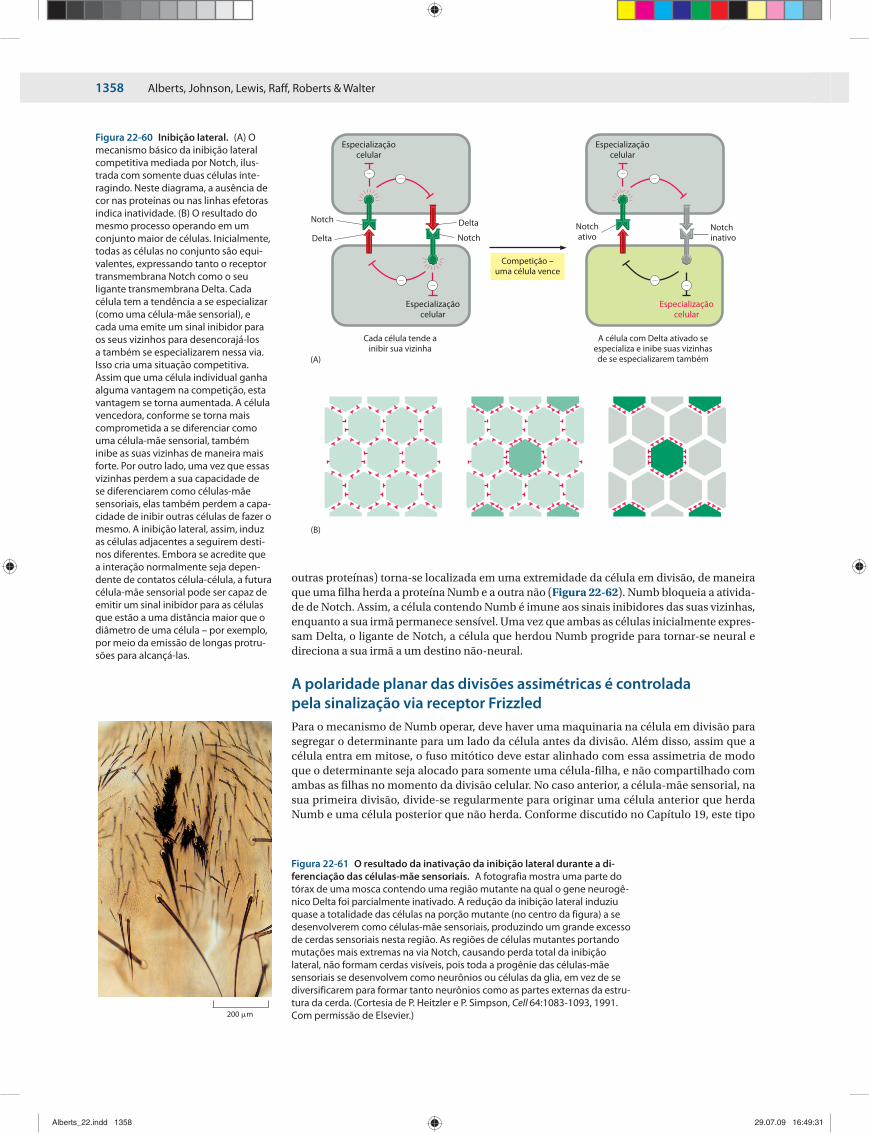
1358 Alberts, Johnson, Lewis, Raff, Roberts & Walter
outras proteínas) torna-se localizada em uma extremidade da célula em divisão, de maneira que uma filha herda a proteína Numb e a outra não (Figura 22-62). Numb bloqueia a ativida-de de Notch. Assim, a célula contendo Numb é imune aos sinais inibidores das suas vizinhas, enquanto a sua irmã permanece sensível. Uma vez que ambas as células inicialmente expres-sam Delta, o ligante de Notch, a célula que herdou Numb progride para tornar-se neural e direciona a sua irmã a um destino não-neural.
A polaridade planar das divisões assimétricas é controlada pela sinalização via receptor FrizzledPara o mecanismo de Numb operar, deve haver uma maquinaria na célula em divisão para segregar o determinante para um lado da célula antes da divisão. Além disso, assim que a célula entra em mitose, o fuso mitótico deve estar alinhado com essa assimetria de modo que o determinante seja alocado para somente uma célula-filha, e não compartilhado com ambas as filhas no momento da divisão celular. No caso anterior, a célula-mãe sensorial, na sua primeira divisão, divide-se regularmente para originar uma célula anterior que herda Numb e uma célula posterior que não herda. Conforme discutido no Capítulo 19, este tipo
Figura 22-60 Inibição lateral. (A) O meca nismo básico da inibição lateral competitiva mediada por Notch, ilus-trada com somente duas células inte-ragindo. Neste diagrama, a ausência de cor nas proteínas ou nas linhas efetoras indi ca inatividade. (B) O resultado do mesmo pro cesso operando em um conjunto maior de célu las. Inicialmente, todas as células no conjunto são equi-valentes, expressando tanto o receptor transmembrana Notch como o seu ligante transmembrana Delta. Cada célula tem a tendência a se especializar (como uma célula-mãe sensorial), e cada uma emite um sinal inibidor para os seus vizinhos para desencorajá-los a também se es pecializarem nessa via. Isso cria uma situação competitiva. Assim que uma célula individual ga nha alguma vantagem na competição, esta vanta gem se torna aumentada. A célula vencedora, conforme se torna mais comprometida a se dife renciar como uma célula-mãe sensorial, também ini be as suas vizinhas de maneira mais forte. Por outro lado, uma vez que essas vizinhas perdem a sua capacidade de se diferenciarem como células-mãe sensoriais, elas também perdem a capa-cidade de inibir outras células de fazer o mesmo. A inibição late ral, assim, induz as células adjacentes a seguirem desti-nos diferentes. Embora se acredite que a interação normalmente seja depen-dente de con tatos célula-célula, a futura célula-mãe sensorial pode ser capaz de emitir um sinal inibidor para as células que estão a uma distância maior que o diâmetro de uma célula – por exemplo, por meio da emissão de longas protru-sões para alcançá-las.
Figura 22-61 O resultado da inativação da inibição lateral durante a di-ferenciação das células-mãe sensoriais. A fotografia mostra uma parte do tórax de uma mosca contendo uma região mutante na qual o gene neurogê-nico Delta foi parcialmente inativado. A redução da inibição lateral indu ziu quase a totalidade das células na porção mutante (no centro da figura) a se desenvolverem como células-mãe sensoriais, produzindo um grande excesso de cerdas sensoriais nesta região. As regiões de células mutantes portando mutações mais extremas na via Notch, causando perda total da inibição lateral, não formam cerdas visíveis, pois toda a progênie das células-mãe sensoriais se desenvolvem como neurônios ou células da glia, em vez de se diversificarem para formar tanto neurônios como as partes externas da estru-tura da cerda. (Cortesia de P. Heitzler e P. Simpson, Cell 64:1083-1093, 1991. Com permissão de Elsevier.)
Especializaçãocelular
Especializaçãocelular
Notch
Delta Notch
Delta
Especializaçãocelular
Especializaçãocelular
Notchativo
Notchinativo
Competição –uma célula vence
Cada célula tende ainibir sua vizinha
A célula com Delta ativado seespecializa e inibe suas vizinhasde se especializarem também(A)
(B)
200 �m
Alberts_22.indd 1358Alberts_22.indd 1358 29.07.09 16:49:3129.07.09 16:49:31
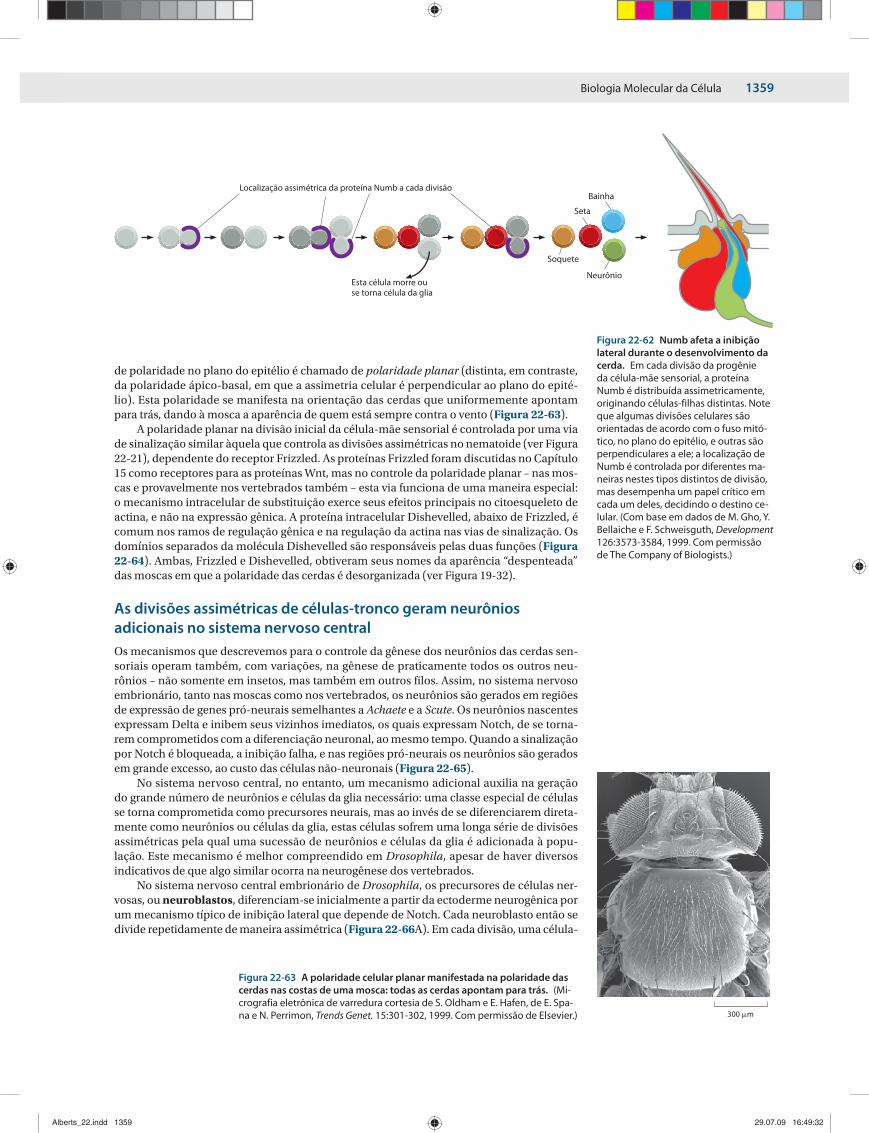
Biologia Molecular da Célula 1359
de polaridade no plano do epitélio é chamado de polaridade planar (distinta, em contraste, da polaridade ápico-basal, em que a assimetria celular é perpendicular ao plano do epité-lio). Esta polaridade se manifesta na orienta ção das cerdas que uniformemente apontam para trás, dando à mosca a aparência de quem está sempre contra o vento (Figura 22-63).
A polaridade planar na divisão inicial da célula-mãe sensorial é controlada por uma via de sinalização similar àquela que controla as divisões assimétricas no nematoide (ver Figura 22-21), dependente do receptor Frizzled. As proteínas Frizzled foram discutidas no Capítulo 15 como receptores para as proteínas Wnt, mas no controle da polaridade planar – nas mos-cas e provavelmente nos vertebrados também – esta via funciona de uma manei ra especial: o mecanismo intracelular de substituição exerce seus efeitos principais no citoesqueleto de actina, e não na expressão gênica. A proteína intracelular Dishevel led, abaixo de Frizzled, é comum nos ramos de regulação gênica e na regulação da actina nas vias de sinalização. Os domínios separados da molécula Dishevelled são responsáveis pelas duas funções (Figura 22-64). Ambas, Frizzled e Dishevelled, obtiveram seus nomes da aparên cia “despenteada” das moscas em que a polaridade das cerdas é desorganizada (ver Figura 19-32).
As divisões assimétricas de células-tronco geram neurônios adicionais no sistema nervoso centralOs mecanismos que descrevemos para o controle da gênese dos neurônios das cerdas sen-soriais operam também, com variações, na gênese de praticamente todos os outros neu-rônios – não somente em insetos, mas também em outros filos. Assim, no siste ma nervoso embrionário, tanto nas moscas como nos vertebrados, os neurônios são gera dos em regiões de expressão de genes pró-neurais semelhantes a Achaete e a Scute. Os neurônios nascentes expressam Delta e inibem seus vizinhos imediatos, os quais expressam Notch, de se torna-rem comprometidos com a diferencia ção neuronal, ao mesmo tempo. Quando a sinalização por Notch é bloqueada, a inibição falha, e nas regiões pró-neurais os neurônios são gerados em grande excesso, ao custo das células não-neuronais (Figura 22-65).
No sistema nervoso central, no entanto, um mecanismo adicional auxilia na geração do grande número de neurônios e células da glia necessário: uma classe especial de células se torna comprometida como precursores neurais, mas ao invés de se diferenciarem direta-mente como neurônios ou células da glia, estas células sofrem uma longa série de divisões assimétricas pela qual uma sucessão de neurônios e células da glia é adicionada à popu-lação. Este mecanismo é melhor compreendido em Drosophila, apesar de haver diversos indicativos de que algo similar ocorra na neurogênese dos vertebrados.
No sistema nervoso central embrionário de Drosophila, os precursores de células ner-vosas, ou neuroblastos, diferenciam-se inicialmente a partir da ectoderme neurogênica por um mecanismo típico de inibição lateral que depende de Notch. Cada neuroblasto então se divide repetidamente de maneira assimétrica (Figura 22-66A). Em cada divisão, uma célula-
Figura 22-62 Numb afeta a inibição lateral durante o desenvolvimento da cerda. Em cada divisão da progênie da célula-mãe sensorial, a proteína Numb é distribuída assimetricamente, originando células-filhas distintas. Note que algumas divisões celulares são orientadas de acordo com o fuso mitó-tico, no plano do epitélio, e outras são perpendiculares a ele; a localização de Numb é controlada por diferentes ma-neiras nestes tipos distintos de divisão, mas desempenha um papel crítico em cada um deles, decidindo o destino ce-lular. (Com base em dados de M. Gho, Y. Bellaiche e F. Schweisguth, Development 126:3573-3584, 1999. Com permissão de The Company of Biologists.)
BainhaLocalização assimétrica da proteína Numb a cada divisão
Neurônio
Seta
Soquete
Esta célula morre ouse torna célula da glia
300 �m
Figura 22-63 A polaridade celular planar manifestada na polaridade das cerdas nas costas de uma mosca: todas as cerdas apontam para trás. (Mi-crografia eletrônica de varredura cortesia de S. Oldham e E. Hafen, de E. Spa-na e N. Perrimon, Trends Genet. 15:301-302, 1999. Com permissão de Elsevier.)
Alberts_22.indd 1359Alberts_22.indd 1359 29.07.09 16:49:3229.07.09 16:49:32
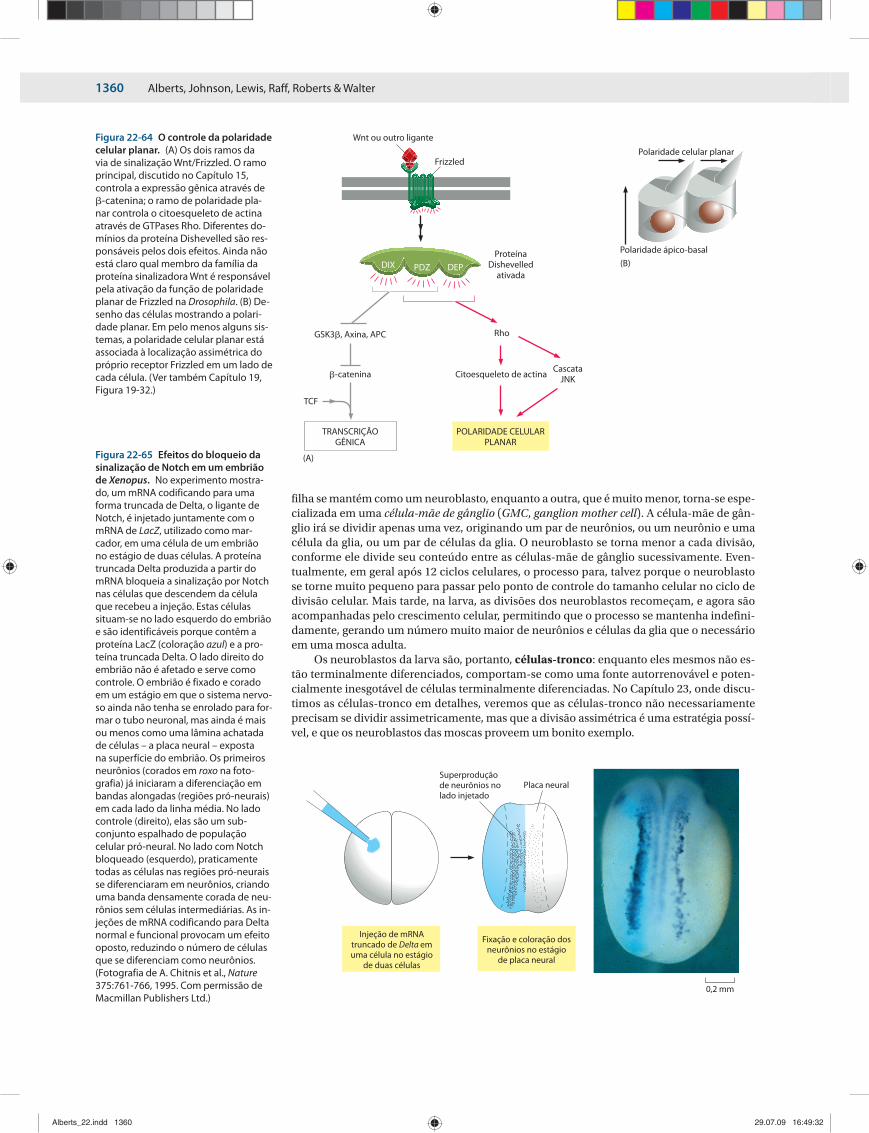
1360 Alberts, Johnson, Lewis, Raff, Roberts & Walter
filha se mantém como um neuroblasto, enquanto a outra, que é muito menor, torna-se espe-cializada em uma célula-mãe de gânglio (GMC, ganglion mother cell). A célula-mãe de gân-glio irá se dividir apenas uma vez, originando um par de neurônios, ou um neurônio e uma célula da glia, ou um par de células da glia. O neuroblasto se torna menor a cada divisão, conforme ele divide seu conteúdo entre as células-mãe de gânglio sucessivamente. Even-tualmente, em geral após 12 ciclos celulares, o processo para, talvez porque o neuroblasto se torne muito pequeno para passar pelo ponto de controle do tamanho celular no ciclo de divisão celular. Mais tarde, na larva, as divisões dos neuroblastos recomeçam, e agora são acompanhadas pelo crescimento celular, permitindo que o processo se mantenha indefini-damente, gerando um número muito maior de neurônios e células da glia que o necessário em uma mosca adulta.
Os neuroblastos da larva são, portanto, células-tronco: enquanto eles mesmos não es-tão terminalmente diferenciados, comportam-se como uma fonte autorrenovável e poten-cialmente inesgotável de células terminalmente diferenciadas. No Capítulo 23, onde discu-timos as células-tronco em detalhes, veremos que as células-tronco não necessariamente precisam se dividir assimetricamente, mas que a divisão assimétrica é uma estratégia possí-vel, e que os neuroblastos das moscas proveem um bonito exemplo.
Figura 22-64 O controle da polaridade celular pla nar. (A) Os dois ramos da via de sinali zação Wnt/Frizzled. O ramo principal, discutido no Capítulo 15, controla a expressão gênica através de �-catenina; o ramo de polaridade pla-nar contro la o citoesqueleto de actina através de GTPases Rho. Diferentes do-mínios da proteína Dishevelled são res-ponsáveis pelos dois efeitos. Ainda não está claro qual membro da família da pro teína sinalizadora Wnt é responsável pela ativa ção da função de polaridade planar de Frizzled na Drosophila. (B) De-senho das células mostran do a polari-dade planar. Em pelo menos alguns sis-temas, a polaridade celular planar está associada à localização assimétrica do próprio receptor Frizzled em um lado de cada célula. (Ver também Capítulo 19, Figura 19-32.)
DIX PDZ DEP
Wnt ou outro ligante
Frizzled
ProteínaDishevelled
ativada
Rho
CascataJNKCitoesqueleto de actina�-catenina
GSK3�, Axina, APC
TCF
TRANSCRIÇÃOGÊNICA
POLARIDADE CELULARPLANAR
(A)
(B)
Polaridade ápico-basal
Polaridade celular planar
Figura 22-65 Efeitos do bloqueio da sinali zação de Notch em um embrião de Xeno pus. No experimento mostra-do, um mRNA co dificando para uma forma truncada de Delta, o ligante de Notch, é injetado juntamente com o mRNA de LacZ, utilizado como mar-cador, em uma célula de um embrião no estágio de duas células. A proteína truncada Delta produzida a partir do mRNA bloqueia a sinalização por No tch nas células que descendem da célula que re cebeu a injeção. Estas células situam-se no lado esquerdo do embrião e são identificáveis porque contêm a proteína LacZ (coloração azul) e a pro-teína truncada Delta. O lado direito do embrião não é afetado e serve como controle. O em brião é fixado e corado em um estágio em que o sistema nervo-so ainda não tenha se enrolado para for-mar o tubo neuronal, mas ainda é mais ou menos como uma lâmina achatada de células – a placa neural – exposta na superfície do embrião. Os primeiros neurônios (corados em roxo na foto-grafia) já iniciaram a diferenciação em bandas alongadas (regiões pró-neurais) em cada lado da linha média. No lado controle (direito), elas são um sub-conjunto espalhado de população celular pró-neural. No lado com Notch bloqueado (esquerdo), praticamente todas as células nas regiões pró-neurais se diferencia ram em neurônios, criando uma banda den samente corada de neu-rônios sem células intermediárias. As in-jeções de mRNA codificando para Delta normal e funcional provocam um efeito oposto, reduzindo o número de células que se diferenciam como neurônios. (Fotografia de A. Chitnis et al., Nature 375:761-766, 1995. Com permissão de Mac millan Publishers Ltd.)
Placa neuralSuperproduçãode neurônios nolado injetado
Injeção de mRNAtruncado de Delta emuma célula no estágio
de duas células
Fixação e coloração dosneurônios no estágio
de placa neural
0,2 mm
Alberts_22.indd 1360Alberts_22.indd 1360 29.07.09 16:49:3229.07.09 16:49:32
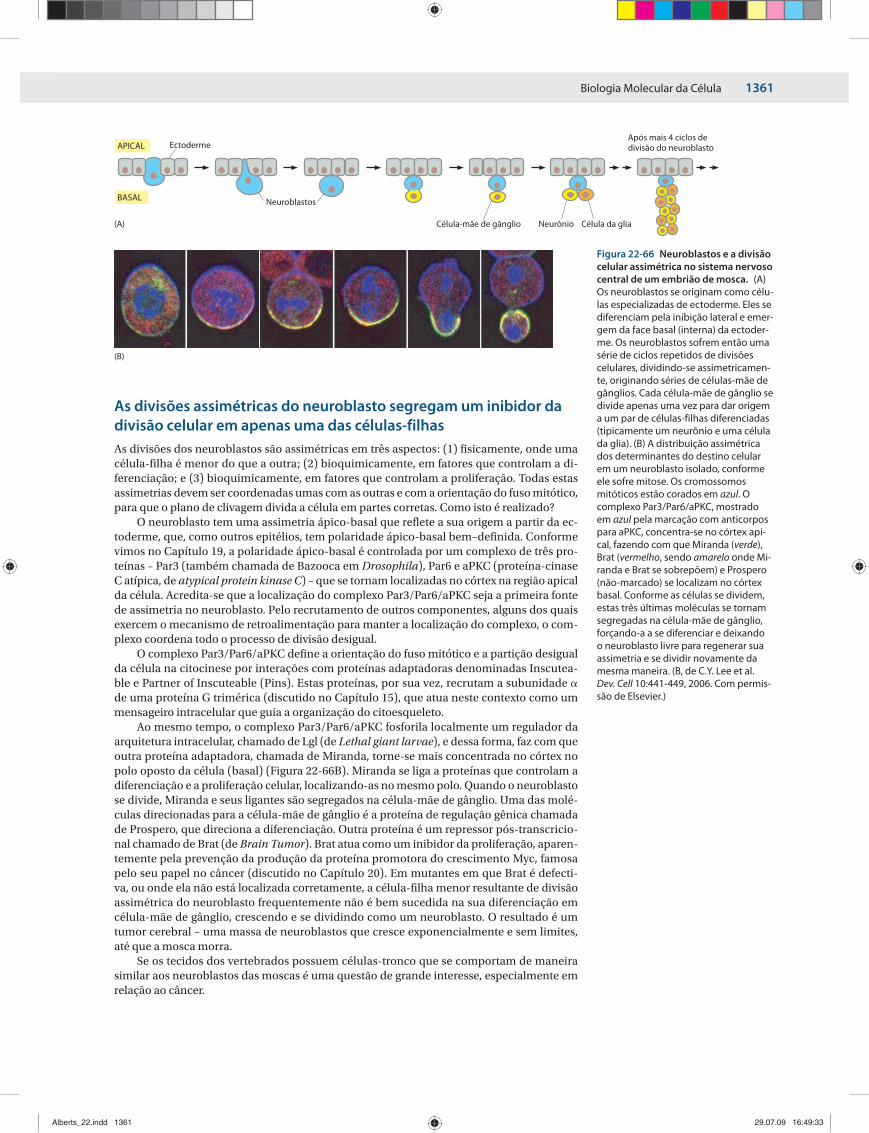
Biologia Molecular da Célula 1361
As divisões assimétricas do neuroblasto segregam um inibidor da divisão celular em apenas uma das células-filhasAs divisões dos neuroblastos são assimétricas em três aspectos: (1) fisicamente, onde uma célula-filha é menor do que a outra; (2) bioquimicamente, em fatores que controlam a di-ferenciação; e (3) bioquimicamente, em fatores que controlam a proliferação. Todas estas assimetrias devem ser coordenadas umas com as outras e com a orientação do fuso mitótico, para que o plano de clivagem divida a célula em partes corretas. Como isto é realizado?
O neuroblasto tem uma assimetria ápico-basal que reflete a sua origem a partir da ec-toderme, que, como outros epitélios, tem polaridade ápico-basal bem–definida. Conforme vimos no Capítulo 19, a polaridade ápico-basal é controlada por um complexo de três pro-teínas – Par3 (também chamada de Bazooca em Drosophila), Par6 e aPKC (proteína-cinase C atípica, de atypical protein kinase C) – que se tornam localizadas no córtex na região apical da célula. Acredita-se que a localização do complexo Par3/Par6/aPKC seja a primeira fonte de assimetria no neuroblasto. Pelo recrutamento de outros componentes, alguns dos quais exercem o mecanismo de retroalimentação para manter a localização do complexo, o com-plexo coordena todo o processo de divisão desigual.
O complexo Par3/Par6/aPKC define a orientação do fuso mitótico e a partição desigual da célula na citocinese por interações com proteínas adaptadoras denominadas Inscutea-ble e Partner of Inscuteable (Pins). Estas proteínas, por sua vez, recrutam a subunidade � de uma proteína G trimérica (discutido no Capítulo 15), que atua neste contexto como um mensageiro intracelular que guia a organização do citoesqueleto.
Ao mesmo tempo, o complexo Par3/Par6/aPKC fosforila localmente um regulador da arquitetura intracelular, chamado de Lgl (de Lethal giant larvae), e dessa forma, faz com que outra proteína adaptadora, chamada de Miranda, torne-se mais concentrada no córtex no polo oposto da célula (basal) (Figura 22-66B). Miranda se liga a proteínas que controlam a diferenciação e a proliferação celular, localizando-as no mesmo polo. Quando o neuroblasto se divide, Miranda e seus ligantes são segregados na célula-mãe de gânglio. Uma das molé-culas direcionadas para a célula-mãe de gânglio é a proteína de regulação gênica chamada de Prospero, que direciona a diferenciação. Outra proteína é um repressor pós-transcricio-nal chamado de Brat (de Brain Tumor). Brat atua como um inibidor da proliferação, aparen-temente pela prevenção da produção da proteína promotora do crescimento Myc, famosa pelo seu papel no câncer (discutido no Capítulo 20). Em mutantes em que Brat é defecti-va, ou onde ela não está localizada corretamente, a célula-filha menor resultante de divisão assimétrica do neuroblasto frequentemente não é bem sucedida na sua diferenciação em célula-mãe de gânglio, crescendo e se dividindo como um neuroblasto. O resultado é um tumor cerebral – uma massa de neuroblastos que cresce exponencialmente e sem limites, até que a mosca morra.
Se os tecidos dos vertebrados possuem células-tronco que se comportam de maneira similar aos neuroblastos das moscas é uma questão de grande interesse, especialmente em relação ao câncer.
Figura 22-66 Neuroblastos e a divisão celular assimétrica no sistema nervoso central de um embrião de mosca. (A) Os neuroblastos se originam como célu-las especializadas de ectoderme. Eles se diferenciam pela inibição lateral e emer-gem da face basal (interna) da ectoder-me. Os neuroblastos sofrem então uma série de ciclos repetidos de divisões celulares, dividindo-se assimetricamen-te, originando séries de células-mãe de gânglios. Cada célula-mãe de gânglio se divide apenas uma vez para dar origem a um par de células-filhas diferenciadas (tipicamente um neurônio e uma célula da glia). (B) A distribuição assimétrica dos determinantes do destino celular em um neuroblasto isolado, conforme ele sofre mitose. Os cromossomos mitóticos estão corados em azul. O complexo Par3/Par6/aPKC, mostrado em azul pela marcação com anticorpos para aPKC, concentra-se no córtex api-cal, fazendo com que Miranda (verde), Brat (vermelho, sendo amarelo onde Mi-randa e Brat se sobrepõem) e Prospero (não-marcado) se localizam no córtex basal. Conforme as células se dividem, estas três últimas moléculas se tornam segregadas na célula-mãe de gânglio, forçando-a a se diferenciar e deixando o neuroblasto livre para regenerar sua assimetria e se dividir novamente da mesma maneira. (B, de C.Y. Lee et al. Dev. Cell 10:441-449, 2006. Com permis-são de Elsevier.)
APICAL
BASAL
Ectoderme
Neuroblastos
Célula-mãe de gânglio(A)
(B)
Célula da gliaNeurônio
Após mais 4 ciclos dedivisão do neuroblasto
Alberts_22.indd 1361Alberts_22.indd 1361 29.07.09 16:49:3329.07.09 16:49:33

1362 Alberts, Johnson, Lewis, Raff, Roberts & Walter
A sinalização por Notch regula o padrão mais refinado dos tipos celulares diferenciados em diversos tipos de tecidosCada célula-filha e uma célula-mãe de gânglio normal podem se tornar tanto um neurônio quanto uma célula da glia. A escolha final, assim como a escolha do destino celular para a progênie de uma célula-mãe sensorial no sistema nervoso periférico, é controlada pela via de sinalização Notch e por inibição lateral. O processo de inibição lateral mediado por Notch mostrou-se crucial para a diversificação celular e para a formação de padrões mais refinados em uma enorme variedade de tecidos diferentes. Na mosca, ele controla a produção não somente dos neurônios, mas também de muitos outros tipos celulares diferenciados – por exemplo, no músculo, no revestimento do intestino, no sistema excretor, na traqueia, no olho e em outros órgãos sensoriais. Nos vertebrados, os homólogos de Notch e seus ligantes são expressos em tecidos correspondentes e desem penham funções semelhantes: as mutações na via de Notch alteram o equilíbrio dos neurônios e das células não-neuronais no sistema nervoso central e dos diferentes tipos celulares espe cializados no revestimento do intestino, das células endócrinas e exócrinas no pâncreas e das células sensoriais e auxiliares em ór-gãos sensoriais como o ouvido, para citar apenas alguns exemplos.
Em todos esses tecidos, é necessária uma mistura balanceada de diferentes tipos celu-lares. A sinalização por Notch fornece os meios para gerar a mistura, possibilitando que cé-lulas individuais expressem um conjunto de genes, direcionando seus vizinhos imedia tos a expressarem outro conjunto.
Alguns genes reguladores chave definem um tipo celular; outros podem ativar o programa para a criação de um órgão inteiroConforme mencionamos no início deste capítulo, existem alguns genes cujos produtos agem como disparadores para o desenvolvimento de um órgão específico, iniciando e co-ordenando todo o complexo programa de expressão gênica necessário para isso. Assim, por exemplo, quando o gene Eyeless é expresso artificialmente em um grupo de células no disco imaginal da pata, uma porção de células bem-organizadas de tecido ocular, com todos os seus tipos celulares corretamente arranjados, se desenvolverá na pata (ver Figura 22-2). De maneira similar, mas muito mais tarde, quando a célula faz a escolha final de um modo particular de diferenciação, como consequência das interações mediadas por Notch, ela deve seguir um complexo programa envolvendo a expressão de um conjunto inteiro de genes, e este programa de diferenciação é iniciado e coordenado por um con-junto muito menor de reguladores de nível superior. Estes reguladores são algumas vezes chamados de “proteínas reguladoras mestras” (mesmo que elas somente possam exercer seus efeitos específicos em combinação com os parceiros corretos, em uma célula que está adequada mente preparada).
Um exemplo é a família MyoD/miogenina de proteínas de regulação gênica. Estas pro-teínas direcio nam as células para diferenciarem-se como músculos, expressando actinas e miosinas musculares específicas e todas as outras proteínas do citoesqueleto, metabólicas e de mem brana necessárias à célula muscular (ver Figura 7-75). As proteínas de regulação gênica que definem tipos particulares de células frequentemente pertencem (assim como MyoD e seus correlatos) à família básica hélice-alça-hélice, codificada por genes homólogos e, em alguns casos, aparentemente idênticos aos genes pró -neurais que já mencionamos. A sua expressão frequentemente é controlada pela via Notch por meio de complicados circui-tos de retroalimentação.
A diferenciação celular terminal trouxe-nos ao final do nosso esboço de como os ge nes controlam a produção de uma mosca. Nossa narrativa foi necessariamente simplifica da. Um número muito maior de genes do que aquele aqui mencionado está envolvido em cada um dos processos do desenvolvimento que descrevemos. Os circuitos de retroa limentação, os mecanismos alternativos operando em paralelo, as redundâncias genéticas e outros fenô-menos complicam o quadro como um todo. Apesar disso, a mensagem prin cipal da genética do desenvolvimento é de uma simplicidade inesperada. Um número limitado de genes e de mecanismos, utilizados repetidamente em diferentes circunstâncias e combinações, é res-ponsável pelo controle das principais características do desenvolvimento de todos os ani-mais multicelulares.
A seguir, abordaremos um aspecto essencial do desenvolvimento animal que até agora temos negligenciado: os movimentos celulares.
Alberts_22.indd 1362Alberts_22.indd 1362 29.07.09 16:49:3329.07.09 16:49:33

Biologia Molecular da Célula 1363
Girino capaz de se alimentar110 horas, 106 células
32 horas, 170.000 células
19 horas, 80.000 células
Nêurula
10 horas, 30.000 células
Gástrula
6 horas, 10.000 células
Blástula
4 horas, 64 células
½ hora, 1 célula
Óvulo fertilizado 1 mm
ResumoAs partes externas de uma mosca adulta desenvolvem-se a partir de estruturas epiteliais chamadas de discos imaginais. Cada disco imaginal está dividido em um pequeno número de domínios que expressam diferentes proteínas de regulação gênica como resultado de processos de formação de pa-drões embrionários iniciais. Estes domí nios são chamados de compartimentos, pois suas células não se misturam. Nos limites dos compartimentos, as células que expressam genes diferentes confrontam-se umas com as outras e interagem, induzindo a produção localizada de morfógenos que governam o crescimento adicional e a formação dos padrões internos de cada compartimento. Assim, no disco da asa, as células dorsais e ventrais interagem pelo mecanismo de sinalização Notch para criar uma fonte da proteína Wingless (Wnt) ao longo dos limites do com partimento dorso-ventral, enquanto as células anteriores e posteriores interagem por meio da sinalização de curto alcance Hedgehog para criar uma fonte da proteína Dpp (um membro da família TGF�) ao longo dos limites do compar-timento ântero-posterior. Todas estas moléculas de sinalização possuem homólogos que desempe-nham papéis se melhantes na formação de padrões dos membros em vertebrados.
Cada compartimento de um disco imaginal, e cada subestrutura dentro dele, cresce até um tamanho precisamente previsível, mesmo em face de distúrbios aparentemente drásticos, como as mutações que alteram a taxa de divisão celular. Embora os gradi entes de morfógenos no disco claramente estejam envolvidos, os mecanismos reguladores críticos que controlam o tamanho dos órgãos não são entendidos.
Dentro de cada compartimento, os gradientes de morfógenos controlam os locais de expressão de conjuntos adicionais de genes, definindo conjuntos de células que intera gem umas com as outras ainda mais uma vez para criarem os detalhes mais refinados dos padrões finais de diferenciação ce-lular. Assim, a expressão gênica pró-neural define os locais onde as cerdas sensoriais irão se formar, e as interações mediadas por Notch entre as células do agrupamento pró-neural, juntamente com as divisões celulares assimé tricas, forçam as células individuais das cerdas a seguirem caminhos distintos para a diferenciação terminal. No sistema nervoso central, neuroblastos se diferenciam da ectoderme por inibição lateral de uma maneira similar, sofrendo uma longa série de divisões assimétricas como células-tronco para originar neurônios e células da glia. Falhas na distribuição assimétrica de moléculas que controlam a diferenciação e a proliferação celular podem converter células-tronco de neuroblastos em células de tumorais.
Acredita-se que muitos destes mecanismos também atuem em tecidos de vertebrados.
MOVIMENTOS CELULARES E A DETERMINAÇÃO DA FORMA DO CORPO DOS VERTEBRADOSA maioria das células do corpo de um animal é móvel, e no embrião em desenvolvimento seus movimentos muitas vezes são extensos, dramáticos e surpreendentes. Mudanças controladas na expressão gênica criam arranjos ordenados de células em diferentes estados; movimen-tos celulares rearranjam esses blocos de construção celulares, colocando-os em seus devidos lugares. Os genes que as células expressam determinam como eles se movem; nesse sentido, o controle da expressão gênica é o fenômeno primordial. Contudo, os movimentos celulares também são cruciais, e maiores explicações não são necessárias se quisermos entender como é formada a arquitetura do corpo. Nesta seção, examinaremos esse tópico no contexto do de-senvolvimento de vertebrados. Tomaremos como nosso exemplo principal a rã Xenopus laevis (Figura 22-67), na qual os movimentos celulares têm sido bem estudados; embora também serão analisadas evidências da galinha, do peixe-zebra e do camundongo.
Figura 22-67 Sinopse do desenvolvimento de Xenopus laevis a partir do óvulo recém-fertiliza-do até o girino capaz de se alimentar. A rã adulta é mostrada na fotografia superior. Os estágios de desenvolvimento são vistos lateralmente, com exceção dos embriões de 10 horas e 19 horas, os quais são vistos de baixo e de cima, respectivamente. Todos os estágios, exceto o de adulto, são mostrados na mesma escala. (Fotografia cortesia de Jonathan Slack; desenhos segundo P. D. Nieuwkoop e J. Faber, Normal Table of Xenopus laevis [Daudin]. Amsterdam: North-Holland, 1956.)
Alberts_22.indd 1363Alberts_22.indd 1363 29.07.09 16:49:3329.07.09 16:49:33
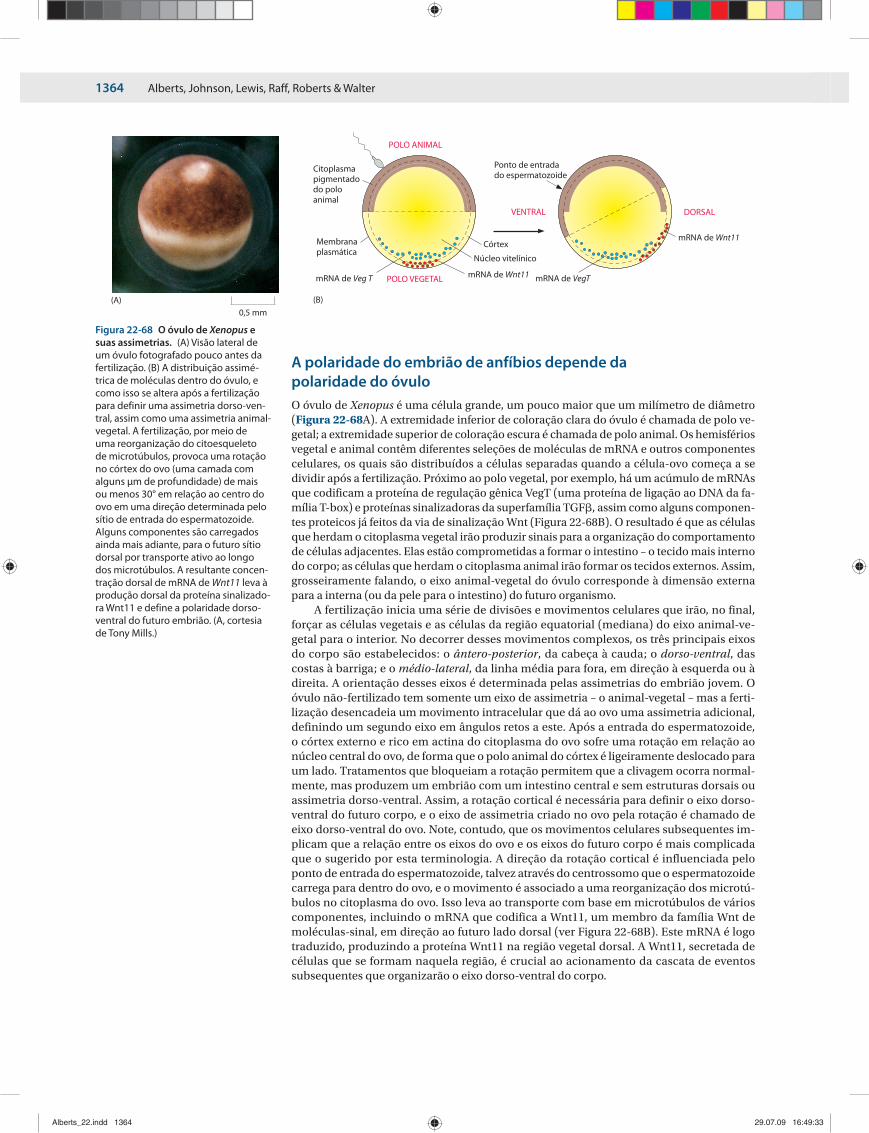
1364 Alberts, Johnson, Lewis, Raff, Roberts & Walter
A polaridade do embrião de anfíbios depende da polaridade do óvuloO óvulo de Xenopus é uma célula grande, um pouco maior que um milímetro de diâmetro (Figura 22-68A). A extremidade inferior de coloração clara do óvulo é chamada de polo ve-getal; a extremidade superior de coloração escura é chamada de polo animal. Os hemisférios vegetal e animal contêm diferentes seleções de moléculas de mRNA e outros componentes celulares, os quais são distribuídos a células separadas quando a célula-ovo começa a se dividir após a fertilização. Próximo ao polo vegetal, por exemplo, há um acúmulo de mRNAs que codificam a proteína de regulação gênica VegT (uma proteína de ligação ao DNA da fa-mília T-box) e proteínas sinalizadoras da superfamília TGF�, assim como alguns componen-tes proteicos já feitos da via de sinalização Wnt (Figura 22-68B). O resultado é que as células que herdam o citoplasma vegetal irão produzir sinais para a organização do comportamento de células adjacentes. Elas estão comprometidas a formar o intestino – o tecido mais interno do corpo; as células que herdam o citoplasma animal irão formar os tecidos externos. Assim, grosseiramente falando, o eixo animal-vegetal do óvulo corresponde à dimensão externa para a interna (ou da pele para o intestino) do futuro organismo.
A fertilização inicia uma série de divisões e movimentos celulares que irão, no final, forçar as células vegetais e as células da região equatorial (mediana) do eixo animal-ve-getal para o interior. No decorrer desses movimentos complexos, os três principais eixos do corpo são estabelecidos: o ântero-posterior, da cabeça à cauda; o dorso-ventral, das costas à barriga; e o médio-lateral, da linha média para fora, em direção à esquerda ou à direita. A orientação desses eixos é determinada pelas assimetrias do embrião jovem. O óvulo não-fertilizado tem somente um eixo de assimetria – o animal-vegetal – mas a ferti-lização desencadeia um movimento intracelular que dá ao ovo uma assimetria adicional, definindo um segundo eixo em ângulos retos a este. Após a entrada do espermatozoide, o córtex externo e rico em actina do citoplasma do ovo sofre uma rotação em relação ao núcleo central do ovo, de forma que o polo animal do córtex é ligeiramente deslocado para um lado. Tratamentos que bloqueiam a rotação permitem que a clivagem ocorra normal-mente, mas produzem um embrião com um intestino central e sem estruturas dorsais ou assimetria dorso-ventral. Assim, a rotação cortical é necessária para definir o eixo dorso-ventral do futuro corpo, e o eixo de assimetria criado no ovo pela rotação é chamado de eixo dorso-ventral do ovo. Note, contudo, que os movimentos celulares subsequentes im-plicam que a relação entre os eixos do ovo e os eixos do futuro corpo é mais complicada que o sugerido por esta terminologia. A direção da rotação cortical é influenciada pelo ponto de entrada do espermatozoide, talvez através do centrossomo que o espermatozoide carrega para dentro do ovo, e o movimento é associado a uma reorganização dos microtú-bulos no citoplasma do ovo. Isso leva ao transporte com base em microtúbulos de vários componentes, incluindo o mRNA que codifica a Wnt11, um membro da família Wnt de moléculas-sinal, em direção ao futuro lado dorsal (ver Figura 22-68B). Este mRNA é logo traduzido, produzindo a proteína Wnt11 na região vegetal dorsal. A Wnt11, secretada de células que se formam naquela região, é crucial ao acionamento da cascata de eventos subsequentes que organizarão o eixo dorso-ventral do corpo.
(A)
DORSAL
(B)
0,5 mm
Citoplasmapigmentadodo poloanimal
Membranaplasmática
mRNA de Veg T mRNA de VegT
Ponto de entradado espermatozoide
Córtex
Núcleo vitelínico
mRNA de Wnt11
mRNA de Wnt11
POLO ANIMAL
POLO VEGETAL
VENTRAL
Figura 22-68 O óvulo de Xenopus e suas assimetrias. (A) Visão lateral de um óvulo fotografado pouco antes da fertilização. (B) A distribuição assimé-trica de moléculas dentro do óvulo, e como isso se altera após a fertilização para definir uma assimetria dorso-ven-tral, assim como uma assimetria animal-vegetal. A fertilização, por meio de uma reorganização do citoesqueleto de microtúbulos, provoca uma rotação no córtex do ovo (uma camada com alguns μm de profundidade) de mais ou menos 30° em relação ao centro do ovo em uma direção determinada pelo sítio de entrada do espermatozoide. Alguns componentes são carregados ainda mais adiante, para o futuro sítio dorsal por transporte ativo ao longo dos microtúbulos. A resultante concen-tração dorsal de mRNA de Wnt11 leva à produção dorsal da proteína sinalizado-ra Wnt11 e define a polaridade dorso-ventral do futuro embrião. (A, cortesia de Tony Mills.)
Alberts_22.indd 1364Alberts_22.indd 1364 29.07.09 16:49:3329.07.09 16:49:33

