Embed Size (px)
Citation preview

Análise da resposta antioxidativa em plantas de
Ricinus communis submetidas ao estresse por metil
jasmonato.
ALEXANDRA MARTINS DOS SANTOS SOARES
Dissertação apresentada à Universidade
Estadual do Norte Fluminense Darcy
Ribeiro, para a obtenção do título de
Mestre em Biociências e Biotecnologia.
Área de concentração: Biologia Celular.
Campos dos Goytacazes
Estado do Rio de Janeiro – Brasil
Abril – 2006

Análise da resposta antioxidativa em plantas de
Ricinus communis submetidas ao estresse por metil
jasmonato.
ALEXANDRA MARTINS DOS SANTOS SOARES
ORIENTAÇÃO: PROFa.DRa. OLGA LIMA TAVARES MACHADO
Dissertação apresentada à Universidade
Estadual do Norte Fluminense Darcy
Ribeiro, para a obtenção do título de
Mestre em Biociências e Biotecnologia.
Área de concentração: Biologia Celular.
Campos dos Goytacazes
Estado do Rio de Janeiro – Brasil
Abril – 2006

OFEREÇO
A Solange Maria e José Jorge, meus pais,
pelo apoio, amor e dedicação irrestritos.
In memoriam
dedico a Alexandrina Duarte Martins.

AGRADECIMENTOS Agradeço a Deus pela honra de viver e por ter estado em todos os
acontecimentos de minha vida. Sou inteiramente feliz na profissão que escolhi
com Vossa ajuda.
Agradeço ao meu grande pai pela sabedoria nos ensinamentos, pelo
investimento em minha educação e pelos conselhos tão valiosos. Sou
imensamente orgulhosa por sua garra.
Agradeço a minha amada mãe por nunca ter desistido de mim e sempre
ter respeitado, mesmo sem aceitar, qualquer decisão que eu tomasse. Por
estar sempre presente, mesmo nos meus piores momentos e muitas vezes me
presenteando com um silêncio necessário acompanhado de um olhar de “pode
contar comigo”. Apesar de sua aposentadoria, gostaria que soubesse que a
semente que plantou, germinou e que uma das minhas maiores aptidões é o
magistério.
Um abraço a minha irmã, futura advogada. Espero que um dia
possamos ter um convívio mais civilizado. Obrigada por me julgar mais
inteligente do que realmente sou.
Durante os seis anos que pude participar das atividades do laboratório,
observei que a minha orientadora tem um brilho e uma essência raros,
acompanhados de uma grande competência e profissionalismo. Agradeço,
então, a minha querida Dra. Olga Lima Tavares, por todos os ensinamentos
compartilhados.
Ao meu, namorado João Cláudio, pela amizade e respeito dedicados a
mim, tão necessários e nobres em dias atuais. Pelo caráter invejável, pela
conduta fiel e pelo amor demonstrado em cada pequeno ato, nesses 1570 dias
de namoro.
A todos os meus familiares, presentes ou distantes, pela simples
existência e pela acolhida em qualquer momento. Em especial, agradeço a
minha prima, Alessandra, e ao meu tio, Lenimar, pelo suporte por de trás das
câmeras desta tese e por todo apoio!
A Rachel e Carol, pela amizade ininterrupta e verdadeira.
Aos companheiros de laboratório: Thiago, Viviane, Shayanny, Hélio,
Natália, Camila, Fábio, Jucélia, Keyson e Cristiane, agradeço por fazer dos dias

de trabalho, uma grande satisfação e pelas pequenas ajudas, que final se
somam e tomam dimensões enormes. Que Deus e a ciência nos mantenham
sempre juntos.
Agradeço em especial ao Igor e ao Renato, que foram mais do que
simples bolsistas de trabalho, me ajudando diretamente nos experimentos.
Agradeço a Leandro Hespanhol pela paciência e gentileza de me
acompanhar em alguns experimentos e ao Eduardo Aragão por me ceder
reagentes e sempre estar a disposição com excelente humor.
Agradeço ao André “Pink” por sempre nos contemplar com sua simpatia
e competência. Você é um cara muito especial!
Agradeço a Willian Cabral pela presença constante e pelo apoio nos
últimos oito anos da minha vida.
Ao engenheiro agrônomo Angelo Savy Filho pela boa vontade em me
auxiliar mesmo a distância e sem me conhecer.
Agradeço a todos os professores e pós-graduandos do LQFPP pela
orientação na execução de diversos experimentos e utilização de
equipamentos.
Aos professores Ricardo, Tânia, Valdirene e Denise, por terem aceitado
fazer parte da banca de avaliadores da minha dissertação e por toda a
contribuição no decorrer do trabalho.
Por fim, agradeço ao grande amor da minha vida, Sra Alexandrina
Duarte Martins, que infelizmente não pôde ver o término deste trabalho, apesar
de estar presente na realização de cada experimento. Vozinha, como ela
sempre era chamada por netos e congêneres, sempre foi o que eu queria ser.
Sem mestrado, foi a maior mestre; sem doutorado, a maior doutora. Com a sua
partida, muita coisa perdeu o sentido e com sua presença consegui finalizar
metas que já não tinha mais coragem nem vontade de finalizar.

SUMÁRIO Lista de figuras .................................................................................................... X
Lista de tabelas .................................................................................................... XII
Lista de siglas, abreviaturas e símbolos .............................................................. XIII
Resumo ............................................................................................................... XVI
Abstract ............................................................................................................... XVII
1.0 – Introdução ................................................................................................... 01
2.0 – Revisão bibliográfica ................................................................................... 03
2.1 – Ricinus communis ....................................................................................... 03
2.2 - Defesa vegetal ............................................................................................. 05
2.2.1 - Rota octadecanóide como mecanismo de resposta de defesa ................ 07
2.2.2 - Variações das expressões protéicas em plantas e participação
hormonal .............................................................................................................. 10
2.2.3 – Rota octadecanóide e sua relação com a produção de peróxido de
hidrogênio (H2O2) ................................................................................................ 13
2.3 - Estresse oxidativo ....................................................................................... 15
2.4 – Antioxidantes enzimáticos .......................................................................... 19
2.4.1 – Superóxido dismutase (SOD) .................................................................. 19
2.4.2 – Catalase (CAT) ........................................................................................ 20
2.4.3 – Ascorbato peroxidase (APX) .................................................................... 21
2.4.4 – Guaiacol peroxidase (GPX) ..................................................................... 22
2.4.5 – Peroxiredoxina (Prx) ................................................................................ 23
2.4.6 – Polifenol oxidase (PPO) ........................................................................... 25
2.5 – Resposta às Espécies Reativas de Oxigênio (ERO) .................................. 27
3.0 – Objetivos gerais .......................................................................................... 29
3.1 – Objetivos específicos .................................................................................. 29
4.0 – Material e Métodos ...................................................................................... 30
4.1 - Equipamentos ............................................................................................. 30
4.2 – Reagentes .................................................................................................. 30

4.3 - Material vegetal e condições de crescimento .............................................. 31
4.4 - Indução de estresse .................................................................................... 31
4.5 - Obtenção de extratos protéicos ................................................................... 32
4.6 - Determinação do teor de proteínas nos extratos brutos .............................. 33
4.7 - Análise do perfil protéico em gel de poliacrilamida descontínua ................. 33
4.7.1 - Na presença de dodecil sulfato de sódio (SDS-PAGE) ............................ 33
4.7.2 - Na ausência de dodecil sulfato de sódio (NATIVE-PAGE) ....................... 33
4.7.3 – Bidimensional .......................................................................................... 34
4.8 - Determinação das atividades antioxidantes por espectofotometria ............. 34
4.8.1 - Determinação da atividade catalase (CAT) .............................................. 34
4.8.2 - Determinação da atividade ascorbato peroxidase (APX) ......................... 34
4.8.3 - Determinação da atividade guaiacol – peroxidase (GPX) ........................ 35
4.8.4 - Determinação da atividade polifenol oxidase (PPO) ................................ 35
4.9 – Determinação das atividades antioxidantes por PAGE não desnaturante .. 36
4.9.1 - Atividade da superóxido dismutase (SOD) ............................................... 36
4.9.1.2 - Determinação das isoformas da superóxido dismutase (SOD) .............. 36
4.9.2 - Atividade da catalase (CAT) ..................................................................... 37
4.9.3 - Atividade da ascorbato peroxidase (APX) ................................................ 37
4.9.4 - Atividade da guaiacol peroxidase (GPX) .................................................. 38
4.10 – Caracterização da GPX ............................................................................ 38
4.10.1 – Determinação do pH ideal da GPX ........................................................ 38
4.10.2 – SDS-PAGE da enzima homogênea ....................................................... 38
4.10.3 – Determinação do ponto isoelétrico da GPX por eletroforese
bidimensional ....................................................................................................... 39
4.11 - Detecção in vivo do peróxido de hidrogênio .............................................. 39
4.12 – Varredura de radicais livres pelo método do DPPH .................................. 40
4.13 - Analise estatística dos resultados ............................................................. 40
5.0 – Resultados .................................................................................................. 41
5.1 – Determinação do perfil protéico por SDS-PAGE ........................................ 41
5.2 – Determinação das atividades enzimáticas .................................................. 42
5.2.1 – Determinação da atividade superóxido dismutase (SOD) após
separação das proteínas por eletroforese não desnaturante .............................. 42

5.2.1.1 – Determinação das isoformas da superóxido dismutase (SOD) ............ 42
5.2.2 – Determinação da atividade catalase (CAT) no extrato bruto ................... 43
5.2.2.1 – Determinação da atividade catalase (CAT) após separação das
proteínas por eletroforese não desnaturante ....................................................... 43
5.2.3 – Determinação da atividade ascorbato peroxidase (APX) no extrato
bruto .................................................................................................................... 45
5.2.3.1 – Determinação da atividade ascorbato peroxidase (APX) após
separação das proteínas por eletroforese não desnaturante .............................. 45
5.2.4 – Determinação da atividade polifenol oxidase (PPO) no extrato bruto ...... 45
5.2.5 – Determinação da atividade guaiacol peroxidase (GPX) no extrato bruto . 47
5.2.5.1 – Determinação da atividade guaiacol peroxidase (GPX) por
eletroforese não desnaturante ............................................................................. 47
5.3 – Caracterização da enzima guaiacol peroxidase (GPX) .............................. 49
5.3.1 – Determinação do pH ideal ....................................................................... 49
5.3.2 – SDS-PAGE da enzima homegênea ......................................................... 50
5.3.3 – Determinação do ponto isoelétrico da GPX por eletroforese
bidimensional ....................................................................................................... 50
5.4 - Detecção in vivo do peróxido de hidrogênio ................................................ 52
5.5 - Varredura de radicais livres pelo método do DPPH .................................... 52
6.0 – Discussão ................................................................................................... 55
7.0 – Conclusão ................................................................................................... 64
8.0 – Referências Bibliográficas .......................................................................... 65

LISTA DE FIGURAS
Fig.1: Lavoura de mamona. Monteiro-PB ........................................................... 4
Fig.2: Estrutura química do metil jasmonato (MeJa) e do ácido jasmônico (AJ)..
Fig.3: Representação esquemática da produção de ácido jasmônico e seu
metil éster, metil jasmonato, pela via octadecanóide. .......................................... 8
Fig.4: Modelo para a regulação diferencial de genes da via de sinalização e
genes defensivos em tomateiros em resposta à injúria ........................................ 14
Fig.5: Etapas de redução univalente do O2 ......................................................... 16
Fig.6: Reação de Fenton ..................................................................................... 16
Fig.7: Sistema antioxidante de defesa composto por componentes enzimáticos
e não enzimáticos ................................................................................................ 17
Fig.8: Dismutação do superóxido catalisada pela SOD ...................................... 18
Fig.9: Degradação do peróxido de hidrogênio catalisada pela CAT .................... 19
Fig.10: Degradação do peróxido de hidrogênio catalisada pela APX ................. 21
Fig.11: Redução do H2O2 e tetramerização do guaiacol catalisada GPX ............ 22
Fig.12: Reação catalisada pela Prx .....................................................................
Fig.13: Reações catalisadas pela PPO ............................................................... 25
Fig.14: Representação esquemática do tratamento com metil jasmonato e
extração protéica em folhas de Ricinus communis .............................................. 31
Fig.15: Eletroforese em gel de poliacrilamida 12% na presença de SDS 0,1%
dos extratos protéicos de folhas de Ricinus communis, obtidos a partir das
plantas controle e tratadas com metil jasmonato.................................................. 40
Fig.16: Atividade da superóxido dismutase de plantas de Ricinus communis
submetidas ao tratamento com metil jasmonato após separação das proteínas
por eletroforese nativa 8%.................................................................................... 41
Fig.17: Isoformas da SOD determinada por eletroforese nativa em plantas de
Ricinus communis................................................................................................. 42

Fig.18: Atividade específica da catalase (µmol peróxido min-1 mg-1 de
proteína) nos extratos de plantas de Ricinus communis submetidas ao
tratamento com metil jasmonato em diferentes intervalos de tempo.................... 43
Fig.19: Atividade catalase determinada em gel após separação das proteínas
por eletroforese nativa 8% ................................................................................... 43
Fig.20: Atividade específica da ascorbato peroxidase (mM min-1 mg-1 de
proteína) nos extratos de plantas de Ricinus communis submetidas ao
tratamento com metil jasmonato em diferentes intervalos de tempo.................... 45
Fig.21: Atividade ascorbato peroxidase determinada em gel após separação
das proteínas por eletroforese nativa 8% . ........................................................... 45
Fig.22: Atividade polifenol oxidase (∆Abs mg -1 min -1) determinada em gel
após separação das proteínas por eletroforese nativa 8%................................... 46
Fig.23: Atividade específica da guaiacol peroxidase (µmol min-1 mg-1 de
proteína) nos extratos de plantas de Ricinus communis submetidas ao
tratamento com metil jasmonato em diferentes intervalos de tempo.................... 47
Fig.24: Atividade guaiacol peroxidase determinada em gel após separação das
proteínas por eletroforese nativa 8% . .................................................................. 47
Fig.25: SDS-PAGE 12% das bandas G1 e G2 obtidas após eletroforese nativa
8%, na qual as proteínas foram ensaiadas para revelação da atividade GPX ..... 50
Fig.26: Eletroforese bidimensional das bandas G1 e G2. .................................... 50
Fig.27: Visualização da formação de peróxido de hidrogênio nas folhas de
Ricinus communis tratadas com metil jasmonato em diferentes intervalos de
tempo.................................................................................................................... 52
Fig.28: Fotografias tiradas das imagens obtidas na lupa (aumento 2X) nas
folhas de Ricinus communis. ................................................................................ 53
Fig.29: Detecção de radicais livres nos extratos foliares de plantas de Ricinus
communis expostas ao tratamento com MeJa ..................................................... 53

LISTA DE TABELAS
TABELA I: Genes que respondem ao ácido jasmônico e suas expressões sob
diversas condições de estresse ........................................................................... 11
TABELA II: Listagem de reagentes e sua procedência ....................................... 29
TABELA III: Preparo das amostras e dos controles para a verificação do
conteúdo de radicais livres nas folhas de Ricinus communis ..............................39
TABELA IV: Determinação das isoformas da SOD ............................................. 42
TABELA V: Determinação da atividade guaiacol peroxidase em diferentes pHs . 48

LISTA DE SIGLAS, ABREVIATURAS E SÍMBOLOS
13-HPOT .......................... Ácido 13-hidroperoxido-octadecatrienóico
2 – CP ..............................2-cys peroxiredoxina
1 – CP...............................1-cys peroxiredoxina
ABA .................................Ácido abscísico
AJ ....................................Ácido jasmônico
AL ....................................Ácido linolênico
AOS .................................Aleno óxido sintase
APX .................................Ascorbato peroxidase
AS ....................................Ácido salicílico
ATP ..................................Adenosina trifosfato
CAT .................................Catalase
KCN- ................................Cianeto de potássio
CuZn-SOD .......................Cobre-zinco superóxido dismutase
DAB .................................3,3 diaminobenzidina
DNA .................................Ácido Desoxirribonucléico
DPI ...................................Cloreto de difenil iodo
DPPH ...............................1,1-difenil-2-picrilhidrazil
DTT ..................................Ditiotreitol
EDTA ............................... Ácido etileno diamino tetracético
EGTA................................Ácido bis (2-aminoetil) etilenoglicol-N,N,N',N'-
tetracético
ERO .................................Espécies reativas de oxigênio
Fe-SOD ...........................Ferro superóxido dismutase
FeCl3 ................................Cloreto férrico

GPX .................................Guaiacol peroxidase
H2O2 .................................Peróxido de hidrogênio
HR ...................................Resposta de hipersensibilidade
K3 [ Fe (CN)6 ] ..................Ferricianeto de potássio
KCN .................................Cianeto de potássio
LOX ................................13-lipoxigenase
LSU ..................................Cadeia pesada de ribulose 1,5- bisfosfato-
carboxilase/ oxigenase
MDA .................................Monodehidroascorbato
MDAR ..............................Monodeidroascorbato redutase
MeJa ................................Metil jasmonato
MJRC21 ...........................Metil Jasmonato Ricinus communis 21 kDa
Mn-SOD ...........................Manganês superóxido dismutase
mRNA ..............................Ácido Ribonucléico Mensageiro
NaCl .................................Cloreto de Sódio
NADPH ............................Nicotinamida adenina di-nucleotídeo fosfato
NBT ..................................Nitro blue tetrazolio
o-metoxi fenol ..................guaiacol
O2 .....................................Oxigênio molecular
O2.- ...................................ânion superóxido
OGA .................................Ácido oligogalacturônico
OH. ...................................Radical hidroxila
OPC .................................Oxo-pentenil-ciclopentanos
OPDA ..............................ácido oxo-fitodienoico
OVA .................................Ovalbumina
PAGE ............................... Eletroforese em gel de poliacrilamida

PAL .................................. Fenilalanina amônia liase
PG ................................... Poligalacturonases
PIIF .................................. Proteinase Inhibitor Inducing Factor
PIs ...................................Inibidores de proteinases
PLA2 ................................Fosfolipase A2
PMSF ...............................Fenil Metil sulfonilfluoride
PPO .................................Polifenol oxidase
PR ....................................Proteínas relacionadas à patogênese
Prx ...................................Peroxiredoxina
PS I .................................. Fotossistema 1
PS II ................................. Fotossistema 2
PVPP ...............................Polivinilpolipirrolidona
RIPs .................................Proteínas inativadoras de ribossomos
SDS .................................Dodecil sulfato de sódio
SOD .................................Superóxido dismutase
SSU .................................Cadeia leve de ribulose 1,5- bisfosfato-carboxilase/
oxigenase
TEMED ............................N,N,N’,N’ Tetrametilenodiamina
TRX .................................Tioredoxina
UV ....................................Ultra violeta

RESUMO
As plantas frequentemente estão expostas à estresses bióticos e abióticos que
prejudicam o seu crescimento, desenvolvimento ou a sua produtividade. Estes
estresses disparam várias respostas, desde alteração na expressão gênica e
metabolismo celular até variação na taxa de crescimento e produção de biomassa. O
estresse oxidativo estimula a biossíntese de componentes antioxidantes e aumenta a
atividade de enzimas antioxidantes. Contudo, no início do estresse, uma redução
transiente de algumas atividades antioxidantes pode ser observada, contribuindo para
um aumento de espécies reativas de oxigênio. Sabe-se que a superóxido dismutase
(SOD), catalase (CAT), ascorbato peroxidase (APX), peroxidase (Prx) e polifenol
oxidase (PPO) influenciam diversas reações envolvidas na sinalização de defesa
vegetal. Suas concentrações e atividades orquestrarão muitos processos fisiológicos
envolvidos no estresse oxidativo. O objetivo do nosso trabalho foi verificar o
comportamento dessas enzimas em plantas Ricinus communis tratadas com metil
jasmonato (MeJa) após 1, 4, 6, 12, 24 e 48 horas. Para isto, duas folhas de cada
planta foram congeladas em nitrogênio líquido. As proteínas foram extraídas com
tampão fosfato e o extrato foi centrifugado. O perfil protéico foi analisado por SDS-
PAGE de acordo com a metodologia proposta por Laemmli (1970) e as atividades
antioxidantes foram verificadas utilizando-se os respectivos protocolos. O tratamento
com MeJa causou uma redução transiente nos níveis da proteína 2-cys
peroxiredoxina. A atividade da isoforma Mn-SOD da enzima superóxido dismutase
decresceu após 24 e 48h de tratamento. A atividade catalase decresceu com uma
hora de tratamento a aumentou até 12h. Após este período, a atividade retornou aos
níveis de controle. A atividade ascorbato peroxidase aumentou após 4 e 24h de
tratamento. A atividade das duas guaiacol peroxidases detectadas decresceram com
1, 4 e 6h. Com 12h retornaram aos níveis de controle e após esse período cresceram
significativamente. O pH ideal para esta enzima foi em torno de 6,0. As bandas com
atividade guaiacol peroxidase foram extraídas do gel e tiveram a massa molecular e pI
determinados. Ambas apresentaram duas bandas protéicas com massa molecular
próxima de 39 kDa e pI entre 8.5 e 8,7. A atividade PPO não mostrou alterações
significativas. O MeJa induziu o acúmulo tanto de peróxido de hidrogênio quanto de
radicais livres com 1, 4, 6 e 12h de tratamento. Após este período não se observou
peróxido de hidrogênio ou radicais livres nas folhas. Esses dados reforçam a idéia de
que o MeJa pode estar relacionado com alterações no estresse oxidativo, alterando as
atividades de algumas enzimas chave que controlam este processo, promovendo o
acúmulo de ERO nos estágios iniciais de estresse.

ABSTRACT
Plants frequently encounter biotic and abiotic stress that adversely affect growth,
development, or productivity. These stresses trigger a wide range of plant responses,
from altered gene expression and cellular metabolism to changes in growth rates and
crop yields. Oxidative stress stimulates synthesis of antioxidant metabolites and
enhances antioxidant enzyme activities that could protect plant tissues. However, in the
early stages of stress a transient reduction of some antioxidant activities were
observed contributing to an increase of oxygen reactive species. It is known that
superoxide dismutase (SOD), catalase (CAT), ascorbate peroxidase (APX), peroxidase
(Prx) and polyphenol oxidase (PPO) influence several reactions involved in plant
defence systems. Their concentrations and activities will orchestrate many
physiological processes involved in oxidative stress. The intention of our work was to
verify the behavior of the enzymes in Ricinus communis plants treated with methyl
jasmonate after 1, 4, 6, 12, 24 and 48 hours. For this, two leaves of each plant were
harvested and frozen in liquid nitrogen. The proteins were extracted with phosphate
buffer and the slurry was centrifuged. The protein pattern was analyzed by SDS-PAGE
according to Laemmli and antioxidant activities were verified by their respective
protocols. Methyl jasmonate treatment caused a transient reduction in the levels of 2-
Cys peroxiredoxin protein. The activity of Mn-SOD isoform of superoxide dismutase
decreased after 24 and 48h of treatment. Catalase activity decrease with 1h of
treatement and then increased even 12h. After this period, the activity remains to
control levels. Ascorbate peroxidase activity increased after 4 and 24h of treatment.
The activity of the two guaiacol peroxidase activities detected decreased with 1, 4 and
6h. With 12 h it returned remains to control levels and after that increased significantly.
The ideal pH for this enzyme was about 6,0. The bands with peroxidase activity were
extracted from the gel and its mass and pI were determined. Both of them presented
two proteic bands after SDS-PAGE near 39kDa and a pI near 8,5-8,7. PPO activity
does not show significant changes. Methyl jasmonate induced both hydrogen peroxide
and free radicals accumulation with 1, 4, 6 and 12h of treatment. After that, hydrogen
peroxide and free radicals were not observed in the leaves. This data supports the idea
that methyl jasmonate can be related to changes in oxidative stress, altering the
activities of some key enzymes that control this process, promoting the accumulation of
ERO at early stages of stress.

