Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DOS VALES DO JEQUITINHONHA E MUCURI
Programa de Pós-Graduação em Zootecnia
Mariane Rodrigues ferreira
ANÁLISE DE REPETIBILIDADE E AGRUPAMENTO EM GENÓTIPOS
DE Panicum maximum Jacq.
Diamantina
2017


MARIANE RODRIGUES FERREIRA
ANÁLISE DE REPETIBILIDADE E AGRUPAMENTO EM GENÓTIPOS
DE Panicum maximum Jacq.
Dissertação apresentada ao programa de Pós-Graduação
em Zootecnia da Universidade Federal dos Vales do
Jequitinhonha e Mucuri, como requisito parcial para
obtenção do título de Mestre.
Orientador: Prof. Dr. Thiago Gomes dos Santos Braz
Coorientadora: Profa. Dra. Janaina Azevedo Martuscello
Coorientadora: Profa. Dra. Márcia Vitória Santos
Diamantina
2017




AGRADECIMENTOS
Agradeço a Deus, por guiar meu caminho, por sempre atender meus pedidos e me
proporcionar calma, tranquilidade e paciência quando mais precisei.
Aos meus pais, José Airto e Rosária, por todo amor, dedicação, incentivo, por
acreditarem e confiarem em mim.
A minha irmã, Ariane, pela cumplicidade e amizade.
A minha sobrinha, Lorena, pelos momentos de descontração, estar ao seu lado faz com
que todos as preocupações entrem em esquecimento. A “titiá” te ama muito!
Aos meus avós, pelas orações e por estarem presentes. Aos meus tios, tias e primos pelo
carinho e apoio.
Ao meu orientador, Professor Thiago Braz, por todo conhecimento transmitido,
paciência e apoio na execução deste trabalho e por sempre me mostrar que sou capaz.
A minha coorientadora, Professora Janaina Martuscello, pela orientação, por todos os
ensinamentos profissionais e pessoais, pela confiança, paciência, carinho e apoio incondicional.
Pela amizade que se fortaleceu e por todos os conselhos. Por sempre me incentivar a seguir em
frente e nunca desistir.
Ao meu amigo de longa data, Artur, pela amizade, cumplicidade e afeto. Pela dedicação
durante o período experimental e por sempre estar presente. Te desejo sucesso!
Aos membros do GEFOR pela ajuda prestada. Agradeço especialmente a Juliana (Ju) e ao
Gabriel (Patrãozinho), pela amizade, descontração e disposição para ajudar sempre que foi
necessário. A ajuda de vocês foi essencial para a realização deste trabalho, vocês tornaram os
dias árduos e estressantes mais leves e divertidos.
As minhas amigas e companheiras de república em Diamantina, Kamilla, Natalia, Sabrina e a
pequena Sofia. Conviver com vocês foi sinônimo de crescimento.
A República Cafundó por me receber mais uma vez de braços abertos em São João de-
Rei. Em especial a Brenda, com quem mais convivi nos últimos meses, por sempre me mostrar
o lado bom dos acontecimentos ruins e por sempre me esperar com um café quente depois de
um dia cansativo.
Ao Kárito e a Natalia (Jararaca) pela amizade estabelecida, companheirismo, momentos
de descontração, troca de informações, ajuda e apoio em qualquer situação. Que nossa amizade
perdure e que a distância não atrapalhe nosso reencontro.

Às minhas amigas da graduação Alessandra e Nahyara, que mesmo com o tempo nunca
se fizeram ausentes e continuaram sempre me apoiando e incentivando.
Aos meus amigos de Candeias, que apesar de estarmos distantes na maior parte do
tempo, a amizade sempre continuou a mesma.
Aos colegas da pós-graduação, e todos que convivi em Diamantina, que de alguma
forma, contribuíram para o meu crescimento.
A Universidade Federal dos Vales do Jequitinhonha e Mucuri, ao Programa de Pós-
Graduação em Zootecnia, pela oportunidade de cursar o mestrado. A todos os professores do
PPGZOO pelos ensinamentos. E a secretaria Elizângela por toda atenção, disposição e
paciência.
A CAPES, pela bolsa de estudos.
A Universidade federal de São João del-Rei, por ceder a área para a condução do
experimento. Aos técnicos Caio e Elson e as funcionárias Lili, Romilda e Meirinha.
Aos membros da banca de defesa pelas sugestões que foram propostas para a melhoria
deste trabalho.

RESUMO
Objetivou-se com este trabalho determinar o melhor método de estimação dos coeficientes de
repetibilidade e as melhores combinações entre cortes de acordo com a estabilização genotípica
para características agronômicas, estimar parâmetros genéticos e formar grupos
morfofuncionais com base nas características morfogênicas e estruturais por meio do
agrupamento de Otimização de Tocher em genótipos de Panicum maximum. Os coeficientes
de repetibilidade para produção de massa seca total (MST), massa seca foliar (MSF), massa
seca do colmo (MSC), porcentagem de folhas (%F), porcentagem de colmo (%C), foram
estimados por meio de quatro métodos: análise de variância (ANOVA), análise estrutural com
base na média dos coeficientes de correlação (AECOR), análise de componentes principais
com base na matriz de covariância (CPCOV) e na matriz de correlações (CPCOR). Para o
estudo da estabilização genotípica, utilizaram-se os coeficientes estimados pela ANOVA e
CPCOR. Para a avaliação das características morfogênicas foram estimadas: taxa de
aparecimento foliar (TAPF), filocrono (FIL), taxa de alongamento foliar (TALF), taxa de
senescência foliar (TSF), comprimento final da lâmina (CFL), número de folhas vivas (NFV),
duração de vida das folhas (DVF), taxa de alongamento de pseudocolmo (TALC), número
médio de perfilhos (NMP), relação lâmina:colmo (RLC). Para MST, foram observados
coeficientes de repetibilidade variando entre 0,3500 e 0,4300 pelos métodos da ANOVA e
CPCOR, respectivamente. Altos coeficientes de repetibilidade também foram encontrados
para a característica MSF. Baixos coeficientes de repetibilidade foram observados para %F e
%Ce relação lâmina:colmo. Para estabilização genotípica da MST, os melhores coeficientes
foram observados para a combinação entre os cortes 6 a 7 e entre os cortes 5 a 8, enquanto os
menores coeficientes foram observados quando se utilizaram apenas os cortes 3 a 4 e de 1 a 2,
em ambos os métodos. Para a relação lâmina:colmo, os melhores coeficientes foram
registrados para os cortes 6 a 7 pelo método da ANOVA, e 1 a 2 pelo método CPCOR. De
maneira geral, a combinação entre os cortes de 6 a 7, também proporcionou maior
repetibilidade e determinação, otimizando a estabilização dos genótipos para as massas e
porcentagens de folha e de colmo. No estudo dos parâmetros genéticos e agrupamento, foi
observado que somente as características TALF, CFL e RLC tiveram o componente variância
genética significativo. Apesar disto, as características TALC, NFV, apresentaram coeficientes
de variação genotípicos (CVg) superiores aos coeficientes de variação residual ou ambiental
(CVe). As características TAPF, FIL, DVF e TSF apresentaram valores abaixo da unidade para

a razão CVg/CVe. Alta razão CVg/CVe foi observada para as características RLC, NFV,
TALC, TALF, CFL, sendo os maiores coeficientes foram registrados para RLC. Após o
agrupamento, constatou-se a formação de cinco grupos morfofuncionais. Os grupos que
apresentaram maiores valores de TALF foram os grupos 3, 5 e 1 com valores superiores à
média geral de todos os genótipos avaliados. Enquanto o grupo 4 obteve menor desempenho
para esta característica. Para a TALC o grupo 2 se destacou, seguido pelo grupo 5. Dentre
todos os grupos a maior RLC constada foi para o grupo 4 e para CFL o grupo 3. Conclui-se
que os métodos que proporcionaram os melhores coeficientes de repetibilidade de
determinação foram os dos componentes principais com base na matriz de correlação e de
covariância. Para a estabilização genotípica, os melhores coeficientes de repetibilidade e
determinação são observados para os cortes realizados no segundo período das águas. As
características TALF, CFL e RLC apresentam variabilidade genética significativa, e as
características TAPF, FIL, DVF e TSF apresentam baixa razão CVg/CVe e necessitam de
maior controle ambiental. Os grupos 3, 5 e 1, apresentam altas taxas de alongamento de folha
como mecanismo de acúmulo de forragem. Já o grupo 4 se destaca pela capacidade de
perfilhamento.
Palavras chave: Forrageiras. Medidas repetidas. Morfogênese. Melhoramento. Valor genético.

ASTRACT
The objective was to evaluate the coefficient of repeatability of the agronomic traits and to
estimate the breeding value of morphogenic characteristics to stablish morphofunctional groups
in the clustering analysis of Tocher in Panicum maximum genotypes. The repeatability
coefficients were estimated by four methods: analysis of variance (ANOVA), structural
analysis based on the correlation matrix (EACOR), principal component analysis based on the
variance and covariance’s matrix (PCCOV) and principal component analysis based on the
correlation matrix (PCCOR). To the genotypic stabilization study, the ANOVA and PCCOR
were used. To the morphogenic evaluation, the characteristics were estimated: leaf appearance
rate (LAR), phillochron (PHC), leaf elongation rate (LER), leaf senescence rate (LSR), number
of live leaves (NLL), leaf life spam (LLS), leaf final length (LFL), stem elongation rate (SER),
average number of tillers (ANT) and leaf:stem ratio (LSR). To total dry matter were observed
repeatability coefficients ranging from 0.3500 to 0.4300 by the ANOVA and PCCOR methods,
respectively. High coefficients of repeatability were estimated to leaf dry mass too. Low
coefficients of repeatability were observed to percentage of leaves, stems and leaf:stem ratio.
In the genotypic stabilization of total dry mass the higher coefficients were observed between
the 6 and 7th harvest and between the 5 and 8th harvest, while the lowest coefficients were
observed when the harvests 3 to 4 and 1 to 2 were considered in both methods. To leaf:stem
ratio the higher coefficients were observed to harvests between 6 and 7 in the ANOVA method
and 1 to 2 in the PCCOR method. In general, the combination between the 6 and 7th harvests
also improves the repeatability and determination coefficients, optimizing the stabilization of
the genotypes to dry mass and percentage of leaf and stems. In the study of genetic parameters
and clustering, were observed significant effect to genetic variance component only to LER,
LFL and LSR. In spite of this the characteristic SER and NLL had CVg higher than CVr. The
characteristics LAR. PHC, LLS and LSR had CVg/CVr above the unity. High CVg/CVr ratio
were observed to LSR, NLL, SER, LER and LFL, so that the highest coefficient observed to
LSR. After the clustering analysis was found five morphofunctional groups. The groups with
higher LER were 3, 5 and 1 with breeding values above the general mean. The group 4 had
lowest LER. The groups 2 and 5 had the highest SER and the highest LSR was observed to the
group 4. The group 3 showed high LFL. It was possible to conclude that the total and leaf dry
mass have higher coefficient of repeatability, indicating better accuracy in the identification of
the superior genotypes of P. maximum. In the genotypic stabilization, the higher coefficient and
determination were observed to the harvests realized in the rainy period. The morphogenic

characteristics that have the highest genetic variance have more possibility of gains with the
selection and the traits with low CVg/CVr ratio need more environmental control. The groups
3, 5 and 1 have high potential to forage production duly its high leaf elongation rate.
Keywords: Breeding value. Forages. Genetic breeding. Morphogenesis. Repeated measures
SUMÁRIO

1. INTRODUÇÃO GERAL ................................................................................................ 9
2. REVISÃO DE LITERATURA ..................................................................................... 11
2.1 Histórico ....................................................................................................................... 11
2.2 Melhoramento genético em forrageiras tropicais............................................................ 12
2.3 Repetibilidade ............................................................................................................... 14
2.4 Morfogênese ................................................................................................................. 15
2.4.1 Características morfogênicas ...................................................................................... 16
2.4.2 Características Estruturais ........................................................................................... 18
3. REFERÊNCIAS BIBLIOGRÁFICA............................................................................ 20
CAPÍTULO I - REPETIBILIDADE E ESTABILIZAÇÃO GENOTÍPICA DE
CARACTERÍSTICAS AGRONÔMICAS EM Panicum maximum .... 24
RESUMO ........... ................................................................................................................ 24
ABSTRACT ....... ....................................................................Erro! Indicador não definido.
1. INTRODUÇÃO .............................................................................................................. 26
2. MATERIAL E MÉTODOS ............................................................................................. 28
3. RESULTADOS E DISCUSSÃO ..................................................................................... 31
4. CONSIDERAÇÕES FINAIS........................................................................................... 41
5. REFERÊNCIAS BIBLIOGRÁFICAS ............................................................................. 43
CAPÍTULO II - ANÁLISE DE AGRUPAMENTO COM BASE EM
CARACTERÍSTICAS MORFOGÊNICAS EM GENÓTIPOS DE
Panicum maximum ................................................................................. 46
RESUMO ........... ................................................................................................................ 46
ABSTRACT ....... ....................................................................Erro! Indicador não definido.
1. INTRODUÇÃO .............................................................................................................. 48
2. MATERIAL E MÉTODOS ............................................................................................. 50
3. RESULTADOS E DISCUSSÃO ..................................................................................... 53
4. CONSIDERAÇÕES FINAIS .......................................................................................... 61
5. REFERENCIAS BIBLIOGRÁFICAS ............................................................................. 62


9
1. INTRODUÇÃO GERAL
O Brasil possui uma vasta extensão territorial destinada para pastagens, assim, a
pecuária nacional é quase que exclusivamente realizada à pasto, sendo uma forma prática e
barata a alimentação dos ruminantes. Contudo, grande parte destas áreas é caracterizada pelo
monocultivo de forrageiras, principalmente com gramíneas do gênero Brachiaria e da espécie
Panicum maximum. Sendo, basicamente as pastagens a sustentação da cadeia produtiva de leite
e carne no Brasil, é fundamental a disponibilização de forrageiras diversificadas para que
atendam às demandas dos produtores rurais.
Em consequência do monocultivo e do modo de reprodução por apomixia, as plantas
utilizadas nas pastagens brasileiras apresentam baixa variabilidade, o que as torna vulneráveis
do ponto de vista genético, podendo ser seriamente prejudicadas pelo surgimento de pragas,
doenças ou pelas mudanças climáticas. Esses fatores representam grande risco para a cadeia
produtiva nacional.
O lançamento de novas cultivares forrageiras resulta de uma demanda por plantas mais
competitivas, menos exigentes em fertilidade do solo, com menor sazonalidade de produção
e maior resistência a pragas e doenças, bom valor nutritivo, elevada produção de sementes entre
outros. Nesse sentido, estudos com avalições de genótipos podem contribuir para o
aperfeiçoamento destas características, além de colaborar para o aumento da diversificação das
pastagens, diminuindo o impacto causado pelo monocultivo (MARTUSCELLO, 2007).
O processo de lançamento de novas cultivares forrageiras é complexo e demorado,
podendo levar até 10 anos e é dividido em três fases de avaliações. Primeiramente, é necessária
a obtenção da variabilidade genética a partir do germoplasma disponível ou híbridos gerados
pelo melhoramento, para dar-se início a seleção, que primeiro ocorre somente em um local e
em seguida em locais diferentes. Na segunda etapa, os genótipos selecionados são avaliados em
rede nacional de ensaios, em pequenos piquetes com animais para se avaliar a tolerância ao
pastejo, além da sua adaptação a diferentes solos e condições climáticas. Na última etapa é
avaliado o potencial de produção animal pelas forrageiras selecionadas nas etapas anteriores.
Somente após todo este período de avalições uma nova cultivar é lançada no mercado.
A espécie P. maximum destaca-se por ser a uma das forrageiras mais produtivas
propagadas por sementes utilizadas nos sistemas de produção a pasto no Brasil. A Empresa
Brasileira de Pesquisa Agropecuária (EMBRAPA) mantem um programa de melhoramento
genético da espécie e vem lançando cultivares promissoras no mercado. Entretanto,
considerando-se a dimensão continental do Brasil e a necessidade de intensificação dos sistemas

10
de produção, a busca por forrageiras mais produtivas, competitivas, de fácil propagação e
adaptadas as diversas regiões do país, está cada vez maior. Assim, o melhoramento genético de
P. maximum tem grande potencial para atender esta demanda.
Objetivou-se com este trabalho determinar o melhor método de estimação dos
coeficientes de repetibilidade e as melhores combinações entre cortes de acordo com a
estabilização genotípica para características agronômicas, estimar parâmetros genéticos e
formar grupos morfofuncionais com base nas características morfogênicas e estruturais por
meio do agrupamento de Otimização de Tocher em genótipos de Panicum maximum.

11
2. REVISÃO DE LITERATURA
2.1 Histórico
Panicum maximum pertence à família Poaceae, subfamília Panicoideae e tribo Paniceae.
O principal centro de origem da espécie é a África do Leste, sendo encontradas formas nativas
até a África do Sul, em margens de florestas como planta pioneira, ocupando solos recém-
desmatados e em pastagens sob sombra (BOGDAN, 1977).
A primeira cultivar de P. maximum (cv. Colonião) foi introduzida no Brasil durante o
período de colonização por meio dos navios negreiros vindos da África, no qual era utilizada
como cama para os escravos (PARSONS, 1972). Devido à semelhança das condições
edafoclimáticas entre a África e o Brasil, o capim-colonião se adaptação muito bem no país e
se disseminou, sendo em algumas regiões considerado nativo (MARTUSCELLO, 2007). No
Brasil, o auge da produção do capim-colonião ocorreu até os anos 80, quando era considerado
como a salvação da pecuária, quando a partir daí se introduziu o germoplasma coletado no seu
centro de origem (JANK, 2003).
Algumas cultivares de P. maximum foram diretamente trazidas da África, sem terem
sido avaliadas no Brasil, como é o caso da cultivar Aruana, lançada pelo Instituto de Zootecnia
de Nova Odessa. Cita-se também a cultivar Tobiatã que foi avaliada pelo ORSTOM, na Costa
do Marfim, e lançada pelo IAC (Instituto Agronômico de Campinas) no Brasil (USBERTI
FILHO, 1982). Outras cultivares chegaram ao Brasil, após terem sido avaliadas na Austrália,
como é o caso das cultivares Gatton Panic e Green Panic. Em 1986 e 1988, respectivamente,
foram lançados pelo IAC as cultivares Centenário e Centauro (MARTUSCELLO, 2007).
Plantas da espécie P. maximum foram coletadas em 1967 e 1969 no Quênia e Tanzânia
na África pelo Institut Français de Recherche Scientifique pour le Développment en
Coopération (ORSTOM) (COMBES & PERNÈS, 1970). Com a finalidade de aumentar a
variabilidade disponível para seleção, a Empresa Brasileira de Pesquisa Agropecuária
(EMBRAPA), firmou um convênio em 1982 com ORSTOM, atualmente Institut de Recherche
pour le Développment (IRD), que viabilizou a transferência do germoplasma de P. maximum,
por eles reunido, para o Centro Nacional de Pesquisa de Gado de Corte (CNPGC) em Campo
Grande, MS, Brasil (JANK et al., 1997). Nesta ocasião a CNPGC recebeu uma coleção de
germoplasma com 426 genótipos apomíticos e 417 plantas sexuais e iniciou-se assim o
programa de melhoramento genético de P. maximum na EMBRAPA (JANK et al., 2008;
SILVA, 2013).

