Embed Size (px)
Citation preview

INSTITUTO DE ZOOTECNIA PROGRAMA DE PÓS-GRADUAÇÃO EM PRODUÇÃO ANIMAL SUSTENTÁVEL
COMPLEXO ENZIMÁTICO PARA SUÍNOS NAS FASES DE
CRESCIMENTO E TERMINAÇÃO
Natália Yoko Sitanaka
Nova Odessa
Fevereiro - 2016

GOVERNO DO ESTADO DE SÃO PAULO
SECRETARIA DE AGRICULTURA E ABASTECIMENTO
AGÊNCIA PAULISTA DE TECNOLOGIA DOS AGRONEGÓCIOS
INSTITUTO DE ZOOTECNIA PROGRAMA DE PÓS-GRADUAÇÃO EM PRODUÇÃO ANIMAL SUSTENTÁVEL
Complexo enzimático para suínos nas fases de crescimento e terminação
Natália Yoko Sitanaka
Orientador: Dr. Fábio Enrique Lemos Budiño
Nova Odessa
Fevereiro -2016
Dissertação apresentada ao Programa de
Pós-graduação do Instituto de Zootecnia,
APTA/SAA, como parte dos requisitos
para obtenção do título de Mestre em
Produção Animal Sustentável.

Ficha Catalográfica elaborada pelo
Núcleo de Documentação e Informação do Instituto de Zootecnia
Bibliotecária: Tatiane Helena Borges de Salles – CRB 8/8946
S623c Sitanaka, Natália Yoko
Complexo enzimático para suínos na fase de crescimento e terminação /
Natália Yoko Sitanaka.
Nova Odessa, SP: [s.n.], 2016.
72p.; il.
Dissertação (mestrado) – Instituto de Zootecnia. APTA/SAA,
Nova Odessa.
Orientador: Dr. Fábio Enrique Lemos Budinõ
1. Aditivos alimentos para animais. 2. Eficiência. 3. Enzimas. 4. Nutrição I.
Budinõ, Fábio Enrique Lemos. II. Título.
CDD – 636.4085

GOVERNO DO ESTADO DE SÃO PAULO
SECRETARIA DA AGRICULTURA E ABASTECIMENTO
AGÊNCIA PAULISTA DE TECNOLOGIA DOS AGRONEGÓCIOS
INSTITUTO DE ZOOTECNIA
PROGRAMA DE PÓS-GRADUAÇÃO EM PRODUÇÃO ANIMAL SUSTENTÁVEL
CERTIFICADO DE APROVAÇÃO
TÍTULO: COMPLEXO ENZIMÁTICO PARA SUÍNOS NA FASE DE
CRESCIMENTO E TERMINAÇÃO
AUTOR: NATÁLIA YOKO SITANAKA
Orientador: Dr. Fábio Enrique Lemos Budiño
Aprovado como parte das exigências para obtenção de título de MESTRE em Produção
Animal Sustentável, pela Comissão Examinadora:
Dr. Fábio Enrique Lemos Budiño
Dra. Keila Maria Roncato Duarte
(Instituto de Zootecnia)
Dr, Dirlei Antônio Berto
(Faculdade de Medicina Veterinária e Zootecnia- UNESP)
Data da realização: 18 de fevereiro de 2016
Presidente da Comissão Examinadora
Prof. Dr. Fábio Enrique Lemos Budiño


DEDICO
Pai e Mãe: minhas inspirações e espelhos;
Vó Nair: meu porto seguro;
Irmãos Fernanda, Yuri e Nicholas: meus pedaços de céu aqui na Terra;
Guilherme: namorado, amigo e parceiro de vida;
Tampiko: a melhor companhia durante essa realização:


AGRADECIMENTOS
À Deus pela dádiva da vida, saúde e proteção; por me guiar para o caminho do bem ao
lado de pessoas abençoadas.
Ao meu orientador Fábio Enrique Lemos Budinõ, pelos ensinamentos transmitidos,
confiança depositada no meu trabalho, paciência e apoio. Muito obrigada por tudo!
À CAPES pela concessão da bolsa de estudos.
À Empresa Comércio e Indústria Uniquímica Ltda, especialmente à Andréia Chagas
pela oportunidade do projeto de pesquisa e auxílio durante todo experimento.
As funcionárias da APTA, Unidade Tanquinho, Simone, Lurdes e Isabel: pessoas
incríveis que além de me ajudarem durante o experimento, fizeram a minha estadia em
Tanquinho mais feliz.
Ao funcionário “Maquina”, ao pesquisador Evandro Moraes e ao amigo Daniel
Malagoli pela ajuda durante o período experimental.
Á Dra. Luciana Katiki e à Dra. Rosana Possenti pelo aceite do convite para
participação da banca de qualificação e pela colaboração na melhora do trabalho.
Ao Dr Dirlei Antônio Berto e Dra Keila Roncato pela participação na banca de Defesa
e pela valiosa contribuição no trabalho.
Aos demais pesquisadores e funcionários do Instituto de Zootecnia que contribuíram
para a minha formação pessoal e profissional, especialmente a Sandra Liepkaln dos Reis,
pela amizade e carinho conquistados.
Aos amigos conquistados Natália Trevizan, Heloisa Fidelis, André Lasmar e Daniela
Fagotti, pela união durante os estudos, parceria nas dificuldades e compartilhamento de
momentos inesquecíveis.
Aos amigos da Casa dos Estudantes, Camila Chaves, Guadalupe, Carol e Thomas:
obrigada pelos bons momentos vividos.
Além disso, minha gratidão àqueles que não foram citados, mas contribuíram de
alguma forma para que eu alcançasse mais esse objetivo.


RESUMO
COMPLEXO ENZIMÁTICO PARA SUÍNOS NA FASE DE CRESCIMENTO E
TERMINAÇÃO
O estudo teve como objetivo de avaliar o uso de um complexo enzimático contendo -
amilase, -glucanase, fitase, celulase, xilanase e protease, sobre o desempenho,
digestibilidade e análise econômica de suínos nas fases de crescimento e terminação.
Foram utilizados 80 suínos, 42 machos e 38 fêmeas, com aproximadamente 63 dias de
idade, num delineamento em blocos ao caso. Foram testados cinco tratamentos com oito
repetições/tratamento, sendo: CP: Dieta controle positivo, com nível energético
recomendado para a fase; CN85: Dieta controle negativo com redução energética (85
kcal/kg EM); CE85: Dieta CN85 com adição de complexo enzimático; CN100: Dieta
controle negativo com redução energética (100 kcal/kg EM); CE100: Dieta CN100 com
adição de complexo enzimático,. Foi utilizado o programa alimentar com 3 dietas: -
Crescimento I (CI) – dos 63 aos 90 dias de idade, Crescimento II (CII) – dos 91 aos 118
dias de idade e Terminação (T) - dos 119 aos 145 dias de idade. Durante cada fase do
período experimental, foi avaliado o desempenho, através das variáveis: consumo diário de
ração (CDR), ganho de peso diário (GPD) e conversão alimentar (CA). A digestibilidade
das dietas foram analisadas nas três fases, através do uso do marcador Celite ® e,
posteriormente, análise de cinza insolúvel em ácido. A análise de viabilidade econômica
foi realizada através da determinação do custo da ração (em R$), por quilograma de peso
vivo ganho e cálculo do Índice de Eficiência Econômica. Os resultados mostraram que para
machos, recomenda-se o uso de complexo enzimático em dietas com redução de 85 Kcal/
kg de EM, durante os períodos de crescimento e terminação, pois melhora a conversão
alimentar e é economicamente mais eficiente durante o Crescimento I. Para fêmeas, o uso
do complexo enzimático não se justifica. Nas dietas com redução energética de 100
Kcal/kg de EM, o uso do complexo enzimático elevou o teor de proteína digestível durante
o Crescimento II; A adição de complexo enzimático nas dietas com redução energética de
85 e 100 Kcal/kg de EM aumentou os teores de energia digestível nas dietas de
Terminação.
Palavras-chave: aditivos de alimentos para animais, eficiência, enzimas, nutrição.


ABSTRACT
ENZYME COMPLEX FOR SWINE DURING GROWING AND FININSHING
The study aimed to evaluate the use of an enzyme complex containing amylase -, -
glucanase, phytase, cellulase, xylanase and protease, on the performance, digestibility and
economic analysis of pigs during growing and finishing. 80 pigs were used, 42 males and 38
females, with approximately 63 days of age in a randomized complete block design case. Five
treatments with eight replicates / treatment were tested, as follows: CP: Diet positive control,
with energy level recommended for phase; CN85: negative control diet with reduced energy
(85 kcal / kg MS); CE85: CN85 diet with addition of enzyme complex; CN100: negative
control diet with energy reduction (100 kcal / kg MS); CE100: CN100 diet with addition of
enzyme complex. The feeding program with three diets were used: - Growth I - of 63 to 90
days of age, Growth II- from 91 to 118 days old and termination (T) - of 119 to 145 days of
age . During each phase of the trial period, the performance was evaluated through the
variables: daily feed intake (DFI), daily weight gain (ADG) and feed conversion (FC).
Digestibility of Diets were analyzed in three stages by the use of Celite ® marker and then
analyzing gray-insoluble acid. The economic viability analysis was performed by determining
the cost of feed (in R $) per kilogram of live weight gain and calculation of Economic
Efficiency Ratio. The results showed that for the males , we recommend the use of enzyme
complex in diets with a reduction of 85 Kcal / kg on during period of growing and finishing ,
it improves the feed conversion and economically more efficient during Growth I. For
females, the use of the enzyme complex is not justified. All diets reducing energy 100 Kcal /
kg of MS, the use of complex enzyme increased digestible protein during growth II; The
addition of enzyme complex in diets with energy reduction of 85 and 100 Kcal / kg of MS
increased the digestible energy content in diets finishing..
Keywords: efficiency, enzymes, feed additives, nutrition.


SUMÁRIO
1 INTRODUÇÃO ................................................................................................................... 15
2 REVISÃO DE LITERATURA ........................................................................................... 17
2.1 Polissacarídeos não amiláceos ..................................................................................... 17
2.1.1 Polissacarídeos não amiláceos do milho e da soja ................................................. 19
2.2 Fitato .............................................................................................................................. 21
2.3 Aditivos utilizados na nutrição animal ....................................................................... 22
2.4 Enzimas ......................................................................................................................... 23
2.5 Enzimas utilizadas na nutrição animal ....................................................................... 25
2.5.1 Fitase ........................................................................................................................ 25
2.5.2 -glucanase .............................................................................................................. 26
2.5.3 -amilase ................................................................................................................. 28
2.5.4 Xilanase ................................................................................................................... 28
2.5.5 Celulase ................................................................................................................... 30
2.5.6 Protease .................................................................................................................... 30
2.6 Complexos enzimáticos ................................................................................................ 31
3 MATERIAL E MÉTODOS ................................................................................................... 34
3.1 Desempenho .................................................................................................................. 34
3.2 Digestibilidade ............................................................................................................... 40
3.3 Análise econômica ........................................................................................................ 42
4 RESULTADOS E DISCUSSÃO ........................................................................................ 44
4.1 Desempenho .................................................................................................................. 44
4.2 Digestibilidade ............................................................................................................... 49
4.3 Análise econômica ........................................................................................................ 52
5 CONCLUSÃO ...................................................................................................................... 57
REFERÊNCIAS ..................................................................................................................... 58


15
1 INTRODUÇÃO
A produção mundial da carne suína, em 2014, foi de 110, 606 milhões ton. O Brasil
ocupa o quarto lugar no ranking de produção de carne suína, com 3.344 milhões ton,
perdendo apenas para China (56.500 milhões ton), a União Européia (22.400 milhões ton) e
os EUA (10.329 milhões ton) .Na exportação, em 2014, o EUA lideram o ranking, com 2.321
milhões ton, seguido pela União Europeia com 2.150 milhões ton e pelo Canadá , com
1.180 milhões ton de carne exportada. O Brasil se manteve na quarta posição, tendo
exportado 505 mil ton (ABIPECS, 2015).
O destaque da suinocultura brasileira no cenário mundial se deve aos avanços na
produção, devido a uma boa nutrição, sanidade, bom manejo da granja, produção integrada e,
principalmente, aprimoramento gerencial dos produtores (MAPA, 2015).
A nutrição é o fator que mais onera a produção suína, por isso, pesquisas têm sido
desenvolvidas com o intuito de buscar ingredientes e aditivos que melhorem o desempenho
animal, e assim, reduzam custos. Além disso, há uma grande preocupação ambiental, pois o
excesso de nutrientes nas rações é um dos agravantes da poluição do ambiente. Portanto, é
necessário que as dietas atendam todas as exigências do animal e apresentem alta
digestibilidade, garantindo, assim, menor concentração dos nutrientes nas excretas.
Os aditivos são produtos destinados à alimentação animal que podem ter ou não valor
nutritivo, e com capacidade de melhorar as características de ingredientes utilizados na dieta,
o desempenho dos animais sadios, atender às necessidades nutricionais ou ainda ter efeito
anticoccidiano. Entre os aditivos utilizados, atualmente, estão as enzimas exógenas,
responsáveis por potencializar a digestão de alguns substratos presentes nas dietas. (MAPA,
2006)
No processo digestivo, os suínos utilizam as enzimas para digestão do alimento,
contudo, não são eficientes na digestão da fibra, portanto o uso de enzimas exógenas que
degradam a fibra pode romper a parede celular, permitindo que as enzimas produzidas pelo
animal tenham acesso ao interior das células dos grãos e, consequentemente, há liberação de
nutrientes, passíveis de absorção, melhorando a metabolização da energia e o desempenho
produtivo dos animais (Graham, 1996).
A utilização de enzimas exógenas na alimentação de suínos e aves tem sido
pesquisada com o objetivo de melhorar o aproveitamento dos alimentos, favorecendo a
hidrólise de compostos resistentes ou de baixa digestibilidade, como é o caso dos
polissacarídeos não amiláceos (Graham, 1996).

16
Para melhorar o valor nutritivo das dietas formuladas com cereais de baixa viscosidade
Carvalho et. al, (2009), sugere que o uso de complexos enzimáticos seja mais efetivo por
atuar sobre polissacarídeos da parede celular dos grãos, levando a um maior aproveitamento
da dieta.
As enzimas exógenas vêm sendo utilizadas com êxito em países da Europa. Nesses
países, as principais fontes de energia, para rações de monogástricos, são cereais, como trigo,
cevada, centeio e aveia, e grãos que possuem baixa disponibilidade de energia e são ricos em
polissacarídeos não amiláceos.(Freitas et al., 2000)
No Brasil, a maioria das dietas para monogástricos são formuladas a base de milho e
farelo de soja, alimentos de alta digestibilidade. Entretanto, pesquisas realizadas que
envolvem o uso de enzimas para suínos, são feitas com base no uso em rações contendo
ingredientes como aveia, cevada, farelo de trigo e farelo de arroz (Fireman et al., 2000, Ludke
et al., 2000;; Mathlouthi et al., 2002), realidade contrária à observada no Brasil.
Além disso, o milho e farelo de soja, apresentam elevada variabilidade nutricional. A
composição do milho pode variar dento de uma mesma região e entre regiões (Soto-Salanova
et al., 1996). O farelo de soja também é um ingrediente de composição variável e de qualidade
altamente dependente do seu processamento (Parsons et al., 2000). Consequentemente ocorre
variação nos seus constituintes considerados antinutricionais, o que compromete uma possível
atuação das enzimas no organismo animal.
O principal objetivo da utilização de um complexo enzimático em dietas à base de
milho e soja é aproveitar ao máximo os nutrientes que se incluem na dieta e, com isso,
melhorar os resultados produtivos (Fuente e Soto-Salanova, 1997). O benefício esperado
seria a atuação sinérgica das emzimas em componentes específicos, permitindo assim uma
resposta mais expressiva no desempenho animal. (Cardoso et al., 2010)
O objetivo do trabalho foi avaliar o efeito da inclusão de um complexo enzimático
composto por alfa-amilase, beta-glucanase, fitase, celulase, xilanase e protease, sobre o
desempenho e digestibilidade dos nutrientes de suínos na fase de crescimento e terminação,
além de verificar a viabilidade econômica do uso desse complexo.

17
2 REVISÃO DE LITERATURA
2.1 Polissacarídeos não amiláceos
Utilizados na nutrição animal como fonte de energia e proteína, alguns alimentos de
origem vegetal trazem consigo fatores antinutricionais e constituintes de baixa digestibilidade
(Charlton, 1996).
Estes alimentos, normalmente, contêm os polissacarídeos não amiláceos (PNAs),
polímeros de monossacarídeos ou açucares que não podem ser digeridos por monogástricos,
devido a natureza de suas ligações (Rosa e Uttapatel, 2007), o que afeta negativamente o
crescimento do animal e a digestibilidade do alimento (Leenhouwers et al., 2006). Deste
modo, quando alimentos que contém PNAs são consumidos, ocorre o aumento da viscosidade
do conteúdo intestinal, dificultando a ação das enzimas endógenas (Brito et al., 2008).
A interferência dos PNA’s na digestibilidade dos nutrientes e da energia de dietas para
suínos depende de inúmeros fatores e concentração: origem botânica dos PNAs que compõem
a fibra, proporção dos tecidos que recobrem o endosperma no cereal e da solubilidade destes
polissacarídeos, bem como da espécie e idade animal, e das propriedades físico-químicas dos
alimentos. (Montagne et al. 2003).
Os PNAs são classificados como solúveis e insolúveis através da sua capacidade em
formar solução homogênea ou não com a água, contudo, muitas das atividades antinutritivas
são atribuídas diretamente aos polissacarídeos solúveis, apesar de os polissacarídeos
insolúveis também apresentarem efeito na taxa de passagem da digesta e na retenção de água
(Lima e Viola, 2001).
Os PNAs solúveis são, basicamente, as pectinas, gomas e a maioria das hemiceluloses.
São os principais responsáveis pela formação de gel e aumento na viscosidade do conteúdo
intestinal, características antinutritivas que impedem a digestão e absorção dos nutrientes
pelos animais (Torres et al., 2003).
Os PNAs insolúveis compreendem o grupo formado por: celuloses, ligninas e parte
das hemiceluloses. Essas fibras possuem baixa digestibilidade para animais monogástricos, e
são capazes de reduzir a digestibilidade de outros nutrientes (Andriguetto et al., 2002).
A figura 1 ilustra a classificação dos polissacarídeos não amiláceos.

