Embed Size (px)
Citation preview

CCRROOSSSS--TTAALLKK NNAA SSIINNAALLIIZZAAÇÇÃÃOO EENNTTRREE RREECCEEPPTTOORREESS TTOOLLLL--LLIIKKEE EE RREECCEEPPTTOORREESS DDEE MMAANNOOSSEE,, NNAA AATTIIVVAAÇÇÃÃOO DDOOSS MMAACCRRÓÓFFAAGGOOSS
PPOORR MMIICCOOBBAACCTTÉÉRRIIAA
MARCELLE RIBEIRO MANHÃES DE ANDRADE
UNIVERSIDADE ESTADUAL DO NORTE FLUMINENSE DARCY RIBEIRO CAMPOS DOS GOYTACAZES – RJ
AGOSTO 2007

CCRROOSSSS--TTAALLKK NNAA SSIINNAALLIIZZAAÇÇÃÃOO EENNTTRREE RREECCEEPPTTOORREESS TTOOLLLL--LLIIKKEE EE RREECCEEPPTTOORREESS DDEE MMAANNOOSSEE,, NNAA AATTIIVVAAÇÇÃÃOO DDOOSS MMAACCRRÓÓFFAAGGOOSS
PPOORR MMIICCOOBBAACCTTÉÉRRIIAA
MARCELLE RIBEIRO MANHÃES DE ANDRADE
Dissertação de Mestrado apresentada ao Centro de Biociências e Biotecnologia da Universidade Estadual do Norte Fluminense Darcy Ribeiro como parte das exigências para obtenção do Título de Mestre em Biociências e Biotecnologia, com ênfase em Biologia Celular.
CAMPOS DOS GOYTACAZES – RJ AGOSTO 200

CROSS-TALK NA SINALIZAÇÃO ENTRE RECEPTORES TOLL-LIKE E RECEPTORES DE MANOSE, NA ATIVAÇÃO DOS MACRÓFAGOS
POR MICOBACTÉRIA
MARCELLE RIBEIRO MANHÃES DE ANDRADE
Dissertação de Mestrado apresentada ao Centro de Biociências e Biotecnologia da Universidade Estadual do Norte Fluminense Darcy Ribeiro como parte das exigências para obtenção do Título de Mestre em Biociências e Biotecnologia, com ênfase em Biologia Celular.
Aprovada em 03 de agosto de 2007
Comissão Examinadora:
___________________________________________________ Martha Maria Oliveira (Dra em Ciências de Clínicas Médicas) – UFRJ
_____________________________________ Milton M. Kanashiro (Dr em Imunologia) – UENF
_____________________________________ Wilmar Dias da Silva (Dr em Imunologia) – UENF
________ __ Elena B. Lassounskaia (Dra em Imunologia) – UENF
(orientador)

AGRADECIMENTOS...
Concedei-me senhor, a serenidade necessária para aceitar as coisas que não posso modificar...
coragem para modificar às que posso... e sabedoria para distiguir umas das outras... Palavras sábias, mas
nem sempre tão fáceis de serem seguidas... Errar tentando acertar... querer e muitas vezes não conseguir...
conseguir e não ser suficiente! Aprendi que todo esforço é válido e que tentar é o começo de tudo, mesmo
que não alcancemos a vitória. Agradeço a DEUS pela serenidade necessária em determinadas ocasiões,
pela coragem para enfrentar as dificuldades e pelas oportunidades de acertos e erros... À minha mãe
Sandra, ao meu pai José Afonso e meu irmão Fábio agradeço por todo apoio, carinho e incentivo que me
fortalecem a cada dia! Agradeço à minha avó Déa pelo amor e toda dedicação... Ao meu namorado
Bruno, agradeço pelo carinho, companheirismo e compreensão nas horas em que mais necessito... Quero
agradecer às minhas amigas Aline, Mariana, Simone, Verônica, Lígia e Marina pelo carinho... pelo apoio
nos momentos difíceis e também pelos belos momentos de diversão!! Afinal, nem só de pão vive o homem...
Agradeço a todos os meus companheiros do LBR pelo excelente convívio que tornaram meu dia-a-dia
muito mais prazeroso. À minha querida orientadora Elena dedico agradecimentos especiais... Muito
abrigada pela paciência, pelos ensinamentos que me proporciona, pelo incentivo nas horas em que acho
que não vai dar nada certo! Sem sua ajuda não conseguiria chegar até aqui! Agradeço aos professores
Wilmar, Milton, Martha e Andréa por terem aceitado meu convite e pela disposição de avaliar meu
trabalho... Quero muito agradecer a professora Thereza pela pronta disposição em revisar minha tese,
apesar de estar passando por momentos delicados em sua vida! Agradeço aos técnicos do LBR pelo
suporte aos meus experimentos... sem vocês nada disso seria possível! Enfim, agradeço a todos que de
certa forma contribuíram para minha vitória. Tenho sorte por fazerem parte da minha vida ...

SUMÁRIO
ÍNDICE DE FIGURAS E TABELAS.....................................................................
ABREVIATURAS..................................................................................................
RESUMO .............................................................................................................
iv
vi
viii
ABSTRACT ......................................................................................................... ix
I. INTRODUÇÃO.................................................................................................
1
1.1. Incidência da tuberculose no mundo...................................................... 1
1.2. O gênero Mycobacterium ...................................................................... 2
1.3. Tuberculose- A doença .......................................................................... 4
1.4.Tratamento daTuberculose .................................................................... 4
1.5. Transmissão da Tuberculose.................................................................. 6
1.6. Resposta imune contra M. tuberculosis................................................. 8
1.6.1 Mecanismos de modulação da resposta imune mediados por
M.tuberculosis............................................................................................
13
1.7. Receptores dos macrófagos ................................................................... 14
1.7.1. Receptores de fagocitose........................................................ 15
1.7.1.1. Fagocitose................................................................... 17
1.7.2.Principais vias de sinalização acionadas pelas micobactérias... 18
1.7.2.1. Receptores Toll-Like (TLRs)........................................ 18
1.7.2.2. Ativação de NF-κB mediada por TLR 2....................... 19
1.7.2.3. Ativação de MAPK por micobactéria........................... 20
1.7.2.4. Ativação de PI3-K/Akt................................................. 22
1.7.2.5. Ativação de PI3-K/Akt mediada por TLR 2.................. 23
1.8. Receptores de Manose.......................................................................... 23
1.8.1. Sinalização celular mediada por MR.............................. 25
1.9. Fenótipo migratório dos macrófagos..................................................... 27
1.9.1. Participação de TLR2 e PI3K nas alterações morfológicas
sofridas pelos macrófagos..........................................................................
28

II. OBJETIVOS.....................................................................................................
31
3.1. Objetivos específicos.................................................................... 31
III. JUSTIFICATIVA..............................................................................................
32
IV. METODOLOGIA.............................................................................................. 33
4.1. Lisados e componentes micobacterianos ............................................... 33
4.2. Cultura Celular ............................................................................................ 33
4.2.1. Extração de células precursoras de medula óssea de
camundongos..................................................................................
34
4.3. Tratamento dos macrófagos................................................................... 34
4.4. Preparação dos lisados Celulares......................................................... 35
4.5. Imunoprecipitação .................................................................................. 35
4.6. Anticorpos ............................................................................................. 36
4.7. SDS PAGE 10%..................................................................................... 36
4.7.1. Western Blotting........................................................……….. 37
4.7.2. Imunodetecção de quinases e fosfatases.............................. 37
4.8. Quantificação das citocinas..................................................................... 37
4.8.1. ELISA..................................................................................... 37
4.8.2. Bioensaio de L929.................................................................. 38
4.8.2.1. Teste de MTT........................................................... 39
4.8.3. Quantificação de óxido nítrico (NO) pela reação de Griess... 39
4.9. Imunofuorescência................................................................................... 39
4.9.1. Ensaio para F-actina.............................................................. 40
4.10. Análises estatísticas............................................................................... 40
V- RESULTADOS................................................................................................. 41
5.1. Efeito de Man-LAM na produção dos mediadores pró-inflamatórios
pelos macrófagos ativados pelas micobactérias...................................................
43

5.1.1. A inibição de IL12p40 induzida por Man-LAM e
independente de IL-10.....................................................................
44
5.2. Efeito modulador de Man-LAM nas vias de sinalização pró-
inflamatórias acionadas pelas micobactérias......................................................
45
5.2.1. O efeito de Man-LAM na ativação de NF-κB induzida por
BCG .................................................................................................
46
5.3. A ativação de PI3-K pode ser mediada por TLR 2................................... 47
5.3.1. O efeito de Man-LAM na ativação da via PI3-K/Akt por
BCG..................................................................................................
48
5.4. Efeito de Man-LAM na ativação das MAPK por BCG............................... 49
5.5. Efeito de Man-LAM na ativação da fosfatase SHP-1 ............................... 50
5.6. Mycobacterium bovis BCG estimula a formação de filapódios,
espalhamento e polarização celular em macrófagos RAW 264.7.........................
51
5.6.1. O rearranjo do citoesqueleto induzido por BCG é mediado
por TLR 2.........................................................................................
52
5.7. Efeito de Man-LAM na regulação dos rearranjos citoesqueléticos dos
macrófagos induzido por BCG..............................................................................
54
5.8. Análise da importância de PI3-K na regulação dos rearranjos no
citoesqueleto dos macrófagos, induzido por BCG................................................
55
VI- DISCUSSÃO....................................................................................................
58
VII- CONCLUSÕES...............................................................................................
65
VIII- REFERÊNCIAS BIBLIOGRÁFICAS.............................................................
67

ÍNDICE DE FIGURAS E TABELAS
Figura 1- Incidência da tuberculose no mundo em 2000........................................ 1
Figura 2- Representação esquemática de PAMPs em diferentes bactérias.............. 3
Figura 3- Transmissão da tuberculose em humanos............................................. 6
Figura 4- Resposta imune contra M. tuberculosis.................................................. 12
Figura 5- Receptores dos macrófagos.................................................................. 15
Figura 6- Ativação de TLR 2 por M. tuberculosis................................................. 20
Figura 7- Representação esquemática das moléculas implicadas na ativação de
MAPK, seguido ao estímulo de LPS......................................................................
21
Figura 8- Estrutura do Receptor de Manose ........................................................ 24
Figura 9- Estrutura das três famílias representativas de moléculas LAM isoladas
de diferentes espécies micobacterianas................................................................
25
Figura 10 A- Produção de IL-12 por macrófagos RAW 264.7 ativados................ 42
Figura 10 B- Produção de TNF-α por macrófagos RAW 264.7 ativados............. 42
Figura 10 C- Produção de NO por macrófagos RAW 264.7 ativados.................. 43
Figura 11- Produção de IL-10 por macrófagos RAW 264.7 ativados................... 44
Figura 12- Verificação da ativação de IκB-α......................................................... 45
Figura 13- Imunoprecipitação de TLR 2................................................................. 46
Figura 14- Inibição da ativação de PI-3K (p85) mediada por Man-LAM.............. 47
Figura 15 A- Avaliação da ativação de Akt após o pré-tratamento dos
macrófagos RAW 264.7 com Man-LAM Mtb e sua posterior ativação com BCG..
48
Figura 15 B- Avaliação da ativação de Akt, nos macrófagos de medula óssea,
após o tratamento destes com Man-LAM de Mtb, LAM de Mav ou BCG WCL......
49
Figura 16- Ativação das MAPKs por BCG e inibição da ativação destas por
Man-LAM...............................................................................................................
50
Figura 17- Aumento da expressão de SHP-1 induzido por Man-LAM.................. 51
Figura 18- Indução dos pseudopódios nos macrófagos ativados com BCG.......... 52
Figura 19- Indução do espalhamento dos macrófagos derivados de medula
óssea (BMM) pela lipoproteína micobactreriana p19.............................................
53

Figura 20- Características morfológicas dos macrófagos tratados com Man-
LAM.......................................................................................................................
54
Figura 21- O inibidor de PI3-K reduziu o teor de F-actina e protusões celulares
mas não o enrugamento da membrana dos macrófagos RAW 264.7.
estimulados por BCG..............................................................................................
55
Figura 22- Ativação de Rac-1 nos macrófagos estimulados com BCG................ 57
Figura 23- Ativação de Rac1 pelos componentes da parede celular de M.
tuberculosis.............................................................................................................
57
Figura 24- Mecanismo de modulação da resposta pró-inflamatória induzida por
M. tuberculosis, mediado por IL-10 induzido por Man-LAM...................................
59
Figura 25- Participação de TLR2 na ativação de PI3-K, mediado por M.
tuberculosis.............................................................................................................
64
Tabela 1- Fatores de transcrição envolvidos na transcrição de genes pró-
inflamatórios............................................................................................................
60