1.0 – Introdução
Nas últimas duas décadas, houve um avanço sem antecedentes em
relação ao estudo da defesa de plantas e seus mecanismos de sinalização
para prevenir ou cessar tanto o desenvolvimento de doenças como o ataque de
patógenos. Muitos são os tipos de estresses que os vegetais podem enfrentar,
como oscilações drásticas de temperatura, umidade, radiação solar, ataque de
pestes ou patógenos, dentre outros (Malick & Raí, 1999). A sobrevivência sob
estas condições de estresse depende da habilidade da planta em perceber os
estímulos, gerar e transmitir sinais e induzir alterações bioquímicas que
ajustem o metabolismo de acordo com a situação de estresse em questão
(Enyedi et al., 1992; Mehdy, 1994). Vários agentes, como por exemplo, Ca+2,
ácido jasmônico, etileno e ácido salicílico têm sido identificados como
transdutores de sinais em plantas (Enyedi et al., 1992; Klessig & Malamy,
1994).
A indução de proteínas de defesa é um processo complexo que segue
um determinado cronograma. Estudos feitos com células de tomate
(lycopersicon esculentum L.) mostram que o ferimento inicial induz à liberação
de um peptídeo altamente móvel denominado sistemina (Pearce et al., 1991) e
que essa liberação ocorre logo nos primeiros minutos (Ryan, 2000). Este
hormônio migra pelo floema até ligar-se ao seu respectivo receptor em células
vegetais em várias regiões da planta, induzindo a biossíntese de metil
jasmonato. Este, por sua vez, induz à produção de proteínas relacionadas com
defesa, tais como inibidores de proteinases (Farmer & Ryan, 1992).
As espécies reativas de oxigênio (ERO) são geralmente vistas como um
metabólito celular tóxico, por reagir avidamente com moléculas biológicas e
levar a uma morte celular programada, controlada geneticamente. Porém, em
baixas concentrações, ERO induzem genes de defesa e resposta adaptativa
(Breusegem et al., 2001). O peróxido de hidrogênio regula diretamente a
expressão de numerosos genes, alguns dos quais, envolvidos em defesa de
plantas e resposta hipersensitiva (Levine et al., 1994). Experimentos de
Nürnberger et al. (1994), indicaram a presença, tanto o superóxido, quanto o
produto de sua dismutação (H2O2), durante interações incompatíveis entre

plantas resistentes e patógenos avirulentos. O H2O2, produzido durante
situações de estresse, é um dos responsáveis pela indução de enzimas
antioxidantes envolvidas na atenuação do estresse oxidativo (Pastori & Trippi,
1992).
Orozco-Cardenas & Clarence Ryan demonstraram em 1999 que
sistemina, ácido oligogalacturônico (OGA), quitosana e metil jasmonato,
induzem o acúmulo de peróxido de hidrogênio (H2O2) em folhas de tomate.
Posteriormente, o mesmo grupo verificou que o H2O2 é gerado em feixes
vasculares de folhas de tomate em resposta à injúria e se comporta como um
sinal difusível para a expressão de genes de defesa em células do mesofilo.
Dentre as enzimas que protegem as células e compartimentos sub
celulares vegetais dos efeitos citotóxicos das ERO, pode-se citar: superóxido
dismutase, ascorbato peroxidase, glutationa redutase, peroxiredoxina, catalase
e polifenol oxidase. Além da ação enzimática, os organismos contam com a
ação de metabólitos, como a glutationa, ácido ascórbico, α-tocoferol e
carotenóides para detoxificar seus ambientes celulares (Scandalios, 1993; Inzé
& Montagu, 1995; Mittler, 2002).
Neste trabalho a atividade de algumas enzimas antioxidantes e o
conteúdo de ERO foram investigados. O estudo das alterações das atividades
de enzimas antioxidantes pode fornecer, de forma direta, evidências sobre o
estresse oxidativo. Indiretamente, pode fornecer dados importantes no
esclarecimento de rotas de sinalização de defesa em plantas. Além disso,
alterações provocadas pela indução de estresses fisiológicos podem contribuir
para uma melhor compreensão da ação específica de cada enzima envolvida
na resposta antioxidativa em plantas.

2.0 – Revisão bibliográfica 2.1 – Ricinus communis
Ricinus communis L. (mamona) é uma Euphorbiacea de origem indiana
ou africana. Apesar da dúvida sobre sua procedência, esta planta é conhecida
desde a antigüidade pelos egípcios. A mamoneira atinge até dez metros de
altura em estado silvestre e floresce ao final do verão. É também conhecida
como rícino, palma – cristo e carrapateira em nosso país. Os espanhóis a
identificam como rícino ou “figuera del diablo”; já os ingleses e norte
americanos a identificam como “castor bean” ou “castor seed” (Fornazieri-
Júnior, 1986). A mamoneira se apresenta na forma de arbusto quando
cultivada e pode crescer em diferentes condições, em vários tipos de solos e
condições ambientais, podendo assim, ser um modelo útil para análise de
mecanismos de sinalização e defesa .
A mamoneira é uma oleaginosa de grande importância econômica e
social, sendo encontrada em estado selvagem em várias regiões do Brasil.
Suas sementes, depois de industrializadas, dão origem à torta e ao óleo de
mamona o qual, entre as diversas utilidades, é empregado na indústria de
plástico, siderurgia, saboaria, perfumaria, curtume, tintas e vernizes. Além
disso, é um excelente óleo lubrificante para motores de alta rotação e
carburante de motores a diesel.
O Brasil foi, durante décadas, o maior produtor mundial de bagas de
mamona e, é ainda, o maior exportador do seu óleo. O estado da Bahia é o
responsável pelas maiores produções no Brasil, seguido pelo estado de São
Paulo. A tecnologia utilizada no cultivo desta espécie tem evoluído
sensivelmente. Pode-se definir, basicamente, dois tipos de sistema de
produção; no primeiro, o qual a cultura assume papel social de grande
relevância, a força de trabalho familiar explora pequenas áreas, geralmente em
regime de consórcio com o feijão e o milho. Neste sistema não existe
mecanização nem utilização de insumos modernos, como sementes
melhoradas, defensivos, fertilizantes, etc. No segundo sistema, o cultivo
assume um caráter mais comercial, com a utilização de tração mecânica e de
insumos modernos (Azevedo & Lima, 2001). Desde 1936, o Instituto

Agronômico de Campinas vem desenvolvendo o projeto “melhoramento da
mamoneira” que visa obter cultivares mais produtivos.
A cultura da mamona pode se tornar em curto prazo, um dos principais
componentes de um programa nacional de biodiesel (fig.1). A estimativa é de
que cerca de 40% do biodiesel produzido no Brasil nos próximos anos, sejam
obtidos a partir dessa oleaginosa (Azevedo & Lima, 2001).
Dentre os insetos que atacam a mamoneira, destacam-se: o percevejo-
verde (Nezara viridula), que provoca murcha e secamento de folhas e frutos; a
cigarrinha (Agallia spp), que produz manchas cloróticas nas folhas, que com o
tempo ficam necrosadas. Também causam danos às folhas de mamona o
ácaro rajado (Tetranychus urticae) e o ácaro vermelho (Tetranychus ludeni).
Estes deixam manchas esbranquiçadas na parte abaxial das folhas e
pontuações cloróticas na parte adaxial, levando ao secamento e queda das
folhas com o passar do tempo. Como representantes dos lepidópteros,
destacam-se duas espécies de larvas conhecidas como lagartas-das-folhas
(Spodoptera latifascia e Thaelesa citrina). Ambas as espécies provocam
ataques severos à mamoneira que podem levar ao desfolhamento completo da
planta. A cultura da mamoneira pode ainda ser afetada por diferentes
patógenos causadores de importantes doenças que podem afetar o seu
desenvolvimento e sua produção, como os fungos: Macrophomina phaseolina e
Botrytis ricini, que causam respectivamente, podridão-de-macrophomina e
mofo cinzento (Azevedo & Lima, 2001).
A mamona (fig. 1) se classifica da seguinte forma (disponível em: < http:
//www.ncbi.nih.gov/ > Acesso em: 03/04/06).

Super Reino: Eucaryota
Reino: Viridiplantae
Filo: Streptophyta
Superdivisão: Spermatophyta
Divisão: Magnoliophyta
Classe: Magnoliopside
Subclasse: Rosidae
Ordem: Malpighiales
Família: Euphorbiacea
Sub família: Acalyphoideae Fig.1: Lavoura de mamona. Monteiro-PB
Tribo: Acalypheae
Gênero: Ricinus
Espécie: Ricinus communis L.
2.2 - Defesa vegetal
Quando feridas, as plantas conseguem mudar a constituição de
compostos moleculares, como um mecanismo de resposta a esse tipo de
estresse. Muitas dessas alterações podem estar diretamente relacionadas com
defesa e proteção. Para sobreviver, durante sua evolução, elas desenvolveram
mecanismos de resposta a danos e doenças que, quando acionados,
reconhecem a agressão. Saber como os vegetais se protegem é essencial para
obter, por meio da bioengenharia, variedades agrícolas mais resistentes, o que
poderia aumentar a produção e a qualidade da planta em questão. Por isso,
diversos grupos de pesquisa buscam definir o papel de cada substância
participante dos processos bioquímicos de defesa das plantas (Shewry &
Lucas, 1997).
Uma das principais revelações das pesquisas sobre defesa vegetal é
que a resistência a pestes e patógenos é usualmente complexa e tem como
base, a ação combinada de diversos fatores e não um componente somente.
Uma distinção fundamental é geralmente feita entre defesas pré-existentes ou
constitutivas das plantas e sistemas induzidos pela agressão da peste ou
patógeno. As defesas vegetais podem ser também classificadas como
estruturais, baseadas em características anatômicas, e químicas, baseadas em

compostos biologicamente ativos de massa molecular variada (Shewry &
Lucas, 1997).
A defesa estrutural é encontrada nas mais diversas partes da planta. A
presença de pêlos, espinhos, tricomas e ceras recobrindo principalmente a
superfície de caules e frutos, são exemplos típicos dessas estruturas de defesa
(Bowles, 1990).
A importância de proteínas como forma de defesas constitutivas de
plantas é um assunto estudado há muito tempo. Como exemplo, o estudo de
Ryan & Balls (1962) constatou, pela primeira vez, a presença de inibidores de
proteinases em solanáceas. Posteriormente, inibidores de proteinases também
foram relacionados a processos de defesa induzidos por ferimento em folhas
de tomate e batata, tanto no local da agressão, quanto em locais afastados
(Green & Ryan, 1972). Os inibidores de proteinases interagem com as
proteinases do intestino de insetos e afetam negativamente a proteólise do
alimento ingerido, reduzindo a disponibilidade de aminoácidos essenciais e
retardando o crescimento e o desenvolvimento dos mesmos (Constabel &
Ryan, 1995). As proteínas protetoras têm expressão significativa e podem ser
descritas como constitutivas ou tecido específicas, sendo restrita a órgãos,
tecidos ou tipos celulares específicos. Esse tipo é particularmente distribuído
em sementes e em alguns tecidos vegetativos (Shewry & Lucas, 1997).
Dentre as substâncias relacionadas à defesa química, destacam-se
aminoácidos não protéicos, alcalóides, fenóis, saponinas, lectinas, RIPs
(proteínas inativadoras de ribossomos) quitinases, glucanases, flavonóides,
inibidores de proteases, e alérgenos (Bowles, 1990; Xavier-Filho, 1993). As
proteínas protetoras, citadas anteriormente, também podem ser induzidas em
resposta à infecção, dano ou predação. Entre elas estão as proteínas
relacionadas à patogênese (PR), que por sua vez são subdivididas em diversos
grupos (β-1,3-glucanases, quitinases, peroxidases, etc) (Shewry & Lucas,
1997).
Fatores anatômicos constitutivos e químicos, como cutículas, parede
celular e inibidores pré-formados podem ser suficientes para prevenir a
colonização de tecidos vegetais. Se a penetração ocorrer, o sistema de defesa
induzido é ativado. Este inclui a rápida geração de espécies reativas de

oxigênio, alterações em polímero da parede celular, síntese de metabólitos de
baixo peso molecular como as fitoalexinas, produção de novas classes de
proteínas relacionadas à defesa e uma resposta de hipersensibilidade seguida
por morte celular programada. Coletivamente, esses sistemas primeiro inibem
e depois impedem o potencial colonizador (Shewry & Lucas, 1997).
2.2.1 - Rota octadecanóide como mecanismo de resposta de defesa.
Por muito tempo, a questão de como ocorre o processo de sinalização
da agressão da área lesada de uma folha até outras folhas de uma planta
permaneceu indefinida. O grupo liderado pelo Dr. Clarence Ryan, pioneiro de
tais investigações, acreditava que um fator molecular móvel era responsável
por esta indução, e o definiu como PIIF – “Proteinase Inhibitor Inducing Factor”
(Walker Simmons & Ryan, 1977). Em 1990, foi descoberto que um metabólito
secundário comum em plantas, o metil jasmonato (MeJa), induzia, por via
aérea, a biossíntese de inibidores de proteinases em diferentes plantas (das
famílias fabacea e solanacea), mesmo estando estas fisicamente separadas
(Farmer & Ryan, 1990). Este grupo observou também que células de tomate
(Lycopersicon esculentum L.) respondem à injúria e a herbivoria, liberando um
peptídeo octadecamérico altamente móvel denominado sistemina, que é um
milhão de vezes mais poderoso que oligossacarídeos (que atuam apenas no
sítio de ataque) na indução da biosíntese de inibidores de proteinases serínicas
em folhas de tomate feridas (Pearce et al., 1991).
A sistemina é originalmente sintetizada como uma proteína precursora
de 200 aminoácidos chamada pró–sistemina (McGurl et al., 1994). Depois da
ativação proteolítica e liberação, o oligopeptídeo é rapidamente translocado
para tecidos não injuriados, onde se liga em um receptor de membrana que
possui 50kDa (Pearce et al., 1993; Schaller & Ryan, 1994). Subseqüente à
ativação do receptor, o caminho de sinalização se dá, hipoteticamente, via
estimulação de uma fosfolipase A2 (PLA2), resultando na liberação intracelular
do ácido linolênico (AL), o qual é convertido, em uma reação catalisada por
uma 13-lipoxigenase (LOX), a ácido 13-hidroperoxido-octadecatrienóico (13-
HPOT) nos cloroplastos. O 13-HPOT é convertido, pela catálise da aleno óxido
sintase (AOS), a uma ciclopentanona, o ácido oxo-fitodienoico (OPDA). A maior

parte do OPDA parece estar esterificada a lipídios de membrana. Estas
ciclopentanonas deixam o cloroplasto e funcionam como moléculas
sinalizadoras no citossol ou se convertem a oxo-pentenil-ciclopentanos (OPC),
que também podem agir como moléculas sinalizadoras no citossol. Os OPC
são metabolizados nos peroxissomos, nos quais ocorre a redução e a β-
oxidação com formação do ácido jasmônico (AJ). O jasmonato produzido deixa
o peroxissomo e pode ser metilado no citossol para formação de metil
jasmonato (fig. 2) e estes podem agir como moléculas sinalizadoras no citossol,
ativar a transcrição de genes relacionados a defesa no núcleo, agir em células
vizinhas ou serem inativados (fig. 3) (Farmer & Ryan, 1992; Weber, 2002).
Fig.2: Estrutura química do metil jasmonato (MeJA) e do ácido jasmônico (JA).
JA MeJA
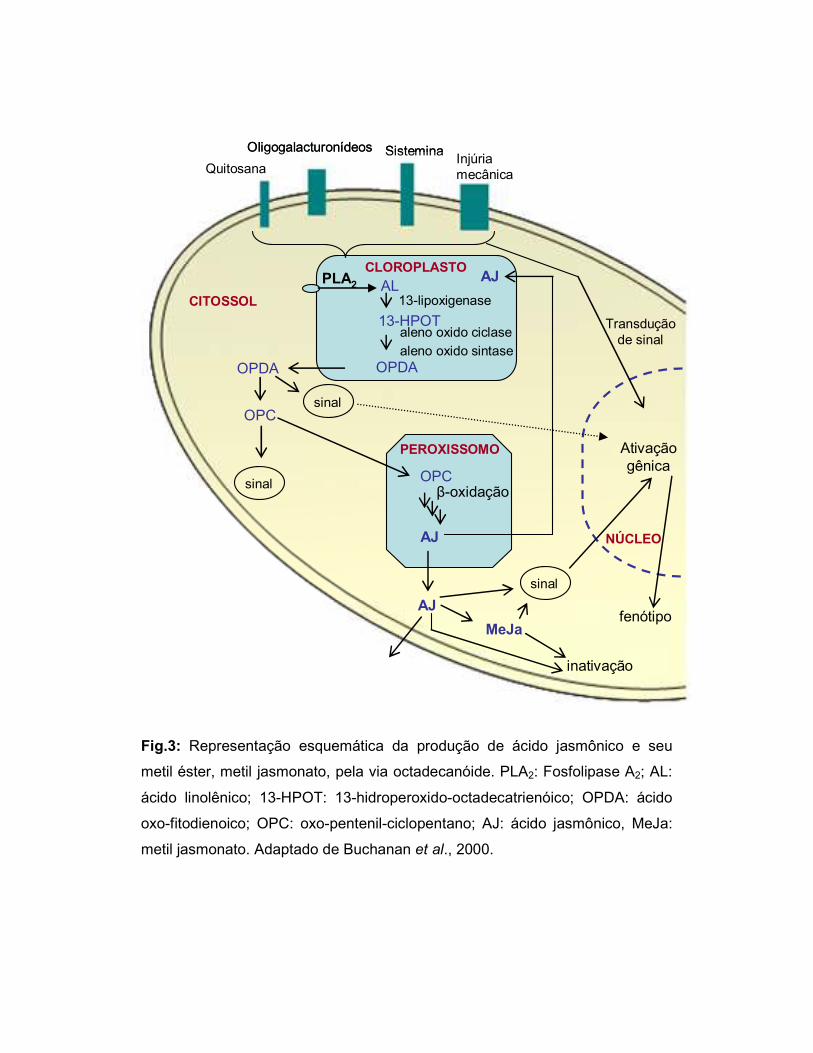
Fig.3: Representação esquemática da produção de ácido jasmônico e seu
metil éster, metil jasmonato, pela via octadecanóide. PLA2: Fosfolipase A2; AL:
ácido linolênico; 13-HPOT: 13-hidroperoxido-octadecatrienóico; OPDA: ácido
oxo-fitodienoico; OPC: oxo-pentenil-ciclopentano; AJ: ácido jasmônico, MeJa:
metil jasmonato. Adaptado de Buchanan et al., 2000.
CLOROPLASTO
PEROXISSOMO
NÚCLEO
Quitosana
Oligogalacturonídeos SisteminaInjúriamecânica
PLA2 AL
13-HPOT
OPDA
13-lipoxigenase
aleno oxido sintase
aleno oxido ciclase
CITOSSOL
OPDA
sinalOPC
sinal OPCβ-oxidação
AJ
AJ
AJ
MeJa
sinal
Ativaçãogênica
inativação
Transduçãode sinal
fenótipo
CLOROPLASTO
PEROXISSOMO
NÚCLEO
Quitosana
Oligogalacturonídeos SisteminaInjúriamecânica
PLA2 AL
13-HPOT
OPDA
13-lipoxigenase
aleno oxido sintase
aleno oxido ciclase
CITOSSOL
OPDA
sinalOPC
sinal OPCβ-oxidação
AJ
AJ
AJ
MeJa
sinal
Ativaçãogênica
inativação
Transduçãode sinal
fenótipo

2.2.2 - Variações das expressões protéicas em plantas e participação
hormonal
Várias pesquisas constataram que estresses bióticos ou abióticos levam
a alteração no padrão de expressão de proteínas das plantas, podendo ocorrer
tanto a inibição quanto à indução da biossíntese de determinados constituintes
protéicos. Em 1972, por exemplo, Green & Ryan verificaram que há indução de
inibidores de proteinases em tomate, como um mecanismo possível de defesa
contra insetos. Cavalcante e colaboradores relataram, em 1999, que o metil
jasmonato altera os níveis da enzima rubisco e de outras proteínas. Ghosh e
colaboradores (2001) correlacionaram alterações nos teores de rubisco com
senescência em Brassica napus.
Diferentes partes da planta, que passaram por injúria, podem responder
de maneiras diferentes a uma dada lesão. Além disso, plantas de idades
diferentes podem apresentar diferentes níveis de expressão protéica em partes
vegetais semelhantes (Buchanan et al., 2000). Estudos de Alarcon & Malone
(1995) relataram que a idade da planta influencia a indução de inibidores de
proteinase em resposta a injúria em tomate. Esses dados evidenciam que a
expressão de proteínas em plantas feridas não é um processo somente
dependente da lesão, mas também do estado fisiológico do vegetal.
Um trabalho feito com plantas mutantes de tomate que apresentava
deleções em genes codificadores de enzimas da rota biossintética do MeJa,
mostrou que estas plantas, quando feridas, foram incapazes de produzir
proteínas relacionadas com defesa (Howe et al., 1996).
O MeJa é um composto de fórmula similar a das prostaglandinas,
hormônios de origem animal envolvidos com respostas inflamatórias (Farmer &
Ryan. 1992). Em plantas, foi descoberto pela primeira vez em jasmim, planta
que deu origem ao seu nome. A atividade biológica do MeJa é altamente
variável e dependente de sua concentração nos tecidos ou em meio de cultura
de células vegetais. Em concentrações maiores que 50 µM, causa senescência
em culturas celulares, induzindo à morte celular. Em concentrações que variam
de 1 a 10 µM, atua na indução da expressão de genes relacionados com
defesa, dentre outros genes, sem causar senescência (Mason & Mullet, 1990).

