Embed Size (px)
Citation preview

UNIVERSIDADE DE SÃO PAULO
FACULDADE DE CIÊNCIAS FARMACÊUTICAS
Curso de Graduação em Farmácia-Bioquímica
Comparação da perda cognitiva e neuroinflamação na
encefalomielite autoimune experimental após tratamento com
dexametasona e fingolimode
Guilherme Dragunas
Trabalho de Conclusão do Curso de
Farmácia-Bioquímica da Faculdade de
Ciências Farmacêuticas da Universidade
de São Paulo.
Orientadora:
Profa. Dra Carolina Demarchi Munhoz
São Paulo
2017

2
SUMÁRIO
1 INTRODUÇÃO ...........................................................................................7
1.1 Fisiopatologia da Esclerose Múltipla ........................................................... 7
1.2 Encefalomielite Autoimune Experimental ................................................... 9
1.3 Glicocorticoides na EAE .............................................................................. 10
1.4 Fingolimode ................................................................................................... 12
2 OBJETIVOS ............................................................................................. 13
3 MATERIAIS E MÉTODOS ....................................................................... 14
3.1 Animais ........................................................................................................... 14
3.2 Indução da EAE ............................................................................................ 14
3.3 Tratamento farmacológico e delineamento experimental ...................... 15
3.4 Testes comportamentais ............................................................................. 16
3.4.1 Campo aberto ................................................................................................ 16
3.4.2 Teste de reconhecimento de novo objeto ................................................. 16
3.4.3 Labirinto em cruz elevado ........................................................................... 17
3.5 Eutanásia ....................................................................................................... 17
3.6 Preparação do extrato proteico nuclear e citosólico ............................... 18
3.7 Determinação da concentração de proteínas .......................................... 18
3.8 Western-Blot .................................................................................................. 18
3.9 Análise dos Resultados ............................................................................... 19
4 RESULTADOS ......................................................................................... 20
4.1 Score clínico e massa corporal .................................................................. 20
4.2 Campo Aberto ............................................................................................... 21
4.3 Reconhecimento de novo objeto ................................................................ 23
4.4 Labirinto em cruz elevado ........................................................................... 26

3
4.5 Análise Bioquímica ....................................................................................... 27
5 DISCUSSÃO ............................................................................................ 31
6 CONCLUSÃO .......................................................................................... 35
7 BIBLIOGRAFIA ........................................................................................ 36
8 ANEXOS .................................................................................................. 39

4
LISTA DE ABREVIATURAS
ANOVA Analise de Variância
AP-1 Proteína Ativadora 1
BCG Bacillus Calmette-Guérin
BHE Barreira Hemato-Encefálica
CAA Células Apresentadoras de Antígenos
CFA Complete Freund’s Adjuvant
EAE Encefalomielite autoimune experimental
EM Esclerose Múltipla
EM-PS Esclerose Múltipla Progressiva Secundária
EM-RR Esclerose Múltipla Recorrente Remitente
EMSA Ensaio de Desvio de Mobilidade Eletroforética
ERK1/2 Quinase Reguladora de Sinal Extracelular
FoxP3 Forkhead Box P3
GC Glicocorticoide
GR Receptor de Glicocorticoide
GREs Elementos Responsivos a Glicocorticoides
HPA Eixo Hipotálamo-Hipófise-Adrenal
IL Interleucina
LCE Labirinto em Cruz Elevado LPS Lipopolissacarídeo
MHC Complexo Principal de Histocompatibilidade
NO Óxido Nítrico
MOG Glicoproteína da Mielina de Oligodendrócitos
MR Receptor de Mineralocorticoides
NFkB Fator Nuclear Kappa B
PLP Proteína de Protolipídeo
PMB Proteína Mielínica Básica
S1P Esfingosina-1-fosfato
SNC Sistema Nervoso Central
TGF-β Fator de Crescimento Transformador Beta
TLRs Receptores do Tipo Toll
TNF-α Fator de Necrose Tumoral alfa

5
RESUMO
DRAGUNAS, G. Comparação da perda cognitiva e neuroinflamação após
tratamento com fingolimode e com o glicocorticoide sintético dexametasona
em encefalomielite autoimune experimental. 2017. Trabalho de Conclusão de
Curso de Farmácia-Bioquímica – Faculdade de Ciências Farmacêuticas –
Universidade de São Paulo, São Paulo, 2017.
Palavras-chave: Glicocorticoide, Fingolimode, EAE
INTRODUÇÃO: A Esclerose Múltipla (EM) é uma doença inflamatória crônica que
atinge o Sistema Nervoso Central (SNC) e é caracterizada por neurodegeneração e
destruição da bainha de mielina por células imunes autoreativas. Na forma
Recorrente Remitente (RR), a EM apresenta surtos agudos de incapacidade
neurológica, os quais são tratados na clínica com altas doses de glicocorticoides
(GC) sintéticos. Estudos recentes e resultados de nosso grupo apontam efeitos
controversos dos GCs no SNC, mostrando que esta classe de fármacos pode
apresentar efeitos pró-inflamatórios, tempo e contexto dependentes. Em
camundongos com encefalomielite autoimune experimental (EAE), o modelo animal
para o estudo da EM, o tratamento com dexametasona antes do aparecimento dos
sintomas motores acarreta em aumento da ativação de células gliais, perda cognitiva
e alterações comportamentais. Outros fármacos também são utilizados para o
tratamento da EM, dentre eles, o fingolimode, o primeiro fármaco para administração
oral aprovado para esse fim. O fingolimode controla a infiltração de células imunes
no SNC e possui efeitos neuroprotetores diretos em todas as células do SNC.
OBJETIVO: Comparar os efeitos da dexametasona e do fingolimode na
neuroinflamação e déficit cognitivo em camundongos com EAE. MATERIAIS E
MÉTODOS: O tratamento com fingolimode foi avaliado em um modelo de EAE
induzida com glicoproteína da mielina de oligodendrócitos (MOG) em camundongos
C57BL/6 fêmeas. Foram realizados os testes comportamentais de campo aberto e
labirinto em cruz elevado (LCE) para análise do comportamento do tipo ansioso e
teste de reconhecimento de objeto novo para análise de memória declarativa. Por
Western Blot, foi analisada a expressão das proteínas Akt, ERK, TNF-α, NF-κB,
MHC classe II e CREB em extratos proteicos hipocampais. RESULTADOS: O

6
tratamento preventivo com fingolimode foi capaz de reduzir o escore clínico motor de
animais com EAE e também impediu o aparecimento de déficit de memória
declarativa, medido pelo índice de discriminação no teste de objeto novo. Em nível
molecular, o tratamento com fingolimode reduziu a expressão de proteínas pró-
inflamatórias no hipocampo. A imunização com EAE e tratamento com fingolimode
não resultaram em diferenças significativas no teste de LCE. CONCLUSÃO: O
fingolimode é capaz de reduzir os sintomas motores, a expressão de proteínas pró-
inflamatórias e déficit cognitivo da EAE. Esses resultados se contrapõe aqueles
obtidos com o tratamento com dexametasona e reforçam a hipótese da presença de
efeitos deletérios com o uso de glicocorticoides sintéticos na EAE.

7
1 INTRODUÇÃO
1.1 Fisiopatologia da Esclerose Múltipla
A Esclerose Múltipla (EM) é uma doença autoimune crônica do Sistema Nervoso
Central (SNC) caracterizada por danos à bainha de mielina e degeneração
neuroaxonal [Compston, 2008]. A EM afeta 2,3 milhões de pessoas ao redor do
mundo com uma prevalência média de 30/100.000 habitantes [World Health
Organization, 2008], atingindo mais mulheres que homens em uma proporção de
2:1. Os sintomas da EM começam geralmente no início da vida adulta,
aproximadamente aos 30 anos de idade. No Brasil, estudos epidemiológicos
apontam uma prevalência que varia de 1,36/100.000 a 27,2/100.000 habitantes [da
Gama Pereira, 2015]. A EM possui heterogeneidade de apresentações clínicas,
sendo a forma Recorrente Remitente (EM-RR) a mais comum da doença, ocorrendo
em 85% dos casos. Na EM-RR, os pacientes apresentam episódios de incapacidade
neurológica que remitem espontaneamente. Grande parte destes desenvolverá,
após alguns anos do início dos episódios agudos, a forma Progressiva Secundária
(PS), na qual a incapacidade neurológica aumenta com o tempo, acompanhada de
degeneração neuronal e grande atrofia cerebral [Dendrou, 2015].
Nos episódios agudos, a EM é caracterizada por ativação microglial e astrocística e
infiltração linfocitária periférica na região perivascular encefálica e na medula
espinhal, principalmente por linfócitos T e B autorreativos contra constituintes da
bainha de mielina. Os principais linfócitos T presentes nos focos de lesão
desmielinizante da matéria cinzenta e branca são do tipo T CD8+, sendo seu número
relacionado ao dano axonal dessas lesões. Os linfócitos CD4+ Th1/Th17 também
são importantes na fisiopatologia das lesões da EM. Eles secretam citocinas pró-
inflamatórias como o IL-17A, GM-CSF e IFN-γ e quimiocinas como CCL-2 e
promovem infiltração de mais células do sistema imune inato e adaptativo para o
SNC [Dendrou, 2015; Simmons, 2013]. Os linfócitos que migram para o SNC são
reativados por células apresentadoras de antígenos (CAA), incluindo CAA derivados
de monócitos meningeais, perivasculares e ventriculares. Uma característica
importante da EM é o espalhamento de epítopo, onde CAAs apresentam outros
peptídeos de mielina que possuem epítopos similares ao antígeno original, gerando

8
uma grande amplificação da resposta auto-imune. A microglia contribui de forma
decisiva, juntamente com os astrócitos, para a inflamação e consequente
desmielinização na EM. A microglia e os astrócitos respondem a insultos
inflamatórios e se tornam ativados, através de receptores como os TLRs e dos
fatores de transcrição NF-kB e AP-1. Estas células ativadas secretam citocinas,
quimiocinas, espécies reativas de oxigênio e nitrogênio, óxido nítrico e aumentam a
expressão de MHC classe II, as quais propagam a inflamação, causam
desmielinização e morte de oligodendrócitos. Através da secreção de citocinas
inflamatórias como o IL-1β e TNF-α, promovem a diferenciação de células T naive
para os subtipos Th1/Th17 [Dendrou, 2015]. Os oligodendrócitos são o alvo celular
na EM, sendo que também a diferenciação de progenitores de oligodendrócitos está
comprometida, resultando em remielinização deficiente [Chamberlain, 2016].
Na forma PS, a infiltração periférica cessa, no entanto, a inflamação prossegue
intensamente por meio de células da glia ativadas e por estruturas linfoides terciárias
presentes na região meningeal, as quais promovem intensa neurodegeneração e
desmielinização secundária pela continua produção de espécies reativas de oxigênio
e nitrogênio. O estresse oxidativo gerado resulta em danos a biomoléculas e
mutações no DNA mitocondrial dos neurônios, que resulta estresse metabólico e do
retículo endoplasmático, déficit energético e subsequente ativação de vias de morte
celular e neurodegeneração anterógrada e retrógada. O acúmulo de glutamato
causado pela lesão axonal resulta em excitotoxicidade e também contribui para a
morte neuronal [Dendrou, 2015].
Apesar da maior compreensão atual da EM, sua patogênese ainda permanece, em
boa parte, desconhecida. Isso gera dificuldades para o desenvolvimento de terapias
novas e mais eficazes, sendo a melhor compreensão da progressão e fisiopatologia
da doença, portanto, de extrema importância. As estratégias atuais de tratamento
da EM-RR são baseadas nos episódios agudos e tentam controlar a infiltração de
linfócitos no SNC e subsequente inflamação. Dentre os fármacos utilizados podemos
citar o recentemente aprovado fingolimode, um fármaco imunomodulador, e os
glicocorticoides sintéticos como dexametasona e metilprednisolona, os quais atuam

9
controlando a inflamação durante as fases de remissão da EM [Burton, 2012; Garris,
2013; Bhise, 2016].
1.2 Encefalomielite Autoimune Experimental
O modelo animal de EM mais comumente utilizado, a encefalomielite autoimune
experimental (EAE) pode ser induzida por meio da aplicação subcutânea de
proteínas componentes da bainha de mielina de roedores, como a glicoproteína da
mielina de oligodendrócitos (MOG), a proteína mielínica básica (PMB) e a proteína
de proteolipídeo (PLP), juntamente com adjuvante CFA contendo BCG e toxina
pertussis intraperitoneal [Mendel, 1995; Lassmann, 2016]. Neste modelo, é gerado
uma resposta celular adaptativa contra a proteína do oligodendrócito, causando
inflamação aguda e crônica, desmielinização e degeneração axonal, mediadas por
macrófagos e células T CD4+ autorreativas, que possui similaridades com o padrão
RR da EM [Simmons, 2013]. A inflamação se concentra, principalmente, na medula
espinhal dos animais com menor afecção cerebral e não há mediação de células T
CD8+ e B, característicos da doença em humanos. Entretanto, se trata de um modelo
muito útil para o estudo de terapias anti-inflamatórias para a EM, uma vez que
reproduz características da doença como desmielinização, alterações na
plasticidade e transmissão sinápticas, perda neuronal e deficiência cognitiva
[Lassmann, 2016].
A EM é marcada por pronunciadas alterações emocionais, neuropsiquiátricas e
cognitivas nos portadores, as quais frequentemente não são relacionadas a surtos
ou progressão da doença. Cerca de 50% dos pacientes com EM sofrem de
depressão e distúrbios cognitivos como déficits de atenção, no processamento de
informações, memória de longo prazo e aprendizado visual [Paparrigopoulos, 2010,
Chiaravalloti, 2008]. Nestes pacientes, a atrofia do hipocampo e dos núcleos da
base são fatores importantes associados ao déficit cognitivo [Damjanovic, 2016].
Interessantemente, em camundongos portadores de EAE, essas alterações
emocionais e cognitivas também podem ser observadas. Camundongos C57Bl/6 na
fase tardia de EAE induzida por MOG35-55 mostraram um aumento de
comportamento ansioso e depressivo comparados ao controle [Peruga, 2011]. Estas
alterações foram correlacionadas a um aumento na quantidade de TNF-α e perda