ABREVIATURAS
AIDS – Síndrome da Imunodeficiência Adquirida
BCG- Bacilo de Calmette Guerrin
CD40 – Cluster of Differentiation 40
CR- região extracelular do receptor de manose que contém um domínio rico em
cisteína.
CR1- Receptor para o complemento tipo 1
CTLs – células T citotóxicas
CTLDs - múltiplos domínios de reconhecimento de carboidratos nos receptores de
manose.
COX2- cicloxigenase-2
DC – células dendríticas
DC-SIGN- receptores de manose das células dendríticas
DD – domínio de morte
DTH – Reação de Hipersensibilidade Tardia
FNII- domínio do receptor de manose com repetições de dois tipos de fibronectina
IκκκκB – complexo quinase inibidor de κB
IFN-γγγγ - interferon gama
IL-10 – interleucina 10
IL-12 – interleucina 12
IL-2 – interleucina 2
iNOS – indutor de óxido nítrico sintase
IFNs- interferons
IRF- Fator Responsável por Interferon
IRAKs – quinases associadas a receptores de IL-1
LAM – lipoarabinomanana
LM – lipomanana
LPS – lipopolissacarídeo
MDR- bacilos multidrogaresistentes
MR- Receptores de Manose

Man-LAM – lipoarabinomanana-manosilada
MAPK – Proteína Quinase Ativada por Mitógeno
MHC – Complexo de Histocompatibilidade Principal
Mo – macrófagos
Mtb – Mycobacterium tuberculosis
Mav- Micobacterium avium
MyD88 – proteína 88 de diferenciação de resposta primária mielóide
NF-κκκκB – fator nuclear κB
NO- óxido nítrico
NIK – quinase indutora de NF-κB
NK – células killer
OMS – Organização Mundial de Saúde
PGN – peptideoglicano
PI3-K – fosfatidilinositol 3 quinase
PRR – reconhecimento de padrão molecular
PS – fosfatidilserina
PTKs- Proteínas tirosina quinase
PTP – tirosinafosfatase
SH2 – domínio 2 de homologia do Src
SHP-1- fosfotirosina
TH1- Célula T helper 1
TB – Tuberculose
TCR- Receptor de célula T
TGF-ββββ - Fator de transformação de crescimento β
TLR – receptor Toll-Like
TNF-αααα - Fator de Necrose Tumoral-α
TRAF-6 – Fator-6 associado ao receptor para TNF-αααα

RESUMO
A parede celular de M. tuberculosis é composta por diversos antígenos que
ativam macrófagos principalmente através de sua ligação com o receptor Toll-like
:TLR 2. Entretanto, Man-LAM um fator de virulência predominante na parede
celular de M.tuberculosis, se liga aos macrófagos através de Receptores de
Manose (MR), os quais a literatura sugere possuir propriedades
imunossupressoras da resposta imune inflamatória, induzida pela micobactéria.
Componentes bacterianos podem se ligar com diversos receptores e ativar várias
vias de sinalização com eventos de interferência (cross-talk) de uma para outra.
Verificamos a hipótese de que Man-LAM de Mtb, através de sua ligação com MR,
pode interferir na sinalização pró-inflamatória, dependente de TLR 2, induzida nos
macrófagos, por BCG. O pré-tratamento de macrófagos RAW 264.7 com Man-
LAM de Mtb diminui a produção dos mediadores pró-inflamatórios IL-12 e NO,
induzidos pelos produtos bacterianos. Entretanto, este efeito de atenuação não foi
observado para TNF-α. IL-10 provavelmente não interfere neste mecanismo de
modulação porque Man-LAM não alterou os níveis de produção desta citocina.
Verificamos o efeito modulador de Man-LAM nas vias de sinalização (NF-κB, PI3-
K e MAPK) acionadas pelas micobactérias através de TLR 2. Demonstramos que
o pré-tratamento dos macrófagos com Man-LAM inibe a ativação de PI3-K,
MAPKs e não altera o perfil de ativação de NF-κB induzidos por BCG. A
combinação de Man-LAM e BCG levou à forte ativação da tirosinafosfatase SHP-1
que pode contribuir com os eventos de desfosforilação de MAPK e suprimir a
ativação de PI3-K. BCG estimula a formação de filapódios, espalhamento e
polarização celular em macrófagos RAW 264.7. A via PI3-K que leva a ativação de
Rac-1, medeia mecanismos de regulação dos rearranjos do citoesqueleto celular,
induzido por BCG, pois a supressão de PI3-K pelo inibidor LY294002 impede a
protusão dos filapódios. Entretanto, não foi observada a alteração da polarização
celular induzida por BCG, nos macrófagos pré-tratados com Man-LAM.

ABSTRACT
Macrophages are activated by M. tuberculosis cell wall ligands mainly
binding withTLR 2. However, Man-LAM a dominant Mycobacterial virulent cell wall
component binds with macrophage through Mannose Receptors (MR), which was
suggested suppress mycobacterial inflammatory response. Severely macrophages
receptors can interact with many bacterial components at same time and it
activates differents signaling pathway and cross-talk between them. We suggest
the cross-talk between MR signaling pathway mediated by Man-LAM and TLR 2
pro-inflammatory signaling, induced by BCG. The macrophages RAW 264.7 Man-
LAM pre-treatment decreases IL-12 and NO pro-inflammatory mediators, induced
by bacterial components. Nevertheless, the lacking effect was not observed to
TNF-α. Probably, IL-10 citokyne doesn´t interfere in the Man-LAM modulation
mechanism because was not observed alteration in IL-10 level. We verified the
modulatory effect of Man-LAM in the (NF-κB, PI3-K e MAPK) signaling pathway
activated by mycobacteria, through TLR 2. We demonstrated that Man-LAM
macrophages pre-treatment inhibit PI3-K and MAPKs activation and it has no effect
to NF-κB activation, induced by BCG. Man-LAM and BCG association induces
strong tyrosinephosphatase (SHP-1) activation which can contributes with MAPK
dephosphorilation events and suppress PI3-K activation. BCG can estimulate
filapodias induction, spreading and cellular polarization in RAW 264.7
macrophages. PI-3K signaling pathway which induces Rac-1 activation mediate
cellular cytoskeleton rearrangement induced by BCG because the protusion
filapodias is suppressed by PI3-K inhibitor, LY 294002. However, it had not been
observed polarization interference in the macrophages pre-treated by Man-LAM.

Figura 1- Incidência da tuberculose no mundo em 2000. (Nature Rev Micro, 2004)Figura 1- Incidência da tuberculose no mundo em 2000. (Nature Rev Micro, 2004)
I- INTRODUÇÃO
1.1. Incidência da tuberculose no mundo
A tuberculose (TB) é
uma doença de alta
prevalência em todo mundo.
A cada ano são notificados 8
milhões de novos casos e
2,4 milhões de mortes (Gao
Q, et al; 2005). O sudoeste
da Ásia e a África são
consideradas as regiões que
apresentam o maior número
de casos, notificando-se 700 casos a cada 100.000 indivíduos, em determinadas
áreas (figura 1). O maior número de mortalidade ocorre na África, chegando a
alcançar um sinergismo com a AIDS, doença que mais mata nesta região (Nature
Reviews, 2004).
A tuberculose é um dos principais problemas de saúde pública na América
Latina. O Brasil é 150 país em número de notificações desta doença, com uma
taxa anual de aproximadamente 114.000 novos casos. O número de mortes por
tuberculose no país é de 4 a 5 mil anualmente. O estado do Rio de Janeiro tem
uma incidência de 110 casos em 100.000 habitantes, uma das maiores taxas do
país (Corbet et al, 2003).
Com o surgimento, em 1981, da Síndrome de Imunodeficiência Adquirida
(SIDA/Aids), vem-se observando, tanto em países desenvolvidos, como nos
países em desenvolvimento, um crescente número de casos notificados de
tuberculose em pessoas infectadas pelo vírus da imunodeficiência humana (HIV).
A associação HIV/TB constitui, nos dias atuais, um sério problema de saúde
pública, podendo levar ao aumento da morbidade e mortalidade pela TB, em
muitos países (OMS).

A análise da mortalidade deve considerar a distribuição geográfica, os
grupos etários e a associação com o HIV (FUNASA 2006).
1.2. O gênero Mycobacterium
Mycobacterium tuberculosis (Mtb), espécie pertencente ao gênero
Mycobaterium, é o principal causador da tuberculose em humanos, embora outras
espécies como M. bovis, M. africanum, M. microti, M. kansasii e M. canetti também
possam promover esta doença. Estes patógenos são caracterizados por serem
organismos aeróbicos, intracelulares facultativos, que crescem lentamente nos
meios de cultura, podendo sobreviver e se multiplicar dentro de macrófagos ou
outras células (Flynn J, et al; 2001).
Outra característica das micobactérias é a presença de uma parede celular
de composição rara devido à presença predominante de ácidos micólicos, que
compõem mais de 50% de seu peso seco, responsável pela sua propriedade de
álcool-ácido resistência, diferenciando-as de bactérias Gram-positivas e -negativas
e micoplasmas (figura 2).
M. tuberculosis é identificável ao microscópio após tratamento com solução
álcool-ácida e coloração pelo método de Ziehl-Neelsen. Nesta técnica, os bacilos
absorvem um corante vermelho brilhante, o qual fica bem contrastado com um
fundo azul. Os bacilos álcool-ácido resistentes também podem ser visualizados
por microscopia fluorescente após coloração com auramina-rodamina (Ibanga H,
et al; 2006).

Figura 2 – Representação esquemática de PAMPs em diferentes bactérias. As
bactérias Gram-positivas apresentam uma camada espessa de peptideoglicano
(PGN) na parede celular. Ácido lipoteicóico, ácidos teicóicos e lipoproteínas estão
também embebidas nesta parede. Bactérias gram-negativas possuem uma camada
mais fina de PGN, porém a parede celular destas bactérias é mais intensificada pela
presença de Lipopolissacarídeo (LPS) em sua superfície. LPS é um componente ativo
derivado do lipídeo A e do O-polissacarídeo. O último é exposto fora da superfície da
célula. Micoplasma não tem uma parede celular, mas as lipoproteínas e os
lipopeptídeos estão encaixados em sua membrana citoplasmática. Mycobacterium
tuberculosis possui uma cada hidrofóbica característica que contém arabinogalactana,
além de uma membrana citoplasmática e uma camada de PGN. Lipoarabinomanana é
um glicolipídeo associado à parede celular. Algumas destas PAMPs mostraram ter
forte atividade imunoestimulatória através da ativação de membros da família TLR.
(Kaisho et al, 2002).

1.3. Tuberculose - A doença
A tuberculose é uma doença infectocontagiosa que pode ser fatal se não
tratada, e que progride silenciosamente podendo se desenvolver em qualquer
parte do corpo, mas geralmente apresenta-se na forma de uma infecção
pulmonar, evoluindo de uma infiltração branda até uma forma crônica, cavitária e
doença severamente destrutiva. Diante de um diagnóstico precoce e tratamento
corretamente administrado, a tuberculose é curável em praticamente 100% dos
casos. O tratamento dos bacilíferos é a atividade prioritária de controle da doença,
uma vez que permite anular rapidamente as maiores fontes de infecção (Flynn O,
et al; 2001).
Estão mais sujeitos à doença, indivíduos que convivam com o doente
bacilífero, determinados grupos de pessoas imunodeficientes, como os silicóticos,
e pessoas que estejam em uso de corticosteróides ou infectados pelo HIV. Outro
fator que contribui para o aparecimento de novos casos de tuberculose é o
aparecimento de cepas multidrogaresistentes (MDR e XDR). A alarmante
emergência destas cepas nos países desenvolvidos dificulta o combate à doença
com o arsenal terapêutico existente (Manabe et.al; 2000).
Antes do advento quimioterapia com antibióticos, a mortalidade era o
indicador utilizado tanto para avaliar a tendência da endemia, como para fazer
estimativas de morbidade. Atualmente, a incidência da mortalidade é descrita
considerando-se a distribuição geográfica, os grupos etários e a associação com o
HIV. (FUNASA 2002).
1.4. Tratamento da Tuberculose
M. tuberculosis foi descrito pela primeira vez em 24 de março de 1882 por
Robert Koch (Chan et al, 2002), que recebeu o Prêmio Nobel de Medicina por sua
descoberta em 1905. Koch não acreditava que as tuberculoses bovina e humana
fossem similares, o que impediu o reconhecimento do leite infectado como fonte
da doença. Mais tarde, esta fonte foi eliminada graças à pasteurização. Koch

apresentou o extrato do bacilo em glicerina, como um remédio para a doença, em
1890, chamando-o de tuberculina (ATS/CDC). A tuberculina não teve eficácia,
porém mais tarde foi adaptada por Von Pirquet para um teste para tuberculose
pré-sintomática (Chan e Iseman et al, 2002).
Antes disso, somente a intervenção cirúrgica era utilizada como tratamento
(além dos sanatórios), incluindo a técnica do pneumotórax: provocar o colapso de
um pulmão infectado para deixá-lo em repouso para permitir que as lesões
cicatrizassem. Esta técnica executada, porém de pouco benefício, foi posta de
lado após 1946. Somente após este período, com o desenvolvimento do
antibiótico estreptomicina, que o tratamento, ao invés de apenas a prevenção,
tornou-se possível (Chan e Iseman et al, 2000).
A descoberta da quimioterapia utilizando antibióticos específicos, a partir da
década de 1940, alterou mundialmente de forma bastante radical o perfil
epidemiológico, a ação institucional e o conhecimento científico em relação à
tuberculose. Paralelamente com a descoberta da quimioterapia, veio o surgimento
de bacilos resistentes às drogas (www.coc.fiocruz.br/tuberculose).
Segundo as normas do Ministério da Saúde, o tratamento básico da
tuberculose inclui a administração de três quimioterápicos associados
(Pirazinamida, Ioniazida e Rifampicina) por um período de seis meses
consecutivos. A utilização de três drogas distintas destina-se a inativação das
micobactérias por diferentes mecanismos. Isto porque, algumas micobactérias
desenvolvem resistência (droga resistentes-DR) à determinados tipos de
medicamentos, tornando-os ineficientes para o combate ao patógeno (Colditz
et.al;1994).
Algumas cepas se tornam resistentes à pelo menos duas ou três drogas de
primeira linha (MDR), devendo-se neste caso ser utilizado o arsenal de antibióticos
reserva. O tratamento da tuberculose MDR (MDRTB) é longo e requer drogas
mais tóxicas, as quais freqüentemente apresentam limitada resolução em
determinados locais e geralmente são menos eficazes em pessoas infectadas pelo
HIV (Naomi B, et al; 1999).

