Embed Size (px)
Citation preview

i
INSTITUTO AGRONÔMICO
CURSO DE PÓS-GRADUAÇÃO EM AGRICULTURA
TROPICAL E SUBTROPICAL
AVALIAÇÃO DE METIL JASMONATO E DE ÁCIDO
SALICÍLICO NO CONTROLE PÓS-COLHEITA DE
PODRIDÕES EM MORANGO ‘OSO GRANDE’
ABIKEYLA DE SOUZA ROBAINA
Orientadora: Dr.ª Patrícia Cia
Dissertação submetida como requisito parcial
para obtenção do grau de Mestre em
Agricultura Tropical e Subtropical, Área de
concentração em Tecnologia da Produção
Agrícola
Campinas, SP
Maio 2013

ii

iii
DEDICATÓRIA
Aos meus pais Adilson e Nilza, pelo apoio,
encorajamento, incentivo, amor, carinho e
pelos constantes ensinamentos que formaram
os alicerces da minha vida.
Ao meu marido Carlos Cesar, por todo apoio,
amor, incentivo, paciência, compreensão e
pela companhia ao longo deste sonho.
As minhas irmãs Isabella e Kasmirra pelo amor,
carinho, apoio e compreensão.
Aos meus avós Paulo (in memorian) e Benedita,
Alfredo e Maria (in memorian).
Ao lado de vocês aprendi o significado da palavra
família e a importância de sempre seguir o
caminho da honestidade e persistência,
DEDIDO
Ao meu pai querido
Adilson de Medeiros Robaina,
pelo esforço de sempre,
pelo incondicional apoio e incentivo
durante todo meu trabalho
pelo amor de pai a mim dedicado,
OFEREÇO

iv
AGRADECIMENTOS
- A Deus pelas oportunidades que Ele semeia em meu caminho, por guiar meus passos,
pela chance de mais uma conquista, pela minha vida, pela vida dos meus familiares e
amigos e por todas as lições que aprendo por meio dos obstáculos que enfrento.
- Ao Instituto Agronômico de Campinas (IAC) pela oportunidade de realização deste
trabalho e pelos conhecimentos adquiridos.
- Ao Centro de Engenharia e Automação, localizado em Jundiaí/SP, pela cooperação e
apoio institucional.
- Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) pela bolsa
de mestrado concedida no primeiro ano do curso para a realização desde projeto de
pesquisa.
- À minha orientadora Dr.ª Patrícia Cia pela orientação, pelos ensinamentos, pela
disponibilidade e pelo exemplo de trabalho e dedicação à pesquisa.
- Aos funcionários do Centro de Engenharia e Automação, pela ajuda no laboratório, em
especial a Maria do Carmo, pela amizade e carinho.
- Aos pesquisadores que ministraram aulas nas disciplinas que eu cursei durante o
mestrado, pelos ensinamentos e disponibilidade.
- Aos meus pais Adilson de Medeiros Robaina e Nilza Ap. de Souza Robaina, pelo amor,
exemplo e apoio incondicional dedicado a mim.
- As minhas irmãs Isabella de Souza Robaina e Kasmirra de Souza Robaina, pelo amor,
pela ajuda e incentivo.
- Ao meu marido Carlos Cesar Silva Alves, pela paciência, companheirismo, amor,
incentivo e apoio de todos estes anos juntos.
- À amiga Francine Scolfaro Ponzo, pela amizade, conversas, risadas, paciência, ajuda nos
experimentos e conhecimentos compartilhados durante este trabalho.
- Aos amigos da Pós-graduação, pelo companheirismo, ajuda sempre que necessário, pelas
conversas e risadas, em especial à Franciele, Paula, Flávia e Allan.
- À Dr.ª Marise Cagnin Martins Parisi pela doação dos patógenos utilizados neste trabalho,
pela atenção e disponibilidade de sempre.
- Ao produtor de morango Daniel Juliato e sua família pelo fornecimento dos frutos.
- Aos meus queridos amigos Mariana Sacchi Armelim e Rony Nunes, pela grande
amizade, por me compreenderem e apoiarem sempre.
- À secretaria da Pós-graduação IAC, pela atenção e ajuda

v
SUMÁRIO
LISTA DE TABELAS ................................................................................................. viii
LISTA DE FIGURAS ................................................................................................. xv
RESUMO .................................................................................................................... xvii
ABSTRACT ................................................................................................................ xix
1 INTRODUÇÃO ........................................................................................................ 01
2 REVISÃO BIBLIOGRÁFICA ................................................................................. 03
2.1 Classificação e Características do Morangueiro .................................................... 03
2.2 Aspectos Socioeconômicos do Morangueiro ......................................................... 05
2.3 Principais Doenças Pós-colheita ............................................................................ 06
2.4 Conservação e Métodos de Controle de Podridões ............................................... 09
2.4.1 Ácido salicílico .................................................................................................. 12
2.4.2 Metil jasmonato ................................................................................................. 19
2.5 Indução de Resistência .......................................................................................... 22
3 MATERIAL E MÉTODOS ..................................................................................... 25
3.1 Avaliação in vivo de Metil Jasmonato e Ácido Salicílico no Controle do Mofo
Cinzento, da Podridão Mole e da Antracnose em Morangos ...................................... 25
3.1.1 Inoculação dos frutos ......................................................................................... 25
3.1.2 Efeito de metil jasmonato no controle in vivo da antracnose, do mofo cinzento
e da podridão mole em morangos ................................................................................ 26
3.1.3 Efeito do ácido salicílico no controle in vivo da antracnose, do mofo cinzento
e da podridão mole em morangos ................................................................................ 26
3.1.4 Efeito do metil jasmonato e ácido salicílico no controle de podridões em
morangos não inoculados artificialmente ................................................................... 27
3.1.5 Armazenamento e avaliação ............................................................................... 27
3.1.6 Análises fisíco-químicas .................................................................................... 28
3.2 Avaliação de Metil Jasmonato e de Ácido Salicílico sobre o Desenvolvimento
In Vitro de Colletotrichum acutatum, Botrytis cinerea e Rhizopus stolonifer............. 29
3.2.1 Metil jasmonato .................................................................................................. 29
3.2.2 Ácido salicílico ................................................................................................... 29
3.2.3 Delineamento experimental e análise dos resultados ......................................... 29

vi
3.3 Avaliação In Vivo da Possibilidade de Indução de Resistência pelo Metil
Jasmonato e Ácido Salicílico em Morangos „Oso Grande‟ contra os Patógenos
Colletotrichum acutatum, Botrytis cinerea e Rhizopus stolonifer............................... 30
3.3.1 Avaliação da possibilidade de indução de resistência em morangos pelo metil
jasmonato ................................................................................................................... 30
3.3.2 Avaliação da possibilidade de indução de resistência em morangos pelo ácido
salicílico ...................................................................................................................... 31
3.4 Análises Estatísticas .............................................................................................. 31
4 RESULTADOS ........................................................................................................ 32
4.1 Avaliação de Metil Jasmonato no Controle In Vivo e In Vitro dos Patógenos
Botrytis cinerea, Rhizopus stolonifer e Colletotrichum acutatum em Morango „Oso
Grande‟ ....................................................................................................................... 32
4.1.1 Avaliação in vivo do metil jasmonato no controle do mofo cinzento em
morangos „Oso Grande‟ inoculados artificialmente ................................................... 32
4.1.2 Avaliação da possibilidade de indução de resistência pelo metil jasmonato em
morangos „Oso Grande‟ contra o mofo cinzento ....................................................... 32
4.1.3 Avaliação de metil jasmonato no desenvolvimento in vitro de Botrytis cinerea 35
4.1.4 Avaliação in vivo do metil jasmonato no controle da podridão mole em
morangos „Oso Grande‟ inoculados artificialmente .................................................... 36
4.1.5 Avaliação da possibilidade de indução de resistência pelo metil jasmonato em
morangos „Oso Grande‟ contra a podridão mole ........................................................ 38
4.1.6 Avaliação de metil jasmonato no desenvolvimento in vitro de Rhizopus
stolonifer ..................................................................................................................... 38
4.1.7 Avaliação in vivo do metil jasmonato no controle da antracnose em morangos
„Oso Grande‟ inoculados artificialmente ................................................................... 39
4.1.8 Avaliação da possibilidade de indução de resistência pelo metil jasmonato em
morangos „Oso Grande‟ contra a antracnose .............................................................. 42
4.1.9 Avaliação de metil jasmonato no desenvolvimento in vitro de Colletotrichum
acutatum ...................................................................................................................... 42
4.1.10 Avaliação de metil jasmonato sobre os atributos de qualidade de morangos
„Oso Grande‟ ............................................................................................................... 44
4.1.11 Avaliação do metil jasmonato no controle de podridões em morangos „Oso
Grande‟ não inoculados artificialmente ....................................................................... 44

vii
4.2 Avaliação de Ácido Salicílico no Controle In Vivo e In Vitro dos Patógenos
Botrytis cinerea, Rhizopus stolonifer e Colletotrichum acutatum em Morango „Oso
Grande‟ ....................................................................................................................... 44
4.2.1 Avaliação in vivo de ácido salicílico no controle do mofo cinzento em
morangos „Oso Grande‟ inoculados artificialmente ...................................................... 44
4.2.2 Avaliação da possibilidade de indução de resistência pelo ácido salicílico em
morangos „Oso Grande‟ contra o mofo cinzento ........................................................... 48
4.2.3 Avaliação de ácido salicílico no desenvolvimento in vitro de Botrytis cinerea .... 48
4.2.4 Avaliação in vivo de ácido salicílico no controle da podridão mole em morangos
„Oso Grande‟ inoculados artificialmente ....................................................................... 49
4.2.5 Avaliação da possibilidade de indução de resistência pelo ácido salicílico em
morangos „Oso Grande‟ contra a podridão mole ........................................................... 50
4.2.6 Avaliação de ácido salicílico no desenvolvimento in vitro de Rhizopus stolonifer 52
4.2.7 Avaliação in vivo de ácido salicílico no controle da antracnose em morangos
„Oso Grande‟ inoculados artificialmente ...................................................................... 52
4.2.8 Avaliação da possibilidade de indução de resistência pelo ácido salicílico em
morangos „Oso Grande‟ contra a antracnose ................................................................ 54
4.2.9 Avaliação de ácido salicílico no desenvolvimento in vitro de Colletotrichum
acutatum ...................................................................................................................... 56
4.2.10 Avaliação in vivo de ácido salicílico sobre os atributos de qualidade de
morangos „Oso Grande‟ ............................................................................................... 57
4.2.11 Avaliação de ácido salicílico no controle de podridões em morangos „Oso
Grande‟ não inoculados artificialmente ........................................................................ 59
5 DISCUSSÃO ............................................................................................................ 60
6 CONCLUSÕES ........................................................................................................ 64
7 REFERÊNCIAS BIBLIOGRÁFICAS...................................................................... 65

viii
LISTA DE TABELA
Tabela 1 - Fungicidas registrados para a aplicação em frutos pós-colheita no
Brasil .................................................................................................. 12
Tabela 2 - Área abaixo da curva de progresso da doença (AACPD) para
incidência (%) e severidade (cm) do mofo cinzento em morangos
„Oso Grande‟, inoculados com Botrytis cinerea, tratados com
diferentes concentrações de metil jasmonato (MeJa) por 24 h e
armazenados a 25°C / 80% UR durante sete dias ou a 5°C / 90% UR
por cinco dias seguido por mais quatro dias a 25°C / 80% UR
(Experimento I) .................................................................................... 33
Tabela 3 - Área abaixo da curva de progresso da doença (AACPD) para
incidência (%) e severidade (cm) do mofo cinzento em morangos
„Oso Grande‟, inoculados com Botrytis cinerea, tratados com
diferentes concentrações de metil jasmonato (MeJa) por 24 h e
armazenados a 25°C / 80% UR durante sete dias ou a 5°C / 90% UR
por cinco dias seguido por mais quatro dias a 25°C / 80% UR
(Experimento II) .................................................................................. 33
Tabela 4 - Incidência (%) do mofo cinzento em morangos „Oso Grande‟
inoculados com Botrytis cinerea, tratados com diferentes
concentrações de metil jasmonato (MeJa) por 24 h, e armazenados a
25°C / 80% UR por sete dias ou a 5°C / 90% UR por cinco dias
seguido por mais quatro dias a 25°C / 80% UR ................................... 34
Tabela 5 - Área abaixo da curva de progresso da doença (AACPD) para
incidência (% de ocorrência) e severidade (cm) do mofo cinzento
(Botrytis cinerea) em morangos „Oso Grande‟ tratados com metil
jasmonato (0,01 mM), inoculados em diferentes intervalos de tempo
(0, 12 e 24 h) após tratamento e armazenados por sete dias a 25 °C /
80% UR................................................................................................. 34

ix
Tabela 6 - Área abaixo da curva de progresso da doença (AACPD) para
incidência (%) e índice de doença (%) da podridão mole em
morangos „Oso Grande‟, inoculados com Rhizopus stolonifer,
tratados com diferentes concentrações de metil jasmonato (MeJa)
por 24 h e armazenados a 25°C / 80% UR durante três dias ou a 5°C
/ 90% UR por cinco dias seguido por mais três dias a 25°C / 80%
UR (Experimento I) .............................................................................. 36
Tabela 7 - Área abaixo da curva de progresso da doença (AACPD) para
incidência (%) e índice de doença (%) da podridão mole em
morangos „Oso Grande‟, inoculados com Rhizopus stolonifer,
tratados com diferentes concentrações de metil jasmonato (MeJa)
por 24 h e armazenados a 25°C / 80% UR durante três dias ou a 5°C
/ 90% UR por cinco dias seguido por mais três dias a 25°C / 80%
UR (Experimento II) ........................................................................... 37
Tabela 8 - Incidência (%) da podridão mole em morangos „Oso Grande‟
inoculados com Rhizopus stolonifer, tratados com diferentes
concentrações de metil jasmonato (MeJa) por 24 h, e armazenados a
25°C / 80% UR por três dias ou a 5°C / 90% UR por cinco dias
seguido por mais quatro dias a 25°C / 80% UR ................................... 37
Tabela 9 - Área abaixo da curva de progresso da doença (AACPD) para
incidência (% de ocorrência) e severidade (cm) da podridão mole
(Rhizopus stolonifer) em morangos „Oso Grande‟ tratados com metil
jasmonato (0,01 mM), inoculados em diferentes intervalos de tempo
(0, 12 e 24 h) após tratamento e armazenados por três dias a 25 °C /
80% UR................................................................................................ 38

x
Tabela 10 - Área abaixo da curva de progresso da doença (AACPD) para
incidência (%) e severidade (cm) da antracnose em morangos „Oso
Grande‟, inoculados com Colletotrichum acutatum, tratados com
diferentes concentrações de metil jasmonato (MeJa) por 24 h e
armazenados a 25°C / 80% UR durante sete dias ou a 5°C / 90% UR
por cinco dias seguido por mais quatro dias a 25°C / 80% UR
(Experimento I) ................................................................................... 40
Tabela 11 - Área abaixo da curva de progresso da doença (AACPD) para
incidência (%) e severidade (cm) da antracnose em morangos „Oso
Grande‟, inoculados com Colletotrichum acutatum, tratados com
diferentes concentrações de metil jasmonato (MeJa) por 24 h e
armazenados a 25°C / 80% UR durante sete dias ou a 5°C / 90% UR
por cinco dias seguido por mais quatro dias a 25°C / 80% UR
(Experimento II) .................................................................................. 41
Tabela 12 - Incidência (%) da antracnose em morangos „Oso Grande‟ inoculados
com Colletotrichum acutatum, tratados com diferentes
concentrações de metil jasmonato (MeJa) por 24 h, e armazenados a
25°C / 80% UR por sete dias ou a 5°C / 90% UR por cinco dias
seguido por mais quatro dias a 25°C / 80% UR .................................. 41
Tabela 13 - Área abaixo da curva de progresso da doença (AACPD) para
incidência (% de ocorrência) e severidade (cm) da antracnose
(Colletotrichum acutatum) em morangos „Oso Grande‟ tratados com
metil jasmonato (0,01 mM), inoculados em diferentes intervalos de
tempo (0, 12 e 24 h) após o tratamento e armazenados por sete dias a
25 °C / 80% UR.................................................................................... 42
Tabela 14 - Atributos de qualidade de morangos „Oso Grande‟ tratados com metil
jasmonato (MeJa) por 24 h e armazenados a 25°C / 80% UR durante
sete dias ou a 5°C / 90% UR por cinco dias seguido por mais quatro
dias a 25°C / 80% UR ........................................................................ 45

xi
Tabela 15 - Área abaixo da curva de progresso da doença (AACPD) para
incidência (%) do mofo cinzento, da podridão mole e da antracnose
em morangos „Oso Grande‟ não inoculados artificialmente, tratados
com diferentes concentrações de metil jasmonato (MeJa) por 24 h e
armazenados a 25°C / 80% UR durante sete dias ou a 5°C / 90% UR
por cinco dias seguido por mais quatro dias a 25°C / 80% UR ........... 46
Tabela 16 - Área abaixo da curva de progresso da doença (AACPD) para
incidência (%) e severidade (cm) do mofo cinzento em morangos
„Oso Grande‟, inoculados com Botrytis cinerea, tratados com
diferentes concentrações de ácido salicílico por dois minutos e
armazenados a 25°C / 80% UR durante sete dias ou a 5°C / 90% UR
por cinco dias seguido por mais quatro dias a 25°C / 80% UR
(Experimento I) .................................................................................... 46
Tabela 17 - Área abaixo da curva de progresso da doença (AACPD) para
incidência (%) e severidade (cm) do mofo cinzento em morangos
„Oso Grande‟, inoculados com Botrytis cinerea, tratados com
diferentes concentrações de ácido salicílico por dois minutos e
armazenados a 25°C / 80% UR durante sete dias ou a 5°C / 90% UR
por cinco dias seguido por mais quatro dias a 25°C / 80% UR
(Experimento II) ................................................................................... 47
Tabela 18 - Incidência (%) do mofo cinzento em morangos „Oso Grande‟
inoculados com Botrytis cinerea, tratados com diferentes
concentrações de ácido salicílico (AS) por dois minutos, e
armazenados a 25°C / 80% UR por sete dias ou a 5°C / 90% UR por
cinco dias seguido por mais quatro dias a 25°C / 80% UR ................ 47

xii
Tabela 19 - Área abaixo da curva de progresso da doença (AACPD) para
incidência (% de ocorrência) e severidade (cm) do mofo cinzento
(Botrytis cinerea) em morangos „Oso Grande‟ tratados com ácido
salicílico (1 mM), inoculados em diferentes intervalos de tempo (0,
12 e 24 h) após o tratamento e armazenados por sete dias a 25 °C /
80% UR................................................................................................. 48
Tabela 20 - Área abaixo da curva de progresso da doença (AACPD) para
incidência (%) e índice de doença (%) da podridão mole em
morangos „Oso Grande‟, inoculado com Rhizopus stolonifer,
tratados com diferentes concentrações de ácido salicílico por dois
minutos e armazenados a 25°C / 80% UR durante três dias ou a 5°C
/ 90% UR por cinco dias seguido por mais três dias a 25°C / 80%
UR (Experimento I) .............................................................................. 50
Tabela 21 - Área abaixo da curva de progresso da doença (AACPD) para
incidência (%) e índice de doença (%) da podridão mole em
morangos „Oso Grande‟, inoculado com Rhizopus stolonifer,
tratados com ácido salicílico por dois minutos e armazenados a 25°C
/ 80% UR durante três dias ou a 5°C / 90% UR por cinco dias
seguido por mais três dias a 25°C / 80% UR (Experimento II) .......... 51
Tabela 22 - Incidência (%) da podridão mole em morangos „Oso Grande‟
inoculados com Rhizopus stolonifer, tratados com diferentes
concentrações de metil jasmonato (MeJa) por 24 h, e armazenados a
25°C / 80% UR por três dias ou a 5°C / 90% UR por cinco dias
seguido por mais quatro dias a 25°C / 80% UR ................................... 51
Tabela 23 - Área abaixo da curva de progresso da doença (AACPD) para
incidência (% de ocorrência) e severidade (cm) da podridão mole
(Rhizopus stolonifer) em morangos „Oso Grande‟ tratados com ácido
salicílcio (1 mM), inoculados em diferentes intervalos de tempo (0,
12 e 24 h) após o tratamento e armazenados por três dias a 25 °C /
80% UR................................................................................................ 52

xiii
Tabela 24 - Área abaixo da curva de progresso da doença (AACPD) para
incidência (%) e severidade (cm) da antracnose em morangos „Oso
Grande‟, inoculado com Colletotrichum acutatum, tratados com
diferentes concentrações de ácido salicílico por dois minutos e
armazenados a 25°C / 80% UR durante sete dias ou a 5°C / 90% UR
por cinco dias seguido por mais quatro dias a 25°C / 80% UR
(Experimento I) ................................................................................... 54
Tabela 25 - Área abaixo da curva de progresso da doença (AACPD) para
incidência (%) e severidade (cm) da antracnose em morangos „Oso
Grande‟, inoculado com Colletotrichum acutatum, tratados com
diferentes concentrações de ácido salicílico por dois minutos e
armazenados a 25°C / 80% UR durante sete dias ou a 5°C / 90% UR
por cinco dias seguido por mais quatro 4 dias a 25°C / 80% UR
(Experimento II) ................................................................................... 55
Tabela 26 - Incidência (%) da antracnose em morangos „Oso Grande‟ inoculados
com Colletotrichum acutatum, tratados com diferentes
concentrações de metil jasmonato (MeJa) por 24 h, e armazenados a
25°C / 80% UR por sete dias ou a 5°C / 90% UR por cinco dias
seguido por mais quatro dias a 25°C / 80% UR ................................... 55
Tabela 27 - Área abaixo da curva de progresso da doença (AACPD) para os
valores de incidência (% de ocorrência) e severidade (cm) da
antracnose (Colletotrichum acutatum) em morangos „Oso Grande‟
tratados com ácido salicílico (1 mM) e inoculados em diferentes
intervalos de tempo (0, 12 e 24 h) após o tratamento e armazenados
por sete dias a 25 °C / 80% UR .......................................................... 56
Tabela 28 – Atributos de qualidade de morangos „Oso Grande‟ tratados com ácido
salicílico por dois minutos e armazenados a 25°C / 80% UR durante
sete dias, a 5°C / 90% UR durante cinco dias ou a 5°C / 90%UR por
cinco dias seguido por mais quatro dias a 25°C / 80% UR................... 58

xiv
Tabela 29 - Área abaixo da curva de progresso da doença (AACPD) para
incidência (%) de doenças vindas do campo em morangos „Oso
Grande‟ tratados com diferentes concentrações de ácido salicílico
(AS) por dois minutos e armazenados a 25°C / 80% UR durante sete
dias ou a 5°C / 90% UR por cinco dias seguido por mais quatro dias
a 25°C / 80% UR (Experimento I) ..................................................... 59
Tabela 30 - Área abaixo da curva de progresso da doença (AACPD) para
incidência (%) de doenças vindas do campo em morangos „Oso
Grande‟ tratados com diferentes concentrações de ácido salicílico
(AS) por dois minutos e armazenados a 25°C / 80% UR durante sete
dias ou a 5°C / 90% UR por cinco dias seguido por mais quatro dias
a 25°C / 80% UR (Experimento II) ..................................................... 60

xv
LISTA DE FIGURAS
Figura 1 - Sintoma de mofo cinzento (Botrytis cinerea) em morango ................ 07
Figura 2 - Sintoma de podridão mole (Rhizopus stolonifer) em morango ........... 08
Figura 3 - Sintoma de antracnose (Colletotrichum acutatum) em morango ........ 09
Figura 4 - Estrutura química do ácido salicílico .................................................. 13
Figura 5 - Esquema simplificado das vias de biossíntese do ácido salicílico (AS)
(PAL – fenilalanina-amonialiase, ICS – isocorismato sintase, IPL –
isocorismato piruvato liase, SEG – saliciloil éster glucosa, SAG – SA
O-β-glucosídeo, MeSa – metil salicilato, MeSAG – metil salicilato
O-β-glucosídeo) (modificado de VLOT et al., 2009) .......................... 13
Figura 6 - Mecanismo de ação do ácido salicílico ............................................... 16
Figura 7 - Aplicação do metil jasmonato (MeJa) em morangos „Oso Grande‟ em
tambores herméticos ............................................................................. 26
Figura 8 - Aplicação de ácido salicílico (AS) em morangos „Oso Grande‟ ........... 27
Figura 9 - Índice de crescimento micelial de Botrytis cinerea cultivado em meio
batata-dextrose-ágar (BDA), após exposição ao metil jasmonato por
24 h. Experimentos I (A) e II (B). Valores seguidos pela mesma letra
não diferem entre si pelo teste de Tukey (P ≤ 0,05) ............................ 35
Figura 10 - Índice de crescimento micelial de Rhizopus stolonifer cultivado em
meio batata-dextrose-ágar (BDA), após exposição ao metil
jasmonato por 24 h. Experimentos I (A) e II (B). Valores seguidos
pela mesma letra não diferem entre si pelo teste de Tukey (P ≤ 0,05) . 39

xvi
Figura 11 - Índice de crescimento micelial de Colletotrichum acutatum cultivado
em meio batata-dextrose-ágar (BDA), após exposição ao metil
jasmonato por 24 h. Experimentos I (A) e II (B). Valores seguidos
pela mesma letra não diferem entre si pelo teste de Tukey (P ≤ 0,05) .. 43
Figura 12 - Índice de crescimento micelial de Botrytis cinerea cultivado em meio
batata-dextrose-ágar (BDA), incorporado com ácido salicílico em
diferentes concentrações. Experimentos I (A) e II (B). Valores
seguidos pela mesma letra não diferem entre si pelo teste de Tukey
(P ≤ 0,05) ............................................................................................. 49
Figura 13 - Índice de crescimento micelial de Rhizopus stolonifer cultivado em
meio batata-dextrose-ágar (BDA), incorporado com ácido salicílico
em diferentes concentrações. Experimentos I (A) e II (B). Valores
seguidos pela mesma letra não diferem entre si pelo teste de
Tukey (P ≤ 0,05) .................................................................................. 53
Figura 14 - Índice de crescimento micelial de Colletotrichum acutatum cultivado
em meio batata-dextrose-ágar (BDA), incorporado com ácido
salicílico em diferentes concentrações. Experimento I (A) e II (B).
Valores seguidos pela mesma letra não diferem entre si pelo teste de
Tukey (P ≤ 0,05) .................................................................................. 57

xvii
Avaliação de metil jasmonato e de ácido salicílico no controle pós-colheita de
podridões em morango ‘Oso Grande’
RESUMO
O morango, embora muito comercializado, possui vida pós-colheita limitada,
apresentando elevada suscetibilidade a patógenos. As podridões são responsáveis por elevadas
perdas em pós-colheita, destacando-se a podridão mole (Rhizopus stolonifer), o mofo cinzento
(Botrytis cinerea) e a antracnose (Colletotrichum acutatum). Atualmente, no Brasil, não há
fungicidas registrados para o controle destas doenças na pós-colheita de morangos, contudo
métodos alternativos aos fungicidas têm apresentado resultados promissores no controle de
podridões e no aumento do período de conservação de frutos, como o metil jasmonato (MeJa)
e o ácido salicílico (AS). Assim, os objetivos deste trabalho foram avaliar os efeitos do MeJa
e do AS no controle pós-colheita de podridões (mofo cinzento, podridão mole e antracnose) e
sobre os atributos de qualidade de morangos „Oso Grande‟, mantidos sob refrigeração e
condições ambiente. Para tanto, morangos „Oso Grande‟ foram artificialmente inoculados
com suspensão de esporos (105 esporos mL
-1) dos patógenos B. cinerea, R. stolonifer e C.
acutatum e, após 2 h, tratados com o MeJa, em tambores herméticos, com circulação forçada
de ar, onde foram expostos ao produto (0,01 e 0,02 mM), durante 24 h ou, com AS, através de
imersão dos frutos nas concentrações de 1, 2 e 4 mM, durante o período de 2 min. Após os
tratamentos, os frutos foram mantidos a 25° C/80% UR ou sob refrigeração (5 °C/90% UR) e
avaliados quanto à incidência e severidade das podridões, bem como quanto aos atributos de
qualidade (coloração, firmeza, teor de sólidos solúveis, acidez titulável e ratio). Os produtos
também foram avaliados no controle das podridões em frutos não inoculados artificialmente,
ou seja, sobre infecções provenientes do campo. A possibilidade de indução de resistência em
morangos contra os patógenos pelo MeJa e AS foi investigada inoculando-se os frutos após
diferentes intervalos de tempo do tratamento. Ensaios in vitro foram realizados para se avaliar
os efeitos de MeJa e AS sobre o crescimento micelial dos patógenos. Os resultados
demonstraram que o metil jasmonato (0,01 mM) reduz a incidência do mofo cinzento e a
severidade da antracnose em morangos „Oso Grande‟, enquanto o ácido salicílico diminui a
ocorrência de Botrytis cinerea, quando os patógenos são inoculados após o tratamento dos
frutos, possivelmente pela ativação de mecanismos de defesa. In vitro, o ácido salicílico reduz
o desenvolvimento de Botrytis cinerea, Colletotrichum acutatum e Rhizopus stolonifer,
enquanto o metil jasmonato não influencia o crescimento micelial dos patógenos. O MeJa e o
AS não reduz a incidência e/ou a severidade das podridões em morangos inoculados antes da

xviii
aplicação dos produtos e não alteram significativamente os atributos de qualidade dos frutos.
Contudo, o tratamento com AS (1 mM) reduz significativamente a incidência de mofo
cinzento em frutos com infecções provenientes do campo e armazenados a 25 °C.
Palavras-Chave: Botrytis cinerea, Rhizopus stolonifer, Colletotrichum acutatum; indução de
resistência.

xix
Evaluation of methyl jasmonate and salicylic acid on the postharvest control of rot in
‘Oso Grande’ strawberry
ABSTRACT
Although very commercialized, strawberry has a limited post-harvest life, showing a high
susceptibility to pathogens. Rots are responsible for large postharvest losses and, among
them is the gray mold (Botrytis cinerea), the soft rot (Rhizopus stolonifer), and anthracnose
(Colletotrichum acutatum). Actually, in Brazil, there are no fungicides registered for the
postharvest control of these diseases in strawberries. However, alternative methods to the
fungicides have shown promising results on the rot control and on the shelf-life increase,
such as the methyl jasmonate (MeJa) and salicylic acid (SA). Thus, the objectives of this
study were to evaluate the effects of MeJa and AS on the postharvest control of gray mold,
soft rot, and anthracnose and on the quality attributes of 'Oso Grande' strawberries kept
under refrigeration and at room condition. 'Oso Grande' strawberries were artificially
inoculated with spore suspension (105 spores mL
-1) of the pathogens B. cinerea, R.
stolonifer and C. acutatum, and, after 2 h, treated with MeJa, in hermetic chamber, with
forced air circulation, where they were exposed to the product (0,01 and 0,02 mM), for 24
h. SA treatment was carried out by dipping fruit in 1, 2, and, 4 mM, for 2 min. After
treatment, fruit were kept at 25 °C/ 80% RH or under refrigeration (5 °C/ 90% RH) and
assessed as to rot incidence and severity and quality attributes (color, firmness, soluble
solids content, titratable acidity and ratio). The products were also evaluated in the control
of non-inoculated artificially fruit, i.e. on the infections from the field. The possibility of
inducing resistance in strawberries against the pathogens by MeJa and SA was investigated
by fruit inoculation after different time intervals of the treatment. In vitro assays were
performed to evaluate the effects of MeJa and SA on the pathogens mycelial growth. The
results showed that MeJa (0,01 mM) reduces the gray mold incidence and anthracnose
severity in „Oso Grande‟ strawberry, and SA reduces Botrytis cinerea occurrence, when
the pathogens are inoculated after treatments. These results show that, probably, MeJa and
SA induces defense response in strawberry. In vitro, SA reduces Botrytis cinerea,
Colletotrichum acutatum and Rhizopus stolonifer development, and MeJa do not reduce
pathogen mycelial growth. MeJa and SA does not reduce the rot incidence and/or severity
on strawberries inoculated before the treatment, and do not significantly change the fruit
quality attributes. However, SA (1 mM) in not-artificially inoculated strawberries reduces
gray mold incidence in fruit kept at 25 °C.

xx
Keywords: Botrytis cinerea, Rhizopus stolonifer, Colletotrichum acutatum; resistance
induction.

