Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DO PARÁ
INSTITUTO DE CIÊNCIAS BIOLÓGICAS
PROGRAMA DE PÓS – GRADUAÇÃO EM BIOLOGIA DE AGENTES
INFECCIOSOS E PARASITÁRIOS
ANÁLISE MORFOLÓGICA, MOLECULAR, HISTOPATOLÓGICA
E PROTEÔMICA DA RELAÇÃO PARASITO-HOSPEDEIRO
Ortleppascaris sp. - Rhinella marina
JEFFERSON PEREIRA E SILVA
BELÉM – Pará
2013

JEFFERSON PEREIRA E SILVA
ANÁLISE MORFOLÓGICA, MOLECULAR, HISTOPATOLÓGICA
E PROTEÔMICA DA RELAÇÃO PARASITO-HOSPEDEIRO
Ortleppascaris sp. - Rhinella marina
Orientadora: Profa. Dra. Jeannie Nascimento dos Santos
Co-Orientador: Prof. Dr. Cláudio Lísias Mafra de Siqueira
BELÉM – Pará
2013
Dissertação apresentada para o Programa de Pós-
Graduação em Biologia de Agentes Infecciosos e
Parasitários do Instituto de Ciências Biológicas da
Universidade Federal do Pará como requisito para a
obtenção do grau de Mestre em Biologia de
Agentes Infecciosos e Parasitários.
Orientadora: Profa. Dra. Jeannie Nascimento dos
Santos.

Dados Internacionais de Catalogação-na-Publicação (CIP) Sistema de Bibliotecas
da UFPA
Silva, Jefferson Pereira e, 1989-
Análise morfológica, histopatológica, molecular e proteômica da relação
parasito-hospedeiro Ortleppascaris sp.-Rhinella marina / Jefferson Pereira e Silva.
- 2013.
Orientadora: Jeannie Nascimento dos Santos;
Coorientador: Cláudio Lísias Mafra de Siqueira.
Dissertação (Mestrado) - Universidade Federal do Pará, Instituto de
Ciências Biológicas, Programa de Pós-Graduação em Biologia de Agentes
Infecciosos e Parasitários, Belém, 2013.
1. Helminto. 2. Nematoda. 3. Relação hospedeiro-parasito. 4. Anuro. 5. Anfíbio. I.
Título.
CDD 22. ed. 592.57

1
JEFFERSON PEREIRA E SILVA
ANÁLISE MORFOLÓGICA, MOLECULAR, HISTOPATOLÓGICA E
PROTEÔMICA DA RELAÇÃO PARASITO-HOSPEDEIRO
Ortleppascaris sp. - Rhinella marina
Dissertação apresentada ao Programa de Pós-Graduação em Biologia de Agentes
Infecciosos e Parasitários, do Instituto de Ciências Biológicas da Universidade Federal
do Pará, como requisito para a obtenção do grau de Mestre em Biologia de Agentes
Infecciosos e Parasitários.
Orientadora: Profa. Dra. Jeannie Nascimento dos Santos (Orientadora)
Laboratório de Biologia Celular e Helmintologia Dra Reinalda
Marisa Lanfredi, ICB, UFPA
Co-orientador: Prof. Dr. Cláudio Lísias Mafra de Siqueira
Laboratório de Parasitologia e Epidemiologia Molecular, UFV
Banca examinadora: Prof. Dr Evonnildo Gonçalves (Titular)
Laboratório de Tecnologia Biomolecular, ICB, UFPA
Prof. Dr Marcio Roberto Teixeira Nunes (Titular)
Centro de Inovações Tecnológicas, IEC
Prof. Dr Leonardo dos Santos Sena (Titular)
Instituto de Ciências Biológicas, UFPA
Prof. Dr Adriano Penha Furtado (Suplente)
LBCH Dra Reinalda Marisa Lanfredi, ICB, UFPA
Data da Defesa: ____/_____/______

2
DEDICATÓRIA
Dedico de mais sincero coração este trabalho e esforço à minha mãe, Maria
Ivoneide Barbosa Pereira, que por sua generosidade e amor esquece os seus próprios
méritos empenhados na minha felicidade.

3
AGRADECIMENTOS
À Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES),
PROCAD-NOVAS FRONTEIRAS E PROPESP/FADESP, pelo auxílio financeiro para
a realização deste trabalho.
Ao Laboratório de Bioquímica e Biologia Molecular de Agentes Infecciosos e
Parasitários do Departamento de Bioquímica e Biologia Molecular da Universidade
Federal de Viçosa, com o apoio do Núcleo de Análise de Biomoléculas (NUBIOMOL)
do Departamento de Bioquímica e Biologia Molecular da Universidade Federal de
Viçosa, pela infraestrturura.
Ao Professor Humberto Ramos pelas sugestões, amizade, incentivo e pelo
exemplo de competência e simplicidade em ajudar.
Aos amigos da Casa 21: Rose, Carla, Leandro, Ronnie pela agradável
convivência.
Às amigas queridas Ana Clara (irmã) e Fernanda, sem as quais não seria
possível a realização deste trabalho. Gostaria de registrar minha admiração, amizade e
meu infinito obrigado a vocês duas de modo especial.
À República “Doidimais”: Carol, Karen, Camila. Agregados: Rafael, Bruno,
Claudinha, Larissa e Flávia por terem feito dos meus dias em Viçosa dias inesquecíveis.
Aos melhores amigos que eu poderia ter: Lorena, Lázaro, Loriane, Tamyris,
Flávia e Rose, que sempre fazem da minha vida mais leve e feliz.
Adriano Furtado, que não poupou esforços em me apoiar durante toda a
realização desta dissertação, devo minha gratidão.
À Sandra e Dante, por terem sido uma família para mim.
À minha família do LBCH, em especial Elane, pelos conselhos. Djane e Tiago
pela grande amizade e companheirismo construídos.
Minha família, pelo apoio incondicional, em particular minha avó (mãezinha) e
Maria pelos ensinametos que só a sabedoria de um coração puro pode trazer.
À minha orientadora e amiga, Jeannie, por me permitir crescer como profissional
sem nunca esquecer de mim como pessoa, compreendendo minhas grandes limitações e
me fazendo superá-las. Este agradecimento é muito pouco frente a dimensão do que
sempre fez por mim. Meu carinho e admiração.
Por fim, a todos aqueles que se fizeram presentes, mesmo em pensamento, e
contribuíram para realização deste trabalho devo minha gratidão.

4
SUMÁRIO
RESUMO .............................................................................................................. 5
ABSTRACT ......................................................................................................... 6
1. INTRODUÇÃO ............................................................................................. 7
1.1. O PARASITO Ortleppascaris sp. .............................................................. 7
1.2. O HOSPEDEIRO Rhinella marina (Linnaeus, 1758) ............................... 7
1.3. HISTOPATOLOGIA HEPÁTICA E HELMINTOLOGIA ................... 8
1.4. A BIOLOGIA MOLECULAR E HELMINTOLOGIA ........................... 10
1.5. PROTEÔMICA E SECRETOMA ........................................................... 12
1.6. ANÁLISE DE PRODUTOS HELMÍNTICOS ........................................ 14
1.7. HELMINTOS E IMUNOMODULAÇÃO ............................................... 15
1.8. POTENCIAIS MOLÉCULAS NA BIOLOGIA E PATOGENICIDADE
DOS HELMINTOS .................................................................................................... 18
1.9. Ortleppascaris x Rhinella marina: MODELO PARA ESTUDO DE
RELAÇÃO HELMINTO-HOSPEDEIRO ................................................................. 18
1.10. OBJETIVOS .......................................................................................... 20
1.10.1 Objetivo Geral: ................................................................................. 20
1.10.2. Objetivos Específicos: ..................................................................... 20
2. ARTIGOS..................................................................................................... 21
2.1. ARTIGO 1 ................................................................................................ 21
2.2 ARTIGO 2 ................................................................................................. 30
2.3. ARTIGO 3 ................................................................................................ 39
2.4. ARTIGO 4 ................................................................................................ 72
3. CONCLUSÃO .............................................................................................. 102
3.1 TAXONOMIA E BIOLOGIA MOLECULAR DE Ortleppascaris sp. . 102
3.2. HISTOPATOLOGIA DA INTERAÇÃO Ortleppascaris sp. x R. marina
.................................................................................................................................. 102
3.3. ANÁLISE PROTEÔMICA DA INTERAÇÃO Ortleppascaris sp. x R.
marina ....................................................................................................................... 103
REFERÊNCIAS BIBLIOGRÁFICAS ............................................................. 105

5
RESUMO
Helmintos são organismos complexos, que podem causar infecções em seres
humanos e animais, as quais são frequentemente negligenciadas. Contudo, estudos
sobre a morfologia e biologia de helmintos parasitos são necessários para o melhor
entendimento das relações parasito-hospedeiro. Neste trabalho é descrita a ocorrência
do nematódeo Ortleppascaris sp., parasito encontrado em fígado de Rhinella marina,
abordando aspectos morfológicos, genéticos, histológicos e proteômicos visando o
entendimento da relação desenvolvida pelo anfíbio hospedeiro e o helminto parasito. A
metodologia utilizada inclui taxonomia alfa, análise ultraestrutural e biologia molecular
para fornecer dados taxonômicos sobre o nematódeo e contribuir para sua sistemática.
Análises histopatológicas identificaram as alterações morfo-fisiológicas no órgão-alvo a
partir da infecção e o impacto causado pelo parasito no hospedeiro bem como a reação
deste frente ao invasor. A análise proteômica, através de eletroforese bi-dimensional e
espectrometria de massas, foi realizada a fim de identificar derivados helmínticos
constituintes de Ortleppascaris sp. além de outras moléculas envolvidas nesta complexa
relação parasito-hospedeiro, permitindo a triagem de marcadores proteicos relativos a
este modelo de estudo. Esta pesquisa adiciona portanto dados sobre a morfologia, status
filogenético e distribuição geográfica de Ortleppascaris sp., permite a avaliação da
dinâmica do processo infeccioso causado pelo helminto e aponta possíveis moléculas
importantes para o campo de estudo de produtos e biomarcadores helmínticos de
interesse humano e veterinário.

6
ABSTRACT
Helminths are complex organisms, that can be responsible for human and animal
infections, which are frequently neglected. However, studies about morphology and
biology of helminthes parasites on are necessary for a better understanding of thehost-
parasite relationship.. The present work describes the occurrence of nematode
Ortleppascaris sp. parasiting the liver of Rhinella marina using morphological, genetic,
proteomic and histological approaches to understand the relationship developed by
amphibian host and helminth parasite. The methodology include alpha taxonomy,
molecular biology and ultrastructural analysis were published, discussingto provide
data about the nematode taxonomy contributing for the systematic of this group.
Histopathology studies, identified the morpho-physiological alterations in the infected
organ assessing the impact caused by the parasite in the host as well as the reactions of
the parasite. Proteomic approaches, using two-dimensional electrophoresis and mass
spectrometry, were performed to identify derivatives constituents helmintics of
Ortleppascaris sp. and other molecules involved in this complex relationship between
parasite-host, allowing the screening of protein markers related to these helminthes. The
present work added new data on the morphology, geographical distribution and
phylogenetic status of the Ortleppascaris sp., allows the evaluation of the dynamics of
infection caused by those helminthes and shows important molecules for the field of
study of anthelmintic products and biomarkers of human and veterinary interest.

7
1. INTRODUÇÃO
1.1. O PARASITO Ortleppascaris sp.
Os nematódeos do gênero Ortleppascaris Sprent, 1978 foram registrados, em
sua forma larval, parasitando fígados de anfíbios como Eleutherodactylus cf.
terraebolivaris, em Tobago por Moravec e Kaiser (1995); Leptodactylus bufonius
Boulenger, 1894, na Argentina, por Gonzáles e Hamann (2006) e Chaunus fernandezae
Gallardo, 1957 por Gonzáles e Hamann (2007) no mesmo país.
Sua forma adulta é encontrada no trato gastrintestinal de animais da Ordem
Crocodylia (Goldberg et al., 1991), tais como Caiman crocodilus (no Brasil), Caiman
sclerops e Crocodylus intermedius (na Venezuela), Crocodylus rhombifer, Caiman
yacare, Alligator mississippiensis, distribuídos pela América do Sul, Cuba, e no sul dos
Estados Unidos (Sprent, 1978; Catto, 1991; Moravec & Kaiser, 1995; Gonzáles &
Hamann, 2007; Wadle et al., 2009).
Entretanto, apesar destes parasitos terem sido registrados em fígados de anfíbios,
o ciclo de vida e modo de infecção desse gênero é ainda desconhecido, pois vale
destacar que anfíbios não são parte significante da dieta de crocodilianos (Sprent, 1978;
Wadle et al., 2009).
As características morfológicas mais relevantes para definição taxonômica de
larvas no gênero Ortleppascaris, são: posição das aletas laterais, as quais se estendem
desde as proximidades do anel nervoso até a região do ânus; o esôfago estreito,
consideravelmente longo; a presença de ventrículo e de ceco intestinal. São também
considerados importantes para a Taxonomia de larvas os dados do comprimento e
largura do corpo do helminto, posição do poro excretor, posição do anel nervoso,
comprimento e largura de esôfago, do ventrículo, do ceco intestinal e comprimento da
porção posterior de acordo com Moravec & Kaiser (1995) e Gonzáles & Hamann (2006,
2007).
1.2. O HOSPEDEIRO Rhinella marina (Linnaeus, 1758)
Os anfíbios constituem uma classe de animais vertebrados, pecilotérmicos e que
não possuem bolsa amniótica, cuja característica mais marcante é o ciclo de vida
dividido em duas fases: uma aquática e outra terrestre (Young et al., 2004).

8
Rhinella marina (Fig. 1) é um anfíbio comum no Brasil. Possuem porte variando
de 2 a 20 cm de comprimento; protuberância acima dos olhos até o focinho; uma grande
glândula paratóide atrás de cada olho; ausência de dentes e membrana interdigital nas
patas dianteiras; pele seca e verrugosa; pupilas horizontais e as íris douradas. A espécie
não apresenta pálpebra inferior, mas uma membrana nictitante (Frost et al., 2006).
São animais sinantrópicos, encontrados na América Central, América do Sul e
Sul do Texas (América do Norte), tendo sido introduzido em muitas regiões tropicais,
para o controle de pragas na agricultura, como por exemplo, na agricultura de cana-de-
açúcar (Zug & Zug, 1979; Brandt & Mazzotti, 2002; Ragoo & Omahmaharaj, 2003).
Assim, a partir da década de 90, o homem auxiliou a dispersão desta espécie de tal
forma que, hoje, podem ser encontrados em várias regiões do planeta e, em muitas
delas, é considerado como praga (Barton, 1997; 1999).
Figura 1: Rhinella marina. Fonte: Arquivo pessoal de Lilian Cristina Macedo (2012).
1.3. HISTOPATOLOGIA HEPÁTICA E HELMINTOLOGIA
A histopatologia caracteriza-se pelo estudo de tecidos ou de fragmentos de
tecidos obtidos através de necropsias/biopsias ou de outra técnica de colheita. Estas

9
amostras passam por processamento histológico que envolve fixação, desidratação,
clarificação, geralmente, por produtos químicos, com posterior infiltração em resinas,
microtomia e coloração para fins de visualização microscópicas de alterações da
estrutura tecidual e celular normais, causadas por agentes externos ou não (Horobin,
1982; Lemos et al., 2003).
Análises morfológicas em conjunto com estudos histológicos são abordagens
que se complementam e auxiliam no diagnóstico de parasitos teciduais além de
possibilitar o entendimento das alterações induzidas pelo parasito no hospedeiro.
De acordo com Eiras & Rego (1989), a repercussão do parasitismo sobre a
condição do hospedeiro pode ser vital e depende de vários fatores, tais como: a natureza
do parasito, a intensidade de infecção e o órgão afetado.
Segundo Rodrigues et al. (2002) helmintos, de maneira geral, podem induzir
respostas imunes com alterações patológicas nocivas, mas não letais, ao seu hospedeiro
e nos estudos sobre infecções por endoparasitos, observam-se várias alterações em
órgãos e tecidos dos hospedeiros.
Geralmente, os tecidos reagem encapsulando os parasitos por hiperplasia celular,
resultando em nódulos, que podem ser macroscópicos (Rodrigues et al., 2002) ou
resultando na formação de granulomas, frequentemente no fígado na esquistossomose
mansônica humana (Andrade, 2005) ou na capilaríase hepática, na qual os parasitos
podem produzir um quadro de hepatite aguda ou sub-aguda com forte eosinofilia e
ainda afetar outros órgãos (Orihuela et al., 2006).
Ratos infectados por Capillaria hepatica Bancroft, 1983, nematódeo zoonótico
de distribuição global presente no fígado, apresentam alterações histopatológicas de
bases imunológicas como, por exemplo, nódulos branco-amarelados contendo ovos e/ou
vermes, caracterizando granulomas, além de fibrose septal (Lemos et al., 2003).
A fibrose hepática é caracterizada pela presença de miofibroblastos e mastócitos;
já os granulomas são apenas focais com características de inflamação, rodeada por
tecido conjuntivo, macrófagos, eosinófilos, linfócitos e células plasmáticas, além de
outras alterações, como vermes e ovos circundados por fibras colágenas (Akiyoshi &
Terada, 1998; Jeong et al., 2008).
Barros (2008) também descreve alterações hepáticas em murinos desnutridos e
eutróficos submetidos à infecção experimental por Schistosoma mansoni Sambon, 1907.
Em alguns casos as alterações histopatológicas são geradas pelos estímulos de citocinas

10
do tipo Th2, em virtude da presença de ovos do parasito no órgão, causando, inclusive,
grande produção de imunoglobulinas.
Estes parasitos também induzem a formação de granulomas pequenos e esparsos
no parênquima hepático, com ausência de fibrose periportal no grupo desnutrido. Outro
tipo de lesão também se caracteriza por infiltração de células mononucleares e de alguns
linfócitos nos espaços-porta e em torno de veias centrolobulares, além de numerosos
granulomas em torno de ovos de S. mansoni degenerados ou isolando fragmentos de
cascas de ovos (Barros, 2008).
A maioria das lesões possuem predominantemente colágeno, às vezes exibindo
componente exsudativo, particularmente eosinófilos. Ocorre ainda a presença de
pigmento pardo-amarelado em aglomerados granulosos, sobretudo nos espaços-porta
mais calibrosos, além de vascularização aumentada, hiperplasia de ductos biliares e
infiltração inflamatória crônica (Barros, 2008).
Em fígado de Salminus maxillosus Valenciennes, 1840 (peixe conhecido como
“dourado”) foram observados os nematóides da espécie Neocucullanus neocucullanus
encerrados em cistos esbranquiçados, situados no parênquima hepático próximo à
superfície do órgão, produzindo infecção tanto nas regiões centrais, como periféricas,
sendo que algumas vezes, realiza perfuração do parênquima do órgão, permitindo uma
visão do cisto na superfície do órgão. Microscopicamente o tecido torna-se
desorganizado e com células típicas de processo inflamatório ativo (Rodrigues et al.,
2002).
1.4. A BIOLOGIA MOLECULAR E HELMINTOLOGIA
Biologia molecular é uma importante ferramenta na área da Parasitologia,
especificamente na Helmintologia, pois contribui com a identificação e a sistemática de
espécies, o diagnóstico de infecções, epidemiologia de parasitos, análises de estruturas
genéticas de uma população, expressão gênica, estudo de resistência a drogas e
desenvolvimento de vacinas (Gasser et al., 1999; Genchi et al., 2007),
Apesar de alguns entraves entre o método molecular e a taxonomia alfa, a
caracterização molecular de helmintos tem se mostrado uma ferramenta de extrema
importância para os estudos em parasitologia, pois além de auxiliar na identificação de
parasitas, pode ainda auxiliar no entendimento de questões ainda não elucidadas, como

11
o mecanismo de ação e resistência de alguns fármacos utilizados no tratamento destas
infecções (Genchi et al., 2007).
Atualmente, os estudos de descrições morfológicas de espécies de helmintos têm
incluído em seus resultados a descrição de uma ou mais sequências gênicas, com
informações que podem ser usadas em estudos sobre filogenia molecular como em
trabalhos desenvolvidos por Nadler et al. (2007), Furtado et al. (2009), Casiraghi et al.
(2006), Melo et al. (2011), Otranto et al., (2011).
Mitreva et al. (2005) realizou um estudo comparativo entre sequências genéticas
de nematódeos baseado em sequências do DNAr encontradas em bancos de dados e o
genoma sequenciado de Caenorhabtidis elegans. Neste trabalho, estes autores
agruparam o Filo Nematoda em cinco clados definidos, que podem ser distintos por
características morfológicas e especificidade parasitária.
Folmer et al. (1994), desenvolveram um par de iniciadores (LCO1490 e
HCO2198) amplificadores de uma região conservada de fragmento de DNA com 710
pb, do gene codificador da subunidade I da citocromo oxidase C (COI). Deste modo,
estes iniciadores puderam gerar análises filogenéticas contemplando estudos em
sistemática de organismos inclusos em 11 filos de invertebrados metazoários,
contabilizando mais de 80 espécies, dentre estas, espécies de nematódeos.
Mar et al. (2002) sugeriram os espaçadores transcritos 1 e 2 mais o gene 5.8S do
DNAr como região de escolha na identificação molecular de helmintos por preservar
similaridades dentro deste grupo de seres vivos, mas também diferenças específicas, as
quais podem ser levadas em consideração para estudos moleculares. De fato, esta região
vem sendo pesquisada por diversos autores que desenvolvem estudos filogenéticos por
seqüenciamento nucleotídico e, mais recentemente, por análise de sua estrutura
secundária em diversos seres vivos, como Dassanayake et al. (2008) e Miao et al.
(2008).
Outro aspecto importante da Biologia Molecular na Helmintologia no que tange
a identificação acurada da biodiversidade desses organismos é a proposta do “DNA
barcoding – código de barras”, caracterizado como um método de identificação, o qual
utiliza um pequeno marcador molecular genético como ferramenta para esta
identificação. O código de barras de DNA tem o sentido de contornar possíveis
problemas na identificação de espécies e sistemática realizadas através da tradicional

12
abordagem morfológica, o que pode gerar identificações incorretas causadas pela
plasticidade fenotípica (Elsasser et al., 2009).
Este método tem como padrão para esta identificação no Reino animal o uso de
uma região gênica codificante da enzima mitocondrial citocromo c oxidase I (COI),
contendo 658 pares de base (Hebert et al., 2003a, b). Estes aspectos conferem a este
DNA um potencial para se tornar uma ferramenta universal, padronizada e econômica
para uma identificação massiva (Ferri et al., 2009).
1.5. PROTEÔMICA E SECRETOMA
Proteínas são polímeros de aminoácidos resultantes da tradução das informações
genéticas contidas no DNA de um genoma. Estão presentes em grande diversidade, nas
membranas, no citoplasma e no interior do núcleo, agindo como enzimas, anticorpos,
componentes estruturais, receptores celulares, agentes de sinalização, transporte e
armazenamento de substâncias entre outras funções variadas. Devido a essa diversidade
funcional, as proteínas exercem papéis fundamentais em quase todos os fenômenos
biológicos, como produção de energia, comunicação celular, defesa, contração
muscular, atividade neuroquímica, reprodução, dentre outros (Souza et al., 1999).
A tecnologia de sequenciamento nucleotídico em larga escala pode proporcionar
uma grande quantidade de genomas inteiros de muitos organismos. Desse modo,
durante muitos anos, o conhecimento genômico propagou a ideia de que não seriam
necessários os estudos em níveis proteicos para se obter dados relacionados ao aumento
ou redução da sua expressão e que somente seria necessário a determinação dos níveis
de mRNA, para avaliar as interações entre as proteínas e suas funções no
desenvolvimento de determinada condição fisiológica (Monti et al., 2005).
Ao serem sintetizadas, as proteínas podem sofrer importantes modificações para
se tornarem funcionais, portanto tais transformações devem ser consideradas juntamente
com informações sobre a sequência genética (Baggerman et al., 2004).
Desse modo o proteoma é, sem dúvida, muito mais complexo que o genoma
pois, enquanto o DNA possui somente quatro bases nitrogenadas, as proteínas estão
compostas de aminoácidos, dos quais existem 20 tipos diferentes, originando em uma
célula, tecido ou organismo milhares de proteínas e apresentando, portanto, uma alta
complexidade. A análise proteômica refere-se à avaliação sistemática dessas proteínas,

13
de forma a comparar a sua expressão celular tecidual ou orgânica em distintos
momentos fisiológicos (Wilkins et al., 1995; Soares et al., 2007).
A proteômica envolve, portanto, uma análise total ou mesmo em larga escala das
proteínas de um organismo, gerando quantidade massiva de dados experimentais
(Cagney et al., 2003). O proteoma, termo cunhado pela primeira vez por Wilkins &
Williams em 1995 é, por definição, o conjunto de todas as proteínas expressas por um
genoma em um determinado tempo (Lanças et al., 2003).
Estudos nessa área são, portanto, capazes de fornecer importantes informações
sobre as modificações pós-traducionais de proteínas, contribuindo para o entendimento
do funcionamento celular (Pandey & Mann, 2000) enquanto que o genoma representa,
através do DNA, um possível código desse dinamismo das transformações protéicas.
A proteômica está subdivida em duas grandes áreas: proteômica estrutural
(determinação estrutural em larga escala) e proteômica funcional (expressão).Os estudos
estruturais avaliam os níveis de inibição e de superexpressão, investigando modelos
celulares de expressão protéica submetidos a duas ou mais situações fisiológicas
diferentes.
Essa pesquisa comparativa é usualmente utilizada para identificar proteínas e
para relacionar o perfil de expressão do proteoma com alguma situação fisiológica
específica (Monti et al., 2005).
O aumento do número de genomas seqüenciados fez crescer exponencialmente o
número de seqüências de proteínas com funções desconhecidas. Como nem sempre a
seqüência genômica é suficiente para elucidar essas funções, a proteômica é uma área
do conhecimento científico altamente dinâmica e competente para tal (Antunes, 2008).
Os dados de seqüenciamento gênico associados a estudos proteômicos e
peptidômicos estruturais e funcionais buscam estabelecer o melhor entendimento do
estado metabólico induzido por situações fisiológicas específicas, assim estudos
proteômicos podem ser valiosos para a identificação de novos alvos terapêuticos e o
entendimento da dinâmica protéica (Antunes, 2008; Sun & Cavalli, 2009).
Neste ensejo, sabe-se que patógenos podem liberar um número amplo de
diversificadas proteínas, as quais podem modificar e adaptar o ambiente hospedeiro,
influenciando a seu favor modificações em vias de sinalização imune, invocando um
processo conhecido como imunomodulação helmíntica (Hewitson et al., 2009).

