Embed Size (px)
DESCRIPTION
Asignatura Citologia. Grado en Medicina. Santiago de Compostela
Citation preview

BIOLOGIA HUMANA
CITOLOGIA. BIBLIOGRAFIA
Biología molecular de la célula. Omega. Alberts B Brae, D. Lewis
Paniagua B Nistal, M sesma P, Álvarez. Biología celular, Mc Graw Hill
Kierzarbraum a l. Histología e Biología Celular. Introducción a la Antonia patológica
GENETICA. BIBLIOGRAFIA
Jorde lb, Genética medica. Elsevier
Nussbaum. Genética en Medicina
Tunpenne. Elementos de xenetica medica. Elservier
Solari. Genética medica. Fundamentos…

BIOLOGIA HUMANA
CITOLOGÍATEMA 1: LA CELULA
1.- Concepto y definición2.- Caracteres generales y tipos de células3.- Relación de las células entre sí y con otras formas de vida4.- Morfología general de la célula eucariota y procariota
1.- LA CÉLULA COMO UNIDAD DE ORGANIZACIÓN BIOLÓGICA
Existen varas definiciones para la célula: Unidad morfológica, fisiológica y de origen de todos los seres vivos (TEORIA CELULAR):
o Morfológica porque todos los seres vivos están constituidos por una o más células
o Fisiológicas porque es capaz de realizar todos los procesos metabólicos necesarios para vivir (unidad estructural de la vida)
o De origen porque las células sol pueden originarse por división de una célula preexistente (omnis cellula ex cellula).
Las propiedades de un organismo son el resultado de las propiedades de sus células individuales
Unidad vital más pequeña
Sin embargo, hoy en día se recurre a la definición:
1.1.- CONCEPTO DE CELULA
CELULA: Ser completo, altamente organizado e independiente capaz de realizar labores metabólicas captando materia y energía del medio y capaz de poseer proteínas y ácidos nucleicos que pueden ser sintetizados por ella misma
-Condiciones para considerarla como célula:
a) Existencia de membrana plasmática y reguladora (barrera que regula intercambios Na+ (materia)/E (energía) con el medio
b) Poseer Ácidos Nucleicos (ADN o ARN) capaces de especificar aminoácidos y dar lugar a proteínas
c) Capacidad de producir energía, maquinaria y enzimas para mantener los procesos metabólicos

2.- TIPOS DE CELULAS
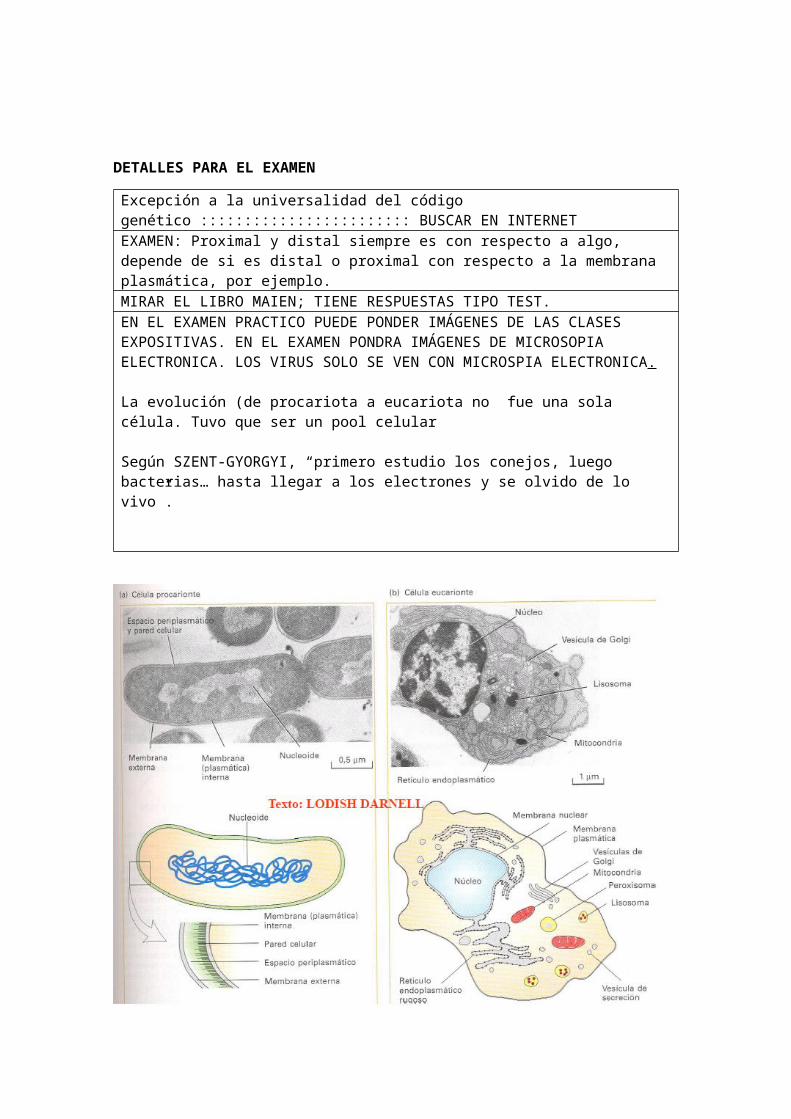
A) PROCARIOTAS:
No tienen núcleo aislado del citoplasma (por lo que no puede hacer reacciones sin que afecte al citoplasma dentro de lo que sería el núcleo). Los organismos son bacterias, cianobacterias y micoplasmas; un tamaño menor (de 1 a 10 micrómetros); no tiene envoltura nuclear (EXAMEN, es envoltura y no membrana);En cuanto al ADN es circular, sin proteínas; No presenta nucléolos; ribosomas 70S; No tiene sistema endomembrana; sus orgánulos son los ribosomas; tienen pared celular (excepto micoplasmas); no hacen endocitosis y exocitosis; no presentan corrientes citoplasmáticas; su locomoción es por flagelo bacteriano; metabolismo aeróbico y anaeróbico; Enzimas respiratorios en mesosomas; división celular por simple bipartición; son formas mayormente unicelularesADN procariota: menos electrodenso y estructura fibrilar. Carece de nucleosoma. Las células procariotas tienen una histona HU que repara el ADN de la procariota cuando se produce un fallo en su replicación
B) EUCARIOTAS:
Características Generales:
TAMAÑO: se refiere al soma (oocitos) no a las prolongaciones. Cajal:o Enanas (3 a 12 micras)o Medianas (12-30 micras)o Gigantes (30-120 micras) o Hay algunas que miden mucho más como el huevo de avestruz o de
gallina aunque el núcleo de la célula requerimos el microscópio FORMA: R/función. Ej: neuronas: morfología estrellada (andan información
lejos). La forma está condicionada por la presión que ejercen las células vecinas,
POLARIDAD: morfo-funcional de la célula: polo basal/polo apical. ; diferenciando la free Surface, y la connective tissue
NUMERO DE NUCLEOS:o Lo normal es unoo Muchos: por ejemplo célula de fibra muscular estriadao Sin núcleo: sincitio (sincitio, trofoblasto de la placenta)
COLOR: Son incoloras (tono: pigmentos endógenos y exógenos) LIBRES/ASOCIADAS: los seres unicelulares son libres, aunque lo normal es el
sistema de asociación celular OTROS DETALLES: Son protistas, hongos, planta y animales; de 10 a 100
micrómetros; Si tiene envoltura nuclear; El ADN son varias moléculas lineales de DNA asociadas a proteínas; tienen nucléolos; ribosomas 80S; Si tienen

sistema endomembrana; Los orgánulos son: núcleo, ribosomas, RE, Golgi, mitocondrias, cloroplastos…; tienen citoesqueleto; solo tienen pared celular las células vegetales; si hacen endocitosis y exocitosis; si tienen corrientes citoplasmáticas; locomoción por cilios y flagelos; el metabolismo es aeróbico; Las enzimas respiratorias son las mitocondrias; división ceular por mitosis; las formas son principalmente pluricelular.
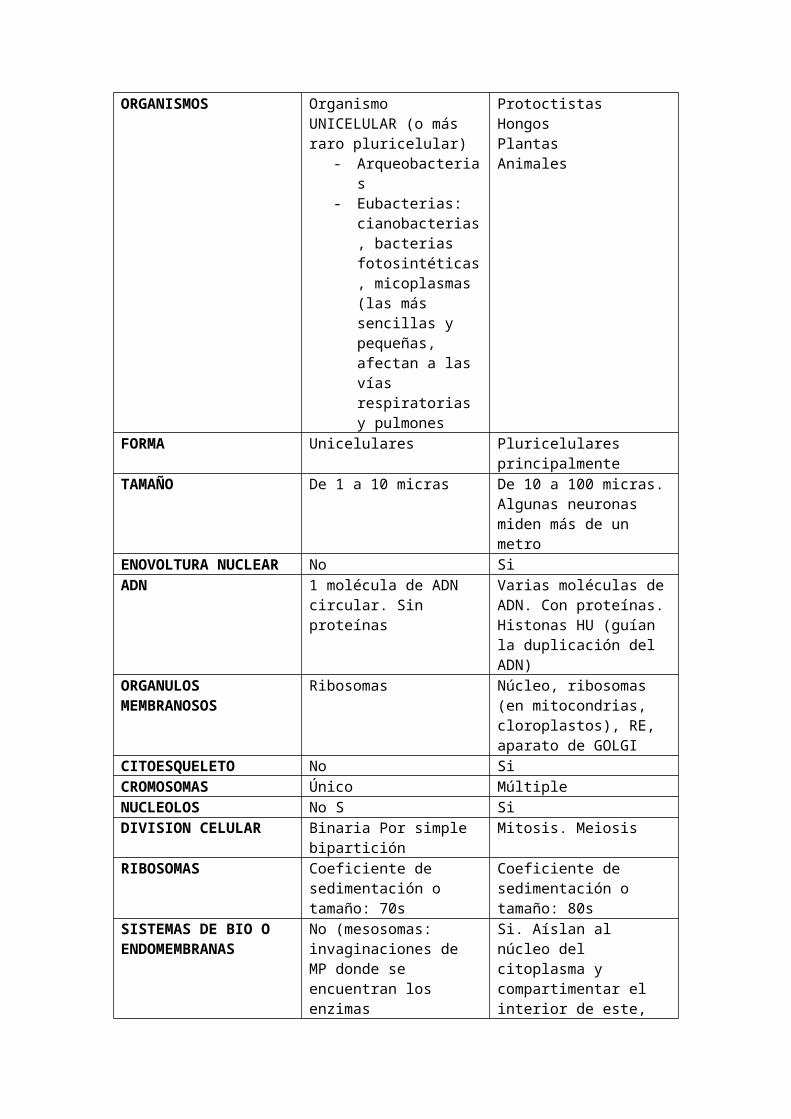
CARACTERISTICAS DE PROCARIOTAS Y EUCARIOTAS EN TABLA:
PROCARIOTA EUCARIOTAORGANISMOS Organismo UNICELULAR (o
más raro pluricelular)- Arqueobacterias- Eubacterias:
cianobacterias, bacterias fotosintéticas, micoplasmas (las más sencillas y pequeñas, afectan a las vías respiratorias y pulmones
ProtoctistasHongosPlantasAnimales
FORMA Unicelulares Pluricelulares principalmenteTAMAÑO De 1 a 10 micras De 10 a 100 micras. Algunas
neuronas miden más de un metro
ENOVOLTURA NUCLEAR No SiADN 1 molécula de ADN circular.
Sin proteínasVarias moléculas de ADN. Con proteínas. Histonas HU (guían la duplicación del ADN)
ORGANULOS MEMBRANOSOS
Ribosomas Núcleo, ribosomas (en mitocondrias, cloroplastos), RE, aparato de GOLGI
CITOESQUELETO No SiCROMOSOMAS Único MúltipleNUCLEOLOS No S SiDIVISION CELULAR Binaria Por simple
biparticiónMitosis. Meiosis
RIBOSOMAS Coeficiente de sedimentación o tamaño: 70s
Coeficiente de sedimentación o tamaño: 80s
SISTEMAS DE BIO O ENDOMEMBRANAS
No (mesosomas: invaginaciones de MP donde se encuentran los enzimas energéticos, pues no tienen mitocondrias)
Si. Aíslan al núcleo del citoplasma y compartimentar el interior de este, se llevan a cabo reacciones químicas diferentes simultáneamente
MITOCONDRIAS No SiENZIMAS RESPIRATORIOS Membrana plasmática
(mesosomas)Si

RESPIRACION. METABOLISMO
Aerobia/anaerobia Aerobia
PARED CELULAR Si (mureina, peptidoglucanos). Excepción: micoplasma, reducida y muy fina
Solo células vegetales (de naturaleza celulósica)
CLOROPLASTOS No Si (vegetales)MOVIMIENTOS INTERNOS No Endocitosis/ExocitosisCORRIENTES CITOPLASMATICOS
No (porque carecen de citoesqueleto)
Si
LOCOMOCIÓN Flagelo bacteriano (Muy primitivo)
Cilio/flagelo
DETALLES PARA EL EXAMEN
Excepción a la universalidad del código genético :::::::::::::::::::::::: BUSCAR EN INTERNETEXAMEN: Proximal y distal siempre es con respecto a algo, depende de si es distal o proximal con respecto a la membrana plasmática, por ejemplo.MIRAR EL LIBRO MAIEN; TIENE RESPUESTAS TIPO TEST.EN EL EXAMEN PRACTICO PUEDE PONDER IMÁGENES DE LAS CLASES EXPOSITIVAS. EN EL EXAMEN PONDRA IMÁGENES DE MICROSOPIA ELECTRONICA. LOS VIRUS SOLO SE VEN CON MICROSPIA ELECTRONICA.
La evolución (de procariota a eucariota no fue una sola célula. Tuvo que ser un pool celular
Según SZENT-GYORGYI, “primero estudio los conejos, luego bacterias… hasta llegar a los electrones y se olvido de lo vivo”.
3.- CONCEPTO DE SER VIVO
Son seres que posee completa estructura material y son capaces de nutrirse, relacionarse y reproducirse (las tres funciones vitales)
4.- SISTEMA ACELULAR
No responde al concepto de unidad vital más pequeña. Mayoritariamente son acido nucleicos que pueden asociarse a otro ácido nucleico y necesita de la maquinaria de replicación de otra célula para crear sus propias proteínas. No pueden ser las células más pequeñas porque necesitan de otra célula para reproducirse=
VIRUS: AN (ADN O ARN) más proteínas. Poseen proteínas pero son incapaces de sintetizarlas por si mismos
VIRION: Virus extracelular. Es inerte, carece del metabolismo de la célula parasitada. Algunos autores los consideran genes móviles que se trasladan de célula en célula (30-300nm) (250-2000A)
VIRUSOIDES: ARN/virus de virus
PRIONES: glicoproteínas sin AN (Síndrome de Geutzfeld-Jacob, encefalitis bovina o mal de las vacas locas, Kuru…) Los priones son capaces de autoreplicarse dentro de las células, debido a la posibilidad de que activaran algún gen del ADN huésped para codificaran las proteínas del prion.
Su tamaño es 100 veces más reducido que los virus más pequeños
Son proteínas con una secuencia de aminoácidos igual o casi igual que la de una proteína normal pero tienen una forma espacial distinta. Los priones son capaces de inducir que las proteínas normales de la célula adopten la forma del prion. Generalmente, los priones son proteínas de membrana de las neuronas, por lo que suelen provocar enfermedades neurodegenerativas
VIRINOS: Son ácidos nucleicos con afinidad a proteínas del sistema nervioso
VIROIDES: fragmento de ARN circular (atacan al núcleo de células vegetales)PLASMIDOS: fragmentos de ADN bicatenario independientes del ADN bacteriano+
TRASNSPOSONES: fragmentos de ADN o genes que a diferencia de los provirus se integran en el ADN celular en lugares bien determinados
PROVIRUS: Genes móviles (fragmentos de ADN.
PROVIRUS: Genes móviles (fragmentos de ADN. Pasan
VIRUS ONCOGÉNICOS: Pedir a la otra clase
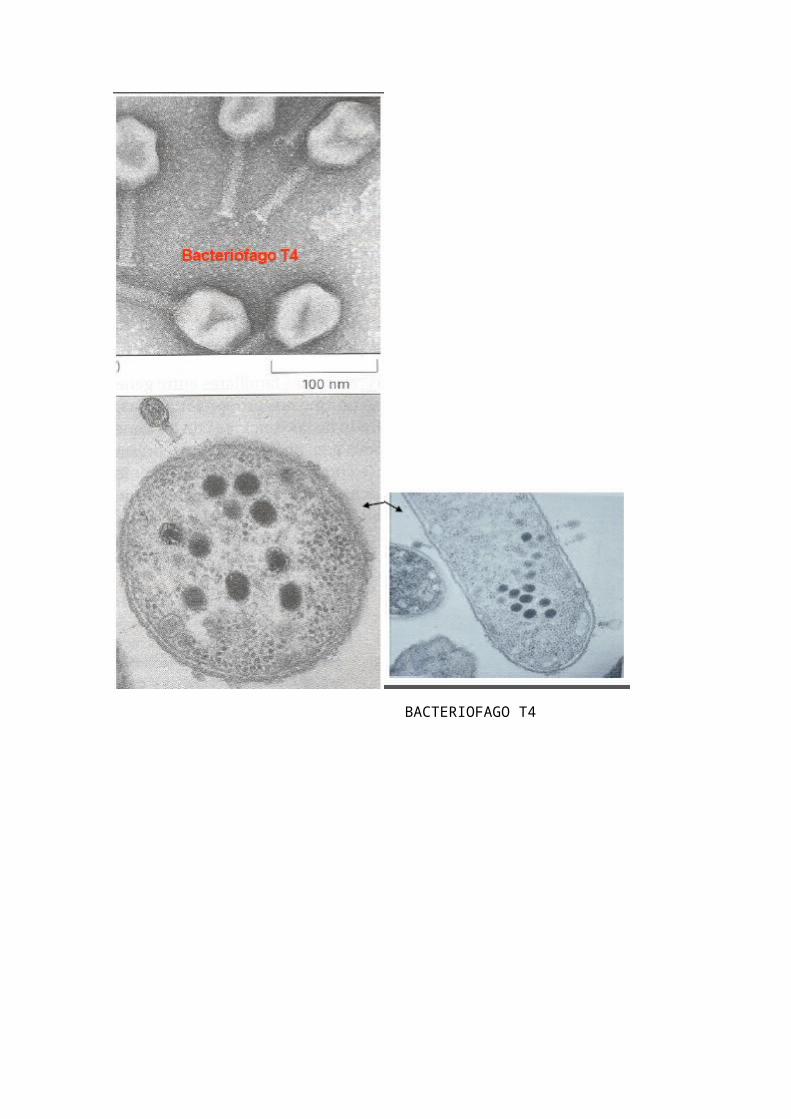
BACTERIOFAGO T4

UNIDAD TEMATICA 2: MEDIO AMBIENTE Y SUPERFICIE CELULAR
MEDIO AMBIENTE DE LA CELULA
MEDIO EXTERNO A LA CELULA. INTRODUCCIÓN
Cuando pertenece a un tejido, la célula sufre un proceso de especialización. Existen elementos que crean la matriz de la célula, es decir, de crear un ambiente alrededor de la célula para que pueda desarrollar sus funciones vitales. Este elemento es esencialmente el fibroblasto. El fibroblasto produce el medio extracelular, la matriz, las cuales les interesa tener a las células que las produce. En el caso de la sangre se trata de un medio extracelular de plasma sanguíneo, por eso es un tejido. Por otro lado, cabe destacar los GLUCOAMINOGLUCANOS, los cuales son espacios donde podemos encontrar gran cantidad de agua, rodeado a su vez de proteínas fibrosas que crean la matriz de este tejido
Existen tres factores clave para entender la asociación entre células y por tanto la formación de tejidos:
1.- Comunicación celular: Se reconocen mutuamente entre ellas asi como la matriz extracelular, solo se asocian células que se reconocen. Es decir una célula hepática solo le da la mano a una hepática, y así todos los tejidos. Los que no tiene células cerca le da la mano a la matriz extracelular.
2.- Adhesión selectiva célula - célula o célula-matriz
3.- Memoria celular. Solo se une con las células que reconoce.
MATRIZ EXTRACELULAR
Se trata del medio altamente hidratado que rodea a la célula, estabilizando su estructura, facilitando su adherencia y regulando su comportamiento.
Los fibroblastos son los encargados de crear esta matriz. Ayudando a los fibroblastos en la síntesis de esta matriz están los ostoblastos, los cuales aportan sales, y los condroblastos. Las proteínas de la matriz extracelular pueden ser muy diferentes.
Según la cantidad de matriz extracelular, puede ser mucha cantidad (dermis, hueso, cartílago) o muy poca matriz como las células nerviosas, cuya matriz la médula espinal.
Sea cual sea el tejido y la cantidad: la composición de la matriz es siempre la misma: proteínas y polisacáridos. Tanto las proteínas como la naturaleza bioquímica de los polisacáridos difieren según el tipo de tejidos.

Dentro de la matriz extracelular, y estando formados por las proteinas y polisacáridos antes nombrados, podemos destacar los AMINOGLUCOSAMINOGLUCANOS (BUSCAR COMPOSICIÓN BIOMOLECULAR) están formados por un aminoazucar que se repite estando unido a proteínas (proteoglucanos). Los proteoglucanos están formando muchas proteínas de la matriz celular formando también parte de la membrana plasmática.
Como ejemplo de proteínas y polisacáridos que podemos encontrar en la matriz, tenemos:
Proteínas : Refuerzan y organizan la matriz. Cabe destacar: la colágena (la encontramos en los tendones ya que tiene que ser muy elásticos), elastina, laminina, fibrilina, fibronectina (esta proteínas une fibras) (BUSCAR PARA QUE VALE CADA UNO DE ESTE TIPO)
Polisacáridos : son glucosaminoglucanos siempre. Hacen que retengan mucha agua. Contribuye a que tengan aspecto de gel o moco
Organización: Variable (de si forman parte de un hueso, de la córnea, tendón…)
Según la composición (proteínas y polisacáridos) forman la lámina basal, estroma (están in vivo) o cultivo (in vitro); al estar en cultivo también necesitan una matriz.
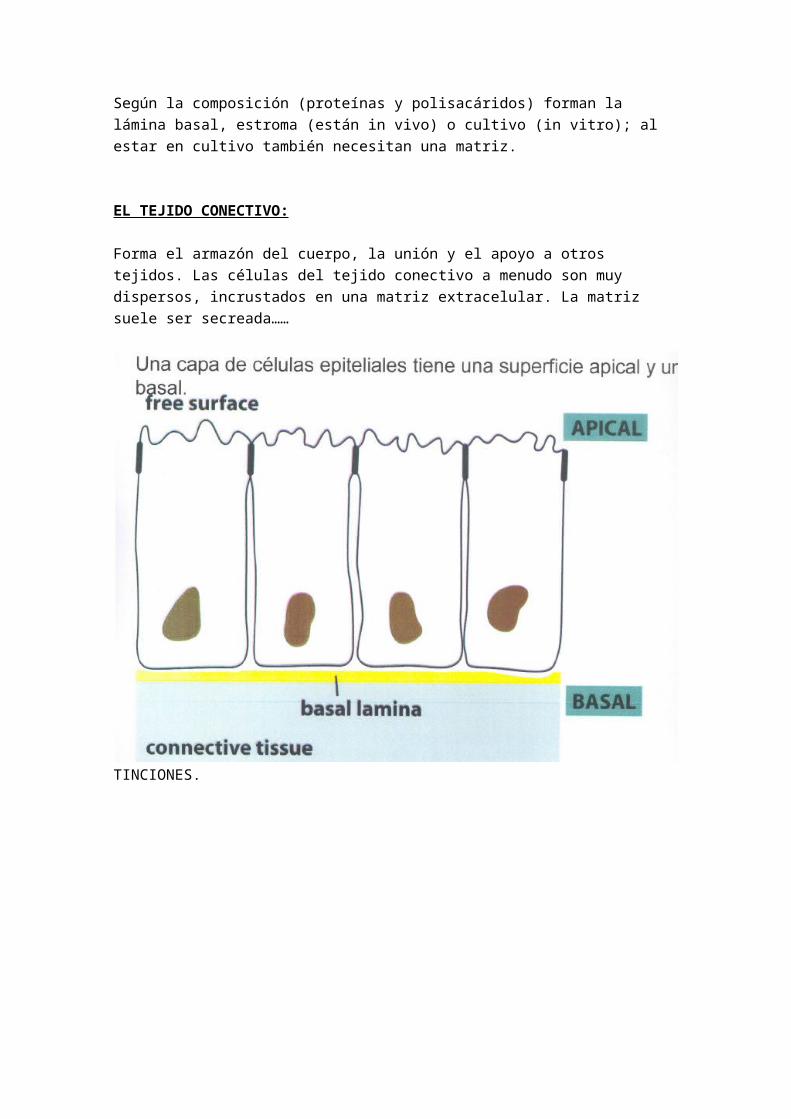
EL TEJIDO CONECTIVO:
Forma el armazón del cuerpo, la unión y el apoyo a otros tejidos. Las células del tejido conectivo a menudo son muy dispersos, incrustados en una matriz extracelular. La matriz suele ser secreada……
TINCIONES.