1
1 INTRODUÇÃO
O morango (Fragaria x ananassa Duch.) é produzido e apreciado em diversas regiões
do mundo, com produção mundial de aproximadamente 4,6 milhões de toneladas no ano de
2011, sendo considerado o fruto de maior destaque entre os pequenos frutos. No Brasil a
produção em 2011 foi de 3.016 toneladas, sendo os Estados de São Paulo, Minas Gerais, Rio
Grande do Sul, Paraná, Espírito Santo, Goiás e Distrito Federal considerados os maiores
produtores (FAO, 2012; HOFFMANN et al., 2008). A maior parte da produção brasileira
destina-se ao consumo in natura, mas o consumo industrial, nas mais diversas formas, cresce
continuamente.
O morango é um pseudofruto muito comercializado, principalmente, devido às suas
qualidades nutricionais, aroma e sabor, no entanto é um produto muito delicado e perecível,
apresentando alta atividade respiratória e alta sensibilidade aos danos mecânicos,
desenvolvimento de patógenos, desidratação e perda de firmeza. Essas alterações causam
redução na qualidade do fruto, diminuindo assim a sua aceitação no mercado e limitando a
vida pós-colheita. Por serem muito perecíveis, os morangos devem ser submetidos a um
manejo rápido e cuidadoso em pós-colheita, para que sua qualidade seja preservada
(HENRIQUE & CEREDA, 1999; FLORES-CANTILLANO et al., 2003).
Para manter a qualidade pós-colheita e prolongar a vida útil dos frutos, em geral, é
necessária a utilização de diversas técnicas, estando entre elas o resfriamento rápido, o
armazenamento refrigerado, e o armazenamento sob atmosfera modificada e/ou controlada
(FLORES-CANTILLANO et al., 2003). Para morangos, a refrigeração é a principal forma de
conservação empregada, auxiliando, principalmente, na conservação do fruto através da
redução da atividade respiratória e metabólica e, portanto, retardando sua senescência.
Entre os principais problemas da cultura do morango estão as doenças, que podem
atingir tanto a parte aérea, como as raízes. As doenças que atacam a parte aérea são mais
importantes, pois interferem na frutificação, reduzindo o tamanho e comprometendo o aspecto
visual dos frutos, além de provocar redução na produtividade e na conservação pós-colheita
(OSÓRIO & FORTES, 2003). Em pós-colheita, as doenças conhecidas como antracnose,
mofo cinzento e podridão mole, são consideradas as principais podridões dos frutos (COSTA
et al., 2003).
A produção de alimentos mais saudáveis, isentos de resíduos tóxicos, é um tema que
vem sendo bastante enfatizado nos últimos anos, e com isso, a busca por métodos alternativos
ao uso de defensivos agrícolas para o controle de fitopatógenos vem aumentando. Vários

2
estudos estão sendo realizados visando a redução da incidência de podridões pós-colheita
através de agentes de controle alternativos aos defensivos agrícolas, podendo-se citar a
quitosana, o tratamento térmico, os sanificantes, a radiação ionizante, a luz pulsante, a
radiação ultravioleta (UV-C), o biocontrole, o metil jasmonato (MeJa) e o ácido salicílico
(AS) (BENATO & CIA, 2009). A utilização do MeJa e do AS em pós-colheita está ganhando
importância, principalmente, pela ação como indutores de resistência em plantas à diversas
espécies de fitopatógenos. Cabe ressaltar que existem defensivos agrícolas registrados no
Brasil para o controle de doenças pós-colheita em diversas frutas, contudo não para o
morango (AGROFIT, 2012).
O AS e o MeJa são moléculas endógenas que regulam o crescimento das plantas,
provocando uma ampla variedade de respostas metabólicas e fisiológicas, além de
desempenharem papel fundamental na defesa da planta em resposta a estresses abióticos e
bióticos. Tanto o MeJa, quanto o AS, vem sendo bastante estudados, pois possuem ação
benéfica para manter e prolongar a vida pós-colheita de diversos produtos hortícolas (TAIZ &
ZEIGER, 2006; TRIPATHI & DUBEY, 2004; ASGHARI & AGHDAM, 2010). O efeito
destes compostos no controle de fitopatógenos tem demonstrado resultados promissores,
como, por exemplo, na redução do desenvolvimento de Botrytis cinerea e Colletotrichum
gloesporioides pelo MeJa e AS, respectivamente (ZHU & TIAN, 2012; LEE et al., 2009).
Porém trabalhos envolvendo a ação direta do MeJa e AS sobre microrganismos são limitados.
O controle de podridões em plantas através da resistência sistêmica adquirida
(“Systemic Acquired Resistance” - SAR), pela aplicação de AS ou MeJa, é um tema bastante
estudado. A SAR é um mecanismo de defesa induzido por agentes bióticos, abióticos ou
infecção localizada por patógeno, que confere proteção de longa duração contra um vasto
espectro de microrganismos. O grau desta proteção varia em função da concentração do
indutor, do inoculo utilizado, do intervalo de tempo entre o tratamento inicial com o indutor e
o contato com o patógeno, e das condições de temperatura e luminosidade. A molécula de AS
e do MeJa estão associadas ao acúmulo de proteínas relacionadas à patogênese, bem como a
inibição e/ou estimulação de diversas enzimas relacionadas com a defesa das plantas,
contribuindo assim para conferir resistência contra estresses abióticos ou bióticos
(DURRANT & DONG, 2004; GUZZO, 2004; TAIZ & ZEIGER, 2006; ASGHARI &
AGHDAM, 2010; ROHWER & ERWIN, 2008).
Neste contexto, o objetivo deste trabalho foi testar a hipótese de que o AS e o MeJa
controlam o mofo cinzento, a antracnose e a podridão mole em morangos „Oso Grande‟,

3
devido a atuação direta sobre os patógenos ou indireta, através de indução de mecanismos de
defesa.
2 REVISÃO BIBLIOGRÁFICA
2.1 Classificação e Características do Morangueiro
O morangueiro (Fragaria x ananassa Duch.), de acordo com o Sistema de
Classificação Vegetal de Cronquist (1988), está classificado da seguinte maneira: Divisão
Magnoliophyta (Angiosperma), Classe Magnoliopsida (Dicotiledônea), Subclasse Rosidae,
Ordem Rosales, Família Rosaceae e Gênero Fragaria L. A principal espécie cultivada,
Fragaria x ananassa Duch., é um híbrido entre espécies originárias do continente americano
(Fragaria virginiana x Fragaria chiloensis) (OLIVEIRA & SANTOS, 2003). O morango é
um pseudofruto de procedência da América do Norte e do Chile e é apreciado no mundo
inteiro, principalmente pelos seus aspectos nutritivos e sabor, sendo consumido in natura e
por diversas maneiras de processamento (REICHERT, 2003; SANTOS & MEDEIROS,
2003).
Espécies de frutas que possuem como características o pequeno tamanho (0,5 cm a 5
cm), porte arbustivo ou rasteiro, plantio em alta densidade, necessidade de manejo intensivo
desde o plantio, elevada exigência de mão-de-obra e conseqüente áreas pequenas para o
cultivo, são denominadas de pequenas frutas. No Brasil este termo é recente, sendo
representada pelo morango, pela amora-preta, pelo mirtilo e pela framboesa, respectivamente
em ordem de importância comercial. Pelo tempo de produção, área cultivada e expansão do
cultivo, o morango é a principal espécie deste grupo (HOFFMANN et al., 2008).
O morangueiro é considerado uma espécie olerícola; é uma planta herbácea
estolonífera, perene e possui caule semi-subterrâneo, chamado de coroa. As folhas se
originam da coroa de forma helicoidal, variando em cor e forma de acordo com a cultivar; são
trifoliadas com um par de espículas triangulares na base. Os folíolos são denteados, com
coloração verde escuro na face superior; já na face inferior possui pilosidade e coloração
acinzentada (MADAIL et al., 2005; SANHUEZA et al., 2005).
O morango é um pseudofruto, ou seja, é resultado do desenvolvimento dos
receptáculos de um conjunto floral compacto fertilizado. Os frutos verdadeiros são as
pequenas pontuações que ficam sob a superfície do morango, chamados de aquênios ou
semente (PBMH, 2009). O morango é considerado um fruto não climatérico, ocorrendo

4
diminuição gradual da respiração, por isso não ocorre amadurecimento nem alterações
expressivas nas condições organolépticas em pós-colheita (MELOTTI, 2011; CHITARRA &
CHITARRA, 2005).
O índice de maturação é baseado na coloração; morangos com menos da metade da
superfície vermelha são considerados impróprios para o consumo ou para o processamento
porque possuem pouco aroma e conservam elevado teor de acidez e adstringência em pós-
colheita. Morangos com mais da metade e até ¾ da superfície vermelha apresentam boas
condições para consumo in natura ou para processamento por alguns dias, dependendo da
variedade, da temperatura e da umidade atmosférica (CHITARRA & CHITARRA, 2005).
Este fruto possui baixo valor calórico, cerca de 40 calorias a cada 100 g, possui ação
antitérmica, diurética, antiartrítica, mineralizante, antiviral e anticancerígeno. Possui na sua
constituição: fósforo, sódio, carboidratos, ferro e vitaminas A, B e C (MELOTTI, 2011). Os
morangos possuem elevada concentração de ácido ascórbico (100 a 300 mg 100g-1
); as frutas
vermelhas, de maneira geral são ricas também em antocianinas (SANTINI, 2006 apud
ROMBALDI et al., 2009).
Os atributos sensoriais, como a cor, a textura, o aroma e o balanço açúcar/acidez, são
características importantes na qualidade do morango. O sabor do morango é um dos aspectos
mais exigidos pelo consumidor, sendo dependente, em parte, do balanço entre os sólidos
solúveis e a acidez titulável do fruto. A coloração do morango é atrativa devido à presença de
antocianinas, que indicam a maturação do fruto. Polissacarídeos, como substâncias pécticas,
determinam a textura deste fruto, sendo que a qualidade e a vida pós-colheita do morango são
influenciadas diretamente pela perda da firmeza que ocorre durante a senescência (FLORES-
CANTILLANO et al., 2003).
As principais cultivares de morango cultivadas no Brasil originam-se dos Estados
Unidos. Cabe ressaltar a cultivar que é importada da Espanha, Milsei-Tudla (OLIVEIRA et
al., 2005). De acordo com REICHERT & MADAIL (2003), as cultivares nacionais,
Campinas, Monte Alegre, Konvoy, Konvoy Cascata e as cultivares estrangeiras, Oso Grande,
Chandler, Dover, Camarosa, Sweet Charlie, Kabarla, Aromas, Tudla, Toyonoca, Seascape,
Selva e Fern. são as mais cultivadas, pois apresentam frutos maiores, polpa mais firme,
melhor sabor, maior produtividade e, sobretudo, melhor condição fitossanitária. ANTUNES
& REISSER (2008) destacam que entre as principais cultivares produzidas no Brasil estão: a
Oso Grande, a Camarosa, a Dover e a Aromas.
A cultivar Oso Grande foi introduzida da Universidade da Califórnia (EUA), sendo
considerada de dias curtos e grande adaptabilidade; é uma planta vigorosa, com folhas

5
grandes de coloração verde escura, possui ciclo mediano e elevada capacidade produtiva. Os
frutos são de tamanho grande com coloração de epiderme vermelho-clara, textura de polpa
firme no início da produção e mediana no final, coloração interna vermelho-claro e aromática,
com sabor subácido, conveniente para o consumo in natura. É suscetível a Mycospharella
fragarieae, Colletotrichum fragarieae e C. acutatum, contudo apresenta tolerância ao fungo
Botrytis cinerea Pers. & F. (SANTOS, 2003; SANTOS, 2005).
2.2 Aspectos Socioeconômicos do Morangueiro
O interesse comercial pelo cultivo do morango é grande em diversos países. A
coloração, o aroma e o sabor da fruta, assim como suas propriedades nutritivas, fazem do
morango um produto muito apreciado pelos consumidores. O maior mercado desta cultura é o
consumo in natura, porém o mercado é muito amplo em produtos processados, tais como em
geléias, caldas, sucos e polpa congelada (ALMEIDA et al., 1999 apud GIMÉNEZ et al.,
2008).
O cultivo do morango é uma atividade de grande importância socioeconômica, pois
emprega grande número de pessoas durante sua condução, sendo realizado principalmente em
propriedades familiares (COSTA & VENTURA, 2006). Mesmo que restrita a pequenas áreas,
esta cultura é uma atividade agrícola que ocorre em muitas regiões do Brasil (TANAKA et al.,
2005).
No Brasil, o morangueiro é cultivado nos Estados do Rio Grande do Sul, São Paulo e
Minas Gerais, bem como em regiões com diferentes solos e climas, como Santa Catarina,
Paraná, Espírito Santo, Goiás e Distrito Federal. No mundo e no Brasil, o morango vem se
destacando como uma das principais frutas produzidas, devido a grande rentabilidade (224%)
quando comparada com outros cultivos (HOFFMANN et al., 2008; REICHERT & MADAIL,
2003).
De acordo com a FAO (2012), a produção nacional de morango em 2011 foi estimada
em 3.016 toneladas, sendo explorada numa área de aproximadamente 376 ha. O Estado de
Minas Gerais é o maior produtor, contribuindo com cerca de 40% do total produzido, seguido
por São Paulo e Rio Grande do Sul, sendo os três Estados responsáveis por 80% da produção
nacional (CARVALHO, 2006). A produção em Minas Gerais é destinada tanto para a
industrialização, como para o consumo in natura; já em São Paulo, a maior parte é destina
para o consumo in natura. No Rio Grande do Sul, há cidades que se destacam pela produção

6
de morangos de mesa, contudo em Pelotas a produção é destinada ao processamento
(HOFFMANN et al., 2008).
O aumento da incidência e da severidade de doenças tem sido um dos grandes
responsáveis por perdas na produtividade. Inúmeras doenças têm se agravado ano após ano,
devido ao clima favorável que permite o desenvolvimento de doenças durante todo o período
de cultivo. Doenças consideradas como secundárias e de ocorrência esporádica vêm se
tornando cada vez mais um problema para várias regiões produtoras (TANAKA et al., 2005).
2.3 Principais Doenças Pós-colheita
A cultura do morango pode ser afetada por inúmeras doenças infecciosas de natureza
fúngica, bacteriana ou viral. Contudo, a maioria é causada por fungos, gerando grandes
perdas, pois afetam o desenvolvimento e a produtividade das plantas. As doenças podem
incidir em diferentes partes aéreas (folhas, frutos, estolões e flores), nas raízes e no colo. A
incidência e a severidade das doenças são dependentes, principalmente, do estado nutricional
da planta, da cultivar, das condições de clima e solo e do manejo da cultura (COSTA et al.,
2003; FORTES, 2003; TANAKA et al., 2005).
Como os morangos possuem como principal mercado o consumo in natura, as
doenças que ocorrem em pós-colheita são extremamente importantes, pois afetam diretamente
a comercialização (TANAKA et al., 2005). As podridões dos frutos são favorecidas por
condições de elevada umidade, pelas injúrias mecânicas, irrigação, colheita, embalagem,
manuseio e transporte inadequados; por isso são consideradas as grandes responsáveis pelas
perdas que ocorrem na produção de morango. A antracnose, o mofo-cinzento e a podridão
mole são consideradas as principais podridões dos frutos, pois se manifestam tanto no campo,
como em pós-colheita (TANAKA, 2003; COSTA et al., 2003; TANAKA et al., 2005).
Segundo BENATO & CIA (2009), em frutos maduros as podridões são principalmente
provenientes de infecções quiescentes estabelecidas no período de produção, mas podem
também ser ocasionadas por patógenos que penetram por ferimento, pela manipulação ou
exposição a ambientes contaminados. Para a prevenção das doenças pós-colheita, recomenda-
se evitar danos mecânicos, já que os ferimentos permitem a penetração de todos os patógenos.
Dentre as podridões dos frutos na cultura do morango, o mofo-cinzento, causado pelo
fungo Botrytis cinerea Pers. Fr., é uma das mais importantes, pois está presente em todas as
regiões de cultivo. Esta doença pode afetar o fruto em qualquer estádio do seu
desenvolvimento, ocasionando grandes prejuízos, seja em condições de campo, ou durante o

7
armazenamento e comercialização (TANAKA, 2002; COSTA et al., 2003; TANAKA et al.,
2005; FORTES & COUTO, 2003).
O fungo B. cinerea não é específico da cultura do morango, podendo ocorrer em
diferentes plantas. O mesmo é considerado um parasita facultativo, vivendo saprofiticamente
na matéria orgânica e formando escleródios e micélio dormente, em restos de cultura. Em
meio de cultura, possui uma coloração acinzentada, com conídios unicelulares, que são
facilmente disseminados no campo pelo vento ou pela água (TANAKA, 2002; TANAKA et
al., 2005).
Segundo TANAKA (2002) e COSTA et al. (2003), os sintomas iniciais no fruto são
manchas de tamanhos variáveis, com cor marrom-clara e consistência mole, não tendo
delimitação entre o tecido doente e o sadio. O fruto apodrece rapidamente e adquire aparência
seca e firme, recoberto por estruturas do fungo, formando uma massa de cor cinza. Quando a
umidade é elevada, o micélio se desenvolve mais denso, apresentando aspecto cotonoso
(Figura 1). Esta doença é favorecida por elevada umidade e temperatura ao redor de 20 °C,
geralmente nas estações de inverno e primavera. De acordo com TANAKA et al. (2005) e
FORTES & COUTO (2003), se as condições foram ideais, os pecíolos, folhas, botões florais,
pétalas e pedúnculos podem apresentar sintomas, sendo que em frutos maduros a doença
avança rapidamente.
Figura 1 - Sintoma de mofo cinzento (Botrytis cinerea) em morango.
Provocada por mais de uma espécie de Rhizopus, geralmente R. stolonifer ou R.
nigricans, a podridão mole ou podridão de Rhizopus é observada com freqüência em pós-
colheita. Em embalagens com frutos maduros, a incidência desta podridão é elevada. No
campo raramente esta doença é observada, porém, os frutos colhidos podem possuir, na sua

8
superfície, estruturas do fungo, que é o inoculo (TANAKA et al., 2005; COSTA et al., 2003;
FORTES, 2003).
Os sintomas são caracterizados por uma área aquosa e mole, podendo estar recoberta
por micélio de aspecto cotonoso de coloração branca a acinzentada, sobre o qual podem
ocorrer pontuações escuras (esporangiósforos e esporângios) (Figura 2). Geralmente esta
doença é acompanhada de odor característico, resultado de infecções de bactérias e leveduras
(BENATO & CIA, 2009). De acordo com BORESZTEIN et al. (2009), R. stolonifer pode
causar infecção em frutos intactos de morango, mas a incidência da doença é menor quando
comparada com frutos feridos.
Devido a penetração do R. stolonifer ocorrer, principalmente, através de ferimentos, a
infecção é pouco afetada pela umidade relativa. Já a temperatura é um fator limitante para o
desenvolvimento da doença e para a germinação dos esporos. A temperatura mínima
requerida é 6°C e a produção de esporângios é inibida por temperaturas inferiores a 8-10° C
(TANAKA et al., 2005; TANAKA, 2002).
Figura 2 - Sintoma de podridão mole (Rhizopus stolonifer) em morango.
De acordo com AGRIOS (2005) e FORTES (2005), o desenvolvimento da antracnose
na cultura do morango pode ser causado por três espécies, Colletotrichum fragariae, C.
acutatum e C. gloeosporioides, produzindo lesões e estrangulamento em estolões, pecíolo,
pedúnculo, fruto e coroa do morango. O C. acutatum provoca a podridão dos frutos
(antracnose) e a flor preta, ao passo que C. gloeosporioides e C. fragariae infectam
principalmente a coroa das plantas e causa podridão da coroa e murcha.
A antracnose é favorecida por elevada temperatura e umidade, porém podem ocorrer
sintomas severos no campo em períodos de clima frio, com alta umidade. O inoculo presente

9
em restos culturais, como frutos mumificados, pode desencadear a doença. Lesões em
estolhos ou folhas, de modo geral, carregam o inoculo, que atinge os frutos por ação da
irrigação, água da chuva, vento, insetos ou pelo manuseio das plantas no cultivo (TANAKA,
2002; TANAKA et al., 2005). Segundo TANAKA et al. (2005), este patógeno pode causar
perdas superiores a 50% em variedades muito suscetíveis e sob condições favoráveis, sendo
um fator limitante para a produção e a comercialização de morangos.
De acordo com BENATO & CIA (2009) e FORTES (2005), em pós-colheita, a
antracnose, causada pelo fungo Colletotrichum spp., possui grande importância. Os sintomas
característicos são pequenas lesões deprimidas, que sob condições de temperatura e umidade
adequadas aumentam de tamanho. As lesões são firmes, no início, com coloração branco-
acinzentada e, posteriormente, marrom. No centro das lesões ocorre uma massa de esporos de
coloração alaranjada ou rosada (Figura 3). Em frutos maduros a podridão é mais comum,
porém em surtos severos, frutos verdes podem ser atacados. Os sintomas de deficiência de
cálcio/boro ou fitotoxidez causados por produtos químicos podem ser confundidos com esta
doença (COSTA et al., 2003).
Figura 3 - Sintoma de antracnose (Colletotrichum acutatum) em morango.
2.4 Conservação e Métodos de Controle de Podridões
O aumento da produção de etileno e a elevada atividade respiratória são os principais
responsáveis pelos rápidos danos causados após a colheita em morangos. Porém existem
ainda outros fatores que também influenciam no processo de deterioração destes frutos, como
a alta suscetibilidade a lesão mecânica, a perda de água e a ação de patógenos, principalmente
fungos (LUNARDI, 2009).

10
Um dos fatores de maior influência na conservação pós-colheita de frutos e hortaliças
é a respiração. Diversos fatores influenciam a intensidade da respiração em um fruto, entre
eles estão o tipo de tecido (jovem ou adulto) e a quantidade de água. O morango apresenta
alta atividade respiratória após a sua colheita (LUNARDI et al., 2009; VIEITES et al., 2006).
A temperatura pode ser considerada como o principal fator externo na conservação de
produtos vegetais, pois influencia na respiração, transpiração, entre outros aspectos da
fisiologia das plantas. A velocidade de deterioração de frutos e hortaliças aumenta cerca de
duas a três vezes, com o aumento de 10 °C na temperatura. Já quando a temperatura mínima
de segurança é ultrapassada, pode ocorrer perda de sabor, aroma, escurecimento da casca e
perda da capacidade de maturação (LUNARDI et al., 2009). A umidade relativa do ar (UR)
também influência na conservação pós-colheita, atuando diretamente na transpiração.
Quando a UR está em níveis adequados, promove o declínio da desidratação, afetando as
interações patógeno-hospedeiro (LUNARDI et al., 2009).
Nos frutos não climatéricos, como o morango, as respostas típicas à ação do etileno
não são tão acentuadas, embora possam ser observadas reduções no teor de clorofilas e ácidos
orgânico, com aumentos temporários na atividade respiratória. Em pós-colheita são
observados alterações de cor, firmeza da polpa e perda do brilho natural dos frutos, e os
procedimentos adotados após a colheita, de modo geral, podem evitar, parcialmente, a perda
da qualidade (ROMBALDI et al., 2009; LUNARDI, 2009).
Por ser um fruto muito sensível, o morango é muito suscetível ao desenvolvimento de
doenças pós-colheita. Nos frutos, em geral, o processo de infecção geralmente ocorre durante
o seu desenvolvimento no campo. Embora alguns frutos possam ser descartados antes de
serem armazenados ou transportados, por apresentarem sintomas visíveis, muitos escapam
dessa seleção, principalmente aqueles frutos cujos sintomas estão nos estádios iniciais de
desenvolvimento. Portanto, algumas infecções acontecem no campo, mas permanecem
quiescentes e os sintomas se desenvolvem somente quando o fruto amadurece (BENATO &
CIA, 2009).
De maneira geral a incidência de doenças pós-colheita é influenciada pelas condições
climáticas da região produtora, espécies e cultivares, tratos culturais e fitossanitários
incorretos. Processos físicos são estudados e aplicados comercialmente para a conservação
pós-colheita de frutos, tais como: termoterapia, resfriamento, radiação UV-C e a atmosfera
modificada/controlada. Nos últimos anos, há uma crescente busca por processos alternativos
aos agroquímicos para o controle de doenças pós-colheita (BENATO & CIA, 2009).