O diagnóstico certo para a identificação do patógeno é fundamental para o
tratamento do paciente. A administração da associação medicamentosa
adequada, assim como a administração de doses corretas e no período
necessário, sob supervisão médica, são os meios designados para evitar a
persistência bacteriana e desenvolvimento de resistência às drogas, assegurando-
se a cura do paciente (FUNASA 2002).
Os bacilos da tuberculose praticamente perdem sua infectividade em poucos
dias após o início da administração da quimioterapia. Adotando-se estes
procedimentos, os doentes portadores da doença pulmonar não necessitam, nem
devem ser segregados do convívio familiar e da comunidade (FUNASA 2002).
1.5. Transmissão da Tuberculose
A transmissão da tuberculose realiza-se através da inalação de aerossóis
oriundos de tosse, espirro ou fala de pacientes portadores da doença pulmonar
(figura 3). A disseminação da bactéria é facilitada devido ao estágio
Figura 3 - Transmissão da Tuberculose em humanos. A transmissão da tuberculose é mediada principalmente pelas vias aéreas, através da inalação de gotículas de aerossóis, oriundas da fala, tosse ou espirro de pacientes com a doença pulmonar. (FUNASA, 2002).

imunopatológico de destruição pulmonar que resulta no desenvolvimento de
cavidades pulmonares portadoras de um elevado número de bacilos que
alcançam os brônquios e podem ser eliminados juntamente com as gotículas de
aerossóis (Kaufmann S. et al, 2001).
As gotículas mais pesadas depositam-se rapidamente no solo, enquanto que
as mais leves podem permanecer em suspensão por diversas horas. Somente os
núcleos secos das gotículas (Núcleo de Wells), com diâmetro de até 5µ e com 1 a
2 bacilos em suspensão, podem atingir os bronquíolos e alvéolos, e aí iniciar a
multiplicação. As gotículas médias são retidas pelas mucosas do trato respiratório
superior, e removidas dos brônquios através do mecanismo muco-ciliar (FUNASA
2002). Bacilos assim removidos são deglutidos, inativados pelo suco gástrico e
eliminados nas fezes. Os que se depositam nas roupas, lençóis, copos e outros
objetos, dificilmente se dispersarão em aerossóis e, por isso, não desempenham
papel importante na transmissão da doença.
A transmissão bacilífera varia de acordo com o grau de infecção do organismo
e quantidade de bacilos expelidos, forma e duração da exposição ao bacilo,
virulência e a competência imunológica do paciente (Fartazzi C. et al; 1997). A
inalação de poucos microrganismos pode causar a infecção, que na maioria dos
indivíduos permanece latente. Estima-se que um terço da população mundial
esteja infectada pelo M. tuberculosis, sem sintomas aparentes, e que 5-10% dos
infectados desenvolvem a doença em algum período da sua vida. Sem tratamento,
fatalmente as mortes chegam a 50% dos casos (Nature Reviews, 2004).
Subsequente a infecção pelo M. tuberculosis, transcorrem em média 4 a 12
semanas para a detecção das lesões primárias. Após a infecção inicial (infecção
primária), transcorrem 12 meses para o estabelecimento da doença pulmonar. A
probabilidade de infecção e progressão para um estado patológico é influenciada
por diversos fatores, dentre os quais destacam-se as condições sócio-econômicas
e certas condições médicas como diabetes mellitus, silicose, uso prolongado de
corticosteróides ou outros imunossupressores, neoplasias, uso de drogas e
infecção pelo HIV (Flynn J, et al; 2001).

A evolução do quadro clínico dependerá do indivíduo estar sendo infectado
pela primeira vez (primo-infecção), ou re-infectado (re-infecção exógena). A
probabilidade de adoecer em uma primo-infecção depende da virulência do bacilo,
da fonte infectante e das características genéticas dos indivíduos infectados. Em
novo contato, após uma infecção natural ou induzida pela BCG, a resistência
dependerá da resposta imunológica.
1.6. Resposta Imune contra M. tuberculosis
As micobactérias primeiramente infectam macrófagos alveolares (Mo) do
hospedeiro, os quais representam a primeira linha de defesa celular contra
invasões de patógenos. Fagócitos mononucleares como Mo e células dendríticas
(DC) são responsáveis pela destruição das micobactérias através de degradação
por hidrolases ácidas, após a fusão dos fagolisossomos. A destruição das
micobactérias depende da capacidade microbicida intrínseca dos fagócitos do
hospedeiro e dos fatores de virulência das micobactérias internalizadas (Van
Crevel, et al; 2002).
Espécies patogênicas de micobactérias desenvolveram mecanismos para se
replicarem e se tornarem resistentes à degradação dentro dos fagócitos. A
multiplicação intracelular continuada destes microrganismos promove a lise destas
células com conseqüente liberação dos patógenos (Schaible et al; 1998).
O desenvolvimento de uma resposta inflamatória local bem sucedida, pelo
hospedeiro, é de fundamental importância para a eliminação das micobactérias. O
processo inflamatório é iniciado pelo recrutamento de células imune
intravasculares (leucócitos) para as proximidades do foco infeccioso. Este
processo é regulado por citocinas, moléculas de adesão (integrinas e selectinas) e
quimiocinas (RANTES, MIP1-α, MIP2, MCP-1, MCP-3, MCP-5, e IP10).
M. tuberculosis induz a secreção de TNF-α, prostaglandinas E1 e IFNs por
macrófagos, DC e células T. O TNF-α é uma citocina produzida principalmente por
Mo, requerida principalmente para o controle da infecção na fase aguda e
desempenha um papel fundamental para o recrutamento dos leucócitos. Atua no

processo de migração e localização celular dentro do tecido infeccioso, bem como
influencia na expressão de moléculas de adesão, quimiocinas (moléculas
quimioatraentes) e receptores de quimiocinas. Foi demonstrado que camundongos
deficientes em TNF-α ou receptores para TNF-α, infectados pelo M. tuberculosis,
desenvolvem uma carga bacteriana substancialmente alta e apresentam uma
meia vida curta em comparação com camundongos normais (Flynn J. et al, 1995).
A secreção de IL-12 pelos macrófagos e pelas células dendríticas (DC) após
a fagocitose do bacilo leva ao desenvolvimento de uma resposta celular tipo 1
(TH1) com a produção de altos níveis de INF-γ (Figura 4). A IL-12 biologicamente
ativa é um heterodímero composto de duas sub-unidades (p40 e p35) ligadas por
pontes de enxofre. O gene p35 está constitutivamente expresso na molécula de
IL-12. Entretanto, o gene p40 somente está expresso em células que produzem IL-
12 bioativas (Pathak S.K et al, 2005).
A deficiência de IL-12 em camundongos induz uma pré-disposição à infecção
causada por M. bovis BCG, assim como organismos que apresentam mutações
que afetam genes correspondentes à IL-12 ou seus receptores, apresentam
suscetibilidade ao desenvolvimento de infecções micobacterianas (Wakeham J.J.
et al, 1998). Entretanto, a administração de IL-12, in vivo, em camundongos,
aumenta a resistência destes à infecção pelo M. tuberculosis, sugerindo que IL-12
seja crítica para o controle de infecções micobacterianas (Flynn, J. L et al; 1995).
Os peptídeos antigênicos, processados a partir dos microrganismos
fagocitados, se associam às moléculas do Complexo de Histocompatibilidade
Principal II (MHC II), expressas pelas células apresentadoras de antígeno (APC) e
são apresentados às células T CD4 (naives), nos linfonodos e no local da
infecção, dando início ao desenvolvimento de uma resposta imune
específica/adquirida mediada por célula.
A ativação celular é proporcionada pela interação do Receptor de Célula T
(TCR) e CD4 com o complexo MHC-II, juntamente com sinais estimuladores
concomitantes, proporcionados pela interação de moléculas CD28 sobre as
células T, com as moléculas B7 existentes nos macrófagos e também por IL-1 e
IL-12 (Nigou J, Zelle-Rieser C, et al, 2001).

A primeira função efetuadora das células TH1 é a produção de IFN-γ e outras
citocinas pró-inflamatórias capazes de potencializar a ativação de macrófagos,
tornando-os mais eficientes no controle ou eliminação das micobactérias. O
principal mecanismo responsável pela atividade antimicobacteriana do IFN-γ e
TNF-α é a regulação positiva de genes envolvidos em respostas de estresses
nitrosativos e
oxidativos que levam a produção, por Mo e DC, de potentes moléculas
microbicidas como o Óxido Nítrico (NO) e superóxidos e peróxido de hidrogênio
(Abbas A; et al, 1996).
Os macrófagos alveolares infectados com M. tuberculosis produzem
intermediários reativos do nitrogênio (RNI) através da ativação da forma induzida
de óxido nítrico sintase (iNOS), utilizando L-arginina como substrato (Nigou J,
Zelle-Rieser C, et al, 2001). O crescimento intracelular dos M. tuberculosis
depende de sua capacidade para evitar sua destruição por enzimas lisossomais,
RNI e ROI. Camundongos geneticamente deficientes em IFN-γ, TNF-α e iNOS,
são altamente susceptíveis a infecção e sucumbem rapidamente de tuberculose
aguda (Raupach B, et al; 2001).
Células T CD4+ participam na ativação de linfócitos T CD8+ citotóxicos (CTLs)
que atuam cooperativamente na defesa contra as bactérias intracelulares através
da lise das células infectadas e produção de IFN-γ (Van Crevel et al; 2002).
A resposta imune humoral é ineficiente em infecções causadas por patógenos
intracelulares. Devido a sua capacidade de reprodução dentro das células
hospedeiras, estes patógenos não podem ser atacados por anticorpos. Entretanto,
as CTLs destroem as células-alvo no momento do contato com a célula infectada.
CTLs ligam-se ao complexo MHCI-antígeno na superfície da célula e então
liberam uma substância denominada perforina. As perforinas formam um poro na
membrana da célula alvo, levando à lise por desequilíbrio osmótico. A lise das
células alvo por células CTLs também podem ser mediadas por granzimas ou
através da via de fas/Fas L (Gerard D et al 2002).
O M. tuberculosis estimula células T que expressam a forma γδ do receptor de
antígeno. Essas células T podem diretamente reconhecer pequenas proteínas

