Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE – UFRN
CENTRO DE TECNOLOGIA – CT
DEPARTAMENTO DE ENGENHARIA QUÍMICA - DEQ
PROGRAMA DE PÓS-GRADUAÇÃO EM ENGENHARIA QUÍMICA - PPGEQ
DISSERTAÇÃO DE MESTRADO
AVALIAÇÃO PRELIMINAR DA VIABILIDADE DE PRODUÇÃO IN VITRO DE UM ISOLADO
BRASILEIRO DE BACULOVÍRUS SPODOPTERA FRUGIPERDA MNPV
Andréa Farias de Almeida
Orientadora: Profª Drª Gorete Ribeiro de Macedo Co-Orientadora: Drª Márcia Regina da Silva Pedrini
NATAL/RN ABRIL/2005

ALMEIDA, Andréa Farias – Avaliação Preliminar da Viabilidade de Produção in vitro de um Isolado Brasileiro de Baculovírus Spodoptera frugiperda MNPV. Dissertação de Mestrado, UFRN, Programa de Pós-Graduação em Engenharia Química. Área de Concentração: Engenharia de Processos, Natal/RN, Brasil. Orientadora: Profª Drª Gorete Ribeiro de Macedo Co-Orientadora: Drª Márcia Regina da Silva Pedrini ___________________________________________________________________________
RESUMO:A produção in vivo de biopesticidas virais é a maior fonte destes inseticidas
presentes no mercado atualmente. Entretanto, o sistema in vivo apresenta limitação em
relação ao escalonamento de produção. No caso do vírus Spodoptera frugiperda
nucleopoliedrovirus (SfMNPV), o inseto usado para sua multiplicação tem características
canibais, o que dificulta ainda mais a sua produção in vivo. As células de inseto são
comumente utilizadas para produção in vitro de baculovírus. Várias linhagens destas células
são derivadas principalmente da espécie Lepidoptera. A linhagem Sf21 é derivada do tecido
ovariano da lagarta Spodoptera frugiperda e um clone isolado da linhagem original,
denominado Sf9, tem sido utilizado para produção de biopesticidas, por apresentar fácil
crescimento em cultivo em suspensão. Neste trabalho, foi testada a viabilidade de produção in
vitro de um isolado viral brasileiro de SfMNPV obtido em lavouras de milho. A produção de
poliedros, em células Sf21 e Sf9, foi determinada com base na avaliação comparativa da
produtividade volumétrica e específica destes poliedros. Ambas as linhagens celulares foram
cultivadas, em suspensão, em meio HyClone suplementado com diferentes concentrações de
soro fetal bovino (2,5 e 5%). A célula Sf9 suplementada com 5% de soro apresentou os
melhores resultados de produção. Os resultados foram confirmados quantitativamente através
dos parâmetros cinéticos estimados para as duas linhagens e diferentes concentrações de soro.
O sistema demonstrou, após sete passagens sucessivas, uma alta produção específica de
poliedros.
Palavras-chaves: Produção in vitro; baculovírus; Spodoptera frugiperda
BANCA EXAMINADORA:
Presidente:
Profª Drª Gorete Ribeiro de Macedo (UFRN)
Membros:
Drª Márcia Regina da Silva Pedrini (DCR/UFRN)
Profª Drª Magarida Maria dos Anjos Magalhães (UFRN)
Prof. Dr. José Luiz Caldas Wolff (UMC)
DATA DA DEFESA: 29 de abril de 2005
ii

Abstract
In vivo production of viral biopesticides is the major source of viral insecticides
currently in the marketplace. However, this system presents limitations during production
scale-up. For the Spodoptera frugiperda nucleopolyhedrovirus (SfMNPV), the insect used for
replication has cannibalistic characteristics, thus production is even more difficult. Insect cells
are commonly used for in vitro baculovirus production. Most of these cell lines are derived
from Lepidoptera species. The Sf21 cell line is derived from Spodoptera frugiperda
caterpillar ovarian tissue, and its clonal isolate Sf9 has been used for biopesticide production
due to its ease of growth in suspension cultures. In this work, the in vitro production
capabilities of a Brazilian SfMNPV isolate obtained from cornfields was evaluated.
Comparison of polyhedra production was carried out using both Sf21 and Sf9 cells, based on
volumetric and specific yields. Both cell lines were cultivated in Hyclone medium
supplemented with different fetal bovine serum concentrations (2,5 and 5%). The best results
were obtained using Sf9 cells supplemented with 5% serum. These results were further
confirmed quantitively through kinetic parameter estimation for both cells lines and different
serum concentrations. After seven successive passages, this system still presented high
specific polyhedra production.
Key-words: in vitro production; baculovirus; Spodoptera frugiperda
iii

Dedico esta dissertação à minha família
pelo amor, apoio e a compreensão de
sempre.
iv