11
De acordo com TANAKA (2002), o controle das podridões deve iniciar-se com os
frutos ainda no campo, tendo continuidade durante e após a colheita. No campo é preciso
realizar práticas culturais como, por exemplo, evitar condições de elevada umidade e realizar
a colheita nos períodos mais secos e frescos do dia, evitando ferimentos e manuseio
excessivo. Outras medidas ainda podem ser tomadas, como a eliminação dos restos de cultura.
TANAKA et al. (2005) destacam que o controle químico, além de aumentar o custo de
produção, esbarra nas poucas opções de produtos registrados para a cultura e na ineficácia de
muitos princípios ativos.
Após a colheita, o principal método utilizado para a preservação dos frutos é o
armazenamento refrigerado ou convencional, que consiste em controlar a temperatura e a
umidade relativa do ar em câmaras frigoríficas. De maneira geral, as condições para a
conservação de morangos são: temperatura de 0 °C, com cerca de 90 a 95% de umidade
relativa (LUNARDI et al., 2009; FLORES-CANTILLANO et al., 2003). Entretanto diversos
trabalhos (COSTA, 2009; SILVA et al., 2010; PINELI et al., 2008) vêm apresentando bons
resultados, quanto aos atributos de qualidade, utilizando a temperatura de 5 °C para a
refrigeração de morangos.
A utilização de substâncias naturais, para induzir resistência ou controlar diretamente
os fitopatógenos em pós-colheita, é um estudo em expansão, e vem apresentando resultados
promissores nos últimos anos (BENATO, 2003b). Agentes abióticos e bióticos podem
desencadear respostas de resistência induzida em frutas e hortaliças. Os elicitores são
classificados como abióticos e bióticos, sendo que os bióticos podem ser de procedência dos
microrganismos (elicitor exógeno) ou pode ocorrer na própria planta (elicitor endógeno).
Quando ativado por elicitores, o metabolismo secundário das plantas pode sintetizar
fitoalexinas, que são compostos antimicrobianos, de baixa massa molecular, que se acumulam
nas células em resposta às infecções (TAIZ & ZEIGER, 2006; PASCHOLATI & LEITE,
1995). Segundo DANNER et al. (2008), o uso de elicitores gera um aumento no teor de
proteínas totais, açúcares totais e redutores, e fenóis totais em frutos.
Os métodos alternativos de controle de doenças pós-colheita, que possuam a
capacidade de induzir respostas de defesa em frutos contra patógenos (compostos naturais,
tratamentos físicos e biológicos), têm chamado a atenção dos pesquisadores, pois atendem as
necessidades de diminuição do uso de fungicidas e os requisitos do mercado internacional.
Contudo, apesar da crescente busca por processos alternativos de controle de doenças em pós-
colheita que sejam economicamente viáveis, a área de fitopatologia pós-colheita no Brasil
ainda precisa expandir, buscar informações e novas tecnologias que possam contribuir para

12
minimizar as perdas e agregar valor aos produtos, aumentando assim a competitividade
(BENATO, 1999; BENATO & CIA, 2009). Vale ressaltar que para o tratamento de doenças
pós-colheita em morango não existem produtos químicos registrados (Tabela 1) (AGROFIT,
2012).
Tabela 1 - Fungicidas registrados para a aplicação em frutos pós-colheita no Brasil
(AGROFIT, 2012).
Ingrediente
ativo Grupo químico Frutos
Dicloran Cloroaromático pêssego
Imazalil Imidazole banana, citros, maçã, mamão, manga e melão
Iprodione Dicarboximida maçã
Tiabendazol Benzimidazole abacate, banana, citros, mamão, manga e melão
Procloraz Imidazolilcarboxamida mamão e manga
Alguns agentes abióticos vêm apresentando um bom resultado no controle de doenças
de plantas e indução de resistência, como o acibenzolar-S-metil (ASM), o AS, o ácido β-
aminobutírico (BABA), o ácido jasmônico, o MeJa e a quitosana (BENATO, 2003a). Estudos
envolvendo tratamento térmico (GARCÍA et al., 1996), atmosfera controlada (JÚNIOR,
2011), quitosana (EL GHAOUTH et al., 1991), ultra-som (CAO et al., 2010), aplicação de
resveratrol (ZAICOVSKI et al., 2006; MALGARIM et al., 2006), ozônio (PÉREZ et al.,
1999), luz ultravioleta (TIBOLA et al., 2007), ASM e a proteína harpina (TOMAZELI, 2010),
vem demonstrando que são eficientes na prevenção de podridões pós-colheita em morangos.
2.4.1 Ácido salicílico
O AS é considerado uma molécula endógena de sinalização comum em todo o reino
vegetal, pertencente ao grupo dos compostos fenólicos definidos como substâncias formadas
por um anel aromático ligado a um grupo hidroxil ou ao seu derivado funcional (Figura 4)
(KERBAUY, 2004; MÉTRAUX, 2002; ASGHARI & AGHDAM, 2010). A origem desse
nome está associada a ter sido encontrado na casca de uma árvore do gênero Salix, sendo
distribuído nas plantas, tanto nas folhas, quanto nas estruturas de reprodução (KERBAUY,
2004).

13
Figura 4 - Estrutura química do ácido salicílico (SOBRINHO et al., 2005).
A biossíntese do AS pode ser através de duas vias enzimáticas diferentes: pela via dos
fenilpropanóides, a partir da L-fenilalanina que por ação da enzima fenilalanina-amonialiase
(FAL), é convertida em ácido trans-cinâmico que irá formar o ácido benzóico e será
convertido em AS por ação da enzima ácido benzóico-2-hidroxilase; e pela via do
isicorismato, sendo que o corismato é convertido em isocorismato por ação da enzima
isocorismato sintase (ICS) e transforma-se em AS por ação da enzima isocorismato piruvato
liase (IPL). O AS produzido nas plantas pode ser convertido em SA O-β-glucosídeo (SAG),
saliciloil éster glucosa SEG, metil salicilato (MeSA) e metil salicilato O-β-glucosídeo
(MeSAG) (Figura 5) (KERBAUY, 2004; SOUZA, 2007; VLOT et al., 2009).
Figura 5 - Esquema simplificado das vias de biossíntese do ácido salicílico (AS) (PAL –
fenilalanina-amonialiase, ICS – isocorismato sintase, IPL – isocorismato piruvato liase, SEG
– saliciloil éster glucosa, SAG – SA O-β-glucosídeo, MeSa – metil salicilato, MeSAG – metil
salicilato O-β-glucosídeo) (modificado de VLOT et al., 2009).

14
Segundo SOUZA (2007), estudos indicam que a via dos fenilpropanóides está
relacionada com a resposta de hipersensibilidade, através da rápida produção de AS, o que
leva a morte celular. Já a via do isocorismato atuaria no aumento da síntese de AS,
aumentando assim a resistência sistêmica adquirida e, consequentemente, conferindo maior
resistência da planta contra o ataque de patógenos.
Em 1874, na Alemanha, iniciou-se a comercialização do AS. Por ser amplamente
utilizado na medicina humana (alívio de dores, prevenindo tromboses cerebrais e acidentes
vasculares), o AS é uma molécula bastante conhecida (SOBRINHO et al., 2005).
O estudo do AS teve início a partir de observações de um análogo (aspirina) que
prolongava a vida pós-colheita de flores, possivelmente por interferir na biossíntese de etileno
(SOBRINHO et al., 2005). Embora o conhecimento dos diversos efeitos fisiológicos e
bioquímicos da aplicação de AS em plantas ser antigo, o seu desempenho como regulador
endógeno só foi estabelecido em um estudo de termogênese em plantas em 1987 (RASKIN,
1992). O uso de AS no controle de perdas pós-colheita de produtos hortícolas vêm sendo
considerado uma alternativa com grande potencial. Recentemente, o estudo do AS tem sido
amplamente explorado, já que é uma molécula chave para a expressão de resistência a
estresses nas plantas, principalmente a estresses abióticos (luz, seca, salinidade, frio, UV e
choque térmico) e ao ataque de patógenos. A ação antifúngica do ácido também vem sendo
descrita (ASGHARI & AGHDAM, 2010).
Os trabalhos envolvendo compostos fenólicos já comprovaram que os mesmos
desempenham papel fundamental na regulação de diversos processos fisiológicos, dentre eles
o crescimento e desenvolvimento das plantas, a fotossíntese e a captação de íons (POPOVA et
al., 1997). De acordo com LEE et al., (1995) e ASGHARI & AGHDAM, (2010), o AS é
essencial na regulação das respostas ao estresse e nos processos de desenvolvimento da
planta, provocando efeitos fisiológicos e bioquímicos, incluindo a indução da floração, a
produção de calor (termogênese), a fotossíntese, a condutância estomática, a transpiração, a
absorção e o transporte de íons (inibição de fosfato e captação de potássio), a germinação de
sementes, a inibição da biossíntese / ação do etileno e a resistência a doenças. O AS pode agir
de modo isolado, associado ou controlar os efeitos de outros hormônios. Os hormônios MeJa
e etileno induzem a ativação genética de diversas proteínas, que por sua vez são inibidas pelo
AS (SOARES & MACHADO, 2007).
O controle de podridões em plantas pode ser realizado por meio de mecanismos de
defesa, conhecidos como resistência sistêmica adquirida (RSA). O mecanismo da RSA inicia-
se no momento da interação planta/patógeno ou do tratamento com fatores abióticos, o

15
mesmo envolve uma cascata de eventos e sinais, que culmina em alterações no seu
metabolismo celular, bem como na emissão de sinais moleculares dirigidos para outras partes
da planta, promovendo a diminuição da severidade da doença. A planta, então, sobrevive à
infecção e induz uma proteção de longa duração contra um vasto espectro de microrganismos,
pois em resposta à distribuição dos sinais, esta seria induzida a sintetizar agentes de defesa,
incluindo as proteínas relacionadas à patogênese (proteínas-RP), além da formação de
barreiras estruturais, como lignificação, papilas e tiloses (FERNANDES et al., 2009;
DURRANT & DONG, 2004; TAIZ & ZEIGER, 2006; RASKIN, 1992). O AS é uma
molécula essencial na sinalização de respostas de resistência, induzindo a biossíntese de
enzimas que atuam na formação de compostos de defesa vegetal, como polifenóis, alcalóides
e também as proteínas-RP, principalmente no local da aplicação, contribuindo assim para
conferir resistência (RASKIN, 1992; ASGHARI & AGHDAM 2010).
O envolvimento do AS na resistência sistêmica adquirida foi evidenciado em 1993, em
plantas de fumo, quando transferiu-se o gene nahG (isolado de Pseudomonas putida), o qual
codifica a enzima, salicilato hidroxilase, que converte o AS em catecol (não possui a
capacidade de induzir respostas de defesa) (CAVALCANTI et al., 2005). De acordo com
ASGHARI & AGHDAM (2010), quando o AS é aplicado exogenamente em plantas, induz a
expressão de genes de proteínas-RP e também confere resistência contra patógenos. TERRY
& JOYCE (2004) destacam que o AS vem mostrando um aumento na resistência em várias
culturas hortícolas, tanto no campo, como em pós-colheita, por exemplo, em kiwi contra o
fungo B. cinerea (POOLE & McLEOD, 1994), em pêra contra o patógeno Penicillium
expansum (CAO et al., 2006), em manga contra C. gloeosporioides (ZENG et al., 2006) e em
caqui a diferentes doenças (KHADEMI et al., 2012).
O modelo de ação do AS foi inicialmente baseado na constatação de que este
composto poderia se ligar a enzima catalase (CAT) e inibir a sua ação, o que levaria a um
aumento na concentração de peróxido de hidrogênio (H2O2) ou de espécies reativas de
oxigênio (ERO) derivadas do H2O2. O H2O2 pode atuar como uma molécula antimicrobiana
contra patógenos, assim como seus derivados que atuam como intermediários na cascata de
sinalização para a defesa vegetal (Figura 6) (CIA et al., 2007; SOARES & MACHADO,
2007).

16
Figura 6 - Mecanismo de ação do ácido salicílico (SOBRINHO et al., 2005).
Contudo o AS não possui somente a função de atuar bloqueando a atividade da
catalase, ele pode interferir na atividade da FAL e da peroxidase (POD) que estão envolvidas
no processo de lignificação da parede celular e também na atividade da quitinase e β-1,3-
glucanase, que promovem a desorganização da parede celular dos patógenos (CIA et al.,
2007; SOBRINHO et al., 2005; PASCHOLATI & LEITE, 1995), bem como agir direta ou
indiretamente estimulando e/ou inibindo a atividade de enzimas antioxidantes [superóxido
desmutase (SOD), CAT e glutationa redutase (GSH), polifenol oxidase (PPO) e ascorbato
peroxidase (APX)] (ASGHARI & AGHDAM, 2010). ZENG et al. (2006) verificaram que
frutos tratados com 1 mM de AS durante 2 min sob baixa pressão (- 80 kPa) e por mais 10
min a pressão normal, e armazenados a 13° C, apresentaram as atividades de enzimas de
defesa (PAL e β-1,3-glucanase) aumentadas e maiores níveis de H2O2 e radicais superóxidos
(O2-
), aumentando assim a resistência de mangas contra a antracnose.
O efeito positivo do AS no controle de podridões tem sido relatado por diversos
autores. De acordo com ZAINURI et al. (2001), a aplicação pré e pós-colheita, 1000 mg/L e
2000 mg/L respectivamente, de AS em mangas diminuiu a severidade da antracnose, causada
pelo fungo C. gloeosporioides, pois promoveu inibição do amadurecimento dos frutos. A
aplicação pós-colheita de 1 mM de AS em mangas promoveu redução de 37,5% na incidência
e 20,9% no diâmetro das lesões de antracnose, segundo ZENG et al. (2006). KHADEMI et al.
(2012) verificaram que caquis cv. Karaj tratados com 2 mM de AS por imersão, durante 10
min. e armazenados sob refrigeração (1 °C e 85% UR) apresentaram redução na incidência de
doenças pós-colheita. O uso de AS (1, 2 e 4 mM) em morangos cv. „Selva‟ reduziu a

17
incidência de doenças pós-colheita (BABALAR et al., 2007). Outros autores também
demonstram resultados positivos no controle das doenças pós-colheita, como em tomate
(MANDAL et al., 2009), cereja (XU & TIAN, 2008) e pêssego (ZHANG et al., 2008; YANG
et al., 2011).
Além de ser um ativador de mecanismos de defesa, o AS também pode exercer
atividade antifúngica direta in vitro, porém os trabalhos que envolvem a ação direta do AS
sobre microrganismos são limitados (CIA et al., 2007; ANAND et al., 2008). YAO & TIAN
(2005a) observaram que o AS na concentração de 2 mM foi eficaz na inibição do crescimento
micelial e na germinação de esporos de Monilinia fructicola, quando avaliado in vitro, o que
demonstra a capacidade de fungitoxicidade do AS. Ao testar o efeito do AS na germinação de
esporos de R. stolonifer, ZHANG et al. (2010) constataram que houve redução de 51,2 e
68,2% na germinação dos esporos nas concentrações de 100 e 1000 µg mL-1
, respectivamente.
Contudo, o crescimento micelial do patógeno Alternaria solani não foi inibido quando
exposto ao AS em concentrações que variaram de 0 a 200 µM, assim como não houve
inibição do crescimento micelial de P. expansum quando exposto a 2 mM (SPLETZER &
ENYEDI, 1999; XU & TIAN, 2008).
Segundo CIA et al. (2007), o AS apresenta resultados contraditórios para diversos
patógenos e em diferentes frutos, podendo causar fitotoxidade. Portanto, estudos para
constatar a eficácia do tratamento com AS em diferentes culturas e patógenos são de extrema
importância.
Os processos de amadurecimento e senescência das frutas são responsáveis por
diversas mudanças nos aspectos de qualidade, tais como amolecimento, diminuição na acidez
total, desenvolvimento da cor, produção de aroma e aumento no teor de açúcares
(CHITARRA & CHITARRA, 2005). O AS, por sua vez, pode promover alterações nos
atributos de qualidade das frutas. O tratamento pré-colheita de laranjas com AS, aumentou
significativamente o teor de carotenóides (licopeno e α-caroteno), ácido ascórbico, glutationa,
compostos fenólicos totais e flavonóides totais na polpa e na casca durante o armazenamento
(HUANG et al., 2008). VALERO et al. (2011) verificaram que a aplicação pós-colheita de AS
(1 mM) em cereja, atrasou o processo de maturação, que se manifesta pela diminuição da
acidez, mudanças de cor e pela perda de firmeza. Entretanto, DING et al. (2007) verificaram
que a aplicação de AS em manga não alterou a firmeza dos frutos, o teor de sólidos solúveis e
a acidez titulável. Pêssegos tratados com AS e armazenados a 20 °C por 7 dias, também não
apresentaram efeito negativo sobre os parâmetros de qualidade (ZHANG et al., 2008). MO et

18
al. (2008) observaram que o tratamento de maçãs com AS não reduziu significativamente o
teor de sólidos solúveis totais.
Mangas armazenadas a 20 °C ou a 13 °C e imersas em solução contendo 0.5, 1 e 5
mmol L-1
de AS apresentaram aumento no teor de sólidos solúveis totais e menor perda de
firmeza, teor de ácido ascórbico e acidez titulável (FONG, 2005). Segundo IMRAN et al.
(2007) o tratamento com AS (0,002 e 0,02 mM) por imersão retardou a senescência em peras.
BAL & CELIK (2010) afirmaram que o tratamento com AS retardou o amadurecimento de
kiwi, sendo um método eficaz para aumentar a vida de prateleira durante o armazenamento
dos frutos. O AS tem notável capacidade de manter a qualidade dos frutos durante o período
de armazenamento, principalmente controlando a perda de firmeza (TAREEN et al., 2010).
Além de atuar no controle de doenças e na manutenção da qualidade dos frutos, o AS
pode atuar também na redução da produção autocatalítica de etileno e parece diminuir a
produção de etileno causada por estresses, bloqueando a passagem ou evitando o acúmulo de
síntese do ácido 1-carboxílico-1-aminociclopropano (ACC), como observado em tecidos
feridos de tomate (JOMORI, 2005; KERBAUY, 2004). Segundo BARBALAR et al. (2007), a
produção de etileno em morangos cv. „Selva‟ foi reduzida com a aplicação de AS.
Além de atuar sobre o etileno, tem sido demonstrado que o AS inibe as enzimas que
degradam a parede celular, tais como a poligalacturonase (PG), a lipoxigenase (LOX),
celulase e a pectinametilesterase (PME), levando a uma redução do processo de amolecimento
de frutos. Os efeitos negativos do AS sobre o ACC, PG, PME, celulose e enzimas
antioxidantes, resultam na redução da produção e ação do etileno, o que leva a diminuição da
respiração dos frutos (ASGHARI & AGHDAM, 2010). Em um estudo com bananas,
SRIVASTAVA & DWIVEDI (2000) verificaram que o tratamento com AS (500 e 1000 µM)
resultou na diminuição da atividade respiratória, bem como no atraso do aparecimento do pico
climatérico, quando comparado com o controle de maneira depende da concentração.
Alguns estudos mostraram também que o AS pode induzir a biossíntese de proteínas
de choque térmico (HSPs), que conferem proteção contra o estresse térmico. O acúmulo de
HSPs permite o armazenamento de frutos em baixas temperaturas sem o desenvolvimento de
dano pelo frio (ASGHARI & AGHDAM, 2010). Foi verificado que a aplicação de AS (1
mM) em melancias promoveu indução de resistência a danos pelo frio (JING-HUA et al.,
2008). Segundo LIU et al. (2009), após a pulverização foliar de AS (1 mM) houve um
aumento na atividade das enzimas POD, SOD e CAT, aumentando assim a resistência de
plântulas de pepino contra baixa temperatura e intensidade de luz.

19
2.4.2 Metil jasmonato
O MeJa, assim com o ácido jasmômico (AJ) e seus derivados, são conhecidos como
jasmonatos, ou seja, uma classe particular de reguladores vegetais que geralmente auxiliam na
defesa de uma planta (TAIZ & ZEIGER, 2006; KERBAUY, 2004).
Em 1962, o MeJa foi descoberto pela primeira vez em plantas, como um composto de
cheiro doce em jasmim, o que deu origem ao seu nome. Por ser volátil, foi isolado a partir do
extrato do óleo essencial de pétalas de Jasminum grandiflorum. Inicialmente, foram
realizados apenas estudos focando a sua síntese e estrutura, devido ao interesse da indústria de
perfumes. Somente após 10 anos a sua atividade biológica foi relatada, como promotor da
senescência em plantas de absinto e inibição do crescimento em favas (ROHWER & ERWIN,
2008; SOARES & MACHADO, 2007; AVANCI et al., 2010).
O AJ e o MeJa, são considerados importantes reguladores do desenvolvimento das
plantas, bem como do mecanismo de resposta das plantas a estresses bióticos (ataque de
patógenos e pragas) e abióticos (seca, baixa temperatura e salinidade). Devido a capacidade
de difundir-se através das membranas e da sua natureza volátil, o MeJa é uma molécula de
sinalização que pode mediar respostas de defesa nas plantas. A presença do MeJa pode
influenciar na germinação de sementes, fertilidade, amadurecimento dos frutos, senescência,
formação de órgãos de armazenamento, floração, regulação da abertura e fechamento de
estômatos e no crescimento radicular (CHEONG & DO CHOI, 2003; ROHWER & ERWIN,
2008; AVANCI et al., 2010; SOBRINHO et al., 2005). Além disso, pode ser caracterizado
como um regulador chave na resposta da planta a ferimento ou ataque de patógenos e insetos,
pois ativa genes envolvidos com a defesa (JUNIOR, 2009; COUTO, 2006).
Em plantas, o MeJa ocorre naturalmente, sendo sua produção dependente tanto de
estímulos externos como de genes de expressão (COUTO, 2006). A sua resposta fisiológica é
altamente variável e dependente da sua concentração, tanto nos tecidos como em meio de
cultura de células vegetais. Em concentrações menores (1 – 10 µM), causa indução da
expressão de diversos genes, entre eles os relacionados com a defesa da planta, sem causar
senescência, já em concentrações maiores (> que 50 µM) atua na indução de morte celular,
levando à senescência (SOARES & MACHADO, 2007).
O MeJa é uma molécula lipídica com enorme capacidade de regular diversos
processos fisiológicos; sua biossíntese ocorre através da via do octadecanóide ou via LOX. O
início desta via ocorre a partir do ácido α-linolênico (α-Lea), presente nas membranas dos
cloroplastos, composto este que é semelhante à prostaglandina (molécula encontrada em

20
animais que atuam na resposta a estresses, infecção e alergias). A formação de α-Lea acontece
devido a estímulos externos, como ferimentos e ataque de patógenos (AVANCI et al., 2010;
JUNIOR, 2009; ROHWER & ERWIN, 2008) .
Após a formação do ácido α-linolênico, ocorre a oxigenação do mesmo, numa reação
catalisada pela enzima 13-lipoxigenase (LOX), gerando o ácido 13-hidroperoxi-9,11,15-
octadecatrienóico (13-HPOT). O ácido 13-HPOT é oxidado pela enzima chave da via, a
Allene Oxide Synthase (AOS), formando um óxido de aleno que é instável, o qual é
convertido em 12,13-epoxilinolênico. Subseqüentemente no citosol este composto é
convertido ao ácido 12-oxofitodienóico (12-OPDA), através da enzima Allene Oxide Cyclase
(AOC). A molécula 12-OPDA é a precursora do AJ, e é o produto final da via de biossíntese
dos jasmonatos que ocorre no cloroplasto. No peroxissomo o ácido 12-ODPA, por meio da
ação da enzima OPDA reductase (OPR), sofre uma redução e após três reações de β-
oxidações, pelas enzimas Acyl-CoA Oxidase (ACX), proteína multifuncional (PMF) e Keto-
acyl-CoA thiolase (KAT), formando o AJ (AVANCI et al., 2010; JUNIOR, 2009, COUTO,
2006).
Uma vez formado no peroxissomo, o AJ pode sofrer diferentes conversões
metabólicas, produzindo assim os seus derivados. O MeJa é formado por meio de reações
catalisadas por metilação, ou seja, por ação da enzima carboxila metil-transferase o AJ é
convertido ao volátil MeJa. Acredita-se que esta reação ocorra no citoplasma (AVANCI et al.,
2010; JUNIOR, 2009; CHEONG & DO CHOI, 2003).
A enzima lipoxigenase, que está envolvida na biossíntese dos jasmonatos, pode ter sua
atividade induzida com a aplicação de jasmonatos, fazendo com que uma planta que seja
considerada suscetível, além de aumentar sua capacidade de produzir outros compostos
lipídicos usados na defesa, suporte melhor o ataque de patógenos (COUTO, 2006).
O MeJa vem sendo muito utilizado pelos pesquisadores, tanto em pré como em pós-
colheita, pois possui ação benéfica para manter, prolongar a vida pós-colheita e proteger os
produtos hortícolas contra o desenvolvimento de doenças. Em baixas concentrações, os
jasmonatos são indicados para reduzir as deteriorações que ocorrem nos frutos, bem como
aumentar a resistência natural contra patógenos (TRIPATHI & DUBEY, 2004; ROHWER &
ERWIN, 2008). A aplicação de MeJa em plantas deve ser feita como gás em meio fechado,
sendo o produto comercial um líquido volátil (ROHWER & ERWIN, 2008).
Alguns trabalhos têm demonstrado que a aplicação exógena de MeJa induz a síntese
e/ou acúmulo de proteínas antifúngicas (defensinas, osmotinas e tioninas), inibidores de
proteinases (PIs), enzimas de síntese de fitoalexinas, a defesa estrutural (presença de pelos,

21
espinhos, tricomas e ceras na superfície de caules e frutos), várias proteínas relacionadas à
patogênese (proteínas-RP) e a biossíntese de flavanóides, terpenos e compostos fenólicos
(ROHWER & ERWIN, 2008; DI PIERO et al., 2005; JUNIOR, 2009; COUTO, 2006). CAO
et al. (2008a) postularam que o controle de doenças pelo MeJa pode ser direto, devido ao
efeito inibitório sobre o crescimento dos patógenos e, indiretamente, pela indução de
resistência provocada por elevar níveis de H2O2. O tratamento com MeJa também pode
influenciar a atividade das enzimas FAL e PPO, aumentar a produção de H2O2 e vem
mostrando efeito positivo na produção de antocianinas em culturas de células (SOARES &
MACHADO, 2007; COUTO, 2006).
Em um estudo com C. acutatum (isolado de nêsperas) foi observado que o tratamento
com MeJa (10 µM) inibiu a germinação de esporos, o crescimento dos tubos germinativos e o
crescimento micelial (CAO et al., 2008a), no entanto, o fungo M. fructicola não apresentou
inibição do crescimento micelial e da germinação com o uso de 0,2 mM de MeJa (YAO &
TIAN, 2005a). Segundo YAO & TIAN (2005b), o MeJa (200 µM) promoveu inibição direta
em P. expansum, fungo causador do mofo azul em pêssego. ZHU & TIAN (2012) afirmaram
que o MeJa (10 mM) afeta negativamente a integridade da membrana plasmática de esporos
do fungo B. cinerea e ainda possui ação direta na atividade antimicrobiana in vitro, o que
sugere que o MeJa seja fungitóxico para este patógeno. Os resultados distintos podem estar
relacionados com a sensibilidade dos patógenos ao MeJa.
O MeJa pode atuar tanto na indução de resistência a pragas, em tomate (Myzus
persicae e Tetranychus urticae) e videiras (T. pacificus), como na defesa das plantas contra
patógenos (ROHWER & ERWIN, 2008). Segundo SOBRINHO et al., (2005), a aplicação de
AS antes da inoculação de Blumeria graminis f. sp. tritici em trigo, foi possível observar um
alto grau de indução de resistência, assim como em batata e tomate com o patógeno
Phytophthora infesntans. Em uvas, armazenadas a 2°C durante seis semanas, o tratamento
com MeJa (50 µM) provocou indução de resistência contra o patógeno P. digitatum, após
inoculação natural ou artificial (DROBY et al., 1999). CAO et al. (2008a) verificou que o
tratamento pós-colheita de nêsperas com 10 µM de MeJa, por 24 h, promoveu indução de
resistência contra C. acutatum.
Segundo JIN et al. (2009), o tratamento de pêssegos em pós-colheita com MeJa (1
mM) por 24 h, promoveu redução da incidência e severidade e aumentou a resistência dos
frutos inoculados com os patógenos P. expansum, B. cinerea e R. stolonifer. YU et al. (2009)
observaram redução dos sintomas de doença causados por B. cinerea em tomates tratados
com MeJa (100 µM). Segundo ZHU & TIAN (2012), a aplicação de MeJA na concentração

22
de 10 mM, acarretou na inibição do tamanho das lesões de podridão cinzenta em tomate. A
manutenção da qualidade dos frutos pelo MeJa também é relatada, como em pêssegos
armazenados a 5°C e sob temperatura ambiente (MENG et al., 2009; JIN et al., 2006), além
de nêsperas (CAO et al., 2008b) e tomates (TZORTZAKIS & ECONOMAKIS, 2007).
Alguns estudos realizados em Arabidopsis thaliana, indicaram que tanto a sinalização
dos jasmonatos, quanto a do etileno estão envolvidos e / ou seriam necessárias para atuar na
defesa contra patógenos necrotróficos. Já a via do AS atuaria na defesa contra patógenos
biotróficos (ROHWER & ERWIN, 2008).
O papel do MeJa não está totalmente elucidado no contexto da indução de resistência.
Alguns autores relataram que esta molécula pode atuar como mensageiro secundário na
sinalização para a resistência sistêmica adquirida, de forma semelhante à resposta por injúria
mecânica, entretanto os resultados de pesquisas sobre o envolvimento do MeJa no processo de
sinalização ainda são muito contraditórios (SOBRINHO et al., 2005; CAVALCANTI et al.,
2005).
O uso do MeJa também pode reduzir o dano pelo frio, como foi visto em estudos com
goiabas (GONZÁLEZ-AGUILAR, et al., 2007), limão (SIBOZA et al., 2010), uva e laranja
(GHASEMNEZHAD et al., 2008). Em um estudo com pêssegos tratados com 1 mM de MeJa,
foi observado um aumento na atividade da enzima PG, o que pode levar a uma redução de
lanosidade em frutos armazenados sob baixas temperaturas. O aumento do teor de açúcar
solúvel e a diminuição de ácidos orgânicos podem também ser conseqüência do uso de MeJa,
como visto em framboesas. Contudo o uso do MeJa também pode promover efeito
desvantajoso, pois pode reduzir a formação de voláteis, por exemplo em maçãs. De maneira
geral, portanto, a utilização do MeJa na horticultura pode representar um ganho para a
fisiologia pós-colheita (ROHWER & ERWIN, 2008).
2.5 Indução de Resistência
Os vegetais, embora não apresentem um sistema imunológico, conseguem
surpreendentemente resistir a diversas doenças provocadas por fungos, bactérias, vírus e
nematóides (TAIZ & ZEIGER, 2006). O sistema de defesa das plantas é multicomponente,
resultando em mecanismos que operam de maneira dinâmica e coordenada, atuando no local
da infecção no momento e magnitude adequada (PASCHOLATI & LEITE, 1995).
A resistência natural das plantas ao ataque de patógenos está relacionada com os
mecanismos de defesa estruturais e bioquímicos, sendo que ambos podem ser pré e /ou pós-