micobacterianas e antígenos micobacterianos homólogos às proteínas de choque
térmico, na ausência de moléculas apresentadoras de antígeno. Devido ao
estresse da infecção, as células infectadas assim como os parasitas produzem
proteínas de choque térmico (Abbas et al; 1996).
Assim como as células T γδ, as células T restritas ao CD1 não reagem com
antígenos protéicos micobacterianos no contexto de moléculas de MHC de classe
I ou II. Contudo, reconhecem lipídios micobacterianos e antígenos glicolipídicos
ligados às moléculas CD1 não polimórficas nas células apresentadoras de
antígeno. Estas células apresentam atividades citotóxicas e são capazes de
produzir IFN-γ (Van crevel, et al; 2002).
As citocinas anti-iflamatórias, IL-4, IL-10 e TGF-β (Fator de transformação de
crescimento β) podem inibir a produção ou efeitos das citocinas pró-inflamatórios
produzidas na tuberculose. Estas citocinas são produzidas pelos macrófagos após
a fagocitose do M. tuberculosis e ligação com LAM micobacteriano. São
antagonistas da resposta pró-inflamatória porque regulam negativamente a
produção de IFN-γ, TNF-α e IL-12 (Mckinney J, et al; 2003).
A infecção causada pelo M. tuberculosis é inicialmente controlada e uma
infecção latente, sem sintomas clínicos de doença é estabelecida, uma vez que
um baixo número de bacilos persiste em dormência, dentro de compartimentos
granulomatosos. A inflamação granulomatosa é uma forma de reação de
hipersensibilidade tardia (DTH) contra os bacilos que previne a disseminação das
micobactérias.
Os granulomas consistem de macrófagos e células gigantes, células T e
fibroblastos recrutados para o sítio da infecção (Raupach B, et al; 2001). Na
infecção latente, o estado da micobactéria dentro do granuloma não é bem
conhecido: o microrganismo pode estar em um estado dormente e não replicativo,
ativamente replicativo, mas sendo eliminado pelo sistema imune ou
metabolicamente alterado com ciclos de replicação limitados ou não freqüentes.
Uma falha na resposta imune designada para conter a infecção pode resultar na
reativação e replicação do bacilo, com necrose e dano no tecido pulmonar
(Jérôme N, et al, 2004).
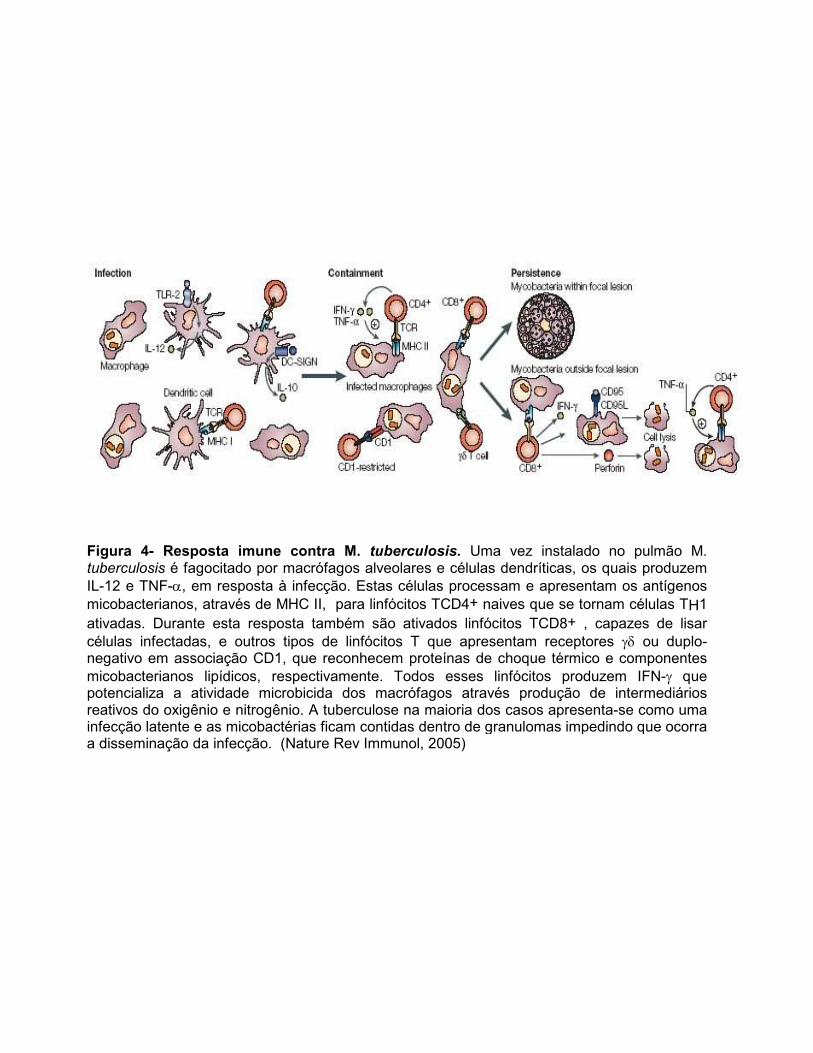
Figura 4- Resposta imune contra M. tuberculosis. Uma vez instalado no pulmão M. tuberculosis é fagocitado por macrófagos alveolares e células dendríticas, os quais produzem IL-12 e TNF-α, em resposta à infecção. Estas células processam e apresentam os antígenos micobacterianos, através de MHC II, para linfócitos TCD4+ naives que se tornam células TH1 ativadas. Durante esta resposta também são ativados linfócitos TCD8+ , capazes de lisar células infectadas, e outros tipos de linfócitos T que apresentam receptores γδ ou duplo-negativo em associação CD1, que reconhecem proteínas de choque térmico e componentes micobacterianos lipídicos, respectivamente. Todos esses linfócitos produzem IFN-γ que potencializa a atividade microbicida dos macrófagos através produção de intermediários reativos do oxigênio e nitrogênio. A tuberculose na maioria dos casos apresenta-se como uma infecção latente e as micobactérias ficam contidas dentro de granulomas impedindo que ocorra a disseminação da infecção. (Nature Rev Immunol, 2005)

12, consideradas serem importantes para imunidade protetora contra tuberculose
do tipo TH1. Mas, provocaram a expressão dos genes IL-13, IL-11 e IL-4 favorável
para a resposta imune humoral (Lopez et al., 2003; Manca et al., 2004).
A variabilidade genética do M. tuberculosis determina o seu grau de
virulência e pode influenciar no caráter da resposta imune contra a bactéria e a
severidade da doença (Gao.Q, et al, 2005). Isso significa que as cepas de Mtb que
possuem diferentes genótipos podem apresentar diferenças na estrutura da
parede celular que são capazes induzir ativação dos macrófagos clássica ou
alternativa e através desta, determinar o caráter do desenvolvimento da infecção
(Gordon S. 2003; Goerdt S. et al, 1999).
1.7. Receptores dos macrófagos
Os macrófagos apresentam um vasto número de receptores em sua
superfície e suas respectivas ligações podem resultar em diversas respostas as
quais variam de simples fagocitose de debris celulares à degradação de
patógenos potenciais e subseqüente ativação de uma variedade de mecanismos
antibacterianos (Jeffrey S., et al; 2003) (figura 5).
Diversos receptores (receptores para complemento, scavanger receptores,
receptores lectina-similar, CD14) estão envolvidos na ligação e internalização da
micobactértia, mas a maioria destes não traduz o sinal para núcleo celular. O
reconhecimento da micobactéria que leva a sinalização celular é função dos
receptores determinados: receptores Toll-like (TLR 2 e TLR 4) e MR que
transduzem sinais até o núcleo e induzem a produção de citocinas pró- e anti-
inflamatórias (Schorey J, et al; 2003).
M. tuberculosis e M. bovis expressam uma variedade de moléculas
agonistas expressas na parede celular que são capazes de ativar macrófagos
através de sua ligação com receptores presentes na superfície celular,
principalmente TLR 2 (liporoteina p19, PIM), TLR 4 (proteína termolábil) e MR
(Man-LAM, lipoarabinomanana- manosilada) (Van Crevel et al, 2002).

A ligação micobacteriana com os diferentes receptores do macrófago pode
induzir diferentes vias de sinalização com fenômeno de interferência de uma para
outra. O efeito final depende do nível da expressão das moléculas agonistas na
parede celular e/ ou secreção destas pela bactéria (Nguyen L,et al, 2005).
Figura 5- Receptores dos macrófagos. As micobactérias são reconhecidas pelos macrófagos através de receptores celulares. Uma classe de receptores dos fagócitos são responsáveis pela internalização das micobactérias outros participam de eventos de sinalização celular e ativam uma resposta inflamatória. (Nature Rer Microbiol, 2004).
Figura 5- Receptores dos macrófagos. As micobactérias são reconhecidas pelos macrófagos através de receptores celulares. Uma classe de receptores dos fagócitos são responsáveis pela internalização das micobactérias outros participam de eventos de sinalização celular e ativam uma resposta inflamatória. (Nature Rer Microbiol, 2004).

1.6.1 Mecanismos de modulação da resposta imune mediados pelo M.
tuberculosis
A sobrevivencia micobacteriana no interior dos macrófagos é um dos
principais focos de pesquisa atualmente. Micobactérias não apenas são capazes
de se adaptarem ao sistema do hospedeiro, mas também interferem ativamente
na maquinaria de sinalização da célula hospedeira, subvertendo ou inibindo parte
do aparato de morte empregado pelos macrófagos (Hestvik A.L, et al; 2005).
Uma variedade de mecanismos é proposta para explicar a sobrevivência do
M. tuberculosis dentro do macrófago, incluindo a fusão do fagossoma com o
lisossoma, inibição da acidificação dos fagossomas e resistência à morte pelos
metabólitos oxigenados. Macrófagos infectados apresentam um processamento
antigênico diminuído, reduzida responsividade ao IFN-γ, e reduzida produção de
citocinas, assim como intermediários do oxigênio e nitrogênio (Jérôme N, et al,
2004).
Uma das estratégias desenvolvidas por M tuberculosis para subverterem o
sistema imune do hospedeiro é a manipulação de diferentes Receptores de
Reconhecimento Padrão (PRR’s) desviando-os dos seus papéis de defesa
originais (Hestvik A.L, et al; 2005).
M. tuberculosis desenvolveram mecanismos moleculares para limitar a
ativação dos macrófagos pelo INF-γ (ativador proeminente dos Mφ). Foi descrito
que indivíduos funcionalmente deficientes na sinalização para INF-γ, seja como
resultado de mutações nos genes de INF-γ ou em seus receptores, eram mais
susceptíveis às infecções causadas por M. Bovis BCG ou micobactérias
oportunistas (Flynn J. et al; 1993).
Um dos mais importantes mecanismos de evasão desenvolvidos por estes
patógenos, tem como alvo a supressão da produção de IL-12, através da
manipulação de sua via de sinalização (Pathak S, et al., 2005). Dados recentes,
obtidos no modelo de monócitos humanos in vitro, demonstraram que isolados
clínicos da cepa mais virulenta de Mtb, do genótipo W/Beijing, em contraste com

cepas non-Beijing inibiram a secreção das citocinas pró-inflamatórias TNF-α, IL-
12.
1.7.1. Receptores de fagocitose
Patógenos alvo como M. tuberculosis que apresentam numerosos e
diversos ligantes em sua superfície são caracterizados por apresentarem múltiplos
tipos de receptores e simultaneamente. Além disso, in vivo, M. tuberculosis
provavelmente não é internalizado por macrófagos utilizando apenas um receptor.
De qualquer modo, em alguns contextos, a utilização de receptores por M.
tuberculosis pode ser baseada no estado de diferenciação ou ativação do
macrófago (Schorey J, et al; 2003).
Por exemplo, durante a diferenciação de monócitos para macrófagos CR3
diminui em abundância, enquanto CR4, MR e receptores scavenger aumentam. O
estímulo dos macrófagos pelo INF-γ regula negativamente a expressão de MR.
Entretanto, distintos tipos de receptores podem cooperar para otimizar a ligação e
internalização de partículas modelo (Ernest J. D, 1998).
A cooperação de entre CR1 e CR3 e receptores para Fc marcadamente
facilitam a fagocitose de alvos opsonizados por IgG . A cooperação pode
acontecer através da fagocitose de partículas ligadas aos receptores que perdem
domínios transmembrana e citoplasmáticos, como os CD14 (Ernest J. et al; 1998).
Receptores do complemento em fagócitos ocorre em duas formas
estruturais distintas. Receptor para o complemento tipo 1 (CR1) que é uma
proteína transmembrana monomérica que liga c3b e c4b, mas não liga c3bi. CR1
possui uma atividade reguladora do complemento e pode mediar a fagocitose de
diversas partículas, mas sua capacidade para transdução de sinal e ativação
celular não foi caracterizada (Ernest J, et al ; 1998).
Assim como bactérias e fungos, M. tuberculosis podem ativar a via
alternativa do complememto, resultando na opsonização com c3b e c3bi. As
bactérias que são suficientemente revestidas com estes ligantes derivados do soro

ligam com CR1, CR3 e CR4 e são conseqüentemente fagocitados (Sendide K, et
al; 2005).
Receptor para o complemento tipo 3 e 4 (CR3 e CR4) são proteínas
heterodiméricas da super família das integrinas. São heterodímeros que contêm
subunidades β idênticas (CD18 ou β 2 integrina) e subunidades α distintas (CD11b
ou αM e CD11c ou αX). CR3 e CR4 ligam c3bi e CR3 também contém um sítio de
ligação de glicano (Schlesinger, et al; 1996).
CD14, uma proteína de membrana ligada ao glicano fosfatidilinisitol (PI) é
melhor conhecido e caracterizado como receptor de alta afinidade por
lipopolissacarídeos (LPS) de bactérias Gram- negativas. De qualquer modo, CD14
também liga lipoarabinomanana (LAM) de M. tuberculosis; e esta ligação induz o
macrófago a secretar IL-8 (Sendide K, et al; 2005).
Receptores Scavenger de macrófagos ligam macromoléculas e partículas
polianiônicas, incluindo LPS de bactéria Gram-negativa e ácido lipoteicóico de
bactérias Gram-positivas. Experimentos usando inibidores competitivos têm
implicado receptores scavenger de classe A como quantitavivamnete importantes
para fagocitose de M. tuberculosis em macrófagos derivados de monócitos
humanos (Zimmerli S, et al; 1996).
Além disso, receptores scavenger de classe A purificados ligam M.
tuberculosis e sulfolipídeos destes competem com outros ligantes pela ligação
com este receptor. Porém ainda não está bem esclarecido se receptores
scavenger podem ativar o citoesqueleto para internalizar a bactéria ou,
alternativamente, se estes receptores atuam na ligação da bactéria, mas a
fagocitose é executada por outros receptores (Ernest J. D, 1998).
Alguns indivíduos infectados possuem anticorpos circulantes contra M.
tuberculosis. Imunoglobulina G (IgG) recobrem as micobactérias e são
internalizadas por macrófagos em vesícolas que rapidamente são fundidas com
lisossomos carregados de ferritina, através de receptores macrofágicos que
reconhecem a porção Fcγ dos anticorpos (Koul A, et al; 2004).