Biologia Molecular da Célula 1365
A clivagem produz muitas células a partir de umaA rotação cortical é completada cerca de uma hora após a fertilização, sendo seguida por cli-vagem, na qual a grande célula-ovo única rapidamente se subdivide, por mitoses repetidas, em muitas células menores, ou blastômeros, sem nenhuma alteração na massa total (Figura 22-69). Dessa maneira, os determinantes assimetricamente distribuídos no ovo ficam distri-buídos em células separadas, com destinos diferentes (Figura 22-70).
Essas primeiras divisões celulares em Xenopus têm um tempo de ciclo de cerca de 30 minutos, com uma alternância direta de fases M e S, como discutido no Capítulo 17. As taxas bastante altas de replicação do DNA e da mitose parecem impedir quase toda a transcrição gênica (embora ocorra a síntese de proteínas), e o embrião em processo de clivagem é quase inteiramente dependente de reservas de RNA, proteína, membrana e outros materiais que se acumularam no ovo enquanto se desenvolvia como oócito na mãe. Após cerca de 12 ciclos de clivagem (7 horas), a taxa de divisão celular diminui, os ciclos celulares começam a seguir o padrão normal com fases G1 e G2 intervenientes entre as fases S e M, e começa a transcrição extensiva do genoma do embrião. Este evento é chamado de transição da blástula média, e ocorre com velocidade aproximadamente similar na maioria das espécies animais (sendo os mamíferos uma exceção). Estudos no peixe-zebra mostram que os transcritos recém-sin-tetizados incluem microRNAs que reconhecem muitos dos transcritos depositados no ovo pela mãe e que controlam a degradação rápida destes. Assim, a transição da blástula média marca o ponto em que o genoma do próprio embrião assume a maior parte do controle do desenvolvimento.
A gastrulação transforma uma bola côncava de células em uma estrutura de três camadas com um intestino primitivoDurante o período de clivagem, o embrião de rã se transforma de uma esfera sólida de cé-lulas em algo semelhante a uma bola côncava, com uma cavidade interna cheia de fluido e cercada por células que se aderem, formando um folheto epitelial. O embrião agora é deno-minado blástula (Figura 22-71).
Logo após, começam os movimentos coordenados da gastrulação. Esse processo dra-mático transforma a simples bola côncava de células em uma estrutura de camadas múlti-plas com um tubo intestinal central e simetria bilateral: por meio de uma versão mais ela-borada do processo anteriormente descrito para o ouriço-do-mar (ver Figura 22-3), muitas das células no exterior do embrião são movidas para dentro dele. O desenvolvimento sub-sequente depende das interações entre as camadas interna, externa e mediana de células assim formadas: a endoderme na parte interna, consistindo nas células que se moveram para o interior a fim de formar o intestino primitivo; a ectoderme na parte externa, consistindo em
Figura 22-69 Os estágios da clivagem em Xenopus. As divisões por clivagem rapidamente subdividem o ovo em muitas células menores. Todas as células se dividem sincronicamente até as pri-meiras 12 clivagens, mas as divisões são assimétricas, de forma que as células vegetais inferiores, carregadas de vitelo, são menos numerosas e maiores.
1 mm
1 hora, 1 célula 3 horas, 8 células 4 horas, 64 células
Figura 22-70 As origens das três camadas germinativas remontam a blastômeros distintos do embrião em seus estágios iniciais de clivagem. A endoderme deriva da maioria dos blastômeros vegetais, a ectoderme, da maioria dos animais, e a mesoderme, de um conjunto intermediário que tam-bém contribui à endoderme e à ecto-derme. A coloração em cada ilustração indica que, quanto maior a intensidade, maior a proporção de progênie celular que irá contribuir a uma dada camada germinativa. (Segundo L. Dale, Curr. Biol. 9:R812-R815, 1999. Com permissão de Elsevier.)ECTODERME MESODERME ENDODERME
Blastômerosanimais
Blastômerosvegetais
Alberts_22.indd 1365Alberts_22.indd 1365 29.07.09 16:49:3429.07.09 16:49:34
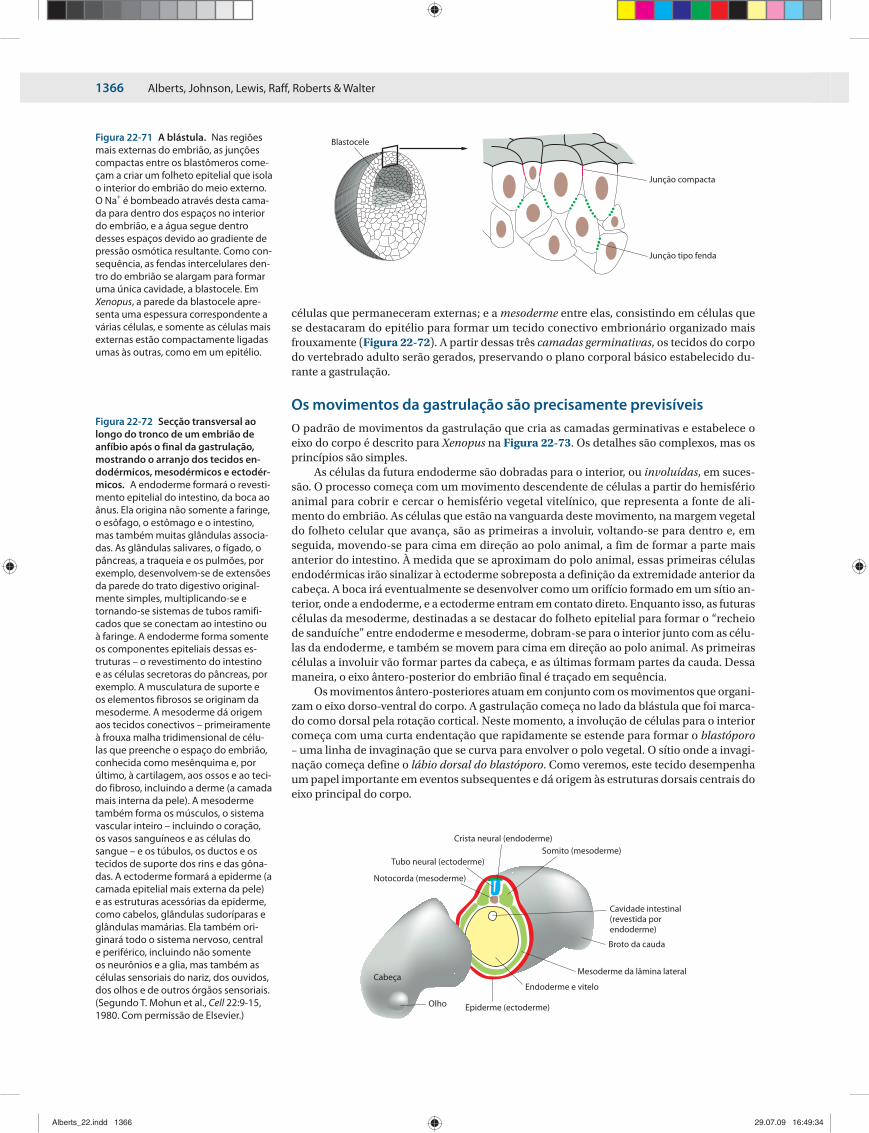
1366 Alberts, Johnson, Lewis, Raff, Roberts & Walter
células que permaneceram externas; e a mesoderme entre elas, consistindo em células que se destacaram do epitélio para formar um tecido conectivo embrionário organizado mais frouxamente (Figura 22-72). A partir dessas três camadas germinativas, os tecidos do corpo do vertebrado adulto serão gerados, preservando o plano corporal básico estabelecido du-rante a gastrulação.
Os movimentos da gastrulação são precisamente previsíveisO padrão de movimentos da gastrulação que cria as camadas germinativas e estabelece o eixo do corpo é descrito para Xenopus na Figura 22-73. Os detalhes são complexos, mas os princípios são simples.
As células da futura endoderme são dobradas para o interior, ou involuídas, em suces-são. O processo começa com um movimento descendente de células a partir do hemisfério animal para cobrir e cercar o hemisfério vegetal vitelínico, que representa a fonte de ali-mento do embrião. As células que estão na vanguarda deste movimento, na margem vegetal do folheto celular que avança, são as primeiras a involuir, voltando-se para dentro e, em seguida, movendo-se para cima em direção ao polo animal, a fim de formar a parte mais anterior do intestino. À medida que se aproximam do polo animal, essas primeiras células endodérmicas irão sinalizar à ectoderme sobreposta a definição da extremidade anterior da cabeça. A boca irá eventualmente se desenvolver como um orifício formado em um sítio an-terior, onde a endoderme, e a ectoderme entram em contato direto. Enquanto isso, as futuras células da mesoderme, destinadas a se destacar do folheto epitelial para formar o “recheio de sanduíche” entre endoderme e mesoderme, dobram-se para o interior junto com as célu-las da endoderme, e também se movem para cima em direção ao polo animal. As primeiras células a involuir vão formar partes da cabeça, e as últimas formam partes da cauda. Dessa maneira, o eixo ântero-posterior do embrião final é traçado em sequência.
Os movimentos ântero-posteriores atuam em conjunto com os movimentos que organi-zam o eixo dorso-ventral do corpo. A gastrulação começa no lado da blástula que foi marca-do como dorsal pela rotação cortical. Neste momento, a involução de células para o interior começa com uma curta endentação que rapidamente se estende para formar o blastóporo – uma linha de invaginação que se curva para envolver o polo vegetal. O sítio onde a invagi-nação começa define o lábio dorsal do blastóporo. Como veremos, este tecido desempenha um papel importante em eventos subsequentes e dá origem às estruturas dorsais centrais do eixo principal do corpo.
Figura 22-71 A blástula. Nas regiões mais externas do embrião, as junções compactas entre os blastômeros come-çam a criar um folheto epitelial que isola o interior do embrião do meio externo. O Na+ é bombeado através desta cama-da para dentro dos espaços no interior do embrião, e a água segue dentro desses espaços devido ao gradiente de pressão osmótica resultante. Como con-sequência, as fendas intercelulares den-tro do embrião se alargam para formar uma única cavidade, a blastocele. Em Xenopus, a parede da blastocele apre-senta uma espessura correspondente a várias células, e somente as células mais externas estão compactamente ligadas umas às outras, como em um epitélio.
Blastocele
Junção compacta
Junção tipo fenda
Tubo neural (ectoderme)
Notocorda (mesoderme)
Crista neural (endoderme)Somito (mesoderme)
Cavidade intestinal(revestida por endoderme)
Broto da cauda
Mesoderme da lâmina lateral
Endoderme e vitelo
Epiderme (ectoderme)Olho
Cabeça
Figura 22-72 Secção transversal ao longo do tronco de um embrião de anfíbio após o final da gastrulação, mostrando o arranjo dos tecidos en-dodérmicos, mesodérmicos e ectodér-micos. A endoderme formará o revesti-mento epitelial do intestino, da boca ao ânus. Ela origina não somente a faringe, o esôfago, o estômago e o intestino, mas também muitas glândulas associa-das. As glândulas salivares, o fígado, o pâncreas, a traqueia e os pulmões, por exemplo, desenvolvem-se de extensões da parede do trato digestivo original-mente simples, multiplicando-se e tornando-se sistemas de tubos ramifi-cados que se conectam ao intestino ou à faringe. A endoderme forma somente os componentes epiteliais dessas es-truturas – o revestimento do intestino e as células secretoras do pâncreas, por exemplo. A musculatura de suporte e os elementos fibrosos se originam da mesoderme. A mesoderme dá origem aos tecidos conectivos – primeiramente à frouxa malha tridimensional de célu-las que preenche o espaço do embrião, conhecida como mesênquima e, por último, à cartilagem, aos ossos e ao teci-do fibroso, incluindo a derme (a camada mais interna da pele). A mesoderme também forma os músculos, o sistema vascular inteiro – incluindo o coração, os vasos sanguíneos e as células do sangue – e os túbulos, os ductos e os tecidos de suporte dos rins e das gôna-das. A ectoderme formará a epiderme (a camada epitelial mais externa da pele) e as estruturas acessórias da epiderme, como cabelos, glândulas sudoríparas e glândulas mamárias. Ela também ori-ginará todo o sistema nervoso, central e periférico, incluindo não somente os neurônios e a glia, mas também as células sensoriais do nariz, dos ouvidos, dos olhos e de outros órgãos sensoriais. (Segundo T. Mohun et al., Cell 22:9-15, 1980. Com permissão de Elsevier.)
Alberts_22.indd 1366Alberts_22.indd 1366 29.07.09 16:49:3429.07.09 16:49:34
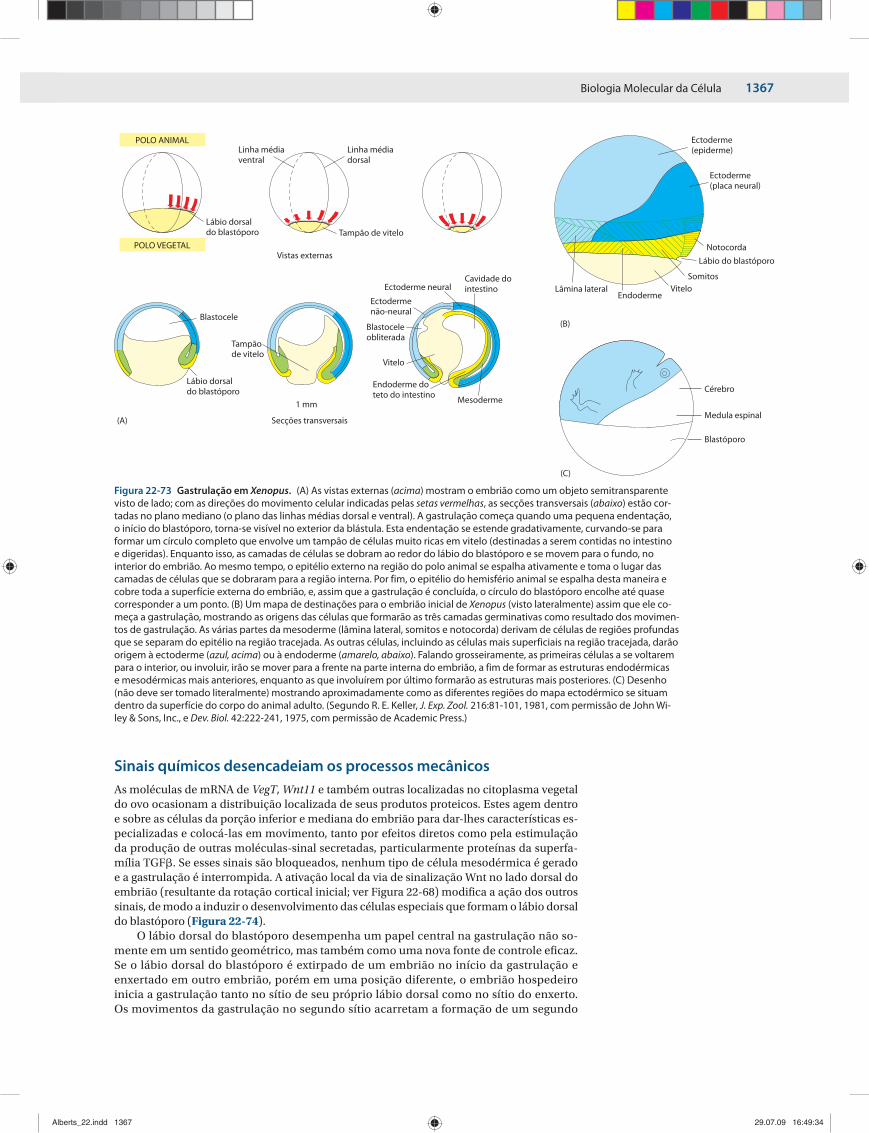
Biologia Molecular da Célula 1367
Sinais químicos desencadeiam os processos mecânicosAs moléculas de mRNA de VegT, Wnt11 e também outras localizadas no citoplasma vegetal do ovo ocasionam a distribuição localizada de seus produtos proteicos. Estes agem dentro e sobre as células da porção inferior e mediana do embrião para dar-lhes características es-pecializadas e colocá-las em movimento, tanto por efeitos diretos como pela estimulação da produção de outras moléculas-sinal secretadas, particularmente proteínas da superfa-mília TGF�. Se esses sinais são bloqueados, nenhum tipo de célula mesodérmica é gerado e a gastrulação é interrompida. A ativação local da via de sinalização Wnt no lado dorsal do embrião (resultante da rotação cortical inicial; ver Figura 22-68) modifica a ação dos outros sinais, de modo a induzir o desenvolvimento das células especiais que formam o lábio dorsal do blastóporo (Figura 22-74).
O lábio dorsal do blastóporo desempenha um papel central na gastrulação não so-mente em um sentido geométrico, mas também como uma nova fonte de controle eficaz. Se o lábio dorsal do blastóporo é extirpado de um embrião no início da gastrulação e enxertado em outro embrião, porém em uma posição diferente, o embrião hospedeiro inicia a gastrulação tanto no sítio de seu próprio lábio dorsal como no sítio do enxerto. Os movimentos da gastrulação no segundo sítio acarretam a formação de um segundo
Figura 22-73 Gastrulação em Xenopus. (A) As vistas externas (acima) mostram o embrião como um objeto semitransparente visto de lado; com as direções do movimento celular indicadas pelas setas vermelhas, as secções transversais (abaixo) estão cor-tadas no plano mediano (o plano das linhas médias dorsal e ventral). A gastrulação começa quando uma pequena endentação, o início do blastóporo, torna-se visível no exterior da blástula. Esta endentação se estende gradativamente, curvando-se para formar um círculo completo que envolve um tampão de células muito ricas em vitelo (destinadas a serem contidas no intestino e digeridas). Enquanto isso, as camadas de células se dobram ao redor do lábio do blastóporo e se movem para o fundo, no interior do embrião. Ao mesmo tempo, o epitélio externo na região do polo animal se espalha ativamente e toma o lugar das camadas de células que se dobraram para a região interna. Por fim, o epitélio do hemisfério animal se espalha desta maneira e cobre toda a superfície externa do embrião, e, assim que a gastrulação é concluída, o círculo do blastóporo encolhe até quase corresponder a um ponto. (B) Um mapa de destinações para o embrião inicial de Xenopus (visto lateralmente) assim que ele co-meça a gastrulação, mostrando as origens das células que formarão as três camadas germinativas como resultado dos movimen-tos de gastrulação. As várias partes da mesoderme (lâmina lateral, somitos e notocorda) derivam de células de regiões profundas que se separam do epitélio na região tracejada. As outras células, incluindo as células mais superficiais na região tracejada, darão origem à ectoderme (azul, acima) ou à endoderme (amarelo, abaixo). Falando grosseiramente, as primeiras células a se voltarem para o interior, ou involuir, irão se mover para a frente na parte interna do embrião, a fim de formar as estruturas endodérmicas e mesodérmicas mais anteriores, enquanto as que involuírem por último formarão as estruturas mais posteriores. (C) Desenho (não deve ser tomado literalmente) mostrando aproximadamente como as diferentes regiões do mapa ectodérmico se situam dentro da superfície do corpo do animal adulto. (Segundo R. E. Keller, J. Exp. Zool. 216:81-101, 1981, com permissão de John Wi-ley & Sons, Inc., e Dev. Biol. 42:222-241, 1975, com permissão de Academic Press.)
POLO ANIMAL
POLO VEGETAL
Lábio dorsaldo blastóporo
Linha médiaventral
Linha médiadorsal
Tampão de vitelo
Blastocele
Lábio dorsaldo blastóporo
Tampãode vitelo
Vistas externas
Secções transversais
1 mm
Ectodermenão-neural
Blastoceleobliterada
Vitelo
Endoderme do teto do intestino
Mesoderme
Ectoderme neuralCavidade dointestino
(A)
Ectoderme(epiderme)
Lábio do blastóporo
Notocorda
SomitosLâmina lateral
Endoderme
Ectoderme(placa neural)
(B)
Vitelo
(C)
Cérebro
Medula espinal
Blastóporo
Alberts_22.indd 1367Alberts_22.indd 1367 29.07.09 16:49:3429.07.09 16:49:34
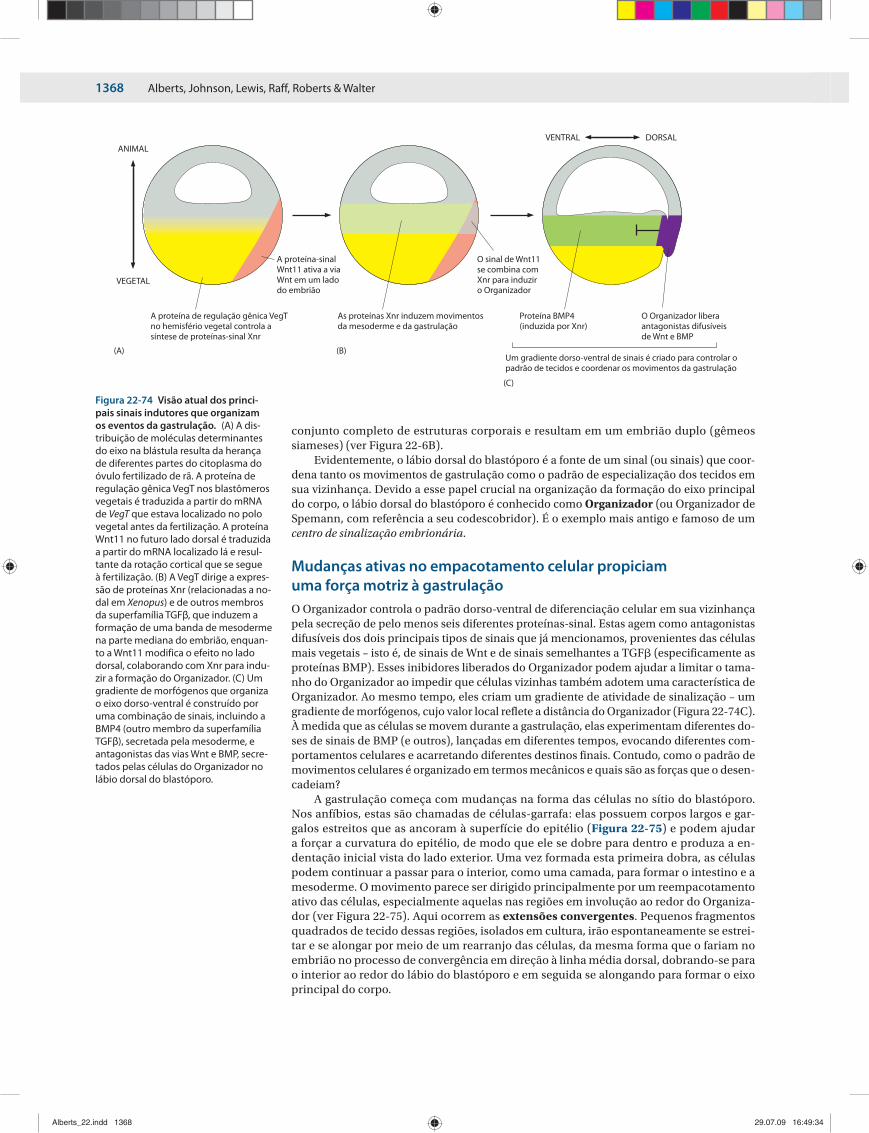
1368 Alberts, Johnson, Lewis, Raff, Roberts & Walter
conjunto completo de estruturas corporais e resultam em um embrião duplo (gêmeos siameses) (ver Figura 22-6B).
Evidentemente, o lábio dorsal do blastóporo é a fonte de um sinal (ou sinais) que coor-dena tanto os movimentos de gastrulação como o padrão de especialização dos tecidos em sua vizinhança. Devido a esse papel crucial na organização da formação do eixo principal do corpo, o lábio dorsal do blastóporo é conhecido como Organizador (ou Organizador de Spemann, com referência a seu codescobridor). É o exemplo mais antigo e famoso de um centro de sinalização embrionária.
Mudanças ativas no empacotamento celular propiciam uma força motriz à gastrulaçãoO Organizador controla o padrão dorso-ventral de diferenciação celular em sua vizinhança pela secreção de pelo menos seis diferentes proteínas-sinal. Estas agem como antagonistas difusíveis dos dois principais tipos de sinais que já mencionamos, provenientes das células mais vegetais – isto é, de sinais de Wnt e de sinais semelhantes a TGF� (especificamente as proteínas BMP). Esses inibidores liberados do Organizador podem ajudar a limitar o tama-nho do Organizador ao impedir que células vizinhas também adotem uma característica de Organizador. Ao mesmo tempo, eles criam um gradiente de atividade de sinalização – um gradiente de morfógenos, cujo valor local reflete a distância do Organizador (Figura 22-74C). À medida que as células se movem durante a gastrulação, elas experimentam diferentes do-ses de sinais de BMP (e outros), lançadas em diferentes tempos, evocando diferentes com-portamentos celulares e acarretando diferentes destinos finais. Contudo, como o padrão de movimentos celulares é organizado em termos mecânicos e quais são as forças que o desen-cadeiam?
A gastrulação começa com mudanças na forma das células no sítio do blastóporo. Nos anfíbios, estas são chamadas de células-garrafa: elas possuem corpos largos e gar-galos estreitos que as ancoram à superfície do epitélio (Figura 22-75) e podem ajudar a forçar a curvatura do epitélio, de modo que ele se dobre para dentro e produza a en-dentação inicial vista do lado exterior. Uma vez formada esta primeira dobra, as células podem continuar a passar para o interior, como uma camada, para formar o intestino e a mesoderme. O movimento parece ser dirigido principalmente por um reempacotamento ativo das células, especialmente aquelas nas regiões em involução ao redor do Organiza-dor (ver Figura 22-75). Aqui ocorrem as extensões convergentes. Pequenos fragmentos quadrados de tecido dessas regiões, isolados em cultura, irão espontaneamente se estrei-tar e se alongar por meio de um rearranjo das células, da mesma forma que o fariam no embrião no processo de convergência em direção à linha média dorsal, dobrando-se para o interior ao redor do lábio do blastóporo e em seguida se alongando para formar o eixo principal do corpo.
Figura 22-74 Visão atual dos princi-pais sinais indutores que organizam os eventos da gastrulação. (A) A dis-tribuição de moléculas determinantes do eixo na blástula resulta da herança de diferentes partes do citoplasma do óvulo fertilizado de rã. A proteína de regulação gênica VegT nos blastômeros vegetais é traduzida a partir do mRNA de VegT que estava localizado no polo vegetal antes da fertilização. A proteína Wnt11 no futuro lado dorsal é traduzida a partir do mRNA localizado lá e resul-tante da rotação cortical que se segue à fertilização. (B) A VegT dirige a expres-são de proteínas Xnr (relacionadas a no-dal em Xenopus) e de outros membros da superfamília TGFβ, que induzem a formação de uma banda de mesoderme na parte mediana do embrião, enquan-to a Wnt11 modifica o efeito no lado dorsal, colaborando com Xnr para indu-zir a formação do Organizador. (C) Um gradiente de morfógenos que organiza o eixo dorso-ventral é construído por uma combinação de sinais, incluindo a BMP4 (outro membro da superfamília TGFβ), secretada pela mesoderme, e antagonistas das vias Wnt e BMP, secre-tados pelas células do Organizador no lábio dorsal do blastóporo.
VENTRAL DORSALANIMAL
VEGETAL
A proteína de regulação gênica VegTno hemisfério vegetal controla a síntese de proteínas-sinal Xnr
As proteínas Xnr induzem movimentosda mesoderme e da gastrulação
Proteína BMP4 (induzida por Xnr)
O Organizador liberaantagonistas difusíveisde Wnt e BMP
(B)
(C)
(A)Um gradiente dorso-ventral de sinais é criado para controlar opadrão de tecidos e coordenar os movimentos da gastrulação
A proteína-sinal Wnt11 ativa a viaWnt em um ladodo embrião
O sinal de Wnt11se combina comXnr para induziro Organizador
Alberts_22.indd 1368Alberts_22.indd 1368 29.07.09 16:49:3429.07.09 16:49:34

Biologia Molecular da Célula 1369
Para que esta transformação extraordinária seja efetuada, as células individuais têm de se arrastar umas sobre as outras de forma coordenada (Figura 22-76). O alinhamento de seus movimentos parece depender da mesma maquinaria que, no verme e na mosca, encontramos controlando a polaridade planar celular: a via de sinalização de polaridade Frizzled/Dishevelled. Quando esta via é bloqueada – por uma forma dominante negativa de Dishevelled, por exemplo – a extensão convergente não ocorre.
Padrões variáveis de moléculas de adesão celular forçam a formação de novos arranjos de célulasOs padrões de expressão gênica governam os movimentos celulares embrionários de muitos modos diferentes. Eles regulam a mobilidade celular, a forma celular e a produção de sinais para seu direcionamento. Também têm a importante função de determinar os conjuntos de moléculas de adesão que as células apresentam em suas superfícies. Por meio de alterações em suas moléculas de superfície, uma célula pode romper ligações antigas e produzir novas. As células em uma região podem desenvolver propriedades de superfície que fazem-nas se aderirem umas às outras e se segregarem de um grupo de células vizinhas cuja superfície química é diferente.
Experimentos feitos há meio século em embriões jovens de anfíbios mostraram que os efeitos de adesão seletiva célula-célula podem ser tão eficazes a ponto de resultar em uma
Figura 22-75 Movimentos celulares na gastrulação. Secção transversal de um embrião de Xenopus em gastrulação, feita no mesmo plano da Figura 22-73, indicando os quatro principais tipos de movimentos envolvidos na gastrulação. O epitélio do polo animal se expande por rearranjo celular, tornando-se mais fino à medida que se espalha. A migra-ção de células mesodérmicas sobre uma matriz rica em fibronectina que reveste a parte superior da blastocele pode ajudar a empurrar os tecidos invagina-dos para a frente. Contudo, a principal força motriz à gastrulação em Xenopus é a extensão convergente na zona mar-ginal. (Segundo R. E. Keller, J. Exp. Zool. 216:81-101, 1981. Com permissão de Wiley-Liss.)
EXTENSÃO
EXTENSÃO
CONVERGÊNCIA CONVERGÊNCIA
(A) (B)Linha média dorsal
Os lamelipódios tentamse arrastar sobre a superfície de células vizinhas, puxando-aspara dentro em direção às setas
Figura 22-76 Extensão convergente e suas bases celulares. (A) Padrão de extensão convergente na zona marginal de uma gástrula visto a partir do lado dorsal. As setas azuis representam a convergência em direção à linha média dorsal, as setas vermelhas representam a extensão do eixo ânte-ro-posterior. O diagrama simplificado não pretende mostrar o movimento acompanhado de involução, pelo qual as células estão se dobrando para o interior do embrião. (B) Diagrama esquemático do comportamento celular que forma a base da extensão convergente. As células formam lameli-pódios, com os quais tentam se arrastar umas sobre as outras. O alinhamento dos movimentos dos lamelipódios ao longo de um eixo comum leva à extensão convergente. O processo depende da via de sinalização de polaridade Frizzled/Dishevelled e é presumivelmente cooperativo, porque as células que já estão alinhadas exercem forças que tendem a alinhar suas vizinhas da mesma maneira. (B, segundo J. Shih e R. Keller, Development 116:901-914, 1992. Com permissão de The Company of Biologists.)
Camada rica em fibronectina
Espaço preenchidopor fluido
O epitélio do poloanimal se expande
As células mesodérmicasmigram sobre a fibronectina
As células-garrafa ajudama forçar a curvatura doepitélio invaginante
A zona marginal sofreextensão convergente
1
2
3
4
Vitelo
Lábio dorsal do blastóporo
Alberts_22.indd 1369Alberts_22.indd 1369 29.07.09 16:49:3529.07.09 16:49:35