12
A seleção a partir de banco de germoplasma é indispensável para o melhoramento em
P. maximum. Desta forma, o convênio criado entre Embrapa e ORSTOM, em 1982, contribuiu
muito para o avanço do melhoramento genético dessa espécie forrageira A caracterização da
coleção de P. maximum do Orstom na Embrapa Gado de Corte demonstrou uma ampla
variabilidade disponível para o melhoramento de todas as características agronômicas e
morfológicas avaliadas. Foi observado variação quanto à porcentagem de proteína nas folhas e
nos colmos, tanto na época das águas quanto na seca, forma das inflorescências, pilosidade no
verticilo e espiguetas, dispersão das espiguetas, porte da planta, porte das folhas e tamanho das
ramificações primárias e secundárias (JANK, 1995; JANK et al., 1997; JANK et al., 2008).
A partir do início do programa de melhoramento em P. maximum as cultivares lançadas
pela Embrapa Gado de Corte até o momento foram: Tanzânia-1 – em 1990, Mombaça – em
1993, Massai – em 2000, BRS Zuri – em 2014, BRS Tamani – 2015 como a primeira cultivar
híbrida de P. maximum desenvolvida pela EMBRAPA, e mais recentemente, em 2017, BRS
Quênia.
2.2 Melhoramento genético em forrageiras tropicais
No Brasil, o interesse em genética de plantas forrageiras deu-se início a partir da década
de 1980. Com o pensamento de introdução de novos genes, seleção de genes de maior produção,
qualidade, adaptação e, finalmente, manipulação desses genes por meio de cruzamentos (JANK
et al., 2005).
Cerca de 40% dos capins se reproduzem sexuadamente, onde os gametas masculinos se
unem com os femininos e produzem híbridos. Entretanto, a outra parte desses capins se
reproduz assexuadamente, ou seja, por apomixia, na qual não acorre meiose, ou, a mesma não
é funcional, os gametas masculinos não se unem com os femininos, e híbridos não são formados
(JANK et al., 2011). Dentre as gramíneas C4 utilizadas em pastagens, a grande maioria se
reproduz por apomixia, produzindo sementes clones da planta-mãe.
A espécie P. maximum não é diferente, sendo considerada modelo de modo de
reprodução apomítica (apomixia do tipo Panicum). A apomixia é um modo de propagação
vegetativa por sementes, uma vez que o embrião não é derivado de uma fecundação. Nesta
espécie, como em outras gramíneas forrageiras tropicais, a apomixia é uma combinação de uma
aposporia (não redução gamética) seguida de uma partenogênese (não fecundação)
(WARMKE, 1954; COMBES, 1975; SAVIDAN, 1982; JANK et al, 2008). Porém, tipos
sexuais são encontrados na natureza, as quais possibilitam o cruzamento com genótipos
apomíticos e, assim, a obtenção de híbridos.

13
Nessas espécies, para que ocorra a fecundação é necessário encontrar plantas da mesma
espécie que permitam tal união (plantas sexuais). Portanto, para todas as espécies de capins é
necessário trazer recursos genéticos do centro de origem para se ter variabilidade para seleção
das espécies apomíticas e, mais ainda, para poder encontrar as plantas que permitem a
realização de cruzamentos (JANK et al., 2005).
De acordo com Valle et al. (2009) a herança da apomixia em P. maximum indicam que
um único gene dominante está envolvido, ou seja, as progênies híbridas incluem plantas sexuais
e apomíticas, na razão 1:1. Portanto, o melhoramento, realizado pelo cruzamento entre uma
planta sexual e uma apomítica, dará uma progênie em que 50% dos híbridos serão apomíticos
e 50% sexuais.
Desta forma, as novas cultivares forrageiras de P. maximum podem ser desenvolvidas
de duas formas: pela seleção dos melhores genótipos apomíticos a partir do germoplasma ou
pela geração de nova variabilidade por cruzamentos, fazendo-se em seguida a seleção para as
características de interesse (JANK et al., 2008).
Após a obtenção dos híbridos, dá-se início as fases de avaliação até o lançamento.
Primeiramente, os híbridos gerados por cruzamento entre plantas sexuais e apomíticas são
comparados em parcelas pequenas de 2 m2, com repetições, geralmente nesta fase há grande
número de plantas a serem avaliadas. Nesses experimentos são avaliadas produção forrageira,
relação folha:colmo, produção de sementes, densidade da rebrota após os cortes, intensidade e
época de florescimento, ocorrência de insetos e doenças e qualidade da forragem. Os híbridos
também são caracterizados morfologicamente nessa fase, isto é, são computados tamanho e
porte das plantas e folhas, pilosidade nas folhas, colmos ou hastes e sementes, tipo e cor da flor
quando leguminosa e tipo da inflorescência quando gramínea, entre outros (JANK et al., 2005).
Essa fase resulta na seleção dos melhores genótipos ou híbridos que, em seguida, são
avaliados em uma rede nacional de ensaios em diferentes locais, em parcelas pouco maiores
que as anteriores, para testar sua adaptação a diferentes solos e condições climáticas. São
avaliadas as mesmas características da primeira fase. Esses ensaios levam à seleção de até oito
genótipos que passam para a fase seguinte de avaliação, em piquetes pequenos (em torno de
1.000 m²) sob pastejo animal, para se avaliar o efeito do animal sobre as plantas, pisoteio e
pastejo. Posteriormente, os melhores genótipos são avaliados quanto ao desempenho animal
sob pastejo (produção de carne ou leite), em áreas grandes de até 10 ha (JANK et al., 2005).
Vale ressaltar que o primeiro objetivo da maioria dos novos programas de
melhoramento é a produção forrageira, para elevar diretamente os índices de produtividade da
pastagem, e consequentemente, da pecuária. Apenas quando o mercado se apresenta saturado

14
de opções de cultivares de alta produtividade de uma dada espécie e/ou gênero é que os
objetivos do melhoramento passam a ser a obtenção de cultivares de maior qualidade da
forragem, resistentes a patógenos, ou para atender demandas especificas (JANK et al., 2008).
2.3 Repetibilidade
Uma das dificuldades encontradas para o melhoramento de forrageiras está relacionada
à determinação do número de avaliações necessárias para estimar, com acurácia, as diferenças
entre os genótipos avaliados. Em plantas perenes, como é o caso da espécie P. maximum, são
necessárias avaliações periódicas, por meio de cortes, de modo a mensurar o comportamento
das principais características fenotípicas, portanto, deve-se considerar todos os dados das
diferentes avaliações para reunir informações satisfatórias para selecionar quais genótipos
obtiveram melhores resultados.
A partir das análises de sucessivas medições de uma característica, em um grupo de
indivíduos, é possível apontar se a superioridade ou inferioridade inicial de um indivíduo em
relação aos demais, se mantem ao longo das medições (MARTUSCELLO et al., 2007). Essas
avaliações sucessivas permitem a estimativa do coeficiente de repetibilidade das características
avaliadas, ou seja, permitem saber qual a probabilidade daquele resultado se repetir em
avaliações futuras. Determina-se, também, o número de avaliações necessárias para se predizer,
com confiabilidade, o comportamento dos materiais avaliados (SOUZA-SOBRINHO et al.,
2004).
A repetibilidade é uma correlação fenotípica entre medidas repetidas sobre o mesmo
grupo de indivíduos. Assim, observa-se uma correlação pela qual se mede a capacidade dos
organismos em repetir a expressão do caráter no tempo ou no espaço (RESENDE, 2002;
SOUZA SOBRINHO et al., 2010). Além disso, a repetibilidade demonstra o valor máximo que
a herdabilidade pode atingir, pois representa a proporção da variância fenotípica, que é atribuída
às diferenças genéticas, misturadas aos efeitos de ambiente permanente. O número de medições
necessárias para a previsão do valor real do indivíduo é aquele em que os efeitos temporários
do ambiente tendem a ser nulos ou minimizados a um valor aceitável (CARGNELUTTI FILHO
et al., 2004). Assim, a repetibilidade, à semelhança da herdabilidade, constitui instrumento
indispensável para orientar os trabalhos de melhoramento (FERREIRA et al., 1999).
Quando uma determinada característica apresenta alta repetibilidade, indica que um
pequeno número de medições já é suficiente para predizer seu valor real, sendo um indício que,
em avaliações sucessivas, haverá pouco ganho em acurácia. Em contrapartida, quando a

15
repetibilidade apresenta valores baixos, maior número de medições serão necessárias para
alcançar resultados satisfatórios.
Ao aumentar o número de medidas tomadas para determinada característica, diminui-se
a variância temporária causada pelo ambiente e, evidentemente, reduz-se também a variância
fenotípica, melhorando a acurácia do coeficiente de repetibilidade. Ressalta-se que, quando a
variância ambiental temporária é baixa e a repetibilidade é alta, o aumento no número de tomada
de dados pouco acrescentará na melhoria da inferência do valor genotípico do indivíduo. No
entanto, se o valor da repetibilidade é baixo, aumentando o número de avaliações haverá
aumento de ganho (MARTUSCELLO et al., 2007).
Um notável benefício do conhecimento do coeficiente de repetibilidade é possibilitar o
avanço da pesquisa de forma mais rápida e com menor demanda por mão de obra, fazendo que
o processo de melhoramento de plantas forrageiras ocorra de forma mais eficiente. Tornando-
se uma ferramenta fundamental para avaliações em plantas que possuem vários ciclos durante
o ano, para indicar, assim, qual o número de medições necessárias para que a seleção dos
melhores genótipos seja feita com confiabilidade.
Deve-se levar em consideração que o genótipo sofre influências pelo meio, e que o
fenótipo apresentado pelo acesso pode ser fruto desta interação (PEREIRA et al., 2002).
Portanto, a utilização de genótipos já estabilizados quando da obtenção de estimativa de
repetibilidade é de suma importância (MARTUSCELLO, 2007). Durante a fase jovem da
planta, antes da estabilização, é provável que apenas parte dos genes responsáveis pelos
caracteres de interesse estejam se expressando, enquanto na idade adulta todo o potencial da
planta manifesta-se, ocasionando mudanças consideráveis no fenótipo. Assim, quando o estudo
de repetibilidade é feito com genótipos ainda não estabilizados, pode-se encontrar baixa
repetibilidade, sem que, necessariamente, a solução do problema esteja no aumento do número
de repetições (PEREIRA et al., 2002).
Em uma visão prática, a maior vantagem da estimativa do coeficiente de repetibilidade
é o fato de poder possibilitar a determinação do número de medições, para avaliar com acurácia
os valores genéticos aditivos, genótipos ou fenótipos dos indivíduos (MARTUSCELLO, 2007).
2.4 Morfogênese
O sucesso na utilização de pastagens não depende apenas da disponibilidade de
nutrientes ou da escolha da espécie forrageira, mas também da compreensão dos mecanismos
morfofisiológicos e de sua interação com o ambiente (FAGUNDES et al., 2006). Assim, o
estudo das características morfofisiológicas em gramíneas forrageiras é imprescindível.

16
O estudo da origem e desenvolvimento dos diferentes órgãos de um organismo e das
transformações que determinam a produção e a mudança na forma e estrutura da planta no
espaço ao longo do tempo é dominado morfogênese (CHAPMAN; LEMAIRE, 1993;
SILVEIRA, 2006). A partir deste estudo é possível determinar as características morfogênicas
e estruturais, que por sua vez, influenciam no índice de área foliar (IAF). Tais características
tornam-se uma ferramenta poderosa para auxiliar nos processos de melhoramento genético em
plantas, por serem baseadas no acompanhamento da dinâmica do crescimento e
desenvolvimento de lâminas foliares e de colmos dos perfilhos.
Folhas e perfilhos constituem as unidades básicas consumidas pelos animais em
sistemas de produção a pasto. Conhecer a dinâmica de crescimento e desenvolvimento dessas
frações da planta constitui o alvo da morfogênese (PORTO, 2009). As características
morfogênicas são inerentes ao genótipo e influenciadas pelas condições ambientais, como
temperatura, disponibilidade hídrica e nutrientes (PORTO, 2009). Já as características
estruturais sofrem influência direta pelas características morfogênicas Apesar de não haver
estudos que comprovem a relação benéfica entre a morfogênese e o melhoramento genético de
plantas forrageiras, existe grande potencial para tal relação. Pois, com o estudo da morfogênese
é possível acompanhar o desenvolvimento da planta forrageira e sua interação com o ambiente,
que são informações consideradas ponto chave na seleção dos melhores genótipos em estudo.
2.4.1 Características morfogênicas
Taxa de Aparecimento Foliar (TApF)
A taxa de aparecimento foliar é expressa em número médio de folhas surgidas por
perfilho, em determinado período de tempo, representada geralmente em número de
folha/dia/perfilho. Seu inverso é o intervalo de aparecimento de folhas, o filocrono, estimado
pelo número de dias entre o aparecimento de duas folhas sucessivas (PORTO, 2009).
Esta característica desempenha papel central na morfogênese, e, por consequência, no IAF,
pois influencia diretamente a estrutura da pastagem. Existe uma relação direta da TApF com a
densidade de perfilhos que determina o potencial de perfilhamento para um dado genótipo,
porque cada folha formada sobre uma haste representa o surgimento de um novo fitômero, isto
é, a geração de novas gemas axilares. Portanto, a TApF determina grandes diferenças na
estrutura da pastagem devido ao seu efeito sobre o tamanho e a densidade de perfilhos
(NABINGER & PONTES, 2001; PORTO, 2009).
A taxa de aparecimento varia dentro de espécies, quando a planta forrageira se encontra
em ambiente uniforme por longos períodos a TApF é considerada constante, porém é

17
amplamente influenciada por mudanças estacionais, causadas não apenas por mudanças na
temperatura, mas também por mudanças na disponibilidade de água, fotoperíodo, intensidade
luminosa e nutrientes no solo.
Taxa de alongamento foliar (TAlF)
O processo de alongamento foliar em gramíneas ocorre na base da folha em expansão (zona
de alongamento), que estão envoltas por bainhas de folhas mais velhas formando o
pseudocolmo, sendo expressa geralmente em mm/dia (SKINER & NELSON, 1995; PORTO,
2009).
O alongamento de folhas apresenta grande variação de respostas às condições do meio como
temperatura, luz e disponibilidade de nutrientes, qualquer efeito sobre a TAlF afeta a taxa de
aparecimento de folhas e surgimento de perfilhos. Ressalta-se também que esta é uma
característica sensível à adubação nitrogenada, sendo que doses crescentes de nitrogênio podem
resultar em aumentos lineares na sua resposta (BRAZ, 2008).
A taxa de alongamento foliar é a variável morfogênica que mais se relaciona com o rendimento
da forragem (massa seca). Portanto, esta é uma variável importante quando se faz uma
abordagem da morfogênese relacionada ao processo de melhoramento e seleção de plantas
forrageiras (SILVEIRA, 2006).
Taxa de Alongamento de Colmo (TAlC)
Gramíneas tropicais, em particular aquelas de crescimento ereto, apresentam
componente de grande relevância e que pode interferir na estrutura do pasto e no equilíbrio do
processo de competição por luz, que é o alongamento do colmo (SBRISSIA & DA SILVA,
2001). Esta característica foi adicionada ao diagrama de Chapman & Lemaire (1996) por
Sbrissia & Da Silva (2001), com a finalidade de melhor descrever a resposta morfogênica das
gramíneas forrageiras tropicais sob influência dos principais fatores do ecossistema pastagem
(PORTO, 2009).
O desenvolvimento de colmos pode favorecer o aumento da produção de massa seca,
porém, pode apresentar efeitos negativos sobre o aproveitamento e a qualidade da forragem
produzida, alterando o comportamento ingestivo dos animais e, consequentemente, o consumo
de forragem, devido à redução na relação lâmina/colmo.
Mesmo que essa redução não afete diretamente o IAF, a distribuição de folhas no perfil
do dossel é alterada pelo alongamento do colmo, o que, por sua vez, modifica a quantidade e a
qualidade da luz no interior do dossel, repercutindo diretamente nas características

18
morfogênicas e no perfilhamento. Apesar disso, os efeitos negativos podem compensar parcial
ou totalmente os benefícios do incremento em produção de biomassa (NASCIMENTO
JÚNIOR & ADESE, 2004).
Duração de Vida da Folha (DVF)
A duração de vida da folha (DVF) representa o período durante o qual há acúmulo de
folhas no perfilho, sem que seja detectada qualquer perda por senescência. Esta característica
determina o número máximo de folhas vivas por perfilho e é influenciada pela temperatura da
mesma forma que a TApF.
O conhecimento da duração de vida das folhas é importante para o manejo do pastejo,
pois indica o potencial de produção da espécie (máxima quantidade de material vivo por área).
Nesse contexto, torna-se possível estabelecer um elo entre a DVF e a avaliação de cultivares,
pois com base nessa característica seria possível obter uma indicação da frequência ideal de
desfolhação das plantas, servindo esse dado como referência para o planejamento das três fases
do protocolo utilizado atualmente na seleção e melhoramento de cultivares forrageiros
(SILVEIRA, 2006).
2.4.2 Características Estruturais
Comprimento Final da Lâmina (CFL)
Os fatores que determinam o comprimento da folha são a TAlF e TApF, uma vez, que
para dado genótipo, o período de alongamento da folha é uma fração constante no intervalo de
folhas sucessivas (DALE, 1982). Por consequência, as variações na TAlF e TApF, por meio de
práticas de manejo (intensidade de pastejo, frequência de desfolhação, adubação) ou variações
climáticas, podem ocasionar variações no comprimento final da folha (PORTO, 2009).
O comprimento da lâmina foliar é uma característica vegetal plástica à intensidade de
desfolha, sendo considerada uma estratégia morfológica de escape da planta ao pastejo
(LEMAIRE & CHAPMAN, 1996), portanto, devido a esse mecanismo, ocorre a diminuição do
comprimento das laminas em pastagens sujeitas à maior intensidade de desfolha (PORTO,
2009).
Número de Folhas Vivas por Perfilho (NFV)

19
A variável número de folhas vivas por perfilho (NFV) sofre influência direta de TApF e de
duração de vida da folha (DVF), por isso, qualquer mudança em uma destas características
morfogênicas afetará o número de folhas vivas por perfilho (NABINGER & PONTES, 2001).
O número de folhas vivas por perfilho é razoavelmente constante conforme o genótipo,
condições de meio de manejo. Por ser resultado da combinação de características morfogênicas,
sofre influência direta e indireta de fatores relacionados ao ambiente e às práticas de manejo
utilizadas.
O progressivo aumento no número de folhas por perfilho e de perfilho, por planta determina o
aumento do IAF do relvado e, então, o rendimento forrageiro, pois aumenta o percentual de
interceptação de captura da radiação luminosa (PORTO, 2009).
Número Médio de Perfilhos (NMP)
O perfilho é a unidade básica de crescimento do pasto e o balanço entre perfilhos vivos e mortos
a cada ciclo de pastejo é de grande importância para a manutenção do crescimento estabilidade
dos mesmos. O perfilhamento é um dos mecanismos utilizados pela forrageira para assegurar o
rápido crescimento da planta após um evento de pastejo. Esse tipo de resposta caracteriza o
processo de tolerância ao pastejo e é fundamental para assegurar a sobrevivência da forrageira
em situações de desfolhação (CARVALHO, 2000).
O perfilhamento é influenciado por vários fatores relacionados ao ambiente e ao manejo
adotado. Logo, a forma como determinada espécie e, ou, cultivar demonstra sua capacidade de
perfilhar serve como indicativo de resposta adaptativa a determinada situação ou região
(SILVEIRA, 2006).
Relação Lâmina Foliar/Colmo (RLC)
A importância da RLC está associada à espécie forrageira, sendo maior naquelas de
crescimento entouceirado e colmo mais lignificado. Essa relação pode ser utilizada como um
indicador do valor nutritivo da forragem e, ao lado da altura do pasto e da disponibilidade de
biomassa, condiciona a facilidade de apreensão da forragem pelo animal e, portanto, seu
comportamento durante o pastejo (NASCIMENTO JÚNIOR & ADESE, 2004). Esta relação
está intimamente ligada aos valores de massa seca de lâminas verdes e massa seca de colmos
verdes estimados em uma pastagem (PORTO, 2009).
Portanto, é importante que a RLC seja alta para conferir à gramínea melhor adaptação
ao pastejo ou tolerância ao corte, por representar um fenótipo, em que meristemas apicais se
apresentam mais próximos ao solo e, portanto, menos vulneráveis destruição (PINTO et al.,

20
1994). Em contrapartida, a baixa RLC compromete o processo de pastejo e, consequentemente,
o desempenho, pois diminui o consumo voluntário de forragem devido a baixa digestibilidade,
menor taxa de passagem e limitações de ordem física na ingestão.
3. REFERÊNCIAS BIBLIOGRÁFICA
BOGDAN, A. V. Panicum maximum. In: BOGDAN. A. V. (Ed.) Tropical pasture and
fodder plants: grasses and legumes. 1997, p.181-191.
BRAZ, T. G. S. Características morfogênicas e estruturais do Capim-Tanzânia sob doses
de nitrogênio e densidade de plantas. 2008. p. 81. Dissertação (Mestrado em Zootecnia) -
Universidade Federal de Viçosa, Viçosa, 2008.

