Embed Size (px)
Citation preview

0
UNIVERSIDADE CEUMA PRÓ-REITORIA DE PÓS-GRADUAÇÃO, PESQUISA E EXTENSÃO
PROGRAMA DE PÓS GRADUAÇAO EM BIOLOGIA PARASITÁRIA MESTRADO ACADÊMICO
CAROLINA XAVIER LIMA BRITO
Avaliação da participação do receptor TRPV1 na infecção murina por Plasmodium
berghei ANKA
São Luís
2014

1
CAROLINA XAVIER LIMA BRITO
Avaliação da participação do receptor TRPV1 na infecção murina por Plasmodium
berghei ANKA
Dissertação apresentada ao Programa de Pós Graduação em Biologia Parasitária da Universidade CEUMA, para obtenção do título de Mestre. Área de concentração: Resposta imune nas doenças infecciosas e parasitárias. Orientador: Profa. Dra. Elizabeth Soares Fernandes Co-orientador: Prof. Dr. Marcos Augusto Grigolin Grisotto
São Luís
2014

2
Brito, Carolina Xavier Lima.
Avaliação da participação do receptor TRPV1 na infecção murina por Plasmodium
berghei ANKA./ Carolina Xavier Lima Brito. São Luís: UNICEUMA, 2014.
73 p.:il.
Dissertação (Mestrado) – Mestrado em Biologia Parasitária. Universidade CEUMA,
2014.
1. Malária. 2. Capsazepina. 3. Receptor TRPV1. 4. C57BL/6.5. Plasmodium berghei ANKA. I. Fernandes, Elizabeth Soares (Orientador) II. Grisotto, Marcos Augusto
Grigolin (Co-orientador). III.Título.
CDU: 576.89
B574a

3
CAROLINA XAVIER LIMA BRITO
Avaliação da participação do receptor TRPV1 na malária
A Comissão julgadora da Defesa do Trabalho Final de Mestrado em
Biologia Parasitária, em sessão pública realizada no dia / / , considerou a
candidata:
( ) APROVADO ( ) REPROVADO
1. Examinador ________________________________________________
2. Examinador ________________________________________________
3. Examinador ________________________________________________

4
Dedico este trabalho à minha
amada família, meu marido Luis,
minha filha Luíse e minhas mães
Ilis e Zezé, pelo amor e
companheirismo.

5
AGRADECIMENTOS
É com grande alegria que concluo essa etapa tão importante na minha vida profissional, mas tenho convicção que não consegui sozinha e gostaria de compartilhar essa vitória com todos que me ensinaram, me incentivaram e me acolheram em prol desta conquista.
Agradeço a Deus por ser o meu sustento, pela força e fé que me deu durante toda esta jornada, que por vezes o fim parecia tão distante, devido aos obstáculos que surgiram. Enfim, agradeço a Ele por não ter permitido que eu desanimasse!!!
Ao meu marido, Luis Henrique, porque sem você, nada disso seria possível. Pelo apoio incondicional que você me deu, principalmente nesse momento final, em que se tornou pai e mãe da nossa filha, colocando-a para dormir, lendo histórias, acompanhando nas tarefas da escola, levando pra cá e levando pra lá, indos aos aniversários... Enfim, tentando suprir ao máximo a minha ausência; pela ajuda com a informática, dúvidas de formatação...; pelos jantares, que tantas vezes você fez para que eu não ficasse sem comer na loucura desses últimos meses;
À minha filha, Luíse, pelo carinho e amor que faziam a renovação das minhas forças e pela compreensão da minha ausência, por saber o quanto este mestrado é importante, que você exemplificava quando dizia, “quando a mamãe terminar o mestrado, ela vai ter mais tempo pra mim”;
Às minhas mães, Ilis e Zezé, que sempre estiveram presentes, prontas a ajudar a qualquer hora, momento ou lugar; pelas orações que tanto me consolaram e me sustentaram;
À minha orientadora, Profa. Dra. Elizabeth Soares Fernandes, pela oportunidade de concluir este sonho, por ter acreditado em mim, quando nem eu mesma acreditava; pelos ensinamentos, paciência e incentivo constantes; muito obrigada mesmo, você foi essencial para a conclusão deste trabalho!! Tenho uma enorme admiração por você!!
Ao meu co-orientador, Prof. Dr. Marcos Augusto Grigolin Grisotto, pelo conhecimento ofertado deste o início do mestrado, pelo encorajamento constante e por não ter desistido de mim. Minha gratidão será eterna!!
Ao prof. Dr. Eduardo Martins de Sousa, componente da banca examinadora de qualificação, pela atenção, disponibilidade e sugestões que muito me ajudaram a concluir esta dissertação;
A todos os professores do Programa de Pós Graduação em Biologia Parasitária da Universidade CEUMA pelo conhecimento repassado e disponibilidade constante, despertando em mim o desejo de sempre querer aprender mais e mais;
Aos meus companheiros da turma IV, Claudia, Fernanda, Fran, Iven, Jéssica, Kátia, Lisiane e Reutoni (in memorian), formamos uma turma incrível!! Foram momentos inesquecíveis que construímos juntos!! Agradeço, em especial, às divas que sempre estavam disponíveis e com uma palavra de carinho e incentivo.

6
À minha amiga Ericka Mesquita que se tornou muito mais que minha companheira de laboratório, sendo hoje minha amiga de todas as horas;
Ao prof. Dr. Ivan Figueiredo, que acima de tudo é um verdadeiro amigo, uma referência de pessoa, pela disponibilidade, porque em meio a tantos afazeres sempre está pronto a me ajudar, pelas orações e palavras de carinho. Muito obrigada!!!
A todos os meus amigos que torceram, oraram e me incentivaram a concluir esta etapa importantíssima da minha vida. Amo vocês!!!
Aos profs. Drs. Claudio Marinho, Renato Barboza e Aramys Reis pelo excelente acolhimento e por terem aberto as portas do laboratório no Departamento de Parasitologia do Instituto de Ciências Biomédicas da USP, disponibilizando equipamentos e material. Sempre seria grata por tudo!!
À Universidade Ceuma pela oportunidade de realização do mestrado em Biologia Parasitária;
Aos camundongos que possibilitaram a execução deste projeto;
A todos que de forma direta ou indireta contribuíram para a realização deste sonho, afinal o sucesso é nosso porque ninguém vence sozinho!!

7
“Aqueles que confiam no Senhor
são como o monte de Sião, que não
pode ser abalado, mas permanece
para sempre.” Salmos 125:1

8
RESUMO
A malária é uma doença infecciosa, com elevada morbidade, mortalidade e elevado impacto
socioeconômico, representando assim, um grave problema de saúde pública global,
responsável por mais de 200 milhões de novos casos anualmente. Destes, os mais acometidos
são mulheres grávidas, adultos imunocomprometidos e crianças com até 5 anos de idade; os
quais podem desenvolver malária grave, com uma série de prejuízos para o paciente, inclusive
o óbito. Embora existam medicamentos eficazes no tratamento da malária, estes não são
capazes de reverter os danos decorrentes da malária grave. Ainda, algumas cepas de
Plasmodium são resistentes às medicações existentes. Assim, faz-se necessário o estudo de
novas vias associadas à malária, e consequentemente novos fármacos para o tratamento desta
doença. Recentemente, um papel protetor foi sugerido para o receptor TRPV1 (do inglês:
Transient Receptor Potential Vanilloid 1), localizado em células neuronais e não neuronais
(como as células endoteliais e macrófagos), na sepse. Este receptor pode ser ativado por
produtos do estresse oxidativo, como o H2O2, além de modular a liberação de espécies
reativas de oxigênio. Ainda, a ausência da ativação do TRPV1 leva a uma disfunção do
macrófago. Ressalta-se que monócitos e macrófagos, assim como o estresse oxidativo gerado
pelas interações destas células com o hospedeiro, são determinantes no prognóstico da
malária. Assim, o presente trabalho avaliou a participação do TRPV1 na malária, através do
tratamento repetido com o antagonista para o TRPV1, a capsazepina (50 µg/animal, 2 x ao
dia, 6 dias), em modelo murino de malária induzida por Plasmodium berghei ANKA. A
infecção foi induzida com uma injeção peritoneal contendo 1,0 x 105 células vermelhas
infectadas com P. berghei ANKA por animal. A parasitemia foi avaliada diariamente por
exame de gota espessa, e os animais foram sacrificados 7 dias após indução da infecção. O
sangue e o baço foram coletados e utilizados para a análise de fenotipagem das células e
também para quantificação de aldeídos e citocinas plasmáticas. A capsazepina mostrou-se
capaz de modular a resposta imune à malária em diferentes níveis. Os resultados demonstram
que a capsazepina promove diminuição da do número de células F4/80+/Ly6G+ e da ativação
de células F4/80+ e células F4/80+/Ly6G+ circulantes. Vale ressaltar que este estudo descreve
pela primeira vez a importância da população de monócitos F4/80+/Ly6G+ circulantes na
malária. Ainda, a capsazepina foi capaz de reduzir o número de células CD19+ e aumentar o
número de células CD3-NK1.1+ circulantes, sem no entanto, interferir com a ativação destas
células. Ainda, o tratamento com capsazepina inibiu a ativação de células CD19+, CD3+CD4+
e CD3+NK1.1+, via o marcador de ativação precoce, CD69. Ainda, a capsazepina promoveu

9
redução dos níveis plasmáticos de aldeídos e TNFα liberados por monócitos circulantes, sem,
no entanto, alterar a produção de IFNγ, em animais infectados. Ressalta-se que a capsazepina
não apresentou efeitos em nenhuma das populações de células avaliadas e nem na produção
de mediadores inflamatórios obtidos de animais não infectados. Em conjunto, estes resultados
demonstram pela primeira vez, que o TRPV1 é um importante modulador da resposta imune à
malária. No entanto, o impacto final da inibição deste receptor no prognóstico da malaria,
permanece por ser melhor investigado.
Palavras-chave: malária; capsazepina; receptor TRPV1; C57BL/6; Plasmodium berghei
ANKA.

10
ABSTRACT
Malaria is an infectious disease with high morbidity, mortality and socio-economic impact,
representing thus a serious public health problem, responsible for more than 200 million new
cases per year. Among these, the most affected are pregnant women, immunocompromised
adultsand children under 5 years old, that may develo grave malaria, with a series of
impairments for the patient, including death. Although there are effective medications for
malaria treatment, they are not able to reverse injuries caused by deep malaria. Also,
some Plasmodium stock are resistant to the available medication. Thus, it is necessary to
study new ways connected to malaria, and consequently new drugs for this disease's
treatment. Recently, a protector role has been suggested for the TRPV1 (from Transient
Receptor Potential Vanilloid 1) receptor, found in neural and non-neural cells (like
endothelial cells and macrophages) in sepsis. This receptor may be activated by oxidative
stress products, like H2O2, and also modulate the liberation of oxygen reactive species. Also,
absence of TRPV1 activation leads to a macrophage disfunction. It is empasyzed that
monocytes and macrophages, as well as oxidative stress generated by these cells interaction
with the host, are determinants for malaria prognosis. As such, the present work evaluated
TRPV1 participation in malaria, through repeated treatment with TRPV1 antagonist,
capsazepine (50 mg/animal, twice a day, 6 days) in Plasmodium berghei ANKA induced
malaria murine model. Infection was induced by a peritoneal injection containing 1,0 x 105
red cells infected with P. berghei ANKA per animal. Parasitemy was evaluated daily by thick
drop exam, and the animals were sacrificed 7 days after infection induction. Blood and spleen
were collected and used for cell phenotype analysis and also for plamatic aldehyde and
cytokine quantification. Capsazepine was found able to modulate immune response to malaria
in different levels. Results demonstrate that capsazepine enhances lowering of F4/80+/Ly6G+
cell numbers and of F4/80+ cell activation and circulating F4/80+/Ly6G+. It is worthy of note
that this study describes for the first time the importance of the F4/80+/Ly6G+ monocyte
circulating population in malaria. Also, capsazepine was able to reduce CD19+ cell number
and raise the number of circulating CD3+NK1.1+ cells, without interfering with these cells
activation. Also, capsazepine treatment inhibited CD19+, CD3+CD4+ and CD3+NK1.1+
activation, through the early activation marker CD69. Likewise, capsazepine provoked
lowering of plasma levels of aldehyde and TNFα liberated by circulating monocytes without
affecting INFγ production in the infected animals. It is empasyzed that capsazepine didn't
show effects in none of the populations of evaluated cells neither in the production of

11
inflammatory mediators obtained from non-infected animals. Together, such results show for
the first time that TRPV1 is an important immune response modulator in malaria. Even
though, the final impact of this receptor inhibition in malaria prognosis remains to be better
investigated.
Key words: malaria; capsazepina; TRPV1 receptor; C57BL/6; Plasmodium berghei ANKA.

12
SUMÁRIO
1. INTRODUÇÃO 17
1.1. A Malária 17
1.2. Aspectos gerais 19
1.3. Ciclo biológico
1.4. A resposta imune na Malária
1.4.1 Resposta imune inata
1.4.2 Resposta imune adquirida
1.5 Modelos experimentais de Malária
1.5.1 Modelo de Malária Cerebral
1.5.2 Modelo de Malária placentária
1.6 Tratamento de malária
1.7 Receptor TRP
1.7.1 TRPV1
1.7.2 O papel do TRPV1 nas infecções
23
24
25
27
28
29
31
33
35
36
37
2. OBJETIVOS
2.1 Geral
2.2 Específicos
40
40
40
3. METODOLOGIA 41
3.1. Animais utilizados no estudo 41
3.2. Indução da malária
3.3 Obtenção de células sanguíneas e esplênicas
41
42
3.4. Fenotipagem por citometria de fluxo em leucócitos de baço e de sangue 43
3.5. Dosagem de aldeídos plasmáticos (TBARS)
3.6. Dosagem de citocinas plasmáticas
3.7. Tratamento farmacológico
3.8. Análise estatística
44
44
45
45
4. RESULTADOS 46
4.1. Efeito do tratamento com o antagonista do receptor TRPV1, capsazepina,
sobre a parasitemia
46
4.2. Participação do receptor TRPV1 na resposta imune celular inata à
Malária
47

13
4.3. Participação do receptor TRPV1 na resposta imune celular adquirida à
Malária
4.4 Efeito do antagonista de receptores TRPV1 na produção sistêmica de
mediadores inflamatórios
56
63
5. DISCUSSÃO 65
6. CONCLUSÃO 81
REFERÊNCIAS 82

14
LISTA DE FIGURAS
Figura 1: Áreas de risco de transmissão da Malária. 20
Figura 2: Ciclo biológico do Plasmodium. 24
Figura 3: Efeito do tratamento com o antagonista do receptor TRPV1, capsazepina, sobre a parasitemia 46
Figura 4: (A) Painel representativo sobre a expressão de marcadores F4/80 e Ly6G no sangue de animais
sadios e infectados. (B) Histograma representativo demonstrando a contagem de eventos relacionada à
expressão de F4/80 e Ly6G em animais sadios e infectados tratados com veículo ou capsazepina.
48
Figura 5: (A) Painel representativo sobre a expressão de marcadores F4/80 e Ly6G no baço de animais
sadios e infectados. (B) Histograma representativo demonstrando a contagem de eventos relacionada à
expressão de F4/80 e Ly6G em animais sadios e infectados tratados com veículo ou capsazepina.
49
Figura 6: Efeito da capsazepina sobre as populações de células F4/80+ e F4/80+Ly6G+ no sangue e no baço
de animais sadios e infectados.
50
Figura 7: Efeito da capsazepina sobre a fluorescência média de IAb em células sanguíneas e esplênicas
F4/80+ e F4/80+Ly6G+em animais sadios e infectados.
51
Figura 8: Efeito da capsazepina sobre a população de células GR1+ no sangue e no baço de animais sadios e
infectados.
52
Figura 9: Painel representativo de células NK e NKT no sangue de animais sadios e infectados tratados
com veículo ou capsazepina.
53
Figura 10: Painel representativo de células NK e NKT esplênicas de animais sadios e infectados tratados
com veículo ou capsazepina.
54
Figura 11: Efeito da capsazepina sobre a população de células NK e NKT circulantes e esplênicas. 55
Figura 12: Efeito da capsazepina sobre a ativação de células NK e NKT circulantes e esplênicas. 56
Figura 13: (A) Painel representativo de células CD3+ e CD19+ em sangue de animais sadios e infectados
tratados com veículo ou capsazepina. (B) Painel representativo de células CD4+ e CD8+ em sangue de
animais sadios e infectados tratados com veículo ou capsazepina.
58
Figura 14: (A) Painel representativo de células CD3+ e CD19+ no baço de animais sadios e infectados
tratados com veículo ou capsazepina. (B) Painel representativo de células CD4+ e CD8+ no baço de animais
sadios e infectados tratados com veículo ou capsazepina.
59
Figura 15: Efeito da capsazepina sobre as células CD3+ e CD19+ no sangue e no baço de animais sadios e
infectados tratados com veículo ou capsazepina.
60
Figura 16: Efeito da capsazepina sobre as células CD4+ e CD8+ no sangue e no baço de animais sadios e
infectados tratados com veículo ou capsazepina.
61
Figura 17: (A) Efeito da capsazepina sobre os níveis de aldeído plasmático em animais infectados tratados
com veículo ou capsazepina. (B) Efeito da capsazepina sobre os níveis de TNF-α plasmático em animais
infectados tratados com veículo ou capsazepina. (C) Efeito da capsazepina sobre os níveis de TNF-γ
plasmático em animais infectados tratados com veículo ou capsazepina.
64

15
LISTA DE TABELAS
Tabela 1: Especificação do anticorpos monoclonais utilizados 43
Tabela 2: Efeito da capsazepina sobre a distribuição da população de células circulantes
62
Tabela 3: Efeito da capsazepina sobre a distribuição de células esplênicas 62

16
LISTA DE ABREVIATURAS E SIGLAS
a.C. – Antes de Cristo
ANOVA – Análise de variância
APC – Célula apresentadora de antígeno
AST – Aspartato-amino transferase
Ca – Cálcio
CBA – Cytometric bead array
CEUA – Comissão de ética no uso de animais
CGRP – Calcitonin Gene-Related Peptide
d.C. – Depois de Cristo
DMSO – Dimetilsulfóxido
FITC – Isotiocianato de fluoresceína
GPI – Glicosilfosfatidilinositol
HNE – 4-hidroxi-2-nonenal
IFN-γ – Interferon-gama
Ig – Imunoglobulina
IL – Interleucina
LPS – Lipopolissacarídeos
MHC – Complexo principal de histocompatibilidade
NaCl – Cloreto de sódio
NADH – Nicotinamida adenina de nucleotídeo
NK – Natural killer
NO – Óxido Nítrico
OMS – Organização Mundial de Saúde
PAMP – Padrões moleculares associados a patógenos
PBS – Tampão fosfato salina

17
PE – Ficoeritrina
ROS – Espécies reativas de oxigênio
SBCAL – Sociedade Brasileira de Ciência de Animais de Laboratório
SD – Desvio Padrão
SP – Substancia P
TBARS – Thiobarbituric acid reactive substances
TNF-α – Fator de necrose tumoral- alfa
Treg – Células T reguladoras
TRP – Transient Receptor Potential
TRPA – Transient Receptor Potential Ankyrin
TRPC - Transient Receptor Potential Canonica
TRPM – Transient Receptor Potential Melastatin
TRPML – Transient Receptor Potential Mucolipin
TRPN – Transient Receptor Potential No mechanopotential
TRPP – Transient Receptor Potential Polycystin
TRPV – Transient Receptor Potential Vanilloid
TRPV1KO – Transient Receptor Potential 1 knockout
USP – Universidade de São Paulo
WT – Camundongos selvagens (do inglês “wild-tipe mice”)

18
1. INTRODUÇÃO
1.1 Malária
A malária é uma doença antiga que acompanha a humanidade desde os primórdios, há
dezenas de milhares de anos. Por apresentar características clínicas bem definidas, como por
exemplo, intensos calafrios que antecedem febre alta que podem durar de três a quatro horas e
se repetirem com intervalos de um ou dois dias até que o paciente se recupere ou chegue ao
óbito, é possível diferenciá-la de outras patologias febris descritas na história (CAMARGO,
2003).
Acredita-se que a principal causa de morte entre os primatas precursores do Homo
sapiens (Australopithecus) foi a malária (BRUCE-CHWATT, 1988). Porém, os primeiros
relatos que podem referenciar esta patologia foram documentados pelos chineses em 3000
a.C., pelos egípcios em 1570 a.C. e em tábuas cuneiformes da Mesopotâmia (hoje Iraque) de
2000 a.C. (COX, 2010).
Os filósofos da antiguidade, como por exemplo, Homero, Aristóteles, Platão, Sócrates,
Horácio e Shakespeare, também descreveram febres relacionadas à malária (BRUCE-
CHWATT, 1988). O fisiologista Hipócrates foi o primeiro a destacar que a presença de água
estagnada (pântanos, várzeas e alagadiços) parecia estar relacionada com o surgimento de
febres na população. E por mais de 2500 anos, a malária foi tida como uma doença decorrente
dos vapores venenosos provenientes dos pântanos, dando origem ao termo “ar ruim” ou “mal
aria” para os italianos, enquanto que para os franceses era conhecida como “paludismo”
(termo relacionado a pântano). Assim, tais termos deram origem aos nomes hoje utilizados
para esta doença, malária/paludismo (COX, 2010).

19
Porém, somente depois da descoberta das bactérias, em 1676, com o surgimento da
teoria dos germes causadores de infecção em 1878-1879, que o questionamento sobre a
origem da malária foi intensificado. O agente causador da malária foi descoberto no ano de
1880, quando o sangue fresco de um soldado doente foi analisado e encontraram-se grânulos
pigmentados e formas móveis que foram chamadas de Oscillaria maláriae. Cinco anos
depois, patologistas italianos identificaram formas ameboides em forma de anel no interior
das hemácias de pacientes doentes e deram-lhes o nome de Plasmodium (DAVES, 1988). Em
1898, foi descoberto o vetor da malária, o mosquito hematófago do tipo Anopheles, porém,
somente em 1950, o ciclo do parasita foi totalmente elucidado (DAVES, 1988).
A malária sempre foi um grande algoz da humanidade, afetando todo o planeta, com
exceção das regiões polares e subpolares. De maneira que grandes ícones da história mundial
podem ter padecido de malária, como pode ter sido o caso do arcebisbo Santo Agostinho em
597 a.C., do imperador Alexandre o Grande em 323 a.C. e do poeta italiano Dante Alighiere
em 1321 d.C. (CAMARGO, 1995).
Durante a colonização da América do Norte, os ingleses trouxeram para este
continente, duas espécies de agentes causadores da malária (Plasmodium vivax e Plasmodium
maláriae) e o tráfico de escravos foi responsável pela importação de outra espécie, o
Plasmodium falciparum (FRANÇA et al, 2008). Assim, sugeriu-se que a malária impediu
grandemente o desenvolvimento da região sul da colônia americana, impactando assim, de
forma negativa a economia da época (FRANÇA et al, 2008).
No Brasil, a malária teve destaque no contexto nacional na década de 1980, quando se
iniciou uma ocupação desordenada da região amazônica, a construção de usinas
hidroelétricas, a instalação de garimpos e o desenvolvimento de projetos agropecuários. Estes
eventos favoreceram a exposição do indivíduo ao mosquito, sendo a região amazônica

20
responsável por cerca de 97,5% dos casos da doença (TAUIL, 1982). Fora da região
conhecida por Amazônia Legal (Acre, Amapá, Amazonas, Maranhão, Mato Grosso, Pará,
Rondônia, Roraima e Tocantins), o maior número de casos foi registrado em Goiás, Paraná,
São Paulo e Mato Grosso do Sul, devido a casos oriundos da região endêmica (MARQUES e
PINHEIRO, 1982).
Na década de 90, houve um aumento alarmante de casos de malária sendo que 99,7%
notificados na Amazônia Legal, de maneira que o governo iniciou um conjunto de ações com
o objetivo de controlar tal patologia (FRANÇA et al, 2008). A partir de 2002, foi possível
observar um declínio no número de casos, embora até os dias de hoje, o Brasil ainda apresente
números alarmantes de casos anuais confirmados de malária (FRANÇA et al, 2008; OMS,
2012).
1.2 Aspectos Gerais
A malária é uma doença infecciosa aguda, não contagiosa, considerada como
parasitose de importância mundial, causada pelo protozoário do gênero Plasmodium, o qual é
transmitido ao homem a partir da picada da fêmea de insetos dípteros do gênero Anopheles
(CROMPTON et al, 2014; OMS, 2010). Porém, nem todas as espécies de Anopheles são
capazes de transmitir tal patógeno, sendo que, somente cerca de 100 das 400 descritas na
literatura funcionam como vetor da malária (FORATTINI, 2002).
Apesar das medidas adotadas após a Segunda Guerra Mundial no intuito de controlar a
transmissão da malária, metade da população mundial está em área de risco de infecção em
países pobres de regiões tropicais e subtropicais (FEACHEM et al, 2010). A maioria dos
casos está concentrada nos países africanos (85% dos casos), nos latino-americanos e nos da
região sudoeste da Ásia (OMS, 2010; 2012).

