Embed Size (px)
Citation preview

7/22/2019 Coprocultura Protocolos Parte A
http://slidepdf.com/reader/full/coprocultura-protocolos-parte-a 1/10
NewsLab - edição 86 - 2008 58
Protocolos de Microbiologia Clínica
CoproculturaParte 1 - Salmonella e Shigella
Carlos Henrique Pessôade Menezes e SilvaDoutor em Microbiologia
Microbiologista do CentroTecnológico de Análises(CETAN), Vila Velha-ESConsultor em Microbiologia doLaboratório Landsteiner, Vitória-ES
Quadro 1. Agentes etiológicos mais relacionados com as diarréias infecciosasBactérias Vírus Protozoários
Vibrio cholerae e outrosvibriões, Shigella spp., Sal-
monella spp. (não tifóide),E. coli (enterotoxigênica,enteropatogênica clássica,enteroinvasora, enterohe-morrágica, enteroagregati-va), Campylobacter jejuni ,Yersinia enterocolitica,Clostridium difficile, Bac-
teroides fragilis (entero-toxigênico)
Rotavírus, Adenovírus en-térico, Calicivírus, Astro-
vírus
Entamoeba histolytica,Giardia lamblia, Cryptos-
poridium, Cyclospora
Introdução
As doenças diarréicas continuamsendo uma freqüente causa de morteainda nos dias atuais, especialmente empaíses em desenvolvimento. A incidênciaanual e o perfil etiológico das diarréias emdiferentes populações podem variar deacordo com os diversos fatores de risco,tais como a idade muito jovem, deficiên-cias nutricionais, higiene inadequada dealimentos e do próprio corpo, ausência
de cuidados sanitários básicos, acesso asuprimentos de água contaminados e atémesmo o verão, estação do ano que sem-pre registra o maior número de casos deinternações devido às diarréias. Nos paísesindustrializados, a freqüência de casos dediarréia por criança é de somente 0,5 a 2episódios/criança/ano, enquanto que empaíses em desenvolvimento este númeropode alcançar facilmente os 10 episódios/
criança/ano. Nos países industrializadosas diarréias por rotavírus predominam,enquanto que nos países em desenvolvi-mento as bactérias são comumente encon-tradas nos casos de diarréia (18).
Os agentes etiológicos envolvidosnas diarréias infecciosas incluem muitasespécies de bactérias, vírus e protozoá-rios, todos possuindo grande número desorogrupos e biotipos e sendo também
A
convite da Revista Newslab, preparamos uma série denominada “Protocolos de Microbiologia Clínica”,
abordando os principais temas de interesse àqueles que militam na área, objetivando a educação conti-
nuada, o aprimoramento científico e a aplicabilidade imediata das diretrizes sugeridas para que os seto-
res de Microbiologia Clínica dos laboratórios brasileiros possam realizar seus exames com acurácia e qualidade.
Proporcionaremos, ainda, uma visão crítica, com sugestões construtivas, para a melhoria da qualidade neste setor,
tanto para aqueles que já possuem um setor ativo, mas que desejam incrementá-lo com informações precisas e
coesas, quanto para os laboratórios de pequeno porte, os quais terão a oportunidade de implementar rotinas de
fácil execução com toda informação científica necessária. Inicialmente, abordaremos o tema Coprocultura, dividi-
do em duas partes para melhor interpretação. Bimestralmente, a Revista Newslab trará novos protocolos, a saber:
Urocultura, Microbiologia dos Líquidos Corporais Estéreis, Microbiologia das Secreções, Microbiologia do Sistema
Genital Humano e Micologia Clínica.
7/22/2019 Coprocultura Protocolos Parte A
http://slidepdf.com/reader/full/coprocultura-protocolos-parte-a 2/10
NewsLab - edição 86 - 2008 59
todos transmitidos pela via fecal-oral.Pelo fato da grande diversidade demicrorganismos que podem estar envol-vidos, é virtualmente impossível paraqualquer laboratório clínico realizar umcompleto exame de fezes diarréicas. Por-tanto, os patógenos mais freqüentes sãoinvestigados, dependendo da localizaçãogeográfica e da estação do ano (13).
Shigella
O homem é o reservatório natural e asinfecções ocorrem via fecal-oral atravésda ingestão de 20 a 200 células viáveisem alimentos e água contaminados. Nashigelose, corre uma invasão e destruiçãoda camada epitelial da mucosa com in-
tensa reação inflamatória, gerando leucó-citos, muco e sangue nas fezes. Deve-sesuspeitar de espécies de Shigella quandoobservamos colônias lactose-negativas,bioquimicamente inertes em muitasprovas bioquímicas de identificação. Elastipicamente não produzem gás a partirde carboidratos, com exceção de algunsbiogrupos de S. flexneri que são aerogê-nicos. Raras cepas de S. sonnei podem
fermentar tardiamente a lactose (2%) ea sacarose (1%) e a maioria das cepasdesta espécie descarboxila a ornitina,característica não observada em outrasespécies de Shigella.