1370 Alberts, Johnson, Lewis, Raff, Roberts & Walter
reconstrução aproximada da estrutura normal de um embrião jovem pós-gastrulação, mes-mo após as células terem sido artificialmente dissociadas. Quando essas células são reagre-gadas em uma mistura aleatória, elas se separam espontaneamente, de acordo com suas características originais (Figura 22-77). Como discutido no Capítulo 19, um papel central em tais fenômenos é desempenhado pelas caderinas – uma grande e variada família de pro-teínas de adesão célula-célula dependentes de Ca2+ evolutivamente relacionadas. Estas e outras moléculas de adesão célula-célula são diferencialmente expressas nos vários tecidos do embrião jovem, sendo que anticorpos contra elas interferem na adesão seletiva normal entre células de um tipo similar.
As mudanças nos padrões de expressão das várias caderinas se correlacionam intima-mente com as alterações dos padrões de associação entre as células durante a gastrulação, a neurulação e a formação dos somitos (ver Figura 19-25). Esses rearranjos são provavelmente regulados e direcionados, em parte, pelo padrão de caderinas. Em particular, as caderinas parecem desempenhar uma função importante no controle da formação e da dissolução de folhetos epiteliais e de agrupamentos de células. Elas não somente colam uma célula à outra, mas também propiciam ancoramento para os filamentos intracelulares de actina nos sítios de adesão célula-célula. Desta maneira, o padrão de tensões e movimentos no tecido em desenvolvimento é regulado de acordo com o padrão de adesões.
A notocorda se alonga, enquanto a placa neural se enrola para formar o tubo neuralA gastrulação é somente a primeira – embora talvez a mais dramática – de uma estonteante variedade de movimentos celulares que moldam as partes do corpo. Temos espaço para dis-cutir somente alguns deles aqui.
No embrião que se segue à gastrulação, a camada de mesoderme é dividida em placas separadas nos lados esquerdo e direito do corpo. Definindo o eixo central do corpo e efe-tuando esta separação, está a especialização mais inicial da mesoderme, conhecida como notocorda. Este delgado bastão de células, com ectoderme acima dele, endoderme abaixo e mesoderme de cada lado (ver Figura 22-72), deriva das células do próprio Organizador. As células da notocorda são caracterizadas pela expressão de uma proteína de regulação gênica denominada Brachyury (palavra grega para “cauda curta”, tomada a partir de um fenótipo mutante) que pertence à mesma família T-box da proteína VegT dos blastômeros vegetais.
Assim que as células da notocorda passam ao redor do lábio dorsal do blastóporo e se movem para o interior do embrião, elas formam uma coluna de tecido que se alonga dra-maticamente por meio de extensão convergente. As células da notocorda também ficam in-chadas com vacúolos, de modo que o bastão se alonga ainda mais e distende o embrião. A notocorda é a peculiaridade definidora dos cordados – o filo ao qual pertencem os vertebra-dos. É uma das principais características dos vertebrados, não tendo nenhum equivalente aparente em Drosophila. Nos cordados mais primitivos, os quais não possuem vértebras, a notocorda persiste como um substituto primitivo para a coluna vertebral. Nos vertebrados, ela funciona como um núcleo ao redor do qual outras células mesodérmicas irão finalmente se reunir para formar as vértebras.
Enquanto isso, outros movimentos estão ocorrendo na camada sobreposta de ectoder-me, a fim de formar os rudimentos do sistema nervoso. Em um processo conhecido como neurulação, uma ampla região central de ectoderme, denominada placa neural, torna-se espessa, se enrola em um tubo e se desprende do resto da camada de células. O tubo assim criado a partir da ectoderme é chamado de tubo neural; ele irá formar o cérebro e a medula espinal (Figura 22-78).
Os mecanismos de neurulação dependem de alterações no empacotamento das cé-lulas e na forma das células que fazem com que o epitélio se enrole em um tubo (Figura
Figura 22-77 Separação por tipos celulares. As células de diferentes partes de um embrião jovem de anfíbio irão se separar por tipos, de acordo com suas origens. No experimento clássico aqui mostrado, as células da mesoderme (verde), as células da placa neural (azul) e as células epidérmicas (vermelho) foram desagregadas e em seguida reagregadas em uma mistura aleatória. Elas se separam por tipos em um arranjo reminiscente ao de um embrião normal, com um “tubo neural” interno, uma epiderme externa e uma mesoderme no meio. (Modificada de P. L. Townes e J. Holtfreter, J. Exp. Zool. 128:53-120, 1955. Com permissão de Wiley-Liss.)
Alberts_22.indd 1370Alberts_22.indd 1370 29.07.09 16:49:3529.07.09 16:49:35
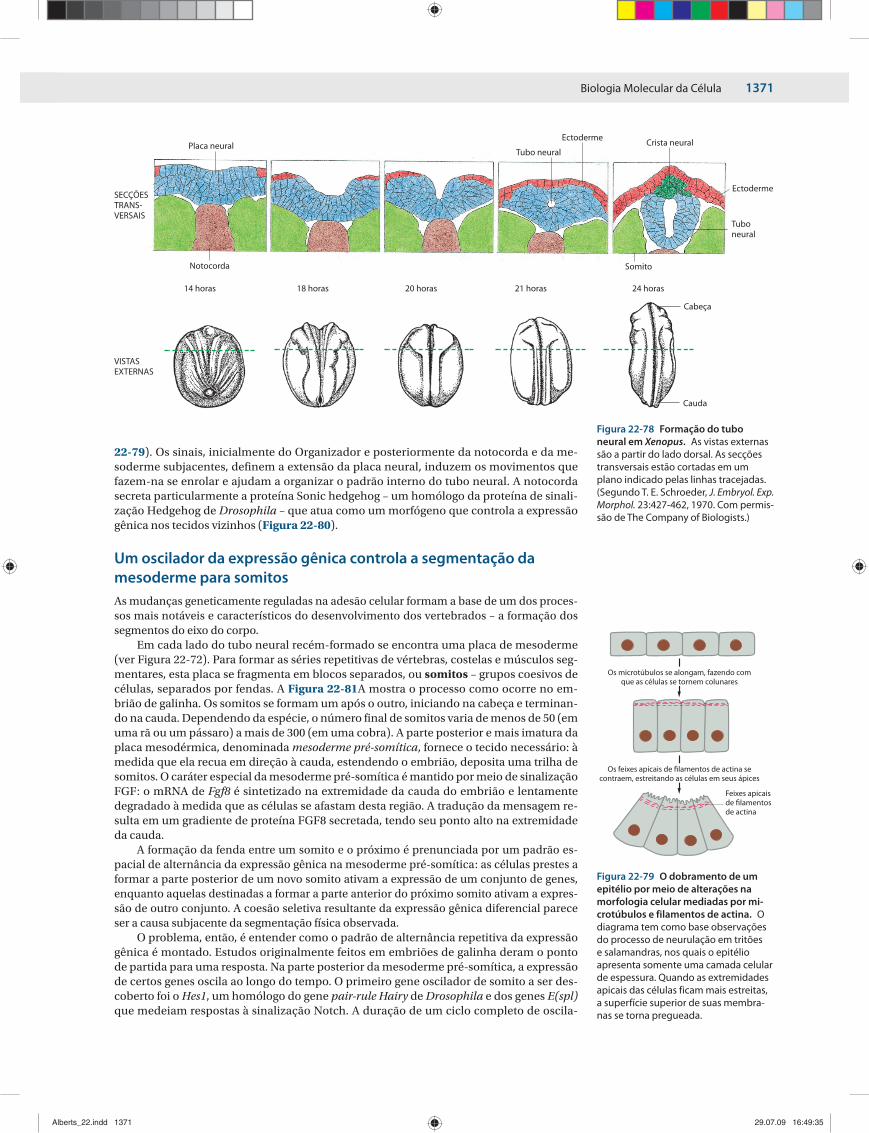
Biologia Molecular da Célula 1371
22-79). Os sinais, inicialmente do Organizador e posteriormente da notocorda e da me-soderme subjacentes, definem a extensão da placa neural, induzem os movimentos que fazem-na se enrolar e ajudam a organizar o padrão interno do tubo neural. A notocorda secreta particularmente a proteína Sonic hedgehog – um homólogo da proteína de sinali-zação Hedgehog de Drosophila – que atua como um morfógeno que controla a expressão gênica nos tecidos vizinhos (Figura 22-80).
Um oscilador da expressão gênica controla a segmentação da mesoderme para somitosAs mudanças geneticamente reguladas na adesão celular formam a base de um dos proces-sos mais notáveis e característicos do desenvolvimento dos vertebrados – a formação dos segmentos do eixo do corpo.
Em cada lado do tubo neural recém-formado se encontra uma placa de mesoderme (ver Figura 22-72). Para formar as séries repetitivas de vértebras, costelas e músculos seg-mentares, esta placa se fragmenta em blocos separados, ou somitos – grupos coesivos de células, separados por fendas. A Figura 22-81A mostra o processo como ocorre no em-brião de galinha. Os somitos se formam um após o outro, iniciando na cabeça e terminan-do na cauda. Dependendo da espécie, o número final de somitos varia de menos de 50 (em uma rã ou um pássaro) a mais de 300 (em uma cobra). A parte posterior e mais imatura da placa mesodérmica, denominada mesoderme pré-somítica, fornece o tecido necessário: à medida que ela recua em direção à cauda, estendendo o embrião, deposita uma trilha de somitos. O caráter especial da mesoderme pré-somítica é mantido por meio de sinalização FGF: o mRNA de Fgf8 é sintetizado na extremidade da cauda do embrião e lentamente degradado à medida que as células se afastam desta região. A tradução da mensagem re-sulta em um gradiente de proteína FGF8 secretada, tendo seu ponto alto na extremidade da cauda.
A formação da fenda entre um somito e o próximo é prenunciada por um padrão es-pacial de alternância da expressão gênica na mesoderme pré-somítica: as células prestes a formar a parte posterior de um novo somito ativam a expressão de um conjunto de genes, enquanto aquelas destinadas a formar a parte anterior do próximo somito ativam a expres-são de outro conjunto. A coesão seletiva resultante da expressão gênica diferencial parece ser a causa subjacente da segmentação física observada.
O problema, então, é entender como o padrão de alternância repetitiva da expressão gênica é montado. Estudos originalmente feitos em embriões de galinha deram o ponto de partida para uma resposta. Na parte posterior da mesoderme pré-somítica, a expressão de certos genes oscila ao longo do tempo. O primeiro gene oscilador de somito a ser des-coberto foi o Hes1, um homólogo do gene pair-rule Hairy de Drosophila e dos genes E(spl) que medeiam respostas à sinalização Notch. A duração de um ciclo completo de oscila-
SECÇÕESTRANS-VERSAIS
Placa neural
Notocorda
14 horas
VISTASEXTERNAS
18 horas 20 horas 21 horas
Tubo neural
EctodermeCrista neural
Ectoderme
Tuboneural
Somito
Cabeça
Cauda
24 horas
Figura 22-78 Formação do tubo neural em Xenopus. As vistas externas são a partir do lado dorsal. As secções transversais estão cortadas em um plano indicado pelas linhas tracejadas. (Segundo T. E. Schroeder, J. Embryol. Exp. Morphol. 23:427-462, 1970. Com permis-são de The Company of Biologists.)
Os microtúbulos se alongam, fazendo comque as células se tornem colunares
Os feixes apicais de filamentos de actina secontraem, estreitando as células em seus ápices
Feixes apicaisde filamentosde actina
Figura 22-79 O dobramento de um epitélio por meio de alterações na morfologia celular mediadas por mi-crotúbulos e filamentos de actina. O diagrama tem como base observações do processo de neurulação em tritões e salamandras, nos quais o epitélio apresenta somente uma camada celular de espessura. Quando as extremidades apicais das células ficam mais estreitas, a superfície superior de suas membra-nas se torna pregueada.
Alberts_22.indd 1371Alberts_22.indd 1371 29.07.09 16:49:3529.07.09 16:49:35
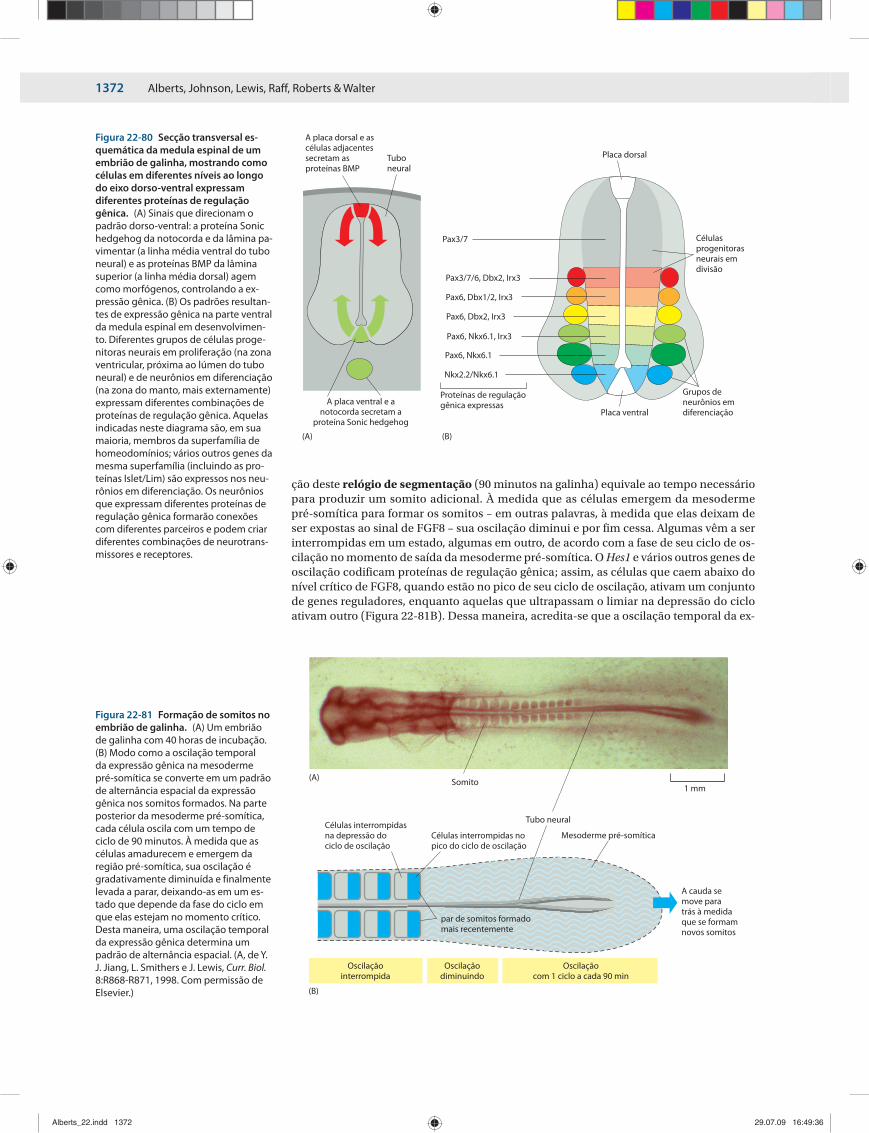
1372 Alberts, Johnson, Lewis, Raff, Roberts & Walter
ção deste relógio de segmentação (90 minutos na galinha) equivale ao tempo necessário para produzir um somito adicional. À medida que as células emergem da mesoderme pré-somítica para formar os somitos – em outras palavras, à medida que elas deixam de ser expostas ao sinal de FGF8 – sua oscilação diminui e por fim cessa. Algumas vêm a ser interrompidas em um estado, algumas em outro, de acordo com a fase de seu ciclo de os-cilação no momento de saída da mesoderme pré-somítica. O Hes1 e vários outros genes de oscilação codificam proteínas de regulação gênica; assim, as células que caem abaixo do nível crítico de FGF8, quando estão no pico de seu ciclo de oscilação, ativam um conjunto de genes reguladores, enquanto aquelas que ultrapassam o limiar na depressão do ciclo ativam outro (Figura 22-81B). Dessa maneira, acredita-se que a oscilação temporal da ex-
Figura 22-80 Secção transversal es-quemática da medula espinal de um embrião de galinha, mostrando como células em diferentes níveis ao longo do eixo dorso-ventral expressam diferentes proteínas de regulação gênica. (A) Sinais que direcionam o padrão dorso-ventral: a proteína Sonic hedgehog da notocorda e da lâmina pa-vimentar (a linha média ventral do tubo neural) e as proteínas BMP da lâmina superior (a linha média dorsal) agem como morfógenos, controlando a ex-pressão gênica. (B) Os padrões resultan-tes de expressão gênica na parte ventral da medula espinal em desenvolvimen-to. Diferentes grupos de células proge-nitoras neurais em proliferação (na zona ventricular, próxima ao lúmen do tubo neural) e de neurônios em diferenciação (na zona do manto, mais externamente) expressam diferentes combinações de proteínas de regulação gênica. Aquelas indicadas neste diagrama são, em sua maioria, membros da superfamília de homeodomínios; vários outros genes da mesma superfamília (incluindo as pro-teínas Islet/Lim) são expressos nos neu-rônios em diferenciação. Os neurônios que expressam diferentes proteínas de regulação gênica formarão conexões com diferentes parceiros e podem criar diferentes combinações de neurotrans-missores e receptores.
A placa dorsal e ascélulas adjacentessecretam as proteínas BMP
A placa ventral e anotocorda secretam a
proteína Sonic hedgehog
(A) (B)
Tuboneural
Proteínas de regulaçãogênica expressas
Placa dorsal
Células progenitorasneurais emdivisão
Grupos deneurônios emdiferenciaçãoPlaca ventral
Pax3/7/6, Dbx2, Irx3
Pax6, Dbx1/2, Irx3
Pax6, Dbx2, Irx3
Pax6, Nkx6.1, Irx3
Pax6, Nkx6.1
Nkx2.2/Nkx6.1
Pax3/7
Somito(A)
Mesoderme pré-somítica
(B)
Tubo neuralCélulas interrompidasna depressão do ciclo de oscilação
Células interrompidas nopico do ciclo de oscilação
par de somitos formadomais recentemente
A cauda semove paratrás à medidaque se formamnovos somitos
1 mm
Oscilaçãointerrompida
Oscilaçãodiminuindo
Oscilaçãocom 1 ciclo a cada 90 min
Figura 22-81 Formação de somitos no embrião de galinha. (A) Um embrião de galinha com 40 horas de incubação. (B) Modo como a oscilação temporal da expressão gênica na mesoderme pré-somítica se converte em um padrão de alternância espacial da expressão gênica nos somitos formados. Na parte posterior da mesoderme pré-somítica, cada célula oscila com um tempo de ciclo de 90 minutos. À medida que as células amadurecem e emergem da região pré-somítica, sua oscilação é gradativamente diminuída e finalmente levada a parar, deixando-as em um es-tado que depende da fase do ciclo em que elas estejam no momento crítico. Desta maneira, uma oscilação temporal da expressão gênica determina um padrão de alternância espacial. (A, de Y. J. Jiang, L. Smithers e J. Lewis, Curr. Biol. 8:R868-R871, 1998. Com permissão de Elsevier.)
Alberts_22.indd 1372Alberts_22.indd 1372 29.07.09 16:49:3629.07.09 16:49:36

Biologia Molecular da Célula 1373
pressão gênica na mesoderme pré-somítica deixe seu rastro em um padrão espacialmente periódico de expressão gênica na mesoderme em maturação, e isso, por sua vez, dita como o tecido irá se fragmentar em blocos fisicamente separados.
A retroalimentação negativa retardada pode gerar as oscilações do relógio de segmentaçãoQual é, então, o mecanismo que gera a oscilação temporal? Como funciona o relógio? No camundongo, descobriu-se que pelo menos três classes de genes apresentam um padrão de expressão oscilante na mesoderme pré-somítica, codificando componentes da via Notch, da via Wnt e da via Fgf, respectivamente; porém, sabe-se que a maioria das mu-tações que interrompem o relógio e desorganizam a segmentação dos somitos está lo-calizada em componentes da via Notch. Aí estão incluídos genes (como o Hes1 e, mais significativamente, seu parente Hes7) que são regulados por Notch e codificam proteínas de regulação gênica inibidoras. Algumas dessas proteínas agem diretamente no DNA re-gulador de seu próprio gene, de modo a inibir sua própria expressão. De acordo com uma teoria, este circuito de retroalimentação negativa simples poderia ser o gerador básico das oscilações (Figura 22-82): quando o gene é transcrito, a quantidade de seu produto pro-teico cessa; a proteína, então, decai, permitindo que a transcrição comece de novo, e assim por diante. Há um intervalo entre o início de um novo evento de transcrição e o primeiro aparecimento, no núcleo, das moléculas de proteínas reguladoras resultantes, porque leva um certo tempo para que a RNA-polimerase percorra o gene, para que o transcrito de RNA resultante amadureça, deixe o núcleo e direcione a síntese de uma molécula proteica, e para que a proteína entre no núcleo a fim de controlar a transcrição. Propõe-se que este atraso no circuito de retroalimentação seja o principal determinante do período de oscila-ção do relógio e, assim, do tamanho de cada somito.
A maioria das células de cada somito recém-formado irá rapidamente se diferenciar para formar um bloco de músculo, correspondente a um segmento de músculo do eixo prin-cipal do corpo. O embrião pode agora (e o faz) começar a se torcer. Os subconjuntos sepa-rados das células dos somitos irão formar as vértebras e outros tecidos conectivos, como a derme. Um subconjunto adicional se desprende do somito e migra para a mesoderme lateral não-segmentada, arrastando-se pelos espaços entre outras células: esses emigrantes irão dar origem a praticamente todas as outras células musculares esqueléticas do corpo, incluindo as dos membros.
Os tecidos embrionários são invadidos de modo estritamente controlado por células migratóriasOs precursores das células musculares, ou mioblastos, que emigram dos somitos estão de-terminados, mas não completamente diferenciados. Nos tecidos que colonizam, eles irão se misturar com outras classes de células das quais parecem praticamente indistintos; contudo, irão manter a expressão de proteínas de regulação gênica específicas de mioblastos (como a Pax3 e os membros da família MyoD) e, quando chegar o momento da diferenciação, eles, e somente eles, irão se transformar em células musculares (Figura 22-83).
INIBIÇÃO
mRNA
Proteínainibidora
200 400 600 minutos
40
80
120
400
800
1.200
Con
cent
raçã
ode
pro
teín
a
Con
cent
raçã
ode
mRN
A
(B)(A)
ATRASO
ATRASOATRASO
Gene que codifica umaproteína inibidora
Figura 22-82 A retroalimentação ne-gativa atrasada dá origem à expressão gênica oscilante. (A) Um único gene, codificando uma proteína de regulação gênica que inibe sua própria expressão, pode se comportar como um oscilador. Para que a oscilação ocorra, deve haver um atraso (ou vários atrasos) no circuito de retroalimentação, e os tempos de vida das moléculas de mRNA e proteína devem ser breves em comparação ao atraso total. O atraso determina o pe-ríodo de oscilação. De acordo com uma teoria, um circuito de retroalimentação como este, com base em um gene cha-mado de Her7 no peixe-zebra, ou Hes7 no camundongo (um parente do gene Hes1), é o marca-passo do relógio de segmentação que governa a formação dos somitos. (B) A oscilação prevista do mRNA de Her7 e da proteína correspon-dente, computada usando-se estima-tivas aproximadas dos parâmetros do circuito de retroalimentação apropria-dos para este gene no peixe-zebra. As concentrações são medidas em número de moléculas por célula. O período pre-visto é próximo ao período observado, que é de 30 minutos por somito no peixe-zebra.
Alberts_22.indd 1373Alberts_22.indd 1373 29.07.09 16:49:3629.07.09 16:49:36
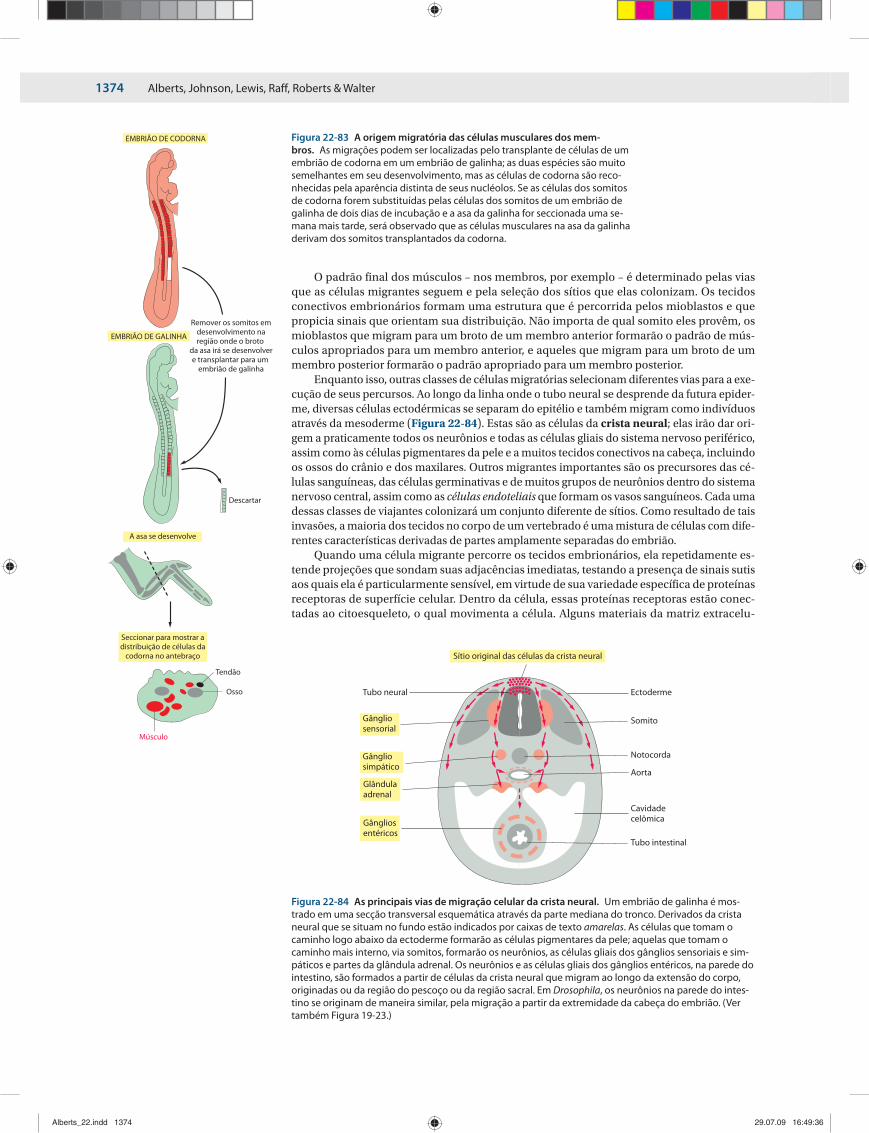
1374 Alberts, Johnson, Lewis, Raff, Roberts & Walter
O padrão final dos músculos – nos membros, por exemplo – é determinado pelas vias que as células migrantes seguem e pela seleção dos sítios que elas colonizam. Os tecidos conectivos embrionários formam uma estrutura que é percorrida pelos mioblastos e que propicia sinais que orientam sua distribuição. Não importa de qual somito eles provêm, os mioblastos que migram para um broto de um membro anterior formarão o padrão de mús-culos apropriados para um membro anterior, e aqueles que migram para um broto de um membro posterior formarão o padrão apropriado para um membro posterior.
Enquanto isso, outras classes de células migratórias selecionam diferentes vias para a exe-cução de seus percursos. Ao longo da linha onde o tubo neural se desprende da futura epider-me, diversas células ectodérmicas se separam do epitélio e também migram como indivíduos através da mesoderme (Figura 22-84). Estas são as células da crista neural; elas irão dar ori-gem a praticamente todos os neurônios e todas as células gliais do sistema nervoso periférico, assim como às células pigmentares da pele e a muitos tecidos conectivos na cabeça, incluindo os ossos do crânio e dos maxilares. Outros migrantes importantes são os precursores das cé-lulas sanguíneas, das células germinativas e de muitos grupos de neurônios dentro do sistema nervoso central, assim como as células endoteliais que formam os vasos sanguíneos. Cada uma dessas classes de viajantes colonizará um conjunto diferente de sítios. Como resultado de tais invasões, a maioria dos tecidos no corpo de um vertebrado é uma mistura de células com dife-rentes características derivadas de partes amplamente separadas do embrião.
Quando uma célula migrante percorre os tecidos embrionários, ela repetidamente es-tende projeções que sondam suas adjacências imediatas, testando a presença de sinais sutis aos quais ela é particularmente sensível, em virtude de sua variedade específica de proteínas receptoras de superfície celular. Dentro da célula, essas proteínas receptoras estão conec-tadas ao citoesqueleto, o qual movimenta a célula. Alguns materiais da matriz extracelu-
Figura 22-83 A origem migratória das células musculares dos mem-bros. As migrações podem ser localizadas pelo transplante de células de um embrião de codorna em um embrião de galinha; as duas espécies são muito semelhantes em seu desenvolvimento, mas as células de codorna são reco-nhecidas pela aparência distinta de seus nucléolos. Se as células dos somitos de codorna forem substituídas pelas células dos somitos de um embrião de galinha de dois dias de incubação e a asa da galinha for seccionada uma se-mana mais tarde, será observado que as células musculares na asa da galinha derivam dos somitos transplantados da codorna.
EMBRIÃO DE CODORNA
EMBRIÃO DE GALINHA
A asa se desenvolve
Remover os somitos emdesenvolvimento naregião onde o broto
da asa irá se desenvolvere transplantar para um
embrião de galinha
Descartar
Tendão
Músculo
Osso
Seccionar para mostrar adistribuição de células da
codorna no antebraço
Tubo neural
Gângliosensorial
Gângliosimpático
Glândulaadrenal
Sítio original das células da crista neural
Ectoderme
Somito
Notocorda
Aorta
Cavidadecelômica
Tubo intestinal
Gângliosentéricos
Figura 22-84 As principais vias de migração celular da crista neural. Um embrião de galinha é mos-trado em uma secção transversal esquemática através da parte mediana do tronco. Derivados da crista neural que se situam no fundo estão indicados por caixas de texto amarelas. As células que tomam o caminho logo abaixo da ectoderme formarão as células pigmentares da pele; aquelas que tomam o caminho mais interno, via somitos, formarão os neurônios, as células gliais dos gânglios sensoriais e sim-páticos e partes da glândula adrenal. Os neurônios e as células gliais dos gânglios entéricos, na parede do intestino, são formados a partir de células da crista neural que migram ao longo da extensão do corpo, originadas ou da região do pescoço ou da região sacral. Em Drosophila, os neurônios na parede do intes-tino se originam de maneira similar, pela migração a partir da extremidade da cabeça do embrião. (Ver também Figura 19-23.)
Alberts_22.indd 1374Alberts_22.indd 1374 29.07.09 16:49:3629.07.09 16:49:36
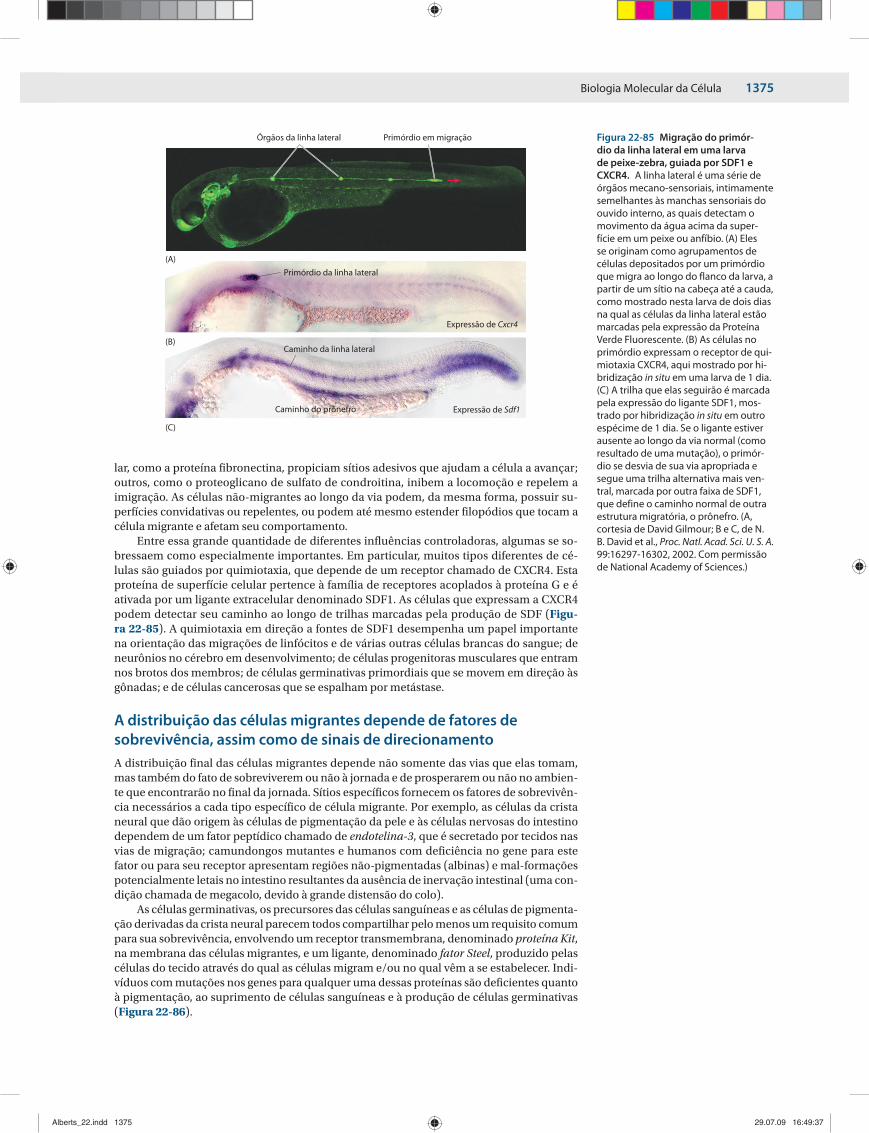
Biologia Molecular da Célula 1375
lar, como a proteína fibronectina, propiciam sítios adesivos que ajudam a célula a avançar; outros, como o proteoglicano de sulfato de condroitina, inibem a locomoção e repelem a imigração. As células não-migrantes ao longo da via podem, da mesma forma, possuir su-perfícies convidativas ou repelentes, ou podem até mesmo estender filopódios que tocam a célula migrante e afetam seu comportamento.
Entre essa grande quantidade de diferentes influências controladoras, algumas se so-bressaem como especialmente importantes. Em particular, muitos tipos diferentes de cé-lulas são guiados por quimiotaxia, que depende de um receptor chamado de CXCR4. Esta proteína de superfície celular pertence à família de receptores acoplados à proteína G e é ativada por um ligante extracelular denominado SDF1. As células que expressam a CXCR4 podem detectar seu caminho ao longo de trilhas marcadas pela produção de SDF (Figu-ra 22-85). A quimiotaxia em direção a fontes de SDF1 desempenha um papel importante na orientação das migrações de linfócitos e de várias outras células brancas do sangue; de neurônios no cérebro em desenvolvimento; de células progenitoras musculares que entram nos brotos dos membros; de células germinativas primordiais que se movem em direção às gônadas; e de células cancerosas que se espalham por metástase.
A distribuição das células migrantes depende de fatores de sobrevivência, assim como de sinais de direcionamentoA distribuição final das células migrantes depende não somente das vias que elas tomam, mas também do fato de sobreviverem ou não à jornada e de prosperarem ou não no ambien-te que encontrarão no final da jornada. Sítios específicos fornecem os fatores de sobrevivên-cia necessários a cada tipo específico de célula migrante. Por exemplo, as células da crista neural que dão origem às células de pigmentação da pele e às células nervosas do intestino dependem de um fator peptídico chamado de endotelina-3, que é secretado por tecidos nas vias de migração; camundongos mutantes e humanos com deficiência no gene para este fator ou para seu receptor apresentam regiões não-pigmentadas (albinas) e mal-formações potencialmente letais no intestino resultantes da ausência de inervação intestinal (uma con-dição chamada de megacolo, devido à grande distensão do colo).
As células germinativas, os precursores das células sanguíneas e as células de pigmenta-ção derivadas da crista neural parecem todos compartilhar pelo menos um requisito comum para sua sobrevivência, envolvendo um receptor transmembrana, denominado proteína Kit, na membrana das células migrantes, e um ligante, denominado fator Steel, produzido pelas células do tecido através do qual as células migram e/ou no qual vêm a se estabelecer. Indi-víduos com mutações nos genes para qualquer uma dessas proteínas são deficientes quanto à pigmentação, ao suprimento de células sanguíneas e à produção de células germinativas (Figura 22-86).
Figura 22-85 Migração do primór-dio da linha lateral em uma larva de peixe-zebra, guiada por SDF1 e CXCR4. A linha lateral é uma série de órgãos mecano-sensoriais, intimamente semelhantes às manchas sensoriais do ouvido interno, as quais detectam o movimento da água acima da super-fície em um peixe ou anfíbio. (A) Eles se originam como agrupamentos de células depositados por um primórdio que migra ao longo do flanco da larva, a partir de um sítio na cabeça até a cauda, como mostrado nesta larva de dois dias na qual as células da linha lateral estão marcadas pela expressão da Proteína Verde Fluorescente. (B) As células no primórdio expressam o receptor de qui-miotaxia CXCR4, aqui mostrado por hi-bridização in situ em uma larva de 1 dia. (C) A trilha que elas seguirão é marcada pela expressão do ligante SDF1, mos-trado por hibridização in situ em outro espécime de 1 dia. Se o ligante estiver ausente ao longo da via normal (como resultado de uma mutação), o primór-dio se desvia de sua via apropriada e segue uma trilha alternativa mais ven-tral, marcada por outra faixa de SDF1, que define o caminho normal de outra estrutura migratória, o prônefro. (A, cortesia de David Gilmour; B e C, de N. B. David et al., Proc. Natl. Acad. Sci. U. S. A. 99:16297-16302, 2002. Com permissão de National Academy of Sciences.)
Órgãos da linha lateral Primórdio em migração
(C)
(B)
(A)
Expressão de Cxcr4
Expressão de Sdf1
Primórdio da linha lateral
Caminho da linha lateral
Caminho do prônefro
Alberts_22.indd 1375Alberts_22.indd 1375 29.07.09 16:49:3729.07.09 16:49:37
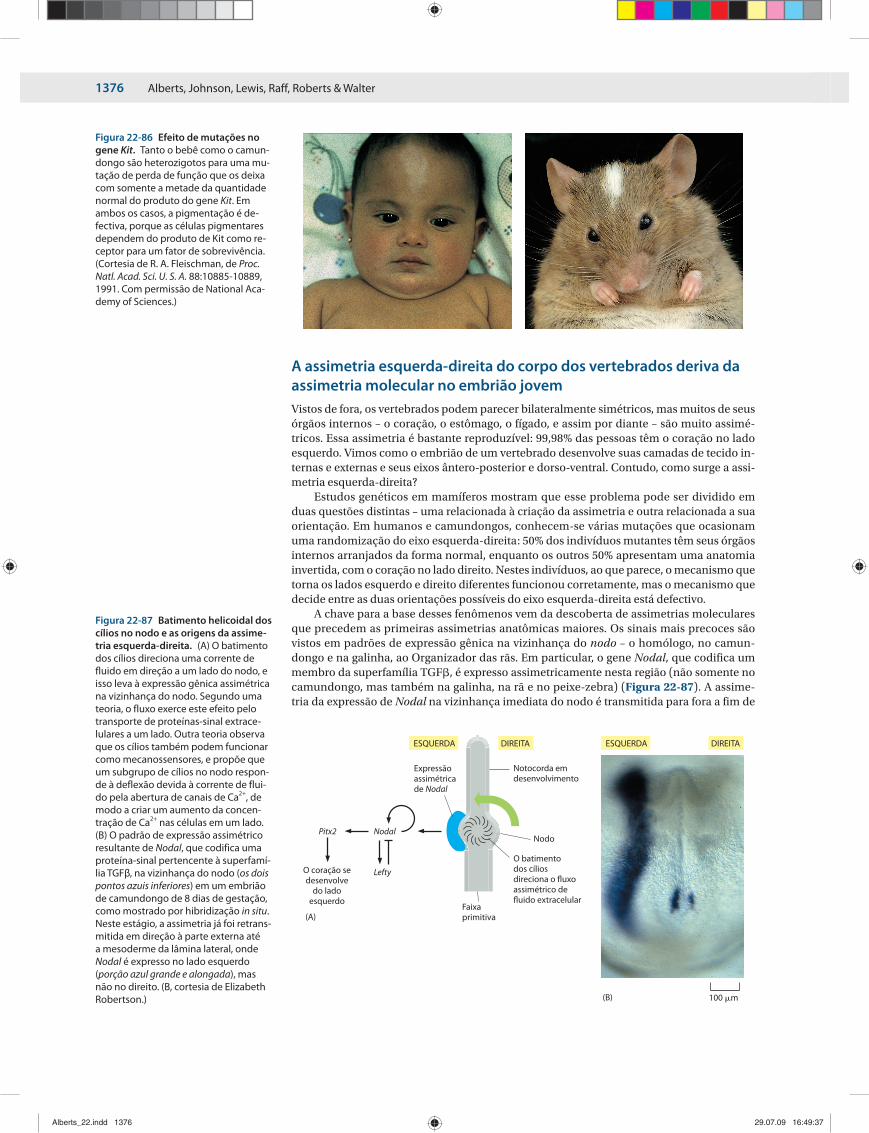
1376 Alberts, Johnson, Lewis, Raff, Roberts & Walter
A assimetria esquerda-direita do corpo dos vertebrados deriva da assimetria molecular no embrião jovemVistos de fora, os vertebrados podem parecer bilateralmente simétricos, mas muitos de seus órgãos internos – o coração, o estômago, o fígado, e assim por diante – são muito assimé-tricos. Essa assimetria é bastante reproduzível: 99,98% das pessoas têm o coração no lado esquerdo. Vimos como o embrião de um vertebrado desenvolve suas camadas de tecido in-ternas e externas e seus eixos ântero-posterior e dorso-ventral. Contudo, como surge a assi-metria esquerda-direita?
Estudos genéticos em mamíferos mostram que esse problema pode ser dividido em duas questões distintas – uma relacionada à criação da assimetria e outra relacionada a sua orientação. Em humanos e camundongos, conhecem-se várias mutações que ocasionam uma randomização do eixo esquerda-direita: 50% dos indivíduos mutantes têm seus órgãos internos arranjados da forma normal, enquanto os outros 50% apresentam uma anatomia invertida, com o coração no lado direito. Nestes indivíduos, ao que parece, o mecanismo que torna os lados esquerdo e direito diferentes funcionou corretamente, mas o mecanismo que decide entre as duas orientações possíveis do eixo esquerda-direita está defectivo.
A chave para a base desses fenômenos vem da descoberta de assimetrias moleculares que precedem as primeiras assimetrias anatômicas maiores. Os sinais mais precoces são vistos em padrões de expressão gênica na vizinhança do nodo – o homólogo, no camun-dongo e na galinha, ao Organizador das rãs. Em particular, o gene Nodal, que codifica um membro da superfamília TGF�, é expresso assimetricamente nesta região (não somente no camundongo, mas também na galinha, na rã e no peixe-zebra) (Figura 22-87). A assime-tria da expressão de Nodal na vizinhança imediata do nodo é transmitida para fora a fim de
Figura 22-86 Efeito de mutações no gene Kit. Tanto o bebê como o camun-dongo são heterozigotos para uma mu-tação de perda de função que os deixa com somente a metade da quantidade normal do produto do gene Kit. Em ambos os casos, a pigmentação é de-fectiva, porque as células pigmentares dependem do produto de Kit como re-ceptor para um fator de sobrevivência. (Cortesia de R. A. Fleischman, de Proc. Natl. Acad. Sci. U. S. A. 88:10885-10889, 1991. Com permissão de National Aca-demy of Sciences.)
ESQUERDA ESQUERDADIREITA DIREITA
Nodo
(A)
(B)
Expressãoassimétricade Nodal
Notocorda emdesenvolvimento
O batimento dos cílios direciona o fluxoassimétrico defluido extracelular
Faixaprimitiva
100 �m
NodalPitx2
LeftyO coração sedesenvolve
do ladoesquerdo
Figura 22-87 Batimento helicoidal dos cílios no nodo e as origens da assime-tria esquerda-direita. (A) O batimento dos cílios direciona uma corrente de fluido em direção a um lado do nodo, e isso leva à expressão gênica assimétrica na vizinhança do nodo. Segundo uma teoria, o fluxo exerce este efeito pelo transporte de proteínas-sinal extrace-lulares a um lado. Outra teoria observa que os cílios também podem funcionar como mecanossensores, e propõe que um subgrupo de cílios no nodo respon-de à deflexão devida à corrente de flui-do pela abertura de canais de Ca2+, de modo a criar um aumento da concen-tração de Ca2+ nas células em um lado. (B) O padrão de expressão assimétrico resultante de Nodal, que codifica uma proteína-sinal pertencente à superfamí-lia TGFβ, na vizinhança do nodo (os dois pontos azuis inferiores) em um embrião de camundongo de 8 dias de gestação, como mostrado por hibridização in situ. Neste estágio, a assimetria já foi retrans-mitida em direção à parte externa até a mesoderme da lâmina lateral, onde Nodal é expresso no lado esquerdo (porção azul grande e alongada), mas não no direito. (B, cortesia de Elizabeth Robertson.)
Alberts_22.indd 1376Alberts_22.indd 1376 29.07.09 16:49:3729.07.09 16:49:37

Biologia Molecular da Célula 1377
criar uma ampla faixa de expressão de Nodal na mesoderme ao longo do lado esquerdo – e somente do lado esquerdo – do corpo do embrião. O mecanismo que transmite a assime-tria a partir do nodo e localiza a expressão de Nodal não é entendido e pode variar entre as diferentes classes de vertebrados. Em todas as espécies, contudo, ele parece depender de circuitos de retroalimentação que envolvem, além de Nodal, um segundo conjunto de genes, os genes Lefty. Estes, como o próprio Nodal, são diretamente regulados pela via de sinalização Nodal, e seus produtos, as proteínas Lefty, são relacionados a Nodal. Porém, as proteínas Lefty se difundem mais amplamente e agem contrariamente, como antagonistas de Nodal. Camundongos com uma mutação nocaute no gene Lefty1 frequentemente apre-sentam o lado direito convertido em uma imagem especular do esquerdo, de forma que a simetria esquerda-direita se perde.
Outro gene diretamente regulado pela via Nodal, Pitx2, que codifica uma proteína de regulação gênica, vincula o resultado das interações entre Nodal/Lefty ao desenvolvimento anatômico subsequente. A Nodal controla a expressão de Pitx2 no lado esquerdo do corpo e, com isso, confere assimetria ao coração e a outros órgãos internos.
Isso nos deixa com o quebra-cabeça de como a assimetria inicial da expressão de No-dal se origina. Seja qual for o mecanismo, o resultado dos eventos no nodo em um animal normal deve ser influenciado de tal forma que os genes específicos do lado esquerdo sejam regularmente expressos no lado esquerdo: deve haver uma ligação entre o mecanismo que cria a assimetria e o mecanismo que a orienta. Um indício para o mecanismo de orientação foi primeiramente observado em uma clínica de infertilidade da Suécia. Descobriu-se que um pequeno subconjunto de homens inférteis possuía espermatozoides sem mobilidade devido a um defeito nas moléculas de dineína necessárias ao batimento dos cílios e flagelos. Esses homens também sofriam de bronquite crônica e sinusite, porque os cílios em seus tratos respiratórios eram defeituosos. E, surpreendentemente, 50% deles tinham os órgãos internos esquerda-direita invertidos, com o coração à direita. As descobertas originalmente pareceram completamente misteriosas; porém, efeitos similares são vistos em mamíferos com outras mutações que resultam em cílios defeituosos. Isso sugere que o batimento ciliar, de algum modo, controla como o eixo esquerda-direita é orientado.
A videomicroscopia em tempo real em um embrião vivo de camundongo revela que as células do nodo, em sua face interna, são dotadas de cílios que batem de modo helicoidal: como uma rosca de parafuso, essas células têm um sentido definido e, no nodo, elas estão arranjadas em uma pequena concavidade que é moldada de forma que seus batimentos direcionam uma corrente de fluido ao lado esquerdo (ver Figura 22-87A). Segundo uma teoria, acredita-se que as proteínas-sinal carregadas nesta corrente produzam a tendência que orienta o eixo esquerda-direita do corpo do camundongo. Outra teoria propõe que os cílios, neste sistema, como em outros contextos, agem não somente como direcionadores do fluxo de fluido, mas também como sensores mecânicos, respondendo à deflexão pela geração de uma corrente assimétrica de íons Ca2+ que atravessa o nodo e influencia o teci-do adjacente.
O sentido do batimento ciliar reflete o sentido – a assimetria esquerda-direita – das mo-léculas orgânicas das quais todos os seres vivos são feitos. Parece que isso, portanto, é a dire-triz final da assimetria esquerda-direita de nossa anatomia.
ResumoO desenvolvimento animal envolve movimentos celulares dramáticos. Assim, na gastrulação, as células do exterior do embrião jovem se dobram para o interior para formar a cavidade intestinal e criar as três camadas germinativas – endoderme, mesoderme e ectoderme – a partir das quais são construídos os animais superiores. Nos vertebrados, os movimentos da gastrulação são organizados por sinais do Organizador (o lábio dorsal do blastóporo de anfíbios, correspondente ao nodo em um embrião de galinha ou camundongo). Esses sinais especificam o eixo dorso-ventral do corpo e governam a extensão convergente, na qual o folheto de células que se move para o interior do corpo se estende ao longo do eixo cabeça-cauda enquanto se estreita em ângulos retos a este eixo. Os movi-mentos ativos de re-empacotamento de células individuais que dirigem a extensão convergente são coordenados pela via de sinalização de polaridade planar Frizzled/Dishevelled – um ramo da via de sinalização Wnt que regula o citoesqueleto de actina.
O desenvolvimento subsequente envolve muitos movimentos celulares adicionais. Parte da ectoderme se torna espessa, enrolando-se e se desprendendo para formar o tubo neural e a crista neural. Na linha média, um bastão de células especializadas denominado notocorda se
Alberts_22.indd 1377Alberts_22.indd 1377 29.07.09 16:49:3829.07.09 16:49:38