Agradecimentos
Agradeço antes de tudo à minha mãe, presente em todos os momentos da minha vida,
sempre.
Pela realização deste trabalho, agradeço imensamente:
Às minhas orientadoras, professora Gorete pela orientação e apoio que vêm desde a
minha graduação e, principalmente, pelo incentivo constante às minhas atividades. Agradeço
ainda, pela parceria com Márcia Pedrini que teve participação importantíssima desde
elaboração do plano de trabalho até os momentos finais desta dissertação. Em especial,
agradeço a Márcia Pedrini pela oportunidade de trabalhar com cultivo de células animais, que
me proporcionou um enorme aprendizado na área.
Aos companheiros do LEB que de alguma forma ajudou-me na realização da parte
experimental do meu trabalho, em especial, agradeço a Sanderson pela ajuda incondicional e
Danielle pela ajuda na parte final dos experimentos.
Às pesquisadoras Maria Elita B. de Castro e Zilda Maria B. Ribeiro, ambas do
Laboratório de Genética Molecular de Microorganismos e Invertebrados (LGM) - Núcleo de
Controle Biológico da Embrapa/Cernagen, por terem cedido as linhagens de Spodoptera
frugiperda.
Ao pesquisador Fernando Valicente por ter cedido o Isolado 18 do baculovírus de
Spodoptera frugiperda (SfMNPV).
Aos professores Everaldo Silvino e Paulo Marinho pela colaboração na escrita da
dissertação.
Ao Laboratório de Metrologia do departamento de Engenharia Mecânica da UFRN,
em especial, a Henrique e Marcelo pela manipulação e utilização do microscópio na fase de
adaptação das células.
Aos colegas das disciplinas da Pós-graduação sempre compartilhando os momentos de
estudos e descontração.
Agradeço ainda aos funcionários Mazinha e Medeiros pelo apoio administrativo e
burocrático.
Finalmente, agradeço ao Programa de Pós-Graduação em Engenharia Química e ao
CNPq pela bolsa concedida no mestrado.
v

SUMÁRIO
Lista de figuras ...................................................................................................viii
Lista de tabelas ..................................................................................................... ix
Nomenclatura ........................................................................................................ x
Lista de variáveis.................................................................................................xii
Capítulo 1. Introdução Geral................................................................................. 1
Capítulo 2. Aspectos teóricos................................................................................ 5
2.1. Cultura de células de inseto............................................................................ 6
2.1.1. Cinética para o cultivo de células de inseto............................................. 7
2.2. Baculovírus..................................................................................................... 8
2.3. Modo de ação dos baculovírus..................................................................... 10
2.4. Produção de baculovírus .............................................................................. 10
2.4.1. Produção in vivo..................................................................................... 10
2.4.2. Produção in vitro.................................................................................... 11
2.4.2.1. Processo de infecção in vitro........................................................... 13
2.4.2.2. Parâmetros de infecção: MOI e TOI ............................................... 14
2.4.3. Passagem seriada ................................................................................... 14
Capítulo 3. Estado da arte ................................................................................... 16
Capítulo 4. Metodologia experimental................................................................ 21
4.1. Lavagem das vidrarias.................................................................................. 22
4.2. Meio de cultura de células............................................................................ 22
4.2.1. Preparação do meio de cultura de células (HyClone) ........................... 23
4.3.Linhagens de células ..................................................................................... 23
4.3.1. Adaptação e manutenção das células..................................................... 24
vi

4.3.2. Contagem de células .............................................................................. 25
4.3.2.1. Cálculos da contagem...................................................................... 25
4.4. Cinética de crescimento celular, consumo de substrato e produção de poliedros .............................................................................................................. 26
4.5. Vírus ............................................................................................................. 27
4.5.1. Obtenção da hemolinfa .......................................................................... 27
4.5.2. Multiplicação viral in vivo do SfMNPV................................................ 27
4.5.3. Multiplicação viral in vitro do SfMNPV............................................... 28
4.6. Infecção in vitro com SfMNPV ................................................................... 29
4.6.1. Preparação do estoque de vírus.............................................................. 29
4.6.2. Infecção com vírus derivado de poliedro (VDP)................................... 29
4.7. Liberação e quantificação de poliedros das células infectadas.................... 30
4.8. Passagem seriada com SfMNPV.................................................................. 30
4.9. Bioensaios para teste de virulência .............................................................. 31
Capítulo 5. Resultados e discussão ..................................................................... 32
5.1. Adaptação das células Sf9 e Sf21 ................................................................ 33
5.2. Teste preliminar de infecção ........................................................................ 38
5.3. Produção de poliedros em células Sf9 e Sf21 .............................................. 45
5.4. Produção de poliedros em diferentes concentrações de SFB....................... 49
5.5. Cinética de crescimento das células Sf9 e Sf21........................................... 53
5.6. Passagem seriada com SfMNPV.................................................................. 58
Capítulo 6. Conclusões........................................................................................ 61
Capítulo 7. Referências bibliográficas ................................................................ 63
Anexos................................................................................................................. 74
vii

Lista de figuras
Figura 2.1. Taxonomia dos baculovírus (Kalmakoff & Ward, 2004) ........................................ 8
Figura 2.2. Infecção de um inseto hospedeiro (Kalmakoff & Ward, 2004)............................. 11
Figura 2.3. Estrutura do virion (Kalmakoff & Ward, 2003) .................................................... 12
Figura 2.4. Ciclo de infecção dos baculovírus (Rohrmann, 1999). .......................................... 13
Figura 4.1. Câmara de Neubauer – hemocitômetro (Invitrogen, 2002). .................................. 25
Figura 5.1. Células Sf9 adaptadas ao cultivo em suspensão .................................................... 34
Figura 5.2. Adaptação e manutenção das células Sf9 .............................................................. 35
Figura 5.3. Adaptação e manutenção das células Sf21 ............................................................ 37
Figura 5.4. Teste preliminar de infecção com VDP em células Sf9 (HyClone + 5%SFB)...... 39
Figura 5.5. Células Sf9 infectadas com Isolado SfMNPV a 72 h.p.i ....................................... 39
Figura 5.6. Produção de poliedros em células Sf9 (HyClone + 5% SFB) ............................... 40
Figura 5.7. Infecção, em células Sf9, com VDP (SfMNPV) de poliedros multiplicados in vivo.
.................................................................................................................................................. 42
Figura 5.8. Infecção, em células Sf9, com VDP (SfMNPV) de poliedros multiplicados in vitro
.................................................................................................................................................. 44
Figura 5.9. Produção de poliedros com VEC (Passagem 2)..................................................... 46
Figura 5.10. Produção de poliedros com VEC (Passagem 3) em células Sf9 .......................... 51
Figura 5.11. Curva de crescimento e consumo de glicose das células Sf9 (HyClone + 5%SFB)
.................................................................................................................................................. 53
Figura 5.12. Curva de crescimento e consumo de glicose das células Sf9 (HyClone +
2,5%SFB) ................................................................................................................................. 55
Figura 5.13. Curva de crescimento e concentração de glicose das células Sf21 em meio
HyClone + 5%SFB................................................................................................................... 56
Figura 5.14. Passagem seriada em células Sf9 (HyClone + 5%SFB) ...................................... 58
Figura 5.15. Produção específica de poliedros em células Sf9 (HyClone + 5%SFB) ............. 60
viii

