Embed Size (px)
Citation preview

0
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
CEPPE
Centro de Pós-Graduação e Pesquisa
MESTRADO EM ANÁLISE GEOAMBIENTAL
CARLOS HUMBERTO BIAGOLINI
ALGUNS COMPONENTES DA MACROFLORA DA FORMAÇÃO
ITAQUAQUECETUBA, PALEÓGENO DA BACIA DE SÃO PAULO E
SUAS EVIDÊNCIAS PALEOCLIMÁTICAS”
Guarulhos
2012

1
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
CARLOS HUMBERTO BIAGOLINI
ALGUNS COMPONENTES DA MACROFLORA DA FORMAÇÃO
ITAQUAQUECETUBA, PALEÓGENO DA BACIA DE SÃO PAULO E
SUAS EVIDÊNCIAS PALEOCLIMÁTICAS”
Dissertação de Mestrado apresentado à
Universidade Guarulhos para obtenção do título de Mestre em Análise Geoambiental
Orientadora: Profa. Dra. Mary E.C. Bernardes-de-Oliveira
Guarulhos
2012

2
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Biagolini, Carlos Humberto
B576a Alguns componentes da macroflora da formação itaquaquecetuba, paleógeno da bacia
de São Paulo e suas evidências paleoclimática / Carlos Humberto Carlos Humberto.
Guarulhos, 2012.
151 f.: il.; 31 cm
Dissertação (Mestrado em Análise Geoambiental) - Centro de Pós-Graduação e
Pesquisa, Universidade Guarulhos, 2012.
Orientador: Profa. Dra. Mary Elizabeth C. Bernardes-de-Oliveira
Referências: f. 107-128
1. Paleógeno 2. Itaquaquecetuba 3. Bacia de São Paulo 4. Microgramma 5.
Podocarpus 6. Bauhinia 7. Leandra
CDD 22 st 550
Ficha catalográfica elaborada pela Biblioteca Fernando Gay da Fonseca

3
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
CEPPE
Centro de Pós-Graduação e Pesquisa
MESTRADO EM ANÁLISE GEOAMBIENTAL
A Comissão Julgadora dos trabalhos de Defesa de Dissertação de MESTRADO,
intitulada “ALGUNS COMPONENTES DA MACROFLORA DA FORMAÇÃO
ITAQUAQUECETUBA, PALEÓGENO DA BACIA DE SÃO PAULO E SUAS
EVIDÊNCIAS PALEOCLIMÁTICAS” em sessão pública, realizada em 29 de maio de
2012, considerou o candidato CARLOS HUMBERTO BIAGOLINI aprovado.
COMISSÃO EXAMINADORA:
1. Prof. Dra. Mary E. C. Bernardes-de-Oliveira – UnG/USP____________
2. Prof. Dra. Maria Judite Garcia - UnG____________________________
3. Prof. Dr. Carlos Alberto Bistrichi – PUC-SP______________________
Guarulhos, 29 de maio de 2012.

4
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
DEDICATÓRIA:
Dedico esta Dissertação de Mestrado à minha querida mãe (in memorian)
Yolanda Gonçalves Biagolini pela oportunidade que me deu de existir e pelo incentivo
constante aos estudos.
À minha irmã (in memorian) Sonia Regina Biagolini da qual guardo na memória
muitos momentos alegres e divertidos e que provavelmente nunca irei esquecer. A você
minha querida irmã dedico também este trabalho.
Ao meu amado pai Ernesto Biagolini, querido e companheiro que, por muitas
vezes, deixei de visitar para empenho total na elaboração e conclusão deste trabalho.
À minha maravilhosa esposa, Rosangela, minha adorada filha Patrícia e meu
precioso filho Carlos Jr. pelo apoio, incentivo e pelos diversos “empurrões” dados, que
ajudaram a superar as dificuldades naturais que surgem diante de um curso de
Mestrado.
À minha querida sobrinha e afilhada Cristiane, pelo apoio e incentivo no decorrer
do curso.
Ao meu querido irmão João Biagolini, pelo apóio e incentivo não só no decorrer
do curso, mas principalmente nos momentos difíceis de minha vida.
Às minhas tias Prof. Dra. Maria Esmeralda Zanllutti e Maria Zélia Minéo, por tanta
ajuda oferecida durante a jornada,sejam no incentivo ou no apoio, para que fosse
possível o cumprimento de todos os compromissos assumidos.
Aos meus amigos professores da Rede Estadual de Ensino de São Paulo que
também torceram pelo sucesso e conclusão deste trabalho. A estes grandes
companheiros e companheiras professores e guerreiros, o meu muito obrigado. Espero
que a conclusão desta caminhada sirva de inspiração para que outros professores
busquem, no aperfeiçoamento profissional, um futuro melhor não só financeiro, mas
pessoal refletindo na forma de transmitir conhecimentos e formando melhor à nova
geração que chega a cada ano nas escolas brasileiras tão carentes de profissionais
realmente apaixonados pelo que fazem.

5
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
AGRADECIMENTOS
À minha querida orientadora e Professora Dra. Mary E. C. Bernardes-de-Oliveira,
que durante todo o período de desenvolvimento deste trabalho, sempre esteve ao meu lado,
ensinando-me e orientando-me com carinho e precisão em tudo que foi necessário para que
fosse possível a conclusão desta obra. Sua dedicação, carinho e competência, serviram de
inspiração e exemplo, resultando em muitos outros trabalhos que realizei no exercício de
minha profissão de professor, ajudando na educação de jovens do ensino médio e
fundamental durante o período em que fiz o curso de mestrado e que com toda certeza, irá
também me acompanhar no exercício da profissão de professor do ensino superior que está
por iniciar.
À querida Isa Mendes que, ao lado de minha orientadora, torceu e me ajudou muito
tornando-se parte do elenco de pessoas as quais serei sempre grato. A você Isa, meu muito
obrigado.
Aos professores coordenadores do curso de Mestrado em Análise Geoambiental,
Professores Dr. Antonio Roberto Saad e Dra. Maria Judite Garcia, pelo carinho, apoio,
amizade e preocupação em fazer com que este trabalho pudesse ser realizado dentro dos
prazos determinados. A vocês o meu muito obrigado por toda ajuda oferecida.
Aos amigos e professores Profa Dra. Ana Olívia B. F. Magalhães; Profo Dr. Mario
Lincoln E. Etchebehere; Profo. Dr. Marcio de Andrade Magalhães, Profo Dr. Antonio Manoel
dos Santos Oliveira e ProfoDr.KenitiroSuguio, pela amizade e paciência na condução das
aulas e na preocupação em “ensinar”.
Aos grandes amigos do laboratório de Geociências que sempre com sorriso no rosto,
não pouparam esforços para que eu pudesse cumprir com minhas obrigações. Daniely dos
Santos; Andréia Barbieri Resende; Júnia Faria; Ricardo Seiffer; MScFabio da Costa Casado
e Professora MSc Rosana Saraiva Fernandes, meu muito obrigado a vocês por toda ajuda
oferecida no decorrer do curso de mestrado. Todos ajudaram muito e são responsáveis
também pela realização deste trabalho. Cada informação fornecida foi uma peça
fundamental a mais na elaboração deste trabalho.
À Gisele Dueñas secretária do curso de Mestrado muito obrigado por toda ajuda
fornecida durante o curso.

6
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Aos amigos e companheiros de turma que, em 2010, iniciaram comigo esta jornada e
que foram responsáveis por momentos alegres, que ficarão para sempre em minha
memória. Sou muito grato a todos vocês pelo carinho e amizade.
Aos botânicos do Horto Florestal, Dr. Osny Tadeu de Aguiar e Dr. João Aurélio
Pastore pela ajuda no fornecimento de material e informações a respeito de vegetais
pesquisados.
À Professora Dra Olga Yano, do Instituto de Botânica, pela grande ajuda no
esclarecimento de dúvidas no decorrer das pesquisas.
Às Funcionárias da USP, Jordana e Elaine do Laboratório de Petrografia Sedimentar
pela grande ajuda na realização dos testes de granulometria, realizados durante a
confecção desta dissertação.
À Secretaria da Educação do Estado de São Paulo, pela ajuda mais do que
importante através do programa “Bolsa Mestrado”, amparado no Decreto no 48.298 de
03/12/2003 e na Resolução SE 131 de 04/12/2003 e Resolução 105 de 01/12/2004, e aos
professores coordenadores da Delegacia de Ensino Leste 01 Professora Mariléa e Professor
Ivan pelo apoio e credibilidade ao projeto por mim desenvolvido, que possibilitaram
financeiramente o desenvolvimento desse trabalho.
Ao Prof.Dr.Carlos Alberto Bistrichi, por valiosas informações transmitidas na área de
coleta dos fitofósseis.
À Itaquareia – Indústria Extrativa de Minérios Ltda, pela facilidade no acesso à área
de estudo.
A todos que contribuíram de alguma forma para a realização desta dissertação, meu
muito obrigado por tudo.

7
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
"Todo aquele que se dedica ao estudo da ciência chega a convencer-se de que nas
leis do Universo se manifesta um Espírito sumamente superior ao do homem, e
perante o qual nós, com os nossos poderes limitados, devemos humilhar-nos."
Albert Einstein

8
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
RESUMO
Esse documento versa sobre um estudo taxonômico de novos elementos componentes da
macroflora Itaquaquecetuba, ocorrente no município homônimo (do estado de São Paulo) e
suas evidências paleoclimáticas. O material estudado corresponde ao registro fitofossilífero
preservado em camadas argilosas da Formação Itaquaquecetuba, documentando uma
vegetação que medrou durante o Paleógeno, não só na área de Itaquaquecetuba, mas por
toda a área da bacia sedimentar de São Paulo e, provavelmente, também, por grande parte
do Planalto Atlântico. Essa tafoflora foi preservada na forma de impressões, compressões e
piritizações ou marcassitizações foliares e caulinares envolvidas por folhelhos ou argilitos de
cor cinza, com tons mais claros ou mais escuros. A boa preservação permite observar, além
do contorno, detalhes de arquitetura foliar, de estruturas caulinares e alguns vestígios de
frutos ou sementes. Seu estudo foi feito através de observação em estereomicroscópio
óptico, com câmara clara, sobre fotomacrografias, fotomicrografias e em microscopia
eletrônica de varredura (MEV). O material foi analisado, do ponto de vista morfográfico da
fisiognomia foliar e taxonômico. Além dos macrofitofósseis já conhecidos de estudos
anteriores, foram aqui identificados novos elementos: Microgramma cf. M.persicariifolia,
Podocarpus cf. P.dubius, Bauhinia sp. e Leandra sp. Os resultados obtidos com a pesquisa
sugerem a presença de uma vegetação adaptada para um clima muito quente e úmido da
Floresta Ombrófila Densa (na época, com temperatura média anual ao redor de 27ºC a
28ºC). O ambiente de deposição sugerido pela tafonomia dos fitofósseis seria calmo e
redutor, possivelmente em meandros abandonados da planície de inundação de um sistema
fluvial meandrante ocorrente na bacia hidrográfica do paleorio Tietê, durante o Paleógeno.
Palavras-chave: Paleógeno, Itaquaquecetuba, Bacia de São Paulo, Microgramma,
Podocarpus, Bauhinia, Leandra.

9
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
ABSTRACT
This work reports about a taxonomic study of new elements belonging to the
Itaquaquecetuba macroflora, occurring in the namesake municipality (state of São Paulo)
and their paleoclimatic evidences. The studied material corresponds to the plant fossil
record preserved in the Itaquaquecetuba Formation clay layers, registering a vegetation that
thrieved during the Paleogene, not only in the area of Itaquaquecetuba, but for the whole
area of the sedimentary São Paulo basin and probably also by a larger part of the Atlantic
Plateau. This taphoflora was preserved as leaf and stem impressions, compressions and
pyritizations or marcassitizations sorrounded by lighter or darker grayish shale or claystones.
The preservation allows to
denot, beyond the shape, foliar architecture details, caulinar structures and some fruits and
seeds evidences. This study was done through the observation in optical stereomicroscope
with lucida camera, on photomacrographies, photomicrographies and scanner electronic
microscope (SEM). The material was analyzed after its foliar physiognomy and taxonomy.
Beyond the already known macrofossil plants from previous studies, new
elements are identified herein: Microgramma cf. M.persicariifolia, Podocarpus cf. P.dubius,
Bauhinia sp. e Leandra sp. The research gotten results suggest a flora with adaptation for a
hotter and wetter climate of the Dense Ombrophilus Forest (at that time under 27ºC to 28ºC).
The depositional environment suggested by the fossil plants taphonomy would be calm and
euxinic environment possibly in oxbow-lakes of a floodplain as part of a meandering fluvial
system occurring in the Tietê paleo-river hydrographic basin during the Paleogene.
Key-words: Paleogene, Itaquaquecetuba, São Paulo basin, Microgramma. Podocarpus,
Bauhinia, Leandra.

10
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
LISTA DE ILUSTRAÇÕES
Figura 2.1 – Tabela de Tempo Geológico 18
Figura 2.2 – Tendências de temperatura Paleógeno e Neógeno 19
Figura 2.3 – (A e B) PaleomapasPaleoceno / Oligoceno 20
Figura 2.4 – Reconstituição da Terra – Paleoceno 21
Figura 2.5 – Reconstituição da Terra – Oligoceno 21
Figura 2.6 – Principais biomas do Eoceno 22
Figura 2.7 – Principais biomas do Oligoceno 23
Figura 2.8 – Mapa com principais ocorrências de tafoflorasPaleógenas 25
Figura 3.1 – Região metropolitana de São Paulo/Área de Estudos 30
Figura 3.2 – Área de Coleta de Fósseis 31
Figura 3.3 – Quadro geológico e tectônico do RCSB 32
Figura 3.4 – Mapa Geológico da Bacia de São Paulo 33
Figura 3.5 – Seção Geológica Bacia de São Paulo 34
Figura 3.6 – Afloramento Itaquareia I em destaque 37
Figura 3.7 – Seção de Correlação de perfis Form. Itaquaquecetuba 39
Figura 3.8 – Idades propostas para a Formação Itaquaquecetuba 42
Figura 4.1 – Perfil do afloramento Itaquareia I 44
Figura 4.2 – Imagem do Afloramento com indicação do local de coleta 45
Figura 4.3 – Classificação de folhas forma laminar 48
Figura 4.4 – Classificação da lâmina foliar 49
Figura 4.5 – Classificação do ângulo da base 49
Figura 4.6 – Avaliação do ângulo do ápice 50
Figura 4.7 – Formas de base 51
Figura 4.8 – Formas de conexão do pecíolo 52
Figura 4.9 – Formas de ápices 52
Figura 4.10 – Diferentes formas de ápices 53
Figura 4.11 – Formas de margens 54
Figura 4.12 – Formas de margens conforme LAWG, (1999) 54
Figura 4.13 – Formas de lobação 55
Figura 4.14 – Tipos de venação 57
Figura 5.1 – Taxa superiores de Filicophyta segundo Taylor et al. (2009) 63
Figura 5.2 – Relações filogenéticas entre angiospermas 64

11
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Figura 5.3 – Lista de componentes da tafoflora Itaquaquecetuba 66
Figura 5.4 – Classificação de subfamílias de Polypodiaceae 68
Figura 5.5 – Cladograma filogenético 69
Figura 5.6 – Fitofóssil Microgramma sp 71
Figura 5.7 – Esquema de venação de Microgramma sp 71
Figura 5.8 – Mapa com distribuição de Microgramma sp 72
Figura 5.9 – Ilustração de ramos folhosos de podocarpácea 74
Figura 5.10 – Grãos de polén de Podocarpus sp 77
Figura 5.11 – Micro folhas de Podocarpus sp 79
Figura 5.12 – Fitofóssil Bauhinia sp 86
Figura 5.13 – Esquema de venação fitofóssil Bauhinia sp 86
Figura 5.14 – Formas de diferentes espécies de Bauhinia 87
Figura 5.15 – Fitofóssil Leandra sp 90
Figura 5.16 – Esquema de venação de fitofóssil Leandra sp 91
Figura 6.1 – Esquema de Duparque 95
Figuras 6.3 – Gráfico de avaliação margem foliar 97
Figura 6.4 – Gráfico relação margem x temperatura 98
Figura 6.5 – Gráfico de avaliação de textura das folhas 99
Figura 6.6 – Gráfico de avaliação das formas dos ápices 100
Figura 6.7 – Gráfico de avaliação de ângulo de ápice 100
Figura 6.8 – Gráfico de avaliação de ângulo de base 101
Figura 6.9 – Gráfico de avaliação de formas de base 101
Figura 6.10 – Componentes identificados na tafoflora Itaquaquecetuba 102
TABELA DE CLASSIFICAÇÃO DE FOLHAS POR ÁREA 56

12
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
SUMÁRIO
TEMA PÁGINA
DEDICATÓRIA IV
AGRADECIMENTOS V
EPÍGRAFE VII
RESUMO VIII
ABSTRAT IX
LISTA DE ILUSTRAÇÕES X
SUMÁRIO XII
CAPÍTULO 1 - INTRODUÇÃO
1.1 – Generalidades 15
1.2 – Objetivos 17
1.3 – Justificativas 17
CAPÍTULO 2 – A PALEOBOTÂNICA PALEÓGENA BRASILEIRA
2.1 – O que é Paleógeno? 18
2.2 – Aspectos Gerais e Climáticos do Período Paleógeno 19
2.3 – A Paleogeografia Paleógena 21
2.4 – Floras e Fauna Paleógenas 22
2.5 – Floras Paleógenas do Brasil 23

13
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
CAPÍTULO 3 – ÁREA DE ESTUDO: LOCALIZAÇÃO GEOGRÁFICA E GEOLOGIA REGIONAL
3.1 – Localização Geográfica da Área de Estudos 30
3.2 – Geologia Regional 31
3.2.1 – Formação Itaquaquecetuba 35
CAPÍTULO 4 – MATERIAL E MÉTODOS
4.1 – Material 43
4.2 – Métodos 45
4.2.1 – Métodos de Campo 45
4.2.2 – Métodos de Laboratório 46
4.2.2.1 – Preparação Mecânica e Codificação de Amostras 46
4.2.2.2 – Preparação Química 46
4.2.2.3 – Documentação Gráfica 46
4.2.2.4 – Análise da Arquitetura Foliar 47
4.2.2.5 – Identificação Taxonômica 59
4.2.3 – Métodos de Gabinete 60
4.2.3.1 – Levantamento Bibliográfico 60
4.2.3.2 – Elaboração de Mapas 60
4.2.3.3 – Integração de Dados 60
CAPÍTULO 5 – SISTEMÁTICA
5.1 – Taxonomia 61
5.2 – Lista Taxonômica da tafoflora da Formação Itaquaquecetuba 66

14
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
5.3 – Novas Identificações e descrições de elementos da tafoflora de
Itaquaquecetuba. 67
5.3.1 – Classe Filicopsida ou Polydiopsida
Ordem Polypodiales 67
5.3.2 - Gimnospermas
Ordem Coniferales 72
5.3.3 – Angiospermas
Generalidades 80
Ordem Fabales 82
Ordem Myrtales 88
CAPÍTULO 6 – ANÁLISE E INTERPRETAÇÃO DE DADOS
6.1 – Análise Tafonômica e sua Interpretação 93
6.2 – Análise morfográfica dos fitofósseis foliares e sua interpretação 96
6.3 – Interpretação dos dados taxonômicos encontrados, associados
a aspectos ecológicos e distribuição geográfica de formas atuais
afins 102
CAPÍTULO 7 – CONSIDERAÇÕES FINAIS 104
REFERÊNCIAS BIBLIOGRÁFICAS 107
ESTAMPAS 129
ANEXOS

15
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
ALGUNS COMPONENTES DA MACROFLORA DA FORMAÇÃO ITAQUAQUECETUBA, PALEÓGENO DA BACIA DE SÃO PAULO E SUAS
EVIDÊNCIAS PALEOCLIMÁTICAS.
Carlos Humberto Biagolini
1.1 – Generalidades
Os fósseis são instrumentos fundamentais para a compreensão do
aparecimento e evolução da vida em nosso planeta, durante sua longa história
geológica, uma vez que fornecem informações acerca do ambiente no qual
habitaram, sua ecologia e época, possibilitando estabelecer idades relativas e
relações estratigráficas (CASSAB, 2000; RICARDI-BRANCO et al., 2008) .
“Paleobotânica é a área da ciência que estuda os organismos vegetais sob todas as formas conhecidas de fossilização ou de vestígios nas rochas considerando a interação sinergística dos organismos com os seus ambientes de vida, no curso de tempo geológico” (MUSSA, 2004).
As adaptações morfoanatômicas necessárias à sobrevivência de um
organismo vegetal atual sob determinadas condições ambientais podem, com base
no princípio do atualismo, ser consideradas semelhantes às vigentes no passado,
tornando os fitofósseis importantes ferramentas para as ciências geológicas na
obtenção de dados paleoecológicos, paleoclimáticos e paleogeográficos (IANNUZZI;
VIEIRA, 2005; TUFANO, 2009).
Todo conhecimento que se possa obter sobre as floras paleógenas, neógenas
e pleistocenas é muito importante, pois através deste conhecimento se
compreenderá a diversidade floral atual existente no planeta, como e porque se
formou. Relacionando com suas características geomorfológicas, biogeográficas e
climáticas, pode-se também obter dados importantes sobre o comportamento do
planeta, ao longo de sua história geológica.
O estudo da paleobotânica fornece subsídios para melhor compreensão sobre
as floras que viveram em um determinado lugar, no passado. Através dos fitofósseis
é possível realizar a análise de clima, composição do solo e ciclos de chuva. Seus
CAPÍTULO 1 – INTRODUÇÃO

16
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
processos adaptativos podem assumir o papel de bioindicadores na análise de
eventos geológicos do passado.
Os principais itens verificados no estudo da Paleobotânica são:
a) Restos ou impressões de folhas ou fragmentos de plantas deixados entre os
sedimentos, durante o processo de tafonomia (transporte, deposição e
fossildiagênese);
b) Venação da folha e a morfologia e\ou anatomia de outros órgãos que
permitem identificar, em muitos casos, a classe, a família e o gênero da
planta-mãe e também estabelecer correlações com diferentes tafofloras;
c) Dados paleoecológicos relacionados à interação planta com animal e com
outros organismos. Através da análise de marcas de danos deixadas nas
folhas, pode-se identificar a ação de diferentes organismos que se
relacionavam com o vegetal no passado: insetos, fungos, etc;
d) Dados paleoclimáticos, uma vez que a morfologia adaptativa foliar e de outros
órgãos vegetais podem indicar a temperatura e a umidade do ambiente a que
estão ou foram submetidos, e
e) Identificação de tipos de solo quanto a umidade e composição química, uma
vez que certos vegetais, só se desenvolvem em tipos específicos de solo.
Cada detalhe observado em uma folha ou fragmento vegetal, como também
marcas deixadas por outros organismos podem representar novas informações que
permitem compreender melhor as etapas pelas quais passou uma determinada
região, no curso do tempo geológico.
A Formação Itaquaquecetuba, objeto de estudo paleobotânico desta
dissertação de mestrado, é uma unidade litoestratigráfica presente na bacia
sedimentar de São Paulo. Apresenta diversos aspectos que facilitam a coleta de
amostras e seu estudo, tanto sedimentológico como paleontológico e que permitem
uma análise morfoanatômica mais precisa de seus bem-preservados fitofósseis. A

17
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
atividade mineradora desta formação para fins de exploração de areia para
construção civil também facilita a retirada de material para estudo.
As idades desta formação obtidas sobre diferentes bases tem gerado muita
controvérsia, no entanto a idade mais aceita é a paleógena, obtida por dados
palinológicos (SANTOS, 2009).
1.2 - Objetivos
Esta pesquisa busca: Identificar, taxonomicamente, os componentes
florísticos preservados na Formação Itaquaquecetuba, aflorantes na área do
município paulista homônimo; correlacionar com outras tafofloras cenozóicas
do Estado de São Paulo, buscando identificar fitofósseis marcadores de
tempo; identificar o bioma do qual os fitofósseis fizeram parte; identificar os
paleoambientes de deposição e aspectos paleoecológicos e entender as
adaptações da vegetação, diante de variações dos níveis pluviométricos e de
temperatura relacionados com o paleoclima.
1.2 – Justificativas
O estudo desse tema justifica-se pelas seguintes razões:
A Formação Itaquaquecetuba representa uma unidade litoestratigráfica ainda
pouco compreendida, que aflora em esparsas áreas da bacia de São Paulo.
Assim, este trabalho surge e justifica-se pela área geográfica que a Formação
Itaquaquecetuba ocupa, pela importância e riqueza de detalhes de seus fitofósseis
e por muitas informações não disponíveis ainda.
Grande parte do conhecimento sobre formações do Estado de São Paulo foi
adquirido nas décadas de 1960 e 1970 em trabalhos de cunho geológico e
geomorfológico, impulsionado pelo interesse nos recursos minerais destas
formações. Na unidade litoestratigráfica considerada, há muita informação ainda a
ser obtida com as pesquisas da tafoflora nela existente, razão principal desta
pesquisa.

18
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
2.1 – O que é o Paleógeno?
Paleógeno é o primeiro período geológico da era Cenozóica, incluído no éon
Fanerozóico, compreendendo o intervalo de tempo entre 65,5 e 23,0 Ma, segundo
Gradstein et al. (2004) e Ogg et al. (2008).
O período Paleógeno sucede ao período Cretáceo da era Mesozóica e
precede o período Neógeno de sua era. Divide-se nas épocas paleocena, eocena e
oligocena, da mais antiga para a mais nova (Figura 2.1).
Figura 2.1: Tabela do Tempo Geológico Cenozóico, incluindo o Paleógeno com suas subdivisões em
épocas e andares. (Extraído da tradução para o Português de SALLUM, 2010, a partir de GRADSTEIN et al.,
2004 e OGG et al. 2008, conforme ICS, 2010)
CAPÍTULO 2 – A PALEOBOTÂNICA PALEÓGENA BRASILEIRA

19
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
2.2 – Aspectos Gerais e Paleoclimáticos do Período Paleógeno
Atualmente, estudos relacionados às mudanças climáticas têm recebido
grande atenção, ao considerar a ocorrência do aquecimento global provocado pelo
aumento nas concentrações atmosféricas de CO2, resultantes principalmente de
atividade humana.
Entretanto, mudanças climáticas importantes também aconteceram de forma
cíclica durante o passado do planeta (independentemente da ação antrópica), sendo
as ocorridas entre o Pleistoceno e Holoceno (Período Quaternário) as mais
conhecidas e discutidas. Nesse período, importantes eventos ocorreram no clima e
na biota brasileira. As alternâncias entre as fases glaciais e interglaciais
(principalmente das áreas nórdicas) foram acompanhadas por fases secas e úmidas,
respectivamente, com mudanças profundas no tipo de vegetação e biomassa
causando extinção, diferenciação e mudanças na distribuição geográfica de plantas
e animais, nas zonas continentais tropicais (ADAMS et al.,1990), principalmente do
hemisfério sul.
No final do Paleógeno (época oligocena), sob um clima mais fresco e mais
seco, a Terra teria alcançado uma “fase de transição” entre o mundo quase
pantropical (neopaleoceno – eoceno) com um máximo térmico (efeito greenhouse) e
o mundo de clima ameno do início do Neógeno até o glacial/ interglacial Quaternário
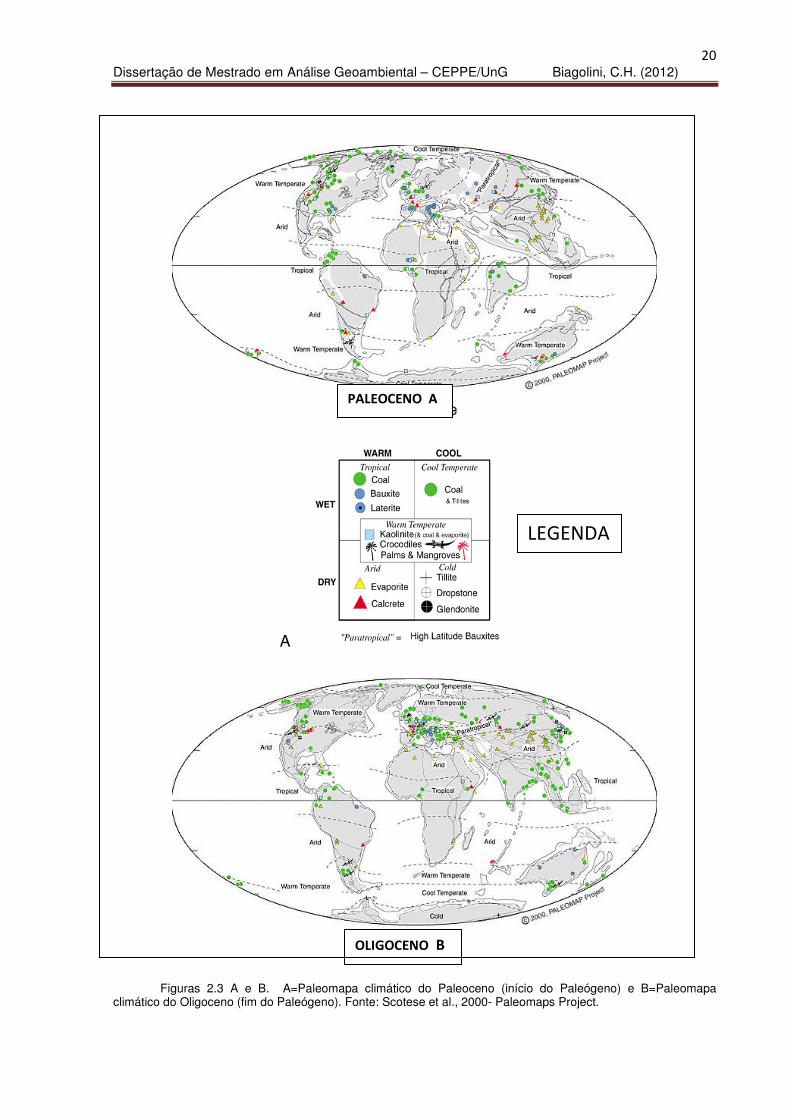
(Figuras 2.2.; 2.3 A e B).
Figura 2.2 – Tendências da temperatura nos períodos Paleógeno e Neógeno, fornecidas a partir da
análise isotópica do oxigênio de registros foraminíferos bentônicos de locais do Atlântico onde a temperatura está em oC, acima dos valores atuais (WILLIS; Mc ELWAIN, 2002).
20
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
A
Figuras 2.3 A e B. A=Paleomapa climático do Paleoceno (início do Paleógeno) e B=Paleomapa climático do Oligoceno (fim do Paleógeno). Fonte: Scotese et al., 2000- Paleomaps Project.
PALEOCENO A
LEGENDA
OLIGOCENO B

21
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
2.3. A Paleogeografia Paleógena
No início do Paleógeno, a paleogeografia mundial ainda era muito semelhante
à do Neocretáceo e os continentes, paulatinamente, moviam-se em direção às suas
posições atuais. A América do Sul, a Antártica e a Austrália permaneciam unidas e a
Índia não havia cruzado ainda a linha do Equador, em seu deslocamento rumo à
Ásia. A Europa meridional e a África setentrional constituíam uma porção de ilhas
banhadas pelo Mar de Tétis, num corredor marinho entre os oceanos Atlântico e
Índico (Figura 2.3 A e 2.4).
Figura 2.4. Reconstituição do mundo no início do Paleoceno (65 milhões de anos atrás). (Fonte: http://celestia.albacete.org/celestia/deriva/paleoce.htm).
No final do Paleógeno, a América do Sul e a Antártica separaram-se com a
abertura do estreito de Drake e consequentemente, a corrente marinha Circumpolar
e a glaciação antártica instalaram-se (Figura 2.5).
Figura 2.5. Reconstituição do mundo no início do Oligoceno (34 milhões de anos atrás).
(Fonte: http://www2.nau.edu/rcb7/035Marect.jpg.

