Embed Size (px)
Citation preview

LIGNINA DE PLÂNTULAS DE Lophantera lactescens
DUCKE
Jairo de Oliveira Tenorio
Rio de Janeiro
Janeiro de 2010

ii
JAIRO DE OLIVEIRA TENORIO
Aluno do Curso de Biotecnologia
Matrícula 0623800230
LIGNINA DE PLÂNTULAS DE Lophantera lactescens
DUCKE
Trabalho de Conclusão de Curso,
TCC, apresentado ao Curso de
Graduação em Biotecnologia da
UEZO como parte dos requisitos
para a obtenção de grau de
Tecnólogo em Biotecnologia, sob a
orientação do Profº. Heber dos
Santos Abreu.
Rio de Janeiro
Janeiro de 2010

Tenorio, Jairo de Oliveira.
Lignina de Plântulas de Lophantera lactescens DUCKE/ Jairo de Oliveira Tenorio. – Rio de Janeiro, 2010.
Monografia (Graduação em Biotecnologia) – Centro Universitário Estadual da Zona Oeste (UEZO).
Orientador: Heber dos Santos Abreu

iii
LIGNINA DE PLÂNTULAS DE Lophantera lactescens
DUCKE
Elaborado por Jairo de Oliveira Tenorio
Aluno do curso de Biotecnologia da UEZO
Este trabalho foi analisado e aprovado com
Grau:____________
Rio de Janeiro, ____ de janeiro de 2010.
______________________________
Max Valério Dória Barbosa, Drº.
______________________________
Tereza Cristina Jesus Rocha, Drª.
______________________________
Heber dos Santos Abreu, Drº, Presidente.
RIO DE JANEIRO, RJ – BRASIL JANEIRO DE 2010

iv
Dedico este trabalho a todos que desejam minha vitória.

v
Inicialmente agradeço aos meus pais, pelo apoio incondicional que me deram desde o início desta jornada, dos risos pelas notas boas, dos abraços e conselhos de momentos que precisei de um colo, e principalmente por estas duas pessoas terem feito de tudo para que pudesse alcançar meus objetivos.
A minha namorada, por ter aguentado meu mau humor, como também pelo apoio, e companheirismo incondicional durante todo o tempo de graduação.
Agradeço também minha madrinha pelo que por mais que pra ela tenha sido pouco, para mim foi de extrema importância nos momentos difíceis, assim como minha tia Cláudia.
Aos eternos e sinceros amigos que fiz ao longo da graduação, estes foram de extrema importância não só para meu aprendizado, como também para que eu crescesse como ser humano.
A minha co-orientadora, pelo total apoio e por ter assumido esta grande responsabilidade num dos momentos mais ocupados de sua vida.
A meu orientador pelo apoio, e pelo fornecimento de ferramentas que me levaram a acreditar que na ciência encontra-se meu futuro.

vi
Resumo
Este trabalho teve o objetivo de estudar o processo de lignificação da
plântula de uma árvore da espécie Lophantera lactescens Ducke, da família
Malpighiaceae endêmica da Amazônia Brasileira, desenvolvida em meio de
cultura sólido (MS). O material caule e raiz de partes diferentes foram estudados
sob o aspecto anatômicos e bioquímicos para observação da lignificação, usando
material pré-tratado e in loco. Foram realizadas análises no infravermelho com
material pré-extraído e sem extração e através de microscopia no infravermelho
IFTR usando, respectivamente o modo de transmitância e reflectância. Os
espectros foram registrados em pastilha de KBr e através de cortes da seção
transversal do tecido sem pré-tratamento com aproximadamente 18 micra de
espessura sob lâmina de KBr. Foram realizados testes de cor usando safrabalu,
teste de Wiesner, observados em microscópio de campo claro e fluorescência
com a aplicação de auramina. As observações permitiram concluir que tanto nas
raízes e no caule, e principalmente no segundo a lignificação é abrangente e
dominante no xilema primário, sem comparação com outras espécies no estado
semelhante de desenvolvimento.
Palavras-chave : Lophantera lactescens Ducke, lignina, lignificação.

vii
Abstract
This study aimed to explore the process of lignification of the seedling of a
tree species Lophantera lactescens Ducke, Malpighiaceae family endemic from
Brazilian Amazon, developed in solid medium (MS). The stem and root material of
different parts were studied under anatomical and biochemical aspect with
observation of lignification, using pre-treated material and in situ. Analyses were
carried out in the infrared from pre and not extracted material using infrared
spectrometry and microscopy FTIR by of method of transmittance and reflectance,
respectively. The spectra were recorded in KBr pellets and by cutting the cross
section of tissue without pretreatment, approximately with 18 micra of thickness on
KBr laminates. Tests were performed using stain technique with safrablau and
Wiesner reagent. It was observed a brightfield and fluorescence with the
application of auramine stain. The observations showed that both the roots and
stem, and especially in the second one the lignification is dominant in the primary
xylem, without comparison with other species in similar state of development.
Keywords: Lophantera lactescens Ducke, lignin, lignification.

viii
Aprender é a única coisa de que a mente nunca se cansa, nunca tem
medo e nunca se arrepende. Leonardo da Vinci

ix
SUMÁRIO
Páginas
Resumo...............................................................................................................vi
Abstract..............................................................................................................vii
1. Introdução........................................................................................................1
1.2 Revisão de Literatura................................................................................2
1.3 Lignina.......................................................................................................2
2. Justificativa......................................................................................................7
3. Objetivos..........................................................................................................9
3.1. Objetivos específicos................................................................................9
4. Material e Métodos........................................................................................10
4.1. Coleta e armazenagem das sementes................................................13
4.2. Desinfestação das sementes e condições de germinação ..................13
4.3. Preparo de material livre de extratos solúveis......................................14
4.4. Infravermelho........................................................................................15
4.5. Preparo do material para microscopia..................................................15
4.6. Reagente de Wiesner...........................................................................15
4.7. Microscopia com fluorescência.............................................................16
5. Resultados e Discussão.................................................................................17
5.1. Microscopia no infravermelho...............................................................20
5.2. Análise anatômica com enfoque a lignificação.....................................23
6. Conclusão.......................................................................................................35
7. Referências Bibliográficas..............................................................................36

1
1 – INTRODUÇÃO
Lignina é um termo que se aplica as macromoléculas que apresentam
composição básica formada por unidades cumarílica, guaiacílica e sinapílica de
origem fenilpropanoídica com estrutura molecular tridimensional. Lignina vem do
latim, da palavra “lignun” que significa madeira, entretanto nem todo material
vegetal que contém lignina é considerado madeira. Mas, as madeiras por outro
lado apresentam essa substância como sendo importante para sua estrutura
física. A maioria das plantas superiores possuem lignina.
O interesse despertado pela espécie Lophantera lactescens Ducke,
decorre por alguns aspectos. Primeiro por ser uma espécie endêmica da
Amazônia, com importância na medicina popular local (Amazônia) para baixar a
febre da malária. Dela foi isolado um nor-triterpeno com atividades de interesse
farmacológico notável (ABREU et al., 1990). Por outro lado a árvore apresenta um
fuste ereto cilíndrico, resistente a broca e com madeira de densidade de média
para alta, podendo ser usada na indústria de madeira. O conhecimento de sua
biologia, no que diz respeito a métodos biotecnológicos e de lignificação tem sido
realizado pelo grupo do Laboratório de Biotecnologia da Madeira.
Outros estudos tem sido realizados como a descrição anatômica, estudos
de cristais, formação de calos e desenvolvimento de células em suspensão.
Visando o aproveitamento da biodiversidade florestal brasileira, a madeira
se destaca para indústria de base florestal e, portanto o estudo sobre sua
composição química, sobretudo a lignina mostra-se de imensa importância,
tendo em vista, insuficiente informações descrita na literatura (ABREU et al.
1990).entretanto o conhecimento mais amplo sobre o crescimento das plântulas,
os parâmetros de desenvolvimento celular e o estudo químico e bioquímico em
todas as fases de crescimento da Lophantera lactescens, tornam-se necessários.
A madeira da Lophantera lactescens é moderadamente pesada, compacta,
medianamente dura, e resistente ao ataque de organismos xilófagos (LORENZI,
2008). TREVISAN (2003) ao estudar a durabilidade natural e a ocorrência de
térmitas em madeiras de cinco espécies em contato com o solo por um período

2
de dois anos constataram que a madeira desta espécie foi a mais durável, em
termos de resistência, dureza e ataque de térmitas. Pesquisadores também
atestaram a resistência da madeira de Lophantera, ao expor toras de madeira
desta espécie ao ataque de C. gestroi, e verificaram a notável resistência da
madeira de L. lactescens ao ataque de escolitídios.
1.2 – REVISÃO DE LITERATURA
1.3 – Lignina
A lignina é considerada um poderoso agente cimentante da parede celular,
característico de tecidos de plantas lenhosas, funciona como barreira química e
física, responsável pela condução de água nos tubos condutores, se
apresentando com alto peso molecular (LATORRACA & ABREU, 1997) e é
proveniente da oxidação desidrogenerativa de três monolignóis álcoois
precursores: p-cumarílico, trans-coniferílico e trans-sinapílico (Figura 1). Quando
estes monolignóis são incorporados à lignina, são chamados trivialmente de p-
hidroxifenil, guaiacil e siringilpropano, respectivamente (RAES et al., 2003).
Figura 1 – Precursores da lignina e respectivos sítios ativos

3
Durante o desenvolvimento celular, a lignina é incorporada como último
componente da parede celular, interpenetrando as fibrilas e fortalecendo a parede
(FENGEL & WEGENER, 1984), ocorrendo assim a lignificação. A deposição de
lignina proporciona à parede celular considerável resistência e rigidez. Essa
lignificação também pode ser induzida por uma infecção ou ferida, para proteger
os tecidos contra o ataque de patógenos (RAES et al., 2003). A lignina também é
indispensável em diversos processos biológicos dos quais os mais importantes
são, assegurar a existência de vias rápidas de circulação de água, minerais e
conferir rigidez necessária à manutenção da verticalidade do caule principal a
vários metros de altura, não apresentando mobilidade entre paredes celulares, o
que torna compreensível à preservação desse material ao longo de milhares de
anos sob a face da terra, em condições rigorosas de alteração ambiental (ABREU
et al., 1999).
A lignificação é um processo bioquímico altamente regulado, que começa
na parede primária das células e na lamela média, se estendendo sobre a parede
secundária em direção ao lúmen (DONALDSON, 2001). Segundo ABREU et. al
(1999), é um processo bioquímico que abrange a formação de monolignóis, seu
transporte e a polimerização na parede celular, que em primeiro estádio é
altamente mediado por enzimas intrínsecas à formação dos precursores nos
compartimentos citoplasmáticos. O segundo estádio de formação da lignina
ocorre na parede celular e caracteriza-se pela reação de oxidação
desidrogenativa dos monolignóis disponíveis. As enzimas oxi-redutoras tais como
peroxidases, e isoenzimas correspondentes, atuam na polimerização da lignina,
dentro da parede celular, formando um complexo coordenado com peróxido de
hidrogênio. A lacase entre outras oxidases também promovem oxidação
desidrogenativa dos monolignóis na parede celular (WHETTEN et al., 1998;
RANOCHA et al., 2002). Além disso, desempenham um importante papel no
crescimento e desenvolvimento da planta (ROGERS & CAMPBELL, 2004), que
provavelmente permitiu a adaptação das plantas aquáticas à vida terrestre
(MONTIES, 1989, BARCELO, 1997; INOUE et al., 1998, NORTHCOTE, 1989).