Lista de tabelas
Tabela 2.1. Produtos comerciais produzidos a partir de baculovírus. ........................................9
Tabela 5.1. Resultados dos ajustes a curva de produção de poliedros (células Sf9 e Sf21).....47
Tabela 5.2. Resultado da ANOVA para a produção volumétrica de poliedros........................47
Tabela 5.3. Resultado da ANOVA para a produção especifica de poliedros...........................48
Tabela 5.4. Resultados dos ajustes a curva de produção de poliedros (5% e 2,5%SFB) .........52
Tabela 5.5. Resultado da ANOVA para a produção volumétrica de poliedros........................52
Tabela 5.6. Resultado da ANOVA para a produção específica de poliedros...........................52
Tabela 5.7. Resultado da regressão linear para as células Sf9 controle (HyClone + 5% SFB)54
Tabela 5.8. Resultado da regressão linear para as células Sf9 infectadas (HyClone + 5% SFB)
..................................................................................................................................................54
Tabela 5.9. Resultado da regressão linear para as células Sf9 controle (HyClone + 2,5% SFB)
..................................................................................................................................................55
Tabela 5.10. Resultado da regressão linear para as células Sf9 infectadas (HyClone + 2,5%
SFB)..........................................................................................................................................55
Tabela 5.11. Resultado do ajuste linear para as células Sf21 controle (HyClone + 5% SFB) .56
Tabela 5.12. Resultado do ajuste linear para as células Sf21 infectadas (HyClone + 5% SFB)
..................................................................................................................................................57
Tabela 5.13. Parâmetros cinéticos para as células Sf9 e Sf21..................................................57
ix

Nomenclatura
AcMNPV Nucleopoliedrovirus de múltiplos capsídeos de Autographa californica
AfMNPV Nucleopoliedrovirus de múltiplos capsídeos de Anagrafa falcifera
AgMNPV Nucleopoliedrovirus de múltiplos capsídeos de Anticarsia gemmatalis
AsMNPV Nucleopoliedrovirus de múltiplos capsídeos de Agrotis segetum
ANOVA Análise de variância
BCIRL-HA-AM1 Linhagem de célula de Helicoverpa armigera
BCIRL-HA-AM3 Linhagem de célula de Helicoverpa zea
BCIRL-HV-AM1 Linhagem de célula de Heliothis virescens
BEVS Sistemas de expressão que utilizam baculovírus como vetor
BmNPV Nucleopoliedrovirus de Bombyx mori
CENARGEN Recursos Genéticos e Biotecnologia
CNPMS Centro Nacional de Pesquisa de Milho e Sorgo
CvMNPV Nucleopoliedrovirus de múltiplos capsídeos de Condylorrhiza vestigialis
DIP Partículas interferentes defectivas
DjjNPV Nucleopoliedrovirus de Dione juno juno
DNS Ácido dinitro-salicílico
dpi Dias de pós-infecção
DSS Dodecil sulfato de sódio
EeGV Granulovírus de Erinnyis ello
EMBRAPA Empresa brasileira de pesquisa agropecuária
FP Mutantes poucos poliedros (few polyhedra)
GmMNPV Nucleopoliedrovirus de múltiplos capsídeos de Galleria mellonella
GV Granulovírus
HaSNPV Nucleopoliedrovirus de simples capsídeo de Helicoverpa armigera
hpi Horas de pós-infecção
IPLB-LdEp Linhagem de célula Lymantria dispar
LdMNPV Nucleopoliedrovirus de múltiplos capsídeos de Lymantria dispar
LGM Laboratório de Genética Molecular de Microorganismos e Invertebrados
MbMNPV Nucleopoliedrovirus de múltiplos capsídeos de Mamestra brassicae
MET Microscopia eletrônica de transmissão
MNPV Nucleopoliedrovirus de múltiplos capsídeos
________________________________________________________________________ xAndréa Farias de Almeida, abril/2005