21
CARGNELUTTI FILHO, A.; CASTILHOS, Z. M.; STORCK, L.; SAVINA, J. F. Análise de
repetibilidade de caracteres forrageiros de genótipos de Panicum maximum, avaliados com e
sem restrição solar. Ciência Rural, Santa Maria, v.34, n.3, p.723-729, 2004.
CARVALHO, C. A. B. Padrões demográficos de perfilhamento e acúmulo de forragem
em pastagens de Cynodon spp. Manejadas em 4 intensidades de pastejo. 2000. 96p.
Dissertação (Mestrado)- Escola Superior de Agricultura de Queiroz, Piracicaba, 2000.
CHAPMAN, D.; LEMAIRE, G. Tissue flows in grazed plant communities. In: HODGSON,
J.; ILLIUS, A.W. (Ed.). The ecology and management of grazing systems. Guilford: CAB
International, 1996. p. 3-36.
CHAPMAN, D.F.; LEMAIRE, G. Morphogenetic and structural determinants of plant
regrowth after defoliation. In: BAKER, M.J. (Ed.) Grasslands for our world.
Wellington:SIR Publishing, 1993. p.55-64.
COMBES, D. Polymorphisme et modes de reproduction dans la section des Maximae du
genre Panicum (Graminées) en Afrique. Paris: ORSTOM, 1975. 99 p. (Mémoires
ORSTOM, 77).
COMBES, D.; PÉRNES, J. Variation ans les nombres chromosomiques du Panicum
maximum em relation avec le mode de reproduction. Compts Rendues Academie Science,
v.270, p.783-785, 1970.
DALE, J.E. Some effects of temperature and irradiance on growth of the first four leaves of
wheat, triticum aestivum. Annals of Botany, v.50, p.851-858, 1982.
FAGUNDES, J.L.; FONSECA, D.M.; MISTURA, C. et al. Características morfogênicas e
estruturais do capim-braquiária em pastagem adubada com nitrogênio avaliadas nas quatro
estações do ano. Revista Brasileira de Zootecnia, v.35, n.1, p.21-29. 2006.
FERREIRA, R. P.; BOTREL, M. A.; PEREIRA, A. V.; CRUZ. C. D. Avaliação de cultivares
de alfafa e estimativas de repetibilidade de caracteres forrageiros. Pesquisa Agropecuária
Brasileira, v.34, n.6, p.995-1002, 1999.

22
JANK, L ; RESENDE, R M S ; VALLE, C. B. DO . Genética em Pastagem. REVISTA USP,
São Paulo, v. 22, p. 87 - 93, 2005.
JANK, L. A história do Panicum maximum no Brasil. Revista JC Maschietto, n.1, p. 14,
2003.
JANK, L. Melhoramento e seleção de variedades de Panicum maximum. In: SIMPÓSIO
SOBRE MANEJO DA PASTAGEM, 12. 1995, Piracicaba. Anais... Piracicaba: FEALQ, p.
21-58, 1995.
JANK, L.; CALIXTO, S.; COSTA, J. C. G.; SAVIDAN, Y. H.; CURVO, J. B. E. Catalog of
the characterization and evaluation of the Panicum maximum germplasm:
morphological description and agronomical performance. Campo Grande, MS: Embrapa
Gado de Corte, p. 53, 1997.
JANK, L.; RESENDE, R. M. S.; VALLE, C. B; RESENDE, M. D. V.; CHIARI, L.;
CANÇADO, L. J.; SIMIONI, C. Melhoramento genético de Panicum maximum. In:
RESENDE, R. M. S.; VALLE, C. B. do; JANK, L. (Ed.) Melhoramento de forrageiras
tropicais. 1.ed. Campo Grande, MS: Embrapa Gado de Corte, 2008, p. 55-87.
JANK, L.; VALLE, C. B.; RESENDE, R. M. S. Breeding tropical forages. Crop Breeding
and Applied Biotechnology. v. 11, p. 27-34. 2011.
MARTUSCELLO, J. A. Repetibilidade e seleção em Panicum maximum Jacq.. 2007. p.
111. Tese (Doutorado em Zootecnia) - Universidade Federal de Viçosa, Viçosa, 2007.
MARTUSCELLO, J. A.; JANK, L.; FONSECA, D. M.; CRUZ, C. D.; CUNHA, D. N. F. A.
Repetibilidade de caracteres agronômicos em Panicum maximum Jacq. Revista Brasileira de
Zootecnia, v.36, n.6, p.1975-1981, 2007.
NABINGER, C & PONTES, L. da S. Morfogênese de plantas forrageiras e estrutura do pasto.
In: REUNIÃO DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 38, 2001. Piracicaba.
Anais... Piracicaba: SBZ, 2001. p. 755-771.

23
NASCIMENTO JUNIOR, D. do; ADESE, B. Acúmulo de biomassa na pastagem. In:
SIMPÓSIO SOBRE MANEJO ESTRATÉGICO DA PASTAGEM, 2., 2004, Viçosa, MG.
Anais... Viçosa, MG: Sociedade Brasileira de Zootecnia, 2004. v. 1, p. 289-346.
PARSONS, J. J. Spread of African pasture grasses of the American tropics. Journal of
Range Management, v.25, n. 1, p. 12-17, 1972.
PEREIRA, A. V.; CRUZ, C. D.; FERREIRA, R. P.; BOTREL, M. A.; OLIVEIRA, J. S.
influência da estabilização de genótipos de capim-elefante (Pennisetum purpureum Schum.)
sobre a estimativa da repetibilidade de características forrageiras. Ciência e Agrotecnologia,
v.26, n.4, p.762-767, 2002.
PINTO, J.C., GOMIDE, J.A., MAESTRI, M. Produção de matéria seca e relação folha/caule
de gramíneas forrageiras tropicais cultivadas em vaso, com duas doses de nitrogênio. Revista
Brasileira de Zootecnia, v.23, n.3, p.313-326, 1994.
PORTO, E. M. V. Morfogênese e rendimento forrageiro de cultivares de Cenchrus
ciliares L. submetidos à adubação nitrogenada. 2009. 107p. Dissertação (Magister
Scientiae) – Universidade Estadual de Montes Claros, Janaúba, 2009.
RESENDE, M.D.V. de. Genética biométrica e estatística no melhoramento de plantas
perenes. Brasília: Embrapa Informação Tecnológica; Colombo: Embrapa Florestas, 2002.
975p.
SAVIDAN, Y. H. Nature et hérédité de l'apomixie chez Panicum maximum Jacq. Paris:
ORSTOM, 1982. 159 p. (ORSTOM. Travaux et documentos, 153).
SBRISSIA, A.F., Da SILVA, S.C. O ecossistema de pastagens e a produção animal In:
REUNIÃO ANUAL DA SOCIDADE BRASILEIRA DE ZOOTECNIA, 38, Piracicaba,
2001. Anais... Piracicaba : SBZ, 2001, p.731-754.
SILVA, P. M. P. Tolerância ao déficit hídrico em Panicum maximum .2013. p. 50.
Dissertação (Mestrado em Ciência Animal) - Universidade Federal do Mato Grosso do Sul,
Campo Grande, 2013.

24
SILVEIRA, M. C. T. Caracterização morfogênica de oito cultivares do gênero Brachiaria
e dois do gênero Panicum maximum. 2006. p. 111. Dissertação (Mestrado em Zootecnia) -
Universidade Federal de Viçosa, Viçosa, 2006.
SKINNER, R.H.; NELSON, C.J. Elongation of the grass leaf and its relationship to the
phyllochron. Crop Science, v.35, n.1, p.4-10, 1995.
SOUZA SOBRINHO, F. de; LÉDO, F.J. da S.; PEREIRA, A.V.; BOTREL, M.A.;
EVANGELISTA, A.R.; VIANA, M.C.M. Estimativas de repetibilidade para produção de
matéria seca em alfafa. Ciência Rural, v.34, p.531-537, 2004.
SOUZA SOBRINHO, F.; BORGES, V.; LÉDO, F. J. S.; KOPP, M. M. Repetibilidade de
características agronômicas e número de cortes necessários para seleção de Urochloa
ruziziensis. Pesquisa Agropecuária Brasileira, v.45, n.6, p.579-584, 2010.
USBERT FILHO, J. A. O Agronômico. Piracicaba, v.34, 1982. 7p. (Edição Especial).
VALLE, C. B.; JANK, L.; RESENDE, R. M. S. O melhoramento de forrageiras tropicais no
Brasil. Revista Ceres. V. 56, n. 4, p. 460-472, 2009.
VALLE, C. B.; JANK, L.; RESENDE, R. M. S. O melhoramento de forrageiras tropicais no
Brasil. Revista Ceres. V. 56, n. 4, p. 460-472, 2009.
WARMKE, H.E. Apomixis in Panicum maximum. American Journal of Botany, v.41, n.1,
p. 5-11, 1954.
CAPÍTULO I - REPETIBILIDADE E ESTABILIZAÇÃO GENOTÍPICA DE
CARACTERÍSTICAS AGRONÔMICAS EM Panicum maximum
RESUMO
Objetivou-se com esse trabalho determinar o melhor método de estimação dos coeficientes de
repetibilidade e as melhores combinações entre cortes de acordo com a estabilização genotípica
para características agronômicas em genótipos de P. maximum. Foram avaliados 17 genótipos

25
em delineamento em blocos ao acaso com três repetições. Os coeficientes de repetibilidade
foram estimados por meio de quatro métodos: análise de variância (ANOVA), análise estrutural
com base na média dos coeficientes de correlação (AECOR), análise de componentes principais
com base na matriz de covariância (CPCOV) e na matriz de correlações (CPCOR). Para o
estudo da estabilização genotípica, utilizaram-se os coeficientes estimados pela ANOVA e
CPCOR. Para massa seca total, foram observados coeficientes de repetibilidade variando entre
0,3500 e 0,4300 pelos métodos da ANOVA e CPCOR, respectivamente. Altos coeficientes de
repetibilidade também foram encontrados para a característica massa seca de folha. Baixos
coeficientes de repetibilidade foram observados para a porcentagem de folhas e colmo e relação
lâmina:colmo. Para estabilização genotípica da massa seca total, os melhores coeficientes foram
observados para a combinação entre os cortes 6 a 7 e entre os cortes 5 a 8, enquanto os menores
coeficientes foram observados quando se utilizaram apenas os cortes 3 a 4 e de 1 a 2, em ambos
os métodos. Para a relação lâmina:colmo, os melhores coeficientes foram registrados para os
cortes 6 a 7 pelo método da ANOVA, e 1 a 2 pelo método CPCOR. De maneira geral, a
combinação entre os cortes de 6 a 7, também proporcionou maior repetibilidade e determinação,
otimizando a estabilização dos genótipos para as massas e porcentagens de folha e de colmo.
Conclui-se que os métodos que proporcionaram os melhores coeficientes de repetibilidade de
determinação foram os dos componentes principais com base na matriz de correlação e de
covariância. Para a estabilização genotípica, os melhores coeficientes de repetibilidade e
determinação são observados para os cortes realizados no segundo período das águas. Os cortes
realizados antes das plantas adquirirem maturidade fisiológica e durante a seca são prejudiciais
no processo de seleção.
Palavras-chave: Relação lâmina:colmo. Folha. Forragem. Medidas repetidas.

26
REPEATABILITY AND GENOTYPIC STABILIZATION OF AGRONOMIC
CHARACTERISTICS IN Panicum maximum
ABSTRACT
The objective was to identify the better estimator of repeatability coefficient and the better
harvest combinations according to genotypic stabilization in Panicum maximum genotypes.
Were evaluated 17 genotypes in completely randomized block design with three replicates. The
coefficient of repeatability were estimated by the methods: variance analysis (ANOVA),
structural analysis based on the mean of the correlation coefficients (SACOR), principal
component analysis based on the matrix of covariance (PCCOV) and the correlation matrix
(PCCOR). The ANOVA and PCCOR were used to study the genotypic stabilization. The total
dry mass had repeatability coefficients ranging from 0.3500 to 0.4300 by the ANOVA and
PCCOR, respectively. The leaf dry mass also showed high repeatability coefficients. The
leaves percentage and leaf:stem ration had the lowest repeatabilities in the study. To the
genotypic stabilization of total dry mass, the highest coefficiets were estimated to 6 to 7
harvests and 5 to 8, while the lowest coefficients were observed in the evaluation of the harvests
ranging from 3 to 4 and 1 to 2 in both methods. To the leaf:stem ratio the repeatability was
improved using the harvests 6 to 7 by ANOVA and 1 to 2 by PCCOR. In general, the
combination the harvests 6 to 7 also improve the repeatability and determination coefficients to
the mass and percentages of leaf and stems. The method of principal component based on the
correlation matrix presented the highest coefficients of repeatability and determination. In the
genotypic stabilization, the highest coefficients were estimated to harvests in the second rainy
season. The harvests made before the physiological maturity of the Panicum maximum
genotypes and during the dry season should be avoided in the evaluation and selection process.
Key words: Leaf:stem ratio. Relação lâmina:colmo. Folha. Forragem. Medidas repetidas.
1. INTRODUÇÃO
A produção animal em pastagens corresponde a boa parte de agronegócio brasileiro,
gerando renda e empregos. Isso se deve à vasta extensão territorial estabelecida com pastagens

27
e às condições de solo e clima favoráveis à utilização de forrageiras de clima tropical. Dentre
as plantas mais utilizadas na formação de pastagens, destaca-se a espécie Panicum maximum,
caracteriza-se por apresentar elevada produção de forragem com bom valor nutritivo, além de
boa resposta a adubação (JANK et al., 2008).
A maioria das plantas de P. maximum, sobretudo as cultivares registradas, se reproduz
por apomixia, modo no qual a nova planta gerada tem as mesmas características da planta-mãe.
Este tipo de reprodução faz com que as pastagens brasileiras sejam consideradas extensos
monocultivos clonais de alta vulnerabilidade genética (VALLE et al., 2009). Neste cenário, o
processo de diversificação é indispensável para o aumento da variabilidade genética, além de
proporcionar forrageiras mais adaptadas para determinada região, melhorando ainda mais seu
potencial de produção.
Para a seleção de genótipos de P. maximum, planta de ciclo perene, são necessárias
avaliações regulares, por meio de sucessivos cortes, podendo-se estimar a variabilidade
genotípica e a repetibilidade da resposta de determinada característica. Ao realizar sucessivos
cortes é possível estimar o coeficiente de repetibilidade das características avaliadas, isto é,
aferir a probabilidade ou a capacidade destas características se repetirem ao longo das
avaliações. A repetibilidade é um parâmetro genético importante para estudos de melhoramento
e para o processo de avaliação de plantas forrageiras (RESENDE et al., 2008).
O coeficiente de repetibilidade é uma ferramenta que pode auxiliar a pesquisa por meio
da mensuração do número de avaliações necessárias para obtenção de coeficientes de
determinação predefinidos (CRUZ et al.,2004) e desta forma, reduzir o período experimental,
a mão-de-obra e, por conseguinte, o gasto financeiro, otimizando o processo no lançamento de
novas cultivares, dado o seu longo período de avaliação. Nesse sentido, o conhecimento do
coeficiente de repetibilidade é indispensável para que os melhoristas avaliem a melhor
estratégia para conduzir o melhoramento de característica de interesse na população.
Quando uma característica possui alta repetibilidade, há indício de pouco ganho em
acurácia com o aumento do número de avaliações. Todavia, quando a repetibilidade apresenta
baixos valores, é necessário grande número de avaliações para se alcançar valor de
determinação satisfatório. Assim, ao aumentar o número avaliações, diminui-se a variância
temporária causada pelo ambiente e, evidentemente, reduz-se também a variância fenotípica,
melhorando a acurácia do coeficiente de repetibilidade. Ressalta-se que, quando a variância
ambiental temporária é baixa e a repetibilidade é alta, o aumento no número de tomada de dados
pouco acrescentará na melhoria da inferência do valor genotípico do indivíduo. No entanto, se

