Embed Size (px)
Citation preview

Avaliação da cadeia trófica em dois ecossistemas de grande profundidade do Oceano Pacífico
Andreia Filipa Campos Alves
Mestrado de Recursos Biológicos Aquáticos Departamento de Biologia 2015
Orientador Doutora Teresa Amaro, Investigadora pós-doutoramento, HCMR
Coorientador Doutora Sandra Ramos, Investigadora pós-doutoramento, CIIMAR

Todas as correções determinadas pelo júri, e só essas, foram efetuadas.
O Presidente do Júri,
Porto, ______/______/_________

“Costumávamos pensar que o nosso destino estava escrito nas estrelas. Hoje
sabemos que em grande parte, ele está nos nossos genes.”
James Watson

i
Agradecimentos
A dissertação de mestrado aqui apresentado é o culminar de um trabalho que só
foi possível devido ao empenho e dedicação de inúmeras pessoas, que de uma forma
ou outra, tiveram um papel muito importante. Como tal, gostaria de agradecer:
aos meus orientadores, Doutora Teresa Amaro e Doutora Sandra Ramos, que me
acompanharam ao longo deste trabalho contribuindo de forma exemplar para a minha
formação cientifica e pessoal, permitindo-me adquirir as mais variadas competências.
Pelo seu profissionalismo, espirito crítico e bom humor, um muito obrigado,
ao Professor Doutor Aires Teles, coordenador de Mestrado de Recursos Biológicos
Aquáticos, por sempre ter facilitado as meios necessários para realização deste
mestrado,
à Doutora Ascensão Ravara, pela sua colaboração que foi fundamental para a
identificação da macrofauna,
ao Doutor Hidetaka Nomaki por possibilitar a realização deste Projeto, pela sua
disponibilidade e colaboração,
ao projeto Pest-C/MAR/LA0015/20, SFRH/BPD/26782/2006 e Marie Curie Actions
através do projeto CEFMED (nº 327488),
a todos os membros do laboratório CIIMAR, que de alguma maneira contribuíram
para a realização deste estudo,
à minha avó, que já não se encontra entre nós, mas que foi uma fonte de
inspiração constante ao longo de todo este trabalho, por todas as qualidades que a
caracterizavam,

ii
aos meus pais, a minha irmã, ao André e aos restantes familiares pela
compreensão, pelo carinho e pelo apoio incondicional que demostraram para comigo
ao longo deste tempo, sem eles este percurso teria sido mais difícil,
aos meus amigos, Nita, Mi e Di, agradeço pelo apoio e carinho, paciência e
amizade, principalmente nos momentos mais difíceis,
ao Gui por fazer parte da minha vida, me fazer sorrir e nunca me deixar desistir,
A todos um muito obrigado!!!

iii
Até ao momento, deste trabalho resultaram duas comunicações (poster) em
reuniões científicas: 1. no Projeto Pluridisciplinares e 2. no Congresso Internacional de
Aveiro “14th Deep-Sea Biology Symposium”.
Póster científico:
Alves, A., Ramos, S., Ravara, A., Amaro, T. (2015). Benthic Macrofaunal
Communities Differences Between Oligotrophic and Eutrophic Abyssal Habitats: A
Case Study of the West Pacific. IJUP – Investigação Jovem na Universidade do Porto
– Projeto Pluridisciplinar, Maio 2015.
Alves, A., Ramos, S., Nomaki, H., Ravara, A., Amaro, T. (2015). Feeding
Preferences of Abyssal Macrofauna Inferred from in situ Pulse Chase Experiments in
the West Pacific. 14th DSBS. Aveiro, Setembro 2015. (comunicação em poster).

iv
Resumo
O ecossistema do mar profundo é o mais extenso ecossistema na terra e cobre
60% da superfície do planeta. O biota associada aos substratos consolidados e não
consolidados, também conhecido como bentos, é altamente diverso e complexo, e tem
um papel fundamental no fluxo de energia, em diferentes níveis tróficos das cadeias
alimentares. Assim, as comunidades bentónicas alimentam-se de matéria orgânica em
suspensão ou depositada no sedimento. Neste contexto, o presente estudo teve como
objetivos caracterizar as comunidades da macrofauna bentónica de dois habitats do
mar profundo do Pacifico ocidental e verificar a resposta destas comunidades face à
presença de diferentes alimentos (cianobactérias e diatomáceas). Para tal, foram
analisadas organismos de dois habitats diferentes do Pacifico ocidental: eutrófico (alto
teor de matéria orgânica - 39⁰00N, 146⁰00E, profundidade da água: 5256 metros) e
oligotrófico (baixo teor de matéria orgânica - 1⁰15N, 163⁰15E, profundidade da água:
4277 metros), recolhidos pelo submarino Shinkai 6500. A diversidade da macrofauna
foi estudada através das abundâncias de seis frações de profundidades (0-1; 1-2; 2-3;
3-5; 5-10 e 10-15 cm). Após identificação até ao nível taxonómico mais baixo possível,
a abundância, diversidade, riqueza específica foram calculadas e atribuída a posição
trófica na cadeia alimentar de cada taxa identificado. A posição trófica da cadeia
alimentar foi inicialmente determinada através da classificação do grupo trófico
funcional de acordo com a informação disponível na literatura e, posteriormente
comprovada através da análise de isótopos estáveis. Os resultados evidenciaram que
no habitat eutrófico a abundância média (1320±277 ind./m2), foi mais elevada com 23
diferentes taxa sendo os Nematoda o grupo mais abundante (74%). Em contraste, no
habitat oligotrófico, a abundância foi em média inferior (83±61 ind./m2) com apenas 6
diferentes taxa identificados, sendo o filo Artropoda o mais abundante (38%). Para
avaliar a resposta da macrofauna a dois tipos de alimentos foi realizada uma
experiencia in situ de modo a simular um bloom de fitoplâncton. Assim, as
comunidades de cada habitat foram injetadas com dois tipos distintos de alimento
(cianobactérias e diatomáceas) marcados radioactivamente, durante um período curto
e longo de incubação. Através da análise de isótopos estáveis, os resultados não
foram conclusivos para o habitat oligotrófico. No entanto, para o habitat eutrófico
verificou-se existir preferência pelas diatomáceas, tanto no período curto, como no
longo de incubação. Este estudo contribuiu para o aumento do conhecimento científico

v
do funcionamento trófico das comunidades de macrofauna bentónica das planícies
abissais do Pacifico profundo.

vi
Abstract
The deep-sea floor ecosystem is the largest ecosystem on Earth, covering about
60% of the globe surface. The biota associated with consolidated and unconsolidated
substrates is highly diverse and complex, and has a fundamental role for the energy
flow at different trophic levels of the food web. Deep benthic and pelagic communities
feed of organic matter sinking through the ocean interior from the euphotic zone. The
content of available organic matter controls the benthic communities, working as a
major environmental driver. To examine the response of abyssal benthic communities
to different food resources we simulated a food sedimentation event containing
diatoms and cyanobacteria at two stations in the Western Pacific. In this context, the
benthic macrofaunal communities at two different habitats of western Pacific were
analysed: one eutrophic (high organic matter content; 39˚00N, 146 ˚00E, water depth:
5256 m) and another oligotrophic (low organic matter content; 1˚15N, 163˚15E, water
depth: 4277 m), collected within Shinkai 6500 submarine. The macrofauna abundance
was studied along six depth layers, namely 0-1, 1-2, 2-3, 3-5, 5-10 and 10-15 cm. After
identification until the possible lowest taxonomic level, abundance, diversity, richness
and trophic position in the food web were calculated. The trophic position in the food
web was initially determined by classification of the functional trophic group according
to the information available in the literature, and later by analysis of stable isotopes.
Mean abundance ranged between 83±61 ind./m2 in the oligotrophic station and
1320±277 ind./m2 in the eutrophic station. Species richness also varied, with 23 taxa
observed at the eutrophic habitat and only 6 taxa in the oligotrophic habitat. At each
sampling station we simulated a food sedimentation event containing labeled
cyanobacteria and diatoms during two incubation periods (short and long). We found
no evidence for selective ingestion of the different food at the oligotrophic habitat,
whereas at the eutrophic habitat, the community seems to have preference for diatoms
during both short and long incubation periods. This study contributed to increase our
scientific knowledge of deep Pacific benthic communities, and understanding of their
trophic functioning.

vii
Índice
CAPÍTULO I – INTRODUÇÃO .......................................................................................................... 1
1.1 – O mar profundo ........................................................................................................... 2
Planícies abissais ....................................................................................................................... 4
Fontes hidrotermais ................................................................................................................... 4
Canhões Submarinos ................................................................................................................ 5
Montes submarinos .................................................................................................................... 7
Fontes Frias ................................................................................................................................. 7
1.2 – O estudo das comunidades bentónicas do mar profundo na planície
abissal ..................................................................................................................................... 8
1.2.1 – Métodos de amostragem em mar profundo ................................................. 9
1.3 – Objetivo ....................................................................................................................... 14
CAPÍTULO II – MATERIAL E MÉTODOS ........................................................................................ 14
2.1 – Locais de amostragem ............................................................................................ 15
2.2 – Experiências in situ ................................................................................................. 17
2.3 – Amostragem .............................................................................................................. 18
2.4 – Tratamento laboratorial .......................................................................................... 19
2.5 – Análise de dados ...................................................................................................... 21
CAPÍTULO III – RESULTADOS ....................................................................................................... 24
3.1 – Estrutura da comunidade de macrofauna ......................................................... 25
3.2 – Posição trófica .......................................................................................................... 29
3.3 Estudo do alimento preferencial da macrofauna ................................................ 31
CAPÍTULO IV – DISCUSSÃO ......................................................................................................... 33
CAPÍTULO V – CONSIDERAÇÕES FINAIS ...................................................................................... 37
CAPÍTULO VI – BIBLIOGRAFIA ..................................................................................................... 39

viii
Lista de Figuras
Figura 1 - Esquema representativo das diferentes zonas e regiões que constituem os
oceanos adaptado de http://www.geoprof.org/mar_prof.php .............................................. 3
Figura 2 - Fontes hidrotermais mais exploradas .................................................................. 5
Figura 3 - Principais equipamentos utilizados na amostragem de organismos
bentónicos A – Multicorer; B – Submersível; C – Boxcorer .............................................. 10
Figura 4 - Cadeia alimentar do meio aquático (modificado de Andréa et al., 2007) ..... 12
Figura 5 - Topografia do Oceano Pacífico – adaptado (Wishner et al., 1995) .............. 15
Figura 6 - Representação dos dois locais de amostragem - campanhas YK 13-09/12 e
YK 14-06/12. ............................................................................................................................. 16
Figura 7 - Desenho Experimental ......................................................................................... 18
Figura 8 - Corer fracionado em seis frações de profundidade ......................................... 19
Figura 9 - Exemplares de macrofauna bentónica encontrados no Oceano Pacifico
entre Julho e Setembro 2013 e Maio e Julho 2014. A – Ostracoda; B – Isopoda; C –
Cumacea; D- Copepoda; E – Sipuncula; F – Nematoda; G – Bivalve; H –
Chrysopetalidae; I – Capitellida; J - Phyllodocidae; K – Fauveliopsidae; L –
Sabelliriidae; M – Cirratulidae; N – Spionidae; O – Terebellidae. ............................ 20
Figura 10 - Percentagem da abundância total dos grupos funcionais para cada local
de amostragem (eutrófico versus oligotrófico) ..................................................................... 25
Figura 11 - Percentagem da abundância relativa da família de poliquetas para cada
local de amostragem (eutrófico versus oligotrófico) ........................................................... 26
Figura 12 - Abundância de poliquetas por grupos funcionais para o local eutrófico e
oligotrófico A classificação das poliquetas em grupos funcionais foi baseada em
Fauchald e Jumars (1979). ..................................................................................................... 27
Figura 13 - Distribuição vertical da abundância total da comunidade de macrofauna no
controlo, nº de cores=4. ........................................................................................................... 28
Figura 14 - Índices de diversidade da macrofaua bentónica do Oceano Pacífico, entre
Julho e Setembro 2013 e Maio e Julho de 2014. H´- Shannon Wiener; J´-
Equitabilidade; d - Riqueza Específica ................................................................................. 28
Figura 15 - Análise isotópica (13C, 15N) do controlo do A - local oligotrófico, B - local
eutrófico. .................................................................................................................................... 30

ix
Figura 16 - Incorporação de cianobactérias (A,B) e diatomáceas (C) durante o período
curto (A,C) e longo (B) no local oligotrófico. ........................................................................ 31
Figura 17 - Incorporação de cianobactérias (A,B) e diatomáceas (C,D) durante o
período curto (A,C) e longo (B,D) no local eutrófico........................................................... 32

x
Lista de Tabelas
Tabela 1 - Resultados das análises ANOSIM e SIMPER entre os dois locais de
amostragem (eutrófico vs oligotrófico). ................................................................................. 29