MOI Multiplicidade de infecção (ufp/mL)
MP Muitos poliedros
NaCl Cloreto de sódio
NaHCO3 Bicarbonato de sódio
NaOH Hidróxido de sódio
NPV Nucleopoliedrovirus
NsSNPV Nucleopoliedrovirus de simples capsídeo de Neodiprion sertifer
OpMNPV Nucleopoliedrovirus de múltiplos capsídeos de Orgyia pseudotsugata
PfMNPV Nucleopoliedrovirus de múltiplos capsídeos de Panolis flammea
SeMNPV Nucleopoliedrovirus de múltiplos capsídeos de Spodoptera exigua
SFB Soro fetal bovino
Sf9 Clone 9 da linhagem de célula Spodoptera frugiperda
Sf21 Clone 21 da linhagem de célula Spodoptera frugiperda
SfMNPV Nucleopoliedrovirus de múltiplos capsídeos de Spodoptera frugiperda
SNPV Nucleopoliedrovirus de simples capsídeo
TN-CL1 Linhagem de célula Trichoplusia ni
TN-368 Linhagem de célula Trichoplusia ni
TOI Tempo de infecção (célula/mL)
ufp Unidades formadoras de placa
VEC Vírus extracelular
VDP Vírus derivado de poliedro
________________________________________________________________________ xiAndréa Farias de Almeida, abril/2005

Lista de variáveis
C Concentração de células viáveis (células/mL)
D Fator de diluição
N Número de células
µxmáx Velocidade específica máxima de crescimento de células não infectadas (h-1)
µxmáx* Velocidade específica máxima de crescimento de células infectadas (h-1)
µsmáx Velocidade específica máxima de consumo de substrato de células não infectadas (h-1)
µsmáx* Velocidade específica máxima de consumo de substrato de células infectadas (h-1)
µpmáx Velocidade específica máxima de produção de poliedros (h-1)
td Tempo de duplicação celular (h)
R Coeficiente de correlação
________________________________________________________________________ xiiAndréa Farias de Almeida, abril/2005

Capítulo 1
Introdução Geral
________________________________________________________________________ xiiiAndréa Farias de Almeida, abril/2005

__________________________________________________________________________________________ Introdução geral
O Brasil, por sua grande extensão territorial, é um país com uma grande vocação
agrícola, por isto, a preocupação com o controle de pragas. O uso constante e muitas vezes
indiscriminado dos agrotóxicos provoca uma redução da população de organismos benéficos e
cada vez mais se cria dependência de produtos químicos, causando assim, resistência da praga
em relação ao pesticida químico utilizado para o seu controle. A redução do uso destes
agrotóxicos na agricultura é interesse de países produtores e exportadores de alimentos devido
a questões comerciais e ambientais.
Por outro lado, os biopesticidas, por serem altamente seletivos, mais seguros ao
aplicador e por não poluírem o ambiente, como os agrotóxicos, são uma alternativa
verdadeiramente ecológica e sustentável, que contribuem para o equilíbrio ecológico. Além
das vantagens ecológicas, evidenciou-se uma vantagem econômica ao redor de 70% nos
custos de utilização de baculovírus, quando compara-se com pesticidas químicos
convencionais (Embrapa, 2004).
A lagarta-do-cartucho do milho, Spodoptera frugiperda J.E. Smith (Lepidoptera:
Noctuidae), também conhecida por lagarta do milharal ou lagarta militar, é a principal praga
da cultura do milho, responsável por grandes prejuízos. No Brasil, onde as perdas de produção
giram em torno de 30%, em nenhuma época do ano as plantações estão a salvo do ataque do
inseto. A lagarta-do-cartucho afeta as culturas de milho não só do Brasil, mas em todo o
continente americano (Guia Rural, 1990).
A preferência da Spodoptera frugiperda pelo cartucho do milho, não exclui o ataque
às folhas e aos grãos nas espigas, o que explica o tamanho do prejuízo causado às plantações.
O controle desta praga é realizado mediante aplicações freqüentes de pesticidas químicos, que
apesar de eficientes e apresentarem baixos custos, o uso destes produtos de alta toxicidade e
de amplo espectro pode provocar graves danos ambientais.
A partir de uma lagarta-do-campo, um baculovírus foi identificado como
nucleopoliedrovirus de Spodoptera frugiperda (SfMNPV) e purificado (Valicente et al.,
1989). Conseqüentemente, metodologias para o controle biológico da lagarta-do-cartucho
pelo SfMNPV foram estabelecidas (Valicente & Cruz, 1991), e estudos de caracterização
biológica e bioquímica foram conduzidos (Gerk; Kitajima; Souza, 1997). Até o ano de 1999,
aproximadamente 5.000 hectares de milho foram tratados com esse vírus produzido em
lagartas criadas com dieta artificial e formulado como pó molhável para distribuição aos
agricultores (Castro et al., 1999).
A alternativa do biopesticida, produzido a partir do baculovírus (extraído de lagartas
infectadas), é recebida com grande entusiasmo, já que os agrotóxicos têm sido, até o
________________________________________________________________________ 14Andréa Farias de Almeida, abril/2005