14
Convencionou-se denominar, portanto, como “Excreção/Secreção” (E/S) os
produtos ativamente exportados através de vias secretórias ou mesmo àquelas que
podem se difundir ao meio por “extravasamento” a partir dos tecidos dos parasitos
(Hewitson et al., 2009).
In vivo, antígenos “secretados” incluem enzimas digestivas que emanam do
intestino de vermes adultos, bem como conteúdo uterino que fêmeas liberam juntamente
com os ovos ou larvas de transmissão estágio. No entanto, os parasitos podem muito
bem adaptar tais "secreções" para cumprir um novo papel no hospedeiro, uma vez que
elas são liberadas a partir do interior do corpo do helminto. Por isso, é necessário para
analisar todos os produtos E/S, submetê-los a uma gama completa de ensaios
bioquímicos, imunológicos e análises proteômicas para que se saiba qual a sua real
origem fisiológica (Hewitson et al., 2009)
Este conjunto de moléculas protéicas e não-protéicas, o secretoma, fornece as
ferramentas necessárias para a sobrevivência do parasita no seu nicho hospedeiro, no
entanto, ao passo que essas moléculas são a chave para os efeitos deletérios e
patogênicos, o secretoma da célula infectada é igualmente importante para o
entendimento dos mecanismos de defesa hospedeira diante do ataque do patógeno (Lee
et al., 2009).
Dados sobre o secretoma de organismos patogênicos são escassos e limitados a
específicos tipos de amostras e experimentos, mesmo àqueles relacionados a agentes
infecciosos causadores de “doenças tropicais negligenciadas”, como no caso dos
helmintos. Nesse sentido, análises da parte protéica de secretomas de helmintos pela
proteômica tem sido realizadas para diminuir essa deficiência e assim, desenvolver
novas soluções terapêuticas para erradicar estas infecções (Hotez, 2009).
1.6. ANÁLISE DE PRODUTOS HELMÍNTICOS
As moléculas expressas ou liberadas por patógenos podem ser classificadas
como proteícas, glicoprotéicas ou lipídicas e são estudadas, em helmintos, a partir da
obtenção de extratos brutos/somáticos desses parasitos e de seus produtos de
excreção/secreção (Northern & Grove, 1987).
Segundo Ehigiator et al. (2000), o extrato de helmintos pode ser obtido através
da maceração do helminto e posterior centrifugação do homogenato. A obtenção de
produtos de excreção/secreção, conforme descrição de Craig et al. (2006), consiste no

15
cultivo in vitro do helminto e retiradas de alíquotas do meio em determinado intervalo
de tempo, assim como, de helmintos para verificar se a estrutura do mesmo não foi
modificada durante cultivo, alterando a composição das substâncias obtidas.
A análise proteômica é o principal método atualmente escolhido para este tipo
de pesquisas (Sickmann et al., 2002) e segundo Cagney et al. (2003), a maioria dos
experimentos proteômicos envolvem quatro etapas:
a) Isolamento proteico a partir de uma amostra biológica (como por exemplo
um extrato celular), seguindo algum tratamento experimental;
b) Fracionamento das proteínas resultantes (ou peptídeos, produtos oriundos de
digestão proteômica) por métodos tradicionais tais como eletroforese
bidimensional em gel de poliacrilamida (2D PAGE) ou Cromatografia
Líquida (LC), por exemplo;
c) Caracterização da proteína ou peptídeo através de Espectrometria de Massas
(MS);
d) Identificação da proteína por meio de interpretação manual ou correlação em
base de dados de acordo com o espectro de massa obtido.
Esta ferramenta facilita na identificação de produtos helmínticos que já estão no
hall de potenciais vacinas ou alvos de intervenção terapêutica (Ranganathan & Garg,
2009).
No entanto, mesmo com alguns trabalhos empregando uma abordagem
proteômica para o estudo da biologia desses animais, uma das principais limitações para
a realização de alguns estudos é a escassez de informações genômicas disponíveis.
Atualmente, além dos modelos helmínticos Caenorhabditis elegans e C. briggsae
apenas cinco genomas nucleares de nematodas foram publicados e são de Brugia
malayi, Meloidogyne incógnita e M. hapla, Pristionchus pacificus e Trichinella spiralis
(Rebello et al., 2011).
1.7. HELMINTOS E IMUNOMODULAÇÃO
O parasitismo é uma condição desvantajosa por definição, embora os helmintos
parasitas sejam profícuos na regulação imune e na supressão de mecanismos
imunopatológicos (Maizels et al., 2004), uma vez que fugir ou amortecer a resposta

16
imune de seus hospedeiros, pode lhes proporcionar oportunidade de atingir a
maturidade, reproduzir-se e completar seu ciclo de vida. Conseqüentemente, muito pode
ser aprendido a partir das relações parasito-hospedeiro, particularmente quando o
parasita é usado como uma ferramenta para avaliar imunidade do hospedeiro (Wilson &
Maizels, 2006).
Embora a base molecular da supressão imune induzida por helmintos ainda
esteja por ser definida completamente, a caracterização dessas moléculas é uma
importante abordagem para essa investigação, pois são elas a razão da subversão das
células imune do hospedeiro (Kwan-Lim et al., 1989).
Numerosos estudos demonstram que moléculas bioativas presentes em extratos
de helmintos e produtos de excreção/secreção são capazes de modular o sistema
imunológico do hospedeiro, favorecendo a sobrevivência do parasito, mas trazendo
também benefícios para o hospedeiro (Johnston et al., 2009).
Recentemente, estes estudos têm evoluído a um ponto onde os dados a partir de
modelos animais e, em menor número, de observações clínicas, sugerem que a infecção
com helmintos parasitas podem reduzir a severidade de doenças como esclerose
múltipla (La Flamme et al., 2003), diabetes mellitus tipo I (Zaccone et al. 2003), asma
(Wilson et al., 2005), doença intestinal inflamatória e, talvez, distúrbios
cardiovasculares (Elliott et al., 2005).
Esses produtos de Excreção/Secreção são expostos para o meio extra-celular do
parasita através das vias secretoras clássica e não clássica, ficando expostos na
superfície do corpo ou mesmo liberados, fazendo-as acessíveis ao sistema imune ou
mesmo à ação de drogas. A via clássica é mediada pela presença de sinais peptídicos
com curtas sequencias amino-terminais, enquanto as proteínas secretadas pela via não-
clássica são de difícil previsão devido estas serem normalmente secretadas por via
independente de Retículo endoplasmático e Complexo de Golgi, além de não necessitar
de sequências de peptídeo sinal (Nickel, 2003; Bendtsen et al., 2004).
A eficácia de anti-helmínticos têm diminuído gradativamente com seu repetido
uso, além disso, uma rápida reinfestação tem ocorrido após a administração desses
medicamentos, requerindo o desenvolvimento de vacinas como uma prioridade para a
melhoria da saúde pública, como no caso da admistração de benzimidazóis em
infestação por Necator americanus e Ancylostoma caninum (Albonico et al., 1995).

17
Parasitoses helmínticas de interesse veterinário também tem sido alvo para
estudos proteômicos abordando mecanismos de sobrevivência dos helmintos em
questão. A maioria desses estudos tem utilizado murinos como modelo animal para
infecções helmínticas. Porém, é importante ressaltar que muitos resultados diferentes
têm sido obtidos usando diferentes modelos animais e helmínticos, fazendo disto uma
dificuldade em extrapolar as informações obtidas aos helmintos e respectivos
hospedeiros naturais (Meeusen, 1999).
A respeito dessas descobertas, o parasitismo de helmintos em modelos animais
tem contribuído diretamente para o conhecimento e compreensão de como o sistema
imune de mamíferos é mobilizado para combater parasitas metazoários (Johnston et al.,
2010).
Em seres humanos e alguns modelos animais, os antígenos ou alérgenos
helmínticos estimulam os linfócitos T a produzirem citocinas Th2, como a interleucina
(IL)-4 e a IL-5. A IL-4 induz os linfócitos B a produzirem imunoglobulina E (IgE) e a
IL-5 atrai e ativa os eosinófilos além de diminuir os níveis de interferon – γ (IFN- γ)
(Tezuka et al., 2003).
Além disso, Th2 associada às citocinas IL-4, IL-9, IL-13, IL-25 e IL-33 são
conhecidas por desempenhar um importante papel na mediação dos mecanismos
efetores que contribuem para expulsão do verme do interior do intestino, tais como:
hiperplasia de células caliciformes, produção de muco, eosinofilia, mastocitose,
recrutamento de macrófagos e hipercontração muscular (Nair et al., 2006; Anthony et
al., 2007; Díaz & Allen, 2007).
Substâncias liberadas em diferentes estágios de sensibilização do modelo animal
(infecções primárias, secundárias e múltiplas), diferentes estágios dos parasitos (larva e
adultos), e sexo dos parasitos (macho e fêmea) podem evocar diferentes perfis de
respostas imunes (Balic et al., 2000).
Segundo Craig et al. (2006), essas moléculas são derivadas da superfície do
helminto e/ou de glândulas excretoras/secretoras especializadas e freqüentemente
liberadas em estágios específicos. Isto foi observado em estudos proteômicos realizados
por Soblik et al. (2011) em Strongyloides ratti, o qual apresentou diferentes
composições nos perfis proteicos entre machos, fêmeas e estágios larvais, comprovando
que o complexo ciclo biológico de helmintos, com estágios de vida livre e separação

18
sexual, fazem com que cada um desses estágios tenha importância única na interação
com seu hospedeiro.
1.8. POTENCIAIS MOLÉCULAS NA BIOLOGIA E PATOGENICIDADE DOS
HELMINTOS
Baseados na abundância de proteínas encontradas em produtos de excreção e
secreção, essas moléculas são classificadas quanto a sua importância para a viabilidade
de vermes, tanto na sua patogenicidade quanto imunossupressão, reprodução adesão,
dentre outros aspectos biológicos condizentes com cada organismo. A partir desses
estudos, começam a surgir a possibilidade de produção de vacinas e de novas
perspectivas a combate desse modelo de infecção (Bethony, 2005).
Os principais antígenos vacinais já determinados são enzimas hidrolíticas,
incluindo proteases e acetilcolinesterases, a partir do estágio larval infeccioso 3 (L3) e
estágios adultos de Ancylostoma sp. As principais moléculas candidatas encontrados em
L3 são as proteínas ASP (Ancylostoma-secreted proteins), astacin-like metaloprotease
(MTP), a acetilcolinesterase (ACH) e transtirretina (TTR). A partir da fase adulta, os
antígenos principais encontradas são o inibidor tecidual das metaloproteinases (como
Ac-TMP), proteases aspárticas e cisteinil proteases (Ranganathan & Garg, 2009).
ASP-1 e ASP-2 são proteínas liberadas em maior abundância no período de
penetração no hospedeiro pelos estágios L3 de helmintos de ciclo geográfico. Estas
moléculas são ricas em cisteína e pertencem à superfamília das proteínas relacionadas à
patogenicidade (PRP) (Hawdon & Hotez, 1996).
ASP-2, que estão localizadas no esôfago glandular e cutícula do estágio L3 de A.
caninum, mostrou-se um antígeno de caráter protetor em cães vacinados em laboratório,
devido à redução na contagem de ovos, carga parasitária e aumento in vitro da
habilidade do soro humano em inibir a migração desse estágio através de tecido da pele
em comparação a amostras controle (Bethony, 2005).
1.9. ORTLEPPASCARIS X RHINELLA MARINA: MODELO PARA ESTUDO DE
RELAÇÃO HELMINTO-HOSPEDEIRO
A sobrevivência dos parasitos em seus hospedeiros depende da relação de
adaptação estabelecida entre os mesmos, pois dentre outros fatores limitantes para essa

19
co-existência, o sistema imunológico do hospedeiro desempenha estratégias de
detecção, inativação e/ou destruição do organismo invasor. Por consequência, este
necessita desenvolver mecanismos para o escape, mimetização e manipulação do
sistema imune, adaptando a resposta do hospedeiro ao seu benefício (McKay, 2006).
Na helmintologia, o estabelecimento dessas relações pode ser elucidado através
de diversos modelos de parasitos e hospedeiros, tendo como ferramentas diversas áreas
do conhecimento, como análises morfológicas, histopatológicas, genômicas e
proteômicas. Neste estudo, temos como exemplo o nematódeo Ortleppascaris sp. e seu
hospedeiro R. marina.
As informações sobre os parâmetros morfológicos em estudos por microscopia
de luz são escarças nas descrições de larvas de Ortleppascaris sp. e análises ultra
estruturais dessas larvas não haviam sido realizadas. A Taxonomia Alfa se utiliza de
estudos morfológicos de helmintos por técnicas de microscopia de luz, no entanto, cada
vez mais tem sido importante a adição de informações por meio de análises
ultraestruturais.
Neste trabalho, aspectos histopatológicos da infecção hepática por
Ortleppascaris sp. em de R. marina são mostradas em tecidos incluídos em parafina
além de técnicas que utilizam método de fixação e processamento de amostra para
resina hidrofílica como suporte para os cortes histológicos, que, segundo Pannocchia et
al., (2008), preservam de melhor maneira as estruturas teciduais, possibilitando melhor
visualização das estruturas do que as técnicas convencionais em parafina.
A caracterização molecular de helmintos tem sido realizada para parasitos de
interesse médico, veterinário e econômico. No entanto, estudos referentes a parasitas
que não estão inclusos nessas categorias se fazem necessários, pois geram sequências
nucleotídicas aos bancos de dados, como o Genbank, com informações que podem ser
usadas em análises filogenéticas.
A caracterização proteômica e identificação de moléculas envolvidas no
parasitismo de Ortleppascaris sp. contribui para o avanço do entendimento da relação
deste parasito com seu hospedeiro, gerando informações e perspectivas no campo de
estudo de biomoléculas.

20
1.10. OBJETIVOS
1.10.1 Objetivo Geral:
- Realizar estudo morfológico, molecular, histopatológico e proteômico de
Ortleppascaris sp. parasito de fígado de Rhinella marina.
1.10.2. Objetivos Específicos:
- Caracterizar as estruturas somáticas de larvas de Ortleppascaris sp. por
microscopia de luz e eletrônica de varredura;
- Identificar as alterações histopatológicas provocadas por este parasita em
fígado de Rhinella marina;
- Caracterizar sequencias gênicas de larvas de Ortleppascaris sp.;
- Construir um mapa 2D de referência de proteínas presentes no extrato somático
e fluido cístico de larvas de Ortleppascaris sp., utilizando espectrometria de massas
como referência;
- Realizar análise comparativa dos perfis proteômicos do extrato somático e
fluido cístico desse parasito.

21
2. ARTIGOS
2.1. ARTIGO 1
Título: Morphological and molecular characterization of Ortleppascaris sp.
larvae parasite of cane toad Rhinella marina from eastern Amazonia. 2013.
Revista: The Journal of Parasitology (ISSN).
Online ISSN: 1937-2345.
Print ISSN: 0022-3395.
Frequência: Bimensal.
Fator de impacto: 1.405.
Volume: 99.
Número: 1.
Páginas: 118-123.
Ano de publicação: 2013.

30
2.2 ARTIGO 2
Título: Liver histopathological alterations in the cane toad, Rhinella marina
(AMPHIBIA: BUFONIDAE), induced by Ortleppascaris sp. larvae (NEMATODA:
ASCARIDIDAE).
Revista: The Journal of Parasitology.
Online ISSN: 1937-2345.
Print ISSN: 0022-3395.
Frequência: Bimensal.
Fator de impacto: 1.405.
Ano de publicação: 2013, no prelo.

39
2.3. ARTIGO 3
Título: Proteomic profile of Ortleppascaris sp. larvae in Rhinella marina.
Revista a ser submetido: International Journal for Parasitology
ISSN: 0020-7519
Frequência: Bimensal
Fator de impacto: 3.393.
Ano de submissão: 2013.

40
PROTEOMIC PROFILE OF ORTLEPPASCARIS SP. LARVAE IN RHINELLA
MARINA
Jefferson Pereira e Silva*, Adriano Penha Furtado *, Humberto Josué de Oliveira
Ramos†, Ana Clara Rosa Salvador†, Claudio Lísias Mafra de Siqueira‡ and
Jeannie Nascimento dos Santos *
*Laboratory of Cell Biology and Helmithology (Laboratório de Biologia Celular e
Helmintologia) “Profa. Dra. Reinalda Marisa Lanfredi”, Biological Sciences Institute
(Instituto de Ciências Biológicas), Federal University of Pará (Universidade Federal do
Pará), Belém, Pará, Brazil. e-mail: [email protected]
†Laboratory of Molecular Biology of Plants (Laboratório de Biologia Molecular de
Plantas)
‡Laboratory of Parasitology and Molecular Epidemiology (Laboratório de Parasitologia
e Epidemiologia Molecular)
ABSTRACT: Ortleppascaris sp. is a helminth that, in its larval stage, infects the liver
parenchyma of the amphibian Rhinella marina, resulting in severe physiological and
pathological changes. This study used a proteomic approach to determine the overall
profile of proteins expressed in a somatic extract from the nematodes to investigate the
relationship between the parasite and its host. A total of 60 abundant proteins were
selected from the two-dimensional electrophoresis, identified by MS, and grouped
based on their Gene Ontology by the biological processes in which they are possibly
involved. Through homology, important helminthic derivatives were identified, such as
the immunoreactive As37 antigen, guanylyl cyclases, proteolytic enzymes, and other
proteins conserved among different parasites. This study represents a new approach to
helminth-related proteomic studies using an amphibian animal model. Furthermore, this
study identified protein markers that are important to the host-parasite relationship and
the viability, development, infectivity, and virulence of helminths.

41
INTRODUCTION
Helminths are complex, often dangerous organisms wich are neglected, even in
this era of health system globalisation [1]. However, with the emergence of "omics"
tools, particularly proteomics, these organisms have been investigated at the molecular
level [2]. Proteomics has provided much information and promising results, identifying
a repertoire of helminth derivatives that may reveal their biology, mode of infection,
and the pathology associated with infection [3-7].
Molecular studies are needed for the design of future interventions, such as
antihelminthic vaccines, which may directly influence the quality of life of billions of
human beings. In addition, the occurrence of infection in animals and plants, the
organisms most affected by parasitic infections, represents an important public health
problem and is of global economic significance [8, 9].
Along with their hosts, helminths have co-evolved for thousands of years,
resulting in a favourable environment for the survival and co-existence of both parties in
a complex adaptive mechanism [10-12]. This host-parasite adaptation is closely related
to the host immune system, which detects and attempts to inactivate and/or destroy the
invader. The parasite, in turn, develops strategies for escape by mimicking and
manipulating the host immune system, thereby adapting the host response to parasitism
[13].
Research shows that by manipulating their immune systems, the hosts can
become vulnerable to other infections or, conversely, suppress their
immunopathological reactions. These processes also depend, in evolutionary terms, on
the duration of the relationship between helminth parasites and their hosts [13, 14].
Johnston et al. [15] suggest that the bioactive molecules present in extracts of helminths
or their excretion/secretion products are responsible for modulating the host immune
response in favour of the parasites.
Helminth biomolecules can be classified as glycoproteins, lipids, and proteins,
and have been studied using extracts of helminths or their excretion/secretion products.
Each protein class contains structural and functional diversity, including cytokines,
chemokines, hormones, digestive enzymes, antibodies, extracellular proteinases, toxins,
and antimicrobial peptides. Some of these proteins are involved in vital biological
processes, such as oxidation-reduction processes, cell adhesion and migration, cell-cell
communication, differentiation, proliferation, morphogenesis, defence, virulence

42
factors, and immune responses [16-18]. According to Craig et al. [6], these substances
are derived from the surface of the helminth or from specialised excretory/secretory
glands and are often released at specific stages. Among the group of helminths that
release immunomodulating substances, the Nematoda phylum is important because
members of this phylum have been used in clinical trials for the treatment of
immunological diseases and vaccine development [13, 19, 20].
In the present study, helminths of the genus Ortleppascaris were used. As adults,
these parasites are found in the gastrointestinal tract of Crocodilians [21], and as larval
forms, they are found in the liver of amphibians [22-25].
According to Silva et al. [25], the nematode infects the liver of R. marina in its
larval parasitic form and develops inside fibrous cysts, which are apparently filled with
amorphous material and cellular debris. Infection by these larvae induces a significant
increase in the area occupied by melanomacrophages, which are phagocytic immune
system cells in amphibians, and other pathological changes, such as the presence of
inflammatory infiltrates distributed throughout the liver parenchyma. However, even
with severe physiological changes due to parasitism, the defence system of the anuran
is, in most cases, unable to curb the presence of the parasite because the larvae remain
alive and develop in these organs from their elementary stages [26].
The identification and proteomic analysis of biomolecules of Ortleppascaris sp.
larval parasites of R. marina contribute to the understanding of the host-parasite
relationship. Further, the screening of molecular markers generates information for the
study of immunomodulatory products, which are targets of interest for the control of
helminth infections in humans and animals.
MATERIAL AND METHODS
Parasites
Helminths were harvested from the interior of cysts present in the hepatic
parenchyma of the host. The parasites were subjected to three washing steps with PBS
(pH 7.4) and then stored in extraction buffer (7 M urea - 2 M thiourea - 2% CHAPS) at
-20ºC.
Preparation of somatic extract of Ortleppascaris sp.

43
Somatic protein extracts were obtained by macerating approximately 200 larvae
(L3 stage) in extraction solution (7 M urea - 2 M thiourea - 2% CHAPS) under cooling
with liquid nitrogen, then centrifuging the extracts at 13,000x g for 15 min at 4°C. The
supernatant was used directly for protein analysis.
Two-dimensional electrophoresis
The total protein concentration was determined by the Bradford method [27],
and samples were stored at -80°C until use. Aliquots of protein extract containing 100
µg of sample were diluted to a final volume of 125 µL in Destreak solution (GE
healthcare) and 2% IPG buffer (pH 3 – 10) (GE healthcare). Seven-centimetre strips
(Immobiline, GE healthcare) with an immobilised pH gradient in the range of 3-10 were
rehydrated with the protein extract for 17 h using IPGBox (GE Healthcare). Isoelectric
focusing was initiated immediately after rehydration.
The isoelectric focusing was performed with an automated system (Ettan
IPGphor III GE Healthcare) at 20°C with a constant current of 50 μA/strip and a total of
5.0 to 6.5 kVh following the four-step program: 300 V for 4 h; linear gradient to 1000 V
for 30 min; linear gradient to 5000 V for 1:20 h; 5000 V for 30 min.
After the isoelectric focusing, the strips were reduced in equilibration buffer (6
M urea, 0.075 M TrisHCl (pH 8.8), 29% glycerol, 2% SDS, 0.02% bromophenol blue)
containing 2% dithiothreitol (DTT) for 30 min and then alkylated for 30 min in the
equilibration buffer containing 2.5% iodoacetamide.
For the second dimension, the strips were placed on a 12.5% polyacrylamide gel
in a Mini Protean Cell system (Bio-Rad). The electrophoresis ran at a constant 80 V for
2 h. The gels were stained with Coomassie Blue G-250 solution overnight with stirring
and scanned with ImageScanner III (GE Healthcare) using Labscan software (GE
Healthcare).
In-gel tryptic digestion and mass spectrometry
The spots detected by ImageMaster 2D Platinum 7.0 software (GE Healthcare)
and observed with the naked eye were manually excised, treated with washing solution
(50% methanol, 5% acetic acid), and then dehydrated in 100% acetonitrile in a vacuum
centrifuge at room temperature. The proteins were subsequently subjected to reduction
(10 mM DTT) and alkylation (100 mM iodoacetamide).

44
The samples were digested at 37°C overnight with proteomic-grade trypsin
(Promega, Madison, WI, USA) in 50 mM ammonium bicarbonate (final concentration
20 ng/μL). Tryptic peptides were extracted from the gel solution with 50% acetonitrile
in 5% formic acid.
The extracted peptides were transferred to a sterile tube and treated with 100
mM ammonium bicarbonate, dried in a vacuum centrifuge, and resuspended in a
solution of 50% acetonitrile, 0.05% formic acid, and 0.1% trifluoroacetic acid.
Aliquots of 0.5 µL of each sample were applied to a steel plate at a 1:1 ratio with
2,5-dihydroxybenzoic acid matrix (Sigma). After crystallisation, the plate was inserted
into the mass spectrometer for analysis. All MS spectra were obtained in the m/z range
of 800 - 4000 Da using a MALDI-TOF Ultraflex III spectrometer (Bruker Daltonics).
The spectra were analysed using FlexAnalysis 3.3 software (Bruker Daltonics) for the
determination of peaks.
The search for protein homology by peptide mass fingerprinting (PMF) was
performed using the Blaxter Lab database (NEMBASE4) and the Mascot Daemon
software (Matrix Science). The search parameters were set as follows: up to two missed
cleavage sites; 0.1 Da error for the identification of peptides; carbamidomethylation of
cysteines as a fixed modification and oxidation of methionine as a variable
modification. PMF data analysis was supplemented by linking with Gene Ontology
through the UniProt database to infer the biological processes in which the identified
molecules are involved. The identified proteins were grouped into functional categories
and subcategories according to their ontology.
RESULTS
Identification of the Somatic Extract Proteome of Ortleppascaris sp.
To characterise the protein profile of the nematode’s somatic extract, the most
abundant proteins detected with the Image Master 2D Platinum software, shown in
Figure 1 and indicated as spots 1-60, were analysed by mass spectrometry.