Vários pesquisadores têm avaliado as respostas de plantas em diversas
condições de estresse. Reinbothe e colaboradores (1994) e Reymond &
Farmer (1998) publicaram revisões sobre estes estudos e a tabela 1 apresenta
um resumo das alterações provocadas pelo ácido jasmônico em plantas.
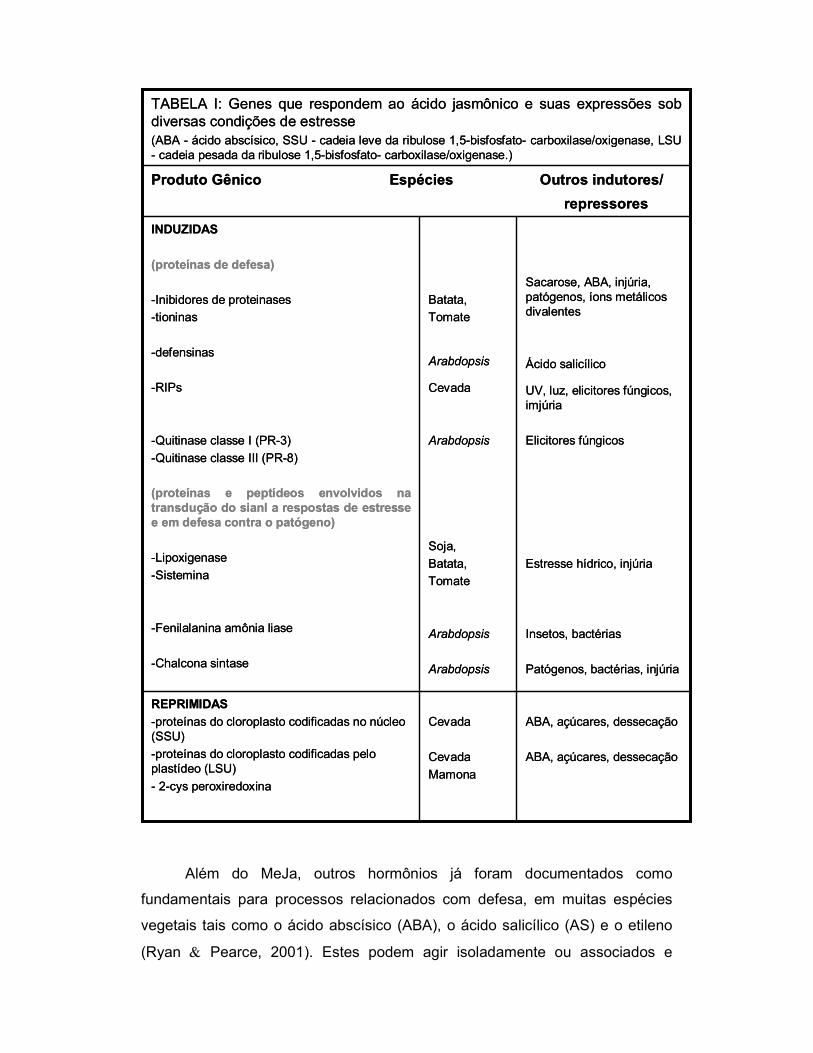
Além do MeJa, outros hormônios já foram documentados como
fundamentais para processos relacionados com defesa, em muitas espécies
vegetais tais como o ácido abscísico (ABA), o ácido salicílico (AS) e o etileno
(Ryan & Pearce, 2001). Estes podem agir isoladamente ou associados e
ABA, açúcares, dessecação
ABA, açúcares, dessecação
Cevada
Cevada
Mamona
REPRIMIDAS
-proteínas do cloroplasto codificadas no núcleo(SSU)
-proteínas do cloroplasto codificadas peloplastídeo (LSU)
- 2-cys peroxiredoxina
Sacarose, ABA, injúria, patógenos, íons metálicosdivalentes
Ácido salicílico
UV, luz, elicitores fúngicos, imjúria
Elicitores fúngicos
Estresse hídrico, injúria
Insetos, bactérias
Patógenos, bactérias, injúria
Batata,
Tomate
Arabdopsis
Cevada
Arabdopsis
Soja,
Batata,
Tomate
Arabdopsis
Arabdopsis
INDUZIDAS
(proteínas de defesa)
-Inibidores de proteinases
-tioninas
-defensinas
-RIPs
-Quitinase classe I (PR-3)
-Quitinase classe III (PR-8)
(proteínas e peptídeos envolvidos natransdução do sianl a respostas de estressee em defesa contra o patógeno)
-Lipoxigenase
-Sistemina
-Fenilalanina amônia liase
-Chalcona sintase
Produto Gênico Espécies Outros indutores/
repressores
TABELA I: Genes que respondem ao ácido jasmônico e suas expressões sob diversas condições de estresse(ABA - ácido abscísico, SSU - cadeia leve da ribulose 1,5-bisfosfato- carboxilase/oxigenase, LSU - cadeia pesada da ribulose 1,5-bisfosfato- carboxilase/oxigenase.)
ABA, açúcares, dessecação
ABA, açúcares, dessecação
Cevada
Cevada
Mamona
REPRIMIDAS
-proteínas do cloroplasto codificadas no núcleo(SSU)
-proteínas do cloroplasto codificadas peloplastídeo (LSU)
- 2-cys peroxiredoxina
Sacarose, ABA, injúria, patógenos, íons metálicosdivalentes
Ácido salicílico
UV, luz, elicitores fúngicos, imjúria
Elicitores fúngicos
Estresse hídrico, injúria
Insetos, bactérias
Patógenos, bactérias, injúria
Batata,
Tomate
Arabdopsis
Cevada
Arabdopsis
Soja,
Batata,
Tomate
Arabdopsis
Arabdopsis
INDUZIDAS
(proteínas de defesa)
-Inibidores de proteinases
-tioninas
-defensinas
-RIPs
-Quitinase classe I (PR-3)
-Quitinase classe III (PR-8)
(proteínas e peptídeos envolvidos natransdução do sianl a respostas de estressee em defesa contra o patógeno)
-Lipoxigenase
-Sistemina
-Fenilalanina amônia liase
-Chalcona sintase
Produto Gênico Espécies Outros indutores/
repressores
TABELA I: Genes que respondem ao ácido jasmônico e suas expressões sob diversas condições de estresse(ABA - ácido abscísico, SSU - cadeia leve da ribulose 1,5-bisfosfato- carboxilase/oxigenase, LSU - cadeia pesada da ribulose 1,5-bisfosfato- carboxilase/oxigenase.)

controlar os efeitos uns dos outros (Finch-Savage et al., 1996). O etileno
também é liberado após a agressão e, junto ao MeJa, atua na expressão de
proteínas de defesa (Pieterse & Van Loon, 1999). O ácido salicílico (AS),
contudo, inibe as vias de ativação de proteínas induzidas por MeJa e etileno
(Fidantsef et al., 1999). Porém, proteínas induzidas por AS podem ser
estimuladas por MeJa e etileno. O ácido abscíssico (ABA) é um hormônio
vegetal relacionado com a proteção de plantas a estresse hídrico, podendo
atuar potencializando a indução de biossíntese protéica relacionada com MeJa
(Finch-Savage et al., 1996). O tratamento de células vegetais, tanto com MeJa
(Kauss et al., 1994), quanto com AS (Kauss & Jeblick, 1995), leva a um
aumento na produção de H2O2. Além disso, a combinação de AS e MeJa leva a
uma super indução de genes relacionados à patogênese (PR1), resultado não
observado quando se usa o MeJa isoladamente (Xu et al., 1994). Bi &
colaboradores (1995) demonstraram que células de plantas agredidas por
injúria mecânica, por patógenos ou tratadas com AS ou MeJa, têm produção de
H2O2 e de genes PR1 aumentados em resposta a H2O2 exógeno.
2.2.3 – Rota octadecanóide e sua relação com a produção de peróxido de
hidrogênio (H2O2).
Sistemina, ácido oligogalacturônico (OGA), quitosana e metil jasmonato,
induzem o acúmulo de peróxido de hidrogênio (H2O2) em folhas de tomate. O
padrão de resposta corresponde a um aumento no mRNA para
poligalacturonases (PG) injúria-induzidas e a um aumento na atividade de tal
enzima. Isto sugere que os fragmentos do OGA, produzidos por PG, levam a
uma resposta de aumento na produção de H2O2 (Orozco-Cardenas & Ryan,
1999). Uma linhagem mutante de tomate com a rota octadecanóide
comprometida não exibe atividade PG ou aumento de H2O2 em resposta à
injúria, sistemina, OGA, ou quitosana, mas responde a metil jasmonato,
indicando que a geração de H2O2 requer uma rota octadecanóide funcional
(Orozco-Cardenas & Ryan, 1999).
Orozco-Cárdenas e colaboradores propuseram um modelo em 2001 (fig.
4), em que o ácido jasmônico, produzido através da rota octadecanóide, ativa
genes sinalizadores (genes iniciais) nos feixes vasculares, que são expressos

0,5h após a injúria (Ryan, 2000), enquanto o H2O2 produzido por derivados de
oligogalacturonideos (OGA) liberados por ação da enzima poligalacturonase
(PG), é um segundo mensageiro que ativa genes defensivos (genes tardios)
que são expressos 4h após a injúria (Ryan, 2000) nas células do mesofilo. Foi
verificado, com auxílio de um inibidor da expressão de genes de defesa tardios
(DPI), que o H2O2 gerado em feixes vasculares de folhas de tomate em
resposta à injúria se comportaria como um sinal difusível para a expressão de
genes de defesa em células do mesofilo, mas não como sinalizador nas células
dos feixes vasculares.
O gene que codifica a PG está entre os genes expressos inicialmente.
Porém, a poligalacturonase libera fragmentos de OGA da parede celular
vegetal que ativa, ambos os tipos de defesa (inicial e tardia) e a produção de
H2O2 em plantas de tomate, o que levanta questionamentos acerca do seu
papel no caminho de trasdução de sinal. (Orozco-Cárdenas et al., 2001).

Herbivoria
(injúria)
Pró sistemina
Sistemina
RMP
Ácido linolênico
Rota octadecanóide
Ácidojasmônico
Feixes vasculares
“Genes Iniciais”
Genes sinalizadores
(PG)
Células do mesofilo
“Genes Tardios”
Genes de defesa
(Pis, PPO)DPI
H2O2
Herbivoria
(injúria)
Pró sistemina
Sistemina
RMP
Ácido linolênico
Rota octadecanóide
Ácidojasmônico
Feixes vasculares
“Genes Iniciais”
Genes sinalizadores
(PG)
Células do mesofilo
“Genes Tardios”
Genes de defesa
(Pis, PPO)DPI
Herbivoria
(injúria)
Pró sistemina
Sistemina
RMP RMP
Ácido linolênico
Rota octadecanóide
Ácidojasmônico
Feixes vasculares
“Genes Iniciais”
Genes sinalizadores
(PG)
Células do mesofilo
“Genes Tardios”
Genes de defesa
(Pis, PPO)DPI
H2O2
Fig.4: Modelo para a regulação diferencial de genes da via de sinalização e
genes defensivos em tomateiros em resposta à injúria. DPI, Cloreto de difenil
iodo; PG, poligalacturonase; PIs, inibidores de proteinases ; MP, membrana
plasmática; PPO, polifenol oxidase; R, receptor. Adaptado de Orozco-Cárdenas
et al. (2001).
2.3 - Estresse oxidativo
O acúmulo de oxigênio molecular (O2) na Terra tornou possível a
evolução de organismos aeróbicos que utilizam o O2 como aceptor final de
elétrons. O O2 é pouco reativo, mas tem a capacidade de originar estados
excitados reativos como radicais livres e derivados (Scandalios, 1993). Com
dois átomos de oxigênio, o O2 é completamente reduzido por quatro elétrons
transportados ao longo da cadeia respiratória, gerando duas moléculas de
água. No entanto, uma pequena parcela dos elétrons escapa da cadeia
respiratória, resultando em uma redução parcial do oxigênio molecular, levando
à produção de espécies reativas de oxigênio (ERO) na forma de oxigênio
singleto (1O2), peróxido de hidrogênio (H2O2), radical hidroxila (OH.) e anion
superóxido (O2.-) (Mittler, 2002). Essas moléculas tóxicas são formadas durante
funções metabólicas normais nos peroxisomos ou induzidas por estímulos
ambientais aos quais as plantas estão constantemente expostas.

ERO são, sobretudo, subprodutos do metabolismo celular regular, mas
podem ser gerados com a destruição do sistema de transporte de elétrons
durante condições de estresse. O principal ponto de produção de ERO na
célula durante o estresse são as organelas com alta atividade de oxidação
metabólica ou com fluxo de elétrons sustentado: cloroplastos e mitocôndrias.
Nos cloroplastos, a formação de ERO está relacionada com eventos da
fotossíntese. O fenômeno de fotorrespiração nos peroxissomos é outra forma
de produção de H2O2. A produção de ERO em mitocôndrias de plantas recebeu
pouca atenção no passado, mas dados recentes sugerem que tais organelas
podem ser fontes de ERO sobre condições de estresse específicas
(Breusegem et al., 2001).
Sob condições adequadas de desenvolvimento, a produção de ERO na
célula é baixa (240 mM s-1 O2.- e um nível ''steady-state'' de 0,5 mM s-1 H2O2
nos cloroplastos), enquanto muitos estresses que alteram a homeostase
celular acentuam a sua produção (240 a 720 mM s-1 O2.- e 5 a 15 mM de H2O2)
(Mittler, 2002). Em plantas, a produção de ERO é favorecida por vários fatores
ambientais de estresse como a exposição a níveis elevados de luminosidade,
seca, metais pesados, elevada concentração de sais, extremos de
temperatura, radiação UV, poluição do ar, herbicidas, estresse físico e
mecânico e também como resposta ao estresses bióticos tais como o ataque
de patógenos (Malick & Raí, 1999).
Algumas ERO são classificadas como radicais livres por apresentarem
elétrons desemparelhados na sua estrutura, fazendo com que reajam
avidamente com moléculas biológicas, como DNA, proteínas e lipídeos,
podendo alterar suas funções. O efeito final gerado depende não só do
compartimento que está sendo afetado, mas do tipo de ERO que está reagindo
(Dröge, 2002).
As ERO são formadas em estapas de redução univalente a partir do
oxigênio molecular (fig. 5). O primeiro passo na redução de O2 produz radicais
de vida relativamente curta, os superóxidos. Esses radicais de oxigênio não
conseguem atravessar membranas biológicas, ficando confinados no
compartimento onde foram gerados. Os superóxidos formam hidroxiperóxidos
com duplas ligações (enos) ou duplas ligações alternadas (dienos), Além de
oxidar aminoácidos específicos, como metionina, histidina e triptofano. O

superóxido também pode causar peroxidação de lipídeos no ambiente celular e
nas membranas celulares (Breusegem et al., 2001). Posteriormente, a redução
do oxigênio gera peróxido de hidrogênio (H2O2), que, apesar de não ser um
radical livre, atravessa as biomembranas e se distribui a partir do local de sua
produção (Breusegem et al., 2001). A última e mais reativa espécie a ser
formada nessa reação é o radical hidroxil (OH.). Esse radical é formado pela
redução do H2O2 por íons metálicos (Fe2+ e Cu+) na reação de Fenton (fig. 6) e
tem grande afinidade por moléculas biológicas em seu sítio de produção. O OH.
Apresenta uma meia-vida muito curta, pois reage muito rapidamente com
moléculas biológicas, seqüestrando aleatoriamente um átomo de hidrogênio
(Breusegem et al., 2001; Nordberg & Arnér, 2001).
Fig.5: Etapas de redução univalente do O2.
Fig.6: Reação de Fenton.
As plantas protegem suas células e compartimentos sub celulares dos
efeitos citotóxicos das ERO com o auxilio de enzimas antioxidantes, como
superóxido dismutase, ascorbato peroxidase, glutationa redutase,
peroxiredoxina, catalase, polifenol oxidase e metabólitos, como a glutationa,
ácido ascórbico, α-tocoferol e carotenóides (Scandalios, 1993; Inzé & Montagu,
1995; Mittler, 2002) (Fig. 7).
O2 � O2.- � H2O2 � .OH + OH- � 2 H2O
e- e- e- e-
2H+ 2H+
H2O2 + Cu+ / Fe2+ � .OH + OH- + Cu2+ / Fe3+
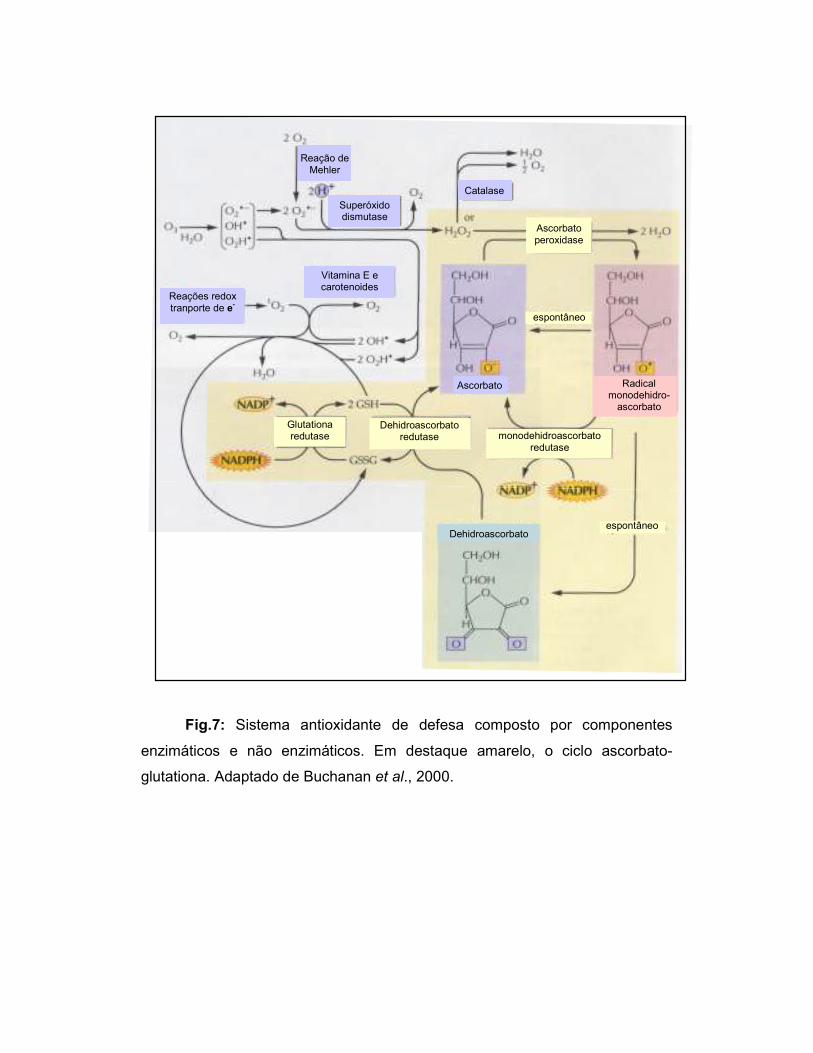
Fig.7: Sistema antioxidante de defesa composto por componentes
enzimáticos e não enzimáticos. Em destaque amarelo, o ciclo ascorbato-
glutationa. Adaptado de Buchanan et al., 2000.
Reação de Mehler
Superóxido dismutase
Catalase
Ascorbato peroxidase
espontâneo
Radical monodehidro- ascorbato
Ascorbato
espontâneo Dehidroascorbato
monodehidroascorbato redutase
Dehidroascorbato redutase
Glutationa redutase
Reações redox tranporte de e-
Vitamina E e carotenoides

2.4 – Antioxidantes enzimáticos
2.4.1 – Superóxido dismutase (SOD)
A enzima superóxido dismutase (SOD) tem um papel importante na
proteção dos organismos contra os efeitos do estresse oxidativo. Participa de
uma via crucial na defesa antioxidativa, pois catalisa a dismutação de O2.- para
H2O2 (fig. 8) (Scandalios, 1993). Quatro diferentes classes de SOD foram
identificadas, dependendo do metal presente no sítio ativo: Manganês, ferro,
cobre e zinco ou níquel, sendo a forma CuZn-SOD, a mais abundante em
vegetais. Elas se apresentam como homodímero no citossol, núcleo,
mitocôndria (membrana interna) e peroxissomo. Nos cloroplastos, a SOD existe
como um homotetrâmero em plantas superiores e como monômero no
periplasma de algumas bactérias gram – negativas (Dupeyrat et al., 2004).
A isoforma Cu/Zn-SOD é, em geral, sensível a CN- e H2O2, e ocorre
geralmente no citoplasma e no cloroplasto; a Fe-SOD é sensível a H2O2, e
resiste a CN-, ocorrendo no cloroplasto; e a Mn-SOD é resistente a H2O2 e CN-,
e está presente nas mitocôndrias (Bowler et al., 1992; Del Rio et al., 1998).
Vários trabalhos têm relatado aumento nas concentrações de anion superóxido
e, conseqüentemente, da atividade SOD, para amenizar os efeitos destrutivos
do radical, sobre diferentes condições de estresse (Larkindale & Huang, 2004;
Vierling, 1991; Dat et al., 2000).
Fig.8: Dismutação do superóxido catalisada pela SOD.
Análises detalhadas da expressão em tabaco e tomate mostram que os
genes da SOD são regulados diferenciadamente tanto no desenvolvimento
como na resposta vegetal a diversos tipos de estresses (Tsang et al., 1991;
Perl-Treves & Galun, 1991). Em Nicotiana plumbaginifolia (tabaco), a
expressão da Cu/Zn-SOD é aumentada preferencialmente pelo choque térmico
e congelamento, enquanto que, o mRNA da isoforma Fe-SOD é induzido
2 O2.- + 2 H+ ���� H2O2 + O2

preferencialmente por congelamento (Tsang et al. 1991). O elevado aumento
de transcritos de SOD sob condições de estresse corresponde, geralmente, a
um aumento bem mais moderado da atividade da enzima (Inzé & Montagu,
1995).
2.4.2 – Catalase (CAT)
Uma das enzimas responsáveis pela degradação de H2O2 é a catalase
(Stryer, 1995 ; Maté et al., 1999). Atualmente, é bem estabelecido que existam
três genes, não relacionados, de famílias de catalase capazes de degradar o
H2O2: a manganês catalase, reconhecida apenas em procariotos (Barnynin et
al., 1986); a catalase peroxidase, que é largamente distribuída em procariotos e
encontrada em eucariotos inferiores (Fraaije et al., 1996) e a família que tem
sido chamada de catalase verdadeira, correspondendo a hemoproteínas com
quatro cadeias (de aproximadamente 60 kDa cada uma) e um grupo heme em
cada uma, encontrada ubiquamente em eucariotos e também em muitos
procariotos (Stryer, 1995). São encontradas no citoplasma, nas mitocôndrias e
nos peroxissomos de células animais, vegetais e microrganismos aeróbicos
(Scandalios, 1993).
A degradação do H2O2 pela catalase se realiza em dois passos (fig. 9). A
primeira molécula de substrato é reduzida a água, deixando a enzima como um
intermediário oxoferril, chamado componente 1 (reação 1). Um dos dois
elétrons transferidos é doado a porfirina (Chance & Herbert, 1950). A forma
resultante da enzima passa pelo segundo passo do ciclo catalítico (reação 2),
pela transferência dos dois elétrons para uma segunda molécula de H2O2
(Dolphin et al., 1971).
Fig.9: Degradação do peróxido de hidrogênio catalisada pela CAT. 1, 2: reação
1 e reação 2, respectivamente. (Maté et al., 1999).

Existem três tipos distintos de catalases identificadas em plantas: CAT-1,
CAT-2 e CAT-3 (Scandalios, 1993). A CAT-1 é responsável por 80% da
atividade total da enzima e se localiza nos peroxissomos, sendo responsável
pela degradação do H2O2 gerado na fotorrespiração. A CAT-2 é encontrada em
tecidos vasculares e a CAT-3 está localizada no mesófilo das folhas
(Scandalios, 1990).
O peróxido de hidrogênio, além de ser tóxico para as células por inativar
enzimas do ciclo de Calvin, pode gerar outros radicais livres. Nos tecidos
infectados, uma redução da atividade da catalase levaria a um aumento de
peróxido de hidrogênio, que poderia provocar a morte celular, que caracteriza a
resposta de hipersensibilidade (HR), (Margis – Pinheiro et al., 1999).
2.4.3 – Ascorbato peroxidase (APX)
Segundo Foyer et al. (1994), a catalase está ausente no cloroplasto e a
degradação do peróxido de hidrogênio nesse compartimento é feita pela APX
ligada ao tilacóide. As moléculas de O2.- e H2O2, que escapam da destruição no
tilacóide, são destruídos no estroma pela SOD ou APX. Várias isoenzimas de
APX são encontradas no citossol, peroxissomo e cloroplasto (estroma e
tilacóides) e sua função é degradar o H2O2 via oxidação do ascorbato
(Karyotou & Donaldson. 2005), (fig. 10). O primeiro produto da reação
catalisada pela APX é o radical monodehidroascorbato (MDA), que pode ser
convertido a ascorbato espontaneamente ou enzimaticamente pela
monodehidroascorbato redutase (MDAR). O MDA também pode se converter
espontaneamente a dehidroascorbato e este pode, pela dehidroascorbato
redutase, regenerar o ascorbato (fig. 10) (Foyer et al., 1994; Karyotou &
Donaldson, 2005).
APX são hemoproteínas contendo o grupo prostético protoporfinia IX,
similar a guaiacol peroxidase em plantas. As propriedades enzimáticas da APX,
contudo, são bem diferentes de outras heme – peroxidases. As isoenzimas
variam em uma faixa de massa molecular entre 27 e 40 kDa. Geralmente, a
APX está presente como um monômero, ou como um homodímero (Asada.
1992).