21
Segundo a Organização Mundial de Saúde (OMS), a malária é causa de grande
morbidade e mortalidade, com 210 milhões de casos/ano em todo o mundo, alcançando 660
mil óbitos/ano (OMS, 2012). Ressalta-se que tais dados podem ser ainda mais expressivos,
uma vez que a subnotificação ainda ocupa papel de destaque nesses países. Ainda, grávidas,
adultos imuno comprometidos e crianças com idade abaixo de cinco anos tendem a ser mais
acometidos pela malária, sendo que nas crianças é frequente a associação da malária a outras
patologias, como infecções intestinais e respiratórias, ocasionando complicações clínicas e
levando muitas vezes ao óbito de cerca de um milhão de crianças por ano (RIDLEY, 2002).
Assim, a malária é considerada uma das três principais doenças transmissíveis causadoras de
morte no mundo (SACHS e MALANEY, 2002).
Figura 1: Áreas de risco de transmissão da malária
Ilustração evidenciando as áreas de risco de transmissão da malária no mundo. Fonte: modificado de OMS, 2012.
Na América Latina, o Brasil e a Colômbia concentraram cerca de 68% dos casos de
malária notificados no ano de 2011 (OMS, 2012). No Brasil, os casos de malária encontram-
se localizados, principalmente, na região conhecida como Amazônia Legal a qual abriga cerca
Área de transmissão de malária Área de transmissão controlada de malária Não aplicável

22
de 20% da população brasileira, e na qual se concentram 99,8% dos casos notificados de
malária no Brasil, que em 2009, corresponderam a 306 mil (SILVEIRA e REZENDE, 2001;
OMS, 2012).
A grande ocorrência de malária nessa região pode ser justificada nesta área pela
presença de floresta tropical úmida, a qual favorece a proliferação do mosquito vetor; pela
presença de grupos humanos com atividade ocupacional, como por exemplo, agricultores,
garimpeiros e madeireiros, os quais se encontram vulneráveis à picada do inseto; pelas
condições climáticas favoráveis ao desenvolvimento do vetor (umidade e temperatura); pela
carência de infraestrutura física, social e de atenção básica de saúde; e pelo uso irracional de
drogas antimaláricas (LOIOLA et al, 2002).
O gênero Plasmodium inclui cinco espécies causadoras de malária no homem: vivax,
falciparum, ovale, maláriae e knowlesi; sendo o P. vivax o de maior incidência no Brasil,
responsável por 87%, seguido do P. falciparum com 13% dos casos novos registrados de
malária (OLIVEIRA-FERREIRA et al, 2010; OMS, 2012).
O P. falciparum é capaz de originar acessos febris em intervalos que variam de 36 a
48 horas, caracterizando assim um quadro conhecido como febre terçã maligna, com
comprometimento, até mesmo, do sistema nervoso central e, por tanto, está associado a
maioria dos óbitos causados pela malária grave (GARCIA et al, 2001). O dano tecidual
característico da infecção causada por esta espécie é consequência do sequestro de eritrócitos
infectados pelo endotélio vascular cerebral e/ou placentário, originando uma diminuição do
fluxo sanguíneo, migração de células inflamatórias e liberação de mediadores inflamatórios,
causando assim a lesão endotelial propriamente dita e a retenção das formas assexuadas do
Plasmodium impedindo que estas sejam destruídas pelo baço (GREENWOOD et al, 2008).

23
Já o ciclo febril do P. vivax é de 48 horas, o que faz com que a infecção por este
protozoário seja conhecida por febre terçã benigna. Assim, a malária causada por P. vivax é
relatada na literatura, normalmente, como uma patogenia relativamente benigna, se
comparada à malária por P. falciparum, podendo não se manifestar, caracterizando pacientes
clinicamente assintomáticos, os quais se tornam reservatórios humanos de Plasmodium
(OLIVEIRA-FILHO e MARTINELLI, 2009). Sua evolução clínica caracteriza,
principalmente, uma patologia menos grave e com baixa letalidade, mas elevada morbidade,
ocasionando graves problemas de ordem sócio-econômicos (MENDIS et al, 2001).
Entretanto, recentemente, foi observado o aparecimento de 57 casos graves e inclusive
fatais de malária por P. vivax, sendo 11 destes ocorridos no Maranhão (RAPOSO, 2006).
Ainda, de 426 pacientes com diagnóstico prévio de malária por P. vivax no Estado do
Amazonas acompanhados periodicamente, 20% evoluíram para um quadro clínico grave,
tanto em aspectos clínicos como laboratoriais (ALECRIM, 2000). É possível que a
complicação do quadro de malária por P. vivax seja decorrente da imediata presença de
formas sexuadas do parasita no sangue periféricos de indivíduos infectados, da existência de
hipnozoítos, dos efeitos colaterais das drogas utilizadas no tratamento e de cepas resistentes
ao tratamento habitual (OLIVEIRA-FERREIRA et al, 2010).
O termo febre terçã benigna também é designado para a infecção por Plasmodium
ovale, sendo este mais frequente no continente africano, porém, recentemente, foi descrito o
primeiro caso notificado de malária causada por ele no Brasil (LIMONGI et al, 2014). Uma
característica comum ao P. vivax e ao P. ovale é a capacidade de gerar formas latentes no
fígado, os hipnozoítos, capazes de promover uma recaída da doença. Este período de latência
pode se estender por 15 dias ou até mesmo por anos (WARRELL, 1998). Outra forma de
manifestação da malária é através da febre quartã, na qual os acessos febris ocorrem a cada 72
horas e tem como agente etiológico o Plasmodium malarie. (DAILY, 2006). O Plasmodium

24
knowlesi tem sido identificado como o principal agente causador da malária no sudeste
asiático e tem sua morfologia muito semelhante ao P. maláriae (FOSTER et al, 2014)
1.3 Ciclo Biológico do Plasmodium
A transmissão da malária acontece a partir da inoculação da forma infectante do
Plasmodium, o esporozoíto, pela fêmea do mosquito Anopheles nos capilares subcutâneos do
hospedeiro. Apesar de esta ser a forma usual de contaminação, é importante ressaltar que
existem outras formas de contágio, como por exemplo, através da via materno-fetal, da
transfusão de sangue e derivados, e mediante o compartilhamento de seringas por usuários de
drogas (GILLES, 1998).
No Brasil, existem cinco espécies responsáveis pela transmissão da malária, sendo que
o Anopheles darlingi é o mais representativo, uma vez que é comum na região amazônica e
em regiões litorâneas (TADEI e THATCHER, 2000). O ciclo de vida do Plasmodium
apresenta duas etapas, uma sexuada e outra assexuada, que diferem também quanto ao local
de ocorrência, uma vez que a sexuada ocorre no interior da fêmea do mosquito e a assexuada,
no hospedeiro humano (CORNEJO, ESCALANTE, 2006).
Em pouco tempo, o esporozoíto atinge os hepatócitos, iniciando, então, um processo
de replicação intracelular chamado de ciclo exo-eritrocítico, o qual dura aproximadamente
oito dias, dando origem à forma do protozoário que penetra nas hemácias, o merozoíto. Este
sofre multiplicação assexuada nos eritrócitos, instalando assim, a malária. Ressalta-se que os
sintomas da malária, episódios repetidos de febre e anemia progressiva, são decorrentes da
liberação de toxinas (como por exemplo, a hemozoína) após o desenvolvimento dos parasitas
no interior dos eritrócitos (SCHOFIELD e GRAU, 2005; NADJIM e BEHRENS, 2012).

25
Ainda no hospedeiro humano, alguns parasitas se diferenciam em suas formas
sexuadas, gametócitos masculinos e femininos, que permanecem no sangue e são ingeridos
pela fêmea do mosquito Anopheles durante a picada. No estômago do mosquito, o zigoto é
formado a partir da penetração do gametócito masculino no feminino. Por ser uma forma
móvel, o zigoto invade a parede intestinal, originando o oocisto que ao crescer se rompe e
libera os esporozoítos, os quais migram para as glândulas salivares do mosquito para que, no
momento da picada, sejam inoculados no homem, dando origem a um novo ciclo (RICHIE e
SAUL, 2002).
Figura 2: Ciclo biológico do Plasmodium
Ilustração evidenciando as fases do ciclo biológico do Plasmodium. Fonte: www.imm.ul.pt
1.4 A Resposta Imune na Malária
A resposta imune do paciente exerce um papel primordial na evolução da malária.
Assim, o estudo do padrão de resposta imune envolvido na evolução do quadro clínico do

26
paciente é essencial para a busca de novos tratamentos para a malária (GALINSKI e
BARNWELL, 2008). A resposta imune do hospedeiro humano ao parasita envolve tanto a
imunidade inata quanto adquirida, sendo o funcionamento adequado de cada uma destas
respostas, determinante para o prognóstico da doença. Assim, os aspectos imunológicos desta
patologia serão descritos a seguir.
1.4.1 Resposta Imune Inata
A resposta imune inata na malária é extremamente importante na patogenia da doença,
ainda que, relativamente, não tenha sido objeto constante de estudo. A literatura descreve
repetidamente que mediadores proinflamatórios, especialmente, interferon-gama (IFN-γ) e
fator de necrose tumoral (TNF-α), são mediadores de proteção imunológica essenciais para a
destruição do parasita no interior dos eritrócitos contaminados, caracterizando assim, uma
resposta imune de caráter pró-inflamatório (STEVENSON et al, 1995).
Estas citocinas podem ser produzidas tanto pela resposta imune inata quanto pela
resposta imune adquirida, de maneira que o IFN-γ é normalmente produzido pelas células
natural killer (NK), NKT e linfócitos T (FAVRE et al, 1997); enquanto que o TNF-α é
produzido por monócitos/macrófagos e linfócitos T (JACOBS et al, 1996). Assim, quando os
eritrócitos infectados pelo Plasmodium aderem no endotélio vascular, ativam
monócitos/macrófagos e ambas citocinas são liberadas (JACOBS et al., 1996).
O IFN-γ e o TNF-α estão relacionados à produção de óxido nítrico (NO) e de espécies
reativas de oxigênio (ROS), que, por serem produtos altamente reativos, destroem o parasita
(PARK et al, 2004). Os monócitos/macrófagos ativados são os maiores responsáveis pela
produção de NO e ROS que, quando em excesso, produzem o estresse oxidativo (RAY e
SHAH, 2005), acarretando em um dano tecidual, e favorecendo, assim, complicações no
quadro clínico da malária. Assim, a liberação sistêmica de altos níveis de citocinas pró-

27
inflamatórias pode ser prejudicial ao hospedeiro (homem), já que muitas vezes o incremento
sérico de IFN-γ e TNF-α, no caso da infecção por Plasmodium, está relacionado com o
aparecimento da malária grave, até mesmo com comprometimento cerebral (NEBL et al,
2005).
Ainda, a ativação destas células e consequente liberação de mediadores inflamatórios
determinam o quadro febril e o controle da parasitemia característico da malária
(KREMSNER et al, 1996), além de controlar a transmissão da malária, uma vez que os
gametócitos também são alvo de destruição pelo hospedeiro e, desta maneira, o ciclo é
interrompido (NAOTUNNE et al, 1991).
As células do sistema imune inato do hospedeiro possuem receptores que reconhecem
estruturas conservadas presentes nos patógenos, os padrões moleculares associados a
patógenos (PAMPs), o que facilita a interação hospedeiro/patógeno, e sendo o
glicosilfosfatidilinositol - GPI - (glicolipídeo presente na membrana do merozoíto do
Plasmodium) um destes padrões. Esta interação funciona como excelente ativador da resposta
imune inata, capaz de induzir a produção de TNF-α e IL-12, e, consequentemente,
desencadear uma cascata de liberação de outros mediadores pró-inflamatórios, incluindo
estimulação da produção de óxido nítrico por macrófagos, ativação do endotélio vascular com
expressão de moléculas de adesão de leucócitos, caracterizando a resposta inicial do
hospedeiro à malária (SCHOFIELD, 2007).
De maneira que, após infecção com esporozoítos, existe a intensa produção de IL-12 e
IFN-γ pelos macrófagos ativados e células NK, respectivamente, no intuito de neutralizar o
Plasmodium antes que ocorra a infecção dos hepatócitos. As células de kupffer, ao
reconhecerem a invasão hepática, induzem a produção de IFNγ, a fim de impedir o
crescimento do parasita dentro do órgão, gerando assim, uma resposta inflamatória no fígado

28
(SCRAGG et al, 1999). Assim, nas primeiras horas de infecção, os parasitas localizados no
fígado induzem a liberação de IFNγ e TNFα, enquanto que monócitos ativados liberam TNFα.
Assim, destaca-se que a liberação de citocinas pró-inflamatórias é dependente de monócitos e
linfócitos ativados (SCRAGG et al, 1999). As células NK ocupam papel de destaque uma vez
que são responsáveis por grande parte do IFNγ produzido e pela lise de eritrócitos infectados
pelo Plasmodium. De fato, experimentos in vitro, demonstraram que células NK quando
cultivadas na presença de eritrócitos infectados, são capazes de gerar grandes quantidades de
IFNγ em um período de 24 horas (ARTAVANIS-TSAKONAS e RILEY, 2002). No entanto,
o papel das células NK na malária permanece por ser melhor esclarecido.
1.4.2 Resposta Imune Adquirida
Ainda hoje, muitos fatores que favorecem o desenvolvimento de uma resposta imune
adquirida eficaz contra a malária, não estão totalmente elucidados, uma vez que pessoas
oriundas de uma área endêmica estão susceptíveis a apresentarem infecções recorrentes, ainda
que existam indícios de que tal proteção é dada através da inter-relação entre a imunidade
humoral e a imunidade celular (WYKES e GOOD, 2008).
Durante o ciclo hepático, os linfócitos T CD8+ são ativados pela presença de IFN-γ e
ocupam papel de destaque devido à sua ação frente à replicação do Plasmodium nas células
hepáticas. No ciclo eritrocítico, para que o parasita seja eliminado na corrente sanguínea são
necessárias células T CD4+, que consequentemente ativam linfócitos B promovendo assim, a
síntese de anticorpos na tentativa de controlar a parasitemia (HAFALLA et al, 2011).
A imunidade produzida pela infecção da malária é parcial, sendo mais efetiva caso a
carga parasitária permaneça alta para a manutenção das células de memória (STEVENSON e
RILEY, 2004). Assim, no processo de reinfecção, o Plasmodium invade o hospedeiro e ocorre
o deslocamento do protozoário desde o local da picada até o fígado. Uma das maneiras de se

29
evitar o estabelecimento da reinfecção é a presença de anticorpos específicos contra os
esporozoítos e no fígado a ativação de linfócitos T citotóxicos (CD8⁺). Já os linfócitos T
auxiliares (CD4⁺) são estimulados pela apresentação de antígenos parasitários pelas células
apresentadoras de antígenos profissionais (APCs), incluindo células dendríticas, macrófagos e
linfócitos B, favorecendo o estabelecimento da resposta imunológica de padrão Th1,
caracterizada pela produção de IFN-γ e TNF-α (BUENO et al. 2010).
É fundamental que exista um equilíbrio entre as células T efetoras (CD4⁺ e CD8⁺) e as
células T reguladoras (CD4⁺CD25⁺FoxP3⁺), uma vez que as células T reguladoras também
estão associadas à resposta imune natural do hospedeiro frente ao patógeno, ainda que seu
mecanismo não esteja totalmente esclarecido (WALTHER et al, 2005). Assim, no momento
em que a parasitemia é controlada, mecanismos supressores são ativados, a fim de controlar o
dano tecidual decorrente da própria ativação do sistema imune que pode ocasionar a
instalação da malária grave. A alta produção de TNF-α também está relacionada às
complicações da malária, uma vez que esta citocina induz febre, e esta relacionada à
interrupção da gravidez, hipotensão, acidose láctica, hipoglicemia e ao coma característicos
da malária grave (POOVASSERY e MOORE, 2009; KARUNAWEERA et al, 1992).
1.5 Modelos Experimentais de Malária
Os modelos animais de infecção por Plasmodium em animais de laboratório têm sido
bastante úteis para uma melhor compreensão da relação parasita-hospedeiro. De fato, diversos
aspectos da malária humana podem ser mimetizados em camundongos (LAMB et al., 2006).
Ainda, as diferentes linhagens de camundongos associadas às diferentes espécies de
Plasmodium favorecem o esclarecimento da heterogeneidade peculiar da malária (DE
SOUZA e RILEY, 2002). A utilização de camundongos geneticamente modificados vem

30
facilitando a compreensão das vias de sinalização associadas a esta patologia (DESSENS et
al, 2001; NITCHEU et al, 2003;). No entanto, a correlação entre o modelo animal e o quadro
malárico humano deve ser sempre realizada levando em consideração as diferenças inerentes
a cada hospedeiro. Assim, serão discutidos a seguir, os modelos murinos de malária
experimental.
1.5.1 Modelo de Malária Cerebral
A malária cerebral é uma complicação da malária, sendo característica de uma malária
grave, acarretando uma série de déficits neurológicos a pacientes que sobrevivem a malária
(MARSH et al, 1996). Estudos mecanísticos da malária cerebral em humanos são difíceis,
sendo as alterações neurológicas monitoradas através do acompanhamento de um quadro
marcado pela rigidez na nuca, delírios, desorientação, sonolência ou excitação, convulsões,
vômitos e estado de coma e as alterações teciduais presentes no cérebro avaliadas post-
mortem (BREWSTER et al, 1990). Assim, modelos experimentais são utilizados como
ferramentas para esclarecer os mecanismos associados à malária cerebral. A infecção pelo P.
berghei ANKA em camundongos vem sendo utilizada como um modelo de malária cerebral,
e, portanto, uma ferramenta valiosa no entendimento desta complicação (RODRIGUES-
DUARTE et al, 2012; VAN DER HEYDE et al, 2001; MACKEY et al, 1980). O
desenvolvimento da malária cerebral em camundongos, no entanto, depende da linhagem de
camundongo utilizada, sendo camundongos C57BL/6 mais susceptíveis e camundongos
BALB-c, resistentes (GRAU et al, 1987). Assim, é possível desenvolver um quadro
semelhante ao da malária causada por P. falciparum quando camundongos C57BL/6 são
infectados com P. berghei ANKA (RODRIGUES-DUARTE et al, 2012; VAN DER HEYDE
et al, 2001; MACKEY et al, 1980). Os camundongos infectados susceptíveis podem
desenvolver anemia, acidose láctica, nefrite e sinais neurológicos (coma, paralisia e
convulsão) compatíveis com a malária cerebral, enquanto que os animais resistentes morrem

31
em, aproximadamente, 20 dias após o início da infecção, em decorrência da anemia grave e da
hiperparasitemia, mas não desenvolvem nenhuma lesão cerebral (MEDANA et al, 1997;
RODRIGUES-DUARTE et al, 2012; VAN DER HEYDE et al, 2001; MACKEY et al, 1980).
O ciclo de vida do P. berghei é semelhante às outras espécies que infectam mamíferos,
variando somente com relação ao tempo referente aos diferentes estágios. No hospedeiro
vertebrado (roedores murinos), este parasita tem preferência por invadir reticulócitos, porém
também pode infectar eritrócitos maduros (GARNKAN, 1996). A duração do ciclo assexuado
varia entre 22 e 25 horas, assim a infecção de hemácias não é sincrônica, coexistindo
diferentes formas parasitárias (LANDAU e BOULARD, 1978).
Assim como na malária humana, a resposta imune do camundongo à infecção pelo
Plasmodium, conta com a ativação de vias similares (DAY et al, 1999; GRAU et al, 1987). A
presença do parasita induz a ativação de células da resposta imune inata, como
monócitos/macrófagos e células NK, com consequente liberação de mediadores pró-
inflamatórios (TNF-α e IFN-γ) e ROS, com a finalidade de provocar a morte do parasita
(JACOBS et al, 1996). Em camundongos, a diferença entre uma infecção letal ou não, no
entanto, depende da maneira como o animal responde à malária, ou seja, da quantidade de
estresse oxidativo gerado pelo animal (CLARK et al, 1992; JACOBS et al, 1996).
Vale ressaltar que existe um importante sinergismo entre a resposta imune inata e a
adaptativa, como por exemplo, durante o ciclo hepático. Recentemente foi demonstrado que
com a chegada do parasita ocorre uma intensa resposta inflamatória caracterizada por um
infiltrado rico em células NK, macrófagos e linfócitos T no fígado (DOOLAN e HOFFMAN,
1999). Existem poucos relatos sobre os mecanismos de controle do estágio eritrocítico do
ciclo do Plasmodium em camundongos, porém a diminuição das células NK em
camundongos resulta em uma piora no curso da infecção, um aumento da parasitemia e da
mortalidade (MOHAN et al, 1997). Ainda, através da apresentação de antígenos que ocorre,

32
os linfócitos T são ativados e assumem um importante papel na produção de IFN-γ em
animais (DICK et al, 1996).
A infecção induzida pelo P. berghei ANKA contribui para a alteração na
permeabilidade vascular da barreira hemato-encefálica, favorecendo o sequestro de eritrócitos
infectados, macrófagos e plaquetas na região nos vasos do córtex cerebral do animal e a
hemorragias (FAVRE et al, 1999). Assim, em semelhança ao que acontece na infecção
humana, esta retenção de células ativa monócitos e macrófagos que passam a secretar
citocinas e a produzir ROS com o intuito de eliminar o patógeno da região cerebral, porém,
em contrapartida, também pode causar lesão tecidual (SHARMA et al, 2012; NEBL et al,
2005).
1.5.2 Modelo de Malária Placentária
As mulheres grávidas são mais susceptíveis às complicações decorrentes da malária e
esta infecção é caracterizada pelo sequestro de eritrócitos infectados pelo Plasmodium na
região placentária com consequências tanto para a mãe quanto para o feto, como por exemplo,
baixo peso ao nascer, retardo no crescimento, parto prematuro, natimorto, anemia e
mortalidade materna (BARDAJI et al, 2011; UMBERS, et al, 2011). No modelo animal,
também foi observada essa maior vulnerabilidade (VAN ZON e ELING, 1980).
De fato, assim como em humanos, camundongos fêmeas primíparas previamente
contaminadas com P. berghei, o Plasmodium é capaz de aderir à placenta, e
consequentemente causar danos ao endotélio vascular placentário, acarretando as
consequências da malária placentária, incluindo aborto (BARDAJI et al, 2011; NERES et al,
2008). Ainda, a recrudesccência é comum em primigrávidas (MARINHO et al, 2009; VAN
ZON et al, 1985). Com a formação de uma imunidade inata adquirida contra o Plasmodium,
no evento de uma gravidez subsequente, os efeitos clínicos da malária placentária estão