4
Durante a formação dos precursores da lignina algumas dessas enzimas
atuam sobre o núcleo aromático dos fenilpropanóides, introduzindo hidroxilas em
C-3 e C-5 e as metilando em seguida enquanto outras desempenham importante
papel na redução da cadeia lateral. A biossíntese da lignina envolve uma série de
enzimas desde a Fenilalanina Amônia-liase (PAL), Cinamato-4-Hidroxilase (C4H),
Hidroxicinamoil-COA-Ligase (4CL), 4-Hidroxicinamato 3-Hidroxilase (C3H), 5-
Adenosil-Metionine:Cafeato/5-Hidroxi (OMT), Ferulato-5-Hidroxilase (F5H),
Hidroxicinamoil COA Redutase (CCR) até a Cinamil Álcool Desidrogenase (CAD).
A estrutura da lignina tem sido também objeto de estudo em inúmeros trabalhos
científicos, aos quais estabelecem propostas estruturais e formação biossíntética
(BOERJAN, 2003). Considera-se para os mesmos a existência de um provável
controle biológico diferenciado segundo os aspectos inerentes a classe botânica a
que o vegetal pertence, a influência genética, aos aspectos espaciais da
subestrutura da parede celular, entre outros aspectos. Dois estádios de formação
da lignina são preconizados, a formação enzimática e a formação semi-
enzimática. Sendo a primeira considerada como oxidação horizontal ocorrendo no
citoplasma e a segunda oxidação vertical que acontece na parede celular
(ABREU et al., 1999). Para formar os precursores terminais (ésteres de ácidos
fenilpropanóides) sucessivas oxidações e metilações são descritas
(CHOINOWSKI et al., 1999). O éster p-cumarato, por exemplo, é hidroxilado na
posição meta a cadeia lateral, formando o éster cafeato. Este precursor
intermediário é determinante na síntese dos demais precursores monolignóis.
Três reações mediadas por cinamil redutase levam aos aldeídos correspondentes
(cumaraldeído, coniferaldeído e sinapaldeído), os quais são reduzidos a álcoois
através da enzima cinamil álcool desidrogenase (CAD). Todo este processo
enzimático ocorre no compartimento citoplasmático. Oxidação dos três
monolignóis na condição de precursores terminais. Esses precursores terminais
(álcoois p-cumarílico, coniferílico e sinapílico) apresentam vários sítios reativos
capazes de constituírem ligações cruzadas entre si, preferencialmente sobre a
cadeia lateral (HIGUCHI, 1997). O aumento do grau de metoxilação a partir dos
precursores: ácido cumárico, ácido coniferílico e ácido sinápico correspondem à
diminuição do número de ligações intermoleculares, possibilitando a formação de

5
uma rede de ligações intermonoméricas na maioria envolvendo ligações C-C.
Isso, portanto constitui, base na composição química e conseqüentemente nas
formas estruturais adquiridas após a polimerização na parede celular.
A composição da lignina varia significativamente entre as espécies, dentro
da espécie e também na mesma planta, pois há variações de célula para célula
de acordo com a localização da parede celular, conforme o estágio de
desenvolvimento da célula e do tecido, e ainda com a influência de estresses
ambientais (CAMPBELL E SEDEROFF, 1996, apud STUDART-GUIMARÃES et
al., 2003). Sua arquitetura molecular difere segundo a origem botânica dos
táxons, entre células e até mesmo dentro da parede celular, respondendo aos
efeitos abióticos e bióticos do ambiente (LARROQUE & PLANCHON, 1990;
KITAYAMA et al., 2004).
Em madeiras de coníferas por exemplos, predominam, basicamente,
subestruturas do tipo guaiacila, provenientes do álcool trans-coniferílico; em
madeiras de folhosas, existe uma mistura de quantidades significativas de
subestruturas guaiacila e siringila resultantes da polimerização dos precursores
trans-coniferílico e trans-sinapílico, respectivamente. Já em palhas e gramíneas,
além de unidades guaiacílicas e siringílicas, encontram-se subestruturas do tipo
cumaríla proveniente da polimerização do álcool p-cumarílico, todas em
quantidades relevantes. A proporção molar dessas subestruturas na planta
depende, principalmente, da espécie (CHEN, 1991). Ligninas de plantas
herbáceas são do tipo H-S-G, sendo mais parecidas com as ligninas de
angiospermas do que de gimnospermas. Portanto, em uma conceituação mais
precisa, as ligninas são classificadas nos seguintes grupos: Tipo G; Tipo G-S; e
Tipo H-G-S (CHEN, 1991; PILÓ-VELOSO et al., 1993).
Um das dificuldades para se estudar lignina é distância existente entre a
protologina e a lignina isolada. Distância essa caracterizada que dificulta uma
diagnose precisa sobre a estrutura da lignina. Existem diferentes tipos de
extração de ligninas, e nenhum deles permite obtê-la como ela se encontra

6
estruturalmente no vegetal, pois haverá sempre interferência entre os
procedimentos de extração química e a estrutura da lignina in situ.

7
2 – JUSTIFICATIVA
Desde 1990 o estudo com a espécie Lophantera lactescens tem sido
desafiante. O interesse a princípio deve-se ao isolamento do nor-triterpeno
(Figura 2) com atividades já descrita na literatura (ABREU et al., 1990).
Recentemente teste in vitro mostrou atividade contra Leishmania amazonensis
(DANELLI, et al., 2009). Também seu interesse aumentou pela possibilidade
dessa substância se apresentar como cristais rombóides nos tecidos do
parênquima radial. Além disso, há interesse muito grande de empresas
multinacionais como a Merck por exemplo, que publicou e registrou patente tendo
como referência o trabalho de Abreu (1990). No Laboratório de Biotecnologia da
Madeira do Departamento de produtos florestais tem-se desenvolvido a formação
de calos, células em suspensão e análise de plântulas. A exploração e o uso mais
adequado, a produção in vitro, e o desenvolvimento de plantas dando enfoque
florestal, tem sido meta do laboratório.
Figura 2 – Estrutura do Nor-triterpeno isolado da madeira de Lophantera lactescens [6α,7α,15β,16β,24-pentacetoxi-22α-carbometoxi-21β,22β-epoxi-18β-hidroxi-27,30-bisnor-3,4-
secofriedela-1,20(29)-dien-3,4R-lideo]
No sentido de apoiar a indústria de base florestal o processo de
lignificação da Lophantera tem uma vertente interessante que é o estudo de
plantas brasileiras para sua utilização na indústria de base florestal. Atualmente
no setor florestal a caracterização de madeiras segundo sua composição físico,
química e anatômica é de extrema importância para se utilizá-las nos setores
23
14
13
1211
10
4
3
29
28
2625
24
22
212019
1817
16
15
98
76
5
2 1
o
CH2OAc
OAc
OAcOAc
OAc
O
COOMeOH
o= A B
C D
E

8
tecnológico e industrial. Pois, a composição e o teor de lignina em madeiras têm
conseqüências de natureza técnica e econômica (industriais, ambientais, e
agrícolas profundas). Atualmente há uma grande porcentagem de espécies não
nativas sendo utilizadas no setor tecnológico, o estudo do processo de lignificação
de Lophantera se torna importante, pois comprovando-se que esta tem uma
madeira de alta qualidade, tal espécie pode então substituir outras como as do
gênero Eucalyptus, utilizadas no setor de base florestal.

9
3 – OBJETIVOS
O objetivo deste trabalho foi verificar a ocorrência de lignina em segmentos
caulinares e radiculares e sua caracterização química em plântulas de Lophantera
lactescens cultivadas in vitro.
3.1 – OBJETIVOS ESPECÍFICOS
• Cultivo de plântulas de Lophantera lactescens em meio nutritivo artificial em
ambiente controlado de temperatura, luminosidade e humidade;
• Verificar a presença de lignina em plântulas de Lophantera lactescens;
• Evidenciar a lignina por uso de corante específico em cortes transversais de
segmentos caulinares e radiculares, por meio de microscopia de campo claro e
fluorescência;
• Evidenciar o processo de lignificação em plântula de Lophantera lactescens
que ainda não foi realizado.

10
4 – MATERIAL E MÉTODOS
A espécie Lophantera lactescens pertence a família Malpighiaceae. Esta
família é de distribuição tropical e subtropical (LORENZI & SOUZA, 2008),
apresentando sua maior diversidade na América do Sul (Judd et al., 1999 apud
SILVA, S. L., 2007). Apresenta cerca de 1.300 espécies, organizadas em 75
gêneros (LORENZI et al., 2008), sendo referida como uma das dez mais bem
representadas no bioma cerrado (MENDONÇA et al., 1998 apud SILVA, S. L.,
2007).
Dentre os gêneros desta família, Lophantera lactescens Ducke (Figura 3),
conhecida vulgarmente, como Lofantera-da-amazônia, Chuva-de-ouro e
Lanterneira, é uma espécie arbórea de 10-20m de altura, com tronco colunar ou
bifurcada próximo à base, de 30-40cm de diâmetro, revestido por casca
pardacenta fina com ritidoma lenticelado (Figura 4).
Figura 3 - Lanterneira (Lophantera lactescens) no campus da Universidade Federal Rural do Rio de Janeiro, Seropédica, com aproximadamente 10m de altura

11
Figura 4 – Tronco colunar
As folhas (Figura 5) são opostas cruzadas, dotadas de estípulas
intrapeciolares, simples, elípticas a frequentemente obovais, de base aguda a
atenuada e ápice arredondado a retuso, cartáceas, glabras, com nervuras
primárias e secundárias imersas na face superior e proeminentes na inferior,
lactescentes quando jovens, de 16-22cm de comprimento por 8-11cm de largura.
Figura 5 – Folhas de Lophantera lactescens
As flores são dispostas em racemos terminais pendentes (Figura 6), são
amarelas, vistosas, bissexuadas e pentâmeras; com disco nectarífero ausente.
Floresce durante os meses de fevereiro-maio.