Há quatro subgrupos principais e 43sorotipos reconhecidos de Shigella (Qua-dro 3). A classificação do CDC combinaS. dysenteriae (grupo A), S. flexneri (grupo
B) e S. boydii (grupo C) como “Shigella sorogrupos A, B e C” por conta de suas si-milaridades bioquímicas. A presença deornitina-descarboxilase e a atividade debeta-galactosidase fazem com que as ce-pas de S. sonnei sejam bioquimicamentedistintas de outras espécies de Shigella. Aincapacidade de fermentar o manitol dis-tingue a S. dysenteriae. Todos os isoladosclínicos identificados bioquimicamente
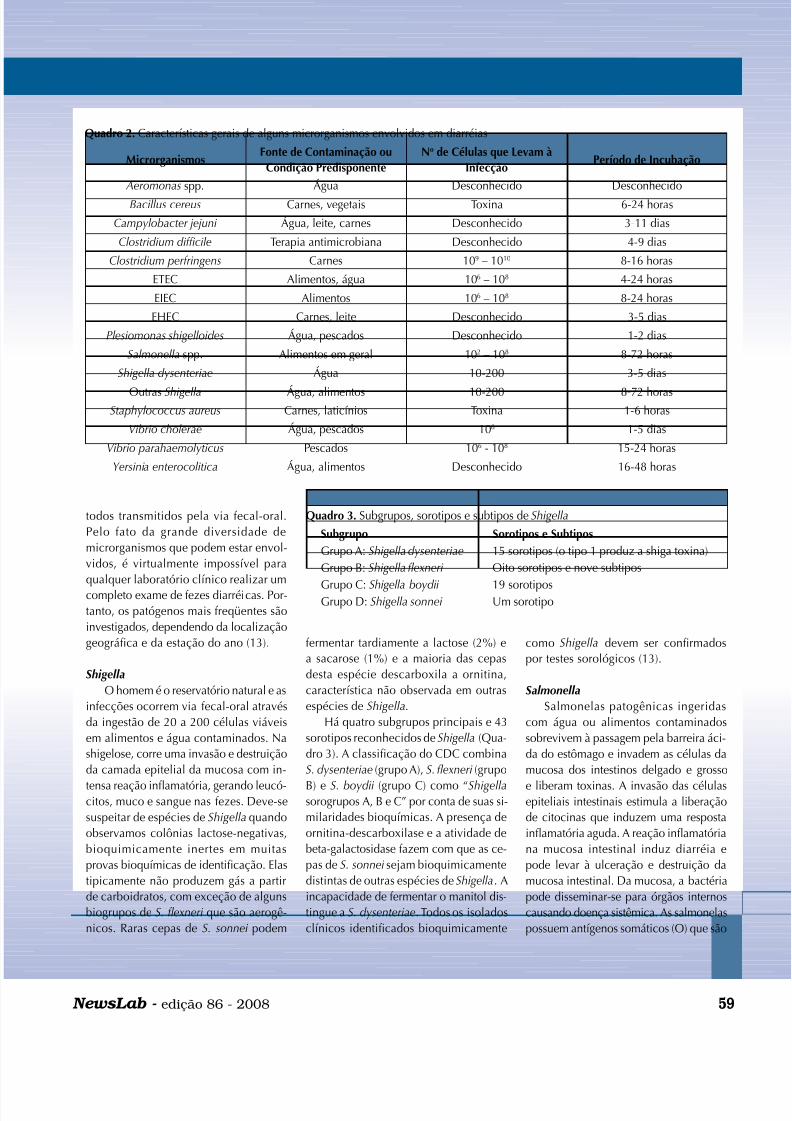
Quadro 2. Características gerais de alguns microrganismos envolvidos em diarréias
MicrorganismosFonte de Contaminação ou
Condição Predisponente
No de Células que Levam à
InfecçãoPeríodo de Incubação
Aeromonas spp. Água Desconhecido Desconhecido
Bacillus cereus Carnes, vegetais Toxina 6-24 horas
Campylobacter jejuni Água, leite, carnes Desconhecido 3-11 dias
Clostridium difficile Terapia antimicrobiana Desconhecido 4-9 dias
Clostridium perfringens Carnes 109 – 1010 8-16 horas
ETEC Alimentos, água 106 – 108 4-24 horas
EIEC Alimentos 106 – 108 8-24 horas
EHEC Carnes, leite Desconhecido 3-5 dias
Plesiomonas shigelloides Água, pescados Desconhecido 1-2 dias
Salmonella spp. Alimentos em geral 102 – 108 8-72 horas
Shigella dysenteriae Água 10-200 3-5 dias
Outras Shigella Água, alimentos 10-200 8-72 horasStaphylococcus aureus Carnes, laticínios Toxina 1-6 horas
Vibrio cholerae Água, pescados 108 1-5 dias
Vibrio parahaemolyticus Pescados 106 - 108 15-24 horas
Yersinia enterocolitica Água, alimentos Desconhecido 16-48 horas
como Shigella devem ser confirmadospor testes sorológicos (13).
Salmonella
Salmonelas patogênicas ingeridascom água ou alimentos contaminadossobrevivem à passagem pela barreira áci-da do estômago e invadem as células damucosa dos intestinos delgado e grosso
e liberam toxinas. A invasão das célulasepiteliais intestinais estimula a liberaçãode citocinas que induzem uma respostainflamatória aguda. A reação inflamatóriana mucosa intestinal induz diarréia epode levar à ulceração e destruição damucosa intestinal. Da mucosa, a bactériapode disseminar-se para órgãos internoscausando doença sistêmica. As salmonelaspossuem antígenos somáticos (O) que são
Quadro 3. Subgrupos, sorotipos e subtipos de Shigella
Subgrupo Sorotipos e Subtipos
Grupo A: Shigella dysenteriae 15 sorotipos (o tipo 1 produz a shiga toxina)Grupo B: Shigella flexneri Oito sorotipos e nove subtipos
Grupo C: Shigella boydii 19 sorotiposGrupo D: Shigella sonnei Um sorotipo

7/22/2019 Coprocultura Protocolos Parte A
http://slidepdf.com/reader/full/coprocultura-protocolos-parte-a 3/10
NewsLab - edição 86 - 2008 60
Protocolos de Microbiologia Clínica
lipopolissacarídeos e antígenos flagelares(H) que são proteínas. S. typhi também pos-sui um antígeno capsular ou de virulência(Vi). Bioquimicamente elas geralmente são
lactose e sacarose-negativas.Desde a época do primeiro isolamento
de Salmonella, reportado em 1884 por Ga-ffky (Bacterium typhosum) e em 1886 porSalmon e Smith (Salmonella choleraesuis),o desenvolvimento da nomenclatura paraeste gênero tem sido variado e complexo.O gênero Salmonella possui mais de 2.400sorotipos descritos no esquema atual deKauffmann-White. Antes de 01 de julhode 1983, três espécies de Salmonella foramusadas para reportar os resultados positivos
dos exames: S. choleraesuis, S. typhi e S.enteritidis (com a maioria dos sorotipospertencendo a este último sorotipo).
Atualmente, todas as antigas denomi-nações das espécies e subgrupos de Sal-
monella e Arizona são consideradas comopertencentes à mesma espécie, mas podemser separadas em sete grupos taxonômicos,representando seis subgrupos distintos. Aúnica exceção é S. bongori , anteriormenteconhecida como subgênero V, sendo umaespécie totalmente distinta através deestudos de homologia de DNA. Portanto,há duas espécies e seis subespécies de S.
enterica no atual sistema de classificaçãodo CDC (Quadro 4).
