Embed Size (px)
Citation preview

Curso de Campo Ecologia da Floresta Amazônica - 2001 1
Dadão (palhaço Goiabada)
Tonhão (Maníaco do Igarapé)
Angelita (epicentro da zoeira)
Gargamel Andrea Cachorrinho
Prof. Jansen Glaucólise Duca... Juruna (Pit Bitoka)
Cris Gaúcha (A Realeza)
Chris Gringa (OK! OK!)
Betsa (Shakira Gandi)
Caê (Caretando Veloso)
Katchaça BaladaFinoGuga Edgar (Rambo)

2 Curso de Campo Ecologia da Floresta Amazônica - 2001
Paula CabelinhoNatalie (lousinha da Xuxa) Leandro Wolverine Maurício Semi-aleatório
Thiago Gorfão Rosinha Time-lag Paula Pé de Cana Rodrigo Pitangui (bisturi de ouro)

Curso de Campo Ecologia da Floresta Amazônica - 2001 3
ÍndiceReserva Florestal Adolpho Ducke - Floresta de terra firmeAbundância e riqueza de plântulas sob a copa de duas espécies de palmeiras (Arecaceae) em função da presença ou ausência deespinhos (Reserva Ducke, Manaus, AM) ................................................................................................................................................... 5Influência do habitat e da temperatura sobre o início da atividade de libélulas (Odonata) ....................................................................... 6Estimativa da biomassa vegetal em floresta de terra firme na Amazônia Central ..................................................................................... 9Variações no tamanho de girinos de Hyla geographica (Hylidae, Anura) na área de influência do barramento do igarapé Barro Branco,Manaus – AM ........................................................................................................................................................................................... 11Madeirera MIL - Itacoatiara - Floresta manejadaComunidade de briófitas epífilas de quatro espécies de palmeiras em uma floresta de terra firme da Amazônia Central ...................... 13Efeito do corte seletivo na regeneração da vegetação de clareiras na Amazônia Central ......................................................................... 15Ocorrência e ritmo de atividade de duas espécies de lagartos (Squamata: Lacertilia: Teiidae) em área de preservação e corte seletivo nafloresta amazônica (Itacoatiara, AM) ........................................................................................................................................................ 17Influência da extração seletiva de madeira sobre as comunidades de aves e morcegos em uma área de floresta da Amazônia Central . 19Influência da extração seletiva de madeira em parâmetros físicos, serapilheira e na biomassa de artrópodes de sub-bosque em umafloresta de terra firme da Amazônia central .............................................................................................................................................. 22Perturbações no habitat afetam a riqueza e abundância de herbívoros galhadores? ................................................................................ 24Efeito do manejo e de clareiras sobre a regeneração natural de Miconia tetraspermoides (Melastomataceae) e Minquartia guianensis(Olacaceae) ............................................................................................................................................................................................... 26Influência da exploração seletiva de madeira sobre a fauna de anuros de serapilheira em área de baixio na Amazônia Central ............ 28Ilha da Marchantaria - VárzeaEfeito da inundação sobre o padrão reprodutivo de Vitex cymosa (Verbenaceae) em uma área de várzea no Rio Solimões, AM ......... 30Estrutura da ictiofauna associada a bancos de macrófitas flutuantes em uma área de várzea do Rio Solimões, AM .............................. 31Estudo da dieta de duas espécies de aves granívoras na várzea da Ilha da Marchantaria, AM ................................................................ 35Análise comparativa da riqueza e distribuição de espécies de plantas herbáceas aquáticas de três ambientes de várzea da Ilha deMarchantaria (Iranduba, AM) ................................................................................................................................................................... 36Um experimento sobre a predação de cupins por gerrídeos (Hemiptera: Gerridae) ................................................................................ 38Análise da dieta e uso da várzea pela população ribeirinha da Ilha da Marchantaria (Rio Solimões): uma breve descrição dascomunidades ribeirinhas de São Lázaro II e São Francisco, Ilha da Marchantaria (Iranduba, AM) ........................................................ 40Zonação de herbáceas aquáticas no gradiente topográfico de um sistema de várzea da Ilha da Marchantaria, Amazônia Central ......... 42Composição funcional trófica de insetos bentônicos emergentes no gradiente topográfico de uma área de várzea na Ilha daMarchantaria, Iranduba, AM .................................................................................................................................................................... 44Arquipélago de Anavilhanas - IgapóEfeito do isolamento em poças sobre a estrutura da comunidade de macroinvertebrados bentônicos no arquipélago das Anavilhanas,AM ............................................................................................................................................................................................................ 46Efeito da inundação da planta hospedeira sobre insetos galhadores em um igapó da Amazônia Central ............................................... 47Efeitos da inundação na mortalidade das árvores adultas de uma floresta de igapó no Arquipélago das Anavilhanas, AM ................... 49Aporte de nutrientes e herbivoria em florestas alagadas da Amazônia: uma abordagem ecológico-evolutiva no arquipélago dasAnavilhanas, AM ...................................................................................................................................................................................... 51Utilização de uma ilha do Arquipélago de Anavilhanas, Baixo Rio Negro (AM) por mamíferos de médio e grande porte .................... 54Distribuição espacial de três famílias de insetos predadores em diferentes micro-habitats em uma poça temporária em floresta de igapóna Amazônia Central ................................................................................................................................................................................ 56Efeito da presença de látex na herbivoria em plantas da floresta de igapó ............................................................................................... 57Atributos ecomorfológicos para determinação de guildas tróficas da ictiofauna de igapó no arquipélago das Anavilhanas, AmazôniaCentral ...................................................................................................................................................................................................... 59A ictiofauna de uma poça sazonal nas ilhas Anavilhanas, rio Negro, AM ............................................................................................... 62Reserva do Km 41 - Floresta de terra firmeFatores estruturais que influenciam o tamanho dos jardins de formiga ................................................................................................... 64Um teste da hipótese de disponibilidade de recursos em clareiras da Reserva do Km 41 ........................................................................ 66Defesa química de duas espécies de hilídeos (Anura) contra um peixe predador ................................................................................... 68Seleção intra e inter individual para sítios de oviposição em Maieta guianensis (Melastomataceae) pela aranha Argyrodes sp. (Araneaea:Theridiidae) ............................................................................................................................................................................................... 69Efeito do ápice foliar e da epifilia no tempo de dessecamento de folhas do subosque de uma floresta de terra firme ............................ 71Reações anti-herbívoros da formiga Pheidole minutula Mayr (Formicidae: Myrmicinae) em uma planta mirmecófita, Maieta guianensisAubl. (Melastomataceae). ......................................................................................................................................................................... 73Ocorrência de galhas em relação à esclerofilia de folhas numa floresta de terra firme da Amazônia Central, Reserva do Km 41, AM .. 75Relação entre a forma da folha e o grau de herbivoria em Iriartella setigera (Arecaceae) ........................................................................ 77Aspectos estruturais e populacionais de agregados de Leucauge sp. (Araneae: Tetragnathidae) ............................................................. 79Observações subaquáticas de Iguanodectes variatus Gerý, 1993 (Characidae) ........................................................................................ 80Caracterização dos jardins de formiga em Conceveiba martiana (Euphorbiaceae) na Reserva do Km 41, Amazônia Central ............... 82Guaribas (Alouatta seniculus, Primates Cebidae) influenciam a distribuição de espécies por eles dispersadas? ..................................... 84Descrição da Estrutura e padrão espacial de inflorescências de Rapatea ulei (Rapataeceae), uma planta herbácea de baixio em mata de

4 Curso de Campo Ecologia da Floresta Amazônica - 2001
terra firme, AM ......................................................................................................................................................................................... 86Composição, riqueza e abundância da ictiofauna de um igarapés de terra firme da Amazônia Central, AM .......................................... 87Respostas a herbívoros simulados na mirmecófita Maieta poepigii (Melastomataceae): Pheidole minutula Mayr (Formicidae) é que eraa formiga de verdade! ............................................................................................................................................................................... 90Influência do tamanho e tipo de presa no recrutamento de indivíduos da aranha social Anelosimus eximius (Araneae: Theridiidae) ... 94Papel do dimorfismo sexual na reprodução de uma espécie de aranha da família Pholcidae .................................................................. 97Estratificação vertical na ocorrência de galhas em uma Floresta de Terra Firme da Amazônia Central ................................................. 100Defensas alternativas en plantas mirmecófitas y en función de la ontogenia ......................................................................................... 102Efeitos da estrutura física de habitats sobre a composição e abundância da ictiofauna em igarapés da floresta de terra firme naAmazônia Central ................................................................................................................................................................................... 106Intensidade de herbivoria por invertebrados ao longo de um gradiente vertical em uma floresta de terra firme na Reserva do Km 41,Manaus, AM ........................................................................................................................................................................................... 110Interações planta-animal na história natural de uma herbácea do baixio, Rapatea ulei (Rapateaceae) na Reserva do KM 41, Manaus,Amazonas ............................................................................................................................................................................................... 113Diferenças de estratégias reprodutivas em duas espécies de palmeiras clonais, Geonoma deversa e Geonoma stricta (Arecaceae) ..... 116Estudo das relações tróficas dos peixes de um igarapé da Reserva do Km 41,Manaus, AM ................................................................. 119Influência do tamanho corpóreo no comportamento defensivo de mariposas: uma abordagem fisiológica ......................................... 122Distribuição longitudinal e estrutura trófica da ictiofauna de igarapés de terra firme da Amazônia Central .......................................... 125Efeito de borda sobre a intensidade de predação de sementes artificiais em uma floresta de terra firme na Amazônia Central ........... 130Fatores que influenciam o tamanho de clareiras e seus impactos em uma floresta de terra firme na Amazônia Central ...................... 133Avaliação de dois índices de integridade ecológica através da comunidade íctica em um igarapé de terra firme da Amazônia Central 136Levantamento da riqueza e composição de espécies de mamíferos terrestres com o auxílio de uma nova metodologia para identificaçãode pegadas .............................................................................................................................................................................................. 141Influência de características ambientais sobre o tamanho de grupos de gerrídeos (Hemiptera, Gerridae) em igarapés na AmazôniaCentral .................................................................................................................................................................................................... 145Interação entre formigas e Cecropia purpurascens (Cecropiaceae) na Amazônia Central: diferentes espécies protegem a planta com amesma eficiência? ................................................................................................................................................................................... 149Interação entre formigas e Cecropia purpurascens (Cecropiaceae) na Amazônia Central: diferentes espécies protegem a planta com amesma eficiência? ................................................................................................................................................................................... 152Influência da abertura de estradas nos padrões do uso de áreas e composição da comunidade de mamíferos de médio a grande portenuma área de floresta de terra-firme na Amazônia Central .................................................................................................................... 155

Curso de Campo Ecologia da Floresta Amazônica - 2001 5
Abundância e riqueza de plântulas sob a copa de duas espéciesde palmeiras (Arecaceae) em função da presença ou ausência de espinhos
(Reserva Ducke, Manaus, AM)Edgar F. Prieto, Angela M. F. Pacheco, Antonio J. C. Aguiar, Paula S. Pinheiro & Thiago Timo
IntroduçãoPalmeiras são elementos característicos das florestas daAmazônia Central, apresentando uma grande variedade dehábitos e ocorrências (Kahn et al. 1988). Nas áreas de solodrenado as palmeiras Attalea attaleoides e Astrocaryumsciophilum representam as espécies dominantes do sub-bosque (Kahn & Castro 1985). Vários fatores podeminfluenciar a presença de plântulas, entre eles podemos citara estocagem de sementes por mamíferos e o uso de refúgiostemporários por dispersores em locais protegidos contrapredadores (Smythe 1989 apud Cintra 1997). Da mesmaforma, a utilização de plantas como poleiros por avesfrugívoras pode resultar em acúmulo de fezes e conseqüenteagrupamento de plântulas sob a copa.Este estudo se baseia na hipótese que a presença de espinhosna raque das folhas de Astrocaryum sciophilum poderiainfluenciar o estabelecimento de plântulas sob a copa. Oobjetivo do presente estudo foi avaliar o efeito da ocorrênciade espinhos em A. sciophilum em relação a Attaleaattaleoides, sobre a riqueza e abundância de plântulas naárea de sombra destas palmeiras.
Material e MétodosO estudo foi realizado na Reserva Florestal Adolpho
Ducke (2o55’ S; 59o59’ W), em uma área de vertente pertoda trilha de acesso a norte-sul 1, próximo a nascente doIgarapé Barro Branco, no dia 09 de agosto de 2001.
Foram coletadas todas as plântulas com no máximo 30cmde altura, localizadas sob a copa de indivíduos de Attaleaattaleoides e Astrocaryum sciophilum a partir de um raio de1m da base do caule das palmeiras. As palmeiras vistoriadasapresentavam pelo menos 3 folhas e 1m de raio de copa, eeram distantes no mínimo de 5m entre si e 5m da trilha. Asplântulas foram contadas e separadas em morfotipos paraobtenção de estimativas de riqueza. Não foi determinado onúmero total de morfoespécies entre as amostras.
A estimativa de tamanho das palmeiras foi realizado deduas formas: (1) contagem direta do número de folhas e (2)área da copa, inferida pela média do comprimento das trêsmaiores folhas. Esta medida foi utilizada como raio paraposterior cálculo da área da copa. Neste procedimento usamosa fórmula da área do circulo assumido que as copas daspalmeiras têm forma circular. Também foi calculada adensidade média do número e morfoespécies de plântulaspor metro quadrado.
Foram amostrados 36 indivíduos, entre jovens e adultos,sendo 18 de Attalea attaleoides e 18 de Astrocaryumsciophilum.
A abundância e a riqueza de morfoespécies foram testadascontra os dois índices de tamanho para palmeiras com e semespinhos. Os dados foram analisados através de ANCOVA(Zar 1984).
ResultadosForam coletadas 994 plântulas, sendo 564 sob a copa de
Astrocaryum sciophilum e 430 sob Attalea attaleoides. Onúmero de plântulas sob a copa de cada indivíduo variou de12 a 97 em A. sciophilum, e 2 a 50 em A. attaleoides, sendoque o primeiro apresentou um número médio de plântulasmaior que o segundo (Tabela 1). O número médio demorfoespécies foi maior em A. sciophilum (Tabela 1), comnúmero de plântulas variando de 8 a 31e em A. attaleoidesde 2 a 30.
Attalea attaleoides apresentou indivíduos com maior áreamédia (Tabela 1), com área de copa variando de 3,0 a 74,6m2 e número de folhas variando de 4 a 16, em relação a A.sciophilum que variou de 1,9 a 49,4 m2 e 4 a 16 folhas.
Não houve correlação entre o número de plântulas e aárea da copa (N = 36; r2 = 0,088; g.l. = 1; p = 0,240), comotambém entre o densidade de morfoespécies e o tamanho dacopa (N = 36; g.l. = 1; r2 = 0,022, p = 0,937).
O número de folhas e o número de plântulas nãoapresentou correlação (N = 36; r2 = 0,325; g.l. = 1; p = 0,197).Também não houve correlação entre o número de folhas e adensidade de morfoespécies (N = 36; r2 = 0,022; g.l. = 1; p =0,785).
Tabela 1. Médias e desvio padrão de índices de tamanho e densidadede plântulas sob a copa de Attalea attaleoides e Astrocaryumsciophilum, em área de vertente Próximo ao igarapé Barro Branco,Reserva Florestal Adolpho Ducke, AM (N = 36).
DiscussãoA presença de espinhos na raque das folhas de
Astrocaryum sciophilum não interferiu no estabelecimentode plântulas no presente estudo. O resultado encontrado porAzevedo-Ramos et al. (1994) e Vicentini et al. (1995a), queestudaram as mesmas espécies apontam para uma maiorabundância de plântulas sob a copa de A. sciophilum. Ambosos autores atribuem este resultado a um possível efeito dedefesa contra herbivoria. Entretanto, o estudo de Vicentiniet al tem um volume de dados pequeno, e o resultado docoeficiente de regressão foi baixo, o que diminui aconfiabilidade dos seus resultados (N = 10; r2 = 0,22).
Em contrapartida, Tabarelli et al. (1994) estudaram o
Attalea attaleoides Astrocaryum sciophilum
No de pl ntulas 29,7 21,8 31,3 22,7
No de morfo-espécies/palmeira 4,5 2,4 4,9 1,9
Densidade de pl ntulas/m2 7,6 4,2 9,9 7,2
No de folhas 9,0 3,0 7,7 3,1
Área da copa (m 2) 29,7 21,8 19,2 16,2

6 Curso de Campo Ecologia da Floresta Amazônica - 2001
padrão de riqueza entre A. sciophilum. e A. attaleoides masnão encontraram maior número de morfoespécies de plântulassob a copa de A. sciophilum.. Além disso, Vicentini et al.(1995b) encontraram uma maior abundância de plântulas emáreas sem influência da copa das palmeiras, o que sugereque não está havendo uma proteção efetiva de plântulas soba copa de A. sciophilum, e que é grande a probabilidade deque os resultados devam-se à fatores aleatórios.
O estabelecimento de plântulas sob a copa destas duasespécies de palmeiras pode estar sendo influenciado poroutros fatores ainda não considerados, como por exemplo achuva de sementes das árvores acima dos indivíduosamostrados, ou a distância da planta-mãe.
Novos estudos enfocando o estabelecimento de plântulassob palmeiras com espinhos devem considerar os fatores quepossam estar influenciando a chegada de sementes e oestabelecimento de plântulas isoladamente. Estudos commétodos de exclusão de aves, de invertebrados ou demamíferos, para um acompanhamento a longo prazo, e aidentificação da síndrome de dispersão das sementes podemgerar um panorama mais interessante.
Agradecimentos
Agradecemos ao Juruna Jurubeba pelo apoio na coleta
dos dados.
Referências BibliográficasAzevedo-Ramos, C.; Tabarelli, M.; Simões-Filho, F.F.L.;
Faria, M.L. & Takahasi, A. 1994. Efeito da palmeiracom espinhos (Astrocaryum: Arecaceae) sobre a
estrutura da comunidade de plântulas abaixo de sua copaem uma mata de terra firme, Amazônia, Brasil: I -Abundância de plântulas. Livro do Curso de Campo“Ecologia da Floresta Amazônica”. INPA/PDBFF,Manaus, AM.
Cintra, R. 1997. Sobrevivência por dispersão de sementes eplântulas de três espécies de palmeiras em relação apresença de objetos naturais na Floresta Amazônia. In:Regeneração Florestal na Amazônia, Moutinho, P. &Gascon, C. (ed.).
Kahn, F.; Meija, K. & Castro, A. 1988. Species richness anddensity of palms in terrafirme forests of Amazônia.Biotropica 20: 266-269.
Kanh, F. & Castro, A. 1985. The palm community in a forestof Central Amazônia. Biotropica 17: 210-216.
Smythe, N. 1989. Seed survival in the palm Astrocaryumstandleyanum: evidence for dependence upon its seeddispersers. Biotropica 21: 50-56.
Tabarelli, M.; Simões-Filho, F.F.L. & Azevedo-Ramos, C.1994. Efeito da palmeira com espinhos (Astrocaryum,Arecaceae) sobre a estrutura da comunidade de plântulasabaixo de sua copa em uma mata de terra firme,Amazônia, Brasil: II – Riqueza de espécies de plântulas.Livro do Curso de Campo “Ecologia da FlorestaAmazônica”. INPA/PDBFF, Manaus, AM.
Zar, J.H. 1984. Biostatistical Analysis. Prentice-Hall, NewJersey.
OrientadoresEduardo Venticinque e Jansen Zuanon
Influência do habitat e da temperatura sobre o início da atividadede libélulas (Odonata)
Leandro M. Scoss, Daniel L. M. Vieira, Katiane M. Ferreira, Paula C. Ceotto & Betsabé Ruiz
IntroduçãoAs libélulas, insetos pertencentes à ordem Odonata, estão
divididas em duas subordens, Zigoptera e Anisoptera. Osadultos, assim como as ninfas, são predadores e estão ativosdurante o dia. Durante o período de reprodução, as espéciesse mantém predominantemente próximas a corpos d’água,onde as fêmeas fazem suas posturas. Nos períodos entre aemergência do adulto e a maturidade sexual, elas podem serencontradas em locais relativamente distantes da água. Ocomportamento da maioria das espécies durante a fase dereprodução é territorial (Borror et al. 1981).
Baseado no comportamento de termorregulação, Corbet(1962 apud De Marco 1998) classificou as libélulas emespécies voadoras (fliers), que se mantém voando a maiorparte do tempo, e as espécies pousadoras (perchers), queficam mais frequentemente pousadas. As espécies pousadoraspodem regular sua temperatura através da irradiação e daconvecção. A irradiação é o recebimento de calor pela
incidência solar direta e a convecção está ligada à troca decalor entre o animal e o meio. Os animais pousadores podemser divididos em dois grupos: aqueles que dependem do calorde convecção, e os heliotérmicos, que dependemprincipalmente da irradiação solar. As diferenças demecanismos termorreguladores entre as espécies podemexercer influências os horários de atividade e sobre oshabitats utilizados pelas libélulas (De Marco 1998).
Neste trabalho, comparamos o horário de início deatividade e espécies de Odonata em duas áreas adjacentes,uma aberta e outra área de mata, com objetivo de avaliar oefeito da temperatura sobre o horário de atividade dessasespécies. Uma vez que as espécies maiores (com menorrelação superfície/volume) demoram mais tempo paraabsorver calor (P. De Macro com. pess.), nossa hipótese erade que as menores iniciariam suas atividades mais cedo, jáque ganhariam calor mais rapidamente. Analisamos tambémo efeito das diferenças de temperatura entre as áreas e o efeito

Curso de Campo Ecologia da Floresta Amazônica - 2001 7
do tamanho corporal sobre a preferência de habitats por estegrupo de insetos.
Material e MétodosO estudo foi realizado na Reserva Florestal Adolpho
Ducke, gerenciada pelo Instituto Nacional de Pesquisas daAmazônia (INPA), um fragmento florestal de terra firme de10.000 ha, localizado a 26 Km do centro de Manaus, AM. Oclima da região é quente e úmido, com temperatura média de26,7 oC e precipitação anual em torno de 2.186 mm (Lovejoy& Bierregaard 1990). O local escolhido para o estudo foi oigarapé Barro Branco, próximo a sede administrativa dareserva.
Dois trechos de coleta foram delimitados, sendo um nosubosque da floresta e outro em uma área aberta, ambos àsmargens do igarapé Barro Branco. As coletas foram realizadaspela manhã, entre 7:30 e 11:30 horas. Entre 9:00 e 10:00horas, as atividades foram interrompidas devido à chuva e,em seguida, reiniciadas.
Para testar a hipótese de que a temperatura influencia ohorário de início de atividade de espécies de libélulas, foramcomparados dois tipos de ambiente: uma área aberta e osubosque de um trecho de floresta. As libélulas foramcoletadas utilizando puças, e para cada indivíduo foramtomados os seguintes dados biométricos: comprimento daasa, largura e comprimento do tórax. A temperatura ambientefoi aferida sempre que um espécime era capturado, e aobservação do primeiro indivíduo de cada espécie foiutilizada para analisar o início da atividade da espécie nosdois locais.
Verificamos se o tamanho das espécies (largura do tórax)influencia o horário de inicio das suas atividades em cadaárea estudada através de regressão linear. Para verificar adiferença na temperatura de início das atividades das espéciesde subosque e área aberta, utilizamos o teste t-student (Zar1984). Da mesma forma, utilizamos o teste t-student paratestar o efeito do habitat sobre o tamanho das espécies delibélulas.
ResultadosRegistramos 8 espécies de libélulas na área aberta e 10
espécies no subosque num total de 34 indivíduos (Tabela 1).Apenas Argia sp.1 e a Protoneuridae sp. 1 foram comunsnas duas áreas. As 18 espécies de libélulas amostradas nestetrabalho são consideradas pousadoras (De Marco 1998). Amaioria das espécies encontradas no subosque pertenciam àsubordem Zigoptera, enquanto a subordem Anisoptera foimais frequente na área aberta.
Na área aberta, a temperatura alcançou valores maiores evariou mais (25 a 34oC) comparado com o subosque (23 a27oC). A Figura 1 ilustra a variação de temperatura duranteo período de amostragem (7:50 a 11:30 h) e o início deatividade das espécies de libélulas nos dois ambientes.
Subosque Area abertaZigoptera
Lestidae (não identificado) 3Argia sp.1 4 1Argia sp.2 1Chalcopteryx scintilans 1Fylgia amazonica 1Hetaerina amazonica 2Hetaerina sempronia 1Lestes sp.1 2Lestes sp.2 1Mecistogaster jocaste 1
AnisopteraErythrodiplax basalis 2Erythrodiplax fusca 1Erythrodiplax cf. funerea 1Micrathyria sp. 1Orthemis discolor 1Protoneuridae sp.2 4Protoneuridae sp.1 1Zenithoptera spp. 5
N. de IndivíduosSubordem / Espécies
Tabela 1. Lista das espécies de libélulas encontradas em um dia deamostragem em dois tipos de habitat do Igarapé do Barro Branco,Reserva Ducke, Manaus, AM.
A temperatura média em que as espécies de libélulasiniciaram suas atividades foi maior na área aberta (X ± DP =29,5oC ± 4.31) do que no subosque (X ± DP = 25,1oC ±1,21)(t = 2,81; g.l. = 7,8; p = 0,024). Contudo, quando utilizamosna análise apenas a variação da temperatura comum às duasáreas (23 a 26oC), não foi observada diferença em relação aoinício da atividade de libélulas (t= 0,854; g.l. = 11,5; p =0,4).
O tamanho das espécies não influenciou a temperaturaem que estas iniciaram suas atividades tanto na área aberta(r2 = 0,12; F = 0,55; p = 0,55), como no subosque (r2 = 0,13;F = 1,08; p = 0,33). As espécies de área aberta apresentaramlargura média do tórax de 4,7 mm (DP = 2,5), enquanto queas espécies do subosque mediram em média 2,54 mm (DP =0,52), não havendo diferença significativa (t = 2,06; g.l. =5,3; p = 0,091) (Figura 2). Por outro lado, as espécies deáreas abertas apresentam o tórax mais comprido (X ± DP =6,52 ±2,22) que espécies de subosque (X ± DP = 3,88 ±0,83)(t = 2,78; g.l. = 5,9; p = 0,03). A Figura 2 representa adistribuição das espécies em relação à largura do tórax paraas duas áreas amostradas.
Figura 1. Horário de início de atividade de espécies de libélulas –área aberta (direita) e subosque no igarapé Barro Branco, ReservaDucke, Manaus, AM. Cada ponto representa o primeiro registro deatividade de uma espécie.

8 Curso de Campo Ecologia da Floresta Amazônica - 2001
DiscussãoOs resultados deste trabalho mostram que os dois habitats
analisados tiveram diferentes composições de espécies, comsobreposição apenas de um gênero e uma família, indicandoo alto grau de especialização das espécies amostradas emfunção do ambiente analisado. A temperatura de início deatividade das diferentes espécies registradas para cada tipode habitat também diferiu entre as áreas. Esta diferença podeestar relacionada à características fisiológicas ecomportamentais envolvidas na regulação da temperaturacorporal de cada espécie.
Figura 2. Comprimento médio do tórax em espécies de libélulas deárea aberta e subosque na área do igarapé Barro Branco, ReservaFlorestal Ducke, Manaus, AM. Os pontos são a média do comprimentodo tórax para cada espécie da área aberta (o) e do subosque (x).
De Marco (1998) sugere que diferenças entre osmecanismos de termorregulação entre voadores e pousadorespodem determinar o uso de diferentes microhabitats. Osdados obtidos neste trabalho, não permitem a discussão dediferenças entre voadores e pousadores, pois apenaspousadoras foram amostrados (18 espécies). Desta forma,nossa discussão é restrita à variação entre comunidades depousadoras.
Nossa hipótese previa que as libélulas menorescomeçariam suas atividades antes das maiores, já queganhariam calor mais rapidamente. No entanto, nãoencontramos relação entre o horário de início de atividade eo tamanho das libélulas dentro de cada ambiente. É possívelque o tamanho do corpo das espécies encontradas não variea ponto de refletir respostas comportamentais associadas àscaracterísticas fisiológicas das mesmas.
Os resultados deste trabalho sustentam a hipótese de quehá diferença entre o tamanho das espécies de libélulas emrelação ao habitat que foram amostrados. O fato de espéciesmaiores de libélulas terem sido mais frequentes na área abertae que as menores mais observadas no subosque, não significa
30
35
Tem
pert
ura
(oC
)necessariamente que estas são exclusivas de cada uma destasáreas.
Uma outra explicação para termos encontrado as espéciesmaiores na área aberta e as menores no subosque seria queas pequenas suportam uma amplitude térmica menor,refletindo uma limitação fisiológica em relação à temperatura.O subosque é uma zona mais estável, principalmente pelascaracterísticas estruturais da vegetação e da temperatura daágua do igarapé, que oscila pouco ao longo do ano (J. Zuanoncom. pess.). Devido ao tamanho, as espécies menores sofremuma influência maior da temperatura ambiente, o que poderiatorná-las mais sensíveis a ambientes com grande amplitudetérmica.
O maior tamanho das espécies encontradas na área abertapode estar relacionado à fisiologia delas. Na área de mata,as espécies contam principalmente com a temperaturaambiente para termorregular. Já na área aberta o aquecimentopor irradiação direta torna-se tão ou mais importante que aconvecção para manter o animal em atividade. O tamanhomaior das espécies da área aberta pode ser explicado pelamaior dificuldade de atingirem o limiar de temperaturanecessário ao início de atividade simplesmente porconvecção, o que explicaria o investimento em habitar umaárea aberta, onde ganhariam calor mais rapidamente porirradiação.
No subosque, a presença de espécies menores poderia serexplicada pela hipótese de que estas espécies não seriamcapazes de competir com espécies maiores em ambientes maisabertos. Outra razão para acreditarmos que espécies menorespreferem ambientes mais fechados é a maior susceptibilidadeà predação. As espécies de Zigoptera não possuem a mesmaeficiência de vôo que as Anisoptera, o que as tornaria presasmais fáceis (De Marco com. pess.). Por outro lado, éimportante considerar que as diferenças de tamanhoencontradas entre as áreas podem estar associadas à filogeniado grupo, já que os Zigoptera são animais mais delgados emenores que os Anisoptera.
AgradecimentosGostaríamos de agradecer especialmente os préstimos de
Paulo De Marco Jr. pela idealização deste projeto e pelaidentificação das espécies de Odonata. Agradecemos tambéma todos os colegas, professores e coordenadores do cursoque contribuiram para melhoria da discussão dos resultadosdeste trabalho. Ao INPA, agradecemos a disponibilizaçãodas instalações da Reserva Ducke para o desenvolvimentodo curso “Ecologia da Floresta Amazônica - 2001”.
Referências BibliográficasBorror, J.D.; De Long, M.D.; & Triplehorn, A. 1981. An
Introduction to the Study of Insects. Saunders CollegePublishing, Philadelphia.
De Marco, P.J. 1998. The amazonian campina dragonflyassemblage: patterns in microhabitat use and behaviourin a foraging habitat (Anisoptera). Odonatologica. 27(2):239-248.
Lovejoy, T.E. & R.O. Bierregaard. 1990. Central Amazonian

Curso de Campo Ecologia da Floresta Amazônica - 2001 9
forests and the minimum critical size of the ecossystemsproject. In: .. Four Neotropical Rainforest, Gentry, A.H. (ed.), pp. 60-71. Yale University Press, Haven.
Zar, H.J. 1984. Biostatisitcal Analysis. Prentice Hall, NewJersey.
OrientadorPaulo De Marco Jr.
Estimativa da biomassa vegetal em floresta de terra firmena Amazônia Central
Cristiane F. Jurinitz, Felipe B. Oliveira, Maria R. Darrigo, Mauricio Camargo & Rodrigo Cogni
IntroduçãoProcessos naturais e artificiais, como a respiração e a
queima de combustíveis fósseis, desprendem gás carbônicopara a atmosfera. Especialistas em ciclos biogeoquímicostêm detectado uma diferença significativa entre a quantidadedesse gás que é liberada e a concentração que é medida naatmosfera. Dessa forma, de acordo com as taxas de emissão,esperaria-se que sua concentração na atmosfera fosse maior(A. Nobre com. pess.). Uma hipótese para o destino dessegás carbônico sugere que as florestas tropicais atuam comosumidouros, incorporando esse CO
2 à sua biomassa (Brown
& Lugo 1992).Os dados obtidos para o fluxo de gás carbônico na
“atmosfera” das florestas tropicais são cada vez mais precisose mostram que, de forma geral, a floresta consome maiscarbono do que libera (A. Nobre com. pess.). Emcontrapartida, a estimativa de biomassa vegetal da floresta,outro elemento necessário para se testar hipóteses sobre adinâmica do carbono nesses ecossistemas, ainda é feita demaneira muito imprecisa. Brown et al. (1995), em umlevantamento dos estudos feitos até o momento, relatadiferenças nas estimativas encontradas para a biomassa dasflorestas tropicais brasileiras de até 230%. Essa variaçãodeve-se principalmente à diferenças no tamanhos dasamostras, nos critérios de inclusão dos indivíduos amostradose às diferentes fisionomias florestais. Por esse motivo, até omomento, não existe um consenso sobre qual é a estimativade biomassa mais adequada para a Amazônia brasileira(Brown et al. 1995).
Recentes acordos internacionais, como o Protocolo deKyoto, sugerem que os processos de fixação de carbonopossam de alguma maneira serem comercializados, o queaumenta a necessidade de maior confiabilidade nasestimativas de biomassa da floresta (Fearnside 1999).
O objetivo deste estudo é comparar quatro métodosdiferentes para estimar a biomassa acima do solo em umtrecho de terra firme na Reserva Ducke (Manaus, AM),discutindo as vantagens e desvantagens de cada um.
Material e MétodosRealizamos o estudo na Reserva Adolpho Ducke, área de
100 km2 próxima a Manaus (AM), predominantemente
ocupada por floresta de terra firme (Ribeiro et al. 1999). Areserva possui uma torre de aproximadamente 45 metros dealtura construída com a finalidade de realizar medidas da“atmosfera” da floresta. Há alguns anos essas medidas foraminterrompidas devido a alterações provocadas pelaproximidade da área urbana (A. Nobre com. pess.). Dequalquer forma, a torre foi construída em um localrepresentativo das matas de terra firme da Amazônia central,motivo que nos levou a escolher essa área para realizar essasestimativas.
Ao redor da torre, delimitamos uma área circular de 1 ha,medindo as árvores vivas de diâmetro à altura do peito (DAP)igual ou maior de 45 cm e estimamos a altura total. Obtivemoso DAP medindo o diâmetro do tronco a 1,30 m de altura eno caso de árvores com sapopemas ou troncos irregulares,estimamos o diâmetro projetando sua extensão sobre umatrena ou medindo-o um pouco acima das sapopemas. Issofoi necessário porque estudos indicam que a rigidez em mediro DAP a 1,30 m em árvores com sapopemas podesuperestimar a biomassa das árvores em até 40% (Clark &Clark 2000). Utilizando essas medidas, estimamos abiomassa acima do solo (BAS) para cada indivíduo, atravésde quatro equações:
1) Biomassa = 0,62 x 0,67 x área basal x altura ou BAS =0,0326 x (DAP)2 x altura (Brown et al. 1995)
2) Biomassa = 21,297 - 6,953 x DAP + 0,74 (DAP)2 (Brown1997)
3) Biomassa = 38,49 – 11,79 x DAP + 1,193 (DAP)2 (Brownet al. 1989)
4) Biomassa = exp {3,323 + [2,546 x ln (DAP/100)]} x 600(Santos, 1997)
Como árvores com DAP maior que 45cm representamaproximadamente 50% da BAS de uma área (Brown et al.1995), a soma da biomassa dos indivíduos foi multiplicadapor dois para se obter a BAS total da área.
ResultadosEncontramos um total de 30 indivíduos com DAP maior
que 45 cm no hectare estudado. As estimativas de BASvariaram entre 154 e 325 toneladas por hectare (Tabela 1).

10 Curso de Campo Ecologia da Floresta Amazônica - 2001
Tabela 1. Estimativas da biomassa (viva sobre o solo) para cadaequação aplicada nos dados obtidos em um hectare de Floresta deTerra Firme na Reserva Ducke, Manaus (AM).
DiscussãoDos resultados obtidos para a biomassa com as quatro
equações, três deles (1, 2 e 3) estão compreendidos nointervalo de estimativas já feitas para a biomassa da FlorestaAmazônica brasileira (Brown et al. 1995). O valor maisdiscrepante foi obtido com a equação 2, derivada por Brownet al. (1989) a partir de medidas realizadas em florestas daCosta Rica, sujeitas à uma precipitação anual de 4000 mm(Clark & Clark 2000), muito maior que a observada naReserva Ducke (2100mm - M. Hopkins com. pess.). Assim,a discrepância entre as estimativas pode ser explicada peladiferença na estrutura da vegetação, resultante das diferençasde precipitação entre os dois locais (Ribeiro et al. 1999).Assim, o resultado da equação 2 parece subestimar a biomassada floresta quando comparado ao obtido com outrasequações, pelo menos no que diz respeito às áreasrelativamente mais secas da Amazônia Central.
As equações 1 e 3 foram derivadas de dadosindependentes, sendo que uma delas incorpora na análiseuma outra variável (altura, no caso da equação 1). Portanto,o fato dessas duas equações fornecerem estimativassemelhantes aumenta a confiabilidade do nosso resultado.No entanto, a utilização de cada equação exige esforçosdiferentes e implica em níveis distintos de incerteza. Issoporque a equação 1 incorpora a altura das árvores, umparâmetro difícil de medir diretamente e com imprecisãorazoável associada à sua estimativa. Assim, nos parece que épossível que uma estimativa de biomassa baseada apenas noDAP possa ser mais adequada.
A equação 4, desenvolvida a partir de dados de uma áreapróxima a Manaus, muito parecida com a Reserva Ducke,forneceu resultado similar ao obtido por Laurance et al.(1999) para a biomassa nas reservas do PDBFF à cerca de60 km de Manaus.. Dessa forma, consideramos que a áreaescolhida para esse estudo é representativa das florestas deterra firme da Amazônia Central, pelo menos no que dizrespeito à biomassa.
Cálculos de biomassa de florestas são extremamentedifíceis de serem realizados, motivo pelo qual se constroemmodelos para estimar esse valor. Assim, por se tratarem deestimativas, existe um erro inerente, que geralmente não émencionado nos estudos realizados. Dos trabalhos
Equação Biomassa (ton./ha)
1 - Brownet al.1995 (DAP e altura) 250
2 – Brown 1997 (DAP) 154
3 – Brown 1989 (DAP) 246
4 - Santos 1997apud
Laurance et al. 1997 (DAP)
325
consultados neste estudo, apenas Brown et al. (1995) discuteessa incerteza, concluindo que ela ainda é muito grande. Poresse motivo, com base nos estudos realizados até o momento,valores encontrados entre 150 e 450 ton/ha podem serconsiderados razoáveis para a Amazônia Central, visto quenão são apresentados intervalos de confiança para essasestimativas.
O foco de maior interesse atual é a questão do sumidourode CO
2, principalmente devido às pressões geradas peos
problemas de aquecimento global (Fearnside 1999). Esseproblema exige que estimativas muito mais precisas sejamfeitas, principalmente em uma escala regional. Taisestimativas precisam incorporar, além da biomassa acima dosolo, os dados para serapilheira, árvores mortas e biomassaradicular, que por natureza são muito mais difíceis de seremobtidos, mas necessários para aumentar a confiança nasestimativas de biomassa.
AgradecimentosAgradecemos ao Dr. Foster Brown pela orientação e ao
Dr. Leandro Ferreira pelos comentários sobre o trabalho.
Referências BibliográficasBrown, I.F.; Martinelli, L.A; Thomas, W.W.; Moreira, M.Z.;
Ferreira, C.A.C. & Victoria, R.A. 1995. Uncertainty inthe biomass of Amazonian forests: an exemple fromRondonia , Brazil. For. Ecol. Manag. 75: 175-189.
Brown, S. & Lugo, A.E. 1992. Above ground biomassestimates for tropical moist forests of the BrazilianAmazon. Interciencia 17: 8-18.
Brown, S. 1997. Estimating biomass and biomass change oftropical rain forest: a primer. Forestry Paper 134, FAO.Roma.
Brown, S.; Gillespie, A.J.R. & Lugo, A.E. 1989. Biomassestimation methods for tropical forests with applicationsto forestry inventory data. For. Sci. 35: 881-904.
Brown, S.; Gillespie, A.J.R. & Lugo, A.E. 1991. Biomassof tropical forests of south and southeast of Asia. Can.J. For. Res. 21:111-117.
Clark, D.B. & Clark, D.A. 2000. Landscape-scale variationin forest structure and biomass in a tropical rain forest.For. Ecol. Manag. 137: 185-198.
Fearnside, P.M. 1999. Como o efeito estufa pode renderdinheiro para o Brasil. Ciência Hoje 26: 41-44.
Gaston, G.; Brown, S.; Lorenzini, M. & Singh, K. D. 1998.State and change in carbon pools in the forest of AfricaTropical. Global Change Biol. 4: 97-114.
Laurance, W.F.; Fearnside, P.M.; Laurance, S.G; Delamonica,P.; Lovejoy, T.E.; Rankin-de Merona, J.M;. Chambers,J.Q. & Gascon, C. 1999. Relationship between soils andforest biomass: a landscape-scale study. For. Ecol.Manag.118: 127-138.

Curso de Campo Ecologia da Floresta Amazônica - 2001 11
Laurance, W.F.; Laurance, S.G.; Ferreira, L.V.; Rankin-deMerona, J.M.; Gascon, C. & Lovejoy, T.E. 1997.Biomass collapse in Amazonian forest fragments.Science 278: 1117-1118.
Ribeiro, J.E.; Hopkins, M.J.G.; Vincentini, A.; Sothers, C.;Costa, M.A.S.; Brito, J.M.; Souza, M.A.D.; Martins,L.H.P.; Lohmann, L.G.; Assunção, P.A.C.L.; Pereira, E.;
Silva, C.F.; Mesquita, M.R.; Procopio L.C. 1999. Florada Reserva Ducke: Guia de Identificação das PlantasVasculares de uma Floresta de Terra-Firme na AmazôniaCentral. INPA, Manaus.
OrientadorI. Foster Brown
Variações no tamanho de girinos de Hyla geographica (Hylidae, Anura) naárea de influência do barramento do igarapé Barro Branco, Manaus – AM
Natalie Olifiers, Andrea H. Lamberts, Christina McCain, Carlos E. Marinelli, Alexander V. Christianini
IntroduçãoA presença de predadores pode afetar não apenas a
distribuição de suas presas naturais (Schmidt 1995), comotambém a composição e estrutura dessas populações,resultando na adoção de comportamentos de defesa (Begon1990).
Entre as estratégias apresentadas pelas formas larvais deanuros, observa-se a tendência dos jovens de algumasespécies manterem-se agregados, locomovendo-se eforrageando em grupos (Wasserburg 1973 apud Azevedo-Ramos 1995). Este comportamento inibe o ataque depredadores, uma vez que a imagem de um grupo tem umtamanho maior do que a maioria dos predadores de anurosnesta fase de seu desenvolvimento (Oliveira 1996).
Estudos sobre o comportamento de agregação sãodifundidos para alguns gêneros (Hyla e Bufo), sendo queestes grupos podem ser formados por indivíduos nãodiretamente aparentados, embora esta suposição ainda nãotenha sido confirmada (Costa et al. 1998).
Hyla geographica tem ocorrência restrita a ambientesflorestais e corpos d’água associados (Duellman 1978). Osgirinos de H. geographica, apresentam comportamento deagregação logo após a fase de eclosão, aliado à produção desecreções tóxicas que os tornam impalatáveis, ampliando aeficiência do aposematismo (Wassersug 1973 apud Azevedo-Ramos 1995).
O objetivo deste estudo foi testar se o tamanho de girinosde H. geographica é determinante na agregação dosindivíduos em cardumes em um igarapé na Amazônia Central.
Material e MétodosVariações no tamanho do corpo de girinos de H.
geographica (Hylidae, Anura) foram analisadas paracardumes ocorrentes na área de influência do barramento doigarapé Barro Branco, na Reserva Adolpho Ducke (2°55’ S;59°59’W), Manaus, AM.
Para obtenção dos espécimes, empregamos técnicasintensivas de captura direta com puçá (35 x 35cm), até quefossem capturados o maior número de exemplares possível.
Os cardumes foram separados em recipientes, anotando-se onúmero de indivíduos presentes em cada um. Foram retiradosao acaso 30 indivíduos de cada cardume, dos quais foi tiradaa medida de comprimento do corpo.
Análise de variância simples (ANOVA) foi empregada paratestar possíveis diferenças no comprimento do corpo degirinos entre os cardumes. Foi também utilizado o teste deTukey para verificar quais cardumes eram formados porindivíduos de tamanhos semelhantes. Para cada cardumeforam ainda calculados os coeficientes de variação dotamanho do corpo dos indivíduos. A partir dos resultadosencontrados empregamos um teste F para verificar eventuaisdiferenças nos coeficientes de variação entre os grupos degirinos ao longo de diferentes fases de desenvolvimento.
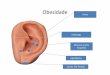
ResultadosForam capturados quatro grupos de girinos, compostos
por indivíduos de tamanhos diferentes (ANOVA: F = 160,4;n = 120; p < 0,001) (Tabela 1). A partir do teste de Tukey foipossível verificar que os grupos amostrados dividiam-se emduas categorias de tamanho: girinos “grandes” (grupos 1 e3) e “pequenos” (grupos 2 e 4) (Tabela 1, Figura 1). Dentrodestas duas categorias de tamanho, os grupos compostos porgirinos em estágios de desenvolvimento mais avançados(grupos 1 e 3) apresentaram maior variação no tamanho decorpo do que grupos formados por girinos mais jovens(grupos 2 e 4) (F = 11.9; g.l. =30; p < 0.05).
Tabela 1. Número total de girinos em cada grupo capturado,
mostrando o tamanho médio do corpo (mm), desvio padrão (DP) e
coeficiente de variação (CV).
Grupos No. Indivíduos X ± DP CV
1 708 12,2 ± 2,7 0.22
2 431 5,3 ± 0,4 0.07
3 520 13,0 ± 2,8 0.22
4 548 4,3 ± 0,3 0.08
12 Curso de Campo Ecologia da Floresta Amazônica - 2001
Figura 1. Distribuição do tamanho do corpo dos girinos dentro decada grupo de Hyla geographica amostrado (N = 30 para cada grupo).
DiscussãoNo presente estudo, os girinos de H. geographica
demonstraram um padrão não casual de agregação, compondogrupos formados por indivíduos de tamanhos semelhantes,confirmando os padrões anteriormente relatados por Costaet al. (1998) para esta espécie.
A partir dos padrões observados, duas hipóteses podemser sugeridas neste estudo:
1) cada grupo seria formado por indivíduos que possuemafinidade de parentesco (mesma ninhada), com a variaçãono tamanho dos indivíduos dentro dos cardumes sendoresultante de taxas de desenvolvimento diferentes após operíodo de eclosão dos ovos (M. Gordo com. pess.). Nestecaso, os cardumes de girinos “pequenos” apresentam variaçãode tamanho relativamente menor que os cardumes de girinos“grandes”, devido ao menor tempo decorrido após a eclosãodos ovos. 2) os indivíduos de diferentes ninhadas agregariam-se a partir de características morfológicas comuns, já que emestágios de desenvolvimento semelhantes possuemcapacidades natatórias e velocidade de escape semelhantes(Azevedo-Ramos 1995). É possível ainda, que mecanismosde competição ou partição de recursos influenciem adistribuição de girinos em grupos (Begon et al. 1990).
Duellman (1978) discute que o número de ovosproduzidos em uma única desova de H. geographyca oscilaentre 2000 e 4000, evidenciando o contraste com os dadosobservados neste trabalho, já que os grupos amostradosapresentaram entre 431 e 708 girinos. As diferençasencontradas poderiam ser devidas a predação ou a divisãodos indivíduos originários de uma desova em vários grupos.Técnicas de marcação e acompanhamento de grupos degirinos em diferentes fases de desenvolvimento poderiamesclarecer estas questões.
Por outro lado, observamos que os agrupamentos deindivíduos maiores não necessariamente apresentam menorabundância que os grupos de menor tamanho corporal, oque seria esperado se os girinos fossem exclusivamente damesma ninhada, uma vez que existe uma probabilidade donúmero de indivíduos em um grupo diminuir com o tempoem função da mortalidade ou simples dispersão.
Dessa maneira, os indivíduos se agrupariam portamanho e não por parentesco. Este agrupamento nãonecessariamente implicaria em algum mecanismo dereconhecimento de tamanho entre os indivíduos, uma vezque existe uma variação natural no desenvolvimento dosgirinos implicando em diferentes eficiências de locomoçãoe forrageio.
AgradecimentosAgradecemos ao Marcelo Gordo pela orientação e apoio
no trabalho e a todos os alunos e professores do cursoEcologia da Floresta Amazônica 2001 pelos comentários esugestões.
Referências BibliográficasAzevedo-Ramos, C. 1995. Ecologia de comunidades de
girinos as margens do Rio Tapajós em uma região desavana Amazônica. Tese de Doutorado, Unicamp,Campinas, SP.
Begon, M.; Harper, J.L. & Townsend, C.R. 1990. Ecology:Individuals, Populations, and Communities. BlackwellScientific Publications, Boston.
Costa, C.P.A.; Boege, K.; Silveira, M.; Fonseca, M.A. &Fáveri, S.B. 1998. Distribuição de tamanho ecomportamento de agregação de girinos de Hyla
geographica (Hylidae). Livro do Curso de Campo“Ecologia da Floresta Amazônica”. INPA/PDBFF,Manaus, AM.
Duellman W.E. 1978. The biology of an EquatorialHerpetofauna in Amazonian Ecuador. MiscellaneousPublications of the University of Kansas Museum ofNatural History, 65: 146-148.
Oliveira, S.N. 1996. Efeitos diretos e indiretos da predaçãosobre três espécies de girinos (Amphibia-Anura) naAmazônia Central. Dissertação de Mestrado, INPA,Manaus, AM.
Schmidt, B. 1995. Predation by belostomids and aeshnids(Insecta: Hemiptera and Odonata) on tadpoles ofPhyllomedusa tarsius (Amphibia: Hylidae).
OrientadorMarcelo Gordo
0
2
4
6
8
10
12
14
16
18
20
0 1 2 3 4Grupos
Co
mp
rim
en
tod
oco
rpo
(mm
)

Curso de Campo Ecologia da Floresta Amazônica - 2001 13
Comunidade de briófitas epífilas de quatro espécies de palmeiras em umafloresta de terra firme da Amazônia Central
Andrea H. Lamberts, Natalie Olifiers, Christina McCain, Carlos E. Marinelli & Alexander V. Christianini
IntroduçãoSucessões ecológicas geralmente definem a dinâmica das
populações que colonizam um hábitat (Tilman 1988). Muitasdas propriedades e funcionamento de comunidades sãoresultantes de sua história de colonização (Begon 1990). Adensidade inicial de colonizadores e suas probabilidades deocorrência para diferentes espécies são únicas em cada fasesucessional.
Embora o fenômeno de colonização seja um eventoprobabilístico, seu sucesso está intrinsecamente ligado aopotencial de ocupação do hábitat pela espécie . Apesar dessaimportância, os padrões sucessionais em comunidades nãosão consensualmente aceitos entre os ecologistas (Tilman1988).
A ocupação de folhas por briófitas, em sua maioria daFamília Lejeuneaceae (Hepaticae) (Lücking 1995), pode sercaracterizada como uma sucessão ecológica, uma vez que deacordo com Begon (1990) uma folha jovem constitui umasuperfície nua que é gradualmente colonizada por diferentesespécies.
Briófitas epífilas podem ser um bom objeto para o estudode comunidades, devido à facilidade de amostragem epossibilidade de acompanhá-las durante todo o período decolonização, inclusive até a fase de queda da folha hospedeira(C. Zartman com. pess.).
Apesar disso, há poucos estudos enfocando suascomunidades. A maioria deles refere-se a levantamentos ecompilações sobre a ocorrência de espécies (e.g. Allen &Grandstein 1996), mas mesmo a distribuição dessas espéciesainda é pouco conhecida. Os relatos de ocorrência de espéciesdeste grupo para a zona neotropical restringem-se a um únicoestudo, onde foram registradas 67 espécies numa única folha(Lücking 1995).
O objetivo do presente estudo foi caracterizar a flora debriófitas epífilas que ocorre sobre folhas de quatro espéciesde palmeiras em uma floresta de terra firme da AmazôniaCentral. Concomitantemente, buscamos investigarassociações específicas e substituição de espécies de briófitasao longo de um gradiente temporal, relacionado à idade dasfolhas amostradas.
Material e MétodosEste estudo foi conduzido ao longo de uma transecção de
cerca de 300 m de extensão em uma floresta de terra firme,localizada na área de preservação da Mil Madeireira ItacotiaraLtda, no município de Itacotiara, AM. As amostragens foramrealizadas em um único dia, durante a estação seca no anode 2001.
Para analisar a estrutura das comunidades, foramamostrados exemplares de folhas das seguintes espécies depalmeiras hospedeiras: Astrocaryum sciophilum, Socratea
exorrhiza, Syagrus inaja, e Geonoma maxima. Para obtençãodas folhas, percorremos uma trilha no gradiente ambientalplatô-baixio, onde a cada 30 metros - no mínimo- era marcadoum ponto onde uma palmeira de cada gênero que se localizavamais próxima deste ponto era amostrada. A amostragemconsistia na coleta de uma folha jovem, uma de idadeintermediária e uma velha de cada planta. Quando osindivíduos apresentavam folhas muito grandes, coletávamosum folíolo de cada categoria de idade de folha (nova,intermediária e velha).
Para cada folha (ou folíolo) de palmeira, foramdeterminadas a riqueza de espécies, área foliar total, e a áreafoliar colonizada pelas espécies de briófitas. A área foliarcolonizada foi quantificada da seguinte forma: um griddividido a cada 1 cm2, confeccionado em uma transparência,foi colocado sobre a folha. Quando a área coberta porbriófitas atingia pelo menos 50% do quadrado de 1cm2, esteera quantificado como preenchido. A soma dos quadradospreenchidos gerou uma estimativa da área total de cada folhacoberta por briófitas. Esta estimativa foi dividida pela áreatotal foliar para gerar uma estimativa de cobertura de briófitasindependente do tamanho da folha. No total, foramamostrados 44 folhas ou folíolos de palmeiras. Contamoscom o auxílio de Charles Zartman e de consultas a Lücking(1995) para auxiliar nas identificações das briófitas.
Calculamos a média e o desvio padrão da riquezade espécies de briófitas em relação à idade da folhahospedeira e o índice de correlação de Spearman parainvestigar relações entre o número de espécies de briófitas,sua cobertura por folha e idade.
ResultadosIdentificamos um total de 23 espécies de briófitas, o que
corresponde a 30% da ocorrência esperada no local, baseadonuma comparação com um levantamento detalhado realizadonas áreas do Projeto de Dinâmica Biológica de FragmentosFlorestais (PDBFF), localizada há cerca de 100 km da áreadeste estudo (C. Zartman com. pess.).
Embora não tenhamos verificado uma grandeespecificidade das briófitas em relação às palmeirashospedeiras, algumas espécies ocorreram preferencialmentenas folhas de algumas espécies de hospedeiras em detrimentode outras (Tabela 1).
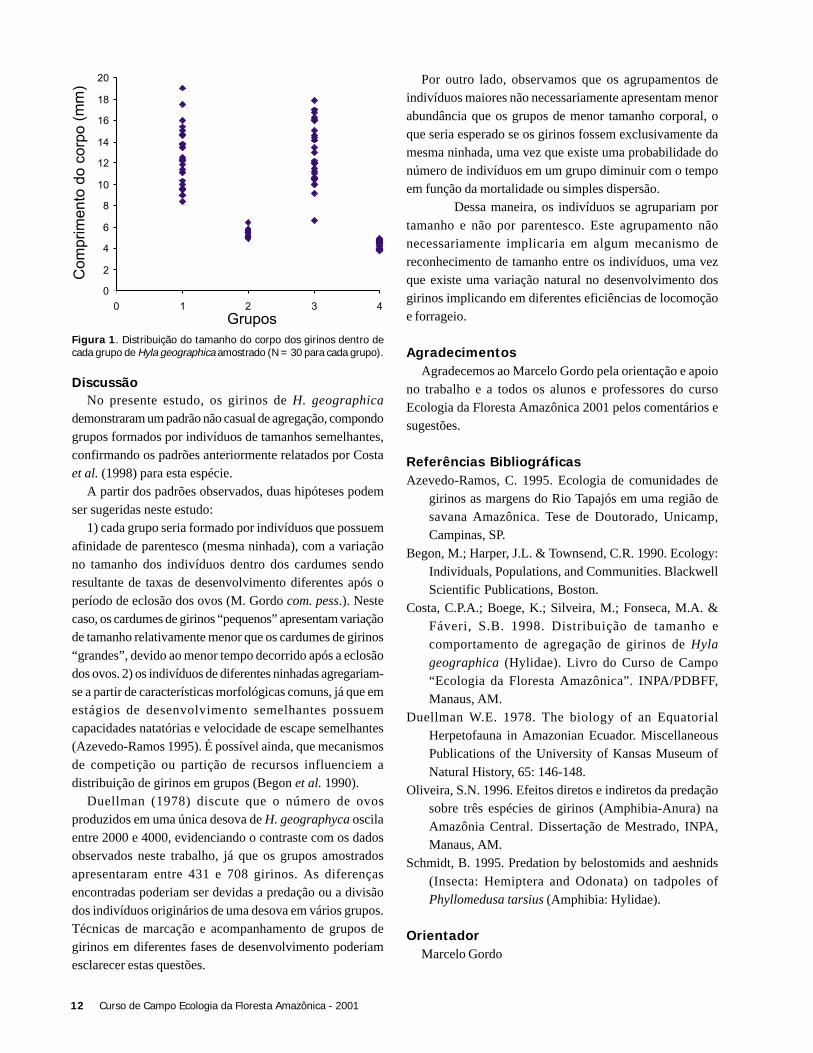
A espécie Cololejeunea thiersana foi encontrada emapenas uma folha, em uma das palmeiras hospedeiras (A.sciophilum). Por outro lado, Cyclolejeunea convexistipaapresentou alta freqüência de ocorrência em todas as espéciesde palmeiras amostrados (Tabela 1). Estas diferenças nasfreqüências de ocorrência evidenciam a existência de algumasespécies raras, pelo menos localmente –e outras mais comuns,que seriam dominantes na área estudada. Entretanto, a Figura
14 Curso de Campo Ecologia da Floresta Amazônica - 2001
1 mostra que esta dominância relativa não seria muitoacentuada, já que a diminuição na frequência de ocorrênciadas diferentes espécies ocorre de forma relativamente“gradual”.
Nenhum padrão quanto a riqueza de espécies de briófitasfoi observado em relação a idade das folhas (r
s = 0,134; N =
44; p > 0,05), nem em relação às espécies de palmeirasestudadas ou à superfície relativa da folha ocupada porbriófitas (r
s = 0,026; N = 44; p > 0,05) (Figura 2).
Algumas espécies ocorreram mais freqüentemente emdeterminados estágios de desenvolvimento das folhas depalmeiras, evidenciando uma variação temporal nacomposição e abundância de espécies, embora esta variaçãonão apresente um padrão direcional de aumento com a idadeda folha. Por exemplo, Lejeunea flava foi encontrada apenasem folhas intermediárias ou velhas, enquanto Ceratolejeuneacoarina foi mais freqüente em folhas velhas (66%) do queem folhas novas (22%) ou intermediárias (11%).
Figura 1. Freqüência de ocorrência das espécies de briófitasamostradas em 44 folhas de palmeiras das espécies Syagrus inaja,Socratea exorrhiza, Geonoma maxima e Astrocaryum sciophilum, naárea de preservação da Madeireira Mil, Itacoatiara, AM.
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
Cyc
lole
jeun
eaco
nvex
istip
a
Col
olej
eune
asu
rina
men
sis
Aph
anol
ejeu
nea
spp.
Lep
ttole
jeun
eael
liptic
a
Cer
atol
ejeu
nea
coar
ina
Cyc
lole
jeun
eape
ruva
na
Aph
anol
ejeu
nea
grac
ilis
Rad
ula
flac
ida
Dra
pano
leje
unea
cruc
iane
lla
Lej
unia
flav
a
Odo
ntol
ejeu
nea
lunu
lata
Dip
lasi
olej
eune
ape
lluci
da
Rha
pido
leje
unea
polr
rhiz
ans.
Col
ura
sp.
Cer
atol
ejeu
nea
cube
nsis
Har
pale
juni
a
Col
olej
eune
ath
iers
ana
Arc
hile
jeun
eafu
scec
ens
Oct
olej
eune
aal
bidu
m
Dra
pole
jeun
ealic
heni
cola
Rap
hale
june
asp
.
Lep
tole
jeun
eatr
iden
tata
Plag
iole
jeun
easp
.
Freq
uênc
ia
0
0,020,04
0,060,08
0,1
0,120,14
0,16
Ast
roca
ryum
scio
philu
m
Geo
nom
am
axim
a
Soc
rate
aex
orrh
iza
Sya
grus
inaj
a
Den
sida
de
Para uma das espécies de briófitas identificadas nopresente trabalho - Cololeujeunea thiersana Tixier – estefoi o segundo registrado no Brasil, evidenciando a escassezde informações sobre o grupo no Brasil (C. Zartman com.pess.).
Figura 2. Densidade média de briófitas sobre folhas de quatroespécies de palmeiras.
DiscussãoA ausência de um padrão nítido de variação na riqueza
total de briófitas com a idade das folhas pode significar queinterações entre as espécies, como facilitação, inibição e/oucompetição podem estar estruturando a comunidade durantea sucessão. Se a variável tempo, representada pelas diferentesidades das folhas de hospedeiras amostradas fosse a únicadeterminante na sucessão de espécies, esperaríamos encontrarmaior riqueza de briófitas nas folhas hospedeiras mais velhas,o que não necessariamente ocorreu.
A cobertura (abundância) de briófitas sobre as folhashospedeiras poderia estar relacionada à localização dasmesmas, ou seja, palmeiras que estivessem mais próximas acursos de água e áreas úmidas deveriam apresentarabundância maior, pois as briófitas dependem da água parase reproduzirem. Infelizmente, o parâmetro umidade não foi
Briófitas/Hospedeiras Syagrus inaja Socratea exorrhiza Geonoma maxima Astrocaryum sciophilum Total
M V J M V J M V J M V
3 4 1 1 0 3 3 3 3 4 5 21
3 4 0 0 1 3 3 3 2 3 3 28
4 2 0 0 0 2 3 3 2 2 3 22
2 2 0 1 0 3 3 1 1 3 1 18
1 1 1 1 4 2 4 15
2 2 2 1 1 1 1 1 1 14
1 1 1 1 1 2 1 1 1 1 11
1 1 1 1 2 3 9
1 1 1 1 1 1 6
2 1 1 3 7
1 1 2 1 1 6
1 1 2 5
1 1 3
1 1 1 1 1 1 6
1 1 1 3
1 1 2
1 1
1 1
1 1
1 1
1 1
1 1
1 1
19 22 9 5 8 14 25 21 10 22 27
Idade das folhas ?
C. convexistipa
C. surinamensis
Aphanolejeunea spp.
L. elliptica
C. coarina
C. peruvana
A. gracilis
R. flacida
D. crucianella
L. flava
O. lunulata
D. pellucida
R. polrrhizans.
Colura sp.
C. cubensis
H. traviscuspis
C. thiersana
A. fuscecens
O. albidum
D. lichenicola
Raphalejunea sp.
L. tridentata
Plagiolejeunea sp.
Total
Tabela 1: Ocorrência das espécies de briófitas em folhas de diferentes idades, nas quatro espécies de palmeiras hospedeiras. J = folhajovem; M = folha de idade intermediária; V = folha velha.

Curso de Campo Ecologia da Floresta Amazônica - 2001 15
mensurado. Apesar de uma aparente diferença na coberturatotal de briófitas nas diferentes espécies de palmeira, adiferença não foi significativa. Nota-se que Syagrus tem umacobertura bem menor que Astrocaryum. É possível que estadiferença se tornasse significativa caso tivéssemos maisamostras.
A variação na composição e na freqüência de ocorrênciadas espécies com a idade das folhas seria o reflexo do poderde colonização de cada espécie - em termos de dispersão ouvelocidade de colonização da folha – e das possíveisinterações entre as diversas espécies durante a sucessão. Paradeterminar quais interações estariam ocorrendo entre asespécies de briófitas, precisaríamos realizar experimentosmais detalhados.
AgradecimentosAgradecemos a Charles Zartman pela orientação,
identificação das briófitas e apoio no trabalho de campo, e atodos os alunos e professores do curso Ecologia da FlorestaAmazônica 2001 pelos comentários e sugestões.
Referências BibliográficasAllen, N.S. & Grandstein, S.R. 1996. A report on the
bryoflora of Perú. In: Manu: La Biodiversidad delSureste del Perú, Wilson, D.E. & Sandoval, A. (eds.),pp. 201-207. Smithsonian Institution, USA.
Begon, M.; Harper, J.L. & Townsend, C.R. 1990. Ecology:Individuals, Populations and Communities. BlackwellScientific Publications, Boston.
Lücking, A. 1995. Diversität und Mikrohabitatpräferenzenepiphyller Moose in einem tropischen Regenwald inCosta Rica. Dissertation zur Erlangung des DoktorgradesDr. Rer. Nat. Der Fakultät für Naturwissenschaften derUniversität Ulm. Ulm, Deutschland.
Tilman, D. 1988. Plant Strategies and the Dynamics andStructure of Plant Communities. Princeton, New Jersey.
OrientadorCharles Zartman
Efeito do corte seletivo na regeneração da vegetação de clareiras naAmazônia Central
Katiane M. Ferreira, Leandro M. Scoss, Daniel L. M. Vieira, Paula C. Ceotto & Betsabé Ruiz
IntroduçãoO corte de madeira para fins comerciais é uma das
alternativas ao desflorestamento de florestas tropicais(Anderson 1990). Contudo, poucas madeireiras aplicammétodos de manejo para efetuar a extração. Atualmentemenos que 0,12% das florestas tropicais têm uma propostade exploração de baixo impacto (Poore 1989). A certificaçãoao contrário da legislação inerente à exploração madeireiraé recente, e poucos são os indícios de que haja efetividadenos planejamentos de baixo impacto deste tipo de exploração(C. Rittl com. pess.).
Uma das primeiras consequências do desmatamento é aabertura do dossel e criação de clareiras, modificandocondições microclimáticas como, incidência de luz e aumentoda temperatura, gerando assim uma diminuição na umidade.Além das consequências relacionadas diretamente ao corte,são provocados impactos por máquinas, que aumentam acompactação e a erosão do solo, que podem afetar acicatrização da floresta.
As clareiras também ocorrem naturalmente, e sãoconsideradas a maneira pela qual a floresta se regenera e serenova (Brokaw 1982). A dinâmica de clareiras é consideradafator preponderante na determinação da composição eestrutura das florestas tropicais e na manutenção da altadiversidade de espécies nestes ecossistemas (Denslow 1980;Hartshorn 1980; Brokaw 1982; Brokaw 1985a,b).
Luz, temperatura, umidade, dimensões das clareiras,distâncias das fontes de sementes, dispersão e banco desementes, relações planta-herbívoros, dentre outros, são
fatores que atuam de forma integrada na regeneração dasclareiras. O tamanho da clareira afeta diretamente a capacidadede regeneração da floresta e a riqueza de espécies, visto quea relação entre intensidade luminosa e o tamanho da clareiraatuará na seleção e proporção de colonização de cada grupoecológico.
Como uma das conseqüências mais evidentes do corteseletivo de madeira é o aumento da densidade de clareiras,nós partimos do pressuposto que o aumento nadisponibilidade de recursos, principalmente luminosidade,aumente a probabilidade de um maior número de espécies seregenerarem nos estágios mais iniciais da sucessão ecológica,em áreas onde há exploração madeireira. Por outro lado, éesperado que em áreas onde há maior densidade de indivíduosde plantas, a competição por recursos e espaço seja maior,favorecendo um número menor de espécies. Sendo assim, nósquestionamos se clareiras em florestas intactas e exploradasdiferem quanto a densidade de indivíduos e riqueza deespécies regenerantes.
Material e MétodosO estudo foi realizado na área da empresa Mil Madereira
Itacoatiara Ltda., localizada no município de Itacoatiara, AM.O clima da região é quente e úmido, com temperatura médiade 26oC, precipitação em torno de 2.200 mm e solo amarelodistrófico argiloso (Mil Madeireira Ltda., Planejo de Manejo1997). Neste trabalho foram propostos dois tratamentos: umtalhão não explorado (controle) e em um talhão que foisubmetido à extração de madeira, sob normas de certificação

16 Curso de Campo Ecologia da Floresta Amazônica - 2001
florestal (explorado) em 1999.Em cada talhão (explorado e controle) foram selecionadas
visualmente quatro clareiras com estágios sucessionais etamanhos semelhantes. No centro de cada clareira foiestabelecida uma parcela de 2x2m, onde todos os indivíduosabaixo de 1m de altura foram amostrados e identificados.Contudo, se o centro da clareira ainda estava ocupado porgalhos da árvore caída (que abafa a regeneração), foiselecionado outro local dentro da clareira, evitando-se asbordas. Utilizamos teste t-student (Zar 1984) para comparara densidade e riqueza de morfoespécies entre as clareiras daárea controle e da área controle e da explorada.
Para testar o grau de similaridade entre as clareiras dasduas áreas, utilizamos a análise de cluster, com o algorítimo(porcentagem de similaridade) e tipo de ligação (vizinho maispróximo), baseado na presença e ausência das morfoespécies(Ferreira com. pess.).
ResultadosAmostramos 231 indivíduos distribuídos em 135
morfoespécies nas 8 clareiras amostradas. O número médiode indivíduos na área explorada foi aproximadamente 1,5vezes maior do que o número de indivíduos observado naárea controle, mas não diferiram significativamente (t = -1,729; df = 3,9; p = 0,16). A variação de densidade e riquezafoi maior na área explorada (Tabela 1), indicando maiorvariabilidade entre as clareiras da área explorada em relaçãoàs clareiras da área de preservação (controle).
Tabela 1. Número de indivíduos, riqueza de espéceis, média e desviopadrão de plantas por tipo de tratamento; clareiras de áreasexploradas (desde de 1999) e em áreas de preservação da MilMadeireira Itacoatiara Ltda (n=4).
A média do número de morfoespécies da área exploradafoi duas vezes maior do que a média de morfotipos observadosna área controle. No entanto, esta diferença não foisignificativa (t = -1,691; df = 3,5; p = 0,17).
O grau de similaridade florística entre clareiras nos doistipos de tratamento (áreas alteradas e de preservação) nãoformam grupos definidos. O resultado da análise mostrouque existe grande variação tanto dentro de cada tratamento,como entre eles (Figura 1).
Área Clareira No de Indivíduos No de Espécies
Controle 1 49 23Controle 2 39 18Controle 3 41 13Controle 4 26 20Média 38,75 18,50Desvio padrão 9,54 4,20Explorada 1 68 39Explorada 2 27 19Explorada 3 66 46Explorada 4 83 19Média 61,00 30,75Desvio padrão 23,90 13,87
Figura 1. Dendograma da análise de agrupamento de morfotiposde plantas em clareiras de uma área preservada (Pres.) e outraperturbada pela extração seletiva de madeira (Exp.) na Mil MadeireiraLtda.
DiscussãoA variação no número de indivíduos entre as parcelas
dentro do mesmo tratamento foi muito alta, tanto para áreaexplorada quanto para a área de preservação, a ponto de nãoser detectada diferença entre as mesmas, apesar de termosobservado mais indivíduos nas clareiras da área explorada.O maior número e variação da densidade de indivíduosencontrados em clareiras da área explorada, pode serexplicada pelo acréscimo de variáveis causais, resultantesda exploração, como maior luminosidade, mudanças naestrutura da vegetação e variações microclimáticas. Dequalquer forma, conforme o hipotetizado, o número deindivíduos foi maior na área explorada, podendo estarrelacionado à maior abertura de dossel encontrada neste tipode ambiente.
Verificamos o mesmo para a riqueza de espécies. Avariabilidade das unidades amostrais foi alta e não foisuficiente para retratar o efeito do manejo florestal sobre aregeneração de comunidades de plantas, ou a exploraçãorealmente não causa impactos significativos sobre a riquezade espécies regenerantes. A área explorada apresentou maiornúmero médio de espécies, mas não diferiu estatisticamenteda área de preservação. Isto pode ser devido ao baixo esforçoamostral. Mais informações sobre os fatores ambientais queatuam sobre clareiras são necessárias para explicar estespadrões.
O mesmo padrão poderia ser aplicado à pouca similaridadeda composição florística entre as clareiras amostrada, tantodentro como entre os tratamentos. Dentre todas as clareiras,as clareiras 2 e 4 da área explorada foram as mais diferentes.Estes resultados suportam que áreas alteradas, mesmo emníveis de exploração de baixo impacto, podem resultar emmaiores aberturas do dossel não só pela derrubada de árvores,mas também pela infra-estrutura necessária para a exploraçãomadeireira.
As clareiras da área de preservação e da área de exploraçãonão foram diferentes, tanto em relação a densidade quanto ariqueza de espécies. Três hipóteses podem ajudar a respondero fenômeno observado: i) a amostra foi muito pequena paradetectar diferenças significativas entre as clareiras das duasáreas; ii) é possível que o tempo desde o início da exploração(2 anos) não tenha seja ainda suficiente para gerar mudanças

Curso de Campo Ecologia da Floresta Amazônica - 2001 17
significativas na composição e abundância dasmorfoespécies; iii) a exploração madeireira da empresa naárea estudada não exerce efeito significativo sobre aregeneração natural de clareiras, já que uma das técnicas demanejo proposta é o direcionamento da queda das árvoresextraídas, evitando a derrubada de outras árvores em grandequantidade.
AgradecimentosSomos gratos a Leandro Valle Ferreira (Massaranduba)
pela colaboração e idealização da proposta deste trabalho.
Referências BibliográficasAnderson, A.B. 1990. Alternatives to Deforestation: Steps
Toward Sustainable Use of the Amazon Rain Forest.Columbia University Press, New York.
Brokaw, N.V.L. 1982. The definition of treefall gap and itseffects on measures of forest dynamics. Biotropica11:158-160.
Brokaw, N.V.L. 1985a. Gap-phase regeneration in a tropicalforest. Ecology 66:682-687.
Brokaw, N.V.L. 1985b. Treefalls, regrowth and communitystructure in tropical forest. In: The Ecology of NaturalDisturbance and Patch Dynamics, Pickett, S.T.A. &White, P.S. (eds.), pp. 53-69. Academic Press, New York.
Denslow, J.S. 1980. Gap partioning among tropical rainforest. Biotropica 12: 47-55.
Hartshorn, G.S. 1980. Neotropical forest dynamics.Biotropica 12: 23-30.
Poore, D. 1989. Conclusions. In: No Timber Without Trees:Sustainability in the Tropical Forest, Poore, D.,Burguess, P., Palmer, J., Rietbergen, S. & Synnott, T.(eds.), pp. 190-209. Earthscan, London.
Zar, H.J. 1984. Biostatisitcal Analysis. Prentice Hall, NewJersey.
OrientadorLeandro Valle Ferreira
Ocorrência e ritmo de atividade de duas espécies de lagartos (Squamata:Lacertilia: Teiidae) em área de preservação e corte seletivo na floresta
amazônica (Itacoatiara, AM)Angela M. F. Pacheco, Antonio J. C. Aguiar, Edgar F. Prieto, Paula S. Pinheiro & Thiago Timo
IntroduçãoLagartos são animais ectotérmicos que podem apresentar
adaptações anatômicas, fisiológicas e comportamentais paramanutenção da temperatura corpórea. Esta estratégia demanutenção térmica é muito econômica metabolicamente emrelação à endotermia dos mamíferos, entretanto implica emlimitações comportamentais, como por exemplo, o tempodiário dispendido em termorregulação e o tempo de insolaçãonecessário antes que se iniciem as atividades diárias(Pianka1994).
Espécies de lagartos heliófilas ou heliotérmicas,apresentam uma alta taxa metabólica em relação a lagartosumbrófilos, e necessitam de alta taxa de insolação para atingire manter uma temperatura corpórea proporcionalmente alta(Schieseri 1994). Estes lagartos periodicamente locomovem-se do sol para sombra para manter-se em amplitudesrelativamente estreitas de temperatura (Castilla & Bawens1991 apud Bawens et al. 1995). Ameiva ameiva e Kentropyxcalcarata são lagartos teídeos heliófilos de porte médio (5 a18 cm e 8 a 15 cm, respectivamente) e freqüentementeencontrados em clareiras no chão da floresta e na borda damata (Schieseri 1994).
A retirada seletiva de madeira, ainda que certificada porórgãos ambientais, promove alterações na dinâmica declareiras de uma floresta, o que pode estar afetando o ritmode atividades e a ocorrência de algumas espécies de lagartosheliófilos. Este estudo teve como objetivo testar a hipótese
de que a ocorrência e o ritmo de atividade de A. ameiva e K.calcarata diferem entre uma área florestal preservada e outraperturbada por corte seletiva de madeira.
Material e MétodosEste estudo foi conduzido em uma área de exploração
comercial de madeira, baseada em corte seletivo, emItacoatiara, AM (2o43’ S; 58o31’ W). A vegetação caracteriza-se por floresta de terra firme com manchas de campinarana eigapó. A temperatura média é 26o C e a pluviosidade é de2000 mm/ano (Anônimo 1997). As amostragens foramrealizadas em duas áreas separadas por uma estrada, uma depreservação permanente e outra que foi explorada em 1999.Esta última apresenta diversas clareiras, fomadas pela quedadas árvores, por trilhas grandes, de cerca de 5m de largura,para o arrasto de troncos, e trilhas menores, de acesso asárvores derrubadas.
Foram utilizadas 12 trilhas de aproximadamente 200metros para observação dos lagartos das espécies Ameivaameiva e Kentropyx calcarata, sendo seis na área alterada eseis na preservada. Cada trilha foi percorrida diversas vezespor um observador, durante intervalos de 30 minutos, das10:30 às 16:00 h no primeiro dia, e das 9:30 às 12:00 h nosegundo dia. Seis observadores percorreram as trilhas, cadaobservador amostrou uma trilha por dia, alternando entreáreas alteradas e preservadas entre os dois dias. O númerototal de horas de observação foi de 48 horas, (soma dos
18 Curso de Campo Ecologia da Floresta Amazônica - 2001
períodos de amostragem dos seis observadores).O número máximo de indivíduos em cada transecto foi
computado como o maior número de avistamentos de lagartosem um turno. Os indivíduos observados entre os turnos nãonecessariamente representavam indivíduos distintos, já quenão foi realizado nenhum método de marcação. O total delagartos avistados foi comparado entre as áreas através doteste de Qui-quadrado de aderência (Zar 1984).
Foram realizadas estimativas da frequência de ocorrênciae horário de atividade de cada espécie nos dois ambientes. Onúmero máximo de indivíduos observados entre as áreas foicomparado separadamente para cada espécie através do testet pareado (Zar 1984). Neste teste cada par foi formado pelomesmo observador que realizou a amostragem nas duas áreas.O ritmo de atividade de K. calcarata entre as áreas foicomparado a partir da freqüência total de indivíduos em cadaintervalo de tempo, através do teste de Kolmogorov-Smirnov(Zar 1984). O nível de significância considerado em todosos testes foi de 0,05. No teste Kolmogorov-Smirnov foramconsideradas apenas as amostras de K. calcarata, pois A.ameiva não apresentou número de ocorrências suficiente.
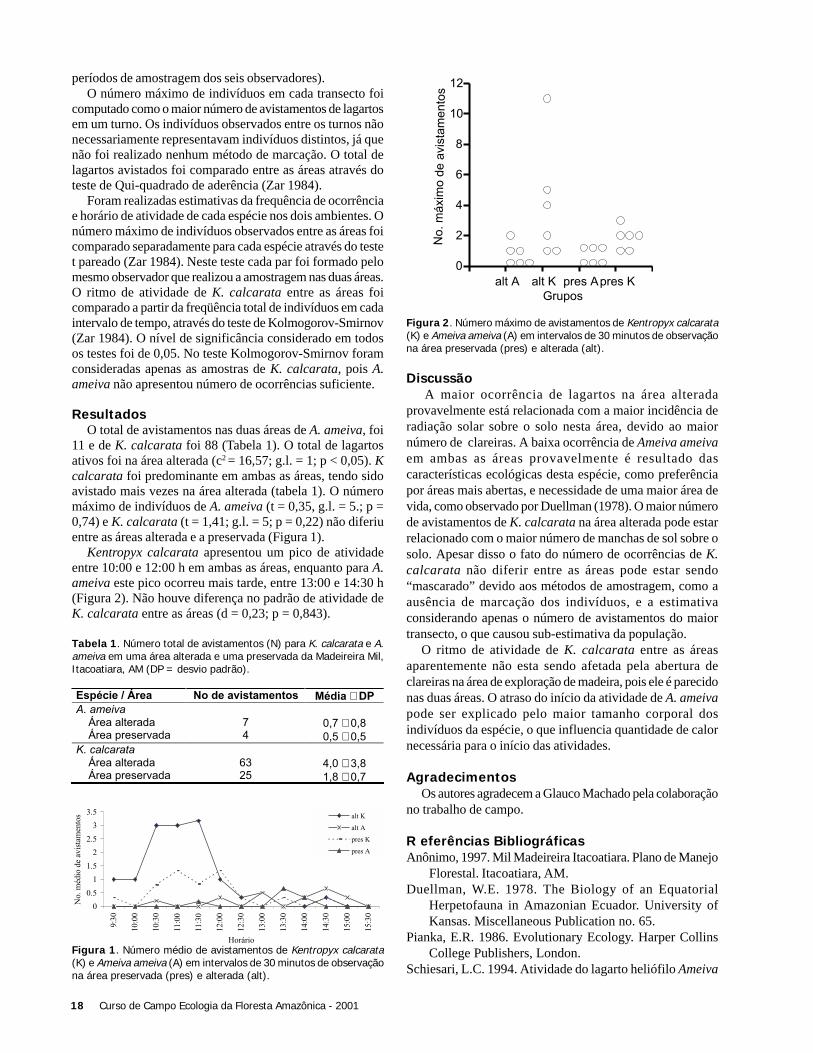
ResultadosO total de avistamentos nas duas áreas de A. ameiva, foi
11 e de K. calcarata foi 88 (Tabela 1). O total de lagartosativos foi na área alterada (c2 = 16,57; g.l. = 1; p < 0,05). Kcalcarata foi predominante em ambas as áreas, tendo sidoavistado mais vezes na área alterada (tabela 1). O númeromáximo de indivíduos de A. ameiva (t = 0,35, g.l. = 5.; p =0,74) e K. calcarata (t = 1,41; g.l. = 5; p = 0,22) não diferiuentre as áreas alterada e a preservada (Figura 1).
Kentropyx calcarata apresentou um pico de atividadeentre 10:00 e 12:00 h em ambas as áreas, enquanto para A.ameiva este pico ocorreu mais tarde, entre 13:00 e 14:30 h(Figura 2). Não houve diferença no padrão de atividade deK. calcarata entre as áreas (d = 0,23; p = 0,843).
Tabela 1. Número total de avistamentos (N) para K. calcarata e A.ameiva em uma área alterada e uma preservada da Madeireira Mil,Itacoatiara, AM (DP = desvio padrão).
Figura 1. Número médio de avistamentos de Kentropyx calcarata(K) e Ameiva ameiva (A) em intervalos de 30 minutos de observaçãona área preservada (pres) e alterada (alt).
Espécie / Área No de avistamentos Média � DPA. ameiva
Área alteradaÁrea preservada
74
0,7 � 0,80,5 � 0,5
K. calcarataÁrea alteradaÁrea preservada
6325
4,0 � 3,81,8 � 0,7
0
0.5
1
1.5
2
2.5
3
3.5
9:30
10:0
0
10:3
0
11:0
0
11:3
0
12:0
0
12:3
0
13:0
0
13:3
0
14:0
0
14:3
0
15:0
0
15:3
0
Horário
No.
méd
iode
avis
tam
ento
s alt K
alt A
pres K
pres A
Figura 2. Número máximo de avistamentos de Kentropyx calcarata(K) e Ameiva ameiva (A) em intervalos de 30 minutos de observaçãona área preservada (pres) e alterada (alt).
Discussão A maior ocorrência de lagartos na área alterada
provavelmente está relacionada com a maior incidência deradiação solar sobre o solo nesta área, devido ao maiornúmero de clareiras. A baixa ocorrência de Ameiva ameivaem ambas as áreas provavelmente é resultado dascaracterísticas ecológicas desta espécie, como preferênciapor áreas mais abertas, e necessidade de uma maior área devida, como observado por Duellman (1978). O maior númerode avistamentos de K. calcarata na área alterada pode estarrelacionado com o maior número de manchas de sol sobre osolo. Apesar disso o fato do número de ocorrências de K.calcarata não diferir entre as áreas pode estar sendo“mascarado” devido aos métodos de amostragem, como aausência de marcação dos indivíduos, e a estimativaconsiderando apenas o número de avistamentos do maiortransecto, o que causou sub-estimativa da população.
O ritmo de atividade de K. calcarata entre as áreasaparentemente não esta sendo afetada pela abertura declareiras na área de exploração de madeira, pois ele é parecidonas duas áreas. O atraso do início da atividade de A. ameivapode ser explicado pelo maior tamanho corporal dosindivíduos da espécie, o que influencia quantidade de calornecessária para o início das atividades.
AgradecimentosOs autores agradecem a Glauco Machado pela colaboração
no trabalho de campo.
R eferências BibliográficasAnônimo, 1997. Mil Madeireira Itacoatiara. Plano de Manejo
Florestal. Itacoatiara, AM.Duellman, W.E. 1978. The Biology of an Equatorial
Herpetofauna in Amazonian Ecuador. University ofKansas. Miscellaneous Publication no. 65.
Pianka, E.R. 1986. Evolutionary Ecology. Harper CollinsCollege Publishers, London.
Schiesari, L.C. 1994. Atividade do lagarto heliófilo Ameiva
alt A alt K pres Apres KGrupos
0
2
4
6
8
10
12
No
.má
xim
od
ea
vist
am
en
tos

Curso de Campo Ecologia da Floresta Amazônica - 2001 19
ameiva (Sauria; Teiidae). Livro do Curso de Campo“Ecologia da Floresta Amazônica”. INPA/PDBFF,Manaus, AM.
Zar, J.H. 1984. Biostatistical Analysis. Prentice Hall, NewJersey.
OrientadorMarcelo Gordo
Influência da extração seletiva de madeira sobre as comunidades de aves emorcegos em uma área de floresta da Amazônia Central
Mauricio Camargo, Cristiane F. Jurinitz, Felipe B. Oliveira, Maria R. Darrigo & Rodrigo Cogni
IntroduçãoInterferências antrópicas no ambiente como a extração
seletiva de madeira têm causado mudanças na estrutura dasflorestas tropicais (Anderson 1990). Estimativas apontampara o fato de que a Amazônia brasileira, que representa umterço das florestas tropicais do mundo, produz 75% damadeira em tora do Brasil (Uhl et al. 1998). Mesmo com autilização de modelos de extração de madeira consideradosde baixo impacto, essa atividade causa perda de biomassa,aberturas no dossel, e perda de fontes de sementes e de pólen,elementos importantes para a manutenção da floresta(Anderson 1990). Todas essas alterações provavelmenteafetam os organismos que habitam esse ambiente emdiferentes graus.
Uma abordagem interessante para averiguar o efeito dessasalterações consiste em estudar os grupos que possuem papel-chave na manutenção da estrutura do ecossistema. Desseponto de vista, as aves e morcegos destacam-se por suaimportância como polinizadores e dispersores de muitasespécies de plantas (Begon et al. 1986).
Uma guilda pode ser definida como um conjunto deespécies que têm em comum um determinado hábitat, tipode forrageio e dieta (Kalko 1996). Desse modo, em cadaguilda podem ser identificados padrões morfológicos comunsentre as espécies (Boege et al. 1998), que podem serrelacionados como táticas alimentares e padrões de vôo. Jáforam relatadas mudanças na estrutura das comunidades dediversos grupos de animais em florestas onde há atividademadeireira (Rittl 1998). Esse fato reflete-se em alteraçõesna proporção desses grupos dentro de diferentes guildas. Demaneira geral, os animais mais especialistas são mais afetados(Rittl 1998).
O presente estudo teve como objetivo estudar as relaçõesentre a estrutura da floresta e as comunidades de aves emorcegos em uma área que sofreu extração seletiva de madeirae uma área preservada. Além disso, realizamos uma análisedas características morfológicas das aves e morcegos quehabitam essas áreas.
Material e MétodosÁrea de estudoRealizamos este estudo em uma área de propriedade da
Mil Madeireira Itacoatiara Ltda., situada no município de
Itacoatiara, AM (2°43’S; 58°31’W). O clima é quente e úmidocom médias anuais de temperatura de 26°C e precipitaçãode 2000mm. A vegetação predominante é a floresta de terrafirme, com algumas manchas de campinarana e igapó. Namaior parte da área, essa empresa realiza a extração seletivade madeira, retirando apenas os indivíduos de maior porte(DAP maior que 50cm) das espécies de maior interessecomercial, além de seguir alguns procedimentos no momentodo corte na tentiva de diminuir os efeitos da extração sobrea floresta. Para o presente estudo, selecionamos dois setores,sendo que um é caracterizado pela recente extração demadeira (aproximadamente 1 ano) e o outro é localizadodentro da área de preservação permanente. Estes dois setoressão contíguos e separados por uma estrada de 20m de largura.
Estimativas de biomassa e de abertura do dosselEm cada uma das áreas, a uma distância de
aproximadamente 100 metros da estrada, delimitamos umtransecto de 100m perpendicular à borda da mata. Nestesdois transectos estabelecemos quatro parcelas não contíguasde 10 x 10m. Nestas parcelas, medimos o diâmetro à alturade 1,30m de todas as árvores com diâmetro maior ou igual a5 cm e de todas as palmeiras com caule. Estimamos abiomassa de cada área com equações utilizadas em outrosestudos na mesma área (Rittl 1998).
Equação A – (árvores de 5cm < DAP < 20cm):(e -1,754 + 2,65 x ln DAP) x 0,604 / 103
Equação B – (árvores de DAP > 20cm): (e -0,151 + 2,17 x ln DAP) x 0,604 / 103
Equação C – (palmeiras):(e 0,9285 x ln DAP + 5,7236) x 1,05 / 106
Para estimar a cobertura vegetal de cada área estudada,fizemos medidas com um densiômetro em dois vértices daparcela. O valor médio dessas medidas foi considerado umindicador da abertura do dossel de cada parcela.
Captura dos animaisPara amostrar a fauna de vertebrados voadores, armamos
ao longo dos transectos sete redes de neblina de 12 x 3mcom malha de 3,6cm, totalizando uma área de 252m2 de redepor transecto.
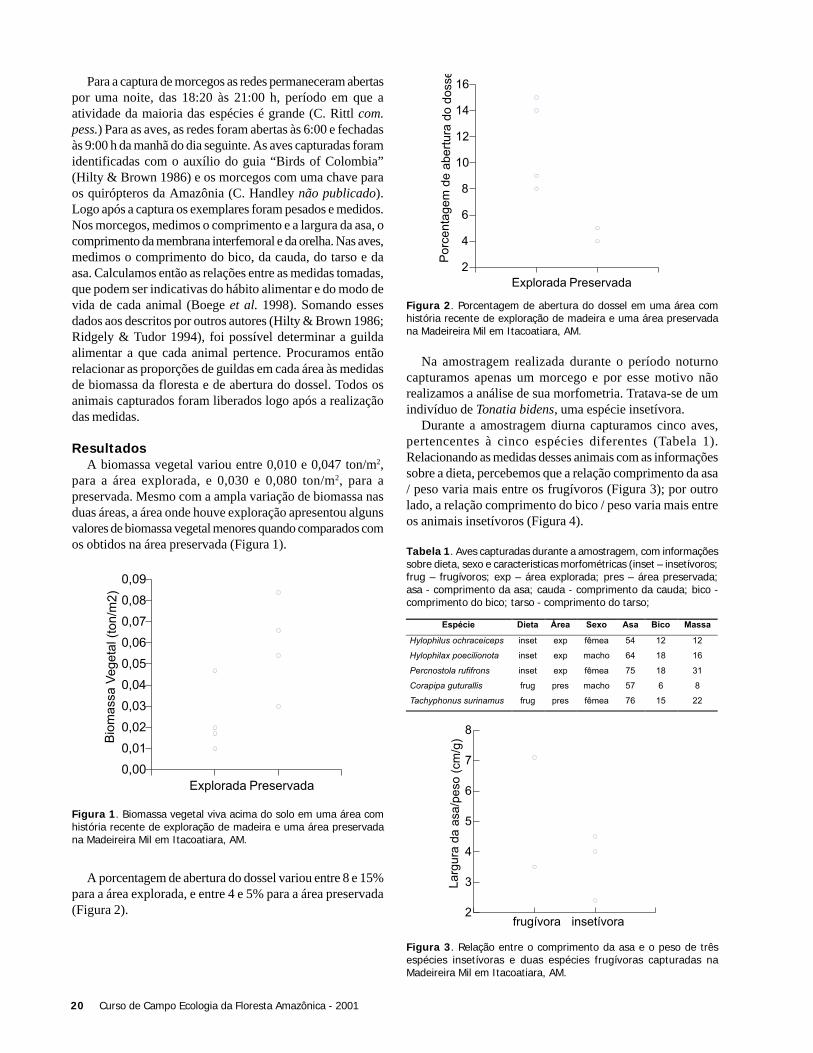
20 Curso de Campo Ecologia da Floresta Amazônica - 2001
Para a captura de morcegos as redes permaneceram abertaspor uma noite, das 18:20 às 21:00 h, período em que aatividade da maioria das espécies é grande (C. Rittl com.pess.) Para as aves, as redes foram abertas às 6:00 e fechadasàs 9:00 h da manhã do dia seguinte. As aves capturadas foramidentificadas com o auxílio do guia “Birds of Colombia”(Hilty & Brown 1986) e os morcegos com uma chave paraos quirópteros da Amazônia (C. Handley não publicado).Logo após a captura os exemplares foram pesados e medidos.Nos morcegos, medimos o comprimento e a largura da asa, ocomprimento da membrana interfemoral e da orelha. Nas aves,medimos o comprimento do bico, da cauda, do tarso e daasa. Calculamos então as relações entre as medidas tomadas,que podem ser indicativas do hábito alimentar e do modo devida de cada animal (Boege et al. 1998). Somando essesdados aos descritos por outros autores (Hilty & Brown 1986;Ridgely & Tudor 1994), foi possível determinar a guildaalimentar a que cada animal pertence. Procuramos entãorelacionar as proporções de guildas em cada área às medidasde biomassa da floresta e de abertura do dossel. Todos osanimais capturados foram liberados logo após a realizaçãodas medidas.
ResultadosA biomassa vegetal variou entre 0,010 e 0,047 ton/m2,
para a área explorada, e 0,030 e 0,080 ton/m2, para apreservada. Mesmo com a ampla variação de biomassa nasduas áreas, a área onde houve exploração apresentou algunsvalores de biomassa vegetal menores quando comparados comos obtidos na área preservada (Figura 1).
Figura 1. Biomassa vegetal viva acima do solo em uma área comhistória recente de exploração de madeira e uma área preservadana Madeireira Mil em Itacoatiara, AM.
A porcentagem de abertura do dossel variou entre 8 e 15%para a área explorada, e entre 4 e 5% para a área preservada(Figura 2).
Explorada Preservada0,00
0,01
0,02
0,03
0,04
0,05
0,06
0,07
0,08
0,09
Bio
ma
ssa
Ve
ge
tal (
ton
/m2
)
Figura 2. Porcentagem de abertura do dossel em uma área comhistória recente de exploração de madeira e uma área preservadana Madeireira Mil em Itacoatiara, AM.
Na amostragem realizada durante o período noturnocapturamos apenas um morcego e por esse motivo nãorealizamos a análise de sua morfometria. Tratava-se de umindivíduo de Tonatia bidens, uma espécie insetívora.
Durante a amostragem diurna capturamos cinco aves,pertencentes à cinco espécies diferentes (Tabela 1).Relacionando as medidas desses animais com as informaçõessobre a dieta, percebemos que a relação comprimento da asa/ peso varia mais entre os frugívoros (Figura 3); por outrolado, a relação comprimento do bico / peso varia mais entreos animais insetívoros (Figura 4).
Tabela 1. Aves capturadas durante a amostragem, com informaçõessobre dieta, sexo e caracteristicas morfométricas (inset – insetívoros;frug – frugívoros; exp – área explorada; pres – área preservada;asa - comprimento da asa; cauda - comprimento da cauda; bico -comprimento do bico; tarso - comprimento do tarso;
Figura 3. Relação entre o comprimento da asa e o peso de trêsespécies insetívoras e duas espécies frugívoras capturadas naMadeireira Mil em Itacoatiara, AM.
Explorada Preservada2
4
6
8
10
12
14
16
Espécie Dieta Área Sexo Asa Bico Massa
Hylophilus ochraceiceps inset exp fêmea 54 12 12
Hylophilax poecilionota inset exp macho 64 18 16
Percnostola rufifrons inset exp fêmea 75 18 31
Corapipa guturallis frug pres macho 57 6 8
Tachyphonus surinamus frug pres fêmea 76 15 22
frugívora insetívora2
3
4
5
6
7
8
La
rgu
rad
aa
sa/p
eso
(cm
/g)

Curso de Campo Ecologia da Floresta Amazônica - 2001 21
Figura 4. Relação entre o comprimento do bico dividido pelo pesode três espécies insetívoras e duas espécies frugívoras capturadasna Madeireira Mil em Itacoatiara, AM.
DiscussãoConforme o esperado, as medidas de biomassa e de
abertura de dossel mostram que a atividade madeireiraefetivamente altera a estrutura da floresta, com possíveisinfluências sobre as comunidades de animais que nelahabitam. Esse resultado também foi obtido por outros autores(Rittl 1998; Johns 1985).
Outras amostragens de morcegos que estão sendorealizadas na área da Mil Madeireira obtêm taxas de capturaque, utilizando o mesmo número de redes de neblina, emdiferentes noites variam de 0 a 35 morcegos, ou seja, emcertas noites a taxa de captura é igual ou até menor que aobtida neste estudo (C. Rittl com. pess.). Outros trabalhosque utilizaram a mesma metodologia de captura sugerem quea atividade dos animais está muito relacionada às condiçõesambientais (Koege et al. 1998); em dias chuvosos, porexemplo, a atividade dos morcegos é bastante reduzida (C.Rittl com. pess.). Aliando essas informações aos resultadosobtidos, ficou claro que para obter dados mais representativossão necessárias muitas horas de amostragem, distribuídasem dias diferentes para minimizar o efeito das condiçõesambientais momentâneas. Outro fator que pode contribuirpara a baixa taxa de captura é a presença de uma estradarelativamente larga (20m) próxima aos locais onde foramcolocadas as redes (aproximadamente 100m). Nessa estradaexiste circulação freqüente de veículos pesados que fazembarulho e jogam poeira no ar, principalmente em dias maissecos (obs. pess.), o que pode afetar negativamente ambasas áreas. Embora em menor escala, a grande circulação depessoas nos dias em que realizamos as capturas também podeter alguma influência.
Analisando as medidas dos animais capturados,percebemos a existência de uma maior variação na relaçãocomprimento do bico / peso entre os insetívoros que entreos frugívoros, embora não tenha sido possível realizarnenhuma análise estatística sobre esses dados. Essa variaçãopode indicar uma maior diversidade de formas de bico entreos insetívoros. Como o tamanho do bico é um atributo muitoligado à dieta de cada animal (Stotz et al. 1996), é possívelque cada insetívoro seja especializado em um tipo de presa
frugívora insetívora0,5
0,6
0,7
0,8
0,9
1,0
1,1
1,2
Co
mp
rim
en
tod
ob
ico
/pe
so(c
m/g
) diferente, aumentando a diversidade morfológica. No entanto,dados da literatura mostram que os frugívoros tambémpossuem vários tipos de bico (Stotz et al. 1996), o que nãofoi detectado na nossa amostragem.
A relação comprimento da asa / peso também é importantepara o hábito de vida de cada animal. Um animal insetívoroprovavelmente precisa manobrar muito quando persegue suaspresas na mata e para isso uma asa relativamente curta pareceser mais adequada (Stotz et al. 1996). Os frugívorosalimentam-se de um recurso estático, e a morfologia da asaparece não ser tão importante em relação a seu modo deforrageio (Boege et al. 1998). No entanto, nossos dados sónos permitem especular nesse campo, pois com os poucosanimais que medimos não temos idéia da real variação damorfologia entre as guildas de aves presentes na áreaestudada.
O baixo número de capturas de aves e morcegos obtidoneste estudo não permitiu investigar a existência dediferenças nos padrões morfológicos desses gruposassociadas às diferenças físicas identificadas entre as duasáreas estudadas. Sugerimos a realização de estudos com maioresforço amostral para averiguar a existência dessas diferenças.
Referências BibliográficasAnderson, A.B. 1990. Alternatives to Deforestation: Steps
Toward Sustainable Use of the Amazon Rain Forest.Columbia University Press, New York.
Begon, M.; Harper, J.L. & Townsend, C.R. 1986. Ecology:Individuals, Populations and Communities. BlackwellScientific Publications, Cambridge.
Boege, K; Ruggiero, P.G.C.; Sá, F.N.; Baptista, S. & Silveira,M. 1998. Variaciones morfométricas en cinco gremiosde murciélagos neotropicales. Livro do Curso de Campo“Ecologia da Floresta Amazônica”. INPA/PDBFF,Manaus, AM.
Hilty, S.L. & Brown, W.L. 1986. A Guide to the Birds ofColombia. Princeton University Press, New Jersey.
Johns, A.G. 1985. Selective logging and wildlife conservationin tropical rainforests: problems and recommendations.Biol. Conserv. 31: 335-375.
Kalko, E.K.; Handley, C.O. & Handley, D. 1996.Organization, Diversity, and Long-Term Dynamics of aNeotropical Bat Community.
Ridgely, R.S. & Tudor, G. 1994. The Birds of South America.Vol II. University of Texas Press, Austin.
Rittl, C.E. 1998. Efeitos da extração seletiva de madeira sobrea comunidade de pequenos mamíferos de uma florestade terra firme na Amazônia Central. Dissertação deMestrado, INPA, Manaus, AM.
Stotz, D.F.; Fitzpatrick, J.W.; Parker, T.A. & Moskovits, D.K.1996. Neotropical Birds: Ecology and Conservation.University of Chicago Press, Chicago.
Uhl, C.; Barreto, P.; Veríssimo, A.; Barros, A.; Amaral, P.;Vidal, E. & Souza Jr. 1998. Uma abordagem integradade pesquisa sobre o manejo dos recursos florestais naAmazônia brasileira. In: Floresta Amazônica: Dinâmica,Regeneração e Manejo, Gascon, C. & Moutinho,P.(eds.). MCT/INPA, Manaus.