28
o valor da repetibilidade é baixo, aumentando o número de avaliações haverá aumento em
acurácia (MARTUSCELLO et al., 2007).
Estudos de estabilização genotípica também são ferramentas importantes na avaliação
de culturas perenes, uma vez que, durante as primeiras colheitas, quando a planta se encontra
na fase jovem, pode haver baixa expressão de genes importantes para características críticas
para a fase produtiva (PEREIRA et al., 2002), o que pode prejudicar o processo de avaliação
das mesmas. Além disso, o estabelecimento das forrageiras no campo e a mudança do manejo
comum nas fases inicias da pesquisa também pode resultar em expressão diferenciada de genes
que modifica a resposta produtiva das mesmas e leva a erros no processo de avaliação (BRAZ
et al., 2015a).
Nos trabalhos de estabilização genotípica, são estimados os coeficientes de
repetibilidade de grupos de colheitas sucessivos, que indicam períodos onde houve maior
correlação entre as medidas repetidas e, consequentemente, períodos onde há maior
confiabilidade no processo de seleção dos genótipos.
Assim, objetivou-se com esse trabalho determinar o melhor método de estimação dos
coeficientes de repetibilidade e as melhores combinações entre cortes de acordo com a
estabilização genotípica para características agronômicas em genótipos de P. maximum.
2. MATERIAL E MÉTODOS
O experimento foi conduzido na Universidade Federal de São João del-Rei, Campus
Tancredo Neves (CTAN), localizada no município de São João del-Rei no Campo das
29
Vertentes, em Minas Gerais, Brasil. O município encontra-se a 904 m de altitude e está situado
nas coordenadas geográficas 21º08ʼ11ʼʼ de latitude Sul e 40º15ʼ43ʼʼ de longitude Oeste. O
clima, pela classificação de Köppen (1948), é tipo Cwa, com estação seca (maio a outubro) e
chuvosa (novembro a abril) bem definidas. Os dados referentes às temperaturas média, máxima,
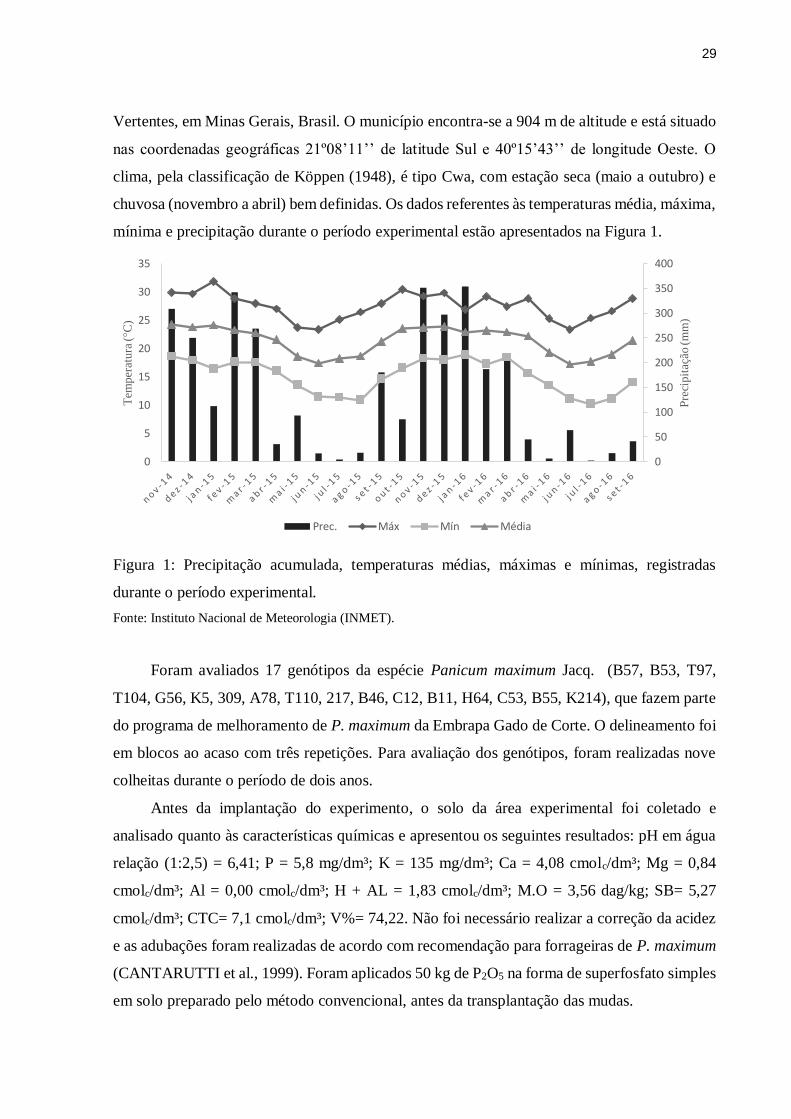
mínima e precipitação durante o período experimental estão apresentados na Figura 1.
Figura 1: Precipitação acumulada, temperaturas médias, máximas e mínimas, registradas
durante o período experimental.
Fonte: Instituto Nacional de Meteorologia (INMET).
Foram avaliados 17 genótipos da espécie Panicum maximum Jacq. (B57, B53, T97,
T104, G56, K5, 309, A78, T110, 217, B46, C12, B11, H64, C53, B55, K214), que fazem parte
do programa de melhoramento de P. maximum da Embrapa Gado de Corte. O delineamento foi
em blocos ao acaso com três repetições. Para avaliação dos genótipos, foram realizadas nove
colheitas durante o período de dois anos.
Antes da implantação do experimento, o solo da área experimental foi coletado e
analisado quanto às características químicas e apresentou os seguintes resultados: pH em água
relação (1:2,5) = 6,41; P = 5,8 mg/dm³; K = 135 mg/dm³; Ca = 4,08 cmolc/dm³; Mg = 0,84
cmolc/dm³; Al = 0,00 cmolc/dm³; H + AL = 1,83 cmolc/dm³; M.O = 3,56 dag/kg; SB= 5,27
cmolc/dm³; CTC= 7,1 cmolc/dm³; V%= 74,22. Não foi necessário realizar a correção da acidez
e as adubações foram realizadas de acordo com recomendação para forrageiras de P. maximum
(CANTARUTTI et al., 1999). Foram aplicados 50 kg de P2O5 na forma de superfosfato simples
em solo preparado pelo método convencional, antes da transplantação das mudas.
0
50
100
150
200
250
300
350
400
0
5
10
15
20
25
30
35
Pre
cipit
ação
(m
m)
Tem
per
atu
ra (
°C)
Prec. Máx Mín Média

30
Os genótipos foram avaliados em parcelas de 4 m². Para garantir uniformidade do estande
no momento do estabelecimento, os genótipos foram previamente semeados em casa de
vegetação no dia 17/10/14, em substrato agrícola comercial e transplantados para a área
experimental 35 dias após a germinação (21/11/14) de modo a assegurar densidade de 30
plântulas/m². Sete dias após a transplantação realizou-se adubação correspondente a 100 kg/há-
1 de N na forma de ureia. Quando as plantas completaram 31 dias no campo foi realizado um
corte de uniformização (22/12/14) e depois de 44 dias efetuou-se o primeiro corte (04/02/15).
No total, foram realizados sete cortes para avaliação de crescimento nas águas e outros dois
cortes no período seco. Os cortes foram realizados a 20 cm do solo em toda área útil da parcela,
descartando as bordaduras, realizados nos dias 04/02/2015, 14/03/2015, 14/05/2015,
29/10/2015, 23/12/2 015, 02/02/2016, 14/03/2016, 20/04/2016, 23/09/2016. Com intervalo
médio de 42 dias entre cortes no período das águas e 90 dias durante a seca.
Imediatamente após os cortes, a forragem foi pesada para obtenção da produção de massa
verde (MV). Subamostras foram retiradas e separadas em lâmina, colmo + bainha e forragem
senescente, secas em estufa de circulação forçada a 55ºC por 72 horas e pesadas para estimativa
da produção de massa seca total (MST), massa seca foliar (MSF), massa seca do colmo (MSC),
porcentagem de folhas, porcentagem de colmo e relação lâmina:colmo. Após os cortes as
parcelas foram adubadas com nitrogênio e potássio, em doses equivalentes a 50 kg /ha-1 de N e
20 kg/ha-1 de KCl com 45g de ureia e 13 g de KCl, respectivamente, em cada parcela.
Os coeficientes de repetibilidade foram estimados por meio de quatro métodos: análise
de variância (ANOVA), análise de componentes principais com base na matriz de covariância
(CPCOV), análise de componentes principais com base na matriz de correlações (CPCOR),
análise estrutural com base na média dos coeficientes de correlação (AECOR), conforme
metodologias descritas em Cruz et al. (2004).
Após a estimativa do coeficiente de repetibilidade pelos diferentes métodos, foi realizada
análise de estabilização genotípica. Utilizaram-se os coeficientes estimados pela ANOVA e
CPCOR para as sucessivas medições, considerando-se 2, 3, ... até todas as n avaliações efetuadas.
Portanto, foram realizadas n-1 análises de duas medições sucessivas; n-2 análises de três avaliações
sucessivas até que fossem obtidas nove medições (CRUZ et al., 2006).
Todas as análises foram realizadas no programa GENES - Aplicativo computacional em
genética e estatística, versão 2016.6.0 (CRUZ, 2013).

31
3. RESULTADOS E DISCUSSÃO
Os métodos dos componentes principais proporcionaram os melhores resultados com os
maiores coeficientes de repetibilidade, sendo que o método de componentes principais obtido
da matriz de variância e covariância (CPCOV) se destacou sobre o método de componentes
principais obtidos da matriz de correlação (CPCOR) (Tabela 1). Independente da característica
avaliada, o método de análise de variância (ANOVA), seguido do método de análise estrutural

32
com base na matriz de correlação (AECOR) foram os que proporcionaram menores valores
dos coeficientes de repetibilidade.
Tabela 1: Estimativas do coeficiente de repetibilidade e seus respectivos coeficientes de
determinação (entre parênteses) das características de produção de massa seca total (MST),
massa seca de folha (MSF), porcentagem de folha (%F), massa seca de colmo (MSC),
porcentagem de colmo (%C), e relação folha/colmo (RFC)
Método Características
MST MSF %F MSC %C RFC
Análise de Variância 0,3500
(82,89)
0,3496
(82,87)
0,0852
(45,60)
0,1954
(68,61)
0,1052
(51,41)
0,1127
(53,33)
Componentes principais
(Covariância)
0,4196
(86,67)
0,5083
(90,29)
0,4407
(87,64)
0,4573
(88,35)
0,4528
(88,16)
0,4477
(87,94)
Componentes principais
(Correlação)
0,4300
(87,16)
0,4527
(88,15)
0,3122
(80,33)
0,3509
(82,94)
0,4329
(87,29)
0,3703
(84,10)
Análise estrutural
(Correlação)
0,4085
(86,13)
0,4347
(87,37)
0,1568
(62,60)
0,2195
(71,68)
0,2456
(74,55)
0,2217
(71,93)
Variações observadas entre os métodos podem estar associadas à metodologia utilizada
no cálculo do coeficiente. Segundo Abeywardena (1972) o coeficiente de repetibilidade pode
ser mais eficientemente estimado por meio do método dos componentes principais, pois esta
metodologia enfatiza situações em que os genótipos avaliados apresentam comportamento
cíclico em relação ao caráter estudado. Nesse sentido, a espécie P. maximum é caracterizada
por apresentar comportamento cíclico, com picos de produção durante o período das águas, e
queda ao longo do período seco do ano, caracterizando resposta cíclica, quando o coeficiente
de repetibilidade é estimado com base nas medidas de cada corte. Este efeito pode variar de
maneira e intensidade diferentes em cada genótipo. De fato, a estacionalidade de produção de
forrageiras tropicais é bastante conhecida e segundo Jank et al. (2008) algumas cultivares de
P. maximum podem apresentar cerca de 80% de produção no período das águas.
O método da ANOVA, por sua vez, pode não eliminar este componente adicional do erro
experimental e, consequente, subestimar o coeficiente de repetibilidade (CRUZ, et al., 2004).
Já a análise estrutural (AECOR) apresenta apenas diferenças conceituais da análise de
componentes principais, e as estimativas desses métodos tendem a ser próximas (CRUZ et al.,

33
2004; NEGREIROS et al., 2014).
Martuscello et al. (2007), avaliando diferentes métodos para estimativa do coeficiente de
repetibilidade de características agronômica de P. maximum, observaram que o método CPCOV
apresentou melhores resultados. Segundo os autores, em geral, as variações na estimativa do
coeficiente de repetibilidade apontam a necessidade de utilização de vários métodos disponíveis
para se obter um intervalo preciso dentro do qual, com maior probabilidade, seja encontrado o
valor real para esse parâmetro. A mesma resposta também foi observada por Cargnelutti Filho
et al. (2004) e Braz et al. (2015b), ambos trabalhando com plantas de P. maximum. Nos
resultados de Braz et al. (2015b), foi notada grande discrepância entre os métodos, que foi
atribuída ao viés dos métodos baseados na análise de componentes principais, quando o valor
real do coeficiente se aproxima de zero (MANSOUR et al.,1981).
Para massa seca total (MST), observaram-se coeficientes de repetibilidade variando
entre 0,3500 e 0,4300 pelos métodos da ANOVA e CPCOR, respectivamente.
Independentemente do método utilizado, o coeficiente de determinação foi maior que 82,89,
indicando confiabilidade na identificação dos melhores genótipos de P. maximum para
produção de massa seca (Tabela 1). Braz et al. (2015b), avaliando características agronômicas
de diferentes progênies de P. maximum, em seis cortes, encontraram resultados semelhantes
aos deste estudo, e independentemente do método utilizado, os autores constataram coeficiente
de determinação acima 82,55. Estes resultados indicam que a mensuração da MST demanda
menor número de medidas repetidas para proporcionar bom nível de confiabilidade na predição
da resposta e identificação dos genótipos superiores. Já Souza Sobrinho et al. (2010),
encontraram valores inferiores aos deste trabalho para estimativas de repetibilidade em
Urochloa ruziziensis em relação a massa de massa seca, no qual os valores observados
variaram de 0,23 a 0,25 para os métodos da análise de variância e de componentes principais
a partir da matriz de correlação, respectivamente.
Maiores coeficientes de repetibilidade também foram encontrados para a característica
massa seca de folha (MSF), sendo que o método CPCOV apresentou maior repetibilidade
(0,5083) e determinação (90,29) (Tabela 1). As características MST e MSF apresentaram
valores muito próximos. De acordo com Braz et al. (2015b) este comportamento pode ser
explicado pela grande quantidade de folhas que usualmente é encontrada na massa de forragem
amostrada em genótipos de P. maximum. Ainda, ressalta-se o fato dos genótipos avaliados
neste estudo serem considerados de bom desempenho para características de interesse para o
melhoramento, como a MSF, o que também contribui para tal resultado. Isso porque, os
genótipos avaliados já foram preciamente selecionados no programa de melhoramento

34
genético de P. maximum da EMBRAPA. Os coeficientes observados neste estudo foram
menores que os registrados na literatura para a espécie P. maximum (CARGNELUTTI FILHO
et al., 2004; MARTUSCELLO et al. 2007; LEDO et al., 2008), onde a amplitude variou de
0,542 (93,90) a 0,869 (98,15). Nesse sentido, a repetibilidade para MSF deste estudo pode ser
considerada baixa em relação aos registros da literatura.
A MSF é indispensável no processo de seleção de novas forrageiras, pois as folhas
apresentam maior digestibilidade em relação ao colmo (FREITAS et al., 2012) e favorecem o
consumo de forragem dos animais em pastejo (BRÂNCIO et al., 2003). Nesse sentido, a
seleção com base nessa característica tende a tornar o processo de melhoramento genético mais
eficiente, principalmente na etapa final, quando a avaliação do ganho de peso é o principal
indicador do potencial de utilização da planta (MARTUSCELLO et al., 2007).
Baixos coeficientes de repetibilidade foram estimados para a porcentagem de folhas
(%F), que apresentou valores que variaram de 0,0852, pelo método da ANOVA a 0,4407, pelo
método CPCOV (Tabela 1), que pode ser justificado pelo fato do experimento ter tido duração
de dois anos consecutivos, incluindo períodos de seca e transições, períodos nos quais a
produção de colmo pode variar muito e, consequentemente, afetar a %F. Coeficientes de
repetibilidade mais baixos para %F também foram observados por Braz et al. (2015b), que
encontraram valores médios de 0,2498 para progênies de irmãos completos de P. maximum.
Segundo estes autores, estes resultados podem estar associados ao baixo grau de controle
genético da característica %F, que fica sujeita a variação ambientais e tende a apresentar
elevado coeficiente de variação. Já Martuscello et al. (2007), trabalhando com progênies de
meios irmãos de P. maximum observaram repetibilidades substancialmente maiores para %F
(0,7067). Essas variações ressaltam a importância de mais estudos acerca deste parâmetro
genético para dar mais subsídios para trabalhos com medidas repetidas e avaliação de
genótipos de forrageiras.
A massa seca de colmos (MSC) apresentou coeficientes que variaram de 0,1954 pelo
método da ANOVA a 0,4573 pelo método CPCOV. Nota-se, assim como para %F, maior
amplitude de variação entre os métodos de estimação, diferente do que foi observado para MST
e MSF. Provavelmente isso se deve ao viés causado pela ocorrência de coeficientes de
repetibilidade mais baixos sobre os métodos baseados em análises de componentes principais,
que tendem a superestimar o valor do parâmetro nestas condições (MANSOUR et al., 1981).
A importância da MSC no melhoramento é baixa, uma vez que este componente não é visado
para melhoria da produção, mas pode ser reduzido por meio da seleção de plantas que possuam
melhor porcentagem de folhas.

