Embed Size (px)
Citation preview

UNIVERSIDADE ESTADUAL DO CENTRO-OESTE, UNICENTRO-PR
Aspectos ecofisiológicos e morfometria da Curitiba
prismatica (D. Legrand) Salywon & Landrum
TESE DE DOUTORADO
EDSON GARDIN
GUARAPUAVA - PR
2017

EDSON GARDIN
Aspectos ecofisiológicos e morfometria da Curitiba prismatica (D. Legrand)
Salywon & Landrum
Tese apresentada à Universidade Estadual do Centro-Oeste, como parte das exigências do Programa de Pós-Graduação em Agronomia, área de concentração em Produção Vegetal, para obtenção do título de Doutor.
Orientador:
Prof. Dr. Luciano Farinha Watzlawick
Co-orientadora: Profª. Drª. Patrícia Carla Giloni de Lima
GUARAPUAVA - PR
2017

Catalogação na Publicação
Biblioteca Central da Unicentro, Campus Santa Cruz
Gardin, Edson
G219a Aspectos ecofisiológicos e morfometria da Curitiba prismatica (D. Legrand) Salywon & Landrum / Edson Gardin. – – Guarapuava, 2017.
xiii, 113 f. : il. ; 28 cm Tese (doutorado) - Universidade Estadual do Centro-Oeste, Programa
de Pós-Graduação em Agronomia, área de concentração em Produção Vegetal, 2017
Orientador: Luciano Farinha Watzlavick Coorientadora: Patrícia Carla Giloni de Lima Banca examinadora: Luciano Farinha Watzlavick, Patricia Carla Giloni de Lima, Vanderlei Aparecido de Lima, Vitor Hugo Zanette, Eleandro Jose Brun, Flavia Gizele Konig Brun
Bibliografia 1. Agronomia. 2. Produção vegetal. 3. Fluorescência da clorofila a. 4.
Pigmentos fotossintéticos. 5. Irradiância. 6. Sazonalidade. 7. Sistema faxinal. 8. Floresta ombrófila mista. 9. Murta. 10. Biometria. 11. Myrtaceae. 12. Efeitos abióticos. 13. Plasticidade. 14. Terpenos. I. Título. II. Programa de Pós-Graduação em Agronomia.
CDD 630


A natureza é o único livro que oferece um conteúdo valioso em todas as suas folhas.
Johann Wolfgang Von Goethe

AGRADECIMENTOS
Ao meu orientador Prof. Dr. Luciano Farinha Watzlawick, pelas oportunidades,
incentivos e pelo auxílio na minha formação acadêmica. Suas observações foram de
fundamental valia para o desenvolvimento desse trabalho.
À Profª. Drª. Patrícia Carla Giloni de Lima, que gentilmente aceitou o convite para co-
orientar, sendo de fundamental importância os seus ensinamentos.
Ao Prof. Dr. Vanderlei Aparecido de Lima por todos os incentivos e ajuda nas análises
estatísticas e laboratoriais.
Aos professores que direta ou indiretamente colaboraram nessa caminhada, desde o
início dos créditos até a conclusão do trabalho.
À minha esposa Daniela do Amaral Oliveira, que me auxiliou e incentivou nas
diversas etapas da realização deste trabalho.
A todos os amigos e colegas que contribuíram de alguma forma para esse trabalho,
entre eles, Francisco Putini, Marcos Leandro Garcia, Mariane Dal Comune, Regina Lopes dos
Santos, Richeliel Rodrigues Silva, Suélen Hellmann, e especialmente a Jey Marinho de
Albuquerque que gentilmente se dispôs a ajudar nas medições nas unidades amostrais do
Faxinal Marmeleiro de Baixo.
A todas as pessoas que direta ou indiretamente colaboraram para a conclusão desse
trabalho.
À UNICENTRO e ao Programa de Pós-graduação em Agronomia pela oportunidade
de realizar o doutorado.

SUMÁRIO
LISTA DE FIGURAS ............................................................................................................. i
LISTA DE TABELAS .......................................................................................................... iv
LISTA DE SÍMBOLOS E ABREVIATURAS ....................................................................... v
1. INTRODUÇÃO GERAL ................................................................................................... 1
2. OBJETIVO GERAL .......................................................................................................... 3
2.1. Objetivos Específicos ...................................................................................................... 3
3. REFERENCIAL TEÓRICO ............................................................................................... 4
3.1. A espécie Curitiba prismatica (D. Legrand) Salywon & Landrum ................................... 4
3.2. Fotossíntese e pigmentos fotossintéticos .......................................................................... 7
3.3. Fluorescência da clorofila a ........................................................................................... 11
3.4. Óleo essencial ............................................................................................................... 15
3.5. Morfometria e dinâmica de crescimento de árvores ....................................................... 21
3.7. Referências Bibliográficas ............................................................................................. 25
4. Efeitos da sazonalidade climática e de diferentes irradiâncias nos parâmetros fotossintéticos
de Curitiba primastica (D. Legrand) Salywon and Landrum ................................................ 36
4.1. Introdução ..................................................................................................................... 38
4.2. Material e métodos ........................................................................................................ 40
4.3. Resultados e Discussão ................................................................................................. 44
4.4. Conclusão ..................................................................................................................... 55
4.5. Referências Bibliográficas ............................................................................................. 56
5. Morfometria e dinâmica de crescimento de Curitiba primastica (D. Legrand) Salywon e
Landrum .............................................................................................................................. 63
5.1 Introdução ...................................................................................................................... 65
5.2. Material e Métodos ....................................................................................................... 67

5.3. Resultados e Discussão ................................................................................................. 71
5.4. Conclusão ..................................................................................................................... 87
6. Composição química e rendimento de óleos essenciais de Curitiba prismatica (D. Legrand)
Salywon e Landrum sob efeito de sazonalidade climática e diferentes irradiâncias ............... 91
6.1. Introdução ..................................................................................................................... 93
6.2. Material e Métodos ....................................................................................................... 95
6.3. Resultados e Discussão ................................................................................................. 98
6.4. Conclusão ................................................................................................................... 109
6.5. Referências Bibliográficas ........................................................................................... 110
7. Considerações finais....................................................................................................... 113

i
LISTA DE FIGURAS
Figura 1. Frutos (A), flores (B), botões florais (C) e exemplar da espécie Curitiba prismatica
(D) no município de Rebouças-PR. ........................................................................................ 5
Figura 2. Local de ocorrência da espécie Curitiba prismatica no Brasil. ................................ 6
Figura 3. Espectro de absorção da luz pelas clorofilas a e b, em diferentes comprimentos de
onda. ...................................................................................................................................... 9
Figura 4. Cinética de emissão da fluorescência ou curva Kautsky registrada de uma
suspensão de Chlorella adaptada ao escuro. (A) cinética rápida; (B) cinética lenta. .............. 14
Figura 5. Representação das variáveis morfométricas, sendo: cc - comprimento de copa , d -
diâmetro a 1,30m do solo (DAP), dc - diâmetro de copa, h - altura total, hf - altura do fuste, hl
- altura da copa exposta a luz e hs - altura da copa sombreada. ............................................. 22
Figura 6. Localização da área de estudo no munícipio de Rebouças-PR. ............................. 40
Figura 7. Curva OJIP da fluorescência da clorofila a da espécie Curitiba prismatica em
ambiente sombreado e a pleno sol, durante as estações da primavera (A), verão (B), outono
(C) e inverno (D) de 2015 a 2016. ........................................................................................ 44
Figura 8. Parâmetros Fo (A), Fm (B), Fv/Fm (C), ABS/RC (D) e PIABS (E) da fluorescência da
cinética rápida da clorofila a obtidos da curva OJIP de folhas de Curitiba prismatica, em
diferentes ambientes de irradiação luminosa, durante as estações da primavera, verão, outono
e inverno de 2015 a 2016. .................................................................................................... 46
Figura 9. Parâmetros ETR (A), Y(II) (B) da fluorescência da cinética lenta da clorofila a da
espécie Curitiba prismatica em ambiente sombreado e a pleno sol, durante as estações da
primavera, verão, outono e inverno de 2015 a 2016. ............................................................. 49
Figura 10. Teores de clorofila a (A), clorofila b (B), clorofila total (C), relação clorofila a/b
(D) e carotenoides totais (E) da espécie Curitiba prismatica em ambiente sombreado e a pleno
sol, durante as estações da primavera, verão, outono e inverno de 2015 a 2016. ................... 51
Figura 11. Localização da área de estudo no munícipio de Rebouças, PR. ........................... 67
Figura 12. Modelos ajustados entre as variáveis morfométricas diâmetro a altura do peito –
DAP com grau de esbeltez – GE (A) e altura total – H com comprimento de copa – CC (B)
para Curitiba prismatica. ..................................................................................................... 76

ii
Figura 13. Resíduos dos modelos ajustados entre as variáveis morfométricas diâmetro a
altura do peito – DAP com grau de esbeltez – GE (A) e altura total – H com comprimento de
copa – CC (B) para Curitiba prismatica. .............................................................................. 76
Figura 14. Histograma das frequências de diâmetros à altura do peito de Curitiba prismatica
avaliadas em 2017, em Rebouças-PR. .................................................................................. 78
Figura 15. Variáveis morfométricas: altura total (A), altura de inserção da copa (B),
comprimento da copa (C), diâmetro da copa (D) e proporção copa (E), da Curitiba prismatica
por classes diamétricas. ........................................................................................................ 79
Figura 16. Variáveis morfométricas: (A) grau de esbeltez, (B) índice de saliência, (C) índice
de abrangência e (D) formal da copa, da Curitiba prismatica por classes diamétricas. .......... 80
Figura 17. Incremento periódico diamétrico relativo no período de 2012 a 2017, por classe
diamétrica. ........................................................................................................................... 84
Figura 18. Variação da área basal, por classes diamétricas, entre 2012 e 2017. .................... 85
Figura 19. Hidrodestilação do óleo essencial de folhas frescas de Curitiba prismatica
coletada no município de Rebouças-PR., utilizando conjunto com aparelho do tipo Clevenger,
balão de vidro e manta aquecedora.. ..................................................................................... 96
Figura 20. Rendimento do óleo essencial (mL.kg-1) das folhas frescas de Curitiba prismatica
coletadas em Rebouças-PR. ................................................................................................. 98
Figura 21. Variações da radiação solar incidente e precipitação média registradas nas
estações da primavera - 2015 e inverno - 2016. .................................................................... 99
Figura 22. Distribuição de compostos terpênicos, agrupados por classes, dos óleos essenciais
das folhas de Curitiba prismatica coletadas em Rebouças-PR, na primavera e inverno, e nos
ambientes de pleno sol e sombreado em sub-bosque........................................................... 102
Figura 23. Dendrograma para o agrupamento das amostras de óleos essenciais das folhas de
Curitiba prismatica coletadas em Rebouças-PR, na primavera e inverno, nos ambientes de
pleno sol e sombreado. ....................................................................................................... 104
Figura 24. Gráfico de distância de ligação das amostras de óleos essenciais das folhas de
Curitiba prismatica coletadas em Rebouças-PR. ................................................................ 105
Figura 25. Dispersão das amostras para os óleos essenciais de folhas das folhas de Curitiba
prismatica coletadas em Rebouças-PR, na primavera e inverno, nos ambientes de pleno sol e
sombreado em sub-bosque. ................................................................................................ 106

iii
Figura 26. Dispersão dos compostos químicos presentes nos óleos essenciais de folhas das
folhas de Curitiba prismatica coletadas em Rebouças-PR, na primavera e inverno, nos
ambientes de pleno sol e sombreado em sub-bosque........................................................... 107

iv
LISTA DE TABELAS
Tabela 1. Caracterização das variáveis morfométricas das árvores de Curitiba prismatica,
avaliadas em 2017. ............................................................................................................... 71
Tabela 2. Matriz de correlação linear de Pearson entre o diâmetro à altura do peito - DAP e a
altura total - H e as variáveis morfométricas. ........................................................................ 75
Tabela 3. Matriz de progressão por classe de diâmetro (cm) das árvores de Curitiba
prismatica avaliadas em 2012 e 2017. .................................................................................. 81
Tabela 4. Composição química dos óleos essenciais das folhas de C. prismatica coletadas em
Rebouças-PR, na primavera e inverno, nos ambientes de pleno sol e de sub-bosque. .......... 100

v
LISTA DE SÍMBOLOS E ABREVIATURAS
ABS/RC Fluxo de absorção por centro de reação ACP Análise de componentes principais
ATP Adenosina trifosfato
CCL Complexo coletor de luz CEO Complexo de evolução de oxigênio
CG/EM Cromatografia gasosa acoplada à espectrometria de massa
Chl a Clorofila a
Chl b Clorofila b CIM Concentração inibitória mínima
CR Centro de reação
CTE Cadeia de transporte de elétrons DAP Diâmetro a 1,30 m do solo
DCMU 3-(3,4-Diclorofenil)-1,1-dimetilurea
DMAPP Dimetilalil difosfato
DXP 1-deoxi-D-xilulose 5-fosfato DXR DXP redutoisomerase
ETR Taxa de transporte de elétrons
FEO Feofetina Fo Fluorescência mínima
Fm Fluorescência máxima
FOM Floresta ombrófila mista
FSI Fotossistema I FSII Fotossistema II
Fv Fluorescência variável
Fv/Fm Rendimento quântico máximo do FSII IK Índice de Kovatz
IPP Isopentenil difosfato
IPd Incremento periódico diamétrico IPAd Incremento periódico anual diamétrico
IPg Incremento periódico em área basal
IPAg Incremento periódico anual em área basal
IVI Índice de Valor de Importância

vi
MEP Metileritritol fosfato
MG Magnésio
NADPH Nicotinamida adenina dinucleótideo fosfato
NADPH+ Nicotinamida adenina dinucleótideo fosfato oxidada OE Óleo essencial
PAM Pulso de amplitude modulada
PIABS Desempenho fotossintético QA Plastoquinona A
qP Coeficiente de dissipação de fluorescência fotoquímica
qN Coeficiente de dissipação de fluorescência não-fotoquímica RFA Radiação fotossinteticamente ativa
RFD Taxa de decréscimo na fluorescência da clorofila a
UV Ultravioleta
VAM Via do ácido mevalônico Y(II) Rendimento fotoquímico do fotossistema II

1
1. INTRODUÇÃO GERAL
No Brasil as florestas nativas foram historicamente exploradas de forma desordenada e
com pouca ou nenhuma preocupação com sua sustentabilidade. As técnicas silviculturais e de
manejo florestal têm sido escassamente empregadas nessas florestas. Um exemplo que se tem
observado ultimamente é o da Floresta Ombrófila Mista (FOM), que caracteriza um dos
ecossistemas florestais mais importantes do Sul do Brasil.
Essa tipologia florestal, antes mesmo de ser adequadamente conhecida, foi
severamente reduzida sua dimensão original. O processo de colonização, exploração
madeireira e expansão agrícola conduziram à progressiva redução deste bioma. Atualmente
grandes partes das áreas remanescentes da FOM se restringem a fragmentos relictuais
alterados ou descaracterizados. No estado do Paraná a área de cobertura da FOM está
reduzida a menos de 30% de sua cobertura original (ACCIOLY, 2013).
A significativa redução da cobertura florestal da FOM ocorrida no Estado do Paraná,
evidenciam que muitas espécies endêmicas de regiões restritas desapareceram. Com elas
podem ter desaparecido informações importantes, relacionadas à estrutura das comunidades
florestais e sua capacidade natural de proteção e fornecimento de produtos benéficos às mais
diversas formas de vida (SILVESTRE, 2009).
O interesse sobre a perda da biodiversidade tropical em função do desflorestamento
conduziu ao reconhecimento do valor dos produtos florestais não-madeireiros – PFNM, que
podem ser extraídos das florestas tropicais (MYERS, 1988; DE BEER e MCDERMOTT,
1989; PETERS et al., 1989). Como exemplos da exploração de PFNM estão os consórcios de
árvores, culturas agrícolas e animais (sistemas agroflorestais em suas diversas conformações),
a prestação de serviços ambientais (neutralização de gases de efeito estufa, conservação da
biodiversidade, conservação do solo e água, conexão de fragmentos florestais e aumento da
permeabilidade da matriz de paisagens alteradas), óleos essenciais, entre outros. Segundo
Zamora (2001), na América Latina, os usos mais importantes dos PFNM são: medicinal e
farmacêuticos (óleos e essências), alimentício e industrial (gomas e resinas).

2
Dessa forma, os PFNM passaram a ser vistos como uma alternativa viável para se
explorar a riqueza biológica de florestas tropicais sem prejudicá-las e, ao mesmo tempo,
estimular o desenvolvimento das comunidades rurais (GENTRY e BLANEY, 1990;
PRANCE et al., 1987).
Ainda, conforme Nepstad e Schwartzman (1992), quando se realiza a extração de
PFNM, a estrutura e a função da floresta não são alteradas e, na maioria dos casos, essa
operação não envolve a degradação dos recursos naturais.
Considerando tais fatos, o uso desses produtos foi introduzido como uma estratégia de
conservação e de desenvolvimento florestal. Diante destes fatos surge a necessidade de que,
para serem sustentáveis, os níveis da colheita associados aos PFNM necessitam ser baseados
no bom conhecimento da floresta, das espécies que ali se desenvolvem e do potencial dos
produtos que podem ser extraídos (WICKENS, 1991).
Atualmente não existem pesquisas da espécie C. prismatica que visem analisar o seu
potencial para produção de PFNM e sua preservação. Nesse contexto a espécie Curitiba
prismatica (D. Legrand) Salywon e Landrum ganha destaque para o desenvolvimento de
pesquisas, visto que, além de ser uma espécie endêmica (SALYWON e LANDRUM, 2007),
estudos ecofisiológicos e de crescimento de C. prismatica são inexistentes, não havendo
registros das respostas adaptativas desta espécie sob diferentes níveis de irradiância, bem
como os efeitos da sazonalidade climática sobre seu desenvolvimento.

3
2. OBJETIVO GERAL
Avaliar a ecofisiologia e a morfometria da Curitiba prismatica (D. Legrand) Salywon
& Landrum
2.1. Objetivos Específicos
Avaliar os efeitos da sazonalidade e da irradiância, em relação à fluorescência da
clorofila a, em folhas de C. prismatica;
Associar a emissão de fluorescência da clorofila a aos teores de pigmentos
fotossintéticos das folhas de C. prismatica;
Analisar a morfometria e biometria de C. prismatica, bem como a sua dinâmica de
crescimento;
Avaliar a influência da irradiância e da sazonalidade na composição química dos óleos
essenciais de C. prismatica.

4
3. REFERENCIAL TEÓRICO
3.1. A espécie Curitiba prismatica (D. Legrand) Salywon & Landrum
A C. prismatica (D. Legrand) Salywon & Landrum é uma espécie arbórea, a qual
também pode ocorrer com porte arbustivo, pertencente à família Myrtaceae. É uma das
inúmeras espécies que compõem a Floresta Ombrófila Mista (FOM). Inicialmente classificada
como Eugenia prismatica (D. Legrand), foi transferida, em 2007, para um novo gênero,
Curitiba Salywon & Landrum (SALYWON e LANDRUM, 2007).
O nome do gênero foi uma homenagem à cidade de Curitiba - PR, região em que pode
ser encontrado crescendo em parques e remanescentes florestais, e também em homenagem
ao Museu Botânico Municipal de Curitiba. A denominação, prismatica refere-se à forma de
prisma que o seu fruto apresenta (SALYWON e LANDRUM, 2007) (Figura 1A).
A criação e classificação do gênero monotípico Curitiba só foi possível após várias
análises e estudos realizados com o DNA da planta. Em 1969, o pesquisador uruguaio Diego
Legrand a classificou como sendo do gênero Eugenia L., o mesmo da pitangueira, conclusão
obtida após perceber as semelhanças entre as flores das duas espécies. No ano de 1990, outros
estudos foram realizados pelo botânico brasileiro Marcos Sobral, especialista no gênero
Eugenia, e este demostrou que a planta tinha características muito diferentes daquele gênero.
Em 1997, o pesquisador americano Leslie Landrum classificou a planta como Mosiera
prismatica (D. Legrand) Landrum. E, finalmente em 2007, estudos genéticos demonstraram
que a classificação não estava adequada e se tratava, na verdade, de uma nova espécie.
Levando, assim, à criação do novo gênero Curitiba (SALYWON e LANDRUM, 2007).
É uma espécie endêmica encontrada no sul do Brasil, ocorrendo no primeiro e
segundo planaltos do Paraná e no planalto norte de Santa Catarina, (Figura 2), fato que
justifica atenção maior em sua preservação devido sua área de ocorrência restrita. Embora não
tenha dispersão geográfica muito ampla, essa espécie coloniza facilmente áreas degradadas.
Frequentemente forma populações densas no sub-bosque da FOM, em áreas com vegetação
mais esparsa, e também em áreas campestres (LORENZI, 2014; SALYMON e LANDRUM,
2007).

5
Figura 1. Frutos (A), flores (B), botões florais (C) e exemplares da espécie Curitiba
prismatica (D e E) no município de Rebouças-PR.
Fonte: Autor (2016).
A semente de C. prismatica é exalbuminosa, espiralada com tegumento coriáceo,
castanho-escuro. O embrião é do tipo cotiledonar pimentoide, axial, cilíndrico, curvado e de
coloração branca. Sua plântula é fanerocotiledonar, epígea e foliácea.

6
Segundo a resposta das sementes de C. prismatica à luz, são classificadas como
fotoblásticas positivas preferenciais. Os maiores valores de porcentagem e índice de
velocidade de germinação são obtidos à temperatura de 25ºC, nos substratos papel toalha e
areia (REGO et al., 2011).
Figura 2. Local de ocorrência da espécie Curitiba prismatica no Brasil.
Fonte: Salywon e Landrum (2007).
A C. prismatica pode atingir de 1 a 8 metros de altura. A floração ocorre durante os
meses de dezembro a fevereiro e os frutos amadurecem de janeiro a março. Sua frutificação
ocorre todos os anos, com produção abundante de sementes viáveis, dispersadas pela
avifauna. Possui fuste reto e casca acinzentada, fissurada, descamando em placas. Folhas
simples, opostas, com até 4 cm de comprimento e 2 cm de largura, não caducifólia. Flores
hermafroditas e de coloração branca. Frutos do tipo baga prismáticos, roxo-escuros,
alongados, com aproximadamente 2 cm de comprimento. Prefere luz solar direta e solos bem
drenados. Popularmente é conhecida por vários nomes, como, guamirim, cambuí ou murta.
(BACKES e IRGANG, 2009; LORENZI, 2014).

7
Estudo realizado por Watzlawick et al. (2011) em um fragmento de FOM no Faxinal
do Salto, município de Rebouças-PR, local endêmico da espécie, a C. prismatica foi a
segunda espécie com maior índice de Valor de Importância (IVI), 11,9%, e densidade de 90
ind.ha-1. Resultados semelhantes também foram encontrados por Albuquerque (2015) no
Faxinal Marmeleiro de Baixo, pertencente àquele mesmo município, onde o autor encontrou a
C. prismatica com maior IVI, 10,15%, e densidade de 111 ind.ha-1.
3.2. Fotossíntese e pigmentos fotossintéticos
O processo fotossintético consiste em duas etapas interdependentes e concomitantes.
Primeiro uma fase fotoquímica, a qual ocorre nos tilacoides, onde há conversão da energia
luminosa em energia química (ATP e NADPH) e o oxigênio é liberado. A outra fase, a
bioquímica, ocorre no estroma, onde o dióxido de carbono (CO2) é fixado e convertido em
carboidratos com a utilização dos produtos da etapa fotoquímica (ATP e NADPH). Essa
energia estocada nas moléculas orgânicas é utilizada nos processos celulares da planta e serve
como fonte de energia para todas as formas de vida (LAWLOR, 2009; DULAI et al., 2011).
O aparato fotossintético localiza-se dentro dos cloroplastos, nas membranas dos
tilacoides. São constituídos por dois tipos de fotossistemas, fotossistema I, FSI e fotossistema
II, FSII. Cada um destes fotossistemas é formado por um complexo coletor de luz (CCL) e
por um centro de reação (CR), interligados pelas proteínas da cadeia de transporte de elétrons
(CTE). Os fótons são absorvidos principalmente nos CCLs, os quais são formados por
pigmentos fotossintéticos, sendo os principais, a clorofila a e b, e os carotenoides (NOBEL,
1991; LAMBERS et al., 2008; TAIZ e ZEIGER, 2013).
O CR do FSII, seus pigmentos antenas do CCL e as proteínas da CTE associadas estão
localizadas predominantemente nas lamelas grana. A enzima conhecida como fator de ligação
que catalisa a formação do ATP, também é encontrada quase exclusivamente nas lamelas do
estroma e nas margens das lamelas granais. O citocromo b6f, pertencente à CTE, está
uniformemente distribuído entre o estroma e o grana. Dessa forma, ambos os eventos
fotoquímicos que ocorrem na fotossíntese estão espacialmente separados, necessitando assim,
que um ou mais carreadores de elétrons, que funcionam entre os fotossistemas, difundam-se
do grana da membrana até a região do estroma, onde os elétrons são entregues ao FSI
(ALLEN e FORSBERG, 2001).

8
Os carotenoides são pigmentos de coloração vermelha, laranja e amarela, encontrados
em todos os cloroplastos e nas cianobactérias. Da mesma forma que as clorofilas, os
carotenoides dos cloroplastos estão embebidos nas membranas dos tilacoides. Nas folhas
verdes, a cor dos carotenoides é mascarada, devido à maior abundância das clorofilas. As
funções dos carotenoides na fotossíntese são a absorção de luz, que será transferida para os
CR, e a fotoproteção contra danos fotoxidativos (TAIZ e ZEIGER, 2013).
Dois grupos de carotenoides, carotenos e xantofilas encontram-se normalmente
presentes nos cloroplastos em plantas. Os carotenoides, contendo apenas hidrocarbonetos são
chamados carotenos, e aqueles que contêm átomos de oxigênio, de xantofilas
(YAROSHEVICH et al., 2015).
A molécula de clorofila é formada por complexos derivados da porfirina, tendo um
átomo de magnésio (Mg) no centro de quatro anéis pirrólicos, ligados a uma cadeia fitol. Na
clorofila a, o anel de porfirina contém um grupo metil (-CH3) no carbono C-3, e na clorofila
b, que é considerada um pigmento acessório, contém um grupo aldeído (-CHO), que substitui
o grupo metil (SCHOEFS, 2002; TANAKA e TANAKA, 2007; VON ELBE, 2000).
As clorofilas a e b, além dos carotenoides, são pigmentos-chave das reações
luminosas. Deste modo, a determinação do conteúdo de clorofilas e carotenoides foliares e o
conhecimento de como ele varia entre espécies é uma característica de interesse dos
ecofisiologistas (BACARIN e MOSQUIM, 2002). Além disso, o estado nutricional das
plantas, geralmente, está associado com a quantidade e qualidade de clorofila a e b
(ZOTARELLI et al., 2003).
A energia dos fótons, que é denominada de quantum, uma vez absorvida pelos
pigmentos do CCL é transferida, por ressonância, para os CRs dos fotossistemas, levando à
excitação das clorofilas a. A faixa de radiação fotossinteticamente ativa (RFA) situa-se no
espectro visível entre 400 a 700 nm (NOBEL, 1991).
Os dois tipos de fotossistemas, FSI e FSII, que trabalham de forma simultânea e
contínua, diferem quanto à faixa de absorção de luz. No FSI, as moléculas de clorofila a do
CR possuem pico ótimo de absorção em 700 nm (P700), enquanto que no FSII essas
moléculas possuem o pico máximo de absorção em 680 nm (P680). Em torno de 95 a 99% de
fótons absorvidos pelos pigmentos CCL são transferidos para os CR (ODUM e BARRET,
2007; FALKOWSKI e RAVEN, 2007).

9
As clorofilas a e b diferem quanto à faixa do espectro da luz visível na qual captam luz
com mais eficiência. Ambas as clorofilas possuem dois picos de absorção em diferentes faixas
do espectro de luz, conforme Figura 3, sendo que para clorofila a são em 420 e 660 nm,
regiões do azul e do vermelho, respectivamente. Para a clorofila b, os máximos de absorção
correspondem, respectivamente, a 435 e 643 nm, também nas regiões azul e vermelho. Os
carotenoides, pigmentos mais envolvidos com outras reações fotoquímicas, têm picos de
absorção ao redor de 425, 450 e 480 nm. A quantidade de cada tipo de pigmento varia entre
os dois fotossistemas (TAIZ e ZEIGER, 2013).
Figura 3. Espectro de absorção da luz pelas clorofilas a e b, em diferentes comprimentos de
onda.
Fonte: Govindjee (2004).
O processo de fotossíntese inicia com a absorção de energia luminosa pelos CCLs e
suas subunidades antena, que estão ligados ao FSI e FSII. Depois de capturada, essa energia é
transferida por excitação para os CR, dos quais fazem parte moléculas de clorofila a,
desencadeando um conjunto de reações fotoquímicas que promovem, a partir da fotólise da
água, o transporte de elétrons entre os dois fotossistemas.
O transporte de elétrons será responsável pela produção da molécula redutora NADPH
e pela formação da molécula de energia ATP (KRAUSE e WEIS, 1991, NOBEL, 1991). Esse
processo, como um todo, forma a cadeia de transporte de elétrons (CTE), e é chamado de
esquema Z, sendo inicialmente proposto por Hill e Bendall (1960).

10
A CTE inicia com a clorofila a do CR do FSII sendo excitada pela luz, P680*, que
então transfere um elétron para a feofitina (Feo). O P680 oxidado é re-reduzido por um
elétron, que foi extraído pela oxidação de uma molécula de água no complexo de evolução de
oxigênio (CEO), liberando H+ e O2 no lúmen do tilacoide. No lado redutor do FSII, a feofitina
transfere elétron para os aceptores primários QA e QB, que são dois tipos de plastoquinonas. O
complexo citocromo b6f é o próximo carreador, que transfere o elétron para a plastocianina
(PC), reduzindo o P700*. Acredita-se que o aceptor de elétrons do P700* seja um tipo de
clorofila, seguido por uma quinona. Os carreadores seguintes são uma sequência de proteínas
ferro-sulfurosas ligadas à membrana (FeSx, FeSA e FeSB), a qual transfere elétrons para uma
ferredoxina solúvel (Fd), e que por fim irá reduzir o NADP+ à NADPH, completando o
transporte acíclico de elétrons da fotossíntese (FAN et al., 2009, JOLIOT e JOLIOT, 2006,
BANIULIS et al., 2008).
Além do transporte de elétrons acíclico, o sistema pode também realizar o cíclico e
pseudocíclico. No transporte cíclico, a ferredoxina reduzida transfere seus elétrons de volta
para o pool de plastoquinona ou diretamente para o citocromo b6f (JOLIOT e JOLIOT, 2006),
originando uma circulação de elétrons ao redor do FSI, sendo assim independente do FSII.
Esse processo causa um bombeamento de prótons para o lúmen, gerando um gradiente
eletroquímico, o qual é útil para síntese extra de ATP (FAN et al., 2009).
No pseudocíclico, em condições de super-redução da cadeia de transporte de elétrons,
a ferredoxina doa elétrons para o oxigênio molecular, não reduzindo NADP+ (ROCHAIX,
2011).
A luz, além de ser fonte de energia para a planta, pode atuar como fator estressante
quando em excesso, gerando um efeito fotodestrutivo (ORT, 2001). Quando a quantidade de
energia luminosa recebida é maior que a capacidade de utilização pela fotossíntese, o aparelho
fotossintético sofre o processo de fotoinibição (ADIR et al., 2003). Alguns fatores,
combinados aos altos níveis de irradiância, podem contribuir para a diminuição da
fotossíntese. Além da luz, o déficit hídrico reduz a fotossíntese porque afeta e limita a
utilização da energia luminosa pelas plantas (CLOSE e DAVIDSON, 2003; PORTES et al.,
2006).
A temperatura também pode afetar o processo fotossintético de diferentes formas,
principalmente na quantidade e ativação das enzimas que participam da fase bioquímica da
fotossíntese (PIMENTEL et al., 2007).