MEMBRANA PLASMATICA I
o Concepto y generalidades de las biomembranas.o Evolución de los modelos de membrana.o Imagen microscópica o Técnicas de aislamientoo Composición química y anatomía molecular de la membranao Las cubiertas de la membrana plasmática
MEMBRANA PLASMATICA I: CONCEPTO Y GANERALIDADES DE LAS BIOMEMBRANAS
1.- CONCEPTO DE MEBRANA PLASMATICA
La membrana se trata de la biomembrana de la superficie celular:
- Presenta: “cell coat” (cubierta celular) glucocalix- Definición de contorno celular- Es una barrera selectiva y dinámica (desigual concentración de iones, carga desigual, presión osmótica interna constante)- Se encarga del reconocimiento y comunicación intercelular
La membrana plasmática es una barrera que se interpone entre el medio externo y el medio interno celular (medios extracelular e intracelular)La membrana plasmática se sitúa en la superficie celular y también tabicando el citoplasma celular.Composición bioquímica variable debido a las moléculas que la constituyen
1.1 CARACTERISTICAS DE LAS BIOMEMBRANAS

o Estructuras BIFACIALES: se enfrentan con 2 medios (externo o interno). En el caso de la membrana plasmática una se enfrenta a la matriz (cara exoplasmatica) y otra al citoplasma
o Estructuras UNITARIAS: tienen el mismo patrón estructural las biomembranas del interior (biomembranas de Golgi, mitocondrias…) que la superficial.
o Estructuras DIVERSIFICADAS: no puede tener la misma función la biomembrana del aparato de GOLGI que la membrana plasmática. Tiene distinta composición química.
1.2 BIOMEMBRANA: FUNCION
o COMPARTIMENTACION: Las biomembranas intervienen en los mecanismos de transporte (limita la entrada y la salida)
o RESPONSABLE DE LAS DIFERENTES (IONES), CARGAo REGULAR LA BIOLOGÍA CITOCELULAR: reciben y procesan la información que llega al
exterior y la que circula dentro. Luego puede emitir una respuestao REGULA LA PRESION OSMOTICAo Es CONTINUA y es responsable de la FORMA DE LA CELULA: Debido a la polaridad de la
celua tenemos membrana basal, membrana apical y membrana lateralo La membrana es la que reconoce a la célula vecina o la rechaza y no se asocia con ellao Locomoción: en muchos casos responsables del movimiento.
La célula debe protegerse del frio. Cell coat (un abrigo de naturaleza glucídica)Es una cubierta celular o GLUCOCALIX:
o Aparece en la car externa de la biomembrana celularo No se suele encontrar en las biomembranas del interior del citoplasma
Superficie celular es igual a la biomembrana + el glicocalix
1.3.- ANATOMIA DE LAS MEMBRANAS (evolución del conocimiento de la MP)
o MICROSCOPIA OPTICA (MO) : No se ve (menor de 0´2 micras). Pero se sabía qe estaba porque si se teñía el medio extracelular no entraba el tinte en la célula y viceversa. Se observa le glucocalix (que se tienen con PAS)
o ME: Trilaminar (20A- 35A- 20A): 75-80A: dos capas electrodensas/una intermedia entre ambas más clara. Asimetría (glucocalix) cubierta externa más gruesa por la presencia del glucocálix
o ME barrido: bilaminar (caras E y P). Asimétrica (partículas intramembrana)
o MICROSCOPIA OPTICA (MO): el estudio de la celula comieza en el siglo XIX con los primeros microscopios. De la observación de celdillas nace la teoría celular. Estas celdillas parecina tener unos limites entre ellas. Pero la escas resolución del MO (0´2micras) no permitía ir mas alla. Se procedio entonces a la tinción de las células, es decir, a inyectar dentro de las células un colorante, observando que éste no se difundia por las otras deduciendo asi que debía haber algún tipo de barrera. Tambien se recurrio a una tecnia de tinción de glúcidos, la TINCION DE PASS (acido periódico de Shiff) lo que producia una parte coloreada en el exterior de la celula (el glucocalix).
CURIOSIDAD: Las células del epitelio intestinal presentan un glucocalix desarrollado (puede alcanzar un grosor de 300 micras y se puede ver con MO
o MICROSCOPIA ELECTONICA CONVENCIONAL (DE TRANSMISION): con esta tecnología en los años 40 se pudo apreicar de forma sencilla la membrana plasmatica.
La biomembrana se definio como una estructura TRILAMINAR:o Una capa electrodensa que se relaciona con la matriz extracelular: capa
OSMIÓFILA (afinidad por el osmio)o Una capa clara intermedia (no electodensa). Capa OSMIÓFOBAo Una capa electrodensa interior (OSMIOFILA)
El grosor de las tres capas unidas varía entre 70A y 80A:o Capa exterior: 35ªo Capa intermedia e interior: 20A cada una
La capa exterior es más gruesa que la interna debido al glucocalix
o ME DE BARRIDO: mediante técnicas de criogenización y fractura (criofractura). Esta nueva técnica permitió una mayor resolución. Se realizó a la congelación de la célula y su fractura con un golpe de cuchillo (para evitar el exceso de agua se recurría a la sublimación). Se observó que la membrana se fracturaba en dos capas: ESTRUCTURA BILAMINR:
o Cara E (Exoplasmatica): Se relaciona con la matriz extracelularo Cara P (Protoplasamatica): más en contacto con el citoplasma.
Al fracturar se observaban unas partículas intramembrana más presentes en una cara que en otra (asimetría). Pero las partículas presentes en una delas caras encajaban en las otras. La cara P tiene abundantes partículas intramembrana.
1.4.- COMPOSICION QUIMICA DE LAS MEMBRANAS
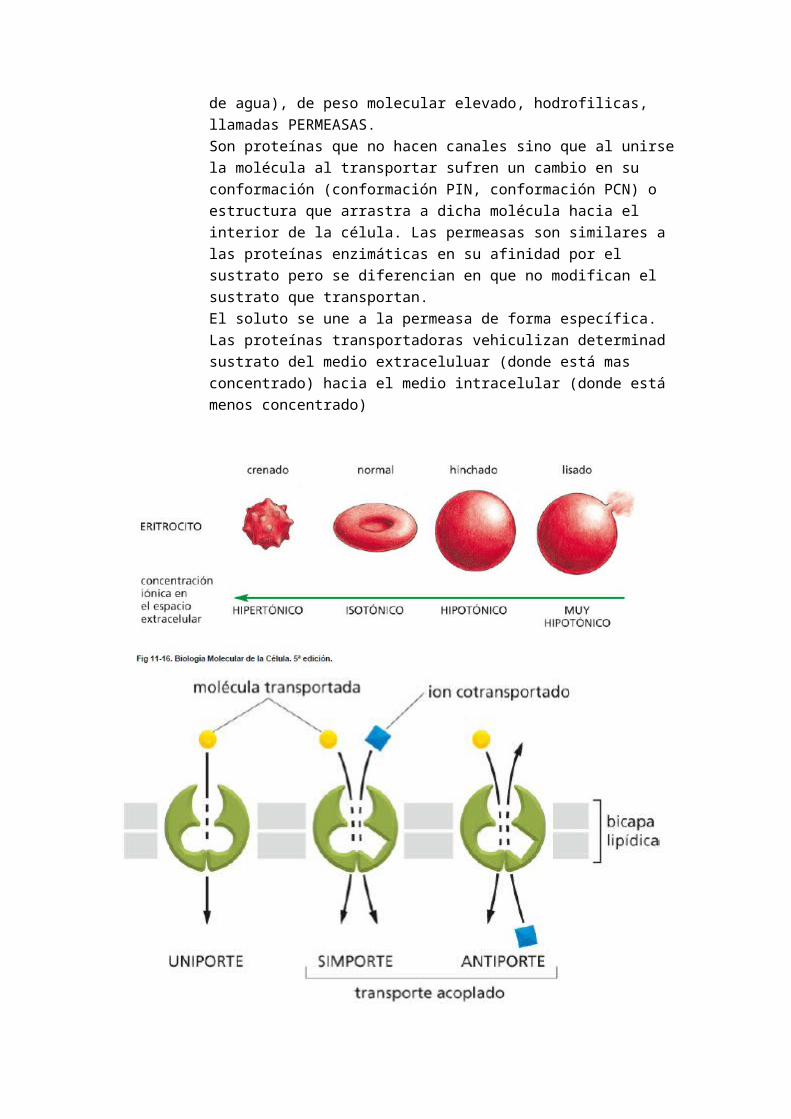
Para el estudio de la composición química de la membrana, se recurrió a un simple experimento. Los eritrocitos o hematíes son el material de elección para el estudio de la membrana plasmática.Se eligió el eritrocito humano su fácil obtención, bajo coste, abundancia y es la membrana menos contaminada (no hay restos de orgánulos) ya que carece de otras biomembranas internas (sin núcleo ni orgánulos) y solo posee hemoglobina.Se procedió al aislamiento de las membranas de los eritrocitos. Para ello se recurrió a someterlos a distintos medios tónicos:
o En medio hipertónico: perdía agua y se arrugabao En medio hipotónico: captaba agua, se hinchaba y se rompía (hemolisis)
La hemolisis hipotónica se produce en un tubo de ensayo mediante técnicas de centrifugación y da lugar a la hemoglobina y a los fantasmas de membrana (restos de la membrana celular del eritrocito).
o Si la lisis es SUAVE, los fantasmas pueden volver a sellarse formando nuevas vesículas de dos tipos:
o Rectos: conformes con la estructura celularo Invertidos: con la estructura invertida, capa invertida hacia el exterior.
o El procedimiento con hemolisis DRASTICA con formación de fantasmas blancos de membrana será el utilizado para el estudio de la membrana celular. Los fantasmas blancos no se pueden volver a unir. Estos fantasmas blancos nos indican la composición de la membrana.Fantasmas blancos de membrana: COMPOSICION QUIMICA

En los fantasmas blancos se encontraron: lípidos y proteínas:o Lípidos: eran las sustancias más abundantes. Tres tipos:
o Lípidos: FOSFOLIPIDOS (los mas abundantes)o Glucolipidoso Colesterol
o Proteínas: podían ser simples o:o Asociadas a glúcidos (glucoproteínas)o Asociadas a lípidos
A) LIPIDOS
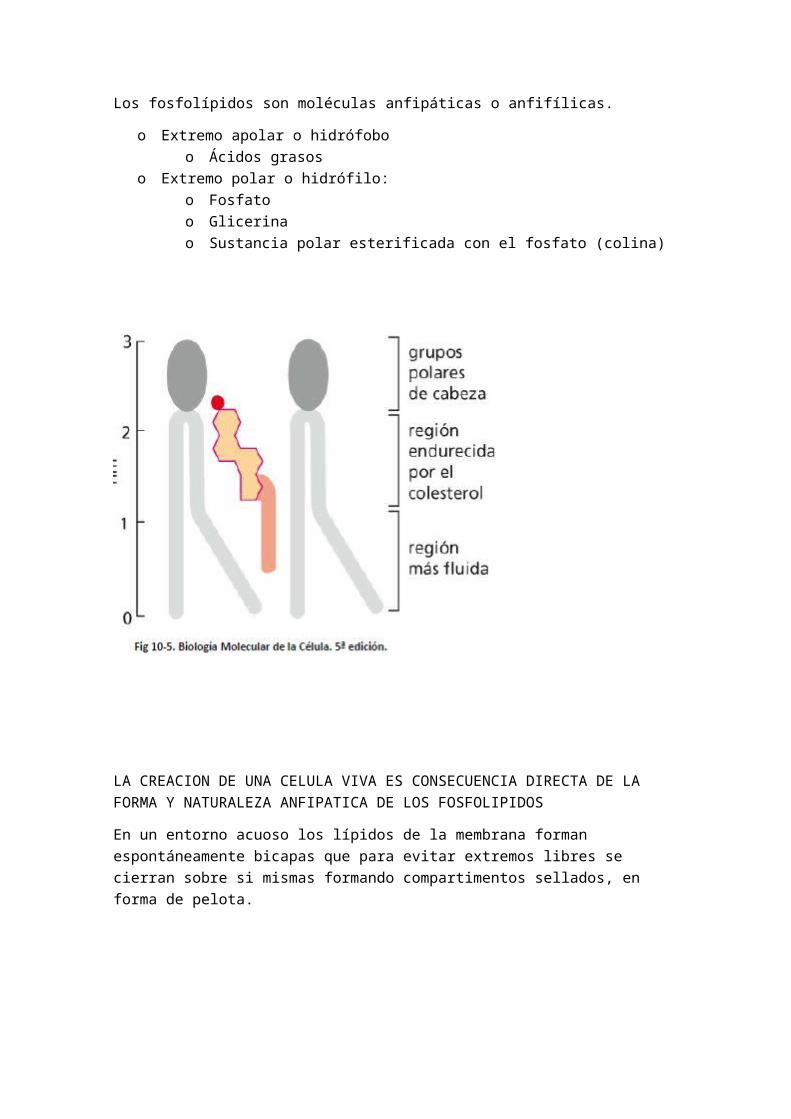
Se trata del elemento que le confiere mayormente la cualidad de fluidez del modelo, ya que están en continuo movimiento. Se tratan de moléculas antipáticas. Cada lípido posee una cabeza hidrófila y una o dos colas hidrófobas.
Los lípidos de las membranas son:
o Fosfolípidoso Colesterolo Glucolipidos
A.1.- FOSFOLIPIDOS
Son los lípidos más abundantes en las membranas celulares. Presentan en la cabeza hidrófila en GRUPO FOSFATO. Hay dos tipos:
o Fosfogliceridoso Esfingolipidos
Fosfoglicéridos
Contienen una molécula de glicerol esterificado con dos ácidos grasos. El tercer grupo hidroxilo (OH) del glicerol se une a un grupo fosfato formando el ácido fosfatidico. El grupo fosfato se une a otros radicales que dan nombre a los fosfogliceridos. La fosfatidiserina está en la cara interna por eso va a tener potencial de membrana. Es importante en la comunicación intracelular fostatidilinsitol, porque envía señales al interior de la célula. La fosfatidiletanolamina tiene función estructural.
Los esfingolipidos tienen también un grupo fosfato, pero se forman a partir de la esfinfosina a la que se une un ácido graso siendo dos cadenas hidrófobas. La esfingomielina es la más importante y tiene también colina.

Colesterol
Es un esterol, presente en las membranas de células animales. Es importante porque contribuye a la rigidez de la membrana, tiene por tanto función estructural. Es precursor de (BUSCAR). Tiene un grupo polar pequeño. Presenta una estructura rígida (esteroide) y una pequeña cadena hidrocarbonada.

Glucolípidos
Se forman a partir de la esfingosina a la que se añade una cadena de ácido graso y grupos de carbohidratos. Los más comunes son los cerebrosidos y los gangliosidos. En la mielina es especialmente abundante en la galactocerebrósidos.

CONFORMACIÓN ESTRUCTURAL DE LOS FOSFOLÍPIDOS
En cuanto a la conformación de las los Fosfolípidos, podemos determinar que: los lípidos forman espontáneamente monocapas o bicapas cuando están en suspensión acuosas. Estas capas pueden ser planas o esféricas.
o MONOCAPAS:o Planas: superficie
(DIBUJO AQUÍ)
o Esféricas: micelaso (DIBUJO AQUÍ)
o BICAPASo Planas: membranas negras
(DIBUJO AQUÍ)
o Liposomas: vesículas esférica formada por una doble capa de fosfolípidos(DIBUJO AQUÍ)
Los fosfolípidos son moléculas anfipáticas o anfifílicas.
o Extremo apolar o hidrófoboo Ácidos grasos
o Extremo polar o hidrófilo:

o Fosfatoo Glicerinao Sustancia polar esterificada con el fosfato (colina)
LA CREACION DE UNA CELULA VIVA ES CONSECUENCIA DIRECTA DE LA FORMA Y NATURALEZA ANFIPATICA DE LOS FOSFOLIPIDOS
En un entorno acuoso los lípidos de la membrana forman espontáneamente bicapas que para evitar extremos libres se cierran sobre si mismas formando compartimentos sellados, en forma de pelota.

B) PROTEINAS DE MEMBRANA
La parte mosaico del modelo. Las proteínas confieren a las membranas sus propiedades funcionales características por ello la cantidad y el tipo de proteínas es muy variable en las diferentes biomembranas. La carga proteica depende de las funciones de la célula.
Las proteínas se asocian con la bicapa lipídica de diferentes maneras:
o Proteínas integrales de membrana (1-4): Poseen una o más regiones incluidas en la bicapa lipídica
o Proteínas periféricas: ancladas a lípidos (5-6) o asociadas con otras proteínas (7-8)
Todas estas tendrán una disposición hélice alfa, que es la más características de las eucariotas, excepto al de tipo 3, que es lamina beta, pero este tipo es más propio de procariotas. En eucariotas más típico es la alfa.
La 5 y la 6, tienen enlaces covalentes, el 7 y 8 están enlazados con enlaces débiles (de hidrogeno); la 7 es una proteína transmembrana unida a otra.
CARACTERÍSTICAS:
o La bicapa es FLUIDA: Esencial para la función de la membrana. La fluidez de la membrana depende de la temperatura y su composición. Las membranas ricas en ácidos grasos con cadenas cortas tienden a ser más fluidas, las membranas son ácidos grasos insaturados son más fluidas.
La fluidez de la membrana está regulada por el colesterol, una molécula corta y rígida especialmente abundante en la membrana plasmática. La membrana es más fluida ya que se trata de fosfolípidos insaturadas. El colesterol también influye en la fluidez, a más colesterol, más rigidez, ya que la célula presenta ciertas regiones más rígidos, necesita esta región para que las proteínas que aquí habrá no se muevan continuamente.

o La mayoría de los lípidos se distribuye de forma desigual entre las dos monocapas, LA MEMBRANA ES ASIMETRICA. La simetría se mantiene de tal forma que la monocapa protoplasmática es aquella capa de las dos que hay de pega la citosol, siendo su estructura siempre igual. La APOPTOSIS es un proceso de muerte programada, por ejemplo, si está infecta con un virus; siempre se muere desde la capa interna hasta la capa externa, al ver esto los monocitos se dan cuenta, al morir de esta forma, y hay que fagocitarla antes de que vierta el virus al medio.

1.- EVOLUCION DE LOS MODELOS DE MEMBRANA
La membrana plasmática envuelve la célula, definiendo sus límites y manteniendo las diferencias entre su contenido y el entorno. Dentro de la célula la membrana delimita orgánulos y mantiene la diferente composición entre el contenido de cada orgánulo y el citosol.
a) En 1895 Overton estableció que las sustancias no polares si podían atravesar la membrana y las polares si, por lo que estableció que debía de ser de tipo lipídico
b) En 1897 Langmuir estableció que tiene dos regiones, una parte hidrófoba y otra hidrófila (fosfolípidos). Al mezclarlo con benceno, consiguió una monocapa de fosfolípidos
c) En 1925 Gorter y Grendel; estableció que existían dos membranas. Realizaron estudios con glóbulos rojos ya que carecen de núcleo, ellos repitieron el experimento de Langmuir, y consiguieron el doble de la superficie de la monocapa de lo que debería ser, por lo que determinaron que era una bicapa.
d) En 1935 Davson y Danielli establecieron la bicapa lipídica con proteínas globulares en sus laterales, ya que se dieron cuenta de que había elementos que no debían atravesar la membrana cuando realmente si acontecía; dedujeron que había proteínas a ambos lados de la bicapa (se le conoce por eso como sándwich). Supuestamente las proteínas era lamina beta
e) 1959 Robertson. Aparece la microscopia electrónica. Observo que exiistia la estructura trilaminar, es una región electrodensa formado por dos capas que rodean a una tercera. Establecio que todos los orgánulos tienen esta estructura. Creian que esto se debía al modelo sándwich anterior
f) 1964 Lucy y Glaubert: determinaron la presencia de micelas envueltas por capa glicoproteica. Ni Robertson ni Davson y Danielli explicaban la transmisión del impulso eléctrico
g) 1972 Singer y Nicolson: Modelo del mosaico fluido. Cambia ya que dice que hay proteínas no solo en la superficie como el modelo sándwich si no que están inmersas en la bicapa.

MODELO MOSAICO FLUIDO
Según este modelo la membrana plasmática está constituida por una doble capa de lípidos a la que se asocian moléculas proteínas, que pueden situarse en ambas caras de la superficie de dicha doble capa o englobadas en la misma formado una estructura denominada “mosaico fluido”
Bicapa lipídica es igual a Fosfolípidos + colesterol + glucolipidos
Con la técnica de criofractura se divide la membrana en dos. La hemimembrana interna o protoplasmática (P) y la Externa o exoplasmatica (E)
La superficie de las hemimembrana nos es lisa. En un lado resaltan partículas que se corresponden con cavidades en el lado complementario.
El modelo de mosaico fluido determina que es: Fluida (ya que sus componentes se mueven a lo largo de la membrana), es Asimétrica (la capa exterior es distinta de la interior), es en Mosaico (es un mosaico formado por biomoléculas unidas entre sí)
Demostración de la teoría del mosaico Fluido
o Demostración microscópica:
o ME convencional: esta teoría explicaba las tres capas de la observación electrónica (capas osmiofilas y capa osmiofoba). Ahora se identifican esas tres capas con la doble capa lipídica
o ME de barrido: el experimento de la criofracutra y la consolidación de las dos bicapas cobrada forma al identificarlas y explicar la composición fosfolipidica de esta bicapa. Además, explicaba por qué se rompía la membrana por el medio: era la zona de enlaces más débiles: uniones entre las colas.
CARACTERISTICAS DE LA MEMBRANA PLASMÁTICA
A) FluidezB) Asimetría
A) FLUIDEZ: MOVIMIENTO DE LAS BIOMOLECULAS EN LA MEMBRANA PLASMATICA
Cada una de las biomoléculas que conforman la membrana plasmatica realiza una serie de movimientos en el interior de la estructura de esta membrana:
A.1.- Fosfolípidos
Realiza tres tipos de movimientos:
o Difusion lateralo Rrotacion sobre el eje mayoro Flexion y flip-flop
DEMOSTRACION DE LA FLUIDEZ FOSFOLIPIDICA
o Cubo agujereado: tenemos un recipiente con una lamina intermedia agujereada y disolvemos fosfolipidos en el agua. Tras un tiempo estos forman una dobel membrana en el agujero.
o Platina de apalstamiento: se podía demostrar la teoría si se formaba una monocapa de fosfolipidos y se comprimia hasta el máximo. El resultado era una concentración de fosfolipidos igual a la mitad de la que podíamos calcular en una membrana. Esto se debe a que el experimento se realiza con una monocapa. Monocapa + monocapa es igual a la bicapa.
o Otra forma de estudiar esa fluidez era estudiar como se movían los fosfolipidos. Los fosofolipidos (y glucolipidos) de la bicapa tienen tendencia a girar sobre si mismo y desplazarse laterlamente por si monocapa. Solo ocasionalmente pueden cambiar de capa lipídica. Esta movilidad de las moléculas origina una fluidez en la membrana, tal que se permite adaptarse a las condiciones variables del medio.
Tipos de movimientos de los fosfolípidos:
1.- ROTACION: giro sobre su propio eje2.- FLEXION: movimiento de sus colas hidrocarbonatada3.- TRASLOCACION: difusión lateral por la monocapa4.- FLIP-FLOP: (raro, ocasional). Cambio de hemicapa. Pueden saltar de una hemicapa a otras (necesita ayuda de otro). En las biomembrana del REL resulta ser un movimiento habitual. Presencia de las enzimas flipasas.
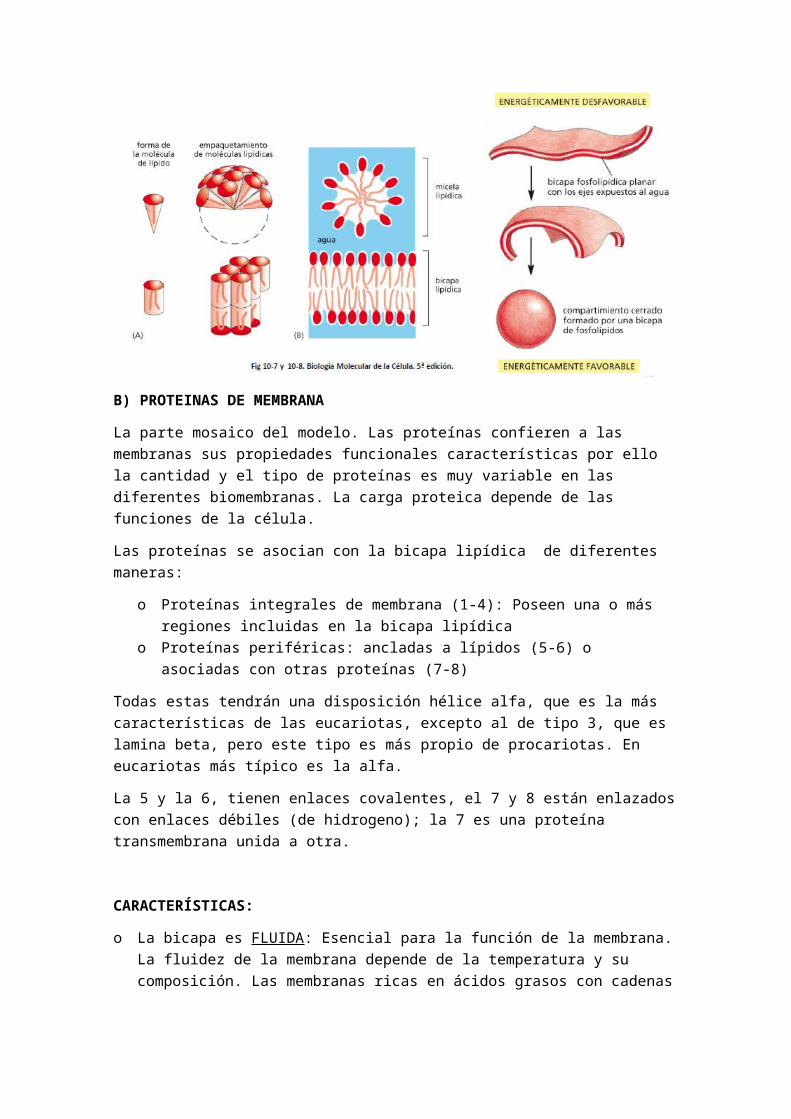
Tipos de movimientos de las proteínas:
1.- Movimientos de difusión lateral: (heterocarionte marcado con anticuerpos fluorescentes). Son muy variadas y debido a su mayor tamaño tiene mayor dificultad de movimiento.
Demostración de la fluidez de las proteínas:
Heterocarionte marcado con anticuerpo fluorescentes: para demostrar la difusión lateral de las proteínas se recurrió a un experimento. Consistió en fusionar una célula humana (con proteínas de membrana marcadas) y otra célula de rata (con sus proteínas también marcadas). Se fusionaron con ayuda de un virus obteniendo una célula con dos núcleos (heterocarionte) y proteínas de ambos tipos en su membrana plasmática. Primero estas proteínas se situaron las humanas en un polo y las de rata en otro. Tras un tiempo las proteínas de ambos tipos aparecieron mezcladas, lo que demostraba su movimiento (su difusión lateral por la membrana)