22 Curso de Campo Ecologia da Floresta Amazônica - 2001
Influência da extração seletiva de madeira em parâmetros físicos, serapilheira ena biomassa de artrópodes de sub-bosque em uma floresta de terra firme da
Amazônia centralFelipe B. Oliveira, Alexander V. Christianini, Thiago P. C. Timo
IntroduçãoA grande riqueza e abundância de árvores de grande porte
na floresta amazônica faz dela um local importante para aextração de madeira. De fato, a atividade madeireira é umadas principais causas de desmatamento na Amazônia(Anderson 1990). A atividade madeireira de baixo impactoé uma prática que se iniciou recentemente e ainda é utilizadapor poucos empreendedores (De Graaf & Poels 1990). Noentanto, trata-se de um sistema de exploração menos danosoà floresta que o sistema tradicional, por explorar espéciescomerciais de forma planejada e a partir de um tamanhomínimo, deixando alguns indivíduos adultos como portasementes. Alguns autores sugerem que este sistema deexploração pode ser uma alternativa para a exploraçãosustentável da floresta amazônica (De Graaf & Poels 1990).
A dinâmica das florestas tropicais abrange processoscomplexos (Rankin-de-Merona et al. 1990), que podem serinfluenciados pela extração de madeira. Perturbações nodossel, por exemplo, aumentam a incidência luminosa,temperatura do ar e penetração de ventos, diminuindo aumidade relativa do ar (Denslow & Hathshorn 1994). Essasmudanças microclimáticas influenciam as plantas dediferentes modos, aumentando a mortalidade para algumasespécies e o recrutamento de outras (Clark & Clark 1991;Denslow & Hartshorn 1994). Assim, espera-se queperturbações físicas promovidas pela retirada de árvores e aabertura de estradas para escoamento de madeira alterem adinâmica da floresta. O aumento na quantidade de plântulase adultos de espécies vegetais secundárias, que em condiçõesnaturais estariam em menor número no interior da floresta,pode causar um incremento na abundância dos herbívorosque delas se alimentam (Coley & Kursar 1996). Emconseqüência, espera-se um incremento no número depredadores desses herbívoros.
Os objetivos deste trabalho são testar se parâmetrosambientais, como a abertura do dossel e a temperatura do ar,são modificados pela extração seletiva de madeira e se essaperturbação tem efeitos na biomassa de insetos herbívoros ede alguns de seus predadores, as aranhas de teia orbicular.
Material e MétodosRealizamos este estudo em uma área de propriedade da
Mil Madeireira Itacoatiara Ltda., situada no município deItacoatiara, AM (2°43’S; 58°31’W). O clima local é quentee úmido com médias anuais de temperatura de 26°C eprecipitação de 2000mm/ano. A vegetação predominante é afloresta de terra firme, com algumas manchas de campinaranae igapó. Na maior parte da área, ocorre extração seletiva demadeiras de interesse comercial, retirando indivíduos de DAP
maior que 50cm. A derrubada também segue algunsprocedimentos no momento do corte na tentativa de diminuiros efeitos da extração sobre a floresta.
Para o presente estudo, selecionamos dois setores, umcaracterizado pela recente extração de madeira(aproximadamente 1 ano) e outro localizado dentro da áreade preservação permanente. Estes dois setores são contíguose separados por uma estrada de 20m de largura.
Em cada setor delimitamos um transecto de 300 mperpendicular à estrada, que se iniciava a 30m da mesma natentativa de reduzir sua influência sobre a amostragem. Otransecto foi dividido a cada 10m, num total de 30 segmentospor transecto, com os segmentos amostrados por sorteio. Atemperatura foi medida em 10 segmentos por transecto, comno máximo 30 minutos de defasagem entre os dois transectospara minimizar o efeito da hora do dia. A biomassa de insetosde sub-bosque foi amostrada em cinco destes segmentossorteados, por transecto. Coletamos insetos na folhagem dosub-bosque com o uso de um batedor entomológico, ao longode metade do segmento sorteado. Medindo os animaiscoletados com um paquímetro, estimamos a biomassa deartrópodes em cada área com a equação:
W = 0,0305 x L 2,62 (Rogers et al. 1976 apud Venticinque 1994),onde W é a biomassa em mg e L é o comprimento dos animais emmm.
Estimamos a porcentagem de abertura do dossel com umdensiômetro em dez pontos por área, escolhidos da seguintemaneira: a partir dos dez pontos escolhidos em cada transecto,sorteamos novamente o lado e a distância perpendicular aoponto de amostragem, até o máximo de 50m de distância dotransecto. Em cada um destes pontos medimos ainda aespessura da serapilheira, espetando quatro locais adjacentesao acaso com uma estaca e contando o número de folhas queficavam presas.
Para estimar a quantidade de aranhas de teia orbicularpercorremos os dois transectos, registrando cada teiaencontrada numa faixa de até 0,5 m de largura de ambos oslados e até dois metros de altura. Medimos o raio maior e oraio menor de cada teia e calculamos a área da teia, assumindoque sua forma é elíptica (A = ë r
1 r
2).
ResultadosNão obtivemos diferenças significativas entre a cobertura
do dossel (teste t = 0,75; df = 18; p = 0,47) e a temperaturado ar (teste t = 0,67, df = 18; p = 0,51) entre a área preservadae a área explorada por corte seletivo de madeira (Fig. 1). Nãodetectamos diferenças na espessura da serapilheira (Mann-Whitney; U = 37; p = 0,32) e na biomassa de artrópodes (U
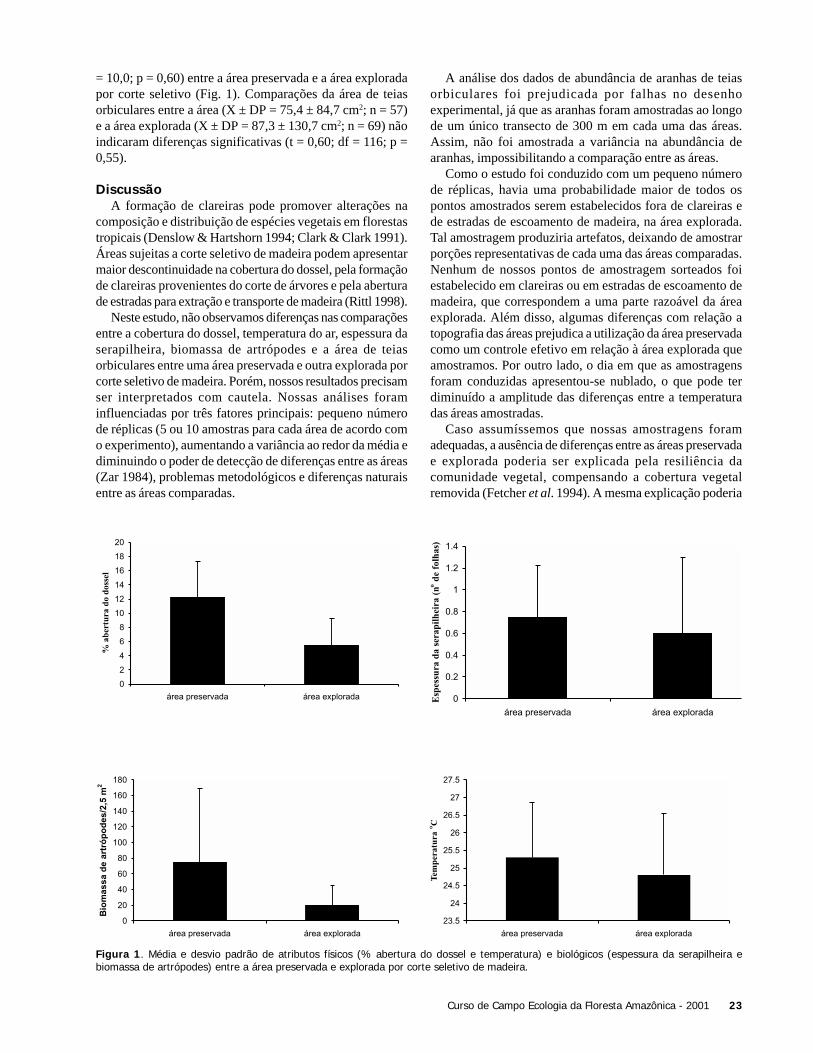
Curso de Campo Ecologia da Floresta Amazônica - 2001 23
= 10,0; p = 0,60) entre a área preservada e a área exploradapor corte seletivo (Fig. 1). Comparações da área de teiasorbiculares entre a área (X ± DP = 75,4 ± 84,7 cm2; n = 57)e a área explorada (X ± DP = 87,3 ± 130,7 cm2; n = 69) nãoindicaram diferenças significativas (t = 0,60; df = 116; p =0,55).
DiscussãoA formação de clareiras pode promover alterações na
composição e distribuição de espécies vegetais em florestastropicais (Denslow & Hartshorn 1994; Clark & Clark 1991).Áreas sujeitas a corte seletivo de madeira podem apresentarmaior descontinuidade na cobertura do dossel, pela formaçãode clareiras provenientes do corte de árvores e pela aberturade estradas para extração e transporte de madeira (Rittl 1998).
Neste estudo, não observamos diferenças nas comparaçõesentre a cobertura do dossel, temperatura do ar, espessura daserapilheira, biomassa de artrópodes e a área de teiasorbiculares entre uma área preservada e outra explorada porcorte seletivo de madeira. Porém, nossos resultados precisamser interpretados com cautela. Nossas análises foraminfluenciadas por três fatores principais: pequeno númerode réplicas (5 ou 10 amostras para cada área de acordo como experimento), aumentando a variância ao redor da média ediminuindo o poder de detecção de diferenças entre as áreas(Zar 1984), problemas metodológicos e diferenças naturaisentre as áreas comparadas.
Figura 1. Média e desvio padrão de atributos físicos (% abertura do dossel e temperatura) e biológicos (espessura da serapilheira ebiomassa de artrópodes) entre a área preservada e explorada por corte seletivo de madeira.
0
2
4
6
8
10
12
14
16
18
20
área preservada área explorada
%a
ber
tura
do
do
ssel
0
0.2
0.4
0.6
0.8
1
1.2
1.4
área preservada área explorada
Esp
essu
rad
ase
rap
ilh
eira
(no
de
folh
as)
0
20
40
60
80
100
120
140
160
180
área preservada área explorada
Bio
ma
ss
ad
ea
rtró
po
de
s/2
,5m
2
23.5
24
24.5
25
25.5
26
26.5
27
27.5
área preservada área explorada
Tem
per
atu
raoC
A análise dos dados de abundância de aranhas de teiasorbiculares foi prejudicada por falhas no desenhoexperimental, já que as aranhas foram amostradas ao longode um único transecto de 300 m em cada uma das áreas.Assim, não foi amostrada a variância na abundância dearanhas, impossibilitando a comparação entre as áreas.
Como o estudo foi conduzido com um pequeno númerode réplicas, havia uma probabilidade maior de todos ospontos amostrados serem estabelecidos fora de clareiras ede estradas de escoamento de madeira, na área explorada.Tal amostragem produziria artefatos, deixando de amostrarporções representativas de cada uma das áreas comparadas.Nenhum de nossos pontos de amostragem sorteados foiestabelecido em clareiras ou em estradas de escoamento demadeira, que correspondem a uma parte razoável da áreaexplorada. Além disso, algumas diferenças com relação atopografia das áreas prejudica a utilização da área preservadacomo um controle efetivo em relação à área explorada queamostramos. Por outro lado, o dia em que as amostragensforam conduzidas apresentou-se nublado, o que pode terdiminuído a amplitude das diferenças entre a temperaturadas áreas amostradas.
Caso assumíssemos que nossas amostragens foramadequadas, a ausência de diferenças entre as áreas preservadae explorada poderia ser explicada pela resiliência dacomunidade vegetal, compensando a cobertura vegetalremovida (Fetcher et al. 1994). A mesma explicação poderia

24 Curso de Campo Ecologia da Floresta Amazônica - 2001
ser aplicada à comunidade de artrópodes, que poderiarecuperar populações afetadas pela retirada da vegetação oumesmo não responder às alterações promovidas pela retiradade madeira, dado seu ciclo de vida curto e áreas de imigraçãopróximas (Janzen 1983), já que a área estudada está conectadacom mata contínua. É possível ainda que o tempo decorridoapós a extração de madeira tenha sido suficientemente longopara que as populações de herbívoros e de aranhas tenhamse estabilizado, diminuindo a amplitude do efeito queesperaríamos detectar (Janzen 1983).
Estudos com melhor desenho experimental podem abordarmelhor estas questões. Contudo, vale lembrar que outrosestudos tem demonstrado efeitos biológicos promovidos pelaextração seletiva de madeira, como diminuição davariabilidade genética de espécies vegetais exploradas(Buchert et al. 1997). Longos tempos de pousio parecem sernecessários para que haja sustentabilidade da retirada seletivade madeira (Condit et al. 1995).
Referências BibliográficasAnderson, A.B. 1990. Alternatives to Deforestation: Steps
Toward Sustainable Use of the Amazon Rain Forest.Columbia University Press, New York.
Buchert, G.P.; Rajora, O.M.P.; James V.H. & Dancik, B.1997. Effects of harvesting on genetic diversity in old-growth Eastern white pine in Ontario, Canada. Cons.Biol. 11: 747-758.
Clark, D.B. & Clark, D.A. 1991. The impact of physicaldamage on canopy tree regeneration in tropicalrainforest. J. Ecol. 79: 447-457.
Coley, P.D. & Kursar, T.A.. 1996. Anti-herbivore defensesof young tropical leaves: physiological constraints andecological trade-offs. In: Tropical Forest PlantEcophysiology, Hulley, S.S., Charson R.L. & Smith, A.P.(eds.) Chapman & Hall, New York.
Condit, R.; Hubbell, S.P. & Foster, R.B. 1995. Demographyand harvest potential of Latin American timber species:data from a large, permanent plot in Panama. J. Trop.Ecol. 7: 599-622.
De Graaf, N.R. & Poels, L.H. 1990. The Celos ManagementSystem: a polyciclic method for sustained timberproduction in South American rainforest. In: Alternativesto Deforestation: Steps Toward Susteinable Use of theAmazon Rainforest, Anderson, A.B. (ed.). ColumbiaUniversity Press, New York.
Denslow, J.S. & Hartshorn, G.S. 1994. Tree-fall gaps:environments and forest dynamic processes. In: La Selva:Ecology and Natural History of a Neotropical Rainforest,McDade, L.A., Bawa, K.S., Hespenheide, H.A. &Hartshorn, G.S. (eds.), pp. 121-127. The University ofChicago Press, Chicago.
Fetcher, N.; Operbauer, S.F. & Chazdon, R.L. 1994.Physiological ecology of plants. In: La Selva: Ecologyand Natural History of a Neotropical Rainforest,McDade, L.A., Bawa, K.S., Hespenheide, H.A. &Hartshorn, G.S. (eds.), pp. 121-127. The University ofChicago Press, Chicago.
Janzen, D.H. 1983. Insects. In: Costa Rican Natural History,pp. 619-779. The University of Chicago Press, Chicago.
Rankin-de-Merona J.M.; Hutchings, R.W.; Lovejoy, T.E.1990. Tree mortality and recruitment over a five-yearperiod in undisturbed upland rainforest of the CentralAmazon. In: Four Neotropical Rainforests, Gentry, A.H.(ed.), pp. 573-584. Yale University Press, New Haven.
Rittl, C.E. 1998. Efeitos da extração seletiva de madeira sobrea comunidade de pequenos mamíferos de uma florestade Terra Firme na Amazônia Central. Dissertação deMestrado, INPA, Manaus, AM.
Venticinque, E.M. 1994. Colonialidade, seleção de micro-hábitat e comportamento de uma aranha tropicalconstrutora de teias orbiculares. Livro do Curso deCampo “Ecologia da Floresta Amazônica”. INPA/PDBFF, Manaus, AM.
Zar, J.H. 1984. Biostatistical Analysis. Prentice Hall, NewJersey.
Perturbações no habitat afetam a riqueza e abundância de herbívorosgalhadores?
Paula C. Ceotto, Andrea H. Lamberts, Antônio J. C. Aguiar, Natalie Olifiers & Paula S. Pinheiro
IntroduçãoA abertura de clareiras altera o microclima das florestas
através do aumento da incidência solar, seguida de elevaçãoda temperatura e da diminuição da umidade, entre outros.Estas alterações geram diferentes respostas da comunidadevegetal. Por exemplo, a maior luminosidade favorece ocrescimento de árvores pioneiras que teriam baixaprobabilidade de germinar no solo sombreado da florestamadura (Uhl 1982; Denslow & Hartshorn 1994). Ainda,plantas pioneiras investem proporcionalmente mais energia
em crescimento do que em defesas químicas, como compostossecundários (Pavón 1999).
O aumento na densidade de plantas com poucas defesascontra herbivoria e patógenos poderia induzir um aumentona riqueza e abundância de insetos fitófagos (Gonçalves-Alvin & Fernandes no prelo). Segundo da Silva et al. (1999),existe uma associação positiva entre o grau de herbivoria e aincidência de patógenos, provavelmente porque os danoscausados pela herbivoria facilitariam a entrada dos patógenos.
Galhas são respostas morfogenéticas das plantas à

Curso de Campo Ecologia da Floresta Amazônica - 2001 25
infestação por organismos fitopatogênicos, os quais podemser insetos, vírus, bactérias ou fungos (Begon et al. 1990;Mani 1964). As galhas de insetos apresentam alta diversidadee especificidade em relação às espécies vegetais hospedeiras,sendo facilmente distinguidas no campo e podendo ser usadaspara estimar a riqueza e abundância de insetos galhadoresem uma comunidade (Fernades & Price 1991; Gonçalves-Alvin & Fernandes no prelo). Por exemplo, Floate et al.(1996) utilizou com sucesso a comunidade de galhas deinsetos para separar genótipos de Chrysothamnus nauseosus.
Neste trabalho, comparamos a riqueza e abundância degalhas em uma área perturbada pela extração seletiva demadeira e uma área adjacente considerada preservada. Nossahipótese é que a riqueza e a abundância de galhas seja maiorna área perturbada devido ao aumento da densidade e riquezade plantas, associadas à abertura de clareiras e trilhas dearraste de árvores.
Material e MétodosO estudo foi desenvolvido em uma área florestal
pertencente a Mil Madeireira Itacoatiara Ltda., em Itacoatiara,AM. A área é cortada por uma estrada que separa a florestaexplorada para corte seletivo de árvores da florestapreservada, que neste estudo serve de controle. O clima daregião é quente e úmido, com temperatura média anual de26o C e precipitação em torno de 2200 mm.
Com o objetivo de comparar a área preservada com aperturbada, foram estabelecidos três transectos em cada umadelas, aproximadamente perpendiculares à estrada principalque separa estas duas áreas, totalizando 500m vistoriadospor área. Estes transectos acompanhavam trilhas já existentesnas áreas. Todas as plantas contendo galhas que estivessemlocalizadas a uma distância de até 1m de ambos os lados datrilha foram amostradas, retirando-se uma folha queapresentasse tal tipo de infestação. As galhas foramposteriormente classificadas em morfotipos de acordo comsuas morfologias (eg. formato e quantidade de galhas) e asda folha na qual estavam presentes. Considerou-seespecificidade máxima entre hospedeiro e parasita, de modoque galhas similares em espécies vegetais distintas foramcomputadas como morfotipos diferentes (veja Floate et al.1996).
Para verificar a possível existência de outrasmorfoespécies de galhas que poderiam ser originárias deárvores do dossel da floresta, foi coletado todo o folhiçocontido em uma circunferência de 0,08 m2, estabelecidaaleatoriamente a cada 50 metros na trilha. As amostras dofolhiço foram triadas e as galhas encontradas foramclassificadas conforme descrito acima.
A riqueza e abundância de galhas em cada área foramcomparadas com uso do teste de Mann –Whitney. Asimilaridade na composição de morfoespécies de galhas paracada par de trilhas foi comparada pelo índice de similaridadede Jaccard (Zar 1984).
ResultadosForam identificadas um total de 73 morfoespécies de
galhas, sendo 47 presentes na área preservada e 31 na áreaalterada. As Figuras 1 e 2 ilustram, respectivamente, a riquezae abundância de morfoespécies de galhas em cada trilhaamostrada.
Não houve diferença significativa entre a área perturbadae a preservada quanto à riqueza (U = 6,0; g.l. = 2; p=0,507),e também quanto à abundância (U = 7,0; g.l. = 2; p = 0,268)de morfotipos de galhas. A composição de morfotipos degalhas, por outro lado, foi muito diferente não só entre asduas áreas, mas também entre cada uma das seis trilhasamostradas (Tabela 1). Somente uma morfoespécie ocorreuem todas as trilhas.
Os índices de similaridade de morfotipos de galhas,calculados para todas as comparações entre pares de trilhasapresentaram valores muito baixos (Tabela 1). A maiorsimilaridade ocorreu entre as trilhas alteradas 2 e 3 (12,5%),e a menor entre a trilha perturbada 1 e a preservada 3 (2,3%).
Tabela 1. Similaridade de composição de morfotipos de galhas entreas trilhas das áreas preservada e perturada (índice de Jaccard), naMil Madeireira, Itacoatiara, AM.
Tabela 2. Riqueza e abundância de galhas em cada trilha nas áreaspreservada (Pres) e perturbada (Pert).
DiscussãoEsperava-se encontrar uma maior riqueza e/ou abundância
de morfotipos de galhas na área perturbada devido aoaumento na riqueza e abundância de espécies hospedeiras,com menor resistência a agentes patógenos. Entretanto, nãohouve diferença entre as duas áreas amostradas com relaçãoa esses dois parâmetros. Desta forma, tanto a riqueza quantoa abundância de organismos indutores de galhaaparentemente não foram afetadas pelo corte seletivo demadeira.
Embora não tenhamos encontrado diferença na riquezade morfotipos de galhas entre as duas áreas, a composiçãode espécies variou muito, mostrando pouca sobreposiçãoentre a área preservada e a alterada. Um estudo mais detalhado
Pert 1 Pert 2 Pert 3 Pres 1 Pres 2 Pres 3
Pert1 0 0,06 0,06 0,04 0,06 0,03
Pert 2 0 0,13 0,04 0,09 0,07
Pert 3 0 0,04 0,04 0,07
Pres 1 0 0,09 0,09
Pres 2 0 0,04
Pres 3 0
Trilha Riqueza Abundância
Pert 1 15 37
Pert 2 20 70b
Pert 3 20 37
Pres 1 8 14
Pres 2 3 3
Pres 3 25 69

26 Curso de Campo Ecologia da Floresta Amazônica - 2001
desta composição de espécies diferenciada talvez revelasseum efeito do corte seletivo de madeira não sobre a riquezaou abundância de galhas, mas sim na composição das espéciesdestas duas áreas.
Neste trabalho, encontramos uma riqueza relativamentemenor de morfotipos de galhas que a encontrada em umestudo no cerrado, onde foram encontrados 92 morfotipos(Gonçalves-Alvim & Fernandes 2001).
De maneira geral, ambientes úmidos como as florestastropicais, possuem uma menor diversidade de morfotipos degalhas, quando comparado a ambientes mais xéricos, comoo cerrado. Isto porque os ambientes úmidos apresentam maiorabundância de fungos e parasitas que pode causar uma grandemortalidade de insetos galhadores (Fernandes & Price 1992).
Fernandes & Price (1988) afirmam que a riqueza deplantas de uma dada área não explicaria uma maior riquezade espécies de galhas, embora seja correlacionada com esta.Por outro lado, o efeito do corte seletivo de madeira naAmazônia pode ter conseqüências em maiores escalas detempo e espaço, impossíveis de serem observadas em estudoscurtos como este. Os efeitos de alterações menos abruptasna paisagem (como é o caso deste tipo de atividade) podemser muito sutis, de forma que só seriam percebidos comestudos mais detalhados. Sugerimos que sejam feitos estudosa longo prazo e em diversas áreas de extração seletiva demadeira para responder a esta questão.
Referências BibliográficasDenslow, J.S. & Hartshorn, G. S. 1994. Tree-fall gap
environments and forest dynamic processes. In: La Selva,Ecology and Natural History of a Neotropical RainForest, McDade, L.A., Bawa, K.S., Hespenhreide, H.A.& Hartshorn, G.S. (eds). University of Chicago Press,London.
Gonçalves-Alvim, S.J. & Fernandes, G.W. 2001.Comunidades de insetos galhadores (Insecta) em
diferentes fisionomias do Cerrado em Minas Gerais,Brasil. Revta bras. Zool. 18: 289-305.
Gonçalves-Alvim, S.J. & Fernandes, G.W. (no prelo).Biodiversity of galling insects: historical, communityand habitat effects in four neotropical savannas. Biodiv.Conserv.
Fernandes, G.W. & Price, P.W. 1988 Biogeopgraphicalgradients in galling species richness tests of hypotheses.Oecologia 76: 161-167.
Fernandes, G.W. & Price, P.W. 1991. Comparisons of tropicaland temperate galling species richness: the roles ofenviromental harshness and plant nutrient status. In:Plant-Animal Interactions: Evolutionary Ecology inTropical and Temperate Regions, Price, P.W.,Lewinsohn, T.M.; Fernandes, G.W. & Benson, W.W.(eds.), pp. 91-115. New York.
Fernandes, G.W. & Price, P.W. 1992. The adaptativesignificance of insect gall distribution: survivorship ofspecies in xeric and mesic habitats . Oecologia 90: 14-20.
Fernandes, G. W.; Silva de Paula, A. & Loyola, R. 1995.Distribuição diferencial de insetos galhadores entrehabitats entre habitats e seu possível uso comobioindicadores. Vida Silv. Neot. 4(2): 133-139.
Floate, K.; Fernandes, G.W. & Nilsson, J. 1996.Distinguishing intrapopulational categories ofplants bytheir insectfauna: galls on rabbitbrush. Oecologia 105:221-229.
Mani, M.S. 1964. The Ecology of Plant Galls. The Hague,The Netherlands.
Uhl, C. 1982. Recovery following disturbances of differentintensities in the Amazon rain forest of Venezuela.Interciência 7: 19-24.
Zar, J.H. 1984. Biostatistical Analysis. Prentice Hall, NewJersey.
Efeito do manejo e de clareiras sobre a regeneração natural de Miconiatetraspermoides (Melastomataceae) e Minquartia guianensis (Olacaceae)
Betsabé Ruiz, Leandro M. Scoss & Daniel L.M. Vieira
IntroduçãoO corte de madeira para fins comerciais é uma das
alternativas ao desflorestamento de florestas tropicais(Anderson 1990). Contudo, poucas madeireiras aplicam osprincípios de manejo florestal para efetuar a extração.Atualmente menos que 0,12% das florestas tropicais têm umaproposta de exploração de baixo impacto (Poore 1989). Acertificação ao contrário da legislação inerente à exploraçãomadeireira, é recente, e poucos são os indícios de que hajacontrole do planejamentos de baixo impacto proposto poreste tipo de exploração (C. Rittl com. pess.).
Uma das primeiras consequências do desmatamento é a
abertura do dossel e criação de clareiras, modificandocondições microclimáticas como, incidência de luz e aumentoda temperatura, gerando assim uma diminuição na umidade.Além das consequências relacionadas diretamente ao corte,são provocados impactos por máquinas, que aumentam acompactação e a erosão do solo, que podem afetar aregeneração da floresta.
Apesar da formação de clareiras potencializar aregeneração natural, em áreas manejadas, a alteração defatores como maior abertura de dossel e luminosidade, podenão permitir que espécies desejáveis regenerem, resultandona colonização de outros grupos de espécies, como por

Curso de Campo Ecologia da Floresta Amazônica - 2001 27
exemplo pioneiras e lianas (Uhl 1990). A regeneração deuma clareira é dependente dos propágulos existentes na áreaantes da sua abertura, daqueles que entram após o distúrbioe da brotação de raízes e troncos. Neste contexto, o bancode sementes e de plântulas existentes no momento da aberturada clareira constitui uma fonte primária de recursos para aregeneração da floresta, desempenhando grande importânciana dinâmica sucessional.
Vianna (1990) propõe que um manejo florestal baseadoem regeneração natural deve considerar disponibilidade desementes e plântulas de espécies desejáveis, a fim depromover a sustentabilidade econômica da floresta. Um fatorque pode retardar a regeneração natural de espécies é acolonização em clareiras de espécies pioneiras “posseiras”,que são caracterizadas pelo crescimento mais lento do queoutras pioneiras, mas que apresentam uma vantagemcompetitiva com espécies tardias, sombreando fortemente ochão da floresta (Williansom 1998).
Neste estudo quantificamos a regeneração natural de umaespécie pioneira “posseira” e uma espécie de interessecomercial para extração de madeira, em habitats de florestapreservada e explorada, a fim de averiguarmos indícios desustentabilidade do manejo florestal realizado pela empresaMil Madeireira Itacoatiara Ltda. Testamos o efeito do manejoflorestal e de clareiras sobre a regeneração natural de duasespécies, Miconia tetraspermoides e Minquartia guianensis.Para testar as diferenças entre o número de indivíduos deambas as espécies entre tratamentos, foram comparadas asdiferenças entre o número de indivíduos que ocorreram nasclareiras e o número de indivíduos que ocorreram nosubosque. Esta diferença é uma medida de efeito do tipo dehabitat que é mais ocupado pelas espécies regenerantesanalisadas neste estudo.
Material e MétodosEste estudo foi realizado nas áreas da empresa Mil
Madereira Itacoatiara Ltda., localizada no município deItacoatiara, AM O clima da região é quente e úmido, comtemperatura média de 26 oC, precipitação em torno de 2.200mm e solo amarelo distrófico argiloso (Mil Madeireira Ltda.,Plano de Manejo 1997). O trabalho foi realizado em umtalhão não explorado (controle) e em um talhão exploradosob normas de certificação florestal de manejo sustentável.
Em cada talhão (explorado e controle) foram selecionadascinco clareiras com estágios sucessionais semelhantes, sendoque as clareiras na área explorada foram resultantes de cortede árvores, e, 30m de distância de cada clareira foi amostradauma área de subosque. No centro de cada clareira e na áreade subosque adjascente foi estabelecida uma parcela de 5x2m(10 m2) e todos os indivíduos de Miconia tetraspermoides eMinquartia guianensis até 1m de altura foram contados. Aabertura do dossel foi medida com densiômetro em doispontos nas extremidades do comprimento de cada parcela,cuja média deles constituiu o valor de abertura de dosselpara cada ponto.
ResultadosOs valores de abertura de dossel e número de espécies de
Miconia tetraspermoides e Minquartia guianensis obtidospara cada parcela e tipo de habitat são apresentados na Tabela1.
Tabela 1. Porcentagem de abertura de dossel e número de indivíduosdas duas espécies, Miconia tetraspermosides e Minquartia guianensis,para áreas de clareiras e subosque de áreas de preservaçãopermanente e de corte seletivo da Mil Madeireira Itacoatiara Ltda.
Não houve diferença significativa da abertura do dosseldo subosque das duas áreas estudadas (t = 0,681; g.l. = 5,7;p = 0,523) (Figura 1). Da mesma forma, as clareiras da áreaexplorada e da área controle não apresentaram diferenças (t= 1,957; df = 6,9; p = 0,092). Mas clareiras e subosque dasduas áreas, apresentaram nítida diferença no percentual deabertura do dossel, onde clareiras estão mais expostas àluminosidade do que o subosque.
Figura 1. Porcentagem média de abertura de dossel na floresta emáreas de preservação (mata preservada) e de exploração (mataexplorada) em clareiras e subosque na área da Mil MadeireiraItacoatiara Ltda. O desvio padrão é indicado por barras.
Miconia tetraspermoides não ocorreu no subosque; nasclareiras das duas áreas esta espécie foi observada, mas nãohouve diferença na densidade de indivíduos entre elas (Mann-Whitney, p = 0,292) (Tabela 2).
As clareiras e as áreas de subosque foram pareadas,portanto utilizou-se a diferença de densidade de Minquartiaguianensis (clareira – subosque = efeito) em cada floresta
0
5
10
15
20
25
Clareira Exp. Clareira Pres. Mata Exp. Mata Pres.
Ab
ert
ura
do
do
ss
el
(%)

28 Curso de Campo Ecologia da Floresta Amazônica - 2001
para comparar as duas florestas através do teste t nãoparamétrico. A subtração clareira – subosque foi negativapara a floresta explorada e 0 para a floresta preservada. Estesvalores diferiram significativamente (Mann-Whitney, p =0,018). Os resultados desta análise indicam que os indivíduosde M. guianensis foram mais freqüentes no subosque da áreaalterada do que nas clareiras. Já na área de preservação, onúmero de indivíduos de M. guianensis encontrados nosubosque e nas clareiras foram muito semelhantes.
Tabela 2. Resultados médios do número de indivíduos para Miconiatetraspermosides e Minquartia guianensis, para clareiras e subosquede áreas de preservação permanente e de corte seletivo da MilMadeireira Itacoatiara Ltda.
Não identificamos diferença significativa entre aabundância de M. guianensis e abertura do dossel (r2 = 0,007;p = 0,73). Já para Miconia tetraspermoides houve diferençasignificativa entre a abertura do dossel e abundância (r2 =0,383; p < 0,01).
DiscussãoA medida de abertura de dossel, utilizada para caracterizar
clareiras e subosque das duas áreas estudadas, não apresentoudiferença significativa entre as áreas, mas podem existircaracterísticas distintas entre as áreas que não foramabordadas neste estudo. Por exemplo Minquartia guianensisrespondeu diferentemente aos tratamentos. Para esta espécie,as clareiras da área explorada provavelmente não são um sítiode regeneração propício para o seu estabelecimento. Osnossos resultados indicam que Minquartia guianensisprovavelmente é uma espécie secundária de estágiosintermediários da sucessão ecológica. Sendo pouco tolerantesà luminosidade, o que pode significar que os indivíduosamostrados neste estudo, mais frequentes no subosque doque em clareiras, poderiam ser os remanescentes do bancode plântulas anteriores à formação das clareiras que aindanão sentiram os efeitos do corte seletivo da madeireira.
A densidade de M. tetraspermoides foi relacionada com
a abertura de dossel e, como pioneira típica, só ocorreu emclareiras, independente da área. Para esta espécie pioneira,as clareiras devem ser o sítio mais propício para a regeneração,mesmo em áreas sob impacto da exploração madeireira.Outras espécies de pioneiras, como as dos gêneros Vismia eCecropia, que necessitam de clareiras para regenerarem(Denslow 1980), foram observadas crescendo apenas emclareiras da área explorada, sendo interessante enfocar estasespécies em estudos futuros que pretendam avaliar o impactodo manejo florestal, visto que a densidade estas espéciespodem indicar o grau de perturbação de uma área e em qualestágio sucessional a área se encontra.
Recomendamos que estudos sobre a regeneração deespécies comerciais e pioneiras facilitadoras da sucessãoflorestal, sejam intensificados, de forma a propiciar amanutenção ou o incremento dessas espécies em sistemasalterados pela exploração florestal. Informações básicas sobrehistória natural e ecologia de espécies com potencialeconômico, bem como de outras espécies que mantêm oequilíbrio de sistemas naturais, são fundamentais paragarantir a sustentabilidade de uso do potencial madeireiroda floresta Amazônica.
Referências BibliográficasDenslow, J.S. 1980. Gap partioning amont tropical rain
forest. Biotropica 12: 47-55.Uhl, C. 1990. Studies of ecosystem response to natural and
anthropogenic disturbances provide guidelines fordesigning sustaintable land-use systems in Amazonia.In: Alternatives to Deforestation: Steps TowardSustainable Use of the Amazon Rain Forest, Anderson,A.B. (ed.). pp.24-37.Columbia University Press.
Vianna, V.M. 1990. Natural seed and seedling avaibility oftropical trees In: Alternatives to Deforestation: StepsToward Sustainable Use of the Amazon Rain Forest,Anderson, A.B. (ed.). pp. 99-111.Columbia UniversityPress.
Willianson, B.; Mesquita, R.C.G.; Kalan, I. & Ganade, G.1998. Estratégias de árvores pioneiras nos trópicos. In:Floresta Amazônica: Dinâmica, Regeneração e Manejo,Gascon, C. & Moutinho (eds.). MCT-INPA.
Influência da exploração seletiva de madeira sobre a fauna de anuros deserapilheira em área de baixio na Amazônia Central
Maria R. Darrigo, Cristiane F. Jurinitz, Angela M. F. Pacheco, Christina McCaine & Rodrigo Cogni
IntroduçãoPerturbações antrópicas resultam em alterações nos
componentes bióticos e abióticos em ecossistemas tropicais(Myers 1986). A exploração seletiva de madeira em florestaspode alterar a estrutura da vegetação, gerando mudanças naumidade e temperatura do solo, além de alterações no volume
da serapilheira, devido ao incremento no número de clareiras(Kapos et al. 1997).
As condições ambientais em florestas tropicais sãorelativamente constantes, apesar de não necessariamenteuniformes (Myers 1986). May (1975), afirma que sistemascom arranjo complexo de espécies e ampla rede de interações,

Curso de Campo Ecologia da Floresta Amazônica - 2001 29
como encontrado em florestas tropicais, tendem a ser maisfrágeis que sistemas mais simples. Assim, podemos suporque perturbações em florestas tropicais, mesmo não sendode grandes proporções, podem causar impacto negativo empopulações animais (Myers 1986).
Um grupo interessante para verificar alterações empopulações animais resultantes de mudanças na estrutura davegetação são os anuros, já que são afetados por alteraçõesmicroclimáticas tais como variações na umidade e volumede serapilheira (Allmon 1991). Além disso, anfíbiosconstituem um grupo de fácil coleta e identificação em campo,podendo assim ser utilizado como indicadores biológicos.
Neste estudo avaliamos diferenças na composição deespécies e densidade de anuros de serapilheira de áreas debaixio, em uma área preservada e uma área onde ocorre corteseletivo de madeira na Amazônia Central.
Material e MétodosRealizamos as coletas na área da Madeireira Mil,
Município de Itacoatiara, AM. (2o 43’S; 58o 31’W). O climado local é quente e úmido, com precipitação média anual de2000 mm e temperatura média de 26o C (C. Rittl com. pess.).Utilizamos como local de amostragem a faixa de 30 m deproteção ambiental, a partir da margem do igarapé,estabelecida pela legislação.
Coletamos as amostras em áreas de baixio de seis igarapésde primeira ordem, sendo três em área de preservação e trêsem área de corte seletivo de madeira, com menos de um anode atividade de extração. Em cada igarapé amostramos trêsparcelas de 9 m2, distantes 15m entre si e localizadas a 2mdo curso d’água. As parcelas foram delimitadas por barbantes,e toda a serapilheira era vasculhada e removida por seispessoas, na direção da borda para o centro, evitando assim afuga dos anuros. Os anuros encontrados foram identificadose em seguida liberados.
ResultadosColetamos um total de 62 indivíduos de três espécies de
anuros. A densidade encontrada foi semelhante nas duas áreas(Figura 1). Colostethus stepheni foi a espécie dominante emambas as áreas (Tabela 1). Não constatamos diferenças nacomposição de espécies entre as duas áreas .
Figura 1. Densidade de anuros de serapilheira em igarapés de umaárea com exploração seletiva de madeira e uma área preservada naMil madeireira, Itacoatiara, AM.
Explorado1
Explorado2
Explorado3
Preserva
do1
Preserva
do2
Preserva
do3
Igarapé
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
Ind
ivíd
uo
sp
or
m2
Tabela 1. Composição de espécies e abundância de anuros deserapilheira em igarapés de uma área com exploração seletiva demadeira e uma área preservada na Mil madeireira, Itacoatiara, AM.
DiscussãoEm uma área similar à deste estudo, Allmon (1991), ao
longo de 15 meses, encontrou 23 espécies de sapos deserapilheira, registrando maior densidade e riqueza nos meseschuvosos. Deste modo, o baixo número de espécieencontrados por nós pode ser explicado pelo fato do trabalhoter sido realizado na época seca e curto período deamostragem. Com nosso esforço amostral, encontramosapenas as espécies mais comuns em serapilheira (Allmon1991). Estas espécies são generalistas e reconhecidamentetolerantes a alterações ambientais resultantes de processosantrópicos (Allmon 1991). Desta maneira, um maior esforçoamostral pode ser necessário para coleta de um maior númerode espécies, e verificar como estas são afetadas pela extraçãode madeira.
Outra possível causa para o fato de não termos encontradodiferenças na composição de espécies e abundância deindivíduos entre as duas áreas pode estar relacionado ao fatodas coletas terem sido conduzidas apenas em área de baixios,onde a exploração de madeira é proibida em uma faixa de 30m de cada lado do igarapé. Deste modo, a extração de madeiranão estaria afetando os anuros da serapilheira no baixio.Assim, amostragens em outros locais, como platô e vertentespodem esclarecer se os anuros são afetados nestes ambientes.Também temos que considerar que as populações de anurospresentes no platô devem ser mais sensíveis a variações deumidade decorrente da ação antrópica que as populaçõespresentes no baixio.
Uma outra hipótese possível é que os anuros de umamaneira geral não são afetados por processos antrópicos,como demostrado no projeto de dinâmica de fragmentosflorestais (H. Vasconcelos com. pess.).
AgradecimentosAgradecemos ao Deputado Caçula Juruna, pela ajuda no
campo e identificação dos anuros. Agradecemos tambémMarcelo Gordo e Carlos Rittl pela orientação.
Referências BibliográficasAllmon, W.D. 1991. A plot study of forest floor litter frogs,
Central Amazon, Brazil. J. Trop. Ecol. 7: 503-522.
Área explorada Área preservada
No total de
indivíduos
No de indivíduos por
igarapé (X� DP)
No total de
indivíduos
No de indivíduos por
igarapé (X� DP)
Colostethus stepheni 20 6,6� 5,5 32 10, 7 � 4,0
C. marchesianus 1 0,3 � 0,6 1 0,3 � 0,6
Adenomera andreae 5 1,1 � 1,1 2 0,7 � 0,6
Não identificado 1 - 0 -
Total 27 9,0 � 6,6 35 11,7 � 3,0

30 Curso de Campo Ecologia da Floresta Amazônica - 2001
Kapos, V.; Wandelli, E.; Camargo, J.L. & Ganade, G. 1997.Edge related changes in environment and plant responsesdue to forest fragmentation in Central Amazon. In:Tropical Forest Remnants, Laurance, W.F. &Bierregaard, R.O. (eds.). Chicago Press, Chicago.
May, R.M. 1975. The tropical rain forest. Nature 257: 737-738.
Myers, N. 1991. Tropical deforestation and mega-extinction spasm. In: Conservation Biology, Soulé, M.(ed.).
Efeito da inundação sobre o padrão reprodutivo de Vitex cymosa (Verbenaceae)em uma área de várzea no Rio Solimões, AM
Paula S. Pinheiro, Angela M. F. Pacheco, Leandro M. Scoss, Mauricio Camargo & Paula C. Ceotto
IntroduçãoAs várzeas são florestas de planície inundáveis
periodicamente, sujeitas a drásticas variações sazonais donível das águas, que podem alcançar até 14m de amplitude(Ferreira 1997). Esta variação da coluna d’água por sua vezé regida pela dinâmica de precipitação e escoamento ao longoda bacia do Amazonas (Sioli 1984). A origem andina dosvários rios de águas brancas na bacia define processos detransporte e deposição de sedimentos recentes nas florestasde várzea. Como conseqüência, tem-se áreas muito ricas emnutrientes nestas formações vegetais (Junk 1997).
A periodicidade e a duração do alagamento podem exercergrande influência em processos ecológicos, padrões deriqueza de espécies, além de padrões reprodutivos comofloração e dispersão de frutos e sementes (Ferreira 1998).Worbes (1997) relatou uma sincronia nos períodos defloração e frutificação de plantas lenhosas nas florestas devárzea ligadas à flutuação sazonal do nível do rio. Duaspopulações de Eschweilera parvifolia localizadas emdiferentes cotas em uma floresta de várzea apresentaramvariações no período de floração e frutificação relacionadascom diferenças no tempo de inundação (Ferreira 1998).
Vitex cymosa é uma espécie arbórea abundante na ilha deMarchantaria, cujos indivíduos estão distribuídos ao longode um gradiente de inundação, porém com maior abundâncianas cotas baixas. Esta planta sofre inundação parcial duranteas cheias e se mantém fora d’água por cerca de apenas trêsmeses ao ano. Os indivíduos de V. cymosa perdem suas folhasno pico da enchente, e a produção das novas folhas ocorreno início da vazante, assim como a floração. O período defloração e frutificação é de aproximadamente dois meses(Worbes 1997). Os frutos são flutuantes e são dispersosprincipalmente através da hidrocoria, mas a ictiocoriaaparentemente também é efetiva para dispersão nessa espécie(L. V. Ferreira com. pess.).
Neste trabalho, estudamos a influência do nível deinundação na presença ou ausência de frutos de Vitex cymosa(Verbenaceae) ao longo de um gradiente batimétrico de umafloresta de várzea na ilha da Marchantaria.
Material e MétodosA área de estudo localiza-se em uma floresta de várzea,
que se encontra na Ilha da Marchantaria (03o 14’S; 59o 57’W),
no Rio Solimões, aproximadamente a 15km da confluênciado rio Solimões com o Rio Negro. A temperatura média anualé de 26,7oC e a pluviosidade varia em torno de 2,186mm porano (RadamBrasil 1978). Nossa coleta de dados ocorreudurante o período de vazante (17/08/2001).
Amostramos 40 indivíduos de Vitex cymosa, metade emcada uma das duas categorias de cota de profundidade, sendoa primeira de 2 a 3 m e a segunda de profundidade entre 3 e4m. Anotamos o diâmetro próximo à lamina d’água, aexposição da copa à luminosidade e o estado fenológico daplanta (presença ou ausência de frutos). Com as informaçõesobtidas para o diâmetro por cota de profundidade, foicalculada a média e o desvio padrão. Simultaneamente,comparamos a presença ou ausência de frutos para cadaprofundidade.
Como forma de explorar uma possível relação daprofundidade da coluna e o diâmetro da planta, plotamos osdados para verificar a existência de alguma relação entre estasduas variáveis.
ResultadosAs observações em campo nos indicaram que, comparada
com outras espécies arbóreas, Vitex cymosa ocorre com muitafreqüência na área alagada da zona de estudo. Do total de 40plantas medidas em seu diâmetro à altura da superfície daágua, 39 apresentaram frutos. O valor médio do diâmetro dotronco à altura da água foi igual para as duas cotas estudadas,com o desvio padrão também similar (Tabela 1).
Tabela 1. Variação do diâmetro das plantas de Vitex cymosa emfunção da cota batimétrica na área de várzea da Ilha da Marchantaria,Rio Solimões, AM.
Não verificamos uma correlação entre as diferentesprofundidades e o respectivo diâmetro de cada um dosindivíduos amostrados (r = 0,11; N = 40; p > 0,05) (Figura1).
Média ± DP Mínimo Máximo
2-3 m 29 ± 9 cm 12 47
3-4 m 29 ± 10 cm 11 47

Curso de Campo Ecologia da Floresta Amazônica - 2001 31
Figura 1. Distribuição diamétrica dos indivíduos de Vitex cymosaem duas categorias de profundidade, na várzea da Ilha daMarchantaria, AM. Cotas maiores significam menor profundidade etempo de inundação.
DiscussãoO grande número de plantas de Vitex cymosa com frutos
nas duas cotas de profundidade nos indica que não existemdiferenças evidentes no período de frutificação entre estas.A planta parece não ser afetada por uma variação de um metrono nível de inundação. Entretanto, algumas limitações dométodo devem ser consideradas.
As medidas de diâmetro não foram realizadas em relaçãoa altura da base das plantas, mas sim em relação ao nível daágua. Consequentemente, medimos as plantas em alturasdiferentes, inviabilizando a comparação do tamanho dasplantas entre as cotas já que elas estavam em profundidadesdistintas. A alternativa mais simples para padronizar asinformações seria realizar a medição do diâmetro a um metrode altura da lâmina da água para o grupo localizado na cotamais alta.
Apesar de não ocorrer uma correlação entre as diferentescotas batimétricas e o diâmetro das árvores, acreditamos que
através de um delineamento mais criterioso em relação aodiâmetro e a uma maior amplitude de cotas, poderíamosdetectar padrões de frutificação mais confiáveis em relaçãoaos pulsos de inundação. Além disso, um estudo como estedeveria abranger uma escala temporal maior, acompanhandotodo o ciclo reprodutivo de Vitex cymosa.
Referências BibliográficasFerreira, L.V. 1997. Effects of the duration of flooding on
species richness and floristic composition in threehectares in the Jaú National Park in floodplain forestsin central Amazônia. Biodiv. Conserv. 6(10): 1353-1363.
Ferreira, L.V. 1998. Intraspecific variation in phenology inrelation to flooding duration in Eschweilera parvifolia(Lecythidaceae) in Central Amazonian flooplain forest.An. Acad. Bras. Ci. 70(3): 677-680.
Junk, W.J. 1997. General Aspects of Floodplain Ecologywith Special Reference to Amazon Floodplains. In: TheCentral Amazon Floodplain Ecology of a PulsingSystem, Junk, W (ed). Springer-Verlag, Berlin.
RadamBrasil. 1978. Levantamento dos Recursos Naturais.Ministério de Minas e Energia. Departamento Nacionalde produção Mineral, Rio de Janeiro.
Sioli, H. 1984. The Amazon and its main affluents:hydrography, morphology of the river courses and rivertypes. In: The Amazon: Limnology and LandscapeEcology of a Mighty Tropical River and its Bbasin, Sioli,H (ed), pp 127-165. Monographiae Biologicae, Junk,Dordrech.
Worbes, M. 1997. The forest ecosystem of the floodplains.In: The Central Amazon Floodplain Ecology of a PulsingSystem, Junk, W (ed). Springer-Verlag, Berlin.
OrientadorLeandro V. Ferreira
Estrutura da ictiofauna associada a bancos de macrófitas flutuantes em umaárea de várzea do Rio Solimões, AM
Angela M. F. Pacheco, Paula S. Pinheiro, Leandro M. Scoss, Mauricio Camargo & Paula C. Ceotto
IntroduçãoAs flutuações sazonais do nível da água e a baixa
declividade da planície Amazônica proporcionam a formaçãode grandes extensões de áreas de várzea, igapós, igarapés,lagos e paranás (Walker 1995). O rio Solimões, que banhaas florestas de várzea, origina-se em bacias sedimentares quedatam do Quaternário, contendo sedimentos ricos emnutrientes e pH quase neutro, o que propicia odesenvolvimento de ilhas de macrófitas flutuantes (Conserva& Piedade 2001).
Cerca de 50 espécies podem compor essas ilhas flutuantes,
sendo as mais comuns em grandes bancos Eichorniacrassipes, Paspalum repens, Salvinia sp., Pistia stratioidese Mimosa sp. Paspalum repens é uma gramínea que possuiraízes com cerca de 2m na coluna d’água. Suas raízesproporcionam um substrato adequado para o estabelecimentode uma rica ictiofauna, que se alimenta basicamente deorganismos planctônicos e insetos aquáticos (Junk 1997).
Os bancos de macrófitas são ambientes temporários queaumentam de tamanho à medida que o nível da água do rioaumenta, iniciando a senescência com o período de vazante.Neste período os bancos podem se desprender da margem
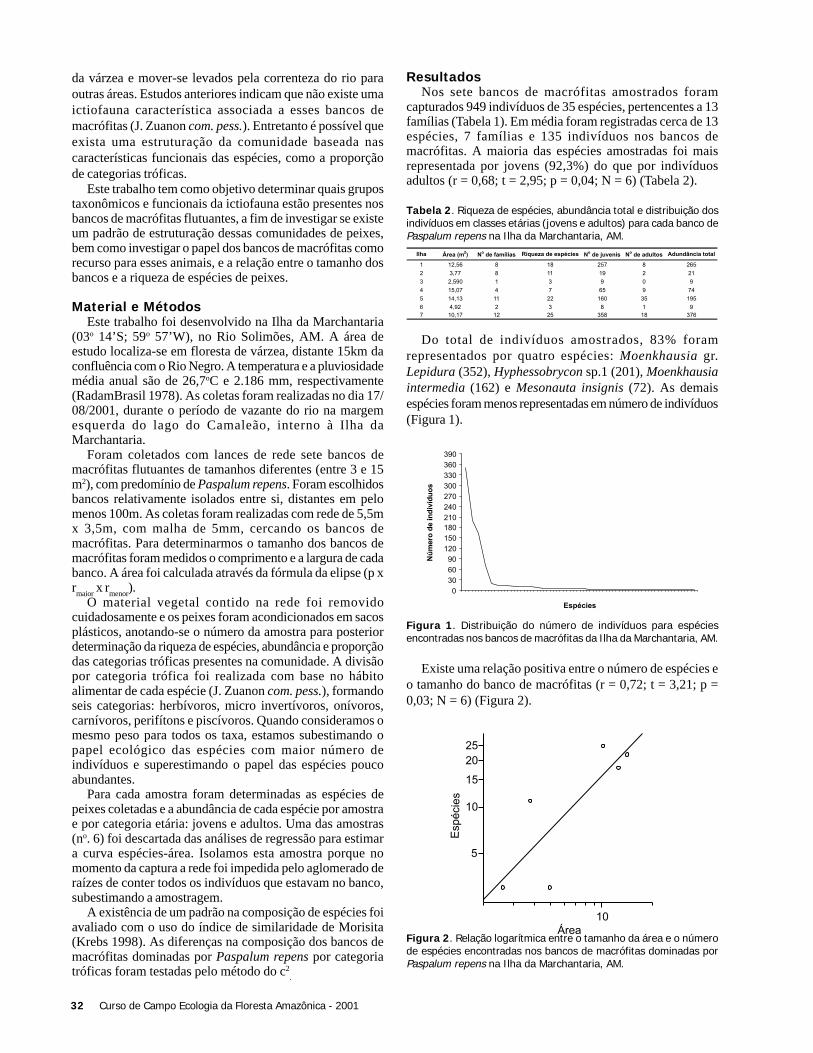
32 Curso de Campo Ecologia da Floresta Amazônica - 2001
da várzea e mover-se levados pela correnteza do rio paraoutras áreas. Estudos anteriores indicam que não existe umaictiofauna característica associada a esses bancos demacrófitas (J. Zuanon com. pess.). Entretanto é possível queexista uma estruturação da comunidade baseada nascaracterísticas funcionais das espécies, como a proporçãode categorias tróficas.
Este trabalho tem como objetivo determinar quais grupostaxonômicos e funcionais da ictiofauna estão presentes nosbancos de macrófitas flutuantes, a fim de investigar se existeum padrão de estruturação dessas comunidades de peixes,bem como investigar o papel dos bancos de macrófitas comorecurso para esses animais, e a relação entre o tamanho dosbancos e a riqueza de espécies de peixes.
Material e MétodosEste trabalho foi desenvolvido na Ilha da Marchantaria
(03o 14’S; 59o 57’W), no Rio Solimões, AM. A área deestudo localiza-se em floresta de várzea, distante 15km daconfluência com o Rio Negro. A temperatura e a pluviosidademédia anual são de 26,7oC e 2.186 mm, respectivamente(RadamBrasil 1978). As coletas foram realizadas no dia 17/08/2001, durante o período de vazante do rio na margemesquerda do lago do Camaleão, interno à Ilha daMarchantaria.
Foram coletados com lances de rede sete bancos demacrófitas flutuantes de tamanhos diferentes (entre 3 e 15m2), com predomínio de Paspalum repens. Foram escolhidosbancos relativamente isolados entre si, distantes em pelomenos 100m. As coletas foram realizadas com rede de 5,5mx 3,5m, com malha de 5mm, cercando os bancos demacrófitas. Para determinarmos o tamanho dos bancos demacrófitas foram medidos o comprimento e a largura de cadabanco. A área foi calculada através da fórmula da elipse (p xr
maior x r
menor).
O material vegetal contido na rede foi removidocuidadosamente e os peixes foram acondicionados em sacosplásticos, anotando-se o número da amostra para posteriordeterminação da riqueza de espécies, abundância e proporçãodas categorias tróficas presentes na comunidade. A divisãopor categoria trófica foi realizada com base no hábitoalimentar de cada espécie (J. Zuanon com. pess.), formandoseis categorias: herbívoros, micro invertívoros, onívoros,carnívoros, perifítons e piscívoros. Quando consideramos omesmo peso para todos os taxa, estamos subestimando opapel ecológico das espécies com maior número deindivíduos e superestimando o papel das espécies poucoabundantes.
Para cada amostra foram determinadas as espécies depeixes coletadas e a abundância de cada espécie por amostrae por categoria etária: jovens e adultos. Uma das amostras(no. 6) foi descartada das análises de regressão para estimara curva espécies-área. Isolamos esta amostra porque nomomento da captura a rede foi impedida pelo aglomerado deraízes de conter todos os indivíduos que estavam no banco,subestimando a amostragem.
A existência de um padrão na composição de espécies foiavaliado com o uso do índice de similaridade de Morisita(Krebs 1998). As diferenças na composição dos bancos demacrófitas dominadas por Paspalum repens por categoriatróficas foram testadas pelo método do c2
.
ResultadosNos sete bancos de macrófitas amostrados foram
capturados 949 indivíduos de 35 espécies, pertencentes a 13famílias (Tabela 1). Em média foram registradas cerca de 13espécies, 7 famílias e 135 indivíduos nos bancos demacrófitas. A maioria das espécies amostradas foi maisrepresentada por jovens (92,3%) do que por indivíduosadultos (r = 0,68; t = 2,95; p = 0,04; N = 6) (Tabela 2).
Tabela 2. Riqueza de espécies, abundância total e distribuição dosindivíduos em classes etárias (jovens e adultos) para cada banco dePaspalum repens na Ilha da Marchantaria, AM.
Do total de indivíduos amostrados, 83% foramrepresentados por quatro espécies: Moenkhausia gr.Lepidura (352), Hyphessobrycon sp.1 (201), Moenkhausiaintermedia (162) e Mesonauta insignis (72). As demaisespécies foram menos representadas em número de indivíduos(Figura 1).
Figura 1. Distribuição do número de indivíduos para espéciesencontradas nos bancos de macrófitas da Ilha da Marchantaria, AM.
Existe uma relação positiva entre o número de espécies eo tamanho do banco de macrófitas (r = 0,72; t = 3,21; p =0,03; N = 6) (Figura 2).
Figura 2. Relação logarítmica entre o tamanho da área e o númerode espécies encontradas nos bancos de macrófitas dominadas porPaspalum repens na Ilha da Marchantaria, AM.
Ilha Área (m2) No de famílias Riqueza de espécies No de juvenis No de adultos Adundância total
1 12,56 8 18 257 8 265
2 3,77 8 11 19 2 21
3 2,590 1 3 9 0 9
4 15,07 4 7 65 9 74
5 14,13 11 22 160 35 195
6 4,92 2 3 8 1 97 10,17 12 25 358 18 376
0
30
60
90
120
150
180210
240
270
300
330
360
390
Espécies
Nú
mer
od
ein
div
ídu
os
10Área
5
10
15
2025
Esp
éci
es

Curso de Campo Ecologia da Floresta Amazônica - 2001 33
Utilizamos o índice de similaridade de Morisita (Krebs1998) para testar se os bancos de macrófitas apresentampadrões de composição de espécies e abundância (Figura 3).A partir do resultado de similaridade, calculamos a diferençaentre o tamanho das áreas dos bancos e relacionamos com oíndice de similaridade (Figura 4) para testar se o tamanhodos bancos está ou não estruturando as comunidades depeixes.
Figura 3. Valores de similaridade (Morisita) entre os bancos demacrófitas de Paspalum repens da área de várzea da Ilha daMarchantaria, AM.
1 2 3 4 5 612 0.113 0.24 0.764 0.45 0.38 0.565 0.66 0.43 0.24 0.326 0.99 0.16 0.25 0.45 0.72
-20 -10 0 10
Diferença entre o tamanho das áreas
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
Sim
ilarid
ad
e
Tabela 3. Espécies e famílias identificadas nos bancos de macrófita da várzea da Ilha da Marchantaria, AM. As categorias tróficas sãorepresentadas por microinvertívoros (1), perifíton (2), carnívoros (3), piscívoros (4), onívoros (5) e herbívoros (6). Os bancos de macrófitasforam numerados de 1 a 7.
Figura 4. Valores de similaridade (Morisita) e diferença entre otamanho das áreas dos bancos Paspalum repens da área de várzeada Ilha da Marchantaria, AM.
No total Ocorrência
de indivíduos (bancos)
Adontosternarchus "anal clara" Apteronotidae 7 5, 7 3Adontosternarchus "anal preta" Apteronotidae 1 5 3Ageneiosus brevis Ageneiosidae 2 5 3Anadoras sp. Doradidae 1 7 5Aphyocharax aff. erythurus Characidae 14 1, 4, 5,7 1Apistogramma agassizi Cichlidae 3 1, 2, 5 1Apistogramma sp. Cichlidae 1 7 1Apteronotus hasemani Apteronotidae 2 2, 5 3Brachyhypopomus pinnicaudatus Hypopomidae 4 1, 5, 7 3Brachyhypopomus brevirostris Hypopomidae 1 5 3Cichla monoculus Cichlidae 1 2 4Cichlasoma amazonarum Cichlidae 1 1 5Crenicichla sp.1 Cichlidae 6 1, 2, 5 3Ctenobrycon hauxwelliannus Characidae 6 5, 7 5Eigenmannia sp. Sternopygidae 5 5, 7 3Hemigrammus levis Characidae 12 1, 4 1Hoplias gr. malabaricus Erythrinidae 12 1, 2, 5, 7 4Hyphessobrycon eques Characidae 25 1, 5 7 1Hyphessobrycon sp.1 Characidae 201 1, 3, 4, 5, 7 1Mesonauta insignis Cichlidae 72 1, 2, 3, 4, 5, 7 2Moenkhausia gr. lepidura Characidae 352 1, 3, 4, 5, 6, 7 5Moenkhausia intermedia Characidae 162 1, 2, 5, 6, 7 5Odontostilbe sp. Characidae 2 5, 7 1Odontostilbe sp.2 "cauda vermelha" Characidae 2 7 1Parauchenipterus cf. galeatus Auchenipteridae 14 1, 2, 5, 7 3Parauchenipterus sp. "malhado" Auchenipteridae 1 7 3Petalodoras cf. eigenmanni Doradidae 11 1, 2, 6, 7 5Prionobrama filigera Characidae 1 7 1Pterophyllum scalare Cichlidae 1 1 3Rivulus ornatus Rivulidae 6 4, 5, 7 1Schizodon fasciatus Anostomidae 3 1, 7Serrasalmus rhombeus Serrasalmidae 1 7 3Serrasalmus spilopleura Serrasalmidae 18 1, 2, 4, 5, 7 3Synbranchus sp. Synbranchidae 4 2, 5, 7 4Triportheus flavus Characidae 1 1 4
Nível tróficoFamíliaEspécies

34 Curso de Campo Ecologia da Floresta Amazônica - 2001
Na Tabela 3 é apresentada a lista de espécies de peixesregistradas neste trabalho e as principais informações queforam utilizadas na análise dos resultados deste trabalho.
Não houve uma relação significativa entre o número deindivíduos (R2 = 0,62; t = 2,59; p = 0,06; N = 6) e de famílias(R2 = 0,56; t = 2,26; p = 0,08; N=6) e o bancos de macrófitas.Os bancos de macrófitas de Paspalum repens apresentaramconjuntos de peixes significativamente diferentes quanto acomposição por categoria trófica (c2 = 322,86; g.l. = 25.; p <0,01). As categorias tróficas mais abundantes foramcarnívoros e microinvertívoros (Tabela 4).
Tabela 4. Proporção de cada categoria trófica em relação ao númerode indivíduos e número de espécies entre os bancos de macrófitasda Ilha da Marchantaria, AM.
DiscussãoAs abordagens através das quais podem ser estudadas as
comunidades, compreendem desde associações espaciais,funcionais ou taxonômicas, até interações dinâmicas dentrodas teias tróficas (MacArthur 1972). Cada uma destas afetaráa compreensão da estrutura da comunidade de formadiferente.
Os nossos resultados indicam que há um aumento nonúmero de espécies quando há um incremento de área.Contudo, de acordo com Westoby (1993), nossa estimativada curva espécies-área é restrita aos bancos de macrófitasque foram amostrados (diversidade alfa), e podem não serrepresentativa do padrão regional de riqueza de espécies(diversidade beta) neste ambiente, em função do tamanho danossa amostra.
Um fator que poderia estar afetando nossos resultados é apossível ocorrência de descontinuidades (Schluter & Ricklefs1993), ou seja, alteração da composição e abundância deespécies de peixes, que podem estar ocorrendo dentro dosdiferentes bancos estudados. Um outro aspecto que de fatorefere-se às diferenças de importância ecológica das espéciescomuns e raras. Neste estudo, a distribuição do número deindivíduos entre as espécies registradas suporta o padrão maisfrequentemente observado na natureza, ou seja, muitasespécies raras e poucas abundantes. A densidade de umaespécie pode ser diretamente influenciada pela abundânciade outras espécies que apresentem hábitos alimentaressemelhantes, uma vez que estas podem estar competindo porrecursos.
Os bancos de Paspalum repens amostrados mostraram-
Categoria trófica No de indivíduos No de espécies
Herbívoro 3 1
Micro invertívoro 255 15
Onívoro 524 1
Carnívoro 63 14
Perifíton 72 4
Piscívoro 18 1
se muito heterogêneos quanto a composição de espécies depeixes, o que significa que além da composição florísticados bancos de macrófitas e da estrutura submersa destesbancos (atuando como abrigo e substrato para outrosorganismos do perifíton), outros fatores devem estarinfluenciando a composição da comunidade de peixes. Comoexemplo podemos citar a composição original de espéciesde peixes nos bancos maiores, de onde estas manchas sedesprendem podendo levar consigo conjuntos menores deespécies de peixes formados ao acaso, em funçãoprincipalmente do tamanho do banco de macrófita.
Em relação à similaridade entre os bancos de macrófitas,estes foram pouco similares. O que observamos é que bancoscom tamanhos mais parecidos têm uma ictiofauna maissimilar. A relação entre a diferença do tamanho da área dosbancos de Paspalum repens e a similaridade entre as áreasindica que o tamanho dos bancos não está estruturando ascomunidades de peixes.
AgradecimentosAgredecemos a Jansen Zuanon pela orientação e auxílio
no trabalho de campo e ao Juruna pelo apoio logístico.
Referências BibliográficasConserva, A.S. & Piedade, M.T.F. 2001. Ciclo de vida e
ecologia de Paspalum fasciculatum Willd. Ex. Fluegge(Poaceae), na Várzea da Amazônia Central. ActaAmazonica 31(2): 205-220.
Junk, W.J. & Robertson, A. 1997. Aquatic invertebrates. In:The Central Amazon Floodplain, Junk, W.J. (ed.).
Krebs, C.J. 1998. Ecological Methodology. Benjamin/Cummings, Menlo Park.
MacArthur, R.H. 1972. Geographical Ecology Patterns inthe Distribution of Species. Princeton University Press.
RadamBrasil. 1978. Levantamento dos Recursos Naturais.Ministério de Minas e Energia. Departamento Nacionalde produção Mineral, Rio de Janeiro.
Schluter, D. & Ricklefs, R. 1993. Species diversity andintroduction to the problem. In: Species Diversity inEcological Communities: Historical and GeographicalPerspectives, Ricklefs, R. & Schluter, D. (eds). TheUniversity of Chicago Press, Chicago and London.
Walker, I. 1995. Amazonian streams and small rivers. In:Limnology in Brazil. Tundisi, J.G.; Bicudo, C.E.M &Matsumura, T. (eds.). Brasilian Academy of Sciences.
Westoby, M. 1993. Biodiversity inAustralia compared withother continents. In: Species Diversity in EcologicalCommunities: Historical and Geographical Perspectives,Ricklefs, R. & Schluter, D. (eds), pp. 170–177. TheUniversity of Chicago Press, Chicago and London.
OrientadorJansen Zuanon

Curso de Campo Ecologia da Floresta Amazônica - 2001 35
Estudo da dieta de duas espécies de aves granívoras na várzea da Ilha daMarchantaria, AM
Natalie Olifiers, Christina McCain, Edgar Prieto, Felipe B. Oliveira & Maria R. Darrigo
IntroduçãoOs rios amazônicos apresentam variações sazonais e
previsíveis no nível de suas águas, com períodos bemmarcados de cheia e vazante (Junk 1997). Esta flutuaçãocíclica produz uma grande diversidade de hábitats,colonizados por diversas espécies de herbáceas aquáticas(Conserva & Piedade 2001).
Entre as herbáceas aquáticas encontradas na várzea, pode-se destacar a presença de gramíneas anuais como Paspalumfasciculatum (Conserva & Piedade 2001), que são resistentesàs perturbações sazonais, apresentando ciclo de vida curto egrande potencial para a colonização desse tipo de hábitat(Conserva & Piedade 2001). A grande produção de sementesencontrada em gramíneas faz delas uma importante fonte derecurso alimentar para determinados pássaros, como osgranívoros (S. Borges com. pess.).
Sabe-se que as espécies de aves encontradas nas florestasalagáveis compõem 15% da avifauna de florestas daAmazônia (Borges & Carvalhaes 2000), sendo muito distintada encontrada em florestas de terra firme. No entanto, poucose sabe sobre a composição de espécies e biologia dessesanimais (Borges & Carvalhaes 2000).
Na Ilha da Marchantaria, uma área de várzea na regiãoAmazônica, é conhecida a presença de sete espécies de avesgranívoras (S. Borges com. pess.), para as quais os frutos degramíneas podem ser de grande importância. Neste estudorelatamos o comportamento de forrageamento de algumasespécies de aves granívoras em uma área de várzea, paraaveriguar a existência de sobreposição na utilização deespécies de gramíneas como recurso alimentar.
Material e MétodosEste estudo foi realizado na várzea da Ilha da Marchantaria
(Iranduba, AM), no Rio Solimões, a aproximadamente 15Km ao sul de Manaus (3°14’S; 59°57’W). Durante duas horase meia (6:00 - 8:30h) procuramos localizar as aves granívorasnos bancos de herbáceas flutuantes. Em cada grupoencontrado, observamos o comportamento de alimentaçãodas aves, registrando as espécies de gramíneas consumidas.A identificação das aves e gramíneas foi realizada com a ajudado professor Sérgio Borges da Fundação Vitória Amazônica.
Uma panícula de cada espécie de gramínea foi coletadapara calcularmos o tamanho médio da semente utilizada pelasaves, a fim de testarmos possíveis relações entre tamanho doalimento e o porte do animal.
Utilizamos o Teste Exato de Fisher para averiguarmosdiferenças na frequência de utilização das espécies gramíneaspelas aves granívoras.
ResultadosObservamos indivíduos ou grupos de indivíduos de quatro
espécies de aves granívoras: Agelaius icterocefalus(Icterideae - 10 observações), Sicalis columbiana(Catamblyrhynchidae - cinco), Sporophila lineola(Fringillideae - 14) e Sporophila americana (Fringillideae -um). Três espécies de gramíneas estavam sendo utilizadascomo item alimentar pelas aves estudadas: Echinochloapolystachya, Paspalum repens e Oryza sp. Como observamospoucos indivíduos de S. columbiana, S. americana e deOryza sp., excluímos esta gramínea e as duas espécies deaves das análises realizadas.
Notamos uma diferença significativa na utilização das duasespécies de gramíneas pelas aves granívoras (c2 = 10,48; g.l.= 1; p = 0,001). A. icterocephalus apenas se alimentou dassementes de E. polystachya, enquanto S. lineola mostroumaior frequência de utilização das sementes de P repens (67%dos registros), embora também tenha utilizado E. polystachya(33% dos registros).
Figura 1. Frequência de utilização das espécies de gramíneas pelasaves granívoras estudadas.
DiscussãoA diferença na frequência de utilização das gramíneas
indica que a sobreposição na dieta das aves granívoras foirelativamente baixa. A. icterocephalus é a espécie de maiortamanho corporal (18 cm de comprimento total),apresentando um bico maior e mais afilado (Hilty & Brown1986). Esta morfologia de bico pode não ser muito eficientena utilização de P. repens, uma vez que as sementes destagramínea são muito pequenas (X ± DP = 2.1 ± 0,02mm decomprimento). Por outro lado, S. lineola, por ser menor (11cm de comprimento total) e possuir um bico mais curto (Hilty& Brown 1986), pode ser mais eficiente quando alimenta-sede E. polystachya (X ± DP = 5,8 ± 0,29mm de comprimento)e portanto apresenta maior freqüência de visitas a essagramínea.
Outra hipótese com respeito à utilização das gramíneaspelas aves pode estar relacionada à morfologia das plantas.
0
2
4
6
8
10
12
Sporophila lineola Agelaius
icterocephalus
Núm
ero
deob
serv
açõe
s
Echinochloa polystechya
Paspalum ripens

36 Curso de Campo Ecologia da Floresta Amazônica - 2001
O ramo que sustenta a panícula de P. repens é mais frágil emaleável que o de E. polystachya. Por este motivo, A.icterocephalus não conseguiria sustentar-se nos ramos dessaespécie para manipular as sementes. Por outro lado, S. lineolautilizaria com maior frequência P. repens devido ao seu bico(e peso corporal) relativamente menor, que seria maisadequado para a manipulação das sementes desta espécie.
As duas hipóteses apresentadas se adequam à observaçãode que S. lineola alimenta-se das duas espécies de gramíneas– embora com diferentes freqüências – enquanto A.icterocephalus não utiliza P. repens, devido ao seu maiortamanho corporal e à fragilidade do ramo desta gramínea e/ou ao seu maior tamanho de bico.
Essas hipóteses não necessariamente são exclusivas eembora sejam especulativas, apresentam um bomembasamento teórico. Características morfológicas corporaisrelacionadas à alimentação, como o tamanho do bico de umaave, definem o conjunto de itens alimentares que o animalpode explorar (Norberg 1981). Nos tentilhões das IlhasGalápagos, por exemplo, observou-se que as dimensões dobico determinam o tamanho do alimento utilizado por eles(Pianka 1994). Por outro lado, Pineschi (1990) comenta quea variação no tamanho do fruto e da semente de capororocas(Rapanea, Myrsinaceae) é muito pequena e que portanto nãodetermina quais espécies de aves estudadas por ele poderiamse alimentar dos frutos destas espécies.
AgradecimentosAgradecemos ao professor Sérgio Borges pela orientação
e identificação das espécies e aos professores Dadão eLeandro pelas sugestões interessantes que enriqueceram otrabalho.
Referências BibliográficasBorges, S.H. & Carvalhaes, A. 2000. Bird species of black
water inundation in the Jaú National Park (Amazonasstate, Brazil): their contribution to regional speciesrichness. Biodiv. Conserv. 9: 201-214.
Conserva, A.S. & Piedade, M.T.F. 2001. Ciclo de vida eecologia de Paspalum fasciculatum Willd. Ex. Fluegge(Poaceae), na várzea da Amazonia Central. ActaAmazonica 31: 205-220.
Junk, W.J 1997. General aspects of foodplain ecology withspecial reference to Amazonian foodplains In: TheCentral Amazonian Foodplain: Ecology of a PulsingSystem. Springer-Verlag, Berlin.
Norberg, V.M. 1981. Flight, morphology, and the ecologicalniche in some birds and bats. Simp. Zool. Soc. Lond.48: 173-197.
Pianka, E.R. 1994. Evolutionary Ecology. Harper CollinsCollege Publishers, New York.
Pineschi, R.B. 1990. Aves como dispersores de sete espéciesde Rapanea (Myrsinaceae) no maciço de Itatiaia, estadosdo Rio de janeiro e Minas Gerais. Ararajuba 1: 73-78.
OrientadorSérgio Borges
Análise comparativa da riqueza e distribuição de espécies de plantas herbáceasaquáticas de três ambientes de várzea da Ilha de Marchantaria (Iranduba, AM)
Alexander V. Christianini, Andrea Lambert, Thiago Timo, Katiane Ferreira & Antônio J. C. Aguiar
IntroduçãoCerca de 300.000 km2, ou 3% de toda superfície da região
Amazônica, são compostos por vegetações de várzea (Junk& Piedade 1993). Nestes ambientes, três espécies degramíneas, Echinochloa polystachia, Paspalum repens ePaspalum fasciculatum destacam-se pela produção debiomassa, sendo responsáveis pela cobertura deaproximadamente 10% de todos os sistemas de várzeas(Piedade et al. 1994).
Em uma área de várzea próxima à cidade de Manaus, Junk& Piedade (1993) registraram 388 espécies herbáceas,distribuídas em diversos habitats. Esses autores relacionarama grande riqueza de espécies na várzea com atributos comoresistência das sementes e esporos à imundação e dessecação;curtos períodos e altas taxas de reprodução; alta produçãoprimária e tolerância das plantas a diferentes níveis deinundação.
O objetivo deste estudo é verificar se existem diferençasentre a riqueza e distribuição de espécies de plantas herbáceas
aquáticas em três ambientes na várzea da Ilha deMarchantaria.
Material e MétodosEste estudo foi realizado no Lago Camaleão, na Ilha de
Marchantaria, município de Iranduba a 13 km de Manaus(03o15‘S; 60o00W). A maior parte da área da ilha fica alagadadurante o período de cheias (dezembro-junho), com umpadrão de alagamento regular de amplitude em torno de 10metros (Piedade 1993).
As três áreas escolhidas para amostragem foram: umamargem da vegetação flutuante de várzea, em uma área centraldo lago, uma área junto à margem da floresta de várzea euma no interior da floresta de várzea, citadas no texto comomargem, borda e floresta, daqui em diante. Em cada localforam amostrados 5 parcelas de 1 m2.
Em cada parcela foram coletados pelo menos um exemplarde cada espécie presente. Foi determinada a composiçãoflorística e o número total de espécies em cada parcela,
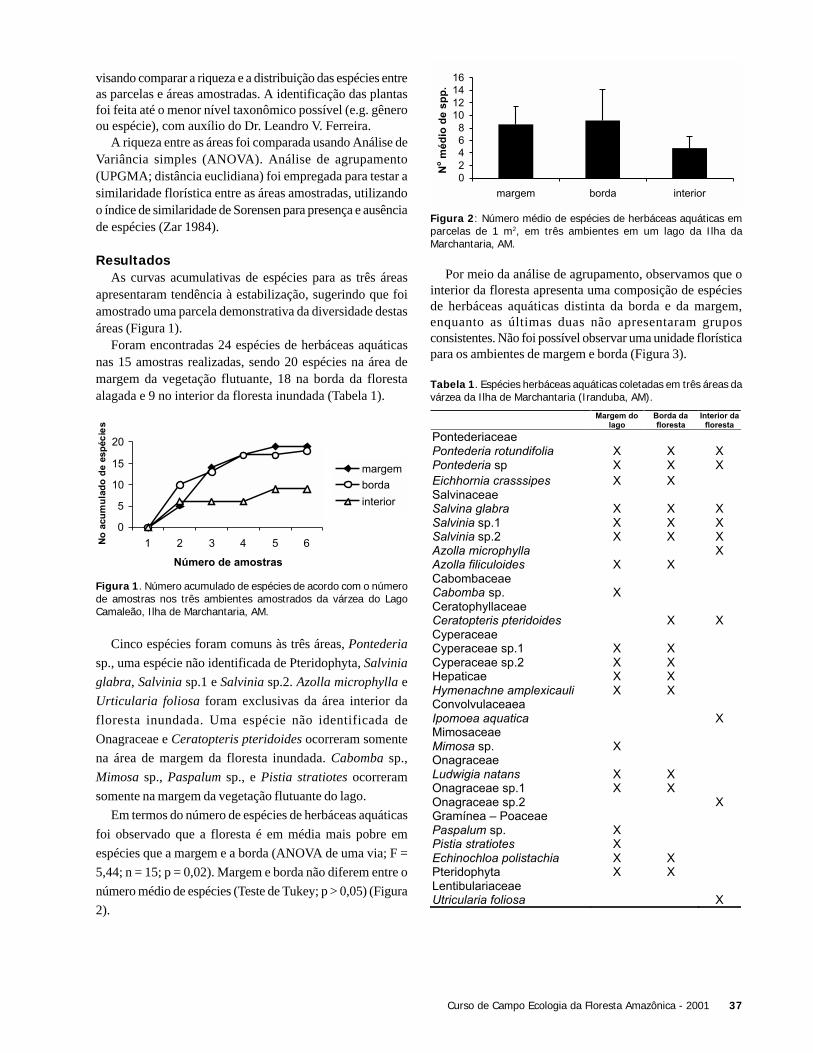
Curso de Campo Ecologia da Floresta Amazônica - 2001 37
visando comparar a riqueza e a distribuição das espécies entreas parcelas e áreas amostradas. A identificação das plantasfoi feita até o menor nível taxonômico possível (e.g. gêneroou espécie), com auxílio do Dr. Leandro V. Ferreira.
A riqueza entre as áreas foi comparada usando Análise deVariância simples (ANOVA). Análise de agrupamento(UPGMA; distância euclidiana) foi empregada para testar asimilaridade florística entre as áreas amostradas, utilizandoo índice de similaridade de Sorensen para presença e ausênciade espécies (Zar 1984).
ResultadosAs curvas acumulativas de espécies para as três áreas
apresentaram tendência à estabilização, sugerindo que foiamostrado uma parcela demonstrativa da diversidade destasáreas (Figura 1).
Foram encontradas 24 espécies de herbáceas aquáticasnas 15 amostras realizadas, sendo 20 espécies na área demargem da vegetação flutuante, 18 na borda da florestaalagada e 9 no interior da floresta inundada (Tabela 1).
Figura 1. Número acumulado de espécies de acordo com o númerode amostras nos três ambientes amostrados da várzea do LagoCamaleão, Ilha de Marchantaria, AM.
Cinco espécies foram comuns às três áreas, Pontederia
sp., uma espécie não identificada de Pteridophyta, Salvinia
glabra, Salvinia sp.1 e Salvinia sp.2. Azolla microphylla e
Urticularia foliosa foram exclusivas da área interior da
floresta inundada. Uma espécie não identificada de
Onagraceae e Ceratopteris pteridoides ocorreram somente
na área de margem da floresta inundada. Cabomba sp.,
Mimosa sp., Paspalum sp., e Pistia stratiotes ocorreram
somente na margem da vegetação flutuante do lago.
Em termos do número de espécies de herbáceas aquáticas
foi observado que a floresta é em média mais pobre em
espécies que a margem e a borda (ANOVA de uma via; F =
5,44; n = 15; p = 0,02). Margem e borda não diferem entre o
número médio de espécies (Teste de Tukey; p > 0,05) (Figura
2).
0
5
10
15
20
1 2 3 4 5 6
Número de amostras
No
acu
mu
lad
od
ees
péc
ies
margem
borda
interior
Figura 2: Número médio de espécies de herbáceas aquáticas emparcelas de 1 m2, em três ambientes em um lago da Ilha daMarchantaria, AM.
Por meio da análise de agrupamento, observamos que ointerior da floresta apresenta uma composição de espéciesde herbáceas aquáticas distinta da borda e da margem,enquanto as últimas duas não apresentaram gruposconsistentes. Não foi possível observar uma unidade florísticapara os ambientes de margem e borda (Figura 3).
Tabela 1. Espécies herbáceas aquáticas coletadas em três áreas davárzea da Ilha de Marchantaria (Iranduba, AM).
02468
10121416
margem borda interior
No
mé
dio
de
sp
p.
Margem dolago
Borda dafloresta
Interior dafloresta
PontederiaceaePontederia rotundifolia X X XPontederia sp X X XEichhornia crasssipes X XSalvinaceaeSalvina glabra X X XSalvinia sp.1 X X XSalvinia sp.2 X X XAzolla microphylla XAzolla filiculoides X XCabombaceaeCabomba sp. XCeratophyllaceaeCeratopteris pteridoides X XCyperaceaeCyperaceae sp.1 X XCyperaceae sp.2 X XHepaticae X XHymenachne amplexicauli X XConvolvulaceaeaIpomoea aquatica XMimosaceaeMimosa sp. XOnagraceaeLudwigia natans X XOnagraceae sp.1 X XOnagraceae sp.2 XGramínea – PoaceaePaspalum sp. XPistia stratiotes XEchinochloa polistachia X XPteridophyta X XLentibulariaceaeUtricularia foliosa X

38 Curso de Campo Ecologia da Floresta Amazônica - 2001
.006 .282 .558 .834 1.110|_______+_______+_______+_______+_______+_______+_______+______
B1__________________________________|
I1___________ |___________| | |
I2________ |______________________| || | |
I5________|__| |_______________________| | |
I4________| | || |
I3______________________________________________| ||
B2_____________ ||______ |
M4_____________| |________ |__| | |
M3____________________| |_______________________ || | |
B3________________ | | ||____________| |_______ |
M1________________| | | || | |
B5________________________ | | ||____________________________| |________|
M2________________________| ||
B4________________________________________ ||____________________|
M5________________________________________|
Figura 3. Dendrograma de similaridade (UPGMA), gerado a partirdo Índice de similaridade de Sorensen e distâncias euclidianas paraa ocorrência de espécies herbáceas aquáticas nas parcelas das áreasde várzea da Ilha da Marchantaria: margem do lago (M), borda dafloresta (B) e interior da floresta (I).
DiscussãoAs diferenças entre a composição e o número de espécies
de herbáceas aquáticas do interior da floresta, margem e bordapodem ser devidas a fatores físicos destes ambientes. Dentreestes, a luminosidade que atinge superfície da água pareceser o fator mais evidente. Junk & Piedade (1993) inferemque em condições de pouca luminosidade há um baixonúmero de espécies de herbáceas aquáticas. Em áreas defloresta alagada, a interceptação da luz do sol pela copa dasárvores pode estar restringindo a possibilidade de ocorrênciade algumas espécies, principalmente as gramíneas.
Plantas do tipo C4, como a maioria das gramíneas e
ciperáceas, necessitam de um grande aporte de luz paraatingirem seu ponto de compensação, o que restringe aocorrência destas plantas a áreas a sol pleno, fora da
influência do sombreamento produzido pela floresta (Junk& Piedade 1997). Plantas do tipo C
3, por outro lado, podem
atingir seu ponto de compensação com cerca de 25% menosluz que as plantas C
4 (Junk & Piedade 1997). De fato,
gramíneas e algumas ciperáceas aquáticas estiveram ausentesno interior da floresta.
O grande número de espécies observados em áreas daborda e da margem da floresta aparentemente deve-se àpossibilidade de colonização destas áreas por plantas C
4.
Por outro lado, a grande biomassa vegetal advinda dasespécies de Poaceae (Piedade et al. 1994) pode gerar algunsmicrohabitats que permitem a fixação e colonização poroutras herbáceas aquáticas, que não se fixam ao solo porrizomas, como Pistia e Azolla, formando um mosaico deespécies nessas áreas.
Referências BibliográficasJunk, W.J. & Piedade, M.T.F. 1993. Herbaceous plants of
the Amazon floodplain near Manaus: species diversityand adaptations to the flood pulse. Amazoniana XII(3/4):467-484.
Junk, W.J. & Piedade, M.T.F. 1997. Plant life in thefloodpplain with special reference to herbaceous plants.In: The Central Amazon Floodplain: Ecology of a PulsingSystem, Junk, W.J. (ed), pp. 147-184. Ecological Studies.Vol. 126. Springer-Verlag, Berlin.
Piedade, M.T.F.; Long, S.P. & Junk, J.W. 1994. Leaf andcanopy uptake of stand of Echinochloa polystachia onthe Central Amazon floodplain. Oecologia 97: 159-174
Zar, J.H. 1984. Biostatistical Analysis. Prentice Hall, NewJersey.
OrientadoraMaria Teresa Piedade
Um experimento sobre a predação de cupins por gerrídeos (Hemiptera:Gerridae)
Christina McCain, Edgar Prieto, Felipe B. Oliveira, Maria R. Darrigo & Natalie Olifiers.
IntroduçãoOs predadores, como todos os animais, tentam maximizar
a eficiência de forrageamento, minimizando seu gasto deenergia. Como os diversos tipos de presa conferem diferentesvalores em termos de ganho energético e implicam emdiferentes custos em termos de tempo de busca e demanipulação, predadores tomam decisões sobre os tipos depresa que caçam (Begon et al. 1986). Estudos sobrepreferências alimentares dos animais são comuns e o interessenesse assunto deve-se à grande significância ecológica queas escolhas dos animais representam para sua sobrevivência.
Um grupo de animais interessante para estudos sobre
predação são os insetos da família Gerridae. Os gerrídeossão hemípteros que se locomovem sobre a superfície da água,caçando os pequenos animais que passam à deriva. Demaneira geral, estão limitados a águas calmas ou áreasprotegidas (Borror et al. 1981). Neste trabalho, testamos ahipótese de que os gerrídeos conseguem avaliar a qualidadede presas potenciais, capturando algumas e rejeitando outras,exibindo preferências por certos grupos.
Material e MétodosRealizamos o estudo em uma área de várzea na Ilha da
Marchantaria, situada 20 km ao sul de Manaus. Observamos

Curso de Campo Ecologia da Floresta Amazônica - 2001 39
dois grupos de Cylindrostebus sp. (Gerridae), sendo que umdeles continha cerca de 35 indivíduos e o outro mais de 160indivíduos.
Em observações anteriores, os gerrídeos pareciam rejeitarcupins soldados (G. Machado com. pess.). Supôs-se que talrejeição poderia ser devido à existência de uma glândulalocalizada na cabeça destes animais que secreta umasubstância pegajosa, utilizada na defesa da colônia (Borroret al. 1981).
Para verificar se os gerrídeos realmente tendem a rejeitaros soldados devido a alguma característica da cabeça destesanimais, oferecemos quatro categorias de cupins para cadagrupo de gerrídeos: operárias intactas, soldados intactos,operárias sem cabeça e soldados sem cabeça. Oferecemosalternadamente os diferentes tipos de presa, lançando-as naágua de maneira a serem carregadas pela correnteza até ogrupo de predadores enquanto observávamos seucomportamento. Caso capturassem a presa, registrávamos otempo de manipulação gasto por cada indivíduo. Para cadagrupo de gerrídeos estudado, oferecemos dez presas de cadatipo, totalizando 80 amostras.
Comparamos as freqüências de captura de cada tipo depresa utilizando o teste de c2, e as diferenças no tempo demanipulação usando o teste t de Student (Zar 1984).
ResultadosObservamos uma taxa de predação três vezes maior sobre
as operárias em relação a soldados intactos (Tabela 1 e Figura1). No entanto, quando removemos a cabeça dos soldadosencontramos um aumento significativo em sua freqüênciade predação, que se tornou muito semelhante a das operárias.A remoção da cabeça não alterou significativamente afrequência de predação sobre as operárias (Tabela 1 e Figura1).O tempo de manipulação da maior presa (operária intacta)em relação a menor (soldado sem cabeça) não foisignificativamente diferente (t = 0,715; g.l. = 9; p = 0,493).
Tabela 1. Comparações realizadas entre as freqüências de predaçãosobre os diferentes tipos de presa (n = 10 por cada tratamento); occ= operária com cabeça, scc = soldado com cabeça, osc = operáriasem cabeça, ssc = soldado sem cabeça.
Figura 1. Frequência de captura para cada categoria de presa; occ= operária com cabeça, scc = soldado com cabeça, osc = operáriasem cabeça, ssc = soldado sem cabeça.
Valor de ÷2 P
Scc x occ 10,1 0,001
Ssc x occ 0,44 0,507
Ssc x osc 1,64 0,200
Scc x ssc 6,67 0,010
0
0.2
0.4
0.6
0.8
occ scc osc ssc
Categoria de presa
Fre
qüên
cia
deca
ptur
a
DiscussãoDurante a época de cheia na várzea, as árvores ficam
parcialmente inundadas e existe a possibilidade de que oscupins arborícolas caiam na água, tornando-se, assim, presaspotenciais dos gerrídeos. Os grupos de gerrídeos exibiramuma preferência significativa pela casta dos operários emrelação aos soldados. A remoção das cabeças dos soldadosindica que alguma característica ali presente inibe a predaçãopor parte dos gerrídeos, mas a causa dessa inibição permanecenão esclarecida. Duas hipóteses são igualmente possíveis:os soldados eram mais difíceis de serem manipulados pelosgerrídeos, devido as características morfológicas de suacabeça, que é mais rígida; além disso, ela apresenta um rostroem forma de espinho, que poderia eventualmente ferir opredador. No caso dos gerrídeos, isso é especialmenteimportante, pois certos tipos de ferimento poderiamcomprometer a capacidade de manutenção sobre a superfícieda água, podendo resultar na morte do animal porincapacidade de forrageamento.
A segunda hipótese é que os gerrídeos evitam os soldadosporque a glândula presente em sua cabeça libera umasubstância repugnante ou adesiva, tornando a presaimpalatável ou dificultando sua manipulação. Uma nova sériede experimentos mais específicos seria necessária paracomprovar a natureza dos fatores que determinam essa menorpreferência da predação dos gerrídeos sobre os soldados decupins.
AgradecimentosAgradecemos ao professor Jorge Nessimean pela
orientação e a Glauco e Juruna pela ajuda na execução doprojeto.
Referências BibliográficasBegon, M.; Harper, J.L. & Townsend, C.R. 1986. Ecology:
Individuals, Populations, and Communities. BlackwellScientific Publications, Oxford, UK.
Borror, D.J.; De Long, D.M. & Triplhorn, C.A. 1981. AnIntroduction to the Study of Insects. Saunders CollegePublishing, New York.
Zar, J.H. 1984. Biostatistical Analysis. Ed. Prentice Hall,New Jersey.
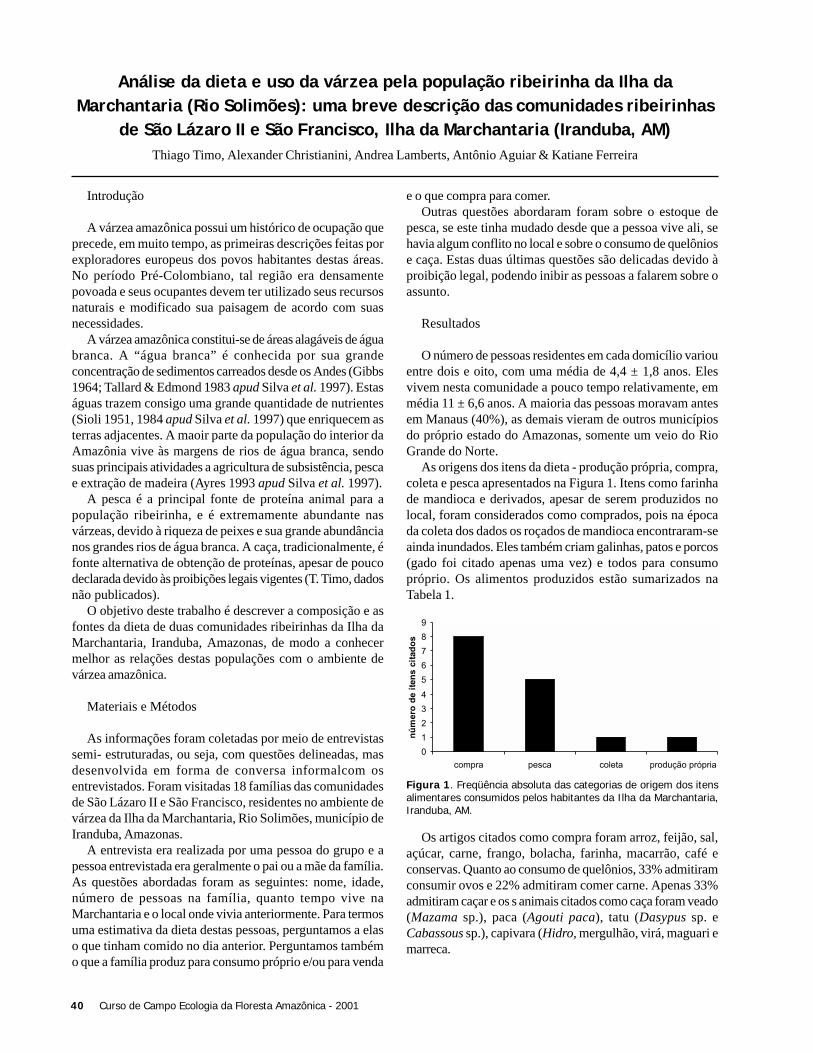
40 Curso de Campo Ecologia da Floresta Amazônica - 2001
Análise da dieta e uso da várzea pela população ribeirinha da Ilha daMarchantaria (Rio Solimões): uma breve descrição das comunidades ribeirinhas
de São Lázaro II e São Francisco, Ilha da Marchantaria (Iranduba, AM)Thiago Timo, Alexander Christianini, Andrea Lamberts, Antônio Aguiar & Katiane Ferreira
Introdução
A várzea amazônica possui um histórico de ocupação queprecede, em muito tempo, as primeiras descrições feitas porexploradores europeus dos povos habitantes destas áreas.No período Pré-Colombiano, tal região era densamentepovoada e seus ocupantes devem ter utilizado seus recursosnaturais e modificado sua paisagem de acordo com suasnecessidades.
A várzea amazônica constitui-se de áreas alagáveis de águabranca. A “água branca” é conhecida por sua grandeconcentração de sedimentos carreados desde os Andes (Gibbs1964; Tallard & Edmond 1983 apud Silva et al. 1997). Estaságuas trazem consigo uma grande quantidade de nutrientes(Sioli 1951, 1984 apud Silva et al. 1997) que enriquecem asterras adjacentes. A maoir parte da população do interior daAmazônia vive às margens de rios de água branca, sendosuas principais atividades a agricultura de subsistência, pescae extração de madeira (Ayres 1993 apud Silva et al. 1997).
A pesca é a principal fonte de proteína animal para apopulação ribeirinha, e é extremamente abundante nasvárzeas, devido à riqueza de peixes e sua grande abundâncianos grandes rios de água branca. A caça, tradicionalmente, éfonte alternativa de obtenção de proteínas, apesar de poucodeclarada devido às proibições legais vigentes (T. Timo, dadosnão publicados).
O objetivo deste trabalho é descrever a composição e asfontes da dieta de duas comunidades ribeirinhas da Ilha daMarchantaria, Iranduba, Amazonas, de modo a conhecermelhor as relações destas populações com o ambiente devárzea amazônica.
Materiais e Métodos
As informações foram coletadas por meio de entrevistassemi- estruturadas, ou seja, com questões delineadas, masdesenvolvida em forma de conversa informalcom osentrevistados. Foram visitadas 18 famílias das comunidadesde São Lázaro II e São Francisco, residentes no ambiente devárzea da Ilha da Marchantaria, Rio Solimões, município deIranduba, Amazonas.
A entrevista era realizada por uma pessoa do grupo e apessoa entrevistada era geralmente o pai ou a mãe da família.As questões abordadas foram as seguintes: nome, idade,número de pessoas na família, quanto tempo vive naMarchantaria e o local onde vivia anteriormente. Para termosuma estimativa da dieta destas pessoas, perguntamos a elaso que tinham comido no dia anterior. Perguntamos tambémo que a família produz para consumo próprio e/ou para venda
e o que compra para comer.Outras questões abordaram foram sobre o estoque de
pesca, se este tinha mudado desde que a pessoa vive ali, sehavia algum conflito no local e sobre o consumo de quelôniose caça. Estas duas últimas questões são delicadas devido àproibição legal, podendo inibir as pessoas a falarem sobre oassunto.
Resultados
O número de pessoas residentes em cada domicílio variouentre dois e oito, com uma média de 4,4 ± 1,8 anos. Elesvivem nesta comunidade a pouco tempo relativamente, emmédia 11 ± 6,6 anos. A maioria das pessoas moravam antesem Manaus (40%), as demais vieram de outros municípiosdo próprio estado do Amazonas, somente um veio do RioGrande do Norte.
As origens dos itens da dieta - produção própria, compra,coleta e pesca apresentados na Figura 1. Itens como farinhade mandioca e derivados, apesar de serem produzidos nolocal, foram considerados como comprados, pois na épocada coleta dos dados os roçados de mandioca encontraram-seainda inundados. Eles também criam galinhas, patos e porcos(gado foi citado apenas uma vez) e todos para consumopróprio. Os alimentos produzidos estão sumarizados naTabela 1.
Figura 1. Freqüência absoluta das categorias de origem dos itensalimentares consumidos pelos habitantes da Ilha da Marchantaria,Iranduba, AM.
Os artigos citados como compra foram arroz, feijão, sal,açúcar, carne, frango, bolacha, farinha, macarrão, café econservas. Quanto ao consumo de quelônios, 33% admitiramconsumir ovos e 22% admitiram comer carne. Apenas 33%admitiram caçar e os s animais citados como caça foram veado(Mazama sp.), paca (Agouti paca), tatu (Dasypus sp. eCabassous sp.), capivara (Hidro, mergulhão, virá, maguari emarreca.
0
1
2
3
4
5
6
7
8
9
compra pesca coleta produção própria
nú
mer
od
eít
ens
cita
do
s

Curso de Campo Ecologia da Floresta Amazônica - 2001 41
Tabela 1. Itens alimentares produzidos pelas comunidades estudadasda Ilha da Marchantaria, Iranduba, AM.
Os artigos citados como compra foram arroz, feijão, sal,açúcar, carne, frango, bolacha, farinha, macarrão, café econservas. Quanto ao consumo de quelônios, 33% admitiramconsumir ovos e 22% admitiram comer carne. Apenas 33%admitiram caçar e os s animais citados como caça foram veado(Mazama sp.), paca (Agouti paca), tatu (Dasypus sp. eCabassous sp.), capivara (Hidro, mergulhão, virá, maguari emarreca.
Os conflitos citados envolvem a presença de barcos depesca comercial apontada como predatória pela populaçãoresidente das comunidades estudadas e o preço baixo pagopelos produtos agrícolas vendidos a intermediários(marreteiros), que pagam apenas cerca de 10% do preço finaldos produtos comercializados em mercados e feiras deManaus (Silva et al. 1997).
DiscussãoA várzea é um ambiente com solo muito fértil mas, por
causa das cheias, não há produção agrícola durante grandeparte do ano. Isto é refletido pelo modo de vida destascomunidades. Grande parte das culturas são anuais, comexceção de algumas árvores frutíferas, como goiabeiras.Porém, tais culturas anuais não suprem de todo asnecessidades básicas das famílias residentes deste ambientee, portanto, são necessárias aquisições de não apenasprodutos que não tem como ser produzidos no local, comosal, açúcar, macarrão, mas também de produtos que sãoproduzidos no local, mas devido à cheia, não estãodisponíveis no momento, como farinha e feijão.
Silva et al. (1997), em um estudo com a comunidade deSão Lázaro II também constataram o pouco tempo deresidência dos atuais moradores na Ilha. Alguns reportaramque nasceram em locais de várzea e igapó, apesar de grandeparte deles ter vindo da Manaus e outras cidades do estadodo Amazonas, como Itacoatiara. Portanto, seus hábitosrelacionados ao consumo de produtos industrializados aindasão muito arraigados.
Quanto à pesca, recurso de extrema importância comofonte de proteína animal, é realizada apenas como atividadede subsistência e, devido à exploração predatória (segundoos moradores) feita por barcos pesqueiros de Manaus, éconsiderada por parte dos residentes como recurso emdeclínio.
No que tange à caça dos referidos animais de pelo e pena,
Itens produzidos Porcentagem de citação
Feijão 11%
Jerimum 8%
Verdura, macaxeira, cebolinha e pepino 7%
Cebolinha, tomate e alface 6%
Banana 4%
milho, melancia, couve e pimentão 3%
Hortaliças, pimenta, abobrinha, batata, repolho e
goiaba
2%
Legumes, alecrim, quiabo, plantas medicinais. 1%
pouco é declarado devido ao conhecimento das proibiçõeslegais à esta atividade. Em função da abundância de peixes epequena de mamíferos e aves terrestres na várzea, a caça émais comum em terra-firme, e mais expressiva como fontede proteína nos igapós, onde o pescado é mais escasso. Empovos indígenas o uso da caça como atividade de subsistênciaé mais importante, resultado de suas tradições, tendo poucosanimais domésticos e menos acesso à carne em conserva(Robinson & Redford 1991). Contudo, animais como tatus,veados, pacas, capivaras, mergulhão, virá, maguari e marrecasão citados como consumidos, estando de acordo com dadosnão publicados sobre a comunidade de Boas Novas no LagoJanauacá, município de Careiro Castanho (Timo, dados nãopublicados).
Em relação aos “bichos de casco” (quelônios), durante aépoca de desova são consumidos carne e ovos dos mesmos.Os quelônios são bastante apreciados, mas foi alegado que émuito difícil encontrá-los. Outros ninhos assaltados são osde marreca.
Os conflitos existentes na Ilha são em grande parte devidoà falta de organização de associações ou cooperativas de pescae agricultura. Na comunidade São Lázaro II há umaassociação que recebe e distribui sementes, materiaisagrícolas, mas conta com pouca participação dos moradores,segundo “Seu” Chico, presidente da associação. Osprincipais conflitos, que são com os barcos pesqueiros e comos “marreteiros”, poderiam ser solucionados caso houvessemais organização.
O tipo de uso do solo pelos moradores, a agriculturapraticada por eles e a pesca para subsistência aparentementesão sustentáveis, devido à pequena densidade de moradores.Estudos mais detalhados tanto sobre a fauna local, comodos costumes destas comunidades são necessários paraelucidar esta questão. Estudos sobre outras fontes deimpactos sobre o ecossistema de várzea, como os barcospesqueiros, também urgem, já que este tipo de exploraçãocomercial dos recursos naturais é muito mais impactante doque a de subsistência praticada pelos moradores locais.
AgradecimentosAgradecemos ao Sérgio Borges pela orientação, à Maria
Teresa Piedade e ao Leandro Ferreira pelas dicas e ao INPApela oportunidade e infra estrutura.
Refererências BibliográficasRobinson, J.G. & Redford, K.H. 1996. Neotropical Wildlife
Use and Conservation. The University of Chicago Press,Chicago and London.
Silva, A.L.; Oliveira, R. & Ritll, C. 1997. Estratégias deadaptação das comunidades ribeirinhas na época dascheias no Baixo Solimões, Ilha da Marchantaria, AM.Livro do Curso de Campo “Ecologia da FlorestaAmazônica”. INPA/PDBFF, Manaus, AM.
OrientadorSérgio Borges