45
Figure 1: 2D gel containing the somatic extract of Ortleppascaris sp. larvae
The list of marked spots on the two-dimensional gel (Figure 1) with their
proteins and corresponding helminth according to the Nembase4 database are
summarised in Table 1. Of these, 26 spots were identified, 24 spots were identified as
hypothetical proteins, and six spots were not characterised (Table 1). Four spots did not
produce spectra of sufficient quality to allow their identification, having insufficient
homology with any protein from the database.

46
Table 1 - Summary of somatic extract proteins of Ortleppascaris sp. identified by
MS using Nembase4 database. (See Supplemental Table 1 for details)
Spot Protein [helminth]
01 hypothetical protein CAEBREN_23080 [Caenorhabditis
brenneri]
02 CRE-ATH-1 protein [Caenorhabditis remanei]
03 hypothetical protein CAEBREN_12726 [Caenorhabditis
brenneri]
04 As37 [Ascaris suum]
05 Sulphotransferase domain containing protein [Brugia malayi]
06, 33, 34, 37,
38, 39,40 Cell division protein kinase 7 [Ascaris suum]
07 CBN-CDC-37 protein [Caenorhabditis brenneri]
08 Dolichyl-diphosphooligosaccharide--protein
glycosyltransferase 48 kDa subunit [Ascaris suum]
09, 29, 48, 60 hypothetical protein CAEBREN_00296 [Caenorhabditis
brenneri]
10 hypothetical protein CAEBREN_00853 [Caenorhabditis
brenneri]
11, 16, 41, 42 LONg family member (lon-3) [Caenorhabditis elegans]
12 hypothetical protein F46F3.3 [Caenorhabditis elegans]
13, 50 guanylyl cyclase [Heterodera glycines]
14 CRE-NSY-4 protein [Caenorhabditis remanei]
15 cleavage and polyadenylation specificity factor subunit 2
[Trichinella spiralis]
17, 19, 44, 49 Uncharacterised protein family UPF0005 containing protein
[Brugia malayi]
18 hypothetical protein LOAG_04901 [Loa loa]
21 hypothetical protein LOAG_06031 [Loa loa]
22 GTP-binding protein SAR1 [Brugia malayi]
23 hypothetical protein CRE_11672 [Caenorhabditis remanei]
25 hypothetical protein CAEBREN_23609 [Caenorhabditis
brenneri]

47
26 hypothetical protein LOAG_09802 [Loa loa]
27 hypothetical protein CRE_25790 [Caenorhabditis remanei]
28, 31, 35, 55, 56 actin variant 1 [Dictyocaulus viviparus]
30, 59 [Pyruvate dehydrogenase acetyl-transferring-phosphatase 1
[Ascaris suum]
32 formyl-CoA transferase [Micromonospora aurantiaca ATCC
27029]
36 hypothetical protein LMOh7858_0493 [Listeria
monocytogenes str. 4b H7858]
43 Hypothetical protein CBG09068 [Caenorhabditis briggsae]
46, 47, 52 hypothetical protein AbauAB_10162 [Acinetobacter
baumannii AB900]
51 rnf126-prov protein [Brugia malayi]
53 Nodal modulator 1 [Ascaris suum]
57 hypothetical protein CBG_14574 [Caenorhabditis briggsae
AF16]
58 hypothetical protein LOAG_01253 [Loa loa]
The products detected in the somatic extract include enzymes involved in
metabolic processes and energy production (spots 1, 2, 3, 15, 30, 59), such as
hydrolases of the PP2C family (protein phosphatase 2C), sulphotransferases (spot 5)
and transferases from the CoA transferase family III (spot 32), which are evolutionarily
conserved and related to energy production in helminths and protozoa.
Some of the abundant proteins were associated with body morphogenesis, such
as spots 11, 16, 41, and 42, which corresponds to the collagen present in the cuticle of
helminths. Similarly, other structural components, such as cytoskeletal proteins, were
identified among the hypothetical proteins (spots 10, 57) or positively identified with
significant scores as proteins from the actin family (spots 28, 31, 35, 55, 56).
Regarding the development process of the larvae, seven proteins (spots 06, 33,
34, 37, 38, 39, 40) related to cell division and protein phosphorylation processes were
found.

48
Guanylyl cyclases (spots 13, 50) were also detected; these molecules are of great
importance to the biology of helminths, as they are involved in cell signalling, neural
circuits, and host recognition.
Endoplasmic reticulum (ER) membrane proteins involved in the process of
asparagine glycosylation (spot 8) and SAR1 protein (spot 22), which is related to
vesicular trafficking between ER and Golgi, were also identified.
Surprisingly, some other relevant proteins were identified: As37 (spot 4), which
is a highly reactive antigen in Ascaris suum and a member of the immunoglobulin
family, and proteolytic enzymes (spot 43), which are considered virulence factors for
the establishment and maintenance of helminthic infection.
Proteomic Analysis of the Somatic Extract
Most of the proteins found in this study had nearly significant scores when
ranked with proteins that are homologous to those found in the database. However,
other proteins, such as those related to the cytoskeleton, exhibited values with
considerable significance.
Table 2 shows the list of identified proteins with their respective spot numbers.
They are grouped into functional categories and subcategories according to their GO, as
described in Supplemental Table 1. Also shown in Supplemental Table 1 are the
following parameters: the identification of each spot from the gel of somatic extract
shown in Figure 1; the protein name and its corresponding helminth; the GI accession
numbers from the National Centre for Biotechnology Information (NCBI) database
corresponding to the protein sequence with the highest match to the peptides; and the
scores, match, coverage, theoretical isoelectric point and molecular weight, and the
amino acid sequences of corresponding peptides obtained by mass spectrometry.

49
Table 2. Summary of the identified proteins from the gel of the somatic extract and the category of biological process in GO
terms. See Supplemental Table 1 for additional information
Molecules of metabolic processes and energy production
GO term Process/Biological function Spot Protein
GO: 0006535 Biosynthesis of cysteine from serine 01 hypothetical protein CAEBREN_23080
GO: 0016787 Hydrolase 02 CRE-ATH-1 protein
GO: 0008152 Metabolic process 03 hypothetical protein CAEBREN_12726
32 formyl-CoA transferase
GO: 0008146 Sulphotransferase 05 Sulphotransferase domain containing protein
GO: 0019901 Protein kinase ligand 07 CBN-CDC-37 protein
GO: 0016787/
GO: 0000104
Hydrolase/succinate dehydrogenase
action 15 cleavage and polyadenylation specificity factor subunit 2
GO: 0016787/
GO: 0006470 Hydrolase/protein dephosphorylation 30, 59 Pyruvate dehydrogenase acetyl-transferring-phosphatase 1
Structural molecules and organic morphogenesis
GO term Process/Biological function Spot Protein
GO: 0051301/
GO: 0004674 Cell division/protein phosphorylation
06, 33, 34,
37, 38,
39,40
Cell division protein kinase 7
GO: 0005856 Cytoskeleton
10 hypothetical protein CAEBREN_00853
57 hypothetical protein CBG_14574
28, 31, 35,
55, 56 actin variant 1 [Dictyocaulus viviparus]
GO: 0010171/
GO: 0042329
Body morphogenesis/constituents of
cuticle structure
11, 16, 41,
42 LONg family member (lon-3)

50
GO: 0016021 Membrane protein
14 CRE-NSY-4 protein
17, 19, 44,
49
Uncharacterised protein family UPF0005 containing
protein
Immunoreactive molecules and virulence factors
GO term Process/Biological function Spot Protein
GO: 0005515 Protein binding 04 As37
GO: 0006508 Proteolysis 43 Hypothetical protein CBG09068
Molecules of the biosynthetic/secretory pathway
GO term Process/Biological function Spot Protein
GO: 0004579/
GO: 0018279
Glycotransferase/N-terminal
Glycosylation in asparagine 08
Dolichyl-diphosphooligosaccharide--protein
glycosyltransferase 48 kDa subunit
GO: 0016192 Transport mediated by vesicles 22 GTP-binding protein SAR1
Molecules of the chemosensory process
GO term Process/Biological function Spot Protein
GO: 0035556 Intracellular signal transduction 13, 50 guanylyl cyclase

51
DISCUSSION
Protein Identification and Interpretation
Problems with the identification of sequences in the proteomic analysis are not
related to the production of significant scores because many of these databases are
incomplete or not well established [28]. According to Antunes [29], these data should
be considered even when they present low scores due to the lack of a complete and
specific database for the organism in question. The scarcity of database entries for each
sequence is responsible for the low scores. However, the expectation is that detecting
these molecules is a strong indication of their presence in the systems studied.
Molecules of the Metabolic Process and Energy Production
This group of molecules includes a large group of enzymes, such as hydrolases
that catalyse the breakage of several covalent bonds important for fighting aging in
nematodes [30]. Succinate dehydrogenase was also identified in our spots and is
involved in the mitochondrial electron transport chain, cell signalling, and apoptosis
[31, 32].
In proteomic studies of Brugya malayi, Hewitson et al. [4] suggested that these
molecules are essential for resisting the oxidative stress caused by reactive oxygen
species and nitrogen produced by immune cells. Thus, these molecules may be involved
in the evasion of the host immune system.
The CoA-transferase family of enzymes is widely found in helminths
parasitising their hosts in hypoxic environments or anaerobic conditions, such as
Ortleppascaris sp. According to Tielens et al. [33], these proteins are responsible for
acetate production from acetyl-CoA, which is directly associated with the production of
metabolic energy in various helminths and protozoa.
Phosphatases from the protein phosphatase 2C (PP2C) family, one of the largest
families of serine/threonine phosphatases, are also involved in the metabolism of
Ortleppascaris sp. According to Wenk et al. [34], these enzymes have a broad spectrum
of specific substrates, including enzymes that regulate many metabolic pathways and
energy production and are likely involved in the metabolic processes of the helminths in
the present study.

52
Homologous proteins of this family have been found in Caenorhabditis elegans
(C. elegans) and are encoded by the FEM-2, F42G9.1, T23F11.1 genes, which may be
related to gender determination in helminths. It is believed that the FEM-2 gene encodes
a protein with phosphatase activity that leads to somatic male development and sperm
production [35, 36]. Because protein phosphorylation is a major mechanism of cellular
regulation and these proteins are highly evolutionarily conserved [37]. In addition to
participating in the metabolic processes of Ortleppascaris sp., these molecules must be
involved in the sexual differentiation of the larvae.
Structural Molecules and Organic Morphogenesis
Similar to proteomic studies performed with other parasitic nematodes, many of
the abundant proteins from the somatic extract of Ortleppascaris sp. are associated with
cell division, the cytoskeleton, or even the structural constitution of the helminth; these
proteins are used in the intense morphological changes that occur during the larval
stage. Evidence of high larval mitotic activity was observed based on the identification
of seven proteins (spots 6, 33, 34, 37, 38, 39, 40) related to cell division and protein
phosphorylation, which are processes required for larval viability during fully active
development.
Structural molecules encoded by the gene lon-3 were identified; these molecules
have been previously studied in C. elegans. This gene encodes cuticular collagen, which
is required during larval development in hypodermal cells and specifies and controls
body length. Expression occurs on the surface of helminths and plays an important role
in the formation of the cuticle [38, 39]. Cox et al. [40, 41] state that cuticular collagens
are synthesised and secreted by hypodermal cells and polymerise in the apical surface of
the epithelium, forming a complex structure of six layers. Furthermore, according to
Johnstone & Barry [42], different genes encoding collagen are expressed in discrete
time periods during nematode larval stages.
The identification of these homologs in the somatic extract from Ortleppascaris
sp. suggests that the helminth undergoes a developmental process in the host, as shown
by Silva et al. [25].
These findings suggest the use of Ortleppascaris sp. and other nematodes as an
experimental model to study the processes regulating body size and shape and other
aspects of development. Many nematode processes remain poorly understood, as

53
indicated by Conlon & Raff [43] and Suzuki et al. [38]. According to Van der Eycken
[44], genes encoding collagen appear to be highly evolutionarily conserved, even
among free-living nematodes and parasites, such as C. elegans, Ascaris suum,
Haemonchus contortus, and Meloidogyne incognita.
The spots 28, 31, 35, 55, 56 were identified by their significant scores as
homologous to actin, a major component of the cytoskeleton that is important to cell
shape maintenance, endocytosis, organelle movement, and cell division. These proteins
interact with myosin, and in invertebrates, they are structural constituents of the
contractile machinery.
Immunoreactive Molecules and Virulence Factors
A database search indicated the presence of As37, a highly immunoreactive 37
kDa antigen belonging to the immunoglobulin superfamily. As37 was previously
identified in the larval stage of Ascaris suum by Tsuji et al. [45], who also suggested its
presence in other nematodes, such as A. lumbricoides and Toxocara canis. In addition,
this antigen was recently identified in Angiostrongylus costaricensis [46] and A.
cantonensis [47]. In the latter study, the authors also described peptide molecules with
high homology to As37 in Baylisascaris schroederi and Brugia malayi.
This important antigen was identified among the molecules expressed by
Ortleppascaris sp. and is a possible candidate target for an antihelminth vaccine. The
existence of As37 homologs in other nematodes suggests that As37 is specific to
nematodes. The helminth used in this study belongs to the order Ascaroidea, the same
as A. suum.
In addition to identification of immunoreactive antigens, homologs of
proteolytic enzymes, such as the hypothetical protein CBG09068, are present in the
somatic extract of Ortleppascaris sp. This group of proteins is considered by Dalton
[48] as a potential candidate for anti-parasitic interventions. They are possibly expressed
in the intestines of parasites; alternatively, they may be constituents of the products of
excretion and secretion.
According to Hong et al. [49], Rhoads et al. [50], Dresden & Xu [51] and Chung
et al. [52], these enzyme homologs have been implicated in many of the developmental
processes of helminths, such as moulting, hatching, and encystment. This finding is
consistent with the larval-stage, encysted parasitism studied in this report.

54
In addition, according to McKerrow et al. [53] and Rebello et al. [54], these are
key molecules for the invasion of the host by the helminth in the infective larval stage;
they are considered the major virulence factors for the establishment and maintenance
of an infection. The proteolytic enzymes assist in the evasion of the host immune
system by degrading the mucus barrier of the gastrointestinal tract, preventing blood
coagulation, facilitating the feeding process, and aiding in the invasion of connective
tissue [55-57]. These factors were observed in the liver of R. marina parasitised by
Ortleppascaris sp. [26], where they were shown to be encysted or free in the
parenchyma with apparent destruction of connective tissue.
Another important factor that should be noted is the association of these
proteolytic enzymes with the host haemoglobin, which is considered the main substrate
in haematophagous nematodes [58]. This correlation was observed in the parasitism of
this study, where nematode infestation caused an exacerbation of melanomacrophage
centres in the parenchyma of R. marina. Melanomacrophages are host defence cells that
have a characteristic pigment formed by the degradation of haemoglobin in the
cytoplasmic compartment.
Molecules of the Secretory Biosynthetic Pathway
Constituents of the vesicular formation and recruitment machinery are present
among the abundant proteins of the somatic extract. In particular, there are several
proteins involved in glycosylation and the export of proteins and lipids from the ER.
According to Varki [59] and Berninsone [60], complex carbohydrates are
involved in multiple biological processes, including the immune response and parasite-
host interaction. A membrane glycotranferase of the ER was detected in both
Ortleppascaris sp. and Ascaris suum; wich is responsible for the production of glycans
with N-terminal oligosaccharides linked to asparagine.
Kelleher & Gilmore [61] reported that glycosylation is one of the most common
protein modifications in eukaryotes. The biosynthetic pathway of these glycans is
highly conserved in most eukaryotes, such as C. elegans [60, 62], and is essential for the
synthesis and transport of the glycolipid by ER vesicles.
Traffic along the biosynthetic secretory pathway occurs through packaging in
vesicles that bud from the ER [63]. COPII vesicles are key components of this pathway,
and the formation of such vesicles is initiated by the GTPase SAR1, which was found in

55
the present study. SAR1 is responsible for recruiting additional proteins and coating the
vesicles.
Glycoproteins are located in subcellular compartments, at the cell surface, and in
the cell matrix and may function as surface antigens of helminths. According to Thomas
& Harn [64], glycoproteins can direct the host immune system toward a typical Th2
response or modulate the system for the maintenance of infection by dendritic cells,
which may recognise these molecules as pathogen-associated molecular patterns.
The presence of proteins involved in production machinery and glycoprotein
trafficking may contribute to the increased cellular immune response described by Silva
et al. [26]. In addition, the glycoproteins may contribute to the export of sugars and
lipoproteins to the larvae cell surface during its full development because many proteins
have been identified as being part of the surface/cuticle composition of the nematode.
For example, collagen trafficking is mediated by "mega vesicles" formed by COPII [63]
and may cause a strong immune response against the nematode.
Molecules of the Chemosensory Process
Much has been speculated about the sensory abilities of helminths, for example,
their ability to sense odours or flavours, memorise temperature, and detect and respond
to different oxygen levels. To maintain vital functions, the nematode must be able to
recognise the "demand" for nutrients and initiate chemo- and thermotaxic processes to
respond to environmental changes and host tropism. Little is known about these
processes in helminths, particularly at the molecular level [65]. In C. elegans, studies
have identified the molecular basis of the genetic and behavioural plasticity of
signalling proteins, including the guanylyl cyclase enzymes [66-68] that are present in
the somatic protein group of Ortleppascaris sp.
Guanylyl cyclases are ubiquitous enzymes found in organisms ranging from
bacteria to humans. They catalyse the conversion of GTP to cGMP and are involved in
cell signalling and other physiological processes, including smelling, muscle
contractility, electrolyte homeostasis, and retinal phosphotransduction [69, 70]. Studies
have shown that the genes encoding guanylyl cyclases are coincidentally expressed in
sensory regions of the helminth body, such as in the nerve ring, amphid, and tail of the
Heterodera glycines nematode [70] and in the gustatory neurons of C. elegans [71].

56
Holden-Dye & Walker [72] believe that such molecules have a significant role in the
neural circuits involved in host recognition by chemotaxis and thermotaxis.
In helminths, it is hypothesised that guanylyl cyclases increase their expression
at low oxygen concentrations [73]. Enzyme upregulation likely occurs at the sites of
Ortleppascaris sp. infection, which correspond to areas with low amounts of free
oxygen. Moreover, according to Hallem et al. [66], enzyme expression is closely related
to CO2 concentrations, which in turn appear to be an important chemotactic component
to attract nematodes. It is believed that the presence of CO2 in C. elegans is mediated by
evolutionarily conserved receptor-like guanylyl cyclases. This receptor, GCY-9, is well
known in C. elegans and has orthologs in other nematodes, such as Heterodera
glycines, Meloidogyne incognita, and the human nematode Brugia malayi, indicating
that CO2 is a determinant of the affinity of helminths for their respective hosts.
Thus, Ortleppascaris sp. may be attracted to its amphibian host because,
according to Taylor et al. [74], this animal has pulmonary and skin respiration that
releases carbon dioxide predominantly through the skin. This study provides new
perspectives regarding the chemotaxis, host recognition, and life cycle of this nematode.
CONCLUSION
Key molecules of helminth biology were found in the present study, such as
guanylyl cyclases that are involved in the chemosensory and neurobiological processes
of parasitic nematodes. Furthermore, immunoreactive antigens specific to nematodes
and virulence factors that are considered potential antihelminthic vaccine candidates and
protein markers for helminth infection, respectively.
Other proteins identified in Ortleppascaris sp., such as actin, proteins related to
lon-3, and PP2C, are highly evolutionary conserved. Thus, this organism has potential
for use in structural studies on the development and sexual differentiation of nematodes.
The set of molecules identified shows the diversity of processes that occur in
these nematodes regarding development, redox metabolism, infectivity, and virulence
and is a further indicator of the physiological complexity of these organisms.
The proteomic approach used here provides information about the host-parasite
relationship at the molecular level and is relevant to co-evolutionary studies, the choice
of antigen for vaccine development, and the duality of the physiological relationship
between these two organisms.

57

58
References
[1] Hotez PJ. One world health: neglected tropical diseases in a flat world. PLoS Negl
Trop Dis. 2009;3:e405.
[2] Loukas A, Gaze S, Mulvenna JP, Gasser RB, Brindley PJ, Doolan DL, et al.
Vaccinomics for the major blood feeding helminths of humans. Omics : a journal of
integrative biology. 2011;15:567-77.
[3] Mutapi F. Helminth parasite proteomics: from experimental models to human
infections. Parasitology. 2012;139:1195-204.
[4] Hewitson JP, Harcus YM, Curwen RS, Dowle AA, Atmadja AK, Ashton PD, et al.
The secretome of the filarial parasite, Brugia malayi: proteomic profile of adult
excretory-secretory products. Mol Biochem Parasitol. 2008;160:8-21.
[5] Jenkins SJ, Hewitson JP, Jenkins GR, Mountford AP. Modulation of the host's
immune response by schistosome larvae. Parasite Immunol. 2005;27:385-93.
[6] Craig H, Wastling JM, Knox DP. A preliminary proteomic survey of the in vitro
excretory/secretory products of fourth-stage larval and adult Teladorsagia circumcincta.
Parasitology. 2006;132:535-43.
[7] Northern C, Grove DI. Antigenic analysis of Strongyloides ratti infective larvae and
adult worms. Immunol Cell Biol. 1987;65 (Pt 3):231-9.
[8] Garg G, Ranganathan S. Helminth secretome database (HSD): a collection of
helminth excretory/secretory proteins predicted from expressed sequence tags (ESTs).
BMC Genomics. 2012;13 Suppl 7:S8.
[9] Nagaraj SH, Gasser RB, Ranganathan S. Needles in the EST haystack: large-scale
identification and analysis of excretory-secretory (ES) proteins in parasitic nematodes
using expressed sequence tags (ESTs). PLoS Negl Trop Dis. 2008;2:301.

59
[10] Allen JE, Maizels RM. Diversity and dialogue in immunity to helminths. Nature
reviews Immunology. 2011;11:375-88.
[11] Jackson JA, Friberg IM, Little S, Bradley JE. Review series on helminths, immune
modulation and the hygiene hypothesis: immunity against helminthes and
immunological phenomena in modern human populations: coevolutionary legacies?
Immunology. 2009;126:18-27.
[12] Brindley PJ, Mitreva M, Ghedin E, Lustigman S. Helminth genomics: The
implications for human health. PLoS Negl Trop Dis. 2009;3:e538.
[13] Mckay DM. The therapeutic helminth? Trends Parasitol. 2006;25:109-14.
[14] Soblik H, Younis AE, Mitreva M, Renard BY, Kirchner M, Geisinger F, et al. Life
cycle stage-resolved proteomic analysis of the excretome secretome from Strongyloides
ratti identification of stage-specific protease. Mol Cell Proteomics. 2011;10:doi:
10.1074/mcp.M111.010157. Epub 2011 Sep 30.
[15] Johnston MJG, Macdonald JA, Mckay DM. Parasitic helminths: a pharmacopeia of
anti-inflammatory molecules. Parasitology. 2009;136:125-47.
[16] Geary J, Satti M, Moreno Y, Madrill N, Whitten D, Headley SA, et al. First
analysis of the secretome of the canine heartworm, Dirofilaria immitis. Parasit Vectors.
2012;5:140.
[17] Bonin-Debs AL, Boche I, Gille H, Brinkmann U. Development of secreted proteins
as biotherapeutic agents. Expert Opin Biol Ther. 2004;4:551-8.
[18] Maizels RM, Yazdanbakhsh M. Immune regulation by helminth parasites: cellular
and molecular mechanisms. Nature reviews Immunology. 2003;3:733-44.
[19] Pearson MS, Tribolet L, Cantacessi C, Periago MV, Valero MA, Jariwala AR, et
al. Molecular mechanisms of hookworm disease: stealth, virulence, and vaccines. The
Journal of allergy and clinical immunology. 2012;130:13-21.