22
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
2.4. Floras e Fauna Paleógenas
Como pode ser visto em Willis e McEwain (2002) e Garcia et al. (2007), a
flora inicial do Paleógeno apresentava semelhança com a flora predominante do
Cretáceo, embora deva ser ressaltado o maior desenvolvimento das angiospermas,
no período Paleógeno. As vegetações dominantes eram as florestas tropicais
úmidas, nas latitudes baixas a médias, e os bosques temperados de coníferas e
árvores decíduas, nas latitudes mais altas (Figura 2.6).
Figura 2.6- Principais biomas do Eoceno (60 – 50 Ma) Reconstrução paleogeográfica com os principais biomas para o Eoceno: 1) Temperado Fresco; 2) Temperado Quente; 3)Subtropical de Verão Úmido; 4) Tropical Sempre Úmido; 5) Subtropical Desértico 6) Subtropical de Verão Úmido Semi-Desertico (Modifi cado de WILLIS; MCELWAIN, 2002) extraído de Garcia et al., 2007.
No final do Paleógeno (Oligoceno), com o crescimento da calota polar
Antártica, o clima tornou-se mais frio e mais seco e os mares desceram seus níveis.
Devido a isto, as florestas tropicais retraíram-se dando lugar a uma extensa

23
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
vegetação mais aberta, constituída de árvores e campinas. Nas proximidades dos
pólos, instalam-se as taigas e tundras (Figura 2.7).
Figura 2.7- Principais biomas do Oligoceno (~ 30 Ma) Reconstrução Paleogeográfica com os principais biomas para o Oligoceno: 1) Temperado Frio; 2) Temperado Fresco; 3) Subtropical de Verão Úmido Semi-Desertico; 4) Subtropical de Verão Úmido; 5) Tropical Sempre Úmido 6) Ártico; 7) Glacial (Modificado de WILLIS; MCELWAIN, 2002) extraído de Garcia et al., 2007.
A fauna paleógena se caracterizou pelos mamíferos primitivos (com
parentesco distante dos atuais) e por aves carnívoras de grande porte, que foram as
primeiras a ocupar o topo da cadeia alimentar após a extinção dos dinossauros
(COWEN, 2005).
2.5. Floras Paleógenas do Brasil
Cada época do Paleógeno apresenta características climáticas próprias que
interferiram diretamente na constituição da paleofloras cenozóicas brasileiras
inclusive na da Formação Itaquaquecetuba. A temperatura acima dos níveis de hoje
e a umidade acentuada são registradas e detectadas na forma e taxonomia dos
fitofósseis.
A figura 2.8 exibe as tafofloras paleógenas brasileiras conhecidas até o
momento; todas ocorrentes no sudeste brasileiro à exceção das paleocenas de
Ipixuna (PA) e Maria Farinha (PE).

24
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
As paleofloras paleocenas brasileiras, além das duas mencionadas acima,
estão documentadas também na bacia de São José de Itaboraí (RJ).
Na Formação Ipixuna, no Pará, apareceu um registro de lenho fóssil de
Malpighiales da Família Humiriaceae (Sacoglottis paraensis), cujo gênero é,
atualmente, característico da mata amazônica de terra firme ou de formações
abertas do Baixo Amazonas (JAPIASSÚ, 1969). Fósseis de frutos de Palmae (Nypa
pernambucencis) foram achados na Formação Maria Farinha, na costa de
Pernambuco (DOLIANITI, 1955). Na bacia de São José do Itaboraí, Rio de Janeiro,
foram descritas sementes de Ulmaceae (Celtis santosi) e Myrtaceae (Psidium sp) (
MAGALHÃES, 1948; BEURLEN; SOMMER, 1954).
As três ocorrências paleocenas fazem parte do bioma tropical sempre úmido
que deve ter medrado não só por todo o território nacional, como também se
estendido pela Bolívia e por toda a área setentrional da Argentina (bacia de Salta)
(GARCIA et al., 2007).
É provável que todas estas ocorrências brasileiras estejam incluídas na
Província de Ulmaceae, associada principalmente às famílias tropicais e
subtropicais (QUATTROCCHIO, 2006).
As paleofloras eocenas brasileiras têm registro apenas no Eoceno médio a
superior, nas formações Fonseca e Gandarela (MG) e, no Eoceno superior, na bacia
de Bonfim (SP), na Bacia de Resende (Formação Resende), na Bacia de Volta
Redonda e no Graben Casa de Pedra (RJ). De idade transicional mesoeocena a
neoeocena, as tafofloras brasileiras das bacias de Fonseca e de Gandarela são as
que melhor caracterizam o intervalo de tempo.
O clima do Eoeoceno a Mesoeoceno (55,8 a 40,4 Ma., conforme
GRADSTEIN et al., 2004 e OGG et al., 2008), continuou o mesmo do Paleoceno,
tendo sido um dos mais quentes da história da Terra. O bioma tropical sempre
úmido, provavelmente, continuou cobrindo todo o país e atingindo até latitudes de
40º S. Todavia, ainda não há registros brasileiros conhecidos dessa idade.

25
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)

26
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Como bem ressaltaram Garcia et al. (2007), a bacia de Fonseca (MG) possui
uma rica associação de lenhos e folhas de Magnoliales (Annonaceae), Lamiales
(Bignoniaceae), Fabales (Fabaceae), Laurales (Lauraceae, Monimiaceae),
Sapindales (Meliaceae, Rutaceae, Sapindaceae), Myrtales (Combretaceae,
Myrtaceae, Melastomataceae), Malpighiales (Euphorbiaceae, Malpighiaceae),
Ranunculales (Menispermaceae), Ericales (Myrsinaceae, Sapotaceae, Theaceae),
Malvales ( Malvaceae, dentre as quais foi detectada uma flor de Eriotheca prima
Duarte, (1974) além de Filicófitas tais como Schizeaceae e Aspleniaceae
(GORCEIX, 1884; BERRY, 1935; DOLIANITI, 1949, 1950; CURVELLO, 1955;
DUARTE, 1956, 1974 e 1985; OLIVEIRA; SILVA, 1982; MELO et al., 2002;
FANTON et al., 2009). Na bacia de Gandarela, foram identificadas as famílias
Blechnaceae, Chrysobalanaceae, Fabaceae, Rutaceae, Vochysiaceae,
Sapindaceae e Myrtaceae (DUARTE; MELLO FILHA, 1980). Lima e Salard-
Cheboldaeff (1981) e Maizzato (2001) assinalaram, para ambas as bacias,
palinomorfos relacionados às Schizaeaceae, Polypodiaceae, Ephedraceae,
Cycadales, Palmae (Nypa), Caesalpinoidea, Guttiferae, Ulmaceae, Caprifoliaceae,
Polygalaceae, Rubiaceae, Proteaceae, Malpighiaceae, Bombacaceae e
Podocarpaceae, que indicam ambiente de vida em áreas alagáveis de um sistema
fluvial meandrante, sob clima quente e úmido.
No Eoceno superior brasileiro (~40 a 33 Ma.), há várias ocorrências de mega
e microfósseis. Na bacia de Bonfim, foram registrados por Paes Leme (1918)
fitofósseis de Podocarpus lambertii, de Rutales (Zanthoxylon) e Ebenales. Oliveira
(1937) e Oliveira; Leonardos, (1978), identificaram poáceas e sementes, nessa
bacia. Também foram registrados palinomorfos de Schizaeaceae, Polypodiaceae,
Ephedraceae, Cycadales, Palmae, Fabaceae, Guttiferae, Caprifoliaceae,
Polygalaceae, Rubiaceae, Proteaceae, Malpighiaceae, Ulmaceae, Bombacaceae e
Podocarpaceae. Essa associação sugere um ambiente pantanoso sob clima úmido,
conforme Lima e Dino (1984).
Ainda entre as associações palinológicas neoeocenas brasileiras citam-se
aquelas descritas por Lima e Amador (1985), para a bacia de Resende (Formação
Resende) e para o Graben Casa de Pedra por Lima e Melo (1994), onde estão
presentes grãos de pólen de Faboideae, Proteaceae, Mimosoideae,

27
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Convolvulaceae, Thymeleaceae, Myrtaceae, Aquifoliaceae, Malpighiaceae,
Plamaceae e grande abundância de esporos de Pteridófitas, que sugerem a
existência de matas em condições climáticas quentes e úmidas, além de
representantes de Onograceae que indicam a existência de prováveis corpos
lacustres. Estas evidências associadas a ausência de palinomorfos de
gimnospermas, sugerem para a região uma vegetação neoeocena de florestas
quentes e úmidas sob um clima tropical.
As macro e microfloras, de idade neoeoceno-oligocena do sudeste brasileiro
documentam uma flora de caráter tropical a sub-tropical úmido. Dentre elas citam-se
a da Formação Macacu (RJ) com palinomorfos identificados por Lima et al. (1996) e
a da bacia do Tanque (SP) e depósitos correlatos com o seu conteúdo palinológico
apresentado por Bistrichi (2001) e Garcia et al. (2004, 2008). Outra ocorrência de
mesma idade corresponde a tafoflora Entre-Córregos da bacia de Aiuruoca (MG),
datada por seu espectro esporo-polínico constituído de pteridófitas, podocarpáceas,
malpiguiáceas, mirtáceas, proteáceas, ulmáceas, euforbiáceas, faboídeas e
mimosóideas, sapindáceas, foi descrita por Garcia et al (2000). Seus macrofósseis
representados por impressões e compressões foliares de Annona, Nectandra,
Apeiba, Luehea, Passiflora, Caesalpinia, Machaerium, Campomanesia, Eugenia,
Myrciaria, Psidium e Sapindus, foram identificados por Franco-Delgado (2002),
Franco-Delgado e Bernardes-de-Oliveira (2004).
O resfriamento climático global e o crescimento da aridez, as grandes
mudanças na circulação oceânica e o início do gelo sobre a Antártica, típicos da
transição eoceno para o oligoceno (~34 Ma), afetaram também os biomas
brasileiros. O bioma tropical sempre úmido restringiu-se a uma faixa equatorial
estreita e, ligeiramente, oblíqua desde ~ 10º N até 10º S, cobrindo a região norte-
nordeste brasileira. O bioma sub-tropical de verão úmido estendeu-se desde o Peru
cobrindo todo o sul do Brasil (WILLIS; McELWAIN, 2002).

28
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
O Oligoceno está bem representado no sudeste brasileiro na bacia de
Resende (Formação Itatiaia), na bacia de Taubaté (Formação Tremembé) e bacia
de São Paulo (Formações Itaquaquecetuba e São Paulo), que atestam clima sub-
tropical de verão úmido. A Formação Itatiaia teve seu conteúdo palinológico
estudado por Lima e Melo, (1994), que detectaram a presença de pteridófitas,
proteáceas, além de espécies de euforbiáceas, nictagináceas, solanáceas,
fagáceas, meliácias, sapotáceas, mirtáceas, palmas, ulmáceas, bombacáceas,
timeleáceas, onagráceas, malpiguiáceas, fabáceas, malváceas e convol-vuláceas.
Junto aos grãos de pólen dessas famílias vegetais ocorreram também grãos de
pólen gimnospérmicos de taxodiáceas, podocarpáceas e efedráceas que sugeriam
condições climáticas mais frias. Na bacia de Taubaté, as formações Tremembé e
São Paulo (= Caçapava), tiveram sua palinologia estudada por Lima et al. (1983).,
Lima et al. (1991)., Yamamoto (1995)., Merces (2005) e Merces et al. (2006). A
palinoflora mostrou-se constituída de esporos de fungos, algas e pteridófitas,
diversidade de grãos de pólen de gimnospermas (taxodiáceas, podocarpáceas e
efedráceas), e de angiospermas: poáceas, liliáceas, tifáceas, bombacáceas,
asteráceas, fabáceas, ulmáceas, clorantáceas, proteáceas, fitolacáceas, malváceas,
palmas, timeleáceas, malpiguiáceas, euforbiáceas, mirtáceas, sapotáceas,
poligaláceas, onagrácias, nictagináceas e meliáceas. Tanto seus registros
palinoflorísticos como megaflorísticos são concordantes com a existência de um
bioma sub-tropical de verão úmido do tipo floresta paratropical.
Na bacia de São Paulo, as formações São Paulo e Itaquaquecetuba tiveram
sua palinoflora documentada por Melo et al. (1985) e Lima et al. (1991). Yamamoto
(1995), Santos (2005), Santos et al (2006) e Santos (2009). Esta última autora, em
seu extenso trabalho de mestrado, identificou a palinoflora e sua evolução, definiu a
idade, interpretou o paleoclima e o paleoambiente e reconstruiu a
paleopalinofitogeografia da região no intervalo de tempo neoeoceno-eo-oligoceno.
As análises indicaram diferentes idades para a deposição dos sedimentos da
Formação Itaquaquecetuba. Foram reconhecidas 4 ecozonas, sendo as três
primeiras de idade neoeocena e a quarta de idade eo-oligocena.

29
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Enquanto os palinomorfos registrados no intervalo neoeoceno sugeriram a
vigência de um clima sub-tropical úmido, com uma palinoflora diversificada, os
palinomorfos da ecozona IV, de idade eo- oligocena sugeriram um paleoclima
subtropical a temperado seco, com significativo aumento de gimnospermas e
diminuição de diversidade das angiospermas.
30
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
3.1 – Localização Geográfica da Área de Estudo
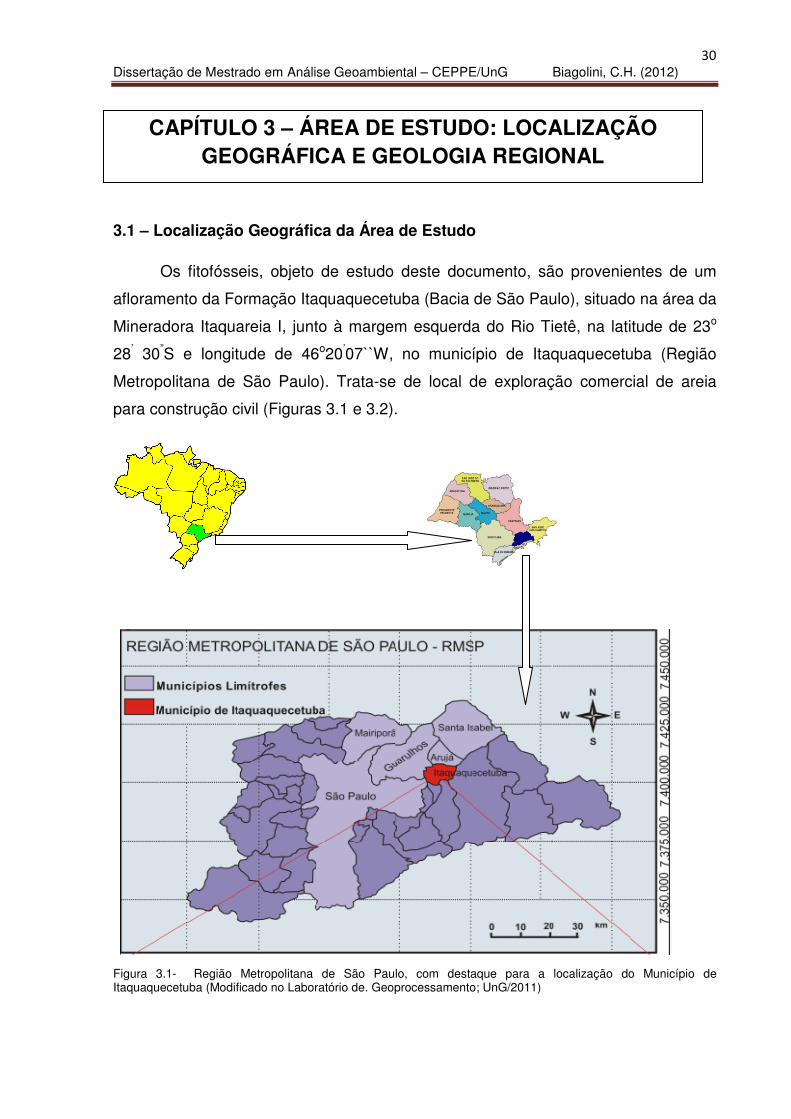
Os fitofósseis, objeto de estudo deste documento, são provenientes de um
afloramento da Formação Itaquaquecetuba (Bacia de São Paulo), situado na área da
Mineradora Itaquareia I, junto à margem esquerda do Rio Tietê, na latitude de 23o
28’ 30”S e longitude de 46o20’07``W, no município de Itaquaquecetuba (Região
Metropolitana de São Paulo). Trata-se de local de exploração comercial de areia
para construção civil (Figuras 3.1 e 3.2).
Figura 3.1- Região Metropolitana de São Paulo, com destaque para a localização do Município de Itaquaquecetuba (Modificado no Laboratório de. Geoprocessamento; UnG/2011)
CAPÍTULO 3 – ÁREA DE ESTUDO: LOCALIZAÇÃO GEOGRÁFICA E GEOLOGIA REGIONAL

31
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Figura 3.2 - Área de coleta de material de pesquisa, na Mineradora Itaquareia I, junto à margem esquerda do Rio Tietê, (fonte: Google Maps /2010 adaptado por CASADO /2011, para este trabalho)
A área de estudo dista 34 km da capital paulista e seu principal acesso é
realizado pela rodovia Airton Senna, designada SP-70. Também pode ser feito pela
SP-66 (Antiga Estrada São Paulo - Rio) ou pela SP-56 (Estrada de Santa Isabel).
A região de Itaquaquecetuba, antes da ação destruidora antropogênica, mas
já durante o Holoceno, era coberta por um ecótono (= uma região de transição entre
diferentes biomas). Havia um mosaico de formações e não um tapete contínuo do
bioma Mata Atlântica como aquele que se vê na Serra do Mar: a Mata de Araucária
originária do sul, o Cerrado do Centro-Oeste, a Mata Atlântica da Encosta da Serra
do Mar e a Mata Semidecídua do interior paulista, encontravam-se nessa área em
um fluxo de diferentes floras, cobrindo variadas paisagens (CARDIM, 2009).
3.2- Geologia Regional
A bacia de São Paulo, situada na Província Geomorfológica Planalto Atlântico
(ALMEIDA, 1964; PONÇANO, 1981), no cenário geológico cenozóico do Sudeste do
Brasil, é uma bacia sedimentar tafrogênica que se encontra inserida no Sistema de
Riftes da Serra do Mar, (ALMEIDA, 1976), posteriormente renomeado Rifte
Continental do Sudeste do Brasil (RCSB) por Riccomini (1989).

32
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Esta província estrutural corresponde a uma feição tectônica paralela à da
faixa litorânea, com cerca de 900 km de extensão, alongada e deprimida, de direção
ENE. Deste sistema de rift fazem parte ainda, as Bacias de Curitiba, Taubaté,
Resende, Volta Redonda, Itaboraí e os grábens de Sete Barras, Cananéia,
Guaraqueçaba e Barra de São João (Figura 3.3).
Figura 3.3 – Quadro geológico e tectônico regional do Rift Continental do Sudeste do Brasil (RCSB) (MELO et al. 1985; RICCOMINI et al. 1996). I) Embasamento pré-cambriano; II) Sedimentos paleozóicos e mesozóicos da Bacia do Paraná; III) Formação Serra Geral; IV) Rochas relacionadas ao inagmalismo Mesozóico-Cenozóico; (Tanguá, Itatiaia, Passa-Quatro). V) Bacias do rift contendo os sedimentos tercidrios do RCSB (1-Bacia de Itaboraí, 2-Grabem de Barra de São João, 3-Bacia de Macaca, 4- Bacia de Volta Redonda, 5- Bacia de Resende, 6- Bacia de Taubaté, 7- Bacia de São Paulo, 8- Graben de Sete Barras, 9- Formação Pariquera-Açu, 10 – Formação Alexandra e Graben de Guaraqueçaba, 11- Bacia de Curitiba, 12 – Graben de Cananéia); VI) Zonas de cisalhamento pré-cambrianas, em parte reativadas durante o Mesozóico e Cenozóico.
A Bacia Sedimentar de São Paulo encontra-se assentada sobre o
embasamento pré-cambriano cristalino (MELO; CAETANO; COIMBRA, 1986). Essa
bacia caracteriza-se por possuir um substrato com soerguimentos e abatimentos
localizados, formados pela ação de falhas pré e pós-sedimentares, e um formato
irregular de 60 X 30 km, com duas ramificações principais: uma de Itaquaquecetuba
para Poá e Mogi das Cruzes, e outra do bairro da Lapa, na cidade de São Paulo,
para Barueri; ambas acompanham o vale do Rio Tietê.
33
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
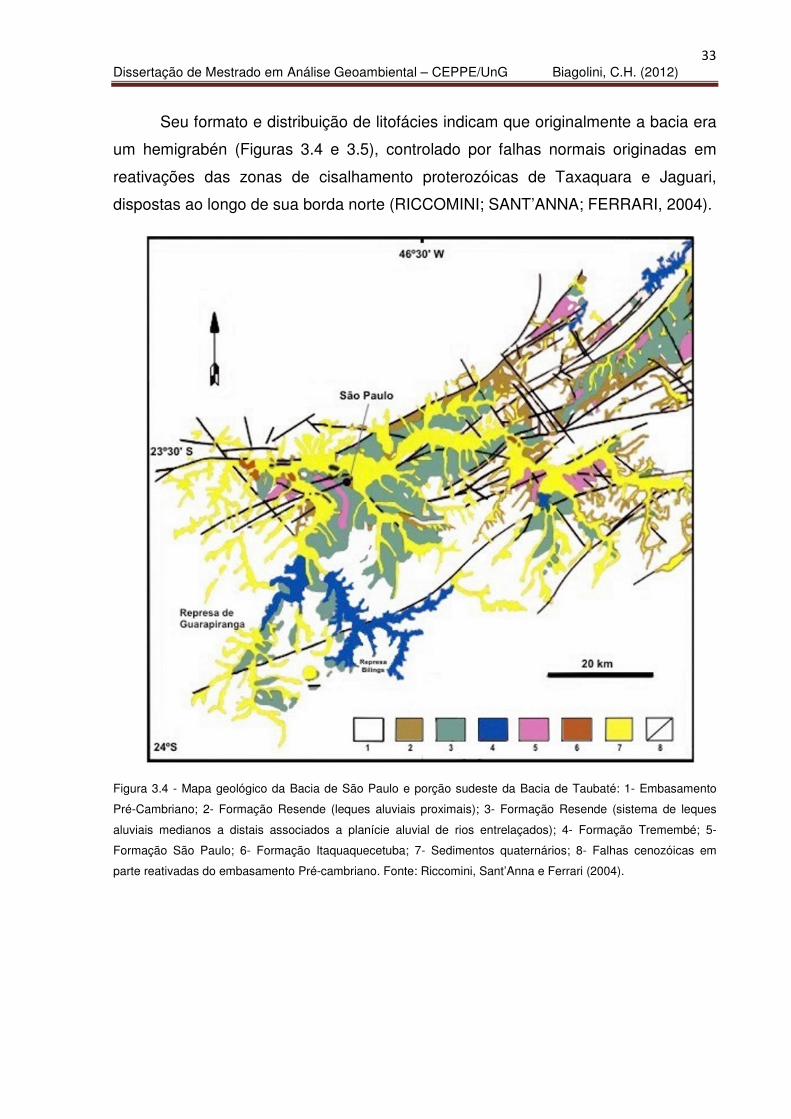
Seu formato e distribuição de litofácies indicam que originalmente a bacia era
um hemigrabén (Figuras 3.4 e 3.5), controlado por falhas normais originadas em
reativações das zonas de cisalhamento proterozóicas de Taxaquara e Jaguari,
dispostas ao longo de sua borda norte (RICCOMINI; SANT’ANNA; FERRARI, 2004).
Figura 3.4 - Mapa geológico da Bacia de São Paulo e porção sudeste da Bacia de Taubaté: 1- Embasamento
Pré-Cambriano; 2- Formação Resende (leques aluviais proximais); 3- Formação Resende (sistema de leques
aluviais medianos a distais associados a planície aluvial de rios entrelaçados); 4- Formação Tremembé; 5-
Formação São Paulo; 6- Formação Itaquaquecetuba; 7- Sedimentos quaternários; 8- Falhas cenozóicas em
parte reativadas do embasamento Pré-cambriano. Fonte: Riccomini, Sant’Anna e Ferrari (2004).
34
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)

35
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Os depósitos sedimentares da Bacia de São Paulo estão irregularmente
distribuídos em uma área de aproximadamente 1000 km2, com o eixo maior
localizado entre os municípios de Arujá e Embu-Guaçu, medindo aproximadamente
75 km, e com o eixo menor localizado entre o bairro de Santana, em São Paulo, e o
município de Santo André, medindo aproximadamente 25 km. O pacote sedimentar
possui espessura altamente variável, o que pode ser explicado pelas diversas
irregularidades do embasamento (HASUI; CARNEIRO, 1980). A maior espessura de
sedimentos foi encontrada no bairro da Mooca e atinge, conforme dados de
sondagens, 290 m (TAKIA, 1991).
O sistema de falhas denominadas Taxaquara-Jaguari é evidenciado na bacia
em sua borda norte retilínea, enquanto que sua borda sul apresenta-se marcada por
contatos irregulares com o embasamento pré-cambriano (TAKYIA, 1997;
RICCOMINI; SANT’ANNA; FERRARI, 2004).
Os sedimentos que compõem a Bacia de São Paulo tiveram como área-fonte
rochas pré-cambrianas provenientes das áreas circundantes (SUGUIO; COIMBRA;
CATTO, 1972) e foram depositados por processos em ambientes lacustres e de
leques aluviais, com rios associados. As unidades litoestratigráficas presentes são
as formações Resende, São Paulo e Tremembé, que constituem a seqüência basal
paleógena denominada Grupo Taubaté; essa unidade é recoberta de maneira
discordante pela Formação Itaquaquecetuba, de acordo com Riccomini, Sant’Anna e
Ferrari (2004).
Quanto aos recursos minerais explorados na Bacia de São Paulo destacam-
se as areias para construção civil e água subterrânea para abastecimento público e
industrial (BEDANI, 2008).
3.2.1 - Formação Itaquaquecetuba
A Formação Itaquaquecetuba teve seus estudos pioneiros realizados por
Junqueira (1969), e posteriormente seus sedimentos foram denominados de
“Aluviões Antigos dos Rios Tietê e Pinheiros” (SUGUIO; TAKAHASHI, 1970;
SUGUIO, 1971) e “Camadas Cruzadas da Fase Butantã” (AB’SABER, 1978, 1980).