18
Figura 1- Esquema de classificação dos carboidratos na célula vegetal (Luchesi, 2013, adaptado de
Hall, 2003)
FDA = fibra em detergente ácido, FDN = fibra em detergente neutro, FSND = fibra solúvel em
detergente neutro (incluindo todos os polissacarídeos não-amiláceos ausentes na FDN). CNF =
Carboidratos não fibrosos.
Dentre os PNAs mais abundantes da parede celular estão a celulose, hemiceluloses e
pectinas. O menor grupo de PNAs compreende os frutanos, glucomananos e galactomananos,
que são utilizados como polissacarídeos internos de reserva (Moreira et al., 2006). As
mucilagens, glucanos e gomas também são exemplos de PNAs, no entanto, as frações
solúveis dos -glucanos e arabinoxilanas nos cereais interferem mais na capacidade de
utilização dos nutrientes da dieta (Classen, 1996).
As pectinas se encontram principalmente na lamela média e parede primária da célula
vegetal, atuando como elemento “cimentante” entre membranas. Encontram-se mais
abundantemente em leguminosas do que em gramíneas, e estão presentes em concentrações
maiores em certos subprodutos ou resíduos agro-industriais como as polpas cítricas e de
beterraba (Van Soest, 1994).
A celulose é um homopolissacarídeo de alto peso molecular, cadeia linear e elevado
grau de polimerização das unidades D-glicose unidas por ligações do tipo -1,4 e -1,6. A
relação lignina/celulose determina a intensidade de degradação microbiana da parede celular,
igualmente condicionada pela presença de outras substâncias incrustantes, como a sílica e a
cutina. Outros fatores macromoleculares intrínsecos da própria celulose também interferem na
intensidade de degradação da parede celular, como a cristalinidade e especificidade de suas
ligações químicas, tanto para ruminantes como em não-ruminantes (Van Soest, 1994).
A lignina está presente em pequenas quantidades em forragens jovens, mas tende a

19
aumentar em função do estado de maturação das plantas e do ambiente em que se
desenvolvem ( Jung, 1989). De acordo com Arruda et al (2003), ela influencia negativamente
a extensão da atividade fermentativa devido ao fato de impedir que as enzimas atuem nos
polissacarídeos e de se ligar com os polissacarídeos.
As hemiceluloses também são classificadas como hexanos, contendo
predominantemente glicose e manose unidas por ligações -1,4 (mananos), polímeros
compostos de resíduos de glicose unidos por ligações -1,3 e -1,4 (β-glicanos) que se
diferenciam da celulose pela solubilidade em meio alcalino, e os polímeros compostos por
unidades de glicopiranose unidas por ligações -1,4 contendo cadeias laterais de
xilanopiranose unidas por ligações α-1,6 (xiloglicanos). Em leguminosas foram verificadas
ramificações com galactose e frutose (Van Soest, 1994).
O consumo de alimentos com elevados teores de PNAs, pode causar o aumento do
tamanho dos órgãos gastrintestinais (Arija et al., 2006), e estimulação da produção demasiada
de secreções digestivas (Sakomura et al., 2004). Isso pode estar associado a redução nos
níveis plasmáticos de glicose, porque os PNAs inibem a produção do polipeptídeo inibitório
gástrico, aumentando a produção de gastrina, que favorece a produção de secreções gástricas.
Substâncias ácidas atingem o duodeno e a entrada destas substâncias no intestino delgado
estimula ainda mais a liberação de secretina que, por sua vez, promove a liberação de suco
pancreático e eletrólitos. Todos estes processos podem representar alto custo metabólico para
animais monogástricos, gerando efeitos negativos na eficácia do aproveitamento energético
das rações (Low, 1989).
Por outro lado, PNAs em baixas concentrações podem ser considerados
imunoestimulantes, é o caso dos glucanos, polissacarídeo não amiláceo, geralmente presente
na parede celular dos grãos ou leveduras (Pereira, 2015), que tem a capacidade de aumentar as
defesas do animal contra ataque de bactérias, vírus e fungos, porém em altas concentrações
provoca efeito antinutricional aos animais. Em suínos, Xiaou et al. (2004) observaram que os
-glucanos solúveis foram capazes de aumentar a imunidade antiviral própria contra a
síndrome viral reprodutiva e respiratória suína.
2.1.1 Polissacarídeos não amiláceos do milho e da soja
O milho e o farelo de soja são os principais ingredientes utilizados, no Brasil, para a
dieta de monogástricos, porém ocorrem grandes variações em suas composições nutricionais.

20
O milho é a principal fonte de energia nas dietas para aves e suínos, e apesar do seu
uso em larga escala, a proporção e conteúdo de PNAs presentes podem variar, dependendo da
variedade ou cultivar, das condições de desenvolvimento diante o cultivo, das temperaturas de
secagem, da estrutura química do amido, entre outros (Cowieson e Adeola, 2005).
Na soja também ocorre essa variabilidade, dependendo da origem do grão, de sua
variedade, do grau de processamento e da proporção da casca presente no farelo, a qual é rica
em PNAs (Nagashio, 2007).
A Tabela 1 mostra as diferenças encontradas por alguns autores na porcentagem de
PNAs no milho e no farelo de soja.
Tabela 1- Polissacarídeos não amiláceos totais (%) presentes no milho e no farelo de soja
Milho Farelo de Soja Autores
9,32 29,02 Malathi e Dvegowda (2001)
9,7 10,3 Ruiz et al. (2008)
8,1 30,3 Tavernari et al (2008)
Fonte: Cardoso (2010).
O farelo de soja possui polissacarídeos não amiláceos (PNAs) (Ward e Fodge, 1996)
na forma de pectinas, hemiceluloses e oligassacarídeos (rafinose e estaquiose) (Charlton,
1996). A estaquiose é formada por duas moléculas de galactose, glicose e frutose, enquanto
que a rafinose é formada por frutose, galactose e glicose unidas por ligações glicosídicas do
tipo alfa 1-6 (Lan et al., 2007). Esses oligossacarídeos dificultam a absorção dos nutrientes e
reduzem o valor da energia metabolizável das rações, porque não podem ser degradados por
animais que não secretam a enzima a-1,6-galactosidase. (Vinjamooi et al., 2004, Vahjen et al.,
2005). Além desses PNAs, fatores antinutricionais como inibidores de proteases e lectinas
estão amplamente distribuídos na soja e não podem ser degradados pelo sistema digestório
dos monogástricos, principalmente aves e suínos. Para Sorbara et al.(2009), a maioria dos
PNAs presentes no farelo de soja é encontrado na forma insolúvel, e consequentemente, não
atua aumentando a viscosidade intestinal, ou seja, interfere pouco na digestão quando
comparado com outros alimentos vegetais.
No entanto, mesmo não sendo considerados grãos viscosos, os componentes insolúveis
dos PNAs presentes no milho e na soja podem encapsular nutrientes, por isso, o uso de

21
enzimas exógenas tem sido proposto para dietas à base destes ingredientes. (Brum et al.,
2006).
2.2 Fitato
O fitato é um composto orgânico de ocorrência natural nos vegetais (Sebastian et al.,
1997) e está presente na forma de ester myo-inositol hexafosfato, ligado a íons de magnésio,
sódio, potássio, cálcio e zinco. Outras duas nomenclaturas utilizadas são: fitina e ácido fítico.
A fitina refere-se ao ester myo-inositol hexafosfato ligado a potássio, magnésio e cálcio, como
acontece nas plantas. O ácido fítico é a forma livre do ester myo-inositol hexafosfato.
(Sakomura et al. 2014).
A molécula de fitato apresenta alto teor de fósforo (28,2%) com alto potencial de
quelação (Keshavarz, 1999). Sua função fisiológica na semente do vegetal é servir de estoque
de fósforo e outros minerais, além da energia, que são liberados pela ação da fitase endógena
à medida que ocorre a germinação (Borges, 1997).
Em média, dois terços do fósforo dos ingredientes de origem vegetal encontra-se na
forma de fitato (Simons & Versteegh, 1990) , e em especial o milho e o farelo de soja. Esses
ingredientes possuem, cerca de 50 a 85% do porcentual de fósforo complexado em forma de
ácido fítico, como mostra a Tabela 2.
Tabela 2- Porcentagem de fósforo fítico em relação ao fósforo total de alguns ingredientes
Ingredientes P fítico/P total (%)
Kornegay (2001) Rostagno et al (2011)
Trigo 69 66
Cevada 64 -
Farelo de Soja 60 61
Farelo de Girassol 77 67
Adaptado de Sakomura et al (2014)
O grupo ortofosfato da molécula de fitato é altamente ionizado e se complexa com
uma variedade de cátions e com fração protéica do alimento, o que pode interferir
negativamente na digestibilidade das dietas (Kasim e Edwards, 1998), pois animais
monogástricos não podem aproveitar o fósforo contido no ácido fítico porque estes não
sintetizam a enzima fitase, necessária para hidrolisar o referido complexo.

22
Esse fato inclui o ácido fítico como um fator antinutricional, pois diminui a
disponibilidade dos minerais (Ca, P, Zn, Mn e Mg) e também a das proteínas e moléculas de
glicose conjugadas (Costa et al., 2006). Além disso, é possível que interfira negativamente na
utilização da energia da dieta, possivelmente por inibir a atividade da enzima α-amilase ou
pela diminuição da digestibilidade da proteína da dieta (Selle e Ravindran, 2008).
Por isso, torna-se necessária a suplementação de fósforo nas dietas de monogástricos.
Um fonte inorgânica é inserida, geralmente em quantidades acima da exigência do animal,
com o objetivo de se ter uma margem de segurança em relação à este elemento na dieta. Com
isso, o fósforo fítico de baixa disponibilidade para estes animais, juntamente com este excesso
de fósforo inorgânico adicionado às rações é eliminado nas fezes dos animais, ocasionando
assim, problemas ambientais.
A quantidade de fósforo e cálcio absorvidos é dependente principalmente dos seus
níveis na dieta e de suas fontes (Ludke, 2002). O excesso desse nutriente no solo traz sérios
problemas para o meio ambiente, devido a eutrofização e nitrificação, que diminuem a
quantidade de oxigênio existente nas águas dos rios e lagos e contaminam o solo e águas
subterrâneas.
2.3 Aditivos utilizados na nutrição animal
Aditivos são substâncias capazes de melhorar o desempenho animal ou as
características físicas dos alimentos, sem prejudicar seu valor nutritivo. Para a FDA (Food and
Drug Administration - órgão governamental dos EUA responsável pelo controle dos
alimentos), aditivo é a substância adicionada com a finalidade de melhorar o desempenho
animal, passível de ser utilizada sob determinadas normas desde que não deixe resíduo no
produto de consumo (FDA,2015).
O uso de aditivos antimicrobianos passou a ser visto como fator de risco para a saúde
humana, principalmente em decorrência de duas contestações: a presença de resíduos dos
antimicrobianos na carne, nos ovos e no leite e a indução de resistência cruzada para bactérias
patógenas para humanos (Menten, 2001).
Apesar de ainda haver controvérsias, do ponto de vista científico sobre possíveis
relações entre o uso de antimicrobianos em animais e o aumento de resistência em bactérias
isoladas do ser humano, a União Européia proibiu o uso dessas substâncias como aditivos
melhoradores de desempenho zootécnico desde 1º de janeiro de 2006 (MAPA,2004).

23
No Brasil, foram estruturadas pesquisas, realizadas pelo MAPA, que avaliaram a
proibição de moléculas como bacitracina de zinco, fosfato de tilosina, avilamicina,
monensina, dentre outras. Acredita-se que estes princípios ativos não deveriam ser excluídos
no Brasil, por não apresentarem riscos à saúde do homem, conforme os conhecimentos atuais
sobre a toxicidade e resistência bacteriana (MAPA, 2006).
Ainda assim, visando se adequar a mercados internacionais, é preciso que o Brasil
busque novas alternativas para minimizar o impacto da retirada dos antimicrobianos como
promotores de crescimento de suínos. As alternativas incluem os probióticos, os prebióticos,
os ácidos orgânicos, os extratos vegetais e as enzimas exógenas, que são aditivos zootécnicos,
responsáveis por modificarem o crescimento animal, a eficiência alimentar, o metabolismo e
o desempenho, sem comprometer a saúde humana.
As enzimas exógenas são responsáveis por amenizar os efeitos negativos provocados
pelos fatores antinutricionais presentes nos diversos ingredientes, aumentando a
disponibilidade de polissacarídeos de reserva, gorduras e proteínas, protegidas da atividade
digestória. Além disso, otimizam a atividade enzimática endógena, principalmente em
animais jovens que possuem um sistema enzimático imaturo (Campestrini et al., 2005).
2.4 Enzimas
Enzimas são proteínas globulares, de estrutura terciária e quaternária, que funcionam
como catalisadores biológicos, ou seja, aumentam a velocidade das reações químicas no
organismo, sendo altamente específicas para os substratos (Champe e Harvery, 1989).
No entanto, existem enzimas que não são secretadas, mesmo na presença de
substratos. Em monogástricos, isso acontece, porque o código genético não dispõe da
indicação para sua síntese. Para um organismo ter em seu código genético a possibilidade da
produção de determinada substância é necessário a real necessidade de produzi-la. Seja
porque o meio não a proporciona ou porque não há substrato disponível para ser utilizado
(Penz Junior, 1998). No caso, esta teoria é questionada, pois os polissacarídeos não amíláceos
e o fitato estão disponíveis em vários grãos ingeridos pelos monogástricos, os quais não
produzem enzimas para digerir estes componentes vegetais.
Por isso, enzimas exógenas são comercialmente produzidas a fim de aumentar a
digestibilidade de nutrientes e melhorar a sua utilização. Estas são provenientes, geralmente
de bactérias do gênero Bacillus ou fungos do gênero Aspergills (Ferket, 1996), e sob

24
condições favoráveis de temperatura, pH e umidade, estas enzimas têm um sitio ativo que
permite a ruptura de uma determinada ligação química (Penz Júnior, 1998).
Para Soto-Salanova, (1996), as enzimas têm quatro formas de ação: rompendo as
paredes celulares das fibras, reduzindo a viscosidade, degradando proteínas e
complementando as enzimas endógenas. No caso da última, é uma ação de grande
importância para animais jovens, pois a produção de enzimas endógenas ainda ocorre em
quantidade insuficiente.
Após a suplementação nas dietas, a ação catalítica das enzimas depende de uma série
de fatores, tais como: concentrações do subtrato e da enzima, temperatura, variação do pH,
umidade, presença de co-enzimas, resistência à atividade proteolítica e inibidores no local em
que ocorrerá a reação (Barbosa et al., 2008), pois as enzimas apresentam estruturas bastante
frágeis, podendo ser desarranjadas, tornando-as ineficazes. Vários processos podem contribuir
para desnaturação enzimática, como por exemplo, em situação de calor excessivo, presença de
ácidos ou agentes oxidantes (Osera et al., 2008).
A termoestabilidade da enzima é um fator que afeta sua ação catalítica e depende do
tipo de microorganismo que produz a enzima, sendo menos resistentes (até 75ºC) aquelas
produzidas por fungos e mais resistentes (80 a 90ºC) as produzidas por bactérias (Officer,
2000).
Uma enzima adicionada ao alimento seco só é ativada no trato digestivo quando
misturada aos fluidos digestivos e sob a temperatura corporal (Rotter, 1990). A ação máxima
ocorre no estômago e na porção inicial do intestino delgado, especificamente no duodeno
(Jongbloed et al., 1992).
Para conseguir o efeito positivo desejado, as enzimas alimentares devem funcionar
cataliticamente dentro do trato digestório. A maioria das enzimas alimentares de origem
bacteriana tem exposição ótima em pH neutro, enquanto as enzimas provenientes de fungos
apresentam atividade máxima em condições ácidas (pH 4,0-6,0). Por isso, enzimas
microbianas atuam mais eficientemente no intestino delgado, onde o pH é neutro. Misturas
de enzimas de fontes de fungos e bactérias são frequentemente incluídas na alimentação para
assegurar a atividade ao longo de um amplo espectro de pH (Walsh, 1993).
Strada et al. (2005) citam os fatores que influenciam a atuação das enzimas no
organismo animal, destacando-se aqueles relacionados ao processamento da ração, pH do
meio, comprimento do trato gastrointestinal, grau de hidratação, temperatura corporal,
susceptibilidade da enzima exógena ao ataque da endógena, concentração do produto e tipo de

25
ingrediente utilizado na ração. O tipo de ingrediente utilizado na dieta pode afetar a utilização
de enzimas pelos animais, pois alguns alimentos podem apresentar fatores antinutritivos que
variam de acordo com as condições de produção da planta, colheita, secagem e
armazenamento, além disso, podem ocorrer variações na quantidade de substrato.
Além da idade e espécie animal, alguns outros fatores secundários também podem
influenciar a ação das enzimas, como o estado sanitário dos animais, porém fatores que
afetam o consumo de ração, também estão diretamente relacionados a interferir na quantidade
de nutrientes ingeridos, o que pode favorecer ou não a ação das enzimas (Sakomura et al.,
2014).
Há três maneiras de utilizar a suplementação enzimática: a “over the top” (por cima) é
a aplicação mais simples e provavelmente mais prática, e consiste em adicionar as enzimas
com uma formulação padrão, sem alterar os níveis nutricionais. Outra alternativa seria alterar
a formulação da ração, por meio da redução dos nutrientes, e adição de enzimas exógenas
para restaurar o valor nutricional da dieta-padrão, visando o mesmo desempenho de uma dieta
com os níveis nutricionais recomendados (Barbosa et al., 2008). A terceira considera
superestimação de algum nutriente, porém mais comumente de energia metabolizável de um
ou mais ingredientes da ração para adição da enzima exógena (Brum et al., 2007)
É comum a utilização de enzimas em dietas de suínos com o intuito de aumentar a
disponibilidade de polissacarídeos de reserva, gorduras e proteínas, pois estes ficam
protegidos pelos polissacarídeos da parede celular, impedindo de realizar a digestão
(Campestrini, 2005). Além disso, é possível minimizar os efeitos indesejáveis dos fatores
antinutricionais e aperfeiçoar a ação das enzimas endógenas, porque disponibilizam certos
nutrientes para absorção e também aumentam o valor energético de ingredientes mais baratos
(Walsh et al., 1993).
A utilização de enzimas tem demonstrado bons resultados quando adicionadas a
alimentos com alto grau de PNA podendo aumentar o valor nutritivo dos alimentos (Bedford,
1995). Para Guenter (2002), além de diminuir os fatores antinutricionais e aumentar a
digestibilidade da dieta, as enzimas exógenas também contribuem para uma menor poluição
ambiental, pois diminuem os nutrientes poluentes excretados nas fezes.
2.5 Enzimas utilizadas na nutrição animal
2.5.1 Fitase

26
Selle e Ravindran (2008), acreditam que o fitato interfira negativamente na utilização
da energia da dieta, possivelmente por inibir a atividade da enzima α-amilase ou pela
diminuição da digestibilidade da proteína da dieta.
A fitase, de um modo prático, tem a capacidade de descomplexar o fitato presente nas
vegetais, para que haja aproveitamento do fósforo, e assim, ele possa ser absorvido pelos
animais. Assim, adicionar fitase às dietas a base de milho e farelo de soja pode ser tão
eficiente quanto suplementá-las com fosfato bicálcico. (Ludke et al., 2002)
Além de aumentar o valor nutricional das rações, há um interesse econômico na
utilização da fitase em dietas de monogástricos, visto que o fósforo torna-se cada dia mais
caro para o custo das rações. Há também a preocupação ecológica em diminuir a
contaminação do meio ambiente com as excretas dos animais. A fitase, melhora a retenção do
fósforo por parte do animal, reduzindo a eliminação e, consequentemente, a contaminação do
ambiente.
De acordo com Ludke et al, (2000) a adição de fitase em dietas propicia maior
disponibilidade de fósforo e cálcio nos alimentos orgânicos, propiciando desempenho dos
animais semelhante, mas com redução na quantidade de cálcio e fósforo excretados nas fezes,
em relação às mesmas dietas suplementadas com fosfato inorgânico.
A resposta da fitase é influenciada pelo nível de fósforo total e disponível (incluindo o
fósforo fítico) na dieta, pela quantidade de fitase suplementada e pela relação cálcio e fósforo
(Kornegay 1996), pois altas concentrações de cálcio na dieta aumentam o pH do conteúdo
intestinal, diminuindo a atividade da fitase (Sandberg et al., 1993). Além disso, o cálcio em
excesso pode competir diretamente com o sítio ativo da fitase (Quian et al., 1996).
Liu et al (1998), sugerem a utilização de Ca: P na proporção de 1:1, pois concluíram
que dietas para suínos em crescimento e terminação à base de milho e farelo de soja, com
baixo teor de fósforo e suplementadas com fitase microbiana têm a utilização do fósforo
otimizada.
2.5.2 -glucanase
Alguns cereais utilizados nas dietas de aves e suínos, como a cevada, possuem
limitações na digestibilidade da energia, devido à presença de -glucanos (Ward &
Marquardt, 1987). Estes componentes reduzem muito a digestibilidade de vários nutrientes
presentes. O uso de -glucanases em dietas com cevada é uma prática comum na Europa.