1378 Alberts, Johnson, Lewis, Raff, Roberts & Walter
alonga para formar o eixo central do embrião. As longas placas de mesoderme em cada lado da notocorda vêm a ser segmentadas em somitos. Células migrantes, como as da crista neural, des-prendem-se de seus vizinhos originais e percorrem o embrião para colonizar novos sítios. Células germinativas primordiais e muitas outras migrantes são guiadas por quimiotaxia dependente do receptor CXCR4 e de seu ligante SDF1. Moléculas específicas de adesão celular, como as caderinas e integrinas, auxiliam no direcionamento das migrações e controlam a coesão seletiva das células em novos arranjos.
Finalmente, o padrão de movimentos celulares é direcionado pelo padrão de expressão gêni-ca, o qual determina as propriedades da superfície celular e a mobilidade. Assim, a formação dos somitos depende de um padrão periódico de expressão gênica, que é estabelecido por um oscilador bioquímico – o relógio de segmentação – na mesoderme e que dita a maneira pela qual a massa de células irá se dividir em blocos separados. Similarmente, a assimetria anatômica esquerda-direita do corpo dos vertebrados é prenunciada pela assimetria esquerda-direita no padrão de expressão gênica no embrião jovem. Acredita-se que essa assimetria, pelo menos em mamíferos, seja em últi-ma análise direcionada pelo sentido do batimento ciliar na vizinhança do nodo.
O CAMUNDONGOO embrião de camundongo – minúsculo e inacessível no útero de sua mãe – representa um desafio difícil para os biólogos do desenvolvimento. Contudo, ele tem dois atrativos ime-diatos. Primeiro, o camundongo é um mamífero, e os mamíferos são os animais que mais interessam a nós, humanos. Segundo, entre os mamíferos, ele é um dos mais convenientes para estudos genéticos, porque é pequeno e se reproduz rapidamente. Esses dois fatores têm estimulado um enorme esforço de pesquisa, resultando no desenvolvimento de algu-mas ferramentas extraordinariamente potentes. Desta maneira, o camundongo tornou-se o principal organismo-modelo para a experimentação em genética de mamíferos e o mais intensivamente estudado substituto para humanos. Ele está evolutivamente separado dos humanos por apenas cerca de cem milhões de anos. Seu genoma tem o mesmo tamanho que o nosso e existe uma correspondência muito próxima de um para um entre os genes de camundongos e os de humanos. Nossas proteínas são, tipicamente, de 80 a 90% idênticas quanto à sequência de aminoácidos, e grandes blocos de estreita similaridade quanto à se-quência de nucleotídeos também são evidentes, quando as sequências reguladoras de DNA são comparadas.
Por meio de engenhosidade e perseverança, os biólogos do desenvolvimento encon-traram meios para ter acesso ao embrião jovem de camundongo sem matá-lo e para gerar camundongos com mutações em qualquer gene selecionado. Quase todas as modificações genéticas que podem ser feitas em um verme, uma mosca ou um peixe-zebra agora tam-bém podem ser feitas no camundongo e, em alguns casos, até de forma melhor. Os custos com pesquisa em camundongos são muito maiores, mas também são maiores os incentivos. Como resultado, o camundongo se tornou uma fonte rica de informação sobre todos os as-pectos da genética molecular do desenvolvimento – um sistema modelo-chave não somente para mamíferos, mas também para outros animais. Ele proporcionou, por exemplo, muito do que sabemos sobre os genes Hox, a simetria esquerda-direita, os controles da morte celu-lar, o papel da sinalização Notch e uma série de outros tópicos.
Já fizemos repetidamente uso de dados do camundongo. Iremos utilizá-los ainda mais no próximo capítulo, em que discutiremos os tecidos adultos e os processos de desenvolvi-mento que ocorrem neles. Nesta seção, examinaremos as características especiais do desen-volvimento do camundongo que têm sido exploradas para possibilitar a manipulação ge-nética. Por meio de exemplos, delinearemos também como o camundongo tem sido usado para esclarecer um importante processo adicional do desenvolvimento – a criação de órgãos, como pulmões e glândulas, por interações entre tecido conectivo embrionário e epitélio.
O desenvolvimento de mamíferos começa com um preâmbulo especializadoO embrião dos mamíferos começa seu desenvolvimento de uma maneira excepcional. Pro-tegido dentro do útero, ele não tem a mesma necessidade que os embriões da maioria das outras espécies têm de completar as etapas iniciais do desenvolvimento rapidamente. Além disso, o desenvolvimento de uma placenta prontamente fornece nutrição a partir da mãe, de
Alberts_22.indd 1378Alberts_22.indd 1378 29.07.09 16:49:3829.07.09 16:49:38
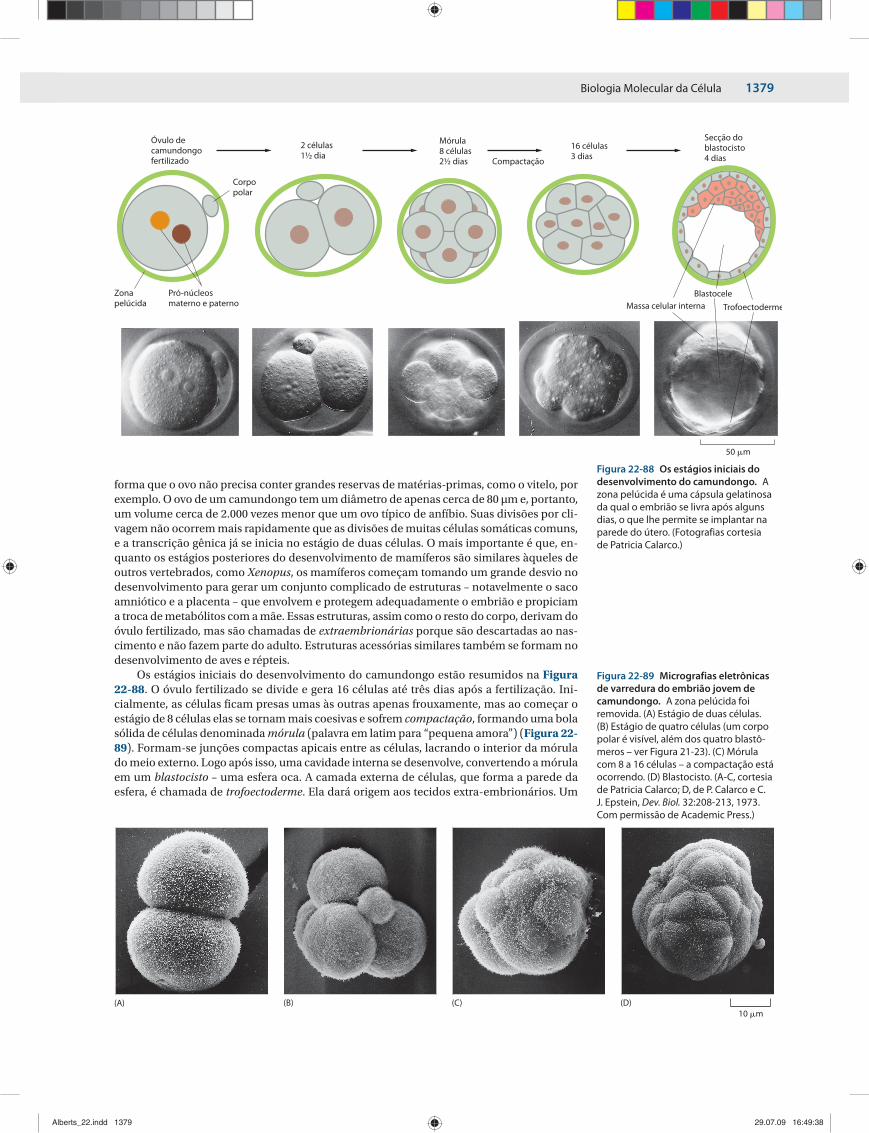
Biologia Molecular da Célula 1379
forma que o ovo não precisa conter grandes reservas de matérias-primas, como o vitelo, por exemplo. O ovo de um camundongo tem um diâmetro de apenas cerca de 80 μm e, portanto, um volume cerca de 2.000 vezes menor que um ovo típico de anfíbio. Suas divisões por cli-vagem não ocorrem mais rapidamente que as divisões de muitas células somáticas comuns, e a transcrição gênica já se inicia no estágio de duas células. O mais importante é que, en-quanto os estágios posteriores do desenvolvimento de mamíferos são similares àqueles de outros vertebrados, como Xenopus, os mamíferos começam tomando um grande desvio no desenvolvimento para gerar um conjunto complicado de estruturas – notavelmente o saco amniótico e a placenta – que envolvem e protegem adequadamente o embrião e propiciam a troca de metabólitos com a mãe. Essas estruturas, assim como o resto do corpo, derivam do óvulo fertilizado, mas são chamadas de extraembrionárias porque são descartadas ao nas-cimento e não fazem parte do adulto. Estruturas acessórias similares também se formam no desenvolvimento de aves e répteis.
Os estágios iniciais do desenvolvimento do camundongo estão resumidos na Figura 22-88. O óvulo fertilizado se divide e gera 16 células até três dias após a fertilização. Ini-cialmente, as células ficam presas umas às outras apenas frouxamente, mas ao começar o estágio de 8 células elas se tornam mais coesivas e sofrem compactação, formando uma bola sólida de células denominada mórula (palavra em latim para “pequena amora”) (Figura 22-89). Formam-se junções compactas apicais entre as células, lacrando o interior da mórula do meio externo. Logo após isso, uma cavidade interna se desenvolve, convertendo a mórula em um blastocisto – uma esfera oca. A camada externa de células, que forma a parede da esfera, é chamada de trofoectoderme. Ela dará origem aos tecidos extra-embrionários. Um
Figura 22-88 Os estágios iniciais do desenvolvimento do camundongo. A zona pelúcida é uma cápsula gelatinosa da qual o embrião se livra após alguns dias, o que lhe permite se implantar na parede do útero. (Fotografias cortesia de Patricia Calarco.)
2 células1½ dia
Mórula8 células2½ dias Compactação
16 células3 dias
Secção doblastocisto4 dias
Massa celular internaBlastocele
Trofoectoderme
Óvulo decamundongofertilizado
Corpopolar
Zonapelúcida
Pró-núcleosmaterno e paterno
50 �m
(A) (B) (C) (D)10 �m
Figura 22-89 Micrografias eletrônicas de varredura do embrião jovem de camundongo. A zona pelúcida foi removida. (A) Estágio de duas células. (B) Estágio de quatro células (um corpo polar é visível, além dos quatro blastô-meros – ver Figura 21-23). (C) Mórula com 8 a 16 células – a compactação está ocorrendo. (D) Blastocisto. (A-C, cortesia de Patricia Calarco; D, de P. Calarco e C. J. Epstein, Dev. Biol. 32:208-213, 1973. Com permissão de Academic Press.)
Alberts_22.indd 1379Alberts_22.indd 1379 29.07.09 16:49:3829.07.09 16:49:38

1380 Alberts, Johnson, Lewis, Raff, Roberts & Walter
agrupamento interno de células, chamado de massa celular interna, está localizado em um lado da cavidade. Ele dará origem à totalidade do embrião propriamente dito.
Após o embrião ter se livrado de sua cápsula gelatinosa (em cerca de quatro dias), as cé-lulas da trofoectoderme estabelecem um estreito contato com a parede do útero, iniciando o processo de implantação que levará à formação da placenta. Enquanto isso, a massa celular interna cresce e começa a se diferenciar. Parte dela dá origem a algumas estruturas extraem-brionárias adicionais, como o saco vitelínico, enquanto o resto segue em frente para formar o embrião propriamente dito pelos processos de gastrulação, neurulação, e assim por diante, que são fundamentalmente similares àqueles vistos em outros vertebrados, embora distorções da geometria muitas vezes tornem a homologia difícil de ser discernida à primeira vista.
O embrião jovem de mamíferos é altamente reguladorOs determinantes intracelulares localizados têm apenas um pequeno papel no desenvolvi-mento inicial dos mamíferos, e os blastômeros produzidos pelas primeiras poucas divisões celulares são extraordinariamente adaptáveis. Se o embrião jovem for dividido em dois, um par de gêmeos idênticos pode ser produzido – dois indivíduos normais completos a partir de uma única célula. Similarmente, se uma das células em um embrião de camundongo com duas células for destruída perfurando-a com uma agulha e o “meio embrião” resultante for colocado no útero de uma mãe adotiva para se desenvolver, em muitos casos um camun-dongo perfeitamente normal irá se formar.
Inversamente, dois embriões de camundongo com 8 células podem ser combinados para formar uma única mórula gigante que, então, se desenvolve em um camundongo de estrutura e tamanho normais (Figura 22-90). Tais criaturas, formadas a partir de agrega-dos de grupos de células geneticamente diferentes, são chamadas de quimeras. As quimeras também podem ser obtidas por meio da injeção de células de um embrião jovem de um genótipo em um blastocisto de outro genótipo. As células injetadas vêm a ser incorporadas na massa celular interna do blastocisto hospedeiro, e um animal quimérico se desenvolve. Desta maneira, uma única célula coletada de um embrião de 8 células ou da massa celular interna de outro blastocisto jovem pode dar origem a qualquer combinação de tipos celu-lares na quimera. Onde quer que a célula adicionada venha a ser encontrada, ela responde corretamente a estímulos de seus vizinhos e segue o caminho apropriado de desenvolvi-mento.
Essas descobertas têm duas implicações. Primeiro, durante os estágios iniciais, o sis-tema de desenvolvimento está se autoajustando, de forma que uma estrutura normal se desenvolve mesmo se as condições iniciais forem perturbadas. Embriões ou partes de em-briões que têm esta propriedade são denominados reguladores. Segundo, as células indivi-duais da massa celular interna são inicialmente totipotentes, ou quase totalmente: embora elas não possam formar o trofoblasto, podem dar origem a qualquer parte do corpo adulto, incluindo as células germinativas.
As células-tronco embrionárias totipotentes podem ser obtidas a partir de um embrião de mamíferoSe um embrião jovem de camundongo normal é enxertado no rim ou testículo de um adulto, seu desenvolvimento é perturbado além de qualquer possibilidade de regulação adequada, mas não é interrompido. O resultado é um crescimento tumoroso bizarro conhecido como teratoma, que consiste em uma massa desorganizada de células contendo muitos tipos de tecidos diferenciados – pele, osso, epitélio glandular, e assim por diante – misturados com células-tronco não-diferenciadas que continuam a se dividir e gerar ainda mais esses tecidos diferenciados.
A investigação das células-tronco em teratomas e tipos relacionados de tumores levou à descoberta de que seu comportamento reflete uma propriedade notável das células normais da massa celular interna: em um ambiente adequado, elas podem ser induzidas a se proliferar indefinidamente enquanto retiverem seu caráter totipotente. Células cultivadas que possuem esta propriedade são chamadas de células-tronco embrionárias ou células ES (de embryo-nic stem cells). Elas podem ser derivadas colocando-se uma massa celular interna normal em cultura e dispersando as células tão logo proliferem. Separar as células de suas vizinhas nor-mais e colocá-las no meio de cultura apropriado evidentemente interrompe o programa nor-mal de mudança de características celulares e, assim, permite que as células continuem a se
Embrião de camundongo no estágio de 8 célulascujos pais são camundongos brancos
Embrião de camundongono estágio de 8 célulascujos pais sãocamundongos pretos
A zona pelúcida de cada ovo é removidapor tratamento com protease
Os embriões são colocados juntos e sefundem quando incubados a 37°C
O desenvolvimento dos embriõesfusionados continua in vitro atéo estágio de blastocisto
O blastocisto é transferido para uma fêmeade camundongo pseudográvida, que atuacomo mãe adotiva
O filhote de camundongo tem quatroprogenitores (mas a mãe adotivanão é nenhum deles)
Figura 22-90 Procedimento para criar um camundongo quimérico. Duas mórulas de diferentes genótipos são combinadas.
Alberts_22.indd 1380Alberts_22.indd 1380 29.07.09 16:49:3929.07.09 16:49:39

Biologia Molecular da Célula 1381
dividir indefinidamente sem diferenciação. Muitos tecidos do corpo adulto também contêm células-tronco que podem se dividir indefinidamente sem diferenciação terminal, como ve-remos no próximo capítulo; porém, estas células-tronco adultas, quando se diferenciam, nor-malmente dão origem somente a uma série bastante restrita de tipos celulares diferenciados.
O estado no qual as células ES são interrompidas parece ser equivalente àquele de cé-lulas normais da massa celular interna. Pode-se mostrar isso coletando células ES da placa de cultura e injetando-as em um blastocisto normal (Figura 22-91). As células injetadas se incorporam à massa celular interna do blastocisto e podem contribuir para a formação de um camundongo quimérico aparentemente normal. As descendentes das células-tronco in-jetadas podem ser encontradas em praticamente qualquer tecido deste camundongo, onde elas se diferenciam de uma maneira bem-comportada e apropriada para o local, poden-do até mesmo formar células germinativas viáveis. O comportamento extraordinariamente adaptável das células ES mostra que sinais dos vizinhos de uma célula não somente orien-tam escolhas entre diferentes vias de diferenciação, mas também podem parar ou iniciar o relógio do desenvolvimento – os processos que impelem uma célula a progredir do estado embrionário para o adulto.
Em nível prático, as células ES têm uma dupla importância. Primeiro, do ponto de vis-ta médico, elas oferecem a perspectiva de uma fonte versátil de células para o reparo de tecidos danificados e defectivos no corpo adulto, como discutiremos no final do próximo capítulo. Segundo, as células ES tornam possíveis as formas mais precisamente controladas de modificação genética, permitindo que animais sejam criados com praticamente qual-quer alteração desejada introduzida em seu genoma. Como discutido no Capítulo 8, a téc-nica usa a recombinação genética para substituir uma sequência de DNA normal por um segmento de DNA construído artificialmente em um local selecionado do genoma de uma célula ES. Embora apenas uma rara célula incorpore a construção de DNA corretamente, foram elaborados procedimentos de seleção para encontrar esta célula entre as milhares de outras nas quais a construção de DNA foi transfectada. Uma vez selecionadas, as células ES geneticamente modificadas podem ser injetadas em um blastocisto para formar um ca-mundongo quimérico. Este camundongo irá, com sorte, ter algumas células germinativas derivadas de ES, capazes de atuar como fundadoras de uma nova geração de camundongos que consistem inteiramente em células carregando a mutação cuidadosamente projetada. Desta maneira, um camundongo mutante completo pode ser ressuscitado a partir da placa de cultura (ver Figura 8-65).
Interações entre o epitélio e o mesênquima geram estruturas tubulares ramificadasOs vertebrados são animais comparativamente grandes, devendo muito de seu tamanho aos tecidos conectivos. Contudo, para a excreção, a absorção de nutrientes e as trocas gasosas, eles também requerem grandes quantidades de vários tipos especializados de superfícies epiteliais. Muitos desses tipos tomam a forma de estruturas tubulares criadas por meio de morfogênese por ramificação, na qual um epitélio invade o tecido conectivo embrionário (mesênquima) para formar um órgão composto. O pulmão é um exemplo típico. Ele se ori-gina da endoderme que reveste a parte basal da porção anterior do intestino. Este epitélio brota e se expande para dentro do mesênquima vizinho para formar a árvore brônquica, um sistema de tubos que se ramificam repetidamente à medida que se estendem (Figura 22-92). O mesmo mesênquima também é invadido por células endoteliais – as células que revestem os vasos sanguíneos – para criar o sistema de vias aéreas e vasos sanguíneos inti-mamente justapostos necessário às trocas gasosas nos pulmões (discutido no Capítulo 23).
Todo o processo depende de trocas de sinais em ambas as direções entre os brotos em crescimento do epitélio e o mesênquima que estão invadindo. Esses sinais podem ser anali-sados por meio de manipulações genéticas no camundongo. Um papel central é desempe-nhado por proteínas-sinal da família do fator de crescimento de fibroblastos (FGF, fibroblast growth factor) e por receptores de tirosina-cinases sobre os quais eles atuam. Essas vias de sinalização têm vários papéis no desenvolvimento, mas parecem ser especialmente impor-tantes nas muitas interações que ocorrem entre o epitélio e o mesênquima.
Os mamíferos têm cerca de 20 diferentes genes Fgf, em comparação com três em Droso-phila e dois em C. elegans. O Fgf mais importante nos pulmões é o Fgf10. Este é expresso em agrupamentos de células mesenquimais próximas às extremidades dos tubos epiteliais em crescimento, enquanto seu receptor é expresso nas próprias células epiteliais. O FGF10 ou
Células ES derivadas delinhagens geneticamente
distintas de camundongos
Blastocisto receptor
Agrupamento de célulasES na micropipetaPipeta de
sucção dandosuporte
Células ES injetadasno blastocisto
As células injetadas seincorporam à massacelular interna doblastocisto hospedeiro
O blastocisto sedesenvolve na mãe adotiva emum camundongoquimérico saudável;as células ES podemcontribuir para qualquer tecido
Figura 22-91 Fazendo um camun-dongo quimérico com células ES. As células ES em cultura podem se com-binar com as células de um blastocisto normal para formar um camundongo quimérico saudável e podem contribuir para qualquer um de seus tecidos, incluindo a linhagem germinativa. Por-tanto, as células ES são totipotentes.
Alberts_22.indd 1381Alberts_22.indd 1381 29.07.09 16:49:3929.07.09 16:49:39

1382 Alberts, Johnson, Lewis, Raff, Roberts & Walter
seu receptor podem ser nocauteados (pelas técnicas-padrão com base na recombinação de células ES). No camundongo mutante nocauteado resultante, todo o processo de morfogê-nese por ramificação então falha – um broto primário de epitélio pulmonar é formado, mas não se expande para dentro do mesênquima para criar uma árvore brônquica. Inversamente, uma esfera microscópica embebida em FGF10 e colocada próxima a um epitélio pulmonar embrionário em cultura induzirá a formação de um broto, que se expandirá em direção a ela. Evidentemente, o epitélio invade o mesênquima apenas por convite, em resposta a FGF10.
Contudo, o que faz com que os tubos epiteliais em crescimento se ramifiquem repetida-mente, à medida que efetuam a invasão? Isso parece depender de um sinal Sonic hedgehog que é enviado na direção oposta, a partir das células epiteliais nas extremidades dos brotos, de volta para o mesênquima. Em camundongos sem Sonic hedgehog, o epitélio dos pulmões cresce e se diferencia, mas forma um saco em vez de uma árvore ramificada de túbulos. En-quanto isso, o FGF10, em vez de estar restrito a pequenos agrupamentos de células mesen-quimais, com cada agrupamento atuando como um sinal para direcionar o crescimento de um broto epitelial separado, é expresso em amplas bandas de células imediatamente adja-centes ao epitélio. Essa descoberta sugere que o sinal Sonic hedgehog pode servir para cortar a expressão de FGF10 nas células mesenquimais mais próximas à extremidade crescente de um broto, dividindo o agrupamento secretor de FGF10 em dois agrupamentos separados, os quais, por sua vez, fazem com que o broto se ramifique em dois (ver Figura 22-92A).
O crescimento por ramificação tanto do epitélio como do mesênquima tem de ser coor-denado com o desenvolvimento dos vasos sanguíneos associados, e todo o processo envolve um grande número de sinais adicionais. Muitos aspectos do sistema ainda não são entendi-dos. Sabe-se, contudo, que Drosophila utiliza mecanismos intimamente relacionados para governar a morfogênese por ramificação de seu sistema traqueal – os túbulos que formam as vias aéreas de um inseto. Novamente, o processo depende da proteína FGF de Drosophi-la, codificada pelo gene Branchless, e do receptor FGF de Drosophila, codificado pelo gene Breathless, ambos operando de forma muito semelhante à observada no camundongo. Na verdade, estudos genéticos do desenvolvimento traqueal em Drosophila também identifica-ram outros componentes da maquinaria de controle, e os genes de Drosophila nos levaram a seus homólogos em vertebrados. As manipulações genéticas no camundongo nos deram os meios para testar se esses genes também possuem funções similares nos mamíferos; e, em um grau extraordinário, eles o fazem.
ResumoO camundongo tem um papel central como organismo-modelo para o estudo da genética molecu-lar do desenvolvimento de mamíferos. O desenvolvimento do camundongo é essencialmente similar ao de outros vertebrados, mas começa com um preâmbulo especializado para formar estruturas extraembrionárias, como o âmnio e a placenta. Técnicas potentes foram elaboradas para a criação de nocautes gênicos e outras alterações genéticas marcadas pela exploração de propriedades alta-mente reguladoras das células da massa celular interna do embrião de camundongo. Essas células podem ser colocadas em cultura e mantidas como células-tronco embrionárias (células ES). Sob condições corretas de cultura, as células ES podem se proliferar indefinidamente sem diferenciação,
(A)(B)
FGF10 feito poragrupamento de
células mesenquimais
Receptor de FGF10nas células
epiteliais do broto
Produção de FGF10inibida por Shh
Criação de dois novos centrosde produção de FGF10
Sonic hedgehog(Shh) produzida
por células epiteliaisna extremidade do
broto em crescimento
Dois novos brotos são formadose todo o processo se repete
Figura 22-92 Morfogênese por ra-mificação nos pulmões. (A) Como se acredita que FGF10 e Sonic hedgehog induzam o crescimento e a ramificação dos brotos da árvore brônquica. Muitas outras moléculas-sinal, como BMP4, também são expressas neste sistema, e o mecanismo de ramificação sugerido é apenas uma das várias possibilidades. (B) Molde da árvore brônquica de um humano adulto, preparada pela injeção de resina nas vias aéreas; resinas de diferentes cores foram injetadas nas di-ferentes ramificações da árvore. (B, de R. Warwick e P. L. Williams, Gray’s Anatomy, 35th ed. Edinburgh: Longman, 1973.)
Alberts_22.indd 1382Alberts_22.indd 1382 29.07.09 16:49:4029.07.09 16:49:40

Biologia Molecular da Célula 1383
enquanto retêm a capacidade de dar origem a qualquer parte do corpo quando injetadas de volta em um embrião jovem de camundongo.
Muitos processos gerais do desenvolvimento, incluindo a maioria daqueles discutidos neste capítulo, foram esclarecidos por estudos em camundongos. Apenas como um exemplo, o camun-dongo tem sido utilizado para investigar o controle da morfogênese por ramificação. Este processo dá origem a estruturas como, por exemplo, pulmões e glândulas, e é governado por trocas de sinais entre as células mesenquimais e o epitélio invasor. As funções desses sinais podem ser analisadas por experimentos de nocaute gênico.
DESENVOLVIMENTO NEURALAs células nervosas, ou neurônios, estão entre os mais antigos de todos os tipos celula-res especializados de animais. Sua estrutura não é comparável com a de nenhuma outra classe de células, e o desenvolvimento do sistema nervoso apresenta problemas que não têm paralelo real em outros tecidos. Um neurônio é sobretudo extraordinário por sua forma bastante estendida, com um axônio longo e dendritos ramificados que o conectam por meio de sinapses a outras células (Figura 22-93). O desafio central do desenvolvimento neural é explicar como os axônios e dendritos crescem, encontram seus parceiros corretos e esta-belecem sinapses com os mesmos de forma seletiva, criando uma rede funcional (Figura 22-94). O problema é formidável: o cérebro humano contém mais de 1011 neurônios, cada um dos quais, em média, deve fazer conexões com milhares de outros, de acordo com um plano de ligação previsível e regular. A precisão necessária não é tão grande como a existente em um computador artificial, pois o cérebro realiza suas computações de maneira diferente e é mais tolerante aos caprichos dos componentes individuais; no entanto, o cérebro supera todas as outras estruturas biológicas em sua complexidade organizada.
Os componentes de um sistema nervoso típico – as várias classes de neurônios, células gliais, células sensoriais e músculos – originam-se em diversos locais extensamente separa-dos no embrião e, inicialmente, não estão conectados. Assim, na primeira fase do desenvol-vimento neural (Figura 22-95), as diferentes partes se desenvolvem de acordo com seus pró-prios programas locais: os neurônios nascem e características específicas lhes são atribuídas de acordo com o local e o momento de seu nascimento, sob o controle de sinais indutivos e mecanismos de regulação gênica similares àqueles que já discutimos para outros tecidos do corpo. A próxima fase envolve um tipo de morfogênese única ao sistema nervoso: os axônios e dendritos crescem ao longo de vias específicas, estabelecendo uma rede provisória, mas organizada, de conexões entre as partes separadas do sistema. Na terceira fase e final, que continua pela vida adulta, as conexões são ajustadas e aperfeiçoadas por meio de interações entre os componentes amplamente distribuídos de uma maneira que depende dos sinais elétricos que passam entre eles.
São designadas diferentes características aos neurônios de acordo com o momento e o local onde nasceramOs neurônios são quase sempre produzidos em associação com as células gliais, que pro-piciam uma estrutura de suporte e criam um ambiente fechado e protegido no qual os neu-rônios podem realizar suas funções. Ambos os tipos de células, em todos os animais, desen-
Figura 22-93 Um neurônio típico de um vertebrado. As setas indicam a di-reção em que os sinais são transmitidos. O neurônio mostrado é da retina de um macaco. Os maiores e mais longos neu-rônios humanos se estendem por cerca de 1 milhão de μm e têm um axônio com 15 μm de diâmetro. (Desenho de neurônio de B. B. Boycott, em Essays on the Nervous System [R. Bellairs e E. G. Gray, eds.]. Oxford, UK: Clarendon Press, 1974.)
25 �m
Corpo celular
Os dendritos recebemsinais sinápticos
Axônio (menos de 1 mm até mais de 1 m de comprimento)
As ramificações terminaisdo axônio formamsinapses nas células-alvo
Figura 22-94 A complexa organiza-ção das conexões das células nervo-sas. Este desenho representa uma secção transversal de uma pequena parte do cérebro de um mamífero – o bulbo olfatório de um cão, corado pela técnica de Golgi. Os objetos pretos são neurônios; as linhas finas são axônios e dendritos, por meio dos quais os vários grupos de neurônios são interconecta-dos de acordo com regras precisas. (De C. Golgi, Riv. sper. freniat. Reggio-Emilia 1:405-425, 1875; reproduzida em M. Ja-cobson, Developmental Neurobiology, 3rd ed. New York: Plenum, 1992.)
Alberts_22.indd 1383Alberts_22.indd 1383 29.07.09 16:49:4029.07.09 16:49:40
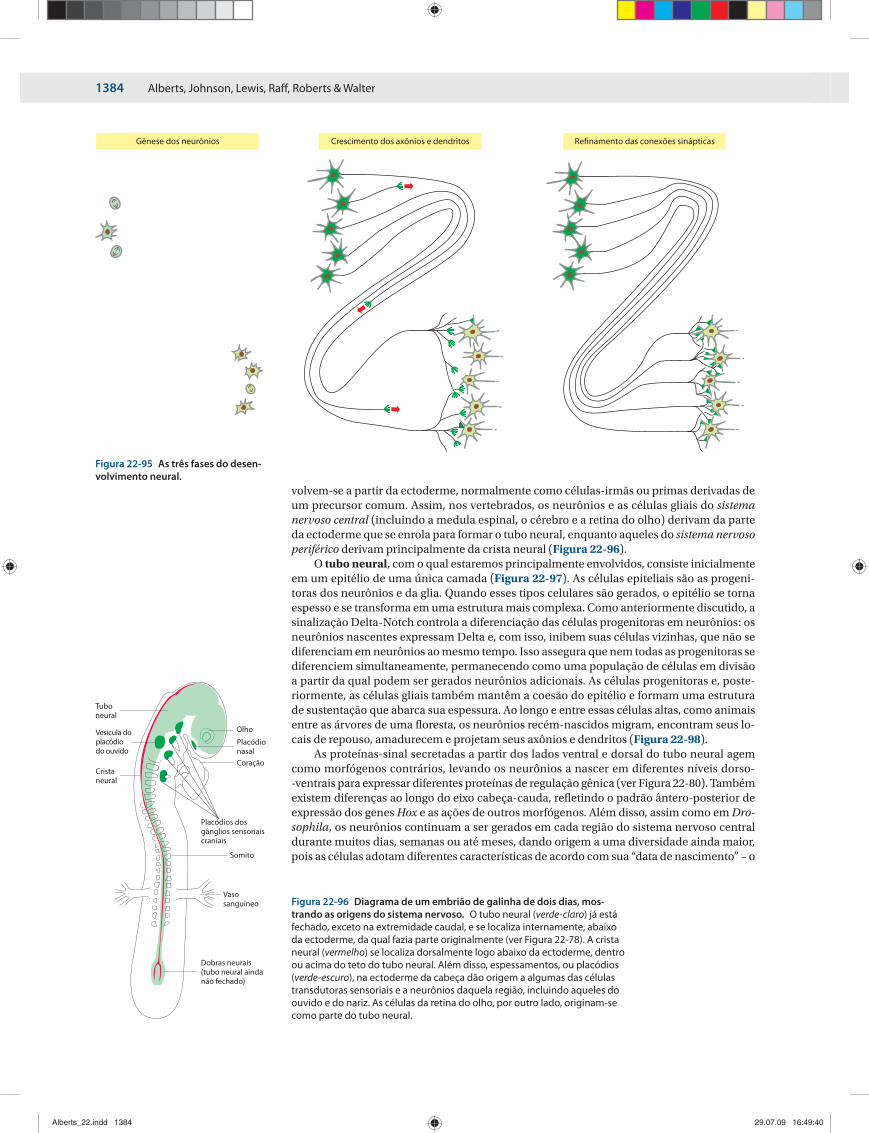
1384 Alberts, Johnson, Lewis, Raff, Roberts & Walter
volvem-se a partir da ectoderme, normalmente como células-irmãs ou primas derivadas de um precursor comum. Assim, nos vertebrados, os neurônios e as células gliais do sistema nervoso central (incluindo a medula espinal, o cérebro e a retina do olho) derivam da parte da ectoderme que se enrola para formar o tubo neural, enquanto aqueles do sistema nervoso periférico derivam principalmente da crista neural (Figura 22-96).
O tubo neural, com o qual estaremos principalmente envolvidos, consiste inicialmente em um epitélio de uma única camada (Figura 22-97). As células epiteliais são as progeni-toras dos neurônios e da glia. Quando esses tipos celulares são gerados, o epitélio se torna espesso e se transforma em uma estrutura mais complexa. Como anteriormente discutido, a sinalização Delta-Notch controla a diferenciação das células progenitoras em neurônios: os neurônios nascentes expressam Delta e, com isso, inibem suas células vizinhas, que não se diferenciam em neurônios ao mesmo tempo. Isso assegura que nem todas as progenitoras se diferenciem simultaneamente, permanecendo como uma população de células em divisão a partir da qual podem ser gerados neurônios adicionais. As células progenitoras e, poste-riormente, as células gliais também mantêm a coesão do epitélio e formam uma estrutura de sustentação que abarca sua espessura. Ao longo e entre essas células altas, como animais entre as árvores de uma floresta, os neurônios recém-nascidos migram, encontram seus lo-cais de repouso, amadurecem e projetam seus axônios e dendritos (Figura 22-98).
As proteínas-sinal secretadas a partir dos lados ventral e dorsal do tubo neural agem como morfógenos contrários, levando os neurônios a nascer em diferentes níveis dorso--ventrais para expressar diferentes proteínas de regulação gênica (ver Figura 22-80). Também existem diferenças ao longo do eixo cabeça-cauda, refletindo o padrão ântero-posterior de expressão dos genes Hox e as ações de outros morfógenos. Além disso, assim como em Dro-sophila, os neurônios continuam a ser gerados em cada região do sistema nervoso central durante muitos dias, semanas ou até meses, dando origem a uma diversidade ainda maior, pois as células adotam diferentes características de acordo com sua “data de nascimento” – o
Figura 22-95 As três fases do desen-volvimento neural.
Crescimento dos axônios e dendritosGênese dos neurônios Refinamento das conexões sinápticas
Dobras neurais(tubo neural aindanão fechado)
Vasosanguíneo
Somito
Placódios dosgânglios sensoriaiscraniais
Olho
Placódionasal
Coração
Vesícula doplacódiodo ouvido
Tuboneural
Cristaneural
Figura 22-96 Diagrama de um embrião de galinha de dois dias, mos-trando as origens do sistema nervoso. O tubo neural (verde-claro) já está fechado, exceto na extremidade caudal, e se localiza internamente, abaixo da ectoderme, da qual fazia parte originalmente (ver Figura 22-78). A crista neural (vermelho) se localiza dorsalmente logo abaixo da ectoderme, dentro ou acima do teto do tubo neural. Além disso, espessamentos, ou placódios (verde-escuro), na ectoderme da cabeça dão origem a algumas das células transdutoras sensoriais e a neurônios daquela região, incluindo aqueles do ouvido e do nariz. As células da retina do olho, por outro lado, originam-se como parte do tubo neural.
Alberts_22.indd 1384Alberts_22.indd 1384 29.07.09 16:49:4029.07.09 16:49:40
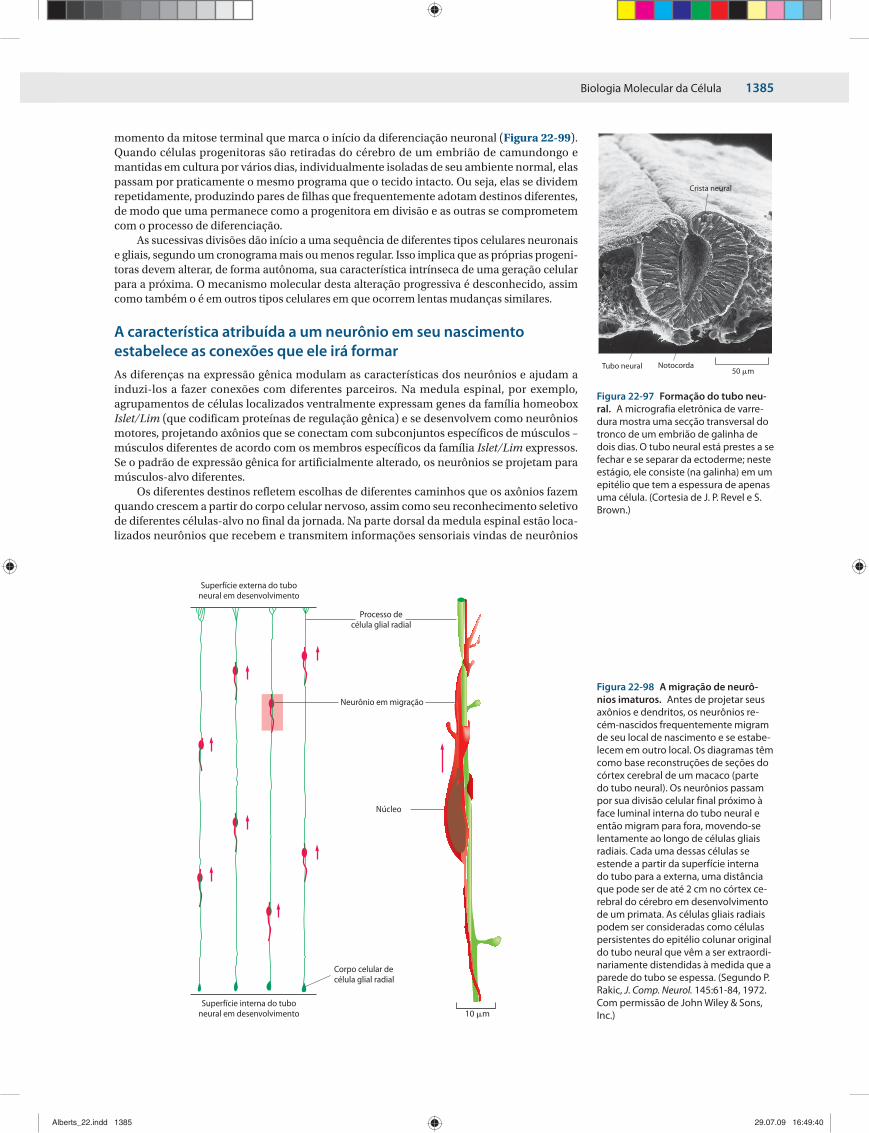
Biologia Molecular da Célula 1385
momento da mitose terminal que marca o início da diferenciação neuronal (Figura 22-99). Quando células progenitoras são retiradas do cérebro de um embrião de camundongo e mantidas em cultura por vários dias, individualmente isoladas de seu ambiente normal, elas passam por praticamente o mesmo programa que o tecido intacto. Ou seja, elas se dividem repetidamente, produzindo pares de filhas que frequentemente adotam destinos diferentes, de modo que uma permanece como a progenitora em divisão e as outras se comprometem com o processo de diferenciação.
As sucessivas divisões dão início a uma sequência de diferentes tipos celulares neuronais e gliais, segundo um cronograma mais ou menos regular. Isso implica que as próprias progeni-toras devem alterar, de forma autônoma, sua característica intrínseca de uma geração celular para a próxima. O mecanismo molecular desta alteração progressiva é desconhecido, assim como também o é em outros tipos celulares em que ocorrem lentas mudanças similares.
A característica atribuída a um neurônio em seu nascimento estabelece as conexões que ele irá formarAs diferenças na expressão gênica modulam as características dos neurônios e ajudam a induzi-los a fazer conexões com diferentes parceiros. Na medula espinal, por exemplo, agrupamentos de células localizados ventralmente expressam genes da família homeobox Islet/Lim (que codificam proteínas de regulação gênica) e se desenvolvem como neurônios motores, projetando axônios que se conectam com subconjuntos específicos de músculos – músculos diferentes de acordo com os membros específicos da família Islet/Lim expressos. Se o padrão de expressão gênica for artificialmente alterado, os neurônios se projetam para músculos-alvo diferentes.
Os diferentes destinos refletem escolhas de diferentes caminhos que os axônios fazem quando crescem a partir do corpo celular nervoso, assim como seu reconhecimento seletivo de diferentes células-alvo no final da jornada. Na parte dorsal da medula espinal estão loca-lizados neurônios que recebem e transmitem informações sensoriais vindas de neurônios
Crista neural
Tubo neural Notocorda50 �m
Figura 22-97 Formação do tubo neu-ral. A micrografia eletrônica de varre-dura mostra uma secção transversal do tronco de um embrião de galinha de dois dias. O tubo neural está prestes a se fechar e se separar da ectoderme; neste estágio, ele consiste (na galinha) em um epitélio que tem a espessura de apenas uma célula. (Cortesia de J. P. Revel e S. Brown.)
Figura 22-98 A migração de neurô-nios imaturos. Antes de projetar seus axônios e dendritos, os neurônios re-cém-nascidos frequentemente migram de seu local de nascimento e se estabe-lecem em outro local. Os diagramas têm como base reconstruções de seções do córtex cerebral de um macaco (parte do tubo neural). Os neurônios passam por sua divisão celular final próximo à face luminal interna do tubo neural e então migram para fora, movendo-se lentamente ao longo de células gliais radiais. Cada uma dessas células se estende a partir da superfície interna do tubo para a externa, uma distância que pode ser de até 2 cm no córtex ce-rebral do cérebro em desenvolvimento de um primata. As células gliais radiais podem ser consideradas como células persistentes do epitélio colunar original do tubo neural que vêm a ser extraordi-nariamente distendidas à medida que a parede do tubo se espessa. (Segundo P. Rakic, J. Comp. Neurol. 145:61-84, 1972. Com permissão de John Wiley & Sons, Inc.)
Superfície externa do tuboneural em desenvolvimento
Superfície interna do tuboneural em desenvolvimento
Corpo celular decélula glial radial
10 �m
Núcleo
Neurônio em migração
Processo decélula glial radial
Alberts_22.indd 1385Alberts_22.indd 1385 29.07.09 16:49:4029.07.09 16:49:40
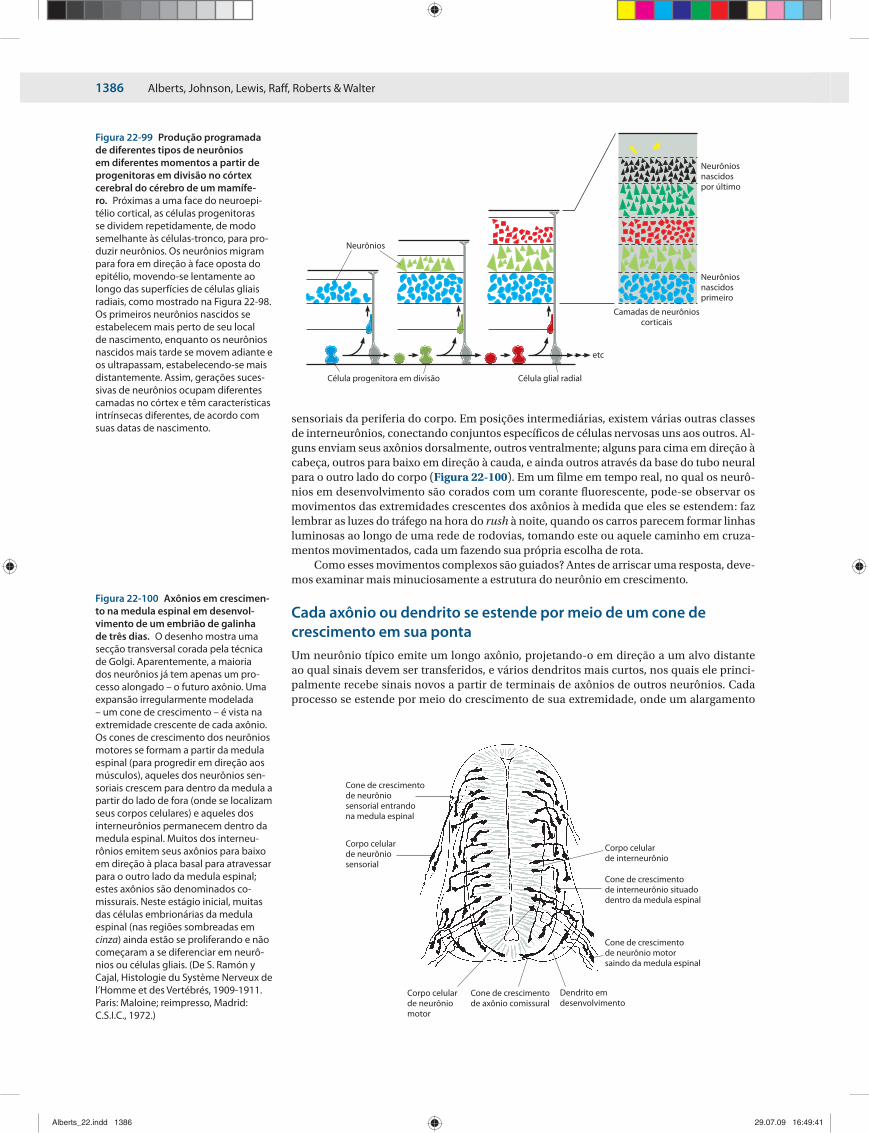
1386 Alberts, Johnson, Lewis, Raff, Roberts & Walter
sensoriais da periferia do corpo. Em posições intermediárias, existem várias outras classes de interneurônios, conectando conjuntos específicos de células nervosas uns aos outros. Al-guns enviam seus axônios dorsalmente, outros ventralmente; alguns para cima em direção à cabeça, outros para baixo em direção à cauda, e ainda outros através da base do tubo neural para o outro lado do corpo (Figura 22-100). Em um filme em tempo real, no qual os neurô-nios em desenvolvimento são corados com um corante fluorescente, pode-se observar os movimentos das extremidades crescentes dos axônios à medida que eles se estendem: faz lembrar as luzes do tráfego na hora do rush à noite, quando os carros parecem formar linhas luminosas ao longo de uma rede de rodovias, tomando este ou aquele caminho em cruza-mentos movimentados, cada um fazendo sua própria escolha de rota.
Como esses movimentos complexos são guiados? Antes de arriscar uma resposta, deve-mos examinar mais minuciosamente a estrutura do neurônio em crescimento.
Cada axônio ou dendrito se estende por meio de um cone de crescimento em sua pontaUm neurônio típico emite um longo axônio, projetando-o em direção a um alvo distante ao qual sinais devem ser transferidos, e vários dendritos mais curtos, nos quais ele princi-palmente recebe sinais novos a partir de terminais de axônios de outros neurônios. Cada processo se estende por meio do crescimento de sua extremidade, onde um alargamento
Figura 22-99 Produção programada de diferentes tipos de neurônios em diferentes momentos a partir de progenitoras em divisão no córtex cerebral do cérebro de um mamífe-ro. Próximas a uma face do neuroepi-télio cortical, as células progenitoras se dividem repetidamente, de modo semelhante às células-tronco, para pro-duzir neurônios. Os neurônios migram para fora em direção à face oposta do epitélio, movendo-se lentamente ao longo das superfícies de células gliais radiais, como mostrado na Figura 22-98. Os primeiros neurônios nascidos se estabelecem mais perto de seu local de nascimento, enquanto os neurônios nascidos mais tarde se movem adiante e os ultrapassam, estabelecendo-se mais distantemente. Assim, gerações suces-sivas de neurônios ocupam diferentes camadas no córtex e têm características intrínsecas diferentes, de acordo com suas datas de nascimento.
Cone de crescimentode neurôniosensorial entrandona medula espinal
Corpo celularde neurôniosensorial
Corpo celularde interneurônio
Cone de crescimentode interneurônio situadodentro da medula espinal
Cone de crescimentode neurônio motorsaindo da medula espinal
Corpo celularde neurôniomotor
Dendrito emdesenvolvimento
Cone de crescimentode axônio comissural
Neurônios
Célula progenitora em divisão Célula glial radial
etc
Camadas de neurônioscorticais
Neurôniosnascidospor último
Neurôniosnascidosprimeiro
Figura 22-100 Axônios em crescimen-to na medula espinal em desenvol-vimento de um embrião de galinha de três dias. O desenho mostra uma secção transversal corada pela técnica de Golgi. Aparentemente, a maioria dos neurônios já tem apenas um pro-cesso alongado – o futuro axônio. Uma expansão irregularmente modelada – um cone de crescimento – é vista na extremidade crescente de cada axônio. Os cones de crescimento dos neurônios motores se formam a partir da medula espinal (para progredir em direção aos músculos), aqueles dos neurônios sen-soriais crescem para dentro da medula a partir do lado de fora (onde se localizam seus corpos celulares) e aqueles dos interneurônios permanecem dentro da medula espinal. Muitos dos interneu-rônios emitem seus axônios para baixo em direção à placa basal para atravessar para o outro lado da medula espinal; estes axônios são denominados co-missurais. Neste estágio inicial, muitas das células embrionárias da medula espinal (nas regiões sombreadas em cinza) ainda estão se proliferando e não começaram a se diferenciar em neurô-nios ou células gliais. (De S. Ramón y Cajal, Histologie du Système Nerveux de l’Homme et des Vertébrés, 1909-1911. Paris: Maloine; reimpresso, Madrid: C.S.I.C., 1972.)
Alberts_22.indd 1386Alberts_22.indd 1386 29.07.09 16:49:4129.07.09 16:49:41

