Embed Size (px)
Citation preview

Biossíntese e deposição de hemicelulose nas paredes celulares Elisson Romanel Departamento de Biotecnologia Escola de Engenharia de Lorena Universidade de São Paulo Lorena, SP

Outline -‐ Revisando sobre a estrutura das hemiceluloses
-‐ Síntese no complexo de Golgi
-‐ Enzimas envolvidas na síntese
-‐ Glucurono(arabino)xylan Synthase Complex (discussão de 1 arBgo sobre o assunto -‐ seminário de aluno )
Ref. básicas: 1. Anita Teleman. Hemicelluloses and Pectin, item 5.3 In: Pulp and Paper Chemistry and Technology Vol 1: Wood Chemistry and Wood Biotechnology. Edited by Monica Ek, Göran Gellerstedt, Gunnar Henriksson , 2009 2. Emilie A Rennie and Henrik Vibe Scheller. Xylan biosynthesis. Current Opinion in Biotechnology 2014, 26:100–107 3. Wei Zeng et al. A Glucurono(arabino)xylan Synthase Complex from Wheat Contains Members of the GT43, GT47, and GT75 Families and Functions Cooperatively. Plant Physiology 154:78–97, 2010.
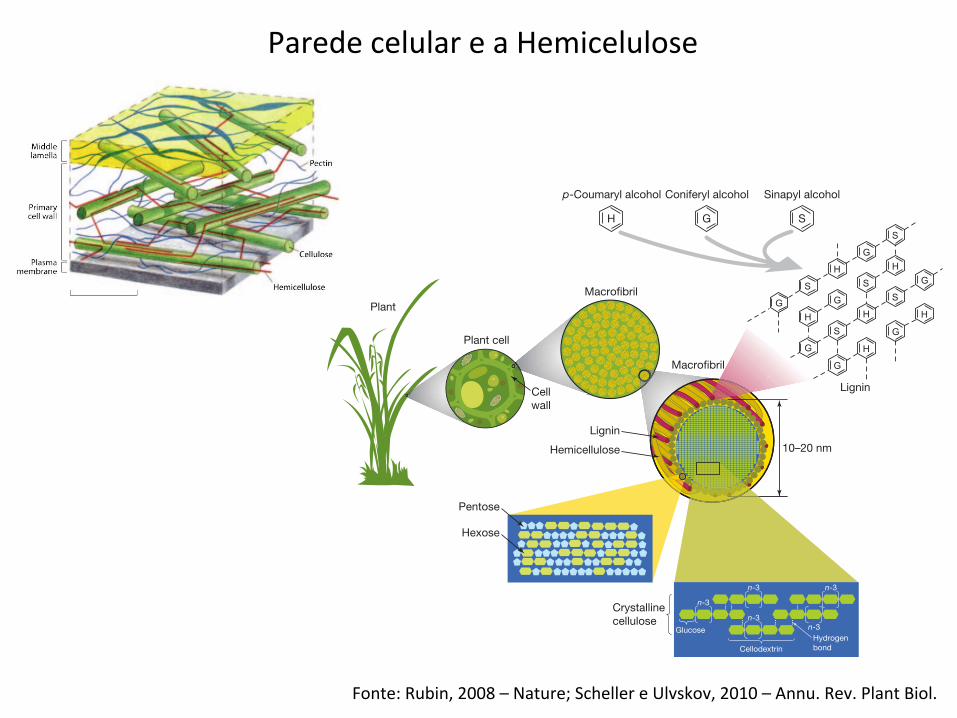
Fonte: Rubin, 2008 – Nature; Scheller e Ulvskov, 2010 – Annu. Rev. Plant Biol.
Parede celular e a Hemicelulose
compared to that of teosinte. Some of the most rapid increases haveoccurred in the past 40 years, both from advances in agronomicpractices and, importantly, from the application of modern genetics.The optimization of bioenergy crops as feedstocks for transportationfuels is in its infancy, but already genomic information and resourcesare being developed that will be essential for accelerating theirdomestication. Many of the traits targeted for optimization in poten-tial cellulosic energy crops are those that would improve growth onpoor agricultural lands, to minimize competition with food cropsover land use.
Populus trichocarpa (poplar), the first tree and potential bioenergycrop to have its genome sequenced (Table 1)9, illustrates some of theissues and potential of applying genomics to the challenge of optim-izing energy crops. The traits for which the genetic underpinningswill be sought in the genomes of bioenergy-relevant plants, such aspoplar, include those affecting growth rates, response to competitionfor light, branching habit, stem thickness and cell wall chemistry.Significant effort will go into maximizing biomass yield per unit landarea, because this more than any other factor will minimize theimpact on overall land use. One can imagine trees optimized to haveshort stature to increase light access and enable dense growth, largestem diameter, and reduced branch count to maximize energy den-sity for transport and processing. Trees have evolved with highly rigidand stable cell walls due to heavy selective pressure for long life and anupright habit. Plants domesticated for energy production, with a
crop cycle time of only a few years, would have less need for a rigidcell wall than wild plants with lifetimes of a hundred years or more.Alterations in the ratios and structures of the various macromole-cules forming the cell wall are a major target in energy crop domest-ication to facilitate post-harvest deconstruction at the cost of a lessrigid plant.
Already, by comparing several of the presently available plant gen-omes (poplar9, rice10,11, Arabidopsis12; see Table 1) coupled with large-scale plant gene function and expression studies, a number of can-didate genes for domestication traits have been identified13,14. Theseinclude many genes involved in cellulose and hemicellulose synthesisas well as those believed to influence various morphological growthcharacteristics such as height, branch number and stem thickness15.In addition to homology-based strategies, other genome-enabledstrategies for identifying domestication candidate genes are beingused. These include quantitative trait analysis of natural variationand genome-wide mutagenesis coupled with phenotypic screensfor traits such as recalcitrance to sugar release, acid digestibilityand general cell wall composition. The availability of high-through-put transgenesis in several plant systems16 will facilitate functionalstudies to determine the in vivo activities of the large number ofdomestication candidate genes. Using these strategies, genes affectingfeatures such as plant height, stem elongation and trunk radialgrowth, drought tolerance, and cell wall stability are but a few ofthe features that are likely to be identified as targets for domestication
OH
Macrofibril
Plant cell
Plant
Cellwall
Lignin
Lignin
Hemicellulose
Pentose
Crystallinecellulose
HydrogenbondCellodextrin
n-3
n-3
n-3
n-3
n-3
Glucose
Hexose
10–20 nm
Macrofibril
OH
OH
OH
OH
p-Coumaryl alcohol Coniferyl alcohol Sinapyl alcohol
OH
O
H G S
OO
Figure 2 | Structure of lignocellulose. The main component oflignocellulose is cellulose, a b(1–4)-linked chain of glucose molecules.Hydrogen bonds between different layers of the polysaccharides contributeto the resistance of crystalline cellulose to degradation. Hemicellulose, thesecond most abundant component of lignocellulose, is composed of various5- and 6-carbon sugars such as arabinose, galactose, glucose, mannose andxylose. Lignin is composed of three major phenolic components, namely
p-coumaryl alcohol (H), coniferyl alcohol (G) and sinapyl alcohol (S). Ligninis synthesized by polymerization of these components and their ratio withinthe polymer varies between different plants, wood tissues and cell wall layers.Cellulose, hemicellulose and lignin form structures called microfibrils,which are organized into macrofibrils that mediate structural stability in theplant cell wall.
NATUREjVol 454j14 August 2008 REVIEWS
843 ©2008 Macmillan Publishers Limited. All rights reserved
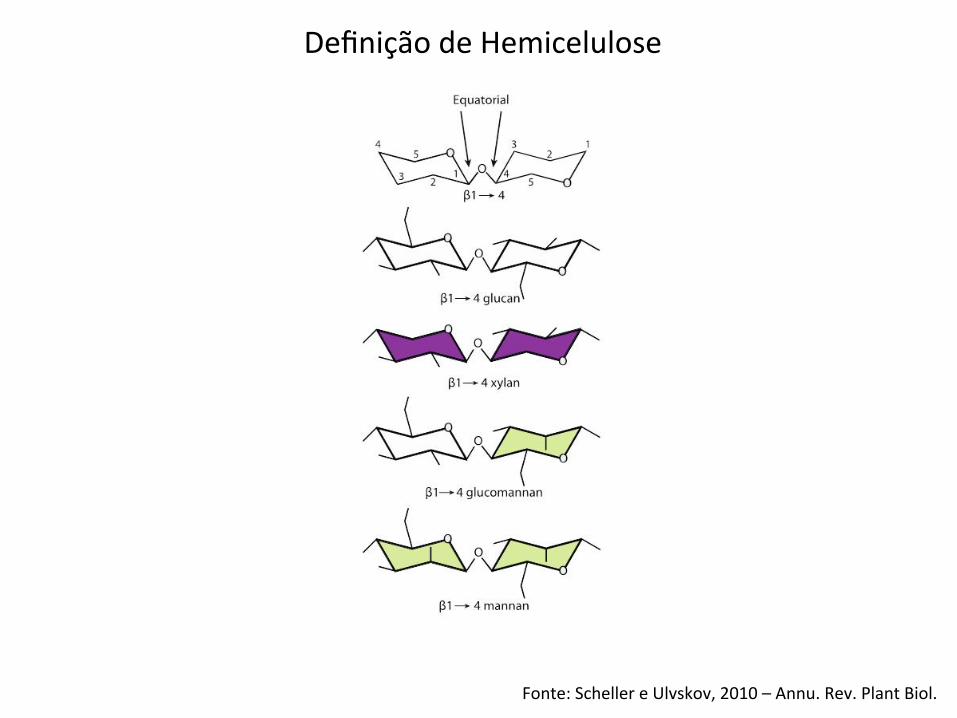
Fonte: Scheller e Ulvskov, 2010 – Annu. Rev. Plant Biol.