23
formado à tentativa de penetração do patógeno. Os mecanismos estruturais atuam como
barreiras físicas, promovendo o atraso na penetração e / ou colonização do patógeno; cutícula,
estômatos, tricomas e vasos condutores são exemplos de defesa estrutural pré-formadas e,
papilas, halos, tiloses e lignificação são pós-formados. Os mecanismos bioquímicos, por sua
vez, atuam através da produção de substâncias tóxicas (fitoantecipinas, catecol, saponinas,
espécies reativas de oxigênio e fitoalexinas), inibindo o crescimento do patógeno ou criando
condições adversas ao estabelecimento do mesmo na planta (SCHWAN-ESTRADA et al.,
2008; PASCHOLATI & LEITE, 1995).
Quando a planta é infectada por um patógeno, ocorre a disponibilização de um vasto
espectro de defesa que irá atuar contra o microrganismo invasor. A reação de
hipersensibilidade (RH) é uma resposta de defesa comum nas plantas. Esta reação mostra-se
como uma resposta celular extrema por parte da planta, na qual um número limitado de
células adjacentes ao local da infecção morre repentinamente, privando assim o acesso do
patógeno aos nutrientes e impedindo a sua propagação, podendo levar a um alto grau de
resistência. Os aspectos fisiológicos da RH, também incluem o aumento acelerado e
transitório de agentes oxidantes, além de síntese de toxinas (fitoalexinas) e proteínas-RP
(PASCHOLATI & LEITE, 1995; TAIZ & ZEIGER, 2006; BARROS et al., 2010).
A reação de hipersensibilidade é muitas vezes precedida pela produção de ERO. Estas
EROs [anion superóxido (O2-), H2O2 e radical hidroxila (OH
-)] podem contribuir para causar a
morte celular como parte da RH ou agir diretamente sobre o patógeno. Em baixas
concentrações, as EROs também podem induzir genes de defesa e resposta adaptativa da
planta (TAIZ & ZEIGER, 2006; SOARES & MACHADO, 2007). As plantas, entretanto,
protegem as suas células e compartimentos sub-celulares dos efeitos citotóxicos das ERO,
através das enzimas antioxidantes, tais como, SOD, CAT, GSH, APX, PPO e metabólitos
(glutationa, carotenóides e ácido ascórbico) (SOARES & MACHADO, 2007).
Outra maneira da planta se defender é através da resistência induzida, que é um
fenômeno estimulado por indutores químicos e biológicos, caracterizado pelo aumento da
resistência das plantas contra doenças (MACAGNAN, 2007). O processo de indução de
resistência baseia-se na interação entre receptores, geralmente localizados na membrana
plasmática vegetal e moléculas elicitoras capazes de sinalizar os diferentes mecanismos
envolvidos no processo de defesa vegetal. Assim, os elicitores são componentes iniciais e
fundamentais no desencadeamento do processo de resistência (CAVALCANTI et al., 2005).
Diversos agentes (indutores) bióticos e abióticos vêm sendo utilizado em pós-colheita para
promover indução de resistência, podendo-se citar a irradiação UV-C, altas concentrações de

24
CO2, o tratamento térmico, a quitosana, o AS, o acibenzolar-S-metil (ASM), o ácido β-
aminobutírico (BABA), os agentes de controle biológico e os jasmonatos (CIA et al., 2007;
GUZZO & HARAKAVA, 2007).
O uso de agentes externos (indutores) gera um aumento do nível de resistência, ou
seja, a resistência induzida. Os indutores, não alteram o genoma da planta, atuam de maneira
não-específica, ativando genes que expressão diversas respostas de defesa, como por
exemplo, as proteínas-RP, dentre elas a quitinase e a β-1,3 glucanase, enzimas com ação de
degradar a parede celular, principalmente de fungos. A enzima FAL também pode ser
destacada, pois promove o acúmulo de lignina em tecidos próximos ao local da infecção por
microrganismos (BONALDO et al., 2005; SOBRINHO et al., 2005; CAMILI et al., 2005).
O estudo de enzimas-chaves envolvidas na defesa das plantas contra o ataque de
patógenos, como as peroxidases (POs) e PPOs, podem confirmar o estado de indução de
resistência, já que a mesma é caracterizada pelo aumento da atividade dessas enzimas. As
peroxidases fazem parte de um conjunto de isoenzimas capazes de catalisar a oxidação de
vários substratos (substâncias aromáticas, ácido ascórbico e compostos fenólicos) na presença
de H2O2, gerando quinonas e água. A ação das POs gera produtos que estão envolvidos na
formação da parede celular e lignificação. As POs também estão envolvidas na formação de
H2O2, que apresenta atividade antimicrobiana direta. Já as PPOs possuem importância em
plantas resistentes ou induzidas devido a sua capacidade de oxidar fenóis (MACAGNAN,
2007).
Os termos “resistência induzida” (RI) e “resistência adquirida” (RA) são utilizados
como sinônimos por alguns autores (EDREVA, 2004). Segundo BONALDO (2005), a
resistência induzida é dependente do intervalo de tempo que ocorre entre o tratamento com o
indutor e a subseqüente inoculação da planta. O fenômeno também exibe outras
características, como a proteção local, em células próximas às tratadas, ou sistêmica, que
ocorre distante do sítio de aplicação do indutor.
Alguns pesquisadores por sua vez, separam a resistência sistêmica adquirida da
resistência sistêmica induzida, pois a RSA é geralmente induzida por patógenos ou ativadores
químicos e utiliza como mecanismo induzido de defesa da planta o acúmulo de proteínas-RP,
sendo esta salicilato-dependente, podendo resultar em alterações visuais na planta. Já na RSI,
a planta não apresenta alterações, não ocorre acúmulo de proteínas-RP e não é salicilato-
dependente, parece haver outra rota de sinalização, provavelmente associada à jasmonatos e
etileno. Porém fenotipicamente são processos semelhantes, já que após a exposição da planta

25
a um agente indutor, a mesma ativa os seus mecanismos de defesa (BONALDO et al., 2005;
BARROS et al., 2010).
A RSA é um processo que envolve a ativação de mecanismos latentes de resistência,
por meio de tratamentos com agentes elicitores, associada à expressão coordenada de um
conjunto de genes de defesa, a fim de formar uma barreira ou atrasar a entrada do patógeno,
restringindo assim o crescimento do patógeno e conseqüentemente, a supressão ou diminuição
dos sintomas de doença (SOBRINHO et al., 2005; GUZZO & HARAKAVA, 2007).
A indução de resistência tem sido uma ferramenta muito utilizada para o controle de
doenças pós-colheita, pois é um método alternativo ao uso efetivo de produtos químicos e
pode controlar diversas doenças. Contudo a indução de resistência no controle de doenças
pós-colheita, segundo CAMILI et al. (2005), é um tema que necessita de pesquisas que
busquem esclarecer questões sobre o efeito na interação patógeno-hospedeiro, bem como,
conhecer quais os impactos dessa nova tecnologia sobre os atributos de qualidade dos frutos.
3 MATERIAL E MÉTODOS
3.1 Avaliação In Vivo de Metil Jasmonato e Ácido Salicílico no Controle do Mofo
Cinzento, da Podridão Mole e da Antracnose em Morangos
3.1.1 Inoculação dos frutos
Foram utilizados para a pesquisa, morangos da cultivar „Oso Grande‟, no estádio de
maturação ¾ vermelho, provenientes de Valinhos-SP. Depois de colhidos, os frutos foram
levados para o Laboratório de Tecnologia Pós-colheita do Centro de Engenharia e
Automação, onde foram selecionados quanto à uniformidade de cor, ausência de defeitos e
podridões, visando à homogeneização dos mesmos. Em seguida, os morangos foram
inoculados com os agentes causais do mofo cinzento (Botrytis cinerea), da antracnose
(Colletotrichum acutatum) e da podridão mole (Rhipozus stolonifer), através de injeção
subcuticular, utilizando-se seringa de cromatografia de 100 µL (Hamilton®), de 10 μL da
suspensão de esporos, ajustada para 105 esporos mL
-1, com o auxílio da câmara de Neubauer.
O inoculo foi obtido da cultura de cada patógeno (C. acutatum, B. cinerea e R. stolofiner),
cultivados em meio de cultura batata-dextrose-ágar (BDA), por oito dias, para C. acutatum e
B. cinerea e dois dias para R. stolofiner, em incubadora BOD, a 25 ºC, com fotoperíodo de 12
h.

26
A área inoculada foi previamente demarcada com caneta para confirmação de que os
sintomas observados foram decorrentes da inoculação e não de infecção prévia (proveniente
do campo). Após 2 h da inoculação, os frutos foram tratados com metil jasmonato (MeJa) ou
ácido salicílico (AS), em diferentes concentrações, conforme descrito abaixo.
3.1.2 Efeito de metil jasmonato no controle in vivo da antracnose, do mofo cinzento e da
podridão mole em morangos
Para a aplicação do MeJa (Sigma®), os morangos foram colocados em tambores
herméticos de 200 L, com circulação forçada de ar, sob temperatura de 25 °C, onde foram
expostos ao produto (0,01 e 0,02 mM) durante 24 horas. O MeJa foi aplicado em recipiente
contendo papel filtro, sendo as concentrações determinadas considerando-se o volume dos
tambores herméticos. Como testemunha utilizou-se frutos inoculados, os quais também foram
mantidos em tambores herméticos, mas não foram expostos aos voláteis do produto (Figura
7).
Figura 7. Aplicação do metil jasmonato em morangos „Oso Grande‟ em tambores herméticos.
3.1.3 Efeito do ácido salicílico no controle in vivo da antracnose, do mofo cinzento e da
podridão mole em morangos
O AS (Synth®) foi aplicado através de imersão dos frutos nas concentrações de 1, 2 e
4 mM, durante o período de 2 min. Os frutos testemunha foram imersos pelo mesmo período
em água sob temperatura ambiente (Figura 8).

27
Figura 8. Aplicação de ácido salicílico (AS) em morangos „Oso Grande‟.
3.1.4 Efeito do metil jasmonato e do ácido salicílico no controle de podridões em
morangos não inoculados artificialmente
Frutos não inoculados artificialmente foram tratados com MeJa (0; 0,01 e 0,02 mM) e
AS (0, 1, 2 e 4 mM) para a avaliação da incidência de infecções provenientes do campo,
sendo a aplicação dos produtos realizada conforme descrito em 3.1.2 e 3.1.3.
3.1.5 Armazenamento e avaliação
Após os tratamentos os frutos foram mantidos a 25 ± 2 °C/80±5 %UR e sob
refrigeração (5±1 °C/90±5% UR) e avaliados diariamente quanto à incidência (porcentagem
de frutos com sintomas da doença) e severidade (diâmetro da lesão – calculando-se a média
de duas medidas perpendiculares). Para avaliação da severidade da podridão mole foi
utilizada escala de notas (0 = ausência de sintomas; 1 = 1 a 25% de área lesionada; 2 = 26 a
50%; 3 = 51 a 75% e 4 = 76 a 100%) e calculou-se o índice de doença (ID), conforme fórmula
abaixo descrita (CIA et al., 2010).
ID (%) = [(1 x n1) + (2 x n2) + (3 x n3) + (4 x n4)] x 100
4 x N
Sendo: ID: Índice de doença; ni: o número de frutos infectados na respectiva escala de notas;
N: o número total de frutos.

28
O delineamento experimental foi inteiramente casualizado com cinco repetições
contendo seis frutos como unidade experimental. Os experimentos foram repetidos duas
vezes, com exceção do experimento que avaliou o efeito do MeJa em frutos não inoculados,
que foi realizado apenas uma vez. Os dados (incidência e severidade das podridões ou índice
de doença) obtidos nos experimentos foram utilizados para o cálculo da área abaixo da curva
de progresso da doença (AACPD), conforme equação (CARDOSO et al., 2005):
Sendo: n, o número de observações; Yi, a incidência da doença na “i”-ésima observação e, Ti,
o tempo em dias na “i”-ésima observação.
3.1.6 Análises físico-químicas
Para as análises físico-químicas foram utilizados frutos não inoculados artificialmente.
Ao final do período de armazenamento foram realizadas análises de coloração, firmeza, teor
de sólidos solúveis, acidez titulável e ratio conforme os métodos abaixo descritos:
Coloração: foi determinada por meio de colorímetro Hunter Lab – MiniScan XE Plus,
sistema L*a*b, sendo L* correspondente à luminosidade [0 (cor preta) a 100 (cor branca)], a*
[-60 (verde) a +60 (vermelho)] e b* [-60 (azul) a +60 (amarelo)], relativos à cromaticidade;
Firmeza: foi determinada com penetrômetro digital PCE - PTR, cujas perfurações
foram feitas na região equatorial de cada fruto, com ponteira de 8 mm de diâmetro. Os
resultados foram expressos em Newton (N);
Sólidos solúveis: foi determinado no suco da fruta, obtido pela centrifugação de
quatro frutos por repetição, realizando-se leitura direta em refratômetro digital Pal - 1, marca
Atago, sendo os resultados expressos em °Brix;
Acidez titulável: foi determinada por titulação, com solução de hidróxido de sódio
(NaOH) 0,2 N até pH 8,10 (pHmetro Digimed), utilizando-se 10 g do suco, diluídos em 90 ml
de água destilada. O resultado foi expresso em porcentagem de ácido cítrico;
Ratio: foi obtido através do cálculo da razão entre sólidos solúveis e a acidez titulável.
O delineamento experimental foi inteiramente casualizado com cinco repetições
compostas por quatro frutos como unidade experimental.

29
3.2 Avaliação de Metil Jasmonato e de Ácido Salicílico sobre o Desenvolvimento In Vitro
de Colletotrichum acutatum, Botrytis cinerea e Rhizopus stolonifer
3.2.1 Metil jasmonato
Para a avaliação do MeJa sobre o desenvolvimento in vitro de C. acutatum, B. cinerea
e R. stolonifer, discos de micélio (7 mm de diâmetro), retirados da borda de colônias dos
patógenos cultivados em BDA por aproximadamente oito dias para C. acutatum e B. cinerea
e, dois dias para R. stolonifer, foram transferidos para o centro de placas de petri (9 cm de
diâmetro) contendo BDA, sendo as mesmas abertas e colocadas em tambores herméticos de
200 L, com circulação de ar, onde foram expostas às diferentes concentrações de MeJa (0,01 e
0,02 mM), por volatilização, durante 24 horas, sob a temperatura de 25 °C. Como testemunha,
placas foram depositadas em tambores herméticos, mas não expostas ao produto. Após o
tratamento as placas foram mantidas em incubadora BOD a 25 °C, com fotoperíodo de 12 h.
As medidas dos diâmetros das colônias foram efetuadas diariamente, sendo realizadas duas
medidas perpendiculares entre si até que o patógeno alcançasse a borda da placa.
3.2.2 Ácido salicílico
Para a avaliação do AS sobre o desenvolvimento in vitro de C. acutatum, B. cinerea e
R. stolonifer, adicionou-se o produto ao meio de cultura BDA nas concentrações de 1, 2 e 4
mM. Após a solidificação do meio, discos de micélio (7 mm de diâmetro), retirados da borda
de colônias dos patógenos cultivados em BDA por aproximadamente oito dias para C.
acutatum e B. cinerea e, dois dias para R. stolonifer, foram transferidos para o centro das
placas de petri (9 cm de diâmetro), as quais foram mantidas em BOD a 25 °C, com
fotoperíodo de 12 h. As medidas dos diâmetros das colônias foram efetuadas diariamente,
sendo efetuadas duas medidas perpendiculares entre si até que o patógeno alcançasse a borda
da placa.
3.2.3 Delineamento experimental e análise dos resultados
Os experimentos foram repetidos duas vezes, utilizando-se oito placas por tratamento,
com uma placa por parcela e os experimentos foram conduzidos em delineamento

30
inteiramente casualizado. Os dados obtidos nos experimentos foram utilizados para o cálculo
do índice de crescimento micelial (ICM), conforme equação (REY et al., 2005):
Sendo: C1, C2 e Cn - crescimento micelial das colônias na primeira, segunda e última
avaliação, respectivamente; N1, N2 e Nn - número de dias.
3.3 Avaliação In Vivo da Possibilidade de Indução de Resistência pelo Metil Jasmonato e
Ácido Salicílico em Morangos ‘Oso Grande’ contra os Patógenos Colletotrichum
acutatum, Botrytis cinerea e Rhizopus stolonifer
Além da avaliação dos efeitos do MeJa e do AS no controle direto de podridões em
morangos (itens 3.1.2, 3.1.3 e 3.2), foi investigada a possibilidade de os produtos atuarem
indiretamente no controle dos patógenos, através, por exemplo, da ativação de mecanismos de
defesa dos frutos.
3.3.1 Avaliação da possibilidade de indução de resistência em morangos pelo metil
jasmonato
Para se avaliar a possibilidade de o MeJa atuar como indutor de resistência contra a
antracnose, o mofo cinzento e a podridão mole, morangos „Oso Grande‟ foram colocados em
tambores herméticos de 200 L, com circulação forçada de ar, sob temperatura de 25° C, onde
foram expostos ao produto (0,01 mM), durante 24 horas. Como testemunha os frutos foram
colocados em tambores herméticos e expostos à água. Os frutos foram inoculados com os
patógenos (C. acutatum, B. cinerea e R. stolonifer) após diferentes tempos (24 h – logo após o
período de exposição ao produto, 36 h e 48 h) de exposição ao volátil.
Os frutos foram mantidos a 25 ± 2 °C/80 ± 5% UR, sendo avaliados diariamente
quanto à incidência (porcentagem de frutos com sintomas da doença) e a severidade (diâmetro
da lesão – duas medidas perpendiculares entre si) da podridão ou índice de doença. O
delineamento experimental foi inteiramente casualizado com cinco repetições contendo seis

31
frutos como unidade experimental. Os dados (incidência e severidade das podridões ou índice
de doença) obtidos nos experimentos foram utilizados para o cálculo da área abaixo da curva
de progresso da doença (AACPD), conforme descrito em 3.1.5. Para análise utilizou-se
esquema fatorial com dois tratamentos e três tempos de inoculação.
3.3.2 Avaliação da possibilidade de indução de resistência em morangos pelo ácido
salicílico
Para se avaliar a possibilidade do AS atuar como indutor de resistência contra a
antracnose, o mofo cinzento e a podridão mole, morangos foram tratados com AS (1 mM),
por imersão, durante 2 min. A utilização da concentração de 1 mM, foi baseada nos resultados
dos experimentos anteriores. Os frutos testemunha foram imersos em água sob temperatura de
25° C, pelo mesmo período. Os frutos foram inoculados com os patógenos (C. acutatum, B.
cinerea e R. stolonifer) após diferentes tempos (0, 12 e 24 h) do tratamento dos frutos. Os
frutos foram mantidos a 25 ± 2 °C/80 ± 5% UR, sendo avaliados diariamente quanto à
incidência (porcentagem de frutos com sintomas da doença) e a severidade (diâmetro da lesão
– duas medidas perpendiculares entre si) da podridão ou índice de doença. O delineamento
experimental foi inteiramente casualizado com cinco repetições contendo seis frutos como
unidade experimental. Os dados (incidência e severidade das podridões ou índice de doença)
obtidos nos experimentos foram utilizados para o cálculo da área abaixo da curva de
progresso da doença (AACPD), conforme descrito em 3.1.5. Para análise utilizou-se esquema
fatorial com dois tratamentos e três tempos de inoculação.
3.4 Análises Estatísticas
Os resultados obtidos nos experimentos foram submetidos à análise de variância
(ANOVA) e comparados pelo teste de Tukey, em delineamento inteiramente casualizado,
sendo discutido a 5% de probabilidade. Para tanto, foi empregado o programa estatístico
ASSISTAT (Assistência Estatística) versão 7.6 beta (2012).

32
4 RESULTADOS
4.1 Avaliação de Metil Jasmonato no Controle In Vivo e In Vitro dos Patógenos Botrytis
cinerea, Rhizopus stolonifer e Colletotrichum acutatum em Morango ‘Oso Grande’
4.1.1 Avaliação in vivo do metil jasmonato no controle do mofo cinzento em morangos
‘Oso Grande’ inoculados artificialmente
O MeJa, nas concentrações de 0; 0,01 e 0,02 mM, aplicado por volatilização durante
24 h, não foi efetivo no controle do mofo cinzento em nenhuma das condições de
armazenamento avaliadas. Observou-se que tanto no experimento I, como no II, não houve
redução da incidência e da severidade da doença nos frutos (Tabelas 2 e 3). Verificou-se no
experimento I, que os frutos armazenados sob refrigeração apresentaram menores valores de
AACPD, tanto para incidência, como para severidade da doença, quando comparados aos
frutos armazenados a 25 °C (Tabela 2). No último dia de avaliação, de maneira geral, os
frutos apresentaram valores de incidência da doença acima de 50%, principalmente no
experimento II (Tabela 4).
4.1.2 Avaliação da possibilidade de indução de resistência pelo metil jasmonato em
morangos ‘Oso Grande’ contra o mofo cinzento
Não houve interação significativa entre o MeJa e o intervalo de tempo entre o
tratamento e a inoculação, com relação à incidência e severidade de B. cinerea. Verificou-se
que o tratamento com MeJa (0,01 mM) exerceu efeito significativo sobre a incidência do
mofo cinzento em morangos „Oso Grande‟, quando o patógeno foi inoculado logo após o
tratamento (24 h), promovendo redução de 49% no número de frutos com sintomas, quando
comparado com os frutos testemunhas. A severidade não foi influência pelo tratamento com
MeJa. O fator tempo influenciou tanto a incidência, quanto o desenvolvimento das lesões,
sendo que as menores AACPD foram observadas para os frutos inoculados logo após o
tratamento (Tabela 5).

33
Tabela 2 - Área abaixo da curva de progresso da doença (AACPD) para incidência (%) e
severidade (cm) do mofo cinzento em morangos „Oso Grande‟, inoculados com Botrytis
cinerea, tratados com diferentes concentrações de metil jasmonato (MeJa) por 24 h e
armazenados a 25 °C / 80% UR durante sete dias ou a 5 °C / 90% UR por cinco dias seguido
por mais quatro dias a 25 °C / 80% UR (Experimento I).
25 °C
MeJa AACPD1
(mM) Incidência (%) Severidade (cm)
0,00 143,34 a 2,78 a
0,01 103,34 a 2,98 a
0,02 86,84 a 3,38 a
CV (%) 17,77 7,32
5 °C + 25 °C
MeJa AACPD1
(mM) Incidência (%) Severidade (cm)
0,00 101,68 a 1,56 a
0,01 93,00 a 1,10 a
0,02 81,66 a 0,92 a
CV (%) 23,55 10,90 1 Valores seguidos pela mesma letra, na coluna, não diferem entre si pelo teste de Tukey (P ≤ 0,05). Os dados
foram transformados em √ x + 1.
Tabela 3 - Área abaixo da curva de progresso da doença (AACPD) para incidência (%) e
severidade (cm) do mofo cinzento em morangos „Oso Grande‟, inoculados com Botrytis
cinerea, tratados com diferentes concentrações de metil jasmonato (MeJa) por 24 h e
armazenados a 25 °C / 80% UR durante sete dias ou a 5 °C / 90% UR por cinco dias seguido
por mais quatro dias a 25 °C / 80% UR (Experimento II).
25 °C
MeJa AACPD1
(mM) Incidência (%) Severidade (cm)
0,00 195,50 a 2,02 a
0,01 170,02 a 1,74 a
0,02 184,84 a 2,22 a
CV (%) 14,65 15,11
5 °C + 25 °C
MeJa AACPD1
(mM) Incidência (%) Severidade (cm)
0,00 196,66 a 2,38 a
0,01 185,66 a 2,50 a
0,02 187,68 a 2,40 a
CV (%) 7,53 17,02 1 Valores seguidos pela mesma letra, na coluna, não diferem entre si pelo teste de Tukey (P ≤ 0,05).
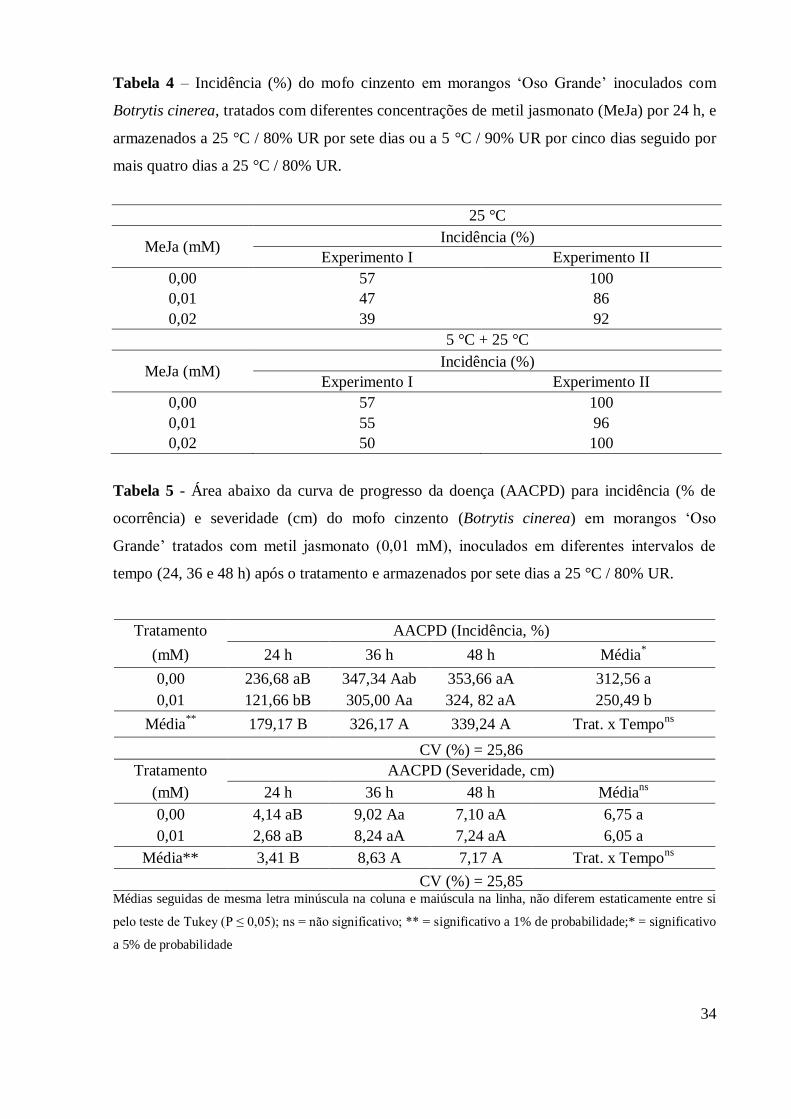
34
Tabela 4 – Incidência (%) do mofo cinzento em morangos „Oso Grande‟ inoculados com
Botrytis cinerea, tratados com diferentes concentrações de metil jasmonato (MeJa) por 24 h, e
armazenados a 25 °C / 80% UR por sete dias ou a 5 °C / 90% UR por cinco dias seguido por
mais quatro dias a 25 °C / 80% UR.
25 °C
MeJa (mM) Incidência (%)
Experimento I Experimento II
0,00 57 100
0,01 47 86
0,02 39 92
5 °C + 25 °C
MeJa (mM) Incidência (%)
Experimento I Experimento II
0,00 57 100
0,01 55 96
0,02 50 100
Tabela 5 - Área abaixo da curva de progresso da doença (AACPD) para incidência (% de
ocorrência) e severidade (cm) do mofo cinzento (Botrytis cinerea) em morangos „Oso
Grande‟ tratados com metil jasmonato (0,01 mM), inoculados em diferentes intervalos de
tempo (24, 36 e 48 h) após o tratamento e armazenados por sete dias a 25 °C / 80% UR.
Tratamento AACPD (Incidência, %)
(mM) 24 h 36 h 48 h Média*
0,00 236,68 aB 347,34 Aab 353,66 aA 312,56 a
0,01 121,66 bB 305,00 Aa 324, 82 aA 250,49 b
Média**
179,17 B 326,17 A 339,24 A Trat. x Tempons
CV (%) = 25,86
Tratamento AACPD (Severidade, cm)
(mM) 24 h 36 h 48 h Médians
0,00 4,14 aB 9,02 Aa 7,10 aA 6,75 a
0,01 2,68 aB 8,24 aA 7,24 aA 6,05 a
Média** 3,41 B 8,63 A 7,17 A Trat. x Tempons
CV (%) = 25,85 Médias seguidas de mesma letra minúscula na coluna e maiúscula na linha, não diferem estaticamente entre si
pelo teste de Tukey (P ≤ 0,05); ns = não significativo; ** = significativo a 1% de probabilidade;* = significativo
a 5% de probabilidade

35
4.1.3 Avaliação de metil jasmonato no desenvolvimento in vitro de Botrytis cinerea
Todas as concentrações do MeJa, aplicadas por volatilização, não reduziram o
crescimento micelial de B. cinerea, tanto no experimento I, como no II (Figura 9).
7,77 a 8,04 a 9,97 a
0
1
2
3
4
5
6
7
8
9
10
0 mM 0,01 mM 0,02 mM
Índ
ice
de
cres
cim
ento
mic
elia
l (c
m)
Tratamento
5,65 a 5,82 a 5,67 a
0
1
2
3
4
5
6
7
8
9
10
0 mM 0,01 mM 0,02 mM
Índ
ice
de
cres
cim
ento
mic
elia
l (c
m)
Tratamento
Figura 9 – Índice de crescimento micelial de Botrytis cinerea cultivado em meio batata-
dextrose-ágar (BDA), após exposição ao metil jasmonato por 24 h. Experimentos I (A) e II
(B). Valores seguidos pela mesma letra não diferem entre si pelo teste de Tukey (P ≤ 0,05).
A
B

36
4.1.4 Avaliação in vivo do metil jasmonato no controle da podridão mole em morangos
‘Oso Grande’ inoculados artificialmente
Observou-se no experimento I, que o MeJa, nas concentrações de 0; 0,01 e 0,02 mM,
não foi efetivo no controle da podridão mole em morangos armazenados a 25 °C, não
reduzindo a incidência e o índice da doença nos frutos (Tabela 6). Resultados semelhantes
foram obtidos no segundo experimento; o MeJa não reduziu a incidência e o índice da doença
nos frutos (Tabela 7). Em ambos os experimentos, foi possível observar que frutos mantidos
sob armazenamento refrigerado apresentaram menor incidência da podridão mole, quando
comparado com os frutos armazenados constantemente a 25 °C (Tabela 8). Após três dias da
inoculação do patógeno constatou-se que a ocorrência da doença não atingiu 100% nas
condições de armazenamento avaliadas, tanto no experimento I, como no II (Tabela 8).
Tabela 6 - Área abaixo da curva de progresso da doença (AACPD) para incidência (%) e
índice de doença (%) da podridão mole em morangos „Oso Grande‟, inoculados com
Rhizopus stolonifer, tratados com diferentes concentrações de metil jasmonato (MeJa) por 24
h e armazenados a 25 °C / 80% UR durante três dias ou a 5 °C / 90% UR por cinco dias
seguido por mais três dias a 25 °C / 80% UR (Experimento I).
25 °C
MeJa AACPD1
(mM) Incidência (%) Índice de doença (%)
0,00 54,98 a 48,74 a
0,01 60,00 a 52,52 a
0,02 76,68 a 65,00 a
CV (%) 16,99 17,27
5 °C + 25 °C
MeJa AACPD1
(mM) Incidência (%) Índice de doença (%)
0,00 23,34 b 20,02 b
0,01 45,00 ab 36,68 ab
0,02 65,00 a 57,92 a
CV (%) 25,96 24,45 1 Valores seguidos pela mesma letra, na coluna, não diferem entre si pelo teste de Tukey (P ≤ 0,05). Os dados
foram transformados em √ x + 1.

