Embed Size (px)
Citation preview

Universidade Federal de Goiás
Escola de Veterinária e Zootecnia
Programa de Pós-Graduação em Ciência Animal
Disciplina: SEMINÁRIOS APLICADOS
ASPECTOS BIOLÓGICOS, EPIDEMIOLÓGICOS, CLÍNICOS E DE
DIAGNÓSTICO DO Toxoplasma gondii
Nome:
Rebeka Cristine de Bastos Costa
Orientadora:
Valéria de Sá Jayme
GOIÂNIA
2013

ii
REBEKA CRISTINE DE BASTOS COSTA
ASPECTOS BIOLÓGICOS, EPIDEMIOLÓGICOS, CLÍNICOS E DE
DIAGNÓSTICO DO Toxoplasma gondii
Seminário apresentado junto à Disciplina
de Seminários aplicados do Programa de
Pós-graduação em Ciência Animal da
Escola de Veterinária e Zootecnia da
Universidade Federal de Goiás.
Nível: Mestrado
Área de Concentração:
Sanidade Animal, Higiene e Tecnologia de Alimentos
Linha de Pesquisa:
Enfermidades de importância em saúde pública
Orientadora:
Prof. Dr. Valéria de Sá Jayme
Comitê de orientação
Prof. Dr. Guido Fontgalland Coelho Linhares
Prof. Dr. Andréa Caetano da Silva
GOIÂNIA
2013

iii
LISTA DE FIGURAS
FIGURA 1 - Morfologia geral da forma taquizoíta do Toxoplasma gondii, protozoário do Filo Apicomplexa, o qual possui as estruturas características do
complexo apical, como róptrias, micronemas, anéis polares. ..................................... 3 FIGURA 2 - Ciclo biológico do Toxoplasma gondii e suas três fases de vida: taquizoítos, bradizoítos (cistos tissulares) e esporozoítos (oocistos), com os diferentes tipos de hospedeiros intermediários (mamíferos e aves) e o hospedeiro
definitivo, felino. .................................................................................................................. 4 FIGURA 3 - Corte de cérebro de paciente com neurotoxoplasmose, mostrando
duas lesões necróticas extensas (setas). ....................................................................... 6 FIGURA 4 - Recém-nascidos com as formas clínicas da toxoplasmose congênita.
A. Hidrocefalia; B. Microcefalia. ........................................................................................ 7 FIGURA 5 - Cisto tecidual de Toxoplasma gondii identificado no material cerebral
de camundongo, inoculado com material cerebral de ovino. Ampliação de 400x. .. 8 FIGURA 6 - Esquema demonstrativo das três etapas da PCR convencional: A. No primeiro passo, ocorre a desnaturação, então a dupla fita de DNA separa-se, a uma temperatura de 94ºC por um minuto, dando origem a duas fitas simples de DNA. B. A uma temperatura de 30 a 60ºC por cerca de um minuto, ocorre o segundo passo e tem-se o anelamento dos primers em cada uma das fitas simples, em posições específicas. C. O terceiro passo ou extensão dá-se pela ação da DNA polimerase, a uma temperatura de 72ºC por dois minutos, a qual
realiza o processo de extensão da sequência a ser amplificada. ............................ 11 FIGURA 7 – Amplificação da sequência de 529 pb (pares de base) e detecção pela PCR do DNA de Toxoplasma gondii em água contaminada
experimentalmente. .......................................................................................................... 12 FIGURA 8 - Oocistos não esporulados de Isospora felis (A), Isospora rivolta (B), e Toxoplasma gondii (C), em comparação com esporocisto esporulado de
Sarcocystis muris (D). Ampliação de 1600 x. .............................................................. 17 FIGURA 9 - Mapa do Brasil demonstrando a distribuição dos isolados do Toxoplasma gondii: Pará, linhagem BrI, em frangos; Espírito Santo, tipo BrI em frangos; São Paulo, BrI, BrII e BrIII em cães e gatos e tipo II, em ovinos; Paraná, tipos BrI, BrII e BrIII, em cães e gatos; Rio Grande do Sul, tipos BrIII e BrIV, em
frangos. ............................................................................................................................... 19

SUMÁRIO
1 INTRODUÇÃO .................................................................................................... 1
2 REVISÃO DE LITERATURA ............................................................................... 2
2.1 Toxoplasma gondii ........................................................................................... 2
2.2 Métodos de diagnóstico e caracterização molecular ....................................... 9
2.2.1 Reação em cadeia da polimerase ................................................................. 9
2.2.2 Teste de polimorfismo de comprimento de fragmentos de DNA gerados por
enzimas de restrição sobre produtos amplificados pela reação em cadeia da
polimerase ............................................................................................................ 13
2.3 Diagnóstico e caracterização molecular do Toxoplasma gondii ..................... 14
2.3.1 Diagnóstico molecular ................................................................................. 14
2.3.2 Caracterização molecular ............................................................................ 17
3 CONSIDERAÇÕES FINAIS .............................................................................. 22
4 REFERÊNCIAS ................................................................................................. 23

1 INTRODUÇÃO
Apesar dos avanços no conhecimento e na prática médica, as
doenças parasitárias continuam a ser um importante problema de saúde global
(VASOO & PRITT, 2013). Entre as principais enfermidades de importância
mundial, estão incluídas as causadas por protozoários, os quais podem atingir
tanto os seres humanos como os animais (THEKISOE & INOUE, 2011), um
exemplo é o caso do Toxoplasma gondii, o qual é capaz de infectar mamíferos
e aves (SILVEIRA, 2009).
O desenvolvimento de métodos simples, rápidos e sensíveis para a
detecção e identificação desse protozoário é importante para o
aperfeiçoamento dos diagnósticos e dos estudos epidemiológicos da
toxoplasmose (SU et al., 2010). Assim, com o crescimento da biologia
molecular foram desenvolvidas ferramentas baseadas na amplificação do DNA,
as quais aprimoraram e aperfeiçoaram o diagnóstico do parasita em questão
(WAAL, 2012).
Inicialmente, a aplicação de técnicas moleculares para o diagnóstico
do T. gondii foram concentradas na detecção do mesmo. Porém, com o
desenvolvimento de métodos baseados em uma variedade de marcadores
genéticos para as populações deste protozoário proporcionou a identificação
de diferentes linhagens genotípicas, o que aprimorou as informações já
conhecidas, em relação à epidemiologia e à patogenicidade para a
toxoplasmose (SU et al., 2010).
Por ser um potencial causador de zoonose e de distribuição
cosmopolita, o T. gondii tem sido um dos protozoários mais amplamente
estudados. Os recentes desenvolvimentos na área da biologia molecular
forneceram ferramentas, as quais foram utilizadas para o aprofundamento do
conhecimento dessa espécie, de elevada importância na clínica médica e
veterinária. Diante disto, objetivou-se com esta revisão de literatura, abordar a
biologia do T. gondii e a aplicação dos métodos moleculares utilizados para a
detecção e caracterização das diferentes linhagens genotípicas do T. gondii.

2
2 REVISÃO DE LITERATURA
2.1 Toxoplasma gondii
Os protozoários pertencem ao Reino Protista, sub-reino Protozoa
(grego, protos=primeiro e zoon=animal) e são classificados em sete filos,
destacando-se o Apicomplexa, o qual representa um dos maiores grupos de
protozoários parasitas, compreendendo muitas espécies de importância
veterinária e médica (TENTER et al., 2002).
Esse filo é composto por parasitas intracelulares obrigatórios que se
caracterizam por possuírem, em determinadas fases de sua vida, uma
estrutura chamada de complexo apical, o qual é composto por organelas
secretórias especializadas, entre elas as róptrias, os micronemas e os
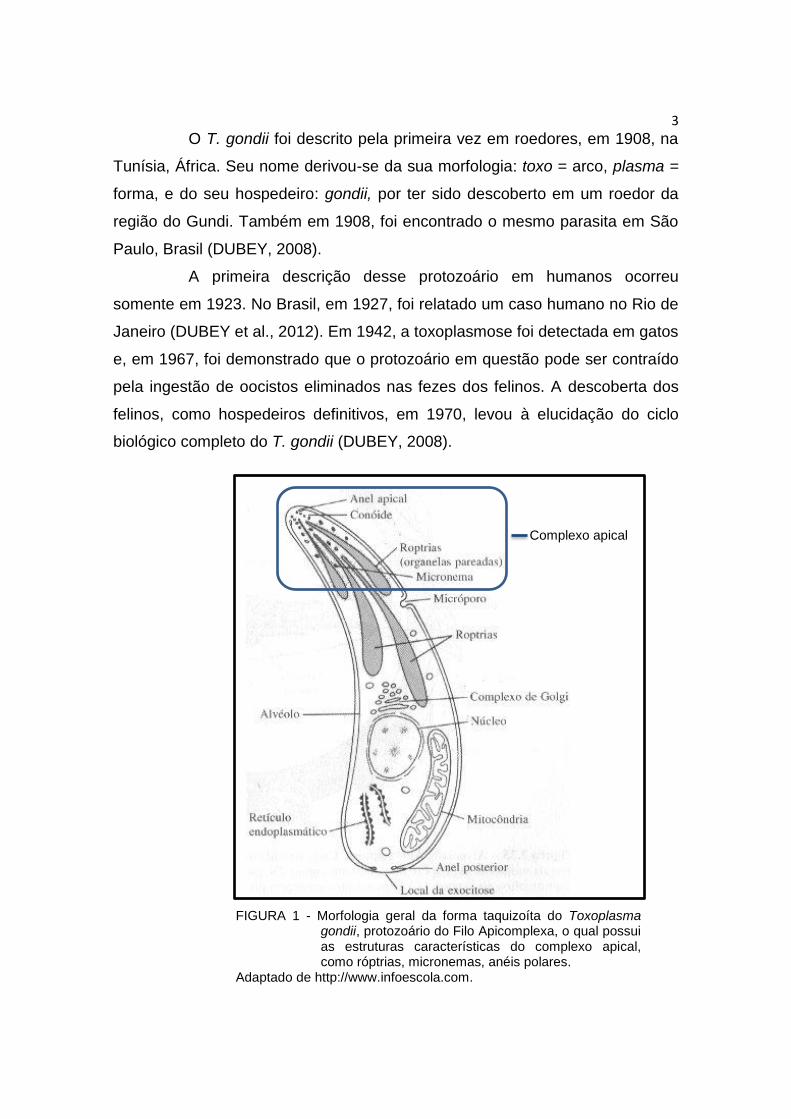
elementos do citoesqueleto, como os anéis apicais (Figura 1) (MONTEIRO,
2006). Este conjunto de organelas está localizado na região anterior da célula
parasitária, pois possui como função principal facilitar a entrada dos parasitas
nas células hospedeiras (TENTER et al., 2002).
Dentro do Filo Apicomplexa destaca-se a família Sarcocystidae por
conter cerca de 200 espécies. Ainda, essa se subdivide em subfamílias, sendo
uma delas a Toxoplasmatinae, na qual estão alocados vários gêneros e entre
eles, o Toxoplasma, no qual se tem uma única espécie: o T. gondii
(SERCONDES, 2010).
Família Sarcocystidae (Poche,1913)
Subfamília Toxoplasmatinae (Biocca de 1957)
Gênero Besnoitia (Henry, 1913)
Gênero Hammondia (Frenkel, 1974)
Género Neospora (Dubey et al., 1988)
Gênero Toxoplasma (Nicolle & Manceaux, 1909)
Toxoplasma gondii (Nicolle & Manceaux, 1909)
3
O T. gondii foi descrito pela primeira vez em roedores, em 1908, na
Tunísia, África. Seu nome derivou-se da sua morfologia: toxo = arco, plasma =
forma, e do seu hospedeiro: gondii, por ter sido descoberto em um roedor da
região do Gundi. Também em 1908, foi encontrado o mesmo parasita em São
Paulo, Brasil (DUBEY, 2008).
A primeira descrição desse protozoário em humanos ocorreu
somente em 1923. No Brasil, em 1927, foi relatado um caso humano no Rio de
Janeiro (DUBEY et al., 2012). Em 1942, a toxoplasmose foi detectada em gatos
e, em 1967, foi demonstrado que o protozoário em questão pode ser contraído
pela ingestão de oocistos eliminados nas fezes dos felinos. A descoberta dos
felinos, como hospedeiros definitivos, em 1970, levou à elucidação do ciclo
biológico completo do T. gondii (DUBEY, 2008).
FIGURA 1 - Morfologia geral da forma taquizoíta do Toxoplasma gondii, protozoário do Filo Apicomplexa, o qual possui as estruturas características do complexo apical, como róptrias, micronemas, anéis polares.
Adaptado de http://www.infoescola.com.
Complexo apical