Na prática do dia-a-dia, os isoladosdesconhecidos provenientes de espécimesclínicos que são sugestivos de perten-ceram ao gênero Salmonella, devem serconfirmados por sorologia. Subcultivodos isolados confirmados devem ser en-caminhados para laboratórios de saúdepública, onde as designações de sorotipos(ex: Salmonella sorotipo Typhimurium)
serão realizadas. Répteis, particularmente
cobras, são o reservatório natural de S. en-
terica subespécie arizonae, mas o homem,aves e outros animais também podem serinfectados por esta bactéria. As infecçõeshumanas podem ser originadas quando damanipulação de aves, répteis e produtos àbase de ovos (13).
Microscopia
A análise microscópica direta do ma-terial fecal é muito útil, pois pode se teruma noção do que será encontrado na co-procultura. A presença de numerosos leu-cócitos polimorfonucleares (PMN) sugereum processo inflamatório invasivo (Figura1) envolvendo o cólon (como a exemplo deinfecções por Shigella ouCampylobacter ).Colites ulcerativas e a colite associada àantibioticoterapia (relacionada ao C. diffi-
cile) estão quase sempre associadas a umexsudato fecal contendo leucócitos. No
entanto, deve-se ter em mente que nemtodos os processos diarréicos causadospor bactérias irão produzir leucócitos nosespécimes fecais. Para a realização do exa-me, coloca-se uma pequena quantidadedo material fecal em uma lâmina de vidrolimpa e desengordurada, misturando umagota de água destilada estéril (ou salina) euma gota de azul de metileno sob lamínu-
la. Observar ao microscópio com aumentode 400x (13).
Isolamento
Grande parte dos métodos de identifica-ção no laboratório ainda é demorada e re-quer uma bateria de meios de cultura, apesardo constante progresso no desenvolvimentode novos métodos rápidos de diagnóstico.Assim, o período decorrido entre a entradado material no laboratório de Microbiologiae a identificação e antibiograma dos agentesisolados pode demorar vários dias para umacompleta análise (13).
A qualidade dos insumos utilizadospor muitos fabricantes é questionável emuitas vezes ruim, levando a um alto graude inconsistência de resultados lote a lotede um mesmo meio de cultura (24). Portan-to, deve-se dar preferência para meios decultura de fabricantes idôneos, seja em póou em placas/tubos prontos para uso. Des-confie de meios muito baratos, oferecidoscomumente no mercado. Geralmente sãode baixa qualidade e colocarão em cheque
a qualidade dos serviços microbiológicosofertados pelo seu laboratório.
A metodologia tradicional para a detec-ção de patógenos entéricos invariavelmenteemprega a combinação de meios de cultura,geralmente um meio seletivo em placa eum caldo de pré-enriquecimento. A neces-sidade de mais de uma placa é defendidapor alguns autores, motivados pelo conhe-cimento de que se um meio é altamente
Quadro 4. Classificação das espécies e subespécies de Salmonella
Salmonella enterica subespécie enterica (I): inclui a maioria dos sorotiposS. enterica subespécie salamae (II)S. enterica subespécie arizonae (IIIa)S. enterica subespécie diarizonae (IIIb)S. enterica subespécie houtenate (IV)S. enterica subespécie indica
Salmonella bongori (antigamente denominada subespécie V)
Quadro 5. Leucócitos PMN em infecções intestinaisPresentes Variáveis Ausentes
Shigella,Campylobacter ,E.coli invasora (EIEC)
Salmonella, Yersinia,Vibrio parahaemolyticus,Clostridium difficile
Vibrio cholerae, E.coli toxigênica(ETEC), E.coli enteropatogênica
(EPEC), Staphylococcus aureus [toxina], Bacillus cereus [toxina]
Figura 1. Gram: Diarréia muco-sanguinolentapor Shigella (numerosas hemácias, leucócitosPMN e acentuada presença de bacilos Gram-negativos; microbiota intestinal normal
praticamente inexistente).

7/22/2019 Coprocultura Protocolos Parte A
http://slidepdf.com/reader/full/coprocultura-protocolos-parte-a 4/10
NewsLab - edição 86 - 2008 61
inibitório para alguns membros da famíliaEnterobacteriaceae (isto é, os coliformes damicrobiota normal intestinal), poderá haveruma concomitante perda de sensibilidade
na detecção de patógenos verdadeiros, taiscomo a Shigella. Os meios de cultura para oisolamento de patógenos entéricos utilizadosatualmente ou são muito inibitórios para Shi-
gella ou não suficientemente inibitórios paraa microbiota intestinal normal. Além disso, adiferenciação das colônias não é tão boa osuficiente para evitar o subcultivo de muitosnão patógenos os quais não fermentam a lac-tose, tornando-se um trabalho que demandatempo e maior custo, especialmente emlaboratórios que analisam muitos espécimes
diariamente (10, 13).Os meios de cultura em placa maisutilizados para o isolamento de Salmonella e Shigella em laboratórios clínicos são:
• Eosina-Azul de Metileno (EMB),
MacConkey (baixa seletividade)• Xilose-Lisina-Desoxicolato (XLD),
Salmonella-Shigella (SS), Hektoen Enteric,Citrato-Desoxicolato (média seletividade)(Figuras 2, 3, 4, 5, 6)
• Verde Brilhante, Bismuto-Sulto (alta
seletividade)
No entanto, os meios de alta sele-tividade só servem para o isolamentoseletivo de Salmonella e foram adaptadosde forma incorreta da microbiologia de
alimentos para a microbiologia clínicahumana. Infelizmente, também, muitossão os laboratórios clínicos brasileiros queutilizam de forma completamente equi-vocada combinações de meios de culturaque nada acrescentam para a realizaçãode uma coprocultura de boa qualidade.Exemplo disso é a utilização de meioscomo ágar manitol hipertônico, ágar CLED,ágar sangue, dentre outros.