Fig.10: Degradação do peróxido de hidrogênio catalisada pela APX.
2.4.4 – Guaiacol peroxidase (GPX)
Peroxidases geralmente reagem com compostos contendo grupos
hidroxila ligados a anéis aromáticos e atuam na lignificação, suberização e em
outros pontos do metabolismo da parede celular vegetal. O guaiacol (o-metoxi
fenol) é comumente usado como substrato para a medição da atividade
peroxidásica (Hiraga et al., 2001).
A enzima guaiacol peroxidase (GPX) oxida uma molécula de guaiacol
para cada molécula de H2O2 que é reduzida. A reação global ocorre com a
utilização de quatro moléculas de guaiacol que se tetramerizam, levando a
redução de quatro moléculas de H2O2, com formação de água, oxigênio
molecular e um composto formado pelas 4 moléculas de guaiacol (fig. 11).
Esse composto posteriormente se dissocia em radicais fenoxi, que levam a
polimerização não enzimática de monômeros. De um modo similar o ácido
hidroxicinâmico e o álcool hidroxicinamílico e seus derivados, são convertidos a
radicais fenoxi pela peroxidase. O ácido hidroxicinâmico contém porções
alifáticas que são incorporados à suberina, enquanto que o álcool
hidroxicinamílico é polimerizado a lignina (Hiraga et al., 2001).
Nas espécies vegetais e em particular no linho (Linum usitatissimum), a
guaiacol peroxidase apresenta formas ácidas e básicas. A isoforma ácida está
envolvida numa variedade de processos relacionada com a biossíntese da
parede celular, incluindo a formação de lignina. A isoforma básica participa da
regulação da degradação do ácido indol-acético e da síntese do etileno
(Fieldes & Gerhardt, 1998).
A GPX utiliza preferencialmente o guaiacol como substrato, mas
catalisa, in vitro, a oxidação de doadores de oxigênio inespecíficos. In vivo, a
GPX pode utilizar ascorbato como substrato para suas reações. Neste caso, a
desintoxicação pode se tornar a função principal de algumas isoformas (Fieldes
& Gerhardt, 1998).
2 Ascorbato + H2O2 � 2 Monodehidroascorbato + H2O

Mika & Lüthje (2003) detectaram, em raiz de milho, a presença de três
isoformas de guaiacol peroxidases com 70, 155 e 38 kDa respectivamente. As
isoformas não possuíam atividade ascorbato peroxidase, provavelmente
perdida com a purificação.
Jung demonstrou em 2004 que a atividade GPX aumenta cerca de 100
vezes, sete dias após o tratamento de folhas de Arabidopsis thaliana com metil
jasmonato. No mesmo ano, Soares e colaboradores verificaram que até 6
horas após a indução do estresse com metil jasmonato, há uma redução da
atividade GPX em Ricinus communis. Contudo, na maioria dos organismos, as
principais funções das GPX não foram bem esclarecidas. O estudo das
alterações provocadas pela indução de estresses fisiológicos pode contribuir
para uma melhor compreensão da ação específica das isoformas da GPX
(Campa, 1991).
Fig.11: Redução do H2O2 e tetramerização do guaiacol catalisada pela GPX.
2.4.5 – Peroxiredoxina (Prx)
Peroxiredoxinas (Prx) são enzimas que catalisam a reação de redução
de peróxido de hidrogênio (H2O2) ou outros alquil hidroperóxidos em água e
álcool correspondente na presença de um doador de hidrogênio, o qual é,
então convertido à forma oxidada (fig. 12). Tal doador de hidrogênio pode ser
uma tioredoxina, uma glutaredoxina ou ambas. Pertencem à família das
peroxidases, porém não possuem grupos prostéticos, contendo um íon metal,
normalmente requerido para a reação catalítica ocorrer. A peroxiredoxina
0 0
OCH3 OCH3
OCH3
0 0
OCH3
OH
OCH3
Peroxidase
+ 4 H2O2
Guaiacol
+ 6 H2O+ O2 4

supera este problema usando uma cisteína catalítica na região N- terminal que
é convertida a ácido sulfênico e regenerada via doador de próton. Seu peso
molecular é de aproximadamente 20kDa, (Rouhier et al., 2001).
A eficiência catalítica moderada, se comparada à catalases, torna a
importância das peroxiredoxinas questionável. No entanto, a alta abundância
de Prxs em uma grande variedade de células e um dado recente de que uma
Prx de bactéria, e não a catalase, é responsável pela redução do H2O2
endógeno, demonstram a importância das Prxs na desintoxicação de células
(Wood et al., 2003).
Segundo Rouhier et al. (2001), peroxiredoxinas se classificam em três
tipos; A, B e C. Tal classificação deve-se às diferenças na organização
molecular, seqüência de aminoácidos, doador de prótons e localização
subcelular.
A elucidação do genoma de Arabidopsis indica que há muitos genes de
Prx em plantas. A expressão de alguns desses genes é regulada temporal e
espacialmente. Em plantas, foram identificadas três tipos de Prx (Rouhier et al.,
2001).
A enzima 2-cys peroxiredoxina (2-CP) se localiza em cloroplastos e
parece ser expressa na maioria dos tecidos vegetais e age na desintoxicação
de produtos do transporte de elétrons no tilacóide, o lugar de maior
metabolismo de oxigênio ativo em células vegetais. Plantas com 2-CP
suprimida compensam com a super expressão de ascorbato peroxidase,
sugerindo que existe um equilíbrio entre diversas enzimas antioxidantes em
cloroplastos (Baier & Dietz, 1997). Para esse grupo de peroxiredoxinas, o
doador de prótons é claramente a tioredoxina (Trx). As 2-cys peroxiredoxinas
exibem estrutura dimérica com duas subunidades idênticas ligadas por ponte
dissulfeto (Choi & Cheng, 1999). Soares et al. (2004) verificaram que o metil
jasmonato promove uma redução transiente nos níveis da enzima 2-CP
peroxiredoxina em folhas de Ricinus communis. A 1-cys peroxiredoxina (1-CP)
é codificada aparentemente por um único gene expresso em sementes de
dicotiledôneas e monocotiledôneas em desenvolvimento. Como função, foi
proposta a proteção desse tecido contra espécies reativas de oxigênio
produzidas durante a dessecação ou como um produto da respiração durante a

embebição das sementes (Schröder & Ponting, 1998). As Peroxiredoxinas do
tipo C foram isoladas recentemente (1999) de Brassica rapa e Arabidopsis. Das
três, é a Prx de menor seqüência de aminoácidos. Ambas as enzimas reduzem
H2O2 na presença de NADPH, Trx e Trx redutase. A organização molecular da
Prx tipo C é muito pouco conhecida. Sugere-se que tenha um enovelamento
similar ao da Trx com uma folha beta pregueada central, rodeada por alfa
hélices, (Schröder & Ponting, 1998).
Fig.12: Reação catalisada pela Prx.
2.4.6 – Polifenol oxidase (PPO)
Polifenol oxidase é uma enzima encontrada em muitas plantas e fungos.
Em plantas superiores se localiza no tilacóide dos cloroplastos ou em outros
plastídeos. Em sua forma ativa, a PPO catalisa a hidroxilação de monofenóis a
o-difenóis (cresolase) e a oxidação de o-difenóis a o-diquinonas (catecolase),
como ilustrado na figura 13. As o-quinonas, reagem então com o oxigênio do
ar, formando um composto marrom escuro chamado melanina. Com a
oxidação de compostos fenólicos, a PPO reduz as agressões em moléculas
biológicas. A enzima pode existir também, parcial ou completamente, em uma
forma latente, dependendo da espécie de planta ou de fungo. Pode ser
convertida a forma ativada com a adição de tripsina (Tolbert, 1973) ou
detergente ao tampão de ensaio. Geralmente possui massa molecular entre 59
e 68KDa (Shin et al., 1997).
2 R'-SH + ROOH � R'-SSR' + H2O + ROH

Fig.13: Reações catalisadas pela PPO.
Durante a alimentação de insetos com folhas, a enzima se mistura aos
substratos fenólicos e as quinonas resultantes alquilam aminoácidos essenciais
da dieta protéica, tornando-os nutricionalmente inviáveis aos insetos
(Constabel & Ryan. 1998).
Em adição, PPO de cloroplastos do mesofilo funcionam nas reações de
Mehler, fotorredução do oxigênio molecular pelo fotosistema Ι da fotossíntese e
regulação dos níveis de oxigênio (Thipyapong et al., 2004).
Uma outra função de PPO está em aumentar a resistência da parede
celular dos tecidos vegetais, uma vez que colabora com a biossíntese de
lignina (Aquino-Bolaños & Mercado-Silva, 2004). Todos estes atributos aqui
descritos conferem a essas proteínas uma ampla ação protetora, atuando não
somente contra pragas, mas também contra patógenos (Yuan et al., 2002).
Polifenol oxidases são as proteínas de defesa mais abundantemente
induzidas em plantas de tomate submetidas a ferimento, ou estimuladas com
MeJa, ou ainda com a sistemina (Constabel et al., 1995). Além disso, o
aumento dos níveis de PPO em plantas causado por lesão ou MeJa,
representa a indução de proteínas de defesa mais amplamente distribuída
entre as espécies vegetais, como exemplos vistos em plantas de tomate,
monofenol o-difenol
o-difenol o-diquinona

tabaco, soja, ervilha dentre outros cultivares. A intensidade de indução da PPO
varia grandemente entre diferentes famílias e espécies de plantas (Constabel &
Ryan. 1998).
2.5 – Resposta às Espécies Reativas de Oxigênio (ERO)
Respostas adequadas às mudanças ambientais são cruciais para o
crescimento e sobrevivência da planta, contudo, os mecanismos moleculares e
bioquímicos que orquestram estas respostas ainda são pouco compreendidos.
Uma função atribuída para as espécies ativas de oxigênio, durante respostas a
estresses bióticos e abióticos, é bem documentada. As ERO podem agir
causando danos ou agir como moléculas sinais que ativam múltiplas respostas
de defesa. Essa dualidade pode ser obtida apenas quando os níveis celulares
de ERO são bem controlados tanto na produção quanto no consumo,
(Breusegem et al. 2001; Neto et al. 2005).
O nível e o tipo das ERO são fatores determinantes para o tipo de
resposta. Peróxido de hidrogênio (H2O2) e superóxido (O2*-) podem induzir
genes diferentes, em conjunto ou separadamente, dando mais flexibilidade a
sinalização de ERO. Em baixas concentrações, ERO induz genes de defesa e
resposta adaptativa. Níveis subletais, contudo, podem levar plantas a
condições de estresse biótico e abiótico e reduzir seu crescimento,
provavelmente como parte de uma resposta adaptacional. Porém, respostas
genômicas substanciais e atividades enzimáticas são afetatadas por ERO e os
mecanismos moleculares de adaptação ainda são pouco conhecidos. Em altas
concentrações, as ERO levam a um programa de morte celular controlado
geneticamente. Também comunicam-se com outras moléculas sinalizadoras e
vias formando uma grande rede de resposta vegetal, (Breusegem et al. 2001).
O acúmulo de H2O2 em tecidos específicos e em quantidades
apropriadas beneficia as plantas mediando aclimatação e tolerância cruzada a
estresses bióticos e abióticos (Bowler & Fluhr, 2000). Tem sido evidenciado
que a adição de H2O2 em tecidos foliares ou sua indução endógena age como
um sinal de indução para a expressão de genes referentes a catalase
(Polidoros & Scandalios, 1999), ascorbato peroxidase (Breusegen et al., 2001),
guaiacol peroxidase e glutationa peroxidase (Janda et al., 1999).

Existem na literatura algumas evidências da participação de H2O2 em
resposta ao estresse. Como exemplo, podemos citar os experimentos de
Orozco–Cardenas & Ryan (1999). Estes autores demonstraram que sistemina,
OGA (oligogalacturonídeos), quitosan e metil jasmonato induzem o acúmulo de
peróxido de hidrogênio em plantas de tomate, como citado anteriormente. Rao
e colaboradores investigaram em 1997 como o ácido salicílico (AS) aumenta o
conteúdo de H2O2 em Arabidopsis thaliana. O tratamento com AS, aumenta a
produção de H2O2, a peroxidação lipídica e o dano oxidativo a proteínas. O
aumento dos níveis de H2O2 foi relacionado a um aumento na atividade da
isoforma Cu/Zn-SOD e foi independente de alterações nas atividades catalase
e ascorbato peroxidase. Contudo não é claro se o ácido salicílico ou os sinais
derivados do H2O2 são percebidos e propagados. Ambos AS e H2O2 parecem
ter um papel regulatório no desenvolvimento de resistência a patógenos (Wu et
al., 1995; Hammond-Kosack & Jones, 1996). Cho & Seo (2005) observaram
um aumento no conteúdo de H2O2, bem como em todas as atividades
antioxidantes testadas (SOD, CAT, APX) em Arabidopsis thaliana exposta ao
Cádmio. O H2O2 gerado em resposta ao tratamento com ABA foi detectado
com 0,5h nas nervuras principais das folhas de milho (Zea mays L.) e
maximizado em torno de 2-4h. O H2O2 foi detectado em células do mesofilo e
em feixes vasculares. O tratamento com ABA levou a aumentos significativos
nas atividades de enzimas antioxidantes presentes em diversos
compartimentos sub-celulares: SOD, APX e GR. Além disso, o estresse
oxidativo induzido por paraquate, que gera O2.- e então H2O2 nos cloroplastos,
também induziu um aumento na atividade das enzimas. Esse dado sugere que
o H2O2 produzido em um local celular específico pode coordenar a atividade de
enzimas antioxidantes em diferentes compartimentos subcelulares.

3.0 – Objetivos gerais:
O nosso estudo tem como objetivos gerais, a detecção e a análise do
comportamento das diversas enzimas relacionadas com o metabolismo de
espécies reativas de oxigênio, em folhas de Ricinus communis tratadas com
metil jasmonato, bem como, relacionar a atividade das mesmas com o
conteúdo de peróxido de hidrogênio “in vivo” nas folhas em diferentes tempos
de tratamento.
3.1 – Objetivos específicos:
− Avaliar, por SDS-PAGE, as modificações protéicas provocadas por
MeJa em Ricinus communis;
− Padronizar e determinar a atividade das enzimas: superóxido dismutase
(SOD), catalase (CAT), ascorbato peroxidase (APX), guaiacol
peroxidase (GPX) e polifenol oxidase (PPO) em extrato bruto de folhas
de Ricinus communis tratadas com metil jasmonato e após separação
das proteínas por PAGE nativo;
− Caracterizar parcialmente a enzima guaiacol peroxidase;
− Detectar o acúmulo de peróxido de hidrogênio e de radicais livres nas
folhas de Ricinus communis em diferentes tempos de tratamento com
MeJa.

4.0 – Material e Métodos
O trabalho foi desenvolvido no Laboratório de Química e Função de
Proteínas e Peptídeos, no Centro de Biociências e Biotecnologia da
Universidade Estadual do Norte Fluminense Darcy Ribeiro durante dois anos.
4.1 – Equipamentos
− Centrífuga Eppendorf - Centrifuge / 5402
− Seqüenciador - SHIMADZU / PPSQ-10
− Leitor de microplacas - labsitems / Multiskan
− Sistema de eletroforese - Bio Rad
− IPG phor - GE Healthcare
− Balança semi analítica e analítica - Sartorius
− Sistema de eletrotransferência - Bio Rad
− pHmetro - Procyon / 310
− Fonte de alta de tensão - Desaga
− Agitador magnético com aquecimento - NT /103
− Espectrofotômetro - Spekol
− Sonicador - Fisher-cientific / Sonic dismembrator-60
− Lupa - Tecnival / SQF-F
− Máquina fotográfica - Sony - Cyber-shot / DSC-S60
4.2 – Reagentes
A seguir, a lista dos principais reagentes necessários à execução do
trabalho. Todos os outros reagentes eram de alto grau analítico.
TABELA II: Listagem de reagentes e sua procedência.
Reagente Sigla Fonte
Polivinilpolipirrolidona PVPP Sigma- Aldrich
Ácido etileno diamino tetracético EDTA VETEC
Ditiotreitol DTT Sigma- Aldrich
Ovalbumina OVA Sigma- Aldrich
Dodecil sulfato de sódio SDS Sigma- Aldrich
3,3 diaminobenzidina DAB Sigma- Aldrich
Cianeto de potássio KCN Merck

Nitro blue tetrazolio NBT USB
N,N,N’,N’ Tetrametilenodiamina TEMED Sigma- Aldrich
Guaiacol - Merck
Peróxido de hidrogênio - Nuclear
Triton - Calbiochem
Ácido ascórbico - Sigma- Aldrich
Cloreto férrico - Sigma- Aldrich
Ferricianeto de potássio - Sigma- Aldrich
Catecol - Sigma- Aldrich
Riboflavina - Sigma- Aldrich
4.3 - Material vegetal e condições de crescimento
As sementes de Ricinus communis, cultivar IAC 226 (Tarabay) foram
fornecidas pelo Instituto Agronômico de Campinas. Esse cultivar foi originado
de linhagem pura, obtida do cruzamento controlado entre o “Pindorama” e o
“Campinas”. Como características principais, possui ramificação com inserção
baixa, conferindo à arquitetura da planta o formato de taça, frutos indeiscentes,
porte alto (2,50 a 3,50 metros) e elevado potencial produtivo (2.681 kg/ha de
sementes e 1.233 kg/ha de óleo) (Savy Filho et al., 1990). As sementes deste
cultivar foram semeadas em potes plásticos contendo o substrato orgânico
Plantmax®, na casa de vegetação. Durante sua germinação e crescimento as
plantas foram irrigadas uma vez ao dia, até atingir o estágio trifoliar.
4.4 - Indução de estresse
Quando as plantas atingiram o estágio trifoliar, foram levadas ao
laboratório para que se efetuasse o tratamento com metil jasmonato. As folhas
foram borrifadas manualmente com uma solução de metil jasmonato (0,025%).
Após a indução de resposta, as plantas foram mantidas em câmaras de vidro
fechadas (30 x 30 x 50 cm) em diferentes intervalos de tempo (1, 4, 6, 12, 24,
48 horas).
As plantas não tratadas foram mantidas em outra câmara de vidro com
condições similares de temperatura e luminosidade. Cada câmara continha em
média seis plantas.

4.5 - Obtenção de extratos protéicos
As plantas foram utilizadas no estágio trifoliar. As duas folhas principais
foram cortadas com o auxílio de uma lâmina de barbear e pesadas, o que
equivalia, em geral, a uma massa de aproximadamente 0,15g por folha. Em
seguida foram congeladas em nitrogênio líquido, trituradas na presença de
polivinilpolipirrolidona insolúvel (PVPP) até a consistência de pó fino, numa
proporção de 10% do valor da massa foliar pesada. Posteriormente foi
acrescentado o tampão de extração (100 mM tampão fosfato de potássio pH
7,5, 1 mM EDTA e 3 mM DTT) na proporção de 1:4 (p/v) ao pó fino obtido. A
extração foi realizada durante 1 hora em gelo. O homogenato foi transferido
para tubos tipo eppendorf de 1,5mL e centrifugado a 15.000 x g por 20 minutos
a 4ºC. Logo após, o sobrenadante foi coletado, redistribuído em tubos tipo
eppendorf e congelados em -20ºC para análises posteriores. A maioria dos
passos da extração estão ilustrados na figura 14.
Fig.14: Representação esquemática do tratamento com metil jasmonato e
extração protéica em folhas de Ricinus communis.
MACERAR
EXTRAÇÃO (1: 4)
Centrifugação a 15.000 x g por 20´
SOBRENADANTESEDIMENTO
ANÁLISES POSTERIORES
Tratamento das folhas com metil jasmonato
Tampão fosfato de potássio 100mM, pH 7,5; 1mM EDTA, 3mM
DTT
MACERARMACERAR
EXTRAÇÃO (1: 4)EXTRAÇÃO (1: 4)
Centrifugação a 15.000 x g por 20´
SOBRENADANTESOBRENADANTESEDIMENTOSEDIMENTO
ANÁLISES POSTERIORESANÁLISES POSTERIORES
Tratamento das folhas com metil jasmonatoTratamento das folhas com metil jasmonato
Tampão fosfato de potássio 100mM, pH 7,5; 1mM EDTA, 3mM
DTT

4.6 - Determinação do teor de proteínas nos extratos brutos
A curva padrão foi preparada com ovalbumina (2 mg/mL) em diversas
diluições. Alíquotas de amostras foram diluídas 20 vezes, tratadas com o
reagente de Bradford e a seguir lidas a 540 nm (Bradford,1976). Todos os
ensaios foram feitos em triplicata.
Os volumes usados representam uma pequena modificação do método.
O volume máximo em cada poço da placa (de 96 poços) foi de 10 µL, tanto
para a curva quanto para as amostras. A esse valor, adicionou-se 200 µL do
reagente de Bradford e a leitura foi realizada em leitor de micro placas.
4.7 - Análise do perfil protéico em gel de poliacrilamida descontínua
4.7.1 - Na presença de dodecil sulfato de sódio (SDS-PAGE)
O perfil protéico dos extratos das plantas controle e das plantas tratadas
foram avaliados por SDS-PAGE empregando a metodologia descrita por
Laemmli (1970). Foram utilizados géis fracionadores e concentradores com,
respectivamente, 12% e 3% em acrilamida.
As amostras foram misturadas ao tampão de amostra contendo SDS e
aquecidas a 100º C por 5 minutos. Foram aplicados 100 µg de proteína em
cada canaleta em géis de 1mm de espessura e foi aplicada uma corrente
elétrica de 35 mA por gel, por aproximadamente uma hora. Para revelação das
proteínas, foi usada a solução corante (0,1% comassie blue, 40% metanol e
10% ácido acético) para revelação das bandas protéicas, após a corrida
eletroforética. A visualização das proteínas coradas foi feita com a imersão do
gel em solução descorante (40% metanol e 10% ácido acético).
4.7.2 - Na ausência de dodecil sulfato de sódio (NATIVE-PAGE)
A avaliação da maioria das atividades enzimáticas em gel de
poliacrilamida é limitada se efetuada na presença do agente desnaturante SDS.
Para realizar nossos experimentos, empregamos a mesma metodologia
descrita no item anterior, porém sem SDS e sem aquecimento (Davis, 1964).
Os géis fracionadores utilizados para este fim eram 8% e também possuíam
1mm de espessura. As amostras foram acondicionadas em banho de gelo e a
eletroforese aconteceu em ambiente refrigerado (4ºC).