CAPÍTULO I – INTRODUÇÃO

Introdução
2
1.1 – O mar profundo
Os ambientes marinhos são os maiores habitats na Terra, sendo os oceanos
responsáveis por cobrirem cerca de 70% da superfície do planeta. São extremamente
diversos com diferentes teores de salinidade e grandes variações de pressão e
temperatura (Kennedy et al., 2010).
O oceano pode ser dividido verticalmente e horizontalmente, de acordo com as
características físicas e biológicas de cada secção. No meio marinho, a luz solar
consegue penetrar até aproximadamente 200 metros (Galand et al., 2010), enquanto
que a profundidades maiores, não há praticamente luz. Assim, a topografia dos
oceanos é tipicamente dividida em várias zonas: (1) região fótica que cobre os
oceanos a partir do nível da superfície até 200 metros de profundidade – zona
epipelágica (Galand et al., 2010); (2) região disfótica caracterizada por fraca
iluminação e, embora ainda haja alguma luz esta não é suficiente para a realização da
fotossíntese - zona mesopelágica (200-1000 metros de profundidade) (Barange et al.,
2010)) e (3) região afótica camada profunda dos ecossistemas aquáticos onde não há
ação direta da luz solar – zona batipelágica (1000-3000 metros de profundidade),
abisopelágica ou abissal (3000-6000 metros de profundidade) e zona hadopelágica ou
hadal com mais de 6000 metros de profundidade (Barange et al., 2010; Galand et al.,
2010) (Figura 1).
Gage & Tyler, (1991), descrevem que o mar profundo começa no declive abrupto
da plataforma, aproximadamente nos 200 metros de profundidade (transição da fauna
de águas superficiais para o mar profundo) e, é caracterizado por um vasto ambiente,
que cobre mais de 65% da superfície terrestre (Sverdrup et al., 1942; Sanders et al.,
1965; Hessler & Jumars, 1974; Merrett, 1989). Apesar das condições neste ambiente
serem extremas, com altas pressões, baixas temperaturas e pequena disponibilidade
de alimento, o mar profundo pode ser caracterizado como um ambiente fisicamente
estável (Sanders, 1968). Embora a área total estudada seja ainda muito pequena, o
conhecimento do mar profundo tem aumentado significativamente durante as últimas
décadas (Cochonat et al., 2007).

Introdução
3
Os primeiros estudos dos ecossistemas marinhos de grande profundidade
(Sanders et al., 1965; Sanders & Hessler, 1969) tiveram como objetivo principal
descrever e explicar a biodiversidade existente nessas regiões (Brandt et al., 2004).
No século XX observaram-se desenvolvimentos notáveis no domínio da
investigação dos oceanos, nas tecnologias submarinas e na acústica, ocorridas
durante a 2ª Guerra Mundial. Multiplicaram-se os cruzeiros científicos e nos anos 60,
com o submersível americano Alvin, iniciou-se uma nova era da investigação do
oceano profundo com acesso a sistemas de visualização. Nos anos 80 surgem outros
submersíveis com capacidades para mergulhar abaixo dos 4000 metros de
profundidade, como o Nautile da França, os MIR da União Soviética e o Shinkai do
Japão. Entretanto com o desenvolvimento das tecnologias robóticas várias outras
plataformas começaram a estar disponíveis para a investigação oceanográfica.
Desta forma, um grande número de habitats foi descoberto, como canhões, montes
submarinos, recifes, fontes hidrotermais e fontes frias, que suportam comunidades
microbianas e faunísticas únicas (Van Dover, 2000; Barry et al., 2002; Brandt et al.,
2007; Roberts, 2009; Tyler et al., 2009).
Figura 1 - Esquema representativo das diferentes zonas e regiões que constituem os oceanos adaptado de http://www.geoprof.org/mar_prof.php

Introdução
4
Planícies abissais
A planície abissal é uma área extensa dos fundos oceânicos com topografia suave
e plana, normalmente coberta por sedimentos pelágicos, situadas entre os 4000 e os
6000 metros de profundidade (Tyler, 2003). Nas planícies abissais podem existir
depressões designadas por fossa, que apresentam grandes profundidades, podendo
mesmo ultrapassar os 11000 metros. Podem ainda existir ilhas e colinas formadas
pela acumulação de materiais vulcânicos emitidos por vulcões submarinos (Smith &
Demopoulos, 2003).
Até aos cinco mil metros de profundidade, a fina textura dos sedimentos é formada
pela deposição das carapaças de microrganismos calcários, como os de foraminíferos
e de certas algas. Abaixo dessa profundidade, o sedimento mais comum é a de
natureza silicosa, formada a partir das carapaças de diatomáceas que afundam desde
a superfície. A grande parte do sedimento nas planícies abissais é depositado a partir
das margens continentais ao longo dos canhões submarinos
Fontes hidrotermais
As fontes hidrotermais são uma espécie de vulcões submarinos, que podem atingir
temperaturas na ordem dos 300ºC proporcionando inúmeras formas de vida, e
aparecem geralmente associadas aos rifts oceânicos ou a zonas de subducção.
Foram descobertas em 1977 e que estão normalmente situadas abaixo de dois mil
metros de profundidade em regiões de fundo oceânico com atividade vulcânica. A
biologia e a fisiologia das comunidades bentónicas em torno das fontes hidrotermais
profundas estão relativamente bem estudadas, no entanto, existe pouca informação
sobre o efeito de fontes hidrotermais de baixa profundidade no ecossistema
(Thiermann et al., 1997).
Atualmente estas áreas tem sido exploradas ativamente e cada vez mais fontes
hidrotermais são descobertas. O submersível Alvin desde de 1977 tem sido utilizado
como meio para as descobrir e catalogar e, já efetuou mais de 1000 mergulhos.
Devido ao cariz provisório das fontes hidrotermais torna-se difícil identificar
concretamente o local onde estas áreas se encontram, pois muitas das que já foram
identificadas já colapsaram e muitas estarão a surgir neste momento. Alguns dos

Introdução
5
sítios mais estudados situam-se no mar Egeu (Dando et al., 1995), na Baía de Plenty,
Nova Zelândia (Pichler & Dix, 1996), na cumeeira de Kolbeinsey, na Islândia (Botz et
al., 1996), em Kraternaya Bight, nas Ilhas Kurile (Tarasov et al., 1990), no porto de
Matupi, na Baía de Tatum, na Papua Nova Guiné (Pichler et al., 1999; Tarasov et al.,
1999) e a região subpolar de baixa profundidade na cumeeira do médio Atlântico
(Fricke et al., 1989) (Figura 2).
A alteração do ambiente por influência das fontes hidrotermais ocorrem tanto em
fontes hidrotermais de elevada profundidade, como nas de baixa profundidade, e têm
um efeito significativo no habitat bentónico. No entanto, o seu efeito na coluna de água
é bastante diferente. Normalmente, o efeito das fontes hidrotermais profundas é
restrito a uma zona muito estreita em proximidade imediata com a descarga ou com a
pluma, ocorrendo um efeito semelhante em áreas abertas com fontes hidrotermais de
baixa profundidade. Em contraste, fontes hidrotermais de baixa profundidade semi-
isoladas ou em baías, têm a camada de água perto do fundo e/ou da superfície entre
0-0,5 m de profundidade afetados pelos fluidos termais (Tarasov et al., 1999).
Canhões Submarinos
Os canhões submarinos são sistemas complexos e heterogéneos. Apresentam
uma forma típica em V, com gargantas estreitas e vertentes muito inclinadas. O
Figura 2 - Fontes hidrotermais mais exploradas

Introdução
6
comprimento médio é de 50 km e, muito embora a maioria tenha poucos quilómetros
de comprimento, existem alguns com cerca de 200 a 300 km de extensão. Assim, até
ao presente o canhão submarino mais extenso possui cerca de 442 km. Estas
estruturas atingem grandes profundidades, estando muitas vezes abaixo dos 3000
metros de profundidade (Shepard & Milliman, 1978).
Os canhões submarinos embutidos na margem continental funcionam como vias
de drenagem de materiais (sedimento, lixo, etc.) do continente para a planície abissal,
e a sua eficácia depende, entre outros fatores, da distância ao litoral a que estão
definidas as suas cabeceiras e da área de plataforma que os influenciam diretamente.
Este tipo de incisões está frequentemente associado a diversos condicionalismos
energéticos (upwelling e acumulação de energia das ondas internas) com capacidade
de transferir os sedimentos acumulados na plataforma, bordo e vertente continental
superior, para a planície abissal. No caso de canhões cuja cabeceira esteja muito
próxima da orla litoral, a sua morfologia tem uma influência significativa sobre os
processos costeiros, como a hidrodinâmica da zona costeira (ex. ondas gigantes,
tsunamis) e o transporte sedimentar (ex. o Canhão Submarino da Nazaré) (Masson et
al., 2011). Além disso, a presença de canhões submarinos afeta fortemente a
circulação e a produtividade oceânica, bem como a distribuição da cobertura
sedimentar de fundo, sendo estas estruturas um dos principais mecanismos de
transferência de massa, entre a zona costeira e o oceano profundo (grandes
transferências catastróficas de massa são despoletadas, por exemplo, pela atividade
neotectónica, sendo que estas depressões submarinas tendem a guiar as correntes de
massa descendentes até às planícies abissais).
Recentemente foram feitos vários estudos focados nos habitats bentónicos
associados aos canhões submarinos e foi descoberto que devido às características
oceanográficas, geológicas anteriormente descritas, estas estruturam contém uma
diversidade elevada e por conseguinte são considerados, muitas vezes, áreas de
hotspot de biodiversidade, não só ao nível de organismos bentónicos (Tyler et al.,
2009; Amaro et al., 2010; De Leo et al., 2010), como populações de peixes
comercialmente importantes ao homem (Company et al., 2008; Würtz, 2012).

Introdução
7
Montes submarinos
Os montes submarinos são estruturas conspícuas, normalmente com flancos
abruptos e uma forma cónica de base circular, elíptica ou mais alongada. Do ponto de
vista geológico são formações que emergem do leito do oceano, geralmente de origem
vulcânica e frequentemente associados como hotspots de biodiversidade do fundo do
mar. Apesar de existirem milhares de montanhas submarinas estes sistemas são
pouco estudados (Gubbay & Deutschland, 2003). Atualmente são conhecidos cerca de
100 000 montes submarinos, dos quais 14 000 são de grandes dimensões (Pitcher et
al., 2008) e, em apenas 200 destes 14 000 montes submarinos foram recolhidas
amostras (Stocks et al., 2004). As correntes que circundam os montes submarinos e
as superfícies rochosas expostas dos mesmos oferecem condições ideias para
animais filtradores, que tendem a dominar a zona bentónica. No Oceano Atlântico
existem pelo menos 800 montes submarinos de grandes dimensões, a maioria deles,
associada à Crista Média Atlântica mas também na zona a oeste de Portugal
Continental (Gubbay & Deutschland, 2003).
Os montes são de natureza rochosa e podem possuir fissuras, falhas, canhões,
mas é frequente a acumulação de sedimentos biogénicos (Gubbay & Deutschland,
2003). O relevo dos montes submarinos tem efeitos profundos sobre a circulação
oceânica circundante, como a formação de ondas retidas, jatos, turbilhões e as
circulações fechadas conhecidas como colunas de Taylor (Rogers, 2004). Os montes
podem ser divididos (a) pouco profundos, quando o cume se encontra na zona fótica;
(b) intermédio, quando o cume se encontra entre o limite inferior da zona fótica e a
profundidade de migração diurna do zooplâncton (aproximadamente 400m) e (c)
profundos, quando o cume está abaixo dos 400 m sem efeito da luz (Pitcher et al.,
2008). Os montes são caracterizados por uma elevada biodiversidade e pensa-se que
os que se aproximam mais da superfície são importantes por facilitarem a dispersão
das espécies nos oceanos agindo como pontos de passagem (stepping stones) para
espécies nas suas rotas de colonização (Gubbay & Deutschland, 2003).
Fontes Frias
As fontes frias encontram-se entre os ambientes redutores mais geologicamente
diversos explorados até à data. Estão dispersos globalmente, ocorrendo em todas as

