Embed Size (px)
Citation preview

UNIVERSIDADE DO VALE DO ITAJAI
ISAAC PITTIGLIANI JORGE
ANÁLISE DO COMPORTAMENTO NOCICEPTIVO DE CAMUNDONGOS SUBMETIDOS À AVULSÃO DO PLEXO BRAQUIAL
Biguaçu 2011

1
ISAAC PITTIGLIANI JORGE
ANÁLISE DO COMPORTAMENTO NOCICEPTIVO DE CAMUNDONGOS SUBMETIDOS À AVULSÃO DO PLEXO BRAQUIAL
Monografia apresentada como requisito parcial para obtenção do título de Bacharel em Fisioterapia na Universidade do Vale do Itajaí, Centro de Ciências da Saúde. Orientadora: Profa. Dra. Nara Lins Meira Quintão
Biguaçu, novembro de 2011.

ISAAC PITTIGLIANI JORGE
ANÁLISE DO COMPORTAMENTO NOCICEPTIVO DE CAMUNDONGOS SUBMETIDOS À AVULSÃO DO PLEXO BRAQUIAL
Esta monografia foi julgada adequada para a obtenção do título de Bacharel em Fisioterapia e aprovada pelo Curso de Fisioterapia da Universidade do Vale do Itajaí, Centro de Ciências da Saúde. Área de Concentração: Farmacologia da dor.
Biguaçu, 21 de novembro de 2011.
______________________________________
Profa. Dra. Nara Lins Meira Quintão UNIVALI – Centro de Ciências da Saúde
Orientador
______________________________________ Profa. Dra. Kathryn Ana Bortolini Simão da Silva
UNIVALI – Centro de Ciências da Saúde Membro
______________________________________
Profa. Ma. Gislaine Franciele da Silva Membro

Este trabalho é dedicado a minha família.
Meus pais, CarlosCarlosCarlosCarlos e GicéliaGicéliaGicéliaGicélia que me ensinaram o valor de um sonho,
e compreenderam minha ausência.
Os meus irmãos IaraIaraIaraIara e IgorIgorIgorIgor, que mesmo longe sempre estiveram ao
meu lado em todas as etapas da minha vida.
À minha esposa MarivaneMarivaneMarivaneMarivane que de forma especial e carinhosa me deu
força e coragem, me apoiando nos momentos mais difíceis desta
minha jornada acadêmica.
E ao meu filho Luiz MiguelLuiz MiguelLuiz MiguelLuiz Miguel que com sua inocência de criança me
fez ser pai.

AGRADECIMENTOS
Agradeço primeiramente à Deus, pela vida, por estar sempre no meu
caminho, iluminando-me e guiando-me às escolhas certas, e por me privilegiar de
exercer uma profissão magnífica.
Aos meus pais, Carlos e Gicélia, que me deram toda a estrutura para que me
tornasse a pessoa que sou hoje. Pela confiança e pelo amor que me fortalece todos
os dias. Que mesmo longe sempre me incentivaram a continuar nesta luta e
compreenderam minha ausência durante todos estes anos fora de casa.
Aos meus irmãos, Iara e Igor, meus companheiros e amigos verdadeiros, os
quais eu deixei em Imbituba para eu poder levar a diante este meu sonho de ser
fisioterapeuta, deixando de participar do crescimento de vocês, e quando me dei
conta já eram adultos e não mais crianças, saudade é o que eu posso descrever,
amo vocês.
A professora Dra. Nara Lins Meira Quintão, meu agradecimento em especial
pela orientação, por seus ensinamentos a mim transmitidos que me acompanharão
por toda a vida, por todos estes anos de estudo e dedicação a pesquisa que
passamos juntos, nas horas que passei cronometrando meus animais e você estava
ali para me ajudar a dar continuidade ao trabalho, por acreditar que hoje eu estaria
chegando aonde cheguei.
Ao Professor Me. Marcelo Dias, por acreditar em meu potencial, transmitindo
seus ensinamentos incansavelmente, mostrando o caminho certo a seguir,
corrigindo meus erros, incentivando meus acertos, estando você junto a nossa turma
desde o início de minha vida acadêmica.
Ao Professor Cristiano Coelho, um grande amigo, pelas suas caronas quando
eu morava em Tijucas, tirando dúvidas em sala de aula, argumentando o porquê das
minhas respostas esperando sempre mais de mim. Deixando saudades quando saiu
do quadro de docentes.
Aos professores, de uma forma geral que me deixavam dormindo até o ultimo
minuto antes das aulas começarem e entenderam minhas horas sonolentas no
período em que eu trabalhava de madrugada, estudava pela manhã e dormia
somente à tarde, repassando a matéria em momentos de dúvidas e incertezas

5
ocasionadas pelo sono.
Aos professores: Aline Moreira, Ana Paula, Angelise Mozerle, Cladis Moraes,
Claudia Thofehrn, Clarice Pamplona, Cristiani Burguer, Daniela Simoni, Eliana
Trevisan, Elaine Farina, Fabiane Dell, Isabela Sell, Juliana Vidotto, Mariem El Saya,
Rita de Cássia, Adriano de Sousa, Alceu Furtado, Felipe Lacerda... entre outros,
pelos ensinamentos durante estes longos seis anos de estudo, que sempre se
propuseram a esclarecer dúvidas, sempre mostrando o melhor caminho e a serem
seguidos, pelos momentos de distração, conversas e seriedade, transmitindo sábios
conhecimentos durante todo esse tempo.
A técnica do laboratório de farmacologia de Itajaí, Maria Angélica, a Maggie,
pelos momentos de ajuda com o cuidado de meus animais após as longas cirurgias,
as horas de conversas, risadas e cafezinhos para descontrair e passar o tempo, meu
imenso carinho.
Aos meus colegas, amigos de laboratório que me ajudaram durante as horas
de pesquisa e experimento, no laboratório de farmacologia, as viagens para
congressos e o companheirismo... são vocês: Gislaine, Isabel, Zeca, Ana Paula,
Juliana, Philipe, Nicole, Lilian, Rosana, Luiz Carlos, Iandra, Maurício, Ana Roseli,
Gabriel, Diogo... amigos que jamais vou esquecer.
Ao Douglas, meu companheiro de quarto nos congressos, pelas horas de
descontração e estudo, pelos momentos de troca de conhecimento e ansiedade
pelas apresentações dos banners.
As minhas amigas e companheiras de jornada, meninas, mulheres em
especial, por me aturarem durante todos estes anos juntos de formação e
aprendizado, pelas horas de descontração e divertimento com nossos pacientes e
nas horas sérias durante as aulas e atendimentos... são vocês: Célia, Fabiane,
Letícia, Mariana, Regiani, Suely e Vanessa... amigas as quais estarão sempre em
meu coração, porque eu sei o quanto é difícil suportar minha presença.
Em especial a minha companheira e amiga de jornada Célia Regina Caetano
Fernandes, pelos vários e incontáveis momentos de risadas e descontração pelos
ensinamentos que dela eu absorvi, pelos momentos de atendimento que não foram
poucos, pela nossa sincronia durante todos estes anos que com você eu convivi,
pelo carinho com o qual você tem e sabe passar aos outros, por me apoiar todo

6
esse tempo que passamos juntos, pessoa igual a você é raridade, que esta nossa
amizade flua eternamente, amo você.
Ao meu amigo Maurício Milanese pelas horas de “happy hour” fora do
cotidiano acadêmico.
Aos meus amigos de jornada na Portobello, que sempre acreditaram que um
dia eu chegaria até aqui, me incentivando, me encobertando enquanto eu estudava
durante o horário de trabalho, me deixando dormir uns minutos a mais durante os
poucos minutos de café, pelas horas incansáveis de trabalho, conversas e risadas...
São vocês: Tonho, Dioclei, Dione, Moisés, Pacheco entre muitos outros que não me
recordo no momento... vocês sempre estarão marcados em minha vida.
Aos meus primos... Ellen, André, Arthur, Lúcia e Clau... por me ajudarem
durante minha morada em Tijucas, sem vocês eu não estaria hoje onde estou e em
especial a minha afilhada Letícia, um amor de menina.
À Marivane... por hoje fazer parte da minha história, esposa e amiga, pelas
inúmeras horas de dedicação ao meu lado, durante as horas extras de experimento,
ficando ao meu lado, pelo seu jeito alegre, seu sorriso encantador, pelos momentos
de descontração e conversas intermináveis ao final dos experimentos aos arredores
de Itajaí e por ter dado a mim a dádiva de ser pai. Amo você e o nosso filho Luiz
Miguel.
À Universidade do Vale do Itajaí, por disponibilizar instalações e materiais
para a realização deste trabalho.
E por fim, agradeço ao PROBIC/PIBIC pelo apoio financeiro.
Muito obrigado...

““““Não despreze Não despreze Não despreze Não despreze oooo filhote fraco.filhote fraco.filhote fraco.filhote fraco.
Ele podeEle podeEle podeEle pode sesesese tornar tornar tornar tornar um tigre feroz”.um tigre feroz”.um tigre feroz”.um tigre feroz”.
(Provérbio Mongol) (Provérbio Mongol) (Provérbio Mongol) (Provérbio Mongol)

ANÁLISE DO COMPORTAMENTO NOCICEPTIVO DE CAMUNDONGOS SUBMETIDOS À AVULSÃO DO PLEXO BRAQUIAL
Isaac Pittigliani Jorge
Resumo
A dor crônica, causada por lesões ou doenças que superam a capacidade do organismo de reverter o quadro, envolve alterações adaptativas, como neuroplasticidade, em diferentes níveis do sistema nervoso. A lesão por avulsão do plexo braquial (APB) é um evento comum em humanos, podendo levar a importantes alterações patológicas responsáveis pela indução e estabelecimento de dor neuropática. O objetivo deste estudo é investigar as alterações nociceptivas agudas em animais submetidos à APB, com a finalidade de expandir o conhecimento dos mecanismos envolvidos nos quadros de dor persistente e auxiliar na busca de tratamentos eficazes. Diferentes grupos de animais foram submetidos ao procedimento cirúrgico de APB e no 6º ou 30º dia após a cirurgia foram desafiados em modelos de nocicepção aguda induzida por formalina, capsaicina, glutamato e bradicinina, tornando-se necessário verificar principalmente a participação do sistema cininérgico e glutamatérgico através de ensaios in vivo utilizando agonistas e antagonistas seletivos em animais operados e falso-operados. Procurou-se também avaliar a nocicepção na pata traseira contralateral à cirurgia, verificando se as alterações nas respostas nociceptivas estariam limitadas ao hemisfério corporal do animal que sofreu APB. Foi demonstrado que a resposta nociceptiva induzida pela formalina em camundongos submetidos à APB foi significativamente diferente quando comparada a resposta dos animais falso-operados. Estas alterações foram revertidas pelo pré-tratamento com o anti-inflamatório indometacina ou dexametasona, ou com baixas doses de morfina. Animais submetidos à APB injetados com glutamato ou bradicinina apresentaram redução do comportamento nociceptivo comparado com o grupo falso-operado. Quando camundongos operados receberam injeção i. pl. de capsaicina, não se observou diferença significativa. Os resultados do presente trabalho demonstram que a APB produz alterações na resposta nociceptiva aguda, sugerindo que a APB desenvolve alterações na atividade do sistema glutamatérgico e de cininas e não interfere com a ação de receptores vaniloides periféricos. Demonstrou-se também o envolvimento de componentes inflamatórios nestas alterações. A redução da resposta nociceptiva após a injeção de formalina foi atribuída ao comportamento de freezing, uma vez que quando administrado um anti-inflamatório não esteroidal ou mesmo doses subliminares de morfina esta redução na resposta foi revertida e o animal passou a responder de maneira semelhante aos animais falso-operados. No modelo de nocicepção induzida por formalina, a administração do HOE-140 e da DALBK (antagonistas dos receptores B2 e B1 para cininas, respectivamente) promoveu um aumento da resposta dolorosa na segunda fase do teste, sugerindo que a redução na resposta frente à injeção de 2,5% de formalina possa estar relacionada a uma maior expressão dos receptores B1 e/ou B2. No modelo de nocicepção induzida por glutamato, os animais submetidos à ABP apresentaram uma redução na resposta nociceptiva em todas as concentrações testadas,

9
sugerindo que a expressão dos receptores glutamatérgicos possa estar diminuída, ou a resposta nociceptiva possa estar bloqueada em nível de controle central da dor, visto que as alterações na neurobiologia causadas pela APB possam estar interferindo nos processos de sinalização das respostas nociceptivas envolvidas. Palavras-chave: Dor. Dor neuropática. Avulsão do plexo braquial.

ANALYSIS OF BEHAVIOR NOCICEPTIVE MICE SUBMITTED TO BRACHIAL PLEXUS AVULSION
Isaac Pittigliani Jorge
Abstract Chronic pain caused by injury or illness that exceed the body's ability to reverse the situation involves adaptive changes, such as neuroplasticity at different levels of the nervous system. The injury of the brachial plexus avulsion (BPA) is a common event in humans and may lead to important pathological changes responsible for the induction and establishment of neuropathic pain. The aim of this study is to investigate the acute nociceptive changes in animals submitted to BPA for the purpose of expanding knowledge of the mechanisms involved in the frames of persistent pain and assist in the search for effective treatments. Different animals groups underwent the surgical procedure of BPA and 6 or 30 days after surgery were challenged in models of nociception-induced acute formalin, capsaicin, glutamate, and bradykinin, making it necessary to check mainly the shareholding cininergic and glutamatergic system through in vivo using selective agonists and antagonists in animals operated and false-operated. We also sought to evaluate the process nociceptive hind paw contralateral to the surgery, making sure that the changes in nociceptive responses were limited to the hemisphere, the animal's body that has BPA.It was shown that formalin-induced nociceptive response in mice submitted to BPA was significantly different when compared the response of false-operated animals. These changes were reversed by pretreatment with anti-inflammatory drug indomethacin or dexamethasone, or with low doses of morphine. Animals treated with glutamate or BPA injected with bradykinin showed a reduction in nociceptive behavior compared to the false-operated group. When operated mice were injected i.pl. of capsaicin, there was no significant difference. The present results demonstrate that the BPA produces changes in acute nociceptive response, suggesting that the BPA develops changes in the activity of glutamatergic system and kinin and does not interfere with the action of peripheral vanilloid receptors. It also demonstrated the involvement of inflammatory components in these changes. The reduction of the nociceptive response after formalin injection was attributed to the freezing behavior, when administered as a non-steroidal anti-inflammatory or even subthreshold doses of morphine reduced this response was reversed and the animal began to respond in a similar way to animals false-operated. In the model of formalin-induced nociception, the administration of HOE-140 and DALBK (receptor antagonists for kinin B1 and B2, respectively) promoted an increase in pain response in the second phase of the trial, suggesting that the reduction in response to the injection front 2.5% formalin may be related to increased expression of B1 receptors and / or B2. In the model of glutamate-induced nociception, the animals treated with BPA showed a reduction in nociceptive response at all concentrations tested, suggesting that the expression of glutamate can be reduced, or the nociceptive response can be blocked at the level of central control of pain since the changes caused by the BPA in neurobiology might be interfering in signaling processes involved in nociceptive responses. Keywords: Pain. Neuropathic pain. Brachial plexus avulsion.

LISTA DE FIGURAS
Figura 1. Tabela dos vários tamanhos de axônios aferentes primários. O diâmetro do axônio está correlacionado com a sua velocidade de condução, e com o tipo de receptor sensorial ao qual está conectado. ............................................................... 27
Figura 2. Esquema representativo do percurso do estímulo doloroso até o córtex cerebral. .................................................................................................................... 28
Figura 3. Ilustração demonstrando os procedimentos cirúrgicos da APB. (A) Região da incisão, (B e C) anatomia do plexo braquial, (D) separação de nervos e vasos, (E) preensão do tronco inferior, (F) avulsão do tronco inferior. Tronco inferior do plexo braquial (setas pretas); vasos subclávios (setas brancas). ....................................... 34
Figura 4. Resposta nociceptiva induzida pela injeção de formalina (0,5 – 2,5 %; A – primeira fase; B – segunda fase) em animais submetidos à APB ou falso-operados, avaliados 6 dias após a cirurgia. Cada grupo representa a média de 6 a 8 animais e as linhas verticais indicam o E.P.M. Significativamente diferente dos valores do grupo falso-operado *p < 0,05 (ANOVA de uma via seguida pelo teste post-hoc de Dunnett)..................................................................................................................... 39
Figura 5. Resposta nociceptiva induzida pela injeção de formalina (0,5 – 2,5 %; A – primeira fase; B – segunda fase) em animais submetidos à APB ou falso-operados, avaliados 30 dias após a cirurgia. Cada grupo representa a média de 6 a 8 animais e as linhas verticais indicam o E.P.M. Significativamente diferente dos valores do grupo falso-operado *p < 0,05 (ANOVA de uma via seguida pelo teste post-hoc de Dunnett)..................................................................................................................... 40
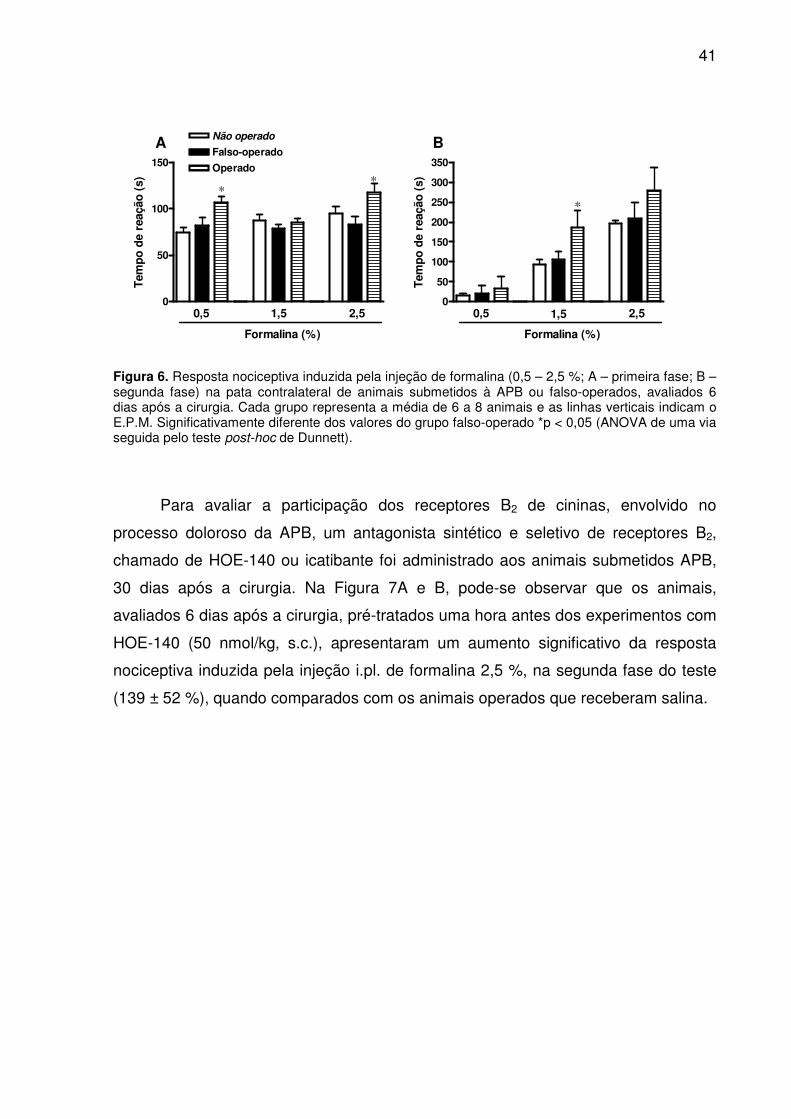
Figura 6. Resposta nociceptiva induzida pela injeção de formalina (0,5 – 2,5 %; A – primeira fase; B – segunda fase) na pata contralateral de animais submetidos à APB ou falso-operados, avaliados 6 dias após a cirurgia. Cada grupo representa a média de 6 a 8 animais e as linhas verticais indicam o E.P.M. Significativamente diferente dos valores do grupo falso-operado *p < 0,05 (ANOVA de uma via seguida pelo teste post-hoc de Dunnett). ................................................................................................ 41
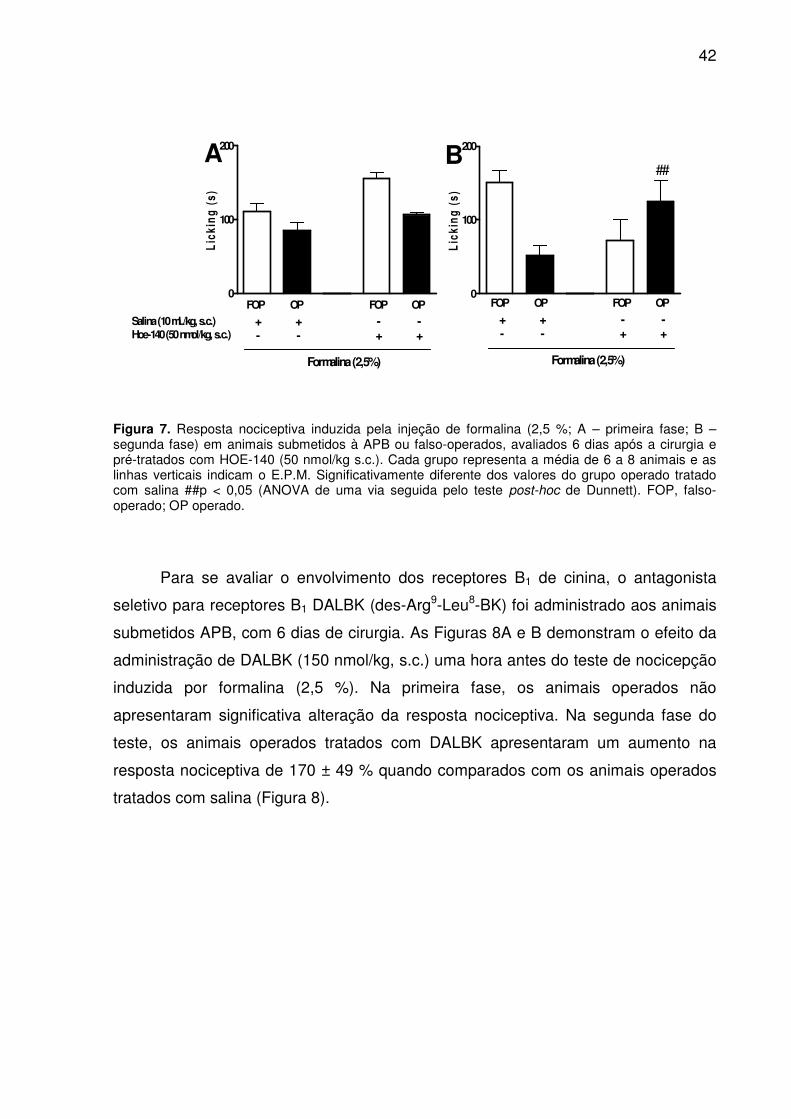
Figura 7. Resposta nociceptiva induzida pela injeção de formalina (2,5 %; A – primeira fase; B – segunda fase) em animais submetidos à APB ou falso-operados, avaliados 6 dias após a cirurgia e pré-tratados com HOE-140 (50 nmol/kg s.c.). Cada grupo representa a média de 6 a 8 animais e as linhas verticais indicam o E.P.M. Significativamente diferente dos valores do grupo operado tratado com salina ##p < 0,05 (ANOVA de uma via seguida pelo teste post-hoc de Dunnett). FOP, falso-operado; OP operado. ............................................................................................... 42
Figura 8. Resposta nociceptiva induzida pela injeção de formalina (2,5 %; A – primeira fase; B – segunda fase) em animais submetidos à APB ou falso-operados, avaliados 6 dias após a cirurgia e pré-tratados. Cada grupo representa a média de 6 a 8 animais e as linhas verticais indicam o E.P.M. Significativamente diferente dos valores do grupo operado tratado com salina ##p < 0,01 (ANOVA de uma via seguida pelo teste post-hoc de Dunnett). FOP, falso-operado; OP operado. ........... 43