35
De maneira semelhante à %F, a porcentagem de colmos (%C) também apresentou
baixos valores estimados pelos métodos da ANOVA (0,1052) e elevado valor pelo método da
CPCOV (0,4528). Provavelmente, os baixos resultados se devem ao efeito marcante do nível
de maturidade das plantas sobre a participação deste componente na composição morfológica
da forragem. Nesse sentido, o controle ambiental exerceria maior efeito sobre a %C, que o
controle genético, o que dificultaria o processo de seleção. Apesar disso, ganhos simultâneos
podem ser obtidos por meio da %F, que apresenta correlação elevada e negativa com a %C
(BRAZ, 2012).
A relação lâmina:colmo (RLC) apresentou coeficientes de repetibilidade próximos a
MSC, que variaram de 0,1127 pelo método da ANOVA e 0,4477 pelo método CPCOV, valores
que são considerados baixos (Tabela 1). Resultados semelhantes foram observados por
Cargnelutti Filho et al. (2004) avaliando cinco genótipos de P.maximum com restrição solar,
no qual os autores encontraram valores de 0,080 e 0,528 pelos métodos da ANOVA e CPCOV,
respectivamente. Este fato pode ser explicado pelas variações climáticas ao longo dos anos,
considerando que as avaliações foram realizadas durante as quatro estações, em que ocorre
diferenças no clima, como altas temperaturas e precipitação durante o verão e redução das
mesmas durante o período da seca e transição água-seca. Além disso, a RLC também influência
é influenciada pelo do estágio fenológico que, por sua vez, pode causar interferência nos
resultados quando as avaliações são realizadas em períodos fixos de colheita como neste
estudo.
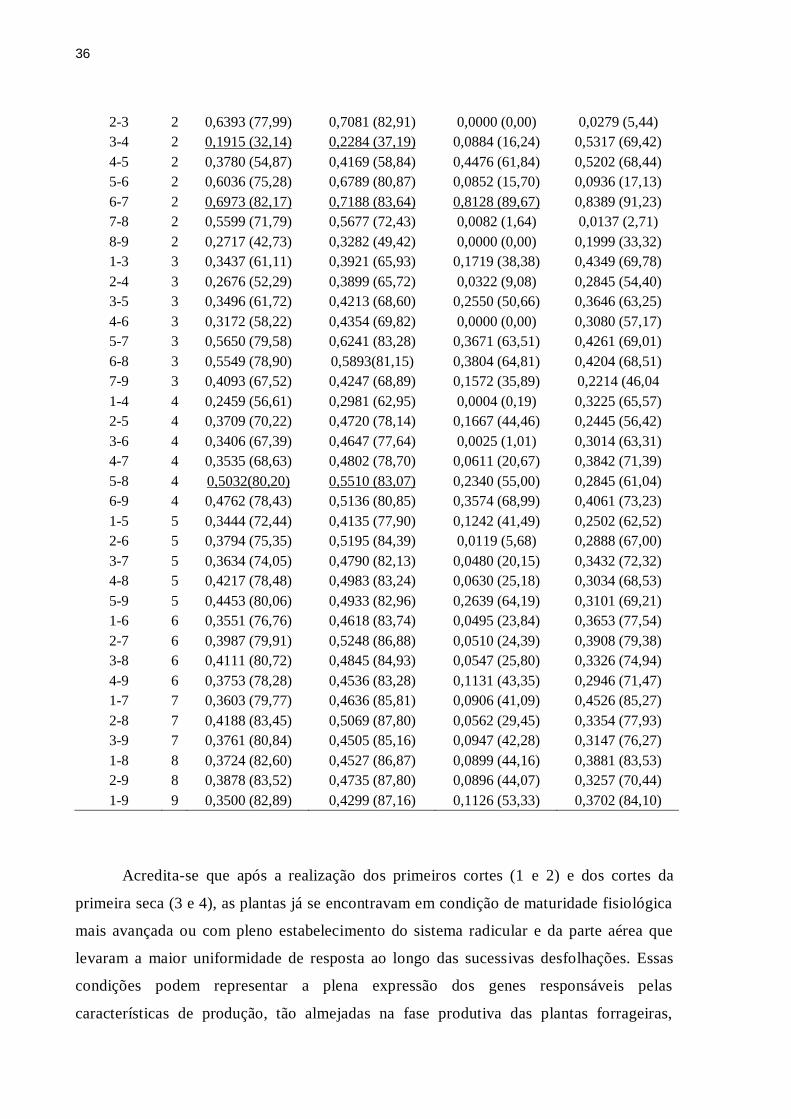
No estudo da estabilização genotípica, os melhores coeficientes de repetibilidade e
determinação para a variável MST foram observados para a combinação entre os cortes 6
a 7 e entre os cortes 5 a 8, ao passo que os menores coeficientes foram observados quando
se utilizaram apenas os cortes 3 a 4 e de 1 a 2, em ambos os métodos (Tabela 2). A melhoria
dos coeficientes pela combinação entre os cortes 6 a 7 e também, pela combinação dos
cortes de 5 a 8 representa as avaliações feitas durante o segundo período das águas após o
estabelecimento.
Tabela 2: Estimativa de número de cortes (N), coeficiente de repetibilidade e coeficiente de
determinação (entre parênteses) da produção de massa seca total (MST) e relação folha/colmo
(RFC), considerando-se diferentes combinações entre as avaliações realizadas em genótipos
de Panicum maximum, segundo as metodologias ANOVA e CPCOR
Avaliação N MST RFC
ANOVA CPCOR ANOVA CPCOR
1-2 2 0,2683 (42,31) 0,2749 (43,12) 0,3794 (55,01) 0,8691 (92,99)
36
2-3 2 0,6393 (77,99) 0,7081 (82,91) 0,0000 (0,00) 0,0279 (5,44)
3-4 2 0,1915 (32,14) 0,2284 (37,19) 0,0884 (16,24) 0,5317 (69,42)
4-5 2 0,3780 (54,87) 0,4169 (58,84) 0,4476 (61,84) 0,5202 (68,44)
5-6 2 0,6036 (75,28) 0,6789 (80,87) 0,0852 (15,70) 0,0936 (17,13)
6-7 2 0,6973 (82,17) 0,7188 (83,64) 0,8128 (89,67) 0,8389 (91,23)
7-8 2 0,5599 (71,79) 0,5677 (72,43) 0,0082 (1,64) 0,0137 (2,71)
8-9 2 0,2717 (42,73) 0,3282 (49,42) 0,0000 (0,00) 0,1999 (33,32)
1-3 3 0,3437 (61,11) 0,3921 (65,93) 0,1719 (38,38) 0,4349 (69,78)
2-4 3 0,2676 (52,29) 0,3899 (65,72) 0,0322 (9,08) 0,2845 (54,40)
3-5 3 0,3496 (61,72) 0,4213 (68,60) 0,2550 (50,66) 0,3646 (63,25)
4-6 3 0,3172 (58,22) 0,4354 (69,82) 0,0000 (0,00) 0,3080 (57,17)
5-7 3 0,5650 (79,58) 0,6241 (83,28) 0,3671 (63,51) 0,4261 (69,01)
6-8 3 0,5549 (78,90) 0,5893(81,15) 0,3804 (64,81) 0,4204 (68,51)
7-9 3 0,4093 (67,52) 0,4247 (68,89) 0,1572 (35,89) 0,2214 (46,04
1-4 4 0,2459 (56,61) 0,2981 (62,95) 0,0004 (0,19) 0,3225 (65,57)
2-5 4 0,3709 (70,22) 0,4720 (78,14) 0,1667 (44,46) 0,2445 (56,42)
3-6 4 0,3406 (67,39) 0,4647 (77,64) 0,0025 (1,01) 0,3014 (63,31)
4-7 4 0,3535 (68,63) 0,4802 (78,70) 0,0611 (20,67) 0,3842 (71,39)
5-8 4 0,5032(80,20) 0,5510 (83,07) 0,2340 (55,00) 0,2845 (61,04)
6-9 4 0,4762 (78,43) 0,5136 (80,85) 0,3574 (68,99) 0,4061 (73,23)
1-5 5 0,3444 (72,44) 0,4135 (77,90) 0,1242 (41,49) 0,2502 (62,52)
2-6 5 0,3794 (75,35) 0,5195 (84,39) 0,0119 (5,68) 0,2888 (67,00)
3-7 5 0,3634 (74,05) 0,4790 (82,13) 0,0480 (20,15) 0,3432 (72,32)
4-8 5 0,4217 (78,48) 0,4983 (83,24) 0,0630 (25,18) 0,3034 (68,53)
5-9 5 0,4453 (80,06) 0,4933 (82,96) 0,2639 (64,19) 0,3101 (69,21)
1-6 6 0,3551 (76,76) 0,4618 (83,74) 0,0495 (23,84) 0,3653 (77,54)
2-7 6 0,3987 (79,91) 0,5248 (86,88) 0,0510 (24,39) 0,3908 (79,38)
3-8 6 0,4111 (80,72) 0,4845 (84,93) 0,0547 (25,80) 0,3326 (74,94)
4-9 6 0,3753 (78,28) 0,4536 (83,28) 0,1131 (43,35) 0,2946 (71,47)
1-7 7 0,3603 (79,77) 0,4636 (85,81) 0,0906 (41,09) 0,4526 (85,27)
2-8 7 0,4188 (83,45) 0,5069 (87,80) 0,0562 (29,45) 0,3354 (77,93)
3-9 7 0,3761 (80,84) 0,4505 (85,16) 0,0947 (42,28) 0,3147 (76,27)
1-8 8 0,3724 (82,60) 0,4527 (86,87) 0,0899 (44,16) 0,3881 (83,53)
2-9 8 0,3878 (83,52) 0,4735 (87,80) 0,0896 (44,07) 0,3257 (70,44)
1-9 9 0,3500 (82,89) 0,4299 (87,16) 0,1126 (53,33) 0,3702 (84,10)
Acredita-se que após a realização dos primeiros cortes (1 e 2) e dos cortes da
primeira seca (3 e 4), as plantas já se encontravam em condição de maturidade fisiológica
mais avançada ou com pleno estabelecimento do sistema radicular e da parte aérea que
levaram a maior uniformidade de resposta ao longo das sucessivas desfolhações. Essas
condições podem representar a plena expressão dos genes responsáveis pelas
características de produção, tão almejadas na fase produtiva das plantas forrageiras,

37
conforme discutido por Pereira et al. (2002) e Braz et al. (2015a).
De maneira geral, nota-se que a inclusão do primeiro corte e dos cortes realizados
no período da seca foram prejudiciais para a estimativa da repetibilidade da MST, levando
a baixos coeficientes e necessidade de maior número de medidas repetidas para se melhorar
a eficiência na seleção e predição da resposta agronômica das plantas. A baixa
repetibilidade se deve ao fato dos genótipos se comportarem de forma distinta durante a
realização destes cortes (antes da maturidade fisiológica e durante o período seco),
diminuindo a correlação entre os cortes, e, consequentemente a repetibilidade.
Para a característica RFC, assim como observado para MST, os melhores
coeficientes de repetibilidade (0,8128) e determinação (89,67) foram registrados para os
cortes 6 a 7 pelo método da ANOVA (Tabela 2). Tais resultados podem ser explicados pelo
fato dos cortes terem ocorridos no segundo ano de experimento durante o período das
águas, com condições ambientais homogêneas em relação a temperatura e precipitação
(Figura 1). Ressalta-se, que a queda de precipitação causada pelo veranico foi muito menos
expressiva no segundo ano, o que pode ter contribuído para maior uniformidade das
condições ambientais.
Pelo método CPCOR, o maior coeficiente de repetibilidade e determinação para RFC
foram observados para as combinações entre os cortes 1 a 2 (Tabela 2). Durante a fase de
formação as plantas apresentam maior quantidade de folhas em relação ao colmo quando
comparadas com pastos já estabelecidos, os cortes 1 e 2 foram realizados em fevereiro e março,
meses com altas temperaturas e precipitação, condições favoráveis para o melhor
desenvolvimento das plantas, logo, o conjunto destes fatores podem ter contribuído para os
melhores resultados encontrados estarem relacionados a estas combinações de cortes. Os
maiores coeficientes de repetibilidade e determinação para os cortes 1 e 2 pelo método
CPCOR, também podem ser explicados pelo fato de que cortes de dois a dois são mais
semelhantes uns aos outros em relação a grupos maiores, além deste método levar em
consideração o comportamento cíclico apresentado pelos genótipos.
A análise da estabilização genotípica da porcentagem de folha (%F) mostrou que os
melhores coeficientes de repetibilidade observados também estão relacionados aos cortes 6 e
7 pelo método da ANOVA, o mesmo comportamento foi observado na CPCOR (Tabela 3). As
avaliações que incluíram do primeiro ao quarto corte, em sua maioria, reduziram os valores de
repetibilidade, possivelmente as plantas não estavam estabilizadas nos primeiros cortes,
evidenciando a necessidade de fazer avaliações durante todos os ciclos de desenvolvimento da
planta, pois há variação nas respostas da fase juvenil para a fase adulta (PEREIRA et al., 2002).

38
Em relação a MSF as combinações que apresentaram maior coeficiente de
repetibilidade e determinação também incluiram os cortes 6 a 7 (Tabela 3). Resultados
satisfatórios também foram observados quando considerada a combinação dos cortes 5 a 7 pelo
método da ANOVA, para o coeficiente de repetibilidade e determinação, com valores de
0,5996 e 81,79, respectivamente. Assim como pelo método CPCOR com as combinações 5 e
6 com valores de 0,6918 de repetibilidade e 81,78 de determinação (Tabela 3). Os melhores
resultados observados durante estes cortes podem ser explicados pelo período de avaliação ter
ocorrido durante a época das águas.
Tabela 3: Estimativa de número de cortes (N), coeficiente de repetibilidade e coeficiente de
determinação (entre parênteses) da porcentagem de folha (%F) e massa seca de folha (MSF),
considerando-se diferentes combinações entre as avaliações realizadas em genótipos de
Panicum maximum, segundo as metodologias ANOVA e CPCOR
Avaliação N %F MSF
ANOVA CPCOR ANOVA CPCOR
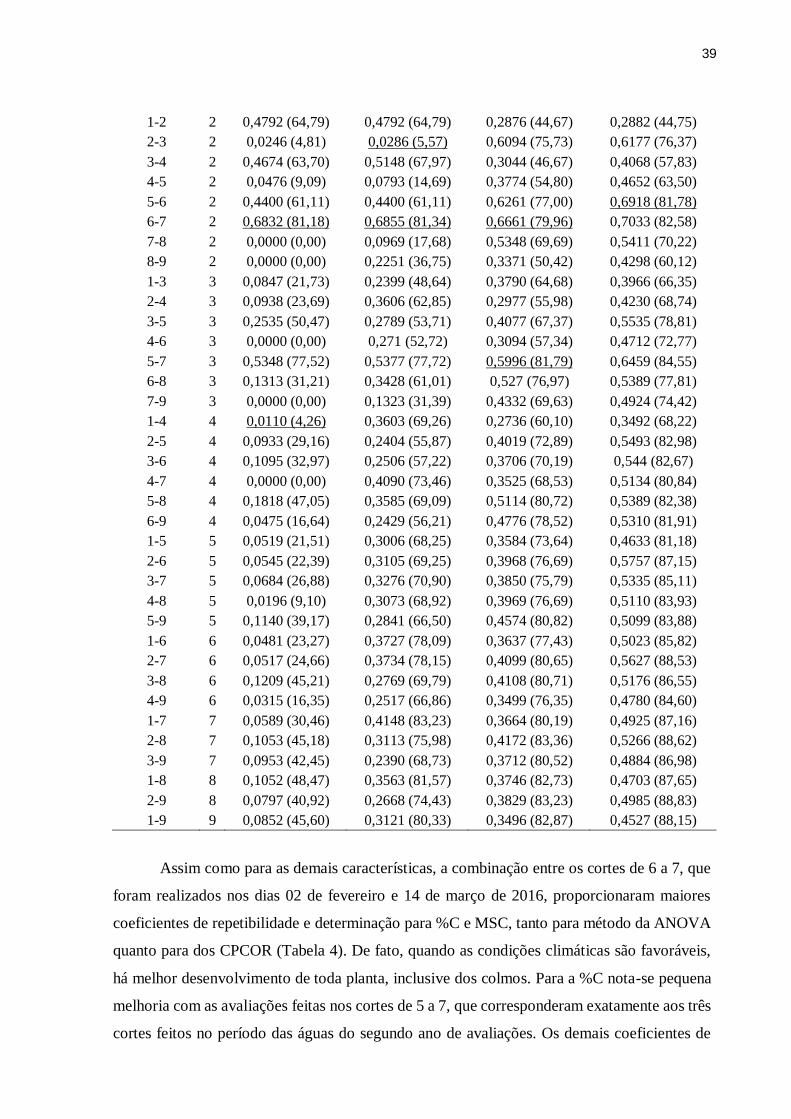
39
1-2 2 0,4792 (64,79) 0,4792 (64,79) 0,2876 (44,67) 0,2882 (44,75)
2-3 2 0,0246 (4,81) 0,0286 (5,57) 0,6094 (75,73) 0,6177 (76,37)
3-4 2 0,4674 (63,70) 0,5148 (67,97) 0,3044 (46,67) 0,4068 (57,83)
4-5 2 0,0476 (9,09) 0,0793 (14,69) 0,3774 (54,80) 0,4652 (63,50)
5-6 2 0,4400 (61,11) 0,4400 (61,11) 0,6261 (77,00) 0,6918 (81,78)
6-7 2 0,6832 (81,18) 0,6855 (81,34) 0,6661 (79,96) 0,7033 (82,58)
7-8 2 0,0000 (0,00) 0,0969 (17,68) 0,5348 (69,69) 0,5411 (70,22)
8-9 2 0,0000 (0,00) 0,2251 (36,75) 0,3371 (50,42) 0,4298 (60,12)
1-3 3 0,0847 (21,73) 0,2399 (48,64) 0,3790 (64,68) 0,3966 (66,35)
2-4 3 0,0938 (23,69) 0,3606 (62,85) 0,2977 (55,98) 0,4230 (68,74)
3-5 3 0,2535 (50,47) 0,2789 (53,71) 0,4077 (67,37) 0,5535 (78,81)
4-6 3 0,0000 (0,00) 0,271 (52,72) 0,3094 (57,34) 0,4712 (72,77)
5-7 3 0,5348 (77,52) 0,5377 (77,72) 0,5996 (81,79) 0,6459 (84,55)
6-8 3 0,1313 (31,21) 0,3428 (61,01) 0,527 (76,97) 0,5389 (77,81)
7-9 3 0,0000 (0,00) 0,1323 (31,39) 0,4332 (69,63) 0,4924 (74,42)
1-4 4 0,0110 (4,26) 0,3603 (69,26) 0,2736 (60,10) 0,3492 (68,22)
2-5 4 0,0933 (29,16) 0,2404 (55,87) 0,4019 (72,89) 0,5493 (82,98)
3-6 4 0,1095 (32,97) 0,2506 (57,22) 0,3706 (70,19) 0,544 (82,67)
4-7 4 0,0000 (0,00) 0,4090 (73,46) 0,3525 (68,53) 0,5134 (80,84)
5-8 4 0,1818 (47,05) 0,3585 (69,09) 0,5114 (80,72) 0,5389 (82,38)
6-9 4 0,0475 (16,64) 0,2429 (56,21) 0,4776 (78,52) 0,5310 (81,91)
1-5 5 0,0519 (21,51) 0,3006 (68,25) 0,3584 (73,64) 0,4633 (81,18)
2-6 5 0,0545 (22,39) 0,3105 (69,25) 0,3968 (76,69) 0,5757 (87,15)
3-7 5 0,0684 (26,88) 0,3276 (70,90) 0,3850 (75,79) 0,5335 (85,11)
4-8 5 0,0196 (9,10) 0,3073 (68,92) 0,3969 (76,69) 0,5110 (83,93)
5-9 5 0,1140 (39,17) 0,2841 (66,50) 0,4574 (80,82) 0,5099 (83,88)
1-6 6 0,0481 (23,27) 0,3727 (78,09) 0,3637 (77,43) 0,5023 (85,82)
2-7 6 0,0517 (24,66) 0,3734 (78,15) 0,4099 (80,65) 0,5627 (88,53)
3-8 6 0,1209 (45,21) 0,2769 (69,79) 0,4108 (80,71) 0,5176 (86,55)
4-9 6 0,0315 (16,35) 0,2517 (66,86) 0,3499 (76,35) 0,4780 (84,60)
1-7 7 0,0589 (30,46) 0,4148 (83,23) 0,3664 (80,19) 0,4925 (87,16)
2-8 7 0,1053 (45,18) 0,3113 (75,98) 0,4172 (83,36) 0,5266 (88,62)
3-9 7 0,0953 (42,45) 0,2390 (68,73) 0,3712 (80,52) 0,4884 (86,98)
1-8 8 0,1052 (48,47) 0,3563 (81,57) 0,3746 (82,73) 0,4703 (87,65)
2-9 8 0,0797 (40,92) 0,2668 (74,43) 0,3829 (83,23) 0,4985 (88,83)
1-9 9 0,0852 (45,60) 0,3121 (80,33) 0,3496 (82,87) 0,4527 (88,15)
Assim como para as demais características, a combinação entre os cortes de 6 a 7, que
foram realizados nos dias 02 de fevereiro e 14 de março de 2016, proporcionaram maiores
coeficientes de repetibilidade e determinação para %C e MSC, tanto para método da ANOVA
quanto para dos CPCOR (Tabela 4). De fato, quando as condições climáticas são favoráveis,
há melhor desenvolvimento de toda planta, inclusive dos colmos. Para a %C nota-se pequena
melhoria com as avaliações feitas nos cortes de 5 a 7, que corresponderam exatamente aos três
cortes feitos no período das águas do segundo ano de avaliações. Os demais coeficientes de