4
Segundo DUBEY (2008), em seu ciclo de vida o T. gondii,
desenvolve organelas características do Filo Apicomplexa, apresentando o
complexo apical e as organelas mencionadas anteriormente, porém é o único
que apresenta o conóide, como demonstrado na Figura 1.
Tal protozoário possui três fases distintas: taquizoítos, bradizoítos
(cistos tissulares) e esporozoítos (oocistos). Ainda, possui ciclo de vida
heteroxeno (Figura 2), sendo que todos os animais de sangue quente
(mamíferos e aves) podem participar do ciclo como hospedeiros intermediários
e os felinos são seus hospedeiros definitivos (TENTER et al., 2000).
FIGURA 2 - Ciclo biológico do Toxoplasma gondii e suas três fases de vida: taquizoítos, bradizoítos (cistos tissulares) e esporozoítos (oocistos), com os diferentes tipos de hospedeiros intermediários (mamíferos e aves) e o hospedeiro definitivo, felino.
Fonte: http://www.icb.ufmg.br/biq/prodap/2000/toxo/ciclo.gif.
Todas as fases de desenvolvimento são infecciosas para ambos
hospedeiros e estes podem adquirir a toxoplasmose por diferentes vias, como
a horizontal, por ingestão oral de oocistos esporulados no ambiente (Figura 2)
e por ingestão de cistos tissulares na carne crua ou mal cozida (Figura 2); ou
verticalmente, pela transmissão transplacentária de taquizoítos (Figura 2).

5
Estes também podem ser transmitidos pelo leite, da mãe para o filho, ou por
transplante de sangue ou órgãos (TENTER et al., 2000).
Os gatos podem eliminar oocistos no ambiente (Figura 2) após a
ingestão de qualquer um dos estágios de desenvolvimento do T. gondii, mas,
somente 30% dos felinos eliminam essa fase no ambiente após ingerir
taquizoítos ou oocistos, enquanto, praticamente todos os gatos que ingerem
bradizoítos eliminam-nos (DUBEY et al., 1998).
O contato com o protozoário promove a formação de anticorpos
contra o parasita em questão, sendo que TENTER et al. (2000) verificaram que
74% da população de gatos adultos, dependendo do tipo de alimentação e
acesso à rua ou não, apresentam anticorpos contra o T. gondii.
Como o homem pode participar do ciclo, sendo um hospedeiro, a
toxoplasmose é considerada uma zoonose, a qual se destaca na medicina
veterinária e humana, pois pode causar aborto ou doença congênita em seus
hospedeiros intermediários (TENTER et al., 2000).
Tal enfermidade apresenta distribuição cosmopolita e estima-se que
um terço, ou mais, da população mundial esteja cronicamente infectada,
apresentando anticorpos para o parasita (MONTOYA & LIESENFELD, 2004),
chegando a índices de soropositividade variando de 23 a 83%, dependendo de
fatores climáticos, socioeconômicos e culturais (FIALHO et al., 2009). No
Brasil, a infecção pelo T. gondii está amplamente prevalente em humanos,
sendo que 50% das crianças e 80% das mulheres em idade fértil têm
anticorpos para esse protozoário (DUBEY et al., 2012).
Infecções primárias em adultos são geralmente assintomáticas, mas
linfadenopatia ou toxoplasmose ocular podem ocorrer em alguns pacientes (SU
et al., 2010). Já para os imunossuprimidos a toxoplasmose apresenta-se de
forma aguda, podendo em alguns casos levar a encefalite fatal, ou
neurotoxoplasmose, e provocar lesões necróticas no cérebro (MONTOYA &
LIESENFELD, 2004), como demonstrado na Figura 3.