1.7.1.1 Fagocitose
Após a entrada no macrófago, as micobactérias patogênicas como M.
avium e M. tuberculosis residem e replicam dentro de fagossomos. Diversas
pesquisas demonstraram que fagossomos micobacterianos são capazes de se
fundirem com endossomos primários ou lisossomos e estão associados com
proteínas como os receptores de transferrina. Estes fagossomos também perdem
a ATPase e falham em acidificar (Koul A, et al; 2004).
As falhas dos fagossomos em adquirirem ou perderem marcadores
associados com a maturação dos fagolisossomos indicam que estes fagossomos
não participam dos eventos normais de fusão, necessários para gerar um
ambiente antimicobacteriano dentro do fagolisossomo.
Alguns dos mecanismos utilizados pelas micobactérias para evadir estes
eventos é o bloqueio da maturação do fagossomo pela manipulação da
maquinaria endossomal do hospedeiro. Foi demonstrado que alguns fagossomos
contendo micobactérias ou produtos micobacterianos falharam em recrutar
moléculas efetoras importantes na biogênese fagossomal (Fratti. et al 2001). Um
proeminente lipoglicano de parede celular de Mtb, lipoarabinomanana coberta com
manose terminal (Man-LAM), tem sido diretamente implicada na regulação de
diversos processos (Strohmeier G, et al; 1999).
A presença de Man-LAM na superfície micobacteriana coloca esta molécula
em uma posição ideal para mediar interações iniciais entre Mtb e macrófagos.
Man-LAM é considerado um regulador crítico na maturação de fagossomos em
macrófagos murinos e monócitos humanos. Ele é capaz de bloquear o aumento
de Ca2 citosólico no macrófago e inibir a interação do fosfatidilinositol 3 quinase
(PI3-K), hVPS4, com a calmodulina, um passo necessário para maturação do
fagolisossoma (Peter B. et al; 2007).

1.7.2. Principais vias de sinalização acionadas pelas micobactérias
A ativação de macrófagos e a produção de citocinas pró-inflamatórias são
cruciais para o desenvolvimento de uma resposta imune bem sucedida contra as
M. tuberculosis. A produção de citocinas pró-inflamatórias pode ser mediada por
receptores Toll-Like 2 e 4 (TLR 2/4), Proteína Quinase Ativada por Mitógeno
(MAPK) e fosfatidilinositol 3 quinase (PI3-K)(Heldwein and Fenton, 2002).
Os eventos de sinalização que levam para transcrição de genes pró-
inflamatórios durante a infecção micobacteriana já foram bem elucidados. As
principais vias de sinalização acionadas pelas micobactérias (NFκB, MAPK e PI3-
K) são principalmente ativadas após a ligação de agonistas micobacterianos com
TLR 2, (Quesniaux V, et al; 2004).
1.7.2.1 Receptores Toll-Like (TLRs)
Os TLRs são mediadores filogenéticamente conservados na imunidade
inata, os quais são essenciais para o reconhecimento dos patógenos. Os
membros da família TLR são proteínas transmembrana que possuem seqüências
ricas em leucina em seus domínios extracelulares, similares para outras proteínas
de reconhecimento padrão do sistema imune inato. O domínio citoplasmático dos
TLR é homólogo aos domínios de sinalização do receptor de IL-1 (IL1R) que liga
IRAK (quinase associada ao IL1R), uma serina quinase que ativa fatores de
transcrição como NFκB (fator nuclear -κ B) para sinalizar a produção de citocinas
(Krutzik S, et al; 2004).
Pelo menos dez TLRs foram identificados, destes, TLR 2, TLR4 e TLR9 são
responsáveis por respostas celulares aos peptídeoglicanos e lipopeptídeos
bacterianos, endotoxina de bactéria Gram-negativa e DNA bacteriano,
respectivamente (Van Crevel, et al; 2002).
Os TLRs também estão envolvidos no reconhecimento micobacteriano.
Através de TLRs, lisado de M. tuberculosis ou lipoproteínas associadas à parede
celular de micobactérias solúveis induzem a produção de IL-12, uma forte citocina

pró-inflamatória. MyD88 (proteína 88 de diferenciação mielóide), um componente
comum da sinalização liga todos os TLRs ao IRAK, é essencial para ativação de
macrófagos induzido por M. tuberculosis (Bulut, Y; et al; 2001).
1.7.2.2. Ativação de NF-κκκκB mediada por TLR 2
A principal via de sinalização mediadas por TLR 2 e TLR 4 já está bem
estabelecida. Ela leva à ativação do NF-κB através da ligação da proteína
adaptadora citosólica MyD88, quinase IRAK, Fator 6 associado com TNFR
(TRAF6) e quinase indutora de NF-κB (NIK). A ativação do complexo da quinase
IκB (IKKα e IKKβ) pela NIK provoca a fosforilação das IκBs, ubiquitinação e
degradação pelo proteossomo, levando a liberação do NF-κB (p65/p50 dímero),
translocação do fator para o núcleo e transativação dos genes pró-inflamatórios
(Kopp, Medzitov, 1999; Underhill et al., 1999) (figura 6).
Figura 6- Ativação de TLR 2 por M. tuberculosis. A ligação de M. tuberculosis com TLR leva a ativação de NF-kB e consequente produção de citocinas imunoestimulatórias que ativam células T e medeiam a morte de micobactérias patogênicas. TLRs iniciam a sinalização pela ligação da proteína adaptadora, MyD88, a qual recruta uma quinase serina/treonina IRAK (Interleukin-1 Receptor-associated- Kinase). IRAK então se associa com TRAF-6 (TNF Receptor- associated –Factor 6). Este permite a ativação do complexo IKK (Inhibtor of kB kinase), o qual fosforila IkB, um inibidor de NF- IkB. A fosforilação de IkB causa sua degradação e permite a translocação de NF-IkB para o núcleo. (Koul A, et al; 2004)

1.7.2.3. Ativação de MAPK por micobactéria
As MAPKs são ativadas nos macrófagos após a ligação de fatores de
crescimento e citocinas inflamatórias com receptores específicos e participam da
ativação da transcrição de genes de citocinas inflamatórias (figura 7). A família das
MAPKs é composta das vias de ERK1/2, p38 e SAPK/JNK. Embora distintas em
sua ativação, existe uma considerável cooperação entre estas quinases, e muitos
substratos são divididos entre estas vias (Schorey J, et al; 2003).
Esta família de quinases é importante em um largo espectro de funções
celulares, incluindo proliferação, apoptose, biosíntese de citocinas e reorganização
do ciotoesqueleto. Todas as MAPKs possuem regiões tirosina-treonina altamente
conservadas, ativadas por regulação positiva de quinases de MAPK (MAPKK)
através da fosforilação do motivo Thr-xxx-Tyr (Martin. B et al, 2000).
Em geral a via ERK1/2 é ativada por fatores de crescimento, estímulos
mitogênicos e promotores de tumor. Estresse ambiental e citocinas inflamatórias
estimulam as vias p38 e JNK. As MAPKs ativadas, são responsáveis pela
fosforilação e ativação de numerosos fatores de transcrição cuja função é
estimular a síntese de várias proteínas inflamatórias incluindo TNF-α, IL-1e IL-6.
Assim como estão envolvidas na regulação transcricional de NOS 2 e
cicloxigenase-2 (COX-2) (Koul A, et al; 2004).
Estudos primários demonstraram que Ara-LAM isolado de micobactéria
avirulenta, e PIM isolados de M. tuberculosis, estimularam a fosforilação de
ERK1/2 e ativação dos fatores de transcrição NFκB e AP-1 em uma linhagem de
macrófagos murinos. Esta estimulação foi aparentemente mediada por TLR2
(Jones B. et al; 2001).

1.7.2.4. Ativação de PI3-K/Akt
As enzimas envolvidas na geração dos distintos fosfoinositídeos são
denominadas famílias de quinases lipídicas, coletivamente conhecidas como PI3-
K. Membros da família do fosfoinositídeo-3 quinase (PI3-K) controlam diversas
respostas, incluindo crescimento celular, sobrevivência, remodelamento do
citoesqueleto e tráfico para vários tipos de organelas diferentes (Koyasu et al;
2003).
A PI3-K exerce importantes funções no sistema imune. Os fosfoinositídeos
são ubiquitinados na membrana celular de vários tipos celulares e tecidos e
Figura 7- Representação esquemática das moléculas implicadas na ativação de MAPK, seguido ao estímulo por LPS. A sinalização mediada por LPS tem sido mostrada ativar diversas pequenas GTPases, incluindo Rac, CDC42 e Ras. Também ocorre na ativação primária com LPS a ativação de PI3-K e subsequente ativação de PKC. Os passos subsequentes a ativação de MAPK inclui MEKK1/4, Raf1, ASK1 e TAK1 e a ativação de MAPK como como as MKKs e MEK1/2. As MAPK podem diretamente fosforilar vários fatores transcricionais ou podem trabalhar através da ativação de quinases como MSK1. (Schorey J, et al; 2003)

funcionam como segundos mensageiros em diversos processos de sinalização
intracelular (Koyasu S. et al; 2003).
As PI3-K são divididas em 4 classes, referidas como IA, IB, II e III, baseado em
sua característica estrutural e especificidade para substrato. Cada tipo de PI3-K
contém um domínio C2 e um domínio catalítico conectado por um domínio
helicoidal que é encontrado em quinases lipídicas, mas não em proteínas quinase.
O domínio C2 liga fosfolipídeo de uma maneira dependente de cálcio e pode estar
envolvida no recrutamento de PI3-K para frações da membrana (Li, X. et al, 2003).
PI3-K de classe IA pode ser ativada durante a sinalização do TLR, mediada
pela associação com proteínas tirosinaquinase (PTK). As enzimas da classe IA são
heterodímeros que apresentam uma subunidade reguladora (p85) e uma catalítica
(p110) e são ativadas através de receptores envolvidos na sinalização, por PTK
(Schlesinger L, et al, 1996).
Neste processo de ativação, ocorre uma interação direta da subunidade p85
reguladora de PI3-K com o receptor (Koyasu S. et al, 2003). Esta interação
envolve um domínio de homologia Src (SH2) da subunidade p85 e um domínio
SH2 no receptor que contém um motivo Tyr-Xaa-Xaa-Met, onde Xaa representa
qualquer aminoácido e quando fosforilado na tirosina é capaz de ativar PI3-K (Li X;
2003).
A subseqüente associação da subunidade catalítica p110 de PI3-K resulta na
sua completa ativação enzimática que fosforila fosfoinositídeos de membrana na
posição D3 do anel de inositol, liberando para a formação de mensageiros
lipídicos secundários (Shizuo et al; 2004).
1.7.2.5 Ativação de PI3-K/Akt mediada por TLR 2
S. aureus foi o primeiro agonista de TLR 2 demonstrado induzir a ativação
de PI3-K através da ligação com TLR2 (Ardibe et al, 2000). Entretanto, foi
demonstrada a associação física de TLR 2 com a subunidade reguladora de PI3-K
(p85-PI3-K) (Lassounskaia E, et al; 2006).

O papel de CD14 tem se revelado essencial para a ativação de PI3-K
através de TLR2 induzida por BCG. Foi demonstrado que BCG ou Man-LAM
purificada, induziu uma significante ativação de PI3-K, somente após a
opsonização por soro ou proteína ligadora de lipopolissacarídeo (LBP) que
aumentou a ligação da micobactéria com CD14 de monócitos THP 1 promovendo
a sinalização através de TLR 2 (Sendide K et a; 2005).
1.8. Receptores de Manose
Os receptores de manose reconhecem polissacarídeos de diversos patógenos
como bactérias Gram-negativas, Gram-positivas, fungos e micobactérias. Além de
sua atividade endocítica, eles são os únicos membros da família das lectinas tipo
C com propriedades fagocíticas e são considerados receptores de reconhecimento
padrão, envolvidos na defesa do hospedeiro e imunidade inata (Taylor P, et al,
2005).
Os MR possuem uma região extracelular que contém um domínio rico em
cisteína (CR), um domínio com repetições de dois tipos de fibronectina (FNII) e
múltiplos domínios de reconhecimento de carboidratos (lectina-like tipo C)
(CTLDs), possuem também um domínio transmembrana e uma pequena cauda
citoplasmática. Os domínios CTDLs medeiam a ligação do MR com açúcares
terminados em manose, fucose e n-acetilglicosamina; na presença de duas
moléculas de Ca2+, por açúcar ligado (Taylor P, et al, 2005).

Figura 8- Estrutura do Receptor de Manose (MR). Estrutura conformacional do domínio de glicosilação ligado ao N-demonstrado na protusão estendida (sialização indicada pelos losangos vermelhos). Domínio CR (vermenlho), segmentos de fibronectina tipo II (marrom) e CTLD (verde). O CTLD 4, o CTLD são responsáveis pela ligação dos açúcares (mostrado em verde escuro). (Taylor P, et al; 2005)
Lipoarabinomananas (LAMs), são lipoglicanos de parede celular e tem sido
consideradas modulinas, por sua capacidade de manipular o sistema imune do
hospedeiro. De fato, foi estabelecido que lipoarabinomananas de Mycobacterium
smegmatis e Mycobacterium species, que são envoltas por fosfoinositóis
(PILAMs), ativaram macrófagos murinos de uma maneira dependente do TLR-2
levando a produção de citocinas como TNF-α e IL-12.
Lipoarabinomananas manosiladas (Man-LAM) são membros do extenso
repertório de moléculas presentes na parede celular de M. tuberculosis e M. bovis
BCG e se ligam aos receptores de manose (macrófagos) ou DC-SIGN (células
dendríticas), murinas ou humanas. Entretanto, poucos estudos demonstram a
participação dos receptores de manose e outros membros da família das lectinas
tipo-C nos eventos de transdução de sinal (Zamze S, et al, 2002).