O surgimento de meios cromogênicose fluorogênicos tem permitido avanços
significativos na formulação dos meiosde cultura diferenciais, os quais detectamcaracteres fenotípicos bacterianos, mani-festos pela ação de enzimas característicasde certos grupos taxonômicos, orientandoa sua identificação presuntiva. Estes meiosincluem em sua composição compostoscromogênicos e/ou fluorogênicos, habitu-almente incolores, os quais servem comosubstrato de enzimas específicas. Quandoa enzima atua sobre o substrato, este sofreuma mudança estrutural, formando um
novo composto colorido (cromóforo) oufluorescente (12). Nos últimos anos sur-giram diversos meios cromogênicos úteispara o isolamento e identificação presunti-
va de Salmonella spp. (CHROMagar, SMID,Rambach, ABC, etc.); no entanto, até omomento, nenhum meio cromogênicofoi desenvolvido para o isolamento e aidentificação presuntiva de Shigella. O ágarRambach (Figura 7) foi o primeiro meiocromogênico disponível comercialmente,seguido pelo SMID.
Muitos estudos demonstraram altosíndices de sensibilidade e especificidade,mas as investigações foram baseadas es-sencialmente no uso de culturas puras (es-
toque) de Salmonella. Estudos posterioresindicaram que a presença da microbiotaintestinal normal, não inibida previamentepor caldos de pré-enriquecimento, atrapa-lhava sobremaneira a performance destesmeios (3). O ágar Rambach permite aidentificação de salmonelas não-tifóides,gerando colônias de cor vermelha no meio.Os coliformes aparecem como colôniasazuis, verdes, violetas ou incolores. Umavez que as características bioquímicas usa-das neste meio são altamente específicas
Figura 5. Ágar Hektoen: Aspecto colonial deSalmonella spp. (ausência de fermentação doscarboidratos do meio e produção de H2S).
Figura 2. Ágar Hektoen: Aspecto colonial deShigella sonnei .
Figura 6. Ágar SS: Aspecto colonial deSalmonella spp. (lactose-negativa e produçãode H2S).
Figura 3. Ágar SS: Aspecto colonial deShigella sonnei .
Figura 7. Ágar Rambach: Aspecto colonialde Salmonella não-tifóide (fermentação dopropilenoglicol).
Figura 4. Ágar XLD: Aspecto colonial deShigella sonnei .

7/22/2019 Coprocultura Protocolos Parte A
http://slidepdf.com/reader/full/coprocultura-protocolos-parte-a 5/10
NewsLab - edição 86 - 2008 62
Protocolos de Microbiologia Clínica
para salmonelas não-tifóides (fermentaçãodo propilenoglicol e ausência de beta-galactosidase), somente poucos resultadosfalsos-positivos são encontrados.
O maior problema deste meio é a in-capacidade de isolamento de Salmonella
enterica sorotipo Typhi, pois estas nãofermentam o propilenoglicol (17). No meioSMID, as colônias de Salmonella são tam-bém distintas das demais pela formaçãode coloração vermelha intensa (devido àformação de ácido a partir do glicuronato eausência de beta-galactosidase), enquantooutras bactérias acompanhantes aparecemcomo colônias azuis, violetas ou incolo-res. Este meio possibilita o isolamento e a
identificação presuntiva de todos os soro-grupos de Salmonella. Já o ágar ABC (ágaralfa-beta-cromogênico) explora o fato dassalmonelas poderem ser diferenciadas dosmembros da família Enterobacteriaceae pela presença da enzima alfa-galactosi-dase, na ausência da atividade da enzimabeta-galactosidase (10).
Um total de 1.022 cepas de Salmonella spp. e 300 outros bacilos Gram-negativosforam inoculados neste meio. Destas,99,7% das cepas de Salmonella produzi-ram colônias com uma coloração verdecaracterística, ao passo que somente umacepa de Escherichia coli atípica (0,33%)também produziu colônias verdes (100%de sensibilidade e 90,5% de especifi-cidade) (10). Atualmente, outros meioscromogênicos para Salmonella já estãodisponíveis comercialmente e quase todoseles (com exceção dos meios Rambach eABC) detectam a atividade de esterase (Fi-gura 8), positiva para todas as salmonelas.
Porém, estes meios possuem custo elevadoe são específicos somente para Salmonella.Outros meios em ágar, invariavelmente,devem ser usados em conjunto para o iso-
lamento de outros enteropatógenos, one-rando muito o custo final da coprocultura.Estes meios obtiveram excelente aceitaçãoem laboratórios de controle de qualidade,os quais pesquisam somente Salmonella (em amostras de águas e alimentos).
Quando qualquer meio de culturadesidrata, o potencial de oxi-redução (eH)é alterado, as concentrações dos agentesinibidores são aumentadas e o efeitomais visível disso é o pouco ou nenhumcrescimento de colônias. Nos laboratórios
clínicos, onde é freqüente a prática de seconfeccionar meios de cultura para umaou mais semanas de trabalho, este efeitopode ser minimizado embalando as placasem sacolas plásticas ou, preferencialmen-te, envoltas por plástico-filme de PVC (omesmo utilizado em cozinha). Mesmoum tempo razoavelmente curto de duassemanas sob refrigeração é suficiente parapromover efeitos deletérios nos meios emplaca sem proteção, proporcionando resul-tados aberrantes nas análises (13).
Isenberg et al . (6) estudaram váriosmeios de cultura seletivos (ágares SS, XLD,
Hektoen) na tentativa de avaliar o grau desupressão da microbiota intestinal normalem espécimes fecais, permitindo o cresci-mento dos reais patógenos significativos.Salmonella e Shigella, isolados de espéci-mes clínicos humanos, foram misturadoscom Escherichia, Klebsiella, Enterobacter ,Serratia e Proteus. Foram feitas diferentesdiluições destas misturas e então seme-adas nas placas de ágar. Os resultadosdemonstraram um maior grau de inibição
de Shigella utilizando o ágar SS, ao passoque os ágares XLD e Hektoen permitiram
o isolamento normal desta bactéria (bemcomo de Salmonella), suprimindo conside-ravelmente a microbiota intestinal normalacompanhante.
Vários estudos (5, 25, 26) comprova-ram a inibição de Shigella por microrga-nismos pertencentes à microbiota intestinalnormal. Os primeiros estudos mostraram
que Klebsiella / Enterobacter inibia o cres-cimento de Shigella em culturas mistas,indicando que uma possível competiçãopor fontes de carboidratos fermentáveis
seria a causa principal. Outros estudoscomprovaram parcialmente esta teoria,uma vez que também foram relatadas ini-bições do crescimento de Shigella quandoem presença de Klebsiella / Enterobacter ,mesmo em meios aerados e com excessode glicose. No entanto, estes estudos tam-bém provaram que a produção de ácidosacético e fórmico por estas bactérias (etambém por E. coli ) fazia com que asShigella entrasse em fase logarítmica demorte, comprovando que a produção de
ácidos voláteis com a concomitante redu-ção acentuada do meio eram os principaismecanismos de inibição do crescimento deShigella em culturas mistas.