Definição de Hemicelulose
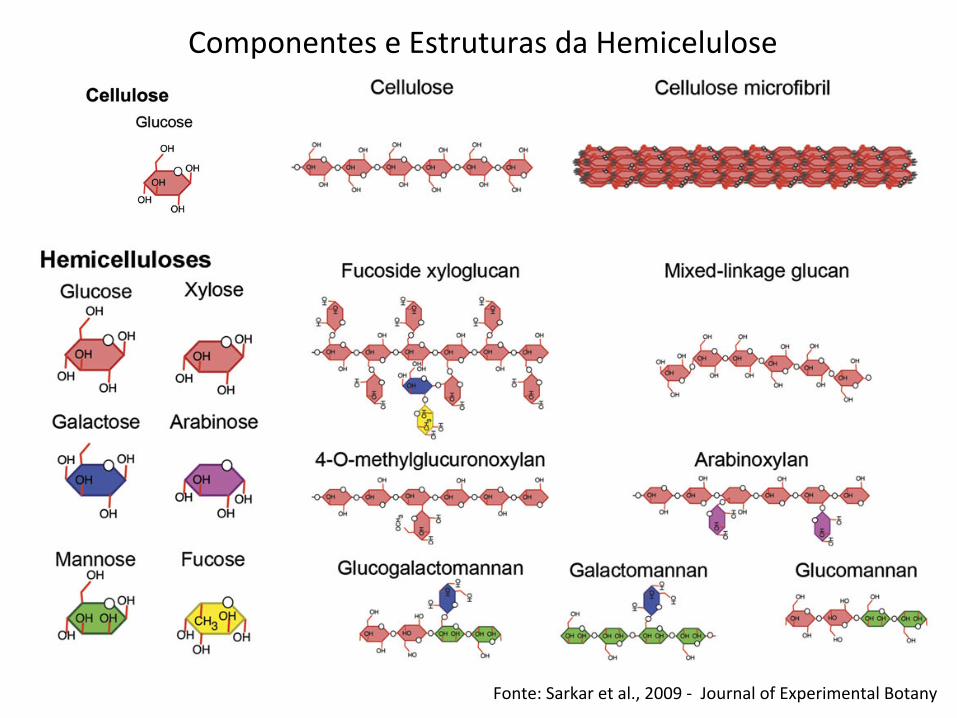
Fonte: Sarkar et al., 2009 -‐ Journal of Experimental Botany
Componentes e Estruturas da Hemicelulose
nature ‘invented’ the protective properties of cell walls morethan once, coming up with vastly different solutions toa common problem. It is widely believed that the cell walldesigns in the two prokaryotic domains have evolved
independently from a common wall-less ancestor, whereascell walls in eukaryotes have evolved by lateral gene transferfrom previously established cell wall-producing organismsduring primary or secondary endosymbiosis (Niklas, 2004).
Fig. 1. Chemical structure of the predominant building blocks of plant cell walls. Left panel: monomers. Right panel: subunit of therespective polymers.
Towards an understanding of plant cell walls | 3617
at FMRP/U
SP/BIBLIOTECA
CENTRA
L on September 19, 2013
http://jxb.oxfordjournals.org/D
ownloaded from
nature ‘invented’ the protective properties of cell walls morethan once, coming up with vastly different solutions toa common problem. It is widely believed that the cell walldesigns in the two prokaryotic domains have evolved
independently from a common wall-less ancestor, whereascell walls in eukaryotes have evolved by lateral gene transferfrom previously established cell wall-producing organismsduring primary or secondary endosymbiosis (Niklas, 2004).
Fig. 1. Chemical structure of the predominant building blocks of plant cell walls. Left panel: monomers. Right panel: subunit of therespective polymers.
Towards an understanding of plant cell walls | 3617
at FMRP/U
SP/BIBLIOTECA
CENTRA
L on September 19, 2013
http://jxb.oxfordjournals.org/D
ownloaded from
nature ‘invented’ the protective properties of cell walls morethan once, coming up with vastly different solutions toa common problem. It is widely believed that the cell walldesigns in the two prokaryotic domains have evolved
independently from a common wall-less ancestor, whereascell walls in eukaryotes have evolved by lateral gene transferfrom previously established cell wall-producing organismsduring primary or secondary endosymbiosis (Niklas, 2004).
Fig. 1. Chemical structure of the predominant building blocks of plant cell walls. Left panel: monomers. Right panel: subunit of therespective polymers.