60
[20] Babayan SA, Luo H, Gray N, Taylor DW, Allen JE. Deletion of parasite immune
modulatory sequences combined with immune activating signals enhances vaccine
mediated protection against filarial nematodes. PLoS Negl Trop Dis. 2012;6:e1968.
[21] Goldberg SR, Bursey CR, Aquino-Shuster AL. Gastric nematodes of the
Paraguayan caiman, Caiman yacare (Alligatoridae). J Parasitol. 1991;77:1009-11.
[22] González CE, Hamann MI. Nematode parasites of two species of Chaunus (Anura:
Bufonidae) from Corrientes, Argentina. Zootaxa. 2007;1393:27-34.
[23] González CE, Hamann MI. Helmintos parásitos de Leptodactylus bufonius
Boulenger, 1894 (Anura: Leptodactylidae) de Corrientes, Argentina. Rev Esp Herpetol.
2006;20:39-46.
[24] Moravec F, Kaiser H. Helminth parasites from west Indian frogs, with descriptions
of two new species. Caribb J Sci. 1995;31:252-68.
[25] Silva JP, Silva DC, Melo VT, Giese EG, Furtado AP, Santos JN. Liver
histopathological alterations in the cane toad, Rhinella marina (AMPHIBIA:
BUFONIDAE), induced by Ortleppascaris sp larvae (NEMATODA: ASCARIDIDAE).
J Parasitol. 2012:18 Sep [Epub ahead of print].
[26] Silva JP, Melo VTV, Silva LCN, Gonçalves EC, Giese EG, Furtado AP, et al.
Morphological and molecular characterization of Ortleppascaris sp. larvae parasite of
cane toad Rhinella marina from eastern Amazonia. J Parasitol. 2013;99:118-23.
[27] Bradford, MM. A rapid and sensitive method of a rapid and sensitive method for
the quantitation of microgram quantities of protein utilizing the principle of protein-dye
binding. or the quantitation of microgram quantities of protein utilizing the principle of
protein-dye binding. Anal Biochem. 1976;72: 248-254.
[28] Medina, ML, Haynes, PA, Breci, L, Franscisco, WA. Analysis of secreted proteins
from Aspergillus flavus. Proteomics. 2005;5:3153-3161,

61
[29] Antunes. 2008. Proteômica e peptidômica aplicadas ao estudo da defesa de plantas
de tomate à injúria mecânica (Mestrado em Bioquímica Agrícola) - Viçosa,
Universidade Federal de Viçosa, 2008. 99p.
[30] Ishii N, Fujii M, Hartman PS, Tsuda M, Yasuda K, Senoo-Matsuda N, et al.
Amutation insuccinate dehydrogenase cytochrome b causes oxidative stress and ageing
in nematodes. Nature.1998,394: 694-7.
[31] Huang J, Lemire BD. Mutations in the C. elegans succinate dehydrogenase iron-
sulfur subunit promote superoxide generation and premature aging. J Mol Biol.
2009;387:559-69.
[32] Schilling B, Murray J, Yoo CB, Row RH, Cusack MP, Capaldi RA, et al.
Proteomic analysis of succinate dehydrogenase and ubiquinol-cytochrome c reductase
(Complex II and III) isolated by immunoprecipitation from bovine and mouse heart
mitochondria. Biochim Biophys Acta. 2006;1762:213-22.
[33] Tielens AG, Van Grinsven KW, Henze K, Van Hellemond JJ, Martin W. Acetate
formation in the energy metabolism of parasitic helminths and protists. Int J Parasitol.
2010;40:387-97.
[34] Wenk J, Trompeter HI, Pettrich KG, Cohen PT, Campbell DG, Mieskes G.
Molecular cloning and primary structure of a protein phosphatase 2C isoform. FEBS
Lett. 1992;297:135-8.
[35] Kuwabara PE. A complex solution to a sexual dilemma. Dev Cell. 2007;13:6-8.
[36] Stothard P, Hansen D, Pilgrim D. Evolution of the PP2C family in Caenorhabditis:
rapid divergence of the sex-determining protein FEM-2. J Mol Evol. 2002;54:267-82.
[37] Rodriguez PL. Protein phosphatase 2C (PP2C) function in higher plants. Plant Mol
Biol. 1998;38:919-27.

62
[38] Suzuki Y, Morris GA, Han M, Wood WB. A cuticle collagen encoded by the lon-3
gene may be a target of TGF-beta signalling in determining Caenorhabditis elegans
body shape. Genetics. 2002;162:1631-9.
[39] Nystrom J, Shen ZZ, Aili M, Flemming AJ, Leroi A, Tuck S. Increased or
decreased levels of Caenorhabditis elegans lon-3, a gene encoding a collagen, cause
reciprocal changes in body length. Genetics. 2002;161:83-97.
[40] Cox GN, Staprans S, Edgar RS. The cuticle of Caenorhabditis elegans II Stage-
specific changes in ultrastructure and protein composition during postembryonic
development. Dev Biol. 1981b;86:456-70.
[41] Cox GN, Kusch M, Edgar RS. Cuticle of Caenorhabditis elegans: its isolation and
partial characterisation. J Cell Biol. 1981a;90:7-17.
[42] Johnstone IL, Barry JD. Temporal reiteration of a precise gene expression pattern
during nematode development. The EMBO journal. 1996;15:3633-9.
[43] Conlon I, Raff M. Size control in animal development. Cell. 1999;96:235–44.
[44] Van, Der, Eycken W, Engler JA, Van, Montagu M, et al. Identification and
analysis of a cuticular collagen-encoding gene from the plant-parasitic nematode
Meloidogyne incognita. Gene. 1994;151:237-42.
[45] Tsuji N, Kasuga-Aoki H, Isobe T, Arakawa T, Matsumoto Y. Cloning and
characterisation of a highly immunoreactive 37 kDa antigen with multi-immunoglobulin
domains from the swine roundworm Ascaris suum. Int J Parasitol. 2002;32:1739-46.
[46] Rebello KM, Barros JS, Mota EM, Carvalho PC, Perales J, Lenzi HL, et al.
Comprehensive proteomic profiling of adult Angiostrongylus costaricensis, a human
parasitic nematode. J Proteomics. 2011;74:1545-59.

63
[47] Morassutti AL, Levert K, Pinto PM, Silva AJ, Wilkins P, Graeff-Teixeira C.
Characterization of Angiostrongylus cantonensis excretory–secretory proteins as
potential diagnostic targets. Exp Parasitol. 2012;130:26-31.
[48] Dalton J. Helminth vaccines: from mining genomic information for vaccine targets
to systems used for protein expression. In J Parasitol. 2003;33:621-40.
[49] Hong X, Bouvier J, Wong MM, Yamagata GY, Mckerrow JH. Brugia pahangi:
identification and characterization of an aminopeptidase associated with larval molting.
Exp Parasitol. 1993;76:127-33.
[50] Rhoads ML, Fetterer RH, Urban JF, Jr. Effect of protease class-specific inhibitors
on in vitro development of the third- to fourth-stage larvae of Ascaris suum. J Parasitol.
1998;84:686-90.
[51] Xu YZ, Dresden MH. Leucine aminopeptidase and hatching of Schistosoma
mansoni eggs. J Parasitol. 1986;72:507-11.
[52] Chung YB, Kong Y, Joo IJ, Cho SY, Kang SY. Excystment of Paragonimus
westermani metacercariae by endogenous cysteine protease. J Parasitol. 1995;81:137-
42.
[53] Mckerrow JH, Caffrey C, Kelly B, Loke P, Sajid M. Proteases in parasitic diseases.
Annu Rev Pathol. 2006;1:497-536.
[54] Rebello KM, De Siqueira CR, Ribeiro EL, Valente RH, Mota EM, Perales J, et al.
Proteolytic activity in the adult and larval stages of the human roundworm parasite
Angiostrongylus costaricensis. Mem Inst Oswaldo Cruz. 2012;107:752-9.
[55] Hasnain SZ, Mcguckin MA, Grencis RK, Thornton DJ. Serine protease(s) secreted
by the nematode Trichuris muris degrade the mucus barrier. PLoS Negl Trop Dis.
2012;6:e1856.

64
[56] Dzik JM. Molecules released by helminth parasites involved in host colonization.
Acta Biochim Pol. 2006;53:33-64.
[57] Mckerrow JH. Parasite proteases. Exp Parasitol. 1989;68:111-5.
[58] Williamson AL, Brindley PJ, Knox DP, Hotez PJ, Loukas A. Digestive proteases
of blood-feeding nematodes. Trends Parasitol. 2003;19:417-23.
[59] Varki AA. Biological roles of oligosaccharides: all of the theories are correct.
Glycobiology. 1993;3:97–130.
[60] Berninsone PM. Carbohydrates and glycosylation. Wormbook. 2006;18:1-22.
[61] Kelleher DJ, Gilmore R. An evolving view of the eukaryotic
oligosaccharyltransferase. Glycobiology. 2006;16:47R-62R.
[62] Haslam SM, Dell A. Hallmarks of Caenorhabditis elegans N-glycosylation:
complexity and controversy. Biochimie. 2003;85:25-32.
[63] Gillon AD, Latham CF, Miller EA. Vesicle-mediated ER export of proteins and
lipids. Biochim Biophys Acta. 2012;1821:1040-9.
[64] Thomas PG, Harn DA, Jr. Immune biasing by helminth glycans. Cell Microbiol.
2004;6:13-22.
[65] Rankin CH. Nematode memory: Now, where was I? Curr Biol. 2005;15:R374-R5.
[66] Hallem EA, Spencer WC, Mcwhirter RD, Zeller G, Henz SR, Rätsch G, et al.
Receptor-type guanylate cyclase is required for carbon dioxide sensation by
Caenorhabditis elegans. PNAS. 2011;108:254-9.
[67] Mori I. Genetics of chemotaxis and thermotaxis in the nematode Caenorhabditis
elegans. Annual review of genetics. 1999;33:399-422.
[68] Baude EJ, Arora VK, Yu S, Garbers DL, Wedel BJ. The cloning of a
Caenorhabditis elegans guanylyl cyclase and the construction of a ligand-sensitive
mammalian/nematode chimeric receptor. J Biol Chem. 1997;272:16035-9.

65
[69] Potter LP. Guanylyl cyclase structure, function and regulation. Cell Signal
2011;23: 1921-1926.
[70] Yan Y, Davis EL. Characterisation of guanylyl cyclase genes in the soybean cyst
nematode, Heterodera glycines. Int J Parasitol. 2002;32:65-72.
[71] Ortiz CO, Faumont S, Takayama J, Ahmed HK, Goldsmith AD, Pocock R, et al.
Lateralized gustatory behavior of C elegans is controlled by specific receptor-type
guanylyl cyclases. Curr Biol. 2009;19:996-1004.
[72] Holden-Dye L, Walker RJ. Neurobiology of plant parasitic nematodes. Invertebrate
neuroscience : IN. 2011;11:9-19.
[73] Gray JM, Karow DS, Lu H, Chang AJ, Chang JS, Ellis RE, et al. Oxygen sensation
and social feeding mediated by a C. elegans guanylate cyclase homologue. Nature.
2004;430:317-22.
[74] Taylor EW, Leite CA, Mckenzie DJ, Wang T. Control of respiration in fish,
amphibians and reptiles. Brazilian journal of medical and biological research = Revista
brasileira de pesquisas medicas e biologicas / Sociedade Brasileira de Biofisica [et al].
2010;43:409-24.

66
Supplemental Table 1 – Expressed proteins of the somatic extract of the Ortleppascaris sp. Identified by PMF using the Nembase4 database.
Spots Proteins GI accession * Score Match Coverage (%) PI/MW
theoretical Peptides
01
hypothetical protein
CAEBREN_23080
[Caenorhabditis brenneri]
341903378 53 4 19 6.18/33828
GFPGTIAAK
LVITMPASMSVER
MALEEGLLCGISSGANVHAACK
DEAEGMGVTTLEADLSR
02 CRE-ATH-1 protein
[Caenorhabditis remanei] 308474210 53 3 43 6.38/15633
ILLGGFSQGAAVSLFSGLTYHR
TPVFFGCGDSDSIVPFGNLMK
SFDYIDSFQEGVEFHVYR
03
hypothetical protein
CAEBREN_12726
[Caenorhabditis brenneri]
341883050 54 5 30 5.80/22246
SDDINCENPK
DDDGMLGGFAHLHKPDITQSR
DDDGMLGGFAHLHKPDITQSR
VDMVELVK
ENLSDSGKPLFDDEDHSWTK
04 As37 [Ascaris suum] 22036079 53 5 17 5.17/34804
APHFPQQPVAR
DLADSDAGAYR
DDGQVMVMEFR
GDEIVAESDR
FEVPQGAPTFTR
05
Sulfotransferase domain
containing protein [Brugia
malayi]
170593967 55 4 26 6.28/37955
EENDILPYLNDSSCHYTICIHEEDGK
FDTFFK
LAFGNYFEHLKPWLPHIYDDNVLFLK
EQSDIIDNMYAQFFSGTEAEFLLTSDMA
06 Cell division protein kinase
7 [Ascaris suum] 324507876 46 3 32 6.54/29952
LLTELYHDNVIALLDVIGHR
TSIQLVFDFMETDLENLIK
NILLQVFLGLEFLHLNFILHR
33, 34, 37 Cell division protein kinase
7 [Ascaris suum] 324507876 50 9 44 4.96/34918
AGFAGDDAPR
AVFPSIVGRPR
IWHHTFYNELR
VAPEEHPVLLTEAPLNPK
LCYVALDFEQEMATAASSSSLEK
38, 39,40 Cell division protein kinase 324507876 48 3 22 6.54/29952 LLTELYHDNVIALLDVIGHR

67
7 [Ascaris suum] TSIQLVFDFMETDLENLIK
NILLQVFLGLEFLHLNFILHR
07 CBN-CDC-37 protein
[Caenorhabditis brenneri] 341896809 44 3 50 6.82/4694
MSHDEDDTYHNIDTPSLFR
MSHDEDDTYHNIDTPSLFR
SHDEDDTYHNIDTPSLFR
08
Dolichyl-
diphosphooligosaccharide--
protein glycosyltransferase
48 kDa subunit [Ascaris
suum]
324509341 41 4 16 5.64/39496
GIALVADELGPLQMILMTASSTAYSHN
PNK
TVEEHPSAVGK
SPPAEYTIMDDVR
WVPFQGK
09
hypothetical protein
CAEBREN_00296
[Caenorhabditis brenneri]
341874383 44 2 58 6.92/5221
MAQVVVAACLLSAIAFALGSHK
GDNAATAEVEIEDRPIHIGR
29, 48, 60
hypothetical protein
CAEBREN_00296
[Caenorhabditis brenneri]
341874383 44 2 85 6.92/5221
MAQVVVAACLLSAIAFALGSHK
GDNAATAEVEIEDRPIHIGR
10
hypothetical protein
CAEBREN_00853
[Caenorhabditis brenneri]
341899417 55 4 52 5.45/18354
LCYVALDFEQEMATAASSSSLEK
CPEALFQPSFLGMESAGIHETSYNSIMK
DLYANTVLSGGSTMYPGIADR
SEYDESGPSIVHR
11 LONg family member (lon-
3) [Caenorhabditis elegans] 17566918 47 3 60 6.78/10985
MGAGTPSNR
ENYGGYGILQQVHSNLFTAK
VALGSLCSCIPQNDCPTGPPGVQGNSG
QPGE
16 LONg family member (lon-
3) [Caenorhabditis elegans] 17566918 44 3 60 6.78/10985
MGAGTPSNR
ENYGGYGILQQVHSNLFTAK
VALGSLCSCIPQNDCPTGPPGVQGNSG
QPGE
41,42 LONg family member (lon-
3) [Caenorhabditis elegans] 17566918 46 3 60 6.78/10985
MGAGTPSNR
ENYGGYGILQQVHSNLFTAK
VALGSLCSCIPQNDCPTGPPGVQGNSG
QPGE
12
hypothetical protein
F46F3.3 [Caenorhabditis
elegans]
115534662 45 3 70 5.58/6972
QGMGSYLHVLPDSK
EGDEPIYTLGASFYGGLEPAGAK
MGYMTRPFG

68
13 guanylyl cyclase
[Heterodera glycines] 4337130 48 3 38 6.67/20988
IGESVEPESFQMATIFFSDVVSFTTLAGK
CSPLLVVNLLNGLYTAFDGIIDTHDCYK
VETIGDGYLVCSGIPK
50 guanylyl cyclase
[Heterodera glycines] 4337130 44 4 38 6.67/20988
IGESVEPESFQMATIFFSDVVSFTTLAGK
IGESVEPESFQMATIFFSDVVSFTTLAGK
CSPLLVVNLLNGLYTAFDGIIDTHDCYK
VETIGDGYLVCSGIPK
14 CRE-NSY-4 protein
[Caenorhabditis remanei] 308465131 58 5 27 8.90/20304
TNITNMGR
SMEFEHSVFVQQAPPPARPR
SMEFEHSVFVQQAPPPARPR
SVADSMPELSR
SYPSDIAMGGR
15
cleavage and
polyadenylation specificity
factor subunit 2 [Trichinella
spiralis]
339247939 52 4 22 6.49/23105
HVIVVGSGLAGLSAALAAFNQGSR
VNGVQDNAK
LFYSDTMK
ANSVGSDR
17
Uncharacterized protein
family UPF0005 containing
protein [Brugia malayi]
170584338 50 3 27 7.93/23630
DHLVNVYATVAMGVGVAAGGASVHL
FTDILR
LGFFFAFCGLSGLGMGPLLER
YIIVLLA
19, 44
Uncharacterized protein
family UPF0005 containing
protein [Brugia malayi]
170584338 48 3 27 7.93/23630
DHLVNVYATVAMGVGVAAGGASVHL
FTDILR
LGFFFAFCGLSGLGMGPLLER
YIIVLLA
49
Uncharacterized protein
family UPF0005 containing
protein [Brugia malayi]
170584338 59 4 37 7.93/23630
DHLVNVYATVAMGVGVAAGGASVHL
FTDILR
ANLLLSLVSIGLMFALISTPHSK
LGFFFAFCGLSGLGMGPLLER
YIIVLLA
18 hypothetical protein
LOAG_04901 [Loa loa] 312075594 46 3 25 8.93/24945
CVIVSQWTILLNILEYHLK
DNLANFGGTICPSK
FISGSTSSDINTGSLIANAVVEK
21 hypothetical protein
LOAG_06031 [Loa loa] 312078154 44 3 25 4.87/18912
IPAAFSGTCHISSELDNGCFK
AGDELNVEFPTGGK
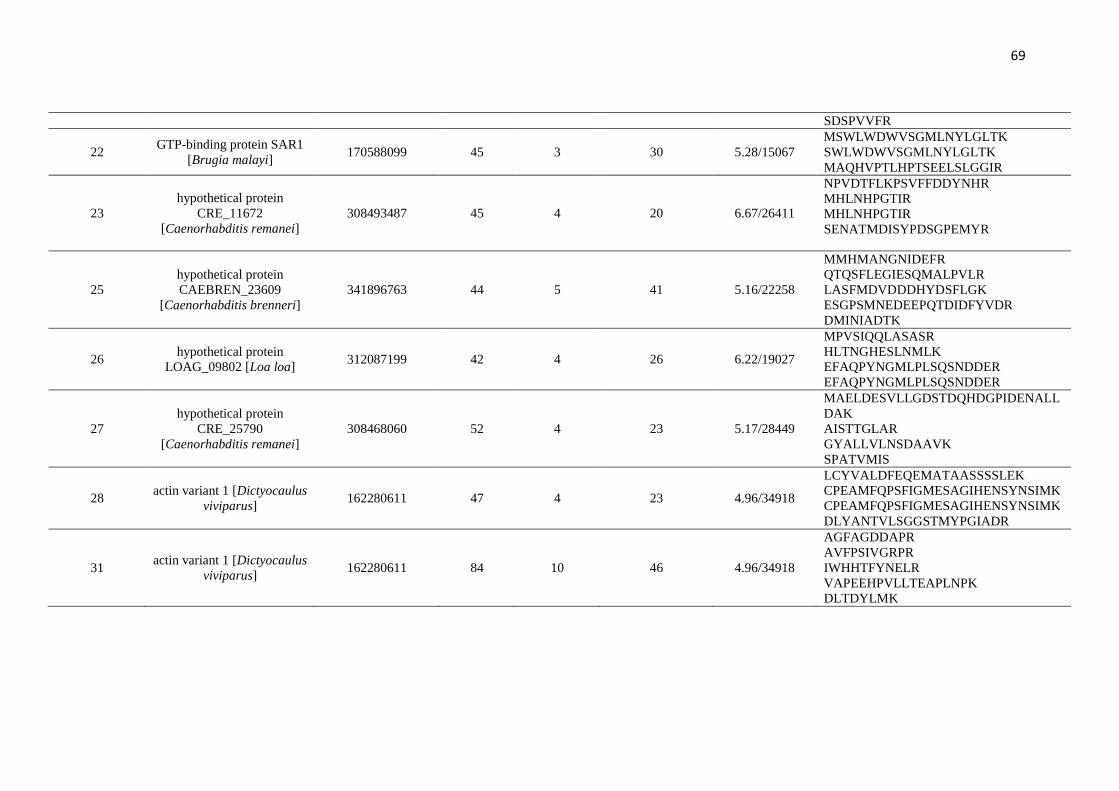
69
SDSPVVFR
22 GTP-binding protein SAR1
[Brugia malayi] 170588099 45 3 30 5.28/15067
MSWLWDWVSGMLNYLGLTK
SWLWDWVSGMLNYLGLTK
MAQHVPTLHPTSEELSLGGIR
23
hypothetical protein
CRE_11672
[Caenorhabditis remanei]
308493487 45 4 20 6.67/26411
NPVDTFLKPSVFFDDYNHR
MHLNHPGTIR
MHLNHPGTIR
SENATMDISYPDSGPEMYR
25
hypothetical protein
CAEBREN_23609
[Caenorhabditis brenneri]
341896763 44 5 41 5.16/22258
MMHMANGNIDEFR
QTQSFLEGIESQMALPVLR
LASFMDVDDDHYDSFLGK
ESGPSMNEDEEPQTDIDFYVDR
DMINIADTK
26 hypothetical protein
LOAG_09802 [Loa loa] 312087199 42 4 26 6.22/19027
MPVSIQQLASASR
HLTNGHESLNMLK
EFAQPYNGMLPLSQSNDDER
EFAQPYNGMLPLSQSNDDER
27
hypothetical protein
CRE_25790
[Caenorhabditis remanei]
308468060 52 4 23 5.17/28449
MAELDESVLLGDSTDQHDGPIDENALL
DAK
AISTTGLAR
GYALLVLNSDAAVK
SPATVMIS
28 actin variant 1 [Dictyocaulus
viviparus] 162280611 47 4 23 4.96/34918
LCYVALDFEQEMATAASSSSLEK
CPEAMFQPSFIGMESAGIHENSYNSIMK
CPEAMFQPSFIGMESAGIHENSYNSIMK
DLYANTVLSGGSTMYPGIADR
31 actin variant 1 [Dictyocaulus
viviparus] 162280611 84 10 46 4.96/34918
AGFAGDDAPR
AVFPSIVGRPR
IWHHTFYNELR
VAPEEHPVLLTEAPLNPK
DLTDYLMK

70
35 actin variant 1 [Dictyocaulus
viviparus] 162280611 104 9 44 4.96/34918
AGFAGDDAPR
AVFPSIVGRPR
IWHHTFYNELR
VAPEEHPVLLTEAPLNPK
LCYVALDFEQEMATAASSSSLEK
55 actin variant 1 [Dictyocaulus
viviparus] 162280611 97 9 44 4.96/34918
AGFAGDDAPR
AVFPSIVGRPR
IWHHTFYNELR
VAPEEHPVLLTEAPLNPK
LCYVALDFEQEMATAASSSSLEK
56 actin variant 1 [Dictyocaulus
viviparus] 162280611 99 9 44 4.96/34918
AGFAGDDAPR
AVFPSIVGRPR
IWHHTFYNELR
VAPEEHPVLLTEAPLNPK
LCYVALDFEQEMATAASSSSLEK
30
[Pyruvate dehydrogenase
acetyl-transferring-
phosphatase 1 [Ascaris
suum]
324508573 46 3 35 6.02/14778
FLIVASDGLWEWFDADTAVR
RPLDTNSATHLLR
LHDQLQLPDGAAR
59
[Pyruvate dehydrogenase
acetyl-transferring-
phosphatase 1 [Ascaris
suum]
324508573 51 3 35 6.02/14778
FLIVASDGLWEWFDADTAVR
RPLDTNSATHLLR
LHDQLQLPDGAAR
32
formyl-CoA transferase
[Micromonospora
aurantiaca ATCC 27029]
302868224 43 3 62 8.76/7336
CQISSNFDGFPPSPLTYDR
HFAGTGFEVVK
VGIFAQHLAR
36
hypothetical protein
LMOh7858_0493 [Listeria
monocytogenes str. 4b
H7858]
47092550 55 3 58 9.47/9612
EEPNNEVLTWNGFKPYKPAK
IDVFLAK
DISDTTFLGLVTVFALVMSCR

71
43
Hypothetical protein
CBG09068 [Caenorhabditis
briggsae]
268572001 46 3 72 4.33/8114
LYDTQPANLSGLNEEFISGAR
LDNLVSTYTAIK
GVVESLEDTDAFDADPNVGSR
46
hypothetical protein
AbauAB_10162
[Acinetobacter baumannii
AB900]
239502665 44 3 26 4.66/19907
NNEEEDGPVIIMSRPDEEK
NSPDDDLINDYVADEQYYR
NCPTSVK
47, 52
hypothetical protein
AbauAB_10162
[Acinetobacter baumannii
AB900]
239502665 47 3 26 4.66/19907
NNEEEDGPVIIMSRPDEEK
NSPDDDLINDYVADEQYYR
NCPTSVK
51 rnf126-prov protein [Brugia
malayi] 170590155 45 3 38 6.58/17389
GGSGSNTPSNGR
AQGPQAIAMQMHPILGQVLSSLGDPHM
QVR
DGTMHGPFGDYVWGENGMDR
53 Nodal modulator 1 [Ascaris
suum] 324501240 44 4 25 6.23/22345
EYFEFDK
IIFFPNR
YLLKPISPMFLPSQIQIDLNNGPVTNLK
SGPVSFLR
57
hypothetical protein
CBG_14574
[Caenorhabditis briggsae
AF16]
309359371 56 6 30 5.02/34059
AGFAGDDAPR
DSYVGDEAQSK
IWHHTFYNELR
VAPEEHPVLLTEAPLNPK
SYELPDGQVITVGNER
58 hypothetical protein
LOAG_01253 [Loa loa] 312067645 44 3 33 6.81/17326
FSQQLKPIFEEASNR
GSAIAWAIVDCDR
QLIPSIQNFTTNVELNNQIDLK
*GenInfo Identifier (GI) – número de acesso no GenBank – NCBI

72
2.4. ARTIGO 4
Título: Abordagem proteômica da relação parasito hospedeiro entre
Ortleppascaris sp. e Rhinella marina.
Revista a ser submetido: International Journal for Parasitology.
ISSN: 0020-7519.
Frequência: Bimensal.
Fator de impacto: 3.393.
Ano de submissão: 2013.