12
Figura 6 – Inflorescência em detalhe
Os frutos são esquizocárpicos, secos (Figura 7). A maturação dos frutos
verifica-se em setembro-outubro (LORENZI et al., 2008).
Figura 7 – Frutos ainda presos aos racemos nas árvores

13
É uma espécie de ocorrência da região Amazônica, na mata de várzea alta,
principalmente na parte central (LORENZI, 2008). Planta semidecídua, heliófita ou
esciófita, seletiva higrófita, característica da floresta pluvial da região amazônica.
Apresenta dispersão bastante restrita, tendo sido encontrada, até o momento,
apenas no baixo Tapajós, em mata de várzea alta. Ocorre tanto no interior da
mata primária densa como em formações secundárias (LORENZI, 2008).
O presente trabalho foi conduzido no Laboratório de Biotecnologia da Madeira
(LBM) do Departamento de Produtos Florestais do Instituto de Florestas da
Universidade Federal Rural do Rio de Janeiro (DPF/IF/UFRRJ).
4.1 – COLETA E ARMAZENAGEM DAS SEMENTES
As sementes para a produção dos explantes foram coletadas de uma única
matriz do próprio campus da UFRRJ, sendo posteriormente armazenadas à
temperatura de aproximadamente 25°C e umidade relat iva do ar 62%.
4.2 – DESINFESTAÇÃO DAS SEMENTES E CONDIÇÕES DE GERMINAÇÃO
As sementes de Lophantera lactescens foram submetidas à seguinte condição
de desinfestação: 20 minutos em presença de hipoclorito de sódio 2,5% contendo
0,2 mL/L de detergente tween 20. Em seguida as sementes foram escorridas em
peneira inox forradas com papel filtro e lavadas 5 vezes em água bidesionizada
estéril. Após a última lavagem as sementes foram imersas em peróxido de
hidrogênio 1% por 30 minutos. Decorrido esse período as sementes foram
novamente escorridas e em capela de fluxo laminar, foram inoculadas no meio de
cultura MS (MURASHIGE & SKOOG, 1962), contendo a seguinte formulação:
1,65g de (NH4)NO3, 1,9g de KNO3, 0,44g de CaCl2, 0,17g de KH2PO4, 0,37g
MgSO4, 0,002g de glicina, 30g de sacarose, 7g de Agar-Agar, 0,1g de Benlate,
1mL de solução de ferro + EDTA, 5mL de solução de micronutriente, 1mL de
solução de vitaminas. O meio de cultura MS foi produzido com 24 horas de
antecedência à desinfestação. Os procedimentos seguidos para produzir o meio
de cultura foram: Colocar 300mL de água bidesionizada (DDH2O) em um Becker

14
de 1L, manter sob agitação e acrescentar cada componente do meio MS
excluindo o Agar-Agar. Completar o volume do Becker até 1000mL e manter sob
agitação até que todos os componentes estejam dissolvidos. Aferir o pH, e, se
necessário, corrigir para 5.7 utilizando solução de KOH ou HCl, então acrescentar
o Agar-Agar e ligar a placa de aquecimento em aproximadamente 200ºC,
mantendo o meio em constante agitação. Cobrir o becker com papel alumínio e
aguardar a fervura do meio. Distribuir o meio nos recipientes colocando-se
aproximadamente 50mL de meio de cultura MS por recipiente, posteriormente o
meio de cultura foi esterilizado em autoclave a 120°C e 1,0 atm por 20 minutos.
Para otimizar as atividades, alguns componentes do meio de cultura foram
otimizados em soluções estoque. Em capela de Fluxo laminar as sementes foram
acondicionadas em recipientes próprios para germinação contendo meio de
cultura, 5 sementes por recipiente, sendo mantidas em sala de germinação com
fotoperíodo de 16 horas, temperatura de 25°C +/- 1º C, e umidade relativa do ar
62%.
O cultivo de plântulas de Lophantera lactescens in vitro foi alcançado (Figura 8).
Figura 8 – Plântulas germinadas no LBM
4.3 – PREPARO DE MATERIAL LIVRE DE EXTRATOS SOLÚVEIS
O material foi pré-extraído com etanol 95%. Em seguida, após seco, foi
efetuado a maceração dos tecidos de caule e raiz (separadamente) em moinho de
facas do tipo Wiley, após extração o tecido foi seco. Em seguida o material sólido

15
foi colocado no Moinho de Bolas, para que este reduzisse a granulométrica das
partículas.
4.4 – INFRAVERMELHO
Os espectros no infravermelho foram registrados dos materiais
lignocelulósicos, (plântulas) dos caules e das raízes. Material pré-extraído e não
pré-extraído foram moídos em moinho de bolas e em seguida foram preparados
pastilhas de KBr com 1 mg de lignina e 100mg de KBr. Os espectros foram
registrados em um espctrômetro VARIAN 640-IRFT-IR no modo transmitância.
A microscopia no infravermelho foi feita no equipamento VARIAN 610-IRFT-IR
no modo reflectância. Os cortes foram liofilizados durante 12 horas e colocados
no equipamento para a obtenção de imagens e os respectivos espectros.
4.5 – PREPARO DO MATERIAL PARA MICROSCOPIA
Cortes transversais foram feitos a mão livre em micrótomo de Ranvier e
corados com Safranina e Azul de Astra (Safrablau), e observados em microscópio
modelo Olympus trinocular com software cell*.
4.6 – REAGENTE DE WIESNER
O reagente foi preparado por uma combinação de 50 mL de uma solução de
floroglucinol 2% em etanol 95% e 25 mL de ácido clorídrico (HCl) concentrado
(LIN & DENCE, 1992).
Como este reagente não é muito estável, a solução de floroglucinol foi
armazenada em vidro âmbar (com rolha de vidro), sendo misturado ao HCl
somente na hora de usar.