37
Tabela 7 - Área abaixo da curva de progresso da doença (AACPD) para incidência (%) e
índice de doença (%) da podridão mole em morangos „Oso Grande‟, inoculados com
Rhizopus stolonifer, tratados com diferentes concentrações de metil jasmonato (MeJa) por 24
h e armazenados a 25 °C / 80% UR durante três dias ou a 5 °C / 90% UR por cinco dias
seguido por mais três dias a 25 °C / 80% UR (Experimento II).
25 °C
MeJa AACPD1
(mM) Incidência (%) Índice de doença (%)
0,00 43,34 a 33,74 a
0,01 53,34 a 44,58 a
0,02 40,00 a 33,75 a
CV (%) 19,58 18,23
5 °C + 25 °C
MeJa AACPD1
(mM) Incidência (%) Índice de doença (%)
0,00 58,32 a 52,50 a
0,01 56,64 a 47,50 a
0,02 53,32 a 49,16 a
CV (%) 29,17 29,52 1 Valores seguidos pela mesma letra, na coluna, não diferem entre si pelo teste de Tukey (P ≤ 0,05). Os dados
foram transformados em √ x + 1.
Tabela 8 – Incidência (%) da podridão mole em morangos „Oso Grande‟ inoculados com
Rhizopus stolonifer, tratados com diferentes concentrações de metil jasmonato (MeJa) por 24
h, e armazenados a 25 °C / 80% UR por três dias ou a 5 °C / 90% UR por cinco dias seguido
por mais três dias a 25 °C / 80% UR.
25 °C
MeJa (mM) Incidência (%)
Experimento I Experimento II
0,00 57 60
0,01 73 67
0,02 53 80
5 °C + 25 °C
MeJa (mM) Incidência (%)
Experimento I Experimento II
0,00 40 30
0,01 40 47
0,02 37 73

38
4.1.5 Avaliação da possibilidade de indução de resistência pelo metil jasmonato em
morangos ‘Oso Grande’ contra a podridão mole
A análise em esquema fatorial demonstrou que não houve interação entre os fatores
MeJa e o tempo de inoculação de R. stolonifer em morangos „Oso Grande‟. A incidência e a
severidade da podridão mole também não foram influenciadas pelo tratamento com MeJa na
concentração de 0,01 mM. Verificou-se que o tratamento com MeJa promoveu aumento no
desenvolvimento da doença quando comparado com frutos não tratados (Tabela 9).
Tabela 9 - Área abaixo da curva de progresso da doença (AACPD) para incidência (% de
ocorrência) e severidade (cm) da podridão mole (Rhizopus stolonifer) em morangos „Oso
Grande‟ tratados com metil jasmonato (0,01 mM), inoculados em diferentes intervalos de
tempo (24, 36 e 48 h) após tratamento e armazenados por três dias a 25 °C / 80% UR.
Tratamento AACPD (Incidência, %)
(mM) 24 h 36 h 48 h Médians
0,00 98,34 aA 90,00 bB 100,00 aA 96,11 a
0,01 100,00 aA 96,66 aA 100,00 aA 98,88 a
Média**
99,17 A 93,33 B 100,00 A Trat. x Tempons
CV (%) = 4,56
Tratamento AACPD (Índice de doença, %)
(mM) 24 h 36 h 48 h Média*
0,00 92,49 aA 69,99 bB 79,74 aB 80,74 b
0,01 94,16 aA 81,84 aA 85,24 aA 87,08 a
Média**
93,33 A 75,92 B 82,49 B Trat. x Tempons
CV (%) = 9,43 Médias seguidas de mesma letra minúscula na coluna e maiúscula na linha, não diferem estaticamente entre si
pelo teste de Tukey (P ≤ 0,05); ns = não significativo; ** = significativo a 1% de probabilidade;* = significativo
a 5% de probabilidade
4.1.6 Avaliação de metil jasmonato no desenvolvimento in vitro de Rhizopus stolonifer
Constatou-se que todas as concentrações de MeJa, aplicadas por volatilização, não
reduziram o crescimento micelial de R. stolonifer, tanto no experimento I, como no II (Figura
10).
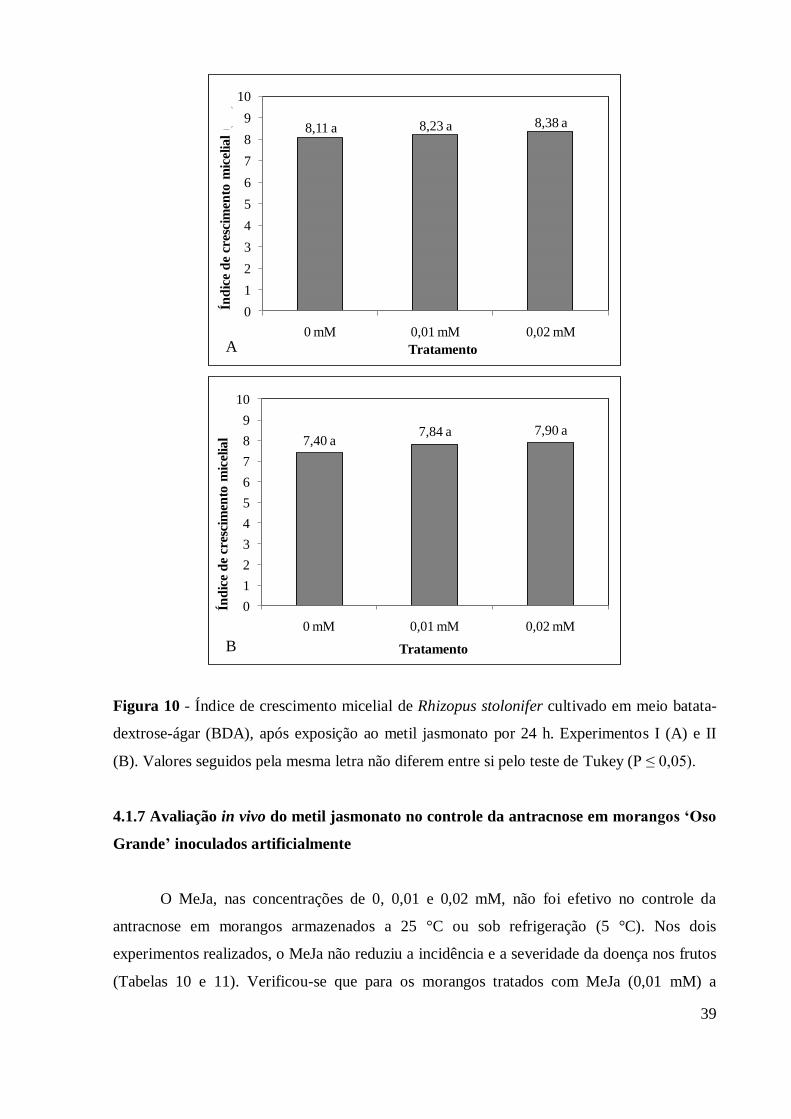
39
8,11 a 8,23 a 8,38 a
0
1
2
3
4
5
6
7
8
9
10
0 mM 0,01 mM 0,02 mM
Índ
ice
de
cres
cim
ento
mic
elia
l (c
m)
Tratamento
7,40 a7,84 a 7,90 a
0
1
2
3
4
5
6
7
8
9
10
0 mM 0,01 mM 0,02 mM
Índ
ice
de
cres
cim
ento
mic
elia
l (c
m)
Tratamento
Figura 10 - Índice de crescimento micelial de Rhizopus stolonifer cultivado em meio batata-
dextrose-ágar (BDA), após exposição ao metil jasmonato por 24 h. Experimentos I (A) e II
(B). Valores seguidos pela mesma letra não diferem entre si pelo teste de Tukey (P ≤ 0,05).
4.1.7 Avaliação in vivo do metil jasmonato no controle da antracnose em morangos ‘Oso
Grande’ inoculados artificialmente
O MeJa, nas concentrações de 0, 0,01 e 0,02 mM, não foi efetivo no controle da
antracnose em morangos armazenados a 25 °C ou sob refrigeração (5 °C). Nos dois
experimentos realizados, o MeJa não reduziu a incidência e a severidade da doença nos frutos
(Tabelas 10 e 11). Verificou-se que para os morangos tratados com MeJa (0,01 mM) a
B
A
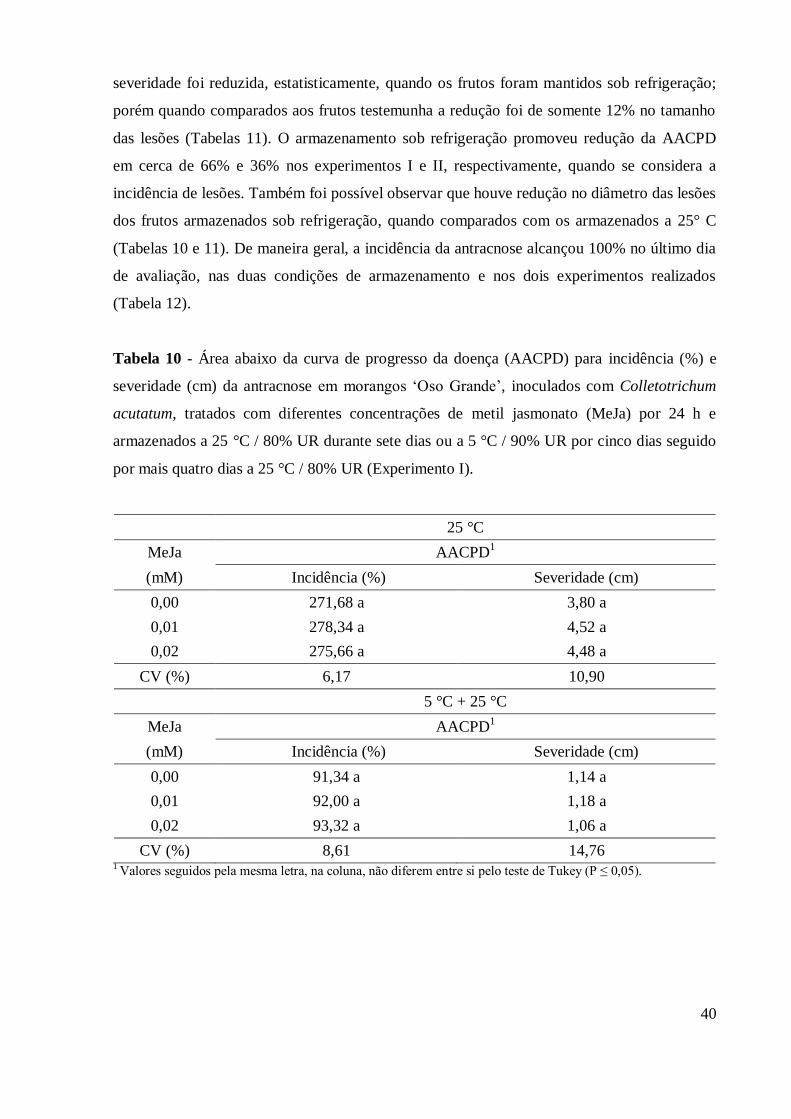
40
severidade foi reduzida, estatisticamente, quando os frutos foram mantidos sob refrigeração;
porém quando comparados aos frutos testemunha a redução foi de somente 12% no tamanho
das lesões (Tabelas 11). O armazenamento sob refrigeração promoveu redução da AACPD
em cerca de 66% e 36% nos experimentos I e II, respectivamente, quando se considera a
incidência de lesões. Também foi possível observar que houve redução no diâmetro das lesões
dos frutos armazenados sob refrigeração, quando comparados com os armazenados a 25° C
(Tabelas 10 e 11). De maneira geral, a incidência da antracnose alcançou 100% no último dia
de avaliação, nas duas condições de armazenamento e nos dois experimentos realizados
(Tabela 12).
Tabela 10 - Área abaixo da curva de progresso da doença (AACPD) para incidência (%) e
severidade (cm) da antracnose em morangos „Oso Grande‟, inoculados com Colletotrichum
acutatum, tratados com diferentes concentrações de metil jasmonato (MeJa) por 24 h e
armazenados a 25 °C / 80% UR durante sete dias ou a 5 °C / 90% UR por cinco dias seguido
por mais quatro dias a 25 °C / 80% UR (Experimento I).
25 °C
MeJa AACPD1
(mM) Incidência (%) Severidade (cm)
0,00 271,68 a 3,80 a
0,01 278,34 a 4,52 a
0,02 275,66 a 4,48 a
CV (%) 6,17 10,90
5 °C + 25 °C
MeJa AACPD1
(mM) Incidência (%) Severidade (cm)
0,00 91,34 a 1,14 a
0,01 92,00 a 1,18 a
0,02 93,32 a 1,06 a
CV (%) 8,61 14,76 1 Valores seguidos pela mesma letra, na coluna, não diferem entre si pelo teste de Tukey (P ≤ 0,05).

41
Tabela 11 - Área abaixo da curva de progresso da doença (AACPD) para incidência (%) e
severidade (cm) da antracnose em morangos „Oso Grande‟, inoculados com Colletotrichum
acutatum, tratados com diferentes concentrações de metil jasmonato (MeJa) por 24 h e
armazenados a 25 °C / 80% UR durante sete dias ou a 5 °C / 90% UR por cinco dias seguido
por mais quatro dias a 25 °C / 80% UR (Experimento II).
25 °C
MeJa AACPD1
(mM) Incidência (%) Severidade (cm)
0,00 300,00 a 5,24 a
0,01 291,34 a 5,16 a
0,02 300,00 a 5,16 a
CV (%) 1,99 12,76
5 °C + 25 °C
MeJa AACPD1
(mM) Incidência (%) Severidade (cm)
0,00 193,50 a 2,68 a
0,01 178,00 a 2,36 b
0,02 192,16 a 2,46 ab
CV (%) 6,25 7,41 1 Valores seguidos pela mesma letra, na coluna, não diferem entre si pelo teste de Tukey (P ≤ 0,05).
Tabela 12 – Incidência (%) da antracnose em morangos „Oso Grande‟ inoculados com
Colletotrichum acutatum, tratados com diferentes concentrações de metil jasmonato (MeJa)
por 24 h, e armazenados a 25 °C / 80% UR por sete dias ou a 5 °C / 90% UR por cinco dias
seguido por mais quatro dias a 25 °C / 80% UR.
25 °C
MeJa (mM) Incidência (%)
Experimento I Experimento II
0,00 100 100
0,01 100 100
0,02 100 100
5 °C + 25 °C
MeJa (mM) Incidência (%)
Experimento I Experimento II
0,00 100 100
0,01 96 100
0,02 97 100

42
4.1.8 Avaliação da possibilidade de indução de resistência pelo metil jasmonato em
morangos ‘Oso Grande’ contra a antracnose
Para a incidência de antracnose em morangos „Oso Grande‟, não houve interação
significativa entre o MeJa e o intervalo de tempo entre o tratamento e a inoculação. Observou-
se que, tanto o tratamento com MeJa (0,01 mM), quanto o tempo de inoculação, não
exerceram efeito significativo sobre a incidência de C. acutatum (Tabela 13).
Contudo, para a severidade das lesões constatou-se que tanto o tratamento com MeJa
(0,01 mM) quanto o fator tempo, exerceram efeito significativo sobre o diâmetro das lesões
de antracnose. Os frutos inoculados após 48 h do tratamento com MeJa apresentaram redução
de 31% na severidade da antracnose, quando comparado com os frutos testemunha (Tabela
13).
Tabela 13 - Área abaixo da curva de progresso da doença (AACPD) para incidência (% de
ocorrência) e severidade (cm) da antracnose (Colletotrichum acutatum) em morangos „Oso
Grande‟ tratados com metil jasmonato (0,01 mM), inoculados em diferentes intervalos de
tempo (24, 36 e 48 h) após o tratamento e armazenados por sete dias a 25 °C / 80% UR.
Tratamento AACPD (Incidência, %)
(mM) 24 h 36 h 48 h Médians
0,00 386,66 aA 378,76 aA 391,50 aA 385,64 a
0,01 394,16 aA 382,50 aA 344,16 aA 373,60 a
Médians
390,41 A 380,63 A 367,83 A Trat. x Tempons
CV (%) = 7,63
Tratamento AACPD (Severidade, cm)
(mM) 24 h 36 h 48 h Média*
0,00 8,24 aA 5,58 aB 7,14 aAB 6,98 a
0,01 7,02 aA 6,10 aAB 4,88 bB 6,00 b
Média** 7,63 A 5,84 B 6,01 B Trat. x Tempo*
CV (%) = 17,29 Médias seguidas de mesma letra minúscula na coluna e maiúscula na linha, não diferem estaticamente entre si
pelo teste de Tukey (P ≤ 0,05); ns = não significativo; ** = significativo a 1% de probabilidade;* = significativo a 5% de probabilidade
4.1.9 Avaliação de metil jasmonato no desenvolvimento in vitro de Colletotrichum
acutatum
Quanto aos efeitos do MeJa sobre o índice de crescimento micelial de C. acutatum,
nota-se que no primeiro experimento, houve redução (30%) do crescimento micelial deste

43
patógeno apenas na concentração de 0,01 mM. Já no segundo experimento, a aplicação do
produto causou aumento no índice de crescimento de C. acutatum, 17% na concentração de
0,01 e 9% quando aplicado 0,02 mM do produto (Figura 11).
2,72 a
1,90 b
2,74 a
0
1
2
3
4
5
6
7
8
9
10
0 mM 0,01 mM 0,02 mM
Índ
ice
de
cres
cim
ento
mic
elia
l (c
m)
Tratamento
2,24 c2,70 a 2,44 b
0
1
2
3
4
5
6
7
8
9
10
0 mM 0,01 mM 0,02 mM
Índ
ice
de
cres
cim
ento
mic
elia
l (c
m)
Tratamento
Figura 11 - Índice de crescimento micelial de Colletotrichum acutatum cultivado em meio
batata-dextrose-ágar (BDA), após exposição ao metil jasmonato por 24 h. Experimentos I (A)
e II (B). Valores seguidos pela mesma letra não diferem entre si pelo teste de Tukey (P ≤
0,05).
A
B

44
4.1.10 Avaliação de metil jasmonato sobre os atributos de qualidade de morangos ‘Oso
Grande’
A avaliação dos frutos quanto aos atributos físico-químicos revelou que, de maneira
geral, o MeJa não exerceu efeito significativo sobre os parâmetros avaliados; o teor de sólidos
solúveis, em frutos mantidos a 25 °C e tratados com 0,02 mM apresentou redução de 13%,
quando comparado com os frutos testemunha. A luminosidade (L*), para os frutos
armazenados sob refrigeração (5 °C), seguido de transferência para 25 °C e tratados com 0,02
mM, estava menor quando comparada aos frutos testemunha, indicando coloração mais escura
(Tabela 14).
4.1.11 Avaliação do metil jasmonato no controle de podridões em morangos ‘Oso
Grande’ não inoculados artificialmente
A avaliação dos efeitos do MeJa no controle de podridões em morangos „Oso Grande‟
não inoculados artificialmente (infecções provenientes do campo) revelou que o produto não
reduziu significativamente a incidência do mofo cinzento, da antracnose e da podridão mole,
tanto quando avaliadas isoladamente, quanto quando avaliadas em conjunto. Observou-se
também que os frutos armazenados sob refrigeração apresentaram menores valores de
AACPD, quando comparados aos frutos armazenados sob condições ambientes (Tabela 15).
4.2 Avaliação de Ácido Salicílico no Controle In Vivo e In Vitro dos Patógenos Botrytis
cinerea, Rhizopus stolonifer e Colletotrichum acutatum em Morango ‘Oso Grande’
4.2.1 Avaliação in vivo de ácido salicílico no controle do mofo cinzento em morangos
‘Oso Grande’ inoculados artificialmente
O AS, nas concentrações de 1, 2 e 4 mM, aplicado por imersão de morangos por 2
minutos, não foi efetivo no controle do mofo cinzento, pois não reduziu a severidade e a
incidência da doença nos frutos (Tabelas 16 e 17). Observou-se, no segundo experimento
(Tabela 17), que a incidência da podridão em frutos armazenados sob condições ambiente, foi
reduzida, estatisticamente, para frutos tratados com AS a 1 mM, mas de maneira não
expressiva, já que quando comparado à testemunha, a redução foi de somente 2,8%. Pôde-se
observar ainda que os frutos armazenados sob refrigeração, apresentaram menor incidência e
severidade da doença, quando comparado com os frutos armazenados a 25 °C (Tabelas 16 e

45
17). Observou-se que a incidência de mofo cinzento no último dia de avaliação atingiu, em
média, 80 e 100%, no experimento I e II, respectivamente (Tabela 18).
Tabela 14 – Atributos de qualidade de morangos „Oso Grande‟ tratados com metil jasmonato
(MeJa) por 24 h e armazenados a 25 °C / 80% UR durante sete dias ou a 5 °C / 90% UR por
cinco dias seguido por mais quatro dias a 25 °C / 80% UR.
25 °C (sete dias)
MeJa Cor Firmeza (N) SS (%) AT (%) Ratio (SS/AT)
(mM) L* a*
Dia 0 34,86 33,12 6,42 7,1 0,9 7,8
0,00 31,08 ab 31,54 ab 6,07 a 8,50 a 1,08 a 7,92 a
0,01 30,79 b 29,87 b 5,87 a 8,14 ab 1,01 a 7,99 a
0,02 32,95 a 33,01 a 7,15 a 7,40 b 1,00 a 7,43 a
CV % 3,95 4,41 20,56 5,75 7,71 8,93
5 °C (quatro dias)
MeJa Cor Firmeza (N) SS (%) AT (%) Ratio (SS/AT)
(mM) L* a*
Dia 0 34,86 33,12 6,42 7,1 0,9 7,8
0,00 38,69 a 36,69 a 8,67 a 7,82 a 1,13 a 7,74 a
0,01 39,04 a 36,61 a 8,58 a 8,04 a 0,98 a 8,17 a
0,02 38,67 a 35,36 a 8,49 a 7,66 a 1,02 a 7,49 a
CV % 5,56 8,25 13,29 7,36 18,02 6,50
5 °C (quatro dias) + 25 °C (quatro dias)
MeJa Cor Firmeza (N) SS (%) AT (%) Ratio (SS/AT)
(mM) L* a*
Dia 0 34,86 33,12 6,42 7,1 0,9 7,8
0,00 50,20 a 47,62 a 7,79 a 8,88 a 1,10 ab 8,02 a
0,01 49,61 ab 46,57 a 7,21 a 7,88 a 0,99 b 7,99 a
0,02 47,82 b 44,98 a 7,48 a 8,48 a 1,15 a 7,38 a
CV % 2,15 6,57 12,76 9,40 8,04 7,12
Cor: em colorímetro Hunter Lab, sistema L*a*b, onde L corresponde à medida de luminosidade (0=preto a
100=branco), a (-60=verde a +60=vermelho) e b (-60=azul a +60=amarelo), significam cromaticidade; SS (%):
sólidos solúveis; AT (%): acidez titulável em % de ácido cítrico; Ratio (SS/AT): relação entre sólidos solúveis e
acidez titulável. Médias seguidas de mesma letra, na coluna, não diferem significativamente entre si pelo teste de
Tukey (P ≤ 0,05).

46
Tabela 15 - Área abaixo da curva de progresso da doença (AACPD) para incidência (%) do
mofo cinzento, da podridão mole e da antracnose em morangos „Oso Grande‟ não inoculados
artificialmente, tratados com diferentes concentrações de metil jasmonato (MeJa) por 24 h e
armazenados a 25 °C / 80% UR durante sete dias ou a 5 °C / 90% UR por cinco dias seguido
por mais quatro dias a 25 °C / 80% UR.
25 °C
MeJa AACPD (Incidência, %)1
(mM) Geral Mofo cinzento Antracnose Podridão mole
0,00 81,66 a 13,32 a 0,00 a 68,34 a
0,01 161,70 a 95,00 a 0,00 a 71,68 a
0,02 111,70 a 45,02 a 5,00 a 83,36 a
5 °C + 25 °C
MeJa AACPD (Incidência, %)1
(mM) Geral Mofo cinzento Antracnose Podridão mole
0,00 33,32 a 13,32 a 0,00 a 20,00 a
0,01 33,32 a 20,00 a 0,00 a 13,32 a
0,02 36,66 a 20,00 a 10,00 a 6,66 a 1 Valores seguidos pela mesma letra, na coluna, não diferem entre si pelo teste de Tukey (P ≤ 0,05).
Tabela 16 - Área abaixo da curva de progresso da doença (AACPD) para incidência (%) e
severidade (cm) do mofo cinzento em morangos „Oso Grande‟, inoculados com Botrytis
cinerea, tratados com diferentes concentrações de ácido salicílico por dois minutos e
armazenados a 25 °C / 80% UR durante sete dias ou a 5 °C / 90% UR por cinco dias seguido
por mais quatro dias a 25 °C / 80% UR (Experimento I).
25 °C
AS AACPD1
(mM) Incidência (%) Severidade (cm)
0 154,00 a 3,42 a
1 153,00 a 3,70 a
2 149,16 a 3,14 a
4 148,50 a 3,22 a
CV (%) 20,88 17,15
5 °C + 25 °C
AS AACPD1
(mM) Incidência (%) Severidade (cm)
0 113,66 a 1,06 ab
1 110,00 a 1,14 ab
2 107,00 a 0,70 b
4 142,32 a 1,22 a
CV (%) 14,62 6,27 1 Valores seguidos pela mesma letra, na coluna, não diferem entre si pelo teste de Tukey (P ≤ 0,05). Os dados
foram transformados em √ x + 1.

