Embed Size (px)
Citation preview

INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA - INPA PROGRAMA DE PÓS-GRADUAÇÃO EM GENÉTICA, CONSERVAÇÃO E BIOLOGIA
EVOLUTIVA – PPG GCBEv
Citogenética clássica e molecular de espécies neotropicais de
serpentes da família Boidae (Gray, 1825)
Patrik Ferreira Viana
Manaus, AM - Fevereiro de 2015

ii
INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA - INPA PROGRAMA DE PÓS-GRADUAÇÃO EM GENÉTICA, CONSERVAÇÃO E BIOLOGIA
EVOLUTIVA – PPG GCBEv
Citogenética clássica e molecular de espécies neotropicais de
serpentes da família Boidae (Gray, 1825)
Patrik Ferreira Viana
Dissertação apresentada ao Programa de Pós Graduação do Instituto Nacional de Pesquisas da Amazônia como parte dos requisitos para obtenção do título de Mestre em Genética, Conservação e Biologia Evolutiva.
Orientadora: Eliana Feldberg, Dra. – INPA
Coorientadora: Maria Claudia Gross, Dra. – UFAM
Manaus, AM - Fevereiro de 2015

iii
FICHA CATALOGRÁFICA
Sinopse
Com intuito de investigar os mecanismos de evolução cariotípica em serpentes da Amazônia, uma caracterização citogenética foi realizada em diferentes espécies de Boidae da região Neotropical. Para isso, foram estudadas três espécies e três subespécies, usando diferentes marcadores citogenéticos clássicos e moleculares, como a detecção das regiões organizadoras de nucléolo e DNAr 18S, detecção das regiões de heterocromatina constitutiva e sequências teloméricas. As três subespécies de Boa constrictor bem como as duas espécies de Eunectes mostraram um cariótipo similar com 2n=36 cromossomos, com diferenças no número de braços cromossômicos. Corallus hortulanus, por sua vez, mostrou uma composição cariotípica totalmente diferenciada para a superfamília Booidea com 2n=40 cromossomos e um possível sistema heteromórfico de determinação sexual. A composição cromossômica dessas serpentes de linhagem basal não é tão conservada quanto se imaginava e futuramente algumas subespécies poderão ser elevadas à categoria de espécie.
Palavras-chave: Booidea, Mapeamento cromossômico, FISH, Cariótipos espécie-específicos.
Viana Patrik Ferreira
Citogenética clássica e molecular de espécies neotropicais de serpentes da família Boidae (Gray,
1825) / Patrik Ferreira Viana. --- Manaus: [s.n.], 2015.
xvi, 67 f. : il. color.
Dissertação (Mestrado) --- INPA, Manaus, 2015. Orientadora: Eliana Feldberg. Coorientadora: Maria Claudia Gross. Área de concentração: Genética, Conservação e Biologia Evolutiva.
1. Booidea. 2. Heterocromatina. 3. Sequências repetitivas. 4. Mapeamento
cromossômico. I. Título.

iv
Dedico aos meus pais, às serpentes
que tiveram que morrer e ao professor
Willy Beçak pela grandiosa contribuição
à citotaxonomia da Herpetofauna
Brasileira.

v
“Se você irá se render por tão pouco, as razões que guiam os seus atos são igualmente insignificantes.”
Asmita de Virgem
“Você pode até encontrar as coisas que perdeu, mas nunca as que abandonou.”
Mithrandir

vi
A realização deste estudo foi possível devido:
Ao Instituto Nacional de Pesquisas da Amazônia (INPA), ao programa de pós-
graduação em Genética, Conservação e Biologia Evolutiva (GCBEV) e aos laboratórios
de Genética Animal (INPA) e de Citogenômica Animal (UFAM).
Aos financiamentos fornecidos pelo Centro de Estudos de Adaptações da Biota
Aquática da Amazônia – ADAPTA (INCT/CNPq/FAPEAM 573976/2008-2); Projetos–
(Pronex/FAPEAM/CNPq 003/2009 - Processo: 653:2009);
MCT/CNPq/MEC/CAPES/FNDCT – Ação Transversal/FAPs Nº 47/2010 - Rede
“Genômica comparativa de peixes amazônicos frente a diferentes desafios ambientais”
e pesquisa para ampliação do conhecimento sobre a biodiversidade de vertebrados da
Amazônia brasileira com aplicações sobre seu uso e conservação – Rede BioPHAM;
Programa Pró-Amazônia: Biodiversidade e Sustentabilidade, Edital Nº 047/2012, Área
Temática Contemplada: Recursos (produtos) naturais – “Estudos citogenéticos e
citogenômicos da biodiversidade da Amazônia, com implementação de avanços
técnicos”. À Fundação de Amparo à Pesquisa do estado do Amazonas (FAPEAM) pela
concessão da bolsa de estudo durante a realização deste estudo.

vii
Agradecimentos
Em especial para minha mãe, que caminhou comigo ao longo dessa jornada. Ela
que foi herpetóloga, mateira, barqueira, cozinheira, psicóloga, diplomata e acima de
tudo mãe.
A todos os colegas de laboratório: Marajozin; Lucas; KK; Ramão; Masseo;
Carlão; Natália; Thatá pela companhia na madrugada; Eduardo; minha prima Milena;
Maelin pela companhia noturna, dicas gerais e troca de experiências; Leilinha que tanto
me ajudou e ensinou nessa caminhada; professor Jorge Porto pelo apoio logístico em
algumas ocasiões; Celeste por adivinhar os momentos em que estava precisando de
uma palavra de apoio e pelas diversas bandejas de salgadinhos; Arlindo pelos
conselhos, conversas e experiência; minhas chefas Eliana Feldberg e Cláudia Gross
por tudo .
Aos colegas da minha divertida turma de pós: Camila; Jéssica; Nayara; Érico
pelas conversas produtivas; Elciomar (Pará) pela vivência em campo e de forma
indireta mostrar como lidar com gente que não presta, é muito gente boa e tenho um
grande apreço; Shizuka e Valdirley pelas risadas espontâneas; Martinha que me
ajudou nos primeiros passos do laboratório; Fabrício (Gaúcho) pelas conversas
divertidas em campo. Em especial Erika, Vanessa e Mayara, foi muito bom ter a
companhia de vocês.
À minha prima Deyla que tanto gosto; Priscila pela alegria de sempre; aos
pescadores Astro e Mario; ao Dick-Vogt pela primeira “permiso” que me concedeu para
realizar minhas pesquisas; Dona Lucia por arrumar minha bagunça o ano inteiro; os
motoristas João Bosco e Orlando que sempre me levaram para minhas coletas e nunca
reclamaram de nada; seu Juruna pelas idas divertidas ao campo; Dona Elci que
sempre me ajudou com a parte burocrática que tive que resolver na secretaria do
GCBEv, a Coleção de Herpeto do INPA.
À prefeitura de Manaus; CETAS; CIGS; Fundação Zoobotânica de Belo
Horizonte; Jibóias Brasil; soldados do CIGS que me ajudaram a conter as anacondas; a

viii
coleção de herpeto do INPA e seus componentes; professor Chalkidis da FIT; professor
Ronis, por me ajudar em momentos iniciais; Laérzio por me passar tanta experiência;
George Myller por me ajudar muito em Belo Horizonte; Camila, Kayan e Ronaldo por
me receberem de portas abertas em sua casa.
À professora Marta Svartman que cedeu seu laboratório com tanta boa vontade;
Mathias, Carla e demais que me ajudaram no laboratório da UFMG; Luiz; Fernanda
Blandy e Herlandes pela ajuda com as serpentes em BH; Raquel por me ajudar com a
papelada no Zoo. “Povo mineiro é bão demais da conta!”
Aos amigos George; Diego; Camila Tekila; Little Sister pela companhia nos
primeiros meses da pós-graduação. Ao mestre Obed por tanto estimular meu senso
crítico; ao meu eterno mestre Rato de Fraga por ensinar quase tudo que sei sobre a
herpetofauna amazônica. A professora Maria José pelas dicas iniciais e a professora
Jeniffer Marshal A. Graves por sempre me mandar os artigos que não tinha como
pagar.
Em especial aos prezados professores Willy Beçak da Universidade de São
Paulo no Brasil, José Maria Gutiérrez do Instituto Clodomiro Picado na Costa Rica e
Ettore Olmo da Università Politecnica delle Marche na Itália, pioneiros na citotaxonomia
da herpetofauna Neotropical e mundial, pelas palavras e cumprimentos de sucesso.
Sei que devo ter esquecido de alguns nomes, mas deixo aqui o meu muitíssimo
obrigado a todos que me ajudaram. E um muitíssimo obrigado às serpentes que foram
material deste.

ix
Resumo
A família Boidae (Gray, 1825) compreende serpentes de uma linhagem basal
que podem ser encontradas nas Américas do Norte, Central e do Sul. Muitas espécies
são divergentes tanto morfologicamente quanto biogeograficamente e mesmo com
muitos estudos evolutivos sendo propostos para algumas espécies ao longo dos
últimos anos, muitos aspectos ainda permanecem desconhecidos. Para uma melhor
compreensão das relações evolutivas dessas serpentes, uma caracterização
citogenética clássica e molecular foi realizada em 44 exemplares pertencentes a três
gêneros, sendo três subespécies de “Jiboias” (Boa contrictor constrictor, Boa contrictor
amarali e Boa contrictor occidentalis), duas espécies de “Anacondas” (Eunectes
murinus e Eunectes notaeus) e uma espécie de “Suaçubóia – Amazon Tree Boa”
(Corallus hortulanus), que além de evidenciar um novo panorama para o entendimento
da evolução cromossômica dos Boideos Neotropicais, revelou três cariótipos inéditos e
uma técnica diferenciada para a obtenção de cromossomos em répteis na Amazônia.
As três subespécies de Jiboias, bem como as duas espécies de Anacondas, mostraram
um cariótipo muito similar com 2n=36 cromossomos, no entanto apresentaram
diferenças de tamanho e no número de braços cromossômicos (NF). As subespécies
de Jibóias apresentaram um NF=48, enquanto as duas espécies de Anacondas
apresentaram um NF=52. A Suaçubóia (Amazon Tree Boa), por sua vez, mostrou uma
composição cariotípica totalmente diferenciada para a superfamília Booidea com 2n=40
♀ com NF=62 e 2n=39 ♂ com NF=60, o que além de revelar um heteromorfismo
cromossômico entre fêmeas e machos, evidenciou uma dupla heterogametia. Os
padrões de distribuição da heterocromatina constitutiva e a localização de sequências
teloméricas, além de revelarem novos insights sobre a evolução cromossômica de
boídeos Neotropicais, mostraram-se como eficientes marcadores espécie/específicos
para os táxons de Boidae analisados. Isso também revelou que a composição
cromossômica dessas serpentes de linhagem basal não é tão conservada quanto se
presumia e que, futuramente, algumas subespécies de “Jibóias” poderão de fato serem
elevadas à categoria específica.

x
Abstract
Boidae (Gray, 1825) family comprehends snakes from a basal lineage, which
may be found in North, Central and South America. Many of these species are both
morphologically and biogeographically divergent, and even with the several evolutionary
studies being proposed for some species along the last years, many aspects remain
unknown. For a better understanding of the evolutionary relationships of these snakes,
a classic and molecular cytogenetic characterization was performed in 44 exemplars
belonging to three genera. It has included three Jiboia subspecies (Boa constrictor
constrictor, Boa constrictor amarali and Boa constrictor occidentalis), two Anaconda
species (Eunectes murinus and Eunectes notaeus) and one species of Amazon Tree
Boa (Corallus hortulanus). The three Jiboia subspecies and the two Anaconda species
presented a very similar karyotype, with 2n=36 chromosomes, with subtle differences in
the number of chromosomes arms. Amazon Tree Boa, however, presented a totally
different karyotype composition for the Booidea superfamily, with 2n=40 chromosomes
♀ and FN=62 and 2n=39 ♂ and FN=60. The constitutive heterochromatin patterns and
the telomeric sequences location, besides revealing new insights on the chromosome
evolution of Neotropical Boidae, were efficient markers for the analyzed taxa. It has also
shown that the chromosome composition of these basal lineage snakes is not as
conserved as it was supposed and that, eventually, some of the subspecies might be
elevated to the species category, as well as the description of new species.

xi
Sumário
1.0 Introdução ................................................................................................................. 1
1.1 Considerações sobre Boa, Corallus e Eunectes .................................................. 3
1.2 Citogenética em Serpentes .................................................................................. 7
1.3 Citogenética molecular em Reptilia..................................................................... 12
2.0 Objetivos ................................................................................................................. 14
2.1 Objetivo geral ..................................................................................................... 14
2.2 Objetivos específicos .......................................................................................... 14
3.0 Material e Métodos.................................................................................................. 15
3.1 Material ............................................................................................................... 15
3.2 Métodos ...............................................................................................................16
3.2.1 Coleta dos Animais .......................................................................................... 16
3.2.2 Obtenção de cromossomos mitóticos.............................................................. 19
3.2.3 Detecção das regiões organizadoras de nucléolo (Ag-RON) .......................... 19
3.2.4 Detecção das regiões de heterocromatina constitutiva (Banda C) ..................20
3.2.5 Extração de DNA ............................................................................................. 20
3.2.6 Isolamento de sequências repetitivas ..............................................................21
3.2.7 Hibridização in situ fluorescente (FISH) .......................................................... 22
3.2.8 Análise cariotípica ........................................................................................... 24
4.0 Resultados .............................................................................................................. 25
4.1 Boa constrictor .................................................................................................... 25
4.2 Eunectes ............................................................................................................. 31
4.3 Corallus hortulanus ............................................................................................. 34
5.0 Discussão ............................................................................................................... 37
5.1 A macroestrutura cromossômica em Boideos Neotropicais ............................... 37
5.2 Evolução das famílias gênicas ........................................................................... 41
5.3 Cromossomos sexuais ....................................................................................... 44
5.4 Evolução cromossômica em Booidea ................................................................ 48
5.3 Estrutura cromossômica em Corallus hortulanus e Corallus grenadensis.......... 54
6.0 Conclusões ............................................................................................................. 56

xii
7.0 Referências Bibliográficas ...................................................................................... 57

xiii
Lista de Figuras
Figura 1. Origem da diversidade em Boídeos na região Neotropical (Colston et al.
2013) .................................................................................................................. 1
Figura 2. Árvore filogenética para Booidea baseada em caracteres moleculares
proposta por Reynolds et al. (2014) ................................................................... 2
Figura 3. Diversidade dos padrões de manchas da pele de Corallus hortulanus .......... 5
Figura 4. Mapa de coleta ............................................................................................. 18
Figura 5. Prancha de cariótipos de Boa constrictor em coloração convencional, região
organizadora de nucléolo e localização de DNAr 18S ..................................... 26
Figura 6. Prancha de cariótipos de Boa constrictor evidenciando as regiões de
heterocromatina constitutiva ............................................................................ 28
Figura 7. Prancha de cariótipos de Boa constrictor evidenciado o padrão de
distribuição de sequências teloméricas ........................................................... 30
Figura 8. Prancha de cariótipos de Eunectes em coloração convencional, região
organizadora de nucléolo e localização de DNAr 18S ..................................... 32
Figura 9. Prancha de cariótipos de Eunectes evidenciado as regiões de
heterocromatina constitutiva ............................................................................ 33
Figura 10. Prancha de cariótipos de Eunectes evidenciado o padrão de distribuição de
sequências teloméricas ................................................................................... 34
Figura 11. Prancha de cariótipos de Corallus hortulanus em coloração convencional,
região organizadora de nucléolo e localização de DNAr 18S .......................... 35
Figura 12. Cariótipo de fêmea de Corallus hortulanus evidenciando as regiões de
heterocromatina constitutiva ............................................................................ 36
Figura 13. Cariótipo de fêmea de Corallus hortulanus evidenciando o padrão de
distribuição de sequências teloméricas ........................................................... 37
Figura 14. Idiogramas representando alguns tipos de cromossomos sexuais em
serpentes ......................................................................................................... 46
Figura 15. Hipótese de evolução cromossômica em boideos Neotropicais, proposta por
Beçak (1965) .................................................................................................... 48

xiv
Figura 16. Hipótese de evolução cromossômica em boideos Neotropicais, proposta por
Gorman e Gress (1970) ................................................................................... 49
Figura 17. Compilação de dados cromossômicos em filogenia de Booidea proposta por
Reynolds et al. (2014) ..................................................................................... 51
Figura 18. Idiograma representando hipótese de cariótipo ancestral para Pythonoidea e
Booidea ........................................................................................................... 53
Figura 19. Cariótipo de Corallus grenadensis conforme Gorman e Gress (1970)
......................................................................................................................... 55
Lista de Tabelas
Tabela 1. Dados cromossômicos para Booidea ............................................................. 8
Tabela 2. Espécies, localidade, sexo e número de indivíduos analisados .................. 16
Tabela 3. Material testemunho, número tombo e coleção ........................................... 19

xv
Lista de Abreviações e Termos
µL – Microlitros
2n – Número Diplóide
Ag-NOR – Região organizadora de nucléolo

xvi
Água mili-Q – Água ultrapura (deionizada e dessalinizada)
Banda C – Técnica de detecção de heterocromatina constitutiva
DNA – Ácido desoxirribonucleico
DNAr – DNA ribossomal
dNTP – Desoxirribonucleotídeo
EDTA – Ácido etilenodiaminotetracético
FISH – Hibridização fluorescente in situ
HCl – Ácido clorídrico
Homeólogo – Mesmo cromossomo em espécies distintas
KCl – Cloreto de potássio
MgCl2 – Cloreto de magnésio
mL – Mililitro
NaCl – Cloreto de Sódio
Nick translation – Deslocamento de corte
PBS – Tampão de Bloqueio
PCR – Reação em cadeia da polimerase
Primer – Oligonucleotídeos iniciadores
RNAse – Enzina que degrada o ácido ribonucleico
Rpm – Rotações por minuto
SSC – Solução Salina
Taq – Enzima termoestável derivada da bactéria Thermus aquaticus
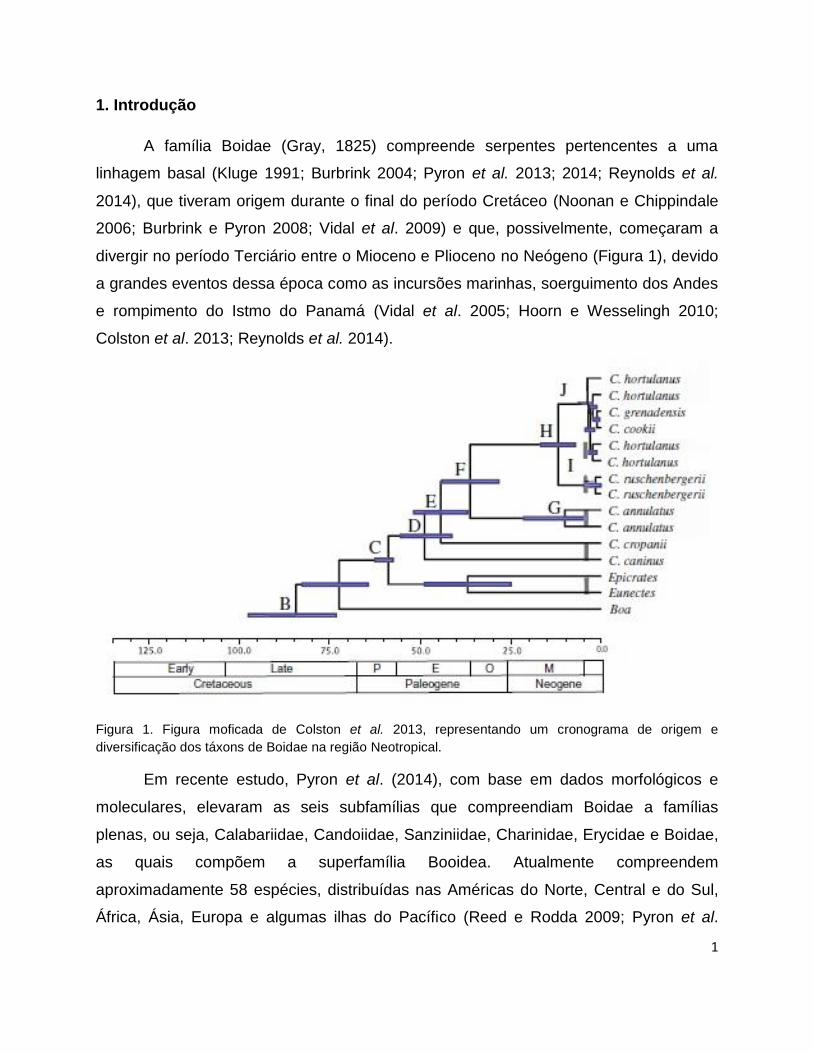
1
1. Introdução
A família Boidae (Gray, 1825) compreende serpentes pertencentes a uma
linhagem basal (Kluge 1991; Burbrink 2004; Pyron et al. 2013; 2014; Reynolds et al.
2014), que tiveram origem durante o final do período Cretáceo (Noonan e Chippindale
2006; Burbrink e Pyron 2008; Vidal et al. 2009) e que, possivelmente, começaram a
divergir no período Terciário entre o Mioceno e Plioceno no Neógeno (Figura 1), devido
a grandes eventos dessa época como as incursões marinhas, soerguimento dos Andes
e rompimento do Istmo do Panamá (Vidal et al. 2005; Hoorn e Wesselingh 2010;
Colston et al. 2013; Reynolds et al. 2014).
Figura 1. Figura moficada de Colston et al. 2013, representando um cronograma de origem e
diversificação dos táxons de Boidae na região Neotropical.
Em recente estudo, Pyron et al. (2014), com base em dados morfológicos e
moleculares, elevaram as seis subfamílias que compreendiam Boidae a famílias
plenas, ou seja, Calabariidae, Candoiidae, Sanziniidae, Charinidae, Erycidae e Boidae,
as quais compõem a superfamília Booidea. Atualmente compreendem
aproximadamente 58 espécies, distribuídas nas Américas do Norte, Central e do Sul,
África, Ásia, Europa e algumas ilhas do Pacífico (Reed e Rodda 2009; Pyron et al.