__________________________________________________________________________________________ Introdução geral
momento, o principal recurso utilizado. Segundo testes realizados por pesquisadores do
Centro Nacional de Pesquisa de Milho e Sorgo, da Embrapa, o biopesticida mostrou-se 100%
eficiente no controle da praga (Embrapa, 2004).
A produção in vivo de biopesticidas virais é a maior fonte desses pesticidas atualmente
presentes no mercado. Neste sistema, insetos criados em laboratório e alimentados com dieta
artificial, geralmente resultam em um produto final de custo elevado, muitas vezes não
competitivo com o custo dos produtos químicos disponíveis (Moscardi, 1999; Moscardi &
Souza, 2002). Além disso, existe uma limitação em relação à ampliação de produção. No caso
do vírus SfMNPV, o inseto usado para a multiplicação deste vírus tem características
canibais, o que dificulta ainda mais a sua produção in vivo.
A produção de baculovírus pode ser aumentada utilizando-se processo in vitro de
infecção de células de inseto. Em tal abordagem, podem-se obter metodologias de maior
facilidade de ampliação de escala, controle de produção e, principalmente, um custo mais
baixo que as adotadas atualmente.
O objetivo principal de um processo in vitro, para produzir baculovírus como
biopesticida, é gerar uma alta produção de poliedros com alta patogenicidade a insetos e que
ofereça baixo custo, por exemplo, minimizando custos do meio de cultura. O produto também
deverá ser eficaz quando aplicado no campo contra insetos específicos (Chakraborty et al.,
1999).
A maior parte dos estudos de produção in vitro é realizada em cultivo de
monocamadas de células dificultando assim a obtenção de baculovírus em escala industrial. Já
o cultivo de células em suspensão facilita a produção de baculovírus em grande escala e como
conseqüência sua produção em biorreatores (Weiss et al., 1994; Chakraborty et al., 1999)
A produção in vitro de baculovírus por cultura de células é realizada normalmente
com o vírus extracelular da hemolinfa de insetos infectados ou do sobrenadante das células
previamente infectadas (King & Possee, 1992).
A necessidade de um controle seguro da praga Spodoptera frugiperda, despertou o
interesse na produção in vitro deste biopesticida viral. O tema desta dissertação de mestrado
foi trabalho inicial na implantação de uma nova linha de pesquisa, cultura de células animais,
especificamente células de inseto, no Laboratório de Engenharia Bioquímica (LEB) da
Universidade Federal do Rio Grande do Norte (UFRN). O sucesso na cultura de células de
inseto requer o estabelecimento de práticas e métodos adequados para seu cultivo.
Considerando que culturas de células de invertebrados são extremamente sensíveis às
condições ambientais de cultivo, cada laboratório de cultura de células desenvolve métodos e
________________________________________________________________________ 15Andréa Farias de Almeida, abril/2005

__________________________________________________________________________________________ Introdução geral
protocolos de acordo com as instalações e ambiente disponíveis. Os métodos aqui
estabelecidos foram foco para o desenvolvimento de técnicas na produção in vitro de
baculovírus como biopesticida.
Neste trabalho propõe-se a avaliação do potencial de produção de poliedros através da
infecção de células Spodoptera frugiperda com o baculovírus SfMNPV. Na etapa inicial,
desenvolveu-se a adaptação e manutenção das células Sf9 e Sf21 em cultivos em suspensão
utilizando o meio HyClone (HyQ SFX-InsectTM). A adaptação inicial tinha o objetivo de
proporcionar às linhagens de células Sf9 e Sf21 condições de crescimento em suspensão, já
que as mesmas estavam adaptadas ao cultivo de monocamadas. Em seguida estudou-se a
adaptação das células ao meio de cultura HyClone (HyQ SFX-InsectTM) em diferentes
concentrações de soro fetal bovino (SFB).
Realizada a adaptação das células, os experimentos de infecção celular foram
conduzidos em cultivo submerso, onde se utilizou como inóculo vírus derivado de poliedro
(VDP) liberado do SfMNPV (Isolado 18) por um combinado de solução alcalina e tripsina.
As duas linhagens de célula, Sf9 e Sf21, foram comparadas quanto à produção de
poliedros. A melhor linhagem foi escolhida para realizar os demais experimentos de infecção
celular. A partir destas infecções foram estimadas as produções volumétrica e específica do
baculovírus SfMNPV em diferentes concentrações de soro (2,5 e 5%).
Alguns experimentos foram realizados para obtenção dos seguintes parâmetros
cinéticos: velocidade específica máxima de crescimento celular (µxmáx); velocidade específica
máxima de consumo de substrato (µsmáx); velocidade específica máxima de produção de
poliedros (µpmáx) e o tempo de duplicação (td), tanto do crescimento celular como da
replicação viral das linhagens citadas.
Na produção de SfMNPV, como biopesticida, é necessário um certo número de
passagens deste vírus em cultura de células de inseto para obter inóculo suficiente para
produção em larga escala. Portanto, a passagem seriada do SfMNPV foi realizada para
verificar a produtividade deste baculovírus quando passado sucessivamente em cultura de
células.
________________________________________________________________________ 16Andréa Farias de Almeida, abril/2005