o Factores que influyen en la fluidez:o Instauración (a mas doble enlace, más fluidez)o Colesterol (regula la fluidez y estabilidad, a más colesterol menos fluidez)o Temperatura ( a menos temperatura menos fluidez)
La fluidez de la membrana plasmática depende de la instauración del os fosfolípidos, la cantidad de colesterol y la temperatura.La instauración de las cadenas hidrocarbontadas aumenta la fluidez de la doble capa lipídica. Por el contrario, la saturación de las cadenas las vuelve viscosasLa cantidad de dobles enlaces determina el grado de fluidez de la membrana plasmática.
El colesterol refuerza la estabilidad de la membrana:
o Grupo hidrofilico: se dispone entre los grupos polares de la cabeza del os fosfolípidoso Grupo hidrofóbico: se intercala en la zona de las cadenas antipáticas de los fosfolípidos

El colesterol inmoviliza las cadenas de fosfolípidos vecinos y hace que la membrana sea rigida. El colesterol se intercala las cadenas hidrocarbonadas. Si baja la temperatura el colesterol evita que se peguen unas colas a otras.
Temperatura: una bajada en la temperatura provoca la síntesis de lípidos insaturadas de membrana – aumenta la fluidez de la membrana. En condiciones normales si baja la temperatura las células se defienden sintetizando dichos lípidos insaturados- aumentando la fluidez.Ej: mantequilla: frio, mas rígida (baja la temperatura), calor, más fluida (sube la temperatura). Para evitarlo las células sintetizan lípidos insaturados
B) ASIMETRIA DE LAS MEMBRANAS
o Fosfolípidos:o Hoja externa: esfingomielina y fosfatidilcolinao Hoja interna: fosfatidiletanolamina y fosfatidilserina. Capa con carga global
negativa
La distribución de los lípidos entre las dos capas es asimétrica: esfingolipidos, fosfatidilcolinas y gluocolipidos son más abundantes en la cara externa de la bicapa lipídica; fosfatidletanolamina y fosfatidilerinas son más abundantes en la cara interna.También se encontró una diferencia de carga eléctrica entre ambas capas. Esto se debe a que la fofatidilserina está cargada eléctricamente (-) y como es más abundante en la cara interna esta es negativa. La cara externa tiene una carga neta global neutra.
o Glucolipidos (glucoesfingolipidos):o Hoja externao Cerebrósidos (sencillos: 1M de azúcar)o Gangliósidos (complejos: oligosacáridos)
Tay-Sach-GMIo Proteínas (antipáticas)
o Estructura hélice, hoja B, P. multipasoo R/lípidos: transmembrana parcial, transmembrana periféricao R/extracción: intrínsecos (SDS-detergente), extrínsecas (s. sal)o R/naturaleza: lipoproteínas, glucoproteínaso R/función: receptores, integrinas, puente, enzimáticas, transportadoras
(bombas Na-k) Glóbulo rojo: Espectrina; periférica, dímero, forma bicóncava
Glucoforina: intrínseca hélice alfa, antígenos MN Banda III: intrínseca, multipaso, T. O2 Y CO2

Las moléculas proteicas son menos numerosas que los lípidos (100 lípidos por cada molécula proteica) pero entre 30 y 50 veces más voluminosas que estas.Además, son un grupo mucho más variado y heterogéneoLas proteínas son moléculas ANFIPATICAS. A lo largo de la proteína se alternan regiones hidrofilicas con regiones hidrofóbicas (en función de la naturaleza de los aminoácidos que las constituyen).Sus radicales polares (o extremos hidrofilicos NH2, COCH) quedan fuera de la membrana y sus radicales lipófilo contactan con los lípidos de la membrana.Estructura:
o Helice alfao Hoja betao Proteínas multipaso (atraviesan la membrana varias veces)(alfa+beta)
TIPOS DE PROTEINAS
Podemos establecer distintas clasificaciones:
o Según las posición de la proteína en relación a la los lípidos:o Transmembrana : atraviesan las dos capas lipídicaso Parcialmente transmembrana : atraviesan una sola capa lipídicao Periféricas : se sitúan en el exterior de la membrana interactuando de forma
débil con lípidos o proteínas de la membrana
o Según la dificultad o modo de extracción de las proteínaso Intrínsecas : están dentro de la doble capa lipídica fuertemente unidas. Es
necesario el uso de un detergente (SDS) para separarlaso Extrínsecas : se separan mediante un lavado en una solución salina. Se
encuentran “adosadas” a la bicapa
o Según la naturaleza de la molécula no proteica que la forma (heteroproteinas):o Lipoproteínas : grupo prostéticos es igual a los ácidos grasoso Glucoproteínas : grupo prostético es igual a la glucosa
o Según su función:o Receptoras : responsables de la biología citosocial de la célula (envío y
recepción de mensajes, reconocimiento celular…)o Integrinas : comunican al medio ambiente interior con el exterior. Interactúan
con el citoesqueleto y con moléculas de la membrana extracelularo Puente : median las conexiones de una célula con la célula vecina, formando
parte de los complejos de unióno Enzimáticas

o Transportadoras : regulan el paso de sustancias a través de la membrana celular (ejemplos: permeasas, bomba Na-K)
Ejemplos de proteínas de membrana del GLOBULO ROJO:o Espectrina:
o Periférica orientada hacia el citoplasmao Es un dímero formado por dos alfa-héliceso Función: mantener la forma bicóncava
o Glucoforina:o Intrínsecao Estructura alfa-héliceo Antígenos MN (responsable de los determinados antigénicos de los eritrocitos)
o Banda IIIo Intrínsecao Multipasoo Responsable del intercambio de O2 y CO2 de la sangre con los tejidos
o Fibronectina:
Características:
o Periféricao Interacciona con proteínas del grupo de las integrinas de al membrana
plasmática y con las fibras de colágeno (en relación con la membrana extracelular, elementos de la m. extracelular)
o Dimérica
La fibronectina interviene en los mecanismos de la coagulación sanguínea y cicatrización. Proteína aislada a partir de los fibroblastos, se encuentra también en el plasma donde interviene en los mecanismos de la coagulación… y en la m. extracelular (formas fibrillas).

DOMINIOS LIPÍDICOS O BALSAS LIPIDAS
o Los lípidos y las proteínas no difunden libremente y no se atribuyen al alzaro Existe una regionalidad de las dos hemimembranas: se forman micro y macrodominioso La membrana plasmática es un mosaico de microdominios lipídicos denominados
dominios enriquecidos en glucolipidos insolubles en detergentes: DIG (detergentes insoluble glycolipid-enriched domains) separados por regiones no DIG (lípidos en fase fluida)
o Esfingolipidos + colesterol (región DIG)--------fosfatidilcolina (No DIG)o Sirven para el anclaje de proteínas de membrana. Intervienen también en la expresión
de los genes, en la biogénesis de la membrana plasmática ya división célulao Las moléculas lipídicas de la membrana pueden ensamblarse transitoriamente
formando dominios especializados (dominios de membrana o BALSAS LIPIDIAS) donde quedan confinadas determinadas proteínas implicadas en la señalización celular.
o Están presentes tanto en el monocapa externa como en la interna, se caracterizan por ser regiones ricas en colesterol y esfingomielina (con ácidos grasos largos y saturados) lo que las hace menos fluidas.
GLUCOCALIX (Cubierta celular o “cell coat”)
Es un recubrimiento fibrilar de carbohidratos
Consiste en la parte más externa de la membrana plasmática (la cara externa)
Está compuesta por PROYECCIONES OLIGOSACARIDAS de:
- Glucoproteinas- Proteoglucanos: los más abundantes (20% de fracción proteica, 80% de polisacáridos)- Glucolípidos
Las cadenas oligosacaridas se unen covalentemente a lípidos y proteínas y por GLUCOPROTEINAS EXTERNAS extracelulares débilmente unidas a las anteriores (fibronectinas, laminina…). Más simple: es el conjunto de cadenas de oligosacáridos pertenecientes a los glucolipidos y glucoproteínas de la membrana plasmática.
- Glucosaminglucanos: largas cadenas de disacáridos que presentan: n-acetilglucosamina o n-acetilgalatosamina en el disacárido.
El glucocalix puede variar con:
- El tipo de célula. Ej.: está más desarrollado en las células que necesitan protección (células del epitelio intestinal) pero el aspecto (estructura) es igual
- Envejecimiento: la célula sintetiza menos glucocalix a medida que pasa el tiempo.
- Patología: una infección en las células por el virus de la gripe. La célula sintetiza un glucocalix anómalo. La célula es más sensible a infecciones bacterianas.
FUNCION DEL GLUCOCALIX:
1.- PROTECCIÓN: La cubierta asegura:
o La protección química de la membrana plasmática (de la célula), porque la hace resistentes a enzimas mucolíticos y proteolíticos
o La protección mecánica al evitar el contacto con moléculas muy grandes, previene roturas en la membrana apical.
2.- RECONOCIMIENTO: entre células vecinas similares pueden reconocerse y asociarse para formar tejidos. Las cadenas oligosacáridos actúan como señales que deben ser reconocidas par que la célula pueda interrelacionarse. Así, moléculas, virus, bacterias y otra células solo pueden penetrar o relacionarse con la célula sí reconoce a estos receptores de membrana
3.- ANTIGENOS: (antígeno es igual a la proteína que el sistema inmune no reconoce). Los receptores de membrana actúan como antígenos específicos para cada célula. Por ello, cuando se producen trasplantes, estos receptores son reconocidos por el sistema inmune de la persona que recibe el órgano como moléculas extrañas y se produce el rechazo.
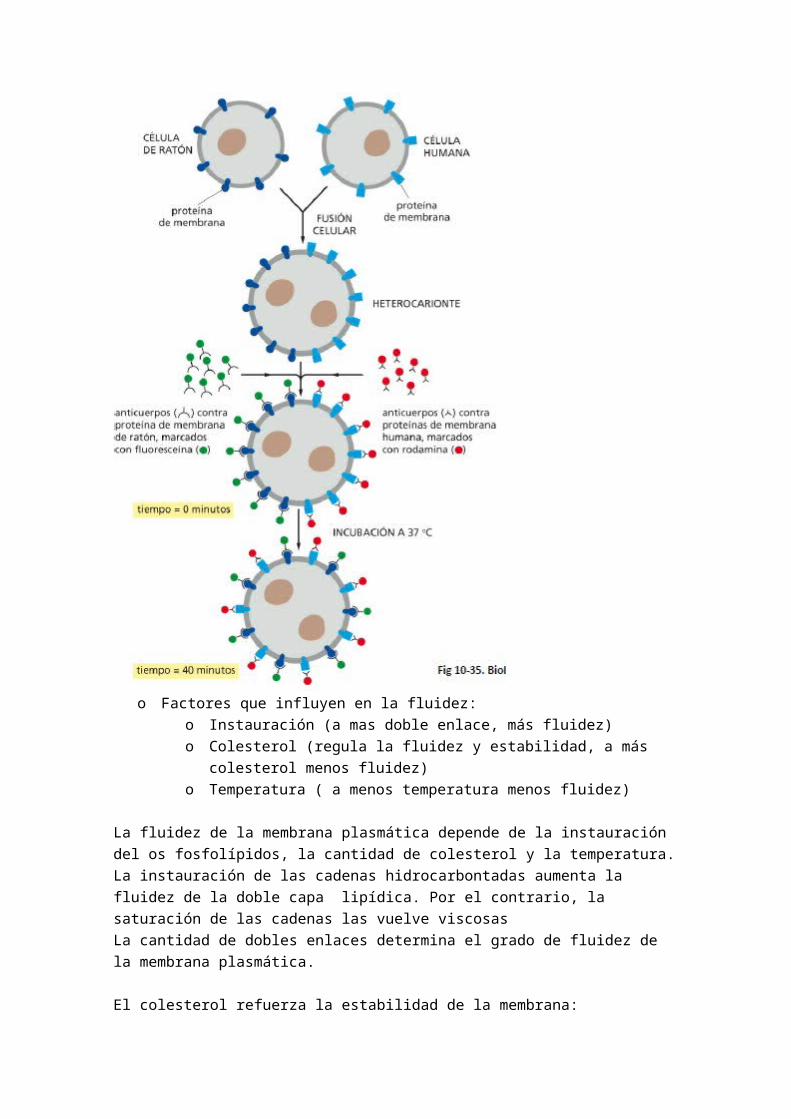
4.- ANCLAJE Y ORIENTACION DE GLUCOPROTEINAS
5.- ESTABILIZACION DE LA ESTRUCTURA PLEGADA DE LAS PROTEINAS
3.- BIOGENESIS DE LAS MEMBRANAS CELULARES
La síntesis de los componentes de las membranas se realiza en el RETICULO ENDOPLASMATICO (RE) LISO Y RUGOSO.
3.1 SINTESIS DE LIPIDOS
Los fosfotolipidos y el colesterol presentes en todas las membranas celulares se sintetizan en el RE liso.La ceramida (esfingosina + ácido graso) se sintetizan en el RE y es exportado al aparato de Golgi.La biosíntesis de fosfolípidos tiene lugar únicamente en al monocapa citosolica del RE. Las translocadoras de fosfolípidos de RE (Escramblasa) transfieren los fosfolípidos de una monocapa a la otras (crecimiento SIMETRICO en ambas monocapas).El translocador (flipasa) presente en la membrana plasmática transfiere la fosfatidiletanolmina y la fosfatidilserina a la cara citosolica contribuyendo a la asimetría de la membrana.

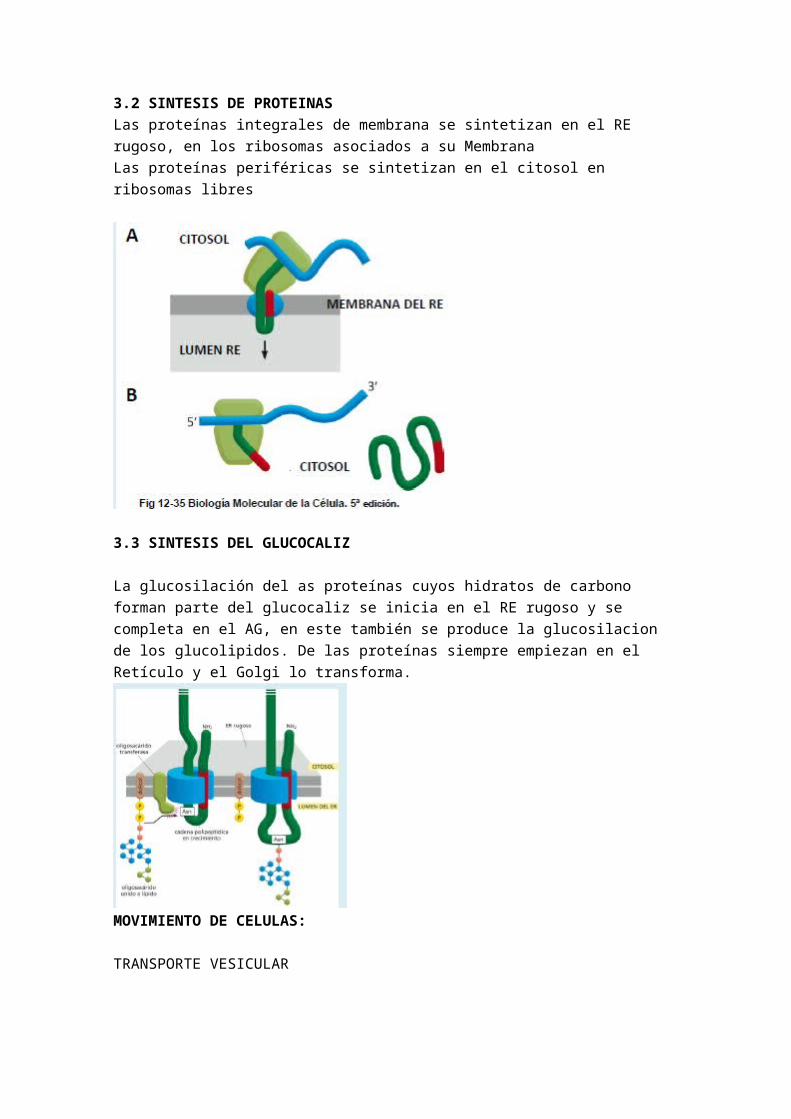
3.2 SINTESIS DE PROTEINASLas proteínas integrales de membrana se sintetizan en el RE rugoso, en los ribosomas asociados a su Membrana Las proteínas periféricas se sintetizan en el citosol en ribosomas libres

3.3 SINTESIS DEL GLUCOCALIZ
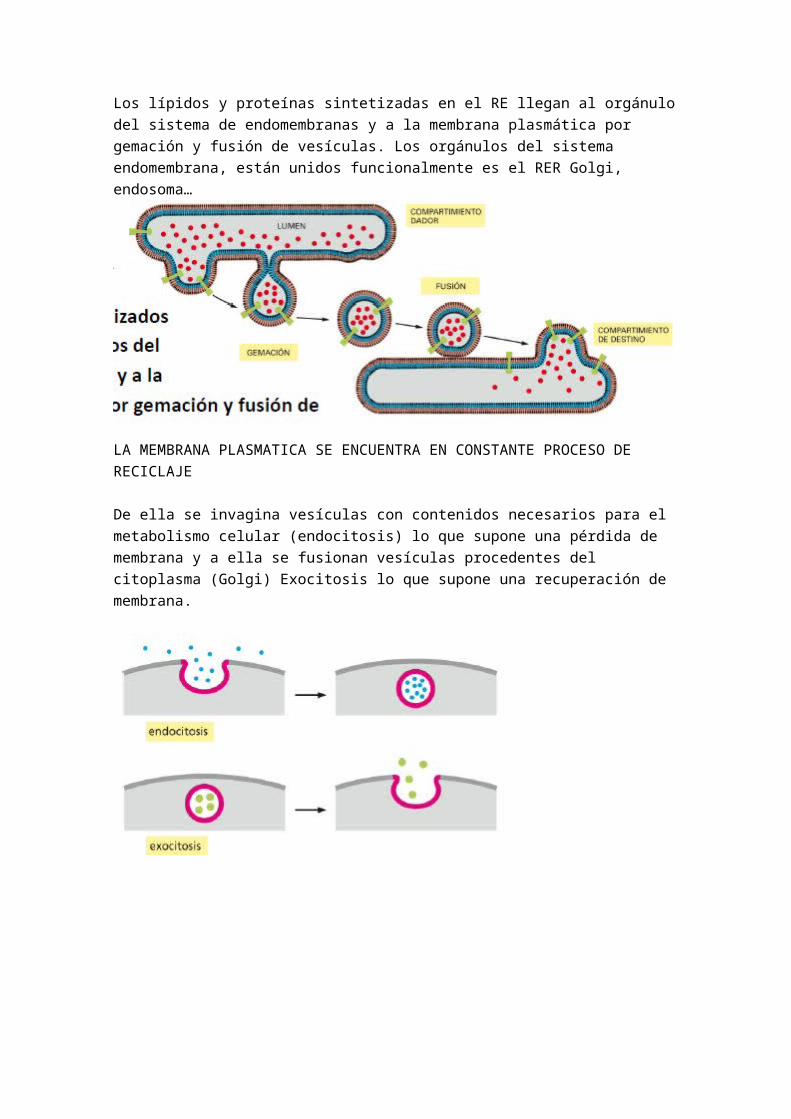
La glucosilación del as proteínas cuyos hidratos de carbono forman parte del glucocaliz se inicia en el RE rugoso y se completa en el AG, en este también se produce la glucosilacion de los glucolipidos. De las proteínas siempre empiezan en el Retículo y el Golgi lo transforma.
MOVIMIENTO DE CELULAS:
TRANSPORTE VESICULAR
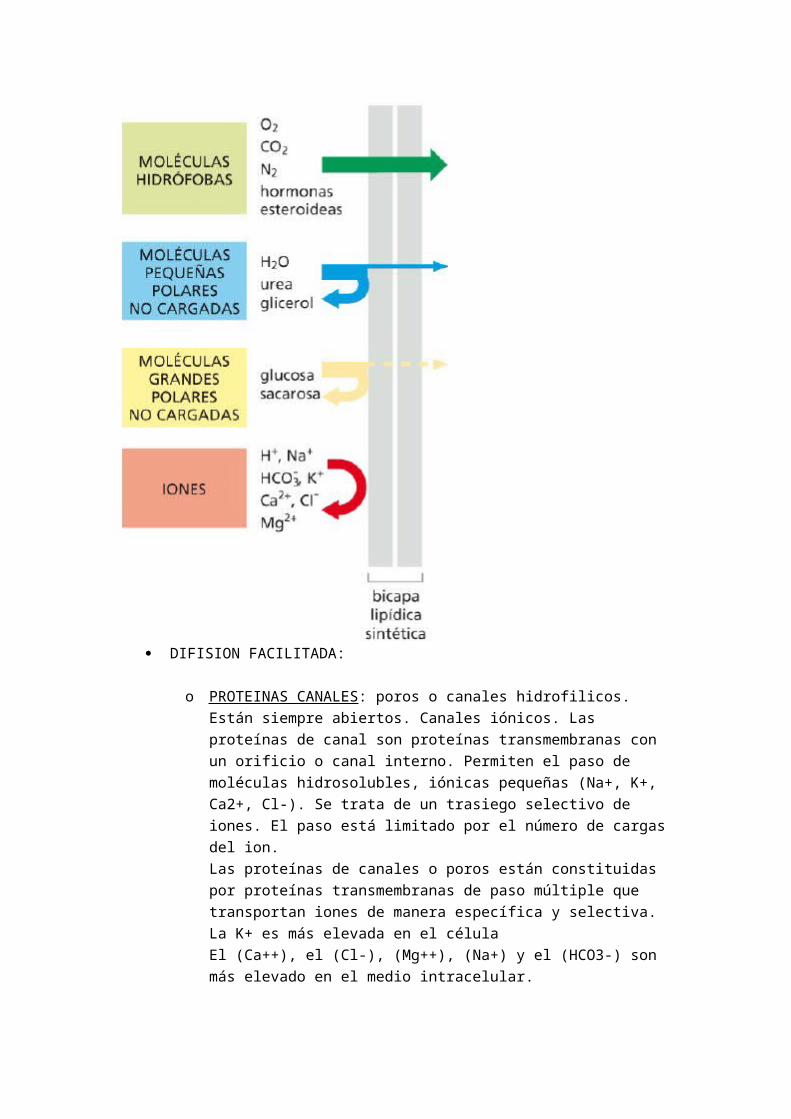
Los lípidos y proteínas sintetizadas en el RE llegan al orgánulo del sistema de endomembranas y a la membrana plasmática por gemación y fusión de vesículas. Los orgánulos del sistema endomembrana, están unidos funcionalmente es el RER Golgi, endosoma…
LA MEMBRANA PLASMATICA SE ENCUENTRA EN CONSTANTE PROCESO DE RECICLAJE
De ella se invagina vesículas con contenidos necesarios para el metabolismo celular (endocitosis) lo que supone una pérdida de membrana y a ella se fusionan vesículas procedentes del citoplasma (Golgi) Exocitosis lo que supone una recuperación de membrana.