Towards an understanding of plant cell walls | 3617
at FMRP/U
SP/BIBLIOTECA
CENTRA
L on September 19, 2013
http://jxb.oxfordjournals.org/D
ownloaded from
nature ‘invented’ the protective properties of cell walls morethan once, coming up with vastly different solutions toa common problem. It is widely believed that the cell walldesigns in the two prokaryotic domains have evolved
independently from a common wall-less ancestor, whereascell walls in eukaryotes have evolved by lateral gene transferfrom previously established cell wall-producing organismsduring primary or secondary endosymbiosis (Niklas, 2004).
Fig. 1. Chemical structure of the predominant building blocks of plant cell walls. Left panel: monomers. Right panel: subunit of therespective polymers.
Towards an understanding of plant cell walls | 3617
at FMRP/U
SP/BIBLIOTECA
CENTRA
L on September 19, 2013
http://jxb.oxfordjournals.org/D
ownloaded from
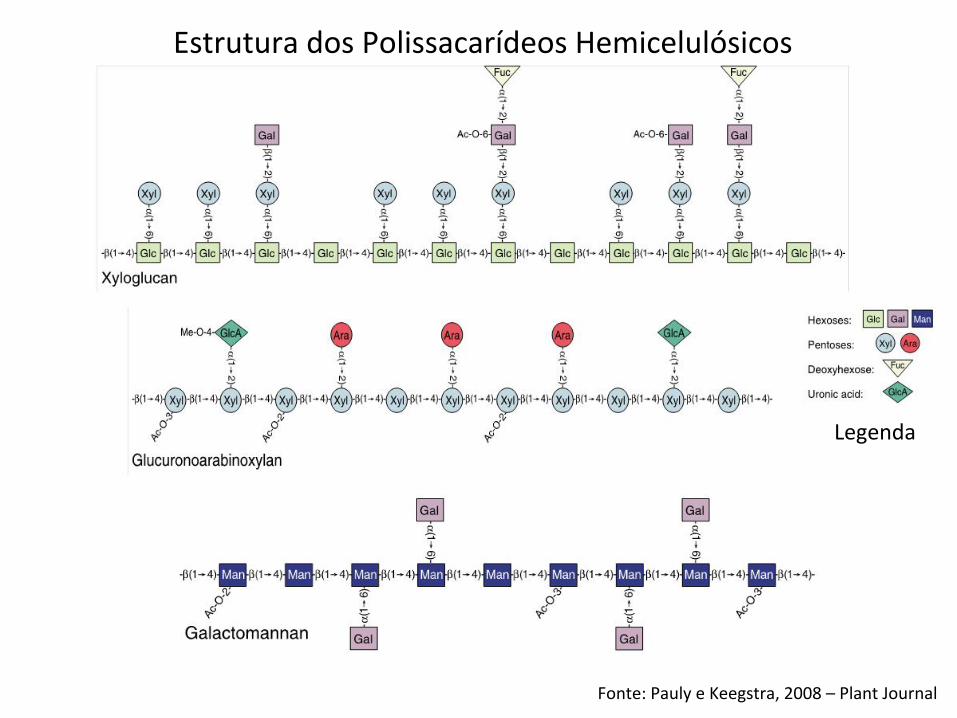
Fonte: Pauly e Keegstra, 2008 – Plant Journal
Legenda
Estrutura dos Polissacarídeos Hemicelulósicos
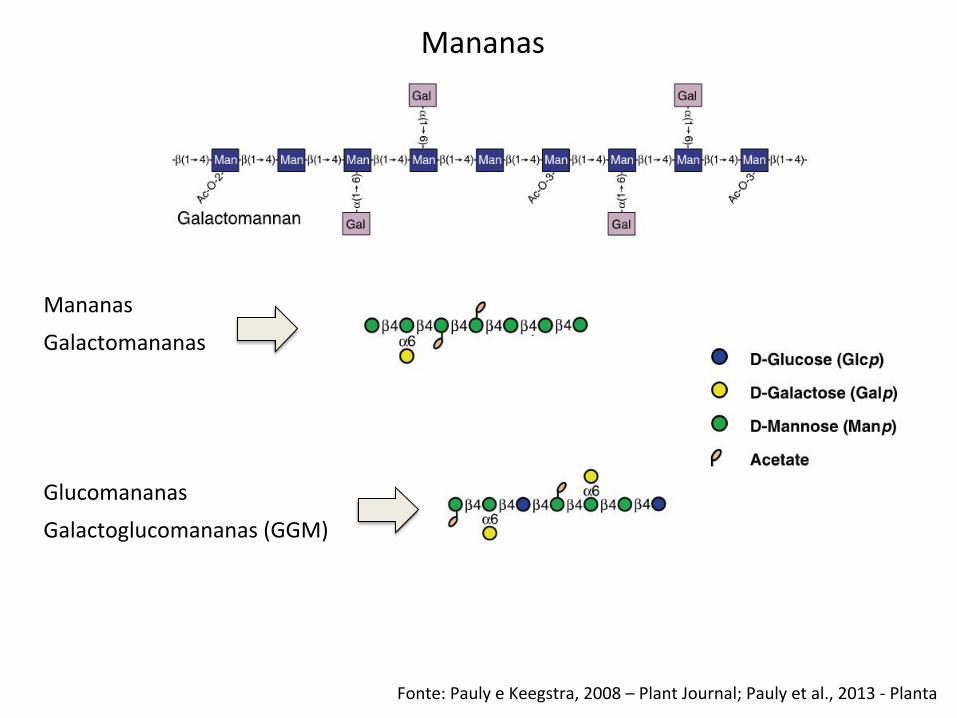
Fonte: Pauly e Keegstra, 2008 – Plant Journal; Pauly et al., 2013 -‐ Planta
Mananas
Mananas
Glucomananas
Galactomananas
Galactoglucomananas (GGM)
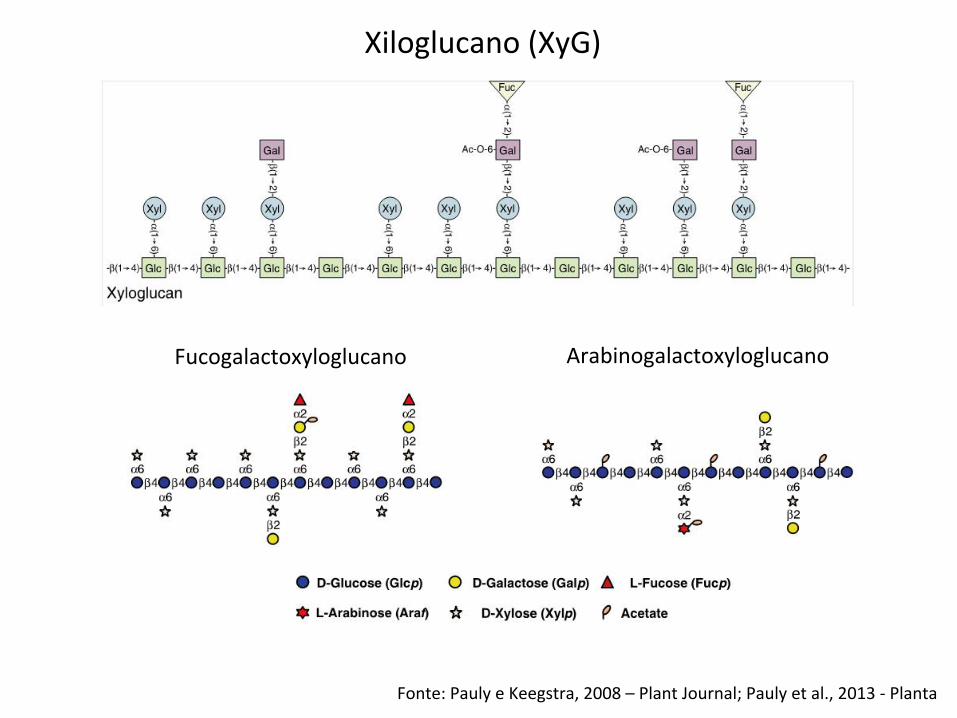
Xiloglucano (XyG)
Fucogalactoxyloglucano Arabinogalactoxyloglucano
Fonte: Pauly e Keegstra, 2008 – Plant Journal; Pauly et al., 2013 -‐ Planta
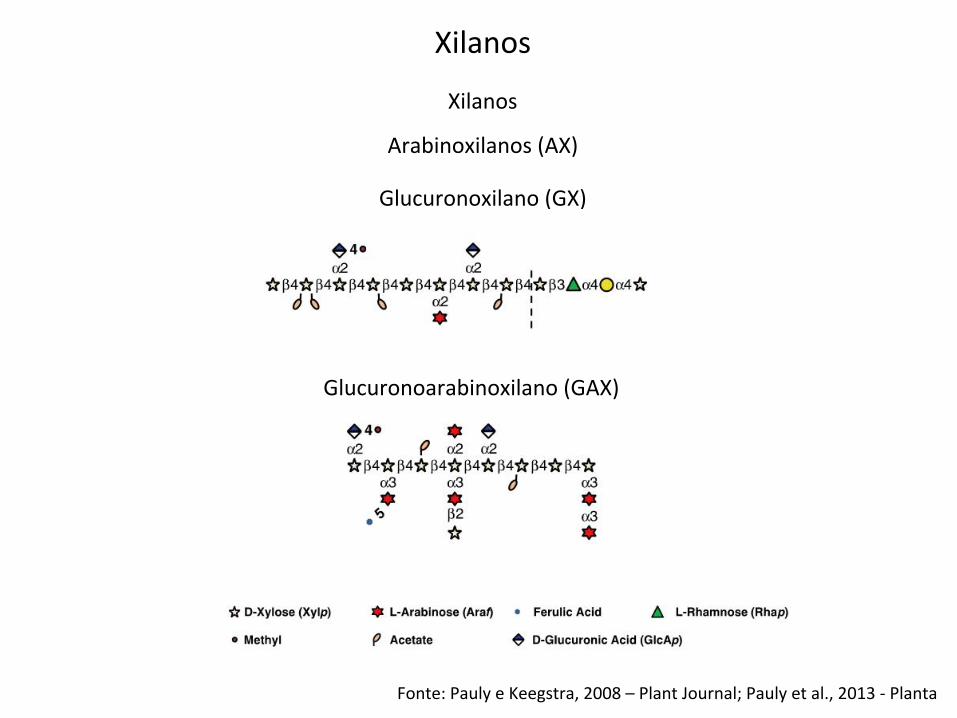
Xilanos
Fonte: Pauly e Keegstra, 2008 – Plant Journal; Pauly et al., 2013 -‐ Planta
Glucuronoxilano (GX)
Glucuronoarabinoxilano (GAX)
Arabinoxilanos (AX)
Xilanos
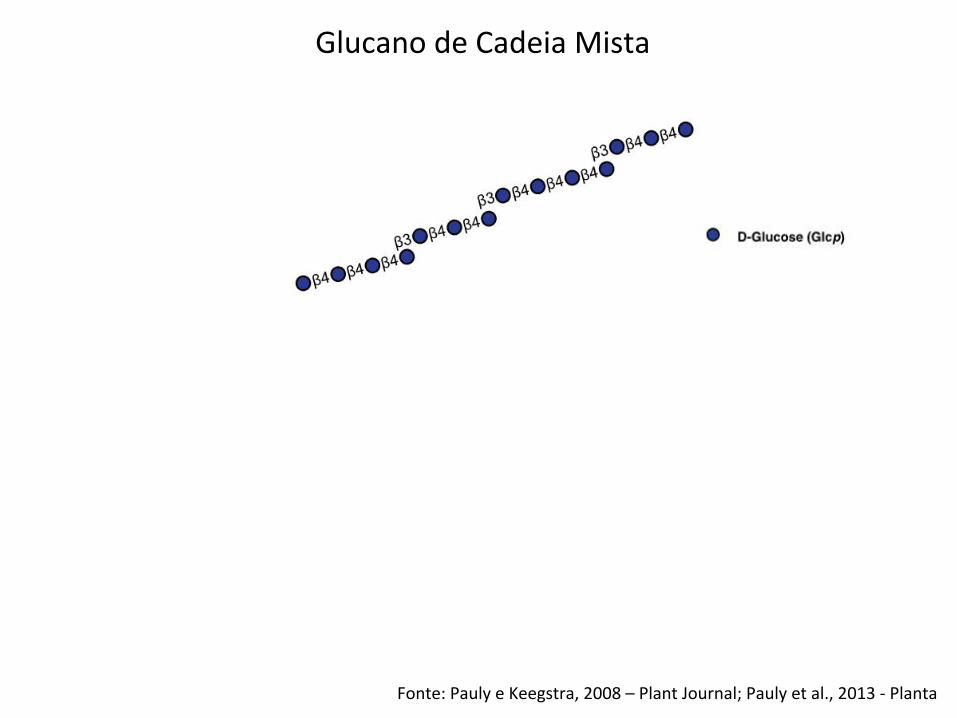
Glucano de Cadeia Mista
Fonte: Pauly e Keegstra, 2008 – Plant Journal; Pauly et al., 2013 -‐ Planta
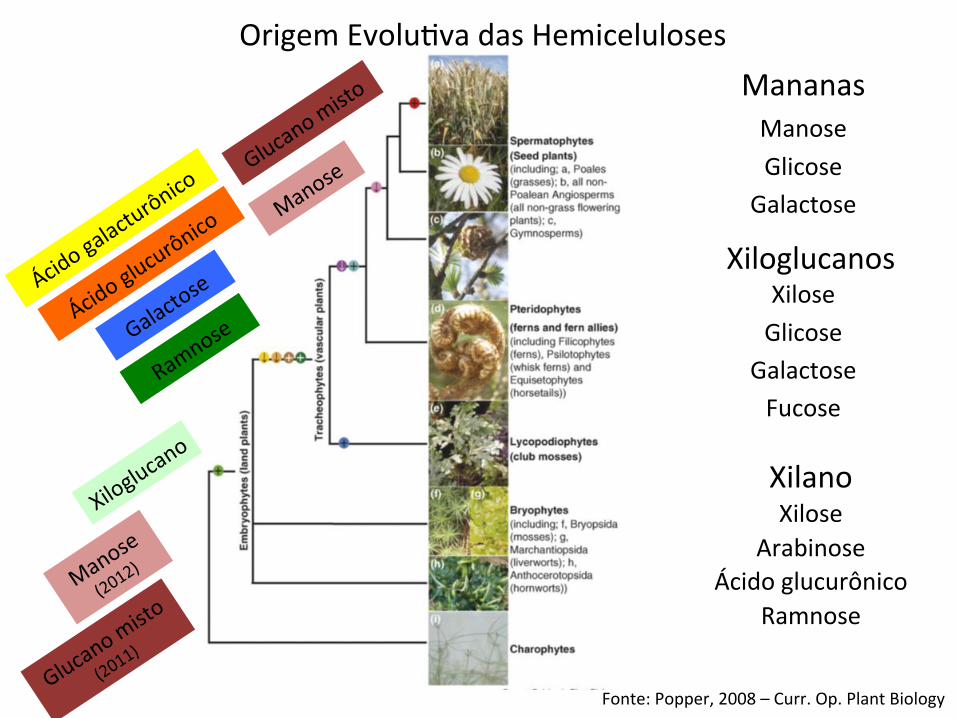
Origem EvoluBva das Hemiceluloses
Fonte: Popper, 2008 – Curr. Op. Plant Biology
Mananas
Xiloglucanos
Xilano
Manose Glicose
Galactose
Xilose Glicose
Galactose Fucose
Xilose Arabinose
Ácido glucurônico Ramnose
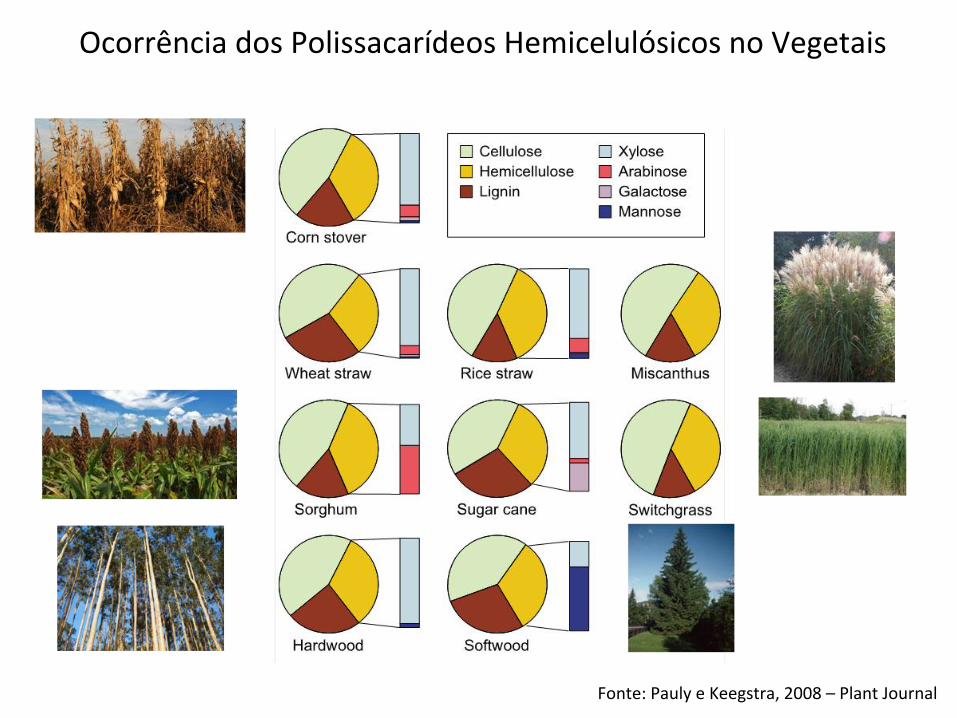
Fonte: Pauly e Keegstra, 2008 – Plant Journal
Ocorrência dos Polissacarídeos Hemicelulósicos no Vegetais
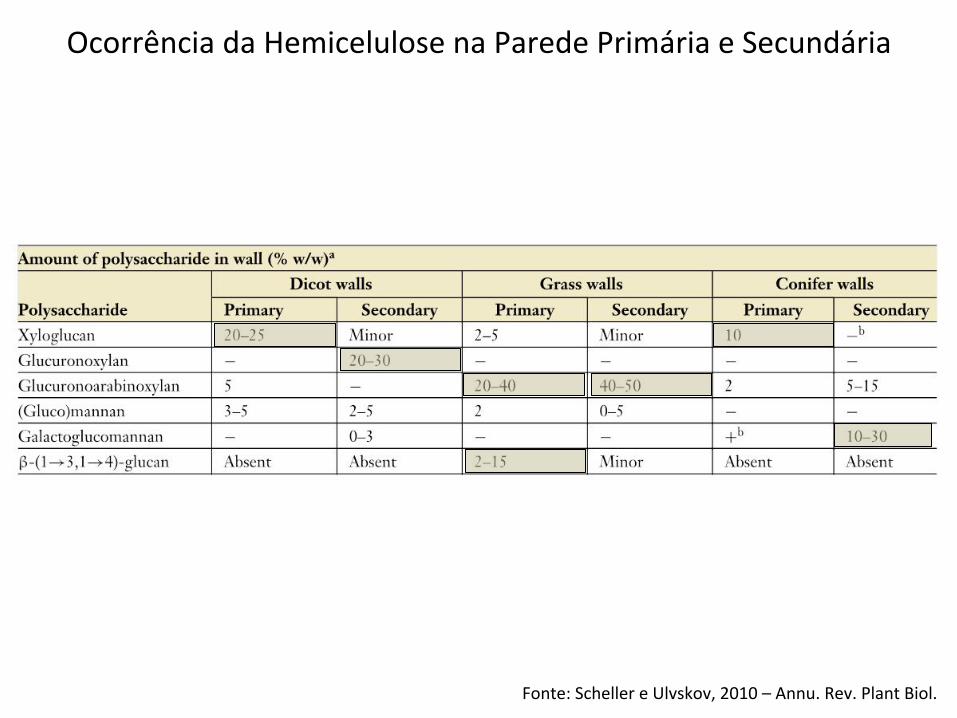
Fonte: Scheller e Ulvskov, 2010 – Annu. Rev. Plant Biol.