4.7.3 – Eletroforese bidimensional
Uma alíquota da amostra a ser analisada foi colodada em 120µL de
tampão de rehidratação (8M de uréia; 2% chaps; 0,5% anfólitos; 0,3% DTT,
0,002% azul d bromo fenol). A solução resultante foi colocada no centro de um
“strip holder”. O “strip” de 7 cm, com faixa de pH entre 3 e 10, foi colocado
sobre a amostra no “strip holder” e coberto com um óleo mineral e, em seguida,
trasferido para a plataforma do aparelho de focalização, IPG Phor, o qual foi
programado para proceder com a focalização até a marcação de 25000 Vh.
O “strip” com as proteínas focalizadas foi colocado por 15 minutos em
um tampão de equilíbrio (0,05M tris-HCl pH8,8; 6M uréia; 30% glicerol; 2%
SDS; 0,0002% azul de bromo fenol). O “strip” foi, então, colocado no mesmo
tampão por mais 15 minutos, porém sem DTT e com 2,5% iodoacetamida
(O’Farel, 1975).
O “strip” equilibrado foi colocado sobre um gel desnaturante, como
descrito no item 4.7.1, para que fosse realizada a segunda dimensão e foi
aplicada uma corrente de 25 mA durante a corrida eletroforética.
4.8 - Determinação das atividades de enzimas antioxidantes por
espectofotometria
4.8.1 - Determinação da atividade da catalase (CAT)
A atividade catalase (EC 1.11.1.6) foi medida em uma mistura reacional
(3mL) composta de 50 mM tampão fosfato de sódio pH 7,0 na qual o peróxido
de hidrogênio foi adicionado numa concentração final de 10 mM (Barka, 2001).
A reação foi iniciada adicionando 30 µL de extrato e a atividade foi monitorada
pelo do declínio da densidade óptica a 240 nm (ε = 36 mM-1 cm-1) durante 1
min, após o início da reação, tendo, como branco, o meio de reação livre de
H2O2. A atividade da CAT foi expressa em µmol min-1 mg-1 de proteína.
4.8.2 - Determinação da atividade da ascorbato peroxidase (APX)
A atividade ascorbato peroxidase (EC 1.11.1.11) foi medida pelo
monitoramento da oxidação do ascorbato pela diminuição da absorbância a
290 nm (ε=2,8 mM-1 cm-1), em uma cubeta de quartzo de 3 mL. O meio

reacional (2 mL) continha 0,5 mM ácido ascórbico, 0,1 mM EDTA, 1 mM H2O2
em 50 mM tampão fosfato de potássio pH 7,0 , (Barka, 2001). A reação foi
iniciada com a adição de 30 µL do extrato enzimático tendo, como branco, o
meio de reação livre de H2O2 e extrato enzimático. A atividade da APX foi
expressa em mmol min-1 mg-1 de proteína.
4.8.3 - Determinação da atividade da guaiacol - peroxidase (GPX)
A determinação da atividade da GPX (EC 1.111.7) foi baseada em
estudos desenvolvidos por Cakmak & Horst (1991), com modificações. O meio
de reação continha 2,58 mM guaiacol e 10 mM água oxigenada em 25 mM
tampão fosfato de sódio, pH 6.0, preparado no escuro. A reação se iniciou com
adição de 30 µL extrato enzimático a 2 mL da solução de guaiacol. Durante
cinco minutos, a tetramerização do guaiacol foi acompanhada em
espectrofotômetro, a 470 nm (ε = 26,6 mM-1 cm-1), contra um branco contendo
o mesmo volume de reação livre do extrato enzimático. A atividade GPX foi
expressa em µmol min-1 mg-1 de proteína.
4.8.4 - Determinação da atividade da polifenol oxidase (PPO)
Para a execução da atividade PPO (EC 1.14.18.1), foi necessária uma
extração direcionada. As folhas de Ricinus communis tratadas e não tratadas
com MeJa, foram maceradas separadamente com auxílio de nitrogênio líquido
e extraídas com tampão 100 mM fosfato de sódio, pH 7,0, contendo 0,1% (v/v)
SDS em banho de gelo. O extrato foi clarificado por centrifugação, após 20
minutos de extração e o sobrenadante foi utilizado para o ensaio da PPO.
A atividade PPO foi monitorada seguindo a oxidação de uma solução
100 mM catecol, 2% SDS em tampão 100 mM fosfato de sódio, pH 6,5 a 490
nm, durante um minuto. A reação se iniciou com adição de 50 µL extrato
enzimático a 1mL da solução de ensaio, tendo como branco o meio de reação
livre de catecol segundo Constabel & Ryan (1998), com modificações. Os
resultados foram expressos como variação de absorvância min-1 mg-1 de
proteína.

4.9 - Determinação das atividades antioxidantes por PAGE não
desnaturante
Como descrito no item 4.7.2, as atividades enzimáticas foram
visualizadas em géis de eletroforese nativa 8%, após separação protéica. A
detecção de cada enzima varia com o substrato usado e com o pH no meio
reacional. Vale ressaltar que todos os extratos foram manipulados em banho
de gelo e que os ensaios foram realizados em temperatura ambiente, 18-30ºC.
A seguir, uma descrição dessas metodologias.
4.9.1 - Atividade da superóxido dismutase (SOD)
A enzima foi ensaiada após separação das proteínas do extrato em gel
não desnaturante. A atividade superóxido dismutase (EC 1.15.1.1) foi
ensaiada medindo a inibição da redução do nitro blue tetrazolio (NBT) na
presença de riboflavina a 560nm de acordo com Azevedo et al. (1998). O meio
reacional, foi preparado, no escuro, da seguinte forma: 30 mL de uma solução
0,005% (p/v) riboflavina em água e 70 mL de 50 mM tampão fosfato de
potássio pH 7,8, acrescido de 0,02 g de NBT, 0,02 g de EDTA e 300 µL de
TEMED. Ambas as soluções foram preparadas imediatamente antes do seu
uso e armazenadas em recipientes cobertos com papel alumínio.
Após a corrida eletroforética, o gel foi lavado com água destilada e
encubado com o meio reacional durante trinta minutos, sob agitação constante,
na ausência de luz. Após esse período, o gel foi levado à luz, onde ocorreu a
fotoxidação do formando uma coloração púrpura, exceto nos locais
correspondentes às bandas com atividades de SOD, que permaneceram sem
se fotoxidar, promovendo uma revelação negativa. A revelação levou em média
de 1 a 3 minutos e a fotoxidação foi interrompida mergulhando-se o gel em
uma solução 6% (v/v) ácido acético.

4.9.1.2 - Determinação das isoformas da superóxido dismutase (SOD)
A determinação das isoformas da SOD foi realizada após eletroforese
não desnaturante 8%, em que foi aplicado 400 µL de uma única amostra
controle em todas as canaletas do gel, nas condições descritas anteriormente.
Após a eletroforese, o gel foi dividido verticalmente em três partes. Uma
das partes foi mantida a 4ºC em 100 mL de tampão fosfato de potássio 100
mM, pH 7,8 (solução controle). A outra foi imersa em 100 mL do mesmo
tampão, contendo 0,0292 g de EDTA e 0,0130 g de KCN (esta solução inibe a
isoforma Cu/Zn-SOD) e a terceira parte foi imersa em 100 mL de tampão,
acrescido de 0,0292 g de EDTA e 70 µL de H2O2 (esta solução inibe as
isoformas Cu/Zn-SOD e Fe-SOD). Todos os passos descritos foram realizados
no escuro. Após 20 min de incubação nestas soluções, os géis foram
submetidos à revelação com NBT e riboflavina, como descrito anteriormente.
Ao final da revelação, foi analisada a presença ou ausência de bandas no
controle e nos tratamentos com KCN e H2O2, de modo a classificá-las, Azevedo
et al. (1998).
4.9.2 - Atividade da catalase (CAT)
O ensaio descrito para a CAT (EC1.11.1.6) foi seguido com
modificações. Após a eletroforese, o gel foi encubado em 100 mL de água
destilada contendo 20 mL de 50% H2O2 por 25 minutos em temperatura
ambiente e então, brevemente lavado com água. O gel foi encubado
subsequentemente em uma solução 2% (m/v) cloreto férrico e 2% (m/v)
ferricianeto de potássio em água. O desenvolvimento da cor amarela continuou
em média por 2 minutos e a reação foi encerrada com água (Rao et al.,1997).
4.9.3 - Atividade da ascorbato peroxidase (APX)
Subseqüente a separação eletroforética, o gel foi equilibrado com
tampão 50mM fosfato de sódio (pH:7,0) e 2 mM ácido ascórbico por 30
minutos, sendo o tampão de equilíbrio trocado a cada 10 minutos. O gel foi
então encubado com tampão 50 mM fosfato de sódio (pH 7,0) e 4 mM ácido
ascórbico e 2 mM de H2O2 por 20 minutos. O H2O2 foi adicionado à solução
imediatamente antes da encubação. O gel foi subsequentemente lavado com
tampão 50mM fosfato de sódio (pH 7,0) por 1 minuto e submergido em uma

solução de 50mM fosfato de sódio (pH 7,8), 28 mM TEMED e 2,45 mM NBT,
com agitação suave.
A atividade APX (EC 1.11.1.11) foi observada como uma banda
acromática em um fundo de cor púrpura (geralmente entre 2 e 5 minutos). A
reação foi interrompida com água destilada. Uma longa encubação com a
solução TEMED/NBT, resulta em um fundo muito escurecido, o que deixa as
bandas acromáticas de atividade nubladas e difusas (Mittler & Zilinskas, 1993).
4.9.4 - Atividade da guaiacol peroxidase (GPX)
Através de modificações no método desenvolvido por Cakmak & Horst
(1991), detectamos a atividade GPX (EC 1.11.1.7) em géis de eletroforese
após separação protéica do extrato bruto. O meio de reação, preparado no
escuro, continha 2,58 mM guaiacol e 10 mM água oxigenada em 25 mM
tampão fosfato (pH 6,0), como descrito anteriormente na avaliação da enzima
por espectofotometria.
Após eletroforese, o gel foi lavado em água destilada e encubado na luz
e sob agitação com o meio reacional. As bandas de cor marrom, referentes a
tetramerização do guaiacol, apareceram em média entre 1 e 3 minutos. Após a
revelação o gel é fotografado.
4.10 – Caracterização da GPX
A enzima guaiacol peroxidase, por sofrer alterações mais severas com o
tratamento com MeJa, passou por alguns passos de purificação.
4.10.1 – Determinação do pH ideal da GPX
A atividade da guaiacol – peroxidase foi avaliada em diferentes pHs. Os
valores de pH avaliados foram: 4,6; 5,0; 5,4; 5,8; 6,8; 7,0; 9,0. Para isso
utilizou-se tampão fosfato de sódio 100 mM.
4.10.2 – SDS-PAGE da enzima homogênea
Após a corrida eletroforética em PAGE nativo 8% (ítem 4.7.2), as bandas
com atividade GPX foram recortadas dos géis para a subseqüente extração.
Para a obtenção do material protéico necessário ao procedimento, foram
usados 10 géis.

As tiras recortadas foram lavadas com tampão 250 mM tris, 250 mM
EDTA, 1 mM PMSF, 1 mM benzamidina, pH 7,4 em um tubo falcon e lavadas 5
vezes com água destilada. A água foi removida com uma pipeta Pasteur e os
pedaços de gel foram partidos em pedaços minúsculos (2-5mm) com uma
espátula. Foi adicionado, 2mL de tampão 20mM tris, pH 7,4, contendo 0,1%v/v
SDS (a razão entre volume de tampão de e o volume de pedaços de gel foi de
aproximadamente 2:1). As amostras foram sonicadas 1 vez por 3 minutos e 6
vezes por 30 segundos em gelo. Após esse processo, as amostras foram
centrifugadas por 10000g por 20 minutos (Retamal et al., 1999). O
sobrenadante foi retirado com uma pipeta Pasteur, dialisado e seco, misturado
em tampão de amostra desnaturante e fervido por 5 minutos a 100ºC. Essa
solução (aproximadamente 30 µL) foi então aplicada no topo de um gel
desnaturante 12% (item 4.7.1). Procedeu-se a eletroforese e a massa
molecular da GPX foi estimada após revelação das bandas protéicas com o
corante comassie, se comparando com a massa dos marcadores de massa
molecular.
4.10.3 – Determinação do ponto isoelétrico da GPX por eletroforese
bidimensional
Para a realização da eletroforese bidimensional, as bandas com
atividade GPX foram recortadas de dez géis nativos e encubadas em tampão
de extração, como descrito no item 4.10.2.
Após passar por diálise e por secagem, a proteína, teoricamente
homogênea, passou pelos passos descritos no item 4.7.3. No final da corrida
eletroforética da segunda dimensão, o gel foi corado com solução comassie e
descorado para visualização da banda protéica.
4.11 - Detecção “in vivo” do peróxido de hidrogênio
O peróxido de hidrogênio (H2O2) foi visualizado e detectado nas folhas
das plantas usando o 3,3 - diaminobenzidina (DAB) como substrato, utilizando
o método descrito por (Orozco-Cardenas & Ryan, 1999), com modificações. As
plantas foram infiltradas a vácuo com uma solução de DAB 1 mg/mL, pH: 3,8 e
triton (0,1% v/v) por meia hora sob luz à 25ºC, após o tratamento com metil
jasmonato por 1, 4, 6, 12, 24 e 48 horas.

Após a infiltração, as folhas foram imersas em etanol fervente 96% por 5
minutos. Este tratamento descolorara as folhas, exceto nos locais onde o H2O2
reage com o DAB. Nesses locais, aparece um produto de polimerização
marrom. Após esfriar, as folhas foram preservadas em etanol na temperatura
ambiente e fotografadas.
4.12 – Varredura de radicais livres pelo método do DPPH.
Para avaliar o conteúdo de radicais livres nas folhas de Ricinus
communis tratadas com metil jasmonato, foi necessária uma extração alcoólica
desses componentes, na proporção 1:5 (1 parte de massa foliar para 5 partes
de etanol absoluto). Os extratos foram centrifugados a 15000g por 10 minutos
e os sobrenadantes obtidos foram utilizados na análise dos radicais livres pelo
método do DPPH. (1,1-difenil-2-picrilhidrazil) adaptado de Yepez et al. (2001).
O ensaio foi preparado como ilustrado na tabela III e as absorvâncias foram
medidas no comprimento de onda de 515 nm imediatamente após o preparo
das amostras e 30 minutos depois. Utilizou-se rutina no lugar da amostra como
controle positivo. O DPPH. foi utilizado numa concentração de 6,5 x 10-5 M.
TABELA III: Preparo das amostras e dos controles para a verificação do conteúdo de radicais livres nas folhas de Ricinus communis.
Teste Controle- Controle+ Branco
amostra 33 µL - - 33 µL
Etanol 799 µL 833 µL 799 µL 1132 µL
DPPH.
333 µL 333 µL 333 µL -
Rutina - - 33 µL -
Por fim, mediu-se o Índice Varredor da amostra em percentual (IV%)
usando a fórmula: IV% = (1 – A/A0 ) x 100. Onde A0 é a absorbância inicial, ou
seja, no tempo zero e A é a absorbância final, ao final dos 30 minutos.
4.13 - Analise estatística dos resultados
Para cada tratamento, foram realizadas três repetições individuais em
todas as avaliações. Os dados de cada avaliação foram analisados
estatisticamente e os resultados expressos como média (± erro-padrão) de três
repetições.

5.0 – Resultados 5.1 – Determinação do perfil protéico por SDS-PAGE Antes de iniciar as investigações acerca do status antioxidante em
Ricinus communis, realizamos um mapeamento das alterações protéicas,
numa faixa de massa molecular entre 14 e 66 Kda, que ocorriam quando as
plantas eram tratadas com o agente estressante, o metil jasmonato (MeJa).
Podemos observar, por eletroforese desnaturante, que o teor protéico
dos extratos de folhas de Ricinus communis é alterado quando as plantas são
tratadas com MeJa, fig.15. As modificações mais notórias referem-se à redução
drástica dos níveis das proteínas com massa molecular superior a 24kDa com
48h de tratamento e a alteração dos níveis de uma proteína de 21Kda, que
teve seus níveis reduzidos nos momentos iniciais de tratamento com MeJa (1-
6h).
A proteína de 21 kDa foi denominada MJRC21. Esta proteína, quando
submetida ao sequenciamento automático, apresentou alto grau de homologia
com 2cys-peroxiredoxinas de várias fontes, segundo Soares et al., 2004.
Fig.15: Eletroforese em gel de poliacrilamida 12% na presença de SDS 0,1%
dos extratos protéicos de folhas de Ricinus communis, obtidos a partir das
plantas controle e tratadas com metil jasmonato. Linhas: C- controle; 01, 04,
06, 12, 24 e 48h – tempo de exposição e M – marcador de peso molecular em
KDa. As setas indicam proteínas cujos níveis foram modificadas pelo metil
jasmonato. As setas indicam as principais alterações observadas.

5.2 – Determinação das atividades enzimáticas 5.2.1 – Determinação da atividade superóxido dismutase (SOD) após
separação das proteínas por eletroforese não desnaturante.
A avaliação da atividade SOD foi feita após eletroforese não
desnaturante 8%. Detectamos quatro isoformas diferentes. Ao contrário das
demais enzimas, a SOD não foi alterada nos momentos iniciais de estresse.
Contudo, uma isoforma, de atividade tênue, teve sua atividade reduzida com 24
e 48h, como indicam as setas na fig. 16.
Fig.16: Atividade da superóxido dismutase de plantas de Ricinus communis
submetidas ao tratamento com metil jasmonato após separação das proteínas
por eletroforese nativa 8%. Linhas: C - controle; 01, 04, 06, 12, 24 e 48h –
tempo de exposição ao metil jasmonato. As setas indicam as alterações
observadas.
5.2.1.1 – Determinação das isoformas da superóxido dismutase (SOD).
As isoformas da SOD foram visualizados após separação por
eletroforese não desnaturante de extratos de plantas controle, como descrito
antreriormente (fig. 17). Com o intuito de identificar as isoformas presentes no
gel, foram usados inibidores específicos para cada uma. Observando a tab IV,
obtida com base nos dados da figura 15, observamos três isoformas: Mn-SOD,
Fe-SOD e Cu/Zn-SOD sendo a isoforma Mn-SOD (mitocondrial) reduzida com
24 e 48h de tratamento.

Fig.17: Isoformas da SOD determinada por eletroforese nativa em plantas de
Ricinus communis. A - Géis pré-incubados em tampão sem inibidor; B - tampão
+ KCN (inibidor da isoforma Cu/Zn-SOD) e C - tampão + H2O2 (inibidor das
isoformas Cu/Zn-SOD e Fe-SOD). 1, 2, 3: isoformas da SOD observadas.
TABELA IV: Determinação das isoformas da SOD Banda KCN H2O2 Tipo Localização
1 resistente resistente Mn-SOD Mitocondria 2 resistente sensível Fe-SOD Cloroplasto
3 sensível sensível Cu/Zn-SOD Citosol e
Cloroplasto
5.2.2 – Determinação da atividade catalase (CAT) no extrato bruto.
O tratamento com MeJa afetou a atividade catalase até 12h de
exposição. Com 1h, ocorreu uma redução de cerca de 30% da atividade,
seguida de um aumento gradativo, no qual, em 4h a atividade era similar a do
controle e com 6h, um pouco maior. Houve um máximo de atividade em 12h. A
partir deste momento, o comportamento transiente da enzima se encerra e com
24 e 48h não há alterações significativas em sua atividade (fig. 18).
5.2.2.1 – Determinação da atividade catalase (CAT) após separação das
proteínas por eletroforese não desnaturante.
Não foram observadas alterações na atividade catalase em eletroforese
não desnaturante após o tratamento com metil jasmonato (fig. 19).
1 2 3
A B C
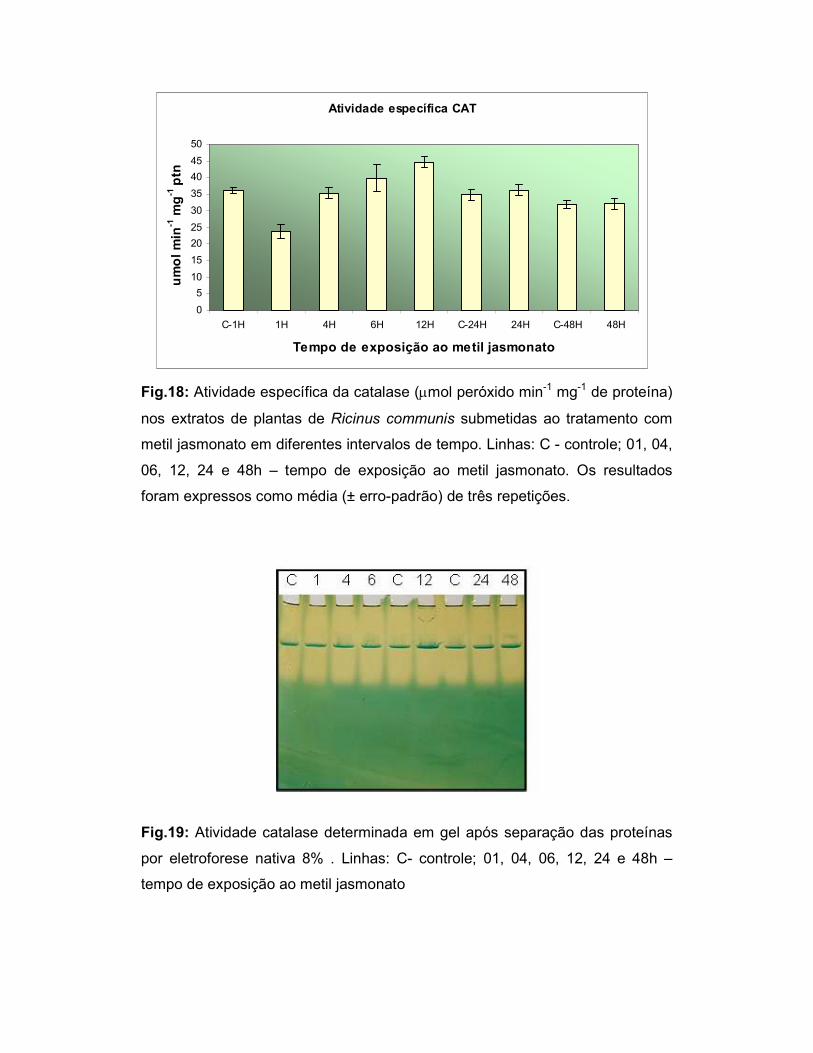
Atividade específica CAT
0
5
10
15
20
25
30
35
40
45
50
C-1H 1H 4H 6H 12H C-24H 24H C-48H 48H
Tempo de exposição ao metil jasmonato
umol min-1 mg-1 ptn
Fig.18: Atividade específica da catalase (µmol peróxido min-1 mg-1 de proteína)
nos extratos de plantas de Ricinus communis submetidas ao tratamento com
metil jasmonato em diferentes intervalos de tempo. Linhas: C - controle; 01, 04,
06, 12, 24 e 48h – tempo de exposição ao metil jasmonato. Os resultados
foram expressos como média (± erro-padrão) de três repetições.
Fig.19: Atividade catalase determinada em gel após separação das proteínas
por eletroforese nativa 8% . Linhas: C- controle; 01, 04, 06, 12, 24 e 48h –
tempo de exposição ao metil jasmonato