6
FIGURA 3 - Corte de cérebro de paciente com neurotoxoplasmose, mostrando duas lesões necróticas extensas (setas).
Fonte: http://www.icb.usp.br/~livropar/img/capitulo3/2.html.
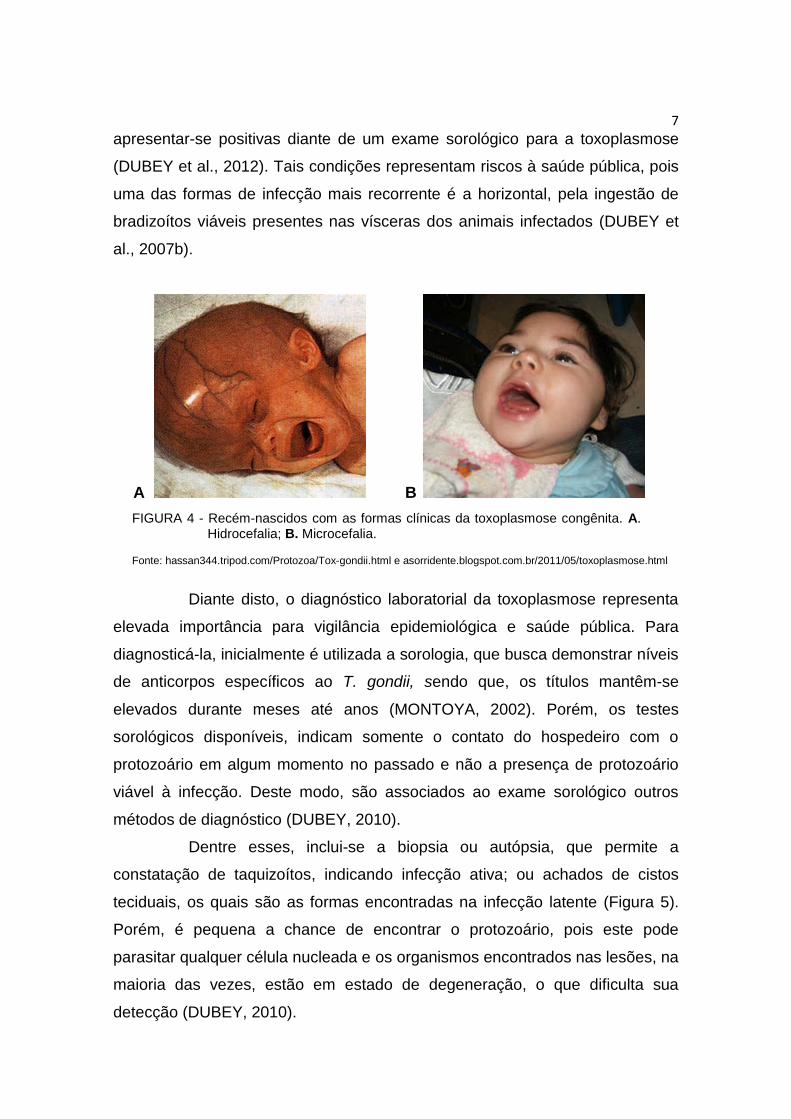
Quando ocorre a transmissão vertical, a enfermidade é conhecida
como toxoplasmose congênita (SU et al., 2010). Porém, se a infecção materna
for adquirida antes da gestação, aquela representa pouco ou nenhum risco
para o feto. Mas quando mulheres, já gestantes, são infectas pela primeira vez
pelo T. gondii, a chance do filho desenvolver a toxoplasmose congênita clínica
é aumentada. Já a infecção subclínica dessa enfermidade, em recém-nascidos,
pode passar despercebida ou o bebê desenvolverá a hidrocefalia ou
microcefalia (MONTOYA & LIESENFELD, 2004), como demonstrado na
Figura4.
Da mesma forma que ocorre nos homens, a infecção pelo T. gondii
está amplamente difundida em animais em todo o mundo. Inquéritos
epidemiológicos relevam que 90% dos animais domésticos e silvestres
possuem anticorpos para aquele protozoário. Desta forma, ressalta-se a
importância da toxoplasmose, pois animais de produção, como suínos, ovinos
e caprinos podem apresentar sorologia positiva para o T. gondii (DUBEY et al.,
2012).
No Brasil, a maioria dos suínos apresenta-se sorologicamente
positiva para o protozoário em questão, e de forma semelhante, este está
largamente distribuído entre ovinos, destacando-se ainda que as aves possam
7
apresentar-se positivas diante de um exame sorológico para a toxoplasmose
(DUBEY et al., 2012). Tais condições representam riscos à saúde pública, pois
uma das formas de infecção mais recorrente é a horizontal, pela ingestão de
bradizoítos viáveis presentes nas vísceras dos animais infectados (DUBEY et
al., 2007b).
A B
FIGURA 4 - Recém-nascidos com as formas clínicas da toxoplasmose congênita. A. Hidrocefalia; B. Microcefalia.
Fonte: hassan344.tripod.com/Protozoa/Tox-gondii.html e asorridente.blogspot.com.br/2011/05/toxoplasmose.html
Diante disto, o diagnóstico laboratorial da toxoplasmose representa
elevada importância para vigilância epidemiológica e saúde pública. Para
diagnosticá-la, inicialmente é utilizada a sorologia, que busca demonstrar níveis
de anticorpos específicos ao T. gondii, sendo que, os títulos mantêm-se
elevados durante meses até anos (MONTOYA, 2002). Porém, os testes
sorológicos disponíveis, indicam somente o contato do hospedeiro com o
protozoário em algum momento no passado e não a presença de protozoário
viável à infecção. Deste modo, são associados ao exame sorológico outros
métodos de diagnóstico (DUBEY, 2010).
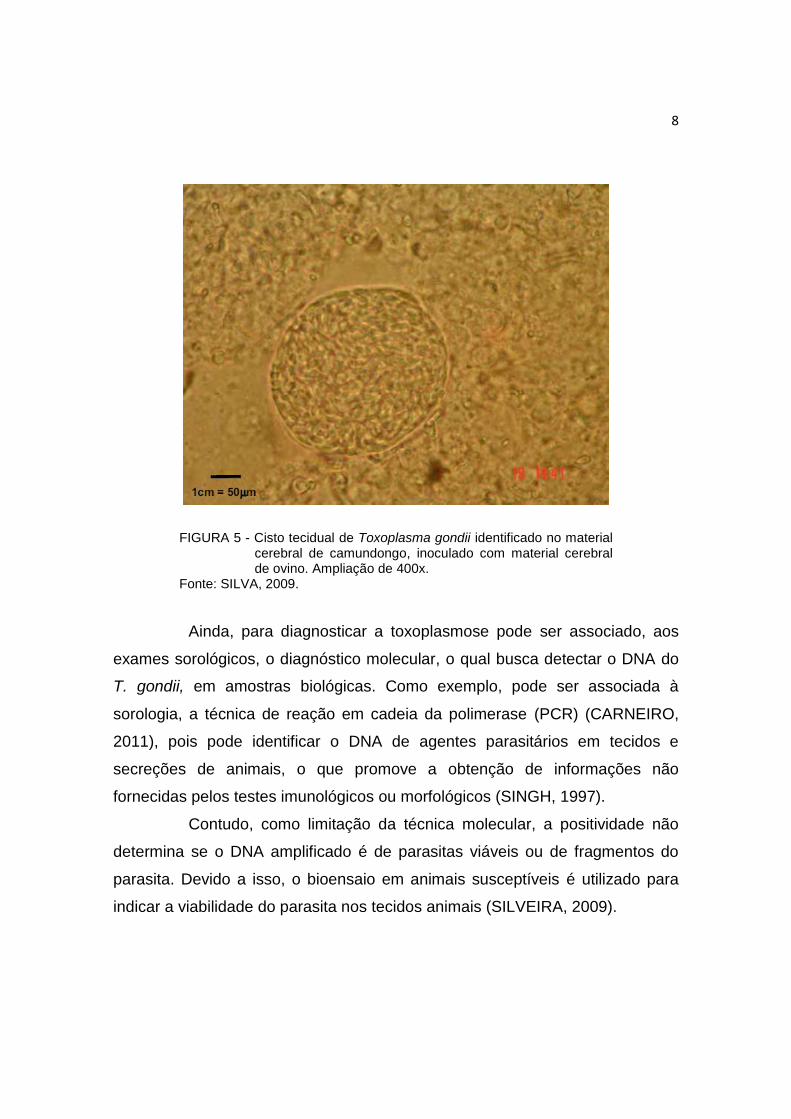
Dentre esses, inclui-se a biopsia ou autópsia, que permite a
constatação de taquizoítos, indicando infecção ativa; ou achados de cistos
teciduais, os quais são as formas encontradas na infecção latente (Figura 5).
Porém, é pequena a chance de encontrar o protozoário, pois este pode
parasitar qualquer célula nucleada e os organismos encontrados nas lesões, na
maioria das vezes, estão em estado de degeneração, o que dificulta sua
detecção (DUBEY, 2010).
8
FIGURA 5 - Cisto tecidual de Toxoplasma gondii identificado no material
cerebral de camundongo, inoculado com material cerebral de ovino. Ampliação de 400x.
Fonte: SILVA, 2009.
Ainda, para diagnosticar a toxoplasmose pode ser associado, aos
exames sorológicos, o diagnóstico molecular, o qual busca detectar o DNA do
T. gondii, em amostras biológicas. Como exemplo, pode ser associada à
sorologia, a técnica de reação em cadeia da polimerase (PCR) (CARNEIRO,
2011), pois pode identificar o DNA de agentes parasitários em tecidos e
secreções de animais, o que promove a obtenção de informações não
fornecidas pelos testes imunológicos ou morfológicos (SINGH, 1997).
Contudo, como limitação da técnica molecular, a positividade não
determina se o DNA amplificado é de parasitas viáveis ou de fragmentos do
parasita. Devido a isso, o bioensaio em animais susceptíveis é utilizado para
indicar a viabilidade do parasita nos tecidos animais (SILVEIRA, 2009).

9
2.2 Métodos de diagnóstico e caracterização molecular
O diagnóstico molecular é baseado na detecção, identificação e
quantificação de ácidos nucleicos, auxiliando no diagnóstico e no prognóstico
de enfermidades (BARRA et al., 2011). Ainda, possui a capacidade de
diferenciar agentes parasitários taxonomicamente muito próximos
(THOMPSON et al., 1998), o que facilita estudos das populações de parasitas,
bem como a identificação de possíveis relações entre um determinado genótipo
e sua virulência (YERA et al., 2003).
Os métodos caracterizados como moleculares dependem da reação
em cadeia da polimerase (PCR) para a detecção específica ou a análise de
DNA do T. gondii (SU et al., 2010).
Tais métodos podem ser divididos em dois grupos: no primeiro grupo
estão as técnicas capazes de detectar especificamente o DNA em amostras
biológicas, como a PCR convencional (cn-PCR) e a nested-PCR (n-PCR). Já
no segundo, tem-se o teste de polimorfismo de comprimento de fragmentos de
DNA gerados por enzimas de restrição sobre produtos amplificados pela PCR
(PCR-RFLP), o qual é capaz de identificar os fragmentos do DNA do T. gondii,
realizando assim, a caracterização genotípica do protozoário (SU et al., 2010).
2.2.1 Reação em cadeia da polimerase
Para conseguir uma boa sensibilidade na detecção do DNA do
parasita, o resultado depende do método utilizado, como também, da técnica
realizada para extração do DNA (SILVA et al., 1999). Tradicionalmente, no
laboratório clínico, os ácidos nucleicos são extraídos de amostras como sangue
total, soro, líquido amniótico e restos placentários (BARRA et al., 2011).
Para realizar a extração do DNA, na maioria das vezes são
utilizados kits comerciais, os quais facilitam o manuseio das amostras. Ainda,
possibilitam a remoção de possíveis inibidores de PCR, como por exemplo, o
grupo heme do sangue e a heparina, que é frequentemente utilizada na coleta