16
4.7 – MICROSCOPIA COM FLUORESCÊNCIA
As análises foram realizadas em um microscópio Olympus trinocular com
software cell*, provido de fluorescência. As ligninas que são auto fluorescentes
foram observadas no caule e raiz. Os cortes foram imersos em auramina (1%) em
solução aquosa para aumentar a intensidade da fluorescência.
17
5 – RESULTADOS E DISCUSSÃO
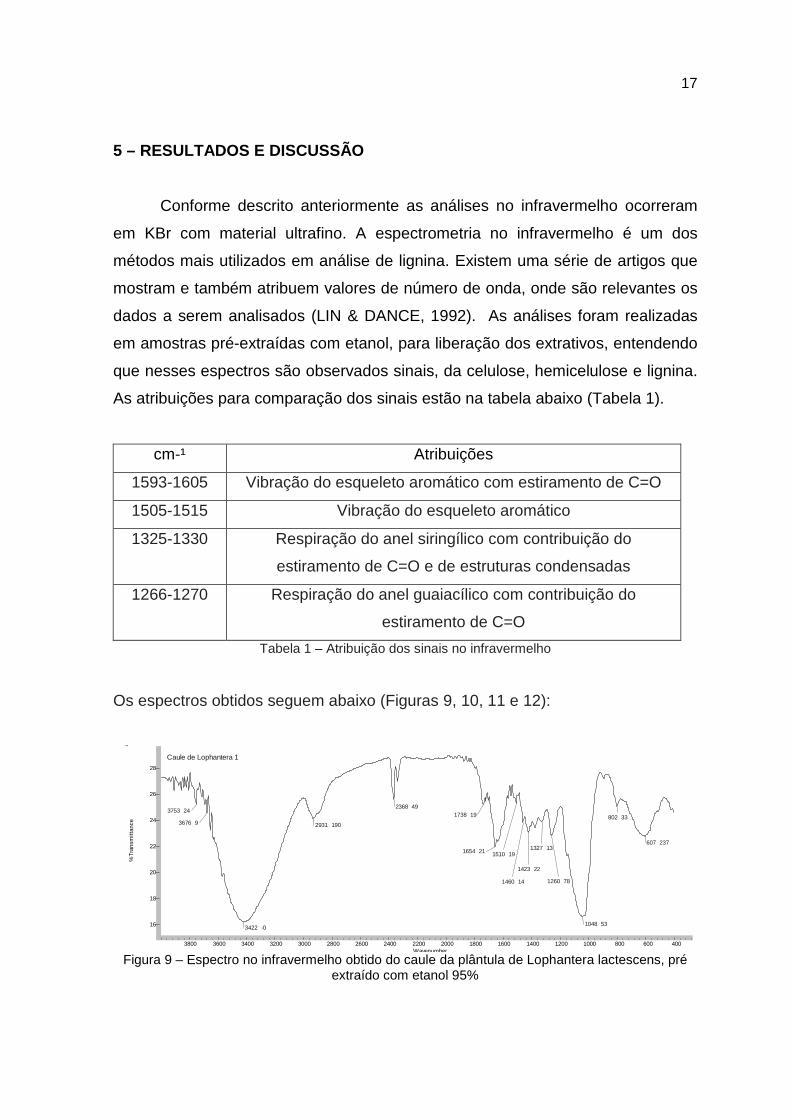
Conforme descrito anteriormente as análises no infravermelho ocorreram
em KBr com material ultrafino. A espectrometria no infravermelho é um dos
métodos mais utilizados em análise de lignina. Existem uma série de artigos que
mostram e também atribuem valores de número de onda, onde são relevantes os
dados a serem analisados (LIN & DANCE, 1992). As análises foram realizadas
em amostras pré-extraídas com etanol, para liberação dos extrativos, entendendo
que nesses espectros são observados sinais, da celulose, hemicelulose e lignina.
As atribuições para comparação dos sinais estão na tabela abaixo (Tabela 1).
cm-¹ Atribuições
1593-1605 Vibração do esqueleto aromático com estiramento de C=O
1505-1515 Vibração do esqueleto aromático
1325-1330 Respiração do anel siringílico com contribuição do
estiramento de C=O e de estruturas condensadas
1266-1270 Respiração do anel guaiacílico com contribuição do
estiramento de C=O
Tabela 1 – Atribuição dos sinais no infravermelho
Os espectros obtidos seguem abaixo (Figuras 9, 10, 11 e 12):
3753 24
3676 9
3422 -0
2931 190
2368 491738 19
1654 21 1510 19
1460 14
1423 22
1327 13
1260 78
1048 53
802 33
607 237
Caule de Lophantera 1
3800 3600 3400 3200 3000 2800 2600 2400 2200 2000 1800 1600 1400 1200 1000 800 600 400
28
26
24
22
20
18
16
Wavenumber
%T
rans
mitt
ance
Figura 9 – Espectro no infravermelho obtido do caule da plântula de Lophantera lactescens, pré extraído com etanol 95%
18
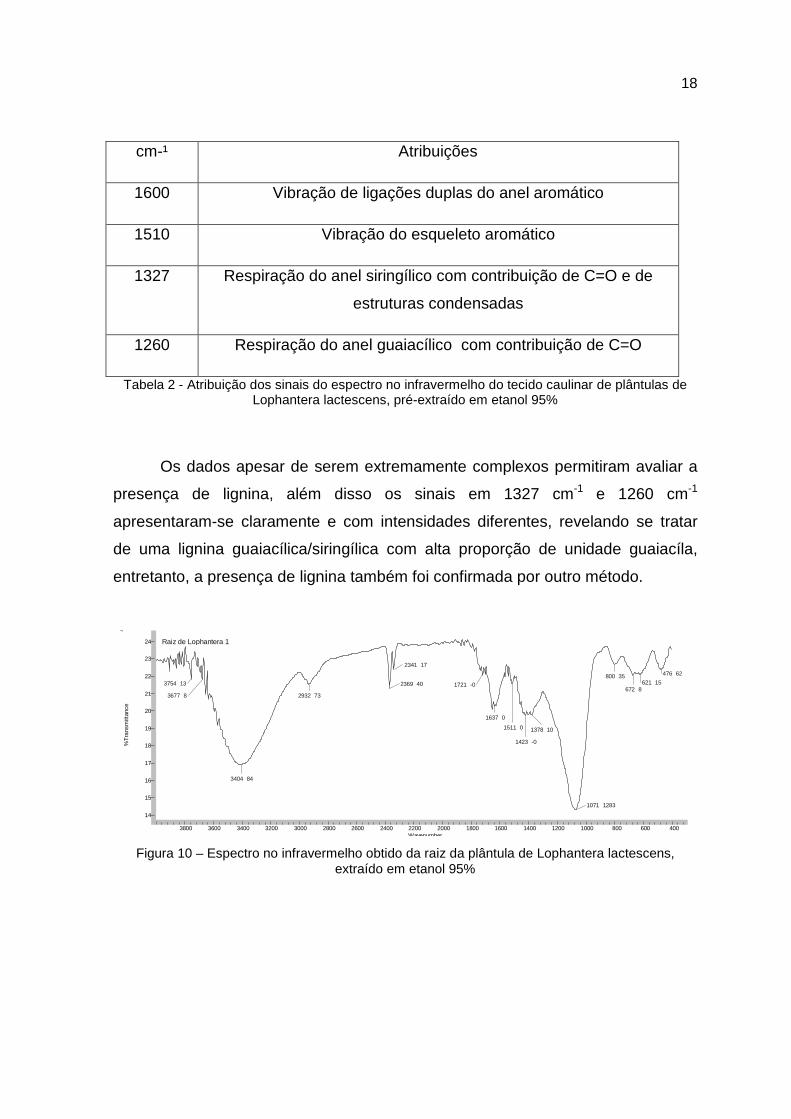
cm-¹ Atribuições
1600 Vibração de ligações duplas do anel aromático
1510 Vibração do esqueleto aromático
1327 Respiração do anel siringílico com contribuição de C=O e de
estruturas condensadas
1260 Respiração do anel guaiacílico com contribuição de C=O
Tabela 2 - Atribuição dos sinais do espectro no infravermelho do tecido caulinar de plântulas de Lophantera lactescens, pré-extraído em etanol 95%
Os dados apesar de serem extremamente complexos permitiram avaliar a
presença de lignina, além disso os sinais em 1327 cm-1 e 1260 cm-1
apresentaram-se claramente e com intensidades diferentes, revelando se tratar
de uma lignina guaiacílica/siringílica com alta proporção de unidade guaiacíla,
entretanto, a presença de lignina também foi confirmada por outro método.
3754 13
3677 8
3404 84
2932 73
2369 40
2341 17
1721 -0
1637 0
1511 0
1423 -0
1378 10
1071 1283
800 35
672 8621 15
476 62
Raiz de Lophantera 1
3800 3600 3400 3200 3000 2800 2600 2400 2200 2000 1800 1600 1400 1200 1000 800 600 400
24
23
22
21
20
19
18
17
16
15
14
Wavenumber
%T
rans
mitt
ance
Figura 10 – Espectro no infravermelho obtido da raiz da plântula de Lophantera lactescens, extraído em etanol 95%
19
cm-¹ Atribuições
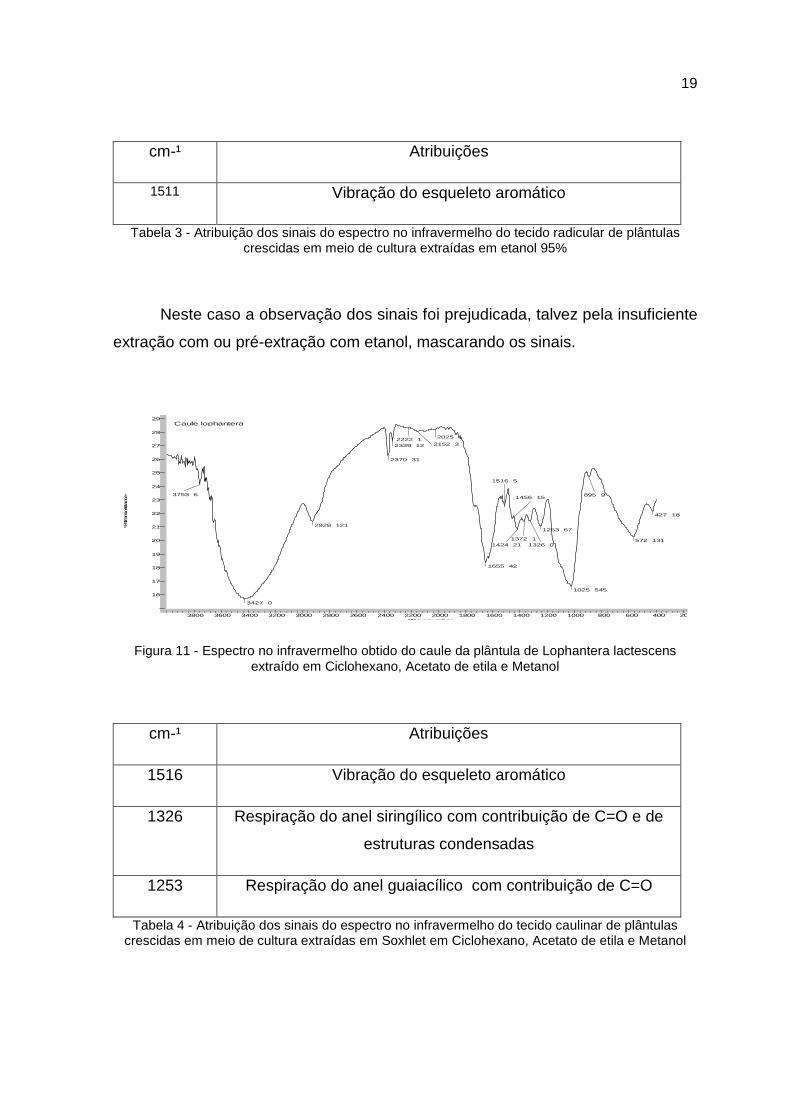
1511 Vibração do esqueleto aromático
Tabela 3 - Atribuição dos sinais do espectro no infravermelho do tecido radicular de plântulas crescidas em meio de cultura extraídas em etanol 95%
Neste caso a observação dos sinais foi prejudicada, talvez pela insuficiente
extração com ou pré-extração com etanol, mascarando os sinais.
3753 6
3427 0
2928 121
2370 31
2339 122222 1
2152 3
2025 1
1655 42
1516 5
1456 15
1424 211372 1
1326 0
1253 67
1025 545
895 9
572 131
427 18
Caule lophantera
3800 3600 3400 3200 3000 2800 2600 2400 2200 2000 1800 1600 1400 1200 1000 800 600 400 200
29
28
27
26
25
24
23
22
21
20
19
18
17
16
Wavenumber
%Trans
mittan
ce
Figura 11 - Espectro no infravermelho obtido do caule da plântula de Lophantera lactescens extraído em Ciclohexano, Acetato de etila e Metanol
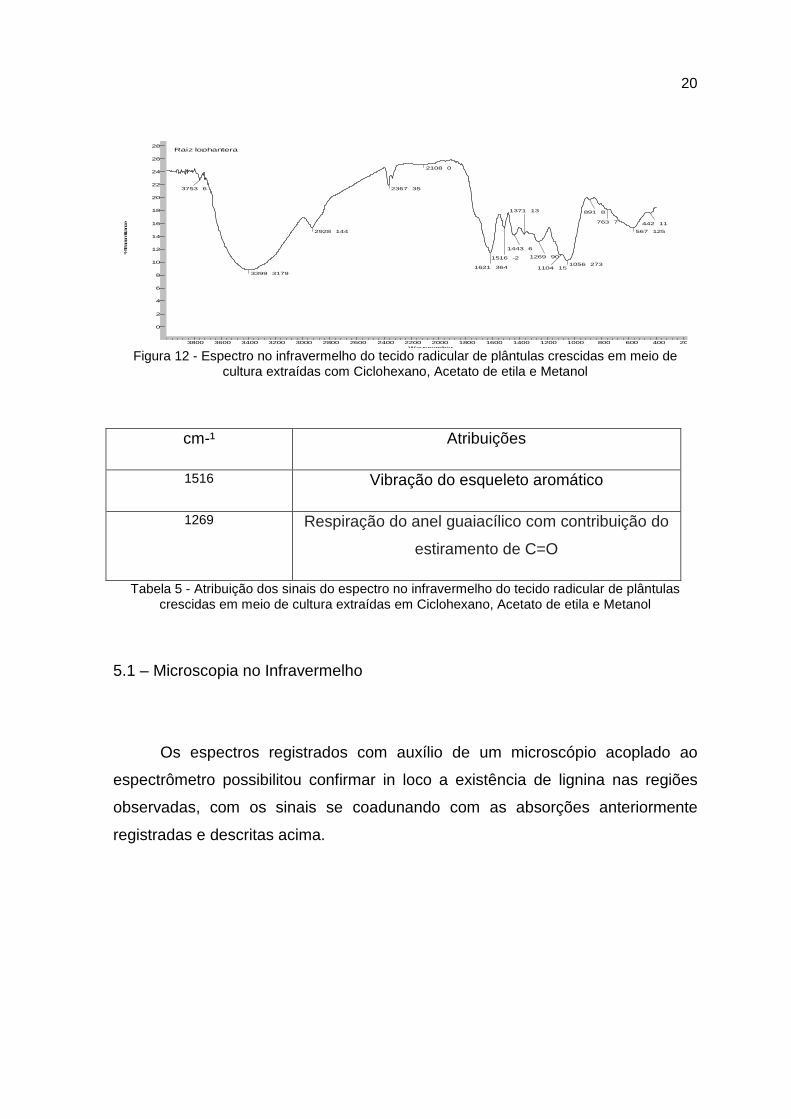
cm-¹ Atribuições
1516 Vibração do esqueleto aromático
1326 Respiração do anel siringílico com contribuição de C=O e de
estruturas condensadas
1253 Respiração do anel guaiacílico com contribuição de C=O
Tabela 4 - Atribuição dos sinais do espectro no infravermelho do tecido caulinar de plântulas crescidas em meio de cultura extraídas em Soxhlet em Ciclohexano, Acetato de etila e Metanol
20
3753 6
3399 3179
2928 144
2367 35
2108 0
1621 364
1516 -2
1443 6
1371 13
1269 90
1104 151056 273
891 8
763 7
567 125
442 11
Raiz lophantera
3800 3600 3400 3200 3000 2800 2600 2400 2200 2000 1800 1600 1400 1200 1000 800 600 400 200
28
26
24
22
20
18
16
14
12
10
8
6
4
2
0
Wavenumber
%Trans
mittan
ce
Figura 12 - Espectro no infravermelho do tecido radicular de plântulas crescidas em meio de cultura extraídas com Ciclohexano, Acetato de etila e Metanol
cm-¹ Atribuições
1516 Vibração do esqueleto aromático
1269 Respiração do anel guaiacílico com contribuição do
estiramento de C=O
Tabela 5 - Atribuição dos sinais do espectro no infravermelho do tecido radicular de plântulas crescidas em meio de cultura extraídas em Ciclohexano, Acetato de etila e Metanol
5.1 – Microscopia no Infravermelho
Os espectros registrados com auxílio de um microscópio acoplado ao
espectrômetro possibilitou confirmar in loco a existência de lignina nas regiões
observadas, com os sinais se coadunando com as absorções anteriormente
registradas e descritas acima.
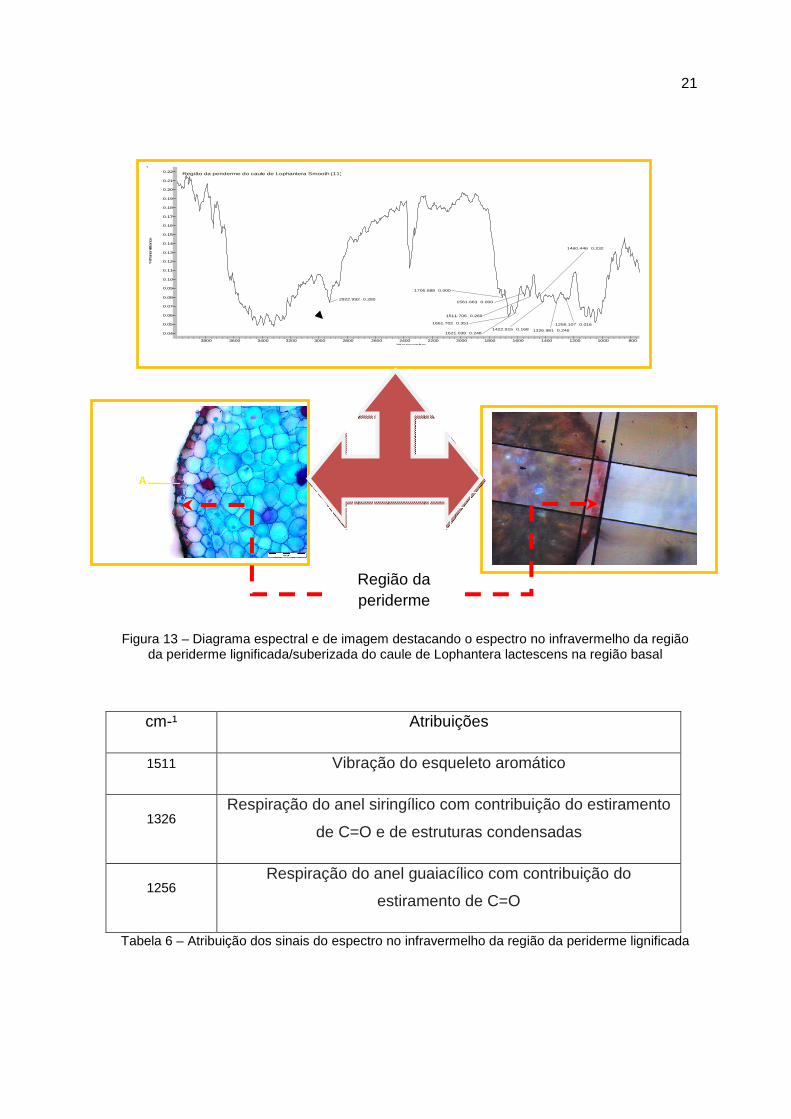
21
Figura 13 – Diagrama espectral e de imagem destacando o espectro no infravermelho da região da periderme lignificada/suberizada do caule de Lophantera lactescens na região basal
cm-¹ Atribuições
1511 Vibração do esqueleto aromático
1326 Respiração do anel siringílico com contribuição do estiramento
de C=O e de estruturas condensadas
1256 Respiração do anel guaiacílico com contribuição do
estiramento de C=O
Tabela 6 – Atribuição dos sinais do espectro no infravermelho da região da periderme lignificada
2922.992 0.280
1706.688 0.000
1661.702 0.351
1621.038 0.248
1561.663 0.000
1511.706 0.269
1460.446 0.232
1422.915 0.168 1326.981 0.246
1256.107 0.016
Região da periderme do caule de Lophantera Smooth (11)
3800 3600 3400 3200 3000 2800 2600 2400 2200 2000 1800 1600 1400 1200 1000 800
0.22
0.21
0.20
0.19
0.18
0.17
0.16
0.15
0.14
0.13
0.12
0.11
0.10
0.09
0.08
0.07
0.06
0.05
0.04
Wavenumber
%Tra
nsm
ittan
ce
Região da periderme
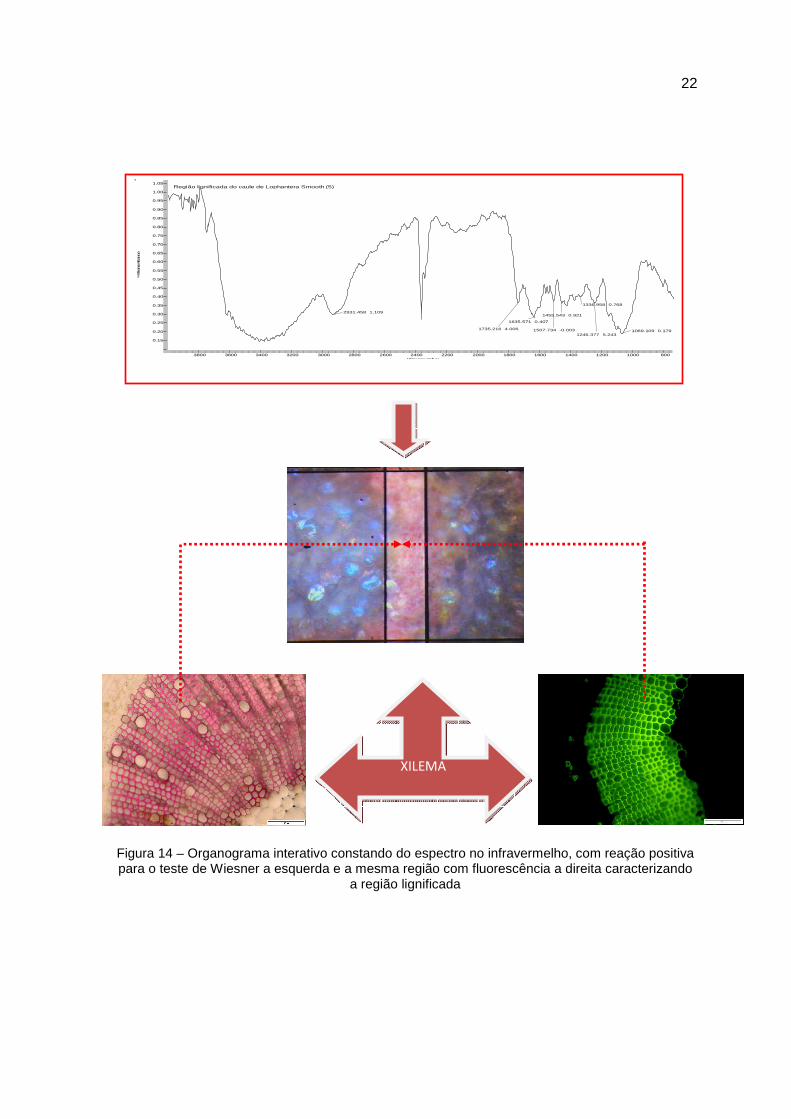
22
Figura 14 – Organograma interativo constando do espectro no infravermelho, com reação positiva para o teste de Wiesner a esquerda e a mesma região com fluorescência a direita caracterizando
a região lignificada
2931.458 1.109
1735.216 4.006
1635.571 0.407
1507.734 -0.003
1455.543 0.921
1336.958 0.768
1246.377 5.2431069.109 0.179
Região lignificada do caule de Lophantera Smooth (5)
3800 3600 3400 3200 3000 2800 2600 2400 2200 2000 1800 1600 1400 1200 1000 800
1.05
1.00
0.95
0.90
0.85
0.80
0.75
0.70
0.65
0.60
0.55
0.50
0.45
0.40
0.35
0.30
0.25
0.20
0.15
Wavenumber
%Trans
mittan
ce
XILEMA