5.2.3 – Determinação da atividade ascorbato peroxidase (APX) no extrato
bruto
Em nosso estudo, o monitoramento da atividade APX foi feito através da
oxidação do substrato ascorbato, o que leva a um decréscimo da absorvância a
290nm.
Pode-se observar na figura 20 que a atividade se mantém até 1 hora de
tratamento sem alterações e que há, com 4h, um aumento de cerca de 20% da
atividade. Com 6 e 12h, a atividade parece retornar a níveis de controle e com
24h há um novo aumento, voltando a se igualar estatisticamente com o
controle em 48h.
5.2.3.1 – Determinação da atividade ascorbato peroxidase (APX) após
separação das proteínas por eletroforese não desnaturante.
A atividade APX verificada em gel de eletroforese nativo 8% foi
visualizada como uma banda acromática em um fundo de cor púrpura após 2
minutos de reação. Como podemos observar na figura 21, os dados obtidos
pela eletroforese foram condizentes com os resultados da atividade em extrato
para a APX, na qual há um aumento discreto da atividade nos períodos de 4 e
24h após o tratamento com MeJa.
5.2.4– Determinação da atividade polifenol oxidase (PPO) no extrato bruto
A atividade polifenol oxidase não variou, no período estimado, em
Ricinus communis exposta ao estresse por metil jasmonato (fig. 22).
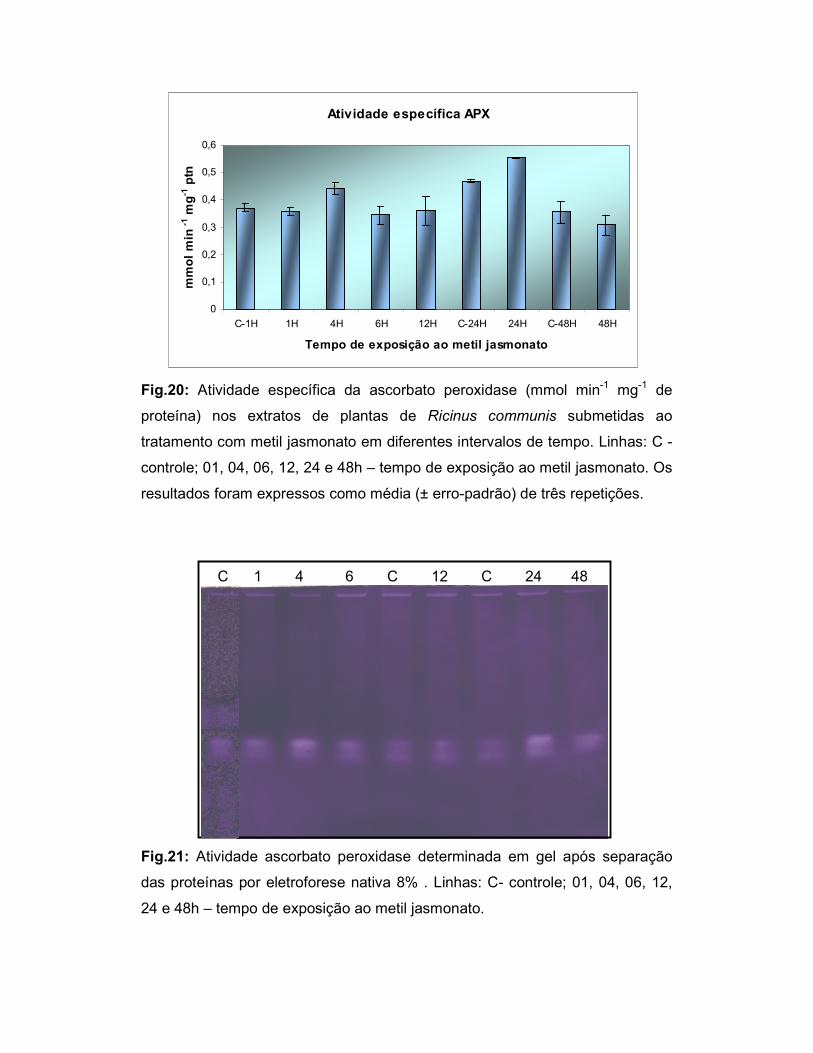
Atividade específica APX
0
0,1
0,2
0,3
0,4
0,5
0,6
C-1H 1H 4H 6H 12H C-24H 24H C-48H 48H
Tempo de exposição ao metil jasmonato
mmol min -1 mg-1 ptn
Fig.20: Atividade específica da ascorbato peroxidase (mmol min-1 mg-1 de
proteína) nos extratos de plantas de Ricinus communis submetidas ao
tratamento com metil jasmonato em diferentes intervalos de tempo. Linhas: C -
controle; 01, 04, 06, 12, 24 e 48h – tempo de exposição ao metil jasmonato. Os
resultados foram expressos como média (± erro-padrão) de três repetições.
Fig.21: Atividade ascorbato peroxidase determinada em gel após separação
das proteínas por eletroforese nativa 8% . Linhas: C- controle; 01, 04, 06, 12,
24 e 48h – tempo de exposição ao metil jasmonato.
C 1 4 6 C 12 C 24 48

Atividade específica PPO
0,00
0,01
0,01
0,02
0,02
0,03
0,03
C-1H 1H C-6H 6H 12H C-24H 24H C-48H 48H
Tempo de exposição ao metil jasmonato
Variação Abs . mg -1 . min
-1
Fig.22: Atividade polifenol oxidase (∆Abs mg -1 min -1) em gel após separação
das proteínas por eletroforese nativa 8% . Linhas: C- controle; 01, 04, 06, 12,
24 e 48h – tempo de exposição ao metil jasmonato. Os resultados foram
expressos como média (± erro-padrão) de três repetições.
5.2.5 – Determinação da atividade guaiacol peroxidase (GPX) no extrato
bruto
Ao analisar a influência do tratamento com MeJa nas folhas de mamona
sobre a atividade GPX, pudemos observar a existência de duas isoformas com
atividade majoritária, onde houve uma redução gradual da atividade em 1, 4 e
6h, sendo o declínio mais pronunciado em 6h. Com doze horas de tratamento,
as atividades tendem a retornar aos valores verificados para o controle. Após
24 e 48h, há um aumento significativo das atividades (fig. 23).
5.2.5.1 – Determinação da atividade guaiacol peroxidase (GPX) após
separação das proteínas por eletroforese não desnaturante.
Foram observadas duas bandas majoritárias com atividade GPX (G1 e
G2) com comportamento análogo, após eletroforese nativa 8% (fig. 24). A
atividade GPX é reduzida até 6h após o tratamento com MeJa. Em 12h de
tratamento a atividade alcança valores similares aos do controle e com 24 e
48h, há um grande aumento relativo para a atividade GPX.
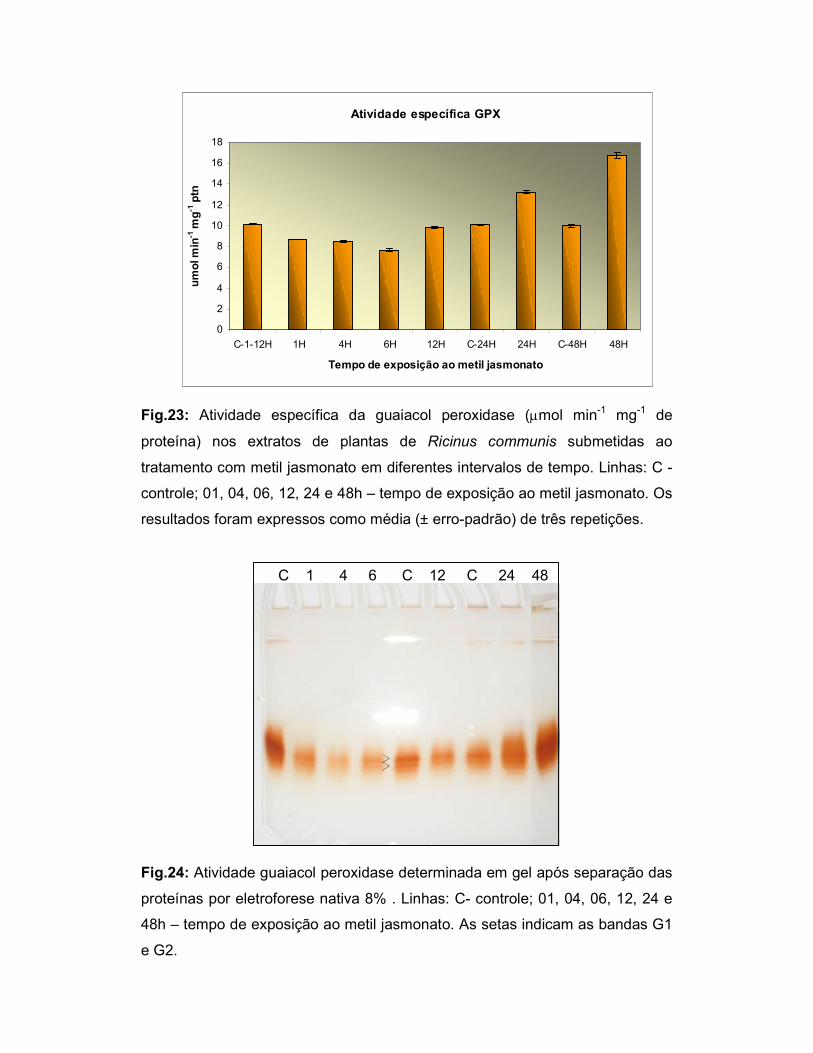
Atividade específica GPX
0
2
4
6
8
10
12
14
16
18
C-1-12H 1H 4H 6H 12H C-24H 24H C-48H 48H
Tempo de exposição ao metil jasmonato
umol min-1 mg-1 ptn
Fig.23: Atividade específica da guaiacol peroxidase (µmol min-1 mg-1 de
proteína) nos extratos de plantas de Ricinus communis submetidas ao
tratamento com metil jasmonato em diferentes intervalos de tempo. Linhas: C -
controle; 01, 04, 06, 12, 24 e 48h – tempo de exposição ao metil jasmonato. Os
resultados foram expressos como média (± erro-padrão) de três repetições.
Fig.24: Atividade guaiacol peroxidase determinada em gel após separação das
proteínas por eletroforese nativa 8% . Linhas: C- controle; 01, 04, 06, 12, 24 e
48h – tempo de exposição ao metil jasmonato. As setas indicam as bandas G1
e G2.
C 1 4 6 C 12 C 24 48

5.3 – Caracterização da enzima guaiacol peroxidase (GPX) 5.3.1 – Determinação do pH ideal
Para verificar em qual pH seria melhor visualizada a(s) atividade(s)
peroxidásica(s), foi realizado um ensaio com vários pHs, como mostra a tabela
IV. Observamos que a enzima tem mais eficiência na catálise em valores de pH
entre 5,4 e 6,8. Os experimentos foram feitos em triplicata.
Tabela V: Determinação da atividade
específica da guaiacol peroxidase
em diferentes pHs.
pH Atividade
(mmol min-1 mg-1 proteína)
4,6 0,52
5,4 5,15
5,8 5,42
6,8 5,58
7,0 4,60
9,0 3,18

5.3.2 – SDS-PAGE da enzima homegênea
Após eletroforese nativa dos extratos de plantas controle, os géis foram
revelados para a atividade GPX e o conteúdo protéico contido nas bandas foi
extraido do géis e homogeneizado em tampão de amostra desnaturante para
que se efetuasse um SDS-PAGE, como mencionado no item 4.10.2.
Com a revelação de proteínas subseqüente à corrida eletroforética,
pode-se observar que as duas bandas com atividade GPX, G1 e G2,
apresentam-se como um dímero com massa molecular entre 36 e 40kDa (fig.
25).
5.3.3 – Determinação do ponto isoelétrico da GPX por eletroforese
bidimensional
Para determinar o ponto isoelétrico das bandas G1 e G2, as mesmas
foram recortadas dos géis de eletroforese nativa e tiveram as proteínas
extraídas, como mencionado no item 4.10.2. Subsequentemente foram
processadas para eletroforese bidimensional, como citado no item 4.10.3.
G1 e G2 apresentaram a massa molecular muito próxima de 39kDa,
confirmando dados obtidos por SDS-PAGE. Em G1 observamos um “spot” não
homogêneo de massa molecular em torno de 39kDa, que talvez represente
dois spots, com um pI próximo de 8,5-8,7 (fig. 26). No gel bidimensional
referente à banda G2, foram observados 3 “spots”: Um deles, também não
uniforme com massa molecular próxima a 39kDa e pI 8,5-8,7 (banda G2); e
dois “spots” minoritários de pI em torno de 6, com massas moleculares de
60kDa e 20kDa, aproximadamente (fig:26).
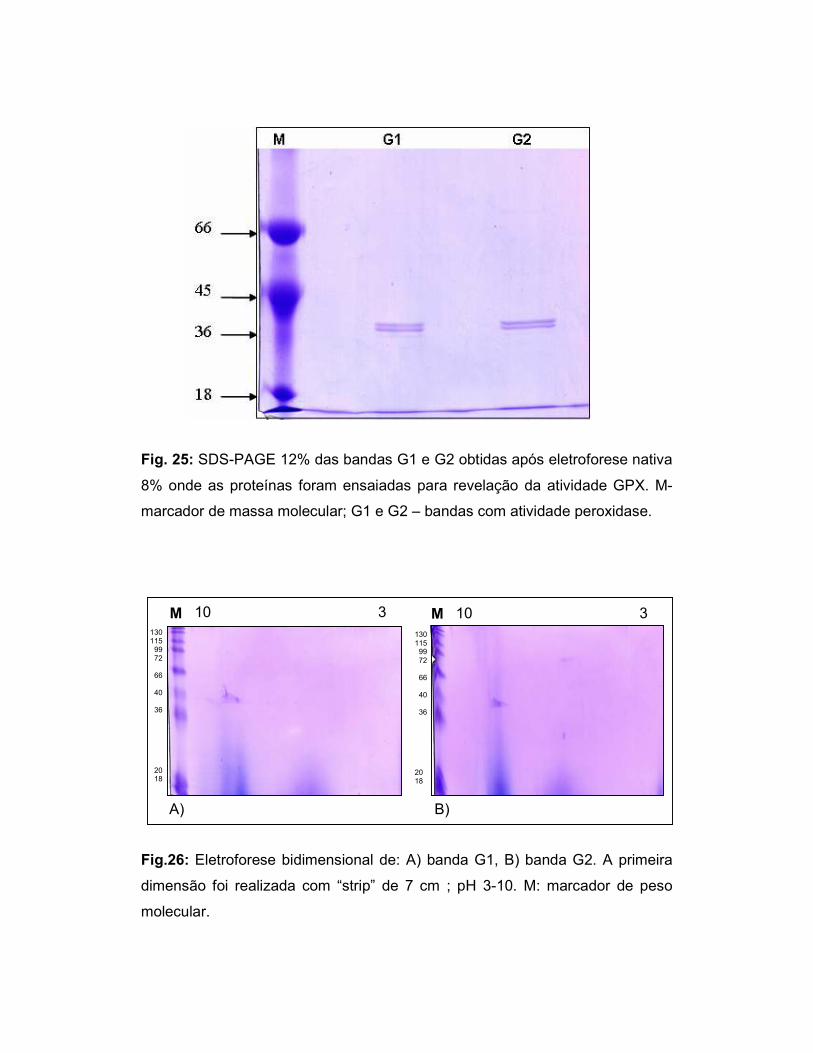
Fig. 25: SDS-PAGE 12% das bandas G1 e G2 obtidas após eletroforese nativa
8% onde as proteínas foram ensaiadas para revelação da atividade GPX. M-
marcador de massa molecular; G1 e G2 – bandas com atividade peroxidase.
Fig.26: Eletroforese bidimensional de: A) banda G1, B) banda G2. A primeira
dimensão foi realizada com “strip” de 7 cm ; pH 3-10. M: marcador de peso
molecular.
A) B)
M M 10 3 10 3 130 115 99 72 66 40 36 20 18
130 115 99 72 66 40 36 20 18

5.4 - Detecção in vivo do peróxido de hidrogênio
Utilizando o substrato DAB, pudemos visualizar o peróxido de hidrogênio
(H2O2) nas folhas das plantas. Com 1h de tratamento, a formação de H2O2 é
quase imperceptível e passa a ser mais bem notada com 4h. As folhas tratadas
por 6h tiveram uma alta produção de H2O2, quando comparadas às folhas não
tratadas (fig. 27).
Com relação às plantas tratadas por 12, 24 e 48h, não podemos
observar pontos de polimerização do DAB. Contudo, essas folhas mantiveram
a nervura central mais escurecida, fig: 27.
Essas folhas foram observadas em lupa e fotografadas, (fig. 28). Vale
ressaltar que as plantas de 12, 24 e 48h foram fotografadas na região da
nervura principal, por ter sido o único diferencial causado pelo MeJa.
5.5 - Varredura de radicais livres pelo método do DPPH Para avaliar o conteúdo de radicais livres nos extratos foliares de Ricinus
communis tratadas com MeJa, foi utilizado o método do DPPH modificado de
Yepez et al., 2001. O aumento do índice de remoção do DPPH em relação ao
controle foi expresso como percentual, tendo como controle positivo a rutina.
Detectamos um aumento de 33% no índice de remoção de DPPH após
1h de tratamento e de 25% após 4, 6 e 12h. Houve um aumento de 13% e com
24 e 48h de tratamento com MeJa. (fig. 29). A rutina teve índice de remoção de
DPPH de 100%.
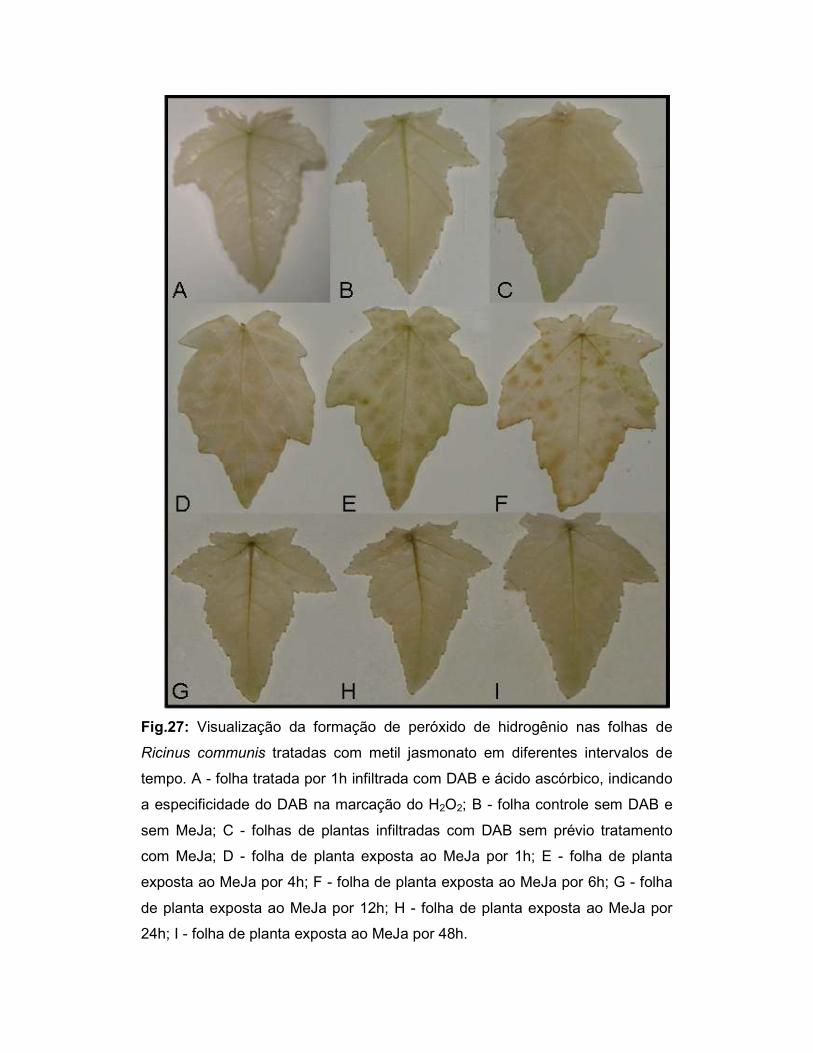
Fig.27: Visualização da formação de peróxido de hidrogênio nas folhas de
Ricinus communis tratadas com metil jasmonato em diferentes intervalos de
tempo. A - folha tratada por 1h infiltrada com DAB e ácido ascórbico, indicando
a especificidade do DAB na marcação do H2O2; B - folha controle sem DAB e
sem MeJa; C - folhas de plantas infiltradas com DAB sem prévio tratamento
com MeJa; D - folha de planta exposta ao MeJa por 1h; E - folha de planta
exposta ao MeJa por 4h; F - folha de planta exposta ao MeJa por 6h; G - folha
de planta exposta ao MeJa por 12h; H - folha de planta exposta ao MeJa por
24h; I - folha de planta exposta ao MeJa por 48h.
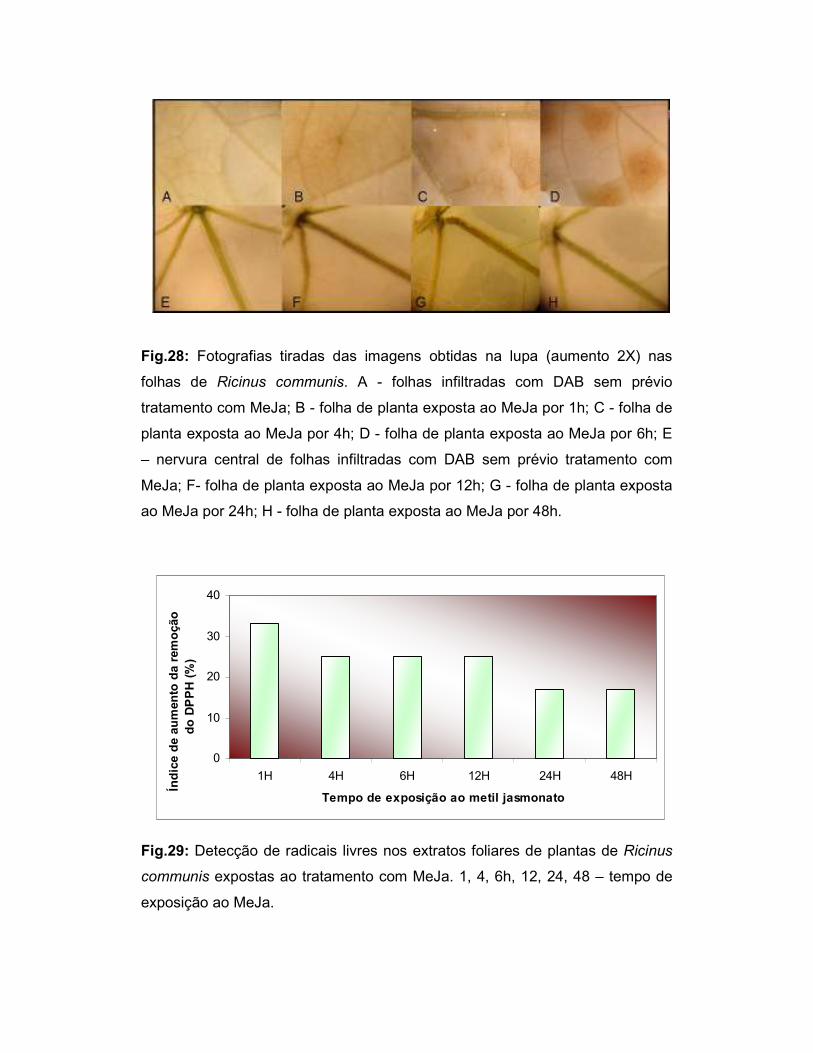
Fig.28: Fotografias tiradas das imagens obtidas na lupa (aumento 2X) nas
folhas de Ricinus communis. A - folhas infiltradas com DAB sem prévio
tratamento com MeJa; B - folha de planta exposta ao MeJa por 1h; C - folha de
planta exposta ao MeJa por 4h; D - folha de planta exposta ao MeJa por 6h; E
– nervura central de folhas infiltradas com DAB sem prévio tratamento com
MeJa; F- folha de planta exposta ao MeJa por 12h; G - folha de planta exposta
ao MeJa por 24h; H - folha de planta exposta ao MeJa por 48h.
0
10
20
30
40
1H 4H 6H 12H 24H 48H
Tempo de exposição ao metil jasmonato
Índice de aumento da remoção
do DPPH (%)
Fig.29: Detecção de radicais livres nos extratos foliares de plantas de Ricinus
communis expostas ao tratamento com MeJa. 1, 4, 6h, 12, 24, 48 – tempo de
exposição ao MeJa.