2
2014; Reynolds et al. 2014; Uetz e Hosek 2014). Essas famílias são distintas tanto
morfologicamente quanto biogeograficamente (Underwood 1967; Kluge 1991), sendo
que Candoiidae está restrita a países da Oceania; Sanziniidae está restrita a
Madagascar; Charinidae tem ocorrência predominantemente nas Américas Central e
do Norte (exceto uma única espécie ocorrente na Colômbia); Erycidae ocorre na
Europa, Oriente Médio; norte da África, parte central e sul da Ásia; e Boidae tem sua
distribuição na região neotropical (Figura 2) (Uetz e Hosek 2014; Pyron et al. 2014;
Reynolds et al. 2014).
Figura 2. Figura adaptada de Reynolds et al. 2014, representando filogenia que compreende os atuais
táxons para Booidea, baseada em genes mitocondriais e nucleares.

3
Aparentemente, as seis famílias que compõem Booidea, constituem um grupo
monofilético (Pyron et al. 2014), sendo a família Boidae a mais numerosa, com 29
espécies, em cinco gêneros (Boa, Corallus, Chilabothrus, Epicrates e Eunectes),
distribuídas principalmente nas Américas Central e do Sul (McDiarmid et al. 1999;
Burbrink 2004; Noonan e Chippindale 2006; Uetz e Hosek 2014; Pyron et al. 2014;
Reynolds et al. 2014), como sendo parte da radiação sul-americana das serpentes
desta superfamília (Reed e Rodda 2009; Pyron et al. 2013).
1.1 Considerações gerais sobre Boa, Corallus e Eunectes
As espécies de Boa (Jiboia) e Corallus (Amazon Tree Boa) apresentam hábitos
arborícolas e terrestres e são mais ativas no crepúsculo e à noite. Possuem uma dieta
variada, consumindo aves, mamíferos, lagartos e anfíbios, que geralmente predam por
constrição (Henderson 1993a; b; Tolson e Henderson 1993; Henderson et al. 1995;
Stafford e Henderson 1996; Monroy-Vilchis et al. 2011). Devido à diversidade de
padrões de coloração e manchas na pele, tamanho, resistência e docilidade relativa, as
espécies destes gêneros têm sido muito difundidas no comércio de “pets” exóticos
(Fogel 1997; Bonny 2007; Russo 2007).
O gênero Boa (Linnaeus, 1758) é monotípico (Boa constrictor) e até
recentemente era composto por nove subespécies, que divergem em diversas
características como dieta, ecologia, comportamento, tamanho, padrões de manchas e
coloração (Reed e Rodda 2009; Boback e Lynn 2010).
Hynková et al. (2009), a partir da análise do gene Citocromo b em seis
subespécies de Boa constrictor, detectaram 67 haplótipos em dois clados, sendo um
para a América Central e outro para a America do Sul e sugeriram a elevação da
subespécie Boa constrictor imperator à espécie plena. Reynolds et al. (2013; 2014)
também encontraram apoio para o reconhecimento de Boa imperator, bem como para
a realocação de Boa constrictor sabogae, como subespécie de Boa imperator (Figura
2).
Esses animais apresentam uma ampla distribuição desde o norte do México até
o sul da América do Sul (Ernest e Zug 1996; Reed e Rodda 2009).

4
Boa constrictor compreende animais de médio a grande porte, podendo
ultrapassar 4m de comprimento. Seu corpo cilíndrico é comprimido nas laterais, o que
evidencia sua forte musculatura constritora (Reed e Rodda 2009). Para o Brasil são
conhecidas duas subespécies: Boa constrictor constrictor, com ampla distribuição na
região amazônica, e Boa constrictor amarali, distribuída na região nordeste, sudeste e
sul (Reed e Rodda 2009).
Corallus (Daudin, 1803) é composto por nove espécies com ampla distribuição
ao longo das Américas Central e do Sul (Henderson et al. 2009). As relações evolutivas
para esse gênero têm sido bem abordadas ao longo dos últimos anos, sendo elas
baseadas em caracteres morfológicos e moleculares (Kluge 1991; Henderson e
Hedges 1995; Henderson 1997; Vi dal et al. 2005; Colston et al. 2013; Pyron et al.
2014; Reynolds et al. 2014). Três espécies podem ser encontradas tanto na América
Central quanto na América do Sul (Corallus blombergi, Corallus annulatus e Corallus
ruschenbergerii); duas endêmicas às Antilhas (Corallus cookii e Corallus grenadensis)
e quatro ao longo da América do Sul (Corallus cropanii, Corallus caninus, Corallus
batesii e Corallus hortulanus), sendo essas últimas encontradas no Brasil. Com
exceção da raríssima Corallus cropanii, as outras espécies ocorrentes no Brasil são
simpátricas e encontram-se amplamente distribuídas na região amazônica (Henderson
et al. 2009; Colston et al. 2013; Pyron et al. 2014). Apresentam o corpo levemente
comprimido lateralmente, dentes bem desenvolvidos na região anterior da boca e
fossas labiais bem visíveis (Henderson 1993b).
As espécies desse gênero apresentam uma infinidade de variações de padrões
de manchas da pele (Figura 3) e coloração com tons que podem ou não ser mistos de
amarelo, vermelho, laranjado, marrom, preto, caramelo, cinza, bege, entre outras.
Esses padrões de coloração e manchas na pele podem estar relacionados a mutações
em vários genes responsáveis pela coloração em vertebrados (Bennett e Lamoreux
2003; Hoekstra et al. 2006; Hubbard et al. 2010; Johnson 2012), porém, mais
frequentemente estão associados a mutações nucleotídicas na sequência do
codificador de melanocortina (Mc1r), podendo essas variações, ser intra e
interespecificas (Rosenblum et al. 2004; 2010). Embora essas variações nucleotídicas

5
já tenham sido bem documentadas em alguns grupos de vertebrados (Ritland et al.
2001; Eizirik et al. 2003; Rosenblum et al. 2004; Mundy et al. 2004; Corso et al. 2012;
Cox et al. 2013), as razões pelas quais há a ocorrência desses polimorfismos de
coloração ainda não estão muito claras, sendo que os mesmos podem estar
associados a eventos epigenéticos ou à presença de elementos transponíveis. Essas
variações de coloração também podem estar associadas à dinâmica de seleção sexual
em diferentes grupos de vertebrados, incluindo os répteis (Sinervo e Lively 1996; Gray
e McKinnon 2006), mimetismo ou adaptabilidade ao meio, como “background” (Mallet e
Joron 1999; Hoekstra et al. 2006; Rosenblum et al. 2010).
Figura 03. Figura retirada de Henderson (1997), representando alguns exemplos das variações
de padrões de manchas da pele de Corallus.

6
Espécies de Corallus normalmente ocorrem em uma ampla variedade de
habitats, podendo ser comumente encontradas na floresta amazônica, em regiões
arbóreas com alta taxa de umidade, como ambientes de várzea, florestas de terra firme
e em zonas secas como o cerrado (Henderson et al. 1995; Martins e Oliveira 1999;
Huang 2006).
Eunectes (Wagler, 1830) é composto por quatro espécies de serpentes
conhecidas popularmente como Anacondas: Eunectes notaeus e Eunectes
deschauenseei consideradas anacondas amarelas; Eunectes murinus e Eunectes
beniensis, as anacondas verdes (Dirksen e Böhme 1998). São serpentes
predominantemente aquáticas, que apresentam ampla distribuição na América do Sul
(Strüssmann e Sazima 1993; Dirksen 2002; Bisplinghoff e Bellosa 2007; Waller et al.
2007). São consideradas as maiores serpentes do mundo em massa, podendo
ultrapassar os 300 quilos (Murphy 1997). Possuem tanto os olhos quanto as narinas
voltadas para a parte dorsal do corpo, o que lhes permite uma melhor eficiência em
suas estratégias de forrageamento em lugares alagados, como pântanos e lagos
(Thorbjarnarson 1995; Murphy e Campbell 1997; Rivas e Owens 2001).
No Brasil são encontradas as espécies E. notaeus, E. deschauenseei e E.
murinus. Apenas a última é simpátrica em relação às demais (Reed e Rodda 2009;
Uetz e Hosek 2014). As anacondas são animais de grande porte e possuem uma dieta
eurifágica (Beebe 1946; Henderson et al. 1995; Murphy e Campbell 1997; Martins e
Oliveira 1999), podendo incluir peixes, tartarugas, jacarés, serpentes aquáticas, aves
aquáticas, roedores e até mesmo carniça (Strussmann e Sazima 1991; Strüssmann
1997; Dirksen 2002; Waller et al. 2007). Foram muito impactadas por causa do
comércio de seu couro (Strimple 1993) e utilização de sua gordura na medicina
tradicional (Rivas 1999). O contato com o ser humano quase sempre resulta na morte
desses animais, por serem capturados em anzóis (Boos 2001) e em redes de pesca
(Martins e Oliveira 1999; Duellman 2005).

7
1.2 Citogenética de Serpentes
Os dados citogenéticos são de grande importância para a compreensão da
evolução de um grupo, podendo atuar em conjunto ou de forma complementar na
sistemática e taxonomia. Em serpentes, a citogenética e evolução cromossômica não
têm sido abordadas de forma integral como em outros grupos de vertebrados, sendo
que a maioria dos estudos restringe-se a descrições cariotípicas em coloração
convencional, existindo poucos estudos com o emprego de bandeamentos
cromossômicos (Mengden e Stock 1980; Moreno et al. 1987; Porter et al. 1991; 1994;
Camper e Hanks 1995; Olmo e Signorino 2005; Aprea et al. 2006; Garcia e Hernando
2007; Pinthong et al. 2013), os quais são fundamentais para se discutir a evolução
cromossômica de um grupo.
Os cariótipos, para a grande maioria das espécies de lagartos, aves, tartarugas
e serpentes são assimétricos, separados em macrocromossomos e
microcromossomos, os quais diferem em relação ao tamanho físico e morfologia
(Beçak e Beçak 1969; Singh 1972a; Bickham e Carr 1983; Sasaki et al. 1984;
Trajtengertz et al. 1995; Matsuda et al. 2005; Matsubara et al. 2006; Ledesma et al.
2006; Fantin e Monjeló 2011; Pokorná et al. 2011; Pinthong et al. 2013).
Os dados citogenéticos para a superfamília Booidea, para as seis famílias,
limitam-se à descrição do número diploide em coloração convencional para algumas
espécies, descrição da região organizadora de nucléolo apenas para a subespécie B.
constrictor constrictor e descrição do padrão de heterocromatina constitutiva para
Sanzinia madagascariensis (Tabela 1).

8
Tabela 1. Dados citogenéticos para a superfamília Booidea compilados da literatura (2n=número diploide;
FC=Fórmula cromossômica; M=macrocromossomos; mi=microcromossomos; Banda C=pares com
heterocromatina; RON=região organizadora do nucléolo).
Família Espécie 2n/FC Cromossomos
Sexuais
Banda
C
RON
Localidade Referência
Sanziniidae Sanzinia
madagascariensis
34/18 M-
16 mi
- 4, 7, 10 - Madagascar Mengden e Stock 1980
Acrantophis dumerilii 34/16 M-
18 mi
ZZ/ZW - Madagascar
Charinidae Lichanura trivirgata 36/16 M-
20 mi
- - El Cajon -
Califórnia
Gorman e Gress 1970
Charina bottae 36/16 M-
20 mi
- - Pinecrest -
Califórnia
Erycidae Eryx conicus 34/16 M-
18 mi
- - Índia Singh et al. 1970
Eryx johnii 34/16 M-
18 mi
- - Índia

9
Eryx jaculus 34/16 M-
18 mi
- - Israel Werner 1959
Candoiidae Candoia bibroni 36/16 M-
20 mi
- - Austrália Mengden 1982
Candoia paulsoni 36/16 M-
20 mi
- - Austrália
Boidae
Boa constrictor
amarali
36/16 M-
20 mi
- Um par
micro
São Paulo -
Brasil
Beçak 1965; De Smet
1978; Camper e Hanks
1995
Boa constrictor
constrictor
36/16 M-
20 mi
- - Rondônia -
Brasil
Beçak et al. 1962;
Bianchi et al. 1969
Corallus caninus 44/24 M-
20 mi
- - Pará - Brasil Beçak 1966
Corallus grenadensis 40/20 M-
20 mi
- - Grenada - West
Indies
Gorman e Gress 1970
Eunectes murinus 36/16 M-
20 mi
- - São Paulo -
Brasil
Beçak 1965; De Smet
1978

10
Epicrates crassus 36/16 M-
20 mi
- Um par
micro
Brasil Beçak 1965; Viana,
comunicação pessoal
Epicrates assisi 36/16 M-
20 mi
Um par
micro
Minas Gerais -
Brasil
Viana, comunicação
pessoal
Epicrates cenchria 36/16 M-
20 mi
Um par
micro
Pará - Brasil Viana, comunicação
pessoal
Chilabothrus striatus 36/16 M-
20 mi
- - Leogane - Haiti Gorman e Gress 1970

11
Camper e Hanks (1995) descreveram a localização e o número de regiões
organizadoras de nucléolo (Ag-RON) para 21 espécies de serpentes, pertencentes a
cinco famílias, dentre essas, a serpente Boa constrictor constrictor, representante da
família Boidae, que apresentou Ag-RON em um par de microcromossomos, sendo este
muito conservado no genoma das serpentes (Burt et al. 1999; Smith et al. 2000).
Embora Ag-RON simples seja considerada uma característica plesiomórfica dentro da
subordem Serpentes, rearranjos como fusões e translocações podem ocorrer entre
macrocromossomos e microcromossomos envolvendo essa região (Baker et al. 1971;
Olmo 2008).
Além do número diploide, fórmula cariotípica e Ag-RON, a descrição do padrão
de heterocromatina constitutiva em serpentes pode contribuir de forma significativa
para o entendimento dos processos evolutivos no grupo. Mengden e Stock (1980)
descreveram o padrão de banda C para oito espécies de serpentes pertencentes às
famílias Colubridae e Boidae, onde verificaram que inúmeros rearranjos
cromossômicos em macrocromossomos e microcromossomos, bem como diferenças
significativas de blocos heterocromáticos e variações na morfologia do cromossomo W
estão presentes, tanto em linhagens primitivas, quanto em linhagens derivadas de
serpentes. Também verificaram que rearranjos cromossômicos e adição de
heterocromatina podem ter sido responsáveis pela diferenciação cromossômica sexual
em Acrantophis dumerili, sendo o primeiro registro de cromossomos sexuais
diferenciados na superfamília Booidea. Estes rearranjos cromossômicos também foram
revelados com a análise do bandeamento G, evidenciando que o cromossomo W é
uma das partes mais variáveis do genoma das serpentes (Beçak 1965; Mengden e
Stock 1980; Aprea et al. 2006; Oguiura et al. 2009; Pinthong et al. 2013).
Estudos cromossômicos com técnicas citogenéticas mais avançadas mostram-
se importantes para a compreensão da taxonomia das espécies de Boidae, bem como
para o entendimento dos mecanismos de evolução cariotípica em serpentes. Tais
mecanismos foram bem documentados para a família Elapidae, onde Micrurus
apresenta inúmeros rearranjos cromossômicos, resultando em diferentes cariótipos,

12
como fusões em microcromossomos para espécies da América Central e fissões para
as espécies da América do Sul, segundo hipótese de Serafim et al. (2007). Para as
espécies que ocorrem na América Central, o número diploide variou entre 2n=26
(Micrurus nigrocinctus, Micrurus ruatanus e Micrurus browni), 2n=30 (Micrurus
mosquitensis, Micrurus diastema, Micrurus elegans, Micrurus hippocrepis), 2n=32
(Micrurus fulvius) e 2n=34 (Micrurus mipartitus, Micrurus alleni) (Graham 1977;
Gutiérrez e Bolaños 1979; Gutiérrez et al. 1988; Luykx et al. 1992) e as da América do
Sul variou entre 2n=38 (Micrurus surinamensis), 2n=40 (Micrurus corallinus), 2n=42
(Micrurus ibiboboca) (Beçak e Beçak 1969; Gutiérrez et al. 1988; Serafim et al. 2007).
Essas grandes variações do número diploide também são descritas para outros
gêneros como Naja e Ophiophagus (Singh 1972a; Ray-Chaudhuri e Singh 1972; Olmo
e Signorino 2005), as quais também podem ser evidenciadas com técnicas de
citogenética molecular.
1.3 Citogenética Molecular em Reptilia
A citogenética molecular, que envolve mapeamentos cromossômicos de
sequências de DNA, é útil no esclarecimento de questões relacionadas à evolução
cromossômica (Bickham e Rogers 1985; King et al. 1986; Amemiya et al. 1993; Porter
et al. 1991; 1994; Srikulnath et al. 2011), à origem de cromossomos sexuais e acima de
tudo sobre a organização genômica, fornecendo novos insights sobre a evolução dos
vertebrados (Matsuda et al. 2005; Ezaz et al. 2005; 2006; Matsubara et al. 2006; 2014
Kawai et al. 2007; Rhen e Schroeder 2010; Pokorná et al. 2011).
Em serpentes, genes ribossomais (DNAr) foram evidenciados em
microcromossomos de diferentes populações de Bothrops neuwiedi (Viperidae) e
também em macrocromossomos dessa serpente, evidenciando um possível evento de
translocação de microcromossomo para macrocromossomo (Trajtengertz et al. 1995).
Ainda, o mapeamento de sequências do gene ribossomal 28S em quatro espécies de
serpentes pertencentes às famílias Viperidae e Colubridae mostrou variabilidade
cariotípica com relação à localização do DNAr nestas serpentes. Crotalus viridis
(Viperidae) apresentou marcações de DNAr em dois pares de microcromossomos;

13
Masticophis flagellum (Colubridae) apresentou marcações em apenas um par de
microcromossomos; Nerodia fasciata e Tamnophis marcianus (Colubridae)
apresentaram marcações em um par de macrocromossomos (Porter et al. 1991).
Abordagens mais avançadas como pintura cromossômica e mapeamento de
genes de cópia única também têm se mostrado como ótimas ferramentas para a
compreensão dos mecanismos de determinação do sexo em répteis e evolução de
cromossomos sexuais. Ezaz et al. (2005) conseguiram identificar em Pogona vitticeps a
existência de microcromossomos sexuais através da hibridização in situ com sondas a
partir de um cromossomo W microdissectado. Matsubara et al. (2006) a partir do
mapeamento de genes responsáveis pela determinação do sexo (RAB5A, DMRT1 e
SOX9) conseguiram identificar os cromossomos sexuais em três espécies de
serpentes, dentre elas Python molurus (Pythonidae), serpente de linhagem basal que
apresentou sequências do RAB5A no braço longo em um par “homomórfico” de
macrocromossomos (número 4) como sendo o par sexual do tipo ZW.
Assim, o mapeamento de sequências de DNA, repetitivas ou não, em
cromossomos de serpentes, tais como DNAr e sequências teloméricas, pode oferecer
importantes insights à cerca da organização genômica desses animais, sendo esta
abordagem ainda insipiente para espécies da família Boidae.