27
Acredita-se que as glucanases aumentem o valor nutritivo de algumas dietas a base de
milho e soja por alterarem a arquitetura das paredes celulares dos grãos (Cowieson e Adeola
2005). Além disso, a -glucanase melhora a digestibilidade da dieta, reduzindo a viscosidade
do conteúdo do intestino e impedindo o crescimento de bactérias (Mathlouthi et al, 2002). Em
frangos de corte, a suplementação com glucanase provoca aumento da digestão das gorduras
durante o crescimento e aumento do conteúdo em ácidos graxos voláteis no ceco durante o
acabamento. A suplementação da enzima também proporcionou maior digestibilidade dos
nutrientes e diminuiu o peso relativo de alguns órgãos do sistema digestivo (Yu et al., 2002).
Camiruaga et al. (2001) trabalhando com rações de triticale suplementadas com -
glucanase para frangos de corte, observaram aumento do peso vivo aos 21 dias e redução da
conversão alimentar. Porém, Yu et al. (1998) não verificaram alterações no peso relativo dos
órgãos do sistema digestivo, ao suplementar uma dieta à base de cevada com glucanase, para
frangos de corte,
Em suínos na fase de crescimento e terminação, foi observado que o uso de -
glucanase em dietas a base de cevada e farelo de soja, aumenta a digestibilidade ileal da
energia bruta, proteína, –glucanas e da maioria dos aminoácido utilizados. Quando as dietas
foram a base de trigo e soja verificou-se efeitos da -glucanase sobre a digestibilidade ileal
das β-glucanas e sobre a digestibilidade fecal da energia bruta ( Li et al., 1996).
Jensen et al. (1998) utilizaram a enzima β-glucanase em grãos de duas cultivares de
cevada, e concluíram que a enzima contribui para a redução na viscosidade do alimento, sem
modificar a digestibilidade do amido, nitrogênio e o desempenho dos leitões.
Para leitões desmamados alimentados com dietas com diferentes teores de PNA, a
adição da enzima β-glucanase aumentou a produção de ácido lático e reduziu a de ácido
acético, evidenciando que o aumento da concentração de PNA e da suplementação enzimática
influencia a proporção dos ácidos graxos voláteis no íleo, indicando alteração na flora
bacteriana (Hogberg et al. 2004).
Para suínos, em dietas a base de milho e farelo de soja, não há estudos que evidenciem
o uso da - glucanase de forma isolada, pois é mais comum a utilização associada a outras
enzimas, em forma de complexos enzimáticos.

28
2.5.3 -amilase
No trato digestório, a α-amilase é responsável por hidrolisar polissacarídeos a
dissacarídeos e oligossacarídeos. Quando essa hidrólise não ocorre eficientemente devido a
deficiência das enzimas pancreáticas, provoca diminuição da digestibilidade aparente dos
componentes da dieta e reduz o crescimento do animal (Nitsan et al., 1991). Por isso, a
suplementação da enzima -amilase é capaz de melhorar o desempenho animal,
principalmente, em períodos em que a produção de enzimas endógenas é prejudicada, por
exemplo, no período pós desmame de leitões.
Brum et al. (2006) testaram o uso da -amilase para frangos de corte e verificaram
uma melhora significativa no ganho de peso e peso corporal das aves até os 23 dias de idade,
contudo não encontraram diferenças na conversão alimentar, consumo de ração e na energia
metabolizável das dietas. Em outras pesquisas, Brum et al. (2007) não verificaram diferenças
no desempenho de frangos de corte ao utilizarem a mesma enzima e nas mesmas quantidades.
Oliveira et al. (2012) observaram que para frangos de cortes, dietas com níveis de
energia reduzidos e adição de - amilase, tiveram boas respostas, suprindo os incrementos
energéticos.
Em leitões, o uso da enzima amilase exógena justifica-se pela baixa atividade da
amilase pancreática no período pós desmame. A amilase atua na hidrólise do amido, principal
fonte energética nas dietas de leitões recém desmamados (Makkink et al., 1994). A -amilase
pode aumentar a energia da dieta de leitões na fase de creche. De acordo com Piovesan et al,
(2011), aumento da energia digestível e metabolizável foi observado ao utilizar a enzima em
dieta a base de milho duro, porém, não houve alterações no desempenho dos animais. Barros
et al., (2014) também testou o uso de amilase em dietas com milho duro, porém não
observaram diferença estatística comparando os resultados aos da dieta controle, sem adição
de enzimas.
2.5.4 Xilanase
A xilanase é obtida a partir de fungos e bactérias e pertence a classe das carboidrases,
que têm por finalidade hidrolisar os arabinoxilanos. A hidrólise dos arabinoxilanos pela
xilanase resulta na diminuição da viscosidade do conteúdo digestivo (Gao et al., 2008). Por
isso, o efeito da xilanase tem sido alvo de vários estudos na nutrição de monogástricos.

29
Em frangos de corte, consumindo dietas com 15% de farelo de arroz, a xilanase
melhorou a conversão alimentar de 1,56 para 1,47 (Conte et al., 2003).
A adição de xilanase também pode reduzir a viscosidade do conteúdo digestivo no
proventrículo e no jejuno aos 21 dias e no cólon aos 49 dias de idade do frango e alterar o pH
do conteúdo digestivo em vários órgãos, como verificado por Gao et. al (2008). Além disso, a
suplementação de xilanase inibe a fermentação no intestino delgado e aumenta no ceco,
estimulando uma maior produção de ácidos graxos voláteis no ceco. Em relação à energia, a
xilanase também pode favorecer a disponibilidade da energia metabolizável aparente (Yang et
al., 2010).
Viana et al., (2014), verificaram que a adição de xilanase melhorou a utilização da
energia e a produção de ovos em galinhas poedeiras. As dietas controle negativas, com
redução de níveis nutricionais, mas com adição de enzimas, determinam o mesmo resultado
que as dietas controle positivas, ou seja, formuladas de acordo com a exigência.
Em suínos, as rações contendo cevada, trigo e centeio, que possuem grande quantidade
de PNA, a digestibilidade da matéria seca e nitrogênio foi melhor na presença de 0,1% de
xilanase derivada de Trichoderma longibrachiatum (Dersjant-Li et al., 2001).
No entanto, Diebold et al. (2005) não observaram efeito positivo na digestibilidade
ileal e cecal quando incluiu-se xilanase em dietas com 60% de trigo para suínos. Este fato
pode ser explicado pelo baixo consumo de dieta pelos animais e, conseqüentemente,
diminuição da taxa de passagem da digesta pelo intestino, além de uma maior eficiência das
enzimas endógenas.
Mavromichalis et al. (2000), estudando aspectos relacionados à adição da enzima
xilanase e diferentes tamanhos de partículas em dietas à base de farelo de trigo para leitões
nas fases de creche, crescimento e terminação, observaram que a adição das enzimas não
influenciou o ganho de peso e o consumo de ração, porém, a digestibilidade dos nutrientes foi
melhorada.
Hauschild et al. (2004) objetivando avaliar a digestibilidade o balanço de nitrogênio e
fósforo de dietas contendo diferentes níveis de triguilho em substituição ao milho, com o uso
ou não de enzimas, observaram que a adição de xilanase não afetou os processos digestivos e
metabólicos dos suínos.

30
2.5.5 Celulase
A celulose é um polissacarídeo formado por várias unidades de glicose unidas entre si
através de ligações químicas e que pode ser convertido a açúcares simples por ação da
celulase (Tasker et al., 1994).
As celulases realizam a quebra das ligações químicas existentes entre as unidades de
glicose que formam a celulose. No caso das celulases, três enzimas fazem parte desse grupo,
que recebem os nomes de endoglucanases, exoglucanases e beta-glicosidases. As
endoglucanases agem na região interna da fibra de celulose e liberam compostos menores
formados por poucas unidades de glicose, os chamados oligossacarídeos (açúcares pequenos).
As exoglucanases agem nas extremidades das fibras de celulose e liberam unidades de glicose
ou celobiose, que são compostos menores, formados por duas unidades de glicose. As beta-
glicosidases quebram a ligação química existente entre as duas unidades de glicose que
formam a celobiose, liberando unidades de glicose livres (Wright et al., 1988).
Em estudo conduzido por Esonu et al. (2004) com a inclusão de celulase em dietas
para frangos de 28 a 35 dias de idade, os autores constataram que o ganho de peso diminuiu e
o consumo de ração aumentou com a inclusão da enzima, não havendo diferença no peso final
e na conversão alimentar.
Tenorio Fireman et al.(2000), observaram que a enzima celulase não apresentou efeito
no desempenho de suínos durante as fases de crescimento e terminação. Este fato pode ser
justificado, no caso da celulase, talvez por causa do baixo nível de fibra das dietas. Se a fibra
bruta na dieta fosse acima de 7% (Kass et al., 1980) ou o FDN fosse superior a 15%
(Cromwell, 1986) possivelmente teria prejudicado o desempenho dos animais.
Na literatura, não há muitos estudos em que tenha sido utilizada apenas a celulase na
nutrição de suínos, pois comumente, ela está contida em complexos enzimáticos. Entretanto,
Pinheiro et al. (2004) estudaram o uso de protease, α-amilase e celulase em dieta à base de
milho e de farelo de soja para frangos de corte e observaram maior peso corporal para as aves
que receberam a suplementação enzimática.
2.5.6 Protease
Sendo a proteína o ingrediente de preço mais elevado nas rações para suínos e aves, a
adição de proteases em dietas para monogástricos possibilita a redução de níveis de inclusão

31
de ingredientes proteicos e diminuição da excreção de nitrogênio, pois melhora o
aproveitamento das proteínas com a liberação de peptídios e aminoácidos (Wang et al., 2006).
O benefício do aumento da digestibilidade da proteína, promovida pela suplementação
enzimática, está mais relacionado à redução da produção de aminoácidos endógenos, do que à
melhor digestão dos aminoácidos da dieta, (Wyatt e Bedford 1998). Tal benefício permite
reduzir o gasto energético, pois o animal gasta menos energia para realizar os processos de
digetão, o que resulta em mais energia disponível para os processos produtivos (Lima et al.,
2007).
Além disso, embora a indústria tente eliminar fatores antinutricionais em alimentos
protéicos por meio de tratamento térmico, ainda são encontrados elevados níveis de princípios
inibidores de proteases, principalmente no farelo de soja. No entanto, Sheppy (2001) observou
que o uso de protease adicionadas em dietas elimina fatores antinutricionais existentes, e
Fireman e Fireman (1998), verificaram que em dietas que tenham a soja como ingrediente, a
suplementação de proteases pode melhorar o valor nutricional desse alimento, pois essas
enzimas degradam inibidores da tripsina e da lecitina presentes na soja mal processada.
Ao ser utilizada no período pós desmame de suínos, a protease pode incrementar a
digestão de proteínas vegetais, como por exemplo, do farelo de soja. Isso porque a capacidade
de digestão protéica fica comprometida devido à aos danos instestinais provocados pelo
desmame (Hedemann; Jensen, 2004).
Odetallah et al. (2002) adicionaram protease em dietas para frangos de corte e
observaram melhorias significativas no crescimento. De acordo com Wang et al. (2006), a
adição de proteases na dieta de aves melhora o desempenho produtivo e o rendimento de
carcaça, sendo as observações mais evidentes em aves alimentadas com rações com baixos
níveis de aminoácidos essenciais ou de proteína total. A explicação para isso é o fato de a
suplementação enzimática disponibilizar maior quantidade de aminoácidos para a síntese
proteica no organismo da ave.
2.6 Complexos enzimáticos
As enzimas são específicas em suas reações, isso determina que produtos que
contenham só uma enzima sejam insuficientes para produzir o máximo benefício, quando
utilizadas em dietas para aves e suínos. Por isso, misturas enzimáticas, também denominadas
de complexos enzimáticos, podem ser mais efetivas no aproveitamento dos nutrientes pelos
animais (Tejedor et al., 2001).

32
Hertado Ney et al. (2000), testaram o uso de três enzimas, amilase, lípase e protease,
de forma isolada e combinadas, na dieta de leitões de 10 a 30 kg. Ao utilizar a amilase e a
lípase isoladamente, não obsevaram efeitos significativos em relação a dieta controle. O uso
isolado da protease determinou melhor conversão alimentar, porém, o uso de complexo
enzimático de amilase, lípase e protease apresentaram melhores resultados na digestibilidade
da proteína.
Yin et al. (2001) conduziram um experimento testando grãos de cinco cultivares de
cevada nas dietas de suínos, enriquecidas com as enzimas β-glucanase, xilanase e o complexo
β-glucanase, xilanase e protease. Foi constatada maior digestibilidade de aminoácidos, fibra
detergente neutro, energia total, proteína bruta e PNA, e redução na fermentação do intestino
grosso, devido, provavelmente, a uma melhor absorção dos nutrientes.
Porém, Amorim et al (2014), concluíam que o uso de 5g/kg de complexo enzimático (
xilanase, - glucanase, celulase, pectinase, protease) numa dieta contendo polpa cítrica não
foi eficaz para melhorar o desempenho ou qualidade de carcaça de suínos na fase de
crescimento e terminação. Os autores justificam a falta de efeito das enzimas pela asssociação
de níveis inadequados das enzimas no complexo ou de níveis de inclusão na dieta. Thacker
(2005), também não verificou qualquer efeito da utilização de enzimas em dietas à base de
milho e farelo de canola sobre as características de carcaça de suínos, quer utilizando
enzimas dietéticas individualmente ou complexos enzimáticos.
Ruiz et al. (2008) também não observaram efeito da amilase, pentosanas, celulases,
protease e -galactase nas dietas de suínos em crescimento e terminação, quando foram
avaliados o desempenho, digestibilidade e impacto ambiental. Porém, a falta de resultados
positivos pode ter ocorridos devido aos baixos níveis de PNAs e oligassacarídeos dietéticos,
pois por se tratar de uma dieta com milho e farelo de soja, tais fatores antinutricionais podem
ter sido insuficientes para causarem efeito negativo. Além disso, a atividade enzimática pode
ter sido prejudicada pelo baixo pH estomacal e pelo baixo nível de inclusão das enzimas na
dieta.
Rodrigues (2002) avaliou o efeito da suplementação de um complexo enzimático
contendo xilanase, amilase, ß-glucanase e pectinase sobre a digestibilidade de nutrientes e o
desempenho de suínos alimentados com rações formuladas com milho ou sorgo. Conclui-se
que a suplementação com complexos enzimáticos melhorou a digestibilidade e os valores
energéticos, independentemente do alimento utilizado. O desempenho dos animais, que
receberam rações formuladas com milho, melhorou com a adição de enzimas. Porém, nas

33
dietas formuladas com sorgo, a suplementação enzimática não obteve respostas significativas
comparadas a dietas sem enzimas. Isso demonstra que o uso de enzimas nas dietas com sorgo
é tecnicamente inviável pra suínos na fase de crescimento e terminação.
Em frangos de corte, apesar de alguns experimentos utilizando complexo enzimático
não apresentarem efeitos significativos no desempenho das aves (Torres et al., 2003; Cardoso
et al., 2011), há estudos que demonstram melhora no desempenho de frangos de corte,
(Opalinski et al.2010) e na digestibilidade (Tejedor et al. 2001; Rodrigues et al., 2003;
Barbosa et. al, 2008).