Os caldos de enriquecimento seletivosão utilizados para diminuir a quantidadede bactérias da microbiota intestinal nor-mal no espécime fecal, inibindo de algumaforma seu metabolismo e propiciando odesenvolvimento franco dos verdadeirosenteropatógenos. Estes caldos devem sersempre utilizados como uma etapa prévia,uma vez que a semeadura direta das fezesem meios sólidos geralmente leva a umsupercrescimento da microbiota normal,em detrimento às poucas colônias quepodem ser formadas pelos verdadeirospatógenos (13).
Deve ser observado que cada caldopossui suas próprias características e seuuso deve ser criterioso, seguindo todas asinformações dadas pelos fabricantes parauma melhor performance. No entanto, emmuitos laboratórios clínicos estes caldossão usados de forma equivocada, muitas
vezes sem o real conhecimento científicode sua utilidade. Muitos são os caldos parao pré-enriquecimento seletivo utilizados emmicrobiologia: tetrationato segundo Muller-Kauffmann, Rappaport-Vassiliadis, selenitosegundo Leifson, selenito-cistina e GNsegundo Hajna. No entanto, com exceçãodos dois últimos, estes meios foram adap-tados, incorretamente, da microbiologiade alimentos para a microbiologia clínica
Figura 8. Ágar cromogênico: Aspecto colonialde Salmonella spp. (utilização do substratocromogênico através da enzima esterase)
7/22/2019 Coprocultura Protocolos Parte A
http://slidepdf.com/reader/full/coprocultura-protocolos-parte-a 6/10
NewsLab - edição 86 - 2008 63
humana, pois são utilizados somente parao isolamento de Salmonella. Eles possuemdiversas substâncias tóxicas para as outrasbactérias (especialmente E. coli e Shigella)
e, portanto, não devem ser utilizados emespécimes fecais (9, 11, 14, 15).Mesmo muito utilizado em micro-
biologia clínica, o caldo selenito-cistinatambém é bastante inibidor para o de-senvolvimento de Shigella, especialmentepelo contato prolongado das células como selenito de sódio (este caldo deve serincubado por 24 horas antes do repiqueem meios sólidos). Já o caldo GN (Gram-Negative), idealizado por Hajna em 1955(4), continua sendo o melhor meio para o
pré-enriquecimento seletivo tanto para oisolamento de Salmonella quanto de Shi-
gella, a partir de espécimes fecais humanos(Figura-9). O meio deve ser inoculado
com pequena porção de fezes, incubadopor até 8 horas em estufa bacteriológica erepicado para meios em placa (o tempoideal para a incubação é de 6 a 8 horas).O citrato e o desoxicolato de sódio pre-sentes em sua formulação impedem o rá-pido crescimento da microbiota intestinalnormal, possibilitando a multiplicaçãodos verdadeiros enteropatógenos. Noentanto, após 8 horas de incubação, estacapacidade inibitória começa a se perdere as bactérias intestinais acompanhantes
começam a se multiplicar normalmente.Portanto, nunca devemos utilizar o cal-do GN após 8 horas de incubação pois,dessa forma, é praticamente como seestivéssemos semeando o espécime fecaldiretamente nas placas (10, 13).
Taylor e Schelhart (21) compararam cal-dos de pré-enriquecimento e três meios emplaca durante a análise de 1.405 espécimesfecais com o intuito de escolherem a melhorcombinação caldo/placa para o isolamentode Shigella. Os caldos GN e selenito foram
semeados em ágar EMB, SS e XLD. Com
o uso dos caldos de pré-enriquecimento,obteve-se o dobro de isolamentos de Sal-
monella e Shigella em comparação com a
semeadura direta em placas das amostrasfecais. Todos os três caldos obtiveramresultados ótimos no pré-enriquecimentoseletivo de Salmonella; no entanto, o caldoGN possibilitou o isolamento de quase trêsvezes mais Shigella do que o caldo selenito(Tabela 1). A comparação entre os meios emplaca mostrou que o ágar XLD foi muito
mais eficiente que os ágares SS e EMB parao isolamento de ambos os gêneros bacteria-nos (Tabela 2), indicando que a combinaçãoGN/XLD foi a mais eciente.
Quatro meios de cultura em placa(MacConkey, citrato-desoxicolato, XLD e
verde brilhante) foram inoculados direta-mente com amostras fecais e também apóso pré-enriquecimento seletivo usando oscaldos selenito, GN e tetrationato, emum estudo conduzido por Taylor e Sche-lhart (22) com 1.117 espécimes fecais natentativa de isolamento de Salmonella eShigella. O ágar XLD foi claramente supe-rior na detecção destas bactérias (Tabela3) e o uso do caldo GN mostrou-se de
Figura 9. Caldo GN Hajna: 1) meio inoculadocom espécime fecal; 2) meio não inoculado
Tabela 1. Performance dos caldos de pré-enriquecimento no isolamento de Salmonella e Shigella
Bactéria Semeadura direta Caldo GN* Caldo Selenito*
Salmonella 160 332 260
Shigella 57 118 38
Total 217 450 298
* pré-enriquecimento com posterior semeadura em placa
Tabela 2. Performance dos meios em placa no isolamento de Salmonella e Shigella
Bactéria EMB XLD SS
Salmonella 235 (47%) 497 (99%) 343 (68%)
Shigella 103 (73%) 137 (96%) 75 (53%)Total 338 (52%) 634 (98%) 418 (65%)
Tabela 3. Total de amostras positivas para Salmonella e Shigella usando caldos de pré-enriquecimento seletivo
Bactéria Semeadura direta Caldo GN* Caldo Selenito* Caldo Tetrationato*
Salmonella 157 250 257 164
Shigella 53 77 66 27
Total 210 327 323 191
* pré-enriquecimento com posterior semeadura em placa

7/22/2019 Coprocultura Protocolos Parte A
http://slidepdf.com/reader/full/coprocultura-protocolos-parte-a 7/10
NewsLab - edição 86 - 2008 64
Protocolos de Microbiologia Clínica
fundamental importância para a detecçãode ambos os patógenos, especialmente
Shigella (Tabela 4).Vassiliadis et al. (24) reportaram que
105 Shigella foram isoladas em ágar citrato-desoxicolato quando semeadas diretamen-te mas somente 16 foram recuperadas apóso uso do pré-enriquecimento com caldoselenito, das quais 13 de 25 foram S. sonnei e somente três de 78 foram S. flexneri .