Biologia Molecular da Célula 1387
irregular e pontiagudo é observado. Esta estrutura, denominada cone de crescimento, mo-vimenta-se lentamente através do tecido adjacente, arrastando um axônio ou dendrito fino atrás de si (ver Figura 22-100). O cone de crescimento compreende tanto o mecanismo que produz o movimento como o aparato de direção que conduz a extremidade de cada proces-so ao longo do caminho adequado (ver Figura 16-105).
Muito do que sabemos a respeito das propriedades dos cones de crescimento veio de estudos em cultura de células ou tecidos. Pode-se observar quando um neurônio começa a estender seus prolongamentos, todos semelhantes no começo, até que um dos cones de cres-cimento realiza uma alteração súbita em sua velocidade, identificando seu prolongamento como axônio, com seu próprio conjunto axônio-específico de proteínas (Figura 22-101). O contraste entre axônio e dendrito estabelecido neste estágio envolve o transporte intracelular polarizado de diferentes materiais para dentro de dois tipos de prolongamento. O resultado é que eles irão se expandir por distâncias diferentes, seguir caminhos diferentes e desempe-nhar diferentes papéis na formação de sinapses.
O cone de crescimento na extremidade de um típico prolongamento em crescimento de uma célula nervosa – ou axônio ou dendrito – move-se para a frente a uma velocidade de cerca de 1 mm por dia, continuamente explorando as regiões que estão adiante e de cada lado por meio da extensão de seus filopódios e lamelipódios. Quando tal protrusão faz con-tato com uma superfície desfavorável, ela se retrai; quando faz contato com uma superfície mais favorável, ela persiste por mais tempo, guiando o cone de crescimento como um todo a se mover naquela direção. Desta maneira, o cone de crescimento pode ser guiado por va-riações sutis nas propriedades de superfície dos substratos sobre os quais se move. Ao mes-mo tempo, ele é sensível a fatores quimiotáticos que se difundem no meio circundante, os quais também podem estimular ou impedir seu avanço. Esses comportamentos dependem da maquinaria citoesquelética dentro do cone de crescimento, como discutido no Capítulo 16. Uma grande quantidade de receptores na membrana do cone de crescimento detectam os sinais externos e, pela ação de reguladores intracelulares, como as GTPases monoméricas Rho e Rac, controlam o agrupamento e o desagrupamento dos filamentos de actina e de outros componentes da maquinaria de movimento celular.
O cone de crescimento guia o neurito em desenvolvimento ao longo de um caminho precisamente definido in vivoEm animais vivos, os cones de crescimento geralmente se deslocam em direção a seus alvos ao longo de vias previsíveis e estereotipadas, explorando uma grande quantidade de diferen-tes sinais para encontrar seu caminho, mas sempre requerendo um substrato de matriz ex-tracelular ou de superfície celular sobre o qual possa deslizar. Frequentemente, os cones de crescimento tomam vias que foram inauguradas por outros neuritos, os quais eles seguem por meio de orientação por contato. Como resultado, as fibras nervosas em um animal ma-duro se encontram normalmente agrupadas em feixes paralelos compactos (denominados fascículos ou sistemas de fibras). Acredita-se que esse rastejamento dos cones de crescimen-to ao longo dos axônios seja mediado por moléculas homofílicas de adesão célula-célula – glicoproteínas de membrana que auxiliam uma célula que as exibe a se ligar a qualquer outra célula que também as apresenta. Como discutido no Capítulo 19, duas das classes
Figura 22-101 Formação de axônios e dendritos em cultura. Um neurônio jovem foi isolado do cérebro de um ma-mífero e colocado para se desenvolver em cultura, onde ele emite seus prolon-gamentos. Um destes prolongamentos, o futuro axônio, começou a crescer mais rápido que o resto (os futuros dendri-tos) e se bifurcou. (A) Uma fotografia de contraste de fase; (B) o padrão de colo-ração com faloidina fluorescente, que se liga a filamentos de actina. A actina está concentrada nos cones de crescimento nas extremidades dos prolongamentos que estão se estendendo ativamente e em alguns outros locais de atividade lamelipodial. (Cortesia de Kimberly Goslin.)
(B)(A)
dendrito corpo celular axônio cone de crescimento
10 �m
Alberts_22.indd 1387Alberts_22.indd 1387 29.07.09 16:49:4229.07.09 16:49:42

1388 Alberts, Johnson, Lewis, Raff, Roberts & Walter
mais importantes dessas moléculas são aquelas que pertencem à superfamília das imuno-globulinas, como as N-CAM, e aquelas da família de caderinas dependentes de Ca2+, como a N-caderina. Os membros de ambas as famílias geralmente estão presentes nas superfícies dos cones de crescimento, dos axônios e de vários outros tipos celulares sobre os quais os cones de crescimento se arrastam, incluindo as células gliais do sistema nervoso central e as células musculares da periferia do corpo. O genoma humano contém mais de 100 genes de caderinas, por exemplo, e a maior parte deles é expressa no cérebro (ver Figura 19-6). Diferentes conjuntos de moléculas de adesão célula-célula, atuando em combinações varia-das, propiciam um mecanismo para a orientação e o reconhecimento neuronal seletivo. Os cones de crescimento também migram sobre componentes da matriz extracelular. Algumas dessas moléculas de matriz, como a laminina, favorecem o crescimento dos axônios, en-quanto outras, como os proteoglicanos de sulfato de condroitina, o inibem.
Os cones de crescimento são guiados por uma sucessão de diferentes sinais em dife-rentes estágios de sua jornada, e não é somente a adesividade do substrato que importa. Outro papel importante é desempenhado por fatores quimiotáticos, secretados a partir de células que agem como guias em pontos estratégicos ao longo do caminho – alguns atraindo e outros repelindo. A trajetória dos axônios comissurais – aqueles que atravessam de um lado do corpo para o outro – constitui um belo exemplo de como uma combinação de sinais de orientação pode especificar um caminho complexo. Os axônios comissurais são uma carac-terística geral de animais bilateralmente simétricos, pois os dois lados do corpo têm de ser coordenados por meio de neurônios. Os vermes, as moscas e os vertebrados utilizam meca-nismos intimamente relacionados para guiar seu crescimento.
Na medula espinal em desenvolvimento de um vertebrado, por exemplo, um grande número de neurônios envia seus cones de crescimento axônicos ventralmente em direção à lâmina pavimentar – uma faixa especializada de células que forma a linha média ventral do tubo neural (ver Figura 22-100). Os cones de crescimento cruzam a lâmina pavimentar e então mudam de direção abruptamente, em ângulo reto, para seguir um caminho longi-tudinal para cima, em direção ao cérebro, paralelamente à lâmina pavimentar, mas nunca cruzando-a novamente (Figura 22-102A). O primeiro estágio da jornada depende de um gradiente de concentração da proteína netrina, secretada pelas células da lâmina pavimen-tar: os cones de crescimento comissurais farejam o caminho em direção a sua fonte. A ne-trina foi purificada de embriões de galinha testando-se extratos de tecido neural quanto a uma atividade que atrairia os cones de crescimento comissurais em uma placa de cultu-ra. Sua sequência revelou que a netrina dos vertebrados era homóloga a uma proteína já identificada em C. elegans por meio de triagens genéticas de vermes mutantes com axônios com orientação alterada – denominados mutantes Unc (de uncoordinated), pois se movem de forma desordenada. Um dos genes Unc, Unc6, codifica o homólogo da netrina. Outro, Unc40, codifica seu receptor transmembrana; este também tem um homólogo vertebrado, denominado DCC, que é expresso nos neurônios comissurais e serve como mediador em sua resposta ao gradiente de netrina.
A ativação localizada de DCC pela netrina leva à abertura de uma classe especializada de canais iônicos na membrana plasmática. Esses canais, denominados canais TRPC (po-
Figura 22-102 A orientação de axô-nios comissurais. (A) O caminho toma-do pelos axônios comissurais na medula espinal embrionária de um vertebrado. (B) Os sinais que os orientam. Os cones de crescimento são primeiramente atraídos para a lâmina pavimentar pela netrina, que é secretada pelas células da lâmina pavimentar e age sobre o recep-tor DCC na membrana axônica. Quando atravessam a lâmina basal, os cones de crescimento aumentam a expressão de Roundabout, o receptor para uma proteína repelente, Slit, que também é secretada pela lâmina basal. A proteína Slit, ligando-se ao receptor Rounda-bout, não somente age como repelente para impedir que as células entrem no-vamente na lâmina basal, mas também bloqueia a receptividade ao atraente netrina. Ao mesmo tempo, os cones de crescimento ativam a expressão de receptores para outra proteína repelen-te, a semaforina, que é secretada pelas células nas paredes laterais do tubo neural. Capturados entre dois territórios repelentes, os cones de crescimento, tendo atravessado a linha média, deslo-cam-se em um fascículo compacto para cima em direção ao cérebro.
Placa dorsal
Neurôniocomissural
Axôniocomissural
Linha médiaPlaca ventral
EM DIREÇÃO AO CÉREBRO
(A) (B)
Neurônio comissuralse aproximando dalinha média Parede do tubo neural
Placa ventralna linha média ventral
Cone de crescimentoexpressando receptor (DCC)para netrina
Atraente(netrina)
Cone de crescimento expressandoreceptor (Roundabout) para Slite receptores para semaforina
Repelente(semaforina)
Repelente(Slit)
PARA O CÉREBRO
Alberts_22.indd 1388Alberts_22.indd 1388 29.07.09 16:49:4229.07.09 16:49:42

Biologia Molecular da Célula 1389
tencial transiente de receptor C, de transient receptor potential C), pertencem a uma grande família (a família TRP), que é responsável por muitos outros processos de transdução sen-sorial, desde sensações mecânicas até a percepção de calor e frio. Quando abertos, os canais TRPC permitem que o Ca2+ (e outros cátions) entre na célula. O aumento localizado de Ca2+ ativa, então, a maquinaria para a extensão de filopódios e o movimento do cone de cresci-mento em direção à fonte de netrina.
Os receptores em cada cone de crescimento determinam a via que ele irá tomar: os neu-rônios não-comissurais no tubo neural, sem DCC, não são atraídos para a lâmina pavimen-tar, e os neurônios que expressam um receptor diferente de netrina – denominado Unc5H nos vertebrados (com o equivalente Unc5 no verme) – são ativamente repelidos pela lâmina pavimentar e enviam seus axônios em direção à lâmina superior.
Os cones de crescimento podem alterar sua sensibilidade à medida que se deslocamSe os cones de crescimento comissurais são atraídos para a lâmina pavimentar, por que eles a atravessam e emergem no outro lado, em vez de permanecer no território atrativo? E, depois de a terem cruzado, por que eles nunca voltam? A provável resposta se encontra em outro con-junto de moléculas, várias das quais também são conservadas entre vertebrados e invertebra-dos. Estudos em mutantes de Drosophila com axônios comissurais com orientação alterada identificaram, primeiramente, três das proteínas-chave: Slit, Roundabout e Commissureless.
A Slit, assim como a netrina, é produzida por células da linha média da mosca em de-senvolvimento, enquanto seu receptor, Roundabout, é expresso nos neurônios comissurais. A Slit, que age sobre Roundabout, tem um efeito exatamente oposto ao da netrina: ela repele os cones de crescimento, bloqueando a entrada no território da linha média. Entretanto, a Commissureless interfere na entrega de Roundabout à superfície celular e, com isso, faz com que os cones de crescimento fiquem inicialmente cegos para este sinal de “mantenha-se afastado”. Os cones de crescimento comissurais neste estado avançam para a linha média; quando eles a atravessam, parecem perder, por um mecanismo que ainda não compreende-mos, sua venda da proteína Commissureless e começam a ser repelidos. Emergindo do outro lado, eles agora têm o receptor Roundabout funcional em suas superfícies e são, desse modo, proibidos de entrar novamente.
Nos vertebrados, opera um mecanismo similar, envolvendo homólogos de Slit e Roun-dabout. Os cones de crescimento comissurais são primeiramente atraídos para a linha média e, então, de alguma maneira alteram suas proteínas receptoras de superfície quando a atra-vessam; desse modo, eles mudam suas sensibilidades, ganhando sensibilidade para a repul-são por Slit – que é expressa na lâmina pavimentar – e perdendo sensibilidade para a atração por netrina. A sensibilidade a Slit na aproximação inicial à linha média não é bloqueada por nenhum homólogo de Commissureless, mas por um membro divergente da família do re-ceptor Roundabout denominado Rig1, que se situa na membrana plasmática e interfere na recepção de sinais por seus primos. Uma vez que os cones de crescimento tenham atraves-sado a linha média, o bloqueio de Rig1 é interrompido por um mecanismo desconhecido. A repulsão a partir da linha média evita que eles se percam e voltem pelo mesmo caminho. Ao mesmo tempo, os cones de crescimento aparentemente se tornam sensíveis a outro grupo de sinais repulsivos, na forma de proteínas denominadas semaforinas, que os impedem de voltar para as regiões dorsais da medula espinal. Estando presos entre os dois conjuntos de sinais repulsivos, os cones de crescimento não têm outra escolha a não ser seguir em frente por um caminho estreito, correndo em paralelo à lâmina pavimentar, mas nunca entrando novamente nela (Figura 22-102B).
Os tecidos-alvo liberam fatores neurotróficos que controlam o crescimento e a sobrevivência das células nervosasFinalmente, os cones de crescimento dos axônios alcançam a região-alvo em que devem parar e fazer sinapses. Os neurônios que emitiram os axônios podem agora começar a se co-municar com suas células-alvo. Embora as sinapses geralmente transmitam sinais em uma direção, do axônio para ou o dendrito ou o músculo, as comunicações que ocorrem durante o desenvolvimento são uma via de mão dupla. Os sinais oriundos do tecido-alvo não só re-gulam quais cones de crescimento devem fazer sinapse (como discutiremos a seguir), mas também quantos dos neurônios em inervação devem sobreviver.
Alberts_22.indd 1389Alberts_22.indd 1389 29.07.09 16:49:4229.07.09 16:49:42

1390 Alberts, Johnson, Lewis, Raff, Roberts & Walter
A maior parte dos tipos de neurônios do sistema nervoso central e periférico dos verte-brados é produzida em excesso; até 50% ou mais deles morrem logo após ter alcançado seu alvo, mesmo que pareçam perfeitamente normais e saudáveis até o momento de sua morte. Cerca de metade de todos os neurônios motores que enviam axônios para os músculos es-queléticos, por exemplo, morre dentro de alguns dias após ter feito contato com suas células musculares-alvo. Uma proporção similar dos neurônios sensoriais que fazem a inervação da pele morre depois que seus cones de crescimento chegaram lá.
Acredita-se que essa morte em grande escala de neurônios reflita o resultado de uma competição. Cada tipo de célula-alvo libera uma quantidade limitada de um fator neuro-trófico específico que os neurônios que inervam aquele alvo precisam para sobreviver. Apa-rentemente, os neurônios competem pelo fator e aqueles que não conseguem o suficiente morrem por morte celular programada. Se a quantidade de tecido-alvo é aumentada – en-xertando um broto de um membro extra em um lado do embrião, por exemplo – mais neu-rônios inervando o membro sobrevivem; inversamente, se o broto do membro é cortado, todos os neurônios inervando o membro morrem. Desta maneira, embora os indivíduos possam variar quanto às suas proporções corporais, eles sempre manterão o número correto de neurônios motores para inervar todos os seus músculos e o número correto de neurônios sensoriais para inervar toda a sua superfície corporal. A estratégia aparentemente dispen-diosa de superprodução seguida pela morte de células excedentes funciona em quase todas as regiões do sistema nervoso. Ela funciona como um meio simples e efetivo de ajustar cada população de neurônios que fazem a inervação, de acordo com a quantidade de tecido que necessita de inervação.
O primeiro fator neurotrófico a ser identificado, que ainda continua sendo o melhor caracterizado, é simplesmente conhecido como fator de crescimento neuronal (NGF, nerve growth factor) – o membro fundador da família das neurotrofinas das proteínas de sinali-zação. Ele promove a sobrevivência de classes específicas de neurônios sensoriais deriva-dos da crista neural e de neurônios simpáticos (uma subclasse de neurônios periféricos que controlam as contrações dos músculos lisos e a secreção das glândulas exócrinas). O NGF é produzido pelos tecidos que estes neurônios inervam. Quando o NGF extra é fornecido, os neurônios sensoriais e simpáticos adicionais sobrevivem, como se o tecido-alvo extra esti-vesse presente. Inversamente, em um camundongo com uma mutação que inativa o gene NGF ou o gene de seu receptor (uma tirosina-cinase transmembrana denominada TrkA), quase todos os neurônios simpáticos e os neurônios sensoriais dependentes de NGF são perdidos. Existem muitos fatores neurotróficos, mas apenas alguns pertencem à família das neurotrofinas, atuando em diferentes combinações para promover a sobrevivência de dife-rentes classes de neurônios.
O NGF e seus assemelhados têm um papel adicional: além de agirem na célula nervo-sa como um todo, controlando sua sobrevivência, eles regulam o crescimento de axônios e dendritos (Figura 22-103). Podem até mesmo agir localmente em somente uma parte da árvore de prolongamentos das células nervosas, promovendo ou podando o crescimento de ramificações individuais: um cone de crescimento exposto ao NGF mostra um aumento imediato de mobilidade. Inversamente, uma ramificação de um axônio que é privado de NGF morre, enquanto o resto do neurônio continua a ser banhado pelo fator.
A ação periférica do NGF continua a ser importante depois da fase de morte neuronal. Na pele, por exemplo, o NGF controla a ramificação das fibras nervosas sensoriais, assegu-rando não só que toda a superfície do corpo fique inervada durante o desenvolvimento, mas também que ela recupere sua inervação após alguma lesão.
Figura 22-103 Efeitos do NGF no desenvolvimento de neuritos. Fotomicro-grafias de campo escuro de um gânglio simpático cultivado por 48 horas com (acima) e sem (abaixo) o NGF. Os neuritos se desenvolvem a partir dos neurônios simpáticos apenas quando o NGF está presente no meio. Cada cultura também contém células de Schwann (gliais) que migraram para fora do gânglio; elas não são afetadas por NGF. A sobrevivência neuronal e a manutenção dos cones de crescimento para a extensão dos neuritos representam dois efeitos distintos do NGF. O efeito nos cones de crescimento é local, direto, rápido e independente de comunicações com o corpo celular; quando o NGF é removido, os cones de crescimento prejudicados interrompem seus movimentos dentro de um ou dois minutos. O efeito do NGF sobre a sobrevivência celular é menos imediato e está associado à absorção de NGF por endocitose e a seu transporte intracelular de volta para o corpo celular. (Cortesia de Naomi Kleitman.)
NGF
Controle
Alberts_22.indd 1390Alberts_22.indd 1390 29.07.09 16:49:4229.07.09 16:49:42
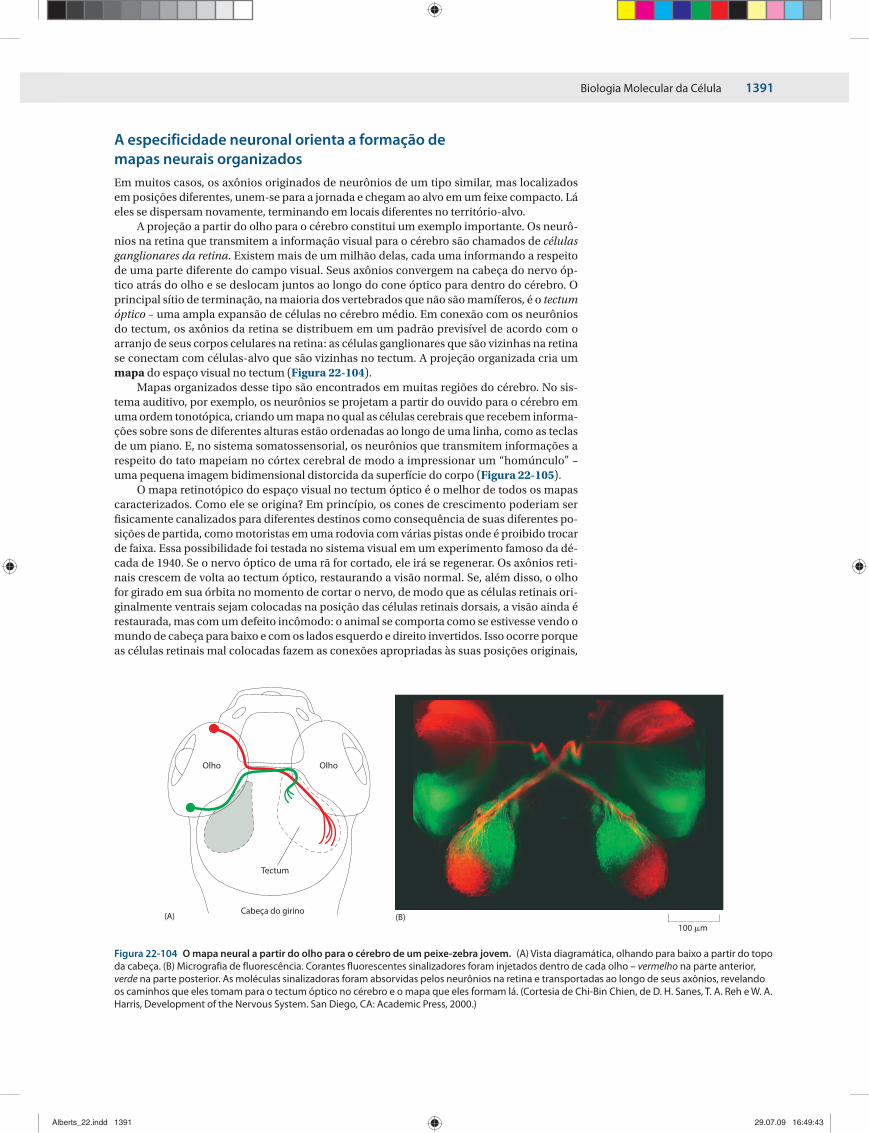
Biologia Molecular da Célula 1391
A especificidade neuronal orienta a formação de mapas neurais organizadosEm muitos casos, os axônios originados de neurônios de um tipo similar, mas localizados em posições diferentes, unem-se para a jornada e chegam ao alvo em um feixe compacto. Lá eles se dispersam novamente, terminando em locais diferentes no território-alvo.
A projeção a partir do olho para o cérebro constitui um exemplo importante. Os neurô-nios na retina que transmitem a informação visual para o cérebro são chamados de células ganglionares da retina. Existem mais de um milhão delas, cada uma informando a respeito de uma parte diferente do campo visual. Seus axônios convergem na cabeça do nervo óp-tico atrás do olho e se deslocam juntos ao longo do cone óptico para dentro do cérebro. O principal sítio de terminação, na maioria dos vertebrados que não são mamíferos, é o tectum óptico – uma ampla expansão de células no cérebro médio. Em conexão com os neurônios do tectum, os axônios da retina se distribuem em um padrão previsível de acordo com o arranjo de seus corpos celulares na retina: as células ganglionares que são vizinhas na retina se conectam com células-alvo que são vizinhas no tectum. A projeção organizada cria um mapa do espaço visual no tectum (Figura 22-104).
Mapas organizados desse tipo são encontrados em muitas regiões do cérebro. No sis-tema auditivo, por exemplo, os neurônios se projetam a partir do ouvido para o cérebro em uma ordem tonotópica, criando um mapa no qual as células cerebrais que recebem informa-ções sobre sons de diferentes alturas estão ordenadas ao longo de uma linha, como as teclas de um piano. E, no sistema somatossensorial, os neurônios que transmitem informações a respeito do tato mapeiam no córtex cerebral de modo a impressionar um “homúnculo” – uma pequena imagem bidimensional distorcida da superfície do corpo (Figura 22-105).
O mapa retinotópico do espaço visual no tectum óptico é o melhor de todos os mapas caracterizados. Como ele se origina? Em princípio, os cones de crescimento poderiam ser fisicamente canalizados para diferentes destinos como consequência de suas diferentes po-sições de partida, como motoristas em uma rodovia com várias pistas onde é proibido trocar de faixa. Essa possibilidade foi testada no sistema visual em um experimento famoso da dé-cada de 1940. Se o nervo óptico de uma rã for cortado, ele irá se regenerar. Os axônios reti-nais crescem de volta ao tectum óptico, restaurando a visão normal. Se, além disso, o olho for girado em sua órbita no momento de cortar o nervo, de modo que as células retinais ori-ginalmente ventrais sejam colocadas na posição das células retinais dorsais, a visão ainda é restaurada, mas com um defeito incômodo: o animal se comporta como se estivesse vendo o mundo de cabeça para baixo e com os lados esquerdo e direito invertidos. Isso ocorre porque as células retinais mal colocadas fazem as conexões apropriadas às suas posições originais,
(B)100 �m
Tectum
Olho Olho
Cabeça do girino(A)
Figura 22-104 O mapa neural a partir do olho para o cérebro de um peixe-zebra jovem. (A) Vista diagramática, olhando para baixo a partir do topo da cabeça. (B) Micrografia de fluorescência. Corantes fluorescentes sinalizadores foram injetados dentro de cada olho – vermelho na parte anterior, verde na parte posterior. As moléculas sinalizadoras foram absorvidas pelos neurônios na retina e transportadas ao longo de seus axônios, revelando os caminhos que eles tomam para o tectum óptico no cérebro e o mapa que eles formam lá. (Cortesia de Chi-Bin Chien, de D. H. Sanes, T. A. Reh e W. A. Harris, Development of the Nervous System. San Diego, CA: Academic Press, 2000.)
Alberts_22.indd 1391Alberts_22.indd 1391 29.07.09 16:49:4329.07.09 16:49:43

1392 Alberts, Johnson, Lewis, Raff, Roberts & Walter
e não às suas posições reais. Parece que as células têm valores posicionais – propriedades bioquímicas específicas para posição que representam registros de sua localização original. Como resultado, as células em lados opostos da retina são intrinsecamente diferentes, assim como os neurônios motores da medula espinal que se projetam para diferentes músculos são intrinsecamente diferentes.
Tal não-equivalência entre neurônios é referida como especificidade neuronal. É esta característica intrínseca que orienta os axônios retinais a seus sítios-alvo adequados no tectum. Os próprios sítios-alvo são distinguíveis pelos axônios retinais, pois as células do tectum também carregam marcas posicionais. Assim, o mapa neuronal depende de uma correspondência entre dois sistemas de marcadores posicionais, um na retina e outro no tectum.
Os axônios de diferentes regiões da retina respondem diferentemente a um gradiente de moléculas repulsivas no tectumOs axônios da retina nasal (o lado mais próximo do nariz) se projetam para o tectum pos-terior, e os axônios da retina temporal (o lado mais distante do nariz) se projetam para o tectum anterior, com regiões intermediárias de retina projetando-se para regiões interme-diárias do tectum. Quando se permite que axônios nasais e temporais cresçam sobre um tapete de membranas do tectum anterior e posterior em uma placa de cultura, eles também mostram seletividade (Figura 22-106). Os axônios temporais preferem substancialmente as membranas do tectum anterior, como in vivo, ao passo que os axônios nasais ou preferem as membranas do tectum posterior ou não têm preferência (dependendo da espécie animal). A diferença-chave entre o tectum anterior e o posterior parece ser um fator repulsivo no tectum posterior, ao qual os axônios retinais temporais são sensíveis, mas os axônios retinais nasais não o são: se um cone de crescimento retinal temporal tocar a membrana do tectum posterior, ele desfaz seus filopódios e se retrai.
Experimentos com base nesses fenômenos in vitro identificaram algumas das molécu-las responsáveis. O fator repulsivo na membrana do tectum posterior parece ser parcial ou inteiramente constituído de proteínas efrina A, um subconjunto da família de proteínas li-gadas a GPI que atuam como ligantes para a família EphA de receptores de tirosina-cinases. No camundongo, duas efrinas diferentes são expressas para formar um gradiente ântero--posterior nas células do tectum. As células anteriores têm pouca ou nenhuma quantidade
Figura 22-105 Mapa da superfície do corpo no cérebro humano. A super-fície do corpo está mapeada na região somatossensorial do córtex cerebral por um sistema organizado de conexões de células nervosas, de modo que a informação sensorial vinda de sítios corporais vizinhos é entregue a sítios vizinhos no cérebro. Isso significa que o mapa no cérebro é em grande parte fiel à topologia da superfície do corpo, mesmo que diferentes regiões do corpo estejam representadas em diferentes proporções, de acordo com sua densi-dade de inervação. O homúnculo (o “pe-queno homem” no cérebro) tem lábios grandes, por exemplo, porque os lábios são uma fonte particularmente grande e importante de informações sensoriais. O mapa foi determinado estimulando-se diferentes pontos no córtex de pa-cientes conscientes durante cirurgias de cérebro e gravando o que eles diziam estar sentindo. (Segundo W. Penfield e T. Rasmussen, The Cerebral Cortex of Man. New York: Macmillan, 1950.)
Genitália
Pern
a
Dedos do pé
Pé
Intra-abdominalFaringe
Língua
Dentes, gengivas e maxilar
Lábio inferior
Lábios
Lábio superior
Face
Nariz
Olho
Polegar
Indicador
Dedo médio
AnularDedo m
indinho
Mão
PulsoA
ntebraçoCotovelo
BraçoO
mb
ro Cab
eça
Pesc
oço
Tron
coQ
uadr
il
Alberts_22.indd 1392Alberts_22.indd 1392 29.07.09 16:49:4329.07.09 16:49:43
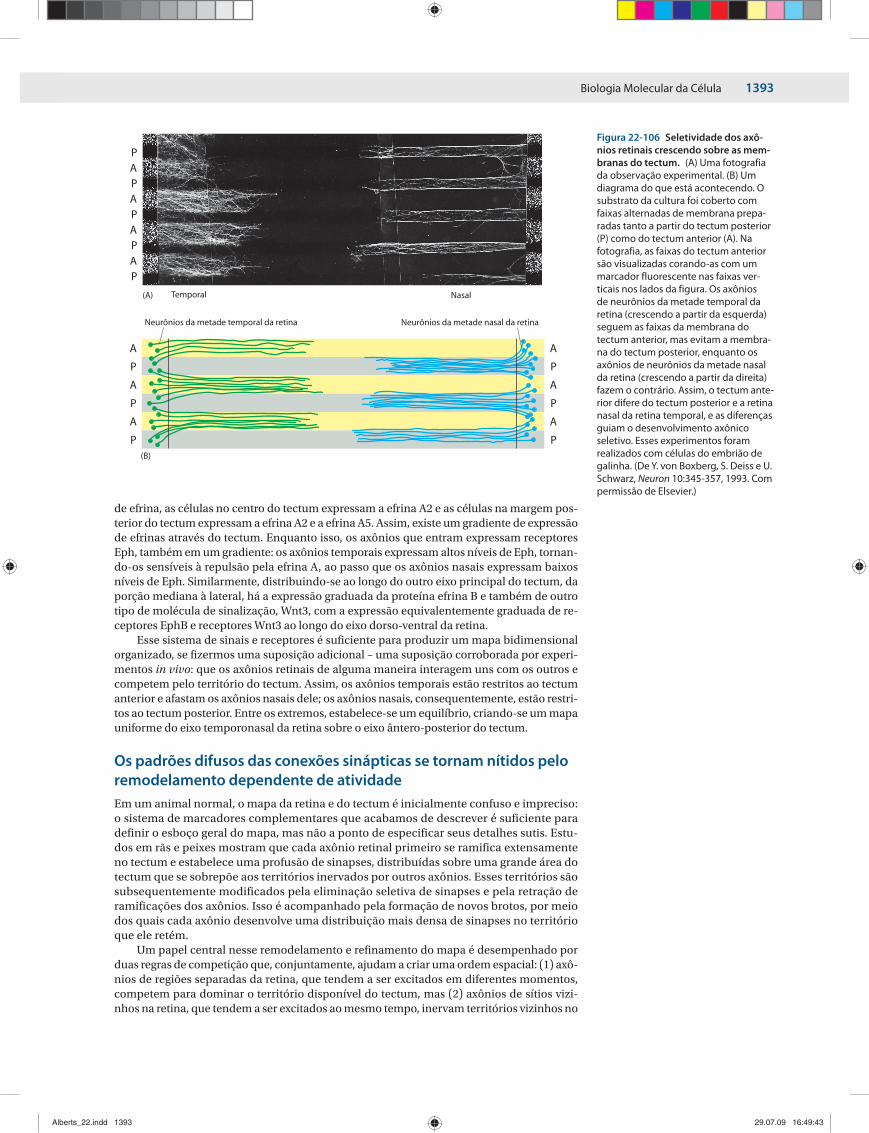
Biologia Molecular da Célula 1393
de efrina, as células no centro do tectum expressam a efrina A2 e as células na margem pos-terior do tectum expressam a efrina A2 e a efrina A5. Assim, existe um gradiente de expressão de efrinas através do tectum. Enquanto isso, os axônios que entram expressam receptores Eph, também em um gradiente: os axônios temporais expressam altos níveis de Eph, tornan-do-os sensíveis à repulsão pela efrina A, ao passo que os axônios nasais expressam baixos níveis de Eph. Similarmente, distribuindo-se ao longo do outro eixo principal do tectum, da porção mediana à lateral, há a expressão graduada da proteína efrina B e também de outro tipo de molécula de sinalização, Wnt3, com a expressão equivalentemente graduada de re-ceptores EphB e receptores Wnt3 ao longo do eixo dorso-ventral da retina.
Esse sistema de sinais e receptores é suficiente para produzir um mapa bidimensional organizado, se fizermos uma suposição adicional – uma suposição corroborada por experi-mentos in vivo: que os axônios retinais de alguma maneira interagem uns com os outros e competem pelo território do tectum. Assim, os axônios temporais estão restritos ao tectum anterior e afastam os axônios nasais dele; os axônios nasais, consequentemente, estão restri-tos ao tectum posterior. Entre os extremos, estabelece-se um equilíbrio, criando-se um mapa uniforme do eixo temporonasal da retina sobre o eixo ântero-posterior do tectum.
Os padrões difusos das conexões sinápticas se tornam nítidos pelo remodelamento dependente de atividadeEm um animal normal, o mapa da retina e do tectum é inicialmente confuso e impreciso: o sistema de marcadores complementares que acabamos de descrever é suficiente para definir o esboço geral do mapa, mas não a ponto de especificar seus detalhes sutis. Estu-dos em rãs e peixes mostram que cada axônio retinal primeiro se ramifica extensamente no tectum e estabelece uma profusão de sinapses, distribuídas sobre uma grande área do tectum que se sobrepõe aos territórios inervados por outros axônios. Esses territórios são subsequentemente modificados pela eliminação seletiva de sinapses e pela retração de ramificações dos axônios. Isso é acompanhado pela formação de novos brotos, por meio dos quais cada axônio desenvolve uma distribuição mais densa de sinapses no território que ele retém.
Um papel central nesse remodelamento e refinamento do mapa é desempenhado por duas regras de competição que, conjuntamente, ajudam a criar uma ordem espacial: (1) axô-nios de regiões separadas da retina, que tendem a ser excitados em diferentes momentos, competem para dominar o território disponível do tectum, mas (2) axônios de sítios vizi-nhos na retina, que tendem a ser excitados ao mesmo tempo, inervam territórios vizinhos no
Figura 22-106 Seletividade dos axô-nios retinais crescendo sobre as mem-branas do tectum. (A) Uma fotografia da observação experimental. (B) Um diagrama do que está acontecendo. O substrato da cultura foi coberto com faixas alternadas de membrana prepa-radas tanto a partir do tectum posterior (P) como do tectum anterior (A). Na fotografia, as faixas do tectum anterior são visualizadas corando-as com um marcador fluorescente nas faixas ver-ticais nos lados da figura. Os axônios de neurônios da metade temporal da retina (crescendo a partir da esquerda) seguem as faixas da membrana do tectum anterior, mas evitam a membra-na do tectum posterior, enquanto os axônios de neurônios da metade nasal da retina (crescendo a partir da direita) fazem o contrário. Assim, o tectum ante-rior difere do tectum posterior e a retina nasal da retina temporal, e as diferenças guiam o desenvolvimento axônico seletivo. Esses experimentos foram realizados com células do embrião de galinha. (De Y. von Boxberg, S. Deiss e U. Schwarz, Neuron 10:345-357, 1993. Com permissão de Elsevier.)
A
P
A
P
A
P
A
P
A
P
A
P
Neurônios da metade nasal da retina
(B)
PAPAPAPAP
(A) Temporal Nasal
Neurônios da metade temporal da retina
Alberts_22.indd 1393Alberts_22.indd 1393 29.07.09 16:49:4329.07.09 16:49:43