33
menos presentes (VAN ZON et al, 1985). Isso se deve ao fato de que existe um acúmulo de
anticorpos específicos contra o parasita, os quais previvem a adesão de eritrócitos infectados à
placenta (VAN ZON at al, 1985; MEGNEKOU et al, 2009; MARINHO et al, 2009). Ainda, a
elevação da carga parasitária, encontra-se relacionada à redução do espaço intravascular
(GOEL e GOWDA, 2011; MARINHO et al, 2009).
Apesar das similaridades entre a malária placentária humana e murina, não existe um
consenso na literatura sobre a utilização de modelos murinos para o esclarecimento da malária
placentária, uma vez que algumas proteínas expressas nas superfícies do eritrócito infectado
humano são distintas daquelas encontradas nos eritrócitos dos roedores (HVIID et al, 2010).
No entanto, muitas características patogênicas e imunológicas são semelhantes, o que justifica
sua utilização (NERES et al, 2008). O modelo mais adequado para o estudo experimental da
recrudescência é através da infecção de camundongos BALB/c infectado com Plasmodium
berghei ANKA (PEREIRA, 2012).
Assim como na malária humana, a resposta imune do camundongo à infecção pelo
Plasmodium, conta com a ativação de vias similares resultantes do acúmulo de eritrócitos
infectados associados à intensa resposta inflamatória monocítica. Tendo como consequência a
liberação de citocinas pró-inflamatórias a indução da apoptose e de fatores relacionados ao
estresse oxidativo, lesionando assim, a placenta (BERENDT et al, 1990; SHARMA et al,
2012). Como decorrência desta ativação, linfócitos iniciam a resposta imune adaptativa com
produção de anticorpos capazes de prevenir o acúmulo de hemácias no endotélio placentário e
neutralizar a infectividade dos que já estão aderidos (STAALSOE et al, 2004). Esta resposta
específica torna-se cada vez mais eficiente à medida que a gestação progride com o intuito de
limitar a replicação e possibilitar a destruição do parasita (MCGREGOR, 1984).
Pereira (2012) observou que, na análise, por citometria de fluxo, de macrófagos e
linfócitos placentários e esplênicos, prevaleceu um aumento na população de células CD4+,

34
CD19+ e GR1+ no baço de animais infectados com malária, porém sem alteração de leucócitos
na região da placenta. Enquanto que no ambiente esplênico, foi encontrado uma maior
ativação de células CD4+ e CD8+, ainda que na placenta este aumento observado somente em
células CD4+ (PEREIRA, 2012). Tais achados sugerem que, na malária placentária murina
exista uma preferência pela resposta TH1, conforme acontece na malária placentária humana
(FRIED et al, 1998)
1.6 Tratamento da Malária
Ao longo dos anos, o tratamento para a malária evoluiu muito pouco. Após a Segunda
Guerra Mundial, a malária passou a ser controlada principalmente, através de medidas de
melhorias socioeconômicas (melhorias na habitação, drenagem de água estagnada, uso de
inseticidas) e drogas antimaláricas (SACHS e MALANEY, 2002).
A Organização Mundial de Saúde recomenda que todas as pessoas, de todas as idades
que tenham confirmado o diagnóstico de malária iniciem o tratamento imediatamente (OMS,
2012). No Brasil, o Ministério da Saúde, seguindo a recomendação, estimula e orienta a
terapêutica à medida que disponibiliza gratuitamente os medicamentos antimaláricos, sob a
supervisão do Programa Nacional de Controle da Malária (BRASIL, 2010). Incluem-se no
grupo de anti-malaricos: cloroquina, primaquina, artemeter, lumefantrina, artesunato,
mefloquina, quinina e doxiciclina (BRASIL, 2010).
O mecanismo de ação das drogas antimaláricas tem como alvo três pontos chaves do
ciclo do Plasmodium: a interrupção da esquizogonia sanguínea (responsável pelas
manisfestações clínicas); destruição dos hipnozoítos (formas latentes) e destruição dos
gametócitos (interrupção da transmissão). Assim, o esquema terapêutico básico utilizado deve

35
atuar em todas as fases do ciclo e neste contexto, duas drogas ocupam papel de destaque:
cloroquina e primaquina, em uso desde a Segunda Guerra Mundial (TEIXEIRA, 2011).
Na malária grave, o tratamento com os antimaláricos deve ser iniciado o mais
precocemente possível, para minimizar as complicações, e por via intravenosa, para que sua
ação inicie em menor tempo. Existem duas drogas disponíveis para uso intravenoso, os
alcalóides de cinchona (quinino e quinidina) e os derivados de artemisinina (artesunato),
sendo que este último reduz o risco de morte em até 39% (RIBEIRO et al, 2013). O uso
destas drogas parenterais deve ser de 24 horas ou até que o paciente tolere a terapia por via
oral, com as drogas habituais, uma vez que estas podem ocasionar náuseas, dor abdominal,
disforia, síndrome neuropsiquiátrica transitória, disfunção cerebelar, hemólise oxidante com
metahemoglobinemia, anemia e hemoglobinuria (WHITE, 1996). Como ainda não existe um
tratamento que reduza a mortalidade da malária grave, buscam-se associações com outras
drogas com ação adjuvante, como por exemplo, dexametasona, anticorpos anti-TNF-α,
pentoxifilina, desferroxamina, N-acetilcisteína, heparina, aspirina e manitol, porém, nenhuma
dessas associações mostrou-se eficaz (NADJIM e BEHRENS, 2012).
Outra problemática existente também relacionada ao tratamento é o surgimento de
casos em que o parasita é resistente às drogas utilizadas. A cloroquina, droga que está no
esquema terapêuticos há décadas, já está resistente (MAY e MEYER, 2003). Fato já
observado também com o artesunato (componente da maioria dos esquemas terapêuticos) em
quatro países da região do sudeste asiático. Desta maneira, são necessárias intervenções que
viabilizem a prevenção e o tratamento de surtos epidêmicos para que aconteça uma redução
do número de casos e de mortes por malária (OMS, 2012).
Esta resistência às drogas antimaláricas em paralelo à resistência dos mosquitos aos
inseticidas é perigosa nas áreas endêmicas porque as ações de profilaxia ficam prejudicadas e

36
seria difícil controlar um possível surto epidêmico com as drogas usuais, devidos aos
mecanismos de resistência permanentes desenvolvidos pelo Plasmodium e pelo mosquito
(SHARMA et al, 2013). Nesse contexto, a relevância dos TRPs para o tratamento da malária
será abordada ao longo desta dissertação.
1.7 Receptor TRP
De importância para este projeto, encontram-se os receptores de potencial transitório
(Transient Receptor Potential), conhecidos como receptores TRP. Estes receptores encontram-
se expressos nas fibras sensoriais to tipo C e Aδ e, quando estimulados, promovem a liberação
dos neuropeptídeos substância P (SP) e peptídeo relacionado ao gene da calcitonina
(Calcitonin Gene-Related Peptide; CGRP) (MAGGI et al, 1990; BRAIN e GRANT, 2004).
A família de receptores TRP engloba mais de 30 membros distribuídos em 7
subfamílias: Canônica (TRPC; inclui TRPC1-7), Melastatina (TRPM; inclui TRPM1-8),
Vanilóide (TRPV; inclui TRPV1-6), Policistina (TRPP; inclui TRPP1-3), Mucolipina
(TRPML; inclui TRPML1-3), Anquirina (TRPA; inclui TRPA1) e NOMPC (TRPN;
encontrados em Drosophila sp, sapos, truta e vermes) (PEDERSEN et al, 2005). Como
característica comum, todos os receptores TRP são permeáveis a cátions, particularmente ao
Ca+2 e podem ser ativados por diversos agonistas endógenos (incluindo mudanças na
temperatura (ZIMMERMANN et al, 2011; VAY et al, 2012), pH (LEFFLER et al, 2006;
SEMTNER et al, 2007; WANG et al, 2011), estímulos mecânicos (GOMIS et al, 2008),
hormônios (QIU et al, 2010), espécies reativas de oxigênio (BESSAC et al, 2008; NAYLOR
et al, 2010) e outros mediadores inflamatórios (FERNANDES et al, 2012a); e exógenos
(como produtos derivados de plantas, fumaça de cigarro, poluição, substâncias químicas, etc
(BAHNASI et al, 2008; FERNANDES et al, 2012a). Recentemente, a importância dos

37
receptores TRP na sepse, começou a ser especulado e no decorrer desta seção, serão
discutidas as diversas evidências de como estes receptores participam na e/ou delimitam a
progressão da sepse em um prognóstico letal. Assim, é importante ressaltar que além de serem
expressos em neurônios, os receptores TRP são também encontrados numa série de células e
tecidos não-neuronais, incluindo células inflamatórias (BASU e SRIVASTAVA, 2005;
SCHWARZ et al, 2007; FERNANDES et al, 2012b) e células da musculatura lisa da aorta e
microvasculatura (YIP et al, 2004; KARK et al, 2008).
1.7.1 TRPV1
Dentre as subfamílias termossensíveis dos TRPs, destaca-se a conhecida como TRPV,
composta por 6 membros que são ativados pelo calor e pela dor; não seletivos para cátions e
moderadamente permeável para o cálcio, foram isolados inicialmente em Drosophilas. O
receptor vanilóide de potencial transitório do tipo 1 (TRPV1) é um canal catiônico não-
seletivo localizado nos nervos sensoriais (CATERINA et al, 1997) e é relativamente
considerado um novo e importante componente da inflamação neurogênica e sua ativação
vem sendo relacionada a diferentes patologias como doenças respiratórias, artrite, prurido,
pancreatite, colite, dor, dentre outras (SCHWARTZ et al, 2013; KEEBLE e BRAIN, 2006;
GEPPETTI et al, 2006; LIDDLE, 2007; STORR, 2007; SHIM et al, 2007; MA e QUIRION,
2007). Além disso, o TRPV1 encontra-se localizado em diversos tipos celulares, incluindo
células endoteliais (POBLETE et al, 2005) através das quais, eles poderiam influenciar
eventos vasculares. Estudos moleculares e celulares, assim como alguns poucos estudos in
vivo têm demonstrado a existência de um número crescente de mecanismos que possuem
relevância endógena como, por exemplo, o calor nocivo e prótons (RANG et al, 1991;
CESARE et al, 1999; FERNANDES et al, 2013) e uma lista crescente de mediadores capazes
de ativar o TRPV1 em vários sistemas, incluindo canabinóides (CHU et al, 2003, HUANG et
al, 2002, OLAH et al, 2001, RALEVIC et al, 2002), metabólitos do ácido araquidônico (ex:

38
20-HETE, SCOTLAND et al, 2004, ROUSSEAU et al, 2005; e vários produtos da via da
lipoxigenase, HWANG et al, 2000). Ademais, estudos celulares e experimentos de nocicepção
in vivo têm sugerido que mecanismos de sinalização comuns podem aumentar a ativação do
TRPV1, como, por exemplo, substâncias inflamatórias que ativam receptores acoplados à
proteína G (ex: prostaglandina E2, MORIYAMA et al, 2005; bradicinina, TANG et al, 2004; e
PAR-2, AMADESI et al, 2006).
Recentemente, um papel protetor foi sugerido para o TRPV1 na sepse. Neste contexto,
a relevância deste receptor na regulação da resposta imune à infecção será discutida à seguir.
1.7.2 O papel do TRPV1 nas infecções
Recentemente, o TRPV1 foi sugerido como um modulador da resposta imune à
infecção, como sendo um componente essencial para o desenvolvimento da resposta
inflamatória contra a infecção bacteriana (CLARK et al, 2007). O primeiro estudo sugerindo o
envolvimento deste receptor na sepse demonstrou que ratos com sepse abdominal pré-tratados
com o agonista TRPV1 capsaícina apresentavam uma maior taxa de sobrevivência (BRYANT
et al, 2003).
Além disso, demonstrou-se que ratos com sepse induzida por LPS não apresentavam
febre quando pré-tratados com capsaícina (ROMANOVSKY, 2004). No entanto, estes
experimentos apresentavam limitações considerando-se que além de ativar o receptor TRPV1,
a capsaícina também provoca a denervação das fibras sensoriais, ocasionando a perda de
outros componentes sensoriais além do TRPV1.
Não muito depois, as evidências obtidas com o uso da capsaícina foram confirmadas
em experimentos usando camundongos com deleção gênica do sítio de ativação do receptor
TRPV1 (TRPV1KO). Foi observado que animais TRPV1KO com sepse induzida pelo LPS
apresentavam menos febre do que camundongos selvagens (WT) (IIDA et al, 2005). Em

39
conjunto, essas evidências iniciais sugeriam que a ativação do receptor TRPV1 era capaz de
mediar uma “proteção” contra a sepse, embora os mecanismos envolvidos nessa proteção
ainda não estavam estabelecidos.
Mais recentemente, foi sugerido que o receptor TRPV1 é um componente essencial
para o controle da resposta inflamatória e imune no combate a infecção bacteriana (CLARK
et al, 2007; FERNANDES et al, 2012b) e desta forma, representa uma importante via de
sinalização no combate à letalidade relacionada a sepse. Camundongos TRPV1KO com sepse
causada por LPS (intraperitoneal) apresentam hipotensão e hipotermia exacerbadas quando
comparados a camundongos WT sépticos (CLARK et al, 2007).
O mesmo estudo demonstrou que TRPV1KOs com sepse apresentavam níveis
aumentados de TNFα e NO no lavado peritoneal em comparação aos WTs. Esses mesmos
TRPV1KOs apresentavam níveis plasmáticos aumentados de aspartato-amino transferase
(AST) sugerindo falência do fígado 4h após o tratamento com LPS, quando comparados a
camundongos WTs. Além do mais, Guptill e colaboradores (2011) demonstraram que
camundongos TRPV1KO com sepse induzida pela ligação e punctura do ceccum apresentam
uma maior taxa de mortalidade quando comparados a seus respectivos WTs. Similarmente,
estudos com antagonistas confirmaram as evidências obtidas através de estudos com
camundongos TRPVKO. O tratamento com o antagonista TRPV1 capsazepina exacerbou a
hipotensão e reduziu a taxa de sobrevivência em ratos com sepse induzida por LPS (WANG
et al, 2008). Da mesma forma, o tratamento prolongado com capsazepina ou o antagonista
seletivo para o TRPV1 SB366791, causou acentuada falência de órgãos (FERNANDES et al,
2012b) e mortalidade em camundongos WT (GUPTILL et al, 2011).
Embora os mecanismos associados à proteção mediada pelo TRPV1 na sepse não
estejam completamente esclarecidos, evidências recentes obtidas de experimentos em animais

40
com sepse causada pela ligação e punctura do ceccum, sugerem que a interrupção da
sinalização mediada pelo receptor TRPV1 resulta em uma resposta imune deficiente e
consequente aumento de proliferação bacteriana (GUPTILL et al, 2011; FERNANDES et al,
2012b). De fato, dados mais recentes (FERNANDES et al, 2012b) demonstram que na
ausência de receptores TRPV1 funcionais no sítio inicial de infecção, existe perda de função
das células inflamatórias, mais especificamente, macrófagos, que tornam-se deficientes no
combate a infecção, uma vez que estas células perdem a capacidade de realizar fagocitose e
gerar mediadores como NO e espécies reativas de oxigênio. Além disso, a deficiência da
sinalização via o receptor TRPV1 promove aumento da apoptose de macrófagos
(FERNANDES et al, 2012b). Essa deficiente resposta imune celular facilita a transição da
resposta inflamatória local em resposta a infecção bacteriana, em uma resposta inflamatória
sistêmica com prognóstico letal.
Tendo em vista os componentes imunológicos da malária e a crescente descoberta do
envolvimento do TRPV1 em diferentes patologias também com forte carater imunológico
como doencas respiratorias e do trato gastrointestinal e a sepse, o presente trabalho procurou
investigar o possivel envolvimento do TRPV1 na malária experimental em camundongos.

41
2 OBJETIVOS
2.1 Geral
Avaliar a participação de receptor TRPV1 na infecção murina por P. berghei ANKA
2.2 Específicos
- Avaliar a expressão de marcadores característicos de monócitos/macrófagos e células
NK em camundongos infectados por P. berghei ANKA tratados e não tratados com a
capsazepina;
- Avaliar a expressão de marcadores característicos de linfócitos e células NKT em
camundongos infectados por P. berghei ANKA tratados e não tratados com a capsazepina;
- Quantificar as citocinas circulantes nos camundongos infectados por P. berghei
ANKA tratados e não tratados com a capsazepina;
- Verificar o efeito imunomodulatório do receptor TRPV1 na resposta imune causada
pelo P. berghei ANKA.

42
3 METODOLOGIA
3.1 Animais utilizados no estudo
Foram utilizados camundongos C57/BL6 machos (6-8 semanas) mantidos em
ambiente com temperatura e umidade controlada (22 ± 2ºC e 60-80%, respectivamente), em
ciclo claro/escuro e com livre acesso à água e ração. Os animais foram fornecidos pelo
Biotério de Parasitologia – BCE_BMP – USP. Para a realização dos experimentos, os animais
foram aclimatizados no laboratório durante um período de pelo menos 1 hora e os mesmos
foram realizados entre 6 e 18 horas. Todos os experimentos foram conduzidos de acordo com
os Princípios Éticos de Experimentação Animal adotado pela Sociedade Brasileira de Ciência
de Animais de Laboratório (SBCAL) e aprovados pela Comissão de Ética no Uso de Animais
(CEUA) da Universidade de São Paulo.
3.2 Indução da Malária
Para indução da malária, os animais receberam uma injeção intraperitoneal contendo 1 x
105 hemácias infectadas com Plasmodium berghei ANKA conforme metodologia descrita por
Elias et al (2012). O nível de parasitemia foi verificado diariamente, do quinto ao sétimo dia
pós-infecção, a partir de amostras de sangue coletada por uma incisão na ponta da cauda do
animal infectado pelo método da gota espessa. Os esfregaços foram corados com panótico
rápido e a contagem dos parasitas foi realizada através da microscopia óptica em objetiva de
imersão (100 x). No sétimo dia pós-infecção, período pré-mortalidade (WRIGHT, 1968;
WHITE e HO, 1992), os animais foram anestesiados terminalmente com uma mistura de
ketamina e xylazina (75mg/kg e 1 mg/kg, respectivamente), e o sangue total foi obtido através
de punção cardíaca com o auxílio de uma seringa heparinizada. O sangue total fresco foi

43
utilizado para os experimentos de citometria de fluxo e plasma foi separado, congelado e
armazenado em freezer -70oC para análise posterior dos níveis de citocinas e aldeídos
plasmáticos. O baço foi removido, pesado e processado imediatamente para análise de
citometria de fluxo. A tíbia foi coletada e removida de todo o tecido adjacente. O peso do
baço foi normalizado em relação ao comprimento da tíbia e este índice, utilizado como
indicativo proliferação celular esplência decorrente da infecção.
3.3 Obtenção de células sanguíneas e esplênicas
Os baços dos animais experimentais foram retirados e pesados, e posteriormente
transferidos para placas de petri contendo 5 mL de solução de PBS contendo 3% de soro
bovino fetal. Os baços foram macerados e dissociados individualmente, e as células foram
lavadas por 2 vezes em PBS contendo 3% de soro bovino fetal (5 minutos , 300g). As células
esplênicas foram mantidas a 4o C até a marcação com anticorpos específicos para marcadores
celulares.
O sangue dos animais foi obtido através de punção cardíaca com os animais
anestesiados e diluídos n proporção de 1:1 (v/v) com PBS contendo 3% de soro bovino fetal.
Após a diluição, os leucócitos foram isolados por separação por gradiente de percoll (60% em
PBS) por centrifugação por 10 minutos a 500g. Após a separação, a fase contendo os
leucócitos foi coletada e lavada por 2 vezes com PBS contendo 3% de soro bovino fetal. Os
leucócitos sanguíneos foram mantidos a 4o C até a marcação com anticorpos específicos para
marcadores celulares.

44
3.4 Fenotipagem por citometria de fluxo em leucócitos de baço e de sangue
Após o isolamento dos leucócitos esplênicos e de sangue periférico, 1 X 106 células
foram transferidas para uma poços de placa de fundo redondo (96 poços), e marcadas com
1µg/amostra dos seguintes anticorpos monoclonais (BD Bioscciences) (Tabela 1)
Tabela 1: Especificação dos anticorpos monoclonais utilizados no presente estudo
FITC PE PerCP APC PACIFIC
BLUE PE-Cy 7 APC-Cy 7 Marcação A F4/80 IA b Ly6C CD3 CD4 CD8 CD19
Marcação B NK1.1 CD3 CD69 FoxP3 CD4 Gr1 CD25
Descrição dos marcadores: F4/80, marcador de monócitos/macrófagos; Ly6G,
marcador de neutrófilos; GR1, marcador de células precursoras monócitos e granulócitos;
IAb, MHC II, utilizado como marcador de ativação de monócitos/macrófagos; CD19,
marcador de linfócitos B; CD3, marcador de linfócitos T; CD8, marcador de linfócitos T
citotóxicos; CD4, marcador de linfócitos T auxiliares; CD69 e CD25, marcadores de ativação
de linfócitos T; FOXP3, marcador de linfócitos T reguladores; e NK1.1, marcador de células
NK. Em seguida, as amostras foram incubadas a 4˚C por 30 minutos e então lavadas com com
200 µl de tampão fosfato pH 7.4 (PBS), transferidas para tubos de polietileno com 400 µl de
PBS e 100 µL de paraformaldeído a 2% para fixação para leitura em citômetro de fluxo
FACSCanto – BD Biosciences. As análises foram realizadas utilizando-se o programa FlowJo
(TreeStar).

45
3.5 Dosagem de aldeídos plasmáticos (TBARS)
Os níveis de aldeídos plasmáticos foram quantificados e tomados como índice de
peroxidação lipídica, indicativa de estresse oxidativo (BIRD e DRAPER, 1984; KEEBLE et
al, 2009). Para isto, 100 µl de plasma foram incubados com 100 µl de PBS e 400 µl de ácido
tiobarbitúrico (0,67%), a 90˚C por 45 minutos. Em seguida, as amostras foram centrifugadas a
1000 x g por 10 minutos, e 300 µl do sobrenadante foi incubado com 300 µl de butanol e 30µl
de solução saturada de NaCl. As amostras foram então homogeizadas em vórtex e
centrifugadas a 1000 x g por 2 minutos. Duzentos microlitros de cada amostra foram então
pipetados em uma placa de 96 poços e a absorbância lida em 535 e 572 nm. A diferença entre
as absorbâncias foi tomada como índice de produção de aldeídos plasmáticos. Os resultados
foram calculados em aldeídos totais por mililitro e expressos como aumento relativo ao
controle, em número de vezes.
3.6 Dosagem de citocinas plasmáticas
Os níveis circulantes de citocinas (IL-2, IL-4, IL-6, IL-10, TNF, IFN-γ e IL-17) foram
quantificados em amostras de plasma, através do uso de um kit CBA (Cytometric Bead
Array) utilizando citometria de fluxo, conforme as especificações do fabricante. Sete
populações de beads com diferentes intensidades de fluorescência foram conjugadas com um
anticorpo de captura específico para cada citocina. As populações de beads foram
visualizadas de acordo com as suas intensidades de fluorescência, da menos brilhante para a
mais brilhante. Nesta metodologia, as beads de captura de citocina foram misturadas com o
anticorpo de detecção conjugado com o fluorocromo PE e depois incubadas com as amostras
para formar o ensaio em sanduíche. Os resultados foram expressos em pg/ml e expressos
como aumento relativo ao controle, em número de vezes.

46
3.7 Tratamento farmacológico
A participação do receptor TRPV1 na malária foi avaliado através do tratamento com
o antagonista de receptores TRPV1, capsazepina de acordo com protocolo descrito por
Fernandes et al (2012). Para isto, 24 h após a infecção com P. berghei ANKA, os animais
receberam uma injeção intraperitoneal contendo capsazepina (50 µg/animal; n = 8), 2 x ao
dia, por 6 dias ou contendo veículo (10% DMSO em salina; n = 8). Animais não infectados
receberam capsazepina (50 µg/animal; n = 5) ou veículo (10% DMSO em salina; n = 5) e
foram utilizados como controle.
3.8 Analise estatística
Os resultados foram expressos como a média ± desvio padrão (S.D.). As porcentagens
de inibição foram calculadas como a média das inibições obtidas para cada experimento
individual. A avaliação estatística dos resultados foi realizada por meio de análise de
variância de uma via (ANOVA), seguida pelo teste de Bonferroni, ou teste "t" não-pareado,
quando apropriado. Valores de P menores do que 0,05 (*P ou #P < 0,05) foram considerados
como indicativos de significância.