10
neuronal no hipocampo desses animais [Peruga, 2011]. Em camundongos C57Bl/6
na fase pré-simtomática da EAE/ MOG35-55, sem apresentar ainda sintomas motores
característicos, foram observadas alterações emocionais e cognitivas. Os
camundongos permanecerem mais tempo na área próxima às paredes no teste de
campo aberto e nos braços fechados do labirinto em cruz elevado, além de reduzida
latência para imobilidade nos testes de nado forçado e suspensão pela cauda,
sugestivos de um comportamento do tipo ansioso e depressivo, respectivamente
[Acharjee, 2013]. Apresentaram também menor tempo no quadrante alvo no teste de
Morris, indicativo de déficit de memória espacial [Acharjee, 2013]. Além disso, foi
observada elevação na concentração de corticosterona sérica nos animais do grupo
EAE, indicando uma modulação do eixo HPA [Acharjee, 2013].
1.3 Glicocorticoides na EAE
Os glicocorticoides (GC) são amplamente conhecidos por suas capacidades anti-
inflamatórias e imunossupressoras; nos humanos, o cortisol (corticosterona nos
roedores) é liberado pelo córtex da glândula suprarrenal em situações de estresse e
liga-se a receptores nucleares de glicocorticoides (GR) e mineralocorticoides (MR),
presentes em virtualmente todas as células do corpo. Os glicocorticoides são
imunossupressores potentes, pois, além de diminuir a expressão de genes pró-
inflamatórios, inibem a ação e proliferação de linfócitos T e B e ativação de
macrófagos e neutrófilos, além de promover aumento de apoptose nessas células
[Cruz-Topete, 2015; Schweingruber, 2012]. Os glicocorticoides sintéticos, ao
contrário dos glicocorticoides endógenos, possuem maior seletividade para o GR e,
portanto, possuem atividade glicocorticoide com baixa atividade mineralocorticoide.
O GR possui duas isoformas em humanos, o hGRα e o hGRβ, os quais geram
respostas diferentes aos glicocorticoides, sendo que, geralmente, o GRα é
responsável pelos efeitos clássicos dos glicocorticoides e o GRβ atua antagonizando
estas ações [Cruz-Topete, 2015; Schweingruber, 2012].
A ligação dos glicocorticoides no receptor GR induz alterações conformacionais e
hiperfosforilação do mesmo, o que o libera da ligação de chaperonas acessórias e
este migra para o núcleo, atuando de 3 maneiras documentadas: 1) dimerização do

11
GR e ligação no DNA em sequências GRE (elementos responsivos a
glicocorticoides), o que causa indução da expressão de genes anti-inflamatórios
como o da β-arrestina1, Anexina-1 e TTP (o qual age desestabilizando mRNAs de
genes pró-inflamatórios) e inibição da expressão de genes pró-inflamatórios como
TNF-α e IL-1β. 2) interação com outros fatores de transcrição para a modulação de
genes anti/pró-inflamatórios GRE-independentes (composição); e 3) ação anti-
inflamatória indireta pela ligação a fatores de transcrição pró-inflamatórios como o
NF-kB e AP-1 (tethering). O GR também possui um mecanismo de ação rápido, não
genômico, por meio da ligação de GR de membrana e interação do GR citosólico
com outras vias de sinalização intracelulares [Cruz-Topete, 2015; Schweingruber,
2012].
O hipocampo e o córtex cerebral são regiões que apresentam grande densidade de
GR e MR, e são suscetíveis a efeitos deletérios quando grandes quantidades de
GCs estão presentes, causando prejuízo na plasticidade sináptica e cognição,
diminuição da neurogênese, diminuição da densidade dendrítica e atrofia dessas
áreas [Sorrells, 2009].
Dentre as patologias que apresentam desregulação no eixo HPA e quantidade
aumentada de GCs, podemos citar o estresse, depressão e a EM [Sorrells, 2009]. A
relação entre estresse crônico e agudo e a presença de sintomas de ansiedade e
depressão já está estabelecida na literatura. O estresse causa inibição da
neurogênese no giro dentado e atrofia de dendritos na região CA3 do hipocampo, ao
passo que pacientes portadores de depressão também apresentam atrofia
hipocampal [McEwen, 2001]. A EAE, como a EM, apresenta inflamação hipocampal
e em outras estruturas relacionadas à cognição e comportamento, além de
desregulação no eixo HPA com concentração aumentada de corticosterona.
Apesar de sua capacidade em reduzir os sinais clínicos da EM e da EAE, em alguns
contextos, os GCs podem ter um efeito controverso, por induzir um aumento da
secreção de mediadores inflamatórios e a diminuição da secreção de mediadores
anti-inflamatórios [Sorrells, 2009; Duque, 2016].
Resultados de nosso grupo mostram que a administração de alta dose preventiva do
glicocorticoide sintético dexametasona (50mg/kg) melhora o escore clínico motor dos

12
animais com EAE (ANEXO 1). Entretanto, a administração da dexametasona
também promove efeitos negativos na neuroinflamação e em resultados de testes
comportamentais. Os animais tratados com a dexametasona mostraram aumento no
comportamento do tipo ansioso, evidenciado pelo teste do Labirinto em Cruz
Elevado (ANEXO 2), déficit cognitivo, através do teste de reconhecimento de objetos
(ANEXO 3) e aumento da expressão de proteínas pró-inflamatórias e diminuição da
expressão de proteínas de expressão rápida no hipocampo (ANEXO 4).
Os resultados obtidos mostram alinhamento com a literatura, a qual aponta efeito
controverso dos glicocorticoides na modulação de respostas inflamatórias
específicas [Duque, 2016]. Entretanto, ainda é preciso utilizar outro fármaco
imunomodulador de outra classe e que seja utilizado na EM como comparador, a fim
de fortalecer mais ainda a hipótese de que os efeitos observados com o tratamento
com a dexametasona na EAE sejam causados por ação específica destes fármacos
e não uma consequência natural da doença.
1.4 Fingolimode
Outro fármaco utilizado na clínica para diminuir a frequência de surtos e retardar a
progressão da doença, o fingolimode (FTY720), foi aprovado em 2010 pelo FDA e é
o primeiro tratamento oral para a EM-RR. O fármaco é derivado de um metabólito
encontrado no fungo Isaria sinclairii, sendo estruturalmente análogo à esfingosina,
um membro da família dos esfingolipídeos. O fingolimode, quando fosforilado por
esfingosina quinases, possui ação moduladora do receptor de esfingosina-1-fosfato
(S1P1,3,4,5) em linfócitos T, promovendo sua internalização, ativando a via de
sinalização da MAPK/ERK, sequestrando estas células nos linfonodos e impedindo
assim seu aporte para o SNC e subsequente exacerbação da inflamação e relapsos
na EM [Chun, 2010]. Este, entretanto, não é o único mecanismo de ação pelo qual o
fingolimode age, possuindo também ação direta protetora sobre o SNC, atuando em
neurônios, microglia, astrócitos e oligodendrócitos. O fingolimode promove
diferenciação de oligodendrócitos e recuperação da remielinização, diminuição da
ativação de células microgliais e astrócitos, com diminuição da produção de
mediadores inflamatórios e restauração da função neuronal com redução da perda
de espinhos dendríticos e proteção contra excitotoxicidade [Hunter, 2016].

13
Em camundongos SJL/J com EAE induzida por PLP139-151, o fingolimode, dado por 7
dias, foi capaz de induzir a proliferação e diferenciação de progenitores de
oligodendrócitos [Zhang, 2015]. Na EAE induzida com MOG33-55 em camundongos
C57BL/6, o fingolimode (0,2mg/kg) dado após a imunização dos animais melhorou o
desenvolvimento da doença, reduzindo a neurodegeneração e o escore clínico. Foi
observado maior número de células Treg e maior expressão dos genes FoxP3 e tgf-
b no grupo tratado com fingolimode [Hou, 2016].
Diversos estudos apontam a diminuição do fator neurotrófico derivado do cérebro
(BDNF) no hipocampo e no sangue periférico de pacientes com depressão e em
animais submetidos a estresse crônico e agudo [Castrén, 2016]. O fingolimode
promove aumento da quantidade de BDNF circulante e no hipocampo de
camundongos em modelos de estresse crônico e agudo através da sua ação
inibidora de histonas desacetilases e gera um efeito antidepressivo nesses animais
[Nuzzo, 2015].
Diante da complexidade da doença e das contradições observadas com os
tratamentos disponíveis, principalmente àquelas relacionadas aos transtornos de
humor e aos déficits cognitivos observados em pacientes com EM ou em animais
com EAE, através deste projeto se espera entender mais sobre os diversos efeitos
dose e contexto dependentes que os glicocorticoides podem exercer em uma
doença neuroinflamatória, comparando-os com o fingolimode. A comparação será
realizada por meio da avaliação comportamental, cognitiva e da neuroinflamação no
hipocampo de animais doentes, que desenvolveram EAE, submetidos ao tratamento
com fingolimode.
2 OBJETIVOS
Avaliar a neuroinflamação e capacidade cognitiva de animais com EAE tratados
preventivamente com fingolimode e comparar os resultados obtidos com o
tratamento com alta dose de dexametasona.

14
3 MATERIAIS E MÉTODOS
3.1 Animais
Foram utilizados 42 camundongos C57BL/6, fêmeas, adultas, entre 6 e 8 semanas e
pesando entre 15 g e 20 g, provenientes do Biotério de Camundongos Isogênicos do
Instituto de Ciências Biomédicas da Universidade de São Paulo. Os animais foram
mantidos em temperatura controlada (20-22 °C) e ciclo claro/escuro de 12 horas,
com início do ciclo de claro as 7:00 horas. Foram alimentados com ração
balanceada para camundongos e água ad libitum.
Todos os procedimentos experimentais, que incluem a imunização, perfusão e
decapitação dos animais, foram aprovados e realizados de acordo com as normas
da Comissão de Ética em Experimentação Animal do Instituto de Ciências
Biomédicas da Universidade de São Paulo (CEUA/ICB-USP), registrados sob o
número 03, na folha 03 do livro 03 e seguiram todas as exigências descritas no
Colégio Brasileiro de Experimentação Animal (COBEA).
3.2 Indução da EAE
A EAE foi induzida por meio de imunização por via subcutânea com 150 µg de
MOG35-55 emulsificados em CFA (complete Freund’s adjuvant) (v/v), com um total de
400 μg de BCG (Bacillus Calmette-Guérin). Além disso, foram feitas duas
administrações, por via intraperitoneal, de 200 ng de toxina de Bordetella pertussis,
0 e 48 horas após a imunização. Os animais foram avaliados diariamente com uma
escala clínica (descrita abaixo) e sofreram eutanásia no 9o e 26º dia pós-imunização
(d.p.i.), que corresponde ao estágio pré-clínico e a fase de recuperação da doença,
respectivamente.
Os animais foram avaliados diariamente e o grau de apresentação clínica distribuído
da seguinte forma: 0) sem doença; 0.5) perda parcial do tônus de cauda; 1) perda
total do tônus da cauda e fraqueza de membros posteriores; 2) hemiplegia; 3)
arrastamento parcial dos membros posteriores; 4) paraplegia e 5) tetraplegia ou
morte.

15
3.3 Tratamento farmacológico e delineamento experimental
Os animais foram divididos em 4 grupos experimentais (Controle, EAE, Fingo e
EAE+Fingo). Foram realizados dois esquemas de administração do fingolimode,
diluído em PBS/ 5% EtOH: a) Subcrônico: 0,5 mg/kg do dia 0 ao dia 9º d.p.i. por via
intraperitoneal (I.P.). e b) Agudo: 10 mg/kg no dia imunização.
Figura 1. Delineamento experimental com administração sub-crônica de fingolimode 0,5 mg/kg/dia em
animais C57BL/6 femeas imunizados com MOG35-55 acrescido de CFA.
Figura 2. Delineamento experimental com administração aguda de fingolimode 10 mg/kg no dia da
imunização com MOG35-55 acrescido de CFA em animais C57BL/6 femeas.