12
Figura 9. Resposta nociceptiva induzida pela injeção de capsaicina (0,016 – 1,6 µg/pata) em animais submetidos à APB ou falso-operados, avaliados 6 dias após a cirurgia. Cada grupo representa a média de 6 a 8 animais e as linhas verticais indicam o E.P.M. Significativamente diferente dos valores do grupo falso-operado *p < 0,05 (ANOVA de uma via seguida pelo teste post-hoc de Dunnett). ...................... 43
Figura 10. Resposta nociceptiva induzida pela injeção de glutamato (0,3 – 30 µmol/pata) em animais submetidos à APB ou falso-operados, avaliados 6 dias após a cirurgia. Cada grupo representa a média de 6 a 8 animais e as linhas verticais indicam o E.P.M. Significativamente diferente dos valores do grupo falso-operado **p < 0,05 (ANOVA de uma via seguida pelo teste post-hoc de Dunnett). ...................... 44
Figura 11. Resposta nociceptiva induzida pela injeção de bradicinina (0,1 – 10 nmol/pata) em animais submetidos à APB ou falso-operados, avaliados 6 dias após a cirurgia. Cada grupo representa a média de 6 a 8 animais e as linhas verticais indicam o E.P.M. Significativamente diferente dos valores do grupo falso-operado **p < 0,05 (ANOVA de uma via seguida pelo teste post-hoc de Dunnett). FOP, falso-operado; OP operado. ............................................................................................... 45
Figura 12. Resposta nociceptiva induzida pela injeção de BK (10 nmol/pata) em animais submetidos à APB ou falso-operados, avaliados 6 dias após a cirurgia. Cada grupo representa a média de 6 a 8 animais e as linhas verticais indicam o E.P.M. Significativamente diferente dos valores do grupo falso-operado **p < 0,05 (ANOVA de uma via seguida pelo teste post-hoc de Dunnett). FOP, falso-operado; OP operado. .................................................................................................................... 46
Figura 13. Resposta nociceptiva induzida pela injeção de bradicinina (1 – 10 nmol/pata) em animais submetidos à APB ou falso-operados, avaliados 6 dias após a cirurgia. Cada grupo representa a média de 6 a 8 animais e as linhas verticais indicam o E.P.M. Significativamente diferente dos valores do grupo falso-operado **p < 0,05 (ANOVA de uma via seguida pelo teste post-hoc de Dunnett). FOP, falso-operado; OP operado. ............................................................................................... 47
Figura 14. Efeito do pré-tratamento com indometacina (10 mg/kg, i.p.) sobre a resposta nociceptiva induzida pela injeção de formalina (1,5 %) em animais submetidos à APB ou falso-operados, avaliados 6 dias após a cirurgia. Cada grupo representa a média de 6 a 8 animais e as linhas verticais indicam o E.P.M. Significativamente diferente dos valores do grupo falso-operado *p < 0,05 e ***p < 0,001. (#) difere significativamente do grupo operado tratado com salina (ANOVA de uma via seguida pelo teste post-hoc de Dunnett). .................................................... 48
Figura 15. Efeito do pré-tratamento com dexametasona (0,5 mg/kg, s.c.) sobre a resposta nociceptiva induzida pela injeção de formalina (1,5 %) em animais submetidos à APB ou falso-operados, avaliados 6 dias após a cirurgia. Cada grupo representa a média de 6 a 8 animais e as linhas verticais indicam o E.P.M. Significativamente diferente dos valores do grupo falso-operado *p < 0,05 e ***p < 0,001. (#) difere significativamente do grupo operado tratado com salina (ANOVA de uma via seguida pelo teste post-hoc de Dunnett). .................................................... 48
Figura 16. Efeito do pré-tratamento com morfina (0,5 mg/kg, s.c.) sobre as respostas nociceptivas (A) neurogênica e (B) inflamatória induzidas pela injeção de formalina

13
(2,5 %) em animais submetidos à APB ou falso-operados, avaliados 6 dias após a cirurgia. Cada grupo representa a média de 6 a 8 animais e as linhas verticais indicam o E.P.M. Significativamente diferente dos valores do grupo falso-operado *p < 0,05 (ANOVA de uma via seguida pelo teste post-hoc de Dunnett). ...................... 49
Figura 17. Resposta hiperalgesica induzida pela injeção de (A) PGE2, (0,1 nmol/pata) (B) epinefrina (100 ng/pata) e (C) BK (0,1 nmol/pata) em animais submetidos à APB ou falso-operados, avaliados 6 dias após a cirurgia. Cada grupo representa a média de 6 a 8 animais e as linhas verticais indicam o E.P.M. Significativamente diferente dos valores do grupo falso-operado #p < 0,05 (ANOVA de duas vias seguida pelo teste post-hoc de Bonferroni). ......................................... 50

LISTA DE ABREVIATURAS
Aαααα - Fibras do tipo A subtipo alfa
Aββββ - Fibras do tipo A subtipo beta
Aδδδδ - Fibras do tipo A subtipo delta
AAE - Aminoácidos excitatórios
AINEs - Anti-inflamatórios não-esteroidais
AMPA - Ácido alfa-amino-3-hidroxi-5-metil-4-isoxazol-propiônico
AMPc - Adenosina monofosfato cíclico
ANOVA - Análise de variância
APB - Avulsão do plexo braquial
ATP - Adenosina trifosfato
B1 - Receptor de bradicinina do tipo 1
B2 - Receptor de bradicinina do tipo 2
BDNF - Fator neurotrófico derivado do cérebro
BK - Bradicinina
C - Fibras do tipo C
CCDV - Canais de cálcio dependentes de voltagem
CFA - Completo adjuvante de Freund
CGRP - Peptídeo relacionado ao gene da calcitonina
COX1 - Enzima ciclo-oxigenase do tipo 1
COX2 - Enzima ciclo-oxigenase do tipo 2
DABK - Des-Arg-BK
DALBK - Des-Arg9-Leu8-BK
DRG - Gânglio da raiz dorsal
E. P. M. - Erro padrão da média
FOP - Falso operado
i. p. - intra peritonial
i. pl. - intra plantar
IASP - International Association for the Study of Pain
NGF - Fator de crescimento neuronal
NMDA - N-metil-D-aspartato
NO - Óxido nítrico

15
OP - Operado
PGE2 - Prostaglandina do tipo E2
PGs - Prostaglandinas
PKA - Proteína quinase do tipo A
PKC - Proteína quinase do tipo C
PKCγ - Proteína quinase do tipo C subtipo gama
PKCε - Proteína quinase do tipo C subtipo episilon
RNAm - Ácido ribonucleico mensageiro
RVM - Medula ventromedial rostral
s. c. - subcutâneo
SNC - Sistema nervoso central
SP - Substância P
TNFα - Fator de necrose tumoral alfa
TrkA - Tirosina quinase do tipo A

SUMÁRIO
1 INTRODUÇÃO ........................................................................................ 18
2 OBJETIVOS ............................................................................................ 20
2.1 Objetivo geral .......................................................................................... 20
2.2 Objetivos específicos ............................................................................... 20
3 REVISÃO DA LITERATURA .................................................................. 21
3.1 Uma breve história sobre a dor ............................................................... 21
3.2 Definição da dor ...................................................................................... 23
3.3 As vias da dor: mecanismos de indução, transmissão e processamento
da dor ................................................................................................................ 25
3.4 Sinalização celular e neuroplasticidade na dor crônica ........................... 29
4 MATERIAIS E MÉTODOS ...................................................................... 33
4.1 Animais .................................................................................................... 33
4.1.1 Delineamento experimental ..................................................................... 33
4.2 Procedimento cirúrgico para a avulsão do plexo braquial ....................... 34
4.3 Análise do limiar mecânico através do filamento de von Frey ................. 35
4.4 Avaliação da Atividade Antinociceptiva ................................................... 35
4.4.1 Modelo de nocicepção espontânea induzida pela formalina ................... 35
4.4.2 Modelo de nocicepção espontânea induzida pela capsaicina ................. 35
4.4.3 Modelo de nocicepção espontânea induzida pelo glutamato .................. 36
4.4.4 Modelo de nocicepção espontânea induzida pela bradicinina ................. 36
4.4.5 Hipernocicepção mecânica induzida pela BK .......................................... 36
4.4.6 Hipernocicepção mecânica induzida pela PGE2...................................... 37
4.4.7 Hipernocicepção mecânica induzida pela epinefrina ............................... 37
4.5 Estudo farmacológico .............................................................................. 37
4.5.1 Envolvimento do sistema de cininas........................................................ 37
4.5.2 Envolvimento de componentes inflamatórios e sistema opióide ............. 37
4.6 Drogas e reagentes ................................................................................. 38
4.7 Análise estatística.................................................................................... 38
5 RESULTADOS ........................................................................................ 39

17
5.1 Resposta nociceptiva induzida pela formalina ......................................... 39
5.2 Resposta nociceptiva induzida pela capsaicina ...................................... 43
5.3 Resposta nociceptiva induzida pelo glutamato ........................................ 44
5.4 Resposta nociceptiva induzida pela bradicinina ...................................... 45
5.5 Participação de componentes inflamatórios na alteração da resposta
nociceptiva de animais submetidos à APB ....................................................................
................................................................................................................ 47
5.6 Efeito do tratamento com morfina sobre a resposta nociceptiva induzida
pela formalina ............................................................................................................ 49
5.7 Resposta hipernociceptiva induzida pela PGE2, epinefrina e BK. ........... 49
6 DISCUSSÃO ........................................................................................... 51
7 CONCLUSÕES ....................................................................................... 60
REFERÊNCIAS ......................................................................................................... 61

18
1 INTRODUÇÃO
Tanto a dor aguda quanto a dor crônica estão frequentemente associadas a
processos inflamatórios, como resultado da lesão tecidual, reatividade imune
anormal ou lesão nervosa (STEIN et al., 2003). Na dor crônica, muitas alterações
ocorrem em associação com os eventos básicos da nocicepção, que modificam a
relação entre o estímulo e a resposta nociceptiva, afetando a modulação do estado
doloroso resultante. Além disso, alterações centrais crônicas na neuroquímica da
sinalização da dor produzem hipersensibilidade, aumentando a frequência e a
duração dos impulsos aferentes. Ademais, mudanças estruturais secundárias à
lesão nervosa periférica incluem a perda de interneurônios espinhais, rearranjos não
apropriados de processos neurais aferentes na medula espinhal e a proliferação de
fibras simpáticas no gânglio sensorial (WATKINS, MAIER, 2002; DOGRUL et al.,
2003).
O que dificulta o desenvolvimento de terapias racionais e efetivas para o
tratamento de processos dolorosos crônicos é a falta de conhecimento do quanto os
mecanismos envolvidos na dor crônica diferem daqueles implicados na dor aguda
(ALEY et al., 2000). O entendimento dos mecanismos envolvidos na transmissão do
sinal doloroso tem progredido muito nos últimos anos, em grande parte devido a um
aprimoramento na compreensão dos mecanismos envolvidos na fisiologia das fibras
aferentes e no processo de neurotransmissão no corno dorsal da medula espinhal
(GRUBB, 1998; FÜRST, 1999; MILLAN, 1999). Este progresso foi só possível com o
uso de múltiplos protocolos experimentais, incluindo estudos comportamentais,
eletrofisiológicos in vivo e in vitro, estudos anatômicos e principalmente através de
técnicas de biologia molecular (GRUBB, 1998; MILLAN, 1999; JULIUS; BASBAUM,
2001).
A dor induzida por lesão nervosa tem sido muito descrita como aumento na
sensibilidade de fibras aferentes primárias com consequente sensibilização do
circuito nociceptivo do corno dorsal da medula espinhal (CAMPBELL; MEYER,
2006). Entretanto, estudos mais recentes revelam que sistemas modulatórios do
tronco cerebral como a medula ventromedial rostral (RVM) também tem um papel
crítico, podendo facilitar ou suprimir a resposta nociceptiva (PORRECA, OSSIPOV,
GEBHART, 2002; FIELDS et al., 2005).
Diante deste contexto, este projeto teve como objetivo investigar os

19
mecanismos envolvidos nas alterações das respostas nociceptivas agudas de
camundongos submetidos à avulsão do plexo braquial (APB), com seis e trinta dias
após a cirurgia, frente a modelos de nocicepção aguda, amplamente descritos na
literatura, na tentativa de melhor conhecer a amplitude das alterações sensoriais
decorrentes da lesão nervosa, avaliando também a participação do sistema
cininérgico e glutamatérgico nos processos de nocicepção induzida por formalina,
capsaicina, bradicinina e glutamato.
Além disso, sabe-se que a lesão por avulsão do plexo braquial (APB) é um
evento comum em humanos, podendo levar a importantes alterações patológicas
responsáveis pela indução e estabelecimento de dor neuropática (CARVALHO,
NIKKHAH, SAMII, 1997). Segundo Wynn Parry (1980), a dor por deaferentação é
uma das mais significantes razões de incapacidade. Em virtude da carência de
conhecimento a respeito dos mecanismos envolvidos nas alterações
hipernociceptivas observadas no modelo de APB, a investigação das alterações no
comportamento nociceptivo agudo destes animais poderá auxiliar no maior
entendimento deste quadro patológico contribuindo para a busca de terapias
clinicamente mais eficazes.

20
2 OBJETIVOS
2.1 Objetivo geral
O presente estudo teve como objetivo investigar os mecanismos envolvidos
nas alterações das respostas nociceptivas agudas de camundongos submetidos à
avulsão do plexo braquial (APB) na tentativa de melhor conhecer a amplitude das
alterações sensoriais decorrentes da lesão nervosa, bem como, identificar melhor os
mecanismos envolvidos nos diferentes tipos de dor.
2.2 Objetivos específicos
Os objetivos específicos do presente estudo foram:
1. Avaliar a resposta nociceptiva de animais submetidos à APB através do
modelo de nocicepção induzida quimicamente por capsaicina, formalina, glutamato,
ou bradicinina.
2. Verificar a resposta hiperalgésica de animais submetidos à APB
através dos modelos de hiperalgesia mecânica induzida pela PGE2 e Epinefrina.
3. Verificar o papel dos receptores de cininas nas alterações da resposta
nociceptiva induzida pela formalina através da administração de antagonistas
seletivos para os receptores B1 e B2.
4. Avaliar a participação do sistema opioide, bem como dos processos
inflamatórios nas alterações da resposta nociceptiva induzida pela APB através do
modelo de formalina.

21
3 REVISÃO DA LITERATURA
3.1 Uma breve história sobre a dor
A dor é uma experiência embutida de inúmeros significados, não podendo
estar limitado somente ao aspecto físico associado à lesão, mas também a aspectos
cognitivos, não tão bem explorados pela neurofarmacologia moderna. Desde o início
dos tempos pré-históricos encontram-se registros de que eventos de dor e seu
controle recebem atenção especial. A interpretação da dimensão da dor varia a cada
sociedade, e talvez não seja sentida de forma idêntica por todos os indivíduos. O
limiar da dor varia de indivíduo para indivíduo e de sociedade para sociedade,
independentemente de suas bases anatômicas e fisiológicas (TEIXEIRA, OKADA,
2001).
Em todas as sociedades primitivas, sejam mesopotâmicos, gregos ou
egípcios, encontramos relatos das consequências da dor e de como proceder em
seu controle. As lendas em torno de heróis e deuses fazem referencias à ocorrência
da dor como forma de punição e controle de atitudes humanas. Ainda nos dias
atuais, podemos encontrar povos com este mesmo conceito, ligando a dor a atos
divinos ou demoníacos. Assim, a medicina era exercida por sacerdotes a serviço dos
deuses, que empregavam diversos métodos para proporcionar o alívio da dor. De
acordo com Hipócrates, pai da medicina, “Divinunstest opus sedare dolorien” (Sedar
a dor é obra divina) (TEIXEIRA, OKADA, 2001).
No início do século XIX, surgiram os primeiros trabalhos relacionando a dor
com informações sensoriais provenientes das raízes ventrais e dorsais da medula
espinhal. O anatomista escocês Charles Bell (1774-1842) defendia a idéia de que os
nervos sensoriais possuíam funções especializadas para detectar informações
somáticas incluindo a dor (PERL, 2007). Em 1826, Johannes Müller (1801-1858),
psicólogo e fisiologista alemão, propôs sua teoria da "energia nervosa específica", a
qual defendia que os diferentes nervos (ótico, auditivo, etc.) transmitiam uma
espécie de "código", que identificava sua origem ao cérebro (MÜLLER, 1840). Já
Mortiz Schiff (1823-1896) fisiologista alemão, aluno do célebre médico neurologista e
fisiologista François Magendie (1783-1855), demostrou que lesões específicas na
medula espinhal resultavam na perda de sensações táteis e dolorosas, propondo

22
que a dor é uma sensação independente das demais (SCHIFF, 1858).
Nas décadas seguintes, foram identificadas as vias ascendentes com origem
nos neurônios espinhais (via espinotalâmica) e que o quadrante antero-lateral da
medula espinhal era a via anatômica que leva a informação dolorosa da periferia até
o cérebro. Anos mais tarde, o tálamo foi considerado a estrutura encefálica
fundamental no reconhecimento da dor (EDINGER, 1890; BROWN-SÉQUARD,
1868; DEJERINE, ROUSSY, 1906; HEAD; HOLMES, 1911).
No entanto, o neurologista Wilhelm Erb (1840-1921) postulou que a dor é
resultado da ativação vigorosa das vias nervosas comuns, contradizendo o conceito
de especificidade da dor, originando assim, uma nova teoria sobre a dor, onde
propunha que a intensidade do estímulo gerador, e que determinaria uma resposta
dolorosa ou não (ERB, 1874). Anos mais tarde, descobriu-se que não apenas a
intensidade do estímulo era necessária, mas que o padrão neuronal gerado também
determinava a resposta de dor (NAFE, 1929). Assim, os médicos e fisiologistas do
início do século XX estavam divididos sobre as origens e natureza da dor.
As observações feitas pelo fisiologista Maximiliano Ruppert Franz von Frey
(1852-1932) concluíram que a estimulação de distintas áreas da pele estavam
relacionadas à estrutura dos terminais nervosos, reforçando a idéia da
especificidade da dor, porém essa sensação específica era dada por estímulos não
específicos (mecânicos, térmicos ou químicos). O histologista, microbiologista e
patologista britânico Sir Charles Scott Sherrington (1857-1952) foi um histologista,
microbiologista e patologista britânico, percebeu que a dor era gerada por estímulos
lesivos (estímulos nocivos) independente de sua natureza física. Assim cunhou o
termo nociceptor como sendo a estrutura responsável por tal detecção dos estímulos
nocivos, embora não tivesse identificado nenhuma estrutura (SHERRINGTON,
1906).
Entre o início e a metade do século XX, os registros de sinais nervosos
levaram os pesquisadores a sugerir que as fibras mielinizadas Aδ e fibras não-
mielinizadas C eram ativadas por estímulos nocivos. Embora estudos apoiassem a
Teoria da Especificidade, a ausência de estudos sobre a existência de receptores
específicos fez com que o Departamento de Anatomia de Oxford repudiasse tal
teoria. Somente em 1965, Patrick Wall e Ronald Melzack, estudante de medicina e
psicólogo respectivamente, postularam a Teoria do Portão, onde o balanço entre a
ativação das fibras de pequeno (C) e de grande (A) diâmetro localizadas na medula

23
espinhal é que determinariam a condução ou não do estímulo nocivo: quando a
ativação das fibras C se sobressai a ativação das fibras A, o “portão” se abre e o
estímulo é conduzido. Se o contrário acontece, o “portão” se fecha e o estímulo
nocivo não é levado aos centros supra-espinhais. Esta teoria se tornou precursora
no conceito de modulação da dor, sendo confirmada em anos mais tarde
(HEINBECKER, BISHOP, O'LEARY, 1933; ZOTTERMAN, 1933; CLARK, HUGHES,
GASSER, 1935; LANDAU, BISHOP, 1953; COLLINS, NULSEN, RANDT, 1960;
MELZACK, WALL, 1965).
Em 1970, os pesquisadores demonstraram que os neurônios presentes na
lamina I do corno dorsal da medula espinhal eram ativados por estímulos térmicos
nocivos, e em 1975 foram identificados neurônios presentes em lâminas espinhais
profundas (V-VI) capazes de responder tanto a estímulos mecânicos e térmicos,
nocivos e inócuos. Estes neurônios foram denominados neurônios polimodais.
Atualmente sabe-se que a medula espinhal apresenta lâminas compostas por
neurônios nociceptivos e polimodais que estão particularmente envolvidas na
condução do estímulo nocivo (CHRISTENSEN, PERL, 1970; PRICE, MAYER, 1975;
CRAIG, 2003).
A detecção de estímulos dolorosos depende de estruturas especializadas,
mas, no entanto, a medula espinhal está sujeita a alterações plásticas quando
submetidas a contínuas estimulações, levando a respostas alteradas. Os
mecanismos envolvidos na condução e modulação da dor não estão completamente
elucidados, sendo que a idéia de dor deve ser estendida não apenas a experiência
física, delimitada apenas por características anatômicas e fisiologias, mas também
por um processo dinâmico sujeito a diversas alterações e modulações ligadas a
processos neuronais complexos (MELZACK, WALL, 1965; WOOLF, 1983).
3.2 Definição da dor
Existe uma grande variedade de definições para a dor, mas nenhuma delas é
capaz de agrupar todos os aspectos desta sensação tão complexa e subjetiva.
Sendo assim, sua definição é difícil, apesar da transposição de seu significado. O
termo dor, como definido pela IASP (International Association for the Study of Pain),
descreve não somente uma sensação, mas também uma emoção desagradável