Ocorrência da Hemicelulose na Parede Primária e Secundária
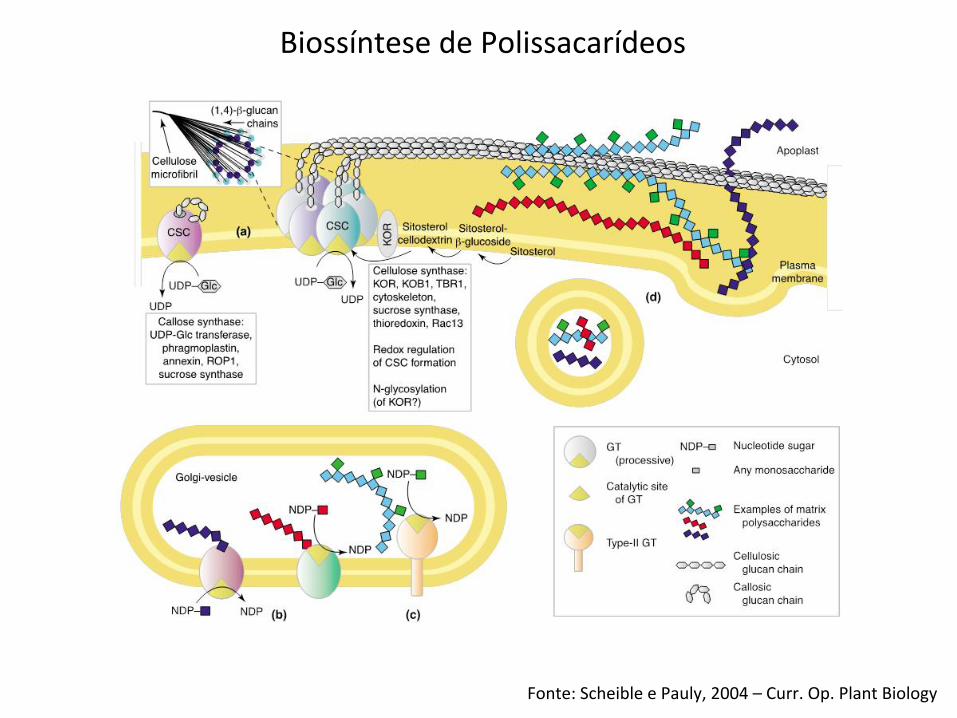
Fonte: Scheible e Pauly, 2004 – Curr. Op. Plant Biology
Biossíntese de Polissacarídeos
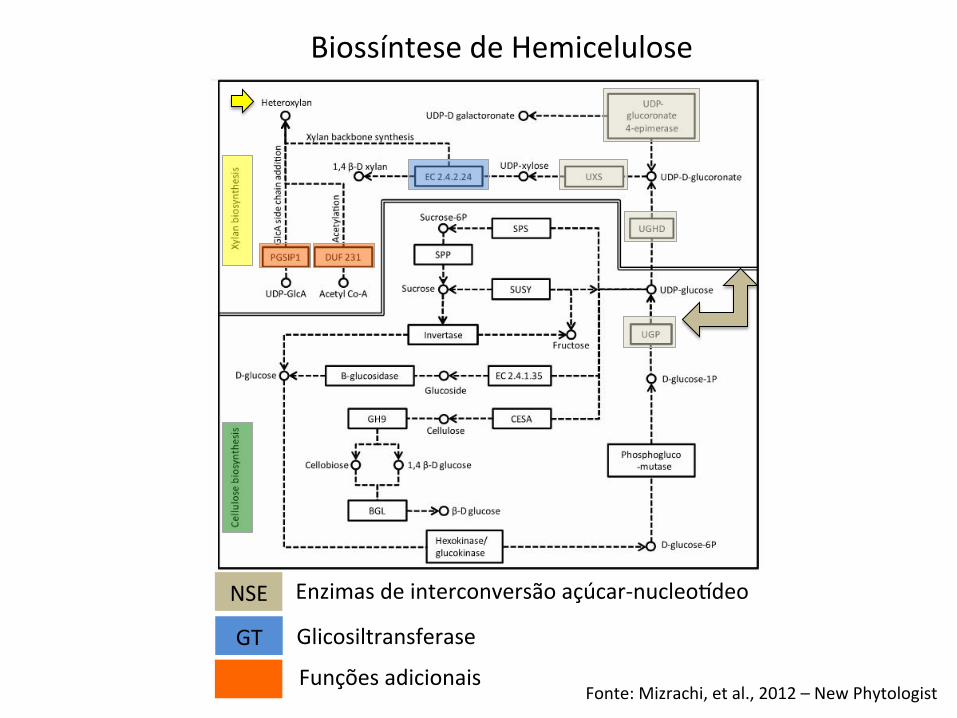
Fonte: Mizrachi, et al., 2012 – New Phytologist
Biossíntese de Hemicelulose
NSE
GT
Enzimas de interconversão açúcar-‐nucleoideo
Glicosiltransferase
Funções adicionais
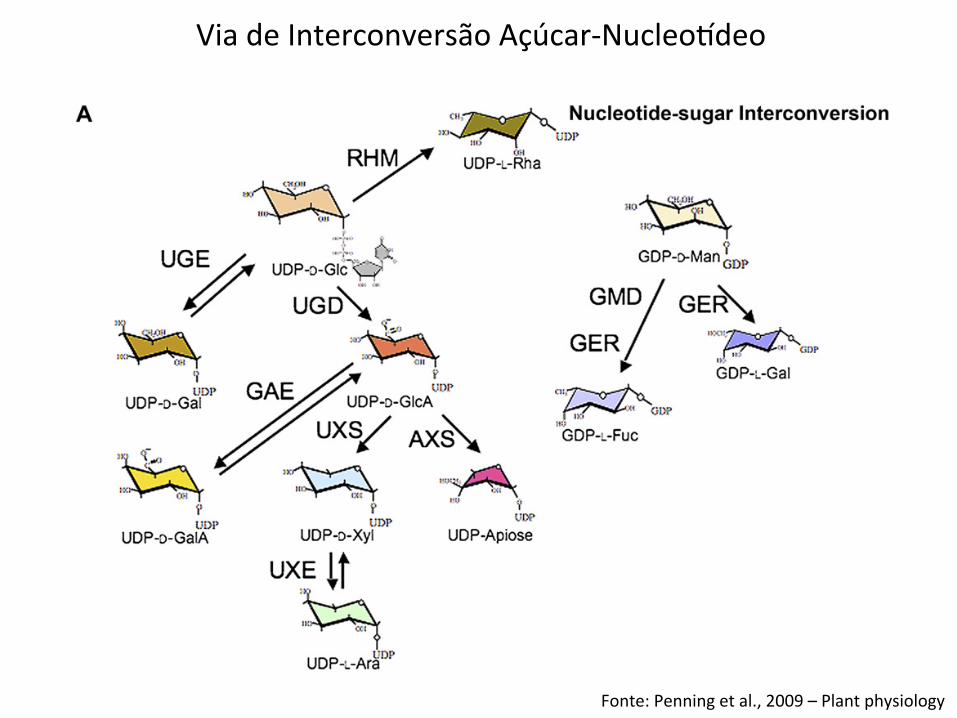
Figure 1. Genes of the nucleotide-sugar interconversion pathways. A, Schematic of pathways for plant nucleotide-sugarinterconversion. The committed step to synthesis of uronic acids and pentoses is catalyzed by UDP-Glc dehydrogenase (UGD);isoforms exhibit different catalytic activities that indicate varied functions (Karkonen et al., 2005). The function of the UDP-GlcAdecarboxylase (carboxyl-lyase) was established for the UXS family in barley (Zhang et al., 2005), with homology to the SUD/AUD group proposed for Arabidopsis (Reiter and Vanzin, 2001). Apiose, the essential monosaccharide in the boron didiestercross-linking of RG II, is synthesized by enzymes encoded by members of the AXS group, which converts irreversibly UDP-GlcAto a mixture of UDP-apiose and UDP-Xyl. A reduction in the levels of these synthases results in an RG II deficiency and cell wallabnormalities (Ahn et al., 2006). Although pectins are a minor component of the walls of grasses, an apiose-containing RG II withonly slightly modified side groups is present (Thomas et al., 1989). B, These evolutionarily distinct families are combined forconvenience into one dendrogram; evolutionary relationships are relevant only within a single family. Three groups of C-4epimerases have been annotated: the UDP-Glc 4-epimerases (UGEs), including REB1, that interconvert UDP-Glc and UDP-Gal(Seifert et al., 2002; Nguema-Ona et al., 2006); the UDP-GlcA 4-epimerases (GAEs) that interconvert UDP-GlcA and UDP-GalA
Penning et al.