Introdução
8
margens continentais (tectonicamente ativas e passivas), e mesmo em lagos e mares
interiores. São conhecidas fontes frias desde profundidades inferiores a 15 m até
profundidades superiores a 7400 m (no Fosso do Japão). As fontes frias encontram-se
com frequência ao longo de fraturas na crista de colinas, nas escarpas baixas e nos
afloramentos planos, além de zonas de falha associadas a tectónicas salinas em
margens continentais passivas (Gibson et al., 2005).
Estes locais são semelhantes às fontes hidrotermais (excetuando no que respeita à
temperatura), com comunidades caracterizadas por uma produção primária quimio-
autotrófica e com associações simbióticas entre bactérias eucariotas. O metano e o
sulfureto são as principais fontes de energia, são oxidados por espécies bacterianas
na presença de oxigénio (ou possivelmente nitrato). As altas concentrações de
metano, hidrocarbonetos ou sulfureto podem estar presentes no fluido, sendo que as
concentrações e o fluxo têm grandes efeitos na composição da comunidade quimio-
autotrófica adjacente (Solem, 1997). O metano existente nas fontes frias pode ser
biogénico (microbiano) ou termogénico na sua origem. As taxas de 13C/12C diferem
entre os mecanismos, com o metano biogénico a ter menores quantidades de 13C na
sua assinatura. Em sedimentos anóxicos controlados por difusão, todo o metano
produzido por metanogénese é oxidado na zona de transição metano/sulfato e nunca
alcança as águas do fundo. Nas fontes frias, a água saturada de metano existente nos
poros do sedimento é transportada na direção da superfície do sedimento e a grande
disponibilidade de metano leva a taxas mais altas de Oxidação Anaeróbica de Metano
na superfície dos sedimentos (Gibson et al., 2005).
1.2 – O estudo das comunidades bentónicas do mar profundo na planície abissal
As comunidades bentónicas são compostas por organismos que têm contato com
o substrato em pelo menos uma etapa do seu ciclo de vida, ou apresentam uma íntima
relação com ele. A estrutura das comunidades é influenciada pela competição entre as
espécies que a compõem, sendo o espaço e o alimento os recursos mais limitantes.
Deste modo, desempenham um papel importante e fundamental na estrutura física
dos seus habitats e nos processos ecológicos e biogeoquímicos destes ecossistemas
(Thrush & Dayton, 2002).

Introdução
9
As comunidades bentónicas podem ser classificadas ou catalogadas em diferentes
grupos funcionais, em função das suas dimensões, biótopos, distribuição vertical e
nutrição (Levinton, 2001). De acordo com o habitat preferencial os organismos
bentónicos podem ser classificados em: infauna (ou endofauna) que incluem todos os
organismos bentónicos que escavam substrato ou encontram-se enterrados no
sedimento ou rochas; e epifauna que é composta por espécies que vivem ou
deslocam-se sobre o substrato (Gray & Elliott, 2009). Relativamente ao tamanho,
podem ser identificados em várias categorias: (1) megafauna - organismos maiores
que 1 cm; (2) macrobentos – organismos maiores do que 0,5 milímetros; (3)
meiobentos ou mesobentos - organismos maiores do que 0,062 milímetros; (4)
microbentos - organismos menores do que 0,062 milímetros (Eleftheriou & Holme,
1984).
1.2.1 – Métodos de amostragem em mar profundo
Os métodos de estudo das comunidades bentónicas do oceano profundo são muito
variados e adaptados ao tipo de substrato e profundidade do local. Alguns
equipamentos são apropriados para realizar amostragens quantitativas, enquanto que
outros servem somente para amostragem qualitativa (tipo de engenho a utilizar,
estratégia de amostragem, evitar os organismos a amostrar, migrações verticais,
microdistribuição, evitamento, extrusão, colmatagem, etc.) (Nybakken, 1993). Nas
últimas décadas, com o avanço da tecnologia oceânica e robótica, têm sido
desenvolvidos aparelhos, com ou sem tripulação, capazes de alcançar grandes
profundidades (Gage & Tyler, 1991).
No entanto, o uso de submersíveis ou câmaras controladas para descer a milhares
de metros de profundidade é uma metodologia muito dispendiosa o que limita os
estudos do fundo marinho. Outra limitação prende-se com as densidades dos
organismos bentónicos, sendo por isso necessário recolher um grande número de
amostras para quantificar de forma representativa a biodiversidade existente. As
técnicas mais utilizadas para a amostrar organismos bentónicos são os corers
(boxcorer, multicorer, megacorer), dragas e redes de arrasto. A técnica de filmagem

Introdução
10
também é um método bastante utilizada para pontos com elevadas profundidades
(Figura 3).
Os corers são normalmente usados para amostrar sedimentos para serem
analisados a nível químico, geo-quimico e biológico e permitem amostrar os
sedimentos de uma forma quantitativa. O boxcorer permite amostrar uma área maior
(600 a 2500 cm2) e a camadas mais profundas (penetração de 50 cm), mas a
qualidade da amostra é menor, especialmente nas camadas mais superficiais. Os
multicorers podem ter 4 a 8 tubos (Diâmetro 6 cm, comprimento 80 cm). Enquanto que
a qualidade da amostra é excelente, a área é bem menor e permite amostrar a uma
profundidade tão grande no sedimento. Embora, tenham tubos maiores em
comprimento do que as boxcorers, a penetração máxima não vai além dos 20 cm. São
mais usados para amostragens dos meiobentos e microbentos, enquanto que os
megacorers (tubos diâmetro 11 cm) são usados para a macrofauna. As dragas são
armações pesadas de metal que são arrastadas sobre o fundo marinho, enquanto os
materiais vão sendo depositados em sacos de forma cónica.
Figura 3 - Principais equipamentos utilizados na amostragem de organismos bentónicos A – Multicorer; B – Submersível; C – Boxcorer

Introdução
11
A composição específica dos ecossistemas varia entre regiões, mas os princípios
de transferência energética entre os diferentes componentes da cadeia, desde os
organismos fotossintéticos e os sucessivos níveis tróficos, são similares em todos os
oceanos (Pope & Knights, 1982; Daan, 1986). A macrofauna é uma componente
chave dentro dos ecossistemas aquáticos, uma vez que participa intensamente nas
atividades de manutenção e regulação dos processos do ecossistema, como
decomposição da matéria orgânica e reciclagem de nutrientes (por bioturbação).
Adicionalmente, ocupa uma posição essencial nas cadeias alimentares aquáticas, por
serem os principais consumidores secundários (Nybakken, 1993). No entanto, podem
também incluir espécies detritívoras e, assim, representam um elo fundamental entre
as cadeias de detritos e de cadeia trófica mais elevada. Devido à ação destes
invertebrados, os detritos são transformados em partículas menores, aumentando a
área disponível para colonização por microrganismos decompositores (Pomeroy,
1980). Deste modo, indiretamente aceleram a decomposição dos detritos e, portanto,
a disponibilização de nutrientes minerais para os produtores (Anderson & Cargill,
1987). Devido à sua grande dependência dos detritos, não apenas como alimento,
mas também como habitat, a disponibilidade e a qualidade da matéria orgânica estão
entre os principais fatores que interferem na estrutura das comunidades
macrobentónicas (Rich & Wetzel, 1978), juntamente com a concentração de oxigênio
dissolvido na água e a granulometria do substrato (Brinkhurst, 1970).
As cadeias alimentares não são sequências isoladas, estão interligadas, e o
padrão de interconexões denomina-se rede alimentar ou rede trófica. Em
comunidades naturais complexas diz-se que os organismos que obtêm o seu alimento
através do mesmo número de estágios pertencem ao mesmo nível trófico (Figura 4).
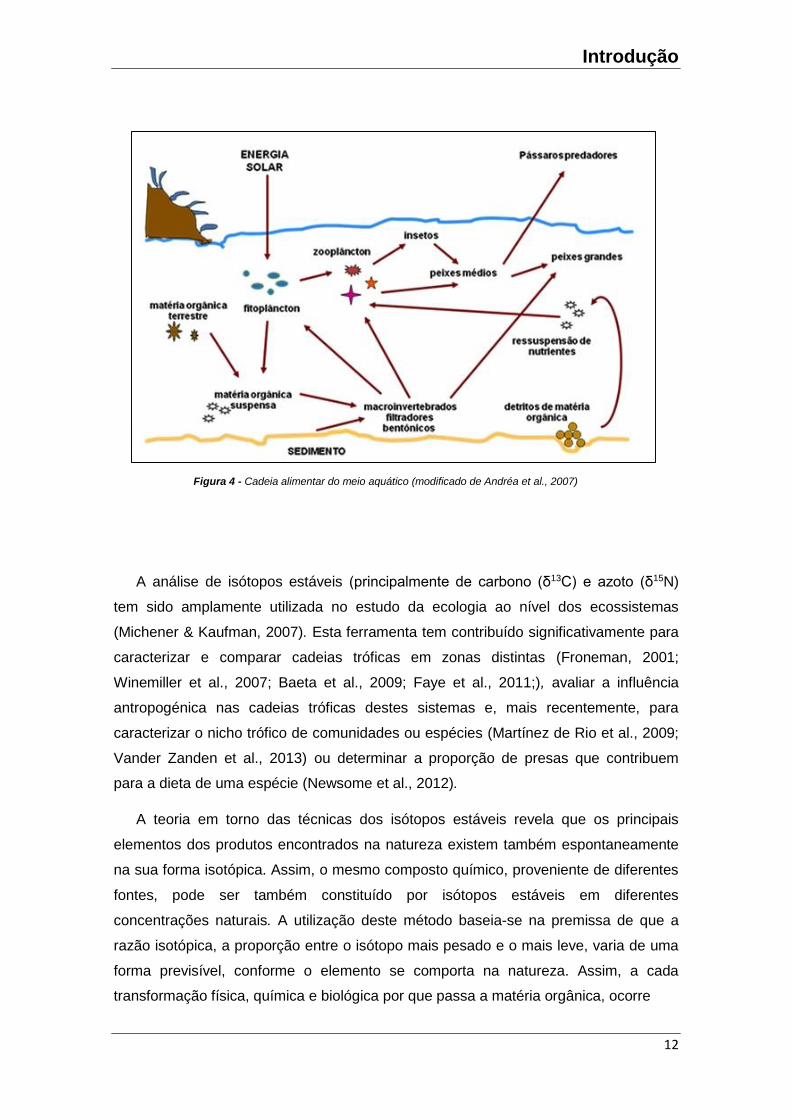
Introdução
12
A análise de isótopos estáveis (principalmente de carbono (δ13C) e azoto (δ15N)
tem sido amplamente utilizada no estudo da ecologia ao nível dos ecossistemas
(Michener & Kaufman, 2007). Esta ferramenta tem contribuído significativamente para
caracterizar e comparar cadeias tróficas em zonas distintas (Froneman, 2001;
Winemiller et al., 2007; Baeta et al., 2009; Faye et al., 2011;), avaliar a influência
antropogénica nas cadeias tróficas destes sistemas e, mais recentemente, para
caracterizar o nicho trófico de comunidades ou espécies (Martínez de Rio et al., 2009;
Vander Zanden et al., 2013) ou determinar a proporção de presas que contribuem
para a dieta de uma espécie (Newsome et al., 2012).
A teoria em torno das técnicas dos isótopos estáveis revela que os principais
elementos dos produtos encontrados na natureza existem também espontaneamente
na sua forma isotópica. Assim, o mesmo composto químico, proveniente de diferentes
fontes, pode ser também constituído por isótopos estáveis em diferentes
concentrações naturais. A utilização deste método baseia-se na premissa de que a
razão isotópica, a proporção entre o isótopo mais pesado e o mais leve, varia de uma
forma previsível, conforme o elemento se comporta na natureza. Assim, a cada
transformação física, química e biológica por que passa a matéria orgânica, ocorre
Figura 4 - Cadeia alimentar do meio aquático (modificado de Andréa et al., 2007)