73
ABORDAGEM PROTEÔMICA DA RELAÇÃO PARASITO HOSPEDEIRO
ENTRE ORTLEPPASCARIS SP. E RHINELLA MARINA
Jefferson Pereira e Silva*, Adriano Penha Furtado *, Humberto Josué de Oliveira
Ramos†, Ana Clara Rosa Salvador†, Claudio Lísias Mafra de Siqueira‡ and
Jeannie Nascimento dos Santos *
*Laboratório de Biologia Celular e Helmintologia “Profa. Dra. Reinalda Marisa
Lanfredi”, Instituto de Ciências Biológicas, Universidade Federal do Pará, Belém, Pará,
Brazil. e-mail: [email protected]
†Laboratório de Biologia Molecular de Plantas
‡ Laboratório de Parasitologia e Epidemiologia Molecular
ABSTRACT: Entre microrganismos e seus hábitats faz-se necessária uma gama de
moléculas indispensáveis para o bom funcionamento deste ambiente, no mesmo sentido,
a relação entre helmintos e seus hospedeiros também carrega um arsenal molecular
bioquímico. Dentre estas moléculas, o proteoma constitui-se talvez no maior e mais
importante conjunto substâncias ativas. Proteínas desempenham papéis imprescindíveis
em processos biológicos vitais na relação parasito/hospedeiro, desde metabolismo
básico e produção de energia até respostas imunes complexas. Em helmintos
representam um importante foco para estudos imunológicos e análises bioquímicas. A
proteômica tem fornecido informações valiosas quanto a estes agentes infecciosos e,
então, porque não se voltar para o agente infectado? Neste estudo foi traçado o perfil
proteômico de fluido presente pela encapsulação de larvas de Ortleppascaris sp. no
parênquima hepático do anfíbio Rhinella marina. Proteínas foram separadas por
eletroforese bidimensional e identificadas por MS a partir de Peptide Mass Fingerprint
protéinas. 54 moléculas puderam ser analisadas neste fluido, revelando uma complexa
relação entre parasito-hospedeiro. Este trabalho contribui para estudos proteômicos
relacionados a marcadores proteicos quanto ao desenvolvimento, infectividade,
virulência e co-existência de helmintos e seus albergadores.

74
INTRODUÇÃO
A sobrevivência dos parasitos em determinado organismo depende da relação
parasito-hospedeiro estabelecida entre os mesmos (MCKAY, 2006). O secretoma é o
conjunto completo de produtos excretados/secretados por uma célula ou organismo
(RANGANATHAN & GARG, 2009), por meio do qual podemos entender como um
organismo se comporta, incluindo informações de como um parasita realiza sua funções
no “meio” hospedeiro (HEWITSON, 2008; NAGARAJ et al., 2008).
Johnston et al. (2009) relatam que moléculas bioativas presentes em extratos de
helmintos ou em seus produtos de excreção/secreção são responsáveis por modular a
resposta imune do hospedeiro a seu favor em termos de co-existência.
Estas biomoléculas podem estar envolvidas nos mais diversos níveis fisiológicos
e segundo Craig et al. (2006) e Soblik, et al., (2011), em helmintos, essas substâncias
são derivadas da superfície ou de glândulas excretoras/secretoras especializadas e
freqüentemente liberadas em estágios específicos.
Este elenco de produtos, principalmente as proteínas, tem sido intensamente
estudado, revelando que a liberação destas moléculas desempenha um papel importante
na infecção pelo patógeno e nas interações Hospedeiro-Patógeno. Esta
Excreção/Secreção de proteínas pelos patógenos pode, muitas vezes, suprimir o sistema
imunológico, colaborando para o fortalecimento da infecção ou mesmo auxiliar na
sobrevivência de patógenos em seus hospedeiros definitivos (RANGANATHAN &
GARG, 2009; LIAO et al., 2011).
Para estudos proteômicos, modelos de praxe tem sido utilizados como C.
elegans e ratos (BOLKER, 2012), no entanto, helmintos de interesse humano e
veterinário também são alvos de promissoras investigações (SOTILLO et al.,2012,
ROBINSON et al., 2013).
Aqui destacamos o parasitismo de Ortleppascaris sp. em fígado de Rhinella
marina, no qual larvas deste nematódeo são encontradas no interior de cistos fibrosos,
preenchidos por fluido viscoso (Silva et al., 2013). Durante o parasitismo, o helminto
em questão permanece vivo e se desenvolve no órgão desde estágios elementares,
comunicando-se com o ambiente hospedeiro.
Neste trabalho identificamos por ferramentas proteômicas a composição proteica
do fluido gerado pelo encistamento da larva, parasita do anfíbio Rhinella marina.

75
A análise proteômica e identificação dessas proteínas contribuirá para o avanço
do entendimento da relação parasito-hospedeiro e seus possíveis marcadores
moleculares, expandindo o campo de análise de biomoléculas em novos modelos de
estudo, para o entendimento de infecção helmínticas de interesse humano e veterinário.
MATERIAL E MÉTODOS
Fluido cístico
Foram coletados por sucção fluidos de aproximadamente 200 cistos por sucção,
utilizando seringa estéril descartável e armazenados em tampão de extração (Uréia 7 M
– Tiouréia 2 M – CHAPS 2%) a -20ºC.
Preparação de Proteínas do Fluido cístico
Os fluidos coletados em tampão de extração foram centrifugados a 13000 × g
por 15 min a 4° C para obtenção do sobrenadante, utilizado diretamente para análise de
proteínas.
Eletroforese bidimensional
A concentração de proteínas totais foi determinada pelo método de Bradford
(1976), em seguida as amostras foram armazenadas a - 80° C até uso.
Alíquotas de extrato proteico contendo 100 µg de amostra foram diluídas ao
volume final de 125 µl em solução Destreak (GE healthcare) e 2% de tampão IPG pH 3
- 10 (GE healthcare). Tiras de 07 cm (Immobiline, GE healthcare) de gradiente
imobilizado de pH na faixa 3-10 foram reidratadas com o extrato proteico por 17 h
utilizando IPGBox (GE Healthcare). Iniciou-se a focalização isoelétrica logo após o
término da reidratação.
A focalização isoelétrica foi realizada a 20°C, corrente constante de 50 µA/strip
com um total de 5,0 - 6,5 KVh, seguindo os steps: 300V-4h; gradiente de 1000V-30min;
gradiente 5000V-1:20h; step 5000V-30min, em sistema automatizado (ETTAN
IPGPhor III GE Healthcare).
Após a focalização isoelétrica, as tiras foram reduzidas em tampão de equilíbrio
(6M uréia, 0,075M TrisHCl pH 8,8; glicerol 29%; SDS 2%; azul de bromofenol 0,02%)
contendo 2% DTT (ditiotreitol) durante 30 minutos e em seguida, alquiladas por 30
minutos em mesmo tampão de equilíbrio mencionado, contendo iodacetamida 2,5%.

76
Para a segunda dimensão, as tiras foram colocadas sobre géis de poliacrilamida
12,5% em sistema Mini Protean Cell (Bio-Rad). A corrida de eletroforese seguiu
voltagem constante de 80V por 2 horas. Os géis foram corados em solução Coomassie
Blue G 250, overnight, sob agitação e escaneados em ImageScanner III (GE
Healthcare), software Labscan (GE Healthcare).
Digestão tríptica in gel e espectrometria de massas
Os spots detectados pelo programa ImageMaster 2D Platinum 7.0 (GE
Healthcare) e observados a olho nu foram manualmente excisados, tratados com uma
solução de lavagem (Metanol 50%, Ácido Acético 5%), em seguida desidratados em
acetonitrila 100% em centrífuga a vácuo à temperatura ambiente. Proteínas foram
submetidas a processo de redução (Ditiotreitol 10 mM) e alquilação (Iodacetamida 100
mM).
Amostras foram digeridas a 37 °C, overnight, com Tripsina grau proteômico
(Promega, Madison, WI, USA) em Bicarbonato de Amônio 50 mM, (concentração final
20ng/µL). Peptídeos trípticos foram extraídos do gel com solução acetonitrila 50% em
ácido fórmico 5%.
Os peptídeos extraídos foram transferidos para um tubo estéril, tratados com
Bicarbonato de Amônio 100 mM, secos em centrífuga a vácuo e resuspendidos em
solução de Acetronila 50%, Ácido Fórmico 0,05% e Ácido Trifluoroacético 0,1%.
Alíquotas de 0,5 µL de cada amostra foram aplicadas sobre uma placa de aço,
juntamente com a matriz de ácido 2,5 di-hidróxi-benzóico (Sigma), numa proporção
1:1. Após a cristalização a placa foi inserida no espectrômetro e a análise iniciada.
Todos os espectros MS foram obtidos no intervalo de m/z 800 – 4000 Da,
usando MALDI - TOF em espectrômetro Ultraflex III (Bruker Daltonics). Os espectros
foram submetidos à análise em FlexAnalisys 3.3 (Bruker-Daltonics) para a formação da
peaklist.
A busca de proteínas por homologia de PMF (peptide mass fingerprint) foi
realizada em pesquisa no banco de dados público do NCBI [Nacional Center for
Biotechnology Information (NCBInr)], limitando-se a busca por taxonomia em Xenopus
laevis nos espectros de amostras de fluido, usando a versão on line do Mascot (Matrix
Science). Como parâmetros de busca, foram utilizados até dois sítios de clivagem
perdidos; erro de 0,1 Da para identificação de peptídeos, além de carbamidometilação

77
de cisteínas como modificação fixa e oxidação de metionina como modificação
variável. No sentido de inferir o processo biológico no qual as moléculas identificadas
estão envolvidas, a análise dos dados de PMF foram enriquecidas confrontando-a junto
aos termos do Gene Ontology (GO) por meio da base de dados on line UniProt. As
proteínas identificadas foram agrupadas em categorias e subcategorias funcionais de
acordo com a ontologia.
RESULTADOS
Para caracterizar o perfil proteico do fluido, utilizamos os 54 spots mais
abundantes detectados com o programa Image Master 2D Platinum (Fig. 1).
Figura 1: Gel bidimensional de proteínas provenientes de fluido cístico presente no
encistamento de Ortleppascaris sp.
A identificação das proteínas foi realizada com auxílio do software Mascot, na
qual foram considerados scores significativos acima de 55 (p < 0,05), indicando
homologia significativa com sequência disponível no banco de dados usado na análise.

78
Buscas na base de dados GO (Gene Ontology) foram realizadas em associação
com nossos dados de PMF. A lista de spots assinalados no gel bi-dimensional (Fig. 1)
com suas respectivas proteínas estão sumarizadas no Quadro 1 a seguir segundo a base
de dados utilizada. Destes spots, 47 foram identificados, 4 como não-caracterizadas (03,
13, 34, 36) e 3 spots (35, 37, 49) não originaram espectros de boa qualidade para
identificação, não possuindo homologia suficiente com nenhuma proteína do banco de
dados utilizado.
As proteínas encontradas no fluido compõem um conjunto de moléculas que se
agrupam em categorias gerais para o funcionamento celular, como balanço redox,
tráfego vesicular, etc, mas também revela um grupo de moléculas presentes em
processos específicos sumarizadas a seguir:
Quadro 1. Sumário de proteínas do fluido cístico de Ortleppascaris sp.
identificadas a partir de PMF na base de dados NCBI, busca por taxonomia
em Xenopus laevis. (Ver Tabela 1 material suplementar para mais detalhes)
Spot Proteína
01 prolyl endopeptidase-like
02 CDKN2A interacting protein N-terminal like
03 uncharacterized protein LOC779198
04 MGC84728 protein
05, 16 Cytoglobin
06 cysteine-rich with EGF-like domain protein 2-B precursor
07 Chain A, Crystal Structure Of X. Laevis T-Cadherin Ec1
08 LOC733224 protein, partial
09, 10, 11, 12, 22 TraB domain containing
13, 34 uncharacterized protein LOC379167
14 regulation of nuclear pre-mRNA domain containing 1B
15 proteasome subunit beta type-7
17, 25 phosphopantothenoylcysteine synthetase
18 peroxisome proliferator-activated receptor alpha
19 fidgetin-like protein 1
20 LOC446946 protein, partial

79
21 LOC733282 protein
23 microtubule-associated protein 1 light chain 3 gamma
24 cell division cycle 73, Paf1/RNA polymerase II complex
component, homolog
26 Cas/HEF-like protein 1
27 structure-specific endonuclease subunit slx1
28 actin, cytoplasmic type 5
29 Unknown (protein for MGC:181785)
30 gamma-crystallin-3
31 autophagy protein 5
32 nitrogen fixation cluster-like
33 vacuolar protein sorting 33 homolog A
36 uncharacterized protein LOC734830
38 LOC398774 protein
39 Pro-opiomelanocortin B (unnamed protein product)
40 LOC443709 protein, partial
41 LOC443713 protein, partial
42, 52 copper chaperone for superoxide dismutase
43, 50 ATPase, H+ transporting, lysosomal 50/57kDa, V1 subunit H
44 RAB3A, member RAS oncogene Family
45 aspartoacylase-2
46 tubulin tyrosine ligase-like family, member 7
47 MARK3 protein
48 5-aminoimidazole-4-carboxamide ribonucleotide
formyltransferase/IMP cyclohydrolase
51, 53 LOC414512 protein, partial
54 methylthioadenosine phosphorylase
Prolyl endopeptidase-like (spot 01), da subfamília S9A, com provável atividade
de endopeptidade em serina. Subunidade beta de proteossomo (spot 15), também com
atividade de endopeptidadase, envolvida em via proteolítica não lisossomal dependente
de ATP/ubiquitina.

80
Proteína similar a inibidor de quinase dependente de ciclina (spot 02), cujos
genes correspondentes são candidatos a supressão de tumores.
Moléculas responsáveis por processos bioquímicos básicos tais como
citoglobina (spots 05, 16) atuante no transporte de oxigênio; caderina (spot 07),
relacionada em processos de adesão intracelular. Proteína fidgetin-like 1 (spot 19),
integrante da ampla família AAA ATPases.
Hidrolases responsáveis pela quebra ATP e transporte de prótons foram
encontradas nos spots 43 e 50 e Aspartociclase no spot 45.
Neste mesmo repertório de moléculas ubíquas expressas em eucariotos,
identificamos actina citoplasmática tipo 5 (spot 28), proteínas referentes ao tráfego
vesicular (spots 33 ,44); moléculas com função de ligação às proteínas (spot 38), ao
zinco (spot 40), modificação proteica (spot 46), fosforilação (spot 47).
Estão presentes moléculas do processo de oxi-redução de ácidos graxos, como
acil-CoA desidrogenase (spot 08) e PPAR-alpha (spot 18) (Peroxisome proliferator-
activated receptor alpha). Além dessas foram detectadas a proteína copper chaperone
for superoxide dismutase (CCS) em 2 spots (42, 52) e proteína nitrogen fixation cluster-
like (Nifu) (spot 32) operante no sistema de montagem de compostos “iron-sulphur”,
importantes para a transferência de elétrons.
Proteínas com função ainda não elucidada estão presentes neste conjunto de
proteínas tais como domínio proteico TraB , identificadas em 05 spots (9, 10, 11, 12,
22) moléculas de ligação a ácidos nucléicos como LOC733282 protein (spot 21) e
cdc73 (spot 24) (cell division cycle 73, Paf1/RNA polymerase II complex component,
homolog) acreditadas como fator acessório da RNA polimerase II. Além de
endonucleases responsáveis por reparo e recombinação de DNA como structure-specific
endonuclease subunit slx1 (spot 27) e LOC443713 protein (spot 41). Em associação aos
ácidos nucléicos foram determinadas moléculas envolvidas na biossíntese de purinas
(spot 48), ligação a ácidos nucléicos (spots 51, 53) e metabolismo de nucleosídeos (spot
54).
Moléculas associadas ao processo de autofagia foram detectadas: Microtubule-
associated protein 1 light chain 3 gamma, também denominada map1lc3b ou LC3 (spot
23), membro da família de proteínas ATg8 ubiquitin-like (autophagy protein) e
autophagy protein 5 (spot 31).

81
Pro-opiomelanocortin B (spot 39), importante hormônio neurotransmissor
precursor de outros neuropeptídeos foi identificado com score significativo dentre este
conjunto de proteínas do fluido.
Análise Proteômica do Fluido Cístico
A pesquisa no banco de dados dos resultados foi realizada delimitando-se o
parâmetro de busca taxonômica para o organismo Xenopus laevis, uma vez que este é o
organismo taxonomicamente mais próximo ao hospedeiro R. marina no banco de dados
utilizado, ademais, nenhum spot referente ao extrato somático do parasito foi detectado
no gel bi-dimensional do fluido, sugestivo de que estas moléculas abundantes são do
hospedeiro e não do parasito.
O Quadro 2 a seguir mostra lista de proteínas identificadas com seus respectivos
spots identificados agrupadas em categorias e subcategorias funcionais de acordo com a
ontologia (Gene Ontology), minuciados na Tabela 2 (material suplementar), a qual
contém os dados referentes identificação pontual de cada spot do gel de fluido cístico
indicados na figura 1, o nome da proteína e seu respectivo helminto, os números de
acesso no GI indicado corresponde a sequencia de proteína do banco de dados do
National Center for Biotechnology Information (NCBI) com maior identidade com os
peptídeos, scores, match (número de peptídeos encontrados), coverage
(representabilidade dos peptídeos para a determinada proteína), ponto isoelétrico e
massa molecular teóricos e as sequências de aminoácidos dos peptídeos correspondentes
obtidas por espectrometria de massa.

82
Quadro 2. Sumário das proteínas identificadas a partir do gel de fluido cístico e processo biológico envolvido em termos
GO (categoria de processo biológico). Veja Tabela 2 com informações adicionais.
Moléculas do processo metabólico básico e produção de energia
Termo GO Processo/função biológico(a) Spot Proteína
GO: 0006508/
GO: 0004252
Proteólise/ Endopeptidase em
serina 01 Prolyl endopeptidade-like
GO: 0015671 Transporte de oxigênio 05, 16 Cytoglobin
GO: 0051603/GO:
0004252
Proteólise de processo
catabólico/ Endopeptidase 15 Proteasome subunit beta type
GO: 0046034 Processo metabólico de ATP 19 fidgetin-like protein 1
GO: 0005856 Citoesqueleto 28 actin, cytoplasmic type 5
GO: 0005515 Ligação a proteínas 38 LOC398774 protein
GO: 0016192 vesicle-mediated transport 33 vacuolar protein sorting 33 homolog A
GO: 0008270 Ligação com íon zinco 40 LOC443709 protein, partial
GO: 0015991 hidrólise de ATP e transporte de
prótons 43, 50 ATPase, H+ transporting, lysosomal 50/57kDa, V1 subunit H
GO: 0015031 Transporte de proteínas 44 RAB3A, member RAS oncogene Family
GO: 0016787 Hidrolase 45 Aspartoacylase-2
GO: 0006464 Modificação proteica celular 46 tubulin tyrosine ligase-like family, member 7

83
GO: 0006468 Fosforilação protéica 47 MARK3 protein
Moléculas envolvidas em processo de oxi-redução e stress oxidativo
Termo GO Processo/função biológico(a) Spot Proteína
GO:0055114/GO:0
003995
Oxi-redução/Acil-CoA
desidrogenase 08 LOC733224 protein, partial
GO: 0008289 Ligação em lipídeos 18 peroxisome proliferator-activated receptor alpha
GO: 0016226 Síntese de compostos Fe-S 32 nitrogen fixation cluster-like
GO: 0055114/ GO:
0006801
Oxi-redução/ processo
metabólico de superóxido 42, 52 copper chaperone for superoxide dismutase
Moléculas relacionadas a ácidos nucléicos
Termo GO Processo/função biológico(a) Spot Proteína
GO: 0003676 Ligação a ácidos nucléicos 21 LOC733282
GO: 0006281 Reparo de DNA 27 structure-specific endonuclease subunit slx1
41 LOC443713
GO: 0006164 Biossíntese de nucleotídeos de
purina 48
5-aminoimidazole-4-carboxamide ribonucleotide
formyltransferase/IMP cyclohydrolase
GO: 0000166 Ligação a ácido nucléico 51, 53 LOC414512 protein, partial
GO: 0009116 Metabolismo de nucleosídeos 54 methylthioadenosine phosphorylase
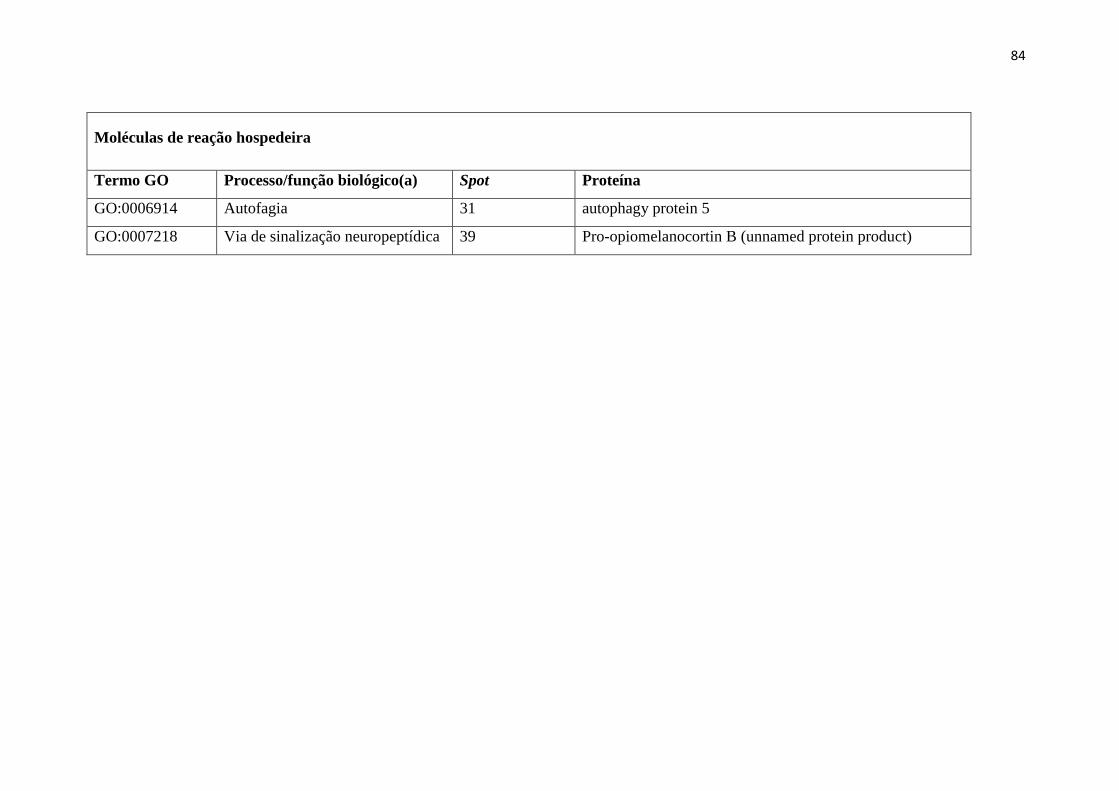
84
Moléculas de reação hospedeira
Termo GO Processo/função biológico(a) Spot Proteína
GO:0006914 Autofagia 31 autophagy protein 5
GO:0007218 Via de sinalização neuropeptídica 39 Pro-opiomelanocortin B (unnamed protein product)

85
DISCUSSÃO
Identificação de Proteínas e Interpretação
Problemas na identificação de seqüências, em análise proteômica estão
relacionados a não produção de scores significantes, já que vários desses bancos estão
incompletos ou não bem estabelecidos (Medina et al., 2005). Segundo Antunes (2008),
esses dados devem ser considerados mesmo em os valores baixos de scores,
considerando a inexistência de um banco de dados completo e específico para os
organismos em questão. A escassez do banco de dados mais apropriado para cada
análise é responsável por redução nos valores dos score.
Apesar deste entrave, a perspectiva é de que a detecção destas moléculas sejam
um forte indicativo de sua presença nos sistemas de estudo.
Moléculas do Processo Metabólico básico e Produção de Energia
Clivagem peptídica é um processo muito comum e requer um número bastante
grande de famílias de enzimas. Dentre essas famílias encontra-se a Prolyl-peptidase,
uma dipeptil peptidase (DPP), com especificidade de clivagem em sítios ricos em
prolina, desempenhando uma função central na produção, regulação e degradação de
peptídeos in vitro (Lone et al., 2010).
Segundo Rosenblum & Kozarich (2003) estas proteínas desempenham um
importante papel na regulação de sinalização por hormônios peptídicos e talvez estejam
envolvidas na sinalização a partir do hormônio neuroendócrino Pro-opiomelanocortin
B, o qual está relacionado a respostas em situações de stress em alguns anuros.
São também indicadores moleculares para doenças oncogênicas, assim como
CDKN2A (Cyclin-dependent kinase inhibitor 2A), também denominada p16 (Drexler,
1998), importante proteína para regulação do ciclo celular e tumoral.
Sua detecção no fluido pode ser justificada pela presença de moléculas de
colágeno, proteína rica em prolina, encontrado na cutícula de Ortleppascaris sp. ou
mesmo na matriz celular da face interna do cisto fibroso desencadeando, de acordo com
Lone et al., (2010), aumento da produção de FAP (fibroblast activation protein)
membro desta família de enzimas, cuja função é degradação dessa matriz.