Capítulo 2
Aspectos teóricos

___________________________________________________________________________ Aspectos teóricos
2.1. Cultura de células de inseto
A utilização de células de insetos visando a produção in vitro de proteínas
recombinantes e de biopesticidas intensificou-se nas últimas décadas. Os sistemas de
expressão tendo baculovírus como vetores (BEVS) constituem-se em uma tecnologia
relativamente nova quando comparada aos sistemas de expressão em células de mamíferos
(Chan, 1998). A maioria das pesquisas é voltada à expressão de uma grande variedade de
proteínas (Chan; Greenfield; Reid, 1998). Avanços tecnológicos originaram meios sintéticos,
completos e padronizados, para cultivo de células animais que suprem as necessidades
nutricionais proporcionando um elevado crescimento celular (Batista et al., 2003).
Várias linhagens de células de inseto são derivadas principalmente da espécie
Lepidoptera. A linhagem de célula Sf21 é derivada do tecido ovariano da lagarta Spodoptera
frugiperda, um clone isolado da linhagem original, Sf9, tem sido comumente utilizado para
produção de biopesticidas, devido ao fato de facilmente crescer em cultivo submerso (King &
Possee, 1992).
O estabelecimento de culturas de células de inseto permitiu o estudo detalhado in vitro
do ciclo infeccioso de vários baculovírus. Estas células de inseto são facilmente cultivadas in
vitro e sua manutenção é relativamente simples. A maioria delas pode ser cultivada em uma
faixa de temperatura de 25-30°C, todavia, a temperatura ótima durante o crescimento celular e
infecção para células Sf9 é considerada ao redor de 27-28°C (King & Possee, 1992).
Para se obter um bom crescimento das células de inseto é necessário que o meio
escolhido satisfaça às exigências, ambientais e nutricionais, da linhagem em questão. Os
pontos principais na seleção deste meio são pH, osmolaridade, quantidade e razão dos sais
inorgânicos. Componentes como aminoácidos, vitaminas e uma fonte de carbono tipo glicose
são comumente encontrados nos meios clássicos de formulação (Lynn, 1996). Os meios de
cultura que não contêm glicose, nutrientes alternativos podem ser consumidos, resultando no
entanto, na redução na taxa de crescimento celular (Drews et al., 2000).
Estas formulações básicas regularmente não favorecem o crescimento celular, mas a
suplementação com soro animal, normalmente soro fetal bovino (SFB) a uma concentração de
5–20%, favorecem o crescimento celular (Lynn, 1996). Embora o SFB tenha importância para
o crescimento das células, sua utilização apresenta desvantagens, por ser caro, sua adição
eleva substancialmente o custo total do meio de cultura. Além disso, a qualidade deste soro
pode variar de lote para lote, provocando alteração dos seus componentes o que pode afetar o
crescimento celular e replicação do vírus (Schlaeger, 1996). Conseqüentemente, o
_______________________________________________________________________ 6Andréa Farias de Almeida, abril/2005

___________________________________________________________________________ Aspectos teóricos
crescimento celular e a produção de baculovírus são dificultados em um processo de
ampliação de escala.
Outra estratégia é utilizar o meio de cultura com uma quantidade reduzida de SFB. A
redução da concentração de soro no sistema de cultivo pode ser desejável por razões técnicas
ou econômicas (Invitrogen, 2004).
O desenvolvimento de meio de cultura sem soro é de grande valor para o processo de
produção em larga escala. Novas formulações não usam mais soro, como é o caso dos meios
SF900II e ExCell 401 que são os mais comumente utilizados (Schlaeger, 1996).
A adição da mistura complexa de aminoácidos e lipídeos ao meio sem soro pode
recuperar o pico de produtividade atingindo altas concentrações de célula (Chan; Greenfield;
Reid, 1998), a utilização de extratos obtidos pelo processamento de leveduras oriundas de
processos de fermentação alcoólica e de concentrados protéicos resultantes da produção de
derivados de leite também são alternativas de suplementação de meios de cultura para células
animais (Batista et al., 2003).
2.1.1. Cinética para o cultivo de células de inseto
O estudo cinético do cultivo de células de inseto consiste inicialmente na análise dos
valores de concentração celular. A concentração celular é a variável mais importante em todo
o cultivo de células de inseto, porque dela depende a estimativa das velocidades especificas de
crescimento, consumo de substrato e produção.
As células de inseto possuem velocidade de crescimento muito baixa variando entre
0,01 e 0,05h-1 (Tonso, 2000). Em particular, as velocidades de crescimento para as células Sf9
e Sf21 estão na faixa de 0,029 – 0,034 h-1, que correspondem ao tempo de duplicação de 20 a
25h (Schmid, 1996).
Os meios de cultura são constituídos por vários nutrientes: aminoácidos, carboidratos,
vitaminas e o carboidrato mais frequentemente consumido pelas células de inseto é a glicose.
A concentração de glicose pode ser determinada a fim de estimar a velocidade específica de
consumo de substrato.
_______________________________________________________________________ 7Andréa Farias de Almeida, abril/2005

___________________________________________________________________________ Aspectos teóricos
2.2. Baculovírus
Baculovírus são vírus da família Baculoviridae, Figura 2.1, que contêm DNA circular
de fita dupla como material genético e são caracterizados pelos nucleocapsídeos de vírus
infeccioso que se encontram dentro de uma matriz protéica ajudando este vírus a sobreviver
fora de seu hospedeiro (King & Possee, 1992).
Há dois gêneros distintos na família dos baculovírus; os nucleopoliedrovirus (NPVs) e
os granulovírus (GVs), que são diferenciados pelo tamanho das partículas de vírus e a
natureza da proteína do corpo de oclusão (King & Possee, 1992).
Os nucleopoliedrovirus apresentam forma poliédrica e podem ser do tipo múltiplo
(MNPV) ou simples (SNPV), de acordo com o número de capsídeos por virion, esta diferença
está ilustrada na Figura 2.1. (Souza et al., 2002).
Figura 2.1. Taxonomia dos baculovírus (Kalmakoff & Ward, 2004)
Uma divisão mais recente tem sido proposta para distinguir os baculovírus em dois
grupos, grupo I e II, que é uma subdivisão dos NPVs pela análise filogenética das proteínas.
Estas proteínas são poliedrina/granulina (Zanotto; Kessing; Maruniak, 1993), Ecdisteróide
UDP-glicosil transferase (EGT) (Chen et al., 1997), fator de expressão tardia 2 (LEF-2) (Chen
et al., 1999), DNA polimerase (Bulach et al., 1999) e do envelope de fusão gp64/67 (Pearson;
Groten; Rohrmann, 2000). Os baculovírus Autographa californica MNPV (Ayres et al.,
1994), Bomyx mori MNPV (Gomi; Majima; Maeda, 1999), Orgyia pseudotsugata MNPV _______________________________________________________________________ 8Andréa Farias de Almeida, abril/2005