10
sanguínea. No caso de amostras fecais, os kits devem extrair o DNA do
parasita a partir dos oocistos, evitar adsorção do material obtido pelos
constituintes das fezes e remover os inibidores da PCR (SILVA et al., 1999).
Após a extração do DNA, é iniciada a cn-PCR e para a realização
desta, são necessários três segmentos de ácidos nucléicos: a dupla fita de
DNA, a qual servirá de molde para amplificação e dois oligonucleotídeos
específicos, também conhecidos como primers ou iniciadores. Além destes,
são necessários: DNA polimerase, dNTPs (trifosfatos desoxinucleótidos),
tampão e sais de cloreto de magnésio (LIMA, 2008).
A DNA polimerase necessita do magnésio para realizar sua
atividade e este geralmente é fornecido sob a forma de cloreto de magnésio.
Os dNTPs são de quatro tipos de nucleotídeos, os quais são utilizados pela
polimerase para estender o iniciador anelado e o tampão garante adequado
ambiente à ação da polimerase (STEPHENSON & ABILOCK, 2012). Já os
primers são pequenas sequências de nucleotídeos que se anelam no início da
sequência alvo, sendo então complementares à fita de DNA (LIMA, 2008).
Assim, o método padrão requer um modelo de DNA, que contenha a
região a ser amplificada pelos dois primers e para realizar a amplificação é
utilizada a Taq DNA polimerase. Ao reconhecer os iniciadores, a polimerase
sintetiza uma cópia complementar, obedecendo à informação contida na
sequência de DNA que será replicada (LIMA, 2008). Com a ação da Taq DNA
polimerase, ocorre a síntese da nova fita complementar e as regiões
específicas do DNA de interesse, são amplificadas enzimaticamente, em
sucessivos ciclos (MONIS & ANDREWS, 1998).
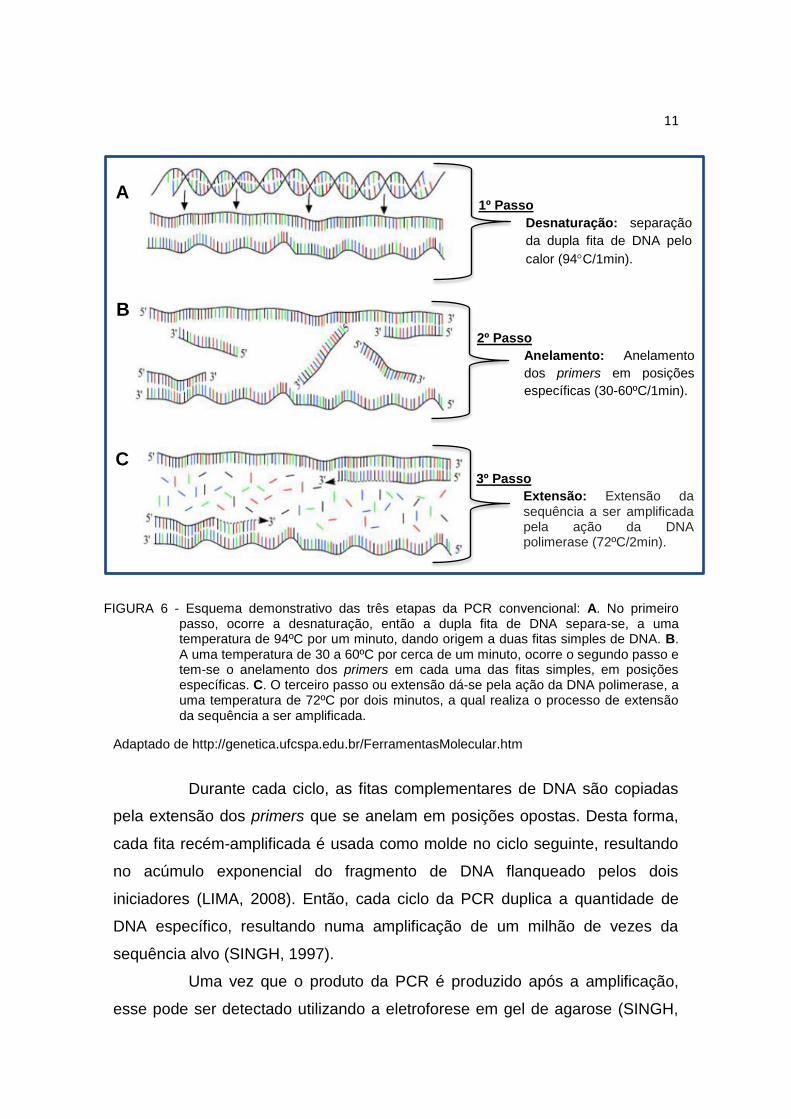
Cada ciclo da PCR é repetido por cerca de 30 vezes e deve ser
realizado em três etapas (Figura 6): desnaturação, anelamento e extensão.
Cada uma das três etapas é realizada em temperaturas diferentes, sendo que
geralmente na desnaturação a etapa ocorre a 94ºC (Figura 6 A), já no segundo
passo, ou anelamento, a temperatura pode variar de acordo com o tamanho e
constituição dos primers, podendo variar de 30 a 60ºC (Figura 6 B) e a
extensão ocorre a 72ºC (Figura 6 C), sendo que essas diferenças na
temperatura são conseguidas com uso do termociclador (LIMA, 2008).
11
FIGURA 6 - Esquema demonstrativo das três etapas da PCR convencional: A. No primeiro passo, ocorre a desnaturação, então a dupla fita de DNA separa-se, a uma temperatura de 94ºC por um minuto, dando origem a duas fitas simples de DNA. B. A uma temperatura de 30 a 60ºC por cerca de um minuto, ocorre o segundo passo e tem-se o anelamento dos primers em cada uma das fitas simples, em posições específicas. C. O terceiro passo ou extensão dá-se pela ação da DNA polimerase, a uma temperatura de 72ºC por dois minutos, a qual realiza o processo de extensão da sequência a ser amplificada.
Adaptado de http://genetica.ufcspa.edu.br/FerramentasMolecular.htm
Durante cada ciclo, as fitas complementares de DNA são copiadas
pela extensão dos primers que se anelam em posições opostas. Desta forma,
cada fita recém-amplificada é usada como molde no ciclo seguinte, resultando
no acúmulo exponencial do fragmento de DNA flanqueado pelos dois
iniciadores (LIMA, 2008). Então, cada ciclo da PCR duplica a quantidade de
DNA específico, resultando numa amplificação de um milhão de vezes da
sequência alvo (SINGH, 1997).
Uma vez que o produto da PCR é produzido após a amplificação,
esse pode ser detectado utilizando a eletroforese em gel de agarose (SINGH,
2º Passo
Anelamento: Anelamento
dos primers em posições
específicas (30-60ºC/1min).
C
B
A
3º Passo
Extensão: Extensão da sequência a ser amplificada pela ação da DNA polimerase (72ºC/2min).
1º Passo
Desnaturação: separação
da dupla fita de DNA pelo
calor (94C/1min).
12
1997). A agarose utilizada no gel é um polissacarídeo que forma uma rede e
permite regular a velocidade da migração das moléculas durante a separação.
Esta técnica faz uso da diferença de potencial elétrico, sendo que as moléculas
de menor massa migram mais rapidamente (LIMA, 2008).
Após a separação dos produtos, ocorre a adição de corante
fluorescente (brometo de etídio) que se intercala entre as bases do DNA e o
uso de radiação ultravioleta permite a visualização dos produtos gerados
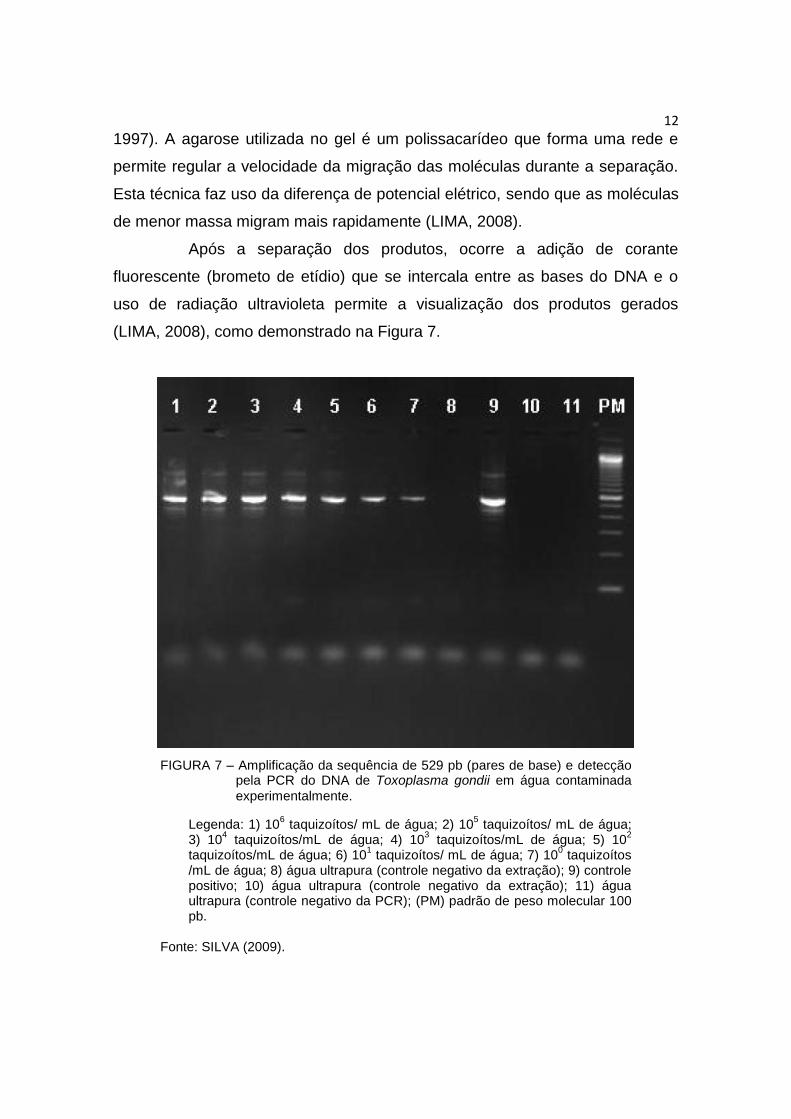
(LIMA, 2008), como demonstrado na Figura 7.
FIGURA 7 – Amplificação da sequência de 529 pb (pares de base) e detecção pela PCR do DNA de Toxoplasma gondii em água contaminada
experimentalmente.
Legenda: 1) 106 taquizoítos/ mL de água; 2) 10
5 taquizoítos/ mL de água;
3) 104 taquizoítos/mL de água; 4) 10
3 taquizoítos/mL de água; 5) 10
2
taquizoítos/mL de água; 6) 101 taquizoítos/ mL de água; 7) 10
0 taquizoítos
/mL de água; 8) água ultrapura (controle negativo da extração); 9) controle positivo; 10) água ultrapura (controle negativo da extração); 11) água ultrapura (controle negativo da PCR); (PM) padrão de peso molecular 100 pb.
Fonte: SILVA (2009).