Introdução
13
uma discriminação entre os seus isótopos, possibilitando a sua utilização como
marcadores naturais (Boutton, 1991). Por exemplo, um animal ao ingerir e assimilar
um determinado alimento reflete-o no sinal isotópico dos seus tecidos. O uso de
isótopos estáveis além de determinar as fontes alimentares, pode também prover uma
contínua mensuração da posição trófica que um organismo integra, a assimilação de
energia ou fluxo de massa através das diferentes vias tróficas de um organismo (Post,
2002). A determinação da composição isotópica do tecido e as suas prováveis fontes
(alimento) fornecem informações quantitativas sobre as contribuições relativas de
cada uma dessas fontes (DeNiro & Epstein, 1978).
A assinatura de carbono é geralmente utilizada para determinar as fontes de
carbono orgânico que suportam as cadeias tróficas, ocorrendo um fracionamento de
0.4‰ ± 1.3‰ da presa para o predador (Post, 2002). Através deste isótopo é possível
avaliar a dependência da cadeia trófica em relação a fontes de energia marinhas ou
terrestres (Paterson & Whitfield, 1997), dado que os produtores primários terrestres
têm, geralmente, valores inferiores de δ13C relativamente aos produtores primários
marinhos (Michener & Kaufman, 2007).
A assinatura isotópica de azoto, cujo fator de fracionamento é de cerca de 3.4‰ ±
1‰, proporciona informação sobre a posição de uma determinada espécie na cadeia
trófica (Post, 2002). No entanto, pode também contribuir para determinar a origem da
matéria orgânica em sistemas costeiros, dado que organismos marinhos são mais
enriquecidos em δ15N que os terrestres (Michener & Kaufman, 2007), ou ainda avaliar
a influência de compostos inorgânicos à base de azoto uma vez que estes tendem a
apresentar valores de δ15N mais elevados (Finlay & Kendall, 2007). O fracionamento
isotópico do azoto pode ser influenciado por diversos fatores, como o conteúdo de
azoto da dieta, condições ambientais, grupo taxonómico e o grupo trófico (McCutchan
et al., 2003; Vanderklift & Ponsard, 2003).
Os valores ᵹ13C (13C/12C) apresentam variação quantitativa ao longo da cadeia
alimentar, sofrendo incremento de 1‰ a 2‰ a cada nova posição em relação à cadeia
trófica de uma comunidade (Fry & Sherr, 1984). No entanto, os valores ᵹ15N (15N/14N)
são maiores que o da dieta, com o enriquecimento do valor isotópico entre 3‰ a 4‰
por nível trófico, permitindo estimar a posição trófica. Isótopos de azoto oferecem um
forte sinal do nível trófico em que se encontra o individuo num certo período de tempo,

Introdução
14
porque é um valor influenciado pela fonte do alimento da proteína ingerida (Minagawa
& Wada, 1984). Deste modo, são necessários cerca de três meses para que se detete
a composição isotópica da nova presa no tecido do consumidor.
1.3 – Objetivo
O presente trabalho esteve inserido num projeto de investigação CEFMED –
Climate effects on the deep-sea ecosystem functioning of the Mediterranean Sea, cujo
objetivo principal visa a criação de um modelo matemático para prever o impacto das
alterações climáticas nas comunidades bentónicas no Oceano Pacifico. Esta
investigação focou uma parte do projeto CEFMED, com o objetivo principal de avaliar
a cadeia trófica das comunidades de macrofauna de dois ecossistemas diferentes do
Oceano Pacifico, mais concretamente:
Caracterizar as comunidades de macrofauna dos dois habitats de mar
profundo, ao nível da abundância, riqueza específica, diversidade e posição
trófica na cadeia alimentar;
Verificar a resposta de ambas as comunidades face à injeção de dois
alimentos (cianobactérias e diatomáceas) radiactivamente marcados.

CAPÍTULO II – MATERIAL E MÉTODOS

Material e Métodos
15
De modo a cumprir os objetivos deste trabalho foram realizadas campanhas
oceanográficas no Pacifico Oeste e experiencias in situ em dois locais diferentes um
oligotrófico e outro eutrófico.
2.1 – Locais de amostragem
O Pacifico é o mais extenso e mais antigo oceano do mundo, com cerca de 200
milhões de anos, envolvendo mais de um terço da superfície da Terra. A crista do
Pacifico estende-se a uma latitude de 15 000 quilómetros das Filipinas ao Panamá e a
uma longitude de 10 000 quilómetros do Antártico ao Alasca (Figura 5).
Excluindo os mares adjacentes, o Oceano Pacífico abrange cerca de 166x106 km2,
tendo uma profundidade média de 4190 metros. Assim, a sua morfologia difere dos
outros oceanos devido a vasta extensão contínua da planície abissal e grande
abundância de ilhas e montes submarinos. Sendo grande parte envolvida por fossas
oceânicas profundas adjacentes a cadeias de montanhas lineares (Andes na América
Figura 5 - Topografia do Oceano Pacífico – adaptado (Wishner et al., 1995)

Material e Métodos
16
do Sul) ou arcos insulares (Ilhas Aleutas e das Marianas), com profundidade de 6700-
11000 metros (Berner, 1982; Smith & Demopoulos, 2003).
Para o presente estudo, foram realizadas quatro campanhas oceanográficas a
bordo do navio Japonês Yokosuka, onde duas estações foram visitadas. A escolha
das estações for feita de acordo com a produtividade do ecossistema do local. Deste
modo, foi escolhido um local oligotrófico (pobre em nutrientes, com baixa
produtividade), situado a 1⁰15N, 163⁰15E e com a profundidade de 4277 m e outro
eutrófico (elevada concentração de nutrientes e alta produtividade), situado a 39⁰00N,
146⁰00E e com a profundidade de 5256 m. Ambas as estações foram visitadas duas
vezes: 1. Local oligotrófico: Julho e em Setembro 2013 nas campanhas
oceanográficas denominadas YK13-09/12, respetivamente, 2. Local eutrófico: Maio e
Julho de 2014 nas campanhas oceanográficas denominadas YK14-06/12,
respetivamente (Figura 6).
Figura 6 - Representação dos dois locais de amostragem - campanhas YK 13-09/12 e YK 14-06/12.

Material e Métodos
17
2.2 – Experiências in situ
Para cada local de amostragem, foram realizadas duas experiências in situ com o
objetivo de cumprir com os objetivos propostos. Em cada local, foram enterradas 6
caixas (25x25 cm) no sedimento com o uso do submarino tripulado Shinkai 6500.
Para simular um evento de bloom de algas no mar profundo com dois alimentos
diferentes, 60 mg de Synechococcus sp. (Cianobactérias) e 60 mg de Chaetoceros
Sociale Lauler (Diatomáceas) foram injetados em cada três caixas de incubação.
Ambos os tipos de alimento foram marcados radioactivamente para posterior análise
laboratorial. A diatomácea Chaetoceros Sociale Lauler e a cianobactéria
Synechococcus sp., usadas na experiência foram cultivadas e etiquetadas no Japan
Agency for Marine – Earth Science and Technology (JAMSTEC), segundo o método
desenvolvido por (Nomaki et al., 2008). Antes de serem injetadas, as duas culturas
foram suspensas em água do mar filtrada a 0,2 mm.
A incubação foi avaliada em dois períodos diferentes curto e longo, mais
concretamente 1 e 2 dias (curto) e 51 e 58 dias (longo) para os locais oligotrófico
eutrófico, respetivamente (Figura 7).

Material e Métodos
18
2.3 – Amostragem
Em ambos os locais, a amostragem da macrofauna (3 cores recolhidos
independentes) foi obtida através do submarino tripulado Shinkai 6500 e usando push
cores (ɸ = 8.2 cm, comprimento = 32 cm). O processamento da amostra foi iniciado a
bordo do navio, dividindo o core em seis frações de profundidade 0-1 cm, 1-2 cm, 2-3
cm, 3-5 cm, 5-10 cm e 10-15 cm, seguindo técnicas standard adotadas para o mar
profundo (Cunha et al., 2011). Cada fração do sedimento foi lavado com água do mar
e crivado com uma malha de 300 µm. O material crivado foi fixado em formaldeído
tamponado, diluído com água do mar a 4% e corado com rosa Bengala (0.5 g/l) para
posterior análise laboratorial (Figura 8).
Figura 7 - Desenho Experimental

Material e Métodos
19
Os seis cores foram processados imediatamente após chegarem a bordo do navio
e as amostras de macrofauna foram usadas para determinar a abundância natural de
C e N dos organismos (referidas aqui como amostras controlo). O sedimento dos
cores usados nas experiências foram processados depois do período curto e longo de
incubação.
2.4 – Tratamento laboratorial
Cada amostra foi novamente crivada, lavada em água doce e transferida para uma
placa de Petri. Todos os organismos foram cuidadosamente identificados à lupa
binocular e microscópio ótico (quando necessário) até à família, com base nos
caracteres morfológicos, recorrendo a chaves taxonómicas e monografias
especializadas ( Saldanha, 1995; Campbell et al., 1994; Fauchald, 1977; Jumars et al.,
2015). Para efeitos de contagem dos indivíduos, apenas foram consideradas as
cabeças. A abundância de cada amostra foi determinada para cada fração de
sedimento de cada réplica e para cada tratamento.
As famílias das poliquetas foram identificadas como filtradores (F, família
Sabeliidae), decompositores de superfície (SDF, famílias Ampharetidae, Cirratulidae,
Oweniidae, Spionidae, Terebellidae), decompositores sub-superficiais (SSDF, famílias
Figura 8 - Corer fracionado em seis frações de profundidade

Material e Métodos
20
Capitellidae, Maldanidae, Paraonidae, Scallibregmatidae), predadores–detritívoros (P–
D, famílias Amphinomidae, Glyceridae, Phyllodocidae, Polynoidae) e omnívoros
(OMN, família Onuphidae) (Fauchald & Jumars, 1979).
Relativamente aos crustáceos e aos bivalves não foi possível classificar os seus
grupos tróficos, porque a resolução de taxonomia foi baixa e como tal foi impossível de
o fazer (Figura 9).
Figura 9 - Exemplares de macrofauna bentónica encontrados no Oceano Pacifico entre Julho e Setembro 2013 e Maio e Julho 2014. A – Ostracoda; B – Isopoda; C – Cumacea; D- Copepoda; E – Sipuncula; F – Nematoda; G – Bivalve; H – Chrysopetalidae; I – Capitellida; J - Phyllodocidae; K – Fauveliopsidae; L – Sabelliriidae; M – Cirratulidae; N – Spionidae; O – Terebellidae.