___________________________________________________________________________ Aspectos teóricos
(Ahrens et al., 1997), Anticarsia gemmatalis MNPV (Zanotto; Kessing; Maruniak, 1993) e o
Lonomia obliqua MNPV (Wolff et al., 2002) são membros do grupo I, enquanto que
Spodoptera frugiperda MNPV (Zanotto; Kessing; Maruniak, 1993), Spodoptera exigua
MNPV (Ijkel et al., 1999), Lymantria dispar MNPV (Kuzio et al., 1999), Helicoverpa
armigera SNPV (Chen et al., 2001), Plutella xyslostella GV (Yoshifumi et al., 2000), e a
Xestia c-nigrum GV (Hayakawa et al., 1999) pertencem ao grupo II.
O uso de baculovírus como alternativa aos inseticidas químicos é devido às suas
características que conferem total segurança à saúde humana e ao meio ambiente. Eles
compreendem o maior grupo de vírus de inseto e possuem grande potencial como agentes de
controle biológico de insetos-praga em agricultura e áreas florestais (Martignoni, 1984;
Moscardi, 1999). Sua proteção em cristais protéicos (poliedros) permite a formulação de
biopesticidas com fácil tecnologia de aplicação, representando economia e biossegurança em
relação aos inseticidas químicos.
A Tabela 2.1 mostra os produtos feitos com base em baculovírus, os insetos
hospedeiros e os cultivos atacados pela praga.
Tabela 2.1. Produtos comerciais produzidos a partir de baculovírus.
Cultivos Praga
dos Insetos Vírus usados Nome
comercial Maçã, pêra,
ameixa e noz Cydia pomonella Granulovírus de C. pomonella. Cyd-X
Algodão, milho e tomate Spodoptera littoralis Nucleopoliedrovirus
de S. littoralis. Spodopterin
Algodão e hortaliças
Helicoverpa zea e Heliothis virescens
Nucleopoliedrovirus de H. zea.
Gemstar LC, Biotrol,
Hortaliças e flores Spodoptera exigua
Nucleopoliedrovirus de S. exigua. Spod-X
Alfafa e outros cultivos Autographa californica Nucleopoliedrovirus de
A. californica.
Gusano Inseticida Biológico
Espécies florestais
Orgyia pseudotsugata
Nucleopoliedrovirus de O. pseudotsugata.
TM Biocontrole
Espécies florestais Lymantria dispar Nucleopoliedrovirus de L.
dispar. Gypchek
Soja Anticarsia gemmatalis Nucleopoliedrovirus de A. gemmatalis
Baculoviron, Baculovirus
Nitral Fonte: Tabela adaptada (Moscardi, 1999)
_______________________________________________________________________ 9Andréa Farias de Almeida, abril/2005

___________________________________________________________________________ Aspectos teóricos
2.3. Modo de ação dos baculovírus
A larva é a única fase susceptível (quanto mais precoce o ínstar mais susceptível) à
infecção pelo vírus. Em condições naturais a praga pode ser contaminada através dos ovos,
dos orifícios de respiração do corpo (espiráculos), através de insetos contendo o vírus ou mais
comumente pela via oral, ingerindo o vírus juntamente com o alimento. Uma vez ingerido, o
vírus começa a se multiplicar, espalhando-se por todo o corpo do inseto e provocando sua
morte, que ocorre geralmente de 6 a 8 dias após a ingestão (Valicente & Cruz, 1991).
O tempo de aparecimento dos primeiros sintomas da doença, bem como para a morte
do inseto infectado é influenciado por diferentes fatores, como a espécie do inseto, o ínstar em
que ocorreu a infecção, a quantidade ingerida, a virulência e as condições climáticas durante o
período em que o inseto ficou infectado. Como consequência, esses fatores têm efeitos
marcantes sobre a rapidez da ação do vírus, quando ele é aplicado no campo. Além disso,
outros fatores também influenciam a eficiência e a estabilidade do vírus antes de ser ingerido
pela praga. Entre eles estão: a irradiação solar, a temperatura, a umidade, o hábito da praga, os
equipamentos e a tecnologia para a sua aplicação (Valicente & Cruz, 1991).
2.4. Produção de baculovírus
A produção de baculovírus pode ser realizada por dois sistemas: produção in vivo que
é a forma mais empregada e a produção in vitro onde se concentra a maioria dos estudos,
estes relacionados com a biologia do baculovírus e o processo de infecção em células de
inseto.
2.4.1. Produção in vivo
No sistema de produção in vivo, insetos são criados em laboratório e alimentados com
dieta artificial, geralmente resultando em um produto final de custo elevado, muitas vezes não
competitivo com o custo dos produtos químicos disponíveis (Moscardi, 1999; Moscardi &
Souza, 2002). Os baculovírus produzidos in vivo são a única fonte de biopesticida viral
atualmente encontrada no mercado (Moscardi & Souza, 2002).
_______________________________________________________________________ 10Andréa Farias de Almeida, abril/2005