36
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Posteriormente, Coimbra; Riccomini; Mello (1983) propuseram a designação
“Formação Itaquaquecetuba”.
Os sedimentos da Formação Itaquaquecetuba apresentam-se em cotas
inferiores a 710 m, preenchendo canais escavados diretamente nas rochas pré-
cambrianas do embasamento (ALMEIDA et al., 1984). Os sedimentos são de origem
clástica e de maneira geral constituem arenitos grossos arcoseanos, com grãos
angulosos a sub-angulosos, mal selecionados, indicativos de curto transporte e
proximidade da área-fonte. Zanão, Castro e Saad (2006 a, b), constataram, ainda, a
presença de diamictitos e lamitos/folhelhos na seção aflorante da mineração
Itaquareia I.
Assim como outras unidades que compõem a Bacia de São Paulo, os
sedimentos da Formação Itaquaquecetuba também tiveram como área-fonte as
rochas pré-cambrianas do embasamento cristalino.
Mineralogicamente, quartzos e feldspatos constituem o arcabouço das rochas
desta unidade, que também apresentam frações de minerais pesados representados
por: zircões, monazitas, turmalinas, sillimanitas, estaurolitas, granadas, cianitas,
espinélios, andaluzitas e hornblendas. A cimentação por marcassita é
freqüentemente constatada, sendo um indicativo de ambiente redutor (COIMBRA;
ATENCIO; BRANDT NETO, 1980; COIMBRA; RICCOMINI; MELO, 1983).
As estruturas sedimentares predominantes são estratificações cruzadas
tabulares e acanaladas, que podem ou não conter níveis argilosos, com coloração
castanha-escura e ricos em matéria orgânica. Níveis conglomeráticos também são
abundantes, sendo compostos majoritariamente por seixos de quartzo e quartzito.
Estudos de paleocorrentes nas estratificações cruzadas constatam mergulhos
preferenciais para NW e SW, indicativos, portanto, de paleocorrentes oriundas de
NE e SE, direções também constatadas nos estudos em seixos imbricados
(COIMBRA; RICCOMINI; MELO, 1983) (Figura 3.6).

37
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Figura 3.6 - Afloramento na Mineradora Itaquareia – Porto 1, Município de Itaquaquecetuba; (a) frente de
exploração; (b) estratificações cruzadas; e (c) níveis orgânicos (cores mais escuras). Fotos cedidas pela Prof.
M.J.Garcia.
De acordo com ALMEIDA (1984) e RICCOMINI (1989), a deposição dessa
unidade foi controlada por falhas de direções ENE e NNW, em cujas bordas
formaram-se cunhas clásticas, que contém entre outras litologias megaclastos ricos
em vegetais oriundos provavelmente da antiga Formação São Paulo. Nas porções
distais verificam-se lamitos arenosos de coloração esverdeada com seixos
arredondados de quartzo e quartzitos, bem como com intercalações de arenitos
maciços médios/finos. Associadas a essas cunhas clásticas, têm-se depósitos
fluviais entrelaçados, que predominam na maior parte dessa unidade (RICCOMINI;
SANT’ANNA; FERRARI, 2004).
Do ponto de vista de sistema deposicional, os estudos realizados por Zanão;
Castro e Saad, (2006a) e Santos (2008) conduziram a uma nova interpretação
estratigráfica para esta unidade. Os primeiros autores, por meio de levantamento de

38
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
seções estratigráficas, definiram nove litofácies distintas e oito ciclos sedimentares.
As litofácies verificadas foram as seguintes: conglomerado com estratificação
cruzada acanalada; arenito grosso/conglomerático com estratificação cruzada
acanalada; arenito fino/grosso com estratificação cruzada acanalada; arenito
fino/médio com laminação horizontal; arenito moderadamente argiloso; ritmito
arenito-siltito carbonoso; lamito mal selecionado, com grãos e seixos; e lamito síltico
bem-selecionado. Os oito ciclos sedimentares reconhecidos caracterizam-se por
apresentarem fining-upward, com a base de cada ciclo sendo constituída por
arenitos conglomeráticos/grossos e o topo por lamitos (Figura 3.7).
Zanão, Castro e Saad (2006a) propuseram para essa área um sistema
deposicional de leques aluviais, onde reconheceram fácies fluviais meandrantes
associadas a canais do tipo ribbon. Este é um tipo de canal fixo e estreito com razão
largura/profundidade menor que 15. De acordo com os autores, um canal do tipo
ribbon ocorre quando há soerguimento da área fonte e normalmente encontram-se
associados a leques aluviais.
Segundo Santos (2009), na Formação Itaquaquecetuba, encontram-se
preservados macrofitofósseis (lenhos, folhas e frutos) e microfitofósseis
(palinomorfos). Tolentino (1965) noticiou a presença de um carvão fóssil piritizado,
nos sedimentos da bacia de São Paulo, próximo à Cidade Universitária. Tratava-se
de fragmentos de troncos vegetais carbonizados no estágio de linhito xilóide
dispersos dentro da areia a cerca de 10 m de profundidade. Os primeiros estudos
paleontológicos da formação foram desenvolvidos por Suguio e Takahashi (1970) e
Suguio (1971), com lenhos fósseis, linhitizados com substituição parcial por
marcassita, provenientes dos depósitos da raia olímpica da Cidade Universitária, às
margens do rio Pinheiros, em São Paulo. Foram identificados gêneros da família
Fabaceae (Myrocarpus sp., Centrolobium sp. e Piptadenia sp.), das Elaeocarpaceae
(Sloania sp.) e das Melastomataceae (Miconia sp.), que sugerem clima tropical
pluvial. As datações pelo método do C14 mostraram idades superiores a 40.000
anos (A.P.), portanto superiores à precisão do método.

39
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Fig
ura:
3.7
: Seç
ão d
e co
rrel
ação
de
perf
is fa
ciol
ógic
os d
a F
orm
ação
Itaq
uaqu
ecet
uba.
Mod
ifica
do d
e Z
anão
; Cas
tro
; Saa
d (2
006)
.

40
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Suguio e Mussa (1978) estudaram lenhos fósseis silicificados, procedentes da
mineradora Itaquareia 1, no município de Itaquaquecetuba e descreveram
Astronioxylon mainieri (Anacardiaceae), Piptadenioxylon chimeloi (Fabaceae-
Mimesoídeae), Myrocarpoxylon sanpaulense (Fabaceae-Faboídeae), Matayboxylon
tietense (Sapindaceae) e Qualeoxylon itaquaquecetubense (Vochysiaceae). Todos
esses gêneros são viventes das florestas úmidas da América tropical. Fittipaldi e
Simões (1989a,1990), Fittipaldi e Simões (1989b), Fittipaldi (1990), Fittipaldi (1993)
estudaram macrofósseis de folhas e frutos, provenientes também da mineradora
Itaquareia 1. Desses estudos foram descritos os seguintes táxons de folhas:
Bryophyta (Isotachis simonesi), Filicopsida (Lindsaea pradoi), Alismataceae
(Echinodorus rossiae), Aquifoliaceae (Aquifoliphyllum ilicioides), Araceae (Monstera
marginata) Fabaceae-Mimosoídeae (Piptadenia terciaria), Fabaceae-
Caesalpinóideae (Cassia rosleri, Schizolobium inaequilaterum), Fabaceae-
Faboídeae (Machaerium piranii), Flacourtiaceae (Casearia serrata), Lauraceae
(Ocotea puchelliformis), Malpighiaceae (Byrsonima bullata), Melastomataceae
(Bertolonia coimbrae), Myrtaceae (Myrcia cf. rostrataformis, Psidium paulense)
Rhamnaceae (Rhamniphyllum caseariformis), Rubiaceae (Tocoyena riccominii),
Rutaceae (Zanthoxyllum gladuliferum), Sapindaceae (Serjania lancifolia, Serjania
itaquaquecetubensis), Tiliaceae (Luehea divaricatiformis), e frutos de Fabaceae
(Sophora giuliettiae).
Na base da seção-tipo da mineradora Itaquareia 1, a palinoflora foi estudada
por Melo et al (1985), que sugeriram a idade neoeocena; Lima et al (1991) também
analisaram amostras dessa seção-tipo, além de outras provenientes do município de
Barueri. Esses autores atribuíram às seções estudadas idade oligocena, justificando
que a presença de certos palinomorfos deve-se ao retrabalhamento de unidades
mais antigas. Posteriormente, Yamamoto (1995) ao estudar amostras da Formação
Itaquaquecetuba provenientes da mesma mineradora, atribuiu a essa unidade idade
oligocena e eo-miocena. Santos (2005) apresentou o resultado da análise de
diversas amostras coletadas na seção-tipo dessa formação: para as amostras da
base, determinou idade neoeocena-oligocena; para as amostras do meio e do topo,
atribuiu idade eomiocena, com retrabalhamento de palinomorfos do Neoeoceno-
Oligoceno.

41
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Santos (2009) ao estudar, do ponto de vista palinológico, as seções
estratigráficas levantadas por Zanão, Castro e Saad (2006), constatou que a maior
parte da seção apresenta idade neoeocena, enquanto o terço final é de idade eo-
oligocena. Cabe citar que as formas mais importantes do ponto de vista
bioestratigráfico são as seguintes: para o intervalo neoeoceno destacam-+se
Perisyncolporites pokornii (Malpighiaceae), Margocolporites vanwijhei
(Fabaceae- Caesalpinoideae), Perfotricolpites digitatus (Convolvulaceae),
Cicatricosisporites dorogensis (Anemiaceae-Schizaeaceae), Echitriletes muelleri
(Selaginella-Selaginellaceae), Spinizonocolpites echinatus (Arecaceae)
Ulmoideipites krempii (Ulmaceae), Catinipollis geiseltalensis (Martyniaceae),
Echiperiporites akanthos (Malvaceae), Striatipollis catatumbus (Fabaceae),
Quadraplanus sp., Polyadopollenites sp. e Acaciapollenites sp. (Mimosaceae); para
o intervalo oligoceno destaca-se o aumento nas quantidades de grãos de
gimnospermas como Podocarpidites (Podocarpus-Podocarpaceae) e Dacrydiumites
florinii (Dacrydium-Podocarpaceae), além da redução na diversidade e quantidade
de angiospermas, com destaque para Proteacidites dehanii (Proteaceae),
Magnaperiporites spinosus (Malvaceae), Tricolpites reticulatus (Gunnera),
Psilatricolpites operculatus (Euphorbiaceae).
Com base no conteúdo palinológico e nas relações estratigráficas, as idades
admitidas para a Formação Itaquaquecetuba podem ser visualizadas na Figura 3.8.

42
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Figura 3.8 - Idades propostas para a Formação Itaquaquecetuba. Fonte: Santos, 2009.

43
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
4.1 – Material
O material analisado foi coletado em seção aflorante da Formação
Itaquaquecetuba, hoje completamente destruída pela exploração de areia na área da
Mineradora Itaquareia, Porto I, no município de Itaquaquecetuba, SP. Um perfil
colunar foi levantado na época da coleta, e encontra-se reproduzido na figura 4.1.
A assembléia tafoflorística é constituída de folhas em disposição caótica de
distribuição esparsa. Muitas folhas sofreram fragmentação durante o transporte até o
sítio de deposição (causas bioestratinômicas), enquanto outros, por serem
extremamente frágeis, na matriz intemperizada, fragmentaram-se durante a coleta,
transporte do campo para o laboratório e na manipulação durante a fase de estudos.
Apesar da fragmentação pela qual algumas folhas passaram, preservam
ainda feições morfológicas como margem, forma da base e/ou ápice embora muitas
vezes com prejuízo para a avaliação da venação secundária e de ordem superior.
CAPÍTULO 4 – MATERIAL E MÉTODOS

44
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Figura 4.1. Perfil do afloramento Itaquareia I, com indicação do nível de coleta dos fitofósseis.
(Fonte: Comunicação Pessoal)

45
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
4.2 – Métodos
O estudo paleobotânico envolve métodos de campo, de laboratório e de
gabinete.
4.2.1 – Métodos de Campo
Os métodos de campo envolveram levantamento de seção colunar do
afloramento fossilífero, conforme já referido anteriormente e registrado nas figuras
4.1 e 4.2. Também aspectos tafonômicos foram aí analisados.
Quanto a esses aspectos, observou-se que o material está fossilizado na
forma de impressão e compressão com cutícula preservada. Há folhas praticamente
inteiras e com todas as suas partes bem preservadas e outras apenas na forma de
fragmentos. A presença de camadas superficiais ferruginosas, em alguns casos
ressalta ou camufla determinadas estruturas das folhas como, por exemplo, as
venações secundária e terciária. Alguns materiais fitofossilíferos aparecem também
piritizados.
Figura 4.2: Afloramento Itaquareia I com indicação do local de coleta do material de estudo. (Foto: Biagolini/2010).
Local da coleta dos fitofósseis no
interior da empresa Itaquareia I em
Itaquaquecetuba

46
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
4.2.2 – Método de Laboratório
As atividades laboratoriais foram desenvolvidas nos Laboratórios de
Geociências (Palinologia e Paleobotânica) da Universidade Guarulhos – UnG, e
consistiram em: preparação mecânica das amostras, preparação química para
clarificação de folhas de espécies atuais, codificação dos exemplares preparados,
documentação gráfica dos espécimes analisados, análise da arquitetura foliar,
identificação taxonômica após mensuração e descrição dos espécimes e integração
de dados obtidos.
4.2.2.1 – Preparação Mecânica e Codificação das Amostras
Através do uso de ferramentas (martelos, talhadeiras, agulhas, estiletes e
outros), a matriz fossilífera foi desbastada para livrar os fósseis de coberturas de
sedimentos.
O material coligido foi incorporado à coleção de estudo do Laboratório de
Palinologia e Paleobotânica da Universidade Guarulhos – UnG, onde recebeu
numeração preliminar de ITQpb 001/11 a ITQpb 111/11 atribuída durante a
preparação mecânica e análise do material para desenvolvimento desta dissertação.
As siglas ITQ referem-se a material proveniente de Itaquaquecetuba, e pb refere-se
a material paleobotânico ou fitofossilífero. Após o término do estudo, receberão a
numeração definitiva para publicação e inclusão na Coleção Científica dos Tipos do
Laboratório de Geociências da UnG.
4.2.2.2 – Preparação Química
Espécimes foliares coletados em herbários ou in natura foram clarificados
através de processo químico, segundo método de Foster, (1952) descrito por Dutra
e Stranz, (2002), para servir de comparação no exercício de identificação
taxonômica dos fitofósseis foliares analisados.
4.2.2.3 – Documentação Gráfica
A documentação gráfica dos espécimes estudados foi obtida por:

47
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
a) Fotos em magnitude natural (fotomacrografia) e ampliadas por
estereomicroscópio (fotomicrografia)
b) Desenhos em câmara clara ou sobre as fotos ampliadas, utilizando mesa de
luz.
Os espécimes foram fotografados utilizando iluminação natural ou artificial.
As fotomacrografias, foram obtidas com câmera Fuji modelo Finepix modelo S-1000.
As fotomicrografias foram obtidas através do fotoestereomicroscópio Zeiss modelo
Stemi CV6 (Laboratório de Palinologia e Paleobotânica da UnG).
Os fitofósseis foram desenhados sob iluminação por fibra ótica (oblíqua ou
rasante), no mesmo estereomicroscópio Zeiss, acoplado à câmara clara para se
obter o detalhamento dos padrões de venação de ordens superiores à secundária e
outros pormenores morfológicos.
4.2.2.4. – Análise da Arquitetura Foliar.
Nesta análise foram utilizados os equipamentos disponíveis no Laboratório de
Palinologia e Paleobotânica da UnG, tais como lupas, estereomicroscópios e
comparação com guia de identificação de arquitetura foliar (LAWG, 1999).
Foram realizadas análises da arquitetura foliar com mensuração e descrição
das impressões estudadas, a fim de permitir uma posterior interpretação
paleoclimática e paleoambiental da vegetação representada por esta assembléia
fitofossilífera.
O termo arquitetura foliar, usado por Hickey (1973), refere-se a localização e
forma dos elementos constituintes da expressão externa da estrutura foliar, incluindo
padrão de venação, configuração marginal, forma foliar e posição de glândulas.
Neste sentido, a arquitetura é o aspecto da morfologia que se aplica à configuração
espacial e coordenação dos elementos que fazem parte de uma planta sem
considerar a histologia, função, origem ou homologia (HICKEY e WOLFE, 1975).

48
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
A arquitetura foliar dos espécimes foi analisada conforme o manual de LAWG
(1999), considerando-se as suas dimensões lineares e angulares, obtidas com o
paquímetro digital marca Mitutoyo e com transferidor sobre o próprio fóssil ou sobre
seu desenho em escala, obtido com o uso da câmara clara e observação através de
estereomicroscópio.
Os aspectos morfográficos relativos à forma do contorno foliar, forma do ápice
e da base e ainda os relacionamentos ao tipo de margem foliar, que refletem com
maior evidência as adaptações das plantas fósseis ao meio ambiente, foram os
primeiros a serem analisados.
Quanto à forma ou contorno laminar, as impressões foliares foram avaliadas
segundo a posição do eixo ou zona de maior largura da folha, perpendicularmente
ao eixo de maior comprimento, conforme LAWG (1999) (Figura 4.3).
Forma Elíptica: A parte mais larga da folha está
sobre uma reta perpendicular, situada no
terceiro quinto do eixo longo da folha
Forma Ovada: A parte mais
larga da folha está sobre uma reta perpendicular situada nos dois quintos
basais do eixo longo da folha
Forma Ovada: A parte mais
larga da folha está sobre uma reta perpendicular, situada nos dois quintos basais do eixo longo da
folha.
Forma Oblonga: A parte
mais larga da folha corresponde ao terço médio
do eixo longo, onde as margens opostas tornam-se aproximadamente, paralelas.
Folha Lobada: Forma de folha com lobação: a folha apresenta aberturas com
venações ramificadas para cada divisão da folha
Figura 4.3: Classificação de folhas conforme ‘forma laminar (LAWG, 1999).

49
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Quanto à simetria, as lâminas são também avaliadas seguindo os parâmetros
da figura 4.4:
Simétrica: A lamina é
praticamente idêntica em ambos os lados, considerando venação
central como referência.
Assimétrica: Os lados da folha, considerando a venação central como divisão, são diferentes.
Base Assimétrica: Embora os lados sejam semelhantes,
somente na área da base os lados são diferentes.
Figura 4.4: Classificação da lâmina foliar quanto a simetria (LAWG, 1999).
Quanto ao Ângulo da Base:
A avaliação de lâminas foliares pode também considerar a base como
parâmetro de condições ambientais em que viveu o vegetal (figura 6.4).
Ângulo agudo: menor que
90o
Ângulo obtuso: Maior que 90o
Caso especial de ângulo obtuso, onde base apresenta
ângulo superior a 180o
Figura 4.5: Forma de ângulos na base de folhas (LAWG, 1999).

50
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Quanto ao Ângulo do Ápice:
O ângulo do ápice das folhas é avaliado conforme o ângulo formado entre as
laterais da folha (Figura 4.6), considerando o ápice como referência principal.
Obtuso-ângulo do ápice
90o-180o
Grande obtuso ângulo de ápice>180o
Ápice lobado obtuso
Ápice lobado agudo
Agudo – ângulo do
ápice < 90o
Figura 4.6 : Avaliação de ângulo dos ápices (LAWG, 1999).
Quanto a Forma da Base da Folha:
A base da folha é avaliada de acordo com a terminação entre lâmina e
pecíolo. A forma como a lâmina foliar se conecta com pecíolo dá à folha a
classificação quanto à base (Figura 4.7).
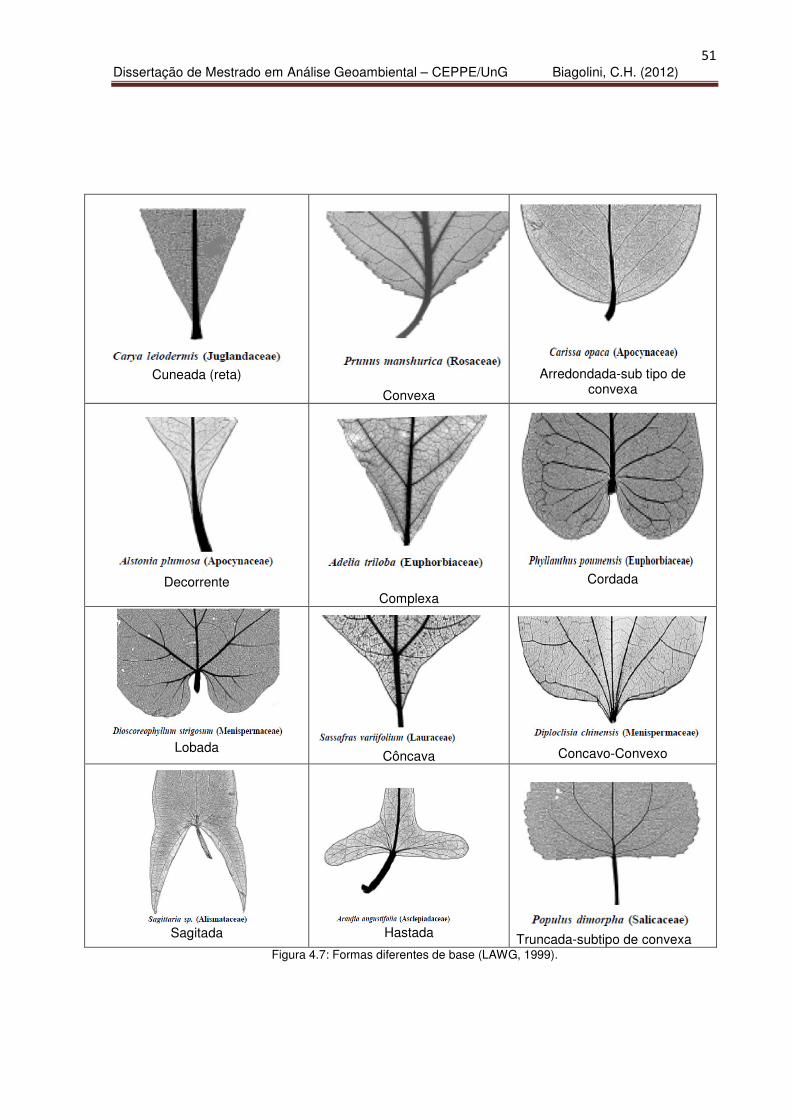
51
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Cuneada (reta)
Convexa
Arredondada-sub tipo de
convexa
Decorrente
Complexa
Cordada
Lobada
Côncava
Concavo-Convexo
Sagitada
Hastada
Truncada-subtipo de convexa Figura 4.7: Formas diferentes de base (LAWG, 1999).

52
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Quanto à Posição de Conexão do Pecíolo:
As folhas apresentam variações quanto à conexão do pecíolo com a lâmina
(Figura 4.8). Estas variações ocorrem em função do ambiente de vida dos vegetais.
Marginal Peltada central
Peltada excêntrica
Figura 4.8: Formas de conexão do pecíolo á lâmina (LAWG, 1999).
Quanto à Forma do Ápice:
Estas condições são aplicáveis aos 25% apicais das lâminas foliares (Figuras e 4.9 e 4.10).
Reto
Convexo
Arredondado
Truncado
Acuminado
Complexo
Figura 4.9: Formas diferentes de ápices foliares (LAWG, 1999).

53
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Figura 4.10: Diferentes formas de ápices foliares (LAWG, 1999)
Quanto ao Tipo de Margem:
As margens de uma folha podem apresentar características diferentes,
variando de lisa a serrilhada ou crenada. Esta característica das folhas, podem
indicar paleoambientes aos quais pertencia o vegetal e por esta razão precisam ser
consideradas.
Segundo Gonçalves e Lorenzi (2007), (Figura 4.11) as margens são
classificadas como: Inteira, Repanda, Crenada, Dentada, Serrada, Erosa, Crespa,
Sinuada e variam em função de adaptações ocorridas ao longo do tempo.
Se La > 0La > 0La > 0La > 0, então a
folha será
considerada retusa,
ou emarginada ou
lobada
Retusa < 5%
Emarginado 5 – 25%
Lobado >25%

54
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Inteira Repanda Crenada Dentada Serrada Erosa Crespa Sinuada
Figura 4.11: Formas de margens de lâminas conforme (GONÇALVES; LORENZI, 2007)
O catálogo Leaf Architecture Working Group traz como definição para folhas
não lisas o seguinte critério: “Os dentes apresentados em alguns tipos de folhas são
projeções marginais”. As projeções podem ser: dentada, serrada ou crenada. As
projeções observadas nas folhas analisadas, foram consideradas como dente.
O conceito de margem dentada, segundo Gonçalves e Lorenzi (2011), é
usado quando a margem é dividida em pequenos lobos agudos e direcionados
perpendicularmente a uma tangente traçada ao longo da margem e não deve ser
confundida com margem serrada ou serreada, onde os dentes estão voltados para o
ápice (Figura 4.12).
Dentada
Serrada
Crenada
Inteiro
Revoluta
Erose (Margem irregular)
Figura 4.12: Formas ou tipos de margens encontradas em lâminas diversas. (LAWG, 1999).

55
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Quanto a Lobação:
Serão consideradas folhas lobadas quando esta ou qualquer outra estrutura
laminar encontrar-se parcialmente dividida em um número determinado de
segmentos, que podem ser iguais entre si ou não, e o lobo, é considerado toda e
qualquer porção ou segmento de um órgão foliáceo (folha, sépala, pétala, etc) com
uma incisão qualquer a partir das margens (GONÇALVES; LORENZI, 2011).
Algumas folhas podem apresentar formas diferenciadas com aberturas nas
bordas também chamadas de lobos. Estas aberturas estão relacionadas a
adaptações ao clima do local de origem da planta. Para LAWG (1999), lobos são
projeções marginais que podem chegar a ¼ ou mais de distância da veia mediana
medida paralelamente em relação ao eixo de simetria do lobo. Na figura 4.13, são
apresentadas diferentes formas de lobação.
Não lobada
Bilobada
Lobada (Palmeada)
Lobada pinada Figura 4.13: Diferentes formas de lobação em folhas (LAWG, 1999).
Cálculo de Área da Superfície Foliar:
Entre os diversos dados obtidos por intermédio da análise das folhas
fossilizadas em argilitos da Formação Itaquaquecetuba, o cálculo de área se
apresenta conforme métodos matemáticos de LAWG (1999), segundo os quais a
superfície foliar corresponde a 2/3 da área retangular obtida pela multiplicação do
comprimento foliar pela largura. As lâminas foliares foram classificadas segundo
suas áreas, conforme a tabela 4.1.
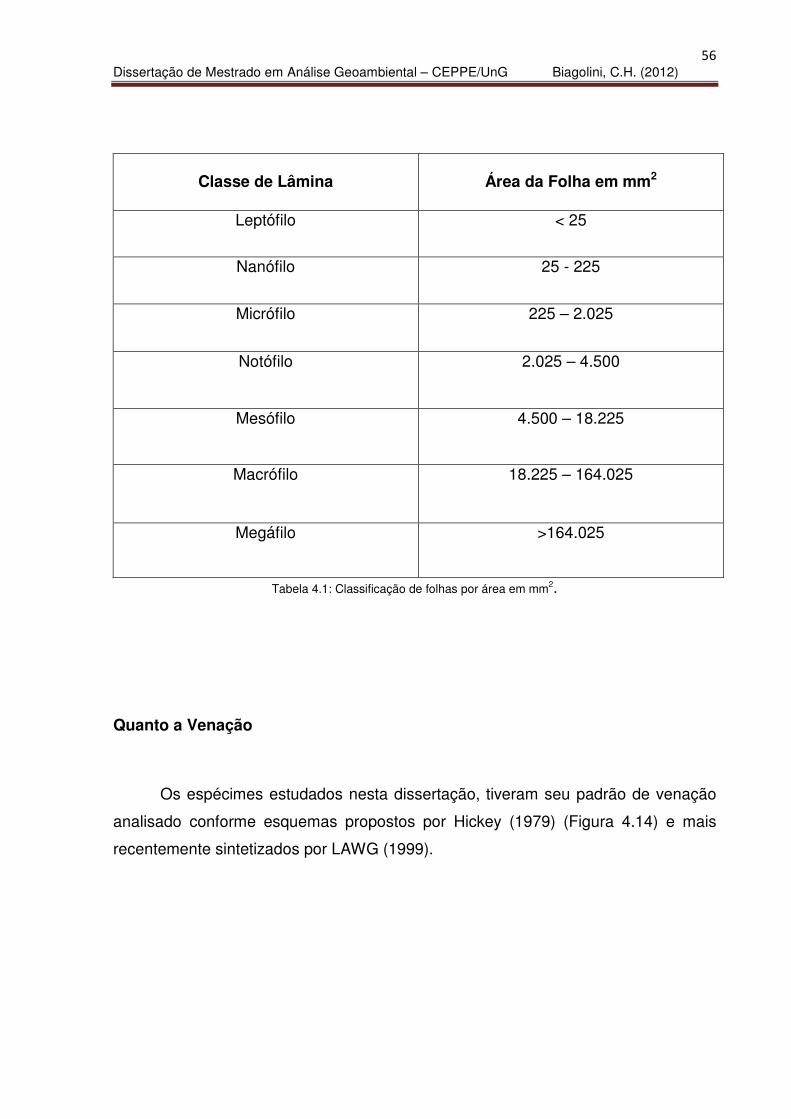
56
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Classe de Lâmina
Área da Folha em mm2
Leptófilo
< 25
Nanófilo
25 - 225
Micrófilo
225 – 2.025
Notófilo
2.025 – 4.500
Mesófilo
4.500 – 18.225
Macrófilo
18.225 – 164.025
Megáfilo
>164.025
Tabela 4.1: Classificação de folhas por área em mm2.
Quanto a Venação
Os espécimes estudados nesta dissertação, tiveram seu padrão de venação
analisado conforme esquemas propostos por Hickey (1979) (Figura 4.14) e mais
recentemente sintetizados por LAWG (1999).