11
3.3. Fluorescência da clorofila a
A fotossíntese se inicia com a absorção de um fóton pelas moléculas de clorofilas do
CCL. Esse evento ocorre num tempo muito curto, em torno de dois fentosegundos (2x10-15s),
levando essas moléculas de clorofila de um estado eletrônico fundamental (S0) para um estado
excitado singleto, rico em energia (FLEMING e GRONDELLE, 1997).
Os estados excitados singletos podem ser S1 e S2, dependendo do comprimento de
onda absorvido. Quando é absorvido um fóton de luz vermelha, a clorofila passa ao estado
excitado singleto S1, e se for um fóton de luz azul, passa ao estado excitado singleto S2. A
diferença de energia entre S2 e S1 é dissipada como calor. A transição do estado excitado S1
para o estado fundamental S0 é lenta e pode ocorrer pelos seguintes caminhos alternativos e
concorrentes (MAXWELL e JOHNSON, 2000; PAPAGEORGIOU e GOVINDJEE, 2011;
TAIZ e ZEIGER, 2013):
- Fluorescência: processo de reemissão do fóton pela clorofila a excitada e desta
maneira voltando para o estado basal. Quando este evento ocorre, o comprimento de onda da
fluorescência é ligeiramente mais longo, e, dessa forma, com menor energia que o
comprimento de onda inicialmente absorvido, devido à parte da energia de excitação ser
convertida em calor antes dos fótons da fluorescência serem emitidos. A fluorescência da
clorofila pertence à região vermelha do espectro.
- Perda por calor: a energia da clorofila excitada pode ser perdida, levando-a para seu
estado não excitado, pela conversão direta da energia absorvida em calor, sem a emissão de
fluorescência;
- Transferência de energia: a clorofila pode transferir a energia recebida pela luz para
outra molécula. Uma vez absorvida pelos pigmentos do CCL é então transferida, por
ressonância, para os CRs dos fotossistemas;
- Reações fotoquímicas: a energia do estado excitado provoca as reações fotoquímicas
da fotossíntese, que estão entre as mais rápidas reações conhecidas. Essa velocidade extrema é
necessária para que as reações fotoquímicas possam competir com as outras três possíveis
vias de dissipação de energia anteriormente descritas.

12
Essas vias de dissipação da energia luminosa pelas moléculas de clorofilas são
competitivas, sendo que alterações nas taxas fotossintéticas e na dissipação de calor causam
alterações complementares na emissão da fluorescência. Dessa forma, alterações na
fluorescência podem revelar alterações e comprometimentos no processo fotossintético
(PAPAGEORGIOU e GOVINDJEE, 2011; TAIZ e ZEIGER, 2013).
A emissão da fluorescência da clorofila a pela luz é um processo espontâneo que
ocorre durante a fase clara da fotossíntese e foi observado pela primeira vez por Kautsky et al.
(1960). As moléculas de clorofila a ao absorverem a energia luminosa atingem um estado
transitório de excitação e repassam elétrons por meio da sequência de reações de oxi-redução
do FSII em direção ao FSI, ocorrendo a conversão da energia luminosa absorvida em energia
fotoquímica (MAXWELL e JOHNSON, 2000; MÜLLER et al., 2001; BAKER e
ROSENQVIST, 2004).
A fluorescência está relacionada principalmente à emissão pelo FSII, pois a sua
emissão pelo FSI é praticamente constante, além de contribuir pouco para o sinal de
fluorescência total (BYRDIN et al., 2000; RAPPAPORT et al., 2007; MURCHIE e LAWSON,
2013). De acordo com Lichtentlaler et al. (2005), mais de 90% da fluorescência da clorofila a
emana do FSII.
A fluorescência da clorofila a tem sido utilizada principalmente no estudo da
capacidade fotossintética das plantas. É um método não destrutivo que permite a análise
qualitativa e quantitativa da absorção e aproveitamento da energia luminosa pelo aparelho
fotossintético. Com o auxílio dessa técnica permitiu-se conhecer o funcionamento dos
processos fotoquímicos e não fotoquímicos que ocorrem na membrana dos tilacoides, além de
possibilitar o estudo de características relacionadas à capacidade de absorção e transferência
da energia luminosa na cadeia de transporte de elétrons em plantas (KRAUSE e WEIS, 1991).
A fluorescência emitida pela clorofila a pode ser utilizada como uma ferramenta muito
eficiente para medições de diversos parâmetros fotossintéticos, principalmente a realização de
diversas análises de plantas em condições de estresse.
As metodologias atuais para a realização das medições da emissão de fluorescência da
clorofila a podem fornecer informações rápidas a respeito da transferência de elétrons,
conversão e dissipação da energia luminosa no FSII, sempre mantendo a integridade do
material vegetal utilizado nas análises (CHRISTEN et al., 2007; WANG et al., 2013).

13
Originalmente as técnicas de medição da fluorescência da clorofila a, foram
desenvolvidas utilizando fluorímetros convencionais e a substância 3-(3,4-Diclorofenil)-1,1-
dimetilurea (DCMU) para a indução da fluorescência máxima da clorofila a. O DCMU liga-se
ao primeiro aceptor de elétrons do FSII, a plastoquinona (QA), bloqueando o fluxo de elétrons
por meio da sequência de transportadores, o que inibe totalmente o processo fotoquímico e
causa a indução da fluorescência máxima da clorofila a (HSU et al., 1986).
Progressos relacionados à pesquisa da fluorescência têm sido obtidos após construção
dos fluorímetros de pulso de amplitude modulada (PAM - Pulse-Amplitude-Modulation
fluorometer), (ÖGREN e BAKER. 1985; SCHREIBER 1986; SCHREIBER et al., 1986),
tornando-se a técnica de fluorescência da clorofila mais frequentemente utilizada e geralmente
aceita (SCHREIBER, 2004).
Os fluorímetros do tipo PAM utilizam uma fonte luminosa de excitação modulada (1 a
100 kHz), em conjunto com um sistema de detecção de fluorescência, permitindo o
monitoramento da fluorescência na frequência e fase da luz modulada de excitação. No
momento em que o pulso de luz é disparado, ocorre a saturação total dos transportadores de
elétrons do FSII, ocasionando o seu bloqueio e, consequentemente, impedindo a ocorrência do
processo fotoquímico e induzindo a máxima fluorescência da clorofila a (SCHREIBER,
2002).
De acordo com Kautsky (1960), quando iluminadas, as folhas emitem fluorescência de
forma contínua, na faixa vermelha do espectro de luz. Essa fluorescência contínua fornece
pouca informação em si, pois muitos fatores físico-químicos e metabólicos podem influenciar
o nível de fluorescência absoluta.
Diante disso, uma das grandes vantagens do fluorímetro PAM, é a possibilidade de se
obter medições de fluorescência da clorofila em amostras adaptadas ao escuro, para impedir
que a luz ambiente possa interferir na medição.
Segundo Schreiber et al. (1998), durante a aplicação do pulso saturante de luz e após o
tecido ter sido mantido no escuro, o rendimento quântico do processo fotoquímico reduz para
zero e a emissão da fluorescência bem como a dissipação do calor tornam-se máximos.
Dessa forma, depois de mantido o tecido fotossintetizante no escuro (em tecidos
saudáveis, uma adaptação de 15 a 30 minutos é normalmente considerada suficiente) e então a
folha ser subitamente iluminada, irá ocorrer o que é chamado de indução de fluorescência,
fluorescência transiente ou simplesmente efeito Kautsky (KAUTSKY e HIRSCH, 1931).

14
A cinética dessa resposta é denominada curva Kautsky, como pode ser visto na Figura
4, sendo formada por duas fases, uma rápida e outra lenta, convencionalmente denominadas
de OIJPSMT (LARCHER e CERNUSCA, 1985; PAPAGEORGIOU, 1975).
Figura 4. Cinética de emissão da fluorescência ou curva Kautsky registrada de uma
suspensão de Chlorella adaptada ao escuro. (A) cinética rápida; (B) cinética lenta.
Fonte: Bolhàr-Nordenkampf e Öquist (1993).
Na fase rápida, conhecida como OJIP, a fluorescência mínima (Fo) ocorre
aproximadamente em 50 μs após a iluminação (ponto O). Os pontos J e I são passos
intermediários que ocorrem a 2 e 30 ms após a iluminação, respectivamente, e P corresponde
a máxima fluorescência (Fm), que é alcançada em torno de 300 ms (STRASSER et al., 2010;
TSIMILLI-MICHAEL e STRASSER, 2008).
A fase lenta é chamada de PSMT, onde S representa um estado semi-estável de
decaimento e subsequente elevação da fluorescência até o ponto máximo M; e T é um nível
de estado estável terminal (PAPAGEORGIOU e GOVINDJEE, 1968).
Com as informações obtidas pela curva de Kautsky, além da determinação de Fo e Fm,
é possível determinar os coeficientes de dissipação fotoquímico (qP) e não fotoquímico (qN e
NPQ). Essa curva possibilita a determinação de outros parâmetros fluorimétricos de
importante significado biológico como fluorescência variável (Fv), rendimento quântico
máximo do FSII (Fv/Fm), além da taxa de decréscimo na fluorescência da clorofila a (RFd),
entre outros (KRAUSE e WEIS, 1991; OXBOROUGH e BAKER, 1997).

15
Estes parâmetros são utilizados de forma integrada em diversos estudos buscando a
compreensão da ação dos fatores ambientais sobre a fisiologia do FSII e a identificação dos
parâmetros fluorimétricos de maior sensibilidade para cada situação (LICHTENTHALER et
al., 2005).
A fluorescência variável, Fv é uma medida aproximada da quantidade de energia
luminosa que foi absorvida pelo FSII e que seria utilizada no processo fotoquímico. A Fv/Fm
indica a eficiência em que a energia luminosa, captada pelo complexo antena e pelas
moléculas de clorofila a do centro de reação P680, é encaminhada em direção ao restante do
desenvolvimento do processo fotoquímico (MAXWELL e JOHNSON, 2000; SCHREIBER et
al., 2002; LICHTENTHALER et al., 2005).
Os diversos parâmetros obtidos pelo PAM são extensivamente utilizados de forma
integrada em diversos estudos buscando um melhor entendimento da ação dos fatores
ambientais sobre a fotossíntese e a fisiologia do FSII em plantas e algas (LICHTENTHALER
et al., 2005).
3.4. Óleo essencial
Os óleos essenciais (OEs) são misturas complexas de substâncias voláteis, lipofílicas,
geralmente odoríferas, líquidas e com densidade menor à da água. A designação de óleo é
devida a algumas características físico-químicas como a de serem geralmente líquidos, de
aparência oleosa, à temperatura ambiente. Sua principal característica é a volatilidade,
diferenciando-os dos óleos fixos, que são misturas de substâncias lipídicas obtidas
normalmente de sementes (BAKKALI et al., 2008; EDRIS, 2007; SIMÕES e SPITZER,
2007).
São geralmente incolores ou ligeiramente amarelados. Poucos são os óleos que
apresentam outra cor, como o óleo de camomila, de coloração azulada, pelo seu alto teor em
azulenos. Em geral, são muito instáveis, principalmente na presença de ar, luz, calor, umidade
e metais. A maioria dos óleos voláteis é opticamente ativo, propriedade essa usada na sua
identificação e controle da sua qualidade (SIMÕES e SPITZER, 2007; OUSSALAH, 2006).

16
Na constituição dos OEs pode ser encontrado até quase uma centena de diferentes
componentes. Cada um desses componentes apresenta concentrações variadas, caracterizados
por dois ou três compostos principais, que representam entre 20 a 70% de todo conteúdo. Os
elementos que possuem menor porcentagem em sua composição são chamados traços
(BAKKALI et al., 2008).
Os OEs por serem voláteis, atuam como sinais de comunicação química entre as
diversas espécies de plantas, e também agem como armas de defesa contra o reino animal.
Essa característica torna as plantas que os produzem poderosas fontes de agentes biocidas
sendo largamente estudadas por apresentarem atividades bactericidas, inseticidas e fungicidas
(BENCHAAR et al., 2007; SOYLU et al., 2010; WOLFFENBÜTTEL, 2007).
Os OEs são abundantemente encontrados nas Angiospermas, como por exemplo, na
família Lamiaceae, Lauraceae, Myrtaceae, entre outras. A distribuição desses metabólitos
ocorre em diversas partes do vegetal, como folhas, cascas, raízes, flores, frutos, sementes,
entre outros. Desempenham importante papel na interação das plantas no ambiente em que
vivem, sendo produto também da interação com fatores bióticos e abióticos (FABROWSKI,
2002; PERES, 2004; SIMÕES e SPITZER, 2007; SANTOS, 2007; SILVA e CASALI, 2000).
Os componentes dos OEs são sintetizados por rotas biossintéticas do metabolismo
especializado vegetal, sendo que os seus constituintes pertencem à classe química dos
terpenos (PERES, 2004).
Os terpenos ou terpenoides constituem uma classe de compostos com maior variedade
estrutural e funcional. Os elementos estruturais básicos dos terpenos são algumas vezes
chamados de unidades isoprênicas. Alguns compostos terpênicos podem participar de
processos do metabolismo primário, como respiração e desenvolvimento celular, contudo, a
maioria deles tem sua origem no metabolismo especializado, tendo como função intermediar a
relação planta e ambiente (TAIZ e ZEIGER, 2013).
Nas plantas superiores os precursores universais dos terpenos, o isopentenil difosfato
(IPP) e seu isômero, o dimetilalil difosfato (DMAPP), podem ser sintetizados por duas vias.
Na primeira delas, que ocorre no citosol, o IPP é gerado pela via do ácido mevalônico ou
mevalonato (VAM ), e por ação da enzima IPP-isomerase pode originar DMAPP.
Quando a síntese acontece nos cloroplastos e em outros plastídeos, ocorre pela via do
metileritritol fosfato (MEP), cujos precursores são piruvato e gliceraldeído-3-fosfato
(PHILLIPS et al., 2008; ARAÚJO et al., 2011, TAIZ e ZEIGER, 2013).

17
Os terpenos atuam nas plantas em diferentes funções, como na produção de hormônios
(giberelinas), pigmentos fotossintéticos (carotenoides), carreadores de elétrons (ubiquinonas)
e em mecanismos de comunicação e de defesa contra insetos e herbívoros. A maioria dos
compostos apresenta baixo peso molecular, natureza lipofílica, grande variedade de estruturas
e alta pressão de vapor à temperatura ambiente (BAKKALI et al., 2008).
Os terpenos podem ser classificados de acordo com o número de isoprenos que
constituem: hemiterpenoides, monoterpenoides, sesquiterpenoides, diterpenoides,
triterpenoides, tetraterpenoides e politerpenoides. Os monoterpenos e sesquisterpenos são os
compostos terpênicos mais frequentes nos óleos essenciais, estando presentes em cerca de
90% dos OEs conhecidos (OLIVEIRA et al., 2003).
Os monoterpenos são constituintes da essência volátil de flores e óleos essenciais
extraídos de plantas medicinais e ervas aromáticas, justificando sua importância para a
indústria de perfumes e aromatizantes. O geraniol, mentol, linalol e o citral são exemplos de
monoterpenos. Muitos sesquiterpenos podem atuar como compostos antimicrobianos
(fitoalexinas) e antiherbivoria. Fazem parte desta classe o farnesol, o zingibereno e o
cariofileno (SIMÕES et al., 2007).
Para a extração dos OEs existem cinco métodos principais, de acordo com Simões et
al. (2007): enfloração, prensagem, extração com solventes orgânicos, por fluído supercrítico e
por arraste de vapor d'água. De acordo com o método de extração utilizado, a composição dos
óleos essenciais pode sofrer alterações, pois suas propriedades bioativas podem ser
comprometidas. As características físico-químicas podem ser alteradas pelas condições
operacionais empregadas na extração, bem como seus efeitos (ROBBERS et al., 1996).
Ainda de acordo com Simões et al. (2007), existem três tipos de extrações distintas de
arraste por vapor d’água. Essa distinção é feita pela forma na qual se estabelece o contato
entre a amostra e a água, na fase líquida ou de vapor. A primeira é chamada de
hidrodestilação, na qual a amostra fica imersa na água contida numa caldeira. Na segunda,
chamada de destilação pela água e vapor, a amostra permanece contida em um recipiente logo
acima da caldeira, ficando assim separada da água.
Na terceira, chamada propriamente de destilação pelo vapor de água, a amostra é
mantida em recipiente separado e o vapor de água que flui provém de um gerador próprio
independente. A indústria utiliza, preferencialmente, a extração por vapor d’água por ser
reduzido o contato da amostra com a água.

18
A hidrodestilação é um método antigo e versátil, sendo o mais usado comercialmente
no Brasil. O material vegetal permanece em contato com a água em ebulição, o vapor faz com
que as paredes celulares se abram e o óleo que está entre as células evapore junto com a água
que vai para o condensador, onde é resfriado e separado por diferença de densidade. No caso
das produções em pequena escala, emprega-se o aparelho denominado de Clevenger. O óleo
essencial obtido, após separar-se da água, deve ser seco com sulfato de sódio (Na2SO4) anidro
(SIMÕES et al., 2007).
Mattana et al. (2015) comparam métodos de extração de óleos essenciais pelas
técnicas de extração com solventes, hidrodestilação e extração por infusão de folhas de
Pothomorphe umbellata (C. DC.) Trel. e Yunck e constatou que foi possível identificar maior
número de compostos na extração pelo método de hidrodestilação.
A produção de metabólitos secundários de uma planta é determinada por fatores
bióticos e abióticos. Consequentemente, a composição do OE também sofre essa influência,
tanto quantitativamente como qualitativamente. Fatores como sazonalidade, índice
pluviométrico, ritmo circadiano, radiação ultravioleta (UV), composição atmosférica,
herbivoria e ataque de patógenos, idade da planta, disponibilidade de água, teor de
macronutrientes e micronutrientes, tipo de cultivo, duração total de exposição ao sol,
temperatura e altitude, fenologia, bem como sua participação em alelopatias, estão entre os
principais. A temperatura, a umidade relativa e o regime de ventos exercem uma influência
direta, sobretudo sobre nas espécies que possuem estruturas histológicas de estocagem de óleo
essencial na superfície da folha (GOBBO-NETO e LOPES, 2007; SALGADO, 2005;
WILLIAMS e STOCKLEY, 1998).
Segundo Gobbo-Neto e Lopes (2007), dentre esses fatores, a sazonalidade e a
fenologia estão entre os mais importantes para espécies vegetais não domesticadas, presentes
em seu meio natural, floresta ou outro ambiente, utilizando todas as suas estratégias
evolutivas para sua manutenção e perpetuação da espécie.
As variações extremas na temperatura são desfavoráveis ao metabolismo das plantas,
sejam pelos seus valores muito elevados ou muito baixos. De forma geral, temperaturas
amenas são mais produtivas. Dessa forma, a alternância das estações e consequentes variações
de temperatura de cada estação, podem estimular a produção de certos metabólitos em
detrimento de outros, o que favorece a síntese de determinadas classes de compostos
(SANGWAN et al., 2001, HUSSAIN et al., 2008).

19
A sazonalidade tem influência sobre o ciclo circadiano das plantas, devido à alteração
nos períodos de luz e escuro sob o qual as plantas estão submetidas. Dessa forma, todo o
funcionamento do maquinário fotossintético é influenciado e, consequentemente, todo o seu
metabolismo e a síntese de produtos secundários. Além da duração dos períodos de luz e
escuro, os diferentes comprimentos de onda podem influenciar positivamente ou
negativamente a síntese de alguns compostos secundários (SANGWAN et al., 2001;
GOBBO-NETO e LOPES, 2007)
A quantidade de luz UV absorvida pela planta pode incentivar a biossíntese de
compostos secundários com valor medicinal para o ser humano. Particularmente a radiação
UV-B estimula a formação de metabólitos como compostos fenólicos, incluindo flavonoides,
alcaloides e terpenoides, entre outros. Possivelmente, esses compostos têm sua produção
estimulada para agir contra a ação estressora da radiação UV, prevenindo contra possíveis
danos causados por espécies reativas de oxigênio (ZHANG e BJÖRN, 2009).
As plantas ricas em OEs devem ser coletadas pela manhã ou à noite, pois o período de
exposição ao sol pode provocar uma perda quantitativa importante do óleo existente no
vegetal. O grau de hidratação do terreno e os teores de macronutrientes também podem
influenciar a composição dos óleos voláteis. Entretanto, não se pode prever ou estabelecer um
único padrão, pois cada espécie reage de forma diferenciada (CASTRO, 2006).
Além de fatores ambientais, a genética e a fenologia da planta também influenciam a
produção de OEs. Dessa forma a época em que as amostras para extração dos OEs são
coletadas é um dos fatores de maior importância, visto que a quantidade e, em algumas
situações, a natureza dos compostos não são constantes durante o ano. Existem estudos
mostrando que a composição de metabólitos secundários de uma planta pode variar durante o
dia e a noite (BRITO, 2009; MARCO et al., 2011; MARTINS et al., 1995; TAVARES et al.,
2005).
Eventos estressantes aos qual a planta está submetida, como ferimentos, chuva, vento,
ataque de patógenos e de herbívoros, podem influenciar a expressão do metabolismo
especializado (JEONG et al., 2004).
Danos causados a plantas por ferimentos ou ataque de herbívoros ou patógenos
frequentemente levam a uma resposta bioquímica, a qual reduz a aceitabilidade do órgão ou
de todo o organismo a ataques futuros. Por exemplo, a produção de fitoalexinas em resposta à
invasão de patógenos (BURNS et al., 2002; PICHERSKY et. al, 2008).

20
A composição do óleo essencial de uma espécie de planta é, geralmente, específica
para um determinado órgão e característica para o seu estágio de desenvolvimento. Apesar de
apresentarem cor e aspecto semelhantes, podem apresentar composição química,
características físico-químicas e odores diferentes (ROBBERS et al., 1996).
O local de armazenamento e secreção dos OEs depende da família das plantas, e para
tal fim, existem estruturas especializadas tais como, pelos glandulares, células
parenquimáticas diferenciadas, canais oleíferos, bolsas lisígenas e esquizolisígenas
(MARQUES et al., 2008). Os OEs que apresentam reconhecidas propriedades como
repelentes de insetos são frequentemente encontrados em tricomas glandulares que se
projetam da epiderme e agem como advertência sobre a toxicidade do vegetal. Nos tricomas
glandulares, os terpenos são armazenados em um espaço extracelular modificado na parede
celular (WOLFFENBÜTTEL, 2007).
Os OEs são raramente encontrados em Gimnospermas (exceto coníferas). Em
Angiospermas Monocotiledôneas, a ocorrência é relativamente rara, com exceção de
Gramíneas e Zingiberáceas. No entanto, plantas ricas em óleos essenciais são abundantes em
Angiospermas Dicotiledôneas, tais como nas famílias Asteraceae, Apiaceae, Lamiaceae,
Lauraceae, Myrtaceae, Myristicaceae, Piperaceae e Rutaceae, entre outras. As espécies da
família Myrtaceae são particularmente ricas em óleos essenciais (CERQUEIRA et al., 2009).
Os OEs podem causar diversos efeitos sobre os insetos, tais como repelência, inibição
da oviposição e da alimentação e alterações no sistema hormonal. Consequentemente causam
distúrbios no desenvolvimento, deformações, infertilidade e mortalidade nas diversas fases,
alterações no crescimento e na emergência de adultos, alterações morfogenéticas, entre outros
(BOEKE et al., 2004; GALLO et al., 2002; KETOH et al., 2005; ROEL, 2001).
O controle de pragas com inseticidas de origem vegetal tem despertado um grande
interesse da sociedade, devido também à segurança e à seletividade que apresentam para tal
função, menor probabilidade de desenvolvimento de resistência pelos insetos e, menor
toxicidade para mamíferos (BENHALIMA et al., 2004; GALLO et al., 2002).

21
3.5. Morfometria e dinâmica de crescimento de árvores
O crescimento das árvores é influenciado por fatores genéticos da espécie, que
interagem com fatores ambientais, tais como temperatura, precipitação, insolação;
características físicas, químicas e biológicas do solo; inclinação, altitude e exposição do
terreno, e também com fatores biológicos, como a competição (FINGER, 1992).
O estudo das características morfométricas ou relações morfométricas, assim como o
acompanhamento da dinâmica das formas de crescimentos das árvores são ferramentas que
fornecem importantes informações para orientar e aprimorar as intervenções silviculturais,
Atividades como desbaste e desrama, principalmente quando se utiliza espécies nativas em
reflorestamentos, podem ser melhores planejados utilizando essas informações (DURLO et
al., 2004).
O conhecimento das características morfométricas de uma árvore como, o DAP e a
altura fornecem subsídio para determinar o manejo correto de acordo com o local em que a
espécie está inserida visando o baixo impacto na floresta nativa, promovendo assim a sua
conservação e expansão da espécie (ORELLANA e KOEHLER, 2008).
A competição talvez seja o fator biológico mais importante no manejo florestal, pois
quase todas as intervenções estão relacionadas com a manipulação desse fator ou de
condições que o alteram. A competição pode ser descrita em função do comprimento e
diâmetro de copa, posição sociológica e tendências de crescimento, originando diferentes
índices, que também são influenciados, principalmente, pela luz e pelo vento (SCHNEIDER e
FINGER, 2000; SCHNEIDER e SCHNEIDER, 2008).
A morfometria das árvores e suas variáveis derivadas permitem entender as relações
interdimensionais e reconstruir o espaço ocupado por cada árvore, podendo-se determinar o
grau de concorrência a que uma espécie está sujeita dentro de um povoamento, assim como se
torna possível inferir sobre a estabilidade, a vitalidade e a produtividade de cada indivíduo
(HASENAUER, 1997; DURLO e DENARDI, 1998).
Para uma melhor compreensão das relações morfométricas, Burguer (1939)
desenvolveu um modelo de árvore e por meio dela, Durlo e Denardi (1998) apresentaram
claramente as definições das principais variáveis morfométricas, Figura 5, descritas a seguir:

22
Figura 5. Representação das variáveis morfométricas, sendo: cc - comprimento de copa , d -
diâmetro a 1,30m do solo (DAP), dc - diâmetro de copa, h - altura total, hf -
altura do fuste, hl - altura da copa exposta a luz e hs - altura da copa sombreada.
Fonte: Wink et al. (2012).
- Índice de abrangência: é calculado pela relação entre o diâmetro de copa e a altura
total da árvore. Havendo uma correlação entre o referido índice e a altura das árvores, eles
podem ser usados também como indicador de desbaste ao longo da vida do povoamento;
- Formal de copa: o formal de copa (FC) expressa a relação entre o diâmetro e o
comprimento da copa. Com a análise de FC é possível indicar a forma geométrica que a
árvore se assemelha (ZIMMERMANN et al., 2012). O valor do formal de copa indica o grau
de achatamento das copas. Caso o índice encontrado esteja próximo a 1 indica que a copa é
redonda, valores acima de 1 indicam copas achatadas e valores abaixo de 1 caracterizam
copas alongadas (DURLO, 2001; ORELLANA et al., 2008; ROMÁN et al., 2009);
- Grau de Esbeltez: é a relação de H e DAP, sendo uma variável que caracteriza a
estabilidade das árvores, podendo servir como indicativo de desbastes em atraso,
especialmente em povoamentos puros e equiâneos. Quanto mais alto o Grau de Esbeltez, tanto
mais instável é a árvore. Tonini e Arco-Verde (2005) mencionam que uma relação superior a
100 indica que o crescimento em diâmetro é reduzido em relação à altura;

23
- Índice de saliência: relação entre o diâmetro de copa e o DAP, pode ser utilizada
como indicador de desbaste, podendo-se determinar o espaço a ser liberado ao redor de uma
árvore selecionada, para que ela cresça sem concorrência. Esse índice expressa quantas vezes
o diâmetro de copa é maior que o DAP. Quando o Índice de Saliência é elevado ao quadrado,
chama-se de índice de espaço vital, sendo que quanto menor a média desse índice em um
povoamento, maior será sua área basal e, provavelmente, também seu volume por hectare.
-Proporção de copa: relação entre o comprimento da copa e a altura total da árvore.
Está variável é um indicador da vitalidade, quanto maior a porcentagem de copa, tanto mais
vital e produtiva é a árvore. A proporção de copa atual de uma árvore dá indicativos do grau
de concorrência por ela sofrido no passado, podendo ser utilizada em modelos de
concorrência (HASENAUER, 1994; DURLO, 1996).
-Projeção da copa: é a área que corresponde à superfície coberta pela projeção vertical
da copa de uma árvore. Através da área de projeção da copa é possível conhecer o espaço
ocupado por uma árvore e, quando se dispõe de dados de incremento, pode-se correlacionar a
produção e a produtividade da mesma.
Quando se prevê o manejo de um povoamento não pela sua idade, mas pelo diâmetro
atingido por seus componentes, o número máximo de árvores por unidade de área, à medida
que elas forem crescendo, pode ser calculado pelo índice de saliência. Para sua utilização
deve existir uma correlação significativa entre este e o DAP (SILVEIRA et. al., 2015).
De acordo com Machado e Figueiredo Filho (2006) entre as variáveis descritas em
uma árvore, o DAP é umas das mais importantes. O DAP representa uma medida básica
necessária para calcular a área transversal e também utilizada na estimativa do diâmetro da
copa. Além deste parâmetro, a altura é indispensável, pois é uma das variáveis que define as
relações dendrométricas.
A medição das variáveis morfométricas, principalmente a área de copa, consome
muito tempo. Por serem de fácil obtenção, e quando existe alta correlação, pode-se utilizar
DAP e a altura total, como variáveis independentes em equações de regressão. A partir desses
modelos pode-se predizer as demais variáveis (TONINI e ARCO-VERDE, 2005).
Juntamente com a morfometria, estudos sobre a dinâmica de florestas que se baseiam
nos parâmetros de ingresso, crescimento e mortalidade constituem em informações que
podem fornecer importantes subsídios para a gestão sustentável de florestas naturais (SHEIL
et al., 2000).

24
O ingresso refere-se às árvores que atingiram um limite mínimo de diâmetro ao longo
de um determinado período de tempo, como resultado de diferentes processos, como o
estabelecimento, crescimento e mortalidade de árvores (LEXERØD e EID, 2005;
VANCLAY, 1994). O crescimento é definido como o aumento de dimensões de um ou mais
indivíduos em uma floresta em um determinado período de tempo, o qual está influenciado
pelas características da espécie interagindo com o ambiente (PRODAN et al., 1997). Tais
dimensões podem ser o diâmetro, altura, área basal, volume, biomassa dentre outros
(SCHAAF et al., 2005).
A mortalidade pode ser compreendida como o número de árvores que foram
mensuradas inicialmente, e morreram durante um período de crescimento determinado
(SANQUETTA et al., 2003).
As informações obtidas pelas variáveis morfométricas e pela dinâmica de crescimento,
juntamente com os dados dos parâmetros fotossintéticos, podem fornecer indicações do
manejo a ser utilizados na C. prismatica visando a alta produção de PFNM, como por
exemplo, extração de OEs.