23
cm-¹ Atribuições
1507 Vibração do esqueleto aromático
1336 Respiração do anel siringílico com contribuição do
estiramento de C=O e de estruturas condensadas
1246 Respiração do anel guaiacílico com contribuição do
estiramento de C=O
Tabela 7 - Atribuição dos sinais do espectro no infravermelho da região lignificada do caule da plântula de Lophantera lactesces da região basal
5.2 – Análise anatômica com enfoque a lignificação
Os materiais de caule e raízes foram preparados manualmente e os cortes
transversais foram feitos em micrótomo manual, com auxílio de lâmina de vidro.
Os tecidos foram coloridos com safrablau e as fotomicrografias mostraram que o
tecido do caule apresenta regiões primárias do floema (B) e do xilema com tecido
lignificado (A) e região de câmbio indicado pela letra (C) (Figura 15, 16 e 17),
mostrando que a medida que os entre nós diminuem em distância da base do
colo da plântula, onde neste entre nó pode ser observado alto processo de
lignificação. A lignificação do tecido radicular fica evidenciado pela coloração
avermelhada, devido a assimilação da safranina à esclereides fibrosos. No caso
do tecido radicular, a lignificação foi detectada na periderme do tecido (A) (Figura
18), como também no tecido xilemático deste (X) (Figura 19 e 20).