16
3.4 Testes comportamentais
3.4.1 Campo aberto
Este teste foi realizado no 4º e 22º d.p.i. para avaliação da atividade locomotora e
comportamento do tipo ansioso. Para tal, utilizou-se aparato circular (400 mm x 600
mm, diâmetro x altura) com as paredes opacas. Os animais foram colocados na
periferia do aparato e submetidos à exploração do ambiente durante 5 minutos. O
número de cruzamentos de quadrantes, velocidade média e velocidade máxima
foram utilizados para avaliação da atividade locomotora, enquanto que o número de
entradas e tempo de permanência no centro do aparato indicaram o nível de
ansiedade desses animais. Todo procedimento foi filmado por uma câmera de vídeo
suspensa sobre o aparelho para posterior análise por meio de software Any Maze
Stoelling.
3.4.2 Teste de reconhecimento de objeto novo
Os animais passaram por treinamento no mesmo aparato de campo aberto durante
3 dias consecutivos no mesmo horário. No 7o d.p.i. e 25º d.p.i, os animais foram
colocados no campo aberto contendo dois objetos idênticos (A e B), para livre
exploração, por 10 minutos. 2 e 24 horas depois do treino (8º d.p.i. e 26 d.p.i.), para
testar a memória de curta duração (MCD) e memória de longa duração (MLD),
respectivamente, os animais foram colocados novamente no campo aberto por 5
minutos para exploração. Entretanto, nestes testes, o objeto B foi substituído por
outro (C, para MCD e D, para MLD). Foi considerada exploração quando os animais
cheiraram ou tocaram o objeto com o nariz ou com as patas dianteiras. O tempo
gasto explorando cada objeto foi anotado e expresso em total de tempo de
exploração computado em segundos, permitindo determinar o índice de
discriminação estabelecido a partir da proporção de tempo de exploração do objeto
novo em relação ao objeto familiar (tnovo-tfamiliar / tnovo + tfamiliar). Todo
procedimento foi filmado por uma câmera de vídeo suspensa sobre o aparelho para
posterior análise manual.

17
3.4.3 Labirinto em cruz elevado (LCE)
O LCE é constituído de dois braços abertos e opostos (20 x 20 cm cada), e outros
dois braços opostos de mesmo tamanho, fechados por paredes laterais de 30 cm de
altura. Os braços abertos e fechados cruzam-se perpendicularmente formando uma
cruz, delimitada por uma pequena área central de 5 x 5 cm, seguindo as
especificações de Pellow e colaboradores [Pellow, 1985]. O aparelho é constituído
de madeira tratada com tinta fosca, suspenso a 40 cm do assoalho por um suporte
central e, durante os testes, é alocado em uma sala de experimentação com
condições de luminosidade e temperatura iguais às do biotério.
No dia do experimento (8 d.p.i.), cada animal foi colocado individualmente na área
central do LCE, com a cabeça voltada para um dos braços abertos, podendo
explorar o aparato durante 5 minutos. Para a análise do comportamento do tipo
ansioso, foram avaliados o número de entradas e o tempo de permanência nos
braços. Todo procedimento foi filmado por uma câmera de vídeo suspensa sobre o
aparelho para posterior análise.
3.5 Eutanásia
Os animais foram submetidos à eutanásia por decapitação para a dissecção do
hipocampo para a realização de ensaios moleculares. Os tecidos retirados foram
armazenados à - 80 °C para análise a posteriori.

18
3.6 Preparação do extrato proteico nuclear e citosólico
Os hipocampos foram homogeneizados em douncer (vidro-vidro) em tampão de lise
(HEPES 100 mM, MgCl2 1,5 mM, KCl 10 mM, PMSF 0,5 mM, DTT 1 mM e coquetel
de proteases e fosfatases [Halt Protease and Phosphatase 100x Cocktail - Thermo
Fisher Scientific]) acrescido de NP40 1:40 (v/v). As amostras foram então
centrifugadas a 4ºC por 30 segundos a 12.000g e o sobrenadante, a fração
citosólica, coletado. Em seguida, é adicionado ao precipitado tampão de extração
(HEPES 20 mM, MgCl2 1,5 mM, NaCl 300 mM, EDTA 0,25 mM, glicerol 25%, PMSF
0,5 mM, DTT 1 mM e coquetel de proteases e fosfatases [Halt Protease and
Phosphatase 100x Cocktail - Thermo Fisher Scientific]) e incubado em gelo sob
agitação. Por fim, é submetido à nova centrifugação por 20 minutos a 12.000g e o
sobrenadante, a fração nuclear, coletado. Os extratos foram mantidos a -80 °C para
análise a posteriori.
3.7 Determinação da concentração de proteínas
A dosagem foi realizada através do método de Bradford [Bradford, 1976] utilizando
reagente da Bio-Rad e subsequente medição da absorbância no comprimento de
onda de 595 nm em um leitor de microplacas Synergy HT Multi-Mode Microplate
Reader (BioTek, Winooski, VT, USA). A comparação com uma curva padrão de
albumina forneceu a concentração de proteínas presente nas amostras.
3.8 Western-Blot
Após dosagem de proteínas pelo método do Bradford, descrito no item 3.7, as
amostras foram diluídas em tampão Laemmli (125 mM de Tris-HCl; 4% de SDS;
20% v/v de glicerol; 200 mM de DTT e 0,02% de azul de bromofenol; pH 6,8)
[Laemmli, 1970]) na concentração de 1 μg/μl e aquecidas a 95 °C por 5 minutos para
que houvesse a denaturação das proteínas. 10-20 μg de cada amostra foram
aplicados em géis de 7,5% a 10% SDS-poliacrilamida (acrilamida/bisacrilamida 5:1,
10% de SDS) e submetidos à eletroforese (100 V, 2 horas), juntamente com um
padrão de peso molecular (Precision Plus Protein™ Dual Color Standards, BioRad)
em tampão de corrida (25 mM de Tris-Base, 192 mM de glicina, 0,1% de SDS em
água destilada). Ao final da corrida, as proteínas separadas foram transferidas