24
associada a um estímulo lesivo ou potencialmente lesivo, sendo de natureza
subjetiva e estabelecendo a necessidade de tratamento para qualquer tipo de dor,
seja acompanhada de lesão tecidual ou não. A dor é uma experiência subjetiva
complexa, cuja percepção é o resultado de processos fisiológicos sensoriais ligados
a características pessoais e sociais sendo um estimulo percebido como doloroso
independente da sua natureza, mas sim, de acordo com o contexto em que o
indivíduo está inserido, ligado a suas emoções e memórias (LOESER, TREEDE,
2008).
Nocicepção é o termo utilizado para descrever os processos neurais na
identificação dos estímulos nocivos, capazes de produzir lesão tecidual. Nocicepção
e dor não devem ser confundidos, pois um pode ocorrer na ausência do outro,
justamente pela complexidade que envolve as definições cognitivas da dor. Em
1979, Merskey definiu a dor como sendo “uma experiência sensorial e emocional
desagradável associada a um dano tecidual real ou potencial”. Esta definição foi
então adotada pela IASP e permanece até os dias atuais. (MOGIL, BASBAUM,
2000; JULIUS, BASBAUM, 2001; PRICE, 2002).
A dor tem como papel fisiológico a função de proteger o organismo de forma a
prevenir e/ou diminuir os danos, avisando o indivíduo de que uma lesão tecidual está
para ocorrer ou já ocorreu. Assim que este mecanismo de alerta é estabelecido em
um indivíduo, a ameaça de dor pode provocar respostas comportamentais
generalizadas, tais como ativação simpática, aumento de corticosterona, taquicardia
e hipertensão arterial. Esses eventos melhoram o desempenho do indivíduo e o
afastam de uma situação de risco (MARKENSON, 1996; MILLAN, 1999; COSTIGAN,
WOOLF, 2000; WOOLF, SALTER, 2000; ALMEIDA, ROIZENBLATT, TUF, 2004).
As manifestações de dor podem ser estendidas com base nos substratos
neurais que medeiam às funções sensoriais e afetivas: enquanto o sistema sensorial
percebe e permite a localização tempo-espacial, a qualificação física e a intensidade
do estímulo nocivo, o componente cognitivo-afetivo atribui emoções as experiências,
sendo responsável pela resposta à dor. Sendo assim, a dor tem uma conotação
individual, sofrendo influência de experiências anteriores e ligadas a fatores sociais e
culturais. Porém, a dor pode perder a sua conotação de sobrevivência em um
ambiente hostil e perigoso, tornando-se resultado de mudanças no sistema nervoso,
tornando-se assim patológica, crônica e debilitante. Esta forma de dor é resultante
de um processo tecidual de reparo, estabelecendo um estado de hipersensibilidade

25
localizada dentro e ao redor da região lesada. Nestas condições, um estímulo
normalmente inócuo torna-se uma sensação extremamente desconfortável (WOOLF,
2000; JULIUS, BASBAUM, 2001; ALMEIDA, ROIZENBLATT, TUF, 2004).
Sendo assim, a dor pode ser classificada de acordo com a duração, podendo
ser aguda ou crônica, fisiológica ou não e se possui localização no sistema nervoso
central ou na periferia. A dor aguda é pontual e delimitada, desaparecendo com a
resolução do processo patológico. Já a dor crônica persiste por um longo período de
tempo, sendo associada a processos patológicos crônicos e mudanças no padrão de
transmissão neuronal (LENT, 2004; ALMEIDA, ROIZENBLATT, TUF, 2004).
3.3 As vias da dor: mecanismos de indução, transmissão e processamento
da dor
Estímulos mecânicos, térmicos ou químicos podem ser interpretados como
estímulos nociceptivos quando promovem a estimulação de vias aferentes primárias
denominadas para este caso de nociceptores, ou seja, qualquer que seja o estímulo
que excite o nociceptor, o efeito imediato é a alteração do potencial elétrico da
membrana da fibra nervosa. A dor se inicia pela ativação ou sensibilização de
nociceptores. Este estímulo é levado da periferia para o sistema nervoso central
através de fibras nociceptivas, que são as aferentes primárias de pequeno diâmetro
dos nervos periféricos. Os nociceptores fazem a transdução e codificação do
estímulo nocivo e estão presentes em praticamente todos os tecidos do organismo.
O processamento neural dos estímulos nocivos segue uma sequência de eventos,
sendo que a primeira etapa se inicia com a estimulação, seguida da transdução dos
estímulos nocivos, e posteriormente transmitida à medula espinhal e outras regiões
do SNC, podendo ocorrer à modulação da dor através do processamento do
estímulo nocivo (MEYER et al., 2005; SANDKUHLER, 2009; GUYTON, HALL, 2002,
LENT, 2004).
A ativação dos nociceptores por estímulos nocivos leva à despolarização do
neurônio e gera um potencial de ação que se propaga por toda a fibra. Em uma
inflamação, esta ativação pode ocorrer por mediadores inflamatórios ou provocar
uma lesão que ativa ou sensibilizar os nociceptores. Muitas vezes, estas fibras
sensibilizadas assumem limiares de ativação mais baixos que o normal (WOOLF,

26
SALTER, 2000; JULIUS, BASBAUM, 2001).
As fibras aferentes de primeira ordem são classificadas de acordo com sua
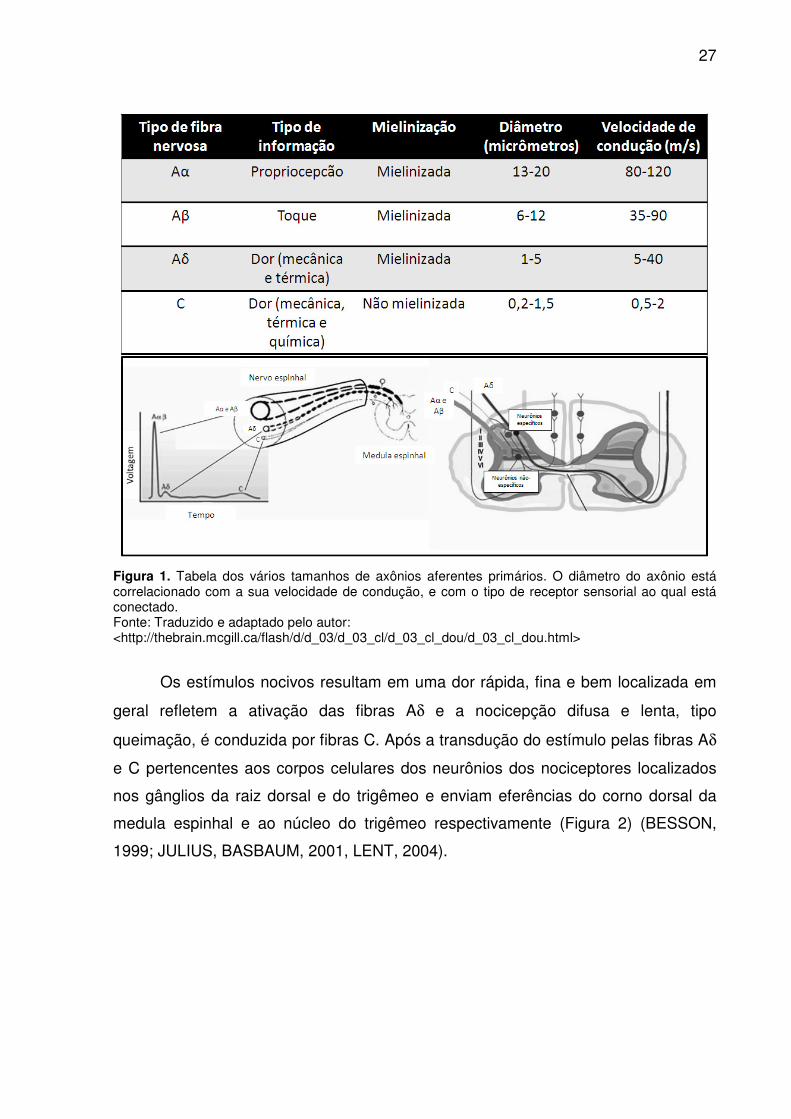
estrutura, diâmetro, mielinização e velocidade de condução: Aα, Aβ, Aδ e C. As
fibras Aα e Aβ são mielinizadas com diâmetro maior que 40 µm e velocidade de
condução de 30-100 m/s. As fibras aferentes Aδ são pouco mielinizadas, variando
seu diâmetro entre 2 e 6 µm e tem velocidade de condução de 12-30 m/s. As fibras
tipo C não são mielinizadas, possuem um diâmetro entre 0,4-1,2 µm e possuem
velocidade de condução de 0,5-2,0 m/s. (Figura 1) (JULIUS, BASBAUM, 2001).
Neurônios que possuem fibras com diâmetro maior (Aα e Aβ) detectam
estímulos inócuos e neurônios com fibras de médio e pequeno diâmetro (Aδ e C)
possuem a maioria dos nociceptores. As fibras Aδ possuem dois tipos de
nociceptores (tipo I e II), que respondem a estímulos mecânicos intensos, mas
podem responder a estímulos de calor intenso. As fibras Aδ do tipo I se ativam em
temperaturas superiores a 53 °C enquanto que as fibras Aδ do tipo II são ativadas
por temperaturas superiores a 43 °C. Já a maioria das fibras C é de característica
polimodal, ou seja, estímulos mecânicos, térmicos e químicos. É difícil identificar
qual o estímulo natural que irá ativar a fibra C. Algumas são insensíveis a estímulos
mecânicos, mas respondem ao calor nocivo. Outras apenas respondem a estímulos
químicos, como ácidos. Estes receptores são chamados de nociceptores silenciosos
ou adormecidos, que passam a ser ativados apenas quando sensibilizados por lesão
tecidual (JULIUS, BASBAUM, 2001).
As fibras C podem expressar o receptor purinérgico do tipo P2X3, cujos
terminais sinápticos localizam-se mais internamente na medula espinhal (lâmina II),
sendo que outro grupo de fibras C sintetiza peptídeos como a substância P e o
peptídeo relacionado ao gene da calcitonina (CGRP) e expressa receptores TrkA
para o fator de crescimento do nervo (NGF), fazendo conexões com neurônios da
lâmina mais externa do corno dorsal da medula (lâmina I) (CUELLO et al.,1993;
AVERIL et al., 1995).
27
Figura 1. Tabela dos vários tamanhos de axônios aferentes primários. O diâmetro do axônio está correlacionado com a sua velocidade de condução, e com o tipo de receptor sensorial ao qual está conectado. Fonte: Traduzido e adaptado pelo autor: <http://thebrain.mcgill.ca/flash/d/d_03/d_03_cl/d_03_cl_dou/d_03_cl_dou.html>
Os estímulos nocivos resultam em uma dor rápida, fina e bem localizada em
geral refletem a ativação das fibras Aδ e a nocicepção difusa e lenta, tipo
queimação, é conduzida por fibras C. Após a transdução do estímulo pelas fibras Aδ
e C pertencentes aos corpos celulares dos neurônios dos nociceptores localizados
nos gânglios da raiz dorsal e do trigêmeo e enviam eferências do corno dorsal da
medula espinhal e ao núcleo do trigêmeo respectivamente (Figura 2) (BESSON,
1999; JULIUS, BASBAUM, 2001, LENT, 2004).
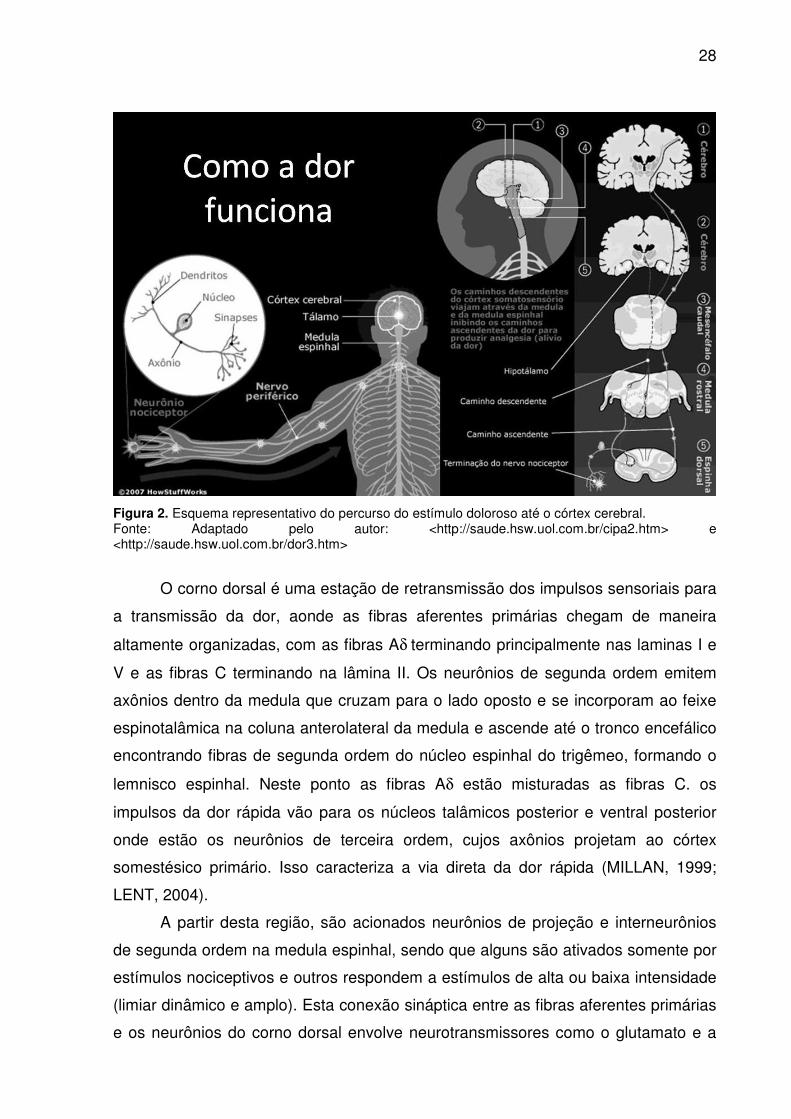
28
Figura 2. Esquema representativo do percurso do estímulo doloroso até o córtex cerebral. Fonte: Adaptado pelo autor: <http://saude.hsw.uol.com.br/cipa2.htm> e <http://saude.hsw.uol.com.br/dor3.htm>
O corno dorsal é uma estação de retransmissão dos impulsos sensoriais para
a transmissão da dor, aonde as fibras aferentes primárias chegam de maneira
altamente organizadas, com as fibras Aδ terminando principalmente nas laminas I e
V e as fibras C terminando na lâmina II. Os neurônios de segunda ordem emitem
axônios dentro da medula que cruzam para o lado oposto e se incorporam ao feixe
espinotalâmica na coluna anterolateral da medula e ascende até o tronco encefálico
encontrando fibras de segunda ordem do núcleo espinhal do trigêmeo, formando o
lemnisco espinhal. Neste ponto as fibras Aδ estão misturadas as fibras C. os
impulsos da dor rápida vão para os núcleos talâmicos posterior e ventral posterior
onde estão os neurônios de terceira ordem, cujos axônios projetam ao córtex
somestésico primário. Isso caracteriza a via direta da dor rápida (MILLAN, 1999;
LENT, 2004).
A partir desta região, são acionados neurônios de projeção e interneurônios
de segunda ordem na medula espinhal, sendo que alguns são ativados somente por
estímulos nociceptivos e outros respondem a estímulos de alta ou baixa intensidade
(limiar dinâmico e amplo). Esta conexão sináptica entre as fibras aferentes primárias
e os neurônios do corno dorsal envolve neurotransmissores como o glutamato e a

29
substância P, que são responsáveis pela produção de potenciais pós-sinápticos
excitatórios rápidos e lentos, respectivamente (MILLAN, 1999).
As fibras C são extensas e ramificadas, que podem se convergir. As fibras
secundárias se dirigem ao tálamo e a área somestésica primária e também para
diversas outras regiões do SNC que controlam fenômenos vegetativos e emocionais.
Algumas fibras de segunda ordem projetam aos neurônios do sistema reticular
ascendente, outras projetam a núcleos do sistema límbico e tálamo. As informações
nociceptivas chegam ao córtex cingulado anterior, além do córtex somestésico.
Quanto à modulação do estimulo nociceptivo, este permite a inibição ou facilitação
seletiva dos sinais nociceptivos que chegam ao corno dorsal da medula espinhal. O
sistema de modulação endógeno constitui-se de interneurônios nas camadas
superficiais da medula espinhal, onde há participação de peptídeos endógenos
(endorfinas, encefalinas, dinorfinas), cuja função é modular as sinapses
nociceptivas, bloqueando a liberação de neurotransmissores endógenos excitatórios
pelo terminal pré-sináptico e hiperpolarizando a membrana pós-sináptica. Todos
esses peptídeos são encontrados na substância cinzenta periaquidural, nos núcleos
da rafe e no corno da medula espinhal (REN, DUBNER, 2002; LENT, 2004).
3.4 Sinalização celular e neuroplasticidade na dor crônica
Primeiramente, uma lesão ativa as fibras nociceptivas produzindo uma dor
aguda, e posteriormente também resulta em uma série de mudanças ao nível de
nociceptores e da medula espinhal, e dependendo do grau dessas mudanças, o
resultado pode ser um estado de dor persistente onde há uma resposta exacerbada
de dor a estímulos nociceptivos (hiperalgesia) ou inócuos (alodínia) ou ainda
produzir a sensação de dor espontânea, isto é, dor sem estímulo externo. Este
estado doloroso persistente é chamado de dor crônica e geralmente persiste por
mais de 12 semanas, ou por um tempo anormal ao da cura. (BESSON, 1999;
ASHBURN; STAATS, 1999; JI, WOOLF, 2001; MERSKEY, BOGDUK, 2002).
A dor crônica é um grave problema de saúde, que afeta centenas de milhões
de pessoas por todo o mundo, diminuindo a qualidade de vida, incapacitando a
pessoa para o trabalho, induzindo a ansiedade e depressão e resultando em
prejuízos para a sociedade (RUSSO, BROSE, 1998; ASHBURN; STAATS, 1999;

30
TANG; CRANE, 2006).
Os tipos de dores crônicas mais comuns são a dor neuropática, que é
decorrente de uma lesão ou doença que afeta ou afetou o sistema somatosensorial,
como por exemplo, a neuropatia diabética; e a dor inflamatória, causada por
doenças inflamatórias crônicas, tais como a artrite reumatoide e a gota. Em qualquer
dos eventos acima citados, o processamento sensorial está alterado, produzindo
mudanças nas propriedades eletrofisiológicas e moleculares dos neurônios dos
nociceptores e da medula espinhal, resultando em hiperalgesia (BESSON, 1999;
WOOLF, SALTER, 2000).
A hiperalgesia é resultado da sensibilização neuronal nos receptores
periféricos, e está relacionada com a diminuição do limiar de estimulação e a uma
resposta aumentada dos nociceptores a estímulos aplicados na região da lesão
(hiperalgesia primária). Já a sensibilização de neurônios na medula espinhal
caracterizada por atividade ectópica ou espontânea aumentada de neurônios
nociceptivos e de neurônios não nociceptivos (hiperalgesia secundária), como
aqueles associados a sensações táteis que adquirem a capacidade de gerar dor
(alodínia). A hiperalgesia primaria está relacionados com processos normais de
proteção e recuperação de tecidos, enquanto que a hiperalgesia secundária está
relacionada com a dor crônica e está intimamente relacionada ao fenômeno de
sensibilização central (WOOLF, 1983; LOESER, TREEDE, 2008).
A sensibilização central envolve uma série de eventos que acabam por alterar
a neurobiologia da dor. Sob circunstâncias patológicas em que a estimulação dos
nociceptores encontra-se aumentada, a transmissão nociceptiva é alterada:
mudanças no potencial de membrana celular levam à ativação de canais iônicos,
como canais de sódio e canais de cálcio dependentes de voltagem (CCDV),
ocorrendo, então, um aumento na liberação de neurotransmissores, como o
glutamato, e a facilitação da transmissão excitatória que, através da ativação de
canais iônicos dependentes de voltagem ou operados por ligantes e da liberação de
estoques intracelulares, produz elevação nos níveis de cálcio. O aumento da
concentração de cálcio intracelular inicia inúmeros processos de sinalização celular.
Dentre eles está à ativação direta ou indireta de uma série de proteínas cinases que
podem em primeira instância, produzir alterações pós-transcricionais, como a
fosforilação de receptores e canais iônicos, e, em última instância, induzir à síntese
de novas proteínas envolvidas na transmissão nociceptiva (WOOLF, THOMPSON,

31
1991; WOOLF, COSTIGAN, 1999; JI, WOOLF, 2000).
Tais eventos ocorrem em intervalos de tempo precisos durante o
desenvolvimento da hiperalgesia e estão diretamente relacionados a alterações
drásticas no fenótipo celular (expressão de neurotransmissores, enzimas, canais
iônicos e receptores) e a mudanças estruturais, como a perda de interneurônios
espinhais, rearranjos inapropriados de neurônios na medula espinhal e a
proliferação de fibras simpáticas nos gânglios sensoriais (WOOLF,
SHORTLAND,COGGESHALL, 1992; MCLACHLAN et al., 1993; MOORE, BABA,
WOOLF, 2000; MOORE et al., 2002).
Embora compartilhem de mecanismos em comum, a dor de origem
inflamatória e neuropática apresenta algumas características únicas. Por exemplo, a
sensibilização decorrente da ação de mediadores inflamatórios como a bradicinina, a
prostaglandina E2 (PGE2), a serotonina e a histamina nos nociceptores, embora
presente na fase inicial da dor neuropática é uma característica que predomina na
dor inflamatória. Por outro lado, a lesão de nervos periféricos produz atividade
ectópica e espontânea de fibras sensoriais e as tornam sensíveis à estimulação pela
noradrenalina (WALL, DEVOR, 1983, TAIWO, LEVINE, 1989; McLACHLAN et al.,
1993; RUEFF, DRAY, 1993; MIZUMURA, 1997; WOOLF, MANNION, 1999).
Ademais, parece haver diferenças quanto à participação de segundos
mensageiros e de cascatas de sinalização celular na geração da dor inflamatória e
da dor neuropática. É amplamente estudada a sinalização via adenosina 3',5'-
monofosfato cíclico (AMPc) e ativação de proteína cinase A (protein kinase A - PKA)
na geração de dor inflamatória. Enquanto que a dor neuropática parece estar mais
atrelada à ativação de determinadas isoformas da proteína cinase C (protein kinase
C - PKC). Por exemplo, em camundongos, a ablação do gene que codifica a
isoforma neuronal da PKA que contém a subunidade regulatória RIβ causa
diminuição na nocicepção evocada por inflamação, mas não altera a dor neuropática
desses animais. Ainda, embora determinadas isoformas de PKC, como a isoforma
epsilon (PKCε), estejam envolvidas em processos de dor inflamatória, evidências
apontam para uma ação quase que exclusiva da isoforma gama (PKCγ) nos
mecanismos de dor neuropática (MAO et al., 1995; ALEY, LEVINE, 1999; OHSAWA
et al., 2001; YAJIMA et al., 2003; HUCHO, LEVINE, 2007).
A dor aguda e crônica representam diferentes condições psicofisiológicas e

32
exigem estratégias de condutas com drogas diferentes. Atualmente existem
múltiplos recursos disponíveis para o tratamento da dor, tais como métodos
fisioterápicos, acupuntura, técnicas de neuroestimulação e bloqueios anestésicos,
porém nenhuma delas é totalmente eficaz (SOBREIRA, ZAMPIER, 1999;
ABRAHAM, MCGINTY, BREWER, 2001).
Tais diferenças justificam o uso de diferentes abordagens terapêuticas no
tratamento da dor crônica inflamatória e neuropática. Por exemplo, a inibição da
síntese de PGE2 pelos anti-inflamatórios não esteirodais (AINES) é efetiva no
tratamento da dor inflamatória, mas não da neuropática. Ademais quadros de dor
inflamatória respondem bem à morfina, que apresenta até mesmo eficácia
analgésica superior nessas situações. Já em situações de dor neuropática a morfina
tem sua eficácia drasticamente reduzida (STANFA, DICKENSON, 1995; OBARA et
al., 2009).
As drogas mais comumente utilizadas no tratamento da dor crônica são os
analgésicos não-opiodes, os anti-inflamatórios não-esteroidais, os analgésicos
narcóticos, os opioides, os anticonvulsivantes, os antidepressivos, os neurolépticos,
os corticoides, vitaminas (principalmente do complexo B) e gangliosídeos. Além
disso, outras drogas tais como a calcitonina e a capsaicina também são utilizadas,
embora não tão frequentes (OLIVEIRA, A. S. B.; GABBAI, 1998)
A farmacoterapia na dor crônica inclui a administração de drogas por via
sistêmica, bem como procedimentos anestésicos locais ou de administração
intratecal. As dificuldades mais críticas surgem nos pacientes jovens, que desejam
levar uma vida social ativa e são impedidos de fazê-lo. O uso à longo prazo de
analgésicos, especialmente os mais potentes, pode levar a vários problemas,
incluindo-se o desenvolvimento de tolerância, ou mesmo ao surgimento de
dependência à droga. O insucesso terapêutico seria evitado através de avaliação
clínica, neurológica e psiquiátrica apropriadas (OLIVEIRA, A. S. B.; GABBAI, 1998).