24
Figura 15 – Micrografia em objetiva de 20x, em corte transversal do 1º entrenó da plântula corado com Safrablau evidenciando a presença de fenóis (lignina) (porção vermelha)
Figura 16 - Micrografia em objetiva de 20x, em corte transversal evidenciando outra região do corte do 1º entrenó da plântula corado com safrablau evidenciando a presença de fenóis (lignina)
(porção vermelha)
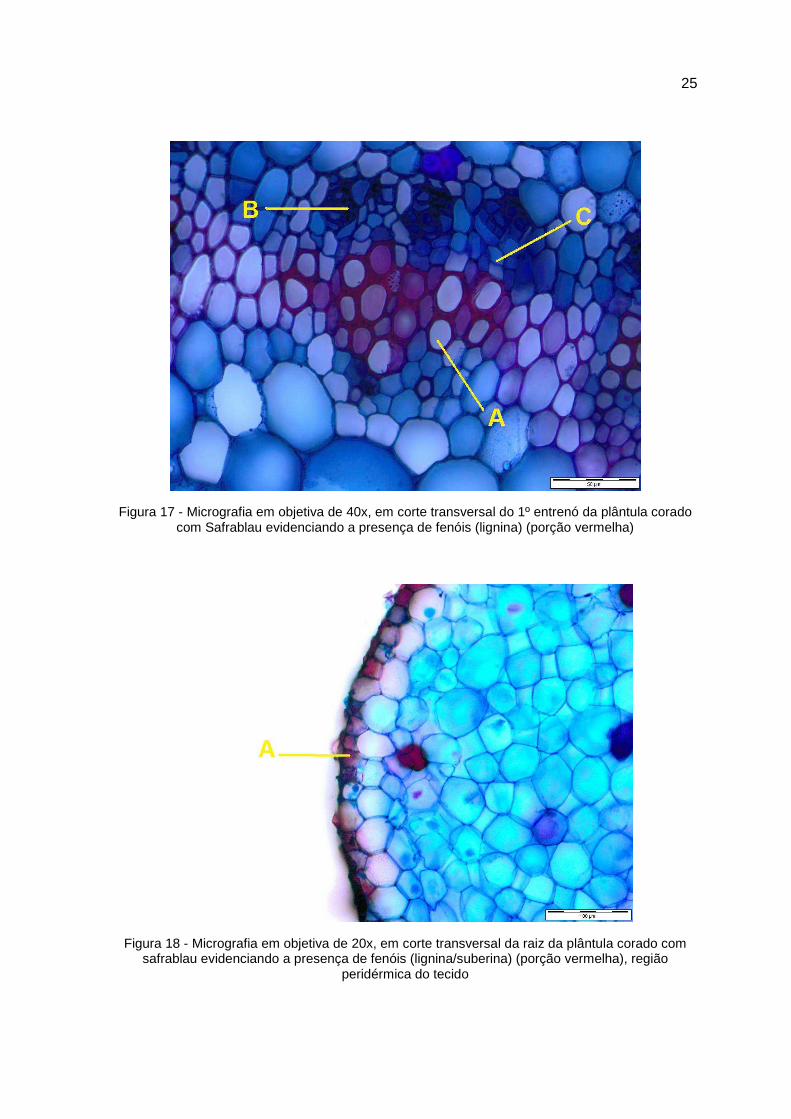
25
Figura 17 - Micrografia em objetiva de 40x, em corte transversal do 1º entrenó da plântula corado com Safrablau evidenciando a presença de fenóis (lignina) (porção vermelha)
Figura 18 - Micrografia em objetiva de 20x, em corte transversal da raiz da plântula corado com safrablau evidenciando a presença de fenóis (lignina/suberina) (porção vermelha), região
peridérmica do tecido
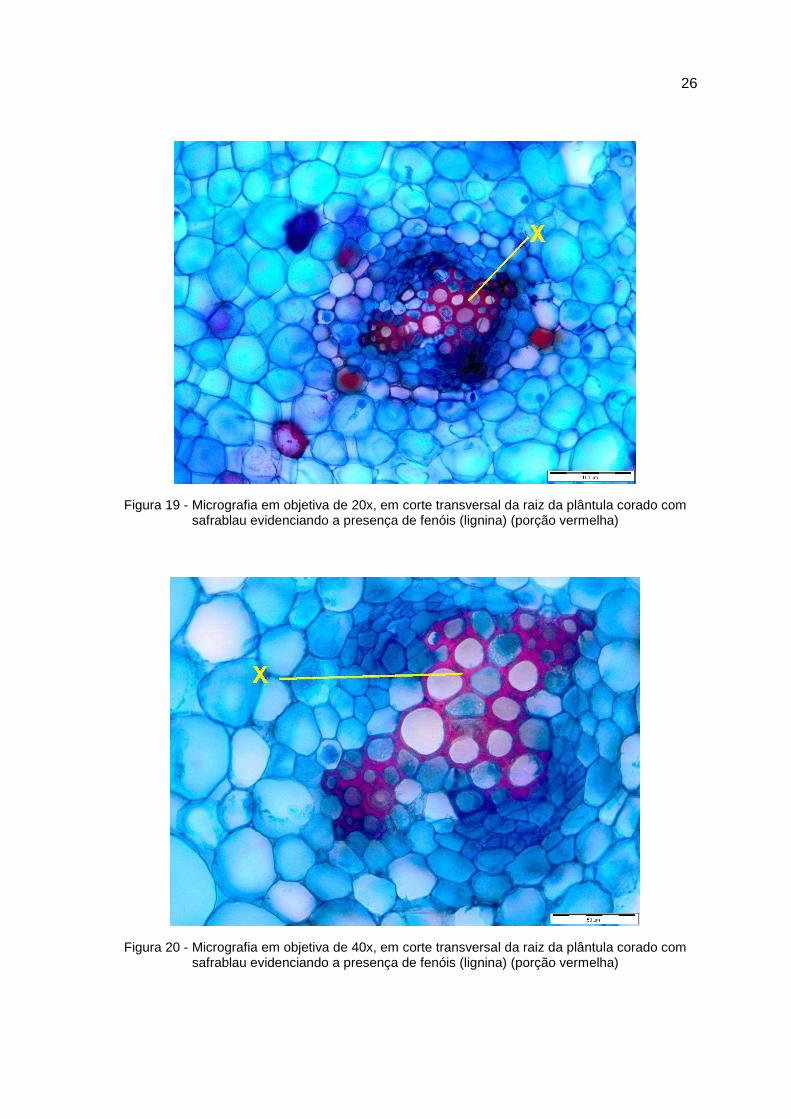
26
Figura 19 - Micrografia em objetiva de 20x, em corte transversal da raiz da plântula corado com safrablau evidenciando a presença de fenóis (lignina) (porção vermelha)
Figura 20 - Micrografia em objetiva de 40x, em corte transversal da raiz da plântula corado com safrablau evidenciando a presença de fenóis (lignina) (porção vermelha)