57
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Figura 4.14: Tipos de venações foliares primárias e secundárias extraído de Hickey (1979),
Nestes esquemas, a venação primária pode ser classificada em pinada,
actinódroma, palinactinódroma, acródroma basal ou suprabasal e campilódroma.

58
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Entende-se que:
a) Venação pinada corresponde a uma única veia primária, normalmente
centralizada na lâmina.
b) Venação actinódroma é encontrada quando três ou mais veias primárias
divergem radialmente a partir de um único ponto.
c) Venação palinactinódroma é divergente com uma série de ramificações
dicotômicas, tanto proximamente ou distantemente espaçadas a distâncias
variáveis.
d) Venação acródroma exibe três ou mais veias primárias que correm em arcos
convergentes rumo ao ápice foliar.
e) Venação campilódroma apresenta várias veias primárias e os seus ramos
originam-se próximos ou em um único ponto e correm em arcos fortemente
recurvados que convergem apicalmente.
No material proveniente da Formação Itaquaquecetuba, estudado nesta
dissertação, registram-se normalmente formas com venação primária pinada. O
estado de conservação e a fragilidade do material, não apresentam na grande
maioria das amostras condições de observação da venação secundária.
A venação secundária, por sua vez, pode ser classificada em broquidódroma,
broquidódroma festonada, eucamptódroma, cladódroma, reticulódroma,
craspedódroma, semicraspedódroma, semicraspedódroma-festonada, acródroma
basal e suprabasal, veia intramarginal e veia intersecundária. Dentre estas
venações, os padrões mais comuns no material estudado são os seguintes:
a) Venação broquidódroma com veias secundárias que se juntam em uma
série de arcos proeminentes:
b) Venação broquidódroma festonada com um ou mais conjuntos de loops
adicionais fora do loop broquidódromo principal.

59
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
c) Venação cladódroma (=marginal) que apresenta veias secundárias
ramificando-se livremente rumo às margens
d) Venação craspedódroma que exibe veias secundárias e terminam nas
margens, normalmente em folhas dentadas.
e) Venação semicraspedódroma com veias secundárias que se ramificam
quase na margem e um dos ramos termina na margem e o outro se junta à
secundária suprajacente.
f) Venação eucamptódroma com veias secundárias que se curvam para cima
e gradualmente diminuem apicalmente no interior da margem, conectadas às
secundárias subrajacentes por uma série de veias terciárias transversais sem
formar quaisquer loops (alças) secundários marginais.
4.2.2.5. – Identificação Taxonômica
Mensuração e descrição dos espécimes
Tendo em vista a descrição e identificação taxonômica dos componentes da
assembléia fitofossilífera, alguns de seus espécimes tiveram suas medidas lineares
mensuradas com paquímetro digital marca Mitutoyo e suas medidas angulares
obtidas com goniômetro.
As identificações taxonômicas das folhas de angiospermas são feitas por
meio da chave de arquitetura foliar (HICKEY, 1973; HICKEY; WOLFE, 1975;
HICKEY, 1979). Esta chave é baseada na localização e forma dos elementos
constituintes da expressão externa foliar, isto é, o padrão de venação, a
configuração marginal, a forma foliar e a posição de glândulas, conforme suas
ordens e famílias.
Ao se identificar a ordem ou família, passou-se à comparação com táxons
menores atuais registrados na literatura botânica produzidas por Lorenzi, (2000;
2002); Souza e Lorenzi, (2008); Gonçalves e Lorenzi, (2007); Simpson, (2006);
Judd (2002) e Judd, et al., (1999) além de outras publicações ou comparações com
material clarificado do Laminário do Laboratório de Geociências da UnG e,
finalmente, com pesquisas de imagens atuais disponíveis em sites acadêmicos.
Quanto à comparação com material fóssil, foram utilizadas fichas de espécies já
descritas na literatura paleobotânica.

60
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
4.2.3. Métodos de Gabinete
Subtendem-se como métodos de gabinete os levantamentos bibliográficos, a
elaboração de mapas e perfis geológicos, bem como a integração de dados obtidos
a partir das observações de campo e de laboratório.
4.2.3.1. Levantamento bibliográfico
As literaturas geológica, botânica e paleobotânica foram consultadas nas
bibliotecas da Universidade Guarulhos, da Universidade de São Paulo e na internet.
Essas pesquisas prosseguiram-se durante todo estudo e envolveram a geologia da
bacia de São Paulo e mais especificamente a Formação Itaquaquecetuba, as
identificações taxonômicas, paleoecológicas, paleoclimáticas e paleofitogeográficas.
Também envolveram a busca na literatura das características diagnósticas de
espécies fósseis paleógenas, já descritas para o Estado de São Paulo e em todo
Brasil, bem como de formas atuais, quando apresentaram semelhanças com as
formas aqui estudadas.
4.2.3.2. Elaboração de mapas
Mapas paleogeográficos foram buscados na internet e mapas de localização
atual foram extraídos com modificações de diferentes fontes.
4.2.3.3. Integração de Dados
Todos os dados obtidos em campo e em laboratório com o material estudado,
foram levados em consideração na interpretação da paleoecologia,
paleofitogeografia, paleoclimatologia, nas correlações estratigráficas e filogenéticas
da paleoflora de Itaquaquecetuba.

61
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
5.1 - Taxonomia
A classificação dos organismos vivos obedece a critérios pré-estabelecidos
onde diversas características são consideradas na determinação de espécie, reino,
família, etc. Assim a nomenclatura científica, tanto dos organismos viventes como
dos elementos do passado muitas vezes, encontrados apenas na forma de fósseis,
obedece às normas de códigos internacionais de nomenclatura. Esses códigos têm
por objetivo a uniformização e padronização da linguagem científica na Zoologia, na
Botânica, na Paleozoologia e na Paleobotânica (RIOS-NETTO, 2010).
A identificação taxonômica é o ato de posicionar um indivíduo em um grupo
de organismos predefinido, com base em similaridade morfológica. Por sua vez o
estudo dos agrupamentos e da diversidade dos organismos e de todas e quaisquer
relações entre eles incluindo sua classificação e aspectos evolutivos, é chamado de
Sistemática. Taxonomia é o conjunto de princípios, procedimentos e regras que
embasam a Classificação e a Sistemática (BLOW, 1979). Em certo sentido, a
Taxonomia inclui a Classificação, a Sistemática e a Nomenclatura (RIOS-NETTO,
2010).
Até um passado recente, as classificações dos seres vivos estabeleciam os
agrupamentos dos organismos baseados em semelhanças, suas inter-relações e
estruturas morfológicas encontradas nesses organismos em comum. Nos últimos
anos têm sido tentadas classificações baseadas na semelhança entre genomas,
com grandes avanços em algumas áreas, especialmente quando se juntam a essas
informações aquelas oriundas dos outros campos da Biologia.
Assim, os principais fatores considerados na classificação sistemática
deveriam ser as semelhanças e diferenças entre os seres vivos, as quais seriam
interpretadas como reflexos das relações de parentesco entre os organismos. Uma
espécie paleontológica agruparia desta maneira, organismos com estreita afinidade
CAPÍTULO 5 – SISTEMÁTICA

62
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
morfológica, fisiológica, filogenética e ecológica, com distribuição estratigráfica
contínua (RIOS-NETTO, 2010).
As informações de caráter taxonômico permitem buscar em vasta literatura já
existente, aspectos morfológicos que indicarão a forma e ambiente de vida de um
organismo. A morfologia vegetal é o ramo da botânica que estuda as formas e
estruturas das plantas, sendo de grande importância em áreas tão diversas como a
sistemática, fisiologia ou ecologia (GONÇALVES e LORENZI, 2011).
As características morfológicas são importantes na identificação de um
vegetal, seja ele atual ou em forma fossilizada. Qualquer característica das plantas
pode ser analisada, desde aquelas relacionadas à morfologia externa, tais como
inserção, formato de folhas, coloração das flores, número de sementes, até à
fitoquímica e composição das macromoléculas tais como DNA ou RNA. (SOUZA e
LORENZI, 2008). As análises químicas em fitofósseis tornam-se, na maioria dos
casos, inviáveis devido aos processos diagenéticos, restando então como
possibilidade de identificação, a análise visual. Observam-se, assim, as
características externas que um vegetal fossilizado tem, principalmente, no que se
refere ao formato da folha e detalhes que podem apresentar como a borda, ápice,
tipo de base, tipo de venação etc. As impressões das feições morfológicas deixadas
na rocha bem como a anatomia preservada nas petrificações ajudam na
identificação se não for da espécie, pelo menos da família ou de alguma categoria
taxonômica maior.
Segundo Souza e Lorenzi (2008): “A Botânica Sistemática é o ramo da
ciência que estuda a diversidade das plantas através da sua organização em
grupos, com base em suas relações evolutivas”. Grandes e rápidos avanços,
especialmente quanto às relações evolutivas entre os táxons vegetais, têm levado à
revisão profunda na sistemática botânica, que se tornou de difícil
acompanhamento.A classificação das filicófitas adotada é aquela apresentada por
Taylor et al. (2009), conforme reproduzida na figura a seguir (Figura 5.1).

63
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Figura 5.1: Taxa superiores de Filicophyta, conforme Taylor et al. (2009)
FILICOPHYTA Cladoxylopsida
Pseudosporochnales (Devonian–Mississippian)
Iridopteridales (Middle–Upper Devonian)
Early Fernlike Plants
Rhacophytales (Middle–Upper Devonian)
[Coenopterids (Devonian–Permian)]
Stauropteridales (Devonian–Pennsylvanian)
Stauropteridaceae
Zygopteridales (Lower Mississippian–Permian)
Zygopteridaceae
True Ferns—Eusporangiates
Marattiales (Carboniferous–recent)
Psaroniaceae
Marattiaceae
Ophioglossales (Jurassic–recent)
Ophioglossaceae
True Ferns—Leptosporangiates
Osmundales
Osmundaceae (Permian–recent)
Guaireaceae (Permian)
Filicales (Carboniferous–recent)
Botryopteridaceae (Carboniferous–Permian)
Anachoropteridaceae (Pennsylvanian–Permian)
Kaplanopteridaceae (Pennsylvanian)
Psalixochlaenaceae (Carboniferous)
Sermayaceae (Pennsylvanian)
Tedeleaceae (Mississippian–Permian)
Skaaripteridaceae (Permian)
Tempskyaceae (Cretaceous)
Schizaeaceae (Jurassic–recent)
Hymenophyllaceae (Triassic–recent)
Gleicheniaceae (Permian–recent)
Dicksoniaceae (Triassic–recent)
Cyatheaceae (Jurassic–recent)
Matoniaceae (Triassic–recent)
Loxsomataceae (Cretaceous–recent)
Dipteridaceae (Triassic–recent)
Polypodiales (Jurassic–recent)
Dennstaediaceae,
Pteridaceae,
Onocleaceae,
Blechnaceae, Polypodiaceae
Salviniales (Cretaceous–recent)
Marsileaceae
Salviniaceae

64
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Ao se levar em consideração esses fatos relacionados à classificação, quanto as
angiospermas, pretende-se adotar na estrutura deste capítulo, o quadro apresentado
pelo APGIII (2009) (Figura 5.2), à medida que os fitofósseis identificados forem
sendo descritos.
Figura 5.2- Relações filogenéticas entre as ordens e algumas famílias de angiospermas de acordo com APGIII
(2009).

65
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
As regras de nomenclatura botânica, relacionadas à hierarquia dos táxons,
não têm sido seguidas na filogenia recente. Acima de gênero, apenas as categorias
taxonômicas de “família” e “ordem” vêm sendo empregadas no sentido mais
tradicional. As categorias taxonômicas acima de ordem estão sendo ignoradas.
Portanto, termos como Rosídeas, Eudicotiledôneas ou Magnoliídeas não devem ser
referidos a determinada posição taxonômica como “classe” ou “subclasse” etc., mas,
informalmente, como grados, clados ou subclados até que uma filogenia mais
acurada seja definida.
As diversas pesquisas, já realizadas na Formação Itaquaquecetuba,
indicaram presença de variadas espécies de briófitas, pteridófitas, gimnospermas e
angiospermas, no entanto, os resultados encontrados, na elaboração desta
dissertação de mestrado, apontam para uma significativa maioria de angiospermas,
embora isto não se reflita neste documento que, por uma questão de urgência na
conclusão, deu ênfase às formas de mais fácil identificação, no momento.
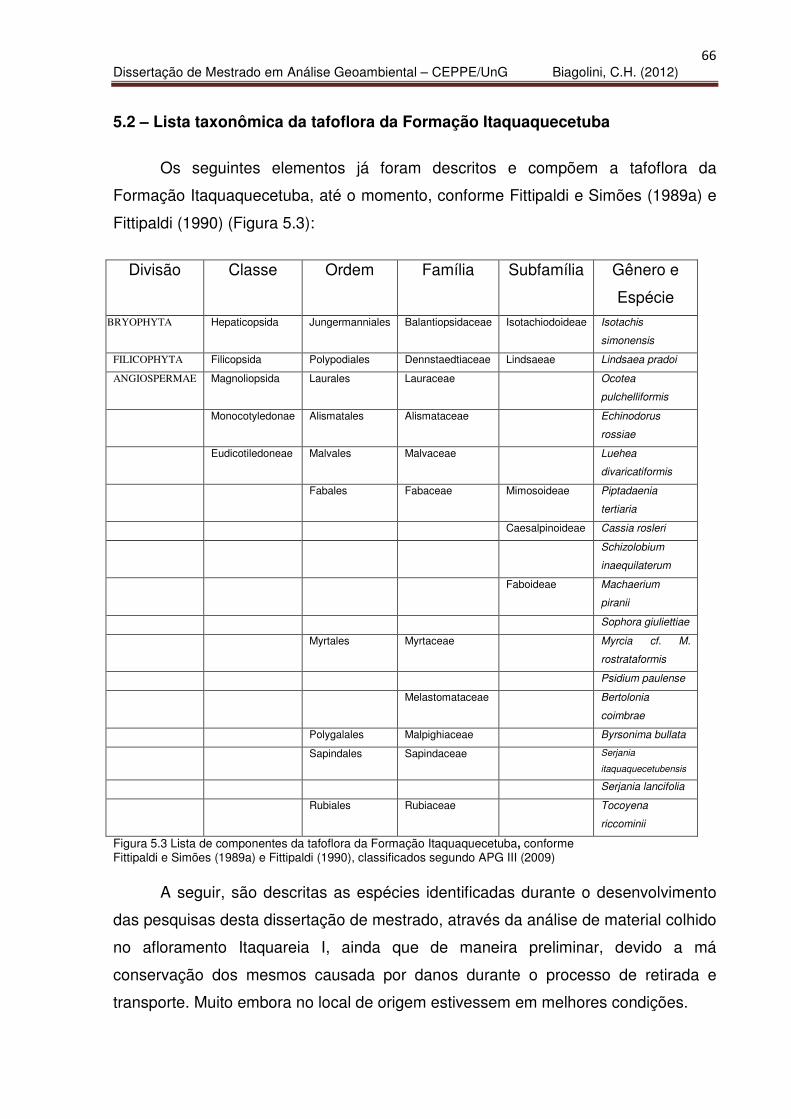
66
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
5.2 – Lista taxonômica da tafoflora da Formação Itaquaquecetuba
Os seguintes elementos já foram descritos e compõem a tafoflora da
Formação Itaquaquecetuba, até o momento, conforme Fittipaldi e Simões (1989a) e
Fittipaldi (1990) (Figura 5.3):
Divisão Classe Ordem Família Subfamília Gênero e
Espécie BRYOPHYTA Hepaticopsida Jungermanniales Balantiopsidaceae Isotachiodoideae Isotachis
simonensis
FILICOPHYTA Filicopsida Polypodiales Dennstaedtiaceae Lindsaeae Lindsaea pradoi
ANGIOSPERMAE Magnoliopsida Laurales Lauraceae Ocotea
pulchelliformis
Monocotyledonae Alismatales Alismataceae Echinodorus
rossiae
Eudicotiledoneae Malvales Malvaceae Luehea
divaricatiformis
Fabales Fabaceae Mimosoideae Piptadaenia
tertiaria
Caesalpinoideae Cassia rosleri
Schizolobium
inaequilaterum
Faboideae Machaerium
piranii
Sophora giuliettiae
Myrtales Myrtaceae Myrcia cf. M.
rostrataformis
Psidium paulense
Melastomataceae Bertolonia
coimbrae
Polygalales Malpighiaceae Byrsonima bullata
Sapindales Sapindaceae Serjania
itaquaquecetubensis
Serjania lancifolia
Rubiales Rubiaceae Tocoyena
riccominii
Figura 5.3 Lista de componentes da tafoflora da Formação Itaquaquecetuba, conforme Fittipaldi e Simões (1989a) e Fittipaldi (1990), classificados segundo APG III (2009)
A seguir, são descritas as espécies identificadas durante o desenvolvimento
das pesquisas desta dissertação de mestrado, através da análise de material colhido
no afloramento Itaquareia I, ainda que de maneira preliminar, devido a má
conservação dos mesmos causada por danos durante o processo de retirada e
transporte. Muito embora no local de origem estivessem em melhores condições.

67
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
5.3- Novas identificações e descrições de elementos da tafoflora de
Itaquaquecetuba
5.3.1 - Classe Filicopsida ou Polypodiopsida (sensu SMITH et al., 2006)
Ordem Polypodiales
As Polypodiales correspondem a 80 % das samambaias atuais. Smith et al.
(2006), consideraram 15 famílias dentro da Ordem Polypodiales. Estima-se que há
cerca de 9.000 espécies de fetos polipoides. Eles distinguem-se dos demais fetos
pela posição e natureza do indúsio, feições dos esporos, tipo de estelo, habitat e
hábito. A posição dos soros é variável e o indúsio é, geralmente, ausente. Seus
esporângios são peltados ou pedunculados, produzem poucos esporos, e possuem
o anulus disposto longitudinalmente. O gênero mais característico é Polypodium,
que inclui folhas que, frequentemente, apresentam veias anastomosantes. A Família
Dennstaedtiaceae é considerada basal entre as Polypodiales. A Pteridaceae
também tem posição basal e as três mais avançadas são eupolipoides:
Onocleaceae, Blechnaceae e Polypodiaceae (TAYLOR et al., 2009).
Família Polypodiaceae
Polypodyaceae apresentam caules variáveis entre eretos e grampantes e
suas frondes podem ser inteiras ou pinatífidas. As raques não possuem estípulas e
rizomas escamosos grampiformes são comuns sobre troncos de árvores. As
espécies de polipodiáceas encontram-se em climas úmidos, principalmente em
florestas tropicais pluviais. Em climas temperados, a maioria tende a ser epífita ou
ruderal.
Por análise filogenética, as Polypodiaceae foram divididas em cinco
subfamílias: Drynariaceae, Grammitidaceae, Gymnogrammitidaceae,
Loxogrammaceae, Platyceriaceae e Pleurisoriopsidaceae, cujos gêneros são citados
no quadro seguinte (Figura 5.4).
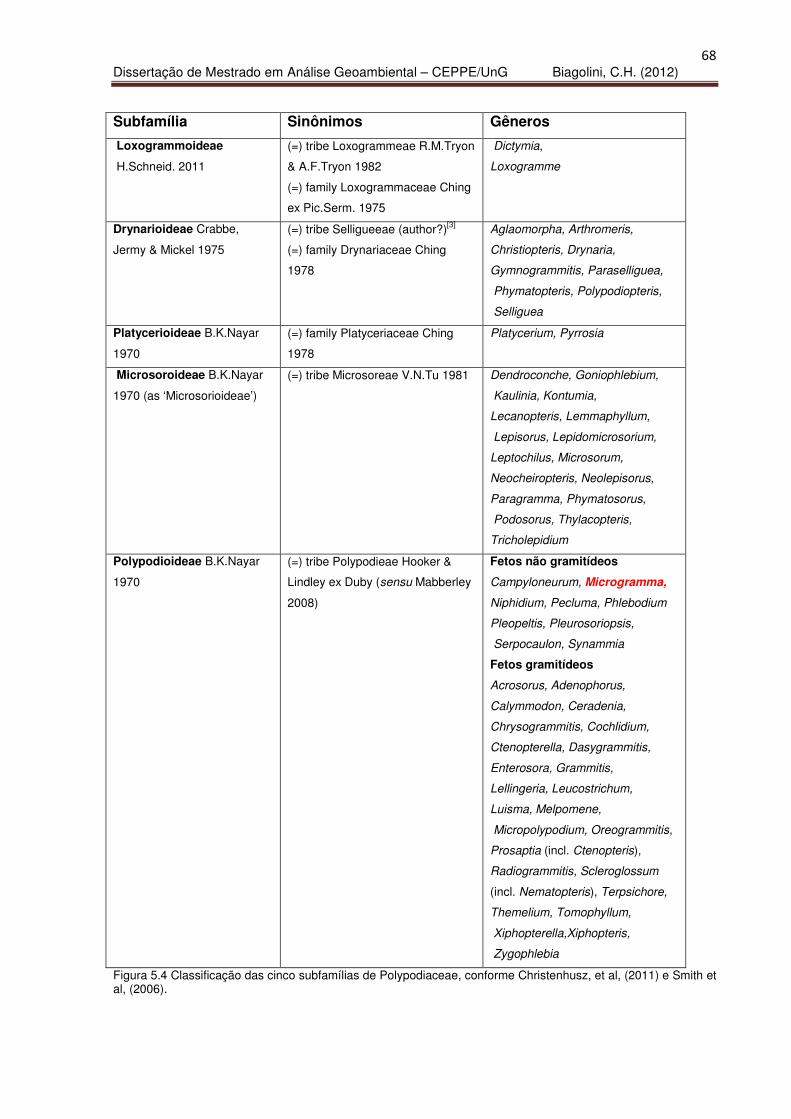
68
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Subfamília Sinônimos Gêneros
Loxogrammoideae
H.Schneid. 2011
(=) tribe Loxogrammeae R.M.Tryon
& A.F.Tryon 1982
(=) family Loxogrammaceae Ching
ex Pic.Serm. 1975
Dictymia,
Loxogramme
Drynarioideae Crabbe,
Jermy & Mickel 1975
(=) tribe Selligueeae (author?)[3]
(=) family Drynariaceae Ching
1978
Aglaomorpha, Arthromeris,
Christiopteris, Drynaria,
Gymnogrammitis, Paraselliguea,
Phymatopteris, Polypodiopteris,
Selliguea
Platycerioideae B.K.Nayar
1970
(=) family Platyceriaceae Ching
1978
Platycerium, Pyrrosia
Microsoroideae B.K.Nayar
1970 (as ‘Microsorioideae’)
(=) tribe Microsoreae V.N.Tu 1981 Dendroconche, Goniophlebium,
Kaulinia, Kontumia,
Lecanopteris, Lemmaphyllum,
Lepisorus, Lepidomicrosorium,
Leptochilus, Microsorum,
Neocheiropteris, Neolepisorus,
Paragramma, Phymatosorus,
Podosorus, Thylacopteris,
Tricholepidium
Polypodioideae B.K.Nayar
1970
(=) tribe Polypodieae Hooker &
Lindley ex Duby (sensu Mabberley
2008)
Fetos não gramitídeos
Campyloneurum, Microgramma,
Niphidium, Pecluma, Phlebodium
Pleopeltis, Pleurosoriopsis,
Serpocaulon, Synammia
Fetos gramitídeos
Acrosorus, Adenophorus,
Calymmodon, Ceradenia,
Chrysogrammitis, Cochlidium,
Ctenopterella, Dasygrammitis,
Enterosora, Grammitis,
Lellingeria, Leucostrichum,
Luisma, Melpomene,
Micropolypodium, Oreogrammitis,
Prosaptia (incl. Ctenopteris),
Radiogrammitis, Scleroglossum
(incl. Nematopteris), Terpsichore,
Themelium, Tomophyllum,
Xiphopterella,Xiphopteris,
Zygophlebia
Figura 5.4 Classificação das cinco subfamílias de Polypodiaceae, conforme Christenhusz, et al, (2011) e Smith et al, (2006).

69
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
O cladograma filogenético (Figura 5.5), demonstra as relações entre as
subfamílias de Polypodiaceae.
Figura 5.5: Cladograma filogenético e relações entre subfamílias Polypodiaceae http://en.wikipedia.org/wiki/polypodiaceae, adaptado por Biagolini (2012).
Apresenta documentário fóssil fidedigno, na forma de compressões ou
rizomas petrificados, desde o Neocretáceo, mas há menção de prováveis rizomas
petrificados envolvidos por raízes de Tempskya a partir do Albiano (SCHNEIDER;
KENRICK, 2001), sugerindo que a radiação de fetos de polipodiáceas foi anterior. A
Formação Clarno (Eoceno do Oregon, EUA) contém rizomas silicificados de, pelo
menos, dois gêneros de polipodiáceas enquanto na Formação Itaquaquecetuba até
o presente momenteo, foram encontrados apenas folhas fossilizadas.

70
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Subfamilia Polypodioideae (NAYAR, 1970)
Gênero Microgramma C. Presl.
Espécie-tipo: M. persicariifolia (Schrad.) C. Presl.
Microgramma C. Presl é um gênero de fetos da Familia Polypodiaceae, cuja
maioria de espécies está restrita à flora neotropical das Américas Central e do Sul.
Possue folhas ou frondes simples, unimórficas ou dimórficas (as férteis são mais
estreitas e mais longas que as estéreis), com ~ 10-350 mm de comprimento; lâminas
herbáceas a coriáceas, elípticas a oblongas, glabras a suavemente pubescentes ou
suavemente escamosas, margem inteira a ondulada; veias anastomosantes,
comumente com uma ou várias vênulas livres em cada aréolas; vênulas excurrentes
ou recurrentes. Soros arredondados a alongados, comumente, numa ou mais séries
sobre cada lado da nervura mediana, abaxial e nascida no final de veias simples ou
na junção de veias; receptáculos suavemente elevados ou não; desprovido de
indúsios. Esporângios de pedúnculos longos (TRYON ; TRYON, 1982).
Microgramma cf. M. persicariifolia (Schrad.) C. Presl.
Estampa II (Figuras 01 e 02) ; Figuras de texto 5.6 e 5.7
Estratigrafia: Porção superior da Formação Itaquaquecetuba, Eo-Oligoceno.
Procedência: Mina Itaquareia I, município de Itaquaquecetuba, SP.
No do Espécime: ITQ pb 026 B/11
Descrição: Compressão de fragmento de fronde simples, provavelmente elíptica,
ligeiramente curva, assimétrica, preservada da porção mediana para o ápice,
comprimento preservado de 40 mm e largura máxima preservada de 11 mm (por
inferência, provavelmente, com área de 590 mm2 = microfila), ápice reto com ângulo
apical de 30o, margem lisa, textura cartácea, veia primária de calibre fino e curvo,
veias secundárias anastomosantes, formando arcos broquidódromos fechados
desde a parte mediana da lâmina, e com vênulas abertas na porção interna do arco.
Comparação: Embora apresente apenas a porção mediano-apical, a comparação
da morfologia e da arquitetura foliar com a atual Microgramma persicariifolia é

71
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
flagrante. Entretanto, devido à sua idade oligocena e à sua preservação parcial,
torna-se mais prudente designá-la como Micrograma cf.
Figura 5.6: Fitofóssil Figura 5.7: Esquema de Micrograma sp (ITQpb 026) venação Micrograma sp (ITQpb 026)
Microgramma persicariifolia (Schrad.) C. Presl., conforme descrição e figura de
Tryon e Tryon (1982, Figura 110.19).
Habitat e Hábito: A espécie atual tem hábito epífito e como habitat, florestas
tropicais pluviais, em matas ciliares. Encontra-se, tipicamente nas altitudes de 0 a
1590 m (TRYON e TRYON, 1982).
Distribuição geográfica: O gênero Microgramma, segundo Tryon e Tryon (1982),
ocorre na América Tropical, desde o México e a Flórida até Buenos Aires. No Brasil,
sua distribuição ocorre por todo território nacional na forma de Microgramma
persicarifolia (Figura. 5.8).
Ocorrência fossilífera no Brasil: Palinomorfos de Microgramma foram encontrados
na Formação Pariquera-Açu (Plioc.- Pleistoc.) da Ilha Comprida, Município de
Iguape, SP, por Sundaram e Suguio, (1985) e em sedimentos neopleistocenos do
vale do rio Embu-Guaçu, SP, por Garcia et al. (1997). Em ambos os casos, a

72
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
associação palinológica sugeria clima quente e úmido. Agora, pelo que se sabe, é
citado, na forma de macrofóssil, pela primeira vez, neste trabalho.
Figura 5.8: Distribuição de Microgramma persicariifolia (Schrad.) Presl.: Bolivia, Brasil, Colombia, Costa Rica,
Ecuador, Guyana, Guyana Francesa, Panamá, Paraguay, Perú, Surinam, Trinidad y Tobago, Venezuela. Fonte:
http://biogeodb.stri.si.edu/herbarium/species/24376/?fam=Polypodiaceae&page=2.
5.3.2 – Gimnospermas
ORDEM CONIFERALES
As coníferas são gimnospermas arbóreas, de até 100 m de altura, de
crescimento monopodial, dióicas ou monóicas, de caule com lenho picnoxílico,
geralmente, disposto em anéis de crescimento e com canais resiníferos. Suas folhas
são simples, uninérveas ou paralelinérveas, lineares, escamiformes ou aciculares,
agrupadas em braquiblastos, helicoidalmente, dispostas (DATTA, 1966; KUBITZKI,
1996; RAPINI, 2008).
Cobrem grandes extensões do globo terrestre, no geral, na forma de plantas
sempre-verdes de florestas temperadas frias e de altitude. Entre as coníferas melhor
conhecidas estão sequoias gigantes das florestas costeiras ocidentais da América
do Norte, cedros do Líbano e as coníferas da Taiga ou Floresta Boreal. No Brasil
meridional, são exemplos de coníferas as araucárias ou “pinheiros do Paraná” e os