47
Tabela 17 - Área abaixo da curva de progresso da doença (AACPD) para incidência (%) e
severidade (cm) do mofo cinzento em morangos „Oso Grande‟, inoculados com Botrytis
cinerea, tratados com diferentes concentrações de ácido salicílico por dois minutos e
armazenados a 25 °C / 80% UR durante sete dias ou a 5 °C / 90% UR por cinco dias seguido
por mais quatro dias a 25 °C / 80% UR (Experimento II).
25 °C
AS AACPD1
(mM) Incidência (%) Severidade (cm)
0 300,00 a 5,50 a
1 291,68 b 5,20 a
2 298,00 ab 6,58 a
4 298,34 ab 5,68 a
CV (%) 1,39 16,50
5 °C + 25 °C
AS AACPD1
(mM) Incidência (%) Severidade (cm)
0 198,00 a 1,56 a
1 183,34 a 1,80 a
2 191,34 a 1,48 a
4 190,00 a 1,58 a
CV (%) 5,78 15,64 1 Valores seguidos pela mesma letra, na coluna, não diferem entre si pelo teste de Tukey (P ≤ 0,05).
Tabela 18 – Incidência (%) do mofo cinzento em morangos „Oso Grande‟ inoculados com
Botrytis cinerea, tratados com diferentes concentrações de ácido salicílico (AS) por 2 min, e
armazenados a 25 °C / 80% UR por sete dias ou a 5 °C / 90% UR por cinco dias seguido por
mais quatro dias a 25 °C / 80% UR.
25 °C
AS (mM) Incidência (%)
Experimento I Experimento II
0 83 100
1 61 100
2 66 100
4 50 100
5 °C + 25 °C
AS (mM) Incidência (%)
Experimento I Experimento II
0 71 100
1 75 93
2 80 100
4 80 100

48
4.2.2 Avaliação da possibilidade de indução de resistência pelo ácido salicílico em
morangos ‘Oso Grande’ contra o mofo cinzento
Não houve interação significativa entre o AS e o intervalo de tempo entre o tratamento
e a inoculação. O tratamento com AS (1 mM) exerceu efeito significativo sobre a incidência
do mofo cinzento em morangos „Oso Grande‟ inoculados 48 h após o tratamento dos frutos,
reduzindo em 47% a AACPD. O diâmetro da lesão não foi influenciado significativamente
pelo AS (Tabela 19).
4.2.3 Avaliação de ácido salicílico no desenvolvimento in vitro de Botrytis cinerea
Os resultados obtidos nos experimentos in vitro mostraram que o AS, na maior
concentração avaliada (4 mM), reduziu significativamente o crescimento micelial de B.
cinerea (Figura 12).
Tabela 19 - Área abaixo da curva de progresso da doença (AACPD) para incidência (% de
ocorrência) e severidade (cm) do mofo cinzento (Botrytis cinerea) em morangos „Oso
Grande‟ tratados com ácido salicílico (1 mM), inoculados em diferentes intervalos de tempo
(24, 36 e 48 h) após o tratamento e armazenados por sete dias a 25 °C / 80% UR.
Tratamento AACPD (Incidência, %)
(mM) 24 h 36 h 48 h Média*
0 332,16 aA 341,66 aA 193,34 aB 289,05 a
1 283,34 aA 342,00 aA 101,66 bB 242,33 b
Média**
307,75 A 341,83 A 147,50 B Trat. x Tempons
CV (%) = 23,31
Tratamento AACPD (Severidade, cm)
(mM) 24 h 36 h 48 h Médians
0 6,20 aAB 7,16 aA 4,48 aB 5,94 a
1 7,08 aAB 7,80 aA 5,68 aB 6,85 a
Média** 6,64 A 7,48 A 5,08 B Trat. x Tempons
CV (%) = 19,94
Médias seguidas de mesma letra minúscula na coluna e maiúscula na linha, não diferem estaticamente entre si
pelo teste de Tukey (P ≤ 0,05); ns = não significativo; ** = significativo a 1% de probabilidade;* = significativo
a 5% de probabilidade

49
6,87 a
5,03 a 4,77 a
2,17 b
0
2
4
6
8
10
12
14
0 mM 1 mM 2 mM 4 mM
Índ
ice
de
cres
cim
ento
mic
elia
l (c
m)
Tratamento
7,14 a6,54 a 6,59 a
3,40 b
0
2
4
6
8
10
12
14
0 mM 1 mM 2 mM 4 mM
Índ
ice
de
cres
cim
ento
mic
elia
l (c
m)
Tratamento
Figura 12 – Índice de crescimento micelial de Botrytis cinerea cultivado em meio batata-
dextrose-ágar (BDA), incorporado com ácido salicílico em diferentes concentrações.
Experimentos I (A) e II (B). Valores seguidos pela mesma letra não diferem entre si pelo teste
de Tukey (P ≤ 0,05).
4.2.4 Avaliação in vivo de ácido salicílico no controle da podridão mole em morangos
‘Oso Grande’ inoculados artificialmente
Todas as concentrações de AS avaliadas não foram eficazes em promover a redução
da incidência e da severidade da podridão mole em morangos „Oso Grande‟. O
armazenamento dos frutos sob refrigeração acarretou em menor ocorrência e desenvolvimento
A
B

50
das lesões, quando comparado com os frutos armazenados a 25 °C (Tabelas 20 e 21). No
último dia de avaliação os frutos armazenados a 25 °C apresentaram incidência de cerca de
100%, enquanto que para os frutos armazenados sob refrigeração a ocorrência da doença foi
de 50% menor, em média (Tabela 22).
4.2.5 Avaliação da possibilidade de indução de resistência pelo ácido salicílico em
morangos ‘Oso Grande’ contra a podridão mole
O tratamento com AS (1 mM) não exerceu efeito significativo sobre a incidência e a
severidade da podridão mole em morangos „Oso Grande‟ inoculados em diferentes intervalos
de tempo após o tratamento. Não houve interação significativa entre o AS e o intervalo de
tempo entre o tratamento e a inoculação (Tabela 23).
Tabela 20 - Área abaixo da curva de progresso da doença (AACPD) para incidência (%) e
índice de doença (%) da podridão mole em morangos „Oso Grande‟, inoculado com Rhizopus
stolonifer, tratados com diferentes concentrações de ácido salicílico por dois minutos e
armazenados a 25 °C / 80% UR durante três dias ou a 5 °C / 90% UR por cinco dias seguido
por mais três dias a 25 °C / 80% UR (Experimento I).
25 °C
AS AACPD1
(mM) Incidência (%) Índice de doença (%)2
0 100,00 a 87,50 a
1 100,00 a 90,83 a
2 100,00 a 91,25 a
4 100,00 a 87,50 a
CV (%) 0,00 2,82
5 °C + 25 °C
AS AACPD1
(mM) Incidência (%) Índice de doença (%)2
0 35,00 a 26,50 a
1 28,35 a 22,50 a
2 18,32 a 13,75 a
4 43,00 a 40,00 a
CV (%) 29,76 33,50 1 Valores seguidos pela mesma letra, na coluna, não diferem entre si pelo teste de Tukey (P ≤ 0,05).
2 Os dados foram transformados em √ x + 1.

51
Tabela 21 - Área abaixo da curva de progresso da doença (AACPD) para incidência (%) e
índice de doença (%) da podridão mole em morangos „Oso Grande‟, inoculado com Rhizopus
stolonifer, tratados com diferentes concentrações de ácido salicílico por dois minutos e
armazenados a 25 °C / 80% UR durante três dias ou a 5 °C / 90% UR por cinco dias seguido
por mais três dias a 25 °C / 80% UR (Experimento II).
25 °C
AS AACPD1
(mM) Incidência (%) Índice de doença (%)
0 100,00 a 97,08 a
1 100,00 a 97,10 a
2 100,00 a 95,82 a
4 100,00 a 97,08 a
CV (%) 0,00 4,48
5 °C + 25 °C
AS AACPD1
(mM) Incidência (%) Índice de doença (%)
0 49,00 a1 39,33 b
1 66,66 a 58,33 a
2 51,66 a 34,58 b
4 46,66 a 35,41 b
CV (%) 27,51 24,69 1 Valores seguidos pela mesma letra, na coluna, não diferem entre si pelo teste de Tukey (P ≤ 0,05).
Tabela 22 – Incidência (%) da podridão mole em morangos „Oso Grande‟ inoculados com
Rhizopus stolonifer, tratados com diferentes concentrações de ácido salicílico (AS) por 24 h, e
armazenados a 25 °C / 80% UR por três dias ou a 5 °C / 90% UR por cinco dias seguido por
mais três dias a 25 °C / 80% UR.
25 °C
AS (mM) Incidência (%)
Experimento I Experimento II
0 100 100
1 100 100
2 100 97
4 100 100
5 °C + 25 °C
AS (mM) Incidência (%)
Experimento I Experimento II
0 37 54
1 33 70
2 20 60
4 48 50

52
Tabela 23 - Área abaixo da curva de progresso da doença (AACPD) para incidência (% de
ocorrência) e severidade (cm) da podridão mole (Rhizopus stolonifer) em morangos „Oso
Grande‟ tratados com ácido salicílcio (1 mM), inoculados em diferentes intervalos de tempo
(24, 36 e 48 h) após o tratamento e armazenados por três dias a 25 °C / 80% UR.
Tratamento AACPD (Incidência, %)
(mM) 24 h 36 h 48 h Médians
0 96,66 aA 90,00 aA 98,34 aA 95,00 a
1 96,68 aA 80,02 bB 93,32 aA 90,00 a
Média**
96,67 A 85,01 B 95,83 A Trat. x Tempons
CV (%) = 8,23
Tratamento AACPD (Índice de doença, %)
(mM) 24 h 36 h 48 h Médians
0 92,08 aA 63,74 aB 91,66 aA 82,49 a
1 87,99 aA 60,41 aB 87,91 aA 78,74 a
Média** 89,99 A 62,07 B 89,78 A Trat. x Tempo*
CV (%) = 6,45
Médias seguidas de mesma letra minúscula na coluna e maiúscula na linha, não diferem estaticamente entre si
pelo teste de Tukey (P ≤ 0,05); ns = não significativo; ** = significativo a 1% de probabilidade;* = significativo
a 5% de probabilidade
4.2.6 Avaliação de ácido salicílico no desenvolvimento in vitro de Rhizopus stolonifer
No primeiro experimento verificou-se que todas as concentrações de AS, incorporadas
ao meio de cultura, reduziram de maneira significativa o crescimento micelial de R. stolofiner,
de maneira dependente da concentração. No entanto, no segundo experimento, somente as
maiores concentrações avaliadas (2 e 4 mM) reduziram significativamente o crescimento
micelial do patógeno (Figura 13).
4.2.7 Avaliação in vivo de ácido salicílico no controle da antracnose em morangos ‘Oso
Grande’ inoculados artificialmente
A imersão de morangos nas diferentes concentrações de AS por 2 minutos não foi
efetiva em reduzir o diâmetro e/ou a incidência de lesões de C. acutatum nos frutos. Pôde-se
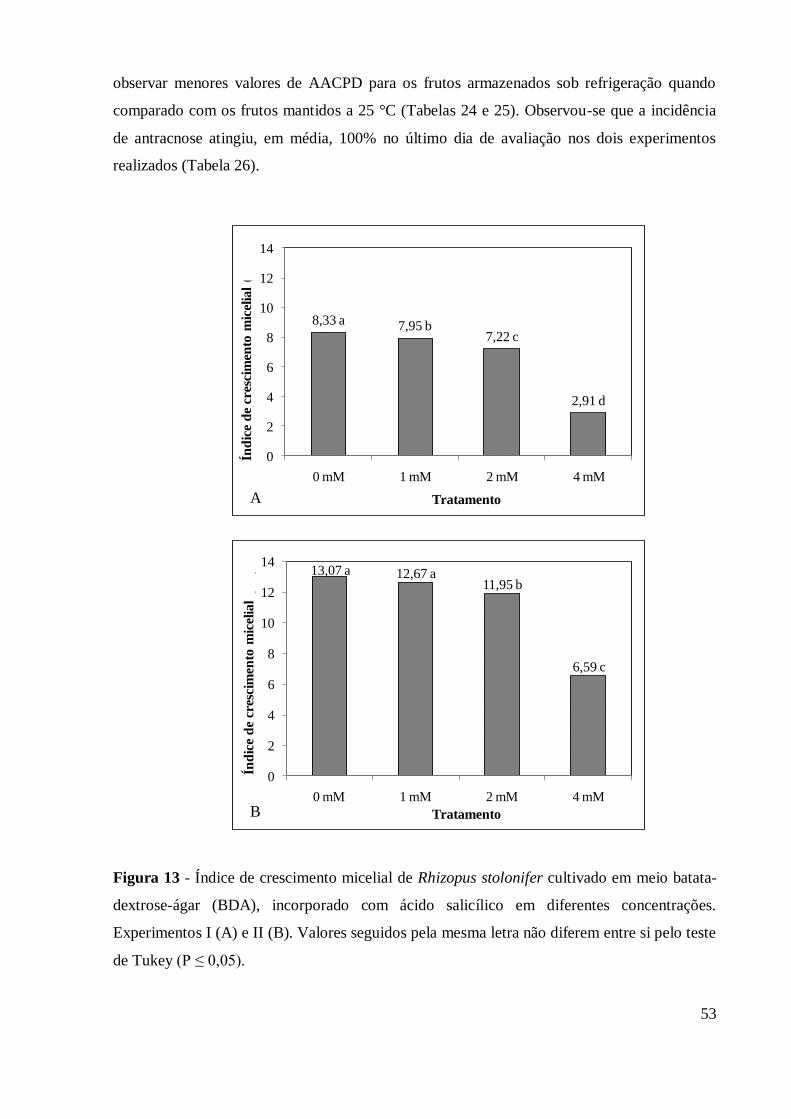
53
observar menores valores de AACPD para os frutos armazenados sob refrigeração quando
comparado com os frutos mantidos a 25 °C (Tabelas 24 e 25). Observou-se que a incidência
de antracnose atingiu, em média, 100% no último dia de avaliação nos dois experimentos
realizados (Tabela 26).
8,33 a 7,95 b7,22 c
2,91 d
0
2
4
6
8
10
12
14
0 mM 1 mM 2 mM 4 mM
Índ
ice
de
cres
cim
ento
mic
elia
l (c
m)
Tratamento
13,07 a 12,67 a11,95 b
6,59 c
0
2
4
6
8
10
12
14
0 mM 1 mM 2 mM 4 mM
Índ
ice
de
cres
cim
ento
mic
elia
l (c
m)
Tratamento
Figura 13 - Índice de crescimento micelial de Rhizopus stolonifer cultivado em meio batata-
dextrose-ágar (BDA), incorporado com ácido salicílico em diferentes concentrações.
Experimentos I (A) e II (B). Valores seguidos pela mesma letra não diferem entre si pelo teste
de Tukey (P ≤ 0,05).
A
B

54
4.2.8 Avaliação da possibilidade de indução de resistência pelo ácido salicílico em
morangos ‘Oso Grande’ contra a antracnose
Não houve interação significativa entre o AS e o intervalo de tempo entre o tratamento
e a inoculação para a incidência de antracnose em morangos „Oso Grande‟. Observou-se
também que tanto o tratamento com AS (1 mM), quanto o tempo de inoculação, não
exerceram efeito significativo sobre a incidência de C. acutatum (Tabela 27).
Entretanto, quando observamos os resultados obtidos para severidade, foi possível
verificar que tanto o tratamento com AS (1 mM), como o fator tempo, exerceram efeito
significativo sobre o diâmetro das lesões de antracnose em morangos „Oso Grande‟. Os frutos
inoculados logo após o tratamento com AS, apresentaram redução de 17% na severidade de
antracnose quando comparado com os frutos testemunhas (Tabela 27).
Tabela 24 - Área abaixo da curva de progresso da doença (AACPD) para incidência (%) e
severidade (cm) da antracnose em morangos „Oso Grande‟, inoculado com Colletotrichum
acutatum, tratados com diferentes concentrações de ácido salicílico por dois minutos e
armazenados a 25 °C / 80% UR durante sete dias ou a 5 °C / 90% UR por cinco dias seguido
por mais quatro dias a 25 °C / 80% UR (Experimento I).
25 °C
AS AACPD1
(mM) Incidência (%) Severidade (cm)
0 338,66 a 6,70 a
1 333,16 a 6,80 a
2 361,82 a 6,62 a
4 378,66 a 6,68 a
CV (%) 10,17 10,26
5 °C + 25 °C
AS AACPD1
(mM) Incidência (%) Severidade (cm)
0 83,34 a1 1,14 a
1 86,34 a 1,14 a
2 79,50 a 1,16 a
4 89,34 a 1,20 a
CV (%) 16,74 13,56 1 Valores seguidos pela mesma letra, na coluna, não diferem entre si pelo teste de Tukey (P ≤ 0,05).

55
Tabela 25 - Área abaixo da curva de progresso da doença (AACPD) para incidência (%) e
severidade (cm) da antracnose em morangos „Oso Grande‟, inoculado com Colletotrichum
acutatum, tratados com diferentes concentrações de ácido salicílico por dois minutos e
armazenados a 25 °C / 80% UR durante sete dias ou a 5 °C / 90% UR por cinco dias seguido
por mais quatro dias a 25 °C / 80% UR (Experimento II).
25 °C
AS AACPD1
(mM) Incidência (%) Severidade (cm)
0 360,84 a 7,90 a
1 381,50 a 7,40 a
2 374,84 a 7,74 a
4 395,00 a 7,94 a
CV (%) 8,89 5,21
5 °C + 25 °C
AS AACPD1
(mM) Incidência (%) Severidade (cm)
0 95,00 a 1,30 a
1 91,00 a 1,22 a
2 96,00 a 1,26 a
4 100,00 a 1,30 a
CV (%) 6,85 6,82 1 Valores seguidos pela mesma letra, na coluna, não diferem entre si pelo teste de Tukey (P ≤ 0,05).
Tabela 26 – Incidência (%) da antracnose em morangos „Oso Grande‟ inoculados com
Colletotrichum acutatum, tratados com diferentes concentrações de ácido salicílico (AS) por
dois minutos e armazenados a 25 °C / 80% UR por sete dias ou a 5 °C / 90% UR por cinco
dias seguido por mais quatro dias a 25 °C / 80% UR.
25 °C
AS (mM) Incidência (%)
Experimento I Experimento II
0 100 100
1 100 100
2 100 95
4 100 100
5 °C + 25 °C
AS (mM) Incidência (%)
Experimento I Experimento II
0 100 97
1 95 100
2 85 96
4 100 100

56
Tabela 27 - Área abaixo da curva de progresso da doença (AACPD) para os valores de
incidência (% de ocorrência) e severidade (cm) da antracnose (Colletotrichum acutatum) em
morangos „Oso Grande‟ tratados com ácido salicílico (1 mM), inoculados em diferentes
intervalos de tempo (24, 36 e 48 h) após o tratamento e armazenados por sete dias a 25 °C /
80% UR.
Tratamento AACPD (Incidência, %)
(mM) 24 h 36 h 48 h Médians
0 384,00 aA 391,00 aA 388,00 aA 387,66 a
1 400,00 aA 379,34 aA 396,00 aA 392,00 a
Médians
392,00 A 385,17 A 392,33 A Trat. x Tempons
CV (%) = 7,63
Tratamento AACPD (Severidade, cm)
(mM) 24 h 36 h 48 h Média*
0 15,50 Aa 9,72 Ba 8,24 Ba 11,15 a
1 12,80 Ab 9,16 Ba 8,86 Ba 10,27 b
Média** 14,15 A 9,44 B 8,55 B Trat. x Tempo**
CV (%) = 17,29 Médias seguidas de mesma letra minúscula na coluna e maiúscula na linha, não diferem estaticamente entre si
pelo teste de Tukey (P ≤ 0,05); ns = não significativo; ** = significativo a 1% de probabilidade;* = significativo
a 5% de probabilidade
4.2.9 Avaliação de ácido salicílico no desenvolvimento in vitro de Colletotrichum
acutatum
A incorporação de AS ao meio de cultura, nas concentrações de 2 e 4 mM inibiu o
crescimento micelial de C. acutatum (Experimento I), demonstrando assim que o ácido
salicílico possui efeito fungistático sobre o desenvolvimento do fungo. Já no experimento II,
observou-se que o AS reduziu o desenvolvimento do patógeno, somente na maior
concentração (4 mM) avaliada (Figura 14).

57
3,70 a 3,82 a3,34 b
2,03 c
0
2
4
6
8
10
12
14
0 mM 1 mM 2 mM 4 mM
Índ
ice
de
cres
cim
ento
mic
elia
l (c
m)
Tratamento
3,67 a 3,64 a 3,23 a
1,57 b
0
2
4
6
8
10
12
14
0 mM 1 mM 2 mM 4 mM
Índ
ice
de
cres
cim
ento
mic
elia
l (c
m)
Tratamento
Figura 14 - Índice de crescimento micelial de Colletotrichum acuctatum cultivado em meio
batata-dextrose-ágar (BDA), incorporado com ácido salicílico em diferentes concentrações.
Experimentos I (A) e II (B). Valores seguidos pela mesma letra não diferem entre si pelo teste
de Tukey (P ≤ 0,05).
4.2.10 Avaliação in vivo de ácido salicílico sobre os atributos de qualidade de morangos
‘Oso Grande’
Os frutos tratados com AS, nas concentrações de 0, 1, 2 e 4 mM, e armazenados a 25
°C ou sob refrigeração, não apresentaram diferença significativa nos atributos de qualidade
avaliados (Tabela 28).
A
B

58
Tabela 28 – Atributos de qualidade de morangos „Oso Grande‟ tratados com ácido salicílico
por dois minutos e armazenados a 25 °C / 80% UR durante sete dias, a 5 °C / 90% UR
durante cinco dias ou a 5 °C / 90%UR por cinco dias seguido por mais quatro dias a 25 °C /
80% UR.
25 °C (sete dias)
MeJa Cor Firmeza (N) SS (%) AT (%) Ratio (SS/AT)
(mM) L* a*
Dia 0 39,47 31,65 8,33 7,8 0,83 9,48
0 28,15 a 27,30 a 7,41 a 7,96 a 1,04 ab 7,64 ab
1 27,36 a 25,56 a 5,65 a 8,22 a 1,03 b 7,94 a
2 27,26 a 26,85 a 7,48 a 8,02 a 1,20 a 6,73 b
4 27,25 a 26,36 a 7,99 a 8,14 a 1,20 a 6,75 b
CV % 4,37 6,35 22,15 8,97 8,10 8,38
5 °C (cinco dias)
MeJa Cor Firmeza (N) SS (%) AT (%) Ratio (SS/AT)
(mM) L* a*
Dia 0 39,47 31,65 8,33 7,8 0,83 9,48
0 34,10 a 38,25 a 11,05 a 8,10 a 0,91 a 8,97 a
1 33,68 a 35,91 ab 9,22 a 8,74 a 0,88 a 9,91 a
2 34,24 a 32,81 b 8,47 a 8,68 a 0,86 a 10,09 a
4 36,46 a 33,41 b 10,38 a 8,70 a 0,92 a 9,45 a
CV % 7,77 6,18 20,06 4,21 6,65 9,52
5 °C (cinco dias) + 25 °C (quatro dias)
MeJa Cor Firmeza (N) SS (%) AT (%) Ratio (SS/AT)
(mM) L* a*
Dia 0 39,47 31,65 8,33 7,8 0,83 9,48
0 29,48 a 27,64 a 8,86 a 8,92 a 0,96 a 9,24 a
1 29,74 a 28,46 a 11,07 a 8,60 ab 1,01 a 8,54 a
2 30,81 a 28,33 a 10,84 a 8,10 b 0,97 a 8,31 a
4 30,50 a 26,73 a 8,04 a 8,46 ab 0,91 a 9,30 a
CV % 3,29 5,00 20,84 4,11 6,06 7,59
Cor: em colorímetro Hunter Lab, sistema L*a*b, onde L corresponde a medida de luminosidade (0=preto a
100=branco), a (-60=verde a +60=vermelho) e b (-60=azul a +60=amarelo), significam cromaticidade; SS (%):
sólidos solúveis; AT (%): acidez titulável em % de ácido cítrico; Ratio (SS/AT): relação entre sólidos solúveis e
acidez titulável. Médias seguidas de mesma letra, na coluna, não diferem significativamente entre si pelo teste de
Tukey (P ≤ 0,05).

59
4.1.11 Avaliação de ácido salicílico no controle de podridões em morangos ‘Oso Grande’
não inoculados artificialmente
No primeiro experimento, constatou-se que o AS não exerceu efeito significativo
sobre a incidência do mofo cinzento, da antracnose e da podridão mole, tanto quando
avaliados individualmente, como quando avaliadas em conjunto (Tabela 29). No entanto, no
segundo experimento, observou-se que o AS (1 mM) reduziu significativamente a incidência
do mofo cinzento em frutos armazenados a 25 °C (Tabela 30). Observou-se ainda que os
valores da AACPD para a incidência de doenças foram menores para os frutos armazenados
sob refrigeração (Tabelas 29 e 30).
Tabela 29 - Área abaixo da curva de progresso da doença (AACPD) para incidência (%) de
doenças vindas do campo em morangos „Oso Grande‟ tratados com diferentes concentrações
de ácido salicílico (AS) por dois minutos e armazenados a 25 °C / 80% UR durante sete dias
ou a 5 °C / 90% UR por cinco dias seguido por mais quatro dias a 25 °C / 80% UR
(Experimento I).
25 °C
AS AACPD (Incidência, %)1
(mM) Geral Mofo cinzento Antracnose Podridão mole
0 193,32 ab 121,66 a 40,07 a 13,34 a
1 205,00 a 125,02 a 36,68 a 0,00 a
2 166,66 ab 95,00 a 28,34 a 8,34 a
4 115,02 b 66,66 a 13,34 a 11,66 a
5 °C + 25 °C
AS AACPD (Incidência, %)1
(mM) Geral Mofo cinzento Antracnose Podridão mole
0 50,00 a 38,33 a 1,00 a 0,00 a
1 41,66 a 38,31 a 1,15 a 1,66 a
2 63,34 a 40,01 a 2,73 a 0,00 a
4 82,32 a 39,00 a 2,57 a 6,66 a 1 Valores seguidos pela mesma letra, na coluna, não diferem entre si pelo teste de Tukey (P ≤ 0,05).
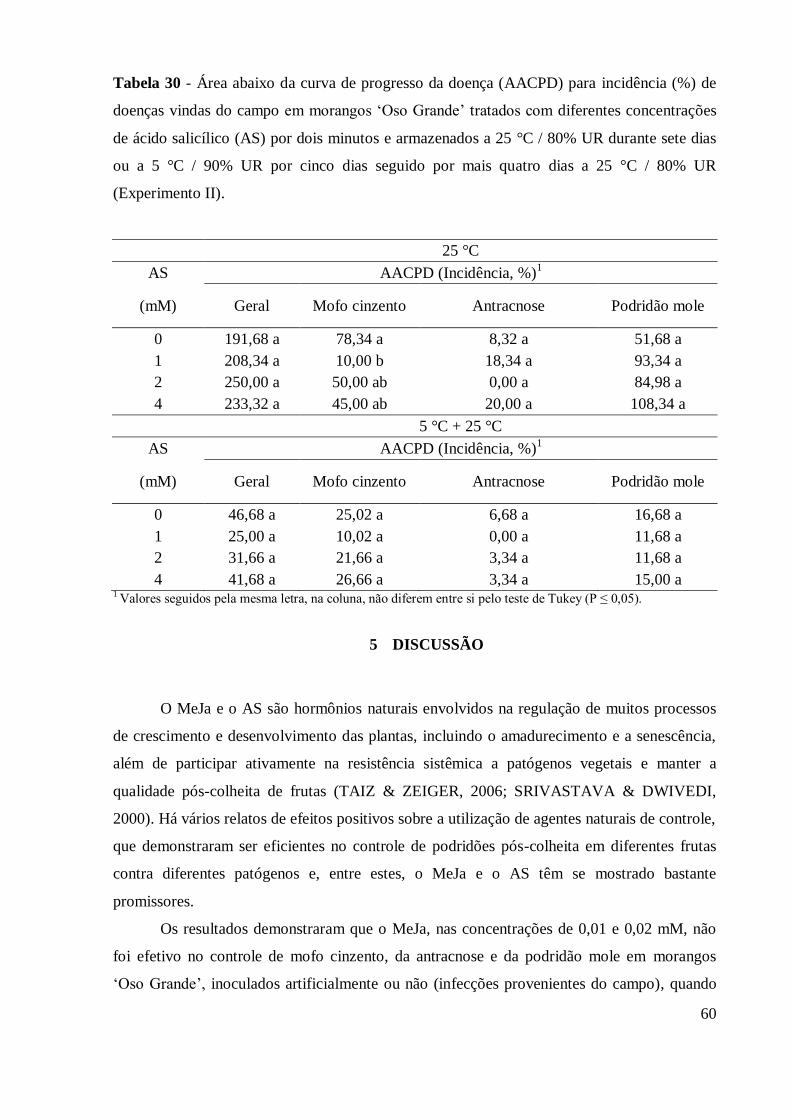
60
Tabela 30 - Área abaixo da curva de progresso da doença (AACPD) para incidência (%) de
doenças vindas do campo em morangos „Oso Grande‟ tratados com diferentes concentrações
de ácido salicílico (AS) por dois minutos e armazenados a 25 °C / 80% UR durante sete dias
ou a 5 °C / 90% UR por cinco dias seguido por mais quatro dias a 25 °C / 80% UR
(Experimento II).
25 °C
AS AACPD (Incidência, %)1
(mM) Geral Mofo cinzento Antracnose Podridão mole
0 191,68 a 78,34 a 8,32 a 51,68 a
1 208,34 a 10,00 b 18,34 a 93,34 a
2 250,00 a 50,00 ab 0,00 a 84,98 a
4 233,32 a 45,00 ab 20,00 a 108,34 a
5 °C + 25 °C
AS AACPD (Incidência, %)1
(mM) Geral Mofo cinzento Antracnose Podridão mole
0 46,68 a 25,02 a 6,68 a 16,68 a
1 25,00 a 10,02 a 0,00 a 11,68 a
2 31,66 a 21,66 a 3,34 a 11,68 a
4 41,68 a 26,66 a 3,34 a 15,00 a 1 Valores seguidos pela mesma letra, na coluna, não diferem entre si pelo teste de Tukey (P ≤ 0,05).
5 DISCUSSÃO
O MeJa e o AS são hormônios naturais envolvidos na regulação de muitos processos
de crescimento e desenvolvimento das plantas, incluindo o amadurecimento e a senescência,
além de participar ativamente na resistência sistêmica a patógenos vegetais e manter a
qualidade pós-colheita de frutas (TAIZ & ZEIGER, 2006; SRIVASTAVA & DWIVEDI,
2000). Há vários relatos de efeitos positivos sobre a utilização de agentes naturais de controle,
que demonstraram ser eficientes no controle de podridões pós-colheita em diferentes frutas
contra diferentes patógenos e, entre estes, o MeJa e o AS têm se mostrado bastante
promissores.
Os resultados demonstraram que o MeJa, nas concentrações de 0,01 e 0,02 mM, não
foi efetivo no controle de mofo cinzento, da antracnose e da podridão mole em morangos
„Oso Grande‟, inoculados artificialmente ou não (infecções provenientes do campo), quando