14
2. Objetivos
2.1 Objetivo Geral
Caracterizar, utilizando marcadores cromossômicos clássicos e moleculares,
espécies de serpentes neotropicais da família Boidae.
2.2 Objetivos Específicos
Investigar a existência de homomorfismo de cromossomos sexuais em espécies
de serpentes neotropicais da família Boidae, pela técnica de banda C.
Mapear sequências repetitivas de DNAr 18S e DNA telomérico em espécies de
serpentes neotropicais da família Boidae.
Investigar se existe variação intraespecífica entre indivíduos provenientes de
diferentes localidades.
15
3. Material e Métodos
3.1 Material
Para o presente estudo foram coletados indivíduos de Boa constrictor constrictor
(ambiente natural), Boa constrictor amarali e Boa constrictor occidentalis (cativeiro),
Eunectes murinus (ambiente natural), Eunectes notaeus (cativeiro) e Corallus
hortulanus (ambiente natural). Os animais foram obtidos de diferentes locais, por meio
de coleta ativa ou de cativeiro (Tabela 2 e Figura 4).
Tabela 2 – Espécies, localidade, sexo e número de indivíduos analisados no presente trabalho.
Espécie Localidade Sexo N° de indivíduos
Boa constrictor constrictor
Manaus – AM 7♂ 8 ♀ 16
FLONA do Tapajós – PA 1 ♀
Boa constrictor amarali
Belo Horizonte – MG (cativeiro) 4 ♀ 6 ♂ 10
Boa constrictor occidentalis
Jibóias Brasil – MG (cativeiro) 1 ♂ 1
Corallus hortulanus
PARNA Anavilhanas – AM 2 ♂ 2 ♀
6 FLONA do Tapajós – PA 1 ♀
ESEC Juami-Japurá – AM 1 ♂
Eunectes murinus
Manaus – AM 3 ♀ 2 ♂
10
Lago Catalão – AM 1♀
Novo Aripuanã – AM 1♀
APA Acariquara – AM 1 ♂
FLONA do Tapajós – PA 1 ♂
Belo Horizonte – MG (cativeiro) 1 ♂
Eunectes notaeus Belo Horizonte – MG (cativeiro) 1 ♀ 1

16
3.2 Métodos
3.2.1 Coleta dos animais
Algumas serpentes foram provenientes de coletas do Projeto
SISBIOTA/BIOPHAM e outras foram coletadas sob a autorização ICMBIO – SISBIO de
número 45275-1 (Tabela 3). O presente estudo foi submetido e aprovado pelo comitê
de ética do Instituto Nacional de Pesquisas da Amazônia, com o protocolo: 013/2014
em 14/07/2014.
O método de coleta ativa consistiu na procura de exemplares durante os
períodos diurno e noturno em florestas de terra firme e em matas ciliares ao longo de
rios e igarapés. Os indivíduos foram capturados de forma manual com o auxílio de
gancho herpetológico, pinção, lanternas e sacos de pano para transporte e trazidos
para uma base laboratorial mais próxima para coleta sanguínea e cultura de linfócitos.
A colheita de sangue deu-se a partir da veia dorsal do animal, utilizando seringas
descartáveis contendo anticoagulante do tipo heparina sódica (hepamax®), sendo
quantidades entre 1ml e 5ml, dependendo do tamanho do animal. Como material
testemunho, os animais provenientes no projeto SISBIOTA foram depositados nas
coleções herpetológicas da Universidade Federal do Amazonas - UFAM ou do Instituto
Nacional de Pesquisas da Amazônia – INPA.
Tabela 3. Material testemunho: espécies, número de registro e coleção.
Espécie Número de Registro Coleção
Boa constrictor constrictor
♀
INPA-H 3096 Coleção de Anfíbios e Répteis / INPA
Boa constrictor constrictor
♀
INPA-H 34712 Coleção de Anfíbios e Répteis / INPA
Boa constrictor constrictor
♂
CTGA-C 84 SISBIOTA / UFAM
Boa constrictor amarali ♀ ------------------------ Zoobotânica - BH
Boa constrictor amarali ♀ ------------------------ Zoobotânica – BH

17
Boa constrictor amarali ♀ ------------------------ Zoobotânica – BH
Boa constrictor amarali ♀ ------------------------ Zoobotânica – BH
Boa constrictor amarali ♂ 9630060000024237 Zoobotânica – BH
Boa constrictor amarali ♂ 963006000024240 Zoobotânica – BH
Boa constrictor amarali ♂ ------------------------ Zoobotânica – BH
Boa constrictor amarali ♂ ------------------------ Zoobotânica – BH
Boa constrictor amarali ♂ ------------------------ Zoobotânica – BH
Boa constrictor amarali ♂ ------------------------ Zoobotânica - BH
Boa constrictor occidentalis
♂
BCOM02 Criadouro - Jiboias Brasil
Corallus hortulanus ♂ CTGA-C 269 SISBIOTA / UFAM
Corallus hortulanus ♀ CTGA-C 95 SISBIOTA / UFAM
Corallus hortulanus ♂ INPA-H 34713 Coleção de Anfíbios e Répteis / INPA
Eunectes murinus ♀ INPA-H 34561 Coleção de Anfíbios e Répteis / INPA
Eunectes murinus ♀ INPA-H 34550 Coleção de Anfíbios e Répteis / INPA
Eunectes murinus ♂ 963006000023750 Zoobotânica - BH
Eunectes murinus ♂ 982009106701490 CIGS - AM
Eunectes murinus ♂ 982009206722122 CIGS - AM
Eunectes murinus ♀ 963006000102214 CIGS – AM
Eunectes murinus ♀ 963006000102023 CIGS – AM
Eunectes murinus ♀ 982009106707295 CIGS - AM
Eunectes notaeus ♀ ------------------------ Zoobotânica - BH

18
Figura 4: Mapa da região Norte do Brasil, indicando os locais de coleta para os animais não provenientes de cativeiro: (1) Manaus, AM; (2) Flona
do Tapajós; (3) Arquipélago de Anavilhanas; (4) Juami-Japurá; (5) Rio Aripuanã. Corallus hortulanus (2,3, 4); Boa constrictor constrictor (1, 2, 3) e
Eunectes murinus (1, 2, 5).

19
Citogenética Clássica
3.2.2 Obtenção de cromossomos mitóticos
Para obtenção das preparações cromossômicas, foi realizada a cultura de
linfócitos de sangue periférico, baseado no protocolo de Beçak et al. 1963, com várias
modificações, que consistiram em:
Adicionar ao frasco de meio de cultura completo para cariótipo com RPMI (5mL) da
marca cultilab®, 0,5 mL de sangue total, misturando-se delicadamente. O frasco foi
incubado em estufa a 29 °C por 96 horas. Após esse período foi adicionado a este
frasco, contendo a cultura de sangue, 0,5 ml de colchicina a 0,025% por 40 minutos.
Em seguida, o material foi transferido para um tubo de centrífuga de 15 ml e
centrifugado por 10 minutos em 1200 rpm. O sobrenadante foi descartado e adicionou-
se uma solução hipotônica de cloreto de potássio (KCL 0,075M). Feito isso, o material
foi homogeneizado e incubado em estufa a 37 °C por 45 minutos. Logo após esse
período, foi acrescentado à solução cerca de 500 µl de fixador Carnoy (metanol 3:1
ácido acético) e homogeneizado novamente. Essa solução foi centrifugada, o
sobrenadante descartado e 8mL de fixador acrescentados, ressuspendida e
centrifugada novamente. Esses passos foram repetidos por mais duas vezes. Após a
última centrifugação, o sobrenadante foi descartado e adicionado 1,5 mL de fixador. O
material foi transferido para um microtubo identificado com o número do indivíduo e
guardado em freezer (-10 °C) até a preparação das lâminas.
As lâminas foram preparadas gotejando-se de duas a três gotas da suspensão
celular em uma lâmina de vidro limpa (aquecida em água destilada a 45 °C), a uma
altura de 40-50 cm, deixando-se secar em temperatura ambiente. Em seguida, as
lâminas foram coradas com solução de Giemsa, diluída a 5% em tampão fosfato pH
6,8, por 7 minutos, lavadas e secas em temperatura ambiente.
3.2.3 Detecção das Regiões Organizadoras de Nucléolo (RON)
As RON foram detectadas utilizando a técnica de impregnação por nitrato de
prata (Ag-RON), descrita por Howell e Black (1980), que consistiu em gotejar sobre a

20
lâmina com a preparação cromossômica uma gota de solução aquosa de gelatina a
2%, acrescida de ácido fórmico na proporção de 1mL/100mL de solução, e duas gotas
de solução aquosa de nitrato de prata (AgNO3) a 50%. A lâmina foi agitada levemente,
coberta com lamínula, colocada em câmara úmida e levada ao banho-maria ou estufa a
60 °C, por um período de dois a quatro minutos, ou até que a lâmina apresentasse uma
coloração marrom-dourada. Em seguida, as lâminas foram lavadas em água destilada,
permitindo que a lamínula e o excesso das soluções fossem removidos, sendo
posteriormente deixadas secar ao ar.
3.2.4 Bandeamento C
Para a caracterização da heterocromatina constitutiva utilizou-se o protocolo
descrito por Sumner (1972), com modificações. A lâmina contendo a preparação
cromossômica foi submetida a um tratamento com solução ácida HCl 0,2 N, 45 °C, por
4 minutos; em seguida foram lavadas com água destilada e deixadas secar ao ar. As
lâminas posteriormente foram incubadas em solução básica de hidróxido de bário
octahidratado (Ba (OH)2.8H2O) 5%, por um período que variou entre 8 e 90 segundos,
a 45 °C. Após este tratamento, as lâminas foram lavadas rapidamente em solução de
HCl 0,2 N, em temperatura ambiente e posteriormente lavadas, várias vezes, em água
destilada e novamente deixadas secar ao ar; em seguida incubadas em solução
2XSSC (cloreto de sódio 0,3M e citrato trissódico 0,06M em pH 6,8) a 60 °C, por 25
minutos. Após isso, as lâminas foram lavadas, secas ao ar e coradas com Giemsa
diluído a 5% em tampão fosfato pH 6,8, por 15 minutos.
Citogenética Molecular
3.2.5 Extração de DNA total
A extração de DNA foi realizada a partir de amostras sanguíneas das serpentes,
utilizando o protocolo de Sambrook et al. (1989), com algumas modificações. Para
tanto, 500 μL de sangue foram transferidos para um tubo de volume de 1,5 mL. Em
seguida, foram adicionados 500 μL de tampão de lise (Tris-HCl 10 mM, pH 8,0, NaCl
0,3M, EDTA 10 mM, SDS 1%). Posteriormente foram acrescentados: 15 μL de

21
proteinase K (10 mg/mL) e 6 μL de RNAse (10 mg/mL). As amostras foram incubadas a
60 ºC por aproximadamente 1 hora para que o tecido fosse totalmente digerido. Foram
adicionados 100 μL de acetato de amônio 3M, 1 volume (600 μL) de fenol-clorofórmio
(1:1) e agitado por inversão durante alguns minutos. Após isso as amostras foram
centrifugadas por 30 minutos a 14.000 rpm em temperatura ambiente. O sobrenadante
foi transferido para um novo tubo. Em seguida adicionou-se 600 μL de clorofórmio e
misturou-se, cuidadosamente, por inversão, durante 1 minuto. A mistura foi então
centrifugada por 20 minutos a 14.000 rpm. O sobrenadante foi transferido para um
novo tubo e acrescentou-se 100 μL de acetato de amônio 3M, 1 volume (600 μL) de
isopropanol gelado e misturado por inversão. O precipitado foi deixado a -20 °C por
cerca de 14 horas. O material foi retirado do freezer e centrifugado por 30 minutos a
14.000 rpm, descartando-se o sobrenadante. O pellet foi lavado com 1 mL de etanol
70% e centrifugado por 20 minutos a 14.000 rpm. O sobrenadante foi descartado
novamente e o pellet foi seco em estufa a 55 °C, ressuspendido em 100 μl de ou água
milli- Q e deixado eluindo por 14 horas. Para possibilitar a análise da quantidade e
integridade do material, o DNA extraído foi quantificado em espectrofotômetro
NanoDrop 2000 (Thermo Scientific).
3.2.6 Isolamento de sequências repetitivas de DNA por PCR (Polymerase Chain
Reaction)
O isolamento das sequências repetitivas de DNAr foi efetuado por meio de
Reação da Polimerase em Cadeia (Saiki et al. 1988), utilizando DNA genômico extraído
do sangue das diferentes espécies e os seguintes pares de primers: a) DNAr 18S: IpF
5’CCGCTTTGGTGACTCTTGAT e IpR 5’CCGAGGACCTCACTAAACCA (Gross et al.
2010). As reações de PCR foram feitas em um termociclador Eppendorf - Mastercycler
Gradient, para volume final de 15 μL (1 μL de DNA genômico (100 ng); 1,5 μL de
Tampão 10X com cloreto de magnésio (1,5 mM); 0,15 μL de Taq DNA polimerase
(5U/µl); 3,0 μL de dNTP (1 mM); 0,6 μL de cada primer (5 mM); água milli-Q para
completar o volume). Para sequências teloméricas, as amplificações foram feitas em
ausência de DNA com os primers (TTAGGG)5x e (CCCTAA)5x (Ijdo et al. 1991).

22
Os ciclos de amplificação seguiram as seguintes etapas:
a) DNAr 18S: 2 minutos a 94 ºC (desnaturação inicial); 35 ciclos de 1 minuto a 95
ºC (desnaturação), 1 minuto a 55 ºC (anelamento dos primers), 1 minuto e 40
segundos a 72 ºC (extensão); 7 minutos a 72 ºC (extensão final);
b) DNA Telomérico: 4 minutos a 94 °C (desnaturação inicial), 12 ciclos de 1 min a
94 °C (desnaturação), 45 segundos a 52 °C (anelamento dos primers) e 1
minuto e 30 segundos a 72 °C (extensão); 1 minuto a 94 °C (desnaturação), 1
minuto e 30 segundos a 60 °C (ligação dos primers) e 1 minuto e 30 segundos a
72 °C(extensão); 7 minutos a 72 ºC (extensão final).
Os produtos de PCR foram utilizados como sondas na hibridização fluorescente in
situ após marcação pelo método de nick translation, utilizando biotina 14-dATP (Bionick
labeling system-Invitrogen) e/ou digoxigenina-11-dUTP (Dig-Nick Translation mix
Roche). Para tanto, em um tubo eppendorf de 1,5 mL, foi preparada uma solução
contendo 1 μL de Mix dNTP 10x; 1 μL de sonda de DNA (200 ng/ μL); 1 μL de Mix de
enzima 10x; 6 μL de água milli-Q, totalizando 9 μL para cada lâmina que foi hibridizada.
Esta solução foi homogeneizada, centrifugada brevemente e incubada a 16 ºC por 90
minutos no termociclador. Em seguida, para interromper a reação foi adicionado 1 μL
de stop buffer contendo 1 μL de acetato de sódio 3M e posteriormente mantida em
freezer para posterior utilização.
3.2.7 Hibridização in situ por fluorescência (FISH)
Foi realizada a técnica de hidridização in situ por fluorescência (FISH) descrita
por Pinkel et al. (1986) com algumas modificações:
Tratamento das lâminas:
As lâminas foram lavadas em tampão PBS 1x por 5 minutos, em temperatura ambiente
e desidratadas em série alcoólica gelada 70, 85 e 100% durante 10 minutos cada. Em
seguida, tratadas com 90 μl de RNase 10 μg/mL (5 μL de RNase 10 mg/mL e 975 μL
de 2XSSC) por 1 hora em câmara úmida a 37 ºC. Após, as lâminas foram lavadas três

23
vezes em 2xSSC por 5 minutos cada e posteriormente lavadas em PBS 1x durante 5
minutos.
Fixação
As lâminas foram fixadas em formaldeído 1% em PBS 1X/50mM MgCl2 durante 10
minutos à temperatura ambiente. Posteriormente, lavadas em PBS 1x por 5 minutos.
Após, as lâminas foram novamente desidratadas em série alcoólica gelada (70, 85 e
100%) durante 5 minutos cada e deixadas secar ao ar.
Pré-hibridização
As lâminas foram desnaturadas em formamida 70% em 2xSSC a 70 ºC por 5 minutos e
novamente desidratadas em etanol gelado 70, 85 e 100% por 5 minutos cada.
Solução de hibridização:
Em um tubo tipo eppendorf foram adicionados 4 μl de sonda 1, 4 μl de sonda 2,
20 μl de formamida, 8 μl de sulfato de dextrano 50% e 4 μl de 20xSSC. As sondas
foram desnaturadas a 99 °C por 10 minutos e transferidas imediatamente ao gelo.
Hibridização:
Foram colocados 40 μl de solução de hibridização sobre uma lamínula e a
lâmina alocada invertida sobre a lamínula. As lâminas foram mantidas com o material
voltado para baixo em câmara úmida (água destilada) a 37 °C por cerca de 14 horas.
Lavagens:
As lamínulas foram removidas das lâminas e, em seguida lavadas uma vez em
formamida 15% a 42 °C durante 5 minutos. Em seguida as lâminas foram lavadas em
solução Tween 0,5% durante 5 minutos à temperatura ambiente.
Detecção:
As lâminas foram incubadas em tampão NFDM (NonFat Dry Milk) por 15
minutos. Após isso foram lavadas duas vezes com solução Tween 5% por 5 minutos à
temperatura ambiente. Foram colocadas sobre cada lâmina 20 µL de anti digoxigenina-
rodamina e 100 µL de solução contendo tampão NFDM e estreptavidina (2 μL de
estreptavidina estoque em 998 μL de NFDM). Em seguida, foram cobertas com
lamínula e deixadas por 60 minutos em câmara úmida com água destilada.
Posteriormente as lamínulas foram removidas e as lâminas lavadas três vezes em

24
solução Tween 5% por 2 minutos cada a temperatura ambiente. As lâminas foram
desidratadas em série alcoólica 70, 85 e 100% durante 5 minutos cada e esperou-se
secar.
Montagem das lâminas:
Foi adicionada, a cada lâmina, solução de DAPI diluído em antifade VectaShield
Vector (20 µL de antifade e 1 µL de DAPI).
3.2.8 Análise cariotípica
As preparações cromossômicas foram analisadas em microscópio óptico
Olympus BX 51 com objetiva de imersão. Contou-se um mínimo de 10 metáfases para
cada indivíduo, com o intuito de se estabelecer o número diploide (2n) e o número
fundamental (número de braços do cariótipo) de cada indivíduo. As melhores
metáfases tiveram suas imagens capturadas com câmera digital acoplada (Olympus
DP71), utilizando o software Image-Pro MC 6.3.
Para a montagem dos cariótipos utilizou-se o software Adobe Photoshop CS6
previamente licenciado, onde os cromossomos metafásicos mitóticos foram recortados,
emparelhados, medidos e colocados em ordem decrescente de tamanho. Os cariótipos
foram divididos em macrocromossomos e microcromossomos e a morfologia para os
macrocromossomos foi determinada de acordo com Levan et al. (1964), com base no
índice de relação de braços (RB= comprimento do braço maior/comprimento do braço
menor), podendo ser metacêntricos (RB= 1,0-1,7), submetacêntricos (RB=1,71-3,0),
subtelocêntricos (RB=3,01-7,0) e acrocêntricos (RB> 7,00). Na determinação do
número de braços (NF) foram considerados os metacêntricos, submetacêntricos e
subtelocêntricos como tendo dois braços e os acrocêntricos e microcromossomos
como tendo apenas um braço.