Dois caldos de enriquecimento e quatromeios em placa foram avaliados por Taylor eSchelhart (23), comparando a eficiência dedetecção de Salmonella e Shigella a partir
de 1.597 espécimes fecais. Todas as 17Shigella foram isoladas no ágar XLD após
o enriquecimento prévio em caldo GN.A semeadura direta das fezes nos quatromeios em placa permitiu o isolamento desomente 92 das 170 Salmonella (54%); damesma forma, somente 10 das 17 Shigella (59%) foram recuperadas. Os caldos depré-enriquecimento se mostraram efetivospara a recuperação de todos os patógenospesquisados, sendo que o caldo GN propor-cionou o isolamento de 87% das Salmonella e 100% das Shigella e o caldo selenito 97 e
76%, respectivamente.Foi observado que o ágar XLD produ-
ziu mais resultados positivos para ambosos patógenos em comparação com osoutros três meios. O ágar XLD recuperou
3%, 12% e 99% mais Salmonella que oságares Hektoen, SS e EMB, respectiva-mente. Para Shigella, o ágar XLD produziu
13%, 33% e 44% mais isolamentos queos ágares Hektoen, SS e EMB, respectiva-
mente. Neste mesmo estudo, os autoresobservaram que o número de resultados
falsos-positivos na observação preliminardas placas (colônias H2S+ que não foramidentificadas como Salmonella) foi maiornos meios SS e Hektoen (Tabela 5).
A vantagem do ágar XLD sobre os ou-tros meios seletivos avaliados neste estudo(SS e Hektoen) provavelmente seja umsistema de diferenciação de colônias commaior poder de discriminação (a xilose,presente no XLD, é fermentada por muito
mais membros da família Enterobacteriace-
ae do que a salicina, presente no Hektoen).
Portanto, todas as colônias salicina(-) de nãofermentadores ou fermentadores tardios delactose/sacarose são uma fonte de possíveisresultados falsos-positivos no ágar Hektoen.No entanto, a maioria destes microrga-nismos fermenta rapidamente a xilose,possibilitando uma maior discriminação noágar XLD. Já o ágar SS, não possuindo nem
xilose nem salicina como características di-ferenciais (somente lactose), é dependentequase que exclusivamente do maior efeitoinibitório de sua formulação.
King e Metzger (7, 8) desenvolveram o
meio de cultura Hektoen Enteric em 1968como um modo de melhorar a performan-ce de isolamento de patógenos intestinais,obtendo bom crescimento de Shigella eSalmonella através da utilização de proteose-peptona e a adição da salicina como umterceiro carboidrato fermentável, além demelhorar o sistema indicador da produçãode H
2S. Durante o estudo, comparando aperformance do recém-criado meio Hektoen
Enteric com os ágares SS e EMB, somente Sal-
monella e Shigella foram pesquisados a partirda semeadura de 2.855 espécimes fecais.Os autores relataram que o meio Hektoenpossibilitou mais isolamentos de Shigella queos demais. Do total de 98 Shigella isoladasde todos os três meios, 97 foram isoladas emHektoen e somente 40 no ágar SS.
Citrobacter freundii , bacilo Gram-negativo pertencente à família Enterobac-
teriaceae, membro da microbiota intestinalnormal da grande maioria das pessoassadias, lactose(-) e produtor de H2S, nãotem o seu crescimento inibido nos ágares
EMB, MacConkey, Hektoen e XLD, mas oágar SS mostra-se bastante inibidor paraesta espécie. Conseqüentemente, C. freun-
dii está entre as causas menos freqüentesde resultados falsos-positivos no ágar SS.No ágar XLD esta espécie geralmente não
causa resultados falsos-positivos pois,devido ao sistema diferencial daquelemeio, as colônias de C. freundii tornam-seamarelas (Figura-10), semelhantes àquelas
Tabela 4. Total de amostras positivas para Salmonella e Shigella utilizando os meios em placa
Bactéria MC XLD CD VB
Salmonella 178 (55%) 305 (94%) 115 (35%) 230 (71%)
Shigella 66 (75%) 78 (89%) 24 (27%) 55 (63%)Total 244 (59%) 383 (93%) 139 (34%) 285 (69%)
MC: MacConkey; XLD: Xilose-lisina-desoxicolato; CD: Citrato-desoxicolato; VB: Verde brilhante.
Tabela 5. Distribuição dos resultados falsos-positivos (H2S+ não Salmonella) nos meios em placa
Bactéria XLD SS Hektoen
Citrobacter freundii 27 (6,7%) 82 (9,5%) 361 (36,5%)
Proteus vulgaris/mirabilis 122 (30,1%) 359 (41,6%) 244 (24,7%)
Total 149 (36,8%) 441 (51,1%) 605 (61,2%)
Figura 10. Ágar XLD: Aspectos coloniais deCitrobacter freundii (Cf), Proteus mirabilis
(Pm) e Escherichia coli (Ec)

7/22/2019 Coprocultura Protocolos Parte A
http://slidepdf.com/reader/full/coprocultura-protocolos-parte-a 8/10
NewsLab - edição 86 - 2008 65
Figura 11. Ágar Hektoen: Aspecto colonial deCitrobacter freundii (ausência de fermentaçãodos carboidratos do meio e produção de H2S).
dos coliformes da microbiota intestinalnormal, mesmo as cepas fermentadorastardias de lactose são xilose(+) e lisina(-).O ágar Hektoen, entretanto, não possui
nenhum sistema inibidor nem diferencialpara distinguir C. freundii de Salmonella,uma vez que ambos podem ser lactose/ sacarose/salicina(-) e H2S(+). Portanto, C.
freundii também é a causa mais freqüentede resultados falsos-positivos quando seusa este meio de cultura em amostras fecais(13) (Figura 11).