34
3 MATERIAL E MÉTODOS
3.1 Desempenho
O experimento foi conduzido na Unidade Experimental de Suínos no Pólo Centro Sul
da Agência Paulista de Tecnologia dos Agronegócios, (APTA/SAA, SP) Tanquinho, São
Paulo, no período de 16 de setembro de 2014 a 10 de dezembro de 2014.
Foram utilizados 80 suínos, 42 machos e 38 fêmeas, com aproximadamente 63 dias de
idade, com peso inicial médio de 20,18 ± 1,98 kg. Os animais foram instalados em baias de
1,00 x 2,00 metros, com piso de concreto liso, divisórias metálicas, comedouros para
abastecimento diário e bebedouros do tipo chupeta. O galpão experimental é de alvenaria,
com pé direito de 3,40 metros, dispondo de janelas laterais basculantes para auxiliar no
controle da ventilação e da temperatura ambiente (Figura 1).
Os animais foram distribuídos em um delineamento em blocos ao acaso, com arranjo
fatorial dos tratamentos, 5 x 2 (dietas x sexo). Para a formação dos blocos foi considerado o
peso inicial dos animais e o sexo.
Foram avaliadas cinco dietas experimentais com adição ou não do complexo
enzimático com oito repetições cada:
CP: Dieta controle positivo, com nível energético recomendado para a fase;
CN85: Dieta controle negativo com redução energética de 85 kcal/kg de EM;
CE85: Dieta CN85 com adição de complexo enzimático (350 g/ton);
CN100: Dieta controle negativo com redução energética de 100 kcal/kg de EM;
CE100: Dieta CN100 com adição de complexo enzimático (500 g/ton).
O complexo enzimático apresentava as seguintes enzimas e respectivas atividades:
Alfa-amilase (400 u/g); Beta-glucanase, (700 u/g); Fitase (1.100 u/g), Celulase (6.000 u/g),
Xilanase (10.000 u/g) e Protease (700u/g). A matriz nutricional do complexo enzimático está
apresentada na tabela 3:
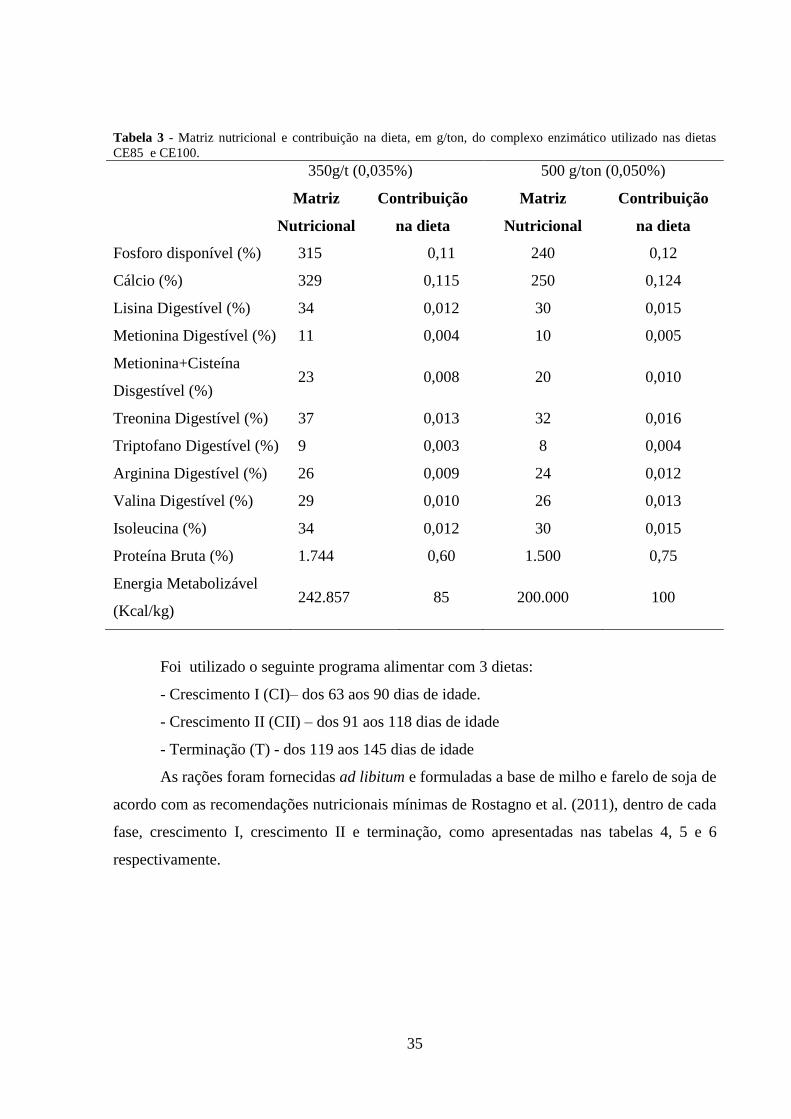
35
Tabela 3 - Matriz nutricional e contribuição na dieta, em g/ton, do complexo enzimático utilizado nas dietas
CE85 e CE100.
350g/t (0,035%) 500 g/ton (0,050%)
Matriz
Nutricional
Contribuição
na dieta
Matriz
Nutricional
Contribuição
na dieta
Fosforo disponível (%) 315 0,11 240 0,12
Cálcio (%) 329 0,115 250 0,124
Lisina Digestível (%) 34 0,012 30 0,015
Metionina Digestível (%) 11 0,004 10 0,005
Metionina+Cisteína
Disgestível (%) 23 0,008 20 0,010
Treonina Digestível (%) 37 0,013 32 0,016
Triptofano Digestível (%) 9 0,003 8 0,004
Arginina Digestível (%) 26 0,009 24 0,012
Valina Digestível (%) 29 0,010 26 0,013
Isoleucina (%) 34 0,012 30 0,015
Proteína Bruta (%) 1.744 0,60 1.500 0,75
Energia Metabolizável
(Kcal/kg) 242.857 85 200.000 100
Foi utilizado o seguinte programa alimentar com 3 dietas:
- Crescimento I (CI)– dos 63 aos 90 dias de idade.
- Crescimento II (CII) – dos 91 aos 118 dias de idade
- Terminação (T) - dos 119 aos 145 dias de idade
As rações foram fornecidas ad libitum e formuladas a base de milho e farelo de soja de
acordo com as recomendações nutricionais mínimas de Rostagno et al. (2011), dentro de cada
fase, crescimento I, crescimento II e terminação, como apresentadas nas tabelas 4, 5 e 6
respectivamente.
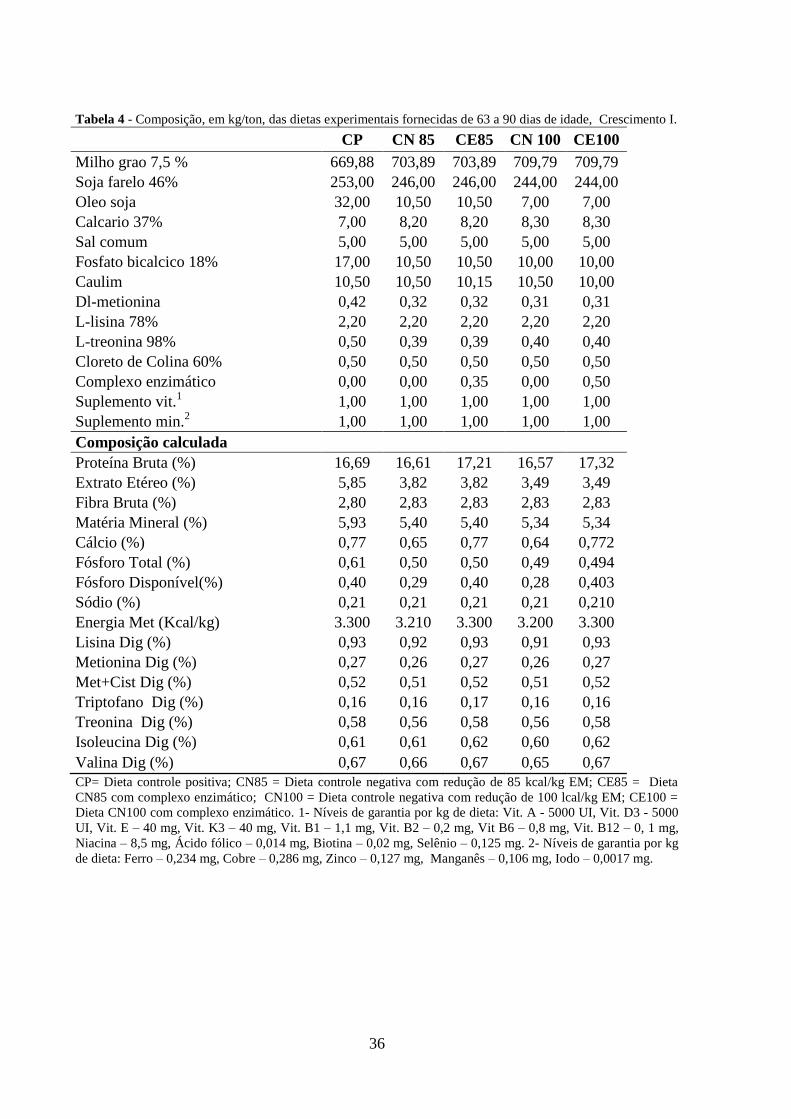
36
Tabela 4 - Composição, em kg/ton, das dietas experimentais fornecidas de 63 a 90 dias de idade, Crescimento I.
CP CN 85 CE85 CN 100 CE100
Milho grao 7,5 % 669,88 703,89 703,89 709,79 709,79
Soja farelo 46% 253,00 246,00 246,00 244,00 244,00
Oleo soja 32,00 10,50 10,50 7,00 7,00
Calcario 37% 7,00 8,20 8,20 8,30 8,30
Sal comum 5,00 5,00 5,00 5,00 5,00
Fosfato bicalcico 18% 17,00 10,50 10,50 10,00 10,00
Caulim 10,50 10,50 10,15 10,50 10,00
Dl-metionina 0,42 0,32 0,32 0,31 0,31
L-lisina 78% 2,20 2,20 2,20 2,20 2,20
L-treonina 98% 0,50 0,39 0,39 0,40 0,40
Cloreto de Colina 60% 0,50 0,50 0,50 0,50 0,50
Complexo enzimático 0,00 0,00 0,35 0,00 0,50
Suplemento vit.1 1,00 1,00 1,00 1,00 1,00
Suplemento min.2 1,00 1,00 1,00 1,00 1,00
Composição calculada
Proteína Bruta (%) 16,69 16,61 17,21 16,57 17,32
Extrato Etéreo (%) 5,85 3,82 3,82 3,49 3,49
Fibra Bruta (%) 2,80 2,83 2,83 2,83 2,83
Matéria Mineral (%) 5,93 5,40 5,40 5,34 5,34
Cálcio (%) 0,77 0,65 0,77 0,64 0,772
Fósforo Total (%) 0,61 0,50 0,50 0,49 0,494
Fósforo Disponível(%) 0,40 0,29 0,40 0,28 0,403
Sódio (%) 0,21 0,21 0,21 0,21 0,210
Energia Met (Kcal/kg) 3.300 3.210 3.300 3.200 3.300
Lisina Dig (%) 0,93 0,92 0,93 0,91 0,93
Metionina Dig (%) 0,27 0,26 0,27 0,26 0,27
Met+Cist Dig (%) 0,52 0,51 0,52 0,51 0,52
Triptofano Dig (%) 0,16 0,16 0,17 0,16 0,16
Treonina Dig (%) 0,58 0,56 0,58 0,56 0,58
Isoleucina Dig (%) 0,61 0,61 0,62 0,60 0,62
Valina Dig (%) 0,67 0,66 0,67 0,65 0,67
CP= Dieta controle positiva; CN85 = Dieta controle negativa com redução de 85 kcal/kg EM; CE85 = Dieta
CN85 com complexo enzimático; CN100 = Dieta controle negativa com redução de 100 lcal/kg EM; CE100 =
Dieta CN100 com complexo enzimático. 1- Níveis de garantia por kg de dieta: Vit. A - 5000 UI, Vit. D3 - 5000
UI, Vit. E – 40 mg, Vit. K3 – 40 mg, Vit. B1 – 1,1 mg, Vit. B2 – 0,2 mg, Vit B6 – 0,8 mg, Vit. B12 – 0, 1 mg,
Niacina – 8,5 mg, Ácido fólico – 0,014 mg, Biotina – 0,02 mg, Selênio – 0,125 mg. 2- Níveis de garantia por kg
de dieta: Ferro – 0,234 mg, Cobre – 0,286 mg, Zinco – 0,127 mg, Manganês – 0,106 mg, Iodo – 0,0017 mg.
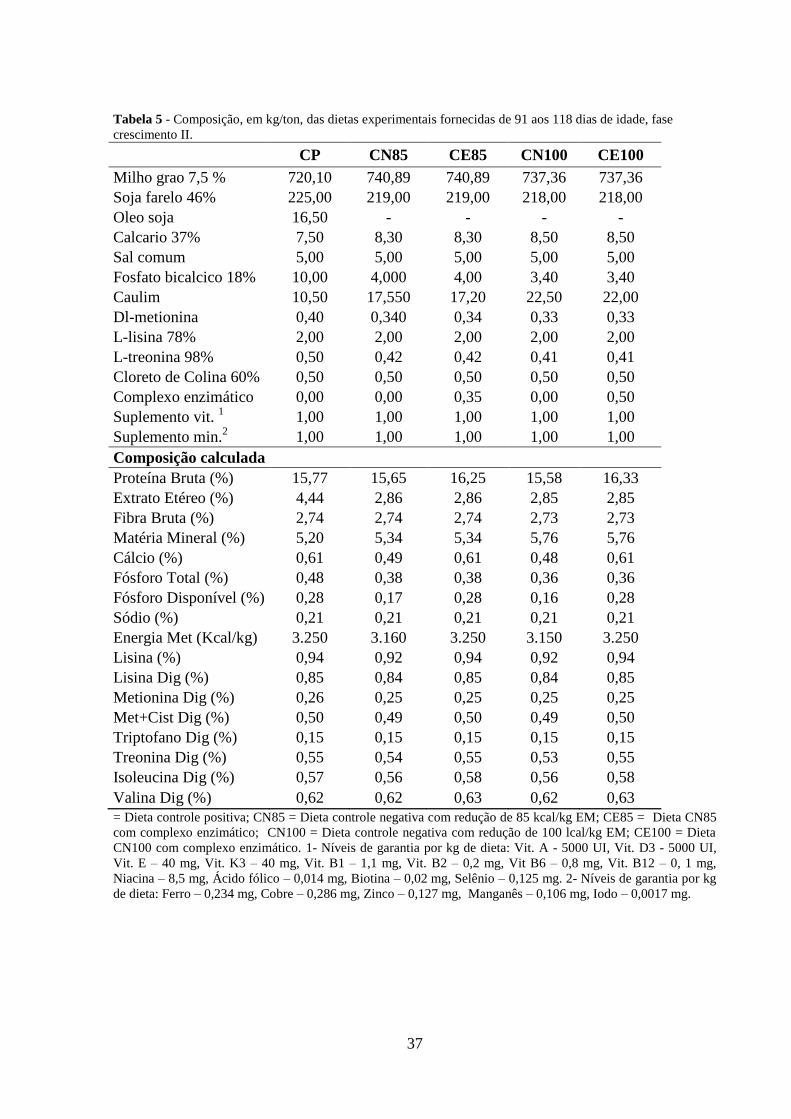
37
Tabela 5 - Composição, em kg/ton, das dietas experimentais fornecidas de 91 aos 118 dias de idade, fase
crescimento II.
CP CN85 CE85 CN100 CE100
Milho grao 7,5 % 720,10 740,89 740,89 737,36 737,36
Soja farelo 46% 225,00 219,00 219,00 218,00 218,00
Oleo soja 16,50 - - - -
Calcario 37% 7,50 8,30 8,30 8,50 8,50
Sal comum 5,00 5,00 5,00 5,00 5,00
Fosfato bicalcico 18% 10,00 4,000 4,00 3,40 3,40
Caulim 10,50 17,550 17,20 22,50 22,00
Dl-metionina 0,40 0,340 0,34 0,33 0,33
L-lisina 78% 2,00 2,00 2,00 2,00 2,00
L-treonina 98% 0,50 0,42 0,42 0,41 0,41
Cloreto de Colina 60% 0,50 0,50 0,50 0,50 0,50
Complexo enzimático 0,00 0,00 0,35 0,00 0,50
Suplemento vit. 1 1,00 1,00 1,00 1,00 1,00
Suplemento min.2 1,00 1,00 1,00 1,00 1,00
Composição calculada
Proteína Bruta (%) 15,77 15,65 16,25 15,58 16,33
Extrato Etéreo (%) 4,44 2,86 2,86 2,85 2,85
Fibra Bruta (%) 2,74 2,74 2,74 2,73 2,73
Matéria Mineral (%) 5,20 5,34 5,34 5,76 5,76
Cálcio (%) 0,61 0,49 0,61 0,48 0,61
Fósforo Total (%) 0,48 0,38 0,38 0,36 0,36
Fósforo Disponível (%) 0,28 0,17 0,28 0,16 0,28
Sódio (%) 0,21 0,21 0,21 0,21 0,21
Energia Met (Kcal/kg) 3.250 3.160 3.250 3.150 3.250
Lisina (%) 0,94 0,92 0,94 0,92 0,94
Lisina Dig (%) 0,85 0,84 0,85 0,84 0,85
Metionina Dig (%) 0,26 0,25 0,25 0,25 0,25
Met+Cist Dig (%) 0,50 0,49 0,50 0,49 0,50
Triptofano Dig (%) 0,15 0,15 0,15 0,15 0,15
Treonina Dig (%) 0,55 0,54 0,55 0,53 0,55
Isoleucina Dig (%) 0,57 0,56 0,58 0,56 0,58
Valina Dig (%) 0,62 0,62 0,63 0,62 0,63
= Dieta controle positiva; CN85 = Dieta controle negativa com redução de 85 kcal/kg EM; CE85 = Dieta CN85
com complexo enzimático; CN100 = Dieta controle negativa com redução de 100 lcal/kg EM; CE100 = Dieta
CN100 com complexo enzimático. 1- Níveis de garantia por kg de dieta: Vit. A - 5000 UI, Vit. D3 - 5000 UI,
Vit. E – 40 mg, Vit. K3 – 40 mg, Vit. B1 – 1,1 mg, Vit. B2 – 0,2 mg, Vit B6 – 0,8 mg, Vit. B12 – 0, 1 mg,
Niacina – 8,5 mg, Ácido fólico – 0,014 mg, Biotina – 0,02 mg, Selênio – 0,125 mg. 2- Níveis de garantia por kg
de dieta: Ferro – 0,234 mg, Cobre – 0,286 mg, Zinco – 0,127 mg, Manganês – 0,106 mg, Iodo – 0,0017 mg.
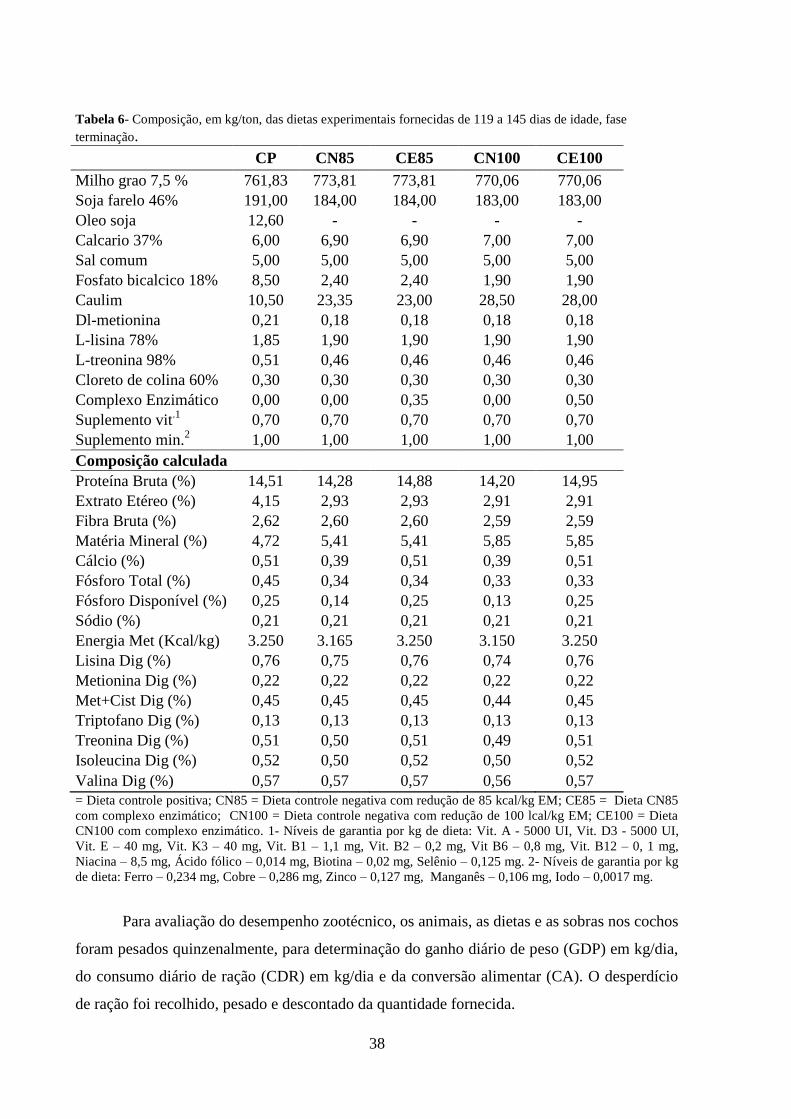
38
Tabela 6- Composição, em kg/ton, das dietas experimentais fornecidas de 119 a 145 dias de idade, fase
terminação.
CP CN85 CE85 CN100 CE100
Milho grao 7,5 % 761,83 773,81 773,81 770,06 770,06
Soja farelo 46% 191,00 184,00 184,00 183,00 183,00
Oleo soja 12,60 - - - -
Calcario 37% 6,00 6,90 6,90 7,00 7,00
Sal comum 5,00 5,00 5,00 5,00 5,00
Fosfato bicalcico 18% 8,50 2,40 2,40 1,90 1,90
Caulim 10,50 23,35 23,00 28,50 28,00
Dl-metionina 0,21 0,18 0,18 0,18 0,18
L-lisina 78% 1,85 1,90 1,90 1,90 1,90
L-treonina 98% 0,51 0,46 0,46 0,46 0,46
Cloreto de colina 60% 0,30 0,30 0,30 0,30 0,30
Complexo Enzimático 0,00 0,00 0,35 0,00 0,50
Suplemento vit.1 0,70 0,70 0,70 0,70 0,70
Suplemento min.2 1,00 1,00 1,00 1,00 1,00
Composição calculada
Proteína Bruta (%) 14,51 14,28 14,88 14,20 14,95
Extrato Etéreo (%) 4,15 2,93 2,93 2,91 2,91
Fibra Bruta (%) 2,62 2,60 2,60 2,59 2,59
Matéria Mineral (%) 4,72 5,41 5,41 5,85 5,85
Cálcio (%) 0,51 0,39 0,51 0,39 0,51
Fósforo Total (%) 0,45 0,34 0,34 0,33 0,33
Fósforo Disponível (%) 0,25 0,14 0,25 0,13 0,25
Sódio (%) 0,21 0,21 0,21 0,21 0,21
Energia Met (Kcal/kg) 3.250 3.165 3.250 3.150 3.250
Lisina Dig (%) 0,76 0,75 0,76 0,74 0,76
Metionina Dig (%) 0,22 0,22 0,22 0,22 0,22
Met+Cist Dig (%) 0,45 0,45 0,45 0,44 0,45
Triptofano Dig (%) 0,13 0,13 0,13 0,13 0,13
Treonina Dig (%) 0,51 0,50 0,51 0,49 0,51
Isoleucina Dig (%) 0,52 0,50 0,52 0,50 0,52
Valina Dig (%) 0,57 0,57 0,57 0,56 0,57
= Dieta controle positiva; CN85 = Dieta controle negativa com redução de 85 kcal/kg EM; CE85 = Dieta CN85
com complexo enzimático; CN100 = Dieta controle negativa com redução de 100 lcal/kg EM; CE100 = Dieta
CN100 com complexo enzimático. 1- Níveis de garantia por kg de dieta: Vit. A - 5000 UI, Vit. D3 - 5000 UI,
Vit. E – 40 mg, Vit. K3 – 40 mg, Vit. B1 – 1,1 mg, Vit. B2 – 0,2 mg, Vit B6 – 0,8 mg, Vit. B12 – 0, 1 mg,
Niacina – 8,5 mg, Ácido fólico – 0,014 mg, Biotina – 0,02 mg, Selênio – 0,125 mg. 2- Níveis de garantia por kg
de dieta: Ferro – 0,234 mg, Cobre – 0,286 mg, Zinco – 0,127 mg, Manganês – 0,106 mg, Iodo – 0,0017 mg.
Para avaliação do desempenho zootécnico, os animais, as dietas e as sobras nos cochos
foram pesados quinzenalmente, para determinação do ganho diário de peso (GDP) em kg/dia,
do consumo diário de ração (CDR) em kg/dia e da conversão alimentar (CA). O desperdício
de ração foi recolhido, pesado e descontado da quantidade fornecida.

39
Os dados consideraram o período acumulado, deste modo: Período I (63-90 dias de
idade), Período II (63-118 dias de idade) e Período III (63-145 dias de idade). Foram
analisados através do programa SISVAR (Ferreira, 2011), e as médias foram comparadas
utilizando teste de Tukey com significância de 5%.
A
B C
Figura 2 - A - Vista externa da Estação de Avaliação de Suínos de Tanquinho; B- Vista do comedouro
e balde de armazenagem de ração; C - Animais alojados em baias experimentais.