25
4. Resultados
4.1 Boa constrictor
Para esta espécie foi possível analisar três subespécies: Boa constrictor
constrictor, Boa constrictor amarali e Boa constrictor occidentalis e todos os
exemplares apresentaram um número diploide igual a 36 cromossomos, com fórmula
cariotípica igual a 6m+2sm+4st+4a+20mi e número de braços (NF) igual a 50, sem
heteromorfismo de cromossomos sexuais e sem diferenças entre as subespécies, em
coloração convencional. Também não foi detectada nenhuma diferença entre os
exemplares de diferentes localidades.
A organização dos cromossomos em ordem decrescente de tamanho evidenciou
que os três primeiros pares de cromossomos são bem maiores, quando comparados
aos outros cinco pares de macrocromossomos. O primeiro, terceiro e quarto pares são
metacêntricos, o segundo par é submetacêntrico, os pares cinco e seis são
subtelocêntricos e os pares sete e oito são acrocêntricos. Com relação aos
microcromossomos, não foi possível determinar com precisão a morfologia para todos
e nem realizar o emparelhamento dos homólogos, devido ao tamanho reduzido dos
mesmos. Portanto, foram organizados em ordem decrescente de tamanho (Figura 5 A,
B, C).
A impregnação com nitrato de prata (Ag-RON) evidenciou a região organizadora
de nucléolo em apenas um par de microcromossomos nas três subespécies. Em
algumas ocasiões, os microcromossomos portadores da região organizadora de
nucléolo foram vistos em associação. O mapeamento da sequência de DNA ribossomal
18S, nas três subespécies, revelou apenas um par de microcromossomos os
portadores da região de DNAr 18S, confirmando as marcações obtidas pela
impregnação por nitrato de prata (Figura 5 A, B, C, em destaque).

26
Figura 5 – Cariótipos de Boa constrictor: (A) Boa constrictor constrictor; (B) Boa constrictor amarali; (C) Boa constrictor occidentalis com suas respectivas regiões organizadoras de nucléolo em um par de microcromossomos, detectada por Ag-NO3 e pela FISH com DNAr 18S (sinais em vermelho).

27
A heterocromatina constitutiva foi evidenciada principalmente nos
macrocromossomos das três subespécies, sendo que alguns microcromossomos
apresentaram marcações difusas, impossibilitando a localização exata desses blocos.
Entretanto, o padrão de banda C foi distinto para cada subespécie.
Boa constrictor constrictor apresentou blocos heterocromáticos principalmente
nos pares 2, 4, 5, 7 e 8, sendo que nos pares 2 e 4 os blocos heterocromáticos
encontram-se na região centromérica; no par 5 a heterocromatina está na região
terminal do braço longo e nos pares 7 e 8, as marcações são biteloméricas (Figura 6
A). Nenhuma diferença foi detectada entre exemplares de diferentes localidades e
entre machos e fêmeas.
Boa constrictor amarali apresentou os blocos nos pares 2, 4, 7 e 8, sendo que
nos pares 2 e 4, assim como em Boa constrictor constrictor, encontram-se em posição
centromérica, já no par de número 7 os blocos estão na região distal do braço curto e
no par de número 8 encontra-se na região terminal do braço longo (Figura 6 B).
Boa constrictor occidentalis apresentou os blocos nos pares 2, 4 e 5, sendo que
nos pares 2 e 4, assim como nas outras duas, encontram-se em posição centromérica,
enquanto o par de número 5 apresenta blocos heterocromáticos biteloméricos (Figura 6
C).
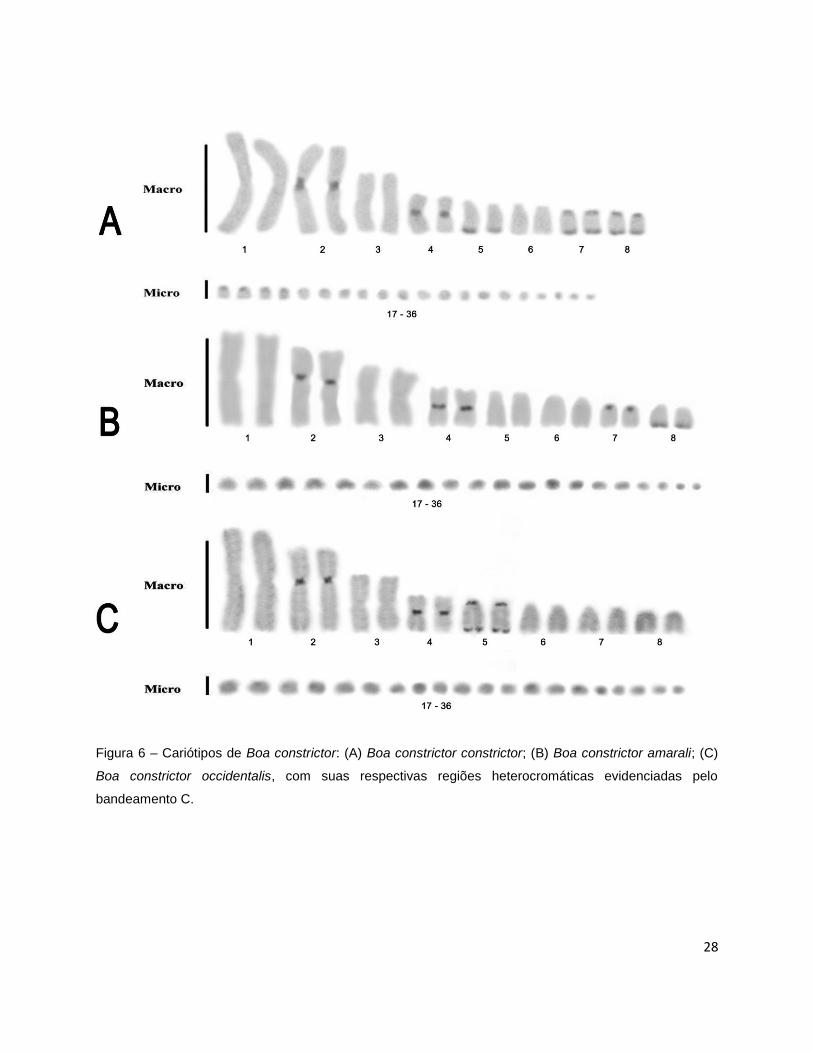
28
Figura 6 – Cariótipos de Boa constrictor: (A) Boa constrictor constrictor; (B) Boa constrictor amarali; (C)
Boa constrictor occidentalis, com suas respectivas regiões heterocromáticas evidenciadas pelo
bandeamento C.

29
O mapeamento cromossômico de sequências teloméricas, além de revelar
marcações terminais em todos os cromossomos do complemento, também evidenciou
sequências teloméricas repetitivas intersticiais (ITS) e padrão distinto para cada
subespécie. O mapeamento foi repetido para confirmação desses padrões e as
marcações foram iguais para animais provenientes de diferentes localidades
Boa constrictor constrictor apresentou ITS nos pares 2 e 4, sendo que no par 2
(sm) as marcações foram observadas nos braços curtos, adjacentes ao bloco de
heterocromatina constitutiva. Já no par 4 (m), a ITS localizou-se na região do
centrômero, coincidindo com o bloco de heterocromatina constitutiva (Figura 7 A).
Nenhuma diferença foi detectada entre os exemplares de diferentes localidades.
Boa constrictor amarali apresentou ITS nos pares 1, 2 e 6, onde o par número 1
apresenta uma ITS na região proximal tanto do braço longo como do curto. No par 2 há
a presença de duas ITS na região intersticial do braço curto e duas ITS na região
proximal do braço longo. Já, o par 6 (st) apresenta uma ITS na região intersticial do
braço longo (Figura 7 B).
Boa constrictor occidentalis apresentou ITS nos pares 2, 4 e 8, sendo que no par
2, a ITS foi proximal no braço curto, adjacente ao bloco de heterocromatina constitutiva,
no para 4 a ITS está na região centromérica coincidente com o bloco de
heterocromatina e no par 8, a ITS foi proximal no braço longo (Figura 7 C).
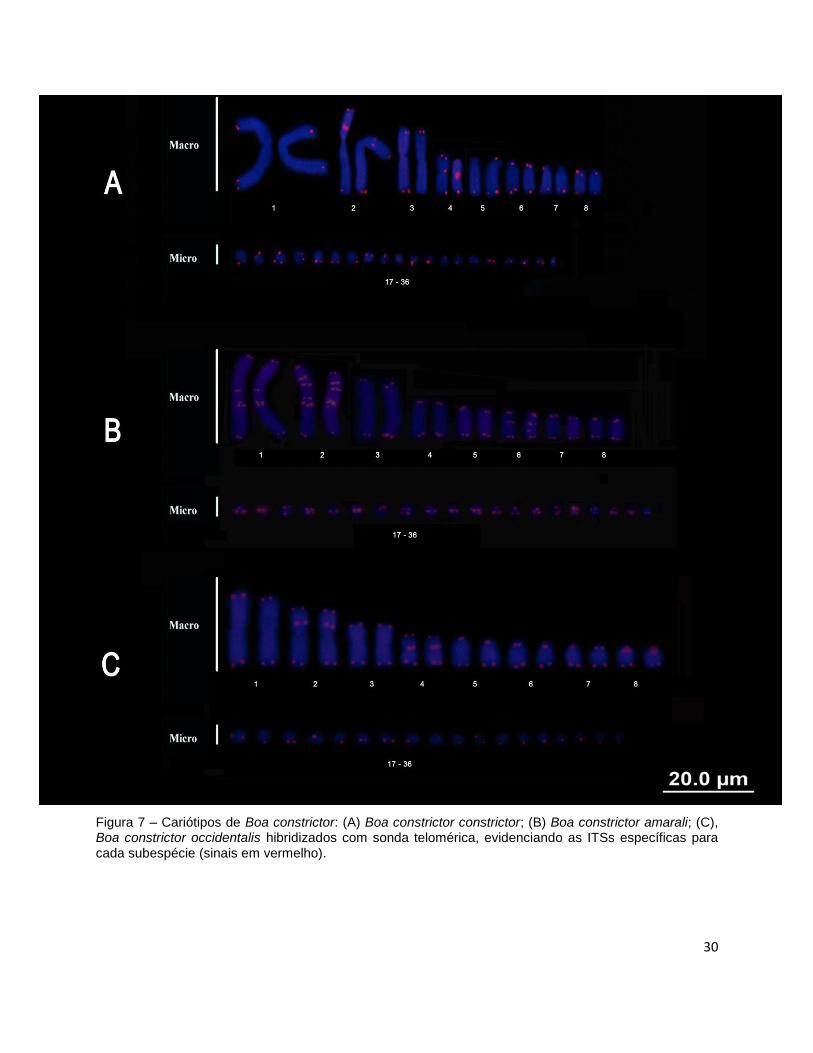
30
Figura 7 – Cariótipos de Boa constrictor: (A) Boa constrictor constrictor; (B) Boa constrictor amarali; (C), Boa constrictor occidentalis hibridizados com sonda telomérica, evidenciando as ITSs específicas para cada subespécie (sinais em vermelho).

31
4.2 Eunectes
Para este gênero foram analisadas duas espécies: Eunectes murinus (anaconda
verde) e Eunectes notaeus (anaconda amarela).
Todos os exemplares de ambas espécies apresentaram um número diploide
igual a 36 cromossomos, com fórmula cariotípica igual a 6m+2sm+8st+20mi e número
de braços (NF) igual a 52, sem heteromorfismo de cromossomos sexuais e sem
diferença entre as espécies, em coloração convencional. Também não foi detectada
nenhuma diferença entre os exemplares das diferentes localidades. A organização dos
cromossomos em ordem decrescente de tamanho evidenciou que, assim como em
Boa, os três primeiros pares de macrocromossomos são bem maiores, em comparação
aos demais pares de macro. Os pares 1, 3 e 4 são do tipo metacêntricos, o par 2 é
submetacêntrico e os pares 5, 6, 7 e 8 são do tipo subtelocêntricos. Os
microcromossomos foram ordenados em ordem decrescente de tamanho devido à
impossibilidade de afirmar com exatidão sua morfologia e emparelhamento dos
homólogos (Figura 8 A, B).
A impregnação com nitrato de prata (Ag-RON) evidenciou a região organizadora
de nucléolo em apenas um par de microcromossomos nas duas espécies. Em algumas
ocasiões, os microcromossomos portadores da região organizadora de nucléolo foram
encontrados em associação para ambas as espécies, porém houve uma diferença
notável de tamanho entre os microcromossomos de Eunectes notaeus. O mapeamento
da sequência de DNA ribossomal 18S, em ambas espécies, revelou apenas um par de
microcromossomos portador da região de DNAr 18S, confirmando as marcações
obtidas pela impregnação por nitrato de prata (Figura 8 A, B em destaque).
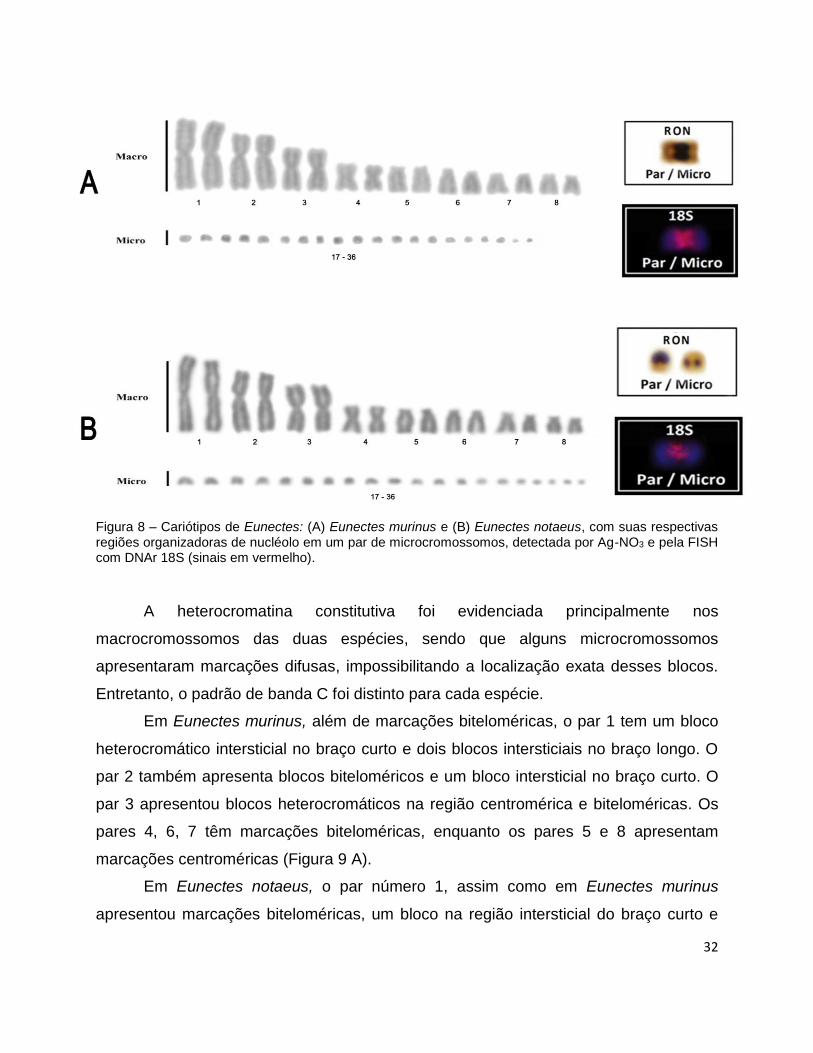
32
Figura 8 – Cariótipos de Eunectes: (A) Eunectes murinus e (B) Eunectes notaeus, com suas respectivas regiões organizadoras de nucléolo em um par de microcromossomos, detectada por Ag-NO3 e pela FISH com DNAr 18S (sinais em vermelho).
A heterocromatina constitutiva foi evidenciada principalmente nos
macrocromossomos das duas espécies, sendo que alguns microcromossomos
apresentaram marcações difusas, impossibilitando a localização exata desses blocos.
Entretanto, o padrão de banda C foi distinto para cada espécie.
Em Eunectes murinus, além de marcações biteloméricas, o par 1 tem um bloco
heterocromático intersticial no braço curto e dois blocos intersticiais no braço longo. O
par 2 também apresenta blocos biteloméricos e um bloco intersticial no braço curto. O
par 3 apresentou blocos heterocromáticos na região centromérica e biteloméricas. Os
pares 4, 6, 7 têm marcações biteloméricas, enquanto os pares 5 e 8 apresentam
marcações centroméricas (Figura 9 A).
Em Eunectes notaeus, o par número 1, assim como em Eunectes murinus
apresentou marcações biteloméricas, um bloco na região intersticial do braço curto e

33
dois blocos na região intersticial do braço longo. O par 2 apresentou blocos
biteloméricos, centromérico e na região intersticial do braço longo. O par 3 apresentou
marcações centromérica e terminal no braço curto. O par 4 apresentou blocos terminais
no braço curto. Os pares 5 e 6 apresentaram marcações apenas centroméricas e os
pares 7 e 8 apresentaram blocos em posição bitelomérica (Figura 9 B).
Figura 9 – Cariótipos de Eunectes: (A) Eunectes murinus e (B) Eunectes notaeus, com suas respectivas regiões heterocromáticas, evidenciadas pelo bandeamento C.
O mapeamento cromossômico de sequências teloméricas, além de revelar
marcações terminais em todos os cromossomos do complemento, também evidenciou
sequências teloméricas repetitivas intersticiais (ITS) em Eunectes murinus, que
apresentou ITS nos pares 1, 2 e 6. No par 1 as marcações foram evidenciadas em
região pericentromérica, do braço curto. No par 2 foram encontradas duas ITS, uma na
região intersticial do braço curto e a outra do braço longo. No par 6 a ITS foi
evidenciada na região intersticial do braço longo (Figura 10 A). Eunectes notaeus, por

34
sua vez não apresentou nenhum vestígio de ITS, o que consequentemente culmina em
dois padrões distintos de sequências teloméricas para Sucuris (Figura 10 B).
Figura 10 – Cariótipos de Eunectes: (A) Eunectes murinus e (B) Eunectes notaeus, hibridizados com
sonda telomérica, evidenciando as ITS em apenas uma das espécies (sinais em vermelho).
4.3 Corallus hortulanus
Para essa espécie, as fêmeas apresentaram número diploide igual a 40
cromossomos com fórmula cariotípica igual a 6m+16st+18mi e número de braços (NF)
igual a 62, enquanto os machos apresentaram número diploide igual a 39
cromossomos com fórmula cariotípica 5m+16st+18mi e número de braços (NF) igual a
60. Os pares 1 e 5 são metacêntricos e os demais subtelocêntricos. Entretanto, as
fêmeas apresentaram mais dois cromossomos metacêntricos, sendo um de tamanho

35
médio (com seu correspondente nos machos) e um pequeno, mas de forma bem
definida, diferenciando assim dos microcromossomos. Esta diferença cromossômica
entre macho e fêmea sugere tratar-se de um sistema heteromórfico de determinação
do sexo. Entretanto, os dois sexos se apresentam heterogaméticos (Figura 11 A, B).
A impregnação com nitrato de prata (Ag-RON) evidenciou a região organizadora
de nucléolo em apenas um par de microcromossomos em ambos os sexos. Em
algumas ocasiões, os microcromossomos portadores da região organizadora de
nucléolo foram encontrados em associação. O mapeamento da sequência de DNA
ribossomal 18S revelou apenas um par de microcromossomos portador da região de
DNAr 18S, confirmando as marcações obtidas pela impregnação por nitrato de prata
(Figura 11, em destaque).
Figura 11 – Cariótipos de Corallus hortulanus: (A) Fêmea e (B) Macho, com suas respectivas regiões organizadoras de nucléolo em um par de microcromossomos, detectada por Ag-NO3 e pela FISH com DNAr 18S (sinal em vermelho).