25
3.7. Referências Bibliográficas
ACCIOLY, P. Mapeamento dos remanescentes vegetais arbóreos do Estado do Paraná e elaboração de um sistema de informações geográficas para fins de análise ambiental do estado. 2013. 127 f. Tese (Doutorado em Engenharia Florestal) – Universidade Federal do Paraná, UFPR, Curitiba, 2013. ADIR, N., ZER, H., SHOCHAT, S., OHAD, I. Photoinhibition – a historical perspective. Photosynthesis Research, v. 76, p. 343-370, 2003. ALBUQUERQUE, J. M. Fatores ecológicos e ocorrência de espécies na floresta ombrófila mista, em sistema faxinal. 2015. 93f. Tese (Doutorado em Engenharia Florestal) - Universidade Federal do Paraná, Curitiba, 2015. ALLEN, J. F.; FORSBERG, J. Molecular recognition in thylakoid structure and function. Trends in Plant Science, v. 6, p.317–326, 2001. ARAÚJO, D. A. M.; FREITAS, C.; CRUZ, J. S. Essential oils components as a new path to understand ion channel molecular pharmacology. Life Sciences, v. 89, p. 540-544, 2011. ASSMAN, E. The Principles of Forest the Yield Study. München: BLV, 1970. 490 p. BACARIN, M. A.; MOSQUIM, P. R. Cinética de emissão de fluorescência das clorofilas de dois genótipos de feijoeiro. Ciência e Agrotecnologia, Lavras, v. 26, n. 4, p. 705-710, 2002. BACKES, P.; IRGANG, B. Árvores do Sul. Guia de identificação & interesse ecológico. As principais espécies nativas Sul-Brasileiras. 2. ed. Santa Cruz do Sul: Instituto Souza Cruz, 2009. 331 p. BAKER, N. R.; ROSENQVIST, E. Applications of chlorophyll fluorescence can improve crop production strategies: an examination of future possibilities. Journal of Experimental Botany, v. 55, n. 403, p. 1607-1621, 2004. BAKKALI, F.; AVERBECK, S.; AVERBECK, D.; IDAOMAR, M. Biological effects of essential oils – A review. Food and Chemical Toxicology, v. 46, n. 2, p. 446-475, 2008. BANIULIS, D.; YAMASHITA, E.; ZHANG, H.; HASAN, S. S.; CRAMER, W. A. Structure-function of the Cytochrome b6f complex. Photochem Photobiol, v. 84, p. 1349–1358, 2008. BENCHAAR, C. et al. Effects of essential oils on digestion, ruminal fermentation, rumen microbial populations, milk production, and milk composition in dairy cows fed alfalfa silage or corn silage. Journal of Dairy Science, v. 90, p. 886–897, 2007. BENHALIMA, H.; CHAUDHRY, M. Q.; MILLS, K. A.; PRICE, N. R. Phosphine resistance in stored-product insects collected from various grain storage facilities in Marocco. Journal Stored Product Research, v. 40, p. 241-249, 2004.

26
BOEKE, S. J.; BOERSMA, M. G.; ALINK, G. M.; VAN LOON, J. J. A, VAN HUIS, A.; Dicke, M.; RIETJENS, I. M. C. M. Safety evaluation of neem (Azadirachta indica) derived pesticides. Journal of. Ethnopharmacology, v. 94, p. 25–41, 2004. BOLHÀR-NORDENKAMPF, H. R.; ÖQUIST, G. Chlorophyll fluorescence as a tool in photosynthesis research. In: HALL, D.; SCURLOCK, J. M. O.; BOLHÀR-NORDENKAMPF, H. R. R.; LEEGOOD, C. R. C.; LONG, S. P. (Org.). Photosynthesis and production in a changing environment: a field and laboratory manual. London: Chapman & Hall, 1993. p. 193-206. BRITO, A. F. R. Análise da variação sazonal e das atividades antifúngica e antimicrobiana em óleos essenciais de Ocotea porosa (Nees) Barroso e Nectandra megapotamica (Spreng.) Mez. 2009. 121f. Dissertação (Mestrado em Química Orgânica) - Instituto de Química da Universidade de São Paulo, São Paulo, 2009. BURGER, H. Baumkrone und zuwachs in zwei hiebsreifen fichtenbeständen. Mitteilungen der Schweizerischen Anstalt für das Forstliche Versuchswesen, v.21, p.147-176, 1939. BURNS, A. E. et al. Light alters the allocation of nitrogen to cyanogenic glycosides in Eucalyptus cladocalyx. Oecologia, 133, p. 288–294, 2002. BYRDIN, M.; RIMKE, I.; SCHLODDER, E.; STEHLIK, D.; ROELOFS, T.A. Decay kinetics and quantum yields of fluorescence in photosystem I from Synechococcus elongates with P700 in the reduced and oxidized state: are the kinetics of excited state decay trap-limited or transfer-limited? Biophysical Journal, v. 79, n. 2, p. 992-1007, 2000. CASTRO, N. E. A. Caracterização e fitoquímica de óleos essenciais de eucalipto e seu efeito sobre o protozoário tripanosomatídeo Herpetomonas samuelpessoai. 2006. 82 f. Tese (Doutorado em Agronomia) – Universidade Federal de Lavras, Lavras, 2006. CERQUEIRA, M. D.; MARQUES, E. J., MARTINS, D.; ROQUE, N. F., CRUZ, F. G.; GUEDES, M. L. S. Variação sazonal da composição do óleo essencial de Myrcia salzmannii Berg. (Myrtaceae). Química Nova, v. 32, n. 6, p. 1544-1548, 2009. CHRISTEN, D.; SCHÖNMANN, S.; JERMINI, M.; STRASSER, R.J.; DÉFAGO, G. Characterization and early detection of grapevine (Vitis vinifera) stress responses to esca disease by in situ chlorophyll fluorescence and comparison with drought stress. Environmental and Experimental Botany, v. 60, p. 504-514, 2007. CLOSE, D. C., DAVIDSON, N. J. Long-term waterlogging: nutrient, gas exchange, photochemical and pigment characteristics of Eucalyptus nitens saplings. Russian Journal of Plant Physiology, v. 50, n. 6, p. 843-847, 2003. DE BEER, J. H.; MCDERMOTT, M. J. Economic value of non-timber forest products in south-east Asia. Amsterdam: The Netherlands Committee for IUCN, 1989.

27
DULAI ,S.; MOLNÁR, I.; MOLNÁR-LÁNG, M. Changes of photosynthetic parameters in wheat/barley introgression lines during salt stress. Acta Biologica Szegdiensis, v. 55, p. 73-75, 2011. DURLO, M. A. Zuwachsuntersuchungen und Einzelbaumwachstumsmodelle für Cabralea glaberrima, Cedrela fissilis und Cordia trichotoma in sekundären Laubmischwälder Südbrasiliens. Wien: Institut für Waldwachstumsforschung, 1996. 175f. Dissertation (Doktorgrades) - Universität für Bodenkultur, 1996. DURLO, M. A.; DENARDI, L. Morfometria de Cabralea canjerana, em mata secundária nativa do Rio Grande do Sul. Ciência Florestal, Santa Maria, v. 8, n. 1, p. 55-66, 1998. DURLO, M. A. Relações morfométricas para Cabralea canjerana (Well.) Mart. Ciência Florestal, Santa Maria, v. 11, n. 1, p. 141-150, jun, 2001. DURLO, M. A.; SUTILI, F. J.; DENARDI, L. Modelagem da copa de Cedrela fissilis Vellozo. Ciência Florestal, v. 14, n. 2, p. 79-89, 2004. EDRIS, A. E. Pharmaceutical and therapeutic potentials of essential oils and their individual volatile constituents: a review. Phytotherapy Research, v. 21, n. 4, p. 308-323, 2007. FABROWSKI, F. J. Eucalyptus smithii R. T. Baker (Myrtaceae) como espécie produtora de óleo essencial no sul do Brasil. 2002. 225 f. Tese (Doutorado em Ciências Florestais) - Universidade Federal do Paraná, Curitiba, 2002. FALKOWSKI, P. G., RAVEN, J. A. Aquatic Photosynthesis. 2. ed. New Jersey: Princeton University Press, 2007. 484 p. FAN, D. Y.; JIA, H.; BARBER, J.; CHOW, W. S. Novel effects of methyl viologen on photosystem II function in spinach leaves. European Biophysics Journal, v. 39, p. 191-199, 2009. FINGER, C. A. G. Fundamentos de Biometria Florestal. Santa Maria: UFSM/CEPEF/FATEC, 1992. 269 p. FLEMING, G. R, GRONDELLE, R. V. Fentosecond spectroscopy of photosynthetic light-harvesting systems. Current Opinion in Structural Biology. v.7, p.738–748. 1997. GALLO, D; NAKANO, O; NETO, S. S; CARVALHO, R. P. L; BAPTISTA, G. C; FILHO, E. B; PARRA, J. R P; ZUCCHI, R. A; ALVES, S. B; VENDRAMIM, J. D; MARCHINI, L. C; LOPES, J. R. S; OMOTO, C. Entomologia Agrícola. Piracicaba: FEALQ, v. 10, 2002. 920p. GENTRY, A. H.; BLANEY, C. L. Tropical forestry. Western-Wildlands, v. 16, n. 1, p. 2-24, 1990. GOBBO-NETO, L. LOPES, N. P. Plantas medicinais: fatores de influência no conteúdo de metabólitos secundários. Química nova, v.30, n. 2, p. 374-381. 2007.

28
GOVINDJEE. Chlorophyll a fluorescence: a bit basics and history. In: PAPAGEORGIOU, G. C.; GOVINDJEE (Org.). Clorophyll a fluorescence: a signature of photosynthesis. Dordrecht:Springer, 2004. p.1-42. HASENAUER, H. Ein Einzelbaumwachtumssimulator für ungleichaltrige Fichten- Kiefern- und Buchen-Fichtenmischbestände. Fostliche Schriftenreihe Universität für Bodenkultur, Wien. Band 8, 152 p. 1994. HASENAUER, H. Dimensional relationships of open-grown trees in Austria. Forest Ecology and Manegement, v. 96, p. 197-206. 1997. HILL, R.; BENDALL, F. Function of the two cytochrome components in chloroplasts: a working hypothesis. Nature, v. 186, v.136–137, 1960. HSU, B; LEE, J., PAN, R. The two binding sites for DCMU in photosystem II. Biochemical and Biophysical Research Communications. v. 141, n. 2, p. 682-688, 1986. HUSSAIN, A. I., ANWAR, F., SHERAZI, S. T. H., PRZYBYLSKI, R. Chemical composition. Antioxidant and antimicrobial activities of basil (Ocimum basilicum) essential oils depends on seasonal variations. Food Chemistry, v. 108, p. 986-995, 2008. JEONG, S. T.; GOTO-YAMAMOTO, N.; KOBAYASHI, S.; ESAKA, M. Effects of plant hormones and shading on the accumulation of anthocyanins and the expression of anthocyanin biosynthetic genes in grape berry skin. Plant Science, v.167, p. 247-252, 2004. JOLIOT, P.; JOLIOT, A. Cyclic electron flow in C3 plants. Biochimica et Biophysica Acta v. 1757, p. 362-368, 2006. KAUTSKY, H.; APPEL, W.; ANAMM, H. Chlorophyllfluorescennz und kohlensaureassimilation. Biochemische Zeitschrift. v. 322, p. 277-292, 1960. KAUTSKY, H.; HIRSCH, A. Neue Versuche zur Kohlensaureassimilation. Naturwissenschaft , v. 19, p. 964, 1931. KETOH, G. K.; KOUMAGLO, H. K.; GLITHO, I. A. Inhibition of Callosobruchus maculates (F.) (Coleoptera: Bruchidae) development with essential oil extracted from Cymbopogon schoenanthus L. Spreng (Poaceae), and the wasp Dinarmus basalis (Rondani)(Hymanoptera: Pteromalidae). Journal of Stored Products Research, v. 41, p. 363-371, 2005. KRAUSE, G. H., WEIS, E. Chlorophyll fluorescence and photosynthesis: The basics. Annual Review of Plant Physiology and Plant Molecular Biology, v. 42, p. 313-49, 1991. KOSUGE, T. The role of phenolics host response to infection. Annual Review of Phytopathology, v. 7, p. 195-222, 1969. LAMBERS, H.; CHAPIM III, F.S.; PONS, T.L. Plant Physiological Ecology. 2. ed. New York: Springer, 2008. 604p.

29
LAKE, J. A.; QUICK, W. P.; BEERLING, D. J.; WOODWARD, E. I. Signals from mature to 388 new leaves. Nature, v. 411, p. 154, 2001. LARCHER, W.; CERNUSCA, A. Bericht der Oesterreichische Akademie der Wissenschaften, v. 194, p. 45-64. 1985. LAWLOR, D. W. Musings about the effects of environment on photosynthesis. Annals of Botany, v. 103, p. 543-549, 2009. LEXERØ, D.; EID, T. Recruitment Models for Norway Spruce, Scots Pine, Birch and Other Broad leaves in Young Growth Forests in Norway. Silva Fennica, v. 39, n. 3. p. 391 - 406. 2005. LICHTENTHALER, H. K.; BUSCHMANN, C.; KNAPP, M. How to correctly determine the different chlorophyll fluorescence parameters and the chlorophyll fluorescence decrease ratio RFd of leaves with the PAM fluorometer. Photosynthetica. v. 43, n. , p. 379-393, 2005. LICHTENTHALER, H. K.; WELLBURN, A. R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochemical Society Transactions, v. 11, p. 591-592, 1983. LORENZI, H. Árvores Brasileiras: manual de identificação e cultivo de plantas arbóreas nativas do Brasil. v.2. Nova Odessa: Plantarum, 2014. 384 p. MACHADO, S. A.; FIGUEIREDO FILHO, A. Dendrometria. 2. ed. Guarapuava: Unicentro. v. 1000. p. 316, 2006. MATTANA, R.S. et al. Efeitos de diferentes tempos de extração no teor e composição química do óleo essencial de folhasde pariparoba [Pothomorphe umbellata (L.) Miq.]. Revista Brasileira de Plantas Medicinais, v.17, n.1, p.150–156, 2015. MARCO, C. A.; SANTOS, H. R.; ROLIM, R. R.; COSTA, J. G. M.; FEITOSA, J. M.; VÁSQUEZ, M. E. F. Influência do estádio fenológico de Vanillosmopsis arborea Baker sobre o rendimento e qualidade do seu óleo essencial. Caderno de Agroecologia, Camaragibe, v. 6, n. 2, 2011. MARQUES, C. A. et al. Considerações anatômicas e análise de óleo essencial do hipanto e do fruto de Hennecartia omphalandra J. Poisson (Monimiaceae). Revista Brasileira Farmacognosia, v. 18, p. 415-429, 2008. MARTINS, E. R.; CASTRO, D. M.; CASTELLANI, D. C.; DIAS, J. E. Plantas medicinais. Viçosa: Universidade Federal de Viçosa, 1995. 220 p. MAXWELL, K.; JOHNSON, N. Chlorophyll Fluorescence – A Pratical Guide. Journal of Experimental Botany, v. 354, p. 659-668, 2000.

30
MÜLLER, P.; LI, X. P.; NIYOGI, K. K. Non-Photochemical Quenching. A Response to Excess Light Energy. Plant Physiology, v. 125, p. 1558–1566, 2001. MURCHIE, E. H.; LAWSON, T. Chlorophyll fluorescence analysis: a guide to good practice and understanding some new applications. Journal Experimental Botany, v. 64, p. 3983-3998, 2013. MYERS, N. Tropical forests: much more than stocks of wood. Journal of Tropical Ecology, v. 4, p. 209-221, 1988. NEPSTAD, D. C.; SCHWARTZMAN, S. Non-timber products from tropical forests: evaluation of a conservation and development strategy. New York : New York Botanical, 1992. 176 p. NOBEL P. S. Physicochemical and environmental plant physiology. San Diego: Academic Press, 1991. 635 p. ODUM, E. P.; BARRET, G. W. Ecologia. São Paulo: Guanabara-Koogan, 2007. 623 p. ÖGREN, E.; BAKER, N.R. Plant, Cell and Environment. v. 8, p. 539-547, 1985. OLIVEIRA, R. B.; GODOY, S. A. P.; COSTA, F. B. Plantas tóxicas: conhecimento e prevenção de acidentes. Ribeirão Preto: Holos, 2003. 64 p. ORELLANA, E.; KOEHLER, A, B. Relações Morfométricas de Ocotea odorífera (vell.) Rohwer. Rev. Acad., Ciênc. Agrár. Ambientais, Curitiba, v. 6, n. 2, p. 229-237, abr./jun. 2008. ORT, D. R. When there is too much light. Plant Physiology, v. 125, p. 29-32, 2001. OUSSALAH, M.; CAILLET, S.; SAUCIER, L.; LACROIX, M. Inhibitory effects of selected plant essential oils on the growth of four pathogenic bacteria: E. coli 0157:H7, Salmonella typhimurium, Staphylococcus aureus e Listerria monocytogenes. Food Control, v. 18, p. 414-420, 2006. OXBOROUGH, K.; BAKER, N. R. Resolving chlorophyll a fluorescence images of photosynthetic efficiency into photochemical and non-photochemical components – calculation of qP and Fv’/Fm’ without measuring Fo’. Photosynthesis Research, v. 54, p. 135–142, 1997. PAPAGEORGIOU, G. C.; GOVINDJEE. Light induced changes in the fluorescence yield of chlorophyll a in vivo. I. Anacystis nidulans. Biophysical Journal, v. 8, p. 299- 1315, 1968. PAPAGEORGIOU, G. In: AMESZ, J.; GOVINDJEE (Org.). Bioenergetics of photosynthesis. New York: Academic Press, 1975. 320 p.

31
PAPAGEORGIOU, G. C.; GOVINDJEE. Photosystem II fluorescence: slow changes—scaling from the past. Journal Photochemestry Photobiology B: Biology, v. 104, p. 258-270, 2011. PERES, L. E. P. Metabolismo Secundário. Piracicaba: Escola Superior de Agricultura Luiz de Queiroz, 2004. 10 p. PETERS, C. M.; GENTRY, A. H.; MENDELSOHN, R. O. Valuation of an Amazonian Rainforest. Nature, v. 339, p.655-656, 1989. PHILLIPS, M. A.; LEON, P; BORONAT, A.; RODRIQUEZ-CONCEPCIÓN, M. The plastidial MEP pathway: unified nomenclature and resources. Trends in Plant Science, v. 13, n. 12, p. 619-23, 2008. PICHERSKY, E.; NOEL, J. P.; DUDAREVA, N. Biosynthesis of plant volatiles:nature’s diversity and ingenuity. Science, v. 311, p. 808–811, 2008. PIMENTEL, C., BERNACCHI, C., LONG, S. Limitations to photosynthesis at different temperatures in the leaves of Citrus limon. Brazilian Journal of Plant Physiology, v. 19, n. 2, p. 141-147, 2007. PORTES, M. T.; ALVES, T. H.; SOUZA, G. M. Water deficits affects photosynthetic induction in Bauhinia forficate Link (Fabaceae) and Esenbeckia leiocarpa Engl. (Rutaceae) growing in understorey and gap conditions. Brazilian Journal of Plant Physiology, v. 18, n. 4, p. 491-502, 2006. PRODAN, M. et al. Mensura Forestal. San Jose, C.R.: Deutsche Gesellschaft fur Technische Zusammenarbeit (GTZ) GmbH: Institute Interamericano de Cooperación para la Agricultura (IICA), 1997, 561 p. RAPPAPORT, F.; BEAL, D.; JOLIOT, A.; JOLIOT, P. On the advantages of using green light to study fluorescence yield changes in leaves. Biochimica et Biophysica Acta, v. 1767, p. 56-65, 2007. REGO, S. S.; COSMO, N. L.; GOGOSZ, A. M.; KUNIYOSHI, Y. S.; NOGUEIRA, A. C. Caracterização morfológica e germinação de sementes de C. prismatica (D. Legrand) Salywon & Landrum. Revista Brasileira de Sementes, v. 33, p. 616-625, 2011. ROBBERS, J. E., TYLER, V. E., SPEEDIE, M. K. Pharmacognosy and pharmacobiotechnology. Nova York: Willian & Wilkim, 1996. 337 p. ROCHAIX, J. D. Reprint of: Regulation of photosynthetic electron transport. Biochimica et Biophysica Acta, v. 1807, p. 878-886, 2011. ROEL, A. R. Utilização de plantas com propriedades inseticidas: uma contribuição para o desenvolvimento rural sustentável. Revista Internacional de Desenvolvimento Local, v. 1, p.43-50, 2001.

32
ROMAN, M.; BRESSAN, D. A.; DURLO, M. A. Variáveis morfométricas e relações interdimensionais para Cordia trichotoma (Vell.) Arráb. Ex Steud. Ciência Florestal, Santa Maria, v. 19, n. 4, p. 473-480, 2009. SALGADO, A. P. S. Efeito da luz na planta e no óleo essencial de tomilho (Thymus vulgaris). 2005. 49 p. Tese (Doutorado em Agronomia) – Universidade Federal de Lavras, Lavras, 2005. SALYWON, A. M.; LANDRUM, L. R. Curitiba (Myrtaceae): A new genus from the planalto of southern Brazil. Brittonia, v. 59, n. 4, p. 301-307, 2007. SANGWAN, N. S.; FAROOQI, A. H. A.; SHABIH, F.; SANGWAN, R. S. Regulation of essential oil production in plants. Plant Growth Regulation, v. 34, p.3-21, 2001. SANQUETTA, C. R. et al. Crescimento, mortalidade e recrutamento em duas florestas de Araucária (Araucaria angustifolia) no estado do Paraná, Brasil. Revista Ciências Exatas e Naturais, v. 5, n. 1, p. 101-112, 2003. SANTOS, R. I. Metabolismo Básico e Origem dos Metabólitos Secundários. In: SIMÕES, C. M. O.; SCHENKEL, E. P.; GOSMANN, G.; MELLO, J. C. P.;MENTZ, L. A.; PETROVICK, P. R (Org.). Farmacognosia: da planta ao medicamento. 6. ed. Porto Alegre: Universidade Federal de Santa Catarina, 2007. 1102 p. SCHAAF, L. B. et al. Incremento diamétrico e em área basal no período de 1979-2000 de espécies arbóreas de uma Floresta Ombrófila Mista localizada no sul do Paraná. Revista Floresta, v.35, n.2, p.271-290, 2005. SCHNEIDER, P. R.; FINGER, C. A. G. Manejo sustentado de florestas inequiâneas heterogêneas. Santa Maria: UFSM, 2000, 195 p. SCHNEIDER, P. R.; SCHNEIDER, P. S. P. Introdução ao manejo florestal. Santa Maria: FACOS/UFSM, 2. ed., 2008, 566 p. SCHOEFS, B. Chlorophyll and carotenoid analysis in food products. Properties of the pigments and methods of analysis. Trends in Food Science & Technology, v. l3, p. 361-371, 2002. SCHREIBER, U. Detection of rapid induction kinetics with a new type of high-frequency modulated chlorophyll fluorometer. Photosynthesis Research, v. 9, p. 261-272, 1986. SCHREIBER, U.; SCHLIWA, U.; BILGER, W. Continuous recording of photochemical and nonphotochemical chlorophyll fluorescence quenching with a new type of modulation fluorometer. Photosynthesis Research, v. 10, p. 51-62, 1986. SCHREIBER, U.; BILGER, W.; HORMANN, H.; NEUBAUER, C. Chlorophyll fluorescence as a diagnostic tool: basics and some aspects of practical relevance. In: Raghavendra, A. S. (Org.). Photosynthesis: a comprehensive treatise. Cambridge: Cambridge University Press, 1998. 376 p.

33
SCHREIBER, U.; MULLER, J. F.; HAUGGL, A.; GADERMANN, R. New type of dual-channel PAM chlorophyll fluorometer for highly sensitive water toxicity biotests. Photosynthesis Research, v. 74, p. 317–330, 2002. SCHREIBER, U. Pulse-amplitude-modulation (PAM) fluorometry and saturation pulse method: an overview. In: PAPAGEORGIOU, G. C.; GOVINDJEE. (Org.). Chlorophyll a fluorescence: a signature of photosynthesis, advances in photosynthesis and respiration, v. 19. Dordrecht: Springer, p. 279–319, 2004. SHEIL, D.; JENNINGS, S.; SAVILL, P. Long-term permanent plot observations of vegetation dynamics in Budongo, a Ugandan rain forest. Journal of Tropical Ecology, v. 16, p. 765-800, 2000. SILVA, F.; CASALI, V. W. D. Plantas medicinais e aromáticas: pós-colheita e óleos essenciais. Viçosa: Universidade Federal de Viçosa, 2000. 135 p. SILVESTRE, R. Comparação da florística, estrutura e padrão espacial em três fragmentos de Floresta Ombrófila Mista no estado do Paraná. 2009. 79f. Dissertação (Mestrado em Engenharia Florestal) - Setor de Ciências Agrárias da Universidade Federal do Paraná, Curitiba, 2009. SILVEIRA, B. D.; FLORIANO, E. P.; NAKAJIMA, N. Y.; HOSOKAWA, R. T.; ROSOT, N. C.; GRACIOLI, C. R. Relações da morfometria e competição com o crescimento de Trichilia claussenii em um fragmento de floresta semidecidual, RS. Floresta, v. 45, n. 2, p. 373-382, 2015. SIMÕES, C. M. O. SPITZER, V. Óleos voláteis. In: SIMÕES, C. M. O.; SCHENKEL, E. P.; GOSMANN, G.; MELLO, J. C. P.;MENTZ, L. A.; PETROVICK, P. R. (Org.). Farmacognosia: da planta ao medicamento. 6. ed. Porto Alegre: Universidade Federal de Santa Catarina, 2007. 1102 p. SOYLU, E. M.;KURT, S.; SOYLU, S. In vitro and in vivo antifungal activities of the essential oils of various plants against tomato grey mould disease agent Botrytis cinerea. International Journal of Food Microbiology, v. 143, p. 183-9, 2010. STRASSER, R. J.; TSIMILLI-MICHAEL, M.; QIANG, S.; GOLTSEV, V. Simultaneous in vivo recording of prompt and delayed fluorescence and 820-nm reflection changes during drying and after rehydration of the resurrection plant Haberlea rhodopensis. Biochimica Biophysica Acta, v. 1797, p. 1313-1326, 2010. TAIZ, L.; ZEIGER, E. Fisiologia vegetal. 5. ed. Porto Alegre: Artmed, 2013. 918 p. TANAKA, R.; TANAKA, A. Tetrapyrrole biosynthesis in higher plants. Annual Review Plant Biology, v. 58, p. 321-46, 2007.

34
TONINI, H.; ARCO-VERDE, M. F. Morfologia da copa para avaliar o espaço vital de quatro espécies nativas da Amazônia. Pesquisa Agropecuária Brasileira, v. 40, n. 7, p. 633-638, 2005. TAVARES, E. S.; JULIÃO, L. S.; LOPES, D.; BIZZO, H. R.; LAGE, C. L. S.; LEITÃO, S. G. Análise do óleo essencial de folhas de três quimiotipos de Lippia alba (Mill.) N. E. Br. (Verbenaceae) cultivados em condições semelhantes. Revista Brasileira de Farmacognosia, v. 15, p.1-5, 2005. TSIMILLI-MICHAEL, M.; STRASSER, R. J. In vivo assessment of plant’s vitality: applications in detecting and evaluating the impact of mycorrhization on host plants. In: VARMA, A. (Org.). Mycorrhiza: State of the Art, Genetics, and Molecular Biology, Eco-Function, Biotechnology, Eco-Physiology, Structure, and Systematics. 3. ed. Dordrecht: Springer, 2008. 797 p. VANCLAY, J. K. Modeling forest growth and yield: aplications to mixed tropical forests. Copenhagen: CAB International, 1994, 312 p. VON ELBE, J. H. Colorantes. In: FENNEMA, O. W. (Org.). Química de los alimentos. 2. ed. Zaragoza : University of Wisconsin - Madison, 2000. 1258 p. WATZLAWICK, L. F.; ALBUQUERQUE, J. M. de; REDIN, C. G.; LONGHI, R. V.; LONGHI, S. J. Estrutura, diversidade e distribuição espacial da vegetação arbórea na Floresta Ombrófila Mista em Sistema Faxinal, Rebouças (PR). Revista Ambiência, v. 7, p. 415-427, 2011. WICKENS, G. E. El desarollo de los productos forestales no madereros: principios de ordenación. Unasylva, v. 42, n. 165, p. 4-8, 1991. WILLIANS, L. R.; STOCKLEY, W. Essential oils with high antimicrobial activity for therapeutic use. The International Journal of Aromatherapy, Amsterdam, v. 8, n. 4. 1998. WINK, C.; MONTEIRO, J. S.; REINERT, D.; LIBERALESSO, E. Parâmetros da copa e a sua relação com o diâmetro e altura das árvores de eucalipto em diferentes idades. Scientia Forestalis, v. 40, n. 93, p. 57-67, 2012. WOLFFENBÜTTEL, A. N. Óleos essenciais. Informativo CRQ-V, ano XI, nov/dez. 2007, n. 105, p. 06-07. YAROSHEVICH, I. A.; KRASILNIKOV, P. M.; RUBIN, A. B. Functional interpretation of the role of cyclic carotenoids in photosynthetic antennas via quantum chemical calculations. Computational and Theoretical Chemistry, v. 1070,p. 27–32, 2015. ZAMORA, M. Análisis de la información sobre productos forestales no madereros en America Latina. San Tiago: FAO, 2001. 88p. ZHANG, W.; BJÖRN, L. The effect of ultraviolet radiation on the accumulation of medicinal compounds in plants. Fitoterapia, v. 80, n. 4, p. 207-218, 2009.

35
ZIMMERMANN, A. P. L.; FLEIG, F. D.; REDIN, C. G.; AGUIAR, A. V. Relações morfométricas para árvores dominantes de Pinus taeda no estado do Paraná. Anais VII Simpósio de Pós-Graduação em Ciências Florestais, Viçosa, MG, 2012. ZOTARELLI, L. CARDOSO, E. G.; PICCININ, J. L.; URQUIAGA, S.; BODDEY, R.M.; TORRES, E.; ALVES, B. J. R. Calibração do medidor de clorofila Minolta SPAD-502 para avaliação do conteúdo de nitrogênio do milho. Pesquisa Agropecuária Brasileira, v. 38, n. 9, p. 1117-1122, 2003.

36
4. Efeitos da sazonalidade climática e de diferentes irradiâncias nos parâmetros
fotossintéticos de Curitiba primastica (D. Legrand) Salywon and Landrum
Resumo
A ecofisiologia das folhas de Curitiba prismatica (D. Legrand) Salywon e Landrum
foram avaliadas em um fragmento de Floresta Ombrófila Mista em sistema faxinal, no
município de Rebouças-PR. O estudo consistiu da análise da indução polifásica (curva OJIP)
e da cinética lenta da fluorescência da clorofila a, e da quantificação do teor de pigmentos
fotossintéticos nas quatro estações do ano, desenvolvidas em ambiente a pleno sol e no sub-
bosque. A análise da fluorescência da clorofila a foi realizada com dados da cinética lenta e
da cinética rápida, em folhas previamente adaptadas ao escuro, realizadas nas quatro estações
entre 2015 e 2016. Foram analisados os parâmetros Fo, Fm, Fv/Fm, ABS/RC, PIABS pelo teste
OJIP, bem como a eficiência fotossintética e a taxa de transporte de elétrons. Os valores
observados de Fo e Fm diferiram significativamente (p<0,05), com os maiores valores nas
árvores que estavam a pleno sol. O efeito da sazonalidade para as árvores de C. prismatica
que estavam a pleno sol demonstrou diferenças significativas (p<0,05) no verão comparado às
demais estações, refletindo em um maior valor de Fm, possivelmente devido a danos causados
por fotoxidação. Para os exemplares de C. prismatica em ambiente de sub-bosque não foi
observado diferenças significativas entre as estações do ano. Durante o inverno, o ambiente a
pleno sol apresentou resultados semelhante ao sub-bosque. O teor de clorofila a, b e total
diferiram significativamente (p<0,05) entre os dois ambientes ao longo de todas as estações
do ano, sendo os maiores valores observados no ambiente de sub-bosque. Os indivíduos de C.
prismatica em sub-bosque foram menos sensíveis às variações sazonais da fluorescência da
clorofila a. Dessa forma, este estudo indica que C. prismatica apresentou mudanças
adaptativas de acordo com o ambiente em que se desenvolveu.
Palavras-chave: Fluorescência da clorofila a, pigmentos fotossintéticos, irradiância, sazonalidade.