47
4. RESULTADOS
4.1 Efeito do tratamento com o antagonista do receptor TRPV1, capsazepina,
sobre a parasitemia
A fim de acompanhar a carga parasitária em relação à evolução da malária, o número a
porcentagem de eritrócitos parasitados com P. berghei ANKA foi avaliado diariamente do 5o
ao 7o dia após a infecção. O número de eritrócitos infectados aumenta de forma crescente com
o passar dos dias em ambos os grupos de animais infectados (veículo e capsazepina) (Figura
1). A capsazepina (50 μg/animal, 2 x ao dia, 6 dias) causou apenas uma pequena redução na
parasitemia (17%), embora esta redução não seja significativa em comparação com o grupo
controle (veículo) (Figura 3).
Figura 3: Efeito do tratamento repetido com capsazepina (50µg/animal, 2 x ao dia, 6 dias) em animais com malária induzida por P. berghei ANKA (105 hemacias infectadas/ animal, i.p.), 7 dias após a indução da malária. (n = 5-8/grupo)

48
4.2 Participação do receptor TRPV1 na resposta imune celular inata à malária
Avaliou-se o efeito do bloqueio do receptor TRPV1, na modulação da imunidade inata
em animais com malária induzida pelo P.berghei ANKA, através do tratamento repetido com
o seu antagonista, capsazepina (50 μg/animal, 2 x ao dia, 6 dias).
Foram avaliados os marcadores para monócitos/macrófagos, F4/80; e neutrófilos,
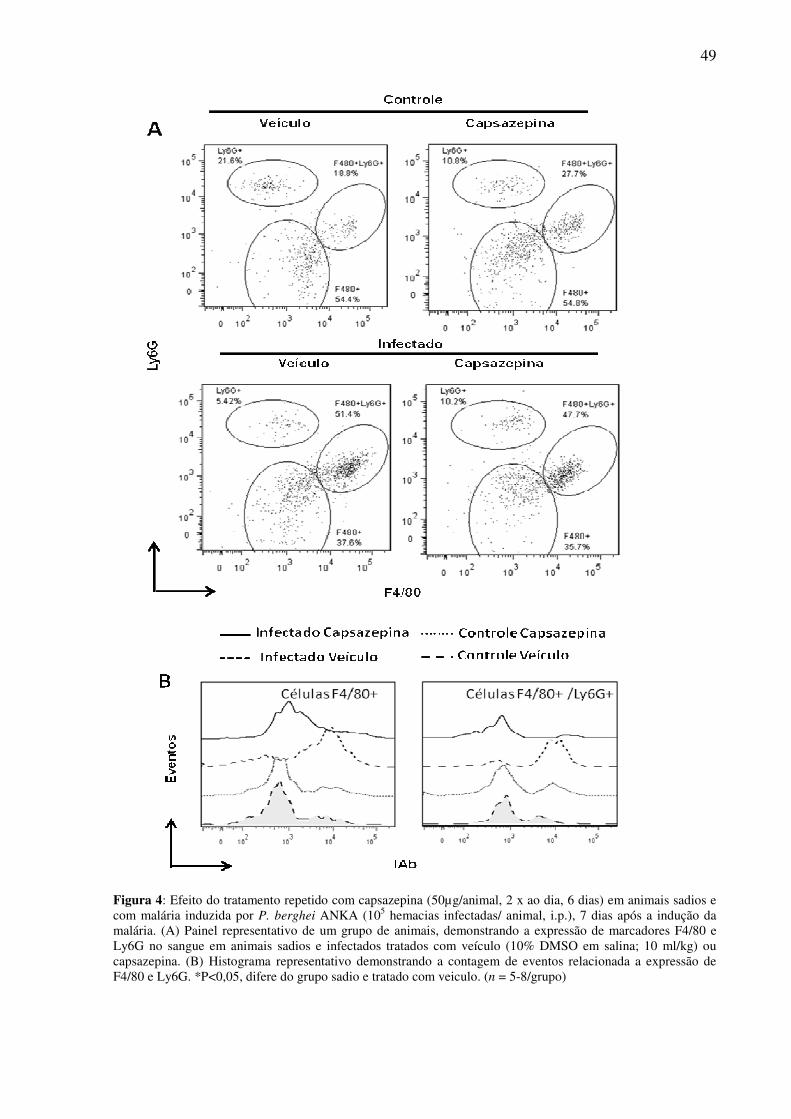
Ly6G. As Figuras 4 e 5 representam as diferentes populações que expressam estes
marcadores. Observou-se a presença de três populações celulares distintas: F4/80+, Ly6G+ e
F4/80+/Ly6G+, tanto no sangue (Figura 4) quanto no baço (Figura 5) de animais infectados ou
não com P. berghei ANKA.
Embora a infecção com P. berghei ANKA não afete o número de células F4/80+
circulantes, observou-se um aumento da população de células F4/80+Ly6G+ (Figura 6 A e B).
Por outro lado, foi observado um aumento de ambas as populações esplênicas, F4/80+ e
F4/80+Ly6G+ (Figura 6 C e D). Ainda, a infecção promoveu o aumento da ativação de ambos
os grupos celulares no sangue (Figura 7 A e B) e no baço (Figura 7 C e D).
O tratamento repetido com capsazepina (50 µg/animal, 2 x dia, 6 dias) reduziu o
número (Figura 6 B) e a expressão de IAb (marcador de ativação; Figura 7 B) de monócitos
sanguíneos do tipo F4/80+Ly6G+, sem no entanto afetar a população de monócitos F4/80+
circulantes ou sua ativação (Figura 6 A e Figura 7 A). Ainda, a capsazepina não teve efeitos
sobre o número (Figura 6 C e D) e a ativação (Figura 7 C e D) de células F4/80+ e
F4/80+Ly6G+ esplênicas. Não foram observadas alterações significantes em relação à
população de células sanguíneas e esplênicas Ly6G+, sendo o número absoluto destas células
similar em todos os grupos avaliados (Tabelas 2 e 3).
49
Figura 4: Efeito do tratamento repetido com capsazepina (50µg/animal, 2 x ao dia, 6 dias) em animais sadios e com malária induzida por P. berghei ANKA (105 hemacias infectadas/ animal, i.p.), 7 dias após a indução da malária. (A) Painel representativo de um grupo de animais, demonstrando a expressão de marcadores F4/80 e Ly6G no sangue em animais sadios e infectados tratados com veículo (10% DMSO em salina; 10 ml/kg) ou capsazepina. (B) Histograma representativo demonstrando a contagem de eventos relacionada a expressão de F4/80 e Ly6G. *P<0,05, difere do grupo sadio e tratado com veiculo. (n = 5-8/grupo)

50
Figura 5: Efeito do tratamento repetido com capsazepina (50µg/animal, 2 x ao dia, 6 dias) em animais sadios e com malária induzida por P. berghei ANKA (105 hemacias infectadas/ animal, i.p.), 7 dias após a indução da malária. (A) Painel representativo de um grupo de animais, demonstrando a expressão de marcadores F4/80 e Ly6G no baço em animais sadios e infectados tratados com veículo (10% DMSO em salina; 10 ml/kg) ou capsazepina. (B) Histograma representativo de um grupo de 5 animais, demonstrando a contagem de eventos relacionada à ativação de células F4/80+ e F4/80+Ly6G+. (n = 5-8/grupo)

51
Figura 6: Efeito do tratamento repetido com capsazepina (50µg/animal, 2 x ao dia, 6 dias) em animais sadios e com malária induzida por P. berghei ANKA (105 hemacias infectadas/ animal, i.p.), 7 dias após a indução da malária. (A) População de células F4/80+ no sangue em animais sadios e infectados tratados com veículo (10% DMSO em salina; 10 ml/kg) ou capsazepina. (B) População de células F4/80+Ly6G+ no sangue em animais sadios e infectados tratados com veículo (10% DMSO em salina; 10 ml/kg) ou capsazepina. (C) População de células esplênicas F4/80+ em animais sadios e infectados tratados com veículo (10% DMSO em salina; 10 ml/kg) ou capsazepina. (D) População de células esplênicas F4/80+Ly6G+ em animais sadios e infectados tratados com veículo (10% DMSO em salina; 10 ml/kg) ou capsazepina.*P<0,05, difere do grupo sadio e tratado com veiculo; #P<0,05, difere do grupo infectado e tratado com veiculo. (n = 5-8/grupo)

52
Figura 7: Efeito do tratamento repetido com capsazepina (50µg/animal, 2 x ao dia, 6 dias) em animais sadios e com malária induzida por P. berghei ANKA (105 hemacias infectadas/ animal, i.p.), 7 dias após a indução da malária. (A) Fluorescência média de IAb em células sanguíneas F4/80+ em animais sadios e infectados tratados com veículo (10% DMSO em salina; 10 ml/kg) ou capsazepina. (B) Fluorescência média de IAb em células sanguíneas F4/80+Ly6G+ em animais sadios e infectados tratados com veículo (10% DMSO em salina; 10 ml/kg) ou capsazepina. (C) Fluorescência média de IAb em células esplênicas F4/80+ em animais sadios e infectados tratados com veículo (10% DMSO em salina; 10 ml/kg) ou capsazepina. (D) Fluorescência média de IAb em células esplênicas F4/80+Ly6G+ em animais sadios e infectados tratados com veículo (10% DMSO em salina; 10 ml/kg) ou capsazepina. *P<0,05, difere do grupo sadio e tratado com veiculo; #P<0,05, difere do grupo infectado e tratado com veiculo. (n = 5-8/grupo)
Avaliou-se também o marcador GR1, presente em células precursoras de monócitos e
granulócitos. Observou-se que, com a instalação da infecção, ocorre uma redução da
população de células sanguíneas GR1+, quando comparado com o grupo controle não
infectado (Figura 8 A). O tratamento com capsazepina por sua vez, promove o aumento desta
população em animais infectados, sem, no entanto, afetá-la em animais não infectados (Figura
8 A). O número de células GR1+ esplênicas não foi alterado pela infecção ou pelo tratamento
com capsazepina (Figura 8 B).
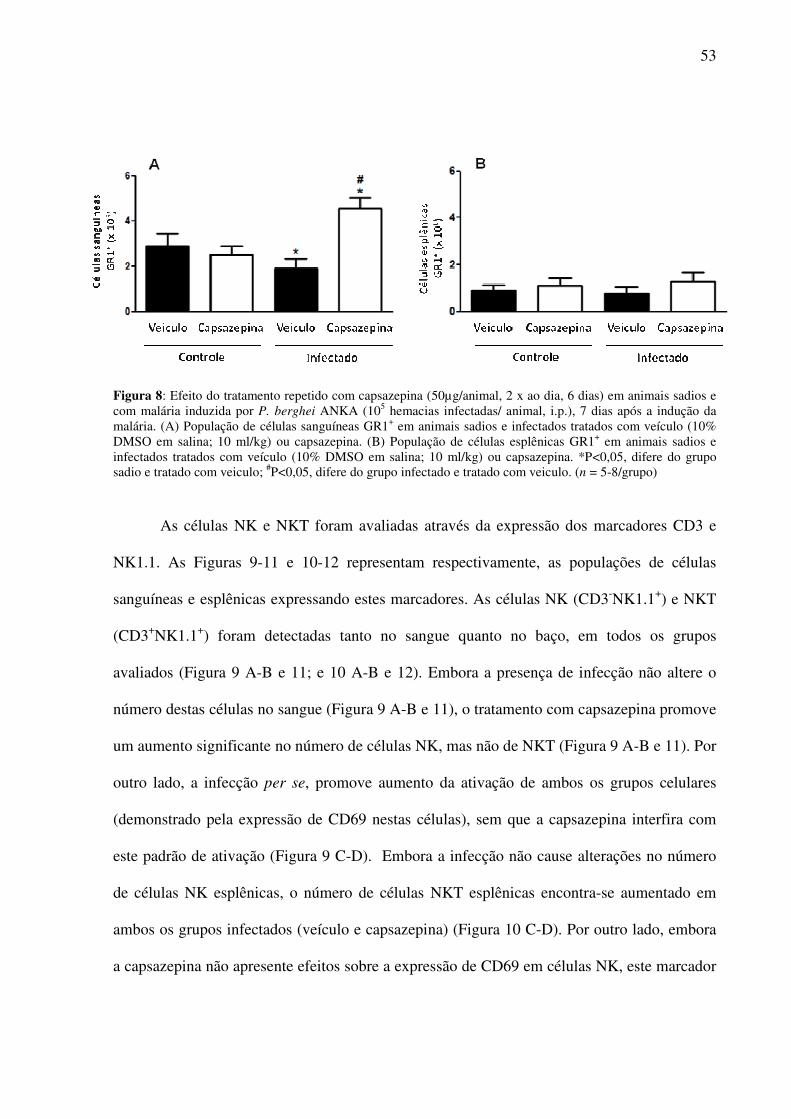
53
Figura 8: Efeito do tratamento repetido com capsazepina (50µg/animal, 2 x ao dia, 6 dias) em animais sadios e com malária induzida por P. berghei ANKA (105 hemacias infectadas/ animal, i.p.), 7 dias após a indução da malária. (A) População de células sanguíneas GR1+ em animais sadios e infectados tratados com veículo (10% DMSO em salina; 10 ml/kg) ou capsazepina. (B) População de células esplênicas GR1+ em animais sadios e infectados tratados com veículo (10% DMSO em salina; 10 ml/kg) ou capsazepina. *P<0,05, difere do grupo sadio e tratado com veiculo; #P<0,05, difere do grupo infectado e tratado com veiculo. (n = 5-8/grupo)
As células NK e NKT foram avaliadas através da expressão dos marcadores CD3 e
NK1.1. As Figuras 9-11 e 10-12 representam respectivamente, as populações de células
sanguíneas e esplênicas expressando estes marcadores. As células NK (CD3-NK1.1+) e NKT
(CD3+NK1.1+) foram detectadas tanto no sangue quanto no baço, em todos os grupos
avaliados (Figura 9 A-B e 11; e 10 A-B e 12). Embora a presença de infecção não altere o
número destas células no sangue (Figura 9 A-B e 11), o tratamento com capsazepina promove
um aumento significante no número de células NK, mas não de NKT (Figura 9 A-B e 11). Por
outro lado, a infecção per se, promove aumento da ativação de ambos os grupos celulares
(demonstrado pela expressão de CD69 nestas células), sem que a capsazepina interfira com
este padrão de ativação (Figura 9 C-D). Embora a infecção não cause alterações no número
de células NK esplênicas, o número de células NKT esplênicas encontra-se aumentado em
ambos os grupos infectados (veículo e capsazepina) (Figura 10 C-D). Por outro lado, embora
a capsazepina não apresente efeitos sobre a expressão de CD69 em células NK, este marcador

54
encontra-se reduzido em células NK obtidas de animais infectados, quando comparado ao
grupo de animais infectados e tratados com veículo (Figura 10 C-D).
Figura 9: Efeito do tratamento repetido com capsazepina (50µg/animal, 2 x ao dia, 6 dias) em animais sadios e com malária induzida por P. berghei ANKA (105 hemacias infectadas/ animal, i.p.), 7 dias após a indução da malária. Painel representativo de células NK e NKT no sangue em animais sadios e infectados tratados com veículo (10% DMSO em salina; 10 ml/kg) ou capsazepina. (n =5-8/grupo)

55
Figura 10: Efeito do tratamento repetido com capsazepina (50µg/animal, 2 x ao dia, 6 dias) em animais sadios e com malária induzida por P. berghei ANKA (105 hemacias infectadas/ animal, i.p.), 7 dias após a indução da malária. Painel representativo de células NK e NKT no baço de animais sadios e infectados tratados com veículo (10% DMSO em salina; 10 ml/kg) ou capsazepina. (n =5-8/grupo)

56
Figura 11: Efeito do tratamento repetido com capsazepina (50µg/animal, 2 x ao dia, 6 dias) em animais sadios e com malária induzida por P. berghei ANKA (105 hemacias infectadas/ animal, i.p.), 7 dias após a indução da malária. (A) População de células NK no sangue em animais sadios e infectados tratados com veículo (10% DMSO em salina; 10 ml/kg) ou capsazepina. (B) População de células NKT no sangue em animais sadios e infectados tratados com veículo (10% DMSO em salina; 10 ml/kg) ou capsazepina. (C) População de células NK ativadas em sangue de animais sadios e infectados tratados com veículo (10% DMSO em salina; 10 ml/kg) ou capsazepina. (D) População de células NKT ativadas em sangue de animais sadios e infectados tratados com veículo (10% DMSO em salina; 10 ml/kg) ou capsazepina.*P<0,05, difere do grupo sadio e tratado com veiculo; #P<0,05, difere do grupo infectado e tratado com veiculo. (n =5-8/grupo)

57
Figura 12: Efeito do tratamento repetido com capsazepina (50µg/animal, 2 x ao dia, 6 dias) em animais sadios e com malária induzida por P. berghei ANKA (105 hemacias infectadas/ animal, i.p.), 7 dias após a indução da malária. (A) População de células NK no baço em animais sadios e infectados tratados com veículo (10% DMSO em salina; 10 ml/kg) ou capsazepina. (B) População de células NKT no baço em animais sadios e infectados tratados com veículo (10% DMSO em salina; 10 ml/kg) ou capsazepina. (C) População de células NK ativadas no baço de animais sadios e infectados tratados com veículo (10% DMSO em salina; 10 ml/kg) ou capsazepina. (D) População de células NKT ativadas no baço de animais sadios e infectados tratados com veículo (10% DMSO em salina; 10 ml/kg) ou capsazepina.*P<0,05, difere do grupo sadio e tratado com veiculo; #P<0,05, difere do grupo infectado e tratado com veiculo. (n = 5-8/grupo)
4.3 Participação do receptor TRPV1 na resposta imune celular adquirida à
malária
Avaliou-se também, o efeito do bloqueio do receptor TRPV1, na modulação da
imunidade adquirida em animais com malária induzida pelo P.berghei ANKA, através do
tratamento repetido com o seu antagonista, capsazepina (50 μg/animal, 2 x ao dia, 6 dias).

58
As Figuras 13 A e 14 A representam as populações CD3+ e CD19+ presentes no
sangue e baço, respectivamente para todos os grupos avaliados. Ainda as populações CD4+ e
CD8+ detectadas no sangue e baço encontram-se demonstradas nas Figuras 13 B e 14 B,
respectivamente.
Com a instalação do processo infeccioso, ocorre uma diminuição de células CD3+ no
sangue, acompanhada de aumento desta população no baço (Figura 15 A e C). Ainda, a
infecção por P. berghei ANKA promove aumento do número de células CD19+ tanto no baço
quanto no sangue (Figura 15 B e C). A capsazepina não apresentou efeitos em células
sanguíneas e esplênicas CD3+ e esplênicas CD19+ (Figura 15 A, C e D). Por outro lado, o
antagonista do receptor TRPV1 reduziu significativamente a população de CD19+ sanguíneas
(Figura 15 C). Avaliaram-se as populações de células sanguíneas e esplênicas CD4+ e CD8+
em linfócitos CD3+ obtidos de todos os grupos estudados. Animais infectados com P. berghei
ANKA apresentam diminuição de células sanguíneas CD8+ (Figura 16 B) e aumento de
células sanguíneas e esplênicas CD4+ e CD8+ (Figura 16 C e D), sem, no entanto,
apresentarem alterações no número de CD4+ sanguíneas (Figura 16 A). O tratamento com
capsazepina não foi capaz de alterar o número de células em nenhuma destas populações
(Figura 16 A-D). Por outro lado, o número absoluto de linfócitos T sanguíneos e esplênicos
CD4+CD25+FOXP3+ foi similar em todos os grupos avaliados tratados ou não com
capsazepina (Tabela 2 e 3).
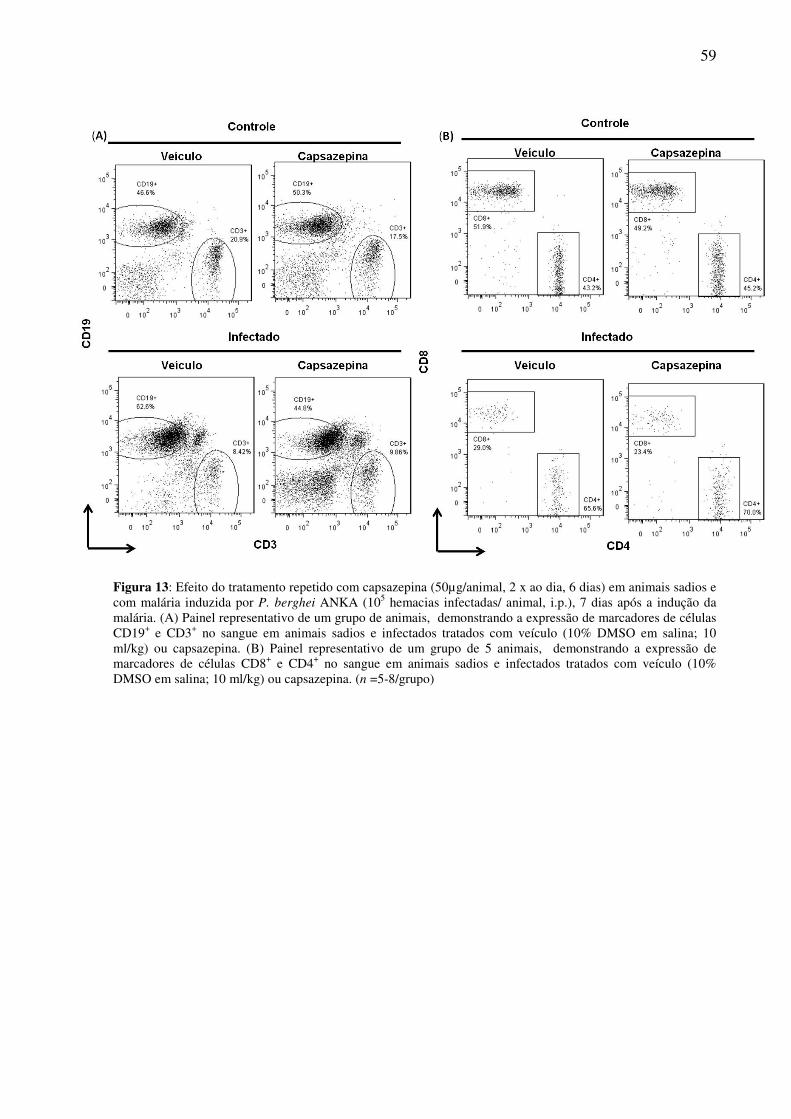
59
Figura 13: Efeito do tratamento repetido com capsazepina (50µg/animal, 2 x ao dia, 6 dias) em animais sadios e com malária induzida por P. berghei ANKA (105 hemacias infectadas/ animal, i.p.), 7 dias após a indução da malária. (A) Painel representativo de um grupo de animais, demonstrando a expressão de marcadores de células CD19+ e CD3+ no sangue em animais sadios e infectados tratados com veículo (10% DMSO em salina; 10 ml/kg) ou capsazepina. (B) Painel representativo de um grupo de 5 animais, demonstrando a expressão de marcadores de células CD8+ e CD4+ no sangue em animais sadios e infectados tratados com veículo (10% DMSO em salina; 10 ml/kg) ou capsazepina. (n =5-8/grupo)