1394 Alberts, Johnson, Lewis, Raff, Roberts & Walter
tectum, pois colaboram para reter e fortalecer suas sinapses em células compartilhadas do tectum (Figura 22-107). O mecanismo subjacente a ambas as regras depende da atividade elétrica e da sinalização nas sinapses que são formadas. Se todos os potenciais de ação forem bloqueados por uma toxina que se liga a canais de Na+ controlados por voltagem, o remode-lamento de sinapses é inibido e o mapa permanece confuso.
O fenômeno de eliminação de sinapses dependente de atividade é encontrado em qua-se todas as partes do sistema nervoso em desenvolvimento dos vertebrados. As sinapses são primeiramente formadas em abundância e distribuídas sobre uma ampla área-alvo; em se-guida, o sistema de conexões é cortado e remodelado por processos competitivos que de-pendem da atividade elétrica e da sinalização sináptica. Desta maneira, a eliminação de si-napses é distinta da eliminação de neurônios excedentes por morte celular, ocorrendo após o período de morte neuronal normal ter acabado.
Muito do que sabemos sobre os mecanismos celulares de formação e eliminação de si-napses vem de experimentos sobre a inervação do músculo esquelético em embriões de ver-tebrados. A troca bidirecional de sinais entre os terminais axônicos dos nervos e as células musculares controla a formação inicial das sinapses. Em locais de contato, os receptores de acetilcolina estão agrupados na membrana da célula muscular, e o aparato para a secreção deste neurotransmissor se organiza nos terminais dos axônios (discutido no Capítulo 11). Cada célula muscular primeiramente recebe sinapses de vários neurônios; contudo, no fi-nal, por um processo que tipicamente leva duas semanas, ela fica inervada por apenas um. A retração de sinapses novamente depende da comunicação sináptica: se a transmissão sináp-tica for bloqueada por uma toxina que se liga aos receptores de acetilcolina na membrana da célula muscular, a célula muscular retém sua inervação múltipla além do tempo normal para a eliminação.
Experimentos com o sistema musculoesquelético, assim como o sistema tectum-reti-nal, sugerem que não só a quantidade de atividade elétrica em uma sinapse é importante a sua manutenção, mas também sua coordenação temporal. O fortalecimento ou o enfra-quecimento de uma sinapse parece depender criticamente do fato de a atividade na célula pré-sináptica ser ou não sincronizada com a atividade das outras células pré-sinápticas que estão fazendo sinapses no mesmo alvo (e assim, também, sincronizadas com a atividade das próprias células-alvo).
Essas e muitas outras descobertas sugeriram uma interpretação simples das regras de competição para a eliminação de sinapses no sistema tectum-retinal (Figura 22-108). Os axônios de diferentes partes da retina disparam em momentos diferentes e, assim, compe-tem. A cada vez que um deles dispara, a sinapse (ou as sinapses) feita pelo outro em uma célula-alvo do tectum é enfraquecida, até que um dos axônios é deixado sozinho no coman-do daquela célula. Os axônios de células retinais vizinhas, por outro lado, tendem a disparar
Figura 22-107 Refinamento do mapa tectum-retinal pela eliminação de si-napses. A princípio o mapa é confuso, porque cada axônio retinal se ramifica extensamente para inervar uma região ampla do tectum que se sobrepõe às regiões inervadas por outros axônios retinais. O mapa é então refinado pela eliminação de sinapses. Onde axônios de partes separadas da retina fazem sinapses com a mesma célula do tec-tum, ocorre a competição, eliminando as conexões feitas por um dos axônios. Porém, axônios de células que são vizinhas próximas na retina cooperam, mantendo suas sinapses em células compartilhadas do tectum. Assim, cada axônio retinal acaba inervando um pe-queno território do tectum, adjacente e parcialmente sobreposto ao território inervado por axônios de sítios vizinhos na retina.
Neurôniosretinais
Axôniosretinais
Neurôniosdo tectum
MAPA INICIAL CONFUSO: CONEXÕES DIFUSAS MAPA FINAL NÍTIDO: CONEXÕES DIFUSAS ELIMINADAS
Alberts_22.indd 1394Alberts_22.indd 1394 29.07.09 16:49:4329.07.09 16:49:43
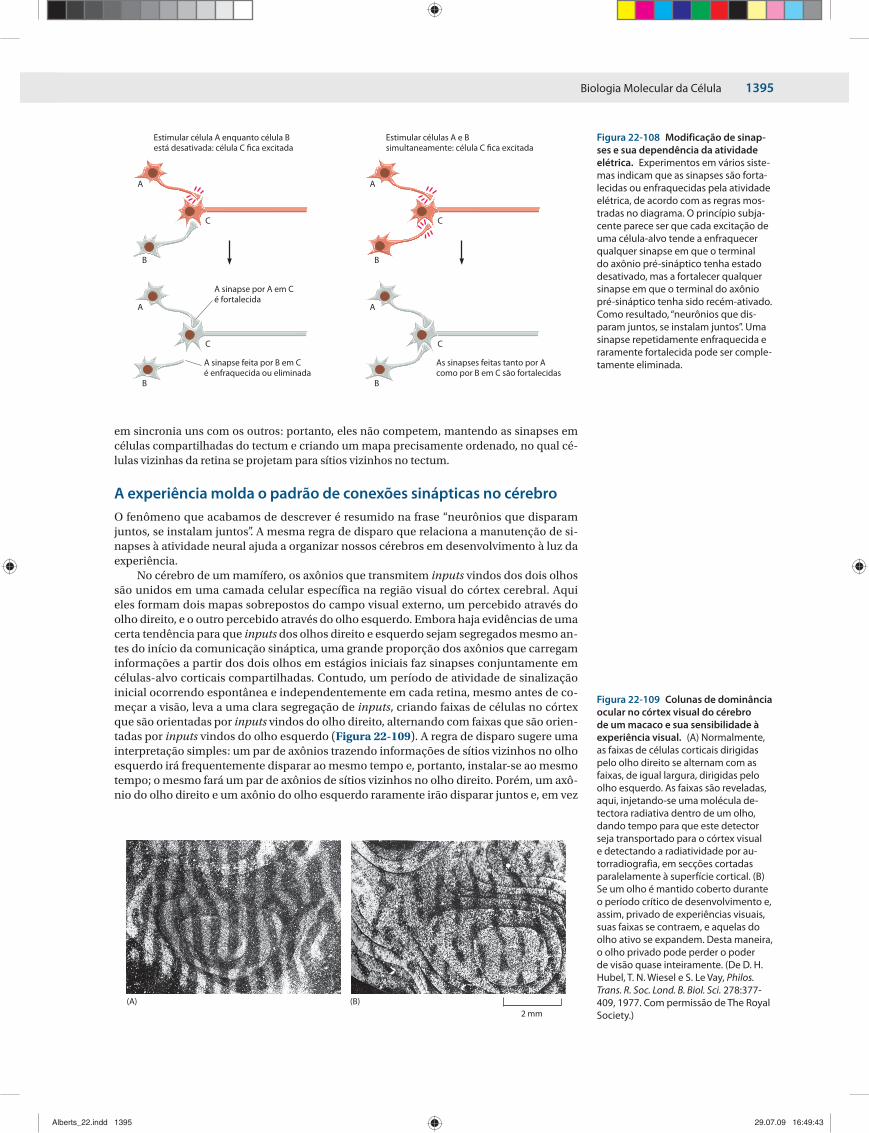
Biologia Molecular da Célula 1395
em sincronia uns com os outros: portanto, eles não competem, mantendo as sinapses em células compartilhadas do tectum e criando um mapa precisamente ordenado, no qual cé-lulas vizinhas da retina se projetam para sítios vizinhos no tectum.
A experiência molda o padrão de conexões sinápticas no cérebroO fenômeno que acabamos de descrever é resumido na frase “neurônios que disparam juntos, se instalam juntos”. A mesma regra de disparo que relaciona a manutenção de si-napses à atividade neural ajuda a organizar nossos cérebros em desenvolvimento à luz da experiência.
No cérebro de um mamífero, os axônios que transmitem inputs vindos dos dois olhos são unidos em uma camada celular específica na região visual do córtex cerebral. Aqui eles formam dois mapas sobrepostos do campo visual externo, um percebido através do olho direito, e o outro percebido através do olho esquerdo. Embora haja evidências de uma certa tendência para que inputs dos olhos direito e esquerdo sejam segregados mesmo an-tes do início da comunicação sináptica, uma grande proporção dos axônios que carregam informações a partir dos dois olhos em estágios iniciais faz sinapses conjuntamente em células-alvo corticais compartilhadas. Contudo, um período de atividade de sinalização inicial ocorrendo espontânea e independentemente em cada retina, mesmo antes de co-meçar a visão, leva a uma clara segregação de inputs, criando faixas de células no córtex que são orientadas por inputs vindos do olho direito, alternando com faixas que são orien-tadas por inputs vindos do olho esquerdo (Figura 22-109). A regra de disparo sugere uma interpretação simples: um par de axônios trazendo informações de sítios vizinhos no olho esquerdo irá frequentemente disparar ao mesmo tempo e, portanto, instalar-se ao mesmo tempo; o mesmo fará um par de axônios de sítios vizinhos no olho direito. Porém, um axô-nio do olho direito e um axônio do olho esquerdo raramente irão disparar juntos e, em vez
Figura 22-108 Modificação de sinap-ses e sua dependência da atividade elétrica. Experimentos em vários siste-mas indicam que as sinapses são forta-lecidas ou enfraquecidas pela atividade elétrica, de acordo com as regras mos-tradas no diagrama. O princípio subja-cente parece ser que cada excitação de uma célula-alvo tende a enfraquecer qualquer sinapse em que o terminal do axônio pré-sináptico tenha estado desativado, mas a fortalecer qualquer sinapse em que o terminal do axônio pré-sináptico tenha sido recém-ativado. Como resultado, “neurônios que dis-param juntos, se instalam juntos”. Uma sinapse repetidamente enfraquecida e raramente fortalecida pode ser comple-tamente eliminada.
A
B
C
A
B
C
Estimular célula A enquanto célula Bestá desativada: célula C fica excitada
A sinapse por A em Cé fortalecida
A sinapse feita por B em Cé enfraquecida ou eliminada
A
B
C
A
B
C
Estimular células A e Bsimultaneamente: célula C fica excitada
As sinapses feitas tanto por Acomo por B em C são fortalecidas
(A) (B)2 mm
Figura 22-109 Colunas de dominância ocular no córtex visual do cérebro de um macaco e sua sensibilidade à experiência visual. (A) Normalmente, as faixas de células corticais dirigidas pelo olho direito se alternam com as faixas, de igual largura, dirigidas pelo olho esquerdo. As faixas são reveladas, aqui, injetando-se uma molécula de-tectora radiativa dentro de um olho, dando tempo para que este detector seja transportado para o córtex visual e detectando a radiatividade por au-torradiografia, em secções cortadas paralelamente à superfície cortical. (B) Se um olho é mantido coberto durante o período crítico de desenvolvimento e, assim, privado de experiências visuais, suas faixas se contraem, e aquelas do olho ativo se expandem. Desta maneira, o olho privado pode perder o poder de visão quase inteiramente. (De D. H. Hubel, T. N. Wiesel e S. Le Vay, Philos. Trans. R. Soc. Lond. B. Biol. Sci. 278:377-409, 1977. Com permissão de The Royal Society.)
Alberts_22.indd 1395Alberts_22.indd 1395 29.07.09 16:49:4329.07.09 16:49:43

1396 Alberts, Johnson, Lewis, Raff, Roberts & Walter
disso, competirão. Na verdade, se a atividade dos dois olhos for silenciada utilizando-se fármacos que bloqueiam potenciais de ação ou a transmissão sináptica, os inputs não se segregam corretamente.
A manutenção do padrão de conexões é extraordinariamente sensível às experiências que ocorrem cedo na vida. Se, durante um certo período crítico (que termina aproximada-mente aos cinco anos de idade em humanos), um olho é mantido coberto por um tempo, de forma a ficar privado de estimulação visual, enquanto o outro olho é estimulado nor-malmente, o olho privado perde suas conexões sinápticas para o córtex e fica quase inteira e irreversivelmente cego. De acordo com o que preveria a regra de disparo, ocorreu uma competição na qual as sinapses no córtex visual feitas por axônios inativos foram elimina-das, enquanto as sinapses feitas por axônios ativos foram consolidadas. Desta maneira, o território cortical é alocado para axônios que carregam informações e não é desperdiçado com aqueles que são silenciosos.
No estabelecimento das conexões nervosas que nos permitem ver, não só a quantidade de estimulação visual é importante, mas também sua coordenação temporal. Por exemplo, a capacidade de ver a profundidade – a visão estérea – depende de células em outras cama-das do córtex visual que recebem inputs transmitidos a partir de ambos os olhos de uma só vez, transportando informações sobre a mesma parte do campo visual como visto de dois ângulos levemente diferentes. Essas células dirigidas de maneira binocular nos permitem comparar o que é visto pelo olho direito com o que é visto pelo olho esquerdo, de forma a deduzir informações sobre as distâncias relativas dos objetos em relação a nós. Contudo, se os dois olhos forem impedidos durante o período crítico de ver a mesma cena ao mesmo tempo – por exemplo, cobrindo primeiro um olho e depois o outro em dias alternados, ou simplesmente como consequência de um estrabismo infantil – quase nenhuma das células dirigidas de maneira binocular é retida no córtex, e a capacidade de percepção estérea é ine-vitavelmente perdida. Evidentemente, de acordo com a regra de disparo, os inputs a partir de cada olho para um neurônio dirigido de maneira binocular são mantidos somente se os dois inputs são frequentemente acionados para disparar em sincronia, como ocorre quando os dois olhos olham juntos para a mesma cena.
A memória adulta e a remodelação das sinapses durante o desenvolvimento podem depender de mecanismos similaresVimos no Capítulo 11 que as alterações sinápticas subjacentes à memória, pelo menos em algumas partes do cérebro adulto, notavelmente o hipocampo, dependem do com-portamento de um tipo particular de receptor para o neurotransmissor glutamato – o re-ceptor NMDA. A inundação de Ca2+ na célula pós-sináptica através de canais abertos por este receptor aciona mudanças duradouras na intensidade das sinapses naquela célula, afetando tanto as estruturas pré-sinápticas como as pós-sinápticas. As alterações que são induzidas pelo mecanismo dependente de NMDA no cérebro adulto obedecem a regras semelhantes à regra de disparo no desenvolvimento: os eventos no mundo exterior que levam dois neurônios a serem ativos ao mesmo tempo, ou em rápida sucessão, favorecem a formação ou o fortalecimento de sinapses entre eles. Tem-se sugerido que esta condi-ção, denominada regra de Hebb, seja o princípio fundamental subjacente ao aprendizado associativo.
É possível, então, que tanto o aprendizado adulto como as formas mais extremas de plasticidade sináptica vistas durante o desenvolvimento dependam da mesma maquina-ria básica de ajuste sináptico? Existem muitos indícios que apontam para tal. Observou-se, por exemplo, que inibidores que bloqueiam especificamente a ativação do receptor NMDA interferem no refinamento e na remodelação de conexões sinápticas no sistema visual em desenvolvimento. Tanto no animal em desenvolvimento como no adulto, as alterações na força das conexões sinápticas correspondem a mudanças na estrutura física. A escala dessas mudanças físicas é, contudo, muito diferente. No organismo em desenvolvimento, a ativi-dade elétrica frequentemente regula a extensão e a regressão de grandes ramificações das árvores axônicas e dendríticas. Porém, no cérebro adulto, os ajustes estruturais que ocorrem em resposta à atividade parecem ser tipicamente muito mais localizados de forma precisa, afetando os tamanhos de espinhos dendríticos individuais – as minúsculas protrusões em forma de maçaneta, com alguns poucos micrômetros de comprimento, nas quais os den-dritos recebem sinapses individuais (Figura 22-110). Parece que o Ca2+ que entra em um espinho por meio dos canais de NMDA, em resposta à excitação da sinapse naquele espinho
Alberts_22.indd 1396Alberts_22.indd 1396 29.07.09 16:49:4429.07.09 16:49:44
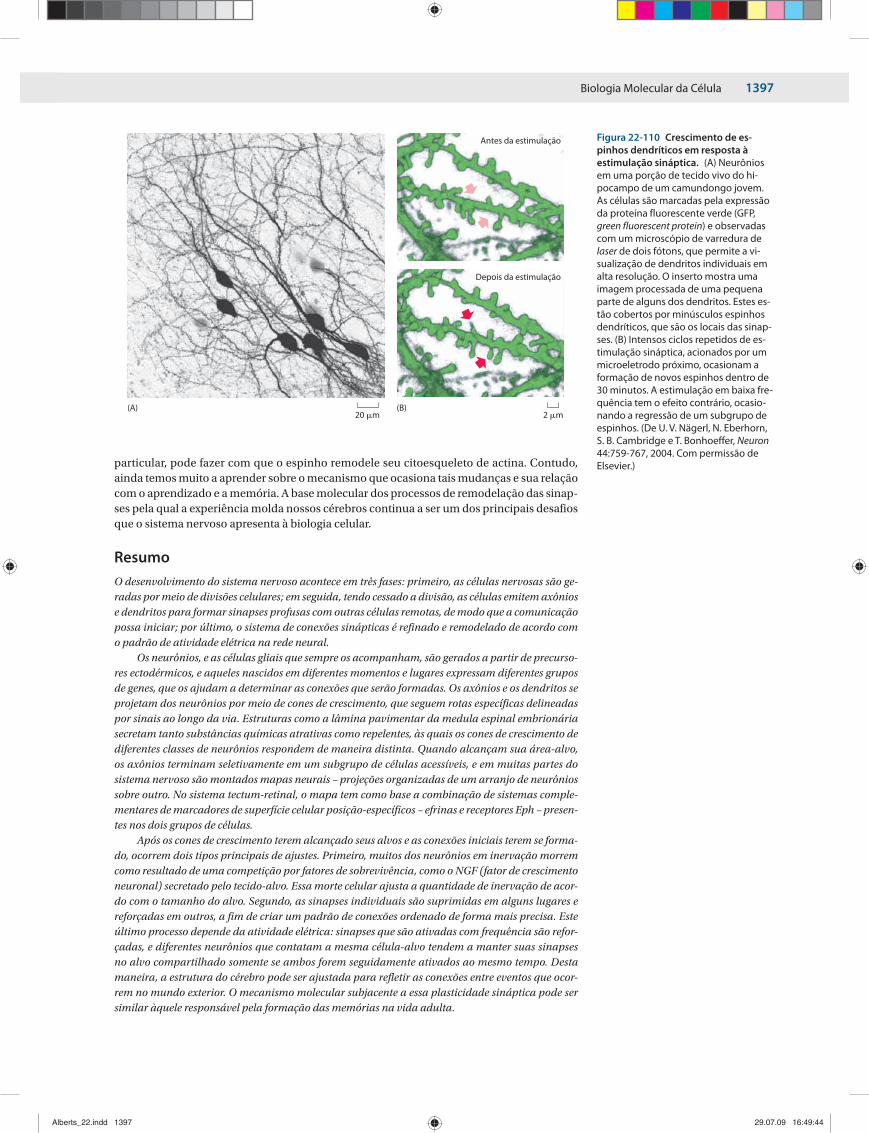
Biologia Molecular da Célula 1397
particular, pode fazer com que o espinho remodele seu citoesqueleto de actina. Contudo, ainda temos muito a aprender sobre o mecanismo que ocasiona tais mudanças e sua relação com o aprendizado e a memória. A base molecular dos processos de remodelação das sinap-ses pela qual a experiência molda nossos cérebros continua a ser um dos principais desafios que o sistema nervoso apresenta à biologia celular.
Resumo
O desenvolvimento do sistema nervoso acontece em três fases: primeiro, as células nervosas são ge-radas por meio de divisões celulares; em seguida, tendo cessado a divisão, as células emitem axônios e dendritos para formar sinapses profusas com outras células remotas, de modo que a comunicação possa iniciar; por último, o sistema de conexões sinápticas é refinado e remodelado de acordo com o padrão de atividade elétrica na rede neural.
Os neurônios, e as células gliais que sempre os acompanham, são gerados a partir de precurso-res ectodérmicos, e aqueles nascidos em diferentes momentos e lugares expressam diferentes grupos de genes, que os ajudam a determinar as conexões que serão formadas. Os axônios e os dendritos se projetam dos neurônios por meio de cones de crescimento, que seguem rotas específicas delineadas por sinais ao longo da via. Estruturas como a lâmina pavimentar da medula espinal embrionária secretam tanto substâncias químicas atrativas como repelentes, às quais os cones de crescimento de diferentes classes de neurônios respondem de maneira distinta. Quando alcançam sua área-alvo, os axônios terminam seletivamente em um subgrupo de células acessíveis, e em muitas partes do sistema nervoso são montados mapas neurais – projeções organizadas de um arranjo de neurônios sobre outro. No sistema tectum-retinal, o mapa tem como base a combinação de sistemas comple-mentares de marcadores de superfície celular posição-específicos – efrinas e receptores Eph – presen-tes nos dois grupos de células.
Após os cones de crescimento terem alcançado seus alvos e as conexões iniciais terem se forma-do, ocorrem dois tipos principais de ajustes. Primeiro, muitos dos neurônios em inervação morrem como resultado de uma competição por fatores de sobrevivência, como o NGF (fator de crescimento neuronal) secretado pelo tecido-alvo. Essa morte celular ajusta a quantidade de inervação de acor-do com o tamanho do alvo. Segundo, as sinapses individuais são suprimidas em alguns lugares e reforçadas em outros, a fim de criar um padrão de conexões ordenado de forma mais precisa. Este último processo depende da atividade elétrica: sinapses que são ativadas com frequência são refor-çadas, e diferentes neurônios que contatam a mesma célula-alvo tendem a manter suas sinapses no alvo compartilhado somente se ambos forem seguidamente ativados ao mesmo tempo. Desta maneira, a estrutura do cérebro pode ser ajustada para refletir as conexões entre eventos que ocor-rem no mundo exterior. O mecanismo molecular subjacente a essa plasticidade sináptica pode ser similar àquele responsável pela formação das memórias na vida adulta.
Figura 22-110 Crescimento de es-pinhos dendríticos em resposta à estimulação sináptica. (A) Neurônios em uma porção de tecido vivo do hi-pocampo de um camundongo jovem. As células são marcadas pela expressão da proteína fluorescente verde (GFP, green fluorescent protein) e observadas com um microscópio de varredura de laser de dois fótons, que permite a vi-sualização de dendritos individuais em alta resolução. O inserto mostra uma imagem processada de uma pequena parte de alguns dos dendritos. Estes es-tão cobertos por minúsculos espinhos dendríticos, que são os locais das sinap-ses. (B) Intensos ciclos repetidos de es-timulação sináptica, acionados por um microeletrodo próximo, ocasionam a formação de novos espinhos dentro de 30 minutos. A estimulação em baixa fre-quência tem o efeito contrário, ocasio-nando a regressão de um subgrupo de espinhos. (De U. V. Nägerl, N. Eberhorn, S. B. Cambridge e T. Bonhoeffer, Neuron 44:759-767, 2004. Com permissão de Elsevier.)
(A) (B)
Antes da estimulação
Depois da estimulação
20 �m 2 �m
Alberts_22.indd 1397Alberts_22.indd 1397 29.07.09 16:49:4429.07.09 16:49:44

1398 Alberts, Johnson, Lewis, Raff, Roberts & Walter
DESENVOLVIMENTO VEGETALAs plantas e os animais estão separados por cerca de 1,5 bilhão de anos de história evoluti-va. Sua organização multicelular evoluiu independentemente, mas utilizando o mesmo jogo inicial de ferramentas – o conjunto de genes herdado de seu ancestral eucariótico unicelu-lar comum. A maioria das diferenças entre suas estratégias de desenvolvimento provém de duas peculiaridades básicas das plantas. Primeiro, elas obtêm sua energia da luz solar, e não pela ingestão de outros organismos. Isso impõe um plano corporal diferente daquele dos animais. Segundo, suas células são envolvidas por paredes celulares semirrígidas e cimen-tadas umas às outras, impedindo-as de se mover como as células animais. Isso impõe um conjunto diferente de mecanismos para a modelagem do corpo e diferentes processos de desenvolvimento para enfrentar um ambiente variável.
O desenvolvimento animal é em grande parte protegido contra mudanças ambientais, e o embrião gera a mesma estrutura corporal, geneticamente determinada, sem ser afetado por condições externas. O desenvolvimento da maioria dos vegetais, por outro lado, é dra-maticamente influenciado pelo ambiente. Como não podem se equiparar a seu ambiente, movendo-se de um lugar para o outro, os vegetais se adaptam, em vez de alterar o curso de seu desenvolvimento. Sua estratégia é oportunista. Um dado tipo de órgão – uma folha, uma flor ou uma raiz, por exemplo – pode ser produzido, a partir de um óvulo fertilizado, por meio de muitos caminhos diferentes, de acordo com sinais do ambiente. De uma folha de begônia fixada ao solo pode crescer uma raiz; a raiz pode formar um broto; o broto, sob luz solar, pode desenvolver folhas e flores.
A planta madura consiste, tipicamente, em muitas cópias de um pequeno conjunto de módulos padronizados, como descrito na Figura 22-111. As posições e os momentos em que cada módulo é gerado são bastante influenciados pelo ambiente, determinando uma ampla variação na estrutura global da planta. As escolhas entre módulos alternativos e sua organi-zação em uma planta inteira dependem de estímulos externos e sinais hormonais de longo alcance que desempenham um papel muito menor no controle do desenvolvimento animal.
Contudo, embora a estrutura global de uma planta – seu padrão de raízes ou ramos, seu número de folhas ou flores – possa ser muito variável, sua organização detalhada em peque-na escala não o é. Uma folha, uma flor ou até mesmo um embrião jovem de planta são espe-cificados de forma tão precisa como qualquer órgão de um animal, possuindo uma estrutura determinada, em contraste com o padrão indeterminado de ramificação e brotamento da planta como um todo. A organização interna de um módulo vegetal gera essencialmente os mesmos problemas relacionados ao controle genético de formação de padrões observados no desenvolvimento animal, e eles são resolvidos de maneiras análogas. Nesta seção, dare-mos enfoque aos mecanismos celulares do desenvolvimento de angiospermas (plantas que florescem). Examinaremos tanto as diferenças como as similaridades com os animais.
A Arabidopsis serve de organismo-modelo para a genética molecular de plantasAs angiospermas, apesar de sua imensa variedade, têm uma origem relativamente recente. Os mais antigos exemplares fósseis conhecidos têm 130 milhões de anos de idade, em com-paração aos 350 milhões ou mais de animais vertebrados. Portanto, subjacente à diversidade de formas, existe um alto grau de similaridade quanto aos mecanismos moleculares. Como veremos, uma pequena mudança genética pode transformar a estrutura de uma planta em grande escala; e, assim como a fisiologia das plantas permite sua sobrevivência em muitos ambientes diferentes, ela também permite a sobrevivência de muitas formas diferencial-mente estruturadas. Uma mutação que origina um animal com duas cabeças geralmente é letal; a mutação que dobra o número de flores ou ramos em uma planta geralmente não é.
Para identificar os genes que controlam o desenvolvimento vegetal e descobrir seu fun-cionamento, os biólogos vegetais selecionaram uma pequena erva daninha, o agrião (ou ára-bis) de parede comum Arabidopsis thaliana (Figura 22-112) como seu organismo-modelo
Meristema apical
Gema axilar
FolhaNó
Nó
Nó
Nó
Entrenó(caule)
Figura 22-111 Um exemplo simples da construção modular nas plantas. Cada módulo (mostrado em diferentes tons de verde) consiste em um caule, uma folha e uma gema contendo um centro de crescimento potencial ou meristema. A gema se forma nos pontos de ramifi-cação ou nós (ou, ainda, nodos), onde a folha se separa do caule. Os módulos se originam sequencialmente a partir da atividade contínua do meristema apical.
Figura 22-112 Arabidopsis thaliana. Esta pequena planta é um membro da família da mostarda (ou das crucíferas, ver também Figura 1-46). É uma erva daninha sem utilidade econômica, mas de grande valor para estudos gené-ticos do desenvolvimento vegetal. (De M. A. Estelle e C. R. Somerville, Trends Genet. 12:89-93, 1986. Com permissão de Elsevier.)15 mm
Alberts_22.indd 1398Alberts_22.indd 1398 29.07.09 16:49:4429.07.09 16:49:44