1708 Plant Physiol. Vol. 151, 2009
Via de Interconversão Açúcar-‐Nucleoideo
Fonte: Penning et al., 2009 – Plant physiology
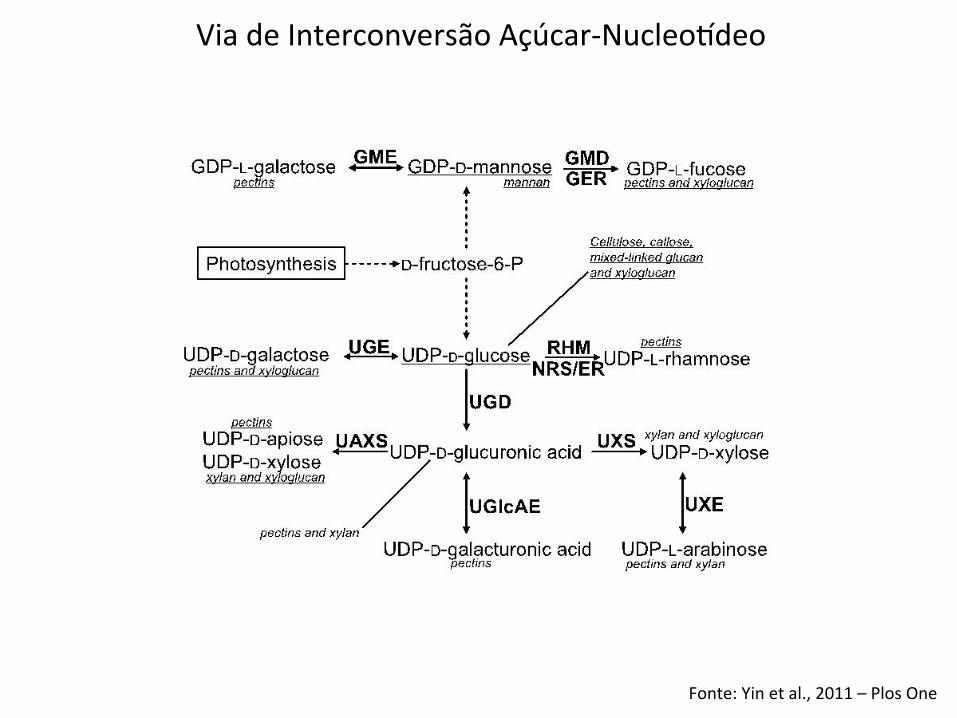
Fonte: Yin et al., 2011 – Plos One
Via de Interconversão Açúcar-‐Nucleoideo
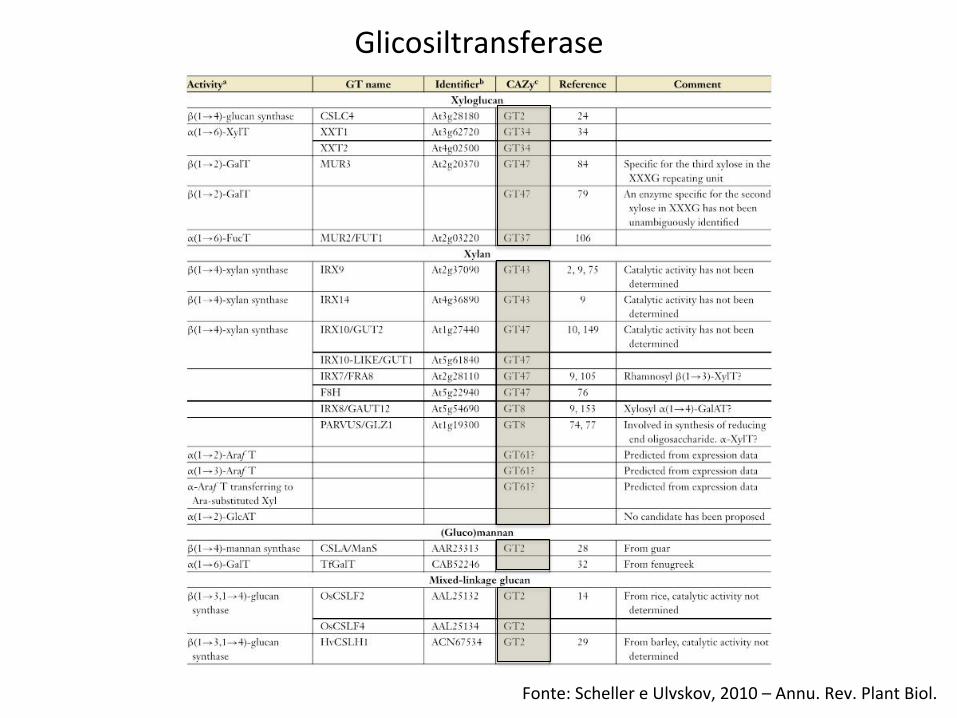
Fonte: Scheller e Ulvskov, 2010 – Annu. Rev. Plant Biol.
Glicosiltransferase
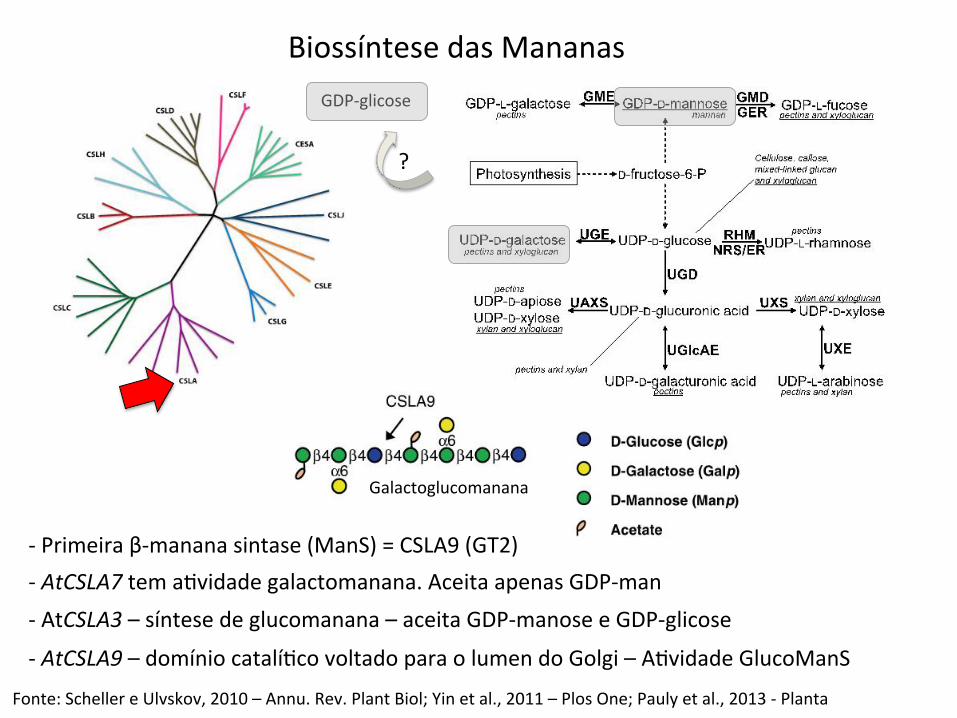
Biossíntese das Mananas GDP-‐glicose
-‐ Primeira β-‐manana sintase (ManS) = CSLA9 (GT2) -‐ AtCSLA7 tem aBvidade galactomanana. Aceita apenas GDP-‐man -‐ AtCSLA3 – síntese de glucomanana – aceita GDP-‐manose e GDP-‐glicose
-‐ AtCSLA9 – domínio catalíBco voltado para o lumen do Golgi – ABvidade GlucoManS
?
Fonte: Scheller e Ulvskov, 2010 – Annu. Rev. Plant Biol; Yin et al., 2011 – Plos One; Pauly et al., 2013 -‐ Planta
Galactoglucomanana
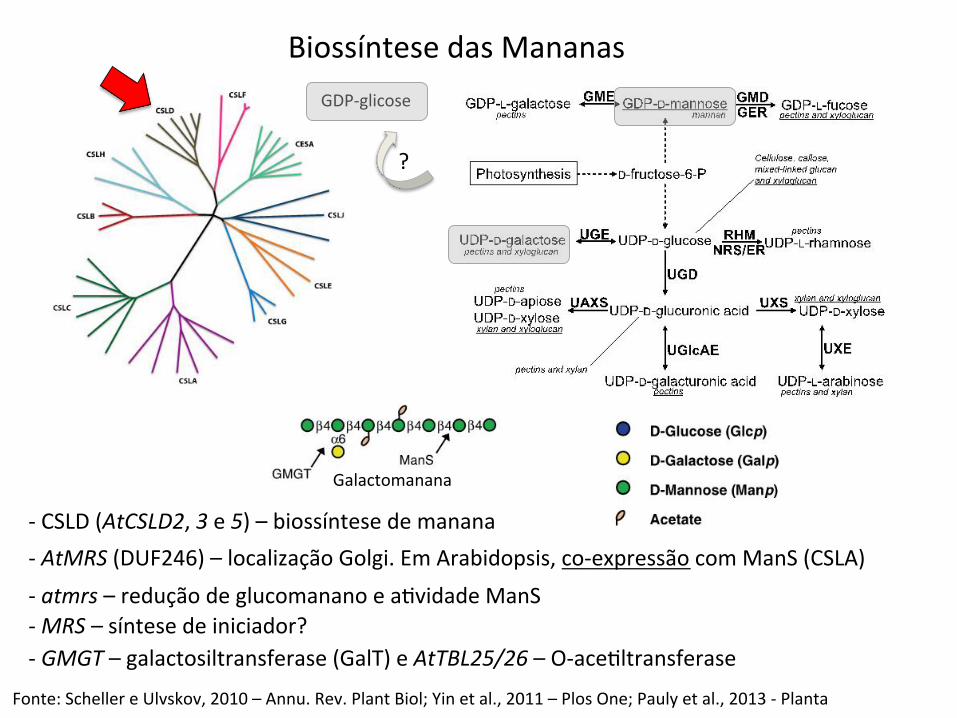
Biossíntese das Mananas
-‐ CSLD (AtCSLD2, 3 e 5) – biossíntese de manana
GDP-‐glicose
?