60
Figura 14: Efeito do tratamento repetido com capsazepina (50µg/animal, 2 x ao dia, 6 dias) em animais sadios e com malária induzida por P. berghei ANKA (105 hemácias infectadas/ animal, i.p.), 7 dias após a indução da malária. (A) Painel representativo de um grupo de animais, demonstrando a população de células CD3+ em linfócitos no baço de animais sadios e infectados tratados com veículo (10% DMSO em salina; 10 ml/kg) ou capsazepina. (B) Painel representativo de um grupo de 5 animais, demonstrando a população de células CD4+ e CD8+ no baço de animais sadios e infectados tratados com veículo (10% DMSO em salina; 10 ml/kg) ou capsazepina. (n =5-8/grupo)
Avaliou-se também o padrão de ativação dos linfócitos T e B. Animais infectados
apresentam aumento da ativação de linfócitos T CD4+ e linfócitos B sanguíneos, como
demonstrado pelo aumento significativo de células T expressando CD25 e linfócitos B
expressando CD69, quando comparados a animais não-infectados (Tabela 2). Além disso,
ocorre também aumento da ativação de linfócitos CD4+ sanguíneos via o marcador CD69,
embora este aumento seja apenas significante em animais infectados e tratados com
capsazepina, quando comparado com o grupo de animais não infectados (Tabela 2). Ainda, a
infecção com P. berghei ANKA em animais tratados com veículo ou capsazepina, causou
aumento da ativação de linfócitos esplênicos T CD4+ e CD8+, tanto via CD25 quanto CD69,
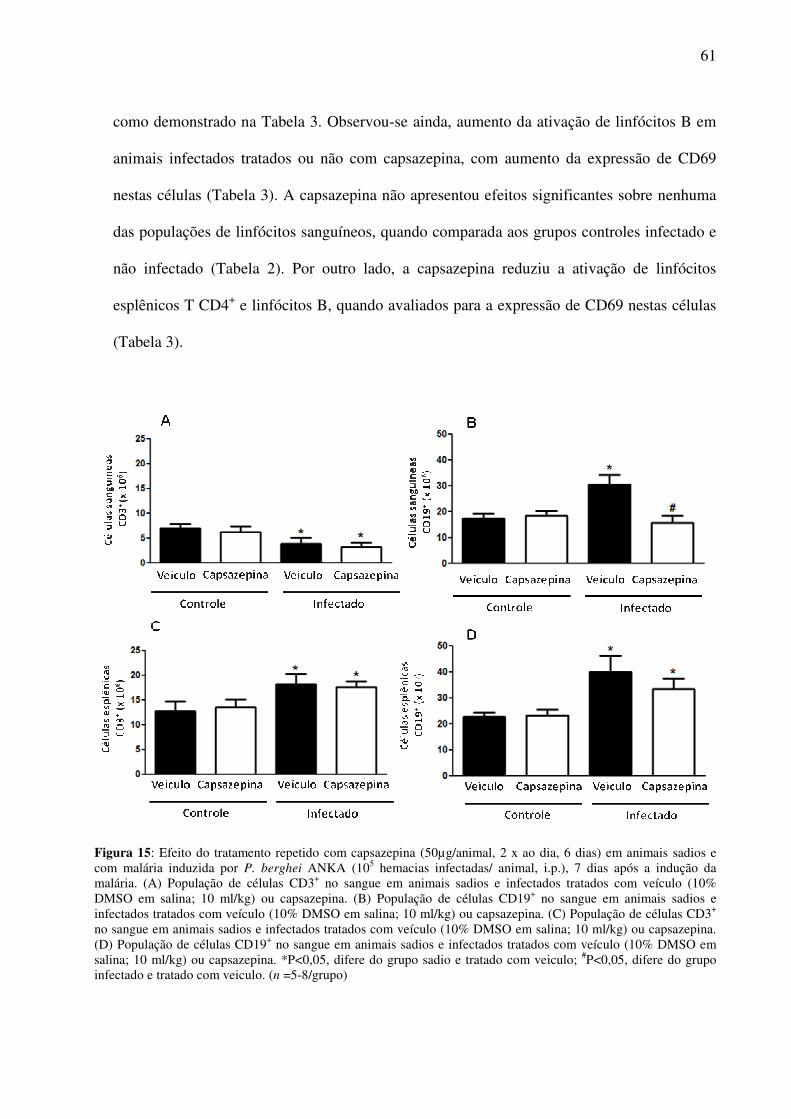
61
como demonstrado na Tabela 3. Observou-se ainda, aumento da ativação de linfócitos B em
animais infectados tratados ou não com capsazepina, com aumento da expressão de CD69
nestas células (Tabela 3). A capsazepina não apresentou efeitos significantes sobre nenhuma
das populações de linfócitos sanguíneos, quando comparada aos grupos controles infectado e
não infectado (Tabela 2). Por outro lado, a capsazepina reduziu a ativação de linfócitos
esplênicos T CD4+ e linfócitos B, quando avaliados para a expressão de CD69 nestas células
(Tabela 3).
Figura 15: Efeito do tratamento repetido com capsazepina (50µg/animal, 2 x ao dia, 6 dias) em animais sadios e com malária induzida por P. berghei ANKA (105 hemacias infectadas/ animal, i.p.), 7 dias após a indução da malária. (A) População de células CD3+ no sangue em animais sadios e infectados tratados com veículo (10% DMSO em salina; 10 ml/kg) ou capsazepina. (B) População de células CD19+ no sangue em animais sadios e infectados tratados com veículo (10% DMSO em salina; 10 ml/kg) ou capsazepina. (C) População de células CD3+ no sangue em animais sadios e infectados tratados com veículo (10% DMSO em salina; 10 ml/kg) ou capsazepina. (D) População de células CD19+ no sangue em animais sadios e infectados tratados com veículo (10% DMSO em salina; 10 ml/kg) ou capsazepina. *P<0,05, difere do grupo sadio e tratado com veiculo; #P<0,05, difere do grupo infectado e tratado com veiculo. (n =5-8/grupo)

62
Figura 16: Efeito do tratamento repetido com capsazepina (50µg/animal, 2 x ao dia, 6 dias) em animais sadios e com malária induzida por P. berghei ANKA (105 hemácias infectadas/ animal, i.p.), 7 dias após a indução da malária. (A) População de células CD3+CD4+ em linfócitos no sangue de animais sadios e infectados tratados com veículo (10% DMSO em salina; 10 ml/kg) ou capsazepina. (B) População de células CD3+CD8+ no sangue de animais sadios e infectados tratados com veículo (10% DMSO em salina; 10 ml/kg) ou capsazepina. (C) População de células CD3+CD4+ em linfócitos no baço de animais sadios e infectados tratados com veículo (10% DMSO em salina; 10 ml/kg) ou capsazepina. (B) População de células CD3+CD8+ no baço de animais sadios e infectados tratados com veículo (10% DMSO em salina; 10 ml/kg) ou capsazepina. *P<0,05, difere do grupo sadio e tratado com veiculo; #P<0,05, difere do grupo infectado e tratado com veiculo. (n =5-8/grupo)

63
Tabela 2: Efeito do tratamento repetido com capsazepina (50µg/animal, 2 x ao dia, 6 dias) em animais sadios e com malária induzida por P. berghei ANKA (105 hemácias infectadas/ animal, i.p.), 7 dias após a indução da malária com relação à distribuição da população de células circulantes. *P<0,05, difere do grupo sadio e tratado com veiculo; #P<0,05, difere do grupo infectado e tratado com veiculo. (n =5-8/grupo)
População Veículo não infectado
Capszepina não infectado
Veículo infectado
Capsazepina infectado
Ly6G+ 2,0 + 0,6 1,3 + 0,4 0,9 + 1,1 1,3 + 0,8
CD3+CD4+CD69+ 0,2 + 0,1 0,2 + 0,1 0,3 + 0,1 0,4 + 0,1 *
CD3+CD4+CD25+FOXP3- 0,2 + 0,04 0,2 + 0,04 0,4 + 0,1 * 0,4 + 0,1 *
CD3+CD4+CD25+FOXP3+ 0,2 + 0,05 0,2 + 0,1 0,2 + 0,04 0,1 + 0,03
CD3+CD8+CD69+ 0,2 + 0,05 0,2 + 0,1 0,2 + 0,1 0,2 + 0,04
CD3+CD8+CD25+ 0,2 + 0,03 0,2 + 0,03 0,15 + 0,04 0,15 + 0,05
CD19+CD69+ 1,1 + 0,2 1,4 + 0,4 4,0 + 0,8 * 3,4 + 0,5 *
Tabela 3: Efeito do tratamento repetido com capsazepina (50µg/animal, 2 x ao dia, 6 dias) em animais sadios e com malária induzida por P. berghei ANKA (105 hemácias infectadas/ animal, i.p.), 7 dias após a indução da malária com relação à distribuição da população de células esplênicas. *P<0,05, difere do grupo sadio e tratado com veiculo; #P<0,05, difere do grupo infectado e tratado com veiculo. (n =5-8/grupo)
População Veículo não- infectado
Capszepina não- infectado
Veículo infectado
Capsazepina infectado
Ly6G+ 0,7 + 0,2 1,1 + 0,4 1,0 + 0,6 0,9 + 0,3
CD3+CD4+CD69+ 0,6 + 0,1 0,5 + 0,1 2,9 + 0,3 * 1,6 + 0,2 * #
CD3+CD4+CD25+FOXP3- 0,4 + 0,06 0,5 + 0,05 1,4 + 0,4 * 1,5 + 0,5 *
CD3+CD4+CD25+FOXP3+ 1,3 + 0,3 1,4 + 0,3 1,4 + 0,4 1,1 + 0,3
CD3+CD8+CD69+ 0,3 + 0,05 0,2 + 0,05 1,7 + 0,2 * 1,5 + 0,1 *
CD3+CD8+CD25+ 0,3 + 0,04 0,4 + 0,1 2,0 + 0,7 * 1,9 + 0,2 *
CD19+CD69+ 1,0 + 0,1 1,2 + 0,5 27,6 + 4,0 * 19,5 + 3,3 * #

64
4.4 Efeito do antagonista de receptores TRPV1 na produção sistêmica de
mediadores inflamatórios
Avaliou-se também, o efeito do bloqueio do receptor TRPV1 na produção sistêmica de
mediadores inflamatórios em animais com malária induzida pelo P.berghei ANKA, através do
tratamento repetido com o seu antagonista, capsazepina (50 μg/animal, 2 x ao dia, 6 dias).
A Figura 17 demonstra o aumento nos níveis plasmáticos de aldeído, produzido como
resultado do estresse oxidativo (A), e os níveis circulantes de TNFα (B) e IFNγ (C) liberados
pela ativação de células inflamatórias, em decorrência da infecção pelo P. berghei ANKA. O
tratamento com capsazepina causa redução dos níveis de aldeído e TNFα, sem, no entanto,
afetar a liberação de IFNγ em animais infectados (Figura 17 A-C). Não foram detectadas IL-2,
IL-4, IL-6, IL-10 nem IL-17, em nenhum dos grupos experimentais avaliados.

65
Figura 17: Efeito do tratamento repetido com capsazepina (50µg/animal, 2 x ao dia, 6 dias) em animais sadios e com malária induzida por P. berghei ANKA (105 hemácias infectadas/ animal, i.p.), 7 dias após a indução da malária. (A) Níveis de aldeído plasmático em animais infectados tratados com veículo (10% DMSO em salina; 10 ml/kg) ou capsazepina. (B) Quantificação de TNF-α plasmático em animais infectados tratados com veículo (10% DMSO em salina; 10 ml/kg) ou capsazepina. (C) Quantificação de IFN-γ plasmático em animais infectados tratados com veículo (10% DMSO em salina; 10 ml/kg) ou capsazepina.*P<0,05, difere do grupo sadio e tratado com veiculo; #P<0,05, difere do grupo infectado e tratado com veiculo. n =5-8/grupo

66
5. DISCUSSÃO
Na tentativa de melhor esclarecer a resposta imunológica e buscar alternativas para um
melhor prognóstico de indivíduos já infectados, modelos em animais têm sido desenvolvidos,
e têm facilitado o entendimento da malária humana. Desta maneira, utilizou-se um modelo
experimental de malária induzida por Plasmodium berghei ANKA em camundongos
C57BL/6, modelo este descrito anteriormente como relevante para o desenvolvimento da
forma neurológica da malária (LOU et al, 2001).
Investigou-se a participação do receptor TRPV1 na malária através do tratamento
repetido com seu antagonista não seletivo, a capsazepina, nesses camundongos. O papel deste
receptor na malária jamais foi descrito na literatura. Assim, avaliou-se a resposta imune dos
animais com malária, por citometria de fluxo, através da análise de populações de células do
sangue e do baço, assim como o processo de ativação destas células.
Os monócitos, população estudada neste trabalho, são células sanguíneas circulantes
derivadas da medula óssea, recrutadas ativamente para locais de inflamação, onde se
diferenciam em macrófagos. Na malária cerebral humana, existe uma alteração na
microcirculação cerebral que justifica os danos ali encontrados. Os eritrócitos infectados
aderem ao endotélio vascular causando vasoconstrição, diminuição do fluxo sanguíneo
cerebral, obstrução vascular e ativação das micróglias, macrófagos localizados no sistema
nervoso central, ocasionando uma alteração na permeabilidade da barreira hematoencefálica e
início da resposta do sistema imune (ADAMS et al, 2000). Com essa adesão endotelial,
células endoteliais e macrófagos liberam espécies reativas de oxigênio e nitrogênio (peróxido
de hidrogênio, superóxido e óxido nítrico) e produtos de peroxidação lipídica
(malonodialdeidos, HNE, dentre outros) que colaboram para o dano tecidual no endotélio
vascular, na tentativa de destruir células infectadas pelo Plasmodium (GRIFFIITHS et al,

67
2001). Ainda, a malária cerebral parece estar relacionada à falência hepática, não só em
animais, mas também em humanos (CASTRO et al, 2004). De fato, sugere-se que a infecção
por P. berghei em camundongos pode levar a um intenso dano hepático devido à presença de
infiltrado de células mononucleares no fígado, aumentando a mortalidade do animal
(YOSHIMOTO et al, 1998; QUEIROZ, 2007).
O presente estudo avaliou a população de monócitos e macrófagos e células
dendríticas esplênicas, através dos marcadores F4/80 e Ly6G. A glicoproteína F4/80 é um
marcador amplamente expresso nestas células (TAYLOR et al, 2003) enquanto que Ly6G
apesar de representar um marcador clássico de neutrófilos (CHOU et al, 2010), foi
recentemente descrita em uma população restrita de macrófagos (FISHER et al, 2011). Ainda,
avaliou-se a ativação de monócitos e macrófagos através da expressão de IAb, utilizado nesse
estudo como um indicador de MHC de classe II, representando assim uma célula em plena
atividade, ou seja, uma célula que esteja apresentando antígenos e consequentemente ativando
linfócitos T (LeGUERN et al, 2010).
Os resultados do presente trabalho demonstram a existência de uma população de
células F4/80+/Ly6G+em camundongos C57BL/6, a qual se encontra aumentada durante o
processo infeccioso por P. berghei ANKA. Esta população celular foi descrita uma única vez
na literatura, em um modelo de infecção viral em camundongos C57BL/6 (FISHER et al,
2011). Este mesmo estudo sugeriu que este grupo de células tem como função primordial a
produção de IFN e espécies reativas de oxigênio. Ainda, foi descrita a existência de uma
população de monócitos Ly6G-, a qual parece estar relacionada principalmente com a
prevenção da disseminação sistêmica da infecção, sendo esta relacionada à produção de TNFα
e óxido nítrico (FISHER et al, 2011). Evidências têm sugerido que a população de células
Ly6G- está relacionada ao momento inicial da infecção, enquanto que a população Ly6G+
compõe o infiltrado existente após a subsequente acumulação de monócitos Ly6G -.

68
O presente estudo demonstrou que o quantitativo das células sanguíneas F4/80+ não
sofre influência com a instalação do quadro da malária, ou seja, dentre os grupos de animais
estudados não houve variação populacional de células F4/80+. Por outro lado, a população
dupla positiva (F4/80+Ly6G+) aumentou com a infecção. Este estudo foi realizado em animais
infectados por 7 dias com P. berghei. Estudos anteriores demonstram que neste período
ocorre o início da mortalidade devido à infecção (QUEIROZ, 2007). De forma similar, neste
período, foi observada uma taxa de mortalidade de 15% em ambos os grupos infectados com
P. berghei. Assim, pode-se sugerir que ao final de 7 dias de infecção a resposta imune é dada
por células efetoras que correspondem a uma resposta não inicial com relação a monócitos, o
que explicaria a predominância da população F4/80+Ly6G+ em animais com malária.
Por outro lado, no baço, foi observado um aumento da quantidade de macrófagos
esplênicos F4/80+ e F4/80+Ly6G+, devido o quadro da malária. O que já era esperado, uma
vez que, durante uma infecção, é para o baço que as células apresentadoras de antígeno
migram a fim de realizar a apresentação de antígenos e ativação de linfócitos (WEISS, 1991).
Ainda, foram observadas alterações importantes no padrão de ativação de ambas as
populações de células F4/80+ e F4/80+Ly6G+. Durante o processo infeccioso por P. berghei
ocorre aumento da ativação destas células quando comparado ao padrão de ativação das
células obtidas de animais não infectados. A ativação de monócitos na malária cerebral é
essencial para a instalação do quadro clínico, devido aos danos teciduais ali existentes,
conforme já citado anteriormente (GRAU e KOSSODO, 1994). Ainda, o aumento da ativação
destas células está relacionado ao aumento da resposta adaptativa a este agente infeccioso
(McLACHALAN et al, 2009).
O tratamento repetido com capsazepina não influenciou a quantidade de monócitos
F4/80+ circulantes. Porém, com relação às células F4/80+Ly6G+, observou-se que apesar do
aumento inerente à infecção, a capsazepina possibilitou um aumento menos intenso destas

69
células. Por outro lado, a capsazepina reduziu a ativação de células sanguíneas F4/80+ e
F4/80+Ly6G+. Estes resultados sugerem pela primeira vez que o receptor TRPV1 é capaz de
modular a resposta imune à outros patógenos além de bactérias. Entretanto, não se sabe ao
certo qual o impacto que o bloqueio da ativação deste receptor pode ter sobre o prognóstico
da malária. Baseado nestes resultados, pode-se sugerir um papel duplo para estes receptores
uma vez que estes podem afetar a ativação de monócitos/macrófagos de fase precoce (Ly6G-)
e tardia (Ly6G+), afetando o controle inicial da infecção, mas também possivelmente os
efeitos deletérios decorrentes da ativação destas células na malária. Assim, a inibição de
funções importantes como a apresentação de antígenos, fagocitose, liberação de mediadores
inflamatórios e ativação da resposta imune adaptativa estariam desreguladas.
De fato, a capsazepina quando utilizada no mesmo esquema de tratamento descrito no
presente estudo, é capaz de modular a resposta imune à infecção bacteriana, ocasionando uma
piora na falência de órgãos característica de quadros de sepse (FERNANDES et al, 2012). Da
mesma forma, a deleção gênica do TRPV1, evoca prognóstico similar, o qual foi associado à
perda de função de macrófagos (fagocitose e liberação de radicais livres). Assim como na
sepse, os presentes resultados demonstram que a capsazepina pode também modular a
infecção pelo P. berghei. O estresse oxidativo gerado pela produção de espécies reativas de
oxigênio está relacionado ao dano tecidual existente na malária cerebral. Como a capsazepina
é capaz de diminuir a ativação de monócitos, consequentemente, espera-se que tais células
produzam menos peróxido de hidrogênio e superóxido e, assim, diminuir o dano ao endotélio.
Investigou-se então, as alterações nos níveis circulantes de produtos decorrentes da
peroxidação lipídica (níveis de malondialdeído ou TBARs) como indicação da presença de
estresse oxidativo. Um aumento de TBARs foi observado em animais infectados, mas não em
animais infectados tratados com capsazepina. Entretanto, o aumento observado em animais
infectados, não foi significativo em comparação ao grupo de camundongos não infectados

70
tratados com veículo, o que sugere a necessidade de um n experimental maior para que este
efeito seja observado de forma significativa. Estes dados sugerem que o bloqueio do TRPV1
pode reduzir o estresse oxidativo na malária por P. berghei, potencialmente afetando de forma
benéfica em relação ao dano vascular caracteristicamente associado à produção excessiva de
produtos deste estresse oxidativo; embora o real impacto deste achado sobre a taxa de
mortalidade, ainda permaneça por ser melhor investigado. Por outro lado, Sanni e
colaboradores (1999) demonstraram que a deleção gênica da NADH oxidase, uma enzima
responsável pela produção de espécies reativas de oxigênio e também pela formação de
fagolisossoma (VAN ASSCHE et al, 2011), resulta em aumento da parasitemia e subsequente
piora do quadro da malária em camundongos. Os dados apresentados no presente estudo
mostram que o tratamento repetido com capsazepina causa redução da parasitemia, sugerindo
que a inibição de espécies reativas de oxigênio no modelo utilizado, não possui um impacto
deletério no que diz respeito à carga parasitária, sugerindo que o bloqueio do receptor TRPV1
pode ser útil em reduzir os danos vasculares, por inibir o estresse oxidativo, sem, no entanto,
exercer efeitos sobre o número de parasitas.
No baço, a capsazepina não influenciou o número de células F4/80+ e F4/80+Ly6G+ e
nem suas ativações. Não se sabe ao certo se a capsazepina realmente não possui efeito sobre
estas células esplênicas, uma vez que existe uma grande diversidade de expressão de
marcadores em células apresentadoras de antígenos do microambiente esplênico. De fato, três
populações diferentes de macrófagos podem ser identificadas no baço, no que diz respeito à
expressão de F4/80: uma com alta expressão de F4/80, uma com mediana expressão e uma
que não expressa este marcador (TAYLOR et al, 2003). Esta variedade populacional pode
mascarar a ação da capsazepina com relação à expressão e ativação de um grupo celular no
baço. Ainda, é possível que a capsazepina tenha sua ação mais direcionada a monócitos que a
macrófagos.