86
Com função semelhante, subunidade beta do proteossomo foi registrada.
Segundo Groll & Huber (2003) é a maior via proteolítica em eucariotos, composta por
um complexo proteico multimérico dependente de ATP/ubiquitina, formado por mais de
31 diferentes subunidades alfas e betas. Esta maquinaria degrada proteínas anormais e,
assim, produz a maior fonte de antígenos peptídicos apresentados para o sistema imune
através do MHC de classe I, mas também regula a degradação em importantes processos
como adaptação metabólica, assim como é observado no interior do cisto, onde o
hospedeiro necessita de uma maquinaria adequada para a apresentação de antígenos,
reconhecimento do helminto, bem como necessita adaptar seu metabolismo frente à
infecção.
A citoglobina ainda é uma incógnita, pois sua função não é bem definida no
metabolismo em anfíbios, porém, estes animais ocupam posição filogenética favorável a
estudos genômicos devido uma conservação evolutiva dessa molécula (Fuchs et al.,
2006). Acredita-se que sua expressão exacerbada esteja relacionada com
hipóxia/isquemia tecidual (Pesce et al., 2002) e/ou síntese de colágeno (Schmidt et al.,
2004).
Estes aspectos estão presentes no fígado do hospedeiro R. marina, onde a
vascularização/oxigenação foi comprometida pelo parasitismo assim como a
organização tecidual mostrados por Silva et al. (2013), justificando, por conseguinte, a
expressão de caderinas, moléculas, as quais segundo Leckband & Prakasam (2006) são
responsáveis, dentre outras funções, pela manutenção da estrutura tecidual.
Quanto a molécula fidgetin-like 1, estudos realizados por Yang et al. (2005)
apontam sua função para o proliferação celular. O que na relação Ortleppascaris sp.
versus R. marina pode estar relacionada a proliferação celular larval assim como ocorre
em C. elegans (Luke-Glaser et al., 2007), sendo liberada para o meio extracorpóreo, e,
portanto presente no fluido.
Moléculas do Processo Oxi-redução e Stress Oxidativo
De acordo com Yang et al. (2005), a molécula fidgetin-like 1 participa, dentre
outras atividade celulares, do transporte mediado por vesículas aos peroxissomos.
Outras moléculas obtidas desta amostra influem nas reações redox, associadas, a
proliferação de peroxissomos, como a proteína PPAR-alpha, indicando um alto
metabolismo e reações beta-oxidação frente a ácidos graxos presentes neste fluido.

87
Além desse conjunto de proteínas outras moléculas-chave para metabolismo
lipídico, reguladoras da via de beta-oxidação ácidos graxos foram encontradas, tais
como, acil-CoA desidrogenase, NifU, envolvida na formação de complexos [Fe-S] e
transporte de elétrons (Johnson et al., 2005) e Copper Chaperone for Superoxide
dismutase (CCS) que segundo Rosenzweig & O’Halloran (2000) é marcador de uma
forte reação antioxidante, presente no fluido.
Moléculas da Reação Hospedeira
LC3 é expressa sob várias condições de stress (Shpilka et al., 2011), como
doenças, infecções microbianas, fase tumoral, e fome, além de ser usado como
marcador molecular de autofagia em mamíferos (Tanida et al., 2004), indicando que R.
marina está sob condições desfavoráveis a partir da infecção por Ortleppascaris sp.,
somado a essas informações foi encontrada autophagy protein 5, a qual assim como
p16, é marcador de processos tumorais e autofagia.
Ainda sim, moléculas de combate ao stress oxidativo, como CSS, aparecem de
modo abundante, sugerindo grande quantidade de radicais livres no fluido cístico, o que
pode certamente contribuir, junto com a injúria celular e hipóxia local, para processos
de autofagia e doenças relacionadas ao câncer, como afirmado por Kongara & Karantza
(2012), uma vez que PPAR-alpha está presente no conjunto de moléculas que podem
aumentar o número de radicais livres.
Todas estas moléculas relatadas podem estar envolvidas em processos de
resposta ao stress celular ou mesmo na resposta imune do anfíbio, uma vez que seu
parênquima sofre alterações na presença dos cistos contendo fases larvais de
Ortleppascaris sp.
A exemplo disto, encontramos com valores significativos de score, a proteína
Pro-opiomelanocortin B, encontrada nos anuros Rana idibunda e X. laevis (Jenks et al.,
2011) precursor de importantes neuropeptídios, como o α-MSH.
A presença dessa molécula está relacionada dentre outros processos, com um
reflexo neuroendócrino de mudança adaptativa de cor de pele em anfíbios, que envolve
a redistribuição e dispersão de pigmentos pelos melanóforos da pele, causando
escurecimento da mesma (Jenks et al., 2003). A Pro-opiomelanocortin B é liberada no
organismo em condições desfavoráveis ou stress intenso, como má nutrição e infecções

88
(Kolk et al., 2002), fatores sob os quais R. marina em questão está certamente
submetido durante a manutenção do parasitismo por Ortleppascaris sp.
CONCLUSÃO
O novo modelo de estudo, anfíbio-helminto, torna a pesquisa relevante, pois até
então, a maioria de estudos têm sido realizados apenas em murinos como modelos
animais, considerando que muitos resultados discrepantes têm sido obtidos usando
diferentes modelos animais e helmínticos, fazendo desta uma oportunidade para
observação da co-evolução destes organismos à luz da variedade e plasticidade de
moléculas protéicas expressas de acordo com diferentes nichos hospedeiros ou ambiente
em que esses helmintos se encontram.
A abordagem molecular de infecções helmínticas em R. marina traz à tona uma
gama de informações sobre o elenco de antígenos proteicos presentes em um animal
sinantrópico, considerado um indicador ambiental e vetor em potencial para zoonoses
helmínticas.
O mapeamento proteico da interface helminto-anfíbio pode demonstrar como o
hospedeiro reage frente ao desequilíbrio causado pelo parasitismo de forma a adaptar-se
a esta condição, contribuindo para elucidação da biologia das relações parasito-
hospedeiro.
REFERÊNCIAS
ANTUNES, P.W.P. Proteômica e peptidômica aplicadas ao estudo da defesa de
plantas de tomate à injúria mecânica (Mestrado em Bioquímica Agrícola) -
Viçosa, Universidade Federal de Viçosa, 2008. 99p.
BOLKER, J. Model organisms: There's more to life than rats and flies. Nature 491
(7422): 31-3, 2012.
CRAIG, H., WASTLING, J.M., KNOX, D.P. A preliminary proteomic survey of the in
vitro excretory/secretory products of fourth-stage larval and adult Teladorsagia
circumcincta, Parasitology 132: 535-543, 2006.
DREXLER, H.G. Review of alterations of the cyclin-dependent kinase inhibitor INK4
family genes p15, p16, p18 and p19 in human leukemia-lymphoma cells.
Leukemia 12: 845-859, 1998.
FUCHS, C., BURMESTER, T. & HANKELN, T. The amphibian globin gene repertoire
as revealed by the Xenopus genome. Cytogenetic and Genome Research 112:
296-306, 2006.

89
GROLL, M. & HUBER, R. Substrate access and processing by the 20S proteasome core
particle. The International Journal of Biochemistry & Cell Biology 35: 606-616,
2003.
HEWITSON, J.P., HARCUS, Y.M., CURWEN, R.S., DOWLE, A.A., ATMADJA,
A.K., ASHTON, P.D., WILSON, A., MAIZELS, R.M. The secretome of the filarial
parasite, Brugia malayi: proteomic profile of adult excretory-secretory products.
Molecular and Biochemical Parasitoly 160: 8-21, 2008.
JENKS, B.G., GALAS, L., KURIBARA, M., DESRUES, L., KIDANE, A.H,
VAUDRY, H., SCHEENEN, W.J., ROUBOS, E.W. & TONON, M.C. Analysis of
the melanotrope cell neuroendocrine interface in two amphibian species, Rana
ridibunda and Xenopus laevis: A celebration of 35 years of collaborative research.
General and Comparative Endocrinology 170: 57-67, 2011.
JENKS, B.G., ROUBOS, E.W., & SCHEENEN, W.J. Ca 2+
oscillations in melanotropes
of Xenopus laevis: their generation, propagation, and function. General and
Comparative Endocrinology 131: 210-219, 2003.
JOHNSON, D.C., DEAN, D.R., SMITH, A.D. & JOHNSON, M.K. Structure, function,
and formation of biological iron-sulfur clusters. Annual Review of
Biochemistry74: 247-8, 2005.
JOHNSTON, M.J.G., MACDONALD, J.A., MCKAY, D.M. Parasitic helminths: a
pharmacopeia of anti-inflammatory molecules. Parasitology 136: 125-147, 2009.
KOLK, S.M., KRAMER, B.M.R., CORNELISSE, L.N., SCHEENEN, W.J.J.M.,
JENKS, B.G. & ROUBOS, E.W. Multiple control and dynamic response of the
Xenopus melanotrope cell. Comparative Biochemistry and Physiology Part B
132 257-268, 2002.
KONGARA, S. & KARANTZA, V. The interplay between autophagy and ROS in
tumorigenesis. Frontiers in Oncology 2: 171, 2012.
LECKBAND, D. & PRAKASAM, A. Mechanism and dynamics of cadherin adhesion.
The Annual Review of Biomedical Engineering 8: 259-87, 2006.
LIAO Q, YUAN X, XIAO H, LIU C, LV Z, et al. Identifying Schistosoma japonicum
Excretory/Secretory Proteins and Their Interactions with Host Immune System.
PLoS ONE 6 (8): e23786, 2011.
LONE, A.M., NOLTE, W.M., TINOCO, A.D. & SAGHATELIAN, A. Peptidomics of
the Prolyl Peptidases. The AAPS Journal 12 (4): 483-91, 2010.
LUKE-GLASER, S., PINTARD, L., TYERS, M. & PETER, M. The AAA-ATPase
FIGL-1 controls mitotic progression, and its levels are regulated by the CUL-
3MEL-26 E3 ligase in the C. elegans germ line. Journal of Cell Science 120:
3179-3187, 2007.

90
MCKAY, D.M. The therapeutic helminth? Trends in Parasitology 25 (3): 109-114,
2006.
MEDINA, M.L., HAYNES, P.A., BRECI, L. & FRANSCISCO, W.A. Analysis of
secreted proteins from Aspergillus flavus. Proteomics 5: 3153-3161, 2005.
NAGARAJ, S.H., GASSER, R.B., RANGANATHAN, S. Needles in the EST haystack:
large-scale identification and analysis of excretory-secretory (ES) proteins in
parasitic nematodes using expressed sequence tags (ESTs). PLoS Neglected
Tropical Diseases 2: 301, 2008.
PESCE, A., BOLOGNESI, M., BOCEDI, A., ASCENZI, P., DEWILDE, S., MOENS,
L., THOMAS HANKELN & BURMESTER, T. Neuroglobin and cytoglobin: Fresh
blood for the vertebrate globin Family. European Molecular Biology
Organization. Reports 3 (12): 1146-1151, 2002.
RANGANATHAN, S. & GARG, G. Secretome: clues into pathogen infection and
clinical applications. Genome Medicine 1: 113. 2009.
ROBINSON MW, DALTON JP, O'BRIEN BA, DONNELLY S. Fasciola hepatica: The
therapeutic potential of a worm secretome. International Journal for
Parasitology 43: 283-291, 2013.
ROSENBLUM, J.S. & KOZARICH, J.W. Prolyl peptidases: a serine protease
subfamily with high potential for drug discovery. Current Opinion in Chemical
Biology 7: 496-504, 2003.
ROSENZWEIG, A.M. & O’HALLORAN, T.V. Structure and chemistry of the copper
chaperone proteins. Current Opinion in Chemical Biology 4: 140-147, 2000.
SCHMIDT, M., GERLACH, F., AVIVI, A., LAUFS, T., WYSTUB, S., SIMPSON,
J.C., NEVO, E., SAALER-REINHARDT, S., REUSS, S., HANKELN, T. &
BURMESTER, T. Cytoglobin is a respiratory protein in connective tissue and
neurons, which is up-regulated by hypoxia. The Journal of Biological Chemistry
279 (9): 8063-8069, 2004.
SHPILKA, T., WEIDBERG, H., PIETROKOVSKI, S. & ELAZAR, Z. Atg8: an
autophagy-related ubiquitin-like protein Family. Genome Biology 12 (7): 226,
2011.
SILVA, J.P., SILVA, D.C.B., MELO, V.T.V., GIESE, E.G., FURTADO, A.P., &
SANTOS, J.N. Liver histopathological alterations in the cane toad, Rhinella marina
(AMPHIBIA: BUFONIDAE), induced by Ortleppascaris sp. larvae
(NEMATODA: ASCARIDIDAE). The Journal of Parasitology, 2013 (no prelo)
SOBLIK, H., YOUNIS, A.E., MITREVA, M., RENARD, B.Y., KIRCHNER, M.,
GEISINGER, F., STEEN, H. & BRATTIG, N.W. Life cycle stage-resolved
proteomic analysis of the excretome secretome from Strongyloides ratti
identification of stage-specific protease. Molecular & Cellular Proteomics 10
(12), 2011.

91
SOTILLO J, TRELIS M, CORTÉS A, VALERO ML, DEL PINO MS, GUILLERMO
ESTEBAN J, MARCILLA A, TOLEDO R. Proteomic analysis of the pinworm
Syphacia muris (Nematoda: Oxyuridae), a parasite of laboratory rats. Parasitology
International 61: 561-564, 2012.
TANIDA, I., UENO, T. & KOMINAMI, E. LC3 conjugation system in mammalian
autophagy. The International Journal of Biochemistry & Cell Biology 36: 2503–
2518, 2004.
YANG, Y., MAHAFFEY, C.L., BÉRUBÉ, N., NYSTUEN, A., FRANKEL. W.N.
Functional characterization of fidgetin, an AAA-family protein mutated in fidget
mice. Experimental Cell Research 304: 50-58, 2005.

92
Tabela suplementar 1 - Proteínas expressas na amostra fluido cístico de Ortleppascaris sp. identificadas por PMF na base de dados NCBI, busca por taxonomia em Xenopus laevis.
Spots Proteína Acesso GI* Score Match Coverage (%) PI/MW
Teórico Peptídeos
01 prolyl endopeptidase-like 291290935 47 8 15 6.81/81428
MRNLAQQVGLFVCR
QKFMAVTLK
RFITINSNSK
IAGVIYYIEHSNGCLYMLRRHGEAAEYK
RHGEAAEYK
HWEPVYEVQERTKLVDMEMLK
DHCLLFLK
ITAYENDQRVPIQGLLGYITRLR
02 CDKN2A interacting
protein N-terminal like 147901853 32 3 43 5.00/11255
FQSFSESDK
AREEFIIR
VCGMAEGIVVEDAPHFTTRDEIMK
03 uncharacterized protein
LOC779198 148235523 43 7 26 6.29/47492
YDPPDPRR
KSRWDELEISGR
LQTLGLKGNPLSQDILGLYQEPDGMR
DQILPSVPFTVMCFNVLCDK
AKIMSDQEK
KHVDGCAIFFRTEK
QLLMVANAHMHWDPEYSDVK
04 MGC84728 protein 148228072 45 6 16 9.93/33217
AQFLVEKAK
HKIVQAEGEATAAKMIGDALSK
IVQAEGEATAAKMIGDALSK
IVQAEGEATAAKMIGDALSKNPGYLK
MIGDALSKNPGYLKLR
RIRAAQSIAK
05 Cytoglobin 147906731 30 3 38 5.90/20708
GVIKETWARVYANCEDVGVSILIR
QHFSQFKHMEDPLEMEGSVQLR
VEPVYFKILTGVMLEVFAEEYAK
16 Cytoglobin 147906731 31 3 38 5.90/20708
GVIKETWARVYANCEDVGVSILIR
QHFSQFKHMEDPLEMEGSVQLR
VEPVYFKILTGVMLEVFAEEYAK
06 cysteine-rich with EGF- 148226184 40 4 13 4.60/41769 QKYPDLLK

93
like domain protein 2-B
precursor
SGDGSCRCK
ECNNGWVK
DCATGFVMLSGKCTDVDECDASEK
07
Chain A, Crystal Structure
Of Xenopus Laevis T-
Cadherin Ec1
290560285 35 3 24 4.97/10779
IPFPKIVGR
LYGKGVDQEPK
GVDQEPKGIFK
08 LOC733224 protein,
partial 62825972 40 4 45 9.54/27128
TDPEAPRHKQQTMILIPMDTAGIK
TRIAFGKPLVEQGTILAGIAQSRAEIEQAR
AAHLMDTVGNKTAALEISLIKMVAPTMAYR
AIQAFGAAGLSSDFPLAQFHAWARALR
09 TraB domain containing 148235771 65 7 37 5.84/42453
TIQEVQPDVVVVELCQYRVSMLK
EASKVPFCK
QKDLLEQMMAEMIGEFPDLHRTIVSER
ASETEPRKSIPSVVVGVVGMGHVPGIER
NWNCELNIQEIMRVPPPSTLSRAVAFAAK
IILSVPATQYCLQRLNQMATDLK
LNQMATDLK
10 TraB domain containing 148235771 42 5 21 5.84/42453
ILWEMKIK
EASKVPFCK
QKDLLEQMMAEMIGEFPDLHRTIVSER
NWNCELNIQEIMRVPPPSTLSRAVAFAAK
LNQMATDLK
11 TraB domain containing 148235771 37 5 16 5.84/42453
ILWEMKIK
TIQEVQPDVVVVELCQYRVSMLK
EASKVPFCK
IILSVPATQYCLQRLNQMATDLK
LNQMATDLK
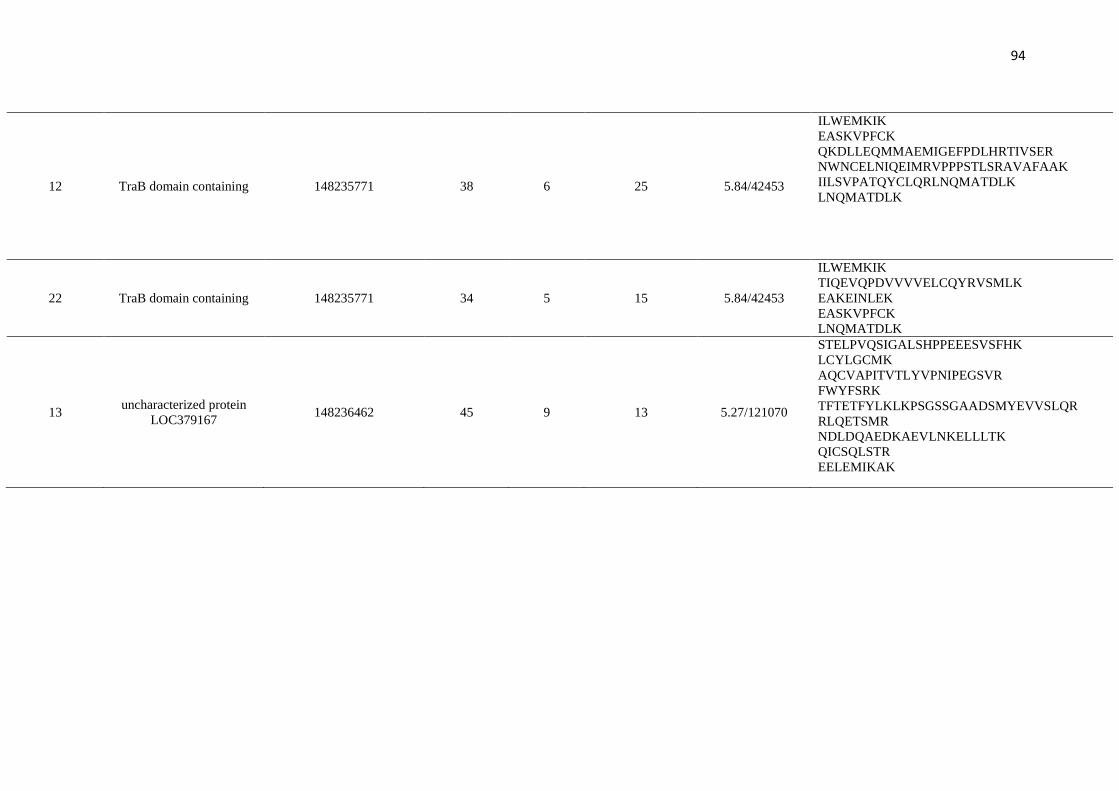
94
12 TraB domain containing 148235771 38 6 25 5.84/42453
ILWEMKIK
EASKVPFCK
QKDLLEQMMAEMIGEFPDLHRTIVSER
NWNCELNIQEIMRVPPPSTLSRAVAFAAK
IILSVPATQYCLQRLNQMATDLK
LNQMATDLK
22 TraB domain containing 148235771 34 5 15 5.84/42453
ILWEMKIK
TIQEVQPDVVVVELCQYRVSMLK
EAKEINLEK
EASKVPFCK
LNQMATDLK
13 uncharacterized protein
LOC379167 148236462 45 9 13 5.27/121070
STELPVQSIGALSHPPEEESVSFHK
LCYLGCMK
AQCVAPITVTLYVPNIPEGSVR
FWYFSRK
TFTETFYLKLKPSGSSGAADSMYEVVSLQR
RLQETSMR
NDLDQAEDKAEVLNKELLLTK
QICSQLSTR
EELEMIKAK

95
34 uncharacterized protein
LOC379167 148236462 53 13 20 5.27/121070
AMEEILRDSEK
STELPVQSIGALSHPPEEESVSFHK
LCYLGCMK
AQCVAPITVTLYVPNIPEGSVR
SEVASFPIYK
NVKNSDMHLLDMESMGKSSDSK
RVYMTVAVDMVVTEVVEPVRFLLETVVR
FWYFSRK
KTFTETFYLK
NTEGDGQESLYK
NDLDQAEDKAEVLNKELLLTK
EELEMIKAK
TLNSIKTATGTQPTSAPSQQGSQASSPVLPAAK
14
regulation of nuclear pre-
mRNA domain containing
1B
147905258 32 4 18 5.41/36911
ASFTEPALEK
LSELSNSQQSVQTLSLWLIHHRK
QLAKMLMEYTQTQKEVLSNK
KLEEYKQK
15 proteasome subunit beta
type-7 147906595 40 5 27 8.64/30389
TGTTIAGIVFK
VATANRMLK
NAIAAGIFNDLGSGSNIDLCVITKNK
KGTTGILSEK
KGTTGILSEKITHFNLDVMEESVQTMDIS
17 phosphopantothenoylcystei
ne synthetase 148234547 39 5 14 7.15/33266
FLDNFSSGR
FLDNFSSGRR
AGYAVIFLHR
HRSMYPYSR
IQSSSGPLQITMK
25 phosphopantothenoylcystei
ne synthetase 148234547 37 5 14 7.15/33266
FLDNFSSGR
FLDNFSSGRR
AGYAVIFLHR
HRSMYPYSR
IQSSSGPLQITMK

96
18 peroxisome proliferator-
activated receptor alpha 148226176 36 6 16 5.58/52966
FEKCLNVGMSHNAIR
FEKCLNVGMSHNAIRFGR
ASTPPFVIHDMETLCMAEKTLVAK
GLRKPIGDMMEPK
LLQKMADLR
LLQKMADLRQLVTEHAQLVQTIK
19 fidgetin-like protein 1 148235265 47 6 14 7.20/72677
YSAILDSDKLEIGLNNYADSILTMAKCQR
WQSSLTTNNVLKLKSVQEMAEAGR
SVQEMAEAGRRAQLSLLNSTDASVR
EQLWVDQQK
NPGPLYGGGKK
QIVVSLMSK
20 LOC446946 protein,
partial 51703613 33 4 13 8.69/32266
HSGSTMAGSR
QQKLLREK
LPEQVTKPLPK
DIVLNVEEK
21 LOC733282 protein 67678022 35 3 8 5.75/31646
IMQKYGFR
YGFREGQGLGK
EECEKYGK
23
microtubule-associated
protein 1 light chain 3
gamma
148227010 39 6 55 9.82/14574
MSQPFKQRK
IPVIVERYRR
EKYLPLLDK
YLPLLDKTK
FLVPKDLSMMQFINIIRNR
DHKDEDGFLYMTYASQEMFG
24
cell division cycle 73,
Paf1/RNA polymerase II
complex component,
homolog
147901047 52 11 29 9.68/ 60701
ADVLSVLRQYNIQK
NVKTNYVIWGTGKEGQPR
VADEISAEAKKPR
EGMVATEQIRSLSEAMSVEKIAAIK
RSTIKTDLDDDITALK
YDQERFKGK
SVTEGASARK
KTQTPAVQPVPRPVSQARPPPNQKK
YDEVRLDPNVQK
FWETLDRYMVK
YMVKNKAHLR