36
A heterocromatina constitutiva foi evidenciada principalmente nos
macrocromossomos, sendo que alguns microcromossomos, assim como as outras
espécies analisadas, apresentaram marcações difusas, impossibilitando a localização
exata desses blocos. Entretanto, não houve diferença significativa entre macho e
fêmea. Nos pares 1 e 5, metacêntricos, as marcações foram no centrômero,
biteloméricas e um bloco intersticial no braço curto. Nos demais pares, as marcações
foram biteloméricas, com exceção do par 3 que também apresentou um bloco
intersticial no braço longo. Para o suposto par sexual, o cromossomo metacêntrico
maior apresenta um bloco heterocromático centromérico, já o pequeno não apresentou
heterocromatina evidente (Figura 12).
Figura 12 – Cariótipo de Corallus hortulanus, com suas respectivas regiões heterocromáticas, evidenciadas pelo bandeamento C.
O mapeamento cromossômico de sequências teloméricas para Corallus
hortulanus, além de revelar marcações terminais em todos os cromossomos do
complemento, também evidenciou sequências teloméricas repetitivas intersticiais (ITS)
em um par de macrocromossomos. O par 2 apresentou uma ITS em posição intersticial
do braço longo que não coincide com heterocromatina constitutiva (Figura 13).

37
Figura 13 – Cariótipo de Corallus hortulanus Fêmea, hibridizado com sonda telomérica, evidenciando
uma ITS (sinais em vermelho).
5. Discussão
5.1 A macroestrutura cromossômica de Boidae
O estudo cromossômico em serpentes não tem ocorrido de forma ampla como
em outros grupos de vertebrados, sendo que a maioria dos estudos restringe-se a
descrições cariotípicas em coloração convencional. Para a Amazônia, em particular,
estes estudos são ainda mais restritos. Como já é sabido, o cariótipo, bem como os
marcadores cromossômicos são de grande utilidade para a sistemática e para a
definição de espécies, principalmente quando características morfológicas não são
suficientes para que problemas taxonômicos sejam esclarecidos. Ainda, com o avanço
da citogenética molecular, que oferece diferentes abordagens, tem sido possível
verificar diferenças na estrutura do genoma e responder perguntas sobre a biologia
evolutiva nos mais diversos grupos de vertebrados (Bickham e Rogers 1985; King et al.
1986; Porter et al. 1991; 1994; Matsuda et al. 2005; Ezaz et al. 2005; 2006; Matsubara
et al. 2006; 2014; Pokorná et al. 2011; Srikulnath et al. 2011).
No presente trabalho, ocorreu uma caracterização citogenética de três espécies
e três subespécies, com a descrição de três cariótipos inéditos de serpentes da família
Boidae, com variações intraespecíficas de coloração e manchas da pele.
Características cromossômicas foram evidenciadas com mais acuracidade, uma vez

38
que, preparações cromossômicas foram obtidas por meio de cultura de sangue, o que
perimitiu um maior alongamento e um melhor espalhamento dos cromossomos.
Os cariótipos foram assimétricos, com macro e microcromossomos, como já
descrito na literatura para serpentes. Para as espécies/subespécies de Boidae, aqui
analisadas, uma característica observada, e mantida, é quanto ao tamanho dos três
primeiros pares de macrocromossomos, que são bem maiores em relação aos outros
macrocromossomos do complemento, à exceção de Corallus hortulanus em que os
mesmos decrescem de forma gradual. Esse padrão já foi observado em outras
espécies de serpentes (Beçak et al. 1964; Beçak e Beçak 1969; Carr e Bickham 1981;
Reed et al. 1991; Garnero et al. 2006), em tartarugas, como a espécie Phrynops
geoffroanus (Viana, comunicação pessoal), em lagartos (King e King 1975) e anfíbios
(Sessions 2008).
A presença e manutenção dos microcromossomos nos Boidae analisados não é
um fato incomum dentro da classe Reptilia, onde a grande maioria apresenta um
complemento cromossômico dividido em macrocromossomos e microcromossomos
(Beçak 1965; Stock 1972; Mengden e Stock 1980; Pellegrino et al. 1999; Srikulnath et
al. 2011) porém, também são encontrados em outras classes de vertebrados, variando
em tamanho e quantidade (Morescalchi 1979; Nanda et al. 2002; Ledesma et al. 2006;
Sessions 2008). Para serpentes, praticamente todas as espécies apresentam a
característica de um cariótipo bimodal, no entando existem algumas exceções como
espécies da família Colubridae, dos gêneros Thamnodynastes, Hydrodnastes,
Erythrolamprus (Beçak e Beçak 1969). O mesmo já foi relatado para quelônios da
família Podocnemididae (Fantin e Monjeló 2011) e em Crocodylia (King et al. 1986).
No presente estudo, foi descrito pela primeira vez o cariótipo de Boa constrictor
occidentalis, que assim como as outras duas subespécies de jiboias pertencentes ao
complexo Boa constrictor, que já tinham seu cariótipo descrito, apresentou o mesmo
complemento cariotípico, ou seja número diploide igual a 36 cromossomos, com 16
macrocromossomos e 20 microcromossomos (Beçak et al. 1963; Beçak 1966; Camper
e Hanks 1995). Entretanto, diferenças quanto às fórmulas cariotípicas foram

39
observadas entre os exemplares das três subespécies de Boa constrictor aqui
analisados e os exemplares de B. c. amarali e B. c. constrictor analisadas por Beçak et
al. (1963). Esta deveu-se à separação dos cromossomos submetacêntricos e
acrocêntricos, onde Beçak et al. (1963) consideraram 6m+2sm+8a+20Mi e no presente
estudo considerou-se 6m+2sm+4st+4a+20mi. Essa diferença dos tipos cromossômicos
foi devida, provavelmente, ao grau de compactação diferencial dos cromossomos e
qualidade da preparação cromossômica.
Interessante notar que, em coloração convencional e pela região organizadora
do nucléolo, os cariótipos das três subespécies são muito semelhantes e nenhuma
diferença em relação a cromossomos sexuais diferenciados foi evidenciada. A família
Boidae ocupa uma posição basal entre as serpentes e heteromorfismo de
cromossomos sexuais é encontrado, mais frequentemente, nos táxons derivados
(Beçak e Beçak 1969; Singh 1972a, 1972b; Gorman 1981; Olmo e Signorino 2005;
Oguiura et al. 2009; Pithong et al. 2013), porém há exceções como Acrantophis
dumerili (Sanziniidae), que apresenta cromossomos sexuais diferenciados (Mengden e
Stock 1980).
No caso de Eunectes, em coloração convencional, as duas espécies de
Anacondas analisadas no presente estudo só se diferenciaram das três subespécies de
Boa pela presença de dois pares a mais de macrocromossomos do tipo
subtelocêntricos que nas subespécies de Boa são do tipo acrocêntricos. Por outro lado,
Corallus hortulanus tem um cariótipo totalmente diferenciado, tanto em relação ao
número diploide como à fórmula cariotípica. O número de microcromossomos é menor
e a diferença no número diploide entre machos (39) e fêmeas (40) sugere a presença
de cromossomos sexuais múltiplos.
O padrão de bandeamento C descrito para B. constrictor constrictor, Boa
constrictor occidentalis e Boa constrictor amarali, mostrou uma baixa quantidade de
heterocromatina constitutiva, principalmente quando comparado ao tamanho dos
macrocromossomos. Pode-se dizer que há uma ampla variação quanto ao padrão de
bandeamento C em serpentes basais, onde por exemplo, Boa apresenta pouca

40
quantidade de heterocromatina constitutiva e Sanzinia apresenta grandes blocos
heterocromáticos (Mengden e Stock 1980).
No presente trabalho, nenhuma das espécies/subespécies de Boa e Eunectes
apresentou diferenças na distribuição dos blocos heterocromáticos entre machos e
fêmeas e nem entre localidades de coleta, descartando assim a presença de
cromossomos sexuais diferenciados nestas serpentes, como descrito para vários
répteis que possuem sistema de determinação sexual do tipo ZZ/ZW, em que o W é
parcialmente ou totalmente heterocromático (Mengden e Stock 1980; Mengden 1981;
Oguiura et al. 2009; Pinthong et al. 2013).
O padrão de banda C foi específico para cada espécie/subespécie, o que o faz
um excelente marcador citogenético e taxonômico para diferenciar todas as serpentes
Boidae aqui analisadas.
Para as subespécies de Boa, os blocos dos pares 2 e 4 apresentam uma alta
homeologia entre os cariótipos, no entanto a variação presente a partir do quinto par de
macrocromossomos encontra forte apoio nos clados recuperados em filogenia proposta
por Reynolds et al. (2014), em que as subespécies analisadas no presente estudo
estão em clados distintos. Hyncová et al. (2009) sugeriram que a subespécie Boa
constrictor imperator fosse elevada à categoria de espécie, devido às divergências
encontradas entre ela e as outras subespécies de Boa constrictor. No entanto, ainda
não existem dados citogenéticos para essa “espécie”, o que seria um grande avanço
para a compreensão da atual taxonomia do gênero.
Em Eunectes, o par número 1 é homeólogo e conservado entre as duas
espécies analisadas, sendo estas, as que apresentam a maior quantidade de
heterocromatina constitutiva, quando comparadas a Boa e Corallus. Contudo, essas
serpentes ainda apresentam uma quantidade escassa de blocos heterocromáticos
quando comparadas a outras serpentes.
Uma característica notável e surpreendente evidenciada aqui é com relação à
distribuição dos blocos heterocromáticos tanto em macrocromossomos quanto em
microcromossomos, que não evidenciou nenhuma diferença entre os indivíduos de
diferentes localidades e nenhuma relação com os padrões de coloração e de manchas

41
da pele dos animais, principalmente em Corallus hortulanus, sendo esta a mais
polimórfica com relação à coloração e padrões de manchas na pele.
Esses blocos heterocromáticos em microcromossomos mostraram-se
indistinguíveis, bem como no cromossomo menor do possível par sexual em fêmas de
Corallus hortulanus.
5.2 Evolução das famílias gênicas
No que refere-se à localização da região organizadora de nucléolo (RON), o
padrão observado foi o mesmo já descrito anteriormente para outras espécies (Camper
e Hanks 1995), ou seja, RON simples em um par de microcromossomos para todas as
espécies analisadas nesse estudo. Os microcromossomos, mesmo sendo
considerados conservados nas serpentes (Mengden e Stock 1980; Nanda et al. 2002),
não estão isentos de rearranjos cromossômicos ou outros eventos como crossing over
desigual.
A RON em um par de microcromossomos é considerada uma característica
plesiomórfica para várias famílias de Serpentes que apresentam 2n=36 cromossomos,
com 16 macrocromossomos e 20 microcromossomos (Moreno et al. 1987; Porter et al.
1991; 1994; Camper e Hanks 1995; Aprea et al. 2006.). Por outro lado, existe uma
grande variação quanto à localização deste marcador, como já descrito para algumas
espécies de serpentes pertencentes a linhagens derivadas, que apresentam a RON em
macrocromossomos e em microcromossomos, caracterizando RON múltipla (Moreno et
al. 1987; Camper e Hanks 1995; Garcia e Hernando 2007). Trajtengertz et al. (1995)
evidenciaram DNAr em microcromossomos e em macrocromossomos de diferentes
populações da serpente Bothrops neuwiedi, pertencente a uma linhagem derivada, o
que corrobora a hipótese de Camper e Hanks (1995), que sugerem a presença de
rearranjos tipo translocação, mudando a região nucleolar de micro para
macrocromossomo, uma vez que o número de microcromossomos se mantém.
Porter et al. (1991), por meio da hibridização in situ fluorescente com sondas de
DNAr 28S em diferentes espécies de serpentes de diferentes famílias, também

42
constataram variabilidade na localização desta sequência em cromossomos, onde
Thamonophis marcianus e Masticophis flagellum (Colubridae) apresentaram
marcações simples em macrocromossomos, enquanto Crotalus viridis (Viperidae)
apresentou marcações múltiplas em dois pares de microcromossomos. Padrão
semelhante foi evidenciado em Bothrocophias hyoprora (Viperidae), que teve
marcações múltiplas em dois pares de microcromossomos, evidenciadas pela
impregnação por nitrato de prata (Carvalho et al. 2013). Também com a impregnação
com nitrato de prata e mapeamento de DNAr 18S foi evidenciado em Spilotes
sulphureus (Colubridae) marcações simples em um único par de macrocromossomos
(Viana, comunicação pessoal), sendo essas características, de RON, DNAr 18S e 28S
com marcações simples em um par de macrocromossomo e múltipla tanto em macro
quanto em microcromossomos, peculiaridades típicas de serpentes pertencentes a
linhagens intermediárias e derivadas.
Inúmeros estudos reforçam que a detecção do marcador RON em Squamata,
bem como estudos comparativos entre diferentes grupos podem fornecer informações
fundamentais para a elucidação do processo evolutivo desses animais (Odierna et al.
1987; Porter et al. 1991; 1994; Pellegrino et al. 1999; Camper e Hanks 1995, Srikulnat
et al. 2009), entretanto para as serpentes estudadas aqui, a RON não se mostrou como
um marcador citogenético e taxonômico eficiente porém, essas marcações simples em
um par de microcromossomos mostraram-se como uma caracteristica conservada
entre serpentes da linhagem basal. Destaca-se então a importância deste e de novos
estudos, usando este marcador em cromossomos de serpentes para estabelecer-se
um padrão evolutivo dentro do grupo.
Um outro marcador, utilizado no presente trabalho e que, sem dúvidas alguma,
abriu um leque de novas perguntas e possibilidades, foi o mapeamento cromossômico
de sequências teloméricas, que no presente estudo evidenciou as marcações terminais
em todos os cromossomos do complemento. Mesmo que de forma indistinta, os
microcromossomos também apresentaram marcações teloméricas evidentes, padrão
semelhante já descrito para microcromossomos de aves, serpentes, lagartos,
tartarugas e crocodilianos, identificando telômeros funcionais (Meyne et al. 1989;

43
Pellegrino et al. 1999; Nanda et al. 2002; Srikulnath et al. 2011, Viana, comunicação
pessoal). Entretanto, além destas marcações, foram observadas sequências
teloméricas intersticiais (ITS) nos macrocromossomos das três subespécies de Boa
constrictor, em Corallus hortulanus e em Eunectes murinus (Figuras 7, 10 e 13). A
intensidade dos sinais teloméricos variou consideravelmente tanto em macro quanto
em microcromossomos em todas as espécies analisadas, sugerindo que as sequências
tenham tamanhos diferenciais (TTAGGG)n entre os pares cromossômicos.
As ITS apresentam diferentes origens entre as várias ordens de vertebrados
(Wurster-Hill et al. 1988; 1992; Meyne 1988; Meyne et al. 1989; Meyne et al. 1990;
Wiley et al. 1992; Lee et al. 1993; Schmid et al. 1994; Svartman e Vianna-Morgante
1998; Pellegrino et al. 1999; Ruiz-Herrera et al. 2002; Metcalfe et al. 2004), como a
ação da telomerase ao reparar quebras cromossômicas, acrescentando sequências
teloméricas em regiões não terminais (Flint et al. 1994; Azzalin et al. 2001, Nergadze et
al. 2007), resquícios da sequência (TTAGGG)n como resultado de fusões, fissões ou
inversões que perderam sua função em posição intersticial ou centromérica, ou mesmo
duplicações desta sequência (Wiley et al. 1992; Meyne et al. 1990), que é o que parece
ter ocorrido no último par de macrocromossomo acrocêntrico de Boa constrictor
occidentalis (Figura 7 C).
Acreditamos que não necessariamente todas essas ITS encontradas nas
serpentes aqui analisadas sejam indicativas de rearranjos cromossômicos, ainda mais
pelo fato de nem todas estarem associadas com heterocromatina constitutiva, mas
existe sim, uma grande possibilidade de estarem associadas a elementos transponíveis
como o “Tel-1”, já documentado em outros grupos de vertebrados (Cherry e Blackburn
1985; Wynen e Blackburn 1991; Nergadze et al. 2007). Além disso, a composição
gênica das ITS não se restringe somente à sequência (TTAGGG)n, podendo
frequentemente estarem associadas a DNA satélite (Lee et al. 1997; Arnason et al.
1988; Garrido-Ramos et al. 1998).
A presença de sequências repetitivas, como DNA satélite e elementos
transponíveis, pode levar a uma inativação temporária ou eliminação física de

44
sequências teloméricas em porções terminais de cromossomos, o que ocasionaria os
rearranjos e consequente presença das mesmas em posições intersticiais e/ou
centroméricas, sendo consideradas como hotspots para recombinação e evolução
cariotípica (Slijepcevic 1998; Cherry e Blackburn 1985; Wyman e Blackburn 1991;
Ashley e Ward 1993; Nergadze et al. 2007). Assim, destaca-se a importância do
mapeamento de sequências teloméricas realizado nas serpentes no presente estudo,
que revelaram um novo panorama sobre a evolução cariotípica para essas serpentes
neotropicais e para a superfamília Booidea.
5.3 Cromossomos Sexuais
A complexidade e diversidade de mecanismos de determinação sexual em
répteis precedem uma evidente história evolutiva de transição entre sistemas de
determinação sexual por temperatura e determinação sexual genética, com
cromossomos sexuais heteromórfios ou homomórficos (Shine et al. 2002; Mank et al.
2006; Radder et al. 2008; Quiin et al. 2011).
Acredita-se que sistemas de cromossomos sexuais heteromórficos simples e
múltiplos, em répteis, evoluíram a partir de uma condição homomórfica, onde
primariamente o sexo era definido por temperatura, sendo progressivamente
diferenciados até uma condição irreversível de heteromorfia (Bull 1983; Charlesworth
1991), em que as mudanças ao nível de DNA estariam principalmente envolvidas com
o cromossomo W ou Y (Matsuda et al. 2005), evoluindo de forma progressiva até
atingir uma condição especializada de determinação do sexo. Porém, o contrário
também pode ocorrer, ou seja, um organismo que possui um sistema genético, definido
e eficiente de determinação do sexo voltar a apresentar a determinação sexual por
temperatura, o que é uma novidade evolutiva sem muitas explicações (Harlow 2001;
Sarre et al. 2004; Ezaz et al. 2005; Radder et al. 2008; Quiin et al. 2011; Sarre et al.
2011).
Quiin et al. (2011) evidenciaram muito bem esse mecanismo, onde ovos de
lagartos que tem um sistema genético para a determinação sexual (SGD), quando
incubados em temperaturas elevadas, nasciam do tipo fêmea, mesmo alguns desses

45
filhotes tendo o genótipo para o sexo masculino. Ou seja, a temperatura age de forma
conjunta ou complementar à determinação sexual em organismos que tenham a
determinação sexual genética, ou influenciando de forma significativa nesse
mecanismo. O que acaba sendo uma preocupação com as constantes elevações de
temperatura, uma vez que isso também ocorre em populações naturais, podendo
alterar drasticamente a razão sexual dessas populações naturais.
A maioria dos estudos acerca dos cromossomos sexuais em serpentes mostra
diferenças significativas no tamanho e na posição do centrômero (Beçak 1965; 1966;
Singh 1972a; 1972b; Mengden e Stock 1980; Singh 2011) (Figura 14-1).
Por apresentar uma alta homeologia, muitos autores consideram o par
homomórfico número 4 em serpentes basais como sendo o par sexual (Beçak 1965;
Beçak 1966; Beçak e Beçak 1969; Mengden e Stock 1980; Singh 1972a). Essa teoria
foi reforçada pela análise do padrão de bandemanto G em espécies de Booidea de
Madagascar, onde o cromossomo Z de Sanzinia, que é homomórfico em relação ao
seu possível W, é presumidamente homeólogo ao de Acrantophis dumerili, que tem o
W diferenciado por uma inversão pericêntrica (Mengden e stock 1980) (Figura 14-2).
Presume-se que o mesmo tenha ocorrido em linhagens intermediárias, como exemplo,
algumas espécies da família Colubridae e Dipsadidae que apresentam o cromossomo
W com o braço curto menor em relação ao Z, sugerindo também uma inversão
pericêntrica (Figura 14-1). Esse mecanismo de inversão em um dos cromossomos do
par 4, quase sempre está associado a um processo de heterocromatinização, o que
pode estar associado à presença de DNA satélite e elementos transponíveis como
previsto por Singh et al. (1976; 1979), Kraemer e Schmidt (1993); Charlesworth et al.
(2005), Ezaz et al. (2013) e evidenciado por Vicoso et al. (2013). Assim, eventos como
esse impedem o crossing over devido a diferenças entre os homólogos, fazendo com
que um dos cromossomos, o W, passe a acumular cada vez mais genes determinantes
do sexo, por supressão de recombinação e por conseguinte a heterocromatinização.
Essa região do braço curto do cromossomo W (par 4), totalmente heterocromatinizada,