13
De acordo com ALMEIDA (2004, vem ocorrendo o desenvolvimento
de técnicas baseadas no princípio da cn-PCR e uma delas é a conhecida n-
PCR, esta é um método sensível, no qual o produto amplificado é submetido a
um segundo processo de amplificação, para isso, são utilizados um conjunto de
iniciadores homólogos às sequências internas do segmento já amplificado.
Este procedimento torna a reação da polimerase mais específica e sensível.
Geralmente, na primeira PCR é utilizado um iniciador gênero-específico e, na
segunda, um iniciador espécie-específico do parasita em estudo.
2.2.2 Teste de polimorfismo de comprimento de fragmentos de DNA
gerados por enzimas de restrição sobre produtos amplificados pela
reação em cadeia da polimerase
Como mencionado anteriormente, a PCR é realizada para a
amplificação de uma região de um gene de interesse que contém uma ou
múltiplas substituições de bases. Quando o produto de tal técnica é submetido
à digestão por uma ou mais enzimas de restrição, ocorre então o PCR-RFLP,
este é um método que detecta variações na região do fragmento alvo que foi
amplificado, sendo que uma única substituição de bases pode originar ou
extinguir um sítio capaz de ser digerido pela enzima de restrição de
endonuclease (ALMEIDA, 2004).
Para isso, tal método busca os marcadores moleculares e baseia-se
na capacidade de que as endonucleases de restrição reconhecem o
polimorfismo do nucleotídeo único dos produtos obtidos pela PCR e
posteriormente, exibe as regiões distintas do fragmento em padrão de bandas
pelo método de eletroforese em gel de agarose (SU et al., 2010).

14
2.3 Diagnóstico e caracterização molecular do Toxoplasma gondii
2.3.1 Diagnóstico molecular
O T. gondii, eucarioto intracelular obrigatório, foi detectado por meio
de técnicas moleculares em vários tecidos vivos e células nucleadas além de
ter sido encontrado em líquidos orgânicos como sangue, linfa, saliva, leite
(colostro), exsudatos, esperma e líquido peritoneal, sendo ainda capaz de se
dividir e multiplicar-se em cultura de células de peixes (SILVEIRA, 2009).
Desta forma, os métodos de diagnósticos moleculares promovem a
detecção do parasito, pois possuem alta sensibilidade e especificidade,
proporcionando a identificação do DNA do protozoário em diferentes amostras,
como fezes e fluidos corpóreos (GONÇALVES, 2010).
Para alcançar elevada sensibilidade pela PCR, deve-se ter
conhecimento prévio da sequência alvo do DNA que se deseja detectar. A
partir disto, dois primers já desenhados são utilizados para amplificar a
sequência desejada (SU et al., 2010).
Cada par de iniciador é específico para um dos marcadores
genéticos existentes para o T. gondii, os quais são frequentemente
pesquisados nas amostras biológicas (SU et al., 2010), exemplo: o gene B1,
inicialmente descrito por BURG et al. (1989); o fragmento de 529 pb (HOMAN
et al., 2000), o espaçado interno transcrito (ITS -1) (SLAPETA et al.,2002),
entre outros.
O gene B1 é um marcador genético com peso molecular de 2,2 Kb
(kilobase) (CARNEIRO, 2011), tal marcador é um dos mais utilizados, pois está
repetido em 35 cópias no genoma do T. gondii, ainda demonstra alta
sensibilidade, sendo capaz de detectar até um parasita em cerca de 100 mil
células hospedeiras. Para a detecção do gene B1 são utilizados os pares de
primers: 5'-GGA ACT GCA TCC GTT CAT GAG-3' e 5'-TCT TTA AAG CGT
TCG TGG TC-3' (BURG et al., 1989).
SPALDING et al. (2002) por meio da cn-PCR utilizou os dois pares
de primers referidos anteriormente, direcionados ao gene B1 do T. gondii e
15
avaliou amostras sanguíneas e placentárias coletadas de mulheres no
momento do parto e a reação foi sensível e específica; evidenciando a
presença de um a dez taquizoítos, no material amostrado.
O marcador genético de 529 pb repete-se 300 vezes no genoma do
parasita em questão, assim, apresenta elevada sensibilidade para a detecção
de tal protozoário e para a amplificação deste marcador podem ser utilizados
os pares de primers: 5’-CGC TGC AGG GAG GAA GAC GAA AGT TG-‘3 e 5’-
CGC TGC AGA CAC AGT GCA TCT GGA TT-3’ (HOMAN et al., 2000).
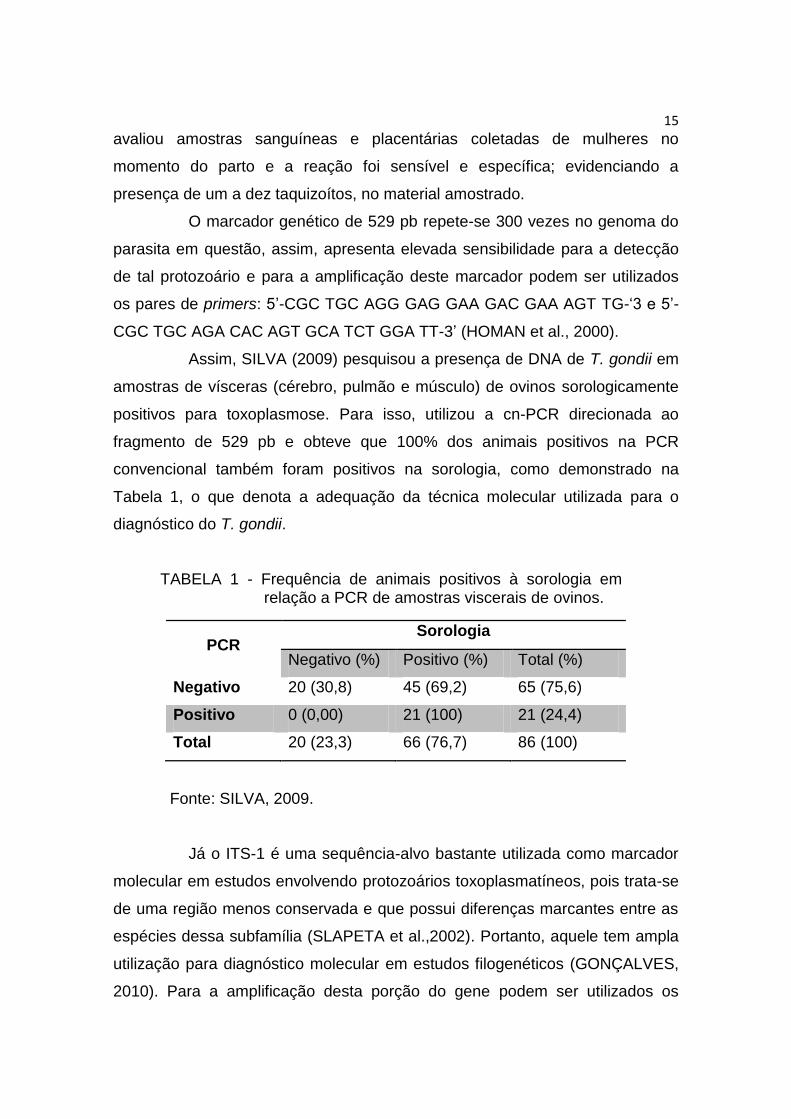
Assim, SILVA (2009) pesquisou a presença de DNA de T. gondii em
amostras de vísceras (cérebro, pulmão e músculo) de ovinos sorologicamente
positivos para toxoplasmose. Para isso, utilizou a cn-PCR direcionada ao
fragmento de 529 pb e obteve que 100% dos animais positivos na PCR
convencional também foram positivos na sorologia, como demonstrado na
Tabela 1, o que denota a adequação da técnica molecular utilizada para o
diagnóstico do T. gondii.
TABELA 1 - Frequência de animais positivos à sorologia em relação a PCR de amostras viscerais de ovinos.
PCR Sorologia
Negativo (%) Positivo (%) Total (%)
Negativo 20 (30,8) 45 (69,2) 65 (75,6)
Positivo 0 (0,00) 21 (100) 21 (24,4)
Total 20 (23,3) 66 (76,7) 86 (100)
Fonte: SILVA, 2009.
Já o ITS-1 é uma sequência-alvo bastante utilizada como marcador
molecular em estudos envolvendo protozoários toxoplasmatíneos, pois trata-se
de uma região menos conservada e que possui diferenças marcantes entre as
espécies dessa subfamília (SLAPETA et al.,2002). Portanto, aquele tem ampla
utilização para diagnóstico molecular em estudos filogenéticos (GONÇALVES,
2010). Para a amplificação desta porção do gene podem ser utilizados os