97
26 Cas/HEF-like protein 1 213623648 45 7 8 8.47/ 96325
ADSPDELTFRK
GLTEIYDIPPSGK
GCYYNTLPPPR
NTSNIWKQR
DSNIQLRIK
QDTSHSPESPK
SSELCGLLK
27 structure-specific
endonuclease subunit slx1 147901424 40 5 13 8.86/ 33182
IQQHNGGKHK
RLTHVPRK
EKGRLGENR
CRVCYER
VCYERVQDK
28 actin, cytoplasmic type 5 288541396 49 8 26 5.30/ 42165
AVFPSIVGRPR
AVFPSIVGRPRHQGVMVGMGQK
DSYVGDEAQSKR
VAPEEHPVLLTEAPLNPK
DLTDYLMK
GYSFTTTAER
SYELPDGQVITIGNER
QEYDESGPSIVHR
29 Unknown (protein for
MGC:181785) 161612103 34 6 20 9.55/39454
KLPQAIIIGVKK
LPQAIIIGVK
GGTRALLEFLR
AVGAEPHFFDRNYDK
VISDYTQTLSK
EFYRPFNMKFYQMTGHDFGWD

98
30 gamma-crystallin-3 285026499 40 6 40 8.18/21842
MGKIFFYEDK
NFQGRSYECNSDCSDLSSYFNR
SCRMSPYHQGQYK
MSPYHQGQYK
FRDIHSCNVFDGNWMFYEEPNYRGR
DIHSCNVFDGNWMFYEEPNYR
31 autophagy protein 5 76150384 47 9 33 5.90/32860
MTDDKDVLR
LLLPRVSYLTLVTDK
VSYLTLVTDKVKK
KHFQKVMR
CQSKDVIEAHFMSSVK
HKSQVINEMQK
QLWMGLQNDK
FEQFWAINRK
FEQFWAINRKLMEFSPEDGGFR
32 nitrogen fixation cluster-
like 147903789 39 3 22 8.18/17373
NVGSLDKNAK
NVGSLDKNAKNVGTGLVGAPACGDVMK
AALADYRLK
33 vacuolar protein sorting 33
homolog A 189217822 37 7 15 6.60/68661
MFTLKSGPLPSSDVKNIIFFVRPR
RSILCEQR
LKDLGVLGSFMHR
LKDLGVLGSFMHR
MKRELGGSQSQITPVFDTLLLLDR
AGLLKLQTSSR
LAQILVRPGWR
36 uncharacterized protein
LOC734830 148234275 38 5 31 4.70/21811
GSGSGAATSLPR
GSGSGAATSLPRMKR
GLATFCLDKEALR
EWEMQFKEK
YEYVGRLLKPGEEPSEYTDEEDVR
38 LOC398774 protein 120577670 35 3 7 5.83/63541 RLQWVSRITNDGDITYYNDSFK
CDDTAITPK

99
ISPGDWENK
39 Pro-opiomelanocortin B
(unnamed protein product) 1334681 37 3 90 10.17/3521
YGGFMTPER
SQTPLMTLFKNAIIK
NAIIKNTHK
40 LOC443709 protein,
partial 49522196 42 5 19 9.53/26306
DLGEPLGGTSK
NSRTNHSSHNDRGPGR
SSSNSNREVVMR
SSSNSNREVVMRVTTV
EVVMRVTTV
41 LOC443713 protein,
partial 49257706 37 7 25 9.54/37426
MPEGPELHR
VCKGLSFAGAVEK
VCKGLSFAGAVEKSAVSK
EVKLILTPLSDESEETHIVFRFGMSGSFK
FTPQDQIPK
LEIPPFTPAR
LEIPPFTPARSVLEAVK
42 copper chaperone for
superoxide dismutase 284520887 45 5 21 5.89/30073
CAHAVKNVLQDVKGVK
AVLKGMGTIK
AVLKGMGTIKSK
ASFRLVDER
QLCTCDGITIWEERNYPIAGPGR
52 copper chaperone for
superoxide dismutase 284520887 40 7 25 5.89/30073
MEELGSSRALSK
MEELGSSRALSKFEFAVQMTCEK
CAHAVKNVLQDVKGVK
AVLKGMGTIK
AVLKGMGTIKSK
ASFRLVDER
LKVYEIIGR

100
43, 50
ATPase, H+ transporting,
lysosomal 50/57kDa, V1
subunit H
148227105 40 7 18 6.01/55745
TFINLMTHISK
VSIFFDYARR
QDLFTVHMAARIIAKLAAWGR
LAAWGRELMEGSDLNYYFNWIK
TQLSSQKLR
NLLEKSTERETR
RVIEQLDGK
44 RAB3A, member RAS
oncogene Family 147901693 45 7 41 4.87/25139
MASATDSRYGQK
ILIIGNSSVGK
ILIIGNSSVGKTSFLFR
TIYRNDKR
YRTITTAYYR
TYSWDNAQVLLVGNK
QTFERLVDIICEKMSESLDTADPAVTGAK
45 aspartoacylase-2 147898393 37 3 14 6.11/34574
SFTADILLANPIAVERCVRYIDR
VAFIATEKK
HCFVPALKLQN
46 tubulin tyrosine ligase-like
family, member 7 148233920 34 10 17 9.37/106393
PSLPNESDHQATCSLSMHSDLPYQPSSSIK
LVIEEMMFTK
FWSDISDLVVK
YEGLLATAFQTFLAGRAASLQR
AASLQREMNNPLK
LSGKPTRPKEPR
NYSSHSSSNSTGSSSDTEEEDEKER
AARPFSNLSSPSSAAPMRRSVSCPR
KVLDIVRTNIR
TNIRSVLQR
47 MARK3 protein 38014795 41 7 13 9.37/67033
AENLLLDADMNIK
FLVLNPSKR
GRSISSSQKQR
GVTPPSPMLGNANNPNK
RNTYVCNER
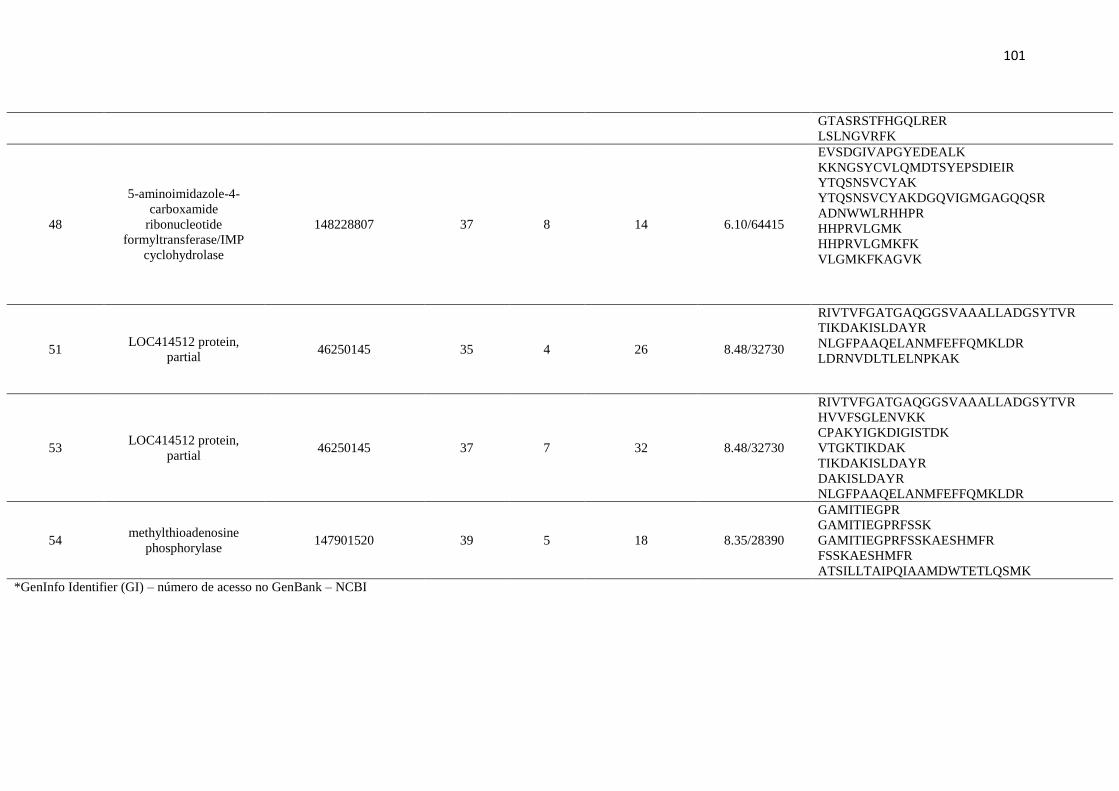
101
GTASRSTFHGQLRER
LSLNGVRFK
48
5-aminoimidazole-4-
carboxamide
ribonucleotide
formyltransferase/IMP
cyclohydrolase
148228807 37 8 14 6.10/64415
EVSDGIVAPGYEDEALK
KKNGSYCVLQMDTSYEPSDIEIR
YTQSNSVCYAK
YTQSNSVCYAKDGQVIGMGAGQQSR
ADNWWLRHHPR
HHPRVLGMK
HHPRVLGMKFK
VLGMKFKAGVK
51 LOC414512 protein,
partial 46250145 35 4 26 8.48/32730
RIVTVFGATGAQGGSVAAALLADGSYTVR
TIKDAKISLDAYR
NLGFPAAQELANMFEFFQMKLDR
LDRNVDLTLELNPKAK
53 LOC414512 protein,
partial 46250145 37 7 32 8.48/32730
RIVTVFGATGAQGGSVAAALLADGSYTVR
HVVFSGLENVKK
CPAKYIGKDIGISTDK
VTGKTIKDAK
TIKDAKISLDAYR
DAKISLDAYR
NLGFPAAQELANMFEFFQMKLDR
54 methylthioadenosine
phosphorylase 147901520 39 5 18 8.35/28390
GAMITIEGPR
GAMITIEGPRFSSK
GAMITIEGPRFSSKAESHMFR
FSSKAESHMFR
ATSILLTAIPQIAAMDWTETLQSMK
*GenInfo Identifier (GI) – número de acesso no GenBank – NCBI

102
3. CONCLUSÃO
3.1 TAXONOMIA E BIOLOGIA MOLECULAR DE ORTLEPPASCARIS SP.
- A utilização de abordagem ultraestrutural gerou novos dados taxonômicos na
descrição do gênero Ortleppascaris, tais como presença de glândula excretora, mucro,
diferença regionalizada das estriações cuticulares, posicionamento de papilas sub-
labiais, anfídios e fasmídios, importante para sistemática desses organismos;
- Apesar de larvas de Ortleppascaris sp. já terem sido registradas previamente
em anuros da América do Sul, este é o primeiro registro deste parasito infectando
anfíbios no Brasil;
- Este é o primeiro registro deste nematódeo em R. marina, adicionando e
ampliando a distribuição geográfica deste helminto;
- A abordagem molecular representa um importante dado para as análises
filogenéticas desses organismos, uma vez que nenhum estudo prévio das características
genéticas do gênero havia sido realizado. Adicionalmente, dados sobre a repetição
dinucleotídica da sequência ITS pode contribuir para futuros estudos em genética de
populações.
3.2. HISTOPATOLOGIA DA INTERAÇÃO ORTLEPPASCARIS SP. X R.
MARINA
- A identificação e observação dos helmintos em seus hospedeiros bem como a
caracterização da resposta imune celular é de fundamental importância para a
compreensão dos processos pelos quais os parasitas podem modular seus hospedeiros;
- A presença dos helmintos em estudo visivelmente influencia na proliferação e
diapedese de melanomacrófagos, processo provavelmente induzido por produtos de
excreção e secreção resultantes do metabolismo das larvas em pleno desenvolvimento;
- Em relação à interação helminto-hospedeiro, acreditamos que o sistema de
defesa deste último não é capaz de coibir, na maioria das vezes, a presença do parasito
uma vez que há grande recrutamento de células de defesa como mostrado nos infiltrados
leucocitários e melanomacrófagos, e, ainda sim o parasito permanece vivo desde
estágios mais elementares;

103
- R. marina caracteriza-se como um hospedeiro adequado para o
desenvolvimento do helminto uma vez que larvas em diferentes estágios foram
observadas pela histologia.
3.3. ANÁLISE PROTEÔMICA DA INTERAÇÃO ORTLEPPASCARIS SP. X R.
MARINA
- São necessários estudos mais aprofundados que contornem os entraves
causados pela falta de banco de dados proteômicos relativos aos organismos em
questão, além de técnicas mais sensíveis para a identificação de proteínas com
parâmetros de identificação extremos, como peso molecular e ponto isoelétrico, não
abrangidos pela técnica de eletroforese bi-dimensional;
- Algumas outras proteínas expressas em Ortleppascaris sp. e seu hospedeiro,
como Actina, proteínas referentes a lon-3, PP2C e citoglobina que apresentam caráter de
conservação evolutiva, conferem a este organismo, potencial para estudos relativos ao
desenvolvimento estrutural e diferenciação sexual de animais, uma vez que a
conservação evolutiva de caracteres em nematódeos têm sido utilizados em estudos e
fornecido informações esclarecedoras;
- Moléculas-chave para a biologia de helmintos foram indicadas aqui, como as
guanilil ciclases, de grande participação em processos quimiosensoriais e neurobiologia
de parasitas nematódeos; antígenos imunorreativos, específicos de nematódeos,
portanto, candidatos vacinais anti-helmínticos; além de fatores de virulência,
considerados marcadores proteicos para infecção helmíntica em geral;
- O modelo de estudo anfíbio-helminto amplia as abordagens de estudo da
relação parasito-hospedeiro, pois até então, a maioria das pesquisas têm sido realizadas
em murinos como modelos animais, considerando que muitos resultados discrepantes
têm sido obtidos usando diferentes modelos animais e helmínticos, fazendo desta uma
oportunidade para observação da co-evolução destes organismos à luz da variedade e
plasticidade de moléculas protéicas expressas de acordo com diferentes nichos
hospedeiros ou ambiente em que esses helmintos se encontram;
- A abordagem molecular de infecções helmínticas em R. marina traz à tona uma
gama de informações sobre o elenco de antígenos protéicos presentes em um animal
sinantrópico, considerado um indicador ambiental e vetor em potencial para zoonoses
helmínticas;

104
- Este conjunto de moléculas encontradas mostra uma diversidade de processos
que ocorrem nesses nematódeos de acordo com o desenvolvimento, metabolismo redox,
infectividade e virulência, afirmando a complexidade fisiológica desses organismos;
- O mapeamento proteico da interface hospedeiro-parasito pode demonstrar
como o hospedeiro reage frente ao desequilíbrio causado pelo parasitismo de forma a
adaptar-se a esta condição. A abordagem proteômica forneceu informações sobre a
relação parasito-hospedeiro a nível molecular, relevantes para estudos co-evolutivos,
escolha de antígenos vacinais e dualidade fisiológica na relação entre estes dois
organismos.

105
REFERÊNCIAS BIBLIOGRÁFICAS
AKIYOSHI, H.; TERADA, T. Mast cell, myofibroblast and nerve terminal complexes
in carbon tetrachloride-induced cirrhotic rat livers. Journal of Hepatology 29 112-
11, 1998.
ALBONICO, M., SMITH, P.G., ERCOLE, E., HALL, A., CHWAYA, H.M.,
ALAWI,K.S. & SAVIOLI, L. Rate of reinfection with intestinal nematodes after
treatment of children with mebendazole or albendazole in a highly endemic area.
Transactions of the Royal Society of Tropical Medicine and Hygiene 89: 538-
541, 1995.
ANDRADE, Z. A. Regressão da fibrose hepática. Revista da Sociedade Brasileira de
Medicina Tropical 38 (6): 514 - 520, 2005.
ANTHONY, R.M, RUTITZKY, L.I., URBAN, J.F.J.R., STADECKER, M.J. e GAUSE,
W.C. Protective immune mechanisms in helminth infection. Nature Reviews
Immunology 7: 975-987, 2007.
ANTUNES, P.W.P. Proteômica e peptidômica aplicadas ao estudo da defesa de
plantas de tomate à injúria mecânica (Mestrado em Bioquímica Agrícola) -
Viçosa, Universidade Federal de Viçosa, 2008. 99p.
BAGGERMAN, G., VERLEYEN, P., CLYNEN, E., HYYBRECHTS, J., DE LOOF, A.
& SCHOOFS, L. Peptidomics. Journal of Chromatography. B. Analytical
Technologies in the Biomedical and Life Sciences 803 (1): 3-16, 2004.
BALIC, A., BOWLES, V.M. & MEEUSEN, E.N. The immunobiology of
gastrointestinal nematode infections in ruminants. Advances in Parasitology
45:181-241, 2000.
BARROS, A. F. Resposta imune humoral e patologia hepática de camundongos
desnutridos, infectados com Schistosoma mansoni. 2008. 124 f. Dissertação
(mestrado) - Centro de Pesquisas Aggeu Magalhães, Fundação Oswaldo Cruz,
2008.
BARTON, D.P. Introduced animals and their parasites: The cane toad, Bufo marinus, in
Australia. Australian Journal of Ecology 22: 316-324, 1997.
BARTON, D.P. Ecology of helminth communities in tropical Australian amphibians.
International Journal of Parasitology 29: 921-926, 1999.
BAUDE, E.J., ARORA, V.K., YU, S., GARBERS, D.L. & WEDEL. B.J. The cloning
of a Caenorhabditis elegans guanylyl cyclase and the construction of a ligand-
sensitive mammalian/nematode chimeric receptor. The Journal of Biological
Chemistry 272 (25): 16035-16039, 1997.

106
BENDTSEN, J.D., JENSEN, L.J., BLOM, N., VON HEIJNE, G. & BRUNAK, S.
Feature-based prediction of non-classical and leaderless protein secretion. Protein
Engineering, Design and Selection 17: 349-356, 2004.
BERNINSONE, P.M. Carbohydrates and glycosylation. Wormbook 18: 1-22, 2006.
BETHONY, J., LOUKAS, A., SMOUT, M., BROOKER, S., MENDEZ, S.,
PLIESKATT, J., GOUD, G., BOTTAZZI, M.E., ZHAN, B., WANG, Y.,
WILLIAMSON, A., LUSTIGMAN, S., CORREA-OLIVEIRA, R., XIAO, S. &
HOTEZ, P.J. Antibodies against a secreted protein from hookworm larvae reduce
the intensity of hookworm infection in humans and vaccinated laboratory animals.
Faseb Journal 19: 1743-5, 2005.
BRANDT, L.A. & MAZZOTTI, F.J. Marine Toads (Bufo marinus). Institute of Food
and Agricultural Sciences Extension, University of Florida, WEC 11, p 1-4,
2002.
CAGNEY, G., SHIVA, A., THANUJA, P., MICHEAL, L. & ANDREW, E. In silico
proteome analysis to facilitate proteomics experiments using mass spectrometry.
Proteome Science 1 (5), 2003.
Casiraghi, M., Bazzocchi, C., Mortarino, M., Ottina, E. & Genchi, C.A simple
molecular method for discriminating common filarial nematodes of dogs (Canis
familiaris). Veterinary Parasitology 141: 368-372. 2006.
CATTO, J.B. Taxonomia e ecologia dos helmintos parasitos de Caiman crocodilus
yacare (CROCODYLIA: ALLIGATORIDAE) no Pantanal Mato-Grossense.
Tese (Doutorado) – Pós Graduação em Medicina Veterinária Parasitologia
Veterinária: Universidade Federal Rural do Rio de Janeiro, 1991. 148p.
CHUNG, Y.B., KONG, Y., JOO, I.J., CHO, S.Y. & KANG, S.Y. Excystment of
Paragonimus westermani metacercariae by endogenous cysteine protease. The
Journal of Parasitology 81: 137-142, 1995.
COX, G.N., KUSCH, M. & EDGAR, R.S. Cuticle of Caenorhabditis elegans: its
isolation and partial characterization. The Journal of Cell Biology 90: 7-17 1981.
(a)
COX, G.N., STAPRANS, S. & EDGAR, R.S. The cuticle of Caenorhabditis elegans. II.
Stage-specific changes in ultrastructure and protein composition during
postembryonic development. Developmental Biology 86: 456-470, 1981. (b)
CONLON, I., & RAFF, M. Size control in animal development. Cell 96: 235–244,
1999.
CRAIG, H., WASTLING, J.M. & KNOX, D.P. A preliminary proteomic survey of the
in vitro excretory/secretory products of fourth-stage larval and adult Teladorsagia
circumcincta, Parasitology 132: 535-543, 2006.

107
DALTON, J. Helminth vaccines: from mining genomic information for vaccine targets
to systems used for protein expression. International Jounal for Parasitology 33:
621-640, 2003.
DASSANAYAKE, R.S., GUNAWARDENE, Y.I. & SILVA, B.D. ITS-2 secondary
structures and phylogeny of Anopheles culicifacies species. Bioinformation 2:
456-560. 2008.
DÍAZ, A. & ALLEN, J.E. Mapping immune response profiles: The emerging scenario
from helminth immunology. European Journal of Immunology 37: 3319-3326,
2007.
DREXLER, H.G. Review of alterations of the cyclin-dependent kinase inhibitor INK4
family genes p15, p16, p18 and p19 in human leukemia-lymphoma cells.
Leukemia 12: 845-859, 1998.
DZIK, J.M. Molecules released by helminth parasites involved in host colonization.
Acta Biochimica Polonica 53: 33-64, 2006.
EHIGIATOR, H.N, STADNYK, A.W. & LEE, T.D.G. Extract of Nippostrongylus
brasiliensis Stimulates Polyclonal Type-2 Immunoglobulin Response by Inducing
De Novo Class Switch. Infection and Immunity 68 (09): 4913-4922, 2000.
EIRAS, J.C. & REGO, A.A. Histologia em peixes resultante de infecções parasitárias.
Publicações do Instituto de Zoologia Dr. Augusto Nobrega 208: 1-12, 1989.
ELSASSER, S.C., FLOYD, R., HEBERT, P.D., SCHULTE-HOSTEDDE, A.I. Species
identification of North American guinea worms (Nematoda: Dracunculus) with
DNA barcoding. Molecular Ecology Resources 9: 707-712, 2009.
ELLIOTT, D.E., SETIAWAN, A., METWALI, BLUM, A., URBAN, J.F, Jr.,
WEINSTOCK, J.V. Heligmosomoides polygyrus inhibits established colitis in IL-
10-deficient mice. European Journal of Immunology 34: 2690–2698, 2004.
FERRI, E., BARBUTO, M., BAIN, O., GALIMBERTI, A., UNI, S., GUERRERO, R.,
FERTÉ, H., BANDI, C., MARTIN, C., CASIRAGHI, M. Integrated taxonomy:
traditional approach and DNA barcoding for the identification of filarioid worms
and related parasites (Nematoda). Frontiers in Zoology 6:1, 2009.
FOLMER, O.; BLACK M.; HOEH, W.; LUTZ, R.; VRIJENHOEK , R. DNA primers
for amplification of mitochondrial cytochrome c oxidase subunit I from diverse
metazoan invertebrates. Molecular Marine Biology and Biotechnology 3 (5):
294-299, 1994.
FROST, D., GRANT, T., FAIVOVICH, J., BAIN, R., HAAS, A., HADDAD, C.,
DESA, R., CHANNING, A., WILKINSON, M., DONNELLAN, S.,
RAXWORTHY, C., CAMPBELL, J., BLOTTO, B., MOLER, P., DREWES, R.C.,
NUSSBAUM, R., LYNCH, J., GREEN, D. & WHELLER, W. The Amphibian
Tree of life. Bulletin of the American Museum of Natural History 297: 1-370,
2006.

108
FUCHS, C., BURMESTER, T. & HANKELN, T. The amphibian globin gene repertoire
as revealed by the Xenopus genome. Cytogenetic and Genome Research 112:
296-306, 2006.
FURTADO, A.P., DO CARMO, E.S., GIESE, E.G., VALLINOTO, A.C.R.,
LANFREDI, R.M. & SANTOS, J.N. Detection of dog filariasis in Marajo Island,
Brazil by classical and molecular methods. Parasitology Research 105: 1509-
1515. 2009.
GASSER, R.B, LEGOFF, L., PETIT, G. & BAIN, O. Rapid delineation of closely-
related filarial parasites using genetic markers in spacer rDNA. Acta Tropica, 62:
143-150. 1999.
GENCHI, C., VENCO, L. & GENCHI, M. Guideline for the laboratory diagnosis of
canine and feline Dirofilaria infections. In: Mappe Parassitologiche 8: Dirofilaria
immitis and D. repens in dog and cat and human infections. Genchi, C., Rinaldi,
L., Cringoli, G. (eds.). Nápoles. Rolando Editore. 2007. p. 137-144.
GILLON, A.D., LATHAM, C.F. & MILLER, E.A. Vesicle-mediated ER export of
proteins and lipids. Biochimica et Biophysica Acta 1821: 1040-1049, 2012.
GOLDBERG, S.R.; BURSEY, C.R. e AQUINOSHUSTER, A.L. Gastric nematodes of
the Paraguayan caiman, Caiman yacare (Alligatoridae). Journal of Parasitology
77: 1009-1011, 1991.
GONZÁLEZ, C.E. & HAMANN, M.I. Helmintos parásitos de Leptodactylus bufonius
Boulenger, 1894 (Anura: Leptodactylidae) de Corrientes, Argentina. Revista
Española de Herpetología 20: 39-46, 2006.
GONZÁLEZ, C.E. & HAMANN, M.I. Nematode parasites of two species of Chaunus
(Anura: Bufonidae) from Corrientes, Argentina. Zootaxa 1393: 27-34, 2007.
GÖRG, A., WEISS, W. & DUNN, M.J. Current two-dimensional electrophoresis
technology for proteomics. Proteomics 4: 3665-3685, 2004.
GRAY, J.M., KAROW, D.S., LU, H., CHANG, A.J., CHANG, J.S., ELLIS, R.E.,
MARLETTA, M.A., & BARGMANN, C.I. Oxygen sensation and social feeding
mediated by a C. elegans guanylate cyclase homologue. Nature 430: 317 - 322,
2004.
GROLL, M. & HUBER, R. Substrate access and processing by the 20S proteasome core
particle. The International Journal of Biochemistry & Cell Biology 35: 606-616,
2003.
HALLEM, E.A., SPENCER, W.C., MCWHIRTER, R.D., ZELLER, G., HENZ, S.R.,
RÄTSCH, G., MILLER III, D.M., HORVITZ, H.R., STERNBERG, P.W. &
RINGSTAD, N. Receptor-type guanylate cyclase is required for carbon dioxide
sensation by Caenorhabditis elegans. PNAS 108 (1): 254-259, 2011.