42 Curso de Campo Ecologia da Floresta Amazônica - 2001
Zonação de herbáceas aquáticas no gradiente topográfico de um sistema devárzea da Ilha da Marchantaria, Amazônia Central
Carlos E. Marinelli, Daniel L. M. Vieira, Cristiane F. Jurinitz, Betsabé R. Guerra & Rodrigo Cogni
IntroduçãoÁreas de várzea compreendem apenas 3% da Amazônia,
entretanto sua representatividade é bastante alta dentro dossistemas inundáveis (Ayres 1993). Essas formações florestaisdatam do período Quaternário, sendo consideradasecossistemas geologicamente recentes (Worbes et al. 1992).Reguladas pelo regime climático, as várzeas sãoperiodicamente inundadas por rios de água branca,caracterizados pela alta quantidade de nutrientes e pHaproximadamente neutro (Worbes et al. 1992). O ciclo daságuas exerce influência na geomorfologia local através deprocessos equilibrados de erosão e sedimentação sazonais,modificando a estrutura da paisagem e influenciando ascaracterísticas estruturais da vegetação (Junk & Piedade1997). Essa sazonalidade gera uma grande variedade dehábitats, como os bancos de herbáceas aquáticas1 , as áreasalagadas e os solos hidromórficos (Conserva & Piedade2001).
Dessa forma, a distribuição da vegetação é influenciadaprincipalmente pela dinâmica do pulso de inundação e aestabilidade física dos hábitats. Em faixas do mesmo nívelde inundação observam-se diferentes estágios da sucessão,com espécies de limitada capacidade de flutuação ocorrendoinicialmente, inseridas em bancos de herbáceas aquáticas(Junk & Piedade 1997).
As herbáceas aquáticas produzem grande biomassa porapresentarem uma alta produtividade primária, representandoum importante recurso nos sistemas alagáveis (Junk &Piedade 1997). Distribuem-se de acordo com a duração desuas fases terrestres e aquáticas, com a estabilidade física doambiente (luz, duração da enchente) e processos sucessionaisrelacionados a seus ciclos de vida (Dantas 1999). Além disso,sua distribuição também resulta de eventos de inundaçãoatuais e passados. Herbáceas anuais são primariamenteinfluenciadas pela hidrografia do respectivo regime deinundação, enquanto as perenes são influenciadas pelo ciclosde inundação mais recentes (Junk & Piedade 1997).
O presente estudo teve como objetivo verificar se (i) acomunidade de herbáceas aquáticas obedece a uma zonaçãoem um gradiente a partir da borda do lago em direção àfloresta inundável; (ii) a composição de espécies estárelacionada com variáveis limnológicas medidas ao longodesse gradiente.
Material e MétodosRealizamos o estudo na várzea do Lago Camaleão na Ilha
da Marchantaria (3o 14 ‘S; 59o 57 O), rio Solimões (AM)(Conserva & Piedade 2001). Estabelecemos três transectos
a partir do início da vegetação na borda do lago até o inícioda floresta inundável, sendo cada um determinadoaleatoriamente pela presença de exemplares dePseudobombax munguba. Essa planta é caracterizada porocorrer em uma faixa restrita da sucessão, indicando o inícioda floresta inundável. Em cada transecto, marcamos trêspontos (borda, intermediário e floresta), sendo que a distânciaentre esses pontos dependia da largura do transecto. O pontoda borda era determinado pela presença das primeirasherbáceas aquáticas, o da floresta pelo P. munguba e ointermediário era o ponto médio entre os demais. Em cadaum desses pontos, colocamos um quadro de 1m2 , onderegistramos a presença das espécies de herbáceas aquáticas.
Para a caracterização do ambiente físico, no centro decada quadro amostrado, a 15 cm de profundidade a partir dasuperfície, medimos as seguintes variáveis liminológicas: pH,temperatura da água, concentração de oxigênio,condutividade. A profundidade foi medida no mesmo pontoque as outras variáveis.
ResultadosEncontramos um total de 18 espécies de herbáceas
aquáticas distribuídas em 13 famílias. Dentre essas espécies,Eichhornia crassipes, Phyllanthus fluitans, Pistia stratiotese Salvinia spp. estiveram presentes nos três pontos dotransecto (Tabela 1).
Tabela 1. Composição florística das herbáceas aquáticas na várzeado lago Camaleão no gradiente de zonação da borda do lago emdireção à floresta inundada, Ilha da Marchantaria, AM (B = borda, I= intermediário, F = floresta).
A temperatura da água manteve-se constante nos diferentespontos amostrados (ca. 29o C). Já a concentração de oxigêniodiminuiu com a aproximação da floresta nos transectos 1 e2, mantendo-se constante no transecto 3. O pH apresentou
1 Adotamos o termo ‘herbáceas aquáticas’ em substituição a ‘macrófitas aquáticas’, pelo fato deste último ser muito amplo, podendo incluiras espécies arbustivo-arbóreas (M. T. F. Piedade com. pess.)
Família Espécie B I FMAGNOLIOPHYTAAraceae Pistia stratiotes Schott X X XCeratophyllaceae Ceratophyllum demersum L. X XConvolvulaceae Ipomoea aquatica Forsk. X XCyperaceae Scirpus cubensis Poepp. & Kunth. X X
Não identificada X XEuphorbiaceae Phyllanthus fluitans Benth. ex Müll. X X XHydrocharitaceae Limnobium laevigatum Heine X X
Limnobium sp. XLeguminosae Não identificada X XLemnaceae Spirodela intermediaW. Koch. XNajadaceae Najas sp. XOnagraceae Ludwigia natans Humb. & Bonpl. XPoaceae Echinochloa polystachya (H. B. K.) Hitch. X X
Leersia hexandra Swartz XPaspalum repens Berg. X
Pontederiaceae Eichornia crassipes (Mart.) Solms. X X XPTERIDOPHYTA X XSalvinaceae Azolla sp. X
Salvinia spp. X X X
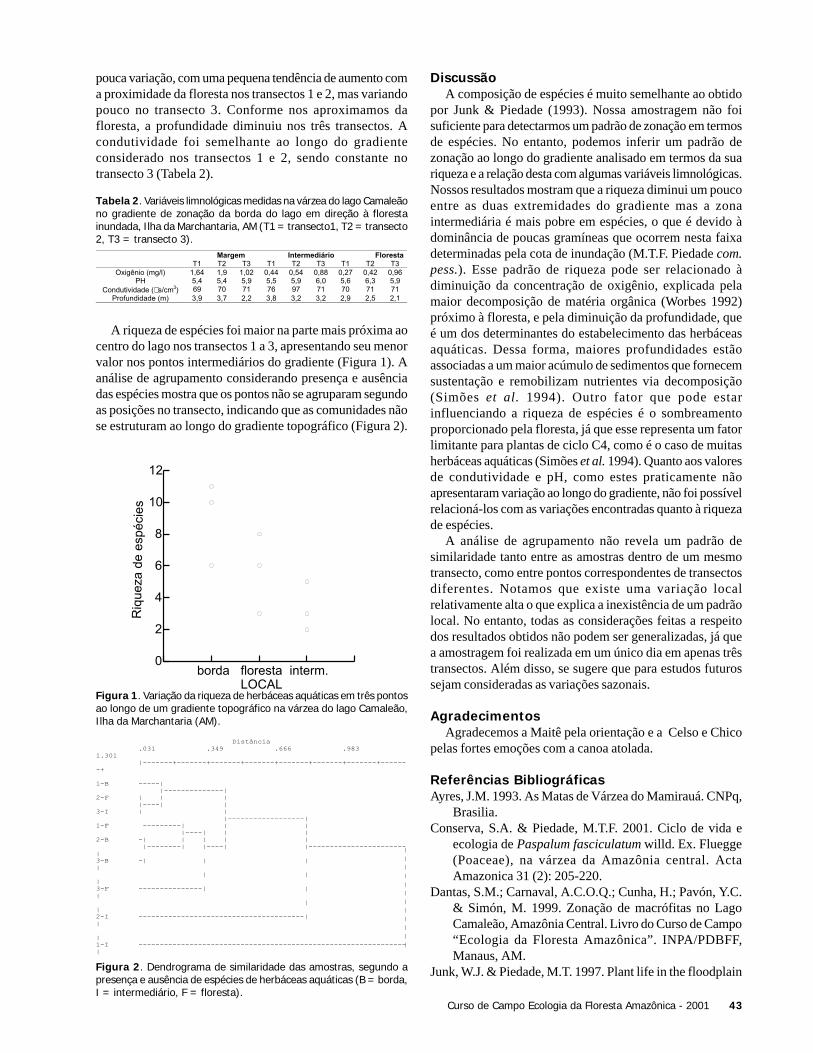
Curso de Campo Ecologia da Floresta Amazônica - 2001 43
pouca variação, com uma pequena tendência de aumento coma proximidade da floresta nos transectos 1 e 2, mas variandopouco no transecto 3. Conforme nos aproximamos dafloresta, a profundidade diminuiu nos três transectos. Acondutividade foi semelhante ao longo do gradienteconsiderado nos transectos 1 e 2, sendo constante notransecto 3 (Tabela 2).
Tabela 2. Variáveis limnológicas medidas na várzea do lago Camaleãono gradiente de zonação da borda do lago em direção à florestainundada, Ilha da Marchantaria, AM (T1 = transecto1, T2 = transecto2, T3 = transecto 3).
A riqueza de espécies foi maior na parte mais próxima aocentro do lago nos transectos 1 a 3, apresentando seu menorvalor nos pontos intermediários do gradiente (Figura 1). Aanálise de agrupamento considerando presença e ausênciadas espécies mostra que os pontos não se agruparam segundoas posições no transecto, indicando que as comunidades nãose estruturam ao longo do gradiente topográfico (Figura 2).
Figura 1. Variação da riqueza de herbáceas aquáticas em três pontosao longo de um gradiente topográfico na várzea do lago Camaleão,Ilha da Marchantaria (AM).
Figura 2. Dendrograma de similaridade das amostras, segundo apresença e ausência de espécies de herbáceas aquáticas (B = borda,I = intermediário, F = floresta).
Margem Intermediário FlorestaT1 T2 T3 T1 T2 T3 T1 T2 T3
Oxigênio (mg/l) 1,64 1,9 1,02 0,44 0,54 0,88 0,27 0,42 0,96PH 5,4 5,4 5,9 5,5 5,9 6,0 5,6 6,3 5,9
Condutividade (� s/cm3) 69 70 71 76 97 71 70 71 71Profundidade (m) 3,9 3,7 2,2 3,8 3,2 3,2 2,9 2,5 2,1
borda floresta interm.LOCAL
0
2
4
6
8
10
12
Riq
ue
zad
ee
spé
cie
s
Distância
.031 .349 .666 .983
1.301
|-------+-------+-------+-------+-------+-------+-------+------
-+
1-B -----|
|--------------|
2-F | | |
|----| |
3-I | |
|------------------|
1-F ---------| | |
|----| | |
2-B -| | | | |
|--------| |----| |-----------------------
|
3-B -| | |
|
| |
|
3-F ---------------| |
|
|
|
2-I ---------------------------------------|
|
|
1-I ---------------------------------------------------------------
|
|
|
|
|
|
|
|
|
|
|
|
|
DiscussãoA composição de espécies é muito semelhante ao obtido
por Junk & Piedade (1993). Nossa amostragem não foisuficiente para detectarmos um padrão de zonação em termosde espécies. No entanto, podemos inferir um padrão dezonação ao longo do gradiente analisado em termos da suariqueza e a relação desta com algumas variáveis limnológicas.Nossos resultados mostram que a riqueza diminui um poucoentre as duas extremidades do gradiente mas a zonaintermediária é mais pobre em espécies, o que é devido àdominância de poucas gramíneas que ocorrem nesta faixadeterminadas pela cota de inundação (M.T.F. Piedade com.pess.). Esse padrão de riqueza pode ser relacionado àdiminuição da concentração de oxigênio, explicada pelamaior decomposição de matéria orgânica (Worbes 1992)próximo à floresta, e pela diminuição da profundidade, queé um dos determinantes do estabelecimento das herbáceasaquáticas. Dessa forma, maiores profundidades estãoassociadas a um maior acúmulo de sedimentos que fornecemsustentação e remobilizam nutrientes via decomposição(Simões et al. 1994). Outro fator que pode estarinfluenciando a riqueza de espécies é o sombreamentoproporcionado pela floresta, já que esse representa um fatorlimitante para plantas de ciclo C4, como é o caso de muitasherbáceas aquáticas (Simões et al. 1994). Quanto aos valoresde condutividade e pH, como estes praticamente nãoapresentaram variação ao longo do gradiente, não foi possívelrelacioná-los com as variações encontradas quanto à riquezade espécies.
A análise de agrupamento não revela um padrão desimilaridade tanto entre as amostras dentro de um mesmotransecto, como entre pontos correspondentes de transectosdiferentes. Notamos que existe uma variação localrelativamente alta o que explica a inexistência de um padrãolocal. No entanto, todas as considerações feitas a respeitodos resultados obtidos não podem ser generalizadas, já quea amostragem foi realizada em um único dia em apenas trêstransectos. Além disso, se sugere que para estudos futurossejam consideradas as variações sazonais.
AgradecimentosAgradecemos a Maitê pela orientação e a Celso e Chico
pelas fortes emoções com a canoa atolada.
Referências BibliográficasAyres, J.M. 1993. As Matas de Várzea do Mamirauá. CNPq,
Brasilia.Conserva, S.A. & Piedade, M.T.F. 2001. Ciclo de vida e
ecologia de Paspalum fasciculatum willd. Ex. Fluegge(Poaceae), na várzea da Amazônia central. ActaAmazonica 31 (2): 205-220.
Dantas, S.M.; Carnaval, A.C.O.Q.; Cunha, H.; Pavón, Y.C.& Simón, M. 1999. Zonação de macrófitas no LagoCamaleão, Amazônia Central. Livro do Curso de Campo“Ecologia da Floresta Amazônica”. INPA/PDBFF,Manaus, AM.
Junk, W.J. & Piedade, M.T. 1997. Plant life in the floodplain

44 Curso de Campo Ecologia da Floresta Amazônica - 2001
with special reference to herbaceous plants. In: TheCentral Amazon Floodplain, Junk, W.J. (ed.), pp. 147-185. Ecological Studies. Springer-Verlag, Berlin.
Simões, F.F.; Takahasi, A.; Aguiar, C.M.L.; Cattanio, J.H.& Prada, M.V. 1994. Zonação de macrófitas aquáticasao longo de um gradiente lago-floresta de várzea (Ilhada Marchantaria, AM). Livro do Curso de Campo“Ecologia da Floresta Amazônica”. INPA/PDBFF,Manaus, AM.
Worbes, M.; Klinge, H.; Revilla, J.D. & Martius, C. 1992.
On the dynamics, floristic subdivision and geographicaldistribution of várzea forest in central Amazonía. J. Veg.Sc. 3: 553-564.
Worbes, M. 1992. The Forest Ecosystem of the Floodplains.In: The Central Amazon Floodplain, Junk, W.J. (ed.),pp. 223-260. Ecological Studies. Springer-Verlag,Berlin.
OrientadoraMaria Teresa Piedade
Composição funcional trófica de insetos bentônicos emergentes no gradientetopográfico de uma área de várzea na Ilha da Marchantaria, Iranduba, AM
Carlos E. Marinelli, Daniel L. M. Vieira, Cristiane F. Jurinitz, Betsabé R. Guerra, Rodrigo Cogni
IntroduçãoÁreas de várzea compreendem apenas 3% da Amazônia,
entretanto sua representatividade é bastante alta dentro dossistemas inundáveis (ca. 250.000 Km2) (Ayres 1993). Essasformações florestais datam do período Quaternário, sendoconsideradas ecossistemas geologicamente recentes (Worbeset al. 1992). Reguladas pelo regime climático, as várzeassão periodicamente inundadas por rios de água branca,caracterizados pela alta quantidade de nutrientes e pHaproximadamente neutro (Worbes et al. 1992). O ciclo daságuas exerce influência na geomorfologia local através deprocessos equilibrados de erosão e sedimentação sazonais,modificando a estrutura da paisagem e influenciando ascaracterísticas estruturais da vegetação (Junk & Piedade1997). Essa sazonalidade gera uma grande variedade dehábitats, como os bancos de herbáceas aquáticas, áreasalagadas e solos hidromórficos (Conserva & Piedade 2001).
Junk et al. (1989 apud Junk & Piedade 1993) afirmamque em grandes rios com planície inundável adjacente, opulso de inundação é o principal fator que regula aprodutividade do sistema rio-planície. Nesse ambiente, afauna bentônica talvez represente o mais efetivo componenteno fluxo de energia entre os compartimentos aquático eterrestre (Nessimian et al. 1998), sendo constituída deorganismos com diversas categorias funcionais tróficas, e,portanto, uma excelente indicadora de alterações ambientaise de disponibilidade de diferentes recursos.
Estudando florestas inundáveis de igapó, Nessimian etal. (1998) verificaram que a fauna bentônica de cortadores éimportante na margem da floresta, pela presença daserapilheira. A fauna de raspadores é mais numerosa na zonaintermediária devido à disponibilidade de perifiton, enquantona margem do lago ocorre um incremento na abundância defiltradores, já que neste ponto a matéria orgânica está bastanteparticulada pela ação de grupos que a utilizaram nas fasesanteriormente descritas. Os demais grupos não sãoaparentemente relacionados com o gradiente topográfico.
Nosso objetivo foi caracterizar a composição das
categorias funcionais tróficas de insetos bentônicosemergentes ao longo do gradiente topográfico de uma áreade várzea do Rio Solimões na Ilha da Marchantaria,Município de Iranduba, AM.
Material e MétodosEstudamos insetos bentônicos, pois estes podem ser
facilmente capturados quando emergem para a fase terrestrecomo adultos. Capturamos insetos bentônicos emergentesem três ambientes da várzea da Ilha da Marchantaria (margemdo lago, borda da mata alagada e interior da mata alagada)utilizando cinco armadilhas para cada ambiente. Cadaarmadilha era constituída de um covo acoplado a um sacoplástico em forma de funil colocado com o lado aberto voltadopara baixo, na altura da lâmina d’água. As armadilhas foramdispostas com pelo menos cinco metros de distância umasdas outras, e permaneceram instaladas durante um períodode 15 horas entre o crepúsculo vespertino e a manhã do diaseguinte. Após o recolhimento das armadilhas, os exemplaresforam sacrificados em câmara etérica e em seguidaidentificados com auxílio do Dr. Jorge L. Nessimian (UFRJ)até os níveis de ordem ou família. Foram totalizadas asabundâncias absolutas de cada taxa, os quais foramclassificados por categorias funcionais tróficas (CFT),conforme sugerido por Nessimian et al. (1998) comocoletores, raspadores, filtradores e predadores. Para aobtenção dos valores de biomassa de cada grupo funcionalforam adotados os valores de peso médio sugeridos porMerritt & Cummins (1984). Como cada taxa podia conterespécies pertencentes a mais de uma categoria, a biomassade cada um foi dividida igualmente entre as CFT possíveispara o grupo. Posteriormente, foi comparada a distribuiçãode cada CFT entre os ambientes estudados.
ResultadosObtivemos 183 exemplares de insetos emergentes, os quais
foram divididos em sete grupos taxonômicos (Tabela 1). Afamília Chironomidae foi predominante com 84,5% da

Curso de Campo Ecologia da Floresta Amazônica - 2001 45
biomassa total amostrada, sendo o único grupo ocorrentenos três ambientes. Os resultados indicaram maiores valoresde riqueza (seis grupos) e biomassa (88,3%) na margem dolago, seguido da borda da mata e interior da mata, comrespectivamente 9,0% e 2,6% da biomassa total. Os valoresde biomassa foram proporcionais aos valores de abundância.
Tabela 1. Ocorrência de insetos bentônicos emergentes e suasrespectivas categorias funcionais tróficas no gradiente topográficode uma área de várzea na Ilha da Marchantaria, Iranduba, AM (CFT= categoria funcional trófica; C = coletores; F = filtradores; P =predadores; R = raspadores; ML = margem do lago; BM = bordo damata; IM = interior da mata).
Abundância (N) Biomassa (mg)Taxa CFT ML BM IM ML BM IMDipteraBrachycera C,P 4 0 0 100 0 0Ceratopogonidae C, F 1 0 0 25 0 0Chironomidae C, F, P 120 5 4 3000 125 100Culicidae C,F 40 0 0 100 0 0Tipulidae C 1 0 0 25 0 0Ephemeroptera C,R 0 2 0 0 100 0Trichoptera C, F 3 3 0 120 120 0Total - 169 10 4 3370 345 100
Com relação às categorias funcionais tróficas, guildas decoletores, filtradores e predadores foram obtidas em todosos ambientes, enquanto que raspadores foram obtidos apenasna borda da mata alagada (Tabela 1). A margem do lago tevemais filtradores, coletores e predadores do que os demaisambientes (Tabela 1), os quais juntos contribuíramaproximadamente com 10% de cada categoria.
DiscussãoO método utilizado para a captura dos espécimes
apresentou baixa eficiência, e a maioria das armadilhas nãocapturou nenhum exemplar. Isso pode ser atribuídoprincipalmente ao colapso do funil de plástico e pequenosfuros utilizados para prender as armadilhas. Além disso, aabundância de indivíduos pode ser baixa nesta época(vazante) (J.L. Nessimian com. pess.). De qualquer maneira,foram encontrados mais indivíduos na margem, que por sermais rasa, e ter o nível da água diminuindo rapidamente (15cm/dia) ocasionará a morte dos indivíduos, se eles nãomigrarem ou emergirem. Portanto, a margem estariacondicionando a emergência antecipada e sincrônica dos
insetos. A identificação dos exemplares em categoriastaxonômicas simples também é uma restrição ao estudo, jáque a mesma ordem ou família pode ocupar diferentescategorias funcionais tróficas.
A maior biomassa de filtradores na margem do lago indicaque nesse ponto há uma maior disponibilidade de matériaorgânica fina, que foi processada nas fases anteriores dogradiente, concordando com o encontrado para a faunabentônica de igapó (Nessimian et al. 1998). Os raspadores,que se alimentam preferencialmente de algas, foramencontrados somente na borda da mata. Isso pode ser devidoà disponibilidade de luz nessa faixa (não tão profunda) e delocais para fixação das algas, como as raízes de herbáceasaquáticas.
Referências BibliográficasAyres, J.M. 1993. As Matas de Várzea do Mamirauá. CNPq,
Brasília.Conserva, S.A. & Piedade, M.T.F. 2001. Ciclo de vida e
ecologia de Paspalum fasciculatum willd. Ex. Fluegge(Poaceae), na várzea da Amazônia central. ActaAmazonica 31 (2): 205-220.
Junk, W.J. & Piedade, M.T. 1997. Plant life in the floodplainwith special reference to herbaceous plants. In:Ecological Studies: The Central Amazon Floodplain,Junk, W.J. (ed.), pp. 147-185. Springer-Verlag, Berlin.
Merritt, R.W. & Cummins, K.W. 1984. An Introduction tothe Aquatic Insects of North America. Kendall/HuntPubl., Dubuque.
Nessimian, J.L.; Dorvillé, L.M.F.; Sanseverino, A.M. &Baptista, D.F. 1998. Relation between flood pulse andfunctional composition of the macroinvertebrate benthicfauna in the lower Rio Negro, Amazonas, Brazil.Amazoniana, XV (1/2): 35-50.
Worbes, M.; Klinge, H.; Revilla, J.D. & Martius, C. 1992.On The dynamics, floristic subdivision and geographicaldistribution of várzea forest in central Amazonía. J. Veg.Sc. 3: 553-564.
OrientadorJ. L. Nessimian

46 Curso de Campo Ecologia da Floresta Amazônica - 2001
Efeito do isolamento em poças sobre a estrutura da comunidade demacroinvertebrados bentônicos no arquipélago das Anavilhanas, AM
Paula C. Ceotto, Angela M. F. Pacheco, Leandro M. Scoss, Maurício Camargo & Paula S. Pinheiro
IntroduçãoFlorestas de igapó sofrem inundações periódicas pelo
aumento do nível de rios de água preta. O rio Negro, principaltributário do rio Amazonas, nasce em terrenos pobres emnutrientes, muito lixiviados e de rápida drenagem. Essascaracterísticas fazem com que sua taxa de decomposição sejabaixa, o que gera uma acumulação de ácidos húmicos efúlvicos (Goulding et al. 1988). Os mesmos autoresconsideram que as inundações periódicas e as característicasfisico-químicas do rio Negro são fatores que limitam acolonização das florestas de igapó pela fauna bentônica.
O rio Negro apresenta uma variação média anual de 10m,submergindo completamente o arquipélago das Anavilhanasno período de cheia. Quando o nível da água sobe, acomunidade bentônica expande sua distribuição para as áreasde floresta, ocupando assim toda a área coberta por água(Nessimian et al. 1998). Durante o período de vazante,formam-se poças em depressões do terreno das nas áreas deflorestas. Nessas poças, são encontrados organismosbentônicos que tinham ampliado sua área de distribuição eficaram presos, perdendo o contato com o rio.
O represamento da água nas ilhas promove alteraçõesfísico-químicas deste ambiente, podendo causar alteraçõesna comunidade bentônica (J. L. Nessimian com. pess.).Nessimian et al. (1998), em um estudo de composiçãofuncional nas margens do rio Negro, encontraram maiorfrequência de predadores no período em que o nível da águaestava mais baixo. Esse padrão provavelmente estárelacionado à concentração dos organismos numa situaçãode diminuição do volume d’água. Neste trabalho testamos oefeito da diminuição do volume devido ao isolamento depoças no interior das ilhas sobre a composição da comunidadebentônica.
Material e MétodosFizemos as coletas no Arquipélago das Anavilhanas
(03005’S e 59059’W) localizado no Rio Negro, distante 80km de Manaus. Coletamos duas amostras em seis pontos,sendo metade em três poças isoladas e a outra metade emmargens de igapó, totalizando 12 amostras. Os pontos decoleta distavam pelo menos 500m entre si. Fizemos asamostragens com peneiras circulares de 0,70 m2 e malha de0,4 cm. Para cada ponto, passamos a peneira na serapilheirasubmersa a uma distância de aproximadamente 1m da margemnos dois. Transferimos o material amostrado para uma baciae, em seguida, o acondicionamos em um coletor de Brundin(malha 180mm) para lavagem do substrato e remoção doexcesso de argila. Posteriormente, triamos as amostras eidentificamos os invertebrados encontrados em nível deordem ou família, quando possível. Com base em Nessimianet al. (1998), categorizamos os grupos taxonômicos
encontrados segundo seu hábito alimentar em predadores enão-predadores. Segundo esta definição, os animaisfiltradores, cortadores, coletores e os parasitas, entre outros,formam o grupo dos não-predadores, enquanto os predadoresincluem os organismos que matam a presa, que é de origemanimal, para se alimentar. Esta definição também levou emconsideração o tamanho das presas, por exemplo, os camarõese caranguejos são onívoros, mas os primeiros foramconsiderados como não predadores porque as presasconsumidas por eles são muito pequenas e a representaçãodelas em sua dieta é reduzida. No caso dos caranguejos, quese alimentam de presas maiores, consideramos metade donúmero de indivíduos em cada categoria.
Calculamos a riqueza e a abundância das famíliasencontradas nas duas áreas e fizemos um teste de c2 entre osdois ambientes para comparar a proporção de indivíduospertencentes a cada categoria trófica.
ResultadosA riqueza de grupos taxonômicos nas poças isoladas e
nas margens de igapó foi similar. Encontramos organismospertencentes a 14 grupos taxonômicos, 12 em áreas demargem de igapó e nove nas poças no interior das ilhas, comdiferentes graus de representatividade entre os grupos (tab.1). Encontramos um total de 38 indivíduos não-predadores(X ± DP = 6,33 ± 6,05 indivíduos/amostra) e 27 indivíduospredadores (X ± DP = 4,5 ± 7,74 ind./amostra) nas áreas demargem de igapó. Nas áreas de poça, coletamos 50 indivíduosnão-predadores (X ± DP = 8,33 ± 6,89 ind./amostra) e 8predadores (X ± DP = 1,50 ± 1,38 ind./amostra).
Tabela 1. Número de indivíduos em cada grupo taxonômico coletadoem poças isoladas e em margens de igapó. P= predador e NP= não-predador.
Grupo taxonômico Hábito alimentar Margens de igapó Poças isoladas
Oligochaeta NP 4 0
Chironomidae NP 2 2
Diplopoda NP 1 0
Elmidae NP 1 0
Ephemeroptera NP 0 1
Trichoptera NP 5 1
Euryrrhynchinae NP 18 30
Paleomonidae NP 6 15
Brachiura NP/P 2 2
Hirudinae P 1 5
Belostomatidae P 0 1
Nepidae P 1 0
Notonectidae P 22 0
Odonata P 1 1
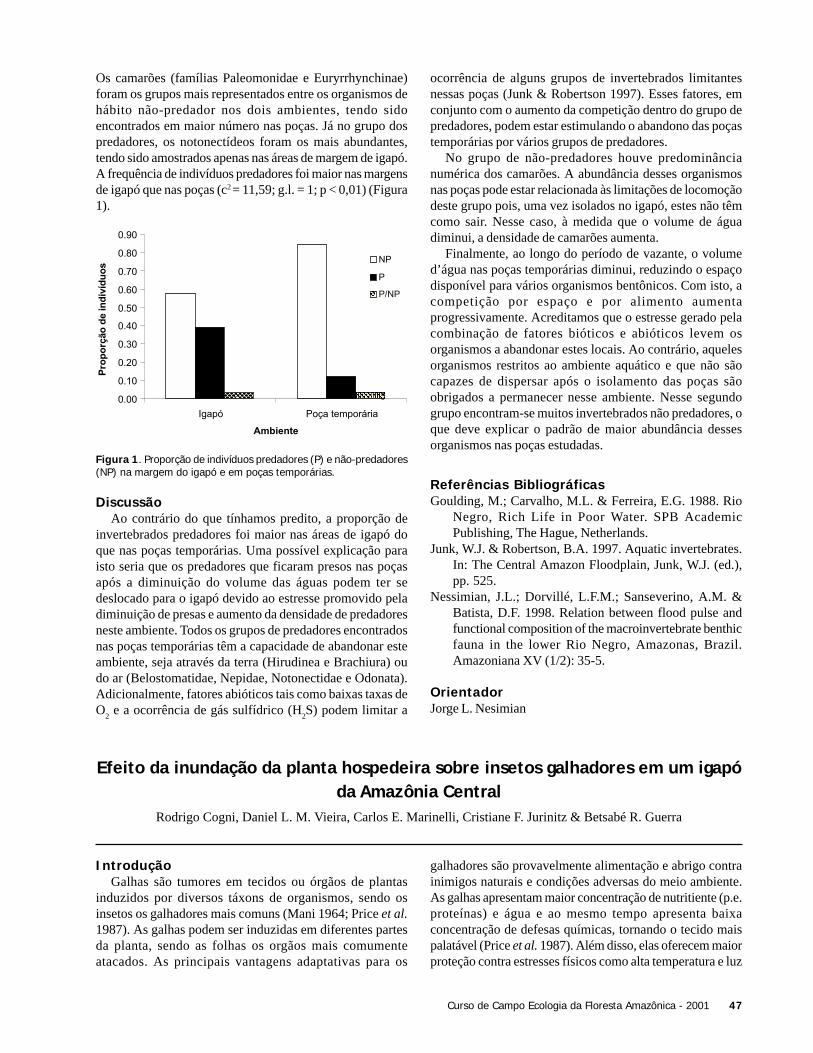
Curso de Campo Ecologia da Floresta Amazônica - 2001 47
Os camarões (famílias Paleomonidae e Euryrrhynchinae)foram os grupos mais representados entre os organismos dehábito não-predador nos dois ambientes, tendo sidoencontrados em maior número nas poças. Já no grupo dospredadores, os notonectídeos foram os mais abundantes,tendo sido amostrados apenas nas áreas de margem de igapó.A frequência de indivíduos predadores foi maior nas margensde igapó que nas poças (c2 = 11,59; g.l. = 1; p < 0,01) (Figura1).
Figura 1. Proporção de indivíduos predadores (P) e não-predadores(NP) na margem do igapó e em poças temporárias.
DiscussãoAo contrário do que tínhamos predito, a proporção de
invertebrados predadores foi maior nas áreas de igapó doque nas poças temporárias. Uma possível explicação paraisto seria que os predadores que ficaram presos nas poçasapós a diminuição do volume das águas podem ter sedeslocado para o igapó devido ao estresse promovido peladiminuição de presas e aumento da densidade de predadoresneste ambiente. Todos os grupos de predadores encontradosnas poças temporárias têm a capacidade de abandonar esteambiente, seja através da terra (Hirudinea e Brachiura) oudo ar (Belostomatidae, Nepidae, Notonectidae e Odonata).Adicionalmente, fatores abióticos tais como baixas taxas deO
2 e a ocorrência de gás sulfídrico (H
2S) podem limitar a
0.00
0.10
0.20
0.30
0.40
0.50
0.60
0.70
0.80
0.90
Igapó Poça temporária
Ambiente
Pro
po
rção
de
ind
ivíd
uo
s
NP
P
P/NP
ocorrência de alguns grupos de invertebrados limitantesnessas poças (Junk & Robertson 1997). Esses fatores, emconjunto com o aumento da competição dentro do grupo depredadores, podem estar estimulando o abandono das poçastemporárias por vários grupos de predadores.
No grupo de não-predadores houve predominâncianumérica dos camarões. A abundância desses organismosnas poças pode estar relacionada às limitações de locomoçãodeste grupo pois, uma vez isolados no igapó, estes não têmcomo sair. Nesse caso, à medida que o volume de águadiminui, a densidade de camarões aumenta.
Finalmente, ao longo do período de vazante, o volumed’água nas poças temporárias diminui, reduzindo o espaçodisponível para vários organismos bentônicos. Com isto, acompetição por espaço e por alimento aumentaprogressivamente. Acreditamos que o estresse gerado pelacombinação de fatores bióticos e abióticos levem osorganismos a abandonar estes locais. Ao contrário, aquelesorganismos restritos ao ambiente aquático e que não sãocapazes de dispersar após o isolamento das poças sãoobrigados a permanecer nesse ambiente. Nesse segundogrupo encontram-se muitos invertebrados não predadores, oque deve explicar o padrão de maior abundância dessesorganismos nas poças estudadas.
Referências BibliográficasGoulding, M.; Carvalho, M.L. & Ferreira, E.G. 1988. Rio
Negro, Rich Life in Poor Water. SPB AcademicPublishing, The Hague, Netherlands.
Junk, W.J. & Robertson, B.A. 1997. Aquatic invertebrates.In: The Central Amazon Floodplain, Junk, W.J. (ed.),pp. 525.
Nessimian, J.L.; Dorvillé, L.F.M.; Sanseverino, A.M. &Batista, D.F. 1998. Relation between flood pulse andfunctional composition of the macroinvertebrate benthicfauna in the lower Rio Negro, Amazonas, Brazil.Amazoniana XV (1/2): 35-5.
OrientadorJorge L. Nesimian
Efeito da inundação da planta hospedeira sobre insetos galhadores em um igapóda Amazônia Central
Rodrigo Cogni, Daniel L. M. Vieira, Carlos E. Marinelli, Cristiane F. Jurinitz & Betsabé R. Guerra
IntroduçãoGalhas são tumores em tecidos ou órgãos de plantas
induzidos por diversos táxons de organismos, sendo osinsetos os galhadores mais comuns (Mani 1964; Price et al.1987). As galhas podem ser induzidas em diferentes partesda planta, sendo as folhas os orgãos mais comumenteatacados. As principais vantagens adaptativas para os
galhadores são provavelmente alimentação e abrigo contrainimigos naturais e condições adversas do meio ambiente.As galhas apresentam maior concentração de nutritiente (p.e.proteínas) e água e ao mesmo tempo apresenta baixaconcentração de defesas químicas, tornando o tecido maispalatável (Price et al. 1987). Além disso, elas oferecem maiorproteção contra estresses físicos como alta temperatura e luz
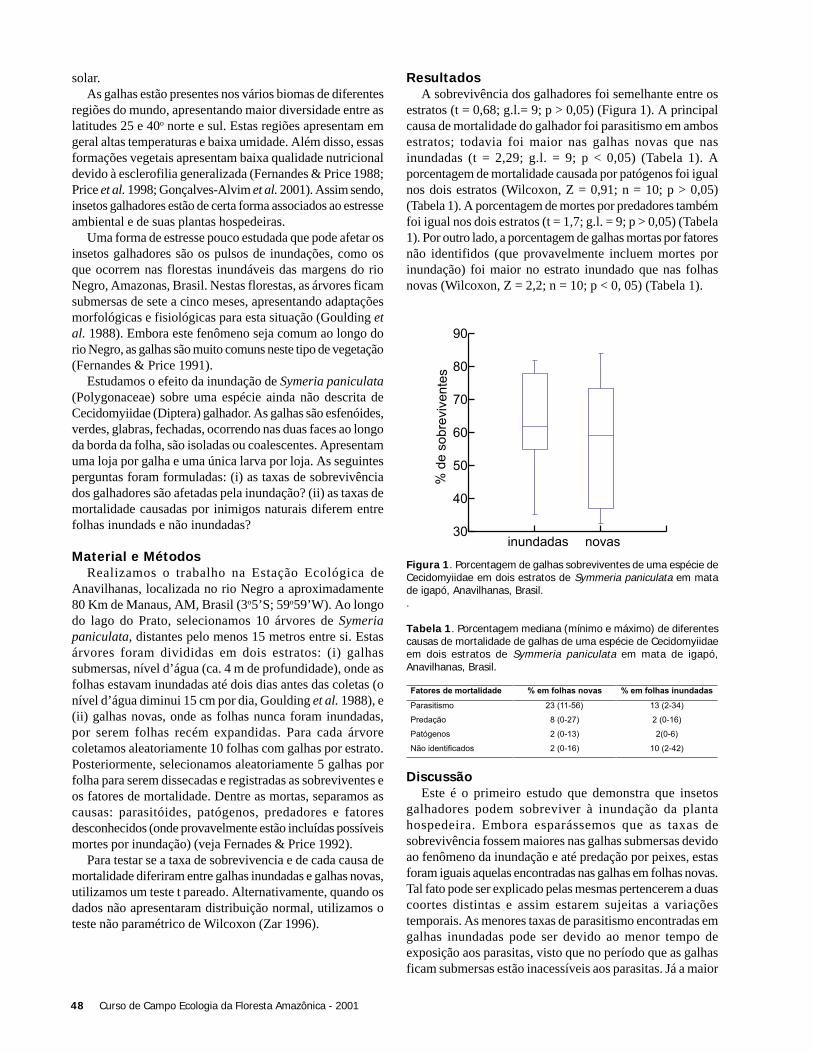
48 Curso de Campo Ecologia da Floresta Amazônica - 2001
solar.As galhas estão presentes nos vários biomas de diferentes
regiões do mundo, apresentando maior diversidade entre aslatitudes 25 e 40o norte e sul. Estas regiões apresentam emgeral altas temperaturas e baixa umidade. Além disso, essasformações vegetais apresentam baixa qualidade nutricionaldevido à esclerofilia generalizada (Fernandes & Price 1988;Price et al. 1998; Gonçalves-Alvim et al. 2001). Assim sendo,insetos galhadores estão de certa forma associados ao estresseambiental e de suas plantas hospedeiras.
Uma forma de estresse pouco estudada que pode afetar osinsetos galhadores são os pulsos de inundações, como osque ocorrem nas florestas inundáveis das margens do rioNegro, Amazonas, Brasil. Nestas florestas, as árvores ficamsubmersas de sete a cinco meses, apresentando adaptaçõesmorfológicas e fisiológicas para esta situação (Goulding etal. 1988). Embora este fenômeno seja comum ao longo dorio Negro, as galhas são muito comuns neste tipo de vegetação(Fernandes & Price 1991).
Estudamos o efeito da inundação de Symeria paniculata(Polygonaceae) sobre uma espécie ainda não descrita deCecidomyiidae (Diptera) galhador. As galhas são esfenóides,verdes, glabras, fechadas, ocorrendo nas duas faces ao longoda borda da folha, são isoladas ou coalescentes. Apresentamuma loja por galha e uma única larva por loja. As seguintesperguntas foram formuladas: (i) as taxas de sobrevivênciados galhadores são afetadas pela inundação? (ii) as taxas demortalidade causadas por inimigos naturais diferem entrefolhas inundads e não inundadas?
Material e MétodosRealizamos o trabalho na Estação Ecológica de
Anavilhanas, localizada no rio Negro a aproximadamente80 Km de Manaus, AM, Brasil (3o5’S; 59o59’W). Ao longodo lago do Prato, selecionamos 10 árvores de Symeriapaniculata, distantes pelo menos 15 metros entre si. Estasárvores foram divididas em dois estratos: (i) galhassubmersas, nível d’água (ca. 4 m de profundidade), onde asfolhas estavam inundadas até dois dias antes das coletas (onível d’água diminui 15 cm por dia, Goulding et al. 1988), e(ii) galhas novas, onde as folhas nunca foram inundadas,por serem folhas recém expandidas. Para cada árvorecoletamos aleatoriamente 10 folhas com galhas por estrato.Posteriormente, selecionamos aleatoriamente 5 galhas porfolha para serem dissecadas e registradas as sobreviventes eos fatores de mortalidade. Dentre as mortas, separamos ascausas: parasitóides, patógenos, predadores e fatoresdesconhecidos (onde provavelmente estão incluídas possíveismortes por inundação) (veja Fernades & Price 1992).
Para testar se a taxa de sobrevivencia e de cada causa demortalidade diferiram entre galhas inundadas e galhas novas,utilizamos um teste t pareado. Alternativamente, quando osdados não apresentaram distribuição normal, utilizamos oteste não paramétrico de Wilcoxon (Zar 1996).
ResultadosA sobrevivência dos galhadores foi semelhante entre os
estratos (t = 0,68; g.l.= 9; p > 0,05) (Figura 1). A principalcausa de mortalidade do galhador foi parasitismo em ambosestratos; todavia foi maior nas galhas novas que nasinundadas (t = 2,29; g.l. = 9; p < 0,05) (Tabela 1). Aporcentagem de mortalidade causada por patógenos foi igualnos dois estratos (Wilcoxon, Z = 0,91; n = 10; p > 0,05)(Tabela 1). A porcentagem de mortes por predadores tambémfoi igual nos dois estratos (t = 1,7; g.l. = 9; p > 0,05) (Tabela1). Por outro lado, a porcentagem de galhas mortas por fatoresnão identifidos (que provavelmente incluem mortes porinundação) foi maior no estrato inundado que nas folhasnovas (Wilcoxon, Z = 2,2; n = 10; p < 0, 05) (Tabela 1).
Figura 1. Porcentagem de galhas sobreviventes de uma espécie deCecidomyiidae em dois estratos de Symmeria paniculata em matade igapó, Anavilhanas, Brasil..
Tabela 1. Porcentagem mediana (mínimo e máximo) de diferentescausas de mortalidade de galhas de uma espécie de Cecidomyiidaeem dois estratos de Symmeria paniculata em mata de igapó,Anavilhanas, Brasil.
DiscussãoEste é o primeiro estudo que demonstra que insetos
galhadores podem sobreviver à inundação da plantahospedeira. Embora esparássemos que as taxas desobrevivência fossem maiores nas galhas submersas devidoao fenômeno da inundação e até predação por peixes, estasforam iguais aquelas encontradas nas galhas em folhas novas.Tal fato pode ser explicado pelas mesmas pertencerem a duascoortes distintas e assim estarem sujeitas a variaçõestemporais. As menores taxas de parasitismo encontradas emgalhas inundadas pode ser devido ao menor tempo deexposição aos parasitas, visto que no período que as galhasficam submersas estão inacessíveis aos parasitas. Já a maior
inundadas novas30
40
50
60
70
80
90
%d
eso
bre
vive
nte
s
Fatores de mortalidade % em folhas novas % em folhas inundadas
Parasitismo 23 (11-56) 13 (2-34)
Predação 8 (0-27) 2 (0-16)
Patógenos 2 (0-13) 2(0-6)
Não identificados 2 (0-16) 10 (2-42)

Curso de Campo Ecologia da Floresta Amazônica - 2001 49
mortalidade causadas por fatores não identificadas podemestar diretamente relacionadas a inundação dos tecidos dagalha.
Visto que as galhas podem sobreviver submersas, torna-se interessante investigar as adaptações morfológicas,bioquímicas e fisiológicas dos galhadores. No igapó, asplantas possuem inúmeras adaptações fisiológicas emorfológicas que as tornam tolerantes a enchentes, comopor exemplo mudanças nas concentrações de etanol,oxigênio, auxina e de taxas fotossintéticas (Hook & Scholtns1978). Essas adaptações devem influenciar enormemente osinsetos galhadores considerando sua alta sintonia com afisiologia da planta (Fernandes & Price 1991). Deste modo,trabalhos futuros deverão investigar de que maneira os insetosgalhadores repondem às mudanças provocadas pelofenômeno de inundação da planta hospedeira.
Referências BibliográficasFernandes, G.W. & Price, P.W. 1988. Biogeographical
gradients in galling species richness: test of hypotheses.Oecologia 76: 161-167.
Fernandes, G.W. & Price, P.W. 1991. Comparations oftropical and temperates galling insects richness: the holesof environmental harshness and plant nutrients status.In: Plant-Animal Interactions: Evolutionary Ecology inTropical and Temperate Regions, Price, P.W.;Lewinsohn, T.M.; Fernandes G.W. & Benson W.W.(eds.)., pp. 237-244.John Willey & Sons, New York.
Fernandes, G.W. & Price, P.W. 1992. The adaptativesignificance of insects gall distributions: survivorship
of species in xeric and mesic habitats. Oecologia 90:14-20.
Fernandes G.W. & Negreiros, D. 2001. The occurrence andeffectiveness of hypersensitive reaction against gallingherbivores across host taxa. Ecol. Entomol. 26: 46-55.
Goulding, M.; Carvalho, M.L. & Ferreira, E.G. 1988. RioNegro: Rich Life in Poor Water. SPB AcademicPublishing, The Hague.
Gonçalves-Alvim, S. J.; Santos, M.C.F.V. & Fernandes, G.W.2001. Leaf gall abundance on Avicennia germinans(Avicenniaceae) along an interstitial salinity gradient.Biotropica 33: 69-77.
Hook, D.D. & Scholtns, J.R. 1978. Adaptation and floodtolerance of tree species. In: Plant life in anaerobicenvironments, Hook, D.D. & Crawford, R.M.M. AnnArbor Science, Michigan.
Mani, M.S. 1964. Ecology of Plants Galls. Dr Junk, TheHague.
Price, P.W.; Fernandes, G.W. & Waring, J.L. 1987.Adaptative nature of insect galls. Environ. Entomol. 16:15-24.
Price, P.W.; Fernandes, G.W.; Lara, A.C F.; Brawn, J.;Barrios, H.; Wright, M.G.; Ribeiro, S.B. & Rothcliff,N. 1998. Global patterns in local number of insect gallingspecies. J. Biogeo. 25: 581-591.
Zar, J.H. 1996. Bioestatistical Analysis. Prentice Hall, NewJersey.
OrientadorGeraldo W. Fernandes
Efeitos da inundação na mortalidade das árvores adultas de uma floresta deigapó no Arquipélago das Anavilhanas, AM
Edgar Prieto, Christina McCain, Felipe de Oliveira, Maria R. Darrigo & Natalie Olifiers
IntroduçãoOs igapós são florestas inundáveis por rios de água preta
ou clara. Suas águas possuem pH baixo, são pobres emnutrientes e ricas em ácidos húmicos e taninos, solubilizadosdurante o processo de decomposição da biomassa da floresta.Esse tipo de vegetação cobre aproximadamente 15.000 km2
da Bacia Amazônica e caracteriza-se pela alta riqueza e baixadensidade por espécie (Pires & Prance 1985). Os picos deinundação dessas florestas apresentam uma sazonalidade bemdefinida, sendo que os níveis mais baixos dos rios ocorremno mês de janeiro e os mais altos em julho, com uma variaçãoanual média de 14 metros e o período de inundação variandoentre 50 e 270 dias (Junk 1989).
A duração do período de inundação e o nível máximoatingido pelas águas são fatores importantes na determinaçãoda riqueza de espécies vegetais, diversidade, distribuição,crescimento, mortalidade, dispersão de frutos e sementes(Junk 1997). A inundação expõe as plantas a um estresse
fisiológico por boa parte do ano, e neste ambienteencontramos árvores com várias adaptações a essas situações,como alterações nas vias metabólicas, queda de folhas, entreoutras (Junk 1989). Apesar dessas adaptações, quando oambiente flutua além do normal, as plantas que se encontramem áreas mais profundas provavelmente têm maior chancede morrer. Neste estudo pretendemos avaliar o efeito daprofundidade da água na taxa de mortalidade das árvoresadultas de uma floresta de Igapó no Arquipélago deAnavilhanas, Rio Negro, AM.
Material e MétodosO estudo foi realizado no Arquipélago das Anavilhanas,
no Rio Negro (03o05’S; 59o59’W), situado a 50 km anoroeste de Manaus. Selecionamos quatro locais comprofundidades diferentes e em cada um deles, escolhemosuma árvore ao acaso, anotando se ela estava morta ou viva.Ao lado desta árvore, medimos a profundidade da coluna
50 Curso de Campo Ecologia da Floresta Amazônica - 2001
d’água. Seguimos o mesmo procedimento para a árvore maispróxima da anterior e assim sucessivamente, até amostrarmos20 indivíduos em cada local. Este método se denomina“Método do Vizinho Mais Próximo” (Krebs 1999).
Utilizamos regressão logística (Harrey 1995) para calculara probabilidade de encontrarmos árvores mortas para cadaprofundidade (a relação entre a quantidade de árvores mortaspresentes nas diferentes profundidades amostradas). Asregressões foram realizadas através do Programa Systat 8.0(Wilkinson 1988).
ResultadosA regressão logística mostrou que não existe uma relação
significativa entre a profundidade em que uma determinadaárvore se encontra e a chance dela sobreviver ou morrer (N =80, p = 0,08). No entanto, quando agrupamos as árvores emclasses de profundidade, observamos que a proporção deárvores mortas em cada classe tende a aumentar com aprofundidade, principalmente depois dos 5m. Na classeintermediária de profundidade que varia de 3 a 4 metros,notamos que a mortalidade de árvores foi extremamentebaixa, quando comparada às outras classes de profundidade(Tabela 1).
Tabela 1. Porcentagem de árvores vivas e mortas em relação àsclasses de profundidade analisadas. N = a número de árvores emcada classe.
DiscussãoÁrvores em profundidades maiores estarão mais sujeitas
a eventos de inundação prolongada, que podem resultar emum aumento da taxa de mortalidade. Sabe-se que, embora amaioria das espécies do igapó sejam bem adaptadas ao ciclode cheia e vazante deste ambiente, existe um limite máximona tolerância das plantas em permanecer submersas (Goulding1988). Eventualmente, podem ocorrer períodos de cheiaprolongados e as árvores situadas em profundidades maiores,sob estresse fisiológico mais prolongado, tenderiam aapresentar taxas de mortalidade maiores. Segundo Goulding(1988), a maioria das espécies não podem suportar umperíodo de inundação de 2 ou 3 anos.
Essa tendência seria reforçada se considerarmos que os
profundidad
e
vivas
(%)
mortas
(%)
N total
0 – 1 m 79 21 29
1 – 2 m 78 22 14
2 – 3 m 75 25 4
3 – 4 m 94 6 16
4 – 5 m 80 20 5
5 – 6 m 62 38 13
6 – 7 m 40 60 5
dados obtidos para as árvores situadas entre três e quatrometros de profundidade são muito discrepantes em relaçãoaos demais. Nessas profundidades encontramos mais árvoresvivas que mortas, o que provavemente causou um aumentodo valor da significância do teste (Tabela 1). Isso pode serdevido a um problema metodológico, já que não foipadronizado um tamanho mínimo para a amostragem dasárvores. Árvores mais jovens têm menores probabilidadesde terem passado por ciclos atípicos e, no ponto onde asprofundidades eram entre três e quatro metros, percebemospor observações que havia muitas árvores mais jovens.
É possível que a mortalidade induzida por períodosprolongados de inundação seja especialmente importante nasprimeiras etapas de vida das plantas e nesse estudoamostramos somente a mortalidade em árvores adultas. Paramelhor compreensão da dinâmica da comunidade em relaçãoaos efeitos da inundação seria necessário uma coleta de dadosa longo prazo, amostrando a comunidade por um ciclocompleto de inundação e seca.
Referências BibliográficasBegon, M., Harper, J.L. & Townsend, C.R. 1986. Ecology:
Individuals, Populations and Communities. BlackwellScientific Publications, Oxford.
Del Val, E.; Fonseca, M.A.; Fáveri, S.; Toppa, R.H. & Gomes-Filho, A. 1998. Largos periodos de inundación y surelación con la mortalidad de árboles en lagos delArchipélago de Anavilhanas. Livro do Curso de Campo“Ecologia da Floresta Amazônica”. INPA/PDBFF,Manaus, AM.
Goulding, M.; Carvalho, M.L. & Ferreira, E.G. 1988. RioNegro: Rich Life in Poor Water. SPB AcademicPublishing, The Netherlands.
Haney, M. 1995. Intuitive Statistics. Oxford University Press,New York
Junk, W.J. 1989. Flood Tolerance and Tree Distribution inCentral Amazonian Floodplains. Academic Press,London, UK.
Junk, W.J. 1997. The physical and chemical environment.In: The Central Amazon Floodplain: Ecology of aPulsing System. Springer, Heidelberg, Germany.
Krebs, C.J. 1999. Ecological Methodology. Addison-WelseyEducational Publishers, Inc.
Pires, J.M. & Prance, G.T. 1985. The vegetation types of theBrazilian Amazon. In: Amazonia, Prance, G.T. &Lovejoy, T.E. (eds.), pp. 192-206. Pergamon Press,Oxford, UK.
Wilkinson, L. 1988. Systat: the system for statistics. Evaston,IL. Systat Inc.
OrientadorLeandro V. Ferreira

Curso de Campo Ecologia da Floresta Amazônica - 2001 51
Aporte de nutrientes e herbivoria em florestas alagadas da Amazônia: umaabordagem ecológico-evolutiva no arquipélago das Anavilhanas, AM
Antônio J. C. Aguiar, Andrea Lamberts, Alexander V. Christianini, Katiane Ferreira & Thiago Timo
IntroduçãoAproximadamente 11% da área foliar anualmente
produzida em florestas tropicais é consumida por herbívorose patógenos (Coley & Aide 1991). Porém, estes danos nãoestão homogeneamente distribuídos entre as espécies deplantas de um ecossistema. Várias espécies de plantasmobilizam recursos para a produção de compostos químicos(taninos, alcalóides) ou estruturas (pelos, espículas,espinhos), que não possuem necessariamente algum papelno metabolismo destas plantas. A hipótese mais aceita paraa evolução destes compostos e estruturas, é a seleção destascaracterísticas em resposta à herbivoria (Coley 1987), já quedanos promovidos por herbívoros podem causar impactosnegativos sobre o crescimento e reprodução de plantas(Marquis 1984).
Plantas que crescem em solos ricos em nutrientes podemtolerar ou compensar a perda de tecido fotossintético paraherbívoros, devido à produção de novas folhas. Contudo,nutrientes são recursos limitantes para o crescimento devegetais que ocorrem em solos pobres. Nestes locais, emgeral, há um maior investimento em defesas de tecidos porparte das plantas, via produção de compostos secundários,como taninos e alcalóides, já que os custos associados àreposição de folhas são mais altos, dado pela escassez denutrientes (Janzen 1974; Coley et al. 1985).
Os rios de água negras drenam solos do terciário equaternário (podzols), pobres em nutrientes. A presença deácidos coloidais (compostos fluviais e húmicos) sãoresponsáveis pela elevada acidez e consequente baixadiversidade da comunidade animal e vegetal (Janzen 1974;Golding 1988). Ao contrário, rios de água branca carregamuma quantidade maior de sedimentos e nutrientes dissolvidos,permitindo sustentar comunidades animais e vegetais maisricas e diversas (Sioli & Klinge 1962).
O Arquipélago das Anavilhanas (aproximadamente59º59’W; 3º05’S) funciona como um divisor natural daságuas do Rio Negro. A margem direita do Rio Negro recebeas águas do rio Jau e outros rios de água preta, enquanto amargem esquerda recebe um grande aporte de nutrientes peladesembocadura dos rios Demini e Branco, afluentes de águasbrancas (Junk, 1984). Devido a estas características ascomunidades vegetais das margens do Rio Negrocompreendem um local ideal para testar a influência do aportede nutrientes sobre o investimento de plantas em defesascontra herbívoros.
O objetivo deste trabalho é verificar o grau de herbivoriaem populações de plantas que crescem em margens opostasdo Rio Negro, distintas com relação ao aporte de nutrientes.
Material e MétodosA vegetação de ambas as margens do Rio Negro é
composta por florestas de igapó, típicas de áreas ribeirinhassazonalmente alagadas durante o período de cheias(dezembro-junho). Na região do Arquipélago dasAnavilhanas, as margens do Rio Negro estão distanciadascerca de 20 Km (Golding et al. 1988).
Na margem direita do Rio Negro foram escolhidas cincoespécies de árvores localmente abundantes: Burchenaviaoxycarpa (Combretaceae), Campsiandra comosa(Mimosoideae), Morfoespécie 1, Escheweilera tenvifolia(Lecythidaceae), Burdachia duckei (Malpighiaceae). Namargem esquerda, que recebe influência de água branca,procurou-se coletar as mesmas espécies. Porém, somente duasespécies foram coincidentes com as coletadas na margemdireita: B. oxycarpa e C. comosa. Outras três espécieslocalmente abundantes foram utilizadas em substituição:Tovomita macrophyla (Clusiaceae), Symeria paniculata(Poligonaceae), Morfoespécie 2.
Foram coletadas ao acaso, cerca de 200 folhas de trêsindivíduos de cada espécie e margem. Destas, foram retiradasao acaso 45-52 folhas para a quantificação de danos causadospor herbivoria. A quantificação dos danos foi feita porinspeção visual, enquadrando cada folha nas seguintescategorias (i): 0- 0% de herbivoria; 1 – 1 a 6%; 2 – 6 a 12%;3 – 12 a 25%; 4 – 25 a 50%; 5 – 50 a 100% (Dirzo &Domínguez 1985). Foi então aplicado um índice deherbivoria (IH), dado por:
IH = S (ni . i)
Nonde, n
i corresponde ao número de folhas de uma espécie
dentro da categoria i e N é o número total de folhas porespécie.
Foi feito um teste de Mann-Whitney (Zar 1984) paracomparar o IH dos conjuntos de espécies de plantas coletadasem margens diferentes. Teste de Qui-quadrado foi empregadopara verificar as freqüências de ocorrência de herbivoria entremargens, segundo cada categoria de dano foliar. Para asespécies coletadas em ambas as margens, procedeu-se àsmesmas análises, separadamente.
ResultadosPlantas encontradas na margem direita do Rio Negro
apresentaram valores de IH significativamente menores queplantas encontradas na margem esquerda. A comparaçãoestatística dos índices de herbivoria das duas margens foialtamente significativa (U = 0; n=10 ; p = 0,009) (Figura 1).Cerca de 60% das folhas coletadas em plantas da margemdireita do Rio Negro não apresentavam qualquer dano, contraapenas 3% das folhas coletadas em plantas da margemesquerda.
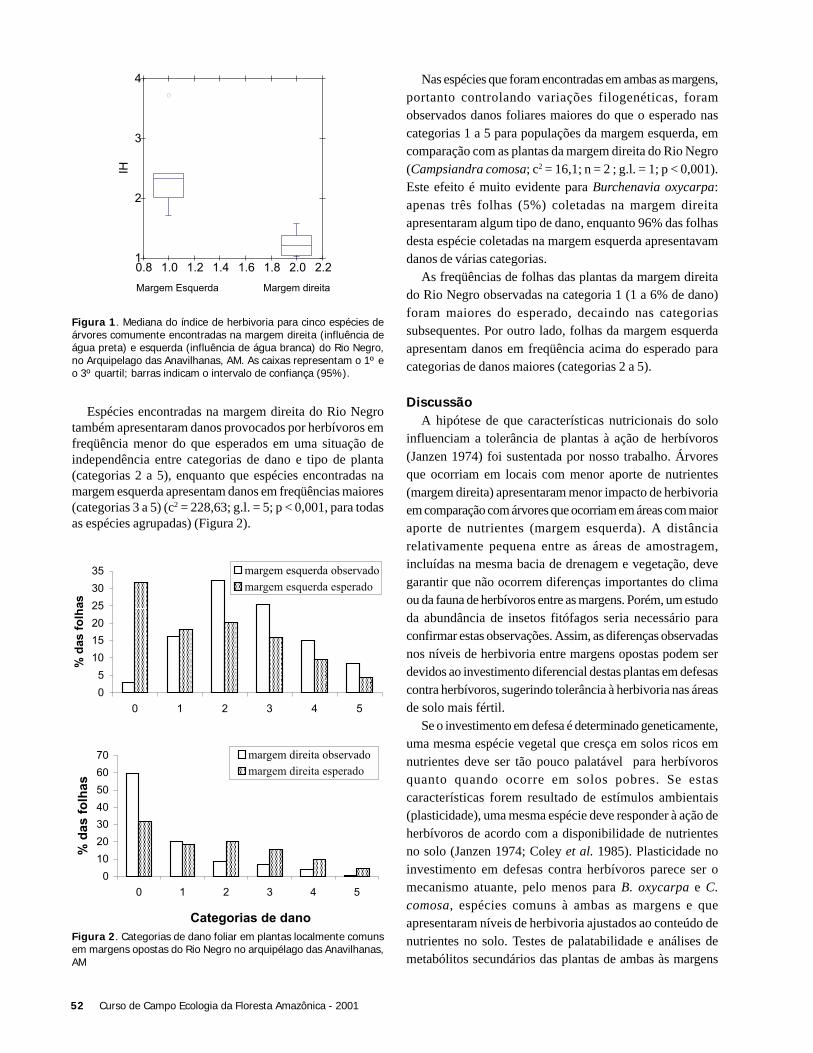
52 Curso de Campo Ecologia da Floresta Amazônica - 2001
0.8 1.0 1.2 1.4 1.6 1.8 2.0 2.21
2
3
4
IH
Margem Esquerda Margem direita
Figura 1. Mediana do índice de herbivoria para cinco espécies deárvores comumente encontradas na margem direita (influência deágua preta) e esquerda (influência de água branca) do Rio Negro,no Arquipelago das Anavilhanas, AM. As caixas representam o 1º eo 3º quartil; barras indicam o intervalo de confiança (95%).
Espécies encontradas na margem direita do Rio Negrotambém apresentaram danos provocados por herbívoros emfreqüência menor do que esperados em uma situação deindependência entre categorias de dano e tipo de planta(categorias 2 a 5), enquanto que espécies encontradas namargem esquerda apresentam danos em freqüências maiores(categorias 3 a 5) (c2 = 228,63; g.l. = 5; p < 0,001, para todasas espécies agrupadas) (Figura 2).
Figura 2. Categorias de dano foliar em plantas localmente comunsem margens opostas do Rio Negro no arquipélago das Anavilhanas,AM
0
5
10
15
20
25
30
35
0 1 2 3 4 5
%d
asfo
lhas
margem esquerda observado
margem esquerda esperado
0
10
20
30
40
50
60
70
0 1 2 3 4 5
%d
as
folh
as
margem direita observado
margem direita esperado
Categorias de dano
Nas espécies que foram encontradas em ambas as margens,portanto controlando variações filogenéticas, foramobservados danos foliares maiores do que o esperado nascategorias 1 a 5 para populações da margem esquerda, emcomparação com as plantas da margem direita do Rio Negro(Campsiandra comosa; c2 = 16,1; n = 2 ; g.l. = 1; p < 0,001).Este efeito é muito evidente para Burchenavia oxycarpa:apenas três folhas (5%) coletadas na margem direitaapresentaram algum tipo de dano, enquanto 96% das folhasdesta espécie coletadas na margem esquerda apresentavamdanos de várias categorias.
As freqüências de folhas das plantas da margem direitado Rio Negro observadas na categoria 1 (1 a 6% de dano)foram maiores do esperado, decaindo nas categoriassubsequentes. Por outro lado, folhas da margem esquerdaapresentam danos em freqüência acima do esperado paracategorias de danos maiores (categorias 2 a 5).
DiscussãoA hipótese de que características nutricionais do solo
influenciam a tolerância de plantas à ação de herbívoros(Janzen 1974) foi sustentada por nosso trabalho. Árvoresque ocorriam em locais com menor aporte de nutrientes(margem direita) apresentaram menor impacto de herbivoriaem comparação com árvores que ocorriam em áreas com maioraporte de nutrientes (margem esquerda). A distânciarelativamente pequena entre as áreas de amostragem,incluídas na mesma bacia de drenagem e vegetação, devegarantir que não ocorrem diferenças importantes do climaou da fauna de herbívoros entre as margens. Porém, um estudoda abundância de insetos fitófagos seria necessário paraconfirmar estas observações. Assim, as diferenças observadasnos níveis de herbivoria entre margens opostas podem serdevidos ao investimento diferencial destas plantas em defesascontra herbívoros, sugerindo tolerância à herbivoria nas áreasde solo mais fértil.
Se o investimento em defesa é determinado geneticamente,uma mesma espécie vegetal que cresça em solos ricos emnutrientes deve ser tão pouco palatável para herbívorosquanto quando ocorre em solos pobres. Se estascaracterísticas forem resultado de estímulos ambientais(plasticidade), uma mesma espécie deve responder à ação deherbívoros de acordo com a disponibilidade de nutrientesno solo (Janzen 1974; Coley et al. 1985). Plasticidade noinvestimento em defesas contra herbívoros parece ser omecanismo atuante, pelo menos para B. oxycarpa e C.comosa, espécies comuns à ambas as margens e queapresentaram níveis de herbivoria ajustados ao conteúdo denutrientes no solo. Testes de palatabilidade e análises demetabólitos secundários das plantas de ambas às margens

Curso de Campo Ecologia da Floresta Amazônica - 2001 53
em relação à herbívoros poderiam auxiliar a esclarecer estasquestões.
AgradecimentosAgradecemos a Rodolfo Dirzo pela abordagem precisa
da hipótese a ser testada e suas implicações biológicas,orientação e auxílio no campo. Agradecemos também aoFlávio (piloteiro) por nos levar em segurança para ambas asmargens do Rio Negro.
Referências BibliográficasColey, P.D. 1987. Patterns of plant defense: why herbivores
prefere certain species. Revta. Biol. Trop. 35: 251-263.Coley, P.D.; Bryant, J.P.; Chapin, F.S. 1985. Resource
availability and plant anti-herbivore defense. Science230: 895-899.
Coley, P.D. & Aide, T.M. 1991. Comparison of herbivoreand plant defenses in temperate and tropical broad-leavedforests. In: Plant-Animal Interactions: Evolution Ecologyof Tropical and Temperate Regions, Price, P.W.,Lewinsohn, T.M., Fernandes, G.W. & Benson, W.W.(eds.), pp. 25-49. John Wiley & Sons, New York.
Dirzo, R. 1984. Herbivory: a phytocentric overview. In:Perspectives on Plant Population Ecology, Dirzo, R. &
Samkam, J. (eds.), pp. 141-165. Sinauer Associates,Semderland, Massachussets.
Golding, M.; Carvalho, M.L. & Ferreira, E.G. 1988. Rich
Life and Poor Water: Amazonian Diversity and
Foodchain Ecology as Seen Through Fish Communities.
The Rague: SPB Academic Publishing, The Netherlands.
Janzen, D.H. 1974. Tropical blackwater rivers, animals, and
mast fruiting by the Dipterocarpaceae. Biotropica 6: 69-
103.
Junk, W.J. 1984. Ecology, fisheries and fish culture in
Amazônia. In: The Amazon Limnology and Landscape
Ecology of a Mighty Tropical River Basin, Sioli, H. (ed.).
Dr. W. Junk Publishers, Dordrecht.
Marquis, R.J. 1984. Leaf herbivores decrease fitness of a
tropical plant. Science 226: 537-539.
Sioli, H. & Klinge, H. 1962. Solos, tipos de vegetação e
águas na Amazônia. Bol. Mus. Paraense E. Goeldi 1:
27-41.
Zar, J.H. 1984. Biostatistical Analysis. Prentice Hall, New
Jersey.
OrientadorRodolfo Dirzo

54 Curso de Campo Ecologia da Floresta Amazônica - 2001
Utilização de uma ilha do Arquipélago de Anavilhanas, Baixo Rio Negro (AM) pormamíferos de médio e grande porte
Andrea Lamberts, Leandro M. Scoss, Natalie Olifiers, Paula S. Pinheiro & Thiago Timo
IntroduçãoGrande parte dos mamíferos silvestres brasileiros não são
facilmente avistados, principalmente porque a maioria delespossui hábitos noturnos e discretos (Becker & Dalponte1991; Wilson et al. 1996). Entretanto, em determinadascondições, podem ser observados indícios da presença destesanimais, tais como fezes, pêlos, rastros, tocas e restosalimentares. Além de possibilitarem em muitos casos aidentificação das espécies (Becker & Dalponte 1991; Wilsonet al. 1996; Piedra & Maffei 2000), tais indícios podemfornecer informações importantes sobre padrões delocomoção e utilização de habitats (Scoss & De Marco 2000),comportamento, idade, e status social dos animais (Wilsonet al. 1996).
As pegadas constituem o registro de interpretação maisconfiável (Becker & Dalponte 1991), sendo bastanteutilizadas em diversos estudos de ecologia e biologia daconservação. Estes estudos incluem censos populacionais(Marten 1972), estimativas de densidade populacional(Caughley 1980) e abundância relativa de espécies (Carey& Witt 1991; Conner et al. 1982), padrões de locomoção(Schaller 1980), utilização de habitats (Scoss & De Marco2000), períodos de atividade (Bider 1962), entre outros.
As ilhas do arquipélago de Anavilhanas podem representarinteressantes sítios para a realização de estudos com pegadas,uma vez que o regime de cheia e vazante da região, cria locaispropícios para a visualização de rastros de mamíferos, soloargiloso úmido.
Este estudo teve como objetivo realizar o levantamentode mamíferos com médio e grande porte que utilizam umadas ilhas do arquipélago de Anavilhanas, avaliando o esforçonecessário para estimar a riqueza de espécies e a freqüênciade observação destes animais, baseada no registro de pegadas.
Material e MétodosRealizamos o estudo em uma ilha do Arquipélago de
Anavilhanas na Estaçã o Ecológica de Anavilhanas (entre2o e 3o S e 60o e 61o W), no Rio Negro, AM, no dia 21 deagosto de 2001, durante o período de vazante do rio Negro.Neste período do ciclo hidrológico (agosto-novembro), o rioapresenta uma diminuição média de aproximadamente 10mdo nível da água (Nessimian et al. 1998), propiciando aexposição dos solos das ilhas mais altas do Arquipélago.
Estabelecemos seis transectos de 100m cada, paralelos àmargem do canal, que foram percorridos em cerca de 30minutos, sendo cada um dos transectos percorrido por doisobservadores que registraram as pegadas de mamíferossituadas até 2m para cada lado do transecto. Os rastros deidentificação duvidosa foram desenhados em folha de acetatoe posteriormente identificados com o auxílio de guia de
campo (Becker & Dalponte 1991). O número de registros decada taxa foi anotado em planilha de campo para posterioranálise. Para obter-se a freqüência de observação de pegadasfoi calculada a razão entre número de transectos em que otaxon ocorreu e o número total de transectos.Resultados
Identificamos um total de 20 rastros pertencentes a cincotáxons, dos quais dois no nível genérico e três no nívelespecífico. Dois registros não foram identificados devido aotipo de substrato ou deformidade das pegadas no terrenoúmido. Dentre os rastros observados, 45% pertenciam aHydrochaeris hydrochaeris (capivara), 15% a Leopardus sp.(gato-do-mato), 15% a Proechymis sp. (rato-toró), 10% aMazama americana (veado mateiro), 5% a Agouti paca(paca) e 10% não foram identificados (Tab. 1).
Tabela 1. Número de registros de rastros de mamíferos para osseis transectos percorridos em uma ilha do Arquipélagos deAnavilhanas, AM, 21 de agosto de 2001.
Calculamos a probabilidade de encontro para cada espécieobservada neste estudo. A probabilidade de encontro é umaestimativa da abundância relativa das espécies para o localestudado, ou seja, a probabilidade de encontrarmos capivarana área estudada é de 25%.
Foram identificadas cerca de uma espécie a cada 343m2
percorridos. A área total percorrida para a obtenção depegadas não foi suficiente para obtermos uma amostrasignificativa da riqueza de espécies da ilha, uma vez que nãoobservamos uma estabilização na curva de número acumuladode espécies por esforço amostral.
A relação entre o esforço amostral e o número acumuladode espécies, onde os transectos foram computados na ordemem que foram amostrados, não foi suficiente para detectar aestabilização do número de espécies que ocorrem na área deestudo (Fig. 1).
Espécies Número de
transectos
Frequência
observada
Leopardus sp. 3 0,5
Hydrochaeris hydrochaeris 4 0,67
Mazama americana 2 0,33
Proechymys sp. 2 0,33
Agouti paca 1 0,16
Não Ident. 1 1 0,16
Não Ident. 2 1 0,16
Curso de Campo Ecologia da Floresta Amazônica - 2001 55
0
1
2
3
4
5
6
7
8
1 2 3 4 5 6
Número de transectos
Nú
mer
oac
um
ula
do
de
esp
écie
s
Figura 1. Número acumulado de espécies de médios e grandesmamíferos em relação ao esforço de amostragem para observaçãode pegadas em uma ilha do Arquipélago de Anavilhanas, AM.
DiscussãoA taxa de observações por área amostrada pode ser
considerada alta, o que contrasta com outras áreas onde osbaixos números de rastros observados podem representar umartefato gerado pelo substrato pouco adequado à visualizaçãode pegadas (solos menos moldáveis e/ou cobertos porserapilheira).
Devido à insuficiência da área amostrada para aestabilização da curva de esforç amostral, concluímos que onúmero de espécies encontrado na ilha foi subestimado.Poderíamos resolver este problema aumentando o esforçoamostral e seguindo o mesmo delineamento aqui proposto.É provável que em uma semana de esforço amostral sejapossível obter-se uma lista de espécies de médios e grandesmamíferos bem representativa da fauna local, percorrendotambém a área seca de outras ilhas do arquipélago.
As maiores freqüências observadas de encontro são paraMazama americana, Hydrochaeris hydrochaeris eLeopardus sp, o que pode relacionar-se à abundância e/oupoder de deslocamento destes animais. É provável que asilhas formadas no Arquipélago de Anavilhanas na época devazante do Rio Negro sejam excelentes fontes alimentarespara espécies herbívoras, por serem sítios apropriados paraa formação de bancos de plântulas e gramíneas (p.e. Oryzasp.). Os felinos, que naturalmente apresentam baixadensidade populacional, estariam utilizando estas ilhas comorefúgio, em razão da pressão de caça das áreas do entorno doarquipélago, oque não descarta a possibilidade de utilizaremestas áreas como fonte de caça (p.e. poças temporárias) ourota de deslocamento.
As espécies encontradas na ilha estudada, provavelmentenão são residentes, uma vez que a ilha está sujeita ao regimeanual de cheia que alaga completamente o ambiente duranteos meses de cheia. Além disso acreditamos que esta pequenaárea não seria capaz de sustentar uma fauna diversificada demamíferos e outros animais. Por outro lado, a ilha podefuncionar como fonte de recursos (ou refúgio) temporáriospara estas espécies, o que faria com que estes animais avisitassem esporadicamente ou em caráter sazonal. O mesmo
poderia ser especulado para as demais ilhas que compõem oArquipélago de Anavilhanas, o que merece ser investigado.
Sugerimos, ainda, que estas ilhas não só funcionem comouma fonte temporária de recursos para algumas espéciescapazes de atravessar os canais e alcançá-las, comoprovavelmente funcionem também como “corredores” de trocade indivíduos entre as duas margens do Rio Negro e ilhas,considerando que algumas destas poderiam não ser capazesde atravessar continuamente de uma margem a outra do rio.
AgradecimentosAgradecemos a Rodolfo Dirzo por ter nos acompanhado
na coleta dos dados e, principalmente, na identificação daspegadas de “BURROS”, que apesar de serem numerosas,muitas vezes a associação delas com seus respectivos donosé difícil. Agradecemos também ao Juruna, por nosacompanhar na água e na lama das Anavilhanas...
Referências BibliográficasBecker, M. & Dalponte, J.C. 1991. Rastros de mamíferos
silvestres brasileiros. Brasília, Ed. Universidade deBrasília, 180pp.
Bider, J.R. 1962. Dynamics and the temporo-spatial relationsof a vertebrate community. Ecology 43(4): 634-646.
Carey, A.B. & Witt, J.W. 1991. Track counts as indices toabundances of arboreal rodents. J. Mammal. 72(1): 192-194.
Caughley, G. 1980. Analysis of Vertebrate Populations. JohnWiley & Sons, London.
Conner, M.C.; Labisky, R.F. & Progulske Jr., D.R. 1983.Scent-station indices as measures of populationabundance for bobcats, raccoons, gray foxes, andopossums. Wildl. Soc. Bull. 11(2): 146-152.
Marten, G.G. 1972. Censusing mouse populations by meansof tracking. Ecology 53(5): 859-867.
Nessimian, J.L.; Dorvillé, L.F.M.; Sanseverino, A.M. &Batista, D.F. 1998. Relation between flood pulse andfunctional composition of the macroinvertebrate benthicfauna in the lower Rio Negro, Amazonas, Brazil.Amazoniana, XV(1/2): 35-5.
Piedra, L. & Maffei, L. 2000. Efecto de las actividadeshumanas sobre la diversidad de mamíferos terrestres enun gradiente altitudinal. Rev. Biol. Trop. 48(1): 263-264.
Schaller, G.B. 1980. Movement patterns of jaguar. Biotropica12(3): 161-168.
Scoss, L.M. & De Marco, P.J. 2000. Estradas no parque:efeitos da fragmentação interna sobre a intensidade deuso do habitat por mamíferos terrestres. Anais IICongresso Brasileiro sobre Unidades de Conservação.Mato Grosso do Sul. p.770-776.
Wilson, D.E.; Cole, F.R.; Nichols, J.D.; Rudran, R. & Foster,M.S. 1996. Measuring and Monitoring BiologicalDiversity: Standard Methods for Mammals. SmithsonianInstitution Press, Washington and London.

56 Curso de Campo Ecologia da Floresta Amazônica - 2001
Distribuição espacial de três famílias de insetos predadores em diferentes micro-habitats em uma poça temporária em floresta de igapó na Amazônia Central
Angela M.i F. Pacheco, Rodrigo Cogni, Paula C. Ceotto & Christina McCain
IntroduçãoO ambiente é usualmente formado por manchas de habitats
favoráveis e desfavoráveis. As condições do habitat podemvariar no tempo e no espaço em relação à quantidade derecursos como alimentos, substrato e reprodução. Ahabilidade de ocupação de uma mancha de habitat favorávelvaria entre espécies e dentro das mesmas, podendo refletir acapacidade competitiva dos organismos (Begon 1996).
Hemípteros são insetos com aparelho bucal picador-sugador com representantes comuns na fauna aquática, dosquais muitos possuem hábito predador. Belostomatídeos,nepídeos e naucorídeos são hemípteros predadores comunsno igapó amazônico que co-ocorrem em diversos ambientes,como nos lagos das ilhas de Anavilhanas, na amazônia central(Nessimian et al. 1998).
As florestas de igapó de Anavilhanas sofrem inundaçõesperiódicas pelo aumento do nível das águas do rio Negro(Goulding et al. 1988). Durante a vazante, a água que cobriacompletamente as ilhas fica represada nas depressões doterreno, formando poças temporárias. Esses ambientesapresentam depósito de lodo e serapilheira no fundo, umaárea marginal com pequena profundidade e uma área centralcom uma coluna d’água relativamente maior, na qualencontra-se uma vegetação submersa. Devido a essasdiferenças, os microhabitats dessas poças podem possuircomposições faunísticas distintas.
Este trabalho objetiva estudar a distribuição de trêshemípteros predadores representantes das famíliasBelostomatidae, Naucoridae e Nepidae, em diferentes micro-habitats em uma poça temporária na Amazônia Central. ParaBelostoma anurum, a espécie de predador mais comum nolocal, estudamos também a distribuição espacial dos adultose juvenis.
Material e MétodosRealizamos o trabalho na Estação Ecológica de
Anavilhanas, localizada no rio Negro a aproximadamente80 Km de Manaus, AM (3o5’S; 59o59’W). Coletamos oshemípteros predadores em uma poça temporária com cercade 20 m largura, 40 m de comprimento e uma profundidadede 1 m no centro. A poça foi dividida em três micro-habitats:(a) margem correspondendo à serrapilheira na faixa de 1m apartir da margem da poça, com menos de 20 cm deprofundidade); (b) fundo, equivalente à serrapilheiradepositada a aproximadamente 1m de profundidade; e (c)vegetação submersa, correspondendo à coluna d’águapróxima a árvores e arbustos total ou parcialmente cobertospor água.
Em cada micro-habitat, realizamos um esforço de coletade uma hora, com cinco pessoas coletando simultaneamentena poça. Amostramos os hemípteros predadores passando
peneiras de 1m de diâmetro, coletando todos os indivíduosdas famílias Belostomatidae, Naucoridae e Nepidae. Onúmero de indivíduos jovens e adultos de belostomatídeosentre os ambientes margem e vegetação foi comparado comteste exato de Fisher (Zar 1994). O ambiente de fundo foidescartado das análises estatísticas devido ao baixo númerode capturas.
ResultadosColetamos 84 indivíduos pertencentes a quatro espécies
de hemípteros aquáticos: uma de Nepidae, uma deNaucoridae e duas de Belastomatidae. A espécie de Nepidaefoi encontrada apenas junto à vegetação submersa (Figura1). Já a espécie de Naucoridae foi encontrada nos trêsmicrohabitats, sendo mais frequente junto à vegetaçãosubmersa (Figura 1). No fundo da poça os Naucoridae foramos predadores mais frequentes. Os Belastomatidae foramencontrados nos três ambientes, sendo também mais comunsna vegetação submersa (Figura 1).
A espécie mais frequentemente encontrada foi Belastomaanurum (Belostomatidae). Nesta espécie, adultos e juvenisutilizam diferentemente os microhabitats (Teste exato deFisher; c2 = 5,38, g.l. = 1; p = 0,03). Enquanto os adultosutilizaram tanto a margem quanto a vegetação, os juvenisforam encontrados quase exclusivamente na vegetaçãosubmersa (Figura 2). Um indivíduo macho com os ovos nascostas foi coletado na vegetação submersa
Figura 1. Número de indivíduos de três famílias de hemípterospredadores capturados em três micro-habitats de uma poçatemporária no arquipélogo de Anavilhanas, AM. Ver texto paradefinição dos micro-habitats.
DiscussãoExiste uma clara preferência pelo micro-habitat da
vegetação submersa em todos os grupos de hemípterospredadores estudados. Provavelmente, a vegetação submersatem uma estrutura mais complexa, oferecendo refúgio e locaisde ancoragem para os indivíduos. A preferência pelavegetação submersa por parte dos naucorídeos e nepídeostambém pode estar associada aos hábitos desses organismos.
0
5
10
15
20
25
30
35
40
45
50
Margem Fundo Vegetação
Micro-habitat
Nú
mer
od
ein
div
ídu
os
Belostomatidae Naucoridae Nepidae
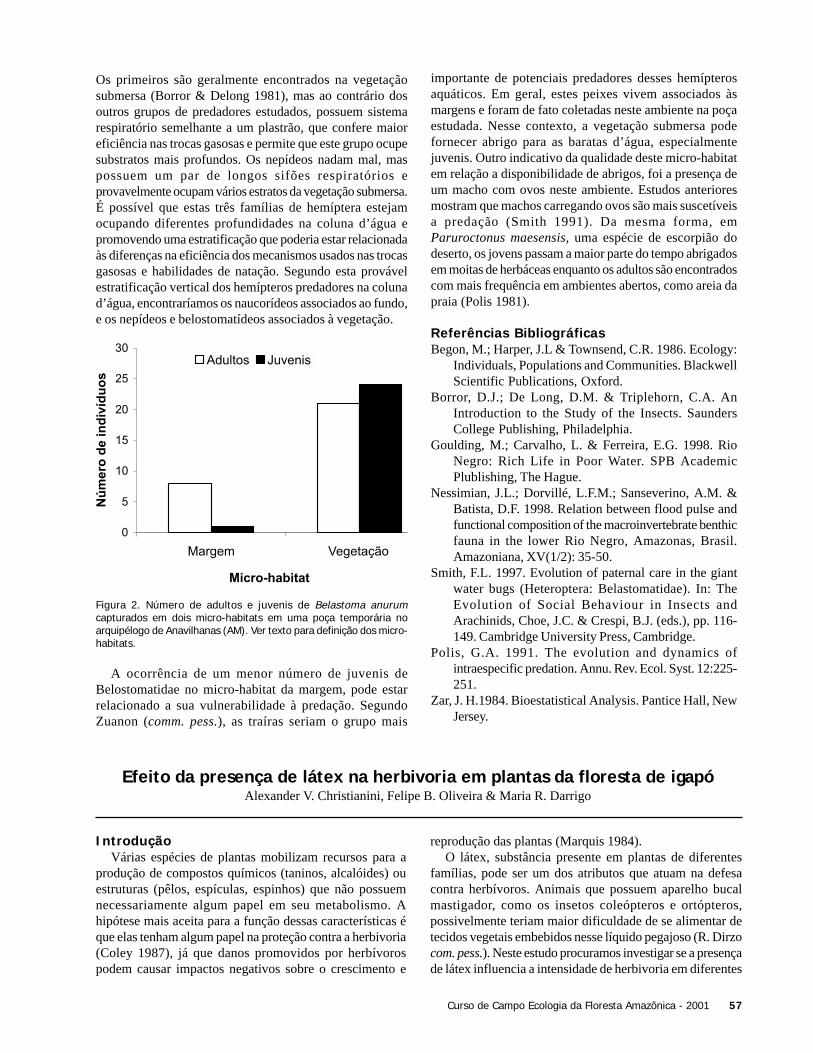
Curso de Campo Ecologia da Floresta Amazônica - 2001 57
Os primeiros são geralmente encontrados na vegetaçãosubmersa (Borror & Delong 1981), mas ao contrário dosoutros grupos de predadores estudados, possuem sistemarespiratório semelhante a um plastrão, que confere maioreficiência nas trocas gasosas e permite que este grupo ocupesubstratos mais profundos. Os nepídeos nadam mal, maspossuem um par de longos sifões respiratórios eprovavelmente ocupam vários estratos da vegetação submersa.É possível que estas três famílias de hemíptera estejamocupando diferentes profundidades na coluna d’água epromovendo uma estratificação que poderia estar relacionadaàs diferenças na eficiência dos mecanismos usados nas trocasgasosas e habilidades de natação. Segundo esta provávelestratificação vertical dos hemípteros predadores na colunad’água, encontraríamos os naucorídeos associados ao fundo,e os nepídeos e belostomatídeos associados à vegetação.
Figura 2. Número de adultos e juvenis de Belastoma anurumcapturados em dois micro-habitats em uma poça temporária noarquipélogo de Anavilhanas (AM). Ver texto para definição dos micro-habitats.
A ocorrência de um menor número de juvenis deBelostomatidae no micro-habitat da margem, pode estarrelacionado a sua vulnerabilidade à predação. SegundoZuanon (comm. pess.), as traíras seriam o grupo mais
0
5
10
15
20
25
30
Margem Vegetação
Micro-habitat
Nú
mer
od
ein
div
ídu
os
Adultos Juvenis
importante de potenciais predadores desses hemípterosaquáticos. Em geral, estes peixes vivem associados àsmargens e foram de fato coletadas neste ambiente na poçaestudada. Nesse contexto, a vegetação submersa podefornecer abrigo para as baratas d’água, especialmentejuvenis. Outro indicativo da qualidade deste micro-habitatem relação a disponibilidade de abrigos, foi a presença deum macho com ovos neste ambiente. Estudos anterioresmostram que machos carregando ovos são mais suscetíveisa predação (Smith 1991). Da mesma forma, emParuroctonus maesensis, uma espécie de escorpião dodeserto, os jovens passam a maior parte do tempo abrigadosem moitas de herbáceas enquanto os adultos são encontradoscom mais frequência em ambientes abertos, como areia dapraia (Polis 1981).
Referências BibliográficasBegon, M.; Harper, J.L & Townsend, C.R. 1986. Ecology:
Individuals, Populations and Communities. BlackwellScientific Publications, Oxford.
Borror, D.J.; De Long, D.M. & Triplehorn, C.A. AnIntroduction to the Study of the Insects. SaundersCollege Publishing, Philadelphia.
Goulding, M.; Carvalho, L. & Ferreira, E.G. 1998. RioNegro: Rich Life in Poor Water. SPB AcademicPlublishing, The Hague.
Nessimian, J.L.; Dorvillé, L.F.M.; Sanseverino, A.M. &Batista, D.F. 1998. Relation between flood pulse andfunctional composition of the macroinvertebrate benthicfauna in the lower Rio Negro, Amazonas, Brasil.Amazoniana, XV(1/2): 35-50.
Smith, F.L. 1997. Evolution of paternal care in the giantwater bugs (Heteroptera: Belastomatidae). In: TheEvolution of Social Behaviour in Insects andArachinids, Choe, J.C. & Crespi, B.J. (eds.), pp. 116-149. Cambridge University Press, Cambridge.
Polis, G.A. 1991. The evolution and dynamics ofintraespecific predation. Annu. Rev. Ecol. Syst. 12:225-251.
Zar, J. H.1984. Bioestatistical Analysis. Pantice Hall, NewJersey.
Efeito da presença de látex na herbivoria em plantas da floresta de igapóAlexander V. Christianini, Felipe B. Oliveira & Maria R. Darrigo
IntroduçãoVárias espécies de plantas mobilizam recursos para a
produção de compostos químicos (taninos, alcalóides) ouestruturas (pêlos, espículas, espinhos) que não possuemnecessariamente algum papel em seu metabolismo. Ahipótese mais aceita para a função dessas características éque elas tenham algum papel na proteção contra a herbivoria(Coley 1987), já que danos promovidos por herbívorospodem causar impactos negativos sobre o crescimento e
reprodução das plantas (Marquis 1984).O látex, substância presente em plantas de diferentes
famílias, pode ser um dos atributos que atuam na defesacontra herbívoros. Animais que possuem aparelho bucalmastigador, como os insetos coleópteros e ortópteros,possivelmente teriam maior dificuldade de se alimentar detecidos vegetais embebidos nesse líquido pegajoso (R. Dirzocom. pess.). Neste estudo procuramos investigar se a presençade látex influencia a intensidade de herbivoria em diferentes
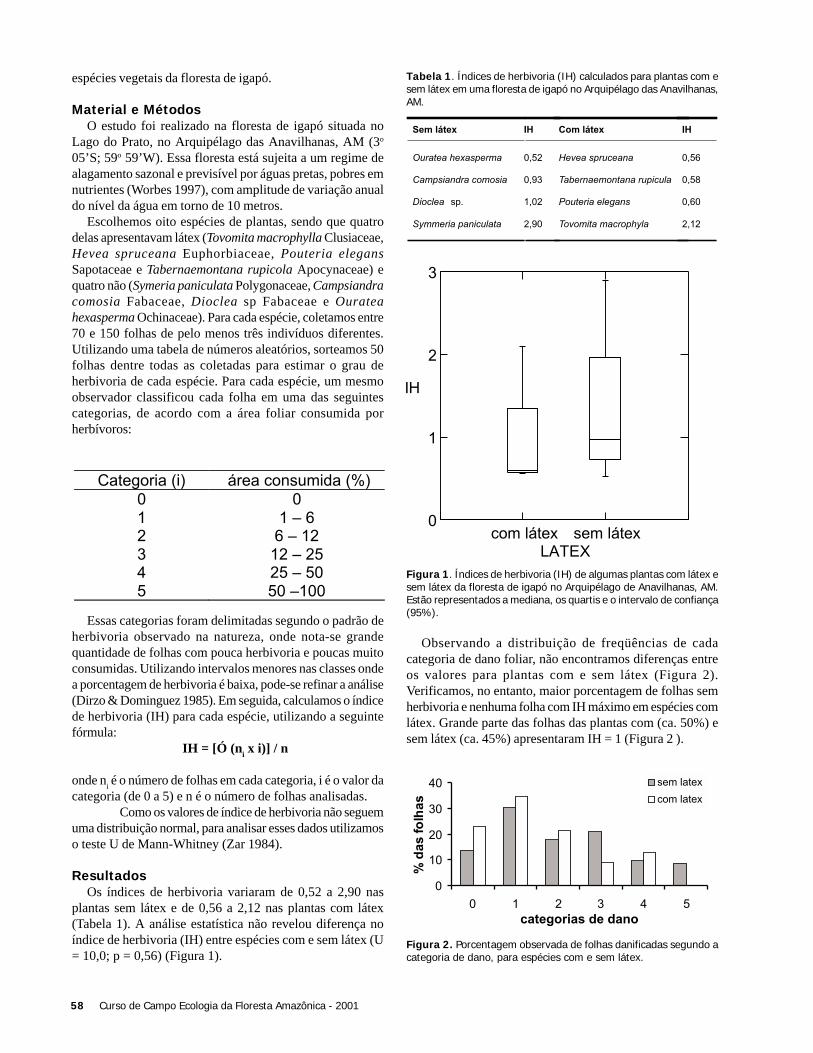
58 Curso de Campo Ecologia da Floresta Amazônica - 2001
espécies vegetais da floresta de igapó.
Material e MétodosO estudo foi realizado na floresta de igapó situada no
Lago do Prato, no Arquipélago das Anavilhanas, AM (3o
05’S; 59o 59’W). Essa floresta está sujeita a um regime dealagamento sazonal e previsível por águas pretas, pobres emnutrientes (Worbes 1997), com amplitude de variação anualdo nível da água em torno de 10 metros.
Escolhemos oito espécies de plantas, sendo que quatrodelas apresentavam látex (Tovomita macrophylla Clusiaceae,Hevea spruceana Euphorbiaceae, Pouteria elegansSapotaceae e Tabernaemontana rupicola Apocynaceae) equatro não (Symeria paniculata Polygonaceae, Campsiandracomosia Fabaceae, Dioclea sp Fabaceae e Ourateahexasperma Ochinaceae). Para cada espécie, coletamos entre70 e 150 folhas de pelo menos três indivíduos diferentes.Utilizando uma tabela de números aleatórios, sorteamos 50folhas dentre todas as coletadas para estimar o grau deherbivoria de cada espécie. Para cada espécie, um mesmoobservador classificou cada folha em uma das seguintescategorias, de acordo com a área foliar consumida porherbívoros:
Essas categorias foram delimitadas segundo o padrão deherbivoria observado na natureza, onde nota-se grandequantidade de folhas com pouca herbivoria e poucas muitoconsumidas. Utilizando intervalos menores nas classes ondea porcentagem de herbivoria é baixa, pode-se refinar a análise(Dirzo & Dominguez 1985). Em seguida, calculamos o índicede herbivoria (IH) para cada espécie, utilizando a seguintefórmula:
IH = [Ó (ni x i)] / n
onde ni é o número de folhas em cada categoria, i é o valor da
categoria (de 0 a 5) e n é o número de folhas analisadas.Como os valores de índice de herbivoria não seguem
uma distribuição normal, para analisar esses dados utilizamoso teste U de Mann-Whitney (Zar 1984).
ResultadosOs índices de herbivoria variaram de 0,52 a 2,90 nas
plantas sem látex e de 0,56 a 2,12 nas plantas com látex(Tabela 1). A análise estatística não revelou diferença noíndice de herbivoria (IH) entre espécies com e sem látex (U= 10,0; p = 0,56) (Figura 1).
Categoria (i) área consumida (%)0 01 1 – 62 6 – 123 12 – 254 25 – 505 50 –100
Tabela 1. Índices de herbivoria (IH) calculados para plantas com esem látex em uma floresta de igapó no Arquipélago das Anavilhanas,AM.
Figura 1. Índices de herbivoria (IH) de algumas plantas com látex esem látex da floresta de igapó no Arquipélago de Anavilhanas, AM.Estão representados a mediana, os quartis e o intervalo de confiança(95%).
Observando a distribuição de freqüências de cadacategoria de dano foliar, não encontramos diferenças entreos valores para plantas com e sem látex (Figura 2).Verificamos, no entanto, maior porcentagem de folhas semherbivoria e nenhuma folha com IH máximo em espécies comlátex. Grande parte das folhas das plantas com (ca. 50%) esem látex (ca. 45%) apresentaram IH = 1 (Figura 2 ).
Figura 2. Porcentagem observada de folhas danificadas segundo acategoria de dano, para espécies com e sem látex.
Sem látex IH Com látex IH
Ouratea hexasperma 0,52 Hevea spruceana 0,56
Campsiandra comosia 0,93 Tabernaemontana rupicula 0,58
Dioclea sp. 1,02 Pouteria elegans 0,60
Symmeria paniculata 2,90 Tovomita macrophyla 2,12
2
3
0
10
20
30
40
0 1 2 3 4 5categorias de dano
%d
as
folh
as
sem latex
com latex

Curso de Campo Ecologia da Floresta Amazônica - 2001 59
DiscussãoEmbora o látex seja visto como um atributo que confere
proteção contra herbívoros (R. Dirzo com. pess.), este estudonão detectou uma associação entre a presença de látex e umamenor taxa de herbivoria nas espécies estudadas na florestade igapó. Várias famílias de plantas não-relacionadaspossuem látex em seus ramos e folhas (p. ex. Euphorbiaceae,Clusiaceae, Sapotaceae), mostrando que esse caráterprovavelmente surgiu mais de uma vez durante a evoluçãodas plantas. Quando comparamos gêneros e famíliasdiferentes quanto à presença de látex e à intensidade deherbivoria, componentes relacionados às histórias evolutivasdiferentes dessas plantas podem influenciar nossasinterpretações. É bastante provável que as plantas queestudamos possuam compostos secundários diferentes, queatuam de maneiras diversas sobre os herbívoros e quepoderiam distorcer nossa análise. Um desenho experimentalinteressante para minimizar esses problemas seria acomparação da intensidade de herbivoria entre espécies come sem látex pertencentes a uma mesma família.
No entanto, algumas características do látex fazem deleuma provável adaptação anti-herbivoria: ainda não se conhecenenhuma função do látex junto ao metabolismo das plantas,e sua consistência pegajosa poderia não ser propriamenteuma defesa química, mas uma barreira física contra herbívorosmastigadores (R. Dirzo com. pess.). Por outro lado, outrascaracterísticas, como esclerofilia das folhas e presença decompostos secundários, podem ser mais importantes eminterações entre plantas e herbívoros (Coley 1987). Plantassubmetidas a condições de estresse edáfico, como as que
ocorrem em regiões de rios de água preta incluindo o RioNegro, investem mais recursos em outros tipos de defesacontra herbívoros (Janzen 1974), o que pode determinar arelativa pouca importância do látex como atributo de defesacontra herbívoros em florestas de igapó.
AgradecimentosAgradecemos ao Zeca (barqueiro) pela grande força no
campo, sem o qual este trabalho teria sido muito mais difícil.A Rodolfo Dirzo, pelas sugestões na concepção do projeto.
Referências BibliográficasColey, P.D. 1987. Patrones en las defensas de las plantas:
porqué los herbívoros prefieren ciertas especies? Rev.Biol. Trop. 35(1):151-164.
Dirzo, R. & Dominguez, C.A. 1985. Plant-herbivoreinteractions in Mesoamerican Tropical Dry Forest. In:Bullock, S.H., Mooney, H.A. & Medina, E. (eds.). JohnWiley & Sons, New York.
Janzen, D.H. 1974. Tropical blackwater rivers, animals, andmast fruiting by the Dipterocarpaceae. Biotropica 6: 69-103.
Marquis, R.J. 1984. Leaf herbivores decrease fitness of atropical plant. Science 226: 537-539.
Worbes, M. 1997. The forest ecosystem of the floodplains.In: The Central Amazonian Floodplain, Junk, W.J. (ed.).Springer, Berlin.
Zar, J.H. 1984. Biostatistical Analysis. Prentice Hall, NewYork.
Atributos ecomorfológicos para determinação de guildas tróficas da ictiofaunade igapó no arquipélago das Anavilhanas, Amazônia Central
Carlos E. Marinelli, Katiane M. Ferreira, Daniel L.M. Vieira, Cristiane F. Jurinitz,
Betsabé R. Guerra & Antônio J.C. Aguiar
IntroduçãoIgapós são áreas florestadas da Amazônia Central que
ficam inundadas pelo transbordamento dos rios de águaspretas (Lowe-McConnell 1999). Apesar da pobreza denutrientes e dos baixos valores de pH, resultantes das altasconcentrações de ácidos húmicos, sistemas de águas pretascomo o rio Negro apresentam uma elevada riqueza biológica.
De modo geral, o ambiente aquático amazônico forneceuma grande variedade de itens alimentares para os peixes,sejam de origem autóctone e alóctone, desde invertebradosaté peixes e frutos (Junk et al. 1997). Essa plasticidadealimentar é limitada por caracteres morfológicos de cadaespécie e pela disponibilidade de itens, ditando não apenascomo as espécies obtêm o seu alimento, mas tambémtornando-as dependentes dos produtos disponíveis desistemas específicos como as florestas de igapó.
Vários estudos têm demonstrando a existência de relaçõesentre as características morfológicas de peixes e a forma como
as espécies utilizam os hábitats e e os recursos alimentares.Neste estudo analisamos as relações entre as característicasmorfológicas da ictiofauna do igapó do arquipélago dasAnavilhanas e seus hábitos alimentares.
Material e MétodosEste trabalho foi realizado na Estação Ecológica das
Anavilhanas, no rio Negro, a aproximadamente 80 Km dacidade de Manaus, AM. O clima da região é caracterizadopor apresentar temperatura média anual de 26,7 °C eprecipitação média anual de 2.186 mm (RADAMBRASIL1978), com o nível d’água variando dez metros em média,anualmente.
As coletas foram realizadas na floresta de igapó do Lagodo Prato, onde utilizamos dez redes de espera, com dez metrosde comprimento cada e aberturas de 24, 30, 40, 50, 60, 70,80, 100, 120 e 140 mm entre nós opostos. As redes foramdispostas as 16h30, sendo monitoradas as 22h, quando os

60 Curso de Campo Ecologia da Floresta Amazônica - 2001
espécimes capturados foram recolhidos e separados em sacosplásticos de acordo com as malhagens. As malhadeiras foramrecolhidas as 7h do dia seguinte, totalizando 15 horas deesforço amostral.
As espécies foram identificadas com auxílio do Dr. JansenZuanon (INPA) e em seguida todos os exemplares forampesados e com paquímetro analógico (precisão de 1mm)tiveram tomadas as seguintes características biométricas:comprimento total (CT): medido da ponta do focinho até aextremidade da nadeira caudal; altura e largura da boca (ABe LB): medida entre o ponto mediano da mandíbula e maxila(com a boca do peixe totalmente aberta); altura do corpo(AC): medida da distância vertical entre a base da nadadeiradorsal e a linha ventral do corpo; largura do corpo (LC):medida da altura da inserção da nadadeira dorsal;comprimento da cabeça (CC): medida da ponta do focinhoaté a extremidade óssea posterior do opérculo. Essas seismedidas morfológicas foram combinadas entre si resultandoem quatro atributos: tamanho relativo da cabeça (TRC),largura relativa do corpo (LRC), tamanho relativo da cabeça(TRC) e tamanho relativo da boca (TRB). Para analisar aimportância relativa das variáveis morfológicas nadiferenciação das espécies no espaço ecomorfológico foirealizada uma análise de componentes principais (SYSTAT8.0).
Todos os exemplares coletados tiveram os seus conteúdosestomacais analisados em estereomicroscópio e suas dietascaracterizadas em categorias tróficas amplas: herbívoros,invertívoros, piscívoros e onívoros. Os resultados da dietaforam comparados com as informações apresentadas porGoulding et al. (1988) para os peixes do rio Negro.
ResultadosForam capturados um total de 20 exemplares pertencentes
a 13 espécies e seis famílias de peixes. A Família Characidae(Characinae, Tetragonopterinae e Acestrorhynchinae) foi amelhor representada com quatro espécies, seguida deAnostomidae com três espécies e Cichlidae com duas. Asoutras três famílias apresentaram apenas uma espécie cada.Os anostomídeos foram o grupo com maior abundância:Anostomoides laticepis (n = 5), Laemolyta proxima (n = 5)e Laemolyta taeniata (n = 1) e totalizando 55% dosexemplares amostrados. Com exceção de Ageneiosusucayalensis (n = 2), capturamos apenas um indivíduo dasoutras nove espécies.
Quanto à composição por guilda trófica, foram registradasseis espécies invertívoras, quatro piscívoras, duas onívorase uma herbívora (Tabela 1), com destaque para a FamíliaAnostomidae que com 11 exemplares foi constituída por duasespécies onívoras (Laemolyta proxima e Laemolyta taeniata)e uma herbívora (Anostomoides laticeps).
Tabela 1. Hábito alimentar das espécies de peixes (sigla para o plotda PCA Figura 1) capturadas na área de igapó da Estação Ecológicadas Anavilhanas e comparação com os dados apresentados porGoulding et al. (1988).
Onde: (*) espécies capturadas na floresta de igapó.
Com relação aos atributos ecomorfológicos, o tamanhorelativo da cabeça e a largura relativa do corpo foram asvariáveis com maior carga noprimeiro eixo, contendo 46,26%da variação, e sendo explicado pelas guildas tróficas. Osegundo eixo foi principalmente correlacionado com a alturae a largura relativas do corpo, contendo 30,7% da variação(Tabela 2). O primeiro eixo ordenou quatro guildas tróficas,sendo elas: onívoros = LAT e LAP; herbívoros = AN;invertívoros = AGE, AG, ANC, CH, TE e ASP e piscívoros= CM, AF e PS.
Figura. 1. Análise de componentes principais para os dados deecomorfométria de peixes do igapó da Estação Ecológica deAnavilhanas, rio Negro, AM.
Espécie Hábito alimentar Goulding et al. (1988)CharacidaeAcestrorhynchus falcirostris (AF) Piscívoro piscívoro*Acestrorhynchus grandoculis (AG) Invertívoro piscívoroAcestrorhynchus microlepis (AM) Piscívoro/invertívoro piscívoroTetragonopterus argenteus (TE) Invertívoro invertívoro*Charax sp. (CH) Invertívoro piscívoro*
AnostomidaeLaemolita proxima (LAP) Onívoro invertívoroAnostomoides laticeps (AN) Herbívoro sem informaçãoLaemolita taeniata (LAT) Onívoro detritívoro/herbívoro
CichlidaeCichla monoculus (CM) Estômago vazio piscívoroPlagioscion squamosissimus (PS) piscívoro invertívoro
AuchenipteridaeAuchenipterichthys thoracatus
(ASP)invertívoro invertívoro
AgeneiosidaeAgeneiosus ucayalensis (AGE) invertívoro piscívoro*
EngraulididaeAnchoviella sp.(ANC) invertívoro invertívoro
-2 -1 1 2EIXO1
-2
-1
1
2
EIXO2
LAT
LAP
AN
TE
AG
ANC
AF
PSCM
ASP
CH
AGE

Curso de Campo Ecologia da Floresta Amazônica - 2001 61
Tabela 2. Cargas dos atributos ecomorfológicos obtidos pela análisede componentes principais para morfologia do corpo de espécies depeixes de uma área de igapó da Estação Ecológica de Anavilhanas,rio Negro, AM.
DiscussãoLowe-McConnell (1999) afirma que ambientes aquáticos
tropicais sazonais e não sazonais demonstram atributosecológicos diferentes quanto a estrutura de suascomunidades de peixes. Como a oferta de itens alimentaresvaria conforme o pulso de inundação, a ictiofauna dessessistemas apresenta algumas estratégias alimentares, como,o pequeno número de espécies especialistas, com o mesmoalimento sendo compartilhado em guildas. Florestas de igapósão caracterizadas por porcentagens relativamente altas deconsumidores de alimentos com origem na floresta alagada(Goulding et al. 1988). Analisando as dietas das dozeespécies obtidas por nós, encontramos números bemparecidos de espécies que compunham cada uma das guildasde hábitos alimentares (i.e. piscívoras, invertívoras, onívorase herbívoras), explicitando de certa forma a diversidade deitens alimentares disponíveis no igapó.
Insetos são os principais organismos na alimentação daictiofauna amazônica, fazendo parte da dieta de muitasespécies em pelo menos uma das fases de seudesenvolvimento ontogenético. Em ambientes de igapó, essegrupo constitui-se de um recurso extremamente importantena sustentação da trama trófica, com as formas aquáticas eas larvas aquáticas de formas terrestres sendo os itens maisconsumidos (Lowe-McConnell 1999). Crustáceos tambémsão itens comuns da alimentação das comunidades de peixesdo rio Negro, fazendo parte do elo de ligação entre detritos,fungos e algas com os peixes, sendo em florestas de igapórepresentados principalmente por camarões e cladóceros(Goulding et al.1988). Com relação à guilda de invertívoros,Acestrorhynchus grandoculis foi a única espécie queconsumiu exclusivamente insetos, enquanto Ageneiosusucayalensis, Charax sp., Tetragonopterus argenteus,Anchoviella sp. e Auchenipterichthys thoracatusconsumiram tanto insetos quanto camarões. Apesar desteúltimo ser um típico consumidor de insetos arbóreos quecaem na água (Lowe-McConnell 1999) a plasticidadealimentar já apontada anteriormente não impede que essaespécie aproveite outros itens altamente disponíveis, comoos camarões em florestas de igapós, onde este grupo encontramatéria orgânica partículada em abundância devido adecomposição da vegetação inundada.
A inundação de matas de igapó disponibiliza uma grandeabundância de recursos alimentares de origem vegetal eanimal, como resultado da submerção do sub-bosque dafloresta. A Família Anostomidae foi o único taxon queapresentou tanto espécies herbívoras (Anostomoideslaticeps) quanto onívoras (Laemolyta taeniata e Laemolyta
Medidas do corpo Eixo 1 Eixo 2
Altura relativa do corpo 0,230 0,749Largura relativa do corpo 0,490 0,647Tamanho relativo da cabeça -0,951 0,100Tamanho relativo da boca (altura e largura) -0,808 0,489Variância explicada pelo eixo 46,26% 30,70%
proxima).A sub-bacia do rio Negro agrupa o maior número de
espécies piscívoras da bacia Amazônica, com a maioria dasespécies desta guilda sendo peixes que engolem as presasinteiras ou as mordiscam (Goulding et al. 1988). Das 450espécies de peixes da bacia do rio Negro, 87 são piscívoros,e para as florestas de igapó são registradas 34 espécies. Aparticipação dos piscívoros só é maior nas praias da baciado rio Negro. Foram amostradas três espécies de peixespiscívoras (Plagiocion squamosissimus, Acestrorhynchusfalcirostris e Acestrorhyncus microlepis), além de Cichlamonoculus, espécie de hábitos preferencialmente piscívoros,mais que no presente estudo apresentava o estômago vazio.
Estudos ecomorfológicos fornecem em sua maior parteinferências com respeito ao uso de micro-hábitats e dieta(Peres-Neto 1999), além da posição do peixe na colunad’água e manobrabilidade natatória. No presente estudo, aanálise de componentes principais ordenou apenas osanostomídeos como umb grupo mais homogêneo e conspícuo,constituído por espécies onívoras com cabeça pequena ecorpo relativamente largo (LAT = Laemolyta taeniata, LAP= Laemolyta proxima e NA = Anostomoides laticeps). Parao restante da comunidade, não obtivemos padrõesconsistentes. No caso dos Siluriformes (ASP =Auchenipterichthys thoracatus e AGE = Ageneiosusucayalensis) por exemplo, apesar das duas espécies seremde hábitos invertívoros e de terem o corpo baixo e alongado,ficaram em lados opostos com relação ao eixo 2, com asegunda espécie se agrupando a outras de corpo curto e alto(p.e. Tetragonopterus argenteus).
Estudando a ecomorfometria das espécies da ictiofaunada Lagoa do Prato, Smith et al. (1998) também nãoencontraram um padrão morfológico que explicasse o hábitoalimentar das espécies de maneira geral. Podemos inferir quetrês podem ter sido os fatores determinantes para obtençãode resultados diferentes do inicialmente esperado: (i) o baixonúmero de exemplares por espécie que tiveram suas dietasanalisadas; (ii) a tendência a captura de um maior número deespécies piscívoras, já que as coletas foram realizadas apenasnos períodos crepuscular e noturno e; (iii) o pequeno númerode variáveis morfológicas tomadas para cada espécie.
AgradecimentosAo Dr. Jansen Zuanon (INPA) pelas sugestões, auxílio
na identificação dos exemplares de peixes e constantedisposição na orientação deste e dos outros trabalhosenvolvendo a ictiofauna de sistemas da Amazônia Centraldurante o Curso de Campo 2001. Ao Dr. EduardoVenticinque pelas valiosas sugestões e, ao Juruna (parceiro)pelo apoio e toda a força nos trabalhos de campo, dicas esua experiência singular.
Referências BibliográficasGoulding, M.; Carvalho, M.L. & Ferreira, E.G. 1988. Rio
Negro: Rich Life in Poor Water. SPB Academic Pub.Lowe-McConnell, R. 1999. Estudos Ecológicos de

62 Curso de Campo Ecologia da Floresta Amazônica - 2001
Comunidades de Peixes Tropicais. Tradução: Vazzoler,M.E.A.M., Agostinho, A.A. & Cunnigham, P.T.M.EDUSP, São Paulo.
RADAMBRASIL 1978. Levantamento de RecursosNaturais. Vols 1-18. Ministério de Minas e Energia.Departamento Nacional de Produção Mineral. Rio deJaneiro.
Santos, G.M. & Ferreira, E.J.G. 1999. Peixes da baciaamazônica. Estudos ecológicos de comunidades depeixes tropicais. In: Vazzoler, E.A.M., Agostinho, A.A.& Cunningham, P.T.M. (eds.). EDUSP, São Paulo.
Smith, M; Fáveri, S.B.; Salcedo, N. & Castilho, C.V. 1998.
Morfometria de peixes: relação com hábito alimentar.Livro do Curso de Campo “Ecologia da FlorestaAmazônica”. INPA/PDBFF, Manaus, AM.
Peres-Neto, P.R. 1995. Estrutura de comunidades de peixesao longo de um gradiente lótico de um rio costeiro doleste brasileiro (rio Macacu, RJ). Dissertação deMestrado, UFRJ, Rio de Janeiro, RJ.
Peres-Neto, P.R., 1999. Alguns métodos e estudos emecomorfologia de peixes de riachos. OecologiaBrasiliensis: ecologia de peixes de riachos. In:Caramaschi, E.P., Mazzoni, R & Peres-Neto, P.R. PPGE,UFRJ.
A ictiofauna de uma poça sazonal nas ilhas Anavilhanas, rio Negro, AMEdgar F. Prieto & Mauricio Camargo
IntroduçãoO rio Negro, ao ser comparado com um rio de águas
brancas como o Solimões, apresenta baixa quantidade decarga de sedimentos. Pórem, as baixas diferenças na suaaltitude ao longo de seu curso, e a contribuição dossedimentos aportados pelo rio Branco, definem a formaçãode séries de ilhas (Goulding et al. 1988) muito dinâmicas noque se refere aos processos de erosão e deposição (obs. pess).Devido á baixa altitude, estas ilhas durante o período decheia ficam totalmente encobertas por água (Goulding et al.1988). De forma inversa durante o período de estiagem, adiminuição da vazão, resulta na retração das águas ao canaldo rio., deixando poças de água estagnada, que podemdiminuir até 20cm de profundidade num período de 24 horas,até secar por completo, deixando camadas de argila úmida,em baixo da serapilheira (obs. pess).
Estes pequenos corpos de água, caracterizam-se porapresentar condições de baixos teores de oxigênio, baixopH, e em geral grandes taxas de decomposição da matériaorgânica de origem alóctone, que pela deficiência de oxigênio(Rai & Hill 1981), geram processos de decomposiçãoanaeróbica, que podem produzir ácido sulfídrico e metanoem condições extremas. Estes pequenos corpos de água,apresentam uma ictiofauna especializada, com adaptaçõespara sobreviver sob condições extremas de baixa qualidadeda água (Junk et al. 1997). Alguns autores já tem estudado,as adaptações fisiológicas de vários grupos de peixes, parasuportar condições ambientais de baixa qualidade doambiente aquático (Machado-Allison 1987; LoweMcConnell 1987).
O objetivo do presente estudo foi avaliar se a estrutura dacomunidade de peixes em uma poça temporaria, é modeladapelas eventuais condições estressantes do ambiente físico.Desta forma, apresentamos uma listagem da composiçãoictiofaunistica de uma poça de águas pretas estagnadas,considerando aspectos do habitat, microhabitats, e hábitosalimentares das espécies presentes, com estimativas dabiomassa e esforço de coleta.
Material e MétodosSelecionamos uma poça temporária, localizada no
arquipélago das Anavilhanas, que recebe aporte de águas dorio Negro, durante a enchente. Inicialmente, caracterizamoso ambiente físico quanto as dimensões da poça, profundidade,tipo de substrato, vegetação predominante e medição do pH.As coletas de peixes tiveram uma duração de 2,5 horas. Pararealizar as capturas, utilizamos duas peneiras de formacircular com áreas de 0,38m2 e 0,13m2 respetivamente. Para2 coletores, definimos um esforço de coleta de 70 lancespara cada aparelho. Cobrimos uma área total de 35,7m2. Oslances foram realizados de forma aleatória, tentando cobrira variação em profundidade e diferentes regiões da área totalda poça. Com base no peso úmido do folhiço de 2 amostras,estimamos a biomassa de folhiço por unidade de área.
Os exemplares capturados foram fixados em álcool, parasua posterior identificação taxonômica com ajuda doProfessor J. Zuanon. Uma vez identificados, os exemplaresos separamos de acordo com seu habitat e categoria trófica.Posteriormente, estes foram contados. Com os cálculos deárea total varrida e sua relação com o número e peso dosindivíduos, estimamos a biomassa e a densidade por unidadede área. Também calculamos a captura por unidade de esforço– CPUE: gramas/lance/homem.
ResultadosO ambiente físico corresponde a uma poça isolada com
uma profundidade máxima de 80cm e uma área do espelhode água de 360m2. A coloração característica é de águas pretascom um pH de 4,0 e cheiro típico de um ambiente anóxico,com odor de enxofre. O sedimento é formado por argilaamarela, encoberta por grande biomassa de folhas emdecomposição (1050-1300g/m2). A vegetação predominantedentro do ambiente aquático é a palmeira Astrocaryum jauaricom algumas folhas submersas.
Encontramos um total de 20 espécies de peixes quecorresponderam a 14 famílias e 18 gêneros (Tabela 1). A

Curso de Campo Ecologia da Floresta Amazônica - 2001 63
família Curimatidae foi a melhor representada em númerode espécies. Observamos um predomínio de Cyphocharaxsp. (Curimatidae) e Apistogramma sp. (Cichlidae). O pesoda captura total, para um esforço de dois coletores foi de5,74g. A captura por unidade de esforço foi de 0,02g/lance/coletor. As estimativas de biomassa e densidade foram de0,16g/m2 e 1,5ind/m2 respectivamente.
A maior parte dos exemplares coletados, corresponderama estádios juvenis das diferentes espécies. Em geral,observamos um predomínio de peixes com hábito alimentardetritívoro e consumidores de pequenos invertebrados.Existe uma divisão dos diferentes microhabitats,predominando os organismos associados ao fundo (85%),que exploram principalmente a serapilheira. O restante dasespécies correspondem aos lebiasinídeos e anostomídeosque fazem uso dos ambientes mais superficiais (Tabela 1).
DiscussãoAs características do ambiente aquático, indicam
condições estresantes para a sobrevivência de vários gruposde peixes. O cheiro característico de enxofre produzido pelaabundante biomassa vegetal em decomposição anaeróbica eo baixo pH são fatores que restringem a ocorrência de certospeixes como alguns characídes e loricarídos. Porém, algunasespecies apresentam adaptações fisiológicas, morfológicas,ou comportamentais que lhes permite suportar estas
condições ambientais. Alguns exemplos como Synbranchussp. que apresenta capacidade para respirar oxigênioatmosférico, lebiasínidos como Copella sp. que utilizan ooxigênio da película mais superficial da coluna de água:Hoplias gr. malabaricus, Apistogramma spp., e algunscurimatídos conseguem através da diminuição em suaatividade de deslocamento, suportar condições extremas debaixos teores de oxigênio.(Junk1992).A abundância defolhas da palmeira Astrocarium javari na poça mesmo quecontribui de forma negativa no ambiente físico ao diminuiros teores de oxigênio, oferece microhabitats para as espéciesde Hypopomidae e Hoplias gr. malabaricus.
Os baixos valores que obtivemos para as estimativas deCPUE, biomassa e densidade, indicam que grande parte dosorganismos que poderiam ocorrer saem antes de diminuir oespelho de água, quando a qualidade torna-se baixa. Estesresultados podem ser explicados através de uma relaçãocusto-benefício no sentido de ter se adaptado as condiçõescríticas do ambiente, mas que pode servir de refúgio para osjovens e novos recrutas. A disponibilidade de fonte alimentar,em especial de organismos de hábitos detritívoros econsumidores de microorganismos animais, é um outro fatorque afeita a densidade de peixes no sistema. Assim, baixasdensidades de microorganismos aquáticos (Junk &Robertson 1992) pode ser o principal fator que define uma
baixa densidade e biomassa da ictiofauna.
Espécie Estádio de desenvolvimentojuvenil
AdultoFonte alimentar principal
MicrohabitatChilodontidaeChilodus sp. 1
Detritos Bentônico
LebiasinidaeCopella gr. nattereriNannostomus eques
56
MicroinvertebradosMicroinvertebrados
SuperficialSuperficial
CurimatidaeCurimatopsis sp.Curimatella sp.Cyphocharax sp.
2110
DetritosDetritosDetritos
BentônicoBentônicoBentônico
EleotrididaeMicrophylipnus amazonicus 5 Microinvertebrados BentônicoAnostomidaeAnostomoides laticeps 1 Microinvertebrados SuperfícieCharacidaeKlausewitzia sp.Characidae sp.
16
MicroinvertebradosMicroinvertebrados
SerapilheiraSerapilheira
TrichomycteridaeTrichomycterus hasemanni 1 Microinvertebrados SerapilheiraCichlidaeApistogramma sp.1Apistogramma sp.2
82
MicroinvertebradosMicroinvertebrados
SerapilheiraSerapilheira
LoricariidaeLoricaria sp. 1 Perifiton SerapilheiraHypopomidaeBrachyhypopomus beebeiMicrosternarchus bilineatus
11
InvertebradosInvertebrados
SerapilheiraSerapilheira
ScoloplacidaeScoloplax dolicholophia Microinvertebrados SerapilheiraErythynidaeHoplias gr. malabaricus IctiófagoSynbranchidaeSynbranchus sp. 1 Ictiófago SerapilheiraGen nov. sp. nov. 3 Carnívoro ? Serapilheira
Tabela 1. Listagem de espécies de peixes coletadas em uma poça temporária no arquipiélago das Anavilhanas, rio Negro, AM.