40
3.2 Digestibilidade
Com o objetivo de determinar os coeficientes de digestibilidade da matéria seca (MS),
proteína bruta (PB) e energia bruta (EB), foi empregado o método da coleta parcial de fezes,
utilizando-se cinza ácido insolúvel – CAI, como indicador.
Foram coletadas amostras de fezes somente dos machos, sendo oitoanimais/tratamento
no período intermediário de cada fase experimental (crescimento I, crescimento II e
terminação), totalizando 120 amostras de fezes e 15 amostras de ração.
Foi adicionado às rações 1% de Celite®, cinza ácido insolúvel, sendo os animais
alimentados com essa ração durante três dias para regular o fluxo do indicador no trato
digestório. Nos três dias subsequentes a esta adaptação, amostras de fezes foram colhidas logo
após a defecação pelos animais, duas vezes ao dia. As amostras foram armazenadas em
freezer a - 8° C, até o momento das análises laboratoriais, quando foram submetidas a pré-
secagem em estufa com ventilação forçada a 55 °C por um período de 72 horas (Figura 3),
sendo posteriormente moídas em moinho tipo “faca”, dotado de peneira com crivos de 1 mm.
As amostras das dietas foram apenas moídas, seguindo os mesmos procedimentos descritos
para as de fezes.
Figura 3 - Amostra de fezes após secagem em estufa a
55ºC por 72 horas.
As amostras de fezes processadas e das rações foram analisadas no Laboratório de
Bromatologia do Instituto de Zootecnia, determinando-se os teores de Matéria seca (MS) -
Resíduo da secagem da amostra em estufa a 103 - 105° C, Proteína Bruta (PB) - Método de
DUMAS (combustão da amostra) e Energia Bruta (EB) - Medida das calorias liberadas pela
substância oxidada em Bomba Calorimétrica da marca IKA, modelo C5001.

41
As determinações de CAI nas dietas e nas fezes foram feitas conforme método
adaptado de Van Keulen e Young (1977). Inicialmente, 2 g de cada amostra foi incinerada na
mufla a 600ºC por um período de 3 horas e 30 minutos, para determinação dos valores de
cinzas. Após, foi feita a digestão das amostras em ácido clorídrico diluído em água destilada
na proporção de 50%. As amostras com o ácido foram aquecidas a 465ºC, até que o ácido
volatizasse por completo. Após, o material foi diluído em água destilada, filtrado em papel de
filtro quantitativo da marca Quanty, faixa branca de 18,5 cm de diâmetro. Posteriormente, os
filtros foram lavados 5 vezes com água destilada para retirar os resíduos de ácido. Finalmente,
filtros e resíduos retidos foram incinerados em forno mufla através do seguinte esquema de
temperatura: 30 minutos a 150ºC, 1 hora a 250ºC, 30 minutos a 450ºC e 4 horas a 550ºC.
(Figura 4)
A
B C
Figura 4 - A- placa aquecedora; B – Filtração do resíduo; C- Resíduo no cadinho após a incineração.

42
Para o cálculo do fator de indigestibilidade (FI) foi usada a seguinte equação proposta
por Sakomura e Rostagno (2007):
FI = CAI dieta / CAI fezes
Posteriormente às análises bromatológicas realizadas, os coeficientes de
digestibilidade foram calculados utilizando-se a seguinte equação:
CD(%) =100–((%FI) * (% nutriente fezes /% nutriente dieta))*100
Para o cálculo do nutriente digestível foi utilizado a seguinte fórmula:
Nutriente digestível= (CD(%) * nutriente da dieta)/100
Os dados foram analisados através do programa SISVAR (Ferreira, 2011), utilizando
teste de Tukey com significância de 5%.
3.3 Análise econômica
A análise de viabilidade econômica foi realizada para cada fase e para o período total
do experimento. Inicialmente, foi determinado o custo da ração (em reais R$), por
quilograma de peso vivo ganho (Bellaver et al., 1985). Em seguida, foi calculado o Índice de
Eficiência Econômica (IEE), como sugerido por Tavernari et al. (2009), como segue:
Yi = (Pi * Qi)/ Gi
Onde: Yi = custo da ração por quilograma de peso vivo ganho no i-ésimo tratamento;
Pi = preço por quilograma da ração utilizada no i-ésimo tratamento;
Qi = quantidade de ração consumida no i-ésimo tratamento;
Gi = ganho de peso do i-ésimo tratamento.
Em seguida, foi calculado o índice de eficiência econômica.
IEE = (MCe/ CTei) * 100
Onde:
MCe = menor custo da ração por quilograma de ganho observado entre os tratamentos;
CTei = custo do tratamento i considerado.

43
Os preços dos ingredientes utilizados no cálculo dos custos das dietas foram baseados
nso preços praticados no estado de São Paulo, no dia 13 de Agosto de 2013 e considerando a
cotação do dólar (Taxa Ptax USD) a R$ 2,60.

44
4 RESULTADOS E DISCUSSÃO
4.1 Desempenho
Os resultados do consumo diário de ração (CDR), ganho diário de peso (GDP) e
conversão alimentar (CA) de machos e fêmeas, nos três períodos estudados, estão
apresentados na Tabela 7.
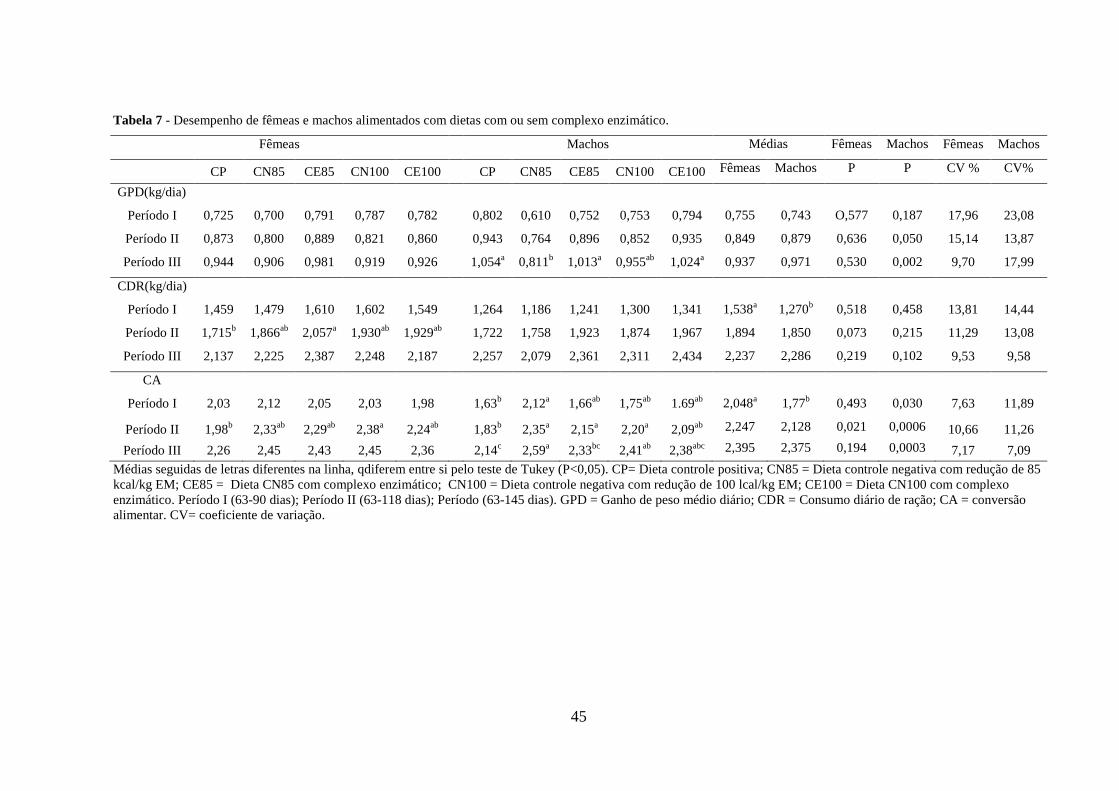
45
Tabela 7 - Desempenho de fêmeas e machos alimentados com dietas com ou sem complexo enzimático.
Fêmeas Machos Médias Fêmeas Machos Fêmeas Machos
CP CN85 CE85 CN100 CE100 CP CN85 CE85 CN100 CE100 Fêmeas Machos P P CV % CV%
GPD(kg/dia)
Período I 0,725 0,700 0,791 0,787 0,782 0,802 0,610 0,752 0,753 0,794 0,755 0,743
O,577 0,187 17,96 23,08
Período II 0,873 0,800 0,889 0,821 0,860 0,943 0,764 0,896 0,852 0,935 0,849
0,879
0,636 0,050 15,14 13,87
Período III 0,944 0,906 0,981 0,919 0,926 1,054a 0,811
b 1,013
a 0,955
ab 1,024
a 0,937 0,971
0,530 0,002 9,70 17,99
CDR(kg/dia)
Período I 1,459 1,479 1,610 1,602 1,549 1,264 1,186 1,241 1,300 1,341 1,538a
1,270b
0,518 0,458 13,81 14,44
Período II 1,715b 1,866
ab 2,057
a 1,930
ab 1,929
ab 1,722 1,758 1,923 1,874 1,967 1,894
1,850
0,073 0,215 11,29 13,08
Período III 2,137 2,225 2,387 2,248 2,187 2,257 2,079 2,361 2,311 2,434 2,237
2,286
0,219 0,102 9,53 9,58
CA
Período I 2,03 2,12 2,05 2,03 1,98 1,63b 2,12
a 1,66
ab 1,75
ab 1.69
ab 2,048
a 1,77
b 0,493 0,030 7,63 11,89
Período II 1,98b 2,33
ab 2,29
ab 2,38
a 2,24
ab 1,83
b 2,35
a 2,15
a 2,20
a 2,09
ab 2,247
2,128
0,021 0,0006 10,66 11,26
Período III 2,26 2,45 2,43 2,45 2,36 2,14c 2,59
a 2,33
bc 2,41
ab 2,38
abc 2,395
2,375
0,194 0,0003 7,17 7,09
Médias seguidas de letras diferentes na linha, qdiferem entre si pelo teste de Tukey (P<0,05). CP= Dieta controle positiva; CN85 = Dieta controle negativa com redução de 85
kcal/kg EM; CE85 = Dieta CN85 com complexo enzimático; CN100 = Dieta controle negativa com redução de 100 lcal/kg EM; CE100 = Dieta CN100 com complexo
enzimático. Período I (63-90 dias); Período II (63-118 dias); Período (63-145 dias). GPD = Ganho de peso médio diário; CDR = Consumo diário de ração; CA = conversão
alimentar. CV= coeficiente de variação.

46
Nas fêmeas, nos períodos I e III, não houve diferença estatística entre as dietas para as
variáveis estudadas.
Os resultados encontrados para essa fase confirmam aqueles obtidos por Pluske et al.
1998 e O’connell et al. 2005, que não obtiveram efeitos significativos no desempenho de
suínos ao utilizar complexos enzimáticos.
Isto demonstra que a falta de efeito para ganho de peso e para conversão alimentar
pode estar relacionada com incrementos muito pequenos na digestibilidade de nutrientes
proporcionados pelas enzimas ou pela inclusão do produto em dietas com níveis nutricionais
adequados (Barrera et al. 2004). Silva et al., 2013, sugere que uso de enzimas exógenas em
rações para suínos em crescimento e terminação sem resultados positivos no desempenho
pode ser decorrente da insuficiente melhora da digestibilidade dos nutrientes das dietas
avaliadas ou ao fato destas apresentarem, independentemente das enzimas empregadas,
níveis nutricionais satisfatórios para atender as exigências das categorias testadas.
Além disso, o uso de milho e farelo de soja podem ter colaborado para a falta de
resultados significativos, pois apesar da grande variabilidade na quantidade de PNA’S, estes
são considerados ingredientes de alta digestibilidade, com baixos teores de oligossacarídeos,
principal substrato para as enzimas presentes no complexo enzimático empregado.
De acordo com Silva et al. (2013), complexos enzimáticos apresentam melhores
resultados no desempenho em dietas com ingredientes de baixa digestibilidade e com níveis
nutricionais e energéticos mais baixos do que em relação às dietas formuladas com milho e
farelo de soja.
No período II, as fêmeas suínas que receberam a dieta CE85, aumentaram seus
consumos. No mesmo período, apresentaram pior conversão alimentar quando consumiram a
dieta CN100.
Segundo Lewis (2001), quando não existem fatores, tais como ambientais, sociais e
animais, que estejam interferindo diretamente o consumo, a concentração de energia da dieta
representa o maior determinante do consumo voluntário de alimento. Quando a alimentação é
a vontade, o consumo de energia geralmente é de 3 a 4 vezes a quantidade de energia exigida
para manutenção (NRC, 1998).
Ruiz et al. (2008) também observaram que fêmeas suínas alimentadas com dietas
contendo complexo enzimáticos aumentaram seus consumos, porém sem diferenças no ganho
diário de peso, interferindo assim, para uma pior conversão alimentar, o que pode demonstrar
que o aumento de consumo é devido as menores concentrações energéticas.

47
Nos machos, durante o Período I, o CDR e GPD não diferiram estatisticamente
(P>0,05), porém é possível notar que a conversão alimentar dos animais que tiveram a dieta
com redução de 85 kcal de energia foi prejudicada em relação aos animais do tratamento CP.
No Período II, apesar de as variáveis CDR e GPD não diferirem (P>0,05), a CA
apresentou diferenças significativas (P<0,05). Os animais que tiveram redução de energia de
85 e 100 Kcal tiveram pior conversão alimentar, mas os que tiveram redução de 100 Kcal e
foram suplementados com o complexo enzimático obtiveram valores estatisticamente iguais
ao da dieta controle.
No período III, o GPD diferiu estatisticamente (P<0,05) e a conversão alimentar dos
animais com redução energética foi prejudicada. A utilização de enzimas melhorou a CA de
machos durante os períodos estudados.
Tais resultados concordam com aqueles obtidos por Rodrigues et al. (2002), em que a
suplementação de enzimas exógenas influenciou significativamente (P<0,05) o GPD e CA
dos animais machos, nas fases analisadas.
Kitchen (1998), verificou melhora no GPD e CA de suínos em crescimento com a
utilização de enzimas, porém em dietas formuladas com cevada, ou seja, cereal de baixa
viscosidade.
No entanto, segundo Lindemann et al. (1997), a adição de um complexo enzimático
contendo protease, celulase, pentosanase, alfa-galactosidase e amilase, para dietas de suínos
em fase de crescimento e terminação, proporciona melhorias no GPD de acordo com o nível
de inclusão do complexo enzimático.
De modo geral, observa-se diferenças entre machos e fêmeas nos resultados das
variáveis estudadas. Geralmente, observa-se que os machos apresentam um maior ganho de
peso com a suplementação enzimática, devido ao maior consumo de ração e à velocidade de
crescimento, porém isso não foi observado em todas as fases nesse estudo.
Ao analisar o período total do experimento, período III, a CA dos machos foi
numericamente melhor do que das fêmeas para o tratamento CE85.
Rodrigues et al, (2002), que verificaram que o sexo dos animais pode ser uma das
variáveis que influenciam a resposta da adição de enzimas e resultados obtidos em seu
experimento evidenciam a necessidade de se preparar uma alimentação específica para
machos e fêmeas durante as fases de crescimento e terminação.
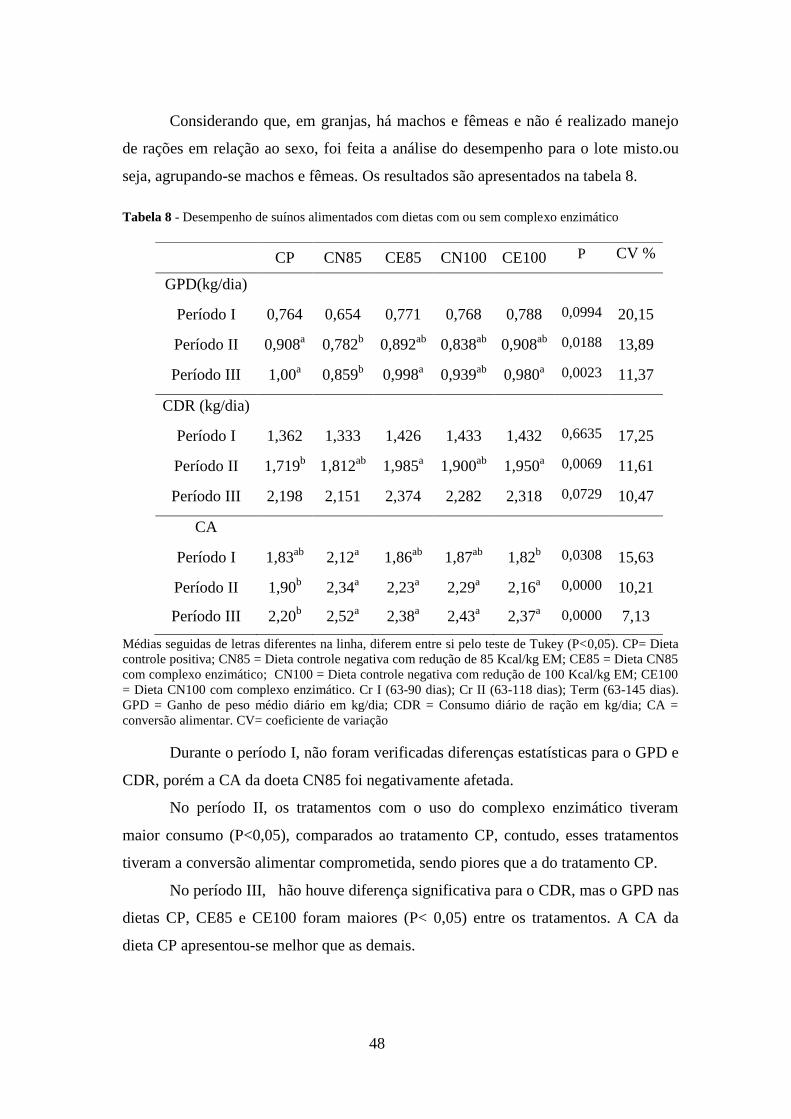
48
Considerando que, em granjas, há machos e fêmeas e não é realizado manejo
de rações em relação ao sexo, foi feita a análise do desempenho para o lote misto.ou
seja, agrupando-se machos e fêmeas. Os resultados são apresentados na tabela 8.
Tabela 8 - Desempenho de suínos alimentados com dietas com ou sem complexo enzimático
Médias seguidas de letras diferentes na linha, diferem entre si pelo teste de Tukey (P<0,05). CP= Dieta
controle positiva; CN85 = Dieta controle negativa com redução de 85 Kcal/kg EM; CE85 = Dieta CN85
com complexo enzimático; CN100 = Dieta controle negativa com redução de 100 Kcal/kg EM; CE100
= Dieta CN100 com complexo enzimático. Cr I (63-90 dias); Cr II (63-118 dias); Term (63-145 dias).
GPD = Ganho de peso médio diário em kg/dia; CDR = Consumo diário de ração em kg/dia; CA =
conversão alimentar. CV= coeficiente de variação
Durante o período I, não foram verificadas diferenças estatísticas para o GPD e
CDR, porém a CA da doeta CN85 foi negativamente afetada.
No período II, os tratamentos com o uso do complexo enzimático tiveram
maior consumo (P<0,05), comparados ao tratamento CP, contudo, esses tratamentos
tiveram a conversão alimentar comprometida, sendo piores que a do tratamento CP.
No período III, hão houve diferença significativa para o CDR, mas o GPD nas
dietas CP, CE85 e CE100 foram maiores (P< 0,05) entre os tratamentos. A CA da
dieta CP apresentou-se melhor que as demais.
CP CN85 CE85 CN100 CE100 P CV %
GPD(kg/dia)
Período I 0,764 0,654 0,771 0,768 0,788 0,0994 20,15
Período II 0,908a 0,782
b 0,892
ab 0,838
ab 0,908
ab 0,0188 13,89
Período III 1,00a 0,859
b 0,998
a 0,939
ab 0,980
a 0,0023 11,37
CDR (kg/dia)
Período I 1,362 1,333 1,426 1,433 1,432 0,6635 17,25
Período II 1,719b 1,812
ab 1,985
a 1,900
ab 1,950
a 0,0069 11,61
Período III 2,198 2,151 2,374 2,282 2,318 0,0729 10,47
CA
Período I 1,83ab
2,12a 1,86
ab 1,87
ab 1,82
b 0,0308 15,63
Período II 1,90b 2,34
a 2,23
a 2,29
a 2,16
a 0,0000 10,21
Período III 2,20b 2,52
a 2,38
a 2,43
a 2,37
a 0,0000 7,13