Material e Métodos
21
Depois da identificação e da contagem, cada indivíduo foi armazenado em
eppendorfs individuais com formaldeído tamponado, diluído com água do mar a 4% e
enviadas para o JAMSTEC (Japão), para posterior análise de isótopos estáveis. Em
resumo, os isótopos C e N foram medidos pelo instrumento Delta plus XP, Thermo -
Finnigan ligado a um analisador de elementos FlashEA1112, CE. Os rácios de
isótopos foram expressos da seguinte forma: δ13C (‰) = [(13C/12C) amostra/(13C/12C)
standard – 1] x 1000, δ15N (‰) =[(15C/14C) amostra/(15C/14C) standard – 1] x 1000
(Nomaki et al., 2011). O enriquecimento em 13C e 15N foram expressos como Δδ13C e
Δδ15N, indicando o aumento relativo em δ13C e δ15N na amostra em comparação com
o valor amostras recolhidas no controlo e foram calculadas como: Δδ13C (‰) =
δ13Csample – δ13Cbackground, Δδ15N (‰) = δ15Nsample – δ15Nbackground, respetivamente (Nomaki
et al., 2008).
2.5 – Análise de dados
A caracterização das comunidades bentónicas de cada local estudado (oligotrófico
versus eutrófico) foi efetuada ao nível da abundância, riqueza especifica, diversidade e
posição trófica na cadeia alimentar. Para tal foram consideradas como amostras os
locais controlo da experiência e, os 4 cores recolhidos em cada controlo de cada
habitat foram utlizados como réplicas para a determinação da abundância, riqueza
especifica e diversidade médias do local eutrófico e oligotrófico.
A distribuição vertical dos organismos foi estudada através da abundância média e
desvio-padrão da macrofauna das seis frações de profundidade (0-1 cm, 1-2 cm, 2-3
cm, 3-5 cm, 5-10 cm e 10-15 cm).
A abundância foi calculada através da soma total de indivíduos de cada réplica
dividida pela área de cada core (0,07 m2) e expressa como número de indivíduos por
m2 (ind.m2).
Para avaliar a diversidade de cada habitat foram calculados três indices de
diversidade, nomeadamente:

Material e Métodos
22
Índice de Riqueza Especifica de Margalef (d)
baseia-se na combinação entre o número de espécies ocorridas (S) e o número
total de indivíduos de todas as espécies (N) (Margalef, 1958) através da equação:
d = (S-1) / log N
Índice de Diversidade de Shannon-Wiener (H´)
baseia-se na proporção das abundâncias das espécies tendo em conta a
equitabilidade e a riqueza específica, assumindo que a amostragem dos indivíduos é
feita aleatoriamente a partir de uma grande população e que todas as espécies se
encontram representadas na amostra (Shannon & Weaver, 2015).
H’= - ∑i pi log (pi)
onde pi é a proporção de indivíduos da espécie i
Índice de Equitabilidade de Pielou (J´)
representa a proporção da diversidade observada pela diversidade máxima,
assumindo que todas as espécies da comunidade estão presentes na amostra,
representando de que forma os indivíduos estão distribuídos entre as diferentes
espécies (Lloyd & Ghelardi, 1964)
J’= H’/Ln (S)
Todos os índices de diversidade foram calculadas recorrendo ao software PRIMER
6 (Clarke & Warwick, 2001).
Foi feita uma análise de variância (ANOVA) para avaliar o efeito do local
(oligotrófico versus eutrófico) na abundância, diversidade (H´) e equitabilidade (J´) da
macrofauna. Todos os dados foram transformados log (x+1) de modo a cumprir os
pressupostos de normalidade e homocesdasticidade da ANOVA (Zar, 1996). As

Material e Métodos
23
análises foram efetuadas a um nível de significância de 0,05 (Zar, 1996) com o
software STATISTICA 12.
Ao nível da estrutura da comunidade de macrofauna, o efeito do local (oligotrófico
versus eutrófico) e tempo (incubação curta versus longa) foi avaliado através análise
ANOSIM (Clarke & Green, 1988). Para tal, os dados de abundância das espécies
foram transformados para raiz quadrada (reduzindo a contribuição das espécies mais
abundantes) e a similaridade entre as amostras foi medida através da matriz de
similaridade utilizando-se o coeficiente de Bray-Curtis. A análise de contribuição
percentual de similaridade (SIMPER) foi efetuada para identificar as espécies que
mais contribuíram para a dissimilaridade entre os grupos. Estas análises estatísticas
multivariadas foram efectuadas através do software PRIMER 6 (Clarke & Warwick,
2001). Mais concretamente, foi efectuado uma forma de caracterizar as comunidades
de acordo com os fatores especiais e temporais.

CAPÍTULO III – RESULTADOS

Resultados
25
3.1 – Estrutura da comunidade de macrofauna
A descrição da estrutura da comunidade baseou-se nos organismos amostrados
nos cores do controlo. No total 375 indivíduos foram identificados, distribuidos por 24
taxa referentes a 8 filos, incluindo Nematoda, Annelida, Artropoda, Mollusca,
Sipuncula, Nemertea, Loricifera e Echinodermata. A percentagem de indivíduos não
identificados (danificados ou em mau estado de conservação) foi de 3% no local
eutrófico e 10% no oligotrófico (Figura 10).
Figura 10 - Percentagem da abundância total dos grupos funcionais para cada local de amostragem (eutrófico versus oligotrófico)
No local eutrófico, os Nematoda foram o filo mais abundante, representando 71%
do total de macrofauna, seguido dos Artropoda com 13% (Figura 10). No habitat
oligotrófico o filo mais abundante foi o Artropoda com 38%, seguido do Nematoda com
36% do total da macrofauna. Os restantes filos apresentaram abundâncias
comparativamente mais baixas em ambos os locais, nomeadamente o filo Annelida,
representados aqui pelas poliquetas, que representaram 5% e 10% do total da
macrofauna no locais eutrófico e oligotrófico, respetivamente (Figura 10).
Resultados
26
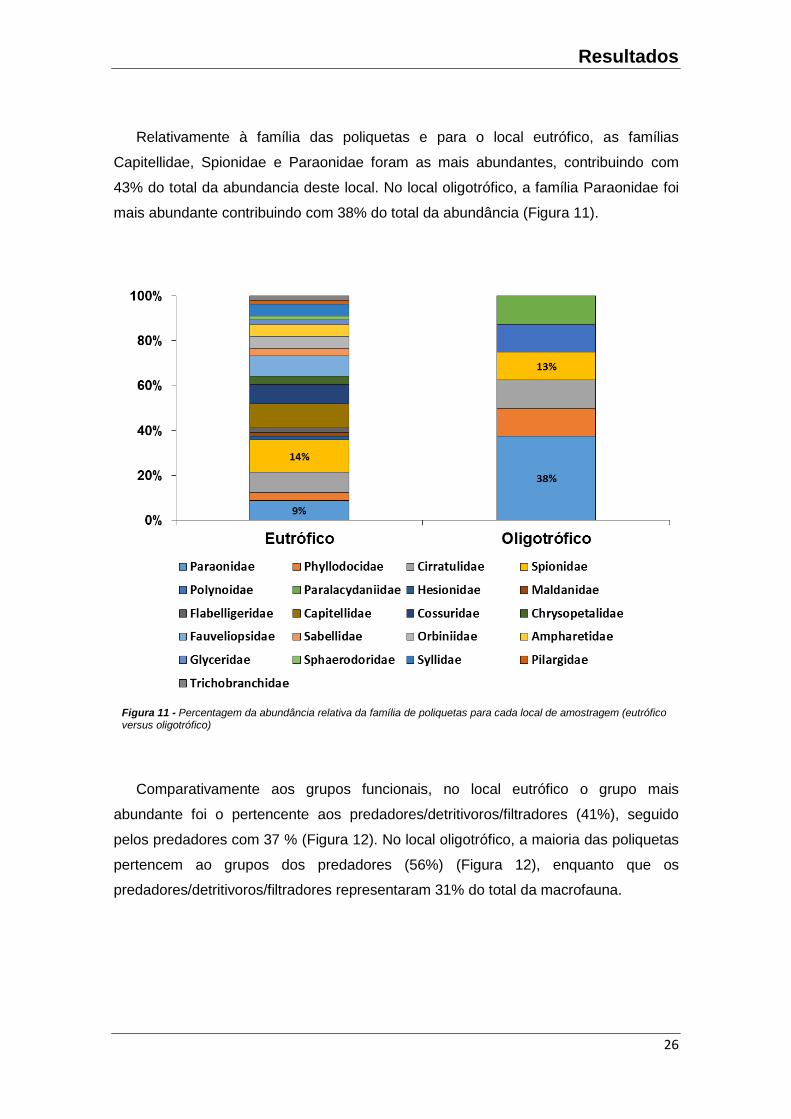
Relativamente à família das poliquetas e para o local eutrófico, as famílias
Capitellidae, Spionidae e Paraonidae foram as mais abundantes, contribuindo com
43% do total da abundancia deste local. No local oligotrófico, a família Paraonidae foi
mais abundante contribuindo com 38% do total da abundância (Figura 11).
Comparativamente aos grupos funcionais, no local eutrófico o grupo mais
abundante foi o pertencente aos predadores/detritivoros/filtradores (41%), seguido
pelos predadores com 37 % (Figura 12). No local oligotrófico, a maioria das poliquetas
pertencem ao grupos dos predadores (56%) (Figura 12), enquanto que os
predadores/detritivoros/filtradores representaram 31% do total da macrofauna.
11%
Figura 11 - Percentagem da abundância relativa da família de poliquetas para cada local de amostragem (eutrófico versus oligotrófico)

Resultados
27
A abundância média do local eutrófico foi de 1320 ± 277 ind.m2 e 83 ± 61 ind.m2 no
local oligotrófico. A macrofauna do local eutrófico foi significativamente mais
abundante do que no local oligotrófico (F= 39,64 p<0.05).
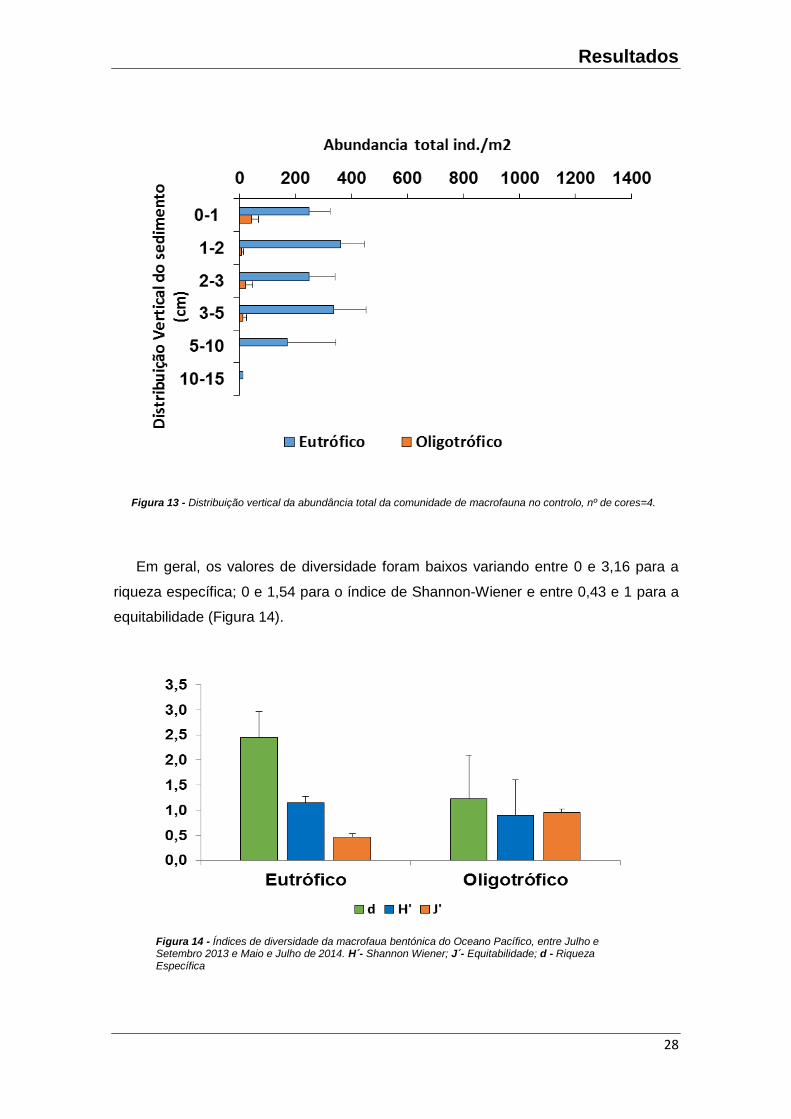
A distribuição vertical dos organismos em ambas os locais mostrou que a
macrofauna comunidade encontrada principalmente nos primeiros 10 cm (Figura 13).
Os organismos amostrados pertencem maioritariamente aos filos Nematoda e
Artropoda. No local eutrófico, as abundâncias estimadas são 4 vezes maiores do que
no local oligotrófico.
Figura 12 - Abundância de poliquetas por grupos funcionais para o local eutrófico e oligotrófico A classificação das poliquetas em grupos funcionais foi baseada em Fauchald e Jumars (1979).
Resultados
28
Em geral, os valores de diversidade foram baixos variando entre 0 e 3,16 para a
riqueza específica; 0 e 1,54 para o índice de Shannon-Wiener e entre 0,43 e 1 para a
equitabilidade (Figura 14).
Figura 13 - Distribuição vertical da abundância total da comunidade de macrofauna no controlo, nº de cores=4.
Figura 14 - Índices de diversidade da macrofaua bentónica do Oceano Pacífico, entre Julho e Setembro 2013 e Maio e Julho de 2014. H´- Shannon Wiener; J´- Equitabilidade; d - Riqueza Específica