71
Avaliou-se ainda, o marcador Ly6G presente em precursores de células dendríticas e
de neutrófilos (GUMLEY et al, 1995). Não foram observadas alterações nas populações
Ly6G+ sanguíneas e esplênicas. Ainda, avaliou-se o marcador GR1, o qual é constituído de
dois epítopos, Ly6C e Ly6G, sendo Ly6C presente somente em células precursoras de
monócitos/macrófagos e granulócitos (FLEMING et al, 1993; GUMLEY et al, 1995). No
presente trabalho, observou-se que a malária diminui a quantidade de células sanguíneas que
expressam este marcador, sem alterar a população de células esplênicas GR1+. Por sua vez,
dados descritos na literatura demonstram que um aumento no percentual de células GR1+
circulantes (sangue) e esplênicas de camundongos infectados por P. berghei ANKA em
modelos de infecção crônica (18 dias) (FRITA et al, 2011; PEREIRA, 2012). Ressalta-se que
enquanto a população de células GR1+ sanguíneas encontra-se em declínio, ocorre aumento
da população F4/80+Ly6G+ enquanto que o número de células F4/80+ permanece inalterado;
sugerindo que a população de células imaturas (GR1+) esteja sendo substituída por células
maduras e ativadas (F4/80+, F4/80+Ly6G+) ao longo dos 7 dias de infecção. Isto foi descrito
de forma similar em um estudo por Randolph e Jakubzick (2008), o qual avaliou monócitos
com capacidade fagocítica e poder para se diferenciar em macrófagos.
O tratamento repetido com capsazepina aumentou o número de células GR1+
sanguíneas em animais infectados, mas, no entanto, diminuiu a população de F4/80+Ly6G+ e
a ativação de ambas as populações F4/80+ e F4/80+Ly6G+. Ainda, a capsazepina não afetou o
número de células Ly6G+. Isto sugere que a capsazepina reduz a maturação e ativação de
monócitos, fazendo com que estes continuem em sua forma imatura, contribuindo para a
disfunção destas células. No entanto, a capsazepina não apresentou efeitos sobre células
apresentadoras de antígeno esplênicas. Sugere-se que isto esteja relacionado ao período do
processo infeccioso, i.e.; 7 dias. Assim, é possível que a ação da capsazepina em diminuir a
atividade fagocítica do monócito/macrófago seja mais notável no sangue, devido,

72
provavelmente, a existência de um maior número células imaturas ou em processo de
maturação no sangue do que no baço. Estes resultados sugerem que a redução da ativação de
monócitos pela capsazepina pode ter um impacto positivo na malária cerebral, uma vez que,
este quadro clínico é consequência de uma resposta imune Th1 (pró-inflamatória) exacerbada,
com ativação de monócitos e macrófagos e consequente produção de TNF e IFN, devido à
intensa resposta inflamatória que é instalada na barreira hematoencefálica (CLARK e
ROCKETT, 1994); além de diminuir a produção de espécies reativas e consequentemente a
lesão tecidual. Ainda, a reduzida capacidade de ativação destas células pode trazer
consequências à ativação dos linfócitos T e B. Assim, avaliou-se a população de linfócitos em
animais infectados e não infectados por P. berghei, tratados e não tratados com capsazepina.
Estudos dos últimos 10 anos enfatizam que as células T CD8+ são as principais células
efetoras da resposta imune adaptativa, enquanto que as células CD4+ assumem um papel mais
regulatório, de acordo com os tipos de citocinas produzidas durante o processo infeccioso
(GORBACHEVe FAIRCHILD, 2001).
As células T possuem um papel de destaque no desenvolvimento da resposta imune,
pois estão relacionadas com a ativação de células B e subsequente produção de anticorpos e
também com a citotoxicidade celular dependente de anticorpos, a qual infelizmente, não é
capaz de erradicar o parasita (WIPASA et al, 2010). A manutenção de antígeno derivados do
Plasmodium no organismo, e favorecendo sua apresentação às células T CD8+,
desencadeando uma melhor imunidade protetora, exercendo sua função citotóxica com
secreção de citocinas inflamatórias incluindo TNF e IFN (COCKBURN et al, 2010).
Recentemente foi sugerido que a ativação de células T na malária pode ser prejudicada devido
a alterações decorrentes de outras células do sistema imune, como por exemplo, a diminuição
de células apresentadoras de antígenos e de sua ativação devido liberação de espécies reativas
de oxigênio, o que pode ocasionar apoptose (GONÇALVES et al 2010). A redução da

73
ativação de células T tem como consequência uma alteração na geração de células de
memória específicas (células T e B) para o Plasmodium (GONÇALVES et al, 2010).
No presente trabalho, foram analisados linfócitos T CD3+CD8+ e observou-se que, em
camundongos C57BL/6 infectados por P. berghei ANKA, há uma diminuição destas células
em sangue. Por outro lado, no baço há um aumento desta população. No caso da malária
cerebral, foi descrito anteriormente que em camundongos infectados por P. berghei ocorre
migração de linfócitos T circulantes, em especial células CD8+, para o cérebro, colaborando
para o dano endotelial ali existente (BELNOUE et al, 2002). Sugere-se que a diminuição de
células CD8+ observadas em animais infectados no presente trabalho ocorra em decorrência
da migração destas células para outros sítios, como o baço e o cérebro, contribuindo tanto
para a apresentação de antígenos quanto para a disfunção endotelial. Ainda, o presente
trabalho avaliou a ativação de linfócitos T CD8+ através da análise da expressão de
marcadores de superfície CD25, receptor da cadeia α da IL-2 e relacionado à etapa final da
ativação, e CD69, marcador de ativação precoce e expresso transitoriamente, nestas células.
Observou-se uma pequena redução da ativação destas células no que diz respeito a CD25,
embora nenhuma alteração tenha sido observada em relação a CD69, em células sanguíneas
obtidas de animais infectados quando estes são comparados a animais tratados com veículo.
Enquanto que no baço, é expressiva a influencia da malária na ativação destas células, as
quais foram altamente positivas para a expressão dos marcadores CD25 e CD69. A ativação
destas células esplênicas sugere que o baço é um órgão de função primordial na malária,
aonde a apresentação de antígenos e ativação de linfócitos T CD8+ são essenciais para que a
quantidade de eritrócitos contaminados seja controlada (MARQUES e PETROIANU, 2003).
O tratamento repetido com a capsazepina em animais infectados, não causou
alterações do número de células CD8+ sanguíneas e esplênicas e nem em sua ativação,
sugerindo que o receptor TRPV1 não está envolvido na regulação das respostas mediadas por

74
estas células. De fato, Takano e colaboradores (2007) demonstraram que, em animais sadios,
o tratamento oral repetido com capsaicina, um agonista de receptores TRPV1, não apresenta
efeitos imunomoduladores sobre células CD8+ isoladas de amostras de placas de Peyer
murinas, embora este agonista module células CD3+ (redução) e CD19+ (aumento) neste
mesmo modelo (TAKANO et al, 2007). No entanto, o tratamento repetido com capsazepina
reverte parcialmente os efeitos imunomoduladores da capsaicina, sugerindo que este agonista
possui ações imunomoduladoras dependentes e independentes de TRPV1 em outros tipos
celulares que não linfócitos CD8+ (TAKANO et al, 2007).
Os linfócitos T CD3+CD4+ são células que auxiliam no desenvolvimento da resposta
imune, uma vez que dependendo dos tipos de citocinas por eles liberados, ocorre a ativação de
vias pró ou anti-inflamatórias, com estas células assumindo características Th1, Th2 ou Th17
(BRADLEY et al, 2011). Na malária, as células T CD4+ são células muito importantes por
auxiliarem no desenvolvimento das respostas imunes imediata e tardia decorrente a exposição
aguda e crônica ao parasita (LANGHORNE et al, 1989). Durante a infecção aguda a resposta
imune ocorre de forma mais intensa e nesta fase, os linfócitos T CD4+ são responsáveis pela
secreção de citocinas pró-inflamatórias (TNFα e IFNγ) e pela ativação de células B com
subsequente secreção de imunoglobulinas. À medida que a infecção progride, grande parte
das células T CD4+ sofre apoptose. Durante a infecção crônica, as células CD4+ auxiliam o
desenvolvimento da resposta imune adaptativa, produzindo IFNγ e estimulando a produção de
anticorpos específicos contra o Plasmodium (FREITAS DO ROSARIO et al, 2008). No
presente estudo, avaliaram-se os linfócitos T CD3+CD4+ circulantes e esplênicos. A indução
da malária não causa alterações na população de células CD4+ circulantes, embora ocorra um
aumento significativo destas células no baço de animais infectados com P. berghei, sugerindo
um aumento populacional destas células em decorrência da apresentação de antígenos neste
órgão linfóide. De acordo com trabalhos anteriores, a principal consequência do aumento de

75
células CD4+ no baço em camundongos infectados por Plasmodium é a intensa secreção de
TNF-α e IFN-γ (SOULARD et al, 2009), e, na malária humana, as células T CD4+ esplênicas
são as principais responsáveis pela produção IFN-γ (ING e STEVENSON, 2009). A produção
destas citocinas está relacionada ao controle da parasitemia porque melhoram a fagocitose e
favorecem a apresentação de antígenos (BASTOS et al, 2002).
Ainda, avaliou-se a ativação das células CD4+ circulantes e esplênicas. Observou-se
que a malária promove ativação de ambas as populações de células, sendo isto demonstrado
pelo aumento de células CD4+ CD25+ e CD4+ CD69+, embora a ativação de células CD4+
circulantes seja significativa somente em relação ao marcador CD25. Estes resultados
demonstram que, conforme descrito na literatura, a existência de células CD4+ ativadas, além
de induzirem a secreção de mediadores inflamatórios, auxiliam a ativação das células B e
consequentemente a produção de IgM e IgG1, principalmente (HANSEN et al, 2003).
Embora o tratamento repetido com a capsazepina não afete o número de células CD4+,
ela causa alterações no padrão de ativação destas células. A capsazepina causou redução da
população de células esplênicas CD4+ CD69+, sem, no entanto, afetar células CD4+ CD25+. O
marcador CD69 é conhecido como um importante sinalizador entre linfócitos CD4+ (NIHEI,
2013) e também para a proliferação dos mesmos, enquanto que o CD25 é necessário para a
proliferação de células CD4+ de forma dependente do receptor para IL-2 (NIHEI, 2013). Isto
sugere que a ação imunmoduladora da capsazepina, e, portanto, do TRPV1 nestas células,
ocorre de forma independente da produção de IL-2 e/ou ativação de seu receptor. Por outro
lado, não foram observadas alterações no padrão de expressão de CD25 ou CD69 em
linfócitos T CD4+ circulantes. Similarmente, Takano e colaboradores (2007) demonstraram
que o tratamento repetido com capsaicina ou capsazepina, não induz alterações no número de
células CD4+ em animais sadios, no entanto, este estudo não avaliou o padrão de ativação
destas células.

76
As células T reguladoras (Treg) são linfócitos T CD4+ que têm como função limitar o
grau de resposta imune efetora. Podendo até mesmo tornar o organismo incapaz de controlar
adequadamente a infecção. Por meio dessa supressão, essas células limitam danos colaterais
que possam ser causados por uma resposta imune vigorosa contra os patógenos (BELKAID e
TARBELL, 2009). A presença das Tregs na malária está relacionada com a carga parasitária e
o aumento desta população celular está associado ao aumento da carga parasitária (MINIGO
et al, 2009) e ao desenvolvimento da malária clínica (TODRYK et al, 2008)
Por outro lado, nem a infecção e nem a capsazepina exerceram efeitos
imunomoduladores sobre os linfócitos T CD3+CD4+CD25+FOXP3+ (células T regulatórias)
circulantes e esplênicos. Por outro lado, estudos têm descrito que a malária desencadeia uma
depleção desta população de células, promovendo uma resposta exclusiva de células efetoras
na tentativa de melhorar a proteção do hospedeiro, ou seja, favorecer o aumento da
capacidade de resposta dos linfócitos T contra os parasitas (HISAEDA et al, 2004).
Os Linfócitos B são as células responsáveis pela produção de anticorpos e,
consequentemente, responsáveis pela resposta humoral. Na malária, os linfócitos B tem papel
fundamental na imunidade do hospedeiro contra a malária, uma vez que os anticorpos
participam ativamente nos estágios pré-eritrocítico e eritrocítico, como por exemplo, os
anticorpos anti-esporozoítos e os anticorpos contra a membrana externa dos esporozoítos,
estão presentes no soro de indivíduos residentes no continente Africano (área com
transmissão estável da malária) e aumentam com a idade e com o desenvolvimento da
resistência (NARDIN et al, 1979).
O marcador CD19 é expresso nas células B, as quais encontram-se aumentadas e
ativamente produzindo anticorpos específicos para o Plasmodium (LEORATTI, 2004). No
presente trabalho, avaliou-se a população de células CD19+. A infecção por P. berghei em
camundongos causou uma elevação no número de células CD19+ circulantes e esplênicas em

77
comparação com o grupo de animais não infectados. Da mesma maneira, a ativação destas
células encontra-se aumentada durante a infecção, demonstrado pelo aumento de células
CD19+ CD69+. Por outro lado, relatos anteriores descrevem que seja possível observar que
crianças são mais susceptíveis à malária que os adultos. E para verificar tal acontecimento, foi
utilizado um modelo experimental com animais semi-imunes, os quais após à terceira
transferência passiva de células CD19+ esplênicas de camundongos já curados já estavam
imunes contra a malária (BAO et al, 2013)
O tratamento repetido com a capsazepina em camundongos infectados por P. berghei
ANKA reduziu a população de CD19+circulantes, porém sem afetar a capacidade de ativação
destas células. Por outro lado, nas células esplênicas CD19+, o tratamento com a capsazepina
não influenciou a população destas células. Entretanto, a ativação deste grupo celular
encontra-se reduzida após o tratamento com capsazepina. Takano e colaboradores (2007)
investigaram os efeitos da capsaícina e capsazepina, administrada por via oral, em animais
sadios e concluíram que a capsaícina é capaz de aumentar a população de células CD19+,
sendo este efeito revertido parcialmente pelo tratamento repetido com capsazepina. Estes
dados sugerem em conjunto com os resultados deste trabalho, que o receptor TRPV1 pode
regular o número ou a ativação de células CD19+, dependendo do microambiente em que
estas células estão inseridas.
Finalmente, avaliou-se o perfil das células NK (CD3-NK1.1+) e NKT (CD3+NK1.1+)
em camundongos infectados por P. berghei ANKA. As células NK são uma linhagem de
células bem caracterizadas as quais reconhecem células infectadas e/ou estressadas
(D’OMBRAIN et al, 2007). Estas células atuam diretamente promovendo a lise dessas células
e secretam citocinas pró-inflamatórias como IFN e IL-4 (D’OMBRAIN et al, 2007). Por outro
lado, as células NKT constituem uma população discreta de células que possuem
características relacionadas ao linfócito T e às células NK (HANSEN et al, 2003), sugeridas

78
como reguladoras da resposta Th1/Th2 decorrentes da malária cerebral em camundongos
(HANSEN et al., 2003). Trabalhos anteriores sugerem que a infecção por malária tem uma
resposta inicial Th1, mas que após 7 dias inicia a resposta tardia. Essa mudança no padrão de
resposta imune é decorrente da ação mecanismos celulares ou mecanismos mediados por
imunoglobulinas, no intuito de controlar a infecção pelo Plasmodium (TAYLOR-ROBINSON
et al, 1993).
Estudos recentes sugerem que as células NK são essenciais para o pleno controle da
malária, pois elas podem produzir citocinas (IFN-γ) ou mediar efeitos citotóxicos diretos.
Existem dados que indicam que a resposta sanguínea inicial à malária em camundongos
infectados por P. chabaudi é marcada pela ação citotóxica das células NK (KIM et al, 2008).
No presente trabalho, não foram observadas alterações no número de células NK circulantes e
esplênicas, com relação a animais infectados ou não com P. berghei. Por outro lado, a
infecção causou um aumento da ativação de células NK. Relatos anteriores demonstraram a
ocorrência de um aumento de células NK circulantes e esplênicas devido a uma proliferação
celular durante o período de até 7 dias após a infecção (KIM et al, 2008). É possível que este
aumento da ativação possa ocasionar aumento dos níveis de IFN-γ durante a infecção por P.
berghei.
As células NKT são grandes produtoras de IFN-γ e podem influenciar tanto as
respostas Th1 e Th2. Apesar das funções deste grupo celular ainda não estarem
completamente elucidadas, um importante papel imunomodulador foi sugerido para as NKT
malária experimental. Hansen e colaboradores (2003) sugeriram que uma população distinta
de células NKT positivas para o marcador CD1d favorecem o desenvolvimento de uma
resposta do tipo Th2 com maior resistência para a malária cerebral, além de induzir altos
níveis de IFN-γ. A população de células NKT circulantes não foi afetada pela infecção por P.
berghei, nem em relação ao número de células nem no que diz respeito à sua ativação (células

79
CD69+). Por outro lado, observou-se um aumento da população e da ativação de células NKT
esplênicas. Sugere-se que, na malária, a população de células esplênicas possa estar em
expansão, conforme comportamento descrito anteriormente para as células NK (KIM et al.,
2008). Da mesma forma, espera-se que alterações no padrão de ativação e no número destas
células possua um impacto na produção de IFN-γ na malária.
Avaliou-se o efeito do tratamento repetido com a capsazepina na ativação e população
de células NK em animais infectados ou não por P. berghei. Observou-se que este antagonista
do TRPV1, causa aumento no número destas células em animais infectados, sem, no entanto,
interferir com a ativação destas células. Sugere-se a capsazepina possa favorecer a migração
de células NK imaturas para a circulação de forma que o aumento populacional não reflita em
uma ativação aumentada. No baço, por sua vez, a capsazepina não afetou nem o número de
células NK nem sua ativação. Assim, sugere-se o envolvimento do receptor TRPV1 se
restringe apenas ao recrutamento de células NK imaturas sem interferir com a ativação de
células NK maduras em animais com malária. Por outro lado, a capsazepina não apresentou
efeitos sobre o número de células NKT, mas reduziu a ativação destas células no baço de
animais infectados. Nenhum efeito significativo foi observado para a capsazepina no que diz
respeito ao número de células NKT ou sua ativação após a instalação do processo infeccioso.
No entanto, não se sabe ao certo qual o impacto que as alterações induzidas pela capsazepina
em células NKT possam ter no prognóstico da malária. Se por um lado, estas células possuam
um perfil pró-inflamatório no início da infecção por Plasmodium, por outro lado, um papel
anti-inflamatório foi sugerido para uma população restrita de NKT CD1d+, numa fase mais
tardia da infecção (HANSEN et al, 2003). Ressalta-se que este efeito anti-inflamatório para as
NKT foi descrito em um modelo experimental de malária em camundongos no qual este papel
foi observado em períodos experimentais mais tardios do que o utilizado no presente trabalho.

80
Assim, é possível que dependendo do momento em que a capsazepina seja administrada, ela
possa modular de forma benéfica ou deletéria a infecção.
Avaliou-se a produção das citocinas pró-inflamatórias IFN e TNFα, em animais
infectados e sadios, tratados ou não com capsazepina. O TNF-α é o principal mediador da
resposta imunológica aguda, sendo responsável por muitas complicações sistêmicas, uma vez
que sua função é estimular o recrutamento e a ativação de neutrófilos e monócitos para os
locais da infecção e sua principal fonte celular são os monócitos/macrófagos e células T
(MEDANA et al, 1997). Além de promover a estimulação de diferentes células, a liberação de
TNFα também leva à apoptose de células inflamatórias e teciduais (RANDALL e
ENGWERDA, 2010). Na malária, a produção de TNFα leva ao aumento da expressão de
moléculas de adesão, favorecendo, portanto, a adesão de células inflamatórias no endotélio
vascular, e subsequente dano tecidual o que leva a disfunção endotelial observada em quadros
de malária cerebral (HEDDINI, 2002; GIMENEZ et al, 2003). Em resposta à infecção, in
vitro, estudos demonstram que o aumento de TNF-α pode destruir gametócitos (NAOTUNNE
et al, 1991). O papel do TNF-α na malária humana e murina é bem descrito na literatura, por
exemplo, está relacionado com a capacidade de induzir sua própria produção e de outras
citocinas pró-inflamatórias, com a ativação de linfócitos e com o estímulo de expressão para o
MHC de classe I e II (GOETZ et al, 2004). Por sua vez, o IFN-γ é a principal citocina
ativadora de macrófagos e exerce funções fundamentais tanto na imunidade inata quanto
adquirida, mediadas por células contra microrganismos intracelulares (BARBALAT et al,
2009). Sua principal fonte celular são células T, NKT e NK (COUPER et al, 2007). Na
malária, o IFN-γ é um potente ativador de monócitos e macrófagos, favorecendo a fagocitose
e subsequente morte do parasita, além de induzir a secreção de citocinas, como IL-1 e IL-6
(HOFFMAN e FRANKE, 1994).

81
No presente estudo, os níveis circulantes de TNFα mostraram-se mais elevados em
animais infectados por P. berghei ANKA quando comparados àqueles de animais sadios. O
mesmo foi observado para os níveis de IFN. O tratamento repetido com a capsazepina por sua
vez, causou a diminuição dos níveis de TNFα circulante. Neste trabalho observou-se que a
capsazepina reduz a ativação de monócitos circulantes. Assim, sugere-se que a redução dos
níveis de TNFα pelo tratamento com capsazepina seja devida à inibição da ativação destes
monócitos. Por outro lado, a capsazepina não afetou a produção de IFNγ em animais
infectados. Como mencionado anteriormente, o IFN pode ser produzido por células T, NK e
NKT. Apesar de aumentar o número de células NK, efeito este provavelmente devido à
presença de células NK imaturas, a capsazepina não afetou a ativação células T, NK e NKT
circulantes. No entanto, este antagonista do receptor TRPV1 foi capaz de reduzir a ativação
de células T CD4+ além de diminuir a ativação de células NKT. Estes resultados sugerem que
a produção de IFN possa ser decorrente da população de células NK esplênicas. Entretanto,
através do modelo experimental empregado, não é possível distinguir qual a população é
realmente a responsável pela produção de IFN. Assim, mais experimentos são necessários
para esclarecer esta hipótese. Sugere-se que com a inibição de células T e NKT produtoras de
IFN pela capsazepina, as células NK possam compensar esta inibição, passando a produzir
mais IFN.

82
6. CONCLUSÃO
Os dados do presente trabalho demonstram que o bloqueio do TRPV1 pela
capsazepina pode modular a resposta imune a malária, embora não se saiba ao certo como
este receptor é capaz de influenciar o prognóstico desta patologia. Ressalta-se que a avaliação
do presente estudo se restringiu à fase aguda à infecção (7 dias de infecção) e portanto, e
portanto, mais estudos são necessários ao entedimento do papel patofisiológico do TRPV1 na
malária. Por outro lado, é importante lembrar que antagonistas do receptor TRPV1
encontram-se em fase clínica para o tratamento da dor crônica e que pacientes com este perfil
farão uso repetido destes medicamentos. Assim, a sua prescrição de antagonistas do TRPV1
deve ser realizada com cautela, uma vez que, como já descrito na literatura, interfere na
resposta imunológica à bacteria e como demonstrado por este trabalho, a resposta à malária.

83
REFERÊNCIAS
ADAMS, S.; TURNER, G. D.; NASH, G. B.; MICKLEM, K.; NEWBOLD, C. I.; CRAIG, A.G. Differential binding of clonal variants of Plasmodium falciparum to allelic forms of intracellular adhesion molecule 1 determined by flow adhesion. Infect Immun. v. 68, p. 264-269, 2000.
ALECRIM, M. G. C. Estudo clínico, resistência e polimorfismo parasitário na malária pelo Plasmodium vivax, Manaus – AM. Brasília: UNB, 2000. Tese de Doutorado, Faculdade de Medicina, Núcleo de Medicina Tropical, Universidade de Brasília, 2000.
AMADESI, S.; COTTRELL, G. S.; DIVINO, L.; CHAPMAN, K.; GRADY, E. F.; BAUTISTA, F.; KARANJIA, R.; BARAJAS-LOPEZ, C.; VANNER, S.; VERGNOLLE, N.;
BUNNETT, N. W. Protease-activated receptor 2 sensitizes TRPV1 by protein kinase Cɛ- and A-dependent mechanisms in rats and mice. The Journal of Physiology. v. 575, n. 2, p. 555-571, 2006.
ARTAVANIS-TSAKONAS, K.; RILEY, E. M. Innate immune response to malária: rapid induction of IFN-γ from human NK cells by live Plasmodium falciparum-infected erythrocytes. The Journal of Immunology. v. 169, n. 6, p. 2956-2963, 2002.
BAHNASI, Y. N.; WRIGHT, H. M.; MILLIGAN, C. J.; DEDMAN, A. M.; ZENG, F.; HOPKINS, P. M.; BATESON, A. N.; BEECH, D. J. Modulation of TRPC5 cation channels by halothane, chloroform and propofol. British Journal of Pharmacology. v. 153, n. 7, p. 1505-1512, 2008.
BAO, L. Q.; TIEN HUY, N. T.; KIKUCHI, M.; YANAGI T.; SENBA, M.; SHUAIBU, M.; N.; HONMA, K.; YUI, K.; HIRAYAMA, K. CD19(+) B Cells Confer Protection against Experimental Cerebral Malaria in Semi-Immune Rodent Model. Plos One. v. 8, n. 5, 2013. BARBALAT, R.; LAU, L.; LOCKSLEY, R. M.; BARTON, G. M. Toll-like receptor 2 on inflammatory monocytes induces type I interferon in response to viral but not bacterial ligands. Nat Immunol. v. 10, p. 1200-1207, 2009.
BARDAJI, A.; SIGAUQUE, B.; SANZ, S.; MAIXENCHS, M.; ORDI, J.; APONTE, J.J.; MABUNDA, S.; ALONSO, P.L.; MENENDEZ, C. Impact of malária at the end of pregnancy on infant mortality and morbidity. J Infect Dis. v. 203, p. 691–699, 2011.
BASTOS, K. R.; BARBOZA, R.; ELIAS, R. M.; SARDINHA, L. R.; GRISOTTO, M. G.; MARINHO, C. R.; AMARANTE-MENDES, G. P.; ALVAREZ, J. M.; LIMA, M. R. Impaired macrophage responses may contribute to exacerbation of blood-stage Plasmodium chabaudi malária in interleukin-12-deficient mice. J Interferon Cytokine Res. v. 22, n. 12, p. 1191-1199, 2002.
BASU, S.; SRIVASTAVA, P. Immunological role of neuronal receptor vanilloid receptor 1 expressed on dendritic cells. Proc Natl Acad Sci USA. v. 102, p. 5120-5125, 2005.
BELKAID, Y.; TARBELL, K. Regulatory T cells in the control of host-microorganism interactions. Annu Rev Immunol., n. 27, p. 551-589, 2009.
BELNOUE, E.; KAYIBANDA, M.; VIGARIO, A. M.; DESCHEMIN, J. C.; VAN ROOIJEN, N.; VIGUIER, M.; SNOUNOU, G.; RÉNIA, L. On the pathogenic role of brain-

84
sequestered αβ CD8+ T cells in experimental cerebral malária. J Immunol. v. 169, p. 6369-6375, 2002.
BERENDT, A. R.; TUMER, G. D.; NEWBOLD, C. I. Cerebral malária: the sequestration hypothesis. Parasitol Today.v. 10, p. 412-414, 1994.
BESSAC, B. F.; JORDT, S. E. Breathtaking TRP channels: TRPA1 and TRPV1 an airway chemosensation and reflex control. Physiology. v. 23, p. 360-370, 2008.
BIRD, R. P.; DRAPER, H. H. Comparative studies on different methods of malonaldehyde determination. Methods Enzymol. v.105, p. 299-305, 1984.
BODKIN, J. V.; FERNANDES, E. S. TRPV1 and SP: key for sepsis outcome. British Journal of Pharmacology. 2013.
BRADLEY, S.; SCHNEIDER, C. M., ROGER P.; SALAHEDDINE, M. Anopheles
stephensi Saliva Enhances Progression of Cerebral Malaria in a Murine Model Vector-Borne and Zoonotic Diseases. vol. 11, n. 4, p. 423-432, 2011. BRAIN, S. D.; GRANT, A. D. Vascular actions of calcitonin gene-related peptide and adrenomedullin. Physiol Rev. v. 84, p. 903-934, 2004.
BRASIL. Ministério da Saúde. Secretaria de Vigilância em Saúde. Departamento de Vigilância Epidemiológica. Guia prático de tratamento da malária no Brasil.. Ministério da Saúde – Brasília, 2010.
BREWSTER, D. R.; KWIATKOWSKI, D.; WHITE, N. J. Neurological sequelae of cerebral malária in children. Lancet. v. 336, p. 1039-1043, 1990.
BRUCE-CHWATT, L. J. Malária: principles and practice of malariology. Edinburg: Churchill Linvingstone. v. 1, p. 1-59, 1988.
BRYANT, P., SHUMATE, M., YUMET, G., LANG, C. H., VARY, T. C., COONEY, R. N. Capsaicin-sensitive nerves regulate the metabolic response to abdominal sepsis. J. Surg. Res. v. 112, p. 152-161, 2003.
BUENO, L. L.; MORAIS, C. G.; ARAÚJO, F. F.; GOMES, J. A. S.; CORRÊA-OLIVEIRA, R.; SOARES, I. S.; LACERDA, M. V.; FUJIWARA, R. T.; BRAGA, E. M. Plasmodium
vivax: induction of CD4⁺CD25⁺FoxP3⁺ regulatory T cells during infection are directly associated with level of circulating parasites. Plos One. v. 5, mar, 2010.
CAMARGO, E. P. A malária encenada no grande teatro social. Estudos Avançados. v. 9, p. 24, 1995.
CAMARGO, E. P. Malária, Malieta, Paludismo. Cienc. Cult. v. 55, n. 1, p. 26-29, 2003.
CASTRO, L.; CARDOSO, A. I.; QUEIRÓS, L.; GONÇALVES, G. Malária na região norte de Portugal (1993-2002): Caracterização Epidemiológica. Acta Méd Port. v. 17, p. 291-298, 2004.