73
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
podocarpos da Floresta Ombrófila Mista ou “Mata dos Pinhais”, que recobrem as
partes altas da Serra da Mantiqueira e os altiplanos da Serra Geral.
Família Podocarpaceae
As podocarpáceas caracterizam-se por serem arbustivas ou arborescentes,
portam folhas escamiformes a lineares, geralmente de filotaxia helicoidal. Todas
apresentam estômatos em criptas, em fileiras descontínuas (STOCKEY; KO, 1988).
Os cones masculinos portam dois esporângios por microesporófilo e seus grãos de
pólen possuem sacos de tamanho reduzido. Cada cone feminino consiste de um
eixo com um pequeno número de brácteas estéreis e um ou dois óvulos (TAYLOR et
al., 2009).
Constitui a família de coníferas viventes com a maior diversidade morfológica
e a segunda maior em número de gêneros. Podocarpus, Dacridium e mais
dezesseis outros gêneros compõem a família, cuja ocorrência se estende desde o
México e América Central, por toda a América do Sul e demais continentes do
Hemisfério Sul. A distribuição geográfica evidencia sua origem gondvânica e
remonta ao Eotriássico (COOPER, 1960; TAYLOR et al., 2009; GUERRA-SOMMER;
PIRES, 2011).
Registros Fossilíferos mesozóicos de Podocarpáceas
Os mais antigos fósseis de podocarpáceas, designados Rissikia, são
compressões de ramos folhosos e cones polínicos e ovulíferos, encontrados no
Triássico Inferior da África do Sul, Austrália, Chile, Argentina, Antártica e
Madagascar (HILL; BRODRIBB, 1999; SUÁRES et al., 2000). Rissikia apresenta
ramos com cicatrizes foliares, que sugerem caráter decíduo. Na base de cada ramo
curto, há várias folhas escamiformes pequenas e no restante do ramo estão
arranjadas folhas de cerca de 10 mm de comprimento. Os estômatos dispõem-se em
quatro fileiras na face abaxial, com células – guardiãs ligeiramente afundadas e
protegidas por papilas ou projeções cuticulares das células subsidiárias, conforme
Taylor et al. (2009) .
Notophytum krauselii inclui caules e folhas do Triássico Médio da Antártica
Ocidental, estudados por Meyer-Berthaud; Taylor (1991). A presença de tecido de
transfusão, em duas porções ao redor dos feixes vasculares, a presença de canais

74
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
resiníferos abaixo dos feixes e de esclerídeos, no mesofilo, são características
utilizadas na comparação de Notophytum com Podocarpáceas (AXSMITH et al.,
1998).
Fósseis jurássicos de podocarpáceas (Nothodacridium), procedentes da
Antártica, foram descritos por Townrow (1967). São ramos vegetativos, em
disposição helicoidal, sem diferenciação de ramos curtos e longos. Suas folhas
divergem dos eixos em várias direções, tem três mm de comprimento e seção
transversal romboide. Cones masculinos de Nothodacridium foram designados na
Antártica como Masculostrobus e, no Jurássico indiano, como Podostrobus (RAO;
BOSE, 1971). Os grãos de pólen desse cone são bi ou trissacados.
Mataia (Figura 5.9) é uma podocarpácea do Jurássico da Austrália e Nova
Zelândia, que exibe folhas de 15 mm de comprimento, helicoidalmente arranjadas e
torcidas na base, a sugerir apenas duas fileiras. Cada folha apresenta estômatos
monocíclicos dispostos em fileiras na superfície abaxial (TOWNROW, 1967).
Figura 5.9- Mataia ramos folhosos de podocarpácea do Jurássico da Austrália e Nova Zelândia. Extraído de
Taylor et al. 2009.
No Cretáceo Inferior da Argentina (Formação Baqueró, Província de Santa
Cruz), ocorrem folhagens e cones de podocarpáceas designados Trisacocladus
tigrensis. Suas folhas possuem arranjo helicoidal, têm 6 mm de comprimento e
deixam uma cicatriz romboide sobre os brotos (ARCHANGELSKY e
GAMERRO,1966). Seus grãos de pólen trissacados são pequenos e suas feições
ultraestruturais assemelham-se às dos gêneros viventes Dacrydium , Podocarpus e
Dacrycarpus (BALDONI; TAYLOR, 1982). Também no Cretáceo Inferior argentino,

75
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
ocorrem ramos de Apterocladus e Podocarpus, cones ovulíferos e polínicos de
Squamastrobus associados a ramos do tipo Brachyphyllum, portando grãos de pólen
que são do tipo Podocarpidites (ARCHANGELSKY; GAMERRO, 1966; GUERRA-
SOMMER; PIRES, 2011).
No Cretáceo Inferior da Chapada do Araripe, Brasil, ocorre um ramo com
folhas alongadas, de filotaxia helicoidal, cuja epiderme exibe fileiras de estômatos,
longitudinalmente dispostos, designado genericamente de Lindleycladus, mas de
posição taxonômica incertae sedis (KUNZMANN et al. 2004).
Registros Fossilíferos cenozóicos de Podocarpáceas
Paleógeno
O documentário fóssil de podocarpáceas, no Hemisfério Norte, é muito
escasso e se restringe a uns poucos grãos de pólen e alguns caules delicados com
folhas aciculares, arranjadas helicoidalmente e anfistomáticas, com anéis espessos
ao redor dos estômatos, feições que indicam claramente sua afinidade com
podocarpáceas. Foram encontrados no Eoceno do Tennessee por Dilcher (1969).
Do Paleógeno da Austrália, Hill e Scriven (1999) descreveram folhas que
relacionaram a podocarpáceas atuais designando-as Sigmaphyllum e Falcatifolium.
Com o uso de análises cuticulares micromorfológicas, evidenciaram que
Falcatifolium viveu, durante o Eoceno, em paleolatitudes altas enquanto hoje está
restrito a latitudes mais baixas. Em lamitos carbonosos paleocenos da Nova
Zelândia, Pole (1998), descreveu uma podocarpácea (Prumnopitys) dentre várias
outras espécies. Seus fósseis eram ramos com folhas dispostas helicoidal e
disticamente achatadas. Para comprovar suas afinidades com podocarpáceas
utilizou o tipo e distribuição dos estômatos.
No Paleógeno do Brasil, há registros de grãos de pólen de Podocarpus, no
Eoceno Inferior, da Bacia de Bonfim, por Lima e Dino (1984).

76
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Outrossim, grãos de pólen relacionados às podocarpáceas foram identificados
nas seguintes ocorrências paleógenas: a) na Bacia de São Paulo, Formação São
Paulo, por Fittipaldi; Simões (1989a), como Dacrydiumites krempsi e Podocarpites
sp. Também identificaram na Formação Itaquaquecetuba, Dacrydiumites florinii,
Podocarpidites sp e P. embrionalis, em Barueri (SP), e Podocarpidites sp cf. P.
sellowiformis, nas áreas de Itaquaquecetuba e Barueri. Lima et al. (1991)
identificaram, na Formação São Paulo (provavelmente Formação Itaquaquecetuba,
no bairro Barra Funda, SP), as espécies de grãos de Podocarpidites embrionalis,
Krutzsch e Podocarpidites sp cf. P. sellowiformis Zaklinskaya; b) Garcia et al. (2000)
identificaram, em argilitos alaranjados de Atibaia, de idade neoeocena a eo-
oligocena, Podocarpidites sp.; c) na Bacia de Taubaté, Lima e Amador (1985)
identificaram em linhito da Formação Caçapava, Neo-oligoceno, grãos de pólen de
Podocarpidites libellus Krutzsch, Podocarpidites sp. 1 e Podocarpidites sp. 2; na
Formação Tremembé (Oligoceno), Wickert (1974) identificou Podocarpus sp ? e
Podocarpidites sp, enquanto Ferreira e Santos (1982) identificaram
Podocarpidites(?) sp 1 e Podocarpidites (?) sp. 2. Lima et al. (1983), em testemunho
de sondagem do CNP no. 42, identificaram na Formação Tremembé: Podocarpidites
libellus Krutzsch, e Podocarpidites sp 1, P. sp 2, P. sp 3, P. sp 4.; d) na Bacia de
Resende, Formação Resende, Lima e Amador (1983) identificaram Podocarpidites
ellipticus Cookson, P. peniverrucatus Weyland; Krieger e Podocarpidites sp.; e) na
Bacia de Gandarella (Mesoeoceno a Oligoceno), Lima e Cheboldaeff (1981)
identificaram Podocarpidites sp..
Dois fragmentos foliares de Podocarpáceas foram mencionados para a
Formação Tremembé (Oligoceno da bacia de Taubaté), por Mandarim-de-Lacerda et
al. (1996). Agora, registram-se ramos folhosos de podocarpáceas entre os fósseis
da eo-oligocena tafoflora de Itaquaquecetuba.
Neógeno
Fósseis miocenos de podocarpáceas, na forma de cones polínicos e
folhagens, ocorrem na Austrália e foram identificados como do gênero vivente
Dacrycarpus (HILL; WHANG, 2000). Microsporófilos arranjados helicoidalmente e
grãos de pólen, cada um com três sacos, são atribuídos aqueles cones.

77
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
No Brasil, Garcia et al (2008) identificaram grãos de Podocarpus em
associações palinológicas extraídas de argilitos de Nazaré Paulista e Pedra Bela, de
idade neomiocena.
Na Província Costeira Paulista, há registros palinológicos de podocarpáceas,
em depósitos plio-pleistocenos da Formação Pariquera-Açu, na Ilha Comprida,
município de Iguape (SP), feitos por Sundaram e Suguio (1985).
Em testemunhos de três sondagens de depósitos pleistocenos-holocenos da
Província Costeira, na área de Santos em testemunhos de três sondagens, na área
de Santos, Itanhaém e Guarujá, Absy e Suguio (1976), identificaram grãos de pólen
de Podocarpus.
Em sedimentos turfosos quaternários do Vale do Paraíba, Garcia (1998)
identificou grãos de pólen (Figura 5.10) do tipo Podocarpus lambertii (Klotsch) Endl.
Também Modenesi (1988) detectou a presença de palinomorfos de Podocarpus sp.
em sedimentos turfosos de Campos de Jordão (Jardim Primavera), SP.
Todos esses registros sugerem que as Podocarpaceae faziam parte da
paisagem paulista desde pelo menos o Eoceno, espalhando-se pelo Vale do Paraíba
e pela área paulistana desde Itaquaquecetuba até Barueri. Indícios de sua
permanência neógeno-quaternária, na região, espalham-se desde a planície costeira
paulista, pelo vale do Paraíba e planalto Atlântico até as áreas montanhosas de
Campos do Jordão.
Figura 5.10 a e b: Grãos de pólen de Podocarpus sp, mostrando sacos
laterais.(MARQUES-SOUZA, 2010)
a b

78
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Podocarpus cf. P. dubius Archangeslky
Estampa III, figuras de 1 a 6, Estampa IV, figuras 1 a 6, Estampa V
figuras 1 e 2. Figuras de texto: 5.11 A,B,C e D.
Estratigrafia: Porção superior da Formação Itaquaquecetuba, Eo-Oligoceno.
Procedência: Mina Itaquareia I, município de Itaquaquecetuba, SP.
No dos Espécimes: ITQ pb 10, ITQ pb 60, ITQ pb 91 A e 91 B, ITQ pb 25.
Descrição: Várias impressões, compressões e/ou piritizações, de ramos folhosos
delicados ou pequenas folhas dispersas na matriz, medindo o maior 25 mm de
comprimento por 10 mm de largura incluindo as folhas. São constituídos de eixos
delicados e flexuosos, com diâmetro da ordem de 0,5 mm, de hábito bifacial. Portam
folhas laminares, longamente decorrentes sobre os eixos caulinares, helicoidal ou
disticamente dispostas. Divergem do eixo caulinar em ângulo ao redor de 30º, são
lineares a lanceoladas longas, suavemente falcadas, de ápice agudo, seção
transversal romboide baixa, de textura coriácea, com nervura mediana marcada e
superfície laminar com duas finas linhas paralelas e longitudinais ou faixas
estomatíferas de cada lado, na superfície abaxial, e mais raros na adaxial. A
margem é lisa ou denteada, com dentes distantes e semelhantes a acúleos. Células
epidérmicas retas e dispostas longitudinalmente. As folhas medem ao redor de 7
mm de comprimento por 1 mm de largura.

79
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
A
B
C
D Figuras 5.11 A e B: ilustração da distribuição de folhas de Podocarpus sp, Figuras 5.11 C e D: Foto
ampliada de possíveis ramos de Podocarpus sp
Comparação e Discussão: Berry (1938) descreveu duas espécies de Podocarpus,
de idade paleógena, provenientes do jazigo Rio Pichileufu, Província de Rio Negro,

80
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Argentina: P. andiniformis Berry (BERRY, 1938; Plate 13, Fig. 4) e P. engelhardti
Berry (BERRY, 1938, Plate 13, Figs. 1-3). Os espécimes aqui analisados diferem de
P.andiniformis Berry por seu tamanho menos e por não apresentar estreitamento
proximal nítido. Por essas feições seria mais semelhante a P.engelhardti Berry, da
qual difere por sua folhagem mais esparsa, isto é, maior distanciamento entre as
folhas. Por outro lado, assemelha-se muito a P.dubius Archangelsky
(ARCHANGELSKY; GAMERRO,1966), Plate 6, Figs. 49-50 e ARCHANGELSKY,
1970, Lam. XIX, Fig. 7), proveniente da Formação Baqueró, Flora de Ticó, ocorrente
em Bajo Grande, Província de Santa Cruz, Argentina, de idade eocretácea, nas
seguintes feições: ramos de última ordem normalmente retos ou suavemente curvos,
medindo aproximadamente 40 mm X 2 mm de diâmetro ou 20mm incluindo as
folhas. Folhas dimórficas, de disposição helicoidal, bilaterais, lineares, de perfil reto
ou falcado, com ângulo de divergência ao redor de 40º e 80º, ápice acuminado ou
agudo e base não contraída e nitidamente decorrente, percorrendo adpressa ao eixo
por até 4 a 5 mm, uninérvias, medindo 13 mm X 1,5 mm na porção livre (setor
decorrente, às vezes, maior que a parte livre), seção transversal tetragonal ou
romboide achatada. Cutículas espessas, células epidérmicas dispostas
longitudinalmente e paralelas à veia mediana. Há que se ressaltar a diferença de
idade e a impossibilidade de uma comparação mais precisa do ponto de vista
cuticular, para uma identificação mais segura. Por isso, considera-se a identificação
como Podocarpus cf. P.dubius.
5.3.3– Angiospermas
5.3.3.1 Generalidades
Como foi ressaltado por Bernardes-de-Oliveira e Mune (2011), as
angiospermas são, atualmente, o grupo dominante do Reino Plantae, incluindo cerca
de 460 famílias e envolvendo, aproximadamente, 260.000 espécies ou mais. Este
domínio iniciou-se há mais de 100 milhões de anos, durante o Neocretáceo, e
ampliou-se na Era Cenozóica até os dias atuais.
Todavia, a origem das angiospermas continua sendo um dos maiores
problemas insolúveis para a paleobiologia moderna. O ponto central de qualquer

81
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
discussão sobre a origem e evolução das angiospermas é a definição precisa do que
é, realmente, uma angiosperma e quais são as características que podem ser
consideradas primitivas nesse grupo (BERNARDES-DE-OLIVEIRA; MUNE, 2011)
Por definição, Angiospermae é um grupo de plantas espermáticas, isto é,
portadoras de sementes e floríferas. Seus óvulos são envolvidos por carpelos
(=folhas modificadas, que se fundem lateralmente constituindo ovário, estilete e
estigma). Uma vez fecundados, esses óvulos tornam-se sementes, que ficarão
abrigadas no interior do fruto.
Constituem um grupo monofilético e entre as características que melhor
delimitam a angiospermia, usadas para separar esse grupo vegetal dos demais
listam-se: um ovário, flores, elementos condutores especializados no xilema (vasos),
no floema (tubos crivados), dupla fecundação, óvulo bitegumentado, endospermia
triploide e grãos de pólen tectados. Seus gametófitos são reduzidos em relação aos
das gimnospermas: o megagametófito feminino (saco embrionário), frequentemente,
é formado por sete células (sendo uma delas constituída de dois núcleos polares,
havendo ainda três antípodas e duas células sinérgides junto à oosfera, o gameta
feminino). O microgametófito masculino é tricelular (duas células gaméticas e uma
célula do tubo polínico). A dupla fecundação envolve os dois gametas masculinos do
microgametófito maduro (grão de pólen), resultando:
a) No zigoto (constituído pela união de um gameta masculino e da oosfera, no
interior do óvulo) que se desenvolverá no embrião;
b) No núcleo endospérmico primário (união do outro gameta masculino com os
dois núcleos polares), que se desenvolverá num tecido nutritivo triplóide
especial – o endosperma.
A diversidade incomparável das angiospermas acaba resultando em algumas
formas desprovidas dessa ou daquela característica. Por outro lado, a maioria
dessas feições não é exclusividade do grupo, podendo ser cada uma delas
compartilhada separadamente com diferentes grupos vegetais. Assim, se constitui
como característica, possivelmente, exclusiva e comum a todas as angiospermas, a
formação de endosperma triploide, conforme (FRIEDMAN 1992).

82
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
A dupla fecundação, que foi, por muito tempo, considerada como uma feição
única da angiosperma, também ocorre nas gimnospermas Ephedraceae. Entretanto,
neste caso, não há desenvolvimento do tecido endospérmico.
Cabe ressaltar que a aquisição de um carpelo e o desenvolvimento de um
fruto a partir dele se constitui em um caráter distintivo para a maioria dos taxa
angiospérmicos.
A adaptação das angiospermas, com relação à fecundação de suas flores,
pode envolver diversos agentes externos tais como: vento, chuva, insetos, aves,
mamíferos. Além desta adaptação, as angiospermas também estão adaptadas a
uma variedade de habitats que vão desde águas salgadas até águas doces e
desertos, desde planícies em nível do mar até altas montanhas e desde regiões
tropicais até regiões polares.
Sua classificação filogenética mais moderna,, com base em dados
moleculares e outros, é aquela apresentada pelo APG III (2009), que pode ser vista
na figura 5.2.
Ordem Fabales
As Fabales contêm cerca de 19.000 espécies, constituindo 9,6 % da
diversidade das eudicotiledôneas (WOJCIECHOWSKIE et al., 2006). É uma das
maiores ordens de plantas angiospérmicas.
A Ordem foi colocada, pelo APG III (2009), dentro do superclado das
Eudicotiledôneas Centrais (Eudicots Core), no clado das Rosídeas e grupo das
Fabídeas (Figura 5.2).
A Ordem Fabales é considerada monofilética com base nas sequências de
rbcL, (JUDD et al, 1999).
Família Fabaceae
Dentre as famílias da Ordem Fabales, a Família Fabaceae/Leguminosae
destaca-se como uma das maiores e mais importantes.

83
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Apresenta-se como: ervas, lianas, arbustos ou árvores, com um metabolismo
de nitrogênio alto, graças às micorrizas (nódulos na raiz, resultantes da simbiose
com fungos), folhas, no geral alternas, compostas pinadas ou bipinadas ou
compostas palmadas, uni ou trifoliadas de margem inteira, com venação pinada,
pulvinos bem desenvolvidos nas folhas ou nos folíolos individuais, eixos foliares ou
folíolos com movimentos retráteis, ocasionalmente, com espinhos.
Tradicionalmente, tem sido reconhecida uma só família (Fabaceae) com três
subfamílias (Papilionoideae ou Faboideae, Caesalpinoideae e Mimosoideae), exceto
por Cronquist (1988) e outros que preferiram mantê-las como famílias distintas
(SOUZA; LORENZI, 2008).
Trata-se de família de distribuição cosmopolita, com 650 gêneros e 18.000
espécies. No Brasil, ocorrem cerca de 180 gêneros e 1.500 espécies.
Sub-família Caesalpinoideae / Cercideae
Conforme Marchiori ( 1997) e Souza; Lorenzi (2008), as cesalpinoídeas,
incluem espécies arbóreas, arbustivas e herbáceas, espinhosas ou inermes de
folhas pinadas, bipinadas ou bifolioladas, com folíolos geralmente fundidos. Assim
como nas Cercideae, suas flores são zigomorfas, a corola dialipétala e a pré-
floração imbricada ascendente (=pétala superior interna). Seus estames, no geral,
são livres entre si e não vistosos, em número duplo ao das pétalas, raramente em
igual ou menor número. Os frutos são legumes de formas variadas. As sementes,
normalmente apresentam pleurograma.
Constituem, provavelmente, a mais primitiva das subfamílias das leguminosas
com cerca de 2.800 espécies, que habitam as regiões tropicais e subtropicais de
todo o planeta. São abundantes no Reino Paleotropical, mormente, na África.
No Brasil, são mais numerosas no centro-norte e mais escassas no sul do
país (MARCHIORI, 1997).
Apareceram durante o Neocretáceo, quando se acredita que haveria fácil e
recíproca dispersão de espécies, dada a proximidade ainda existente entre América
do Sul e África (~800 km, com numerosas ilhas vulcânicas. Após o Paleoceno , por
vicariância, uma diferenciação florística apreciável instalou-se entre os dois

84
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
continentes, determinando inclusive uma grande diferenciação genérica dentre as
cesalpinoídeas.
O gênero Bauhinia L., nativo do Brasil, tem sido colocado na subfamília
informal Cercideae ou na Caesalpinoideae.
Gênero Bauhinia L.
O gênero Bauhinia L. reúne cerca de 250 espécies, ocorrentes nas regiões
tropicais e temperadas quentes do globo (MARCHIORI, 1997).
Conforme Vaz; Tozzi (2005), Bauhinia L. é pantropical, com cerca de 300 spp,
(WUNDERLIN et al., 1987), das quais 200 são brasileiras. O gênero aparece no
Brasil em cinco seções (VAZ; TOZZI, 2003). Dessas, a mais amplamente distribuída
na região neotropical é a Pauletia, que conta com 70 espécies, das quais 53
ocorrem no Brasil.
O gênero se estende por todo Brasil, mas duas espécies são típicas da área
sudeste: B. longifolia (Bong) Stend e B.forficata Link. A primeira espécie distribui-se
pelos estados de São Paulo e Minas Gerais, nos cerradões e na mata latifoliada
semidecídua de altitude. Trata-se de planta semidecídua, heliófita, indiferente às
condições de umidade do solo, característica de mata semidecídua, onde ocorre
dispersa, mas em alta frequência. Ocorre em capoeirões sobre solos argilosos de
boa fertilidade e em terrenos de topografia ondulada.
A segunda espécie (B. forficata), distribui-se pelo Rio de Janeiro e Minas
Gerais até o Rio Grande do Sul, principalmente na floresta pluvial Atlântica. Trata-se
de planta decídua ou semidecídua, também heliófita, mas característica da floresta
atlântica. Ocorre, preferencialmente, em planícies aluviais úmidas ou início de
encostas, quase sempre em formações secundárias, como capoeiras e capoeirões.
É rara sua ocorrência no interior da mata primária densa. Dada sua alta diversidade
em terras brasileiras, o gênero ou tem o Brasil como centro de origem ou de
dispersão ou, pelo menos, é muito antigo por aqui.
Um gênero fóssil (Bauhcis – Cercideae, Caesalpinieae) foi criado para o
Oligoceno do México por Calvillo- Canadell; Cevallos-Ferriz (2002) que detectaram
ali características de arquitetura foliar similares às do gênero Bauhinia misturadas a
outras do gênero Cercis.

85
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
No Brasil, Garcia (1998), documentou a presença do gênero Bauhinia, ao
estudar a palinologia de turfeiras, quaternárias do médio vale do rio Paraíba do Sul,
nas áreas dos municípios de Jacareí, Eugênio de Melo e Taubaté. Grãos de pólen
de Bauhinia também foram observados em sedimentos quaternários do vale do rio
Embu Guaçu, no estado de São Paulo, em palinoflora rica e diversificada típica de
ambiente úmido (GARCIA et al. 1997).
Bauhinia cf. B. divaricata.
Estampa VI (Figuras 1,2) - Texto figuras 5.12 e 5.13
Estratigrafia: Porção superior da Formação Itaquaquecetuba, Eo-Oligoceno.
Procedência: Mina Itaquareia I, município de Itaquaquecetuba, SP.
No do Espécime: ITQ pb 0111/11
Descrição: Fragmento de compressão de folha simples, dividida em duas porções
laminares na posição da veia central ou mediana, pecíolo não-preservado. É uma
microfila, medindo aproximadamente 25 mm de comprimento por 35 mm de largura
máxima, com área de 585 mm2. Sua porção mais larga, na área média, dá-lhe forma
elíptica, bilobada, assimétrica, de base foliar cordada não típica, pecíolo marginal,
ápice não-preservado, margem lisa. A veia mediana apresenta-se rompida até a
base. A venação primária é actinódroma, composta de tres pares de veias laterais e
uma central (total de sete veias primárias), partindo de dois centros de irradiação ou
um só centro partido em dois, na fossildiagênese, mantendo um ângulo de 20o a 30o
entre si. As veias primárias laterais convergem, ligeiramente para o ápice. As veias
secundárias têm percurso suavemente curvo, tendendo para
broquidódromas/eucamptodromas. As veias terciárias emergem a ângulo reto,
dispõem-se em ângulos obtuso em relação à primeira mediana/central, tem trajetória
convexa, delimitando aréolas, aproximadamente retangulares, com as veias

86
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
primárias e secundárias. Há presença de veias agróficas ou fimbriais. A textura da
lâmina seria cartácea.
Figura 5.12: Fitofóssil Bauhinia sp (origem F. Itaquaquecetuba)
Figura 5.13: Esquema de venação
Comparação e discussão: Este novo achado para o Brasil, aproximadamente
coetâneo ao registro mexicano (Oligoceno), tem características muito semelhantes
às de Bauhinia da Seção Paulitea, aproximando-se, pelas características acima
descritas, mais das espécies B. forficata (= B. longiflora), B. longifolia, B.

87
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
subrotundifolia e B. glabra. Dentre essas, as semelhanças maiores recairiam sobre
B. longifólia, B. subrotundifolia e B.divaricata, conforme pode ser visto na figura 5.14.
A B
C
D
E F
Figura 5.14: Formas de diferentes espécies de Bauhinia sp. A) B. divaricata, B) B. glabra, C) B.Subrotundifolia, D e F) B. longiflora, F) B. longifólia (CALVILLO-CANADELL., CEVALLOS-FERRIZ, 2002), Adaptado por (BIAGOLINI, 2012)

88
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Entretanto vale a pena ressaltar, que entre as espécies de Bauhinia que
apresentam dois pontos basais de divergência das veias primárias, conforme pode
ser observado no trabalho de Calvillo- Canadell; Cevallos-Ferriz, (2002), a espécie
que mais se aproximaria da forma aqui analisada (por possuir três pares de veias
primárias de cada lado da primária mediana) é a espécie B. divaricata que é nativa
da região sul do Brasil, Paraguai, Uruguai e Argentina, conforme Alves et al.(2008).
Dado seu caráter fragmentário e por não apresentar feições que permitam
optar por uma ou outra forma é designada Bauhinia sp.
Ordem Myrtales
Myrtales é uma ordem de plantas angiospérmicas pertencentes ao grupo das
Malvideas, clado Rosideas, do clado Eudicotiledoneas centrais (core eudicots).
Caracteriza-se por ter folhas simples, inteiras, flores normalmente tetrâmeras,
sincárpicas e estiletes soldados. Trata-se de uma ordem com nove famílias,
segundo APG III (2009). É um dos principais componentes da flora brasileira, onde
estão presentes as famílias Combretaceae, Onagraceae, Lythraceae, Myrtaceae,
Vochysiaceae e Melastomataceae.
Família Melastomataceae
Dentre as famílias da Ordem Myrtales, a família Melastomastaceae destaca-
se com cerca de 5000 espécies. Tem distribuição subtropical e tropical, sendo
representada por ervas, arbustos ou lianas, notadamente na América do Sul. São
principalmente plantas herbáceas ou arbustivas, menos comumente árvores. Suas
folhas são simples, opostas cruzadas, frequentemente revestidas por tricomas,
margens inteiras ou serreadas e 3 a 9 nervuras curvinérvias. Duas ou mais nervuras
secundárias divergem a partir da base e percorrem paralelas à periferia do limbo
foliar, em arcos convergentes, até unirem-se à nervura central no ápice (venação
acródroma) (MARCHIORI; SOBRAL, 1997).