19
eletroforeticamente para uma membrana de PVDF, em tampão de transferência
gelado (25 mM de Tris-base, 192 mM de glicina, 20% de metanol em água
destilada), a 400 mA por aproximadamente 2 horas. Após a transferência, para
verificar a igualdade da quantidade das proteínas aplicadas ao gel e a eficiência da
transferência, as membranas foram incubadas em reagente de Ponceau S (0,5%
Ponceau-S; 5% ácido tricloro acético e água bidestilada) por 3 a 5 minutos
[Salinovich, 1986]. Após essa etapa, as membranas foram lavadas com água
destilada para retirar o reagente de Ponceau e incubadas por 2 horas em solução de
bloqueio, constituído por 5% leite desnatado (Molico, Nestlé) em TBS-T (100 mM de
Tris-base, 0,9% de NaCl, 0,1% de Tween 20 em água destilada), para bloquear
ligações inespecíficas. Para proteínas fosforiladas, as membranas são incubadas
em solução de 5% BSA (albumina sérica bovina) em TBS-T. Após essa etapa, as
membranas são incubadas com anticorpos específicos: p65 (1:1000, Santa Cruz
Technology), ERK e p-ERK (1:5000, Cell Signaling Technology), TNF-α (1:500,
Santa Cruz Technology), Akt e p-Akt (1:2000, Santa Cruz Technology e EMD
Millipore, respectivamente), MHC classe II (1:3000, Santa Cruz Technology) e CREB
(1:1000, Cell Signaling Technology) overnight a 4 °C, na mesma solução utilizada no
bloqueio. Em seguida, foram incubadas com anticorpos secundários na
concentração adequada durante 2 horas. A detecção do complexo antígeno-
anticorpo foi feita através do kit de quimioluminescência Luminata Forte Western
HRO substrate (Millipore, Billerica, MA, EUA) no fotodocumentador ChemiDoc
Imager (Bio-Rad Laboratories, Inc., Hercules, CA, EUA). A análise da densidade
óptica das bandas foi realizada com o auxílio do programa Image Lab 5.2 (Bio-Rad
Laboratories, Inc.). Vários tempos de exposição foram feitos e analisados para
garantir a linearidade das intensidades das bandas. Os anticorpos β-actina ou
GAPDH (1:20000, Santa Cruz Biotechnology) foram usados como controle interno.
3.9 Análise dos Resultados
Para todos os testes, foi utilizado teste t de Student para comparação de 2 grupos
ou análise de variância (ANOVA) de duas vias (Two-way ANOVA) associada à
análise post hoc de Tukey para comparação de mais grupos. Foram adotadas
diferenças de p<0,05 como sendo estatisticamente significantes.
20
4 RESULTADOS
4.1 Escore clínico e massa corporal
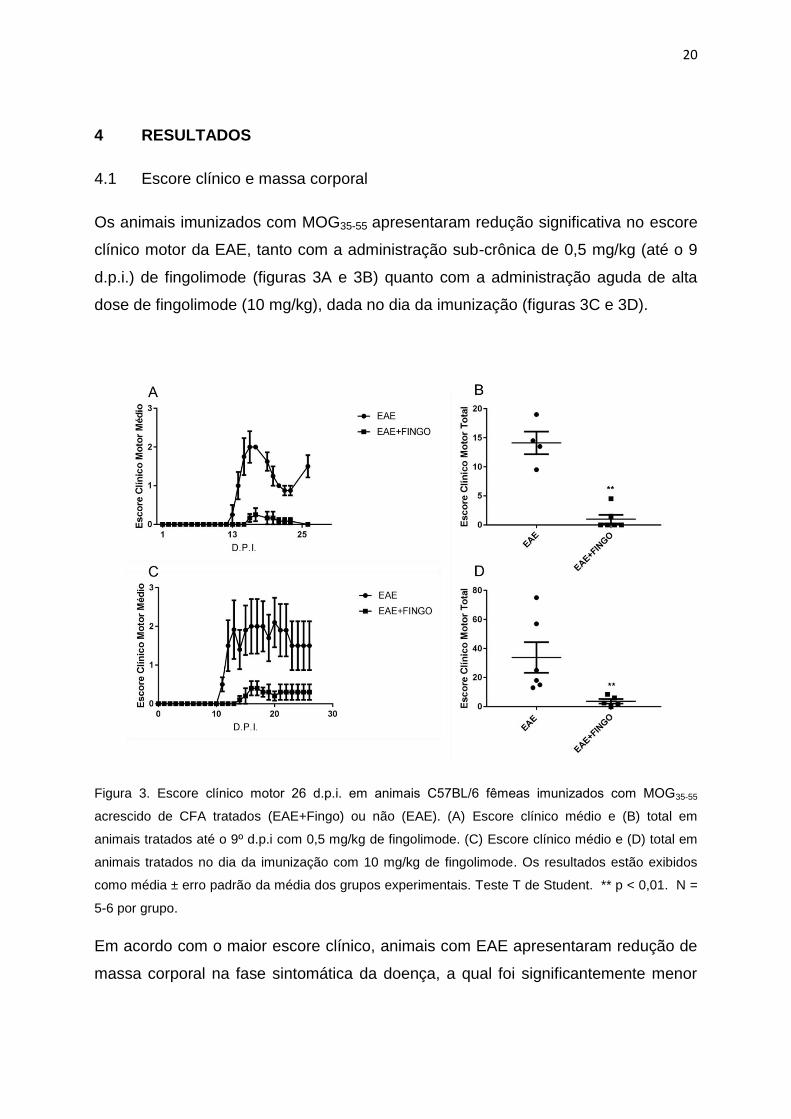
Os animais imunizados com MOG35-55 apresentaram redução significativa no escore
clínico motor da EAE, tanto com a administração sub-crônica de 0,5 mg/kg (até o 9
d.p.i.) de fingolimode (figuras 3A e 3B) quanto com a administração aguda de alta
dose de fingolimode (10 mg/kg), dada no dia da imunização (figuras 3C e 3D).
Figura 3. Escore clínico motor 26 d.p.i. em animais C57BL/6 femeas imunizados com MOG35-55
acrescido de CFA tratados (EAE+Fingo) ou não (EAE). (A) Escore clínico médio e (B) total em
animais tratados até o 9º d.p.i com 0,5 mg/kg de fingolimode. (C) Escore clínico médio e (D) total em
animais tratados no dia da imunização com 10 mg/kg de fingolimode. Os resultados estão exibidos
como média ± erro padrão da média dos grupos experimentais. Teste T de Student. ** p < 0,01. N =
5-6 por grupo.
Em acordo com o maior escore clínico, animais com EAE apresentaram redução de
massa corporal na fase sintomática da doença, a qual foi significantemente menor
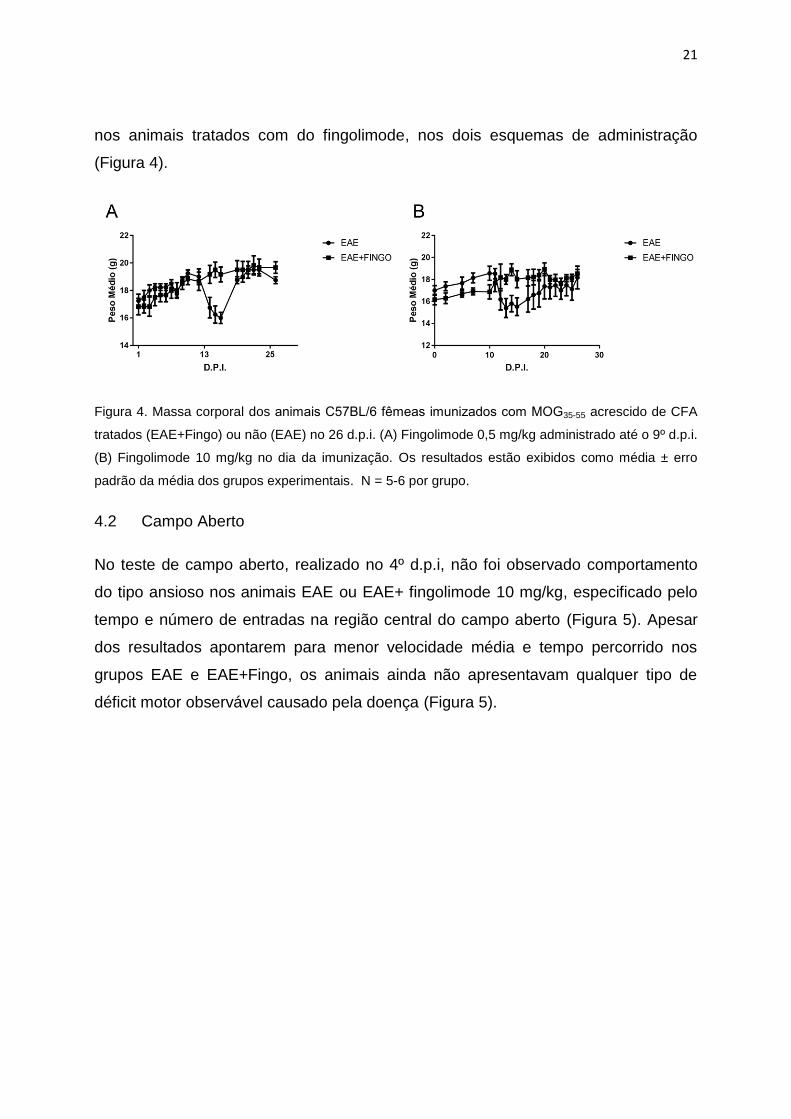
21
nos animais tratados com do fingolimode, nos dois esquemas de administração
(Figura 4).
Figura 4. Massa corporal dos animais C57BL/6 femeas imunizados com MOG35-55 acrescido de CFA
tratados (EAE+Fingo) ou não (EAE) no 26 d.p.i. (A) Fingolimode 0,5 mg/kg administrado até o 9º d.p.i.
(B) Fingolimode 10 mg/kg no dia da imunização. Os resultados estão exibidos como média ± erro
padrão da média dos grupos experimentais. N = 5-6 por grupo.
4.2 Campo Aberto
No teste de campo aberto, realizado no 4º d.p.i, não foi observado comportamento
do tipo ansioso nos animais EAE ou EAE+ fingolimode 10 mg/kg, especificado pelo
tempo e número de entradas na região central do campo aberto (Figura 5). Apesar
dos resultados apontarem para menor velocidade média e tempo percorrido nos
grupos EAE e EAE+Fingo, os animais ainda não apresentavam qualquer tipo de
déficit motor observável causado pela doença (Figura 5).
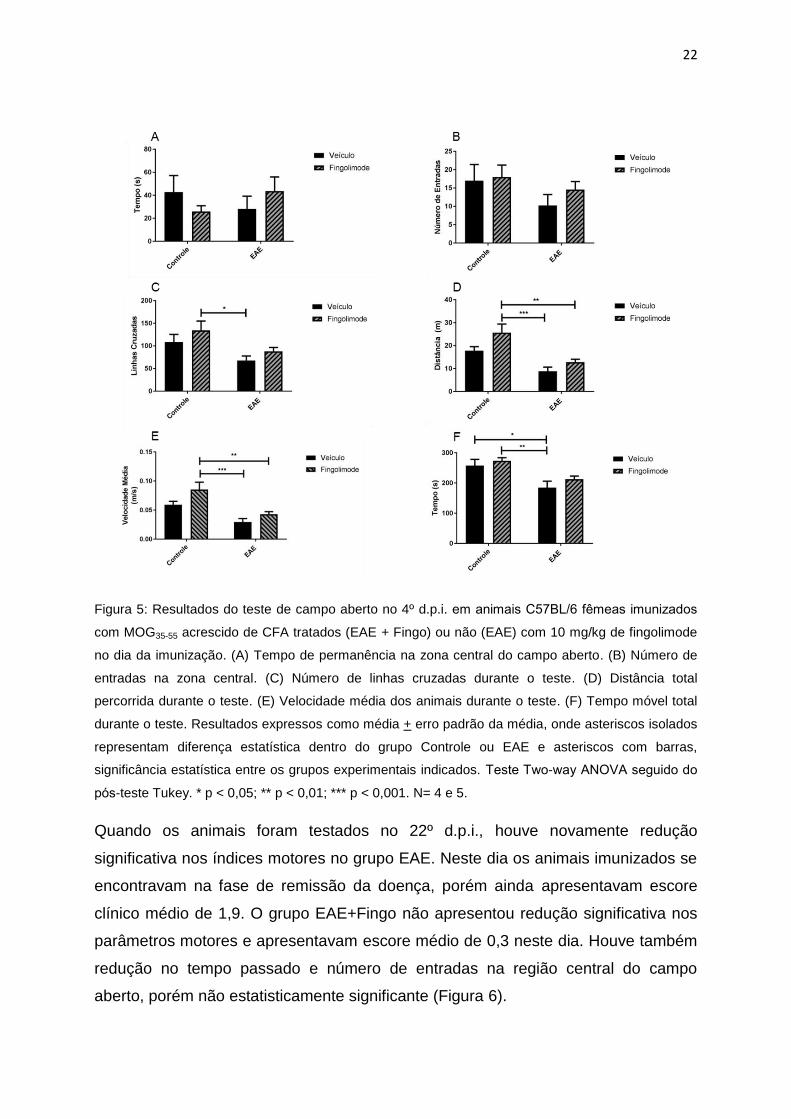
22
Figura 5: Resultados do teste de campo aberto no 4º d.p.i. em animais C57BL/6 femeas imunizados
com MOG35-55 acrescido de CFA tratados (EAE + Fingo) ou não (EAE) com 10 mg/kg de fingolimode
no dia da imunização. (A) Tempo de permanência na zona central do campo aberto. (B) Número de
entradas na zona central. (C) Número de linhas cruzadas durante o teste. (D) Distância total
percorrida durante o teste. (E) Velocidade média dos animais durante o teste. (F) Tempo móvel total
durante o teste. Resultados expressos como média + erro padrão da média, onde asteriscos isolados
representam diferença estatística dentro do grupo Controle ou EAE e asteriscos com barras,
significância estatística entre os grupos experimentais indicados. Teste Two-way ANOVA seguido do
pos-teste Tukey. * p < 0,05; ** p < 0,01; *** p < 0,001. N= 4 e 5.
Quando os animais foram testados no 22º d.p.i., houve novamente redução
significativa nos índices motores no grupo EAE. Neste dia os animais imunizados se
encontravam na fase de remissão da doença, porém ainda apresentavam escore
clínico médio de 1,9. O grupo EAE+Fingo não apresentou redução significativa nos
parâmetros motores e apresentavam escore médio de 0,3 neste dia. Houve também
redução no tempo passado e número de entradas na região central do campo
aberto, porém não estatisticamente significante (Figura 6).

23
Figura 6: Resultados do teste de campo aberto no 22º d.p.i. em animais C57BL/6 femeas imunizados
com MOG35-55 acrescido de CFA tratados (EAE + Fingo) ou não (EAE) com 10 mg/kg de fingolimode
no dia da imunização. (A) Tempo de permanência na zona central do campo aberto. (B) Número de
entradas na zona central. (C) Número de linhas cruzadas durante o teste. (D) Distância total
percorrida durante o teste. (E) Velocidade média dos animais durante o teste. (F) Tempo móvel total
durante o teste. Resultados expressos como média + erro padrão da média, onde asteriscos isolados
representam diferença estatística dentro do grupo Controle ou EAE e asteriscos com barras,
significância estatística entre os grupos experimentais indicados. Teste Two-way ANOVA seguido do
pos-teste Tukey. * p < 0,05; ** p < 0,01. N= 4 e 5.
4.3 Reconhecimento de objeto novo
O teste de reconhecimento de objeto novo foi realizado no 8º d.p.i. em ambos
esquemas de administração do fingolimode, agudo e sub-crônico. No tratamento
sub-crônico, dado até o 9º d.p.i., o fingolimode mostrou aumentar de forma
significativa o índice de discriminação do novo objeto, 2h e 24h após o treinamento,
quando comparado ao grupo EAE (Figura 7).
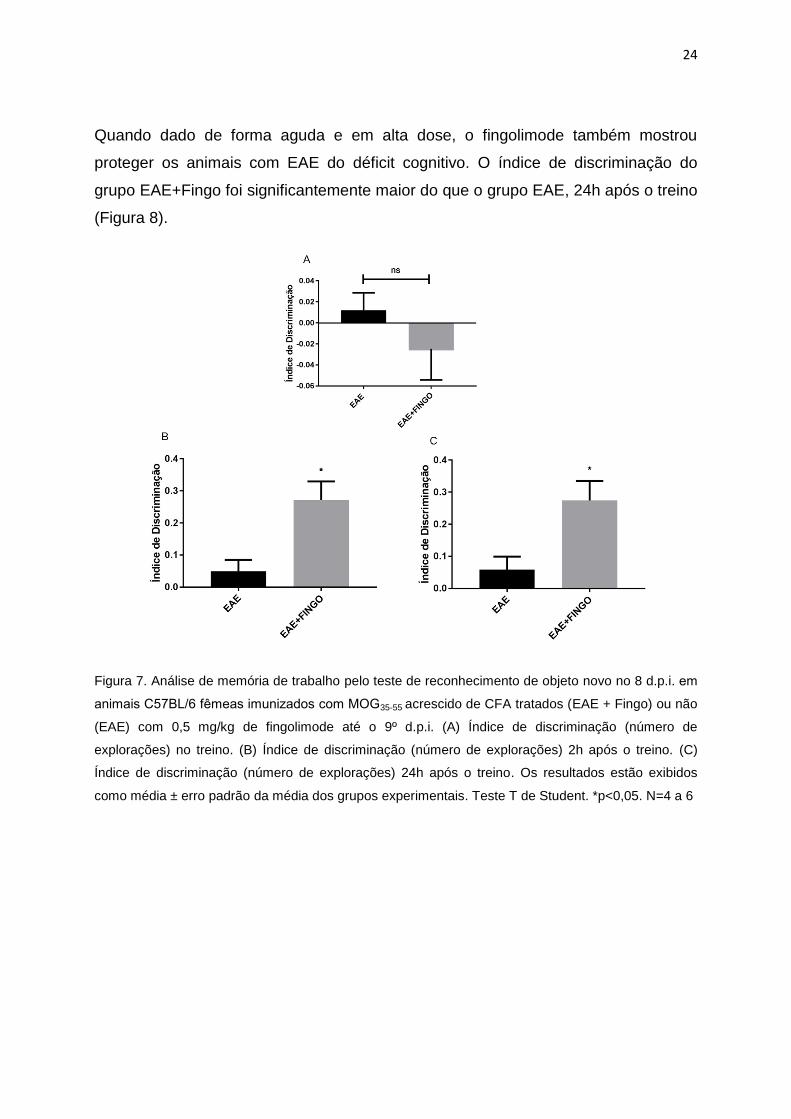
24
Quando dado de forma aguda e em alta dose, o fingolimode também mostrou
proteger os animais com EAE do déficit cognitivo. O índice de discriminação do
grupo EAE+Fingo foi significantemente maior do que o grupo EAE, 24h após o treino
(Figura 8).
Figura 7. Análise de memória de trabalho pelo teste de reconhecimento de objeto novo no 8 d.p.i. em
animais C57BL/6 femeas imunizados com MOG35-55 acrescido de CFA tratados (EAE + Fingo) ou não
(EAE) com 0,5 mg/kg de fingolimode até o 9º d.p.i. (A) Índice de discriminação (número de
explorações) no treino. (B) Índice de discriminação (número de explorações) 2h após o treino. (C)
Índice de discriminação (número de explorações) 24h após o treino. Os resultados estão exibidos
como média ± erro padrão da média dos grupos experimentais. Teste T de Student. *p<0,05. N=4 a 6