85
CATERINA, M. J.; SCHUMACHER, M. A.; TOMINAGA, M.; ROSEN, T. A.; LEVINE, J. D.; JULIUS, D. The capsaicin receptor: a heat-activated ion channel in the pain pathway. Nature. v. 389, p. 816-826, 1997.
CESARE, P., MORIONDO, A., VELLANI, V., MCNAUGHTON, P. A. Ion channels gated by heat. Proc. Natl. Acad. Sci. U. S. A. v. 96, p. 7658-7663, 1999.
CHOU, R. C.; KIM, N. D.; SADIK, C. D.; SEUNG, E.; LAN, Y.; BYME, M. H.; HARIBABU, B.; IWAKURA, Y.; LUSTER, A. D. Lipid-cytokine-chemokine cascade drives neutrophil recruitment in a murine model of inflammatory arthritis. Immunity. v. 33, p. 266-278, 2010.
CHU, C. J., HUANG, S. M., DE PETROCELLIS, L., BISOGNO, T., EWING, S. A., MILLER, J. D., ZIPKIN, R. E., DADDARIO, N., APPENDINO, G., DI MARZO, V., WALKER, J. M. N-oleoyldopamine, a novel endogenous capsaicin-like lipid that produces hyperalgesia. J. Biol. Chem. v. 278, p. 13633-13639, 2003.
CLARK, I. A.; MAcMICKING, J. D.; GRAY, K. M.; ROCKETT, K. A.; COWDEN, W. B. Malária mimicry with tumor necrosis factor. Contrasts between species of murine malária and Plasmodium falciparum. Am J Pathol. v. 140, n. 2, p. 325-336, 1992.
CLARK,, I. A.; ROCKETT, K. A. The cytokine theory of human cerebral malária. Parasitol Today. v. 10, p. 410-412, 1994.
CLARK, N.; KEEBLE, J.; FERNANDES, E. S.; STARR, A.; LIANG, L.; SUGDEN, D.; WINTER, P.; BRAIN, S. D. The transient receptor potential vanilloid 1 (TRPV1) receptor protects against the onset of sepsis after endotoxin. v.21, n. 13, p. 3747-3755, 2007.
COCKBURN, I. A.; CHEN, Y. C.; OVERSTREET, M. G.; LEES, J. R.; VAN, R. N.; FARBER, D. L.; ZAVALA, F. Prolonged antigen presentation is required for optimal CD8+ T cell responses against malária liver stage parasites. Plos Pathogens. v. 6, n. 5, 2010.
CORNEJO, O. E.; ESCALANTE, A. A. The origin and age of Plasmodium vivax. Trends in Parasitology. v. 22, p. 12, 2006.
COUPER, K. N.; BLOUNT, D. G.; HAFALLA, J. C.; VAN ROOIJEN, N.; SOUZA, J. B.; RILEY, E. M. Macrophage-mediated but gamma interferon-independent innate immune responses control the primary wave of Plasmodium yoelli parasitemia. Infect Immun. v. 75, p. 5806-5818, 2007.
COX, F. E. G. History of the discovery of the malária parasites and their vectors. Parasites & Vectors. v. 3, p. 5, 2010.
CROMPTON, P. D.; MOEBIUS, J.; PORTUGAL, S.; WAISBERG, G. H.; GARVER, L. S.; MILLER, L. H.; BARILLAS, C.; PIERCE, S. K. Malária Immunity in man and mosquito: insights into unsolved mysteries of a deadly infectious disease. Annual Review of Immunology. v. 32, p. 157-187, 2014.
DAILY, J. P. Antimalárial Drug Therapy: The Role of Parasite Biology and Drug Resistance. Journal of Clinical Pharmacology. v. 46, p. 1487-1497, 2006.

86
DAVES, B. Advances in parasitoly. London, New York: Academic Press, 1988.
DAY, N. P. J.; HIEN, T. T.; SCHOOLLAARDT, T.; LOC, P. P.; CHUONG, L. V.; CHAU, T. T. H.; MAI, N. T. H.; PHU, N. H.; SINH, D. X.; WHITE, N. J.; HO, M. The prognostic and pathophysiologic role of pro- and anti-inflammatory cytokines in severe malária. The Journal of Infectious Diseases. v. 180, p. 1288-1297, 1999.
DE SOUZA, J. B.; RILEY, E. M. Cerebral malária: the contribution of studies in animal models to our understanding of immunopathogenesis. Microbes Infect. v. 4, n. 3, p. 291-300, 2002. DESSENS, J. T.; MENDONZA, J.; CLAUDIANOS, C.; VINETZ, J. M.; KHATER, E.; HASSARD, S.; RANAWAKA, G. R.; SINDEN, R. E. Knockout of the rodent malária parasite chitinase PbCHT1 reduces infectivity to mosquitoes. Infection and Immunity. v. 69, n. 6, p. 4041-4047, 2001.
DICK, S.; WATERFALL, M.; CURRIE, J.; MADDY, A.; RILEY, E. Naive human alpha beta T cells respond to membrane-associated components of malária-infected erythrocytes by proliferation and production of interferon-gamma. Immunology. v. 88, n. 3, p. 412-420, 1996.
DOOLAN, D. L.; HOFFMAN, S. L. IL-2 and NK cells are required for antigen-specific adaptive immunity against malária initiated by CD8+ T cells in the Plasmodium yoelli model. The Journal of Immunology. v. 163, p. 884-892, 1999.
D’OMBRAIN, M. C.; HANSEN, D. S.; SIMPSON, K. M.; SCHOFIELD, L. Gammadelta-T cells expressing NK receptors predominate over NK cells and conventional T cells in the innate IFN-gamma response to Plasmodium falciparum malária. Eur J Immunol. v. 37, p. 1864-1873, 2007.
ELIAS, R. M.; CORREA-COSTA, M.; BARRETO, C. R.; SILVA, R. C.; HAYASHIDA, C. Y.; CASTOLDI, A.; GONÇALVES, G. M.; BRAGA, T. T.; BARBOZA, R.; RIOS, F. J.; KELLER, A. C.; CENEDEZE, M. A.; HYANE, M. I.; D'IMPÉRIO-LIMA, M. R.; FIGUEIREDO-NETO, A. M.; REIS, M. A.; MARINHO, C. R.; PACHECO-SILVA, A.; CÂMARA, N. O. Oxidative stress and modification of renal vascular permeability are associated with acute kidney injury during P. berghei ANKA infection. PLoS One. v. 7, n. 8, 2012.
FAVRE, N. RYFFEL, B.; BORDMANN, G.; RUDIN, W. The course of Plasmodium chabaudi chadaudi infections in interferon-γ receptor deficient mice. Parasite Immunol. v. 19, n. 375, 1997.
FEACHEM, R. G.A.; PHILLIPS, A. A.; HWANG, J.; COTTER, C.; WIELGOSZ, B.; GREENWOOD, B. M.; SABOT, O. RODRIGUEZ, M. H.; ABEYASINGHE, R. R.; GHEBREYESUS, T. A.; SNOW, R. W. Shrinking the malária map: progress and prospects. The Lancet. v. 376, p. 1566-1578, 2010.
FERNANDES, E. S.; LIANG, L.; SMILLIE, S-J.; KAISER, F.; PURCELL, R.; RIVETT, D. W.; ALAM, S.; HOWAT, S.; COLLINS, H.; THOMPSON, S. J.; KEEBLE, J. E.; RIFFO-VASQUEZ, Y.; BRUCE, K. D.; BRAIN, S. D. TRPV1 deletion enhances local inflammation and accelerates the onset of systemic inflammatory response syndrome. The Journal of Immunology. v. 188, p. 5741-5751, 2012.

87
FERNANDES, E. S.; FERNANDES, M. A.; KEEBLE, J. E. The functions of TRPA1 and TRPV1: moving away from sensory nerves. British Journal of Pharmacology. v. 166, n. 2, p. 510-521, 2012.
FERNANDES, E. S.; VONG, C. T.; QUEK, S.; CHEONG, J.; AWAL, S.; GENTRY, C.; AUBDOOL, A. A.; LIANG, L.; BODKIN, J. V.; BEVAN, S.; HEADS, R.; BRAIN, S. D. Superoxide generation and leukocyte accumulation: key elements in the mediation of leukotriene B4-induced itch by transient receptor potential ankyrin 1 and transient receptor potential vanilloid 1. The FASEB Journal. v. 27, n. 4, p. 1664-1673, 2013. FISCHER, M. A.; DAVIES, M. L.; REIDER, I. E.; HEIPERTZ, E. L.; EPLER, M. R.; SEI, J. J.; INGERSOLL, M. A.; ROOIJEN, N. V.; RANDOLPH, G. J.; NORBURY, C. C. CD11b+, Ly6G+ cells produce type I interferon and exhibit tissue protective properties following peripheral virus infection. Plos Pathgens. v. 7, n. 11, p. 1-13, 2011.
FLEMING, T. J.; FLEMING, M. L.; MALEK, T. R. Selective expression of Ly- 6G on myeloid lineage cells in mouse bone marrow. RB6-8C5 mAb to granulocytedifferentiation antigen (Gr-1) detects members of the Ly-6 family. J. Immunol. v. 151, p. 2399–2408, 1993. FORATTINI, O. P. Culicidologia Médica. v. 2. São Paulo: Editora da Universidade de São Paulo, 2002.
FOSTER, D.; COX-SINGH, J.; MOHAMAD, D. S. A.; KRISHNA, S.; CHIN, P. P.; SINGH, B. Evaluation of three rapid diagnostic tests for the detection of human infections with Plasmodium knowlesi. Malária Journal. v. 13, n. 60, 2014.
FRANÇA, T. C. C.; SANTOS, M. G.; FIGUEROA-VILLAR, J. D. Malária: aspectos históricos e quimioterapia. Quim, Nova. v. 31, n. 5, p. 1271-1278, 2008.
FREITAS DO ROSARIO, A. P.; MUXEL, S.M.; RODRIGUEZ-MALAGA, S. M.; SARDINHA, L. R.; ZAGO, C. A.; CASTILLO-MENDEZ, S. L.; ALVAREZ, J. M.; D’IMPERIO LIMA, M. R. Gradual decline in malária-specific memory T cell responses leads to failure to maintain long-term protective immunity to Plasmodium chabaudi A S despite persistence of B cell memory and circulating antibody. J Immunol. v. 181, n. 12, p. 8344-8355, 2008.
FRIED, M.; MUGA, R. O.; MISORE, A. O.; DUFFY, P. E. Malária elicits type 1 cytokines in the human placenta: IFN-gamma and TNF-alpha associated with pregnancy outcomes. J Immunol. v. 160, p. 2523-2530, 1998.
FRITA, R.; REBELO, M.; PAMPLONA, A.;1 VIGARIO, A. M; MOTA, M. M.; GROBUSCH, M. P.; HÄNSCHEID, T. Simple flow cytometric detection of haemozoin containing leukocytes and erythrocytes for research on diagnosis, immunology and drug sensitivity testing. Malaria Journal. v. 10, p. 74, 2011.
GALINSKI, M. R.; BARNWELL, J. W. Plasmodium vivax: who cares? Malária Journal. v. 7, suppl 1: S9, 2008.
GARCIA, C. R.; MARKUS, R. P.; MADEIRA, L. Tertian and quartan fevers: temporal regulation in malária infection. J Biol Rhythms. v. 16, p. 436-443, 2001.
GARNHAN, P. C. C. Malária parasites and other haemosporidia. Oxford: Blackwell

88
Scientific Publications. 1996.
GEPPETTI, P.; MATERAZZI, S.; NICOLETTI, P. The transient receptor potential vanilloid 1: Role in airway inflammation and disease. European Journal of Pharmacology. v. 533, p. 207-214, 2006.
GIMENEZ, F.; BARRAUD de LAGERIE, S.; FERNANDEZ, C.; PINO, P.; MAZIER, D. Tumor necrosis factor alpha in the pathogenesis of cerebral malária. Cell Mol Life Sci. v. 60, p. 1623-1635, 2003.
GILLES, H; WARRELL, D.; BRUCE-CHWATT. D. Essential Malariology. 3 ed. London: Edward Arnold publishers, p. 12-34, 1998.
GOEL, S.; GOWDA, D. C. How specific is Plasmodium falciparum adherence to chondroitin 4-sulfate? Trends Parasitol. v. 27, p. 375-381, 2011.
GOETZ, F. W.; PLANASB, J. V.; MACKENZIE, S. Tumor necrosis factors. Develop Compar Immunology. v. 28, p. 487-497, 2004.
GOMIS, A.; SORIANO, S.; BELMONTE, C.; VIANA, F. Hypoosmotic- and pressure-induced membrane stretch activate TRPC5 channels. The Journal of Physiology. v. 586, n. 23, p. 5633-5649, 2008.
GONÇALVES, R. M.; SALMAZI, K.; SANTOS, B. A. N.; BASTOS, M. S.; ROCHA, S. C.; BOSCARDIN, S. B.; SILBER, A. M.; KALLAS, E. G.; FERREIRA, M. U.; SCOPLER, KEZIS G. S. CD4+ CD25+ Foxp3+ Regulatory T Cells, Dendritic Cells, and Circulating Cytokines in Uncomplicated Malaria: Do Different Parasite Species Elicit Similar Host Responses? Infection and Immunity. v. 78, n. 11, p. 4763-4772.
GORBACHEV, A. V.; FAIRCHILD, R. L. Regulatory role of CD4+ T cells during the development of contact hypersensitivity responses. Immunol Res. v. 24, p. 69-77, 2001.
GRAU, G. E.; FAJARDO, L. F.; PIGUET, P. F.; ALLET, B.; LAMBERT, P. H.; VASSALLI, P. Tumor necrosis factor (cachectin) as an essential mediator in murine cerebral malária. Science. v. 237, p. 1220-1212, 1987. GRAU, G. E.; KOSSODO, S. Cerebral malária: mediators, mechanical obstruction or more? Parasitol Today. v. 10, 408-409, 1994.
GREENWOOD, B. M.; FIDOCK, D. A.; KYLE, D. E.; KAPPE, S.H.; ALONSO, P. L.; COLLINS, F. H.; DUFFY, P. E. Malária: progress, perils, and prosprects for eradications. J. Clin. Invest. v, 118, p. 1266-1276, 2008.
GRIFFITHS, M. J.; NDUNGU, F.; BAIRD, K. L.; MULLER, D. P.; MARSH, K.; NEWTON, C. R. Oxidative stress and erythrocyte damage in Kenyan children with severe Plasmodium falciparum malária. Br J Haematol. v. 113, n. 2, p. 486-491, 2001.
GUMLEY, T. P.; MCKENZIE, I. F.; SANDRIN, M. S. Tissue expression, structure and function of the murine Ly-6 family of molecules. Immunol Cell Biol. v. 73, p. 277-296, 1995.
GUPTILL, V.; CUI, X.; KHAIBULLINA, A.; KELLER, J. M.; SPORNICK, N.; MANNES, A.; IADAROLA, M.; QUEZADO, Z. M. Disruption of the transient receptor potential

89
vanilloid 1 can affect survival, bacterial clearance, and cytokine gene expression during murine sepsis. Anesthesiology. v. 114, n. 5, p. 1190-1199, 2011.
HAFALLA, J. C.; SILVIE, O. MATUSCHEWSKI, K. Cell biology and immunology of malária. Immunol Rev. v. 240, n. 1, p. 297-316, 2011.
HANSEN, D. S.; SIOMOS, M.; DE KONING-WART, T.; BUCKINGHAM, L.; CRABB, B. S.; SCHOFIELD, L. CD1d-restrict NKT cells contribute to malárial splenomegaly and enhance parasite-specific antibody responses. Eur J Immunol. v. 33, p. 2588-2598, 2003.
HEDINI, A. Malaria pathogenesis: a jigsaw with an increasing number of pieces. Int J Parasitol. v. 32, n. 13, p. 1587-1598, 2002.
HISAEDA, H. MAEKAWA, Y.; IWAKAWA, D.; OKADA, H.; HIMENO, K.; KISHIHARA, K., SHIN-ICHI K.; K.; TSUKUMO, S.; YASUTOMO, K. Escape of malaria parasites from host immunity requires CD4+CD25+ regulatory T cells. Nat Med. v. 10, p. 29-30, 2004.
HUANG, S. M.; BISOGNO, T.; TREVISANI, M.; AL-HAYANI, A.; DE PETROCELLIS, L.; FEZZA, F.; TOGNETTO, M.; PETROS, T. J.; KREY, J. F.; CHU, C. J.; MILLER, J. D.; DAVIES, S. N.; GEPPETTI, P.;WALKER, J. M.; DI MARZO, V. An endogenous capsaicin-like substance with high potency at recombinant and native vanilloid VR1 receptors. Proc. Natl. Acad. Sci. U. S. A.v. 99, p. 8400-8405, 2002.
HVIID, L.; MARINHO, C. R.; STAALSOE, T.; PENHA-GONÇALVES, C. Of mice and women: rodent models of placental malária. Trends Parasitol. v. 26, n. 8, p. 412-419, 2010.
HWANG, S. W.; CHO, H.; KWAK, J.; LEE, S. Y.; KANG, C. J.; JUNG, J.; CHO, S.; MIN, K. H.; SUH, Y. G.; KIM, D.; OH, U. Direct activation of capsaicin receptors by products of lipoxygenases: endogenous capsaicin-like substances. Proc. Natl. Acad. Sci. U.S.A. v. 97, p. 6155-6160, 2000.
IIDA, T., SHIMIZU, I., NEALEN, M. L., CAMPBELL, A., CATERINA, M. Attenuated fever response in mice lacking TRPV1. Neurosci. Lett. v. 378, p. 28-33, 2005.
ING, R.; STEVENSON, M. M. Dendritic cell and NK reciprocal cross talk promotes gamma interferon-dependent immunity to blood-stage Plasmodium chabaudi AS infection in mice. Infect Immun. v. 77, n. 2, p. 770-782, 2009.
JACOBS, P.; RADZIOCH, D.; STEVENSON, M. M. A Th1-associated increase in tumor necrosis factor α expression in the spleen correlates with resistance to blood-stage malária in mice. Infect. Immun. v. 64, p. 535, 1996.
KARK, T.; BAGI, Z.; LIZANECZ, E.; PÁSZTOR, E. T.; ERDEI, N.; CZIKORA, A.; PAPP, Z.; EDES, I. PORSZASZ, R.; TOTH, A. Tissue-specific regulation of microvascular diameter: opposite functional roles of neuronal and smooth muscle located vanilloid receptor-1. Mol Pharmacol. v. 73, p. 1405–1412, 2008.
KARUNAWEERA, N. D.; GRAU, G. E.; GAMAGE, P.; CARTER, R.; MENDIS, K. N. Dynamics of fever and serum levels of tumor necrosis factor are closely associated during clinical paroxysms in Plasmodium vivax malária. Proc Natl Acad Sci. v. 89, n. 8, p. 3200-3203, 1992.

90
KEEBLE, J. E.; BRAIN, S. D. Capsaicin-induced vasoconstriction in the mouse knee joint: a study using TRPV1 knockout mice. Neurosci Lett. v. 401, p. 55–58, 2006.
KEEBLE, J. E.; BODKIN, J. V.; LIANG, L.; WODARSKI, R.; DAVIES, M.; FERNANDES, E. S.; COELHO, C. F.; RUSSELL, F.; GRAEPEL, R.; MUSCARA, M. N.; MALCANGIO, M.; BRAIN, S. D. Hydrogen peroxide is a novel mediator of inflammatory hyperalgesia, acting via transient receptor potential vanilloid 1-dependent and independent mechanisms. Pain. v. 141, p. 135-142, 2009.
KIM, C. C.; PARIKH, S.; SUN, J. C.; MYRICK, A.; LANIER, L. L.; ROSENTHAL, P. J.; DeRISI, J. L. Experimental malária infection triggers early expansion of natural killer cells. Infection and Immunity. v. 76, n. 12, p. 5873-5882, 2008.
KREMSNER, P. G.; WINKLER, X.; WILDING, E.; PRADA, J.; BIENZLE, U.; GRANINGER, W.; NUSSLER, A. High plasma levels of nitrogen oxides are associated with severe disease and correlate with rapid parasitological and clinical cure in Plasmodium
falciparum malária. Trans R Soc Trop Med Hyg. v. 90, p. 44, 1996.
LAMB, T. J.; BROWN, D. E.; POTOCNIK, A. J.; LANGHORNE, J. Insights into the immunopathogenesis of malária using mouse models. Expert Rev. Mol. Med. v.8, n.6, p. 1-22, 2006. LANDAU, I.; BOULARD, Y. Life cycles and Morphology. in: KILLICK-KENDRICK, R. e PETERS, W. Rodent Malária. London: Academic Press, p. 53-84, 1978. LANGHORNE, J.; GILLARD, S.; SIMON, B.; SLADE, S.; EICHMANN, K. Frequencies of CD4+ T cells reactive with Plasmodium chabaudi: distinct response kinectics for cells with Th1 and Th2 characteristics during infection. Int Immunol. v. 1, n. 4, p. 416-424, 1989.
LEFFLER, A.; MONTER, B.; KOLTZENBURG, M. The role of the capsaicin receptor TRPV1 and acid-sensing ion channel (ASICS) in proton sensitivity of subpopulations of primary nociceptive neurons in rates and mices. Neuroscience. v. 139, p. 699-709, 2006.
LeGUERN, C.; AKIYAMA, Y.; GERMANA, S.; TANAKA, K.; FERNANDEZ, L.; IWAMOTO, Y.; HOUSER, S.; BENICHOU, G. Intracellular MHC class II controls regulatory tolerance to allogeneic transplants. J Immunol. v. 184, n. 5, p. 2394-2400, 2010.
LEORATTI, F. M. S. Resposta immune humoral na malária humana: quantidade e qualidade de anticorpos anti-Plasmodium falciparum. Tese de Mestrado. Universidade de São Paulo. 2004.
LIDDLE, R. A. The role of Transient Receptor Potential Vanilloid 1 (TRPV1) channels in pancreatitis. Biochim Biophys Acta. v. 1772, p. 869–878, 2007.
LIMONGI, J. E.; COSTA, D. C.; CARVALHO, L. H.; LOPES, I. S.; SILVA, A.A.; FERREIRA, M. S. Plasmodium ovale malária in Brazil: report of an imported case with a prolonged incubation period. Journal of Infection in Developing Countries. v. 8, n. 4, p. 554-557, 2014.
LOIOLA, C. C. P.; SILVA, C. J. M. D.; TAUIL, P. L. Controle da malária no Brasil: 1965-2011. Revista Panamericana de Salud Publica. v. 11, n. 4, p. 235-244, 2002.