33
4 MATERIAIS E MÉTODOS
4.1 Animais
Foram utilizados camundongos machos Swiss (pesando entre 20 e 28 g),
provenientes do Biotério Central da Universidade do Vale do Itajaí. Os animais foram
mantidos em ambiente com temperatura e umidade controladas (22 ± 1°C, 60 a 80%
de umidade), em ciclo 12 h claro/12 h escuro, com água e ração fornecidos ad
libitum. Os animais permaneceram no laboratório durante um período de adaptação
de pelo menos 1 h antes da realização dos testes farmacológicos, realizados
geralmente entre 8 e 17 h, a temperatura de 22 ± 1 ºC.
Os experimentos descritos foram conduzidos de acordo com as diretrizes
atuais de cuidados com os animais de laboratório e com as diretrizes éticas para
investigações de dor experimental em animais conscientes (ZIMMERMANN, 1983).
O Comitê de Ética da Universidade do Vale do Itajaí aprovou todos os
procedimentos experimentais (com parecer nº 363/07 na data de 25 de julho de
2007). O número de animais e a intensidade do estímulo nocivo utilizado foram os
mínimos necessários para demonstrar os efeitos dos tratamentos com as drogas.
4.1.1 Delineamento experimental
Nos testes de nocicepção espontânea os animais foram divididos em 2
grupos experimentais, sendo um grupo falso-operado e um grupo operado. Cada
grupo foi composto de 10 animais. Oito procedimentos experimentais, com 6 grupos
(3 doses de cada agente) de 10 animais cada (total de 480 animais). Para avaliar o
efeito farmacológico de drogas, os animais foram divididos em 4 grupos, sendo 1
grupo falso-operado tratado com salina, 1 grupo falso-operado tratado com a droga
em questão, 1 grupo operado tratado com salina e 1 grupo operado tratado com a
droga em questão. Como foram testadas 8 drogas diferentes, serão realizados 8
procedimentos experimentais, com 4 grupos de 10 animais cada (total de 480
animais). Ensaios de biologia molecular: 6 grupos de 10 animais cada (total de 60
animais).

34
4.2 Procedimento cirúrgico para a avulsão do plexo braquial
A metodologia utilizada para a realização da APB foi similar àquela descrita
para ratos (RODRIGUES-FILHO et al., 2003) e adaptada para camundongos
(QUINTÃO et al., 2006). Primeiramente, os animais foram anestesiados com hidrato
de cloral 7% (8 ml/kg; i.p.). O plexo braquial direito foi abordado através de uma
incisão longitudinal paralela a clavícula, transcorrendo do esterno à região axilar (1
cm aproximadamente, Figura 3 A). Os vasos subclávios foram localizados e o tronco
inferior foi dissecado (Figura 3 B – D). Um grupo de animais teve o tronco inferior
pinçado e avulsionado por tração utilizando duas pinças cirúrgicas (Figura 3 E e F).
No grupo falso-operado, o plexo braquial foi exposto e dissecado sem sofrer
qualquer lesão. Por fim a pele do animal foi suturada utilizando fio de sutura de seda
4,0 (Ethicon, Edinburgh).
Figura 3. Ilustração demonstrando os procedimentos cirúrgicos da APB. (A) Região da incisão, (B e C) anatomia do plexo braquial, (D) separação de nervos e vasos, (E) preensão do tronco inferior, (F) avulsão do tronco inferior. Tronco inferior do plexo braquial (setas pretas); vasos subclávios (setas brancas). Fonte: Adaptado de QUINTÃO et al., 2006.

35
4.3 Análise do limiar mecânico através do filamento de von Frey
Para avaliar a hipernocicepção mecânica, os animais foram colocados
individualmente em compartimentos de acrílico transparente individuais (9 X 7 X 11
cm) localizados em uma plataforma de arame elevada para permitir o acesso à
superfície ventral das patas traseiras. Os animais foram aclimatizados por pelo
menos 30 min antes dos testes comportamentais. A frequência de resposta de
retirada foi obtida através de 10 aplicações (duração de 1 s cada) do filamento de
von Frey 0,6 g (VFH, Stoelting, Chicago, USA). Os estímulos foram realizados na
superfície plantar da pata traseira direita do animal (QUINTÃO et al., 2005; 2006).
Objetivando determinar o limiar mecânico basal (B), todos os grupos de animais
foram submetidos à avaliação pré-cirúrgica e novamente reavaliados em diferentes
tempos após a APB.
4.4 Avaliação da Atividade Antinociceptiva
4.4.1 Modelo de nocicepção espontânea induzida pela formalina
O procedimento que foi utilizado é similar ao descrito na literatura (MENDES
et al., 2000), sendo composto de duas fases distintas: 1ª fase (resposta fásica 0-5
min) e 2ª fase (resposta tônica 15-30 min). Animais operados e falso-operados, 6
dias e 30 dias após a cirurgia, receberam uma injeção i.pl. de 20 µL de formalina
(0,5 - 2,5 %), na pata traseira direita. Após a injeção de formalina, os animais foram
imediatamente colocados em funis de vidro de 20 cm de diâmetro, e o tempo que o
animal ficou lambendo ou mordendo a pata injetada nas duas fases descritas acima
foi considerado com indicativo de nocicepção.
4.4.2 Modelo de nocicepção espontânea induzida pela capsaicina
Com o objetivo de avaliar o comportamento nociceptivo de animais
submetidos à APB frente à nocicepção neurogênica, camundongos operados e
falso-operados foram submetidos, 6 dias após a cirurgia, ao teste de nocicepção

36
induzida pela capsaicina descrito por Sakurada et al., (1992), com algumas
modificações (CORRÊA et al., 1996). Os animais receberam 20 µL de capsaicina
(0,016 - 1,6 µg/pata), injetado por via i.pl. na pata traseira direita. Imediatamente
após a injeção de capsaicina, os animais foram individualmente colocados em funis
de vidro cilíndricos de 20 cm de diâmetro, e o tempo que o animal lambeu ou
mordeu a pata injetada dentro de um intervalo de 5 min foi considerado como
indicativo de nocicepção.
4.4.3 Modelo de nocicepção espontânea induzida pelo glutamato
Para avaliar o efeito da APB no sistema glutamatérgico, foi utilizado o teste de
nocicepção induzida pelo glutamato descrito por Beirith et al. (2002). Primeiramente,
os animais foram submetidos à APB e transcorridos 6 dias da cirurgia os
camundongos receberam uma injeção i.pl. de 20 µL de glutamato (0,3 - 30
µmol/pata) na para direita traseira. O tempo que o animal ficou lambendo ou
mordendo a pata injetada num intervalo de tempo de 15 min foi considerado como
indicativo de nocicepção.
4.4.4 Modelo de nocicepção espontânea induzida pela bradicinina
Este teste avalia a participação dos receptores B2 para cininas. Os animais
foram submetidos à APB e, após 6 dias receberam uma injeção i.pl. de bradicinina
(BK; 0,1 - 10 nmol/pata) e o tempo que o animal lambeu ou mordeu a pata injetada,
durante 10 min, foi tomado como indicativo de nocicepção (FERREIRA et al., 2004).
4.4.5 Hipernocicepção mecânica induzida pela BK
Os animais foram submetidos à APB 6 dias antes da injeção de 20 µL de BK
(0,1 nmol/pata, i.pl.), e a hipernocicepção mecânica foi avaliada através do
monofilamento de von Frey 0,4 g nos intervalos de tempo de 1/2, 1, 2, 4, 6 e 24 h
após a injeção i.pl. (FERREIRA et al., 2004).

37
4.4.6 Hipernocicepção mecânica induzida pela PGE2
Os animais foram submetidos à APB 6 dias antes da injeção i.pl. de 20 µL de
PGE2 (0,1 nmol/pata, i.pl.), e a hipernocicepção mecânica foi avaliada através do
monofilamento de von Frey 0,4 g nos intervalos de tempo de 1/2, 1, 2, 4, 6 e 24 h
após a injeção i.pl. (KASSUYA et al., 2007).
4.4.7 Hipernocicepção mecânica induzida pela epinefrina
Os animais serão submetidos à APB 6 dias antes da injeção de i.pl. de
Epinefrina (100 ng/pata), e a hipernocicepção mecânica foi avaliada através do
monofilamento de von Frey 0,4 g nos intervalos de tempo de 1/6, 1/2, 1, 2, 4 e 6 h
após a injeção i.pl. (KHASAR et al., 2005).
4.5 Estudo farmacológico
4.5.1 Envolvimento do sistema de cininas
Para o estudo farmacológico foram utilizados animais falso-operados e
animais submetidos à APB. Com base na avaliação das respostas obtidas nos
modelos citados anteriormente foram utilizados antagonistas dos receptores de
cininas [HOE-140, antagonista dos receptores B2 para cininas; DALBK (Des-Arg9-
Leu8-BK) antagonista de receptores B1 frente à nocicepção induzida pela formalina
2,5% ou pela BK (10 nmol/pata) 6 dias após a realização da APB. Em seguida estes
animais foram submetidos a testes de nocicepção aguda.
4.5.2 Envolvimento de componentes inflamatórios e sistema opioide
Para o estudo farmacológico foram utilizados animais falso-operados e
animais submetidos à avulsão do plexo braquial. Com base na avaliação das
respostas obtidas no modelo de nocicepção induzida pela formalina citado