Biologia Molecular da Célula 1399
principal. Assim como a Drosophila ou o Caenorhabditis elegans, ela é pequena, de rápida reprodução e conveniente para a genética. Pode ser crescida dentro de ambientes fechados (como placas de Petri ou minúsculos potes de planta) em grande quantidade, produzindo centenas de sementes por planta após 8 a 10 semanas. Ela tem, em comum com o C. elegans, uma vantagem significativa sobre a Drosophila ou os animais vertebrados para estudos ge-néticos: como muitas angiospermas, ela pode se reproduzir como hermafrodita, pois uma única flor produz tanto óvulos como gametas masculinos que podem fertilizá-la. Portanto, quando uma flor heterozigota para uma mutação letal recessiva é autofertilizada, um quarto de suas sementes irá apresentar o fenótipo embrionário homozigoto. Isso facilita a realiza-ção de triagens genéticas (Figura 22-113) e também a obtenção de um catálogo dos genes necessários a processos específicos do desenvolvimento.
O genoma de Arabidopsis é rico em genes controladores do desenvolvimentoA Arabidopsis tem um dos menores genomas de plantas – 125 milhões de pares de nucleotí-deos, estando no mesmo nível de C. elegans e Drosophila – e a sequência completa de DNA é agora conhecida. Ela contém aproximadamente 26.000 genes. Contudo, este total inclui muitas duplicatas recentemente geradas, de modo que o número de tipos funcionalmente distintos de proteínas representadas pode ser consideravelmente menor. Foram estabeleci-dos métodos de cultura de células e de transformação genética, assim como imensas biblio-tecas de sementes carregando mutações produzidas por inserções aleatórias de elementos genéticos móveis, de forma que plantas com mutações em qualquer gene selecionado po-dem ser obtidas sob medida. Assim, existem ferramentas potentes para analisar as funções de muitos genes. Embora apenas uma pequena fração do conjunto total de genes tenha sido experimentalmente caracterizada até agora, poder-se tentar atribuir funções a vários genes – cerca de 18.000 – com base nas similaridades de sequência com genes bem caracterizados em Arabidopsis e outros organismos.
O genoma da Arabidopsis é ainda mais rico em genes que codificam proteínas de re-gulação gênica que os genomas de muitos animais multicelulares (Tabela 22-2). Algumas importantes famílias de proteínas de regulação gênica de animais (como a família Myb de proteínas que se ligam ao DNA) são bastante difundidas, enquanto outras (como os recepto-res de hormônios nucleares) parecem estar completamente ausentes, e existem grandes fa-mílias de proteínas de regulação gênica nos vegetais que não têm homólogos nos animais.
Embora proteínas de regulação gênica homólogas (como as proteínas homeodomínio) possam ser reconhecidas tanto nos vegetais quanto nos animais, elas têm pouco em comum no que se refere aos genes que regulam ou aos tipos de decisões relacionadas ao desenvol-vimento que controlam, e há pouca conservação das sequências de proteína fora dos domí-nios de ligação ao DNA.
Figura 22-113 Produção de mutan-tes em Arabidopsis. Uma semente, contendo um embrião multicelular, é tratada com uma substância química mutagênica e cultivada até o estágio de planta adulta. Em geral, esta planta será um mosaico de clones de células carre-gando diferentes mutações induzidas. Uma flor individual produzida por esta planta normalmente será composta por células pertencentes ao mesmo clone, todas carregando a mesma mutação, m, na forma heterozigota (m/+). A au-tofertilização de flores individuais por seu próprio pólen resulta em vagens de sementes, cada qual contendo uma família de embriões em que metade dos membros será, em média, hetero-zigota (m/+), um quarto será mutante homozigoto (m/m) e um quarto será o tipo selvagem homozigoto (+/+). Fre-quentemente, a mutação terá um efeito letal recessivo, como indicado aqui pela ausência de uma raiz na plântula m/m. O estoque mutante é então mantido pelo cruzamento dos heterozigotos, que irão produzir vagens de sementes (geração F2) que contêm uma mistura de sementes +/+, m/+ e m/m.
PlântulaSementemutagenizada
Setor de célulasmutantes nomeristema
Setor mutante Setor não-mutanteVagens de sementes do:
As sementes se desenvolvem em plântulas da geração F1
100% +/+25% +/+50% m/+25% m/m
A autofertilização deflores individuaisproduz uma geraçãode sementes F
1
Alberts_22.indd 1399Alberts_22.indd 1399 29.07.09 16:49:4529.07.09 16:49:45
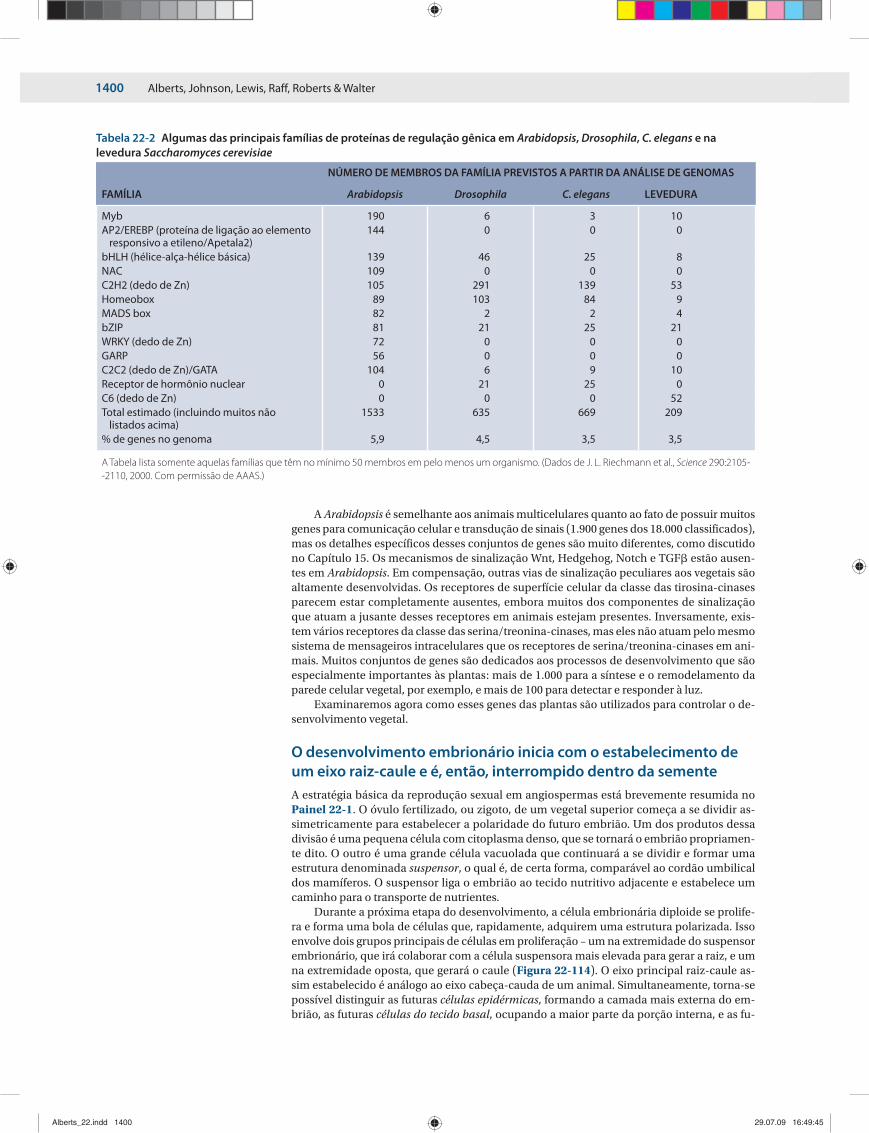
1400 Alberts, Johnson, Lewis, Raff, Roberts & Walter
A Arabidopsis é semelhante aos animais multicelulares quanto ao fato de possuir muitos genes para comunicação celular e transdução de sinais (1.900 genes dos 18.000 classificados), mas os detalhes específicos desses conjuntos de genes são muito diferentes, como discutido no Capítulo 15. Os mecanismos de sinalização Wnt, Hedgehog, Notch e TGF� estão ausen-tes em Arabidopsis. Em compensação, outras vias de sinalização peculiares aos vegetais são altamente desenvolvidas. Os receptores de superfície celular da classe das tirosina-cinases parecem estar completamente ausentes, embora muitos dos componentes de sinalização que atuam a jusante desses receptores em animais estejam presentes. Inversamente, exis-tem vários receptores da classe das serina/treonina-cinases, mas eles não atuam pelo mesmo sistema de mensageiros intracelulares que os receptores de serina/treonina-cinases em ani-mais. Muitos conjuntos de genes são dedicados aos processos de desenvolvimento que são especialmente importantes às plantas: mais de 1.000 para a síntese e o remodelamento da parede celular vegetal, por exemplo, e mais de 100 para detectar e responder à luz.
Examinaremos agora como esses genes das plantas são utilizados para controlar o de-senvolvimento vegetal.
O desenvolvimento embrionário inicia com o estabelecimento de um eixo raiz-caule e é, então, interrompido dentro da sementeA estratégia básica da reprodução sexual em angiospermas está brevemente resumida no Painel 22-1. O óvulo fertilizado, ou zigoto, de um vegetal superior começa a se dividir as-simetricamente para estabelecer a polaridade do futuro embrião. Um dos produtos dessa divisão é uma pequena célula com citoplasma denso, que se tornará o embrião propriamen-te dito. O outro é uma grande célula vacuolada que continuará a se dividir e formar uma estrutura denominada suspensor, o qual é, de certa forma, comparável ao cordão umbilical dos mamíferos. O suspensor liga o embrião ao tecido nutritivo adjacente e estabelece um caminho para o transporte de nutrientes.
Durante a próxima etapa do desenvolvimento, a célula embrionária diploide se prolife-ra e forma uma bola de células que, rapidamente, adquirem uma estrutura polarizada. Isso envolve dois grupos principais de células em proliferação – um na extremidade do suspensor embrionário, que irá colaborar com a célula suspensora mais elevada para gerar a raiz, e um na extremidade oposta, que gerará o caule (Figura 22-114). O eixo principal raiz-caule as-sim estabelecido é análogo ao eixo cabeça-cauda de um animal. Simultaneamente, torna-se possível distinguir as futuras células epidérmicas, formando a camada mais externa do em-brião, as futuras células do tecido basal, ocupando a maior parte da porção interna, e as fu-
Tabela 22-2 Algumas das principais famílias de proteínas de regulação gênica em Arabidopsis, Drosophila, C. elegans e na levedura Saccharomyces cerevisiae
NÚMERO DE MEMBROS DA FAMÍLIA PREVISTOS A PARTIR DA ANÁLISE DE GENOMAS
FAMÍLIA Arabidopsis Drosophila C. elegans LEVEDURA
Myb 190 6 3 10AP2/EREBP (proteína de ligação ao elemento
responsivo a etileno/Apetala2)144 0 0 0
bHLH (hélice-alça-hélice básica) 139 46 25 8NAC 109 0 0 0C2H2 (dedo de Zn) 105 291 139 53Homeobox 89 103 84 9MADS box 82 2 2 4bZIP 81 21 25 21WRKY (dedo de Zn) 72 0 0 0GARP 56 0 0 0C2C2 (dedo de Zn)/GATA 104 6 9 10Receptor de hormônio nuclear 0 21 25 0C6 (dedo de Zn) 0 0 0 52Total estimado (incluindo muitos não
listados acima)1533 635 669 209
% de genes no genoma 5,9 4,5 3,5 3,5
A Tabela lista somente aquelas famílias que têm no mínimo 50 membros em pelo menos um organismo. (Dados de J. L. Riechmann et al., Science 290:2105--2110, 2000. Com permissão de AAAS.)
Alberts_22.indd 1400Alberts_22.indd 1400 29.07.09 16:49:4529.07.09 16:49:45
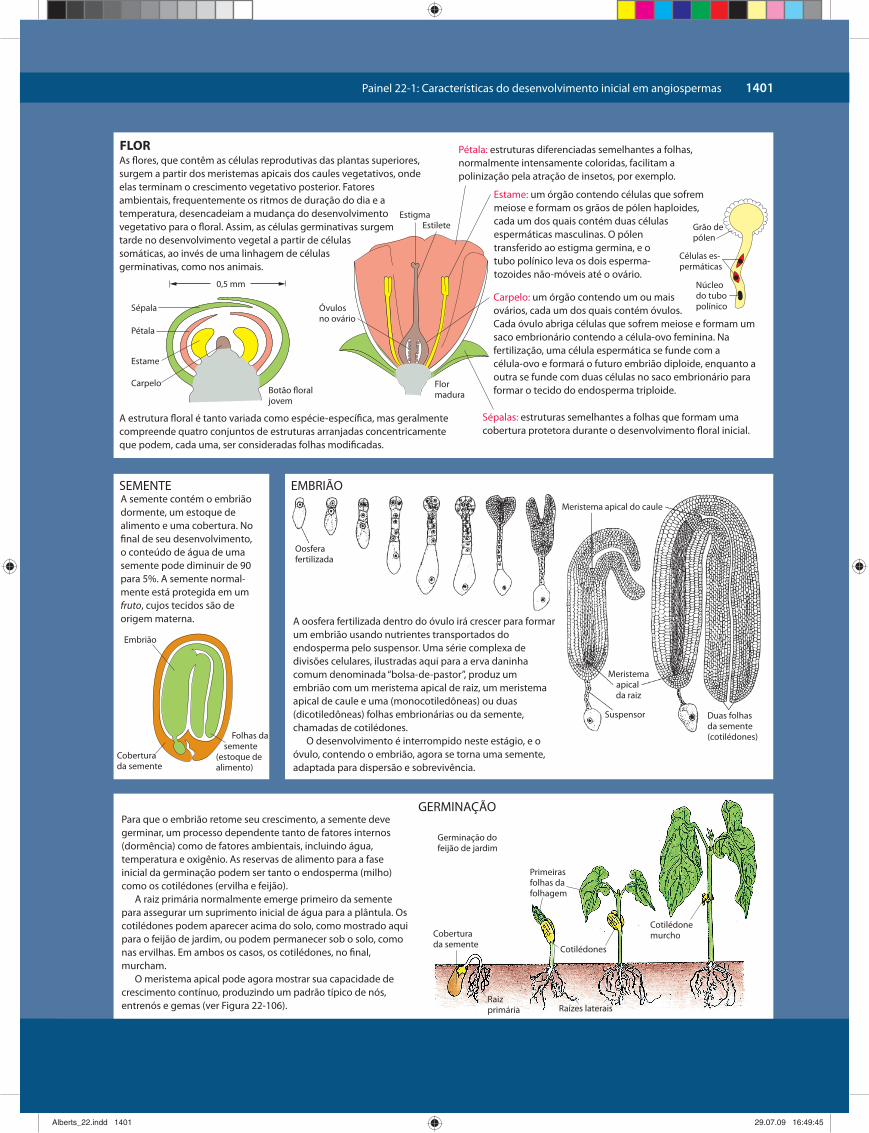
FLOR
A estrutura floral é tanto variada como espécie-específica, mas geralmente compreende quatro conjuntos de estruturas arranjadas concentricamente que podem, cada uma, ser consideradas folhas modificadas.
Pétala: estruturas diferenciadas semelhantes a folhas, normalmente intensamente coloridas, facilitam a polinização pela atração de insetos, por exemplo.
Sépalas: estruturas semelhantes a folhas que formam umacobertura protetora durante o desenvolvimento floral inicial.
0,5 mm
Sépala
Pétala
Estame
CarpeloBotão floraljovem
Óvulosno ovário
EstigmaEstilete
Flormadura
Grão depólen
Células es-permáticas
Núcleodo tubopolínico
SEMENTEA semente contém o embriãodormente, um estoque de alimento e uma cobertura. No final de seu desenvolvimento, o conteúdo de água de uma semente pode diminuir de 90 para 5%. A semente normal-mente está protegida em um fruto, cujos tecidos são de origem materna.
Embrião
Coberturada semente
Folhas da semente(estoque dealimento)
GERMINAÇÃO
Germinação dofeijão de jardim
Primeirasfolhas dafolhagem
Coberturada semente Cotilédones
Raizprimária Raízes laterais
Cotilédonemurcho
As flores, que contêm as células reprodutivas das plantas superiores, surgem a partir dos meristemas apicais dos caules vegetativos, onde elas terminam o crescimento vegetativo posterior. Fatores ambientais, frequentemente os ritmos de duração do dia e a temperatura, desencadeiam a mudança do desenvolvimento vegetativo para o floral. Assim, as células germinativas surgem tarde no desenvolvimento vegetal a partir de célulassomáticas, ao invés de uma linhagem de células germinativas, como nos animais.
A oosfera fertilizada dentro do óvulo irá crescer para formar um embrião usando nutrientes transportados do endosperma pelo suspensor. Uma série complexa de divisões celulares, ilustradas aqui para a erva daninha comum denominada “bolsa-de-pastor”, produz um embrião com um meristema apical de raiz, um meristema apical de caule e uma (monocotiledôneas) ou duas (dicotiledôneas) folhas embrionárias ou da semente, chamadas de cotilédones.
O desenvolvimento é interrompido neste estágio, e o óvulo, contendo o embrião, agora se torna uma semente, adaptada para dispersão e sobrevivência.
Para que o embrião retome seu crescimento, a semente deve germinar, um processo dependente tanto de fatores internos (dormência) como de fatores ambientais, incluindo água, temperatura e oxigênio. As reservas de alimento para a fase inicial da germinação podem ser tanto o endosperma (milho) como os cotilédones (ervilha e feijão).
A raiz primária normalmente emerge primeiro da semente para assegurar um suprimento inicial de água para a plântula. Os cotilédones podem aparecer acima do solo, como mostrado aqui para o feijão de jardim, ou podem permanecer sob o solo, como nas ervilhas. Em ambos os casos, os cotilédones, no final, murcham.
O meristema apical pode agora mostrar sua capacidade de crescimento contínuo, produzindo um padrão típico de nós, entrenós e gemas (ver Figura 22-106).
Estame: um órgão contendo células que sofrem meiose e formam os grãos de pólen haploides, cada um dos quais contém duas células espermáticas masculinas. O pólen transferido ao estigma germina, e o tubo polínico leva os dois esperma-tozoides não-móveis até o ovário.
Carpelo: um órgão contendo um ou mais ovários, cada um dos quais contém óvulos. Cada óvulo abriga células que sofrem meiose e formam um saco embrionário contendo a célula-ovo feminina. Na fertilização, uma célula espermática se funde com a célula-ovo e formará o futuro embrião diploide, enquanto a outra se funde com duas células no saco embrionário para formar o tecido do endosperma triploide.
EMBRIÃO
Oosfera fertilizada
Meristemaapical da raiz
Suspensor Duas folhas da semente(cotilédones)
Meristema apical do caule
Painel 22-1: Características do desenvolvimento inicial em angiospermas 1401
Alberts_22.indd 1401Alberts_22.indd 1401 29.07.09 16:49:4529.07.09 16:49:45
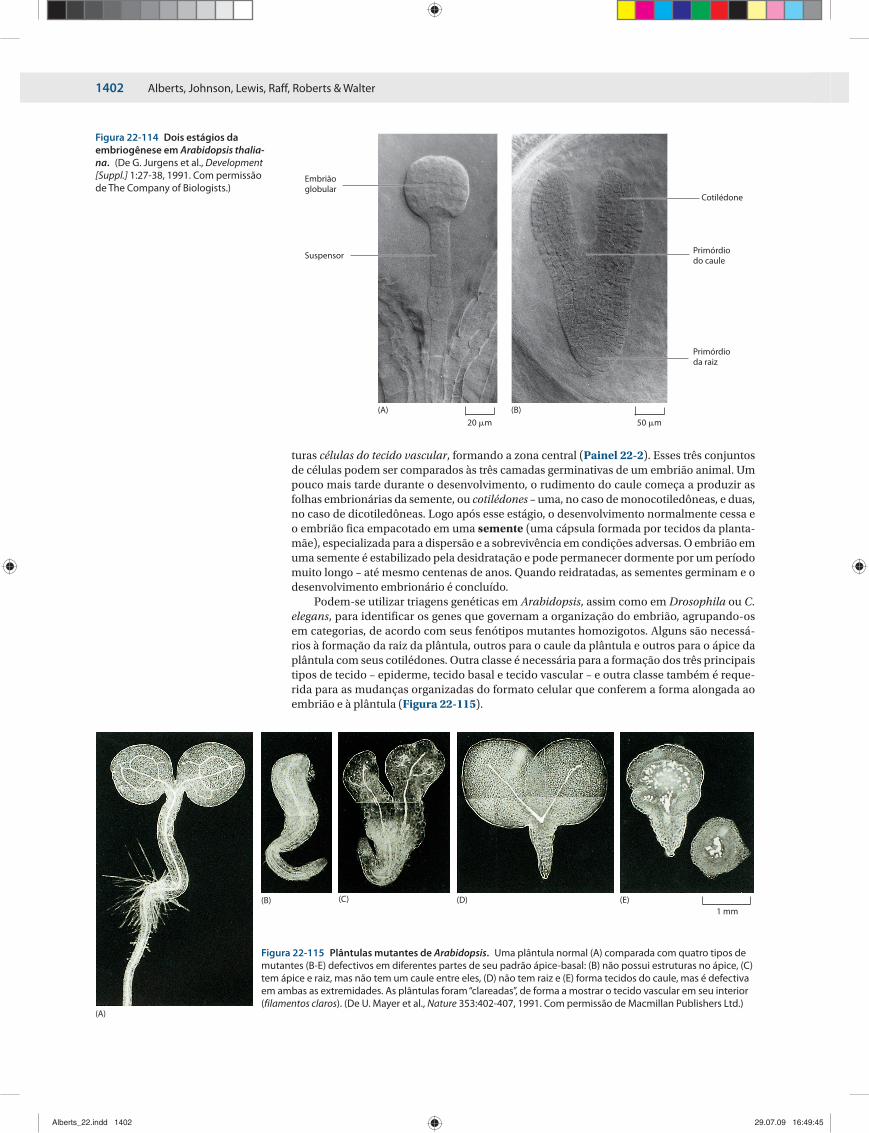
1402 Alberts, Johnson, Lewis, Raff, Roberts & Walter
turas células do tecido vascular, formando a zona central (Painel 22-2). Esses três conjuntos de células podem ser comparados às três camadas germinativas de um embrião animal. Um pouco mais tarde durante o desenvolvimento, o rudimento do caule começa a produzir as folhas embrionárias da semente, ou cotilédones – uma, no caso de monocotiledôneas, e duas, no caso de dicotiledôneas. Logo após esse estágio, o desenvolvimento normalmente cessa e o embrião fica empacotado em uma semente (uma cápsula formada por tecidos da planta-mãe), especializada para a dispersão e a sobrevivência em condições adversas. O embrião em uma semente é estabilizado pela desidratação e pode permanecer dormente por um período muito longo – até mesmo centenas de anos. Quando reidratadas, as sementes germinam e o desenvolvimento embrionário é concluído.
Podem-se utilizar triagens genéticas em Arabidopsis, assim como em Drosophila ou C. elegans, para identificar os genes que governam a organização do embrião, agrupando-os em categorias, de acordo com seus fenótipos mutantes homozigotos. Alguns são necessá-rios à formação da raiz da plântula, outros para o caule da plântula e outros para o ápice da plântula com seus cotilédones. Outra classe é necessária para a formação dos três principais tipos de tecido – epiderme, tecido basal e tecido vascular – e outra classe também é reque-rida para as mudanças organizadas do formato celular que conferem a forma alongada ao embrião e à plântula (Figura 22-115).
Figura 22-114 Dois estágios da embriogênese em Arabidopsis thalia-na. (De G. Jurgens et al., Development [Suppl.] 1:27-38, 1991. Com permissão de The Company of Biologists.)
(A) (B)
Embriãoglobular
Suspensor
20 �m 50 �m
Primórdioda raiz
Primórdiodo caule
Cotilédone
Figura 22-115 Plântulas mutantes de Arabidopsis. Uma plântula normal (A) comparada com quatro tipos de mutantes (B-E) defectivos em diferentes partes de seu padrão ápice-basal: (B) não possui estruturas no ápice, (C) tem ápice e raiz, mas não tem um caule entre eles, (D) não tem raiz e (E) forma tecidos do caule, mas é defectiva em ambas as extremidades. As plântulas foram “clareadas”, de forma a mostrar o tecido vascular em seu interior (filamentos claros). (De U. Mayer et al., Nature 353:402-407, 1991. Com permissão de Macmillan Publishers Ltd.)
(D) (E)
(A)
(B) (C)1 mm
Alberts_22.indd 1402Alberts_22.indd 1402 29.07.09 16:49:4529.07.09 16:49:45

Biologia Molecular da Célula 1403
As partes de uma planta são sequencialmente geradas por meristemasO embrião de um inseto ou um animal vertebrado é um modelo rudimentar, em escala miniatura, do organismo adulto, e os detalhes da estrutura corporal são progressivamente preenchidos, à medida que ele cresce. O embrião de uma planta se torna adulto de uma maneira completamente diferente: as partes da planta adulta são criadas de forma sequen-cial por grupos de células que se proliferam e constroem estruturas adicionais na periferia da planta. Esses importantíssimos grupos de células são denominados meristemas apicais (ver Figura 22-111). Cada meristema consiste em uma população autorrenovável de células do caule. À medida que elas se dividem, deixam para trás uma série de progênies que se deslocam da região do meristema, crescem e, finalmente, diferenciam-se. Embora os meris-temas apicais do caule e da raiz gerem todas as variedades básicas de células que são neces-sárias para construir folhas, raízes e caules, muitas células externas aos meristemas apicais também mantêm a capacidade de proliferação futura e retêm o potencial de meristema. Desta maneira, as árvores e outras plantas perenes, por exemplo, são capazes de aumentar a circunferência dos caules e das raízes à medida que os anos passam e podem produzir novos brotos a partir de regiões dormentes, se a planta é danificada.
Os rudimentos dos meristemas apicais das raízes e dos caules já estão determinados no embrião. Tão logo se rompe a cobertura da semente durante a germinação, ocorre um dramático aumento de tamanho das células não-meristemáticas, direcionando primeiro a emergência de uma raiz para estabelecer uma base de apoio imediata no solo e, então, de um caule (Figura 22-116). Isso é acompanhado por divisões celulares rápidas e contínuas nos meristemas apicais: no meristema apical de uma raiz de milho, por exemplo, as células se dividem a cada 12 horas, produzindo 5 × 105 células por dia. A raiz e o caule em rápido crescimento sondam o ambiente – a raiz aumentando a capacidade da planta de captar água e minerais do solo, e o caule aumentando a capacidade de fotossíntese (ver Painel 22-1).
O desenvolvimento da plântula depende de sinais ambientaisDa germinação em diante, o curso do desenvolvimento dos vegetais é fortemente influencia-do por sinais do ambiente. O caule tem de se desenvolver rapidamente no solo, e deve abrir seu cotilédone e começar a fotossíntese somente após ter atingido a luz. A duração desta transição entre o rápido desenvolvimento subterrâneo e o crescimento com o uso da luz não pode ser geneticamente programada, pois a profundidade em que a semente é enterrada é imprevisível. A ativação do desenvolvimento é controlada pela luz que, entre outros efeitos, atua na plântula inibindo a produção de uma classe de reguladores do crescimento vegetal denominados brassinosteroides, discutidos no Capítulo 15. As mutações nos genes neces-sários à produção ou à recepção de sinais de brassinosteroides tornam o caule da plântula verde, retardam seu alongamento e abrem os cotilédones prematuramente, enquanto as plântulas continuam no escuro.
Sinais hormonais de longo alcance coordenam eventos do desenvolvimento em partes separadas da plantaAs partes separadas de uma planta experimentam ambientes diferentes, reagindo individu-almente a eles por meio de mudanças em seu modo de desenvolvimento. A planta, contudo, deve continuar a funcionar como um todo. Isso demanda que escolhas de desenvolvimento e os eventos daí decorrentes em uma parte da planta afetem escolhas de desenvolvimento em outros locais. Devem existir sinais de longo alcance para efetuar tal coordenação.
Os jardineiros sabem, por exemplo, que quando se arranca a ponta de um ramo po-de-se estimular seu crescimento lateral: a remoção do meristema apical alivia os meriste-mas axilares quiescentes de uma inibição e permite que eles formem novos ramos. Nesse caso, o sinal de longo alcance do meristema apical, ou pelo menos um componente-chave, foi identificado. Trata-se de uma auxina, membro de uma de várias classes de reguladores do crescimento vegetal (às vezes denominados hormônios vegetais), os quais influenciam fortemente o desenvolvimento vegetal. Outras classes conhecidas incluem as giberelinas, as citocininas, o ácido abscísico, o gás etileno e os brassinosteroides. Como mostrado na Figu-ra 22-117, todos são moléculas pequenas que prontamente penetram as paredes celulares. Todos são sintetizados pela maioria das células vegetais e podem ou agir localmente ou ser
Meristema apical docaule (oculto) Cotilédone
(folha da semente)
1 mmMeristema apical da raiz
Figura 22-116 Uma plântula de Ara-bidopsis. Os dois objetos marrons à direita da plântula jovem são as duas metades da cobertura descartada da se-mente. (Cortesia de Catherine Duckett.)
Alberts_22.indd 1403Alberts_22.indd 1403 29.07.09 16:49:4729.07.09 16:49:47

OS TRÊS SISTEMAS TISSULARES A PLANTA
TECIDO BASAL
30 �m
Posições típicas degrupos de suporte decélulas em um caule
Fibras do esclerênquima
Feixe vascular
Colênquima
10 �m
feixe de fibras
100 �m
Esclereídeo
50 �m
Núcleo
Células domesófilo da folha
Vacúolo
Cloroplasto
Células do me-ristema da raiz
Vaso doxilema
Célula detransferência
Gema
NóEntrenó
Nó
Angiospermajovem
Meristema apical da raiz
Meristema apical do caule
CAULE
FOLHA
RAIZ
Nervura central
Nervurada folha
Mesófilo(parênquima)
Colênquima
Epiderme
Epidermesuperior
Estômatosna epidermeinferior
Feixe vascular
Endoderme Periciclo
A divisão celular, o crescimento e a diferenciação dão origem a sistemas tissulares com funções especializadas.
TECIDO DA DERME ( ): Esta é a cobertura externa protetora da planta que está em contato com o ambiente. Ela facilita a captação de água e íons pelas raízes e regula as trocas gasosas nas folhas e nos caules.
TECIDO VASCULAR: Em conjunto, o floema ( ) e o xilema ( ) formam um sistema vascular contínuo em toda a planta. Este tecido conduz água e solutos entre os órgãos e também proporciona suporte mecânico.
TECIDO BASAL ( ): Este tecido de empacotamento e suporte é responsável pela maior parte do volume da planta jovem. Ele também funciona na produção e no armazenamento de alimento.
A angiosperma jovem mostrada à direita é construída a partir de três tipos principais de órgãos: folhas, caules e raízes. Cada órgão vegetal, por sua vez, é constituído de três sistemas tissulares: basal ( ), dérmico ( ) e vascular ( ).
Os três sistemas tissulares são basicamente derivados da atividade proliferativa das células dos meristemas apicais da raiz ou do caule, e cada um contém um número relativamente pequeno de tipos celulares especializados. Estes três sistemas tissulares comuns, e as células que os formam, são descritos neste painel.
O sistema de tecido basal contém três tipos celulares principais denominados parênquima, colênquima e esclerênquima.
O colênquima são células vivas similares às células do parênquima,com exceção das paredes celulares, que são muito mais espessas, normalmente alongadas e empacotadas em longas fibras em forma de corda. Elas são capazes de se estender e proporcionam suporte mecânico ao sistema tissular basal das regiões em alongamento da planta. As células do colênquima são especialmente comuns nas regiões subepidérmicas dos caules.
O esclerênquima, assim como o colênquima, tem funções de reforço e suporte. Contudo, trata-se normalmente de células
mortas com espessas paredes celulares secundárias lignificadas, que as impedem de se alongar à medida que a planta cresce. Os dois tipos comuns são fibras, que frequentemente formam longos feixes, e esclereídeos, que são células ramificadas mais curtas encontradas nas cascas de sementes e frutas.
As células do parênquima são encontradas em todos os sistemas tissulares. São células vivas, geralmente capazes de divisões adicionais, e têm uma fina parede celular primária. Estas células têm várias funções. As células meristemáticas apicais e laterais das gemas e raízes fornecem as novas células necessárias ao crescimento. A produção e o armazenamento de alimento ocorrem nas células fotossintéticas da folha e do caule (denominadas células mesofílicas); as células do parênquima de reserva formam a maior parte do volume da maioria das frutas e verduras. Devido a sua capacidade proliferativa, as células do parênquima também servem de células-tronco para a cicatrização e regeneração.
A célula de transferência, uma forma especializada de célula do parênquima, é prontamente identificável por um crescimento elaborado da parede celular primária. O aumento da área da membrana plasmática abaixo dessas paredes facilita o transporte rápido de solutos para e a partir de células do sistema vascular.
1404 Painel 22-2: Os tipos celulares e tecidos a partir dos quais as plantas superiores são construídas
Alberts_22.indd 1404Alberts_22.indd 1404 29.07.09 16:49:4729.07.09 16:49:47

TECIDO DA DERMEA epiderme é a cobertura protetora externaprimária do corpo da planta. As células daepiderme também são modificadas para formar os estômatos e pelos de vários tipos.
Epiderme Camada cerosa
Cutícula
Epiderme superiorde uma folha
Epiderme deum caule
50 �m
Estômatos Células-guarda
Espaço com ar
5 �m
Epiderme Pelo 100 �m
Epiderme
Pelo da raiz
Um pelo secretormulticelular de uma
folha de gerânioOs pelos unicelulares da raiztêm uma função importantena captação de água e íons
10 �m
Feixes vasculares
Um feixe vascular típico do caulejovem de um ranúnculo
TECIDO VASCULAR
50 �m
Poro deperfuraçãoPlaca de
perfuração
Célulacompanheira
Área deperfuração
Membranaplasmática
Secção longitudinal doelemento do tubode perfuração
Vista externa doelemento do tubode perfuração
Floema
Xilema
Células parenquimais do xilema
Elemento de vaso
Elemento de vasogrande e maduro
Elemento de vasopequeno na ponta
de uma raiz
Bainha doesclerênquima
Floema
Xilema
Parênquima
50 �m
A epiderme (geralmente formada por uma camada de células de espessura) cobre todo o caule, a folha e a raiz da planta jovem. As células são vivas, têm uma parede celular primária espessa e são cobertas, na superfície externa, por uma cutícula especial com uma camada cerosa externa. As células são firmemente entrelaçadas em diferentes padrões.
Os estômatos são aberturas na epiderme, principalmente na superfície inferior da folha, que regulam as trocas gasosas na planta. Eles são formados por duas células epidérmicas especializadas denominadas células-guarda, que regulam o diâmetro do poro. Os estômatos estão distribuídos em um padrão espécie-específico distinto dentro de cada epiderme.
As raízes normalmente têm um único feixe vascular, mas os caules têm vários feixes. Estes estão arranjados em simetria estritamente radial nas dicotiledôneas, mas estão dispersos mais irregularmente nas monocotiledôneas.
Juntos, o floema e o xilema formam um sistema vascular contínuo por toda a planta. Nas plantas jovens, eles normalmente estão associados a vários outros tipos celulares nos feixes vasculares. Tanto o floema como o xilema são tecidos complexos. Seus elementos condutores estão associados a células do parênquima, que mantêm os elementos e trocam materiais com eles. Além disso, os
O floema está envolvido no transporte de solutos orgânicos nas plantas. As principais células condutoras (elementos) estão alinhadas para formar os tubos denominados tubos de filtração. Os elementos dos tubos de filtração, na maturidade, são células vivas interconectadas por perfurações formadas, em suas paredes externas, por plasmodesmata aumentados e modificados (placas de perfuração). Essas células mantêm sua membrana plasmática, mas perderam o núcleo e a maior parte do citoplasma; elas dependem, portanto, de células companheiras associadas para sua manutenção. As células companheiras têm a função adicional de transportar ativamente moléculas de alimento solúvel para dentro e para fora dos elementos do tubo de filtração através de áreas de perfuração porosas na parede.
Os elementos de vaso estão intimamente associados às células parenquimais do xilema, que transportam ativamente solutos selecionados para dentro e para fora dos elementos através da membrana plasmática das células do parênquima.
O xilema transporta água e íons dissolvidos nos vegetais. As principais células condutoras são os elementos de vasos aqui mostrados, os quais, na maturidade, são células mortas que não possuem membrana plasmática. A parede celular
Os pelos (ou tricomas) são apêndices derivados das células epidérmicas. Eles existem em uma grande variedade de formas e normalmente são encontrados em todas as partes da planta. Os pelos funcionam na
proteção, na absorção e na secreção. Por exemplo, quando os pelos unicelulares jovens na epiderme da semente de algodão crescem, as paredes serão secundariamente espessadas com celulose para formar as fibras do algodão.
secundariamente foi espessada e lignificada em grande escala. Como mostrado abaixo, as porções da parede são removidas, permitindo a formação de tubos longos e contínuos.
grupos de células do colênquima e do esclerênquima proporcionam suporte mecânico.
Capítulo 22 Painéis 1405
Alberts_22.indd 1405Alberts_22.indd 1405 29.07.09 16:49:4829.07.09 16:49:48

1406 Alberts, Johnson, Lewis, Raff, Roberts & Walter
transportados para influenciar células-alvo à distância. A auxina, por exemplo, é transporta-da de célula a célula a uma taxa de cerca de 1 cm por hora a partir da ponta de um caule em direção à sua base. Cada regulador do crescimento tem múltiplos efeitos, e esses são modu-lados por outros reguladores do crescimento, assim como por sinais ambientais e pelo esta-do nutricional. Assim, a auxina pode isoladamente promover a formação de raízes, mas em conjunto com uma giberelina pode promover o alongamento do caule, com uma citocinina pode suprimir o crescimento de gemas laterais e, com o etileno, pode estimular o crescimen-to de raízes laterais. Notavelmente, como veremos a seguir, a auxina também controla os padrões detalhados de especialização celular, em escala microscópica, no meristema apical. Os receptores que reconhecem alguns desses reguladores do crescimento são discutidos no Capítulo 15.
A forma de cada nova estrutura depende da divisão e da expansão celular orientadaAs células vegetais, aprisionadas dentro de suas paredes celulares, não podem se mover e se espalhar à medida que a planta cresce; porém, podem se dividir e inchar, esticar e se curvar. A morfogênese de uma planta em desenvolvimento, portanto, depende de divisões celulares organizadas seguidas de expansões celulares estritamente orientadas. A maioria das célu-las produzidas no meristema apical da raiz, por exemplo, passa por três fases distintas de desenvolvimento – divisão, crescimento (alongamento) e diferenciação. Estas três etapas, que se sobrepõem tanto no espaço como no tempo, originam a arquitetura característica da extremidade de uma raiz. Embora o processo de diferenciação celular frequentemente comece enquanto uma célula ainda está se alongando, é fácil distinguir, por comparação, em uma extremidade de raiz, uma zona de divisão celular, uma zona de alongamento celular orientado (que responde pelo crescimento no comprimento da raiz) e uma zona de diferen-ciação celular (Figura 22-118).
Na fase de expansão controlada, que geralmente sucede a divisão celular, as células-fi-lha frequentemente podem aumentar 50 vezes ou mais em volume. Esta expansão é orienta-da por uma pressão osmótica de turgor que pressiona a parede celular da planta para fora, e sua direção é determinada pela orientação das fibrilas de celulose na parede celular, as quais forçam a expansão ao longo de um eixo (ver Figura 19-73). A orientação da celulose, por sua vez, é aparentemente controlada pela orientação dos arranjos de microtúbulos justapostos à membrana plasmática, os quais, acredita-se, guiam a deposição de celulose (discutido no Capítulo 19). Esta orientação pode ser rapidamente alterada por reguladores do crescimento vegetal, como o etileno e o ácido giberélico (Figura 22-119), mas os mecanismos molecula-res subjacentes a esses rearranjos dramáticos no citoesqueleto ainda não são conhecidos.
Figura 22-117 Reguladores do crescimento vegetal. É mostrada a fórmula de uma molécula represen-tativa, que ocorre naturalmente, de cada um dos seis grupos conhecidos de moléculas reguladoras do crescimento vegetal.
C C
H
H H
H
O
O
OH
CH3 CH3
CH3
CH3
COOH
CH2COOH
N
H
N
H
NN
N
NH CH2
OH
C
CH3
CH2CH
COOH
O
CO
HO
CH3
OH
CH2
Etileno
Ácido abscísico (ABA)
Ácido indol-3-acético (IAA) [uma auxina]
Zeatina [uma citocinina] Brassinolídeo [um brassinosteroide]
Ácido giberélico (GA3) [uma giberelina]
H3C
CH3
CH3
CH3
H3C
HO
HO
OH
OH
OH
CH3
Alberts_22.indd 1406Alberts_22.indd 1406 29.07.09 16:49:4829.07.09 16:49:48
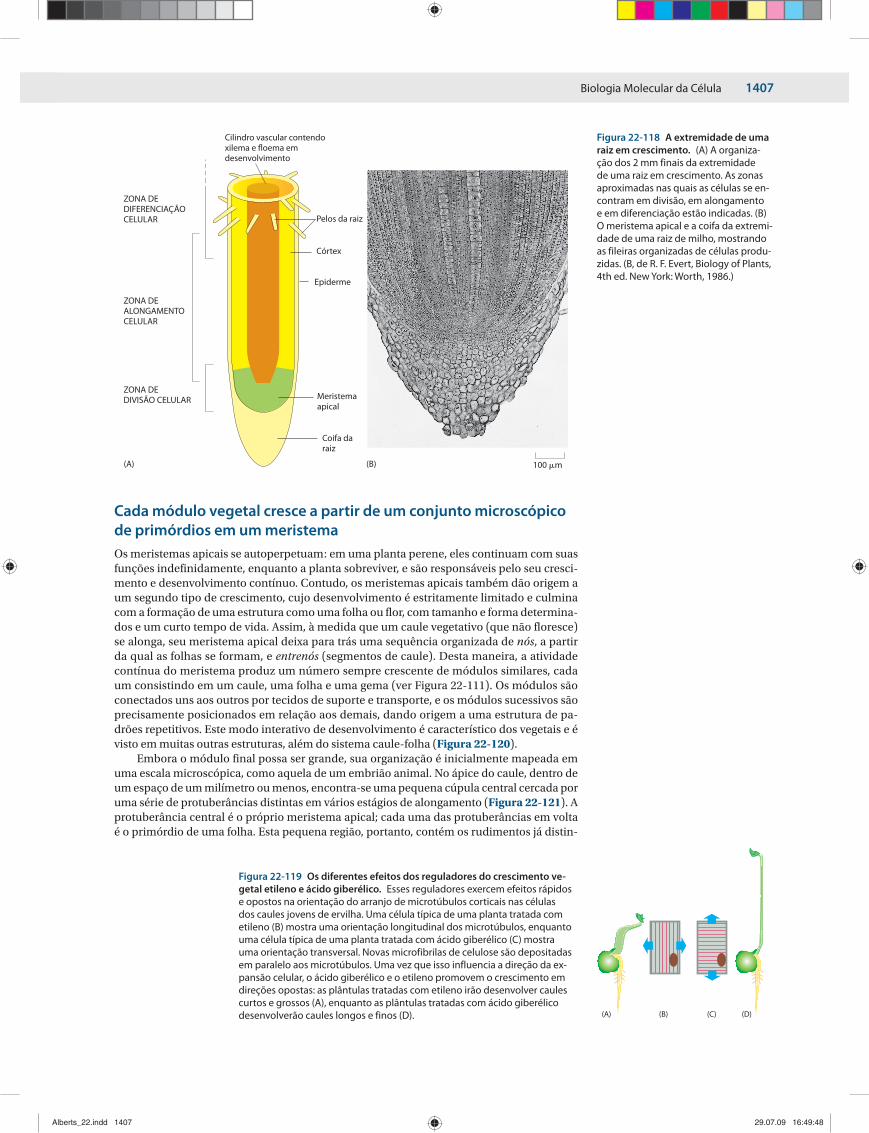
Biologia Molecular da Célula 1407
Cada módulo vegetal cresce a partir de um conjunto microscópico de primórdios em um meristemaOs meristemas apicais se autoperpetuam: em uma planta perene, eles continuam com suas funções indefinidamente, enquanto a planta sobreviver, e são responsáveis pelo seu cresci-mento e desenvolvimento contínuo. Contudo, os meristemas apicais também dão origem a um segundo tipo de crescimento, cujo desenvolvimento é estritamente limitado e culmina com a formação de uma estrutura como uma folha ou flor, com tamanho e forma determina-dos e um curto tempo de vida. Assim, à medida que um caule vegetativo (que não floresce) se alonga, seu meristema apical deixa para trás uma sequência organizada de nós, a partir da qual as folhas se formam, e entrenós (segmentos de caule). Desta maneira, a atividade contínua do meristema produz um número sempre crescente de módulos similares, cada um consistindo em um caule, uma folha e uma gema (ver Figura 22-111). Os módulos são conectados uns aos outros por tecidos de suporte e transporte, e os módulos sucessivos são precisamente posicionados em relação aos demais, dando origem a uma estrutura de pa-drões repetitivos. Este modo interativo de desenvolvimento é característico dos vegetais e é visto em muitas outras estruturas, além do sistema caule-folha (Figura 22-120).
Embora o módulo final possa ser grande, sua organização é inicialmente mapeada em uma escala microscópica, como aquela de um embrião animal. No ápice do caule, dentro de um espaço de um milímetro ou menos, encontra-se uma pequena cúpula central cercada por uma série de protuberâncias distintas em vários estágios de alongamento (Figura 22-121). A protuberância central é o próprio meristema apical; cada uma das protuberâncias em volta é o primórdio de uma folha. Esta pequena região, portanto, contém os rudimentos já distin-
Figura 22-118 A extremidade de uma raiz em crescimento. (A) A organiza-ção dos 2 mm finais da extremidade de uma raiz em crescimento. As zonas aproximadas nas quais as células se en-contram em divisão, em alongamento e em diferenciação estão indicadas. (B) O meristema apical e a coifa da extremi-dade de uma raiz de milho, mostrando as fileiras organizadas de células produ-zidas. (B, de R. F. Evert, Biology of Plants, 4th ed. New York: Worth, 1986.)
(A) (B)
ZONA DEDIFERENCIAÇÃOCELULAR
ZONA DEALONGAMENTOCELULAR
ZONA DEDIVISÃO CELULAR
Cilindro vascular contendoxilema e floema emdesenvolvimento
Pelos da raiz
Córtex
Epiderme
Meristemaapical
Coifa daraiz
100 �m
Figura 22-119 Os diferentes efeitos dos reguladores do crescimento ve-getal etileno e ácido giberélico. Esses reguladores exercem efeitos rápidos e opostos na orientação do arranjo de microtúbulos corticais nas células dos caules jovens de ervilha. Uma célula típica de uma planta tratada com etileno (B) mostra uma orientação longitudinal dos microtúbulos, enquanto uma célula típica de uma planta tratada com ácido giberélico (C) mostra uma orientação transversal. Novas microfibrilas de celulose são depositadas em paralelo aos microtúbulos. Uma vez que isso influencia a direção da ex-pansão celular, o ácido giberélico e o etileno promovem o crescimento em direções opostas: as plântulas tratadas com etileno irão desenvolver caules curtos e grossos (A), enquanto as plântulas tratadas com ácido giberélico desenvolverão caules longos e finos (D). (A) (B) (C) (D)
Alberts_22.indd 1407Alberts_22.indd 1407 29.07.09 16:49:4829.07.09 16:49:48