46
é perdida em serpentes de linhagens derivadas, como por exemplo, animais da família
Colubridae e Viperidae (Beçak 1965; Viana, comunicação pessoal).
Acredita-se, também, que mecanismos como adição de heterocromatina e
eventos epigenéticos também possam estar relacionados com essa diferenciação
gradual, de forma independente em diferentes linhagens de serpentes. Vicoso et al.
(2013) sugerem que essa evolução e diferenciação de cromossomos sexuais não
tenham ocorrido de forma independente, sendo essa supressão de crossing over e
heteromorfia surgido muito antes da divergência de Colubridae e Viperidae. Porém, no
estudo de Vicoso et al. (2013) foram analisadas apenas três espécies de serpentes,
sendo elas da família Boidae, Colubridae e Viperidae. Por outro lado, existe uma ampla
variedade de cromossomos sexuais heteromórficos e presença de homomorfia dentro
de Colubridae e Dipsadidae, que são de linhagens intermediárias (Beçak 1966; Beçak
e Beçak 1969; Beçak et al. 1971; Van Devender e Cole 1977; Mengden e Stock 1980),
o que sugere que essa evolução possa sim ter ocorrido de forma independente para
algumas linhagens. Isso mostra a necessidade de mais estudos comparativos de
espécies da mesma e de linhagens diferentes para a melhor compreensão da evolução
dos cromossomos sexuais em serpentes.
Figura 14. (1) Diversidade de cromossomos sexuais ZZ/ZW em serpentes brasileiras: (A) par 4,
homomórfico em linhagem basal (Boidae); (B; C; D) pares 4, heteromórfico em linhagens derivadas
(Colubridae e Dipsadidae); (E) par 4, heteromórfico de linhagem derivada (Viperidae). (2) Cromossomos
sexuais de serpentes de Madagascar: (A) par 4 homomórfico de Sanzinia; (B) par heteromórfico de
Acrantophis (Sanziniidae) (Beçak 1965; Beçak 1966; Mengden e Stock 1980).

47
Sistemas múltipos de determinação sexual estão presentes em diferentes
classes de vertebrados (Singh et al. 1970; Ray-Chaudhuri e Singh 1972a, 1972b;
Bertolotto et al. 2001; Rens et al. 2007; Oguiura et al. 2009; Veyrunes et al. 2008),
sendo que em serpentes este sistema é encontrado somente em linhagens derivadas
(Singh et al. 1970; Singh 1972a, 1972b; Ray-Chaudhuri e Singh 1972; Singh et al.
1979).
Em lagartos são encontrados sistemas simples de determinação sexual (XX/XY)
e (ZZ/ZW) e também sistemas múltiplos como (X1X1X2X2/X1X2Y e Z1Z1Z2Z2/Z1Z2W)
(Janzen e Paukstis 1991; Olmo e Signorino 2005), enquanto que em serpentes, até o
presente momento, foram encontrados apenas o sistema simples ZZ/ZW e múltiplos do
tipo (ZZ/ZW1W2 e Z1Z1Z2Z2/Z1Z2W), ou seja apenas as fêmeas são heterogaméticas
(Singh et al. 1970; Singh 1972a, 1972b; Ray-Chaudhuri e Singh 1972; Singh et al.
1979).
Em Boa, embora os cromossomos sexuais ZW sejam homomórficos, os
mesmos são tão diferenciados a nível molecular quanto em serpentes de linhagens
derivadas, apresentando uma composição gênica significativamente singular entre Z e
W, porém ainda continuam de forma eficiente a realizar “crossing over” (Hoff 2013;
Vicoso et al 2013). Com isso podemos hipotetizar que talvez esse sistema
homomórfico de determinação sexual, também esteja presente em outras espécies de
boídeos que não têm heteromorfismo cromossômico. Assim, em Corallus hortulanus,
caso o par 5, que é evidentemente homeólogo ao par cromossômico 4 em outras
espécies de Boideos Neotropicais, seja um par de cromomossos sexuais
homomórficos, este seria Z1Z1 no macho e Z1W1 na fêmea, o que não explica a
presença de dois cromossomos que não se emparelham na fêmea, tampouco a
presença de um cromossomo a menos nos machos. Da mesma forma que, se estes
cromossomos heteromórficos forem realmente sexuais, ocorreria a existência de
cromossomos sexuais múltiplos e dupla heterogametia. Portanto, apesar de haver
cromossomos heteromórficos e números diploides diferentes, a determinação sexual

48
em Corallus hortulanus não necessariamente esteja relacionada a isso, fato que
merece mais investigação para ser explicado.
5.4 Evolução cromossômica em Booidea
Beçak (1965), ao analisar o cariótipo de boídeos Neotropicais e se deparar com
um complemento macrocromossômico formado por subtelocêntricos e acrocêntricos
em Corallus caninus com 2n=44, presumiu que esse cariótipo fosse o basal para
Boidae e que o cariótipo com 2n=36 teria se originado a partir de fusões (Figura 15).
Figura 15. Idiograma representando a hipótese formulada por Beçak (1965), de origem dos cariótipos de
Boidae por fusões de macrocromossomos a partir de um cariótipo com 2n=44 (Corallus caninus),
formando um complemento com 2n=40, semelhante ao encontrado em Corallus grenadensis e Corallus
hortulanus (fêmeas) e por mais algumas fusões de macrocromossomos acrocêntricos ou
subtelocêntricos, formando um complemento com 2n=36, semelhante ao encontrado nas outras espécies
de Boidae como Boa, Eunectes, Epicrates e Chilabothrus.

49
Gorman e Gress (1970), por sua vez, ao analisar o cariótipo de uma fêmea de
Corallus grenadensis, sugeriram que o cariótipo dos boídeos Neotropicais, na verdade
teria evoluido a partir de um ancestral com 36 cromossomos, que, a partir de fissões
em macrocromossomos metacêntricos, teria originado o 2n=40, semelhante ao
encontrado em Corallus grenadensis (Figura 16) e por mais algumas fissões em
macrocromossomos metacêntricos teria formado o 2n=44, semelhante ao encontrado
em Corallus caninus.
Figura 16. Idiograma representando a hipótese formulada por Gorman e Gress (1970) para a origem dos
cariótipos de Boidae, por fissões cêntricas em macrocromossômos metacêntricos, originando o cariótipo
com 2n=40, semelhante ao encontrado em Corallus grenadensis.
Entretanto, discordamos da hipótese de Beçak (1965) e discordamos em parte
da hipótese de Gorman e gress (1970), pois ao analisarmos a árvore filogenética
proposta por Reynolds et al. (2014) para a superfamília Booidea (Figura 17), com a
sobreposição do número diploide das espécies já analisadas citogeneticamente,
incluindo as desse estudo, verificamos que o número diploide igual a 36 está presente
em Charinidae e Candoiidae, e que em Boidae não só no clado basal (Boa), mas
também nos clados derivados como Eunectes, Epicrates e Chilabothrus e que
2n=40/39 e 44 só estão presentes em Corallus, que não é o clado basal da família e
que, por sua vez, apresenta não só um número diploide diferente, mas também a
estrutura cariotípica. Além disso, Corallus caninus é clado irmão de Corallus hortulanus
e Corallus grenadensis e não foi evidenciado em Corallus hortulanus a presença de ITS

50
em macrocromossomos, correspondentes às fusões necessárias para obter-se um
cariótipo 2n=40 a partir de um 2n=44, visto em Corallus caninus, muito menos a
presença do possível sistema sexual encontrado em Corallus hortulanus. A ITS
presente no macrocromosssomo, par número 2 (st) de Corallus hortulanus, poderia ser
indicativa de uma inversão pericêntrica seguida de duplicações de fragmentos
(TTAGGG)n, o que seria equivalente ao par de macrocromossomo do tipo
submetacêntrico mantido em Boa, Eunectes, Epicrates e Chilabothrus, no entanto não
há heterocromatina constitutiva associada a essa região no par cromossômico número
2 de Corallus hortulanus, podendo este, ser resultado de um evento recente.
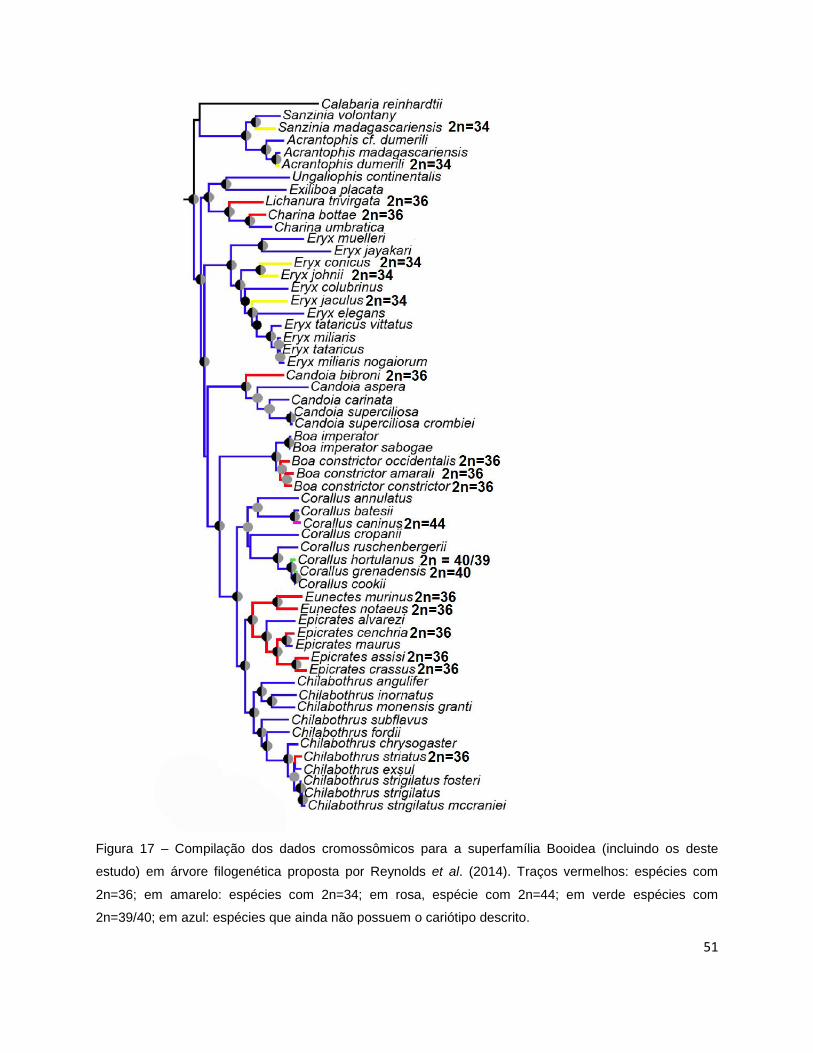
51
Figura 17 – Compilação dos dados cromossômicos para a superfamília Booidea (incluindo os deste
estudo) em árvore filogenética proposta por Reynolds et al. (2014). Traços vermelhos: espécies com
2n=36; em amarelo: espécies com 2n=34; em rosa, espécie com 2n=44; em verde espécies com
2n=39/40; em azul: espécies que ainda não possuem o cariótipo descrito.

52
O número diploide 2n=36 também está presente em grande parte das espécies
que compõem a superfamília Pythonoidea, que é irmã da superfamília Booidea e os
cariótipos com 2n=36 mantidos nos clados recuperados para a superfamília Booidea,
tanto em posição basal quanto em posição derivada, apresentam indícios de rearranjos
cromossômicos. Para as famílias Sanziniidae e Erycidae, o número diploide 2n=34
encontrado, poderia ser resultado de fusões de microcromossomos em
macrocromossomos ou até mesmo de deleções, uma vez que o número de
macrocromossomos se mantém, a exceção de Sanzinia madagascariensis, que
apresenta 9 pares de macrocromossomos (Branch 1980; Mengden e Stock 1980).
Sanziniidae é a família que apresenta o menor número de microcromossomos
dentro de Booidea, que assim como o clado formado por Corallus, formaram uma
macroestrutura cariotípica diferencial, sendo Sanzinia (2n=34 / 18M – 16mi) com um
número maior de macrocromossomos metacêntricos e um número menor de
microcromossomos (Branch 1980; Mengden e Stock 1980) e Achrantophis (2n=34 /
16M – 18mi) com uma composição similar aos outros Booideos com 2n=36, porém,
com uma inversão pericêntrica em um dos homólogos do par 4, que ocasionou a
primeira diferenciação cromossômica sexual entre machos e fêmeas dentro de
Sanziniidae e dentro de Booidea. Uma hipótese para o número menor de
microcromossomos encontrado em Sanziniidae, (16) em Sanzinia e (18) em
Achrantophis, seria rearranjos cariotípicos do tipo fusão entre microcromossomos e
macrocromossomos. Caso esses microcromossomos fusionados fossem os portadores
das regiões de DNAr 18S/28S, as regiões organizadoras de nucléolo estariam agora
em macrocromossomos, onde a confirmação dessas marcações nestes pares poderia
corroborar essa hipótese de fusões, tanto em Sanziniidae quanto em Erycidae. No
entanto, ainda são inexistentes dados cariotípicos sobre o mapeamento de regiões de
DNAr ou de cistrons ribossomais em Sanziniidae e Erycidae. Ainda assim, podemos
supor que esses possíveis eventos de fusão entre macro e microcromossomos teriam
ocorrido de forma independente ao longo da evolução da superfamília Booidea, uma
vez que o clado Sanziniidae é irmão do gênero Calabaria e o clado Erycidae é grupo
irmão de Candoiidae e Boidae (Figura 17).

53
Tendo em vista que a origem de diversificação entre Pytonoidea e Booidea
datam o Cretácio (Noonan e Chippindale 2006, Colston et al. 2013, Reynolds et al.
2014), podemos supor que o cariótipo ancestral para essas superfamílias irmãs teria
sido superior a 2n=36 com a presença de mais macrocromossomos subtelocêntricos ou
acrocêntricos, e que esse cariótipo ancestral já estaria envolvido em rearranjos
cromossômicos onde, por fusões nos macrocromossomos acro ou subtelocêntricos
teriam originado o cariótipo 2n=36, encontrado em grande parte das espécies de
Pythonoidea e Booidea (Figura 18).
Figura 18: Idiograma representando a hipótese de cariótipo ancestral para Pythonoidea e Booidea, onde
por fusões em macrocromossomos teria a formação do cariótipo 2n=36 encontrado em Pythonoidea e
Booidea.
Com relação à variação de número diploide encontrado dentro de Booidea , mais
especificamente em Boidae, que tem sua diversificação e origem mais recente (Colston
et al. 2013), acreditamos que o número diploide 2n=36 ancestral se manteve para o
clado Boa, tendo gradativamente a perda dos braços curtos nos dois últimos pares de
macrocromossomos, e que esse cariótipo ancestral tenha divergido de forma
independente, possivelmente por fissões em macrocromossomos metacêntricos e
submetacêntricos, como previsto por Gorman e Gress (1970), formando uma nova
estrutura cariotipica e composição diferencial para o clado Corallus, com 2n=44
(Corallus caninus), 2n=40 (Corallus grenadensis) e 2n=39/40 para Corallus hortulanus,
sendo esta última com um número menor de microcromossomos (18) e uma
diferenciação cromossômica entre os sexos. Já, para o clado irmão de Corallus,

54
formado por Eunectes, Epicrates e Chilabothrus, manteve-se a composição cariotípica
formada por 2n=36 e fórmula cariotípica 6m+2sm+8st+20mi e NF=52, característica
partilhada com Xenopeltidae, Loxocemidae e Pythonidae (Pythonoidea), sendo o
número de microcromossomos (20) e o par nucleolar simples, como uma característica
conservada para as espécies/subespécies de Boa, Eunectes e Epicrates. Além disso,
as ITS detectadas nos pares 2 e 4 de Boa constrictor occidentalis e de Boa constrictor
constrictor e nos pares 1 e 2 de Eunectes murinus podem ser resultado de fusões ou
inversões paracêntricas, o que reforça a hipótese do possível cariótipo ancestral para
Pythonoidea e Booidea ter sido superior a 2n=36. Entretanto, a partir de dados de
alguns bandeamentos já é possível sugerir que os cariótipos dos boídeos neotropicais
de fato não são tão conservados quanto se imaginava e que sua evolução vem
ocorrendo de forma independente nos gêneros.
5.5 Estrutura cromossômica de Corallus hortulanus e Corallus grenadensis
Gorman e Gress (1970) descreveram o cariótipo de quatro espécies de Boidae
neotropicais, dentre elas uma espécie de Corallus, na época denominada como
Corallus enhydris cooki, que veio a ser sinonimizada com Corallus hortulanus alguns
anos depois. O espécime em questão tratava-se de uma fêmea com 2n=40,
proveniente da ilha de Grenada, nas Índias ocidentais. Porém, na verdade tratava-se
de um espécime de Corallus grenadensis, endêmico da ilha. Corallus grenadensis, por
muito tempo foi mantida como sinonímia de Corallus hortulanus, e em estudo recente,
utilizando técnicas e ferramentas moleculares, Coulston et al. (2013) contestaram a
validade de Corallus grenadensis, porém Reynolds et al. (2014) e Pyron et al. (2014)
encontraram forte apoio e validade para a espécie endêmica da ilha de Grenada.
Portanto, consideramos neste estudo a primeira descrição cariotípica de Corallus
hortulanus da região amazônica.
Em seu estudo, Gorman e Gress (1970) relataram para Corallus grenadensis um
cariótipo diferente ao descrito neste estudo (2n=40, 4m+16a+20mi NF=44 ♀), com
diferenças significativas no número fundamental e fórmula cariotípica (Corallus

55
hortulanus 2n=40, 6m+16st+18mi, NF=62 ♀). Porém, esta diferença é devido à
qualidade da preparação cromossômica, que na época os impediu de visualizar a
morfologia exata de alguns cromossomos (Figura 19).
Figura 19. Cariótipo em coloração convencional de Corallus grenadensis, conforme Gorman e Gress
(1970).
Os autores também não relataram em Corallus grenadensis, o suposto par
sexual, aqui descrito para Corallus hortulanus, o qual os autores descreveram apenas
como microcromossomo, ou seja uma interpretação subestimada do cariótipo, como
revelado por Ezaz et al. (2005), que em muitos estudos os microcromossomos são
omitidos ou subestimados por serem de difícil caracterização, ou também pelo fato de
terem analisado apenas uma fêmea. Vicoso et al. (2013) também ressaltam essa
possibilidade de subestimação na interpretação de dados citogenéticos para a não
identificação de cromossomos sexuais. Assim, com os dados obtidos nesse estudo,
podemos confirmar que de fato Corallus grenadensis e Corallus hortulanus são
espécies diferentes e que a presença desse possível sistema heteromórfico de
cromossomos sexuais, bem como a dupla heterogametia entre machos (2n=39) e
fêmeas (2n=40), até o presente momento, é exclusivo para o clado Coralus hortullanus,
o que representa o primeiro caso de cromossomos sexuais heteromórficos para a
família Boidae.