38
anteriormente foram utilizadas drogas de uso clínico, tais como indometacina (10
mg/kg, i.p.), dexametasona (0,5 mg/kg, s.c.) ou morfina (0,5 mg/kg, s.c.). Estes
ensaios tiveram o objetivo de avaliar o papel dos componentes inflamatórios nas
alterações da resposta nociceptiva apresentadas pelos animais submetidos à APB,
bem como investigar o motivo da redução da resposta nociceptiva apresentada
pelos animais operados quando injetados com formalina 2,5 %.
4.6 Drogas e reagentes
As seguintes drogas e reagentes foram utilizados: Formalina (Merck,
Darmstadt, Alemanha); capsaicina, glutamato, bradicinina e dexametasona (Sigma
Chemical Co.;St. Louis, Missouri, EUA); morfina (Dimorf®, Cristália, Itapina, São
Paulo Brasil); indometacina (DEG Importação de Produtos Químicos Ltda, Brasil),
HOE- 140, DALBK e DABK. (Sigma Chemical Co.; St. Louis, Missouri, EUA);
4.7 Análise estatística
Os resultados foram apresentados como a média ± erro padrão da média
(E.P.M., 95%). As porcentagens de inibição foram citadas como a média ± o erro
padrão da média da diferença (em porcentagem) entre as áreas sob as curvas
obtidas para cada experimento individual em relação ao grupo controle
correspondente ou por cada experimento individual. A análise estatística dos dados
foi realizada por meio de análise de variância (ANOVA) de duas vias seguida pelo
teste de Bonferroni, de análise de variância (ANOVA) de uma via seguida pelos
testes de Dunnett ou Newman-Keuls, ou através do teste t de Student, quando
apropriado. Valores de p menores que 0,05 (P < 0,05) foram considerados como
indicativos de significância. Todas as análises citadas acima foram realizadas
utilizando o programa GraphPad PRISM®.
39
5 RESULTADOS
5.1 Resposta nociceptiva induzida pela formalina
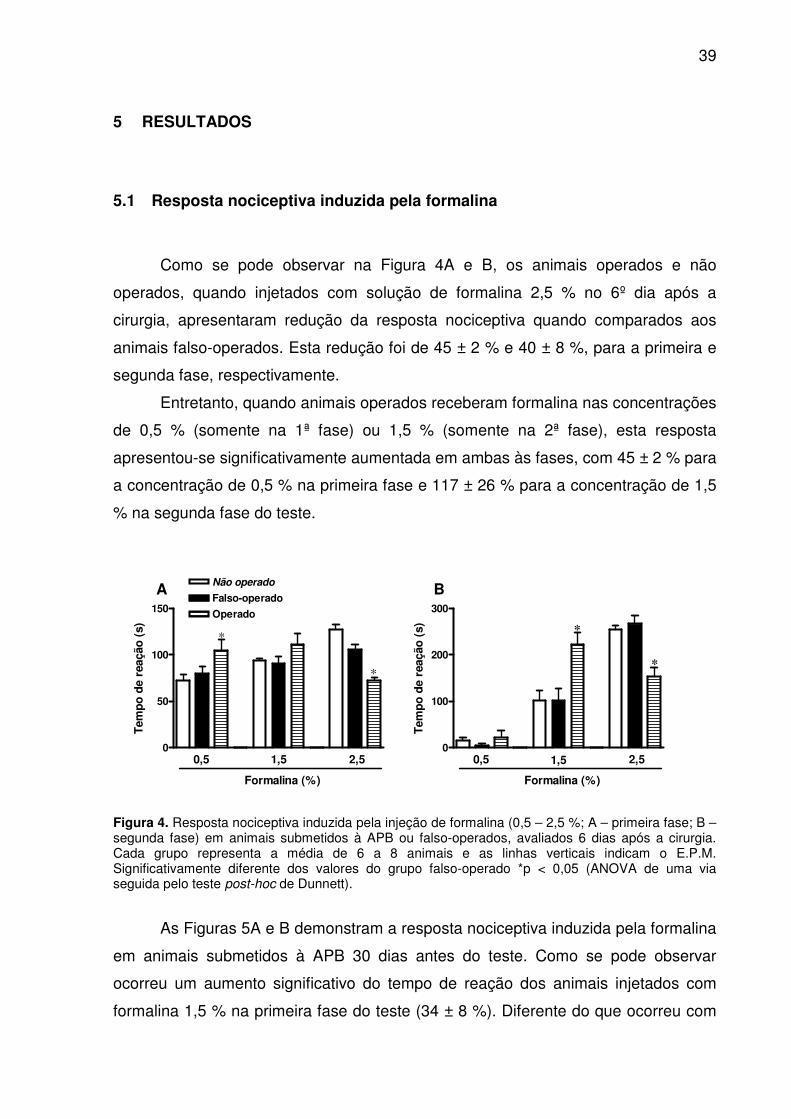
Como se pode observar na Figura 4A e B, os animais operados e não
operados, quando injetados com solução de formalina 2,5 % no 6º dia após a
cirurgia, apresentaram redução da resposta nociceptiva quando comparados aos
animais falso-operados. Esta redução foi de 45 ± 2 % e 40 ± 8 %, para a primeira e
segunda fase, respectivamente.
Entretanto, quando animais operados receberam formalina nas concentrações
de 0,5 % (somente na 1ª fase) ou 1,5 % (somente na 2ª fase), esta resposta
apresentou-se significativamente aumentada em ambas às fases, com 45 ± 2 % para
a concentração de 0,5 % na primeira fase e 117 ± 26 % para a concentração de 1,5
% na segunda fase do teste.
0
50
100
150
Não operado
Falso-operado
Operado
0,5 1,5 2,5
Formalina (%)
*
*
Tem
po
de
reaç
ão (
s)
0
100
200
300
0,5 1,5 2,5
Formalina (%)
*
*
Tem
po
de
reaç
ão (
s)
A B
Figura 4. Resposta nociceptiva induzida pela injeção de formalina (0,5 – 2,5 %; A – primeira fase; B – segunda fase) em animais submetidos à APB ou falso-operados, avaliados 6 dias após a cirurgia. Cada grupo representa a média de 6 a 8 animais e as linhas verticais indicam o E.P.M. Significativamente diferente dos valores do grupo falso-operado *p < 0,05 (ANOVA de uma via seguida pelo teste post-hoc de Dunnett).
As Figuras 5A e B demonstram a resposta nociceptiva induzida pela formalina
em animais submetidos à APB 30 dias antes do teste. Como se pode observar
ocorreu um aumento significativo do tempo de reação dos animais injetados com
formalina 1,5 % na primeira fase do teste (34 ± 8 %). Diferente do que ocorreu com
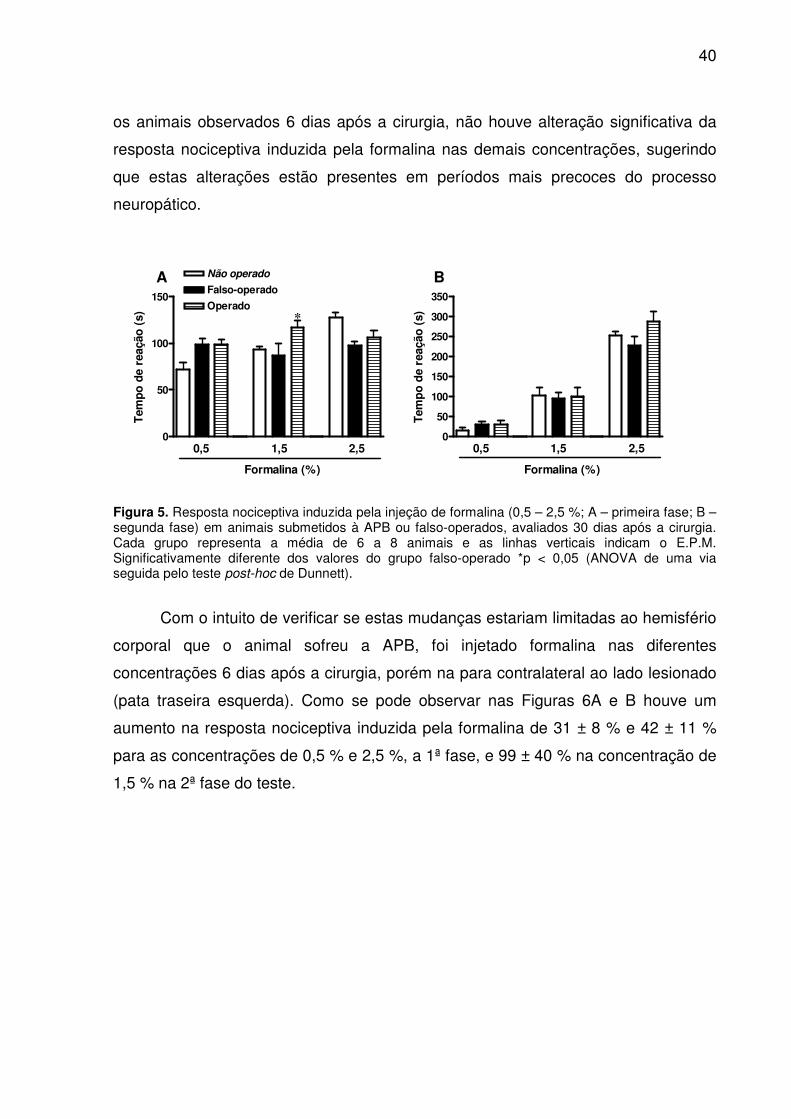
40
os animais observados 6 dias após a cirurgia, não houve alteração significativa da
resposta nociceptiva induzida pela formalina nas demais concentrações, sugerindo
que estas alterações estão presentes em períodos mais precoces do processo
neuropático.
0
50
100
150
Não operado
Falso-operado
Operado
0,5 1,5 2,5
Formalina (%)
*
Tem
po
de
reaç
ão (
s)
0
50
100
150
200
250
300
350
0,5 1,5 2,5
Formalina (%)
Tem
po
de
reaç
ão (
s)
A B
Figura 5. Resposta nociceptiva induzida pela injeção de formalina (0,5 – 2,5 %; A – primeira fase; B – segunda fase) em animais submetidos à APB ou falso-operados, avaliados 30 dias após a cirurgia. Cada grupo representa a média de 6 a 8 animais e as linhas verticais indicam o E.P.M. Significativamente diferente dos valores do grupo falso-operado *p < 0,05 (ANOVA de uma via seguida pelo teste post-hoc de Dunnett).
Com o intuito de verificar se estas mudanças estariam limitadas ao hemisfério
corporal que o animal sofreu a APB, foi injetado formalina nas diferentes
concentrações 6 dias após a cirurgia, porém na para contralateral ao lado lesionado
(pata traseira esquerda). Como se pode observar nas Figuras 6A e B houve um
aumento na resposta nociceptiva induzida pela formalina de 31 ± 8 % e 42 ± 11 %
para as concentrações de 0,5 % e 2,5 %, a 1ª fase, e 99 ± 40 % na concentração de
1,5 % na 2ª fase do teste.
41
0
50
100
150
Não operado
Falso-operado
Operado
0,5 1,5 2,5
Formalina (%)
**
Tem
po
de
reaç
ão (
s)
0
50
100
150
200
250
300
350
0,5 1,5 2,5
Formalina (%)
*
Tem
po
de
reaç
ão (
s)
A B
Figura 6. Resposta nociceptiva induzida pela injeção de formalina (0,5 – 2,5 %; A – primeira fase; B – segunda fase) na pata contralateral de animais submetidos à APB ou falso-operados, avaliados 6 dias após a cirurgia. Cada grupo representa a média de 6 a 8 animais e as linhas verticais indicam o E.P.M. Significativamente diferente dos valores do grupo falso-operado *p < 0,05 (ANOVA de uma via seguida pelo teste post-hoc de Dunnett).
Para avaliar a participação dos receptores B2 de cininas, envolvido no
processo doloroso da APB, um antagonista sintético e seletivo de receptores B2,
chamado de HOE-140 ou icatibante foi administrado aos animais submetidos APB,
30 dias após a cirurgia. Na Figura 7A e B, pode-se observar que os animais,
avaliados 6 dias após a cirurgia, pré-tratados uma hora antes dos experimentos com
HOE-140 (50 nmol/kg, s.c.), apresentaram um aumento significativo da resposta
nociceptiva induzida pela injeção i.pl. de formalina 2,5 %, na segunda fase do teste
(139 ± 52 %), quando comparados com os animais operados que receberam salina.
42
sham salOp sal hoe shamhoe op0
100
200
Salina (10 mL/kg, s.c.)Hoe-140 (50 nmol/kg, s.c.)
+-
+-
-+
-+
FOP FOPOP OP
Formalina (2,5%)
Lic
kin
g (
s)
sham salOp sal hoe shamhoe op0
100
200
##
+-
+-
-+
-+
FOP FOPOP OP
Formalina (2,5%)
Lic
kin
g (
s)
A B
Figura 7. Resposta nociceptiva induzida pela injeção de formalina (2,5 %; A – primeira fase; B – segunda fase) em animais submetidos à APB ou falso-operados, avaliados 6 dias após a cirurgia e pré-tratados com HOE-140 (50 nmol/kg s.c.). Cada grupo representa a média de 6 a 8 animais e as linhas verticais indicam o E.P.M. Significativamente diferente dos valores do grupo operado tratado com salina ##p < 0,05 (ANOVA de uma via seguida pelo teste post-hoc de Dunnett). FOP, falso-operado; OP operado.
Para se avaliar o envolvimento dos receptores B1 de cinina, o antagonista
seletivo para receptores B1 DALBK (des-Arg9-Leu8-BK) foi administrado aos animais
submetidos APB, com 6 dias de cirurgia. As Figuras 8A e B demonstram o efeito da
administração de DALBK (150 nmol/kg, s.c.) uma hora antes do teste de nocicepção
induzida por formalina (2,5 %). Na primeira fase, os animais operados não
apresentaram significativa alteração da resposta nociceptiva. Na segunda fase do
teste, os animais operados tratados com DALBK apresentaram um aumento na
resposta nociceptiva de 170 ± 49 % quando comparados com os animais operados
tratados com salina (Figura 8).
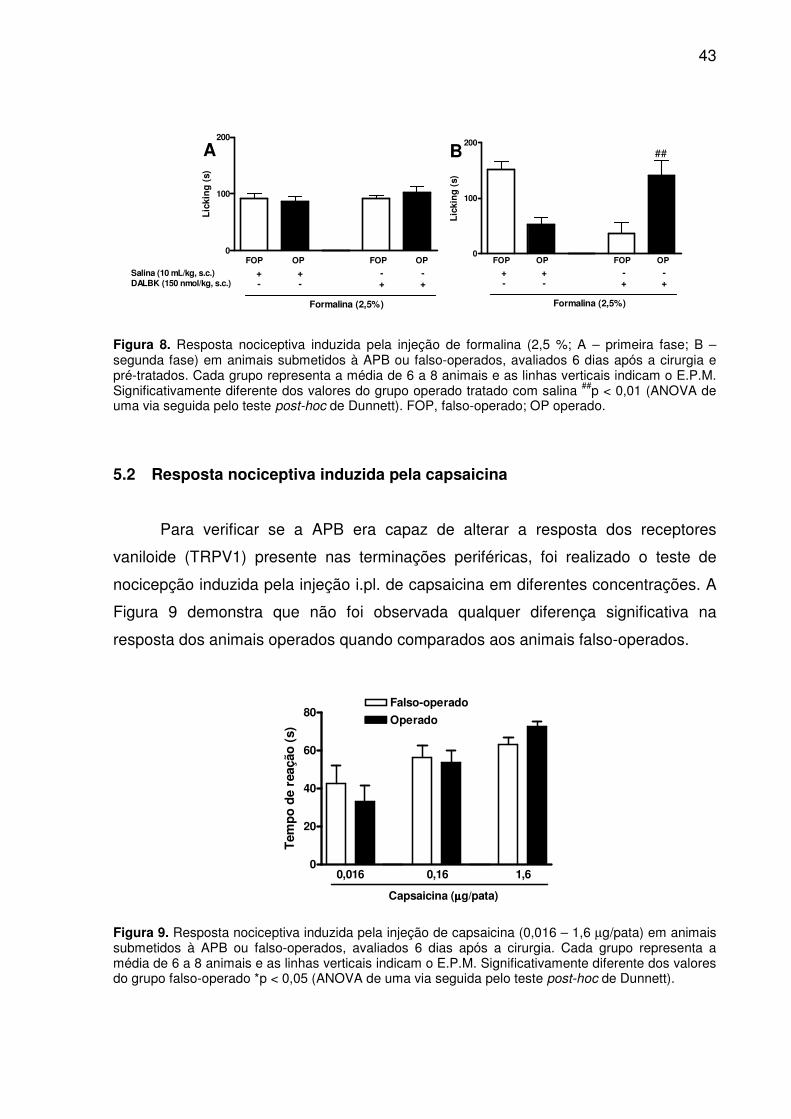
43
Sal Sal DALBK DALBK 0
100
200
Salina (10 mL/kg, s.c.)DALBK (150 nmol/kg, s.c.)
+-
+-
-+
-+
FOP FOPOP OP
Formalina (2,5%)
Lic
kin
g (
s)
sham sal Op sal DALBK hoe op0
100
200##
+-
+-
-+
-+
FOP FOPOP OP
Formalina (2,5%)
Lic
kin
g (
s)
A B
Figura 8. Resposta nociceptiva induzida pela injeção de formalina (2,5 %; A – primeira fase; B – segunda fase) em animais submetidos à APB ou falso-operados, avaliados 6 dias após a cirurgia e pré-tratados. Cada grupo representa a média de 6 a 8 animais e as linhas verticais indicam o E.P.M. Significativamente diferente dos valores do grupo operado tratado com salina ##p < 0,01 (ANOVA de uma via seguida pelo teste post-hoc de Dunnett). FOP, falso-operado; OP operado.
5.2 Resposta nociceptiva induzida pela capsaicina
Para verificar se a APB era capaz de alterar a resposta dos receptores
vaniloide (TRPV1) presente nas terminações periféricas, foi realizado o teste de
nocicepção induzida pela injeção i.pl. de capsaicina em diferentes concentrações. A
Figura 9 demonstra que não foi observada qualquer diferença significativa na
resposta dos animais operados quando comparados aos animais falso-operados.
0
20
40
60
80
0,016 1,6
Capsaicina (µµµµg/pata)
Falso-operado
Operado
0,16
Tem
po
de
reaç
ão (
s)
Figura 9. Resposta nociceptiva induzida pela injeção de capsaicina (0,016 – 1,6 µg/pata) em animais submetidos à APB ou falso-operados, avaliados 6 dias após a cirurgia. Cada grupo representa a média de 6 a 8 animais e as linhas verticais indicam o E.P.M. Significativamente diferente dos valores do grupo falso-operado *p < 0,05 (ANOVA de uma via seguida pelo teste post-hoc de Dunnett).
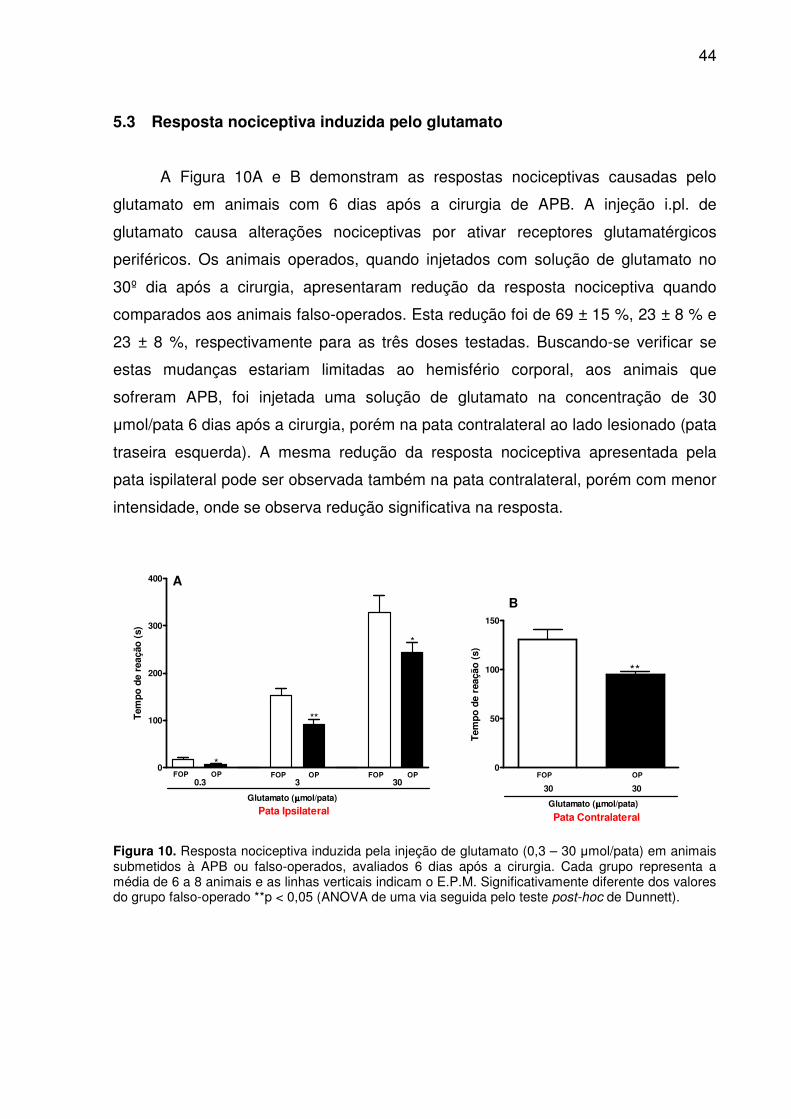
44
5.3 Resposta nociceptiva induzida pelo glutamato
A Figura 10A e B demonstram as respostas nociceptivas causadas pelo
glutamato em animais com 6 dias após a cirurgia de APB. A injeção i.pl. de
glutamato causa alterações nociceptivas por ativar receptores glutamatérgicos
periféricos. Os animais operados, quando injetados com solução de glutamato no
30º dia após a cirurgia, apresentaram redução da resposta nociceptiva quando
comparados aos animais falso-operados. Esta redução foi de 69 ± 15 %, 23 ± 8 % e
23 ± 8 %, respectivamente para as três doses testadas. Buscando-se verificar se
estas mudanças estariam limitadas ao hemisfério corporal, aos animais que
sofreram APB, foi injetada uma solução de glutamato na concentração de 30
µmol/pata 6 dias após a cirurgia, porém na pata contralateral ao lado lesionado (pata
traseira esquerda). A mesma redução da resposta nociceptiva apresentada pela
pata ispilateral pode ser observada também na pata contralateral, porém com menor
intensidade, onde se observa redução significativa na resposta.
0
100
200
300
400
0.3 30
Glutamato (µµµµmol/pata)
**
*
*
3FOP OP FOP OP FOP OP
Tem
po
de
reaç
ão (
s)
Sham OP0
50
100
150
**
30 30
FOP OP
Glutamato (µµµµmol/pata)
Tem
po
de
reaç
ão (
s)
Pata IpsilateralPata Contralateral
A
B
Figura 10. Resposta nociceptiva induzida pela injeção de glutamato (0,3 – 30 µmol/pata) em animais submetidos à APB ou falso-operados, avaliados 6 dias após a cirurgia. Cada grupo representa a média de 6 a 8 animais e as linhas verticais indicam o E.P.M. Significativamente diferente dos valores do grupo falso-operado **p < 0,05 (ANOVA de uma via seguida pelo teste post-hoc de Dunnett).
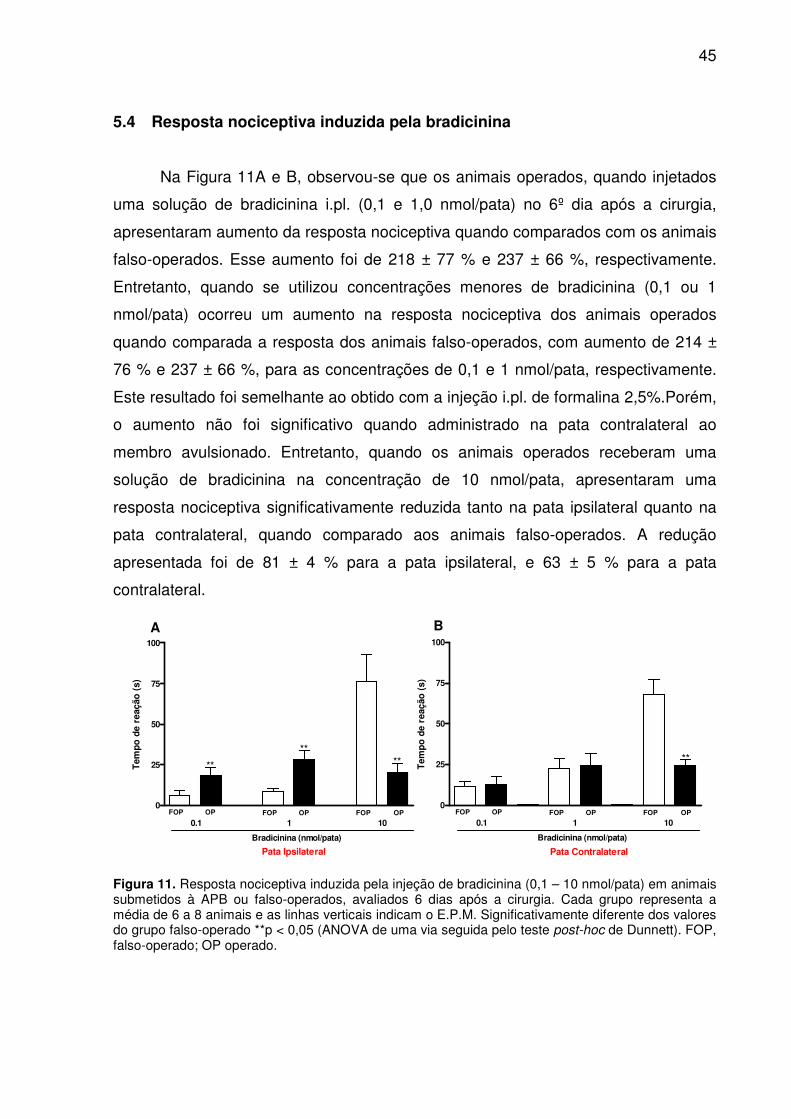
45
5.4 Resposta nociceptiva induzida pela bradicinina
Na Figura 11A e B, observou-se que os animais operados, quando injetados
uma solução de bradicinina i.pl. (0,1 e 1,0 nmol/pata) no 6º dia após a cirurgia,
apresentaram aumento da resposta nociceptiva quando comparados com os animais
falso-operados. Esse aumento foi de 218 ± 77 % e 237 ± 66 %, respectivamente.
Entretanto, quando se utilizou concentrações menores de bradicinina (0,1 ou 1
nmol/pata) ocorreu um aumento na resposta nociceptiva dos animais operados
quando comparada a resposta dos animais falso-operados, com aumento de 214 ±
76 % e 237 ± 66 %, para as concentrações de 0,1 e 1 nmol/pata, respectivamente.
Este resultado foi semelhante ao obtido com a injeção i.pl. de formalina 2,5%.Porém,
o aumento não foi significativo quando administrado na pata contralateral ao
membro avulsionado. Entretanto, quando os animais operados receberam uma
solução de bradicinina na concentração de 10 nmol/pata, apresentaram uma
resposta nociceptiva significativamente reduzida tanto na pata ipsilateral quanto na
pata contralateral, quando comparado aos animais falso-operados. A redução
apresentada foi de 81 ± 4 % para a pata ipsilateral, e 63 ± 5 % para a pata
contralateral.
0
25
50
75
100
0.1 1 10
Bradicinina (nmol/pata)
****
**
FOP OP FOP OP FOP OP
Tem
po
de
reaç
ão (
s)
0
25
50
75
100
**
0.1 1 10
Bradicinina (nmol/pata)
FOP OP FOP OP FOP OP
Tem
po
de
reaç
ão (
s)
Pata Ipsilateral Pata Contralateral
A B
Figura 11. Resposta nociceptiva induzida pela injeção de bradicinina (0,1 – 10 nmol/pata) em animais submetidos à APB ou falso-operados, avaliados 6 dias após a cirurgia. Cada grupo representa a média de 6 a 8 animais e as linhas verticais indicam o E.P.M. Significativamente diferente dos valores do grupo falso-operado **p < 0,05 (ANOVA de uma via seguida pelo teste post-hoc de Dunnett). FOP, falso-operado; OP operado.