61
armazenados a 25° C ou sob refrigeração (Tabelas 2 a 10; Figuras 9, 11). No entanto, a
inoculação dos frutos logo após a exposição ao MeJa (0,01 mM) reduziu a incidência do mofo
cinzento em morangos, bem como a severidade da antracnose em frutos inoculados 48 h após
exposição ao produto. O MeJa é relatado, principalmente, como indutor de resistência em
frutas e hortaliças e, desta forma já era esperado que não atuasse diretamente no controle dos
patógenos. Assim é possível que o MeJa tenha atuado positivamente no controle de B. cinerea
em morangos através da ativação de mecanismos de defesa, como por exemplo, de enzimas
relacionadas à patogênese; no entanto, outros estudos deverão ser realizados com objetivo de
investigar o mecanismo de ação do produto. Sob tal aspecto, ZHANG et al. (2006) obtiveram
resultados distintos àqueles encontrados neste estudo. Os autores verificaram que o tratamento
com 1 µmol L-1
de MeJa em morangos reduziu a podridão causada por B. cinerea quando os
frutos foram mantidos sob refrigeração (5 °C). MARTÍNEZ-FERRER & HARPER (2005)
observaram que a exposição de abacaxis minimamente processados ao MeJa na concentração
de 0,1 mmol L-1
, durante 5 minutos, em um recipiente selado, diminuiu o crescimento
microbiológico depois de 12 dias de armazenamento a 7 °C. A aplicação pós-colheita de
MeJa (10 µmol L-1
) em uva reduziu significativamente o desenvolvimento natural de bolor
verde, causado por P. digitatum, durante o armazenamento a 20 °C e também em frutas
inoculadas artificialmente e armazenadas a 24 °C (DROBY et al., 1999). Segundo os autores
o efeito do MeJa na redução da doença foi obtido pela ativação de respostas de defesa nos
frutos.
YAO & TIAN (2005a) observaram que a aplicação de MeJa (0,2 mM), em pós-
colheita, não foi eficiente no controle da podridão parda em cereja, tanto no armazenamento a
25 °C, quanto a 0 °C. O tratamento de frutas e hortaliças com MeJa para controlar doenças
pós-colheita pode variar de acordo com o fruto, cultivar, época de aplicação, concentração,
tempo e modo de aplicação, forma de armazenamento após o tratamento e o patógeno
avaliado. Portanto, neste estudo, o MeJa pode não ter promovido efeito direto sobre os
patógenos devido à cultivar estudada, ao estádio de maturação utilizado (¾ vermelho), bem
como a concentração aplicada (0,01 e 0,02 mM).
Os resultados obtidos nos ensaios in vitro, que avaliaram o efeito do MeJa sobre o
crescimento micelial de B. cinerea, R. stolonifer e C. acutatum evidenciaram que o produto
não atua diretamente sobre o desenvolvimento dos patógenos (Figuras 10, 12 e 13), indicando
que o MeJa pode realmente atuar no controle do mofo cinzento em morangos através da
ativação de mecanismos de defesa. Resultados semelhantes foram obtidos por DROBY et al.

62
(1999), que verificaram que os jasmonatos (MeJa e JA) não demonstraram efeito inibitório
sobre o desenvolvimento in vitro de P. digitatum.
A imersão de morangos em diferentes concentrações de AS por 2 minutos, não reduziu
o diâmetro e/ou a incidência de lesões de B. cinerea, R. stolonifer e C. acutatum nos frutos
(Tabelas 11 a 17; Figuras 14 e 16). Contudo foi possível observar que o tratamento com 1
mM de AS reduziu significativamente a incidência do mofo cinzento em frutos não
inoculados artificialmente (Tabela 30). De acordo com FONG (2005), o tratamento com 0,1 e
1 mmol L-1
de AS inibiu significativamente a incidência de Colletotrichum gloesporioides em
mangas. Segundo o autor a concentração de 1 mmol L-1
atrasou o processo de senescência
pós-colheita de mangas e ainda inibiu as doenças pós-colheita. A aplicação pré-colheita de AS
(2,5 mM) em peras reduziu efetivamente a incidência de doenças, enquanto a utilização pós-
colheita inibiu o desenvolvimento de mofo azul (P. expansum). O tratamento com 5 mM de
AS promoveu a redução das doenças causadas por P. expansum e Alternaria alternata em
peras (CAO, 2005). Porém, segundo ZHANG et al. (2010), o tratamento com AS não reduziu
a incidência e a severidade da doença causado por R. stolonifer em morangos. YAO & TIAN
(2005a) também observaram que a aplicação de AS (2 mM), em pós-colheita, não foi
eficiente no controle da podridão parda em cereja, tanto no armazenamento a 25 °C, quanto a
0°C.
O AS pode atuar positivamente no controle de patógenos através da ativação de
mecanismos de defesa nos frutos (ASGHARI & AGHDAM, 2010). Constatou-se, neste
estudo, que morangos inoculados com B. cinerea 24 h após a aplicação de AS (1 mM)
apresentaram redução significativa da incidência do mofo cinzento. A proteção dos frutos à
invasão de agentes patogênicos é em grande parte realizada pela ativação coordenada de
respostas de defesa, que inibem a infecção ou a colonização dos tecidos pelos patógenos.
Portanto, provavelmente, o AS promoveu a indução de compostos de defesa nos tecidos dos
frutos que puderam impedir ou reduzir o desenvolvimento do mofo cinzento. Segundo CIA
(2005) a aplicação de AS (2,5, 5, 10 e 40 mM) antes ou após a inoculação dos frutos não
reduziu a incidência e a severidade da antracnose em mamão. LEE et al. (2009) verificaram
efeito positivo do AS na inibição da formação de apressórios por C. gloesporioides por meio
da resistência local adquirida, protegendo assim os frutos verdes de pimenta contra a infecção
pelo fungo.
Nos ensaios in vitro, verificou-se que o AS inibiu de maneira significativa o
desenvolvimento de B. cinerea, R. stolonifer e C. acutatum (Figuras 15, 17 e 18), diferindo
dos resultados obtidos quando o produto foi aplicado em morangos para o controle dos

63
patógenos inoculados antes do tratamento. De maneira semelhante, CIA (2005) verificou que
o AS atua diretamente sobre o crescimento micelial de C. gloesporioides.
A limitação para o uso comercial do AS é devido a sua fitotoxidade em diversas
plantas (ROCHA et al., 2000). FILHO (2003) descreveu que o tratamento de laranjas, com
AS, em concentrações acima de 1 mM, promoveu fitotoxidez. Este efeito também foi
verificado em tomateiros, tratados com AS nas concentrações de 2.5, 5 e 10 mM, de maneira
dependente da concentração; as lesões apresentavam-se irregulares, de áreas variáveis e eram
abundantes (VANETTI, 2001). O AS, neste estudo, não apresentou efeitos visíveis de
fitotoxidez.
Quanto aos atributos físico-químicos, neste estudo, tanto o MeJa, quanto o AS, não
alteraram significativamente a qualidade dos morangos (Tabelas 9 e 18). Sob tal aspecto,
NILPRAPRUCK et al. (2008) relataram que abacaxis tratados com 10-3
, 10-4
e 10-5
M de
MeJA e, em seguida, armazenados a 10 °C, também não apresentaram diferenças
significativas no teor de sólidos solúveis totais, acidez titulável, açúcares totais e açúcares
redutores com relação aos frutos controle. Contudo efeitos contrários já foram relatados,
como no estudo de PÉREZ et al. (1997) que demonstrou efeito significativo do tratamento
com MeJa (50 µM) na evolução da cor de morangos, constatando-se também estímulo da
biossíntese de antocianina, após 2 dias de tratamento, e a degradação acelerada das clorofilas
a e b, e em menor grau, de β-caroteno e luteína.
Com relação ao AS, a aplicação de 1 mM em nêsperas cv. Fukuhara diminuiu a
incidência de escurecimento interno da polpa, enquanto que a concentração de 10 mM causou
fitotoxidez (EDAGI, 2009). Recentemente, KAZEMI et al. (2011) observaram que em maçãs
imersas em solução de AS (0, 1, 3, 5 mM) durante 5 min e armazenadas a 5 °C por até 60
dias, a perda de massa dos frutos diminuiu significativamente em todas as concentrações de
AS em comparação com o controle. Além disso, os resultados mostraram que as frutas
tratadas com AS apresentaram maior firmeza, acidez titulável, atividade da peroxidase e da
superóxido desmutase em relação aos frutos controle. Morangos da cv. Camarosa tratados em
pré-colheita com AS, em solução nutritiva, apresentaram menor perda de massa e
deterioração e maior firmeza, sendo que a aplicação em pós-colheita também manteve as
características de qualidade dos frutos, armazenados durante sete dias a 2 ºC (SHAFIEE et al.,
2010).
Segundo LU et al. (2011) a aplicação de AS em abacaxis armazenados por dez dias a
10 °C, seguido por mais dois dias a 20 °C, tanto em pré-colheita, na forma de spray, como em
pós-colheita, por imersão, promoveu efeito benéfico sobre a qualidade dos frutos e do

64
abacaxizeiro. O AS não afetou o teor de sólidos solúveis totais, de acidez titulável e o
conteúdo de compostos fenólicos totais, mas atrasou o declínio do conteúdo de ácido
ascórbico. Pêssegos cv. Flordaking tratados com AS (0.5, 1, 1.5 e 2 mM) por 5 minutos e
armazenados a 0 °C, não apresentaram alterações nos atributos de qualidade nas
concentrações mais baixas. Entretanto, frutos tratados com a maior concentração
apresentaram menor perda de massa, aumento da luminosidade da casca, diminuição dos
valores de cromaticidade, maior firmeza, níveis mais elevados de acidez titulável, sólidos
solúveis totais, ácido ascórbico e conteúdo de compostos fenólicos. Assim, os autores
sugeriram que o AS a 2 mM pode ser usado comercialmente para preservar pêssegos por até
cinco semanas, sem qualquer deterioração (TAREEN et al., 2012). O AS reduz
significativamente a perda de qualidade em pêssegos (WANG et al., 2006), tomates (DING et
al., 2001) e nêsperas (CAI et al., 2006).
A eficácia do tratamento, com MeJa e AS, na conservação pós-colheita de frutas e
hortaliças, pode variar de acordo com a espécie de fruto, cultivar, concentração, tempo e
modo de aplicação e a forma de armazenamento após o tratamento. LOLAIE et al. (2012)
observaram que a qualidade de morangos cv. Camarosa é mantida com a aplicação de AS,
promovendo assim aumento na vida de prateleira. O resultado encontrado por LOLAIE et al.
(2012) não se assemelha aos encontrados neste estudo. Apesar de ambos os estudos utilizarem
morango, o tempo de aplicação, a concentração utilizada do ácido e a temperatura de
armazenamento foram diferentes, bem como a cultivar utilizada, o que pode explicar a
diferença encontrada nos resultados.
6 CONCLUSÕES
A aplicação de 0,01 mM de metil jasmonato reduz a incidência do mofo cinzento e a
severidade da antracnose em morangos „Oso Grande‟, enquanto o ácido salicílico diminui a
ocorrência de Botrytis cinerea, quando os patógenos são inoculados após o tratamento dos
frutos, possivelmente pela ativação de mecanismos de defesa.
O metil jasmonato e o ácido salicílico não reduzem a incidência e/ou a severidade da
podridão mole em morangos „Oso Grande‟.
In vitro, o ácido salicílico reduz o desenvolvimento de Botrytis cinerea,
Colletotrichum acutatum e Rhizopus stolonifer, enquanto o metil jasmonato não influencia o
crescimento micelial dos patógenos.

65
7 REFERÊNCIAS BIBLIOGRÁFICAS
AGRIOS, G.N. Plant Pathology. 5ª ed. United States of America: Academic Press. 2005.
497p.
AGROFIT – Sistema de Agrotóxicos Fitossanitários. 2012. Disponível em: <
http://agrofit.agricultura.gov.br/agrofit_cons/principal_agrofit_cons>. Acesso em: 26 de
novembro de 2012.
ANAND, A.; UPPALAPATI, S.R.; RYU, C.; ALLEN, S.N.; KANG, L.; TANG, Y.;
MYSORE, K.S. Salicylic Acid and Systemic Acquired Resistance Play a Role in Attenuating
Crown Gall Disease Caused by Agrobacterium tumefaciens. Plant Physiology. v. 146, p. 703
– 715, February 2008.
ANTUNES, L.C; REISSER JUNIOR, C. Produção integrada de morango: oportunidade de
mercado. In: Simpósio Nacional do Morango, 4.; Encontro de Pequenas Frutas e Frutas
Nativas do Mercosul, 3., Pelotas, RS. Anais...Pelotas: [s.n], 2008. p.15 – 20.
ASGHARI, M.; AGHDAM, M.S. Impact of salicylic acid on post-harvest physiology of
horticultural crops. Trends in Food Science & Technology. v. 21, p. 502 – 509, 2010.
AVANCI, N.C.; LUCHE, D.D.; GOLDMAN, G.H.; GOLDMAN, M.H.S. Jasmonates are
phytohormones with multiple functions, including plant defense and reproduction. Genetics
and Molecular Research. v. 9, n. 1, p. 484 – 505, 2010.
BABALAR, M.; ASGHARI, M.; TALAEI, A.; KHOSROSHAHI, A. Effect of pre- and
postharvest salicylic acid treatment on ethylene production, fungal decay and overall quality
of „Selva‟ strawberry fruit. Food Chemistry. v. 105, p. 449 – 453, 2007.
BAL, E.; CELIK, S. The effects of postharvest treatments of salicylic acid and potassium
permanganate on the storage of kiwifruit. Bulgarian Journal of Agricultural Science. v.16,
n. 5, p. 576 – 584, 2010.

66
BARROS, F.C.; SAGATA, E.; FERREIRA, L.C.C.; JULIATTI, F.C. Indução de resistência
em plantas contra fitopatógenos. Bioscience Journal. Uberlândia, v. 26, n. 2, p. 231 – 239,
Mar./Apr. 2010.
BENATO, E.A. Controle de doenças pós-colheita em frutos tropicais. Summa
Phytopathologica. Botucatu, v. 25, n. 1, p. 90 – 93, 1999.
BENATO, E.A. A indução de resistência no controle de doenças pós-colheita: frutas e
hortaliças. Summa Phytopathologica. Botucatu, v. 29, p. 125 – 126, 2003a.
BENATO, E.A. Potencial de indução de resistência em frutas pós-colheita. In: ENFRUTE
Encontro Nacional sobre Fruticultura de Clima Temperado, 5., Fraiburgo. Anais. Caçador,
SC. Epagri, p. 215 – 220, 2003b.
BENATO, E.A.; CIA, P. Doenças de Frutos em Pós-colheita e Controle. In: Manual Pós-
colheita da Fruticultura Brasileira / Leandro Camargo Neves (organizador). Londrina:
EDUEL, 2009. 494p.
BONALDO, S.M.; PASCHOLATI, S.F.; ROMEIRO, R.S. Indução de resistência: noções
básicas e perspectivas. In: Indução de Resistência em plantas a patógenos e insetos.
Editores: CAVALCANTI, L.S.; DI PIERO, R.M.; CIA, P.; PASCHOLATI, S.F.; RESENDE,
M.L.V.; ROMEIRO, R.S. Piracicaba, SP, FEALQ, 2005. 263p.
BORESZTEIN, F.; LOURENÇO, S.A.; AMORIM, L. Incidência de Rhizopus stolonifer em
frutos de morango com ou sem ferimento. Escola Superior de Agricultura “Luiz de Queiroz”
– USP – Piracicaba (SP). In: 17º SIICUSP – Simpósio Internacional de Iniciação
Científica da USP. Resumo n° 4036. Agropecuária – 12 e 13 de Novembro de 2009.
Disponível em: <http://www.usp.br/siicusp/Resumos/17Siicusp/resumos/4036.pdf>. Acesso
em: 18 de Maio de 2011.
CAI, C; LI, X.; CHEN, K.S. Acetylsalicylic acid alleviates chilling injury of postharvest
loquat (Eriobotrya japonica Lindl.) fruit. European Food Research and Technology. v.
223, p. 533 – 539, 2006.

67
CAMILI, E.C.; CIA, P.; BENATO, E.A. Indução de resistência contra doenças pós-colheita.
Cap. 10. In: Indução de Resistência em plantas a patógenos e insetos. Editores:
CAVALCANTI, L.S.; DI PIERO, R.M.; CIA, P.; PASCHOLATI, S.F.; RESENDE, M.L.V.;
ROMEIRO, R.S. Piracicaba, SP, FEALQ, 2005. p. 195 – 218.
CAO, J. K. Effects of SA, ASM, INA and citric acid on postharvest disease resistance and
quality of Yali pear fruit. 2005. Tese (Doutorado em Ciências Agrícolas) – Universidade
Agrícola da China, Pequim.
CAO; J. ZENG, K.; JIANG, W. Enhancement of postharvest disease resistance in Ya Li pear
(Pyrus bretschneideri) fruit by salicylic acid sprays on the trees during fruit growth.
European Journal of Plant Pathology. v. 114, n. 4, p. 363 – 370, April 2006.
CAO, S.; ZHENG, Y.; YANG, Z.; TANG, S.; JIN, P.; WANG, K.; WNAG. X. Effect of
methyl jasmonate on the inhibition of Colletotrichum acutatum infection in loquat fruit and
the possible mechanisms. Postharvest Biology and Technology. v. 49, n. 2, p. 301 – 307,
August 2008a,
CAO, S.; ZHENG, Y.; YANG, Z.; TANG, S.; JIN, P. Control of anthracnose rot and quality
deterioration in loquat fruit with methyl jasmonate. Journal of the Science of Food and
Agriculture. v. 88, n. 9, p. 1598 – 1602, july 2008b.
CAO, S.; HU, Z.; PANG, B. Optimization of postharvest ultrasonic treatment of strawberry
fruit. Postharvest Biology and Technology. v. 55, p. 150-153, março 2010.
CARDOSO, J.E.; SANTOS, A.A.; BEZERRA, M.A.; NETO, J.S.; SOUZA, R.M.N.
Epidemiologia do Mofo-Preto e Danos na Produção do Cajueiro. Fortaleza: Embrapa
Agroindústria Tropical, 16 p. (Boletim de pesquisa e desenvolvimento, 23), 2005.
CARVALHO, S.P. Histórico, importância e socioeconômica e zoneamento da produção no
Estado de Minas Gerais. In: Carvalho, S. P. (Coord.). Boletim do morango: cultivo
convencional, segurança alimentar, cultivo orgânico. Belo Horizonte: FAEMG, 2006. p. 9
– 14.

68
CAVALCANTI, L.S.; BRUNELLI, K.R.; STANGARLIN, J.R. Aspectos bioquímicos e
moleculares da resistência induzida. In: Indução de Resistência em plantas a patógenos e
insetos. Editores: CAVALCANTI, L.S.; DI PIERO, R.M.; CIA, P.; PASCHOLATI, S.F.;
RESENDE, M.L.V.; ROMEIRO, R S. Piracicaba, SP, FEALQ, 2005. p. 81 – 124.
CHITARRA, M I F.; CHITARRA, A. B. Pós-colheita de frutas e hortaliças: fisiologia e
manuseio. 2º ed., Editora: UFLA, 2005. 783p.
CHEONG, J.J.; CHOI, Y.D. Methyl jasmonate as a vital substance in plants. Trends in
Genetics. v. 19, n.7, p. 409 – 413, 2003.
CIA, P. Avaliação de agentes bióticos e abióticos na indução de resistência e no controle
pós-colheita da antracnose (Colletotrichum gloeosporioides) em mamão (Carica papaya).
2005. 197p. Tese (Doutorado em Agronomia) – Escola Superior de Agricultura Luiz de
Queiroz, Piracicaba.
CIA, P. PASCHOLATI, S.F.; BENATO, E.A. Indução de resistência no manejo de doenças
pós-colheita. In: Indução de resistência em plantas a patógenos. Editores: RODRIGUES,
F. A.; ROMEIRO, R. S. Anais da III Reunião Brasileira de Plantas a Patógenos. Viçosa, MG,
2007. p. 245 – 280.
CIA, P.; BENATO, E.A.; PASCHOLATI, S.F.; GARCIA, E.O. Quitosana no controle pós-
colheita da podridão mole em caqui „Rama Forte‟. Bragantia. Campinas, v. 69, n. 3, p 745 –
752, 2010.
COSTA, H.; ZAMBOLIM, L.; VENTURA, J.A. Manejo integrado das doenças do
morangueiro. Cap. 6. In: Manejo integrado – Produção Integrada de Fruterias Tropicais:
Doenças e Pragas. Editor: Laércio Zambolim. Viçosa, MG, 2003. 587p.
COSTA, H.; VENTURA, J.A. Doenças do morangueiro: diagnóstico e manejo. In:
BALBINO, J.M. de S. (ed.) Tecnologias para produção, colheita e pós-colheita de
morangueiro. 2° ed. Vitória: Incaper, 2006. p. 41 – 57.

69
COSTA, F.B. Fisiologia e conservação de cultivares de morangos inteiros e minimamente
processados. 2009. 115p. Tese (Doctor Scientiae em Fisiologia Vegetal) – Universidade
Federal de Viçosa, Viçosa.
COUTO, E.B. Avaliação de metil jasmonato na indução de resistência de plantas de
Eucalyptus spp. ao psilídeo-deconcha Glycaspis brimblecombei moore (hemiptera:
psyllidae). 2006. 52p. Dissertação (Mestrado em Agronomia) – Faculdade de Ciências
Agronômicas – UNESP, Botucatu.
DING, Z.; TIAN, S.; ZHENG, X.; ZHOU, Z.; XU, Y. Responses of reactive oxygen
metabolism and quality in mango fruit to exogenous oxalic acid or salicylic acid under
chilling temperature stress Physiologia Plantarum. v. 130, n. 1, p. 112 – 121, May 2007.
DING, C.K.; WANG, C.Y.; GROSS, K.C.; SMITH, D.L. Reduction of chilling injury and
transcript accumulation of heat shock protein genes in tomatoes by MeJA and MeSA. Plant
Science. v. 161, p. 1153 – 1159, 2001.
DANNER, M.A.; SASSO, S.A.Z.; MEDEIROS, J.G.S.; MARCHESE, J.A.; MAZARO, S.M.
Indução de resistência à podridão-parda em pêssegos pelo uso de eliciadores em pós-colheita.
Pesquisa Agropecuária Brasileira, Brasília, v. 43, n. 7, p.793 – 799, jul. 2008.
DI PIERO, R.M.; KUHN, O.J.; PASCHOLATI, S.F. Indução de resistência e a produtividade
das culturas. Cap. 12. In: Indução de Resistência em plantas a patógenos e insetos.
Editores: CAVALCANTI, L. S.; DI PIERO, R. M.; CIA, P.; PASCHOLATI, S. F.;
RESENDE, M. L. V.; ROMEIRO, R. S. Piracicaba, SP, FEALQ, 2005. p. 239 – 258.
DROBY, S.; PORAT, R.; COHEN, L.; WEISS, B.; SHAPIRO, B.; PHILOSOPH-HADAS,
S.; MEIR, S. Suppressing green mold decay in grapefruit with postharvest jasmonate
application. Journal of the American Society for Horticultural Science, v. 124, p. 184 –
188, 1999.
DURRANT, W.E.; DONG, X. Systemic acquired resistance. Annual Review of
Phytopathology. v. 42, p. 185 – 209, 2004.

70
EDAGI, F.K. Pós-colheita de nêsperas ‘Fukuhara’: efeito de compostos salicilatos, 1-
metilcicloporpeno e tratamentos térmicos. 2009. 86p. Dissertação (Mestrado em
Agronomia) – Escola Superior de Agricultura “Luiz de Queiroz”, Piracicaba.
EDREVA, A. A novel strategy for plant protection: Induced resistance. Journal of Cell and
Molecular Biology. v. 3, p. 61 – 69, 2004.
EL GHAOUTH, A.; ARUL, J.; PONNAMPALAM, R.; BOULET, M. Chitosan coating effect
on storability and quality of fresh strawberries. Journal of Food Science. v. 56, n. 6, p. 1618
– 1620, 1991.
FERNANDES, C.F.; JÚNIOR, J.R.V.; SILVA, D.S.G.; REIS, N.D.; JÚNIOR, H.A.
Mecanismos de defesa de plantas contra o ataque de agentes fitopatogênicos. Porto
Velho/RO: Embrapa Rondônia, 2009. 14 p. – (Documentos / Embrapa Rondonia, 0103-9865;
133).
FILHO, J.A.C. Efeito de extratos de albedo de laranja (Citrus sinensis) e dos indutores de
resistência ácido salicílico, acibenzolar-s-metil e Saccharomyces cerevisiae no controle de
Phyllosticta citricarpa (Teleomorfo: Guignardia citricarpa). 2003. 125p. Tese (Doutorado
em Agronomia) – Escola Superior de Agricultura “Luiz de Queiroz”, Piracicaba.
FLORES-CANTILLANO, F.; BENDER, R.J.; LUCHSINGER, L. Fisiologia e manejo pós-
colheita. In: MORANGO: Pós-colheita. Editor técnico: FLORES-CANTILLANO, F.
Embrapa Clima Temperado (Pelotas, RS). – Brasília, DF, Embrapa Informações
Tecnológicas, 2003. 29p. (Frutas do Brasil, 42).
FONG, Z.K. Effects Of Bagging, Sa And Ina On Postharvest Quality And Disease
Resistance Of Mango (mangifera Indica L.) Fruit. 2005. 119p. Tese (Doutorado em
Ciências Agrícolas) – Universidade Agrícola da China, Pequim.
FOOD AND AGRICULTURE ORGANIZATION OG THE UNITED NATIONS – FAO.
FAOSTAT. Production/Crops – Brazil/Strawberries. 2012. Disponível em: <
http://faostat.fao.org/site/567/default.aspx#ancor>. Acesso em: 29 de nov. de 2012.

71
FORTES, J.F. Doenças Pós-colheita. In: MORANGO: Pós-colheita. Editor técnico:
FLORES-CANTILLANO, F. Embrapa Clima Temperado (Pelotas, RS). – Brasília, DF,
Embrapa Informações Tecnológicas, 2003. 29p. (Frutas do Brasil, 42).
FORTES, J.F.; COUTO, M.E. Doenças Fúngicas e Bacterianas. In: MORANGO:
Fitossanidade. Editores técnicos: FORTES, J. F.; OSÓRIO, V. A. Embrapa Clima
Temperado (Pelotas, RS). – Brasília, DF, Embrapa Informações Tecnológicas, 2003. 36p.
(Frutas do Brasil, 41).
FORTES, J.F. Sistema de Produção do Morango: Doenças do morangueiro. Embrapa
Clima Temperado. Sistemas de Produção, 5, Versão Eletrônica, Nov., 2005. Disponível em:
<http://sistemasdeproducao.cnptia.embrapa.br/FontesHTML/Morango/SistemaProducaoMora
ngo/cap06.htm>. Acesso em: 18 de Maio de 2011.
GARCÍA, J.M.; AGUILERA, C.; JIMÉNEZ, A.M. Gray Mold in and Quality of Strawberry
Fruit following Postharvest Heat Treatment. HortScience, v. 31, n. 2, p. 255 – 257, abril
1996.
GRASEMNEZHAD, M.; BABALAR, M.; MOSTOUFI, Y. Effect of methyl jasmonate and
methyl salicylate in reducing chilling injury and decay in marsh grapefruit and thompsom
navel orange produced in North and south of Iran. Iranian Journal of Agricultural Sciences. v.
39, n. 1, p. 1 – 7, 2008.
GIMÉNEZ, G.; ANDRIOLO, J.; GODOI, R. Cultivo em solo do morangueiro. Ciência
Rural, Santa Maria, v. 38, n. 1, p. 273-279, jan/fev 2008.
GONZÁLEZ-AGUILAR, G.A.; ZAVALETA-GATICA, R.; TIZNADO-HERNÁNDEZ,
M.E. Effect of methyljasmonate on physiological response of guava (Psidium guajava) stored
at low temperatures. Revista Chapingo, Serie Horticultura. v. 13, n. 1, p. 63 – 69, 2007.
GUZZO, S.D. Aspectos bioquímicos e moleculares da resistência sistêmica adquirida em
cafeeiro contra Hemileia vastatrix. 2004. 236p. Tese (Doutorado em Ciências) – Centro de
Energia Nuclear na Agricultura – USP. Piracicaba.