56
6. Conclusões
● O cariótipo de boídeos Neotropicais não é tão conservado quanto era suposto.
● O padrão de bandeamento C de forma geral, mostrou-se um ótimo marcador para
diferenciar os táxons de boídeos Neotropicais.
● A citogenética clássica e molecular apresenta uma contribuição efetiva para a
taxonomia de serpentes.
● Os microcromossomos são de fato conservados no genoma dos boídeos, bem como
a presença da região organizadora de nucléolo em um único par deles.
● O homomomorfismo de cromossomos sexuais foi evidente em Boa e Eunectes, tanto
em coloração convencional quanto com bandeamento C.
● A presença de um possível sistema sexual em Corallus hortulanus é o primeiro caso
de cromossomos sexuais diferenciados em Boidae e o segundo para Booidea.
● Os resultados obtidos no presente trabalho, somados aos já descritos na literatura,
contribuem de maneira significativa para o entendimento da evolução cromossômica,
taxonomia e sistemática de Boideos neotropicais, pois a identificação e a posição
filogenética de algumas espécies ainda não estão bem definidas.

57
7.0 Referências
Amemiya, C.T.; Powers, P.K.; Gold, J.R. 1993. Chromosomal evolution in North
American fishes. In: Mayden, R.L. (ed.), Systematics, Historical Ecology and
North American Fishes, Stanford Univ. Press, Stanford, California. p. 515 – 533.
Aprea, G.; Gentilli, A.; Zuffi, M.A.L.; Odierna, G. 2006. The karyology of Vipera aspis, V.
atra, V. hugyi, and Cerastes vipera. Amphibia-reptilia, 27: 113– 119.
Arnason, U.; Allderdice, P.W.; Lien, J.; Widegren, B. 1988. Highly repetitive DNA in the
baleen whale genera Balaenoptera and Megaptera. Journal of Molecular
Evolution, 27: 217-221.
Ashley, T.; Ward, D.C. 1993. A “hot spot” of recombination coincides with an interstitial
telomeric sequence in the Armenian hamster. Cytogenetics and Cell Genetics,
62: 169–171.
Azzalin, C.M.; Nergadze, S.G.; Giulotto, E. 2001. Human intrachromosomal telomeric-
like repeats: sequence organization and mechanisms of origin. Chromosoma,
110: 75–82.
Baker, R.J.; Bull, J.J.; Mengden, G.A. 1971. Chromosomes of Elaphe subocularis
(Reptilia: Serpentes), with the description of an in vivo technique for preparation
of snake chromosomes. Experientia, 27: 1208-1229.
Beçak, W.; Beçak, M.L.; Nazareth, H.R.S. 1962. Karyotypic studies of two species of
South American snakes (Boa constrictor amarali and Bothrops jararaca).
Cytogenetics, 1: 305–313.
Beçak, W.; Beçak, M.L.; Nazareth, H.R.S. 1963. Chromosomes of snakes in short term
cultures of leucocytes. The American Naturalist, 97(895): 253-256.
Beçak, W.; Beçak, M.L.; Nazareth, H.R.S.; Ohno, S. 1964. Close karyological kinship
between the reptilian suborder Serpentes and the class Aves. Chromosoma, 15:
606–617.
Beçak, W. 1965. Constituição cromossômica e mecanismo de determinação do sexo
em ofídios sul americanos. I. Aspectos cariotípicos. Memórias do Instituto
Butantan, 32: 37-78.
Beçak, W. 1966. Constituição cromossômica e mecanismo de determinação do sexo
em ofídios sul americanos. II. Cromossomos sexuais e Evolução do Cariótipo.
Memórias do Instituto Butantan, 33(3): 775-798.

58
Beçak, W.; Carneiro, S.M.; Beçak, M.L. 1971. Cariologia comparada em seis espécies
de Colubrídeos (Serpentes). In: XXIII Reunião da SBPC e XVIII Reunião da
SBG. Ciência e Cultura, 23: 123.
Beçak, W.; Beçak, M.L. 1969. Cytotaxonomy and chromosomal evolution in Serpentes.
Cytogenetics, 8: 247-262.
Beebe, W. 1946. Field notes on the snakes of Kartabo, British Guyana, and Caripito,
Venezuela. Zoologica, 31: 11–64.
Bennett, D.C.; Lamoreux, M.L. 2003. The color loci of mice a genetic century. Pigment
Cell Research, 16: 333–344.
Bertolotto, C.E.V.; Rodrigues, M.T.; Yonenaga-Yassuda, Y. 2001. Banding patterns,
multiple sex chromosomes system and localization of telomeric (TTAGGG)n
sequences by FISH on two species of Polychrus (Squamata, Polychrotidae).
Caryologia, 54: 217–226.
Bianchi, N.O.; Beçak, W.; De Bianchi, M.S.; Beçak, M.L.; Rabello, M.N. 1969.
Chromosome replication in four species of snakes. Chromosoma, 26: 188–200.
Bickham, J.; W, Carr, J. L. 1983. Taxonomy and phylogeny of the higher categories of
cryptodiran turtles based on a cladistic analysis of chromosomal data. Copeia,
1983: 918-932.
Bickham, J.W.; Rogers, D.S. 1985. Structure and Variation of the nucleolus organizer in
turtles. Genetica, 67: 171-184.
Bisplinghoff, H.; Bellosa, H. 2007. Professional breeder series: Anacondas. Frankfurt
am Main, Germany, Edition Chimaira, 127 pp.
Boback, S.M.; Lynn, M.S. 2010. Variation in color and color change in Island and
Mainland Boas (Boa constrictor). Journal of Herpetology, 44(4): 506-515.
Bonny, K. 2007. Die Gattung Boa: Taxonomie und Fortpflangzung. Rheinstetten,
Germany, KUS-Verlag, 262 pp.
Boos, H.E.A. 2001. The snakes of Trinidad and Tobago. College Station, Texas. Texas
A&M University Press, 270 pp.
Branch, W. R. 1980: Chromosome morphology of the Madagascar tree boa Sanzinia
madagascariensis. South African Journal of Zoology, 152: 79-82.

59
Bull, J.J. 1983. Evolution of Sex Determining Mechanisms. Menlo Park, California:
Benjamin/Cummings 316 pp.
Burbrink, F.T. 2004. Inferring the phylogenetic position of Boa constrictor among the
Boinae. Molecular Phylogenetics and Evolution, 34: 167–180.
Burbrink, F.T.; Pyron, R.A. 2008. The taming of the skew: estimating proper confidence
intervals for divergence dates. Systematic Biology, 57: 317–328.
Burt, D.W.; Bruley, C.; Dunn, I.C.; Jones, C.T.; Ramage, A.; Law, A.S.; Morrice, D.R.;
Paton, I.R.; Smith, J.; Windsor, D.; Sazanov, A.; Fries, R.; Waddington, D. 1999.
The dynamics of chromosome evolution in birds and mammals. Nature, 402:
411-413.
Camper, J.D.; Hanks, B. 1995. Variation in the nucleolus organizer region among New
World snakes. Journal of Herpetology, 29: 468-471.
Carr, J.L.; Bickham, J. 1981: Sex chromosomes of the Asian black pond turtle,
Siebenrockiella crassicollis (Testudines: Emydidae). Cytogenetics and Cell
Genetics, 31: 178-183.
Carvalho, V.T.; Fraga, R.; Eler, E.S.; Kawashita-Ribeiro, R.A.; Feldberg, E.; Vogt, R.;
Carvalho, M.A.; Noronha, J.C.; Condrati, L.H.; Bittencourt, S. 2013. Toad-headed
pitviper Bothrocophias hyoprora (Amaral, 1935) (Serpentes, Viperidae): New
records of geographic range in Brazil, hemipenial morphology, and chromosomal
characterization. Herpetological Review, 44(3): 410-414.
Charlesworth, B. 1991. The evolution of sex chromosomes. Science, 251: 1030-1033.
Charlesworth, D.; Charlesworth, B.; Marais, G. 2005. Steps in the evolution of
heteromorphic sex chromosomes. Heredity, 95: 118–128.
Cherry, E.H.; Blackburn, J.M. 1985. The internally located telomeric sequences in the
germ-line chromosomes of Tetrahymena are at the ends of transposon-like
elements. Cell, 43: 747–758.
Colston, T.J.; Grazziotin, F.G.; Shepard, D.B.; Vitt, L.J.; Colli, G.R.; Henderson, R.W.;
Hedges, S.B.; Bonatto, S.; Zaher, H.; Noonan, B.P.; Burbrink, F.T. 2013.
Molecular systematics and historical biogeography of tree boas (Corallus spp.).
Molecular Phylogenetics and Evolution, 66: 953–959.

60
Corso, J.; Gonçalves, G.L.; de Freitas, T.R. 2012. Sequence variation in the
melanocortin-1 receptor (MC1R) pigmentation gene and its role in the cryptic
coloration of two South American sand lizards. Genetics and Molecular Biology,
35(1): 81–7.
Cox, C.L; Rabosky, A.R; Chippindale, P.T. 2013. Sequence variation in the Mc1r gene
for a group of polymorphic snakes. Gene, 513: 282-286.
De Smet, H.O. 1978. The chromosomes of the 23 species of snakes. Acta Zoologica et
Pathologica, 70: 85-115.
Dirksen, L.; Böhme, W. 1998. Studien an Anakondas 2: Zum taxonomischen Status von
Eunectes murinus gigas (Latreille, 1801) (Serpentes: Boidae), mit neuen
Ergebnissen zur Gattung Eunectes Wagler 1830. Salamandra, 34: 1–16.
Dirksen, L. 2002. Anakondas: Monographische Revision der Gattung Eunectes Wagler,
1830 (Serpentes, Boidae). Münster, Germany, Natur-und-Tier-Verl., 189 pp.
Duellman, W.E. 2005. Cusco Amazonico: the lives of amphibians and reptiles in an
Amazonian rainforest. Ithaca, N.Y., Comstock Publ., 433 pp.
Eizirik, E.N.; Yuhki, W.E.; Johnson, M.; Menotti-Raymond, S.S.; Hannah, S.J.; O’Brien.
2003. Molecular genetics and evolution of melanism in the cat family. Current
Biology, 13: 448–453.
Ernest, C.H.; Zug, G.R. 1996. Snakes in question. Washington: Smithsonian Institution
Press. 203pp.
Ezaz, T.; Quinn, A.E.; Miura, I.; Sarre, S.D.; Georges, A.; et al. 2005. The dragon lizard
Pogona vitticeps has ZZ/ZW micro-sex chromosomes. Chromosome Research,
13: 763–776.
Ezaz, T.; Valenzuela, N.; Grützner, F.; Miura, I.; Georges, A.; Burke, R.L.; Graves,
J.A.M. 2006. An XX/XY sex microchromosome system in a freshwater turtle,
Chelodina longicollis (Testudines: Chelidae) with genetic sex determination.
Chromosome Research, 14: 139-150
Ezaz, T.; Azad, B.; O’Meally, D.; Young, M.J.; Matsubara, K.; et al. 2013. Sequence and
gene content of a large fragment of a lizard sex chromosome and evaluation of
candidate sex differentiating gene R-spondin 1. BMC Genomics, 14: 899.
Fantin, C.; Monjeló, L.A.S 2011. Cytogenetic studies in Podocnemis expansa and
Podocnemis sextuberculata (Testudines, Podocnemididae), turtles of the
Brazilian Amazon. Caryologia, 64: 154-157.

61
Flint, J.; Craddock, C.F.; Villegas, A. et al. 1994. Healing of broken human
chromosomes by the addition of telomeric repeats. The American Journal of
Human Genetics, 55: 505–512.
Fogel, D. 1997. Captive husbandry and propagation of the Boa constrictor and related
boas. Melbourne, Fla., Krieger Publ., 98 pp.
García, J.A.R.; Hernando, A. 2007. Standard karyotype and nucleolus organizer region
of Neotropical blindsnake Typhlops brongersmianus (Serpentes: Typhlopidae).
Acta Herpetologica, 2: 117–120.
Garnero, A.V.; Ledesma, M.A.; Gunski, R.J. 2006. Alta homeologia cariotípica na
família Tinamidae (Aves, Tinamiformes). Revista Brasileira de Ornitologia, 14:
53–58.
Garrido-Ramos, M.A.; Herrán, R.; Ruiz-Rejón, C.; Ruiz-Rejón, M. 1998. A satellite DNA
of the Sparidae family (Pisces, Perciformes) associated with telomeric
sequences. Cytogenetics and Cell Genetics, 83: 3-9.
Gorman, G.C.; Gress, F. 1970. Chromosome cytology of four boid snakes and a varanid
lizard, with comments on the cytosystematics of primitive snakes. Herpetologica,
26(3): 308-317.
Gorman, G. 1981. The chromosomes of Laticauda and a review of karyotypic evolution
in the Elapids. Journal of Zoology, 15: 225-233.
Graham, G. 1977. The karyotype of the Texas coral snake, Micrurus fulvius tenere.
Herpetologica, 33: 345–348.
Gray, S.M.; McKinnon, J.S. 2006. Linking color polymorphism maintenance and
speciation. Trends Ecology and Evolution, 22: 71–79.
Gross, M.C.; Schneider, C.H.; Valente, G.T.; Porto, J.I.R.; Martins, C.; Feldberg, E.
2010. Variability of 18S rDNA locus among Symphysodon fishes: chromosomal
rearrangements. Journal of Fish Biology, 76: 1117–1127.
Gutiérrez, J.M.; Bolaños, R. 1979. Cariótipos de las principales serpientes coral
(Elapidae: Micrurus) de Costa Rica. Revista de Biología Tropical, 27: 57–73.
Gutiérrez, J.M.; Solorzano, A.; Cerdas, L. 1988. Karyotypes of five Species of coral
snakes (Micrurus). Journal of Herpetology, 22: 109-112.

62
Harlow, P.S. 2001. The Ecology of Sex-Determining Mechanisms in Australian Agamid
Lizards. PhD thesis. School of Biological Sciences. Macquarie University,
Sydney.
Henderson, R.W. 1993a. On the diets of some arboreal boids. Herpetological Natural
History, 1: 91-96.
Henderson, R.W. 1993b. Foraging and diet in West Indian Corallus enydris (Serpentes:
Boidae). Journal of Herpetology, 27: 24-28.
Henderson, R. W. 1997. A taxonomic review of the Corallus hortulanus complex of
Neotropical tree boas. Caribbean Journal of Science, 33: 198–221.
Henderson, R.W.; Hedges, S.B. 1995. Origin of West Indian populations of the
geographically widespread boa Corallus enydris inferred from mitochondrial DNA
sequences. Molecular Phylogenetics and Evolution, 4: 88–92.
Henderson, R.W.; Micucci, T.W.P.; Puorto, G.; Bourgeois, R.W. 1995. Ecological
correlates and patterns in the distribution of Neotropical boines (Serpentes:
Boidae): a preliminary assessment. Herpetological Natural History, 1: 15-27.
Henderson, R.W.; Passos, P.; Feitosa, D. 2009. Geographic variation in the Emerald
Treeboa, Corallus caninus (Squamata: Boidae). Copeia, 2009: 572–582.
Hoekstra, H.E.; Hirschmann, R.J.; Bundley, R.A.; Insel, P.A.; Crossland, J.P. 2006. A
single amino acid mutation contributes to adaptive beach mous color pattern.
Science, 313: 101–104.
Hoff, M. 2013. Slithering Toward Clarity: Snakes Shed New Light on the Evolution and
Function of Sex Chromosomes. PLoS Biology, 11(8): e1001644.
Hoorn, C.; Wesselingh, F.P. 2010. Amazonia, landscape and species evolution: a look
into the past. London, UK: Wiley-Blackwell. 464 pp.
Howell, W.M.; Black, D.A. 1980. Controlled silver-staining of nucleolus organizer region
with a protective colloidal developer: a 1-step method. Experientia, 36: 1014-
1015.
Huang, P. 2006. “Introdução” http://www.corallus.com/hortulanus/index.html.
Hubbard, J.K.; Uy, J.A.C.; Hauber, M.E.; Hoekstra, H.E.; Safran, R.J. 2010. Vertebrate
pigmentation: from underlying genes to adaptive function. Trends Genetics, 26:
231–239.

63
Hynková, I.; Starostová, Z.; Frynta, D. 2009. Mitochondrial DNA variation reveals recent
evolutionary history of main Boa constrictor clades. Zoological Science, 26: 623–
631.
Ijdo, J.W.; Wells, R.A.; Baldini, A.; Reeders, S.T. 1991. Improved telomere detection
using a telomere repeat probe (TTAGGG)n generated by PCR. Nucleic Acids
Research, 19(17): 4780.
Janzen, F.J.; Paukstis, G.L. 1991. Environmental sex determination in reptiles: ecology,
evolution, and experimental design. Quarterly Review of Biology, 66: 149-179.
Johnson, J.A. 2012. Genetics of plumage color in the gyrfalcon (Falco rusticolus):
analysis of the melanocortin-1 receptor gene. Journal of Heredity, 103: 315–321.
Kawai, A.; Nishida-Umehara, C.; Ishijima, J.; Tsuda, Y.; Ota, H.; Matsuda, Y. 2007.
Different origins of bird and reptile sex chromosomes inferred from comparative
mapping of chicken Zlinked genes. Cytogenetics Genome Research, 117: 92–
102.
King, M.; Honeycutt, R.; Contreras, N. 1986. Chromosomal repatterning in crocodiles:
C, G, and N-banding and the in situ hybridization of 18S and 26S rRNA cistrons.
Genetica, 70: 191-201.
King, M e King, D. 1975. Chromosomal evolution in the lizard
genus Varanus (Reptilia). Australian Journal BioI. Science. 28: 89–108
Kluge, A.G. 1991. Boine snake phylogeny and research cycles. Miscellaneous
Publications Museum of Zoology University of Michigan, 178: 1–58.
Kraemer, C.; Schmidt, E.R. 1993. The sex determining region of Chironomus thummi is
associated with highly repetitive DNA and transposable elements. Chromosoma,
102: 553–562.
Ledesma, M.A.; Martinez, P.A.; Calderón, O.S.; Boeris, J.M.; Meriles, J.M. 2006.
Descrição do cariótipo e padrões de bandas C e NOR em Pheucticus
aureoventris (Emberizidae, Cardinalinae). Revista Brasileira de Ornitologia, 14:
59-62.
Lee, C.; Sasi, R.; Lin, C.C. 1993. Interstitial localization of telomeric DNA sequences in
the Indian muntjac chromosomes: further evidence for tandem chromosome
fusions in the karyotypic evolution on the Asian muntjacs. Cytogenetics and Cell
Genetics, 63: 156–159.

64
Lee, C.; Wevrick, R.; Fisher, R.B.; Ferguson-Smith, M.A.; Lin, C.C. 1997. Human
centromeric DNAs. Human Genetics, 100: 291–304.
Levan, A.; Fredga, K.; Sandberg, A.A. 1964. Nomenclature for centromeric position on
chromosomes. Hereditas, 52: 201-220.
Luykx, P.; Slowinski, J.B.; McCranie, J.R. 1992. The karyotype of the coral snake
Micrurus ruatanus. Amphib Reptil, 13: 289–292.
Mallet, J.; Joron, M. 1999. Evolution of diversity in warning color and mimicry:
polymorphisms, shifting balance and speciation. The Annual Review of Ecology,
Evolution, and Systematics, 30: 201–233.
Mank, J.E.; Promislow, D.E.L.; Avise, J.C. et al. 2006. Evolution of alternative sex-
determining mechanisms in teleost fishes. Biological Journal of the Linnean
Society, 87: 83–93.
Martins, M.; Oliveira, M. 1999. História natural de serpentes em florestas da região de
Manaus, Amazônia Central, Brasil. Herpetological História Natural, 6: 78-150.
Matsubara, K.; Tarui, H.; Toriba, M.; Yamada, K.; Nishida-Umehara, C.; Agata, K.;
Matsuda, Y. 2006. Evidence for different origin of sex chromosomes in snakes,
birds, and mammals and step-wise differentiation of snake sex chromosomes.
Proceedings of the National Academy of Sciences of the United States of
America, 103: 18190–18195.
Matsubara K, Sarre SD, Georges A, Matsuda Y, Marshall Graves JÁ, Ezaz T. 2014
Highly Differentiated ZW Sex Microchromosomes in the Australian Varanus
Species Evolved through Rapid Amplification of Repetitive Sequences. Plos One,
9(4): e95226.
Matsuda, Y.; Nishida-Umehara, C.; Tarui, H.; Kuroiwa, A.; Yamada, K. 2005. Highly
conserved linkage homology between birds and turtles: Bird and turtle
chromosomes are precise counterparts of each other. Chromosome Research,
13: 601–615.
McDiarmid, R.W.; Campbell, J.A.; Toure, T.A. 1999. Snakes species of the World. A
Taxonomic and Geographic Reference, vol. 1, Herpetol., League, Washington.
Mengden, G.A.; Stock, D. 1980. Chromosomal evolution in Serpentes; a comparision of
G and C chromosome banding patterns of some colubrid and boid genera.
Chromosoma, 79: 53-64.