46
Ainda, buscando-se avaliar a participação dos receptores B2 de cinina, o
antagonista seletivo de receptores B2, HOE-140, foi administrado aos animais
submetidos APB, com 6 dias de cirurgia, uma hora antes do teste de nocicepção
induzida por bradicinina. A Figura 12 apresenta os resultados observados no pré-
tratamento dos animais com HOE-140 (50 nmol/kg), e submetidos à nocicepção
induzida por bradicinina (10 nmol/pata) no 6º dia após a cirurgia de APB. Quando
injetada a bradicinina, tanto os animais operados quanto falso-operados pré-tratados
com HOE-140 apresentaram completa redução da resposta nociceptiva, quando
comparados com os animais tratados com salina.
0
20
40
60
80
100
120
#
** **
Salina (10 mL/kg, s.c.)Hoe-140 (50 nmol/kg, s.c.)
+-
+-
-+
-+
Bradicinina (10 nmol/pata)
FOP FOPOP OP
Tem
po
de
reaç
ão (
s)
Figura 12. Resposta nociceptiva induzida pela injeção de BK (10 nmol/pata) em animais submetidos à APB ou falso-operados, avaliados 6 dias após a cirurgia. Cada grupo representa a média de 6 a 8 animais e as linhas verticais indicam o E.P.M. Significativamente diferente dos valores do grupo falso-operado **p < 0,05 (ANOVA de uma via seguida pelo teste post-hoc de Dunnett). FOP, falso-operado; OP operado.
Como se pode observar na Figura 13, a injeção i.pl. de DABK, em todas as
concentrações testadas não foi capaz de evocar comportamento nociceptivo, tanto
quando injetada na para ipsilateral quanto contralateral em animais submetidos à
cirurgia (6º ou 30º dia PO – dado não mostrado).
47
1 1 3 3 10 100
5
10
15
DABK (nmol/pata)
FOP FOP FOPOP OP OP
1 2 3
Tem
po
de
reaç
ão(s
)
1 1 3 3 10 100
1
2
3
4
5
6
DABK (nmol/pata)
FOP FOP FOPOP OP OP
1 2 3
Tem
po
de
reaç
ão (
s)
Pata Ipsilateral Pata Contralateral
A B
Figura 13. Resposta nociceptiva induzida pela injeção de bradicinina (1 – 10 nmol/pata) em animais submetidos à APB ou falso-operados, avaliados 6 dias após a cirurgia. Cada grupo representa a média de 6 a 8 animais e as linhas verticais indicam o E.P.M. Significativamente diferente dos valores do grupo falso-operado **p < 0,05 (ANOVA de uma via seguida pelo teste post-hoc de Dunnett). FOP, falso-operado; OP operado.
5.5 Participação de componentes inflamatórios na alteração da resposta
nociceptiva de animais submetidos à APB
Na tentativa de determinar os possíveis mecanismos de ação e quais os
processos inflamatórios envolvidos no aumento da resposta nociceptiva presente
nos animais operados injetados com formalina 1,5 %, foi realizado o pré-tratamento
destes animais com dois anti-inflamatórios clássicos, a indometacina (inibidor não
seletivo da enzima COX1 e COX2) ou dexametasona (glicocorticóide, inibidor de
síntese proteica). Os dados apresentados na Figura 14 demonstra que o pré-
tratamento com indometacina (10 mg/kg, i.p.) 30 min antes da administração da
formalina, foi capaz de reverter significativamente o aumento da resposta
nociceptiva apresentado pelos animais operados, quando comparado aos animais
falso-operados.
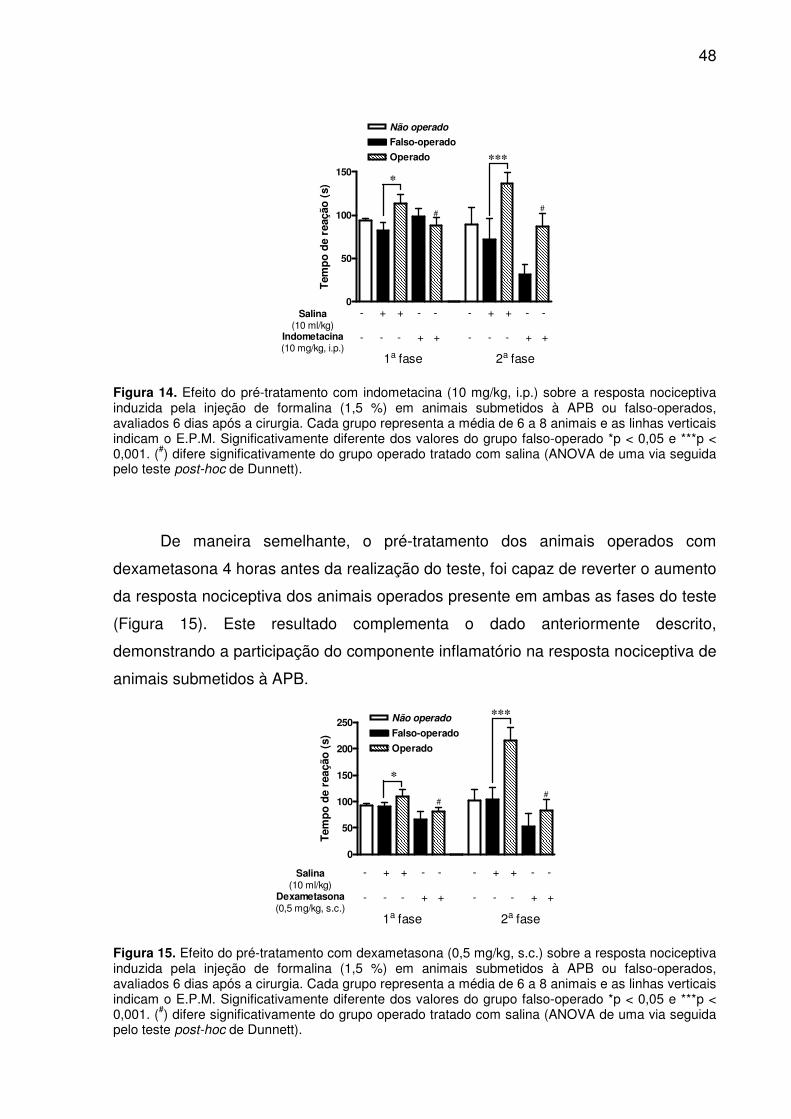
48
0
50
100
150
Não operado
Falso-operado
Operado
*
***
##
2a fase
Salina(10 ml/kg)
Indometacina(10 mg/kg, i.p.)
-
-
+
-
+
-
-
+
-
+
-
-
+
-
+
-
-
+
-
+
1a fase
Tem
po
de
reaç
ão (
s)
Figura 14. Efeito do pré-tratamento com indometacina (10 mg/kg, i.p.) sobre a resposta nociceptiva induzida pela injeção de formalina (1,5 %) em animais submetidos à APB ou falso-operados, avaliados 6 dias após a cirurgia. Cada grupo representa a média de 6 a 8 animais e as linhas verticais indicam o E.P.M. Significativamente diferente dos valores do grupo falso-operado *p < 0,05 e ***p < 0,001. (#) difere significativamente do grupo operado tratado com salina (ANOVA de uma via seguida pelo teste post-hoc de Dunnett).
De maneira semelhante, o pré-tratamento dos animais operados com
dexametasona 4 horas antes da realização do teste, foi capaz de reverter o aumento
da resposta nociceptiva dos animais operados presente em ambas as fases do teste
(Figura 15). Este resultado complementa o dado anteriormente descrito,
demonstrando a participação do componente inflamatório na resposta nociceptiva de
animais submetidos à APB.
0
50
100
150
200
250 Não operado
Falso-operado
Operado
*
***
#
1a fase 2a fase
Salina(10 ml/kg)
Dexametasona(0,5 mg/kg, s.c.)
-
-
+
-
+
-
-
+
-
+
-
-
+
-
+
-
-
+
-
+
#
Tem
po
de
reaç
ão (
s)
Figura 15. Efeito do pré-tratamento com dexametasona (0,5 mg/kg, s.c.) sobre a resposta nociceptiva induzida pela injeção de formalina (1,5 %) em animais submetidos à APB ou falso-operados, avaliados 6 dias após a cirurgia. Cada grupo representa a média de 6 a 8 animais e as linhas verticais indicam o E.P.M. Significativamente diferente dos valores do grupo falso-operado *p < 0,05 e ***p < 0,001. (#) difere significativamente do grupo operado tratado com salina (ANOVA de uma via seguida pelo teste post-hoc de Dunnett).
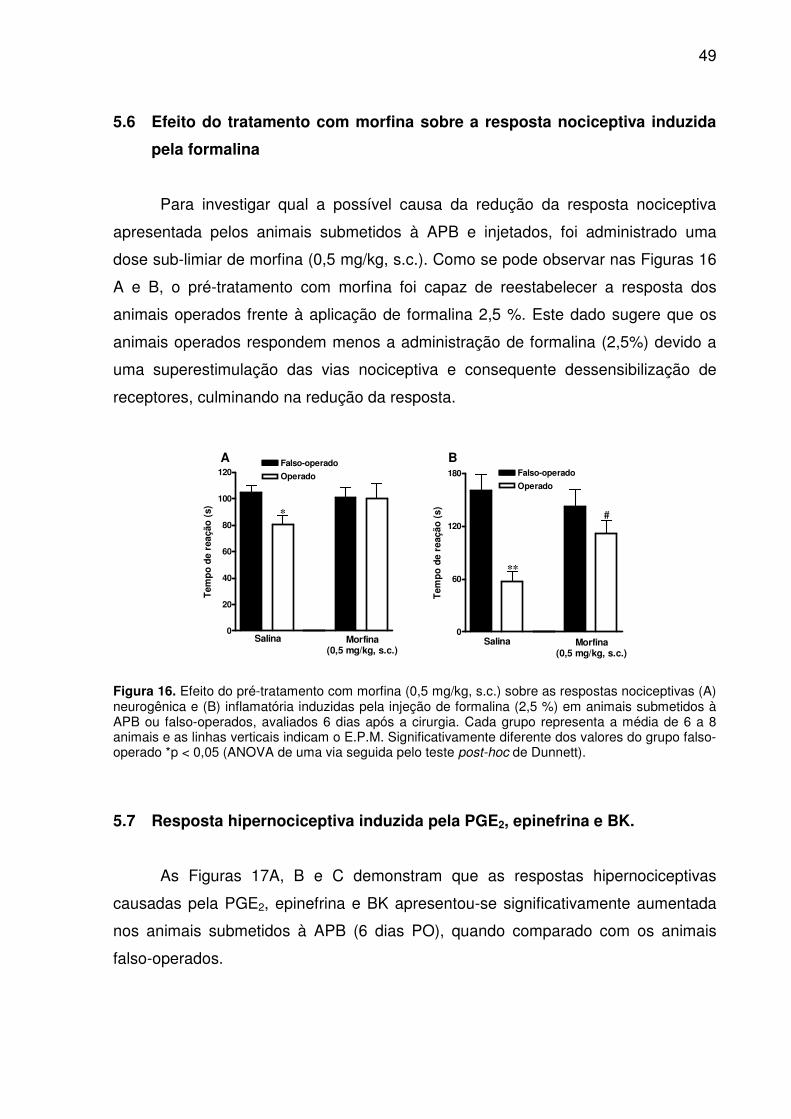
49
5.6 Efeito do tratamento com morfina sobre a resposta nociceptiva induzida
pela formalina
Para investigar qual a possível causa da redução da resposta nociceptiva
apresentada pelos animais submetidos à APB e injetados, foi administrado uma
dose sub-limiar de morfina (0,5 mg/kg, s.c.). Como se pode observar nas Figuras 16
A e B, o pré-tratamento com morfina foi capaz de reestabelecer a resposta dos
animais operados frente à aplicação de formalina 2,5 %. Este dado sugere que os
animais operados respondem menos a administração de formalina (2,5%) devido a
uma superestimulação das vias nociceptiva e consequente dessensibilização de
receptores, culminando na redução da resposta.
0
20
40
60
80
100
120
*
Salina Morfina(0,5 mg/kg, s.c.)
Falso-operado
Operado
Tem
po
de
reaç
ão (
s)
0
60
120
180
**
#
Salina Morfina(0,5 mg/kg, s.c.)
Falso-operado
Operado
Tem
po
de
reaç
ão (
s)
A B
Figura 16. Efeito do pré-tratamento com morfina (0,5 mg/kg, s.c.) sobre as respostas nociceptivas (A) neurogênica e (B) inflamatória induzidas pela injeção de formalina (2,5 %) em animais submetidos à APB ou falso-operados, avaliados 6 dias após a cirurgia. Cada grupo representa a média de 6 a 8 animais e as linhas verticais indicam o E.P.M. Significativamente diferente dos valores do grupo falso-operado *p < 0,05 (ANOVA de uma via seguida pelo teste post-hoc de Dunnett).
5.7 Resposta hipernociceptiva induzida pela PGE2, epinefrina e BK.
As Figuras 17A, B e C demonstram que as respostas hipernociceptivas
causadas pela PGE2, epinefrina e BK apresentou-se significativamente aumentada
nos animais submetidos à APB (6 dias PO), quando comparado com os animais
falso-operados.
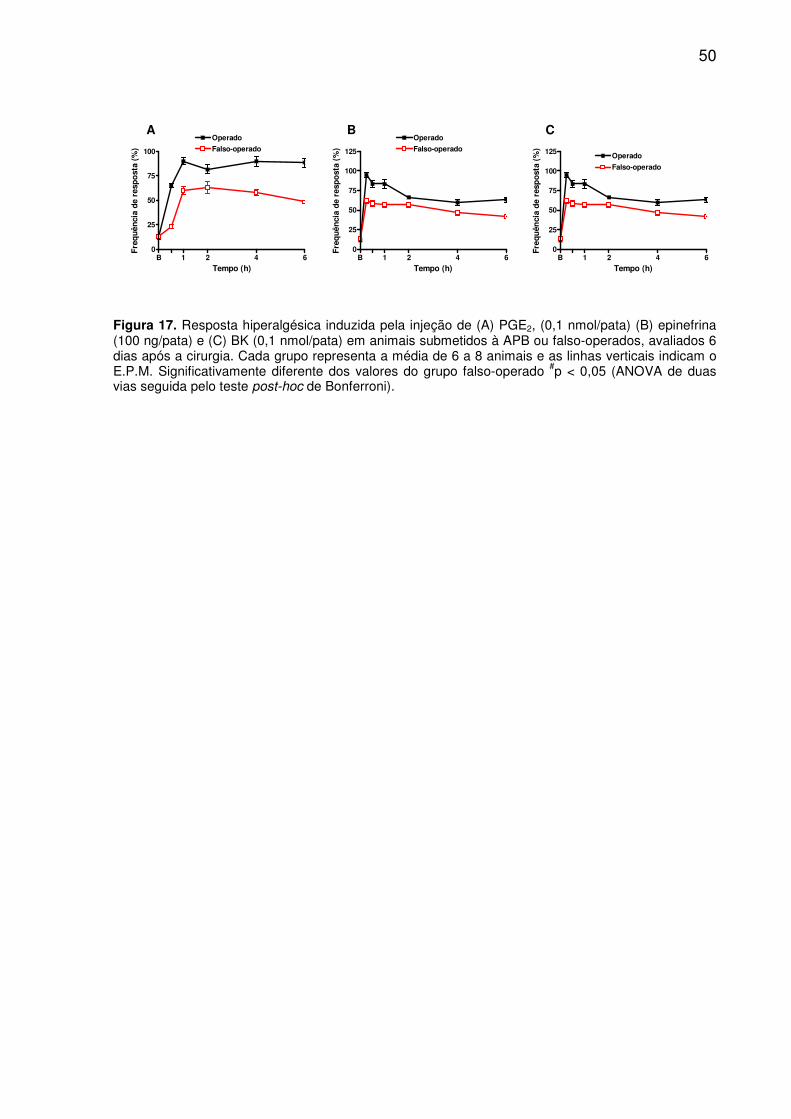
50
B 1 2 4 60
25
50
75
100
Operado
Falso-operado
Tempo (h)
Fre
qu
ênci
a d
e re
spo
sta
(%)
B 1 2 4 60
25
50
75
100
125
Operado
Falso-operado
Tempo (h)
Fre
qu
ênci
a d
e re
spo
sta
(%)
B 1 2 4 60
25
50
75
100
125Operado
Falso-operado
Tempo (h)
Fre
qu
ênci
a d
e re
spo
sta
(%)
A B C
Figura 17. Resposta hiperalgésica induzida pela injeção de (A) PGE2, (0,1 nmol/pata) (B) epinefrina (100 ng/pata) e (C) BK (0,1 nmol/pata) em animais submetidos à APB ou falso-operados, avaliados 6 dias após a cirurgia. Cada grupo representa a média de 6 a 8 animais e as linhas verticais indicam o E.P.M. Significativamente diferente dos valores do grupo falso-operado #p < 0,05 (ANOVA de duas vias seguida pelo teste post-hoc de Bonferroni).

51
6 DISCUSSÃO
A lesão do plexo braquial é causada pela avulsão da raiz medular, produzindo
dor característica constante, intermitente e frequentemente intratável (BERMAN,
BIRCH, ANAND et al., 1998; ANAND; BIRCH, 2002). A dor geralmente acomete a
projeção do ombro e região supra-escapular com irradiação para o membro superior
ipsilateral, dependendo da intensidade e da região do plexo lesionado. A APB foi
primeiramente descrita em ratos como um novo modelo de dor neuropática
persistente (RODRIGUES-FILHO et al., 2003). Recentemente, este modelo foi
adaptado para camundongos (QUINTÃO et al., 2006). Na primeira semana após
uma lesão nervosa periférica, o processo de reorganização dentro dos neurônios
sensoriais lesionados produz um aumento pronunciado no transporte axonal
retrógrado de pequenas proteínas, tais como citocinas e fatores neurotróficos
(CAHILLl et al., 2003). Por outro lado, os fatores neurotróficos são capazes de
modular a expressão de outras proteínas, tais como os receptores B1 (induzido) e B2
(constitutivo) para cininas (LEE et al., 2002).
A dor crônica, diferente da dor aguda, é caracterizada por modificar a relação
entre os estímulos e respostas nociceptivas. Ocorrem alterações neuroquímicas
centrais, resultando em hipersensibilidade, aumento da frequência e duração dos
impulsos aferentes, além de mudanças estruturais secundárias a lesão nervosa
periférica, como a perda de interneurônios espinhais, rearranjos nos processos
neurais aferentes da medula espinhal e proliferação de fibras simpáticas no gânglio
sensorial (WATKINS; MAIER, 2002; DOGRUL et al., 2003).
O modelo de nocicepção induzida por formalina foi descrito por Dubuisson e
Dennis (1977) em ratos e gatos, e tem sido utilizado para o estudo de drogas
analgésicas em roedores, principalmente em camundongos. Este teste permite
avaliar duas fases distintas da sensibilização dolorosa: a primeira fase, que ocorre
nos primeiros 5 min após a injeção de formalina (resposta nociceptiva neurogênica),
caracterizada pela estimulação direta dos nociceptores presentes nas fibras
aferentes tipo C, e, em parte, fibras Aδ (liberação de glutamato e substancia P). A
segunda fase da nocicepção, que ocorre entre 15 e 30 min após à injeção de
formalina (resposta nociceptiva inflamatória), está relacionada com a liberação de
mediadores pró-inflamatórios tais como adenosina, bradicinina, histamina,
prostaglandina e serotonina (REEVE, DICKENSON, 1995). Estudo realizado por

52
Hunskaar e Hole (1987) demonstrou que doses crescentes de formalina, acima de
3%, induzem a uma redução da resposta nociceptiva dos animais, e não ao
aumento.
O passo seguinte foi verificar se concentrações menores de formalina (0,5 %
e 1,5 %) produziriam a mesma resposta da dose maior (2,5 %) em animais
operados. Nas menores doses, a resposta nociceptiva foi maior nos animais
operados do que nos animais falso-operados, sugerindo que a hipótese de hiper-
estimulação na dose de 2,5 % possa estar correta. Os mesmos testes foram
realizados na pata traseira esquerda de animais submetidos à APB (pata
contralateral a cirurgia), e foi observado um aumento da resposta nociceptiva em
todas as dores testadas. O comportamento de freezing não foi observado em
nenhuma das concentrações testadas, sugerindo que as alterações sensoriais
decorrentes da lesão nervosa na periferia unilateral são distintas quando
comparados os dois hemisférios corporais. Quando animais submetidos à APB
foram avaliados 30 dias após a cirurgia não foi observada alteração significativa.
A persistência do quadro nociceptivo resulta em alterações centrais crônicas
na neuroquímica da sinalização da dor produzindo hipersensibilidade, aumento da
frequência e a duração dos impulsos aferentes. Estas alterações estruturais
secundárias à lesão nervosa periférica englobam a perda de interneurônios
espinhais, rearranjos não apropriados de processos neurais aferentes na medula
espinhal e a proliferação de fibras simpáticas no gânglio sensorial. (WATKINS;
MAIER, 2002; DOGRUL et al., 2003).
Dados da literatura demonstram que os mecanismos envolvidos no controle
da dor aguda e crônica são bem distintos e que há um balanço diferente entre a
facilitação e a inibição descendentes da dor nos neurônios espinhais e periféricos,
contribuindo para um balanço distinto de receptores e mediadores envolvidos na dor
aguda e crônica (YASHPAL et al., 2001; ZHAO et al., 2006; HÖSL et a., 2006).
O modelo de nocicepção induzida por capsaicina demonstra o envolvimento
dos componentes neurogênicos na dor. A capsaicina, quando injetada na pata de
camundongos, é capaz de ativar uma subpopulação distinta de fibras aferentes
primárias, as quais transmitem informações nociceptivas ao SNC e induzem a
liberação de neuropeptídeos pró-inflamatórios (HOLZER, 1991). Em 1992, Sakurada
e colaboradores demonstraram que a injeção i.pl. de capsaicina na pata traseira de
camundongos induz um profundo comportamento nociceptivo, caracterizado pela

53
mordida e lambida da pata injetada, relacionado à dor neurogênica. Sabe-se que há
a liberação de alguns mediadores químicos, tais como aminoácidos excitatórios,
substancia P, cininas, CGRP, PGE2 e NO, que contribuem para o aumento do
processo doloroso (WU et al., 1998). Nossos resultados sugerem que a APB não
induz a uma alteração na resposta dos receptores vanilóides periféricos (receptores
TRPV1) frente à injeção de capsaicina, mesmo reduzindo a concentração da
mesma.
Atualmente, sabe-se que dentre os diferentes neurotransmissores envolvidos
nos modelos de nocicepção anteriormente citados, os aminoácidos excitatórios (AAE
- glutamato e aspartato) têm um papel relevante. Perifericamente, demonstra-se o
importante papel fisiológico do glutamato. Alguns pesquisadores têm demonstrado a
intensa liberação de AAE na medula espinhal, principalmente glutamato e aspartato,
após a injeção i.pl. de formalina e capsaicina (OMOTE et al., 1998). Os dados
apresentados neste estudo demonstram que animais submetidos à APB respondem
menos à injeção i.pl. de glutamato do que animais falso-operados, sugerindo que
receptores glutamatérgicos possam estar com sua expressão reduzida ou que a
resposta nociceptiva induzida pelo glutamato possa estar sendo bloqueada por
algum mecanismo central de controle da dor.
As cininas constituem uma importante família de peptídeos biologicamente
ativos envolvidos em diversos efeitos biológicos incluindo a homeostasia
cardiovascular, inflamação e nocicepção (CALIXTO et al., 2000; 2001; 2004;
CAMPOS et al., 2006; LEEB-LUNDBERG et al., 2005; MOREAU et al., 2005). Estes
peptídeos estão entre os primeiros mediadores a serem liberados nos tecidos
lesionados, sendo originados a partir da clivagem dos cininogênios (de alto e baixo
peso molecular) pela calicreína plasmática, a qual é ativada precocemente na
cascata de coagulação ou, pela calicreína tecidual, que por sua vez é ativada por
proteases liberadas no sítio da lesão (REGOLI, BARABÉ, 1980; VAVREK,
STEWART, 1985; MURRAY et al., 1990; REGOLI et al., 1990; BEAUBIEN et al.,
1991; BHOOLA et al., 1992; REGOLI et al., 1994; HALL, MORTON, 1997).
As cininas desempenham seus efeitos através da ativação de dois receptores
de membrana específicos, denominados B1 e B2. A classificação dos dois subtipos
de receptores teve origem em estudos farmacológicos conduzidos no final da
década de 70, que determinaram a ordem de potência dos agonistas cininérgicos
em preparações de órgãos isolados (REGOLI, BARABÉ, 1980). Assim, os

54
receptores do tipo B1 são ativados preferencialmente pela des-Arg9-BK e pela Lys-
des-Arg9-BK, enquanto que os receptores B2 são praticamente insensíveis a estes
metabólitos. Por outro lado, os receptores B2 apresentam alta afinidade pela BK e
pela Lys-BK, ao passo que os peptídeos produzidos pela ação da cininase I são
praticamente inativos neste receptor (MOREAU et al., 2005).
O papel das cininas nas respostas nociceptivas em animais já está bem
estabelecido, há mais ou menos duas décadas (CALIXTO et al., 2000; 2001; 2004;
CAMPOS et al., 2006; LEEB-LUNDBERG et al., 2005). Atualmente, a ativação do
receptor B2 tem sido relacionada com a fase inicial da dor inflamatória, enquanto a
estimulação do receptor B1 estaria associada com processos patológicos mais
persistentes. De fato, o aumento nos níveis de RNAm para o receptor B1, ou da
proteína, foi descrito no DRG após lesão por constrição do nervo ciático em ratos e
camundongos (PETERSEN et al., 1998; ECKERT et al., 1999; LEVY, ZOCHODNE,
2000; YAMAGUCHI-SASE et al., 2003; RASHID et al., 2004).
Walker et al. (1995) já afirmavam a existência de componentes do sistema
calicreína-cininas no SNC, em regiões como córtex, tronco encefálico, cerebelo e
hipotálamo, o que seria condizente com a atuação das cininas no controle central da
dor. Foram evidenciadas também, concentrações razoáveis de BK na medula,
cerebelo e córtex, bem como uma densidade moderada de sítios de ligação para a
BK no hipocampo (PERRY, SNYDER, 1984; KARIYA et al., 1985; FUJIWARA et al.,
1989; COUTURE, LINDSEY, 2000). Petersen e colaboradores (1998) demonstraram
que após a lesão parcial ou axotomia do nervo ciático ocorrem alterações na
expressão de neuropeptídeos e de receptores para cininas em neurônios do DRG.
Ademais, em estudos in vitro, o aumento na expressão de receptores B1 e B2 para
cininas parece estar relacionado com a interação da neurotrofina NGF com o
receptor p75NTR (PETERSEN et al., 1996). Todas estas alterações ocorrem tanto
no lado ipsilateral à lesão, como também no lado contralateral. Estas mudanças
parecem ter como pré-requisito a lesão nervosa, uma vez que em animais falso-
operados não ocorre aumento na expressão destes receptores (PETERSEN et al.,
1998). Levy e Zochodne (2000) demonstraram que a lesão nervosa periférica está
associada com o aumento na expressão do RNAm para os receptores B1 e B2 nos
DRG, apesar da existência de receptores de forma constitutiva. Ainda, os mesmos
autores relataram o aumento da expressão destes receptores no lado contralateral à
lesão.