Fonte: Scheller e Ulvskov, 2010 – Annu. Rev. Plant Biol; Yin et al., 2011 – Plos One; Pauly et al., 2013 -‐ Planta
-‐ AtMRS (DUF246) – localização Golgi. Em Arabidopsis, co-‐expressão com ManS (CSLA) -‐ atmrs – redução de glucomanano e aBvidade ManS -‐ MRS – síntese de iniciador? -‐ GMGT – galactosiltransferase (GalT) e AtTBL25/26 – O-‐aceBltransferase
Galactomanana
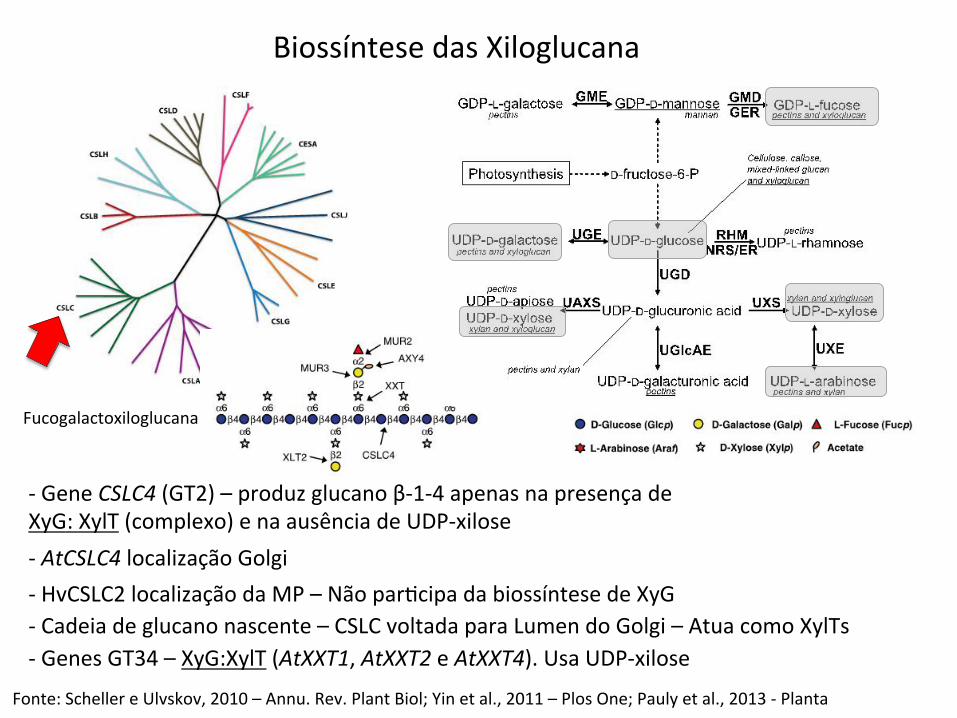
Biossíntese das Xiloglucana
-‐ Gene CSLC4 (GT2) – produz glucano β-‐1-‐4 apenas na presença de XyG: XylT (complexo) e na ausência de UDP-‐xilose
Fonte: Scheller e Ulvskov, 2010 – Annu. Rev. Plant Biol; Yin et al., 2011 – Plos One; Pauly et al., 2013 -‐ Planta
-‐ AtCSLC4 localização Golgi -‐ HvCSLC2 localização da MP – Não parBcipa da biossíntese de XyG -‐ Cadeia de glucano nascente – CSLC voltada para Lumen do Golgi – Atua como XylTs -‐ Genes GT34 – XyG:XylT (AtXXT1, AtXXT2 e AtXXT4). Usa UDP-‐xilose
Fucogalactoxiloglucana
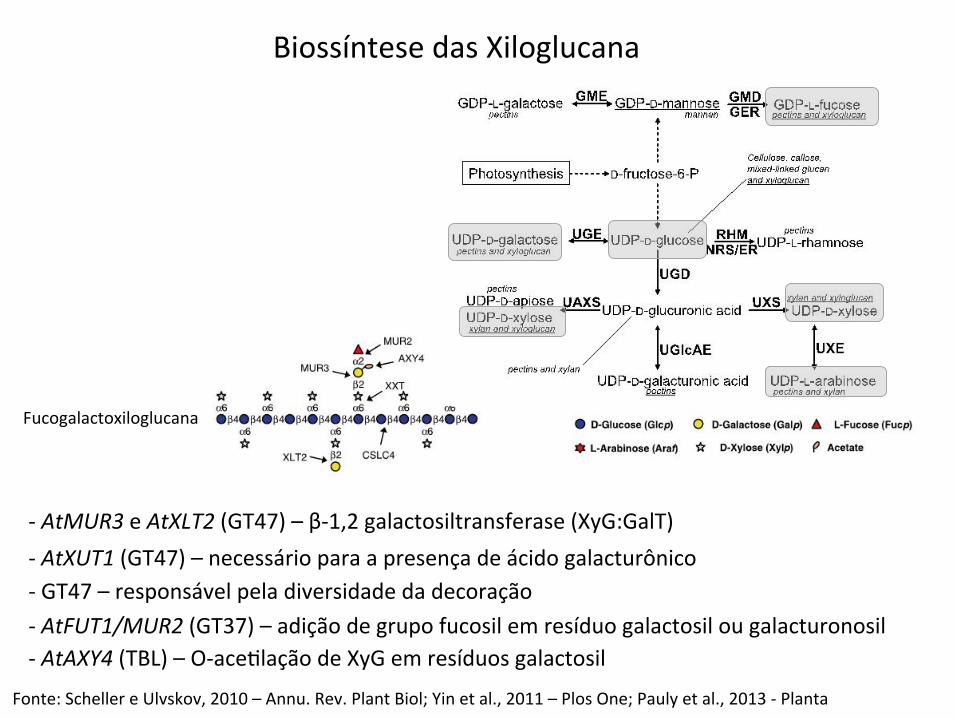
Biossíntese das Xiloglucana
-‐ AtMUR3 e AtXLT2 (GT47) – β-‐1,2 galactosiltransferase (XyG:GalT)
Fonte: Scheller e Ulvskov, 2010 – Annu. Rev. Plant Biol; Yin et al., 2011 – Plos One; Pauly et al., 2013 -‐ Planta
-‐ AtXUT1 (GT47) – necessário para a presença de ácido galacturônico -‐ GT47 – responsável pela diversidade da decoração -‐ AtFUT1/MUR2 (GT37) – adição de grupo fucosil em resíduo galactosil ou galacturonosil -‐ AtAXY4 (TBL) – O-‐aceBlação de XyG em resíduos galactosil
Fucogalactoxiloglucana
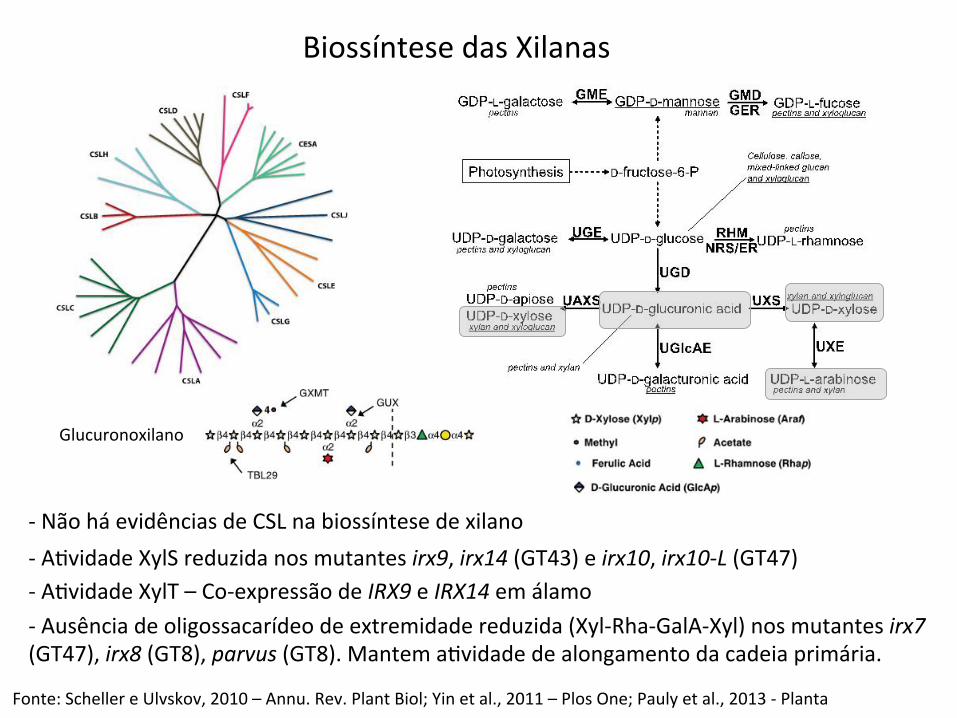
-‐ Não há evidências de CSL na biossíntese de xilano
Fonte: Scheller e Ulvskov, 2010 – Annu. Rev. Plant Biol; Yin et al., 2011 – Plos One; Pauly et al., 2013 -‐ Planta
-‐ ABvidade XylS reduzida nos mutantes irx9, irx14 (GT43) e irx10, irx10-‐L (GT47) -‐ ABvidade XylT – Co-‐expressão de IRX9 e IRX14 em álamo -‐ Ausência de oligossacarídeo de extremidade reduzida (Xyl-‐Rha-‐GalA-‐Xyl) nos mutantes irx7 (GT47), irx8 (GT8), parvus (GT8). Mantem aBvidade de alongamento da cadeia primária.