65
Mengden, G.A. 1981. Linear differentiation of the C-band pattern of the W chromosome
in snakes and birds. Chromosoma, 83(2): 275-287.
Mengden, G.A. 1982. Chromosomal evolution and the phylogeny of elapid snakes. PhD
Thesis, Austr. Nat. Univ. Canberra.
Metcalfe, C.J.; Eldridge, M.D.B.; Johnston, P.G. 2004. Mapping the distribution of the
telomeric sequence (TTAGGG) in the 2n=14 ancestral marsupial complement
and in the macropodines (Marsupialia: Macropodidae) by fluorescence in situ
hybridization. Chromosome Research, 12: 405-414.
Meyne, J. 1988. Fragile sites, telomeric DNA sequences, B chromosomes, and DNA
content in raccoon dogs, Nyctereutes procyonoides, with comparative notes of
foxes, coyote, Wolf and raccoon. Cytogenetics and Cell Genetics, 49: 278–281.
Meyne, J.; Ratliff, R.L.; Moyis, R.K. 1989. Conservation of the human telomere
sequence (TTAGGG)n among vertebrates. Proceedings of the National Academy
of Sciences of the United States of America, 86: 7049–7053.
Meyne, J.; Baker, R.J.; Hobart, H.H.; et al. 1990. Distribution of nontelomeric sites of the
(TTAGGG)n telomeric sequences in vertebrate chromosomes. Chromosoma, 99:
3–10.
Monroy-Vilchis, O.; Sánchez, Ó.; Urios, V. 2011. Consumo de un jaguarundi adulto
Puma yagouaroundi (Felidae) por la serpiente Boa constrictor (Boidae) en el
centro de México. Revista mexicana de biodiversidad, 82(1): 319-321.
Moreno, R.; Navarro, J.; Iturra, P.; Veloso, A. 1987. The karyotype of Philodryas
chamissonis (Colubridae). Identification of nucleolar organizer regions (NOR) and
sex chromosomes by banding methods. Brazilian Journal of Genetics, 10: 497-
506.
Morescalchi, A. 1979. New developments in vertebrate cytotaxonomy. Cytotaxonomy of
the amphibians. Genetica, 50: 179-193.
Mundy, N.I.; Badcock, N.S.; Hart, T.; Scribner, K.; Janssen, K.; Nadeau, N.J. 2004.
Conserved genetic basis of a quantitative plumage trait involved in mate choice.
Science, 303: 1870–1873.
Murphy, J.B.; Campbell, J.A. 1997. Captive maintenance. In: Seigel, R.A.; Collins, J.T.;
Novak, S.S. (Eds.) Snakes: Ecology and Evolutionary Biology. p. 165-181.
Murphy, J.C. 1997. Tales of Giant Snakes: A Historical Natural History of Anacondas
and Pythons. Malabar, Fl.: Krieger Publishing Company.

66
Nanda, I.; Schrama, D.; Feichtinger, W.; Haaf, T.; Schartl, M.; Schmid, M. 2002.
Distribution of telomeric (TTAGGG)n sequences in avian chromosomes.
Chromosoma, 111: 215–227.
Nergadze, S.; Santagostino, M.; Salzano, A.; Mondello, C.; Giulotto, E. 2007.
Contribution of telomerase RNA retrotranscription to DNA double-strand break
repair during mammalian genome evolution. Genome Biology, 8: 260.
Noonan, B.P.; Chippindale, P.T. 2006. Dispersal and vicariance: the complex
evolutionary history of boid snakes. Molecular Phylogenetics and Evolution, 40:
347–358.
Odierna, G.; Olmo, E.; Cobror, O. 1987. Taxonomic implications of NOR-localization in
lacertid lizards. Amphibia-Reptilia, 8: 373-382.
Oguiura, N.; Ferrarezzi, H.; Batistic, R.F. 2009. Cytogenetics and molecular data in
Snakes: A phylogenetic approach. Cytogenetic and Genome Research, 127:
128-142.
Olmo, E.; Signorino, G. 2005. Chromorep: a reptile chromosomes database. Retrieved
from http: //193.206.118.100/ professori/chromorep.pdf
Olmo, E. 2008. Trends in the evolution of reptilian chromosomes. Integrative and
Comparative Biology, 48: 486–493.
Pellegrino, K.C.M.; Rodrigues, M.T.; Yonenaga-Yassuda, Y. 1999. Chromosomal
evolution in the Brazilian lizards of genus Leposoma (Squamata,
Gymnophthalmidae) from Amazon and Atlantic rain forests: Banding patterns and
FISH of telomeric sequences. Hereditas, 131: 15-21.
Pinkel, D.; Straume, T.; Gray, J.W. 1986. Cytogenetic analysis using quantitative,
highsensitivity, fluorescence hybridization. Proceedings of the Natural Academy
of Science, 83: 2934-2938.
Pinthong, K.; Tanomtong, A.; Getlekha, N.; Sangpadee, W.; Sanoamuang, L. 2013.
First cytogenetic study of Puff-Faced Water Snake, Homalopsis buccata
(Squamata, Colubridae) by conventional staing, Ag-NOR banding and GTG –
banding techniques. Cytologia, 78(2): 1-10.
Pokorná, M.; Kratochvíl, L.; Kejnovský, E. 2011. Microsatellite distribution on sex
chromosomes at different stages of heteromorphism and heterochromatinization
in two lizard species (Squamata: Eublepharidae: Coleonyx elegans and
Lacertidae: Eremias velox). BMC Genetics, 12: 90.

67
Porter, C.A.; Hamilton, M.J.; Sites Jr., J.W.; Baker, R.J. 1991. Location of ribosomal
DNA in chromosomes of squamate reptiles: systematic and evolutionary
implications. Herpetologica, 47: 271-280.
Porter, C.A.; Haiduk, M.W.; De Queiroz, K. 1994. Evolution and phylogenetic
significance of ribosomal gene location in chromosomes of squamate reptiles:
systematic and evolutionary implications. Copeia, 1994: 302-313.
Pyron, R.A.; Burbrink, F.T.; Wiens, J.J. 2013. A phylogeny and updated classification of
Squamata, including 4161 species of lizards and snakes. BMC Evolutionary
Biology, 13: 93.
Pyron, R.A.; Reynolds, R.G.; Burbrink, F.T. 2014. A Taxonomic Revision of Boas
(Serpentes: Boidae). Zootaxa, 3846(2): 249-260.
Quinn, A.E.; Sarre, S.D.; Ezaz, T.; et al. 2011. Evolutionary transitions between
mechanisms of sex determination in vertebrates. Biology Letters, 7: 443–448.
Radder, R.S.; Quinn, A.E.; Georges, A.; Sarre, S.D.; Shine, R. 2008. Genetic evidence
for co-occurrence of chromosomal and thermal sex-determining systems in a
lizard. Biology Letters, 4: 176–178.
Ray-Chaudhuri, S.P.; Singh, L. 1972. DNA replication pattern in sex chromosomes of
snakes. Nucleus, 15: 200–210.
Reed, R.N.; Rodda, G.H. 2009. Giant constrictors: biological and management profiles
and an establishment risk assessment for nine large species of pythons,
anacondas, and the Boa constrictor. U.S. Geological Survey Open-File Report.
302 pp.
Reed, K.M.; Hanks, B.G.; Bickham, J.W,; Rhodin,A.G.J.; Greenbaum, I.F.; Mittermeier,
R.A.; Fedullo, L.P. 1991. Cytogenetic analysis of the plurodine turtle Phrynops
hogei and its taxonomic implications. Amphibia-Reptilia, 12: 203-212.
Rens, W.; O’Brien, P.C.; Grutzner, F.; Clarke, O.; Graphodatskaya, D.; Tsend-Ayush,
E.; Ferguson-Smith, M.A. 2007. The multiple sex chromosomes of Platypus and
Echidna are not completely identical and several share homology with the avian
Z. Genome Biology, 8(11): R243.
Reynolds, R.G.; Puente-Rolón, A.R.; Reed, R.N.; Revell, L.J. 2013. Genetic analysis of
a novel invasion of Puerto Rico by an exotic constricting snake. Biological
invasions, 15(5): 953-959.

68
Reynolds, R.G.; Niemiler, M.L.; Revell, L.J. 2014. Toward a Tree-of-Life for the boas
and pythons: multi locus species-level phylogeny with unprecedented taxon
sampling. Molecular Phylogenetics and Evolution, 71: 201–213.
Rhen, T.; Schroeder, A. 2010. Molecular mechanisms of sex determination in reptiles.
Sexual Development, 4: 16-28.
Rivas, J.A. 1999. The life history of the Green Anaconda (Eunectes murinus), with
emphasis on its reproductive biology. Knoxville, Univ. Tennessee, Ph.D.
dissertation, 168 p.
Rivas, J.A.; Owens, R.Y. 2001. Eunectes murinus: juvenile predation.” Herpetological
Review, 32: 107-108.
Ritland, K.; Newton, C.; Marshall, H.D. 2001. Inheritance and population structure of the
white-phased ‘‘Kermode’’ black bear. Current Biology, 11: 1468–1472.
Rosenblum, E.B.; Hoekstra, H.E.; Nachman, M.W. 2004. Adaptive reptile color variation
and the evolution of the MC1R gene. Evolution, 58: 1794–1808.
Rosenblum, E.B.; Rompler, H.; Shoneberg, T.; Hoekstra, H.E. 2010. Molecular and
functional basis of phenotypic convergence in white lizards at White Sands.
Proceedings of the National Academy of Sciences of the United States of
America, 107: 2113–2117.
Ruiz-Herrera, A.; García, F.; Azzalin, C.; Giulotto, E. 2002. Distribution of
intrachromosomal telomeric sequences (ITS) on Macaca fascicularis (Primates)
chromosomes and their implication for chromosome evolution. Human Genetics,
110: 578-586.
Russo, V. 2007. The complete Boa Constrictor: A comprehensive guide to the care,
breeding, and geographic races. ECO Herpetological Publishing, 281 pp.
Saiki, R.K.; Gelfand, D.H.; Stoffel, S.; Scharf, S.J.; Higuchi, R.; Horn, G.T.; Mullis, K.B.;
Erlich, H.A. 1988. Primer-directed enzymatic amplification of DNA with a
thermostable DNA polymerase. Science, 239: 487-491.
Sambrook, J.; Fritsch, E.F.; Maniatis, T. 1989. Molecular Cloning: a Laboratory Manual.
Cold Springs Harbor Laboratory Press, Cold Springs Harbor, NY.
Sarre, S.; Georges, A.; Quinn, A. 2004. The ends of a continuum: Genetic and
temperature-dependent sex determination in reptiles. Bioessays, 26: 639-645.

69
Sarre, S.D.; Ezaz, T.; Georges, A. 2011. Transitions between sex-determining systems
in reptiles and amphibians. Annual Review of Genomics and Human Genetics,
12: 391-406.
Sasaki, M.; Tagaki, N.; Nishida, C.H. 1984.Current profiles of avian cytogenetics, with
notes on chromosomal diagnosis of six birds. Nucleus, 27: 63–73.
Schmid, M.; Feichtinger, W.; Nanda, I.; et al. 1994. An extraordinarily low diploid
chromosome number in the reptile Gonatodes taniae (Squamata, Gekkonidae).
Journal of Heredity, 85: 255–260.
Serafim, H.; Peccinini-Seale, D.M.; Batistic, R.F. 2007. Estudo cariotípico de duas
espécies brasileiras do gênero Micrurus (Ophidia: Elapidae). Biota Neotropica,
7(n1).
Sessions, S.K. 2008. Evolutionary cytogenetics in salamanders. Chromosome
Research, 16: 183–201.
Shine, R.; Elphick, M.; Donnellan, S. 2002. Co-occurrence of multiple, supposedly
incompatible modes of sex determination in a lizard population. Ecology Letters,
5: 486-489.
Sinervo, B.; Lively, C.M. 1996. The rock-paper-scissors game and the evolution of
alternative male strategies. Nature, 380: 240–243.
Singh, L. Sharma, T., Ray-Chaudhuri, S.P. 1970. Multiple sex chromosomes in the
common Indian krait, Bungarus caeruleus Schneider. Chromosoma, 31: 386-391.
Singh, L. 1972a. Evolution of Karyotypes in Snakes. Chromosoma, 38: 185-236.
Singh, L. 1972b. Multiple W chromosome in a sea snake, Enhydrina schistose Daudin.
Experientia, 28: 95-97.
Singh, L.; Purdom, I.F.; Jones, K.W. 1976. Satellite DNA and evolution of sex
chromosomes. Chromosoma, 59: 43-62.
Singh, L.; Purdom, I.F.; Jones, K.W. 1979. Behaviour of sex chromosome associated
satellite DNAs in somatic and germ cells in Snakes. Chromosoma, 71: 167-181.
Singh, L. 2011. The charms of snake sex chromosomes. Journal of Biosciences, 36(1):
17-21.
Slijepcevic, P. 1998. Telomeres and mechanisms of Robertsonian fusion. Chromosoma,
107: 136–140.

70
Smith, J.; Bruley, C.K.; Paton, I.R.; Dunn, I.; Jones, C.T.; Windsor, D.; Morrice, D.R.;
Law, A.S.; Masabanda, J.; Sazanov, A.; Waddington, D.; Fries, R.; Burt, D.W.
2000. Differences in gene density on chicken macrochromosomes and
microchromosomes. Animal Genetics, 31: 96–103.
Srikulnath, K.; Uno, Y.; Matsubara, K.; Thongpan, A.; Suputtitada, S.; Apisitwanich, S.;
Nishida, C.; Matsuda, Y. 2011. Chromosomal localization of 18S-28S and 5S
rRNA genes and (TTAGGG) sequences of butterfly lizards (Leiolepis belliana
belliana and Leiolepis boehmei, Agamidae, Squamata). Genetics and Molecular
Biology, 34(4): 582-586.
Stafford, P.J.; Henderson, R.J. 1996. Kaleidoscopic Tree Boas: the genus Corallus of
Tropical America. Krieger Publishing Company. 120 pp.
Stock, A.D. 1972. Karyological relationships in turtles (Reptilia, Chelonia). Canadian
Journal Genetics Cytology, 14: 859-869.
Strimple, P.D. 1993. Overview of the natural history of the Green Anaconda:
Herpetological Natural History, 1(1): 25–35.
Strussmann, C.; Sazima, I. 1991. Predation on avian eggs by the boid snake, Eunectes
notaeus. Herpetological Review, 22: 118–120.
Strussmann, C.; Sazima, I. 1993. The snake assemblage of the Pantanal at Poconé,
Western Brazil: Faunal composition and ecological summary. Studies on
Neotropical Fauna and Environment, 28: 157–168.
Strussmann, C. 1997. Hábitos alimentares da sucuri-amarela, Eunectes notaeus Cope,
1862, no pantanal Matogrossense. Biociencias, 1: 35–52.
Sumner, A.T. 1972. A simple technique for demonstrating centromeric heterochromatin.
Experimental Cell Research, 75: 304-306.
Svartman, M.; Vianna-Morgante, A.M. 1998. Karyotype evolution of marsupials: from
higher to lower diploid numbers. Cytogenetics and Cell Genetics, 82: 263–266.
Thorbjarnarson, J. 1995. Trailing the Mythical Anaconda. Americas, 47(4): 38.
Tolson, P.J.; Henderson, R.W. 1993. The natural history of West Indian Boas. Tauton,
England: RA Publishing.
Trajtengertz, I.; Beçak, M.L.; Ruiz, I.R.G. 1995. Ribosomal cistrons in Bothrops
neuwiedi (Serpentes) subspecies from Brazil. Genome, 38: 601–606.

71
Uetz, P.; Hosek, J. 2014. The Reptile Database,hhtp://www.reptile-database.org.
Accessed Jul 20.
Underwood, G. 1967. A Contribution to the Classification of Snakes. Trustees of the
British Museum (Natural History), London, 179 pp.
Van Devender, R.W.; Cole, C.J. 1977. Notes on a colubrid snake, Tantilla vermiformis,
from Central America. Apuntes sobre una serpiente colúbrida, Tantilla
vermiformis, de Centroamérica. American Museum Novitates, 2625: 1-12.
Veyrunes, F.; Waters, P.D.; Miethke, P.; Rens, W.; McMillan, D.; Alsop, A.E.; Grützner,
F.; Deakin, J.; Whittington, C.; Kyriena, S.; Colin, K.; Graves, T.; Ferguson-Smith,
M.; Warren, W.; Marshall Graves, J.A. 2008. Bird-like sex chromosomes of
Platypus imply recent origin of mammal sex chromosomes. Genome Research,
18(6): 965–973.
Vicoso, B.; Emerson, J.J.; Zektser, Y.; Mahajan, S.; Bachtrog, D. 2013. Comparative
sex chromosome genomics in snakes: Differentiation, evolutionary strata, and
lack of global dosage compensation. PLoS Biology, 11: e1001643.
Vidal, N.; Henderson, R.W.; Delmas, A.S.; Hedges, S.B. 2005. A phylogeny of the
emerald treeboa (Corallus caninus). Journal of Herpetology, 39: 500–503.
Vidal, N.; Rage, J.C.; Couloux, A.; Hedges, S.B. 2009. Snakes (Serpentes). In: Hedges,
S.B., Kumar, S. (Eds.). The Timetree of Life. Oxford University Univ. Press, New
York, pp. 390–397.
Waller, T.; Micucci, P.A.; Alvarenga, E., 2007. Conservation biology of the Yellow
Anaconda (Eunectes notaeus) in Northeastern Argentina. In: Henderson, R.W.;
Powell, R. (eds.) Biology of the boas and pythons. Eagle Mountain, Utah, Eagle
Mountain Publishing. p. 340–362.
Werner, Y.L. 1959. Chromosomes of primitive snakes from Israel. Bulletin of the
Research Council of Israel, Section B, Zoology, 8B(4): 197-198.
Wiley, J.E.; Meyne, J.; Little, M.L.; Stout, J.C. 1992. Interstitial hybridization sites of the
(TTAGGG)n telomeric sequence on the chromosomes of some North American
hylid frogs. Cytogenetic Cell Genetic, 61: 55–57.
Wurster-Hill, D.H.; Ward, O.G.; Davis, B.H.; Park, J.P.; Moyis, R.K.; Meyne, J. 1988.
Fragile sites, telomeric DNA sequences, B chromosomes, and DNA content in
raccoon dogs, Nyctereutes procyonoides, with comparative notes of foxes,
coyote, Wolf and raccoon. Cytogenetics and Cell Genetics, 49: 278–281.

72
Wurster-Hill, D.H.; Ward, O.G.; Davis, B.H.; Park, J.P.; Moyis, R.K.; Wiley, J.E.; Meyne,
J.; Little, M.L.; Stout, J.C. 1992. Interstitial hybridisation sites of the (TTAGGG)n
telomeric sequence on the chromosomes of some North American hylid frogs.
Cytogenetics and Cell Genetics, 61: 55–57.
Wyman, C.; Blackburn, E.H. 1991. Tel-1 transposon-like elements of Tetrahymena
thermophila are associated with micronuclear genome rearrangements.
Genetics, 129: 57-67.





