89
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
No Brasil, vegetais da Família Melastomataceae são amplamente
encontrados, principalmente, na região sudeste em floresta de mata atlântica,
cerrados ou ainda em áreas devastadas pelo crescimento urbano (RODRIGUES
LIMA; AFFONSO, 2010).
Gênero Leandra Raddi
Este gênero neotropical distingue-se do gênero Miconia Ruiz et Pav. por ter
botões florais agudos, em vez de obtusos (BARROSO, 1984). É constituído
principalmente de arbustos de beira da mata e regiões campestres (MARCHIORI;
SOBRAL, 1997). Aparece também na floresta atlântica juntamente com Miconia e
Tibouchina. Apresenta cerca de 200 espécies que se distribuem desde o México e
Antilhas até o sul do Brasil e Argentina. Os últimos estudos revisionais ou
abrangentes sobre o gênero para o Brasil e o mundo, datam do século 19
(COGNIAUX, 1886; 1891, apud BAUMGRATZ; D’EL REI SOUZA, 2005).
Recentemente duas novas espécies foram descritas por Baumgratz; D’El Rei
Souza (2005) para a flora do estado de São Paulo e em um levantamento do gênero
Leandra Raddi para o Parque Estadual da Serra do Mar (SP), por Rodrigues Lima e
Affonso (2010), onde foram detectadas 14 espécies. No estado do Paraná foram
identificadas sete espécies da seção Leandraria na Região Litorânea, Serra do Mar
e Primeiro Planalto ou de Curitiba (CAMARGO; GOLDENBERG, 2007).
Leandra Raddi caracteriza-se principalmente por apresentar-se na forma de
subarbustos decumbentes, arbustos ou arvoretas, ramos cilíndricos, glabros ou
pilosos; folhas oblongas, elípticas, ovais ou cordiformes; membranáceas, cartáceas
ou coriáceas; margem inteira a crenulada; glabra ou pilosas; longo ou curto
pecioladas; com três a sete nervuras acródromas basais ou supra basais;
inflorescências terminais e ou pseudo-axilares, botões florais de ápice agudo a
acuminado, pétalas atenuado-acuminadas a atenuado-agudas e frutos carnosos
polispérmicos (BAUMGRATZ; D’EL REI SOUZA, 2005; RODRIGUES LIMA;
AFFONSO, 2010).

90
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
No Brasil, não há registros do gênero na forma fossilizada até o presente
momento.
Leandra sp.
Estampa VII (Figuras 1 e 2) e Figuras de texto 5.15 e 5.16
Estratigrafia: Porção superior da Formação Itaquaquecetuba, Eo-Oligoceno.
Procedência: Mina Itaquareia I, município de Itaquaquecetuba, SP.
No do Espécime: ITQPB 012/11.
Descrição: Fragmento basal de compressão de folha simples, pecíolo não
preservado. É uma microfila (cerca de 2000 mm2), medindo 52 mm de comprimento
por 35 mm de largura máxima preservados. Sua porção mais larga, na área média
da folha, dá-lhe forma elíptica, não lobada, simétrica, com conexão de pecíolo
marginal, ápice não preservado, margem lisa e base cuneada/reta. A venação
primária é acródroma, do tipo suprabasal, partindo as duas primárias laterais
aproximadamente a 10 mm da base, em percurso curvo e paralelo as margens do
limbo da folha, aparentemente em direção ao ápice. A venação secundária/terciária
se dispõe de maneira percorrente oposta entre as veias primárias emergindo a um
ângulo obtuso próximo a nervura central e agudo junto as primárias laterais, em um
curso retilíneo-convexo. A textura da lâmina seria cartácea.
Figura 5.15 Fitofóssil Leandra sp – ITQpb 012/11

91
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Figura 5.16: Esquema venação baseado em ITQpb 012/11.
Comparação e discussão:
O fato da folha, apresentar-se provavelmente elíptica, cartácea, de margem
inteira, glabra, com três nervuras acródroma suprabasais e venação secundário-
terciário percorrente oposta de curso retilíneo convexo leva a uma comparação com
Melastomatácea do gênero Leandra. Dentro deste gênero as folhas de nervura
acródroma suprabasal conforme levantamento de Rodrigues Lima e Affonso (2010),
para o Parque Estadual da Serra do Mar, há as seguintes espécies: L. fallax, L.
melastomoides, L. australis, L. acutiflora, L. ribesiaeflora, L. collina, L.
quinquedentata, L. hirtella, L. laxa. Acrescenta-se também com estas características
Leandra mattosii Baumgratz; D’El Rei Souza, descrita para a região de Salezópolis
em torno de 850 metros de altitude, em borda de matas pluviais e capoeiras,
próximos a cursos de rios.
Outro gênero de melastomataceae identificado por Fittipaldi (1990) na flora
fóssil de Itaquaquecetuba é Bertolonia Raddi, do qual se distingue o espécime

92
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
estudado pela nervação acródroma, suprabasal com três nervuras primárias e não
cinco basais, pelo tamanho foliar maior, pelo contorno elíptico e estreito em vez de
elíptico largo e pela base reta/cuneada em vez de convexa.
Hábito: O gênero é caracterizado como sendo arbustos a arvoretas, com
indumento variável ou ausente. O habitat da espécie Leandra mattosii, representada
por subarbustos a arbustos de até 1 metro de altura corresponde a área atual de
ocorrência deste fitofóssil uma vez que foi localizada restrita ao município de
Salesópolis (SP), onde ocorre em torno de 850 metros de altitude, em bordas de
matas pluviais e capoeiras, próximos a cursos de rios.

93
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
A assembléia fitofossilífera da Formação Itaquaquecetuba apresenta-se na
forma de micro e macrorrestos, incarbonizados, piritizados ou através de impressões
bem marcadas em matriz de granulação fina, que permitem, de forma satisfatória,
sua identificação quanto aos aspectos morfológicos. Aspectos tafonômicos,
litológicos, morfográficos, taxonômicos, bioestratigráficos e fitogeográficos são
considerados através de diferentes tipos de análises.
6.1 – Análise Tafonômica e sua Interpretação
Tafonomia (do grego: taphos = sepultamento; nomos = leis, arranjo,
ordenação) é o termo proposto por Efremov (1940) para designar o estudo das “leis”
que governam a transição dos restos orgânicos da biosfera para litosfera. Esta
definição é, no entanto, muito ampla podendo também ser considerada sinônimo de
processos de fossilização e de bioestratinomia. De acordo com Holz e Simões
(2002), a Tafonomia esclarece como um organismo tornou-se fóssil. Explica como os
processos sedimentológicos, pedogênicos, biogênicos e diagenéticos atuam na
história da fossilização de um organismo e de seus vestígios vitais. A tafonomia
compreende duas amplas subdivisões: a) Bioestratinomia, que engloba a história
sedimentar dos restos esqueléticos até o seu soterramento, incluindo as causas da
morte de um organismo, sua decomposição, transporte e soterramento e b)
Fossildiagênese ou processos de preservação. Além disso, alguns autores
consideram ainda na análise tafonômica, a fase do soerguimento tectônico e erosão,
bem como a influência das técnicas e métodos de coleta e preparação dos fósseis.
Nas análises tafonômicas, dois aspectos precisam ser considerados: em
primeiro lugar, a chamada fase curta que considera desde a morte do organismo até
sua cobertura por sedimentos, fase também chamada de sepultamento
(Bioestratinomia de WEIGELT, 1919, apud LOPES, 2000).; em segundo lugar, a
chamada de fase longa, que leva em conta o período desde a cobertura ou
CAPÍTULO 6 - ANÁLISE E INTERPRETAÇÃO DE DADOS

94
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
sepultamento até a coleta dos restos ou vestígios daquele organismo pelo
paleontólogo (Fossildiagênese).
Infelizmente muitos detalhes tafonômicos dos fitofósseis da Formação
Itaquaquecetuba deixaram de ser observados e registrados no momento da coleta,
tais como a orientação paleomagnética, a distância entre os espécimes, etc.
Apesar disso, se pode afirmar que, no nível analisado, os fitofósseis mais
abundantes são folhas de diversos tamanhos, predominando as micrófilas, e
pequenos caules depositados sem orientação preferencial, em alguns casos
sobrepostos e em outros distantes uns dos outros, perfazendo, em média, um total
de 80 pequenos fitofósseis por 100 cm2. Isto denota uma abundante vegetação nas
proximidades da área de deposição.
A presença de folhas sobrepostas, sem um direcionamento preferencial, isto
é, em disposição caótica, pode também evidenciar ambiente sedimentar lento e
calmo, permitindo o depósito sobreposto destes órgãos, ou seja, depósito por
decantação. A grande quantidade de folhas pode evidenciar ainda uma vegetação
do tipo decídua, tipo leguminosas que, em períodos sazonais de redução de água,
perderia suas folhas.
Por vezes, alguns fitofósseis foliares apresentam sinais de herbivoria, tanto
nas bordas, como nas áreas centrais dos limbos foliares, evidenciando a presença
de insetos e outros organismos (fungos, por ex.) que denotam também clima quente
e relações ecológicas a serem estudadas.
A integridade de muitos dos fitofósseis encontrados na Formação
Itaquaquecetuba indica proximidade da área de origem em relação ao local de
depósito e o desprendimento natural da planta-mãe.
Muitas folhas se apresentam inteiras, enquanto outras com fragmentação
principalmente das regiões apical ou basal. A fragmentação é devida a delicadeza
do material, pode ter causas bioestratinômicas, tendo ocorrido durante seu
transporte, ainda que curto, e seu sepultamento, ou durante coleta e transporte da
área fonte até a área de estudo (Laboratórios de Geociências da Universidade
Guarulhos – UnG). Há que se considerar ainda que os danos e fragmentações
ocorreram também durante o manuseio das amostras na abertura dos blocos de

95
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
argilito, quando muitos desses antigos registros foram expostos a atmosfera atual e,
principalmente, os piritizados acabaram se oxidando.
Igualmente, a alta frequência de folhas com textura do tipo cartácea seria
evidência de transporte curto, considerando que tal tipo de folha é de natureza
delicada e sofreria maiores danos, caso o transporte aquoso ou eólico fosse longo e
agitado.
Outrossim, a predominância do número de folhas sobre o de outras partes
vegetais constitui-se evidência de transporte curto e deposição em bordas de corpo
aquoso, considerando-se a fragilidade dos órgãos foliares, seu ataque rápido e
preservação por um filme bacteriano e sua deposição, normalmente, mais rápida
que a de outros órgãos vegetais, como caules, raízes e sementes que podem viajar
com o agente transportador por mais tempo, a flutuar e serem depositados mais
distalmente no ambiente sedimentar, conforme esquema de Duparque (1933, apud
EMBERGER, 1968) exibido na figura 6.1.
Figura 6.1 – Esquema de Duparque (1933) para ambientes lacustres ou lagunares hulheíferos. Extraído e traduzido a partir de Emberger (1968).
A rocha matriz composta de sedimento, no geral, de granulação fina (argilito
ou siltito) e acinzentado (rico em matéria orgânica), pode refletir também uma

96
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
sedimentação calma, em ambiente redutor, rico em acúmulo de matéria orgânica,
cuja evidência é corroborada pelo tipo de fóssildiagênese: incarbonização e
piritização ou marcassitização.
6.2 – A análise morfográfica dos fitofósseis foliares e sua interpretação.
A morfologia foliar está muito relacionada com a adaptação do vegetal ao
ambiente de vida. Portanto, estudos deste tipo permitem chegar a evidências
paleoambientais, paleoclimáticas, paleoecológicas, paleofitogeográficas e de
correlações estratigráficas. Como foi dito logo na introdução deste documento, é
possível realizar a análise de clima, composição do solo e ciclos de chuva, a partir
de seus processos adaptativos que podem assumir o papel de bioindicadores na
análise de eventos geológicos do passado.
As folhas constituem importantes elementos para inferências paleoclimáticas
de áreas restritas, por serem depositadas, relativamente, próximas aos locais de
origem e por apresentarem morfologia adaptativa, como resposta quase imediata às
mudanças das condições ambientais (CHRISTOPHEL; BLACKBUM, 1978;
CRISTALLI, 2006).
Cerca de 150 espécimes de fitofósseis, encontrados no afloramento
Itaquareia I da Formação Itaquaquecetuba (SP), foram analisados e avaliados
numericamente quanto a suas feições morfográficas ou de arquitetura foliar.
As feições morfo-adaptativas analisadas e os resultados foram os seguintes:
A) Tamanho: cerca de 90 % das folhas medidas corresponderam à categoria
de micrófilas (225- 2025 mm2) e poucas seriam nanófilas. Esse resultado
poderia estar relacionado com seleção realizada durante o transporte que,
embora curto, teria depositado as folhas maiores mais próximas à margem
dos corpos aquosos e as menores (embora em situação marginal também)
um pouco mais para dentro do corpo aquoso. Por outro lado, em caso de
depósitos de planície de inundação, as maiores ficariam mais distante da
margem do corpo aquoso, nos níveis mais altos alcançados pela inundação,
quando as águas perdem grande parte de sua força de transporte para, a

97
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
seguir, retornar ao leito normal dos rios trazendo as menores. Tamanho
foliar menor é, muitas vezes, relacionado com escassez de água, isto é,
com climas mais secos, ou com a posição mais alta nos níveis do dossel.
Também pode ser decorrente da composição florística, com predomínio de
plantas com folhas compostas, onde se estaria considerando tamanho de
folíolos e não propriamente, o tamanho total da fronde. Por isso, requer
combinação com outras evidências para melhor interpretação.
B) Margem Foliar: Na avaliação da margem foliar, entre os fitofósseis foram
detectados 94% com margem do tipo lisa. (Figuras 4.11; 4.12 e 6.3).
Figura 6.3: Resultados da avaliação da Margem Foliar.
As correlações entre clima e margem foliar são, suficientemente, fortes para
que os paleobotânicos utilizem rotineiramente, estatísticas de margem foliar a partir
de dados obtidos de floras fósseis como indicadores de climas antigos (BAILEY;
SINNOTT, 1916; WOLFE, 1978; GIVNISH, 1987; BAKER-BROSH; PEET, 1997;
ROYER et al., 2005).
As espécies de florestas tropicais perenifólias e úmidas contêm,
predominantemente, folhas de margem lisa ou inteira. O predomínio de margem
inteira também está relacionado aos ambientes de temperaturas mais altas (BAKER-
BROSH; PEET, 1997 e GREEN, 2006).
Como uma premissa, admite-se que espécies arbóreas de florestas
temperadas e frias contêm, quase que exclusivamente, folhas com margens

98
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
lobadas ou denteadas, enquanto as espécies perenifólias de áreas tropicais e
úmidas contêm, predominantemente, folhas de margem lisa ou inteira. É nítido o
crescimento da proporção de espécies arbóreas, com folhas lobadas ou denteadas,
ao longo do gradiente decrescente da temperatura média anual e crescente de
sazonalidade, desde a região tropical até as temperadas. Adicionalmente, há que
ressaltar, que o gradiente de umidade e sazonalidade de florestas tropicais úmidas
até florestas tropicais secas (isto é, dentre florestas de temperaturas e latitudes
semelhantes, mas de precipitações distintas) correspondem a uma porção também
crescente de espécies de folhas lobadas ou dentadas, apesar de ser menos
pronunciada que no caso anterior (BAKER-BROSH; PEET, 1997). Em síntese,
quanto à margem foliar, o predomínio de margem inteira está relacionado a
ambientes mais úmidos e de temperaturas mais altas (GREEN, 2006).
Figura 6.4 - Relação margem inteira e temperatura para nove amostras florísticas, extraída de Wilf (1997).
Wilf (1997), numa avaliação da aplicação de dados de análise da margem
foliar em relação à temperatura, estabeleceu o gráfico da figura 6.4. Aplicando a
percentagem de margem inteira fornecida pela flora estudada (94%) sobre esse
gráfico, verifica-se que a temperatura média anual correspondente seria de,
aproximadamente, 29ºC.
Ao utilizar o método de Green (2006) e considerando a mesma percentagem
de folhas de margem inteira (94%) chega-se à média anual de 26ºC.
A temperatura média anual atual de Itaquaquecetuba é ~ 23º C, (PORTAL
ALTO TIETÊ, 2012). Ao se comparar com os dados anteriores (26ºC - 29ºC),
verifica-se que a temperatura era em torno de ~5ºC superior aos índices atuais, isto
Temperatura
média anual o C
% de margem inteira

99
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
é, semelhante aos índices da Amazônia de hoje. Temperaturas semelhantes a
essas são estimadas para o tempo Paleógeno do centro-sul da América do Sul,
conforme Hinojosa (2005).
C) Textura da lâmina - A textura refere-se à espessura da lâmina foliar. Esta
espessura é uma feição muito relacionada à abundância de água no ambiente. A
textura cartácea corresponde a espessuras foliares do tipo folha de papel ou
pergaminho. Esse tipo de textura se manifesta mais frequente em ambientes de alta
umidade, isto é, folhas cartáceas estão relacionadas a climas mais úmidos enquanto
folhas coriáceas correspondem a uma adaptação a climas mais secos. Entre os
elementos analisados, 98% apresentaram textura do tipo cartácea, portanto
evidenciaria também um clima muito úmido.(Figura 6.5).
Figura 6.5: Resultado da avaliação da textura foliar.
D) Forma do ápice foliar refere-se ao contorno do quarto mais distal da folha,
oposto ao pecíolo. Na interpretação das relações entre tipos de ápice e fatores
climáticos, a partir da literatura pertinente, sabe-se que o predomínio de formas
apicais agudas está relacionado a climas mais úmidos (WOLFE, UPCHURCH,
1987; FITTIPALDI 1990).
Na tafoflora estudada, há um relativo predomínio de formas de ápice convexo
(60%), seguido de acuminado (24%) e de reto (7%), que somados dão 91% de
predomínio de formas agudas sobre formas mais arredondadas ou retusas. Mais

100
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
uma vez evidencia-se um paleoclima muito úmido, para a região durante a
deposição dos elementos da paleoflora de Itaquaquecetuba. (Figuras 4.6 e 6.6)
Figura 6.6: Resultado da avaliação das formas de ápices
E) O ângulo do ápice das folhas refere-se àquele obtido com vértice no ponto
extremo da folha, com a união das duas linhas que tocam nas margens a 25% do
comprimento foliar total. Cerca de 87% dos ápices são agudos (Figuras 4.6 e
6.7), confirmando o item anteriormente obtido.
Figura 6.7: Resultado da avaliação do ângulo do ápice

101
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
F) O ângulo da base refere-se àquele formado pelas duas linhas que perfazem o
limbo desde o ponto de inserção do pecíolo até 25% do comprimento foliar total.
Há um predomínio de 92% de base com ângulo agudo. .(Figuras 4.5 e 6.8).
Figura 6.8: Resultado da avaliação do ângulo da base
G) Por sua vez, a forma da base está relacionada com o tipo de pecíolo e sua
forma de inserção na lâmina foliar. A base foliar em 46% dos espécimes
analisados é cuneada e em 35% deles, decorrente. As predominantes bases
cuneadas, decorrentes e convexas (total 95%), segundo Green (2006),
possuem forte correlação com margem inteira, o que corroboraria a
associação às temperaturas elevadas (Figuras 4.7 e 6.9)
Figura 6.9: Resultado da avaliação das formas da base
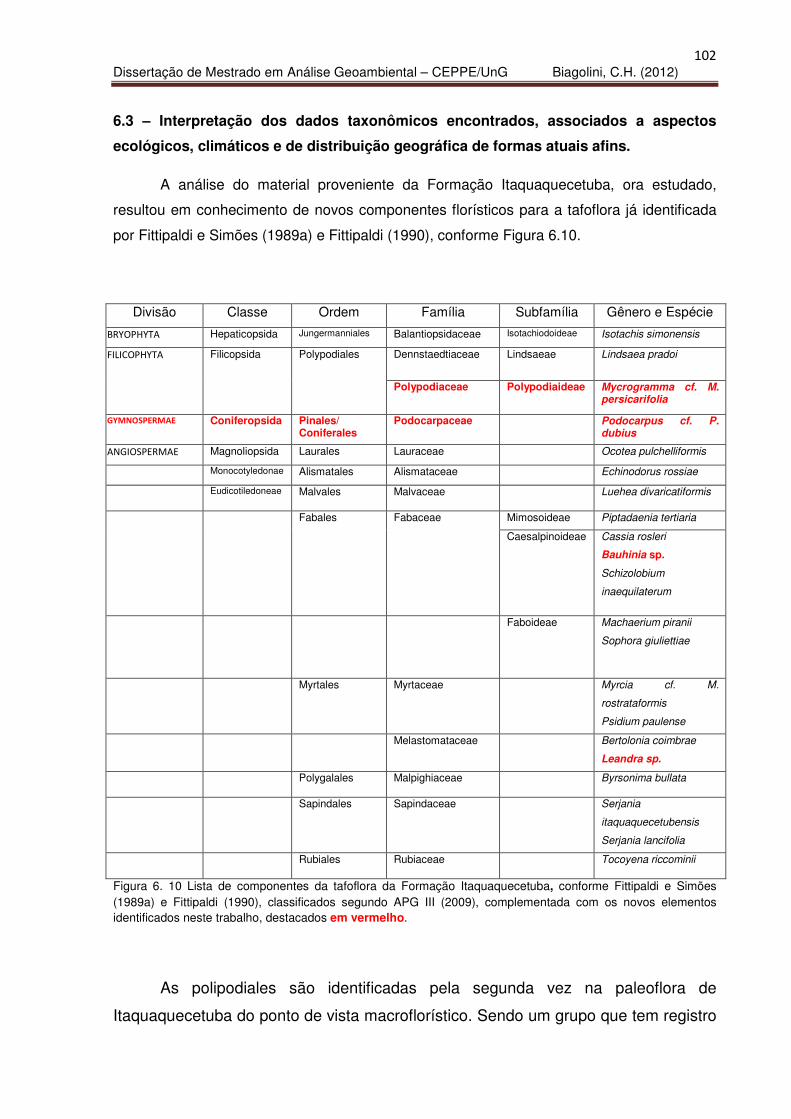
102
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
6.3 – Interpretação dos dados taxonômicos encontrados, associados a aspectos
ecológicos, climáticos e de distribuição geográfica de formas atuais afins.
A análise do material proveniente da Formação Itaquaquecetuba, ora estudado,
resultou em conhecimento de novos componentes florísticos para a tafoflora já identificada
por Fittipaldi e Simões (1989a) e Fittipaldi (1990), conforme Figura 6.10.
Divisão Classe Ordem Família Subfamília Gênero e Espécie
BRYOPHYTA Hepaticopsida Jungermanniales Balantiopsidaceae Isotachiodoideae Isotachis simonensis
Dennstaedtiaceae Lindsaeae Lindsaea pradoi FILICOPHYTA Filicopsida Polypodiales
Polypodiaceae Polypodiaideae Mycrogramma cf. M. persicarifolia
GYMNOSPERMAE Coniferopsida Pinales/ Coniferales
Podocarpaceae Podocarpus cf. P. dubius
ANGIOSPERMAE Magnoliopsida Laurales Lauraceae Ocotea pulchelliformis
Monocotyledonae Alismatales Alismataceae Echinodorus rossiae
Eudicotiledoneae Malvales Malvaceae Luehea divaricatiformis
Mimosoideae Piptadaenia tertiaria Fabales Fabaceae
Caesalpinoideae Cassia rosleri
Bauhinia sp.
Schizolobium
inaequilaterum
Faboideae Machaerium piranii
Sophora giuliettiae
Myrtales Myrtaceae Myrcia cf. M.
rostrataformis
Psidium paulense
Melastomataceae Bertolonia coimbrae
Leandra sp.
Polygalales Malpighiaceae Byrsonima bullata
Sapindales Sapindaceae Serjania
itaquaquecetubensis
Serjania lancifolia
Rubiales Rubiaceae Tocoyena riccominii
Figura 6. 10 Lista de componentes da tafoflora da Formação Itaquaquecetuba, conforme Fittipaldi e Simões (1989a) e Fittipaldi (1990), classificados segundo APG III (2009), complementada com os novos elementos identificados neste trabalho, destacados em vermelho.
As polipodiales são identificadas pela segunda vez na paleoflora de
Itaquaquecetuba do ponto de vista macroflorístico. Sendo um grupo que tem registro

103
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
desde o Neocretáceo, sua presença na flora eoceno-oligocena poderia ser
esperada. A espécie atual Microgramma persicariifolia afim à forma fóssil tem hábito
epífito e como habitat matas ciliares de florestas tropicais pluviais, podendo ocorrer
desde 0 m até 1590 m de altitude. A presença de forma afim na flora Neoeoceno-
Eo-oligocena da Grande São Paulo corrobora a idéia de clima quente e úmido para
a área naquela época.
Por sua vez, a presença de Podocarpaceae como formas remanescentes
desde o Gondwana (Eotriassico no caso) seria esperada. Aliás já era documentada
na forma de grãos de pólen anemófilos em várias as bacias brasileiras, desde a
Bacia de Santos até o Planalto Atlântico. Agora sua presença macroscópica, na
Bacia de São Paulo, é detectada como já havia sido, na Bacia de Taubaté,
anteriormente.
A análise dos fitofósseis do afloramento Itaquareia I, da Formação
Itaquaquecetuba, denotou grande número de fabáceas, embora não tenham sido
identificadas taxonomicamente. Este fato evidencia que, desde longo tempo, a
família Fabaceae é uma das maiores famílias de Angiospermas, de ampla
distribuição geográfica e considerada como a de maior riqueza de espécies arbóreas
nas florestas neotropicais, além de apresentar grande número de táxons endêmicos
(MAKI et al., 2010). Um novo gênero de Fabaceae foi detectado dentro da
composição florística da paleoflora de Itaquaquecetuba: Bauhinia. Trata-se de
gênero pantropical, como já foi dito, cuja seção Pauletia é tipicamente neotropical. A
espécie atual B. forficata com a qual o espécime estudado apresenta a maior
afinidade de carcterísticas, distribui-se desde o Rio de Janeiro e Minas Gerais até o
Rio Grande do Sul, principalmente, na floresta tropical atlântica. Ocorre, segundo
Lorenzi (2002), preferencialmente, em planícies aluviais e início de encostas.
A espécie de melastomatácea identificada aqui (Leandra sp.) apresenta certa
afinidade com L. Mattosii, Baumg. e D”El Rei Souza (2005), que ocorre atualmente
na área da grande São Paulo, restrita à altitude de 850 m, em bordas de matas
pluviais e capoeiras, próximas a cursos de rios, provavelmente em ambiente similar
ao das plantas da paleoflora de Itaquaquecetuba.

104
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Sabe-se que: todo conhecimento que se possa obter sobre as floras paleógenas,
neógenas e pleistocenas é muito importante, pois através deste conhecimento se
compreenderá a diversidade floral atual existente no planeta, como e porque se formou.
Relacionando com suas características geomorfológicas, biogeográficas e climáticas, pode-
se também obter dados importantes sobre o comportamento do planeta, ao longo de sua
história geológica.
A Formação Itaquaquecetuba, objeto de estudo paleobotânico nesta dissertação, é
uma unidade litoestratigráfica presente na Bacia Sedimentar de São Paulo. As idades desta
formação obtidas sobre diferentes bases tem gerado muita controvérsia, no entanto a idade
mais aceita é a paleógena, obtida por dados palinológicos, principalmente na época
Oligoceno.
A Formação Itaquaquecetuba representa uma unidade litoestratigráfica ainda pouco
compreendida, que aflora em esparsas áreas da Bacia de São Paulo.
Os fitofósseis, objeto de estudo deste documento, são provenientes de um
afloramento da Formação Itaquaquecetuba (Bacia de São Paulo), situado na área da
Mineradora Itaquareia I, junto à margem esquerda do Rio Tietê, na latitude de 23o 28’ 30”S e
longitude de 46o20’07``W, no município de Itaquaquecetuba (Região Metropolitana de São
Paulo).
A Bacia Sedimentar de São Paulo encontra-se assentada sobre o embasamento pré-
cambriano cristalino e caracteriza-se por possuir um substrato com soerguimentos e
abatimentos localizados, formados pela ação de falhas pré e pós-sedimentares. Seu formato
e distribuição de litofácies indicam que originalmente a bacia era um hemigrabén, controlado
por falhas normais originadas em reativações das zonas de cisalhamento proterozóicas.
O material fitofossilífero estudado, neste documento, passou por análise
paleobotânica do ponto de vista: morfográfico, taxonômico e tafonômico, com vistas às
interpretrações paleoambiental, paleoclimática, paleofitogográfica e paleoecológica de uma
tafoflora paleógena da Bacia Sedimentar de São Paulo, Formação Itaquaquecetuba.’
CAPÍTULO 7 – CONSIDERAÇÕES FINAIS

105
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
O material tafoflorístico examinado encontra-se preservado na forma de impressões,
contra-impressões, folhas carbonizadas e piritizadas ou marcassitizadas.
A observação da granulometria fina da matriz (analisada a partir dos blocos contendo
fitofósseis), verificada em análises granulométricas no laboratório da Universidade de São
Paulo-USP, confirmam que os depósitos ocorreram em sistema fluvial meandrante, onde
foram encontrados diversos seixos angulosos, em conglomerados, denotando curto
transporte, intercalado às camadas argilosas de deposição calma, no local do afloramento
estudado. Apesar da predominância da cor cinza nos argilitos estudados, alternando entre
claro e escuro, foram encontradas também, manchas de cor amarelo/ferruginoso,
evidenciando ambiente oxidante, alternado com redutor.
Em blocos com grande quantidade de fitofósseis preservados, observou-se que não
há uma direção predominante, ou seja, os fitofósseis preservados, representados em sua
maioria por folhas; apresentam uma distribuição caótica, fato que indica provavelmente
ambiente calmo e de baixa energia.
No início do Paleógeno, a paleogeografia mundial ainda era muito semelhante à do
Neocretáceo e os continentes, paulatinamente, moviam-se em direção às suas posições
atuais. A América do Sul, a Antártica, a Austrália e a África permaneciam quentes e úmidas.
Esse seria, no geral, o clima dominante durante o tempo de vida dos elementos
tafoflorísticos de Itaquaquecetuba.
As evidências de clima quente e úmido na Formação Itaquaquecetuba, característico
do período, são endossadas pelos resultados encontrados com grande número de folhas
com margem lisa, não lobada. As lâminas cartáceas e finas e os ápices tendendo a agudos
são típicos de umidade alta.
Embora não tenha sido o principal objeto de estudo desta dissertação, sementes
aparentemente de Mirtáceas ou ainda vestígios de frutos de leguminosas foram encontradas
durante as pesquisas e confirmam o clima indicado, assemelhando-se a diversas outras
tafofloras estudadas em outras formações e que ocorreram em condições climáticas
semelhantes.
Com estas considerações finais, espera-se que este documento possa contribuir,
fornecendo novos dados para o estudo da Formação Itaquaquecetuba.