37
Effects of climate seasonality and of different irradiances on photosynthetic
parameters of Curitiba primastica (D. Legrand) Salywon and Landrum
Abstract
The ecophysiology of Curitiba prismatica (D. Legrand) Salywon and Landrum leaves
was evaluated in a fragment of Mixed Ombrophilous Forest in faxinal system, in Rebouças
Municipality, PR, Brazil. This study aimed at analyzing polyphase induction (OJIP curve) and
slow kinetics of chlorophyll a fluorescence, besides quantifying photosynthetic pigment
levels in trees developed under full sun and understory environments in the four seasons of
the year. Chlorophyll a fluorescence was analyzed with data of slow and rapid kinetics in
previously dark adapted leaves, between 2015 and 2016. The parameters Fo, Fm, Fv/Fm,
ABS/RC, PIABS by the OJIP test, photosynthetic efficiency, and electron transport rate were
evaluated. The observed Fo and Fm values were significantly different (p<0.05), with the
highest values in trees under full sun. The effect of seasonality on C. prismatica trees under
full sun showed significant differences (p<0.05) in the summer when compared to other
seasons, resulting in a higher Fm value, probably due to damages caused by photooxidation.
For understory C. prismatica trees, no significant differences were observed among seasons.
In the winter, similar results were obtained in the two environments. Chlorophyll a, b and
total levels were significantly different (p<0.05) between the two environments throughout all
seasons, with the highest values observed in understory trees, which were less sensitive to
seasonal variations of chlorophyll a fluorescence. This study indicates that C. prismatica
showed adaptive changes according to the environment.
Keywords: Chlorophyll a fluorescence, photosynthetic pigments, irradiance, seasonality

38
4.1. Introdução
A luz tem sido entendida como o fator abiótico mais determinante no crescimento e
nos processos biológicos das plantas. A disponibilidade e capacidade de uso da irradiância
afeta as características das folhas, e, por conseguinte, a fotossíntese (GONÇALVES et al.,
2007). A intensidade luminosa também regula a sobrevivência das plantas e a distribuição
geográfica de uma determinada espécie. Em outros casos, em que a densidade do fluxo de
radiação fotossinteticamente ativa (RFA) se encontra abaixo do nível de saturação da
fotossíntese, devido ao sombreamento, pode levar à redução do crescimento e de produção de
determinada espécie (BOYER, 1982; SOTO-PINTO et al., 2000; ZHANG et al., 2004;
STEPHAN et al., 2008).
O aparelho fotossintético sob alta irradiância absorve a energia luminosa
excessivamente, resultando na inativação ou a deterioração das clorofilas contidas nos centros
de reação (CRs) dos cloroplastos (BERTAMINIA et al., 2006). Como consequência, a
atividade fotossintética é reduzida pela fotoinibição (OSMOND, 1994).
Em geral, a adaptação da folha desenvolvida sob diferentes níveis de luminosidade
apresenta consideráveis diferenças em sua estrutura, forma e tamanho, no conteúdo de
pigmentos fotossintéticos, nos transportadores de elétrons e nas taxas fotossintéticas, em
comparação com folhas desenvolvidas em pleno sol (TSUKAYA, 2005; VALLADARES e
NIINEMETS, 2008; DAI et al., 2009).
As folhas alocadas na sombra, em que os níveis de luz solar são baixos, normalmente,
a planta investe no aumento da área foliar. Esse aumento visa capturar o máximo de luz para a
realização da fotossíntese (SIMS et al, 1994; NIINEMETS et al., 1998).
Para o crescimento normal, as plantas necessitam equilibrar o fluxo de energia,
elemento determinante em respostas adaptativas que garantem sua sobrevivência, crescimento
e adaptações às constantes mudanças no ambiente (GONÇALVES et al., 2007).
Para avaliar o aproveitamento da energia luminosa e o estado do aparato fotossintético
das plantas pode-se utilizar a fluorescência da clorofila a. A fluorescência da clorofila a têm
sido utilizada na avaliação do efeito de fatores ambientais, como a disponibilidade de energia
luminosa sobre o metabolismo fotossintético, bem como, para medir o grau de tolerância das
plantas frente aos estresses abióticos (STIRBET e GOVINDJEE, 2011).

39
Estudos utilizando a fluorescência da clorofila a indicam que o Fotossistema II (FSII)
é um dos principais alvos do estresse por baixas (BERTAMINI et al., 2007) e altas
temperaturas (BRESTIC et al., 2012).
A análise da fluorescência da clorofila a é uma ferramenta não destrutiva, rápida e
tornou-se um meio eficiente e amplamente utilizado para a obtenção de informações sobre o
estado do FSII (WANG et al., 2013), a funcionalidade do aparato fotossintético, o fluxo de
elétrons e a concentração de clorofila (CHRISTEN et al., 2007). A fluorescência da clorofila a
permite analisar a eficiência fotossintética, fornecendo informações sobre a relação entre
estrutura e função do FSII e CR (ADAMS e DEMMIG-ADAMS, 2004).
Curitiba prismatica é uma espécie endêmica que ocorre na Floresta Ombrófila Mista
(FOM), encontrada no sul do Brasil, ocorrendo no primeiro e segundo planaltos do Paraná e
no planalto norte de Santa Catarina. A ocorrência da espécie se dá tanto em sub-bosque como
em áreas abertas, porém estudos ecofisiológicos de C. prismatica são inexistentes, não
havendo registros das respostas adaptativas desta espécie sob diferentes níveis de irradiância,
bem como a alternância das estações do ano sobre o aparato fotossintético desta espécie.
Diante do exposto acima, esse capítulo teve como objetivo a análise de alguns
aspectos da ecofisiologia de C. prismatica, pela análise fluorescência da clorofila a e dos
teores de pigmentos fotossintéticos e carotenoides. Para as avaliações foram analisados dois
ambientes naturais com diferentes níveis de luminosidade, a pleno sol e sub-bosque, ao longo
de quatro estações climáticas, visando compreender a influência dos efeitos da sazonalidade
climática na adaptação das plantas de C. prismatica desenvolvidas sob diferentes condições
de irradiância luminosa.

40
4.2. Material e métodos
Local do experimento
O estudo foi realizado em um fragmento de FOM no município de Rebouças-PR,
sendo a área localizada a uma latitude sul 25°40'12”, e longitude oeste 50°32'56’’, com uma
altitude média de 815 m acima do nível do mar, conforme apresentada na Figura 6.
Figura 6. Localização da área de estudo no munícipio de Rebouças-PR.
Fonte: Autor (2016).
O clima regional é do tipo Cfb, de acordo com a classificação de Köppen,
apresentando estações climáticas bem definidas, com chuvas distribuídas durante todo o ano,
com geadas severas e umidade relativa do ar acentuada. A precipitação média anual está entre
1500 e 1600 mm (PARANÁ, 2016a). A temperatura média anual é de aproximadamente
18°C, com mínima de -2°C e máxima de 32°C.

41
O substrato geológico da região é formado pelo derrame de trapp da formação
basáltico (EMBRAPA, 1984), apresentando relevo desde plano, ondulado e montanhoso,
sendo o último de maior predominância (EMBRAPA, 1999). Os solos de maior ocorrência
são Neossolos Litólicos, Cambissolos e Argissolos (PARANÁ, 2016).
Com relação à vegetação e a FOM em Sistema Faxinal, na área de estudo, há o
predomínio das famílias Myrtaceae, Lauraceae, Salicaceae, Sapindaceae, Aquifoliaceae. As
espécies com maiores valores de importância são a C. prismatica, Campomanesia
xanthocarpa (Mart.) ex O. Berg, Casearia obliqua Spreng., Casearia sylvestris Sw, Ocotea
puberula (Rich.) Nees, Ilex paraguariensis A. St.-Hil., Ocotea porosa (Nees & Mart.)
Barroso, entre outras (ALBUQUERQUE, 2015).
No local de estudo, a espécie C. prismatica ocorre tanto da forma arbustiva, em locais
a pleno sol que estão desprovidos de floresta, como em forma arbórea, nos sub-bosques e em
bordas florestais. Para as análises realizadas nesse trabalho foram selecionadas árvores de
cada um desses ambientes, sub-bosque e a pleno sol. A identificação botânica e catalogação
da espécie foram realizadas no Herbário da Universidade Estadual do Centro-Oeste (HUCO),
onde as exsicatas foram depositadas e receberam a identificação 6303.
Avaliação da fluorescência da clorofila a
As avaliações da fluorescência da clorofila a, por meio da indução da cinética e da
indução polifásica (OJIP) em C. prismatica, foram realizadas utilizando o delineamento
experimental inteiramente casualizado com medidas repetidas ao longo do tempo. As análises
realizadas avaliaram os efeitos da irradiação luminosa em árvores a pleno sol e de sub-bosque,
com sete repetições em cada condição. O estudo foi realizado durante os anos de 2015 e 2016,
e as análises foram realizadas nas seguintes datas: 17 de novembro de 2015, 20 de fevereiro
de 2016, 28 de abril de 2016 e 30 de julho de 2016, que correspondem às estações da
primavera, verão, outono e inverno, respectivamente, no período das 07h às 11h.

42
Foram selecionadas para as análises de fluorescência folhas de C. prismatica que
estavam completamente expandidas e livres de danos por patógenos ou herbivoria. As folhas
ficaram por um período de 25 minutos de adaptação ao escuro, antes de cada coleta, para que
todos os centros de reação ficassem abertos com capacidade de receber elétrons. Para essa
adaptação foram utilizados clipes específicos para essa finalidade. Para medição da
fluorescência utilizou-se um fluorômetro portátil PAM (PAM-2500, Heinz Walz GmbH ),
sendo avaliados os parâmetros Fo, Fm, Fv/Fm, ABS/RC, PIABS pelo teste JIP, bem como a
rendimento fotossintético (Y(II)) e a taxa de transporte de elétrons (ETR).
Coleta do material vegetal
As coletas das amostras de folhas de C. prismatica para determinação dos teores de
pigmentos fotossintéticos (clorofila a, b e total) e carotenoides totais foram realizadas
utilizando o delineamento experimental inteiramente casualizado com medidas repetidas em
cada estação do ano, nas mesmas datas e horários em que ocorreu a avaliação da fluorescência
da clorofila a. O tratamento utilizado avaliou os efeitos da irradiância em árvores de C.
prismatica a pleno sol e em sub-bosque, sendo realizadas sete repetições em cada ambiente.
Para essas avaliações foram utilizadas folhas completamente expandidas e livres de
danos por patógenos ou herbivoria. No momento da coleta as folhas foram acondicionadas em
envelopes de papel alumínio e refrigeradas em gelo até serem transferidas para ambiente
refrigerado e consequente realização das análises.
Determinação dos teores de pigmentos fotossintéticos
A extração e quantificação das clorofilas e dos carotenoides de folhas de C. prismatica
foram realizadas segundo a metodologia proposta por Lichtenthaler e Wellburn (1983) e Porra
et al. (1989), utilizando amostras foliares de 20 mg de massa fresca da folha de C. prismatica,
evitando-se as nervuras. Cada amostra foi então macerada com carbonato de cálcio e
homogeneizada, com o uso de graal e pistilo, com 8 mL de acetona 80%.
O processo de maceração e homogeneização foi realizado no Laboratório de
Fitopatologia, do departamento de Agronomia, no campus CEDETEG, da Universidade
Estadual do Centro-Oeste do Paraná, UNICENTRO, em ambiente escuro, visando evitar a
degradação dos pigmentos fotossintéticos.

43
Após o material ser totalmente homogeneizado, então, foi centrifugado por 15 minutos
a 4000 rpm. O sobrenadante foi analisado nas absorbâncias de 470 nm, 663,6 nm e 646,6 nm;
no espectrofotômetro UV-VIS (UV-1800, Shimadzu), pertencente ao Laboratório Fruticultura
e Pós-Colheita, do departamento de Agronomia da UNICENTRO.
Para o cálculo dos teores dos pigmentos fotossintéticos foram utilizadas as Equações 1
a 4 propostas por Lichtenthaler e Wellburn (1983) e Porra et al. (1989).
Clorofila a (µg/ml) = 12,21A663,6 – 2,81A646,6 (1)
Clorofila b (µg/ml) = 20,13A646,6 – 5,03A663,6 (2)
Chls a + b (µg/ml) = 17,76A646,6 + 7,34A663,6 (3)
Carotenoides (µg/ml) =
(4)
Em que,
A = absorbância no comprimento de onda indicado;
Chl = clorofila.
Análises Estatísticas
As análises estatísticas foram realizadas com o software SPSS, versão 22. Foi
utilizando o Modelo Linear Generalizado (MLG), pelo método da Equação de Estimativa
Generalizada (EEG), com a distribuição Gama, devido aos dados não serem normais. A
função de ligação utilizada foi identidade. As diferenças nas médias foram avaliadas pelo
teste de Bonferroni, com nível de significância de 5%.
Para o cálculo da área acima da curva OJIP, entre os pontos O e P, utilizou-se o
software Origin, versão 8.5,
1000A470 – 3,27[Chl a] – 104[Chl b] 229

44
4.3. Resultados e Discussão
Análise da fluorescência da clorofila a
As curvas da cinética rápida de emissão da fluorescência transitória da clorofila a em
folhas adaptadas ao escuro de C. prismatica estão apresentadas, em escala logarítmica, na
Figura 7. Nessas curvas estão representados o alcance da fluorescência transitória, partindo da
fluorescência basal Fo até um nível máximo Fm, para cada um dos ambientes, sombreado em
sub-bosque e a pleno sol, durante quatro estações climáticas.
1
2
3
4
5
1
2
3
4
5
1
2
3
4
5
1
2
3
4
5
IP
J
Fluo
resc
ênci
a (µ
.mol
.m-1.s-1
)
Sol Sombra
A
O
B
0,001 0,1 1 10 100 10000,001 0,1 1 10 100 1000
C D
Tempo (ms) Figura 7. Curva OJIP da fluorescência da clorofila a da espécie Curitiba prismatica em
ambiente sombreado e a pleno sol, durante as estações da primavera (A), verão
(B), outono (C) e inverno (D) de 2015 a 2016.
A curva OJIP durante o verão, estação que apresenta maior irradiação solar, foi
relativamente maior nas plantas a pleno sol quando comparada as demais estações. Variação
menor foi observada nas plantas à sombra, independente da estação. A menor variação da
fluorescência, entre os ambientes, foi observada para as curvas correspondentes ao inverno,
em que se observam variações menos intensas no Fo e Fm nas plantas de C. prismatica. Essa
menor variação da fluorescência durante o inverno pode estar associada a menor irradiação
solar incidente e baixa atividade biossintética da planta ocasionada pela estação.

45
A área acima da curva OJIP da fluorescência entre Fo e Fm foi menor para as árvores
que estavam no ambiente sombreado, durante as quatro estações analisadas. Segundo Mehta
et al. (2010) a área acima da curva OJIP é proporcional ao estado de oxidação dos aceptores
de elétrons no lado redutor do FSII e inversamente proporcional ao estado de redução. Quanto
menor é esta área, maior é a transferência de elétrons dos CR para o pool de plastoquinona.
Dessa forma, as menores áreas sobre as curvas OJIP indicam que no ambiente sombreado a C.
prismatica apresentou maior capacidade de absorção de energia luminosa, maior atividade do
lado doador do FSII, refletindo em maior fluxo de elétrons para o FSI.
Nas curvas da Figura 7, a fase O-J (50 µs a 2 ms) refere-se ao comportamento da
fluorescência nos primeiros 2 ms da curva e corresponde à fase fotoquímica primária da
fotossíntese, representando a redução do lado aceptor do FS II (QA para QA-) (STRASSER et
al., 1995; TÓTH et al., 2007). A maior redução da QA é observada na primavera e no verão
em comparação às curvas do outono e do inverno. A fase J-I (2 ms – 30 ms) corresponde à
redução do pool de plastoquinonas (PQ) (TÓTH et al., 2007) e, por último, a fase I-P (30 ms a
300 ms) refere-se a transferência de elétrons do FSII a partir da plastoquinona reduzida
(PQH2) aos aceptores do FSI (YUSUF et al., 2010).
Na Figura 8 estão representados os parâmetros do Teste JIP (STRASSER e
STRASSER, 1995) para os ambientes sombreados e a pleno sol, durante as quatro estações
estudadas. Foram identificadas diferenças significativas nos parâmetros Fo e Fm entre os dois
ambientes (Figura 8A e 8B), nas estações da primavera, verão e outono. As árvores que
estavam a pleno sol sempre apresentaram os maiores valores de fluorescência, porém, no
inverno não houve diferenças significativas entre os dois ambientes, como pode ser visto na
Figura 7D, em que as curvas da fluorescência foram muito próximas.
Os maiores valores para o parâmetro Fo foram observados em C. prismatica que se
desenvolveram a pleno sol, sendo que no verão essas árvores diferiram estatisticamente das
demais estações. Para as árvores do sub-bosque os valores de Fo não diferiram
significativamente entre as estações.

46
0,6
0,8
1,0
1,2
3,0
3,5
4,0
4,5
0,72
0,74
0,76
0,78
0,80
Inverno Outono Primavera Verao0,4
0,6
0,8
1,0
1,2
Inverno Outono Primavera Verao0,2
0,4
0,6
0,8
1,0
1,2
aC aBC aB
aA
aAB
bB bA bA
aB
aAB
aAB aA
aA bAbA bA
aA
bA
aAB
bB
aB
aA
aBaB
bA bAbA
bA
aA aA aA
aA
aA
aA
aB aB
aA
aA aA
aA
Sol SombraA B
Fluo
resc
ênci
a (µ
.mol
.m-1
.s-1)
Estações
C
Estações
D
Estações
E
*Médias seguidas pelas mesmas letras, minúsculas entre os ambientes e maiúsculas entre as estações, não diferem estatisticamente, ao nível de 5 % de probabilidade.
Figura 8. Parâmetros Fo (A), Fm (B), Fv/Fm (C), ABS/RC (D) e PIABS (E) da fluorescência da
cinética rápida da clorofila a obtidos da curva OJIP de folhas de Curitiba
prismatica, em diferentes ambientes de irradiação luminosa, durante as estações da
primavera, verão, outono e inverno de 2015 a 2016.
Essa diminuição da fluorescência em árvores que estavam a pleno sol, durante o
inverno, pode ser devido à redução no suprimento de CO2 para a fotossíntese que pode
ocorrer devido a temperaturas mais baixas observadas nesta estação. Sabe-se que os estômatos
respondem à temperatura foliar, sendo que a transpiração é seu principal mecanismo
envolvido na regulação da temperatura foliar e que menores aberturas estomáticas levam a
redução da transpiração e aumento da temperatura dos tecidos (MACHADO et al., 2005;
NOBEL, 1999). Dessa forma as baixas temperaturas do inverno podem afetar diretamente as
reações fotoquímicas da fotossíntese, entre elas a fluorescência da clorofila a (GEORGIEVA,
1999; VESTE et al., 2000).

47
Aumentos de Fo podem ocorrer, quando há danos no centro de reação do FS II, ou pela
redução na transferência de energia de excitação do sistema coletor de luz para o centro de
reação (MATHIS e PALLOTIN, 1981; BAKER e ROSENQVIST, 2004). No verão a
ocorrência de danos às folhas, e, por conseguinte, no aparelho fotossintético, é maior devido à
alta insolação, podendo levar a fotoxidação das clorofilas.
Segundo Lichtenthaler et al. (1981, 1982ab, 1984) folhas que se desenvolvem em
ambientes sob alta irradiância luminosa possuem um nível muito mais baixo de complexos
coletores de luz (CCL), e um menor grau de empilhamento dos tilacoides, quando comparado
a cloroplastos de plantas adaptadas a menor intensidade de irradiação luminosa. Dessa
maneira, as plantas sujeitas a sombreamento tendem a aproveitar melhor a luz incidente,
apresentando menor valor de Fo, comparativamente com as árvores desenvolvidas a pleno sol.
Diante deste fato, aumentos de Fo podem ocorrer por uma redução na transferência de energia de
excitação do CCL para o CR, visto que os CCL das árvores a pleno sol não são desenvolvidos
para ter um alto aproveitamento da energia luminosa devido à abundância de energia incidente
sobre as folhas (MATHIS e PALLOTIN, 1981; BAKER; ROSENQVIST, 2004).
A Fm manteve perfil semelhante a Fo, entre os ambientes sombreado e a pleno sol,
apresentando os maiores valores nas árvores de C. prismatica que estavam a pleno sol e não
ocorrendo diferenças somente no inverno. Os maiores valores de Fo e Fm em árvores a pleno
sol ocorreram em virtude das folhas que se desenvolvem em ambientes sob alta irradiação
luminosa possuírem um nível muito mais baixo de CCL, tendo um menor aproveitamento da
energia luminosa, comparativamente às folhas de C. prismatica alocadas em sub-bosque
(LICHTENTHALER et al., 1981, 1982ab, 1984).
A relação Fv/Fm, representa o rendimento quântico do FSII, diferiu significativamente
entre os dois ambientes no outono e no verão, bem como revelou diferenças provocadas pela
sazonalidade das estações do ano. Bolhàr-Nordenkampf et al. (1989) afirmam que, a relação
Fv/Fm representa um indicador sensível do desempenho do aparato fotossintético das plantas,
em que, para plantas saudáveis espera-se valores entre 0,75 e 0,85.

48
O menor valor para a relação Fv/Fm para C. prismatica foi obtido no verão, em
ambiente de pleno sol, enquanto que o maior valor ocorreu no sub-bosque durante o outono.
Embora tenha ocorrido diferenças significativas na relação Fv/Fm, todas as árvores analisadas
nos dois ambientes, ao longo das quatro estações do ano, Figura 8C, apresentaram valores
muito próximos do esperado para plantas saudáveis. Isso reflete que o aparato fotossintético
está trabalhando satisfatoriamente e que a espécie apresenta plasticidade adaptativa.
A relação ABS/RC, que estima o fluxo de absorção por centro de reação, também
estima a quantidade de clorofila por centro de reação completamente ativo (redutor de QA)
(STRASSER et. al, 2004, 2010). Este parâmetro (Figura 8D) diferiu significativamente entre
os dois ambientes, em todas as estações. No ambiente sombreado as árvores de C. prismatica
apresentaram os maiores valores.
Esses valores estão de acordo com os relatados por Lichtenthaler et al. (1981, 1982ab,
1984), que conforme mencionado, relataram que folhas que se desenvolvem em ambientes
sob baixa irradiação luminosa possuem um nível muito mais alto de CCL, aproveitando
melhor a luz que incide sobre as folhas. Esse maior aumento de clorofila, tanto clorofila a
quanto b, pôde ser verificado e diferiu significativamente, nas análises de extração de
pigmentos fotossintéticos que serão apresentadas mais adiante nesse capítulo. Nas análises ao
longo das quatro estações do ano não foram verificadas diferenças significativas dentro de
cada ambiente, para a relação ABS/RC.
Para o índice de desempenho fotossintético (PIABS) das plantas de C. prismatica não
ocorreu diferenças entre os ambientes ao longo das estações. Porém dentro de cada ambiente
houve redução à medida que a temperatura e insolação aumentaram, sendo que na primavera e
no verão as árvores que estavam a pleno sol diminuíram os valores deste índice, quando
comparadas as estações inverno e outono. Isso indica um efeito negativo na maquinaria
fotossintética devido à fotoxidação, visto que esses parâmetros estão relacionados com fluxo
de energia em toda a cadeia de transporte de elétrons fotossintética.
Conforme Gonçalves e Santos Junior (2005), a diminuição no PIABS ocorre devido a
maior dissipação da energia na forma de calor e fluorescência. No ambiente a pleno sol as
árvores de C. prismatica estão expostas à alta irradiância, que ocorrem principalmente no
verão, levando-as a dissipar a energia que não pode ser utilizada nas reações fotoquímicas.

49
Os resultados obtidos a partir das avaliações da indução de curva lenta da
fluorescência da clorofila a em folhas de C. prismatica, adaptadas à luz, para os parâmetros
ETR e Y(II) estão representados na Figura 9.
aC
aA
aB aB
aAbA
bA
bAaC
aA
aBaB
aAbA
bA
bA
Inverno Outono Primavera Verao0
5
10
15
20
25
30
35
Fluo
resc
ênci
a (µ
.mol
.m-2
.s-1)
Fluo
resc
ênci
a (µ
.mol
.m-2.s-1
)
Estações
Sol Sombra
A
Inverno Outono Primavera Verao0,0
0,1
0,2
0,3
0,4
Estações
B
*Médias seguidas pelas mesmas letras, minúsculas entre os ambientes e maiúsculas entre as estações, não
diferem estatisticamente, ao nível de 5 % de probabilidade.
Figura 9. Parâmetros ETR (A), Y(II) (B) da fluorescência da cinética lenta da clorofila a da
espécie Curitiba prismatica em ambiente sombreado e a pleno sol, durante as
estações da primavera, verão, outono e inverno de 2015 a 2016.
A estimativa da taxa relativa de transporte de elétrons apresentou os maiores valores
em árvores que estavam a pleno sol, sendo que na primavera, verão e outono os valores do
ETR diferiram significativamente dos encontrados no ambiente sombreado.
Para as árvores que estavam a pleno sol ocorreram diferenças, nessa taxa, entre as
estações do ano, observando-se no outono os maiores valores e no inverno os menores
valores. Na primavera e no verão não ocorreram diferenças significativas nas árvores que
estavam neste ambiente.
Os valores observados para ETR para o ambiente sombreado não demonstrou
variações significativas entre as estações. Isso indica que os efeitos da sazonalidade são
atenuados pela proteção oferecida pelo dossel florestal, tornando esse ambiente de sub-bosque
mais homogêneo durante todo o ano.

50
Os maiores valores de ETR nas árvores de C. prismatica a pleno sol podem indicar
que folhas de sol apresentam maior capacidade fotossintética sobre uma mesma área foliar,
enquanto que folhas de sombra geralmente possuem baixa taxa de assimilação líquida de CO2
e se tornam saturadas em baixa irradiação (TATENO e TANEDA, 2007).
O rendimento fotossintético do FSII, Y(II) seguiu o mesmo padrão da ETR. No
ambiente sombreado não ocorreram variações significativas entre as estações. No ambiente a
pleno sol, no inverno e no outono, ocorreu diferenças significativas das demais estações do
ano, apresentando os maiores e os menores valores, respectivamente.
Os valores encontrados tanto para o ETR quanto para o Y(II) durante o inverno, nas
árvores a pleno sol, não apresentaram diferenças estatísticas com as árvores de sub-bosque.
Em trabalho realizado por Ptushenko et al. (2013), em ambiente sombreado e a pleno sol,
observaram resultados semelhantes a este, em que sobre condições sombreadas Y(II)
apresentou valores inferiores.
As maiores taxas de transporte de elétrons e rendimento fotoquímico observado nas
árvores a pleno sol pode ter sido ocasionada pela adaptação das folhas à alta incidência
luminosa a que estão expostas. Plantas que se desenvolvem nesses ambientes possuem maior
quantidade de CR e menos CCL, levando a uma maior capacidade de transporte de elétrons e
conversão fotoquímica, com consequente aumento em sua produção fotossintética (MEIER e
LICHTENTHALER, 1981; LICHTENTHALER et al., 1981, 1982, 1984).
Teor de pigmentos fotossintéticos
Os resultados na Figura 10 demonstram diferenças significativas nas concentrações de
pigmentos fotossintéticos de C. prismatica entre o ambiente sombreado e a pleno sol, bem
como diferenças ao longo das estações do ano. No ambiente sombreado de sub-bosque foram
verificadas maiores concentrações de clorofilas a e b, bem como a clorofila total, em relação
às plantas que se desenvolveram a pleno sol. Essas maiores concentrações de clorofila em
plantas alocadas no sub-bosque se mantiveram constantes ao longo das quatro estações do ano
analisadas.
Segundo Kramer e Kozlowski (1979), a clorofila é constantemente sintetizada e
destruída em presença de luz, principalmente quando expostas a intensidades luminosas muito
altas. No caso das plantas de C. prismatica a pleno sol, a velocidade de decomposição foi
maior, sendo o equilíbrio estabelecido a uma concentração mais baixa.

51
bD
bA
bCbB
aCaA
aC aB
bBbA bAB
bC
aA aAaA aA
bC
bA
bBC bB
aBaA
aB aB
aC
aBaC
aA
aB
aA
bAB
bAB
bB
bBbB
aA
aB
aAB aA
bB
0
5
10
Clo
rofil
a a
(µg.
ml-1
)
Sol Sombra
A
0
1
2
3
4
Clor
ofila
b (µ
g.m
l-1)
B
0
5
10
15
Clo
rofil
a To
tal (
µg.m
l-1)
C
Inverno Outono Primavera Verao0
1
2
3
4
Rel
ação
a/b
Estações
D
Inverno Outono Primavera Verao0,0
0,5
1,0
1,5
2,0
2,5
Car
oten
oide
s Tot
ais (
µg.m
l-1)
Estações
E
Figura 10. Teores de clorofila a (A), clorofila b (B), clorofila total (C), relação clorofila a/b
(D) e carotenoides totais (E) da espécie Curitiba prismatica em ambiente
sombreado e a pleno sol, durante as estações da primavera, verão, outono e
inverno de 2015 a 2016.
A sazonalidade climática também refletiu em alterações nas concentrações de
clorofila. A clorofila a, Figura 10A, diferiu significativamente em todas as estações no
ambiente a pleno sol, sendo que no inverno foram observadas as menores concentrações e no
outono as maiores concentrações.
Para o ambiente de sub-bosque as alterações foram mais tênues, porém manteve o
mesmo padrão que o ambiente a pleno sol, com a maior concentração no outono e a menor no
verão.
No inverno as baixas temperaturas, ocasionaram alterações mais pronunciadas nas
concentrações de clorofila para os dois ambientes, diminuindo significativamente o conteúdo
de clorofila a. Os baixos conteúdos de clorofila a registrados na primavera sugerem que
devido a grande quantidade de folhas novas, principalmente verificadas nas árvores a pleno
sol, não havia ainda alcançando a sua máxima produção de clorofila.

52
Em relação à clorofila b, Figura 10B, igualmente ao que ocorreu com a clorofila a,
houve diferenças significativas entre os dois ambientes em todas as estações analisadas, sendo
que o ambiente a pleno sol apresentou as menores concentrações deste pigmento em C.
prismatica. No ambiente a pleno sol as concentrações de clorofila b diferiram
significativamente entre o verão, em que foram obtidos os mais baixos teores, e as demais
estações. Para o ambiente de sub-bosque não ocorreu diferenças nos teores de clorofila b ao
longo das estações do ano, demonstrando que as alterações climáticas provocadas pela
sazonalidade tem uma menor influência para as concentrações deste pigmento fotossintético.
No ambiente de sub-bosque, o nível de irradiância reduzido demonstra ser mais
determinante na indução de alterações morfológicas e fisiológicas na espécie. O aumento nos
teores de clorofila b em ambiente com baixas intensidades de luz é uma característica
importante de adaptação da planta em ambientes sombreados, pelo fato desse pigmento captar
fótons com maiores comprimentos de onda (WHATLEY e WHATLEY, 1982). Dessa forma,
pode-se concluir que C. prismatica apresenta adaptação em ambientes com baixos níveis de
luminosidade, como é o ambiente de sub-bosque, em relação aos pigmentos fotossintéticos.
O aumento das concentrações de clorofila b em plantas sombreadas pode ser
considerado como característica importante na adaptabilidade vegetal em ambientes
sombreados. Isto porque a clorofila b absorve energia em comprimento de onda diferentes da
clorofila a e a transfere para os CRs, maximizando, assim, a captura energética que
efetivamente atua nas reações fotoquímicas, estratégia que permite a planta melhorar sua
adaptação em ambientes com baixa disponibilidade luminosa (HUANG et al., 2004;
MITCHELL, 1979; TAIZ e ZEIGER, 2013).
Os resultados obtidos para a relação clorofila a/b, Figura 10D, demonstraram redução
no ambiente sombreado durante a primavera e o verão, diferindo significativamente (p<0,05)
do ambiente a pleno sol. Esses menores valores da relação clorofila a/b refletem uma menor
diferença entre os dois tipos de clorofila como resposta adaptativa de incremento dos
complexos antenas associados aos fotossistemas (LAGE-PINTO et al., 2008). De maneira
geral, vários trabalhos demonstram que a proporção entre clorofila a e b tende a diminuir com
a redução da intensidade luminosa, devido a uma maior proporção relativa de clorofila b em
ambientes sombreados (LEE et al., 2000, CASTRO et al., 2005; LIMA et al., 2010).