MEMBRANA PLASMATICA II
1.- Mecanismos de permeabilidad y transporte2.- Endocitosis y exocitosis3.- Biogénesis de la membrana plasmática
1.- PERMEABILIDAD Y TRANSPORTE
Se puede producir el transporte de:o Pequeñas moléculas e iones: (no es objetivable, no se puede ver…)
o Transporte pasivo: sin aporte energéticoo Transporte activo: con aporte energético
o Grandes macromoléculas y partículas: (se puede observar con MO y ME)o Endocitosis (hacia adentro)o Exocitosis (hacia fuera)
La membrana plasmática es una barrera altamente impermeable a los iones/selectiva en cuanto a lo que entra y sale:
o Es capaz de mantener una diferencia de concentración de iones a ambos lados de la membrana plasmática. Consecuencia: mantiene una distinta concentración de cargas a ambos lados constantemente. (interior: negativo – carga eléctrica interna de signos negativo):Como consecuencia, tiene que mantener una PRESION OSMOTICA INTERNA constante (sin eso no sería posible la vida)
Se han desarrollado dos sistemas para el transporte de sustancias:o Para pequeñas moléculas e iones.o Para macromoléculas y grandes partículas
TRANSPORTE DE PEQUEÑAS MOLECULAS E IONES
TRANSPORTE PASIVO:A favor de gradiente de concentración y/o eléctrico.

Según las leyes físico-químicas que rigen cualquier transporte a través de la biomembrana, no necesita energía.Es un proceso espontaneo de difusión de sustancias a través de la membrana. Se produce siempre a A FAVOR DE GRADIENTE: del medio donde hay más al medio donde hay menos.
o Gradiente de concentración : por simple difusión, las moléculas pasan desde el medio donde están más concentradas hacia donde su concentración es menor
o Gradiente eléctrico: generalmente: medio externo celular se mayor; medio interno de las células es menor. Por simple difusión, los iones positivos entran en la célula, mientras que los iones negativos salen de ella.
Gradiente electroquímico es igual al gradiente de concentración mas eléctrico.
El gradiente electroquímico puede darse por:o DIFUSION SIMPLE: es el paso de pequeñas moléculas a favor de gradiente
electroquímico. La difusión simple depende de:o Coeficiente de permeabilidad de la membrana plasmática : el transporte es
más rápido cuanto más lipófila o apolar sea la sustancia (pasan mejor las liposolubles que las hidrosolubles)
o Coeficiente de reparto de soluto : mejor transporte cuanto mayor sea la diferencia de concentración a ambos lados de la membrana plasmática
o Tamaño y carga del soluto : los solutos que pasan con más dificultad son: Los de mayor tamaño Los de mayor carga (difunden mejor los monovalentes que los
divalentes o trivalentes) : +cargar----+hidratados-----+tamaño
Solutos de pequeño tamaño y sin carga pasan mejor:
1.- Pequeñas moléculas polares sin carga. Hidrosolubles: H2O, CO2, urea, etanol, glicerol2.- Liposolubles apolares hidrófobas: O2, N2, benceno, algún anestésico (éter y cloroformo)

DIFISION FACILITADA:
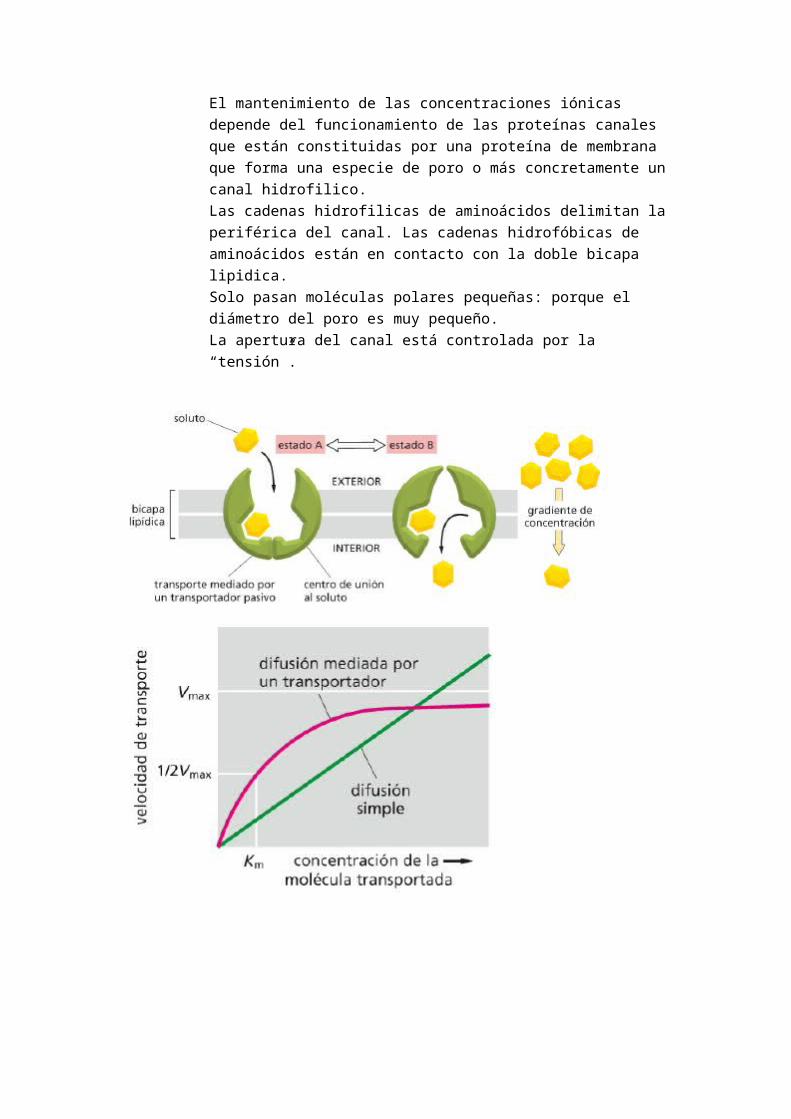
o PROTEINAS CANALES : poros o canales hidrofilicos. Están siempre abiertos. Canales iónicos. Las proteínas de canal son proteínas transmembranas con un orificio o canal interno. Permiten el paso de moléculas hidrosolubles, iónicas pequeñas (Na+, K+, Ca2+, Cl-). Se trata de un trasiego selectivo de iones. El paso está limitado por el número de cargas del ion.Las proteínas de canales o poros están constituidas por proteínas transmembranas de paso múltiple que transportan iones de manera específica y selectiva.La K+ es más elevada en el célulaEl (Ca++), el (Cl-), (Mg++), (Na+) y el (HCO3-) son más elevado en el medio intracelular.El mantenimiento de las concentraciones iónicas depende del funcionamiento de las proteínas canales que están constituidas por una proteína de membrana que forma una especie de poro o más concretamente un canal hidrofilico.Las cadenas hidrofilicas de aminoácidos delimitan la periférica del canal. Las cadenas hidrofóbicas de aminoácidos están en contacto con la doble bicapa lipidica.
Solo pasan moléculas polares pequeñas: porque el diámetro del poro es muy pequeño.La apertura del canal está controlada por la “tensión”.
o PROTEINAS CANALES DE APERTURA CONTROLADA : permiten el paso de moléculas hidrosolubes iónicas pequeñas (Na+, K+, Ca++, Cl-). Es decir, las proteínas canales de apertura controlada permiten el trasiego de lso mismo compuestos (Iones) que los poros o canales hidrofilicos.La apertura puede estar controlada por ligando o por voltaje (onda eléctrica). Es decir, los poros se abren por estímulos del ligando o de onda eléctrica.
Apertura controlada por ligando: proteínas canales cuya apertura depende de un estímulo: cierta sustancias (neurotransmisores u hormas) se unen a un sitio receptor especifico de la proteína canal,

la cual sufre una trasformación estructural que induce la apertura del canal.Ejemplo: neuronas: la acetilcolina (un neurotransmisor) al unirse a su proteína receptora en la membrana postsinatica e induce la apertura del canal de Na+ el cual entra en el célula
Apertura controlada por voltaje: canales de Na+, K+, Ca++, y Cl-. Proteinas de canal cuya apertura depende del potencial de membrana (del gradiente eléctrico, diferencia de cargas a ambos lados de la membrana).Los canales de Ca++ se abren durante la despolarización de la membrana plasmática. Permite, entonces, el paso especifico de Ca++ al interior de la célula.
o PROTEINAS TRANSPORTADORAS : Pasan sustancias polares, hidrosolubles, no tónicas de mayor tamaño (pero aun así pequeñas) con o sin carga. Ej: aminoácidos (polares con carga), glucosa y sacarosa (granes moléculas polares sin carga).Presencia de transportador o “carrier”Afinidad especifica por el sustratoSe lleva a cabo a través de proteínas trasportadoras (ej. Acuaporinas: permeases especificas par el paso de agua), de peso molecular elevado, hodrofilicas, llamadas PERMEASAS.Son proteínas que no hacen canales sino que al unirse la molécula al transportar sufren un cambio en su conformación (conformación PIN, conformación PCN) o estructura que arrastra a dicha molécula hacia el interior de la célula. Las permeasas son similares a las proteínas enzimáticas en su afinidad por el sustrato pero se diferencian en que no modifican el sustrato que transportan.El soluto se une a la permeasa de forma específica. Las proteínas transportadoras vehiculizan determinad sustrato del medio extraceluluar (donde está mas concentrado) hacia el medio intracelular (donde está menos concentrado)
DIFERENCIA CONCEPTUAL ENTRE DIFUSION SIMPLE Y FACILITADA
En la difusión facilitada la velocidad de transporte alcanza su máximo (Vmax) en el momento de saturación del transportador, cuando todos los sitios de unión están ocupada. La Vmax es característica de cada transportador específico. Este tipo de reacción se parece a la reacción enzima-sustrato.Hay divergencias entre autores y a que unos toman todos los mecanismos de transporte pasivo como difusión simple. Esto se debe a la diferenciación que se establece entre transporte por proteínas canal y difusión favorecida. Unos mantienen que la difusión por canal pude llegar a saturarse.Dirían que la difusión simple seria como al facilitada y se saturaría la proteína canal.La difusión simple se distingue de la favorecida porque a mayor (soluto) va aumentando la velocidad de transporte en la simple, pero en la facilitada por mucho que aumente la (soluto) 1º va a aumentar la velocidad de transporte pero luego se estabiliza esa velocidad de transporte y no va a subir porque las proteínas transportadoras están saturadas y ocupadas.En el caso de la simple a mayor soluto, mayor velocidad de transporte.Algunos autores piensan que las proteínas de canal o reguladas por ligando o voltaje también se saturan, es decir, que no permiten el trasiego libre por su configuración especial.En condiciones fisiológicas esto no ocurre – algunos autores la difusión simple mediada por canal las esquematización con la misma curva que la difusión facilitada medida por transportador.Cuando el transporte se produce a la mitad de la Vmax, la concentración de soluto se aproxima a la constante de unión del transportador para el soluto (km) y es análoga a la Km de una enzima para su sustrato.
TIPOS DE TRANSPORTES
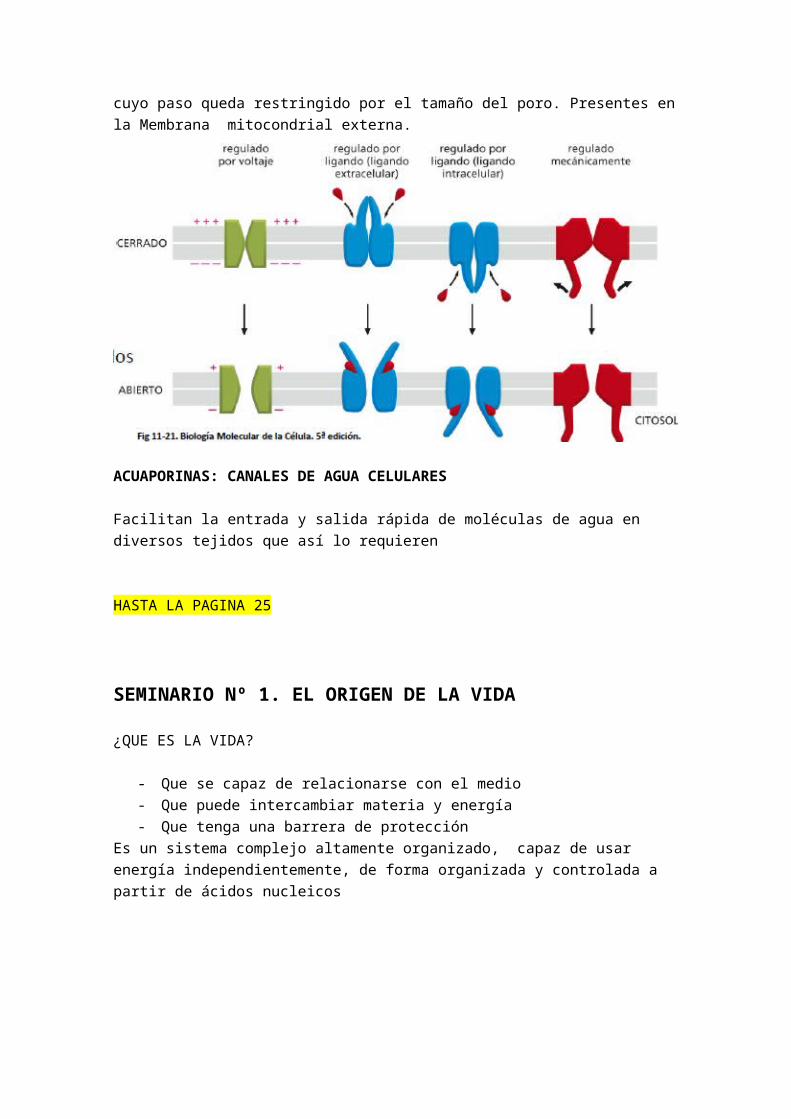
Cuando una permeasa moviliza un solo soluto el proceso se denomina UNIPORTE
Cuando se transportan dos solutos simultáneamente se habla de TRANSPOERTE ACOPLADO O CONTRAPOERTE
o SIMPORTE: Si los dos solutos se mueven en el mismo sentidoo ANTIPORTE: Si los solutos se mueven en sentidos opuestos.
TRANSPORTADORES DE GLUCOSA
La glucosa es la principal fuente corporal de energía directa o la mayoría de las células contienen una proteína que facilita la difusión de la glucosa al interior de la célula
2.2 PROTEINAS CANAL
Facilitan la difusión de sustancias formando túneles acuosos en la membrana. Aunque en algunos canales el flujo de sustancias transportadoras aumenta linealmente siguiendo las leyes de la difusión simple, en la mayoría de ellos el proceso tiende a saturarse a altas concentraciones. La cinética seria por lo tanto intermedia entre la difusión simple y las proteínas transportadoras.
CANALES IONICOS
Los iones presentan carga y por ello no difunden por la membrana.El movimiento de un ion depende de su potencial o gradiente electroquímicoEstos canales son muy selectivos: permiten el paso de una sola especie iónicaEstán regulados: el poro se abre o se cierra en respuesta a determinados estímulos
PORINAS: Proteínas que atraviesan varias veces la membrana formando cilindros cerrados de láminas B, (barriles B) Delimitan un poro lleno de agua permitiendo el paso de solutos hidrófilos cuyo paso queda restringido por el tamaño del poro. Presentes en la Membrana mitocondrial externa.

ACUAPORINAS: CANALES DE AGUA CELULARES
Facilitan la entrada y salida rápida de moléculas de agua en diversos tejidos que así lo requieren
HASTA LA PAGINA 25
SEMINARIO Nº 1. EL ORIGEN DE LA VIDA
¿QUE ES LA VIDA?
- Que se capaz de relacionarse con el medio- Que puede intercambiar materia y energía- Que tenga una barrera de protección
Es un sistema complejo altamente organizado, capaz de usar energía independientemente, de forma organizada y controlada a partir de ácidos nucleicos

IONOFOROS
Difusion facilitada por ionoforos.Los ionoforos son quepñas moeleculas hidorofbicas que se disuelven en las bicapas lipdicas. Incrementan (alteran)la permeabildad de la membrana a determinados iones inorgánicos.La mayoría so nsitentizados por micrrorgansimosExisten dos clses de ionenoforos:
- Transpoertadores móviles- FOrmadres de canal
Ambos tpos actúan roedando la carga del ion de forma que pueda atravesar el interior hidrofóbico de la bicapa lipídica.Dado que no están acoplado, a fuentes de energía solo permiten el moimiento neto de iones a favor de su gradietne electroquímico.
IONOFORO MOVIL
EJ: VALINOMICINA: transportador específico para el potasio (K+)Es un polímero en forma de anillo que transporta potasio a favor de su gradiente electroquímico, tomando potasio de un lado de la membrana, difundiéndolo a través de la membrana y liberándolo al otro lado
IONOFORO FORMADOR DE CANAL.
EJ:GRAMICIDINA
Péptido lineal de 15 residuos de aminoácido, todos ellos con una cadena lateral hidrofóbica, por lo que constituye el canal iónico mayor conocido.Se cree que dos moléculas de gramicidina se unen formado un canal transmembrana que permite selectivamente que los iones monovalente fluyen a favor de sus gradientes electroquímicosSon dimeros proteincos inestables: se están ofmrando y disociandocontinuamnte, de forma que leteimpo medio de apertura de esoso candales es 1 segundo. Es mas rápido, por tanto, que le ionofor móvil.Es sintetizada por ciertas bacterias, quizás para matar otros microorganismos colapsando los gradientes de H+, Na+ y K+ que osn esenciales para la supervivencia de la celula. Ha sido utilizada con éxito como antibiótico.
PROTEINAS TRANSPORTADORAS- Transportadores sencillos o UNIPORTES: transportan un solo soluto de un lado a
otro de la membrana- CONTRAPORTE: (unión de transporte activo y pasivo según Naillet, o trasporte
activo secundario). Se trasportan dos solutos. La trasferencia de un soluto depende de la transferencia simultanea o secuencial de un 2º soluto, ya sea:
o SIMPORTE o transporte unidireccional: en la misma dirección. Ej: transporte

de Na glucosa en células intestinales y renales (penetran juntos en la célula)
o ANTIPORTE: o trasponerte de intercambio: en dirección opuesta: Ej bomba de Na+/K+ que bombea Na+ hacia el exterior en contra de su gradiente electroquímico y K+ hacia el interior
1.- TRANSPORTE ACTIVO
En contra de gradiente de concentración y/o electrquimico. Energía de la hidrolisis del ATP por la ATPasa.La diferencia de potencial de la membrana (gradiente eléctrico o de voltaje) favorece la entrada de iones cargados positivamente y se opone al a entrada de iones cargados negativamente.Si no hubiese transporte activo llegaría un momento en que habría igual número de cargas a los dos lados de la membrana plasmática.Hace posible el movimiento de solutos en contra del equilibrio termodinámico por lo que se requiere siempre el APORTE DE ENERGIA.Las proteínas de membrana implicadas en el transporte activo se denominan BOMBAS. Estas bombas consumen ATP mediante su hidrolisisEs un proceso unidireccional que puede ser:
- Directo: Por ejemplo, el transporte de protones a favor de gradiente electroquímico, transportando al lado contrario moléculas de menor o mayor peso molecular
- Indirecto
Características
- Transporte de moléculas HIDROSOLUBLES, IONICAS Y NO IONICAS- Hace permanecer CONSTANTE la PRESION OSMOTICA INTERNA- Hace permanecer constante la diferencia de concentración de Na+ y K+ a ambos lados
de la membrana plasmática- trasmisión del impulso nervioso- Introduce nutrientes e iones aunque estén en muy bajas concentraciones fuera de la
célula- Hace permanecer constante el reparto desigual de carga a ambos lados de la
membrana
La célula debe tener el mismo número de cargas positivas que negativos, es decir, ha de ser eléctricamente neutra.Estado normal-membrana polarizada
La concentración de K+ en el interior celular es de 10 a 20 veces mayor que en el exterior. La concentración donde Na+ en el interior es menor que en el exterior.
1.3 TRANSPORTE ACTIVO PRIMARIO O DIRECTO
Se realiza mediante 4 tipos de ATPasas: tipo P, tipo V, tipo F y tipo ABC.
Las bombas tipo P . (Phosphorilation). Se fosforilan reversiblemente. Bomba de Na+/K+
Las bombas de protones tipo F (y de tipo V ). V viene de vacuola, forma parte de lisosomas, mitocondrias… ATP sintasas en mitocondrias y bomba de H+ en lisosomas.
Trasnportador ABC . (ATP binding cassette) Tienen dos componentes proteicos transmembrana y dos periféricos. Cabe destacar el transportador MDR (resistencia a multidrogas, significa) con este transportador la célula se hace resistente a los fármacos.
A) BOMBAS TIPO P. BOMBA SODIO POTASIO
La moayorias de las proteínas que realizan este transporte es una ATPasa. Saca tres iones de Na+ de al ceula y a la vez hace entrar en ellas dos iones K+ (por cada molecula de ATP) en contrad e sus gradientes electrquimicos.
El funcionamiento de la bomba electrogénica de Na+/ K+(sodio-potasio) , se debe a un cambio
de conformación en la proteína que se produce cuando es fosforilada por el ATP. Como el
resultado de la catálisis es el movimiento transmembrana de cationes, y se
consume energía en forma de ATP, su función se denomina transporte activo. La demanda
energética es cubierta por la molécula de ATP, que al ser hidrolizada, separa un grupo fosfato,
generando ADP y liberando la energía necesaria para la actividad enzimática. En las
mitocondrias, el ADP es fosforilado durante el proceso de respiración generándose un
reservorio continuo de ATP para los procesos celulares que requieren energía. En este caso, la
energía liberada induce un cambio en la conformación de la proteína una vez unidos los tres
cationes de sodio a sus lugares de unión intracelular, lo que conlleva su expulsión al exterior de
la célula. Esto hace posible la unión de dos iones de potasio en la cara extracelular que provoca
la desfosforilación de la ATP, y la posterior traslocación para recuperar su estado inicial
liberando los dos iones de potasio en el medio intracelular
Los procesos que tienen lugar en el transporte son:
1. Unión de tres Na+ a sus sitios activos en la cara intracelular de la molecula proteica
Na+/K+ ATPasa
2. Fosforilación de la cara citoplasmática de la bomba que induce a un cambio de
conformación en la proteína. Esta fosforilación se produce por la transferencia del
grupo terminal del ATP a un residuo de ácido aspártico de la proteína. Se raliza en
presencia de Ma++ y de 3 Na+
3. El cambio de conformación hace que el Na+ sea liberado al exterior.
4. Una vez liberado el Na+, se unen dos iones de K+ a sus respectivos sitios de unión de la
cara extracelular de las proteínas.
5. La proteína se desfosforila produciéndose un cambio conformacional de ésta, lo que
produce una transferencia de los iones de K+ al citosol.
B) BOMBA DE CA2+
Las células eucariotas baja la concentración de calcio (aproximadamente 10 elevado a 7). El flujo a favor de gradiente de concentración es una forma muy rápida de transmisión de señales extracelulares a través de la membrana plasmática. El mantenimiento de un elevado gradiente de calcio es importante para la célula. El gradiente de calcio se mantiene (en parte) por la acción de bombas de calcio que existen en la membrana plasmática.Bombas de calcio transportan calcio de forma activa hacia el exterior de la célula (en contra de gradiente para mantener la concentración de calcio extracelular elevada).Bombas de calcio: una es una ATPasa. Otra es un transportador de intercambio (antiporte) impulsada por el gradiente electroquímico o de Na+
C) BOMBA DE H+
Es una ATPsintasa o sintetasa, porque sintetiza ATP.En lugar de ser la hidrolisis de ATP en ADP+Pi la que impulsa el tranposrte ionico, en este caso e sun gradiente de H+ a través de la membrana plasmática el que dirige la síntesis de ATP a partir de ADP+Pi.Este gradiente de H+ se genera durante el proceso de transporte de electrones en la fosforilazion oxidativa.La enzima también puede hidrolizar ATP y bombear H+ a través de la membrana plasmática, es decir, puede funcionar como una ATPasa aunque normalmente actua en sentido opuesto.