Os efeitos de temperaturas de incu-bação diferentes (20, 35 e 40 oC), longosperíodos de transporte, caldos de pré-enriquecimento e meios em placa foram
determinados por análises exaustivas,pesquisando Salmonella eShigella, em 132amostras fecais (20). Estes espécimes foramsemeados diretamente em ágares SS, XLD e
EMB, meios de transporte Cary-Blair (CB) esalina e caldos de pré-enriquecimento GNe selenito. Os melhores resultados dos cal-dos na recuperação de Salmonella foramGN > selenito > salina > CB > semeaduradireta. Para Shigella, GN > salina > seme-adura direta > CB > selenito, provandoque o caldo selenito não é adequado parao isolamento de Shigella. A eficiência dosmeios em placa, tanto para Salmonella quanto para Shigella, foi: XLD > EMB > SS.
Além disso, 10 das 39 Shigella não foramisoladas a partir da combinação selenito/ SS. As temperaturas de incubação nãoafetaram as taxas de recuperação de Sal-
monella; no entanto, somente metade dasShigella foi isolada a 40oC e os isolamentosa 20 e 35oC foram equivalentes. A análisedas placas com crescimento de Salmonella
e Shigella revelou que a semeadura diretadas amostras fecais deve ser desencoraja-da, uma vez que o supercrescimento damicrobiota intestinal normal atrapalha o
desenvolvimento daquelas colônias dosverdadeiros patógenos, principalmenteatravés da competição por nutrientes.
Tem sido provado por diferentesautores (2, 10, 13, 14, 16,) que a combi-nação de pré-enriquecimento com caldoselenito e semeadura posterior em ágar SSé excessivamente inibidora para Shigella.Taylor e Schelhart (23) mostraram que 19das 39 cepas de Shigella não cresceram emnenhuma das 78 replicatas por espécimeem placas de ágar SS e 20 não foram iso-
ladas dos 48 subcultivos de caldo selenitocom Shigella. Cinco de seis S. flexneri nãocresceram em ágar SS, quatro de seis nãocresceram em selenito, nenhuma das cincocepas de S. dysenteriae foi detectada nacombinação selenito/SS e somente umacresceu em ágar SS. Desta forma, somen-te 10 das 39 cepas de Shigella puderamser isoladas da combinação selenito/SS,mesmo com 16 replicatas de cada amostrafecal. Os autores concluíram que é alta-mente insatisfatória a combinação destesmeios, embora eles sejam utilizados pormuitos laboratórios clínicos para a reali-zação de coproculturas.
Em um estudo conduzido por Pollocke Dahlgren (16), em um período de 12meses, vários meios em placa (MacConkey,XLD, SS, Hektoen) e caldos de pré-enri-quecimento (selenito, tetrationato) para oisolamento de Salmonella e Shigella foramcomparados, utilizando 455 amostras fe-cais. Destas foram isolados 53 patógenos,dos quais 56% foram S. sonnei e 13%foram S. flexneri . Destes isolados, 90%
foram recuperados em ágar XLD, 87% emágar Hektoen e 80% em ágar MacConkey,mas somente 28% em ágar SS. Menos dametade das Shigella foi isolada a partir dopré-enriquecimento em caldo selenito esomente duas foram isoladas à partir docaldo tetrationato. O ágar XLD mostrou-
se o melhor meio para o isolamento tantode Salmonella quanto de Shigella, seguidopelo ágar Hektoen. Ambos possuem for-
mulações que evitam a combinação deingredientes sabidamente responsáveispela baixa eficiência na recuperação devárias cepas de Shigella, como ocorre no
ágar SS (sais biliares + citrato).Dunn e Martin (2) avaliaram cinco
meios de transporte, oito meios em placae três caldos de enriquecimento seletivopara o isolamento de Salmonella eShigella em oito laboratórios de diferentes regiõesdos Estados Unidos, os quais submeterampara análise 490 espécimes fecais de suasrotinas nos meios de transporte fornecidospelos autores. Os resultados sugerem autilização de mais de um meio de culturapara o isolamento primário aliado ao uso
de um caldo de pré-enriquecimento sele-tivo e a obrigatoriedade de utilização deswabs com meios de transporte nos casosonde as amostras “in natura” não puderemser inoculadas imediatamente. Os autoresconcluíram, também, que o uso de ágarXLD em conjunto com o caldo GN como
pré-enriquecimento seletivo foi a melhorcombinação para o isolamento de Salmo-
nella e Shigella.Embora um número bastante grande
de meios de transporte tenha sido descrito,eles possuem as mesmas funções básicas:manter o status quo da população bacteria-na no espécime clínico, bem como preveniro supercrescimento de uma populaçãobacteriana em particular (como as entero-bactérias da microbiota normal intestinal).Swabs retais podem ser utilizados, desdeque adequada e racionalmente. Deve-se darpreferência para a inoculação das amostras“in natura” mas, nos casos onde a inocu-lação destas amostras no laboratório nãoseja possível (como nos casos de envio deespécimes para laboratórios de referência
localizados em outras cidades), os swabspodem e ser utilizados. No entanto, jamaisutilizar swabs “secos”, ou seja, sem meiosde transporte. Atualmente os swabs comer-ciais possuem preços bastante acessíveis aqualquer laboratório e devem ter preferên-cia. Colocando os custos diretos e indiretosna ponta do lápis, os swabs confeccionadosno próprio laboratório elevam substancial-mente o valor final do teste (13).

7/22/2019 Coprocultura Protocolos Parte A
http://slidepdf.com/reader/full/coprocultura-protocolos-parte-a 9/10
NewsLab - edição 86 - 2008 66
Protocolos de Microbiologia Clínica
Alguns estudos relataram a importânciada utilização de meios conservantes paraespécimes fecais com a finalidade de rea-lização de coproculturas. Stuart (19) foi o
primeiro a publicar um meio quimicamentedefinido para a preservação de células bac-terianas até o momento da inoculação emmeios apropriados no laboratório, inclusivepara amostras destinadas ao cultivo de Sal-
monella e Shigella. Seu estudo concluiu queas evidências sugeriam que os swabs retaispreservados no meio de transporte propostonão possuíam performance inferior aos es-pécimes fecais “in natura” para o isolamen-to de Salmonella eShigella após um ou doisdias de transporte. Posteriormente, Cary e
Blair (1) reportaram o desenvolvimento deum novo meio de transporte para amostrasfecais, indicando que Salmonella eShigella poderiam ser isoladas por até 49 dias, Vibrio
cholerae por até 22 dias e Yersinia pestis por,pelo menos, 75 dias quando armazenadasneste novo meio.