40
repetibilidade, envolvendo outras combinações entre as nove colheitas estudadas, foram em
geral baixos, o que indica a grande variação desta característica e o seu baixo potencial para
avaliação dentro de programas de melhoramento, com foco na seleção negativa.
A MSC, apesar da oscilação, apresentou coeficientes maiores que a %C (Tabela 4), que
deve ser reflexo da maior facilidade de mensuração. Nesse sentido, a variação no peso
acumulado de colmos ao longo de um corte pode ter sido menor que a sua participação relativa
na produção total da forragem.
Foram observados valores iguais a zero para as características RLC, %F, %C e MSC,
todas pelo método da ANOVA (Tabelas 2,3 e 4), sendo que as combinações entre cortes que
mais se destacaram com este resultado foram de 4 a 6, para RLC, %F e %C. Repetibilidade
zero significa ausência de relação entre as medidas repetidas e variância genotípica para o
período.
Tabela 4: Estimativa de número de cortes (N), coeficiente de repetibilidade e coeficiente de
determinação (entre parênteses) da porcentagem de colmo (%C) e massa seca de colmo (MSC),
considerando-se diferentes combinações entre as avaliações realizadas em genótipos de
Panicum maximum, segundo as metodologias ANOVA e CPCOR
Avaliação N %C MSC
ANOVA CPCOR ANOVA CPCOR
1-2 2 0,4490 (61,98) 0,4518 (62,24) 0,1282 (22,73) 0,1437 (25,13)
2-3 2 0,0000 (0,00) 0,0693 (12,96) 0,4740 (74,31) 0,4790 (64,78)
3-4 2 0,5079 (67,36) 0,5309 (69,35) 0,0000 (0,00) 0,0655 (12,29)
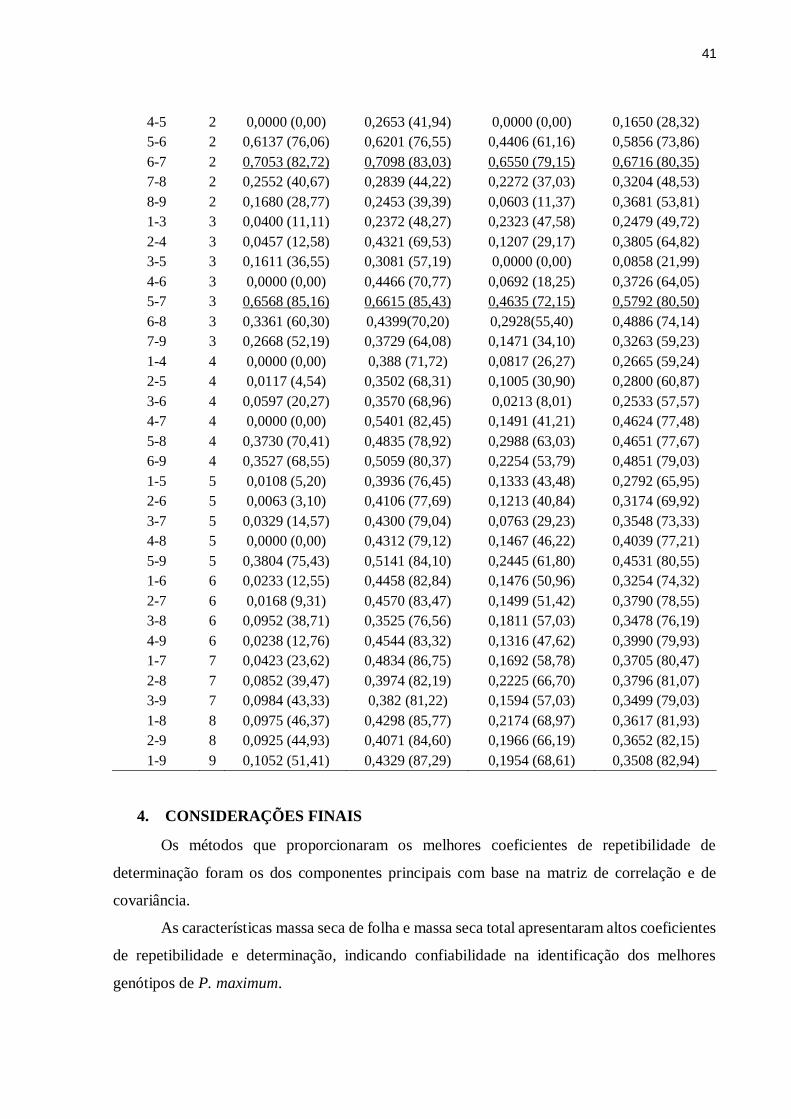
41
4-5 2 0,0000 (0,00) 0,2653 (41,94) 0,0000 (0,00) 0,1650 (28,32)
5-6 2 0,6137 (76,06) 0,6201 (76,55) 0,4406 (61,16) 0,5856 (73,86)
6-7 2 0,7053 (82,72) 0,7098 (83,03) 0,6550 (79,15) 0,6716 (80,35)
7-8 2 0,2552 (40,67) 0,2839 (44,22) 0,2272 (37,03) 0,3204 (48,53)
8-9 2 0,1680 (28,77) 0,2453 (39,39) 0,0603 (11,37) 0,3681 (53,81)
1-3 3 0,0400 (11,11) 0,2372 (48,27) 0,2323 (47,58) 0,2479 (49,72)
2-4 3 0,0457 (12,58) 0,4321 (69,53) 0,1207 (29,17) 0,3805 (64,82)
3-5 3 0,1611 (36,55) 0,3081 (57,19) 0,0000 (0,00) 0,0858 (21,99)
4-6 3 0,0000 (0,00) 0,4466 (70,77) 0,0692 (18,25) 0,3726 (64,05)
5-7 3 0,6568 (85,16) 0,6615 (85,43) 0,4635 (72,15) 0,5792 (80,50)
6-8 3 0,3361 (60,30) 0,4399(70,20) 0,2928(55,40) 0,4886 (74,14)
7-9 3 0,2668 (52,19) 0,3729 (64,08) 0,1471 (34,10) 0,3263 (59,23)
1-4 4 0,0000 (0,00) 0,388 (71,72) 0,0817 (26,27) 0,2665 (59,24)
2-5 4 0,0117 (4,54) 0,3502 (68,31) 0,1005 (30,90) 0,2800 (60,87)
3-6 4 0,0597 (20,27) 0,3570 (68,96) 0,0213 (8,01) 0,2533 (57,57)
4-7 4 0,0000 (0,00) 0,5401 (82,45) 0,1491 (41,21) 0,4624 (77,48)
5-8 4 0,3730 (70,41) 0,4835 (78,92) 0,2988 (63,03) 0,4651 (77,67)
6-9 4 0,3527 (68,55) 0,5059 (80,37) 0,2254 (53,79) 0,4851 (79,03)
1-5 5 0,0108 (5,20) 0,3936 (76,45) 0,1333 (43,48) 0,2792 (65,95)
2-6 5 0,0063 (3,10) 0,4106 (77,69) 0,1213 (40,84) 0,3174 (69,92)
3-7 5 0,0329 (14,57) 0,4300 (79,04) 0,0763 (29,23) 0,3548 (73,33)
4-8 5 0,0000 (0,00) 0,4312 (79,12) 0,1467 (46,22) 0,4039 (77,21)
5-9 5 0,3804 (75,43) 0,5141 (84,10) 0,2445 (61,80) 0,4531 (80,55)
1-6 6 0,0233 (12,55) 0,4458 (82,84) 0,1476 (50,96) 0,3254 (74,32)
2-7 6 0,0168 (9,31) 0,4570 (83,47) 0,1499 (51,42) 0,3790 (78,55)
3-8 6 0,0952 (38,71) 0,3525 (76,56) 0,1811 (57,03) 0,3478 (76,19)
4-9 6 0,0238 (12,76) 0,4544 (83,32) 0,1316 (47,62) 0,3990 (79,93)
1-7 7 0,0423 (23,62) 0,4834 (86,75) 0,1692 (58,78) 0,3705 (80,47)
2-8 7 0,0852 (39,47) 0,3974 (82,19) 0,2225 (66,70) 0,3796 (81,07)
3-9 7 0,0984 (43,33) 0,382 (81,22) 0,1594 (57,03) 0,3499 (79,03)
1-8 8 0,0975 (46,37) 0,4298 (85,77) 0,2174 (68,97) 0,3617 (81,93)
2-9 8 0,0925 (44,93) 0,4071 (84,60) 0,1966 (66,19) 0,3652 (82,15)
1-9 9 0,1052 (51,41) 0,4329 (87,29) 0,1954 (68,61) 0,3508 (82,94)
4. CONSIDERAÇÕES FINAIS
Os métodos que proporcionaram os melhores coeficientes de repetibilidade de
determinação foram os dos componentes principais com base na matriz de correlação e de
covariância.
As características massa seca de folha e massa seca total apresentaram altos coeficientes
de repetibilidade e determinação, indicando confiabilidade na identificação dos melhores
genótipos de P. maximum.

42
As características porcentagem de folha, porcentagem de colmo, massa seca de colmo
e relação lâmina:colmo, apresentam grandes variações entre os métodos avaliados, apesar disso,
os coeficientes de repetibilidade foram considerados baixos.
Para a estabilização genotípica, os melhores coeficientes de repetibilidade e
determinação são observados para os cortes realizados no segundo período das águas (6 e 7).
Os cortes realizados antes das plantas adquirirem maturidade fisiológica e durante a seca são
prejudiciais no processo de seleção, devido à baixa repetibilidade.

43
5. REFERÊNCIAS BIBLIOGRÁFICAS
ABEYWARDENA, V. An application of principal component analysis in genetics. Journal
Genetics, v.16, p.27, 1972.
BRÂNCIO, P. A.; NASCIMENTO, JR. D.; EUCLIDES, V. P. B.; FONSECA, D. M.
Avaliação de três cultivares de Panicum maximum Jacq. sob pastejo: composição da dieta,
consumo de matéria seca e ganho de peso animal. Revista Brasileira de Zootecnia v. 32,
p.1037-1044, 2003.
BRAZ, T. G. S. Parâmetros genéticos, repetibilidade e diversidade genética em híbridos
de Panicum maximum, 2012. p. 117. Tese (Doutorado Zootecnia) – Universidade Federal de
viçosa, Viçosa, 2012.
BRAZ, T. G. S.; FONSECA, D. M.; JANK, L.; CRUZ, C. D.; MARTUSCELLO, J. A.
Genotypic stabilization of agronomic traits in Panicum maximum (Jacq.) hybrids. Revista
Brasileira Zootecnia, v.44 p.377-383, 2015a.
BRAZ, T. G. S.; FONSECA, D. M.; JANK, L.; CRUZ, C. D.; MARTUSCELLO, J. A.
Repeatability of agronomic traits in Panicum maximum (Jacq.) hybrids. Genetics and
Molecular Research, v. 14, p. 19282-19294, 2015b.
CANTARUTTI, R. B.; ALVARES V. V.H.; RIBEIRO, A. C. Amostragem do solo. In:
RIBEIRO, A.C.; GUIMARAES, P.T.G.; ALVAREZ V., V.H. (Ed.). Recomendação para o
uso de corretivos e fertilizantes em Minas Gerais: 5º Aproximação. Viçosa: Comissão de
Fertilidade do Solo do Estado de Minas Gerais. 1999, p. 13-20.
CARGNELUTTI FILHO, A.; CASTILHOS, Z. M.; STORCK, L.; SAVINA, J. F. Análise de
repetibilidade de caracteres forrageiros de genótipos de Panicum maximum, avaliados com e
sem restrição solar. Ciência Rural, v.34, n.3, p.723-729, 2004.
CRUZ, C. D. Programa genes, Biometria. 1 ed. Viçosa: Editora UFV, 2006. 282p.

44
CRUZ, C.D. GENES - a software package for analysis in experimental statistics and
quantitative genetics. Acta Scientiarum Agronomy v.35, n.3, p.271-276, 2013.
CRUZ, C.D.; REGAZZI, A.J.; CARNEIRO, P.C.S. Modelos biométricos aplicados ao
melhoramento genético. 3.ed. Viçosa, MG: Universidade Federal de Viçosa, 2004. v.1.
480p.
DAHER, R. F.; MALDONADO, H.; PEREIRA, A. V.; AMARAL JÚNIOR, A.T.;
PEREIRA, M. G.; FERREIRA, C. F.; RAMOS, S. R. R.; TARDIN, F. D.; SILVA, M. P.
Estimativas de parâmetros genéticos e de coeficientes de repetibilidade de caracteres
forrageiros em clones de capim-elefante (Pennisetum purpureum Schum.). Acta Scientiarum
Agronomy, v. 26, n. 4, p. 483-490, 2004.
FREITAS, F. P.; FONSECA, D. M.; BRAZ, T. G. S.; MARTUSCELLO, J. A.; SANTOS, M.
E. R. Forage yield and nutritive value of Tanzania guinea grass under nitrogen supplies and
plant densities. Revista Brasileira de Zootecnia, v.41, n.4, p.864-872, 2012.
JANK, L.; RESENDE, R. M. S.; VALLE, C. B; RESENDE, M. D. V.; CHIARI, L.; 32
CANÇADO, L. J.; SIMIONI, C. Melhoramento genético de Panicum maximum. In: 33
RESENDE, R. M. S.; VALLE, C. B. do; JANK, L. (Ed.) Melhoramento de forrageiras 34
tropicais. 1.ed. Campo Grande, MS: Embrapa Gado de Corte, 2008. p. 55-87.
KÖPPEN, W. Climatologia: com um estúdio de los climas de la tierra. New Gersey: 1948.
LÉDO, F. J. S; PEREIRA, A. V.; SOBRINHO, F. S.; AUAD, A. M.; JANK, L.; OLIVEIRA,
J. S. Estimativa de repetibilidade para caracteres forrageiros em Panicum maximum. Ciência
e Agrotecnologia. v. 32, n. 4, p. 1299-1303, 2008.
MANSOUR, H.; NORDHEIM, E. V.; RUTLEDGE, J. J. Estimators of repeatability.
Theoretical and Applied Genetics. v. 60, p. 151-156, 1981.
MARTUSCELLO, J. A.; JANK, L.; FONSECA, D. M.; CRUZ, C. D.; CUNHA, D. N. F. A.
Repetibilidade de caracteres agronômicos em Panicum maximum Jacq. Revista Brasileira de
Zootecnia, v.36, n.6, p.1975-1981, 2007.

45
NEGREIROS, J. R. S.; SARAIVA, L.L.; OLIVEIRA, T. K.; ÁLVARES, V. S.;
RONCATTO, G. Estimativas de repetibilidade de caracteres de produção em laranjeiras-
doces no Acre. Pesquisa Agropecuária Brasileira, v.43, n.12, p.1763-1768, 2008.
PEREIRA, A. V.; CRUZ, C. D.; FERREIRA, R. P.; BOTREL, M. A.; OLIVEIRA, J. S.
Influência da estabilização de genótipos de capim elefante (Pennisetum purpureum Schum.)
sobre a estimativa da repetibilidade de características forrageiras. Ciência e Agrotecnologia,
v.26, n.3, p.762-767, 2002.
RESENDE, R. M. S.; VALLE, C. B. do; JANK, L. (Ed.) Melhoramento de forrageiras
tropicais. 1.ed. Campo Grande, MS: Embrapa Gado de Corte, 2008. p. 55-87.
SOUZA SOBRINHO, F.; BORGES, V.; LÉDO, F. J. S.; KOPP, M. M. Repetibilidade de
características agronômicas e número de cortes necessários para seleção de Urochloa
ruziziensis. Pesquisa Agropecuária Brasileira, v.45, n.6, p.579-584, 2010.
VALLE, C. B.; JANK, L.; RESENDE, R. M. S. O melhoramento de forrageiras tropicais no
Brasil. Revista Ceres. V. 56, n. 4, p. 460-472, 2009.

46
CAPÍTULO II - ANÁLISE DE AGRUPAMENTO COM BASE EM
CARACTERÍSTICAS MORFOGÊNICAS EM GENÓTIPOS DE Panicum maximum
RESUMO
Objetivou-se com este trabalho estimar parâmetros genéticos e formar grupos morfofuncionais
com base nas características morfogênicas e estruturais por meio do agrupamento de
Otimização de Tocher em genótipos de Panicum maximum. Foram avaliados 17 genótipos em
delineamento em blocos ao acaso com três repetições. Foram estimadas: taxa de aparecimento
foliar (TAPF), filocrono (FIL), taxa de alongamento foliar (TALF), taxa de senescência foliar
(TSF), número de folhas vivas (NFV), duração de vida das folhas (DVF), taxa de alongamento
de pseudocolmo (TALC), número médio de perfilhos (NMP), relação lâmina:colmo (RLC) e
comprimento final da lâmina (CFL). Foi observado que somente as características TALF, CFL
e RLC tiveram o componente variância genética significativo. Apesar disto, as características
TALC, NFV, apresentaram coeficientes de variação genotípicos (CVg) superiores aos
coeficientes de variação residual ou ambiental (CVe). As características TAPF, FIL, DVF e
TSF apresentaram valores abaixo da unidade para a razão CVg/CVe. Alta razão CVg/CVe
foram observadas para as características RLC, NFV, TALC, TALF, CFL, sendo os maiores
coeficientes foram registrados para RLC. Após o agrupamento, constatou-se a formação de
cinco grupos morfofuncionais. Os grupos que apresentaram maiores valores de TALF foram os
grupos 3, 5 e 1 com valores superiores à média geral de todos os genótipos avaliados. Enquanto
o grupo 4 obteve menor desempenho para esta característica. Para a TALC o grupo 2 se
destacou, seguido pelo grupo 5. Dentre todos os grupos a maior RLC constada foi para o grupo
4 e para CFL o grupo 3. Conclui-se que as características TALF, CFL e RLC apresentam
variabilidade genética significativa, e as características TAPF, FIL, DVF e TSF apresentam
baixa razão CVg/CVe e necessitam de maior controle ambiental. Os grupos 3, 5 e 1, apresentam
altas taxas de alongamento de folha como mecanismo de acúmulo de forragem. Já o grupo 4 se
destaca pela capacidade de perfilhamento
Palavras-chave: Alongamento. Aparecimento. Estrutura. Perfilhos. Multivariada. Tocher.