53
No sub-bosque foram observadas diferenças significativas (p<0,05) na relação
clorofila a/b somente entre o inverno e outono, sendo que no inverno foram obtidos os
menores valores. Já nas árvores a pleno sol houve diferenças entre todas as estações, mas
igualmente, no inverno foram observados os menores valores, e no verão os maiores valores.
Os menores valores da relação clorofila a/b durante o inverno podem indicar adaptação das
folhas para melhor aproveitamento da energia luminosa, visto que durante o inverno, o
fotoperíodo e a intensidade luminosa são reduzidos.
Os valores da relação clorofila a/b, Figura 10D, observados nas árvores de C.
prismatica analisadas estão próximos de 3:1. Segundo Lichtenthaler (1987) a proporção entre
clorofila a e b (relação a/b) é um indicador da adaptação à luz ou de condição de aclimatação
do aparato fotossintético e valores na relação (a/b) de aproximadamente 3:1 indicam plantas
saudáveis.
Os maiores teores de clorofila a, b e total, observados no sub-bosque, indicam que a
espécie consegue se adaptar a essa condição de luminosidade. A maior quantidade de
pigmentos também indica alta síntese de clorofila e uma degradação mais lenta da clorofila b
em relação à clorofila a (ENGEL e POGGIANI, 1991).
O teor de carotenoides, Figura 10E, em C. prismatica foi significativo (p<0,05) nas
avaliações entre os ambientes, sendo consistentes essas diferenças durante todas as estações
do ano. No inverno, outono e primavera os maiores resultados foram encontrados no sub-
bosque. Em relação ao verão, ocorreu o inverso, sendo encontrados os maiores teores de
carotenoides no ambiente a pleno sol.
No ambiente de sub-bosque não ocorreu diferenças significativas entre as estações do
ano nos teores de carotenoides totais. Porém na condição de pleno sol o teor de carotenoides
diferiu significativamente (p<0,05) entre o verão e as demais estações. Os teores mais
elevados de carotenoides no verão pode indicar uma estratégia adaptativa nas árvores a pleno
sol contra danos ocasionados pela fotoxidação.
Os carotenoides são pigmentos acessórios que funcionam como fotoprotetores por
meio da rápida extinção dos estados excitados da clorofila protegendo-a da fotoxidação
(TAIZ e ZEIGER, 2013). Dessa forma, os altos níveis deste pigmento nas árvores a pleno sol
podem estar associados ao desenvolvimento de proteção para os CCL dos fotossistemas, visto
que o verão apresenta as maiores incidências de radiação luminosa.

54
Diversos artigos analisaram as diferenças do conteúdo de pigmentos fotossintéticos de
plantas desenvolvidas sob diferentes luminosidades. Pode-se citar, por exemplo, Atroch et al.
(2001) que estudaram a espécie Bauhinia forficata Link submetidas a diferentes condições de
sombreamento. Avelar et al. (2013) que avaliaram a espécie Piper aduncum L sob diferentes
níveis de irradiação. Engel e Poggiani (1991) que analisaram mudas de quatro espécies
florestais nativas. Lima Júnior et al. (2006) que trabalharam com a espécie Cupania vernalis
Camb. Martins et al. (2009) que avaliaram a espécie Ocimum gratissimum Lineu. Oliveira et
al. (2009) que estudaram os efeitos de telas coloridas em Artemisia vulgaris L. Souza et al.
(2011) que analisaram o efeito de malhas coloridas em plantas de Mikania laevigata Schultz
Bip. ex Baker. Todos esses pesquisadores analisaram o efeito da radiação solar em diferentes
espécies de plantas e encontraram resultados semelhantes a este estudo.

55
4.4. Conclusão
Pelas análises realizadas verificou-se que a C. prismatica possui diferenças no aparato
fotossintético. As árvores desenvolvidas em ambiente sombreado apresentaram melhor
aproveitamento da energia luminosa comparativamente às desenvolvidas em ambiente de
pleno sol.
A espécie C. prismatica possui plasticidade dos fotossistemas, em relação aos teores
de clorofila a, b e carotenoides, em resposta a incidência de energia luminosa disponível no
ambiente. Os maiores teores de clorofila a e b, observados no sub-bosque, indicam que a
espécie desenvolveu alterações fisiológicas para adaptar-se à condição de menor
disponibilidade luminosa. Os teores mais elevados de carotenoides em C. prismatica a pleno
sol durante o verão provavelmente ocorreram para fotoproteção dos fotossistemas.
Os efeitos observados pela sazonalidade foram mais pronunciados em árvores de C.
prismatica que estavam a pleno sol. Nesse ambiente as respostas da fluorescência da clorofila
a diferiram entre o inverno e as demais estações do ano. Em relação às árvores de C.
prismatica de sub-bosque observou-se que a emissão da fluorescência foi mais homogênea
durante as quatro estações analisadas, provavelmente, devido ao extrato arbóreo superior
fornecer um nível de proteção contra os efeitos climáticos, devido a seu sombreamento.

56
4.5. Referências Bibliográficas
ADAMS I. W.W., DEMMIG-ADAMS, B., Chlorophyll fluorescence as a tool to monitor plant response to the environment. In: PAPAGEORGIOU, G.C., GOVINDJEE ((Org.), Chlorophyll Fluorescence: A Signature of Photosynthesis, Advances in Photosynthesis and Respiration Series. Dordrecht: Springer, 2004. 820 p. ATROCH, E. M. A. C.; SOARES, A. M. ALVARENGA, A. A.; CASTRO, E. M. Crescimento, teor de clorofilas, distribuição de biomassa e características anatômicas de plantas jovens de Bauhinia forficata Link. submetidas a diferentes condições de sombreamento. Ciência e Agrotecnologia, v. 25, n. 4, p. 853-862, 2001. AVELAR, R. P.; PACHECO, F. V.; ALVARENGA, A. A.; PINTO, J. E. P. B.; FRANCESCHINI, L. A. Teores de pigmentos fotossintéticos de Piper aduncun cultivadas sob diferentes condições de radiação. Anais do Congresso Nacional de Botanica, 64, Belo Horizonte: SBB, 2013. BAKER, N. R.; ROSENQVIST, E. Applications of chlorophyll fluorescence can improve crop production strategies: an examination of future possibilities. Journal of Experimental Botany, v. 55, n. 403, p. 1607-1621, 2004. BERTAMINIA, M.; MUTHUCHELIANB, K.; RUBINIGGA, M.; ZORERA, R.; VELASCOA, R.; NEDUNCHEZHIANA, N. Low-night temperature increased the photoinhibition of photosynthesis in grapevine (Vitis vinifera L. cv Riesling) leaves. Environmental and Experimental Botany, v. 57, p. 25-31, 2006. BERTAMINI, M.; ZULINI, L.; MUTHUCHELIAN, K.; NEDUNCHEZHIAN, N. Low night temperature effects on photosynthetic performance on two gravepine genotypes. Biologia Plantarum, v.51, p.381-385, 2007. BOARDMAN, N. K. Comparative photosyntesis of sun and shade plants. Annual Review of Plant Physiology, v. 28, p. 355-377, 1977. BOLHÀR-NORDENKAMPF, H. R.; LONG, S. P., BAKER, N. R., et al. Chlorophyll fluorescence as probe of the photosynthetic competence of leaves in the field: a review of current instrument. Functional Ecology, v.3, p.497-514, 1989. BOYER, J. S., Plant productivity and environment. Science, v. 218, p. 443-448, 1982. BRESTIC, M.; ZIVCAK, M.; KALAJI, H. M.; CARPENTIER, R.; ALLAKHVERDIEV, S. I. Photosystem II thermostability in situ: environmentally induced acclimation and genotype-specifi creactions in Triticum aestivum. Plant Physiology and Biochemistry, v. 57, p. 93-105, 2012. CASTRO, E. M.; PINTO, J. E. B. P.; MELO, H. C.; SOARES, A. M.; ALVARENGA, A. A.; LIMA JÚNIOR, E. C. Aspectos anatômicos e fisiológicos de plantas de Guaco submetidas a fotoperíodos. Horticultura Brasileira, v. 23, n. 3, p. 846-850, 2005.

57
CHRISTEN, D.; SCHÖNMANN, S.; JERMINI, M.; STRASSER, R.J.; DÉFAGO, G. Characterization and early detection of grapevine (Vitis vinifera) stress responses to esca disease by in situ chlorophyll fluorescence and comparison with drought stress. Environmental and Experimental Botany, v. 60, p. 504-514, 2007. DAI, Y. J.; SHEN, Z. G.; LIU, Y.; WANG, L. L., HANNAWAY, D.; LU, H. F. Effects of shade treatments on the photosynthetic capacity, chlorophyll fluorescence and chlorophyll content of Tetrastigma hemsleyanum Diels and Gilg. Environmental and Experimental Botany, v. 65, p. 177–182, 2009. EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA - EMBRAPA. Serviço Nacional de Levantamento e Conservação de Solos. Levantamento de reconhecimento dos solos do Estado do Paraná. Londrina: EMBRAPA-SNLCS/SUDESUL /IAPAR, 1984. 2. v. (EMBRAPA-SNLCS. Boletim de Pesquisa, 27; IAPAR. Boletim Técnico, 16). EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA - EMBRAPA. Centro Nacional de Pesquisa de Solos. Sistema brasileiro de classificação de solos. Rio de Janeiro, Brasil, 1999. 412 p ENGEL, V. L.; POGGIANI, F. Estudo da concentração de clorofilas nas folhas e seu espectro de absorção de luz em função do sombreamento em mudas de quatro espécies florestais nativas. Revista Brasileira de Fisiologia Vegetal, v. 3, n. 1, p. 39-45, 1991. GEORGIEVA, K. Some mechanisms of damage and acclimation of the photosynthetic apparatus due to high temperature. Bulgarian Journal of Plant Physiology, v. 25, p. 89-99, 1999. GONÇALVES, J. F. C.; MARENCO, R. A.; VIEIRA, G. Concetration of Photosynthetic Pigments and Chlorophyll Flourescence of Mahogany and Tonka Bean under Two Light Environments. Revista Brasileira de Fisiologia Vegetal, v. 13, n. 2, p. 149-157, 2001. GONÇALVES, J. F. C.; SANTOS JUNIOR, U. M. Utilization of the chlorophyll a fluorescence technique as a tool for selecting tolerant species to environments of high irradiance. Brazilian Journal of Plant Physiology, v.17, p. 307-313, 2005. GONÇALVES, J. F. C.; SANTOS-JUNIOR, U. M.; NINA-JUNIOR, A. R.; CHEVREUIL, L. R. Energetic flux and performance index in copaiba (Copaifera multijuga Hayne) and mahogany (Swietenia macrophylla King) seedlings grown under two irradiance environments. Brazilian Journal of Plant Physiology, v. 19, p. 171-184, 2007. GUBERT FILHO, F. A. O Faxinal – estudo preliminar. Revista do direito Agrário e Meio Ambiente, v. 2, p. 32-40, ago. 1987. HUANG, Z. A.; JIANG, D. A.; YANG, Y; SUN, J. W.; JIN, S. H. Effects of nitrogen deficiency on gas exchange, chlorophyll fluorescence, and antioxidant enzymes in leaves of rice plants. Photosynthetica, v. 3, n. 42, p.357-364, 2004.

58
JOLIOT, P; JOLIOT, A. Cyclic electron transfer in plant leaf. Proceedings of the National Academy of Sciences of the United States of America, v. 99, n. 15, p.10209-10214, 2002. KRAMER, T; KOZLOWSKI, T. Physiology of woody plants. New York: Academic Press, 1979. 811p. KRAUSE, G. H; WEIS, E. Chlorophyll fluorescence and photosynthesis: The basics. Annual Review of Plant Physiology and Plant Molecular Biology, v. 42, p. 313-49, 1991. LAGE-PINTO, F; OLIVEIRA, J. G.; CUNHA, M.; SOUZA, C. M. M.; REZENDE, C. E.; AZEVEDO, R. A.; VITÓRIA, A. P. Chlorophyll a fluorescence and ultrastructural changes in chloroplast of water hyacinth as indicators of environmental stress, Environment and Experimental Botany, v. 64, p. 307-313, 2008. LEE, D. W.; OBERBAUER, F.; JOHNSON, P.; BASKARAN, K.; MANSOR, M.; MOHAMAD, H.; YAP, S. K. Effects of irradiance and spectral quality on leaf structure and function in seedlings of two southeast asian Hopea (Dipterocarpaceae) species. American Journal of Botany, v. 87, n. 4, p. 447-455, 2000. LICHTENTHALER, H. K. Chlorophylls and carotenoids: pigment photosynthetic biomembranes. Methods in Enzymology, v.148, p. 362-385, 1987. LICHTENTHALER, H. K.; BUSCHMANN, C.; DÖLL, M.; FIETZ, H. J.; BACH, T.; KOZEL, U.; MEIER, D.; RAHMSDORF, U. Photosynthetic activity, chloroplast ultrastructure, and leaf characteristics of high-light and low-light plants and of sun and shade leaves. Photosynthesis Research, v. 2, p. 115–141, 1981. LICHTENTHALER, H. K.; KUHN, G.; PRENZEL, U.; MEIER, D. Chlorophyll-protein levels and stacking degree of thylakoids in radish chloroplasts from high-light, low-light and bentazon-treated plants. Physiologia Plantarum, v. 56, p. 183-188, 1982a. LICHTENTHALER, H. K.; PRENZEL, U.; KUHN, G. Carotenoid composition of chlorophyll-carotenoid-proteins from radish chloroplasts. A Journal of Biosciences, v. 37, p. 10–12, 1982b. LICHTENTHALER, H. K.; WELLBURN, A. R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochemical Society Transactions, v. 11, p. 591-592, 1983. LICHTENTHALER, H. K.; MEIER, D.; BUSCHMANN, C. Development of chloroplasts at high and low light quanta fluence rates. Israel Journal of Botany, v. 33, p. 185–194, 1984. LICHTENTHALER, H. K.; RINDERLE, U. The role of chlorophyll fluorescence in the detection of stress conditions in plants. CRC Critical Reviews in Analytical Chemistry, v.19, Suppl. I, p. 29–85, 1988.

59
LICHTENTHALER, H. K. In Vivo Chlorophyll Fluorescence as a Tool for Stress Detection in Plants. In: LICHTENTHALER, H. K (Org.). Applications of chlorophyll fluorescence. Dordrecht: Kluwer, 1988. 262 p. LICHTENTHALER, H. K.; BABANI, F. In: PAPAGEORGIOU, G. C, GOVINDJEE (Org.). Chlorophyll fluorescence: a signature of photosynthesis. Dordrecht: Springer, 2004. 820 p. LIMA, A. L. S.; ZANELLA, F.; CASTRO, L. D. M. Crescimento de Hymenaea courbaril L. var. stilbocarpa (Hayne) Lee et Lang. e Enterolobium contortisiliquum (Vell.) Morong (Leguminosae) sob diferentes níveis de sombreamento. Acta Amazônica, Manaus, v. 40, n. 1, p. 43-48, 2010. LIMA JUNIOR, E. C.; ALVARENGA, A. A.; CASTRO, E. M. Physioanatomy traits of leaves in young plants of Cupania vernalis camb. subjected to different shading levels. Revista Árvore, v. 30, n. 1, p. 33-41, 2006. LORENZI, H. Árvores Brasileiras: manual de identificação e cultivo de plantas arbóreas nativas do Brasil. 2. v. Nova Odessa: Plantarum, 2014. 384p. MACHADO, E. C.; SCHMIDT, P. T.; MEDINA, C. L.; RIBEIRO, R. V. Respostas da fotossíntese de três espécies de citros a fotores ambientais. Pesquisa Agropecuária Brasileira, v. 40, p. 1161-1170, 2005. MATHIS, P.; PALLOTIN, G. Primary process of photosynthesis. In: HATCH, M.D.; BOARDMAN, N. K. (Org.). The biochmistry of plants. New York: Academic Press, 1981. p.97-161. MARTINS, J. R.; ALVARENGA, A. A.; CASTRO, E. M.; SILVA, A. P. O.; OLIVEIRA, C.; ALVES, E. Anatomia foliar de plantas de alfavaca-cravo cultivadas sob malhas coloridas. Ciência Rural, v. 39, n. 1, p. 82-87, 2009. MAXWELL, K.; JOHNSON, N. Chlorophyll Fluorescence – A Pratical Guide. Journal of Experimental Botany. v. 354, p. 659-668, 2000. MEIER, D.; LICHTENTHALER, H. K. Ultrastructural development of chloroplasts in radish seedlings grown at high and low light conditions and in the presence of the herbicide bentazon. Protoplasma, v. 107, p. 195–207. 1981. MEHTA, P.; JAJOO, A.; MATHUR, S.; BHARTI, S. Chlorophyll a fluorescence study revealing effects of high salt stress on Photosystem II in wheat leaves. Plant Physiology and Biochemistry, v. 48, p. 16-20, 2010. MITCHELL, R. L. Crop Growth and Culture. Ames: The Iowa State University Press, 1979, 349p. NIINEMETS, Ü.; KULL, O.; TENHUNEN, J. D. An analysis of light effects on foliar morphology, physiology and light interception in temperate deciduous woody species of contrasting shade tolerance. Tree Physiology, v. 18, p. 681– 696, 1998.

60
NOBEL, P. S. Physicochemical and environmental plant physiology. San Diego: Academic Press, 1999. 474p. OLIVEIRA, M. I.; CASTRO, E. M.; COSTA, L. C. B.; OLIVEIRA, C. Características biométricas, anatômicas e fisiológicas de Artemisia vulgaris L. cultivada sob telas coloridas. Revista Brasileira de Plantas Medicinais, v. 11, n. 1, p. 56-62, 2009. OSMOND, C. B. What is photoinhibition? Some insights from comparisons of shade and sun plants. In: BAKER, N. R.; BOWYER, J. R. (Org.), Photoinhibition of Photosynthesis, from the Molecular Mechanisms to the Field. Oxford: BIOS Scientific Publ., 471p. 1994. PARANÁ. Secretaria do Meio Ambiente e Recursos Hídricos, Instituto das Águas do Paraná. Atlas de Recursos hídricos do estado do Paraná. Curitiba: 2016. Disponível em: <http://www.aguasparana.pr.gov.br/arquivos/ File/mp07.pdf>. Acesso em: 12 mar. 2016a. PARANÁ. Secretaria do Meio Ambiente e Recursos Hídricos, Instituto de Terras, Cartografia e Geociências. Solos – Estado do Paraná. Curitiba: 2016. Disponível em: <http:// www.itcg.pr.gov.br/arquivos/File/Produtos_DGEO/Mapas_ITCG/PDF/Mapa_Solos.pdf>. Acesso em: 03 mar. 2016b. PORRA, R. J.; THOMPSON, W. A.; KRIEDEMANN, P. E. Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: verification of the concentration of chlorophyll standards by atomic absorption spectroscopy. Biochimica et Biophysica Acta, v. 975, p. 384-394, 1989. PTUSHENKO, V. V.; PTUSHENKO, E. A.; SAMOILOVA, O. P; TIKHONOV, A. N. Chlorophyll fluorescence in the leaves of Tradescantia species of different ecological groups: Induction events at different intensities of actinic light. Biosystems, v. 114, n. 2, p. 85–97, 2013. RUBAN, A. V.; HORTON, P. Regulation of non–photochemical quenching of chlorophyll fluorescence in plants. Australian Journal of Plant Physiology, v. 22, p. 221-230, 1995. SCHANSKER, G.; TÓTH, S. Z.; STRASSER, R. J. Methylviologen and dibromothymoquinone treatments of pea leaves reveal the role of photosystem I in the Chl a fluorescence rise OJIP. Biochimica et Biophysica Acta, v. 1706, p. 250– 261, 2005. SCHREIBER, U.; SCHLIWA, U.; BILGER, W. Continuous recording of photochemical and non-photochemical chlorophyll fluorescence quenching with a new type of modulation fluorometer. Photosynthesis Research, v.10, p. 51–62, 1986. SIMS, D. A.; GEBAUER, R.; PEARCY, R. W. Scaling sun and shade photosynthetic acclimation of Alocasia macrorrhiza to whole plant performance. II. Simulation of carbon balance and growth at different photon flux densities. Plant, Cell & Environment, v. 17, p. 889–900, 1994.

61
SOTO-PINTO, L.; PERFECTO, I.; CASTILLO-HERNANDEZ, J.; CABALLERO-NIETO, J. Shade effect on coffee production at the northern Tzeltal zone of the state of Chiapas, Mexico. Agriculture, Ecosystems & Environment, v. 80, p. 61–69, 2000. SOUZA, G. S.; CASTRO, E. M.; SOARES, A. M.; SANTOS, A. R.; ALVES, E. Teores de Pigmentos Fotossintéticos, Taxa de Fotossíntese e Estrutura de Cloroplastos de Plantas Jovens de Mikania laevigata Schultz Bip. ex Baker Cultivadas sob Malhas Coloridas. Ciências Agrárias. 32:1843-1854, 2011. STEPHAN, J.; SINOQUET, H.; DONÈS, N.; HADDAD, N.; TALHOUK, S.; LAURI, P. E. Light interception and partitioning between shoots in apple cultivars influenced by training. Tree Physiology, v. 28, p. 331–342, 2008. STIRBET, A.; GOVINDJEE. On the relation between the Kautsky effect (chlorophyll a fl uorescence induction) and Photosystem II: Basics and applications of the OJIP fluorescence transient. Journal of Photochemistry and Photobiology B: Biology, v.104, p. 236–257, 2011. STRASSER, R. J.; SRIVASATAVA, A.; GOVINDJEE. Polyphasic chlorophyll a fluorescence transient in plants and cyanobacteria. Photochemistry and Photobiology, v. 61, p. 32-34, 1995. STRASSER, B. J.; STRASSER, R. J. Measuring fast fluorescence transients to address environmental question: The JIP test. In: MATHIS, P. (Org.), Photosynthesis: From Light to Biosphere. Dordrecht: Kluwer Academic Publisher, v. 5, 1995. 5168p. STRASSER, R. J.; TSIMILLI-MICHAEL, M.; SRIVASTAVA, A. Analysis of the fluorescence transient. In: PAPAGEORGIOU, G. C.; GOVINDJEE, (Org.). Chlorophyll fluorescence: a signature of photosynthesis. Dordrecht: Springer; 2004. 820 p. STRASSER, R. J.; TSIMILLI-MICHAEL, M.; QIANG, S.; GOLTSEV, V. Simultaneous in vivo recording of prompt and delayed fluorescence and 820-nm reflection changes during drying and after rehydration of the resurrection plant Haberlea rhodopensis. Biochimica et Biophysica Acta, v. 1797, p. 1313-1326, 2010. TAIZ, L.; ZEIGER, E. Fisiologia vegetal. 5. ed. Porto Alegre: Artmed p.918, 2013. TATENO, M.; TANEDA, H. Photosynthetically versatile thin shade leaves: a paradox of irradiance-response curves. Photosynthetica, v. 45, p. 299–302, 2007. TÓTH, S. Z.; SCHANSKER, G.; STRASSER, R. J. A non-invasive assay of the plastoquinone pool redox state based on the OJIP-transient. Photosynthesis Research, v. 93, p. 193–203, 2007.

62
TSIMILLI-MICHAEL, M.; STRASSER, R. In vivo assessment of stress impact on plants' vitality: applications in detecting and evaluating the beneficial role of Mycorrhization on host plants. In: VARMA, A (Org.). Mycorrhiza: state of the art, genetics and molecular biology, ecofunction, biotechnology, eco-physiology, structure and systematic. Uttar Pradesh: Springer, p. 679-703, 2008. TSUKAYA, H. Leaf shape: genetic controls and environmental factors. The International Journal of Developmental Biology, v. 49, p. 547–555, 2005. VALLADARES, F.; NIINEMETS, Ü. Shade tolerance, a key plant feature of complex nature and consequences. Annual Review of Ecology, Evolution, and Systematics, v. 39, p. 237–257, 2008. VESTE, M.; BEN-GAL, A.; SHANI, U. Impact of thermal tress and high VPD on gas exchange and chlorophyll fluorescence of Citrus grandis under desert conditions. Acta Horticulturae, v. 531, p. 143-149, 2000. WANG, P.; SUN, X.; LI, C.; WEI, Z.; LIANG, D.; MA, F. Long-term exogenous application of melatonin delays drought-induced leaf senescence in appleq. Journal of Pineal Research, v. 54, p. 292-302, 2013. WHATLEY, J. M.; WHATLEY, F. R. A luz e a vida das plantas. São Paulo, EPU-EDUSP, 1982. 101p. YUSUF, M. A.; KUMAR, D.; RAJWANSHI, R.; STRASSER, R. J.; TSIMILLI-MICHAEL, M.; GOVINDJEE; SARIN, N. B. Overexpression of γ-tocopherol methyl transferase gene in transgenic Brassica juncea plants alleviates abiotic stress: Physiological and chlorophyll a fluorescence measurements. Biochimica et Biophysica Acta, v. 1797, p. 1428–1438, 2010. ZHANG, J. Z.; SHI, L.; SHI, A. P.; ZHANG, Q. X. Photosynthetic responses of four Hosta cultivars to shade treatments. Photosynthetica, v. 42, p. 213–218, 2004.

63
5. Morfometria e dinâmica de crescimento de Curitiba primastica (D. Legrand) Salywon
e Landrum
Resumo
A morfometria e a dinâmica de crescimento da Curitiba prismatica (D. Legrand)
Salywon e Landrum foram avaliadas em um fragmento de Floresta Ombrófila Mista em
sistema faxinal, no município de Rebouças-PR. Foram incluídas nas análises as árvores com
DAP maior ou igual a 10 cm, as quais foram realizadas de forma agrupada e por classes
diamétricas. Para a análise morfométrica, a coleta dos dados ocorreu em 2017, ocasião em que
foi medido o DAP, a altura total, a altura de inserção da copa e também os quatro raios de
projeção de copa. Com essas informações, foram determinadas as variáveis morfométricas de
diâmetro da copa, comprimento de copa, proporção de copa, grau de esbeltez, índice de
saliência, índice de abrangência e formal de copa. Para estudo da dinâmica de crescimento os
dados foram coletados no período de 2012 e 2017. Os indivíduos de C. prismatica
apresentaram uma altura total média de 8,98 m e a altura média de inserção da copa de 3,60
m, decorrendo que, em média, 60,05% da altura de cada árvore é constituída pela copa.
Devido à amplitude das medidas dos DAP e da altura total, ocorreu alta variabilidade do grau
de esbeltez, do índice de saliência e do índice de abrangência. A variável formal da copa
indica que a espécie tem predomínio de copas arredondadas. A espécie C. prismatica
apresentou alta correlação do DAP com o grau de esbeltez, e da altura total com o
comprimento da copa. Durante o períódo de 2012 a 2017 ingressaram 41,43 árvores ha-1 e
morreram 16,43 árvores ha-1. O DAP apresentou um incremento periódico anual de 0,29
cm.ano-1, em que os indivíduos que apresentaram maior taxa de crescimento foram os que
possuiam os menores DAP. O incremento períodico anual em área basal foi de 0,13 m2.ha-1,
sendo a classe diamétrica que apresentou maior incremento basal foi a de 15-20cm.
Palavras-chave: Sistema faxinal, floresta Ombrófila Mista, murta, biometria.

64
Morphometry and growth dynamics of Curitiba primastica (D. Legrand) Salywon
and Landrum
Abstract
Morphometry and growth dynamics of Curitiba prismatica (D. Legrand) Salywon and
Landrum were evaluated in a fragment of Mixed Ombrophilous Forest in faxinal system, in
Rebouças city, PR, Brazil. Trees with diameter at breast height (DBH) ≥ 10 cm were included
into analyses, grouped and per diameter classes. For morphometry, data were collected in
2017, were measured DBH, total height, canopy insertion height, and the four canopy
projection rays. Based on such information, the following morphometric traits were measured,
i.e. canopy diameter, canopy length, canopy proportion, degree of slenderness, salience index,
canopy coverage index, and crown formal. For growth dynamics data were collected between
2012 and 2017. C. prismatica trees presented 8.98 m mean total height and 3.60 m mean
canopy insertion height, so that, on average, 60.05% height of each tree consisted of canopy.
Due to the range of DBH and total height measurements, a high variability was detected for
the degree of slenderness, salience index and canopy coverage index. The trait crown formal
indicates that such a species has a predominance of rounded canopies. The species C.
prismatica presented a high correlation between DBH and the degree of slenderness, and
between total height and canopy length. From 2012 to 2107, 41.43 trees ha-1 entered and
16.43 trees ha-1 died. DBH had an increase of 0.29 cm year-1, so that trees showing a higher
growth rate were those with the lowest DBH. The annual increase in basal area was 0.13 m2
ha-1, and the 15-20cm diameter class presented the highest basal area increase.
Keywords: Faxinal system, Mixed Ombrophilous Forest, myrtle, biometrics.

65
5.1 Introdução
A fonte de energia das plantas é a luz solar, sendo posteriormente transformada em
energia química pela fotossíntese. Nas árvores, a copa é o órgão onde ocorre esse processo,
visto que nela se localizam as folhas, e em última análise, o sistema fotossintético. Dessa
forma, variáveis como diâmetro de copa e comprimento de copa apresentam relação direta
com o crescimento e produtividade de uma planta (NUTTO et al., 2001; TONINI e ARCO-
VERDE, 2005).
O crescimento de plantas lenhosas está intimamente relacionado com a sua forma e
estruturas. O conhecimento das variações na forma da estrutura é tão essencial para a
compreensão dos processos fisiológicos que regulam o crescimento das plantas como o
conhecimento das reações químicas envolvidas nesses processos. As características da copa
são importantes para estudos da taxa de crescimento da planta, tais como o aumento do
crescimento em diâmetro do fuste e produção de frutos e sementes (KOZLOWSKI e
PALLARDY, 1997).
O conhecimento da morfometria e da dinâmica de crescimento das formas das árvores
torna-se necessário para aperfeiçoar as intervenções silviculturais. A necessidade desse
conhecimento ganha maior importância principalmente quando se deseja utilizar espécies da
flora nativa em programas de reflorestamento com finalidades econômicas (DURLO et al.,
2004, ROMAN et al., 2009).
O estudo das características morfométricas de uma árvore e suas variáveis podem ser
eficazes para transmitir as suas relações interdimensionais. Pode-se assim, reconstituir o
espaço utilizado por cada árvore, pressupor o grau de competição de um povoamento e
permitir, também, mensurar a respeito da estabilidade, vitalidade e produtividade de cada
indivíduo (DURLO e DENARDI, 1998).
Ainda de acordo com Durlo e Denardi (1998), a altura total, altura do fuste, DAP e
diâmetro da copa são as variáveis básicas na dedução das demais variáveis morfométricas da
árvore. As principais variáveis morfométricas são proporção de copa, índice de saliência,
índice de abrangência, formal de copa e grau de esbeltez.