No entanto, estudos mais recentes,comparando a eficiência na manutençãode células viáveis de Shigella em meiosde transporte comumente utilizados emlaboratórios clínicos (Cary-Blair e salinaglicerinada tamponada), questionaram areal utilidade dos swabs com meios detransporte no isolamento de patógenosentéricos mais delicados e que morremrapidamente quando há demora no seuisolamento. Em um estudo (1), 376 es-pécimes fecais de pacientes envolvidosem surtos de diarréia por Shigella foramanalisados utilizando swabs com meio detransporte Cary-Blair e salina glicerinadatamponada. A duração do transporte dasamostras até o laboratório variou de um aseis dias. Os maiores índices de isolamento
foram obtidos em espécimes refrigeradosou congelados, em ambos meios de trans-porte, em comparação com os meios emtemperatura ambiente. Esta diferença foimais evidente na análise dos espécimesdeixados à temperatura ambiente por maisde três dias. Os autores concluíram que asalina glicerinada tamponada é melhorpara o transporte e posterior isolamentode Shigella do que o swab com Cary-Blair,
mas o grau de superioridade é dependenteda temperatura de transporte.
Baseado nas informações anteriorese ainda na nossa experiência prática,
sugerimos a utilização do caldo GN (comincubação a 35-37oC por 6 a 8 horas)
e repique deste para placas de ágaresMacConkey e XLD, para uma ótima recu-peração de Salmonella, Shigella e outrosenteropatógenos bacterianos. No entanto,
a análise destes últimos será tema da Parte2 deste protocolo de coprocultura.
Referências Bibliográficas
1. Cary SG, Blair EB. New transport medium for shipment of clinical specimens. J. Bacteriol.88:96-98, 1964.2. Dunn C; Martin WJ. Comparison of media for isolation of salmonellae and shigellae fromfecal specimens. Appl. Microbiol. 22:17-22, 1971.3. Freydiere AM, Gille Y. Detection of salmonellae by using Rambach agar and by a C8 esterasespot test. J. Clin. Microbiol. 29:2357-2359, 1991.4. Hajna AA. A new enrichment broth medium for gram negative organisms of the intestinalgroup. Publ. Hlth. Lab. 13:83-89, 1955.5. Hentges DJ. Inhibition of Shigella flexneri by the normal intestinal flora. J. Bacteriol. 93:1369-1373, 1967.
6. Isenberg HD et al. Isolation of Salmonella and Shigella from na artificial micture of fecalbacteria. Appl. Microbiol. 18:656-659, 1969.7. King S, Metzger WI. A new plating medium for the isolation of enteric pathogens. I: Hektoenenteric agar. Appl. Microbiol. 16:577-578, 1968.8. King S, Metzger WI. A new plating medium for the isolation of enteric pathogens. II: Comparisonof Hektoen enteric agar with SS and EMB agar. Appl. Microbiol. 16:579-581, 1968.9. Knox R et al. The selective action of tetrathionate in bacteriological media. J. Hyg. 43:147-158,1943.10. Koneman EW et al. Color atlas and textbook of diagnostic microbiology. Baltimore: LippincottWilliams & Wilkins. 6a ed., 2006.11. Leifson E. The effect of sodium selenite on the growth of bacteria and its use as the basisfor a new enrichment medium for the isolation of typhoid bacilli from feces, water, mil, etc. J.Bacteriol. 31:26-27, 1936.12. Manafi M, Sommer R. Comparison of three rapid screening methods for Salmonella spp.:MUCAP test, MicroScreen latex and Rambach agar. Lett. Appl. Microbiol. 14:163-166, 1992.13. Menezes e Silva CHP, Neufeld PM. Bacteriologia e Micologia para o laboratório clínico.Rio de Janeiro: Ed. Revinter, 1a ed., 2006.14. Muller VR, Banwart GJ. Milleu d’enrichment pour la recherche du Bacilie typhique et desParatyphiques. Compt. Rend. Soc. Biol. 89:434, 1923.15. Palumbo AS, Alford JA. Inhibitory action of tetrathionate enrichment broth. Appl. Microbiol.20:970-976, 1970.16. Pollock HM, Dahlgren BJ. Clinical evaluation of enteric media in the primary isolation ofSalmonella and Shigella. Appl. Microbiol. 27:197-201, 1974.17. Rambach A. New plate medium for facillitated differentiation of Salmonella spp. from Proteus spp. and other enteric bacteria. Appl. Environ. Microbiol. 56:301-303, 1990.18. Souza EC et al. Etiologic profile of acute diarrhea in children in São Paulo. J. Pedriatr.78:31-38, 2002.19. Stuart RD. Transport medium for specimens in Public Health bacteriology. Pub. Hlth Rep.74:431-438, 1959.20. Taylor WI, Schelhart D. Effect of temperature on transport and plating media for entericpathogens. J. Clin. Microbiol. 2:281-286, 1975.
21. Taylor WI, Schelhart D. Isolation of Shigellae - V. Comparison of enrichment broths withstools. Appl. Microbiol. 16:1383-1386, 1968.22. Taylor WI, Schelhart D. Isolation of Shigellae - VI. Performance of media with stool specimens.Appl. Microbiol. 16:1387-1393, 1968.23. Taylor WI, Schelhart D. Isolation of Shigellae - VII. Comparison of xylose lysine deoxycholateagar, Hektoen enteric agar, Salmonella-Shigella agar, and Eosin methylene blue agar with stoolspecimens. Appl. Microbiol. 21:32-37, 1971.24. Vassiliadis P, Pateraki E, Politi G. Compertement des Shigella dans le milieu d’enrichissementau selenite. Bull. Soc. Pathol. Exotique 59:31-42, 1966.25. Wynne ES. Antagosnism by Aerobacter strains. J. Bacteriol. 44:209-219, 1947.26. Wynne ES; Norman, JO. On the concept of “direct antagonism” in bacteria. J. Infect. Dis.93:243-246, 1953.

7/22/2019 Coprocultura Protocolos Parte A
http://slidepdf.com/reader/full/coprocultura-protocolos-parte-a 10/10
NewsLab - edição 86 - 2008 67





