25
Figura 8. Teste de reconhecimento de objeto novo no 8 d.p.i. em animais C57BL/6 femeas
imunizados com MOG35-55 acrescido de CFA tratados (EAE + Fingo) ou não (EAE) com 10 mg/kg de
fingolimode no dia da imunização. (A) Índice de discriminação no treino, determinado pelo tempo de
exploração de dois objetos iguais no 7 d.p.i. (B) Índice de discriminação no teste após 2h da troca de
um dos objetos. (C) Índice de discriminação no teste após 24h da troca de um dos objetos. (D)
Tempo de exploração de cada objeto 2h após e (E) 24h após o treino. Resultados expressos como
média ± erro padrão da média, onde asteriscos isolados representam diferença estatística dentro do
grupo Controle ou EAE e asteriscos com barras, significância estatística entre os grupos
experimentais indicados. Teste Two-way ANOVA seguido do pos-teste Tukey. * p < 0,05; ** p < 0,01;
*** p < 0,001, ****p<0,0001. N= 4 e 5.
Quando realizado no 26º d.p.i, o índice de discriminação foi significantemente menor
no grupo EAE quando comparado com o grupo EAE+Fingo no teste 2h após o
treino. 24h após o treino houve também redução do índice de discriminação,
entretanto, este não foi estatisticamente significante (Figura 9).
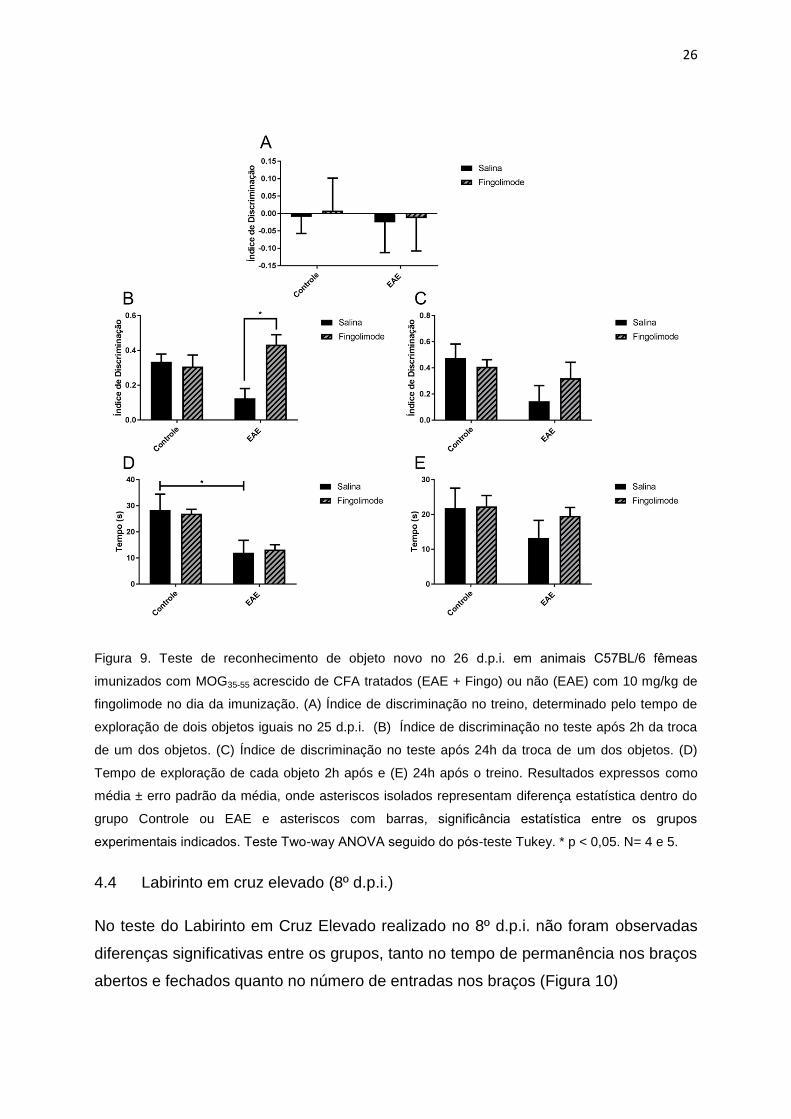
26
Figura 9. Teste de reconhecimento de objeto novo no 26 d.p.i. em animais C57BL/6 femeas
imunizados com MOG35-55 acrescido de CFA tratados (EAE + Fingo) ou não (EAE) com 10 mg/kg de
fingolimode no dia da imunização. (A) Índice de discriminação no treino, determinado pelo tempo de
exploração de dois objetos iguais no 25 d.p.i. (B) Índice de discriminação no teste após 2h da troca
de um dos objetos. (C) Índice de discriminação no teste após 24h da troca de um dos objetos. (D)
Tempo de exploração de cada objeto 2h após e (E) 24h após o treino. Resultados expressos como
média ± erro padrão da média, onde asteriscos isolados representam diferença estatística dentro do
grupo Controle ou EAE e asteriscos com barras, significância estatística entre os grupos
experimentais indicados. Teste Two-way ANOVA seguido do pos-teste Tukey. * p < 0,05. N= 4 e 5.
4.4 Labirinto em cruz elevado (8º d.p.i.)
No teste do Labirinto em Cruz Elevado realizado no 8º d.p.i. não foram observadas
diferenças significativas entre os grupos, tanto no tempo de permanência nos braços
abertos e fechados quanto no número de entradas nos braços (Figura 10)
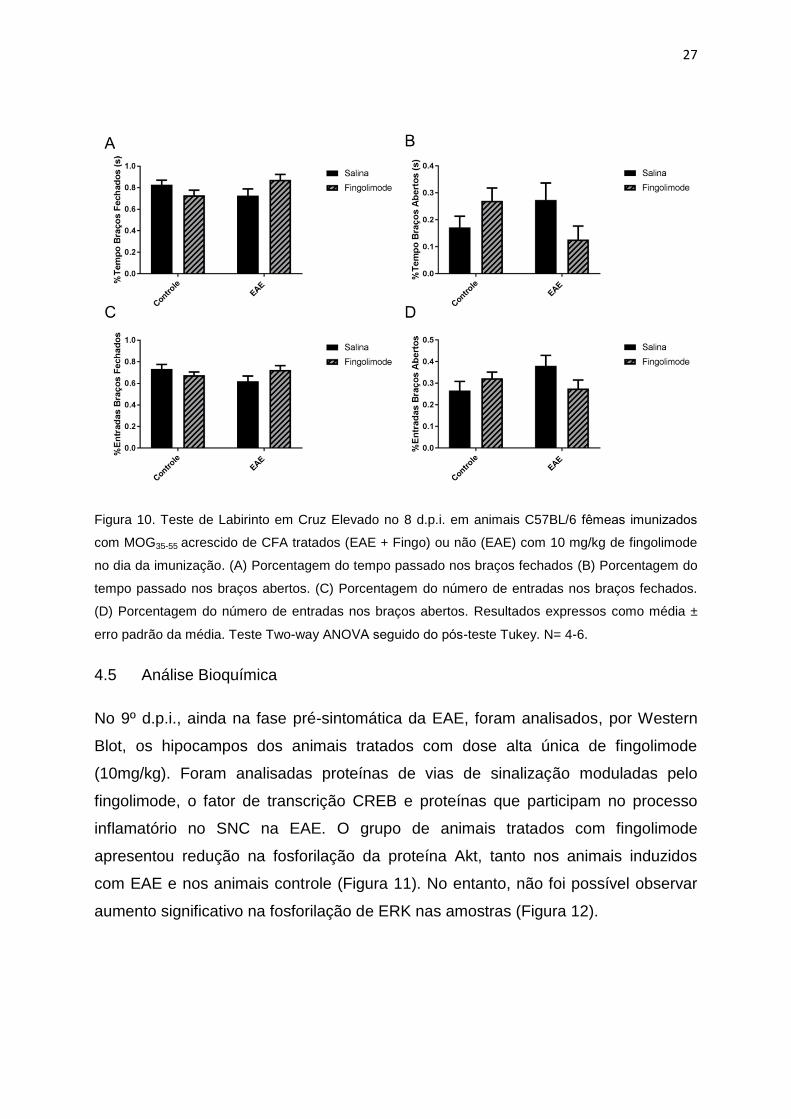
27
Figura 10. Teste de Labirinto em Cruz Elevado no 8 d.p.i. em animais C57BL/6 femeas imunizados
com MOG35-55 acrescido de CFA tratados (EAE + Fingo) ou não (EAE) com 10 mg/kg de fingolimode
no dia da imunização. (A) Porcentagem do tempo passado nos braços fechados (B) Porcentagem do
tempo passado nos braços abertos. (C) Porcentagem do número de entradas nos braços fechados.
(D) Porcentagem do número de entradas nos braços abertos. Resultados expressos como média ±
erro padrão da média. Teste Two-way ANOVA seguido do pos-teste Tukey. N= 4-6.
4.5 Análise Bioquímica
No 9º d.p.i., ainda na fase pré-sintomática da EAE, foram analisados, por Western
Blot, os hipocampos dos animais tratados com dose alta única de fingolimode
(10mg/kg). Foram analisadas proteínas de vias de sinalização moduladas pelo
fingolimode, o fator de transcrição CREB e proteínas que participam no processo
inflamatório no SNC na EAE. O grupo de animais tratados com fingolimode
apresentou redução na fosforilação da proteína Akt, tanto nos animais induzidos
com EAE e nos animais controle (Figura 11). No entanto, não foi possível observar
aumento significativo na fosforilação de ERK nas amostras (Figura 12).
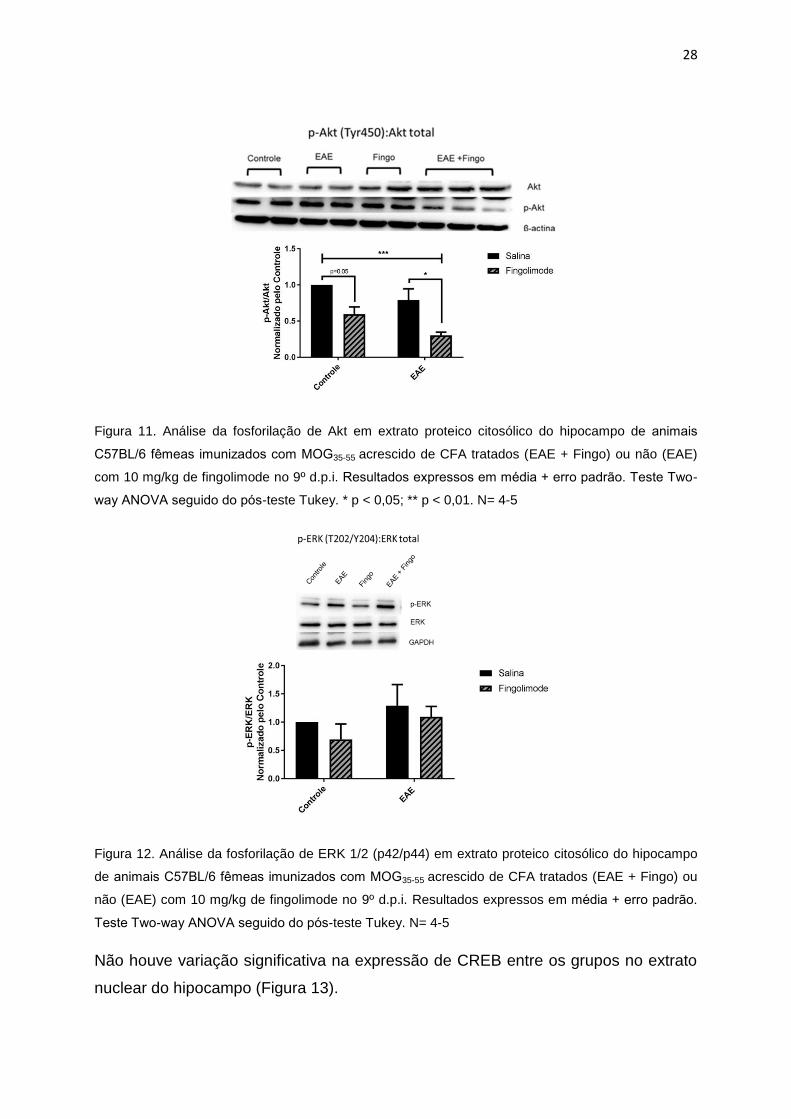
28
Figura 11. Análise da fosforilação de Akt em extrato proteico citosólico do hipocampo de animais
C57BL/6 femeas imunizados com MOG35-55 acrescido de CFA tratados (EAE + Fingo) ou não (EAE)
com 10 mg/kg de fingolimode no 9º d.p.i. Resultados expressos em média + erro padrão. Teste Two-
way ANOVA seguido do pos-teste Tukey. * p < 0,05; ** p < 0,01. N= 4-5
Figura 12. Análise da fosforilação de ERK 1/2 (p42/p44) em extrato proteico citosólico do hipocampo
de animais C57BL/6 femeas imunizados com MOG35-55 acrescido de CFA tratados (EAE + Fingo) ou
não (EAE) com 10 mg/kg de fingolimode no 9º d.p.i. Resultados expressos em média + erro padrão.
Teste Two-way ANOVA seguido do pos-teste Tukey. N= 4-5
Não houve variação significativa na expressão de CREB entre os grupos no extrato
nuclear do hipocampo (Figura 13).