Biossíntese das Xilanas
Glucuronoxilano
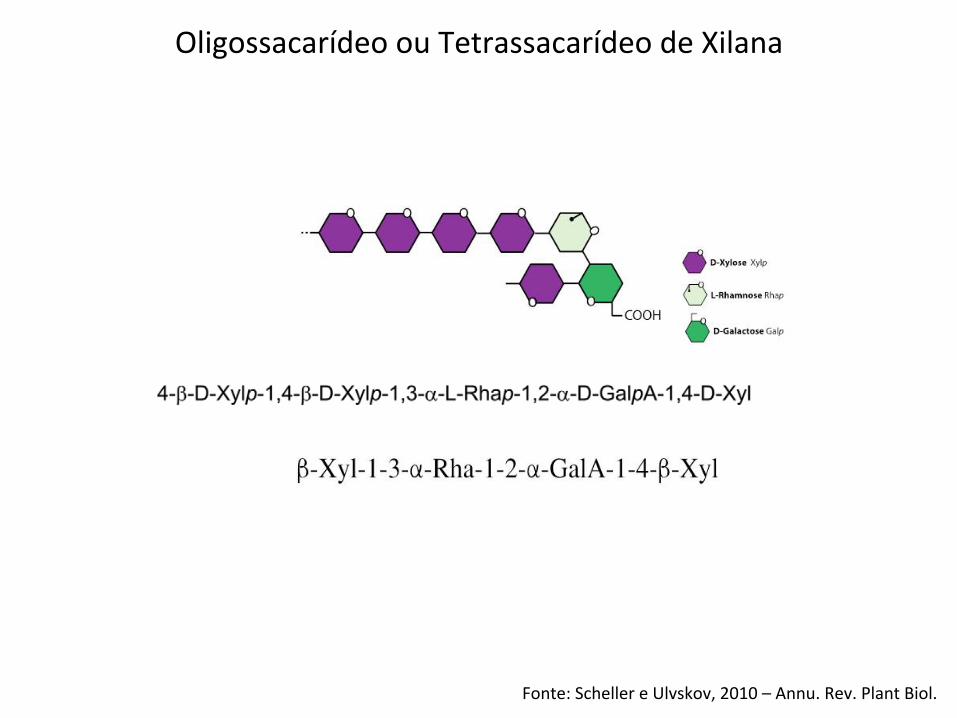
Fonte: Scheller e Ulvskov, 2010 – Annu. Rev. Plant Biol.
Oligossacarídeo ou Tetrassacarídeo de Xilana
bones [9!,15–17]. Two non-CSL genes (IRX9 and IRX14)have been implicated in xylan backbone biosynthesis[14!!,18!,19,20]. Nevertheless, the possibility that CSLgenes also have a direct role in this process cannot bediscounted.
Glucuronoxylan has typically been viewed as a polysac-charide whose synthesis would require a xylan synthasefor backbone formation and one or two glycosyltrans-ferases for the addition of GlcA and 4-O-Me GlcAside chains. This perspective changed when Pena et al.[14!!] demonstrated that Arabidopsis GX contains! 4)-b-D-Xylp-(1! 3)-a-L-Rhap-(1! 2)-a-D-GalpA-(1! 4)-D-Xylp (sequence 1, see Figure 1b) at its reducing end
and that this sequence is required for normal GX syn-thesis in secondary walls. At least three glycosyltrans-ferases are required to form this sequence. At least onemore glycosyltransferase is required if this sequence linksxylan to another polymer [14!!].
To date five genes (FRA8 [At2g28110], IRX8 [At5g54690],IRX9 [At2g37090], PARVUS [At1g19300], and IRX14[At4g36890]) that are likely to have a role in GX synthesishave been identified in Arabidopsis [14!!,19, 21–23]. Thesegenes encode putative glycosyltransferases that may have arole in forming reducing end sequence 1 (FRA8, familyGT47; IRX8, family GT8; PARVUS, family GT8) and inxylan backbone synthesis (IRX9 and IRX14, family GT43).
Biochemical control of xylan biosynthesis York and O’Neill 259
Figure 1
Diversity of xylan structures in land plants. (a) The glucuronoxylans produced by dicots and gymnosperms. (b) Sequence 1 present at thereducing end of dicot and gymnosperm glucuronoxylan. (c) Feruloylated arabinoxylans produced by grasses. (d) Glucuronoarabinoxylanspresent in the secondary walls of soft woods.
www.sciencedirect.com Current Opinion in Plant Biology 2008, 11:258–265
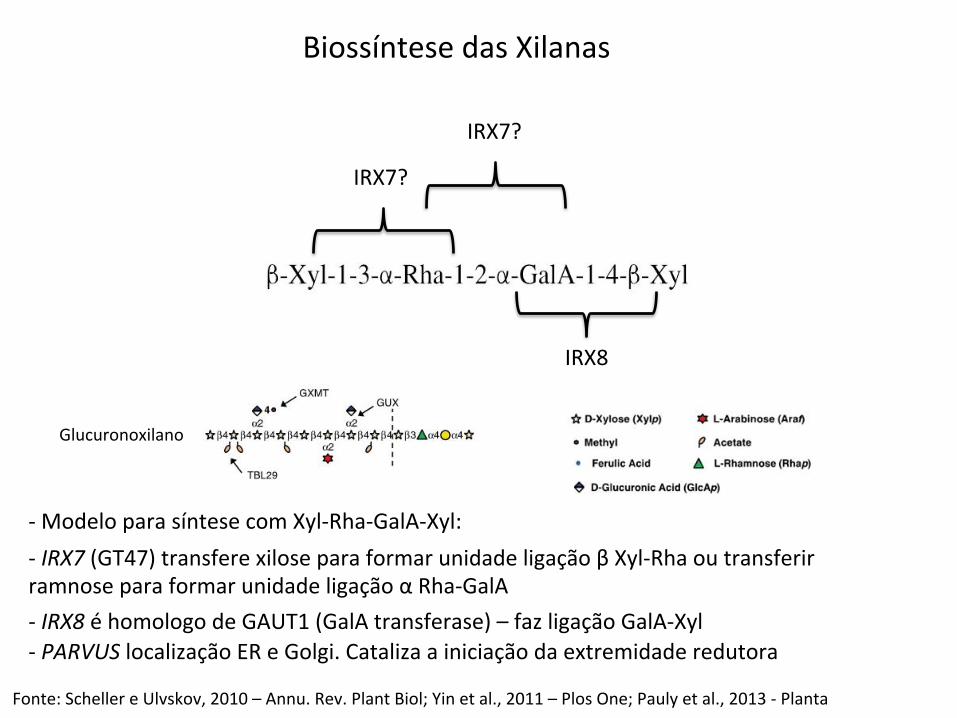
-‐ Modelo para síntese com Xyl-‐Rha-‐GalA-‐Xyl:
Fonte: Scheller e Ulvskov, 2010 – Annu. Rev. Plant Biol; Yin et al., 2011 – Plos One; Pauly et al., 2013 -‐ Planta