D) BOMBA DE TIPO ABC CTFR (regulador de la conductancia transmembrana de la fibrosis quística)
Esta bomba mueve el cloro desde el exterior al interior de la célula, cuando hay un problema en estas proteínas se produce la enfermedad de la fibrosis quística. Es una enfermedad autosómica recesiva, es decir, la patología se desarrolla cuando la persona recibe dos alelos mutados, por lo que el paciente no tiene proteínas funcionales para dicha actividad. Debido a esta anomalía el cloro no va a salir al exterior de la célula, tampoco agua ni Na+. En última instancia, a nivel bronquial, el moco que fisiológicamente debe secretarse en las vías respiratorias queda sin moverse debido a esta anomalía, conllevando por tanto el cumulo del mismo en las vías respiratorias y el desarrollo de las bacterias. La causa de la muerte suele ser fallos respiratorios. Tiene consecuencias también a nivel pancreático o la esterilidad en el caso de los hombres.
1.4.- TRANSPORTADOR ACTIVO INDIRECTO O SECUNDARIO: SODIO/GLUCOSA
No utiliza directamente la hidrolisis del ATP sino la energía potencial almacenada en forma de gradiente ionico (en el transporte activo primario).La energía liberada durante el desplazamiento de unión a favor de su gradiente electroquímico se utiliza como fuerza impulsada para bombear otros solutos en contra de sus gradientes electroquímicos.Asi pues, todas estas proteínas actúan como “cotransporte” (simpote o antiporte).Na+:
- Ion contransportado cuyo gradiente electroquímico proporciona la fuerza impulsora para el transporte activo de una segunda molecula
- El Na+ qu entra en la celula durante este transporte es bombeado hacia el exterior medante la bomba Na+K+, la cual, al mantener el gradiente del Na+, impulsa indrectamtne el trasnpote de las otras moléculas que se cotransportan con el Na+
Es de tipo simporte acoplado; cabe destacar la bomba Na+/glucosa. Este tipo de transportador es importante para la captación de glucosa en el intestino, aprovechando el exceso de Na+ en este órgano. Tras el transporte activo indirecto, el nivel de glucosa será mayor dentro de la célula que fuera. El gradiente se sodio se mantendrá por la bomba sodio/potasio. La glucosa también se consigue por transporte pasivo, este se produce en el dominio basal, no en el dominio apical. A nivel estomacal cabe destacar las células parietales. En este caso se produce la secreción de entre dos a tres litros de ácido al día. Se trata de una bomba de protones. La acción se secreción se inhibe con Omeoprazol, fármaco que anula el transporte final de H+ al lumen gástrico.
En las células epiteliales del intestino y del riñon:
Simporte de glucosa-Na (unidireccional)
- La membrana plasmática del polo apical tiene simportes glucosa-Na+- La glucosa penetra en la célula gracias a la energía obtenida del gradiente
electroquímico/de concentración de Na+-- La glucosa sale de la célula por el polo basal a través de una PERMEASA (difusión
facilitada) y a favor de su gradiente de concentración (si la concentración es más elevada en el polo basal de la célula que en el medio extracelular)
Una bomba de Na+/K+ bombea el sodio al exterior de la célula para mantener baja la concentración intracelular de Na+ y así mantener el gradiente de Na+ que impulsa el transporte unidireccional.
Relación clínica:
- Mala absorción de glucosa-galactosa (la mayoría de origen genético)- Aminoácidos: cistinuria. Mala absorción de la cistina, se acumula y forma piedras en el
riñón- Cl-: fibrosis quística
Vía experimental:- El receptor de la acetilcolina
MUERTE POR INYECCION LETAL: El dr J. Kevorkiw ayuda a suicidirse a la Ddra J.Adkins (54 años y Alzheimer) con un catéter intravenoso con solución de cloruro potásico.
CANALOPATIAS:- Alteración en canales catiónicos- Alteración en canales anionicos
Tanto unas como otras pueden ser adquiridas (reacción inmunitaria, toxina..) o hereditarias (mutaciones en los genes que codifican las proteínas)

TEMA 5. MEMBRANA PLASMATICA III. TRANSPORTE DE MACROMOLECULAS: ENDOCITOSIS Y EXOCITOSIS.
1.- ENDOCITOSIS- FAGOCITOSIS- PINOCITOSIS- ENDOCITOSIS MEDIAD POR RECEPTOR

2.- EXOCITOSIS- VIA CONSTITUTIVA Y REGULAR
3.- TRNSCITOSIS4.- TRAFICO VESCULAR
ENDOCTOSIS Y EXOCITOSIS: INTRODUCCIÓN
Son procesos a través de los cuales la célula transporta moléculas mediante FORMACION Y FUSION DE VESICULAS.El proceso de exocitosis suministra proteínas de nueva síntesis, carbohidratos y lípidos. La membrana plasmática y al medio extracelular. La endocitosis permite a las células eliminar componentes de la membrana plasmática y captar nutrientes. En ambos casos se trata de transportes que conllevan gasto energético.
1.- ENDOCITOSISEs un proceso por el cual las células captan macromoléculas, sustancias y otras células. Los diferentes tipos son:
- Fagocitosis : se ingieren partículas grandes- Pinocitosis : se ingiere fluido y solutos por medio de pequeñas vesículas- Endocitosis medida por receptor
La formación de invaginaciones en la membrana plasmática conduce a la formación de vacuolas o vesículas que contienen el material a importar
FAGOCITOSIS (partículas sólidas)
Tiene lugar solo en unas pocas células especializadas (algunos leucocitos como los macrófagos, neutrófilo o monocitos)Las células extienden proyecciones de la membrana (seudópodos) que se fusionan para formar fagosomas.
Papel defensivo: las células fagocíticas ingieren microrganismos y nos defienden de infecciones. Los macrófagos también desempeñan importante papel al eliminar células moribundas o muertas.Los macrófagos fagocitan más de 10 elevado a 11 eritrocitos a diario.Formaion de fagosoma conlleva la foramcion de vacuolas de gran tamaño (visibles a MO) que contienen partículas solidas o grandes desechos celulares(ESTUDIAR LAS IMÁGENES PARA EL EXAMEN PRACTICO Y TEORICO, PORQUE CAERAN IMÁGENES DE LAS CLASES EXPOSITIVAS).
o MECANISMO DE PRODUCCIONDE LA FAGOCITOSIS
INvaginacion de la membrana plasmática en varias fases. Pare ello utilizamso el ejemplo de sistema de desechos fagocitados por macrófagos1.- OPSONIZACION: (del griego “aderezo”) Las células fagocitados tenen que sentires atraídas por lque vayan a fagocitar. Es to consiste en rodear de anticuerpos (opsoninas) que producen otras células a los cuerpos (bacteria, sustancia de desecho)2.- QUIMIOTACTISMO:El macrófago es atraído por los anticuerpos químicos3.- CONTACTO Y ADHERENCIALos macrófagos forman pseudopodos que contacta con el linfocito (sustancia de desecho).Posteriorente se adhiere y se activa la fagocitosis4.- INVAGINACION: (fase reologica)Los pseudopodos envuelven al lifocito formando una vacuola (fagosoma) y sus bordes librs se fusionan a modo de cierre de cremallera.
La fagocitosis no se produce se el cuerpo a ingerir no se rodea completamente de opsoninas.
El DESTINO para el fagosoma (o también endosoma) se pude unir a una vesicula que contenga enzimas hidroliticoas provenintes del aparato de Golgi con lo que esta estructrua se convierte

en un liposoma- que lo degradará. Si no se fusiona queda en forma de un cuerpo extraño que la celula tratara de expulsar.
LA FAGOCITOSIS ES UN PROCESO REGULADO
Las células fagociticas tienen receptores de superficie especializados que reconocen los microorganismos o células muertas.La unión induce a la célula fagocitica a extender seudópodos que rodean a la partícula y se fusionan por sus extremos para formar fago soma. Es un proceso altamente regulado, implica la aparición de receptores. Para la formación de seudópodos se requiere la actividad de actina. Este proceso solo se da en algunas células, no en la mayoría.
LA PINOCITOSIS.
Es la entrada de líquidos y moléculas a través de vesículas que atrapan de FORMA INDSICRIMINANDA, sustancias presentes en la superficie extracelular. Hay varios mecanismos mediante los cuales las células pueden formar vesículas.Uno de los mecanismos comienza en zonas especializadas de la membrana llamadadas fosas recubiertas de clatrina, estas depresiones se invaginan y se separan de las membrana formando vesículas recubiertas
En general, las pinocitosis no son visibles. Es visible solo a MO la macropinocitosis.- La presentan todos los tipos celulares- Mecanismos: invaginación, lamina ectoplasmatica- Vesiculas: lisas o revestidas (clatrina/trisquelion)

o CLATRINA
Está formada por tres cadenas polipeptidicas grandes y tres pequeñas que forma una estructura denominada TRISQUELION, que se ensambla formando una red de hexágonos y pentágonos.La clatrina no es la única responsable, juega un papel importante otras:
- Adaptina: en el ensamblaje de complejo de adaptadores de moléculas de clatrina en la cara citosólica de la membrana. Genera la fuerza que permite la formación de vesículas.
- La dinamina: Proteínas citosolicas como la dinamina se ensambla formando un anillo alrededor del cuello aproximando membranas que se fusionan. Cuando la vesícula se ha separado de la membrana pierde rápidamente la forma.
- Otra vía alternativa a ésta se inicia en las caveolas, son invaginaciones de la membrana plasmática formadas a partir de las balsas lipídicas. Las caveolas son capaces de invaginarse y concentrar proteínas de transportadoras gracias a la composición lipídica de la membrana y la caveolina, la cual es una proteína integral de la membrana que no se separa de la vesícula.


Tamaño y destino de la PINOCITOSIS:
MICROPINOCITOSIS:o Pinocitosis de vesículas lisas (endocitosis de fase liquida)
Constitutivo y continuo/todas las célulaso Pinocitosis de vesículas recubiertas de clatrina (endocitosis mediada por
receptor) Inducido y discontinuo/abosrcion de colesterol Placas asteroescleroticas/ateroesclerosis juvenil
o Potocitosis (potos_accion de beber, bebida)”caveolae intracellularis” Inducido (ligando-receptores): trnasmision de
señaes/hormonas Microdominios DIG (glucoesfingolidops y colesterol) Revestimiento granular dispuesto en espiral (CAVEOLINA):
proteína intrínseca de la membrana de las vesículas. MACROPINOCITOSIS
Vesiculas no revestidas. Macropinosomas/endsomas tempranos
Mecanismos: invaginación/lamina ectoplasmica:
Formas de pinocitosis:o INVAGINACION: El mecanismo mayoritario es una invaginaciond de la
membrana plasmática. Las invaginaciones pueden ser de dos tipos: Invagiancion sencilla: es el modelo clásico.
Las moléculas de clatrina se agregan espontanemante en un medio acuoso – se inicia un proceso de ENSAMBLAJE. Finamente constituye una malla a base de 8 hexagonos y 12 pentagonos, que rodea la membrana de la vesicula de endocitosis.
Clatrina-subunidad

Union de tres polipéptidos mayores y tres menores
CASO MAS CONOCIDO: MECANISMO DE ABSORCION DEL COLESTEROL
-El colesterol se transporte en la célula por medio de vesículas revestidas de clatrina-El colesterol es insoluble en los líquidos del organismo y solo puede circular con la ayuda de un transportador lipoproteico de baja densidad (LDL: low density lipid): transporta esteres de colesterol- La membrana plasmática posee receptores para LDL y ambos se agrupan en las depresiones u hoyos revestidos de la membrana plasmática- Después de la endocitosis, las LDL, pierden su revestimiento de clatrina y se fusionan con endosomas tempranos- La bomba de H+ en la membrana acidifica el interior del endosoma y se liberan las LDL de las protteinas receptoras.- Vesícula de colesterol: por gemación y estrangulamiento una parte migra a la membrana plasmática donde vuelve a captar colesterol y otra parte que contiene las LDL se fusionan con un lisosoma (gemas del aparato de Golgi) que contiene encimas hidrolíticas específicas que hidrolizan el colesterol liberando colesterol que formara la membrana plasmática y ácidos grasos
Debido a una anomalía genética en la proteína receptora la célula no puede introducir la LDL y por tanto el colesterol queda pegado a los vasos sanguíneos y se forman placas ateroesclerótica y produce ateroesclerosis juvenil
POTOCITOSIS:
- “Caveolar intracellularis”. Se llaman asi para distinguirlas de otros tpios de vesículas de pinocitosis
- Son las vesículas de ponocitosis + PEQUEÑAS- Se trata de un mecanismo inducido por la unión de un ligando a receptores de
membrana y produce una transmisión de señaeles que se relacionan con algunas hormonas
- Esa unión conduce a la fomracion de vesículas (caveolas) que poseen un revestimeinto granular dispuesto en espiral. Este revestimiento está constituido por una proteína intrínseca de la membrana a las vesículas: CAVEOLINA
- La biomembrana se internalizan estas vesículas protocitoicas, además de caveolina se caracteriza por su riqueza en glucoesfingolipcos y colesterol, por lo que las membranas de las vesículas se considera un MICRODOMINIO DIG.
- Las regiones DIG son menos fluidos por su abundancia en colesterol.- Las caveolas contienen, concentración e internalizan numerosas moléculas conocidas
por su papel en al transmisión de señales- Estas vacuola son más pequeñas que las vesículas lisas (diámetro de 0 a 80nm) nose
puede ver con el microscopio electrónico convencional.

- Las vesículas de potocitos se quedan unidas a membrana plasmática o también pueden internalizarse en el citoplasma pero no es frecuente.
TRANSCITOSIS (CITOPEMPSIS)
Mecanismo MIXTO: combina la endocitosis con la exocitosis. Se produce en células polarizadas. Se produce el transporte de macromoléculas en células polarizadas. En uno de los dominios hay endocitosis y exocitosis en el otro. Por ejemplo los anticuerpos de la leche materna que pasan al torrente sanguíneo del bebe. Solo se produce en células polarizadas debido a que hay un complejo de unión, es decir, están fuertemente unidas, por lo que los anticuerpos y otros elementos no pueden ir entre estos elementos.De forma general se puede establecer que:
- Conjunto de fenómenos que permiten que una sustancia o partícula (bacteriana o no) atraviese el citoplasma de una célula y pase de una región extracelular a otra región extracelular.
- Proceso que necesita ATP- Ej: el paso a través de CELULAS ENDOTELIAES (que tapizan los capilares sanguinos) DE
MOLECULAS CONTENIDAS EN AL SANTRE CIRUCLANTE.- Ej: CELULAS EPITELIAES POLARIZADAS.- Afectan a vesículas y a moléculas que atraviesan el citoplasma- Puede existir transcitosis presentan en su membrana proteínas especfias: TAP: su
presencia – que las vesículas se fusionan con el compartimiento diana oa la región de la membrana plasmática adecuada
- Las vesículas se captan por endocitosis en el extremo apical de la célula pero no se fusionan con el endosoma, utilizan el citoplasma como a atajo para atravesarla y fusionarse en la membrana de otro extremo (basal) donde descargaría su contenido por exocitosis hacia la matriz celular

-
ENDOCITOSIS MEDIADA POR RECEPTOR
Es un mecanismo de incorporación SELECTIVO de moléculas a través de los receptores de la membrana plasmática. El colesterol viaja en la sangre en forma de lipoproteínas, o bien LDL (desde la sangre a las células) y la HDL (que se forman para ser eliminados) El HDL es malo porque está en la circulación. La única diferencia es que la pinocitosis es inespecífica, esta es muy específica.

2.- EXOCITOSIS
Es el proceso por el cual las vesículas de secreción se fusionan con la membrana plasmática. Las proteínas de membrana y los lípidos de estas vesículas aportan nuevos componentes a la membrana plasmática y las proteínas son excretadas al espacio extracelular.Las vesículas de exocitosis tiene su origen en las cisternas mas trans del aparato de Golgi por mecanismos de gemación y estrangulamiento.Transporte de macromoléculas y partículas GRANDES.Existen dos modalidades de exocitosis
VIA CONSTITUTIVIA
Es un proceso continuo presentando por todos lso tipos celulares. Presenta vesículas recubiertas de coatomero formado por 7 subunidades proteicas llamadas COP (coat proteins) (diferente a la clatrina), por eso se llaman vesículas COP. Las vesículas pierden su revestimiento antes de fusionarse con la membrana plasmática. Necesitan aporte energético, generalmente en forma de ATP.Todos los tipo celulares mandan al exterior los componentes de su matriz extracelular (el medio ambiente que le interesa tener a la célula lo sintetiza ella). Así mediante vesículas se

envían al exterior. Los componentes que forman la membrana extracelular (diferente y o más o menos abundantes según la membrana externa que rodea a la célula):
- Glucosaminoglucanos- Ciertos iones- Otras proteínas: colágeno, elastina, fibronectina
VIA REGULADA
Las vesículas de secreción esperan cerca de la membrana plasmática hasta que una señal les haga liberar su contenido.En la vía regulada, las proteínas de secreción se concentran a media que se desplazan desde el RE ya través del AG. Las vesículas de secreción van madurando y se fusiona unas con otras y sus contenidos se concentran. El aumento total de la concentración es de entre 200-400 veces superior desde que abandonan el REPor ejemplo, la histamina es liberada mediante la ruta regulada en respuesta de ligandos específicos que se unen a los receptores de la superficie celular

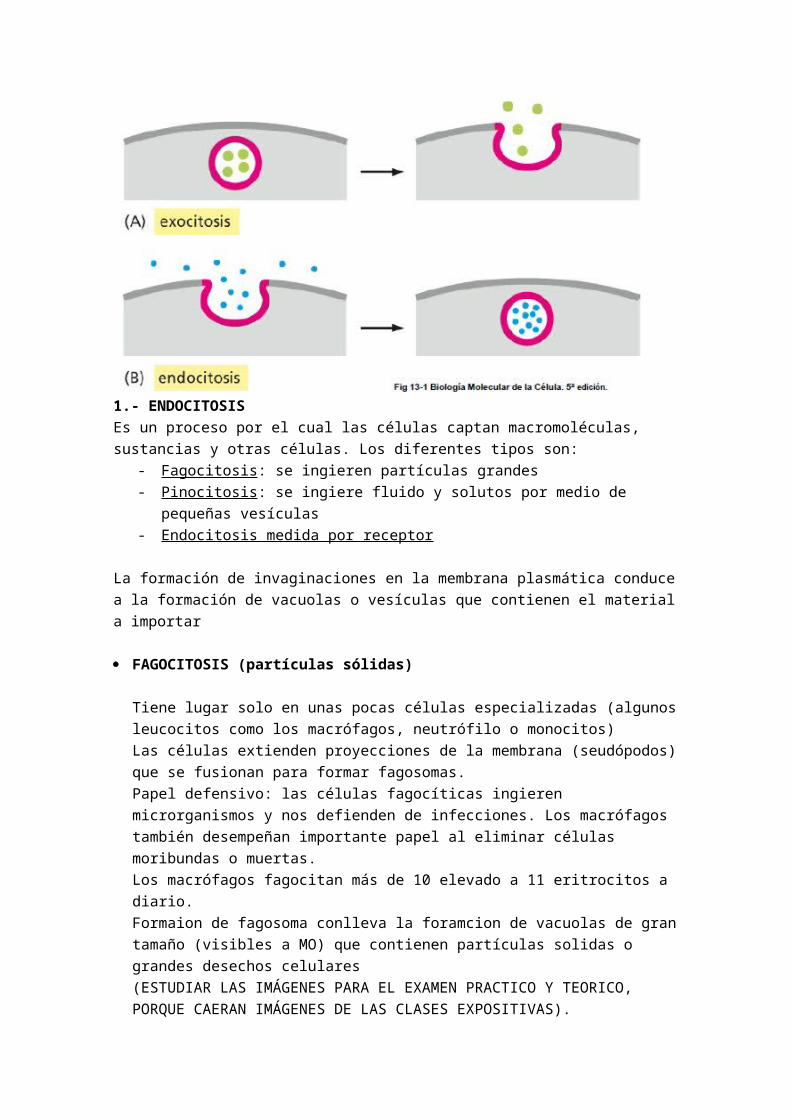
(DIA 24/9 Y 25/9 COMPLEMENTAR CON EL LIBRO BIOLGOIA MOELECULAR DE LA CELULA. 5 EDICION)
VIA INDUCIDA (discontinua): es un proceso discontinuo. Se realiza en respuesta a una estimulo (es especifica). Son vesículas recubiertas de clatrina y que pierden su revestimiento (también) antes de fusionarse con la membrana plasmática.Ejemplo de exocitosis inducida: secreción de insulina en respuesta a los niveles altos de glucosa en sangre. La célula pancreática por exocitosis libera la insulina. La síntesis de la insulina se realiza vía RE- aparato de Golgi. Lo que en principio aparece en vesículas rodeadas de clatrina.Continua su proceso así y después se libera de esa clatrina y es secretada la insulina junto con el péptido C.Los niveles del péptido C en sangre valoran el número de células beta pancráticas que funcionan sintetizando insulina. En la diabetes tipo I los macrófago destruyen las células beta pancreáticas porque no las reconocen.La insulina es el ligando para receptores de células musculares y células adiposas e interaccionan con sus proteínas receptoras y aumenta el número de proteínas transportada de glucosa en la membrana plasmática de esas células – la glucosa entre en la célula a favor de gradiente de concentración. Es la propia célula muscular la qu degrada la glucosa, n la insulina.Ejemplo: de exocitosis inducida cuando el estímulo está en la misma célula: NEUTRASNMISORES
Entrada masiva de Ca2+--exocitosis de actilcolina
Los movimientos de endocitosis y exocitosis no son los únicos que tiene la membrana plasmática también seudópodos y membranas ondulantes.
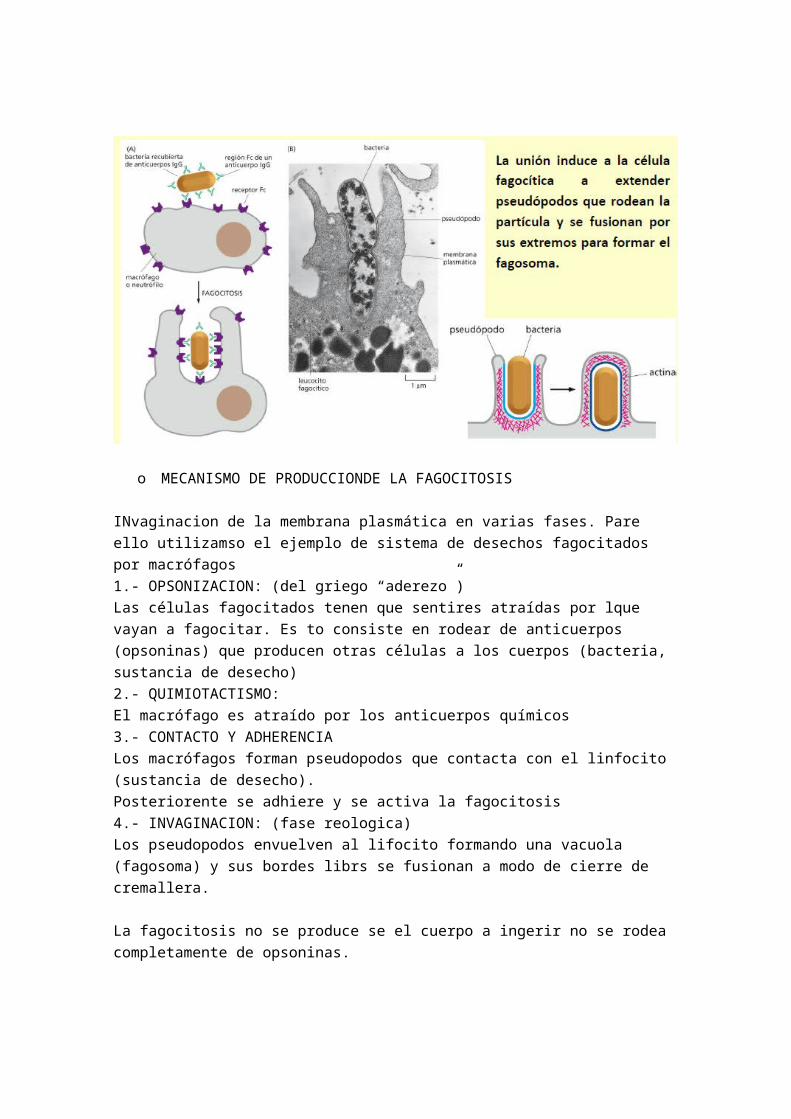
4.- TRAFICO VESICULAR
Existen distintas tipo de vesículas recubiertas. Este revestimiento proporciona una primera clasificación de estas, pero no exhaustiva.No todas las vesículas con el mismo revestimiento tiene el mismo destino. Podmeos destacar la clatrina, la COPI Y LA COP II. Podemos establecer una clasificaron dependiendo del tipo de destino.En la superficie de las vesículas hay moléculas que actúan como marcadores de fusión con el órgano diana. En la membrana de este hay un receptor para este marcador son las denominadas SNARE.Para la fusión se requiere de las proteínas SNARE y también de una RAB-GTP y su efector RAB, este presente en la membrana diana. Tras la fusión de las membranas se hidroliza la RAB-GTP a RAB-GDP y se desacople la pareja SNARE.