29
Figura 13. Análise da expressão de CREB em extrato proteico nuclear do hipocampo de animais
C57BL/6 femeas imunizados com MOG35-55 acrescido de CFA tratados (EAE + Fingo) ou não (EAE)
com 10 mg/kg de fingolimode no 9º d.p.i. Resultados expressos em média + erro padrão. Teste Two-
way ANOVA seguido do pos-teste Tukey. N= 4-5
A neuroinflamação foi avaliada por meio da análise da translocação nuclear da
subunidade p65 do complexo NF-kB e a expressão das proteínas TNF-α e MHC
classe II. A translocação de p65 se encontra significantemente elevada nos animais
com EAE, retornando a níveis similares ao controle no grupo EAE+Fingo (Figura 14).
A expressão da subunidade p34 da molécula de MHC-II apresentou aumento
significativo no grupo EAE, o que retornou a níveis próximos ao controle com o
tratamento com fingolimode. Interessantemente, foi observado elevação da
expressão do MHC-II no grupo tratado apenas com fingolimode (Figura 15). Ainda, o
tratamento com fingolimode reduziu, embora não significantemente, a expressão da
molécula precursora de TNF-α no grupo EAE+Fingo, comparado com o grupo EAE
(Figura 16).
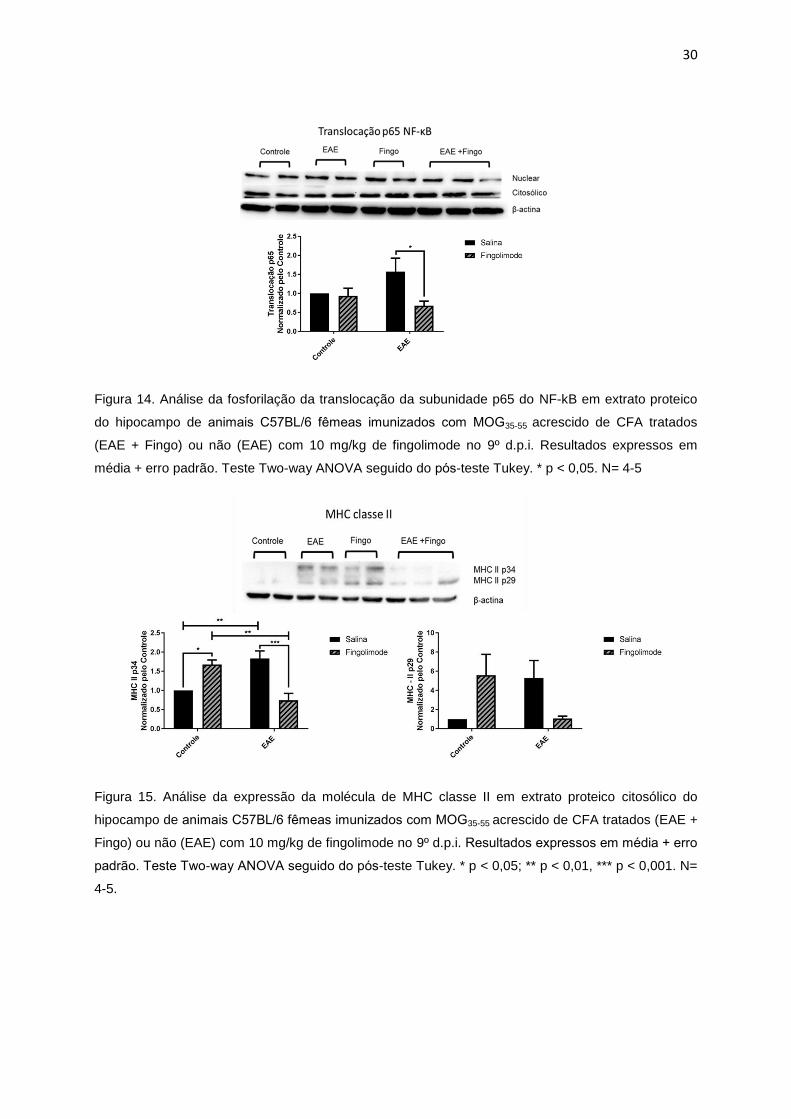
30
Figura 14. Análise da fosforilação da translocação da subunidade p65 do NF-kB em extrato proteico
do hipocampo de animais C57BL/6 femeas imunizados com MOG35-55 acrescido de CFA tratados
(EAE + Fingo) ou não (EAE) com 10 mg/kg de fingolimode no 9º d.p.i. Resultados expressos em
média + erro padrão. Teste Two-way ANOVA seguido do pos-teste Tukey. * p < 0,05. N= 4-5
Figura 15. Análise da expressão da molécula de MHC classe II em extrato proteico citosólico do
hipocampo de animais C57BL/6 femeas imunizados com MOG35-55 acrescido de CFA tratados (EAE +
Fingo) ou não (EAE) com 10 mg/kg de fingolimode no 9º d.p.i. Resultados expressos em média + erro
padrão. Teste Two-way ANOVA seguido do pos-teste Tukey. * p < 0,05; ** p < 0,01, *** p < 0,001. N=
4-5.

31
Figura 16. Análise da expressão da forma precursora do TNF-α em extrato proteico citosólico do
hipocampo de animais C57BL/6 femeas imunizados com MOG35-55 acrescido de CFA tratados (EAE +
Fingo) ou não (EAE) com 10 mg/kg de fingolimode no 9º d.p.i. Resultados expressos em média + erro
padrão. Teste Two-way ANOVA seguido do pos-teste Tukey. N=4-5
5 DISCUSSÃO
No presente estudo, através de testes comportamentais e moleculares, buscou-se
analisar os efeitos do tratamento preventivo com fingolimode na cognição e
neuroinflamação em camundongos com EAE. Ainda, utilizando dados prévios de
nosso grupo, os resultados obtidos com o fingolimode foram comparados aos
observados no tratamento preventivo com dexametasona.
O fingolimode, tanto dado na dose de 0,5mg/kg/dia por 10 dias ou como dose única
de 10 mg/kg no dia da imunização, gerou redução significativa no escore clínico
motor dos animais com EAE (Figura 3). A dose de 10 mg/kg é a maior dose testada
em animais para o tratamento da EM e foi utilizada como comparador para a alta
dose de dexametasona (50mg/kg), utilizada no estudo anterior de nosso grupo [dos
Santos, 2017]. O fingolimode é um agonista dos receptores de S1P, entretanto a
constante ativação desses receptores pelo fármaco acarreta em sua internalização e
degradação pelo sistema proteassoma, consequentemente diminuindo sua
densidade. O receptor S1P1 é um receptor acoplado à proteína Gi importante para o
egresso de linfócitos no SNC, o qual, quando ativado, resulta em ativação da via

32
PI3K/Akt/mTOR, inibindo a expressão do gene FoxP3 e reduzindo
consequentemente a diferenciação de linfócitos Treg. A melhora clínica de animais
com EAE pelo fingolimode é dependente da inibição da via Akt/mTOR [Hou, 2016].
Condizentemente, no 9º d.p.i. em animais tratados com fingolimode, a fosforilação
de Akt está reduzida no hipocampo, indicando que a modulação desta via foi
importante para o efeito protetor do fingolimode neste modelo (Fig. 11). Outra via
também modulada pelo tratamento com fingolimode, a MAPK/ERK, não apresentou
diferença significativa nos grupos tratados com fingolimode no 9 d.p.i. (Figura 12). O
tratamento com fingolimode induz a proliferação de células tronco neurais
hipocampais através da ativação da via de sinalização MAPK/ERK [Sun, 2016].
Pelos testes de campo aberto e LCE não foi observado diferenças sugestivas de
comportamento do tipo ansioso em nenhum grupo experimental. Esta observação
também pode ser encontrada na literatura [Rodrigues, 2011]. A avaliação da
capacidade motora dos animais no campo aberto mostrou que os animais dos
grupos EAE e EAE+Fingo se movimentam menos devido aos sintomas motores da
EAE na fase de remissão (22º d.p.i.), mas também no período pré-simtomático (4º
d.p.i.). Esta observação pode ter sido causada devido a possível efeito inflamatório
subclínico ou a um comportamento do tipo ansioso ou anedônico não detectado pelo
teste de campo aberto ou LCE. Animais com EAE possuem, principalmente nos dias
próximos à imunização e no aparecimento dos sintomas clínicos, síndrome do tipo
sickness behavior [Pollak, 2000]. Será realizado futuramente outro teste
comportamental que exige menor movimentação, como a caixa claro-escuro, no
período pré-sintomático para avaliação do comportamento do tipo ansioso nesses
animais e sua modulação pelo fingolimode.
No 8º e 26º d.p.i., o grupo EAE+Fingo apresentou melhora no índice de
discriminação de objeto novo comparados ao grupo EAE, mostrando que este
fármaco possui efeito protetor na memória declarativa destes animais. Este efeito
pode ter sido causado por efeitos distintos: diminuição da inflamação no hipocampo
e estruturas límbicas responsáveis pela consolidação da memória pela redução de
infiltrado leucocitário, efeito protetor e anti-inflamatório direto do fingolimode nos
diferentes tipos celulares do SNC e secreção aumentada de BDNF no hipocampo.
No entanto, através do ensaio de Western Blot, não foi observada diferença

33
significativa na expressão de CREB entre os grupos no 9º d.p.i., mostrando que não
há modulação quantitativa deste fator de transcrição (Figura 13). Para melhor
comparação, será realizado também análise da expressão de CREB fosforilado e
ensaio de desvio de mobilidade electroforética (EMSA) para CREB. Será realizado
ainda ensaio de Western Blot para animais eutanasiados 60 minutos após a
realização do teste de reconhecimento de objeto novo para a avaliação da
expressão da proteína de expressão rápida EGR-1.
Trabalhos mostraram que camundongos tratados com fingolimode 0,3mg/kg por 15
dias apresentaram neurogênese hipocampal aumentada e melhora na memória
contextual no teste de memória aversiva, indicando um efeito mediado pela
neurogênese hipocampal, dependente do BDNF liberado [Efstathopoulos, 2015].
Após isquemia cerebral focal em ratos pela oclusão da artéria cerebral média, a
administração de 0,5 mg/kg de fingolimode resultou em diminuição da área da lesão
isquêmica e melhora do déficit de memória nesses animais [Nazari, 2016]. O
tratamento de camundongos C57BL/6 adultos por 7 dias com fingolimode também
promoveu neurogênese hipocampal e melhorou a memória contextual e memória
espacial, medidos pelos testes de memória ao contexto e labirinto de Morris,
respectivamente [Sun, 2016].
O grupo EAE+Fingo apresentou menor translocação nuclear da subunidade p65 do
NF-kB no 9º d.p.i., sugerindo que a inflamação hipocampal está diminuída neste
grupo (Figura 14). Em linha com esta observação, a expressão de MHC classe II,
molécula importante para a reapresentação de antígenos no SNC e progressão da
EAE, é menor no grupo EAE+Fingo comparado ao grupo EAE (Figura 15).
Curiosamente, o grupo Controle+Fingo apresentou aumento significativo da
expressão do MHC no hipocampo. Este efeito pode ter sido causado pelo efeito
agonista do fingolimode nos receptores S1P1, o qual, sem uma condição
inflamatória concomitante, gera aumento na expressão dessa molécula. Apesar de
não estatisticamente significante, houve redução na expressão do TNF-α no grupo
EAE+Fingo em relação ao grupo EAE. Como a infiltração periférica no SNC não é
significante no ponto pré-sintomático, provavelmente a quantidade de TNF-α ainda
não é grande o suficiente no grupo EAE para uma diferença ser captada pela técnica
de Western Blot. Um ensaio futuro irá avaliar a expressão dessas proteínas pró-

34
inflamatórias e NF-kB no hipocampo no 26º d.p.i. por Western Blot e EMSA, na fase
de remissão da doença.
O tratamento com dexametasona 50mg/kg dada no dia da imunização, apesar de
gerar melhora no escore motor dos animais com EAE tratados, acarretou também
em efeitos negativos. Dentre eles, a não proteção da piora na razão de
discriminação de novo objeto 24h após o treino no 8º e 26º d.p.i e diminuição do
tempo nos braços abertos do LCE, indicativos de déficit de memória e
comportamento do tipo ansioso, respectivamente. Ainda, no grupo tratado com
dexametasona se observou que a inflamação no SNC é modulada diferentemente
por este fármaco dado nesta dose, em relação à periferia. O grupo EAE+DEX não
apresentou redução significativa de translocação nuclear da subunidade p65 do NF-
kB ou expressão do TNF-α no 8º d.p.i. No 26º d.p.i., a expressão de TNF-α é maior
que a do grupo EAE, há ligação do fator de transcrição NF-kB ao DNA semelhante
ao grupo EAE, medido pelo ensaio de EMSA [Dos Santos, 2017]. Além disso, no 26º
d.p.i, o grupo EAE+DEX apresentou maior expressão do marcador de ativação
astrocitário GFAP e menor expressão de delta-FosB na região CA1 do hipocampo.
Esses resultados fortalecem resultados na literatura que mostram que os
glicocorticoides geram efeitos biológicos complexos e variados, dependendo da
dose, contexto e tempo que são utilizados [Sorrells, 2009; Duque, 2016].
Outro glicocorticoide utilizado na EM, a metilpredinisona, dada de forma terapêutica,
após a aparição dos sintomas clínicos da EAE, em doses até 20 mg/kg, reduz o
escore clínico dos animais imunizados. Entretanto, uma dose alta de 100 mg/kg
dada de forma preventiva, ou seja, antes da aparição dos sintomas motores,
contraditoriamente, promove um aumento do escore clínico em relação ao grupo não
tratado. Isso mostra mais uma vez que os GCs possuem efeitos que podem ser anti-
ou pró- inflamatórios, dependendo da dose e contexto que são utilizados [Wüst,
2012]. O tratamento da EAE com GCs também resulta em diminuição da expressão
do marcador de células T reguladoras (Treg), FoxP3+ [Lühder, 209], deficiência
semelhante ao visto na imunopatologia da EM. Células microgliais hipocampais de
ratos pré-expostas à corticosterona respondem à um segundo insulto com LPS com
expressão aumentada de MHC classe II, TLR-4, IL-1β, TNF-α e NF-κB.