55
Estes dados, juntamente com evidências comportamentais, permitem sugerir
que os receptores cininérgicos estão envolvidos no desenvolvimento e na
manutenção da hipersensibilidade térmica após a lesão nervosa. Mais
recentemente, Rashid e colaboradores (2004) demonstraram a ocorrência da
síntese de novo dos receptores B1 para cininas após a lesão parcial do nervo ciático,
principalmente em neurônios mielinizados do DRG, enquanto que a expressão dos
receptores B2 encontra-se reduzida. Dados da literatura relatam que os antagonistas
para o receptor B2 são capazes de causar redução significativa, porém em menor
grau do que a induzida por antagonistas do receptor B1, no modelo de hiperalgesia
mecânica ipsilateral induzida pela injeção de CFA (PERKINS et al., 1993; BURGESS
et al., 2000). Entretanto, outros autores demonstraram que a hiperalgesia térmica
induzida pela injeção intraplantar de CFA não foi modificada em animais com
deleção gênica para o receptor B2 (RUPNIAK et al., 1997).
Resultados similares foram obtidos por Ferreira et al. (2001), onde animais
com deficiência do receptor B2 apresentaram essencialmente o mesmo
comportamento hipernociceptivo após a injeção de CFA, em relação aos animais
selvagens. Estes resultados foram confirmados através da utilização do antagonista
seletivo para o receptor B2, HOE-140. Outro fato interessante é que a resposta
hiperalgésica induzida pelo agonista seletivo do receptor B1, a des-Arg9-BK, é
potencializada em animais com deleção gênica para o receptor B2, em comparação
com animais selvagens (FERREIRA et al., 2004). Estes dados reforçam a hipótese
de que pode ocorrer um aumento compensatório na expressão do receptor B1 em
animais com deleção gênica para o receptor B2 (MADEDDU et al., 1997; DUKA et
al., 2001).
Ademais, pode-se sugerir que o aumento da resposta presente nos animais
operados e injetados com concentrações menores de formalina (0,5 – 1,5 %) deva-
se a presença de componentes inflamatórios já na primeira fase do teste. Esta
hipótese se deve a reversão do quadro quando estes animais são pré-tratados com
anti-inflamatórios clássicos, como indometacina e dexametasona. Estudos prévios
demonstraram que estes mesmos anti-inflamatórios não tinham efeito algum sobre a
primeira fase do teste de nocicepção induzido pela formalina (HUNSKAAR, HOLE,
1987).
Eventos fisiológicos e o envolvimento dos receptores de cininas podem ser
avaliados e confirmados pelo uso de ferramentas farmacológicas importantes, tais

56
como o uso de antagonistas e agonistas seletivos. No modelo de APB em
camundongos, foi observado que a citocina pró-inflamatória, o fator de necrose
tumoral-α (TNFα), tem um papel relevante na gênese e manutenção da
hipernocicepção mecânica induzida pela APB (QUINTÃO et al., 2006). Ademais,
dados da literatura demonstram que os fatores neurotróficos, principalmente o fator-
neurotrófico-derivado-do-cérebro (BDNF), bem como o aumento na expressão dos
receptores B1 para cininas são fundamentais na indução e na perpetuação do
quadro hipernociceptivo observado no modelo de formalina.
Nos modelos de nocicepção induzida por formalina e bradicinina, avaliou-se a
participação dos receptores de cininas através da administração de antagonistas
seletivos, o HOE-140 (D-Arg0[Hyp3, Thi5, D-Tic7, Oic8]-BK ou icatibante), para
receptores B2, e a DALBK ([Leu8][des-Arg9]-BK), para receptores B1, e administração
i.pl. de um agonista seletivo, a DABK ([des-Arg9]-BK) para receptores B1 (VAVREK,
STEWART, 1985; LEMBECK et al., 1991; WIRTH et al., 1991; TRIFILIEFF, DA
SILVA, GIES, 1993; HALL, MORTON, 1997).
Os receptores B2 são amplamente encontrados nos tecidos (isoforma
constitutiva), e os receptores B1 são regulados a partir de estímulos pró-inflamatórios
(isoforma induzida). (REGOLI, BARABÉ, 1980; MARCEAU, REGOLI, 2004). O
ligante natural dos receptores B1 é a bradicinina, sendo menos potente que seus
metabólitos, as des-Arg-cininas. Os receptores B1 são potentemente ativados pelos
metabólitos des-Arg-cininas, formados pela enzima cininase I a partir da bradicinina,
tais como a Lis-[des-Arg9]-BK e a [des-Arg9]-BK; e são inibidos pelos agonistas
sintéticos [Leu8][des-Arg9]-BK (TRIFILIEFF, DA SILVA, GIES, 1993; MOREAU et al.,
2005).
Os receptores B2 apresentam uma maior afinidade para Lis-BK (calidina) e a
bradicinina do que para seus metabólitos des-Arg-cininas (FARMER, BURCH, 1991).
A bradicinina também é o ligante natural para os receptores B2. O papel das cininas
nas respostas nociceptivas já está bem estabelecido, porém o perfil farmacológico,
fisiológico e patológico destes receptores não está completamente elucidado,
tornando-se necessário o estudo da sua participação nestes processos, buscando-
se a pesquisa de novos fármacos agonistas ou antagonistas que possam ser
utilizados nas terapias de doenças crônicas, tais como em quadros neuropáticos
(REGOLI, RIZZI, CALO, 1997; HESS et al., 2001; CALIXTO et al., 2000; 2001; 2004;
CAMPOS et al., 2006; LEEB-LUNDBERG et al, 2005; MOREAU et al., 2005).

57
As cininas constituem uma importante família de peptídeos biologicamente
ativos, e são os primeiros mediadores a serem liberados em tecidos lesionados. A
calicreína plasmática, por exemplo, é ativada precocemente na cascata de
coagulação. Já a calicreína tecidual é ativada por proteases liberadas no sítio da
lesão. (REGOLI, BARABÉ, 1980; VAVREK, STEWART, 1985; MURRAY et al., 1990;
REGOLI et al., 1990; BEAUBIEN et al., 1991; BHOOLA et al., 1992; REGOLI et al.,
1994; HALL, MORTON, 1997).
Como o quadro nociceptivo é resultante das alterações crônicas na
neuroquímica da sinalização da dor e o envolvimento dos receptores de cininas
pode estar relacionado. No modelo de nocicepção induzida por formalina foi
observado que os animais submetidos à APB com 6 dias de cirurgia apresentaram
um aumento da resposta dolorosa na segunda fase do teste, tanto nos tratamentos
com HOE-140 e com DALBK. Isso pode estar relacionado com a expressão de
receptores B1 para cininas, visto que os receptores B2 estão bloqueados pela ação
do HOE-140. Podemos também sugerir que a redução na resposta dos animais
operados injetados com formalina 2,5% possa representar um comportamento de
freezing desencadeado pela hiper-estimulação das vias nociceptivas decorrentes do
aumento na expressão dos receptores B2 para cininas. O mesmo pode ser
observado com a administração da DALBK, a qual foi capaz de inibir a nocicepção
induzida pela formalina. A ação dos antagonistas cininérgicos também pode ser
observada no modelo de nocicepção induzida pela BK.
A ativação do receptor B2 das cininas tem sido descrita como participante da
fase inicial da dor inflamatória produzida por agentes nociceptivos, tais como a
formalina, enquanto que a estimulação do receptor B1 estaria associada com
processos patológicos mais persistentes, visto que em modelos animais de
constrição do nervo ciático em ratos e camundongos observou-se o aumento nos
níveis de RNAm para a transcrição do receptor B1 (PETERSEN et al.,1998; ECKERT
et al., 1999; LEVY, ZOCHODNE, 2000; YAMAGUCHI-SASE et al., 2003; RASHID et
al., 2004).
Trabalhos na literatura descrevem a existência de componentes do sistema
cininérgico em regiões do sistema nervoso central (córtex, tronco encefálico,
cerebelo e hipotálamo) e a presença de bradicinina na medula, córtex, cerebelo e
hipocampo, demonstrando a atuação das cininas no controle central da dor (PERRY,
SNYDER, 1984; KARIYA, YAMAUCHI, SASAKI, 1985; FUJIWARA et al., 1989;

58
WALKER, PERKINS, DRAY, 1995; COUTURE, LINDSEY, 2000).
Após a lesão do nervo do plexo ocorrem alterações na expressão de
neuropeptídeos e de receptores para cininas em neurônios do gânglio da raiz dorsal.
O aumento na expressão desses receptores parece estar relacionado com a
interação dos fatores de crescimento neuronal (neurotrofina NGF) com o receptor
p75NTR. Essas alterações ocorrem tanto no lado ipsilateral quanto no lado
contralateral a lesão, sendo pré-requisito para o aparecimento da lesão nervosa,
visto que animais falso-operados não apresentam aumento da expressão desses
receptores (PETERSEN et al., 1996; PETERSEN et al., 1998; LEVY, ZOCHONE,
2000).
A participação dos receptores cininérgicos envolvidos no desenvolvimento e
manutenção do quadro nociceptivo em processos onde ocorrem danos nervosos,
como no caso da lesão do nervo ciático ou da avulsão do plexo braquial é, em parte,
determinada pela síntese de “novos” receptores B1, principalmente em neurônios do
gânglio da raiz dorsal, sendo que a expressão de receptores B2 encontra-se
diminuída. Ainda, a participação de antagonistas de receptores B2 causa uma
redução significativa na expressão desses receptores, porém em menor grau do que
os antagonistas de receptores B1. Além disso, o bloqueio do receptor B2 causa um
aumento compensatório na expressão de receptores B1 (PERKINS, CAMPBELL,
DRAY, 1993; MADEDDU et al., 1997; BURGESS et al., 2000; DUKA et al., 2001;
RASHID et al., 2004).
O principal neurotransmissor presente em todos os nociceptores é o
aminoácido excitatório glutamato, que atua principalmente em receptores
ionotrópicos do tipo NMDA, AMPA e cainato, porém, pode atuar também em
receptores metabotrópicos. Perifericamente, o glutamato tem importante papel
fisiológico, atuando em processos nociceptivos induzidos, tais como os observados
no modelo de formalina. Este estudo demonstrou que os animais submetidos à APB
com 30 dias após a cirurgia apresentaram uma reduzida resposta nociceptiva frente
à injeção i.pl. de glutamato quando comparados com os animais falso-operados,
tanto na pata ipsilateral, quanto na contralateral. Esses dados sugerem que a
expressão dos receptores glutamatérgicos possa estar reduzida, ou que a resposta
nociceptiva induzida pelo glutamato possa estar sendo bloqueada por algum
mecanismo de controle central da dor, visto que as alterações na neurobioquímica
causadas pela APB possam estar interferindo nos processos de sinalização

59
mediados por CGRP, SP, ATP, PGs, histamina, neuropeptídeo Y, neurotrofinas,
somatostatinas, serotonina, entre outros (BESSON, 1999; LOESER, MELZACK,
1999; HUNT, JULIUS, BASBAUM, 2001; KIDD, PHOTIOU, INGLIS, 2004).
Além disso, a ativação contínua e sucessiva das fibras pelo glutamato ocorre
através da ativação de canais iônicos e receptores específicos, que sinalizam
através de uma cascata de mediadores intracelulares, pode acarretar em alterações
na expressão gênica, induzindo assim alterações na estrutura e função dos
neurônios sensoriais, resultando no aumento ou diminuição da expressão de canais
iônicos e receptores, na produção de mediadores e na indução de enzimas (KIDD,
PHOTIOU, INGLIS, 2004; SHIBASAKI, 2004, KNYIHAR, CSILLIK, 2006).
Cabe ressaltar ainda o envolvimento do prostanóide PGE2 e a amina
simpatomimética epinefrina, nos papéis de seus respectivos receptores, nas
alterações das vias nociceptivas após a indução de um processo doloroso crônico.
Já está bem estabelecido que a PGE2 participa da sensibilização das vias
nociceptivas, tornando os neurônios muito mais responsivos. Cabe a este sistema o
papel de induzir o processo conhecido como Wind up, juntamente com o aumento
na liberação do glutamato, onde os neurônios ficam incapazes de estabelecer
potencial de repouso e adquirem um comportamento de memória. Ao menor
estímulo, em que normalmente não seria evocada resposta nociceptiva, os
nociceptores despolarizam-se e retomam o estado doloroso anteriormente
vivenciado.

60
7 CONCLUSÕES
Este trabalho demonstra pela primeira vez que uma lesão nervosa periférica é
capaz de causar alteração na resposta nociceptivas frente aos modelos de
nocicepção aguda clássicos. Inicialmente foi demonstrado que animais submetidos à
avulsão do plexo braquial, quando injetados com formalina 2,5% (concentração
utilizada nos testes de nocicepção para avaliar o potencial analgésico de
substâncias), responderam menos quando comparado aos animais falso-operados.
Esta resposta poderia significar ou a redução da resposta nociceptiva dos animais
decorrente da redução da sensibilidade (ausência de nocicepção) ou uma redução
da resposta devido a uma dessensibilização das vias nociceptivas decorrente de
uma estimulação excessiva (resposta conhecida como freezing).
Nossos resultados demonstraram que os animais operados apresentam uma
resposta reduzida frente à injeção i.pl. de bradicinina (agonista B2) quando
comparados aos animais falso-operados. Porém, esta resposta se altera quando a
concentração da bradicinina é reduzida, ocorrendo o mesmo quadro presente nos
experimentos realizados com a formalina. Estes dados nos levam a sugerir que as
alterações presentes nos experimentos realizados com a formalina possam, pelo
menos em parte, ser decorrentes de alterações na expressão de receptores B2 para
cininas. Acredita-se que estes possam sofrer dessensibilização e internalização,
reduzindo a resposta nociceptiva frente ao estímulo.
Estes dados em conjunto nos leva a concluir que a APB é capaz de causar
uma alteração nas vias nociceptivas, porém distintas no que se refere aos
hemisférios corporais e tempo de duração do processo neuropático. Acredita-se que
estas alterações envolvam um componente inflamatório, que levam ao aumento da
resposta nociceptiva e até mesmo a supressão da resposta por hiper-estimulação
dos terminais sensoriais. Estudos adicionais são necessários para que estes
mecanismos de controle da nocicepção sejam mais definidos, podendo colaborar na
busca de estratégias seguras e eficazes no controle da dor neuropática.

61
REFERÊNCIAS
ABRAHAM, K. E.; MCGINTY, J. F.; BREWER, K. L. The role of kainic acid/AMPA and metabotropic glutamate receptors in the regulation of opioid mRNA expression and the onset of pain-related behavior following excitotoxic spinal Cord injury. Neurocience, v. 104, p. 863-74, 2001.
ALEY, K. O.; LEVINE, J. D. Role of protein kinase A in the maintenance of inflammatory pain. Journal of Neuroscience, v. 19, n. 6, p. 2181-2186, 1999.
ALEY, K. O.; MESSING, R. O.; MOCHLY-ROSEN, D.; LEVINE, J. D.. Chronic hypersentivity for inflammatory nociceptor sensitization mediated by the epsilon isoenzyme of protein kinase C. Journal of Neuroscience, v. 20, p. 4680-4685, 2000.
ALMEIDA, T. F., ROIZENBLATT, S.; TUFIK, S. Afferent pain pathways: a neuroanatomical review. Brain Research, v. 1000, p. 40-56, 2004.
ANAND, P. BIRCH, R.. Restoration of sensory function and lack of long-term chronic pain syndromes after brachial plexus injury in human. Brain, v. 125, p. 113-122, 2002.
ASHBURN, M. A.; STAATS, P. S. Management of chronic pain. Lancet, v. 353, n. 9167, p. 1865-1869, 1999.
AVERILL, S.; MCMAHON, S. B.; CLARY, D. O.; REICHARDT, L. F.; PRIESTLEY, J. V. Immunocytochemical localization of trkA receptors in chemically identified subgroups of adult rat sensory neurons. Europan Journal of Neuroscience, v. 7, n. 7, p. 1484-1494, 1995.
BEAUBIEN, G.; ROSINSKI-CHUPIN, I.; MATTEI, M. G.; MBIKAY, M.; CHRETIEN, M.; SAIDAH, N. G.. Gene structure and chromossomal localization of plasmakallikrein. Biochemistry, v. 30, p. 1628-1635, 1991.
BEIRITH, A.; SANTOS, A. R. S.; RODRIGUES, A. L. S.; CRECZYNSKI-PASA, T. B.; CALIXTO, J. B.. Spinal and supraspinal antinociceptive action of dypirone on formalin, capsaicin and glutamate tests. Study of mechanisms of action. European Journal of Pharmacology, v. 345, p. 233-245, 1998.
BERMAN, J. S.; BIRCH, R.; ANAND, P.. Pain following human brachial plexus injury with spinal cord root avulsion and the effect of surgery. Pain, v. 75, p. 199-207, 1998.
BESSON, J. M. The neurobiology of pain. Lancet, v. 353, n. 9164, p. 1610-1615, 1999.
BHOOLA, K. D.; FIGUEROA, C. D.; WORTHY, K.. Bioregulation of kinins: kallikreins, kininogens, and kininases. Pharmacological Reviews. v. 44, p. 1-44, 1992.

62
BROWN-SÉQUARD, C. E. Lectures on the physiology and pathology of the nervous system; and on the treatment of organic nervous affections. Lancet, v. 2, p. 593–596, 1868.
BURGESS, G.M.; PERKINS, M. N.; RANG, H.P.; CAMOBELL, E. A.; BROWN,M. C.; MCINTYRE, P.; URBAN, L.; DZIADULEWCZ, E. K.; RITCHIE, T. J.; HALLET, A.; SNELL, C. R.; WRIGLESWORTH, R.; LEE, W.; DAVIS, C.;PHAGOO, S. B.; DAVIS, A. J.; PHILLIPS, E.; DRAKE, G. S.; HUGHES, G. A.; DUNSTAN, A.; BLOOMFIELD, G. C.. Bradyzide, a potent non-peptide receptor antagonist with long-lasting oral activity in animal models of inflammatory hyperalgesia. British Journal of Pharmacology, v. 129, p. 77-86, 2000.
CAHILL, C. M.; MORINVILLE, A.; HOFFERT, C.; O’DONNELL, D.; BEAUDET, A.. Up-regulation and trafficking of delta opiod receptor in a model of chronic inflammation: implications for pain control. Pain, v. 101, p. 199-208, 2003.
CALIXTO, J. B.; BEIRITH, A.; FERREIRA, J.; SANTOS, A. R.; CECHINEL-FILHO, V.; YUNES, R. A.. Naturally occurring antinociceptive substances from plants. Phytotheraphy Research, v. 14, n. 6, p. 401 – 418, 2000.
CALIXTO, J. B.; MEDEIROS, R.; FERNANDES, E. S.; FERREIRA, J.; CABRINI, D.; CAMPOS, M. M.. Kinin B1 receptors: key G-protein-coupled receptors and their role in inflammatory and painful processes. British Journal of Pharmacology, v. 143, p. 803-818, 2004.
CALIXTO, J. B.; SCHEIDT, C.; OTUKI, M. F.; SANTOS, A. R. S.. Biological activity of plant extracts: novel analgesic drug. Expert Opinion on Emerging Drugs, v. 6, p. 261- 279, 2001.
CAMPBELL, J. N.; MEYER, R .A. Mechanisms of neuropathic pain. Neuron, v. 52, p. 77-92, 2006.
CAMPOS, M. M.; LEAL, P. C.; YUNES, R. A.; CALIXTO, J. B.. Non-peptide antagonists for kinin B(1) receptors: new insights into their therapeutic potential for the management of inflammation and pain. Trends in Pharmacological Sciences, v. 27, n. 12, p. 646-51, 2006.
CARVALHO, G. A.; NIKKHAH, G.; SAMII, M.. Pain management after post-traumatic brachial plexus lesions. Conservative and surgical therapy possibilities. Orthopade, v. 26, p. 621-625, 1997.
CECHINEL FILHO, V; CAMPOS-BUZZI, F; CORREA, R; YUNES,R.A; NUNES, R. J. Aspectos químicos e potencial terapêutico de imidas cíclicas: uma revisão da literatura. Química Nova, v. 26, p. 230-241, 2003.
CHRISTENSEN, B. N.; PERL, E. R. Spinal neurons specifically excited by noxious or thermal stimuli: marginal zone of the dorsal horn. Journal of Neurophysiology, v.33, n. 2, p. 293-307, 1970.

63
CLARK, D.; HUGHES, J.; GASSER, H. S. Afferent function in the group of nerve fibers of slowest conduction velocity. American Journal of Physiology, v. 114, p. 69-76, 1935.
COLLINS, W. R., JR.; NULSEN, F. E.; RANDT, C. T. Relation of peripheral nerve fiber size and sensation in man. Archives of Neurology, v.3, p. 381-385, 1960.
CORRÊA, C. R.; KYLE, D. J.; CHAKRAVARTY, S.; CALIXTO, J. B.. Antinociceptive profile of pseudopeptide B2 bradykinin receptor antagonist NPC 18688 in mice. British Journal of Pharmacology, v. 117, p. 552-558, 1996.
COSTIGAN, M.;WOOLF, C. J. Pain: molecular mechanisms. Journal of Pain, v.1, p. 35-44,2000.
COUTURE, R.; HARRISSON, M.; VIANNA, R. M.; CLOUTIER, F.. Kinin receptors in pain and inflammation. European Journal of Pharmacology, v. 429, p. 161-176, 2001.
CRAIG, A.D. Pain mechanisms: labeled lines versus convergence in central processing. Annual Review of Neuroscience, v. 26, p.1-30, 2003.
CUELLO, A.C.; RIBEIRO-DA-SILVA, A.; MA, W.; DE KONINCK, Y.; HENRY, J. L. Organization of substance P primary sensory neurons: ultrastructural and physiological correlates. Regulatory Peptides, v. 46, n. 1-2, p. 155-164, 1993.
DEJERINE, J.; ROUSSY, G. Le syndrome thalamique. Revue Neurologique, v. 14, p. 521–532, 1906.
DOGRUL, A.; GARDELL, L. R.; OSSIPOV, M. H.; TULUNAY, F. C.; LAI, J.; PORRECA, F. Reversal of experimental neuropathic pain by T-type calcium channel blockers. Pain, v. 105, n. 1-2, p. 159- 168, 2003.
DOGRUL, A.; GARDELL, L. R.; OSSIPOV, M. H.; TULUNAY, F. C.; LAI, J.; PORRECA, F.. Reversal of experimental neuropathic pain by T-type calcium channel blockers. Pain, v. 105, n. 1-2, p. 159-168, 2003.
DUBUISSON, D.; DENNIS, S. G. The formalin test: a quantitative study of the analgesic effects of morphine, meperidine, and brain stem stimulation in rats and cats. Pain, v. 4, p. 161-174, 1977.
DUKA, I.; KINTSURASHVILI, E.; GAVRAS, I.; JOHNS, C.; BRESNAHAN, M.; GAVRAS, H.. Vasoactive potential of the B1 bradykinin receptor in normotension and hypertension. Circulation Research, v. 88, p. 275–281, 2001.
ECKERT, A.; SEGOND VON BANCHET, G.; SOPPER, S.; PETERSEN, M.. Spatiotemporal pattern of induction of bradykinin receptors and inflammation in rat dorsal root ganglia after unilateral nerve ligation. Pain, v. 83, p. 487-497, 1999.
EDINGER, L. Einigles von verlauf der gefühlsbahnen in centralen nerven systeme. Deutsche Med Woch, v. 16, p. 421–426, 1890.