16
primers: 5’CGA AAT GGG AAG TTT TGT GAA C3’ e 5’-CAG CAG CTA CAT
ACG TAG A-3’ (SLAPETA et al.,2002).
GONÇALVES (2010), a fim de detectar parasitas da subfamília
Toxoplasmatinea, presentes em amostras viscerais de galinhas, utilizou dois
pares de primers, para amplificar a porção ITS-1, por meio da n-PCR e foi
observado que 42% das amostras apresentaram resultado com bandas com
cerca de 400pb, equivalentes ao DNA desta subfamília.
O autor realizou ainda a cn-PCR para o T. gondii, sendo que para o
primeiro foi amplificado o gene B1 e obteve que 66% de amostras positivas na
n-PCR também foram positivas para a cn-PCR direcionada ao T. gondii, o que
demonstra que as aves apresentam-se como hospedeiros intermediários
eficientes deste protozoário.
Em geral, a PCR é muito sensível, pois pode detectar o DNA de
apenas um taquizoíto, proporcionando um diagnóstico rápido, e o uso de tal
método é adequado para espécimes clínicos, porém amostras fecais
apresentam inibidores, os quais podem diminuir a sensibilidade da PCR. No
entanto, do ponto de vista da saúde pública, é necessário distinguir os oocistos
de T. gondii dos oocistos de outros protozoários, como Isospora felis. I. rivolta,
Sarcocystis muris (Figura 8), os quais também podem estar presentes nas
fezes do gato, porém aqueles não parasitam o homem (DUBEY, 2010).
Diante disto, foi desenvolvida uma técnica baseada na cn-PCR,
conhecida como copro-CPR na qual procedesse a concentração dos oocistos e
o rompimento da parede desses, antes da extração do DNA. Tal técnica foi
direcionada à sequência de 529 pb e permitiu a detecção de DNA de T. gondii,
a partir de amostras fecais de gatos (SALANT et al., 2010).
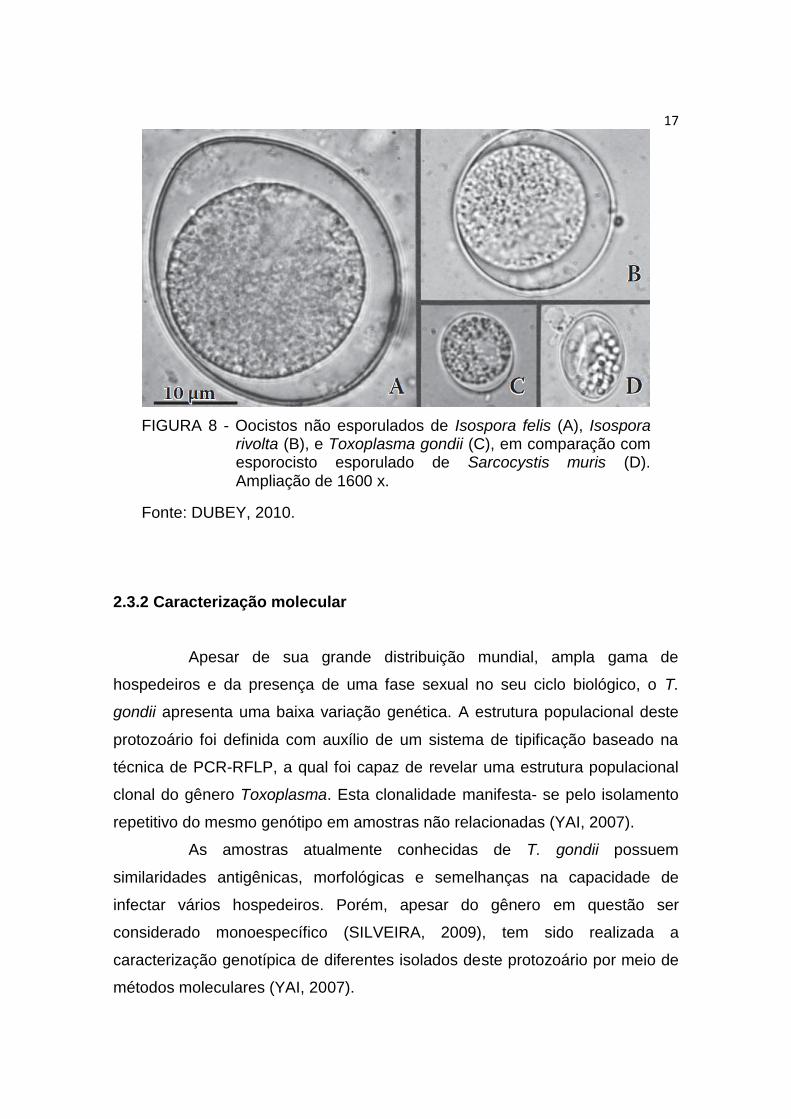
17
FIGURA 8 - Oocistos não esporulados de Isospora felis (A), Isospora rivolta (B), e Toxoplasma gondii (C), em comparação com esporocisto esporulado de Sarcocystis muris (D). Ampliação de 1600 x.
Fonte: DUBEY, 2010.
2.3.2 Caracterização molecular
Apesar de sua grande distribuição mundial, ampla gama de
hospedeiros e da presença de uma fase sexual no seu ciclo biológico, o T.
gondii apresenta uma baixa variação genética. A estrutura populacional deste
protozoário foi definida com auxílio de um sistema de tipificação baseado na
técnica de PCR-RFLP, a qual foi capaz de revelar uma estrutura populacional
clonal do gênero Toxoplasma. Esta clonalidade manifesta- se pelo isolamento
repetitivo do mesmo genótipo em amostras não relacionadas (YAI, 2007).
As amostras atualmente conhecidas de T. gondii possuem
similaridades antigênicas, morfológicas e semelhanças na capacidade de
infectar vários hospedeiros. Porém, apesar do gênero em questão ser
considerado monoespecífico (SILVEIRA, 2009), tem sido realizada a
caracterização genotípica de diferentes isolados deste protozoário por meio de
métodos moleculares (YAI, 2007).

18
Para a realização da genotipagem dos isolados de T. gondii pode
ser utilizada a PCR-RFLP, a qual é direcionada a vários marcadores
moleculares, como por exemplo: SAG1, SAG2, SAG3. O que proporciona a
detecção do polimorfismo genético e possibilita a caracterização genotípica em
diferentes isolados, originados de material humano e animal, demonstrando a
existência de diversidade genética no gênero, ao qual pertence tal protozoário
(CARNEIRO, 2011).
Assim, com a utilização da técnica mencionada foi possível
descrever a subpopulação do T. gondii em várias regiões geográficas do
mundo. Desta forma, a maioria dos isolados deste protozoário, de humanos e
animais, na América do Norte, Europa e África foram agrupadas em uma da
três linhagens clonais conhecidas: tipo I, II e III (SU et al., 2010).
Em humanos, a linhagem clonal de T. gondii classificada como tipo I
é normalmente associada a casos de toxoplasmose aguda, podendo provocar
lesões oculares (VALLOCHI et al., 2005). Já o genótipo tipo II é predominante
encontrado em pacientes imunossuprimidos e em casos de toxoplasmose
congênita e a do tipo III é comumente encontrada em animais (CARNEIRO,
2011).
Assim, de acordo com DUBEY et al. (2004), por meio da PCR-RFLP
e a partir de amostras viscerais de gatos sorologicamente positivos para o T.
gondii, da região do Paraná, Brasil, foram isoladas somente as linhagens tipo I
e tipo III. Outro estudo, com isolados de vísceras de cães, também positivos
para toxoplasmose, provenientes do estado de São Paulo, confirmou somente
a presença da linhagem tipo III (DUBEY et al., 2007a).
Já DUBEY et al. (2007b), demonstraram a partir de amostras
viscerais de galinhas caipiras, sorologicamente positivas para toxoplasmose e
provenientes da região do Pará e Rio grande do Sul, que os genótipos isolados
eram diferentes dos já descritos anteriormente, porém intimamente
relacionados com as linhagens clonais tipos I ou III.
PENA et al. (2008) avaliaram os resultados obtidos por DUBEY et al.
(2004), DUBEY et al. (2007a), DUBEY et al. (2007b) e diante da alta
diversidade genética dos isolados, determinaram, por meio da PCR-RFLP,
19
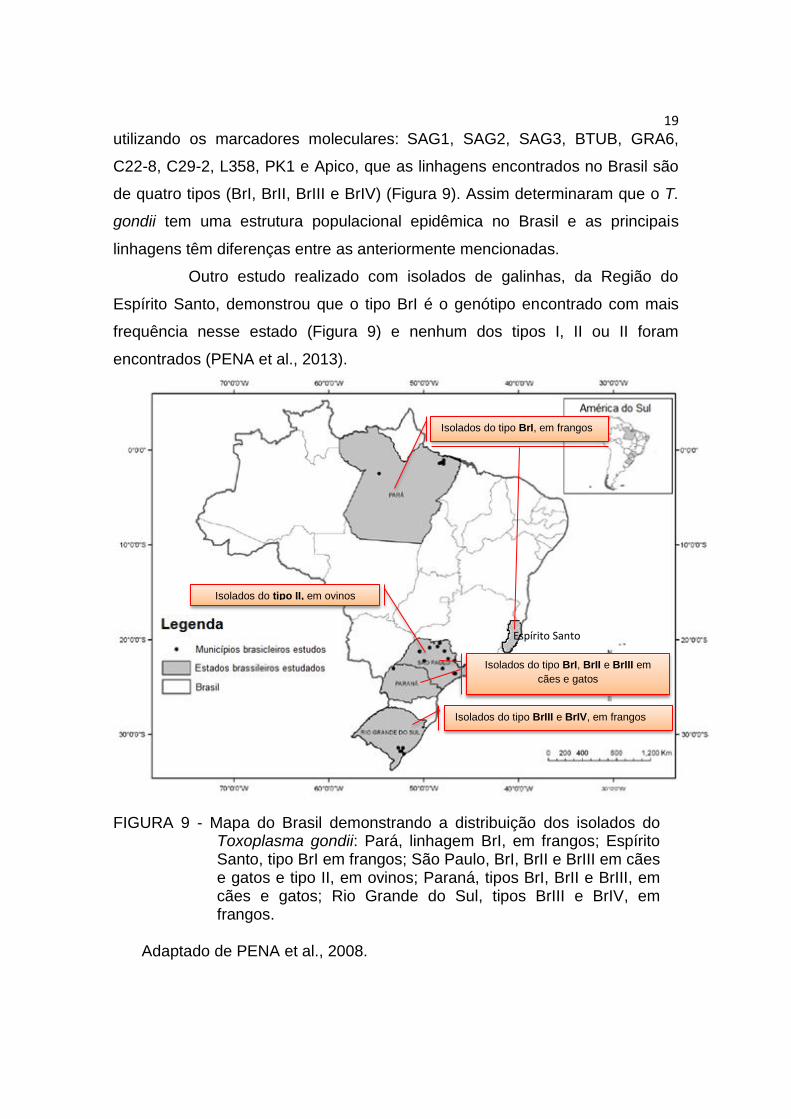
utilizando os marcadores moleculares: SAG1, SAG2, SAG3, BTUB, GRA6,
C22-8, C29-2, L358, PK1 e Apico, que as linhagens encontrados no Brasil são
de quatro tipos (BrI, BrII, BrIII e BrIV) (Figura 9). Assim determinaram que o T.
gondii tem uma estrutura populacional epidêmica no Brasil e as principais
linhagens têm diferenças entre as anteriormente mencionadas.
Outro estudo realizado com isolados de galinhas, da Região do
Espírito Santo, demonstrou que o tipo BrI é o genótipo encontrado com mais
frequência nesse estado (Figura 9) e nenhum dos tipos I, II ou II foram
encontrados (PENA et al., 2013).
FIGURA 9 - Mapa do Brasil demonstrando a distribuição dos isolados do Toxoplasma gondii: Pará, linhagem BrI, em frangos; Espírito Santo, tipo BrI em frangos; São Paulo, BrI, BrII e BrIII em cães e gatos e tipo II, em ovinos; Paraná, tipos BrI, BrII e BrIII, em cães e gatos; Rio Grande do Sul, tipos BrIII e BrIV, em frangos.
Adaptado de PENA et al., 2008.
Espírito Santo
Isolados do tipo BrI, em frangos
Isolados do tipo BrI, BrII e BrIII em
cães e gatos
Isolados do tipo BrIII e BrIV, em frangos
Isolados do tipo II, em ovinos