66
A razão entre o diâmetro da copa e a altura total da árvore é conhecida como índice de
abrangência. Essa relação tende a diminuir com o desenvolvimento em altura, visto que este
crescimento não é acompanhado na mesma proporção pelo aumento em diametro da copa
(TONINI e ARCOVERDE, 2005).
O grau de esbeltez é a relação entre a altura total e o DAP. Valores pequenos indicam
que as árvores estão crescendo mais em diâmetro do que em altura. Quando o grau de esbeltez
apresenta valor superior a 100 é um indicativo que a árvore está suscetível a danos físicos,
causados por ventos e tempestades, entre outros (ROVEDA et al., 2012).
O índice de saliência, que relaciona o diâmetro da copa e o DAP, demonstra quantas
vezes o diâmetro da copa é maior que o DAP da árvore. Essa relação pode então ser utilizada
como indicador de desbaste e pode-se determinar, a qualquer tempo, o espaço a ser liberado
ao redor de uma árvore selecionada, para que ela cresça sem concorrência. O formal de copa
expressa a relação entre o diâmetro da copa e o comprimento da copa, caracterizando a forma
geométrica que a copa possui. A proporção de copa é a relação entre o comprimento da copa e
a altura total da árvore (DURLO e DENARDI, 1998).
C. prismatica é uma espécie arbórea endêmica encontrada no sul do Brasil, conhecida
popularmente como murta. Ocorrendo no primeiro e segundo planaltos do Paraná e no
planalto norte de Santa Catarina. Mesmo não tendo dispersão geográfica muito ampla, essa
espécie coloniza facilmente áreas degradadas. Frequentemente, forma populações densas no
sub-bosque da FOM, em áreas com vegetação mais esparsa, e também em áreas campestres
(LORENZI, 2014; SALYMON e LANDRUM, 2007).
Dessa forma, esse trabalho teve como objetivo avaliar as características morfométricas
de C. prismatica e suas possíveis correlações, bem como a análise da dinâmica de seu
crescimento.

67
5.2. Material e Métodos
Caracterização e localização da área de estudo
Os dados desse trabalho foram coletados em um fragmento de FOM em sistema
faxinal, no município de Rebouças-PR, conforme descrito no capítulo 4. O faxinal é
denominado Faxinal Marmeleiro de Baixo, sendo constituído por uma área de 1.524 ha,
Figura 11.
Figura 11. Localização da área de estudo no munícipio de Rebouças, PR.
Fonte: Albuquerque (2015).
O histórico da área consta de mais de um século de uso da floresta para extrativismo
de produtos madeireiros (lenha, mourões, madeira em tora) e produtos não madeireiros, como
a erva-mate e sementes, principalmente o pinhão. A atividade pastoril de bovinos, equinos,
caprinos, suínos, entre outros, também é bastante difundida nas áreas de floresta
(ALBUQUERQUE, 2015).

68
Essa área encontra-se cadastrada, desde o ano de 1997, no Instituto Ambiental do
Paraná - IAP, como Área de Uso Regulamentado - ARESUR. Após ser reconhecida
formalmente como Unidade de Conservação, foi incluída no Cadastro Estadual de Unidades
de Conservação (CEUC).
Nesse local, em 2012, foram instaladas e inventariadas 50 unidades permanentes, com
o objetivo de avaliar o estado da FOM em Sistema Faxinal. Cada unidade permanente foi
instalada com comprimento de 50 m, traçado no sentido norte-sul, e largura de 10 m, no
sentido leste-oeste, formando um retângulo de 500 m² (ALBUQUERQUE, 2015).
Levantamento dos Dados
Para o levantamento das variáveis morfométricas foram medidos, em abril de 2017,
225 indivíduos de C. prismatica presentes nas unidades permanentes, que apresentaram DAP
igual ou superior a 10 cm. Os dados coletados foram o DAP, altura total, altura de inserção da
copa e também os quatro raios de projeção de copa nos sentidos norte, sul, leste e oeste. A
medida dos quatro raios foi utilizada para calcular o diâmetro da copa.
A identificação botânica e catalogação da espécie foram realizadas no Herbário da
Universidade Estadual do Centro-Oeste (HUCO), onde a exsicata foi depositada, recebendo o
número de tombo 6303.
As alturas das árvores foram medidas com o hipsómetro (Forestor Vertex III,
SoilControl). Para as medições dos DAP e dos raios de copa utilizou-se fita métrica. No caso
da copa foi feita a média dos quatro raios tomados no sentido norte, sul, leste e oeste, do
centro do tronco até o limite da copa.
Análise dos Dados
Para a análise morfométricas foram avaliadas as variáveis: altura total (H), altura de
inserção da copa (HIC), DAP, diâmetro de copa (DC), comprimento de copa (CC), proporção
de copa (PC), grau de esbeltez (GE), índice de saliência (IS), índice de abrangência (IA) e
formal de copa (FC), conforme Burger (1939). Os dados foram analisados tanto de forma
agrupada quanto estratificada por classes de diâmetro.
Os cálculos referentes ao crescimento foram obtidos por meio das Equações 1 a 4, de
acordo com Finger (1992). As taxas de mortalidade e ingresso foram calculadas de acordo
com as Equações 5 e 6, conforme Schaaf (2001).

69
= − (1)
(2)
= − (3)
(4)
% . 100 (5)
% . 100 (6)
Em que: d2012 (cm): DAP medido em 2012, d2017 (cm): DAP medido em 2017, g2012
(m2.ha-1): área basal da espécie medida em 2012, g2017 (m2.ha-1): área basal da espécie medida
em 2017, IPd (cm): incremento periódico diamétrico, IPAd (cm): incremento periódico anual
diamétrico, IPg (m2.ha-1): incremento periódico em área basal, IPAg (m2.ha-1): incremento
periódico anual em área basal, P (anos): intervalo entre as avaliações, I% (%): taxa de
ingresso, M% (%): taxa de Mortalidade, N2012: número de árvores medidas em 2012, Ni:
número de árvores que ingressaram no período e Nm: número de árvores mortas no período.
Para determinação das árvores ingressantes considerou-se as que não estavam
presentes na medição de 2012 e que atingiram o limite de inclusão de 10 cm de diâmetro em
2017. Para a contagem da mortalidade foram consideradas às árvores vivas existentes no
inventário de 2012 e que morreram no período de 2012 a 2017.
Para definição do número de classes diamétrica (nc) utilizou-se a fórmula de Sturges,
Equação 7:
nc = 1 + 3,3.log n (7)
Em que: n = número de indivíduos observados e nc = número de classes.
A determinação do intervalo de classes foi realizada utilizando-se o resultado da razão
entre a amplitude total e o número de classes (nc).

70
As análises estatísticas foram realizadas pela análise de variância, com o software
Statistica, versão 10 e os gráficos com o software Origin, versão 8.5. As diferenças nas
médias foram avaliadas pelo teste de Tukey, com nível de significância de 5%.

71
5.3. Resultados e Discussão
Características Morfométricas
Na área de estudo foram avaliados 225 indivíduos de C. prismatica, sendo os
resultados das variáveis morfométricas apresentados na Tabela 1. Estão dispostos os valores
mínimos, máximos, médios, mediana e coeficiente de variação para as variáveis DAP, H
(altura total), HIC (altura do ponto de inserção da copa), DC (diâmetro de copa), CC
(comprimento de copa), PC (proporção de copa), GE (grau de esbeltez), IS (índice de
saliência), IA (índice de abrangência) e FC (formal de copa).
Tabela 1. Caracterização das variáveis morfométricas das árvores de Curitiba prismatica,
avaliadas em 2017.
Variáveis Mínimo Máximo Médio Mediana CV (%)
DAP (cm) 10,35 46,41 15,91 14,42 32,76 H (m) 3,00 16,50 8,98 8,90 21,88 HIC (m) 1,32 8,50 3,60 3,20 36,90 DC (m) 1,95 12,10 4,80 4,65 33,16 CC (m) 0,70 13,80 5,38 5,30 34,16 PC (%) 8,79 87,89 59,71 61,82 24,27 GE 22,15 95,10 59,71 59,24 26,46 IS 11,32 71,91 31,32 30,30 32,83 IA 0,21 1,40 0,55 0,54 35,03 FC 0,34 2,20 0,98 0,90 43,87 DAP – diâmetro a altura do peito, H – altura total da árvore, HIC – altura de inserção da copa, DC – diâmetro da copa, CC - comprimento da copa, PC – proporção da copa, GE - grau de esbeltez, IS – índice de saliência, IA – índice de abrangência, FC – formal da copa.
Dentre todas as variáveis avaliadas, as que apresentaram os maiores coeficientes de
variação foram a IA (35,03%), HIC (36,90%) e FC (43,87%). Essa alta variabilidade
provavelmente ocorreu devido às diferentes idades e estágios sucessionais, bem como a
diferentes níveis de concorrência em que a C. prismatica estava submetida.
Pela análise da Tabela 1 observa-se que alta variação do DAP, em que o coeficiente de
variação obtido foi de 32,76%. Essa variabilidade possivelmente ocorreu devido a diferentes
idades das árvores inventariadas. O DAP médio foi de 15,91cm, com DAP mínimo e máximo
de 10,35 e 46,41 cm, respectivamente. A frequência de distribuição dos indivíduos da espécie
foi assimétrica em relação à média, apresentando uma mediana de 14,42 cm.

72
A altura total dos indivíduos de C. prismatica apresentaram uma média de 8,98 m e
mediana de 8,90 m, com coeficiente de variação de 21,88%. As alturas mínima e máxima foi
de 3,00 e 16,50 m, respectivamente.
Conforme estudo realizado por Albuquerque (2015) nessa mesma área, foram
encontradas 89 espécies, sendo a média de altura total de 9,76 m. A espécie C. prismatica
ocupava a décima posição das espécies com menor altura, com o extrato arbóreo superior
dominado por espécies como: Ocotea porosa, Araucaria angustifolia, Eugenia uniflora L.,
Campomanesia xanthocarpa, Casearia obliqua, Ocotea puberula, entre outras.
A altura de inserção da copa (HIC) apresentou valores entre 1,32 a 8,50 m, com média
de 3,60 m. A variabilidade entre os indivíduos foi alta, com coeficiente de variação de
36,90%. O valor da mediana foi de 3,20 m, demonstrando que a maioria dos indivíduos
possui HIC abaixo do valor médio. Conforme Durlo et al. (2004), por se tratar de uma floresta
inequiâneas, a grande amplitude de variação da HIC é esperado, visto que foram medidas
árvores em diferentes idades e estágios sucessionais.
Estes valores de HIC observados para C. prismatica são muito próximos aos
resultados obtidos por Hellmann (2016), que encontrou HIC para Eugenia pyriformis
Cambess valores variando de 1,10 a 7,30 m, com média de 3,58 m.
O diâmetro de copa (DC) dos indivíduos de C. prismatica apresentou uma variação,
de 1,95 a 12,10 m com média de 4,80 m e coeficiente de variação de 33,16%. Tendo como
base o valor médio observado de 4,80 m para a variável morfométrica DC, Tabela 1, pode-se
concluir que a espécie C. prismatica necessita de uma área de 20 m2 para seu
desenvolvimento. Com isso, a densidade máxima para o plantio de C. prismatica, para não ter
limitações em seu crescimento seria 500 ind. ha-1.
Homczinski (2017), em trabalho na Floresta Nacional de Irati-PR (Flona de Irati),
obteve para C. xanthocarpa os valores médios de 14 m para H, 7,2 m para HIC e DAP de
32,09 cm. Em relação à copa, a média para DC, CC e IA foram de 9,5 m 7,2 m e 0,66,
respectivamente.
A média para proporção de copa (PC) observada na C. prismatica (Tabela 1) foi de
59,71%, com mínimo de 8,79% e máximo de 87,90 %. O coeficiente de variação para essa
variável morformétrica foi de 24,27%. Segundo Roman et al. (2009), essa amplitude para a
PC é resultado das diferentes idades e graus de concorrência a que as árvores estão
submetidas.

73
Segundo Condé et al., (2013), espécies com elevada proporção de copa, como é o caso
de C. prismatica, apresentam a maior parte de sua estrutura em forma de copa, estando assim
altamente dependente da radiação solar para realizar sua manutenção e desenvolvimento.
Além disso, essa característica pode ser desejável para produtos não madeireiros, derivados de
uma copa exuberante (sementes, frutos, folhas, etc), porém, para produtos com fins
madeireiros, alta PC pode não ser adequado (TONINI et al., 2008; CONDÉ, 2013).
Os valores encontrados nesse trabalho para PC são semelhantes aos dados de Orellana
e Koehler (2008) obtido em um fragmento de FOM. Neste estudo citado foi obtida uma média
de PC igual a 55,2%, em que as árvores analisadas também tinham DAP igual ou superior a
10 cm.
O grau de esbeltez (GE) para a C. prismatica apresentou valor mínimo de 22,15 e
máximo de 95,10, com média de 59,71. A amplitude das medidas dos DAP e da altura total
resultou em uma alta variabilidade do GE, cujo coeficiente de variação foi de 26,46%.
Segundo Durlo e Denardi (1998), o GE caracteriza a instabilidade das árvores, e
quanto maior o grau de esbeltez, mais instável é a árvore. Lanzarin (2016) afirma que valores
maiores que 100, para GE podem ser resultados da falta de luz devido à competição entre as
árvores, ocasionando a inibição do crescimento em diâmetro. Indivíduos com estas
características apresentam troncos finos e altura elevada, o que pode levar a sua instabilidade,
principalmente no caso de fortes ventos. Dessa forma, os valores para o GE observados em C.
prismatica indicam, que na área de estudo, a maioria dos indivíduos da espécie não se
encontra suscetível a danos físicos, causados principalmente por ventos.
O índice de saliência (IS) observado indica que em média as copas de C. prismatica
são 31,32 vezes maiores que o DAP. A amplitude do valor de IS variou entre 11,37 e 71,91,
com coeficiente de variação de 32,83%. Essa grande variabilidade reflete a adaptação de C.
prismatica nos diferentes ambientes inventariados, em virtude, possivelmente, da
concorrência com as outras espécies arbóreas.
O IS também pode ser utilizado como indicador de produção para a espécies, sendo
que as espécies de maior crescimento em termos de volume de madeira apresentam valores
entre 15 e 20 para o IS, espécies com valores superiores a 20 tendem a um baixo crescimento.
Valores acima de 40 indicam baixa produção (TONINI e ARCO-VERDE, 2005). Assim,
conforme os valores observados na Tabela 1 para variável IS, a espécie C. prismatica pode ser
considerada uma espécie de baixo crescimento.

74
O índice de abrangência (IA) apresentou valores mínimo e máximo de 0,21 e 1,40,
respectivamente. A média para os indivíduos inventariados foi de 0,55, com um coeficiente de
variação de 35,67%. Verificou que o IA para a C. prismatica apresentou a tendência em
diminuir com o aumento da altura para todas das árvores. Conforme Tonini e Arco-verde
(2005), isso ocorre devido ao crescimento em altura ser maior que o crescimento do diâmetro
de copa.
Orellana e Koehler (2008) encontraram valores do IA em estudo com Ocotea
odorifera, variando entre 0,30 e 1,00, com média de 0,70. Durlo (2001) observou valores
entre 0,17 e 0,69 para o IA com média 0,33 para a Cabralea canjerana (Vell.) Mart.
O formal de copa (FC) apresentou a maior variabilidade para as variáveis
morfométricas analisadas. Os valores variaram entre 0,34 e 2,20, com média de 0,98 e
coeficiente de variação de 43,87%. De acordo com esses resultados encontrados para FC de
C. prismatica é possível indicar que na comunidade existem os três tipos de copas: copas
alongadas (FC abaixo de 1), copas arredondas (FC próxima a 1) e copas chatas (FC acima de
1). Porém, levando em consideração a média dos resultados, (1,03) a tendência dos indivíduos
de C. prismatica é apresentar copas arredondadas.
Conforme Durlo e Denardi (1998), pode-se determinar o número máximo de
indivíduos em uma área por meio das variáveis morfométricas. Utilizando-se o valor médio
de 31,32 do IS, Tabela 1, e considerando-se que este é invariável com a mudança nas
dimensões das árvores, para o desenvolvimento dee C. prismatica com 15,91 cm de DAP
(DAP médio observado), teria-se um DC de 4,98 m (15,91 cm x 31,32).
Esse valor do DC calculado está muito próximo do observado, que foi de 4,80 m.
Supondo-se também copas redondas, conforme observado para FC, poderia-se fazer um
plantio com aproximadamente 510 árvores.ha-1, sem que houvesse nenhuma concorrência
entre as árvores.

75
Análise de correlação entre as variáveis morfométricas
Segundo Tonini e Arco-Verde (2005), a medição das variáveis morfométricas
consome muito tempo. Em seu lugar podem ser utilizadas variáveis independentes de fácil
medição, quando existe boa correlação, para criação de modelos de estimativas. O DAP e a H
são variáveis de mais fácil medição, e é indicada para serem usadas nesses modelos. Dessa
forma, a correlação das variáveis DAP e a H com as demais variáveis morfométricas foram
analisadas e os resultados apresentados na Tabela 2.
Tabela 2. Matriz de correlação linear de Pearson entre o diâmetro à altura do peito - DAP e a
altura total - H e as variáveis morfométricas.
Variável DAP H HIC DC CC PC GE IS IA FC
DAP - 0,37** 0,15* 0,39** 0,26** 0,05 -0,60** -0,36** 0,10 -0,01
H 0,37** - 0,44** 0,27** 0,67** 0,07 0,45** -0,06 -0,40** -0,30**
DAP – diâmetro a altura do peito, H – altura total da árvore, HIC – altura de inserção da copa, DC – diâmetro da copa, CC - comprimento da copa, PC – proporção da copa, GE - grau de esbeltez, IS – índice de saliência, IA – índice de abrangência, FC – formal da copa. * Correlação é significativa ao nível de 0,05, ** Correlação é significativa ao nível de 0,01.
Os resultados da Tabela 2 demonstram que a variável com a melhor correlação com o
DAP foi o GE. Essa variável apresentou uma correlação significativa de -0,60, indicando que
o GE diminui à medida que as árvores têm seu DAP incrementado.
Para a variável H, a melhor correlação ocorreu com a variável morfométrica CC, em
que se observou uma correlação linear significa de 0,67. Esse resultado indica que a
velocidade do crescimento apical acompanha linearmente o aumento da altura total nas
árvores de C. prismatica.
As Figuras 12 e 13 apresentam os modelos ajustados e a distribuição gráfica dos
resíduos para as variáveis DAP x Ge, e H x CC. O modelo que correlaciona o DAP com a
variável GE embora tenha apresentado um R2 ajustado menor obteve um melhor ajuste, pois
os resíduos apresentaram menor variação e regularmente distribuídos (Figura 13A).

76
8 12 16 20 24 28 32 36 40 440,0
0,2
0,4
0,6
0,8
1,0
1,2
Gra
u de
Esb
elte
z
DAP (cm)
A GE = e (0,13459 - 0,04245.DAP)
R2 Ajustado = 0,4029
Syx = 0,0637
2 4 6 8 10 12 14 16 180
2
4
6
8
10
12
14CC = -0,4146 + 0,6457.H
R2 Ajustado = 0,4492
Syx = 0,48544
B
Com
prim
ento
da
Cop
a (m
)
Altura total (m) Figura 12. Modelos ajustados entre as variáveis morfométricas diâmetro a altura do peito –
DAP com grau de esbeltez – GE (A) e altura total – H com comprimento de copa
– CC (B) para Curitiba prismatica.
Verifica-se também que o GE, Figura 12A, seguiu uma tendência exponencial
negativa, com curva mais íngreme nas árvores de menores diâmetros, tendendo a estabilizar
com o aumento do DAP.
8 12 16 20 24 28 32 36 40 44
-4
-2
0
2
4 A
Res
íduo
s
DAP (cm)2 4 6 8 10 12 14 16 18
-4
-2
0
2
4
Res
íduo
s
B
Altura Total (m) Figura 13. Resíduos dos modelos ajustados entre as variáveis morfométricas diâmetro a
altura do peito – DAP com grau de esbeltez – GE (A) e altura total – H com
comprimento de copa – CC (B) para Curitiba prismatica.

77
A tendência de diminuição do grau de esbeltez com o aumento do DAP foi observada
por Curto (2015) para árvores de plantio superestocado de Araucaria angustifolia. Hellmann
(2016) também observou esta característica para espécie E. pyriformis, em um fragmento de
FOM, em Faxinal do Céu-PR. Lanzarin (2016) estudando as relações morfométricas da
Maclura tinctoria (L.) D. Don Ex. Steud novamente constatou essa mesma tendência de
exponencial negativa.
Homczinski (2017), na Floresta Nacional de Irati-PR (Flona de Irati), observou para a
espécie C. xanthocarpa altas correlação entre as variáveis DAP e H e as variáveis HIC, DC e
CC. A variável DAP obteve correlação de 0,59, 0,90 e 0,95 com as variáveis HIC, DC e CC,
respectivamente. Para a variável H também foi observada correlação positiva, sendo 0,88,
0,79 e 0,89 com as variáveis HIC, DC e CC, respectivamente. Esses resultados encontrados
por Homczinski (2017) demonstram diferenças na arquitetura de copa existente entre
diferentes espécies de Myrtaceas da FOM.
Os resultados apresentados na Tabela 2 também demonstram haver correlação
significativa entre o DAP e as variáveis H, DC, CC e IS. Dessa forma, apesar do coeficiente
de determinação ser relativamente baixo e não permitir uma inferência segura a respeito da
variação dessas variáveis é possível considerar esse comportamento como tendência, uma vez
que a correlação é significativa ao nível de 1%. Essa mesma afirmação pode ser estendida
para a variável H, que apresentou correlação baixa, mas significativa, ao nível de 1%, com as
variáveis DAP, HIC, DC, GE, IA e FC.
Características morfométricas por classes diamétricas
As frequências por distribuição diamétrica dos indivíduos de C. prismatica, bem como
de todas as espécies da área de estudo, estão representadas na Figura 14. As classes
diamétricas acima de 35 cm por apresentarem frequência inferior a um indivíduo de C.
prismatica foram suprimidas das análises estatísticas posteriores.

78
0
50
100
150
200
250
300
350
12,5 17,5 22,5 27,5 32,5 37,5 42,5 47,5 52,5 57,5 62,5 >65
Freq
uênc
ia (i
nd.h
a-1)
Centro de Classe Diamétrica (cm)
Todas Espécies Curitiba prismatica
Figura 14. Histograma das frequências de diâmetros à altura do peito de Curitiba prismatica
avaliadas em 2017, em Rebouças-PR.
De acordo com Figura 14, observa-se que a classe diamétrica de 10-15 cm é formada
por 308,57 ind.ha-1, incluindo todas as espécies encontradas na área de estudo. Desse total, a
C. prismatica, com 90,71 ind.ha-1, representa 29,40% de todas as árvores. Na classe
diamétrica de 15-20 cm a ocorrência de C. prismatica foi relativamente maior, representando
38,82% de todos os indivíduos inventariados. Nas classes diamétricas acima de 20 cm a
participação relativa de C. prismatica sofre gradativa redução, até não mais ocorrer nas
classes acima de 50 cm de DAP.
O maior DAP encontrado para a C. prismatica foi de 46,41 cm, indicando ser uma
característica da espécie o crescimento diamétrico limitado a essa faixa de valores. Embora a
espécie não esteja presente nas classes diamétricas acima de 50 cm, observa-se que a sua
distribuição diamétrica apresentou o característico formato de “J” invertido, típico da
estrutura diamétrica em florestas naturais.
As Figuras 15 e 16 apresentam as variáveis morfométricas observadas de C.
prismatica, divididas por classes diamétricas. Na Figura 15 estão representadas as variáveis
H, HIC, DC, CC e PC, enquanto na Figura 16 as variáveis GE, IS, IA e FC.

79
As variáveis H e HIC apresentaram um comportamento semelhante, aumentando à
medida que ocorreu incremento do DAP. Para as classes diamétricas de 10 a 25 cm não
ocorreram diferenças significativas. Para as classes diamétricas que engloba os diâmetros
acima de 25 cm observaram-se diferenças significativas nessas variáveis, conforme Figura
15A e 15B. Isso pode indicar que a C. prismatica está dividida em dois extratos arbóreos,
sendo as árvores com mais de 10 m pertencente ao primeiro extrato e as abaixo dessa medida,
compondo o segundo extrato.
c c bcab
a*
b bb
aba
cb b b
a
a a a aa
aa a a a
02468
101214 A
Altu
ra T
otal
(m)
01234567
Inse
rção
da
Cop
a (m
)B
0
2
4
6
8
10 C
Diâ
met
ro d
a C
opa
(m)
0
2
4
6
8
10
Com
prim
ento
Cop
a (m
)D
12,5 17,5 22,5 27,5 32,5 Centro de Classe Diamétrica (cm)
010203040506070 E
12,5 17,5 22,5 27,5 32,5
Prop
orçã
o da
Cop
a (%
)
Centro de Classe Diamétrica (cm) * Médias seguidas pela mesma letra não se diferenciam entre si pelo teste de Tukey a 5% de probabilidade.
Figura 15. Variáveis morfométricas: altura total (A), altura de inserção da copa (B), diâmetro
da copa (C), comprimento da copa (D) e proporção copa (E), da Curitiba
prismatica por classes diamétricas.
De acordo com a Figura 15C, a classe diamétrica de 30-35 cm se difere
significativamente das demais classes. Essa classe é a que possui maior H e conforme
observado à campo, elas fazem parte do dossel da floresta. Nessa condição de maior
luminosidade a copa se expande, aumentado em diâmetro. Conforme Wadsworth (2000), as
árvores do dossel superior têm copas horizontais, pouco densas, e as do estrato inferior têm
copas verticais e profundas, devido à busca por recursos luminosos. Após as árvores atingirem
o dossel as copas se expandem para aumentarem a área das folhas em contato com a energia
luminosa.

80
As variáveis morfométricas CC e PC, Figura 15D e 15E, não diferiram em nenhuma
das classes diamétricas analisadas. Isso indica que essas variáveis se mantem constante com o
aumento do DAP, sendo provavelmente, essa uma característica da espécie e pouco afetada
com as condições do ambiente, como luminosidade e competição.
As classes diamétricas, Figura 16A, apresentaram valores decrescentes para o GE,
sendo que as classes de 10-15 cm e de 15-20 cm diferiram significativamente das demais. As
classes acima de 20 cm não se diferenciaram, indicando estabilidade entre o crescimento em
altura e o DAP.
As árvores da classe diamétrica 10-15 cm possuem o maior valor para GE. Conforme
Roveda et al. (2012), isso indica que essas árvores são as mais suscetíveis a danos físicos,
como vento e tempestades. Esses indivíduos estão buscando primeiramente alcançarem o
dossel, em busca de mais energia luminosa, e após isso, aumentarem o DAP.
a*
bc c c
aab
b b
ab
aa a a
a
a a aa
a
0,0
0,2
0,4
0,6
0,8
1,0A
Gra
u de
Esb
elte
z
0
5
10
15
20
25
30
35
40
Índi
ce d
e Sa
liênc
ia
B
0,0
0,2
0,4
0,6
0,8
1,0C
12,5 17,5 22,5 27,5 32,5
Índi
ce d
e A
bran
gênc
ia
Centro de Classe Diamétrica (cm)
0,0
0,5
1,0
1,5Fo
rmal
da
Cop
a
12,5 17,5 22,5 27,5 32,5 Centro de Classe Diamétrica (cm)
D
* Médias seguidas pela mesma letra não se diferenciam entre si pelo teste de Tuke a 5% de probabilidade.
Figura 16. Variáveis morfométricas: (A) grau de esbeltez, (B) índice de saliência, (C) índice
de abrangência e (D) formal da copa, da Curitiba prismatica por classes
diamétricas.

81
Para as variáveis IS, IA e FC, Figura 16B, 16C e 16D respectivamente, não foram
observadas diferenças significativas entre as classes diamétricas. Além da grande
variabilidade dos dados, que contribuiu para não ter ocorrido diferenças significativa, isso
pode demonstrar que essas são também características da própria espécie e são pouco
alteradas pelas características de competição da floresta.
Os valores apresentado pela variável morfométrica FC, devido a grande variabilidde,
não apresentou diferenças significativas entre as classes diamétricas. Dessa forma, conforme
Roman et al. (2009), pode-se classificar a forma das copas de C. prismatica, independente do
diâmetro da árvore, como tendo majoritariamente a forma arredondada.
Crescimento, ingresso e mortalidade
Utilizando os dados de crescimento diamétrico, foi construída a matriz de progressão
(Tabela 3) para avaliar a dinâmica de crescimento de C. prismatica no período de 2012 a
2017. A matriz de progressão apresenta, por classes diamétricas, o ingresso, a mortalidade e a
progressão das árvores de árvores de C. prismatica, sendo representado na horizontal o
número de indivíduos por classe diamétrica observados em 2017, e na vertical o número de
indivíduos observados em 2012, conforme proposto por Sanquetta et al. (1995).
Tabela 3. Matriz de progressão por classe de diâmetro (cm) das árvores de Curitiba prismatica
avaliadas em 2012 e 2017. Classes de
Diâmetro (cm) 10-15 15-20 20-25 25-30 30-35 35-40 40-45 45-50 Mortas Total
Ingresso 58 58
10-15 127 33 15 175
15-20 33 6 5 44
20-25 13 3 3 19
25-30 3 2 5
30-35 2 1 3
35-40 0 0
40-45 1 1 2
45-50 0
Total 185 66 19 6 4 1 1 1 23 306

82
O total de árvores vivas avaliadas em 2012 e 2017 foram 248 e 283, respectivamente.
De acordo com a Tabela 3, observa-se que durante o período de avaliação ingressaram na área
amostrada 58 árvores (41,43 árvores.ha-1), sendo todas com DAP abaixo de 15cm. A taxa de
ingresso no período e a taxa média anual de ingresso foram de 23,39% e 4,67%,
respectivamente.
A mortalidade registrada foi de 23 árvores (16,43 árvores.ha-1), representando uma
taxa de mortalidade no período de 9,27% e anual de 1,85%. Computados os valores da
mortalidade e do ingresso, a espécie apresentou um acréscimo de 14,11% no número de
indivíduos entre 2012 e 2017. Demonstrando que houve um balanço positivo de 25
árvores.ha-1 para a espécie, gerando um ganho líquido de 35 árvores.
Essas taxas de ingresso e mortalidade indicam que a floresta esta em um processo de
evolução, em que o ingresso é mais abundante provavelmente em virtude dessa área ter
sofrido intervenções recentes. Cessando a ocorrência de novas perturbações, a espécie e a
floresta como um todo tenderá ao equilíbrio entre ingressos e mortalidade.
A maioria das mortes (65,22%) ocorreu em árvores presentes na classe diamétrica de
10-15 cm. A mortalidade nas classes superiores foi baixa, não tendo ocorrido mortes nas
classes diamétricas acima de 25 cm. Esses resultados estão de acordo com Longhi (1997),
segundo o qual a maior mortalidade, em florestas naturais, ocorre nas classes diamétricas com
maior frequência de indivíduos.
Homczinski (2017), utilizando dados de 2002 a 2014 e limite de inclusão de 10 cm de
DAP, na Floresta Nacional de Irati-PR (Flona de Irati), obteve para a espécie C. xanthocarpa
uma taxa de 1,53% de ingressos e uma mortalidade de 0,63%. Schaaf et al. (2005), utilizando
dados de 1979 a 2000 e limite de inclusão de 20 cm, em São João do Triunfo-PR, obtiveram
também para a C. xanthocarpa uma taxa de 57,89 e 7,89% para ingressos e mortalidade,
respectivamente, sendo considerada pelos pesquisadores uma das espécies que sofreu menos
competição.
Conforme Tabela 3, observa-se que as duas classes diamétricas inferiores foram as
mais numerosas em ambas as avaliações, abrigando 88,31% e 88,69% de todas as árvores em
2012 e 2017, respectivamente. Na classe diamétrica de 10-15 cm concentrava-se 70,56% dos
indivíduos, no ano de 2012. Em 2017 essa porcentagem diminuiu para 65,37%, embora em
valore absoluto tenha ocorrido aumento de 11 árvores.