106
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Alguns dos resultados obtidos nesta pesquisa foram apresentados na forma painel e
oral, no evento PALEO 2010, ocorrido na UNESP, em promoção do Núcleo Sao Paulo da
Soc. Brasileira de Paleontologia e no 12º Simpósio de Geologia do Sudeste e 16º Simpósio
de Geologia de MG, promovido pela Soc. Brasileira de Geologia, realizado na cidade Nova
Friburgo, entre 8 e 11 de novembro de 2011 (anexos), respectivamente.

107
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
AB’SABER, A.N. A planície Tietê no planalto paulistano, geomorfologia. v.57.
São Paulo: ABGE/SBG, 1978. 24 p.
______. Súmula geomorfológica do planalto paulistano. In: MESA REDONDA
SOBRE ASPECTOS GEOLÓGICOS E GEOTÉCNICOS DA BACIA SEDIMENTAR
DE SÃO PAULO, 1980, São Paulo. Publicação Especial SBG. São Paulo:
ABGE/SBG, 1980. p.33-36.
ABSY, M.L; SUGUIO, K. Palynological content and paleoecological: significance of
the drilled sediment samples from the Baixada Santista, Brazil. Anais da Academia
Brasileira de Ciências, Rio de Janeiro, v.47, p.235-242. 1976.
ADAMS, R.M.; et al. Global climate change and US agriculture. Oregon: Nature
Publishing Group, Oregon.1990. 345 p.
ALMEIDA, F.F.M. The system of Continental Rift bordering the Santos Basin, Brazil.
Anais da Academia Brasileira de Ciências, Rio de Janeiro, 48 (supl.), p.15-26,
1976.
______. Tectônica da Formação Itaquaquecetuba na grande São Paulo. In:
CONGRESSO BRASILEIRO DE GEOLOGIA, 33, 1984, Rio de Janeiro. Anais. Rio
de Janeiro: SBG. p.1794 -1808.
ALVES, F.C.; et al. Ocorrência da espécie exótica Lithobates catesbeianus (rã-touro)
em ambientes naturais nos municípios de Chapecó e Guatambu, Santa Catarina,
Brasil. Acta Amb. Catar. Santa Catarina, v.5, n.1-2, p.35-42, 2008.
APG III. An update of the Angiosperm Phylogeny Group classification for the orders
and families of flowering plants: APG III.121: Botanical Journal of the Linnean
Society, Publisher: Wiley Online Library,v.161, n.2, p.105–121, 2009.
ARCHANGELSKY, S. Fundamentos de Paleobotânica. Argentina: Ed. Universidad
Nacional de La Plata, 1970. 347 p. (Série Técnica y Didactica).
REFERÊNCIAS BIBLIOGRÁFICAS

108
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
ARCHANGELSKY,S.; GAMERRO, J.C. Pollen Grains found in Coniferous Cones
from the Lower Cretaceous of Patagonia Argentina. Review of Palaeobotany and
Palynology. Argentina: Universidad Nacional de La Plata, 1966. v.5, n.1-4, p.179-
182.
ASHAF, A.R.; STINNESBECK, W. Pollen und sporen an der kreide–Tertiärgrenze im
staate Pernambuco, Nordost–Brasikien: Paleontographica. Abt. B, v.208, p.39-51,
1988.
AXSMITH, B.J.; TAYLOR, T.N.; TAYLOR, E.L. Anatomically preserved leaves of the
conifer Notophytum krauseli (Podocarpaceae) from the Triassic of Antarctica.
American Journal of Botany, New York, v.85, p.704-713. 1998.
BAKER-BROSH, K.F.; PEET, R.K. The ecological significance of lobed and toothed
leaves in temperate foresttrees. Ecology v.78, p.1250–1255. Disponível em:
<http://dx.doi.org/10.1890/0012-9658. 078 [1250:TESOLA] 2.0.CO;2. 1997>. Acesso
em novembro/2011.
BALDONI, A.M.; TAYLOR, T.N. The ultrastructure of Trisaccites pollen from the
Cretaceous of southern Argentina. Review of Palaeobotany and Palynology, v.87,
n.81, p.323, 1982.
BARROSO, G.M. Sistemática de angiospermas do Brasil. Viçosa: ed. UFV, 1984.
v.2. 377 p.
BAUMGRATZ, J.F.A.; D’EL REI SOUZA, M.L. Duas novas espécies de Leandra
Raddi (Melastomataceae) para o Estado de São Paulo, Brasil. Acta Botânica
Brasílica, Rio de Janeiro, v.19, n.3, p.573-578. 2005.
BAYLEY, I.W.; SINNOT, E.W. The climate distribution of certain types of
Angiosperms Leaves. American Journal of Botany, New York, n.3 p.24-39, 1916.
BEDANI, E.F. Paisagem natural paleógena da Bacia Sedimentar de São Paulo,
no município de Guarulhos, Estado de São Paulo. 2008. 119 f. Dissertação
(Mestrado em Análise Geoambiental) – Centro de Pós-Graduação e Pesquisa,
Universidade Guarulhos, Guarulhos, 2008.

109
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
BERNARDES-DE-OLIVEIRA, M.E.C.; CASTRO-FERNANDES, M.C.; FRANCO-
DELGADO, S.G. Folhas de Mirtáceas na Formação Entre-Córregos, Paleógeno da
Bacia de Aiuroca, Sudeste de Minas Gerais, Brasil. In: SIMPÓSIO DO CRETÁCEO
DO BRASIL, 7, SIMPÓSIO DO TERCIÁRIO DO BRASIL, 1, 2006, Serra Negra.
Boletim de Resumos, Rio Claro: Ed. UNESP, 2006. p.22.
BERNARDES-DE-OLIVEIRA, M.E.C.; et al. Jazigo Rodovia Quiririm Campos do
Jordão, Km 11, (Tremembé), SP – (Macrofósseis Vegetais do Terciário). In:
SCHOBBENHAUS, C.; et al. (Ed). Sítios Geológicos e Paleontológicos do Brasil.
n.87, v.1, Brasília: Ed.SIGEP, 2002. p.55-62.
BERNARDES-DE-OLIVEIRA, M.E.C.; MUNE, S.E. A Origem e Evolução das
Angiospermas. Cap. 16. In: CARVALHO, I.S. Paleontologia. 3 Ed. Rio de Janeiro:
Editora Interciência, 2011. cap.16, p.347-383.
BERRY, E.W. Terciary plant from Brazil. Philadelphia: American Philosophical
Society, 1935. v.75, p.565-590.
______. Tertiary flora from the Río Pichileufú, Argentina. New York: Geological
Society of America Special Paper, 1938. v.12, p.1-149.
BEURLEN, K.; SOMMER, F.W. Restos de vegetal fóssil e tectônica da bacia
calcárea de Itaboraí Estado do Rio de Janeiro. Boletim Depto. Nacional de
Produção Mineral/Divisão de Geologia e Mineralogia. Rio de Janeiro, Boletim
149, p.7-27. 1954.
BISTRICHI, C.A. Análise estratigráfica e geomorfológica do Cenozóico da
região de Atibaia-Bragança Paulista, Estado de São Paulo. 2001. 184 f. Tese
(Doutorado em Geologia Regional)–Instituto de Geociências e Ciências Exatas,
Univesidade Estadual Paulista, Rio Claro, 2001.
BLOW, W.H. The Cenozoic Glolsigerimida. New York: Ed.E. J. Brill, 1979. 3v.

110
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
CALVILLO-CANADELL, L.; CEVALLOS-FERRIZ, S.R.S.; BAUHCISMORANII, G. et
sp. nov (Cercideae, Caesalpinieae), an Oligoceneplant from Tepexi de Rodríguez,
Puebla,Mex., with leaf architecture similar to Bauhinia and Cercis. Review of
Palaeobotany and Palynology.México, v.122, p.171-184, 2002.
CAMARGO, E.A.; GOLDENBERG, R. Leandra seção Leandraria (Melastomataceae)
no Estado do Paraná, Brasil. IHERINGIA, Sér. Bot., Porto Alegre, v.62, n. 1-2,
p.103-111, jan./dez. 2007.
CARDIM, R. Como era a vegetação original da cidade de São Paulo no
passado? Disponívelem: <Erro! A referência de hiperlink não é válida.. 2009> Acesso em
21/novembro/2011.
CARVALHO, I. S. Paleontologia. Rio de Janeiro: Editora Interciência, 2000. p.628.
CASSAB, R.C.T. Objetivos e Principios da Paleontologia. In: CARVALHO, I.
Paleontologia. Rio de Janeiro: Editora Interciência, 2000. cap.1, p.3.
CHRISTENHUSZ, J.M; ZHANG, X.C.; SCHNEIDER, H. A linear sequence of extant
families and genera of lycophytes and ferns. Phytotaxa, New Zealand, n.19, p.7-54.
2011.
CHRISTOPHEL, D.C., BLACKBURN, D.T. Tertiary megafossilflora of Maslin Bay,
South Australia. A Preliminary. Report. Alcheringa, n.2, p.311-319. 1978.
COIMBRA, A.M.; ATENCIO, D.; BRANDT NETO, M. Sulfatos secundários
associados aos aluviões antigos do Rio Tietê (Itaquaquecetuba-SP). In: XXXII
CONGRESSO BRASILEIRO DE GEOLOGIA, 32. 1980, Camboriú. Resumos.
Camboriú: SBG, p.1970-1981. 1980.
COIMBRA, A.M.; RICCOMINI, C.; MELO, M.S. A Formação Itaquaquecetuba:
Evidências de Tectonismo no Quaternário Paulista. In: SIMPÓSIO REGIONAL DE
GEOLOGIA, 4, 1983, São Paulo, Anais SBG, São Paulo, p.253-266. 1983.
COOPER, J.P. Short-day and low-temperatura induction in Lolium. Ann Botany
London in. press. Oxford, v.24, p.232-246. 1960.

111
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
COWEN, R. History of Life Fourth Edition. Oxford: Blackwell Publishing. 2005.
560p.
CRISTALLI, P.S. Macrofitofósseis em Tufos Calcários Quaternários do Norte da
Bahia como Indicadores Paleoclimáticos. 2006. 195 f. Tese (Doutorado em
Geociência) - Instituto de Geociências, Universidade de São Paulo, São Paulo,
2006.
CRONQUIST, A. The Evolution and Classification of Flowering Plants. New
York: Ed. Botanical Garden, 1988. 555 p.
CURVELLO, W.S. Sobre um Vegetal do Linhito de Fonseca, Minas Gerais. Anais da
Academia Brasileira de Ciências. Rio de Janeiro, v.17, n.3, p.293-296. 1955.
DATTA, S.C. An Introduction to Gymnosperms. London: Asia Publishing House,
1966.168 p.
DILCHER, D.L. Podocarpus from the Eocene of North America. Science, n.164,
p.299-301. 1969.
DOLIANITI, E. Contribuição a Flora Pliocênica de Fonseca, Minas Gerais. Anais da
Academia Brasileira de Ciências. Rio de Janeiro, v.21, n.3, p.239-244. 1949.
______. Contribuição a flora pliocênica de Fonseca, Minas Gerais. Anais da
Academia Brasileira de Ciências. Rio de Janeiro, v.21, n.3, p.303-306. 1950.
______. Frutos de Nypa no Paleoceno de Pernambuco, Brasil. Departamento
Nacional de Produção Mineral/Divisão de Geologia e Mineralogia. Boletim, Rio
de Janeiro, v.158, p.7-36. 1955.
DUARTE, L. Melastomataceae fósseis da Bacia Terciária de Fonseca, MG. Boletim
Departamento Nacional da Produção Mineral/Divisão de Geologia e
Mineralogia. Rio de Janeiro, n.161 p.7-32. 1956.

112
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
______. Pteridophita da Bacia de Fonseca, Minas Gerais. In: CAMPOS, D.A.;
FERREIRA, C. S., BRITO I.M & VIANA, C.F. (org). Coletânea de Trabalhos
Paleontológicos, (27). Brasília: Depto. Nacional da Produção Mineral, Série
Geologia, Seção Paleontológica e Estratigráfica, 1985. p.553-555.
______. Sobre uma flor de Bombacaceae, da Bacia Terciária de Fonseca, MG.
Anais da Academia Brasileira de Ciências, Rio de Janeiro, v.46, n.3-4, p.407-411.
1974.
DUARTE, L.; MANDARIM-DE-LACERDA, A.F. Flora Cenozóica do Brasil, Formação
Tremembé, Bacia de Taubaté; São Paulo. (Nota Preliminar). In: X CONGRESSO
BRASILEIRO DE PALEONTOLOGIA, 10, 1989, Curitiba, PR. Anais..., SBP, São
Paulo, p.879-884, 1987.
______. Flora Cenozóica do Brasil, Formação Tremembé, Bacia de Taubaté; São
Paulo. III. Frutos (Phitolacaceae e Leguminosae). In: CONGRESSO BRASILEIRO
DE PALEONTOLOGIA, 11, 1989, Curitiba, PR. Resumos das Comunicações SBP,
São Paulo, 1, p.395-410, 1989.
______. Flora Cenozóica do Brasil, Formação Tremembé, Bacia de Taubaté, São
Paulo I: Celastraceae, Loganeaceae e Typhaceae. Anais da Academia Brasileira
de Ciências, Rio de Janeiro, v.64, n.1, p.29-41, 1992.
DUARTE, L.; MELLO FILHA, M.C. Flórula Cenozóica de Gandarela, MG. Anais da
Academia Brasileira de Ciências. Rio de Janeiro,v.52, n.1, p.77-91. 1980.
DUTRA, T.; STRANZ, A. Clarificação e Diafanização de Folhas. In: T. L. DUTRA.
Técnicas e Procedimentos de Trabalho com Fósseis e Formas Modernas
Comparativas. São Leopoldo: Ed. Unisinos, 2002. p.27-29.
EFREMOV, J.A. Preliminary description of the new Permian and Triassic Tetrapoda
from USSR. Academie des Sciences de L’Union des Republiques Sovietiques
Socialistes. n.10, p.93-140, 1940.
EMBELGER, l. Les plantes ossiles dans leurs rapports avec les vegétaux
vivants. 2. ed. Paris: Ed. Paris Masson, 1968. 758p.

113
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
FANTON, J.C.M.; et al. O. Afloramento de Canoinhas, SC: Única localidade de
ocorrência de conífera permiana Krauselcladus da Bacia do Paraná. In: WINGE, M.,
et al. Sítios Geológicos e Paleontológicos do Brasil. v.2. Brasília: CPRM
(SIGEP), 2009, p.381-388.
FERREIRA, C.S.; SANTOS, A.S. Novos Dados Sobre a Geocronologia da Formação
Tremembé, Vale do Paraíba, SP, com base Palinológica. Anais da Academia
Brasileira de Ciências, Rio de Janeiro, v.54, n.1, p.264. 1982.
FITTIPALDI, F.C. Vegetais Fósseis da Formação Itaquaquecetuba (Cenozóico,
Bacia de São Paulo). 1990. 146 f. Tese (Doutorado em Geociência) – Instituto de
Geociência, Universidade de São Paulo, São Paulo, 1990.
______. Primeira ocorrência de briófitas na Bacia de São Paulo (Cenozóico). In:
CONGRESSO BRASILEIRO DE PALEONTOLOGIA, SÃO LEOPOLDO, 13, 1993,
São Leopoldo. Boletim de Resumos SBP. Porto Alegre, p.104, 1993.
FITTIPALDI, F.C.; SIMÕES, M.G. Fóssil plants from the Itaquaquecetuba formation
(Cenozoic of the São Paulo Basin) and their possible paleoclimatic significance. São
Paulo: Boletim IGc USP, São Paulo, n.7, p.183-204. 1989a.
______. Estado atual do conhecimento sobre a paleontologia da Bacia de São
Paulo. Workshop “Geologia da Bacia de São Paulo”. In: I WORKSHOP “Geologia
da Bacia de São Paulo”, São Paulo: Ed. Instituto Geológico-SMA e Sociedade
Brasileira de Geologia, 1989b. p.27-34.
______. Restos Vegetais da Formação Pindamonhangaba (Cenozóica, Bacia de
Taubaté). In: REUNIÃO DE PALEOBOTÂNICOS E PALINÓLOGOS, 7, 1990, São
Paulo. Resumos. São Paulo: DPE-IGUSP, 1990, p.18-19.
FOSTER, A. S. Foliar venation in angiosperms from an ontogenetic view point.
American Journal of Botany. n.39, p.752-766. 1952.

114
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
FRANCO-DELGADO, S.G. Tafoflora paleógena da Formação Entre-Córregos,
Bacia de Aiuruoca, Estado de Minas Gerais, Brasil. 2002. 163 f. Tese (Mestrado
em Geociências opção em Paleontologia Estratigráfica), Universidade Guarulhos,
Guarulhos, 2002.
FRANCO-DELGADO, S.G.; BERNARDES-DE-OLIVEIRA, M.E.C. Annonaceae e
Lauraceae da Formação Entre Córregos (Paleógeno) na Bacia de Aiuruoca:
Implicações Paleoclimáticas. Revista Brasileira de Paleontologia. v.7, n.2, p.117-
126. 2004.
FRIEDMAN, W. E. Evidence of a pre-angiosperm origin of endosperm, implications
for the evolution of flowering plants. Science, n.255, p.336-339. 1992.
GARCIA, M.J. Palinologia de Turfeiras Quaternárias do médio vale do Rio Paraíba
do Sul, Estado de São Paulo. Parte II. Gymnospermae e Magnolophyta.
Geociência, Revista Universidade de Guarulhos, Guarulhos-SP, v.3, n.6, p. 84-
107, 1998.
GARCIA, M.J.; et al. Considerações preliminares sobre a palinoflora quaternária dos
depósitos sedimentares do vale do Rio Embu-Guaçu, Estado de São Paulo, Brasil.
In: REUNIÃO DE PALEOBOTÂNICOS E PALINÓLOGOS, 9, 1997, Guarulhos.
Geociências, Revista Universidade Guarulhos. Guarulhos, v.2, número especial,
p.241. Resumos. UnG/SP; IG-SMA/SP,1997.
GARCIA, M.J.; et al. Nova Metodologia na Interpretação Palinoflorística da
Formação Itaquaquecetuba, na área da Mineradora Itaquareia (portos 1, 2 e 3),
Bacia de São Paulo, Brasil. In: REUNIÃO DE PALEOBOÂNICOS E PALINÓLOGOS,
11, 2004, Gramado, Boletim de Resumos UFRGS/UNISINOS. Porto Alegre. V.1,
p.65, 2004.
GARCIA, M.J.; et al. Floras Paleógenas Sul–Americanas no Contexto Mundial. In:
PALEONTOLOGIA: Cenários da Vida. Vol. 1,. Rio de Janeiro: Editora Interciências,
2007. p. 690-723.

115
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
GARCIA, M.J.; et al. Stratigraphy and paleo environments of the Tanque Basin,
southeastern Brazil. Revista Brasileira de Paleontologia, Porto Alegre, v.11, p.147-
168, 2008.
GARCIA, M.J.; SANTOS, M.; HASUI, Y. Palinologia da parte aflorante da Formação
Entre-Córregos, Bacia de Aiuruoca, Terciário de Minas Gerais, Brasil. Geociências
Revista Universidade Guarulhos, Guarulhos, v.5, número especial, p.259. 2000.
GIVNISH. T.J. Comparative Studies of Leaf Form: Assessing the Relative Roles of
Selective Pressures and Phylogenetic Constraints. New Phytologist, v.106, n.1,
p.131-160. 1987.
GONÇALVES, E.G.; LORENZI, H. Morfologia Vegetal: Organografia e dicionário
ilustrado de morfologia das plantas vasculares. Nova Odessa: Instituto Plantarum de
Estudos da Flora, 2007.
______. Morfologia Vegetal. Nova Odessa: Instituto Plantarum de Estudos da
Flora, 2011. 512 p.
GORCEIX, H. Bacia Terciária d’água doce nos arredores de Ouro Preto (Gandarela
e Fonseca) Minas Gerais – Brasil. Anais da Escola Minas.de Ouro Preto, n.3,
p.75-92. 1884.
GRADSTEIN, F.M.; OGG, J.; SMITH, A. A Geologic Time Scale. Cambridge:
Ed.Cambridge University Press, 2004. p.83-100.
GREEN, W.A. Loosening the CLAMP: An Exploratory Graphical Approach to the
Climate Leaf Analysis Multivariate Program. Paleontologia Eletrônica. v.9, n.2,
p.17p. 2006.
GUERRA-SOMMER, M.; PIRES, E.F. Growth ring analysis of fossil coniferous woods
from early cretaceous of Araripe Basin (Brazil). Anais da Academia Brasileira de
Ciências, Rio de Janeiro, v.83, p.409-423. 2011.

116
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
HASUI, Y.; CARNEIRO, C.D.R. Origem e evolução da bacia sedimentar de São
Paulo. In: MESA REDONDA SOBRE ASPECTOS GEOLÓGICOS E GEOTÉCNICOS
DA BACIA SEDIMENTAR DE SÃO PAULO, 1980, São Paulo. Publicação Especial
SBG. São Paulo, ABGE/SBG, p.5-13, 1980.
HICKEY, L.J. Classification of the architecture features of dicotydedonous leaves.
American. Journal Botany, n.60, p.17-33, 1973.
______A revised classification of the architecture of dicotyledonous leaves. In:
C.R.METCALFE e L.W. CHALK (eds). Anatomy of the dicotyledons. Oxford:
Oxford Science Publications-Clarendon Press., 1979. v.1, p.25-39.
HICKEY, L. J.; WOLFE, J. A. The bases of angiosperm phylogeny: vegetative
morphology. An. Missouri Bot. Gard., n.62, p.538-589, 1975.
HILL, R.S.; BRODRIBB, T.J. Southern conifers in time and space (Dedicated to the
memory of L.A.S. Johnson a great Australian botanist). A. Journal of Botany n.47,
p.639-696. 1999.
HILL, R.S.; WHANG, S.S. Dacrycarpus (Podocarpaceae). Macrofossils from Miocene
sediments at Ealands, eastern Australia. Australian Systematic Botany, n.13,
p.395-408. 2000.
HINOJOSA, L.F. Cambios Climáticos y Vegetacionales Inferidos a partir de
Paleofloras Cenozoicas del Sur de Sudamerica. Revista Geológica Del Chile, v.32,
n.1, p.95-115, 2005.
HOLZ, M.; SIMÕES M.G. Elementos fundamentais da Tafonomia. Porto Alegre:
Editora da UFRGS, 2002. 240 p.
IANNUZZI, R.; VIEIRA, C.E.L.L. Paleobotânica. Porto Alegre: Editora UFRGS,
2005. 167p.
JAPIASSU, A.M.S. Madeira Fóssil Humiriaceae de Iritúia. Estado do Pará. Boletim
Emílio Goeldi-Geologia, Belém, n.14, p.1-12. 1969.

117
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
JUDD, W.S. Plant systematic: A Phylogenetic Approach. 2 ed. Sunderland:
Sinauer Associates, 2002. 576p.
JUDD, W.S.; et al. Plant Systematic: A Philogenetic Approach. Sunderland
Massachusetts: Sinauer Associates,1999. 465p.
JUNQUEIRA, C.B. Camadas cruzadas de areias, gravas e cascalhos fluviais do Vale
do Pinheiros (“Campus” da Cidade Universitária). Geomorfologia, São Paulo, v.10,
p.79. 1969.
KUBITZKI, K. The Families and Genera of Vascular Plants. Berlin: Springer
Verlag,1996.
KUNZMANN, L.; MOHR, B.A.R.; BERNARDES-DE-OLIVEIRA, M.E.C.
Gymnosperms from the Lower Cretaceous Crato Formation (Brazil). Araucariaceae
and Lindleycladus (incertae sedis). Mitteilungen aus dem Museum für
Naturkunde Berlin, Geowissenschaftliche Reihe. n.7, p.155-174. 2004.
LAWG, Leaf Architecture Working Group. Manual of leaf architecture:
morphological description and categorization of dicotyledonous and net-veined
monocotyledonous angiosperms. Washington, DC: Smithsonian Institution, 1999.
LIMA, M.R.; AMADOR, E.S. Análise palinolôgica de sedimentos da Formação
Resende, Terciário do Estado do Rio de Janeiro, Brasil. Anais do 8º Congresso
Brasileiro de Paleontologia. Rio de Janeiro: 1983.
______. Análise Palinológica dos sedimentos da Formação Resende, Terciário do
Estado do Rio de Janeiro, Brasil. In: COLETÂNEA DE TRABALHOS
PALEONTOLÓGICOS, Brasília, DNPM. Série Geológica,27. Seção Paleontológica e
Estratigráfica. 2: 371-378. 1985.
LIMA, M.R.; CUNHA, F.L.S. Análise Palinológica de um nível linhítico da Bacia de
São José de Itaboraí, Terciário do Estado do Rio de Janeiro, Brasil. Anais da Acad.
Brasil. de Ciências, Rio de Janeiro, n.58, p.579-588. 1986.

118
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
LIMA, M.R.; DINO, R. Palinologia de amostras da Bacia de Bonfim, Terciário do
Estado de São Paulo, Brasil. Boletim IGUSP. São Paulo, Série Científica. n.15. p.1-
11. 1984.
LIMA, M.R.; MELO, M.S., COIMBRA, A.M. Palinologia de sedimentos da Bacia de
São Paulo, Terciário do Estado de São Paulo, Brasil. Revista do Instituto
Geológico, São Paulo, v.2, n.1. 1991.
LIMA, M.R.; MELO, M.S. Palinologia de depósitos rudáceos da região de Itatiaia,
bacia de Resende, RJ. Geonomos. Rio de Janeiro, v.2, n.1, p.12-21. 1994.
LIMA, M.R.; SALARD-CHEBOLDAEFF, M. Palinologie dês Bassins de Gandarela et
Fonseca (Eocene de L’Etat de Minas Gerais, Brésil). Boletim do Instituto de
Geociências. Universidade de São Paulo, São Paulo, série científica. n.12, p.33-
54.1981.
LIMA, M.R.; SALARD-CHEBOLDAEFF, M.; SUGUIO, K. Étude palynologique de la
Formation Tremembé, Tertiaire du Bassin de Taubaté (État de São Paulo, Brésil).
d'après les èchantillons du sondage nº 42 du CNP. In: CONGR. BRAS.
PALEONTOLOGIA DO RIO DE JANEIRO, 8, 1983, Rio de Janeiro. Anais do 8º
Congresso de Paleontologia. Rio de Janeiro, 1983.
LIMA, M.R.; VESPUCCI, J.B.O.; SUGUIO, K. Estudo Palinológico de uma camada
de linhito da Formação Caçapava, Bacia de Taubaté. Terciário do Estado de São
Paulo, Brasil. Anais da Academia Brasileira de Ciências. São Paulo, v.57, n.2,
p.183-197. 1996.
LOPES, R.F. Temas da Tafonomia. Madrid: Ed.Universidad Complutense de
Madrid-Depto Paleontologia, 2000.
LORENZI, H. Plantas Daninhas do Brasil: Terrestres, aquáticas, parasitas e
tóxicas. 3. ed. Nova Odessa: Ed.Plantarum, 2000. 608b p.
______. Árvores Brasileiras. Manual de identificação e cultivo de plantas arbóreas
do Brasil: Nova Odessa: Ed.Plantarum. 2002. 352 p.

119
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
MAGALHÃES, J. Sobre a ocorrência de vegetais fósseis na Fazenda São José,
Município de Guaxindiba, Rio de Janeiro. Revista Mineração e Metalurgia, Rio de
Janeiro, v.13, n.73, p.36. 1948.
MAIZATTO, J.R. Análise bioestratigráfica, paleoecológica e sedimentológica
das Bacias de Gandarela e Fonseca – Quadrilátero Ferrífero–MG, com base
nos aspectos palinológicos e sedimentares. 2001. 249f. Tese (Doutorado em
Evolução Crustal e Recursos Naturais) – Programa de Pós-graduação e Pesquisa,
Universidade Federal de Ouro Preto, Ouro Preto, 2001.
MAKI, E.S.; et al. Atlas Polínico da Vegetação da Área de Proteção Especial
(APE) Pau-de-Fruta, Diamantina, Minas Gerais – Família Fabaceae – Andira,
Chamaecrista e Zornia. São Paulo: XVIII Congresso da Sociedade Botânica de
São Paulo, 2010.
MANDARIM-DE-LACERDA, A.F. Étue d’une Flore Cenozóique du Bassin de
Tremembé (état de São Paulo, Brésil). 1985.140f. Rapporte de D.E.A de Biologie
Vegetale Tropicale, (Université Pierre et Marie Curie) Paris,1985.
MANDARIM-DE-LACERDA, A.F.; BERNARDES-DE-OLIVEIRA, M.E.C.; PONS, D.
Microscopia Eletrônica de Varredura de Macrofitofósseis da Formação Tremembé,
Eoterciário da Bacia de Taubaté, Estado de São Paulo, Brasil. In: CONGRESSO
NACIONAL DE BOTÂNICA, 47. 1996, Nova Friburgo. Resumos. Rio de Janeiro,
p497-498.1996.
MANDARIM-DE-LACERDA, A.F.; et al. Afloramentos fitofossilíferos da Formação
Tremembé, Município de Tremembé, Eoterciário da Bacia de Taubaté, Leste do
Estado de São Paulo, Brasil. Roteiro para a Excursão de Taubaté, Terciário. In:
REUNIÃO DE PALEOBOTÂNICOS E PALINÓLOGOS, 9, 1997, Guarulhos. Revista
da Universidade Guarulhos Geociências. Guarulhos: Universidade Guarulhos -
UnG, dez/1997. p.1-16.
MARCHIORI, J.N.C. Dendrologia das Angiospermas: Leguminosas. Santa Maria:
Ed. UFSM, 1997. 199 p.