91
LOU, J.; LUCAS, R.; GRAU, G. E. Pathogenesis of cerebral malária: recent experimental data and possible applications for humans. Clin Microbiol. v. 14, p. 810-820, 2001.
MA, W.; QUIRION, R. Inflammatory mediators modulating the transient receptor potential vanilloid 1 receptor: therapeutic targets to treat inflammatory and neuropathic pain. Expert Opin Ther Targets. v. 11, p. 307–320, 2007. MACKEY, L.J.; HOCHMANN, A.; JUNE, C.H.; CONTRERAS, C.E.; LAMBERT, P.H. Immunopathological aspects of Plasmodium berghei infection in five strains of mice. Clinical Experimental Immunology. v.42, p.412-420, 1980.
MAGGI, C. A.; PATACCHINI, R.; TRAMONTANA, M.; AMANN, R.; GIULIANI, S.; SANTICIOLI, P. Similarities and differences in the action of resiniferatoxin and capsaicin on central and peripheral endings of primary sensory neurons. Neuroscience. v. 37, p. 531-539, 1990.
MARINHO, C. R.; EPIPHANIO, S.; GONÇALVES, L. A.; CATARINO, M. B.; PENHA-GONÇALVES, C. Recrudescent Plasmodium berghei from pregnant mice displays enhanced binding to the placenta and induces protection in multigravida. Plos One. 2009.
MARQUES, A. C.; PINHEIRO, E. A. Revista Brasileira de Malariologia e Doenças Tropicais. v. 34, p. 1, 1982.
MARQUES, R. G.; PETROIANU, A. Infecção fulminante pós-esplenectomia. Arq Gastroenterol. v. 40, n. 1, 2003.
MARSH, K.; ENGLISH, M.; CRAWLEY, J.; PESHU, N. The pathogenesis of severe malária in African children. Ann. Trop. Med. Parasitol. v. 90, p. 395-402, 1996. MAY, J.; MEYER, C. G. Chemoresistance in falciparum malária. Trends in Parasitology. v. 19, n. 10, p. 432-435, 2003. McLACHLAN, J. B.; CATRON, D. M.; MOON, J. J.; JENKINS, M. K. Dendritic cell antigen presentation drives simultaneous cytokine production by effector and regulatory T cells in inflamed skin. Immunity. v. 30, p. 277-288, 2009.
MEDANA, I. M.; HUNT, N. H.; CHAUDHRI, G. Tumor necrosis factor alpha expression in the brain during fatal murine cerebral malária: evidence for production by microglia and astrocytes. Am J Pathol. v. 150, p. 1473-1486, 1997.
MEGNEKOU, R.; HVIID, L.; STAALSOE, T. Variant-specific immunity to Plasmodium
berghei in pregnant mice. Infect Immun. v. 77, p. 1827-1834, 2009.
MENDIS, K.; SINA, B. J.; MARCHESINI, P.; CARTER, R. The Negleted Burden of Plasmodium vivax malária. American Journal of Tropical Medicine and Hygiene. v. 64, p. 97-106, 2001.
MINIGO, G.; WOODBERRY, T.; PIERA, K. A.; SALWATI, E.; TJITRA, E.; KENANGALEM, E.; PRINCE, R. N.; ENGWERDA, C. R.; ANSTEY, N. M.; PLEBANSKI, M. Parasite-dependent expansion of TNF receptor II-positive regulatory T cells with enhanced suppressive activity in adults with severe malária. Plos Pathogens. v. 5, n. 4, 2009.

92
MOHAN, K.; MOULIN, P.; STEVENSON, M. M. Natural killer cell cytokine production, not cytotoxicity, contributes to resistance against blood-stage Plasmodium chabaudi AS infection. J Immunol. v. 159, p. 4990-4498, 1997.
MORIYAMA, T.; HIGASHI, T.; TOGASHI, K.; IIDA, T.; SEGI, E.; SUGIMOTO, Y.; TOMINAGA, T.; NARUMIYA, S.; TOMINAGA, M. Sensitization of TRPV1 by EP1 and IP reveals peripheral nociceptive mechanism of prostaglandins. Mol. Pain. v. 1, p. 1-13, 2005.
NADJIM, M.; BEHRENS, R. H. Malária: Na Update for Physicians. Infect Dis Clin N Am. v. 26, p. 243-259, 2012.
NAOTUNNE, T. S.; KARUNAWEERA, N. D.; DEL GIUDICE, G.; KULARATNE, M. U.; GRAU, G. E.; CARTER, R.; MENDIS, K. N. Cytokines kill malária parasites during infections crisis: extracellular complementary factors are assential. J Exp Med. v. 173, n. 3, p. 523-529, 1991.
NARDIN, E. H.; NUSSENZWEIG, R. S.; MCGREGOR, I. A.; BRYAN, J. H. Antibodies to sporozoites: their frequent occurrence in individuals living in an area of hyperendemic malária. Science. v. 206, p. 597-599, 1979.
NAYLOR, J.; LI, J.; MILLIGAN, C. J.; ZENG, F.; SUKUMAR, P.; HOU, B.; SEDO, A.; YULDASHEVA, N.; MAJEED, Y.; BERI, D.; JIANG, S.; SEYMOUR, V. A.; MCKEOWN, L.; KUMAR, B; HARTENECK, C.; O’REGAN, D.; WHEATCROFT, S. B.; KEARNEY, M. T.; JONES, C.; PORTER, K. E.; BEECH, D. J. Pregnenolone sulphate- and cholesterol-regulated TRPM3 channels coupled to vascular smooth muscle secretion and contraction. Circ Res. v. 106, p. 1507–1515, 2010.
NEBL, T.; DE VEER, M. J.; SCHOFIELD, L. Stimulation of innate responses by malárial glycosylphosphatidylinositol via pattern recognition receptors. Parasitology. v. 130, S45-S62, 2005.
NERES, R.; MARINHO, C. R.; GONCALVES, L. A.; CATARINO, M. B.; PENHA-GONCALVES, C. Pregnancy outcome and placenta pathology in Plasmodium berghei ANKA infected mice reproduce the pathogenesis of severe malária in pregnant women. PLoS One,. 2008.
NIHEI, J. S. Participação das células T de memória e linfócitos CD4+CD25+Foxp3+ (reguladores) nos mecanismos de resistência à infecção experimental pelo Trypanosoma
cruzi. Tese de Doutorado. Universidade Federal da Bahia, 2013.
NITCHEU, J. BONDUELLE, O.; COMBADIERE, C.; TEFIT, M.; SEILHEAN, D.; MAZIER, D.; COMBADIERE, B. Perforin-dependent brain-infiltrating cytotoxic CD8+ T lymphocytes mediate experimental cerebral malária pathogenesis. The Journal of Immunology. v. 170, n. 4, p. 2221-2228, 2003.
OLAH, Z.; KARAI, L.; IADAROLA, M. J. Anandamide activates vanilloid receptor 1 (VR1) at acidic pH in dorsal root ganglia neurons and cells ectopically expressing VR1. J. Biol. Chem. v. 276, p. 31163-31170, 2001. OLIVEIRA-FERREIRA, J.; LACERDA, M. V. G.; BRASIL, P.; LADISLAU, J. L. B.; TAUIL, P. L.; DANIEL-RIBEIRO, C. T. Malária in Brazil: na overview. Malária Journal. v. 9, p. 115, 2010.

93
OLIVEIRA-FILHO, A. B.; MARTINELLI, J. M. Casos notificados de malária no Estado do Pará, Amazônia Brasileira, de 1998 a 2006. Epidemiol. Serv. Saúde. Brasília, v. 18, n. 2, p. 277-284, jul-set, 2009.
ORGANIZAÇÃO MUNDIAL DE SAÚDE. World malária report: 2010. Suíça, 2010.
ORGANIZAÇÃO MUNDIAL DE SAÚDE. World malária report: 2012. Suíça, 2012.
PARK, H. S., JUNG, H. Y., PARK, E. Y., KIM, J., LEE, W. J., BAE, Y. S. Direct interaction of TLR4 with NAD(P)H oxidase 4 isozyme is essencial for lipopolysaccharide-induced production of reactive oxygen species and activation of NF-κB. The Journal of Immunology. v. 173, n. 6, p. 3589-3593, 2004. PEDERSEN, S. F.; OWSIANIK, G.; NILIUS, B. TRP channels: an overview. Cell Calcium. v. 38, p. 233-252, 2005. PEREIRA, K. R. B. Estudo dos mecanismos imunológicos envolvidos na recrudescência da malária experimental durante a gravidez. Tese de Mestrado. Universidade de São Paulo, 2012. POBLETE, I. M.; ORLIAC, M. L.; BRIONES, R.; ADLER-GRASCHINSKY, E.;
HUIDOBRO-TORO, J. P. Anandamide elicits an acute release of nitric oxide through endothelial TRPV1 receptor activation in the rat arterial mesenteric bed. The Journal of Physiology. v. 568, v. 2, p. 539-551, 2005.
POOVASSERY, J.; MOORE, J. M. Association of malária-induced murine pregnancy failure with robust peripheral and placental cytokine responses. Infect Immu. v. 77, p. 4998-5006, 2009. QIU, J.; FANG, Y.; RONNEKLEIV, O. K.; KELLY, M. J. Leptin excites proopiomelanocortin neurons via activation of TRPC channels. J Neurosci. v. 30, n. 4, p. 1560-1565, 2010.
QUEIROZ, N. L. Avaliação da resposta inflamatória cerebral em camundongos BALB/c e C57BL/6 infectadoso por Plasmodium berghei cepa NK65. Tese de Mestrado. Universidade Federal de Minas Gerais. 2007.
RALEVIC, V.; KENDALL, D. A.; RANDALL, M. D.; SMART, D. Cannabinoid modulation of sensory neurotransmission via cannabinoid and vanilloid receptors: roles in regulation of cardiovascular function. Life Sci. v. 71, p. 2577-2594, 2002.
RANDALL, L. M.; ENGWERDA, C.R. TNF family members and malária: old observations, new insights and future directions. Exp Parasitol. v. 126, p. 326-331, 2010.
RANDOLPH, G. J.; JAKUBZICH, C.; QU, C. Antigen presentation by monocytes and monocyte-derived cells. Curr Opin Immunol. v. 20, n. 1, p. 52-60, 2008.
RANG, H. P., BEVAN, S., DRAY, A. Chemical activation of nociceptive peripheral neurones. Br. Med. Bull. v. 47, p. 534-548, 1991. RAPOSO, C. C. B. S. Malária vivax na pré Amazônia maranhense: aspectos epidemiológicos e características clínicas. Tese de Mestrato. Universidade de Brasília, 2006.

94
RAY, R.; SHAH, A. M. NADPH oxidase and endothelial cell function. Clinical Science. v. 109, p. 217–226, 2005.
RIBEIRO, E.; OLIVEIRA, E.; MARCO, M.; XAVIER, B. Malária grave secundária a co-infecção por Plasmodium falciparum e Plasmodium ovale. Galicia Clin. v. 74, p. 25-28, 2013.
RICHIE, T. L.; SAUL, A. Progress and challenges for malária vaccines. Nature. v. 415, p. 664-70, 2002.
RIDLEY, R. G. Chemotherapeutic hope on the horizon for Plasmodium vivax malária. Proceedings of National Academy of Science. v. 99, n. 21, p. 13362-13364, 2002.
RODRIGUES-DUARTE, L.; MORAES, L. V.; BARBOZA, R.; MARINHO, C. R. F.; FRANKE-FAYARD, B.; JANSE, C. J.; PENHA-GONÇALVES, C. Distinct placental malária pathology caused by different Plasmodium berghei lines that fail to induce cerebral malária in the C57BL/6 mouse. Malária Journal. v. 11, p. 231, 2012.
ROMANOVSKY, A.A. Signaling the brain in the early sickness syndrome: are sensory nerves involved. Front. Biosci. v. 9, n. 494–504, 2004.
ROUSSEAU, E.; CLOUTIER, M.; MORIN, C.; PROTEAU, S; Capsazepine, a vanilloid antagonist, abolishes tonic responses induced by 20-HETE on guinea pig airway smooth muscle. Am. J. Physiol. v. 288, p. L460-L470, 2005.
SACHS, J.; MALANEY, P. The economic and social burden of malária. Nature. v. 415, p. 680-685, 2002.
SANNI, L. A, FU. S.;, DEAN, R. T.; BLOOMFIELD, G.; STOCKER, R.; CHAUDHRI, G.; DINAUER, M. C.; HUNT, N. H. Are reactive oxygen species involved in the pathogenesis of murine cerebral malaria? J Infect. Dis. v. 179, p. 217-222, 1999.
SCHOFIELD, L. Intravascular infiltrates and organ-specific inflammation in malária pathogenesis. Immunology and Cell Biology. v. 85, p.130-137, 2007.
SCHOFIELD, L.; GRAU, G. E. Immunological processes in malária pathogenesis. Nature Reviews. v. 5, p. 722-735, sept, 2005.
SCHWARTZ, E. S.; LA, J.; SCHEFF, N. N.; DAVIS, B. M.; ALBERS, K. M.; GEBHART, G. F. TRPV1 and TRPA1 antagonists prevent the transition of acute to chronic inflammation and pain in chronic pancreatitis. J Neurosci. v. 33, n. 13, p. 5603-5611, 2013.
SCHWARZ, E. C.; WOLFS, M. J.; TONNER, S.; WENNING, A. S.; QUINTANA, A.; GRIESEMER, D.; HOTH, M. TRP channels in lymphocytes. Handb Exp Pharmacol. v. 179, p. 445-456, 2007.
SCOTLAND, R. S., CHAUHAN, S., DAVIS, C., DE FELIPE, C., HUNT, S., KABIR, J., KOTSONIS, P., OH, U., AHLUWALIA, A. Vanilloid receptor TRPV1, sensory C-fibers, and vascular autoregulation: a novel mechanism involved in myogenic constriction. Circ. Res.
v.95, p. 1027-1034, 2004.

95
SCRAGG, I. G.; HENSMANN, M.; BATE, C. A.; KWIATKOWSKI, D. Early cytokine induction by Plasmodium falciparum is not a classical endotoxin-like process. Eur J Immunol. v. 29, p. 2636-2644, 1999.
SEMTNER, M.; SCHAEFER, M.; PINKENBURG, O.; PLANT, T. D. Potentiation of TRPC5 by protons. J Biol Chem. v. 282, p. 33868–33878, 2007.
SHARMA, L. S. KAUR, J.; SHUKLA, G. Role of oxidative stress and apoptosis in the placental pathology of Plasmodium berghei infected mice. Plos One. v. 7, n. 3, 2012.
SHARMA, M. C.; SHARMA, S.; SHARMA, P.; KUMAR, A. Pharmacophore and QSAR modeling of some structurally diverse azaaurones derivatives as anti-malárial activity. Med Chem Res. 2013.
SHIM, W.; TAK, M.; LEE, M.; KIM, M.; KIM, M.; KOO, J.; LEE, C.; KIM, M.; OH, U. TRPV1 Mediates Histamine-Induced Itching via the Activation of Phospholipase A2 and 12-Lipoxygenase. The Journal of Neuroscience. v. 27, n. 9, p. 2331-2337, 2007.
SILVEIRA, A.C.; REZENDE, D. F. Avaliação da estratégia global de controle integrado da malária no Brasil. Brasília: Organização Pan-Americana de Saúde, 2001.
SOULARD, V.; ROLAND, J.; GORGETTE, O.; BARBIER, E.; CAZENAVE, P. A.; PIED, S. An early burst of IFN-gamma induced by the pre-erythrocytic stage favours Plasmodium
yoelli parasitemia in B6 mice. Malária Journal. v. 8, n. 128, 2009.
STAALSOE, T.; SHULMAN, C. E.; BULMER, J. N.; KAWUONDO, K.; MARSH, K.; HVIID, L. Variant surface antigen-specific IgG and protection against clinical consequences of pregnancy-associated Plasmodium falciparum malária. The Lancet. v. 363, p. 283-289, 2004.
STEVENSON, M. M.; TAM, M. F.; WOLF, S. F.; SHER, A. IL-12-induced protection against blood-stage Plasmodium chabaudi AS requires IFN-γ and TNF-α and occurs via a nitric oxide-dependent mechanism. J Immunol. v. 155, p. 2545, 1995.
STEVENSON, M. M.; RILEY, E. M. Innate immunity to malária. Nature Review Immunology. v. 4, p. 169-180, 2004.
STOOR, M. TRPV1 in colitis: is it a good or a bad receptor? – a viewpoint. Neurogastroenterology & Motility. v. 19, n. 8, p. 625-629.
TADEI, W. P. & THATCHER, B. D. Malária vectors in the Brazilian Amazon: Anopheles of the subgenus Nyssorhynchus. Revista do Instituto de Medicina Tropical de São Paulo. v. 42, p.87-94, 2000.
TAKANO, F.; YAMAGUCHI, M.; TAKADA, S.; SHODA, S.; YAHAGI, N.; TAKAHASHI, T.; OHTA, T. Capsicum ethanol extracts and capsaicin enhance interleukin-2 and interferon-gamma production in cultured murine Peyer’s patch cells ex vivo. v. 80, n. 17, p. 1553-1563, 2007.
TANG H. B.; INOUE A.; OSHITA K.; NAKATA Y. Sensitization of vanilloid receptor 1 induced by bradykinin via the activation of second messenger signaling cascades in rat primary afferent neurons. Eur. J. Pharmacol. v. 498, p. 37-43, 2004.

96
TAUIL, P. L. Resumos do XVII Congresso da Sociedade Brasileira de Medicina Tropical. Ribeirão Preto, 1982.
TAYLOR, P.; BROWN, G.; GELDHOF, A.; MARTINEZ-POMARES, L.; GORDON, S. Pattern recognition receptors and differentiation antigens define murine myeloid cell heterogeneity ex vivo. Eur Journal Immunology. v. 33, n. 8, p. 2090-2097, 2003.
TAYLOR-ROBINSON, A. W.; PHILLIPS, R. S.; SEVEM, A.; MONCADA, S.; LYEW, F. Y. The role of Th1 and Th2 cells in a rodent malária infection. Science. v. 260, p. 1931-1934, 1993.
TEIXEIRA, J. R. M. Resposta terapêutica da malária por Plasmodium vivax a cloroquina e a primaquina. Tese (Doutorado) – Universidade Federal do Pará, Instituto de Ciências Biológicas, Programa de Pós-Graduação em Biologia de Agentes Infecciosos e Parasitários, Belém, 2011.
TODRYK, S.M.; BEJON, P.; MWANGI, T.; PLEBANSKI, M.; URBAN, B.; MARSH, K.; HILL, A. V. S.; FLANAGAN, K. L. Correlation of memory T cell responses against TRAP with protection from clinical malária, and CD4 CD25 high T cells with susceptibility in Kenyans. PLoS One. n. 3, 2008. UMBERS, A. J.; AITKEN, E. H.; ROGERSON, S. J. Malária in pregnancy: small babies, big problem. Trends Parasitol. v. 27, p. 168–175, 2011. VAN ASSCHE, T.; DESSCHACHT, M. DA LUZ, R. A.; MAES, L.; COS, P. Leishmania-macrophage interactions: insights into the redox biology. Free Radic Biol Med. v. 51, n. 2, p. 337-351, 2011.
VAN DER HEYDE, H. C.; BAUER, P.; SUN, G.; CHANG, W. L.; YIN, L.; FUSELER, J.; GRANGER, D. N. Assessing vascular permeability during experimental cerebral malária by a radiolabeled monoclonal antibody technique. Infect Immun. v. 69, n. 5, p. 3460-3465, 2001. VAN ZON, A. A.; ELING, W. M. Depressed malárial immunity in pregnant mice. Infect Immun. v. 28, p. 830-832, 1980. VAN ZON, A. A.; ELING, W. M.; HERMSEN, C. C. Pregnancy-induced recrudescences strengthen malárial immunity in mice infected with Plasmodium berghei. Parasitol. v. 91, p. 9-17, 1985. VAY, L.; GU, C.; McNAUGHTON, P. A. The thermo-TRP ion channel family: properties and therapeutic implications. British Journal of Pharmacology. v. 165, n. 4, p. 787-801, 2012. WALTHER, M.; TONGREN, J. E.; ANDREWS, L.; KORBEL, D.; KING, E.; FLETCHER, H.; ANDERSEN, R. F.; BEJON, P.; THOMPSON, F.; DUNACHIE, S. J.; EDELE, F.; SOUZA, J. B.; SINDEN, R. E.; GILBERT, S. C.; RILEY, E. M.; HILL, A. V. S. Upregulation of TGF-β, FOXP3 and, CD4⁺ ⁺CD25 Regulatory T cells correlates with mora rapid parasite growth in human malária infection. Immunity. v. 23, p. 287-296, sept, 2005
WANG, H.; WANG, D. H.; GALLIGAN, J. J. Interaction between P2Y receptors and TRPV1 on kidney specific sensory neurons. The FASEB Journal. v.22, p. 937-942, 2008.

97
WANG, Y. Y.; CHANG, R. B.; ALLGOOD, S. D.; SILVER, W. L.; LIMAN, E. R. A TRPA1-dependent mechanism for the pungent sensation of weak acids. J Gen Phisiol. v. 137, n. 6, p. 493-505, 2011.
WARRELL, D. A. Clinical features of malária. Bruce-Chwatt´s Essential Malariology. 3º ed. Londres: Edward Arnold publishers, p. 35-45, 1998.
WEISS, L. Barrier cells in the spleen. Immunology Today. v. 12, p. 24-29, 1991.
WHITE, N. J.; HO, M. The pathophysiology of Malaria. Advances in Parasitology. Academic Press, New York. p. 84-173, 1992.
WHITE, N. J. The treatment of malária. The New England Journal of Medicine. v. 335, p. 800-806, 1996.
WIPASA, J.; SUPHAVILAI, C.; OKELL, L. C.; COOK, J.; CORRAN, P. H.; THAIKLA, K. LIEWSAREE, W.; RILEY, E. M.; HAFALLA, J. C. Long-lived antibody and B Cell memory responses to the human malária parasites, Plasmodium falciparum and Plasmodium vivax. Plos pathogens. 2010.
WRIGHT, D.H. The effect of neonatal thymectomy on the survival of golden hamsters infected with Plasmodium berghei. Bristish Journal of Experimental Pathology. v. 49, p. 379-384, 1968.
WYKES, M. N.; GOOD, M. F. What really happens to dendritic cells during malária? Nature Reviews. v. 6, p. 864-870, nov, 2008.
YIP, H.; CHAN, W. Y.; LEUNG, P. C.; KWAN, H. Y.; LIU, C.; HUANG, Y.; MICHEL, V.; YEW, D. T.; YAO, X. Expression of TRPC homologs in endothelial cells and smooth muscle layers of human arteries. Histochem Cell Biol. v. 122, p. 553-561, 2004.
YOSHIMOTO, T.; TAKAHAMA, Y.; WANG, C.; YONETO, T.; WAKI, S.; NARIUCHI, H. A Pathogenic Role of IL-12 in Blood-Stage Murine Malaria Lethal Strain Plasmodium
berghei NK65 Infection. The Journal of Immunology. v. 160, n. 11, p. 5500-5505, 1998. ZIMMERMANN, K.; LENNERZ, J. K.; HEIN, A.; LINK, A. S.; KACZMAREK, J. S.; DELLING, M.; UYSAL, S.; PFEIFER, J. D.; RICCIO, A.; CLAPHAM, D. E. Transient receptor potential cation channel, subfamily C, member 5 (TRPC5) is a cold-transducer in the peripheral nervous system. PNAS. v. 108, n. 44, p. 18114-18119, 2011.





