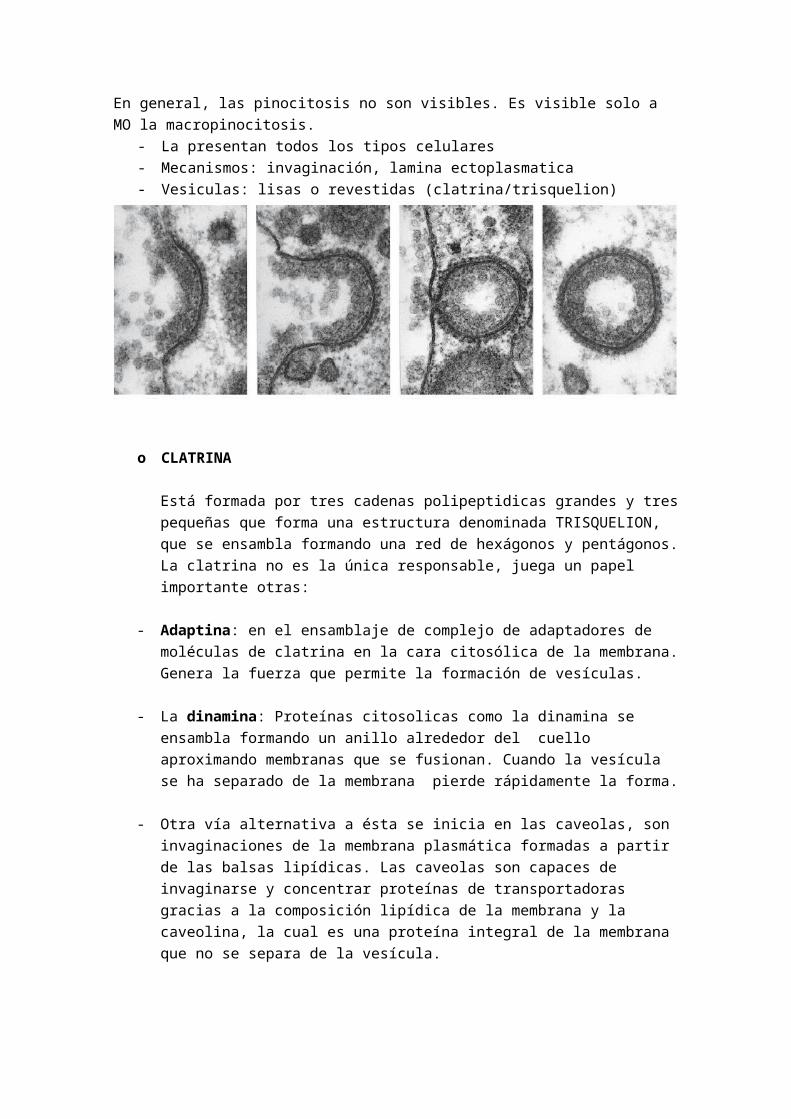
PASADO A LIMPIO HASTA AQUI
TEMA 6: ESPECIALIZACIONES DE LA MEMBRANA CELULUAR
1.- CONTACTOS INTERCELULARES Y COMPLEJOS DE UNION2.- MOCROVELLOSIDADES E INVAGINACIONES3.- COMUNIACION INTERCELULAR A DISTANCIA4.- ALTERACION DEBIDO A LA FALTA DE COMUNICACIÓN CELULUAR
INTRODUCCION
Las células pueden unirse unas a otras a través de uniones intercelulares directa so a través de materiales que ellas mismas secretan.Las células deben cohesionarse para formar una estructura pluricelular organizada.
1.- TIPOS DE UNION
CONTACTOS INTERCELULARES Y COPLEJOS DE UNION.
Podemos destacar:- Complejo de unión : formado por la unión estrecha, la unión adherente, y
desmosomas.- Las uniones de tipo GAP
TEMINOLOGIA/TIPOS DE UNION:
Según la distancia entre las membranas adyacentes:- ADHERENS : (adherente, anclaje) hay cierta distancia entre las células contiguas (20-25
nm)- OCCLUDENS (estrecha o tight en lngles) no hay separación entre las dos células con
fusión de hemimebranas externas- GAP (formadoras de canal, hendidura) no hay fusión entre las membranas de las dos
células, pero están muy próximas (separadas unos 2nm)
Según la extensión de la unión- ZONULA: si es una bada (1 micrometro de ancha) que rodea totlamente la celula- MACULA: uino puntual de uan superficie aproximada de 1micra al cuadrado- FASCIA: mancha o parche irregular de mayor superficie que la macula.
BIOLOGIA CELULAR. PANIAGUA. 3 EDICION
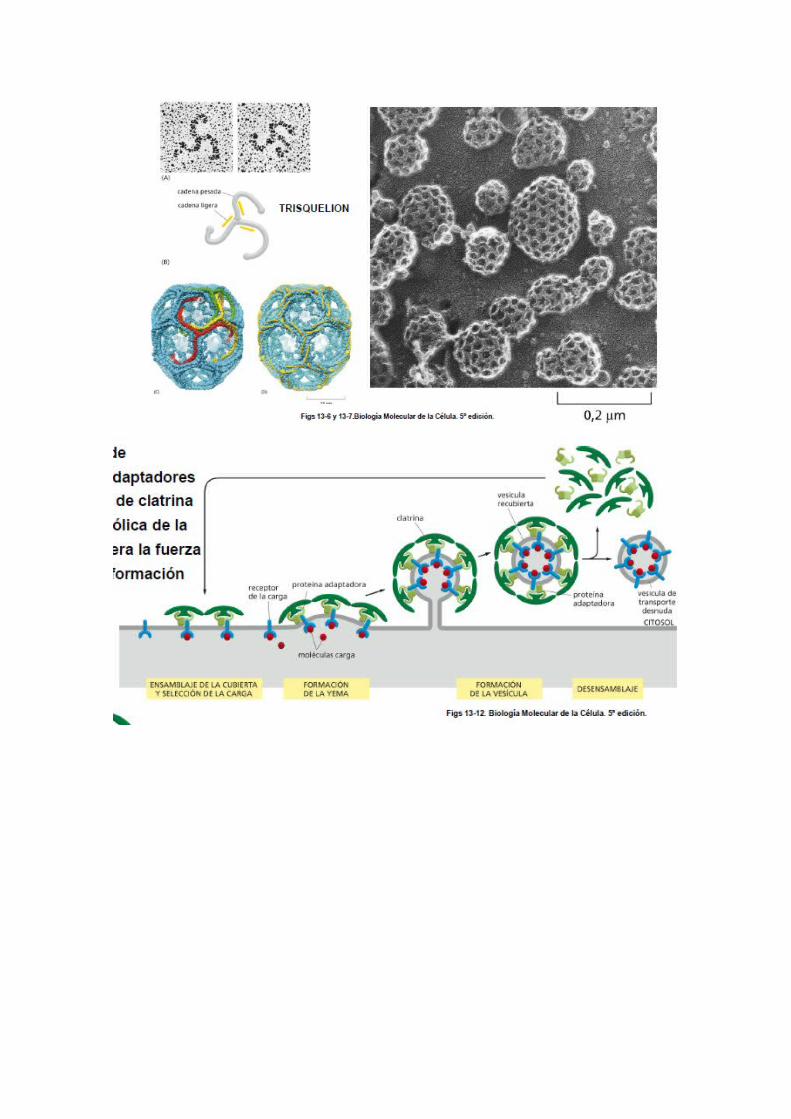
CLASIFICACION FUNCIONAL:
A) Uniones de Anclaje Zonas de anclaje de filamentos de actina
- Uniones célula - célula (adherentes o adherens)- Uniones célula-matriz (adhesiones célula-matriz asociadas a actina)
Zonas de anclaje de filamentos intermedios- Uniones célula - célula (desmossoma)- Uniones célula - matriz (hemidesmosoma)
B) Uniones de oclusión Uniones estrechas (vertebrados) Uniones separadas (invertebrados)
C) Uniones formadores de canal Tipo gap Plasmodesmos (vegetales)
D) Uniones transmisoras de señal
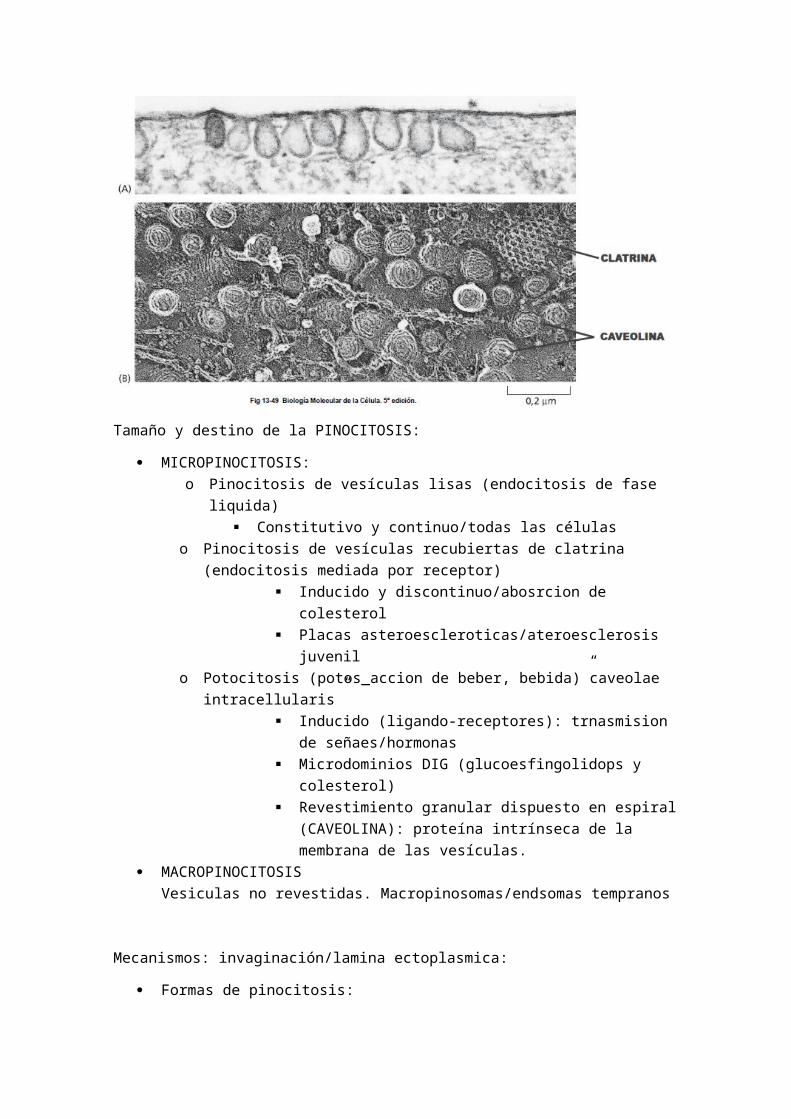
E)
COMPEJO DE UNION
Tres tipos de unión:- La unión estrecha sella los espacios entre las células epiteliales- Unión adherente que conecta los haces de filamentos de actina de una célula con los
de la célula adyacente- Los desmosomas conectan filamentos intermedios de una célula.

a) UNION ESTREA O ZONULA OCCLUDENS
Las membranas de las células adyacentes establecen contacto per on continuo sino a intervalos. Forma un cinturón alrededor de toda la célula e impide el paso de cualquier distancia. En la formación de estas uniones intervienen fundamentalmente dos proteínas, la CLAUDINA y OCLUDINA.Estas uniones, presentes en epitelios, son impermeables para la mayoría de las moléculas. Evitan que receptores y transportadores específicos, ubicados en la superficie apical difundan hacia la superficie basolateral.


b) UNION ADHRENTE/ZONULA ADHRENS/DESOMOSOMA EN BANDA
Son uniones de anclaje a FILAMENTOS DE ACTINA. Forman un cinturón alrededor de la célula. La unión indirecta de los filamentos de una célula a los de sus vecinas permite a las células de un tejido utilizar el esqueleto de actina de forma coordinada.Se forman por la unión adherente en la que interviene las proteínas transmembrana CADHERINAS presentes en las dos células (INTERACCIÓN HOMOFILA)
(BIOLOGIA CELULAR. PANIAGUA. 3 EDICION)

c) DESMOSOMAS/MACULA ADHERENS.
Son uniones PUNTUALES de anclaje a FIMLAMENTOS INTERMEDIOS entre dos células. Intervienen proteínas de la familia de las CADHERINAS mediante INTERACCIÓN HOMOFILA. La placa densa de proteínas de anclaje es propia de esta unión.
(BIOLOGIA MOLECULUAR DE LA CELULA. 5 EDICION)
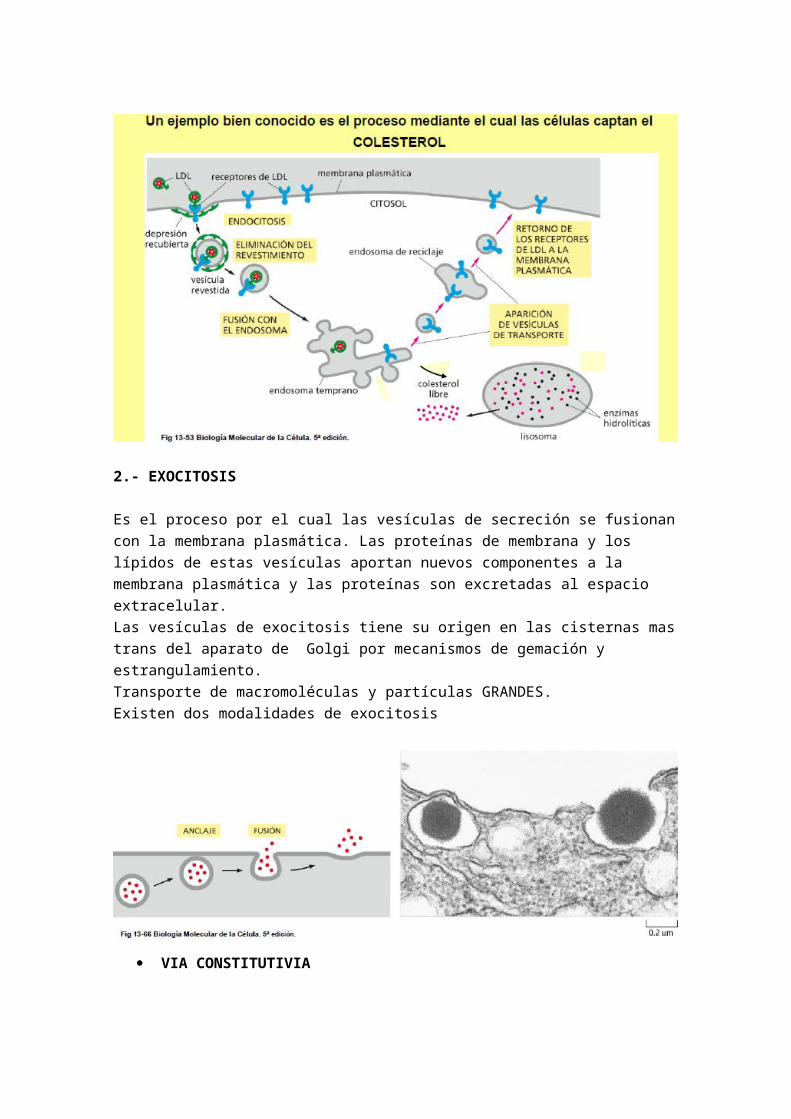
UNIONES DE ANBLAJE CELULA MATRIZ
UNIONES CELULA-MATRIZ: CONTACTORS FOCALES
Forman una placa de adhesión que conecta los filamentos de actina con la matriz extracelular.La mayor diferencia con las uniones adherentes radica en que la glucoproteína transmembrana es una INTEGRINA.Las integrinas sobresalen en el espacio extracelular y se unen a la fibronectina (INTERACCION HETEROGINAL) que a su vez se relaciones con los componentes de la matriz extracelular (colágeno y proteoglicanos)
UNIONES CELULA-MATRIZ: HEMIDESMOSOMAS
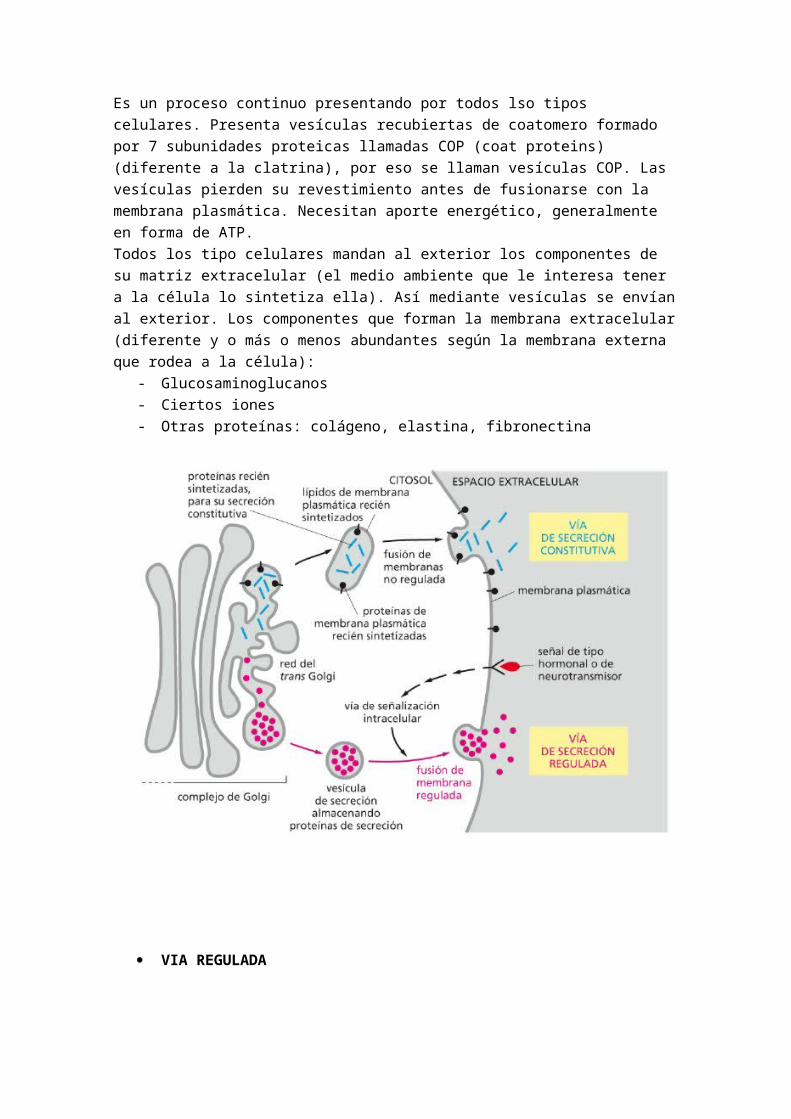
Conecta los filamentos intermedios con la matriz extracelular.La proteína transmembrana es una INTEGRINA, que se une a moléculas de colágeno en una interacción heterofila o heterofilica.En el hemidesmososoma son fialmentes intermedios.
UIONE TIPO GAP/ FORMADORAS DE CANAL/EN HENDIDURA (ES EL MISMO NOMBRE)
Estan presentes ne la moyoria de tipos celulares. La unión se purduce debido del acoplameinto de las células elctrica y metabólicamente (permiten sincronizar las contracciones en el musculo cardicao en el musuclo liso del intestino)La membranas de las células adyacentes separadas por un espacio de 2-4nam, este espcaio esta constituido por proteínas formadoras de canales CONEXINAS. Los canales se pueden regualr por lo que la permeabilidad entre las células puede disminuir. Si una celula se daña se cierran los canales para imperdir la propagación del daño. A través de estos iones entra agua e iones, si este anal no para, entrara agua también en las células vecinas.
CLASIFICION FUNCIONAL DE LAS UNIONES CELULARES:
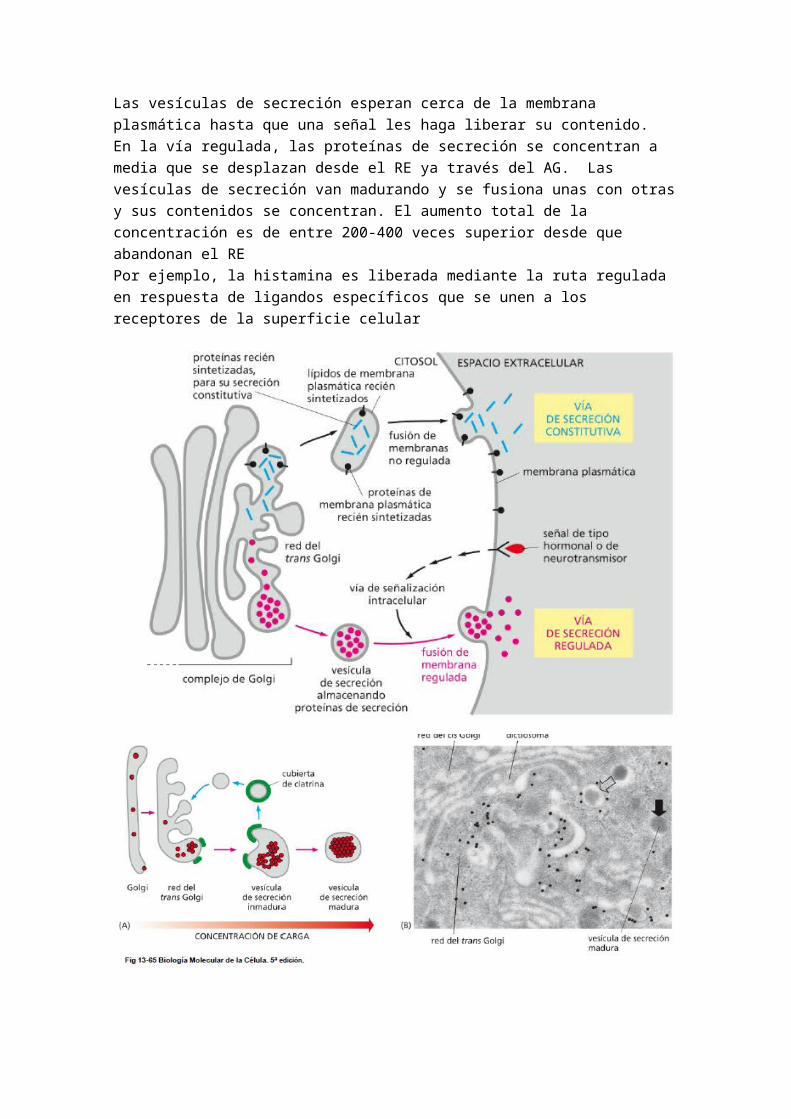

SINAPSIS QUIMICA
En la región sináptica hay muchas proteínas y físicamente hay unión. Esta unión se basan en al formación de proteínas de unión, de la familia de las cadherinas, de la familia de las inmunoglubulinas, proteínas especifivas de las sinapsis. Es decir, las neuronas no solo están cerca sino que también están unidas físicamente, de tal forma que se aumenta la eficacia de la transmisión.