6.0 – Discussão
Espécies reativas de oxigênio (ERO), como H2O2, O2.- e OH- são
detectadas sob diversas condições de estresses (Mehdy, 1994; Kauss &
Jeblick, 1995; Breusegem et al., 2001; Hadrami et al. 2005). Há relatos que as
ERO são liberadas após a exposição ao metil jasmonato em Petrosehum
crispum L. (Kauss et al., 1994), Lycopersicon esculentum L. (Orozco-Cardenas
& Ryan, 1999) e Arabidopsis thaliana (Jung, 2004). Em altas concentrações, as
ERO são tóxicas, porém, em concentrações baixas, podem servir como
potentes sinalizadores (Breusegem et al., 2001). Podem induzir, inclusive, a
rota octadecanóide, caminho de sinalização vegetal mais conhecido, que
culmina com a produção do ácido jasmônico e do metil jasmonato e indução de
genes de defesas (Orozco-Cardenas & Ryan, 1999). Para que a célula possa
resistir aos efeitos tóxicos das ERO, um efetivo sistema antioxidante enzimático
e não enzimático pode orquestrar a concentração das ERO nos mais variados
componentes e compartimentos sub-celulares (Inzé & Montagu, 1995; Mittler,
2002). O excesso de ERO pode resultar em danos no aparelho fotossintético
(fotoinibição), causando danos celulares drásticos e clorose nas folhas
(Hancock et al., 2002).
Nosso estudo demonstrou que os níveis de uma 2-Cys peroxiredoxina
(2-CP), reduzem nos momentos iniciais, 1-6h, após a exposição com metil
jasmonato (fig. 15). As 2-CP constituem um grupo ubíquo de enzimas de
cloroplasto que reduzem alquil hidroperóxidos tóxicos. Evidências de que a 2-
CP tem um papel protetor significativo durante a fotossíntese foram
demonstradas por Baier & Dietz em 1997 e em 2002 por Konig e coloboradoes.
Baier & Dietz (1997) observaram que mutantes de Arabidopsis thaliana que
expressavam quantidades reduzidas de 2-CP, tiveram um decréscimo na
atividade fotossintética e em proteínas associadas ao fotossistema II, bem
como ATP sintase e rubisco. Nós sugerimos que a redução da 2-CP em
Ricinus communis levaria a um acúmulo de H2O2 que, por meio de
mecanismos de sinalização, induziria um decréscimo de moléculas envolvidas
na fotossíntese, como a degradação da enzima rubisco (ribulose 1,5 -
bisfosfato- carboxilase/oxigenase) verificada por Soares e colaboradores em
2004.

A enzima de defesa relacionada com as etapas iniciais de produção de
ERO nas células é a superóxido dismutase (SOD). Presente em organismos
aeróbicos e anaeróbicos facultativos, a SOD caracteriza um grupo de
metaloenzimas que catalisam a formação do peróxido de hidrogênio a partir de
radicais superóxidos (Scandalios, 1993). Existem três isoformas desta enzima
em plantas, as quais se classificam de acordo com os íons metálicos em seu
sítio ativo: cobre/zinco (Cu/Zn-SOD), manganês (Mn-SOD) e ferro (Fe-SOD)
(Bowler et al., 1992; Dupeyrat et al. 2004).
A alteração no padrão de atividade desta enzima é bastante variável e
pode depender do tipo de estresse, da espécie, tecido, fases de
desenvolvimento da planta, além das próprias particularidades das isoformas
(Azevedo et al., 1998). Em nosso estudo detectamos três isoformas da SOD
em folhas de Ricinus communis, porém apenas uma delas apresentou
alteração na atividade mediante tratamento com Meja (fig. 16). A utilização de
inibidores específicos para cada isoforma, leva a crer que a isoforma alterada
se tratava de uma Mn-SOD (fig. 17). Após a exposição das folhas ao MeJa por
24 e 48 horas, a isoforma Mn-SOD teve sua atividade reduzida (fig. 16).
Hernández e colaboradores (1993) estudaram o efeito do estresse salino
em dois cultivares de Pisum sativum L., sendo um tolerante e um sensível ao
estresse. No cultivar sensível a salinidade levou a uma redução significativa da
atividade da Mn-SOD e da Cu/Zn-SOD. Tais atividades foram aumentadas no
cultivar resistente. Os autores concluem que a toxidez sub celular do NaCl em
nível mitocondrial, nas plantas estudadas, se relaciona com um mecanismo de
estresse oxidativo que é mediado por radicais superóxido e Mn-SOD.
Esperávamos um aumento da atividade SOD nos períodos iniciais (1-6h) após
o tratamento com metil jasmonato, para a remoção do superóxido que
geralmente é produzido como resposta inicial ao estresse oxidativo, contudo a
atividade SOD pode ter sido inibida nos tempos iniciais pela concentração
exacerbada de Meja para esta enzima. Lijun & colaboradores (2005)
observaram a inibição desta enzima por altas concentrações de cádmio.
Nossos resultados são concordantes com os de Kochhar & Kochhar
(2004), que observaram redução da atividade SOD em plantas de Vigna mungo
submetidas à temperatura de 40ºC por 2h. Jung verificou, em 2004, que a
atividade total da SOD decresceu, 3 dias após o tratamento com MeJa e então

aumentou cerca de 2 vezes após sete dias de tratamento em folhas de
Arabidopsis thaliana. Um estudo desenvolvido por Jebara et al. (2005) em
nódulos de raízes de Phaseolus vulgaris sob estresse salino, revelou que, nas
raízes, há redução da atividade da SOD, enquanto que, nos nódulos a
atividade da enzima é estimulada. Como um balanço total, foi verificado um
aumento na atividade da enzima.
Catalases são hemo proteínas presentes em procariotos e nos
peroxissomos, citoplasma e mitocôndria das células eucarióticas. Como
mencionado, catalisa a decomposição do H2O2 em oxigênio e água, sem
produzir radicais livres (Fraaije et al. 1996). A atividade catalase apresentou um
comportamento transiente em nossas experimentações, quando avaliada em
extrato bruto (fig. 18). A análise da atividade após separação por eletroforese
não foi conclusiva. Observamos um arraste no gel, decorrente da baixa
mobilidade da enzima nas condições analisadas (fig. 19). Quando revelamos
as proteínas nessa região com comassie, nenhuma banda protéica foi
detectada, indicando que a enzima tem uma atividade específica elevada. A
redução da catalase em 1h pode estar relacionada a um acúmulo de H2O2 nos
peroxissomos, como uma estratégia para que o H2O2 não fosse degradado
totalmente para ser usado em outras partes da célula vegetal como um
sinalizador de defesa. Como tal, ele induziria, inclusive a própria rota
octadecanóide para auxiliar no aumento da produção do AJ e do MeJa.
Portanto, o aumento da atividade CAT, em nossos experimentos, após 12h de
exposição ao MeJa e sua estabilização em 24 e 48h pode refletir um momento
de feedback negativo, no qual a produção do AJ e do MeJa estaria se
estabilizando.
Outros trabalhos têm documentado o decréscimo da atividade catalase,
como em nódulos de raízes de Phaseolus vulgaris sob estresse salino,
enquanto que em outras partes das raízes, não se detectou a atividade CAT
(Jebara et al. 2005). Tratamentos prolongados com ácido salicílico (AS)
inativam a CAT e a APX contribuindo para sintomas fitotóxicos, sugerindo que
a inativação de enzimas metabolizadoras de H2O2 funciona como um indicador
da morte celular hipersensitiva (Rao et al., 1997). Chanomgpol &
colaboradores (1998) relataram a catalase como um mediador de resposta de
defesa uma vez que a redução de sua atividade foi relacionada ao acúmulo de

H2O2 em tabaco. Contudo, a indução do gene CAT ou o aumento da atividade
da enzima também tem sido relatado. Jung (2004) verificou que a atividade
catalase não alterou entre o 3º e o 5º dia após a exposição ao tratamento com
metil jasmonato, mas aumentou 58%, sete dias após o tratamento em
Arabidopsis thaliana. Dependendo da concentação de ácido jasmônico utilizada
na indução de resposta, a atividade das enzimas antioxidantes é alterada de
forma distinta. Foi observado em Morinda elliptica que a atividade catalase é
aumentada após 1 dia e drasticamente reduzida após 3 e 6 dias de tratamento
com 50 µM AJ. Quando se utilizou o AJ numa concentração de 100 µM, não
houve alteração significativa da enzima até o terceiro dia, havendo uma
redução da atividade apenas após 6 dias (Chong et al., 2005). A atividade
catalase reduzida foi documentada também em culturas de Catharanthus
roseus, eliciadas por fungo (Zhao et al., 2001) e por estresse hídrico em
morango (Wang, 1999).
Algumas enzimas envolvidas na atenuação do estresse oxidativo
possuem os mesmos substratos, tornando difícil o entendimento dos
mecanismos antioxidativos. A catalase a ascorbato peroxidase, por exemplo,
tem em comum o substrato H2O2. As concentrações de H2O2 ditarão se a CAT
ou a APX será responsável pela desintoxicação desta ERO. (Karyotou &
Donaldson, 2005). Além disso, como existem diferentes isoformas das enzimas
antioxidantes nos vários compartimentos subcelulares, em um dado momento,
um tipo de estresse interferirá mais pronunciadamente em uma ou outra
enzima. A ascorbato peroxidase tem como função degradar o H2O2 via
oxidação do ascorbato (Karyotou & Donaldson. 2005). Está presente no
citossol, peroxissomo e apresenta-se como a principal enzima responsável pela
degradação do peróxido de hidrogênio no cloroplasto (Foyer et al., 1994). Foi
observado, em nosso trabalho, que a atividade APX é aumentada 4 e 24h após
o tratamento com metil jasmonato e folhas de Ricinus communis. Contudo, a
enzima não sofre alterações em seu padrão de atividade num período de 1, 6,
12 e 48h subseqüente ao tratamento, como verificado nas figuras 20 e 21. O
aumento da atividade da APX pode estar relacionada com a diminuição dos
níveis da 2-CP até 6h após o tratamento com MeJa. Sabe-se que plantas com
2-CP suprimida compensam com a super expressão de ascorbato peroxidase,

sugerindo que existe um equilíbrio entre diversas enzimas antioxidantes em
cloroplastos (Baier & Dietz, 1997). Chong & colaboradores (2005) também
observaram um comportamento transiente da enzima APX em Morinda
elliptica, na qual a atividade ascorbato peroxidase não sofre alteração após 1
dia, é aumentada após 3 dias e drasticamente reduzida após 6 dias de
tratamento com 50 µM AJ. Quando se utilizou o AJ numa concentração de 100
µM, houve um aumento sutil da atividade até o terceiro dia e uma redução
maior que 50% da atividade após 6 dias de tratamento.
O guaiacol é geralmete utilizado como substrato para a medição da
atividade peroxidásica (Hiraga et al. 2001). A enzima guaiacol peroxidase, uma
classe de peroxidases também envolvida em ações protetoras em plantas,
participam da lignificação da parede celular, biossíntese de etileno,
restabelecimento após injúria e defesa contra patógenos (Gazaryan et al 1996).
Nossos experimentos elucidaram o comportamento enzimático da GPX frente
ao tratamento com MeJa. Observamos, pelas análises espectrofotométricas,
um decréscimo gradual da atividade até 6h e um restabelecimento dos valores
referentes ao controle após 12h de tratamento. Após esse período, a atividade
aumenta significativamente, tanto em 24h quanto em 48h, sendo que este
aumento chega a valores próximos a 50% com 48h (fig. 23). A determinação da
atividade GPX por eletroforese nativa, revelou um comportamento similar ao
obtido para análises espectofotométricas (fig. 24). Foi observado por Jung em
2004 que a atividade GPX aumenta cerca de 100 vezes depois de sete dias de
tratamento com metil jasmonato em Arabidopsis thaliana. O autor considera
que a maior parte da proteção contra o estresse oxidativo em plantas tratadas
com MeJa pode resultar de antioxidantes enzimáticos, especialmente GPX.
Nossos resultados indicam um aumento progressivo da atividade GPX com 24
e 48h, porém a atividade reduzida desta enzima nos momentos iniciais após o
tratamento com MeJa pode estar envolvida com a geração de algumas ERO,
que neste momento, poderiam ter um envolvimento com a sinalização de
defesa em resposta ao estresse. Provavelmente o aumento da atividade GPX
em momentos tardios está relacionado com a desintoxicação celular e uma
retomada ao metabolismo normal (fig. 23 e 24). O estudo desenvolvido por
Jebara & colaboradores (2005), citado anteriormente, revelou que há redução

da atividade de três isoformas GPX (uma isoforma ácida e duas básicas) em
raízes, enquanto nos nódulos, ocorre uma redução da atividade GPX em
Phaseolus vulgaris sob estresse salino. Isso sugere que a GPX tem um papel
importante na desintoxicação do H2O2 sob estresse salino.
Conduziu-se experimentos com o objetivo de purificar e fazer algumas
caracterizações da enzima GPX. Foram observadas, por eletroforese não
desnaturane, duas bandas majoritárias (G1 e G2) com o mesmo padrão de
atividade (fig. 24), sendo que a isoforma G1 possuía atividade mais intensa.
Essas bandas de isoformas foram extraídas separadamente dos géis nativos,
ensaiados para GPX, e verificou-se por SDS-PAGE que se tratavam de
dímeros com aproximadamente 39kDa cada (fig. 25). A massa molecular de G1
e G2 foi confirmada pela eletroforese bidimensional, na qual foi detectado um
pI de 8,5 para G1 e G2 (fig. 26). Notamos a presença de dois “spots” em torno
de 39kDa para G1, evidenciando que cada um dos dois monômeros possui pI
ligeiramente diferente, em torno de 8,5-8,7. Quando realizamos uma
eletroforese bidimensional da isoforma G2, percebemos dois “spots” com
aproximadamente 39 kDa, conforme visto por SDS-PAGE, com pI ligeiramente
diferente, também em torno de 8,5-8,7 e mais dois “spots” com massas
moleculares diferentes que podem ter sido extraídas junto com a isoforma G2
(fig. 26). Observamos que uma das duas cadeias polipeptídicas de G1 tem alta
similaridade com uma das cadeias de G2, por apresentarem a massa
molecular próxima e o mesmo pI (fig. 25 e 26). A guaiacol peroxidase de
Ricinus communis, exerce sua função de catálise mais pronunciadamente entre
os pH 5,4 e 6,8, de acordo com nossos dados (Tab. V). Peroxidases com
características similares foram detectadas em brócolis e tabaco. Uma proteína
de 44k Da e pI: 8,7 foi detectada em brócolis com atividade peroxidase e pela
espectrometria de massas foi comprovada a alta identidade da proteína com
uma peroxidase de tabaco de 40kDa (Palomares & Préstamo, 2005).
Karyotou & Donaldson (2005), purificaram uma APX de glioxissomos de
Ricinus communis com aproximadamente 34 KDa. Tal enzima também utiliza
pirogalol e guaiacol como agentes redutores. Mika & Lüthje (2003) isolaram
três GPX de membrana plasmática de milho e avaliaram algumas propriedades
das isoformas. As enzimas nativas revelaram uma massa molecular aparente

de 38, 70 e 155 KDa. Doadores de elétrons artificiais, como o ascorbato,
guaiacol, álcool coniferil, ácido coumarico e ácido ferúlico, foram usados e
observou-se que as isoformas oxidam esses substratos de forma diferenciada.
Todas utilizam o guaiacol, porém outros agentes redutores são utilizados de
forma mais ou menos eficiente pelas isoformas em suas catálises. A atividade
ascorbato peroxidase, provavelmente teria sido perdida durante os passos de
purificação.
Recentemente a pesquisa acerca da variação da atividade antioxidativa
tem se aprofundado e vários eliciadores têm sido testados. Neto et al., (2005)
verificaram que o pré tratamento com H2O2 sob condições normais, induz a
tolerância ao estresse salino em trigo. O pré tratamento com H2O2 aumentou as
atividades SOD e CAT, enquanto que a atividade APX se manteve constante.
O estresse salino aumentou a atividade de todas as enzimas antioxidativas
estudadas, em plantas aclimatadas ou não. Argarwal & colaboradores (2005)
conduziram um estudo com o objetivo de esclarecer o papel regulatório de uma
série de compostos na indução de enzimas antioxidantes em genótipos de
trigo. Os resultados revelaram que AS (1 mM), ABA (0,5 mM), Ca+2 (5 mM) e
H2O2 (0,05 mM) foram mais efetivas no aumento das atividades da SOD, APX
e CAT e no conteúdo de H2O2. EGTA, um quelante de cálcio, reduziu a
atividade de todas as enzimas antioxidantes abaixo dos valores dos controles,
bem como o conteúdo de H2O2. O grupo sugere que o sinal de estresse abiótico
é transduzido via ABA, Ca+2 e H2O2, que seriam responsáveis pela ativação de
alguns fatores de transcrição associados à SOD, APX e CAT.
Estresses bióticos e abióticos podem aumentar a atividade da enzima
fenilalanina amônia liase (PAL), que está situada em um ponto de ramificação
entre o metabolismo primário e secundário. A reação catalisada pela PAL é um
passo regulador importante na formação de muitos compostos fenólicos (Taiz
& Zeiger, 2004). Alguns destes compostos, tais como, flavonóides e
precursores de ligninas, são potentes antioxidantes (Sakihama et al., 2002).
Outra forma de oxidação de fenóis ocorre por meio da enzima polifenol oxidase
(PPO), que catalisa a oxidação de compostos fenólicos para quinonas, usando
oxigênio molecular como aceptor de elétrons.
Em nossos experimentos, avaliamos o envolvimento do metabolismo
secundário com o estresse causado pelo MeJa pela medição da atividade PPO

dos extratos das folhas de Ricinus communis. Nossos resultados indicaram que
os níveis desta enzima permaneceram praticamente constantes ao longo do
tratamento com metil jasmonato (fig. 22). Constabel & Ryan analisaram os
níveis desta enzima em diferentes espécies de cinco famílias de plantas, nas
quais foram detectados valores constitutivos e/ou induzidos muito variáveis. A
atividade, determinada como (∆ A490 min 1 g-t fr. wt x 103), em arroz (Oryza
sativa) foi de 65 e em soja (Glycine max) de 19.500. Em plantas da família
Brassicacea, a atividade enzimática pouco variou 60 minutos após injúria
mecânica, ou exposição aos vapores de MJ, no entanto, em plantas da família
solanaceae, a atividade aumentou em ambos os tratamentos. Em Lycopersicon
esculentum, a atividade aumentou 1630 vezes após a injúria, e 61800 vezes
após MeJa.
A sobrevivência das espécies vegetais varia grandemente com sua
capacidade de responder a injúria e ao MeJa com o aumento da atividade
PPO. A ausência de indução da atividade PPO não impede, contudo, uma
indução potencial em outras condições de estresse, exposições em tempos
variados de um mesmo tipo de estresse ou por diferentes tipos de tecidos
danificados (Felton et al. 1994; Stout et al. 1994; Constabel & Ryan. 1998).
A variação da atividade antioxidante dentro da célula pode alterar o
conteúdo de ERO e desencadear uma resposta de aclimatação ou de morte.
Com o objetivo de verificar se a alteração do arsenal antioxidante em Ricinus
communis se relaciona com variações na formação de ERO, realizamos
experimentos para detectar tanto o conteúdo de H2O2 quanto o de radicais
livres.
Verificamos que há um acúmulo de H2O2 progressivo entre 1 e 6h após o
tratamento com MeJa. Nesse período a marcação com DAB revelou lesões
circulares que se espalharam por toda a superfície da folha. Com 12 e 24h
após a o tratamento, a marcação com DAB foi menos intensa e se restringiu à
nervura central e com 48h, não houve formação aparente de H2O2 (fig. 27).
Esses resultados foram melhor evidenciados quando as folhas foram
fotografadas em lupa com aumento de 2X (fig. 28).
Diversos estudos relataram a produção de peróxido de hidrogênio em
resposta à estresses. Rao & colaboradores investigaram em 1997 como o

ácido salicílico (AS) aumenta o conteúdo de H2O2 em Arabidopsis thaliana. O
tratamento com AS, aumenta a produção de H2O2, a peroxidação lipídica e o
dano oxidativo a proteínas. Cho & Seo (2005) observaram um aumento no
conteúdo de H2O2 em Arabidopsis thaliana exposta a Cádmio.
Nossos dados referentes à detecção do conteúdo de radicais livres em
folhas tratadas com metil jasmonato complementam os resultados obtidos com
relação à produção de H2O2 nas folhas de Ricinus communis. Observamos que
há um aumento de 33% do conteúdo de radicais livres após 1h de tratamento
com MeJa em folhas de Ricinus communis e os valores decrescem, tendendo
aos nívies de controle, até 48h (fig. 29). Aparentemente a formação
pronunciada de ERO nos momentos iniciais de exposição ao estresse com
MeJa em Ricinus communis, está relacionada com o decréscimo dos níveis da
2-CP e das atividades da CAT, e GPX. As demais enzimas orquestram o
estresse oxidativo de forma a contornar o estado fisiológico normal da planta. A
APX, por exemplo, aumentaria em 4 e 24h para balancear os níveis de ERO
nos compartimento celulares de forma a evitar toxidez excessiva. A GPX
parece ser a principal enzima detoxificadora em Ricinus communis após 24h de
tratamento com MeJa.

7.0 – Conclusões Podemos concluir que a maioria das enzimas antioxidativas estudadas
são atenuadas nos momentos iniciais de estresse, período compreendido entre
1 e 6 horas de tratamento com MeJa. Essa redução se relaciona, de forma
inversamente proporcional, ao conteúdo das ERO, que foi contabilizado como a
soma dos teores de peróxido de hidrogênio e de radicais livres. Valores
elevados das ERO perduram até o momento em que o balanço das atividades
antioxidantes deixa de ser favorável a sua formação, ou seja, a partir de 12h de
tratamento. Nesta ocasião, a maioria das enzimas antioxidantes têm sua
atividade elevada e a guaiacol peroxidase comporta-se como principal enzima
detoxificadora em Ricinus communis submetidas ao tratamento com metil
jasmonato.

8.0 – Referências
Alarcon, J.J. & Malone, M. (1995). The influence of plant age on wound
induction of proteinase inhibitors in tomato. Physilogia Plantarum, 95: 423 –
427.
Aquino-Bolaños, E.N. & Mercado-Silva, E. (2004). Effects of polyphenol
oxidase and peroxidase activity, phenolics and lignin content on the browning
of cut jicama. Postharvest Biology and Technology, 33: 275-283.
Asada, K. (1992). Ascorbate peroxidase – a hydrogen peroxide scavenging
enzyme in plants. Physiologia Plantarum, 85: 235-241.
Azevedo, D.M.P. & Lima, E.F. (2001). O agronegócio da mamona no Brasil.
EMBRAPA, p: 213-227.
Azevedo, R.A., Alas, R.M., Smith, R.J., Lea, P.J. (1998). Response of
antioxidant enzymes to transfer from elevated carbon dioxide to air and
ozone fumigation, in the leaves and roots of wild-type and a catalase-
deficient mutant of barley. Physiologia Plantarum, 104: 280 - 292.
Baier, M. & Dietz, K.J. (1997). The plant 2-cys peroxiredoxin BAS1 is a nuclear-
encoded chloroplast protein: its expressional regulation, phylogenetic origin,
and implications for its specific physiological function in plants. Plant Journal,
12: 179 – 190.
Barynin, V.V., Vagin, A.A., Melik-Adamyan, V.R., Grebenko, A.I., Vainshtein,
B.K. (1986). Three-dimensional structure of T-catalase with 3 A resoluction.
Soviet Physics-Crystallography, 31: 457-459. apud Maté, M.J., Zamocky, M.,
Nykyri, L.M., Herzog, C., Alzari, P.M., Betzel, C., Koller, F., Fita, I., (1999).
Structure of catalase-A from saccharomyces cerevisae. Journal of Molecular
Biology, 268: 135-149.
Barka, E.A. (2001). Protective enzymes against reactive oxygen species during
ripening of tomato (Lycopersicon esculentum) fruits in response to low
amounts of UV-C. Australian Journal of Plant Physiololy, 28: 785-791.