27
Os estudos realizados para identificação com tecidos em várias partes do
caule das plântulas de Lophantera lactescens foram realizados com teste de
Wiesner e mostrou que a região visualizada apresenta-se altamente lignificada, e
que no xilema há células fibrosas, células de parênquima radial lignificados,
evidenciando que neste caso o tecido jovem mostrou-se peculiar em relação a
outros tecidos de plantas herbáceas (Figura 21, 22, 23 e 24). No tecido radicular,
o teste de Wiesner evidenciou a porção lignificada (Figura 25 e 26).
Figura 21 - Micrografia em objetiva de 10x, em corte transversal da porção basal do caule da plântula, evidenciando a presença de lignina (campo claro)
28
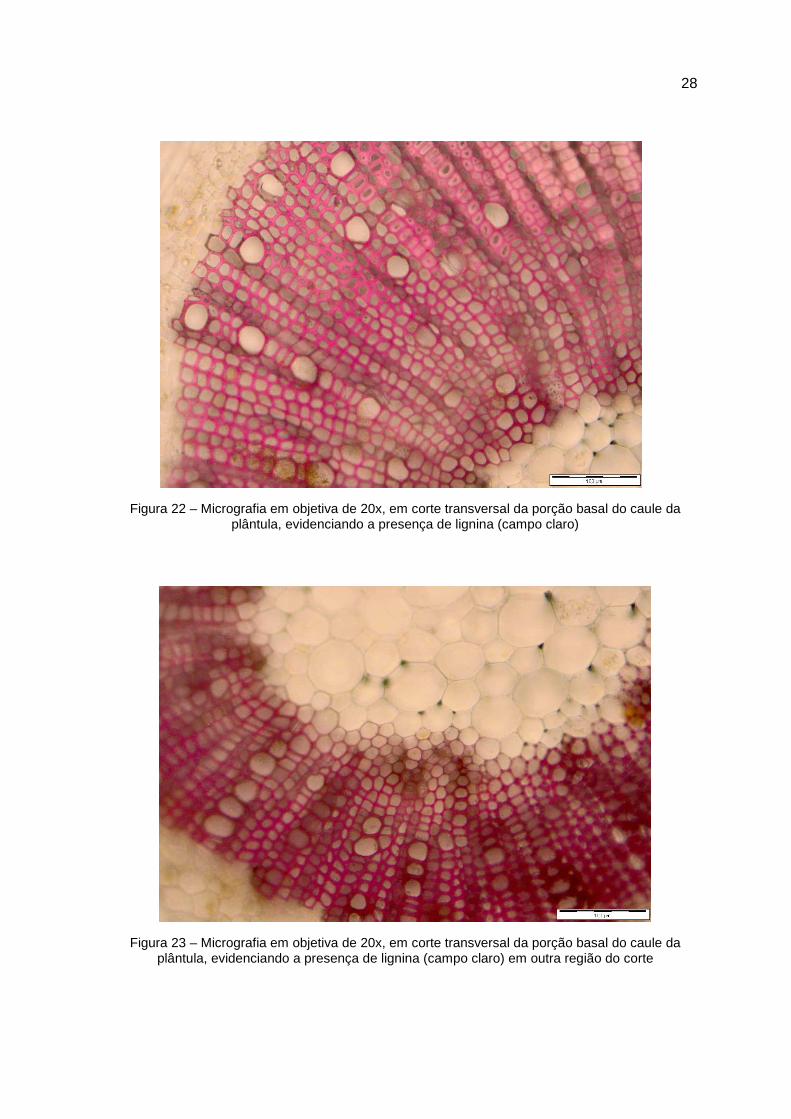
Figura 22 – Micrografia em objetiva de 20x, em corte transversal da porção basal do caule da plântula, evidenciando a presença de lignina (campo claro)
Figura 23 – Micrografia em objetiva de 20x, em corte transversal da porção basal do caule da plântula, evidenciando a presença de lignina (campo claro) em outra região do corte
29
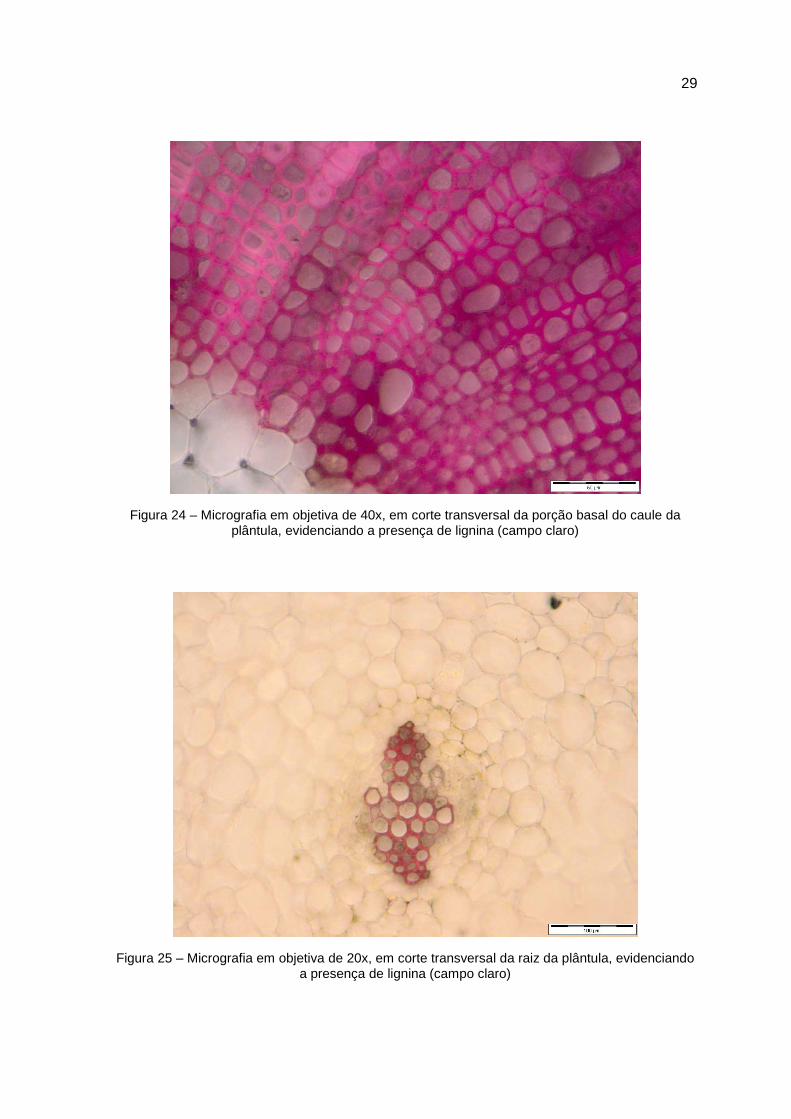
Figura 24 – Micrografia em objetiva de 40x, em corte transversal da porção basal do caule da plântula, evidenciando a presença de lignina (campo claro)
Figura 25 – Micrografia em objetiva de 20x, em corte transversal da raiz da plântula, evidenciando a presença de lignina (campo claro)
30
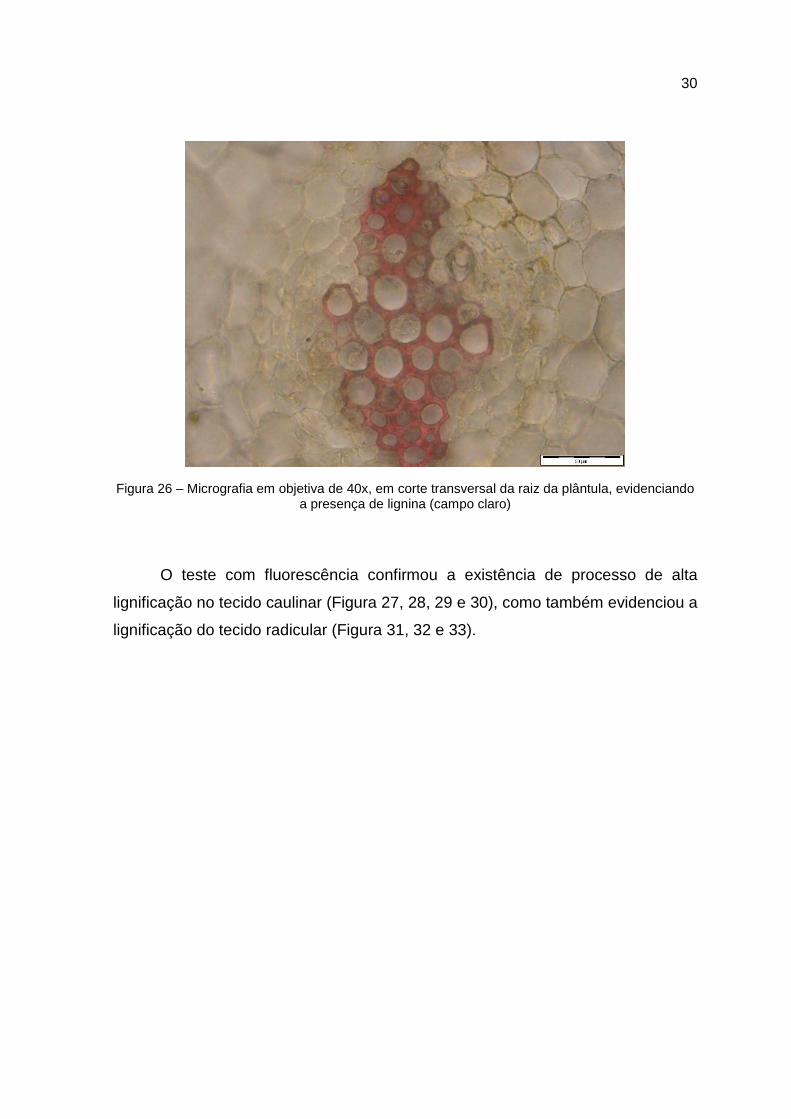
Figura 26 – Micrografia em objetiva de 40x, em corte transversal da raiz da plântula, evidenciando a presença de lignina (campo claro)
O teste com fluorescência confirmou a existência de processo de alta
lignificação no tecido caulinar (Figura 27, 28, 29 e 30), como também evidenciou a
lignificação do tecido radicular (Figura 31, 32 e 33).
31
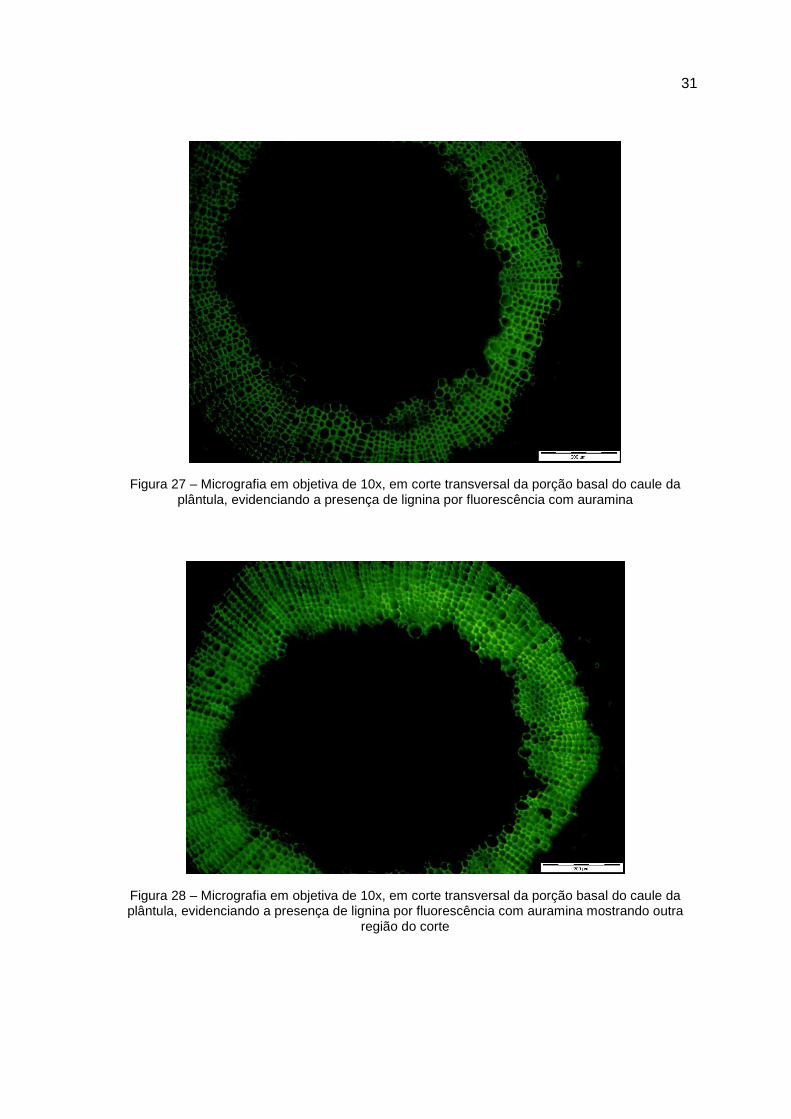
Figura 27 – Micrografia em objetiva de 10x, em corte transversal da porção basal do caule da plântula, evidenciando a presença de lignina por fluorescência com auramina
Figura 28 – Micrografia em objetiva de 10x, em corte transversal da porção basal do caule da plântula, evidenciando a presença de lignina por fluorescência com auramina mostrando outra
região do corte
32
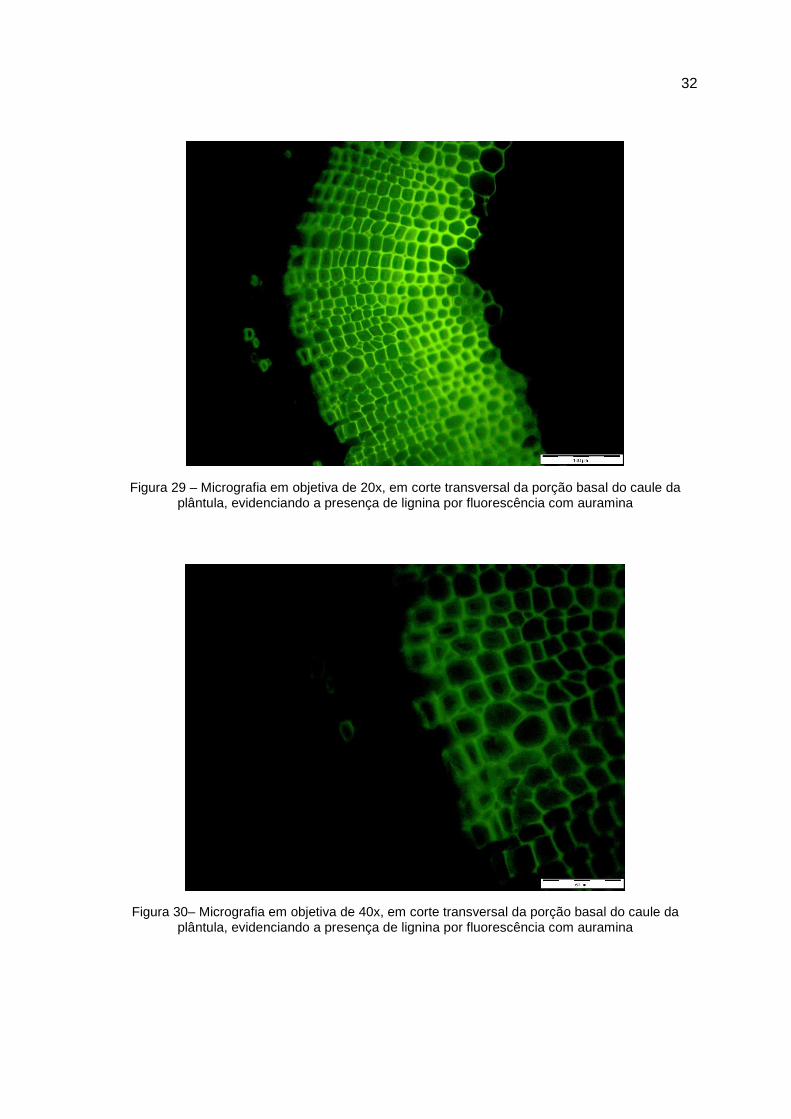
Figura 29 – Micrografia em objetiva de 20x, em corte transversal da porção basal do caule da plântula, evidenciando a presença de lignina por fluorescência com auramina
Figura 30– Micrografia em objetiva de 40x, em corte transversal da porção basal do caule da plântula, evidenciando a presença de lignina por fluorescência com auramina