RESUMEN:- UNIONES CELULARESAS:
o Unión estrecha o zonula occludens (ocludina y Claudina)o Unión adhrente o zonula adherens (cahderinas) conectan con filaments de
actinao Desmosomas o macula adheres (cadherinas) conectan con filamentos
intermedioso Uniones GAP formadoras de canal o en hendidura (conexinas)
- UINOES CELULA MATRIZo Contactos focales (integrinas) conectan con filamentos de actinao Hemidesmosomas (integrinas) conectan con filamentos intermedios
ESPECIALIZACIONES DE LA MEMBRANA PLASMATICA
La membrana plasmática media entre las células, bien mediante las conexiones vistas antres o mediante las especiliazaciones siguientes: microvellosidadse, esterocilios y cilios.
2.1 MICROVELLOSIDADES
Son diferenciaciones muy especializadas de la membrana plasmática para el trnsporte de sustancias hacia el interior de la celula presentes en epitelios absorbentes. El citoplasma de setas prolongocaiones esta constituido principalmente por actina y otras proteína asociadas.Los filamentos están unidso mediante haces de microfilamentos, la vilina y la fimbrina hacen que se mantengan rectos. Las microvellosidades no son móviles, los cilios si.
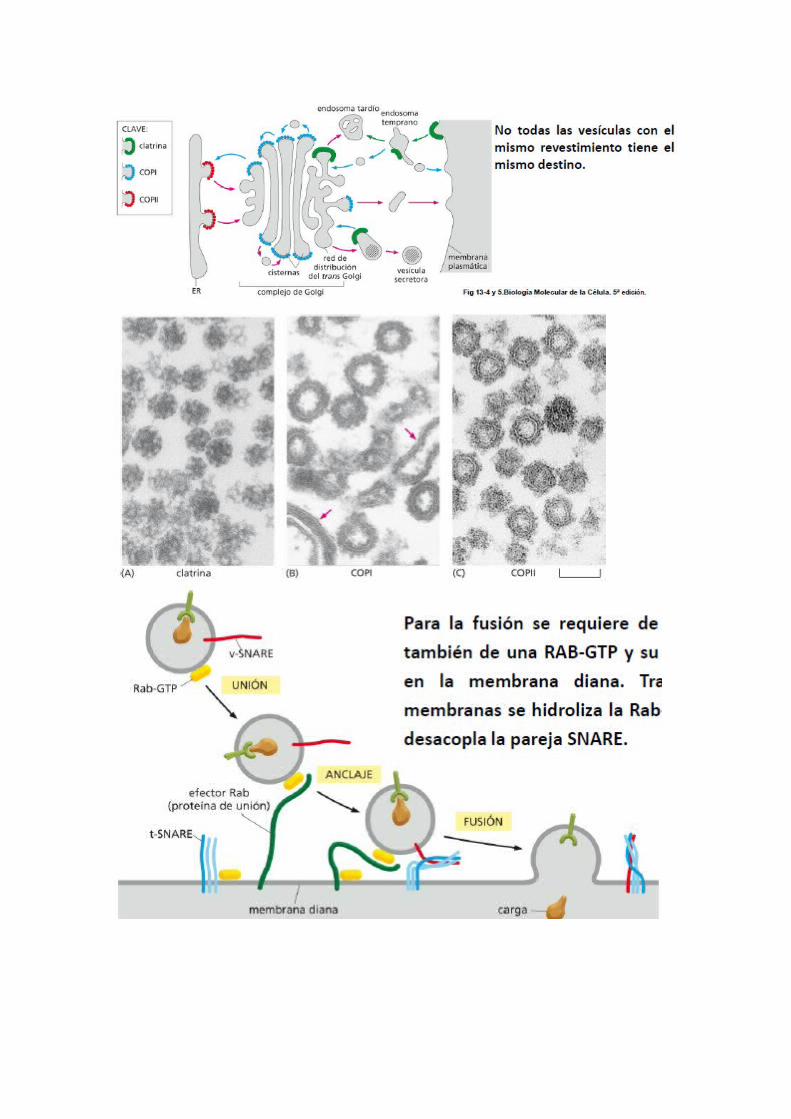
EXAMEN: FOTO DE MICROVELLOSIDAD Y DE CILIOS, DIFERENCIAS (TANTO DE CORTE TRANSVERSAL COMO LONGITUDINAL, MEDAINTE CRIOFRACTURA, POR EJEMPLO, TAMBIEN DE TRANSMISION O COMO SEA).
Las membranas plasmáticas de las microvellosidades intestinales poseen enzimas específicos que terminan la degradación de los compuestos que ingerimos. Disacaridasa (rompoen la sacarosa, maltosa y lactosa)Aminopeptidasa (liberan aminoácidos)Fosfatasa alcalina (hidroliza compuestos fosfatados)
Tambien poseen sistemas de transporte para incorporar a la celula moléculas resltado de la digestión (glucosa, amionacidos, pequeños péptidos… etc).La misoin principal de las microvellosidades consiste en incrementar la velcodidad del proceso de absorción, aumentando para ello la superficie de la membrana plasmática (de 15m2 a 300m2).

2.2 ESTEROCILIOS
Semejantes a las microvellosidades son los ESTEROCILIOS. Son más largas y delgadas que la microvellosidades y se estrechan conforme se acercan a la base. Son uy específicos, por ejemplo están en el epitelio renal, en el epidídimo.a) El EPIDIDIMO: en el domino apical de las células epiteliales presentan esterocilios largos y
ramificados que permiten aumentar la superficie de membrana. Esas células absorben el 90% del líquido testicular y secretan glucoproteínas. La endocitosis y pinocitosis son muy llamativas
b) PELOS SENSORIALES: aquí también hay esterocilios. Estan en las maculas del utirculo y del sáculo en el oído.
Estos pelos no guardan relación con el transpoerte, sion con la transformación del estimulo mecanico en impulso nervioso. Mucho ojo, en los pelos sensoriales, existe el llamado kinocilium, que esta formado por microtubulos, todos los demás son estercilios. El kinocilio es el que tiene capacidad de movimiento, gracias a este movimiento se producirá la apertura de ciertos canales, produciendo una excitación si se mueve a un lado, o hyprpolarizacion (inhibion) si se mueve para el otro lado. Solo hay un kinocilium por penacho (este penacho esta formado por le kinocilio y por los estercilios).

2.2 INVAGINACIONES/INTERDIGITACIONES
Son pliegues generalmente laterales o basales que se observan en algunos epitelios (túbulos contorneados distales del riñón, conductos excretores de glándulas salivales).
Las principales funciones son:A) Reforzar la cohesión entre las célulasB) Proporcionar una reserva de membranaC) Aumentar la superficie de intercambio
(FOTOS DE: BIOLOGIA CELULAR. PANIAGUA. 3º EDICION)
3.- COMUNIACION INTERCELULAR A DISTANCIA

Las ceulas se comunicaran medante la membrana, pero también se peude establecer relación a distancia. Estas relaciones pueden ser:
a) Dependiente de contacto: la información va de una celula a otra porque están al aladob) Paracrina: comunicación a distancia corta. La celula señalizadora emite un mediador
local a las células adyacentesc) Endocrina: La señalizadora emite una horma a la sangre y acutara a larga distanciad) Sinaptcia: mediante neurotransores. A corta distania
3.1 Recepcion de la señal
a) Receptores de superficie celular:La maoria de las de las veces se producirá a través de la membrnan, mediante poreteinas receptores de superficie celular. Por ejemplo la insulina
b) Receptores intracelularesHay elementos liposolubles que atraviesan la membrana plasmática, estando el receptor en el interior de la celula
c) Comunciacion intercelular a distancia: la molecual señal se une a la proteína recepora, se movieran a través de proteínas señalizadoras tnracelulalares, actuando sobre las proteínas efectoras como enzimas metablicas (cambios en el metabolismo), proteínas reguladora de laexpresion génica, o la proteína del citoesqueleto
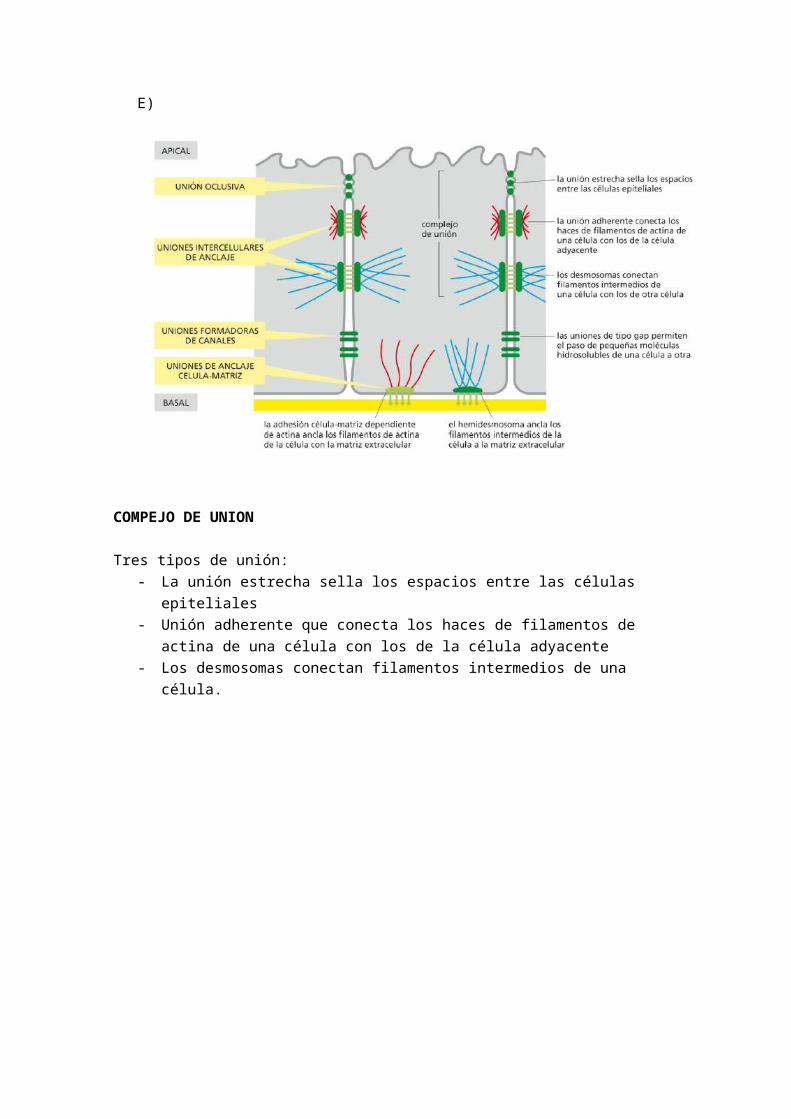
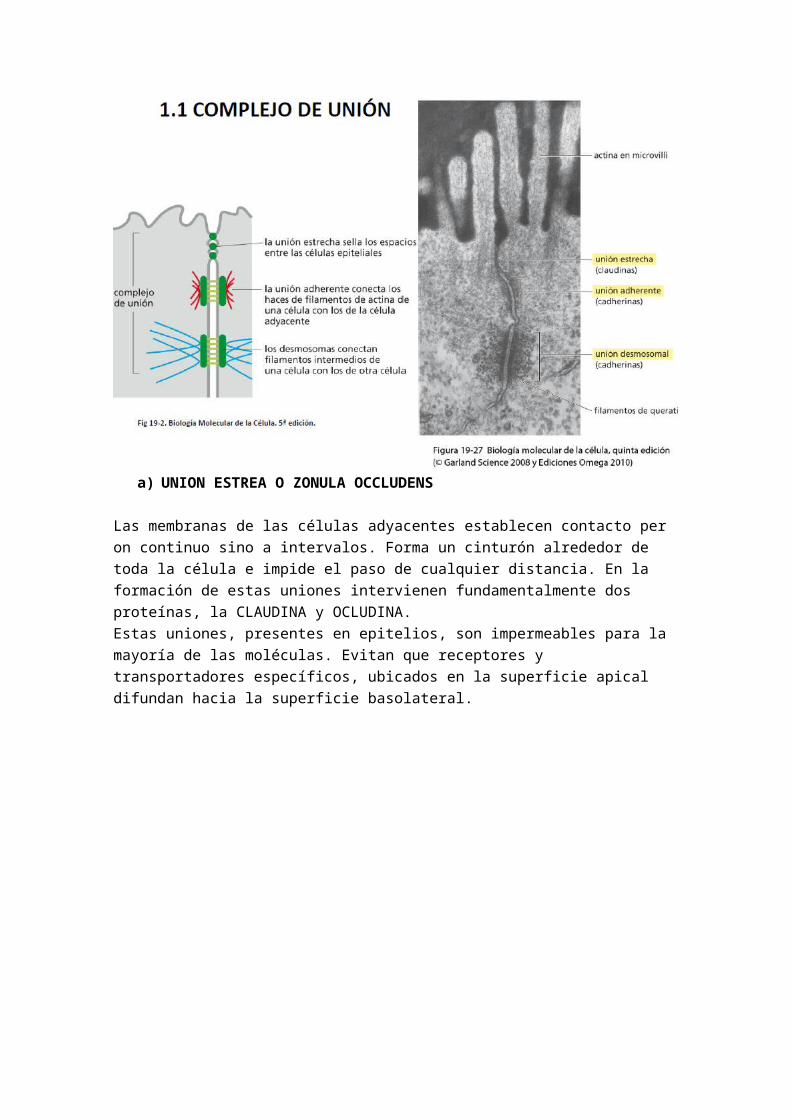
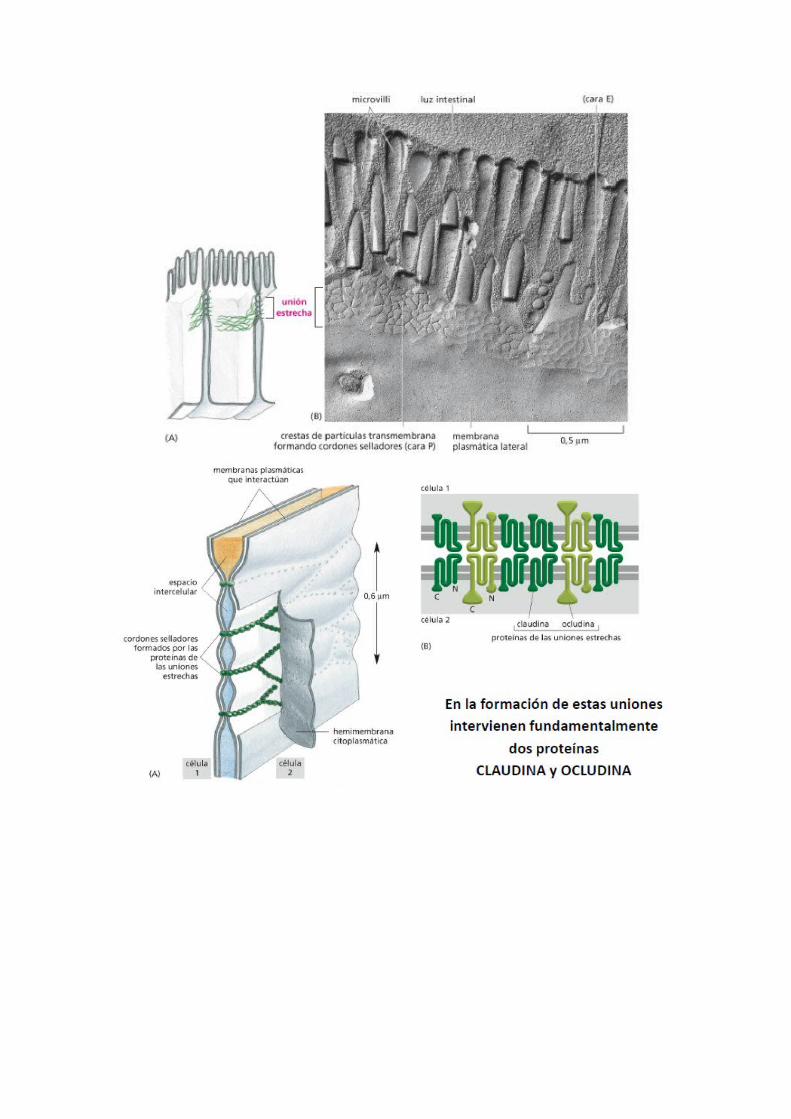
3.2 Tipos de recpetoes
a) Receptores acplados a cnales ionicos:
b) Receptores acoplados a proteínas G. Suelen tener 7 domonios intermembrana, cando llega la señal al receptor, courre la activación de la proteína G, y se crea el segundo mensajero (AMP CICLIOC)
c) Receptores acoplados a enzimasSe produce la activiaciondel domino catalicito o bien la activación del enzima asociada activada.
4.- ALTERACIONES DEBIDAS A LA FALTA DE COMUNICACIÓN CELULAR
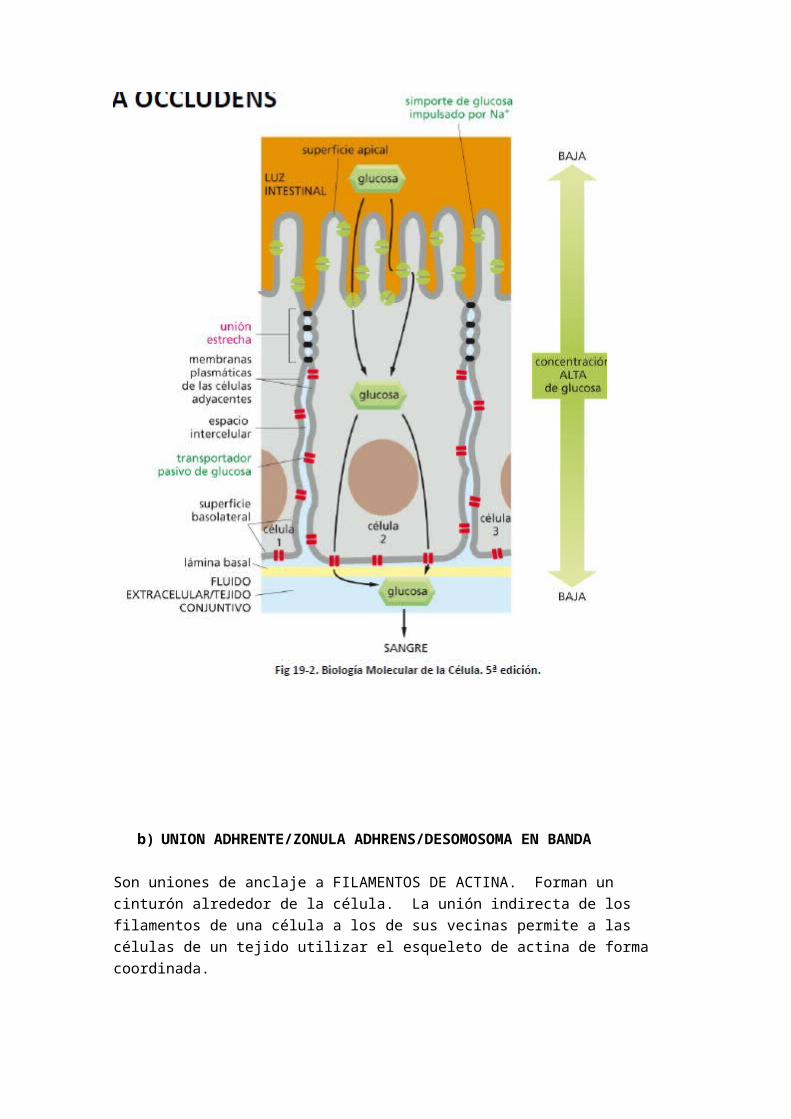
Por ejemplo la epidermólisis bullosa simple producida por una alteración en las unoies celula-matriz. Fallan los hemidesmosomas. Otro ejemplo es la sordera genética por fallo en los transmisores de ifnromacion.
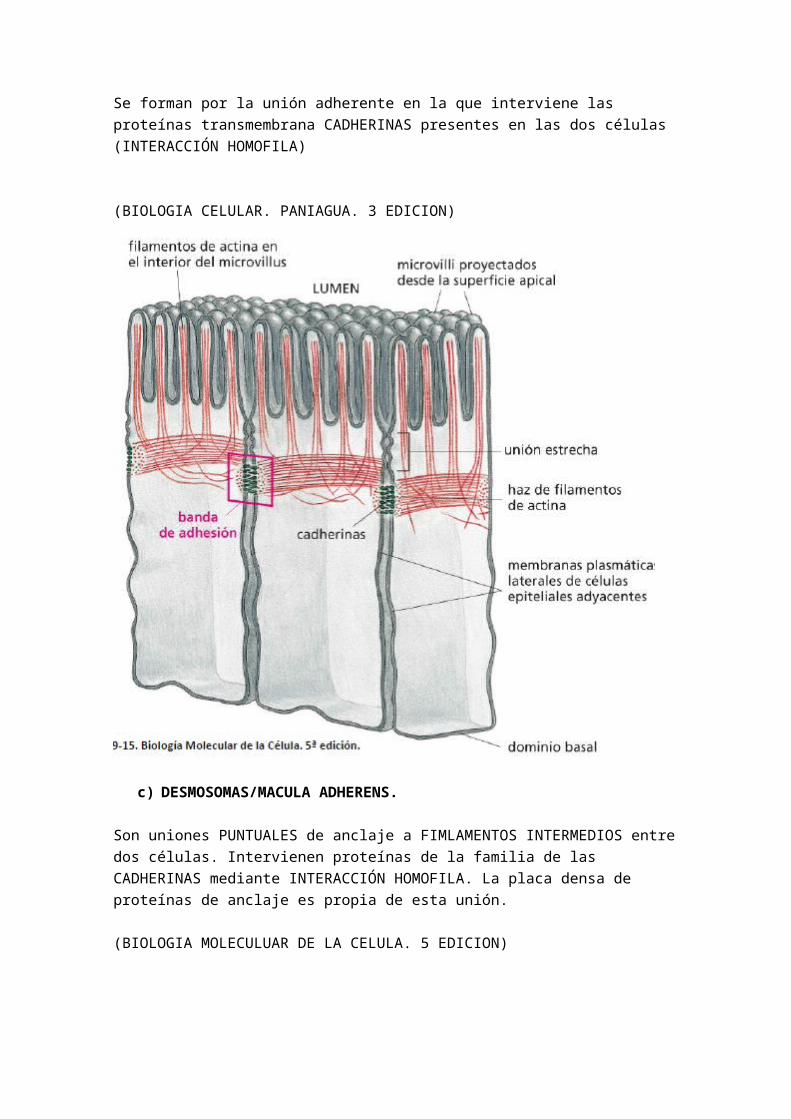
HIALOPLASMA Y CITOESQUELEOT:
CONCEPTO DE HIALOPLASMA
Se entieende por plasma transparente. nO se aprecia ni con MO ni ME. tenemos que recurri a la bioqumica para verlo, y bioquimicamente esto es el sobrenadante una vez realicemos la centrigucaion diferencial.
COMPOSCION QUIMIC Y FUNCION
Esta compuesto por 85% de agua, aminoacidos mayoritameiramete y proteinas (esats s proteinas son el sustrato morfologico para el citoesqueleto celular). Ademas otros mucos como ARNm, ARNt, nucleosidos, nucleoticos azucares, acidos grasos, productos metabolicos en general, ENZIMAS. Dede el punto de vista de la funcion, el hialoplasma es el sustrato donde se realizará interacciones quimicas.en esta lugar se produce:- sintesis proteica, lipidica y de glucogeno- glucolisis anaerobia (ruta de embden meyerhof)- atajo de las pentosas (ruta de bardurg dickens litman)
en el citoesqueleto tambien encontramos diferenciaciones morfologicas de fases o dominios resultado de las reacciones qumicas que se dan en el hialoplasma, estas fases odominios son:
PARAPLASMA
Es un plasma menos fluido que el hialoplasma. Dentro de esta plasma podemos encontrar:- Granulos de almacenamiento:
- GLucogeno: metodo de PAS, se ven cumulos rosados en el hialoplasma. MEdiante ME particulas alfa (1500A), beta (300A), gamma (200x30a). sE ven con MO. Para ponerlo de manifiesto se usa el PAS
- procesos de calcificcacion- sintesis de proteoglucanos
eN la imagen se ve un hepatocito y vemos las organelas citoplasmaticas, en un fragmento como las inclusiones de glucogeno muy electrodensa se aprecia acumulos de roseta (1500 Amstron), a esto se le denomino alfa de glucogeno; por otro lado esta el glucogeno Beta, tiene un diamtro de unos 300 amstron. Si la ampliamos estos puntitos, estan formado por un entramado formado por glucogeno gamma (entre 200a de longitud y 30 de anchura). a la derecha se va la corteza suprarrenal donde se aprecia gotitas lipidcias transparentes (no tienen un oclorate adecuado)
en la segunda imgaen se ven inclusiones lipidicas en celula hepatica humana (en el rincon izquierdo superir) y de una rata es la imagen de una rata (debajo inferior); la fijacion se hizo con glutarldehido y una posfijacion con osmio., lOs puntitos negros (que no electrodeonsos porque es Optica) son incursiones lipidicas. Este fragmento, de la rata, son hepatocitos se aprecian los incursiones lipidicas; las flechas señalan las incuseiones glucogeno intesamente rosadas. Tambien se aprecia en algunos nucleos el nucleolo
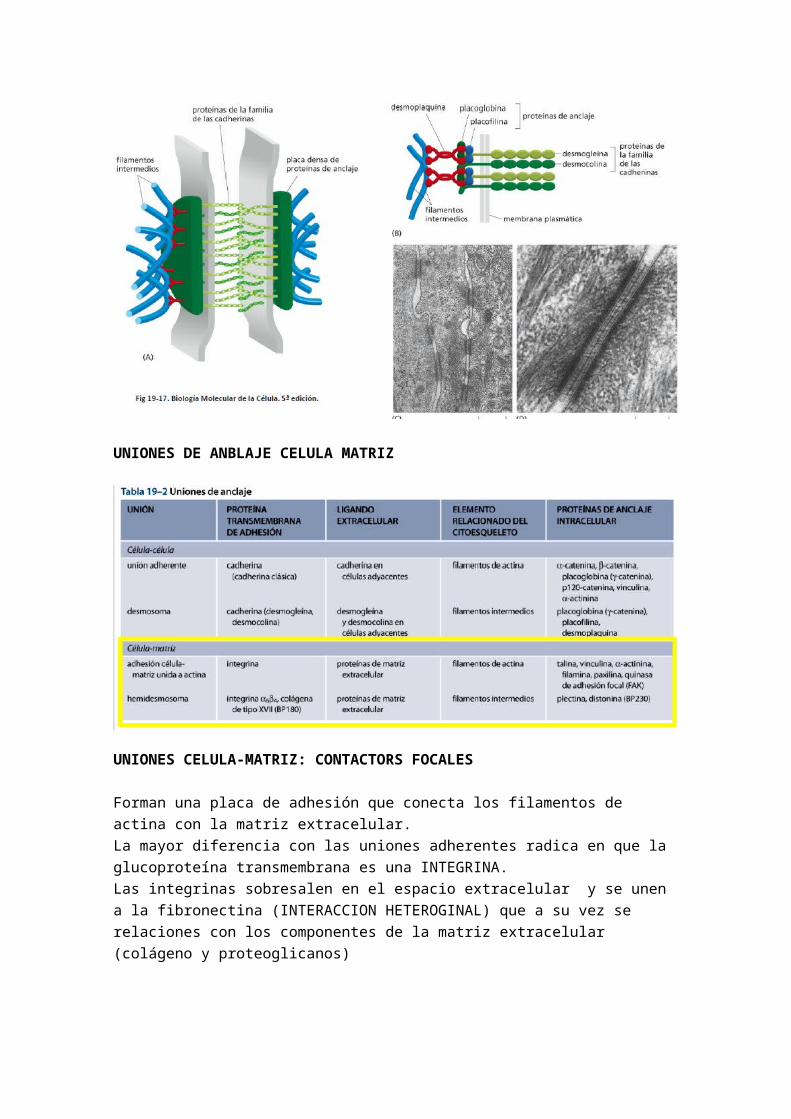
En la tercera imagen (tres fotos) se aprecian a MO, la primera de las tres, se ven cartilagos. la tercera foto esta hecha con hematoxilina, de cartilago, se ven bien separadas las celulas porque esta dentro de la matriz extracelular donde se aloja el condrocito, por eso se aprecia tan bien. Tambie se ve la incursion lipida, en la parte que no se ve nada, porque la grasa se disolvio al meterla en xilol. La primera foto se ve con sudan negro y la segunda con azul nilo; la primera foto se ven las incursiones lipidicas, tanto en la primera foto y la segunda, la primera negra y la segunda rosa. En el sudan no se ve la matriz, por lo que se uso rojo neutro, por eso se ve rojo.En la tercera imagen se ven muy bien los nucleos del condrocito, las dos primeras imagenes se han usado tinciones especiales para las incurisones.
eN la cuarta imagen vemos un adipocito joven, adipoblasto, se ven las gotitas lpidicas, estas gotas se acabaran fusionando gotas mayores, llegara un momento que sera tan grande qe sera una sola y el nucelo por falta de espacio quedara recluido e nal periferria(morfologia en anillo de sello), la gras no se aprecia. En los adipocitos, en la hipodermis, encontramos estas reservas, los adipocitos, en la matriz extracelular estan los adipocitos, que al ser un corte semifino, al ser con glutarladelhido los lipidos se conservan con tonalidad azulada. Con ME se veria el anillo de sello.(EN EL EXAMEN NO TE VA A PREGUNTAR EL TIPO CELULAR, ES DECIR NO TE VA A PEDIR EL ORGANO DE DONDE VIEN SI ES UN FRAGMENTO DE CITOPLASMA) . eN la imagne superrio derecha, vemos gracias a su electrodensidad, las gotas lipidicas, de varios tamaños diferentes. Abajo a la derecha hay una imgaen ampliada deonde se aprecia el reticulo endoplasmatico liso
Otra imgane: Sin hacer postfinaicon con osmio, se aprecia en la foto a la ziquierda, hay un fragmento de un adipocito, es un fragemto de uan iclusion lipidica, se aprecia un vacio histologico, al no usar osmio hay este vacio. Se ven pequeñas gotas lipidicas que se van uniendo a la mayor. Podemos encontrar tambien un neotrofilo, de color negro
- lipidos: adipocitos. Rojo Sudan, Negro S. Azul nIlo- fUente de cadenas de carbono- sumideros de NADH
Tanto el glucogeno como los lipidos se usan para la sintesis de proteoglucanos. las lipidicas en el cartilago pueden ser movilizadas cuando lo necesita la celula, para formar nuevas biomembranas (sobre todo catilaginos). Es decir son fuentes de carbono.Tambien son sumideros de NADH para que no afecten a otras reacciones quimicas