1408 Alberts, Johnson, Lewis, Raff, Roberts & Walter
tos de vários módulos completos. Por meio de um programa bem-definido de proliferação e alongamento celular, cada primórdio foliar e suas células adjacentes irão crescer para formar uma folha, um nó e um entrenó. Enquanto isso, o próprio meristema apical dará origem a novos primórdios foliares, de modo a gerar mais e mais módulos, em uma sucessão poten-cialmente infinita. A organização serial dos módulos dos vegetais é, assim, controlada por eventos no ápice do caule.
O transporte polarizado de auxina controla o padrão de primórdios no meristemaQuais são os sinais que operam na minúscula região apical e determinam o arranjo de pri-mórdios, e como esses sinais são gerados no padrão adequado? Um indício provém de uma mutação em um gene denominado Pin1, cuja perda impede a formação de primórdios fo-
Figura 22-120 Padrões repetitivos nas plantas. A localização precisa dos su-cessivos módulos a partir de um único meristema apical produz esses padrões elaborados, porém regulares, em folhas (A), flores (B) e frutos (C). (A, de John Si-bthorp, Flora Graeca. London: R. Taylor, 1806-1840; B, de Pierre Joseph Redou-té, Les Liliacées. Paris: chez l’Auteur, 1807; C, de Christopher Jacob Trew, Uitgezochte planten. Amsterdam: Jan Christiaan Sepp, 1771 – todas cortesias de John Innes Foundation.) (A)
(B) (C)
Figura 22-121 O ápice de um broto de uma planta jovem de tabaco. (A) Uma micrografia eletrônica de var-redura mostra o ápice do caule com dois primórdios foliares emergindo sequencialmente, vistos aqui como intumescimentos laterais em cada lado da cúpula do meristema apical. (B) Uma secção fina de um ápice similar mostra que o primórdio foliar mais jovem se origina de um pequeno grupo de célu-las (cerca de 100) nas quatro ou cinco camadas externas de células. (C) Um desenho esquemático mostrando que o aparecimento sequencial de primórdios foliares ocorre em um pequeno espaço e muito cedo no desenvolvimento do caule. O crescimento do ápice formará, no devido tempo, entrenós que separa-rão as folhas, de maneira ordenada, ao longo do caule (ver Figura 22-111). (A e B, de R. S. Poethig e I. M. Sussex, Planta 165:158-169, 1985. Com permissão de Springer-Verlag.)
1
1
2
1
3 2
1
3 2
4
(A) (B)
(C) 300 �m
100 �m
Alberts_22.indd 1408Alberts_22.indd 1408 29.07.09 16:49:4829.07.09 16:49:48
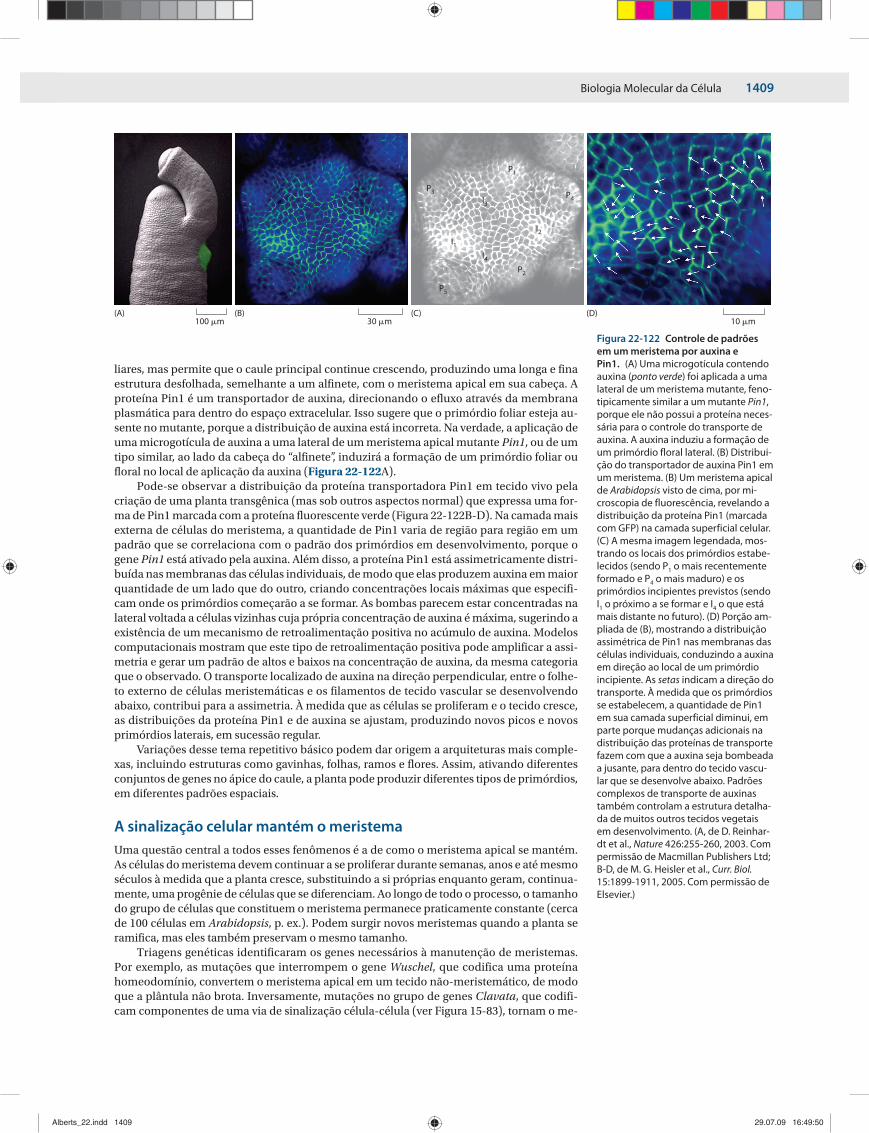
Biologia Molecular da Célula 1409
liares, mas permite que o caule principal continue crescendo, produzindo uma longa e fina estrutura desfolhada, semelhante a um alfinete, com o meristema apical em sua cabeça. A proteína Pin1 é um transportador de auxina, direcionando o efluxo através da membrana plasmática para dentro do espaço extracelular. Isso sugere que o primórdio foliar esteja au-sente no mutante, porque a distribuição de auxina está incorreta. Na verdade, a aplicação de uma microgotícula de auxina a uma lateral de um meristema apical mutante Pin1, ou de um tipo similar, ao lado da cabeça do “alfinete”, induzirá a formação de um primórdio foliar ou floral no local de aplicação da auxina (Figura 22-122A).
Pode-se observar a distribuição da proteína transportadora Pin1 em tecido vivo pela criação de uma planta transgênica (mas sob outros aspectos normal) que expressa uma for-ma de Pin1 marcada com a proteína fluorescente verde (Figura 22-122B-D). Na camada mais externa de células do meristema, a quantidade de Pin1 varia de região para região em um padrão que se correlaciona com o padrão dos primórdios em desenvolvimento, porque o gene Pin1 está ativado pela auxina. Além disso, a proteína Pin1 está assimetricamente distri-buída nas membranas das células individuais, de modo que elas produzem auxina em maior quantidade de um lado que do outro, criando concentrações locais máximas que especifi-cam onde os primórdios começarão a se formar. As bombas parecem estar concentradas na lateral voltada a células vizinhas cuja própria concentração de auxina é máxima, sugerindo a existência de um mecanismo de retroalimentação positiva no acúmulo de auxina. Modelos computacionais mostram que este tipo de retroalimentação positiva pode amplificar a assi-metria e gerar um padrão de altos e baixos na concentração de auxina, da mesma categoria que o observado. O transporte localizado de auxina na direção perpendicular, entre o folhe-to externo de células meristemáticas e os filamentos de tecido vascular se desenvolvendo abaixo, contribui para a assimetria. À medida que as células se proliferam e o tecido cresce, as distribuições da proteína Pin1 e de auxina se ajustam, produzindo novos picos e novos primórdios laterais, em sucessão regular.
Variações desse tema repetitivo básico podem dar origem a arquiteturas mais comple-xas, incluindo estruturas como gavinhas, folhas, ramos e flores. Assim, ativando diferentes conjuntos de genes no ápice do caule, a planta pode produzir diferentes tipos de primórdios, em diferentes padrões espaciais.
A sinalização celular mantém o meristemaUma questão central a todos esses fenômenos é a de como o meristema apical se mantém. As células do meristema devem continuar a se proliferar durante semanas, anos e até mesmo séculos à medida que a planta cresce, substituindo a si próprias enquanto geram, continua-mente, uma progênie de células que se diferenciam. Ao longo de todo o processo, o tamanho do grupo de células que constituem o meristema permanece praticamente constante (cerca de 100 células em Arabidopsis, p. ex.). Podem surgir novos meristemas quando a planta se ramifica, mas eles também preservam o mesmo tamanho.
Triagens genéticas identificaram os genes necessários à manutenção de meristemas. Por exemplo, as mutações que interrompem o gene Wuschel, que codifica uma proteína homeodomínio, convertem o meristema apical em um tecido não-meristemático, de modo que a plântula não brota. Inversamente, mutações no grupo de genes Clavata, que codifi-cam componentes de uma via de sinalização célula-célula (ver Figura 15-83), tornam o me-
Figura 22-122 Controle de padrões em um meristema por auxina e Pin1. (A) Uma microgotícula contendo auxina (ponto verde) foi aplicada a uma lateral de um meristema mutante, feno-tipicamente similar a um mutante Pin1, porque ele não possui a proteína neces-sária para o controle do transporte de auxina. A auxina induziu a formação de um primórdio floral lateral. (B) Distribui-ção do transportador de auxina Pin1 em um meristema. (B) Um meristema apical de Arabidopsis visto de cima, por mi-croscopia de fluorescência, revelando a distribuição da proteína Pin1 (marcada com GFP) na camada superficial celular. (C) A mesma imagem legendada, mos-trando os locais dos primórdios estabe-lecidos (sendo P1 o mais recentemente formado e P4 o mais maduro) e os primórdios incipientes previstos (sendo I1 o próximo a se formar e I4 o que está mais distante no futuro). (D) Porção am-pliada de (B), mostrando a distribuição assimétrica de Pin1 nas membranas das células individuais, conduzindo a auxina em direção ao local de um primórdio incipiente. As setas indicam a direção do transporte. À medida que os primórdios se estabelecem, a quantidade de Pin1 em sua camada superficial diminui, em parte porque mudanças adicionais na distribuição das proteínas de transporte fazem com que a auxina seja bombeada a jusante, para dentro do tecido vascu-lar que se desenvolve abaixo. Padrões complexos de transporte de auxinas também controlam a estrutura detalha-da de muitos outros tecidos vegetais em desenvolvimento. (A, de D. Reinhar-dt et al., Nature 426:255-260, 2003. Com permissão de Macmillan Publishers Ltd; B-D, de M. G. Heisler et al., Curr. Biol. 15:1899-1911, 2005. Com permissão de Elsevier.)
10 �m30 �m100 �m(A) (B) (C) (D)
P3
P1
P4
P2
P5
I2
I4
I1
I3
Alberts_22.indd 1409Alberts_22.indd 1409 29.07.09 16:49:5029.07.09 16:49:50

1410 Alberts, Johnson, Lewis, Raff, Roberts & Walter
ristema anormalmente grande. Esses genes são expressos em diferentes camadas de células na região do meristema (Figura 22-123A). As duas camadas mais superficiais de células, chamadas de camadas L1 e L2, juntamente com a parte mais superior da camada L3, contêm as células do próprio meristema, isto é, as células-tronco, capazes de se dividir indefinida-mente para dar origem a futuras partes da planta. As células meristemáticas das camadas L1 e L2 expressam Clavata3, uma pequena proteína sinalizadora secretada. Logo abaixo, na ca-mada L3, encontra-se um grupo de células que expressam Clavata1 (o receptor de Clavata3). No centro desta porção de Clavata1 estão células que expressam a proteína de regulação gênica Wuschel.
O padrão de divisões celulares implica que as células que expressam Wuschel não são parte do meristema propriamente dito; novas células que expressam Wuschel são, aparente-mente, continuamente recrutadas a partir da parte meristemática (relativa a células-tronco) da população L3, logo acima do domínio Wuschel. No entanto, as células que expressam Wuschel estão no centro do mecanismo que mantém o meristema. Um sinal que elas produzem man-tém o comportamento meristemático nas células acima, estimula a expressão dos genes Cla-vata e, presumivelmente, faz com que novas células recrutadas para o domínio Wuschel ati-vem Wuschel. A retroalimentação negativa das células meristemáticas superiores, distribuída pela via de sinalização Clavata, influencia as regiões abaixo para limitar o tamanho do domínio Wuschel, impedindo, com isso, que o meristema se torne muito grande (Figura 22-123B).
Essa descrição do meristema vegetal, embora incerta em alguns detalhes e certamente bastante simplificada, proporciona um dos exemplos mais claros de uma importante estratégia geral de desenvolvimento: ela mostra como um circuito de retroalimentação envolvendo um sinal de ativação de curto alcance (como aquele produzido pelas células que expressam Wus-chel) e um sinal de inibição de longo alcance (como Clavata3) podem manter, de forma estável, um centro de sinalização de um tamanho bem-definido mesmo quando existe proliferação e circulação contínua das células que formam aquele centro. Como chamamos a atenção no início deste capítulo, acredita-se que sistemas análogos de sinais operem no desenvolvimento animal para manter centros de sinalização localizados – como o Organizador da gástrula de anfíbios ou a zona de atividade polarizadora em um broto de membro. E, assim como essa es-tratégia serve para manter os meristemas de uma planta madura, ela também pode servir nos tecidos de um animal adulto, como, por exemplo, o revestimento do intestino (discutido no Capítulo 23), para manter os importantíssimos grupos de células-tronco adultas.
Mutações reguladoras podem transformar a topologia vegetal pela alteração do comportamento celular no meristemaPara que o caule de uma planta possa se ramificar, novos meristemas apicais do caule devem ser criados, e isso também depende de eventos próximos ao ápice do caule. A cada nó em desenvolvimento, no ângulo agudo (axila) entre o primórdio foliar e o caule, é formada uma gema (Figura 22-124). Esta gema contém um ninho de células, derivadas do meristema api-cal, que mantém um caráter meristemático. Elas têm a capacidade de se tornar o meristema apical de um novo ramo ou o primórdio de uma estrutura como a flor; porém, também têm a opção alternativa de permanecer quiescentes como gemas axilares. O padrão de ramifica-ção dos vegetais é regulado por esta escolha de destinação, e mutações que o afetam podem transformar a estrutura da planta. O milho constitui um belo exemplo.
O milho representa um dos feitos mais extraordinários de engenharia genética produzi-dos pela humanidade. Os indígenas americanos o criaram por meio de cruzamento seletivo, durante um período de vários séculos, ou talvez milênios, entre 5.000 e 10.000 anos atrás.
L3L2L1
(A) (B)
Meristema apical do caule
Epiderme
Primórdiofoliar
Clavata3
Clavata1 Wuschel
Figura 22-123 Acredita-se que os cir-cuitos de retroalimentação mantêm o meristema apical do caule. (A) Arranjo das camadas de células que constituem o meristema apical do caule. (B) Padrão de comunicação célula-célula que mantém o meristema. A superexpressão artificial de Wuschel na região L3 leva a um aumento do número de células nas camadas L1 e L2, que se comportam como células meristemáticas e expres-sam Clavata3; a superexpressão artificial de Clavata3 nas camadas L1 e L2 leva a uma redução da expressão de Wuschel na região L3 abaixo e a uma diminuição do número de células meristemáticas. Clavata3 codifica uma pequena proteí-na-sinal, enquanto Clavata1 codifica seu receptor, uma proteína-cinase trans-membrana. Wuschel, que é expressa na parte central da região que expressa o receptor Clavata1, codifica uma pro-teína de regulação gênica da classe homeodomínio. Acredita-se que o tamanho do meristema seja controlado por um equilíbrio autorregulador entre um sinal estimulador de curto alcance produzido por células que expressam Wuschel (seta amarela) e um sinal inibi-dor de alcance mais longo liberado por Clavata3 (barras vermelhas).
Alberts_22.indd 1410Alberts_22.indd 1410 29.07.09 16:49:5129.07.09 16:49:51

Biologia Molecular da Célula 1411
Eles partiram de uma gramínea selvagem conhecida como teosinte, com caules folhosos muito ramificados e minúsculas espigas carregando sementes duras e incomestíveis. A aná-lise genética detalhada identificou alguns loci genéticos – cerca de cinco – como os sítios das mutações responsáveis pela maior parte das diferenças entre este ancestral não-promissor e o milho moderno. Um desses loci, com um efeito particularmente dramático, corresponde a um gene denominado Teosinte branched-1 (Tb1). No milho com mutações de perda de função em Tb1, o caule simples não-ramificado comum, com algumas folhas grandes em intervalos ao longo do caule, é transformado em uma massa de folhas densa e ramificada re-manescente do teosinte (Figura 22-125A). O padrão de ramificação no mutante implica que as gemas auxiliares, que se originam em posições normais, escaparam de um mecanismo de inibição que as impede, no milho normal, de se desenvolverem em ramos.
No milho normal, o caule único é coroado com um pendão (ou coruto) – uma flor mas-culina – enquanto algumas das gemas axilares ao longo do caule se desenvolvem em flores femininas e, após a fertilização, formam as espigas de milho que comemos. No milho mu-tante, defectivo no gene Tb1, essas gemas axilares frutíferas são transformadas em ramifica-ções carregando pendões. A planta de teosinte selvagem é como o milho defectivo em Tb1 quanto a sua aparência folhosa e altamente ramificada, mas diferentemente desse mutante ela produz espigas em muitos de seus ramos laterais, como se o gene Tb1 estivesse ativo. A análise de DNA revela a explicação. Tanto o teosinte como o milho normal possuem o gene Tb1 funcional, com uma sequência codificante quase idêntica, mas no milho a região regula-dora sofreu uma mutação que elevou o nível da expressão gênica. Assim, no milho normal o gene é expresso em um nível alto em cada gema axilar, inibindo a formação de ramificações, enquanto no teosinte a expressão em muitas gemas axilares é baixa, de modo que ocorre a formação de ramos (Figura 22-125B).
Meristema apical do caule
Primórdios da gema Base da folha
Axila
Figura 22-124 Gemas axilares na vizinhança do ápice de um caule. A fotografia mostra uma secção longitu-dinal de Coleus blumei, uma folhagem comum. (De P. H. Raven, R. F. Evert e S. E. Eichhorn, Biology of Plants, 6th ed. New York: Freeman/Worth, 1999, utilizada com permissão.)
Figura 22-125 Transformação da arquitetura vegetal por mutação: uma comparação entre o teosinte, o milho normal e o milho defectivo em Tb1. (A) Fotografias de três tipos de plantas. (B) Comparação esquemática entre a arquitetura do teosinte, do milho normal e do milho defectivo em Tb1. O produto do gene Tb1 é neces-sário ao desenvolvimento de espigas. Ele está ausente no mutante Tb1 e está presente tanto no teosinte como no milho normal, mas essas duas plantas diferem, porque o gene é diferentemen-te regulado. (A, imagem da esquerda, de J. Doebley e R. L. Wang, Cold Spring Harbor Symp. Quant Biol. 62:361-367, 1997. Com permissão de Cold Spring Harbor Laboratory Press, A, imagem do meio e da direita, de J. Doebley, A. Stec e L. Hubbard, Nature 386:485-488, 1997. Com permissão de Macmillan Publi-shers Ltd.)
(A)
Teosinte Milho normal Milho mutantedefectivo em Tb1
Pendões
Espigas
(B)
Alberts_22.indd 1411Alberts_22.indd 1411 29.07.09 16:49:5129.07.09 16:49:51

1412 Alberts, Johnson, Lewis, Raff, Roberts & Walter
Esse exemplo mostra como mutações simples, ativando o comportamento de células meristemáticas, podem transformar a estrutura vegetal – um princípio de enorme impor-tância no cruzamento de plantas para a alimentação. De forma mais ampla, o caso de Tb1 ilustra como novos planos corporais, sejam vegetais ou animais, podem evoluir por meio de mudanças no DNA regulador sem alterações nas características das proteínas sinteti-zadas.
A ativação para a floração depende de sinais ambientais passados e presentesOs meristemas enfrentam outras escolhas de desenvolvimento além daquela existente entre quiescência e crescimento, como já vimos em nossa discussão sobre o milho, e essas tam-bém frequentemente são reguladas pelo ambiente. A mais importante é a decisão de formar uma flor (Figura 22-126).
A mudança do crescimento meristemático para a formação de flores é desencadeada por uma combinação de sinais. A planta não leva apenas em conta a temperatura, a intensi-dade luminosa e as condições nutricionais atuais; ela também baseia sua decisão de flores-cer em condições passadas. Um sinal importante, para muitas plantas, é a duração do dia. Para detectá-lo, a planta utiliza seu relógio circadiano – um ritmo endógeno de 24 horas da expressão gênica – para gerar um sinal positivo para o florescimento somente quando há luz na parte apropriada do dia. O próprio relógio é influenciado pela luz e, na realidade, a planta utiliza o relógio para comparar as condições luminosas passadas e presentes. Partes importantes da maquinaria genética subjacente a esses fenômenos foram identificadas, dos fitocromos e criptocromos que agem como receptores luminosos (discutido no Capítulo 15) ao gene Constans, cuja expressão nas folhas da planta representa um sinal para a floração. Acredita-se que o sinal seja retransmitido das folhas ao meristema, via vasculatura, pelo pro-duto de outro gene, Flowering locus T (Ft), que é regulado por Constans.
Contudo, este mesmo sinal alcançará o meristema e desencadear a floração somente se a planta estiver em uma condição receptiva, que tipicamente depende de sua história ao longo de um período mais longo. Muitas plantas florescerão somente se tiverem previa-mente passado por um longo período no frio: elas devem atravessar o inverno antes de se comportarem como se fosse primavera – um processo denominado vernalização. O frio prolongado ocasiona mudanças na estrutura da cromatina, dependentes de outro grande conjunto de genes, incluindo homólogos de membros do grupo Polycomb que menciona-mos anteriormente devido a seu papel na perpetuação de padrões de expressão gênica em Drosophila. Essas mudanças epigenéticas (discutido nos Capítulos 4 e 7) resultam no silen-ciamento gradual do gene Flowering locus C (Flc). O efeito é duradouro, persistindo por mui-tos ciclos de divisão celular, mesmo quando o clima fica mais quente. O gene Flc codifica um inibidor da floração, antagonizando a expressão e a ação do gene Ft. Assim, a vernalização, ao bloquear a produção do inibidor, permite que o meristema receba o sinal Ft e responda a ele por mudanças na expressão de um conjunto de genes de identidade de meristema floral no meristema apical.
Mutações que afetam a regulação da expressão de Flc alteram o momento de floração e, assim, a capacidade de uma planta de florescer em um dado clima. Assim, o sistema de con-
Figura 22-126 A estrutura de uma flor de Arabidopsis. (A) Fotografia. (B) Vista esquemática de uma secção longitu-dinal. O plano básico, como mostrado em (B), é comum à maioria das angios-permas dicotiledôneas. (A, cortesia de Leslie Sieburth.)
Pétala CarpeloEstame
Sépala
(B)(A)
Alberts_22.indd 1412Alberts_22.indd 1412 29.07.09 16:49:5129.07.09 16:49:51

Biologia Molecular da Célula 1413
trole global que governa a ativação para a floração é de vital importância para a agricultura, especialmente em uma época de rápidas mudanças climáticas.
Os genes seletores homeóticos especificam as partes de uma florAo ativar os genes de identidade de meristema floral, o meristema apical abandona suas chances de continuar o crescimento vegetativo e arrisca seu futuro na produção de gametas. Suas células se envolvem em um programa rigorosamente finito de crescimento e diferen-ciação: por uma modificação dos mecanismos comuns para a geração de folhas, uma série de verticilos de órgãos acessórios especializados é formada de maneira precisa – tipicamen-te as sépalas primeiro, em seguida as pétalas, depois os estames carregando as anteras que contêm o pólen e, por último, os carpelos que contêm os óvulos (ver Painel 22-1). No final desse processo, o meristema desapareceu, mas junto com sua progênie ele criou as células germinativas.
A série de folhas modificadas que formam uma flor pode ser comparada à série de seg-mentos do corpo que formam uma mosca. Nos vegetais, assim como nas moscas, podem-se encontrar mutações homeóticas que convertem uma parte do padrão em características do outro. Os fenótipos mutantes podem ser agrupados em no mínimo quatro classes, nas quais grupos diferentes, mas sobrepostos, de órgãos estão alterados (Figura 22-127). A primeira, ou classe “A”, exemplificada pelo mutante Apetala2 de Arabidopsis, tem seus dois verticilos mais externos transformados: as sépalas estão convertidas em carpelos e as pétalas em esta-mes. A segunda, ou classe “B”, exemplificada por Apetala3, tem seus dois verticilos do meio transformados: as pétalas estão convertidas em sépalas e os estames em carpelos. A terceira, ou classe “C”, exemplificada por Agamous, tem seus dois verticilos mais internos transfor-mados, com uma consequência mais drástica: os estames estão convertidos em pétalas, os carpelos estão ausentes e, em seu lugar, as células centrais da flor se comportam como um meristema floral, que começa todo o processo de desenvolvimento novamente, gerando ou-tro conjunto anormal de sépalas e pétalas aninhado dentro do primeiro e, potencialmente,
Figura 22-127 Flores de Arabidopsis mostrando uma seleção de mutações homeóticas. (A) Em Apetala2, as sépa-las estão convertidas em carpelos e as pétalas em estames. (B) Em Apetala3, as pétalas estão convertidas em sépalas e os estames em carpelos. (C) Em Aga-mous, os estames estão convertidos em pétalas e os carpelos em meristema floral. (D) Em um mutante triplo, onde estas três funções são defectivas, todos os órgãos da flor estão convertidos em folhas. (A-C, cortesia de Leslie Sieburth; D, cortesia de Mark Running.)
(A) (B)
(D)(C)
Alberts_22.indd 1413Alberts_22.indd 1413 29.07.09 16:49:5229.07.09 16:49:52
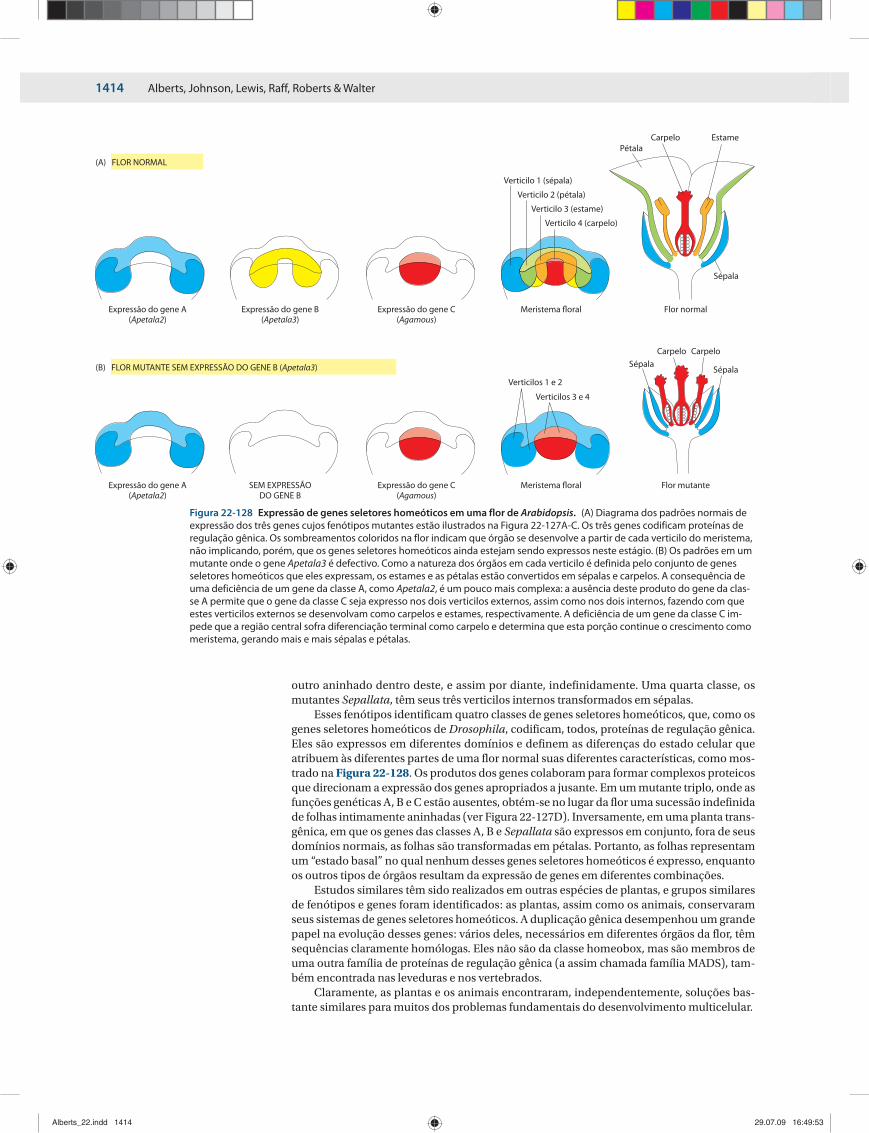
1414 Alberts, Johnson, Lewis, Raff, Roberts & Walter
outro aninhado dentro deste, e assim por diante, indefinidamente. Uma quarta classe, os mutantes Sepallata, têm seus três verticilos internos transformados em sépalas.
Esses fenótipos identificam quatro classes de genes seletores homeóticos, que, como os genes seletores homeóticos de Drosophila, codificam, todos, proteínas de regulação gênica. Eles são expressos em diferentes domínios e definem as diferenças do estado celular que atribuem às diferentes partes de uma flor normal suas diferentes características, como mos-trado na Figura 22-128. Os produtos dos genes colaboram para formar complexos proteicos que direcionam a expressão dos genes apropriados a jusante. Em um mutante triplo, onde as funções genéticas A, B e C estão ausentes, obtém-se no lugar da flor uma sucessão indefinida de folhas intimamente aninhadas (ver Figura 22-127D). Inversamente, em uma planta trans-gênica, em que os genes das classes A, B e Sepallata são expressos em conjunto, fora de seus domínios normais, as folhas são transformadas em pétalas. Portanto, as folhas representam um “estado basal” no qual nenhum desses genes seletores homeóticos é expresso, enquanto os outros tipos de órgãos resultam da expressão de genes em diferentes combinações.
Estudos similares têm sido realizados em outras espécies de plantas, e grupos similares de fenótipos e genes foram identificados: as plantas, assim como os animais, conservaram seus sistemas de genes seletores homeóticos. A duplicação gênica desempenhou um grande papel na evolução desses genes: vários deles, necessários em diferentes órgãos da flor, têm sequências claramente homólogas. Eles não são da classe homeobox, mas são membros de uma outra família de proteínas de regulação gênica (a assim chamada família MADS), tam-bém encontrada nas leveduras e nos vertebrados.
Claramente, as plantas e os animais encontraram, independentemente, soluções bas-tante similares para muitos dos problemas fundamentais do desenvolvimento multicelular.
Figura 22-128 Expressão de genes seletores homeóticos em uma flor de Arabidopsis. (A) Diagrama dos padrões normais de expressão dos três genes cujos fenótipos mutantes estão ilustrados na Figura 22-127A-C. Os três genes codificam proteínas de regulação gênica. Os sombreamentos coloridos na flor indicam que órgão se desenvolve a partir de cada verticilo do meristema, não implicando, porém, que os genes seletores homeóticos ainda estejam sendo expressos neste estágio. (B) Os padrões em um mutante onde o gene Apetala3 é defectivo. Como a natureza dos órgãos em cada verticilo é definida pelo conjunto de genes seletores homeóticos que eles expressam, os estames e as pétalas estão convertidos em sépalas e carpelos. A consequência de uma deficiência de um gene da classe A, como Apetala2, é um pouco mais complexa: a ausência deste produto do gene da clas-se A permite que o gene da classe C seja expresso nos dois verticilos externos, assim como nos dois internos, fazendo com que estes verticilos externos se desenvolvam como carpelos e estames, respectivamente. A deficiência de um gene da classe C im-pede que a região central sofra diferenciação terminal como carpelo e determina que esta porção continue o crescimento como meristema, gerando mais e mais sépalas e pétalas.
(A) FLOR NORMAL
(B) FLOR MUTANTE SEM EXPRESSÃO DO GENE B (Apetala3)
Expressão do gene A(Apetala2)
Expressão do gene B(Apetala3)
Expressão do gene C(Agamous)
Meristema floral Flor normal
Expressão do gene A(Apetala2)
SEM EXPRESSÃODO GENE B
Expressão do gene C(Agamous)
Meristema floral Flor mutante
Verticilo 1 (sépala)
Verticilo 2 (pétala)
Verticilo 3 (estame)
Verticilo 4 (carpelo)
Verticilos 1 e 2
Verticilos 3 e 4
PétalaCarpelo
Carpelo Carpelo
Estame
Sépala
SépalaSépala
Alberts_22.indd 1414Alberts_22.indd 1414 29.07.09 16:49:5329.07.09 16:49:53

Biologia Molecular da Célula 1415
ResumoO desenvolvimento de uma angiosperma, como o de um animal, começa com a divisão de um óvulo fertilizado para formar um embrião com uma organização polarizada: a parte apical do embrião formará a gema, a parte basal, a raiz, e a parte intermediária, o caule. Inicialmente, a divisão celu-lar ocorre ao longo de todo o corpo do embrião. Entretanto, à medida que o embrião cresce, a adição de novas células se torna restrita a pequenas regiões conhecidas como meristemas. Os meristemas apicais, nas pontas das gemas e nas pontas das raízes, persistirão por toda a vida da planta, permi-tindo que ela cresça pela adição sequencial de novas partes corporais em sua periferia. Tipicamen-te, a gema gera uma série repetitiva de módulos, cada qual consistindo em um segmento de caule, uma folha e uma gema axilar. O transporte polarizado de auxina controla o posicionamento dos primórdios dessas estruturas, à medida que elas surgem nas cercanias do meristema. Uma gema axilar é potencialmente um novo meristema, capaz de dar origem a um ramo lateral; o ambiente e os sinais hormonais de longo alcance dentro da planta podem controlar seu desenvolvimento pela regulação da ativação de gemas. Mutações que alteram as regras para a ativação de gemas axilares podem ter um efeito drástico sobre a forma e a estrutura da planta; uma única dessas mutações – uma de cerca de cinco alterações genéticas-chave – é responsável por uma grande parte da diferença dramática entre o milho moderno e seu ancestral selvagem, o teosinte.
A pequena erva daninha Arabidopsis thaliana é amplamente utilizada como organismo-mo-delo para estudos genéticos, sendo a primeira planta a ter tido seu genoma completamente sequen-ciado. Como nos animais, os genes que governam o desenvolvimento vegetal podem ser identifica-dos por meio de triagens genéticas, e suas funções podem ser testadas por manipulações genéticas. Tais estudos começaram a revelar os mecanismos moleculares pelos quais a organização interna de cada módulo da planta é projetada em uma escala microscópica por interações célula-célula nas proximidades do meristema apical. O próprio meristema parece ser mantido por um circuito de retroalimentação local, no qual células que expressam a proteína de regulação gênica Wuschel produzem um estímulo positivo, e por uma retroalimentação negativa, dependente da via de sina-lização célula-célula Clavata, que impede que o meristema se torne muito grande.
Sinais ambientais – especialmente a luz apropriadamente ajustada – podem induzir a ex-pressão de genes que alteram o meristema apical que formaria uma folha para um que formará uma flor. As partes de uma flor – suas sépalas, pétalas, estames e carpelos – são formadas por uma modificação do mecanismo de desenvolvimento das folhas, e as diferenças entre essas partes são controladas por genes seletores homeóticos que são intimamente análogos (mas não homólogos) àqueles dos animais.
REFERÊNCIASGeralCarroll SB, Grenier JK & Weatherbee SD (2004) From DNA to Diversity:
Molecular Genetics and the Evolution of Animal Design, 2nd ed. Malden, MA: Blackwell Science.
Gilbert SF (2006) Developmental Biology, 8th ed. Sunderland, MA: Sinauer.Nusslein-Volhard C (2006) Coming to Life: How Genes Drive Development.
London: Yale University PressWolpert L, Jessell TM, Lawrence P et al (2006) Principles of Development, 3rd
ed. Oxford: Oxford University Press.
Mecanismos universais de desenvolvimento animalGilbert SF & Raunio AM (eds) (1997) Embryology: Constructing The
Organism. Sunderland, MA: Sinauer.Giraldez AJ, Cinalli RM, Glasner ME et al (2005) MicroRNAs regulate brain
morphogenesis in zebrafish. Science 308:833–838.Lander AD (2007) Morpheus unbound: reimagining the morphogen
gradient. Cell 128:245–256.Meinhardt H & Gierer A (2000) Pattern formation by local self-activation and
lateral inhibition. BioEssays 22:753–760.Spemann H (1938) Embryonic Development and Induction. New Haven:
Yale University Press. Reprinted 1988, New York: Garland Publishing.Tyson JJ, Chen KC & Novak B (2003) Sniffers, buzzers, toggles and blinkers:
dynamics of regulatory and signaling pathways in the cell. Curr Opin Cell Biol 15:221–231.
Caenorhabditis elegans: o desenvolvimento a partir da perspectiva de uma célula individualGirard LR, Fiedler TJ, Harris TW et al (2007) WormBook: the online review of
Caenorhabditis elegans biology. Nucleic Acids Res 35:D472–475.Harris WA & Hartenstein V (1991) Neuronal determination without cell
division in Xenopus embryos. Neuron 6:499–515.Kamath RS, Fraser AG, Dong Y et al (2003) Systematic functional analysis of
the Caenorhabditis elegans genome using RNAi. Nature 421:231–237.Lyczak R, Gomes JE & Bowerman B (2002) Heads or tails: cell polarity and axis
formation in the early Caenorhabditis elegans embryo. Dev Cell 3:157–166.Metzstein MM, Stanfield GM & Horvitz HR (1998) Genetics of programmed
cell death in C. elegans: past, present and future. Trends Genet 14:410–416.Pasquinelli AE, Reinhart BJ, Slack F et al (2000) Conservation of the sequence
and temporal expression of let-7 heterochronic regulatory RNA. Nature 408:86–89.
Strome S & Lehmann R (2007) Germ versus soma decisions: lessons from flies and worms. Science 316:392–393.
Drosophila e a genética molecular da formação de padrões: a gênese do plano corporalBate M & Martinez Arias A (eds) (1993) The Development of Drosophila
melanogaster. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press.
Gregor T, Bialek W, de Ruyter van Steveninck RR, Tank DW & Wieschaus EF (2005) Diffusion and scaling during early embryonic pattern formation. Proc Natl Acad Sci U.S.A. 102:18403–18407.
Alberts_22.indd 1415Alberts_22.indd 1415 29.07.09 16:49:5329.07.09 16:49:53

1416 Alberts, Johnson, Lewis, Raff, Roberts & Walter
Huynh JR & St Johnston D (2004) The origin of asymmetry: early polarisation of the Drosophila germline cyst and oocyte. Curr Biol 14:R438–449.
Lawrence PA (1992) The Making of a Fly: The Genetics of Animal Design. Oxford: Blackwell Scientific.
Nusslein-Volhard C & Wieschaus E (1980) Mutations affecting segment number and polarity in Drosophila. Nature 287:795–801.
Peel AD, Chipman AD & Akam M (2005) Arthropod segmentation: beyond the Drosophila paradigm. Nature Rev Genet 6:905–916.
Stathopoulos A & Levine M (2004) Whole-genome analysis of Drosophila gastrulation. Curr Opin Genet Dev 14:477–484.
Genes seletores homeóticos e a formação de padrões do eixo ântero-posteriorLemons D & McGinnis W (2006) Genomic evolution of Hox gene clusters.
Science 313:1918–1922.Lewis EB (1978) A gene complex controlling segmentation in Drosophila.
Nature 276:565–570.McGinnis W, Garber RL,Wirz J et al (1984) A homologous protein-coding
sequence in Drosophila homeotic genes and its conservation in other metazoans. Cell 37:403–408.
Ringrose L & Paro R (2007) Polycomb/Trithorax response elements and epigenetic memory of cell identity. Development 134:223–232.
Wellik DM & Capecchi MR (2003) Hox10 and Hox11 genes are required to globally pattern the mammalian skeleton. Science 301:363–367.
Organogênese e a formação dos padrões dos órgãos acessóriosBellaiche Y & Schweisguth F (2001) Lineage diversity in the Drosophila
nervous system. Curr Opin Genet Dev 11:418–423.Betschinger J & Knoblich JA (2004) Dare to be different: asymmetric cell
division in Drosophila, C. elegans and vertebrates. Curr Biol 14:R674–685.Betschinger J, Mechtler K & Knoblich JA (2006) Asymmetric segregation of
the tumor suppressor brat regulates self-renewal in Drosophila neural stem cells. Cell 124:1241–1253.
Irvine KD & Rauskolb C (2001) Boundaries in development: formation and function. Annu Rev Cell Dev Biol 17:189–214.
Harvey K & Tapon N (2007) The Salvador-Warts-Hippo pathway—an emerging tumour-suppressor network. Nature Rev Cancer 7:182–191.
O’Farrell P (2004). How metazoans reach their full size : the natural history of bigness, In Cell Growth: The Control of Cell Size, MN Hall et al, eds. (Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press), pp. 1–22.
Panganiban G, Irvine SM, Lowe C et al (1997) The origin and evolution of animal appendages. Proc. Natl. Acad. Sci. USA 94:5162–5166.
Rogulja D & Irvine KD (2005) Regulation of cell proliferation by a morphogen gradient. Cell 123:449–461.
Tickle C (2003) Patterning systems—from one end of the limb to the other. Dev Cell 4:449–458.
Movimentos celulares e a determinação da forma do corpo dos vertebradosDavid NB, Sapede D, Saint-Etienne L et al (2002) Molecular basis of cell
migration in the fish lateral line: role of the chemokine receptor CXCR4 and of its ligand, SDF1. Proc Natl Acad Sci U S A 99:16297–16302.
Heasman J (2006) Maternal determinants of embryonic cell fate. Semin Cell Dev Biol 17:93–98.
Hirokawa N, Tanaka Y, Okada Y & Takeda S (2006) Nodal flow and the generation of left-right asymmetry. Cell 125:33–45.
Kalcheim C & Le Douarin NM (1999) The Neural Crest, 2nd ed. Cambridge: Cambridge University Press.
Lewis J (2003) Autoinhibition with transcriptional delay: a simple mechanism for the zebrafish somitogenesis oscillator. Curr Biol 13:1398–1408.
Pourquie O (2003) The segmentation clock: converting embryonic time into spatial pattern. Science 301:328–330.
Raz E (2004) Guidance of primordial germ cell migration. Curr Opin Cell Biol 16:169–173.
Schier AF (2007) The maternal-zygotic transition: death and birth of RNAs. Science 316:406–407.
Tabin CJ (2006) The key to left-right asymmetry. Cell 127:27–32.Takeichi M, Nakagawa S, Aono S et al (2000) Patterning of cell assemblies
regulated by adhesion receptors of the cadherin superfamily. Philos Trans R Soc Lond B Biol Sci 355:885–890.
O camundongoGhabrial A, Luschnig S, Metzstein MM & Krasnow MA (2003) Branching
morphogenesis of the Drosophila tracheal system. Annu Rev Cell Dev Biol 19:623–647.
Hogan BL & Kolodziej PA (2002) Organogenesis: molecular mechanisms of tubulogenesis. Nature Rev Genet 3:513–523.
Larsen WJ (2001) Human Embryology, 3rd ed: Churchill Livingstone.Lu CC, Brennan J & Robertson EJ (2001) From fertilization to gastrulation:
axis formation in the mouse embryo. Curr Opin Genet Dev 11:384–392.Smith AG (2001) Embryo-derived stem cells: of mice and men. Annu Rev Cell
Dev Biol 17:435–462.Weaver M, Dunn NR & Hogan BL (2000) Bmp4 and Fgf10 play opposing
roles during lung bud morphogenesis. Development 127:2695–2704.Zernicka-Goetz M (2006) The first cell-fate decisions in the mouse embryo:
destiny is a matter of both chance and choice. Curr Opin Genet Dev 16:406–412.
Desenvolvimento neuralBi G & Poo M (2001) Synaptic modification by correlated activity: Hebb’s
postulate revisited. Annu Rev Neurosci 24:139–166.Flanagan JG (2006) Neural map specification by gradients. Curr Opin
Neurobiol 16:59–66.Hubel DH & Wiesel TN (1965) Binocular interaction in striate cortex of kittens
reared with artificial squint. J Neurophysiol 28:1041–1059.Kandel ER, Schwartz JH & Jessell TM (2000) Principles of Neural Science, 4th
ed. New York: McGraw-Hill.Nägerl UV, Eberhorn N, Cambridge SB & Bonhoeffer T (2004) Bidirectional
activity-dependent morphological plasticity in hippocampal neurons. Neuron 44:759–767.
Pearson BJ & Doe CQ (2004) Specification of temporal identity in the developing nervous system. Annu Rev Cell Dev Biol 20:619–647.
Sanes DH, Reh TA & Harris WA (2000) Development of the Nervous System. San Diego: Academic Press.
Tessier-Lavigne M (2002) Wiring the brain: the logic and molecular mechanisms of axon guidance and regeneration. Harvey Lect 98:103–143.
Waites CL, Craig AM & Garner CC (2005) Mechanisms of vertebrate synaptogenesis. Annu Rev Neurosci 28:251–274.
Wang GX & Poo MM (2005) Requirement of TRPC channels innetrin-1-induced chemotropic turning of nerve growth cones. Nature 434:898–904.
Zou Y, Stoeckli E, Chen H & Tessier-Lavigne M (2000) Squeezing axons out of the gray matter: a role for slit and semaphorin proteins from midline and ventral spinal cord. Cell 102:363–375.
Desenvolvimento vegetalBäurle I & Dean C (2006) The timing of developmental transitions in plants.
Cell 125:655–664.Doebley J, Stec A & Hubbard L (1997) The evolution of apical dominance in
maize. Nature 386:485–488.Heisler MG, Ohno C, Das P et al (2005) Patterns of auxin transport and gene
expression during primordium development revealed by live imaging of the Arabidopsis inflorescence meristem. Curr Biol 15:1899–1911.
Howell SH (1998) Molecular Genetics of Plant Development. Cambridge: Cambridge University Press.
Jack T (2001) Relearning our ABCs: new twists on an old model. Trends Plant Sci. 6:310–316.
Schoof H, Lenhard M, Haecker A et al (2000) The stem cell population of Arabidopsis shoot meristems is maintained by a regulatory loop between the CLAVATA and WUSCHEL genes. Cell 100:635–644.
Alberts_22.indd 1416Alberts_22.indd 1416 29.07.09 16:49:5329.07.09 16:49:53



















![equilibriopot.ppt [Somente leitura] - Departamento de ...fisfar.ufc.br/v2/graduacao/arquivo_aulas/leticia/equilibriopot.pdf · Potencial de ação Fonte: Alberts et al., 1998. Essential](https://img.document.onl/doc/110x75/5beadea809d3f231188bd9aa/somente-leitura-departamento-de-fisfarufcbrv2graduacaoarquivoaulasleticiaequilibriopotpdf.jpg)