64 Curso de Campo Ecologia da Floresta Amazônica - 2001
Podemos concluir que a ictiofauna observada na poçaestudada é resultado das condições estresantes nesseecossistema. Nos casos extremos de limitação do ambienteaquático, diferentes estratégias podem ser utilizadas pelospeixes, desde se manter em poças de lama ou retornar ao riopor meio de deslocamento sobre vegetação herbácea. Para osgrupos de peixes que não apresentam adaptações a estascondições e não conseguem sair sofrem mortalidade natural.
Referências BibliográficasGoulding, M.; Carvalho, M.L. & Ferreira, E.G. 1988. Rio
Negro: Rich Life in Poor Water: Amazonian Diversityand Foodchain Ecology as Seen through the FishCommunities. Academic Publishing.
Junk, W.J.; Soares, G.M. & Carvalho, F.M. 1983.Distribution of fish species in a lake of the Amazon riverfloodplain near Manaus (Lago Camaleão), with specialreference to extreme oxygen conditions. AmazonianaVII(4): 397-432.
Junk, W.J. & Robertson, B.A. 1992. Aquatic Invertebratesin Ecological Studies. In: The Central Amazon
Floodplain, Junk, W.J. (ed), pp. 279-297.Lowe-McConnell, R.H. 1987. Ecological Studies in Tropical
Fish Communities. Cambridge University Press,London.
Machado-Allison, A. 1987. Los peces de las planiciesinundábles de Venezuela, um estudo de su historianatural. Universidad Central de Venezuela, Caracas.57pp.
Nessimian, J.L.; Dorvillé, L.F.M.; Sanseverino, A.M. &Baptista, D.F. Relation between floodpulse and funtionalcomposition of macroinvertebrate benthic fauna in thelower Rio Negro, Amazonas, Brazil. Amazoniana XV(1/2): 35-50.
Rai, H. & Hill, G.1981. Physical and Chemical studies ofLago Tupí, a Central Amazon Black water ria lake. Int.Revue. Ges. Hydrobiol. 66(1): 37-82.
Walker, I. & Henderson, P.A. 1980. Ecophysiological aspectsof Amazonian blackwater litterbank fish communities.In: Physiology and Biochemistry of the Fishes of theAmazon.
Fatores estruturais que influenciam o tamanho dos jardins de formigaFelipe B. de Oliveira, Christina McCain, Edgar Prieto, Maria R. Darrigo & Natalie Olifiers
IntroduçãoAs formigas desempenham uma série de funções
ecológicas importantes como dispersão de sementes,predação e movimentação do solo, que as tornam animais degrande relevância no estudo das florestas tropicais(Hölldobler & Wilson 1990). Além disso, esses insetosinteragem com várias plantas, em alguns casos de maneirabastante específica; estão descritas as relações de formigascom Cecropia spp Cecropiaceae (Santamaria et al. 1997),com Maieta guianensis Melastomataceae (Vasconcelos1991), entre outras (Hölldobler & Wilson 1990).
Um tipo de interação muito comum em florestasneotropicais e considerada o mais complexo tipo demutualismo entre formigas e plantas são os jardins deformigas (Hölldobler & Wilson 1990), estruturas formadaspor epífitas enraizadas em ninhos desses insetos. As formigasalimentam-se dos frutos de algumas plantas e descartam assementes nas câmaras de lixo no ninho; essas sementes muitasvezes germinam e, nutridas pelos detritos trazidos pelasformigas, crescem sobre a colônia e suas raízes tornam-separte integrante da estrutura do ninho (Hölldobler& Wilson1990). A formação dos jardins de formigas não é ocasional,já que envolvem sempre as mesmas espécies de plantas,principalmente das famílias Araceae e Gesneriaceae (Ribeiroet al. 1999) e de formigas como Crematogaster, Camponotuse Solenopsis (Hölldobler & Wilson 1990).
As plantas que crescem em jardins de formigas sebeneficiam de um ambiente com maior quantidade denutrientes; indivíduos de Codonanthe crassifolia, uma
gesneriácea muito comum em jardins de formigas, apresentamum crescimento maior e mais rápido quando associadas aCrematogaster longispania em relação a indivíduos quecrescem sozinhos (Kleinfeldt 1978). Além disso, as epífitasque nascem em jardins são menos atacadas por herbívoros(Santos 1999). No entanto, os benefícios que as formigasobtém com essa relação, caso existam, ainda sãodesconhecidos. Uma possível hipótese seria a de que asformigas retiram das plantas do jardim algum tipo dealimento: Codonanthe crassifolia, por exemplo, possuinectários extra-florais em suas folhas (Kleinfeldt 1998). Umaoutra hipótese seria a de que as formigas se beneficiam dasraízes das epífitas, que confeririam mais pontos de apoio aoninho e suporte estrutural, permitindo um maior crescimento.
Neste trabalho averiguamos a existência de relações entreo tamanho dos jardins de formigas e o número de plantasque crescem sobre eles. Adicionalmente, verificamos se onúmero de ramos da planta suporte onde o ninho está apoiadoestá relacionado ao tamanho do jardim.
Material e MétodosRealizamos este estudo na Reserva do Km 41 do Projeto
Dinâmica Biológica de Fragmentos Florestais (INPA/SI), naFazenda Esteio, Manaus, AM (2°24’S; 59°44’W). Nessa áreapredomina a floresta de terra firme, que ocupa grande parteda Amazônia Central (Pires & Prance 1985).
Percorremos algumas das trilhas da reserva (trilha L,próxima ao alojamento) procurando por jardins de formiga.Amostramos um total de 34 ninhos, registrando o
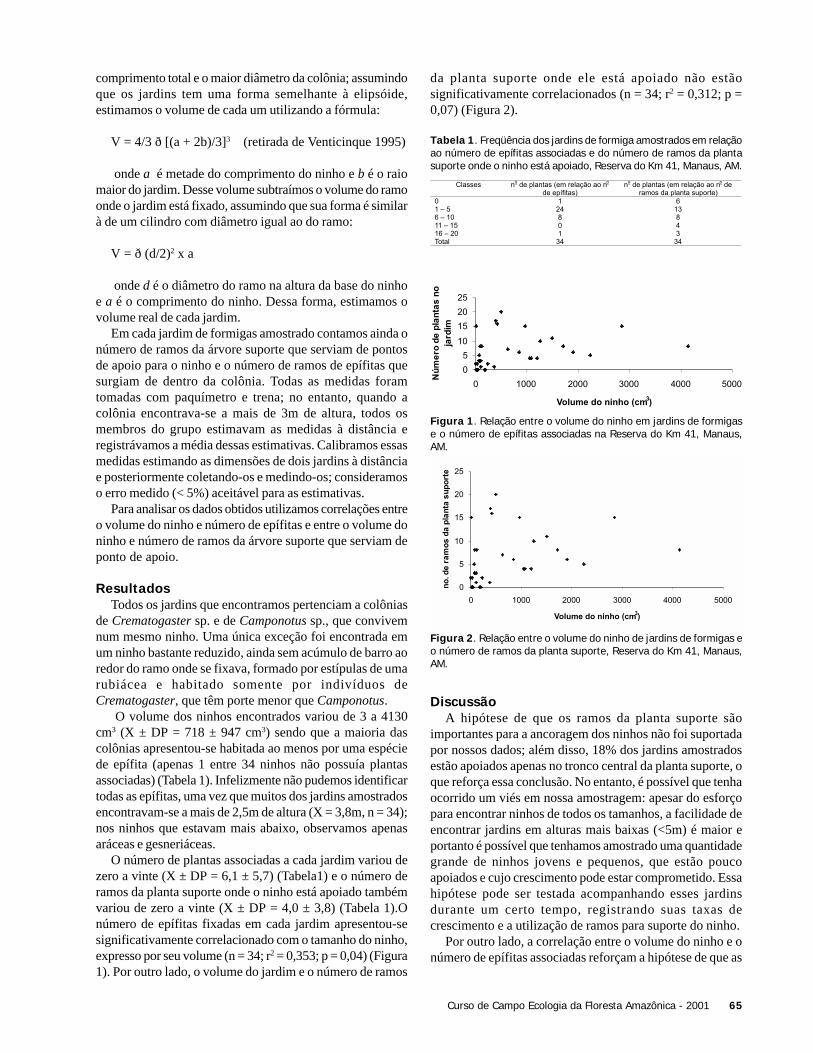
Curso de Campo Ecologia da Floresta Amazônica - 2001 65
comprimento total e o maior diâmetro da colônia; assumindoque os jardins tem uma forma semelhante à elipsóide,estimamos o volume de cada um utilizando a fórmula:
V = 4/3 ð [(a + 2b)/3]3 (retirada de Venticinque 1995)
onde a é metade do comprimento do ninho e b é o raiomaior do jardim. Desse volume subtraímos o volume do ramoonde o jardim está fixado, assumindo que sua forma é similarà de um cilindro com diâmetro igual ao do ramo:
V = ð (d/2)2 x a
onde d é o diâmetro do ramo na altura da base do ninhoe a é o comprimento do ninho. Dessa forma, estimamos ovolume real de cada jardim.
Em cada jardim de formigas amostrado contamos ainda onúmero de ramos da árvore suporte que serviam de pontosde apoio para o ninho e o número de ramos de epífitas quesurgiam de dentro da colônia. Todas as medidas foramtomadas com paquímetro e trena; no entanto, quando acolônia encontrava-se a mais de 3m de altura, todos osmembros do grupo estimavam as medidas à distância eregistrávamos a média dessas estimativas. Calibramos essasmedidas estimando as dimensões de dois jardins à distânciae posteriormente coletando-os e medindo-os; consideramoso erro medido (< 5%) aceitável para as estimativas.
Para analisar os dados obtidos utilizamos correlações entreo volume do ninho e número de epífitas e entre o volume doninho e número de ramos da árvore suporte que serviam deponto de apoio.
ResultadosTodos os jardins que encontramos pertenciam a colônias
de Crematogaster sp. e de Camponotus sp., que convivemnum mesmo ninho. Uma única exceção foi encontrada emum ninho bastante reduzido, ainda sem acúmulo de barro aoredor do ramo onde se fixava, formado por estípulas de umarubiácea e habitado somente por indivíduos deCrematogaster, que têm porte menor que Camponotus.
O volume dos ninhos encontrados variou de 3 a 4130cm3 (X ± DP = 718 ± 947 cm3) sendo que a maioria dascolônias apresentou-se habitada ao menos por uma espéciede epífita (apenas 1 entre 34 ninhos não possuía plantasassociadas) (Tabela 1). Infelizmente não pudemos identificartodas as epífitas, uma vez que muitos dos jardins amostradosencontravam-se a mais de 2,5m de altura (X = 3,8m, n = 34);nos ninhos que estavam mais abaixo, observamos apenasaráceas e gesneriáceas.
O número de plantas associadas a cada jardim variou dezero a vinte (X ± DP = 6,1 ± 5,7) (Tabela1) e o número deramos da planta suporte onde o ninho está apoiado tambémvariou de zero a vinte (X ± DP = 4,0 ± 3,8) (Tabela 1).Onúmero de epífitas fixadas em cada jardim apresentou-sesignificativamente correlacionado com o tamanho do ninho,expresso por seu volume (n = 34; r2 = 0,353; p = 0,04) (Figura1). Por outro lado, o volume do jardim e o número de ramos
da planta suporte onde ele está apoiado não estãosignificativamente correlacionados (n = 34; r2 = 0,312; p =0,07) (Figura 2).
Tabela 1. Freqüência dos jardins de formiga amostrados em relaçãoao número de epífitas associadas e do número de ramos da plantasuporte onde o ninho está apoiado, Reserva do Km 41, Manaus, AM.
Figura 1. Relação entre o volume do ninho em jardins de formigase o número de epífitas associadas na Reserva do Km 41, Manaus,AM.
Figura 2. Relação entre o volume do ninho de jardins de formigas eo número de ramos da planta suporte, Reserva do Km 41, Manaus,AM.
DiscussãoA hipótese de que os ramos da planta suporte são
importantes para a ancoragem dos ninhos não foi suportadapor nossos dados; além disso, 18% dos jardins amostradosestão apoiados apenas no tronco central da planta suporte, oque reforça essa conclusão. No entanto, é possível que tenhaocorrido um viés em nossa amostragem: apesar do esforçopara encontrar ninhos de todos os tamanhos, a facilidade deencontrar jardins em alturas mais baixas (<5m) é maior eportanto é possível que tenhamos amostrado uma quantidadegrande de ninhos jovens e pequenos, que estão poucoapoiados e cujo crescimento pode estar comprometido. Essahipótese pode ser testada acompanhando esses jardinsdurante um certo tempo, registrando suas taxas decrescimento e a utilização de ramos para suporte do ninho.
Por outro lado, a correlação entre o volume do ninho e onúmero de epífitas associadas reforçam a hipótese de que as
Classes no de plantas (em relação ao no
de epífitas)no de plantas (em relação ao no de
ramos da planta suporte)0 1 61 – 5 24 136 – 10 8 811 – 15 0 416 – 20 1 3Total 34 34
0
5
10
15
20
25
0 1000 2000 3000 4000 5000
Volume do ninho (cm3)
Nú
me
rod
ep
lan
tas
no
jard
im
0
5
10
15
20
25
0 1000 2000 3000 4000 5000
Volume do ninho (cm3)
no
.de
ram
os
da
pla
nta
su
po
rte

66 Curso de Campo Ecologia da Floresta Amazônica - 2001
epífitas são realmente importantes para a ancoragem dojardim, conforme já havia sido proposto por alguns autores(Höldobbler & Wilson 1990; Kleinfeldt 1978). Porém, nãoé possível afirmar que essa é uma condição sine qua nonpara o crescimento dos ninhos, ou seja, não se podeestabelecer relações de causa e efeito. É possível que agerminação das plantas favoreça o crescimento do ninho,mas esse aumento no volume por sua vez pode favorecer afixação de eventuais sementes de epífitas trazidas pelo vento.Em outras palavras, os dois parâmetros interagem, tornandodifícil isolar o efeito de cada um deles. Para isso serianecessários que acompanhássemos o desenvolvimento dosjardins e manipulássemos alguns deles, não permitindo queas epífitas se desenvolvessem e observando os efeitos queisso causaria nas colônias.
AgradecimentosAgradecemos a Glauco Machado pelo auxílio no campo.
Referências BibliográficasHöldobbler, B. & Wilson, E.O. 1990. The Ants. Springer-
Verlag, Berlin.Kleinfeldt, S.E. 1978. Antgardens: the interaction of
Codonanthe crassifolia (Gesneriaceae) andCrematogaster longispina (Formicidae). Ecology 59:449-456.
Pires, J.M. & Prance, G.T. 1985. The vegetation types of theBrazilian Amazon. In: Amazonia, Prance, G. & Lovejoy,T.E. (eds.), pp 109-145. Pergamon, New York.
Santamaria, M.; Gomes, F.P.; Dutra, G.F.; Montenegro, S.& Lima, M.G. 1997. Variación interespecifica en ladefensa anti-herbivoros en Cecropia spp. Livro do Cursode Campo “Ecologia da Floresta Amazônica”. INPA/PDBFF, Manaus, AM.
Santos, A.J. 1999. Defesa contra herbivoria por formigasem Philodendron megalophyllum (Araceae), uma epífitade jardins de formiga. Livro do Curso de Campo“Ecologia da Floresta Amazônica”. INPA/PDBFF,Manaus, AM.
Vasconcelos, H.L. 1991. Mutualism between Maietaguianensis Aubl., a myrmecophytic melastome, and oneof its inhabitants: ant protection against insectherbivores. Oecologia 87: 295-298.
Venticinque, E.M. 1995. Dinâmica populacional deAnelosimus eximius (Simom, 1891)(Araneae:Theridiidae) em mosaicos ambientais na AmazôniaCentral. Dissertação de Mestrado, UNESP, Botucatu.
OrientadorSelvino N. Oliveira
Um teste da hipótese de disponibilidade de recursos em clareiras daReserva do Km 41
Christina McCain, Edgar Pietro, Felipe B. Oliveira, Maria R. Darrigo & Natalie Olifiers
IntroduçãoA herbivoria é um processo ecológico de grande
importância. A perda de área foliar implica em menorquantidade de recursos energéticos para o crescimento ereprodução das plantas atacadas, reduzindo assim seu sucessoreprodutivo (Braker & Chazdon 1993). Por esse motivo, osvegetais desenvolveram inúmeras estratégias para proteger-se dos herbívoros, ao longo de sua história evolutiva. Entreos diversos tipos de defesa desenvolvidas pelas plantas,destaca-se a produção de compostos secundários impalatáveispara os herbívoros, uma estratégia amplamente difundidaentre vários grupos vegetais (Coley & Kursar 1996). Noentanto, a produção desses compostos pode envolver custosmetabólicos e, em algumas situações, sua produção não évantajosa para a planta (Coley & Kursar 1996).
Coley & Kursar (1996) sugerem que plantas que vivemem ambientes muito ricos em nutrientes, onde a produção defolhas é relativamente menos custosa, o investimento emdefesas químicas é menor e as plantas tendem a ser maisatacadas por herbívoros. Por outro lado, se o ambiente é maispobre, o custo das folhas é mais alto e a longevidade foliartende a ser maior. Nesses ambientes, é muito difícil recuperar
a perdida de material foliar e o investimento em defesasquímicas torne-se mais vantajoso. Segundo esse hipótese, ograu de herbivoria em tais plantas tende a ser menor (Coley& Kursar 1996). Essa hipótese recebeu o nome de hipóteseda disponibilidade de recursos.
Nas florestas tropicais, a luz é um recurso escasso para asplantas do sub-bosque. Nesse estrato crescem plantasadaptadas a essas condições, geralmente de crescimento lento,taxa de troca foliar baixa e custo de produção de folhas alto.No entanto, quando uma clareira se abre na floresta e a luzpassa a penetrar de maneira mais direta, plantas classificadascomo pioneiras começam a crescer (Hutchinson 1961). Essasplantas apresentam crescimento rápido, alta taxa de trocafoliar e pequeno custo de produção de folhas (Coley & Kursar1996). Em clareiras relativamente antigas, é possível observarplantas pertencentes a essas duas categorias ocorrendo nomesmo local.
Este trabalho tem como objetivo testar a hipótese dadisponibilidade de recursos em clareiras de uma floresta naAmazônia Central, averiguando se o grau de herbivoria émaior em plantas que não experimentam restrição de luz emrelação a plantas com maiores restrições.

Curso de Campo Ecologia da Floresta Amazônica - 2001 67
Material e MétodosRealizamos o presente estudo na Reserva do Km 41 do
Projeto Dinâmica Biológica de Fragmentos Florestais, naFazenda Esteio, Manaus, AM (2°24’S, 59°44’W). Nessaárea predomina a floresta de terra firme na Amazônia Central.
Em clareiras naturais escolhemos dez espécies (Tabela 1),sendo que cinco delas ocorrem apenas em clareiras (espéciespioneiras) e as outras cinco ocorrem na região sombreada dosub-bosque. Amostrando todas as espécies em clareirasevitamos que a presença diferencial ou agregada de herbívorosinfluenciasse os resultados.
Tabela 1: Categorias das espécies utilizados em clareiras.
Para cada espécie, coletamos entre 70 a 150 folhas depelo menos três indivíduos diferentes. Utilizando uma tabelade números aleatórios, sorteamos 50 folhas dentre todas ascoletadas para estimar o grau de herbivoria de cada espécie.Para cada espécie, um mesmo observador classificou cadafolha em uma das seguintes categorias, de acordo com a áreafoliar consumida por herbívoros (Tabela 2).
Tabela2 : Categorias de dano foliar e a porcentagem de áreaconsumida associada a cada uma delas.
Essas categorias foram delimitadas segundo o padrão deherbivoria observado na natureza, onde nota-se grandequantidade de folhas com pouca herbivoria e poucas muitoconsumidas. Utilizando intervalos menores nas classes ondea porcentagem de herbivoria é baixa, pode-se refinar a análise(R. Dirzo com. pess.). Em seguida, calculamos o índice deherbivoria (IH) para cada espécie, utilizando a seguintefórmula:
IH = [Ó (ni x i)] / n
onde ni é o número de folhas em cada categoria, i é o valor
da categoria (de 0 a 5) e n é o número de folhas analisadas (n= 50, no caso deste estudo). Como os valores de índice deherbivoria não seguem uma distribuição normal, utilizamosum teste não-paramétrico Mann-Whitney (Zar 1984) paraanalisar esses dados.
ResultadosOs índices de herbivoria (IH) obtidos para as espécies de
Crescimento Rápido Crescimento Lento
Piper sp. Terminalia sp.
Miconia sp. Combretaceae
Vismia sp. Boraginaceae
Solanum sp. Acariquara sp.
Burseraceae Icacinaceae
Categoria (i) Porcentagem de área consumida0 01 1 – 62 6 – 123 12 – 254 25 – 505 50 –100
crescimento lento foram significativamente diferentes dasespécies de crescimento rápido (U = 25; g.l. = 1; p = 0,009;n = 10) (Figura 1).
Figura 1: Índices de herbivoria das espécies de crescimento lento erápido. Estão representados a mediana, os quartis, o intervalo deconfiança (95%) e o ‘outlier’.
Por outro lado, se observarmos as freqüências de folhasem cada categoria de herbivoria (Tabela 3), notaremos quenas categorias mais baixas (0 e 1), é significativamente maiora presença de plantas de crescimento lento, enquanto que asplantas mais rápidas são bem mais abundantes nas categorias3, 4 e 5.
Tabela 3: Freqüências observadas e esperadas (entre parênteses)de cada categoria de herbivoria para espécies de crescimento rápidoe lento (?2 = 291,5, p < 0,001, cat. 4&5 combinadas).
DiscussãoOs dados apoiam a hipótese inicial segundo a qual as
plantas que crescem rapidamente, isto é, sem a limitação daquantidade de luz, apresentariam índices de herbivoriasignificativamente maiores que as que crescem maislentamente. É improvável que o efeito observado deva-se àabundância maior de herbívoros onde coletamos as espéciesde crescimento rápido, uma vez que as duas categorias deplantas foram coletadas nas mesmas clareiras.
As freqüências observadas em cada categoria de herbivoriaesclarecem ainda mais as diferenças entre as duas categoriasde plantas (Tabela 1). As plantas lentas apresentaram umafreqüência muito maior que a esperada de folhas enquadradasna categoria 1, ou seja, com 1% a 6% de herbivoria. Por
IH Rápidas Lentas Total
0 4 (36.4) 70 (37.6) 74
1 25 (80.6) 139 (83.4) 164
2 29(31.9) 36 (33.1) 65
3 82 (46.2) 12(47.8) 94
4 76 (39.3) 4 (40.7) 80
5 36(17.7) 0(18.3) 36
Total 252 261 513

68 Curso de Campo Ecologia da Floresta Amazônica - 2001
outro lado, as espécies rápidas possuíam muitas folhas nascategorias de maior área foliar consumida (3, 4 e 5). Istopode significar que as plantas de crescimento lento são emmuitos casos “provadas” pelos herbívoros, que a abandonamem seguida devido à presença de substâncias impalatáveisnessas folhas. Em contraste, quando esses animais provamplantas de crescimento rápido, permanecem se alimentandodestas, destruindo uma quantidade significativa da área dafolha, pois não encontram impedimentos grandes ao consumi-la. Essa idéia fica ainda mais reforçada pelo fato deencontrarmos apenas quatro folhas intactas entre as 252analisadas para espécies de crescimento rápido.
Considerando os dados obtidos, pensamos que a hipóteseda disponibilidade de recursos, conforme proposta por Coley& Kursar (1996), pode explicar a herbivoria diferencialobservada entre as plantas de crescimento rápido e lento queestudamos.
Referências BibliográficasBraker, E. & Chazdon, R.L. 1993. Ecological, behavioural
and nutritional factors influencing use of palms as hostplants by a Neotropical forest grasshopper. J. Trop. Ecol.9: 183-197.
Coley, P.D. & Kursar, T.A. 1996. Anti-herbivore defenses ofyoung tropical leaves: physiological constraints andecological trade-offs. In: Tropical Forest PlantEcophysiology, Hulley, S.S., Charson, R.L. & Smith,A.P., pp 305-336. Chapman & Hall, New York.
Hutchinson, G.E. 1961. The paradox of the plankton. Am.Nat. 95: 137-145.
Zar, J.H. 1984. Biostatistical Analisys. Prentice Hall, NewJersey.
OrientadorRodolpho Dirzo
Defesa química de duas espécies de hilídeos (Anura) contra um peixe predadorCarlos E. Marinelli, Betsabé R. Guerra, Cristiane F. Jurinitz, Daniel L. M. Vieira &Rodrigo Cogni
IntroduçãoPeixes predadores exercem um papel fundamental na
estruturação das comunidades de água doce (Hero et al. 1998apud Hero et al. 2001), influenciando a composição temporale espacial de comunidades de girinos (Heyer et al. 1975 apudHero et al. 2001) e conseqüentemente determinando acomunidade de anuros (Gascon 1992). Tamanho,palatabilidade e uso de microhabitats como refúgio sãopossíveis mecanismos que permitem que girinos convivamcom peixes (Hero et al. 1998 apud Hero et al. 2001). Espéciesde girinos mostram diferentes intensidades de defesa química,permitindo às mais tóxicas habitarem um corpo d’águapovoado por peixes.
Hyla minuta (Hylidae) é uma espécie “r” estrategista, temfácil dispersabilidade, produz uma grande quantidade deprole e ocorre em ambientes degradados; em contrapartida,Phyllomedusa tarsius, “k” estrategista, é uma espécie maisexigente, ocorrendo preferencialmente no interior da floresta,apesar de ser encontrada, em baixa freqüência, em ambientespouco alterados (S. N. Oliveira com. pess.). Portanto, éesperado que H. minuta invista menos em defesa química doque P. tarsius, e que essa seja uma das causas para a ausênciada primeira em ambientes preservados onde ocorrem peixespredadores.
Testamos experimentalmente se girinos de H. minuta sãomais predados por peixes que girinos de P. tarsius.
Material e MétodosÁrea de estudoRealizamos este estudo na reserva de mata contínua do
PDBFF, situada no Km 41 da estrada ZF 3, a 80 Km aonorte de Manaus (2° 25’S; 59° 50’W). O clima da região
caracteriza-se por duas estações bem definidas, uma seca(julho a novembro), e uma chuvosa (dezembro a junho). Apluviosidade anual varia de 1900 a 2300 mm, comtemperatura média anual de 26° C (Toppa 1998).
Desenho experimentalRealizamos dois experimentos. (1) Sem escolha de presa
- O tratamento T1 consistiu em colocar em bacias um peixepredador e 12 girinos de H. minuta de tamanhos variados.T2 seguiu os mesmos moldes substituindo-se a presa por P.tarsius. Para cada um dos tratamentos foram feitas trêsréplicas. O experimento teve a duração de sete horas. (2) Emcada bacia foram colocados três indivíduos de cada espéciede presa com um predador, para verificar a escolha da presa.Nesse experimento foram feitas cinco réplicas, tendo aduração de quatro horas.
Coletamos peixes da espécie Hemigrammus sp.(Characidae) que foram utilizados como predadores. Ospeixes foram coletados por meio de bloqueio e cerco comredes em um igarapé no interior da mata. As presas, girinosde Hyla minuta e Phyllomedusa tarsius, foram coletadascom peneira e rapiché em uma poça ao lado da estrada deacesso à reserva. Utilizamos bacias de mesmo tamanho nasquais foram colocados areia (cinco punhados) e folhiço (atécobrir cerca de um terço da bacia), para tentarmos reproduzirda melhor forma possível o ambiente natural das espécies.
Os girinos foram considerados predados quando eramcompletamente consumidos (desaparecidos) ou quandoestavam mortos apresentando mutilações.
ResultadosNo experimento sem escolha de presa, após sete horas os

Curso de Campo Ecologia da Floresta Amazônica - 2001 69
peixes predaram apenas oito girinos de H. minuta dos 36colocados inicialmente nas bacias. P. tarsius teve apenas doisgirinos predados. Não houve diferença na predação das duasespécies de girinos (Mann-Whitney, U = 3; p = 0,49; n = 3).Em três bacias não houve predação (Tabela 1).
Tabela 1. Número de girinos de Hyla minuta e Phyllomedusa tarsiussobreviventes dentre os 12 colocados em cada tratamento no iníciodo 1o experimento (sem escolha), realizado na Reserva do km 41(AM).
No segundo experimento, quando as duas espécies degirinos foram dispostas na mesma bacia, apenas um indivíduode H. minuta foi predado. Novamente não houve diferençana intensidade de predação entre as duas espécies de girinos(Wilcoxon, Z = 1; p = 0,32; n = 5). Os girinos de P. tarsiustiveram apenas a ponta da cauda predada, enquanto os de H.minuta foram inteiramente consumidos.
DiscussãoAlgumas hipóteses podem explicar as intensidades de
predação semelhantes e baixas para as duas espécies. A defesaquímica pode ser eficiente para ambas, mas o fato de que osindivíduos de P. tarsius tenham tido apenas a ponta da caudaconsumida indica um menor grau de toxicidade. Já que ospeixes predadores utilizados no experimento habitamigarapés, eles não teriam memória genética para evitar P.tarsius e, portanto, a predação de apenas a ponta da cauda(que ocasionou a morte dos indivíduos) seria uma evidênciada falta de contato entre populações naturais da presa e dopredador.
Outra explicação para a baixa intensidade de predçãoobservada é o fato de os peixes terem ficado apenas umahora em jejum antes do início do experiemento, bem como o
H. minuta (T1) P. tarsius (T2)
Bacia 1 2 3 4 5 6
N. de sobreviventes 5 11 12 12 12 10
Teste Mann-Whitney; n= 3; U= 3; p= 0,49.
curto tempo de duração do experimento (sete horas). Doisestudos experimentais abordando predação em espécies degirinos isolaram as espécies de predadores em jejum por 12horas e os experimentos duraram pelo menos 24 horas (Heroet al. 2001; Oliveira 1996), obtendo-se assim maiorintensidade de predação. Além disso, o fato da espécie depeixe utilizada no experimento habitar igarapés e, portantonão ser predadora natural desses girinos de poças, pode tê-la levado a investir em presas tóxicas pela sua inexperiência.Por último, os girinos podem ter usado o folhiço comorefúgio, evitando a predação pelos peixes. Experimentosacumulam naturalmente uma variedade de falhas ao tentarsimular ambientes naturais, mas são essenciais paraentendermos processos ecológicos.
Referências BibliográficasGascon, C. 1990. The relative importance of habitat
characteristics in the maintenance of a larval anuranspecies assemblage in the tropics. Ph.D. Thesis, FloridaState University, Tallahassee.
Hero, J.- M.; Magnusson, W.E. & Rocha, C.F.D. 2001.Antipredator defenses influence the distribution ofamphibian prey species in the central amazon rain forest.Biotropica 33(1): 131-141.
Oliveira, S.N. 1996. Efeitos diretos e indiretos da predaçãosobre três espécies de girinos (Amphibia-Anura) naAmazônia Central. Dissertação de Mestrado, INPA/UFAM. Manaus.
Toppa, R.H. 1998. Efeitos da predação em função do tamanhodo corpo, uma interação interespecífica (Osteocephalustaurinus, Hylidae versus náiades de Odonata,Libellulidae). Livro do Curso de Campo “Ecologia daFloresta Amazônica”. INPA/PDBFF, Manaus, AM.
OrientadorSelvino N. Oliveira
Seleção intra e inter individual para sítios de oviposição em Maieta guianensis(Melastomataceae) pela aranha Argyrodes sp. (Araneaea: Theridiidae)
Rodrigo Cogni, Daniel L. M. Vieira, Carlos E. Marinelli, Cristiane F. Jurinitz & Betsabé R. Guerra
IntroduçãoO gênero Argyrodes (Theridiidae) é conhecido como
cleptoparasita (Wise 1993) já que suas espécies habitam asteias de outras espécies de aranhas capturando pequenaspresas que não tenham sido atacadas pela hospedeira ouroubando aquelas que já tenham sido capturadas (Fáveri etal. 1998). Na Amazônia Central ocorre uma espécie dessegênero associada a indivíduos de Maieta guianensis(Melastomataceae), um arbusto pequeno do sub-bosque daFloresta Amazônica que apresenta uma associação com asformigas Pheidole minutula e Crematogaster sp.(Vasconcelos 1993). Essa interação mirmecófita tem sido
bastante estudada (Vasconcelos 1993; Fonseca 1995; Gortari1998; Paré 1998), porém, a possível interação de Argyrodessp. com M. guianensis e as formigas tem recebido poucaatenção, sendo que poucos foram os estudos que avaliaramessa situação (Fáveri 1998; Oliveira 1995).
A seleção de sítios de oviposição é uma das maisimportantes decisões das fêmeas. Para as espécies comcuidado parental, os sítios selecionados deverão apresentarcondições físicas apropriadas para o desenvolvimento dosovos, assim como para os pais (Morse 1985; Resetarits 1996).Um local inadequado pode afetar tanto a prole como os pais,devido a fatores desfavoráveis como desidratação, predação,
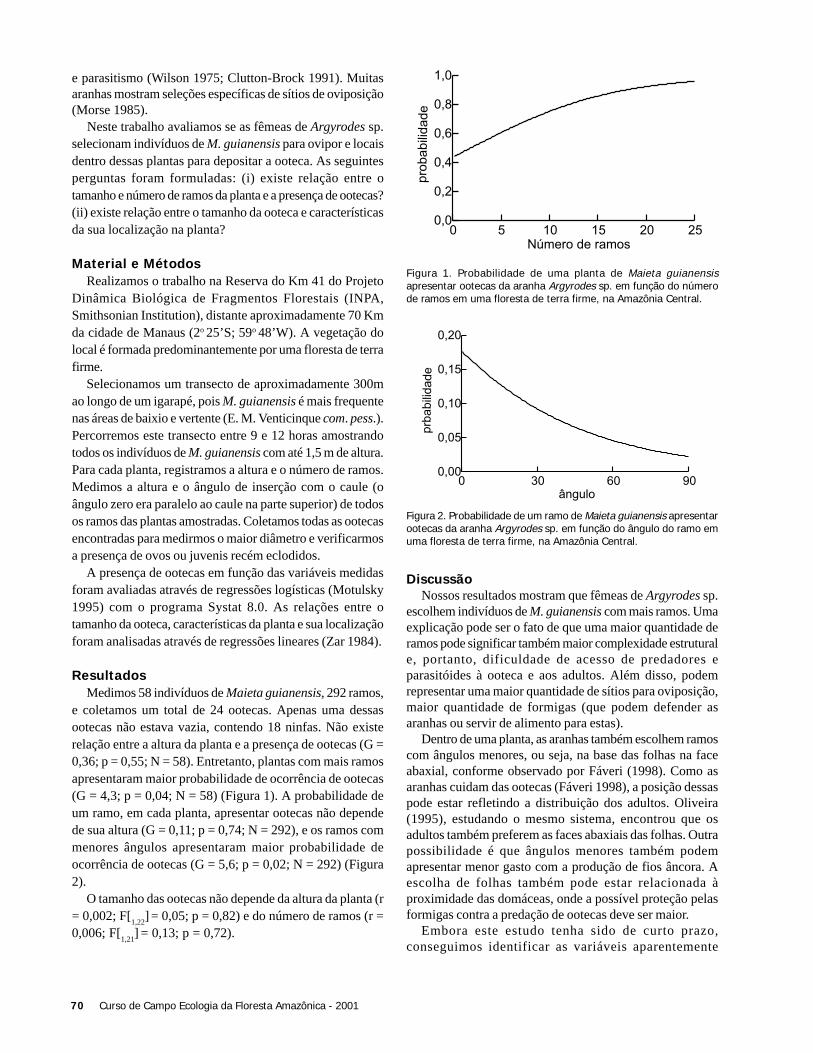
70 Curso de Campo Ecologia da Floresta Amazônica - 2001
e parasitismo (Wilson 1975; Clutton-Brock 1991). Muitasaranhas mostram seleções específicas de sítios de oviposição(Morse 1985).
Neste trabalho avaliamos se as fêmeas de Argyrodes sp.selecionam indivíduos de M. guianensis para ovipor e locaisdentro dessas plantas para depositar a ooteca. As seguintesperguntas foram formuladas: (i) existe relação entre otamanho e número de ramos da planta e a presença de ootecas?(ii) existe relação entre o tamanho da ooteca e característicasda sua localização na planta?
Material e MétodosRealizamos o trabalho na Reserva do Km 41 do Projeto
Dinâmica Biológica de Fragmentos Florestais (INPA,Smithsonian Institution), distante aproximadamente 70 Kmda cidade de Manaus (2o 25’S; 59o 48’W). A vegetação dolocal é formada predominantemente por uma floresta de terrafirme.
Selecionamos um transecto de aproximadamente 300mao longo de um igarapé, pois M. guianensis é mais frequentenas áreas de baixio e vertente (E. M. Venticinque com. pess.).Percorremos este transecto entre 9 e 12 horas amostrandotodos os indivíduos de M. guianensis com até 1,5 m de altura.Para cada planta, registramos a altura e o número de ramos.Medimos a altura e o ângulo de inserção com o caule (oângulo zero era paralelo ao caule na parte superior) de todosos ramos das plantas amostradas. Coletamos todas as ootecasencontradas para medirmos o maior diâmetro e verificarmosa presença de ovos ou juvenis recém eclodidos.
A presença de ootecas em função das variáveis medidasforam avaliadas através de regressões logísticas (Motulsky1995) com o programa Systat 8.0. As relações entre otamanho da ooteca, características da planta e sua localizaçãoforam analisadas através de regressões lineares (Zar 1984).
ResultadosMedimos 58 indivíduos de Maieta guianensis, 292 ramos,
e coletamos um total de 24 ootecas. Apenas uma dessasootecas não estava vazia, contendo 18 ninfas. Não existerelação entre a altura da planta e a presença de ootecas (G =0,36; p = 0,55; N = 58). Entretanto, plantas com mais ramosapresentaram maior probabilidade de ocorrência de ootecas(G = 4,3; p = 0,04; N = 58) (Figura 1). A probabilidade deum ramo, em cada planta, apresentar ootecas não dependede sua altura (G = 0,11; p = 0,74; N = 292), e os ramos commenores ângulos apresentaram maior probabilidade deocorrência de ootecas (G = 5,6; p = 0,02; N = 292) (Figura2).
O tamanho das ootecas não depende da altura da planta (r= 0,002; F[
1,22]
= 0,05; p = 0,82) e do número de ramos (r =
0,006; F[1,21
] = 0,13; p = 0,72).
0 5 10 15 20 25Número de ramos
0,0
0,2
0,4
0,6
0,8
1,0
pro
ba
bili
da
de
Figura 1. Probabilidade de uma planta de Maieta guianensisapresentar ootecas da aranha Argyrodes sp. em função do númerode ramos em uma floresta de terra firme, na Amazônia Central.
Figura 2. Probabilidade de um ramo de Maieta guianensis apresentarootecas da aranha Argyrodes sp. em função do ângulo do ramo emuma floresta de terra firme, na Amazônia Central.
DiscussãoNossos resultados mostram que fêmeas de Argyrodes sp.
escolhem indivíduos de M. guianensis com mais ramos. Umaexplicação pode ser o fato de que uma maior quantidade deramos pode significar também maior complexidade estruturale, portanto, dificuldade de acesso de predadores eparasitóides à ooteca e aos adultos. Além disso, podemrepresentar uma maior quantidade de sítios para oviposição,maior quantidade de formigas (que podem defender asaranhas ou servir de alimento para estas).
Dentro de uma planta, as aranhas também escolhem ramoscom ângulos menores, ou seja, na base das folhas na faceabaxial, conforme observado por Fáveri (1998). Como asaranhas cuidam das ootecas (Fáveri 1998), a posição dessaspode estar refletindo a distribuição dos adultos. Oliveira(1995), estudando o mesmo sistema, encontrou que osadultos também preferem as faces abaxiais das folhas. Outrapossibilidade é que ângulos menores também podemapresentar menor gasto com a produção de fios âncora. Aescolha de folhas também pode estar relacionada àproximidade das domáceas, onde a possível proteção pelasformigas contra a predação de ootecas deve ser maior.
Embora este estudo tenha sido de curto prazo,conseguimos identificar as variáveis aparentemente
0 30 60 90ângulo
0,00
0,05
0,10
0,15
0,20
prb
ab
ilid
ad
e

Curso de Campo Ecologia da Floresta Amazônica - 2001 71
importantes na determinação da escolha de locais para afixação de ootecas de Argyrodes sp. em M. guianensis. Istopode indicar que o padrão de escolha dos sítios é forte. Maspara um melhor entendimento das razões para escolha dossítios de oviposição por Argyrodes sp. precisam-se estudosmais detalhados que demonstrem quais as relações entre asaranhas e as formigas. As seguintes questões poderiam serestudadas: (i) as formigas protegem as aranhas e suas ootecas?(ii) as aranhas predam as formigas? (iii) as formigas predamas aranhas?
Referências BibliográficasClutton-Brock, T.H. 1991. The Evolution of Parental Care.
Princeton University Press, Princeton.Craig, C.L. 1989. Alternative foraging modes of orb web
weaving spiders. Biotropica 21(3): 257 - 264Fáveri, B.S. 1998. Comportamento de Argirodes sp. (
Araneae: Theridiidae) em Maieta guianensis(Melastomataceae). Livro do Curso de Campo “Ecologiada Floresta Amazônica”. INPA/PDBFF, Manaus, AM.
Gortari, E.V. 1998. Sistema defensivo en algumas plantasmirmecófitas de Amazonia Central. Livro do Curso deCampo “Ecologia da Floresta Amazônica”. INPA/PDBFF, Manaus, AM.
Morse, D.H. 1985: Nests and nest-site selection of the crab
spider Misumena vatia (Araneae, Thomisidae) onmilkweed. J. Arachnol. 13: 383-390.
Motulsky, H. 1995. Intuitive Biostatistics. Oxford UniversityPress, New York.
Oliveira, S.N., 1995. Predação e seleção de microhabitat porArgyrodes sp. (Theridiidae) em Maieta guianensis(Melastomataceae) na Amazônia Central. Livro do Cursode Campo “Ecologia da Floresta Amazônica”. INPA/PDBFF, Manaus, AM.
Paré, B.K. 1998. Defensas inducidas en el sistemamirmecófilo Maieta guianensis – Pheidole minutula.Livro do Curso de Campo “Ecologia da FlorestaAmazônica”. INPA/PDBFF, Manaus, AM.
Vasconcelos, H.L. 1993. Ant colonization of Mateiaguianensis seedlings, an Amazonian ant–plant.Oecologia 95: 439 - 443
Resetarits, W.J. 1996: Oviposition site choice and life historyevolution. Amer. Zool. 36: 205-215.
Wise, D.H. 1993. Spiders in Ecological Webs. CambridgeUniversity Press, New York.
Zar, J.H. 1984. Biostatistical Analysis. Prentice Hall, NewJersey.
OrientadorEduardo M. Venticinque
Efeito do ápice foliar e da epifilia no tempo de dessecamento de folhas dosubosque de uma floresta de terra firme
Paula S. Pinheiro, Paula C. Ceotto, Angela M. F. Pacheco, Leandro M. Scoss & Maurício Camargo
IntroduçãoAs adaptações morfológicas que plantas de florestas
tropicais têm desenvolvidas são muito variadas, dependendodos fatores que limitam seu estabelecimento e sobrevivência(Tilman 1988). Um desses fatores é a limitação de luz solar,que diminui consideravelmente do dossel em direçãosubosque. Por esta razão que as plantas do subosque têmque aproveitar ao máximo a luz disponível, geralmente como desenvolvimento de folhas maiores para otimizar asuperfície fotossintética (Richards 1952 apud Boege et al.1998). Por outro lado, as plantas tem que diminuir o excessode umidade provocado pelas altas precipitaçõespluviométricas. Este acúmulo de água pode promover oestabelecimento de epífilas ou o ataque por patógenos, quereduzem o potencial fotossintético das folhas (Richards 1996apud Ivey & De Silva 2001).
Folhas com ápice acuminado são comuns em plantas dosubosque de florestas tropicais (Richards 1996 apud Ivey &De Silva 2001). A hipótese mais aceita para uma possívelfunção adaptativa do ápice acuminado é que este propicia orápido escoamento da água das folhas, reduzindo o tempode secagem da superfície das mesmas (Lightbody 1985 apud
Ivey & De Silva 2001). As plantas com folhas de ápiceacuminado aparentemente ocorrem com frequência maior nosubosque que no dossel, o que pode estar relacionado com ograu de exposição à luz das mesmas (Boege et al. 1988).
O objetivo deste trabalho é testar experimentalmente se otempo de secagem de folhas com e sem ápice acuminado,cobertas ou não por epífilas varia. Partimos do pressupostoque folhas com ápice acuminado e sem epífilas secam maisrápido do que folhas sem ápice acuminado e sem epífilas.
Material e MétodosO presente estudo foi realizado em uma área de platô
coberta por floresta densa de terra firme (Pires & Prance1985), na Reserva do Km 41, do PDBFF/INPA, localizadaaproximadamente a 80 km ao norte de Manaus (2o 24’S; 59o
52’W).Realizamos um experimento com duas espécies do
subosque, uma espécie de palmeira (Astrocaryum) e umaespécie de dicotiledônia (Burseraceae), ambas com ápiceacuminado. Para cada espécie foram coletadas dez folhasque apresentavam epífilas no limbo foliar. Na mesma folha,removemos as epífilas de um lado e não removemos no outro,

72 Curso de Campo Ecologia da Floresta Amazônica - 2001
resultando em dados pareados. As folhas foram dispostasem um varal lado à lado, na sombra, num ângulo de 30o emolhadas com um borrifador. Marcamos o tempo que cadafolha levou para secar com cronômetro. A secagem foiavaliada visualmente, considerando secas as folhas com cercade 80% da superfície sem gotículas de água. Os tempos desecagem de cada tratamento, dez réplicas para cada planta,foram comparados com teste não paramétrico de Wilcoxon(Zar 1984).
Repetimos o processo nas mesmas folhas após cortar oápice do limbo, formando os seguintes tratamentos para cadauma das espécies: (A) com ápice acuminado e com epífilum;(B) com ápice acuminado e sem epífilum; (C) sem ápiceacuminado e com epífilum; (D) sem ápice acuminado e semepífilum.
ResultadosO tempo de secagem das folhas de Astrocaryum foi maior
nos tratamentos sem ápice acuminado que no tratamento comápice acuminado (z = 3,11, N= 13, p < 0,01). O tempo desecagem foi maior para o tratamento sem epifílum, tanto parafolhas com ápice acuminado (z = -3,19, n = 13, p < 0,01),quanto para folhas sem ápice acuminado (z = -3,19, N = 13,p < 0,01) (Tabela 1).
Para Burseraceae sp., o tempo de secagem foi maior nostratamentos com epífilum (z = -2,81, n = 10, p < 0,01), quenos tratamentos sem epífilum (z = -3,19, n = 13, p < 0,01).Existe uma diferença significativa entre os tratamentos come sem ápice acuminado (z = -2,81, n = 10, p < 0,05).Entretanto, o efeito (tempo de secagem das folhas com ápiceacuminado menos o tempo das folhas sem ápice acuminado)foi contrário ao observado para a espécies de palmeira, ouseja, folhas sem epífilum e sem o ápice acuminado, secarammais rápido que a folhas com o ápice acuminado (Tabela 1)
Tabela 1: Médias e desvios padrão do tempo de dessecamento(minutos) em tratamentos com ou sem acúmen, com e sem epífilum,para as espécies Astrocarium sp. e Burseraceae sp.
DiscussãoOs nossos resultados para Astrocaryum, reforçam a
hipótese de que a epifilia aumenta o tempo de retenção deágua sobre as folhas de plantas do subosque, mesmo emfolhas que têm ápice acuminado. O efeito da presença deepífilas na superfície das folhas é ainda pouco conhecido, enão se sabe o quanto a presença destas pode afetar afotossíntese ou a fixação de patógenos. Entretanto, C.
Com ápice acuminado Sem ápice acuminado
Com epífilum (A) Sem epífilum (B) Com epífilum (C) Sem epífilum (D)
Astrocaryum45,8 ± 5,3 17,0 ± 3,9 59,1 ± 6,1 29,4 ± 8,2
Burseraceae sp.57,6 ± 6,5 26,6 ± 9,7 76,03 ± 16,5 14,3 ± 9,6
Zartman (com. pess.) indica que a epifilia pode reternutrientes sobre o limbo foliar e que a associação de briófitascom as algas cianofícias, pode resultar em um efeito benéficopara as plantas “hospedeiras”.
O efeito do ápice acuminado sobre o tempo de secagemde folhas da espécie de dicotiledônea (Burseraceae), écontrário às observações de Ivey & De Silva (2001), queobservaram que o ápice acuminado é uma adaptaçãomorfológica de plantas do subosque para acelerar oescoamento de água da superfície das folhas. Já para as folhasde Astrocaryum (Arecaceae) os resultados encontrados nopresente estudo, reforçam as observações destes autores. Éprovável que o método de amostragem utilizado no nossoestudo, tenha sido falho e subjetivo para a espécie deBurseraceae. A experimentação foi realizada por váriosobservadores sequencialmente, sendo que os tratamentosdesta espécie (Burseraceae) foram realizados por pessoasdiferentes e em horários diferentes. Também é possível queo método proposto para avaliar se as folhas estavam ou nãosecas (visualmente > que 80%), tenha sido inadequado, jáque aumenta a chance de erros durante a experimentação. Ofato dos tratamentos terem sido realizados ao longo de umdia (manhã e início da tarde), pode ter influenciado o tempode secagem para todos os tratamentos, devido a variação datemperatura e umidade.
Sugerimos que os trabalhos experimentais que pretendamtestar o efeito do ápice acuminado em plantas do subosquesejam realizados sob condições climáticas controladas. Damesma forma, indicamos a necessidade de aplicação de outrosmétodos para a avaliação do tempo de secagem das folhas.
Referências BibliográficasBegon, M.; Harper, J.L. & Townsend, C.R. 1990. Ecology:
Individuals, Populations and Communities. BlackwellScientific Publications, Massachusetts.
Ivey, C.T. & De Silva, N. 2001. A test of the function of driptips. Biotropica 33(1): 188-191.
Pires, J.M. & Prance, G.T. 1985. The vegetation types of theBarzilian Amazon. In Amazônia, Prance, G. & Lovejoy,T. (eds.), pp. 109-145. Pergamon, New York.
Tilman, D. 1988. Plant Strategies and the Dynamics andStructure of Plant Communities. Princeton UniversityPress, New Jersey.
Zar, J.H. 1984. Biostatistical Analysis. Prentice Hall, NewJersey.
OrientadorGeraldo W. Fernandes

Curso de Campo Ecologia da Floresta Amazônica - 2001 73
Reações anti-herbívoros da formiga Pheidole minutula Mayr (Formicidae:Myrmicinae) em uma planta mirmecófita, Maieta guianensis Aubl.
(Melastomataceae).Alexander V. Christianini, Andrea Lamberts, Antônio J. C. Aguiar, Katiane Ferreira & Tiago Timo
IntroduçãoEm todo o mundo são conhecidas cerca de 415 espécies
de plantas que apresentam interações estreitas com formigas,chamadas de plantas mirmecófitas (Benson 1985). Estasplantas normalmente desenvolvem estruturas que funcionamcomo abrigo e suporte para construção de ninhos de formigas,como domáceas. Em alguns casos, as plantas fornecemtambém alimento para colônias destes animais (Hölldobler& Wilson 1990). Somente na região Neotropical sãoconhecidas aproximadamente 250 espécies de plantas queapresentam associação com formigas (Fowler1993).
Associações entre formigas e plantas mirmecófitasparecem ter evoluído em resposta a vantagens adaptativaspara ambos organismos, sob a forma de associaçõesmutualísticas (Hölldobler & Wilson 1990). Plantas fornecemabrigo e alimento às formigas, que por sua vez protegem asplantas da ação de herbívoros. Porém, hipóteses alternativasenvolvendo parasitismo, oportunismo e relações assimétricasentre organismos também têm sido levantadas (Hölldobler& Wilson 1990; Fonseca 1995). Embora haja literaturarelativamente abundante sobre associações de plantasmirmecófitas e formigas (Fonseca 1995 e referênciasincluídas), abordagens experimentais ainda são poucoempregadas (e.g. Vasconcelos 1991).
Maieta guianensis (Melastomataceae) é um arbustoabundante no sub-bosque de florestas de terra firme daAmazônia Central (Fowler 1993). Apresenta domáceas queconsistem em duas câmaras adjacentes localizadas na baseda lâmina foliar. Cada uma das câmaras se abre por umapequena passagem na superfície abaxial da folha, na junçãocom a veia central e a nervura lateral principal (Benson 1985).Pheidole minutula está presente em 95% dos indivíduos deM. guianensis que se associam com formigas. Crematogasterlaevis é a outra espécie associada, mas apresenta colôniasmais instáveis e normalmente associadas a plantas menores(Vasconcelos & Davidson 2000). Enquanto P. minutula estáobrigatoriamente associada com M. guianensis e outrasplantas mirmecófilas, C. laevis pode formar colônias no solo,onde forrageiam (Vasconcelos & Davidson 2000; Fonseca1995).
O objetivo deste trabalho foi analisar se as formigashóspedes de M. guianensis podem agir como defensoraseficazes contra herbívoros, respondendo a herbívorossimulados, danos mecânicos e/ou estímulos químicosrelacionados à perda de tecido foliar.
Material e MétodosEste estudo foi realizado na Reserva do Km 41 (2o 24’S;
59o 44’W) pertencente ao projeto Dinâmica Biológica de
Fragmentos Floestais (INPA/Smithsonian Institution),localizada a aproximadamente 80 Km ao norte de Manaus,na Amazônia Central. A vegetação é composta por florestade terra firme. A temperatura média anual é de 26,7o C, commédia de precipitação anual de 2.186 mm. Março e abril sãoos meses mas chuvosos do ano, enquanto julho e setembrocompreendem aos meses mais secos. O solo predominante édo tipo latossolo amarelo (RadamBrasil 1978).
Plantas de M. guianensis foram escolhidas aleatoriamentepara cada experimento, onde foram determinadas uma folhatratamento e uma folha controle, em ramos distintos e alturasequivalentes. Cupins (Isoptera) foram utilizados comoherbívoros simulados. Foram realizados três tipos deexperimentos com o objetivo de testar a reação anti-herbívoros das formigas associadas. Por sua abundância,apenas interações entre M. guianensis e P. minutula foramconsideradas neste estudo.
O primeiro experimento consistiu em depositar um cupimvivo próximo ao ápice de uma das folhas um cupim vivo,enquanto outra folha em um ramo oposto recebiasimultâneamente um pedaço de palito de fósforo de tamanhosemelhante ao cupim, que funcionou como controle. Osegundo experimento quantificou a resposta das formigas adanos físicos provocados pelo observador, simulando a açãode um herbívoro cortador. Em dada planta, duas folhas emramos apicais opostos foram selecionadas. Uma das folhasteve o ápice cortado com o auxílio de uma tesoura, enquantosimultaneamente outra folha foi pressionada levemente comuma pinça (controle). No terceiro experimento, quantificamosa resposta de formigas a uma eventual indução química,promovida por folhas danificadas. Utilizamos um extratoproveniente de uma folha de M. guianenis macerada em água,como tratamento. A seiva diluída em água foi pinceladapróxima ao ápice da folha selecionada, formando um círculode ca. 1 cm de diâmetro. Água foi aplicada simultaneamentecomo controle em uma folha de um ramo oposto. Cadaexperimento foi replicado de nove a dez vezes, de acordocom a disponibilidade de plantas e restrições de tempo.
Dois parâmetros de resposta foram medidos: a) o tempode detecção do herbívoro/dano, representado pelo primeirocontato entre uma formiga e os tratamentos e b) orecrutamento de indivíduos, dado pelo número de formigasem contato com os tratamentos a intervalos de dois minutos,até o máximo de 10 minutos de observação após o contatoinicial.
Comparações entre tratamentos foram realizadas por meiode teste pareado de Wilcoxon e ANOVA de medidasrepetidas, para o tempo de detecção e recrutamento deformigas, respectivamente.
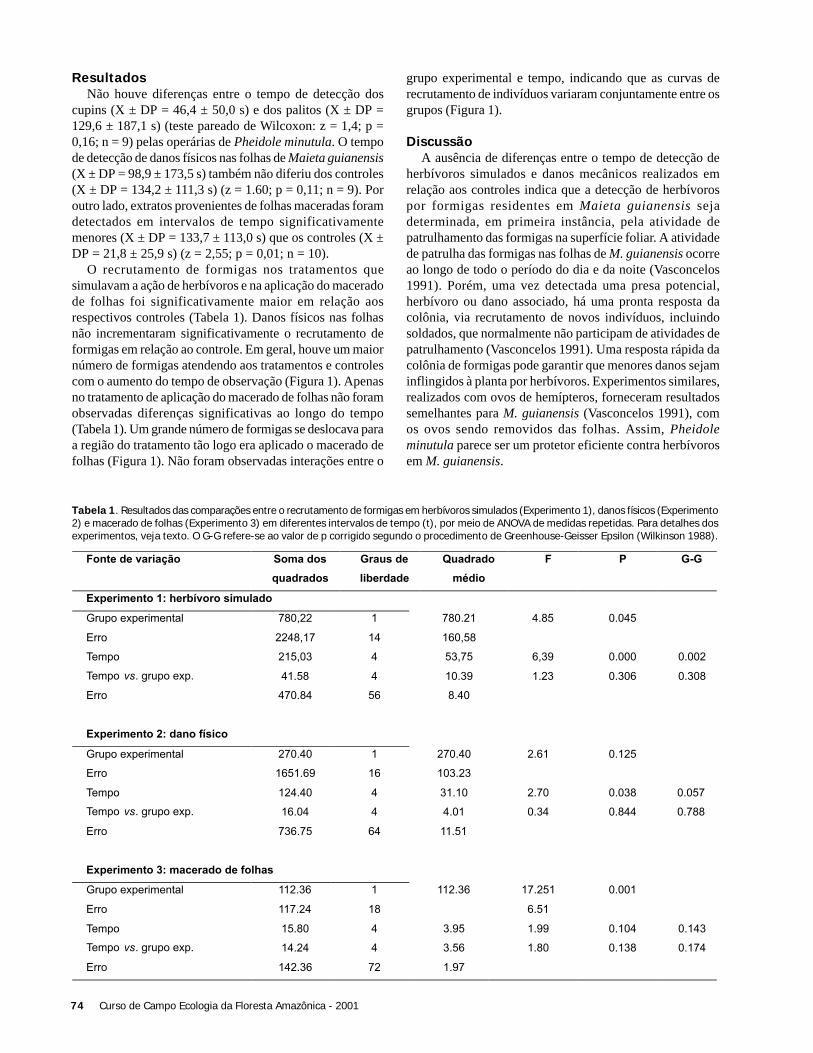
74 Curso de Campo Ecologia da Floresta Amazônica - 2001
ResultadosNão houve diferenças entre o tempo de detecção dos
cupins (X ± DP = 46,4 ± 50,0 s) e dos palitos (X ± DP =129,6 ± 187,1 s) (teste pareado de Wilcoxon: z = 1,4; p =0,16; n = 9) pelas operárias de Pheidole minutula. O tempode detecção de danos físicos nas folhas de Maieta guianensis(X ± DP = 98,9 ± 173,5 s) também não diferiu dos controles(X ± DP = 134,2 ± 111,3 s) (z = 1.60; p = 0,11; n = 9). Poroutro lado, extratos provenientes de folhas maceradas foramdetectados em intervalos de tempo significativamentemenores (X ± DP = 133,7 ± 113,0 s) que os controles (X ±DP = 21,8 ± 25,9 s) (z = 2,55; p = 0,01; n = 10).
O recrutamento de formigas nos tratamentos quesimulavam a ação de herbívoros e na aplicação do maceradode folhas foi significativamente maior em relação aosrespectivos controles (Tabela 1). Danos físicos nas folhasnão incrementaram significativamente o recrutamento deformigas em relação ao controle. Em geral, houve um maiornúmero de formigas atendendo aos tratamentos e controlescom o aumento do tempo de observação (Figura 1). Apenasno tratamento de aplicação do macerado de folhas não foramobservadas diferenças significativas ao longo do tempo(Tabela 1). Um grande número de formigas se deslocava paraa região do tratamento tão logo era aplicado o macerado defolhas (Figura 1). Não foram observadas interações entre o
Fonte de variação Soma dos
quadrados
Graus de
liberdade
Quadrado
médio
F P G-G
Experimento 1: herbívoro simulado
Grupo experimental 780,22 1 780.21 4.85 0.045
Erro 2248,17 14 160,58
Tempo 215,03 4 53,75 6,39 0.000 0.002
Tempo vs. grupo exp. 41.58 4 10.39 1.23 0.306 0.308
Erro 470.84 56 8.40
Experimento 2: dano físico
Grupo experimental 270.40 1 270.40 2.61 0.125
Erro 1651.69 16 103.23
Tempo 124.40 4 31.10 2.70 0.038 0.057
Tempo vs. grupo exp. 16.04 4 4.01 0.34 0.844 0.788
Erro 736.75 64 11.51
Experimento 3: macerado de folhas
Grupo experimental 112.36 1 112.36 17.251 0.001
Erro 117.24 18 6.51
Tempo 15.80 4 3.95 1.99 0.104 0.143
Tempo vs. grupo exp. 14.24 4 3.56 1.80 0.138 0.174
Erro 142.36 72 1.97
Tabela 1. Resultados das comparações entre o recrutamento de formigas em herbívoros simulados (Experimento 1), danos físicos (Experimento2) e macerado de folhas (Experimento 3) em diferentes intervalos de tempo (t), por meio de ANOVA de medidas repetidas. Para detalhes dosexperimentos, veja texto. O G-G refere-se ao valor de p corrigido segundo o procedimento de Greenhouse-Geisser Epsilon (Wilkinson 1988).
grupo experimental e tempo, indicando que as curvas derecrutamento de indivíduos variaram conjuntamente entre osgrupos (Figura 1).
DiscussãoA ausência de diferenças entre o tempo de detecção de
herbívoros simulados e danos mecânicos realizados emrelação aos controles indica que a detecção de herbívorospor formigas residentes em Maieta guianensis sejadeterminada, em primeira instância, pela atividade depatrulhamento das formigas na superfície foliar. A atividadede patrulha das formigas nas folhas de M. guianensis ocorreao longo de todo o período do dia e da noite (Vasconcelos1991). Porém, uma vez detectada uma presa potencial,herbívoro ou dano associado, há uma pronta resposta dacolônia, via recrutamento de novos indivíduos, incluindosoldados, que normalmente não participam de atividades depatrulhamento (Vasconcelos 1991). Uma resposta rápida dacolônia de formigas pode garantir que menores danos sejaminflingidos à planta por herbívoros. Experimentos similares,realizados com ovos de hemípteros, forneceram resultadossemelhantes para M. guianensis (Vasconcelos 1991), comos ovos sendo removidos das folhas. Assim, Pheidoleminutula parece ser um protetor eficiente contra herbívorosem M. guianensis.
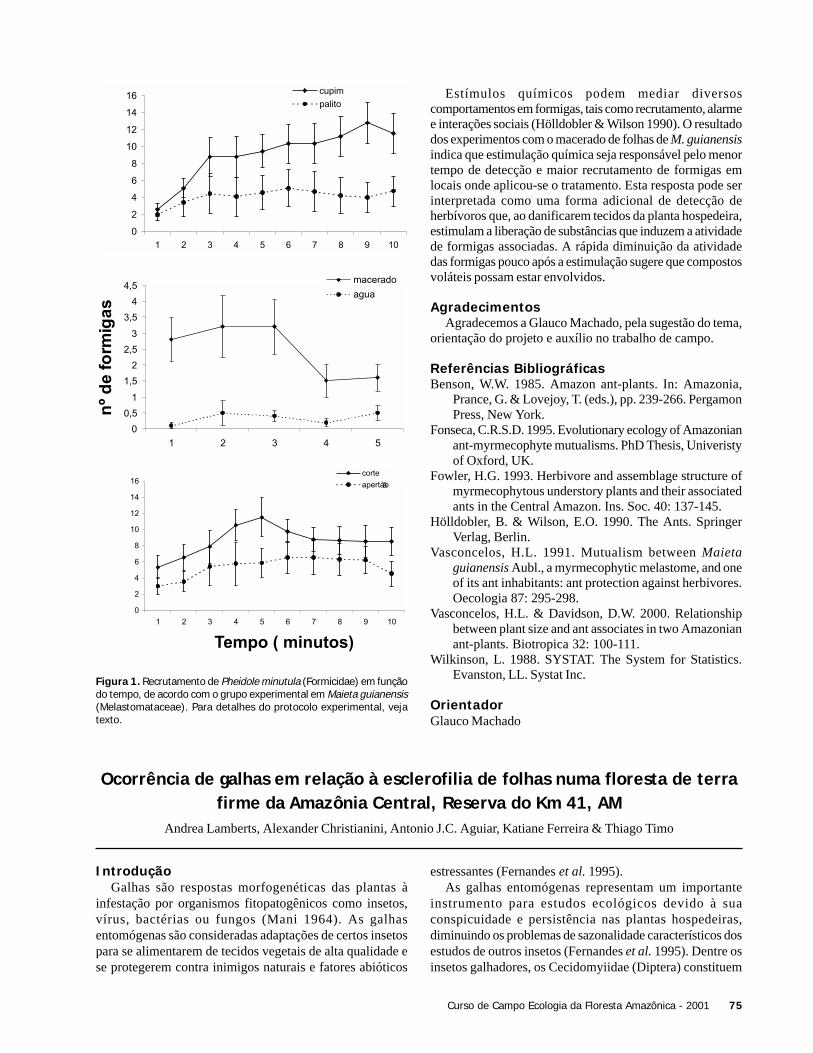
Curso de Campo Ecologia da Floresta Amazônica - 2001 75
0
2
4
6
8
10
12
14
16
1 2 3 4 5 6 7 8 9 10
cupim
palito
0
2
4
6
8
10
12
14
16
1 2 3 4 5 6 7 8 9 10
corte
apertàoã
0
0,5
1
1,5
2
2,5
3
3,5
4
4,5
1 2 3 4 5
nº
de
form
iga
s
macerado
agua
Tempo ( minutos)
Figura 1. Recrutamento de Pheidole minutula (Formicidae) em funçãodo tempo, de acordo com o grupo experimental em Maieta guianensis(Melastomataceae). Para detalhes do protocolo experimental, vejatexto.
Estímulos químicos podem mediar diversoscomportamentos em formigas, tais como recrutamento, alarmee interações sociais (Hölldobler & Wilson 1990). O resultadodos experimentos com o macerado de folhas de M. guianensisindica que estimulação química seja responsável pelo menortempo de detecção e maior recrutamento de formigas emlocais onde aplicou-se o tratamento. Esta resposta pode serinterpretada como uma forma adicional de detecção deherbívoros que, ao danificarem tecidos da planta hospedeira,estimulam a liberação de substâncias que induzem a atividadede formigas associadas. A rápida diminuição da atividadedas formigas pouco após a estimulação sugere que compostosvoláteis possam estar envolvidos.
AgradecimentosAgradecemos a Glauco Machado, pela sugestão do tema,
orientação do projeto e auxílio no trabalho de campo.
Referências BibliográficasBenson, W.W. 1985. Amazon ant-plants. In: Amazonia,
Prance, G. & Lovejoy, T. (eds.), pp. 239-266. PergamonPress, New York.
Fonseca, C.R.S.D. 1995. Evolutionary ecology of Amazonianant-myrmecophyte mutualisms. PhD Thesis, Univeristyof Oxford, UK.
Fowler, H.G. 1993. Herbivore and assemblage structure ofmyrmecophytous understory plants and their associatedants in the Central Amazon. Ins. Soc. 40: 137-145.
Hölldobler, B. & Wilson, E.O. 1990. The Ants. SpringerVerlag, Berlin.
Vasconcelos, H.L. 1991. Mutualism between Maietaguianensis Aubl., a myrmecophytic melastome, and oneof its ant inhabitants: ant protection against herbivores.Oecologia 87: 295-298.
Vasconcelos, H.L. & Davidson, D.W. 2000. Relationshipbetween plant size and ant associates in two Amazonianant-plants. Biotropica 32: 100-111.
Wilkinson, L. 1988. SYSTAT. The System for Statistics.Evanston, LL. Systat Inc.
OrientadorGlauco Machado
Ocorrência de galhas em relação à esclerofilia de folhas numa floresta de terrafirme da Amazônia Central, Reserva do Km 41, AM
Andrea Lamberts, Alexander Christianini, Antonio J.C. Aguiar, Katiane Ferreira & Thiago Timo
IntroduçãoGalhas são respostas morfogenéticas das plantas à
infestação por organismos fitopatogênicos como insetos,vírus, bactérias ou fungos (Mani 1964). As galhasentomógenas são consideradas adaptações de certos insetospara se alimentarem de tecidos vegetais de alta qualidade ese protegerem contra inimigos naturais e fatores abióticos
estressantes (Fernandes et al. 1995).As galhas entomógenas representam um importante
instrumento para estudos ecológicos devido à suaconspicuidade e persistência nas plantas hospedeiras,diminuindo os problemas de sazonalidade característicos dosestudos de outros insetos (Fernandes et al. 1995). Dentre osinsetos galhadores, os Cecidomyiidae (Diptera) constituem
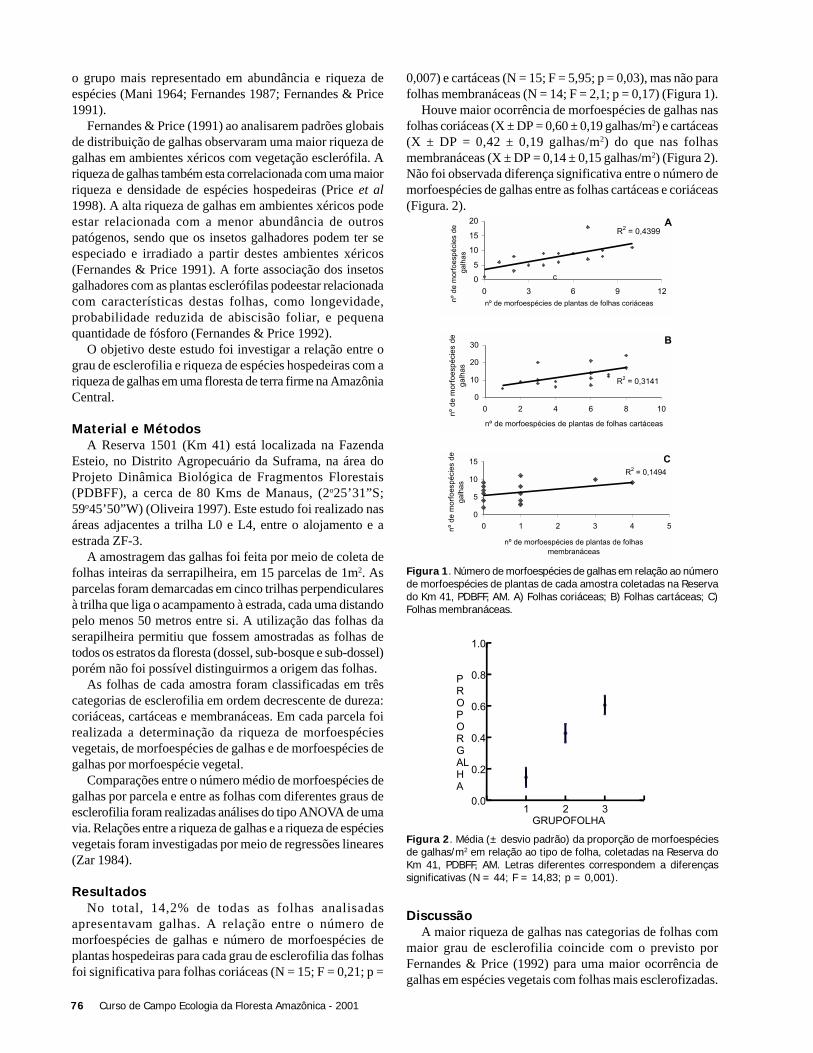
76 Curso de Campo Ecologia da Floresta Amazônica - 2001
o grupo mais representado em abundância e riqueza deespécies (Mani 1964; Fernandes 1987; Fernandes & Price1991).
Fernandes & Price (1991) ao analisarem padrões globaisde distribuição de galhas observaram uma maior riqueza degalhas em ambientes xéricos com vegetação esclerófila. Ariqueza de galhas também esta correlacionada com uma maiorriqueza e densidade de espécies hospedeiras (Price et al1998). A alta riqueza de galhas em ambientes xéricos podeestar relacionada com a menor abundância de outrospatógenos, sendo que os insetos galhadores podem ter seespeciado e irradiado a partir destes ambientes xéricos(Fernandes & Price 1991). A forte associação dos insetosgalhadores com as plantas esclerófilas podeestar relacionadacom características destas folhas, como longevidade,probabilidade reduzida de abiscisão foliar, e pequenaquantidade de fósforo (Fernandes & Price 1992).
O objetivo deste estudo foi investigar a relação entre ograu de esclerofilia e riqueza de espécies hospedeiras com ariqueza de galhas em uma floresta de terra firme na AmazôniaCentral.
Material e MétodosA Reserva 1501 (Km 41) está localizada na Fazenda
Esteio, no Distrito Agropecuário da Suframa, na área doProjeto Dinâmica Biológica de Fragmentos Florestais(PDBFF), a cerca de 80 Kms de Manaus, (2o25’31”S;59o45’50”W) (Oliveira 1997). Este estudo foi realizado nasáreas adjacentes a trilha L0 e L4, entre o alojamento e aestrada ZF-3.
A amostragem das galhas foi feita por meio de coleta defolhas inteiras da serrapilheira, em 15 parcelas de 1m2. Asparcelas foram demarcadas em cinco trilhas perpendicularesà trilha que liga o acampamento à estrada, cada uma distandopelo menos 50 metros entre si. A utilização das folhas daserapilheira permitiu que fossem amostradas as folhas detodos os estratos da floresta (dossel, sub-bosque e sub-dossel)porém não foi possível distinguirmos a origem das folhas.
As folhas de cada amostra foram classificadas em trêscategorias de esclerofilia em ordem decrescente de dureza:coriáceas, cartáceas e membranáceas. Em cada parcela foirealizada a determinação da riqueza de morfoespéciesvegetais, de morfoespécies de galhas e de morfoespécies degalhas por morfoespécie vegetal.
Comparações entre o número médio de morfoespécies degalhas por parcela e entre as folhas com diferentes graus deesclerofilia foram realizadas análises do tipo ANOVA de umavia. Relações entre a riqueza de galhas e a riqueza de espéciesvegetais foram investigadas por meio de regressões lineares(Zar 1984).
ResultadosNo total, 14,2% de todas as folhas analisadas
apresentavam galhas. A relação entre o número demorfoespécies de galhas e número de morfoespécies deplantas hospedeiras para cada grau de esclerofilia das folhasfoi significativa para folhas coriáceas (N = 15; F = 0,21; p =
0,007) e cartáceas (N = 15; F = 5,95; p = 0,03), mas não parafolhas membranáceas (N = 14; F = 2,1; p = 0,17) (Figura 1).
Houve maior ocorrência de morfoespécies de galhas nasfolhas coriáceas (X ± DP = 0,60 ± 0,19 galhas/m2) e cartáceas(X ± DP = 0,42 ± 0,19 galhas/m2) do que nas folhasmembranáceas (X ± DP = 0,14 ± 0,15 galhas/m2) (Figura 2).Não foi observada diferença significativa entre o número demorfoespécies de galhas entre as folhas cartáceas e coriáceas(Figura. 2).
Figura 1. Número de morfoespécies de galhas em relação ao númerode morfoespécies de plantas de cada amostra coletadas na Reservado Km 41, PDBFF, AM. A) Folhas coriáceas; B) Folhas cartáceas; C)Folhas membranáceas.
Figura 2. Média (± desvio padrão) da proporção de morfoespéciesde galhas/m2 em relação ao tipo de folha, coletadas na Reserva doKm 41, PDBFF, AM. Letras diferentes correspondem a diferençassignificativas (N = 44; F = 14,83; p = 0,001).
DiscussãoA maior riqueza de galhas nas categorias de folhas com
maior grau de esclerofilia coincide com o previsto porFernandes & Price (1992) para uma maior ocorrência degalhas em espécies vegetais com folhas mais esclerofizadas.
R2 = 0,4399
0
5
10
15
20
0 3 6 9 12
nº de morfoespécies de plantas de folhas coriáceasnº
de
morf
oesp
éci
es
de
galh
as
c
A
R2 = 0,3141
0
10
20
30
0 2 4 6 8 10
nº de morfoespécies de plantas de folhas cartáceas
nºde
mor
foesp
écie
sde
galh
as
B
R2 = 0,1494
0
5
10
15
0 1 2 3 4 5
nº de morfoespécies de plantas de folhasmembranáceas
nºde
mor
foes
péci
es
de
galh
asC

Curso de Campo Ecologia da Floresta Amazônica - 2001 77
Ambientes submetidos a algum tipo de estresse, comodeficiência de água e de nutrientes essenciais no solo, tendema apresentar uma maior ocorrência de espécies vegetais comfolhas esclerofizadas e compostos secundários (Fernandes& Price 1988, 1991).
Provavelmente a maior parte das folhas que apresentaramgalhas vieram de árvores no dossel e do estrato emergente,onde concentra-se a maior parte da biomassa de folhas dafloresta. O dossel representando um ambiente submetido amaiores graus de estresse que o estrato inferior (McDade etal. 1994). Intensa radiação solar, assim como deficiência denutrientes e de água durante a estação seca normalmente estãorelacionados com esclerofilia em folhas e correspondem aosfatores estressantes aos quais a vegetação do dossel estásubmetida.
Foi também possível observar uma correlação entre ariqueza de espécies hospedeiras e a riqueza de morfoespéciesde galhas. A grande especificidade da relação entre insetosgalhadores e plantas pode explicar este resultado. Ambientescom maior riqueza de espécies vegetais normalmenteapresentam um maior número de espécies de galhas (Price etal. 1998).
Este estudo abre perspectivas para uma melhorcompreensão da distribuição de espécies vegetais de folhasesclerófilas dentro e entre as diversas formações da FlorestaAmazônica e suas relações com a riqueza e abundância degalhas. Os dados indicam a necessidade de estudosdetalhados sobre o porquê das fortes correlações entreesclerofilia e riqueza de galhas, mesmo em formaçõesflorestais. Uma hipótese levantada por G. W. Fernandes (com.pess.) é que as galhas se beneficiariam da condição deesclerofilia, pois elas não sofrem com a falta de água nem denutrientes, pois formam um dreno para estes elementos.Ocorrendo nestes ambientes as galhas também evitamcompetição com outros herbívoros.
AgradecimentosAgradecemos ao professor Geraldo Wilson Fernandes pela
orientação.
Referências BibliográficasFernandes, G.W & Price, P.W. 1991. Comparisons of tropical
and temperate galling species richness: the roles ofenvirovinmental harshness and plant nutrients status. In:Plant-Animal Interactions: Evolutionary Ecology inTropical and Temperate Regions, pp. 91-115. John Wiley& Sons, New York.
Fernandes, G.W. & Price, P.W. 1988. Biogeographicalgradients in galling species richness: tests of hypotheses.Oecologia 76: 161-167.
Fernandes, G.W. & Price, P.W. 1992. The adaptativesignificance of insect gall disribution: Survivorship ofspecies in xeric andmesic habitats. Oecologia 90: 141-20.
Fernandes, G.W.; Paula, A.S. & Loyola Jr, R. 1995.Distribuição de insetos galhadores entre habitats e seupossível uso como bioindicadores. Vida Silv.Neotropical 4(2): 133-139.
Gonçalves–Alvim, S.J.; Vaz dos Santos, M.C.F. & Fernandes,G.W. 2001. Leaf gall abundance on Avicennia germinans(Avicenniaceae) along an intersticial salinity gradient.Biotropica 33(1): 69-77.
McDade, L.A.; Bawa, K.S.; Herpenheide, H.A. & Hartshorn,G.S. 1994. La Selva: Ecology and Natural History of aNeotropical Rainforest. The University of Chicago Press,London.
Mani, M.S. 1964 The Ecology of Plant Galls. The Hague.Oliveira, A.A. 1987. Diversidade, estrutura e dinâmica do
componente arbóreo de uma floresta de terra firme deManaus, AM. Tese de Doutorado, Universidade de SãoPaulo, São Paulo.
Price, P.W.; Fernandes, G.W.; Lara, A.C.F.; Brawn, J.;Barrios, H.; Wright, M.G.; Ribeiro, S.P. & Rothcliff,N. 1998. Global patterns in local number of insect gallingspecies. J. Biogeog. 25: 581-591.
Zar, J.H. 1984. Biostatistical Analysis. Prentice Hall, NewJersey.
OrientadorGeraldo W. Fernandes
Relação entre a forma da folha e o grau de herbivoria emIriartella setigera (Arecaceae)
Maurício Camargo, Angela M. F. Pacheco, Leandro M. Scoss, Paula C. Ceotto &
Paula S. Pinheiro
IntroduçãoA herbivoria é um processo de interação animal-planta de
grande importância na estruturação de comunidades deflorestas tropicais. Os herbívoros são responsáveis porgrandes impactos sobre as plantas, tanto no tempo ecológicoquanto no tempo evolutivo (Coley et al. 1985). Umcomponente importante nesta interação é a defesa das plantas(Loyola Jr. & Fernandes 1993).
Após o ataque de herbívoros, a qualidade nutricional dotecido vegetal atacado pode diminuir, ao mesmo tempo que
a quantidade de compostos secundários e defesas físicaspodem aumentar (Cornelissen & Fernandes 2001). Uma vezque estas substâncias ocorrem com frequência em um grandenúmero de plantas, as respostas induzidas são reconhecidascomo função defensiva devido ao seu forte efeito naperformance dos herbívoros (Fernandes et al. 2000).
Iriartella setigera (Arecaceae) é uma palmeira comfolhas compostas cujos folíolos têm bordas naturalmenterecortadas, semelhantes a marcas deixadas por herbívorosmastigadores. Esta característica pode conferir vantagem a

78 Curso de Campo Ecologia da Floresta Amazônica - 2001
uma planta com pouca ou nenhuma defesa química, já que aocorrência prévia de herbívoria poderia ter elicitado umaresposta induzida por parte da planta, diminuindo seu valorcomo alimento.
Este estudo teve como objetivo testar a hipótese de queIriartella setigera, por apresentar morfologia com traçosaparentes de herbivoria, sofre menos predação que plantasque não possuem tal característica.
Material e MétodosO presente estudo foi realizado em uma área de platô
coberta por floresta densa de terra firme (Pires & Prance1985), na Reserva do Km 41, do PDBFF/INPA, localizadaaproximadamente a 70 km ao norte de Manaus (2o 24’S; 59o
52’W).Ao longo da trilha N4 foram percorridos 320m para
amostrar 22 indivíduos de I. setigera (planta focal). Em cadaplanta estimou-se o grau de herbivoria de seus folíolos pormeio da proporção de área foliar de cada folíolo queapresentava danos por herbívoria. O grau de herbivoria foicategorizado (Tabela 1) e um índice de herbivoria (IH) foicalculado para cada planta, a partir da seguinte fórmula:
onde ni é o número de folíolos referentes a uma dadacategoria, i representa a categoria que especifica a proporçãode área foliar danificada e N representa o número de folíolosamostrados. Para cada planta focal estimamos o grau deherbivoria da palmeira e da dicotiledônea mais próximas.Estas foram escolhidas por sua afinidade filogenética erepresentatividade ecológica dentro da área, respectivamente.Foram coletados 15 folíolos/folhas de cada uma dessasplantas, aleatorizando-se os ramos amostrados e os folíolos/folhas dentro desses ramos. A estimativa da proporção deárea foliar danificada por herbivoria como foi feita como jádescrita para I. setigera.
Tabela 1. Categorias de dano (%) por herbivoria em folhas e folíolosde plantas.
Diferenças nos índices de herbivoria entre I. setigera e aspalmeiras (Arecaceae), e dicotiledôneas, foram avaliadas comuso do teste de Wilcoxon (µ = 5%).
ResultadosTodos os indivíduos de Iriartella setigera apresentaram
IH = n i / Ni * ,
Categorias % de área foliar danificada
0 0
1 1 – 6
2 6 – 12
3 12 – 25
4 25 – 50
5 50 – 100
0
0.5
1
1.5
2
2.5
3
3.5
4
4.5
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22
Amostras
Índi
cede
herb
ivor
ia(I
H)
Iriartella setigeraArecaceaeDicotiledôneas
menores índices de herbivoria quando comparados comoutras palmeiras (n = 22; z = 4,107; p < 0,001) edicotiledôneas (n = 22; z = 4,107; p < 0,001) vizinhas (Figura1).
Figura 1. Comparação dos índices de herbivoria dos indivíduos deIriartella setigera, palmeiras (Arecaceae) e dicotiledôneas.
DiscussãoOs menores índices de herbivoria apresentados por
Iriartella setigera em relação àqueles encontrados para asoutras plantas estudadas indicam que a forma da folha podeexercer um papel importante na redução da herbivoria nestaespécie. Estes resultados confirmam os obtidos por Cachúet al. (1999). Isto pode ser uma resposta adaptativa àspressões seletivas causadas por grandes herbívoros queexistiam no passado, e o padrão recortado da folha nãorepresenta necessariamente uma estratégia de escape contraos herbívoros encontrados atualmente, embora possa estarfuncionando como tal.
Por outro lado, a uma possível defesa mecânica, I. setigerapode apresentar outras formas de resistência à herbivoria,como a produção de compostos secundários (Rosenthal &Janzen 1979) ou defesas induzidas (Fernandes 1990). Aforma como foi conduzido o presente estudo não nos permiteseparar os possíveis efeitos da forma da folha e de eventuaisdefesas químicas apresentadas por I. setigera. Como formade avaliar estes aspectos, podemos propor experimentos cominsetos herbívoros em cativeiro, aos quais seriam oferecidosfolíolos de áreas iguais de I. setigera e de mais duas espéciesde palmeiras, uma apresentando defesa induzida e a outranão. Ao final do experimento, poderíamos comparar o graude herbivoria nos folíolos de I. setigera com aqueleencontrado para as outras duas espécies, e assim constatarse a primeira apresenta algum tipo de defesa química. A partirde nossos resultados, podemos inferir que a morfologia foliarpode ser um possível mecanismo de defesa de plantas paraevitar herbivoria (Lawton 1983).
AgradecimentosAgradecemos ao Rodolpho Dirzo pela orientação e
amizade. Ao Juruna pela ajuda no trabalho de campo.
Referências BibliográficasCachú, Y.P.; Valdivieso, M.A.; Duarte de Souza, M.A.;
Batista, M.A. & Dantas de Melo, S. 1999. La morfologiafoliar de Iriartella setigera: un caso de engaño para los

Curso de Campo Ecologia da Floresta Amazônica - 2001 79
herbívoros?. Livro do Curso de Campo “Ecologia daFloresta Amazônica”, INPA, Smithsonian Institution,PDBFF, Manaus, AM.
Coley, P.D.; Bryant, J.P. & Chapin, S. 1985. Resource andplant antiherbivory defense. Science 230: 895-899.
Cornelissen, T.G. & Fernandes, G.W. 2001. Induced defensesin the neotropical tree Bauhinia breviceps (Vog.) toherbivory: effects of damage-induced changes on leafquality and insect attack. Trees 15: 236-241.
Fernandes, W.G. 1990. Hypersensitivity: a neglected plantresistance mechanism against insect herbivores. Environ.Entomol. 19(5): 1173-1182.
Fernandes, W.G.; Cornelissen, T.G.; Isaias, R.M.S. & Lara,T.A.F. 2000. Plants fight gall formation: hypersensitivity.Ciênc. Cult. 52(1): 49-54.
Lawton, J.H. 1983. Plant architeture and the diversity ofphytophagous insects. Ann. Rev. Entomol. 28: 23-39.
Loyola Jr., R. & Fernandes, G.W. 1993. Herbivoria emKielmeyra coriacea (Guttiferae): efeitos da idade daplanta, desenvolvimento e aspectos qualitativos defolhas. Revta. Bras. Biol. 53(2): 295-304.
Pires, J.M. & Prance, G.T. 1985. The vegetation types of theBarzilian Amazon. In: Amazonia, Prance, G.T. &Lovejoy, T. (eds.), pp. 109-145. Pergamon, New York.
Rosenthal, G.A. & Janzen, D.H. 1979. Herbivores: TheirInteractions with Secondary Plant Metabolites.Academic Press, New York.
OrientadorRodolpho Dirzo
Aspectos estruturais e populacionais de agregados de Leucauge sp.(Araneae: Tetragnathidae)
Felipe B. de Oliveira, Antônio J. C. Aguiar, Paula C. Ceotto & Natalie Olifiers
IntroduçãoA construção de teias é uma eficiente estratégia de captura
de presas utilizada por muitos grupos de aranhas. Paramaximizar esta eficiência é importante que as aranhasdimensionem o tamanho, a disposição e localização das teias,otimizando a captura de presas (Ebehard 1990).
Leucauge sp. (Tetragnathidae) é uma aranha que constróiteias individuais orbiculares, mas vive em agrupamentos comco-específicos (Santos & Silva 1999), onde as teias próximasestão conectadas muitas vezes por fios de sustentação(Venticinque 1994). Os agregados ocorrem freqüentementeacima de pequenos cursos d’água e normalmente sãoassociados a áreas de correnteza (Gonçalves de Lima 1997Venticinque 1994). Os agregados de espécies de Leucaugeapresentam variações nas formas de suas teias relacionadasa fatores como densidade de presas e estrutura da vegetação(Eberhard 1990).
Embora existam alguns estudos enfocando a seleção depresas, alometria das teias (Marques 1992) e estratégias deforrageamento (Costa et al. 1992), pouco se sabe sobreaspectos populacionais e a estrutura física dos agregados deLeucauge sp.
O objetivo deste trabalho foi verificar se existe relaçãoentre a proporção de fêmeas e de indivíduos juvenis noagregado e o tamanho e estrutura deste. Adicionalmente,investigamos se a localização dos agregados (se estão emclareiras ou não) está relacionada a seu tamanho e ao númerode aranhas que o compõem.
Materiais e MétodosA área de estudo localiza-se na Reserva do Km 41, na
área do Projeto Dinâmica Biológica de Fragmentos Florestais
(PDBFF-INPA), a cerca de 70 km ao norte de Manaus(2o24’S; 59o45‘W). Esta área apresenta pluviosidade entre1900-2500 mm/ano, uma estação seca bastante pronunciadaentre os meses de julho e outubro (Rankin-de-Merona et al.1992).
Percorremos as margens de um igarapé próximo aoacampamento em busca de agregados de Leucauge sp.Quando encontrado, medíamos o seu comprimento, largurae altura máximas, a partir dos pontos mais extremos deancoragem das teias. Estimamos o volume do agregado atravésda multiplicação dessas três dimensões.
Medimos também a largura do igarapé abaixo do agregado,avaliando se este localizava-se em uma área de clareira ou dedossel fechado. Registramos o número total de machos,fêmeas e juvenis presentes em cada agregado;
Os machos de Leucauge sp. - assim como em outrasespécies de aranhas - possuem pedipalpos robustos econspícuos, o que permite distingui-los facilmente dasfêmeas, cujos pedipalpos finos e longos. Os juvenis foramdistinguidos dos adultos pela diferença do tamanho do corpo.
Correlacionamos as proporções de fêmeas e de juveniscom as dimensões das teias com o objetivo de investigarpossíveis relações entre estes parâmetros. Para investigar aexistência de possíveis diferenças entre os tamanhos dosagregados dentro e fora de clareiras, utilizamos o teste t deStudent. Utilizamos o programa SYSTAT 8.0 (Wilkinson1987) para realizar todas as análises estatísticas.
ResultadosEncontramos 16 agregados de Leucauge sp., onde o
número de indivíduos variou de 2 a 25 (10,2 ± 7,5). Não

80 Curso de Campo Ecologia da Floresta Amazônica - 2001
observamos correlação entre as dimensões dos agregados e aproporção de fêmeas (R2 = 0,002, p > 0,05) e a proporção deindivíduos juvenis (R2 = 0,03; p > 0,05) (tab. 1).
Aproximadamente a metade dos agregados medidosencontravam-se sob o dossel (56,2%). A média de indivíduosem agregados situados em clareiras foi de 13,1 ± 8,5 (x ±dp), enquanto a dos situados sob o dossel foi de 6,4 ± 5,3,mas não detectamos diferenças entre o número total dearanhas por agregado e o local onde ele estava localizado (t= 1,91, gl = 14, p = 0,07).
Tabela 1: Dimensões dos agregados de Leucauge sp. medidos emum igarapé da Reserva do Km 41, Manaus, AM; máx = valor máximo;mín = valor mínimo.
Das três dimensões da teia, a largura (R2 = 0,97; p =0,04; n = 16) e o comprimento (R2 = 0,98; p = 0,03; n = 16)mostraram-se positivamente correlacionados ao número totalde indivíduos da colônia, enquanto a altura não está (R2 =0,93; p = 0,18; n = 16).
DiscussãoEm muitas das espécies de aranhas é comum a fêmea
construir teias maiores que a dos machos, devido ao seu maiortamanho e sua maior demanda por alimento (Eberhard 1990).Não medimos cada teia individualmente, mas nossos dadosmostram que em agregados a proporção de fêmeas não estácorrelacionada ao volume total da colônia. Isto pode ocorrerdevido a uma pequena diferença de tamanho entre machos efêmeas, em comparação a outras espécies de aranhas (obs.pess.).
Esperávamos que os agregados com maior número deindivíduos juvenis, que são menores que os adultos,apresentassem tamanho menor. Esta hipótese não foicorroborada por este estudo. No entanto, os menoresagregados que observamos apresentaram apenas indivíduosjuvenis.
A presença das teias em clareiras e em locais sombreadosé praticamente igual. Portanto, acreditamos que os fatoresque influenciam mais decisivamente a estrutura da teia são alargura do igarapé e a estrutura da vegetação ao redor,
média máx mín
Altura (cm) 113 ± 30 160 70
Comprimento (cm) 219 ± 168 630 40
Largura (cm) 143 ± 57 305 60
conforme mostram outros estudos (Venticinque 1994).Com relação à estrutura da teia, nossos dados mostram
que os agregados com poucas e muitas aranhas não diferemquanto à altura, mas diferem quanto ao comprimento e àlargura. Isso sugere que para esta espécie de Leucauge é maisimportante maximizar a área de captura sobre o igarapé,sugerindo que sua dieta esteja baseada em insetos queemergem da água. Nossos dados complementam os de Soares(1999) que mostrou que os agregados maiores estão situadosonde a biomassa de insetos emergentes é maior, reforçandoa idéia de que esses insetos constituem fração importante dadieta dessa espécie de aranha.
AgradecimentosAgradecemos a Glauco Machado pela ajuda no campo e
sugestões.
Referências BibliográficasEberhard, W.G. 1990. Functional and phylogeny of spiders
webs. Annu. Rev. Ecol. Syst. 21: 341-372.Gonçalves de Lima, M. 1997. Relação entre tempo de
manuseio de presas e biomassa em Leucauge sp.: umatática de forrageamento ótimo. Livro de Curso de Campo/ 1997, “Ecologia da Floresta Amazônica”. 276-280.
Venticinque, E. M. 1994. Colonialidade, seleção de micro-habitat e comportamento de uma aranha tropicalconstrutora de teias orbiculares. Livro de Curso deCampo / 1994, “Ecologia da Floresta Amazônica”. 401-407.
Rankin-de-Merona, J.M.; G. T. Prance; Hutchings, R.W.;Silva, M.F.; Rodrigues, W.A. & Uehling, M.E. 1992.Preliminar results of large scale tree inventory of uplandrain forest in the Central Amazon. Act. Amaz. 22 (4):493-534.
Santos, A.J. & K. L. Silva. 1999. Distribuição espacial eestado nutricional de uma aranha da Amazônia Central,Leucauge sp. (Tetragnathidae): um teste da hipótese doefeito ricochete. Livro de Curso de Campo / 1999,“Ecologia da Floresta Amazônica”. 23-24.
Soares, M.G. 1999. Distribuição de agregados de Leucagesp. (Araneae; Tetragnathidae) ao longo de um igarapéna Amazônia Central: análise dos fatores determinantesda escolha do habitat. Livro de Curso de Campo / 1999,“Ecologia da Floresta Amazônica”. 165-167.
Wilkinson, L. 1987. SYSTAT: The System for Statistics.Systat inc., Evanston.
Observações subaquáticas de Iguanodectes variatus Gerý, 1993 (Characidae)Mauricio Camargo & Edgar F. Prieto
IntroduçãoA observação das atividades diárias dos organismos tem
sido uma metodologia muito aplicada com o objetivo deentender as relações com o ambiente e aspetos de biologiapopulacional. As informações a respeito da história naturalde uma espécie, são de fundamental importância para a
conservação, pois permite identificar fatores que poderiamcoloca-las em risco de extinção (Gilpin & Soulé 1986).
A informação disponível sobre a história natural dospeixes amazônicos, provém de inferências feitas a partir dospadrões de captura e da análise de amostras coletadas. Amaioria dos estudos realizados para os peixes de pequeno