Resultados
29
A riqueza específica e diversidade de Shannon-Wiener foram superiores no local
eutrófico (Figura 14), apesar de não serem significativamente diferentes (F= 3,93 p≥
0.05) (F= 0,7 p≥ 0.05), respetivamente. No entanto, a equitabilidade da macrofauna do
local oligotrófico foi superior, ainda que não significativa (F= 0,49 p≥ 0.05).
O resultado da análise de similaridade ANOSIM mostrou que a estrutura da
macrofauna variou significativamente entre os dois locais (R global = 0,572 p<0,05).
Segundo a análise SIMPER, os Nematoda (47,34%) juntamente com os copepodes
(13,83%) foram responsáveis por 61,17% da dissimilaridade (83,42%) da estrutura da
macrofauna dos locais oligotrófico e eutrófico (Tabela 1).
Tabela 1 - Resultados das análises ANOSIM e SIMPER entre os dois locais de amostragem (eutrófico vs oligotrófico).
Grupos ANOSIM SIMPER
R p
Dissimilaridade média
Taxa Contrib% Cum.%
Eutrófico vs Oligotrófico 0,572 0,001 83,42% Nematoda 47,34 47,34
Copepoda 13,83 61,17
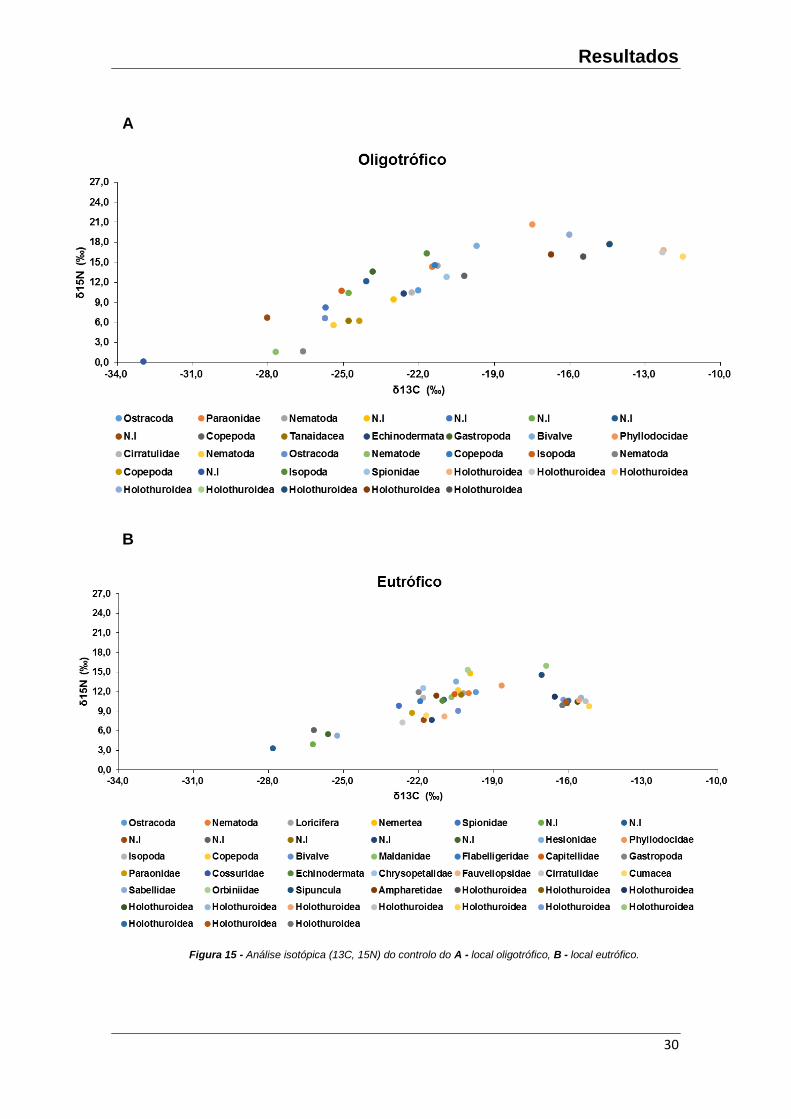
3.2 – Posição trófica
No local eutrófico, o valor mais baixo de 15N/13C ficaram associados aos
organismos a que não se conseguiu atribuir nenhuma identificação (N.I), mais
Sabellidae, encontradas nas camadas mais superficiais do sedimento (0-2 cm) e
sendo classificados como filtradores. Os grupos com uma % em 15N mais elevada
pertencem às classes Holothuroidea e Sipunculidae (detritivoros e predadores). No
local oligotrófico, o valor mais baixo de 15N/13C ficaram associados aos organismos a
que não se consegui atribuir nenhuma identificação (N.I), mais à classe dos nematoda
(maioritariamente filtradores). Os grupos com uma % em 15N mais elevada pertencem
às classes Holothuroidea e Phyllodocidae (detritivoros e predadores) (Figura 15A,B).
Resultados
30
A
B
Figura 15 - Análise isotópica (13C, 15N) do controlo do A - local oligotrófico, B - local eutrófico.
Resultados
31
3.3 Estudo do alimento preferencial da macrofauna
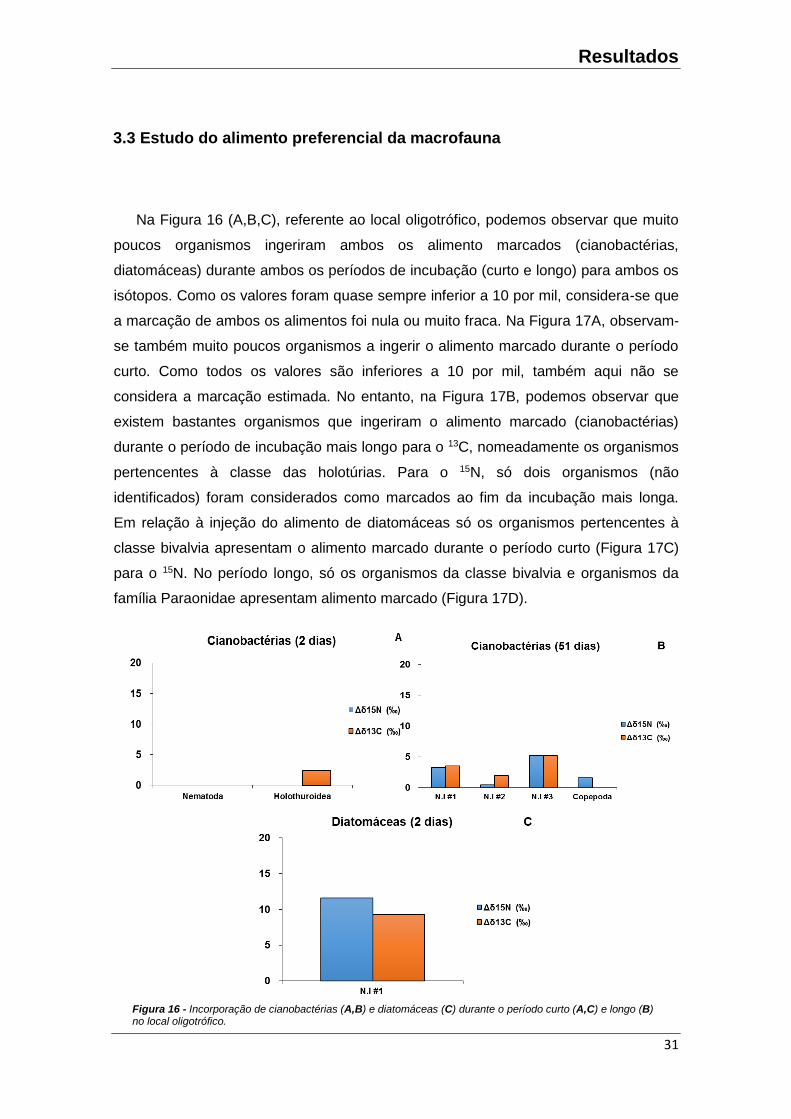
Na Figura 16 (A,B,C), referente ao local oligotrófico, podemos observar que muito
poucos organismos ingeriram ambos os alimento marcados (cianobactérias,
diatomáceas) durante ambos os períodos de incubação (curto e longo) para ambos os
isótopos. Como os valores foram quase sempre inferior a 10 por mil, considera-se que
a marcação de ambos os alimentos foi nula ou muito fraca. Na Figura 17A, observam-
se também muito poucos organismos a ingerir o alimento marcado durante o período
curto. Como todos os valores são inferiores a 10 por mil, também aqui não se
considera a marcação estimada. No entanto, na Figura 17B, podemos observar que
existem bastantes organismos que ingeriram o alimento marcado (cianobactérias)
durante o período de incubação mais longo para o 13C, nomeadamente os organismos
pertencentes à classe das holotúrias. Para o 15N, só dois organismos (não
identificados) foram considerados como marcados ao fim da incubação mais longa.
Em relação à injeção do alimento de diatomáceas só os organismos pertencentes à
classe bivalvia apresentam o alimento marcado durante o período curto (Figura 17C)
para o 15N. No período longo, só os organismos da classe bivalvia e organismos da
família Paraonidae apresentam alimento marcado (Figura 17D).
Figura 16 - Incorporação de cianobactérias (A,B) e diatomáceas (C) durante o período curto (A,C) e longo (B) no local oligotrófico.

Resultados
32
Figura 17 - Incorporação de cianobactérias (A,B) e diatomáceas (C,D) durante o período curto (A,C) e longo (B,D) no local eutrófico

CAPÍTULO IV – DISCUSSÃO

Discussão
34
Nos fundos marinhos a produção biologica está altamente dependente do fluxo de
propagação de materia orgânica a partir das zonas mais à superficie. Este fluxo de
materia orgância é muito baixo, mas constitui uma pequena percentagem da produção
primária nas águas sobrejacentes. Segundo Borowski & Thiel, (1998) na zona
equatorial do Pacifico (local eutrófico), a macrofauna bentónica apresenta grande
diversidade de taxa, incluindo poliquetas, tanaidaceos, isópodes e bivalves, sendo as
poliquetas o grupo mais dominante, representando cerca de 62% da abundância local.
Deste modo, a elevada produção primária, devido à ressurgência de nutrientes
(Berger, 1989; Murray et al., 1994; Lundry et al., 1997) e à alta produtividade perto do
equador produz um maior fluxo de carbono orgânico particulado para o interior (Honjo
et al., 1995). Hessler & Jumars, (1974), no norte do Pacifico (local oligotrófico),
também observaram a ocorrência dos mesmos grupos, mas em menor abundância,
pois a macrofauna bentónica é escassa devido à baixa concentração de nutrientes. Os
resultados deste estudo diferem consideravelmente em relação aos estudos
anteriores, uma vez que o grupo dos nematoda foi o mais abundante, sobretudo no
local eutrófico. A capacidade de adaptação e proliferação destes organismos deve-se
essencialmente ao seu caracter não-seletivo na decomposição da matéria orgânica
existente em cada local.
No local eutrófico, de acordo com Borowski & Thiel (1998), as famílias de
poliquetas que predominaram foram Cirratulidae, Paraonidae, Sabellidae e Spionidae.
Enquanto que, no local oligotrófico, de acordo com Hessler & Jumars, (1974), a família
de poliquetas mais abundantes foram Cirratulidae (25%), Capitellidae (14%),
Fauveliopsidae (11%) e Paraonidae (>6%). Mais uma vez, os resultados deste
trabalho diferem dos estudos anteriores, uma vez que as famílias mais abundantes de
poliquetas no local eutrófico foram Capitellidae, Spionidae e Paraonidae, e no local
oligotrófico foi Paraonidae. No entanto, os nossos resultados devem ser interpretados
com precaução, uma vez que o número de amostras recolhidas e a área total
amostrada ter sido inferior aos trabalhos citados, o que pode ter influenciado as
diferenças encontradas. Mas, atendendo à grande profundidade dos locais
amostrados, os custos associados para recolha destas amostras e a escassez de
estudos relativos à macrofauna bentónica no oceano Pacifico ocidental a
profundidades igual ou superior 5700 metros, os resultados deste trabalho constituem
um avanço para o aumento do conhecimento científico da macrofauna de planícies
abissais do oceano Pacifico.