1.8.1. Sinalização celular mediada por MR
Ainda não foi bem descrito se os MR são requeridos para a ativação das
MAPK. Um engajamento entre MR e a ativação das vias de MAPK foi
demonstrado através do tratamento de monócitos humanos da linhagem THP-1
com Man-LAM, resultando em uma diminuída reação ao estímulo com PMA, LPS
e INF-γ. Esta redução em resposta ao macrófago está associada com a diminuição
da ativação de ERK1/2 nas células tratadas, sugerindo que Man-LAM pode limitar
a ativação do macrófago pela redução da fosforilação de ERK1/2 (knutson K., et
al; 1998).
Man-LAM de M. tubeculosis foi demonstrada atenuar a expressão de TNF-
α e IL-12 em fagócitos mononucleares humanos. Assim como falhou em induzir a
produção de reativos intermediários do O2 nestas células (Astarie-Dequeker et
al.,1999). A ligação de Man-LAM com MR falhou em induzir a produção de
Fig 9- Estrutura das três famílias representativas de moléculas LAM isoladas de difertentes espécies de micobactéria. (Dao, et al. Infect. Immun)

citocinas pró-inflamatórias, por macrófagos, de maneira dependente de TLR
(Quesniaux V.J, et al., 2004).
Todos esses achados sugerem que Man-LAMs de M.tuberculosis e M.bovis
podem contribuir para a persistência das micobacterias dentro de macrófagos.
Provavelmente os MR medeiam a internalização de bactérias patogênicas e não
patogênicas e desviam a resposta micobacteriana nos macrófagos, direcionando
então para ativação ou inibição celular, através da interferência com a sinalização
mediada por TLR (Chieppa M. et al, 2003).
Foi proposto que a indução de IRAK-M, mediada por Man-LAM de Mtb,
regula negativamente a produção de IL-12p40 dependente de TLR por
macrófagos estimulados por LPS, independentemente da modulação de uma
resposta anti-inflamatória mediada por IL-10 (Kobayashi K, et al; 2002).
A família das IRAKs consiste de duas quinases ativas, IRAK e IRAK 4, e
duas quinases inativas, IRAK 2 e IRAK-M. Sendo a expressão desta última,
restrita para monócitos e macrófagos. IRAK-M é um regulador negativo de
sinalização mediada por receptores Toll-like. Ela previne a dissociação de IRAK ou
IRAK-4 com MyD88 e impede formação do complexo IRAK-TRAF6 e posterior
ativação de NFκB, JNK, ERK e p38 (Kobayashi K et al, 2002). Isto sugere que a
repressão da produção de IL-12 pode aumentar a sobrevivência do M.
tuberculosis devido ao impedimento da atuação da resposta imune inata e o
desenvolvimento de uma resposta adaptativa (Pathak S.K. et al, 2005).
Em contrapartida, o cross-talk dos receptores de manose em células
dendríticas derivadas de monócitos imaturos, foi demonstrado ativar um programa
imunosupressor, anti-inflamatório, com a produção de IL-1, IL-10 e TGF-β e inibiu
a produção de IL-12, após o estímulo por LPS (Chieppa M. et al, 2003).
Apesar desses dados, pouco se sabe sobre os efeitos anti-inflamatórios de
Man-LAM. O entendimento dos mecanismos de atenuação da expressão de IL-12
estimuladas pelos agonistas de TLR, por Man-LAM, promove novas introspecções
para a patogênese da tuberculose (Pathak S.K. et al, 2005).
Baseado nos efeitos anti-inflamatórios induzidos por Man-LAM, foi
demonstrado que a via de sinalização mediada por MR é responsável pela

expressão da tirosina fosfatase (SHP-1), que ativa-se e provoca a inibição das
MAPK (Knutson et al., 1997; Nandan D. et al., 2000). Similarmente, sugeriu-se que
o mecanismo de ação do M. tuberculosis, por meio de seu fator de virulência
(Man-LAM), pode estar relacionado com a ativação de SHP-1 (Nandan et al,
2000).
As tirosinas fosfatases são enzimas que removem os radicais fosfato dos
resíduos de tirosina, restringindo as ações das PTKs. A desfosforilação de
proteínas reguladas por tirosina fosfatases é um mecanismo de controle
importante para numerosos processos fisiológicos como: crescimento celular,
motilidade, metabolismo, regulação do ciclo celular e integridade do citoesqueleto
(Knutson et al, 1997).
A tirosina fosfatase SHP-1 participa da alteração de importantes funções
dos macrófagos. Durante a exposição de macrófagos à Yersinia
pseudotuberculosis observou-se a desfosforilação de proteínas celulares, levando
ao impedimento da fagocitose. A infecção de macrófagos por Leishmania
donovani impediu a fosforilação da tirosina e a ativação de JAK quinase 1 e 2 e a
resposta ao IFN-γ (Nandan et al; 2000).
1.9 . Fenótipo migratório dos macrófagos
A migração dos macrófagos em direção ao seu alvo é um importante
mecanismo destas células para desempenharem sua função. As alterações
morfológicas sofridas pelos macrófagos ativados pelas micobactérias são
caracterizadas pela formação de projeções em sua superfície, denominada
filapódios, que são proporcionadas devido a polimerização da actina.
O citoesqueleto de actina é o primeiro determinante para a forma e a
motilidade da célula. O processo de polimerização da actina é relevante para
mudanças morfológicas dos macrófagos, in vivo, durante o extravasamento,
migração e diferenciação destes. Entretanto, os eventos de sinalização que levam
ao espalhamento e adesão dos macrófagos, em resposta às micobactérias, não
estão caracterizadas em detalhes (Claire M.W, et al, 2005).

1.9.1 . Participação de TLR2 e PI3K nas alterações morfológicas sofridas
pelos macrófagos
Os eventos que levam à transcrição de genes pró-inflamatórios, induzidos
pela micobactéria, já estão bem estudados. Além disso, está bem estabelecido o
papel fundamental das vias de sinalização que levam a ativação de NF-κB, MAPK
e PI3-K após a ligação de agonistas micobacterianos com TLRs, principalmente
TLR 2.
A Dissecção das vias que levam ao rearranjo da actina induzida por uma
variedade de fatores extracelulares, tem sido focada naquelas iniciadas por
receptores de tirosinaquinases, para fatores de crescimento, e receptores de
integrinas e associados à proteína G para quimiocinas. De qualquer modo, a
participação das vias de sinalização mediadas pelos TLRs, nos eventos de
remodelamento da actina está começando a ser descoberto.
Micobactérias viáveis ou lisadas, assim como a lipoproteína p19 (agonista
de TLR 2) purificada da parede celular são capazes de ativar PI3-K de uma
maneira dependente de TLR2, levando a reorganização do citoesqueleto e
espalhamento da célula. Sugerindo que TLR2 contribui com os arranjos
morfológicos, envolvidos no processo migratório, induzido por BCG (Lassounskaia
E, et al 2006).
Recentemente, o remodelamento da actina mediado por TLR foi
demonstrado aumentar a captura de antígenos por DCs (West, M. A., et al, 2004).
Além disso, a ativação de TLR2 dependente de CD14, por LAM de micobactéria,
promoveu a fagocitose da micobactéria BCG por macrófagos (Sendide, K et al
2005).
A família Rho das pequenas GTPases inclui as proteínas Rac, Rho e
Cdc42, Estas proteínas apresentam ciclos de troca morfológicos entre um estado
de GTP-ligada (ativa) e GDP-ligada (inativa), para repassar as mensagens
emitidas pelos receptores da superfície celular, para o citoesqueleto (Wells C, et
al; 2004). O ciclo de ligação de GTP/GDP é regulado pela interação de Rho da
família das GTPases com fatores de troca e nucleotídeo-guanina (GEFs) e

proteínas que ativam GTPases (GAPs). GEFs catalisam a troca de GDP para
GTP, enquanto GAPs aumentam a intrínsica atividade das GTPases (Symons
M.,et al, 2000).
As proteínas Rac desempenham um papel central na migração celular pela
indução da extensão de lamelopódios, estruturas finas e espalhadas que
empurram a membrana plasmática para fora, devido à polimerização da actina
(Ridley A. J,et al, 2001).
Existem três isoformas de Rac (1, 2 e 3) em mamíferos, mas pouco se sabe
sobre as relativas contribuições de cada isoforma para as respostas dependentes
de Rac. Todas as três proteínas são altamente homólogas. Porém, apresentam
diferenças primárias concentradas na extremidade carboxi-teminal final dessas
proteínas. Cada isoforma é altamente conservada entre as espécies: Rac1 murino
difere por apenas um aminoácido da RAC1 humana.
Estudos prévios do papel das proteínas Rac na extensão dos lamelopódios
estimulados por citocinas, fator de crescimento e adesão e /ou as dobras na
membrana, tem sido baseados no uso de mutantes constitutivamente ativos ou
negativamente dominantes (Allen W. E, et al, 1997). Microinjeção em fibroblasto
com Rac 1 e Rac 2 ativados, induz o enrugamento da membrana. O contrário do
que acontece com os Rac1 dominantes negativos (N17Rac1), que inibe a
formação dos lamelopódios, dobramento da membrana e migração celular, em
vários tipos celulares, incluindo macrófagos (Allen W. E, et al, 1997).
Macrófagos deficientes em Rac-1 exibem defeitos no espalhamento celular e
dobramento da membrana, mas não na sua migração (Wells C, et al, 2004).
As integrinas são proteínas transmembranas heterodiméricas que consistem
de uma subunidade α e β que funcionam como moléculas de adesão e receptores
de sinalização. Agrupamentos de integrinas na superfície celular produzem
respostas de sinalização intracelulares as quais levam ao rearranjo do
citoesqueleto de actina, espalhamento e adesão celular (Meng F, et al; 1998).
Dentre os eventos de transdução de sinal mais recentes, mediados pelas
integrinas, destaca-se o aumento de fosforilação de tirosina em múltiplas proteínas
associadas ao citoesqueleto: paxilina, tensina, cortactina e p 130. As integrinas

dependem de tirosinaquinases associadas para emitirem seus eventos de
sinalização.
PI3-K tem sido incluída nos eventos de transdução de sinal mediados pelas
integrinas. Foi demonstrado que a inibição farmacológica da atividade quinásica
de PI3-K, bloqueia o espalhamento celular (King et al; 1997). Similarmente, o
tratamento de células com inibidores de PI3-K inibiu o rearranjo normal do
citoesqueleto de actina, dobramento da membrana e formação dos filapódios
(Hartwing, et al; 1996). Em células de carcinoma, PI3-K está envolvida nos
eventos de sinalização induzidos pela ligação das integrinas α4β2, as quais são
requeridas para a invasão celular, em ensaios in vitro (Shaw et al; 1997). Neste
sistema, os fosfoinositídeos D3 produzidos pela ativação de PI3-K atuam na
pequena GTPase Rac 1 para promover o dobramento de membrana, extensão
dos filapódios e migração celular (Meng F, et al; 1998).
Na ausência de quinases da família Src, em células mielóide, a sinalização
via integrina é bloqueada assim como macrófagos são impedidos de migrarem,
em modelo in vivo de peritonite aguda. Estas mutações parecem não afetar a
ativação de MAPK mediada pelas integrinas ou translocação de NF-κB, sugerindo
que as vias de sinalização que regulam as mudanças no citoesqueleto e migração
celular são diferentes das vias que ativam MAPK e NF-κB (Meng F, et al; 1998).

II- OBJETIVO
Verificar o efeito de interferência entre as vias de sinalização mediadas por TLR2,
e receptor de manose, MR, na ativação dos macrófagos pelas micobactérias.
Objetivos específicos:
1. Estudar o caráter da ativação dos macrófagos por BCG e/ou seus
componentes purificados, Man-LAM (agonista de MR), p19 ou PIM
(agonistas de TLR2) e verificar o efeito do pré-tratamento com Man-LAM na
ativação dos macrófagos por BCG, através de :
a) quantificação da produção de citocinas pró-inflamatórias, IL-12, TNF-α e NO, versus anti-inflamatórias, IL-10,
b) análise das mudanças morfológicas pró-migratórias sofridas pelos
macrófagos (espalhamento e polarização).
2. Verificar o efeito da estimulação do MR na modulação das vias de
sinalização acionadas por micobactéria, através TLR2 (ativação do NF-kB,
das vias de MAPK e PI3-K/Akt quinases, ativação da GTPase Rac-1 e
fosfatase SHP-1).

III- JUSTIFICATIVA
O estudo detalhado da patogenia causada por micobactéria serve de
ferramenta para o desenvolvimento de novos tratamentos baseados nos
mecanismos naturais de defesa do hospedeiro. Já está bem estabelecido o papel
dos macrófagos como célula alvo durante a infecção micobacteriana assim como
sua importância para eliminação da micobactéria através de fagocitose e
desenvolvimento de uma resposta pró-inflamatória, mediada principalmente por
TLR 2. O processamento das informações extracelulares requer uma atividade
coordenada de uma ampla gama de vias de sinalização intracelular. Entretanto, a
interferência (cross-talk) entre essas vias, acionadas por diferentes receptores,
consiste em uma nova área de estudo. Foi descrito que os patógenos são capazes
de ativar diferentes vias de sinalização com o fenômeno de interferência de uma
para a outra. A parede celular micobacteriana é complexa e possue agonistas
para vários receptores. A ativação pró-inflamatória do macrófago pela
micobactéria é mediada principalmente por TLR2, que liga lipoproteínas
micobactérianas. O antigeno imunodiminante da parede celular, Man-LAM, liga
com o receptor de manose, MR, do macrófago. Dados recentes sugerem que a
ativação de MR, por Man-LAM, induz eventos de interferência na sinalização
mediada por TLR.