Curso de Campo Ecologia da Floresta Amazônica - 2001 81
porte, tem sido feitos em aquários sob condições não naturais(Sabino & Zuanon 1998). Alguns autores têm enfatizado aimportância de estudos naturalísticos em ambientes tropicais(Sazima 1986; Sabino & Castro 1990).
As características físicas dos pequenos igarapés, comobaixa profundidade, leito bem delimitado e correntezarelativamente acentuada, determinam uma colonizaçãoprincipal por peixes de pequeno porte (Lowe-McConnel1987). Dentro destes cursos se destacam pequenos caracídeoscomo Iguanodectes variatus, um peixe de hábitos diurnosque freqüenta as áreas rasas dos igarapés (Bührheim 1998).
Este estudo tem como objetivo observar algumas variáveiscomportamentais de Iguanodectes variatus, com a finalidadede contribuir para o conhecimento de sua história natural.
Material e MétodosO presente estudo foi realizado no igarapé do
acampamento, que integra as cabeceiras da drenagem da baciado rio Urubu, numa área de platô na Reserva do Km 41 (59o
43’40”W; 2o 24’ 26”S). Para as observações docomportamento da espécie, fizemos 20 mergulhos em sessõesde 5 minutos cada, que totalizaram 100 minutos. Asobservações foram feitas entre as 09:00h e 12:00h.
A metodologia utilizada para fazer as observações foi ado indivíduo focal. Previamente definimos 6 variáveiscomportamentais a serem avaliadas (forrageio, deslocamento,gregarismo, agressividade, comportamento de fuga, e uso dohábitat). Todas as variáveis exceto o uso do habitat, foramcategorizadas em 0 – para a não ocorrência e 1- paraocorrência de atividade comportamental. O uso do hábitatfoi categorizado através de três variáveis (localização nacoluna de água, tipo de sustrato e localização em relação àmargem do igarapé).
Os dados foram preenchidos em uma matriz onde ascolunas corresponderam as variáveis comportamentais e aslinhas a cada uma das observações. Para definir a distribuiçãodo tempo de atividade dentro das cinco variáveis, calculamosas freqüências de ocorrência para cada uma. A soma de estasfreqüências correspondeu a 100% de atividade. Com baseneste total calculamos a porcentagem de tempo investido emcada atividade e os resultados foram plotados em um diagramade barras.
Para definir as preferências de uso do habitat, calculamosa freqüência de ocorrência para cada uma das variáveisconsideradas, as quais foram adicionadas para obter um 100%de tempo de uso. Com este total calculamos a porcentagemde tempo investido em habitat. Os resultados foram plotadosem um diagrama de barras.
ResultadosIguanodectes variatus é um peixe que prefere águas de
correnteza. Esta espécie se agrega em pequenos grupos deindivíduos dentro da coluna, compartilhando o espaço comoutras espécies como Aequidens pallidus e Nannostomusmarginatus. O forrageio e deslocamento foram as atividadesrealizadas com maior frequência. Também, I. variatusapresentou uma alta frequência de interações de agressividade
e de fuga. Por outro lado, o grau de gregarismo foi baixo(Figura 1).
Figura 1. Porcentagem de tempo utilizado nas diferentes atividadescomportamentais de I. variatus em um igarapé da Reserva do Km41.
Com respeito à variável uso do habitat, os indivíduosobservados, ocorreram com maior frequencia emprofundidades menores de 40cm, se localizandofundamentalmente entre o canal central com predomino deareia e a margem do igarapé, com predomino de serapilheira(Figura 2).
Figura 2. Porcentagem do tempo utilizado nos diferentes habitatsde I. variatus
DiscussãoAs caraterísticas fisico-químicas citadas para igarapés da
mesma microbacia hidrográfica, indicam uma pobreza denutrientes dentro de estes sistemas (Bührheim 1998), quepor sua vez definem uma limitação nas fontes de produçãoprimária (Sioli 1968). Esta pobreza de nutrientes determinaque as fontes principais de alimento, sejam de origemalóctone, da floresta circundante (Garutti 1988). Estes fatoressão condicionantes da presença e distribuição das populaçõesde peixes. As diferenças na velocidade da correnteza tambémdefinem a estruturação das comunidades de peixes.
Dentre os characídeos de porte menor, que habitam ospequenos igarapés, é comum uma tendência de agregaçãoenvolvendo diferentes espécies (Lowe-McConnell 1987).Esta estratégia foi observada no presente estudo, no qualIguanodectes variatus compartilha o espaço com outrasespécies como Aequidens pallidus e Nannostomusmarginatus.
30.4
22.5
3.5
2023.6
0
5
10
15
20
25
30
35
Forrageio Deslocamento H. Gregário Comp.Agressivo
Ataque outroindiv.
Atividade
%de
tem
po
Areia Serap.Margem Ca nal <40cm >40cm
0
10
20
30
40
50
60
70
Local Profundidade sustrato
Ocupação do habitat
%de
tem
po

82 Curso de Campo Ecologia da Floresta Amazônica - 2001
O desenho e forma corporal fusiforme de Iguanodectesvariatus, são indícios de sua boa adaptação para habitar locaisde correnteza forte. As nadadeiras pares desenvolvidas ecaudal homocerca são típicas de um nadador veloz. Estedesenho corporal da espécie é determinante para que oindivíduo consiga manter a posição espacial contra a correntee assim aumentar a probabilidade de obter alimento trazidopela correnteza. A alta frequencia de atividade de forrageioe deslocamento confirmam a tendência da espécie a ser ativadurante o horário de observação.
A alternância de uso de diferentes microhabitats por I.variatus (canal central e margem) pode ser uma estratégiapara otimizar a sua capacidade de obtenção de fontesalimentares produzidas no sistema, ou de origem externo.
Existe uma discordância na caracterização da categoriatrófica para o gênero Iguanodectes, o qual segundo Goulding(1988) apresenta um hábito detritívoro, em quanto queSampaio (1994) determinou que a espécie prefere amirmecofagia como dieta principal. As nossas observaçõesindicam que a espécie é de hábitos onívoros, com preferênciapela captura de organismos que caem na superficie da água.
O comportamento agressivo observado para a espécie podeser explicado pela disputa pelos recursos ou por algum tipode territorialidade, que foi muito evidente nas margens. Adesagregação dos indivíduos na coluna d’água, indica quemesmo que estes constituam um grupo, não apresentamrelações duradoras de gregariedade.
Podemos concluir que Iguanodectes variatus utilizagrande parte do tempo das primeiras horas do dia na atividadede forrageio, nadando solitariamente e utilizando a tática decatar itens alimentares à deriva.
AgradecimentosAo Jansen e ao Dadão, pelas dicas e amizade.
Referências BibliográficasBührheim, C.M. 1998. Estrutura de comunidades de peixes
em igarapés de floresta de terra firme na AmazôniaCentral. Dissertação de Mestrado, Universidade doAmazonas, INPA.
Garutti, V. 1988. Distribuição longitudinal da ictiofauna emum córrego da região noroeste do estado de São Paulo,Bacia do rio Paraná. Rev. Brasil. Biol. 48(4): 747-759.
Gilpin, M.E. & Soulé, M.E. 1986. Minimum viablepopulations Processes of species extinction. In: M.E.Soulé (ed.), Conservation Biology -The Science ofScarcity and Diversity pp 19-34. Sinauer Associates,Sunderland, M.A.
Goulding, M.; Carvalho, L.M. & Ferreira, E.G. 1988. RioNegro, Rich Life in Poor Water: Amazonian Diversityand Foodchain Ecology as Seen Through FishCommunities. Academic Publishing, The Netherlands.
Lowe-McConnel, R.H. 1987. Ecological Studies in TropicalFish Communities. Cambridge University Press,London.
Sabino , J. & Castro, R.M.C. 1990. Alimentação, períodode atividade e uso de espaço dos peixes de um riacho dafloresta Atlântica (Sudeste do Brasil). Revta. Bras. Biol.,50: 23-36.
Sabino, J. & Zuanon, J. 1998. A stream fish assemblage inCentral Amazonia: distribution, activity patterns andfeeding behavior. Ichthyol. Explor. Freshwaters 8(3):201-210.
Sampaio, M.M.A. 1994. Estudo comparativo decomunidades de peixes em áreas de remanso e decorrenteza em um igarapé de água clara da regiãoamazônica. Livro do Curso de Campo “Ecologia daFloresta Amazônica”. INPA/PDBFF, Manaus, AM.
Sazima, I. 1986. Similarities in feeding bahaviour betweensome marine and freshwater fishes in two tropicalcommunities. J. Fish. Biol. 49: 325-333.
Sioli, H. 1968. Hydrochemistry and geology in the BrazilianAmazon Region. Amazoniana I: 267-277.
Caracterização dos jardins de formiga em Conceveiba martiana(Euphorbiaceae) na Reserva do Km 41, Amazônia Central
Betsabé R. Guerra, Angela M. F. Pacheco, Rodrigo Cogni, Paula S. Pinhero & Alexander V. Christiani
IntroduçãoDentre as diversas formas de interação entre formigas e
plantas, os jardins de formigas estão entre as mais complexas(Hölldobler & Wilson 1990). Estes consistem em agregadosde epífitas reunidas por formigas. As formigas coletam assementes destas epífitas e as carregam para o ninho, ondeestas germinam e suas raízes tornam-se parte da estrutura doninho (Hölldobler & Wilson 1990).
Os jardins são formados por diferentes espécies deformigas, sendo Crematogaster, Solenopsis, Azteca, Monacise Camponotus os gêneros mais comuns (Davidson 1998).Na Amazônia a principal formiga formadora de jardim éCamponotus femoratus, conhecida como uma das mais
abundantes espécies arborícolas em florestas das regiõestropicais (Wilson 1987). Os jardins são geralmenteelispsoidais e seus diâmetros variam de seis a mais de 60cm. As formigas constróem os ninhos formando câmarasirregulares divididas por paredes cartáceas ao longo das raízesdas epífitas (Hölldobler & Wilson 1990).
Colônias de formigas mirmecófitas são normalmentelimitadas pelo espaço da domácea utilizada como ninho(Fonseca 1993; Fonseca & Ganade 1996). Entretanto, nãoexistem estudos investigando se o tamanho do jardim serelaciona com a demografia da colônia. Este trabalho objetivaestudar o jardim de formigas em Conceveiba martiana(Euphorbiaceae), enfocando a estrutura do ninho e a

Curso de Campo Ecologia da Floresta Amazônica - 2001 83
correlação entre o número de formigas e o tamanho do jardim.
Material e MétodosRealizamos o trabalho na Reserva do Km 41 do Projeto
Dinâmica Biológica de Fragmentos Florestais (INPA,Smithsonian Institution), distante aproximadamente 70 Kmda cidade de Manaus (2o 25’S; 59o 48’W). a vegetação dareserva é formada por uma floresta de terra firme (Pires &Prance 1985).
Coletamos quatro jardins de formigas em indivíduos deConceveiba martiana, uma árvore de pequeno porte comumem áreas de baixio. Essa planta apresenta estípulas foliáceasinterpeciolares muito desenvolvidas no ápice dos ramos, comcerca de 4 cm de comprimento e 3 cm de largura. Estasestípulas formam diversas bolsas ao longo do ramo, quepodem abrigar adultos e imaturos de formigas (Ribeiro et al.1999).
Medimos o comprimento e a largura de cada jardim parao cálculo do volume. Assumimos que estes apresentam formaelipsóide e utilizamos a seguinte fórmula para o cálculo dovolume 4/3ð(a+b)/3)3, onde a = ½ comprimento total, b =largura. Para calcular o volume total do jardim, descontamoso volume do tronco suporte, dado pela fórmula do cilindro
(pr2h), onde r = raio, h = altura. Abrimos os jardins,descrevemos sua estrutura e contamos o número de operáriasde Camponotus femoratus. Para se testar a correlação entreo volume do jardim e o número de operárias utilizamos oteste de correlação de Spearman (Zar 1996).
ResultadosNos quatro jardins encontramos colônias de Camponotus
femoratus e Crematogaster limata parabiotica ocorrendojuntas. Três dos ninhos se dispunham verticalmente, ao longodo caule, em torno da inserção dos pecíolos das folhas. Aspartes mais externas do jardim apresentam câmaras de paredescartáceas que abrigam C. femoratus (Figura 1). O maior ninhocoletado não estava ancorado no ramo principal da planta,mas num ramo secundário, e estava totalmente estruturadopor raízes das epífitas. Os jardins apresentaram volume entre31 e 188 cm3, contendo entre 48 e 2109 operárias de C.femoratus. O número de operárias por colônia não apresentoucorrelação com o volume do jardim (Spearman r
s = 0,4; n =
4; p > 0,05) (Figura 2). Somente no maior ninho foramencontrados imaturos e alados, indicando que somente essacolônia estava reproduzindo.
Os jardins são mais defendidos por Camponotus, que em
Figura 1: Representação externa (a) e interna (b) de jardins-de-formigas junto ao ápice dos ramos de Conceveiba martiana (Euphorbiaceae).Câmara de parede cartácea (A) e estípula com ovos de Crematogaster limata parabiotica (Formicidae) (B).
(entra desenho)

84 Curso de Campo Ecologia da Floresta Amazônica - 2001
todos os casos responderam rapidamente aos danosmecânicos produzidos durante a coleta, enquantoCrematogaster não foi observada defendendo o ninho. Emtrês dos ninhos houve uma separação espacial dentro dojardim, com Crematogaster ocupando o interior das estípulase Camponotus o restante do jardim. Em todos os ninhosforam encontrados imaturos de C. limata.
Figura 2. Relação entre número de operárias da formiga Camponotusfemoratus e o volume do jardim, em indivíduos de Conceveibamartiana na Amazônia Central.
DiscussãoO fato de não verificarmos correlação entre o volume do
jardim e o número de indivíduos da colônia indica que ovolume pode não estar influenciando diretamente o tamanhoda colônia no intervalo de tamanho de ninhos observado.Outros fatores, como a idade do jardim, a disponibilidadede alimento para a colônia e características estruturais doninho, como número de raízes de plantas epífitas, podemestar influenciando o crescimento da colônia. A grandevariação no número de indivíduos indica também que onúmero de ninhos coletados foi insuficiente para o estudoda relação entre tamanho do ninho e tamanho da colônia.Entretanto, parece existir uma relação causal entre a idadedo ninho e o número de indivíduos, pois o único ninhoreprodutivo foi o maior, que apesar de apresentar um volume
0
0.5
1
1.5
2
2.5
3
3.5
0 50 100 150 200
Volume (cm3)
log
do
nú
mer
od
efo
rmig
asligeiramente superior, apresentou um número de indivíduoscomparativamente muito maior que os demais.
Aparentemente Crematogaster se estabelece antes naplanta, pois ocorre nas estípulas de plantas sem jardins deformiga (Ribeiro et al. 1999). A presença de imaturos deCrematogaster em todos os ninhos coletados também apoiaesta hipótese. Segundo Hölldobler & Wilson (1990), écomum a coocorrência das duas espécies e existe uma relaçãode parabiose facultativa entre as espécies. Não existemevidências claras de que esta relação é simétrica.Crematogaster aparentemente se beneficia da proteçãofornecida por Camponotus, mas estas competem peloalimento. Num estudo aprofundado desta relação realizadopor Wheeler (1921 apud Hölldobler & Wilson 1990) foramobservados eventos onde Crematogaster foi excluída doalimento, evidenciando forte competição entre as espécies.
Referências BibliográficasDavidson, D.W. 1998. Ecological studies of the Neotropical
ant gardens. Ecology 69: 1138–1152.Fonseca, C.R. & Ganade, G. 1996 Asymmetries
compartments and null interactions in an Amazonian ant– plat community. J. Anim. Ecol. 65: 339-347.
Holldober, B. & Wilson, O.E. 1990. The Ants. HarvardUniversity Press, Cambrige.
Ribeiro, J.E.L.S.; Hopkins, M.J.G.; Vicentini, A.; Sothers,C.A.; Costa, M.A.S.; Brito, J.M.; Souza, M.A.D.;Martins, L.H.P.; Lohmann, L.G.; Assunção, P.A.C.L.;Pereira, E.C.; Silva, C.F.; Mesquita, M.R. & Procópio,L.C. 1999. Flora da Reserva Ducke: Guia deIdentificação das Plantas Vasculares de uma Floresta deTerra Firme da Amazônia Central. INPA-DFID, Manaus.
Wilson, E.O. 1987. The arboreal ant fauna of PeruvianAmazon Forest: a first assement. Biotropica 19: 245–251.
Zar, J.H. 1984. Biostatistical Analysis. Prentice Hall, NewJersey.
Guaribas (Alouatta seniculus, Primates Cebidae) influenciam a distribuição deespécies por eles dispersadas?
Daniel L. M. Vieira, Cristiane F. Jurinitz, Andrea H. Lamberts, Leandro Scoss, Maria R. Darrigo & Thiago Timo
IntroduçãoA distribuição de espécies arbóreas não ocorre ao acaso,
sendo em grande parte influenciada pelo processo dedispersão de sementes (Janzen 1970). Em florestasneotropicais, muitas espécies arbóreas dependem dadispersão de sementes por primatas para o estabelecimentoe recrutamento de plântulas (Janzen 1970; Howe 1986).Estudos apontam estreita relação entre alguns primatas e adispersão de sementes de determinadas plantas. Forget &Sabatier (1997) quantificaram que o macaco aranha (Atelespaniscus) removia e dispersava cerca de 53% das sementes
de Virola sp. em uma região na Guiana Francesa. Apesar dealguns estudos realizados analisarem a dispersão de sementespor macacos, pouco se sabe sobre a sua influência nadistribuição das espécies arbóreas.
Em um estudo realizado na Amazônia, Neves (1985)acompanhou a alimentação de um bando de guaribas(Alouatta seniculus) que se caracterizam por seremessencialmente folívoros e frugívoros. Dentre os frutosutilizados pelos macacos, podemos destacar os das famíliasSapotaceae, Myrtaceae e Euphorbiaceae. Algumas famíliasforam identificadas pela mesma autora como base da

Curso de Campo Ecologia da Floresta Amazônica - 2001 85
alimentação de guaribas, entre elasNeste trabalho avaliamos se há uma distribuição agregada
de plântulas de epécies conhecidamente dispersas porguaribas embaixo das suas árvores dormitório em uma florestacontínua na Amazônia Central.
Material e MétodosRealizamos o trabalho na Reserva do Km 41 do Projeto
Dinâmica Biológica de Fragmentos Florestais (INPA,Smithsonian Institution), distante aproximadamente 80 Kmda cidade de Manaus (2o 25’S; 59o 48’W). A vegetação dareserva é formada por uma floresta de terra firme.
Selecionamos três árvores “dormitório” de A. seniculus.Para avaliar o efeito da dispersão promovida por essesanimais escolhemos três espécies de árvores cujos frutosreconhecidamente fazem parte da sua dieta: Protium sp.,Pouruma sp. e uma espécier não identificada de Sapotaceae(O. Souza com. pess.).
Marcamos uma parcela de 2 x 5m embaixo da copa decada uma das árvores dormitório e, como controle, umaparcela do mesmo tamanho a 30m de distância. A direção(Norte, Sul, Leste ou Oeste) das parcelas controle foisorteada. Contamos todos os indivíduos com até 30cm dealtura que pertenciam às espécies de interesse. Além disso,procuramos por fezes de guariba embaixo de cada árvoredormitório, afim de saber quais as espécies estavam sendoconsumidas por estes macacos. As fezes foramposteriormente analisadas em laboratório, a fim de identificaras sementes presentes.
ResultadosForam encontradas somente plântulas das espécies em
questão em uma parcela embaixo da árvores dormitório. Nestaparcela foram encontrados. 52 indivíduos de Protium sp, 28de uma espécie não identificada de Sapotaceae e dois dePouruma sp. As demais parcelas embaixo da árvoredormitório e as três parcelas controle não apresentaramnenhum indivíduo das espécies em questão.
Encontramos apenas três amostras de fezes embaixo deuma das árvores dormitório, indicando presença recente deguaribas no local. Nestas amostras encontramos sementesdas espécies Minquartia guianensis (Oleacaceae), umaespécie de Sapotaceae e uma espécie de Leguminosae.
DiscussãoO baixo número e freqüência de plântulas das espécies
arbóreas estudadas, assim como das fezes de A. seniculuspodem ser uma evidência de que o bando utiliza muitasárvores para dormir e/ou que defecam pouco nos sítiosdormitório. Isto favoreceria a regeneração de espécies poreles dipersadas, uma vez que se as fezes poderiam estar maisespalhadas no espaço e as sementes, ao germinarem, poderiamsofrer menor grau de mortalidade devido à densidade. Noestudo realizado por Neves (1985) em um fragmento florestalde 10 ha a poucos quilômetros do local deste estudo, umbando de A. seniculus utilizou 10 árvores para dormir e 18para descansar durante o dia.
Estudos enfocando a distribuição espacial de sementes,plântulas, juvenis e adultos de espécies dispersas por A.seniculus, assim como outras espécies de frugívoros podemajudar a explicar a organização espacial das comunidadesvegetais em florestas tropicais. Nesse sentido, espécies dafamília Sapotaceae e Moraceae, que são as maisfrequentemente utilizadas por A. seniculus em florestas daregião de Manaus (Neves 1985), seriam modelosinteressantes para estudos de interações com seus dispersores.Será que ao longo de séculos os guaribas plantaram seualimento no próprio quintal?
AgradecimentosAgradecemos ao Juruna e ao Osmaildo pela identificação
de árvores dormitório e auxílio na coleta de dados.
Referências BibliográficasForget, P.M. & Sabatier, D. 1997. Dynamics of the seedling
shadow of a frugivore-dispersed tree species in FrenchGuiana. J. Trop. Ecol. 13: 767-773.
Howe, H.F. 1986. Seed dispersal by fruit-eating birds andmammals. In: Seed Dispersal, Murray, D.R. (ed.), pp.123-189. Academic Press, Sydney, Austrália.
Janzen, D.H. 1970. Herbivores and the number of species intropical forest. Am. Nat. 104: 501-528.
Neves, A.M. 1985. Alguns aspectos da ecologia de Alouattaseniculus em reserva isolada na Amazônia Central.Dissertação deMestrado, Universidade de São Paulo,Ribeirão Preto.

86 Curso de Campo Ecologia da Floresta Amazônica - 2001
Descrição da Estrutura e padrão espacial de inflorescências de Rapatea ulei(Rapataeceae), uma planta herbácea de baixio em mata de terra firme, AM
Christina McCain
IntroduçãoA família Rapataeceae é composta de 16 gêneros com 85
espécies de ervas neotropicais, sendo que quatro dessasocorrem na mata de terra firme na região de Manaus, AM,Brasil. Todas as espécies ocorrem em ambientes de baixio,que são definidos por sua proximidade de igarapés. Apesarda sua ocorrência freqüente em nesses habitats e deprovavelmente serem um componente importante nascomunidades de baixio, sua morfologia, ecologia e fenologiasão pouco conhecidas. Segundo a única informaçãopublicada sobre essas plantas na região, em geral florescementre setembro e janeiro (Ribeiro et al. 1999), mas nenhumestudo detalhado investigou isso.
Essas plantas têm uma estrutura floral interessante, commuitas flores (aproximadamente, entre 100-200) em cadainflorescência, dentro de brácteas. Não produzem néctar(Ribero et al. 1999), mas se nota claramente a presença deum exudato transparente que cobre todas as inflorescênciasdentro das brácteas, mas sua função é desconhecida. Umestudo não publicado indica que pelo menos o gêneroRapatea é polinizado por abelhas (Ribeiro et al. 1999).
Para que haja um entendimento básico dessa planta, queprovavelmente têm um papel ecológico importante por suaabundância e função como microhabitat para muitos insetosque vivem dentro do exudato, é preciso descrever suaestrutura floral. Pela grande quantidade de flores em cadaplanta, pode ser desejável utilizar só uma sub-amostra deflores em qualquer estudo. Quais flores serão escolhidas,segundo o arranjo e idade dos múltiplos grupos de flores,constitui um aspecto importante. O objetivo do presenteestudo é descrever a estrutura geral das flores de Rapateaulei e determinar uma metodologia de amostragem dasinflorescências, sem vício de idade ou abundância das floresem uma determinada área.
Material e MétodosO estudo foi realizado na Reserva do Km 41 do Projeto
Dinâmica Biológica de Fragmentos Florestais, na FazendaEsteio, Manaus, AM (2° 24’S; 59° 44’W). Rapatea uleiocorre freqüentemente nas áreas de baixio por toda a reserva(Figura 1). As plantas foram observadas em flor durantetodo o período do estudo (agosto, 2001). Plantas foramescolhidas ao caso, seguindo a trilha “L” da Reserva, correndoao lado de um igarapé. Foram utilizadas apenas plantasadultas com pelo menos uma flor aberta saindo do exudatoque cobre as inflorescências imaturas.
Coletas de inflorescências de 5 plantas (selecionadas aocaso) foram feitas para descrição da morfologia das estruturasflorais. As brácteas foram removidas, para exame dainflorescência e sua distribuição espacial dentro da estrutura.Posteriormente, todas as flores (maduras e imaturas) foram
removidas, separando-as das brácteas na base com cortes nosentido do comprimento das brácteas.
Quinze plantas foram observadas no campo e examinadascom relação à presença, idade e posição de cada flor relativaao comprimento da bráctea. Baseado nos resultados dadissecção da estrutura floral, o comprimento das brácteasfoi dividido em sete compartimentos e a contagem feita porcompartimento. Só flores saindo do exudato ou aparentesforam contadas. Essas estruturas foram categorizadas emtrês grupos: flores abertas, fechadas (antes de abrir) esenescentes. Defino as estruturas florais da seguinte forma:flor, cada unidade dentro de uma inflorescência, que possuientre 100-200 flores, agrupamentos, que são conjuntos deflores dentro de cada inflorescência, e inflorescência. que serefere a todas as flores em conjunto, dentro das duas brácteas.
ResultadosApresento a seguir os dados de 5 das plantas (selecionadas
ao caso) observadas para contagem e distribuição das flores(Tabela 1). Não há um padrão espacial óbvio, mas os dadosnão permitem um teste estatístico.
Tabela 1. Número de flores de Rapatea ulei em cada estadofenológico.
Das quinze plantas amostradas duas tinham 8agrupamentos, mas não foram escolhidas para a amostra dedados apresentada aqui. As brácteas amostradas tinham seteagrupamentos (Figura 2). Todas as plantas tinham duasinflorescências. As flores foram arranjadas mais ou menosem forma de círculos concêntricos em cada agrupamento(Figura 3), com o material floral imaturo no centro e cadacírculo mais externo com estruturas florais mais
Planta 1:Categoria Agrupamento
1 2 3 4 5 6 7Aberta 0 1 0 0 0 0 1Fechada 0 0 0 1 1 0 0Senescente 0 0 0 0 0 0 0Planta 2Categoria Agrupamento
1 2 3 4 5 6 7Aberta 0 0 0 1 0 0 0Fechada 1 0 0 0 0 0 1Senescente 0 1 0 0 0 0 0Planta 3Categoria Agrupamento
1 2 3 4 5 6 7Aberta 0 0 0 0 0 0 1Fechada 1 0 2 0 1 1 0Senescente 0 1 1 1 1 0 0Planta 4Categoria Agrupamento
1 2 3 4 5 6 7Aberta 0 0 0 0 0 0 0Fechada 1 4 0 1 2 2 1Senescente 0 0 1 0 0 1 0Planta 5Categoria Agrupamento
1 2 3 4 5 6 7Aberta 0 1 0 0 0 0 0Fechada 0 0 2 0 1 1 0Senescente 1 0 0 0 0 0 0

Curso de Campo Ecologia da Floresta Amazônica - 2001 87
desenvolvidas.
DiscussãoCada agrupamento aparentemente floresce independente
dos outros e pode ter flores em todas as etapas dedesenvolvimento, independente do número e idade das floresdos outros agrupamentos na mesma inflorescência.Entretanto, não posso concluir isso com certeza, porque nãoé possivel fazer um teste estatístico confiável em função dopequeno número de flores por planta. A variação em termosde número de flores por agrupamento dentro da inflorescênciafoi muita baixa.
Deste modo, concluo que a melhor maneira deamostragem, evitando vício e obtendo uma boa estimativada variabilidade real, seria realizar amostragens deinflorescências correspondentes em cada planta (por
exemplo, amostrando sempre no meio). Outra possibilidadeseria amostrar um grupo do meio e um grupo do lado paraassegurar uma amostra representativa. Assim seria possívelamostrar um agrupamento de cada flor como representativoda população inteira de flores na planta, aumentando aeficiência de amostragem.
Referências BibliográficasRibeiro, J.E.L.S.; Hopkins, M.J.G.; Vicentini, A.; Sothers,
C.A.; Costa, M.A.S.; Brito, J.M.; Souza, M.A.D.;Martins, L.H.P.; Lohmann, L.G.; Assunção, P.A.C.L.;Pereira, E.C.; Silva, C.F.; Mesquita, M.R. & Procópio,L.C. 1999. Flora da Reserva Ducke: Guia deIdentificação das Plantas Vasculares de uma Floresta deTerra Firme da Amazônia Central. INPA-DFID, Manaus.
Composição, riqueza e abundância da ictiofauna de um igarapés de terra firmeda Amazônia Central, AMKatiane Ferreira & Carlos E. Marinelli
IntroduçãoO conhecimento sobre a diversidade da ictiofauna sul-
americana e brasileira é ainda incipiente, particularmente comrespeito às espécies de peixes de água doce (Bölke et al.1978). A América do Sul possui o maior número de espéciesde peixes de água doce entre todas regiões zoogeográficas(Lowe-McConnell 1975), com estimativas variando de 2400a 5000 espécies (Rosa & Menezes 1996).
Pequenos igarapés da Amazônia Central constituembiótopos peculiares para peixes, uma vez que o dossel dafloresta adjacente se fecha sobre os mesmos limitando apenetração de luz na superfície da água, tornando a cadeiaalimentar dependente de aporte de material da florestaadjacente (Lowe-McConnell 1999). Dentre os fatores queregulam a ocorrência e distribuição das espécies de peixesem riachos de pequeno porte, estão a ordem, largura,profundidade, presença de herbáceas aquáticas e estruturada vegetação ripária (Caramaschi 1986), além dadisponibilidade de alimento, intensidade de predação etolerância às condições fisico-químicas da água seconsiderada uma abordagem ecológica mais ampla (Lowe-McConnell 1975).
Estudos sobre a ictiofauna tropical são mais freqüentescom relação a peixes de importância comercial (Lowe-McConnell 1999). Alguns estudos tem sido realizados emigarapés enfocando o análise do recurso alimentar, riqueza,endemismo ou a ampla distribuição geográfica de espécies(Knöppell 1970; Saul 1975; Soares 1979; Silva 1992; Prada-Pedreros 1992). Bührnheim (1998), estudou os igarapés dareserva do km 41 enfocando a estrutura de comunidade depeixes comparando com outros riachos da Amazônia Central.
O presente trabalho teve como objetivo verificar acomposição, riqueza e abundância da ictiofauna de umigarapé de primeira ordem em uma floresta de terra firme.
Material e MétodosO presente trabalho foi desenvolvido em um igarapé de
primeira ordem, localizado na Reserva do Km 41 (ProjetoDinâmica Biológica de Fragmentos Florestais INPA/PDBFF-Smithsonian Institution), na estrada vicinal ZF-3, rodoviaBR 174, 70 km ao norte de Manaus (20º 26’S; 59º 46’W). Areserva é uma área de 10.000 ha de floresta contínua de terrafirme, apresenta temperatura média anual de 26 °C eprecipitação média em torno de 2200 mm/ano (RadamBrasil1978).
As amostragens foram realizadas em três trechos(montante, médio e jusante) de 20 metros longitudinais deum igarapé de primeira ordem. Os três trechos estão situadosmais ou menos eqüidistantes, possibilitando o emprego dametodologia de trabalho de campo semelhantes para os trêslocais. De modo geral, as características ambientais sãohomogêneas, apresentando margem pouco sinuosa, comgalhos e troncos submersos, onde ocorre deposição defolhiço. A correnteza é moderada, sem definição de poços ecorredeiras, com a profundidade variando entre 05 e 37 cm.O substrato é composto por areia fina, sedimento e folhiço.A área delimitada para amostragem foi bloqueada com redesde bloqueio para que não haja evasão dos peixes. Nos trêstrechos dois coletores equipados com peneiras metálicas de50 cm de diâmetro e 3 mm de malhagem, fizeram duaspassagem junto às margens, a coleta em cada trecho teveduração média de uma hora. Os espécimes capturados foram
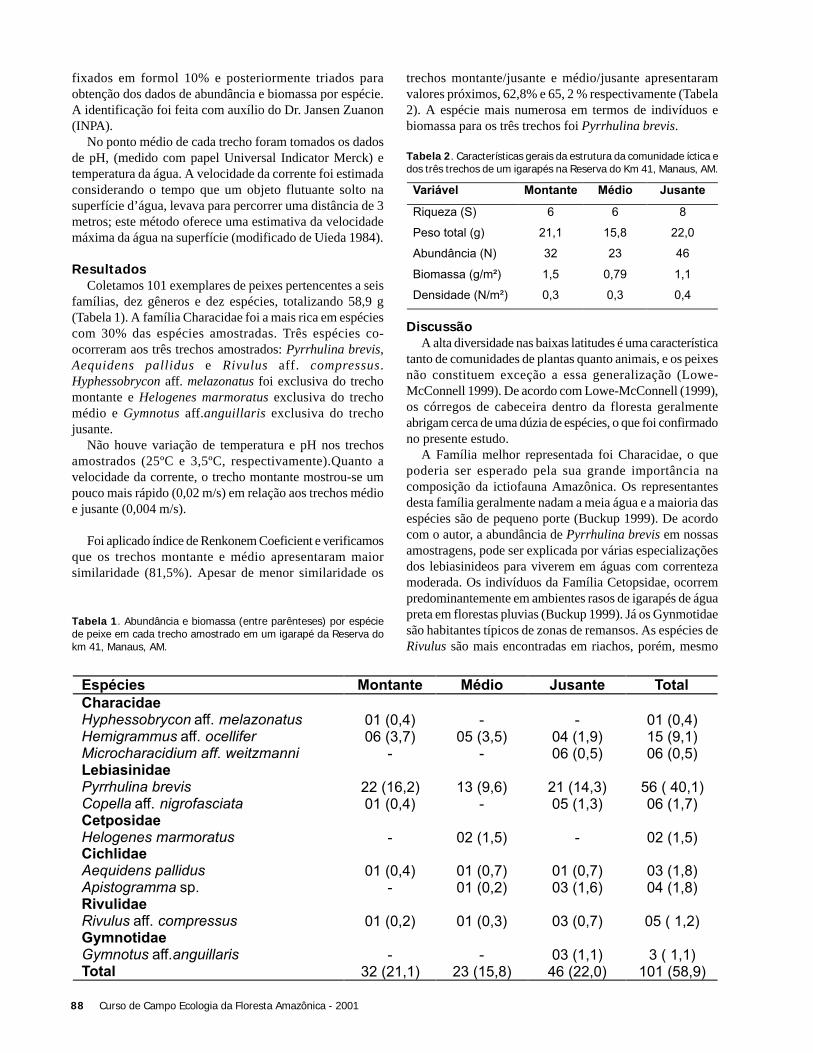
88 Curso de Campo Ecologia da Floresta Amazônica - 2001
fixados em formol 10% e posteriormente triados paraobtenção dos dados de abundância e biomassa por espécie.A identificação foi feita com auxílio do Dr. Jansen Zuanon(INPA).
No ponto médio de cada trecho foram tomados os dadosde pH, (medido com papel Universal Indicator Merck) etemperatura da água. A velocidade da corrente foi estimadaconsiderando o tempo que um objeto flutuante solto nasuperfície d’água, levava para percorrer uma distância de 3metros; este método oferece uma estimativa da velocidademáxima da água na superfície (modificado de Uieda 1984).
ResultadosColetamos 101 exemplares de peixes pertencentes a seis
famílias, dez gêneros e dez espécies, totalizando 58,9 g(Tabela 1). A família Characidae foi a mais rica em espéciescom 30% das espécies amostradas. Três espécies co-ocorreram aos três trechos amostrados: Pyrrhulina brevis,Aequidens pallidus e Rivulus aff. compressus.Hyphessobrycon aff. melazonatus foi exclusiva do trechomontante e Helogenes marmoratus exclusiva do trechomédio e Gymnotus aff.anguillaris exclusiva do trechojusante.
Não houve variação de temperatura e pH nos trechosamostrados (25ºC e 3,5ºC, respectivamente).Quanto avelocidade da corrente, o trecho montante mostrou-se umpouco mais rápido (0,02 m/s) em relação aos trechos médioe jusante (0,004 m/s).
Foi aplicado índice de Renkonem Coeficient e verificamosque os trechos montante e médio apresentaram maiorsimilaridade (81,5%). Apesar de menor similaridade os
Espécies Montante Médio Jusante TotalCharacidaeHyphessobrycon aff. melazonatus 01 (0,4) - - 01 (0,4)Hemigrammus aff. ocellifer 06 (3,7) 05 (3,5) 04 (1,9) 15 (9,1)Microcharacidium aff. weitzmanni - - 06 (0,5) 06 (0,5)LebiasinidaePyrrhulina brevis 22 (16,2) 13 (9,6) 21 (14,3) 56 ( 40,1)Copella aff. nigrofasciata 01 (0,4) - 05 (1,3) 06 (1,7)CetposidaeHelogenes marmoratus - 02 (1,5) - 02 (1,5)CichlidaeAequidens pallidus 01 (0,4) 01 (0,7) 01 (0,7) 03 (1,8)Apistogramma sp. - 01 (0,2) 03 (1,6) 04 (1,8)RivulidaeRivulus aff. compressus 01 (0,2) 01 (0,3) 03 (0,7) 05 ( 1,2)GymnotidaeGymnotus aff.anguillaris - - 03 (1,1) 3 ( 1,1)Total 32 (21,1) 23 (15,8) 46 (22,0) 101 (58,9)
Tabela 1. Abundância e biomassa (entre parênteses) por espéciede peixe em cada trecho amostrado em um igarapé da Reserva dokm 41, Manaus, AM.
trechos montante/jusante e médio/jusante apresentaramvalores próximos, 62,8% e 65, 2 % respectivamente (Tabela2). A espécie mais numerosa em termos de indivíduos ebiomassa para os três trechos foi Pyrrhulina brevis.
Tabela 2. Características gerais da estrutura da comunidade íctica edos três trechos de um igarapés na Reserva do Km 41, Manaus, AM.
DiscussãoA alta diversidade nas baixas latitudes é uma característica
tanto de comunidades de plantas quanto animais, e os peixesnão constituem exceção a essa generalização (Lowe-McConnell 1999). De acordo com Lowe-McConnell (1999),os córregos de cabeceira dentro da floresta geralmenteabrigam cerca de uma dúzia de espécies, o que foi confirmadono presente estudo.
A Família melhor representada foi Characidae, o quepoderia ser esperado pela sua grande importância nacomposição da ictiofauna Amazônica. Os representantesdesta família geralmente nadam a meia água e a maioria dasespécies são de pequeno porte (Buckup 1999). De acordocom o autor, a abundância de Pyrrhulina brevis em nossasamostragens, pode ser explicada por várias especializaçõesdos lebiasinideos para viverem em águas com correntezamoderada. Os indivíduos da Família Cetopsidae, ocorrempredominantemente em ambientes rasos de igarapés de águapreta em florestas pluvias (Buckup 1999). Já os Gynmotidaesão habitantes típicos de zonas de remansos. As espécies deRivulus são mais encontradas em riachos, porém, mesmo
Variável Montante Médio Jusante
Riqueza (S) 6 6 8
Peso total (g) 21,1 15,8 22,0
Abundância (N) 32 23 46
Biomassa (g/m²) 1,5 0,79 1,1
Densidade (N/m²) 0,3 0,3 0,4

Curso de Campo Ecologia da Floresta Amazônica - 2001 89
assim, encontram-se apenas em ambientes periféricos rasos.Os Cichlidae são um dos maiores grupos entre os teleosteosde água doce, incluindo cerca de 1500 espécies, ocorrendoem diferentes ambientes, sendo bem representados em detodas as regiões do Brasil.
As análises mostraram maior similaridade entre os trechosmontante e médio, e menor similaridade entre os trechosmontante e jusante. Esta similaridade entre os trechomontante e o trecho médio, talvez possa ser explicada pelasemelhança entre suas características ambientais, entre elas,deposição de folhiço nas margens do igarapé e pontosisolados de herbáceas aquáticas.
No trecho jusante encontramos a maior riqueza eabundância, este trecho apresentou maior quantidades defolhiço e herbáceas aquáticas, quando comparamas com ostrechos montante e médio. Camadas de folhas arrastadasfornecem cobertura ao substrato formando mosáicos comareias descobertas, restos de árvores e raízes de epífitas caídassob a água, estas servem de esconderijo e cobertura parainvertebrados dos quais os peixes se alimentam (Lowe-McConnell 1999).
Igarapés de pequeno porte, possuem produtividadeprimária in situ relativamente baixa (embora podendo serlocalmente elevada em trechos abertos e ensolarados),dependendo grandemente da importação de matéria orgânicada vegetação ripária para sustentar uma comunidadepredominantemente heterotrófica (Vannote et al. 1980).Assim as fontes principais de alimentação para os peixesneste ambientes são folhas, frutos e artrópodes terrestres quecaem da vegetação ripária, além de larvas aquáticas de insetosque se alimentam, direta ou indiretamente, deste materialorgânico alóctone importado da vegetação adjacente (Castro1999).
AgradecimentosAo Dr. Jansen Zuanon (INPA), pela identificação das
espécies e valiosas sugestões ao trabalho. Ao EduardoVenticinque (Dadão) pela leitura crítica do relatório.
Referências BibliográficasBöhlke, J.E.; Wietzaman, S.H. & Menezes, N.A. 1978.
Estado atual da sistemática dos peixes de água doce daAmerica do sul. Acta Amaz. 8 (4): 657-677.
Buckup, P.A. 1999. Sistemática e biogeografia de peixes deriachos. In: Ecologia de Peixes de Riachos, SérieOecologia Brasiliensis, vol. VI, Caramaschi, E.P.,
Mazzoni, R. & Peres-Neto, P. R. (eds.), pp. 91-138.PPGE-UFRJ, Rio de Janeiro, Brasil.
Bührnheim, C.M. 1998. Estrutura de comunidades de peixesem igarapés floresta de terra firme na Amazônia Central.Dissertação de Mestrado, Universidade do Amazonas,Instituto Nacional de Pesquisa da Amazônia, Manaus.
Castro, R.M.C. 1999. Evolução da ictiofauna de riachos Sul-Americanos: padrões gerais e possíveis processoscasuais. In: . Ecologia de Peixes de Riachos, SérieOecologia Brasiliensis, vol. VI, Caramaschi, E.P.,Mazzoni, R. & Peres-Neto, P. R. (eds), pp. 139-155.PPGE-UFRJ, Rio de Janeiro, Brasil.
Knöppel, H.A. 1970. Food of central Amazonian fishes.Amazoniana 2: 257-352.
Lowe-McConnell, R.H. 1975. Fish Communities in TropicalFreshwaters: Their Distribution, Ecology and Evolution.Longman, London and New York.
Lowe-McConnell, R.H. 1999. Estudos Ecológicos deComunidades de Peixes Tropicais. EDUSP, São Paulo.
Prada-Pedreros, S. 1992. Abundância e distribuição docardinal tetra, Paracheirodon axelrodi (Pisces,Characidae) e diversidade dos peixes nas planíciesinundáveis do médio Rio Negro, Amazonas, Brasil.INPA/FUA. Manaus.
Rosa, R.S.; Menezes, N. A. 1996. Relação preliminar dasespécies de peixes (Pisces: Elasmobranchii:Actinopterygii) ameaçadas no Brasil. Revta. Bras. Zool.13(3): 647-667.
Saul, W.G. 1975. An ecological study of fishes at a site inupper amazonian Ecuador. Proc. Acad. Nat. Sc.Philadelphia 127: 93-34.
Silva, C.P.D. 1992. Influência das modificações ambientaissobre a comunidade de peixes de um igarapé da cidadede Manaus (Amazonas). Dissertação de Mestrado, INPA/FUA, Manaus.
Soares, M.C.M. 1979. Aspectos ecológicos (alimentação ereprodução) dos peixes do igarapé do Porto Aripuanã,MT. Acta Amaz. 2 (3): 257-352.
RadamBrasil. 1978. Levantamento de recursos naturais.Folha SA 20 Manaus 261 p. Ministério de Minas eEnergia: Departamento Nacional de Produção Mineral,Rio de Janeiro, Brasil.
Uieda, V.S. 1984. Ocorrência e distribuição dos peixes emum riacho de água doce. Revta. Bras. Biol. 44(2): 203-213.

90 Curso de Campo Ecologia da Floresta Amazônica - 2001
Respostas a herbívoros simulados na mirmecófita Maieta poepigii(Melastomataceae): Pheidole minutula Mayr (Formicidae) é que era a
formiga de verdade!Alexander Vicente Christianini
IntroduçãoPlantas mirmecófitas são um elemento conspícuo em
diversos ecossistemas Neotropicais (Benson 1985; Fonseca& Ganade 1996). Estas plantas possuem domáceas e nectáriosextraflorais que permitem a instalação de colônias deformigas (Hölldobler & Wilson 1990). Interações entreplantas mirmecófitas e formigas têm sido apontadas comomutualísticas em diversos estudos (Schupp 1986; Fonseca1993, 1999; Fonseca & Ganade 1996; Vasconcelos 1991).Nestas interações, as formigas obtêm locais para ninho erecursos alimentares oferecidos pelas plantas, como néctar ecorpúsculos alimentares (Benson 1985; Fonseca 1993, 1999;Fonseca & Ganade 1996), enquanto as plantas sãobeneficiadas pela proteção oferecida pelas formigas contraherbívoros (Schupp 1986; Oliveira et al. 1987; Vasconcelos1991). A maior parte destes trabalhos se concentra na induçãoda defesa por ação direta dos herbívoros.
Contudo, estudos recentes mostram que as própriasplantas possuem mecanismos de indução de defesa,estimulando a atuação das formigas contra herbívoros pormeio de mediadores químicos (Agrawal 1998; Agrawal &Dubin-Thaler 1999). A efetividade destas outras formas deindução é pouco conhecida.
Neste trabalho eu comparei a efetividade de três formasde defesa induzida em Maieta poepigii (Melastomataceae),uma mirmecófita ocorrente nos baixios de florestas de terrafirme da Amazônia (Ribeiro et al. 1999). Após uma brevecaracterização do sistema, eu investiguei as seguintesquestões:
1) Formigas são agentes bióticos eficientes contraherbívoros em M. poepigii ?
2) M. poepigii possui mecanismos de indução de defesabiótica contra herbívoros (e.g. físico-químicos) ?
3) Qual a efetividade destas outras formas de indução pelaplanta contra herbívoros ?
Material e MétodosÁrea de estudoEu desenvolvi este estudo na Reserva Florestal do Km 41
(2º24’S; 59º44’W), uma área de mata contínua pertencenteao projeto Dinâmica Biológica de Fragmentos Florestais(PDBFF - INPA/Smithsonian Institution), a ca. 70 Km aonorte de Manaus, AM. A vegetação é composta por florestade terra firme. A precipitação anual é de 2.100 mm, comestação chuvosa entre novembro e maio e estação seca entrejunho e outubro. Uma descrição detalhada da área de estudopode ser encontrada em Lovejoy & Bierregard Jr. (1990).
Caracterização do sistemaMaieta poepigii (Melastomataceae) é um arbusto que
ocorre no sub-bosque dos baixios de florestas de terra firmeda Amazônia (Ribeiro et al. 1999). Apresenta domáceas queconsistem em duas câmaras adjacentes localizadas na baseda lâmina foliar. Cada uma das câmaras se abre por umapequena passagem na superfície abaxial da folha, na junçãocom a veia central e a nervura lateral principal (Benson 1985).Colônias de formigas se estabelecem nas domáceas de M.poepigii, mas abordagens experimentais que caracterizem asinterações das formigas com a planta hospedeira não sãoreportadas na literatura. Um estudo com Maieta guianensis,simpátrica e localmente mais abundante, indicou uma quedadrástica no sucesso reprodutivo das plantas, quando daremoção das colônias de formigas (Vasconcelos 1991). Poroutro lado, Pheidole minutula, a formiga presente em 95%dos indivíduos de M. guianensis, é conhecida por sobreviverapenas em plantas mirmecófitas (Vasconcelos 1991).
Eu realizei as observações deste trabalho entre 29/08 e01/09/01, concentrando-me em uma área de 200 m ao longodas margens do igarapé do acampamento (entre trilhas L2 eL4) e seu afluente, junto à represa de captação de água.Visando caracterizar a associação entre formigas e indivíduosde M. poepigii de diferentes tamanhos, foram marcadosalguns indivíduos de M. poepigii (n = 50) encontrados a até15 m da margem dos igarapés. Para cada indivíduo foianotado a altura da planta a partir do solo até a gema apical,o número total de folhas, o número de folhas com domácease a presença de formigas. Eu coletei uma amostra de folhasde indivíduos de diferentes tamanhos para caracterizareventuais relações alométricas da superfície foliar com ovolume das domáceas, utilizada como indicador do espaçodisponível para instalação da colônia de formigas (Fonseca1993)
O padrão de atividade das formigas sobre as folhas de M.poepigii foi determinado por contagens instantâneas dasformigas presentes na superfície adaxial de uma folha novae uma folha velha em cada uma de 11 plantas previamentemarcadas. Como as plantas ocorrem em agrupamentos,buscou-se amostrar uma ou duas plantas por agrupamento,num total de dez agrupamentos amostrados.
ExperimentosEu realizei os experimentos de forma simultânea com a
caracterização do sistema de associação entre formigas e M.poepigii. A resposta das formigas a herbívoros simulados edanos induzidos foi investigada em plantas de M. poepigiiescolhidas aleatoriamente em diferentes agrupamentos.Embora duas espécies de formigas tenham sido registradas

Curso de Campo Ecologia da Floresta Amazônica - 2001 91
em associação com M. poepigii, eu conduzi os experimentossomente em plantas associadas com Pheidole minutula,presentes em ca. 90% das associações. Eu conduzi osexperimentos de forma pareada em folhas de tamanhossimilares e nós distintos de uma mesma planta. Foramrealizados três experimentos, com dez réplicas cada:
a) herbívoros simulados: cupins (Isoptera) foramempregados como herbívoros simulados sobre as folhas(Oliveira et al. 19897). Um operário vivo de cupim (ausênciade defesa química) foi depositado próximo ao ápice de umafolha, enquanto outra folha de um nó próximo recebiasimultaneamente um pedaço de pedaço de palito de dente detamanho similar ao cupim, que funcionou como controle;
b) danos físicos: a resposta de formigas a danosprovocados por um herbívoro cortador foi simulada por danosfísicos provocados pelo observador. Neste segundoexperimento, uma das folhas selecionadas tinha o ápice (1/5da folha) cortado com uma tesoura, enquanto o controle eracomposto por outra folha que foi pressionada levemente comuma pinça, simultaneamente;
c) indução química: visando quantificar uma eventualresposta de formigas a estímulos químicos associados comtecidos da planta hospedeira danificados (Agrawal 1998;Agrawal et al. 1999), uma fração de uma folha (2,5 x 1,5cm) foi macerada em 2 ml de água. Parte do líquido resultante(0,2 ml) foi aplicado cuidadosamente próximo ao ápice deuma das folhas com o uso de uma seringa plástica. Comocontrole, outra folha recebia no mesmo instante um volumeequivalente de água próximo ao ápice.
Observações sobre o comportamento das formigas foramrealizadas ad libitum. A resposta das formigas àsmanipulações experimentais foi quantificada por duasvariáveis: a) o tempo de detecção do herbívoro simulado/dano, representado pelo intervalo de tempo entre o início doexperimento e o primeiro contato físico entre uma formiga eo tratamento em questão; b) o número de formigas recrutadas,dado pelo número de formigas em contato com os tratamentosem contagens instantâneas em intervalos de dois minutos apartir da detecção, até o máximo de dez minutos.
Regressões logísticas foram empregadas para investigaras variáveis individuais de M. poepigii relacionadas com apresença/ausência de formigas associadas. Comparaçõesentre o tempo de detecção dos tratamentos e dos controlesdos experimentos pelas formigas foram realizadas com o testepareado de Wilcoxon. O recrutamento de formigas entretratamentos ao longo do intervalo de tempo das observaçõesfoi comparado com o uso de ANOVA de medidas repetidas,utilizando tratamento e tempo como fatores e o número deformigas em cada intervalo de dois minutos como variáveldependente. Todos os testes seguiram Zar (1984).
ResultadosForam registradas duas espécies de formigas associadas
às domáceas de Maieta poepigii: Pheidole sp. (provavelmenteP. minutula) e Crematogaster sp.(provavelmente C. laevis).Pheidole minutula está presente na maioria dos indivíduos(n = 23 plantas; 88,5%) e agrupamentos de plantas observadas
(n = 10 plantas; 83,5%) com formigas. A presença deformigas está mais fortemente relacionada com o número dedomáceas de uma planta (Tabela 1). O número de domáceaspor sua vez é determinado pelo número total de folhas (r2 =0,84; n =51; F = 248,6; p < 0,001) e pela altura da planta (r2
= 0,68; n = 51; F = 105,8; p < 0,001).Formigas patrulham as folhas de M. poepigii
ininterruptamente durante o dia e a noite (Fig. 1). Operáriasforam predominantes em atividades de patrulha das folhas.Soldados foram registrados em apenas um evento (6:30 h),quando houve o aprisionamento de um inseto e suamanipulação por cerca de 20 formigas sobre a folha.
Tabela 1. Regressões logísticas de características individuais de M.poepigii em relação à presença (n = 22) ou ausência (n = 28) deformigas associadas.
Figura 1. Ciclo diário de atividade de formigas associadas à Maietapoepigii. Valores correspondem à contagens instantâneas do númerode formigas (média ± erro padrão) sobre a face adaxial das folhasde 11 plantas em 28 de agosto de 2001.
Não foram encontradas diferenças entre o tempo dedetecção dos tratamentos em relação aos controles paraherbívoros simulados (teste pareado de Wilcoxon; z = 1,48;n = 10; p = 0,14), danos físicos (z = 1,36; n= 10; p = 0,17)ou indução química (z = -0,059; n = 10; p = 0,95). Contudo,houve um recrutamento significativo de formigas para a áreaafetada pelos tratamentos (Tabela 2, Figura 2). Foramobservadas variações temporais no número de formigas emcontato com os tratamentos para herbívoros simulados edanos físicos, mas não houve diferenças temporais ao longodo intervalo de observações do experimento de induçãoquímica (Tabela 2). Houve uma interação significativa entreo intervalo de observação e o tratamento de herbívorossimulados, com um número crescente de formigas (até setevezes mais) atendendo o tratamento em relação aos controles(Figura 2).
A ausência de efeitos temporais e de interação entre algunstratamentos e o tempo de observação pode ter sido decorrente
Característica Odds
ratio
Intervalo
superior
Intervalo
inferior
G P
Altura da planta 1,059 1,096 1,022 13,404 < 0,001
Nº total de folhas 1,308 1,583 1,081 10,340 0,001
Nº total de
domáceas
1,689 2,357 1,211 14,515 <0,001
0
0,2
0,4
0,6
0,8
1
1,2
6:30
8:30
10:3
012
:30
14:3
016
:30
18:3
020
:30
22:3
00:
302:
304:
30
Horário
Nº
de
form
igas
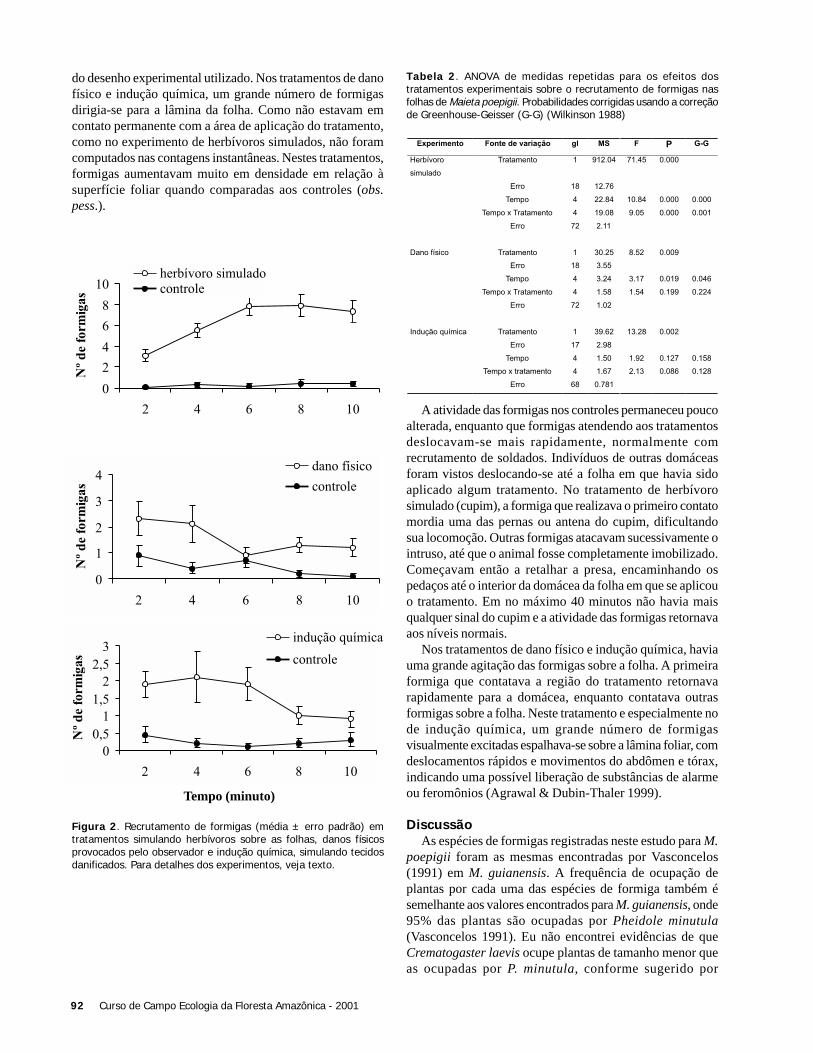
92 Curso de Campo Ecologia da Floresta Amazônica - 2001
do desenho experimental utilizado. Nos tratamentos de danofísico e indução química, um grande número de formigasdirigia-se para a lâmina da folha. Como não estavam emcontato permanente com a área de aplicação do tratamento,como no experimento de herbívoros simulados, não foramcomputados nas contagens instantâneas. Nestes tratamentos,formigas aumentavam muito em densidade em relação àsuperfície foliar quando comparadas aos controles (obs.pess.).
Tempo (minuto)
Figura 2. Recrutamento de formigas (média ± erro padrão) emtratamentos simulando herbívoros sobre as folhas, danos físicosprovocados pelo observador e indução química, simulando tecidosdanificados. Para detalhes dos experimentos, veja texto.
0
2
4
6
8
10
2 4 6 8 10
Nº
de
form
iga
s
herbívoro simuladocontrole
0
1
2
3
4
2 4 6 8 10
Nº
de
form
iga
s
dano físico
controle
00,5
11,5
22,5
3
2 4 6 8 10
Nº
de
form
iga
s
indução química
controle
Tabela 2. ANOVA de medidas repetidas para os efeitos dostratamentos experimentais sobre o recrutamento de formigas nasfolhas de Maieta poepigii. Probabilidades corrigidas usando a correçãode Greenhouse-Geisser (G-G) (Wilkinson 1988)
A atividade das formigas nos controles permaneceu poucoalterada, enquanto que formigas atendendo aos tratamentosdeslocavam-se mais rapidamente, normalmente comrecrutamento de soldados. Indivíduos de outras domáceasforam vistos deslocando-se até a folha em que havia sidoaplicado algum tratamento. No tratamento de herbívorosimulado (cupim), a formiga que realizava o primeiro contatomordia uma das pernas ou antena do cupim, dificultandosua locomoção. Outras formigas atacavam sucessivamente ointruso, até que o animal fosse completamente imobilizado.Começavam então a retalhar a presa, encaminhando ospedaços até o interior da domácea da folha em que se aplicouo tratamento. Em no máximo 40 minutos não havia maisqualquer sinal do cupim e a atividade das formigas retornavaaos níveis normais.
Nos tratamentos de dano físico e indução química, haviauma grande agitação das formigas sobre a folha. A primeiraformiga que contatava a região do tratamento retornavarapidamente para a domácea, enquanto contatava outrasformigas sobre a folha. Neste tratamento e especialmente node indução química, um grande número de formigasvisualmente excitadas espalhava-se sobre a lâmina foliar, comdeslocamentos rápidos e movimentos do abdômen e tórax,indicando uma possível liberação de substâncias de alarmeou feromônios (Agrawal & Dubin-Thaler 1999).
DiscussãoAs espécies de formigas registradas neste estudo para M.
poepigii foram as mesmas encontradas por Vasconcelos(1991) em M. guianensis. A frequência de ocupação deplantas por cada uma das espécies de formiga também ésemelhante aos valores encontrados para M. guianensis, onde95% das plantas são ocupadas por Pheidole minutula(Vasconcelos 1991). Eu não encontrei evidências de queCrematogaster laevis ocupe plantas de tamanho menor queas ocupadas por P. minutula, conforme sugerido por
Experimento Fonte de variação gl MS F P G-G
Herbívoro
simulado
Tratamento 1 912.04 71.45 0.000
Erro 18 12.76
Tempo 4 22.84 10.84 0.000 0.000
Tempo x Tratamento 4 19.08 9.05 0.000 0.001
Erro 72 2.11
Dano físico Tratamento 1 30.25 8.52 0.009
Erro 18 3.55
Tempo 4 3.24 3.17 0.019 0.046
Tempo x Tratamento 4 1.58 1.54 0.199 0.224
Erro 72 1.02
Indução química Tratamento 1 39.62 13.28 0.002
Erro 17 2.98
Tempo 4 1.50 1.92 0.127 0.158
Tempo x tratamento 4 1.67 2.13 0.086 0.128
Erro 68 0.781

Curso de Campo Ecologia da Floresta Amazônica - 2001 93
Vasconcelos & Davidson (2000) para M. guianensis.A distância de outras plantas com formigas é pouco
explicativa para a ausência de formigas em alguns indivíduosde M. poepigii, já que plantas sem formigas foramencontradas nos mesmos agrupamentos de plantas associadascom formigas. A presença de formigas está relacionada maisfortemente com o número de domáceas de uma planta, quepor sua vez está relacionado com a altura e o número total defolhas de uma planta. Assim, plantas maiores e provavelmentemais velhas, tem uma maior probabilidade de estaremassociadas com formigas. Este padrão pode ser resultado defatores históricos e/ou relacionados à biologia das plantas.As plantas menores podem ter sido originadas a partir desementes germinadas na última ou penúltima estaçãochuvosa. Como a colonização por formigas é dependente dachegada de rainhas aladas que dispersam especialmentedurante a estação chuvosa (Vasconcelos 1993), é possívelque estas plantas não tenham sido encontradas ou nãoapresentem condições adequadas que possibilitem ainstalação e desenvolvimento de uma colônia de formigas. Adisponibilidade de espaço para ninho é um recurso limitanteao desenvolvimento de colônias de formigas em plantasmirmecófitas (Fonseca 1993, 1999). Plantas menoresapresentam folhas menores (obs. pess.). O volume dedomáceas está positivamente relacionado com a área foliarem M. poepigii (r = 0,85; n = 12 folhas; p < 0,001), indicandoque a disponibilidade de espaço para ninho esteja limitandoa ocupação de indivíduos menores de M. poepigii.
Formigas são patrulhadores eficientes contra herbívorossobre as folhas de M. poepigii. A atividade ininterrupta deformigas garante que eventuais herbívoros que cheguem àsfolhas possam ser detectados e repelidos antes que promovamdanos significativos (Oliveira et al. 1987). Esta resposta éconfirmada pela ausência de diferenças entre o tempo dedetecção dos tratamentos experimentais em relação aoscontroles, indicando que a detecção de herbívoros sobre asfolhas é determinada em primeira instância pela atividade depatrulhamento de formigas.
Porém, existem diferenças com relação ao comportamentoe recrutamento de formigas que atendem a diferentestratamentos. A presença de um inseto (herbívoro simulado)estimula uma resposta mais intensa da colônia de formigasem comparação a danos físicos e indução química. Esteresultado precisa ser interpretado com cautela, já que ametodologia utilizada não amostrou o número de formigaspresentes na folha como um todo, que aumentavam muitoem abundância quando da aplicação dos tratamentos de danofísico e indução química..
Formigas reconheceram danos físicos associados à perdade tecido foliar, indicando que estímulos físicos e químicospossam estar envolvidos. A resposta positiva ao tratamentode indução química reforça a idéia de que compostosliberados por tecidos danificados estimulem o recrutamentode formigas. Danos físicos e indução química estimulamcomportamento agressivo e recrutamento de formigas Aztecaassociadas a Cecropia obtusifolia (Urticaceae) (Agrawal1998; Agrawal & Dubin-Thaler 1999). Comportamento
semelhante foi observado para Maieta guianensis(Christianini et al. 2001), indicando que estímulos físico-químicos podem estar presentes em associações de plantasmirmecófitas e formigas não relacionadas filogeneticamente.
Estudos recentes têm reforçado o papel da induçãoquímica em detrimento de estímulos visuais e mecânicos emreações anti-herbívoros de formigas associadas a plantasmirmecófitas (Agrawal 1998; Agrawal & Dubin-Thaler1999). Compostos voláteis parecem estar envolvidos,promovendo recrutamento mais intenso de formigas durantepoucos momentos após o dano (Agawal & Dubin-Thaler1999). Queda brusca no recrutamento de formigas poucosminutos após a aplicação de líquidos resultantes de tecidosfoliares danificados foi verificada também para M. guianensis(Christianini et al. 2001). A ausência de diferenças temporaisno número de formigas recrutadas no tratamento de induçãoquímica em M. poepigii pode estar relacionada ao rápidorecrutamento de formigas logo após a detecção do tratamento,seguido pelo pouco tempo de observação subsequente, alémdas restrições do método de contagem empregado.
Em Cecropia o número de formigas está positivamenterelacionado com a resistência da planta contra herbívoros(Rocha & Bergallo 1992). Estímulos físicos e químicos àresposta de formigas devem aumentar o valor adaptativo dasplantas que se utilizam de respostas induzidas contraherbívoros, aumentando rapidamente o número de formigassobre a lâmina foliar (Agrawal 1998; e referências aíindicadas). Danos provocados por herbívoros podemcomprometer seriamente o sucesso reprodutivo de plantas(Marquis 1984). Indivíduos de M. guianensis que tiveramsuas colônias de formigas removidas experimentalmente,apresentaram intensa herbivoria e sucesso reprodutivo 45vezes menor quando comparado a plantas em que as colôniasde formigas foram mantidas (Vasconcelos 1991).
As observações deste trabalho indicam que as interaçõesentre formigas e M. poepigii podem ser encaradas comomutualísticas. Formigas parecem ser protetores eficientescontra herbívoros em M. poepigii, enquanto beneficiam-sedo espaço para ninho, provavelmente consumindo alimentooferecido pela planta sob a forma de corpúsculos alimentares(Vasconcelos 1991). Esta associação é bastante refinada, aponto de possuir mecanismos de resposta anti-herbívorosbaseados em indução química (Agrawal 1998). Observaçõese abordagens experimentais em outras plantas mirmecófitaspodem permitir avaliar a extensão das conclusões destetrabalho para outras plantas mirmecófitas (e.g. Tococa,Duroia, Hirtella, Pouroma, Tachigali e Cecropia) e formigasassociadas (e.g. Azteca, Allomerus, Pseudomyrmex,Camponotus, Pheidole e Crematogaster) ocorrentes na áreade estudo (Fonseca 1999). Na Amazônia Central, plantasmirmecófitas podem atingir densidade de 380 plantas/ha, coma comunidade local de plantas mirmecófitas e formigas quese associam a estas plantas atingindo 16 e 25 espécies,respectivamente (Fonseca & Ganade 1996).
Referências Bibliográficas

94 Curso de Campo Ecologia da Floresta Amazônica - 2001
Agrawal, A.A. & Dubin-Thaler, B.J. 1999. Induced responsesto herbivory in the Neotropical ant-plant associationbetween Azteca ants and Cecropia trees: response ofants to potential inducing cues. Behav. Ecol. Sociol. 45:47-54.
Agrawal, A.A. 1998. Leaf damage and associated cues induceagressive ant recruitment in a Neotropical ant-plant.Ecology 79: 2100-2112.
Benson, W.E. 1985. Amazon ant-plants. In: G. Prance & T.Lovejoy (eds.), Amazônia, Pp. 239-266. Pergamon Press,Elmsford, New York.
Christianini, A.V.; Lamberts, A.; Aguiar, A.J.C.; Ferreira,K. & Timo, T. 2001. Reações anti-herbívoros da formigaPheidole minutula Mayr (Formicidae, Myrmicinae) emuma planta mirmecófita, Maieta guianensis Aubl.(Melastomataceae). Livro do Curso de Campo “Ecologiada Floresta Amazônica”, INPA/Smithsonian Instiution,Manaus, AM.
Fonseca, C.R.S.D. 1993. Nesting space limits colony sizeof the plant-ant Pseudomyrmex concolor. Oikos 67: 473-482.
Hölldobler, B. & Wilson, E.O. 1990. The Ants. SpringerVerlag, Berlin.
Lovejoy, T.E. & Bierregard Jr., R.O. 1990. CentralAmazonian forests and the minimum critical size ofecossystems project. In: Four Neotropical Rainforests,Gentry, A.H. (ed.)., pp. 60-71. Yale University Press,London.
Marquis, R.J. 1984. Leaf herbivores decrease fitness of atropical plant. Science 226: 537-539.
Oliveira, P.S.; Oliveira-Filho, A.T. & Cintra, R. 1987. Antforaging on ant-inhabited Triplaris (Polygonaceae) in
western Brazil: a field experiment using live termite-baits. J. Trop. Ecol. 3:
Ribeiro, J.E.L.S.; Hopkins, M.J.G.; Vicentini, A.; Sothers,C.A.; Costa, M.A.S.; Brito, J.M.; Souza, M.A.D.;Martins, L.H.P.; Lohmann, L.G.; Assunção, P.A.C.L.;Pereira, E.C.; Silva, C.F.; Mesquita, M.R. & Procópio,L.C. 1999. Flora da Reserva Ducke: Guia deIdentificação das Plantas Vasculares de uma Floresta deTerra-Firme na Amazônia Central. INPA, Manaus.
Rocha, C.F.D. & Bergallo, H.G. 1992. Bigger ant coloniesreduce herbivory and herbivore residence time on leavesof na ant-plant: Azteca muelleri vs. Coelomeraruficornis on Cecropia pachystachia. Oecologia 91:249-252.
Schupp, E.W. 1986. Azteca protection of Cecropia: antocupation benefits juvenile trees. Oecologia 70: 379-385.
Vasconcelos, H.L. 1991. Mutualism between Maietaguianensis Aubl., a myrmecophytic melastome, and oneof its ant inhabitants: ant protection against herbivores.Oecologia 87: 295-298.
Vasconcelos, H.L. 1993. Ant colonization of Maietaguianensis seedlings, na Amazonian ant-plant.Oecologia 95: 439-443.
Vasconcelos, H.L. & Davidson, D.W. 2000. Relationshipbetween plant size and ant associates in two Amazonianant-plants. Biotropica 32: 100-111.
Wilkinson, L. 1988. SYSTAT. The System for Statistics.Evanston, LL. Systat Inc.
Zar, J.H. 1984. Biostatistical Analysis. Prentice Hall, NewYork.
Influência do tamanho e tipo de presa no recrutamento de indivíduos da aranhasocial Anelosimus eximius (Araneae: Theridiidae)
Andrea von der Heyde Lamberts
IntroduçãoAranhas em geral são animais estritamente solitários,
porém, algumas espécies desenvolveram a socialidade (Foelix1982). Para que uma espécie de aranha seja consideradasocial, ela deve preencher três requisitos básicos: tolerânciaa indivíduos coespecíficos; tendência de gregarismo ecooperação em atividades como captura de presas e cuidadocom a prole (Foelix 1982). O fenômeno da socialidade emaranhas originou-se diversas vezes, já que as famílias quepossuem espécies sociais são distintas filogeneticamente(Eresidae, Dictynidae, Theridiidae e Agelenidae) (Foelix1982; D’Andrea 1987). Aranhas sociais dividem tarefas comocaptura de presas e cuidado da prole, além de dividir a mesmateia (Pasquet et al. 1997). O comportamento de socialidadetraz vantagens como metabolismo e crescimento mais rápidos,maior expectativa de vida, menor taxa de mortalidade e
redução na fecundidade (Foelix 1982).Anelosimus eximius é uma espécie de aranha social da
família Theridiidae, sendo que sua distribuição geográficavai do Panamá ao Estado de Santa Catarina, Brasil (Levi1963). As colônias são formadas por cerca de 100 a 1000indivíduos e as teias se localizam em árvores ou arbustos aum metro ou mais do solo. Há predomínio de fêmeas e doistipos de reprodução da colônia: 1) haplometrose, na qualfêmeas grávidas dispersam e iniciam novas colônias e 2)sociotomia, na qual ocorre migração de grupos de aranhaspara formar novas colônias em conjunto (Venticinque 1995e ref. cit.).
Por serem aranhas sociais, indivíduos de A. eximius podemsubjugar presas maiores do que espécies solitárias de tamanhoequivalente. Portanto, o esforço individual que seria investidoem uma presa grande é diluído quando se atua em grupo na
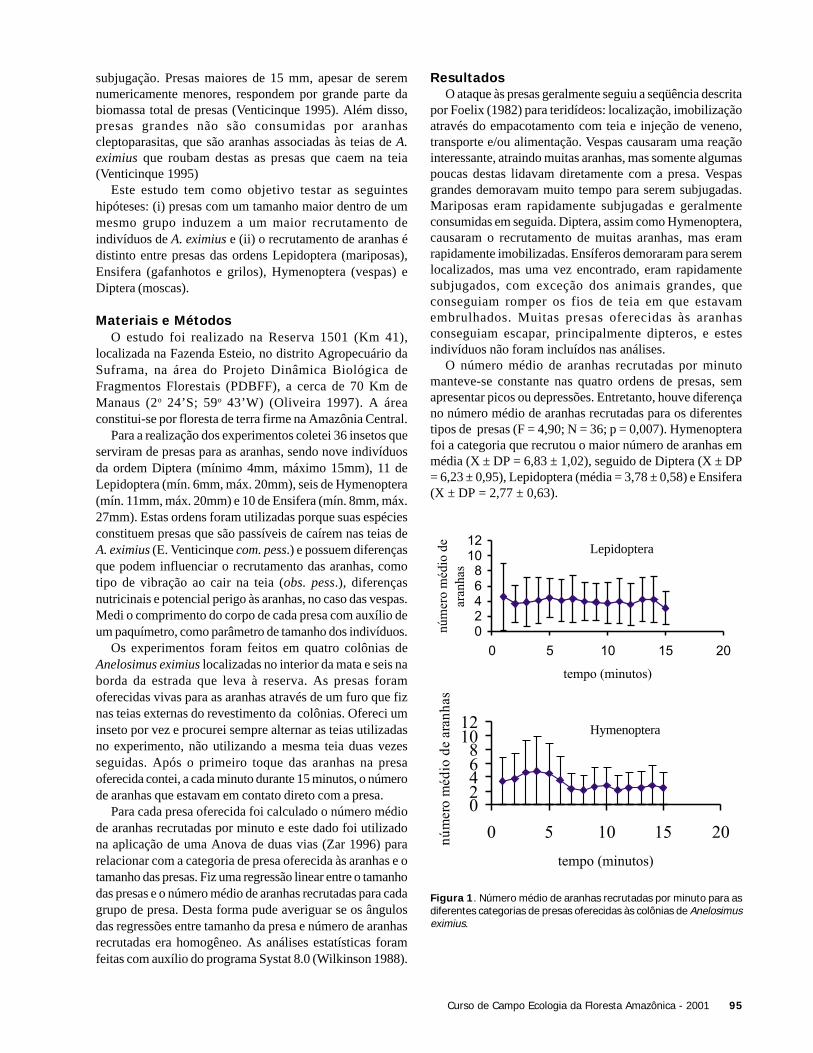
Curso de Campo Ecologia da Floresta Amazônica - 2001 95
subjugação. Presas maiores de 15 mm, apesar de seremnumericamente menores, respondem por grande parte dabiomassa total de presas (Venticinque 1995). Além disso,presas grandes não são consumidas por aranhascleptoparasitas, que são aranhas associadas às teias de A.eximius que roubam destas as presas que caem na teia(Venticinque 1995)
Este estudo tem como objetivo testar as seguinteshipóteses: (i) presas com um tamanho maior dentro de ummesmo grupo induzem a um maior recrutamento deindivíduos de A. eximius e (ii) o recrutamento de aranhas édistinto entre presas das ordens Lepidoptera (mariposas),Ensifera (gafanhotos e grilos), Hymenoptera (vespas) eDiptera (moscas).
Materiais e MétodosO estudo foi realizado na Reserva 1501 (Km 41),
localizada na Fazenda Esteio, no distrito Agropecuário daSuframa, na área do Projeto Dinâmica Biológica deFragmentos Florestais (PDBFF), a cerca de 70 Km deManaus (2o 24’S; 59o 43’W) (Oliveira 1997). A áreaconstitui-se por floresta de terra firme na Amazônia Central.
Para a realização dos experimentos coletei 36 insetos queserviram de presas para as aranhas, sendo nove indivíduosda ordem Diptera (mínimo 4mm, máximo 15mm), 11 deLepidoptera (mín. 6mm, máx. 20mm), seis de Hymenoptera(mín. 11mm, máx. 20mm) e 10 de Ensifera (mín. 8mm, máx.27mm). Estas ordens foram utilizadas porque suas espéciesconstituem presas que são passíveis de caírem nas teias deA. eximius (E. Venticinque com. pess.) e possuem diferençasque podem influenciar o recrutamento das aranhas, comotipo de vibração ao cair na teia (obs. pess.), diferençasnutricinais e potencial perigo às aranhas, no caso das vespas.Medi o comprimento do corpo de cada presa com auxílio deum paquímetro, como parâmetro de tamanho dos indivíduos.
Os experimentos foram feitos em quatro colônias deAnelosimus eximius localizadas no interior da mata e seis naborda da estrada que leva à reserva. As presas foramoferecidas vivas para as aranhas através de um furo que fiznas teias externas do revestimento da colônias. Ofereci uminseto por vez e procurei sempre alternar as teias utilizadasno experimento, não utilizando a mesma teia duas vezesseguidas. Após o primeiro toque das aranhas na presaoferecida contei, a cada minuto durante 15 minutos, o númerode aranhas que estavam em contato direto com a presa.
Para cada presa oferecida foi calculado o número médiode aranhas recrutadas por minuto e este dado foi utilizadona aplicação de uma Anova de duas vias (Zar 1996) pararelacionar com a categoria de presa oferecida às aranhas e otamanho das presas. Fiz uma regressão linear entre o tamanhodas presas e o número médio de aranhas recrutadas para cadagrupo de presa. Desta forma pude averiguar se os ângulosdas regressões entre tamanho da presa e número de aranhasrecrutadas era homogêneo. As análises estatísticas foramfeitas com auxílio do programa Systat 8.0 (Wilkinson 1988).
ResultadosO ataque às presas geralmente seguiu a seqüência descrita
por Foelix (1982) para teridídeos: localização, imobilizaçãoatravés do empacotamento com teia e injeção de veneno,transporte e/ou alimentação. Vespas causaram uma reaçãointeressante, atraindo muitas aranhas, mas somente algumaspoucas destas lidavam diretamente com a presa. Vespasgrandes demoravam muito tempo para serem subjugadas.Mariposas eram rapidamente subjugadas e geralmenteconsumidas em seguida. Diptera, assim como Hymenoptera,causaram o recrutamento de muitas aranhas, mas eramrapidamente imobilizadas. Ensíferos demoraram para seremlocalizados, mas uma vez encontrado, eram rapidamentesubjugados, com exceção dos animais grandes, queconseguiam romper os fios de teia em que estavamembrulhados. Muitas presas oferecidas às aranhasconseguiam escapar, principalmente dipteros, e estesindivíduos não foram incluídos nas análises.
O número médio de aranhas recrutadas por minutomanteve-se constante nas quatro ordens de presas, semapresentar picos ou depressões. Entretanto, houve diferençano número médio de aranhas recrutadas para os diferentestipos de presas (F = 4,90; N = 36; p = 0,007). Hymenopterafoi a categoria que recrutou o maior número de aranhas emmédia (X ± DP = 6,83 ± 1,02), seguido de Diptera (X ± DP= 6,23 ± 0,95), Lepidoptera (média = 3,78 ± 0,58) e Ensifera(X ± DP = 2,77 ± 0,63).
02468
1012
0 5 10 15 20
tempo (minutos)
núm
ero
méd
iode
aran
has
02468
1012
0 5 10 15 20
tempo (minutos)
núm
ero
méd
iode
aran
has
Lepidoptera
Hymenoptera
Figura 1. Número médio de aranhas recrutadas por minuto para asdiferentes categorias de presas oferecidas às colônias de Anelosimuseximius.

96 Curso de Campo Ecologia da Floresta Amazônica - 2001
Há uma relação do número de aranhas recrutadas emfunção do tamanho das presas (F = 5,96; N = 36; p = 0,021).No entanto, quando testei a homogeneidade dos ângulosrepresentada pela interação, notei que o número de aranhasrecrutadas varia em função do tamanho da presa de formadiferente (F = 5,31; N = 36; p = 0,001). Esse resultadodemonstra que os ângulos das regressões entre o número dearanhas recrutadas e o tamanho da presa oferecida não sãohomogêneos (Figura 2, Tabela 1).
Figura 2. Regressão entre o tamanho do corpo de presas das ordensLepidoptera e Diptera e o número médio de aranhas recrutadas porminuto.
O número de aranhas recrutadas em função do tamanhoda presa foi fortemente relacionado para dípteros elepidópteros (Figura 2, Tabela 1). Em Diptera o ângulo daregressão foi maior indicando que existe um acréscimo maiorno número de aranhas para cada milímetro de tamanho daspresas do que em Lepidoptera (Tabela 1). A regressão nãofoi significativa para Ensifera. A relação foi inversa parahimenópteros e, embora não significativa, indica umatendência de que as aranhas recrutam menos para vespasgrandes. Esse resultado provavelmente não foi significativodevido ao pequeno tamanho amostral (Tabela 1).
Tabela 1. Resultados da regressão entre tamanho do corpo e númerode aranhas recrutadas por minuto para cada categoria de presautilizada.
DiscussãoO intervalo de tempo utilizado no experimento não foi
suficiente para acompanhar todo o processo de manipulaçãoda presa, desde a imobilização até o consumo. Isto indicaque o processo de captura e manipulação de presas não é umevento que demanda pouca energia. A relação custo(recrutamento)/benefício (biomassa adquirida) para presasdentro de uma mesma categoria deve ser maior para presasmaiores. Para melhores resultados, seria interessante arealização de estudos comparando o tempo total de
categoria r 2 N constante tamanho F P
Diptera 0,732 9 -0,862 � 1.203 0,583 � 0.133 19,14 0,003
Hymenoptera 0,449 6 12,292 � 3.93 -0,449 � 0.249 3,25 0,146
Ensifera 0,291 10 -0,418 � 2.142 0,262 � 0.145 3,28 0,18
Lepidoptera 0,723 11 -0,836 � 1.031 0,38 � 0.078 23,50 0,001
manipulação da presa em relação à massa e volume, dentrode presas do mesmo grupo taxonômico.
A diferença no número de aranhas recrutadas em relaçãoaos diferentes tipos de presa contraria os resultadosencontrados por Kristoch & Varassin (1995), que nãoobservaram diferença de recrutamento de aranhas parasubjugação de abelhas, formigas e moscas. Já Acosta et al.(1995) observaram que Ensifera provocaram maiorrecrutamento de aranhas, contrariando completamente osresultados do presente estudo.
Diptera e Hymenoptera recrutaram mais aranhaspossivelmente porque seus movimentos são mais fortes,aparentemente vibrando os fios da teia com maior freqüênciae intensidade. Vibrações são utilizadas como indicadores dapresença de presas na teia (Foelix 1982). Hymenoptera,entretanto, são presas potencialmente perigosas, podem picaras aranhs, por isso a relação negativa entre tamanho erecrutamento. O tempo de subjugação de uma vespa grandetambém pode ser um fator que pode ter influenciado opequeno recrutamento para indivíduos grandes de presasdeste grupo. Presas grandes de dípteros, no entanto, nãooferecem riscos às aranhas, apesar de vibrarem muito as asas.
A hipótese de que presas maiores induziriam a umrecrutamento maior foi corroborada para Lepidoptera eDiptera. Presas pequenas grupos freqüentemente foramcapturadas por apenas um ou dois indivíduos, em algunscasos por juvenis. As moscas e mariposas maiores sempreatraíram mais aranhas, seja pelo batimento rápido de suasasas, que poderia se propagar melhor pela teia, seja porrealmente constituírem uma presa mais atrativanutricionalmente.
Ensifera possivelmente causou o menor recrutamento dearanhas por apresentar movimentos mais pontuais, consistiamem tentativas de sair da teia pulando. Ao contrário da vibraçãoconstante das presas das demais categorias, os movimentosdeste grupo eram mais difíceis de serem localizados pelasaranhas.
Este trabalho reforça e idéia de que aranhas sociais têmacesso a uma gama muito maior de presas do que espéciesequivalente do mesmo tamanho. Apesar das aranhas seremconsideradas generalistas (Foelix 1982), esta espécie reagede forma diferente a diferentes categorias de presas.
AgradecimentosAgradeço primeiramente ao Dadão pela orientação em
todas as etapas deste projeto; ao Professor Janzen, colegasPaula “panema”, Gargamel e Fino pela ajuda nas coletas daspresas; ao Juruna e à Rosinha por me ajudarem a encontraras teias; ao Machu Pichu e ao Emílio pelo esforço emrecuperar os arquivos que perdi no prazo final da entregadeste relatório. Às aranhas, à Floresta Amazônica. A todopessoal do curso, ao Inpa, , à Dona Eduarda e Seu Jorge peladeliciosa comida, aos colegas pela convivência.
Referências BibliográficasAcosta, L.A. 1995. Estratégias de ataque da aranha social
Anelosimus eximius (Araneae: Theridiidae) em função

Curso de Campo Ecologia da Floresta Amazônica - 2001 97
do tipo de presa. Curso de Campo “Ecologia da FlorestaAmazônica”, INPA/Smithsonian Institution, Manaus,AM.
Foelix, R.F. 1982. Biology of Spiders. Harvard UniversityPress, Cambridge.
Kristosch, G.C. & Varassin, I.G. 1995. Recrutamento em trêscolônias de Anelosimus eximius (Araneae: Theridiidae).Curso de Campo “Ecologia da Floresta Amazônica”,INPA/Smithsonian Institution, Manaus, AM.
Oliveira, A.A. 1987. Diversidade, estrutura e dinâmica docomponente arbóreo de uma floresta de terra firme deManaus, AM. Tese de Doutorado, Universidade de SãoPaulo, São Paulo.
Pasquet, A.; Trabalon, M.; Bagnères, A.G. & Leborgne, R.1997. Does group closure exist in the social spiderAnelosimus eximius? Behavioral and chemical approach.Ins. Soc. 44: 159-169.
Venticinque, E.M. 1995. Dinâmica populacional deAnelosimus eximius (Simon, 1891) (Araneae:Theridiidae) em mosaicos ambientais na AmazôniaCentral. Dissertação de Mestrado, Botucatu, São Paulo.
Wilkinson, L. Systat: The System for Statistics. Evanston,IL, Systat Inc.
Zar, J.H. 1984. Biostatistical Analysis. Prentice Hall, NewJersey.
Papel do dimorfismo sexual na reprodução de umaespécie de aranha da família Pholcidae
Angela Midori Furuya Pacheco
IntroduçãoO dimorfismo sexual é freqüentemente explicado pela
seleção sexual, um processo onde a escolha da fêmea (seleçãointer-sexual) e a competição entre os machos (seleção intra-sexual) são as forças dominantes. A seleção inter-sexualpromove o desenvolvimento de características morfológicasatrativas para as fêmeas, que possivelmente permitem aavaliação da qualidade do pretendente (Krebs & Davies1981). A seleção intra-sexual promove o desenvolvimentode características utilizadas em interações agonísticas entreindivíduos do mesmo sexo, como processos de avaliação deadversários e características de combate (Anderson & Vitt1990), que conferem ao macho vantagens na capacidade deacesso às fêmeas ou a sítios reprodutivos (Futuyma 1986).
Aranhas são sempre dióicas e muitas vezes apresentamdimorfismo sexual no tamanho corporal e estruturas ligadasa reprodução, sendo que fêmeas geralmente são maiores quemachos (Foelix 1982) pois necessitam de maior quantidadede reservas para reprodução devido ao custo energético daprodução de ovos. O menor tamanho pode conferir maioragilidade aos machos, que no caso de aranhas geralmente éo sexo que busca ativamente por parceiras sexuais. Em algunscasos os machos adultos podem até mesmo praticar“balonismo” para dispersão, como fazem]alguns indivíduosjovens. Além disso, geralmente machos necessitam de menornúmero de mudas para alcançar a maturidade reprodutiva(Foelix 1982).
Na Reserva do Km 41 ocorre uma espécie de aranha dafamília Pholcidae muito abundante, na qual machos adultossão maiores que fêmeas adultas. Indivíduos desta espécieconstróem teias do tipo cúpula, próximas de troncos ebarrancos e ao longo das margens de igarapés. Na mesmaárea, Carnaval et al. (1999), estudaram uma espécie dePholcidae, gênero Mesabolivar (A.J. Santos com. pess.)muito semelhante à espécie do presente trabalho, cujos
machos além de apresentarem tamanho corporal maior quede fêmeas, apresentam coloração conspícua no fêmur do 3o
par de pernas. Registros de dimorfismo sexual com maiortamanho corporal de machos e diferenças na coloração entreos sexos são escassos para a família (Carnaval et al. 1999),além de raros para aranhas em geral (Foelix 1982), indicandoa possibilidade da existência de processos de seleção sexualparticulares neste grupo. Nessa espécie os machos quecompartilham teias com fêmeas defendem a teia de machosinvasores e capturam alimento oferecendo-o como presentenupcial (Carnaval et al. 1999).
O objetivo deste trabalho é buscar padrões na formaçãode pares reprodutivos na espécie, avaliando característicasmorfológicas dos indivíduos, além de característicasestruturais da teia e do ambiente. Esse estudo tem interesseno significado adaptativo do dimorfismo sexual expresso notamanho corporal do macho e relações entre suas medidas,baseando-se na hipótese de que machos maiores têm maiorsucesso na aquisição de parceiras reprodutivas.
Material e MétodosColetei as aranhas em três igarapés de 2o ordem, em área
de baixio, próximo à base de pesquisa da Reserva do Km 41do Projeto de Dinâmica Biológica de Fragmentos Florestais,INPA, localizada à aproximadamente 70 km ao Norte deManaus (2o 24’S; 59o 52’W).
Coletei indivíduos em 74 teias ao longo dos igarapés edas trilhas de acesso, sendo que nas 50 primeiras teiasencontradas todos os indivíduos foram coletados, mas nasúltimas amostras procurei ativamente por teias contendocasais ou fêmeas, que eram elementos mais raros naamostragem.
Anotei o número de teias em agregados, medi ocomprimento e a largura da teia e anotei a altura em queestava fixada. Observei também o tipo de substrato onde a
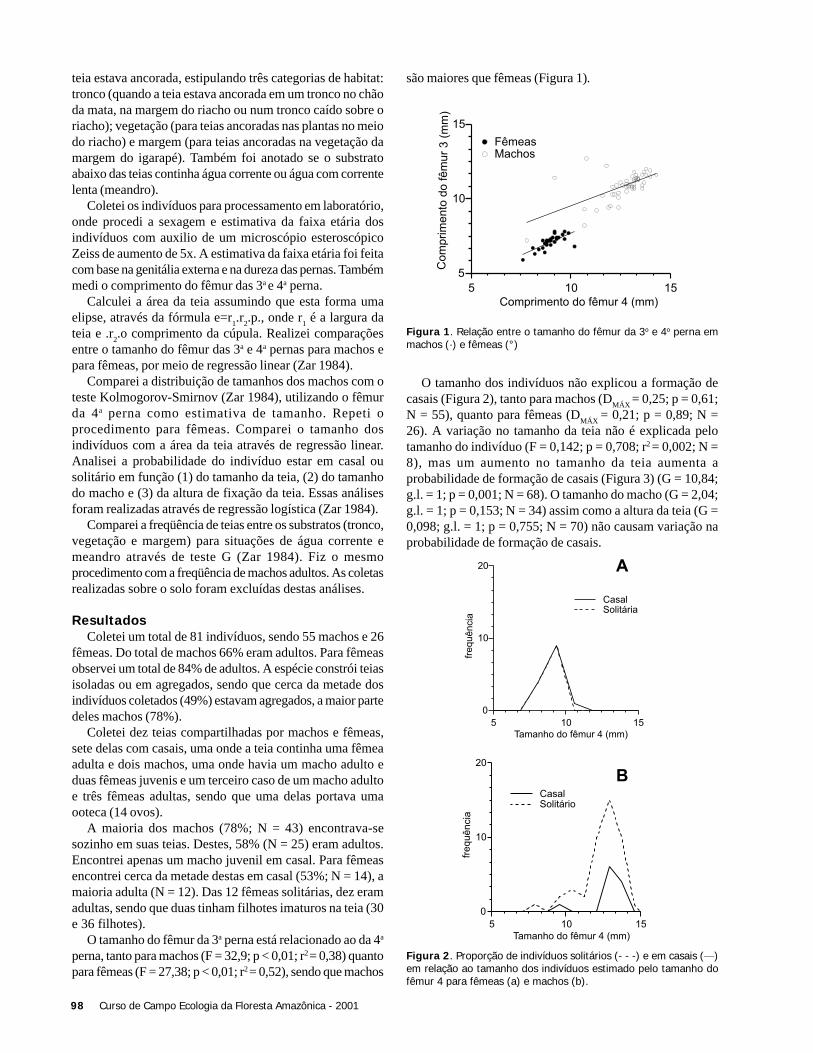
98 Curso de Campo Ecologia da Floresta Amazônica - 2001
teia estava ancorada, estipulando três categorias de habitat:tronco (quando a teia estava ancorada em um tronco no chãoda mata, na margem do riacho ou num tronco caído sobre oriacho); vegetação (para teias ancoradas nas plantas no meiodo riacho) e margem (para teias ancoradas na vegetação damargem do igarapé). Também foi anotado se o substratoabaixo das teias continha água corrente ou água com correntelenta (meandro).
Coletei os indivíduos para processamento em laboratório,onde procedi a sexagem e estimativa da faixa etária dosindivíduos com auxilio de um microscópio esteroscópicoZeiss de aumento de 5x. A estimativa da faixa etária foi feitacom base na genitália externa e na dureza das pernas. Tambémmedi o comprimento do fêmur das 3a e 4a perna.
Calculei a área da teia assumindo que esta forma umaelipse, através da fórmula e=r
1.r
2.p., onde r
1 é a largura da
teia e .r2.o comprimento da cúpula. Realizei comparações
entre o tamanho do fêmur das 3a e 4a pernas para machos epara fêmeas, por meio de regressão linear (Zar 1984).
Comparei a distribuição de tamanhos dos machos com oteste Kolmogorov-Smirnov (Zar 1984), utilizando o fêmurda 4a perna como estimativa de tamanho. Repeti oprocedimento para fêmeas. Comparei o tamanho dosindivíduos com a área da teia através de regressão linear.Analisei a probabilidade do indivíduo estar em casal ousolitário em função (1) do tamanho da teia, (2) do tamanhodo macho e (3) da altura de fixação da teia. Essas análisesforam realizadas através de regressão logística (Zar 1984).
Comparei a freqüência de teias entre os substratos (tronco,vegetação e margem) para situações de água corrente emeandro através de teste G (Zar 1984). Fiz o mesmoprocedimento com a freqüência de machos adultos. As coletasrealizadas sobre o solo foram excluídas destas análises.
ResultadosColetei um total de 81 indivíduos, sendo 55 machos e 26
fêmeas. Do total de machos 66% eram adultos. Para fêmeasobservei um total de 84% de adultos. A espécie constrói teiasisoladas ou em agregados, sendo que cerca da metade dosindivíduos coletados (49%) estavam agregados, a maior partedeles machos (78%).
Coletei dez teias compartilhadas por machos e fêmeas,sete delas com casais, uma onde a teia continha uma fêmeaadulta e dois machos, uma onde havia um macho adulto eduas fêmeas juvenis e um terceiro caso de um macho adultoe três fêmeas adultas, sendo que uma delas portava umaooteca (14 ovos).
A maioria dos machos (78%; N = 43) encontrava-sesozinho em suas teias. Destes, 58% (N = 25) eram adultos.Encontrei apenas um macho juvenil em casal. Para fêmeasencontrei cerca da metade destas em casal (53%; N = 14), amaioria adulta (N = 12). Das 12 fêmeas solitárias, dez eramadultas, sendo que duas tinham filhotes imaturos na teia (30e 36 filhotes).
O tamanho do fêmur da 3a perna está relacionado ao da 4a
perna, tanto para machos (F = 32,9; p < 0,01; r2 = 0,38) quantopara fêmeas (F = 27,38; p < 0,01; r2 = 0,52), sendo que machos
são maiores que fêmeas (Figura 1).
Figura 1. Relação entre o tamanho do fêmur da 3o e 4o perna emmachos (·) e fêmeas (°)
O tamanho dos indivíduos não explicou a formação decasais (Figura 2), tanto para machos (D
MÁX = 0,25; p = 0,61;
N = 55), quanto para fêmeas (DMÁX
= 0,21; p = 0,89; N =26). A variação no tamanho da teia não é explicada pelotamanho do indivíduo (F = 0,142; p = 0,708; r2 = 0,002; N =8), mas um aumento no tamanho da teia aumenta aprobabilidade de formação de casais (Figura 3) (G = 10,84;g.l. = 1; p = 0,001; N = 68). O tamanho do macho (G = 2,04;g.l. = 1; p = 0,153; N = 34) assim como a altura da teia (G =0,098; g.l. = 1; p = 0,755; N = 70) não causam variação naprobabilidade de formação de casais.
Figura 2. Proporção de indivíduos solitários (- - -) e em casais (___)em relação ao tamanho dos indivíduos estimado pelo tamanho dofêmur 4 para fêmeas (a) e machos (b).
5 10 15Comprimento do fêmur 4 (mm)
5
10
15
Co
mp
rim
en
tod
ofê
mu
r3
(mm
)
MachosFêmeas
5 10 15Tamanho do fêmur 4 (mm)
0
10
20
fre
qu
ênci
a
SolitáriaCasal
5 10 15Tamanho do fêmur 4 (mm)
0
10
20
fre
qu
ênci
a
SolitárioCasal
A
B
Curso de Campo Ecologia da Floresta Amazônica - 2001 99
0 200 400 600 800 1000Área da teia (cm2)
0.0
0.2
0.4
0.6
0.8
1.0
pro
ba
bili
da
de
Figura 3. Probabilidade de formação de casais em relação à áreada teia (cm2). Obtida com uso da equação da regressão logística.
Encontrei a maioria das teias na vegetação suspensa sobreo riacho (46%) ou em troncos (39%). A proporção de teiasfoi também maior sobre os meandros e poças (70%). Ainteração entre os fatores ambientais foi marginalmentesignificativa (G = 5,684; g.l. = 2; p = 0,058; N = 74). Afreqüência de machos adultos foi maior em troncos caídossobre água corrente (G = 6,046; g.l. = 2; p = 0,049; N = 36)que nos demais micro-ambientes, não refletindo o padrãopara freqüência de teias.
DiscussãoA espécie de Pholcidae estudada apresenta grande
tolerância à proximidade de indivíduos co-específicos, tantopara machos como para fêmeas. A ocorrência de indivíduosda espécie parece estar associada a locais com vegetaçãoherbácea e acúmulo de troncos para fixação das teia o quepoderia explicar a grande agregação das teias. Entretanto, atolerância à agregação apresenta custos, como por exemploa competição por recursos limitados, como o espaçodisponível para fixação de teias. Teias em locais com grandenúmero de teias circunvizinhas podem ter menor diâmetro(Eberhard 1990), o que interfere na quantidade de alimentodisponível. Outro ônus da agregação é o aumento daprobabilidade de ser detectado por um predador (Krebs &Davies 1981) o que no caso de predadores senta-espera podeser especialmente custoso (Pianka 1994). Como osindivíduos da espécie aparentemente não se beneficiam dasvantagens do gregerismo, uma vez que não apresentamcomportamento de alarme ou forrageio em grupo, acreditoque estejam competindo pelo espaço.
Entretanto, como a proporção de machos adultos solitáriosno ambiente amostrado é muito grande, é possível queagregados possam estar facilitando ou aumentando as chancesde acasalamento, possivelmente refletindo a qualidade domicro-ambiente como sítio reprodutivo. A predominância demachos e a quantidade de machos adultos ao longo dos
igarapés podem estar indicando um desvio na razão sexualna população. Caso isso seja verdade, é esperada uma fortecompetição entre machos por parceiras reprodutivas. Épossível também que os agregados representem retratosposteriores à um evento de reprodução, onde machos seacumularam em torno de um casal. Uma possibilidade,entretanto, é que as coletas, por terem sido realizadas deforma sistemática, não tenham amostrado os dois sexosigualmente, caso fêmeas ocupem preferencialmente outroambiente. Carnaval et al. (1999) coletando ao longo de trilhase barrancos da estrada, encontraram uma maior abundânciade teias com fêmeas para uma espécie muito semelhante a dopresente estudo.
Em contrapartida, é possível que os machos dessa espécieconstruam e defendam teias em sítios de qualidade e nelapermaneçam. Fêmeas apresentariam o padrão inverso aoconhecido para aranhas, realizando a busca ativa porparceiros reprodutivos. Assim, o menor número de fêmeascoletadas poderia ser resultado destas estarem vagando embusca de teias. Essa possibilidade, apesar de inverossímilpara aranhas, é reforçada pela existência na amostra de teiascom um macho e mais de uma fêmea (até 3 fêmeas numateia), e a ausência de teias sem macho e com mais de umafêmea. Além disso, a maior probabilidade de casais em teiasmaiores reforça a hipótese de que fêmeas escolhem osmachos.
A presença de mais de uma fêmea com um macho na teiaprovavelmente é decorrente do fato das fêmeas da famíliaPholcidae serem haplogínicas (Coddington 1991), isto é,possuem o aparelho reprodutivo em forma de canal, com umabolsa copuladora dorsal em fundo cego, que se comunicadiretamente com o útero (Foelix 1982). Essa característicaconfere prioridade de inseminação inversa à ordemcopulatória, ou seja, neste contexto, o estoque de espermado último macho a copular será o primeiro a ser utilizadopara inseminação. Uma implicação importante deste sistemade estocagem de esperma é a predisposição do macho dedefender a fêmea por um período após a cópula. Dois eventosobservados corroboram essa hipótese: uma teia com ummachos, uma fêmea portando ooteca e mais duas fêmeas epela teia contendo uma fêmea com filhotes instalada em áreacontígua a uma teia com casal. Assim, machos poderiam estarmantendo fêmeas de cópulas sucessivas (ou simultâneas) emsuas teias, caso haja alimento suficiente para todos osindivíduos, caso o macho tenha capacidade de defender ateia de machos invasores. A produção de teias nessa espécieaparentemente é muito cara, pois as teias são muito grandesem relação ao tamanho dos indivíduos. Assim, a permanênciade fêmeas nas teias representaria uma economia energéticapara a fêmea.
O fato de machos serem realmente maiores que fêmeasreforça a hipótese de que existem forças de seleção sexualatuando na espécie. Entretanto, o tamanho do machoaparentemente não interfere na escolha da fêmea.Possivelmente o maior tamanho do macho seja efeito deseleção intera-sexual, o que poderia estar sendo acentuadapela distribuição agregada dos indivíduos. Apesar de existir

100 Curso de Campo Ecologia da Floresta Amazônica - 2001
a possibilidade do tamanho da teia ser influenciada pelotamanho do indivíduo (Eberhard 1990) indivíduos maioresnão apresentaram teias maiores, o que indica que outrosfatores, como por exemplo o vigor, o estado fisiológico e ocomportamento de defesa do macho possam estar interferindona habilidade de construção e defesa de teias. Outrapossibilidade é que o aumento do número de inquilinos nateia incremente a habilidade de construção reparo e defesade teias, possibilitando teias maiores em casais.
Estudos que acompanhem o ciclo reprodutivo da espéciee manipulações experimentais, como indução de coorte porintrodução de alimentos e de machos invasores, poderiamtrazer informações adicionais sobre o sistema reprodutivodessa aranha. Esta espécie, bem como a espécie estudadapor Carnaval (1999), por serem muito abundantes, porapresentarem este surpreendente dimorfismo sexual, e porserem aparentadas filogeneticamente podem ser bonsorganismos para o estudo da evolução de sistemasreprodutivos.
AgradecimentosAgradeço Glauco Machado pela companhia bem-
humorada e pelo auxílio nas coletas de campo, pelasdiscussões relevantes ao longo da elaboração do trabalho, epelo companheirismo e sugestões na fase final do trabalho.Agradeço Eduardo Martins Venticinque (Dadão) pela ajudafundamental nas análises estatísticas e pelos comentáriosdurante a elaboração e na fase final desse trabalho.
Referências BibliográficasAnderson, R.A. & Vitt, L.J. 1990. Sexual selection versus
alternative cases of sexual dimorfism in teiid lizards.Oecologia 84: 145-157.
Carnaval, A.C.O.Q.; Cunha, H.F; Simon, M.; Pavón, Y.C.& Dantas, S. 1999. Investigações preliminares sobre osignificado adaptativo de um dimorfismo sexual raro emuma espécie de aranha da família Pholcidae. Livro doCurso “Ecologia da Floresta Amazônica”, INPA/Smithsonian Institution, Manaus, AM.
Coddington, J.A. & Levi, H.W. 1991. Systematics andevolution of spiders (Araneae). Ann. Rev. Ecol. Syst.22: 565-592.
Eberhard, W.G. 1990. Function and phylogeny of spiderwebs. Ann. Rev. Ecol. Syst. 21: 341-347.
Foelix, R.F. 1982. Biology of Spiders. Harvard UniversityPress, Cambridge.
Futuyma, D.J. 1986. Evolutionary Biology. SinauerAssociates, INC., Massachussets.
Krebs, J.R. & Davies, N.B. 1981. Behavioural Ecology: AnEvolutionary Approach. Sinauer Associates, INC.,Massachussets.
Pianka, E.R. 1994. Evolutionary Ecology. Harper CollinsCollege Publishers. New York.
Zar, J.H. 1984. Biostatistical Analysis. Prentice Hall, NewJersey.
Estratificação vertical na ocorrência de galhas em uma Floresta deTerra Firme da Amazônia Central
Antonio José Camilo de Aguiar
IntroduçãoOs insetos galhadores representam um grupo de herbívoros
com grande especificidade quanto à planta hospedeira e comgrande variedade de habitas (Fernandes & Price 1992;Fernandes 1992). Devido a sua fácil identificação em campo,galhas são uma importante ferramenta para estudosecológicos, sendo um bom indicador das condiçõesambientais (Fernandes et al. 1995).
A maior abundância e riqueza de galhas está normalmenterelacionada a vegetações com plantas esclerófilas emambientes estressados higrotermicamente (Fernandes & Price1991; Price et al. 1998; Gonçalves-Alvim & Fernandes2001). Fernandes & Price (1988) observaram que as galhassão mais comuns e diversas em ambientes secos do queúmidos, sendo as áreas de savana consideradas entre as maisbiodiversas em insetos galhadores.
Vários estudos foram realizados sobre padrões dedistribuição geográfica de insetos galhadores (Cornell 1985;Price 1991; Price et al. 1998), porém não existem estudosfocando a estratificação vertical da ocorrência destes nos
ambientes florestados. Dentro deste contexto, objetivou-seneste estudo estudar a ocorrência de galhas no gradientevertical de uma floresta de Terra firme na Amazônia Central.A hipótese deste estudo é que haja uma maior ocorrência degalhas no dossel devido a fatores como a arquitetura dasárvores, maior estresse ambiental e o maior grau deesclerofilia das folhas nesse estrato.
Material e MétodosA área de estudo está dentro da Reserva 1501 (Km 41),
localizada na Fazenda Esteio, no Distrito Agropecuário daSuframa, na área do Projeto Dinâmica Biológica deFragmentos Florestais (PDBFF), a cerca de 70 Km deManaus, (2o24’S; 59o45’W). Esta área apresenta pluviosidadeentre 1900-2500 mm/ano, altitudes variando entre 50-125m acima do nível do mar, e uma estação seca bastantepronunciada entre os meses de julho e outubro (Rankin-de-Merona et al. 1992).
A amostragem foi feita em três parcelas de 20x20 m, dentrode uma área de 100 ha previamente estabelecida para estudo