49
Assim como Silva (2013), que ao avaliar o desempenho de 25 machos e 25 fêmeas,
considerando todo o período experimental (fases de crescimento I, II e terminação), não foram
observadas diferenças entre os tratamentos para a característica CDR (P>0,05). Porém estes
autores observaram que os animais que consumiram a ração com complexo enzimático
(pectinase - 4000µ/g, protease - 700µ/g, fitase - 300µ/g, ß-glucanase - 200µ/g, xilanase -
100µ/g, celulase - 40µ/g, amilase - 30µ/g), adicionado na forma on the top, apresentaram
maior ganho diário de peso (P<0,05) em relação a os animais submetidos ao tratamento cuja
ração foi formulada sem o complexo enzimático e com menores níveis nutricionais.
Foi observado uma melhora de aproximadamente 14% da CA na dieta com complexo
enzimático, CE85, em relação a dieta sem complexo enzimático, CN85, no período I.
Em termos práticos, esperava-se que a suplementação enzimática compensasse a
redução energética das dietas (CN85 e CN100). Deste modo, os suínos que se alimentassem
com a dieta CE85 e CE100 deveriam ter o GPD e Ca semelhantes comparada a dieta sem
redução energéticas e sem o uso de enzimas. Este fato não aconteceu de modo tão evidente,
sendo, portanto parcialmente comprovado, pois a CA e CDR durante o Período II e a CA no
período III foram piores nos tratamentos com enzimas em relação ao tratamento CP.
Isso demonstra respostas diferentes no desempenho para machos, fêmeas e lotes
mistos, indicando que o uso de complexos enzimáticos e manejos de rações em relações ao
sexo devem ser estudados.
4.2 Digestibilidade
Os resultados do ensaio de digestibilidade nos três períodos experimentais são
apresentados na tabela 9.
Tabela 9 - Médias observadas para os coeficientes de digestibilidade e valores digestíveis de dietas com e sem o
uso de complexo enzimático consumidas por machos castrados nas fases de Crescimento I, Crescimento II e
Terminação.
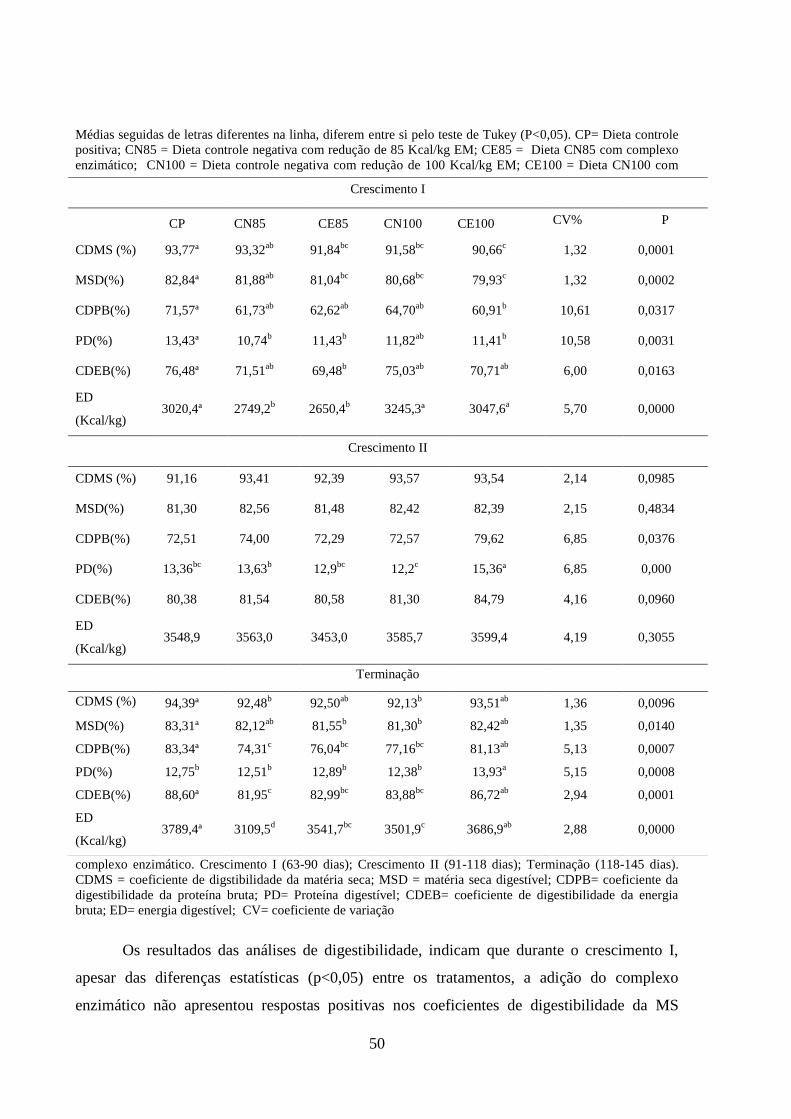
50
Médias seguidas de letras diferentes na linha, diferem entre si pelo teste de Tukey (P<0,05). CP= Dieta controle
positiva; CN85 = Dieta controle negativa com redução de 85 Kcal/kg EM; CE85 = Dieta CN85 com complexo
enzimático; CN100 = Dieta controle negativa com redução de 100 Kcal/kg EM; CE100 = Dieta CN100 com
complexo enzimático. Crescimento I (63-90 dias); Crescimento II (91-118 dias); Terminação (118-145 dias).
CDMS = coeficiente de digstibilidade da matéria seca; MSD = matéria seca digestível; CDPB= coeficiente da
digestibilidade da proteína bruta; PD= Proteína digestível; CDEB= coeficiente de digestibilidade da energia
bruta; ED= energia digestível; CV= coeficiente de variação
Os resultados das análises de digestibilidade, indicam que durante o crescimento I,
apesar das diferenças estatísticas (p<0,05) entre os tratamentos, a adição do complexo
enzimático não apresentou respostas positivas nos coeficientes de digestibilidade da MS
Crescimento I
CP CN85 CE85 CN100 CE100 CV% P
CDMS (%) 93,77ª 93,32ab
91,84bc
91,58bc
90,66c 1,32 0,0001
MSD(%) 82,84ª 81,88ab
81,04bc
80,68bc
79,93c 1,32 0,0002
CDPB(%) 71,57ª 61,73ab
62,62ab
64,70ab
60,91b 10,61 0,0317
PD(%) 13,43ª 10,74b 11,43
b 11,82
ab 11,41
b 10,58 0,0031
CDEB(%) 76,48ª 71,51ab
69,48b 75,03
ab 70,71
ab 6,00 0,0163
ED
(Kcal/kg) 3020,4ª 2749,2
b 2650,4
b 3245,3ª 3047,6
a 5,70 0,0000
Crescimento II
CDMS (%) 91,16 93,41 92,39 93,57 93,54 2,14 0,0985
MSD(%) 81,30 82,56 81,48 82,42 82,39 2,15 0,4834
CDPB(%) 72,51 74,00 72,29 72,57 79,62 6,85 0,0376
PD(%) 13,36bc
13,63b 12,9
bc 12,2
c 15,36ª 6,85 0,000
CDEB(%) 80,38 81,54 80,58 81,30 84,79 4,16 0,0960
ED
(Kcal/kg) 3548,9 3563,0 3453,0 3585,7 3599,4 4,19 0,3055
Terminação
CDMS (%) 94,39ª 92,48b 92,50
ab 92,13
b 93,51
ab 1,36 0,0096
MSD(%) 83,31ª 82,12ab
81,55b 81,30
b 82,42
ab 1,35 0,0140
CDPB(%) 83,34ª 74,31c 76,04
bc 77,16
bc 81,13
ab 5,13 0,0007
PD(%) 12,75b 12,51
b 12,89
b 12,38
b 13,93
a 5,15 0,0008
CDEB(%) 88,60ª 81,95c 82,99
bc 83,88
bc 86,72
ab 2,94 0,0001
ED
(Kcal/kg) 3789,4ª 3109,5
d 3541,7
bc 3501,9
c 3686,9
ab 2,88 0,0000

51
(CDMS), da proteína bruta (CDPB), da energia bruta (CDEB) e nos valores de da matéria
seca digestível (MSD) e da proteína digestível (PD).
A redução de 85 Kcal no teor de energia das rações determinou piores resultados para
a energia digestível (ED) com ou sem o uso de enzimas, em relação a dieta CP, porém a
redução de 100 kcal/ kg de EM com ou sem adição do complexo enzimático resultou em
valores de energia digestível semelhante ao verificado para a dieta CP.
No crescimento II, os valores de CDMS, CDPB, CDEB, MSD e ED não diferiram
entre os tratamentos (p>0,05). O valor de PD no tratamento CE100 foi maior (p<0,05) em
relação aos demais. Isso demonstra o efeito positivo ao uso do complexo enzimático em dietas
com redução de 100 kcal de energia. Entre os tratamentos CN85 e CE85 não houve diferenças
estatísticas ( p>0,05)
Na Terminação, os coeficientes de digestibilidade das dieta CN85 e CN100 foram
negativamente afetados, porém o uso do complexo enzimático na dieta CE100, foi eficaz,
pois os coeficientes de digestibilidade dessas dietas foram e iguais ao da dieta controle
positiva, o que demonstra que na fase da terminação, a dieta com redução energéticas de 100
kcal com o uso de enzimas foi eficiente, trazendo resultados iguais ou melhores que a dieta
sem redução energética.
Hurtado Nery et al., (2000) observaram melhora na digestibilidade da proteína bruta
quando uma ração à base de milho e farelo de soja para suínos dos 10 aos 30 kg foi
suplementada com um complexo enzimático composto por amilase, lípase e protease. No
entanto, o referido autor não observou diferença significativa entre os valores energéticos, o
que discorda dos resultados obtidos no presente trabalho.
O complexo enzimático utilizado possuía as enzimas xilanase e B-glucanase,
responsáveis por hidrolisar as arabinoxilanas ( Hogberg e Lindberg, 2004), agindo nas
paredes celulares dos PNA’s, e consequentemente, melhorando a digestibilidade dos
componentes proteicos e energéticos.
O complexo enzimático utilizado também possuía enzima celulase, porém as dietas
utilizadas continham um baixo nível de fibra. Quando o teor de fibra bruta na dieta for
acima de 7% (Kass et al.,1980) ou o FDN for superior a 15% (Cromwell, 1986) pode
interferir negativamente na digestibilidade e desempenho dos animais. A influência das
enzimas na digestibilidade dos constituintes da dieta, entretanto, depende da
disponibilidade de substrato (Knudsen e Hansen, 1991).

52
Além disso, as dietas experimentais eram compostas por ingredientes de origem
vegetal, ou seja, continham outro fator antinutricional para monogástricos, o fitato. Acredita-
se que ele possa interferir negativamente na utilização da energia, possivelmente por inibir a
atividade da enzima α-amilase ou pela diminuição da digestibilidade da proteína da dieta
(Selle & Ravindran, 2008). Porém, o complexo enzimático possuía a enzima fitase, capaz de
agir sobre o fitato e disponibilizar nutrientes que encontravam-se complexados com moléculas
de ácido fítico (RUIZ et al., 2008) e fornecer melhores resultados na digestibilidade.
A utilização isolada de fitase em dietas para suínos, pode ter sua eficiência
comprometida devido à falta de acesso ao substrato, quando encontra-se complexado aos
PNAs (Barbosa, 2008). Porém, a xilanase é capaz de degradar a camada de PNAs
promovendo a despolimerização de arabinoxilanas (Barbosa, 2012), o que facilita a ação da
fitase sobre fitato armazenado na parede celular (Olukosi et al., 2007) e o acesso das enzimas
endógenas e exógenas (amilase e protease) aos nutrientes encapsulados, o que aumenta a
disponibilidade de nutrientes para o crescimento (Nagashiro, 2007).
Um dos resultados esperados do uso de enzimas seria a diminuição da viscosidade do
conteúdo digestivo, proporcionando maior digestibilidade, porém algumas pesquisas sugerem
que a viscosidade da digesta não é facilmente influenciada em suínos, como é em aves, e, por
conseguinte, não é provavelmente um fator que afeta a digestibilidade (Campbell e Bedford,
1992; Bedford, 1995). A viscosidade da digesta em suínos é consideravelmente menor do que
nas aves por causa do teor de água mais elevado e, por conseguinte, a redução de viscosidade
pelo uso de enzimas é comparativamente baixo (Bedford e Classen, 1992
4.3 Análise econômica
A Tabela 10 apresenta os resultados da análise econômica para dietas sem e com o uso
de complexos enzimáticos para machos castrados e fêmeas.
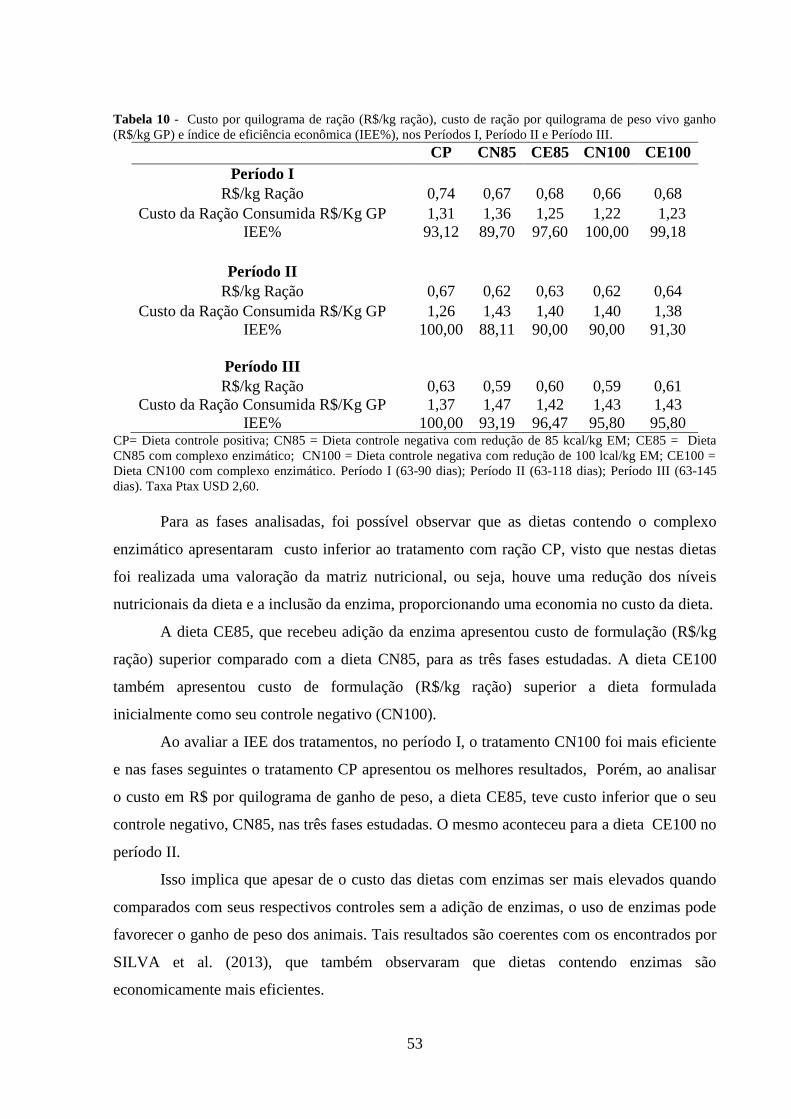
53
Tabela 10 - Custo por quilograma de ração (R$/kg ração), custo de ração por quilograma de peso vivo ganho
(R$/kg GP) e índice de eficiência econômica (IEE%), nos Períodos I, Período II e Período III.
CP CN85 CE85 CN100 CE100
Período I
R$/kg Ração 0,74 0,67 0,68 0,66 0,68
Custo da Ração Consumida R$/Kg GP 1,31 1,36 1,25 1,22 1,23
IEE%
93,12 89,70 97,60 100,00 99,18
Período II
R$/kg Ração 0,67 0,62 0,63 0,62 0,64
Custo da Ração Consumida R$/Kg GP 1,26 1,43 1,40 1,40 1,38
IEE% 100,00 88,11 90,00 90,00 91,30
Período III
R$/kg Ração 0,63 0,59 0,60 0,59 0,61
Custo da Ração Consumida R$/Kg GP 1,37 1,47 1,42 1,43 1,43
IEE% 100,00 93,19 96,47 95,80 95,80 CP= Dieta controle positiva; CN85 = Dieta controle negativa com redução de 85 kcal/kg EM; CE85 = Dieta
CN85 com complexo enzimático; CN100 = Dieta controle negativa com redução de 100 lcal/kg EM; CE100 =
Dieta CN100 com complexo enzimático. Período I (63-90 dias); Período II (63-118 dias); Período III (63-145
dias). Taxa Ptax USD 2,60.
Para as fases analisadas, foi possível observar que as dietas contendo o complexo
enzimático apresentaram custo inferior ao tratamento com ração CP, visto que nestas dietas
foi realizada uma valoração da matriz nutricional, ou seja, houve uma redução dos níveis
nutricionais da dieta e a inclusão da enzima, proporcionando uma economia no custo da dieta.
A dieta CE85, que recebeu adição da enzima apresentou custo de formulação (R$/kg
ração) superior comparado com a dieta CN85, para as três fases estudadas. A dieta CE100
também apresentou custo de formulação (R$/kg ração) superior a dieta formulada
inicialmente como seu controle negativo (CN100).
Ao avaliar a IEE dos tratamentos, no período I, o tratamento CN100 foi mais eficiente
e nas fases seguintes o tratamento CP apresentou os melhores resultados, Porém, ao analisar
o custo em R$ por quilograma de ganho de peso, a dieta CE85, teve custo inferior que o seu
controle negativo, CN85, nas três fases estudadas. O mesmo aconteceu para a dieta CE100 no
período II.
Isso implica que apesar de o custo das dietas com enzimas ser mais elevados quando
comparados com seus respectivos controles sem a adição de enzimas, o uso de enzimas pode
favorecer o ganho de peso dos animais. Tais resultados são coerentes com os encontrados por
SILVA et al. (2013), que também observaram que dietas contendo enzimas são
economicamente mais eficientes.