64
ERB, W. Handbuch der Krankheiten des Nervensystems II. Leipzig: F. C. W. Vogel, 1874.
FERREIRA, J.; DA SILVA, G. L.; CALIXTO, J. B.. Contribution of vanilloid receptors to the overt nociception induced by B2 kinin receptor activation in mice. British Journal of Pharmacology, v. 141, p. 787-794, 2004.
FIELDS, H. L.; BASBAUM, A. I.; HEINRICHER, M. M. Central nervous system mechanisms of pain modulation. In: MCMAHON, S. B.; KOLTZENBURG, M. Wall and Melzack’s textbook of pain, 5 ed., p. 125–142, London: Elsevier, 2005.
FUJIWARA, Y.; MANTIONE, C. R.; VAVREK, R. J.; STEWART, J. M.; YAMAMURA, H. I.. Characterization of (3H) bradykinin binding sites in guinea-pig central nervous system: possible existence of B2 subtypes. Life Science, v. 44, p. 1645-1653, 1989.
FÜRST, S.. Transmitters involved in antinociception in the spinal cord. Brain Research Bulletin, v. 48, p. 129-141, 1999.
OLIVEIRA, A. S. B.; GABBAI, A. A. Abordagem terapêutica da dor neuropática na clínica neurológica. Revista Neurociências, v. 6, n. 2, p. 87-95, 1998.
GRUBB, B. D.. Peripheral and central mechanisms of pain. British Journal of Anaesthesia, v. 81, n. 1, p. 8-11, 1998.
GUYTON, A. C.; HALL, J. E. Tratado de Fisiologia Médica. Guanabara Koogan, Rio de Janeiro, 2002.
HALL, J. M., MORTON, I. K. M.. The pharmacology and immunopharmacology of kinin receptors. In: FARMER, S. G. The handbook of immunopharmacology: The kinin system. Academic Press: London, p. 9-43, 1997.
HEAD, H.; HOLMES, G. Sensory disturbances from cerebral lesions. Brain, v. 34, p. 102–254, 1911.
HEINBECKER, P.; BISHOP, G. H.; O'LEARY, J. Pain and touch fibers in peripheral nerves. Archives of Neurology and Psychiatry, v. 29, p. 771–789, 1933.
HESS, J. F.; HEY, P. J.; CHEN, T. B.; O'BRIEN, J.; OMALLEY, S. S.; PETTIBONE, D. J.;CHANG, R. S.. Molecular cloning and pharmacological characterization of the canine B1 and B2 bradykinin receptors. Biological Chemistry, v. 382, p. 123-129, 2001.
HOLZER, P.. Capsaicin: cellular targets mechanisms of action and selectivity for thin sensory neurons. Pharmacological Reviews, v. 43, p. 144-201, 1991.
HÖSL, K.; REINOLD, H.; HARVEY, R. J.; MÜLLER, U.; NARUMIYA, S.; ZEIHOFER, H. U.. Spinal prostaglandin E receptors of the EP2 subtype and the glycine receptor alpha3 subunit, which mediate central inflammatory hyperalgesia, do not contribute to pain after peripheral nerve injury or formalin injection. Pain, v. 126, p. 46-53, 2006.

65
HUCHO, T.; LEVINE, J. D. Signaling pathways in sensitization: toward a nociceptor cell biology. Neuron, v. 55, n. 3, p. 365-376, 2007.
HUNSKAAR, S.; HOLE, K. The formalin test in mice: dissociation between inflammatory and non-inflammatory pain. Pain, v. 30, p. 103-114, 1987.
JI, R. R.; WOOLF, C. J. Neuronal plasticity and signal transduction in nociceptiveneurons: implications for the initiation and maintenance of pathological pain. Neurobiology of Disease, v. 8, n. 1, p. 1-10, 2001.
JULIUS, D.; BASBAUM, A. I.. Molecular mechanisms of nociception. Nature, v. 413, n. 6852, p. 203-10, 2001.
KARIYA, K.; YAMAUCHI, A.; SASAKI, T.. Regional distribution and characterization of kinin in the CNS of the rat. Journal of Neurochemistry, v. 44, p. 1892-1897, 1985.
KASSUYA, C. A.; FERREIRA, J.; CLAUDINO, R. F.; CALIXTO, J. B.. Intraplantar PGE2 causes nociceptive behaviour and mechanical allodynia: the role of prostanoid E receptors and protein kinases. British Journal of Pharmacology, v. 150, p.727-737, 2007.
KHASAR, S. G.; DINA, O. A.; GREEN, P. G.; LEVINE, J. D.. Estrogen regulates adrenal medullary function producing sexual dimorphism in nociceptive threshold and beta-adrenergic receptor-mediated hyperalgesia in the rat. European Journal of Pharmacology, v. 21, n. 12, p. 3379-3386, 2005.
KIDD, B. L.; PHOTIOU, A.; INGLIS, J. J.. The role of inflammatory mediators on nociception and pain in arthritis. Novartis Foundation Symposium, v. 260, p. 122-33; discussion 133-8, 277-9. 2004.
KNYIHAR, E.; CSILLIK, B.. Plasticity of nociception: recent advances in function- oriented structural pain research. Ideggyóyászati szemle, v. 59, n. 3-4, p. 87-97, 2006.
LANDAU, W.; BISHOP, G. H. Pain from dermal, periosteal, and fascial endings and from inflammation; electrophysiological study employing differential nerve blocks. Archives of Neurology and Psychiatry, v. 69, n. 4, p. 490-504, 1953.
LEE, Y.-J.; ZACHRISSON, O.; TONGE, D. A.; MACNAUGTON, P. A.. Upregulationof bradykinin B2 receptor expression by neurotrophic factors and nerve injury in mouse sensory neurons. Mol. Cell. Neuroscience, v. 19, p. 186-200, 2002.
LEEB-LUNDBERG, L. M.; MARCEAU, F.; MÜLLER-ESTERL, W.; PETTIBONE, D. J.; ZURAW, B. L.. International Union of Pharmacology. XLV. Classification of the kinin receptor family: from molecular mechanisms to pathophysiological consequences. Pharmacological Reviews, v. 57, n. 1, p. 27-77, 2005.
LEMBECK, F.; GRIESBACHER, T.; ECKHARDT, M.; HENKE, S.; BREIPOHL, G.;KNOLLE J.. New, long-acting, potent bradykinin antagonists. British Journal of Pharmacology, v. 102, p.297-304, 1991.

66
LENT, R. Cem Bilhões de Neurônios: Conceitos Fundamentais de Neurociência. Atheneu, São Paulo, 2004.
LEVY, D.; ZOCHODNE, D. W.. Increased mRNA expression of the B1 and B2 bradykinin receptors and antinociceptive effects of their antagonists in an animal model of neuropathic pain. Pain, v. 86, p. 265-271, 2000.
LOESER, J. D.; TREEDE, R. D. The Kyoto protocol of IASP Basic Pain Terminology. Pain, v. 137, n. 3, p. 473-477, 2008.
MADEDDU, P.; VARONI, M. V.; PALOMA, D.; EMANUELI, C.; DEMONSTIS, M. P.;GLORIOSO, N.; DESSI-FULGHERI, P.; SARZANI, R.; ANANIA, V.. Cardiovascular phenotype of a mouse strain with disruption of bradykinin B2-receptor gene. Circulation, v. 96, p. 3570-3578, 1997.
MAO, J.; PRICE, D. D.; PHILLIPS, L. L.; LU, J.; MAYER, D. J. Increases in protein kinase C gamma immunoreactivity in the spinal cord dorsal horn of rats with painful mononeuropathy. Neurosci Letters, v. 198, n. 2, p. 75-78, 1995.
MARCEAU, F.; REGOLI, D.. Bradykinin receptor ligands: therapeutic perspectives. Nature Reviews Drug Discovery, v. 3, p. 845-852, 2004.
MARKENSON, J. A. Mechanisms of chronic pain. American Journal of Medicine, v. 101, p. 6S-18S,1996.
MCLACHLAN, E. M.; JANIG, W.; DEVOR, M.; MICHAELIS, M. Peripheral nerve injury triggers noradrenergic sprouting within dorsal root ganglia. Nature, v. 363, n. 6429, p. 543-546, 1993.
MELZACK, R.; WALL, P. D. Pain mechanisms: a new theory. Science, v. 50, n. 699, p. 971-979, 1965.
MENDES, G. L.; SANTOS, A. R. S.; MALHEIROS, A.; CECHINEL-FILHO, V.; YUNES, R. A.; CALIXTO, J. B.. Assessment of mechanisms involved in antinociception caused by sesquiterpene polygodial. Journal of Pharmacology and Experimental Therapeutics, v. 292, p. 164-172, 2000.
MERSKEY, H; BOGDUK, N. Classification of chronic pain – descriptions of chronic pain syndromes and definitions of pain terms. 2th ed. Seattle, USA: IASP Press, 2002.
MEYER, RA.; RINGKAMP, M.; CAMPBELL, J. N.; RAJA, S.N. Peripheral mechanisms of cutaneous nociception. In: MCMAHON SB, KOLTZENBURG M. M. Wall and Melzack’s textbook of pain, 5 ed., p. 125–142, London: Elsevier, 2005.
MILLAN, M. J. The induction of pain: An integrative review. Progress in Neurobiology, v. 57, p. 1-164, 1999.
MIZUMURA, K. Peripheral mechanism of hyperalgesia--sensitization of nociceptors. Nagoya Journal of Medical Science, v. 60, n. 3-4, p. 69-87, 1997.

67
MOGIL, J. S.; YU, L.; BASBAUM, A. I. Pain genes?: natural variation and transgenic mutants. Annual Review of Neuroscience, v. 23, p. 777-811, 2000.
MOORE, K. A.; BABA, H.; WOOLF, C. J. Synaptic transmission and plasticity in the superficial dorsal horn. Progress in Brain Research, v. 129, p. 63-80, 2000.
MOORE, K. A.; KOHNO, T.; KARCHEWSKI, L. A.; SCHOLZ, J.; BABA, H.; WOOLF, C. J. Partial peripheral nerve injury promotes a selective loss of GABAergic inhibition in the superficial dorsal horn of the spinal cord. Journal of Neuroscience, v. 22, n. 15, p. 6724-6731, 2002.
MOREAU, M. E.; GARBACKI, N.; MOLINARO, G.; BROWN, N. J.; MARCEAU, F.; ADAM, A.. The kallikrein-kinin system: current and future pharmacological targets. Journal of Pharmacological Sciences, v. 99, p. 6-38, 2005.
MÜLLER, J. Handbuch der Physiologie des Menschen für Vorlesungen. Koblenz: J. Hölscher, 1840.
MURRAY, S. R.; CHAO, J.; LIN, F.; CHAO, L.. Kallikrein multigenes families and the regulation of their expression. Journal of Cardiovascular Pharmacology, v. 15, p. S7-S15, 1990.
NAFE, J. P. A quantitative theory of feeling. Journal of General Psychology, v. 2, p. 199–211, 1929.
OBARA, I.; PARKITNA, J. R.; KOROSTYNSKI, M.; MAKUCH, W.; KAMINSKA, D.; PRZEWLOCKA, B.; PRZEWLOCKI, R. Local peripheral opioid effects and expression of opioid genes in the spinal cord and dorsal root ganglia in neuropathic and inflammatory pain. Pain, v. 141, n. 3, p. 283-291, 2009.
OHSAWA, M.; NARITA, M.; MIZOGUCHI, H.; CHENG, E.; TSENG, L. F. Reduced hyperalgesia induced by nerve injury, but not by inflammation in mice lacking protein kinase C gamma isoform. European Journal of Pharmacology, v. 429, n. 1-3, p. 157-160, 2001.
OMOTE, K.; KAWAMATA, T.; KAWAMATA, M.; NAMIKI, A.. Formalin-induced release of excitatory amino acids in the skin of the rat hindpaw. Brain Research, v. 787, p. 161-164, 1998.
PERKINS, M. N.; CAMPBELL, E.; DRAY, A.. Antinociceptive activity of the bradykinin B1 and B2 receptor antagonists, des-arg9,[leu8]-bk and hoe140, in two models of persistent hyperalgesia in the rat. Pain, v. 51, p. 191-197, 1993.
PERL, E. R. Ideas about pain, a historical view. Nature Reviews Neuroscience, v. 8, n. 1, p. 71-80, 2007.
PERRY, D. C.; SNIDER, S. H.. Identification of bradykinin in mammalian brain. Journal of Neurochemistry, v. 43, p. 1072-1080, 1984.

68
PETERSEN, M.; ECKERT, A. S.; SEGOND VON BANCHET, G.; HEPPELMANN, B.; KLUSCH, A.; KNIFFKI, K.-D.. Plasticity in the expression of bradykininbinding sites in the sensory neurons after mechanical nerve injury. Neuroscience, v. 83, n. 3, p. 949-959, 1998.
PETERSEN, M.; SEGOND VON BANCHET, G.; HEPPELMANN, B.; KOLTZENBURG, M.. NGF regulates the expression of bradykinin biding sites onadult sensory neurons via the neurotrophin receptor p75. Neuroscience, v. 83, p. 161-168, 1996.
PORRECA, F.; OSSIPOV, M. H.; GEBHART, G. F. Chronic pain and medullary descending facilitation. Trends in Neurosciences, v. 25, p. 319-325, 2002.
PRICE, D. D. Central neural mechanisms that interrelate sensory and affective dimensions of pain. Molecular Interventions, v. 2, n. 6, p. 392-403, 339, 2002.
PRICE, D. D.; MAYER, D. J. Neurophysiological characterization of the anterolateral quadrant neurons subserving pain in M. mulatta. Pain, v.1, n. 1, p. 59-72, 1975.
QUINTÃO, N. L. M.; MEDEIROS, R.; SANTOS, A. R. S.; CAMPOS, M. M.; CALIXTO, J. B.. The effects of diacerhein on mechanical allodynia in inflammatory and neuropathic models of nociception in mice. Anesthesia & Analgesia, v. 101, n. 6, p. 1763-1769, 2005.
QUINTÃO, N. L. M.; BALZ, D.; SANTOS, A. R. S.; CALIXTO, J. B.. Long-lasting neuropathic pain induced by brachial plexus injury in mice: Role triggered by the pro-inflammatory cytokine, tumour necrosis factor α. Neuropharmacology, v. 50, p. 614-620, 2006.
RASHID, H.; INOUE, M.; MATSUMOTO, M.; UEDA, H.. Switching of bradykinin mediated nociception following partial sciatic nerve injury in mice. Journal of Pharmacology and Experimental Therapeutics, v. 308, p. 1158-1164, 2004.
REEVE, A. J.; DICKENSON, A. H.. The roles of spinal adenosine receptors in the control of acute and more persistent nociceptive response of dorsal horn neuronsin the anesthetized rat. . British Journal of Pharmacology, v. 116, p. 2221-2228, 1995.
REGOLI, D., RIZZI, A.; CALO, G.. Pharmacology of the kallikrein-kinin system. Pharmacological Research, v. 35, p. 513-515, 1997.
REGOLI, D.; BARABÉ, J.. Pharmacology of bradykinin and related kinins. Pharmacological Reviews, v. 32, p. 1 - 46, 1980.
REGOLI, D.; GOBIEL, F.; NGUYEN, Q. T.; JUKIC, D.; SEONE, P. R.; SALVINO,J. M.; SAWUTZ, D. G.. Bradykinin receptor types and B2 subtypes. Life Science, v. 55,p. 735-749, 1994.
REGOLI, D.; RHALEB, N-E.; DRAPEAU, G.; DION, S.. Kinin receptors subtypes. Journal of Cardiovascular Pharmacology, v. 15, p. S30-S38, 1990.

69
REN, K.;DUBNER, R. Descending modulation in persistent pain: an update. Pain, v. 100, p. 1-6,2002.
RODRIGUES-FILHO, R.; SANTOS, A .R. S.; BERTELLI, J. A.; CALIXTO, J. B.. Avulsion injury of the rat brachial plexus triggers hyperalgesia and allodynia in the hindpaws: a new model for the study of neuropathic pain. Brain Research, v. 982, n. 2, p. 186-194, 2003.
RODRIGUES-FILHO, R.; SANTOS, A. R. S.; BERTELLI, J. A.; CALIXTO, J. B.. Avulsion injury of the rat brachial plexus triggers hyperalgesia and allodynia in the hindpaws: a new model for the study of neuropathic pain. Brain Research, v. 982, n. 2, p. 186-194, 2003.
RUEFF, A.; DRAY, A. Sensitization of peripheral afferent fibres in the in vitro neonatal rat spinal cord-tail by bradykinin and prostaglandins. Neuroscience, v. 54, n. 2, p. 527-535, 1993.
RUPNIAK, N .M. J.; BOYCE, S.; WEBB, J. K.; WILLIAMS, A. R.; CARLSON, E. J.;HILL, R. G.; BORKOWSKI, J. A.; HESS, J. F.. Effects of the bradykinin B1
receptor antagonist des-Arg9-[Leu8]-BK and genetic disruption of the B2 receptor on nociception in rats and mice. Pain, v. 71, p. 89-97, 1997.
RUSSO, C. M.; BROSE, W. G. Chronic pain. Annual Review of Medicine, v. 49, p. 123-133, 1998.
SAKURADA, T.; KATSUMATA, K.; TAN-NO, K.; SAKURADA, S.; KISARA, K.. The capsaicin test in mice for evaluating tachykinin antagonists in the spinal cord. Neuropharmacology, v. 31, p. 1279-1285, 1992.
SANDKUHLER, J. Models and mechanisms of hyperalgesia and allodynia. Physiologycal Reviews, v. 89, p. 707-58, 2009.
SCHIFF, J. M. Lehrbuch der physiologie des menschen. Lahr: Schauenberg, 1858.
SHERRINGTON, C. S. The Integrative Action of the Nervous System. Cambridge, UK: Cambridge Univ. Press, 1906.
SHIBASAKI, H. Central mechanisms of pain perception. Supplements of Clinical Neurophysiology, v. 57, p.39-49, 2004.
SOBREIRA, C. R.; ZAMPIER, M. R. Revisão de literatura: Terapia farmacológica nas desordens temporomandibulares. Revista da Universidade de Alfenas, v. 5, p. 239-245, 1999.
STANFA, L.; DICKENSON, A. Spinal opioid systems in inflammation. Inflammation Research,v. 44, n. 6, p. 231-241, 1995.
STEIN, C.; SCHÄFER, M.; MACHELSKA, H.. Attacking pain at its source: new perspectives on opioids. Nature Medicine, v. 9, n. 8, p. 1003 – 1008, 2003.

70
TAIWO, Y. O.; LEVINE, J. D. Prostaglandin effects after elimination of indirect hyperalgesic mechanisms in the skin of the rat. Brain Research, v. 492, n. 1-2, p. 397-399, 1989.
TANG, N. K.; CRANE, C. Suicidality in chronic pain: a review of the prevalence, risk factors and psychological links. Psychological Medicine, v. 36, n. 5, p. 575-586, 2006.
TEIXEIRA, M. J.; OKADA, M. A dor: na antiguidade: punição dos deuses à qualidade sensorial. Revista da Aventis Pharma, p. 55-56, 2001.
TRIFILIEFF, A.; DA SILVA, A.; GIES, J. P. Kinins and respiratory tract diseases. European Respiratory Journal, v. 6, p. 576-87, 1993.
VAVREK, R. J.; STEWART, J. M. Competitive antagonists of bradykinin. Peptides, v. 6, p.161-164, 1985.
WALKER, K.; PERKINS, M.; DRAY, A.. Kinins and kinin receptors in the nervous system. Neurochemistry International, v. 26, p. 1-16, 1995.
WALL, P. D.; DEVOR, M. Sensory afferent impulses originate from dorsal root ganglia as well as from the periphery in normal and nerve injured rats. Pain, v. 17, n. 4, p. 321-339, 1983.
WALL, P. D.; GUTNICK, M. Ongoing activity in peripheral nerves: the physiology and pharmacology of impulses originating from a neuroma. Experimental Neurology, v. 43, n. 3, p. 580-593, 1974.
WATKINS, L. R.; MAIER, S. F.. Beyond neurons: evidence that immune and glial cells contribute to pathological pain states. Physiological Reviews, v. 82, n. 4, p. 981-1011, 2002.
WIRTH, K.; BREIPOHL, G.; STECHL, J.; KNOLLE, J., HENKE, S.; SCHOLKENS, B..DesArg9-D-Arg[Hyp3,Thi5,D-Tic7,Oic8]bradykinin (desArg10-[Hoe140]) is a potente bradykinin B1 receptor antagonist. European Journal of Pharmacology, v. 205, p. 217-218, 1991.
WOOLF, C. J. Evidence for a central component of post-injury pain hypersensitivity. Nature, v. 306, n. 5944, p. 686-688, 1983.
WOOLF, C. J.; COSTIGAN, M. Transcriptional and post translational plasticity and the generation of inflammatory pain. Proceedings of the National Academic of Science USA, v. 96, n. 14, p. 7723-7730, 1999.
WOOLF, C. J.; MANNION, R. J. Neuropathic pain: a etiology, symptoms, mechanisms, and management. Lancet, v. 353, n. 9168, p. 1959-1964, 1999.
WOOLF, C. J.; SALTER, M. W. Neuronal plasticity: increasing the gain in pain. Science, v. 288, p. 1765-9, 2000.
WOOLF, C. J.; SHORTLAND, P.; COGGESHALL, R. E. Peripheral nerve injury triggers central sprouting of myelinated afferents. Nature, v. 355, n. 6355, p. 75-78,

71
1992.
WOOLF, C. J.; THOMPSON, S. W. The induction and maintenance of central sensitization is dependent on N-methyl-D-aspartic acid receptor activation; implications for the treatment of post-injury pain hypersensitivity states. Pain, v. 44, n. 3, p. 293-299, 1991.
WU, J.; LIN, Q.; MCADOO, D. J.; WILLIS, W. D.. Nitric oxide contributes to central sensitization following intradermal injection of capsaicin. Neuroreport, v. 9, p. 589-592, 1998.
WYNN PARRY, C. B.. Pain in avulsion lesions of the brachial plexus. Pain, v. 9, p. 41-53, 1980.
YAJIMA, Y.; NARITA, M.; SHIMAMURA, M.; NARITA, M.; KUBOTA, C.; SUZUKI, T. Differential involvement of spinal protein kinase C and protein kinase A in neuropathic and inflammatory pain in mice. Brain Research, v. 992, n. 2, p. 288-293, 2003.
YAMAGUCHI-SASE, S.; HAYASHI, I.; OKAMOTO, H.; NARA, Y.; MATSUZAKI,S.; HOKA, S.; MAJIMA, M.. Amelioration of hyperalgesia by kinin receptor antagonists or kininogen deficiency in chronic constriction nerve injury in rats. Inflammation Research, v. 52, p. 164-169, 2003.
YASHPAL, K.; FISHER, K.; CHABOT, J. G.; CORDERRE, T. J.. Differential effects of NMDA and group I mGluR antagonists on both nociception and spinal cord protein kinase C translocation in the formalin test and a model of neuropathic pain in rats. Pain.v. 94, p.17-29, 2001.
ZHAO, J.; SEEREERAM, A.; NASSAR, M. A.; LEVATO, A.; PEZET, S.; HATHAWAY, G.; MORENILLA-PALAO, C.; STIRLING, C.; FITZGERALD, M.; MACMAHON, S. B.; RIOS, M.; WOOD, J. N.. Nociceptor-derived brain-derived neurotrophic factor regulates acute and inflammatory but not neuropathic pain. Molecular and Cellular Neuroscience, v. 31, p. 539-548, 2006.
ZIMMERMANN, M.. Ethical guidelines for investigations of experimental pain in conscious animals. Pain. v. 16, p. 109-110, 1983.
ZOTTERMAN, Y. Studies in the peripheral nervous mechanism of pain. Acta Medica Scandinavica, v. 80, p. 1–64, 1933.





