47
ANALYSIS OF GROUPING BASED ON MORPHOGENIC CHARACTERISTICS IN
GENOTYPES OF Panicum maximum
ABSTRACT
The objective was to estimate genetic parameters and form morphofunctional groups based on
the morphogenic and structural characteristics using Tocher clustering in Panicum maximum
genotypes. In the study, were estimated the variables: leaf appearance rate (LAR), phillochron
(PHI), leaf elongation rate (LER), leaf senescence rate (LSR), number of live leaves (NLL),
leaf life spam (LLS), stem elongation rate (SER), leaf:stem ratio (LSR), number of tillers (NTI)
and leaf final length (LFL). In the study of the genetic parameters, only the traits LER, LFL and
LSR had significant genotypic variance. The SER and NLL did not have significant genotypic
variance but have genotypic coefficient of variation (CVg) highest than the residual coefficient
of variation (CVe). The traits LAR, PHI, LLS and LSR presented CVg/CVe ratio lower than 1.
High values of CVg/CVe ratio were also observed to LSR, NLL, SER, LER, being LSR the
variable with the highest coefficient. The Tocher clustering analysis allowed the formation of
five morphofunctional groups. The plants with highest LER were the were in the groups 3, 5
and 1 with values above the average. The group 4 had the the lower SER. The groups 2 and 5
showed high values to SER. The group 4 had high LSR and the group 3 had a high LFL. The
traits LER, LFL and LSR have significant genetic variability and the traits LAR, PHI, LLS and
LSR have low CVg/CVe ratio, demanding more environmental control. The groups 3, 5 and 1
have the elongation of leaves as the mainly mechanism of herbage accumulation and the group
4 stands out by the number of tillers.
Key words: Appearance. Elongation. Multivariate. Structure. Tillers. Tocher.

48
1 . INTRODUÇÃO
As pastagens são uma importante fonte de alimento para os sistemas de produção de
ruminantes situados nas regiões tropicais do planeta, onde as condições de clima e solo são
favoráveis à utilização de gramíneas de metabolismo C4 como o Panicum maximum. Nestes
ambientes, os estudos sobre o melhoramento de forrageiras e a diversificação de pastagens vem
se intensificando nas últimas décadas frente a demanda por plantas mais adaptadas e
competitivas que atendam às demandas dos produtores e também proporcionem boa resposta
produtiva.
O melhoramento de forrageiras de P. maximum no Brasil resultou no lançamento de
plantas como as cultivares Tanzânia e Mombaça, que fazem parte dos sistemas de produção
animal desde a década de 90 (JANK, 1995), da cultivar Massai e de cultivares mais recentes
como Zuri, Tamani e Quênia, entre outras. Estas plantas apresentam grande potencial produtivo
e a sua diversificação tem se intensificado nos últimos anos, o que demanda protocolos de
avaliação mais eficientes e que possam antecipar informações acerca do manejo exigido pelas
plantas. Contudo, durante todo o processo de avaliação de novas cultivares forrageiras, a
definição de protocolos de manejo normalmente está associada as últimas etapas que antecedem
o lançamento (VALLE et al., 2009; JANK et al., 2011) ou até, posteriormente à sua liberação
no mercado. Ressalta-se que, assim como o melhoramento, o manejo tem contribuído para
grandes melhorias na produção animal em pastagens, garantindo maior produtividade,
qualidade e persistência das forrageiras.
No processo de avaliação de estratégias de manejo, o estudo da resposta
morfofisiológica das plantas às condições de meio e à presença dos animais em pastejo, permite
melhor compreensão das condições que podem favorecer características de interesse para a
produção como a massa de folhas, relação folha colmo, entre outras (ALEXANDRINO et al.,
2001; CASAGRANDE, 2010). Muitos destes estudos foram realizados por meio da
morfogênese, que segundo Chapman e Lemaire (1993) pode ser definida como o estudo da
origem e desenvolvimento dos diferentes órgãos de um organismo e das transformações que
determinam a produção e a mudança na forma e estrutura da planta no espaço ao longo do
tempo.
Dentro do melhoramento genético espera-se que a avaliação da morfogênese em
genótipos em processo de seleção possa contribuir para guiar os esquemas de avaliação em
função da aptidão dos genótipos para determinado nível de produtividade ou de crescimento.
Isso se deve ao fato de que em diversos estudos de manejo de pastagens (COSTA et al., 2016;

49
MARTUSCELLO et al., 2015; RESENDE JÚNIOR, 2011; MACEDO et al., 2010), estas
características foram grandes balizadoras da resposta morfofisiológica de plantas de P.
maximum a diversas condições de ambiente e manejo, podendo, então, ser um indicador
morfofisiológico de capacidade produtiva e de crescimento dos diferentes genótipos.
Desta maneira, o estudo da morfogênese permite acompanhar o desenvolvimento e
crescimento da planta forrageira e sua interação com o ambiente, que são informações
importantes na seleção dos melhores genótipos. Assim, antecipar o uso da morfogênese no
processo de avaliação, pode contribuir para o conhecimento de suas exigências de manejo antes
do lançamento de novas cultivares. Contudo, devido à grande quantidade de dados gerados a
partir da avaliação da morfogênese em grande número de genótipos, pode haver dificuldade no
estudo ou identificação do perfil de indivíduos por meio da comparação de cada característica
individualmente. A análise multivariada passa a ter aplicação justamente nestas situações
(MINGOTI, 2005). O uso da análise de agrupamento, por sua vez, possibilita o estudo e a
caracterização dos genótipos através de grupos com características afins, facilitando a

50
interpretação dos resultados e a seleção dos melhores grupos ou indivíduos que obtiveram
melhor desempenho na avaliação.
Diante disso, objetivou-se com este trabalho estimar parâmetros genéticos e formar
grupos morfofuncionais com base nas características morfogênicas e estruturais por meio do
agrupamento de Otimização de Tocher em genótipos de Panicum maximum.
2 . MATERIAL E MÉTODOS
O experimento foi conduzido na Universidade Federal de São João del-Rei, Campus
Tancredo Neves (CTAN), localizada no município de São João del-Rei no Campo das
Vertentes, em Minas Gerais, Brasil. O município encontra-se a 904 m de altitude e está situado
nas coordenadas geográficas 21º08ʼ11ʼʼ de latitude Sul e 40º15ʼ43ʼʼ de longitude Oeste. O
clima, pela classificação de Köppen (1948), é tipo Cwa, com estações secas (maio a outubro) e
chuvosa (novembro a abril) bem definidas. Os dados referentes às temperaturas média, máxima,
mínima e precipitação durante o período experimental estão apresentados na Figura 1.
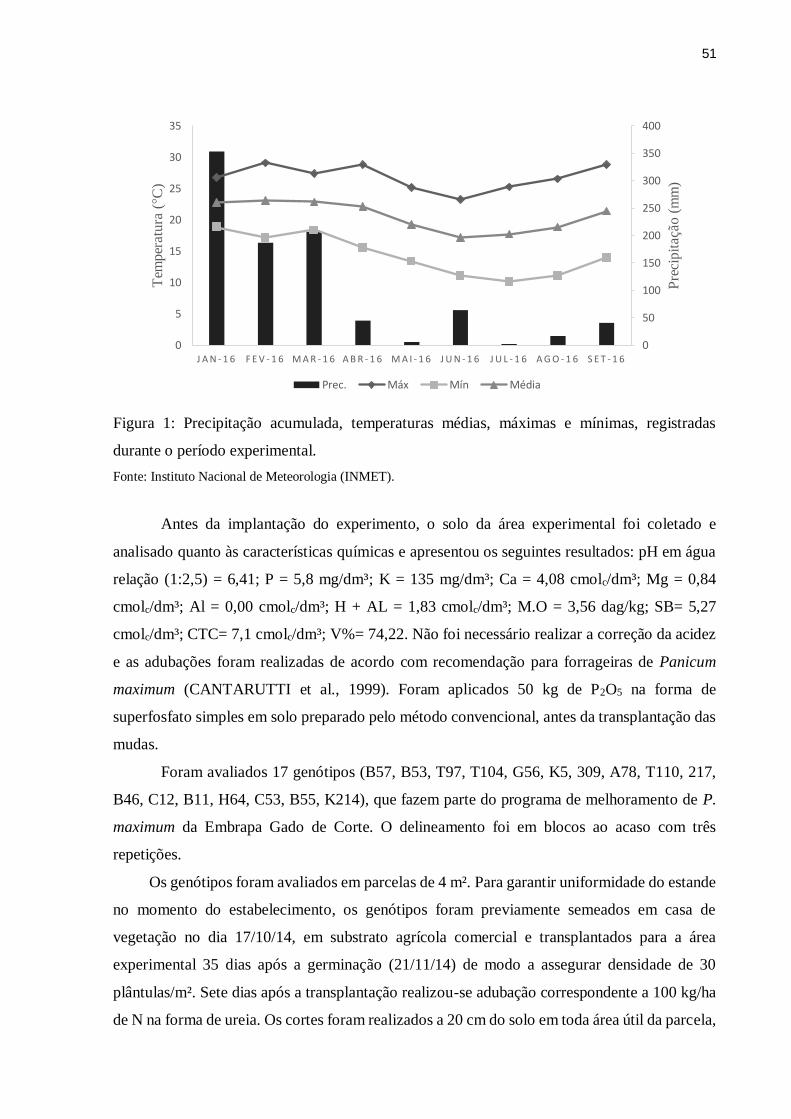
51
Figura 1: Precipitação acumulada, temperaturas médias, máximas e mínimas, registradas
durante o período experimental.
Fonte: Instituto Nacional de Meteorologia (INMET).
Antes da implantação do experimento, o solo da área experimental foi coletado e
analisado quanto às características químicas e apresentou os seguintes resultados: pH em água
relação (1:2,5) = 6,41; P = 5,8 mg/dm³; K = 135 mg/dm³; Ca = 4,08 cmolc/dm³; Mg = 0,84
cmolc/dm³; Al = 0,00 cmolc/dm³; H + AL = 1,83 cmolc/dm³; M.O = 3,56 dag/kg; SB= 5,27
cmolc/dm³; CTC= 7,1 cmolc/dm³; V%= 74,22. Não foi necessário realizar a correção da acidez
e as adubações foram realizadas de acordo com recomendação para forrageiras de Panicum
maximum (CANTARUTTI et al., 1999). Foram aplicados 50 kg de P2O5 na forma de
superfosfato simples em solo preparado pelo método convencional, antes da transplantação das
mudas.
Foram avaliados 17 genótipos (B57, B53, T97, T104, G56, K5, 309, A78, T110, 217,
B46, C12, B11, H64, C53, B55, K214), que fazem parte do programa de melhoramento de P.
maximum da Embrapa Gado de Corte. O delineamento foi em blocos ao acaso com três
repetições.
Os genótipos foram avaliados em parcelas de 4 m². Para garantir uniformidade do estande
no momento do estabelecimento, os genótipos foram previamente semeados em casa de
vegetação no dia 17/10/14, em substrato agrícola comercial e transplantados para a área
experimental 35 dias após a germinação (21/11/14) de modo a assegurar densidade de 30
plântulas/m². Sete dias após a transplantação realizou-se adubação correspondente a 100 kg/ha
de N na forma de ureia. Os cortes foram realizados a 20 cm do solo em toda área útil da parcela,
0
50
100
150
200
250
300
350
400
0
5
10
15
20
25
30
35
J A N - 1 6 F E V - 1 6 M A R - 1 6 A B R - 1 6 M A I - 1 6 J U N - 1 6 J U L - 1 6 A G O - 1 6 S E T - 1 6
Pre
cip
itaçã
o (
mm
)
Tem
per
atu
ra (
°C)
Prec. Máx Mín Média

52
as avaliações morfogênicas foram realizadas a partir do corte do dia 23/12/2015, totalizando
três cortes no período das águas e um corte na seca, |realizados nas seguintes datas: 02/02/2016,
14/03/2016, 20/04/2016, 23/09/2016. Após os cortes as parcelas foram adubadas com
nitrogênio e potássio, em doses equivalentes a 50 kg /ha-1 de N e 20 kg/ha-1 de KCl com 45g de
ureia e 13 g de KCl, respectivamente, em cada parcela.
Para a avaliação das características morfogênicas, foram marcados dois perfilhos por
parcela com fios de cores diferenciadas, de modo haver identificação dos perfilhos que foram
acompanhados. Sempre que houve necessidade, em caso de morte ou remoção do meristema
apical, os perfilhos eram trocados. Para no número médio de perfilhos, foi selecionada uma
touceira representativa dentro de cada parcela, na qual foi realizada a contagem do número de
perfilhos.
As medidas morfogênicas foram tomadas com uso de régua milimétrica duas vezes por
semana no período das águas e apenas uma vez por semana durante o período seco do ano.
Foram efetuadas medidas do comprimento de lâminas foliares, do comprimento da porção
verde das lâminas senescentes e da altura da lígula da última folha expandida além do registro
de novas folhas surgidas em cada um dos perfilhos em cada uma das datas de avaliação. A partir
dessas medidas foram estimadas:
• Taxa de aparecimento foliar (folha/perfilho/dia): quociente entre o número de folhas por
perfilho surgidas no período avaliado e número de dias do período;
• Filocrono (dias/folha/perfilho): número de dias para o aparecimento de duas folhas
consecutivas, inverso da taxa de aparecimento foliar;
• Taxa de alongamento foliar (cm/perfilho/dia): variação média no comprimento da folha em
expansão durante o período de avaliação;
• Taxa de senescência foliar (cm/perfilho/dia): variação média no comprimento da porção
senescente da folha, resultado do produto entre o comprimento da lâmina foliar senescente e
a proporção de tecido senescente correspondente, observada ao longo do período de
avaliação;
• Número de folhas vivas: contagem do número de folhas vivas;
• Duração de vida das folhas (dias): determinado pelo produto do número de folhas vivas pelo
filocrono;
• Taxa de alongamento de pseudocolmo (cm/perfilho/dia): variação média no comprimento do
pseudocolmo durante o período de avaliação;

53
• Número médio de perfilhos: número de médio de perfilhos por touceira dentro de cada
parcela;
• Relação lâmina: colmo: quociente entre massa seca de folha e massa seca de colmo.
Para a análise estatística, foi utilizado o modelo linear misto:
y = Xm + Zg + Wp + e,
em que y é o vetor de dados, m é o vetor dos efeitos das combinações medição-repetição
(assumidos como fixos) somados à média geral, g é o vetor dos efeitos genotípicos (assumidos
como aleatórios), p é vetor dos efeitos de ambiente permanente (assumidos como aleatórios) e
e é o vetor de erros ou resíduos (aleatórios). As letras maiúsculas representam as matrizes de
incidência para os referidos efeitos.
De posse dos valores genotípicos estimados, os dados foram padronizados e utilizados
para estimar a distância de Mahalanobis, utilizada para realizar a análise de agrupamento pelo
método de Otimização de Tocher. Após o grupamento, realizou-se a média dos indivíduos dos
grupos para estudar a caracterização morfofuncional dos mesmos.
Também foram realizados estudos preliminares da significância dos componentes de
variância, dos coeficientes de variação genotípicos, residuais e da razão entre os coeficientes
de variação genotípicos e residuais.
Todas as análises foram realizadas utilizando-se o Software Selegen – REML/BLUP
(Sistema Estatístico e Seleção Genética Computadorizada via Modelos Lineares Mistos)
(RESENDE, 2007).
3 . RESULTADOS E DISCUSSÃO
A partir da análise dos dados, foi observado que as características taxa de alongamento
de folha (TALF), comprimento final da lâmina foliar (CFL) e relação lâmina:colmo (RLC)
tiveram o componente variância genética (Vg) significativo, enquanto a característica número
médio de perfilhos (NMP) e comprimento final da lâmina (CFL) apresentaram diferença
significativas para o componente variância de parcela (Vparc) (Tabela 1). As demais
características avaliadas tiveram componentes de variância estatisticamente iguais a zero
(Tabela 1). Braz et al. (2013), em trabalho sobre parâmetros genéticos para caracteres
agronômicos em híbridos de P. maximum, observaram variância genotípica significativa para

54
todas as características de produção (massa verde total, massa seca total, massa seca de folha e
massa seca de colmo) e para os caracteres morfoagronômicos (altura e rebrota), a única
característica que não apresentou variância genotípica significativa foi porcentagem de folha.
Em um trabalho semelhante, Figueiredo et al. (2012) estimaram os parâmetros genéticos para
características agronômicas de B. humidicola, e também constataram variância genotípica
significativa para todos os caracteres avaliados (massa seca total, porcentagem de folha, massa
seca de folha e relação lâmina:colmo). Segundo estes autores a significância da variância
genética permite inferir sobre a existência da variabilidade genética para os caracteres
avaliados, demonstrando a possibilidade de ganho com a seleção, sendo, então, uma variação
importante para o melhoramento genético.
Tabela 1: Parâmetros genéticos das características morfogênicas e estruturais, taxa de
alongamento de colmo (TALC), taxa de alongamento de folha (TALF), taxa de aparecimento
de folha (TAPF), duração de vida da folha (DVF), filocrono (FIL), taxa de senescência foliar
(TSEF), comprimento final da lâmina (CFL), relação lâmina:colmo (RLC) e número médio de
perfilhos (NMP) em genótipos de P. maximum
Variável Parâmetros
Vg Vparc Média CVg CVe CVg/CVe
TALC 0,0009 ns 0,0004 ns 0,2099 14,5351 9,2616 1,5694
TALF 0,1946** 0,0825 ns 2,9862 14,7731 9,619 1,5358

55
TAPF 0,00008 ns 0,0001 ns 0,1042 2,3514 8,3134 0,2828
FIL 0,0130 ns 0,0663 ns 12,5761 0,9061 2,0472 0,4426
NFV 0,0438 ns 0,0034 ns 4,5172 4,6345 1,2832 3,6116
DVF 0,8821 ns 2,5891 ns 53,738 1,7477 2,9943 0,5837
TSF 0,0002 ns 0,0016 ns 0,2679 5,6231 14,8304 0,3792
CFL 17,2669* 18,1058** 36,2529 11,4621 11,7373 0,9766
RLC 79,3587** 1,5824 ns 15,7153 56,6858 8,0046 7,0816
NMP 87,1709 ns 258,4301** 47,0735 19,8339 34,1503 0,5808
Vg: variância genética; Vparc: variância da parcela; CVg: coeficiente de variação genético; CVe: coeficiente de
variação residual; CVg/CVe: razão entre o coeficiente de variação genético e residual; **, * significativo a 1 e 5%
de probabilidade pelo teste qui-quadrado, respectivamente, ns não significativo.
As características taxa de alongamento de colmo (TALC), e número de folhas vivas
(NFV) não apresentaram diferenças significativas, porém, apresentaram coeficientes de
variação genotípicos (CVg) superiores aos coeficientes de variação residual ou ambiental (CVe)
(Tabela 1). Altos CVg indicam a possibilidade de seleção, uma vez que a variação observada
nessas características é de natureza genética.
Coeficientes de variação residual ou ambiental superiores ao CVg foram observados
para as características taxa de aparecimento de folha (TAPF), filocrono (FIL), duração de vida
das folhas (DVF) (Tabela 1). De certa forma estas características estão interligadas, de modo
que o comportamento de uma pode interferir na resposta da outra, a medida que diminui a TAPF
e aumenta a DVF, a influência do ambiente pode aumentar, já que a temperatura,
disponibilidade de luz, água e nutrientes interferem consideravelmente a taxa metabólica da
planta, que determinará se a duração das folhas será maior ou menor.
Devido aos altos CVe observados para as características TAPF, FIL, DVF e TSF, houve
redução considerável na razão CVg/CVe, que apresentou valores abaixo da unidade, sendo
0,2828; 0,4426; 0,5837; 0,3792, respectivamente (Tabela 1). Baixa razão CVg/CVe também
foi observada para massa seca de colmo em um trabalho realizado por Braz et al. (2013), no
qual o autor ressalta que a ocorrência da razão CVg/CVe abaixo da unidade pode diminuir a
possibilidade de observação de diferenças significativas entre os genótipos avaliados, sendo,
assim, prejudicial ao processo de seleção. Portanto, altos valores de CVe seguido de reduzido
valor CVg demonstram a necessidade de maior controle ambiental visando minimizar a
influência do meio no processo de discriminação dos genótipos (DAHER et al.,2004).
Alta razão CVg/CVe foram observadas para as características RLC, NFV, TALC,
TALF, CFL, sendo os maiores coeficientes foram registrados para RLC, com a razão de 7,0816