54
A tabela 11 apresenta os resultados para a análise econômica de machos e fêmeas
analisados separadamente.
Os dados da tabela 11 demonstram que para fêmeas, no período I, o tratamento
CE100 foi economicamente mais viável e no período II e III, apesar de o tratamento CP
foi mais eficiente.
Nos machos, no período I, o tratamento CE85 apresentou-se economicamente mais
eficiente. Nas fases seguintes, apesar de apresentar-se menos eficiente que o CP, foi mais
eficiente que o seu controle negativo (CN85) e o tratamento que também utilizou enzima
(CE100).
Esses resultados sugerem que, economicamente, para machos e fêmeas, os níveis de
redução energética e suplementação com complexo enzimático trazem respostas diferentes
no período I, visto que para machos foi mais viável o tratamento CE85, enquanto que para
fêmeas o tratamento CE100.
Considerando os dados de desempenho do Período I, não houve diferenças
estatísticas para as variáveis GPD e CDR para machos ou fêmeas. A CA dos machos,
apesar de apresentar diferenças estatística, não demonstra um evidente benefício ao uso do
complexo enzimático, portanto os resultados na análise econômica no período I, não
coincidem com os resultados de desempenho.
Porém, é notável que complexos enzimáticos trazem respostas economicamente
diferentes para machos e fêmeas, provavelmente pelas diferenças no consumo de ração
entre machos e fêmeas no Período I.
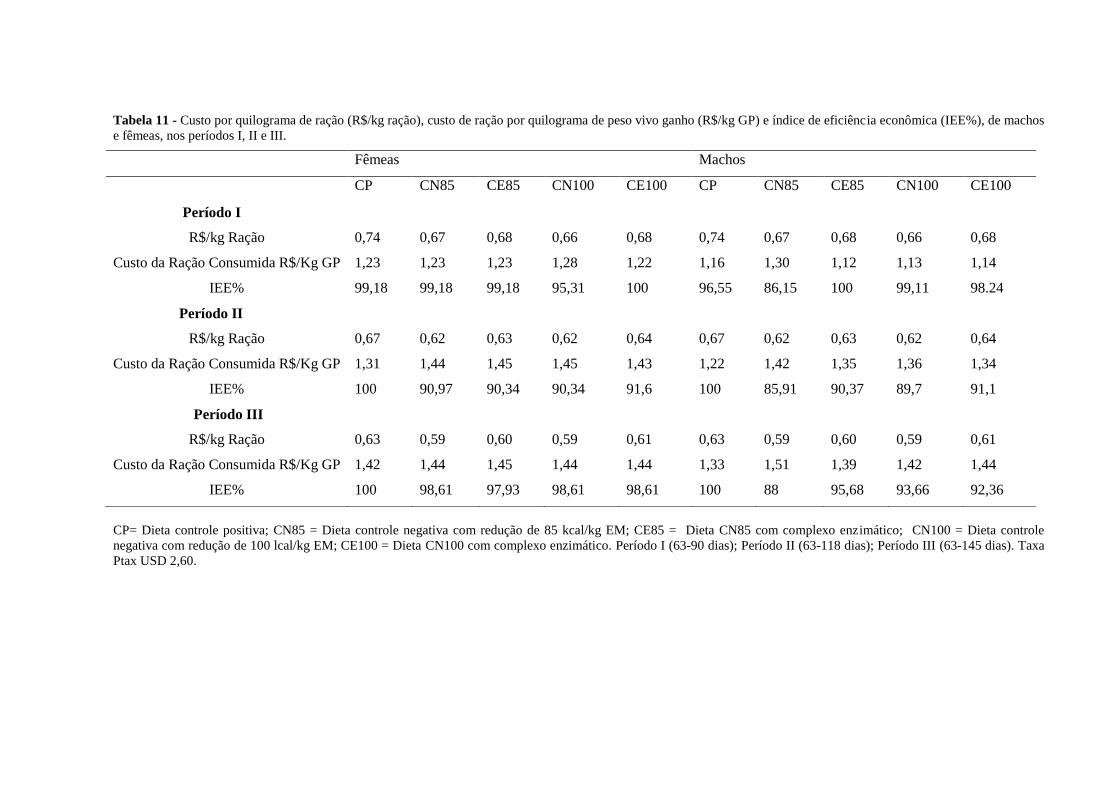
Tabela 11 - Custo por quilograma de ração (R$/kg ração), custo de ração por quilograma de peso vivo ganho (R$/kg GP) e índice de eficiência econômica (IEE%), de machos
e fêmeas, nos períodos I, II e III.
CP= Dieta controle positiva; CN85 = Dieta controle negativa com redução de 85 kcal/kg EM; CE85 = Dieta CN85 com complexo enzimático; CN100 = Dieta controle
negativa com redução de 100 lcal/kg EM; CE100 = Dieta CN100 com complexo enzimático. Período I (63-90 dias); Período II (63-118 dias); Período III (63-145 dias). Taxa
Ptax USD 2,60.
Fêmeas Machos
CP CN85 CE85 CN100 CE100 CP CN85 CE85 CN100 CE100
Período I
R$/kg Ração 0,74 0,67 0,68 0,66 0,68 0,74 0,67 0,68 0,66 0,68
Custo da Ração Consumida R$/Kg GP 1,23 1,23 1,23 1,28 1,22 1,16 1,30 1,12 1,13 1,14
IEE% 99,18 99,18 99,18 95,31 100 96,55 86,15 100 99,11 98.24
Período II
R$/kg Ração 0,67 0,62 0,63 0,62 0,64 0,67 0,62 0,63 0,62 0,64
Custo da Ração Consumida R$/Kg GP 1,31 1,44 1,45 1,45 1,43 1,22 1,42 1,35 1,36 1,34
IEE% 100 90,97 90,34 90,34 91,6 100 85,91 90,37 89,7 91,1
Período III
R$/kg Ração 0,63 0,59 0,60 0,59 0,61 0,63 0,59 0,60 0,59 0,61
Custo da Ração Consumida R$/Kg GP 1,42 1,44 1,45 1,44 1,44 1,33 1,51 1,39 1,42 1,44
IEE% 100 98,61 97,93 98,61 98,61 100 88 95,68 93,66 92,36

56

57
5 CONCLUSÃO
Para machos, recomenda-se o uso de complexo enzimático em dietas com redução
de 85 Kcal/ kg de EM, durante os períodos de crescimento e terminação, pois melhora a
conversão alimentar e é economicamente mais eficiente durante o Crescimento I. Para
fêmeas, o uso do complexo enzimático não se justifica.
O uso do complexo enzimático nas dietas com redução energética de 100 Kcal/kg
de EM elevou o teor de proteína digestível durante o Crescimento II; A adição de
complexo enzimático nas dietas com redução energética de 85 e 100 Kcal/kg de EM
aumentou os teores de energia digestível nas dietas de Terminação.

58

59
REFERÊNCIAS
ABIPECS. Relatórios Anuais da ABIPECS. 2012. Disponível em <http://abpa-
br.com.br/setores/suinocultura>. Acesso em: 31 julho de 2015.
AMORIM, A. B.; THOMAZ, M C.; RUIZ, U. S.; MARTINEZ, J. F.; PASCOAL, L. A.
F.; DANIEL, E.; WATANABE, P.H.; ROSALEN, D. L. Citrus pulp and enzyme complex
for growing and finishing pigs. Revista Brasileira Saúde de Produção e Animal , v.15,
p.369-380, 2014
ANDRIGUETTO, J.M.; PERLY, L.; MINARDI, I.; GEMAEL, A.; FLEMMUNG, L.S.;
SOUZA, G.A.; BONA FILHO, A. Nutrição Animal. São Paulo: Nobel, 2002, v.1, 396p.
ARIJA I.; CENTENO C.; VIVEROS A.;MARZO F; ILERA J.C; SILVAN G.. Poultry
Science v.85, p.635-644, 2006.
ARRUDA A.M.V.; PEREIRA E.S.; MIZUBUTI I.Y.;SILVA L.D.F. Importância da fibra
na nutrição de coelhos. Semina: Ciências Agrárias, v.24, p.181-190, 2003.
BARRERA, M.; CERVANTES, M.; SAUER, W.C.; ARAIZA, A.B.; TORRENTERA,
N.; CERVANTES, M. Ileal amino acid digestibility and performance of growing pigs fed
wheat-based diets supplemented with xylanase. Journal of Animal Science, v. 82, p.
1997-2003, 2004.
BARBOSA, N.A.A.; SAKOMURA, N.K.; FERNANDES, J.B.K.; DOURADO, L.R.B.
Enzimas exógenas no desempenho e na digestibilidade ileal de nutrientes em frangos de
corte. Pesquisa agropecuária brasileira, v.43, p. 755-762, 2008.
BARBOSA,N.A.A.;SAKOMURA,N.K.;BONATO,M. A.; HAUSCHILD, L.; OVIEDO-
RONDON, E. Enzimas exógenas em dietas de frangos de corte: desempenho. Ciência
Rural, v.42, p.1497-1502, 2012.
BARROS, P.C.; OLIVEIRA, V.; GEWEHR C.E.; NUNES, R.V.; Efeito da adição de
enzimas na digestibilidade total aparente de dietas para leitoes desmamados. Semina:
Ciências Agrárias, v.35, p.2211-2218, 2014.
BEDFORD, M.R.; CLASSEN, H.L. Reduction of intestinal viscosity through
manipulation of dietary rye and pentosanase concentration is effected through changes in
the carbohydrate composition of the intestinal aqueous phase and results in improved

60
growt rate and food conversion efficiency of broiler chicks. Journal of Nutrition, v.122,
p.560-569, 1992.
BEDFORD, M.R. Mechanism of action and potential environmental benefits from the use
of feed enzymes. Animal Feed Science Technology, v.53, p.145-155, 1995.
BELLAVER, C.; FIALHO, E.T.; PROTAS, J.F.S.; GOMES, P.C. Radícula de malte na
alimentação de suínos em crescimento e terminação. Pesquisa Agropecuária Brasileira,
v.20, p.969-974, 1985.
BORGES, F.M.O. Utilização de enzimas em dietas avícolas. Cadernos Técnicos da
Escola de Veterinária da UFMG, v.20, p.5-30, 1997.
BRITO M.S.;OLIVEIRA C.F.S.; SILVA T.R.G.; LIMA R.B.; Morais S.N.; Silva J.H.V.;
Polissacarídeos não amiláceos na nutrição de monogástricos. Acta Veterinaria Brasilica,
v.2, p.111-117, 2008.
BRUM, P.A.R.; AVILA,V.S.; LIMA, G.J.M.M.; COLDEBELLA, A.; SHEUERMANN,
G.N. Utilização de a-amilase em dietas à base de milho e farelo de soja de frangos de
corte: efeito da energia metabolizável e no desempenho. In: REUNIÃO ANUAL DA
SOCIEDADE BRASILEIRA DE ZOOTECNIA, 43., 2006. João Pessoa. Anais... João
Pessoa: Sociedade Brasileira de Zootecnia, 2006. 1 CD-ROM.
BRUM, P.A.R.; LIMA, G.J.M.M.; AVILA, V.S.; COLDEBELLA, A.; ZANOTTO, D.L.;
ROSA, P. S. Uso da alfa- amilase em dietas, superestimando ou não a energia
metabolizável do farelo de soja, no desempenho de frangos de corte. In: Reunião ANUAL
DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 44., 2007, Jaboticabal. Anais...
Jaboticabal: UNESP, 2007. 1 CD-ROM.
BRUM, P.A.R.; LIMA, G.J.M.M.; AVILA,V.S.; COLDEBELLA, A.; SCHEUERMANN,
G.N.; USINGLER, F.; TORGO, G.C.; Efeito da utilização de alfa-amilase em dietas a
base de milho e farelo de soja na digestibilidade da energia das rações e no
desempenho de frangos de corte. Concórdia: Embrapa, 2006. 3p. (Comunicado Técnico,
425).
CAMIRUAGA M.; GARCIA, F.; ELERA, R.; SIMONETTI, C. Respuesta productiva de
pollos broilers a la adicion de enzimas exogenas a dietas basadas en maiz o triticale
Ciência e Investigacão Agrária, v.28, p.23-36, 2001.
CAMPESTRINI E.; SILVA, V.T.M.; APPELT, M.D. Utilização de enzimas na
alimentação animal. Revista Eletrônica Nutritime, v.2, p.259-272, 2005.

61
CARDOSO D.M.; LARA L.J.C.; MIRANDA D.J.A.; POMPEU M.A.; MACHADO
A.L.C.; MATIAS C.F.Q.Uso de enzimas exógenas na avicultura: uma visão crítica,
Boletim da Indústria Animal, v.67, p.191-198, 2010.
CARDOSO, D.M.,; MACIEL, M.P.; PASSOS, D.P; SILVA, F.V.; REIS, S.T.; AIURA,
F.S. Efeito do uso de complexo enzimático em rações para frangos de corte. Archivos de
Zootecnia, v.60, p.1053-1064, 2011.
CARVALHO, J.C.C.; BERTECHINI, A.G.; FASSANI, E.J.; RODRIGRUES, P.B.;
PEREIRA, R.A.N. Desempenho e características de carcaça de frangos de corte
alimentados com dietas a base de milho e farelo de soja suplementadas com complexos
enzimáticos. Revista Brasileira de Zootecnia, v.38, p.292-298, 2009.
CHAMPE, P.C.; HARVEY, R.A. Bioquimica Ilustrada. 2.ed. São Paulo: Artes médicas,
1989. 446p.
CHARLTON, P. Expanding enzyme application: higher aminoacid and energy values for
vegetable proteins. In: BIOTECHNOLOGY IN THE FEED INDUSTRY, 12., 1996,
Nottingham. Proceedings...Nottingham: Nottingham University Press, 1996. p.317-326.
CLASSEN, H.L.Cereal grain starch and exogenous enzymes in poultry diets. Animal
Feed Science Technology, v.62, p.21-27, 1996.
CONTE, A.J.; TEIXEIRA, A.S.; FIALHO, E.T.; SHOULTEN, N.A.; BERTECHINI,
A.G.; Efeito da fitase e xilanase sobre o desempenho e as características ósseas de frangos
de corte alimentados com dietas contendo farelo de arroz. Revista Brasileira de
Zootecnia, v.32, p.1147-1156, 2003.
COSTA, F.G.P.; JÁCOME, I.M.T.D.; SILVA, J..H.V.; ARAÚJO, M.J.; CAMPOS,
K.M.F.; BARBOSA, J.G.; PEIXOTO, J.P.N.; SILVA, J.C.A.; NASCIMENTO, G.A.J.;
CLEMENTINO, R.H. Níveis de fósforo disponível e de fitase na dieta de poedeiras de
ovos de casca marrom. Ciência Animal Brasileira, v.5, p.73-81, 2006.
COWIESON, A.J.; ADEOLA, O. Carbohydrase, protease and phytase have an additive
beneficial effect in nutritionanally marginal diets for broiles chicks. Poulty Science, v.84,
p.1860-1867, 2005.

62
CROMWELL, G.L. Fiber Notes of Course ASC 686 - Advanced Non- ruminant
Nutrition. Lexington: University of Kentucky, 1986.
DERSJANT-LI, Y.; SCHULZE, H.; SCHRAMA, J.W.; VERSTEGEN, M.W. Feed
intake, growth, digestibility of dry matter and nitrogen in young pigs as affected by
dietary cation-anion difference and supplementation of xylanase. Journal of Animal
Physiology and Animal Nutrition, v.85, p.101-109, 2001.
DIEBOLD, G.; MOSENTHIN, R.; SAUER, W.C.; DUGAN, M.E.R.; LIEN, K.A.
Supplementation of xylanase and phospholipase to wheat based diets for weaner pigs.
Journal of Animal Physiology and Animal Nutrition, v.89, p.316–325, 2005.
ESONU, B.O.; AZUBUIKE, J.C.; EMENALOM, O.O.; ETUK, I.C.; OKOLI, H.; NNEJI
C.S. Effect of enzyme supplementation on the performance of broiler finisher
fed Microdesmis puberula leaf meal. International Journal of Poultry Science, v.3,
p.112-114, 2004.
FDA- FOOD AND DRUG ADMINISTRATION. Disponível em: <http://
http://www.fda.gov>. Acesso em: 3 jul. de 2015.
FERKET, P. Enzymes offer way to reduce waste, improve perfomance. Feedstuffs, v.01,
p.30-34, 1996.
FERREIRA, D.F. Sisvar: a computer statistical analysis system. Ciência e
Agrotecnologia, v.35, p.1039-1042, 2011.
FIREMAN, F.A.T.; FIREMAN, K.B.A.T. Enzimas na alimentação de suínos. Ciência
Rural, v.28, p.173-178, 1998.
FIREMAN, F.A.T.; LÓPEZ, J.; FIREMAN, A.K.B.T. Desempenho e custos de suínos
alimentados com dietas contendo 50% de farelo de arroz integral suplementados com
fitase e/ou celulase. Archive Latinoam Production Animal, v.1, p.18-23, 2000.
FREITAS, E.R.; FUENTES M.F.F.; ESPINDOLA G.B.; Efeito da Suplementação
Enzimática em Rações à Base de Milho/Farelo de Soja sobre o Desempenho de Poedeiras
Comerciais. Revista Brasileira de Zootecnia, v.29, p.1103-1109, 2000.
FUENTE, J.M.; SOTO-SALANOVA, M.F. Utilización de enzimas para mejorar el valor
nutritivo de las dietas maízsorgo/soja en avicultura. Seleciones Avicolas, v.39, p.271-275,
1997.

63
GAO, F.; JIANG, Y.; ZHOU, G.H.; HAN, Z.K.; The effects of xylanase supplementation
on performance, characteristics of the gastrointestinal tract, blood parameters and gut
microflora in broilers fed on wheat-based diets. Animal Feed Science and Technology,
v.142, p.173-184, 2008.
GRAHAM, H. Mode de action of feed enzymes in diets based on low viscous and viscous
grains, In: SIMPOSIO LATINO AMERICANO DE NUTRIÇÃO DE SUÍNOS E AVES,
1996, Campinas. Anais... Colégio Brasileiro de Nutrição Animal, Campinas, 1996. p.60-
69.
GUENTER, W. Pratical experience with the use of enzymes. 2002. Online. Disponível
em: < http://www.idrc.ca/books/focus/821> Acesso em setembro de 2015.
HALL, M.B. Challenges with nonfiber carbohydrate methods. Journal of Animal
Science, v.81, p.3226-3232, 2003.
HAUSCHILD, L.; LOVATTO, P.A.; GARCIA G.G. Digestibilidade, balanços do
nitrogênio e Fósforo para suínos contendo diferentes níveis de triguilho em substituição ao
milho com ou sem adição de enzimas. Ciência Rural, v.37, p.1557-1562, 2004.
HEDEMANN, M.S.; JENSEN, B.B. Variations in enzyme activity in stomach and
pancreatic tissue and digesta in piglets around weaning. Archives of Animal Nutrition, v.
58, p.47-59, 2004.
HOGBERG, A.; LINDBERG, J.E. Influence of cereal nonstarch polysaccharides and
enzyme supplementation on digestion site and gut environment in weaned pigs. Animal
Feed Science and Technology, v.116, p.113-128. 2004.
HURTADO NERY, V.L.; LIMA J.A.F.; MELO R.C.A.; FIALHO E.T. Adição de
enzimas exôgenas para Leitões dos 10 aos 30 kg de Peso. Revista brasileira de
zootecnia, v.29, p.794-802, 2000
JENSEN, M.S.; BACK KNUDSEN K.E.; INBORR J., JAKOBSEN, K. Effect of â-
glucanase supllementation on pancreatic enzyme activity and nutrient digestibility in
piglets fed diets based on hulled and hulless barley varieties. Animal Feed Science and
Technology, v.72, p.329-345, 1998.

64
JONGBLOED, A.W.; KEMME, P.A.; MROZ, Z. The role of microbial phytases in pig
production. In: Enzymes In ANIMAL NUTRITION, 1., 1992, Switzerland.
Proceedings… Switzerland: [s.n.], 1992. p.173-180.
JUNG, H.G. Forage lignins and their effects on fiber digestibility. Agronmy Journal,
v.81, p.33- 38, 1989.
KASIM, A.B.; EDWARDS, H.M. The analysis for inositol phosphate forms in feed
ingredients. Journal of the Science of Food and Agriculture, v.76, p. 1–9, 1998.
KASS, M.L.; VAN SOEST, P.J.; POND, W.G. Utilization of dietary fiber from alfalfa by
growing swine. I. Apparent digestibility of diet components in specific segments of
gastrointestinal tract. Journal of Animal Science, v.50, p.175 -191, 1980.
KESHAVARZ, K. Por que es necesario emplear La fitasa en la dieta de las ponedoras?.
Industria Avicola, v.46, p.13-14, 1999.
KITCHEN, D.I. Enzimes aplications in pigs’ diets. Feed Compounder, v.2, p.14-18,
1998.
KORNEGAY, E.T. Digestion of phosphorus and other nutrients: the role of phytases and
fsctores influencing their activity. In: BEDFORD, M.R.; PARTRIDGE, G.G. Enzymes in
farm nutrition. Londres : Cab Internacional, 2001, p.237-271.
KORNEGAY, E.T.; QIAN, H. Replacement of inorganic phosphorus by microbial
phytase for young pigs fed on a maize-soybean-meal diet. British Journal of Nutrition,
v.76, p.563-578, 1996.
LAN, Y; WILLIAMS, B.A.; VERTEGEN, M.W.A.; PATTERSON R.; TAMMINGA S.
Soy oligosaccharides in vitro fermentation characteristics and its effect on caecal
microorganisms of young broile chickens. Animal Feed Science and Technology, v.
133, p.286-297, 2007.
LEENHOUWERS, J.I.; ADJEI-BOATENG, J.A.J.; VERRETH; SCHRAMA, J.W.
Digesta viscosity, nutrient digestibility and organ weights in african catfish (Clarias
gariepinus) fed diets supplemented with different levels of a soluble non-starch
polysaccharide. Aquaculture Nutrition, v.12, p.111-116, 2006.
LEWIS A. J., Amino acids in swine nutrition. In: LEWIS, A.J.; SOUTHERN, L.L. (Ed)
Swinw nutrition. 2thed. Boca raton: CRC press, 2001. p.131-141