33
Figura 31 – Micrografia em objetiva de 10x, em corte transversal da raiz da plântula, evidenciando a presença de lignina por fluorescência com auramina
Figura 32 – Micrografia em objetiva de 20x, em corte transversal da raiz da plântula, evidenciando a presença de lignina por fluorescência com auramina

34
Figura 33 – Micrografia em objetiva de 40x, em corte transversal da raiz da plântula, evidenciando a presença de lignina por fluorescência com auramina

35
6 – CONCLUSÃO
Sob diagnóstico do processo de lignificação dos tecidos do caule e da raiz
de plântulas de Lophantera lactescens, com base nas análises por infravermelho,
microscopia de campo claro e de fluorescência ficou evidenciado que o processo
de lignificação em Lophantera lactescens tem início nos primórdios caulinares, e
que o sistema de crescimento apresenta xilema primário totalmente lignificado,
condizente com um sistema adulto. O teste de Wiesner, fluorescência e
infravermelho, serviram como ferramentas indispensáveis à caracterização da
lignina existente, com ênfase a microscopia no infravermelho permitindo a priori
evidenciar uma composição guaiacila/siringila e que nestes termos são tão
evidentes que não deixam dúvidas sobre a sua ocorrência em plântula de
Lophantera lactescens. Os espectros não foram tão claros, devido se tratarem de
material lignocelulósico sem procedimento de isolamento, entretanto os materiais
pré-extraidos com ciclohexano, acetato de etila e metanol, foram considerados
espectros mais típicos de lignina em um sistema multimolecular da parede celular.
Os tecidos xilemáticos conforme fotomicrografia em fluorescência evidenciado
pela auramina mostra que estes tecidos são altamente lignificados. O que permite
torná-la objeto vivo de modelos para novos estudos sobre lignificação.

36
7 – REFERÊNCIAS BIBLIOGRÁFICAS
ABREU, H. S.; FO R. B.; GOTTLIEB H. E.; SHOOLERY J. N. A Nor-triterpenoid from Lophantera Lactescens. Phytochemistry , v. 29, n 7, p. 2257 – 2261, 1990.
ABREU, H. S.; NASCIMENTO, A. M. & MARIA, M. A. Lignin and structure, Wood and Fiber , V.31, p.426-433, 1999.
BARCELO, A. R. Lignification in plant cell walls. Int. Rev. Cytol . 176, p.87–132, 1997.
BOERJAN, W; RALPH, J; BAUCHER, M. Lignin biosynthesis. Annu Rev Plant Biol , V.54, p.519–546, 2003.
CHEN, C.-L. Lignins: occurrence in woody tissues, isolation, reactions, and structure. In: LEWIN, M.; GOLDSTEIN, I. S. Wood structure and composition. New York: Marcel Decker, p. 183-261, 1991.
CHOINOWSKI, T.; BLODIG, W.; WINTERHALTER, K. H.; PIONTEK, K. The crystal structure of lignin peroxidase at 170 Ao resolution reveals a hydroxy group on the Cb of tryptophan 171: A nonel radical site formed during the redox cycle. J Mol Biol , V.286, p.809-827, 1999.
DANELLI, M.G.M.; SOARES, D.C.; ABREU, H.S.; PEÇANHA L.M.T.; SARAIVA E.M. Leishmanicidal effect of LLD-3 (1), a nor-triterpene isolated from Lophanthera lactescens. Phytochemistry . v. 70, p. 608-614, 2009.
DONALDSON, L. A. Lignification and lignin topochemistry – an ultrastructural view. Phytochemistry , v.57, p. 859-873, 2001.
FENGEL, D.; WEGENER, G. Wood chemistry, ultrastructure, reactions. New York. Walter de Gruyter, p. 2-220, 1984.

37
HIGUCHI, T. Biochemistry and molecular biology of wood . Springer, p. 506 1997.
INOUE, K.; SEWALT, V.J.; MURRAY, G.B.; NI, W. STURZER, C.; DIXON, R.A. Developmental expression and substrate specificities of alfalfa caffeic acid 3-Omethyltransferase and caffeoyl coenzyme A 3-Omethyltransferase in relation to lignification. Plant Physiology, V.117, p.761–770, 1998.
KYTAYAMA, K.; SUZUKI, S.; HORI, M.; TAKYU, M.; AIBA, S.I.; MALAJALAP-LEE, N.; KIKUZAWA, K. On the relationships between leaf-litter lignin and net primary productivity in tropical rain forests. O junho ecologia , V.140, p.335-339, 2004.
LARROQUE, C. M. & PLANCHON C. L. Lignification and physiological factors of yield in maize. Published in Crop Sci , V.30, p.1105-1109, 1990.
LATORRACA, J.V.F.; ABREU, H dos S. Extrativos da madeira . Departamento de Produtos Florestais, Imprensa Universitária, Rio de Janeiro, p. 27, 1997.
LIN, S. Y. & DENCE, C.W. Methods in lignin chemistry, Berlin: Springer-Verlag, p. 578 1992.
LORENZI, H. Árvores Brasileiras. Vol 1. 5 Ed. São Paulo: Instituo Plantarum, p. 239. 2008.
LORENZI, H.; SOUZA, C. V. Botânica Sistemática. 2 Ed. São Paulo: Instituto Plantarum, p. 347. 2008.
MONTIES, B. Molecular structure and biochemical properties of lignins in relation with possible selforganization of lignin networks. Annales des Sciences Forestières , V.46, p.846-855, 1989.

38
NORTHCOTE, D. H. Control of plant cell wall biogenesis: an overview. ACS Symp. Ser. V.399, p.1-15, 1989.
PILÓ-VELOSO, D., NASCIMENTO, E.A., MORAIS, S.A.L. Isolamento e análise estrutural de ligninas. Química Nova , v.16, p.435-448, 1993
RAES, J. et al. Genome – Wide Characterization of the lignification toolbox in Arabidopsis. Plant Physiology , vol.133, p.1051-1071, 2003.
RANOCHA, P.; CHABANES, M.; CHAMAYOU, S.; JAUNEAU, A.; BOUDET, A. M.;GOFFNER, D. Laccasse Down-Regulation Causes Alterations in Phenolic Metabolism and cell wall structure in poplar. Plant Physiology , V.129, n.1, p.145-155, 2002.
ROGERS, L. A.; CAMPBELL, M. M. The genetic control of lignin deposition during plant growth and development. New Phytologist . V.164, n.1 p.17, 2004.
SILVA, S. L. MORFOANATOMIA E ONTOGÊNESE DE FRUTOS E SEMENTES DE ESPÉCIES DE BANISTERIOPSIS C. B. ROBINSON E DIPLOPTERYS A. JUSS.(MALPIGHIACEAE) . 2007. 72 p. Dissertação (Doutor em Morfologia e Diversidade Vegetal) – Instituto de Biociências, UNESP, Botucatu – São Paulo. 2007. Orientador: DENISE MARIA TROMBERT DE OLIVEIRA.
STUDART-GUIMARÃES, et al. Transformações genéticas em espécies florestais. Ciência Florestal , v.13, n.1, 2003.
TREVISAN, H. Ocorrência de térmitas subterrâneos (Isoptera: Rhinotermitidae e Termitidae) e durabilidade natural da madeira de cinco essências florestais. Revista Ciência Florestal , Santa Maria, v. 13, n. 2, p.153-158, 2003.

39
WHETTEN, R. W.; MACKAY, J. J. & SEDEROFF, R.R. Recent advances in understanding lignin biosynthesis. Annual Review of plant physiology and plant molecular biology , V.49, p.585-609, 1998.





