56
(Tabela 1), valores iguais ou superiores a 1 indicam que o coeficiente de variação genético
supera o residual ou ambiental, ressaltando o potencial destas características serem melhoradas.
Braz (2012) observou elevado CVe e baixo CVg para RLC em híbridos de P. maximum e
atribuiu os resultados ao efeito do estágio fenológico no desenvolvimento do dossel de
forrageiras.
Daher et al. (2004), com objetivo de obter os parâmetros genéticos do número de
perfilhos por metro linear de clone de capim-elefante em 12 cortes, observaram que a
característica em estudo apresentou alto CVe (38,7505), mas também foi acompanhado de
elevado CVg (15,7470), reduzindo assim a magnitude do efeito ambiental que atua sobre os
genótipos, com valor de 0,4063 para a razão CVg/CVe. Resultados semelhantes foram
observados neste trabalho, no qual, o número médio de perfilhos apresentou alto CVe e CVg,
com valores de 34,1531 e 19,8339, respectivamente, fazendo com que a razão VCg/VCe
apresentasse valor de 0,5808 (Tabela 1).
Após o estudo e caracterização genética das variáveis morfogênicas e estruturais,
realizou-se a análise de agrupamento, de acordo com a metodologia de otimização de Tocher.
Após o agrupamento, constatou-se a formação de cinco grupos, sendo os mesmos constituídos
por G1- (217, A78, B11, B46, B53, B55, B57, C12, G56, K214, K5, T104, T97), G2- (309),
G3- (C53), G4- (T110) e G5- (H64) (Tabela 2). Vale destacar, que o agrupamento tem a
finalidade de reunir indivíduos com base em características afins, de forma a obter
homogeneidade dentro e heterogeneidade entre os grupos (CRUZ et al., 2004). Sendo que o
método de agrupamento por otimização de Tocher possui a particularidade de apresentar a
distância média dentro dos grupos sempre menor que a distância média entre os grupos
(VASCONCELOS et al., 2007).
Os grupos que apresentaram maiores valores de alongamento de folha (TALF) foram
os grupos 3 (3,2257), 5 (3,1249) e 1 (3,0141), com valores superiores à média geral de todos os
genótipos avaliados. Enquanto o grupo 4 obteve menor desempenho para esta característica
(2,2868) (Tabela 2). Costa et al. (2016), avaliando o efeito de diferentes doses de nitrogênio
sobre as características morfogênicas de Panicum maximum cv. Massai em casa-de-vegetação,
encontraram resultados próximos aos grupos que apresentaram alta TALF, quando as doses de
nitrogênio aplicadas foram de 80, 120, 160 mg/kg de solo, com valores de 3,90, 4,38 e 4,47,
respectivamente, as quais não diferiram estatisticamente. Enquanto o menor valor observado
para o grupo 4, foi semelhante nas doses 0 (2,17) e 40 mg/kg de solo (2,55) neste mesmo
trabalho.
57
Tabela 2: Análise de agrupamento de acordo com a metodologia de otimização de Tocher com base na distância de Mahalanobis genéticas de
características morfogênicas estruturais, taxa de alongamento de colmo (TALC), taxa de alongamento de folha (TALF), taxa de aparecimento de
folha (TAPF), duração de vida da folha (DVF), filocrono (FIL), taxa de senescência foliar (TSEF), comprimento final da lâmina (CFL), relação
lâmina:colmo (RLC) e número médio de perfilhos (NMP) em genótipos de P. maximum
Grupo Genótipos TALC TALF TAPF FIL NFV DVF TSEF CFL RFC NMP
1 Maioria 0,2086 3,0141 0,1041 12,5763 4,5452 53,7386 0,2671 36,2685 14,5958 45,6151
2 309 0,2310 2,9452 0,1041 12,5756 4,6763 53,9453 0,2699 36,2827 7,5680 46,3249
3 C53 0,2078 3,2257 0,1041 12,5753 4,3983 53,7053 0,2718 41,1198 19,7889 51,9942
4 T110 0,2020 2,2868 0,1039 12,5906 4,2871 53,7681 0,2733 30,8078 38,9125 63,0436
5 H64 0,2162 3,1249 0,1052 12,5600 4,3427 53,5254 0,2681 36,5984 11,1462 45,8910
Média geral 0,2099 2,9862 0,1042 12,5761 4,5172 53,7380 0,2679 36,2529 15,7153 47,0735
Maioria: 217, A78, B11, B46, B53, B55, B57, C12, G56, K214, K5, T104, T97

58
A TALF é uma característica morfogênica que está diretamente relacionada com a
produção de forragem, pois, à medida que se aumenta a TALF há incremento na proporção de
folhas e o aumento da área foliar fotossinteticamente ativa (MARTUSCELLO et al., 2015).
Portanto, os grupos 3, 5 e 1 que apresentaram altos valores para TALF indicam grande potencial
para o aumento na produção de forragem de qualidade, sendo uma característica de grande
importância para seleção e melhoramento de P. maximum.
Para taxa de alongamento de colmo (TALC) o grupo 2, composto pelo genótipo 309, se
destacou apresentando o maior valor observado para esta característica (0,2310), seguido pelo
grupo 5 (0,2162). Os demais grupos apresentaram valores muito próximos (Tabela 2). Em um
trabalho realizado por Resende Júnior (2011), onde foram avaliadas características
morfogênicas e estruturais de P. maximum cv. Tanzânia submetido a distintos níveis de
desfolhação, constatou que quanto maior a severidade de desfolhação maior foi a taxa de
alongamento de colmo.
A proximidade entre os grupos para TALC se deve à baixa variância genotípica
observada para esta característica, o que pode ser explicado pelo fato destes genótipos terem
sido pré-selecionados e fazerem parte de um grupo de plantas superiores que compõem ensaios
regionais. Provavelmente, isso reduziu a variabilidade genética de determinadas características.
A TALC é uma característica de baixo interesse para o melhoramento genético, pois,
no colmo concentra-se a maior parte de compostos lignificados da planta, pois diminui a
digestibilidade da forragem consumida (LEMPP,2007), além de prejudicar a estrutura do pasto
(Sbrissia e Da Silva, 2001). O P. maximum é caracterizado por apresentar alta TALC, por isso
a avaliação dessa característica é importante, pois um dos principais problemas relacionados ao
manejo, é o aumento da participação do colmo na produção, que além de reduzir o valor
nutricional, pode prejudicar a colheita de forragem (pastejo) pelo animal (SANTOS, 2002).
Outra alternativa que pode ser dada a avaliação do alongamento de colmos em plantas de P.
maximum é a seleção negativa, com intuito de reduzir. Nesse sentido, a busca de genótipos ou
grupos com menor TALC demandaria populações com maior variabilidade para esta
característica.
A relação entre estas duas características (TALF e TALC) influencia diretamente a
relação lâmina:colmo (RLC). De acordo com Sbrissia & Da Silva (2001) altos valores para
TALC podem até melhorar o rendimento forrageiro, contudo reduzem a relação lâmina:colmo,
interferindo significativamente na estrutura do pasto e prejudicando a eficiência de pastejo. De
fato, observou-se que em alguns grupos a TALC influenciou consideravelmente na RLC. O
grupo 2 (genótipo 309) apresentou a maior TALC e valor inferior aos demais grupos para

59
TALF, o que se refletiu diretamente na baixa RLC, proporcionando resultado muito inferior
(7,5680) ao grupo 3, onde a relação lâmina:colmo foi de 19,7889 e as taxas de alongamento de
colmo e folha apresentadas foram inversas às observadas no grupo 2 (Tabela 2).
Dentre todos os grupos, a maior RLC constatada (38,9125) foi para o grupo 4 (T110),
ainda que as taxas de alongamento de colmo e de folha tenham sido baixas, o alto valor para
RLC pode ser explicado pelo maior alongamento de folha. O contrário pode ser observado para
o grupo 5 (H64), no qual as taxas alongamento de colmo e folha foram maiores, porém, a RLC
foi baixa (11,1462) (Tabela 2). A RLC é determinada pelo quociente da massa seca de folha
pela massa seca de colmo, assim, os resultados observados podem ser explicados pela variação
do peso dos colmos dos genótipos avaliados. As variações na RLC também podem estar
associadas ao maior número médio de perfilhos, como foi observado para as plantas de T110.
Uma das características que contribuiu para a diferenciação do grupo 3 foi o
comprimento final da lâmina (CFL), o qual apresentou valor superior aos demais grupos
(Tabela 2). A alta TALF também observada para este grupo, associada ao maior CFL, evidencia
que o grupo tem grande potencial para aumento de produção de folhas, pois quanto maior a área
foliar maior será a capacidade fotossintética da planta e também há aumento da fração de maior
interesse na alimentação de ruminantes, que é a folha.
Assim como o CFL da lâmina contribuiu para diferenciação do grupo 3, o número médio
de perfilhos (NMP) foi fundamental para a separação do grupo 4 (Tabela 2). O NMP pode ser
considerado uma característica com grande potencial para ser inserida nas avaliações do
melhoramento genético de forrageiras, uma vez que a partir da densidade populacional de
perfilhos é possível se estimar o potencial produtivo da forrageira (HODGSON, 1990;
HOESCHL et al., 2007). Ademais, a quantidade de perfilhos existente na pastagem está
diretamente relacionada com a persistência da mesma, sendo que a durabilidade da pastagem
depende da capacidade da planta em emitir novos perfilhos e substituir os que são perdidos no
processo de pastejo ou que morrem durante o florescimento ou entram em senescência. Porém,
deve-se levar em consideração o manejo adotado para plantas com alta capacidade de
perfilhamento, haja vista que maior quantidade de plantas por unidade de área provavelmente
resultara em competição por luz, causando alongamento de colmo e aumentando a taxa de
senescência foliar, além de diminuir a duração de vida das folhas (BRAZ et al., 2011). De fato,
este comportamento foi observado para este grupo, o qual apresentou TALC e taxa de
senescência foliar (TSF) superior aos demais (Tabela 2).
Para as características, taxa de aparecimento de folha (TAPF), filocrono (FIL), número
de folhas viva (NFV), duração de vida da folha (DVF), os valores observados não apresentaram

60
diferenças relevantes entre os grupos formados (Tabela 2). Provavelmente, a baixa variação
observada entre grupos ocorreu porque a influência do ambiente se destacou em relação à
genética, portanto são características com baixo potencial de estudo e inserção no
melhoramento. Contudo, destaca-se que a TAPF, FIL e DVF apresentaram baixa razão
CVg/CVe, o que indica maior influência do ambiente e que a resposta via estratégias de manejo
pode ser otimizada.

61
4 . CONSIDERAÇÕES FINAIS
As características morfogênicas taxa de alongamento de folha, comprimento final da
lâmina e relação lâmina:colmo apresentam variabilidade genética significativa entre os
genótipos de Panicum maximum.
Taxa de alongamento de folha e colmo, e número de folhas vivas e relação
lâmina:colmo possuem alto coeficiente de variação genotípico, apontando possibilidade de
seleção.
As características taxa de aparecimento de folha, filocrono, duração de vida das folhas
e taxa de senescência foliar apresentam baixa razão entre coeficiente de variação genotípico e
ambiental, evidenciando necessidade de maior controle ambiental.
Os genótipos de P.maximum formam 5 grupos morfofuncionais distintos. Sendo que os
grupos 3, 5 e 1, apresentam altas taxas de alongamento de folha como mecanismo de acúmulo
de forragem. Já o grupo 4 se destaca pela capacidade de perfilhamento.
.

62
5 . REFERENCIAS BIBLIOGRÁFICAS
ALEXANDRINO, A.; CANDIDO, M. J. D.; GOMIDE, J. A. Fluxo de biomassa e taxa de
acúmulo de forragem em capim Mombaça mantido sob diferentes alturas. Revista Brasileira
de Saúde e Produção Animal, v.12, n.1, p. 59-71, 2011.
BRAZ, T. G. S. Parâmetros genéticos, repetibilidade e diversidade genética em híbridos
de Panicum maximum, 2012. p. 117. Tese (Doutorado Zootecnia) – Universidade Federal de
viçosa, Viçosa, 2012.
BRAZ, T. G. S.; FONSECA, D. M.; FREITAS, F. P.; MARTUSCELLO, J. A.; SANTOS, M.
E. R.; SANTOS, M. V.; PEREIRA, V. V. Morphogenesis of Tanzania guinea grass under
nitrogen doses and plant densities. Revista Brasileira de Zootecnia, v.40, n. 7, p.1420-1427,
2011.
BRAZ, T. G. S.; FONSECA, D. M.; JANK, L.; RESENDE, D. V.; MARTUSCELLO, J. A.;
SIMEÃO, R. M. Genetic parameters of agronomic characters in Panicum maximum hybrid.
Revista Brasileira de Zootecnia, v. 42, n. 4, p. 231-237, 2013.
CANTARUTTI, R. B.; ALVARES V. V.H.; RIBEIRO, A. C. Amostragem do solo. In:
RIBEIRO, A.C.; GUIMARAES, P.T.G.; ALVAREZ V., V.H. (Ed.). Recomendação para o
uso de corretivos e fertilizantes em Minas Gerais: 5º Aproximação. Viçosa: Comissão de
Fertilidade do Solo do Estado de Minas Gerais. 1999, p. 13-20.
CASAGRANDE, D. R.; RUGGIERI, A. C.; JANUSCKIEWICZ, A. R.; GOMIDE, J. A.;
REIS, R. A.; VALENTE, A. L. S. Características morfogênicas e estruturais do capim-
marandu manejado sob pastejo intermitente com diferentes ofertas de forragem. Revista
Brasileira de Zootecnia, v.39, n.10, p.2108-2115, 2010.
CHAPMAN, D.F.; LEMAIRE, G. Morphogenetic and structural determinants of plant
regrowth after defoliation. In: BAKER, M.J. (Ed.) Grasslands for our world.
Wellington:SIR Publishing, 1993. p.55-64.

63
COSTA, N. L.; PAULINO, V. T.; MAGALHÃES, J. A.; RODRIGUES, B. H. N.; SANTOS,
F. J. S. Eficiência do nitrogênio, produção de forragem e morfogênese do capim-massai sob
adubação. Nucleus, v. 13, n. 2, 2016.
CRUZ, C.D.; REGAZZI, A. J.; CARNEIRO, P. C. S. Modelos biométricos aplicados ao
melhoramento genético. v. 1. 3 ed. Viçosa: UFV. 2004. 480p.
DAHER, R. F.; MALDONADO, H.; PEREIRA, A. V.; AMARAL JÚNIOR, A.T.;
PEREIRA, M. G.; FERREIRA, C. F.; RAMOS, S. R. R.; TARDIN, F. D.; SILVA, M. P.
Estimativas de parâmetros genéticos e de coeficientes de repetibilidade de caracteres
forrageiros em clones de capim-elefante (Pennisetum purpureum Schum.). Acta Scientiarum
Agronomy, v. 26, n. 4, p. 483-490, 2004.
FIGUEIREDO, U. J.; NUNES, J. A. R.; VALLE, C. B. Estimation of genetic parameters and
selection of Brachiaria humidicola progenies using a selection index. Crop breeding and
Applied Biotechnology, v. 12, p. 237-244, 2012.
HODGSON, J. Grazing management – Science into practice. New York: John Wiley &
Sons, Inc., Longman Scientific & Technical. 1990. 203p.
HOESCHL, A. L.; CANTO, M. W.; BONA FILHO, A. et al. Produção de forragem e
perfilhamento em pastos de capim Tanzânia-1 adubados com doses de nitrogênio. Scientia
Agraria, v. 8, n. 1, p. 81-86, 2007.
JANK, L. Melhoramento e seleção de variedade de Panicum maximum. Im. SIMPOSIO
SOBRE MANEJO DA PASTAGEM, 12, Piracicaba, 1995, Anais... Piracicaba: FEALQ,
1995.p. 21-58.
JANK, L.; VALLE, C. B.; RESENDE, R. M. S. Breeding tropical forages. Crop breeding
and Applied Biotechnology. P. 27-34, 2011.
KÖPPEN, W. Climatologia: com um estúdio de los climas de la tierra. New Gersey: 1948.

64
LEMPP, B. Avanços metodológicos da microscopia na avaliação de alimentos. Revista
Brasileira de Zootecnia, v. 36, p.316-329, 2007.
MACEDO, C. H. O.; ALEXANDRINO, E.; KAKELAITS, A.; VAZ, R. G. M. V.; REIS, R.
H. P.; VENDRUSCULO, J. Características agronômicas, morfogênicas e estruturais do capim
Panicum maximum cv. Mombaça sob desfolhação intermitente. Revista Brasileira de Saúde
e Produção animal, v. 11, n. 4, p. 941-952, 2010.
MARTUSCELLO, J. A.; SILVA, L. P.; CUNHA, D. N. F. V.; BATISTA, A. C. S.; BRAZ, T.
G. S.; FERREIRA, P. S. Adubação nitrogenada em capim-massai: morfogênese e produção.
Ciência Animal Brasileira, v. 16, n. 1, 2015.
MINGOTTI, S.A. Análise de Dados Através de Métodos de Estatística Multivariada:
Uma Abordagem Aplicada. Belo Horizonte: Editora UFMG, 2005.
RESENDE JÚNIOR, A. J. Morfogênese, acúmulo de forragem e teores de nutrientes de
Panicum maximum cv. Tanzânia submetidos a diferentes severidades de desfolhação e
fertilidades contrastantes. 2011. p. 104. Dissertação (Ciência Animal e Pastagem) - Escola
Superior de Agricultura “Luiz de Queiroz”, Piracicaba, 2011.
RESENDE, M. D. V. Matemática e estatística na análise de experimentos e no
melhoramento genético. Colombo: Embrapa Florestas, 2007.
SANTOS, P. M. Controle do desenvolvimento das hastes de capim Tanzânia: um desafio.
2002. p. 112. Tese (Doutorado Agronomia) – Escola Superior de Agricultura “Luiz de
Queiroz”, Piracicaba, 2002.
SBRISSIA, A.F., Da SILVA, S.C. O ecossistema de pastagens e a produção animal In:
REUNIÃO ANUAL DA SOCIDADE BRASILEIRA DE ZOOTECNIA, 38, Piracicaba,
2001. Anais... Piracicaba: SBZ, 2001, p.731-754.
VALLE, C. B.; JANK, L.; RESENDE, R. M. S. O melhoramento de forrageiras tropicais no
Brasil. Revista Ceres. V. 56, n. 4, p. 460-472, 2009.

65
VASCONCELOS, E. D.; CRUZ, C. D.; BHERING, L. L.; RESENDE JÚNIOR, M. R.
Método alternativo para análise de agrupamento. Pesquisa Agropecuária Brasileira, v. 42,
n. 10, p. 1421-1428, 2007.





