Curso de Campo Ecologia da Floresta Amazônica - 2001 101
da ecologia e sistemática da família Lecythidaceae (Mory &Becker 1991). Foram coletados ramos de árvores entre 1,5 e25 m de altura. A amostragem das árvores abaixo dos 8 m dealtura foi feita com auxílio do podão manual de cabo longoe acima desta altura, foi necessário subir na árvore comauxílio de corda e mosquetões. A partir desta árvore basecoletou-se os ramos das demais árvores circunvizinhas comauxílio do podão.
Para diminuir efeitos de dependência espacial em relaçãoa infestação por insetos galhadores, foram coletados nomínimo três ramos de cada árvore em galhos diferentes. Paraa amostragem das galhas nas folhas, estas foram colocadasem um saco e misturadas, e destas retiradas cinqüenta folhasao acaso.
A folhas foram classificadas subjetivamente quanto àesclerofilia em três tipos: membranáceas, cartáceas ecoriáceas. O grau de ocorrência das galhas em cada uma dasárvores amostradas foi definida através da porcentagem defolhas com galhas. A ocorrência de galhas foi relacionadacom a altura das árvores através de regressão linear. Estaanálise foi feita para cada tipo de folha e para todas as folhas,independetemente da categoria (Zar 1984).
Os limites de cada estrato da floresta foram definidos doseguinte modo: de 0 a 10 m para o sub-bosque, de 10 a 20 mpara o sub-dossel e acima dos 20 m para o dossel. Francis(1994) e Ribeiro et al. (1999) definem o dossel da florestaAmazônica como acima dos 30 metros, porém é importantesalientar que nas parcelas estudadas as árvores mais altasapresentavam entre 25 a 28 m (obs. pess.).
Para testar as diferenças na ocorrência dos tipos de folhasem cada estrato foi utilizado o teste de c2 e na comparaçãoda ocorrência das galhas entre os estratos foi utilizadaANOVA (Zar 1984).
ResultadosForam amostradas 70 árvores, sendo 41 árvores no sub-
bosque, 20 no sub-dossel e 9 no dossel. O número de árvorescom folhas coriáceas (n = 9) foi menor que o número deárvores com folhas cartáceas (n = 28) e membranáceas (n =33). Quanto à ocorrência dos três tipos foliares nos trêsestratos amostrados, foi observado uma predominância dasfolhas coriáceas no dossel, das folhas cartáceas no sub-dossele das folhas membranaceas no sub-bosque (c2 = 23,83; g.l.=4; p < 0,01) (Figura1).
Figura 1. Número de árvores com diferentes graus de esclerofilianos três estratos arbóreos na Floresta de Terra Firme da AmazôniaCenral, Reserva 1501 (Km 41; PBDFF)
0
5
10
15
20
25
30
SUB-BOSQUE
SUB-DOSSEL
DOSSEL
NÚ
ME
RO
DE
ÁR
VO
RE
S
membranácea
cartácea
coriácea
A ocorrência de galhas nos três tipos de folhas apresentouuma grande variação. As árvores com folhas membranáceasapresentaram em média 10% (DP = 14 %) de suas folhascom galhas; as árvores com folhas cartáceas, 12% (DP =18%); e as com folhas coriáceas, 6% (DP = 14 %).
Os dados de variação da ocorrência de galhas em relaçãocom a altura dos tipos foliares não demonstrou umacorrelação significativa (Figura 2). A maioria das árvoresapresentou abaixo de 20% de suas folhas com galhas, e 28%de todas as árvores amostradas não apresentaram folhas comgalhas.
Os estratos de altura da floresta apresentaram uma grandevariação de folhas com galhas em relação a média: sub-bosque, 11% (DP = 16%); sub-dossel, 15% (DP = 2 %),dossel, 8% (DP=13%). Não foi observada relação entre aporcentagem de folhas com galhas e altura das árvores (r2 =0.005; n = 70; p = 0.54). Também, não foi observada umadiferença na porcentagem de folhas com galhas entre osestratos (F = 0.872; n = 67; p = 0.423).
DiscussãoO crescente grau de esclerofilia das folhas observado para
o gradiente de altura da floresta provavelmente estárelacionado com um aumento do estresse ambiental nosestratos superiores, como aumento da temperatura, radiaçãosolar, baixa umidade, alta evapotranspiração e correntes devento (Kira & Yoda 1988; Fitzjarrald & Moore 1995).
A distribuição de galhas pode estar sendo influenciadapor uma série de fatores independentes do gradiente de alturada floresta, entre eles as características da planta hospedeiracomo idade, fenologia, defesas induzidas, compostossecundários e hipersensibilidade (Lara & Fernandes 1994).
O único estudo realizado que comparou a ocorrência degalhas entre o dossel e os estratos inferiores foi o realizadopor Price et al. (1998) na forma de levantamento rápido emduas áreas de florestas tropicais (uma na Amazônia e outrano Panamá). Price et al. (1998) observaram uma maior riquezade espécies no estrato inferior do que no dossel em ambasáreas estudadas. A comparação deste estudo com o realizadopor Price et al. (1998) fica prejudicada devido aos estudostratarem de questões diferentes, o primeiro a abundância e ooutro a riqueza de galhas. Porém, tanto para este estudo comopara o daqueles autores, o pequeno número de réplicas deáreas amostradas e o curto período de amostragem, fazemcom que essas amostras para o nível de comunidade possamser consideradas pontuais, tanto espacialmente quantotemporalmente.
Este estudo não demonstrou uma diferenciação daabundância de galhas no gradiente de altura na floresta,provavelmente devido ao fato de que foi realizado em umaárea de platô, onde a floresta aparentemente não exibia limitesnítidos entre seus estratos, com um dossel relativamenteaberto (obs. pess.). Outro problema da amostragem que podeter prejudicado o entendimento da distribuição das galhas,foi o número desproporcional de amostras entre os estratos,com um pequeno número de amostras no dossel.
O dossel apresenta uma maior biomassa de recursos para

102 Curso de Campo Ecologia da Floresta Amazônica - 2001
os insetos galhadores, recursos estes representados pelasfolhas. Porém não é possível inferir até o presente momentode que essa maior abundância de recursos associada àcaracterísticas de esclerofilia das folhas do dossel possamestar favorecendo uma maior ocorrência de insetos galhadoresneste estrato.
Para uma abordagem mais completa da estratificaçãovertical da ocorrência de galhas na floresta, sugiro que, paraestudos futuros, sejam caracterizadas com mais detalhes asvariáveis de estressse ambiental como deficiência nutricionaldas plantas, umidade e vento.
AgradecimentosAgradeço a Cristiane pelo auxílio no trabalho de campo,
ao Dadão, Glauco e Jansen pelas sugestões, a Dona Eduardapela maravilhosa comida, ao Geraldinho e ao Leandro Várzeapelo brain storm, ao Celso e a Isabelha por mais essa força,e a todos amigos pela simples e boa amizade.
Referências BibliográficasCornell, H.V. 1985. Local and regional richness of cynipine
gall wasps on California oaks. Ecology 66: 10-16.Fernandes, G.W. & Price, P.W. 1988. Biogeographical
gradients in galling species richness: tests of hypotheses.Oecologia 76: 161-167.
Fernandes, G.W & Price, P.W. 1991. Comparisons of tropicaland temperate galling species richness: the roles ofenvirovinmental harshness and plant nutrients status. In:Plant-Animal Interactions: Evolutionary Ecology inTropical and Temperate Regions, Price, P.W.,Lewinsohn, T.M., Fernandes, G.W. & Benson, W.W.(eds.), pp. 91-115. John Wiley & Sons, New York.
Fernandes, G.W. & Price, P.W. 1992. The adaptativesignificance of insect gall disribution: Survivorship ofspecies in xeric andmesic habitats. Oecologia 90: 141-20.
Fernandes, G.W.; Silva de Paula, A. & Loyola, R. 1995.Distribuição diferencial de insetos galhadores entrehabitats entre habitats e seu possível uso comobioindicadores. Vida Silv. Neotrop. 4(2): 133-139.
Fitzjarrald, D.R. & Moore, K.E. 1995. Physical mechanisms
of heat and mass exclange between forest andatmosphere. In: Forest Canopies, Lowman, M.D. &Nadkarni, N.M. (eds). Academic Press, San Diego.
Francis, C.M. 1994. Vertical stratification of fruit bats(Ptepodidae) in lowland dipterocarp rain forest inMalaysia. J. Trop. Ecol. 10(4): 523-530.
Gonçalves-Alvim, S.J. & Fernandes, G.W. 2001. Biodiversityof galling insects: historical, community and habitateffects in four neotropical savannas. Biodiv. Conserv.no prelo.
Kira, T. & Yoda, K. 1989. Vertical stratification inmicroclimate. In: Tropical Rainforest Ecosystem, Lieth,H. & Werger, M.J.A. (eds.). Elswevier, Amsterdam.
Lara, A.C.F. & Fernandes, G.W. 1994. Padrões dedistribuição de Neopelma baccharidis (Homoptera:Psyllidae) em Bacharis dracunculifolia (Asteraceae).Revta. Bras. Biol. 54: 661-668.
Mori, S.A. & Becker, P. 1991. Flooding affects survival ofLecythidaceae in Terra Firme Forest near Manaus, Brasil.Biotropica 23(1): 87-90.
Price, P. 1991. Patterns in communities along latitudinalgradients. In: Plant-Animal Interactions: EvolutionaryEcology in Tropical and Temperate Regions, Price, P.W.,Lewinsohn, T.M., Fernandes, G.W. & Benson, W.W.(eds.), pp. 51-69. John Wiley & Sons, New York.
Price, P.W.; Fernandes, G.W.; Lara, A.C.F.; Brawn, J.;Barrios, H.; Wright, M.G.; Ribeiro, S.P. & Rothcliff,N. 1998. Global patterns in local number of insect gallingspecies. J. Biog. 25: 581-591.
Rankin-de-Merona, J.M.; Prance, G.T.; Hutchings, R.W.;Silva, M.F.; Rodrigues, W.A. & Uehling, M.E. 1992.Preliminar results of large scale tree inventory of uplandrain forest in the Central Amazon. Acta Amaz. 22(4):493-534.
Ribeiro, J.E.L.S.; Hopkins, M.J.G.; Vicentini, A.; Sothers,C.A.; Costa, M.A.S.; Brito, J.M.; Martins, L.H.P.;Lohmann, L.G.; Assunção, P.A.C.L.; Pereira, E.C.; Silva,C.F.; Mesquita, M.R. & Procopio, L.C. 1999. Flora daReserva Ducke. INPA, Manaus, AM.
Zar, J.H. 1984. Biostatistical Analysis. Prentice Hall, NewJersey.
Defensas alternativas en plantas mirmecófitas y en función de la ontogeniaBetsabé Ruiz Guerra
IntroducciónLas interacciones entre plantas y animales nos permiten
darnos cuenta de que los organismos no evolucionan demanera independiente y únicamente en interacción con elambiente físico, sino que se encuentran inmersos en unaintricada red de fuerzas selectivas bióticas y abióticas. Deesta forma, las presiones de selección ejercidas por uno delos componentes puede originar respuestas adaptativas en elotro, que a su vez pueden convertirse en una presión selectiva
para el primero, y esto puede continuar de manera recurrentepudiendo originar procesos de coevolución y especiación(Del Val 1999). Un claro ejemplo de estos procesosevolutivos lo encontramos en las selvas tropicales donde lasrelaciones entre plantas y herbívoros han permitido laevolución de mecanismos de defensa como lo son latolerancia y la resistencia. La primera involucra la capacidadde las plantas para soportar la herbivoría, con pérdidasmínimas en su crecimiento y reproducción (Rosenthal &

Curso de Campo Ecologia da Floresta Amazônica - 2001 103
Welter 1995). La resistencia, por otro lado, implica eldespliegue de mecanismos que repelen, envenenan oadvierten del peligro (Rosenthal & Kotanen 1994), dandocomo resultado una disminución del daño en la planta. Dentrode esta encontramos las siguientes estrategias: a) resistenciafísica representada con espinas, dureza del tejido, silicatosy tricomas, b) resistencia química conformada principalmentepor metabolitos secundarios así como por derivados decianuro y alcaliodes (Coley 1983), c) resistencia biótica, dadaa partir de una asociación de beneficio mutuo entre plantasy animales, principalmente hormigas. En esta interacción lashormigas protegen a la planta de los ataques de losherbívoros, y en ocasiones evitan la invasión por parte deotras plantas. Como recompensa las plantas ofrecen un pagoconsistente en albergue (estructuras denominadas domacios)y suministro de alimento en forma de nectarios extrafloraleso de otros cuerpos alimenticios, esta asociación se denominamirmecofilia (Beattie 1985). En respuesta a los mecanismosde defensa química por parte de las plantas, los herbívoroshan desarrollado en el curso de su evolución la capacidad dedigerir ciertos compuestos químicos así como evadir lasdefensas físicas. En contraste, poco se ha demostrado de larespuesta de los herbívoros a las defensas bióticas (Price1997), asi como las estrategias de defensa utilizadas por lasplantas mirmecofitas en la fase inicial de su ontogenia, en lacual carcen de los atributos de recompensa para las hormigasy por tanto la consecuente defensa brindada por las mismas.
Este trabajo pretende evaluar la eficiencia de dosmecanismos de defensa en dos especies de Cecropia, unacon hábito mirmecofito (Cecropia purpurascens) y la otrano mirmecofita (Cecropia sciadophylla). En particularpretendo comparar diferencias en los niveles de herbivoríaentre las etapas juveniles y adultas de Cecropia purpurascensy Cecropia sciadophylla.
Material y MétodosRealicé este estudio en la Reserva del Km 41 del Proyecto
Dinámica Biológica de Fragmentos Forestales (PDBFF),localizada a 70 Km al Norte de Manaus (AM). La reservacuenta con 800 hectáreas de bosques primarios contínuosde tierra firme. La temperatura media anual es de 26 o C y lapluviosidad varía de 1900mm a 2400mm por año (Lovejoy& Bierregaard 1990).
En la carretera ZF-3 escogí 15 plantas adultas y 15juveniles de las especies Cecropia sciadophylla y Cecropiapurpurascens, el criterio para determinar a los individuosjóvenes fue la ausencia de colonias de hormigas así comouna altura máxima de 3 m. Procuré que los indivíduos de lasdos especies estuviesen lo más proximo uno del otro(distancia máxima de 10m) y que tuviesen una altura similar.En estos individuos realicé los siguientes experimentos:
1) Experimento de cuantificación de los niveles deherbivoría
Para cada uno de los individuos seleccioné aleatoreamente15 hojas en las cuales se estimó visualmente el porcentajede área folear comida (AFC) utilizando el diseño de
categorías de daño propuesta por Dirzo y Domínguez (1995):0 = 0%, 1=1-6%, 2 = 6-12%, 3 = 12-25%, 4 = 25-50%, 5 =50-100%. A partir de estas categorías se calculó el índice deherbivoría (IH) para cada planta de la siguiente manera : IH= å (nj) i / N , donde nj es el número de hojas en la categoríai de daño y N el número total de as de la planta.
Las diferencias en los niveles de herbivoría fueronevauadas mediante un análisis de Mann-Withney. Devido ala repetición del estadistico los valores de probabilidadfueron corregidos mediante la corrección de Bonferroni (Zar1984), utilizando un valor de a = 0,01 como nivel designificancia.
2) Experimento de evaluación de la prescencia decompuestos secundarios.
Para este experimento realicé una prueba de palatabilidadpara la cual colecté cinco hojas de cinco individuos para lasdos especies, procurando que las hojas fuesen de la mismaedad (posición 4 de la filotaxis). Posteriormente de cada unade las hojas corté muestras de 4 cm2 y coloque en recipientescon herbívoros generalistas (Acrydiidae), a manera que cadarecipiente contenía muestras de follaje de ambas especies.Este experimento fue separado en cuatro tratamientospareados: 1) C. purpurascens de individuos juveniles conC. purpurascens de individuos adultos, 2) C. sciadophyllade individuos juveniles con C. sciadophylla de individuosadultos, 3) C. purpurascens de individuos juveniles con C.sciadophylla de individuos juveniles, 4) C. purpurascensde individuos adultos con C. sciadophylla de individuosadultos. Cada uno de estos tratamientos tuvo ocho réplicas.
Los herbívoros fueron alimentados con lechuga yporteriormente puestos en ayuno por un periodo de 12 horascon el fin de estandarizarlos. El experimento duró 8 horasdespués de las cuales se registró el porcentage de herbivoríade manera visual para cada una de las muestras. Paradeterminar si existían diferencias en la palatabilidad dentroen cada uno de los cuatro tratamientos se realizó un análisisde Wilcoxon.
3) Experimento de determinación de durezaLas medidas de dureza fueron estimadas utilizando el
criterio del experimento anterior para la elección de las hojasy en cada una de ellas se midió la dureza utilizando unpenetrometro manual, en cada hoja se tomaron cinco medidasy después se calculó la media. Para verificar si existendiferencias en la dureza de las hojas de los individuosjuveniles y adultos de ambas especies realicé una prueba deMann-Withney. Todos los análisis fueron realizados en elprograma Systat versión 8.0 (Wilkinson 1988).
ResultadosExperimento 1Los niveles de herbivoría fueron mayores en los individuos
adultos (mediana IH = 1,87) que en los juveniles de C.purpurascens (mediana IH = 1,1), (U = 58; p = 0,009), en elcaso de los individuos juveniles y adultos de C. sciadophyllano hubo diferencias estadisticamente significativas en los

104 Curso de Campo Ecologia da Floresta Amazônica - 2001
niveles de daño (Figura 1).También hubo diferencias marginalmente significativas
(U = 206; p = 0,039) entre los niveles de herbivoría de losindivos adultos de las dos especies, de tal manera que losniveles de herbivoría son mayores en C.purpurascens(mediana IH = 1,87) que en C. sciadophylla(mediana = 1,26) (Figura 1).
Figura1. Grafico mostrando mediana, primer y tercer cuartil, valormáxímo y mínimo de la relación del índice de herbivoría de C.purpuracens en individuos jóvenes (PJ) y adultos (PA) y de C.sciadophylla en individuos jovenes (SJ) y adultos (SA). * representanvalores extremos de la muestra
Experimento 2El estadístico de Wilcoxon realizado para evaluar las
diferencias de palatabilidad mostró que existen diferenciassignificativas entre los individuos jóvenes y adultos de C.sciadophylla (Z = -2,207; P = 0,027), sugiriendo que losindividuos jóvenes son más palatables. No existen diferenciasestadísticamente significativas para los tratamientos:
1) C. purpurascens de individuos juveniles (mediana delporcentaje de herbivoría = 0) con C. purpurascens deindividuos adultos (mediana del porcentaje de herbivoría =0) (Z = -0,36; p = 0,715).
2)FALTA UM PEDAÇO DE TEXTO IGUAL A 1, 3 OU 4
3) C. purpurascens de individuos juveniles (mediana delporcentaje de herbivoría = 0) con C. sciadophylla deindividuos juveniles (mediana del porcentaje de herbivoría= 0) (Z = 0; p = 1).
4) C. purpurascens de individuos adultos (mediana delporcentaje de herbivoría = 4,5) con C. sciadophylla deindividuos adultos (mediana del porcentaje de herbivoría =0) (Z = -1,57;p = 0,116).
Experimento 3No se detectó efecto significativo de los valores de dureza
PA PJ SA SJEspecies
0
1
2
3
4
Ind
ice
de
He
rbiv
orí
a(I
H)
entre: a) individuos jóvenes y adultos de C. purpurascens(U = 6; p = 0,175), b) individuos jóvenes de las dos especies(U = 16; p = 0,46), c) individuos adultos de las dos especies(U = 8; p = 0,35). Para los individuos jóvenes y adultos deC. sciadophylla existen diferencias marginalmentesignificativas (U = 4; p = 0,076).
DiscusiónLas diferencias encontradas en los niveles de daño entre
los individuos jóvenes y adultos de C. purpureans enprincipio sugieren que esta especie presenta diferencias enlas estrategias de defensa a lo largo de su ontogenia, el hechode que los adultos presenten un mayor nivel de daño puedeestar relacionado con las características fisiológicas de laespecie, ya que es una planta pionera que de acuerdo con lateoría de disponibilidad de recursos propuesta por Coley(1985) estas especies optan por asignar mas recursosenergéticos al crecimiento de tal manera que presentan unaalta tasa de recambio foliar, repercutiendo en la cantidad derecursos asignados a la defensa.
Por otra parte estos resultados también sugieren que ladefensa brindada por las hormigas no es tan eficiente hecho,que se refuerza con los resultados obtenidos de lacomparación con la especie no mirmecófita (C. sciadophylla)donde también presentó mayores niveles de daño . Este patronha sido observado en otros estudios (Andrade 1982; Fáveri2000) realizados con plantas mirmecófitas. Longino (1991)observó ausencia de patrullaje por parte de las hormigasAzteca alfari en hojas de Cecropia, asi como que existe unagran cantidad de organismos que utilizan Cecropia comoalimento y habitat. También existe evidencia que sugiere quela respuesta de las hormigas puede ser diferente en relaciónal tamaño de la colonia (Rocha & Bergallo 1992).
Otra posible explicación para estos niveles de daño puedeestar relacionada con el hecho de que existen diferencias enla eficiencia de protección por parte de las hormigas en contrade herbívoros de C. purpurascens dependiente de la especiecolonizadora,siendo más eficiente Azteca alfari queCamponotus balzani (R. Cogni com. pers.), en el presenteestudio este factor no fue considerado para la elección delas plantas por lo que muy probablemente influenció losresultados.
Los niveles de daño en los individuos jóvenes y adultosde C. sciadophylla en contraste con C. purpurascens nofueron diferentes, y los individuos jovenes presentaron unamayor palatabilidad y por consiguiente una menor cantidadde compuestos secundarios, razón por la cual se podría pensarque los individuos de etapa juvenil utilizan una estrategiade defensa basada en la tolerancia en vez de la resistencia,no obstante, para corroborrar este mecanismo de defensa setendrían que hacer estudios a largo plazo tales como elanálisis de crecimiento compensatorio, tasas fotosintética osobrevivencia bajo tratamientos de defoliación experimental.
Los valores no significativos registrados al comparar ladureza de las hojas pueden deberse a la gran variación queexiste dentro de cada hoja asi como a un limitado número demuestras, no obstante se registró un valor marginalmente

Curso de Campo Ecologia da Floresta Amazônica - 2001 105
significativo lo que puede estar indicando que con un estudiomas detallado muy probablemente se encuentren diferenciasestadisticamente significativas.
Respecto a las pruebas de palatabilidad existen dosfactores importantes que pudieron influenciar los resultados:1) Los herbivoros no eran de la misma especie ni de la mismaedad, esto sucedio debido a las limitantes de colecta, 2) Eltiempo y estrategia de manipulación de los herbivorospudieron influenciar su compotamiento.
La inconsistencia de los resultados arrojados pordiferentes trabajos referentes al papel de las hormigas comodefensores contra herbivoros da pie para pensar que quiza laprescencia de las hormigas se trata de un anacronismo , esdecir, que es un mecanismo de defensa que surguío bajo lapresión de selección de otro u otros organismos que afectabanel desempeño de estas plantas y que ahora ya no existe, noobstante, este mecanismo en algunos casos sirve comodefensa antihervívoro asi como para la remoción de lianas yplantas parásitas.
La teoría del mutualismo en mosaico propuesta porThompson (1982) dice que existe una variación geográficaen la intensidad de selección dada por la calidad de reursosdisponibles en el ambiente. Esta teoría puede ser aplicada ael sistema de estudio considerando que los suelos de laamazonía son pobres en nutrientes (Lovejoy & Bierregaard1990), razón por la cual las plantas se enfrentan a uncompromiso en la asignación de recursos destinada a lasdiferentes funciones (Coley et al. 1985). De tal manera quepodríamos pensar que estas plantas están invirtiendo másrecursos en crecimiento y reproducción, repercutiendo en lacalidad de los mecanismos de defensa, asi como la producciónde cuerpos mullerianos lo que asu vez traería comoconsecuencia un déficit en la eficiencia del papel de lashormigas como antiherbívoros reflejandose en altas tasas deherbivoría.
En suma las estrategias de defensa antiherbívoro dependende la calidad de recursos disponible en los diferentes hábitatsprovocando el despliegue de diversos mecanismos derespuesta bajo una determinada presión de selección. Losresultados de este proyecto pueden representar un indiciode las posibles respuestas diferenciales a una misma presiónde selección.
Para futuros estudios realizados con este sistema sugieroque se considere el tamaño de las plantas así como la especiede hormiga colonizadora, para obtener resultados más finosrespecto a la presencia de compuestos secundarios serecomienda hacer dichos análisis en un laboratorio químico.
AgradecimientosAgradezco a Eduardo Venticinque por la discusión del
proyecto, el análisis de datos, y el apoyo en el campo. Alprofesor Jansen por su apoyo incondicional. A Ocírio Pereira(Juruna), Paula S, Felipe, Rodrigo, Glauco y Gargamel por
su ayuda en la colecta de los bichos asi como su compania.A todos los integrantes del curso de campo 2001 por lasdiscusiones, alegrias, y buenos ratos. A los señores EduardaCosta y Raimundo J. por esas deliciosas comidas y porapapacharnos durante la estancia en la Reserva del Km 41.ABrasil.
Referencias BibliograficasAndrade, J.C. & Caruta, J.J.P. 1982. The Cecropia-Azteca
association: a case of mutualism? Biotropica 14:15Beattie, A.J. 1985. The evolutionary ecology of ant–plant
mutualisms. Cambridge University Press, London.Coley, P.D. 1983. Herbivory and defensive characteristics
of tree species in a lowland tropical forest. Ecol. Monog.53: 209–233
Coley, P.D; Bryant, J.P. & Chapin III, F.S. 1985. Resourceavailability and plant antiherbivore defense. Science 230:895–899
Del Val, G.E. 1999. Defensa química y por hormigas durantela ontogénia de Cecropia peltata (Cecrpiaceae) en LaSelva Lacandona. Tesis de Licenciatura. Facultad deCiencias, UNAM, Mexico
Dirzo, R. & Domínguez, C. 1995. Plant–herbivoreinteracctions in Mesoamerican tropical dry forest. In:Seasonally Dry Tropical Forest, Bullock, S.H., Mooney,A. & Medina, E. (eds.), pp.304–309. CambridgeUniversity Press, Cambridge.
Fáveri, S.B. 2000. Riqueza e composição da comunidade deartrópodos associados a Cecropia spp. Dissertação deMestrado. Universidade do Amazonas, InstitutoNacional de Pesquisas da Amazônia.
Lovejoy, E.T. & Bierregaard, J.R. 1990. Central AmazonianForest and the Minimum Critical Size of EcosystemsProyect. In: Four Neotropical Rainforest, Gentry, A.H.(ed). Yale University Press, EUA.
Longino, J.T. 1991. Azteca ants in Cecropia trees: taxonomy,colony structure, and behaviour. In: Ant-PlantInteractions, Huxley, C. & Culver, D.F. (eds.), pp 271-288. Oxford University Press, Oxford.
Rocha, C.F.D. & Bergallo, H.G. 1992. Bigger ant coloniesreduce herbivory and herbivore residence time on leavesof ant-plant: Azteca mulleri vs. Coelomera ruficornison Cecropia pachystachya. Oecologia 91: 249-252
Rosenthal, G.A. & Kotanen, P.M. 1994. Terrestrial planttolerance to herbivory. TREE 9: 145–148.
Rosenthal, G.A. & Welter, S.C. 1995. Tolerance to herbivoryby stemboring caterpillar in architecturally distinctmaizes and wild relatives. Oecologia 102: 146–155.
Wilkinson, L. 1988. Systat 8.0 for Windows. SPSS Inc.,Chicago, Illinois.
Zar, J.H. 1984. Biostatistical Analysis. Prentice–Hall, NewJersey.

106 Curso de Campo Ecologia da Floresta Amazônica - 2001
Efeitos da estrutura física de habitats sobre a composição e abundância daictiofauna em igarapés da floresta de terra firme na Amazônia Central
Carlos Eduardo Marinelli
IntroduçãoDentro da diversidade de ecossistemas aquáticos que
compõem a bacia Amazônica, os igarapés são corpos d’águade pequeno porte, leito bem delimitado e de baixa produçãobiológica, devido ao fluxo d’água e a limitação de luz impostapela floresta de terra firme, que encobre totalmente seuscursos médio e superior (Santos & Ferreira 1999). Corposd’água inseridos em florestas possuem importantes relaçõescom esses ecossistemas, como o aporte de folhas formandosubstratos constituídos de serapilheira que originam habitatse provocam alterações na profundidade e na correnteza doriacho (Barrella et al. 1999).
O principal componente ambiental na constituição dehabitats para a ictiofauna é o espaço (Hayes et al. 1996),sendo que em rios de pequeno porte, características como otipo de substrato, profundidade e velocidade da correntezaatuam como fatores fundamentais na distribuição eorganização das suas comunidades (Gorman & Karr 1978;Richards et al. 1996). Hipóteses sobre a uniformidade físicade igarapés de porte comparável são reforçadas pelasimilaridade na riqueza de espécies de suas ictiofaunas(Sabino 1999). Segundo o autor, este fato sugere a existênciade padrões generalizados de ocupação espacial e de umpossível “limite ecológico” para formação dos conjuntos deespécies nestes ecossistemas, tema que há décadas vêm sendodiscutido pela literatura especializada, devido especialmenteàs suas implicações na biologia da conservação efragmentação de ecossistemas (Angermeier & Schlosser1989).
As características ambientais de igarapés amazônicos,como seu pequeno porte, a vazão reduzida e elevadatransparência, facilitam a realização de estudos naturalísticosbaseados na observação das espécies in situ, permitindoestimar o tamanho das populações, sua distribuição e habitatsespecíficos (Keenleyside 1979; Sabino 1999).
No presente trabalho apresento os resultados de um estudonaturalístico onde analiso a influência da estrutura física dehabitats na composição de espécies e abundância de peixesem igarapés de terra firme da Amazônia Central.
Material e MétodosRealizei o estudo em três igarapés de segunda ordem na
área da Reserva do Km 41, situada na Fazenda Esteio (2º28’S;59º46’W), aproximadamente 70km ao norte de Manaus, AM.A Reserva é drenada pela bacia do rio Urubu, afluente damargem esquerda do médio rio Amazonas e faz parte dasáreas de estudo do PDBFF/INPA-Smithsonian Institution,sendo considerada área controle, já que encontra-se inseridaem floresta contínua de terra firme (Bührnheim 1998). Pelaclassificação de Köppen, o clima local é tropical chuvoso(Amw), com temperatura média anual de 26ºC, precipitação
anual entre 1750 e 2000mm e umidade relativa anual de 80%(Radambrasil 1978).
Determinei aleatoriamente um trecho de 20 metros de cadaum dos igarapés, nos quais primeiramente isolava suassecções a montante e jusante com redes de bloqueio paraevitar a fuga de peixes. Fiz um esboço geral de cada trechoem vista superior, caracterizando sua sinuosidade e adistribuição dos tipos de substrato e de raízes adventícias davegetação ripária. Amostrei locais com substrato formadoapenas de areia, cobertos por serrapilheira, com emaranhadosde raízes, e com serrapilheira associada a raízes. Seguindo omodelo de classificação hierárquica para habitats de riachos,proposto por Hawkins et al. (1993), os locais amostradosforam subdivididos em: 1) áreas inseridas no meio da calhado igarapé, com fundo relativamente uniforme e extensõeslaterais próximas das margens e 2) poções escavados pelaação da água, com paredes laterais mais íngremes e de formatoelíptico, na maioria das vezes dispostos lateralmente na calhado igarapé. Identifiquei os habitats de cada igarapé no mínimocom 4 metros de distância entre um e outro, e para cada umcalculei as médias de comprimento, largura, profundidade evelocidade da correnteza, esta última adotando o método doflutuador. Tomei medidas de largura e o perfil deprofundidade de secções transversais a montante, jusante ena porção intermediária de cada trecho. Os valores de pH(utilizando papel indicador) e da temperatura da água(termômetro com coluna de mercúrio) também foram obtidospara cada igarapé.
Para amostragem da ictiofauna, utilizando máscarasemifacial e respirador, realizei três sessões de mergulho livreem cada igarapé, totalizando seis horas de esforço amostral.As observações foram realizadas no sentido jusante-montante, com o intuito de evitar a ressuspensão dosedimento, o que limitaria a visibilidade. Para o registro dasobservações adotei o método denominado animal-focal,anotando as informações em uma placa de PVC branca (15x 30cm) com uma planilha previamente esquematizada,conforme sugerido por Sabino (1999). Procurei identificarrapidamente a(s) espécie(s), contando o número de indivíduose registrei a posição dos mesmos na coluna d’água.Considerei como habitat preferencial das espécies os locaisonde foram registradas mais de 55% das observações para ogrupo.
ResultadosOs valores de pH (4,0) e de temperatura da água (27ºC)
mantiveram-se estáveis nos três trechos amostrados. A larguramédia dos igarapés foi de 2,5m (1,2-1,3m) e a profundidademédia foi de 0,22m (0,09-0,39m), com as maiores amplitudesde variação das características físicas sendo observadas entreos dois tipos de habitats (Tabela 1).

Curso de Campo Ecologia da Floresta Amazônica - 2001 107
Tabela 1. Valores médios (amplitude) das variáveis físicas dosmesohábitats de três igarapés estudados na área da Reserva do Km41, AM. N = número de habitats amostrados.
Registrei 164 exemplares de peixes pertencentes a noveespécies, três famílias e duas ordens (Tabela 2). As espéciesmais comuns foram Hyphessobrycon aff. melazonatus,Iguanodectes variatus, Hyphessobrycon aff. heteroharbduse Aequidens pallidus. As espécies menos abundantes foramCrenicichla gr. saxatilis e Apistogramma gr. steindachneri,com apenas um exemplar cada.
Tabela 2. Espécies de peixes/abundâncias observadas em trechosde três igarapés na área da Reserva do Km 41, AM.
Registrei a ocorrência das espécies em sete trechos decalha e oito de poções. Os substratos registrados foramarenoso com emaranhados de raízes (três ocorrências),arenoso coberto por folhas (cinco ocorrências), e arenosocoberto por folhas e com raízes (seis ocorrências).
Os valores de riqueza de espécies por habitat variaram dezero à cinco em trechos de calha, onde registrei até 25indivíduos de cinco espécies diferentes. Nannostomus
marginatus, Iguanodectes variatus, Hemigrammus gr.ocellifer e Aequidens pallidus sempre estiveram ausentesnestas áreas. Em poções observei entre três e oito espécies,com a abundância total oscilando entre 03 e 24 indivíduos.As diferenças nas amplitudes de riqueza de espécies eabundância de peixes entre os habitats foram maiscontrastantes quando associados ao tipo de substrato e/ou apresença de raízes (Tabela 3).
Habitats (N) Comprimento (m) Largura (m) Profundidade (m) Correnteza (m/s)
Calha (07) 3,4 (2,1-4,7) 0,91 (0,55-1,4) 0,34 (0,21-0,5) 0,2 (0,12-0,33)
Poção (08) 2,3 (0,67-4,2) 0,86 (0,3-1,5) 0,32 (0,18-0,5) 0,25 (0,12-0,6)
Espécies Igarapé Total %
Q13 M13 L6
CHARACIFORMES
Characidae
Hyphessobrycon aff. melazonatus 02 35 11 48 29,2
Hyphessobrycon aff. heteroharbdus 07 19 02 28 17,1
Iguanodectes variatus 08 13 12 33 20,1
Hemigrammus gr. ocellifer 0 01 08 09 5,5
Moenkhausia gr. lepidura 05 03 0 08 4,9
Lebiasinidae
Nannostomus marginatus 03 06 01 10 6,1
PERCIFORMES
Cichlidae
Aequidens pallidus 02 20 04 26 15,8
Apistogramma gr. steindachneri 01 0 0 01 0,6
Crenicichla gr. saxatilis 01 0 0 01 0,6
Total 29 97 38 164 -
Tabela 3. Abundância total de espécies de peixes observadas emhabitats com diferentes estruturas físicas dos três igarapés estudadosna área da Reserva do Km 41, AM. R = raízes adventícias flutuantes;F = substrato coberto com folhas; A = espécie ausente.
Habitats com substrato constituído apenas de areia, ondeo fluxo d’água era turbulento não registrei a ocorrência depeixes. Os menores valores de riqueza e abundância de peixesforam obtidos em ambientes estruturados apenas com raízesflutuantes, tanto nas áreas de calha como de poções. Poçõescom substrato coberto por serapilheira associada à presençade raízes apresentaram as maiores riquezas. Os maioresvalores de abundância de peixes foram registrados em habitatsde calha onde o substrato era coberto por folhas.
DiscussãoA similaridade na ordem dos igarapés, no número de
habitats por trecho, na proporção entre calhas e poções, e nadimensão dos habitats pareceu-me constituírem-se deevidências suficientes de que essas características sãodeterminantes potenciais da complexidade e heterogeneidadede habitats nos igarapés estudados. Bührnheim (1998) fazconsiderações sobre a relevância da dimensão do sistema naestruturação das comunidades de peixes, atuando como fatordeterminante da composição da ictiofauna em igarapés daAmazônia Central. Estudando riachos da bacia do rio Paraná,Caramaschi (1986) identificou que fatores relacionados aovolume do sistema e também a presença de refúgios(vegetação marginal e tipo de fundo) são os principais fatoresatuantes na distribuição da ictiofauna.
Qualquer sistema biológico é influenciado por um grandenúmero de fatores, que interferem nos padrões de distribuiçãoespacial e temporal de suas espécies (Barrella 1992). Cadaespécie de peixe tem tolerância a características singulares,como a qualidade da água, habitat e outras condições (Araújo1998), em geral com a riqueza de espécies aumentando emfunção do volume e da complexidade ambiental (Lowe-McConnell 1969; Angermeier & Schlosser 1989). Trabalhossobre a influência da estrutura do habitat na composição edinâmica de comunidades de peixes podem ser encontradosem Pusey et al. (1995).
A heterogeneidade de habitats em riachos netropicaisinflui na distribuição espacial das comunidades de peixes
Espécies Habitats
Calha Poção Total %
R F R e F R F R e F R F R e F
Hyphessobrycon aff. melazonatus 1 20 2 1 5 19 4,1 52,1 43,8
Hyphessobrycon aff. heteroharbdus 1 4 3 4 0 16 17,8 14,3 68
Iguanodectes variatus 2 5 2 2 3 19 12,1 21,2 66,7
Hemigrammus gr. ocellifer 0 1 0 0 4 4 A 55,5 44,4
Moenkhausia gr. lepidura 0 2 0 2 0 4 25 25 50
Nannostomus marginatus 0 0 0 0 1 9 A 10 90
Aequidens pallidus 0 5 0 0 3 18 A 30,7 69,2
Apistogramma gr. steindachneri 0 0 0 0 0 1 A A 100
Crenicichla gr. saxatilis 0 0 0 0 0 1 A A 100
Abundância total 4 37 7 9 16 91 7,9 32,3 59,8
Riqueza de espécies 3 6 3 4 5 9 - - -

108 Curso de Campo Ecologia da Floresta Amazônica - 2001
em função das variações espaciais verticais definindomicrohabitats (Silva 1993). Esta heterogeneidade, assimcomo variação da coluna d’água são definidas principalmentepelas variáveis físicas: substrato, profundidade e correnteza(Gorman 1987). Fittkau (1967) afirma que em igarapés deterra firme da Amazônia Central dois biótopos principais serepetem a curtas distâncias, podendo de forma geral serdivididos em: ambientes fundos, pouco correntosos e desubstrato fino e, ambientes mais rasos, correntosos e desubstrato formado de areia grossa. Nesses ambientes, adistribuição espacial das comunidades de peixes parece nãoestar relacionada à competição por alimento (Knöppel 1970)ou a sazonalidade, mas sim as condições locais de cadaigarapé (Bührnheim 1998).
No total dos três igarapés amostrados, observei noveespécies de peixes, números bem inferiores as 19 espéciesregistradas por Bührnheim (1998) também utilizandomergulho livre durante o dia em um igarapé de segunda ordemna área da Reserva do Km 41. Entre as famílias registradaspela autora, não observei três delas: Synbranchidae,Gymnotidae e Rivulidae, espécies conhecidas como dedistribuição mais específica, onde torna-se impraticável seusregistros por meio de observação direta, mas sim por meiode coletas noturnas utilizando peneiras. Segundo Buckup(1999) simbranquídeos e gimnotídeos são comuns de áreasde remanso, com os primeiros normalmente sendoencontrados sob a lama do substrato em condições de anoxia,e o segundo, sob o folhiço em condições de hipoxia, já queambos possuem respiração acessória. Rivulídeos tratam-sede um grupo maior, com o gênero mais comum na baciaamazônica (Rivulus) sendo representado por até 90 espécies(Buckup 1999). Também são de difícil acesso através demergulho por residirem em ambientes periféricos rasos.
Bürhnheim (1998) relata que para os três igarapés de terrafirma que ela investigou, Hyphessobrycon aff. melazonatus,Hyphessobrycon aff. heteroharbdus, Iguanodectes variatuse Aequidens pallidus são as espécies mais abundantes, fatoque confirmei com este estudo, mesmo utilizando um esforçoamostral incomparável. Trabalhando com a distribuição depeixes em riachos de segunda ordem do EUA. Guido et al.(1997) listou 14 espécies de peixes para ambientes de riachos,enquanto Knöppel (1970) registrou 17 e Soares (1979) 20espécies, ambos para igarapés da bacia amazônica.
Embora cada tipo de hábitat tenha condições físicasparticulares, há, especialmente, um efeito local influenciandoa distribuição das comunidades, além de hábitats rasos efundos desempenharem diferentes papéis no comportamentodas espécie nos diferentes períodos do dia, refletindo naestrutura das comunidades (Bührnheim 1998)
Entre as nove espécies estudadas, quatro (Iguanodectesvariatus, Nannostomus marginatus, Moenkhausia gr.lepidura e Aequidens pallidus) seguiram o padrão deaumento na ocorrência seguindo a complexidade eheterogeneidade dos ambientes, com esse grupo, e maisHyphessobrycon aff. melazonatus demonstrando preferênciapor habitats de poções. Poções com raízes e serrapilheiraforam os únicos hábitats onde foram observados indivíduos
de Apistogramma gr. steindachneri e Crenicichla gr.saxatilis, ambas representadas por apenas um indivíduo.Estudando três igarapés da mesma bacia Bührnheim (1998)relata que também só registrou a presença de Crenicichla gr.saxatilis em poções. Apesar do baixo número de registros,esta espécie é reconhecida como sendo típica de poções maisheterogêneos e complexos onde é maior o número de abrigos.
Poças com diferentes formas, profundidade e tipo desubstrato afetam a composição das comuidades (Elso & Giller2001). No caso de Moenkhausia gr. lepidura, apesar dosresultados não permitirem que a espécie seja classificadacomo de ocorrência preferencial em ambientes maiscomplexos, 50% da abundância da espécie foi registradanesses locais.
Duas espécies (Hyphessobrycon melazonatus eHemigrammus gr. ocellifer) tiveram abundâncias similaresregistradas para substratos cobertos por serrapilheira e deserrapilheira associada à raízes adventícias. O valores menosexpressivos para a comnidade em geral foi registrado paraobservações em locais estruturados apenas por raízes, ondesomente Hyphessobrycon aff. heteroharbdus apresentoumaiores valores de abundância.
Não é de surpreender que a riqueza seja correlatada coma diversidade de substrato (Tramer & Rogers 1973 apudHorwitz 1978), concentrando a maioria das espécies empoções. Bührnheim (1998) descreve a freqüência deAequidens pallidus em poções onde a espécie tem o hábitode se forragear especulando o substrato.
Acredito que a associação entre um habitat espacialmenteheterogêneo, abundância de itens alimentares carreados pelacorrenteza e menor fluxo d’água próximo ao substratooferecem as melhores condições possíveis para abrigo eforrageamento com minimização do gasto energético. Amesma explicação valeria para substratos de serapilheira emáreas de calha, com a ressalva de que nesses locais o fluxod’água é maior. Este fato pode ser comprovado pelasabundâncias das duas espécies do gênero Hyphessobryconrelatados por Sazima (1986) como catadores de itensarrastados na coluna d’água e na superfície.
Concluo que, assim como Bührnheim (1998) já haviaespeculado, a heterogeneidade de habitats e microhabitatsem igarapés aparentemente similares em suas condiçõesecológicas detêm composições de peixes diferentes quandoanalisadas a nível de habitats.
AgradecimentosAgradeço ao Dr. Jansen Zuanon pela oportunidade de ter
trabalhado neste e em outros três projetos que participeienvolvendo ecologia de peixes da Amazônia sob suaorientação. Pelas valiosas sugestões, aulas relâmpago deecologia e ictiologia e pela amizade. Ao Dr. EduardoVenticinque (Dado) por todas as sugestões e orientação nodelineamento e análise de dados de diferentes projetos, epor ter com seu pandeiro à tiracólo nos proporcionadomomentos sem iguais de “samba de roda” da garoa nachurrasqueira do 41, emplena floresta Amazônica. AoGlaucão pela disponibilidade integral para socorrer a galera

Curso de Campo Ecologia da Floresta Amazônica - 2001 109
e pelas sugestões e correções destruidoras de vários“relats.doc 1,2,3...”. A Kati, nova amiga das aventurasictiológicas, por todo o apoio, sugestões e oportunidades dediscussões. Aos parceiros Dani (Fisher) e Leandro (Alemão)pela convivência nesses 35 dias, papos, risadas, trocas deidéias, de experiências, sugestões nos projetos, bebedeirasetc. As parceiras Cris (Gaúcha), Natalie, Betsabé e Andréia,com quem tive muita honra de desenvolver vários projetos.Ao Tonhão, Angelita, Paula P., Rosinha, Felipão, Tiago,Machu Pichu, Juruna, Jorjão, Dna. Eduarda que juntostornaram o curso mais prazeroso. A todo pessoal do PDBFF.Aos orientadores M.C. Calijuri e Mauro Ribeiro por toda aforça para minha participação no curso. Ao amigo e mestrePaulo Inácio Prado pelo incentivo nesta e em outrasempreitadas.
Referências BibliográficasAngermeier, P.L. & Schlosser, I.J. 1989. Species-area
relationships for stream fishes. Ecology 70(5): 1450-1462.
Araújo, F.C. 1988. Adaptação do índice de integridadebiótica usando a comunidade de peixes para Rio Paraíbado Sul. Revta. Bras. Biol. 58 (4): 547-558.
Barrella, W. 1992. Princípios básicos da metodologiaaplicada em estudos ambientais. Terra Gasta: a questãoambiental. Educ, São Paulo.
Barrella, W.; Petrere Jr., M.; Smith W.S. & Montag, L.F.A.1999. As relações entre as matas ciliares, os rios e ospeixes. In: Matas Ciliares - Conservação e Recuperação,Rodrigues, R.R. & Leitão Filho, H.F, pp. 187-207.Edusp.
Buckup, P.A. Sistemática e biogeografia de peixes de riachos.In: Ecologia de Peixes de Riachos, Série OecologiaBrasiliensis vol. VI, Caramaschi, E.P., Mazzoni, R &Peres-Neto, P.R. (eds.), pp. 91-138. PPGE-UFRJ, Riode Janeiro, Brasil.
Bürnheim, C.M. 1998. Estrutura de Comunidades de Peixesem Igarapés de Floresta de Terra Firme na AmazôniaCentral. Dissertação de Mestrado, Universidade doAmazonas/INPA, Manaus, AM.
Caramaschi E.P. 1986. Distribuição da ictiofauna de riachosdas bacias do Tietê e do Paranapanema junto ao divisorde águas (Botucatu, SP). Tese de Doutorado,Universidade Federal de São Carlos.
Elso, J.I. & Giller, P.S. 2001. Physical characteristicsinfluencing the utilization of pools by brown trout in anafforested catchment in Southern Ireland. J. Fish. Biol.58: 201-21.
Fittkau, E.J. 1964. Remarks on limnology od central-Amazonrainforest streams. Verh. Intern. Limnol. 15: 1092-1096.
Gido, K.B.; David, L.P. & Molles-Jr., M.C. 1997. Spatialand temporal variation of fish communities in secondarychannels of the San Juan River, New Mexico and Utah.Environ. Biol. Fish. 49: 417-434.
Gorman, O.T & Karr, J.R. 1978. Habitat structure and streamfish communities. Ecology 59(3): 507-515.
Hawkins, C.P.; Kershner, J.L.; Bisson, P.A.; Bryant, M.D.;Decker, L.M.; Gregory, S.V.; McCullough, D.A.;Overton, C.K.; Reeves, G.H.; Steedman, R.J. & Young,M.K. 1993. A hierarchical approach to classifying streamhabitat features. Fisheries Vol 18, No. 6.
Hayes, D.B.; Ferreri, C.P. & Taylor, W.W 1996. Linking fishhabitat to their populations dynamics. Can. J.. Fish.Aquat. Sci. 53: 383-390.
Horwitz, R.T. 1978. Temporal variability patterns and thedistributional patterns of stream fishes. Ecol. Monogr.48: 307-21p.
Keenleyside, M.H.A. 1979. Diversity and Adaptation in FishBehavior. Springer-Verlag, New York.
Knöppel, H.A. 1970. Food of Central Amazonian fishes:contribution to the nutrient ecology of Amazonian rainforest streams. Amazoniana 2: 257-352.
Lowe-McConnell, R.H. 1969. Speciation in tropicalfreshwater fishes. In: Speciation in TropicalEnvironments, Lowe-McConnell, R.H. (ed.), pp. 51-75.Linean Society of London, Academic Press.
Pusey, B.J.; Arthington, A.H.; Read, M.G. 1995. Speciesrichness and special variation in fish assemblagestructure in two rivers of the Wet Tropics of northernQueensland, Australia. Environ. Biol. Fish 42: 181-99.
Radambrasil, 1976. Folha SA.21-Santarém. Levantamentode Recursos Naturais (Geologia, Pedologia, Vegetação,Uso Potencial da Terra). Vol. 10. DNPM, Ministériodas Minas e Energia, Rio de Janeiro, 510p.
Richards, C.; Johnson, L.B. & Host, G.E. 1996. Landscape-scale influence on stream habitats and biota. Can. Jour.Fish. Aquat. Sci. 53: 295-311.
Sabino, J 1999. Comportamento de peixes em riachos:métodos de estudo para uma abordagem naturalística.In: Ecologia de Peixes de Riachos, Série OecologiaBrasiliensis, vol. VI, Caramaschi E.P., Mazzoni, R. &Peres-Neto, P.R. (eds.), pp. 183-208. PPGE-UFRJ, Riode Janeiro, Brasil.
Santos, G.M. & Ferreira, E.J.G. 1999. Peixes da BaciaAmazônica. In: Estudos Ecológicos de Comunidades dePeixes Tropicais, Lowe-McConnell, R.H. (Tradução:Vazzoler, A.E.A.M., Agostinho, A.A. & Cunningham,P.C.), pp. 345-373. Edusp, São Paulo.
Sazima, I. 1986. Similarities in feeding behavior betweensome marine and freshwater fishes in two tropicalcommunities. J. Fish. Biol. 29: 53-65.
Silva, C.P.D. 1993. Alimentação e distribuição espacial dealgumas espécies de peixes do igarapé do Candiru,Amazonas, Brasil. Acta Amaz. 23: 271-285.
Soares, M.G.M. 1979. Aspectos ecológicos (alimentação ereprodução) dos peixes do Igarapé do Porto, Aripuanã,MT. Acta Amaz. 9: 325-352.

110 Curso de Campo Ecologia da Floresta Amazônica - 2001
Intensidade de herbivoria por invertebrados ao longo de um gradiente verticalem uma floresta de terra firme na Reserva do Km 41, Manaus, AM
Cristiane Follmann Jurinitz
IntroduçãoA alta diversidade de plantas característica das florestas
tropicais tem sido atribuída a padrões espaciais demortalidade de sementes e plântulas resultante da herbivoria(Janzen 1970; Connell 1971). Nesses ecossistemas,aproximadamente 11% da área foliar produzida anualmenteé consumida por herbívoros e patógenos (Coley & Aide1991).
A suscetibilidade das plantas ao ataque de herbívoros éfreqüentemente influenciada de forma substancial peloambiente, podendo envolver mudanças na qualidade do tecidofoliar como função do nível de nutrientes do solo (Gershezon1984 apud Marquis & Braker 1994) e de níveis de incidênciade luz (Langenheim et al. 1981 apud Marquis & Braker1994). Esses efeitos podem determinar a probabilidade deataque por herbívoros ao afetarem a qualidade da folha. Domesmo modo, os fatores abióticos exercem influência sobreo forrageamento dos herbívoros e seu padrão de colonização(Moore et al. 1988).
A grande maioria dos estudos sobre herbivoria enfoca aTeoria da Disponibilidade de Recursos, segundo a qualespécies com diferentes taxas intrínsecas de crescimentodiferem no investimento em defesas químicas em respostaao grau de limitação de recursos do ambiente (Coley et al.1985). Existem dois tipos gerais de defesas, representandoextremos de um contínuo: i) defesas quantitativas, queincluem compostos que reduzem digestibilidade, em geralassociadas a espécies de vida longa e de maior tamanho, asquais são mais facilmente encontradas pelos herbívoros; ii)defesas qualitativas (tóxicas a herbívoros especialistas),associadas a espécies pioneiras ou imprevisíveis, de vida maiscurta e porte menor (Marquis & Braker 1994).
A floresta tropical não é organizada em estratos oucamadas horizontais bem definidas. As plantas formam umcontínuo de distribuição vertical conforme suas adaptaçõesà disponibilidade de luz dentro da floresta (Richards 1996).Simplificadamente, as duas condições contrastantes deluminosidade na floresta, dossel e interior, determinam umasérie de adaptações, dentre elas a variação no grau deesclerofilia das folhas como proteção à dessecação (Fetcheret al. 1994), tendo efeito na preferência dos herbívoros (Coley1987).
O estudo dos padrões de herbivoria nas florestas tropicaisnão tem considerado esse gradiente vertical de variação(Marquis & Braker 1994). Wint (1983) avaliou a herbivoriapor insetos no dossel de três florestas tropicais, porém nãoestabelece nenhum tipo de comparação com o que pode serencontrado no interior da floresta.
O objetivo desse trabalho foi verificar como se comporta
a intensidade de herbivoria por invertebrados ao longo deum gradiente vertical e em relação à textura foliar nessegradiente.
Material e MétodosRealizei este estudo entre os dias 28 e 31 de agosto de
2001 na Reserva do Km 41 (2° 24’S; 59° 52’W) a 70 Km aonorte de Manaus. A reserva é constituída de floresta de terrafirme contínua, com temperatura e pluviosidade médiasanuais de 26,7°C e 2186mm, respectivamente (Lovejoy &Bierregaard 1990).
Selecionei três parcelas de 20x20m, distantes pelo menos200m umas das outras, já marcadas em um quadrado de 100ha no qual foi realizado um estudo de longo prazo sobre afamília Lecythidaceae. Em cada parcela, escolhi uma árvorede dossel com possibilidade de escalada com equipamento,da qual coletei ramos de alguns indivíduos mais próximos(lenhosos, exceto lianas) com o auxílio de um podão de cabolongo. Para coleta das árvores com até 8m de altura foiutilizado apenas o podão de cabo longo. Coletei ramos decerca de 20 indivíduos por parcela, buscando contemplartodas as classes de altura. Para cada indivíduo coleteiaproximdamente 100 folhas em uma única determinada classede altura, ou seja, não repetindo outras alturas no mesmoindivíduo. Cada indivíduo foi numerado tendo suas folhasseparadas em sacos plásticos etiquetados com a respectivaaltura de coleta e parcela, sendo levados ao laboratório paraestudo.
Com uso de uma tabela de números aleatórios sorteei 50folhas de cada indivíduo nas quais estimei a porcentagem deherbivoria conforme as categorias a seguir. Além disso, asfolhas foram classificadas de acordo com sua textura emmembranáceas, cartáceas e coriáceas.
Para a estimativa da porcentagem de herbivoria utilizeicinco categorias (0 = 0%, 1 = 1 a 6%, 2 = 6 a 12%, 3 = 12 a25%, 4 = 25-50%, 5 = 50 a 100% de herbivoria) a partir dasquais calculei o índice de herbivoria (IH) para cada planta(Dirzo & Domínguez 1995):
onde ni = número de folhas na categoria de dano foliar,i=categoria de dano; N=número total de folhas.
Para a análise estatística utilizei os softwares Stat View eSystat 8.0. As diferenças na composição por tipos de folhasao longo do gradiente vertical e do IH para cada tipo de folhaforam avaliadas com uso do teste não-paramétrico de Kruskal-Wallis (Zar 1984; Motulsky 1995). Para a análise dacorrelação entre a altura e o IH utilizei a correlação não-paramétrica de Spearman (Zar 1984; Motulsky 1995).
IH = n i / Ni * ,
Curso de Campo Ecologia da Floresta Amazônica - 2001 111
ResultadosColetei 70 indivíduos de árvores (jovens e adultas) e
arbustos entre 1,5m e 24m de altura. O IH apresentou umacorrelação negativa com a altura quando considerado oconjunto total de folhas (r
s = -0,452; p = 0,002; n = 70),
mostrando que, à medida que a altura aumenta, a intensidadede herbivoria diminui (Figuras 1, 2 e 3). A variação em termosde intensidade de herbivoria foi muito maior nas alturasmenores do que nas maiores (Figuras 1, 2 e 3).
Dentre os 70 indivíduos coletados, 33 tiveram suas folhasclassificadas em membranáceas, 28 cartáceas e novecoriáceas. O IH não diferiu entre as classes de textura foliar(H = 0,19; p = 0,90; n = 70). Porém, a ocorrência de folhasmembranáceas, cartáceas e coriáceas ao longo do gradientevertical apresentou uma separação nítida (H = 21,16; p =0,0001; n = 70) (Figura 4). Não encontrei folhas coriáceasem alturas inferiores a 12m, com uma altura mediana de20,5m. A mediana das alturas de ocorrência de folhascartáceas foi de 10,3m, enquanto que para as membranáceasfoi de 6,5m, tendo apenas um indivíduo fora do padrãoobservado, com cerca de 25 m de altura (Figura 4).
Correlacionando os valores de IH com a alturaseparadamente para cada classe foliar, membranáceas ecartáceas exibem o mesmo padrão de correlação negativa (r
s
= -0,433; p = 0,014; n = 33; rs = -0,577; p = 0,0027; n=28;
respectivamente), enquanto que para as folhas coriáceas essepadrão não foi encontrado (r
s = -0,274; p = 0,439; n = 9)
(Figuras 1, 2 e3).
0 10 20 30Altura (m)
0
1
2
3
4
Índ
ice
de
he
rbiv
ori
a
Figura 1. Correlação entre o índice de herbivoria (IH) e a alturaconsiderando-se as folhas membranáceas coletadas na floresta deterra firme Reserva do Km 41, Manaus, AM.
0 5 10 15 20 25Altura (m)
0
1
2
3
4
Índ
ice
de
he
rbiv
ori
a
Figura 2. Correlação entre o índice de herbivoria (IH) e a alturaconsiderando-se as folhas cartáceas coletadas na floresta de terrafirme Reserva do Km 41, Manaus, AM.
10 15 20 25Altura (m)
0.5
1.0
1.5
2.0
2.5
Índ
ice
de
he
rbiv
ori
a
Figura 3. Correlação entre o índice de herbivoria (IH) e a alturaconsiderando-se as folhas coriáceas coletadas na floresta de terrafirme Reserva do Km 41, Manaus, AM.
CART CORI MEMB.
0
10
20
30
Altu
ra(m
)
Figura 4. Distribuição dos tipos de folhas em relação à altura decoleta na floresta de terra firme da Reserva do Km 41, Manaus, AM(CART = cartáceas, CORI = coriáceas, MEMB = membranáceas;linha horizontal equivale à mediana, barras mostram a amplitudeesperada e a caixa os quantis).

112 Curso de Campo Ecologia da Floresta Amazônica - 2001
DiscussãoOs fatores determinantes da distribuição espacial de
herbívoros tropicais não são bem conhecidos. A maioria dostrabalhos têm enfocado essa questão somente no sub-bosqueou no dossel, relatando as preferências dos herbívoros comrelação à idade da folha e diferenças interespecíficas nasdefesas químicas e no status sucessional dessas plantas(Basset 1991). Porém, a floresta é um espaço tridimensional,não havendo estudos que apresentem uma abordagem emtermos da variação da herbivoria ao longo do gradientevertical. Assim, o fato do presente trabalho demonstrar quea intensidade de herbivoria está negativamente correlacionadacom a altura das plantas é algo novo.
Esperava-se que houvesse uma diferença na intensidadede herbivoria entre os três tipos de folhas, já que elas diferemno grau de esclerofilia (Coley 1987), o que não foiencontrado. No entanto, a distribuição desses tipos foliaresmostra uma estratificação nítida quando consideradas asalturas em que ocorrem. Assim, ao correlacionarseparadamente o IH e a altura para cada tipo de folha, o padrãoencontrado foi bastante similar ao obtido para o conjunto defolhas, exceto para as folhas coriáceas. Para essas últimas,uma explicação pode ser o fato delas estarem concentradasnas maiores alturas e portanto apresentarem menor variação.Além disso, deve-se considerar que as diferenças entre asclasses foliares definidas previamente nesse estudo podemnão estar representando uma diferença real para os herbívoros,o que só um teste de palatabilidade poderia responder. Dequalquer forma, essas constatações indicam que é a alturaque está determinando as diferenças no IH por influenciar adistribuição dos tipos de folhas ao longo do gradientevertical.
Estudos afirmam que o dossel apresenta uma maiorabundância de herbívoros em relação ao interior da floresta(Basset 1991; Erwin 1983; Wilson 1991 apud Primack1993). Assim, o fato de ter havido menor herbivoria no dosselpode indicar que a floresta estudada apresenta um padrãodiferente do já descrito para a abundância de herbívoros noestrato superior, ou ainda que as plantas apresentam maisdefesas contra esses herbívoros. Essas duas hipóteses nãosão mutuamente excludentes, e podem estar atuando juntasno padrão encontrado. Além disso, as folhas no dosselapresentam uma textura mais resistente, são mais coriáceas,o que já é reconhecidamente um fator que afeta a herbivoriade forma negativa (Coley 1987). Resta saber se o fato dasfolhas serem coriáceas no dossel é primariamente umaconseqüência do ambiente mais estressante em função dasaltas taxas de irradiação solar e incidência de ventos, ou umaproteção contra o grande número de herbívoros naqueleambiente. Parece mais razoável supor que essa é uma proteçãoselecionada pelo tipo de ambiente, e que adicionalmenteconfere uma resistência a herbívoros de forma econômicapara as plantas.
Mas como esse padrão de variação da intensidade deherbivoria ao longo do gradiente vertical pode ser explicado?Algumas hipóteses especultivas podem ser levantadas. Amaior amplitude de variação na intensidade de herbivoria
em plantas de sub-bosque pode indicar que estas apresentammaior diversidade de defesas químicas e físicas em diferentesgraus do que plantas de dossel. Outra explicação pode ser ofato do dossel ser mais homogêneo em termos de condiçõesambientais em relação ao sub-bosque, que apresenta clareiras,e conseqüentemente, variações na umidade, quantidade deluz, entre outras, o que pode ser uma explicação para a menorvariabilidade de intensidade de herbivoria no dossel. Alémdisso, o tipo de herbívoro, se generalista ou especialista, podeter uma influência grande na intensidade de herbivoria nasdiferentes alturas, uma vez que os especialistas consomemuma maior quantidade da área foliar do que os generalistas(Coley 1987).
São necessárias abordagens complementares queconsiderem as diferenças entre espécies pioneiras e tolerantesà sombra, e ainda que avaliem a intensidade de herbivoriaem uma mesma espécie em diferentes estágios da sua vida, afim de melhor compreender as variações observadas naintensidade de herbivoria. De qualquer forma, o padrãoencontrado parece ser forte, pois foi detectado numaabordagem comunitária, sem considerar essas diferençasintrínsecas de cada espécie e estágio de desenvolvimento.
AgradecimentosAgradeço a todos que contribuíram, de uma forma ou de
outra, para a relaização desse projeto: ao Prof. Geraldinhopela idéia, Tonhão (pelas coletas e companhia no campo),Juruna (ajuda no campo), Daniel (pelas discussões e ajudano campo), Rosinha e Betsa (pela ajuda no sufoco deescrever), Prof. Emílio (pela grande ajuda na análise eestruturação do trabalho), Leandro (ajuda nos gráficos), eainda ao pessoal da correção (Jansen e Dadão, pelassugestões). Agradeço também aos colegas de grupo: Fino,que agüentou meus devaneios com bom humor; Rosinha,companheira de risadas e confidências; Rodrigo e Maurício,quase sempre sérios; Betsa, sempre tranqüila; Caê e Daniel,pelas boas risadas e por me deixarem acreditar quecoordenava o grupo. Aos funcionários que garantiram a boacomida: Osmaildo, Eduarda e Jorge. E por último, mas nãomenos importantes, aos coadjuvantes que tornam esse cursopossível, além dos coordenadores, Jansen e Dadão e doGlauco, sempre de alto astral dispostos a ajudar. Finalizandoagradeço à toda turma pela convivência excelente,brincadeiras, aulas de forró e brega (em especial à Angelita),etc; enfim, por terem transformado esse mês de curso numaexperiência de vida inesquecível.
Referências BibliográficasBasset, Y. 1991. Influence of leaf traits on the spatial
distribution of insect herbivores associted with anoverstory rainforest tree. Oecologia 87: 388-393.
Coley, P.D. 1987. Patrones en las defensas de las plantas:porqué los herbívoros prefieren ciertas especies? Revta.Biol. Trop. 35(1): 151-164.
Coley, P.D. & Aide, T.M. 1991. Comparison of herbivoryand plant defenses in temperate and tropical broad-leavedforests. In: Plant-Animal Interactions: Evolution Ecology

Curso de Campo Ecologia da Floresta Amazônica - 2001 113
in Tropical and Temperate Regions, Price, P.W.,Lewinsohn, T.M., Fernandes, G.W. & Benson, W.W.(eds.), pp. 25-49. .John Wiley & Sons, New York.
Coley, P.D.; Bryant, J..P. & Chapin III, F.S. 1985. Resourceavailability and plant-herbivore defense. Science 230:895-899.
Connell, J.H. 1971. On the role of natural enemies inpreventing competitive exclusion in some marine animalsand rain forest trees. In: Dynamics of Populations, Boer,P.J. & Gradwell, G.R. (eds.), pp. 298-312. Centre forAgricultural Publishing and Documentantion,Wageningen.
Dirzo, R. & Domínguez, C. 1995. Plant-herbivoreinteractions in mesoamerican tropical dry forest. In:Seasonally Dry Tropical Forests, Bullock, S.H., Mooney,A. & Medina, E. (eds.), pp. 304-309. CambridgeUniversity Press, Cambridge.
Erwin, T.L. 1983. Beetles and other insects of tropical forestcanopies at Manaus, Brazil, sampled by insecticidalfogging. In: Tropical Rain Forest: Ecology andManagement, Sutton, S.L., Whitmore, T.C. & Chadwick,A.C. (eds.), pp. 59-75. Blackwell Scientific Publications,Edinburgh.
Fetcher, N.; Oberbauer, S.F. & Chazdon, R.L. 1994.Physiological ecology of plants. In: La Selva: Ecologyand Natural History of a Neotropical Rain Forest,McDade, L.A., Bawa, K.S., Hespenheide, H.A. &Hartshorn, G.S (eds.), pp. 261-281. University ChicagoPress, Chicago.
Janzen, D.H. 1970. Herbivores and the number of tree speciesin tropical forests. Am. Nat. 104: 501-528.
Lovejoy, T.E. & Bierregaard, O. 1990. Central AmazonianForests and the minimal critical size of ecosystemsproject. In: Four Neotropical Rain Forests, Gentry, H.(ed.), pp. 60-74. Yale University Press, London.
Marquis, R.J. & Braker, H.E. 1994. Plant -HerbivoreInteractions: Diversity, Specificity and Impact. In: LaSelva: Ecology and Natural History of a NeotropicalRain Forest, McDade, L.A., Bawa, K.S., Hespenheide,H.A. & Hartshorn, G.S (eds.), pp. 261-28. UniversityChicago Press, Chicago.
Moore, L.V.; Myers, J.H. & Eng., R. 1988. Western tentcaterpillars prefer the sunny side of the tree, but why?Oikos 51: 321-326.
Motulsky, H.M.D. 1995. Intuitive Biostatistics. OxfordUniversity Press, Oxford.
Primack, R.B. 1993. Essentials of Conservation Biology.Sinauer Associates, Massachuusetts.
Richards, P.W. 1996. The Tropical Rain Forest. CambridgeUniversity Press, Cambridge. 575p.
Wint, G.R.W. 1983. Leaf damage in tropical rain forestcanopies. In: Tropical Rain Forest: Ecology andManagement, Sutton, S.L.; Whitmore, T.C. & Chadwick,A.C. (eds.), pp. 229-239. Blackwell ScientificPublications, Edinburgh.
Zar, J.H. 1984. Biostatistical AnalysisPrentice-Hall, NewJersey.
Interações planta-animal na história natural de uma herbácea do baixio,Rapatea ulei (Rapateaceae) na Reserva do KM 41, Manaus, Amazonas
Christina McCain
IntroduçãoInterações entre organismos formam a base fundamental
da idéia de comunidade biológica. As interações planta-animal são um componente importante dessas interações, naforma de predação, herbivoria, parasitismo e mutualismo.Ao contrário das idéias tradicionais de coevolução, variosestudos recentes têm mostrado que a dinâmica dessasinterações pode variar muito, tanto no espaço comotemporalmente (Thompson 1994). Entender os fatores queinfluenciam essas diferenças na dinâmica da comunidade éimportante para a conservação, porque é preciso levar emconta que as comunidades que procuramos preservar, tantocomo o ambiente, não são estáticas.
Rapatea ulei (Commelinales: Rapateaceae), uma herbáceacomum das comunidades de baixio na Amazônia central, temmúltiplas associações com animais, de tipos e intensidadesdesconhecidos. A única referência disponível sobre essasplantas é limitada a uma descrição breve da morfologia daplanta e a flor (Nee 1995). O presente estudo procura em
entender uma parte da história natural dessa planta herbáceae suas interações com a comunidade de insetos associadoscom ela a fim de contribuir para esse conhecimento.
Material e Métodos
O estudo foi realizado na Reserva do Km 41 do ProjetoDinâmica Biológica de Fragmentos Florestais, na FazendaEsteio, Manaus, AM (2°24’S; 59°44’W). Rapatea uleiocorre freqüentemente nas áreas de baixio por toda a reserva.Trechos de seis igarapés na bacia do rio Urubu forampercorridos em busca de áreas de R. ulei em flor.
Um total de 27 inflorescências (uma por planta) foramcoletadas, encontrados ao longo de dois igarapés. Novedessas inflorescências foram dissecadas e examinadas paraobservação da estrutura floral e presença de insetosassociados. Também, coloquei sacos plásticos sobre ainflorescência de cada uma de cinco plantas para captura deinsetos emergindo das flores. Os sacos foram removidos

114 Curso de Campo Ecologia da Floresta Amazônica - 2001
depois de cinco dias para identificação dos insetoscapturados.
Dezoito plantas foram coletadas para quantificar o nívelrelativo de predação em frutos/sementes e herbivoria dasinflorescências. Uma coleta foi realizado em um igarapé(trilhas P- 2 até R-2), em quatro áreas com plantas e floresabundantes. Para cada agregação, amostrei (ao acaso) umaplanta reprodutiva, independente da idade, tamanho,condição de folhas etc. Não escolhi indivíduos com menosde cinco metros de distancia entre si. Amostrei em cadalugar uma planta em cada tipo de solo: “alagado”; “barromolhado”; e “barro seco.” Se uma pegada no solo enchia deágua imediatamente, a área foi definida como “barromolhado”; se não tinha água imediatamente abaixo dasuperfície do solo, foi definida como “barro seco.” Áreasalagadas foram definidas por a posição justo ao lado doigarapé, com a base da planta muito perto, ou parcialmentedentro da água.
Medi a folha mais larga e seu comprimento para umaestimativa de tamanho da planta. Selecionei a inflorescênciamais larga, ou se eram aparentemente iguais, selecionei ainflorescência mais alta (distante do solo). As inflorescênciasforam colocadas em sacos plásticos e levadas para olaboratório. Examinei 18 plantas no total (712 flores).
A amostragem de flores foi realizada segundo a seguintemetodologia: de cada inflorescência, amostrei apenas umagrupamento de flores, sempre no meio da inflorescência(as inflorescências tinham entre cinco e nove agrupamentos).Removi o agrupamento da bráctea, separei todas as flores eabri cada flor individualmente, para determinar sua condição.A condição das flores foi dividido em nove categorias: floraberta, flor intacta (fechada mas madura, sem evidência deherbivoria), flor imatura (fechada e pouca desenvolvida), florcomida (flores fechadas mas maduras, com evidência deherbivoria), frutos intactos (imaturos), frutos intactos(maduros), frutos predados (cheio de material podre,sementes consumidas), frutos abortados (frutos secos comsementes encarquilhadas, impossível de determinar a causado dano) e frutos com sementes soltas (onde as sementesaparentemente já foram dispersadas). Anotei a presença delarvas, pupas, ou adultos de cada inseto encontrado porinflorescência. Dados foram analisados usando umacomparação ANOVA para determinar o efeito de tipo de solo(alagado, molhado, seco) com os dois tipos de dano aos frutos(fruto abortado ou fruto predado) e também com tamanho daplanta. Um teste Kruskal-Wallis foi utilizado para análisedo efeito de solo em herbivoria de flores. A análise de dadosfoi realizado com uso do programa SYSTAT 8.0.
ResultadosHistória NaturalCada planta produz duas inflorescências, cada uma
protegida por duas brácteas grandes e composta de centenasde flores. As flores são perfeitas, hermafroditas com ovárioínfero, sete anteras, e têm três pétalas, três sépalas (Figura1). As flores são agregadas em grupos ao longo das brácteas,arranjadas em círculos concêntricos. O material floral mais
imaturo fica no centro e as flores vão crescendo do centropara as extremidades. Assim a inflorescência cresce emlargura com a idade. Foram observadas em flor durante todoo mês de agosto de 2001 com inflorescências em diferentesestágios, desde senescentes até imaturas. Uma característicasaliente dessa inflorescência é a gelatina entre as duasbrácteas, cobrindo as flores imaturas e os frutos emdesenvolvimento. Quando a flor está madura, ela cresce atéemergir da gelatina e abre. Assim a corola aflora desseexudato e ali pode ser visitada por vários insetos.
A distribuição das plantas foi geralmente agregada, comagregações de plantas em flor e outros grupos seminflorescências. Rapatea ulei foi encontrada em quatro dosseis igarapés amostrados, tanto em igarapés de primeira comode segunda ordem. Na Reserva, foi encontrado do igarapédas trilhas P-2 até R-2, na nascente do igarapé. Outros gruposabundantes também foram encontrados mais a jusante nessemesmo igarapé, mas não estavam em flor durante o períodode estudo. Também encontrei agregações no trecho entre astrilhas L-4 e L-7 no igarapé formado pelos dois igarapésprimários nascentes nas trilhas I-0 e R-2. Outras espéciestambém encontradas ocasionalmente foram R. paludosa, eoutra morfologicamente distinta de R. ulei e R. paludosa,ainda não identificada.
Muitos insetos ocupam a gelatina que cobre asinflorescências. Dentro de aproximadamente 30inflorescências coletadas e examinadas, coletei pelo menos10 espécies de artrópodos de quatro ordens (Diptera,Coleoptera, Acari e Hymenoptera) e um espécie de Annelida(Oligochaeta) não identificada (Tabela 1). Muitas espéciesainda não foram identificadas por falta de chaves para asetapas larvais. Isso inclui somente insetos coletadosdiretamente da inflorescência, dentro da estrutura ou em cima.
Tabela 1. Animais coletados em amostras das inflorescências deRapatea ulei na Reserva do Km 41, Manaus, AM.
Adultos das moscas Cyclorrhapha foram observadosfreqüentemente em cima das inflorescências, particularmenteconcentrados em cima de flores abertas. Larvas e pupas dessaespécie foram identificadas em função de sua presença dentrodo calix com material podre da flor e ovário. As larvas, comoas pupas, têm um tubo para respiração muito comprido e aspupas muito desenvolvidas podem ser relacionados às moscasadultas pelos olhos alaranjados e brilhantes das duas. Apresença dessas larvas ou pupas foi sempre consistente comesse tipo de dano (material pútrido, rosa, ovário descomposto
Classificação Espécie Larva Pupa Adulto
Diptera (Cyclorrhapha) ? esp. 1 X X XDiptera (?) esp. 2 X
Diptera (?) esp. 3 X
Diptera (?) esp. 4 ? X
Diptera (?) esp. 5 ? X
Coleoptera: Phalacridae esp. 1 X X
Coleoptera: Bruchidae esp. 1 X
Acari (?) esp. 1 X?
Annelida (?) esp. 1 X?
Coleoptera (?) esp. 1 X

Curso de Campo Ecologia da Floresta Amazônica - 2001 115
o parcialmente predado).Besouros adultos da família Phalacridae, foram
encontrados em 100 % das inflorescências examinadas e emgrandes quantidades (estimativa casual, 50 –100 indivíduospor inflorescência). Apesar dos adultos serem muitoabundantes nas inflorescências, o registro das larvas foi maisinfreqüente (0-21% das flores, X= 6.6%) e foramcompletamente ausentes em cinco das 12 amostras (Tab. 1).Essas larvas foram observados alimentando-se de materialfloral e foram encontradas dentro de flores fechadas. Emcontraste com os dipteros, eles comem principalmente acorola, e provavelmente anteras. Besouros adultos da famíliaBruchidae foram encontrados em cima das inflorescênciasem vários indivíduos de Rapatea ulei.
Predação e herbivoriaObservei uma relação significativa entre tipo de solo e
nível de predação de frutas/sementes (Figura 1). Plantascrescendo nos solos alagados tinham menos predação desementes. As plantas crescendo no barro seco tinham umnível intermediario, e as crescendo no barro molhado tinhammais (F = 4,769, g.l. = 2, n = 4, p = 0,039). Devido ao baixonúmero de casos de herbivoria das flores, uma análiseparamétrica não foi possível. O resultado do teste Kruskal-Wallis para o efeito de tipo de solo na predação de flores porcoleópteros foi marginalmente significativo (U = 4,744, n =4, p = 0,093) com a maior predação nas inflorescências dossolos alagados. Não encontrei uma relação entre tipo desolo e tamanho da planta.
Tabela 2. Porcentagem do total de inflorescências em cadaagrupamento predados por varias causas.
Planta Agregamento Solo Frutos
abortados
Frutos
predados
Flores
comidas
Total
de perdas
2 A alagado 0,00 0,00 0,18 0,18
4 B alagado 0,24 0,14 0,10 0,48
14 C alagado 0,33 0,19 0,00 0,65
17 D alagado 0,30 0,04 0,13 0,69
1 A seco 0,40 0,23 0,06 0,69
6 B seco 0,00 0,11 0,21 0,73
13 C seco 0,40 0,16 0,12 0,48
15 D seco 0,39 0,26 0,00 0,68
3 A molhado 0.43 0.30 0.00 0.61
18 B molhado 0.63 0.17 0.00 0.32
5 C molhado 0.36 0.25 0.00 0.79
16 D molhado 0.35 0.33 0.00 0.52
0
0.1
0.2
0.3
0.4
alagado molhado seco
Tipo de Solo
%P
red
ado
Figura 1. Predação de sementes por insetos em três tipos de solo(F = 4.769; n = 4; p = 0.039).
DiscussãoA família Rapateaceae é muito pouco conhecida na
Amazônia central. O Guia da Flora da Reserva Ducke(Ribeiro et al 1999), fala que foram observados indivíduosem flor entre setembro e janeiro, mas na Reserva da KM 41,foram observadas florescendo durante todo o mês de agostode 2001.
A estrutura das inflorescências de Rapatea uleirepresentam um microhabitat muito importante para o ciclode vida de muitos insetos que crescem, vivem e reproduzemdentro das inflorescências. A presença da gelatina cobrindoas flores obviamente providencia um meio agrado ounecessário para muitos animais vivendo imersos nela. Issotalvez represente um ambiente mais estável em termos detemperatura, umidade, proteção, ou alimento para os insetos.
A composição dessa gelatina é desconhecida, mas nãoapresenta um cheiro marcado e dissolve em água. Foiobservado que muitas das inflorescências mais velhas já nãotêm essa gelatina, provavelmente por ter sido dissolvidadevido a maior tempo de exposição à chuva. A função queesse exudato teria para a própria planta é ainda desconhecida.A possibilidade de defesa para as flores inicialmente pareceprovável, mas registrei no presente estudo uma perda de até79% de reprodução potencial por inflorescência devido apredação de flores, frutos, e sementes, o que põe a dúvida aeficiência desse mecanismo de proteção.
Esse impacto na reprodução de R. ulei chama atenção paraas pressões ecológicas nessa planta e o balanço das interaçõesque têm com os vários insetos que dependem dela. Até opresente momento, os insetos mais importantes em termosde impacto negativo para a reprodução de Raptea ulei sãouma espécie de Diptera, subordem Cyclorrhapha e umColeoptera da família Phalacridae. Com os níveis depredação e herbivoria encontrados no presente estudo devemrepresentar uma forte pressão negativa na reprodução dessaplanta.
A presença de larvas e pupas em frutos em descomposiçãoé consistente com a deposição de ovos dentro das floresabertas pelos adultos de Diptera Cyclorrhapha. Logo aslarvas consomem o material do ovário, e possivelmente dasestruturas florais, para seu desenvolvimento. O impacto totaldesse espécie foi grande na reprodução potencial da planta,sendo até 33% dos frutos destruídos somente por essas larvas.
A família Phalacridae é uma família de besourosconhecidos por tipicamente comer flores (Borror et al. 1981).O dano às flores foi menos intenso que o dano feito pelosdipteros Cyclorrhapha, mas quando estiveram presentes naplanta, impactaram até 21% das flores, o que também ésignificativo. Nunca encontrei uma larva de coleoptero dentrode uma flor com dano ao ovário, mas o dano ao flor, pode sersuficiente para previr a função dela. Além de todas asinflorescências examinadas terem adultos desse besouro,larvas e evidência de dano por as larvas foi completamenteausente em quase 42% das inflorescências coletadas. Até omomento, a discrepância entre o número de adultos e larvaspresentes não tem explicação plausível, mas é possível que

116 Curso de Campo Ecologia da Floresta Amazônica - 2001
os adultos estejam procurando outros recursos nasinflorescências.
O encontro ocasional de besouros Bruchidae é tambéminteressante, porque a família é especializada em alimentar-se das sementes (Borror et al. 1981), e foi citado como umafonte importante de predação de sementes a florestasneotropicais (Janzen 1971 apud Penny & Arias 1982). Alémdisso, uma grande parte da perda de reprodução total foicontada como “aborto.” No entanto, nessa amostra deplantas, não identifiquei larvas, pupas ou dano a sementesque fosse possível associar com esses besouros.
A relação entre microhabitat (tipo de solo) e predação desementes e flores é um resultado importante, porque relaçõesentre variáveis ambientais e interações ecológicas é um temamuito corrente na ecologia, evolução e biologia deconservação. No momento, não tenho uma explicação paraessa relação mas isso indica que essas interações podeminfluir no sucesso reprodutivo e consequentemente na suadistribuição no ambiente.
Além de serem distribuídas em muitas áreas de baixio eserem relativamente abundantes nos igarapés examinadosnesse estudo, observei que essas plantas se encontram nosbaixios de largura maior e não encaixados entre vertentes.Isso pode ser explicado por muitas variáveis ambientais,principalmente disponibilidade de luz, água, e/ou nutrientes.Igualmente, esses fatores podem influenciar as populaçõesde insetos na área, que também tem um impacto para o
sucesso de R. ulei.As respostas a essas questões demandam um
conhecimento da história natural dessa planta e os animaisassociados, e podem contribuir para o entendimento dainfluência das variações espacias e ambientais das interaçõesplanta-animal.
Referências BibliográficasBorror, D.J.; De Long, D.M., & Triplehorn, C.A. 1981. An
introduction to the study of insects. Saunders CollegePublishing, New York.
Janzen, D.H. 1971. Seed predation by animals. Ann. Rev.Ecol. Syst. 2:465-492.
Nee, M. 1995. Flora Preliminar do Projeto DinâmicaBiológica de Fragmentos Florestais (PDBFF). New YorkBotanical Garden e INPA-PDBFF, Manaus.
Penny, N.D. & Arias, J.R. 1982. Insects of an Amazon forest.Columbia University Press, New York.
Ribeiro, J.E.; Hopkins, M.J.G.; Vicentini, A.; Sothers, C.A.,Costa, M.A.; Brito, J.M.; Souza, M.A.; Martins, L.H.;Lohmann, L.G.; Assunção, P.A.; Pereira, E.; Silva, C.F.;Mesquita, M.R. & Procópio, L.C. 1999. Flora daReserva Ducke: Guia de Identificação das PlantasVasculares de uma Floresta de Terra-firme na AmazôniaCentral. INPA, Manaus, Brasil.
Thompson, J.N. 1994. The Coevolutionary Process. ChicagoUniversity Press, Chicago.
Diferenças de estratégias reprodutivas em duas espécies de palmeiras clonais,Geonoma deversa e Geonoma stricta (Arecaceae)
Daniel L. M. Vieira
IntroduçãoEm plantas com crescimento modular, reprodução pode
ser alcançada tanto sexualmente, pela produção de sementes,ou clonalmente, pela produção de novos rametspotencialmente independentes, mas geneticamente idênticos(Harper 1978 apud Stökclin 1999). Populações de espéciesclonais, que apresentam reprodução sexuada e vegetativa,podem ser analisadas em dois níveis: um considera a touceira(genet) como indivíduo e o outro considera o caule (ramet)como indivíduo, já que cada ramet pode se reproduzir comrelativa independência (Harper 1977).
O crescimento clonal apresenta diversas vantagens, o genetpode aumentar seu espaço e capturar mais recursos, areprodução vegetativa tem menor custo energético que asexuada e genets estabelecidos têm baixa probabilidade demorrer, já que qualquer caule pode regenerá-lo (Silvertown& Doust 1993). Via de regra existe uma forte correlação entreprodução de flores e crescimento clonal (Silvertown & Doust1993). Entretanto o baixo investimento em reproduçãosexuada pode gerar perdas a curto e longo prazos para umaespécie. Deixando de produzir sementes e criando uma
uniformidade genética ela pode responder fracamente àseleção natural e, em caso de competição, a probabilidadede espécies com mais genótipos se estabelecerem é maior,pois um deles deve responder melhor a um novo ambiente(Silvertown & Doust 1993). Além disso, as sementes servemcomo banco de vegetação potencial para áreas já colonizadase como colonizadoras de novas áreas, disseminando a espécie(Harper 1977). Características essenciais da história de vidade uma espécie, como habilidade competitiva ou decolonização, são afetadas de acordo com o balanço dos modosde reprodução (Stökclin 1999).
Nas florestas tropicais úmidas uma variedade de espéciesclonais, que formam moitas, dominam a vegetação de sub-bosque. Entre essas estão as palmeiras de múltiplos estipes,as quais produzem brotos laterais na base da planta (Holttum1955; Hallé et al. 1998 apud Chazdon 1992). Geonoma, umdos maiores gêneros de palmeiras das Américas, é constituídoem grande parte por espécies clonais com alta abundânciano subosque de todos os tipos de florestas úmidas(Henderson 1995).
Com o objetivo principal de estudar o balanço entre

Curso de Campo Ecologia da Floresta Amazônica - 2001 117
reprodução vegetativa e sexuada e suas consequências naspopulações de duas espécies do gênero Geonoma, por meiode comparação entre estrutura de tamanho, distribuiçãoespacial e investimento em reprodução sexuada, postulei asquestões: (i) as espécies diferem no investimento emreprodução sexuada, ou seja, no potencial de produção desementes? Se diferem, existe um balanço entre potencial deprodução de sementes, tamanho das sementes e crescimentoclonal? (ii) espécies com maior investimento em reproduçãosexuada apresentam maior densidade de moitas novas eadultas?
Material e MétodosEspécies estudadasBaseei a descrição das espécies estudadas em Henderson
(1995). Geonoma deversa, “ubim”, possui estipes geralmenteagrupados, de 1-4 m de altura. Suas folhas chegam a 70 cmde comprimento, usualmente divididas em três pinas por ladoe as inflorescências são ramificadas em uma a duas ordens.É largamente distribuída e abundante na América Central enorte da América do Sul. Habita o subosque da floresta úmidade planície ou pré-montana. As folhas são usadas paracobertura de casas e algumas tribos indígenas obtêm salqueimando a planta toda e filtrando as cinzas.
Geonoma stricta var. stricta, “ubim”, possui estipessolitários ou agrupados variando de meio a três metros dealtura. Suas folhas são simples e pequenas (ca. 14 cm decomprimento) e as inflorescências são geralmente solitárias.Sua distribuição é restrita ao norte da América do Sul. Suasfolhas são usadas para cobertura de casas.
Área de estudoO estudo foi realizado na reserva 1501 (Km 41), uma área
de 1000 ha situada aproximadamente a 70 Km ao norte dacidade de Manaus, AM (02o24’S; 59o43’W). A pluviosidadeanual é de aproximadamente 2200 mm, sendo fortementesazonal. A estação seca se estende de junho a outubro, quandoa pluviosidade média mensal é comumente menor que 100mm. O solo é do tipo latossolo amarelo álico distrófico, comalta quantidade de argila. A área é coberta por florestas deterras baixas intactas, em sua maioria sobre terra firme. Odossel da floresta tem cerca de 35 m de altura e árvoresemergentes chegam a 50 m. O sub-bosque é dominado porpalmeiras sem estipe. A reserva pertence ao Projeto DinâmicaBiológica de Fragmentos Florestais (INPA/SmithsonianInstitution).
Coleta de dadosEm uma área de 1000 x 1000 m (100ha) subdividida em
parcelas de 20 x 20 m, selecionei aleatoriamente 30 parcelas.Contei e medi a altura dos estipes de G. deversa e G. strictacom uma vara métrica e registrei as moitas onde estãopresentes os estipes. Moitas com estipes menores de 30 cmforam registradas apenas em sub-parcelas de 10 x 10 mestabelecidas no canto de cada parcela maior.Com isso
adquiri dados de estrutura de tamanho de estipes (ramets),estrutura de tamanho de moitas (genets – número de estipespor moita) e distribuição espacial das moitas. Dado quemoitas podem ser formadas por um único e pequeno estipe,para evitar confusão, o texto que segue tratará apenas degenets e ramets. Registrei a ocorrência de rebrota após quedade ramets, espontânea ou induzida por queda de galhos. Arebrota, nesse caso, é o lançamento de raízes próxima à copado ramet.
Estimei o investimento em reprodução sexuada, potencialde produção de sementes (PPS), para cada espécie a partirde uma amostra de quatro espádices de indivíduos adultos.Contei o número de lóculos de insersão de flores ou frutos.Medi o diâmetro de sete sementes de cada espécie que,associado ao PPS, indica o investimento em reproduçãosexuada de cada espécie. Considero clonalidade, brotaçãona base do estipe, como evento reprodutivo para enfatizarsua importância na manutenção de populações.
Análise de dadosComparei as curvas de frequência de altura dos ramets
das duas espécies através do teste Kolmogorov-Smirnov, poisas medidas de altura são contínuas (Sokal & Rohlf 1981).Comparei as curvas de freqüência de tamanho dos genets(número de ramets) das duas espécies através do teste G, jáque as medidas de tamanho são discretas. Para a análise asclasses 4, 5, 6 e 7 (número de ramets)foram agrupadas porcausa de muitos valores 0 (Sokal & Rohlf 1981). Para asduas análises anteriores o número de genets, econsequentemente os ramets contidos neles, abaixo de 30cm foram multiplicados por quatro para ponderar o tamanhoda área. Analisei a distribuição espacial das moitas dasespécies pela razão variância/média e testei a diferença de 1(distribuição aleatória) através do teste t de uma amostra(Kershaw 1973). Verifiquei a diferença do PPS e do tamanhodas sementes das duas espécies utilizando o teste t (Zar 1999).
ResultadosForam amostrados 210 ramets (175 ramets/ha) de G.
deversa distribuídos em 138 genets (115 genets/ha).Geonoma stricta teve densidade menor, 90 ramets (75ramets/ha) que formaram 50 moitas (42 genets/ha). As duasespécies apresentaram maior frequência de ramets pequenos,diminuindo exponencialmente com o aumento da altura(Figura 1). Entretanto G. deversa apresentou um pico maiorde estipes nas menores alturas, com a freqüência diminuindoabruptamente nas alturas superiores, enquanto que G. strictaapresentou uma distribuição de freqüência menos abrupta(K-S, d
max = 0,17; p = 0,053).
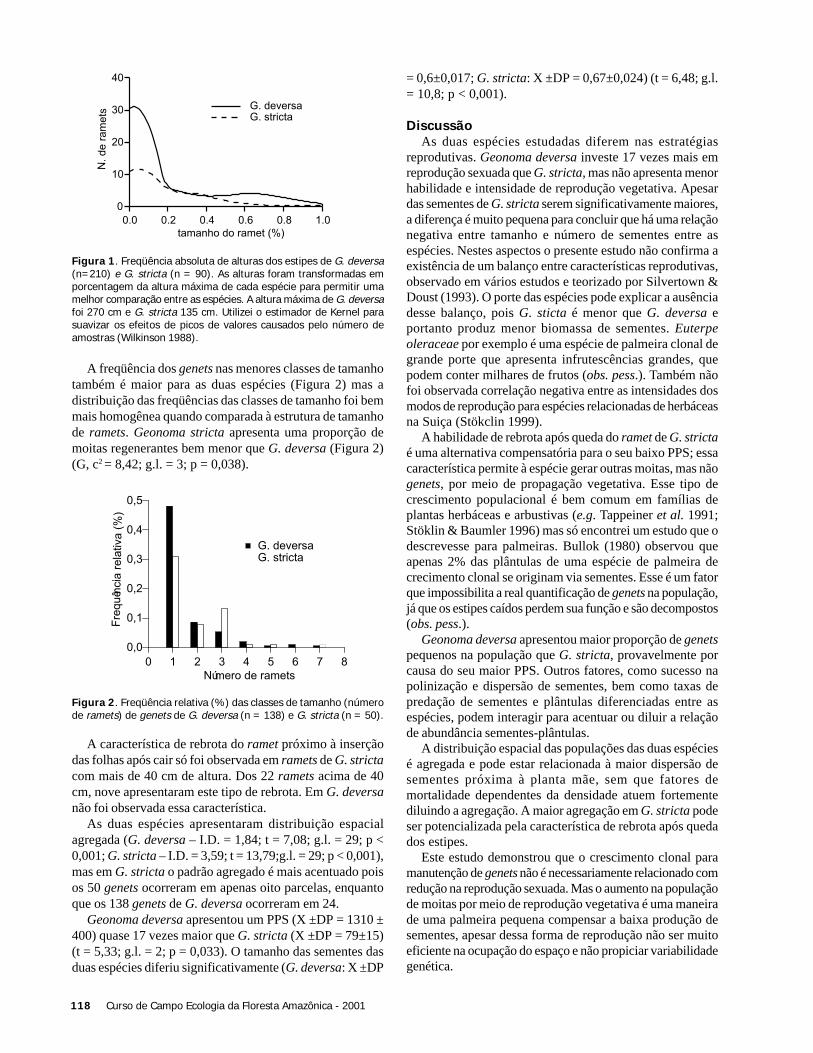
118 Curso de Campo Ecologia da Floresta Amazônica - 2001
0.0 0.2 0.4 0.6 0.8 1.0tamanho do ramet (%)
0
10
20
30
40
N.
de
ram
ets G. stricta
G. deversa
Figura 1. Freqüência absoluta de alturas dos estipes de G. deversa(n=210) e G. stricta (n = 90). As alturas foram transformadas emporcentagem da altura máxima de cada espécie para permitir umamelhor comparação entre as espécies. A altura máxima de G. deversafoi 270 cm e G. stricta 135 cm. Utilizei o estimador de Kernel parasuavizar os efeitos de picos de valores causados pelo número deamostras (Wilkinson 1988).
A freqüência dos genets nas menores classes de tamanhotambém é maior para as duas espécies (Figura 2) mas adistribuição das freqüências das classes de tamanho foi bemmais homogênea quando comparada à estrutura de tamanhode ramets. Geonoma stricta apresenta uma proporção demoitas regenerantes bem menor que G. deversa (Figura 2)(G, c2 = 8,42; g.l. = 3; p = 0,038).
0 1 2 3 4 5 6 7 8Número de ramets
0,0
0,1
0,2
0,3
0,4
0,5
Fre
quên
cia
rela
tiva
(%)
G. strictaG. deversa
Figura 2. Freqüência relativa (%) das classes de tamanho (númerode ramets) de genets de G. deversa (n = 138) e G. stricta (n = 50).
A característica de rebrota do ramet próximo à inserçãodas folhas após cair só foi observada em ramets de G. strictacom mais de 40 cm de altura. Dos 22 ramets acima de 40cm, nove apresentaram este tipo de rebrota. Em G. deversanão foi observada essa característica.
As duas espécies apresentaram distribuição espacialagregada (G. deversa – I.D. = 1,84; t = 7,08; g.l. = 29; p <0,001; G. stricta – I.D. = 3,59; t = 13,79;g.l. = 29; p < 0,001),mas em G. stricta o padrão agregado é mais acentuado poisos 50 genets ocorreram em apenas oito parcelas, enquantoque os 138 genets de G. deversa ocorreram em 24.
Geonoma deversa apresentou um PPS (X ±DP = 1310 ±400) quase 17 vezes maior que G. stricta (X ±DP = 79±15)(t = 5,33; g.l. = 2; p = 0,033). O tamanho das sementes dasduas espécies diferiu significativamente (G. deversa: X ±DP
= 0,6±0,017; G. stricta: X ±DP = 0,67±0,024) (t = 6,48; g.l.= 10,8; p < 0,001).
DiscussãoAs duas espécies estudadas diferem nas estratégias
reprodutivas. Geonoma deversa investe 17 vezes mais emreprodução sexuada que G. stricta, mas não apresenta menorhabilidade e intensidade de reprodução vegetativa. Apesardas sementes de G. stricta serem significativamente maiores,a diferença é muito pequena para concluir que há uma relaçãonegativa entre tamanho e número de sementes entre asespécies. Nestes aspectos o presente estudo não confirma aexistência de um balanço entre características reprodutivas,observado em vários estudos e teorizado por Silvertown &Doust (1993). O porte das espécies pode explicar a ausênciadesse balanço, pois G. sticta é menor que G. deversa eportanto produz menor biomassa de sementes. Euterpeoleraceae por exemplo é uma espécie de palmeira clonal degrande porte que apresenta infrutescências grandes, quepodem conter milhares de frutos (obs. pess.). Também nãofoi observada correlação negativa entre as intensidades dosmodos de reprodução para espécies relacionadas de herbáceasna Suiça (Stökclin 1999).
A habilidade de rebrota após queda do ramet de G. strictaé uma alternativa compensatória para o seu baixo PPS; essacaracterística permite à espécie gerar outras moitas, mas nãogenets, por meio de propagação vegetativa. Esse tipo decrescimento populacional é bem comum em famílias deplantas herbáceas e arbustivas (e.g. Tappeiner et al. 1991;Stöklin & Baumler 1996) mas só encontrei um estudo que odescrevesse para palmeiras. Bullok (1980) observou queapenas 2% das plântulas de uma espécie de palmeira decrecimento clonal se originam via sementes. Esse é um fatorque impossibilita a real quantificação de genets na população,já que os estipes caídos perdem sua função e são decompostos(obs. pess.).
Geonoma deversa apresentou maior proporção de genetspequenos na população que G. stricta, provavelmente porcausa do seu maior PPS. Outros fatores, como sucesso napolinização e dispersão de sementes, bem como taxas depredação de sementes e plântulas diferenciadas entre asespécies, podem interagir para acentuar ou diluir a relaçãode abundância sementes-plântulas.
A distribuição espacial das populações das duas espéciesé agregada e pode estar relacionada à maior dispersão desementes próxima à planta mãe, sem que fatores demortalidade dependentes da densidade atuem fortementediluindo a agregação. A maior agregação em G. stricta podeser potencializada pela característica de rebrota após quedados estipes.
Este estudo demonstrou que o crescimento clonal paramanutenção de genets não é necessariamente relacionado comredução na reprodução sexuada. Mas o aumento na populaçãode moitas por meio de reprodução vegetativa é uma maneirade uma palmeira pequena compensar a baixa produção desementes, apesar dessa forma de reprodução não ser muitoeficiente na ocupação do espaço e não propiciar variabilidadegenética.

Curso de Campo Ecologia da Floresta Amazônica - 2001 119
AgradecimentosSou grato a Ocírio de Souza Pereira (Juruna), Cristiane
Jurinitz, Emílio Bruna e Leandro Scoss pela ajuda na coletade dados e eventuais discussões no campo. EduardoVenticinque e Emílio Bruna ajudaram na análise eapresentação dos resultados. Glauco Machado e JansenZuanon fizeram valiosas sugestões a este trabalho.
Referências BibliográficasBullok, H.S. 1980. Demography of an undergrowth palm in
littoral cameroon. Biotropica 12(4): 247-255.Chazdon, R.L. 1992. Patterns of growth and reproduction
of Geonoma congesta, a cluster understory palm.Biotropica 24(1): 45-51.
Harper, J.L. 1977. Population Biology of Plants. AcademicPress.
Henderson, A.; Galeano, G. & Bernal, R. 1995. Field Guideto the Palms of the Americas. Princeton University Press,Princeton, New Jersey.
Kershaw, K.A. 1973. Quantative and Dynamic Plant Ecology.Edward Arnold Limited, London.
Silvertown, J.W. & Doust, J.L. 1993. Introduction to PlantPopulation Biology. Blackwell Scientific Publications,Oxford.
Sokal, R.R. & Rohlf, J. 1981. Biometry. Freeman andCompany, New York.
Stöcklin, J. 1999. Diferences in life history traits of relatedEpilobium species: clonality, seed size and seed number.Folia Geobot. 34: 7-18.
Stöcklin, J. & Bäumler, E. 1996. Seed rain, seedlingestabelishment and clonal growth strategies on a glacierforeland. J. Veg. Sci. 7: 45-56.
Tappeiner, J.; Zasada, J.; Ryan, P. & Newton, M. 1991.Salmonberry clonal and population structure: the basisfor a persistent cover. Ecology 72(2): 609-618.
Wilkinson, L. 1988. Systat: The System for Statistics.SYSTAT Inc, Evanston, IL.
Zar, J.H. 1999. Biostatistical Analisis. Prentce Hall, NewJersey.
Estudo das relações tróficas dos peixes de umigarapé da Reserva do Km 41,Manaus, AM
Edgar Francisco Prieto Piraquive
IntroduçãoOs igarapés são corpos de água de pequeno porte que se
caracterizam pelo leito bem delimitado, correntezarelativamente acentuada, água com temperatura baixa e poucovariável ao longo do ano. Os pequenos igarapés nas florestastropicais da Amazônia central apresentam biótopos extremospara os peixes, uma vez que as árvores que se fecham sobreos mesmos impedem que a luz atinja a superfície da água,diminuindo a quantidade de nutrientes. São sistemas de baixaprodução biológica e muito dependentes da florestacircundante, que atua como principal elemento de produçãoprimária e consequentemente como base da cadeiatrófica.(Goulding et al 1988,) As mudanças sazonais dosbiótopos aquáticos da Amazônia implicam câmbios naquantidade e qualidade de comida disponível para os peixes(Welcomme 1979).
A disposição dos diferentes alimentos que os peixespodem consumir responde a um processo dinâmico: assim,na época de cheias, os peixes de hábitos herbívoros ecarnívoros que aproveitam os produtos da floresta (sementes,insetos, frutos e flores) têm seu melhor momento, enquantoos peixes piscívoros têm dificuldades para obter alimento(Goulding 1990). Entretanto, a situação da redução doscorpos d’água durante a seca muda a situação porque oscarnívoros passam a ter uma melhor oportunidade paraalimentarem-se (Prieto 2000).
A utilização dos diferentes recursos alimentares pelosmembros das comunidades ícticas, implica em que algumas
espécies podem consumir alimentos de diferentes tipos eorigens, as denominadas generalistas, enquanto outras sófazem uso de um tipo de recurso, as chamadas especialistas.Com base nisso, minha questão tenta descrever qual é atendência de uso dos recursos alimentares, das espéciespredominantes e se estes apresentam sobreposição das dietasna época de seca em um igarapé de águas claras, de terrafirme.
Material e MétodosEste estudo foi realizado em um igarapé de águas claras
de segunda ordem localizado na reserva Km 41 (2o25’S;59o48”W) do projeto Dinâmica Biológica de FragmentosFlorestais (INPA/Smithsonian), Manaus, AM. A temperaturamédia anual é de 27,2 °C (Schroeder 1995) e a precipitaçãomédia anual de 2.127 mm (Junk 1997).
As coletas tiveram uma duração de 30 h, e foram feitas nohorário vespertino conjuntamente com Mauricio Camargodurante três dias. Para cada um dos coletores definimos umesforço de captura de 5 h/dia. Para realizar as capturas utilizeiuma peneira plástica de 0,38 m2. Os lances foram realizadosde forma aleatória, tentando cobrir a variação da profundidadee as diferentes regiões ao longo do igarapé.
Os exemplares capturados foram fixados em formol a 10% para sua posterior identificação Escolhi as cinco espéciesmais abundantes e que tivessem no mínimo 10 indivíduosde comprimento semelhante; de cada um deles foi extraído oestômago e estimado seu grau de repleção segundo as
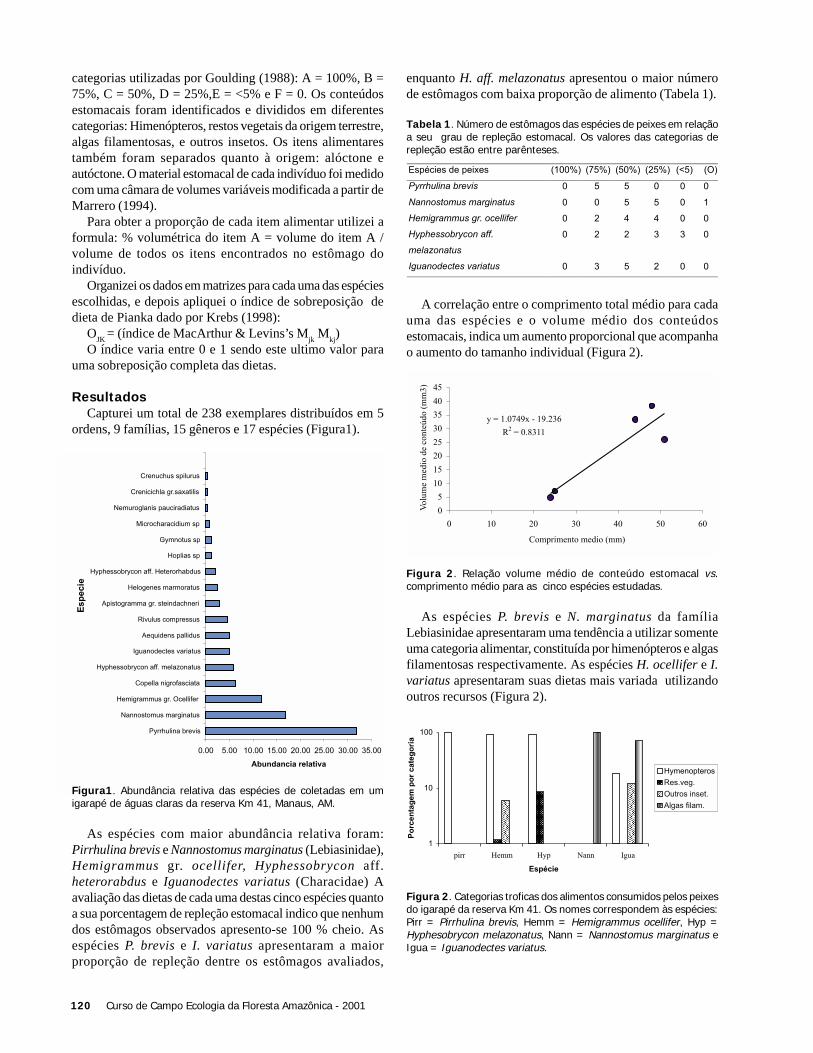
120 Curso de Campo Ecologia da Floresta Amazônica - 2001
categorias utilizadas por Goulding (1988): A = 100%, B =75%, C = 50%, D = 25%,E = <5% e F = 0. Os conteúdosestomacais foram identificados e divididos em diferentescategorias: Himenópteros, restos vegetais da origem terrestre,algas filamentosas, e outros insetos. Os itens alimentarestambém foram separados quanto à origem: alóctone eautóctone. O material estomacal de cada indivíduo foi medidocom uma câmara de volumes variáveis modificada a partir deMarrero (1994).
Para obter a proporção de cada item alimentar utilizei aformula: % volumétrica do item A = volume do item A /volume de todos os itens encontrados no estômago doindivíduo.
Organizei os dados em matrizes para cada uma das espéciesescolhidas, e depois apliquei o índice de sobreposição dedieta de Pianka dado por Krebs (1998):
OJK
= (índice de MacArthur & Levins’s Mjk M
kj)
O índice varia entre 0 e 1 sendo este ultimo valor parauma sobreposição completa das dietas.
ResultadosCapturei um total de 238 exemplares distribuídos em 5
ordens, 9 famílias, 15 gêneros e 17 espécies (Figura1).
0.00 5.00 10.00 15.00 20.00 25.00 30.00 35.00
Pyrrhulina brevis
Nannostomus marginatus
Hemigrammus gr. Ocellifer
Copella nigrofasciata
Hyphessobrycon aff. melazonatus
Iguanodectes variatus
Aequidens pallidus
Rivulus compressus
Apistogramma gr. steindachneri
Helogenes marmoratus
Hyphessobrycon aff. Heterorhabdus
Hoplias sp
Gymnotus sp
Microcharacidium sp
Nemuroglanis pauciradiatus
Crenicichla gr.saxatilis
Crenuchus spilurus
Esp
ecie
Abundancia relativa
Figura1. Abundância relativa das espécies de coletadas em umigarapé de águas claras da reserva Km 41, Manaus, AM.
As espécies com maior abundância relativa foram:Pirrhulina brevis e Nannostomus marginatus (Lebiasinidae),Hemigrammus gr. ocellifer, Hyphessobrycon aff.heterorabdus e Iguanodectes variatus (Characidae) Aavaliação das dietas de cada uma destas cinco espécies quantoa sua porcentagem de repleção estomacal indico que nenhumdos estômagos observados apresento-se 100 % cheio. Asespécies P. brevis e I. variatus apresentaram a maiorproporção de repleção dentre os estômagos avaliados,
enquanto H. aff. melazonatus apresentou o maior númerode estômagos com baixa proporção de alimento (Tabela 1).
Tabela 1. Número de estômagos das espécies de peixes em relaçãoa seu grau de repleção estomacal. Os valores das categorias derepleção estão entre parênteses.
Espécies de peixes (100%) (75%) (50%) (25%) (<5) (O)
Pyrrhulina brevis 0 5 5 0 0 0
Nannostomus marginatus 0 0 5 5 0 1
Hemigrammus gr. ocellifer 0 2 4 4 0 0
Hyphessobrycon aff.
melazonatus
0 2 2 3 3 0
Iguanodectes variatus 0 3 5 2 0 0
A correlação entre o comprimento total médio para cadauma das espécies e o volume médio dos conteúdosestomacais, indica um aumento proporcional que acompanhao aumento do tamanho individual (Figura 2).
y = 1.0749x - 19.236
R2 = 0.8311
0
5
10
15
20
25
30
35
40
45
0 10 20 30 40 50 60
Comprimento medio (mm)
Vol
ume
med
iode
cont
eúdo
(mm
3)
Figura 2. Relação volume médio de conteúdo estomacal vs.comprimento médio para as cinco espécies estudadas.
As espécies P. brevis e N. marginatus da famíliaLebiasinidae apresentaram uma tendência a utilizar somenteuma categoria alimentar, constituída por himenópteros e algasfilamentosas respectivamente. As espécies H. ocellifer e I.variatus apresentaram suas dietas mais variada utilizandooutros recursos (Figura 2).
1
10
100
pirr Hemm Hyp Nann Igua
Espécie
Po
rce
nta
ge
mp
or
ca
teg
ori
a
Hymenopteros
Res.veg.
Outros inset.
Algas filam.
Figura 2. Categorias troficas dos alimentos consumidos pelos peixesdo igarapé da reserva Km 41. Os nomes correspondem às espécies:Pirr = Pirrhulina brevis, Hemm = Hemigrammus ocellifer, Hyp =Hyphesobrycon melazonatus, Nann = Nannostomus marginatus eIgua = Iguanodectes variatus.