83
A classe diamétrica de 15-20 cm continha, em 2012, 17,74% dos indivíduos. Na
avaliação ocorrida em 2017 essa classe teve um incremento, passando a representar 23,32%
do total de árvores vivas. As demais classes diamétricas tiveram pouca variação, juntas
representam 11,69% e 11,31% das árvores avaliadas em 2012 e 2017, respectivamente.
A espécie demonstrou descontinuidade na distribuição diamétrica em classes
sucessivas, em que pouco ou nenhum indivíduo, ingressaram em classes diamétricas
superiores. Isso pode indicar que alguns locais sofreram alterações na sua estrutura por
interferência antrópica.
A análise do incremento diamétrico de C. prismatica indica uma grande variabilidade
no padrão de crescimento da espécie. Durante o período de avaliação, a espécie apresentou a
média de 0,29 cm.ano-1 de incremento periódico anual diamétrico (IPAd) variando de 0,01 a
0,93 cm.ano-1, com coeficiente de variação de 54,22%.
Quando avaliada a evolução do diâmetro médio para todas as árvores da espécie,
observa-se uma taxa de incremento no período de 9,92% (14,48 cm, em 2012, para 15,91 cm,
em 2017). Os indivíduos que apresentaram maiores taxas de incremento diamétrico foram os
que possuem os menores DAP.
De forma geral, observou-se a tendência de diminuição do incremento diamétrico
relativo à medida que o DAP aumentava seus valores, conforme Figura 17. As classes
diamétricas acima de 35 cm, por terem frequência de apenas um indivíduo, foram suprimidas
das análises estatísticas posteriores.
Para as árvores da classe de 10-15 cm foi observado um incremento médio de 12,19%
em relação a 2012, diferindo das demais classes. Para a classe de 15-20 cm o incremento foi
de 9,53%. As classes acima de 20 cm apresentaram uma taxa de incremento variando de
4,27% a 4,96% e não diferiram entre si. A baixa frequência de indivíduos nessas classes
ocasionou um alto erro padrão.
Ebling (2012), observou em fragmento da FOM avaliado no período de 2000 a 2009,
pertencente à Flona de São Francisco de Paula-RS, a média de 0,13 cm.ano-1 de IPAd, com
variação de 0,007 a 0,322 cm.ano-1. As espécies pertencentes à família Myrtaceae obtiveram
um incremento de 0,06 cm.ano-1, sendo a famílias com menor incremento diamétrico e com
maior densidade e frequência relativa.

84
a*
b
c c c
0
2
4
6
8
10
12
14
12,5 17,5 22,5 27,5 32,5
Incr
emen
to D
iam
étric
o R
elat
ivo
(%)
Centro de Classe Diamétrica (cm)
* Médias seguidas pela mesma letra não se diferenciam entre si pelo teste de Tukey a 5% de probabilidade.
Figura 17. Incremento periódico diamétrico relativo no período de 2012 a 2017, por classe
diamétrica.
Em estudo realizado por Cubas et al. (2016), em remanescente da FOM em Três
Barras - SC, encontraram IPAd variando entre 0,09 a 0,43 cm.ano-1. Naquele estudo, espécies
características da FOM, como Ilex paraguariensis, Araucaria angustifolia, Ocotea puberula e
Ocotea porosa, apresentaram incrementos anuais de 0,09, 0,29, 0,32 e 0,43 cm.ano-1,
respectivamente. Para as espécies da família Myrtaceae pertencentes ao gênero Eugenia,
mesmo gênero que a C. prismatica pertencia antes da atual classificação, foram observados
IPAd variando de 0,27 a 0,33 cm.ano-1.
Homczinski (2017), utilizando dados de 2002 a 2014 e limite de inclusão de 10 cm de
DAP, na Floresta Nacional de Irati-PR (Flona de Irati), obteve para a espécie C. xanthocarpa
um IPAd médio de 0,29 cm.ano-1, variando entre 0,01 a 0,72 cm.ano-1.
Em relação à área basal, a espécie C. prismatica apresentou em 2012 um total de 3,29
m2.ha-1. Na avaliação de 2017 observou-se um incremento periódico em área basal (IPg) de
0,63 m2.ha-1, totalizando 3,92 m2.ha-1. O incremento periódico anual em área basal (IPAg) foi
de 0,13 m2.ha-1.
Avaliando o comportamento da área basal por classes diamétrica, Figura 18, observa-
se incremento em todas as classes, com exceção da classe de 40-45 cm. A classe diamétrica
de 10-15 cm apresentou uma taxa de incremento de 9,57%, em relação a 2012. Nessa classe
ocorreu o ingresso de 58 árvores e a morte de 23 indivíduos no período de 2012 a 2017,
resultando assim, em um incremento basal de 0,14 m2.ha-1.

85
A classe de 15-20 cm foi a que apresentou o maior incremento, tanto absoluto (0,34
m2.h-1) quanto relativo (45,99%) . Esse alto incremento ocorreu devido a dois fatores, menor
quantidade de árvores mortas e alto ingresso na classe. De acordo com a Tabela 3, nessa
classe ocorreu a morte de cinco árvores no período, enquanto que outras 33 arvores se
deslocaram da classe diamétrica anterior. Com isso, a área basal passou de 0,73 m2.ha-1, em
2012, para 1,04 m2.ha-1 , em 2017.
Na classe de 20-25 cm, em virtude de ter ocorrido igual quantidade de mortes e
ingresso de classes inferiores, o IPg permaneceu praticamente inalterado. A taxa de
incremento foi de apenas 0,54%.
A classe de 25-30 e a de 30-35 cm apresentaram taxa de incremento de 13,65 e
23,96%, respectivamente. Esse aumento pode ser atribuído tanto ao crescimento das árvores
dentro da classe, quanto à entrada de árvores advindas da classe anterior.
As classes de 35-40 e a de 45-50 cm não continham indivíduos em 2012. Com o
crescimento houve o deslocamento de uma árvore para cada classe, passando a possuírem
área basal de 0,08 e 0,12 m2.ha-1, respectivamente.
De todas as classes avaliadas, somente a classe de 40-45 cm apresentou decremento na
área basal. Na avaliação de 2012 existiam duas árvores nessa classe, porém uma delas, devido
ao crescimento, passou a integrar a classe seguinte. Dessa forma essa classe apresentou uma
taxa de incremento de -52,89%.
0,0
0,5
1,0
1,5
12,5 17,5 22,5 27,5 32,5 37,5 42,5 47,5
Áre
a Ba
sal (
m2 .h
-1)
Centro de Classe Diamétrica (cm)
2012 2017
Figura 18. Variação da área basal, por classes diamétricas, entre 2012 e 2017.

86
Homczinski (2017), utilizando dados de 2002 a 2014 e limite de inclusão de 10 cm de
DAP, na Floresta Nacional de Irati-PR (Flona de Irati), obteve para a espécie C. xanthocarpa
um IPAg médio de 0,007 m2.ha-1, com uma taxa de incremento anual média de 3,45%.
Schaaf et al. (2005), utilizando dados de 1979 a 2000 e limite de inclusão de 20 cm,
em São João do Triunfo-PR, obtiveram para a C. xanthocarpa uma média de 0,20 cm.ano-1,
variando entre -0,08 a 0,74 cm.ano-1.

87
5.4. Conclusão
A grande variabilidade apresentada pelas variáveis biométricas e morfométricas
estudadas, indica que C. prismatica possui plasticidade adaptativa. Isso foi observado,
principalmente em relação à copa, em virtude da ampla variação dos raios da copa
observados.
Essas características observadas pelas variáveis morfométricas, principalmente em
relação a sua copa, podem ser utilizadas para desenvolver planos de manejo visando a
extratação de PFNM, como os OEs.
Pelos dados observados na área de estudo, as variáveis altura total, altura de inserção
da copa, diâmetro da copa e formal da copa podem indicar haver concorrência intra e/ou
interespécie, aumentando ou diminuindo seus valores em virtude dessa disputa por recursos.
A frequência da distribuição diamétrica demonstra que poucos indivíduos estão
migrando para as classes diamétricas superiores. Isso indica que a espécie pode estar sofrendo
alterações na sua estrutura por interferência antrópica.
O aumento de área basal, juntamente com a taxa de ingresso maior que a taxa de
mortes, demonstra que a espécie encontra-se em processo contínuo de sucessão.

88
5.5. Referências Bibliográficas
ALBUQUERQUE, J. M. Fatores ecológicos e ocorrência de espécies na floresta ombrófila mista, em sistema faxinal. 2015. 93f. Tese (Doutorado em Engenharia Florestal) - Universidade Federal do Paraná, Curitiba, 2015. BURGER, H. Baumkrone und zuwachs in zwei hiebsreifen fichtenbeständen. Mitteilungen der Schweizerischen Anstalt für das Forstliche Versuchswesen, v. 21, p.147-176, 1939. CONDÉ, T. M.; LIMA, M. L. M.; LIMA NETO, E. M.; TONINI, H. Morphometric of four species in agroforestry systems in the municipality of Porto Velho, Rondônia. Revista Agroambiente On-line, v. 7, n. 1, p. 18-27, 2013. CUBAS, R.; WATZLAWICK, L. F.; FIGUEIREDO FILHO, A. Incremento, ingresso, mortalidade em um remanescente de Floresta Ombrófila Mista em Três Barras - SC. Ciência Florestal, v. 26, n.3 p. 889-900, 2016. CURTO, R. A. Avaliação do Crescimento e Potencial de Manejo em plantio superestocado de Araucaria angustifolia (Bert.) O. Ktze. 2015. 250 f. Tese (Doutorado em Engenharia Florestal). Universidade Federal do Paraná, 2015. DURLO, M. A.; DENARDI, L. Morfometria de Cabralea canjerana, em mata secundária nativa do Rio Grande do Sul. Ciência Florestal, v. 8, n. 1, p. 55-66, 1998. DURLO, M. A.; SUTILI, F. J.; DENARDI, L. Modelagem da copa de Cedrela fissilis Vellozo. Ciência Florestal, v. 14, n. 2, p. 79-89, 2004. EBLING, A. A. Dinâmica e projeção diamétrica em remanescente de Floresta Ombrófila Mista na Flona de São Francisco de Paula, RS. 2012. 126 f. Dissertação (Mestrado em Engenharia Florestal) - Universidade Estadual do Centro - Oeste, Irati, 2012. FINGER, C. A. G. Fundamentos de biometria florestal. Santa Maria: UFSM – CEPEF-FATEC, 1992. 269 p. HELLMANN, S. S. Ecofisiologia e morfometria como ferramenta para o manejo sustentável de Eugenia pyriformis. 2016. 90 f. Dissertação (Mestrado em Agronomia) - Universidade Estadual do Centro-Oeste, Guarapuava, 2016. HOMCZINSKI, I. Distribuição espacial, dinâmica e biometria de campomanesia xanthocarpa (mart.) O. Berg. Em floresta ombrófila mista, 2017. 88 F. Dissertação (Mestrado em Engenharia Florestal) - Universidade Estadual do Centro - Oeste, Irati, 2017. KOZLOWSKI, T. T.; PALLARDY, S. G. Growth control in woody plants. 2 ed. San Diego: Academic Press, 1997, 411p.

89
LANZARIN, K. Relações Morfométricas e Potencial de Manejo de Maclura tinctoria (L.) D. Don Ex. Steud em Formações Secundárias do Noroeste do rio Grande do Sul. 2016. 77 f. Dissertação (Mestrado). Universidade Federal de Santa Maria, 2016. LONGHI, S. J. Agrupamento e análise fitossociológica em comunidades florestais na subbacia hidrográfica do Rio Passo Fundo - RS. 1997. 198 p. Tese (Doutorado em Engenharia Florestal) – Universidade Federal do Paraná, Curitiba, PR. LORENZI, H. Árvores Brasileiras: manual de identificação e cultivo de plantas arbóreas nativas do Brasil. v.2. Nova Odessa: Plantarum, 2014. 384 p. NUTTO, L.; TONINI, H.; BORSOI, G.A.; MOSCOVICH, F.A.; SPATHELF, P. Utilização dos parâmetros da copa para avaliar o espaço vital em povoamentos de Pinus elliottii Engelm. Boletim de Pesquisa Florestal, v.42, p.110-122, 2001. ORELLANA, E.; KOEHLER, A.B. Relações morfométricas de Ocotea odorifera (Vell.) Rohwer. Revista Acadêmica Ciências Agrárias e Ambientais, v. 6, n. 2, p. 229-237, 2008. ROMAN, M.; BRESSAN, D. A.; DURLO, M. A. Variáveis morfométricas e relações interdimensionais para Cordia trichotoma (Vell.) Arráb. Ex Steud. Ciência Florestal, v. 19, n. 4, p. 473-480, 2009. ROVEDA, M. ; DALGALLO, B. ; DIAS, A. N. ; FIGUEIREDO FILHO, A. ; MULLER, C. S. . Morfometria de Araucaria Angustifolia (Bertol.) Kuntze na Floresta Nacional de São Francisco de Paula-RS. In: Congresso Florestal Paranaense, 4., 2012, Curitiba. Anais eletrônicos... Curitiba: CIETEP, 2012. Disponível em: <http://congressoflorestalpr.sistemasra pidos.com.br>. Acesso em: 03 jul. 2015. SALYWON, A. M.; LANDRUM, L. R. Curitiba (Myrtaceae): A new genus from the planalto of southern Brazil. Brittonia, v. 59, n. 4, p. 301-307, 2007. SANQUETTA, C. R.; ÂNGELO, H.; DOADI, A. B.; MENDES, J. B.. Predição da distribuição diamétrica, mortalidade e recrutamento de floresta natural com matriz Markoviana de potência. Floresta, v. 24, n. 1-2, p. 23-26, 1995. SCHAAF, L. B. Florística, estrutura e dinâmica no período 1979-2000 de uma Floresta Ombrófila Mista localizada no Sul do Paraná. 2001. 131 f. Dissertação (Mestrado em Engenharia Florestal) – Universidade Federal do Parana, Curitiba – PR, 2001. STEPKA, T. F. Modelagem da dinâmica e prognose da estrutura diamétrica de uma Floresta Ombrófila Mista por meio de matriz de transição e razão de movimentação. 2008. 138 f. Dissertação (Mestrado em ciências florestais) – Universidade Estadual do Centro Oeste, Irati – PR, 2008. TONINI, H.; ARCO-VERDE, M. F. Morfologia da copa para avaliar o espaço vital de quatro espécies nativas da Amazônia. Pesquisa Agropecuária Brasileira, v. 40, n. 7, p. 633-638, 2005.

90
TONINI, H.; KAMINSKI, P. E.; COSTA, P. Relação da produção de sementes de castanha-do-brasil com características morfométricas da copa e índice de competição. Pesquisa Agropecuária Brasileira, v. 43, n. 11, p. 1509-1516, 2008. WADSWORTH, F. H. Producción florestal para America Tropical. Washington: USDA, 2000. 602p.

91
6. Composição química e rendimento de óleos essenciais de Curitiba prismatica (D.
Legrand) Salywon e Landrum sob efeito de sazonalidade climática e diferentes
irradiâncias
Resumo
Os óleos essenciais (OE) são compostos sintetizados por rotas biossintéticas do
metabolismo especializado de vegetais, sendo os seus constituintes pertencentes à classe
química dos terpenos. Neste trabalho foi avaliada a composição química e o rendimento de
OE extraído por hidrodestilação de folhas frescas de C. prismatica. O material foi coletado
avaliando-se dois fatores: efeitos da sazonalidade e de diferentes irradiâncias incididas em C.
prismatica. O efeito da sazonalidade foi analisado pelas amostras de OEs extraídos na
primavera e no inverno, enquanto que os efeitos da irradiância foram avaliados por meio de
amostras coletadas em ambientes de sub-bosque e a pleno sol. Observou-se que houve
variações significativas no rendimento entre as estações, sendo o rendimento das amostras do
inverno 15,85% inferiores às amostras da primavera. Em relação aos ambientes, o rendimento
a pleno sol (9,73 mL kg-1 folhas frescas) foi significativamente superior às amostras do
ambiente sombreado (7,09 mL kg-1 folhas frescas). Foram identificados 54 compostos
químicos distribuídos entre as estações primavera e inverno. Os resultados mostraram
diferenças estatísticas significativas nos teores e compostos dos OEs. No inverno, em
ambiente de sub-bosque, os compostos majoritários foram γ-gurjuneno (14,78%), elixeno
(10,95%) e δ-cadineno (9,14%), enquanto que no ambiente a pleno sol, os compostos com
maiores teores foram o β-cariofileno (15,24%), γ-elemeno (14,45%) e elixeno (10,95%). Na
primavera, os principais compostos encontrados em ambiente a pleno sol foi o cariofileno
(22,01%), careno (21,18%) e o 1,5-dimetil-1,5-cyclooctadieno (10,39%), enquanto que no
sub-bosque foi o tau-cadinol (22,50%), cadina-1(2),4-dieno (16,42%) e o eremophileno
(10,26%). Utilizando as técnicas multivariadas de análise de agrupamento e analise de
componentes principais foram identificados quatro grupos distintos de OEs. Diante dos
resultados encontrados, evidenciaram-se os efeitos da sazonalidade (primavera e inverno) e do
ambiente (pleno sol e sombreamento) na alteração do teor e da composição química dos OE
das folhas de C. prismatica.
Palavras-chave: Myrtaceae, efeitos abióticos, plasticidade, terpenos.

92
Chemical composition and yield of essential oils of Prismatic Curitiba (D.
Legrand) Salywon and Landrum under effects of climate seasonality and different
irradiances
Abstract
Essential oils (EO) are compounds synthesized by biosynthetic routes of the
specialized metabolism of vegetables, and its constituents belong to the chemical class of
terpenes. In this work, the chemical composition and yield of EO extracted by
hydrodistillation of fresh leaves of C. prismatica were evaluated. The material was collected
by evaluating two factors: effects of seasonality and different irradiances on C. prismatica.
The effect of seasonality was analyzed by EO samples extracted in spring and winter, while
the effects of irradiance were evaluated by means of samples collected under understory
environments and in full sun. It was observed that there were significant variations in the
yield between seasons, with the winter’s samples yield being 15.85% lower than of the
spring’s samples yield. Regarding the environments, the yield at full sun (9.73 mL.Kg-1 fresh
leaves) was significantly higher than in shaded environment (7.09 mL.Kg-1 fresh leaves). It
was identified 54 chemical compounds distributed between spring and winter seasons. The
resulted showed significant differences on content and in composition of EOs. In the winter,
in the understory environment, the main compounds was γ-gurjunene (14.78%), elixene
(10.95%) and δ-cadinene (9.14%), while in full sun environment, the compounds with
hightest content was β-caryophyllene (15.24%), γ-elemene (14.45%) and elixene (10.95%). In
the spring, the main compounds found in the full sun environment was caryophyllene
(22.01%), carene (21.18%) and 1,5-dimethyl-1,5-cyclooctadiene (10.39%), while in the
understory was tau-cadinol (22.50%), cadina-1(2),4-diene (16.42%) and eremophilene (
10.26%). Using multivariate cluster analysis techniques and principal component analyzes
was identified four distinct groups of EOs. Considering the results found, the effects of
seasonality (spring and winter) and of the environment (full sun and shading) on the alteration
of the content and chemical composition of the C. prismatica leaves EO was evidenced.
Keywords: Myrtaceae, abiotic effects, plasticity, terpenes.

93
6.1. Introdução
A família Myrtaceae é uma família pantropical, com aproximadamente 139 gêneros e
5760 espécies identificadas (GOVAERTS et al., 2017). No Brasil, é constituída por 23
gêneros e 990 espécies, todos pertencentes à subfamília Myrtoideae (SOBRAL et al. 2015),
sendo reconhecida a sua elevada riqueza de espécies e seu importante papel na fitossociologia
das Florestas do Sul e Sudeste do Brasil (ROMAGNOLO e SOUZA, 2004).
Essa é uma das famílias de plantas ricas em OE, sendo que os estudos com Myrtaceae
neotropical, focados em seus compostos voláteis, são relativamente recentes. Das várias
espécies desta família que tiveram seus OEs estudados, diversos resultados de interesse
agrícolas e farmacológicos foram comprovados (STEFANELLO et al., 2011).
Os trabalhos de Becker et al. (2017), por exemplo, revelaram que OE de folhas de E.
uniflora possuem propriedades antifúngicas (COSTA et al., 2010), antibacterianos (BECKER
et al., 2017), antinociceptivas e hipotérmicas (AMORIM et al., 2009) e atividade antioxidante
e antimicrobiana (VICTORIA et al., 2012).
O OE de outra espécie da família de Myrtaceae, Eugenia calycina Cambess,
demonstrou alta atividade bactericida em bactérias anaeróbicas Gram-negative Prevotella
nigrescens e Porphyromonas gingivalis (SOUZA et al., 2015).
De acordo com Stefanello et al. (2011), a composição química da maioria dos OE de
Myrtaceae neotropical estudados são caracterizados por apresentarem grande diversidade
química e pela predominância de sesquiterpenos e monoterpenos.
O gênero Eugenia apresenta predominância de sesquiterpenos, e os monoterpenos
representam uma fração menor. O sesquiterpeno β-cariofileno e o monoterpeno α-pineno são
os mais abundantemente encontrados nesse gênero.
A composição química dos OE do gênero Myrcia é muito semelhante aos da Eugenia,
com predominância de sesquiterpenos cíclicos na maioria das espécies e algumas espécies
apresentando OEs ricos em monoterpenos. O β-cariofileno, como também ocorreu no gênero
Eugenia, é o sesquiterpeno mais comumente encontrado em seus OEs, porém o α-pineno não
foi encontrado em quantidades significantes na maioria das espécies desse gênero. Outras
espécies da família Myrtaceae, pertencentes a gêneros menos numerosos, também apresentam
composição semelhante aos observados para Eugenia spp. (STEFANELLO et al., 2011).

94
Os OEs por serem voláteis, atuam como sinais de comunicação química entre as
diversas espécies de plantas, e também agem na defesa contra patógenos e herbivoria. Essas
características tornam as plantas que os produzem poderosas fontes de agentes biocidas, sendo
largamente estudadas por apresentarem atividades bactericidas, inseticidas, fungicidas, entre
outras (SOYLU et al., 2010). Aproximadamente 60% dos OEs apresentam atividade
antifúngica e 35% mostram propriedades antibacterianas (SANTOS et al., 2010).
Vários fatores bióticos e abióticos podem influenciar a composição química e o
rendimento de OEs. Entre os fatores que podem influenciar sua síntese, a sazonalidade e a
fenologia estão entre os mais importantes para espécies vegetais não domesticadas, presentes
em seu meio natural, floresta ou em outro ambiente (BRITO, 2010; OLIVEIRA et al., 2012).
De acordo com Oliveira et al., (2012), além dos fatores genéticos e ambientais, que
influenciam a composição dos OEs, o método e o tempo de extração também podem alterar
seus teores de forma quantitativa e qualitativa. Mattana et al. (2015), comparou métodos de
extração pelas técnicas que utilizam solventes, por hidrodestilação e extração por infusão, em
folhas de Pothomorphe umbellata (L.) Miq. e constatou que foi possível identificar maior
número de compostos na extração pelo método de hidrodestilação.
C. prismatica pertencente à família Myrtaceae, uma espécie arbórea endêmica, que
também pode apresentar porte arbustivo. Essa é uma das inúmeras espécies que compõem a
FOM. Inicialmente classificada como E. prismatica, foi transferida em 2007, para um novo
gênero, Curitiba Salywon & Landrum (SALYWON e LANDRUM, 2007).
Atualmente não existe na literatura nenhum estudo sobre a composição química dos
OEs de C. prismatica e diante do exposto, o objetivo desse estudo foi analisar a composição
química e o rendimento dos OEs dessa espécie, em indivíduos alocados sob diferentes
irradiâncias e em duas estações do ano; primavera e inverno.

95
6.2. Material e Métodos
Local do experimento
O local da coleta do material vegetal (folhas) utilizado para extração do OE é o mesmo
descrito no capítulo 4, item 4.2.
Coleta do material vegetal para extração do óleo essencial
O material vegetal foi coletado entre os anos de 2015 e 2016, nas seguintes datas: 20
de outubro de 2015 (primavera) e 30 de julho de 2016 (inverno). As folhas de C. prismatica
foram coletadas das 08:00 às 10:00 horas, para evitar influências no rendimento dos OEs.
Além disso, foi observado, antes das coletas, um período de no mínimo três dias sem
ocorrência de chuvas. As coletas das folhas foram realizadas em dois ambientes com distintos
níveis de luminosidade, sub-bosque e a pleno sol, sendo coletado ao acaso material de sete
árvores de cada ambiente.
Depois de realizada cada coleta, as folhas de C. prismatica foram acondicionadas em
sacos plásticos hermeticamente fechados, borrifado água e mantidas refrigeradas em gelo até
serem transferidas para refrigerador para posterior realização das extrações. As extrações
foram realizadas no Laboratório de Ecotoxicologia, do Departamento de Biologia, campus
CEDETEG, da Universidade Estadual do Centro-Oeste do Paraná, UNICENTRO.
Extração do óleo essencial
Foram utilizadas na extração, as folhas de C. prismatica que estavam livres de danos
por fungos ou herbivoria. As folhas secas, amarelas ou com material estranho (deposição de
ovos de insetos, musgo, entre outros) também foram desprezadas. Após essa seleção, as folhas
foram pesadas, lavadas com água destilada, divididas manualmente com tesoura e transferidas
para o balão volumétrico.
As extrações do OE das folhas de C. prismatica foram realizadas em triplicata para
cada ambiente (sub-bosque e a pleno sol) e em cada estação (primavera e verão) avaliada. A
técnica utilizada foi a de arraste a vapor, pelo método de hidrodestilação, com folhas frescas.

96
Para hidrodestilação foi utilizando o conjunto manta aquecedora, balão volumétrico
com junta e aparelho tipo Clevenger, conforme Figura 19. As folhas de C. prismatica foram
aquecidas por um período de 3 horas, contadas a partir do início do refluxo da água
condensada do tubo separador para o balão de destilação.
Figura 19. Hidrodestilação do óleo essencial de folhas frescas de Curitiba prismatica
coletada no município de Rebouças-PR., utilizando conjunto com aparelho do
tipo Clevenger, balão de vidro e manta aquecedora.
Fonte: Autor.
Após a extração, o OE foi recolhido e a água residual foi eliminada por meio de
secagem com sulfato de sódio anidro (Na2SO4). Em seguida, o OE foi transferido para frasco
de vidro, que foi vedado por tampa e envolvido por parafilme e papel alumínio, para evitar
possíveis perdas de constituintes voláteis e impedir a entrada de luz. Após esse procedimento
foi armazenado em ambiente refrigerado até ser submetido à análise química dos seus
constituintes.

97
Análise do óleo essencial e identificação dos constituintes por cromatografia
gasosa hifenada com espectrometria de massa
As amostras de OEs foram analisadas no laboratório Central de Análises, da
Universidade Tecnológica Federal do Paraná, campus Pato Branco. Para esta análise foi
utilizado o cromatógrafo a gás Varian 431/CP-3800 acoplado a um detector de massa Varian
210 íon trap MS, faixa de aquisição: 50-250 u; energia de ionização: 70 eV; modo de
aquisição normal (impacto de elétrons; transferline coluna/MS a 200 ºC; íon trap à 170 ºC. A
identificação dos constituintes foi realizada com base nos índices de retenção (ADAMS,
2007), obtidos da coinjeção de uma mistura de n-alcanos, com número de átomos de carbono
entre 7 e 30, utilizados como padrões, e por comparação de seus espectros de massas, nas
seguintes condições: coluna capilar Rtx-5MS (30 x 0,25 x 0,25), temperatura do injetor 250
°C, rampa de aquecimento 60°C – 3,5°C/min – 240°C, hélio como gás de arraste, fluxo de 1
mL/min).
A identificação individual dos compostos químicos dos OEs foi realizada comparando
os dados dos espectros de massas obtidos com os disponíveis no sistema (NIST, 2002). Para
análise quantitativa, a composição percentual das amostras dos OEs, foi calculada pela
integração da área dos picos dos cromatogramas.
Análises Estatísticas
O perfil da identificação e os teores dos óleos essenciais de C. prismatica foram
submetidos à análise estatística multivariada, por meio das técnicas de agrupamento
hierárquico e análise de componentes principais.
A análise de agrupamentos hierárquicos foi realizada utilizando-se o dendrograma
horizontal para a exibição dos agrupamentos das amostras. Nesta análise, foi utilizado o
método Ward e separados pela distância Euclidiana.
Na análise de componentes principais foram geradas novas variáveis (componentes), a
partir dos compostos químicos dos óleos que contivessem significativa variância original dos
dados. Com estes componentes foram gerados gráficos de dispersão dos casos (amostras) e
das variáveis (compostos químicos). Todas as análises foram realizadas no software Statistica
StatSoft, versão 8.0.

98
6.3. Resultados e Discussão
Rendimento do óleo essencial de Curitiba prismatica
As análises estatísticas demonstraram que houve variações significativas no
rendimento dos OEs, das folhas de C. prismatica entre os ambientes de sub-bosque e a pleno
sol, e também entre as estações do ano, conforme Figura 20.
*bAbA
aB
aA
Inverno Primavera0
2
4
6
8
10
12
Ren
dim
ento
(mL.
kg-1)
Estações
Sombra Sol
* Médias seguidas pelas mesmas letras, minúsculas dentro de cada estação e maiúscula entre os mesmos ambientes, não diferem estatisticamente pelo Teste de Tukey, ao nível de 5 % de probabilidade.
Figura 20. Rendimento do óleo essencial (mL.kg-1) das folhas frescas de Curitiba prismatica
coletadas em Rebouças-PR.
As médias dos rendimentos no OE no inverno (5,61 mL.Kg-1 folhas frescas) foram
15,85% inferiores que o rendimento obtido na primavera (6,67 mL.Kg-1 folhas frescas). Essa
diferença deve-se principalmente ao rendimento superior no ambiente a pleno sol, visto que
não houve diferenças significativas entre as estações no ambiente sombreado, Figura 20.
No ambiente a pleno sol, a maior média de produção de OE (9,73 mL.Kg-1 folhas
frescas) ocorreram durante a primavera. No inverno, neste mesmo ambiente, ocorreu
significativa diminuição do rendimento (27,40%). Essa diminuição, provavelmente está
relacionada com a menor disponibilidade hídrica, menor radiação solar incidente e baixa
atividade biossintética das plantas.

99
De acordo com os dados meteorológicos, referentes à região de Rebouças-PR, obtidos
do Instituto Nacional de Meteorologia (INMET) no período de setembro de 2015 a julho de
2016 (Figura 21) verifica-se diminuição durante o inverno de 17,60% e 23,54% na
precipitação e na radiação solar incidente, respectivamente, em relação à primavera. Essa
menor disponibilidade hídrica, aliada a redução da radiação solar e aos efeitos climáticos da
estação podem justificar as alterações fisiológicas que ocasionaram a diminuição da produção
de OE no inverno.
200
225
250
275
300
325
350
Precipitação Radiação solar
Estação do ano
Prec
ipita
ção
(mm
)
Primavera Inverno10
11
12
13
14
15
Rad
iaçã
o so
lar (
MJ.m
-1.d
ia-1
)
Figura 21. Variações da radiação solar incidente e precipitação média registradas nas
estações da primavera - 2015 e inverno - 2016.
Os dados obtidos nesse trabalho estão de acordo com Castro et al. (2005), os quais
afirmam que devido aos óleos essenciais também possuírem ação protetora em relação a
estresse abiótico, como aqueles associados com mudanças de temperatura, conteúdo de água,
níveis de luz, exposição à UV, entre outros, esses maiores rendimentos podem representar
uma resposta adaptativa a essa condição ambiental em que se desenvolveram os indivíduos de
C. prismatica. Ainda de acordo com Souza (2007), a baixa disponibilidade hídrica e
principalmente as baixas temperaturas podem afetar além do desenvolvimento da planta, e seu
teor de OE.