- proteinas: ovocitos. eL ovocito guarda proteinas, pero las inclusiones proteicas son como respuestas a procesos patologicos (por eso no hay imagenes de estas inclusiones, pero si de glucogeno y de lipgidos)
- CUerpos de russel (plamatica), inclusiones de mallory (si se bebe mucho)
- PIGMENTOS (ENOGENOS Y EXOGENOS)los exogenos son mayoritarimente en patologicas, por ejemplo los mineros tienen dentro de las celulas pigemos abosrbidos.cabe destacar: melanina, hemoglobina (hemosiderina), lipofuschina (resultado de la activdiad de los lisosomas, se acumula en el citoplasma, y se acumula como lipofuschina)Exogenos: Antracosis (carbon), silicosis (silicio), siderosis (fe)
-CRISTALES (Reinke) Cistales que se acumulan en el testiculo, en el citoplasma celular o en el nucleo. Son incursiones de precipitaciones de proteinas.
eN la imagen se aprecia a la derecha la melanina en neurona de la sustancia negra, teñida con Azan. A la izquierda se aprecian son las unicas celulas que sin teñirlas se aprecian la melanina que acumula en su citoplama por su capacidad bioquimica
otra foto. Se ve el cristal de reinke y otros pigmenos debido a la actividad lisosomica. A la izquierda se ve una celula de testiculo, el nucleo electrodenso muy negro y se aprecia la eucromatina con menos electrodensidad.
CITOESQUELETO
En la imagen se ve un fibroblasto en cultvo donde se usaron distintos colorantes fluorescentes y se aprecian los distintos elementos del citoesqueleto. son colorantes especificos del citoesqueleto. en azules esta la ctina, de rojo se ve la pinpurina, y de verde los filmentos intermedios que ofmran parte de los microtubulos. el AMARILLO.....(SALE NE LAS FOTO LOS COLORINES)
CONCEPTO DE CITOESQUELETO.
se trata de un conjunto de estrucutras ....... que poermente el movmiento de las organelas (muy rapido) CON el citoesquelo es una estructura dinamica, es una espcei de andamiaje, se puede formar y deformar segun se necesite. Se trata de polimeros lineales o ramificados de

proteinas, se encuentra en equilibrio entre formas libres y polimericas. Se adaptan a cambois morofuncionales. Es responsable de cualquier tipo de movimiento celuluar.esta integrado por microfilamentos, f intermedios y microtubos
FILAMENTO DE ACTINA
Reciben tambien al nombre de . Son polimero lineales. Existe una ralacion entre su forma libre Actina globular o G y su forma polimerica, actina f. Esta intimmamente con las zonas de adherencia, suele situarse por debajo de la membrana plasmatica, formando el eje de las microvellosidades. Su distribucion en el seno guarda relacion con las proteinas de la matriz extraceuluar. Algunas proteinas de las membrnas son proteoglucanos por lo que a veces discriminar entre el medio extraceuluar y el intrecelular es dificil. La actina por si sola no puede hacer nada, para todo tiene que interaccionar con otras proteinas. Su principal funion es la funcion contractill (endocitosis y exocitosis, participan estas proteinas) hay por tanto proteinas asociadas a las actinas; se les llama Factores de Polimerizacion. CUando la actina forma haces con los ejes necestia la F. formadora de haces; si tiene funcion contractil, necestia de los factores de contraccion; por ultimo tambien necestia
FILAMENTO INTERMEDIO
Su nombre viene dado porque su tamaño es intermedio a los demasTIenen un diametro de 100AQuimicamente heterogeneosmenos labiles y + resistentes a desnaturalizacion

funcion estructural y contribuyen a mantener las organelas en su sitioTIpos: VIMENTINA (ondulados, celula mesodermica-fibroblasto) QUERATINA (celula epitelial-desmosoma) GLIALES (celula Glia) NEUROFILAMENTOS (neuronas)Importancia biomedica: anatomia patologica
NO CONFUNDIR LAS FIBRAS MUSCULARES (QUE SON CELULAS) CON LAS FIBRAS DE LOS TEJIDO QUE SON PROTEINAS (EXAMEN)
Existe una relacion directa entre ciertas proteinas y un cierto tejido:1.- QUERATINA: En el epitelio2.- VIMENTINA: T. conjuntivo3.- DESMINA: Musculo4.- NEUROFILAMENTOS: SOmas y prolongaciones neuronales
ELEMENTOS DE LOS FILAMNETOS INTERMEDIOS
Los fi estan formansod por dos monomeros, en este caso econtramos o bien un dimero o bien un tetramero. eL tetramero es la estructura basica de lso filamentos intermedios. esta estructura tiene estructura antiparalela, es dcir el extermo aminoterminal enfrentado al COOH terminal. Los tetrameros se disponene de una forma desfasada entrealazadas para constituir uno de los subfilamentos que forman parte de lso filamentos intermedios (es decir, los COOH y el NH2 no estan enfrentados unos enfrente de otros, si no que se van alternando primero uno y luego otro). La estrucutra basica al final son 8 subufilamentos que se disponen formando una cuerda enrollados, es decir, una estructura cordiforme. Los expertos disienten en la formacion final, no sabne si es 7 y uno en el centro, o si hay tres arriba, tres abajo y dos entre medias, no habinedo nada justo en el centro.
En a una paotlogia de los filamentos intermedios es la ELA, (stephen hawking), en esta caso los nourofilamentos estan afectados y por tanto hay una incapacidad de movilidad muscular. Se trata de una acumulacion y ensamblaje anormal de neurofilamentos. Sobreexpresion e NF-L y o de NF-H.
MICROTUBULOS
Los microtubulos son las elementos mas grueosos del citoplasma, pero son huecos. Si son huecos tieen que haber una luz y una pared, asi pues 150Amstron mide el hueco y 50A la pared.
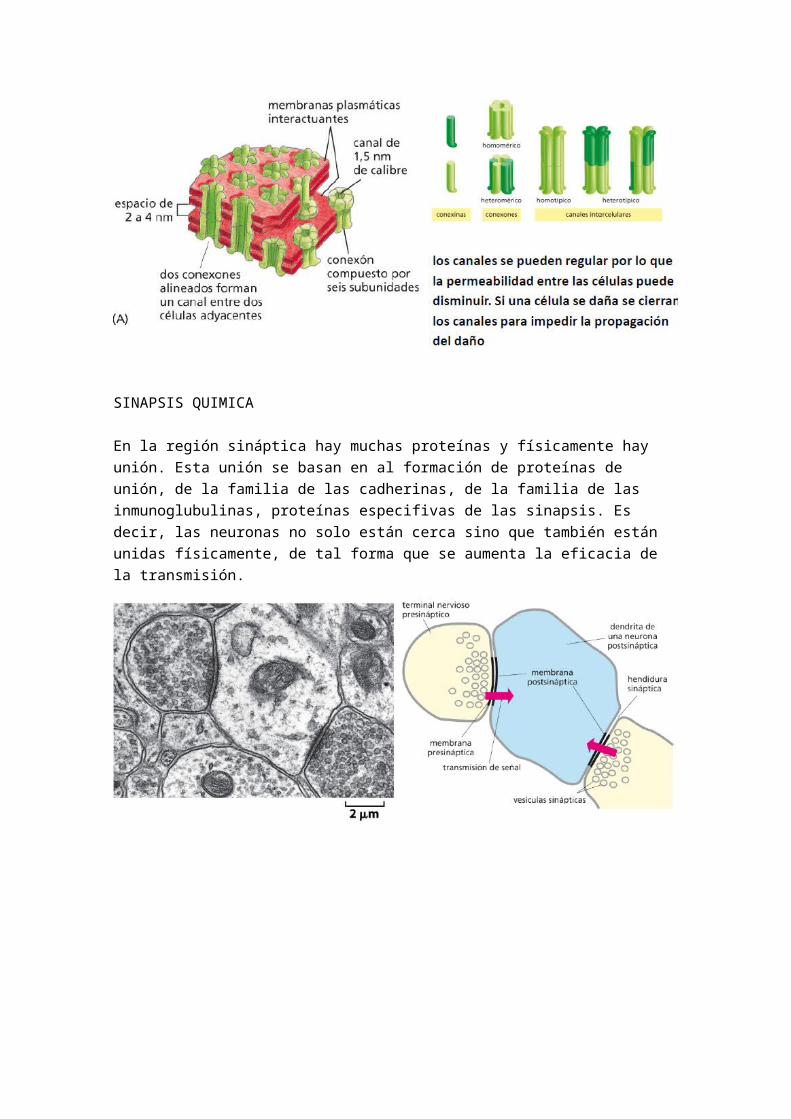
esta formado por 13 protofilamentos (polimeros de tubulina alfa y tubulina Beta)El conocimieto sobre lso microtubulos viene por la patologia de la gota, donde se prodcue una acumulacion de cristales en las articulaciones por problemas metabolicos; el dolor se calma con polvo de azafran. En estos pacientes se encontro una gran cantidad de mitosis, se determino que la colchicina (esta en el azafran) paralizaba la mitosis, se empezo entonces a usar con los tumores. Habia otros alcaloides que hacia los mismo, como la vinblastina, vincristina actuan de forma similar)Los microtubulos estan formados por estructuras tubulina alfa y beta. Son 13 subfilamentos de filamentos, existe un desfase entre los dimeros de tubulina alfa y beta, como si se hubieran dispuesto como forma helicoidal. En total tenemos 13 subufilametes que forma una estrucutra hueca. EN la luz de esta trucutra puede circular elementos de pequeño tamaño. existe una relacion entre la forma tubular y los tubulos libres en el citoplasma, ya que por el extremo + se produce la polimerizacion de la estrucutra y por el extremo - se despolimeriza (es decir se destruye y se libera tubulina epor el mas y se forma de neuvo por el menos). es decir se estan formando y deformando continuamente. SOlo si la celula entre en mitosis los microtubulos cambian su formacion, es decir son tambien autopistas especificas por donde cirucula macromoleculas para que estos se situen en el lugar adecuado en ciertos procesos ( por ejemplo, dirigen donde ir a las vesiculas de exocitosis y endocitosois); los microtuublos les dicen a los cromosomas como ponerse en la mitosis.; es decir responden a las necesidades de la celula, a estos se les llama microtubulos labiles, son los responsables del movimiento tambien del interior de la celulla
Se tratan de labiles porque no soportan cierta temperatura (4grados), son sensibles a fijadores como glutaraldehido, a alcaloides como la colchicina, vinblastina y vincristinaa.lOs enfermos de gota, al tener los microbulos afectados, les llega el urato a las celulas y no lo pueden digerir, el cristal rompe la membrana plasmatica, las enzimas salen fuera y la matriz es degradada y provoca el dolor.El taxol es el que actua de tal fomra que no deja que no se despolarice los microbulos. Los microtubulos cran un canal en los exones de las neuronas (pueden medir hasta dos metros), estas autopistas cricula neurotransmisores desde el soma celular (el extremo menos de los microtubulos, hasta el extremo + en la terminal sinaptica) al reves desde el menoas al mas, esta el transporte restrogrado. Hay dos protteinas asociadoas a los microtubulos, unas proteinas hacen el transporte retrogrado, la dineina y desde el polo menos hasta el polo ma son las quinesina.
MICROTUBULOS ESTABLES
Son los que forman organizaciones como los centriolos y el citoesqueleto interno como cilios y flagelos
- CENTRIOLOS
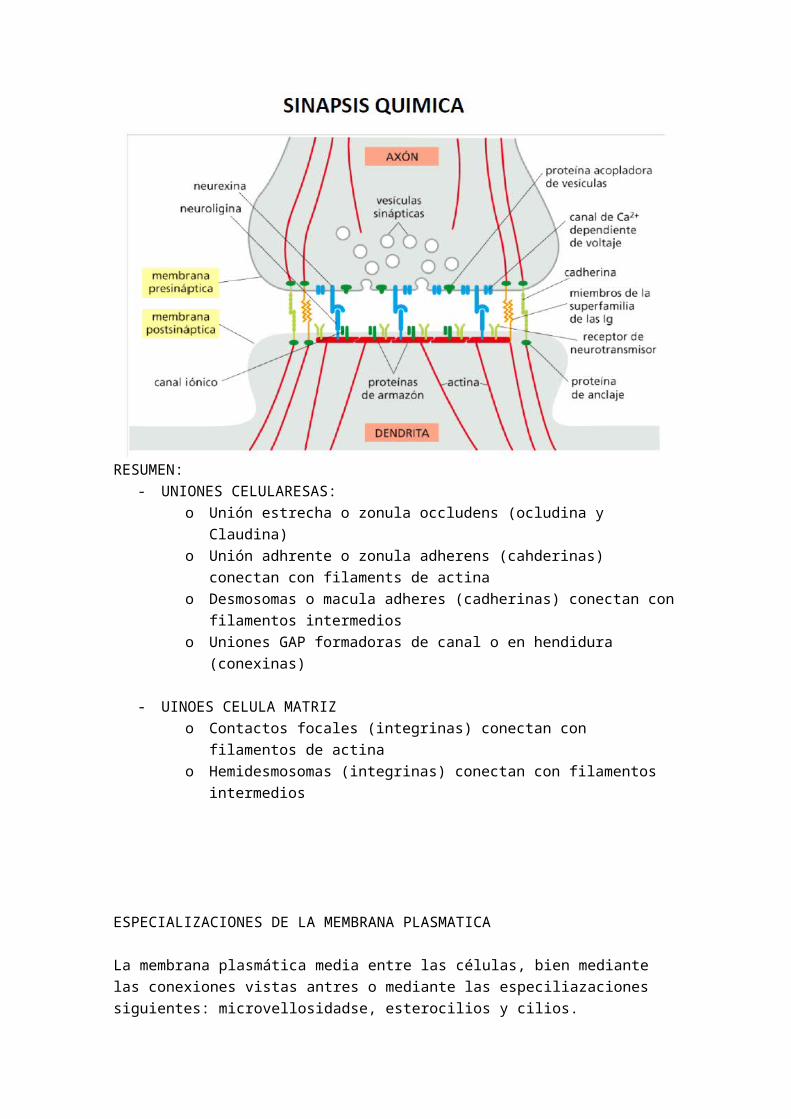
EXAMEN: Es una estructura tubular hueca, y la pared esta formada por microtubulos y lau pared de estos microtubulos es tambien hueca pero esta constituida por protofilamentos. Se puede ver con MO, aquellos sginificativamente grande. los centriolos suelen ir en pareja. en la celula en interfase forman una pareja, recibe esta estructura el nombre de diplosoma, ademas uno de estos centriolos se dispone perpendicularmente al otro centriolo. DEntro del centriolo encontramos la centrosfera, no hay electronegatividad alrededor del centriolo, hay mas electrodensidad en el centro, se le conoco como la esfera atractiva. Al ser una estructura microtubular, necestia un centro donde van a polimerizar los microtubulos que forman parte del citoesquleto norma en estado normal, en mitosis hay que añadirle el aster. Cuando entra en mitosis los microtubulos irradian de los centriolos y al conjunto se le llama centrosfera. Encontramos tambien los satelites pericentriolares.
Estrucutra del centriolo
La pared del centriolo esta formada por microtubulos, estos microtubulos se organizan formando tripletes, cada uno de los cuales tiene 13 protofilamentos. Hay un microtubulo mas proximo al eje central electrondenso, a este microtubulo se le llama microtubulo a y al mas alejado es la llamado microtublo c, y el que esta en medio es el b. Cada triplete se dispone de forma arqueda con respecto a triplete vecino, ademas existen zonas electrodensas entre los tripletes y entre los tripletes y la zona central. esta union se organiza siempre entre el tubulo a de uno y el tubulo c del vecino. esta union viene dada por la proteina nexina y le esta dando estabilidad a la estrucutra.El termino proximal es aquella que tiene la disposcion en rueda de carreta, y por tanto, la otra es la llamada extremo distal.Los satelites pericentrioloraes constituye el centro de nucleacion, y hay globulina alfa. en las plantas para la mitosis hace falta glubulina ganma. eN los microtubulos se polimerizan por el extremo mas y se despolimerizan por el extremo menos.Desde el punto de vista bioquimico, no se ha encontrado ADN, pero tal vez el centriolo tenga en su inicio endosimbiotico tuviera ADN. Ahora no lo hay.LOs centriolos se duplican, lo primero es la aparicio del eje central electrodenso, luego poliermiza el microtubulo A, luego el B y luego el C.
CILIOS
Los cilios son mas pequeños que los flagelos, mientras que los flagelos son de unas 50 micras aunque en ambos casos tienen la misma estrucutra, ademas en abmos la membrana plasmatica se invagina, en su interior habra los microtubulo, esto sera la principal diferencia con el microvellosidades, que no tienen citoesqueleto.
Si cogemos el tallo cilial, y lel damos un corte transversal, vemos los microtubulos se conforman en forma de dobletes, ahy 9 grupos de dobletes y en el centro hay dos centrales. Estos tambien necestian estabilidad, la da las proteinas, por un lado la mesina y por otro la dineina; de dineina hay dos brazos, uno hacia otro doblete y otro brazo de dineina que va hasta el centro del flagelo.lOs cilios tienen movimientos ondluantes, estos movimientos son gracias a la dineina. Si en vez del tallo, el corte lo ahcemos en la punta, desaprece el microtublo b y solo persiste el microtubulo a. Si nos amos a la base del cilio, el contacto con la MB, nos econtramos ante un

corte transversal estara la placa basal, muy electrodensa, los dos microtubulos centrales desaparece para dar paso a esta placa basal. si nos adentramos en el citoplasma y coratmos transversalmente, la base del cilio es un centriolo enteramente, en la extremidad distal estamos hablando (con respecto al nucleo), la distal tendra forma de reueda de carreta.Solo en microtubulo A tiene estructrua en forma redonda, ya que el b comparte protofilamentos con el A, por lo que se quda como media luna. esto es transversal, si lo vemos longitudinal, vemos que surge de vez en cuando desde esta estructura y sin que sea de forma continuidad pero si cierta peridicidad aleatoria aparece el llamado radio. Pasa los mismo con los brazos de dineinaLa funcion ciliar son los movimientos ondulante, gracias a la dienia por su transpte retrogado hasta el extremo menos. la estrucutra de los flagelos se repite en la de los espermatozoides, las mitocondrias le aporta la energi
A PARTIR DE ESTE MOMENTO SOLO SE COPIARA LO QUE NO APARECE EN LA DIAPOSITIVA