109
HASLAM, S.M. & DELL, A. Hallmarks of Caenorhabditis elegans N-glycosylation:
complexity and controversy. Biochimie 85: 25-32, 2003.
HASNAIN, S.Z., MCGUCKIN, M.A., GRENCIS, R.K. & THORNTON, D.J. Serine
Protease(s) Secreted by the nematode Trichuris muris degrade the mucus barrier.
PLoS Negleted Tropical Dieases 6 (10): e1856, 2012.
HAWDON, J.M. & HOTEZ, P.J. Hookworm: developmental biology of the infectious
process. Current Opinion In Genetics & Development 6: 618-623, 1996.
HEBERT, P.D.N., CYWINSKA, A., BALL, S.L., DEWAARD, J.R. Biological
identifications through DNA barcodes. Proceedings of the Royal Society B:
Biological Sciences 270: 313-321, 2003a.
HEBERT, P.D.N., RATNASINGHAM, S., DEWAARD, .J.R. Barcoding animal life:
cytochrome c oxidase subunit 1 divergences among closely related species.
Proceedings of the Royal Society B: Biological Sciences (Supplement) 270:
S96–S99, 2003b.
HEWITSON, J.P., HARCUS, Y.M., CURWEN, R.S., DOWLE, A.A., ATMADJA,
A.K., ASHTON, P.D., WILSON, A. &MAIZELS, R.M. The secretome of the
filarial parasite, Brugia malayi: proteomic profile of adult excretory-secretory
products. Molecular and Biochemical Parasitoly 160: 8-21, 2008.
HOLDEN-DYE, L. & WALKER, R.J. Neurobiology of plant parasitic nematodes.
Invertebrate Neurosciense 11: 9-19, 2011.
HONG, X., BOUVIER, J., WONG, M.M., YAMAGATA, G.Y. & MCKERROW, J.H.
Brugia pahangi: identification and characterization of an aminopeptidase associated
with larval molting. Experimental Parasitology 76: 127-133, 1993.
HOROBIN, R. W. Histochemistry: an explanatory outline of histochemistry and
biophysical staining. 1. ed. Nova Iorque: Gustav Fischer Verlag, Stuttgart, 1982.
311 p.
HOTEZ, P.J. One world health: neglected tropical diseases in a flat world. PLoS
Neglected Tropical Diseases 3: 405, 2009.
HUANG, J. & LEMIRE, B.D. Mutations in the C. elegans succinate dehydrogenase
iron-sulfur subunit promote superoxide generation and premature aging. Journal of
Molecular Biology 387: 559-569, 2009.
ISHII, N., FUJII, M., HARTMAN, P.S, TSUDA, M., YASUDA, K, SENOO-
MATSUDA, N., YANASE, S., AYUSAWA, D. & SUZUKI, K. Amutation
insuccinate dehydrogenase cytochrome b causes oxidative stress and ageing in
nematodes. Nature 394 (13): 694-697, 1998.
JEONG, W. Macrophages, myofibroblasts and mast cells in a rat liver infected with
Capillaria hepatica. Journal of Veterinary Science 9 (2): 211-213, 2008.

110
JENKS, B.G., GALAS, L., KURIBARA, M., DESRUES, L., KIDANE, A.H,
VAUDRY, H., SCHEENEN, W.J., ROUBOS, E.W. & TONON, M.C. Analysis of
the melanotrope cell neuroendocrine interface in two amphibian species, Rana
ridibunda and Xenopus laevis: A celebration of 35 years of collaborative research.
General and Comparative Endocrinology 170: 57-67, 2011.
JENKS, B.G., ROUBOS, E.W., & SCHEENEN, W.J. Ca 2+
oscillations in melanotropes
of Xenopus laevis: their generation, propagation, and function. General and
Comparative Endocrinology 131: 210-219, 2003.
JOHNSON, D.C., DEAN, D.R., SMITH, A.D. & JOHNSON, M.K. Structure, function,
and formation of biological iron-sulfur clusters. Annual Review of
Biochemistry74: 247-8, 2005.
JOHNSTON, M.J.G., MACDONALD, J.A. & MCKAY, D.M. Parasitic helminths: a
pharmacopeia of anti-inflammatory molecules. Parasitology 136:125-147, 2009.
JOHNSTON, M.J., WANG, A., CATARINO, M.E., BALL, L., PHAN, V.C.,
MACDONALD, J.A. & MCKAY, D.M. Extracts of the Rat Tapeworm,
Hymenolepis diminuta, Suppress Macrophage Activation In Vitro and Alleviate
Chemically Induced Colitis in Mice. Infection and Immunity 78 (3): 1364–1375,
2010.
JOHNSTONE, I.L. & BARRY, J.D. Temporal reiteration of a precise gene expression
pattern during nematode development. EMBO Journal 15: 3633-3639, 1996.
KELLEHER, D.J. & GILMORE, R. An evolving view of the eukaryotic
oligosaccharyltransferase. Glycobiology 16 (4): 47R-62R, 2006.
KOLK, S.M., KRAMER, B.M.R., CORNELISSE, L.N., SCHEENEN, W.J.J.M.,
JENKS, B.G. & ROUBOS, E.W. Multiple control and dynamic response of the
Xenopus melanotrope cell. Comparative Biochemistry and Physiology Part B
132 257-268, 2002.
KONGARA, S. & KARANTZA, V. The interplay between autophagy and ROS in
tumorigenesis. Frontiers in Oncology 2: 171, 2012.
KUWABARA, P.E. A complex solution to a sexual dilemma. Developmental Cell 13:
6-8, 2007.
KWAN-LIM, G.E., GREGORY, W.F., SELKIRK, M.E., PARTONO, F. & MAIZELS,
R.M. Secreted antigens of filarial nematodes: survey and characterisation of in vitro
excretory/secretory (E/S) products of adult Brugia malayi filarial parasites.
Parasite Immunology 11: 629–54, 1989.
LA FLAMME, A.C., RUDDENKLAU, K. & BACKSTROM, B.T. Schistosomiasis
decreases central nervous system inflammation and alters the progression of
experimental autoimmune encephalomyelitis. Infection and Immunity 71: 4996-
5004, 2003.

111
LANÇAS, F.M., SILVA, J.C.R., BICUDO, R.C. & NETO, M.B. A química analítica do
proteoma. Revista Analytica 06: 60-66, 2003.
LECKBAND, D. & PRAKASAM, A. Mechanism and dynamics of cadherin adhesion.
The Annual Review of Biomedical Engineering 8: 259-87, 2006.
LEE, D.S., KIM, B.K., KWON, S.J., JIN, H.C. & PARK, O.K. Arabidopsis GDSL
lipase 2 plays a role in pathogen defense via negative regulation of auxin signaling.
Biochemical and Biophysical Research Communications 379:1038-1042, 2009.
LEMOS, Q.T., MAGALHÃES-SANTOS, I.F. & ANDRADE, Z.A. Immunological
basis of septal fibrosis of the liver in Capillaria hepatica-infected rats. Brazilian
Journal of Medical and Biological Research 36: 1201-1207, 2003.
LONE, A.M., NOLTE, W.M., TINOCO, A.D. & SAGHATELIAN, A. Peptidomics of
the Prolyl Peptidases. The AAPS Journal 12 (4): 483-91, 2010.
LUKE-GLASER, S., PINTARD, L., TYERS, M. & PETER, M. The AAA-ATPase
FIGL-1 controls mitotic progression, and its levels are regulated by the CUL-
3MEL-26 E3 ligase in the C. elegans germ line. Journal of Cell Science 120:
3179-3187, 2007.
MAIZELS, R. M., BALIC, A., GOMEZ-ESCOBAR, N., NAIR, M., TAYLOR, M.D. &
ALLEN, J. E. Helminth parasites: masters of regulation. Immunological Reviews
201: 89-116, 2004.
MAR, P.H., YANG, I.C., CHANG, G.N. & FEI, A.C.Y. Specific polymerase chain
reaction for differential diagnosis of Dirofilaria immitis and Dipetalonema
reconditum using primers derived from internal transcribed spacer region 2 (ITS2).
Veterinary Parasitology 106: 243-252. 2002.
MCKAY, D.M. The therapeutic helminth? Trends in Parasitology 25 (3): 109-114,
2006.
MCKERROW, J.H. Parasite proteases. Experimental Parasitology 68: 111-115, 1989.
MCKERROW, J.H., CAFFREY, C., KELLY, B., LOKE, P., SAJID, M. Proteases in
parasitic diseases. Annual Review of Pathology 1: 497-536, 2006.
MEDINA, M.L., HAYNES, P.A., BRECI, L. & FRANSCISCO, W.A. Analysis of
secreted proteins from Aspergillus flavus. Proteomics 5: 3153-3161, 2005.
MEEUSEN, E.N.T. Immunology of helminth infections, with special reference to
immunopathology. Veterinary Parasitology 84: 259-273,1999.
MELO, F.T.V., GIESE, E.G., FURTADO, A.P., SOARES, M.J., GONÇALVES, E.C.,
VALLINOTO, A.C.R. & SANTOS, J.N.S. Lanfrediella amphicirrus gen. nov. sp.
nov. Nematotaeniidae (Cestoda: Cyclophyllidea), a tapeworm parasite of Rhinella
marina (Linnaeus, 1758) (Amphibia: Bufonidae). Memórias do Instituto Oswaldo
Cruz 106 (6): 670-7, 2011.

112
MIAO, M., WARREN, A., SONG, W., WANG, S., SHANG, H. & CHEN, Z. Analysis
of the internal transcribed spacer 2 (ITS2) region of scuticociliates and related taxa
(Ciliophora, Oligohymenophorea) to infer their evolution and phylogeny. Protist
159: 519-533, 2008.
MITREVA, M., BLAXTER, M.L., BIRD, D.M. & MCCARTER, J.P. Comparative
genomics of nematodes. Trends Genetics 21 (10): 573-81. 2005.
MONTI, M., ORRÚ, S., PAGNOZZI, D. & PUCCI, P. Functional proteomics. Clinica
Chimica Acta 357: 140-150, 2005.
MORASSUTTI, A.L., LEVERT, K., PINTO, P.M., SILVA, A. J., WILKINS, P. &
GRAEFF-TEIXEIRA, C. Characterization of Angiostrongylus cantonensis
excretory–secretory proteins as potential diagnostic targets. Experimental
Parasitology 130: 26–31, 2012.
MORAVEC, F. & KAISER, H. Helminth parasites from west Indian frogs, with
descriptions of two new species. Caribbean Journal of Science 31: 252-268,
1995.
MORI, I. Genetics of chemotaxis and thermotaxis in the nematode Caenorhabditis
elegans. Annual Review of Genetic 33: 399-422, 1999.
NADLER,S.A, CARRENO, R.A, MEJIA-MADRID, H., ULLBERG, J., PAGAN, C.,
HOUSTON, R. & HUGOT, J.P. Molecular phylogeny of clade III nematodes
reveals multiple origins of tissue parasitism. Parasitology 134: 1421-1442, 2007.
NAIR, M.G., GUILD, K.J., ARTIS, D. Novel effector molecules in type 2
inflammation: lessons drawn from helminth infection and allergy. The Journal of
Immunology 177: 1393-1399, 2006.
NICKEL, W. The mystery of nonclassical protein secretion. A current view on cargo
proteins and potential export routes. European Journal of Biochemistry 270:
2109-2119, 2003.
NORTHERN, C., GROVE, D.I. Antigenic analysis of Strongyloides ratti infective
larvae and adult worms. Immunology and Cell Biology 65 (3): 231-239, 1987.
NYSTROM, J., SHEN, Z.Z., AILI, M., FLEMMING, A.J., LEROI, A., & TUCK, S.
Increased or decreased levels of Caenorhabditis elegans lon-3, a gene encoding a
collagen, cause reciprocal changes in body length. Genetics 161: 83-97, 2002.
ORIHUELA-CHÁVEZ, O.E, REYNA-FIGUEROA, J., WAKIDA-KUSUNOKI, G.,
LIMÓN-ROJAS, A.E., PASQUEL, P. & NAVA-JÁCOME, R. Capilariasis
hepática en niños: reporte del cuarto caso en México. Enfermedades Infecciosas y
Microbiología 26 (3): 86-88, 2006.
ORTIZ, C.O., FAUMONT, S., TAKAYAMA, J., AHMED, H.K., GOLDSMITH, A.D.,
POCOCK, R. MCCORMICK, K.E., KUNIMOTO, H., IINO, Y., LOCKERY, S. &

113
HOBERT, O. Lateralized gustatory behavior of C. elegans is controlled by specific
receptor-type guanylyl cyclases. Current Biology 19 (12): 996-1004, 2009.
OTRANTO, D., DINIZ, D.G., DANTAS-TORRES, F., CASIRAGHI, M., DE
ALMEIDA, I.N., DE ALMEIDA, L.N., DOS SANTOS, J.N., FURTADO, A.P.,
DE ALMEIDA SOBRINHO, E.F. & BAIN, O. Human intraocular filariasis caused
by Dirofilaria sp. nematode, Brazil. Emerging Infectious Diseases 17: 863-6,
2011.
PANDEY, A. & MANN, M. Proteomics to study genes and genomes. Nature 405
(6788): 837-46, 2000.
PANNOCCHIA, M.A., SILVA, C.A., BORELLA, M.I., CAMARGO, A.C.M. &
GILIO, J.M. Estratégia efetiva de fixação do testículo de ratos Wistar paraavaliar os
parâmetros morfológicos e morfométricos do epitélio seminífero. ConScientiae
Saúde 7 (2): 227-273. 2008.
PESCE, A., BOLOGNESI, M., BOCEDI, A., ASCENZI, P., DEWILDE, S., MOENS,
L., THOMAS HANKELN & BURMESTER, T. Neuroglobin and cytoglobin: Fresh
blood for the vertebrate globin Family. European Molecular Biology
Organization. Reports 3 (12): 1146-1151, 2002.
POTTER, L.P. Guanylyl cyclase structure, function and regulation. Cellular Signalling
23: 1921-1926, 2011.
RAGOO, R.M. & OMAHMAHARAJ, I.R. Helminths of the Cane Toad Bufo marinus
from Trinidad, West Indies. Caribbean Journal of Science 39 (2): 242-245, 2003.
RANGANATHAN, S. & GARG, G. Secretome: clues into pathogen infection and
clinical applications. Genome Medicine 1: 113. 2009.
RANKIN, C.H. Nematode memory: Now, where was I? Current Biology 15 (10):
R374–R375, 2005.
REBELLO, K. M.; BARROS, J. S. L.; MOTA, E. M., CARVALHO, P. C., PERALES,
J., LENZI, H. L., NEVES-FERREIRA, A. G. C. Comprehensive proteomic
profiling of adult Angiostrongylus costaricensis, a human parasitic nematode.
Journal of Proteomics 74: 1545-1559. 2011.
REBELLO, K.M., DE SIQUEIRA, C.R., RIBEIRO, E.L., VALENTE, R.H., MOTA,
E.M., PERALES, J., NEVES-FERREIRA, A.G. & LENZI, H.L. Proteolytic
activity in the adult and larval stages of the human roundworm parasite
Angiostrongylus costaricensis. Memórias do Instituto Oswaldo Cruz 107
(6):752-9, 2012.
RODRIGUES, E.D., RANZANI-PAIVA, M.J.P. & SANTOS, A.A. Alterações
histopatológicas em fígado de dourado Salminus maxillosus Valenciennes, 1840
(Osteichthyes, Characidae) causadas por Neocucullanus neocucullanus Travassos,
Artigas & Pereira 1828 (Nematoda). Acta Scientarium 24 (2): 455-459, 2002.

114
RHOADS, M.L., FETTERER, R.H. & URBAN, J.F. JR. Effect of protease class-
specific inhibitors on in vitro development of the third to fourth-stage larvae of
Ascaris suum. The Journal of Parasitology 84: 686-690, 1998.
ROSENBLUM, J.S. & KOZARICH, J.W. Prolyl peptidases: a serine protease
subfamily with high potential for drug discovery. Current Opinion in Chemical
Biology 7: 496-504, 2003.
ROSENZWEIG, A.M. & O’HALLORAN, T.V. Structure and chemistry of the copper
chaperone proteins. Current Opinion in Chemical Biology 4: 140-147, 2000.
SCHILLING, B., MURRAY, M., YOO, C.B., ROW, R.H., CUSACK, M.P.,
CAPALDI, R.A., GIBSON. B.W. Proteomic analysis of succinate dehydrogenase
and ubiquinol-cytochrome c reductase (Complex II and III) isolated by
immunoprecipitation from bovine and mouse heart mitochondria. Biochimica et
Biophysica Acta 1762: 213-222, 2006.
SCHMIDT, M., GERLACH, F., AVIVI, A., LAUFS, T., WYSTUB, S., SIMPSON,
J.C., NEVO, E., SAALER-REINHARDT, S., REUSS, S., HANKELN, T. &
BURMESTER, T. Cytoglobin is a respiratory protein in connective tissue and
neurons, which is up-regulated by hypoxia. The Journal of Biological Chemistry
279 (9): 8063-8069, 2004.
SOBLIK, H., YOUNIS, A.E., MITREVA, M., RENARD, B.Y., KIRCHNER, M.,
GEISINGER, F., STEEN, H. & BRATTIG, N.W. Life cycle stage-resolved
proteomic analysis of the excretome secretome from Strongyloides ratti
identification of stage-specific protease. Molecular & Cellular Proteomics 10
(12), 2011.
SICKMANN, A., MREYEN, M. & MEYER, H.E. Identification of modified proteins
by mass spectrometry. IUBMB Life 54: 51-57, 2002.
SOUZA, M.V., FONTES. W. & RICART, C.A.O. Análise de Proteomas: O despertar
da era pós genômica. Biotecnologia, Ciência e Desenvolvimento 2: 12-14, 1999.
SILVA, J.P., SILVA, D.C.B., MELO, V.T.V., GIESE, E.G., FURTADO, A.P., &
SANTOS, J.N. Liver histopathological alterations in the cane toad, Rhinella marina
(AMPHIBIA: BUFONIDAE), induced by Ortleppascaris sp. larvae
(NEMATODA: ASCARIDIDAE). The Journal of Parasitology, 2013 (no prelo)
(a).
SILVA, J.P., MELO, V.T.V., SILVA, L.C.N., GONÇALVES, E.C., GIESE, E.G.,
FURTADO, A.P., & SANTOS, J.N. Morphological and molecular characterization
of Ortleppascaris sp. larvae parasite of cane toad Rhinella marina from eastern
Amazonia. The Journal of Parasitology, 2013 99(1): 118-123.
SHPILKA, T., WEIDBERG, H., PIETROKOVSKI, S. & ELAZAR, Z. Atg8: an
autophagy-related ubiquitin-like protein Family. Genome Biology 12 (7): 226,
2011.

115
SOARES, A.J.C., SANTOS, M.F., CHUNG, J., DAVID, C.M.N., DOMONT, G.B.
Proteomica e Sepse. Novas Perspectivas para o Diagnostico. Revista Brasileira de
Terapia Intensiva 19 (1): 14-22, 2007.
SPRENT, J.F.A. Ascaridoid nematodes of amphibians and reptiles: Gedoelstascaris n.
g. and Ortleppascaris n. g. Journal of Helminthology 52: 261-282, 1978.
STOTHARD, P., HANSEN, D. & PILGRIM, D. Evolution of the PP2C Family in
Caenorhabditis: Rapid Divergence of the Sex-Determining Protein FEM-2.
Journal of Molecular Evolution 54: 267-282, 2002.
SUN, F. & CAVALLI, V. Neuroproteomics: towards understanding neuronal
regeneration and degeneration. Molecullar & Cellular Proteomics, 2009.
SUZUKI, Y., MORRIS, G.A., HAN, M. & WOOD, W.B. A cuticle collagen encoded
by the lon-3 gene may be a target of TGF-beta signaling in determining
Caenorhabditis elegans body shape. Genetics 162: 1631-9, 2002.
TANIDA, I., UENO, T. & KOMINAMI, E. LC3 conjugation system in mammalian
autophagy. The International Journal of Biochemistry & Cell Biology 36: 2503–
2518, 2004.
TAYLOR, E.W., LEITE, C.A.C.; MCKENZIE, D.J.; & WANG, T. Control of
respiration in fish, amphibians and reptiles. Brazilian Journal of Medical and
Biological Research 43 (5): 409-424, 2010.
TEZUKA, H., IMAI, S., HIDANO, S., TSUKIDATE, S. & FUJITA, K. Various Types
of Dirofilaria immitis Polyproteins Selectively Induce a Th2-Type Immune
Response. Infection and Immunity 71 (07): 3802–3811, 2003.
THOMAS, P.G. & HARN JR, D.A. Immune biasing by helminth glycans. Cellular
Microbiology: 6 (1): 13-22, 2004.
TIELENS, A.G.M., VAN GRINSVEN, K.W.A., HENZE, K, VAN HELLEMOND, J.J,
& MARTIN, W. Acetate formation in the energy metabolism of parasitic helminths
and protists. International Journal for Parasitology 40: 387-397, 2010.
TSUJI, N., KASUGA-AOKIA, H., ISOBEA, T., ARAKAWAB, T. & MATSUMOTO,
Y. Cloning and characterisation of a highly immunoreactive 37 kDa antigen with
multi-immunoglobulin domains from the swine roundworm Ascaris suum.
International Journal for Parasitology 32: 1739–1746, 2002.
VAN DER EYCKEN, W., ENGLER, J.A., VAN MONTAGU, M. & GHEYSEN, G.
Identification and analysis of a cuticular collagen-encoding gene from the plant-
parasitic nematode Meloidogyne incognita. Gene 151: 237-242, 1994.
VARKI, A. Biological roles of oligosaccharides: all of the theories are correct.
Glycobiology 3: 97–130, 1993.

116
WADDLE, A.R., KINSELLA, J.M., ROSS, J.P., ROJAS-FLORES, E., PERCIVAL,
H.F. & FORRESTER, D.J. Nematodes collected by gastric lavage from live
American alligators, Alligator mississippiensis, in Florida. Journal of Parasitology
95 (5): 1237-1238, 2009.
WENK, J., TROMPETER, H.I., PETTRICH, K.G., COHEN, P.T., CAMPBELL, D.G.
& MIESKES, G. Molecular cloning and primary structure of a protein phosphatase
2C isoform. FEBS Lett 297(1-2):135-8, 1992.
WILKINS, M. R., SANCHES, J.C., GOOLEY, A.A., APPEL, R.D., HUMPHERY-
SMITH, I., HOCHSTRASSER, D.F. & WILLIANS, K.L. Progress whith proteome
projects: Why all proteins expressed by genome should be identified and how to do
it? Biotechnology and Genetic Engineering Reviews 13: 19-50, 1995.
WILSON, M.S., MAIZELS, R.M. Regulatory T cells induced by parasites and the
modulation of allergic responses. Chemical Immunology and Allergy 90: 176–
195, 2006.
WILSON, M.S., TAYLOR, M.D., BALIC, A., FINNEY, C. A., LAMB, J. R. &
MAIZELS, R.M. Suppression of allergic airway inflammation by helminthinduced
regulatory T cells. The Journal of Experimental Medicine 202: 1199–1212,
2005.
WILLIAMSON, A.L., BRINDLEY, P.J., KNOX, D.P., HOTEZ, P.J. & LOUKAS, A.
Digestive proteases of blood-feeding nematodes. Trends in Parasitology 19: 417-
423, 2003.
WILSON, M.S. & MAIZELS, R.M. Regulatory T cells induced by parasites and the
modulation of allergic responses. Chemical Immunology and Allergy 90: 176-
195, 2006.
XU, Y.Z. & DRESDEN MH. Leucine aminopeptidase and hatching of Schistosoma
mansoni eggs. The Jounal of Parasitology 72: 507-511, 1986.
YAN, Y. & DAVIS, E.L. Characterisation of guanylyl cyclase genes in the soybean
cyst nematode, Heterodera glycines. International Journal for Parasitology 32:
65-72, 2002.
YANG, Y., MAHAFFEY, C.L., BÉRUBÉ, N., NYSTUEN, A., FRANKEL. W.N.
Functional characterization of fidgetin, an AAA-family protein mutated in fidget
mice. Experimental Cell Research 304: 50-58, 2005.
YOUNG, B.E, STUART, S.N, CHANSON, J.S, COX, N.A & BOUCHER, T.M.
Disappearing Jewels: The Status of New World Amphibians. Nature Serve,
Arlington Virginia, 2004.
ZACCONE, P., FEHÉRVÁRI, Z., JONES, F.M., SIDOBRE, S., KRONENBERG, M.,
DUNNE, D.W. & COOKE, A. Schistosoma mansoni antigens modulate the activity
of the innate immune response and prevent onset of type 1 diabetes. European
Journal of Immunology 33: 1439-1449, 2003.

117
ZUG, G.R & ZUG, P.B. The Marine Toad, Bufo marinus: A Natural History
Resumé of Native Populations. Smithsonian Institution Press. City of
Washington, 1979.






















![parasito schistosoma[1]](https://img.document.onl/doc/110x75/5571fb6f497959916994e04d/parasito-schistosoma1.jpg)