100
A menor produção de OE em ambiente de sub-bosque pode estar relacionada à menor
atividade metabólica devido a menor irradiância. De acordo com Oliveira et al. (2007), a
planta responde aos ambientes com baixa incidência de radiação diminuindo as atividades
metabólicas de assimilação do carbono e absorção de nutrientes minerais, inibindo seu
crescimento e fazendo com que a produção de OE seja também influenciada, diminuindo
assim o seu rendimento.
Diversos trabalhos, sobre o rendimento de OEs, que analisaram o efeito da
luminosidade encontraram resultados semelhantes aos apresentados nesse estudo. Paulus et al.
(2013), estudando teor de óleo essencial em Aloysia triphylla (L'Hér.) Britton, observaram
maior teor de óleo essencial nas plantas cultivadas em condições de maior espaçamento de
plantas, e constataram que nesta condição houve maior irradiância em relação aos cultivos
mais adensados. Ventrella e Ming (2000) trabalhando com Lippia alba (Mill.) N.E.Br
alocadas a pleno sol verificaram elevação no teor de óleo essencial em relação aos ambientes
sombreados. Silva et al., 2006 verificou que o nível de radiação a pleno sol também aumentou
o rendimento de óleo essencial em Baccharis trimera (Less.) DC (SILVA et al., 2006).
Composição química do óleo essencial da Curitiba prismatica
A análise química das amostras de OEs identificou um total de 54 compostos,
conforme Tabela 4. Desses compostos 51,85%, pertence à classe dos sesquiterpenos
hidrocarbonados. Os demais compostos, com 18,52%, 16,67% e 12,96% dividem-se em
monoterpenos oxigenados, sesquiterpenos oxigenados e monoterpenos hidrogenados,
respectivamente.
Tabela 4. Composição química dos óleos essenciais das folhas de C. prismatica coletadas em
Rebouças-PR, na primavera e inverno, nos ambientes de pleno sol e de sub-bosque.
Constituinte Classe IR Inverno Sol
Inverno Sombra
Primavera Sol
Primavera Sombra
1,5-Dimetil-1,5-cyclooctadieno1 MH 1103 - - 10,39 - 3,7,11,15-Tetrametil-2-hexadecen-1-ol2 SO 2045 2,01 4,45 - - 6-Isopropenil-4,8a-dimetil-1,2,3,5,6,7,8,8a -octahidro-naphthalen-2-ol (3)3 SO 1690 - 0,30 - - Alloaromadendreno4 SH 1386 - 1,31 8,17 1,80
Continua na próxima página

101
Constituinte Classe IR Inverno Sol
Inverno Sombra
Primavera Sol
Primavera Sombra
Aromadendreno5 SH 1386 4,27 3,44 - - Benzil isovalerato6 SH 1394 - - 2,22 - Bicyclosesquiphellandreno7 SH 1386 1,67 1,37 - 4,75 Cadala-1(10),3,8-trieno8 SH 1548 - - - 0,51 Calareno9 SH 1403 3,31 3,14 - - Careno10 MH 919 0,52 - 21,18 - Cariofileno11 SH 1494 - - 22,01 - Carvomenthono12 MO 1148 - - - 0,07 Carvono13 MO 1190 - - 0,36 - Cis-Carveol14 MO 1206 - 0,07 0,94 - Cis-óxido limoneno15 MO 1031 - - 0,05 - Cis-Pinen-3-ol16 MO 1070 - - 0,34 - Cis-p-Mentha-2,8-dien-1-ol17 MO 1140 - - 0,17 - Cis-verbenono18 MO - - 0,44 - Cubenol19 SO 1580 5,35 5,98 - - Dioxido de limoneno20 MO 1127 - - 0,20 - Elixeno21 SH 1431 10,95 10,30 - 0,29 Epizonareno22 SH 1469 3,14 2,39 - - Eremophileno23 SH 1515 - - - 10,26 Germacreno D24 SH 1523 - 2,51 - 0,76 Guaieno25 SH 1018 - 0,93 - - Limoneno26 MH 1018 - 2,59 - 1,03 Cadina-1(2),4-dieno 27 SH 1440 10,12 8,69 - 16,42 Origaneno28 MH 902 - 4,95 - 5,91 Óxido cariofileno29 SO 1507 1,22 - - 2,40 Pineno-epoxide30 MH 961 - - 0,58 - Pirocarvono31 MO 1114 - - 0,22 - Spathulenol32 SO 1536 2,30 2,41 - 2,59 Tau-cadinol33 SO 1580 5,05 - - 22,50 Terpineol34 MO 1143 1,03 0,25 3,94 0,38 Torreyol35 SO 1580 1,00 - - 1,26 Trans-Cariophylleno36 SH 1494 - 7,56 - 9,67 Valenceno37 SH 1474 - - 2,89 0,50 γ-cadineno38 SO 1435 - 5,16 - - γ -terpineno39 MH 998 0,54 - - - α-cadinol40 SO 1580 - 1,01 - - α-calacoreno41 SH 1547 - 0,36 - 0,91 α-cariofileno42 SH 1579 2,24 1,81 4,36 2,45 α-cubebeno43 SH 1314 3,06 0,47 7,96 2,90 α-guaieno44 SH 1490 - - - 0,39 α-gurjuneno45 SH 1419 3,24 2,92 6,90 3,88 α-muuroleno46 SH 1440 - - 6,29 3,13 α-phellandreno47 MH 969 - - 0,39 0,63
Continua na próxima página

102
Constituinte Classe IR Inverno Sol
Inverno Sombra
Primavera Sol
Primavera Sombra
β-cariofileno48 SH 1494 15,24 - - - β-elemeno49 SH 0,66 - - γ-elemeno50 SH 1465 14,45 1,06 - 3,40 γ-gurjuneno51 SH 1461 - 14,78 - 0,65 γ-langeno52 SH 1221 - - - 0,28 γ-muuroleno53 SH 1435 - - - 0,27 δ-cadineno54 SH 1469 9,28 9,14 - - IR - índice de retenção, MH - Monoterpeno hidrocarbonado, MO - Monoterpeno oxigenado, SH - Sesquiterpeno hidrocarbonado, SO – Sesquiterpeno oxigenado.
Observou-se que OEs das folhas de C. prismatica apresentaram variações nos teores e
nos compostos químicos identificados entre as estações e também entre os ambientes
analisados, conforme Figura 22.
0
5
10
15
20
25
30
35
Qua
ntid
ade
de C
ompo
stos
Estações e ambientes avaliados
MH MO SH SO
PrimaveraSol
PrimaveraSombra
InvernoSombra
InvernoSol
MH - Monoterpeno hidrocarbonado, MO - Monoterpeno oxigenado, SH - Sesquiterpeno hidrocarbonado, SO – Sesquiterpenos oxigenado.
Figura 22. Distribuição de compostos terpênicos, agrupados por classes, dos óleos essenciais
das folhas de Curitiba prismatica coletadas em Rebouças-PR, na primavera e
inverno, e nos ambientes de pleno sol e sombreado em sub-bosque.
Nas amostras coletadas no inverno, em ambiente sombreado de sub-bosque, foram
identificados 28 compostos, enquanto que a pleno sol foram 21 compostos. Em ambos
ambientes predominaram os sesquiterpenos hidrogenados, com 57,14% e 64,29%,
respectivamente, nos ambientes a pleno sol e sombreado.
De todos os compostos identificados no inverno, 15 compostos ocorreram em ambos
os ambientes, representando os teores de 76,43% e 57,82% dos OEs extraídos das folhas
coletadas em ambiente a pleno sol e no sub-bosque, respectivamente.

103
No ambiente de sub-bosque, os compostos majoritários foram γ-gurjuneno (14,78%),
elixeno (10,95%) e δ-cadineno (9,14%). Nessa mesma estação, porém no ambiente a pleno
sol, os compostos com maiores teores foram o β-cariofileno (15,24%), γ-elemeno (14,45%) e
elixeno (10,95%).
A análise dos OEs de folhas coletadas na primavera, tanto em ambiente sombreado
como a pleno sol, apresentou o mesmo número de compostos químicos encontrados no
inverno. Porém, ocorreram diferenças nos teores e nos compostos químicos identificados,
conforme Figura 22. Nessa estação no ambiente a pleno sol não foram identificados
compostos pertencentes à classe dos sesquiterpenos oxigenados, sendo os monoterpenos
oxigenados, com 42,86% os principais constituintes. No ambiente sombreado, os
sesquiterpenos hidrogenados (67,86%) formaram a maior classe dos compostos identificados.
A maior produção de monoterpenos (42,86%), moléculas pequenas e de maior
volatilidade, observadas na primavera e somente no ambiente a pleno sol, pode estar ligado
com a sinalização química. Foi observado que essas árvores estavam em pleno
desenvolvimento da brotação, preparando-se para o início da floração e consequente
frutificação.
No ambiente a pleno sol, na primavera, os principais compostos foram o cariofileno
(22,01%), careno (21,18%) e o 1,5-dimetil-1,5-cyclooctadieno (10,39%), enquanto que no
sub-bosque os principais compostos foram o tau-cadinol (22,50%), cadina-1(2),4-dieno
(16,42%) e o eremophileno (10,26%).
Os compostos químicos α-cariofileno, α-cubebeno e α-gurjuneno foram encontrados
em todos os ambientes, em ambas as estações analisadas. Esses compostos apresentaram
maiores rendimentos na primavera, em ambiente a pleno sol.
Carneiro (2016) analisando OEs de Eugenia klotzschiana Berg, encontrou como
compostos químicos majoritários o α-cariofileno (17,38%), γ-elemeno (13,14%) e elixeno
(10,20%). SCALVENZI (2017) encontrou o β-cariofileno, no OE de Myrcia splendens (Sw.)
DC. como sendo o terceiro mais abundante.
Nessa mesma estação, porém em ambiente de sub-bosque, a quantidade de
monoterpenos diminuiu para 7,14%. Essa diminuição pode estar relacionada ao fato que foi
observada baixa atividade de brotação, e consequentes atividades reprodutivas no ambiente de
sub-bosque.

104
Por outro lado, a maior concentração de sesquiterpenos no inverno, momento em que
as árvores estão em estado de repouso vegetativo e mais suscetíveis ao ataque de patógenos
devido ao excesso de umidade e baixas temperaturas, podem indicar funções de defesa para
estas classes de compostos.
A análise dos OEs das folhas de C. prismatica utilizando a técnica multivariada de
análise de agrupamento apresentou a formação de quatro distintos grupos, conforme Figura
23.
0 20 40 60 80 100 120 140
Distância de Ligação
Primavera-Sol-R2
Primavera-Sol-R2
Primavera-Sol-R1
Primavera-Sombra-R3
Primavera-Sombra-R2
Primavera-Sombra-R1
Inverno-Sombra-R3
Inverno-Sombra-R2
Inverno-Sombra-R1
Inverno-Sol-R3
Inverno-Sol-R2
Inverno-Sol-R1
Am
ostra
s
Figura 23. Dendrograma para o agrupamento das amostras de óleos essenciais das folhas de
Curitiba prismatica coletadas em Rebouças-PR, na primavera e inverno, nos
ambientes de pleno sol e sombreado.
Os grupos formados foram definidos pelo traçado de uma linha paralela ao eixo
vertical, denominada “linha fenon”. Definiu-se a distância para traçar essa linha entre as
distâncias euclidianas de ligação 44,03 e 74,86, que representam as maiores diferenças entre
as amostras de OEs.
Essas distâncias foram obtidas pela análise da Figura 24, em que o corte foi realizado
no dendrograma entre as distâncias no qual ocorre a maior distância de ligação, conforme
indicado no gráfico pelo retângulo cinza.
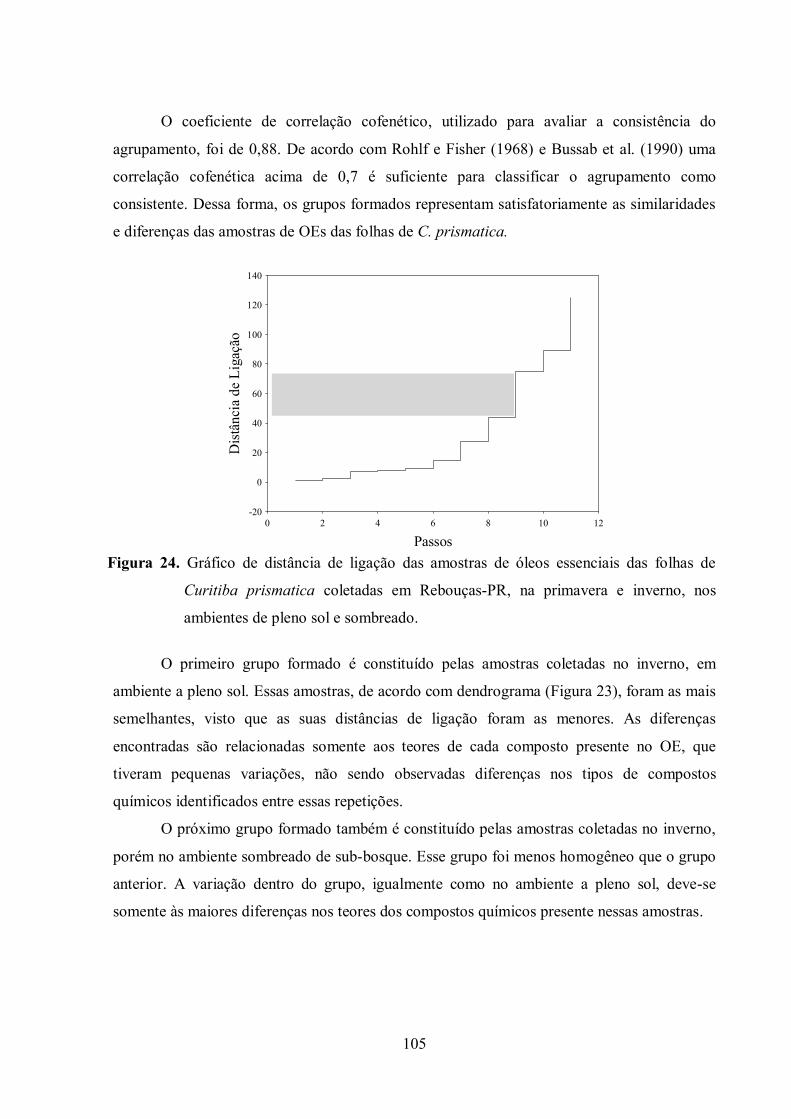
105
O coeficiente de correlação cofenético, utilizado para avaliar a consistência do
agrupamento, foi de 0,88. De acordo com Rohlf e Fisher (1968) e Bussab et al. (1990) uma
correlação cofenética acima de 0,7 é suficiente para classificar o agrupamento como
consistente. Dessa forma, os grupos formados representam satisfatoriamente as similaridades
e diferenças das amostras de OEs das folhas de C. prismatica.
0 2 4 6 8 10 12
Passos
-20
0
20
40
60
80
100
120
140
Dis
tânc
ia d
e Li
gaçã
o
Figura 24. Gráfico de distância de ligação das amostras de óleos essenciais das folhas de
Curitiba prismatica coletadas em Rebouças-PR, na primavera e inverno, nos
ambientes de pleno sol e sombreado.
O primeiro grupo formado é constituído pelas amostras coletadas no inverno, em
ambiente a pleno sol. Essas amostras, de acordo com dendrograma (Figura 23), foram as mais
semelhantes, visto que as suas distâncias de ligação foram as menores. As diferenças
encontradas são relacionadas somente aos teores de cada composto presente no OE, que
tiveram pequenas variações, não sendo observadas diferenças nos tipos de compostos
químicos identificados entre essas repetições.
O próximo grupo formado também é constituído pelas amostras coletadas no inverno,
porém no ambiente sombreado de sub-bosque. Esse grupo foi menos homogêneo que o grupo
anterior. A variação dentro do grupo, igualmente como no ambiente a pleno sol, deve-se
somente às maiores diferenças nos teores dos compostos químicos presente nessas amostras.

106
A similaridade das amostras coletadas no inverno pode ser observada pela análise de
componentes principais (ACP), Figura 25. As amostras de ambos os ambientes estão todas
situadas no primeiro quadrante, formando um grande grupo (círculo tracejado). Essa
proximidade por ser observada também na Figura 23, na qual utilizando como ponto de corte
a distância próxima de 80, tanto o ambiente a pleno sol como o sombreado ficariam no
mesmo grupo.
SOL-INV-R1SOL-INV-R2
SOL-INV-R3
SOM-INV-R1
SOM-INV-R2
SOM-INV-R3
SOL-PRI-R1
SOL-PRI-R2
SOL-PRI-R3
SOM-PRI-R1
SOM-PRI-R2
SOM-PRI-R3
-12 -10 -8 -6 -4 -2 0 2 4 6 8
Componente 1 (40,60%)
-8
-6
-4
-2
0
2
4
6
Com
pone
nte
2 (2
6,87
%)
SOL-INV-R1SOL-INV-R2
SOL-INV-R3
SOM-INV-R1
SOM-INV-R2
SOM-INV-R3
SOL-PRI-R1
SOL-PRI-R2
SOL-PRI-R3
SOM-PRI-R1
SOM-PRI-R2
SOM-PRI-R3
SOL-PRI - folhas em ambiente de pleno sol coletadas na primavera, SOM-PRI - folhas em ambiente de sub-bosque coletadas na primavera, SOL-INV - folhas em ambiente de pleno sol coletadas no inverno, SOM-INV - folhas em ambiente de sub-bosque coletadas no inverno.
Figura 25. Dispersão das amostras para os óleos essenciais de folhas das folhas de Curitiba
prismatica coletadas em Rebouças-PR, na primavera e inverno, nos ambientes de
pleno sol e sombreado em sub-bosque.
Os grupos formados pelas amostras da primavera apresentaram maior dissimilaridade,
tanto entre os ambientes como entre as duas estações analisadas. Essa variação deve-se a
diversidade química e aos teores dos compostos identificados. Os compostos trans-
cariophylleno, eremophileno, cadina-1(2),4-dieno e o tau-cadinol, que constitui 58,85% dos
OEs do ambiente de sub-bosque, não ocorreram à pleno sol. Da mesma forma, no ambiente a
pleno sol os compostos cariofileno, careno e 1,5-Dimetil-1,5-cyclooctadieno, que representam
53,58% do teor de OE, somente ocorreram nas amostras do ambiente a pleno sol.

107
Essas maiores diferenças na primavera podem ser observadas na Figura 23 e 25. Na
Figura 23 a maior dissimilaridade dos OEs do ambiente a pleno sol é representada pela maior
distância de ligação com os demais grupos. Na Figura 25 observa-se que as amostras
pertencentes à primavera ficaram em quadrantes separados, refletindo as diferenças na
composição química do OE.
A análise de ACP das amostras de OEs das folhas de C. prismatica apresentou cinco
componentes com autovalores acima de um, dos quais quatro componentes contêm 92,14% da
variância total acumulada nos dados.
Os gráficos da dispersão dos casos (amostras do OEs) e da dispersão das correlações
das variáveis (compostos químicos dos OEs) estão representados na Figura 25 e 24,
respectivamente. Os componentes 1 e 2 representam 67,47% de toda variância acumulada.
Existe uma relação para as projeções espaciais entre os dois tipos de gráficos (Figura
25 e 26). A dispersão das variáveis (compostos químicos do OE) influencia a dispersão dos
casos (amostras de OEs), formando quatro grupos distintos. Esses grupos formados são os
mesmos identificados na análise de agrupamento, em que ocorreu a separação das amostras
por estação e por tipo de ambiente. Cada grupo formado é influenciado por um conjunto
específico dos compostos químicos identificados e seus respectivos teores (Figura 26).
1
2
3
4
6
7
8
9
10
11
12
13
14
15
1617
18 1920
21
22
23
24
25
26
2728
29
3031
32
33
34
3536
37
38
39
5
40
41
4243
44
4546
47
48 49
50
51
52
53
54
-1,0 -0,5 0,0 0,5 1,0
Componente 1 (40,60%)
-1,0
-0,5
0,0
0,5
1,0
Com
pone
nte 2
(26,
87%
)
1
2
3
4
6
7
8
9
10
11
12
13
14
15
1617
18 1920
21
22
23
24
25
26
2728
29
3031
32
33
34
3536
37
38
39
5
40
41
4243
44
4546
47
48 49
50
51
52
53
54
1-54: constituintes químicos identificados. Figura 26. Dispersão dos compostos químicos presentes nos óleos essenciais de folhas das
folhas de Curitiba prismatica coletadas em Rebouças-PR, na primavera e inverno,
nos ambientes de pleno sol e sombreado em sub-bosque.

108
De acordo, com os dados analisados, pode-se observar que os efeitos da irradiância e
da sazonalidade influenciam a composição química dos OEs em folhas de C. prismatica.
Esses resultados estão de acordo com estudos realizados por diversos autores (GOBBO-
NETO e LOPES, 2007; HESS et al., 2007; BARRA, 2009; BRITO, 2010; KESZEI et al.,
2010; MARCO et al., 2011), os quais também constataram que os fatores abióticos alteram a
morfologia, fisiologia e a produção de OEs em plantas.
Além dos fatores ambientais estudados, outros podem estar correlacionados com as
variações encontradas nos OEs de C. prismatica, como o padrão genético (LAMIEN-MEDA
et al., 2010), expressão gênica diferencial (PAULA et al., 2011), entre outros.

109
6.4. Conclusão
As análises de agrupamentos e de componentes principais demonstram serem
ferramentas adequadas para avaliar a variabilidade de OEs de C. prismatica. Ambas as
técnicas multivariadas evidenciaram os efeitos da sazonalidade (primavera e inverno) e de
diferentes ambientes (pleno sol e sombreado) e sua correlação com a composição química dos
OEs das folhas de C. prismatica.
Pela análise dos OEs pode-se observar a plasticidade de C. prismatica em resposta às
variações sazonais e de irradiância, influenciando a síntese dos compostos terpênicos
provenientes do metabolismo especializado desta espécie. Essa influência pode ser observada
tanto no rendimento total dos OEs como nos diferentes teores e diversidade de compostos
químicos identificados em cada estação e ambiente.
Este estudo demonstrou que cada um dos diferentes grupos de OEs encontrado nas
análises realizadas possui rendimentos e características químicas específicas. Dessa forma, os
dados presentes nesse trabalho podem servir de base para o planejamento de bioensaios que
visem compreender melhor as possíveis aplicações dos OEs de C. prismatica.

110
6.5. Referências Bibliográficas
AMORIM, A. C. L.; LIMA, C. K. F.; HOVELL, A. M. C.; MIRANDA, A. L. P.; REZENDE, C. M. Antinociceptive and hypothermic evaluation of the leaf essential oil and isolated terpenoids from Eugenia uniflora L. (Brazilian Pitanga). Phytomed. v. 16, p. 923–928, 2009. BARRA, A. Factors affecting chemical variability of essential oils: A review of recent developments. Natural Product Communications, v. 4, n. 8, p. 1147-1154, 2009. BECKER, N. A. et al. Biological properties of Eugenia uniflora L. essential oil: phytochemistry composition and antimicrobial activity against gram negative bacteria. Vittalle, v. 29, n. 1, 2017. BRITO, H. R. Caracterização química de óleos essenciais de Spondias mombin L., Spondias purpurea L. e Spondias sp. (canjarana do sertão). 2010. 68 f. Dissertação (Mestrado em Ciências Florestais), Universidade Federal de Campina Grande, Patos, 2010. BUSSAB, W. de O.; MIAZAKI, E. S.; ANDRADE, D. F. Introdução à Análise de Agrupamentos. In: 9º Simpósio Nacional de Probabilidade e Estatística. Associação Brasileira de Estatística, São Paulo, 1990, 105p. CARNEIRO, N. S. Caracterização química e avaliação da atividade antioxidante da polpa e óleo essencial da pera do cerrado (Eugenia klotzschiana Berg.). 63 f. 2016. Dissertação (Mestrado em Agroquímica) – Instituto Federal Goiano, Rio Verde, 2016. CASTRO, P. R. C.; KLUGE, R. A.; PERES, L. E. P. Manual de fisiologia vegetal: teoria e prática. Piracicaba: Editora Agronômica Ceres, 2005. 650 p. COSTA, D. P. et al. Influence of fruit biotypes on the chemical composition and antifungal activity of the essential oils of Eugenia uniflora leaves. Journal of the Brazilian Chemical Society, v. 21, p. 851–858, 2010. GOBBO-NETO, L. LOPES, N. P. Plantas medicinais: fatores de influência no conteúdo de metabólitos secundários. Química nova, v.30, n. 2, p. 374-381, 2007. GOVAERTS, R. et al. World Checklist of Myrtaceae. Facilitated by the Royal Botanic Gardens, Kew. Disponível em <http://www.kew.org/wcsp/>. Acesso em: 12 dez. 2015. HESS, S. C.; PERES, M. T. L. P.; BATISTA, A. L.; RODRIGUES, J. P.; TIVIROLI, S. C.; OLIVEIRA, L. G. L.; SANTOS, C. W. C.; FEDEL, L. E. S.Evaluation of seasonal changes in chemical composition and antibacterial activity of Elyonurus muticus (Sprengel) O. Kuntze (Gramineae). Quimica Nova, v. 30, n. 2, p. 370-373, 2007. KESZEI, A.; HASSAN, Y.; FOLEY, W. J. A biochemical interpretation of terpene chemotypes in Melaleuca alternifolia. Journal of Chemical Ecology, Lexington, v. 36, p.652-661, 2010.

111
LAMIEN-MEDA, A., SCHIMIDERER, C.; LOHWASSER, U.; BÖRNER, A.; FRANZA, C.; NOVAKA, J. Variability of the essential oil composition in the sage collection of the Genebank Gatersleben: a new viridiflorol chemotype. Flavour and Fragrance Journal, Geneva, v. 25, p.75-82, 2010. MARCO, C.A.; SANTOS, H.R.; ROLIM, R.R.; COSTA, J.G.M.; FEITOSA, J.M.; VÁSQUEZ, M.E.F. Influência do estádio fenológico de Vanillosmopsis arborea Baker sobre o rendimento e qualidade do seu óleo essencial. Caderno de Agroecologia, Camaragibe, v. 6, n. 2, 2011. MATTANA, R. S. et al. Efeitos de diferentes tempos de extração no teor e composição química do óleo essencial de folhas de pariparoba [Pothomorphe umbellata (L.) Miq.]. Revista Brasileira de Plantas Medicinais, v.17, n.1, p.150–156, 2015. NIST. NIST/EPA/NIH mass spectral library & search/analysis programs NIST 02 update. Hoboken, NY: John Wiley & Sons, 2002 OLIVEIRA, C. B. A.; FERRI, P. H.; PAULA, J. R.; SERAPHIN, J. C. Composition and chemical variability in the essential oil of Hyptis marrubioides Epl. Journal of Essential Oil Research, v. 19, n. 6, p. 552-556, 2007. OLIVEIRA, A. R. M. F.; JEZLER, C. N.; OLIVEIRA, R. A.; MIELKE. M. S., COSTA, L. C. B.. Determinação do tempo de hidrodestilação e do horário de colheita no óleo essencial de menta. Horticultura Brasileira, v. 30, p. 155–159, 2012. PAULA, J. M. A.; FERRI, P. H.; BARA, M. T. F.; TRESVENZOL, L. M. F.; SÁ, F. A. S.; PAULA, J. R. Infraspecific chemical variability in the essential oils of Pimenta pseudocaryophyllus (Gomes) L.R. Landrum (Myrtaceae). Biochemical Systematics and Ecology, Edimburgo, v. 39, p.643-650, 2011. PAULUS, D. et al. Teor e composição química do óleo essencial e crescimento vegetativo de Aloysia triphylla em diferentes espaçamentos e épocas de colheita, Revista Ceres, Viçosa, v. 60, n.3, p. 372-379 , 2013. ROHLF, F. J.; FISHER, D. R. Test for hierarchical structure in random data sets. Systematic Zoology, v. 17, p. 407-412, 1968. ROMAGNOLO, M. B.; SOUZA, M. C. Os gêneros Calycorectes O. Berg, Hexachlamys O. Berg., Myrciaria O.Berg e Plinia L. (Myrtaceae) na planície alagável do alto rio Paraná, Brasil. Acta Botânica Brasílica, v. 18, n. 3, p. 613- 627, 2004. SALYWON, A. M.; LANDRUM, L. R. Curitiba (Myrtaceae): A new genus from the planalto of southern Brazil. Brittonia, v. 59, n. 4, p. 301-307, 2007.

112
SANTOS, T. T. et al. Efeito do estresse hídrico na produção de massa foliar e teor de óleo essencial em sambacaitá (Hyptis pectinata L.). In: CONGRESSO BRASILEIRO DE OLERICULTURA, 44., 2004. Campo Grande. Anais... Campo Grande: SOB, v. 22, p.1-4. 2004. SANTOS, A. C. A.; ROSSATO, M.; SERAFINI, L. A.; BUENO, M.; CRIPPA, L. B.; SARTORI, V. C.; DELLACASSA, E.; MOYNA, P. Efeito fungicida dos óleos essenciais de Schinus molle L. e Schinus terebinthifolius Raddi, Anacardiaceae, do Rio Grande do Sul. Revista Brasileira de Farmacognosia, v. 20, n. 2, p. 154-159, 2010. SCALVENZI, L. et al. Myrcia splendens (Sw.) DC. (syn. M. fallax (Rich.) DC.) (Myrtaceae) Essential Oil from Amazonian Ecuador: A Chemical Characterization and Bioactivity Profile. Molecules, v. 22, n. 7, p. 1163-1174. 2017. SILVA, F. G. et al. Influência do nível de radiação no crescimento da planta, rendimento e qualidade do óleo essencial de carqueja. Ciência e Agrotecnologia, Lavras, v. 30, n. 1, p. 52-57, 2006. SOBRAL, M., et al. Lista de Espécies da Flora do Brasil. Jardim Botânico do Rio de Janeiro. Disponível em <http://floradobrasil.jbrj.gov.br/>. Acesso em: 03 de maio 2017. SOUZA, M. F. Influência do sombreamento na produção de fitomassa e óleo essencial em Alecrim-pimenta (Lippia sidoides Cham.). Revista Brasileira de Biociências, v. 5, p. 108-110, 2007. SOUZA, R. M. F. et al. Chemical composition, cytotoxic, and antibacterial activity of the essential oil from Eugenia calycina Cambess. leaves against oral bacteria. Industrial Crops and Products, v. 65, p. 71-78, 2015. SOYLU, E. M.; KURT, S.; SOYLU, S. In vitro and in vivo antifungal activities of the essential oils of various plants against tomato grey mould disease agent Botrytis cinerea. International Journal of Food Microbiology, v. 143, p. 183-9, 2010. STEFANELLO, M. E. A.; PASCOAL, A. C. R. F.; SALVADOR, M. J. Essential oils from neotropical Myrtaceae: chemical diversity and biological properties. Chemistry & Biodiversity, v. 8, p. 73–94, 2011. VENTRELLA, M. C.; MING, L. C. Produção de matéria seca e óleo essencial em folhas de erva-cidreira sob diferentes níveis de sombreamento e épocas de colheita. Horticultura Brasileira v.18, p.972-974,2000. VICTORIA, F. N. M. et al. Essential oil of the leaves of Eugenia uniflora L.: Antioxidant and antimicrobial properties. Food and Chemical Toxicology, v. 50, p. 2668-2674, 2012.

113
7. Considerações finais
Estudos com a C. prismatica são muitos escassos, particamente inexistentes. Com
isso, durante a realização deste trabalho verificou-se a necessidades e possibiidade do
desenvolvimento de outras pesquisas complementares, para contribuirem e aperfeiçoarem no
conhecimento dessa espécie, conforme segue:
- Análise dos efeitos da sazonalidade e da irradiância no desempenho fotossintético,
pela avaliação das atividades das enzimas catalase, peroxidase e superóxido dismutase. Essas
enzimas atuam na proteção contra danos oxidativos causados pelas espécies reativas de
oxigênio (EROs) devido a estresse bióticos e abióticos.
- Estudo da fenologia, estrutura vertical e arquitetura de copas, que juntamente com os
disponíveis nesse trabalho, poderão ser usados para definir técnicas de manejo específicas
para espécie.
- A extração dos OEs realizada nesse trabalho utilizou amostras de folhas frescas de C.
prismatica. Futuros trabalhos poderão avaliar os Oes presentes nos frutos, folhas secas e
casca, coletados de árvores de diferentes idades.



























![Aspectos ecofisiológicos no manejo de pastagens_Nabinger[1]](https://img.document.onl/doc/110x75/5873638a1a28ab431c8c3d95/aspectos-ecofisiologicos-no-manejo-de-pastagensnabinger1.jpg)