Curso de Campo Ecologia da Floresta Amazônica - 2001 121
Os maiores valores dos índices de sobreposição das dietascorresponderam às pares de espécies P. brevis vs. H. ocellifer,Hy. melazonatus vs. H. ocellifer e P. brevis vs. Hy.melazonatus. Observei uma baixa sobreposição das dietasde outras espécies (Tabela 3).
Tabela 3. Valores do índice de sobreposição entre as dietas dasespécies de peixes do igarapé do Km 41
P.
brevis
N.
marginatus
H.
ocellifer
I.
variatus
Hy.
melazonatus
P. brevis - 0,24 0,98 0,24 0,90
N. marginatus - - 0 0,17 0
H. ocellifer - - - 0,24 0,98
I.variatus - - - - 0,24
DiscussãoAs diferenças observadas no número de estômagos com
alimento podem ser explicadas por vários fatores: o horáriode consumo alimentar dos peixes, o requerimento energéticoassociado à atividade diária de cada uma das espécies edisponibilidade de alimento no sistema. O maior número deexemplares com estômagos com maior proporção de volumealimentar observado para P. brevis e I. variatus indicam queestas espécies apresentam uma atividade alimentar no ultimohorário do dia. O fato de que espécies como I. variatus semantenham sempre nadando contra a corrente em águas commaior velocidade indica um requerimento energético alto,que pode ser associado a uma contínua procura de alimentobasicamente de origem alóctone, como formigas, mais nocaso de I. variatus tambem se alimenta de fontes autóctonescomo as algas perifíticas. O contrário ocorre com a espécieP. brevis que fica mais associada a águas de remanso, nadandona superfície e aguardando alimento de origem externa aosistema.
Pode-se prever uma relação custo-benefício que consisteem que pela mesma atividade de nado contínuo uma espéciede correnteza precisará de maior consumo diário de alimento,trazido pela correnteza, enquanto uma espécie menos ativa,que habite ambientes de águas paradas vai requerer menortaxa de consumo.
A virtual ausência de produção primária nos igarapés deáguas pretas e claras, implica que a fauna ali existente obtenhaseus nutrientes básicos da mesma floresta, originados daserapilheira caída na água (Walker 1990). Isto implicaria queas poucas fontes de alimento disponíveis para os peixesdeterminariam uma alta taxa de competição, fato este que sereflete nos altos valores obtidos para sobreposição das dietasdurante o período de estudo.
Um forma para diminuir a pressão sobre o mesmo recursoalimentar por parte das espécies de peixes é a utilização deoutros recursos alternativos que implicaria em um aumentodo espectro alimentar da espécie.
Como conclusão deste trabalho, encontrei que entre asespécies, estudadas algumas apresentam especificidade por
uma categoria alimentar, enquanto outras diversificam suadieta com a inclusão de outras categorias. A alta sobreposiçãodas dietas observadas se explica a traves duma tendência afazer uso do mesmo recurso durante este período do ano.
Como recomendação para posteriores estudos da ecologiatrófica dos peixes deve-se levar em consideração aidentificação ao menor nível taxonômico das categoriasalimentares.
AgradecimentosMi eterna gratitud a todas las personas del proyecto
PDBFF que me ayudaron a conocer desde otra dimensionesta nuestra Amazonia; a Mauricio Camargo por su amistady enseñanzas, tambien a Jansen (Prusiano) y Dadão(Dionisiaco!!!) por ese liderazgo y vivir plenamente laBiologia, Para Juruna y Maria do Carmo mi aprecio, a Jorgey el resto de encargados de alimentarnos com esas viandas,para los compas y las bellas con las que comparti el cursotodo mi aprecio y mision cumplida !!!
Referências BibliográficasJunk, W.J. 1997. Ecology of Swamps on the middle Amazon.
In: Mires Swamp, Bog Fen and Moor, Goret, A.J.P (ed.),pp. 269-294. Elsevier Scientific Publishing Company,Ansterdam.
Knöpel, H.A. 1970. Food of Central Amazonian fishes;contribution to the nutrient-ecology of Amazonian rain-forest streams. Amazoniana II: 257-352.
Krebs, C. 1998. Ecological Methodology. Benjamin/Cummins, Menlo Park Ca.
Lowe-McConnell, R.H. 1987. Ecological Studies in TropicalFish Communities. Cambridge University Press,London.
Goulding, M. Amazon, the Flooded Forest. Sterling, NewYork.
Goulding, M.; Leal Carvalho, M. & Ferreira, E.G. 1988. RioNegro: Rich Life in Poor Water: Amazonian Diversityand Foodchain Ecology as Seen Throught the FishCommunities. Academic Publishing, The Hague.
Marrero, C. 1994. Métodos para cuantificar contenidosestomacales en peces. Universidad de los Llanosoccidentales “Ezequiel Zamora” Unellez Caracas.
Prieto, E. 2000. Estudio ictiologico de um caño de aguasnegras de la Amazonia colombiana Leticia-Amazonas.Trabajo de Grado, Departamento de Biologia,Universidad Nacional. Bogotá.
Shroder, R. 1955. Temperat und Nieuderschalp in Brasilien1 Steden-jahbuch. São Paulo, 3: 7-18.
Walker, I.1990.Ecologia e biologia dos igapós e igarapés.Ciência Hoje 64: 44-53.
Welcomme, RL. 1979. Fisheries Ecology of FloodplainRivers. Longman, London.

122 Curso de Campo Ecologia da Floresta Amazônica - 2001
Influência do tamanho corpóreo no comportamento defensivo de mariposas:uma abordagem fisiológica
Felipe Bandoni de Oliveira
IntroduçãoA temperatura pode afetar muitos aspectos da vida de um
animal, desde seu ritmo diário de atividade até suadistribuição geográfica (Pianka 1994). A influência datemperatura é ainda mais marcante em animais ectotermos,na medida em que eles utilizam fontes externas de calor paramanter sua temperatura corpórea (Begon et al. 1986). Deacordo com o tamanho do animal, o grau de influência datemperatura é diferente: animais pequenos, que possuem umaárea de contato com o ambiente grande em relação a seuvolume, aquecem-se e resfriam-se mais rapidamente que osanimais maiores (Schmidt-Nielsen 1984; Bauwens et al.1995).
Em algumas situações, a influência da temperatura podeter implicações diretas na sobrevivência dos animais. Emalguns insetos, como libélulas e mariposas, sabe-se que alocomoção está intimamente relacionada à temperatura (DeMarco 1998; Schmidt-Nielsen 1984). No caso das mariposas,para voar os animais maiores precisam atingir temperaturascorpóreas mais altas que as ambientais (Schmidt-Nielsen1984), o que é feito através de contrações musculares, emum processo semelhante ao tremor observado em mamíferose aves (obs. pess.). Dessa forma, por demorarem mais paraaquecer-se, os animais de maior tamanho passam mais tempoexpostos termorregulando, além de terem maiorprobabilidade de sofrer ataques de predadores enquantoestão resfriados e despreparados para o exercício (De Marco1998). Assim, seria esperado que ao longo de sua históriaevolutiva as espécies maiores desenvolvessem uma maiordiversidade de comportamentos defensivos, uma vez que têma velocidade com que iniciam o comportamento de fugarestringida por suas condições fisiológicas.
O objetivo deste trabalho é testar, através de experimentosque simulam predação, a hipótese de que as mariposasmaiores possuem um repertório maior de comportamentosdefensivos em relação às menores.
Material e MétodosOs animais estudados neste trabalho foram coletados no
período de 27 a 30 de agosto de 2001 na Reserva do Km 41(INPA/PDBFF), situada a aproximadamente 70 km ao nortede Manaus, AM (2°24’S; 59°43’W).
As mariposas, independente do grupo taxonômico à quepertenciam, foram coletadas com rede entomológica earmadilhas de luz entre as 19 e 23h. Os animais forammantidos em recipientes umidificados até o dia seguinte.Trinta minutos antes de cada simulação esses recipientesforam colocados à mesma temperatura na qual o experimentoseria realizado.
No centro de uma arena de 1,80 x 1,20 x 1,00 m cercada
por uma tela, soltei cada mariposa no plano horizontal. Emseguida, com o objetivo de simular um ataque de um predador,apertei levemente uma de suas asas posteriores com uma pinçae registrei sua reação imediata. Todos esses experimentosforam realizados entre 9 e 11h.
Para testar se eventuais diferenças no comportamentodefensivo entre mariposas grandes e pequenas eram devidasà temperatura ambiental, realizei os experimentos a 20 e a25°C, sorteando antes do início das simulações quais animaisiriam para cada experimento. A temperatura do ambiente foimonitorada durante os quatro dias de estudo por umcarregador de dados (StowAway Inc.) colocado em localsombreado próximo ao alojamento da Reserva, com oobjetivo de verificar se as temperaturas em que foramrealizadas as simulações são efetivamente enfrentadas pelasmariposas na natureza.
Após os experimentos medi o comprimento (da cabeçaaté a ponta do abdômen) e a maior largura do corpo de cadaanimal. Assumindo que a forma do corpo das mariposas ésimilar a um cilindro, estimei seu volume utilizando afórmula:
V = p (a/2)2 x bonde a é a largura e b é o comprimento do corpo dos
animais.Em seguida, identifiquei as mariposas até o nível de
família, com o auxílio da chave para identificação delepidópteros presente em Borror et al. (1981). Com a intençãode minimizar o efeito filogenético sobre os comportamentosestudados, analisei isoladamente os indivíduos de Sphingidaee Geometridae, as duas famílias que apresentaram maiorvariação em volume do corpo.
Para testar se as respostas comportamentais das mariposasestão relacionadas ao tamanho corpóreo, utilizei regressõeslogísticas, que quantificam a associação entre uma variáveldependente expressa de forma binária (neste estudo aresposta) e uma variável independente (tamanho) (Motulsky1995). Realizei essas análises com o programa SYSTAT 8.0(Wilkinson, 1988).
ResultadosColetei um total de 76 mariposas, cujo comprimento do
corpo variou de 4,2 a 64 mm (X ± DP = 16,2 ± 11,4 mm) e ovolume de 1 a aproximadamente 11.000 mm3 (X ± DP = 452,7± 1477,0 mm3). No entanto, a maior parte dos animaiscoletados eram relativamente pequenos – 65% deles possuíamcomprimento menor ou igual a 18 mm. As famílias mais bemrepresentadas nas coletas foram Noctuidae, Pyralidae,Geometridae e Sphingidae. Outras famílias e indivíduos nãoidentificados totalizaram 21 animais (Tabela 1).
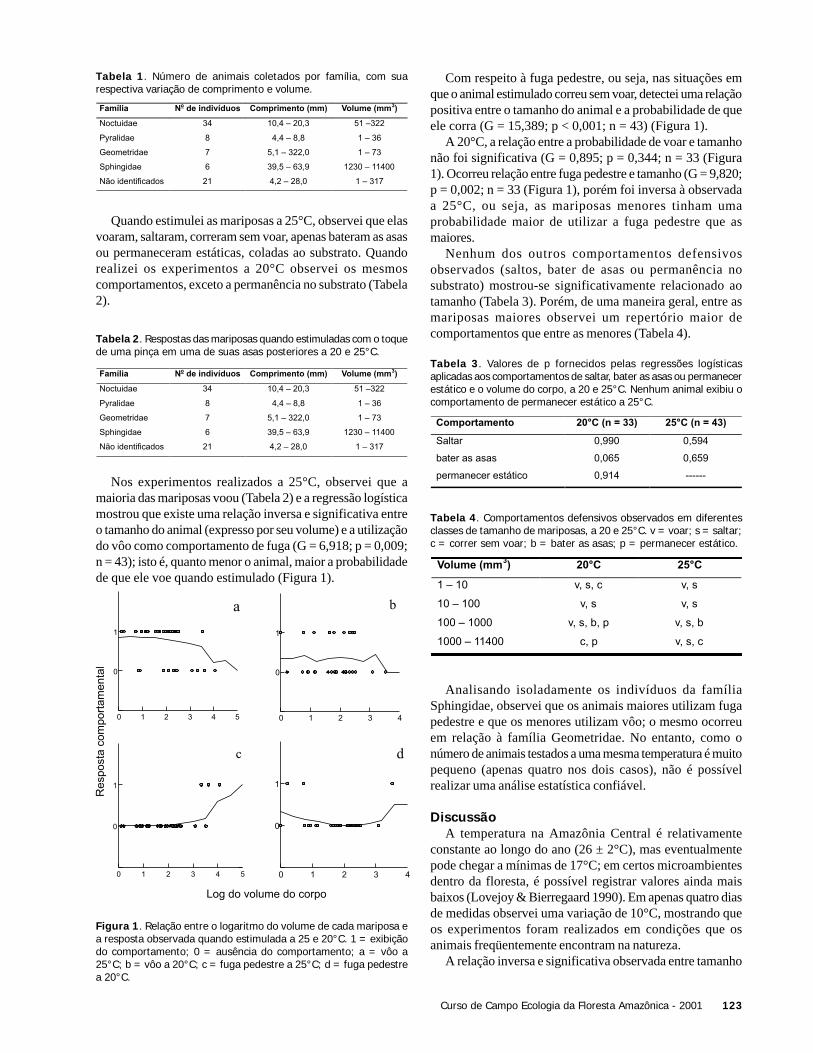
Curso de Campo Ecologia da Floresta Amazônica - 2001 123
Tabela 1. Número de animais coletados por família, com suarespectiva variação de comprimento e volume.
Família No de indivíduos Comprimento (mm) Volume (mm3)
Noctuidae 34 10,4 – 20,3 51 –322
Pyralidae 8 4,4 – 8,8 1 – 36
Geometridae 7 5,1 – 322,0 1 – 73
Sphingidae 6 39,5 – 63,9 1230 – 11400
Não identificados 21 4,2 – 28,0 1 – 317
Quando estimulei as mariposas a 25°C, observei que elasvoaram, saltaram, correram sem voar, apenas bateram as asasou permaneceram estáticas, coladas ao substrato. Quandorealizei os experimentos a 20°C observei os mesmoscomportamentos, exceto a permanência no substrato (Tabela2).
Tabela 2. Respostas das mariposas quando estimuladas com o toquede uma pinça em uma de suas asas posteriores a 20 e 25°C.
Família No de indivíduos Comprimento (mm) Volume (mm3)
Noctuidae 34 10,4 – 20,3 51 –322
Pyralidae 8 4,4 – 8,8 1 – 36
Geometridae 7 5,1 – 322,0 1 – 73
Sphingidae 6 39,5 – 63,9 1230 – 11400
Não identificados 21 4,2 – 28,0 1 – 317
Nos experimentos realizados a 25°C, observei que amaioria das mariposas voou (Tabela 2) e a regressão logísticamostrou que existe uma relação inversa e significativa entreo tamanho do animal (expresso por seu volume) e a utilizaçãodo vôo como comportamento de fuga (G = 6,918; p = 0,009;n = 43); isto é, quanto menor o animal, maior a probabilidadede que ele voe quando estimulado (Figura 1).
Resp
ost
aco
mport
am
enta
l
Log do volume do corpo
Figura 1. Relação entre o logaritmo do volume de cada mariposa ea resposta observada quando estimulada a 25 e 20°C. 1 = exibiçãodo comportamento; 0 = ausência do comportamento; a = vôo a25°C; b = vôo a 20°C; c = fuga pedestre a 25°C; d = fuga pedestrea 20°C.
Com respeito à fuga pedestre, ou seja, nas situações emque o animal estimulado correu sem voar, detectei uma relaçãopositiva entre o tamanho do animal e a probabilidade de queele corra (G = 15,389; p < 0,001; n = 43) (Figura 1).
A 20°C, a relação entre a probabilidade de voar e tamanhonão foi significativa (G = 0,895; p = 0,344; n = 33 (Figura1). Ocorreu relação entre fuga pedestre e tamanho (G = 9,820;p = 0,002; n = 33 (Figura 1), porém foi inversa à observadaa 25°C, ou seja, as mariposas menores tinham umaprobabilidade maior de utilizar a fuga pedestre que asmaiores.
Nenhum dos outros comportamentos defensivosobservados (saltos, bater de asas ou permanência nosubstrato) mostrou-se significativamente relacionado aotamanho (Tabela 3). Porém, de uma maneira geral, entre asmariposas maiores observei um repertório maior decomportamentos que entre as menores (Tabela 4).
Tabela 3. Valores de p fornecidos pelas regressões logísticasaplicadas aos comportamentos de saltar, bater as asas ou permanecerestático e o volume do corpo, a 20 e 25°C. Nenhum animal exibiu ocomportamento de permanecer estático a 25°C.
Comportamento 20°C (n = 33) 25°C (n = 43)
Saltar 0,990 0,594
bater as asas 0,065 0,659
permanecer estático 0,914 ------
Tabela 4. Comportamentos defensivos observados em diferentesclasses de tamanho de mariposas, a 20 e 25°C. v = voar; s = saltar;c = correr sem voar; b = bater as asas; p = permanecer estático.
Volume (mm3) 20°C 25°C
1 – 10 v, s, c v, s
10 – 100 v, s v, s
100 – 1000 v, s, b, p v, s, b
1000 – 11400 c, p v, s, c
Analisando isoladamente os indivíduos da famíliaSphingidae, observei que os animais maiores utilizam fugapedestre e que os menores utilizam vôo; o mesmo ocorreuem relação à família Geometridae. No entanto, como onúmero de animais testados a uma mesma temperatura é muitopequeno (apenas quatro nos dois casos), não é possívelrealizar uma análise estatística confiável.
DiscussãoA temperatura na Amazônia Central é relativamente
constante ao longo do ano (26 ± 2°C), mas eventualmentepode chegar a mínimas de 17°C; em certos microambientesdentro da floresta, é possível registrar valores ainda maisbaixos (Lovejoy & Bierregaard 1990). Em apenas quatro diasde medidas observei uma variação de 10°C, mostrando queos experimentos foram realizados em condições que osanimais freqüentemente encontram na natureza.
A relação inversa e significativa observada entre tamanho

124 Curso de Campo Ecologia da Floresta Amazônica - 2001
do corpo e a probabilidade de voar a 25°C mostra que osanimais maiores não são capazes de voar instantaneamentequando estimulados, apoiando a idéia de que existemrestrições do tamanho sobre este comportamento de fuga. A20°C alguns animais pequenos não voaram, tornando arelação entre resposta de vôo e tamanho não significativaestatisticamente. Provavelmente esses animais que nãovoaram a 20°C possuem um volume de corpo grande emrelação à envergadura de suas asas. Dessa forma, a 20°C suapequena relação área/volume dificulta o aquecimento e essasmariposas não atingem temperaturas suficientes para alçarvôo.
A 25°C apenas as mariposas maiores correram sem voar,mas a 20°C alguns animais menores também fizeram isso. Atemperatura em que os animais foram capturados na noiteanterior ao teste era próxima de 25°C, de modo que o fato deterem sido colocados a 20°C pode ter sido mais crítico paraalguns indivíduos, que quando testados não conseguiramvoar. Além disso, não controlei o grau de atividade dasmariposas durante sua permanência em cativeiro. É possívelque alguns animais tenham se debatido nos recipientes e,quando submetidos a condições menos favoráveis aoexercício não possuíam energia suficiente para voar. Tambémexiste um outro fator, além do tamanho, que pode influenciaro grau de trocas de calor com o ambiente: o tamanho dacamada termoisolante (Pianka 1994). A maioria dasmariposas possui pêlos em seu corpo, que podem ter a funçãode reter calor suficiente para o vôo. Eventuais diferençasnessa camada isolante podem determinar a capacidade devoar em condições mais restritivas como o experimento a20°C, independentemente do tamanho corpóreo.
Apesar dos diferentes comportamentos provavelmenterequererem diferentes níveis de dispêndio de energia, é difícilfazer inferências sobre qual a eficiência de cada um delesem situações naturais, pois isso depende de que tipo depredador ataca as mariposas. Se o predador é um morcegoou um anuro, que detectam movimento, talvez a melhorestratégia seja manter-se estático; caso seja um animal comolfato desenvolvido, como um roedor, talvez saltar ou voarseja mais eficiente. Dessa forma, não existe necessariamenteuma relação entre eficiência do comportamento e a energiagasta para executá-lo.
A hipótese de que os comportamentos observadosdemandam diferentes gastos energéticos explicaria o fato deque a 20°C, uma condição mais restritiva, observa-se umafreqüência alta de comportamentos de “baixa energia” emrelação aos testes realizados a 25°C. Em contrapartida, a 25°Cmais de 75% dos animais voaram (Tabela 1), provavelmenteo tipo de fuga que requer mais energia e é mais freqüente emcondições mais permissivas. Um experimento interessanteseria realizar as mesmas simulações em temperaturassuperiores a 25°C, onde provavelmente uma porcentagemainda maior de animais utilizaria o vôo como estratégia defuga. Apesar do comportamento de permanecer estático poderser interpretado também como uma restrição ao vôo, talvezele seja uma alternativa defensiva já descrita para outrosanimais, conhecida como freeze behavior (Begon et al. 1986).
No entanto, todos os padrões que observei podem serdevidos a um efeito filogenético ao invés de uma restriçãofisiológica ao exercício; ou seja, é possível que animaispertencentes a uma mesma unidade evolutiva possamapresentar um comportamento defensivo parecido não porqueestão submetidos a condições fisiológicas e ambientaissemelhantes, mas porque compartilham uma históriafilogenética comum (Harvey & Pagel 1991). O própriotamanho do corpo, cujo efeito comportamental este trabalhose propôs a investigar, é um caráter utilizado na chave deidentificação das mariposas (Borror et al. 1981). Apesar denão ter sido possível realizar análises estatísticas para osindivíduos Sphingidae e Geometridae isoladamente, aobservação do comportamento dessas mariposas reforça ahipótese de que os animais maiores são mais restringidospelas condições ambientais. Assim, se considerarmos asvariações nas tentativas de fuga como comportamentosdefensivos diferentes, então este estudo corrobora a hipótesede que essa variação é maior entre as mariposas maiores.
Entretanto, a afirmação de que os animais maiores sãomais susceptíveis à predação devido às suas restriçõeslocomotoras apresenta alguns limites. Uma das hipóteses paraexplicar o gigantismo em alguns animais, inclusive insetos,é justamente a redução do número de potenciais predadores(Pianka 1994). Portanto, o mais provável é que cada espéciemantenha-se em um balanço evolutivo entre suas restriçõesfisiológicas e as pressões ecológicas a que está submetida.
AgradecimentosAgradeço a todos os colegas de curso que me ajudaram
neste projeto, desde sua concepção até a discussão dosresultados. A Dadão, Jansen, Glauco, Ana e Leandro pelassugestões interessantes. A Christina, Edgar, Natalie eRosinha por manter o grupo A em harmonia até o final (ufa!).A Dona Eduarda, Jorge, Osmaildo e Antonio pelas refeiçõessensacionais. A Juruna, Everaldo e todos os mateiros portodo o conhecimento sobre a mata que me passaram. A todomundo de São Paulo que fez uma força pra eu poder fazer ocurso: Aninha, Gus, Rodrigo, Tank, Paulo Sano e CarlosNavas. A toda a galera do curso de campo de 2001 pelosbons momentos vividos e a Dadão e Jansen pela coordenaçãoimpecável.
Referências BibliográficasBauwens D.; Garland Jr., T.; Castilla, A.M. & Van Damme,
R. 1995. Evolution of sprint speed in lacertid lizards:morphological, physiological and behavioral covariation.Evolution 49(5): 848-863.
Begon M.; Harper, J.L. & Townsend, C.R. 1986. Ecology:Individuals, Populations and Communities. BlackwellScientific Publications, Boston.
Borror D.J.; De Long, D.M. & Triplehorn, C.A. 1981. AnIntroduction to the Study of Insects. Saunders CollegePublishing, Philadelphia.
De Marco, P.J. 1998. The amazonian campina dragonflyassemblage: patterns in microhabitat use and behaviorin a foraging habitat (Anisoptera). Odonatologica 27(2):

Curso de Campo Ecologia da Floresta Amazônica - 2001 125
239-248.Harvey P.H. & Pagel, M.D. 1991. The Comparative Method
in Evolutionary Biology. Oxford University Press,Oxford.
Motulsky, H. 1995. Intuitive Biostatistics. Oxford UniversityPress, Oxford.
Pianka, E.R. 1994. Evolutionary Ecology. Harper CollinsCollege Publishers, New York.
Schmidt-Nielsen, K. 1984. Scaling: Why Animal Size is soImportant? Cambridge University Press, Cambridge.
Wilkinson, L. 1988. Systat: the System for Statistics.SYSTAT Inc., Evanston, USA.
Distribuição longitudinal e estrutura trófica da ictiofauna deigarapés de terra firme da Amazônia Central
Katiane Ferreira
IntroduçãoA região neotropical possui a maior bacia hidrográfica
do mundo com cerca de 7 x 106 km2, formada pelo rioAmazonas e vários rios e igarapés. O ecossistema aquáticoda Amazônia pode ser dividido, de modo geral, em seteambientes principais: leito de grandes rios, lagos, vegetaçãoflutuante, corredeiras, igapós, praias e igarapés (Lowe-McConnell 1987).
Na América do Sul encontra-se uma mais diversasictiofaunas do mundo, podendo conter mais de 3000 espécies,muitas ainda por serem descritas (Böhlke et al.1978). A baciaamazônica é responsável por grande diversidade, mais de1.300 espécies descritas aproximadamente (Roberts 1972apud Bührnheim 1998; Lowe-McConnell 1987). Menezesressalta que o conhecimento da ampla diversidade de peixesna bacia Amazônica depende de maior esforço de coletavoltado para os sistemas aquáticos ainda pouco estudados,especialmente em regiões de cabeceiras de tributários.Importantes aspectos da ictiofauna de richos neotropicaissão pouco conhecidos, como o endemismo ou a ampladistribuição geográfica de algumas espécies (Lowe-McConnell 1987).
Igarapés são corpos d’água de pequeno porte,caracterizados pelo leito bem delimitado, correntezarelativamente acentuada, água com temperatura baixa e poucovariável ao longo do ano (cerca de 26 °C). Devido à reduçãoda penetração da luz pelo bloqueio da floresta, o reduzidoteor de sais minerais dissolvidos e correnteza moderada, osigarapés são sistemas aquáticos com baixa produçãobiológica e muito dependentes do aporte de nutrientes dafloresta adjacente, que atua como principal elemento deprodução primária e, consequentemente, como base da cadeiatrófica (Lowe-McConnell 1987, 1999).
Minshall et al. (1985) revisaram e sintetizaram diferentesteorias sobre ecologia de riachos, propondo um padrão geralde funcionamento baseado na entrada de matéria orgânica enos grupos tróficos que se alternam longitudinalmente nosistema. Este modelo, desenvolvido originalmente para riosde ambientes temperados, é conhecido como Conceito doRio Contínuo (“River Continuun Concept”, Vannote et al.1980). De acordo com este modelo, em ambientes lóticos,
da cabeceira à foz, pode-se encontrar um gradiente contínuode condições físicas e ajustes bióticos (Vannote et al. 1980).Segundo esses autors, a região das cabeceiras sofre forteinfluência da vegetação circundante, que reduz a produçãoprimária devido ao sombreamento da água, e contribui commaterial alóctone (principalmente vegetais e insetosterrestres). À medida que o riacho aumenta em direção à foz,a importância do aporte de matéria orgânica terrestre éreduzida e aumenta a significância da produção primáriaautóctone. Esse padrão tem origem na matéria orgânicatransportada do trecho superior e provavelmente baseia-sena maior disponibilidade de luz, devido à modificação dacomunidade vegetal de densa para mais rala (Vannote et al.1980).
O presente trabalho teve como objetivo analisar acomposição e estrutura trófica de igarapés da Reserva doKm 41 (INPA/PDBFF), relacionando os resultados obtidoscom o Conceito do Rio Contínuo.
Material e MétodosColetei em igarapés localizados na Reserva do Km 41
(Projeto Dinâmica Biológica de Fragmentos Florestais INPA/PDBFF- Smithsonian Institution), na estrada vicinal ZF-3,rodovia BR 174, 70 km ao norte de Manaus (20° 26’S; 59°46’W). A reserva tem uma área aproximada de 1.000 hainserida em uma área de floresta contínua de terra firme,apresenta temperatura média anual de 26 °C e precipitaçãomédia anual em torno de 2200 mm (Radambrasil 1978).
A partir de uma análise de distribuição dos igarapés naárea de estudo, escolhi como locais de amostragens trechosde igarapés localizados a montante de intersecção destes como sistemas de trilhas. Amostrei nove trechos de igarapés de20 metros de comprimento cada, sendo três em igarapés deprimeira ordem (trilhas P2, L2 e N8), três em igarapés desegunda ordem (trilhas Q13, M13 e N17) e três em igarapéde terceira ordem (trilhas N16 e N17), segundo a classificaçãode Horton-Strahler (Wetzel & Likens 1991). A áreadelimitada para amostragem foi bloqueada com redes de 6mm de malhagem, para que não houvesse evasão dos peixes.Em todos os trechos, dois coletores equipados com peneirasmetálicas de 50 cm de diâmetro e 3 mm de malhagem, fizeram
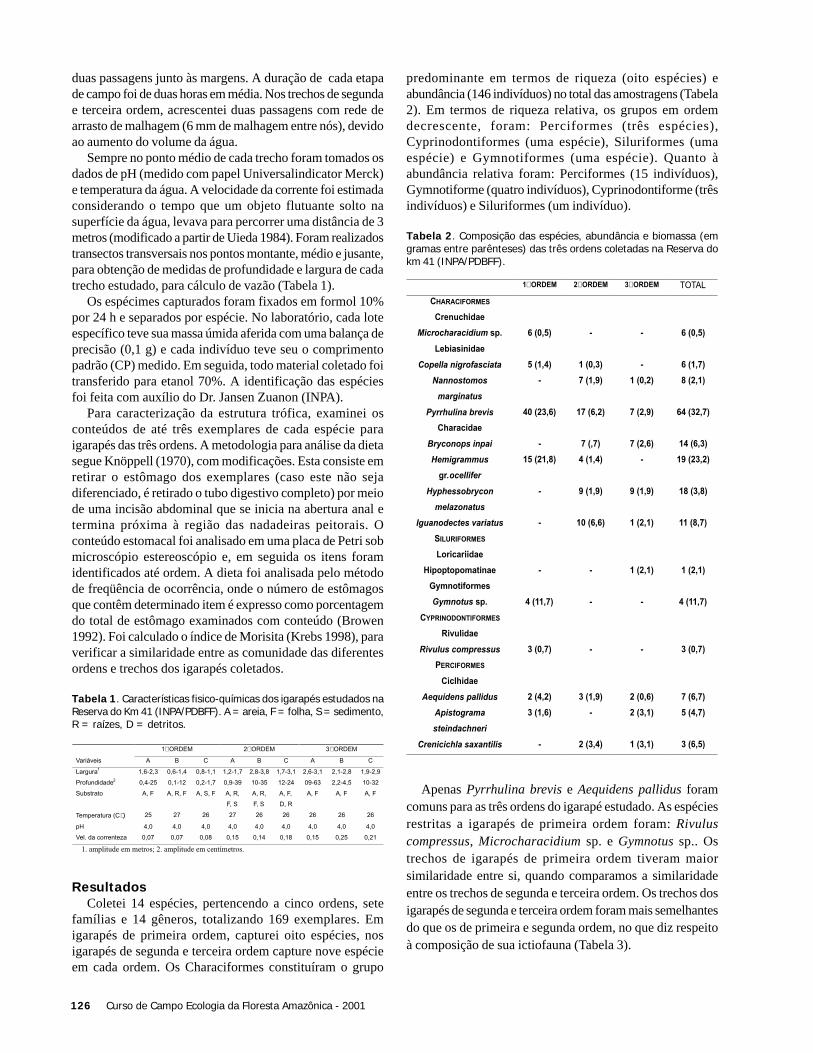
126 Curso de Campo Ecologia da Floresta Amazônica - 2001
duas passagens junto às margens. A duração de cada etapade campo foi de duas horas em média. Nos trechos de segundae terceira ordem, acrescentei duas passagens com rede dearrasto de malhagem (6 mm de malhagem entre nós), devidoao aumento do volume da água.
Sempre no ponto médio de cada trecho foram tomados osdados de pH (medido com papel Universalindicator Merck)e temperatura da água. A velocidade da corrente foi estimadaconsiderando o tempo que um objeto flutuante solto nasuperfície da água, levava para percorrer uma distância de 3metros (modificado a partir de Uieda 1984). Foram realizadostransectos transversais nos pontos montante, médio e jusante,para obtenção de medidas de profundidade e largura de cadatrecho estudado, para cálculo de vazão (Tabela 1).
Os espécimes capturados foram fixados em formol 10%por 24 h e separados por espécie. No laboratório, cada loteespecífico teve sua massa úmida aferida com uma balança deprecisão (0,1 g) e cada indivíduo teve seu o comprimentopadrão (CP) medido. Em seguida, todo material coletado foitransferido para etanol 70%. A identificação das espéciesfoi feita com auxílio do Dr. Jansen Zuanon (INPA).
Para caracterização da estrutura trófica, examinei osconteúdos de até três exemplares de cada espécie paraigarapés das três ordens. A metodologia para análise da dietasegue Knöppell (1970), com modificações. Esta consiste emretirar o estômago dos exemplares (caso este não sejadiferenciado, é retirado o tubo digestivo completo) por meiode uma incisão abdominal que se inicia na abertura anal etermina próxima à região das nadadeiras peitorais. Oconteúdo estomacal foi analisado em uma placa de Petri sobmicroscópio estereoscópio e, em seguida os itens foramidentificados até ordem. A dieta foi analisada pelo métodode freqüência de ocorrência, onde o número de estômagosque contêm determinado item é expresso como porcentagemdo total de estômago examinados com conteúdo (Browen1992). Foi calculado o índice de Morisita (Krebs 1998), paraverificar a similaridade entre as comunidade das diferentesordens e trechos dos igarapés coletados.
Tabela 1. Características fisico-químicas dos igarapés estudados naReserva do Km 41 (INPA/PDBFF). A = areia, F = folha, S = sedimento,R = raízes, D = detritos.
1� ORDEM 2� ORDEM 3� ORDEM
Variáveis A B C A B C A B C
Largura1 1,6-2,3 0,6-1,4 0,8-1,1 1,2-1,7 2,8-3,8 1,7-3,1 2,6-3,1 2,1-2,8 1,9-2,9
Profundidade2 0,4-25 0,1-12 0,2-1,7 0,9-39 10-35 12-24 09-63 2,2-4,5 10-32
Substrato A, F A, R, F A, S, F A, R,
F, S
A, R,
F, S
A, F,
D, R
A, F A, F A, F
Temperatura (C � ) 25 27 26 27 26 26 26 26 26
pH 4,0 4,0 4,0 4,0 4,0 4,0 4,0 4,0 4,0
Vel. da correnteza 0,07 0,07 0,08 0,15 0,14 0,18 0,15 0,25 0,21
1. amplitude em metros; 2. amplitude em centímetros.
ResultadosColetei 14 espécies, pertencendo a cinco ordens, sete
famílias e 14 gêneros, totalizando 169 exemplares. Emigarapés de primeira ordem, capturei oito espécies, nosigarapés de segunda e terceira ordem capture nove espécieem cada ordem. Os Characiformes constituíram o grupo
predominante em termos de riqueza (oito espécies) eabundância (146 indivíduos) no total das amostragens (Tabela2). Em termos de riqueza relativa, os grupos em ordemdecrescente, foram: Perciformes (três espécies),Cyprinodontiformes (uma espécie), Siluriformes (umaespécie) e Gymnotiformes (uma espécie). Quanto àabundância relativa foram: Perciformes (15 indivíduos),Gymnotiforme (quatro indivíduos), Cyprinodontiforme (trêsindivíduos) e Siluriformes (um indivíduo).
Tabela 2. Composição das espécies, abundância e biomassa (emgramas entre parênteses) das três ordens coletadas na Reserva dokm 41 (INPA/PDBFF).
1� ORDEM 2� ORDEM 3� ORDEM TOTAL
CHARACIFORMES
Crenuchidae
Microcharacidium sp. 6 (0,5) - - 6 (0,5)
Lebiasinidae
Copella nigrofasciata 5 (1,4) 1 (0,3) - 6 (1,7)
Nannostomos
marginatus
- 7 (1,9) 1 (0,2) 8 (2,1)
Pyrrhulina brevis 40 (23,6) 17 (6,2) 7 (2,9) 64 (32,7)
Characidae
Bryconops inpai - 7 (,7) 7 (2,6) 14 (6,3)
Hemigrammus
gr.ocellifer
15 (21,8) 4 (1,4) - 19 (23,2)
Hyphessobrycon
melazonatus
- 9 (1,9) 9 (1,9) 18 (3,8)
Iguanodectes variatus - 10 (6,6) 1 (2,1) 11 (8,7)
SILURIFORMES
Loricariidae
Hipoptopomatinae - - 1 (2,1) 1 (2,1)
Gymnotiformes
Gymnotus sp. 4 (11,7) - - 4 (11,7)
CYPRINODONTIFORMES
Rivulidae
Rivulus compressus 3 (0,7) - - 3 (0,7)
PERCIFORMES
Ciclhidae
Aequidens pallidus 2 (4,2) 3 (1,9) 2 (0,6) 7 (6,7)
Apistograma
steindachneri
3 (1,6) - 2 (3,1) 5 (4,7)
Crenicichla saxantilis - 2 (3,4) 1 (3,1) 3 (6,5)
Apenas Pyrrhulina brevis e Aequidens pallidus foramcomuns para as três ordens do igarapé estudado. As espéciesrestritas a igarapés de primeira ordem foram: Rivuluscompressus, Microcharacidium sp. e Gymnotus sp.. Ostrechos de igarapés de primeira ordem tiveram maiorsimilaridade entre si, quando comparamos a similaridadeentre os trechos de segunda e terceira ordem. Os trechos dosigarapés de segunda e terceira ordem foram mais semelhantesdo que os de primeira e segunda ordem, no que diz respeitoà composição de sua ictiofauna (Tabela 3).
Curso de Campo Ecologia da Floresta Amazônica - 2001 127
Tabela 3. Similaridade de Morisita, para os igarapés estudados naReserva do km 41. P = primeira ordem; S = segunda ordem; T =terceira ordem.
Igarapés P1 P2 P3 S1 S2 S3 T1 T2 T3
P1 1.05
P2 0.95 1.08
P3 0.86 0.87 1.07
S1 0.55 0.54 0.78 1.26
S2 0.61 0.61 0.65 0.50 1.23
S3 0.62 0.62 0.69 0.65 1.05 1.25
T1 0.44 0.46 0.53 0.57 0.65 1.08 1.38
T2 0.27 0.31 0.30 0.27 0.52 0.65 0.94 1.20
T3 0.60 0.63 0.89 1.48 0.97 1.14 0.84 0.39 2.36
Quanto às dietas, analisei os conteúdos estomacais de 48espécimes de peixes (Tabela 4). Para igarapés de primeira esegunda ordem, predominaram alimentos de origem alóctone,principalmente formigas (Formicidae: Hymenoptera),enquanto para os igarapés de terceira ordem, os insetosaquáticos predominaram na dieta (Figura 1).
0
10
20
30
40
50
60
FR OIT IA FV FA D CR Al
Itens alimentares
Fre
qü
ên
cia
de
oc
orr
ên
cia
%
1 ordem
2 ordem
3 ordem
Figura 1. Freqüência de ocorrência (%) de itens alimentares nadieta das espécies de peixes coletadas em igarapés da Reserva doKm 41 (INPA/PDBFF). FR = formigas; OIT = outros invertebradosterrestres; IA = insetos aquáticos; FV = fragmentos vegetais; D =detritos; CR = crustáceos e AL = algas.
Tabela 4. Dieta das espécies estudadas em igarapés da Reserva doKm 41 (INPA/PDBFF). NE = número total de estômagos analisadospor espécie. FR = formigas; OIT = outros invertebrados terrestres;IA = insetos aquáticos; FV = fragmentos vegetais; D = detritos; CR= crustáceos e AL = algas.
Espécies Ordem NE 1º Ordem 2º Ordem 3º Ordem
Microcharacidium
sp.
1 3 FV - -
C. nigrofasciata 1 e 2 5 FR; OIT FR; D -
N. marginatus 2 e3 4 - FV; FR; OIT;
IA; D
FV; FR; OIT;
IA; D
P. brevis 1, 2 e 3 8 FR; OIT FR; IA FR; CR; IA
B. inpai 1 e 2 6 FR FR; IA -
H. ocellifer 1 e 2 5 FR FR; OIT -
H. melazonatus 2; 3 6 - IA; FR; OIT IA; FR; OIT
I. viriatus 2; 3 4 - FR; OIT FR; IA
Gymnotus sp. 1 1 D; IA - -
R. compressus 1 2 FR; OIT; D - -
A. pallidus 1; 2; 3 4 D D; FR; CR D; IA
A. steindachneri 1; 3 4 D - D; CR; IA
C. saxantilis 2; 3 3 - IA; FR D; FR
DiscussãoDesde os primeiros trabalhos realizados com a ictiofauna
de igarapés, tem sido apontada a riqueza elevada de peixesem pequenos cursos d’água na Amazônia (Fittakau 1967,Knöppel 1970, Junk 1983 apud Bührnhein 1998). Diversasespécies de peixes pequenos, principalmente Characidae,convivem em um mesmo biótopo, caracterizando uma elevadadiversidade alfa (Lowe-McConnell 1987). Em geral a riquezade espécies encontradas nos igarapés estudados aqui podeser considerada baixa, uma vez que Lowe-McConnell (1987)estimou uma diversidade de 30-50 espécies para igarapésamazônicos. Bürhernhein (1998), estudando os mesmosigarapés na reserva do Km 41, encontrou 24 espécies, umnúmero maior do que o encontrado no presente estudo. Istopode ser explicado pela ineficiência da metodologia aplicadano presente estudo e a diferença do esforço amostral. A maiorabundância encontrada foi em igarapés de primeira ordem,onde utilizamos apenas peneiras. Neste tipo de ambiente, ométodo da peneira é eficiente devido o pequeno volume deágua, já em igarapés de segunda e terceira ordem, o volumeda água aumenta de forma exponencial. Deste modo,aumentamos o esforço de coleta usando redes de cerco, porémestas redes foram consideradas ineficientes, uma vez que amaioria dos biótopos apresentam troncos, galhos e poçascom maior profundidade no meio da calha, dificultando oarraste e o cerco com a rede (ver também Bürhernhein 1998).
A abundância de Perciformes representada pela famíliade Cichlidae, maior do que a de Siluriformes em igarapés deterra firme, também foi encontrada por Knöppel (1970), noigarapé Barro Branco, Amazônia Central. Silva (1992)encontrou percentuais de abundância aproximadas deSiluriformes (22,7%) e Cichlidae (20,5%) em igarapés deterra firme, também observou uma tendência de maiorabundância nos meses chuvosos do que nos meses deestiagem, sugerindo expansão de recursos espacial do fatordeterminante. No presente estudo a composiçãoictiofaunística foi predominantemente representada porCharaciformes (57%) seguidos por Perciformes (21,4%).Quanto a abundância, os Characiformes também foram osmais representativos (87%), seguidos por Perciformes (9%).
Houve uma substituição da composição de espécies dosigarapés de primeira ordem quando comparamos comigarapés de segunda ordem. Já os igarapés de terceira ordem,a substituição de espécie foi pequena quando comparamoscom a composição da ictiofauna de igarapés de segundaordem, porém esperaria que houvesse um acréscimo deespécies nos trechos de terceira ordem, devido à modificaçãoe aumento do número de habitats, substratos, profundidade,correnteza e presença ou ausência da vegetação marginal. Oíndice de similaridade de Morisita mostrou semelhança entrea composição da ictiofauna dos igarapés de segunda e terceiraordem, A heterogeneidade de habitats em riachos neotropicaisinflui na distribuição espacial das comunidades de peixesem função das variações espaciais longitudinais (Uieda 1984;Garutti 1988; Sabino &Castro 1990).
A velocidade da correnteza, quantidade de luz eprofundidade do canal, afetam a disponibilidade e

128 Curso de Campo Ecologia da Floresta Amazônica - 2001
distribuição de recursos como o folhas sob o sedimento queé a base da cadeia trófica desses ecossistemas (Walker 1995).Estudos sobre comunidades de organismos aquáticos emigarapés de mata de terra firme da Amazônia são importantesdo ponto de vista do funcionamento do ecossistema emcondições de baixa ou nenhuma produção primária autóctone,com um suprimento de recursos e nutrientes totalmentedependentes da floresta. No presente estudo podemosobservar a mudança dos itens alimentares em relação a ordemdo igarapé. Esta mudança provavelmente está relacionadacom a variação de um gradiente contínuo de condições físicase ajustes bióticos, que foram encontradas por Vannote etal.(1980).
De acordo com Vannote et al. (1980), a medida que oigarapé vai aumentando em direção a foz, a importância doaporte de matéria orgânica terrestre é reduzida e aumenta asignificância da produção primária autóctone, provavelmenterelacionada com a maior disponibilidade da luz. Isto ficouevidente no presente estudo, uma vez que os peixes deigarapés de primeira ordem estudados, se alimentambasicamente de itens alóctones. De acordo com Vannote etal. (1980), a região de cabeceiras sobre influências navegetação circundante, resultando na redução da produçãoautotrófica devido do sombreamento da água, contribuindoassim para a entrada do material de origem alóctone. A dietada ictiofauna dos igarapés de segunda ordem estudado aindativeram uma forte influência de itens alóctones, pois encontreiitens autóctones com pouca freqüência. Já a dieta dos peixesde igarapés de terceira ordem tiveram influência maior deitens autóctones, com insetos aquáticos, crustáceos e a adiçãode uma espécie de Loricariidae que alimenta-se de algas.
A grande dependência dessas comunidades de peixesestudadas com os itens alimentares provenientes da florestanos mostra a importância da preservação dos fragmentosflorestais. Sérias alterações em sua estrutura poderão ocorrercom a retirada da mata adjacente, mesmo levando emconsideração a plasticidade alimentar dessas espécies.
AgradecimentosAo prof. Jansen Zuanon (INPA), pela amizade e orientação
em todas as etapas deste trabalho, ao Eduardo Venticinque(Dadão) pela leitura crítica e pelas valiosas sugestões, aomeu novo amigo “peixólogo” Carlos E. Marinelli (Caê) pelaindispensável ajuda nos trabalhos de campo e valiosasdiscussões que enriqueceram este trabalho, a todos os amigosdo Curso de Campo Ecologia da Floresta Amazônica que,direta ou indiretamente, participaram da elaboração desteprojeto e ao INPA e PDBFF pela maravilhosa oportunidade.
Referências BibliográficasAngermeier, P.L. & Karr, J.R. 1984. Fish communities along
environmental gradients in a system of tropical streams.In: Evolutionary Ecology of Neotropical FreshwaterFishes, Zaret, T.M. (ed.), pp. 39-57. Dr. W. JunkPublications, The Netherlands.
Bowen, S.H. 1983. Quantitative description of the diet. In:Fisheries Techniques, Nielsen, L.A. & Johnson, D.L.
(eds.), pp 325-336. American Fisheries Society,Bethesda, Maryland, USA.
Bürhnheim, C.M. 1998. Estrutura de comunidades de peixesem igarapés de floresta de terra firme da AmazôniaCentral. Dissertação de Mestrado, INPA/UA, Manaus,AM.
Garutti, V. 1988. Distribuição longitudinal da ictiofauna emum córrego da região noroeste do Estado de São Paulo,Bacia do rio Paraná. Revta. Bras. Biol. 48(4): 747-759.
Goulding, M.; Carvalho, M.L & Ferreira, E. 1988. Rio Negro:Rich Life in Poor Water. The Hague, SPB AcademicPublishing.
Henderson, P.A. & Walker, I. 1986. On the leaf littercommunity of the Amazonian blackwater streamTauruzinho. J. Trop. Ecol. 2:1-17.
Henderson, P.A. & Walker, I. 1986. Spatial organization andpopulation density of the litter banks within a centralAmazonian blackwater. J. Fish Biol. 37: 401-411.
Knöppel, H.A. 1970. Food of central Amazonian fishes:contribution to the nutrient ecology of Amazonian rainforest stream. Amazoniana 2(3): 257-352.
Lowe-McConnell, R.H. 1987. Ecological Studies in TropicalFish Communities. Cambridge University Press,Cambridge.
Lowe-McConnell, R.H. 1999. Estudos Ecológicos deComunidades de Peixes Tropicais. EDUSP, São Paulo.
Minshall, G.W.; Cummins, K.W.; Petersen, R.C.; Cushing,C.E.; Bruns, D.A.; Sedell, J.R.& Vannote, R.L. 1985.Developments in streams ecossystem theory. Can. J. Fish.Aq. Sci. 42: 1045-1055.
Radambrasil, 1978. Levantamento de recursos naturais. FolhaAS 20 Manaus 261 p. Ministério de Minas e Energia:Departamento Nacional de Produção Mineral, Rio DeJaneiro, Brasil.
Sabino, J. & Castro, R.M.C. 1990. Alimentação, período deatividade e distribuição espacial dos peixes de um riachoda Floresta Atlântica (Sudeste do Brasil). Revta. Bras.Biol. 50(1): 23-36.
Silva, C.P.D. 1992. Alimentação e distribuição espacial dealgumas espécies de peixes do igarapé do Candirú,Amazonas, Brasil. Acta Amaz. 23(2-3): 271-285.
Soares, M.C.M. 1979. Aspectos ecológicos (alimentação ereprodução) dos peixes do igarapé do Porto, Aripuanã,MT. Acta Amaz. 9(2): 325-352.
Uieda, V.S. 1984. Ocorrência e distribuição dos peixes emum riacho de água doce. Revta. Bras. Biol. 44(2): 203-213.
Vannote, R.L.; Minshall, G.W.; Cummins, K.W.; Sedell,J.R.& Cushing, C.E. 1980. The river continuum concept.Can. J. Fish. Aq. Sci., 37: 130-137.
Walker, I. 1995. Amazonian streans and small rivers. In:Limnology in Brasil, Tundisi J.G., Bicudo, C.E.M.&Tundisi, T.M. (eds.), pp 167-193. Academy of Sciences& Brazilian Limnological Society, Rio de Janeiro.
Wetzel, R.G. & Likens, G.E., 1991. Limnological Analyses.Springer-Verlang, New York.

Curso de Campo Ecologia da Floresta Amazônica - 2001 129

130 Curso de Campo Ecologia da Floresta Amazônica - 2001
Efeito de borda sobre a intensidade de predação de sementes artificiais emuma floresta de terra firme na Amazônia Central
Leandro Moraes Scoss
IntroduçãoUma borda é definida ecologicamente como o local onde
duas ou mais comunidades de plantas encontram-se ou aindaonde dois ecossistemas se encontram (Hanson 1962; Pianka1974). É na borda que a maior parte dos processos ligados afragmentação geralmente se iniciam. Murcia (1995) descrevetrês tipos de efeito de borda: os abióticos, que envolvemmudanças nas condições ambientais resultante daproximidade de uma matriz dissimilar; os biológicos diretosque envolvem mudanças na distribuição e abundância deespécies causada diretamente pelas condições físicas próximaa borda, e à tolerância fisiológica das espécies; e os biológicosindiretos que manifestam-se nas interações interespecíficasdas populações.
A variação de condições abióticas induz uma modificaçãoda ocupação do espaço, formando também um gradienteflorístico e fitossociológico, alterando a paisagem original.As observações de Lovejoy et al. (1991) reforçam osurgimento de um gradiente entre borda e mata, mostrandoque em poucos anos de isolamento de um fragmento defloresta amazônica, houve uma redução da umidade relativado ar em 5 a 20%, além de alterar a temperatura, a umidadedo solo e a intensidade luminosa.
Algumas estimativas indicam que estas diferenças podemdesaparecer após os primeiros 50 m para o interior da mata(Murcia 1995). Porém outras, permitem concluir que osefeitos de borda podem ser detectados 500m após a bordaem direção ao interior da mata (Laurance 1989). A diferençaentre essas estimativas indica que a estrutura da vegetação éum importante fator para determinar o resultado final causadopela fragmentação.
Os efeitos biológicos indiretos provocados por uma borda,relacionam-se às mudanças nas interações interespecíficas,tais como predação, competição, herbivoria, parasitismo,polinização e dispersão (Murcia 1995). Segundo Angelstam(1986 apud Magro 1988), a intensidade de predação aumentano sentido da vegetação mais natural em direção à borda.Wiens (1989) afirma que as bordas são importantes áreas deinterações entre populações, sugerindo mudanças abruptasnos fluxos de indivíduos e materiais.
Alguns estudos têm demonstrado claramente que emregiões tropicais um grande percentual de sementes sãoconsumidas por predadores (Kiltie 1981; Fragoso 1997;Beck-King, von Helversen & Beck-King 1999). Por outrolado algumas das espécies predadoras também atuamfavoravelmente na dispersão de sementes, como por exemploantas (Tapirus terrestris), porcos-do-mato (Tayassu pecarie T. tajacu), alguns roedores (Agouti paca, Dasyprocta sp.),aves e principalmente formigas (Terborgh et al. 1993;
Holldobler & Wilson 1990).Hubbell & Foster (1986 apud Freitas 1998) sugeriram
que em florestas tropicais as espécies arbóreas sãocompetitivamente equivalentes, e a sobrevivência de espéciestolerantes à sombra deve ser mais afetada por fatores bióticos(predação e herbivoria) do que por fatores abióticos(diferenças físicas entre habitats), já que estas espéciesconseguem sobreviver tanto em clareiras quanto em subosquede mata contínua.
O objetivo deste estudo é testar se sementes de tipos etamanhos diferentes são mais predadas na borda que nointerior da mata, partindo do pressuposto que na borda assementes são menores e mais susceptíveis a predação. A bordaconsiderada neste estudo é a margem de uma estrada, quenão provoca alterações abióticas e bióticas na mesmaintensidade que bordas de fragmentos florestais, mas provocaa fragmentação interna de ambientes naturais e o incrementode habitats de borda.
Material e MétodosO presente estudo foi realizado em uma área de platô
coberta por floresta densa de terra firme (Pires & Prance1985), na Reserva Km 41 (INPA/PDBFF/SI), localizadaaproximadamente a 80 km ao norte de Manaus (2o 24’S; 59o
52’W), Amazônia Central. Os solos pertencem em suamaioria ao grupo dos latossolos amarelos. A temperaturamédia anual em Manaus é 26,7oC, onde a precipitação mediaanual está entre 1.900 e 2.400 mm. A precipitação mensalgeralmente é inferior a 100 mm nos meses mais secos, julho,agosto e setembro. A área de estudo encontra-se dentro dodomínio da floresta perenifolia de terra firme, apresentandoaltura do dossel entre 30 e 37 m (Lovejoy & Bierregard 1990).
O experimento foi instalado às margens da estrada ZF-3único acesso rodoviário a reserva, pela BR-174. Foramutilizados três transectos paralelos e em diferentes distânciasda estrada (10, 70 e 130m), totalizando duas réplicas paracada distância da estrada (Figura 1). Em cada transecto foramdistribuídos nove pontos espaçados 25 m um do outro, ondeforam acondicionadas cinco sementes. As sementes utilizadasforam milho de pipoca, amendoim e castanha do pará. Ostrês primeiros pontos foram aleatorizados (milho, castanhae amendoim) e depois sistematizados, totalizando trêsrepetições com cinco sementes de cada um dos tipos, portransecto. A castanha (Bertholletia excelsa, Lecythidaceae)foi considerada “artificial” porque na área da reserva Km 41apenas um indivíduo foi identificado até o momento. Poresta razão, as sementes
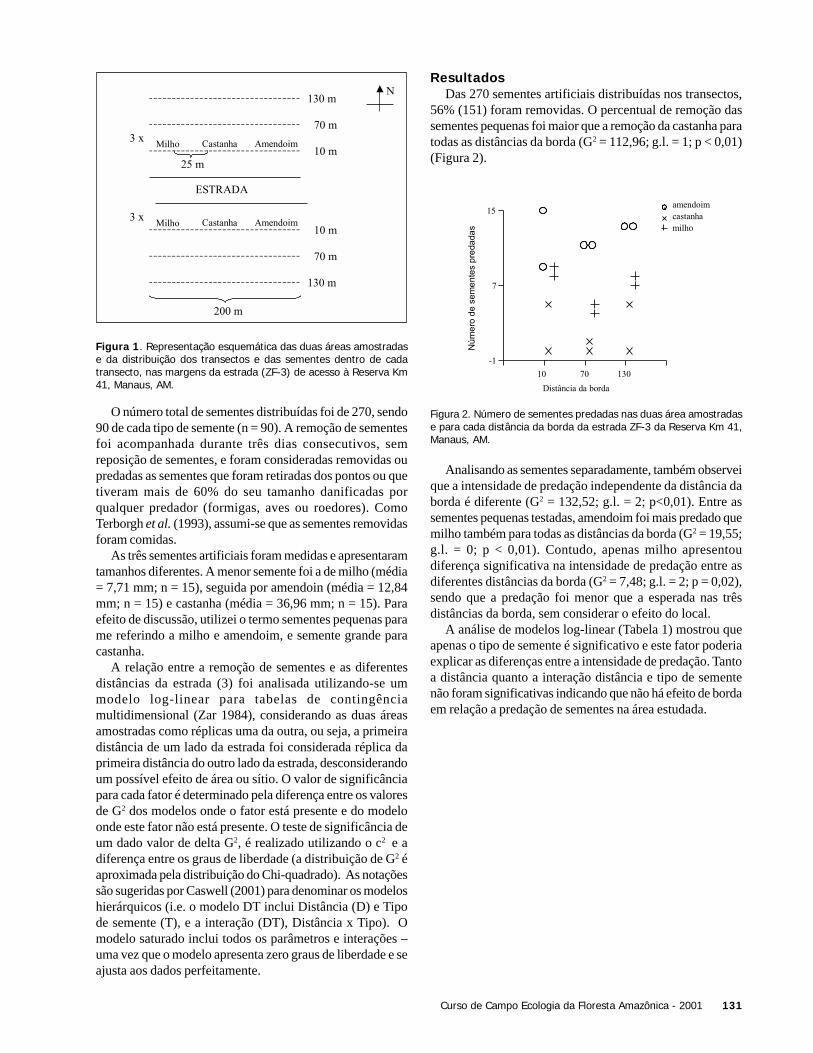
Curso de Campo Ecologia da Floresta Amazônica - 2001 131
Figura 1. Representação esquemática das duas áreas amostradase da distribuição dos transectos e das sementes dentro de cadatransecto, nas margens da estrada (ZF-3) de acesso à Reserva Km41, Manaus, AM.
O número total de sementes distribuídas foi de 270, sendo90 de cada tipo de semente (n = 90). A remoção de sementesfoi acompanhada durante três dias consecutivos, semreposição de sementes, e foram consideradas removidas oupredadas as sementes que foram retiradas dos pontos ou quetiveram mais de 60% do seu tamanho danificadas porqualquer predador (formigas, aves ou roedores). ComoTerborgh et al. (1993), assumi-se que as sementes removidasforam comidas.
As três sementes artificiais foram medidas e apresentaramtamanhos diferentes. A menor semente foi a de milho (média= 7,71 mm; n = 15), seguida por amendoin (média = 12,84mm; n = 15) e castanha (média = 36,96 mm; n = 15). Paraefeito de discussão, utilizei o termo sementes pequenas parame referindo a milho e amendoim, e semente grande paracastanha.
A relação entre a remoção de sementes e as diferentesdistâncias da estrada (3) foi analisada utilizando-se ummodelo log-linear para tabelas de contingênciamultidimensional (Zar 1984), considerando as duas áreasamostradas como réplicas uma da outra, ou seja, a primeiradistância de um lado da estrada foi considerada réplica daprimeira distância do outro lado da estrada, desconsiderandoum possível efeito de área ou sítio. O valor de significânciapara cada fator é determinado pela diferença entre os valoresde G2 dos modelos onde o fator está presente e do modeloonde este fator não está presente. O teste de significância deum dado valor de delta G2, é realizado utilizando o c2 e adiferença entre os graus de liberdade (a distribuição de G2 éaproximada pela distribuição do Chi-quadrado). As notaçõessão sugeridas por Caswell (2001) para denominar os modeloshierárquicos (i.e. o modelo DT inclui Distância (D) e Tipode semente (T), e a interação (DT), Distância x Tipo). Omodelo saturado inclui todos os parâmetros e interações –uma vez que o modelo apresenta zero graus de liberdade e seajusta aos dados perfeitamente.
ResultadosDas 270 sementes artificiais distribuídas nos transectos,
56% (151) foram removidas. O percentual de remoção dassementes pequenas foi maior que a remoção da castanha paratodas as distâncias da borda (G2 = 112,96; g.l. = 1; p < 0,01)(Figura 2).
Figura 2. Número de sementes predadas nas duas área amostradase para cada distância da borda da estrada ZF-3 da Reserva Km 41,Manaus, AM.
Analisando as sementes separadamente, também observeique a intensidade de predação independente da distância daborda é diferente (G2 = 132,52; g.l. = 2; p<0,01). Entre assementes pequenas testadas, amendoim foi mais predado quemilho também para todas as distâncias da borda (G2 = 19,55;g.l. = 0; p < 0,01). Contudo, apenas milho apresentoudiferença significativa na intensidade de predação entre asdiferentes distâncias da borda (G2 = 7,48; g.l. = 2; p = 0,02),sendo que a predação foi menor que a esperada nas trêsdistâncias da borda, sem considerar o efeito do local.
A análise de modelos log-linear (Tabela 1) mostrou queapenas o tipo de semente é significativo e este fator poderiaexplicar as diferenças entre a intensidade de predação. Tantoa distância quanto a interação distância e tipo de sementenão foram significativas indicando que não há efeito de bordaem relação a predação de sementes na área estudada.

132 Curso de Campo Ecologia da Floresta Amazônica - 2001
Tabela 1. Resultados da análise log-linear do efeito da distância daestrada (D) e do tipo de semente (T; milho, amendoim, castanha), eda interação desses dois fatores sobre a predação de sementes (P).Os modelos são significativos (***) ou não (NS). As notações sãosugeridas por Caswell (2001) para denominar os modelos hierárquicos(i.e. o modelo DT inclui Distância (D), Tipo de semente (T) e ainteração (DT) Distância x Tipo).
DiscussãoO modelo log-linear não detectou efeito da distância da
borda sobre a predação das sementes, o que poderia serexplicado por: i) as distâncias utilizadas neste experimentonão foram suficientes para detectar os efeitos de borda, ouporque foram grandes demais ou porque foram pequenas,para refletir um gradiente de predação; ii) o fenômenobiológico estudado não é afetado pela borda da estrada daárea de estudo; iii) o tamanho da amostra não foi suficientepara detectar diferenças significativas; iv) a área de vida euso do habitat dos potenciais predadores das sementes.
Este estudo não sustenta claramente nenhuma dasexplicações acima para a semente de milho, já que a predaçãodesta semente foi menor que o esperado para a primeiradistância e para a última distância. Isto indica que pode haverespecificidade entre esta semente e seus potenciaispredadores e, possivelmente há uma utilização diferenciada(gradiente em relação à borda e interior da mata) dospredadores em relação à proximidade da borda, até mesmoda borda da floresta com uma estrada (Didham 1997). Paraas outras duas sementes (amendoim e castanha), não detecteiefeito de borda sobre a intensidade de predação das sementesquando considerei as áreas nas analises, sustentando asegunda hipótese proposta acima. . Entretanto, há umadiferença entre as primeira distâncias da borda da estrada naintensidade de predação das sementes de amendoim,reforçando a hipótese de área de vida e uso do habitat dealguns predadores (principalmente aves terrestres), quepodem também ser diferentes para cada área.
Os principais predadores e dispersores de sementesgrandes são as antas, porcos-do-mato, roedores e ungulados(Terborgh et al. 1993). Já para as plantas que investem na
produção de sementes pequenas, espera-se que formigassejam os principais predadores e dispersores dessas sementes(Holldobler & Wilson 1990). van der Pijl (1982) descrevealgumas síndromes de dispersão que normalmente estãoassociadas à coloração dos frutos e sementes e oleosidade,indicando alto grau de especificidade de plantas e organismosdispersores/predadores. Das sementes artificiais utilizadasneste estudo, amendoim foi mais predada que milhopossivelmente devido a consistência da semente, maioroleosidade e à coloração mais próxima a encontradanaturalmente no local, visto que a cor vermelha épredominante em florestas tropicais (van der Pijl 1982).
Os resultados observados para milho, indicam que estasemente artificial pode ser útil em futuros estudos para avaliaros efeitos de borda sobre a predação de sementes e respostaa predação em razão da cor. Se utilizado em arranjosexperimentais diferentes do empregado neste estudo, épossível que seja detectado um efeito de borda mais evidentee respostas diferenciadas por predadores diferentes, sendotambém necessário a experimentação levando emconsideração os potenciais predadores deste tipo de semente,por exemplo isolando o efeito de formigas ou de aves emamíferos.
Referências BibliográficasBeck-King, von Helversen, H. O. & Rachel Beck-King. 1999.
Home range, population density, and food resources ofAgouti paca (Rodentia: Agoutidae) in Costa Rica: astudy using alternative methods. Biotropica 31(4): 675-685.
Caswell, H. 2001. Matrix Population Models. Sinauer,Sunderland, MA.
Didham, R.K. 1997. The influence of edge effects and forestfragmentation on leaf litter invertebrates in CentralAmazonia. In: Tropical Forest Remnants, Laurance, W.F. & Bierregaard, R. O. Jr. (eds.), pp. 55-70. TheUniversity of Chicago Press.
Fragoso, J.M.V. 1997b. Tapir-generated seed shadows: scale-dependent patchiness in the Amazon rain forest. J. Ecol.85:519-529.
Freitas, M.A. 1998. O efeito de fatores biótico e abióticosna sobrevivência pós-dispersão de sementes e plântulasde cinco espécies arbóreas na mazônia Central.Dissertação de mestrado, INPA/Universidade doAmazonas, Manaus, AM.
Hanson, H.C. 1962. Dictionary of Ecology. PhilosophicalLibrary, Washington, DC.
Holldobler, B. & Wilson, E.O. 1990. The Ants. The BelknapPress of Harvard University Press, Cambridge, MA.
Kiltie, R.A. 1981. Distribution of palm fruits on a rain forestfloor: why white-lipped peccaries forage near objects.Biotropica 13: 141-145
Laurance, W.F. 1989. Ecological impacts of tropical forestfragmentation on nonflying mammals and their habitats.University of California, Berkeley, California, USA.
Lovejoy, T.E. & Bierregaard, Jr. R.O. 1990. Centralamazonian forest and minimal critical size of ecosystems

Curso de Campo Ecologia da Floresta Amazônica - 2001 133
project. In: Four Neotropical Rain Forests, Gentry, A.H.(ed.), pp. 61-71. Yale University Press, London.
Lovejoy, T.E.; Bierregaard, R.O. & Rylands, A.B. 1991. Edgeand other effects of isolation on Amazon forestfragments. In: Conservation Biology: The Science ofScarcity and Diversity, Soulé, M. E. (ed.), pp 257-85.Sinauer Associates, Sunderland, Massachusetts.
Magro, T.C. 1988. Avaliação da qualidade de habitatfaunístico pela análise de bordas. Dissertação deMestrado. Departamento de Engenharia Florestal, UFV,Viçosa
Murcia, C. 1995. Edge effects in fragmented forests:implications for conservation. TREE 10(2): 58-62.
Pianka, E.R. 1974. Evolutionary Ecology. Harper & Row,
New York.Pires, J.M. & Prance, G.T. 1985. The vegetation types of the
Barzilian Amazon. In: Amazônia, Prance, G. & Lovejoy,T. (eds.), pp. 109-145. Pergamon, New York.
Terborgh, J.; Losos, E.; Riley, M.P. & Bolãnos Ryley, M.B.1993. Predation by vertebrates and invertebrates on theseeds of five canopy tree species of Amazonian forest.Vegetatio 107/108: 375-386.
Van der Pijl, L. 1982. Principles of Dispersal in HigherPlants. Germany.
Wiens, J.A. 1989. Spatial scaling in ecology. Funct. Ecol. 3:385-97.
Zar, J.H. 1984. Biostatistical Analysis. Prentice Hall, NewJersey.
Fatores que influenciam o tamanho de clareiras e seus impactos em umafloresta de terra firme na Amazônia Central
Maria Rosa Darrigo
IntroduçãoFlorestas tropicais têm sido descritas como um mosaico
de manchas composto por vegetações primárias e secundáriasde diferentes tamanhos, originado pela dinâmica sucessionalde clareiras resultantes de quedas de árvores (Oldeman 1978).Nessas florestas, muitas espécies de árvores de dosseldependem, em pelo menos um estágio de sua vida, docrescimento em clareiras para atingir sua maturidade(Hartshorn 1978; Garwood 1982). Sendo assim, fica claro opapel das clareiras na manutenção da dinâmica e diversidadedas florestas tropicais, já que possibilita a permanência deespécies de crescimento rápido ou lento, tolerantes ouintolerantes à sombra, de curta ou longa expectativa de vida(Lieberman et al. 1990; Deslow 1980).
Como parte de um processo dinâmico, vários fatorescondicionam a formação de clareiras como vento, declividadedo terreno, entre outros (Denslow & Hashtorn 1994). Essesfatores, inclusive podem estar relacionados com sua forma,extensão e impacto sobre a vegetação adjacente. Taiscaracterísticas devem influenciar fortemente no tipo desucessão, já que estudos têm indicado que espécies pioneirassó são encontradas em clareiras com mais de 150 m2
(Whithmore 1975; Hashtorn 1980), além de interferir notempo de regeneração (Denslow & Hashtorn 1994). Sabe-seque condições microclimáticas encontradas em uma clareirasão condicionadas pelo seu tamanho (Denslow 1980),orientação, forma, altura do dossel e latitude (MartinezRamos et al. 1988).
Nesse estudo avaliei alguns fatores de fácil e rápidacaracterização, que estariam exercendo grande influência notamanho, impacto e orientação clareiras encontradas em umafloresta de terra firme na Amazônia Central.
Material e MétodosRealizei este estudo em uma área de floresta localizada a
70km ao Norte de Manaus, na Reserva do Km 41, do projetoPDBFF (2o 24’S; 59o 52’W). A floresta tropical de terra firmeé a principal formação vegetal encontrada no local, comdossel variando de 30 a 37 m. O clima na região é quente eúmido, com marcada estação seca e chuvosa (Lovejoy &Bierregaard 1990).
As clareiras foram localizadas por meio de censosrealizados ao longo das trilhas já existentes na reserva (entreas trilhas J4 e E10), distribuídas em uma área de 42 ha. Paracada clareira tomei medidas de comprimento e largura (paraestimar a área através da fórmula da elipse, sendo área =comprimento * altuta * p), orientação da queda da árvorecausadora (quando possível identifica-la), estado das árvoresque caíram (se tinham caído com a raiz, estavam com o troncoquebrado ou continuavam vivas) e número de árvores caídasjunto com a árvore causadora. Também foi medido o DAPda árvore causadora da clareira. Com o uso de regressõeslineares, utilizando o programa Systat 8.0 (Willkinson 1988),verifiquei possíveis relações entre os parâmetros analisados,como área da clareira e número de árvores envolvidas naqueda; DAP da árvore causadora e área da clareira; DAP daárvore causadora e número de árvores envolvidas na queda(Zar 1984). Utilizei análise de distribuição circular a fim decaracterizar a distribuição do sentido de queda das árvorescausadoras (Zar 1984). Para verificar se a topografiainfluenciou o sentido da queda das árvores, avaliei adistribuição das clareiras analisadas em mapa de topografiarealizado para o mesmo local por Oliveira (1997).
A idade das clareiras também foi estimada, através dosseguintes parâmetros:
- árvores causadoras tombadas ainda com folhasindicavam que a clareira havia sido formada a
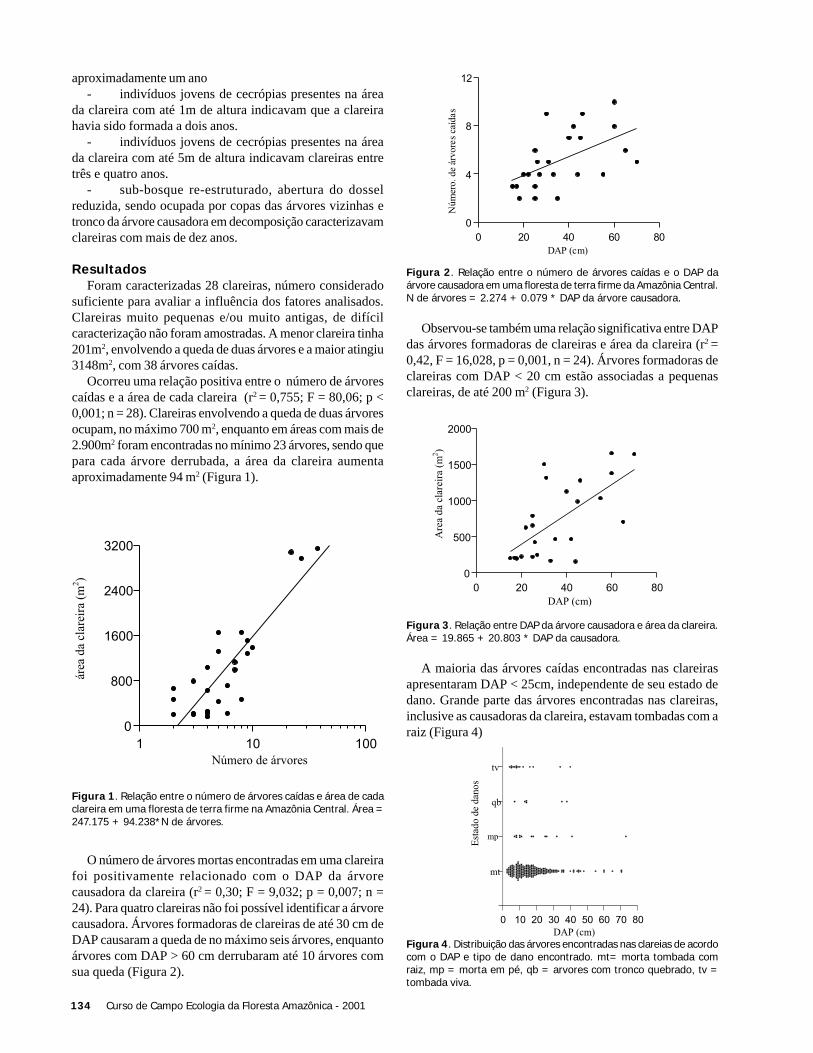
134 Curso de Campo Ecologia da Floresta Amazônica - 2001
aproximadamente um ano- indivíduos jovens de cecrópias presentes na área
da clareira com até 1m de altura indicavam que a clareirahavia sido formada a dois anos.
- indivíduos jovens de cecrópias presentes na áreada clareira com até 5m de altura indicavam clareiras entretrês e quatro anos.
- sub-bosque re-estruturado, abertura do dosselreduzida, sendo ocupada por copas das árvores vizinhas etronco da árvore causadora em decomposição caracterizavamclareiras com mais de dez anos.
ResultadosForam caracterizadas 28 clareiras, número considerado
suficiente para avaliar a influência dos fatores analisados.Clareiras muito pequenas e/ou muito antigas, de difícilcaracterização não foram amostradas. A menor clareira tinha201m2, envolvendo a queda de duas árvores e a maior atingiu3148m2, com 38 árvores caídas.
Ocorreu uma relação positiva entre o número de árvorescaídas e a área de cada clareira (r2 = 0,755; F = 80,06; p <0,001; n = 28). Clareiras envolvendo a queda de duas árvoresocupam, no máximo 700 m2, enquanto em áreas com mais de2.900m2 foram encontradas no mínimo 23 árvores, sendo quepara cada árvore derrubada, a área da clareira aumentaaproximadamente 94 m2 (Figura 1).
Figura 1. Relação entre o número de árvores caídas e área de cadaclareira em uma floresta de terra firme na Amazônia Central. Área =247.175 + 94.238*N de árvores.
O número de árvores mortas encontradas em uma clareirafoi positivamente relacionado com o DAP da árvorecausadora da clareira (r2 = 0,30; F = 9,032; p = 0,007; n =24). Para quatro clareiras não foi possível identificar a árvorecausadora. Árvores formadoras de clareiras de até 30 cm deDAP causaram a queda de no máximo seis árvores, enquantoárvores com DAP > 60 cm derrubaram até 10 árvores comsua queda (Figura 2).
Figura 2. Relação entre o número de árvores caídas e o DAP daárvore causadora em uma floresta de terra firme da Amazônia Central.N de árvores = 2.274 + 0.079 * DAP da árvore causadora.
Observou-se também uma relação significativa entre DAPdas árvores formadoras de clareiras e área da clareira (r2 =0,42, F = 16,028, p = 0,001, n = 24). Árvores formadoras declareiras com DAP < 20 cm estão associadas a pequenasclareiras, de até 200 m2 (Figura 3).
Figura 3. Relação entre DAP da árvore causadora e área da clareira.Área = 19.865 + 20.803 * DAP da causadora.
A maioria das árvores caídas encontradas nas clareirasapresentaram DAP < 25cm, independente de seu estado dedano. Grande parte das árvores encontradas nas clareiras,inclusive as causadoras da clareira, estavam tombadas com araiz (Figura 4)
Figura 4. Distribuição das árvores encontradas nas clareias de acordocom o DAP e tipo de dano encontrado. mt= morta tombada comraiz, mp = morta em pé, qb = arvores com tronco quebrado, tv =tombada viva.

Curso de Campo Ecologia da Floresta Amazônica - 2001 135
As árvores tombadas encontravam-se principalmentedirecionadas entre 600 e 1000 (÷2 = 43,17, g.l. = 17, p <0,001, n = 28) (Figura 5). No entanto, este não coincidiucom a inclinação do terreno em que cada clareira estavalocalizada; já que das dez clareiras localizadas na vertente,apenas duas estavam orientadas no sentido do gradientetopográfico. Entre as árvores localizadas no platô nãoobservei sentido predominante para a queda de árvores.
Foram encontradas clareiras com idades estimadas entreum e dez anos. A maioria (50%) se encontrava em uma faixade dois anos, sendo poucas clareiras antigas, com mais de10 anos (20% das clareiras). Esses dados no entanto nãoserão discutidos devido a falha na metodologia utilizada, jáque em várias clareiras não haviam cecrópias ou a análise daregeneração do sub-bosque foi imprecisa.
0
90
180
270
Ân
gu
lo0 2 4 6
frequência
Figura 5. Freqüência de ocorrência de clareiras ocorrendo de acordocom a sua orientação, em intervalos de ângulos de 200.
DiscussãoAs relações positivas encontradas entre o DAP da árvore
causadora, número de árvores caídas e área da clareira nosindica grande influência do tamanho das árvore principal(formadora) na extensão e impacto da clareira. Esses fatoresafetam a sucessão encontrada na área alterada, já que grandesclareiras apresentam maior potencial para serem colonizadaspor espécies pioneiras, não tolerantes à sombra (comoCecropia, Vismia) que normalmente não são encontradas nointerior da floresta (Harshtorn 1980). O tempo de regeneraçãotambém apresenta relação positiva com o tamanho da clareira,sendo necessário mais tempo para áreas de clareiras maioresvoltarem a seu “estado original” (Denslow & Hashtorn 1994).Tais observações têm importantes implicações para o manejode áreas florestais, principalmente em áreas de extração demadeira, onde árvores com DAP>50cm são preferencialmentecortadas (obs. pess.). Nessas áreas é necessário grandecuidado com o direcionamento da queda das árvores e seuamortecimento, bem como avaliação do entrelaçamento das
copas e presença dos cipós (não avaliados no presenteestudo), a fim de minimizar o impacto causado pela quedade grandes árvores. Grande parte das árvores derrubadas juntocom a causadora da clareira tinham DAP < 20 cm,consequentemente não aproveitáveis comercialmente e coma possibilidade de serem indivíduos jovens de espécies dealto valor comercial. O fato da grande maioria das árvoresderrubadas serem pequenas pode estar relacionado aopredominância de indivíduos na floresta com esse porte(Oliveira 1997) ou à maior suscetibilidade destes à queda .
Grande parte das árvores encontradas nas clareiras estavamtombadas com a raiz, padrão também encontrado porHartshorn (1980) na Costa Rica, que segundo o autor é devidoao solo raso encontrado no local. Na floresta em que serealizou esse estudo o solo é muito profundo (Oliveira 1997),no entanto, grande parte dos nutrientes de encontram nohorizonte A e B do solo e em conseqüência disto, as raízesse mantêm nas camadas mais superficiais, não garantindogrande estabilidade. Também, tal fato pode estar relacionadoa um alto vigor encontrado nas árvores do local estudado,diminuindo assim o risco de quebra no tronco.
Apesar de ter sido encontrada maior freqüência na quedade árvores com orientação entre 600 e 1000, não verifiqueinenhuma relação com os fatores analisados. A direção dosventos alísios (leste-oeste) é contrária ao principal sentidode queda das árvores no local e de qualquer forma, essesventos são fracos e dificilmente poderiam ter ocasionado taisquedas (E. M. Venticinque com. pess.). Alguns estudosapontam como principal fator na formação de clareiras asazonalidade na umidade de solos (Oldeman 1972), ventosfortes decorrentes de tempestades (Sarukha 1978) e forteschuvas (Ghoyne 1968), já que estas encharcam o solodiminuindo a estabilidade das raízes e aumentando o pesodos troncos. As clareiras estudadas devem ser resultantes dacombinação de fortes ventos e chuvas causadas pelas grandestempestades que ocorrem no inverno amazônico. Brokaw(1980) aponta esses fatores como influentes na formação declareiras na Costa Rica. Um fator que tem sido maisrecentemente considerado influente na direção da queda deárvores é a assimetria de copa (Young & Hubbel 1991).Segundo esses autores, árvores (principalmente as menores)crescendo em ambiente com pouca luminosidade crescemassimetricamente, tendendo a aumentar a massa de suas copasna direção de mais luz incidente. Isso resulta em árvoresestruturalmente não estáveis, com tendência a cair de seulado mais pesado. Essa situação ocorre principalmente emambientes com alta variação de luz, como bordas de clareirasantigas ou “sunflex” encontrados no interior de mata.
Sendo assim, a direção da queda da árvore pode estarsendo influenciada por muitos fatores como o sentido dasfortes chuvas e ventos decorrentes das grandes tempestades,assimetria e entrelaçamento das copas e presença de lianas.No entanto, fatores como a inclinação do terreno e ventosconstantes e fracos (ventos alísios) não estão associados àdireção da queda de árvores. Para a certeza de quais dosfatores citados estão influenciando a formação de clareiras equal seria mais importante é necessário um estudo sazonal

136 Curso de Campo Ecologia da Floresta Amazônica - 2001
enfocando a maior freqüência de queda de árvores e direçãodos ventos mais fortes e estrutura das árvores, comacompanhamento da formação de clareiras.
AgradecimentosMuito obrigada ao Dadão e Jansen pela perfeita
coordenação e pela diversão garantida! Agradeço ao Dadão,de novo, pela ajuda nos probleminhas estatísticos, à CrisJurinitz e ao Emílio Bruna pela força moral no meu projeto,à Angelinha pela massagem fundamental no pé docomputador e ao Glauco, por me ensinar a gostar mais dasformigas e opiliões. Ao grupo Força Total : Chris, Edgar,Fino e Natalie. À D. Eduarda pelas suas mãos de fada nacozinha, ao S. Jorge e Osmaildo pela força dada. Muitosagradecimentos ao Juruna, por emprestar um pouco de seuconhecimento da mata e pelo seu incrível bom-humor eprestatividade. Agradeço, é claro, a todos os alunos eprofessores do curso de 2001 pelos fantásticos momentosvividos!
Referências BibliográficasBrokal, N.V.L. 1980. Treefalls: frequency, timing and
consequences. In: The Ecology of a Tropical Forest:Seasonal Rhythms and Long Term Changes, Leigh, E.G.et al. (ES.). Smithsonian Institute Press, Washington.
Denslow, J. 1980.Gap partitioning among tropical rain foresttrees. Biotropica 12: 47-55.
Denslow, J. & Hashtorn, G.H. 1994. Tree fall gapsenvironments and forest dynamics processes. In: LaSelva, Mc Dade, L. et al. (eds.). University of ChicagoPress. Chicago.
Garwood, N.C.1982. Seasonal rhythms of seed germinationin a semi-deciduous Tropical Forest. In: The Ecology ofa Tropical Forest, Leight, E.G. et al. (eds.), pp. 173-185. Smithsonian Institution Press, Washington.
Gloyne, R.W. 1968. The structure of the wind and itsrelevance to forestry. Forestry 41: 7-9.
Hashtorn, G.S. 1978. Treefalls and tropical forest dynamics.In: Tropical Trees as Living Systems, Tomlinson, P.B.& Zimmerman, M.H. (eds.), pp. 617-683. CambrigeUniversity Press, Cambrige.
Hashtorn, T.H. 1980. Neotropical forest dynamics. Biotropica
12: 23-30Lieberman, D.; Hashtorn, G.S.; Lieberman, M. & Peralta,
R. 1990. Forest dynamics at La Selva Biological Station,1969-1985. In: Four Neotropical Rain Forest, Gentry,A.H. (ed.), pp. 509-521. Yale University Press, NewHaven.
Lovejoy, T & Bierregaard, R.O. 1990.Central Amazonianforests and the minimum critical size of ecosystemsprojects. In: Four Neotropical Rain Forest, Gentry, A.H.(ed.). Yale University Press, New Haven.
Martinez Ramos, M.; Sarukhan, J. & Piñero, D. 1988. Thedemography of tropical trees in the context of forest gapsdynamics. In: Plant Population Ecology, 28th Symposiumof the British Ecological Society, Davy, A.J. et al. (eds.).Blackwell Scientific Publications, Oxford.
Oldeman, R.A.A. 1972. L’architeture de la végétationrupicule forestiere des fleuves et criques guyanais.Adansonia 12:253-265.
Oldeman, R.A.A. 1978. Architecture and energy exchangeof dicotiledonious trees in the forest. In: Tropical Treesas Living Systems, Tomlinson, P.B. & Zimmerman, M.H.(eds.), pp. 535-560. Cambrige University Press,Cambrige.
Oliveira, A.A. 1997. Diversidade, estrutura, e dinâmica docomponente arbóreo de uma floresta de terra firme emManaus. Tese de Doutorado, Universidade de São Paulo,São Paulo.
Sarukhan, J. 1978. Studies on the demography of tropicaltrees. In: Tropical Trees as Living Systems, Tomlinson,P.B. & Zimmerman, M.H. (eds.), pp. 163-184. CambrigeUniversity Press, Cambrige.
Whithmore, T.C. 1978. Gaps in the forest canopy. In: TropicalTrees as Living Systems, Tomlinson, P.B. & Zimmerman,M.H. (eds.), pp. 639-655. Cambrige University Press,Cambrige.
Willkinson, L. 1988. Systat: the System for Statistics.SYSTAT Inc., Evanston, IL.
Young, T.P. & Hubbel, S.P. Crown assimetry, treefalls andrepeat disturbance of broad-leaved forest igapó. Ecology72: 1464-1471.
Zar, J.H. 1984. Biostatistical Analysis. Prentice Hall, NewJersey.
Avaliação de dois índices de integridade ecológica através da comunidade ícticaem um igarapé de terra firme da Amazônia Central
Mauricio Camargo-Zorro
IntroduçãoDiferentes abordagens têm sido propostas a fim de
compreender e, consequentemente, apontar formas de manejoe conservação nos vários níveis da biodiversidade,especialmente em ambientes aquáticos tropicais queapresentam acelerado processo de degradação (Christensen1998; Pauly et al. 1998).
A utilização de abordagens parciais, que fundamentam-se na aplicação de modelos empíricos, baseados num únicorecurso do ecossistema e que não contemplam as interaçõesdas espécies entre si e com o meio ambiente, são deficientespara subsidiar cientificamente políticas de manejo que visemmanter a integridade biológica dos ecossistemas. Nestesentido, o estudo de formas de manejo que tenham o

Curso de Campo Ecologia da Floresta Amazônica - 2001 137
ecossistema como unidade básica de gerenciamento, devemser considerados como prioritários (Mangel et al. 1996).
Com essa visão holística, surgiram várias abordagensquantitativas que permitem prever, por meio da integraçãode medições de uma série de atributos biológicos dascomunidades e de sua funcionalidade, possíveis efeitosantrópicos sobre os ecossistemas. Assim, foram propostos oÍndice Multimétrico de Avaliação das Condições Biológicas–IBI (Karr 1981) e recentemente o Índice de Integridade dasAssembléias de Peixes – FAII (Kleynhans 1999). Os doisíndices fundamentam-se na utilização das comunidades depeixes como indicadoras de qualidade do ambiente aquático.
Estes índices foram desenvolvidos para ambientestemperados, é na atualidade não se sabe sobre suasensibilidade e aplicação nas comunidades de peixes quefreqüentam os ambientes de um pequeno sistema aquático,de forma tal de verificar sua capacidade para detectar asmudanças no ecossistema.
O presente trabalho tenta através da caracterização dascomunidades de peixes que ocorrem dois trechos de umigarapé de terra firme amazônico, diagnosticar o estado atualde conservação deste ecossistema. De modo mais específico,pretende-se: (i) avaliar vários atributos das assembléias depeixes, que permitam definir o estado atual de integridadeecológica do ambiente estudado e (ii) avaliar a sensibilidadedestes índices como indicadores, através de mudanças nacomposição ictiofaunística com o objetivo de prever o efeitodas especies com maior distribuição espacial e de suaabundância no ecossistema.
Material e MétodosO presente estudo foi desenvolvido na área de reservas
florestais administradas pelo Projeto Dinâmica Biológica deFragmentos Florestais, do convênio entre INPA eSmithsonian Institution (PDBFF –INPA/SI). A reserva 1501situa-se na Fazenda Esteio e é conhecida como “Reserva doKm 41”. O igarapé estudado faz parte das cabeceiras dedrenagem da bacia do rio Urubu, afluente da margem esquerdado curso médio do rio Amazonas. O clima é tropical chuvoso(Amw) na classificação Köppen, com temperatura médiaanual de 26oC, precipitação anual entre 1750 e 2000mm eumidade relativa anual de 80%.
Inicialmente, como forma de definir um ecossistemadenominado “referência” ou com condições de conservaçãoideais, utilizei informações contidas no estudo de Bührheim(1998) para um igarapé de segunda ordem. Esse igarapé estácontido na mesma microbacia hidrográfica, do igarapé quefoi o objeto do presente estudo, na reserva do km 41. Aseleção dos habitats foi realizada ao longo deaproximadamente 800m do igarapé. Calculei a velocidadeda correnteza pela medição do tempo em que uma placa deisopor percorreu uma distância de 2,0m na superfície d’água.
Realizei as coletas durante três dias, no período de menorregime de precipitação na área (agosto de 2001). Para estafinalidade, defini dos trechos de igarapé, com base naocorrência de uma clareira natural de aproximadamente800m2. Para as capturas, utilizei o puçá, regionalmente
conhecido como rapiché, e a peneira. Amostrei 100m deigarapé, numa área sem evidências de clareiras e 100m deárea com clareiras. Dois coletores participaram das coletas,padronizando um tempo de coleta de 6 horas por trecho.
O material biológico coletado foi colocado em sacosplásticos e fixado em formalina 10% até sua identificaçãotaxonômica e contagem do número de indivíduos por espéciepara calcular das abundâncias relativas. Realizei todas ascoletas em horários vespertinos e noturnos, pois verificou-se maior eficiência na captura durante estes horários.Simultaneamente, com uso de mergulho livre, registrei adistribuição na das espécies coluna d’água e a ocorrência depossíveis espécies não coletadas.
Para a definição dos parâmetros originais do índice deintegridade biótica-IBI construi uma matriz de dados quecontinha nas colunas 12 atributos da comunidade íctica, osquais agrupei de acordo com Karr (1981) em 5 categorias:(i) composição e riqueza de espécies; (ii) espéciesindicadoras; (iii) função trófica; (iv) função reprodutiva e(v) abundância e grau de malformações ou ocorrência deespécies exóticas. Para categorizar o igarapé com condições“ideais”, cada um destes 12 atributos foi ordenado segundoKarr et al. (1986) (Tabela 1). O IBI, proposto por estesautores, foi feito através da soma dos ordenamentosassinalados para cada um dos 12 critérios definidos, os quaisforam comparados com os valores de referência propostospelos mesmos autores (Tabela 2).
Tabela 1. Medições utilizadas para avaliar a integridade biológica
das comunidades, com base no Índice de Integridade Biológica -
IBI (Karr 1981; Karr et al. 1986).
Atributo medido
5
Ordenação
3 1
A. Riqueza e composição das espécies
1. No. total de espécies Os valores esperados para as
categorias 1-5 variam com a
região e tamanho do sistema
2. No. de espécies bentônicas
3. No. de espécies da coluna d’água
4. Espécies k estrategistas
5. No. de espécies intolerantes
6. % de espécies tolerantes <5 5-20 >20
Composição trófica
7. % de espécies onívoras-detritívoras <20 20-45 >45
8. % de espécies insetívoras >45 45-20 <20
9. % de espécies piscívoras >5 5-1 <1
B. Abundância e condições físicas dos
peixes
10. No. de indivíduos na amostra Varia com o sistema e região
11. % de espécies exóticas 0 >0-1 >1
12. % de espécies com anomalias ou
doentes
0-2 >2-5 >5
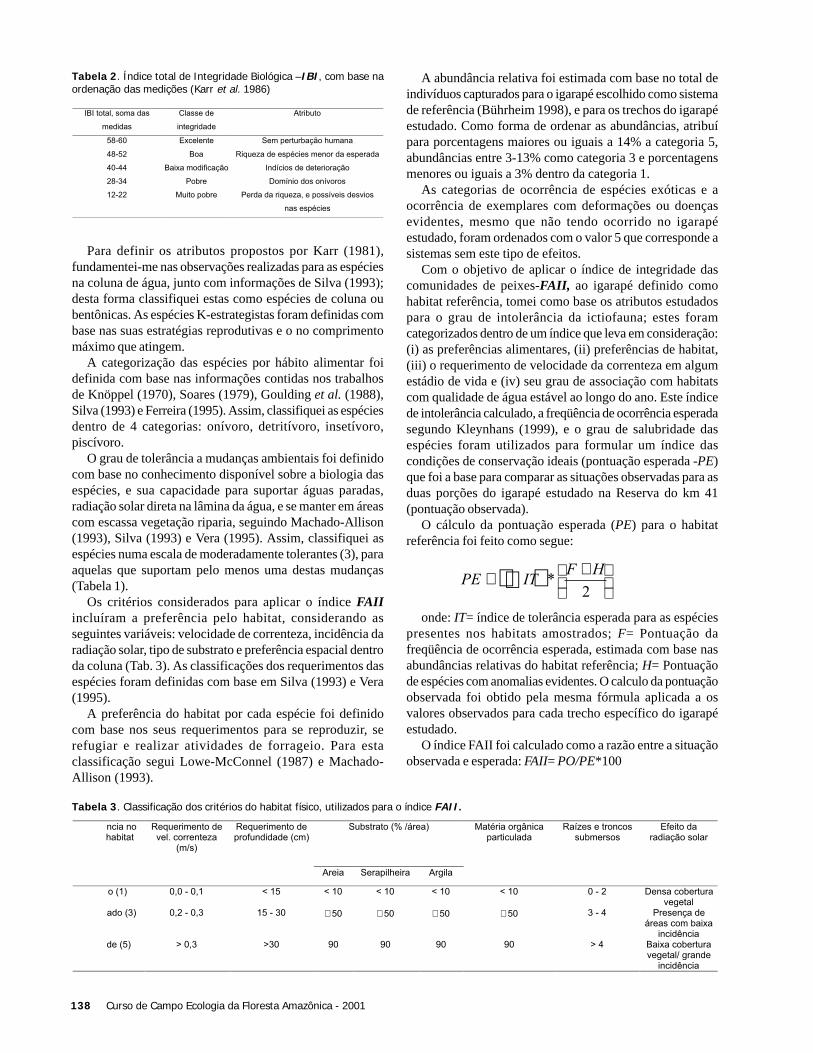
138 Curso de Campo Ecologia da Floresta Amazônica - 2001
Tabela 2. Índice total de Integridade Biológica –IBI, com base naordenação das medições (Karr et al. 1986)
IBI total, soma das
medidas
Classe de
integridade
Atributo
58-60 Excelente Sem perturbação humana
48-52 Boa Riqueza de espécies menor da esperada
40-44 Baixa modificação Indícios de deterioração
28-34 Pobre Domínio dos onívoros
12-22 Muito pobre Perda da riqueza, e possíveis desvios
nas espécies
Para definir os atributos propostos por Karr (1981),fundamentei-me nas observações realizadas para as espéciesna coluna de água, junto com informações de Silva (1993);desta forma classifiquei estas como espécies de coluna oubentônicas. As espécies K-estrategistas foram definidas combase nas suas estratégias reprodutivas e o no comprimentomáximo que atingem.
A categorização das espécies por hábito alimentar foidefinida com base nas informações contidas nos trabalhosde Knöppel (1970), Soares (1979), Goulding et al. (1988),Silva (1993) e Ferreira (1995). Assim, classifiquei as espéciesdentro de 4 categorias: onívoro, detritívoro, insetívoro,piscívoro.
O grau de tolerância a mudanças ambientais foi definidocom base no conhecimento disponível sobre a biologia dasespécies, e sua capacidade para suportar águas paradas,radiação solar direta na lâmina da água, e se manter em áreascom escassa vegetação riparia, seguindo Machado-Allison(1993), Silva (1993) e Vera (1995). Assim, classifiquei asespécies numa escala de moderadamente tolerantes (3), paraaquelas que suportam pelo menos uma destas mudanças(Tabela 1).
Os critérios considerados para aplicar o índice FAIIincluíram a preferência pelo habitat, considerando asseguintes variáveis: velocidade de correnteza, incidência daradiação solar, tipo de substrato e preferência espacial dentroda coluna (Tab. 3). As classificações dos requerimentos dasespécies foram definidas com base em Silva (1993) e Vera(1995).
A preferência do habitat por cada espécie foi definidocom base nos seus requerimentos para se reproduzir, serefugiar e realizar atividades de forrageio. Para estaclassificação segui Lowe-McConnel (1987) e Machado-Allison (1993).
Substrato (% /área)ncia nohabitat
Requerimento devel. correnteza
(m/s)
Requerimento deprofundidade (cm)
Areia Serapilheira Argila
Matéria orgânicaparticulada
Raízes e troncossubmersos
Efeito daradiação solar
o (1) 0,0 - 0,1 < 15 < 10 < 10 < 10 < 10 0 - 2 Densa coberturavegetal
ado (3) 0,2 - 0,3 15 - 30 � 50 � 50 � 50 � 50 3 - 4 Presença deáreas com baixa
incidênciade (5) > 0,3 >30 90 90 90 90 > 4 Baixa cobertura
vegetal/ grandeincidência
Tabela 3. Classificação dos critérios do habitat físico, utilizados para o índice FAII.
A abundância relativa foi estimada com base no total deindivíduos capturados para o igarapé escolhido como sistemade referência (Bührheim 1998), e para os trechos do igarapéestudado. Como forma de ordenar as abundâncias, atribuípara porcentagens maiores ou iguais a 14% a categoria 5,abundâncias entre 3-13% como categoria 3 e porcentagensmenores ou iguais a 3% dentro da categoria 1.
As categorias de ocorrência de espécies exóticas e aocorrência de exemplares com deformações ou doençasevidentes, mesmo que não tendo ocorrido no igarapéestudado, foram ordenados com o valor 5 que corresponde asistemas sem este tipo de efeitos.
Com o objetivo de aplicar o índice de integridade dascomunidades de peixes-FAII, ao igarapé definido comohabitat referência, tomei como base os atributos estudadospara o grau de intolerância da ictiofauna; estes foramcategorizados dentro de um índice que leva em consideração:(i) as preferências alimentares, (ii) preferências de habitat,(iii) o requerimento de velocidade da correnteza em algumestádio de vida e (iv) seu grau de associação com habitatscom qualidade de água estável ao longo do ano. Este índicede intolerância calculado, a freqüência de ocorrência esperadasegundo Kleynhans (1999), e o grau de salubridade dasespécies foram utilizados para formular um índice dascondições de conservação ideais (pontuação esperada -PE)que foi a base para comparar as situações observadas para asduas porções do igarapé estudado na Reserva do km 41(pontuação observada).
O cálculo da pontuação esperada (PE) para o habitatreferência foi feito como segue:
� � *2
F HPE IT
�� � � ��
onde: IT= índice de tolerância esperada para as espéciespresentes nos habitats amostrados; F= Pontuação dafreqüência de ocorrência esperada, estimada com base nasabundâncias relativas do habitat referência; H= Pontuaçãode espécies com anomalias evidentes. O calculo da pontuaçãoobservada foi obtido pela mesma fórmula aplicada a osvalores observados para cada trecho específico do igarapéestudado.
O índice FAII foi calculado como a razão entre a situaçãoobservada e esperada: FAII= PO/PE*100
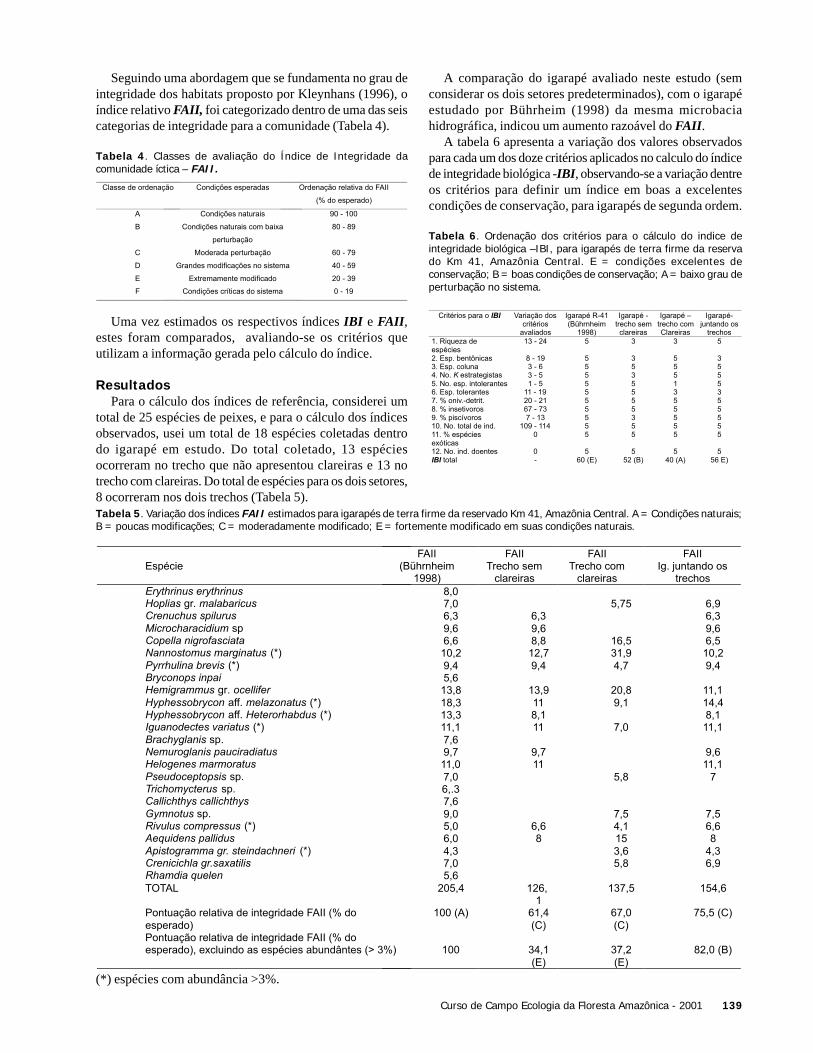
Curso de Campo Ecologia da Floresta Amazônica - 2001 139
Seguindo uma abordagem que se fundamenta no grau deintegridade dos habitats proposto por Kleynhans (1996), oíndice relativo FAII, foi categorizado dentro de uma das seiscategorias de integridade para a comunidade (Tabela 4).
Tabela 4. Classes de avaliação do Índice de Integridade dacomunidade íctica – FAII.
Classe de ordenação Condições esperadas Ordenação relativa do FAII
(% do esperado)
A Condições naturais 90 - 100
B Condições naturais com baixa
perturbação
80 - 89
C Moderada perturbação 60 - 79
D Grandes modificações no sistema 40 - 59
E Extremamente modificado 20 - 39
F Condições críticas do sistema 0 - 19
Uma vez estimados os respectivos índices IBI e FAII,estes foram comparados, avaliando-se os critérios queutilizam a informação gerada pelo cálculo do índice.
ResultadosPara o cálculo dos índices de referência, considerei um
total de 25 espécies de peixes, e para o cálculo dos índicesobservados, usei um total de 18 espécies coletadas dentrodo igarapé em estudo. Do total coletado, 13 espéciesocorreram no trecho que não apresentou clareiras e 13 notrecho com clareiras. Do total de espécies para os dois setores,8 ocorreram nos dois trechos (Tabela 5).
EspécieFAII
(Bührnheim1998)
FAIITrecho sem
clareiras
FAIITrecho com
clareiras
FAIIIg. juntando os
trechosErythrinus erythrinus 8,0Hoplias gr. malabaricus 7,0 5,75 6,9Crenuchus spilurus 6,3 6,3 6,3Microcharacidium sp 9,6 9,6 9,6Copella nigrofasciata 6,6 8,8 16,5 6,5Nannostomus marginatus (*) 10,2 12,7 31,9 10,2Pyrrhulina brevis (*) 9,4 9,4 4,7 9,4Bryconops inpai 5,6Hemigrammus gr. ocellifer 13,8 13,9 20,8 11,1Hyphessobrycon aff. melazonatus (*) 18,3 11 9,1 14,4Hyphessobrycon aff. Heterorhabdus (*) 13,3 8,1 8,1Iguanodectes variatus (*) 11,1 11 7,0 11,1Brachyglanis sp. 7,6Nemuroglanis pauciradiatus 9,7 9,7 9,6Helogenes marmoratus 11,0 11 11,1Pseudoceptopsis sp. 7,0 5,8 7Trichomycterus sp. 6,.3Callichthys callichthys 7,6Gymnotus sp. 9,0 7,5 7,5Rivulus compressus (*) 5,0 6,6 4,1 6,6Aequidens pallidus 6,0 8 15 8Apistogramma gr. steindachneri (*) 4,3 3,6 4,3Crenicichla gr.saxatilis 7,0 5,8 6,9Rhamdia quelen 5,6TOTAL 205,4 126,
1137,5 154,6
Pontuação relativa de integridade FAII (% doesperado)
100 (A) 61,4(C)
67,0(C)
75,5 (C)
Pontuação relativa de integridade FAII (% doesperado), excluindo as espécies abundântes (> 3%) 100 34,1
(E)37,2(E)
82,0 (B)
Tabela 5. Variação dos índices FAII estimados para igarapés de terra firme da reservado Km 41, Amazônia Central. A = Condições naturais;B = poucas modificações; C = moderadamente modificado; E = fortemente modificado em suas condições naturais.
(*) espécies com abundância >3%.
A comparação do igarapé avaliado neste estudo (semconsiderar os dois setores predeterminados), com o igarapéestudado por Bührheim (1998) da mesma microbaciahidrográfica, indicou um aumento razoável do FAII.
A tabela 6 apresenta a variação dos valores observadospara cada um dos doze critérios aplicados no calculo do índicede integridade biológica -IBI, observando-se a variação dentreos critérios para definir um índice em boas a excelentescondições de conservação, para igarapés de segunda ordem.
Tabela 6. Ordenação dos critérios para o cálculo do indice deintegridade biológica –IBI, para igarapés de terra firme da reservado Km 41, Amazônia Central. E = condições excelentes deconservação; B = boas condições de conservação; A = baixo grau deperturbação no sistema.
Critérios para o IBI Variação doscritérios
avaliados
Igarapé R-41(Bührnheim
1998)
Igarapé -trecho sem
clareiras
Igarapé –trecho comClareiras
Igarapé-juntando os
trechos1. Riqueza deespécies
13 - 24 5 3 3 5
2. Esp. bentônicas 8 - 19 5 3 5 33. Esp. coluna 3 - 6 5 5 5 54. No. K estrategistas 3 - 5 5 3 5 55. No. esp. intolerantes 1 - 5 5 5 1 56. Esp. tolerantes 11 - 19 5 5 3 37. % oniv.-detrit. 20 - 21 5 5 5 58. % insetivoros 67 - 73 5 5 5 59. % piscívoros 7 - 13 5 3 5 510. No. total de ind. 109 - 114 5 5 5 511. % espéciesexóticas
0 5 5 5 5
12. No. ind. doentes 0 5 5 5 5IBI total - 60 (E) 52 (B) 40 (A) 56 E)

140 Curso de Campo Ecologia da Floresta Amazônica - 2001
Enquanto que o IBI calculado para os dos trechos doigarapé estudado, indicou boas condições de conservaçãopara o setor sem clareiras e baixo grau de perturbação para osetor de clareiras, já com aplicação do índice FAII, obtivevalores que indicam que estes mesmos setores apresentaramcaracterísticas de um ecossistema moderadamente perturbado,onde o maior efeito foi gerado pela diminuição na riquezade espécies e das abundâncias das espécies intolerantes amudanças no ambiente (Tabelas 5 e 6). A tentativa de diminuirao riqueza ao recalcular os índices com exclusão das espéciescom maior abundância relativa dentro das amostragens,definiu um efeito negativo no FAII, o qual caracterizou ossistemas como seriamente modificados (Tabela 5).
DiscussãoDevido à eficiência diferenciada das artes de coleta para
a comparação dos índices IBI e FAII em ecossistemasaquáticos com uma mesma ordem de classificação, se fazimportante primeiro definir e padronizar as metodologias eo esforço de coleta. Isto se justifica pelo fato de ter-seencontrado para um igarapé da mesma bacia hidrográfica,diferencias nas capturas, que foram atribuídas ao tipo deapetrecho de pesca utilizado (Bührheim 1998). Um outroaspecto de grande relevância para aplicar estas metodologiasé a manutenção de escalas de comparação a nível da ordemdo sistema aquático, considerando o efeito da variaçãotemporal na riqueza e categorias tróficas. Como forma dedefinir um índice padrão que será a base de comparação,este deve ser procedente de amostragens padronizadas, quepossam ser repetidas em posteriores monitoramentos.
A tentativa de exclusão das espécies que apresentaramabundâncias relativas superiores a 3% indicou que o índiceFAII não é aplicável para ecossistemas com baixa riquezade espécies, como foi observado quando consideramos osdois setores do igarapé de forma isolada (trechos com e semclareiras naturais), talvez explicável pela pequena escalaespacial de amostragem. Estes resultados podem serexplicados também pela importância dentro do índice dasespécies raras, sendo que algumas ocorreram em um únicotrecho. O aumento observado no FAII depois de comparar oigarapé avaliado neste estudo (sem considerar os dois trechospredeterminados), com o igarapé estudado por Bührheim(1998) para a mesma microbacia hidrográfica, pode serexplicado pela contribuição das espécies raras para a riquezatotal.
O fato de não ter sido observado com a exclusão dasespécies abundantes de ampla distribuição espacial nosistema em estudo um efeito negativo no valor do FAII, indicaque em ecossistemas com grande riqueza de espécies (maisde 15 espécies), estas não afetam de maneira drástica o cálculodo FAII.
A redução do numero de espécies parece indicar que oFAII não se aplica para pequenos sistemas, com baixariqueza, devido a que não se observou uma boa resposta parao recalculo com diminuição das mesmas. Uma tentativa deaplicação do IBI e do FAII em sistemas com riqueza menorque 5 espécies, indica que estes não fornecem bons
resultados. Neste caso é possível propor um índice que meçaos atributos da população de uma espécie dentro doecossistema, como parâmetro indicador de algum tipo deperturbação. Contudo, Karr & Chu (1998) fazem uma críticaa esta abordagem no sentido de que mesmo sob condiçõesnaturais, as populações mudam muito em sua densidade eoutros aspetos de sua biologia.
Devido ao fato de que os critérios utilizados no cálculodos índices são o resultado de observação e julgamentoprofissional (ausência de parâmetros claros), talvez sejaimportante observar com mas detalhe o grau de tolerânciadas espécies como uma forma de melhorar o índice deintolerância para o FAII. Nesse caso é possível propor talvezmas uma categoria a mais que indique melhor o grau deintolerância, como tem sido proposto por Hughes et al.(1998).
A tentativa de aplicar estes índices a um igarapéamazônico, foi feita pensando em dispor de uma estimativaquantitativa da integridade ambiental aplicável aos pequenossistemas aquáticos de terra firme. Devido ao fato do IBI tomarem consideração atributos mais relacionados com acomunidade e não com o habitat físico, torna-se fácil suaaplicação para avaliações ecológicas rápidas. Talvez emsistemas maiores o FAII possa ser muito informativo,especialmente quando se deseja avaliar algum tipo deperturbação, com efeitos ambientais imediatos, enquanto queo IBI pode ser melhor aplicado quando se deseja avaliarefeitos nas comunidades aquáticas com uma magnitude detempo relativamente grande, após a ocorrência daperturbação.
AgradecimentosAgradeço a todos os professores do curso pelas ricas
palestras e pelos contínuos fluxos de informação, que nosindicaram que somos mais um semiponto na “madrenaturaleza”. Obrigado ao Jansen e ao Dadão pela excelentecoordenação e pelo seu esforço em nos ensinar mas um poucona apreciação do entorno. Aos colegas e novos amigos dosdiferentes cantos do Brasil. Ao Rodrigo e a Ruth pela infinitaforça e paciência nas minhas ausências.
Referências BibliográficasBührnheim, C.M. 1998. Estrutura de comunidades de peixes
em igarapés de floresta de terra firme na AmazôniaCentral. Dissertação de Mestrado, Universidade doAmazonas/INPA, Manaus, AM.
Christensen, V. 1998. Fishery induced changes in a marineecosystem: insight form models of the Gulf of Thailand.J. Fish. Biol. 53:128-142.
Ferreira, E.J.G. 1995. Composição, distribuição e aspectosecológicos da ictiofauna de um trecho do rio Trombetas,na área de influência da futura UHE Cachoeira Porteira,Estado do Pará, Brasil. Acta Amaz. 23(1/4):1-89.
Goulding, M. L. Carvalho & E.G. Ferreira. 1988. Rio Negro:Rich Life in Poor Water: Amazonian Diversity andFoodchain Ecology as Seen Throught the FishCommunities. Academic Pub.