35
Similarmente, a exposição à corticosterona antes do LPS causou, in vivo, o aumento
de citocinas como IL-1β, TNF-α e IL-6 no hipocampo [Frank, 2010] e córtex frontal
[Munhoz, 2010] de ratos, comparados ao controle. A exposição à corticosterona
após o LPS resulta em diminuição da expressão de marcadores pró-inflamatórios
[Frank, 2010; Munhoz, 2010], mostrando um contexto tempo-dependente no tipo de
ação que os GCs podem exercer, pró ou anti-inflamatória. Ainda em linha com os
estudos anteriores, ratos submetidos a estresse crônico imprevisível apresentaram
maior ativação de NF-κB no córtex frontal e hipocampo após injeção de LPS. Este
fenômeno foi revertido com a administração de mifepristona (RU-486), um
antagonista do GR [Munhoz, 2006].
Efeitos distintos observados em testes comportamentais e moleculares quando se
trata animais preventivamente com fingolimode ou dexametasona, fortalece a ideia
que os efeitos deletérios presentes no grupo EAE+DEX são exclusivamente
causados pelos glicocorticóides sintéticos. A futura avaliação da expressão de EGR-
1 e proteínas pró-inflamatórios e a ligação dos fatores de transcrição CREB e NF-kB
no DNA no 26º d.p.i. em animais tratados com fingolimode, irá ajudar a reforçar
ainda mais essa hipótese.
6 CONCLUSÃO
O tratamento preventivo da EAE com fingolimode em dose única de10mg/kg e por
10 dias 0,5 mg/kg/dia resultou na melhora significativa do score clínico motor dos
animais. Os animais tratados com fingolimode não apresentaram, entretanto, os
mesmos resultados comportamentais e expressão de moléculas pró-inflamatórias no
hipocampo como observado no tratamento com dexametasona.
Os resultados de nosso grupo mostram que a utilização de altas doses de
glicocorticoides pode trazer efeitos deletérios que são tempo e contexto
dependentes. Como esta classe farmacológica é profundamente utilizada no
tratamento da EM estes estudos alertam para que a sua utilização seja feita com
cautela, a fim de se evitar possíveis complicações.

36
7 BIBLIOGRAFIA
ACHARJEE, S; et al. Altered cognitive-emotional behavior in early experimental autoimmune
encephalitis - cytokine and hormonal correlates. Brain Behavior Immunity. v. 33, p. 164-72,
2013
BHISE, V.; DHIB-JALBUT, S. Further understanding of the immunopathology of multiple
sclerosis: impact on future treatments. Expert Review of Clinical Immunology. v. 1, p. 21,
2016.
BRADFORD, M.M. A rapid and sensitive method for the quantitation of microgram quantities of
protein utilizing the principle of protein-dye binding. Analytical Biochemistry. v.72, p. 248-54,
1976.
BURTON, J.M.; O'CONNOR, P.W.; HOHOL, M. Oral versus intravenous steroids for treatment of
relapses in multiple sclerosis. Cochrane Database. v. 12, CD006921, 2012
CASTRÉN, E.; KOJIMA, M. Brain-derived neurotrophic factor in mood disorders and
antidepressant treatments. Neurobiology of Disease. v. 97, pt. B p. 119-126, 2017.
CRUZ-TOPETE, D.; CIDLOWSKI, J.A. One hormone, two actions: anti- and pro-inflammatory
effects of glucocorticoids. Neuroimmunomodulation. v. 22 p. 20-32, 2015
CHAMBERLAIN, K.A.; et al. Oligodendrocyte regeneration: Its significance in myelin replacement
and neuroprotection in multiple sclerosis. Neuropharmacology, v. 110, pt. B p. 633-643, 2016.
CHIARAVALLOTI, N.D.; DELUCA, J. Cognitive impairment in multiple sclerosis. Lancet
Neurology. v. 12, p. 1139-51, 2008.
CHUN, J.; HARTUNG, H.P. Mechanism of action of oral fingolimod (FTY720) in multiple
sclerosis. Clinical Neuropharmacology. v. 33, p. 91–101, 2010.
COMPSTON, A.; COLES, A. Multiple sclerosis. Lancet. v. 372, p.1502-17, 2008
DA GAMA PEREIRA, A.B.; et al. Prevalence of multiple sclerosis in Brazil: A systematic review.
Multiple Sclerosis and Related Disorders. v. 6, p. 572-9, 2015
DAMJANOVIC, D.; et al. Hippocampal and Deep Gray Matter Nuclei Atrophy Is Relevant for
Explaining Cognitive Impairment in MS: A Multicenter Study. American Journal of
Neuroradiology, v. 38, n. 1, p.18-24, 2017.
DENDROU, C.A.; FUGGER, L.; FRIESE, M.A. Immunopathology of multiple sclerosis. Nature
Review Immunology. v.15, n. 9, p. 545-58, 2015
DOS SANTOS, N.B. Participação dos glicocorticoides na progressão e no prejuízo cognitivo da
encefalomielite autoimune experimental em camundongos C57BL/6. Tese (Doutorado em
Ciências) – Instituto de Ciências Biomédicas, Universidade de São Paulo, São Paulo, 2017.
DUQUE, E. A.; Munhoz, C.D. The Pro-inflammatory Effects of Glucocorticoids in the Brain.
Frontiers in Endocrinology. v. 7, n. 78, 2016
EFSTATHOPOULOS, P.; et al. Fingolimod induces neurogenesis in adult mouse hippocampus
and improves contextual fear memory. Translational Psychiatry. v. 5, n. 11, 2015

37
FRANK, M.G. Prior exposure to glucocorticoids sensitizes the neuroinflammatory and peripheral
inflammatory responses to E. coli lipopolysaccharide. Brain Behavior Immunity. v. 24, n. 1, p.
19-30, 2010
GARRIS, C.S.; et al. Defective sphingosine 1-phosphate receptor 1 (S1P1) phosphorylation
exacerbates Th17 mediated autoimmune neuroinflammation. Nature Immunology. v. 14, n. 11,
p. 1166-72, 2013
HOU, H.; et al. Fingolimod ameliorates the development of experimental autoimmune
encephalomyelitis by inhibiting Akt–mTOR axis in mice. International Immunopharmacology. v.
30, p. 171-178, 2016.
HUNTER, S. F.; BOWEN, J. D.; REDER, A. T. The Direct Effects of Fingolimod in the Central
Nervous System: Implications for Relapsing Multiple Sclerosis. CNS Drugs. v. 30, n. 2, p. 135-
147, 2016.
LASSMANN, H.; BRADL, M. Multiple sclerosis: experimental models and reality. Acta
Neuropathologica. v.133, n. 2, p. 223-244, 2017
LÜHDER, F.; REICHARDT, H.M. Traditional concepts and future avenues of glucocorticoid action
in experimental autoimmune encephalomyelitis and multiple sclerosis therapy. Critical Reviews
in Immunology. v. 29, n. 3, p. 255-73, 2009
MCEWEN, B.S.; MAGARINOS, A.M. Stress and hippocampal plasticity: implications for the
pathophysiology of affective disorders. Human Psychopharmacology. v. 16, n. S1, p. S7-S19,
2001.
MENDEL, I.; KERLERO DE ROSBO, N.; BEN NUN, A. A myelin oligodendrocyte glycoprotein
peptide induces typical chronic experimental autoimmune encephalomyelitis in H-2b mice: fine
specificity and T cell receptor V beta expression of encephalitogenic cells. European Journal of
Immunology. v. 25, p. 1951–1959, 1995
MUNHOZ, C.D.; et al., Chronic unpredictable stress exacerbates lipopolysaccharide-induced
activation of nuclear factor-kappaB in the frontal cortex and hippocampus via glucocorticoid
secretion. Journal of Neuroscience. v. 26, n. 14, p. 3813-20, 2006.
MUNHOZ, C.D.; et al. Glucocorticoids exacerbate lipopolysaccharide-induced signaling in the
frontal cortex and hippocampus in a dose-dependent manner. Journal of Neuroscience. v. 30,
n. 41, p.13690-8, 2010
NAZARI, M.; et al. Fingolimod (FTY720) improves hippocampal synaptic plasticity and memory
deficit in rats following focal cerebral ischemia. Brain Research Bulletin. v. 124, p. 95-102,
2016.
NUZZO, L.; et al. Antidepressant activity of fingolimod in mice. Pharmacology Research &
Perspectives. v.3, n.3, 2015.
PAPARRIGOPOULOS, T.; et al. The neuropsychiatry of multiple sclerosis: focus on disorders of
mood, affect and behaviour. International Review of Psychiatry. v. 22, n. 11, p. 4-21, 2010