Bi, Y.M., Kenton, P., Murr, L., Darby, R., Draper, J. (1995). H2O2 does not
function downstream of salicylic acid in the induction of PR protein
expression. Plant Journal, 8: 235-245.
Bowler, C. & Fluhr, R. (2000). The role of calcium and activated oxygens as
signals for controlling cross-tolerance. Trends in Plant Science, 5: 241-246.
Bowler, C., Van Montagu, M., Inzé, D. (1992). Superoxide-dismutase and stress
tolerance. Annual Review of Plant Physiology and Plant Molecular Biology,
43: 83-116.
Bowles, D.J. (1990). Defense related proteins in higher plants. Biochemistry,
59: 837-907.
Bradford, M.M. (1976). A rapid and sensitive method for the quantification of
microgram quanties of protein utilizing the principle of protein-dye binding.
Analytical Biochemistry, 72: 248-254.
Breusegem, F. V., Vranová, E., Dat, J.F., Inzé, D. (2001). The role of active
oxygen species in plant signal transduction. Plant Science, 161: 405-414.
Buchanan, B.B., Gruíssem, W., Jones, R.L. (2000). Biochemistry & Molecular
Biology of plants. Editado pelos autores, p: 853.
Cakmak, I. & Horst, W.J. (1991). Effect of aluminum on lipid-peroxidation,
superoxide-dismutase, catalase, and peroxidase-activities in root-tips of
soybean (glycine-max). Physiologia plantarum, 83: 463-468 .
Campa, A. (1991). Biological roles of plant peroxidases: known and potential
function. Peroxidases in Chemistry and Biology, 2: 25-50.
Cavalcante, A.P.R., Jacinto, T., Machado, O.L.T. (1999). Methyl jasmonate
changes the levels of rubisco and other proteins in Ricinus communis. ACTA
Physiologiae Plantarum, 21: 161-166
Chance, B. & Herbert, D. (1950). The enzyme-substrate compounds of
bacterial catalase and peroxides. Biochemical Journal, 46: 402-414. apud
Maté, M.J, Zamocky, M., Nykyri, L.M., Herzog, C., Alzari, P.M., Betzel, C.,

Koller, F., Fita, I., (1999). Structure of catalase-A from saccharomyces
cerevisae. Journal of Molecular Biology, 268: 135-149.
Cho, U.H. & Seo, N.H. (2005). Oxidative stress in Arabidopsis thaliana exposed
to cadmium is due to hydrogen peroxide accumulation. Plant Science,
168:113-120.
Choi, Y.O. & Cheng, N.E. (1999). Cloning and expression of a new isotype of
peroxiredoxin gene of Chinese cabbage and its comparison to 2 cys –
peroxiredoxin isolated from the same plant. Biochemistry Biophysics
Research Communnication, 258: 768-771.
Chong, T.M., Abdullah, M.A., Fadzillah, N.M., Lai, O.M., Lajis, N.H. (2005).
Jasmonic acid elicitation of anthraquinones with some associates enzymic
and non enzymic antioxidant responses in Morinda elliptica. Biochemical
Journal, 36: 469-477.
Constabel, C.P. & Ryan, C.A. (1998). A survey of wound - and methyl
jasmonate - induced leaf polyphenol oxidase in crop plants. Phytochemistry,
47: 507-511.
Constabel, C.P. & Ryan, C.A. (1995). Systemin activates synthesis of wound –
inducible tomato leaf polyphenol oxidase via the octadecanoid defense
signaling pathway. Proceedins of the National Academy of Sciences of the
United States of America, 92: 407-411.
Dat, J.F., Lopez-Delgado, H., Foyer, C.H. Scott, I.M. (2000). Effects of salicylic
acid on oxidative stress and thermotolerance in tobacco. Journal of Plant
Physiology, 156: 659-665.
Davis, B.J. (1964). Disc electrophoresis, part II. Annals New York Academy of
Sciences, 121: 404-427.
Del Rio, L.A., Pastori, G.M., Palma, J.M., Sandalio, L.M., Sevilla, F., Corpas,
F.J., Jimenez, A., Lopez-Huertas, E., Hernandez, J.A. (1998). The activated
oxygen role of peroxisomes in senescence. Plant Physiology, 116: 1195-
1200.

Dolphin, D., Forman, A. Borg, D.C., Fajer, J., Felton, R.H. (1971). Compounds I
of catalase and horse radish peroxide: π-cation radicals. Proceedings of the
National Academy of Sciences of the United States of America, 68: 614-618.
apud Maté, M.J, Zamocky, M., Nykyri, L.M., Herzog, C., Alzari, P.M., Betzel,
C., Koller, F., Fita, I. (1999). Structure of catalase-A from saccharomyces
cerevisae. Journal of Molecular Biology, 268:135-149.
Dröge, W. (2002). Free radicals in the physiological control of cell function.
Physiological Reviews, 82: 47- 85.
Dupeyrat, F., Vidaud, C., Lorphelin, A., Berthomieu, C. (2004). Long distance
charge redistribution upon CuZn-superoxide dismutase reduction. The
Journal of Biological Chemistry, 279: 48091-48101
Edman, P. (1950). Method for determination of amino acid sequence in
peptides. Acta Chemica Scandinavia, 28: 283-293.
Farmer, E.E. & Ryan, C.A. (1992). Octadecanoid precursors of jasmonic acid
activate the synthesis of wound – inducible proteinase inhibitor. Plant Cell, 4:
129-134.
Farmer, E.E. & Ryan, C.A. (1990). Interplant communication: airbone methyl
jasmonate induces syntesis of proteinase inhibitors in plant leaves.
Proceedings of the National Academy of Sciences of the United States of
America, 87: 7713-7716.
Felton, G.W., Summers, G.B., Mueller, A.J. (1994). Oxidative responses in
soybean foliage to herbivory by bean leaf beetle and three-cornered alfalfa
hopper. Journal of Chemistry and Ecology, 20: 639-650.
Fidantsef, A.L., Stout, M.J., Thaler, J.S., Duffey, S.S., Bostock, R.M. (1999).
Signal interactions in pathogen and insect attack: expression of
lipoxygenase, proteinase inhibitor II, and pathogenesis – related protein P4 in
the tomato, Lycopersicon esculentum. Physiological and Molecular Plant
Pathology, 54: 97-114.

Fieldes, M.A. & Gerhard, K.E. (1998). Flax guaiacol peroxidases can be used to
illustrate the possiblility of misinterpreting the effects of stress on the activity
of development regulates enzymes. Plant Science, 132: 89-99.
Finch – Savage, W.E., Blake, P.S., Clay, A.A. (1996). Desiccation stress in
recalcitrnt Quercus robur L. seeds results in lipid peroxidation and Increased
Synthesis of jasmonates and abscisic acid. Journal of experimental Botany,
47: 661-667.
Foyer, C.H., Lelandais, M., Kunert, K.J. (1994). Photoxidative stress in plants.
Physiologia Plantarum, 92: 696-717.
Fornazieri-Júnior, A. (1986). Mamona – uma rica fonte de óleos e de divisas.
Editora Ícone, p: 46.
Fraaije, M.W., Roubroeks, H.P., Hagen, W.R., Van Berkel, W.J.H. (1996).
Purification and characterization of an intracellular catalase-peroxidase from
Penicillium simplicissium. European Journal of Biochemistry, 235: 192-198.
Gazaryan, I.G., Lagrimini, L.M., Ashby, G.A., Thorneley, R.N.F. (1996).
Mechanism of indole-3-acetic acid oxidation by plant peroxidases: anaerobic
stopped-flow spectrophotometric studies on horseradish and tobacco
peroxidases. Biochemical Journal, 313: 841-847.
Ghosh, S., Mahoney, S.R., Penterman, J.N. (2001). Ultastructural and
biochemical changes in chloroplasts during Brassica napus senescence.
Plant Physiology Biochemistry, 39: 777-784.
Green, T.R. & Ryan, C.A. (1972). Wound induced proteinase inhibitors from
tomato leaves: a possible defense mechanism against insects. Science, 175:
776-777.
Hadrami, A.E., Kone, D. Lepoivre, P. (2005). Effect of juglone on active oxygen
species and antioxidant enzymes in susceptible and partially resistant
banana cultivars to Black Leaf Streak Disease. European Journal of Plant
Pathology, 113:241-254.

Hammond-Kossack, K.E. & Jones, J.D.G. (1996). Resistance gene dependent
plant defense responses. Plant Cell, 8: 1773-1807.
Hancock, J.T., Desikan, R., Clarke, A., Hurst, R.D., Neill, S.J. (2002). Cell
signaling following plant–pathogen interactions involves the generation of
reactive oxygen and reactive nitrogen species. Plant Physiology and
Biochemistry, 40: 611-617.
Hernández, J.A., Corpas, F.J., Gómez, M., Rio, L.A., Sevilla, F. (1993). Salt-
induced oxidative stress mediated by activated oxygen species in pea leaf
mitochondria. Physiologia Plantarum, 89: 103-110.
Hiraga, S., Sasaki, K., Ito, H., Ohashi, Y., Matsui, H. (2001). A large family of
class III plant peroxidases. Plant Cell Physiology, 42: 462-468.
Howe, G.A., Lightner, J., Browse, J., Ryan, C.A. (1996). An octadecanoid
pathway
mutant (JL5) of tomato is compromised in signaling for defense against insect
attack. Plant Cell, 8: 2067-2077.
Inzé,D. & Montagu, M.V. (1995). Oxidative stress in plants. Current Opinion in
Biotechnology, 6: 153-158.
Janda, T., Szalai, G., Tari, I., Páldi, E. (1999). Hydroponic treatment with
salicylic acid decreases the effects of chilling injury in maize (Zes maysL.)
plants. Planta, 208: 175-180.
Jebara, S., Jebara, M., Limam, F., Aouani, M.E. (2005). Changes in ascorbate
peroxidase, catalase, guaiacol peroxidase and superoxide dismutase
activities in common bean (Phaseolus vulagis) nodules under salt estress.
Journal of Plant Physiology, 162: 929-936.
Jung, S. (2004). Effect of chlorophyll reduction in Arabidopsis thaliana by methyl
jasmonate or norflurazon on antioxidant systems. Plant Physiology and
Biochemistry, 42: 225-231.

Karyotou, K. & Donaldson, R.P. (2005). Ascorbate peroxidase, a scavenger of
hydrogen peroxide in glyoxysomal membranes. Archives of Biochemistry and
Biophysics, 434: 248-257.
Kauss, H. & Jeblick, W. (1995). Pretreatment of parsley suspension cultures
with salicylic acid enhances spontaneous and elicited production of H2O2.
Plant Physiology, 108: 1171-1178.
Kauss, H., Jeblick, W., Zielger, J., Kraber, W. (1994). Pretreatment of parsley
(Petvoselinum crispum L.) suspension culture with methyl jasmonate
enhances the elicitation of active oxygen species. Plant Physiololy, 105: 89-
94.
Klessig, D.F. & Malamy, J. (1994). The salicylic acid signal in plants. Plant
Molecular Biology, 26: 1439-1458.
Kochhar, S. & Kochhar, V.K. (2004). Expression of antioxidant enzymes and
heat shock proteins in relation to combined stress of cadmium and heat in
Vigna mungo seedlings. Plant Science, 168: 921-929.
Konig, J., Baier, M., Horling, F., Kahmann, U., Harris, G., Schurmann, P., Dietz,
K.J. (2002). The plant-specific function of 2-Cys peroxiredoxinmediated
detoxification of peroxides in the redox-hierarchy of photosynthetic electron
flux. Proceedings of the National Academy of Sciences of the United States
of America, 99: 5738-5743.
Larkindale, J. & Huang, B. (2004). Thermotolerance and antioxidant systems in
Agrostis stolonifera: Involvement of salicylic acid, abscisic acid, calcium,
hydrogen peroxide, and ethylene. Journal of Plant Physiology, 161: 405-413.
Laemmli, U.K. (1970). Cleavage of structural protein during assembly of the
head of becteriophage T4. Nature, 227: 680-685.
Levine, A. Tenhaken, R., Dixon, R. Lamb, C. (1994). H2O2 from de oxidative
burst orchestrates the plant hypersensitive disease resistance response.
Cell, 79: 583-593.

Lijun, L., Xuemei, L., Yaping, G., Enbo, M. (2005). Activity of the enzymes of the
antioxidative system in cadmium-treated Oxya chinesis (Orthoptera
Acridoidae). Environmental Toxicology and Pharmacology, 20:412-416.
Mallick, N. & Rai, L.C. (1999). Response of the antioxidant systems of the
nitrogen fixing cyanobacterium Anabaena doliolum to the copper. Journal of
Plant Physioloy, 155: 146-149.
Margis-pinheiro, M., Sandroni, M., Lummerzeim, M., Oliveira, D. (1999). A
defesa das plantas contra as doenças. Ciência Hoje, 25: 24-31.
Mason, H. S. & Mullet, J.E. (1990). Expression of two soybean vegetative
storage protein genes during development and in response to water deficit,
wound, and jasmonic acid. The Plant Cell, 2: 569-579.
Maté, M.J, Zamocky, M., Nykyri, L.M., Herzog, C., Alzari, P.M., Betzel, C.,
Koller, F., Fita, I. (1999). Structure of catalase-A from saccharomyces
cerevisae. Journal of Molecular Biology, 268: 135-149.
McGurl, B., Orozco-Cardenas, M., Pearce, G., Ryan, C.A. (1994).
Overexpression of the prosystemin gene in transgenic tomato plants
generates a systemic signal that constitutively induces proteinase inhibitor
synthesis. Proceedings of the National Academy of Sciences of the United
States of America, 91: 9799-9802.
Mehdy, M.C. (1994). Active oxygen species in plant defense against pathogens.
Plant Physiology, 105: 467-472.
Mika, A. & Lüthje, S. (2003). Properties of guaiacol peroxidase activities
isolated from corn root plasma membranes. Plant Physiology, 132: 1489-
1498
Mittler, R. (2002). Oxidative stress, antioxidants and stress tolerance. Trends in
Plant in Science, 9: 405-410.

Mitter, R. & Zilinskas, B.A. (1993). Detection of ascorbate peroxidase activity in
native gels by inhibition of the ascorbate-dependent reduction of nitroblue
tetrazolium. Analytycal Biochemistry, 212: 540-546.
Neto, A.D.A., Prisco, J.T., Filho, J.E., Medeiros, J.V.R., Filho, E.G. (2005).
Hydrogen peroxide pre-treatment induces salt-stress acclimation in maize
plants. Journal of Plant Physiology, 162: 1114-1122.
Nordberg, J. & Arner, E.S.J. (2001): Reactive oxygen species, antioxidants, and
the mammalian thioredoxin system. Free Radical Biology & Medicine, 31:
1287-1312.
O’Farel, P. H. (1975). High resolution two dimensional electrophoresis of
proteins. Journal of Biological Chemistry, 250: 4007-4021.
Orozco-Cardenas, M., Narváez-Vásquez, J., Ryan, C.A. (2001). Hydrogen
peroxide acts as a second messenger for the induction of defense genes in
tomato plants in response to wounding, systemin, and methyl jasmonate.
Plant Cell, 13: 179-191.
Orozco-Cardenas, M. & Ryan, C. A. (1999). Hydrogen peroxide is generated
systemically in plant leaves by wounding and systemin via the octadecanoid
pathway. Plant Biology, 96: 6553 – 6557.
Palomares, L. & Préstamo, G. (2005). Detection of peroxidase activity in two-
dimensional gel electrophoresis. European Food Research and Technology,
220: 644-647.
Pastori, G.M. & Trippi, V.S. (1992). Oxidative stress induces high rate of
glutathione reductase synthesis in a drought resistant maize strain. Plant
Cell Physiology, 33: 957-996.
Pearce, G., Johnson, S., Ryan, C.A. (1993). Structure-activity of detected and
substituted acid polypeptide induced of plant defensive genes. Journal of
Biology Chemistry, 268: 212-216.

Pearce, G., Strydom, D., Johnson, S. (1991). A polypeptide from tomato leaves
induces wound-inducible proteinase-inhibitor proteins. Science, 253: 895-
898.
Perl-Treves, R. & Galun, E. (1991). The tomato Cu,Zn superoxide dismutase
gene are developmentally regulated and respond to light and stress. Plant
Molecular Biology, 17: 745-760.
Pieterse, C.M.J. & Van Loon, L.C. (1999). Salicylic acid – independent plant
defense pathways. Trends in Plant Science, 4: 52-58.
Polidoros, A. & Scandalios, J. (1999). Role of hydrogen peroxide and different
classes of antioxidants in the regulation of catalase and glutatione-S-
transferase gene expression in mayze (Zea mays L.). Physiologia Plantarum,
106: 112-120.
Rao, M.V., Paliyath, G., Ormrod, D.P., Murr, D.P., Watkins, C. B. (1997).
Influence of salicylic acid on H2O2 production, oxidative stress, and H2O2 -
metabolizing enzimes. Plant Physiology, 115: 137 – 149.
Reinbothe, S., Mollenhauer, B., Reinbothe, C. (1994 a). JIPs and RIPs: The
regulation of plant gene expression by jasmonates in response to
environmental cues and pathogens. The plant cell, 6: 1197-1209.
Retamal, C.A., Thiebaut, P., Alves, E.A. (1999). Protein purification from
polyacrylamide gels by sonication extraction. Analytical Biochemistry, 268:
15-20.
Reymond, P. & Farmer, E.E. (1998). Jasmonate and salicylate as global signals
for defense gene expression. Current opinion in plant biology, 1: 404-11.
Rouhier, N., Gelhaye, E., Jacquot, J. P. (2001). Isolation and Characterization
of a new Peroxiredoxin from Poplar sieve tubes that uses either glutaredoxin
or thioredoxin as a proton donor. Plant Physiology, 127: 1299-1309.

Ryan, C.A. & Balls, K.A. (1962). An inhibitor of chymotrypsin from Solanum
tuberosum and its behavior toward trypsin. Proceedings of the National
Academy of Sciences of the United States of America, 48: 1839-1844.
Ryan CA & Pearce G.L. (2001). Polypeptide hormones. Plant Physiology, 125:
65-68.
Ryan, C. A. (2000). The systemin signaling pathway: differential activation of
plant defensive genes. Biochimica et Biophysica Acta, 1477: 112-121.
Sakihama, Y., Cohen, M.F., Grace, S.C., Yamasaki, H. (2002). Plant phenolic
antioxidant and prooxidant activities: phenolics-induced oxidative damage
mediated by metals in plants. Toxicology, 177: 67-80.
Savy Filho, A., Banzatto, N.V., Veiga, R.F.A., Campana, M.P., Junior, A.P.
(1990). Novo cultivar de mamona: IAC-226 (TARABAY). Bragantia,
Campinas, 49: 269-280.
Scandalios, J.G. (1990). Response of plant antioxidant defense genes to the
environmental stress. Advances in Genetics, 28: 1-41.
Scandalios, J.G. (1993). Oxygen stress and superoxide dismutase. Plant
Physiology, 101: 7-12.
Schaller A. & Ryan C.A. (1994). Identification of a 50-kda systemin binding
protein in tomato plasma-membranes having kex2p-like properties.
Proceedings of the National Academy of Sciences of the United States of
America, 91: 11802-11806.
Schröder, E. & Ponting, C.P. (1998). Evidence that peroxiredoxins lare novel
members of the thioredoxin fold superfamily. Protein Science, 7: 2465-2468.
Shewry, P.R. & Lucas, J.A. (1997). Plant proteins that confer resistance to pests
and pathogens. Advances In Botanical Research Incorporating Advances In
Plant Pathology, 26: 135-192.
Shin, R., Froderman, T., Flurkey, W. H. (1997). Isolation and characterization
of a mung bean leaf polyphenol oxidase. Phytochemistry, 45: 15-21.

Soares, A.M.S., Souza, T.F., Domingues, S.J.S., Jacinto T., Machado, O.L.T.
(2004). Methyl jasmonate promotes the transient reduction of the levels of 2-
Cys peroxiredoxin in Ricinus communis plants. Plant Physiology and
Biochemistry, 42: 543-547.
Stout, M.J., Workman, J., Duffey, S.S. (1994). Differential induction of tomato
foliar proteins by arthropod herbivores. Journal of Chemical Ecology, 20:
2575-2594.
Stryer ,L. (1995). Biochemistry. Editora W.H. Freeman and Company, p: 554.
Taiz , L. & Zeiger, E. (2004). Fisiologia vegetal. Editora Artemed, p: 32.
Tolbert, N.E. (1973). Activation of polyphenol oxidase of chloroplasts. Plant
Physiology, 51: 234-244.
Thipyapong, P. Melkonian, J., Wolfe, D.W., Steffens, J.C. (2004). Suppression
of polyphenol oxidases increases stress tolerance in tomato. Plant Science,
167: 693-703.
Tsang, E.W.T., Bowler, C., Herouart, D., Cham, V. W., Villarroel, R., Montagu,
M.V., Inzé, D. (1991). Differential regulations of superoxide dismutases in
plants exposed to environmental stress. Plant Cell, 3: 783-792.
Vierling, E. (1991). The roles of heat shock proteins in plants. Annual Review of
Plant Physiology and Plant Molecular Biology, 42: 579-620.
Walker Simmons, M. & Ryan, C. A. (1977). Wound-induced peptidase activity
in tomato leaves. Biochemical and Biophysical Research Communications,
74: 411-416.
Wang, S.Y. (1999). Methyl jasmonate reduces water stress in strawberry. Plant
Growth Regulation, 18: 127-134.
Weber, H. (2002). Fatty acid-derived signals in plants. Trends in Plant Science,
7: 217-224.

Wood, Z. A., Schroder, E., Harris, J. R., Poole, L.B. (2003). Structure,
mechanism and regulation of peroxiredoxins. Trends in Biochemical
Sciences, 28: 32-40.
Wu, G., Short, B.J., Lawrence, E.B., Levine, E.B., Fitzsimmons, K.C., Shah,
D.M. (1995). Disease resistance conferred by expression of a gene
encoding H2O2 - generating glucose oxidase in transgenic potato plants.
Plant Cell, 7: 1357-1368.
Yepez, B., Espinosa, M., López, S., Bolaños, G. (2002). Producing
antioxidant fractions from herbaceous matrices by supercritical fluid
extraction. Fluid Phase Equilibria, 194-197: 879-884.
Yuan, Y.J., Li, C., Hu, Z. D., Wu, J.C. (2002). A double oxidative burst for taxol
production in suspension cultures of Taxus chinensis var. mairei induced by
oligosaccharide from Fusarium oxysporum. Enzyme and Microbial
Technology, 30: 774-778.
Xavier-Filho (1993). Sementes e suas defesas contra insetos. Projeto
Multinacional de Biotecnologia e Alimentos.UFC – Ceará. Organizações dos
Estados Americanos. OEA: 1-31.
Xu, Y., Chang, P.F.L., Lui, D., Narashimhan, M.L., Raghothama, K.G.,
Hasegawa, P.M., Bressan, R.A. (1994). Plant defense genes are
synergistically induced by ethylene and methyl jasmonate. Plant Cell, 6:
1077-1085.























![Esterilização por Peróxido de Hidrogênio [ETEC-CARLOS DE CAMPOS]](https://img.document.onl/doc/110x75/558884f6d8b42a69538b4708/esterilizacao-por-peroxido-de-hidrogenio-etec-carlos-de-campos.jpg)