IV. METODOLOGIA
4.1. Lisados e componentes micobacterianos
Mycobacterium bovis BCG (vacina onco-BCG, linhagem Moreau,
Copenhagem SEED# julho de 1978 produzida no Instituto Butantan (São Paulo,
Brasil) para tratamento de câncer de bexiga). As suspensões da vacina foram
estocadas a 40C e utilizadas em experimentos, antes da expiração da data
recomendada pelo fabricante. Para obtenção do lisado de BCG (BCG WCL) , a
vacina foi lavada duas vezes com tampão salino fosfato (PBS), pH 7.4 e sonicadas
em tampão de lise (10 mM Tris-HCL, 100 mM NaCl, 0,5% SDS e 25 mM EDTA),
pH 7.8.
Os lisados de M bovis (BCG WCL) foram feitos através da coleta das
culturas de micobactérias crescidas em meio Middlebrook 7H9 (8.7 x 1013 CFU),
centrifugação por 12,000 x g por 20 min e ressuspensão em tampão de lise (2 ml
de 0.1% Triton X-100) (Sigma). Em seguida, os sobrenadantes foram misturados
durante 30 min em t.a., e centrifugados por 2 min 10,000 x g. A concentração da
proteína no sobrenadante foi quantificada pelo método de Bradford.
O lisado de M.tuberculosis 7Hrv e os componentes purificados de M.
tuberculosis, lipoproteína 19-kDa (p19) , PIM e Lipoarabinomanana manosilada
(Man-LAM) foram obtidos de Dr. J. Belisle (Colorado State University, Ft Collins,
NIH contract), através de contrato para teste de vacina (Instituto Nacional de
Saúde e Instituto Nacional de Alergia e Doenças Infecciosas NIH –NIAID NO1-A1-
40091). A fração p19 liofilizada foi ressuspensa em Dimetilsulfóxido (DMSO) e
estocado a -800 C.
4.2. Cultura Celular
Os macrófagos murinos RAW 264.7 (ATCC TIB-71; American type Culture
Collection, Mannasas,VA) foram cultivados em meio de cultura Dulbecco’s
modified Eaglee’s medium-F12 (DMEM-F12) suplementado com 10% de soro fetal

bovino. Os monócitos humanos da linhagem THP-1 (ATCC 202-TIB) foram
plaqueados em meio DMEM-F12, 10% soro fetal bovino (FCS) em placas de
cultura de 96 orifícios (3 × 105 células/ l) e cultivados a 37°C , 5% CO2 , por 72h,
na presença de 100 nM phorbol 12-myristate 13-acetate (PMA; Sigma) (Tsuchiya
et al., 1982). Após diferenciação dos macrófagos, a monocamada foi lavada e as
células foram incubadas por 24h com meio DMEM-F12 completo, sem PMA.
4.2.1. Extração de células precursoras de medula óssea de camundongos
Os macrófagos derivados da medula óssea (BMM) foram obtidos da
medula femural de camundongos C57BL/6 TLR2 -/- ou TLR2+/+ (controle) (Doação
de S. Akira, Osaka, Japão). Os fêmures dos camundongos TLR2+/+ e TLR 2 KO
foram cuidadosamente retirados e cortados nas duas extremidades. Em seguida,
foram lavados com 5ml de meio DMEM-F12 completo (10% de SFB),
suplementado com gentamicina (50ug/ml). Após serem retirados, as células
precursoras de medula foram plaqueadas para garrafas grandes. Em 24h, os
sobrenadantes foram retirados e às células foram acrescentado meio
suplementado com 20% de sobrenadante de fibroblasto murino da linhagem L929
por 6 dias, para obtenção da monocamada de macrófagos aderentes. Em seguida
os macrófagos receberam os devidos tratamentos.
4.3. Tratamentos dos macrófagos
Para os experimentos de imunofluorescência e avaliação morfológica,
células RAW 264.7 ou BMM foram crescidas em placas de cultura de 24 poços,
cobertas com lamínulas de vidro, numa concentração de 3x105 células /ml a 370 C
em 5% de CO2, por 24h. As células RAW 264.7 foram lavadas com PBS e
incubadas por 18h adicionais com meio sem soro. A monocamada de macrófagos
resultante foi infectada com BCG viáveis numa proporção de 1:10
macrófagos/bactéria, ou tratado com BCG WCL 2µg/ml, ou 5µg/ml de p19, e
incubados a 370 C e submetidos à diversas análises subseqüentes.

Para análise de proteínas intracelulares ativadas nos macrófagos, as
células (RAW 264.7) foram crescidas em garrafas de 75 cm2, e receberam os
distintos tratamentos: BCG WCL (2µg/ml) ou Man-LAM (5µg/ml) ou Man-LAM
precedida de BCG. Após os períodos de incubação, as células foram lavadas com
PBS estéril e retiradas mecanicamente com scrapers. Estas suspensões foram
posteriormente utilizadas para Imunoprecipitação, ensaios de quinase e
fosfatasess e análises por Western Blotting.
Para quantifição de citocinas, as monocamadas dos macrófagos, crescidas
em placas de cultura de 96 poços foram tratadas com BCG ou MTB WCL ou
Lipopolissacarídeo (LPS) (2µg/ml); Man-LAM (5µg/ml); p19 ou PIM cis 3 (5µg/ml)
a uma temperatura de 37°C, 5% CO2. Após o período de incubação, os
sobrenadantes das culturas foram coletados e congelados à –20oC para
quantificação das citocinas através de ELISA, Bioensaio de L929 para TNF-α ou
Reação de Griess, para NO.
4.4. Preparação dos Lisados Celulares
Os extratos celulares foram obtidos através da ressuspensão dos pellets de
3x105 células em 200µl de tampão de lise Low Ripa (0,5% Nonidet P-40, 3 mM
pirofosfato de sódio, 50mM Tris-HCl pH 7.6, 150 mM NaCl, 0.1 mM EDTA, 50mM
NaF, 1mM Na3VO4, 1mM PMSF, DTT 1mM, 10 µg/ml leupeptina, 1 µg/ml
pepstatina, and 1 µg/ml aprotinina). Os lisados celulares foram centrifugados por
14,000 x g, por 10 min para separação do extrato protéico. A concentração das
proteínas no sobrenadante foi determinada pelo Método de Bradford (Bradford,
1976). As proteínas obtidas foram precipitadas com acetona num volume 3x maior
que o volume protéico, por 1h, -200 C. As amostras foram centrifugadas 14,000 x
g, por 5 min e acetona foi descartada. Os pellets protéicos foram incubados
overnight, 40 C. As proteínas então foram desnaturadas em tampão de amostra 1x
(15mM Tris pH 6,8, 0,5% SDS, 1,25% β-2-mercaptoetanol, 2,5% glicerol, 0,025%

azul bromofenol), fervidas por 5 minutos, e centrifugadas a 3500g por 5 minutos e
analisadas por eletroforese em SDS-PAGE a 10%.
4.5. Imunoprecipitação
Para imunoprecipitação, os lisados celulares, 600 µg, foram incubados com
anticorpo específico, seguido da proteína A/G Plus agarose (Santa Cruz
Biotechnology) a 4oC por 1h cada. Em seguida, foram tratadas com anticorpo de
coelho anti-PI3K anti camundongo ou anticorpo de cabra anti-TLR2 de
camundongo (Santa CRUZ Biotechnology). As imunoprecipitações foram lavadas
com PBS e centrifugadas 2500g, 4min, 4oC, antes da adição de tampão de
amostra 2x. As amostras foram fervidas a 100oC por 5 min, e subsequentemente
analisadas por SDS-PAGE, seguida de transferência eletroforética para
membranas de nitrocelulose (Immobilon-P; Millipore, Bedford, MA). Após a
transferencia eletroforética as membranas foram bloqueadas com leite desnatado
(Molico) 5%, diluído em PBS tween 20, por 18h, 40C. O tratamento da membrana
foi feito com anticorpos anti-TLR2, anti-p85 (subunidade de PI3K) ou anti-Rac-1
(Santa CRUZ Biotechnology). As transferências foram visualizadas através da
marcação com anticorpos anti cabra ou anti coelho conjugados com HRP(1/3000).
(Amersham, Little Chalfont, UK) e reveladas usando Kit ECL ou Diaminobenzidina
(DAB).
4.6. Anticorpos
ERK 1 (k-23)- G, IgG policlonal anti cabra; p-ERK (E-4) IgG monoclonal anti
camundongo; JNK 1(FL) IgG policlonal anti coelho; p-JNK (G-7) IgG monoclonal
anti camundongo; p 38 (C-20)-G IgG policlonal anti cabra; p-p 38 (Thr 180/Tyr
182)-R IgG anti policlonal coelho; Akt ½ (N-19) IgG policlonal anti goat; p-Akt (Ser
473)-R IgG policlonal anti coelho; p-PI3-K p85 α (Tyr 508) IgG policlonal anti
cabra; PIκB-α (B-9) IgG 2b monoclonal anti camundongo e SHPTP-1 (19)-G IgG
policlonal anti cabra- Santa Cruz Biotechnology;

4.7. SDS-PAGE 10%
A eletroforese em gel de poliacrilamida foi realizada segundo a metodologia
de Laemmli (1970), em gel descontínuo constituído de: gel separador (10%
acrilamida, 0,2% bisacrilamida, 0,1% SDS e 0,375M tris pH 8,8), gel de
concentração (5% acrilamida, 0,5% bisacrilamida, 0,1% SDS e 0,125M tris pH 6,8)
e tampão de corrida (25mMTris, 192mM glicina e 0,1% SDS pH 8,3). As corridas
eletroforéticas foram realizadas no sistema de mini-gel (Dual Gel Caster
Amersham Pharmacia Biotech), utilizando-se tampão de corrida sob 60V
constantes no gel de concentração de 10% e 100V constantes no gel separador.
Iguais quantidades de proteína celular das diferentes amostras foram aplicadas nas
canaletas e a massa molecular das amostras de proteínas foi determinada por
comparação com padrões de massas moleculares conhecidas (Amersham Life
Sciences).
4.7.1. Western blotting
Após a corrida eletroforética, as proteínas foram eletrotransferidas para
matriz de nitrocelulose com poros de 0,45µm (Amersham Life Sciences), de
acordo com Towbin et al. (1979), utilizando o equipamento de transferência líquida
Bio Rad em tampão de transferência Tris-Glicina (25mMTris, 192mM glicina e 20%
de metanol) por 45min a 250mA. Após transferência das proteínas, as membranas
foram incubadas com solução bloqueadora (5% de leite desnatado em PBS)
durante 18 horas a 4ºC. Então, foi feita a marcação das proteínas de interesse
através de anticorpos específicos.
A revelação foi realizada tratando-se as membranas com a solução
contendo substrato enzimático, H2O2 e a substância cromógena DAB
(diaminobenzidina) (Sigma) (100µl Tris- HCL 2M pH 7,5, 4,9 ml de H2O destilada,
5mg DAB, 0,3 ml imidazol 0,1M, 5µl H2O2).