Curso de Campo Ecologia da Floresta Amazônica - 2001 141
Hughes R.M.; Kaufmann, P.R.; Herlihy, A.T.; Kincaid, T.M.;Reynolds, T.M. & Larsen, D.P. 1998. A proccess fordeveloping and evaluating indices of fish assemblageintegrity. Can. J. Fish. Aquat. Sci. 55: 1618-1631.
Karr, J.R. 1981. Assessment of biotic integrity using fishcommunities. Fisheries 6(6): 21-27.
Karr, J.R.; Fausch, K.D.; Angermeier, P.L.; Yant, P.R. &Schlosser, I.J. 1986. Assessing Biological Integrity inRunning Waters: A Method and its Rationale. IllinoisNatural History Survey Special Publication 5,Champaign, IL.
Karr, J.R & Chu, E.W. 1998. Biological Monitoring:Essential Foundation for Ecological Risk Assessment.www./salmoweb/conta ct.html.
Kleyhans, C.J. 1996. A qualitative procedure for theassessment of the habitat integrity status of the LuvuvhuRiver (Limpopo System, South Africa). J. Aquat.Ecosystem Healt 5:41-54.
Kleyhans, C.J. 1999. The development of a fish index itassess the biological integrity of South African rivers.Water AS 25(3): 265-278.
Knöppel, H.A. 1970. Food of Central Amazonian fishes;contribution to the nutrient-ecology of Amazonian rain-forest streams. Amazoniana II:257-352.
Lowe-McConnell, R.H. 1987. Ecological Studies in TropicalFish Communities. Cambridge University Press,London.
Machado-Allison, A. 1993. Los peces de las planíciesinundábles de Venezuela, um estudio de su histórianatural. Universidad Central de Venezuela, Caracas.
Mangel, M.; Talbot, L.M. & Meffe, G.K. 1996. Principlesfor the conservation of wild living resources. Ecol. Appl.6(2): 238-362.
Pauly, D; Christensen, V.; Dalsgaard, J. & Froese, R. 1998.Fishing down marine food webs. Science 279: 860-863.
Silva, C.P.D. 1993. Alimentação e distribuição espacial dealgumas espécies de peixes do igarapé do Candirú,Amazonas, Brasil. Acta Amaz. 23(2-3): 271-285.
Soares, M.G.M. 1979. Aspectos ecológicos (alimentação ereprodução) dos peixes do igarapé do Porto, Aripuanã,MT. Acta Amaz. 9:325-352.
Vera, M.C.V. 1995. A diversidade de habitats é responsávelpela riqueza de espécies de uma comunidade? Livro doCurso de Campo “Ecologia da Floresta Amazonica”,INPA/ Smithsonian Institution, Manaus, AM.
Walker, I. 1987. The biology of streams as part of Amazonianforest ecology. Experientia 43: 279-287.
Levantamento da riqueza e composição de espécies de mamíferos terrestrescom o auxílio de uma nova metodologia para identificação de pegadas
Natalie Olifiers
IntroduçãoGrande parte dos mamíferos silvestres brasileiros não são
facilmente avistados, principalmente porque a maioria delespossui hábitos noturnos e discretos (Becker & Dalponte1991; Wilson et al. 1996). Entretanto, em determinadascondições, podem ser observados indícios da presença destesanimais, tais como fezes, pêlos, tocas, restos alimentares epegadas. Além de possibilitarem em muitos casos aidentificação do animal ao nível de espécie (Becker &Dalponte 1991; Wilson et al. 1996; Piedra & Maffei 2000),tais rastros podem fornecer também informações sobre ocomportamento, idade, modo de locomoção e status socialdo animal (Wilson et al. 1996).
As pegadas constituem os rastros de interpretação maisconfiável (Becker & Dalponte 1991) e a sua utilização emtrabalhos científicos com diferentes enfoques tem sidorazoavelmente ampla. Assim, já foram utilizadas comoestimadores de densidade populacional (Caughley 1980), emcensos populacionais (Marten 1972) e mais especificamentena estimativa da abundância relativa de espécies (Carey &Witt 1991; Conner et al. 1982), em estudos de padrões delocomoção (Schaller 1980) e de períodos de atividade (Bider1962).
Várias metodologias específicas para a obtenção de
pegadas já foram desenvolvidas. Freqüentemente utiliza-seareia ou argila umedecida, para que o animal pise e deixe amarca das suas patas. A partir da pegada deixada na areiaumedecida, pode-se obter o molde negativo desta com gessoou parafina.
Também foram propostas outras metodologias, como autilização de papel quimiográfico (Raphael et al. 1986 apudCarey & Witt 1991) ou placas de alumínio enfumaçados(Taylor & Raphael 1988 apud Carey & Witt 1991). Ratz(1997) propôs um método modificado a partir do apresentadopor King & Edgar em 1977 (apud Ratz 1997), envolvendoum tabuleiro com tinta por onde o animal passa e, em seguida,deixa a sua pegada em uma folha de papel adjacente aotabuleiro.
Apesar do esforço dos pesquisadores em desenvolver umaforma eficiente e prática para a obtenção de pegadas demamíferos, todas as metodologias desenvolvidas até opresente apresentam limitações significativas. A maioria delas- como no caso da areia e do papel e alumínio enfumaçados- são suscetíveis à chuva, já que a elaboração de uma estruturaprotetora as tornaria menos práticas. O alumínio é pesado, oque torna o seu transporte para a confecção de “armadilhasde pegada” um trabalho árduo (Carey & Witt 1991). Asarmadilhas feitas com areia apresentam variações na

142 Curso de Campo Ecologia da Floresta Amazônica - 2001
consistência que freqüentemente distorcem as dimensões daspegadas (Ratz 1997). Já a metodologia que envolve otabuleiro de tinta raramente resulta em pegadas perfeitas,além de apresentar variações geradas pela quantidade de tintalevada até o papel pela pata do animal, o que também alterariaas dimensões da pegada; além disso, esta metodologia excluimamíferos de grande porte (Ratz 1997).
O objetivo do presente projeto é realizar o levantamentoda riqueza e composição de espécies de mamíferos terrestresde uma reserva florestal, através da observação de pegadas.Adicionalmente, uma nova metodologia para a obtenção depegadas de mamíferos terrestres será proposta, testada eempregada no auxílio deste levantamento.
Material e MétodosÁrea de estudoO estudo foi realizado entre os dias 28 de agosto e primeiro
de setembro, durante a estação seca (junho a outubro). Aárea estudada localiza-se na Reserva do Km 41 do ProjetoDinâmica Biológica de Fragmentos Florestais (INPA/SI), naFazenda Esteio, Manaus, AM (2°24’S; 59°44’W). Esta regiãocaracteriza-se pela predominância de mata de terra firme(Pires & Prance 1985), sendo recortada por cursos d’água(igarapés) que correm em áreas de baixio.
Descrição da nova metodologiaO material proposto como uma nova metodologia para a
obtenção de pegadas consiste em duas folhas de plásticosuperpostas; a folha superior é um plástico de naturezafluorescente que quando pressionado adere à folha inferiormais fina, emitindo sua fluorescência e permitindo avisualização da área pressionada. Somente a área pressionadaemite fluorescência e é ressaltada, porque apenas esta áreaadere de fato ao plástico inferior. Dessa maneira, a pegadado animal pode ser identificada porque sua “imagem” éformada no plástico devido à pressão imposta pelo peso doanimal.
Teste do material em laboratórioForam feitos testes preliminares em laboratório, onde
mamíferos de diferentes pesos corporais foram induzidos acaminhar sobre o material. O teste foi feito com as espécies:Canis familiares (cão doméstico), Felis catus (gatodoméstico), Didelphis aurita (gambá), Metachirusnudicaudatus (cuíca-marrom-de-quatro-olhos), Philanderfrenata (cuíca-cinza-de-quatro-olhos), Akodon cursor (rato-bolinha), Sphiggurus villosus (ouriço-cacheiro) e Trichomyssp. (roedor).
Teste do material no campoForam estabelecidas estações de armadilhagem em três
locais distintos: numa área de mata fechada e ao longo deduas estradas, sendo uma delas parcialmente desativada e aoutra utilizada regularmente. Em cada uma destas áreas, foramcolocadas cinco armadilhas de pegadas (0,7 x 0,7 m) comespaçamento de 200 m entre si, totalizando quinze estaçõesde armadilhagem. As armadilhas foram cobertas com saco
plástico preto, para uma proteção extra contra a chuva ecolocadas sobre um suporte de plástico rígido que fosse capazde eliminar possíveis irregularidades do terreno, já que estaspoderiam distorcer as dimensões ou a própria imagem daspegadas no material.
As armadilhas foram iscadas alternadamente com bananae toucinho e todas continham um punhado de sal grosso. Aintenção de tal diferenciação foi a obtenção de pegadas tantode animais de hábitos frugívoros quanto carnívoros. Comoo número de estações de armadilhagem cada área era ímpar,uma delas foi iscada tanto com toucinho quanto com banana.A isca sempre era colocada no centro da armadilha. Asestações foram inspecionadas todas as manhãs e iscadas àtarde, permanecendo ativadas por cinco noites consecutivas.
Levantamento das espécies de mamíferos da áreaAliado ao teste da nova metodologia proposta neste
trabalho, foi feito um levantamento da riqueza e composiçãode mamíferos terrestres da área, através da procura ativa porpegadas em áreas de baixio da região.
Foram amostrados 7 trechos de baixios banhados porigarapés, pelo estabelecimento de um transecto de 100 m emcada um deles, disposto paralelamente ao baixio. Todas aspegadas encontradas nestes transectos foram desenhadas emfolha de acetato e as suas dimensões (comprimento total elargura da pegada, comprimento e largura da passada)anotadas e comparadas com as indicações oferecidas no guiade Rastros de Mamíferos Silvestres Brasileiros (Becker &Dalponte 1991). Também foi amostrada uma área de campinaalagada que foi percorrida inteiramente à procura de pegadas.
ResultadosNos testes preliminares desenvolvidos em laboratório, as
pegadas de C. familiares e F. catus mostraram-seperfeitamente visíveis e identificáveis. As demais espéciesdeixaram sinais no material, mas a identificação de tais rastrosnão foi possível. Entretanto, nos testes realizados commarsupiais (D. aurita, P. frenata e M. nudicaudatus), foivisualizado no material o polegar opositor destes animais,característico da família Didelphidae. Ainda, observou-se queo material foi capaz de conservar as marcas das pegadas portempo indeterminado, até que as duas folhas que o compõemfossem separadas manualmente.
A nova metodologia não pôde ser adequadamente testadaem campo porque nenhum animal visitou as estações deamostragem durante o período em que as armadilhaspermaneceram nas diferentes áreas. Entretanto,aparentemente não foi observada nenhuma limitaçãoassociada ao material que o qualificasse como inadequadopara o registro de pegadas, uma vez que mostrou-se resistenteao calor, por exemplo, além de ter minimizado o ataque deformigas à isca, provavelmente por representar um substratonão muito propício para o deslocamento destas (obs. pess.).Além disso, armadilhas de areia colocada nas mesmas árease iscadas da mesma forma também não registraram pegadas(T. Timo com. pess.), o que indica que a ausência de registrosprovavelmente reflete uma baixa abundância de mamíferos

Curso de Campo Ecologia da Floresta Amazônica - 2001 143
naquelas áreas e não um problema associado ao material.A procura ativa por pegadas resultou na identificação de
12 espécies de mamíferos, sendo duas destas identificaçõesduvidosas (Tabela 1). A espécie Tayassu sp. provavelmenteé T. pecari, uma vez que um grupo desta espécie foi avistadodeslocando-se em alguns trechos amostrados (J. Zuanon com.pess.). Ainda, em dois baixios amostrados não foramencontradas pegadas de mamíferos (Tabela 1).
Na área de campina (R24), foram encontradas seteespécies, o que representa cerca de 60% do total de espéciesidentificadas neste estudo. As espécies Tayassu sp. e H.Hidrochaeri foram as que apresentaram maior frequência deocorrência, aparecendo em cinco e quatro áreas,respectivamente (Tabela 1).
DiscussãoA nova metodologiaA nova metodologia proposta para a obtenção de pegadas
apresenta características desejadas para trabalhos em campoporque é leve, apresenta baixo custo, pode ser facilmenteprotegido da chuva, além de poder ser utilizadaindefinidamente, já que basta levantar o plástico fluorescentepara que perca a aderência ao plástico inferior e as marcasde pegadas desapareçam.
As desvantagens deste método são a cor do plástico (rosa),que poderia repelir alguns animais, e o fato de provavelmentenão ser tão eficaz para animais menores, como pequenos
Espécie L10 Q1
3
Q1
7
Q2
0
R2
4
T12 D1
3
O0
8
Ocorrência
(%)
Agouti paca (paca) ** X 12,5
Chironectes minimus (cuíca-
d’água) ***
X 12,5
Dasyprocta sp. (cutia) ** X 12,5
Dusicyon sp. (graxaim) X 12,5
Eira barbara (irara) X* 12,5
Galictis sp. (furão) X* X 25
Hydrochoerus hydrochaeris
capivara) ***
X X X X 50
Mazama sp. (veado-mateiro) ** X 12.5
Panhera onca (onça) X X 25
Tapirus terrestris (anta) ** X X 25
Tayassu sp. (porco-do-mato) ** X X X X X 62.5
Total de espécies 6 3 1 3 7 1 0 0 -
Tabela 1. Espécies identificadas por meio de pegadas em áreas de baixio da Reserva do Km 41 (PDBFF/INPA/SI). As letras associadas anúmeros representam a identificação do local no qual foi realizado o censo – segundo o sistema de coordenadas de trilhas da reserva. Acoluna “ocorrência” representa a frequência de ocorrência das espécies em relação ao total de áreas amostradas.
*identificações duvidosas; **espécies com distribuição associada a cursos d’água ou áreas alagadas; *** espécies com distribuiçãodeterminada pela presença de cursos d’água ou áreas alagadas.
roedores. Entretanto, o primeiro problema pode ser resolvidocobrindo-se as folhas com um plástico fino e escurbo. Osegundo trata-se de um problema que não é restrito a estametodologia, já que a identificação de pequenos roedorespor meio de pegadas freqüentemente é difícil ou impossível,mesmo no caso da obtenção de pegadas perfeitas.
Entretanto, a impossibilidade de testar o materialadequadamente no campo limita a discussão acerca de suaeficiência. O teste em condições reais de trabalho éindispensável para a qualificação do material como adequadoaos fins propostos. Dessa maneira, os testes realizados até omomento ainda são inconclusivos.
A nova metodologia em breve será testada no Zoológicodo Rio de Janeiro – RJ e, em seguida, novamente no campo,sob condições reais de trabalho. O teste a ser realizado nozoológico será de grande importância para a identificaçãocorreta das espécies, uma vez que a imagem da pegada nomaterial aqui apresentado provavelmente não corresponderáexatamente às imagens que são obtidas em areia ou argilaumedecida. Como os guias de identificação existentesbaseiam-se nessa metodologia tradicional (utilizando areiaou argila), talvez haja a necessidade de formular-se um novoguia de identificação de pegadas para a nova metodologiaproposta neste trabalho.
O levantamento das espécies de mamíferos da áreaatravés de pegadas: considerações gerais
A riqueza de espécies encontradas nos baixios amostrados

144 Curso de Campo Ecologia da Floresta Amazônica - 2001
indica que estas áreas, de maneira geral, são locais propíciospara a observação de rastros - durante o período da estaçãoseca - porque o seu terreno mantém-se constantemente úmidopela presença dos igarapés. Entretanto, durante a estaçãochuvosa, quando o volume de água nos igarapés aumenta,estas áreas permanecem alagadas e a visualização de taisrastros é dificultada.
Segundo Emmons (1997), 50% das espécies identificadasneste estudo são comumente associadas a ambientes úmidose, destas, aproximadamente 33% têm sua distribuiçãodeterminada pela presença de cursos d’ água (Tabela 1). Estefato, associado ao terreno propício para a identificação derastros, provavelmente contribuiu para a identificação de umnúmero relativamente alto de espécies nessas áreas.
Por outro lado, o fato de nenhum animal ter visitado asarmadilhas de pegadas estabelecidas em áreas de platô –mesmo contendo iscas atrativas – somado à identificação deespécies não diretamente associadas a cursos d’água em áreasde baixio (Tabela 1), poderia indicar que a densidade destasespécies seria também maior em áreas vizinhas a cursosd’água, pelo menos durante a estação seca, quandodisponibilidade de água em outras áreas diminui.
Sabe-se que a densidade de mamíferos é considerada baixana Amazônia Central (Emmons 1984; de Lima & Gascon1999), mas o número de registros encontrados nos baixiosnão corrobora esta informação. Embora numa escala regionala densidade de mamíferos possa ser considerada baixa, numaescala mais local a densidade de algumas espécies mostraria-se mais variável, em função da presença de igarapés e daépoca do ano (seca ou úmida).
Os baixios – principalmente aqueles banhados porigarapés de segunda e terceira ordem, que tendem a acumularmais matéria orgânica em suas áreas de remanso -representariam áreas de maior concentração de recursos paraas espécies que são comumente encontradas em locaisalagados. A presença destas espécies, muitas delas de médioa grande porte, representaria uma concentração de recursospara outras espécies de topo de cadeia alimentar, como P.onca. Dessa maneira, seria observada uma maior densidadelocal de espécies de mamíferos médio e grande nessas áreas.
Obviamente, a área de vida destes animais –principalmente a de predadores de topo de cadeia – não serialimitada a baixios, mas sugere-se que a frequência de visitaçãodessas áreas por estas espécies seja relativamente maior(principalmente na estação seca),não só por representaremfontes de água como também de concentração de alimento.Sabe-se, por exemplo, que predadores como P. onca e Pumaconcolor alimentam-se preferencialmente de mamíferos demédio e grande porte, como veados, cutias, pacas, capivarase porcos-do-mato (Emmons 1997) e todas estas espéciespossuem algum grau de associação com áreas banhadas porcursos d’água.
É importante ressaltar que, embora essas idéias sejamplausíveis, representam hipóteses a serem devidamentetestadas. Entretanto, este trabalho oferece diretrizes paraestudos posteriores de mamíferos terrestres na Amazônia,principalmente de médio a grande porte.
Sugere-se que estudos de curto prazo com mamíferos naAmazônia utilizem a busca ativa de evidências indiretas, comoa presença de rastros em geral (pegadas, tocas, fezes, entreoutros), uma vez que o esforço de amostragem por capturadireta precisa ser freqüentemente muito grande para obter-se um número razoável de indivíduos.
AgradecimentosAgradeço do fundo do meu estômago – por ser mais
profundo que meu coração – a todas as pessoas que meincentivaram neste meu projeto e se empolgaram como eume empolguei!. Agradeço ao Vitor por ser tão dedicado,prestativo, amoroso e atencioso, ao Américo pela atenção eapoio às minhas idéias, ao pessoal do Laboratório deVertebrados por ter me apoiado nos testes da novametodologia, ao Juruna pelos bregas e caminhadas, ao Jansenpor me acompanhar no “plano B”, quando descobri que osmamíferos daqui não gostam de brincar de “lousa mágica”,ao Dadão por sempre achar uma saída para as coisas que nãoestão dando certo e pelo seu pandeiro sempre bem-vindo, àsPaulas cariocas e empolgadas que me acompanharam pelastrilhas da Amazônia, ao Leandro Scoss por ajudar-me aidentificar as pegadas e, por fim – obviamente sempre nofim– agradeço ao Thiago por me aturar este tempo todoimplicando com ele (e sempre ganhando porque ele é paulistae eu carioca!) e por ter me acompanhado na canoa furadaque a gente entrou, mas conseguiu sair! Agradeço do fundodo meu coração a todos os que estiveram aqui - professores,alunos, mateiros e monitores (porque o Glauco vale porvários) – a ao Jorjão e dona Eduarda pelo apoio e o rangoincrivelmente saboroso!
Referências BibliográficasBecker, M. & Dalponte, J.C. 1991. Rastros de mamíferos
silvestres brasileiros. Ed. Universidade de Brasília,Brasília.
Bider, J.R. 1962. Dynamics and the temporo-spatial relationsof a vertebrate community. Ecology 43(4): 634-646.
Carey, A.B. & Witt, J.W. 1991. Track counts as indices toabundances of arboreal rodents. J. Mammal. 72(1): 192-194.
Caughley, G. 1980. Analysis of Vertebrate Populations. JohnWiley & Sons, London.
Conner, M.C.; Labisky, R.F. & Progulske Jr., D.R. 1983.Scent-station indices as measures of populationabundance for bobcats, raccons, gray foxes, andopossums. Wildl. Soc. Bull. 11(2): 146-152.
de Lima, M.G. & Gascon, C. 1999. The conservation valueof linear forest remnants in central Amazonia. Conserv.Biol. 91(2-3): 241-247.
Emmons, L.H. 1984. Geographic variation in densities anddiversities of non-volant flying mammals in Amazonia.Biotropica 16: 210-222.
Emmons, L.H. 1997. Neotropical Rainforest Mammals: AField Guide. The University Chicago Press, Chicago.
King, C.M. & Edgar, R.L. 1977. Techniques for trappingand tracking stoats (Mustela erminea); a review and a

Curso de Campo Ecologia da Floresta Amazônica - 2001 145
new system. NZ J. Zoology, 4: 193-212.Malcom, S.R. 1990. Estimation of mammalian densities in
continuous forest north of Manaus, In: Four NeotropicalRainforests, Gentry, A.H. (ed.). Yale University Press,New York.
Marten, G.G. 1972. Censusing mouse populations by meansof tracking. Ecology 53(5): 859-867.
Piedra, L. & Maffei, L. 2000. Efecto de las actividadeshumanas sobre la diversidad de mamíferos terrestres enun gradiente altitudinal. Rev. Biol. Trop. 48(1): 263-264.
Pires, J.M. & Prance, G.T. 1985. The vegetation types of theBrazilian Amazon. In: Amazonia, Prance, G. & Lovejoy,T. E. (eds), pp 109-145. Pergamon, New York.
Raphael, M.G.; Taylor, C.A. & Barret, R.H. 1986. Smokedaluminum track stations record flying squirreloccurrence. United States Department of Agriculture,Forest Service, Research Note, PSW 384:1-3.
Ratz, H. 1997 Identification of footprints of some smallmammals. Mammalia 61(3): 431-441.
Schaller, G.B. 1980. Movement patterns of jaguar. Biotropica12(3): 161-168.
Taylor, C.A. & Raphael, M.G. 1988. Identification ofmammal tracks from sooted track stations in the PacificNortwest. California Fish and Game 74(4): 4-15.
Wilson, D.E.; Cole, F.R.; Nichols, J.D.; Rudran, R. & Foster,M.S. 1996. Measuring and monitoring biologicaldiversity: standard methods for mammals. SmithsonianInstitution Press, Washington.
Influência de características ambientais sobre o tamanho de grupos degerrídeos (Hemiptera, Gerridae) em igarapés na Amazônia Central
Paula Ceotto
IntroduçãoA maioria dos ambientes consiste de um mosaico espaço-
temporal de elementos abióticos e bióticos. Devido a estaheterogeneidade ambiental, a localização de um organismono espaço é um importante determinante do seu valoradaptativo. A densidade de indivíduos em uma mancha dehabitat influencia a qualidade deste para os demais indivíduos(Pianka 1994).
Os organismos podem estar distribuídos no espaço deforma regular (geralmente associada à competição), aleatóriaou agregada. A distribuição agregada indica pouco sobre osfatores que influenciam a disposição dos organismosenvolvidos (Pianka 1994). No entanto, estudos acerca darelação entre fatores abióticos e bióticos com os organismospodem ajudar a entender eventuais padrões de distribuição.
Gerrídeos são insetos da ordem Hemiptera que vivem nasuperfície da água onde deslizam alimentando-se de insetos(Borror & Delong 1981). As espécies deste táxon sãofreqüentemente encontradas em grupos, o que as tornaparticularmente interessantes para estudos sobre a relaçãoentre fatores ambientais e mecanismos de agregação.
Os igarapés que atravessam as florestas da AmazôniaCentral apresentam variações quanto à largura, profundidadee velocidade da correnteza que podem influenciar adistribuição espacial das espécies de gerrídeos. Um exemploda influência de fatores abióticos na distribuição deorganismos aquáticos é a agregação do zooplânctonsuperficial, causada principalmente pela movimentação daágua em função do vento (Laugford & Jermolajev 1966 apudDowning 1991).
Com relação aos fatores bióticos, a disponibilidade dealimentos pode exercer forte influência sobre o grau deagregação dos gerrídeos. As larvas de insetos aquáticos aoemergirem como adultos podem servir como fonte de alimento
para organismos deste grupo. Dessa forma, os substratos defundo do igarapé que apresentam maior densidade de larvasde insetos associadas (Rincón 1999) podem ser maispropícios à ocorrência dos grupos de gerrídeos.
Neste trabalho estudei a relação entre o número deindivíduos em grupos de diferentes morfoespécies degerrídeos e variáveis ambientais em igarapés na AmazôniaCentral. Um objetivo secundário deste trabalho foi estimar ariqueza de morfoespécies de gerrídeos encontradas nostrechos de igarapés estudados e relacioná-la com a ordemdos igarapés.
Material e MétodosRealizei este estudo entre os dias 29 de agosto e 01 de
setembro de 2001, na Reserva Florestal do Km 41 do ProjetoDinâmica Biológica de Fragmentos Florestais (INPA-Smithsonian Institution), localizada a 70 km ao norte deManaus (2º24’S; 59º44’W). A reserva possui 10.000ha esitua-se em uma área de mata contínua bem preservada. Atemperatura média anual é de 26,7ºC e a precipitação anualda região é de 2.186 mm, com média mensal de 300 mm,sendo março e abril os meses mais chuvosos e julho, agostoe setembro os mais secos (Radambrasil 1978).
Amostrei grupos de gerrídeos em nove trechos de igarapés(três na calha principal e seis em tributários) e na poçatemporária da campina R24. Nomeei os trechos amostradosdos igarapés com a combinação de letras e números queidentificam o cruzamento de trilhas mais próximo ao pontode coleta. Em cada trecho, procurava por gerrídeos no sentidocontrário à correnteza, a partir do ponto no qual a trilhacortava o igarapé, até conseguir cinco grupos de gerrídeos.Para cada grupo avistado, anotava o número de indivíduos,media a largura e a profundidade do ponto onde estavam ecalculava a velocidade da correnteza. Medi a correnteza

146 Curso de Campo Ecologia da Floresta Amazônica - 2001
soltando um pedaço de isopor na superfície da água, emarquei o tempo que este levava para percorrer uma distânciaque variava de acordo com as condições do trecho amostrado.Em cada ponto, fiz três medidas de velocidade da corrente,de modo que os valores usados neste estudo correspondem àmédia destas três medidas.
Estimei também a porcentagem de cada tipo de substratopresente no fundo do igarapé, nos locais onde encontravam-se os grupos. Esses substratos foram classificados como areia(A), folhedo associado a sedimento fino (F) e areia associadaa sedimento fino (AS). Nos pontos onde havia dois tipos desubstrato, conjuguei os símbolos de modo a informar qualsubstrato ocorria em maior proporção. Por exemplo, osímbolo AF significa que a proporção de areia era maior quea de folhedo associada a sedimento fino. A área usada paraestimar a proporção de cada tipo de substrato variava emfunção de obstáculos naturais (e.g. troncos, galhos, folhedo)que delimitavam as áreas ocupadas pelos grupos. Quandonão havia tais obstáculos, a área usada para as estimativasdo tipo de substrato correspondia à área ocupada pelosgerrídeos.
Assumindo que os grupos são formados por indivíduosde uma única espécie, coletei no mínimo um indivíduo decada grupo, a fim de calcular a riqueza de espécies degerrídeos nos igarapés. Usando uma chave de identificação(Merrit & Cummins 1984), determinei uma das espéciesencontradas no estudo. Como não foi possível determinar asdemais espécies com a chave mencionada, que foi feita paraos insetos norte-americanos, separei-as com base emcaracterísticas morfológicas, observando os animaiscoletados sob lupa binocular. Entre as principaiscaracterísticas diagnósticas usadas na separação incluem-sea forma do abdome, a forma da fronte, presença ou ausênciade asas nos adultos, coloração do corpo, presença ou ausênciade faixas laranjas nos esternitos abdominais e distância dosolhos ao tórax.
Para as duas espécies mais comuns investiguei se o númerode indivíduos nos grupos estava relacionado às variáveisambientais medidas e se os tipos de substratos agrupavam-se em função dessas variáveis e do número de indivíduos.Realizei este procedimento por meio de uma inspeção gráficados dados ordenados.
ResultadosA distância entre os grupos de gerrídeos nos igarapés foi
geralmente curta, de modo que percorria cerca de 10 metrospara amostrar os cinco grupos em cada trecho. A largura médiados trechos onde encontrei os gerrídeos foi de 1,05m (DP =0,07m), com variação de 1m a 5,4m. A profundidade varioude 2,5cm a 32,0cm (X ± DP = 12,11 ± 6,29 cm) e a velocidademédia da corrente foi de 0,09 ± 0,05 m/s, variando de 0 a0,21m/s. O tamanho dos grupos variou de 1 a 26 indivíduos(X ± DP =9,2 ± 4,8). Muitas vezes os grupos amostradosestavam em áreas de remanso ou correnteza fraca, onde osedimento predominante era folhedo associado a sedimentofino. Aparentemente, nessas áreas onde a correntezaaproximava-se de zero, o número de indivíduos era
particularmente baixo.Encontrei a espécie Limnogonus sp. na campina e seis
morfoespécies nos igarapés. Das espécies restritas aosigarapés, quatro ocorreram em apenas uma área, uma ocorreuem sete e outra foi coletada em nove áreas (Tabela 1). Asmorfoespécies encontradas em apenas um trecho de igarapéforam coletadas somente em um dos cinco grupos amostrados.Em duas ocasiões coletei indivíduos pertencentes a diferentesmorfoespécies em um mesmo grupo. Os pontos de coletanos quais encontrei mais de uma morfoespécie foramexcluídos das análises. Não houve relação entre a riquezade morfoespécies e a ordem dos igarapés (Tabela 1).
Tabela 1: Riqueza e composição de morfoespécies de gerrídeosencontradas nos trechos de igarapé (indicado pelo código do localamostrado) e na campina.
Local Ordem Número de
morfoespécies
Composição
de morfoespécies
L3 1a 1 sp.4
Q16 1a 4 sp.1, sp.2, sp.3, sp.4
08 1a 2 sp.1, sp.4
018 2a 3 sp1, sp4, sp5
L6 2a 2 sp.1, sp.4
L12 2a 3 sp.1, sp.4, sp.7
Q13 2a 2 sp.1, sp.4
Q16 1a 4 sp.1, sp.2, sp.3, sp.4
T12 2a 1 Sp.4
O17 3a 2 sp.1, sp.4
Campina - 1 sp.6
A baixa representatividade da maioria das morfoespéciesinviabilizou a avaliação das características ambientaisimportantes para a formação dos grupos nestas espécies,aparentemente raras. Desta forma, os fatores que influenciama formação dos grupos foram analisados apenas para asmorfoespécies sp.1 e sp.4. A inspeção gráfica dos pontosordenados por tipo de substrato não evidenciou relação entreo número de indivíduos por grupo e o tipo de sedimento nofundo do igarapé para ambas as espécies (Figura 1). Alémdisso, a velocidade da correnteza, a profundidade e a largurado igarapé também não mostraram relação evidente com onúmero de indivíduos por grupo, para as duas espécies(Figura 1).
Curso de Campo Ecologia da Floresta Amazônica - 2001 147
0.00 0.05 0.10 0.15 0.20Velocidade da corrente (m/s)
5
10
15202530
FAFASFAF
0 1 2 3 4 5 6Largura (m)
5
10
15202530
0 5 10 15 20Profundidade (cm)
2
4
6
810
0.00 0.05 0.10 0.15 0.20 0.25Velocidade da corrente (m/s)
2
4
6
810
1.0 1.5 2.0 2.5 3.0Largura (m)
2
4
6
810
Núm
ero
de
ind
ivíd
uos
Núm
ero
de
indiv
íduos
0 10 20 30 40Profundidade (cm)
5
10
15202530
Núm
ero
de
indiv
íduos
Figura 1. Ordenação para inspeção gráfica das relações entre o tamanho dos grupos de sp.1 e sp.4 e as variáveis ambientais velocidade dacorrenteza, largura e profundidade do igarapé. As siglas representam as classificações de substrato: AF = predominância de areia fina comrelação ao folhedo, ASF = predominância de areia com sedimento fino com relação ao folhedo, F = folhedo predominante com relação aosedimento fino, FA = predominância de folhedo com sedimento fino com relação à areia.

148 Curso de Campo Ecologia da Floresta Amazônica - 2001
DiscussãoO padrão de riqueza de morfoespécies encontrado neste
estudo, com poucas espécies comuns e várias espécies raras,é comum na natureza (Pianka 1994). O número de espéciesde gerrídeos nos igarapés da reserva provavelmente é aindamaior do que o registrado no presente estudo, uma vez que aárea total amostrada em cada igarapé foi pequena.
No que concerne à ausência de relação entre a ordem doigarapé e o número de morfoespécies de gerrídeos, é possívelque as variações ambientais existentes entre igarapés deprimeira a terceira ordens não sejam grandes o suficientepara gerar padrões distintos de composição de espécies.Dessa forma, as diferenças na riqueza observada demorfoespécies podem estar relacionadas a fatores que nãopuderam ser acessados a partir das variáveis medidas nestetrabalho. Fatores bióticos como pressão de predação ecompetição interespecífica poderiam ser responsáveis pelasdiferenças na riqueza e composição de espécies encontradasneste trabalho.
A ausência de relação entre as variáveis ambientais(largura, profundidade e velocidade da correnteza) e o númerode indivíduos nos grupos das morfoespécies sp.1 e sp.4podem estar relacionadas à acurácia das medidas.Microvariações na correnteza superficial poderiam gerarpadrões de distribuição de gerrídeos ao longo dos igarapés.
Quanto ao tipo de substrato, é possível que esse não exerçainfluência direta sobre os gerrídeos, mas reflitam apenas oresultado da dinâmica de deposição do material em funçãoda velocidade da correnteza. A hipótese de que a medida daproporção de substrato forneceria uma estimativa daquantidade de presas disponíveis para os gerrídeos, pode nãoser verdadeira. Nos trópicos, os recursos de origem terrestre,como insetos que caem na superfície da água, constituemuma das fontes alimentares mais importantes para os animaispresentes nos corpos d’água (Lowe-McConell 1999). Éprovável que o material alóctone, na forma de invertebradosque caem da floresta adjacente, constitua a maior parte dadieta destes gerrídeos. A variação espacial na disponibilidadedeste recurso talvez explique melhor a formação dos grupos.
Segundo Rimón (1999), peixes da coluna d’água devemnadar contra a correnteza para manter sua posição, com umcusto metabólico que aumenta exponencialmente com avelocidade da água. Da mesma maneira, o gasto energéticopara os gerrídeos se manterem em áreas de correnteza fortedeve ser muito alto. Realmente, os grupos não foramencontrados em áreas dos igarapés onde a água corria maisrapidamente.
O baixo número de indivíduos nos grupos encontradosem áreas de remanso pode estar relacionado à baixadisponibilidade de alimento nestes locais. Os animais só terãoacesso às presas que caírem na área de remanso que estãoocupando, enquanto em áreas de correnteza o fluxo dealimento é maior, já que as presas vêm carregadas pela água.Deve haver, portanto, um débito energético associado aobaixo aporte de alimento nas áreas de remanso.
Finalmente, acredito que deve ocorrer uma amplitude decondições adequadas para a ocupação de determinados locais
pelos gerrídeos, condições estas que devem exercer influênciasobre o tamanho dos grupos. Os gerrídeos provavalmenteprocuram áreas onde possam maximizar os benefíciosrelacionados à aquisição de determinado alimento em relaçãoaos custos envolvidos com a captura das presas, como preditopela teoria de forrageamento ótimo (Pianka 1994). É provávelque esta amplitude seja determinada pelo balanço entre ogasto de energia para manutenção da posição espacial eobtenção de alimento.
AgradecimentosAo Jansen pela ajuda no trabalho de campo, pelas
discussões, o cuidado e por não ter rido tanto assim dasminhas atolações. Ao palhaço Dadão goiabada por ter-mefeito rir muito ao longo deste mês e pelas sugestões para arealização deste trabalho, além do empréstimo de seupandeiro nota 10. Ao Juruna, por toda a informação quepassou ao longo deste mês. Ao Glauco, por ter ido comigoatrás das cigarrinhas, pelo cuidado com a correção dosrelatórios e por nos contar um pouco sobre o maravilhosomundo dos opiliões. A Natalie e ao Thiago, por ficaremesperando nos igarapés enquanto eu catava os gerrídeosfujões. A paula e a Natalie por darem sugestões para melhoraro texto. A Angelita, por ser tão empolgada e divertida, e porter-me acompanhado nas madrugadas a escrever relatórios.A dona Eduarda, ao Jorge e ao Osmaildo pela comidamaravilhosa. Ao Jorge por ter paciência conosco e acordarno meio da noite para desligar o gerador. Aos professoresque passaram alguns dias conosco e deixaram muito de si.Por fim, a todos os amigos que fiz ao longo deste mês.Saudades!
Referências BibliográficasBorror, D.J.; DeLong, D.M. & Triplehorn, C.A. 1981. An
Introduction to the Study of Insects. Saunders CollegePublishing, Philadelphia.
Downing, J.A. 1991. The effect of habitat structure on thespatial distribution of freshwater invertebratepopulations. In: Habitat Structure: The PhysicalArrangement of Objects in Space, Beel, S.S., McCoy,E.D. & Mushinsky, H.R, (eds.), pp 87-102. Chapmanand Hall, London.
Lowe-McConnell, R.H. 1999. Estudos ecológicos decomunidades de peixes tropicais. Edusp, São Paulo.
Merrit, R.W. & Cummins, K.W. 1996. An Introduction tothe Aquatic Insects of North America. Kendall/HuntPublishing Company, Iowa.
Pianka, E.R. 1994. Evolutionary Ecology. Harper CollinsCollege Publishers, New York.
Radambrasil, 1978. Levantamento de recursos naturais, vol.1-18. Ministério das Minas e Energia, Departamento deProdução Mineral, Rio de Janeiro.
Rincón, P.A. 1999. Uso do Micro-habitat em Peixes deRiachos: Métodos e Perspectivas. In: OecologiaBrasiliensis, vol. VI, Caramaschi, E.P., Mazzoni, R. &Peres-Neto, P.R. (eds.), pp 23-90. Rio de Janeiro.

Curso de Campo Ecologia da Floresta Amazônica - 2001 149
Interação entre formigas e Cecropia purpurascens (Cecropiaceae) na AmazôniaCentral: diferentes espécies protegem a planta com a mesma eficiência?
Rodrigo Cogni
IntroduçãoÁrvores do gênero Cecropia (Cecropiaceae) são
extremamente comuns em florestas abertas e áreasperturbadas da região neotropical. A maioria das espéciesdeste gênero apresentam interações mutualísticas comformigas. As formigas utilizam as cavidades naturaispresentes no tronco da planta como ninhos, além dealimentarem-se dos corpúsculos müllerianos produzidos naregião axilar do pecíolo (Janzen 1969). Ao forragearem pelaplanta as formigas retiram lianas e epífitas (Janzen 1969;Schupp 1986), além de repelirem herbívoros (Schupp 1986;Rocha & Bergallo 1992; Vasconcelos & Casimiro 1997).
Espécies do gênero Azteca são as mais comumenteencontradas nas associações com Cecropia spp. (Harada &Benson 1988), no entanto, outros gêneros (comoCamponotus, Crematogaster e Pachycondyla) podemtambém ser encontradas associadas (Yu & Davidson 1997).Normalmente uma população de Cecropia apresentaindivíduos colonizados por diferentes espécies de formigas(Yu & Davidson 1997) e, consequentemente, a proteçãoconferida por cada espécie pode diferir, como ocorre emmuitas interações mutualísticas (Bronstein 1994).
Na região de Manaus, a espécie Cecropia purpurascensé frequente (Harada & Benson 1988; Faveri 2000), sendocolonizada principalmente por duas espécies de formigas:Camponotus balzani e Azteca alfari (Bonato 2000). Nestelocal, plantas com altura maior que 1,5 m estão quase semprecolonizadas e a proporção de indivíduos colonizados pelasduas espécies mais comum (C. balzani e A. alfari) é igual.Além disso, estas duas espécies de formigas utilizam plantascom as mesmas características (como altura, volume dadomácea e quantidade de corpúsculos) (Bonato 2000). Estasduas espécies devem apresentar características como tamanhodo corpo, agressividade e intensidade de forrageamento sobrea planta diferentes, que podem causas diferenças na eficiênciada proteção conferida à planta.
O objetivo deste trabalho foi investigar diferenças naproteção à plantas de C. purpurascens por A. alfari e C.balzani. As seguintes perguntas foram formuladas: (i) a taxade herbivoria sobre plantas colonizadas por C. balzani diferedas colonizadas por A. alfari? (ii) as duas espécies deformigas atacam herbívoros simulados nas folhas de C.purpurascens? (iii) o tempo de detecção dos herbívorossimulados difere para as duas espécies de formigas?
Material e MétodosRealizei o trabalho na Reserva do Km 41, uma área de
mata contínua do Projeto Dinâmica Biológica de FragmentosFlorestais (PDBFF/INPA) localizada a 70 Km ao norte deManaus (02º24’S; 59º44’W), com altitude entre 50 e 150 m.A temperatura média no local é de 26,7ºC, e a precipitação
anual é de 2.186 mm (Lovejoy & Bierregaard 1991). Realizeio trabalho no mês de agosto, na estação seca, que vai dejunho a setembro.
Percorri 20 Km ao longo da estrada ZF-3, marcandoindivíduos de C. purpurascens com até 4 metros de alturacolonizados por Azteca alfari (n = 15) e por Camponotusbalzani (n = 15). Para determinar a herbivoria nas folhasdestes indivíduos utilizei o método de Dirzo & Domingues(1995). Para cada indivíduo, determinei aleatoriamente cincofolhas, atribuindo a cada uma destas uma categoria de acordocom a porcentagem da área foliar danificada por herbívoros:0 = folhas sem marcas de herbivoria, 1= folhas com marcasde herbivoria atingindo entre 1 e 5% da superfície da folha,2 = entre 6 e 12%, 3 = entre 13 e 25%, 4 = entre 26 e 50% e5 = entre 51 e 100%. Para determinar o índice de herbivoriapor indivíduo de C. purpurascens, calculei a média das cincofolhas.
Para comparar o comportamento das duas espécies deformiga frente a herbívoros simulados, realizei experimentoscolando um operário vivo de cupim (Nasutitermes sp.) porplanta (veja Oliveira et al. 1987). Colei o cupim pelo dorso,no centro da face adaxial da lâmina da quarta folha basal.Aparentemente, as formigas não foram atraídas ou repelidaspela cola. Observei o cupim por 15 minutos, anotando sehouve ataque pelas formigas e o tempo necessário para aprimeira formiga tocar a isca.
Para comparar a atividade de forrageamento das duasespécies de formigas nas folhas de C. purpurascens, marqueiquatro plantas colonizadas por A. alfari e quatro colonizadospor C. balzani. Escolhi plantas com tamanho e número defolhas semelhantes. Nestes indivíduos realizei quatroamostragens ao longo do dia (8h, 12h, 16h e 20h), nas quaiscontei o número de operárias forrageando em todas as folhasda planta.
Comparei o índice mediano de herbivoria dos indivíduosde C. purpurascens colonizados por A. alfari com oscolonizados por C. balzani através do teste não paramétricode Mann-Whitney. As proporções entre cupins atacados enão atacados pelas duas espécies foram comparadas por umc2 de contingência. O tempo mediano necessário para cadaespécie detectar o cupim foi comparado com o teste nãoparamétrico de Mann-Whitney. Realizei as análises noprograma Systat 8.0, como especificado em Zar (1999),utilizando nível de significância de 5%.
ResultadosTodas as plantas observadas apresentaram marcas de danos
causados por herbívoros. A herbivoria observada em plantascolonizadas por Camponotus balzani (mediana entre 5 e12%) foi maior que em plantas colonizadas por Azteca alfari(mediana entre 1 e 5%) (Mann-Whitney, U = 49; n
1 = n
2=
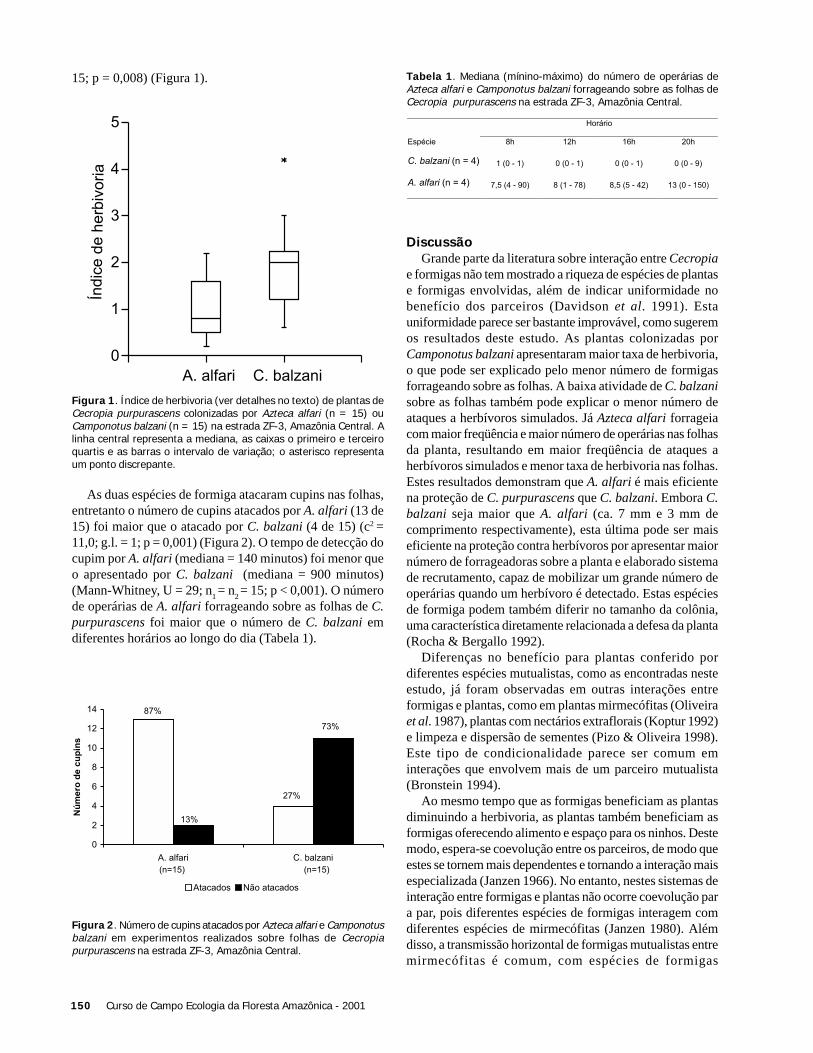
150 Curso de Campo Ecologia da Floresta Amazônica - 2001
15; p = 0,008) (Figura 1).
A. alfari C. balzani0
1
2
3
4
5
Índ
ice
de
he
rbiv
ori
a
Figura 1. Índice de herbivoria (ver detalhes no texto) de plantas deCecropia purpurascens colonizadas por Azteca alfari (n = 15) ouCamponotus balzani (n = 15) na estrada ZF-3, Amazônia Central. Alinha central representa a mediana, as caixas o primeiro e terceiroquartis e as barras o intervalo de variação; o asterisco representaum ponto discrepante.
As duas espécies de formiga atacaram cupins nas folhas,entretanto o número de cupins atacados por A. alfari (13 de15) foi maior que o atacado por C. balzani (4 de 15) (c2 =11,0; g.l. = 1; p = 0,001) (Figura 2). O tempo de detecção docupim por A. alfari (mediana = 140 minutos) foi menor queo apresentado por C. balzani (mediana = 900 minutos)(Mann-Whitney, U = 29; n
1 = n
2 = 15; p < 0,001). O número
de operárias de A. alfari forrageando sobre as folhas de C.purpurascens foi maior que o número de C. balzani emdiferentes horários ao longo do dia (Tabela 1).
0
2
4
6
8
10
12
14
A. alfari C. balzani
Nú
me
rod
ec
up
ins
Atacados Não atacados
(n=15) (n=15)
87%
73%
27%
13%
Figura 2. Número de cupins atacados por Azteca alfari e Camponotusbalzani em experimentos realizados sobre folhas de Cecropiapurpurascens na estrada ZF-3, Amazônia Central.
Tabela 1. Mediana (mínino-máximo) do número de operárias deAzteca alfari e Camponotus balzani forrageando sobre as folhas deCecropia purpurascens na estrada ZF-3, Amazônia Central.
Horário
Espécie 8h 12h 16h 20h
C. balzani (n = 4) 1 (0 - 1) 0 (0 - 1) 0 (0 - 1) 0 (0 - 9)
A. alfari (n = 4) 7,5 (4 - 90) 8 (1 - 78) 8,5 (5 - 42) 13 (0 - 150)
DiscussãoGrande parte da literatura sobre interação entre Cecropia
e formigas não tem mostrado a riqueza de espécies de plantase formigas envolvidas, além de indicar uniformidade nobenefício dos parceiros (Davidson et al. 1991). Estauniformidade parece ser bastante improvável, como sugeremos resultados deste estudo. As plantas colonizadas porCamponotus balzani apresentaram maior taxa de herbivoria,o que pode ser explicado pelo menor número de formigasforrageando sobre as folhas. A baixa atividade de C. balzanisobre as folhas também pode explicar o menor número deataques a herbívoros simulados. Já Azteca alfari forrageiacom maior freqüência e maior número de operárias nas folhasda planta, resultando em maior freqüência de ataques aherbívoros simulados e menor taxa de herbivoria nas folhas.Estes resultados demonstram que A. alfari é mais eficientena proteção de C. purpurascens que C. balzani. Embora C.balzani seja maior que A. alfari (ca. 7 mm e 3 mm decomprimento respectivamente), esta última pode ser maiseficiente na proteção contra herbívoros por apresentar maiornúmero de forrageadoras sobre a planta e elaborado sistemade recrutamento, capaz de mobilizar um grande número deoperárias quando um herbívoro é detectado. Estas espéciesde formiga podem também diferir no tamanho da colônia,uma característica diretamente relacionada a defesa da planta(Rocha & Bergallo 1992).
Diferenças no benefício para plantas conferido pordiferentes espécies mutualistas, como as encontradas nesteestudo, já foram observadas em outras interações entreformigas e plantas, como em plantas mirmecófitas (Oliveiraet al. 1987), plantas com nectários extraflorais (Koptur 1992)e limpeza e dispersão de sementes (Pizo & Oliveira 1998).Este tipo de condicionalidade parece ser comum eminterações que envolvem mais de um parceiro mutualista(Bronstein 1994).
Ao mesmo tempo que as formigas beneficiam as plantasdiminuindo a herbivoria, as plantas também beneficiam asformigas oferecendo alimento e espaço para os ninhos. Destemodo, espera-se coevolução entre os parceiros, de modo queestes se tornem mais dependentes e tornando a interação maisespecializada (Janzen 1966). No entanto, nestes sistemas deinteração entre formigas e plantas não ocorre coevolução para par, pois diferentes espécies de formigas interagem comdiferentes espécies de mirmecófitas (Janzen 1980). Alémdisso, a transmissão horizontal de formigas mutualistas entremirmecófitas é comum, com espécies de formigas

Curso de Campo Ecologia da Floresta Amazônica - 2001 151
colonizando novas espécies de plantas ao longo do tempoevolutivo (Yu & Davidson 1997). Neste contexto, C. balzanipode representar um parceiro de C. purpurascens maisrecente no tempo evolutivo que A. alfari e por isso não sertão eficiente na proteção da planta.
Visto que existem diferenças na proteção por diferentesespécies de formigas, pode-se esperar que as plantasselecionem os parceiros mais eficientes ao longo do tempoevolutivo. Algumas das possíveis formas das plantasselecionarem as espécies de formigas é limitando o tamanhoda entrada e o volume da domácea (Fonseca 1995) eapresentando pêlos e tricomas que limitam o tamanho dasformigas que podem andar sobre as folhas (Davidson et al.1989). Em C. purpurascens colônias maduras das duasespécies de formiga (C. balzani e A. alfari) são encontradasem plantas com mesmas características (Bonato 2000). Noentanto seria interessante investigar a influência decaracterísticas da planta (como tricomas e tamanho da entradae volume da domácea) no sucesso de fundação de colôniasdestas duas espécies.
AgradecimentosAgradeço a Betsabé R. Guerra, Glauco Machado e Dadão
Venticique pela ajuda no trabalho de campo. Tambémagradeço aos colegas e professores do curso Ecologia daFloresta Amazônica 2001 pelas discussões, dicas e sugestões.G. Machado, E. Bruna III, E. M. Venticinque e J. Zuanonrevisaram e melhoraram a primeira versão.
Referências BibliográficasBonato, V. 2000. Colonização de C. purpurascens
(Cecropioaceae) por formigas na reserva do Km 41. Livrodo Curso de Campo “Ecologia da Floresta Amazônica”,INPA/Smithsonian Institution, Manaus, AM.
Bronstein, J.L. 1994. Conditional outcomes in mutualisticinteractions. TREE 9: 214-216.
Davidson, D.W.; Snelling, R.R. & Longino, J.T. 1989.Competition among ants for mirmecophytes and thesignificance of plant trichomes. Biotropica 21: 64-73.
Davidson, D.W.; Foster, R.B.; Snelling, R.R. & Lozada, P.W.1991. Variable composition of some tropical ant-plantsymbioses. In: Plant-Animal Interactions: EvolutionaryEcology in Tropical and Temperate Regions, Price, P.W.,Lewinsohn, T.M., Fernandes, G.W. & Benson, W.W.(eds.), pp. 145-162. John Wiley & Sons, New York.
Dirzo, R. & Dominguez, C. 1995. Plant-animal interactionsin Mesoamerican Tropical dry forest. In: Seasonally DryTropical Forests, Bullock, E., Medina, E. & Mooney,E. (eds). Cambridge University Press, Cambridge.
Faveri, S.B. 2000. Riqueza e composição de comuniadde de
artrópodos associados a Cecropia spp. Dissertação deMestrado, INPA/UA, Manaus, AM.
Fonseca, C.R. 1995. Evolutionary ecology of Amazonianant-myrmecophyte mutualisms. PhD Thesis. Universityof Oxford, Oxford.
Harada, A.Y. & Benson, W.W. 1988. Espécies de Azteca(Hymenoptera: Formicidae) especializadas em Cecropiaspp. (Moraceae): distribuição geográfica e consideraçõesecológicas. Revta. Bras. Entomol. 32: 423-435.
Janzen, D.H. 1966. Coevolution of mutualism between antsand Acacia in Central America. Evolution 20: 249-275.
Janzen, D.H. 1969. Allelopathy by myrmecophytes: the antAzteca as an allelopathic agent of Cecropia. Ecology50: 147-153.
Janzen, D.H. 1980. When is it coevolution? Evolution 34:611-612.
Koptur, S. 1992. Extrafloral nectary-mediated interactionsbetween insects and plants. Insect-Plant Interactions. E.Bernays (ed.), 4: 81-129. CRC Press, Boca Raton.
Lovejoy, T.E. & Bierregaard, R.O. 1991. Central AmazonianForests and minimal critical size of ecossystem project.In: Four Neotropical Rain Forests, Gentry, A. H. (ed.).Yale University Press, New Haven.
Oliveira, P.S.; Silva, A.F. & Martins, A.B. 1987. Ant foragingon extrafloral nectaries of Qualea grandiflora(Vochysiaceae) in cerrado vegetation: ants as potentialantiherbivore agents. Oecologia 74: 228-230.
Oliveira, P.S.; Oliveira-Filho, A.T. & Cintra, R. 1987. Antforaging on ant-inhabited Triplaris (Polygonaceae) inwestern Brazil: a field experiment using live termites-baits. J. Trop. Ecol. 3: 193-200.
Pizo, M.A. & Oliveira, P.S. 1998. Interaction between antsand seeds of a nonmyrmecochorous neotropical tree,Cabralea canjerana (Meliaceae), in the Atlantic forestof southeast Brazil. Am. J. Bot. 85: 669-674.
Rocha, C.F.D. & Bergallo, H.G. 1992. Bigger ant coloniesreduce herbivory and herbivore residence time on leavesof an ant-plant: Azteca muelleri vs. Coelomera ruficornison Cecropia pachystachya. Oecologia 91: 249-252.
Schupp, E.W. 1986. Azteca protection of Cecropia: antoccupation benefits juvenile trees. Oecologia 70: 379-385.
Vasconcelos, H.L. & Casimiro, A.B. 1997. Influence ofAzteca alfari ants on the exploitation of Cecropia treesby a leaf-cutting ant. Biotropica 29: 84-92.
Yu, D.W. & Davidson, D.W. 1997. Experimental studies ofspecies-specifity in Cecropia-ant relationships. Ecol.Monog. 67: 273-294.
Zar, J.H. 1999. Bioestatistical Analysis. Prentice Hall, NewJersey.

152 Curso de Campo Ecologia da Floresta Amazônica - 2001
Interação entre formigas e Cecropia purpurascens (Cecropiaceae) na AmazôniaCentral: diferentes espécies protegem a planta com a mesma eficiência?
Rodrigo Cogni
IntroduçãoÁrvores do gênero Cecropia (Cecropiaceae) são
extremamente comuns em florestas abertas e áreasperturbadas da região neotropical. A maioria das espéciesdeste gênero apresentam interações mutualísticas comformigas. As formigas utilizam as cavidades naturaispresentes no tronco da planta como ninhos, além dealimentarem-se dos corpúsculos müllerianos produzidos naregião axilar do pecíolo (Janzen 1969). Ao forragearem pelaplanta as formigas retiram lianas e epífitas (Janzen 1969;Schupp 1986), além de repelirem herbívoros (Schupp 1986;Rocha & Bergallo 1992; Vasconcelos & Casimiro 1997).
Espécies do gênero Azteca são as mais comumenteencontradas nas associações com Cecropia spp. (Harada &Benson 1988), no entanto, outros gêneros (comoCamponotus, Crematogaster e Pachycondyla) podemtambém ser encontradas associadas (Yu & Davidson 1997).Normalmente uma população de Cecropia apresentaindivíduos colonizados por diferentes espécies de formigas(Yu & Davidson 1997) e, consequentemente, a proteçãoconferida por cada espécie pode diferir, como ocorre emmuitas interações mutualísticas (Bronstein 1994).
Na região de Manaus, a espécie Cecropia purpurascensé frequente (Harada & Benson 1988; Faveri 2000), sendocolonizada principalmente por duas espécies de formigas:Camponotus balzani e Azteca alfari (Bonato 2000). Nestelocal, plantas com altura maior que 1,5 m estão quase semprecolonizadas e a proporção de indivíduos colonizados pelasduas espécies mais comum (C. balzani e A. alfari) é igual.Além disso, estas duas espécies de formigas utilizam plantascom as mesmas características (como altura, volume dadomácea e quantidade de corpúsculos) (Bonato 2000). Estasduas espécies devem apresentar características como tamanhodo corpo, agressividade e intensidade de forrageamento sobrea planta diferentes, que podem causas diferenças na eficiênciada proteção conferida à planta.
O objetivo deste trabalho foi investigar diferenças naproteção à plantas de C. purpurascens por A. alfari e C.balzani. As seguintes perguntas foram formuladas: (i) a taxade herbivoria sobre plantas colonizadas por C. balzani diferedas colonizadas por A. alfari? (ii) as duas espécies deformigas atacam herbívoros simulados nas folhas de C.purpurascens? (iii) o tempo de detecção dos herbívorossimulados difere para as duas espécies de formigas?
Material e MétodosRealizei o trabalho na Reserva do Km 41, uma área de
mata contínua do Projeto Dinâmica Biológica de FragmentosFlorestais (PDBFF/INPA) localizada a 70 Km ao norte deManaus (02º24’S; 59º44’W), com altitude entre 50 e 150 m.A temperatura média no local é de 26,7ºC, e a precipitação
anual é de 2.186 mm (Lovejoy & Bierregaard 1991). Realizeio trabalho no mês de agosto, na estação seca, que vai dejunho a setembro.
Percorri 20 Km ao longo da estrada ZF-3, marcandoindivíduos de C. purpurascens com até 4 metros de alturacolonizados por Azteca alfari (n = 15) e por Camponotusbalzani (n = 15). Para determinar a herbivoria nas folhasdestes indivíduos utilizei o método de Dirzo & Domingues(1995). Para cada indivíduo, determinei aleatoriamente cincofolhas, atribuindo a cada uma destas uma categoria de acordocom a porcentagem da área foliar danificada por herbívoros:0 = folhas sem marcas de herbivoria, 1= folhas com marcasde herbivoria atingindo entre 1 e 5% da superfície da folha,2 = entre 6 e 12%, 3 = entre 13 e 25%, 4 = entre 26 e 50% e5 = entre 51 e 100%. Para determinar o índice de herbivoriapor indivíduo de C. purpurascens, calculei a média das cincofolhas.
Para comparar o comportamento das duas espécies deformiga frente a herbívoros simulados, realizei experimentoscolando um operário vivo de cupim (Nasutitermes sp.) porplanta (veja Oliveira et al. 1987). Colei o cupim pelo dorso,no centro da face adaxial da lâmina da quarta folha basal.Aparentemente, as formigas não foram atraídas ou repelidaspela cola. Observei o cupim por 15 minutos, anotando sehouve ataque pelas formigas e o tempo necessário para aprimeira formiga tocar a isca.
Para comparar a atividade de forrageamento das duasespécies de formigas nas folhas de C. purpurascens, marqueiquatro plantas colonizadas por A. alfari e quatro colonizadospor C. balzani. Escolhi plantas com tamanho e número defolhas semelhantes. Nestes indivíduos realizei quatroamostragens ao longo do dia (8h, 12h, 16h e 20h), nas quaiscontei o número de operárias forrageando em todas as folhasda planta.
Comparei o índice mediano de herbivoria dos indivíduosde C. purpurascens colonizados por A. alfari com oscolonizados por C. balzani através do teste não paramétricode Mann-Whitney. As proporções entre cupins atacados enão atacados pelas duas espécies foram comparadas por umc2 de contingência. O tempo mediano necessário para cadaespécie detectar o cupim foi comparado com o teste nãoparamétrico de Mann-Whitney. Realizei as análises noprograma Systat 8.0, como especificado em Zar (1999),utilizando nível de significância de 5%.
ResultadosTodas as plantas observadas apresentaram marcas de danos
causados por herbívoros. A herbivoria observada em plantascolonizadas por Camponotus balzani (mediana entre 5 e12%) foi maior que em plantas colonizadas por Azteca alfari(mediana entre 1 e 5%) (Mann-Whitney, U = 49; n
1 = n
2=
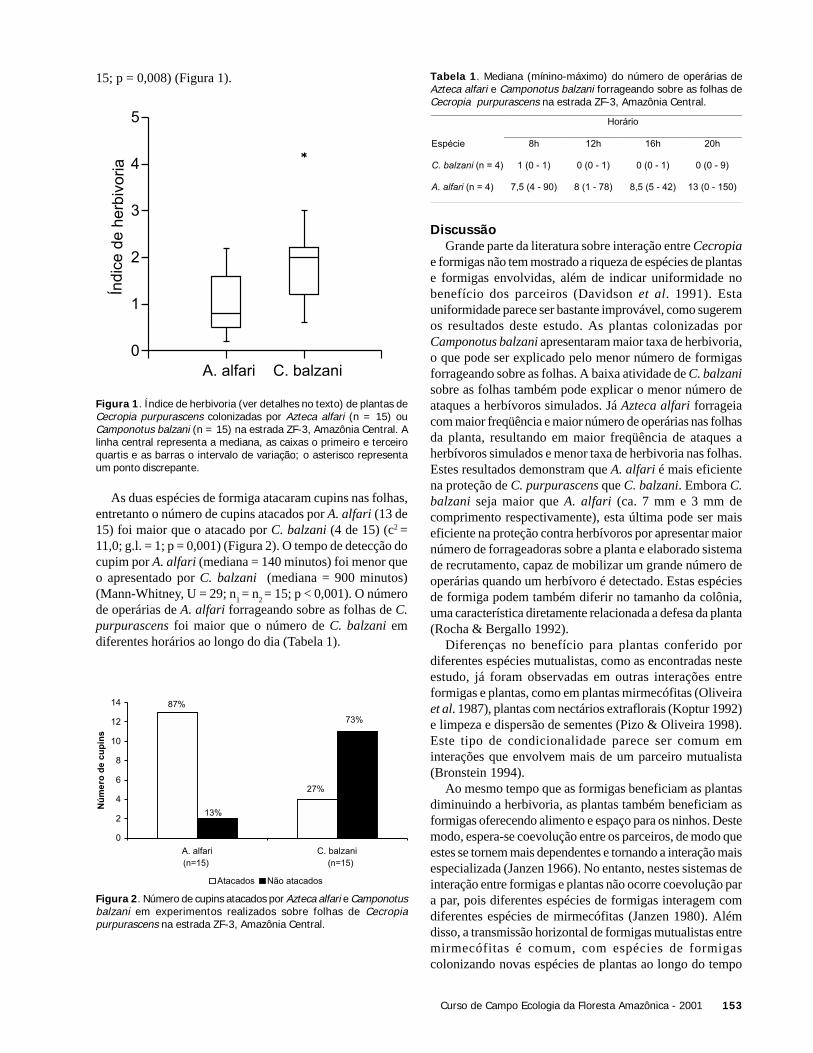
Curso de Campo Ecologia da Floresta Amazônica - 2001 153
15; p = 0,008) (Figura 1).
A. alfari C. balzani0
1
2
3
4
5Ín
dic
ed
eh
erb
ivo
ria
Figura 1. Índice de herbivoria (ver detalhes no texto) de plantas deCecropia purpurascens colonizadas por Azteca alfari (n = 15) ouCamponotus balzani (n = 15) na estrada ZF-3, Amazônia Central. Alinha central representa a mediana, as caixas o primeiro e terceiroquartis e as barras o intervalo de variação; o asterisco representaum ponto discrepante.
As duas espécies de formiga atacaram cupins nas folhas,entretanto o número de cupins atacados por A. alfari (13 de15) foi maior que o atacado por C. balzani (4 de 15) (c2 =11,0; g.l. = 1; p = 0,001) (Figura 2). O tempo de detecção docupim por A. alfari (mediana = 140 minutos) foi menor queo apresentado por C. balzani (mediana = 900 minutos)(Mann-Whitney, U = 29; n
1 = n
2 = 15; p < 0,001). O número
de operárias de A. alfari forrageando sobre as folhas de C.purpurascens foi maior que o número de C. balzani emdiferentes horários ao longo do dia (Tabela 1).
0
2
4
6
8
10
12
14
A. alfari C. balzani
Nú
me
rod
ec
up
ins
Atacados Não atacados
(n=15) (n=15)
87%
73%
27%
13%
Figura 2. Número de cupins atacados por Azteca alfari e Camponotusbalzani em experimentos realizados sobre folhas de Cecropiapurpurascens na estrada ZF-3, Amazônia Central.
Tabela 1. Mediana (mínino-máximo) do número de operárias deAzteca alfari e Camponotus balzani forrageando sobre as folhas deCecropia purpurascens na estrada ZF-3, Amazônia Central.
Horário
Espécie 8h 12h 16h 20h
C. balzani (n = 4) 1 (0 - 1) 0 (0 - 1) 0 (0 - 1) 0 (0 - 9)
A. alfari (n = 4) 7,5 (4 - 90) 8 (1 - 78) 8,5 (5 - 42) 13 (0 - 150)
DiscussãoGrande parte da literatura sobre interação entre Cecropia
e formigas não tem mostrado a riqueza de espécies de plantase formigas envolvidas, além de indicar uniformidade nobenefício dos parceiros (Davidson et al. 1991). Estauniformidade parece ser bastante improvável, como sugeremos resultados deste estudo. As plantas colonizadas porCamponotus balzani apresentaram maior taxa de herbivoria,o que pode ser explicado pelo menor número de formigasforrageando sobre as folhas. A baixa atividade de C. balzanisobre as folhas também pode explicar o menor número deataques a herbívoros simulados. Já Azteca alfari forrageiacom maior freqüência e maior número de operárias nas folhasda planta, resultando em maior freqüência de ataques aherbívoros simulados e menor taxa de herbivoria nas folhas.Estes resultados demonstram que A. alfari é mais eficientena proteção de C. purpurascens que C. balzani. Embora C.balzani seja maior que A. alfari (ca. 7 mm e 3 mm decomprimento respectivamente), esta última pode ser maiseficiente na proteção contra herbívoros por apresentar maiornúmero de forrageadoras sobre a planta e elaborado sistemade recrutamento, capaz de mobilizar um grande número deoperárias quando um herbívoro é detectado. Estas espéciesde formiga podem também diferir no tamanho da colônia,uma característica diretamente relacionada a defesa da planta(Rocha & Bergallo 1992).
Diferenças no benefício para plantas conferido pordiferentes espécies mutualistas, como as encontradas nesteestudo, já foram observadas em outras interações entreformigas e plantas, como em plantas mirmecófitas (Oliveiraet al. 1987), plantas com nectários extraflorais (Koptur 1992)e limpeza e dispersão de sementes (Pizo & Oliveira 1998).Este tipo de condicionalidade parece ser comum eminterações que envolvem mais de um parceiro mutualista(Bronstein 1994).
Ao mesmo tempo que as formigas beneficiam as plantasdiminuindo a herbivoria, as plantas também beneficiam asformigas oferecendo alimento e espaço para os ninhos. Destemodo, espera-se coevolução entre os parceiros, de modo queestes se tornem mais dependentes e tornando a interação maisespecializada (Janzen 1966). No entanto, nestes sistemas deinteração entre formigas e plantas não ocorre coevolução para par, pois diferentes espécies de formigas interagem comdiferentes espécies de mirmecófitas (Janzen 1980). Alémdisso, a transmissão horizontal de formigas mutualistas entremirmecófitas é comum, com espécies de formigascolonizando novas espécies de plantas ao longo do tempo

154 Curso de Campo Ecologia da Floresta Amazônica - 2001
evolutivo (Yu & Davidson 1997). Neste contexto, C. balzanipode representar um parceiro de C. purpurascens maisrecente no tempo evolutivo que A. alfari e por isso não sertão eficiente na proteção da planta.
Visto que existem diferenças na proteção por diferentesespécies de formigas, pode-se esperar que as plantasselecionem os parceiros mais eficientes ao longo do tempoevolutivo. Algumas das possíveis formas das plantasselecionarem as espécies de formigas é limitando o tamanhoda entrada e o volume da domácea (Fonseca 1995) eapresentando pêlos e tricomas que limitam o tamanho dasformigas que podem andar sobre as folhas (Davidson et al.1989). Em C. purpurascens colônias maduras das duasespécies de formiga (C. balzani e A. alfari) são encontradasem plantas com mesmas características (Bonato 2000). Noentanto seria interessante investigar a influência decaracterísticas da planta (como tricomas e tamanho da entradae volume da domácea) no sucesso de fundação de colôniasdestas duas espécies.
AgradecimentosAgradeço a Betsabé R. Guerra, Glauco Machado e Dadão
Venticique pela ajuda no trabalho de campo. Tambémagradeço aos colegas e professores do curso Ecologia daFloresta Amazônica 2001 pelas discussões, dicas e sugestões.G. Machado, E. Bruna III, E. M. Venticinque e J. Zuanonrevisaram e melhoraram a primeira versão.
Referências BibliográficasBonato, V. 2000. Colonização de C. purpurascens
(Cecropioaceae) por formigas na reserva do Km 41. Livrodo Curso de Campo “Ecologia da Floresta Amazônica”,INPA/Smithsonian Institution, Manaus, AM.
Bronstein, J.L. 1994. Conditional outcomes in mutualisticinteractions. TREE 9: 214-216.
Davidson, D.W.; Snelling, R.R. & Longino, J.T. 1989.Competition among ants for mirmecophytes and thesignificance of plant trichomes. Biotropica 21: 64-73.
Davidson, D.W.; Foster, R.B.; Snelling, R.R. & Lozada, P.W.1991. Variable composition of some tropical ant-plantsymbioses. In: Plant-Animal Interactions: EvolutionaryEcology in Tropical and Temperate Regions, Price, P.W.,Lewinsohn, T.M., Fernandes, G.W. & Benson, W.W.(eds.), pp. 145-162. John Wiley & Sons, New York.
Dirzo, R. & Dominguez, C. 1995. Plant-animal interactionsin Mesoamerican Tropical dry forest. In: Seasonally DryTropical Forests, Bullock, E., Medina, E. & Mooney,E. (eds). Cambridge University Press, Cambridge.
Faveri, S.B. 2000. Riqueza e composição de comuniadde deartrópodos associados a Cecropia spp. Dissertação de
Mestrado, INPA/UA, Manaus, AM.Fonseca, C.R. 1995. Evolutionary ecology of Amazonian
ant-myrmecophyte mutualisms. PhD Thesis. Universityof Oxford, Oxford.
Harada, A.Y. & Benson, W.W. 1988. Espécies de Azteca(Hymenoptera: Formicidae) especializadas em Cecropiaspp. (Moraceae): distribuição geográfica e consideraçõesecológicas. Revta. Bras. Entomol. 32: 423-435.
Janzen, D.H. 1966. Coevolution of mutualism between antsand Acacia in Central America. Evolution 20: 249-275.
Janzen, D.H. 1969. Allelopathy by myrmecophytes: the antAzteca as an allelopathic agent of Cecropia. Ecology50: 147-153.
Janzen, D.H. 1980. When is it coevolution? Evolution 34:611-612.
Koptur, S. 1992. Extrafloral nectary-mediated interactionsbetween insects and plants. Insect-Plant Interactions. E.Bernays (ed.), 4: 81-129. CRC Press, Boca Raton.
Lovejoy, T.E. & Bierregaard, R.O. 1991. Central AmazonianForests and minimal critical size of ecossystem project.In: Four Neotropical Rain Forests, Gentry, A. H. (ed.).Yale University Press, New Haven.
Oliveira, P.S.; Silva, A.F. & Martins, A.B. 1987. Ant foragingon extrafloral nectaries of Qualea grandiflora(Vochysiaceae) in cerrado vegetation: ants as potentialantiherbivore agents. Oecologia 74: 228-230.
Oliveira, P.S.; Oliveira-Filho, A.T. & Cintra, R. 1987. Antforaging on ant-inhabited Triplaris (Polygonaceae) inwestern Brazil: a field experiment using live termites-baits. J. Trop. Ecol. 3: 193-200.
Pizo, M.A. & Oliveira, P.S. 1998. Interaction between antsand seeds of a nonmyrmecochorous neotropical tree,Cabralea canjerana (Meliaceae), in the Atlantic forestof southeast Brazil. Am. J. Bot. 85: 669-674.
Rocha, C.F.D. & Bergallo, H.G. 1992. Bigger ant coloniesreduce herbivory and herbivore residence time on leavesof an ant-plant: Azteca muelleri vs. Coelomera ruficornison Cecropia pachystachya. Oecologia 91: 249-252.
Schupp, E.W. 1986. Azteca protection of Cecropia: antoccupation benefits juvenile trees. Oecologia 70: 379-385.
Vasconcelos, H.L. & Casimiro, A.B. 1997. Influence ofAzteca alfari ants on the exploitation of Cecropia treesby a leaf-cutting ant. Biotropica 29: 84-92.
Yu, D.W. & Davidson, D.W. 1997. Experimental studies ofspecies-specifity in Cecropia-ant relationships. Ecol.Monog. 67: 273-294.
Zar, J.H. 1999. Bioestatistical Analysis. Prentice Hall, NewJersey.

Curso de Campo Ecologia da Floresta Amazônica - 2001 155
Influência da abertura de estradas nos padrões do uso de áreas e composição dacomunidade de mamíferos de médio a grande porte numa área de floresta de
terra-firme na Amazônia CentralThiago P. C. Timo
IntroduçãoO impacto da criação de estradas em áreas de florestas é
pouco conhecida, ainda que certamente causadora deperturbações na dinâmica das atividades da fauna local, alémde características da flora e do uso de solos já que estasestradas dão acesso a áreas inexploradas e, no caso daAmazônia brasileira, com grande número de espécies arbóreasde interesse comercial, animais de caça, e solos, os quaisainda que inapropriados para cultivos e pastagens, sãoexplorados para roçado e criação de gado bovino (Fearnside1989).
Este felizmente não é o caso da área em estudo, porém apresença de uma estrada oferece a oportunidade de observar-se o impacto desta alteração na paisagem sobre os padrõesdo uso de área e composição das espécies de mamíferos demédio a grande porte. Estes animais deslocam-se por grandesdistâncias e devem fazer uso oportunista de trilhas e estradasnas suas rotas de deslocamento, como observadoempiricamente através do grande número de pegadas degrandes mamíferos em estradas e da maioria dos trabalhosde coleta de fezes de grandes carnívoros ser realizada nasmesmas (e.g. Leite 1995).
Na realização de estudos com mamíferos de médio egrande porte, a maior dificuldade deve-se aos hábitosreservados, e portanto a baixa frequência em que são vistosem estudos de senso visual (Voss & Emmons 1996). Portantopara a confecção de inventários de mamíferos terrestres demédio e grande porte ficou comprovado nos últimos anosque o método mais eficiente é o uso da contagem de pegadas(Bekcer & Dalponte 1991), preferencialmente utilizando-secamadas finas de areia colocadas em áreas pequenaspreviamente limpas.
A presente proposta tem o objetivo de realizar umaavaliação preliminar das possíveis influências da aberturade estradas em áreas de floresta de terra-firme na AmazôniaCentral sobre a composição da comunidade dos mamíferosterrestres de médio a grande porte em áreas de estrada e demata contínua e seus padrões de uso destas áreas.
Material e MétodosA Reserva do Km 41 do Projeto Dinâmica Biológica de
Fragmentos Florestais (INPA/Smithsonian) situa-se aaproximadamente 70 Km a Norte de Manaus (2o 30’S; 60o
00’W), a aproximadamente 100 m do nível do mar. O climasegundo Köeppen é tropical úmido, com temperatura médiade 26,7oC e pluviosidade média anual de 2186 mm. Os mesesmais chuvosos são março e abril (média de 300 mm/mês) e
os menos chuvosos julho, agosto e setembro com médiapluviométrica de 100 mm/mês (Leite 1995).
Para a observação do uso das estradas pelas diferentesespécies de mamíferos terrestres de médio a grande porte,foram instaladas 15 de armadilhas de pegadas, constituídasde áreas cobertas com camadas finas de areia dispostas aolargo de uma trilha na mata contínua e em dois segmentos daestrada, um em franco uso e outro abandonado. Cincoarmadilhas em cada local a um espaçamento de 200 m entreas mesmas, em plots de 70 cm x 70 cm cada uma, iscadasalternadamente com bacon e sal e banana e sal, sendo aprimeira de uma série de cinco iscada com ambos os tipos deisca.
Devido à falta de registros obtidos nas fases iniciais dopresente estudo, (o que será posteriormente apresentado ediscutido), foi adotada a estratégia de procura ativa deevidências indiretas da presença e atividade de mamíferosna área da Reserva do Km 41. As evidências buscadas foramfezes e restos alimentares deixados em baixios. Para tantoforam percorridos trechos de 100 m em sete igarapés situadosnas trilhas L5, L10, Q13, Q17, Q19, T12 e D13, onde foramrecolhidos frutos com marcas de predação, tais comomordeduras ou roimentos, destes frutos foram medidos ocomprimento, a largura e a espessura da casca, foramidentificados em nível de família. Os baixios tiveram aslarguras medidas e uma análise de regressão linear (Campbell1989) foi efetuada relacinando o número de frutos coletadosà largura do baixio. Também foram recolhidas duas amostrasde fezes de onça (Panthera onca) na estrada próximo àporteira da trilha que leva ao acampamento e duas amostrasde fezes de anta (Tapirus terrestris) de diferentes idades ,uma fresca e outra seca, numa lagoa na trilha R24.
ResultadosNos três dias onde as armadilhas foram visitadas
regularmente e as iscas substituídas a cada visita, não houveregistro de nenhum mamífero de pequeno, médio ou grandeporte, apenas evidências de ataque às iscas por lagartos e apresença constante de grande número de formigas. Em funçãodestes resultados, o uso das armadilhas foi abandonado epassou-se a utilizar as demais evidências recolhidas na estradae nos baixios.
Os frutos coletados nos sete igarapés foram agrupadosem cinco categorias de predação para possível inferênciaquanto ao taxon que o utiliza. As dimensões médias dosfrutos são apresentadas na Tabela 1.
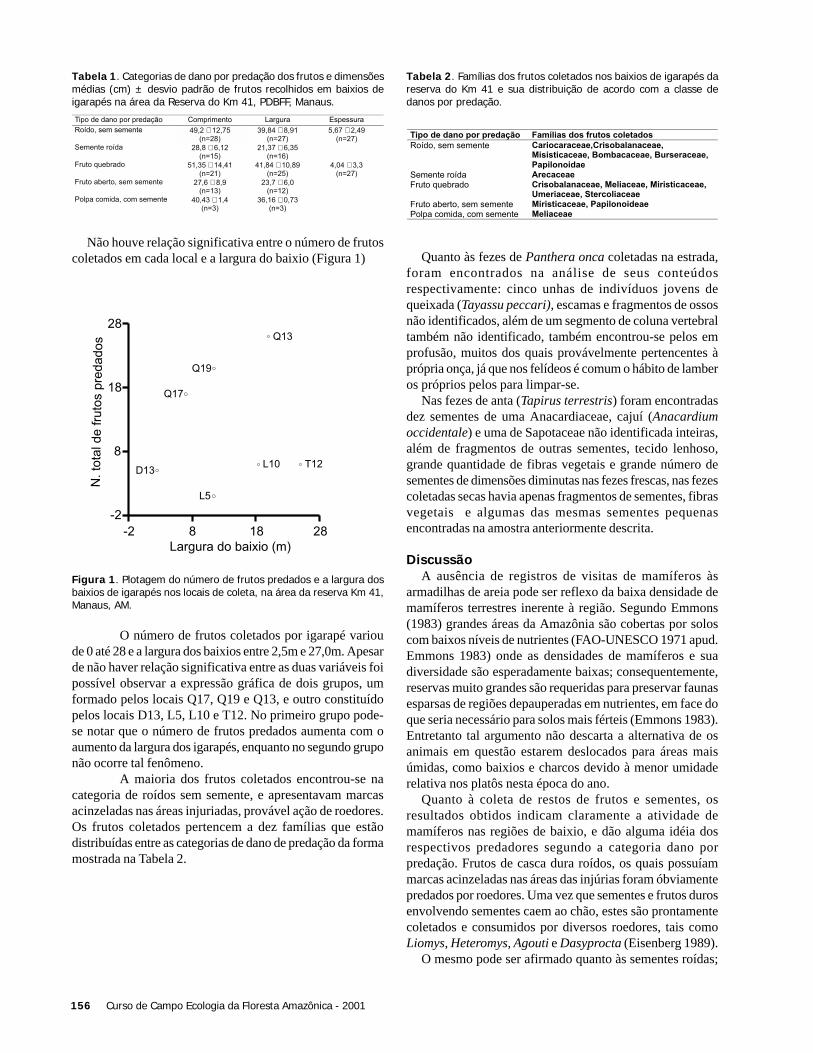
156 Curso de Campo Ecologia da Floresta Amazônica - 2001
Tabela 1. Categorias de dano por predação dos frutos e dimensõesmédias (cm) ± desvio padrão de frutos recolhidos em baixios deigarapés na área da Reserva do Km 41, PDBFF, Manaus.
Tipo de dano por predação Comprimento Largura Espessura
Roído, sem semente 49,2 � 12,75(n=28)
39,84 � 8,91(n=27)
5,67 � 2,49(n=27)
Semente roída 28,8 � 6,12(n=15)
21,37 � 6,35(n=16)
Fruto quebrado 51,35 � 14,41(n=21)
41,84 � 10,89(n=25)
4,04 � 3,3(n=27)
Fruto aberto, sem semente 27,6 � 8,9(n=13)
23,7 � 6,0(n=12)
Polpa comida, com semente 40,43 � 1,4(n=3)
36,16 � 0,73(n=3)
Não houve relação significativa entre o número de frutoscoletados em cada local e a largura do baixio (Figura 1)
-2 8 18 28Largura do baixio (m)
-2
8
18
28
N.t
ota
l de
fru
tos
pre
da
do
s
D13
Q17
Q19
T12
Q13
L10
L5
Figura 1. Plotagem do número de frutos predados e a largura dosbaixios de igarapés nos locais de coleta, na área da reserva Km 41,Manaus, AM.
O número de frutos coletados por igarapé varioude 0 até 28 e a largura dos baixios entre 2,5m e 27,0m. Apesarde não haver relação significativa entre as duas variáveis foipossível observar a expressão gráfica de dois grupos, umformado pelos locais Q17, Q19 e Q13, e outro constituídopelos locais D13, L5, L10 e T12. No primeiro grupo pode-se notar que o número de frutos predados aumenta com oaumento da largura dos igarapés, enquanto no segundo gruponão ocorre tal fenômeno.
A maioria dos frutos coletados encontrou-se nacategoria de roídos sem semente, e apresentavam marcasacinzeladas nas áreas injuriadas, provável ação de roedores.Os frutos coletados pertencem a dez famílias que estãodistribuídas entre as categorias de dano de predação da formamostrada na Tabela 2.
Tabela 2. Famílias dos frutos coletados nos baixios de igarapés dareserva do Km 41 e sua distribuição de acordo com a classe dedanos por predação.
Tipo de dano por predação Famílias dos frutos coletadosRoído, sem semente Cariocaraceae,Crisobalanaceae,
Misisticaceae, Bombacaceae, Burseraceae,Papilonoidae
Semente roída ArecaceaeFruto quebrado Crisobalanaceae, Meliaceae, Miristicaceae,
Umeriaceae, StercoliaceaeFruto aberto, sem semente Miristicaceae, PapilonoideaePolpa comida, com semente Meliaceae
Quanto às fezes de Panthera onca coletadas na estrada,foram encontrados na análise de seus conteúdosrespectivamente: cinco unhas de indivíduos jovens dequeixada (Tayassu peccari), escamas e fragmentos de ossosnão identificados, além de um segmento de coluna vertebraltambém não identificado, também encontrou-se pelos emprofusão, muitos dos quais provávelmente pertencentes àprópria onça, já que nos felídeos é comum o hábito de lamberos próprios pelos para limpar-se.
Nas fezes de anta (Tapirus terrestris) foram encontradasdez sementes de uma Anacardiaceae, cajuí (Anacardiumoccidentale) e uma de Sapotaceae não identificada inteiras,além de fragmentos de outras sementes, tecido lenhoso,grande quantidade de fibras vegetais e grande número desementes de dimensões diminutas nas fezes frescas, nas fezescoletadas secas havia apenas fragmentos de sementes, fibrasvegetais e algumas das mesmas sementes pequenasencontradas na amostra anteriormente descrita.
DiscussãoA ausência de registros de visitas de mamíferos às
armadilhas de areia pode ser reflexo da baixa densidade demamíferos terrestres inerente à região. Segundo Emmons(1983) grandes áreas da Amazônia são cobertas por soloscom baixos níveis de nutrientes (FAO-UNESCO 1971 apud.Emmons 1983) onde as densidades de mamíferos e suadiversidade são esperadamente baixas; consequentemente,reservas muito grandes são requeridas para preservar faunasesparsas de regiões depauperadas em nutrientes, em face doque seria necessário para solos mais férteis (Emmons 1983).Entretanto tal argumento não descarta a alternativa de osanimais em questão estarem deslocados para áreas maisúmidas, como baixios e charcos devido à menor umidaderelativa nos platôs nesta época do ano.
Quanto à coleta de restos de frutos e sementes, osresultados obtidos indicam claramente a atividade demamíferos nas regiões de baixio, e dão alguma idéia dosrespectivos predadores segundo a categoria dano porpredação. Frutos de casca dura roídos, os quais possuíammarcas acinzeladas nas áreas das injúrias foram óbviamentepredados por roedores. Uma vez que sementes e frutos durosenvolvendo sementes caem ao chão, estes são prontamentecoletados e consumidos por diversos roedores, tais comoLiomys, Heteromys, Agouti e Dasyprocta (Eisenberg 1989).
O mesmo pode ser afirmado quanto às sementes roídas;

Curso de Campo Ecologia da Floresta Amazônica - 2001 157
ainda que estas possam ter sido experimentadas por primatase deixadas de lado por sua dureza, ainda há a hipótese deserem restos de alimentação de antas e porcos-do-mato, quesegundo Bodmer (1991), consomem a polpa de Oenocarpusbataua e Mauritia flexuosa (Arecacea) (sendo a primeira dasduas espécies encontrada nas amostras deste estudo) e depoisdescartaram as sementes.
No caso dos frutos encontrados quebrados, pode-se inferirque representam o resultado do forrageamento destrutivo deporcos-do-mato, já que havia em suas cascas marcas de fortesmordidas e segundo Eisenberg (1989) os porcos-do-matosão importantes predadores de sementes e suas fortesmandíbulas podem quebrar muitas das formas semelhantesa nozes para obter suas sementes. Como foi observado porOlifiers (neste volume) pelo menos um bando destes animaisfreqüenta as áreas amostradas no presente estudo.
Os frutos abertos com sementes ausentes e aqueles comapenas a polpa consumida devem ser resultado daforrageamento de primatas que possivelmente têm preferênciapor polpa de algumas espécies e semente de outras. Outrofato curioso a ser comentado é o de que poucas sementes depalmeiras foram encontradas predadas e não se pode afirmarse estas são pouco usadas ou engolidas inteiras ou aindatrituradas pelos artiodáctilos, resultando na quase ausênciade evidências de forrageamento destas plantas por mamíferos.
Devem haver diferenças que agrupariam os locais de coletaem dois diferentes gradientes de largura que teriam comofatores determinantes os tipos de solo, ou talvez a coberturavegetal e sua composição ou ainda a ordem dos igarapés e onível de encharcamento dos solos, já que nos igarapés Q17,Q19, e Q13 os solos dos baixios eram menos encharcados econtinham mais frutos caídos e predados, e no grupo formadopelos locais D13, L5, L10 e T12 os solos eram maisencharcados e o número de frutos coletados foi menor quenos locais do grupo anterior, independentemente da largurado baixio (J. Zuanon com. pess.)
Fezes, de mamíferos silvestres podem ser dispostasaleatóriamente, mas muitas espécies, particularmentecarnívoros, depositam suas fezes em locais previsíveis, comotrilhas, ou ao lado de marcações territoriais proeminentes.Poucas espécies defecam habitualmente em locaisparticulares, ou latrinas que podem ser usadas por um únicoanimal ou vários indivíduos ( Wemmer et al. 1996 apudWilson et al. 1996).
A presença de fezes de Panthera onca nesta área contendounhas de porcos-do-mato são um forte indicativo daintegridade das matas desta região já que onças depende degrandes áreas, superiores a 25 Km² para machos adultos(Emmons 1991; Eisenberg 1989). A ocorrência de pegadasde porcos-do-mato observadas por Olifiers (neste volume)também corroboram esta idéia, pois seus bandos utilizamgrandes áreas durante atividades de forrageamento.
Quanto às sementes encontradas nas fezes frescas de anta,Bodmer (1991) o descreve-o como o ungulado amazônicoque ingere q maior quantidade de sementes intactas e 3,1 %das sementes presentes nos conteúdos estomacais estudadose fezes coletadas pelo esmo autor foram de Anacardiacea, o
que concorda com os resultados da análise do conteúdo dasfezes de anta do presente estudo, onde eram presentes dezsementes de Anacardium occidentale.
Ainda que todos os resultados obtidos tenham sidobaseados em coletas realizadas em mata contínua pode-seinferir que a completa ausência de pegadas de mamíferosnas armadilhas é reflexo da baixa densidade destes animaisnas florestas com solos pobres da Amazônia Central e,finalmente, o pouco tempo disponível para a amostragemsomado ao número provavelmente insuficiente de armadilhasespaçadas a grande distância. Contudo Leite (1995) realizoucoletas de fezes e pegadas de mamíferos de médio e grandeporte na mesma área e época, e percorrendo grandes distânciasde bicicleta na mesma estrada, obteve número satisfatóriode registros.
AgradecimentosAgradeço a todos os componentes do meu grupo,
principalmente a Andréa pela paciência no uso doscomputadores, à Natalie pelo companheirismo em campo,ao Pof. Jansen pelo anti-histamínico, ao Juruna pelaidentificação das famílias de frutos e ao Dr. Venticinque peloapoio e as boas rizadas, além de toda a equipe do curso etodos os colegas.
Referências BibliográficasBecker, M. & Dalponte, J.C. 1991. Rastros de Mamíferos
Silvestres Brasileiros. Editora Universidade de Brasília.Bodmer, R.E. 1991. Strategies of seed dispersal and seed
predation in Amazonian ungulates. Biotropica 23(3):255-261
Eisenberg, J. F. 1989. Mammals of the Neotropics. TheNorthern Neotropics. Vol. 1 The University of ChicagoPress, Chicago.
Emmons L.H. 1997. Neotropical Rainforest Mammals. AField Guide. The University of Chicago Press, Chicago.
Emmons, L. H. 1984. Geographic variation in densities anddiversities of non-flying mammals in Amazonia.Biotropica 16(3): 210-222
Fearnside, P.M. 1989. A Ocupação Humana da Rondônia.Impactos, Limites e planejamento. ProgramaPolonoroeste – Relatório de Pesquisa Nº 5. SCT/PRCNPq.
Leite, M.R.P. 1995. Dieta , uso de habitat e disponibilidadede presas para felinos em uma área de floresta de terra-firme na Amazônia Central. Livro do Curso de Campo“Ecologia da Floresta Amazônica”, INPA/SmithsonianInstitution, Manaus, AM.
Malcom, J.R. 1990. Estimation of mammalian densities incontinuous forest North of Manaus. In: Four NeotropicalRainforests, Gentry, A.H. (ed.). Yale University Press
Pardini, R. no prelo. Nova Metodologia para LevantamentoRápido de Mamíferos de Médio e Grande PorteTerrestres. Projeto RestaUna. Universidade Estadual deSanta Cruz/ Fundação Pau-Brasil.
Voss, R.S. 1996. Mammalian Diversity in NeotropicalLowland Rainforests: a Preliminary Assesment.American Museum of Natural History.
Wilson, D.E.; Cole, F.R, Nichols, J.D., Rudram, R. & Foster,M.E. 1996. Measuring and Monitoring BiologicalDiversity. Standard Methods for Mammals. SmithsonianInstitution Press, Washington and London.



![[Versão corrigida]...todos os dias de nossa vida em comum. À minha família, por apoiarem o meu sonho de ser Doutor em História. Aos meus pais, Tonhão e Elisete, por todo o carinho](https://img.document.onl/doc/110x75/60c5c4d33219b71bc411d823/verso-corrigida-todos-os-dias-de-nossa-vida-em-comum-minha-famlia.jpg)