Discussão
35
Relativamente aos grupos funcionais, no local eutrófico o grupo mais abundante foi
o pertencente aos predadores/detritívoros/filtradores, seguido pelos predadores. No
local eutrófico, a maioria das poliquetas pertenceram aos grupos dos predadores,
enquanto que os predadores/detritívoros/filtradores representaram o total da
macrofauna. Segundo Alba-Tercedor & Sanchez-Ortega, (1988), o estudo do número
de organismos, da sua distribuição e biomassa ou das alterações funcionais nos
ecossistemas são aspetos nos quais se deve debruçar um estudo de caracterização
de uma comunidade. Neste estudo, a caracterização da macrofauna bentónica a nível
funcional ficou bastante incompleta, devido à impossibilidade da identificação dos
organismos até ao nível taxonómico mais baixo (espécie). Por conseguinte, existiram
vários organismos que foram classificados como pertencentes a vários grupos
funcionais, como por exemplo predadores/detritívoros/filtradores.
No estudo da comunidade bentónica dos dois locais de amostragem não foram
identificados, preferências nítidas por determinados estratos da coluna de substrato
(1-2 cm). A abundância média não diferiu muito nas quatro primeiras frações de
sedimento. De acordo com estudos anteriores (Witte, 2000; Enge et al., 2011;
Gontikaki et al., 2011), a maior abundância de organismos tende a estar concentrada
até 2 cm de profundidade (as duas primeiras frações de sedimento), contrariamente
ao que se observou neste estudo. Os nossos resultados devem ser interpretados
como precaução, uma vez que tal como foi anteriormente dito, o número de amostras
recolhidas e a área total amostrada ter sido inferior aos trabalhos citados, o que pode
ter influenciado as diferenças encontradas.
No presente estudo pode-se verificar que as espécies oportunistas foram as que
mais contribuíram para as diferenças entre os dois locais, uma vez que as suas
abundâncias foram notoriamente maiores em zonas onde existe maior concentração
de nutrientes, ou seja o local eutrófico.
Para verificar qual a preferência de alimento das comunidades bentónicas nos dois
locais de amostragem (oligotrófico, eutrófico), foram injetados dois alimentos
diferentes (cianobactérias e diatomáceas). No local oligotrófico foi possível observar
que existiram poucos organismos com alimento radioactivamente marcado durante os
períodos de incubação (curto e longo). A marcação de ambos os alimentos foi
praticamente nula ou muito fraca. Tal pode ter sido devido ao período de experiência,
ou seja, um dia ser tempo insuficiente para observar que ingeriram o alimento, e 51
dias tempo demasiado logo para ainda se observar o sinal radioativo. Como este local

Discussão
36
é caracterizado por ser pobre em nutriente, a escassez de alimento pode ter feito com
que os organismos absorvessem todo o alimento até o final da experiência e como tal,
a marcação de ambos os alimentos ser práticamente nula. No entanto, não é possivel
afimar que a experiencia não resultou, porque não existe uma amostragem de tempo
intermédio para comparação. No local eutrófico observaram-se poucos organismos
que ingeriram o alimento marcado durante o período curto. Em contrapartida e, no
periodo de incubação mais longo foi possivel observar bastantes organismos que
ingeriram o alimento marcado.

CAPÍTULO V – CONSIDERAÇÕES FINAIS

38
No presente estudo foi possível concluir que a macrofauna bentónica diferiu entre o
local eutrófico e o local oligotrófico. No local eutrófico, a macrofauna foi dominada
pelos oportunistas nematoda, associados à maior disponibilidade de matéria orgânica,
enquanto que os artropoda juntamente com os nematoda dominaram a macrofauna do
local oligotrófico. As comunidades foram mais diversas com maior número de taxa no
local eutrófico, mas mais equitativas no local oligotrófico. Em relação aos grupos
tróficos, verificou-se uma maior dominância de predadores no local eutrófico e o grupo
predadores/detritívoros/filtradores teve maior abundância no oligotrófico. Finalmente, a
nível de poliquetas, três famílias foram mais abundantes no habitat eutrófico
(Capitellidae, Spionidae e Paraonidae), em comparação com apenas uma família do
local oligotrófico (Paraonidae).
Relativamente ao alimento preferencial no local oligotrófico observaram-se poucos
organismos que ingeriram o alimento marcado durante o período curto, não tendo sido
possível identificar qual o alimento preferencial. Em contrapartida, verificou-se uma
preferência evidente de diatomáceas como alimento da macrofauna bentónica do local
eutrófico. Em futuros trabalhos será necessário existir um período de incubação
intermédio, para avaliar a preferência dos alimentos pela comunidade de
macroinvertebrados destes locais do Pacifico profundo.

CAPÍTULO VI – BIBLIOGRAFIA

Bibliografia
40
Alba-Tercedor, J., & Sánchez-Ortega, A. (1988). Un método rápido y simple para
evaluar la calidad biológica de las aguas corrientes basado en el de Hellawell
(1978). Limnetica, 4(51-56).
Amaro, T., Bianchelli, S., Billett, D., Cunha, M., Pusceddu, A., & Danovaro, R. (2010).
The trophic biology of the holothurian Molpadia musculus: implications for
organic matter cycling and ecosystem functioning in a deep submarine canyon.
Biogeosciences, 7(8), 2419-2432.
Anderson, N., & Cargill, A. (1987). Nutritional ecology of aquatic detritivorous insects.
Nutritional Ecology of Insects, Mites, Spiders, and Related Invertebrates, 903-
925.
Baeta, A., Valiela, I., Rossi, F., Pinto, R., Richard, P., Niquil, N., & Marques, J. C.
(2009). Eutrophication and trophic structure in response to the presence of the
eelgrass Zostera noltii. Marine Biology, 156(10), 2107-2120.
Barange, M., Field, J. G., Harris, R. P., Hofmann, E. E., Perry, R. I., & Werner, F. E.
(2010). Marine ecosystems and global change: Oxford University Press Oxford,
412 pp.
Barry, J. P., Buck, K. R., Kochevar, R. K., Nelson, D. C., Fujiwara, Y., Goffredi, S. K., &
Hashimoto, J. (2002). Methane‐based symbiosis in a mussel, Bathymodiolus
platifrons, from cold seeps in Sagami Bay, Japan. Invertebrate Biology, 121(1),
47-54.
Berger, W. (1989). Global maps of ocean productivity. Productivity of the Ocean:
Present and Past., 429-455.
Berner, R. A. (1982). Burial of organic carbon and pyrite sulfur in the modern ocean: its
geochemical and environmental significance. Am. J. Sci, 282(4), 451-473.
Borowski, C., & Thiel, H. (1998). Deep-sea macrofaunal impacts of a large-scale
physical disturbance experiment in the Southeast Pacific. Deep Sea Research
Part II: Topical Studies in Oceanography, 45(1), 55-81.
Botz, R., Stüben, D., Winckler, G., Bayer, R., Schmitt, M., & Faber, E. (1996).
Hydrothermal gases offshore Milos Island, Greece. Chemical Geology, 130(3),
161-173.
Boutton, T. W. (1991). Stable carbon isotope ratios of natural materials: I. Sample
preparation and mass spectrometric analysis. Carbon isotope techniques, 155-
171.

Bibliografia
41
Brandt, A., De Broyer, C., Gooday, A. J., Hilbig, B., & Thomson, M. R. (2004).
Introduction to ANDEEP (ANtarctic benthic DEEP-sea biodiversity: colonization
history and recent community patterns)—a tribute to Howard L. Sanders. Deep
Sea Research Part II: Topical Studies in Oceanography, 51(14), 1457-1465.
Brandt, A., Gooday, A. J., Brandão, S. N., Brix, S., Brökeland, W., Cedhagen, T.,
Choudhury, M., Cornelius, N., Danis, B., De Mesel, I., Diaz, R.J., Gillan, D.C.,
Ebbe, B., Howe, J.A., Janussen, D., Kaiser, S., Linse, K., Malyutina, M.,
Pawlowski, J., Raupach, M., Vanreusel, A. (2007). First insights into the
biodiversity and biogeography of the Southern Ocean deep sea. Nature,
447(7142), 307-311.
Brinkhurst, R. (1970). Distribution and abundance of tubificid (Oligochaeta) species in
Toronto Harbour, Lake Ontario. Journal of the Fisheries Board of Canada,
27(11), 1961-1969.
Campbell, A. C., Nicholls, J., Múrias, A., dos Santos, P. T., & Soares, M. (1994). Fauna
e flora do litoral de Portugal e Europa, 320 pp.
Clarke, K., & Green, R. (1988). Statistical design and analysis for a" biological effects"
study. Mar. Ecol. Prog. Ser., 46(1), 213-226.
Clarke, K., & Warwick, R. (2001). An approach to statistical analysis and interpretation.
Change in Marine Communities, 2, 5-176.
Cochonat, P., Dürr, S., & Gunn, V. (2007). The Deep-Sea Frontier: Science challenges
for a sustainable future. Office for Official Publications of the European
Communities, Luxembourg, 53 pp.
Company, J. B., Puig, P., Sarda, F., Palanques, A., Latasa, M., & Scharek, R. (2008).
Climate influence on deep sea populations. PloS one, 3(1):e1431,doi:
10.1371/journal.pone.001431.
Cunha, M. R., Paterson, G. L., Amaro, T., Blackbird, S., de Stigter, H. C., Ferreira, C.,
Glover, A., Hilário, A., Kiriakoulakis, K., Neal, L., Ravara, A., Rodrigues, C.,
Tiago, A., Billett, D. (2011). Biodiversity of macrofaunal assemblages from three
Portuguese submarine canyons (NE Atlantic). Deep Sea Research Part II:
Topical Studies in Oceanography, 58(23), 2433-2447.
Daan, R. (1986). Food intake and growth of Sarsia tubulosa (Sars, 1835), with
quantitative estimates of predation on copepod populations. Netherlands
Journal of Sea Research, 20(1), 67-74.
Dando, P., Hughes, J., & Thiermann, F. (1995). Preliminary observations on biological
communities at shallow hydrothermal vents in the Aegean Sea. Geological
Society, London, Special Publications, 87(1), 303-317.

Bibliografia
42
De Leo, F. C., Smith, C. R., Rowden, A. A., Bowden, D. A., & Clark, M. R. (2010).
Submarine canyons: hotspots of benthic biomass and productivity in the deep
sea. Proceedings Biological sciences/The Royal Society 277: 2783-2792. doi:
10.1098/rspb.2010.0462.
DeNiro, M. J., & Epstein, S. (1978). Influence of diet on the distribution of carbon
isotopes in animals. Geochimica et cosmochimica acta, 42(5), 495-506.
Eleftheriou, A., & Holme, N. (1984). Macrofauna techniques. Methods for the study of
marine benthos, 140-216.
Enge, A. J., Nomaki, H., Ogawa, N. O., Witte, U., Moeseneder, M. M., Lavik, G., . . .
Heinz, P. (2011). Response of the benthic foraminiferal community to a
simulated short-term phytodetritus pulse in the abyssal North Pacific. Marine
Ecology Progress Series, 438, 129-142.
Fauchald, K. (1977). The polychaete worms: definitions and keys to the orders, families
and genera, 188 pp.
Fauchald, K., & Jumars, P. A. (1979). The diet of worms: a study of polychaete feeding
guilds (Vol. 17): Aberdeen University Press, 193-284.
Faye, D., de Morais, L. T., Raffray, J., Sadio, O., Thiaw, O. T., & Le Loc’h, F. (2011).
Structure and seasonal variability of fish food webs in an estuarine tropical
marine protected area (Senegal): Evidence from stable isotope analysis.
Estuarine, Coastal and Shelf Science, 92(4), 607-617.
Finlay, J. C., & Kendall, C. (2007). Stable isotope tracing of temporal and spatial
variability in organic matter sources to freshwater ecosystems. Stable isotopes
in ecology and environmental science, 2, 283-333.
Fricke, H., Giere, O., Stetter, K., Alfredsson, G., Kristjansson, J., Stoffers, P., &
Svavarsson, J. (1989). Hydrothermal vent communities at the shallow subpolar
Mid-Atlantic ridge. Marine Biology, 102(3), 425-429.
Froneman, P. (2001). Feeding ecology of the mysid, Mesopodopsis wooldridgei, in a
temperate estuary along the eastern seaboard of South Africa. Journal of
Plankton Research, 23(9), 999-1008.
Fry, B., & Sherr, E. B. (1984). delta super (13) C measurements as indicators of carbon
flow in marine and freshwater ecosystems. Contributions in Marine Science, 27,
13-47.
Gage, J. D., & Tyler, P. A. (1991). Deep-sea biology: a natural history of organisms at
the deep-sea floor: Cambridge University Press, 509 pp.