4.7.2. Imunodetecção de quinases e fosfatases celulares
A verificação da ativação de IκB-α, MAPKs (ERK1, JNK1/2 e p38), AKT e
PI3K nos macrófagos estimulados pelo lisado de micobactéria e/ou seus
componentes, foi feita através da marcação com anticorpos anti a forma
fosforilada da quinase. A verificação da ativação da fosfatase SHP1 foi feita
através da marcação da sua forma fosforilada com o anticorpo anti pSHP-1
4.8. Quantificação das citocinas
4.8.1. ELISA
As concentrações de IL-10 e IL-12 nos sobrenadantes das culturas celulares
foram medidos através do ensaio imuno enzimático (ELISA- sandwich), utilizando-
se pares de anticorpos monoclonais (BD Pharmingen) anti IL-12, IL-10, adsorvidos
em placas específicas para citocinas (Nunc, Maxi Sorp). Para quantificação de IL-
12, foi utilizado anticorpo monoclonal de rato anti- camundongo IL-12p70-3µg/ml
(captura) e anticorpo monoclonal de rato anti IL-12 (p40/70) de camundongo
biotinilado 1,5µg/ml (detecção). Para quantificação de IL-10, foi utilizado anticorpo
monoclonal de rato, purificado, anti IL-10 de camundongo 3µg/ml (captura) e
anticorpo monoclonal de rato, biotinilado, anti IL-10 de camundongo 1,5µg/ml
(detecção).
A placa foi sensibilizada com anticorpo de captura por 18h, 4oC. Seguindo-se
de lavagem com tampão de lavagem (PBS HCL+ Tween 0,05%) e posteriormente,
bloqueio com solução de bloqueio (PBS HCL+ 1% de BSA) por 1h em temperatura
ambiente. Os sobrenadantes das culturas dos macrófagos foram aplicados para
serem comparados com a curva padrão. A curva padrão foi construída a partir de
diluições seriadas das citocinas recobinantes correspondentes, em tampão
diluente (PBS HCL+ Tween 0,05% +1% BSA), de modo que a curva iniciasse com
4000pg/ml. Posteriormente as amostras foram incubadas por 18h à 40C e os
poços tratados com anticorpos biotinilados correspondentes, por 2h, T.A. Em
seguida foi feita lavagem dos poços com PBS (1x) seguido do tratamento com
estreptavidina HRP por 30 min, T.A. As amostras foram incubadas por 15 minutos

com uma solução reveladora, em T.A. (50% H2O destilada, 28% de ácido cítrico
0,1M, 22% de Na2H2PO4 0,2M, 0,03% de H2O2 30%, 0,1% de ABTS-10mg). A
reação enzimática foi interrompida com a adição de 50µl de uma solução de ácido
cítrico 0,2M. A absorbância das amostras foi medida no leitor de ELISA (Dynatech
MR5000), com filtro de referência de 405nm.
4.8.2. Bioensaio de L929
Os sobrenadantes das culturas dos macrófagos murinos e humanos
estimulados pelos lisados das micobactérias foram descongelados e testados
através do bioensaio utilizando fibroblastos da linhagem L929 para a quantificação
de TNF-α. As células L929 (linhagem de fibroblastos murinos) foram tripsinizadas
com solução de tripsina 0,025% e EDTA 0,2% e plaqueadas na concentração
2,5x105 células/ml de meio DMEM-F12 contendo 10% de SFB e 20µg/ml de
gentamicina (100µl/poço), em placas de cultura com 96 poços (Corning). Após a
incubação das placas a 370C por 24 horas, o meio de cultura foi substituído pelo
meio completo (SFB 10%) acrescido de actinomicina -D (SIGMA). A curva padrão
foi construída a partir de diluições seriadas de TNF-α recombinante com volume
final de 100µl. No final foi acrescentado, por poço, 2µl dos sobrenadantes das
culturas dos macrófagos. A viabilidade celular foi avaliada através da técnica de
MTT.
4.8.3. Teste de MTT
As células foram incubadas à 37oC durante 1 hora com 0,5mg/ml da
solução de corante MTT ("3'-[4,5-dimethylthiazol-2yl] 2,5-diphenyltetrazolium
bromide"). Esta droga, nas células vivas, é convertida em cristais azuis insolúveis
(formazana) pela enzima succinato desidrogenase mitocondrial. Para dissolver os
cristais, foi adicionado às células sem o sobrenadante 0,04M de HCl/Isopropanol.
A densidade óptica foi medida em leitor de ELISA (Dynatech MR5000) com filtro
de teste 570nm e filtro de referência 630nm.

4.8.4. Quantificação de óxido nítrico (NO) pela Reação de Griess
A acumulação de nitrito produzida pelos macrófagos foi determinada pela
adição de 50µl dos sobrenadantes das cultura dos macrófagos, misturadas com
50µl da solução A (2,5ml ácido fosfórico, 0,5g sulfanilamida, 47,5ml H2O milliQ) e
a solução B (0,05g N-1-naphitiletilenodiamina, 50ml H2O milliQ) de Griess (vol/vol).
A absorbância à 570nm foi medida no espectrofotômetro. As concentrações de
nitrito foram calculadas utilizando-se uma curva padrão de NaN3 à 100 µM, e os
resultados foram apresentados em µM de nitrito.
4.9. Imunofluorescência
As células RAW 264.7 ou precursores de medula óssea (BMM) foram
crescidos em placas de cultura de 24 poços, cobertos com lamínulas de vidro,
numa concentração de 3x105 células /ml a 370 C em 5% de CO2, por 24h. As
células foram lavadas com PBS e incubadas por 18h adicionais com meio sem
soro. As monocamadas de macrófagos resultantes foram infectadas com BCG
viáveis numa proporção de 1:10 macrófagos/bactéria, ou tratadas com BCG WCL
2µg/ml, ou 5µg/ml de p19, e incubadas a 370 C e submetidas à diferentes análises
subseqüentes.
4.9.1. Ensaio para f-actina
Para localização dos filamentos de actina e quantificação do espalhamento e
polarização dos macrófagos, as células fixadas e permeabilizadas (0,5% de Triton
X-100 em PBS, por 10 min), foram incubadas com 0,1µg/ml de faloidina conjugada
com TRITC (SIGMA), por 30 min e lavadas com PBS. As lâminas foram montadas
com N- propilgalato e observadas em microscópio confocal com contraste de fase
(Zeiss Axioplan). Cada amostra celular foi primeiramente examinada em
microscópioZeiss Axioplan equipado com contraste de interferência diferencial
(DIC) e então em microscópio de fluorescência. As imagens foram capturadas
através de filmes fotográficos Kodak Tris X (ASA 400). As dimensões celulares

foram medidas usando o programa L33K (Rasband, W.S, Image J, Institutos
Nacionais de Saúde, Bethesda MD, EUA, http:/rsb.info.nih.gov/ij/, 1997-2004). O
espalhamento celular foi estimado através de medidas manuais do perímetro da
célula e medida da área das protusões celulares.
4.10. Dados Estatísticos
Os dados de quantificação das citocinas foram analisados pelo ANOVA e teste
Student t.

V- RESULTADOS
5.1. Efeito de Man-LAM na produção dos mediadores pró-inflamatórios pelos
macrófagos ativados por micobactérias
Para determinar a interferência de Man-LAM na produção das citocinas
pró-inflamatórias, IL-12p40 e TNF-α, induzidas por BCG e Mtb, em macrófagos
murinos RAW 264.7, pré-tratamos as células com Man-LAM (5 µg/ml) por 16 h ou
1h, seguida do desafio com BCG ou Mtb lisada (2µg/ml), LPS (2µg/ml) ou PIM
(2µg/ml) por 24h. Como esperado, todos os produtos bacterianos utilizados,
exceto Man-LAM, induzem alto nível da secreção de IL-12 (Figura 10 A), TNF-α
(Figura 10 B) e NO (Figura 10 C) pelos macrófagos.
A indução da síntese de IL-12 e TNF-α por Man-LAM não foi
significativa em comparação com o controle, sendo que as células RAW
264.7 produzem baixos níveis destas citocinas espontaneamente. O
tratamento com Man-LAM não induziu a produção de NO no período
observado de 24h. O pré-tratamento das células com Man-LAM diminuiu a
produção de IL 12p40 induzida por BCG, Mtb e PIM em uma proporção de
30%, 60% e 50%, respectivamente. A redução da síntese da IL-12 induzida
pelo LPS foi menor, cerca de 15% (Figura 10A).
O pré-tratamento dos macrófagos por Man-LAM não teve efeito
significativo na indução de TNF-αααα pelos produtos bacterianos citados
(Figura 10B). Diferentemente de TNF-αααα, o pré-tratamento por Man-LAM inibiu
completamente a síntese de NO induzida por BCG (Figura 10C).
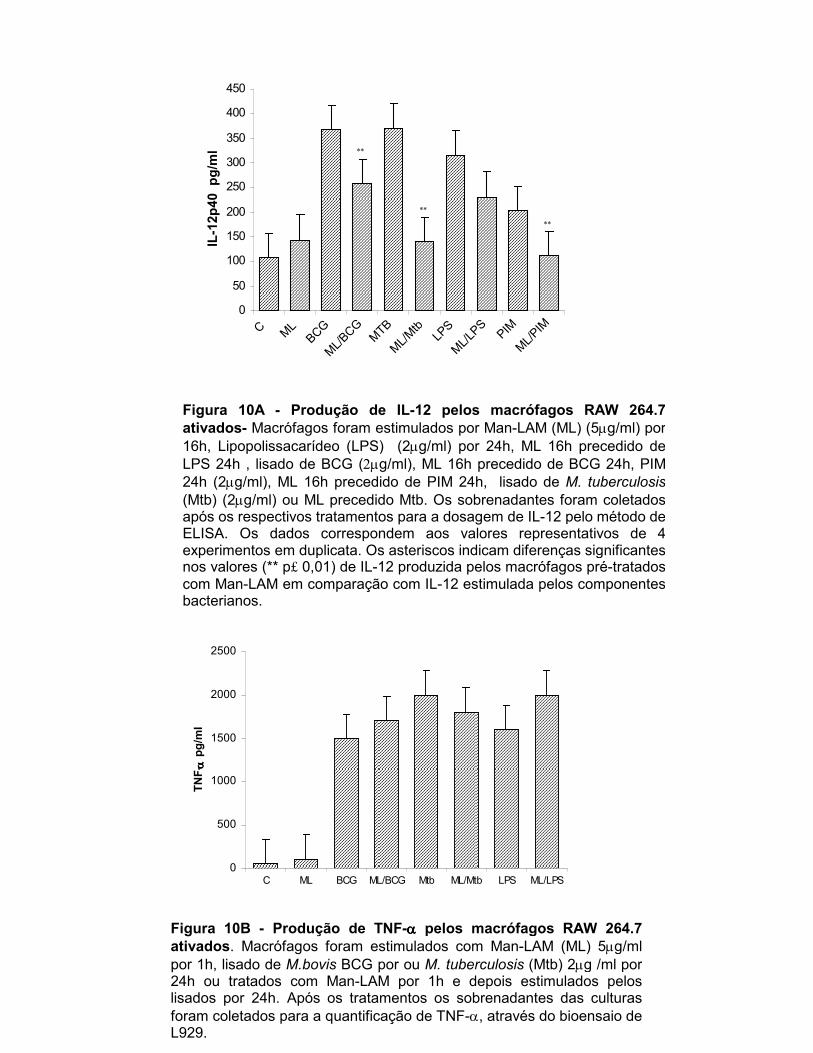
Figura 10A - Produção de IL-12 pelos macrófagos RAW 264.7 ativados- Macrófagos foram estimulados por Man-LAM (ML) (5µg/ml) por 16h, Lipopolissacarídeo (LPS) (2µg/ml) por 24h, ML 16h precedido de LPS 24h , lisado de BCG (2µg/ml), ML 16h precedido de BCG 24h, PIM 24h (2µg/ml), ML 16h precedido de PIM 24h, lisado de M. tuberculosis(Mtb) (2µg/ml) ou ML precedido Mtb. Os sobrenadantes foram coletados após os respectivos tratamentos para a dosagem de IL-12 pelo método de ELISA. Os dados correspondem aos valores representativos de 4 experimentos em duplicata. Os asteriscos indicam diferenças significantes nos valores (** p£ 0,01) de IL-12 produzida pelos macrófagos pré-tratados com Man-LAM em comparação com IL-12 estimulada pelos componentes bacterianos.
0
50
100
150
200
250
300
350
400
450
C ML
BCG
ML/BCG
MTB
ML/Mtb
LPS
ML/LPS
PIM
ML/PIM
IL-12p
40 p
g/m
l **
**
**
0
50
100
150
200
250
300
350
400
450
C ML
BCG
ML/BCG
MTB
ML/Mtb
LPS
ML/LPS
PIM
ML/PIM
IL-12p
40 p
g/m
l **
**
**
Figura 10B - Produção de TNF-αααα pelos macrófagos RAW 264.7 ativados. Macrófagos foram estimulados com Man-LAM (ML) 5µg/ml por 1h, lisado de M.bovis BCG por ou M. tuberculosis (Mtb) 2µg /ml por 24h ou tratados com Man-LAM por 1h e depois estimulados pelos lisados por 24h. Após os tratamentos os sobrenadantes das culturas foram coletados para a quantificação de TNF-α, através do bioensaio de L929.
0
500
1000
1500
2000
2500
C ML BCG ML/BCG Mtb ML/Mtb LPS ML/LPS
TNF- αα αα pg/m
l

5.1.1. A inibição de IL12p40 induzida por Man-LAM é independente de IL-10
IL-10 é um potente inibidor da expressão de genes de IL-12 e da produção
desta proteína. Para verificar se o efeito inibitório de Man-LAM na produção de
IL12p40 e NO é dependente do efeito anti-inflamatório de IL-10, foi analisada a
produção de IL-10 nas células RAW 264.7 tratadas com Man-LAM por 16h na
concentração de 5µg/ml; ou pelos lisados de BCG ou Mtb (2µg/ml), LPS (2µg/ml),
PIM (2µg/ml) por 24h; ou pré-tratadas com Man-LAM e posteriormente
estimuladas. Man-LAM e os outros produtos micobacterianos estudados induzem
a síntese de IL-10 (500-700pg/ml). O pré-tratamento dos macrófagos por Man-
Figura 10C - Produção de Òxido Nítrico (NO) pelos macrófagos RAW 264.7 ativados. Macrófagos foram estimulados com Man-LAM (ML) 5µg/ml por 1h, lisado de M. bovis BCG 1µg/ml por 24h ou tratados com Man-LAM por 1h e depois estimulados pelo lisado. Após os tratamentos os sobrenadantes das culturas celulares foram coletados para a quantificação de NO, através da Reação de Griess. Os dados correspondem aos valores representativos de 2 experimentos em duplicata. Os asteriscos indicam diferenças significantes nos valores (** p£ 0,01) de NO produzido pelos macrófagos pré-tratados com Man-LAM em comparação com os estimulados apenas pelo lisado de BCG.





