72
GUZZO, S.D.; HARAKAVA, R. Mecanismos envolvidos na resistência induzida em plantas
a doenças: sinalização e expressão de genes de defesa. Cap. 12. In: Indução de resistência
em plantas a patógenos. Editores: RODRIGUES, F. A.; ROMEIRO, R. S. Anais da III
Reunião Brasileira de Plantas a Patógenos. Viçosa, MG, 2007. p. 281 – 302.
HENRIQUE, C.M.; CEREDA, M.P. Utilização de biofilmes na conservação pós-colheita de
morango (Fragaria Ananassa Duch) cv. IAC Campinas. Revista Ciência. Tecnologia de
Alimentos, v.19, n. 2, p. 231 – 233, 1999.
HOFFMANN, A., ANTUNES, L.E.C.; RASEIRA, M.C.B.; NAKASU, B.H; BERNARDI, J.
Pequenas frutas. Cap. 3. In: Agricultura Tropical – Quatro décadas de inovações
tecnológicas, institucionais e políticas. Editores Técnicos: ALBUQUERQUE, A.C.S.;
SILVA, A.G. de. Produção e produtividade agrícola. Brasília, DF, Embrapa Informações
Tecnológicas. v. 1, 2008, 1337p.
HUANG, R.; XIA, R.; LU, Y.; HU, L.; XU, Y. Effect of pre-harvest salicylic acid spray
treatment on postharvest antioxidant in the pulp and peel of „Cara cara‟ navel orange (Citrus
sinenisis L. Osbeck). Journal of the Science of Food and Agriculture. v. 88, n. 2, p. 229 –
236, January 2008.
IMRAN, H.; ZHANG, Y.; DU, G.; WANG, G.; ZHANG, J. Effect of salicylic acid (SA) on
delaying fruit senescence of Huang Kum pear. Frontiers of Agriculture in China. v. 1, n. 4,
p. 456 – 459, October, 2007.
JIN, P.; ZHENG, Y.H.; CHENG, C. M.; GAO, H.Y.; CHEN, W.X.; CHEN, H.J. Effect of
methyl jasmonate treatment on fruit decay and quality in peaches during storage at ambient
temperature. Acta Horticulturae. v. 712, p. 711 – 716, 2006.
JIN, P.; ZHENG, Y.; TANG, S.; RUI, H.; WANG, C.Y.; Enhancing disease resistence in
peach fruit with methyl jasmonate. Journal of The Science of Food and Agriculture. v. 89,
n. 5, p. 802 – 808, 2009.
JING-HUA, Y., YUAN, G., YAN-MAN, L., XIAO-HUA, Q.; ZHANG, M.F. Salicylic acid-
induced enhancement of cold tolerance through activation of antioxidative capacity in
watermelon. Scientia Horticulture. v. 118, p. 200 – 205, 2008.

73
JUNIOR, C.G.L. Identificação e expressão de genes de biossíntese do jasmonato na
interação entre Theobroma cacao e Moniliophthora perniciosa. 2009. 89p. Dissertação
(Mestrado em Ciências) - Centro de Energia Nuclear na Agricultura – USP, Piracicaba.
JÚNIOR, L.C.C. Atmosfera controlada na conservação de morango. 2011. 120p. Tese
(Doutorado em Ciências) – USP, Piracicaba.
KERBAUY, G.B. Fisiologia Vegetal. 1ª ed. Editora: Guanabara Koogan S.A.. 2004. 452p.
KHADEMI, O.; ZAMANI, Z.; MOSTOFI, Y.; KALANTARI, S.; AHMADI, A. Extending
Storability of Persimmon Fruit cv. Karaj by Postharvest Application of Salicylic Acid.
Journal of Agricultural Science and Technology. v. 14, p. 1067 – 1074, 2012.
KAZEMI, M.; ARAN, M.; ZAMANI, S. Effect of Salicylic Acid Treatments on Quality
Characteristics of Apple Fruits During Storange. American Journal of Plant Physiology. v.
6, n. 2, p. 113 – 119, 2011.
LEE, H.; LEON, J.; RASKIN, I. Biosynthesis and metabolism of salicylic acid. Proc. Natl.
Acad. Sci. USA. v. 92, p. 4076 – 4079, May 1995.
LEE, S.; HONG, J.; JEON, W.B.; CHUNG, Y.; SUNG, S.; CHOI, D.; JOUNG, Y.H.; OH, B.
The salicylic acid-induced protection of non-climacteric unripe pepper fruit against
Colletotrichum gloeosporioides is similar to the resistance of ripe fruit. Plant Cell Reports.
v. 28, p. 1573 – 1580, 2009.
LI, D.; XU, Y.; SUN, L.; LIU, L.; HU, X.; LI, D.; SHU, H. Salicylic Acid, Ethephon, and
Methyl Jasmonate Enhance Ester Regeneration in 1-MCP-Treated Apple Fruit after Long-
Term Cold Storage. Journal of Agricultural and Food Chemistry. v. 54, n. 11, p. 3887 –
3895, 2006.
LIU, W; AI, X.Z.; LIANG, W.J.; WANG, H.T.; LIU, S.X.; ZHENG, N. Effects of salicylic
acid on the leaf photosynthesis and antioxidant enzyme activities of cucumber seedlings under
low temperature and light intensity. Chinese Journal of Applied Ecology. v. 20, p. 441-445,
2009.

74
LOLAIE, A.; KAVIANI, B.; REZAEI M.A.; RAAD, M.K.; MOHAMMADIPOUR, R. Effect
of Pre- and Postharvest Treatment of Salicylic Acid on Ripening of Fruit and Overall Quality
of Strawberry (Fragaria ananasa Duch cv. Camarosa) Fruit. Annals of Biological Research.
v. 3, n. 10, p. 4680 – 4684, 2012.
LU, X.; SUN, D.; LI, Y.; SHI, W. SUN, G. Pre- and post-harvest salicylic acid treatments
alleviate internal browning and maintain quality of winter pineapple fruit. Scientia
Horticulturae. v. 130, n. 1, p. 97 – 101, August, 2011.
LUNARDI, R. Tecnologia de Armazenamento em Atmosfera Controlada. In: Manual Pós-
colheita da Fruticultura Brasileira / Leandro Camargo Neves (organizador). Londrina:
EDUEL, 2009. 494p.
LUNARDI, R.; TERUEL, B.; NEVES, L.C. Armazenamento Refrigerado e Boas Práticas na
Conservação de Frutos. In: Manual Pós-colheita da Fruticultura Brasileira / Leandro
Camargo Neves (organizador). Londrina: EDUEL, 2009. 494p.
MACAGNAN, D. Indução de resistência e biocontrole em cacaueiro. In: Indução de
resistência em plantas a patógenos. Editores: RODRIGUES, F. A.; ROMEIRO, R. S. Anais
da III Reunião Brasileira de Plantas a Patógenos. Viçosa, MG, 2007. p. 183 – 218.
MADAIL, J.C.M.; REICHERT, L.J.; MARTINS, C.R. Mercado Internacional e Nacional. In:
MORANGO: Pós-colheita. Editor técnico: FLORES-CANTILLANO, F. Embrapa Clima
Temperado (Pelotas, RS). – Brasília, DF, Embrapa Informações Tecnológicas, 2003. 29p.
(Frutas do Brasil, 42).
MADAIL, J.C.M.; REICHERT, L.J.; MIGLIORINI, L.C. Sistema de Produção do
Morango: Coeficientes técnicos para a cultura do morangueiro. Embrapa Clima
Temperado. Sistemas de Produção, 5, Versão Eletrônica, Nov., 2005. Disponível em: <
http://www.cpact.embrapa.br/publicacoes/catalogo/tipo/sistemas/morango/cap15.htm>.
Acesso em: 18 de Maio de 2011.

75
MALGARIM, M.B.; TIBOLA, C.S.; ZAICOWISK, C.; FERRI, V.C.; SILVA, P.R.
Modificação da atmosfera e resveratrol na qualidade pós-colheita de morangos cv. Camarosa.
Revista Brasileira de Agrociência, Pelotas, v. 12, n. 1, p. 67 – 70, jan-mar, 2006.
MANDAL, S.; MALLICK, N.; MITRA, A. Salicylic Acid-induced Resistance to Fusarium
oxysporum f. sp. lycopersici in Tomato. Plant Physiology and Biochemistry. v. 47, p. 642 –
649, 2009.
MARTÍNEZ-FERRER, M.; HARPER, C. Reduction in microbial growth and improvement of
storage quality in fresh-cut pineapple after methyl jasmonate treatment. Journal of Food
Quality. v. 28, n. 1, p. 3 – 12, February, 2005.
MELOTTI, M. Secretaria da Agricultura, Abastecimento, Aqüicultura e Pesca - SEAG, Portal
do Governo do Estado do Espírito Santo. Comercialização de morango na Ceasa/ES cresce
mais de 65% em 21 dias. 2009. Disponível em: <http://www.seag.es.gov.br/?p=1718>.
Acesso em: 19 de Maio de 2011.
MENG, X.; HAN, J.; WANG, Q.; TIAN, S. Changes in physiology and quality of peach
fruits treated by methyl jasmonate under low temperature stress. Food Chemistry. v. 144, n.
3, p. 1028 – 1035, June 2009.
MÉTRAUX, J.P. Recent breakthroughs in the study of salicylic acid biosynthesis. Trends in
Plant Science. v. 7, n. 8, p. 332 – 334, 2002.
MO, Y.; GONG, D.; LIANG, G.; HAN, R.; XIE, J.; LI, W. Enhanced preservation effects of
sugar apple fruits by salicylic acid treatment during post-harvest storage. Journal of the
Science of Food and Agriculture. v. 88, n. 15, p. 2693 – 2699, December 2008.
NILPRAPRUCK, P.; PRADISTHAKARN, N.; AUTHANITHEE, F.; KEEBJAN, P. Effect of
Exogenous Methyl Jasmonate on Chilling Injury and Quality of Pineapple (Ananas comosus
L.) cv. Pattavia. Silpakorn University Science and Technology Journal. v. 2, n. 2, p. 33 –
42, 2008.

76
NILPRAPRUCK, P.; YODMINGKHWAN, P. Effect of Exogenous Methyl Jasmonate on the
Internal Browning of Pineapple Fruit (Ananas comosus L.) cv. Pattavia.
KKU Research Journal. v. 14, n. 6, p. 489 – 498, June, 2009.
OLIVEIRA, R.P.; NINO, A.F.P.; SCIVITTARO, W.B. Mudas certificadas de morangueiro:
maior produção e melhor qualidade da fruta. A Lavoura, Rio de Janeiro, v. 108, n. 655, p. 35
– 38, 2005.
OLIVEIRA, M.A.B. de. Introdução a pós-colheita de frutos. Editora Noryam, 2010. 72p.
OLIVEIRA, M.A.C. de; SANTOS, A. M. de. Classificação Botânica, Origem e Evolução. In:
MORANGO: Produção. Editores técnicos: SANTOS, A. M. dos; MEDEIROS, A. R. M.
Embrapa Clima Temperado (Pelotas, RS). – Brasília, DF, Embrapa Informações
Tecnológicas, 2003. 81p. (Frutas do Brasil, 40).
OSÓRIO, V.A.; FORTES, J.F.; Introdução. In: MORANGO: Fitossanidade. Editores
técnicos: FORTES, J. F.; OSÓRIO, V. A. Embrapa Clima Temperado (Pelotas, RS). –
Brasília, DF, Embrapa Informações Tecnológicas, 2003. 36p. (Frutas do Brasil, 41).
PALLAMIN, M.L. Alternativas no controle fitossanitário em diferentes cultivares de
morangueiro como ferramenta na produção integrada. 2007. 60p. Dissertação (Mestrado
em Agronomia – Horticultura) – UNESP, Botucatu.
PASCHOLATI, S.F.; LEITE, B. Hospedeiro: mecanismos de resistência. In: BERGAMIN
FILHO, A.; KIMATI, H.; AMORIM, L. Manual de fitopatologia: princípios e conceitos. 3ª
ed. Pompéia: Ceres, 1995. p. 417-453.
PBMH – Programa Brasileiro para a modernização da Horticultura e Produção integrada de
morango. Normas de Classificação de Morango. São Paulo: CEAGESP, 2009.
(Documentos, 33).
PÉREZ, A.G.; SANZ, C.; OLÍAS, R.; OLÍAS, J.M. Effect of Methyl Jasmonate on in
Vitro Strawberry Ripening. Journal of Agricultural and Food Chemistry. v. 45, n. 10, p.
3733 – 3737, 1997.

77
PÉREZ, A.G.; SANZ, C.; RÍOS, J.J.; OLÍAS, R.; OLÍAS, J.M. Effects of ozone treatment on
postharvest strawberry quality. Journal of Agricultural and Food Chemistry. v. 47, n. 4, p.
1652 – 1656, 1999.
PINELI, L.L.O.; ROCHA, T.O.; MORETTI, C.L.; CAMPOS, A.B.; BRASILEIRO, A.V.;
GLEICY, G.; SANTANA, M.A.; SANTOS, M.S.; CAMPOS, N.A. Caracterização física,
química e sensorial de morangos „Oso Grande‟ e „Camino Real‟ armazenados a 5 e a 15 °C.
Horticultura Brasileira. v. 26, n. 2, p. 5497 – 5501. 2008.
POOLE, R.R.; McLEOD, L.C. Development of resistance to picking wound entry Botrytis
cinerea storage rots in kiwifruit. New Zealand Journal of Crop and Horticultural Science.
v. 22, p. 387–392, 1994.
POPOVA, L.; PANCHEVA, T.; UZUNOVA, A. Salicylic Acid: Properties, Biosynthesis and
Physiological Role. Bulgarian Journal of Plant Physiology. v. 23, p. 85 – 93, 1997.
RASKIN, I. Role of salicylic acid in plants. Annual Review of Plant Physiology and Plant
Molecular Biology. v. 43, p. 439 – 463, 1992.
REICHERT, L.J. Comercialização. In: MORANGO: Produção. Editores técnicos:
SANTOS, A.M. dos; MEDEIROS, A.R.M. Embrapa Clima Temperado (Pelotas, RS). –
Brasília, DF, Embrapa Informações Tecnológicas, 2003. 81p. (Frutas do Brasil, 40).
REICHERT, L.J.; MADAIL, J.C.M. Aspectos socioeconômicos. In: MORANGO:
Produção. Editores técnicos: SANTOS, A.M. dos; MEDEIROS, A.R.M. Embrapa Clima
Temperado (Pelotas, RS). – Brasília, DF, Embrapa Informações Tecnológicas, 2003. 81p.
(Frutas do Brasil, 40).
REY, M. dos S.; LIMA, N.B.; SANTOS, J. dos; FARIAS, C.R.J. de1; PIEROBOM, C.R.
Crescimento micelial de raças de Colletotrichum lindemuthianum sob diferentes potenciais
hídricos. Revista brasileira de Agrociência, Pelotas, v.11, n. 3, p. 309 – 313, jul-set, 2005.
.

78
ROCHA, M.R.; CASTRO, R.M.; PINA, R.C.; MARTINI, A.L. Efeito do acibenzolar-s-
methyl (benzothiadiazole), como indutor de resistência sistêmica em soja (Glycine max cv.
FT-Cristalina), sobre Heterodera glycines. Pesquisa Agropecuária Tropical, v. 30, n. 2, p.
35 - 38, jul./dez. 2000.
ROHWER, C.L.; ERWIN, J.E. Horticultural applications of jasmonates: A review. Journal
of Horticultural Science & Biotechnology. v. 83, n. 3, p. 283 – 304, 2008.
ROMBALDI, C.V.; SILVA, J.A.; LUCCHETTA, L.; ZANUZZO, M.R.; NEVES, L.C.
Biotecnologia Aplicada à Conservação e à Qualidade de frutos. In: Manual Pós-colheita da
Fruticultura Brasileira / Leandro Camargo Neves (organizador). Londrina: EDUEL, 2009.
494p.
SHAFIEE, M.; TAGHAVI, T.S.; BABALAR, M. Addition of salicylic acid to nutrient
solution combined with postharvest treatments (hot water, salicylic acid, and calcium dipping)
improved postharvest fruit quality of strawberry. Scientia Horticulturae. v. 124, p. 40 – 45,
2010.
SANHUEZA, R.M.V.; HOFFMANN, A.; ANTUNES, L.E.C.; FREIRE, J. de M. Sistema de
Produção de Morango para a Mesa na Região da Serra Gaúcha e Encosta Superior do
Nordeste. Embrapa Uva e Vinho. Sistema de Produção, 6. Versão Eletrônica, Dez., 2005.
Disponível em: <
http://sistemasdeproducao.cnptia.embrapa.br/FontesHTML/Morango/MesaSerraGaucha/impo
rtancia.htm>. Acesso em 30 de Maio de 2011.
SANTOS, A.M. dos. Cultivares. In: MORANGO: Produção. Editores técnicos: SANTOS,
A.M. dos; MEDEIROS, A.R.M. Embrapa Clima Temperado (Pelotas, RS). – Brasília, DF,
Embrapa Informações Tecnológicas, 2003. 81p. (Frutas do Brasil, 40).
SANTOS, P.E.T. dos. Sistema de Produção do Morango: Características básicas das
principais cultivares de morango plantadas no Brasil. Embrapa Clima Temperado.
Sistemas de Produção, 5, Versão Eletrônica, Nov., 2005. Disponível em:
<http://sistemasdeproducao.cnptia.embrapa.br/FontesHTML/Morango/SistemaProducaoMora
ngo/cap02.htm>. Acesso em 18 de Maio de 2011.

79
SCHWAN-ESTRADA, K.R.F.; STANGARLIN, J.R.; PASCHOLATI, S.F. Mecanismos
bioquímicos de defesa vegetal. In: PASCHOLATI, S.F.; LEITE, B.; STANGARLIN, J.R.;
CIA, P. (Ed.). Interação Planta Patógeno – fisiologia, Bioquímica e Biologia Molecular.
Piracicaba: FEALQ, 2008. p.227-248.
SIBOZA, X.I.; BOWER, J.P.; BERTLING, I. Effect of methyl jasmonate and salicylic acid
on chilling injury of 'Eureka' lemons. Acta Horticulturae. v. 911, p. 409 – 414, 2010.
SILVA, P.A.; QUEIROZ, E.R.; ABREU, C.M.P.; CORRÊA, A.D.; SANTOS, C.D.
Avaliação da qualidade de morangos submetidos ao 1-metilciclopropeno e armazenados
sob refrigeração. XIX Congresso de Pós-Graduação da UFLA. Set. / Out. de 2010. 6 p.
SOARES, A.M.S.; MACHADO, O.L.T. Defesa de plantas: Sinalização química e espécies
reativas de oxigênio. Revista Trópica – Ciências Agrárias e Biológicas. v. 1, n. 1, p.9, 2007.
SOBRINHO, C.A.; FERREIRA, P.T.O.; CAVALCANTI, L.S. Indutores Abióticos. In:
Indução de Resistência em plantas a patógenos e insetos. Editores: CAVALCANTI, L.S.;
DI PIERO, R.M.; CIA, P.; PASCHOLATI, S.F.; RESENDE, M.L.V.; ROMEIRO, R.S.
Piracicaba, SP, FEALQ, 2005. p. 51 – 80.
SOUZA, A.P.M. Estudo do efeito de respostas de hipersensibilidade sobre a parede
celular em cultura de células de amora-preta (Rubus fruticosus). 2007. 145p. Dissertação
(Mestrado em Ciências Farmacêuticas) – Faculdade de Ciências Farmacêuticas – USP,
Ribeirão Preto.
SPLETZER, M.E., AND ENYEDI, A.J. Salicylic acid induces resistance to Alternaria solani
in hydroponically grown tomato. Phytopathology. v. 89, nº. 9, p. 722 – 727, 1999.
SRIVASTAVA, M. K.; DWIVEDI, U. N. Delayed ripening of banana fruit by salicylic acid.
Plant Science. v. 158, p. 87 – 96, 2000.
TAIZ, L.; ZEIGER, E. Fisiologia vegetal. 3ª ed. Porto Alegre: Artmed, 2006, 719 p.

80
TANAKA, M.A.S. Controle das doenças causadas por fungos e bactérias em morangueiro.
Cap. 2. In: ZAMBOLIM, L.; VALE, F.X.R do; MONTEIRO, A.J.A.; COSTA, H. Controle
de Doenças de Plantas Fruteiras. Viçosa, MG, v. 1, 2002. 674p.
TANAKA, M.A.S.; BETTI, J.A.; KIMATI, H. Doenças do Morangueiro (fragaria x
ananassa). Cap. 56. In: KIMATI, H.; AMORIM, L.; REZENDE, J.A.M.; FILHO, A.B.;
CAMARGO, L.E.A. Manual de Fitopatologia – Doenças de plantas cultivadas. v. 2. 4ª ed.,
São Paulo, SP, Agronômica Seres, 2005. 663p.
TAREEN, M.J.; ABBASI, N.A.; HAFIZ, I.A. Effect of salicylic acid treatments on storage
life of peach fruits cv. „flordaking‟. Pakistan Journalof Botany. v. 44, n. 1, p. 119 – 124,
2012.
TERRY, L.A.; JOYCE, D.C. Elicitors of induced disease resistance in postharvest
horticultural crops: a brief review. Postharvest Biology and Technology. v. 32, p. 1 – 13,
2004.
TERUEL, B. Tecnologias de Resfriamento Rápido de Frutos. In: Manual Pós-colheita da
Fruticultura Brasileira / Leandro Camargo Neves (organizador). Londrina: EDUEL, 2009.
494p.
TIBOLA, C.S.; MALGARIM, M.B.; ZAICOVSK, C.B.; PEGORARO, C.; CERO, J.D.;
FERRI, V.C. Luz ultravioleta na inibição de podridões pós-colheita de morangos (fragaria
ananassa, duch.) “Camarosa”. Revista Brasileira de Agrociência. Pelotas, v. 13, n. 4, p. 509
– 512, out-dez, 2007.
TRIPATHI, P.; DUBEY, N.K. Exploitation of natural products as an alternative strategy to
control postharvest fungal rotting of fruit and vegetables. Postharvest Biology and
Technology, v. 31, n. 3, p. 235-245, 2004.
TOMAZELI, V.N. Indução de resistência a doenças em morangueiro com o uso de
acibenzolar-S-metil e harpina em pré e pós-colheita. 2010. 102p. Dissertação (Mestrado
em Agronomia) - Universidade Tecnológica Federal do Paraná, Pato branco.

81
TZORTZAKIS, N.G.; ECONOMAKIS, C.D. Maintaining postharvest quality of the tomato
fruit by employing methyl jasmonate and ethanol vapor treatment. Journal of Food Quality.
v. 30, n. 5, p. 567 – 580, October 2007.
VALERO, D.; DÍAZ-MULA, H.M.; ZAPATA, P.J.; CASTILLO, S.; GUILLÉN, F.;
MATÍNEZ-ROMERO, D.; SERRANO, M. Postharvest treatments with salicylic acid,
acetylsalicylic acid or oxalic acid delayed ripening and enhanced bioactive compounds and
antioxidant capacity in sweet cherry. Journal of Agricultural and Food Chemistry. v. 59, n.
10, p. 5483 – 5489, 2011.
VANETTI, C.A. Resistência do Tomateiro a Alternaria solani induzida por quitina,
ácido salicílico, benzotiadiazol e ácido acetil salicílico tamponado. 2001. 65p. Tese
(Doctor Scientiae em Fitopatologia) – Universidade Federal de Viçosa, Viçosa.
VIEITES, R.L.; EVANGELISTA, R.M.; SILVA, C. de S.; MARTINS, M.L. Conservação do
morango armazenado em atmosfera modificada. Semina: Ciências Agrárias, Londrina, v.
27, n. 2, p. 243-252, abr./jun. 2006.
VLOT, A.C.; DEMPSEY, D.M.A.; KLESSIG, D. F. Salicylic Acid, a Multifaceted Hormone
to Combat Disease. Annual Review of Phytopathology. v. 47, p. 177 – 206, 2009.
XU, X.; TIAN, S. Salicylic acid alleviated pathogen-induced oxidative stress in harvested
sweet cherry fruit. Postharvest Biology and Technology. v. 49, p. 379 – 385, 2008.
WANG, L.; CHENA, S.; KONG, W.; LI, S.; ARCHBOLD, D.D. Salicylic acid pretreatment
alleviates chilling injury and affects the antioxidant system and heat shock proteins of peaches
during cold storage. Postharvest Biology and Technology. v. 41, p. 244 – 25, 2006.
YAMASHITA, F. Embalagens Pós-colheita para Frutos. In: Manual Pós-colheita da
Fruticultura Brasileira / Leandro Camargo Neves (organizador). Londrina: EDUEL, 2009.
494p.

82
YANG, Z.; CAO, S.; CAI, Y.; ZHENG, Y. Combination of salicylic acid and ultrasound to
control postharvest blue mold caused by Penicillium expansum in peach fruit. Innovative
Food Science and Emerging Technologies. v. 12, p. 310 – 314, 2011.
YAO, H.; TIAN, S. Effects of pre- and post-harvest application of salicylic acid or methyl
jasmonate on inducing disease resistance of sweet cherry fruit in storage. Postharvest
Biology and Technology. v. 35, p. 253 – 262, 2005a.
YAO, H.J.; TIAN, S.P. Effects of a biocontrol agent and methyl jasmonate on postharvest
diseases of peach fruit and the possible mechanisms involved. Journal of Applied
Microbiology. v. 98, p. 941 – 950, 2005b.
YU, M.; SHEN, L.; FAN, B.; ZHAO, D.; ZHENG, Y.; SHENG, J. The effect of Meja on
ethylene biosynthesis and induced disease resistance to Botrytis cinerea in tomato.
Postharvest Biology and Technology. v. 54, n 3, p. 153 – 158, December 2009.
ZAICOVSKI, C.B.; TIBOLA, C.S.; MALGARIM, M.B; FERRI, V.C.; PEGORARO, C.;
CERO, J.D.; SILVA, P.R. Resveratrol na qualidade pós-colheita de morangos “Camarosa”.
Revista Brasileira de Agrociência. Pelotas, v. 12, n. 4, p. 443 – 446, out-dez, 2006.
ZAINURI, D.C.J.; WEARING, H.; COATES, L.; TERRY, L. Effects of phosphonate and
salicylic acid treatments on anthracnose disease development and ripening of „Kensington
Pride‟ mango fruit. Australian Journal of Experimental Agriculture. v. 41, n. 6, p. 805–
813, 2001.
ZHANG, F.S.; WANG, X.Q.; MA, S.J.; CAO, S.F.; LI, N.; WANG, X.X.; ZHENG, Y.H.
Effects of methyl jasmonate on postharvest decay in strawberry fruit and the possible
mechanisms involved. Acta Horticulturae. v. 712, p. 693 – 698, 2006.
ZHANG, H.; MA, L.; WANG, L.; JIANG, S.; DONG, Y.; ZHENG, X. Biocontrol of Gray
Mold Decay in Peach Fruit by Integration of Antagonistic Yeast with Salicylic Acid and Their
Effects on Postharvest Quality Parameters. Biological control. v. 47, p. 60 – 65, 2008.

83
ZHANG, H.; MA, L.; TURNER, M.; XU, H.; ZHENG, X.; DONG, Y.; JIANG, S. Salicylic
acid enhances biocontrol efficacy of Rhodotorula glutinis against postharvest Rhizopus rot of
strawberries and the possible mechanisms involved. Food Chemistry. v. 22, n. 3, p. 577 –
583, October, 2010.
ZENG, K.; CAO, J.; JIANG, W. Enhancing disease resistance in harvested mango (Mangifera
indica L. cv. „Matisu‟) fruit by salicylic acid. Journal of The Science of Food and
Agriculture. v. 86, n. 5, p. 694 – 698, April 2006.
ZHU, Z. TIAN, S. Resistant responses of tomato fruit treated with exogenous methyl
jasmonate to Botrytis cinerea infection. Scientia Horticulturae. v. 142, p. 38 – 43, july,
2012.

















![At Sb Te Ge As Si B - [DePa] Departamento de Programas ...depa.fquim.unam.mx/amyd/archivero/ManualdeNomenclatura...Ácido hipocloroso Ácido nitroso Ácido sulfúrico Ácido fosfórico](https://img.document.onl/doc/110x75/607b776c072ab702de445b0a/at-sb-te-ge-as-si-b-depa-departamento-de-programas-depafquimunammxamydarchiveromanualdenomenclatura.jpg)