Bibliografia
43
Galand, P. E., Potvin, M., Casamayor, E. O., & Lovejoy, C. (2010). Hydrography
shapes bacterial biogeography of the deep Arctic Ocean. ISME J, 4(4), 564-
576. doi:10.1038/ismej.2009.134
Gibson, R., Atkinson, R., & Gordon, J. (2005). Ecology of cold seep sediments:
interactions of fauna with flow, chemistry and microbes. Oceanography and
Marine Biology: an annual review, 43, 1-46.
Gontikaki, E., Mayor, D., Thornton, B., Black, K., & Witte, U. (2011). Processing of
13C-labelled diatoms by a bathyal community at sub-zero temperatures. Mar
Ecol Prog Ser, 421, 39-50.
Gray, J. S., & Elliott, M. (2009). Ecology of marine sediments: from science to
management: Oxford University Press, 215 pp.
Gubbay, S., & Deutschland, W. (2003). Seamounts of the north-east Atlantic: Oasis,
Hamburg & WWF Germany.
Hessler, R. R., & Jumars, P. A. (1974). Abyssal community analysis from replicate
cores in the central North Pacific. Paper presented at the Deep Sea Research
and Oceanographic Abstracts.
Honjo, S., Dymond, J., Collier, R., & Manganini, S. J. (1995). Export production of
particles to the interior of the equatorial Pacific Ocean during the 1992 EqPac
experiment. Deep Sea Research Part II: Topical Studies in Oceanography,
42(2), 831-870.
Jumars, P. A., Dorgan, K. M., & Lindsay, S. M. (2015). Diet of worms emended: an
update of Polychaete feeding guilds. Marine Science, 7 497-520.
Kennedy, J., Flemer, B., Jackson, S. A., Lejon, D. P., Morrissey, J. P., O’gara, F., &
Dobson, A. D. (2010). Marine metagenomics: new tools for the study and
exploitation of marine microbial metabolism. Marine Drugs, 8(3), 608-628.
Levinton, J. S. (2001). Marine biology: function, biodiversity, ecology: Oxford University
Press New York, 420 pp.
Lloyd, M., & Ghelardi, R. J. (1964). A Table for Calculating theEquitability'Component
of Species Diversity. The Journal of Animal Ecology, 217-225.
Lundry, M. R., Barber, R. T., Bidare, R. R., Chai, F., Coale, K. H., Dam, H. G., Lewis,
M.R., Lindley, S.T., McCarthy J.J., Roman, M. R., Stoecker, D.K., Verity, P.G.,
White, J.R. (1997). Iron and grazing constraints on primary production in the
central equatorial Pacific: an EqPac synthesis. Limnology and Oceanography,
42(3), 405-418.
Margalef, D. R. (1958). Information theory in ecology: Society for General Systems
Research 3:36-71.

Bibliografia
44
Martínez del Rio, C., Wolf, N., Carleton, S. A., & Gannes, L. Z. (2009). Isotopic ecology
ten years after a call for more laboratory experiments. Biological Reviews,
84(1), 91-111.
Masson, D., Huvenne, V., de Stigter, H., Arzola, R., & LeBas, T. (2011). Sedimentary
processes in the middle Nazaré Canyon. Deep Sea Research Part II: Topical
Studies in Oceanography, 58(23), 2369-2387.
McCutchan, J. H., Lewis, W. M., Kendall, C., & McGrath, C. C. (2003). Variation in
trophic shift for stable isotope ratios of carbon, nitrogen, and sulfur. Oikos,
102(2), 378-390.
Merrett, N. R. (1989). The elusive macrourid alevin and its seeming lack of potential in
contributing to intrafamilial systematics. Papers on the systematics of gadiform
fishes. Science Series, 32, 175-151.
Michener, R. H., & Kaufman, L. (2007). Stable isotope ratios as tracers in marine food
webs: an update. Stable isotopes in ecology and environmental science, 238-
282.
Minagawa, M., & Wada, E. (1984). Stepwise enrichment of 15 N along food chains:
further evidence and the relation between δ 15 N and animal age. Geochimica
et cosmochimica acta, 48(5), 1135-1140.
Murray, J. W., Barber, R. T., Roman, M. R., Bacon, M. P., & Feely, R. A. (1994).
Physical and biological controls on carbon cycling in the equatorial Pacific.
Science, 266(5182), 58-65.
Newsome, D., Moore, S. A., & Dowling, R. K. (2012). Natural area tourism: Ecology,
impacts and management (Vol. 58): Channel View Publications, 442 pp.
Nomaki, H., Ogawa, N. O., Ohkouchi, N., Suga, H., Toyofuku, T., Shimanaga, M.,
Nakatsuka, T., Kitazato, H. (2008). Benthic foraminifera as trophic links
between phytodetritus and benthic metazoans: carbon and nitrogen isotopic
evidence. Marine Ecology-progress series, 357, 153.
Nomaki, H., Ogawa, N. O., Takano, Y., Suga, H., Ohkouchi, N., & Kitazato, H. (2011).
Differing utilization of glucose and algal particulate organic matter by deep-sea
benthic organisms of Sagami Bay, Japan. Mar. Ecol.-Prog. Ser, 431, 11-24.
Nybakken, J. W. (1993). Marine biology: an ecological approach.
Paterson, A., & Whitfield, A. (1997). A stable carbon isotope study of the food-web in a
freshwater-deprived South African estuary, with particular emphasis on the
ichthyofauna. Estuarine, Coastal and Shelf Science, 45(6), 705-715.
Pichler, T., & Dix, G. R. (1996). Hydrothermal venting within a coral reef ecosystem,
Ambitle Island, Papua New Guinea. Geology, 24(5), 435-438.

Bibliografia
45
Pichler, T., Veizer, J., & Hall, G. E. (1999). The chemical composition of shallow-water
hydrothermal fluids in Tutum Bay, Ambitle Island, Papua New Guinea and their
effect on ambient seawater. Marine Chemistry, 64(3), 229-252.
Pitcher, T. J., Morato, T., Hart, P. J., Clark, M. R., Haggan, N., & Santos, R. S. (2008).
Seamounts: ecology, fisheries & conservation: John Wiley & Sons, 527 pp.
Pomeroy, L. (1980). Detritus and its role as a food source. Fundamentals of aquatic
ecosystems, 84-102.
Pope, J., & Knights, B. (1982). Comparison of length distributions of combined catches
of all demersal fishes in surveys in the North Sea and at Faroe Bank.
Multispecies approaches to fisheries management advice, 59, 116-118.
Post, D. M. (2002). Using stable isotopes to estimate trophic position: models,
methods, and assumptions. Ecology, 83(3), 703-718.
Rich, P. H., & Wetzel, R. G. (1978). Detritus in the lake ecosystem. American
naturalist, 57-71.
Roberts, J. M. (2009). Cold-water corals: the biology and geology of deep-sea coral
habitats: Cambridge University Press, 334 pp.
Rogers, A. D. (2004). The biology, ecology and vulnerability of seamount communities
(Vol. 12): IUCN.
Saldanha, L. (1995). Fauna Marinha Atlântica. Publicaçoes Europa-America, Mem-
Martins, 364 pp.
Sanders, H., Hessler, R., & Hampson, G. (1965). An introduction to the study of deep-
sea benthic faunal assemblages along the Gay Head-Bermuda transect. Paper
presented at the Deep Sea Research and Oceanographic, 12:845-867.
Sanders, H. L. (1968). Marine benthic diversity: a comparative study. American
naturalist, 243-282.
Sanders, H. L., & Hessler, R. R. (1969). Ecology of the deep-sea benthos. Science,
163(3874), 1419-1424.
Shannon, C. E., & Weaver, W. (2015). The mathematical theory of communication:
University of Illinois press.
Shepard, F. P., & Milliman, J. D. (1978). Sea-floor currents on the foreset slope of the
Fraser River Delta, British Columbia (Canada). Marine Geology, 28(3), 245-
251.
Smith, C. R., & Demopoulos, A. W. (2003). The deep Pacific ocean floor. Ecosystems
of the World, 179-218.
Solem, I. (1997). Analysis of microbial diversity in cold seep sediments by 16 Sr
DNAbased denaturing gradient gel electrophoresis.

Bibliografia
46
Stocks, K. I., Boehlert, G. W., & Dower, J. F. (2004). Towards an international field
program on seamounts within the Census of Marine Life. Archive of Fishery and
Marine Research, 51(1-3), 320-327.
Sverdrup, H. U., Johnson, M. W., & Fleming, R. H. (1942). The Oceans: Their physics,
chemistry, and general biology (Vol. 7): Prentice-Hall New York, 1087 pp.
Tarasov, V., Gebruk, A., Shulkin, V., Kamenev, G., Fadeev, V., Kosmynin, V.,
Malakhov, V., Starynin, D., Obzhirov, A. (1999). Effect of shallow-water
hydrothermal venting on the biota of Matupi Harbour (Rabaul Caldera, New
Britain Island, Papua New Guinea). Continental Shelf Research, 19(1), 79-116.
Tarasov, V., Propp, M., Propp, L., Zhirmunsky, A., Namsakakv, B., Gorlenko, V., &
Starynin, D. (1990). Shallow‐Water Gasohydrothermal Vents of Ushishir
Volcano and the Ecosystem of Kraternaya Bight (The Kurile Islands). Marine
ecology, 11(1), 1-23.
Thiermann, F., Akoumianaki, I., Hughes, J., & Giere, O. (1997). Benthic fauna of a
shallow-water gaseohydrothermal vent area in the Aegean Sea (Milos, Greece).
Marine Biology, 128(1), 149-159.
Thrush, S. F., & Dayton, P. K. (2002). Disturbance to marine benthic habitats by
trawling and dredging: implications for marine biodiversity. Annual review of
ecology and systematics, 449-473.
Tyler, P. (2003). Ecosystems of the world, vol. 28. Ecosystems of the deep ocean:
Amsterdam, The Netherlands: Elsevier, 1-569.
Tyler, P. A., Amaro, T., Arzola, R., Cunha, M. R., Stigter, H., Gooday, A., HUveene, V.,
Ingels, J., Kiriakoulakis, K., Lastras Membrive, G. Masson, D., Oliveira, A.,
Pattenden, A., Vanreusel, A., Weering, T. Van., Vitorino, J., Witte, U., Wolff, G.
(2009). Europe's grand canyon: Nazaré submarine canyon. Oceanography,
2009, vol. 22, num. 1, p. 52-57.
Van Dover, C. (2000). The ecology of deep-sea hydrothermal vents: Princeton
University Press, 425 pp.
Vander Zanden, H. B., Arthur, K. E., Bolten, A. B., Popp, B. N., Lagueux, C. J.,
Harrison, E., Campbell, C.L., Bjorndal, K. A. (2013). Trophic ecology of a green
turtle breeding population. Marine Ecology Progress Series, 476, 237-249.
Vanderklift, M. A., & Ponsard, S. (2003). Sources of variation in consumer-diet δ15N
enrichment: a meta-analysis. Oecologia, 136(2), 169-182.
Winemiller, K. O., Akin, S., & Zeug, S. C. (2007). Production sources and food web
structure of a temperate tidal estuary: integration of dietary and stable isotope
data. Marine Ecology Progress Series, 343(6).

Bibliografia
47
Wishner, K. F., Ashjian, C. J., Gelfman, C., Gowing, M. M., Kann, L., Levin, L. A.,
Mullineaux, L.S., Saltzman, J. (1995). Pelagic and benthic ecology of the lower
interface of the Eastern Tropical Pacific oxygen minimum zone. Deep Sea
Research Part I: Oceanographic Research Papers, 42(1), 93-115.
Witte, U. (2000). Vertical distribution of metazoan macrofauna within the sediment at
four sites with contrasting food supply in the deep Arabian Sea. Deep Sea
Research Part II: Topical Studies in Oceanography, 47(14), 2979-2997.
Würtz, M. (2012). Mediterranean submarine canyons: Ecology and governance: IUCN,
216 pp.
Zar, J. H. (1996). Biostatistical analisys. Biostatistical analisys, 662 pp.





