-‐ IRX7 (GT47) transfere xilose para formar unidade ligação β Xyl-‐Rha ou transferir ramnose para formar unidade ligação α Rha-‐GalA -‐ IRX8 é homologo de GAUT1 (GalA transferase) – faz ligação GalA-‐Xyl -‐ PARVUS localização ER e Golgi. Cataliza a iniciação da extremidade redutora
Biossíntese das Xilanas
Glucuronoxilano
IRX7?
IRX7?
IRX8
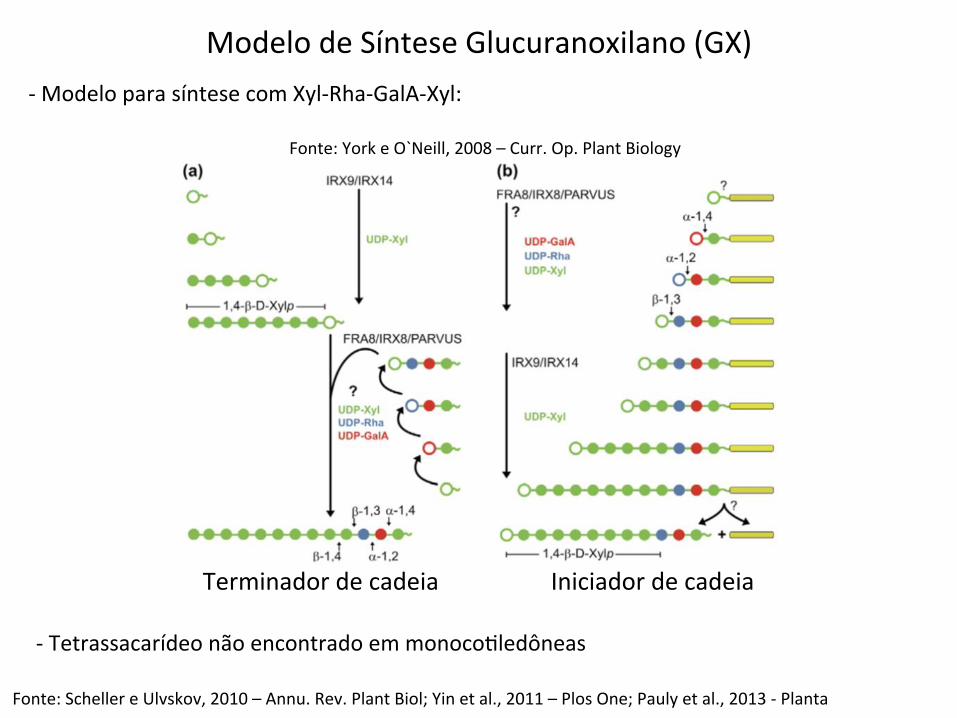
Fonte: York e O`Neill, 2008 – Curr. Op. Plant Biology
Modelo de Síntese Glucuranoxilano (GX) -‐ Modelo para síntese com Xyl-‐Rha-‐GalA-‐Xyl:
Fonte: Scheller e Ulvskov, 2010 – Annu. Rev. Plant Biol; Yin et al., 2011 – Plos One; Pauly et al., 2013 -‐ Planta
-‐ Tetrassacarídeo não encontrado em monocoBledôneas
Terminador de cadeia Iniciador de cadeia
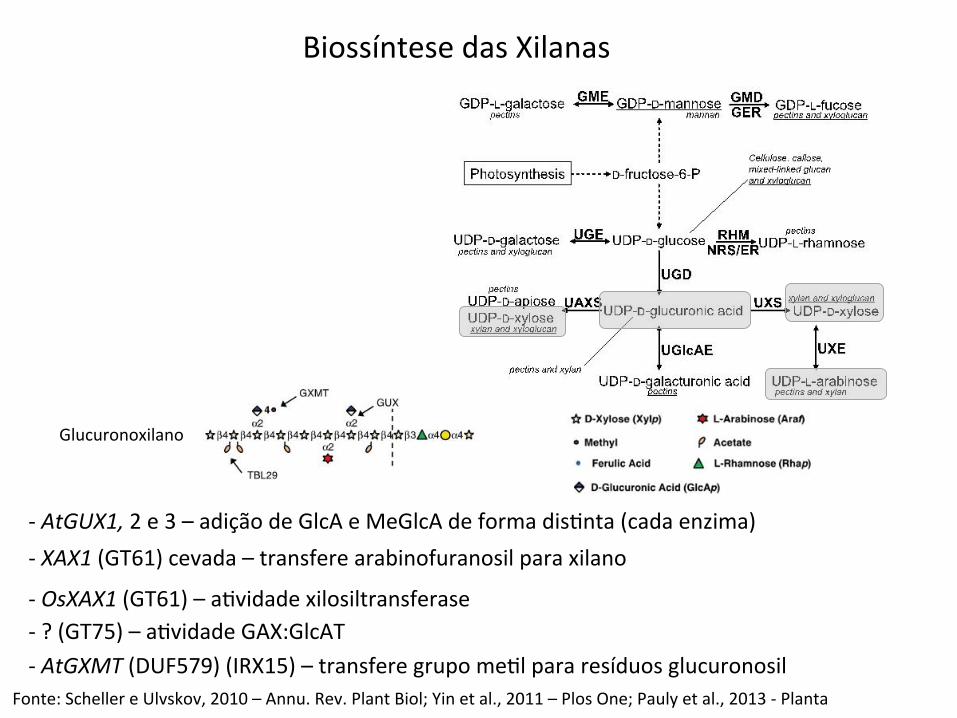
-‐ AtGUX1, 2 e 3 – adição de GlcA e MeGlcA de forma disBnta (cada enzima)
Fonte: Scheller e Ulvskov, 2010 – Annu. Rev. Plant Biol; Yin et al., 2011 – Plos One; Pauly et al., 2013 -‐ Planta
-‐ XAX1 (GT61) cevada – transfere arabinofuranosil para xilano
-‐ OsXAX1 (GT61) – aBvidade xilosiltransferase -‐ ? (GT75) – aBvidade GAX:GlcAT -‐ AtGXMT (DUF579) (IRX15) – transfere grupo meBl para resíduos glucuronosil
Biossíntese das Xilanas
Glucuronoxilano
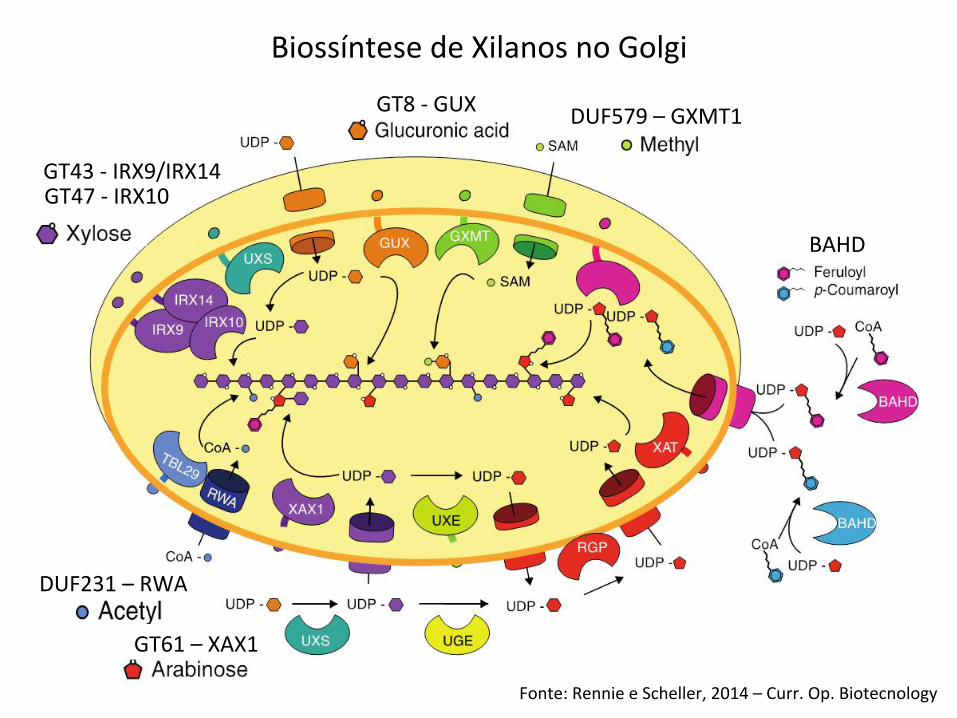
Biossíntese de Xilanos no Golgi
Fonte: Rennie e Scheller, 2014 – Curr. Op. Biotecnology
GT43 -‐ IRX9/IRX14 GT47 -‐ IRX10
GT8 -‐ GUX DUF579 – GXMT1
DUF231 – RWA
BAHD
GT61 – XAX1
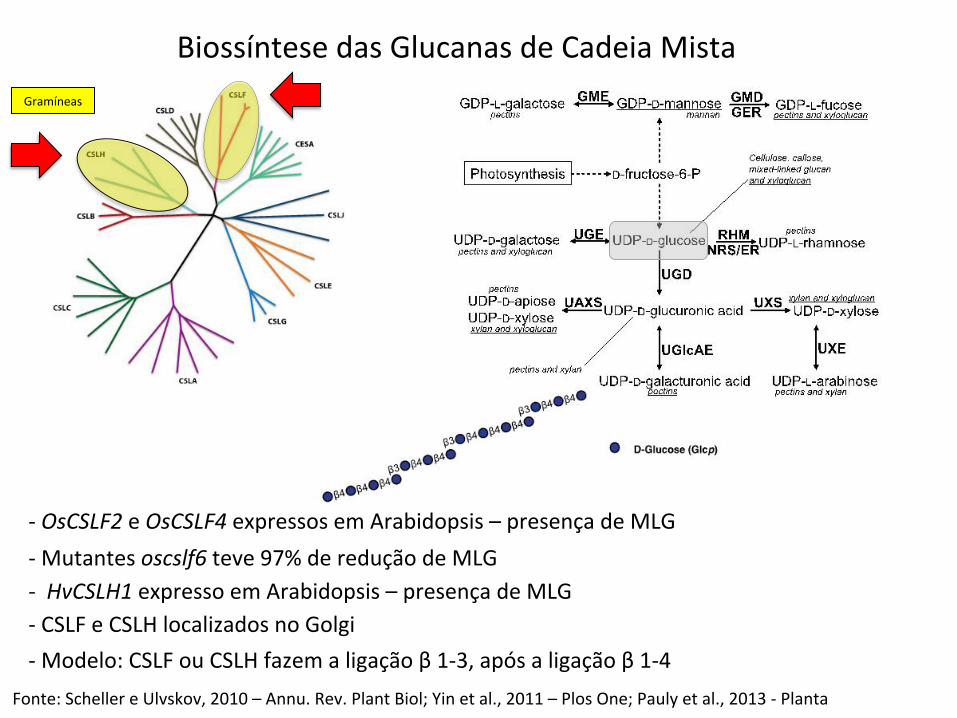
Biossíntese das Glucanas de Cadeia Mista
-‐ OsCSLF2 e OsCSLF4 expressos em Arabidopsis – presença de MLG
Fonte: Scheller e Ulvskov, 2010 – Annu. Rev. Plant Biol; Yin et al., 2011 – Plos One; Pauly et al., 2013 -‐ Planta
-‐ Mutantes oscslf6 teve 97% de redução de MLG -‐ HvCSLH1 expresso em Arabidopsis – presença de MLG -‐ CSLF e CSLH localizados no Golgi -‐ Modelo: CSLF ou CSLH fazem a ligação β 1-‐3, após a ligação β 1-‐4
Gramíneas
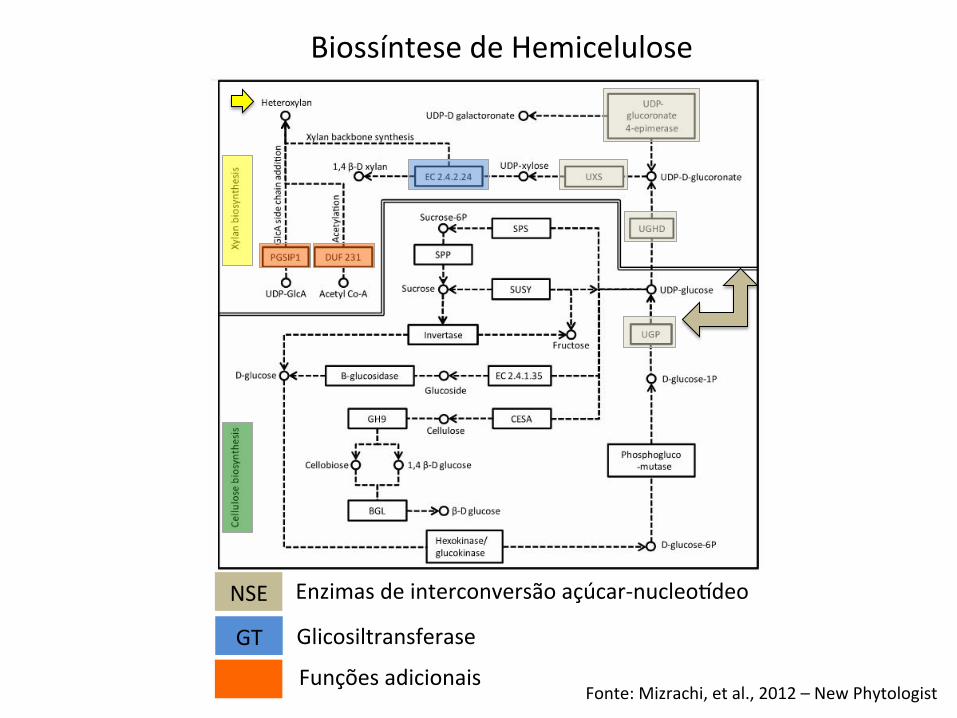
Fonte: Mizrachi, et al., 2012 – New Phytologist
Biossíntese de Hemicelulose
NSE
GT
Enzimas de interconversão açúcar-‐nucleoideo
Glicosiltransferase
Funções adicionais





