20
Porém, SILVA et al. (2011) pesquisaram a taxa de infecção por T.
gondii em ovinos de frigoríficos do estado de São Paulo e os que foram
positivos sorologicamente para toxoplasmose, tiveram suas amostras viscerais
avaliadas pela cn-PCR e PCR-RFLP, sendo que para esta utilizaram 11
marcadores: SAG1, SAG2, SAG3, BTUB, GRA6, C22-8, C29-2, L358, PK1,
Apico e CS3.
A partir disto, os pesquisadores demonstraram que o genótipo tipo II,
anteriormente não encontrado no Brasil, estava presente nas amostras
viscerais de ovinos provenientes do estado de São Paulo. Tais resultados
confirmaram a elevada diversidade de genótipos do protozoário em questão no
território brasileiro (SILVA et al., 2011).
A infecção pelo T. gondii, bem como o desenvolvimento da
toxoplasmose, tem sido detectada com maior sensibilidade através dos
métodos moleculares. Contudo, como limitação da técnica, a positividade não
discrimina se o DNA amplificado provém de parasitas viáveis ou de fragmentos
do parasita. Devido a isso, o isolamento em animais susceptíveis reproduz a
infecção quando há parasitas viáveis nos tecidos animais, refletindo a presença
ativa dos mesmos (SILVEIRA, 2009).
Dentre os animais sensíveis à inoculação, citam-se hamsters,
cobaias, camundongos e coelhos. Destes, os camundongos são os melhores
por serem os mais susceptíveis à infecção experimental, podendo, fornecer
milhões de taquizoítos em apenas três dias depois da infecção experimental
(SILVEIRA, 2009).
Para o isolamento do T. gondii podem ser utilizados camundongos
heterozigotos imunocompetentes, os quais são naturalmente susceptíveis à
infecção pelo parasito; a via de inoculação comumente utilizada é a via
intraperitoneal, que se mostra mais efetiva do que outras vias, como via oral ou
intracerebral (GONÇALVES, 2010).
Os camundongos são inoculados com taquizoítos ou bradizoítos, e
dependendo da linhagem inoculada, se virulenta o animal pode desenvolver
infecção aguda, com a formação de ascite, apresentar taquizoítos no líquido
peritoneal, formar cistos teciduais e desenvolver encefalite, o que causa a

21
sintomatologia neurológica observada em alguns animais (GONÇALVES,
2010), ou quando a linhagem é não-virulenta a infecção pode apresentar de
forma crônica, não desenvolvendo sintomas (YAI, 2007).
Considerando que a patogenicidade do T. gondii está estreitamente
relacionada à virulência da cepa e à espécie hospedeira (YAI, 2007), PENA et
al. (2008) realizaram o isolamento e o bioensaio em camundongos das
linhagens encontradas no Brasil (BrI, BrII, BrIII e BrIV) e constrataram que o
isolado do tipo BrI pode ser considerado virulento, o tipo BrIII não-virulento, e
os tipos BrII e BrIV são considerados virulentos intermediários. Já o tipo II,
encontrado em ovinos no estado de São Paulo, não foi virulento ao bioensaio
em camundongos (SILVA et al., 2011).
Os resultados apresentados demonstram a diferença da distribuição
e das linhagens genotípicas apresentadas no Brasil em relação às
demonstradas anteriormente para a América do Norte, pois nesta são relatadas
apenas a presença de três linhagens: tipo I, II e III (DUBEY et al., 2004).
Diante do que foi exposto, é de extrema importância investigar a
relação do genótipo e as manifestações clínicas da toxoplasmose em animais,
pois estas podem ser relacionadas às linhagens patogênicas aos seres
humanos (SU et al., 2006).

22
3 CONSIDERAÇÕES FINAIS
Os protozoários são parasitas causadores de doenças de
importância médica e veterinária, destacando-se o T. gondii por ser agente
causador de zoonose apresentar distribuição cosmopolita.
O diagnóstico da toxoplasmose baseia-se no exame sorológico, o
qual é capaz de demonstrar o contato com o parasita. Porém, quando a
infecção toma a sua forma crônica, a técnica mencionada, não indica a
presença de parasita viável. O que é importante, por exemplo, quando se
investiga a forma de contaminação: a horizontal, pela presença de bradizoítos
em vísceras, ou a forma vertical, pela detecção de taquizoítos em anexos
placentários.
Para obter tais resultados, as técnicas baseadas no diagnóstico
molecular, especificamente a PCR, são utilizadas juntamente com os métodos
sorológicos, objetivando-se a detecção do T. gondii. Assim, as técnicas
moleculares são ferramentas eficientes, pois podem detectar o DNA do
protozoário em diversos materiais biológicos, o que demonstra a presença do
mesmo, no material analisado.
Porém, somente a identificação do material genético do T. gondii,
não indica a sua viabilidade, desta forma é requisitado o bioensaio, pois este é
capaz de reproduzir a infecção em animais, como por exemplo, em
camundongos. Tal técnica é capaz de demonstrar a presença de parasitas
viáveis e assim, será definido que o T. gondii, na amostra analisada, possui
potencial infeccioso.
Ainda, o estudo da diversidade genética de tal protozoário, por meio
da PCR-RFLP, é importante, pois pode esclarecer dúvidas quanto à
distribuição epidemiológica nos hospedeiros intermediários, pois estes podem
ser importantes fontes de contaminação horizontal. E juntamente com o
bioensaio, as técnicas baseadas na PCR, fornecem subsídios para avaliação
das características biológicas, como a virulência e patogenicidade da linhagem.
Além de elucidar aspectos das distintas manifestações clínicas da doença, o
que resulta em obtenção ou ampliação de conhecimentos importantes para o
controle, diagnóstico e tratamento da toxoplasmose.

23
4 REFERÊNCIAS
1 ALMEIDA, T. T. C. Padronização e avaliação de métodos moleculares para detecção de oocistos de Cryptosporidium spp. (Apicomplexa: Cryptosporiidae) em amostras fecais: extração de DNA genômico e PCR (reação em cadeia pela polimerase). 2004. 143 f. Tese (Doutorado em saúde pública) - Universidade de São Paulo, Faculdade de Saúde Pública, São Paulo.
2 BARRA, G. B.; CAIXETA, M. C. S. A. S. B.; COSTA, P. G. G.; SOUSA, C. F.; VELASCO, L. F. R. Diagnóstico molecular - passado, presente e futuro, Revista Brasileira de Análises Clínicas, Rio de Janeiro, v. 43, n. 3, p.254-260, 2011.
3 BURG, J. L.; GROVER, C. M.; POULETTY, P.; BOOTHROYD, J. C. Direct and sensitive detection of a pathogenic protozoan, Toxoplasma gondii, by polymerase chain reaction. Journal of Clinical Microbiology, Washington, v. 27, n. 8, p. 1787-1792, 1989.
4 CARNEIRO, A. C. A. V. Caracterização molecular de isolados de Toxoplasma gondii obtidos de crianças com toxoplasmose congênita no Estado de Minas Gerais. 2011. 191 f. Tese (Doutorado em Parasitologia). Universidade Federal de Minas Gerais - Instituto de Ciências Biológicas, Belo Horizonte.
5 DUBEY, J. P. Diagnosis. In: ________ Toxoplasmoses of animals and humans. 2º ed. Boca Raton: CRC Press, 2010. Session 1.7, p. 52-71.
6 DUBEY, J. P. The history of Toxoplasma gondii - the first 100 years. Journal of Eukaryotic Microbiology, Lawrence, v. 55, n. 6, p. 467-475, Dec. 2008.
7 DUBEY, J. P.; GENNARI, S. M.; SUNDAR, N. M.; VIANNA, C B.; BANDINI, L. M.; YAI, L. E O.; KWOK, O. C H.; SU, C. Diverse and atypical genotypes identified in Toxoplasma gondii from dogs in São Paulo, Brazil. The Journal of Parasitology, Lawrence, v. 93, n. 1, pp. 60-64, 2007a.
8 DUBEY, J. P.; LAGO, E. G.; GENNARI, S. M., SU, C., JONES, J. L. Review Article. Toxoplasmosis in humans and animals in Brazil: high prevalence, high burden of disease, and epidemiology. Parasitology, Cambridge, v. 139, n. 11, 1375-1424, 2012.
9 DUBEY, J. P.; LINDSAY, D. S.; SPEER, C. A. Structures of Toxoplasma gondii tachyzoites, bradyzoites, and sporozoites and biology and development of tissue cysts. Clinical Microbiology Reviews, Washington, v. 11, n. 2, p. 267-299, 1998.