38
PELLOW, S; et al. Validation of open:closed arm entries in an elevated plus-maze as a measure
of anxiety in the rat. Journal of Neuroscience Methods. v. 14, n. 3, p.149-67, 1985.
PERUGA, I.; et al. Inflammation modulates anxiety in an animal model of multiple sclerosis.
Behavior Brain Research. v. 220, n. 1, p. 20-9, 2011
POLLAK, Y.; et al. Behavioral aspects of experimental autoimmune encephalomyelitis. Journal
of Neuroimmunology. v.104, n. 1, p. 31-6, 2000.
RODRIGUES, D.H.; et al. Behavioral investigation of mice with experimental autoimmune
encephalomyelitis. Arquivos de Neuropsiquiatria. v. 69, n.6, p. 938-42, 2011.
LAEMMLI, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage
T4. Nature. v. 227 n. 259, p.680-5, 1970
SALINOVICH, O; MONTELARO, R.C. Reversible staining and peptide mapping of proteins
transferred to nitrocellulose after separation by sodium dodecylsulfate-polyacrylamide gel
electrophoresis. Analytical Biochemistry. v. 156, n. 2, p. 341-7, 1986.
SCHWEINGRUBER, N.; et al. Mechanisms of glucocorticoids in the control of
neuroinflammation. Journal of Neuroendocrinology. v. 24, n.1, p. 174-82, 2012
SIMMONS, S.B.; et al. Modeling the heterogeneity of multiple sclerosis in animals. Trends in
Immunology. v.34, n.8, p. 410-22, 2013
SORRELLS, S.F.; et al. The stressed CNS: when glucocorticoids aggravate inflammation.
Neuron. v. 64, n.1, p. 33-9, 2009
SUN, Y.; et al. The sphingosine-1-phosphate analogue, FTY-720, promotes the proliferation of
embryonic neural stem cells, enhances hippocampal neurogenesis and learning and memory
abilities in adult mice. British Journal of Pharmacology. v.173, n.18, p. 2793-2807, 2016
WORLD HEALTH ORGANIZATION; MULTIPLE SCLEROSES INTERNATIONAL FEDERATION.
Atlas – Multiple Sclerosis resources in the world, 2008. Disponível em:
<http://www.who.int/mental_health/neurology/Atlas_MS_WEB.pdf>. Acesso em 27 abr. 2017.
WÜST, S.; et al. Preventive treatment with methylprednisolone paradoxically exacerbates
experimental autoimmune encephalomyelitis. International Journal of Endocrinology, v. 2012,
p. 1-8, 2012.
ZHANG, J.; et al. Fingolimod treatment promotes proliferation and differentiation of
oligodendrocyte progenitor cells in mice with experimental autoimmune encephalomyelitis.
Neurobiology of Disease. v. 76, p. 57-66, 2015.
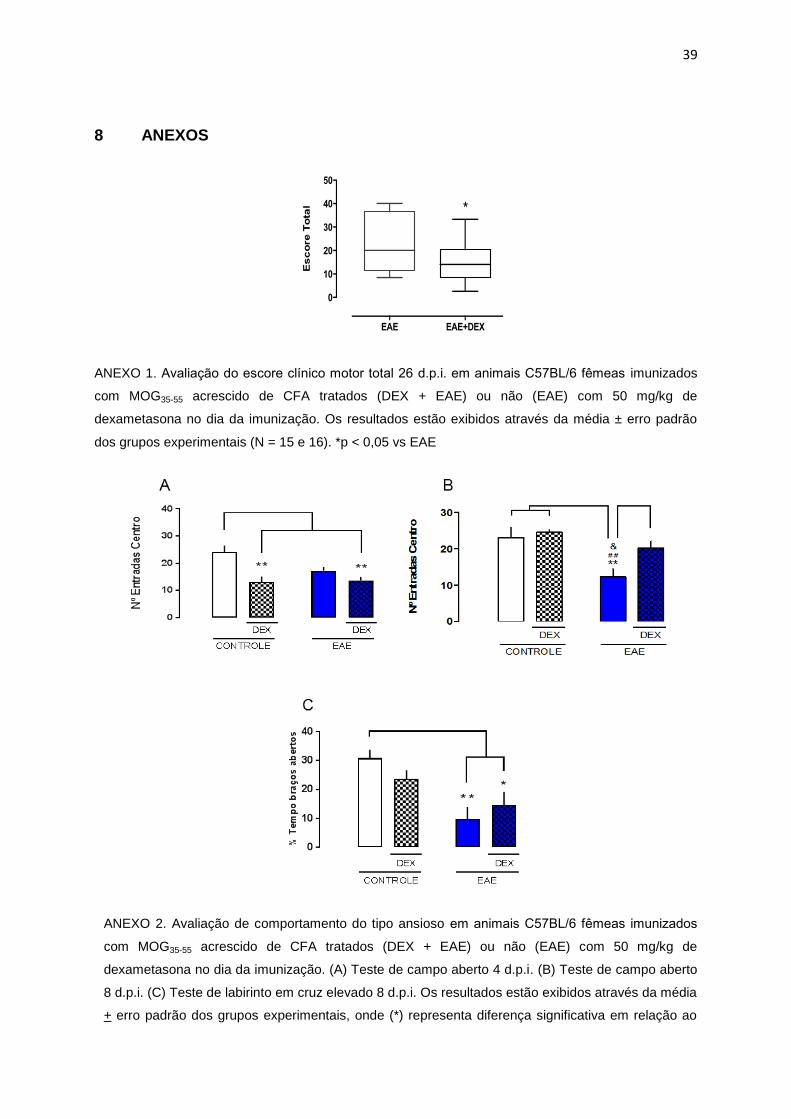
39
8 ANEXOS
ANEXO 1. Avaliação do escore clínico motor total 26 d.p.i. em animais C57BL/6 femeas imunizados
com MOG35-55 acrescido de CFA tratados (DEX + EAE) ou não (EAE) com 50 mg/kg de
dexametasona no dia da imunização. Os resultados estão exibidos através da média ± erro padrão
dos grupos experimentais (N = 15 e 16). *p < 0,05 vs EAE
ANEXO 2. Avaliação de comportamento do tipo ansioso em animais C57BL/6 femeas imunizados
com MOG35-55 acrescido de CFA tratados (DEX + EAE) ou não (EAE) com 50 mg/kg de
dexametasona no dia da imunização. (A) Teste de campo aberto 4 d.p.i. (B) Teste de campo aberto
8 d.p.i. (C) Teste de labirinto em cruz elevado 8 d.p.i. Os resultados estão exibidos através da média
+ erro padrão dos grupos experimentais, onde (*) representa diferença significativa em relação ao
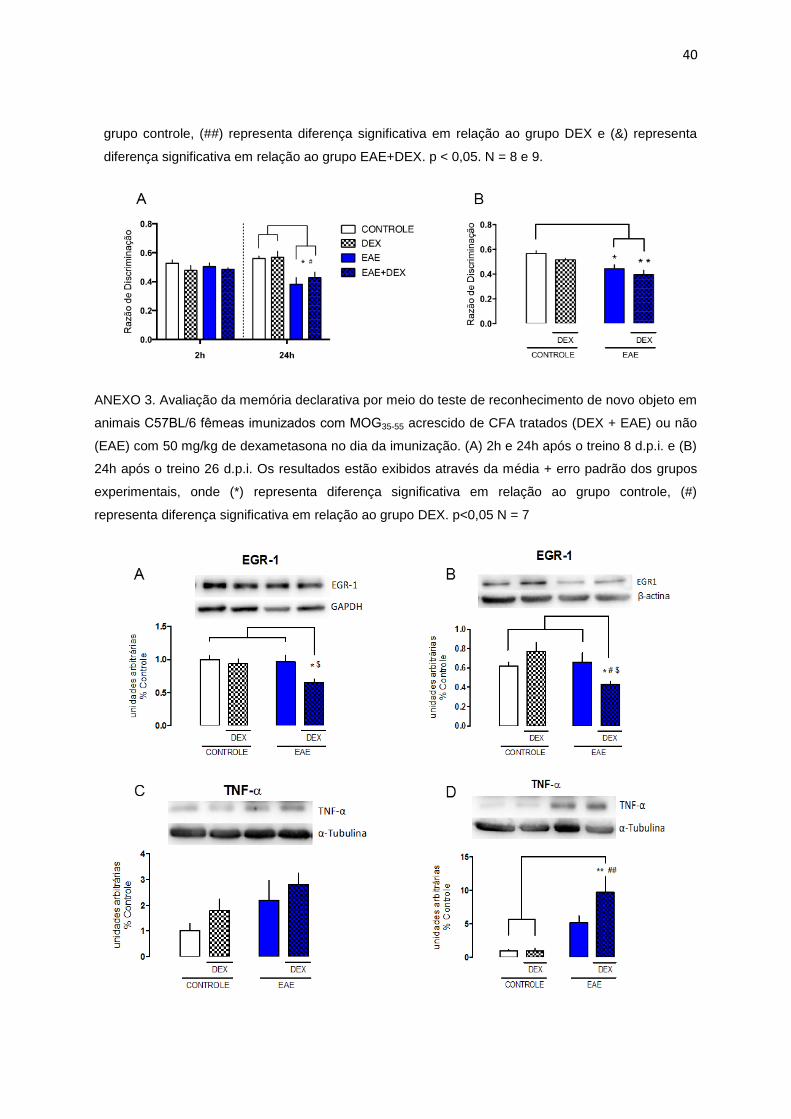
40
grupo controle, (##) representa diferença significativa em relação ao grupo DEX e (&) representa
diferença significativa em relação ao grupo EAE+DEX. p < 0,05. N = 8 e 9.
ANEXO 3. Avaliação da memória declarativa por meio do teste de reconhecimento de novo objeto em
animais C57BL/6 femeas imunizados com MOG35-55 acrescido de CFA tratados (DEX + EAE) ou não
(EAE) com 50 mg/kg de dexametasona no dia da imunização. (A) 2h e 24h após o treino 8 d.p.i. e (B)
24h após o treino 26 d.p.i. Os resultados estão exibidos através da média + erro padrão dos grupos
experimentais, onde (*) representa diferença significativa em relação ao grupo controle, (#)
representa diferença significativa em relação ao grupo DEX. p<0,05 N = 7

41
ANEXO 4. Avaliação por Western Blot das proteínas EGR-1 e TNF-α no hipocampo de animais
C57BL/6 femeas imunizados com MOG35-55 acrescido de CFA tratados (DEX + EAE) ou não (EAE)
com 50 mg/kg de dexametasona no dia da imunização. (A) Proteína de expressão imediata EGR-1 no
hipocampo 8 d.p.i e (B) 26 d.p.i. (C) Forma madura da proteína TNF-α no hipocampo 8 d.p.i e (D) 26
d.p.i. Os resultados estão expressos como média ± erro padrão da média dos grupos
experimentais, onde (*) representa diferença significativa em relação ao grupo controle. p < 0,05
(N = 4).

D~-
UNIVERSIDADE DE SÃO PAULOINSTITUTO DE CIÊNCIAS BIOMÉDICAS
Cidade Universitária "Armando de Salles Oliveira"Av. Prof. lineu Prestes, 2415 - CEPo 05508-000 São Paulo, SP BrasilTelefone :(55) (011) 3091.7733 - e-mail: [email protected]
CERTIFICADO
Certificamos que o protocolo registrado sob nO003 nas fls. 03 do livro 03 para USO
de animais em experimentação, sob a responsabilidade do Prof(a) Or(a)) Carolina
Demarchi Munhoz, Coordenador (a) da Linha de pesquisa "Participação dos
glicocorticoides na progressão e no prejuízo cognitivo de encefalomielite autoimune
experimental em camundongos C57BL/6" do qual participam o(s) aluno(s), Nilton
Barreto dos Santos, Leonardo Santana Novaes, Dielly Catrina Favacho Lopes, Érica
de Almeida Duque, está de acordo com os Princípios Éticos de Experimentação
Animal adotado pela Sociedade Brasileira de Ciência de Animais de Laboratório
(SBCAL) e foi aprovado pela COMISSÃO DE ÉTICA NO USO DE ANIMAIS (CEUA)
em 26.03.2013, com validade de 4 anos.
São Paulo, 26 de março de 2013.
Prof. Dr. WOTHAN TAVARESDELIMACoordenador-CEUA - ICB/USP
Profa. Ora. ANA PAULALEPIQUESecretária- CEUA - ICB/USP

Cidade Universitária "Armando de Salles Oliveira", Butantã, São Paulo, SP . Av. Professor Lineu Prestes, 2415 - ICB III - 05508 000Comissão de Ética no Uso de Animais - Telefone (11) 3091-7733 - e-mail: [email protected]
Of.CEUA.013,2017São Paulo, 20 de março de 2017.
Prezadofa) Professor(a),
Informo que o projeto intitulado "Participação dos glicocorticoides na progressãoe no prejuízo cognitivo de encefalomielite autoimune experimental em camundongosC57BLj6", registrado sob o protocolo nº 003/2013 e aprovado em 26/03/2013 que envolve aprodução, manutenção e/ou utilização de animais pertencentes ao filo Chordata, subfiloVertebrata (exceto o homem), para fins de pesquisa científica, foi prorrogado até 26/03/2021.
Diante desta prorrogação e da declaração de que não houve alteração dametodologia e das técnicas descritas na licença inicial para o uso de animais, autorizo a inclusãodas espécies e quantidades descritas abaixo para continuidade ao referido projeto:
Espécie Linhagem Sexo Idade/Peso Quantidade por anoCamundongo C57BL/6 Fêmea 5 semanas 1º: 120
2º: 803º: 804º: 80
Camundongo C57BL/6 Fêmea Neonatos 1º: 1202º: 1203º: 1004º: 100
Reitero que havendo alteração de metodologia e inserção de novos alunos ao projetode pesquisa vinculado à referida licença a CEUA-ICB deverá ser informada.
Cordialmente,
fJWJJAProf. Dr. Anderson de Sá NunesCoordenador - CEUA-ICB/USP
Prof.(a) Dr.(a) Carolina Demarchi MunhozDepartamento de FarmacologiaInstituto de Ciências Biomédicas - USP

Cidade Universitária "Armando de Salles Oliveira", Butantã, São Paulo, SP . Av. Professor Lineu Prestes, 2415 - ICB 111 - 05508 000Comissão de Ética no Uso de Animais· Telefone (11) 3091-7733 - e-rnail: [email protected]
Decl. CEUA085.2016
DECLARAÇÃO
Em adendo ao Certificado nº 003/2013/CEUA, datado de
26/03/2013 e por solicitação da Profa. Ora. Carolina Demarchi Munhoz, do
Departamento de Farmacologia, responsável pela linha de Pesquisa, autorizo a
inclusão dora) alunoja) Guilherme Dragunas ao Projeto de Pesquisa
"Participação dos glicocorticoides na progressão e no prejuízo cognitivo de
encefalomielite autoimune experimental em camundongos C57BLj6", uma vez
que se trata de utilização da mesma espécie animal e de métodos experimentais
similares ao Projeto.
São Paulo, 19 de setembro de 2016
iJWJJ?AProf. Dr. Anderson de Sá NunesCoordenador da CEUA-ICB/USP

UNIVERSIDADE DE SÃO PAULO
FACULDADE DE CIÊNCIAS FARMACÊUTICASComissão de Ética no Uso de Animais - CEUA
Ofício CEUAlFCF 7.2017
CIÊNCIA
A Comissão de Ética no Uso de Animais da Faculdade de Ciências Farmacêuticas da
Universidade de São Paulo, em 10/0312017, tomou ciência da pesquisa Perda cognitiva e
neuroinflamação causados pelo tratamento com o glicocorticoide sintético dexametasona
em encefalomielite autoimune experimental, conforme aprovada pela CEUA do Instituto
de Ciências Biomédicas da USP (CEUA 00312013, de 26/03/2013), conforme certificado
assinado em 26103/2013.
Orientador(a): Profa. Dra. Carolina Demarchi MunhozPesquisador(a): Guilherme Dragunas
I)
São Pa~~o, !Qie ~~ç1de 20p._ .. '- __. f:(jÇC;2' 1'--;;t'_ ••..• I A
, 'f v{~.. .\r i.i. " -_ .. - .(1Prof. Dr. Jai'son de Oliveira Martins, .
Coordenador da CEUAlFCF/USP
Av. Praf. Lineu Prestes, 580, Bloco 13 A. Cidade Universitária, CEP 05508-900, São Paulo, SPTelefone: (11) 3091-3622 - e-mai!: [email protected]