120
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
MARCHIORI, J.N.C.; SOBRAL, M. Dendrologia das angiospermas: Myrtales.
Santa Maria: Editora UFSM, 1997. 304 p.
MARQUES-SOUZA, A.C. Ocorrência de pólen de Podocarpus sp (Podocarpaceae)
nas coletas de Frieseomelitta varia lepeletier 1836 (Apidae:Meliponinae) em uma
área de Manaus, AM, Brasil. Acta Botânica Brasilica, Manaus, v.24, n.2. jun. 2010.
MELLO, C.L.; SANT’ANNA, L.G.; BERGQVIST, L.P. Fonseca, MG - Vegetais
Fósseis do Terciário Brasileiro. In: SCHOBBENHAUS, C. et al – Sítios Geológicos
e Paleontológicos do Brasil. Brasília: SIGEP, DNPM, CPRM, 2002. p.73-79.
MELO, M.S.; CAETANO, S.L.V.; COIMBRA, A.M. Tectônica e sedimentação na área
das Bacias de São Paulo e Taubaté. In: CONGRESSO BRASILEIRO DE
GEOLOGIA, 1986, Goiânia. Anais. São Paulo: SBG, 1986. p.321-336.
MELO, M.S.; VINCENS, A.; TUCHOLKA, P. Contribuição á cronologia da Formação
Itaquaquecetuba, SP. Anais da Academia Brasileira de Ciências, Rio de Janeiro
v. 57, n.2. 1985.
MENDES, J.C.; PETRI, S. Geologia do Brasil. Rio de Janeiro: Instituto Nacional do
Livro- MEC, 1971. 207 p.
MERCES, M.F.B. Palinologia do Jazigo Quiririm Formação Tremembé, Bacia de
Taubaté: Implicações Palinocronoestratigráficas, Paleovegetacionais e
Paleoclimáticas. 2005. 85 f. TCC (Graduação em Ciências Biológicas) – Curso de
Ciências Biológicas, Universidade Guarulhos, Guarulhos, 2005.
MERCES, M.F.B.; GARCIA, M.J.; BERNARDES-DE-OLIVEIRA, M.E.C. A
Paleopalinologia do Jazigo Quiririm Formação Tremembé, Bacia de Taubaté:
Implicações Paleoambientais. In: SIMPÓSIO DO CRETÁCIO DO BRASIL, 7,
SIMPÓSIO DO TERCIÁRIO DO BRASIL, 1, 2006, Serra Negra. Boletim de
Resumos. Rio Claro: UNESP, 2006. p.85.
MEYER-BERTHAUD, B.; TAYLOR, T.N. A probable conifer with podocarpaceae
affinities from the Triassic of Antarctica. Review of Paleobotany and Palinology.
n.67, p.179-198. 1991.

121
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
MODENESI, M.C. Significado dos depósitos correlativos quaternários em Campos
de Jordão – SP. Implicações paleoclimáticas e paleoecológicas. Boletim do
Instituto Geológico, São Paulo, Boletim 7, p.155. 1988.
MUSSA, D. Paleobotânica: Conceituação Geral e Grupos Fósseis. Rio de Janeiro:
Editora Interciência, 2004. cap.28, p.413-508.
NAYAR, B.K. A Phylogenetic Classification of the Homosporous Ferns. Vienna:
Ed. Intern. Assoc. for. Plant Taxonomy (IAPT), 1970. p.229-236.
OGG, J.G.; OGG, G.; GRADSTEIN, F.M. The concise geologic time scale.
Cambridge: Cambridge University Press, 2008. p.177.
OLIVEIRA, A.I. Linhito no Brasil: Mineração e Metalurgia. Rio de Janeiro, v.2 n.8,
p.105-107. 1937.
OLIVEIRA, A.I.; LEONARDOS, O.H. Geologia do Brasil. 3.ed. Mossoró: Coleção
Mossoroense, 1978. p.685-686.
OLIVEIRA e SILVA, M.I.M.N. Flórula da bacia de Fonseca, Minas Gerais, Brasil.
1982. 199 f. Dissertação (Mestrado em Botânica) – Centro de Pós-Graduação em
Botânica, Universidade Federal do Rio de Janeiro, Rio de Janeiro, 1982.
PAES LEME, A.B. Sobre a Formação de Linhito de Caçapava. Rio de Janeiro:
Pap. Macedo, 1918. 37 p.
PONÇANO, W.L. Mapa Geomorfológico do Estado de São Paulo. São Paulo: IPT,
2v. 1981.
POLE, M. The Proteaceae record in New Zeland. Australian Systematic Botany,
1998. p.343-372.
QUATTROCCHIO, M. Palynology and paleocommunities of the Paleogene of
Argentina. Revista Brasileira de Paleontologia. Porto Alegre, v.9 n.1, p.101-108.
2006.

122
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
RAO, A.N.; BOSE, N.N. Podostrobus Gen. Nov., a Petrified Podocarpaceous
Male Cone from the Rajmahal Hills. India: Palaeobotanist, 1971. p.3.
RAPINI, A. Sistemática Vegetal. Feira de Santana: Universidade Estadual Feira de
Santana, 2008. 200p.
RICARDI-BRANCO, F.; CAIRES, E.T.; SILVA, A.M. Levantamento de Ocorrências
Fósseis nas Pedreiras de Calcário do Subgrupo Irati no Estado de São Paulo, Brasil.
Revista Brasileira de Geociências, São Paulo, 2008.
RICCOMINI, C. Rift Continental do Sudeste do Brasil. 1989. 256 f. Tese
(Doutorado em Geociência) - Instituto de Geociências, Universidade de São Paulo,
São Paulo, 1989.
RICCOMINI, C.; SANT’ANNA, L.G.; FERRARI, A.L. Rift Continental do Sudeste do
Brasil. In: MANTESSO-NETO. Evolução Geológica. São Paulo: Ed. Beca, 2004.
p.383-406.
RIOS-NETTO, A.M. Taxonomia e sistemática. In: CARVALHO, I. Paleontologia.
2.ed. Rio de Janeiro: Interciência, 2010. cap.6, p.83-144.
RODRIGUES LIMA, A.A.; AFFONSO, P. Levantamento de Leandra Raddi no Núcleo
Carucutu, Parque Estadual da Serra do Mar, São Paulo. Revista do Instituto
Florestal de São Paulo. São Paulo, v.22, n.1, p.15-32. 2010.
ROYER, D.L.; et al. Correlations of climate and plant ecology to leaf size and shape:
potential proxies for the fossil record. American Journal of Botany. New York, v.
92. p.1141-1151. 2005.
SALLUM, A. Tabela estratigráfica internacional traduzida. 2010. Disponível em:
<http://www.igeologico.sp.gov.br/downloads/folders/Tabela Estratigráfica
Internacional>. Acesso em: 28/11/2011.

123
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
SANTOS, D.B. Palinologia de amostras da Seção-Tipo da Formação
Itaquaquecetuba, Bacia de São Paulo, na Mineradora Itaquareia I: Implicações
palinocronoestratigráficas e paleoambientais. 2005. 133 f. TCC (Graduação em
Ciências Biológicas) – Curso de Ciências Biológicas -Universidade Guarulhos,
Guarulhos, 2005.
______. A Paleopalinologia na Reconstituição da Paisagem Terciária da Formação
Itaquaquecetuba (Mineradora Itaquareia I), Município de Itaquaquecetuba, Estado de
São Paulo, Brasil. In: SIMPÓSIO ARGENTINO DE PALEOBOTÂNICA Y
PALINOLOGIA BAHIA BLANCA (ARGENTINA), 13, 2006, Argentina. Anais. São
Paulo: SBG, 2006. p.72.
______. A Paleopalinologia na Reconstituição da paisagem Paleógena, na
Formação Itaquaquecetuba (Mineradora Itaquareia 1), Bacia de São Paulo,
Brasil. 2008. 204 f. Dissertação (Mestrado em Análise Geoambiental) – Centro de
Pós-Graduação e Pesquisa, Universidade Guarulhos, Guarulhos, 2008. Disponível
em:<http://tede.ung.br/tde_arquivos/1/TDE20101028T080542Z119/Publico/Danieli%
20Bento%20dos%20Santos%20I.pdf>. Acesso em:18/07/2012.
______. A Paleopalinologia na Reconstituição da Paisagem Paleógena na
Formação Itaquaquecetuba (Mineradora Itaquareia 1), Bacia de São Paulo,
Brasil. 2009. 302 f. Dissertação (Mestrado em Análise Geoambiental) – Centro de
Pós-Graduação e Pesquisa, Universidade Guarulhos, Guarulhos, 2009.
SANTOS, D.B.; et al. Composição Paleopalinoflorística dos Depósitos Terciários da
Formação Itaquaquecetuba (Mineradora Itaquareia I), Município de Itaquaquecetuba,
Estado de São Paulo, Brasil. In: SIMPÓSIO BRASILEIRO DO CRETÁCIO, 7,
SIMPÓSIO DO TERCIÁRIO DO BRASIL, 1, 2006, Serra Negra. Boletim de
Resumos Rio Claro: UNESP, 2006. v.1, p.20-20.
SANTOS, M. Serra da Mantiqueira e Planalto do Alto do Rio Grande: A Bacia de
Terciária de Aiuroca e Evolução Morfotectônica. 1999. 134f. Tese (Doutorado em
Geociência) - Instituto de Geociência e Ciências Exatas – UNESP, Rio Claro, 1999.

124
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
SCHNEIDER, H.; KENRICK, P. An Early Cretaceous root clibing Lindsaeaceae and
its significance for calibrating the diversification of Polypodiaceous ferns. Review of
Paleobotany and Palynology. Department of Botany, Chicago, v.115, n.11, p.33-
41. 2001.
SCOTESE, C.R. Calibrating the Late Ordovician glaciation and mass extinction by
the eccentricity cycles of Earth'sorbit Geology.Geological Society, London, n.28,
p.967-970, nov. 2000.
SIMPSON, M.G. Plant systematics. New York: Ed. Elsevier, 2006. (Academic
Press).
SMITH, A.R.; et al. A classification for Extant Ferns. Taxon. California:
International Association for Plant Taxonomy, 2006. v.55, n.3, p.705-731. 2006.
SOUZA, V.C.; LORENZI, H. Botânica Sistemática: Guia ilustrado para identificação
das famílias de Angiospermas da flora brasileira, baseado em APG II. 2 ed. Nova
Odessa: Ed. Plantarum, 2008. 221 p.
STOCKEY, A.R.; KO, H. Cuticle micromorphology of Dacrydium (Podocarpaceae)
from New Caledonia. Botany Gaz.-The University of Chicago, Chicago,138-
149.1988.
SUÁREZ.M.; DE LA CRUZ, R.; TRONCOSO, A. Tropical/subtropical Upper
Paleocene-Lower Eocene fluvial deposits in eastern central Patagonia. Journal of
South American Earth Sciences, Chile, n.13, p.527-536. 2000.
SUGUIO, K. Estudo dos troncos de árvores lignificadas dos aluviões antigos do rio
Pinheiros (SP). Significados geocronológico e possivelmente paleoclimático. In:
CONGRESSO BRASILEIRO DE GEOLOGIA, 25, 1971. São Paulo, Anais São
Paulo: SBG, 1971. v.1. p.63-69.
SUGUIO. K.; COIMBRA. A.M.; CATTO, A.J. Estudo comparativo dos sedimentos e
rochas cristalinas circundantes da Bacia de São Paulo (granulometria e minerais
pesados). In: XXVI CONGRESSO BRASILEIRO DE GEOLOGIA, 26. 1972, Belém,
PA. Anais... Belém, PA: SBG. 1972. p.141-152.

125
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
SUGUIO, K.; MUSSA, D. Madeiras fósseis dos aluviões antigos do Rio Tietê.
Boletim IG. USP, São Paulo, v.9, 1978.
SUGUIO.K.; TAKAHASHI, L.J. Estudo dos aluviões antigos dos rios Pinheiros e
Tietê, São Paulo, SP. Anais da Academia Brasileira de ciências, Rio de Janeiro,
v.42, p.555-570. 1970.
SUNDARAM, D.; SUGUIO, K. Nota preliminar sobre uma assembléia mioflorística na
Formação Pariquera-Açu, Estado de São Paulo. In: DEPARTAMENTO NACIONAL
DE PRODUÇÃO MINERAL. Coletânea de trabalhos paleontológicos. n.27.
Brasília: Série Geologia-Seção de Paleontologia e Estratigrafia, 1985. p.503-505.
TAKIYA, H. Aplicação de métodos quantitativos espaciais a dados geológicos
da Bacia de São Paulo. 1991. 109 f. Dissertação (Mestrado em Geociências) -
Instituto de Geociências, Universidade de São Paulo, São Paulo, 1991.
______. Estudo da sedimentação Neogênico-Quaternária no município de São
Paulo: Caracterização dos depósitos e suas implicações na geologia urbana. 1997.
152 f. Tese (Doutorado em Geociência) - Instituto de Geociência, Universidade de
São Paulo, São Paulo, 1997.
TAYLOR, T.N.; TAYLOR, E.L.; KRINGS, M. Paleobotany the Biology Evolution of
Fóssil Plants. London: Editora Elsevier-AP(Academic Press), 2009.
TOLENTINO, M. Algumas considerações sobre carvão da Bacia de São Paulo.
São Carlos: Publ. Escola Engenharia de São Carlos, USP. São Carlos, 1965.
TOWNROW, J.A. On Voltziopsis, a southern conifer of Lower Triassic age.
Papers and Proceedings of the Royal Society of Tasmania. 1967. n.101, p.173-188.
TRYON, R.M.; TRYON, A.F. Ferns and allied plants, with special reference to
tropical America. New York: Ed. Springer-Verlag, 1982.
TUFANO, P. Tafoflora Neógena da Formação Pindamonhangaba, Bacia de
Taubaté, em Jacareí, Estado de São Paulo, Brasil. 2009. 200 f. Dissertação
(Mestrado em Análise Geoambiental) – Centro de Pós-Graduação e Pesquisa,
Universidade Guarulhos, Guarulhos, 2009.

126
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
VAZ, A.M.S.F.; TOZZI, A.M.G.A. Bauhinia ser. Cansenia
(Leguminosae:Caesalpinoideae) no Brasil. Rodriguésia, Rio de Janeiro, n.54, p.55-
143. 2003.
______. Sinopse de Bauhinia sect. Pauletia (Cav.) D.C.(Leguminosae,
Caesalpinoideae e Cercideae) no Brasil. Revista Brasileira de Botânica. São
Paulo,n.28, p.477-491. 2005.
WICKERT, W.Q. Contribuição ao Estado de Paleontologia da Bacia de Taubaté,
Estado de São Paulo. 1974. 69 f. Dissertação (Mestrado em Geociência) - Instituto
de Geociências, Universidade Federal do Rio de Janeiro, Rio de Janeiro, 1974.
WILF, P. When are Leaves good Thermometers? A New Case for leaf margin
Analysis. New York, v.23, n.3, p.373-390.1997.
WILLIS, K.J.; MC.ELWAIN, J.C. The Evolution of Plants. Oxford: Ed.University
Press. Oxford. 2002. 378 p.
WOJCIECHOWSKIE, M.F.; JOHANNA M.; BRUCE J. Fabaceae legumes. Version
14 June 2006. Disponível em: http://tolweb.org/Fabaceae/21093/2006.06.14>,
acesso em:18/06/2010.
WOLFE, J.A. A Paleobotanical Interpretation of Tertiary Climates. In: THE
NORTHEM HEMISPHERE. American Scientist, v.66, Ed.Scientific Research
Society, 1978. 694 p.
WOLFE, J.A.; UPCHURCH, G.R. Mid-Cretaceous to Early Tertiary vegetation and
climate: Evidence from fossil leaves and woods. In: FRIIS, E.M., CHALONER, W.G.
AND GRANE, P.H. (eds). The origins of angiosperms and their biological
consequences. Cambridge: Ed. University Press, 1987. p.75-105.
WUNDERLIN, R.P.; LARSEN, K.; LARSEN, S.S. Reorganization of the Cercideae
(Fabaceae: Caesalpinioideae). Det Kongelige Danske Videnskabernes Selskab
Biologiske Skrifter, v.28, p.1–40. 1987.

127
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
YAMAMOTO I. T. Palinologia das Bacias Tafrogênicas do Sudeste (bacias de
Taubaté, São Paulo e Resende): Análise bioestratigráfica integrada e interpretação
paleoambiental. 1995. 217 f. Dissertação (Mestrado em Geociência) - Inst. de
Geociências e Ciências Exatas, Universidade Estadual Paulista, Rio Claro, 1995.
ZANÃO, R.; CASTRO, J.C.; SAAD, A.R. Caracterização Geométrica de um Sistema
Fluvial, Formação Itaquaquecetuba, Terciário da Bacia de São Paulo. Revista
Geociências, Guarulhos, v.25, n.3, p.307 – 315. 2006a.
______. Caracterização geométrica de reservatórios fluviais com base na Formação
Itaquaquecetuba, Terciário da Bacia de São Paulo, SP. In: SIMPÓSIO DO
CRETÁCEO DO BRASIL,7 / SIMPÓSIO DO TERCIÁRIO DO BRASIL.1, 2006, Serra
Negra. Resumos. Serra Negra, SP, p.135, 2006b.
SITES CONSULTADOS PARA OBTENÇÃO DE DADOS E IMAGENS:
<http//celestia.albacete.org/celestial/deriva/paleocene.htm>:(acesso em
fevereiro/2012).
<http://maps.google.com.br/> (acesso outubro/2011).
<http://sigam.ambiente.sp.gov.br/2011> (acesso novembro/2011).
<http//www2.nau.edu/rcb7/035marect..jpg> (acesso em novembro/2011).
<http://en.wikipedia.org/wiki/polypodiaceae> (acesso em março/2012).

128
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
<http://biogeodb.stri.si.edu//herbarium/species/24376/?fam=polypodiaceaeepage=2
(acesso em março/2012)>
<http://www.freewebs.com/rapinibot/embriofitas/parte1.pdf> acesso em 25/06/2010.
http://www.portalaltotiete.com.br/index_cidade_itaqua.asp (acesso em 04/07/2012).

129
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
ESTAMPA I
Figura 01: Martelete utilizado na abertura de blocos de argila
Figura 02: Ferramentas de aço inoxidável, utilizadas na retirada de sedimentos finos
e indesejados sobre o fitofóssil analisado, ou ao redor do mesmo.
Figura 03: Ferramentas para auxilio na limpeza do fitofóssil a ser analisado.

130
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Figura 01 : Martelete
Figura 02: Instrumentos para limpeza e desbaste
Figura 03: Instrumentos para limpeza e desbaste.
ESTAMPA I

131
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
ESTAMPA II
Figura 01: Fitofóssil Microgramma cf., M. persicarifolia ITQ pb 026B/11
Figura 02: Forma atual de Microgramma persicariifolia

132
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Figura 01 Figura 02
ESTAMPA II

133
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
ESTAMPA III
Figura 01: Imagem ampliada de folha de Podocarpus cf. P.dubius., retirada do
material de estudo, obtida através de *M.E.V,
Figura 02: Imagem ampliada em detalhe da região próxima ao limbo, com detalhes
da formação celular e presença de possíveis sinais de estômatos obtida através de
*M.E.V.
Figura 03: Imagem comparativa com folha fossilizada e folha de Podocarpus sp.,
atual.
Figura 04: Fitofóssil, com folhas de Podocarpus cf. P.dubius
Figura 05: Imagem com fragmento de folha de Podocarpus cf. P. dubius, retirado
do material analisado, utilizando-se *M.E.V.
Figura 06: Imagem com detalhes da formação de “espinhos nas margens da folha
fossilizada, obtida utilizando-se *M.E.V.
*Microscopia Eletrônica de Varredura

134
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Figuras 01, 02, 03, 04, 05 e 06.
ESTAMPA III
1 2
3 4
5 6

135
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
ESTAMPA IV
Figura 01 e 02: Imagem em detalhes de folhas do fitofóssil de Podocarpus cf.
P. dubius., retirado da área de estudo.
Figura 03: Imagem ampliada de folha de Podocarpus sp. , fossilizada, obtida
através de *MEV.
Figura 04: Imagem ampliada da região apical de Podocarpus cf. P. dubius. Obtida
através de *MEV.
Figura 05: Imagem em detalhes da região apical de Podocarpus cf. P. dubius,
retirado da área de estudo, obtida através de *MEV.
Figura 06: Imagem em detalhes da formação celular de um fragmento de folha do
fitofóssil estudado, obtida através de *MEV.
*Microscopia Eletrônica de Varredura
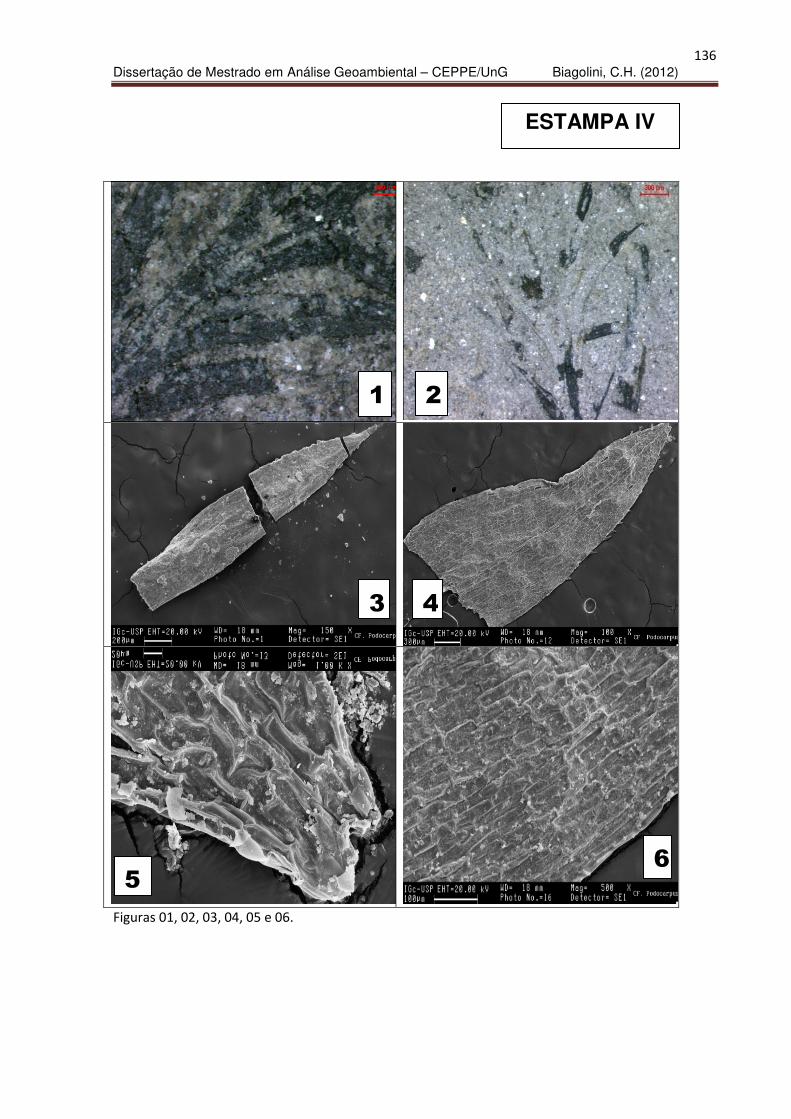
136
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Figuras 01, 02, 03, 04, 05 e 06.
ESTAMPA IV
1 2
3 4
5 6

137
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
ESTAMPA V
Figura 01: Imagem em detalhe de folhas da região apical Podocarpus cf.
P. dubius, fossilizado e encontrado na área de estudo.
Figura 02: Podocarpus sp. forma atual.

138
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Figura 01
Figura 02
ESTAMPA V

139
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
ESTAMPA VI
Figura 01: Imagem em detalhes de fitofóssil Bauhinia sp. ou cf. B. divaricata
encontrado na área de estudo.
Figura 02: Lado inferior da folha Bauhinia sp, forma atual

140
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Figura 01
Figura 02
ESTAMPA VI

141
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
ESTAMPA VII
Figura 01: Imagem em detalhes de fitofóssil Leandra sp. , encontrado na área de
estudo.
Figura 02: Folha de Leandra sp., forma atual

142
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Figura 01 Figura 02
ESTAMPA VII

143
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
ESTAMPA VIII
Figura 01: Tabela de classificação utilizada no estudo e análise dos fitofósseis da
Formação Itaquaquecetuba.

144
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)

145
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
ESTAMPA IX
Figura 01: Lupa com iluminação para observação, manipulação e análise dos fitofósseis.
Figura 02: Equipamento de observação com encaixe de máquina fotográfica para obtenção de imagens do material analisado.
Figura 03: Equipamento de observação que permite a confecção do desenho das estruturas de venação do fitofóssil analisado.

146
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Figura 01
Figura 02
Figura 03
ESTAMPA IX

147
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
ESTAMPA X
Figura 01: Foto do Afloramento Itaquareia 1, com indicação do ponto de retirada dos fitofósseis.
Figura 02: Área de trabalho e as “lagoas” formadas no local com marcas de cor alaranjada indicando a presença de oxidação devido ao material ferruginoso presente entre os minerais.
Figura 03: Pessoal do Laboratório de Geociência da Universidade Guarulhos-UnG, realizando coleta de fitofósseis, ocorrida em março/2010.

148
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Figura 01
Figura 02
Figura 03
ESTAMPA X

149
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
ANEXOS
Apresentação de resultados parciais obtidos durante as pesquisas para dissertação de mestrado no
12o Congresso de Geologia do Sudeste .
Novembro/2011
Local: Nova Friburgo / RJ
Forma de apresentação: Oral e Painel

150
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
ANÁLISE MORFOGRÁFICA DA TAFOFLORA ITAQUAQUECETUBA, PALEÓGENO DA BACIA DE SÃO
PAULO, BRASIL E SUAS IMPLICAÇÕES PALEOCLIMÁTICAS
MORPHOGRAPHIC ANALYSIS OF THE ITAQUAQUECETUBA TAPHOFLORA, PALEOGENE OF THE THE
SÃO PAULO BASIN, BRAZIL AND THEIR PALEOCLIMATIC IMPLICATIONS
CARLOS H. BIAGOLINI 1; MARY E.C. BERNARDES-DE-OLIVEIRA 1, 2
1 Mestrado em Análise Geoambiental do CEPPE/UnG;
2 Programa de Pósgraduação em Geoquímica e
Geotectônica do IGc/USP.
[email protected]; [email protected]
A Formação Itaquaquecetuba, da bacia de São Paulo, apresenta nas camadas silto-argilosas,
fitofósseis como impressões, piritizações e incarbonizações. A análise morfográfica de mais de 100
espécimes foliares pertencentes à sua tafoflora, provenientes de afloramento da Mineradora
Itaquareia I, Município de Itaquaquecetuba (SP), possibilitou extrair informações de caráter
paleoclimático. A orientação caótica de seus eixos longos, sobre as camadas da rocha matriz fina,
permitiu inferir sedimentação em ambiente calmo. A textura cartácea de 99% dos espécimes está
relacionada à alta umidade. Os 95% de folhas de margem lisa, sugere uma temperatura média
anual,de 26 ºC superior a atual (18º) na área. O teor de 90% de ápices agudos ou acuminados leva a
inferir um clima altamente úmido. As bases foliares predominantemente cuneadas, decorrentes ou
convexas (~94%), associadas a margens inteiras corroboram na evidência de temperaturas elevadas.
Essas condições climáticas foram observadas na fase “greenhouse” do Paleógeno, idade sugerida
pela Palinologia.
Palavras-chave: Paleógeno, Paleobotânica, Itaquaquecetuba e greenhouse.

151
Dissertação de Mestrado em Análise Geoambiental – CEPPE/UnG Biagolini, C.H. (2012)
Poster apresentado no 12o Congresso de Geologia do Sudeste,
Nova Friburgo-RJ em novembro/2011. (Forma oral e painel)

![“INICIATIVAS DAS ESCOLAS PÚBLICAS E PARTICULARES NA ...tede.ung.br/bitstream/123456789/225/1/Maria das Gracas.pdf · obesidad infantil del municipio de Amparo-SP [Disertación]](https://img.document.onl/doc/110x75/5bd5d7cd09d3f26c3e8c7f4d/iniciativas-das-escolas-publicas-e-particulares-na-tedeungbrbitstream1234567892251maria.jpg)



