24
10 DUBEY, J. P.; NAVARRO, I. T.; SREEKUMAR, C.; DAHL, E.; FREIRE, R. L.; KAWABATA, H. H.; VIANNA, M. C. B.; KWOK, O. C. H.; SHEN, S. K.; THULLIEZ, P.; LEHMANN, T. Toxoplasma gondii infections in cats from Paraná, Brazil: seroprevalence, tissue distribution, and biologic and genetic characterization of isolates. The Journal of Parasitology, Lawrence, v. 90, n. 4, p. 721-726, 2004.
11 DUBEY, J. P.; SUNDAR, N.; GENNARI, S. M.; MINERVINOC, A. H. H.; FARIAS, N. A.; RUAS, R. J. L.; SANTOS, T. R. B.; CAVALCANTE, G. T.; KWOKA, O.C. H.; SU, C. Biologic and genetic comparison of Toxoplasma gondii isolates in free-range chickens from the northern Pará state and the southern state Rio Grande do Sul, Brazil revealed highly diverse and distinct parasite populations. Veterinary Parasitology, Amsterdam, v. 143, n. 2, p. 182-188, 2007b.
12 FIALHO, C. G.; TEIXEIRA, M. C.; ARAUJO, F. A. P. Toxoplasmose animal no Brasil. Acta Scientiae Veterinariae, Porto Alegre, v. 37, n. 1, p.1-23, 2009.
13 GONÇALVES, I. N. Investigação sorológica, molecular e isolamento de coccídios toxoplasmatíneos em galinhas (Gallus domesticus). 2010. 82 f. Dissertação (Mestrado em Ciência Animal nos trópicos) - Universidade Federal da Bahia, Escola de Medicina Veterinária, Bahia.
14 HOMAN, W. L.; VERCAMMEN, M.; BRAEKELEER, J. D.; VERSCHUEREN, H. Identification of a 200- to 300-fold repetitive 529 bp DNA fragment in Toxoplasma gondii, and its use for diagnostic and quantitative PCR. International Journal for Parasitology, Oxford, v. 30, p. 69-75, 2000.
15 LIMA, L. M. Conceitos básicos de técnicas em biologia molecular, Embrapa Algodão. Documentos, 191, 1ª ed, Embrapa: Campina Grande, 2008. 27 p.
16 MONIS, P. T.; ANDREWS, R. H. Molecular epidemiology: assumptions and limitations of commonly applied methods. International Journal for Parasitology, Oxford, v. 26, n. 6, p. 981-987, 1998.
17 MONTEIRO, R. M. Caracterização de protozoários pertencentes à Subfamília Toxoplasmtinae pela análise molecular do gene codificador de proteína do choque térmico (HSP70) e do espaçador interno transcrito 1 (ITS-1). 2006. 106 f. Dissertação (Mestrado em Epidemiologia experimental aplicada às zoonoses) - Universidade de São Paulo, Faculdade de Medicina Veterinária e Zootecnia, São Paulo.

25
18 MONTOYA, J. G. Laboratory diagnosis of Toxoplasma gondii infection and toxoplasmosis. The Journal of Infectious Diseases, Oxford, v. 15, n. 185 p. 73-82, 2002.
19 MONTOYA, J. G.; LIESENFELD, O. Toxoplasmosis. The Lancet, London, v. 363, n. 9425, p.1965-1976, 2004.
20 PENA, H. F. J.; GENNARI, S. M. DUBEY, J. P.; SU, C. Population structure and mouse-virulence of Toxoplasma gondii in Brazil. International Journal for Parasitology, Oxford, v. 38, n. 5 p. 561-569, 2008.
21 PENA, H. F. J.; VITALIANO, S. N.; M. BELTRAME, A. V.; PEREIRA, F.E.L.; GENNARI, S.M.; SOARES, R. M. PCR-RFLP genotyping of Toxoplasma gondii from chickens from Espírito Santo state, Southeast region, Brazil: New genotypes and a new SAG3 marker allele. Veterinary Parasitology, Amsterdam, 192, n1-2, p. 111-117, 2013.
22 SALANT, H.; SPIRA, D. T.; HAMBURGER, J. A comparative analysis of coprologic ciagnostic methods for detection of Toxoplama gondii in cats. The Americam Journal of Tropical Medicine and Hygiene, Mclean, v. 82, n. 5, p. 865–870, 2010.
23 SERCUNDES, M. K. Filogenia molecular dos protozoários pertencentes à subfamília Toxoplasmatinae pela análise de genes mitocondriais e de apicoplasto. 2010. 94 f. Dissertação (Mestrado em Epidemiologia experimental aplicada às zoonoses) - Universidade de São Paulo, Faculdade de Medicina Veterinária e Zootecnia, São Paulo.
24 SILVA, A. J., BORNAY-LLINARES, F. J., MOURA, N. S., SLEMENDA, SUSAN B., TUTTLE, J. L., PIENIAZEK, NORMAN J. Fast and reliable extraction of protozoan parasite DNA from fecal specimens. Molecular Diagnosis, Naperville, v. 4, n. 1, p. 57-64, 1999.
25 SILVA, R. C. Caracterização genotípica de amostras de Toxoplasma gondii isoladas de ovinos de abatedouro. 2009. 159f. Tese (Doutorado em Medicina Veterinária) - Universidade Estadual Paulista, Faculdade de Medicina Veterinária e Zootecnia, Botucatu.
26 SILVA, R. C.; LANGONI, H.; SU, C.; SILVA, A. V. Genotypic characterization of Toxoplasma gondii in sheep from Brazilian slaughterhouses: new atipical genotypes and the clonal type II strain identified. Veterinary Parasitology, Amsterdam, v. 175, n. 1-2, p. 173-177, 2011.
27 SILVEIRA, L. H. Caracterização biológica e genotípica de isolados de Toxoplasma gondii obtidos de galinhas de criação livre do Pantanal do Mato Grosso do Sul. 2009. 136 f. Tese (Doutorado em Epidemiologia

26
Experimental e Aplicada às Zoonoses) - Faculdade de Medicina Veterinária e Zootecnia da Universidade de São Paulo, São Paulo.
28 SINGH, B. Molecular methods for diagnosis and epidemiological studies for parasitic infections. International Journal for Parasitology, Oxford, v. 27, n. 10, p. 1135-1145, 1997.
29 SLAPETA, J. R.; KOUDELA, B.; VOTYPKA, J.; MODRY, D.; HOREJS, R.; LUKES, J. Coprodiagnosis of Hammondia heydorni in dogs by PCR based amplification of ITS 1 rRNA: differentiation from morphologically indistinguishable oocysts of Neospora caninum. The Veterinary Journal, London, v. 163, n.2, p. 147-154, 2002.
30 SPALDING, S. M.; AMENDOEIRA, M. R. R.; COELHO, J. M. C.; ANGEL, S. O. Otimização da reação de polimerase em cadeia para detecção de Toxoplasma gondii em sangue venoso e placenta de gestantes. Jornal Brasileiro de Patologia e Medicina Laboratorial. Rio de Janeiro, v. 38, n. 2, p. 105-110, 2002.
31 STEPHENSON, F. H.; ABILOCK, M. C. Optimizing the Polymerase Chain Reaction. Bay Area Biotechnology Education Consortium, Hayward, CA, 2012, 33 p.
32 SU, C.; SHWAB, E. K.; ZHOU, P.; ZHU, X. Q.; DUBEY, J. P. Moving towards an integrated approach to molecular detection and identification of Toxoplasma gondii. Parasitology, London, v. 137, p. 1-11, 2010.
33 SU, C.; ZHANG, X.; DUBEY, J. P. Genotyping of Toxoplasma gondii by multilocus PCR-RFLP markers: A high resolution and simple method for identification of parasites. International Journal for Parasitology, Oxford, v. 36, n. 7, p. 841-848, 2006.
34 TENTER, A. M.; BARTA, J. R.; BEVERIDGE, I.; DUSZYNSKI, D. W.; MEHLHORN, H.; MORRISON, D. A.; THOMPSON, R. C. A.; CONRAD, P. A. The conceptual basis for a new classification of the coccidian. International Journal for Parasitology, Oxford, v. 32, n. 5, p. 595-616, 2002.
35 TENTER, A. M.; HECKEROTH, A. R.; WEISS, L.M. Toxoplasma gondii: from animals to humans. International Journal for Parasitology, Oxford, v. 30, n. 12-13, p. 1217-1258, 2000.
36 THEKISOE O. M. M.; INOUE, N. Molecular diagnosis of protozoan infections by loop-mediated isothermal amplification (LAMP): A diagnostic manual. [online] 2011. Research Unit for Advanced Preventive medicine, National Research Center for Protozoan Diseases, Obihiro University of Agriculture and Veterinary Medicine. Disponível em:

27
http://www.ciens.ucv.ve/ciencias/varios/ibe/Oriel_y_Noboru_LAMP_Manual_2011.pdf. Acesso em: 13 de setembro de 2013.
37 THOMPSON, R. C. A.; CONSTANTINE, C. C.; MORGAN, U. M. Overview and significance of molecular methods: what role for molecular epidemiology? Parasitology, Cambridge, v. 117, n. 7, p. 161-175, 1998.
38 VALLOCHI, A. L.; MUCCIOLI, C.; MARTINS, M. C.; SILVEIRA, C.; BELFORT JR., R.; RIZZO, L. V. The genotype of Toxoplasma gondii strains causing ocular toxoplasmosis in humans in Brazil. American journal of ophthalmology, Chicago, v. 139, n. 2, p. 350-351, 2005.
39 VASOO, S.; PRITT, B. S. Molecular diagnostics and parasitic disease. Clinics in Laboratory Medicine, Philadelphia, v. 33, n. 3, p. 461-503, 2013.
40 WAAL, T. Advances in diagnosis of protozoan diseases. Veterinary Parasitology, Amsterdam, v. 189, n. 1, p. 65-74, 2012.
41 YAI, L. E. O. Caracterização biológica e genotípica de isolados de Toxoplasma gondii de capivaras (Hydrochaeris hydrochaeris) do Estado de São Paulo. 2007. 138 f. Tese (Doutorado em Epidemiologia experimental aplicada às zoonoses) - Universidade de São Paulo, Faculdade de Medicina Veterinária e Zootecnia, São Paulo.
42 YERA, H.; TZEN, M.; DUPOUY-CAMET, J. Molecular biology for detection and characterization of protozoan infections in humans. European Journal of Protistology, Stuttgart, v. 39, n. 4, p. 435-443, 2003.





