65
LI, S.; SAUER, W.C.; HUANG, S.X.; GABERT, G.M. Effect of â-glucanase
supplementation to hulless barley or wheat - soybean meal diets on the digestibilities of
energy, protein, b-glucans and amino acids in young pigs. Journal of Animal Science,
v.74, p.1649-1656, 1996.
LIMA G.J.M.M.;VIOLA E.S. Ingredientes energéticos: trigo e triticale na alimentação
animal. In: SIMPÓSIO SOBRE INGREDIENTES NA ALIMENTAÇÃO ANIMAL.
Campinas. Anais... CBNA, Campinas, 2001, p.33-61.
LIMA, M.R; SILVA J,H.V.; , JARAÚJO, J.A.; LIMA, C.B.; OLIVEIRA E.R.A. .
Enzimas exógenas na alimentação de aves. Acta Veterinaria Brasilica, v.1, p.99- 110,
2007.
LINDEMANN, M.D.; GENTRY, J.L.; MONEGUE, H.J.; CROMWELL, G.L.;
JACQUES, K.A. Determination of the contribution of the enzyme combination to the
growth performance of pig. Journal of Animal Science, v.7, p.184, 1997. Supplement.
LIU, J.; BOLLINGER, D.W.; LEDOUX, D.R. et al. Lowering the dietary calcium to total
phosphorus ratio increases phosphorus utilization in low-phosphorus corn-soybean meal
diets supplemental with microbial phytase for growing-finishing pigs. Journal of Animal
Science., v.76, p.808-813, 1998.
LOW, A.G. Secretory responses of the pig gut to no-starch polysaccharides. Animal Feed
Science and Technology, v.23, p.55-65, 1989
LUCCHESI L., Substituição total de leite em pó por soja micronizada e enzima na
desmama precoce de leitões. 2003. 64p. Dissertação (Mestrado) Instituto de Zootecnia,
Nova odessa, 2013.
LUDKE, M.D.C.M.M.; LÓPEZ, J.; LUDKE, J.V.; NICOLAIEWSKY, S. Utilização da
fitase em dietas com ou sem farelo de arroz desengordurado para suínos em
crescimento/terminação. Revista Brasileira de Zootecnia, v.31, p.2002-2010, 2002.
LUDKE, M.D.C.M.M.; LÓPEZ, J.; NICOLAIEWSKY, S.Efeito da fitase em dietas com
ou sem fosfato inorgânico para suínos em crescimento.Revista Brasileira de Zootecnia,
v.29, p.485-494, 2000 .
MAKKINK, C.A.; NEGULESCUS’S, G.P.; GUIXIN, Q.; VERSTEGEN, M.W. A. Effect
of dietary protein source on feed intake, growth, pancreatic enzyme activities and jejunal

66
morphology in newly-weaned piglets. British Journal of Nutrition, v.72, p.353-368,
1994.
MALATHI, V.; DEVEGOWDA, G. In vitro evaluation of nonstarch polysaccharide
digestibility of feed ingredients by enzymes. Poultry Science, v.80, p.302- 305, 2001.
MAPA. MINISTÉRIO DA AGRICULTURA PECUÁRIA E ABASTECIMENTO.
Relatório Técnico. Portaria do Ministério da Agricultura, Pecuária e Abastecimento,
n. 808, de 05 de maio de 2004. Disponível em
<http://www.agricultura.gov.br/animal/especies/suínos> . Acesso em: 4 julho de 2015.
MAPA. MINISTÉRIO DA AGRICULTURA PECUÁRIA E ABASTECIMENTO.
Relatório Técnico. Portaria do Ministério da Agricultura, Pecuária e Abastecimento, n.
40,de 08 de janeiro de 2006. Disponível em
<http://www.agricultura.gov.br/animal/especies/suínos> Acesso em: julho de 2015.
MATHLOUTHI N.; MALLET S.; SAULNIER L.; QUEMENER B.; LARBIER, M.
Effects of xylanase and b-glucanase addition on performance, nutrient digestibility, and
physico-chemical conditions in the small intestine contents and caecal microflora of
broiler chickens fed a wheat and barley-based diet. Animal Reserch, v.51, p.395–406,
2002.
MAVROMICHALIS, I.; HANCOCK, I. D.; SENNE B. W. Enzyme supplementation and
particle size of wheat in diets for nursery and finishing pigs. Journal of Animal Science,
v.78, p.3086-3095, 2000.
MENTEN, J.F.M. Aditivos alternativos na produção de aves: probióticos e prebióticos. In:
REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 38., 2001,
Piracicaba. Anais... Piracicaba: Sociedade Brasileira de Zootecnia, 2001. p.141-157.
MONTAGNE, L.; PLUSKE, J.R.; HAMPSON, D.J. A review of interactions between
dietary fibre and the intestinal mucosa, and their consequences on digestive health in
young non-ruminant animals. Animal Feed Science and Technology, v.108, p.95-117,
2003.
MOREIRA I.; MOURINHO, F.L.; CARVALHO, O.P.L.; PAIANO, D.; PIANO, L.M.;
KURODO JUNIOR, I.S. Avaliação nutricional da casca de soja com ou sem adição de
complexo enzimático para leitões na fase inicial. Revista Brasileira de Zootecnia, v.38,
p.2408-2416, 2009.

67
NAGASHIO,C. Enzimas na nutrição de aves. In: Conferência APINCO DE CIÊNCIAS E
TECNOLOGIA AVÍCOLAS, 2007, Santos. Anais... Santos: FACTA, 2007. p.309-327.
NRC - NATIONAL RESEARCH COUNCIL. Subcommittee on Swine Nutrition.
Committee on Animal Nutrition. Nutrient requeriments of swine. 10thed. Washington:
National Academy Press, 1998.
NITSAN, Z.; BEN-AVRAHAM, G.; ZOREF, Z.; NIR, I. Growth and development of the
digestive organs and some enzymes in broiler chicks after hatching. British Poultry
Science, v.32, p.515-523, 1991.
O’CONNELL, J.M.; CALLAN, J.J.; O’DOHERTY J.V. The effect of dietary crude
protein level, cereal type and exogenous enzyme supplementation on nutrient digestibility,
nitrogen excretion, faecal voltile fatty acid concentration and ammonia emissions from
pigs. Animal Feed Science Technology, v.127, p.73 88, 2005.
ODETALLAH, H.N.; FERKET, P.R.; GRIMES J.L.; MCNAUGHTON, J.L.. Effect of
mannanendo-1,4-_-mannosidade on the growth performance of turkeys fed diets
containing 44 and 48% crude protein soybean meal. Poultry Science, v.81, p.1322-1331,
2002.
OFFICER, D.I. Feed enzymes. IN: D’MELLO, J.P.F. (Ed.). Farm animal metabolism
and nutrition. Edinburgh: The Scottish Agricultural College, 2000. p.405-426.
OLIVEIRA, H. B.; BERTECHINI, A.G.; BRITO J.A.G.; GROPPO, D.P.; OLIVEIRA,
TFB ; MESQUITA, F.R. ; DIEGO REMOLINA ; SALIBA, E.O.S. Uso De Uma Alfa-
Amilase Exógena Em Rações De Frangos De Corte. Arquivo Brasileiro de Medicina
Veterinária e Zootecnia, v.60, p.1053-1064, 2012.
OLUKOSI, O.A.; COWIESON, A.J.; ADEOLA, O. Age-related influence of a cocktail of
xylanase, amylase, and protease or phytase individually or in combination in
broilers. Poultry Science, v.86, p.77-86, 2007.
OPALINSKI M.; MAIORKA A.; CUNHA F.; ROCHA C.; BORGES S.A.dição de
complexo enzimático e da granulometria da soja integral desativada melhora desempenho
de frangos de corte.Ciência Rural, v.40, p.628-632, 2010
OSERA, R.H.; DALANEZI, J.A.; JUNQUEIRA, O.M. Efeito da inclusão de enzimas
digestivas sobre o rendimento de partes nobres de frangos de corte. Pubvet, v.2, 2008.

68
PARSONS, C.M.; ZHANG, Y.; ARABAT, M. Nutritional evaluation of soybean meals
varying in oligosaccharide content. Poultry Science, v.75, p.1127-1131, 2000.
PENZ JUNIOR, A.M. Enzimas em rações para aves e suínos. In: REUNIÃO ANUAL DA
SOCIEDADE BRASILEIRA DE ZOOTECNIA, 25., 1998, Botucatu. Anais... Sociedade
Brasileira de Zootecnia, 1998. p.165-178.
PEREIRA, F.M.A.; ROCHA, A.S.; SILVA, B.C.; BORGES, A.M.; MARTINS, T,P,A,;
NAVARRO, R.D. Uso de polissacarídeos não amiláceos na aquicultura. Revista
Científica de Medicina Veterinária, v.13, p.11-27, 2015.
PINHEIRO, D.F.; CRUZ, V.C.; SARTORI, J.R.; VICENTI PAULIMO, M.L. Effect of
early feed restriction and enzyme supplementation on digestive enzyme activities in
broilers. Poultry Science, v.83, p.1544-1550, 2004.
PIOVESAN, V.; OLIVEIRA, V.; GEWEHR, C.E., Milhos com diferentes texturas de
endosperma e adição de alfa-amilase na dieta de leitões. Ciência Rural , v.41, p.2014-
2019, 2011.
PLUSKE, J.R.; PETHICK, D.W.; MULLAN, B.P. Differential effects of feeding
fermentable carbohydrate to growing pigs on performance, gut size and slaughter
characteristics. Animal Science, v.67, p. 147-156, 1998.
QIAN, H.; VEIT, H.P.; KORNEGAY, E.T.; RAVINDRAN, V.; BENBOW, D.M. Effects
of supplemental phytase and phosphorus on histological and other tibial bone
characteristics and performances of broilers fed semi-purified diets. Poultry Science,
v.75, p.618-626, 1996.
RODRIGUES P.B.; FREITAS R.T.F.; FIALHO E.T.; SILVA H.O.; GONÇALVES T.M.
Digestibilidade dos nutrientes e desempenho de suínos em crescimento e terminação
alimentados com rações à base de milho e sorgo suplementadas com enzimas. Revista
Brasileira de Milho e Sorgo, v.1, p.91-100, 2002.
RODRIGUES P.B.; ROSTAGNO H.S..; ALBINO L.F.T.; GOMES P.C.; BARBOZA
W.A.; TOLEDO R.S. Desempenho de Frangos de Corte, Digestibilidade de Nutrientes e
Valores Energéticos de Rações formuladas com vários milhos, suplementadas com
enzimas. Revista Brasileira de Zootecnia, v.32, p.171-182, 2003.
ROSA, A.P; UTTPATEL, R. Uso de enzimas nas dietas para frango de corte, In: VIII
SIMPÓSIO BRASIL SUL DE AVICULTURA, 2007, Chapecó. Anais... Chapecó, 2007,
p 102-115.

69
ROSTAGNO, H.S.; ALBINO, L.F.T.; DONZELE, J.L.; GOMES, P.G.; OLIVEIRA, R.F.;
LOPES, D.C.; FERREIRA, A.S.; BARRETO, S.L.T.; EUCLIDES, R.F. Tabelas
brasileiras para aves e suínos: composição de alimentos e exigências nutricionais. 3 ed.
Viçosa, MG: UFV, 2011. 252 p.
ROTTER, B.A. The future of crude enzyme supplements in pig nutrition. Pig News
Information, v.11, p.15-17, 1990.
RUIZ, U.S.; THOMAZ, M.C.; HANNAS, M.I.; FRAGA, A.L.; WATANABE, P.H.;
SILVA, S.Z. da. Complexo enzimatico para suinos: digestao, metabolismo, desempenho e
impacto ambiental. Revista Brasileira de Zootecnia, v.37, p.458-468, 2008.
SAKOMURA N.K.; BIANCHI M.D.; PIZAURO JUNIOR, J.M.; CAFÉ M.B.; FREITAS
E.R. Efeito da idade de frangos de corte sobre a atividade enzimática e digestibilidade dos
nutrientes do farelo de soja e da soja integral. Revista Brasileira de Zootecnia, v.33,
p.924-935, 2004.
SAKOMURA N.K., SILVA J.H.V., COSTA F.G.P., FERNANDES J.B.K., HAUSCHILD
L. Nutrição de não-ruminantes. Jaboticabal: Funep, 2014.
SAKOMURA, N.K.; ROSTAGNO, H.S. Métodos de pesquisa em nutrição de
monogástricos. Jaboticabal: Funep, 2007. 283p.
SANDBERG, A.S.; LARSEN, T.; SANDSTRÖM, B. High dietary calcium level
decreases colonic phytate degradation in pigs fed a rapeseed diet. Journal of Nutrition,
v.123, p559-566, 1993.
SEBASTIAN, S.; TOUCHBURN, S.P.; CHAVEZ, E.R.; LAGUE, P.C. Apparent
digestibility of protein and amino acids in broiler chickens fed a cornsoybean diet
supplemented with microbial phytase. Poultry Science, v.76, p.1760-1769, 1997.
SELLE, P.H.; RAVINDRAN, V. Phytate-degrading enzymes in pig nutrition. Livestock
Science, v. 113, p. 99-122, 2008.
SHEPPY, C. The current feed enzyme market and likely trends. In:BEDFORD, M.;
PATRIDGE, G. (Ed.). Enzymes in farm animal nutrition finnfeeds international,
Marlborough:Wiltshire, 2001. p.1-10.

70
SILVA J.A.M.; PEREIRA FILHO M.; CAVERO B.A.S.; OLIVEIRA PEREIRA M.I.
Digestibilidade aparente dos nutrientes e energia de ração suplementada com enzimas
exógenas para juvenis de tambaqui. Acta Amazonica, v.37, p.157 – 164, 2007.
SILVA, C. A.; VINOKUNOVAS, S. L.; BRIDI, A. M.; OBA, A.; PACHECO, G.D.;
LOZANO, A.P.; SILVA, R.A.M.; DALTO, D.B.; AGOSTINI P.S. Utilização de um
complexo enzimático para rações contendo farelo de gérmen de milho desengordurado
para suínos em fase de crescimento e terminação. Semina: Ciências Agrárias, v.34, p.
4065-4082, 2013. Suplemento, 2.
SIMONS, P.C.M.; VERSTEEGH, H.A.J.Improvement of phosphorus availability by
microbial phytase in broilers and pigs. British Journal Nutrition, v.64, p.525-540, 1990
SOARES, E.C.; PEREIRA FILHO, M.; ROUBACH, R.; SOARES E SILVA R.C.
Protease exógena em dietas para juvenis de tucunaré-paca (Cichla sp.) Revista Brasileira
de Zootecnia, v.37, p.971-976, 2008
SOBARA, J.O.B.; MURAKAMI A.E.; NAKAGE E.S.; PIRACES F. Enzymatic
programs for broilers. Brazilian Archives pf Biology and Technology, v.52, p.233-240,
2009.
SOTO-SALANOVA, M.F.; GARCIA, O.; GRAHAM, H.; PACK, M. Uso de enzimas em
dietas de milho e soja para frangos de corte. In: CONFERÊNCIA APINCO DE CIÊNCIA
E TECNOLOGIA AVÍCOLA, Curitiba, PR, 1996. Anais... Curitiba: APINCO, 1996,
p.71-76
STRADA, E.S.O.; ABREU, R.D.; OLIVEIRA, G.J.C.; COSTA, M.C.M.M.;
CARVALHO, G.J.L.; FRANCA, A.S.; CLARTON, L.; AZEVEDO, J.L.M.Uso de
enzimas na alimentação de frangos de corte. Revista Brasileira de Zootecnia, v.34,
p.2369-2375, 2005.
TASKER, S.J.P.S.; BADYAL, S.C.E.; BACKSON, R.W. Hydroxyl accessibility in
celluloses. Polymer, v.35, p.4717-4721, 1994.
TAVERNARI, F.C. ; DUTRA JUNIOR, W.M.; ALBINO, L.F.T.; ROSTAGNO, H.S.;
VIEIRA, R.A.; SILVA, C.R. Efeito da utilização de farelo de girassol na dieta sobre o
desempenho de frangos de corte. Revista Brasileira de Zootecnia, v.38, p.1745-1750,
2009.

71
TAVERNARI, F.C.; CARVALHO, T.A.; ASSIS, A.P.; LIMA, H.J.D. Polissacarídeos
não amiláceo solúvel na dieta de suínos e aves. Revista Eletrônica Nutritime, v.5, p.673-
689, 2008
TEJEDOR, A.A.; ALBINO, L.F.T.; ROSTAGNO, H.S.; LIMA, C.A.R.; VIEITES, F.M.
Efeito da adição de enzimas em dietas e frangos de corte à base de milho e faelo de soja
sobre a digestibilidade ileal de nutrientes. Revista Brasileira de Zootecnia, v.39, p.151-
165. 2001.
TENORIO FIREMAN F.A.; LOPEZ J.; KYONARA BARBOSA A.; TENRIO FIREMN
A. Desempenho e custo de suínos alimentados com dietas contendo 50% de farelo de
arroz integral suplementados com fitase e/ou celulase. Archivos Latinoamericano de
Produção Animal, v.8, p.18-23, 2000.
THACKER, P.A. Effect of xylanase and protease on the performance of growing-
finishing pigs fed corn-based diets. Journal of Applied Animal Research,v.28, p.17-23,
2005.
TORRES, D.M. ; COTTA, J.T.B ; TEIXEIRA, A.S.; MUNIZ, J.A.; FONSECA, R.A.;
SANTOS, E.C.; ALVES, E.L. Dietas à base de milho e soja suplementadas com enzimas
na alimentação de frangos de corte. Ciência e Agrotecnologia , v.27, p.199-205, 2003.
VAHJEN, W.; BUSCH, T.; SIMON, O. Study on the use of soy bean polysaccharide
degrading enzymes in broiler nutrition. Animal Feed Science and Technology, v.120,
p.259-276, 2005.
VAN KEULEN, J.; YOUNG, B.A. Evaluation of acid-insoluble ash as natural marker in
ruminant digestibility studies. Journal of Animal Science, v.44, p.282-287, 1977.
Van Soest P.J. Nutritional ecology of the ruminant. 2thed. New York: Cornell
University Press, 1994.
VIANA, M.T.S.; ALBINO, L.F.T.; ROSTAGNO, H.S.; SILVA, E.A.; VIEIRA, R.A.;
RIBEIRO JUNIOR, V., Utilização de xilanase em dietas compostas por milho e faelo de
soja de poedeiras comerciais em postura. Revista Brasileira de Zootecnia, v.40, p.385-
390, 2011
WALSH, G.A.; POWER, R.F.; HEADON, D.R. Enzymes in the animal feed industry.
Trends in Biotechnology, v.11, p.946-957,1993.

72
WANG, J.J.; GARLICH, J.D.; JSHIH, J.C.H. Beneficial effects of versazyme, a
keratinase feed additive, on body weight, feed conversion, and breast yield of broilers
chickens. Journal of Applied Poultry Research, v.15, p.544-550, 2006.
WARD, A.T.; MARQUARDT, R.R. Antinutritional activity of a water-soluble
pentosanrich fraction from rye grain. Poultry Science, v.66, p.1665-1674, 1987.
WARD, N.E.; FODGE, D. Ingredients to counter antinutritional factors: soybean - based
feeds need enzymes too. Feed Management, v.47, p.13-18, 1996.
WRIGHT, J. D.; WYMAN, C. E.; GROCHMANN, K. Simultaneous saccharification and
fermentation of lignocellulose: Process evaluation. Applied Biochemistry and
Biotechnology, v.17, p.75-90, 1988.
WYATT, C.L.; BEDFORD, M.R. O uso de enzimas nutricionais para maximizar a
utilização de nutrientes pelo frango de corte em dietas à base de milho: recentes
progressos no desenvolvimento e aplicações práticas. In: SEMINÁRIO TÉCNICO
FINNFEEDS, Curitiba. Anais... Curitiba: Finnfeeds, 1998, p.2-12.
XIAO Z.; TRINCADO C.A.; MURTAUGH M.P. Beta-glucan enhancement of T cell IFN
gamma response in swine. Veterinarian Immunology and Immunopathol, v.102,
p.315- 320, 2004.
YANG, Z.B.; YANG, W.R.; JIANG, S.Z.; ZHANG, G.G.; ZHANG, Q.Q.; SIOW, K.C.;
Effects of a thermotolerant multi-enzyme product on nutrient and energy utilization of
broilers fed mash or crumbled corn – soybean meals diets. Journal of Applied Poultry
Research, v.19, p.38-45, 2010.
YIN, Y. L.; BAIDOO, S. K.; JIN, L. Z.; LIU, Y. G.; SHULZE, H.; SIMMINS P.H. The
effect of different carbohydrase and protease supplementation on apparent (ileal and
overall) digestibility of nutrients of five hulless barley varieties in young pig. Livestock
Production Science, v.71, p.109-120, 2001.
YU, B., HSU, J.C., CHIOU, P.W.S. Effects of glucanase supplementation of barley diets
on growth performance of broilers. Animal Feed Science and Technology, v.70, p.353-
361, 1998.
YU, B., SUN, Y.M., CHIOU, P.W.S. Effects of glucanase inclusion in a de-hulled barley
diet on the growth performance and nutrient digestion of broiler chickens. Animal Feed
Science and Technology, v.102, p.35-52, 2002.





