___________________________________________________________________________ Aspectos teóricos
O processo de infecção, utilizando poliedros, é possível devido ao conteúdo do trato
digestivo dos insetos susceptíveis pois possui um pH alto e a matriz protéica destes poliedros
(poliedrina) é solúvel nesta condição. Assim, o processo de infecção é iniciado quando o
inseto susceptível consome os poliedros que contaminam a sua dieta. Ao alcançar o ambiente
alcalino do intestino do inseto, estes poliedros se dissolvem, liberando as partículas virais.
Após o ajuste na parede da membrana do intestino, o vírus então se funde com a membrana
plasmática das células da parede do intestino e libera os nucleocapsídeos (envelope do vírus
que contém uma nucleoproteína no centro) dentro do citoplasma, como mostrado na Figura
2.2. Os nucleocapsídeos servem para transportar o vírus de DNA para o núcleo da célula
(King & Possee, 1992).
O vírus extracelular (VEC) liberado atinge a hemolinfa espalhando-se e provocando
infecção secundária em outras células do inseto, disseminando assim a infecção de célula para
célula do hospedeiro. As células infectadas no segundo estágio da replicação do vírus na larva
do inseto também produzem VEC.
Figura 2.2. Infecção de um inseto hospedeiro (Kalmakoff & Ward, 2004)
2.4.2. Produção in vitro
A produção in vitro é relativamente mais fácil, quando comparada à produção in vivo,
para estabelecer cultivos contínuos de células. As células de inseto mais comumente usadas
são a Sf9 e a Sf21 (King & Possee, 1992).
_______________________________________________________________________ 11Andréa Farias de Almeida, abril/2005

___________________________________________________________________________ Aspectos teóricos
A produção de vírus em cultura de células de inseto oferece várias vantagens: podem
ser selecionadas cepas específicas, preservadas e armazenadas em meios criogênicos para uso
futuro; podem ser testadas para assegurar que estejam livres de contaminantes indesejáveis,
que certamente afetariam o produto final (King & Possee, 1992).
O estudo para replicação de baculovírus in vitro simplificou enormemente os
experimentos para a compreensão da cinética do vírus e sua replicação. A infecção do vírus
deve ser estabelecida in vitro normalmente pela coleta da hemolinfa de larvas infectadas,
contendo partículas infecciosas de vírus (VEC), e utilizada para inocular cultura de células
susceptíveis. As partículas de vírus (virions), Figura 2.3, que subseqüentemente brotam da
membrana plasmática destas células, dentro do meio de cultura, podem ser usadas para
multiplicar o estoque de vírus (Chan; Greenfield; Reid, 1998).
Figura 2.3. Estrutura do virion (Kalmakoff & Ward, 2003)
A Figura 2.3 ilustra, também, a forma morfológica do vírus importante para a
expansão horizontal do baculovírus no ambiente (poliedros).
Com os avanços da tecnologia de cultura de células de insetos e análise ultra-estrutural
por microscopia eletrônica, tornou-se possível um estudo mais detalhado do processo de
infecção in vitro e o desenvolvimento de uma série de investigações sobre os mecanismos
moleculares envolvidos na replicação dos baculovírus (Castro et al., 1999).
_______________________________________________________________________ 12Andréa Farias de Almeida, abril/2005

___________________________________________________________________________ Aspectos teóricos
2.4.2.1. Processo de infecção in vitro
Em cultura de células, a infecção ocorre em três fases: a) nas primeiras seis horas após,
as células de inseto são reprogramadas, com ação dos vírus, para replicar mais vírus; b) entre
seis e 24 horas após a infecção, novos vírus são produzidos e liberados para o meio de cultivo;
c) após 24 horas são produzidos vírus que ficam armazenados no núcleo das células, envoltos
na matriz de poliedrina, esperando a morte e a desintegração celular para se espalhar no meio
de cultivo e infectar outras células (Rohrmann, 1999). Estas fases estão ilustradas na Figura
2.4.
A primeira indicação da infecção por vírus é o inchaço do núcleo celular e a formação
de um material granular no centro do núcleo (Lua & Reid, 2000). Estas indicações de
alterações morfológicas que podem ser visualizadas ao microscópio óptico recebem o nome
de efeito citopático. O efeito citopático in vitro a ser observado fundamenta-se na análise de
formação de poliedros virais.
A próxima fase da infecção (tardia), compreende o período de 6 a 24 h.p.i., quando
uma estrutura elétron-densa (estroma virogênico) é formada e ocorre a intensa produção de
vírus extracelular (VEC) e replicação do DNA viral. A formação de poliedro é detectada por
volta de 18 h.p.i. e é continuada até a sua deposição completa na membrana em 48 h.p.i.
(Knudson & Harrap, 1975)
Na terceira fase da infecção (muito tardia), há a síntese de novo da membrana que vai
interagir com os nucleocapsídeos para a formação do envelope viral e a posterior oclusão de
virions na matriz protéica, no núcleo da célula.
_______________________________________________________________________ 13
Figura 2.4. Ciclo de infecção dos baculovírus (Rohrmann, 1999).
Poliedro VEC
Andréa Farias de Almeida, abril/2005

___________________________________________________________________________ Aspectos teóricos
2.4.2.2. Parâmetros de infecção: MOI e TOI
Toda a infecção de vírus deriva de uma única partícula infecciosa e assim, esta se
replica dando origem a uma população de vírus. Este vírus infeccioso pode ser usado então
como o inóculo para geração de um estoque puro de vírus. O número de partículas de vírus
infecciosas contidas na solução (meio de cultivo) é o título do vírus. A multiplicidade de
infecção (MOI) é a relação das partículas virais infecciosas com as células viáveis, enquanto
que o tempo de infecção (TOI) refere-se à densidade celular a qual as células são infectadas
(células/mL).
A relação da concentração do produto final e a MOI é altamente dependente do
crescimento das células. Desta forma, um fator importante para obtenção de bons vírus é a
condição das células. Elas devem ser saudáveis e crescendo ativamente no tempo de infecção
onde esta concentração de célula é de importância indiscutível (O'Reilly; Miller; Luckow,
1994; Chakraborty et al., 1999).
A MOI usual para infectar cultura de células está entre 5 e 50, para o estudo da
replicação viral, isto é feito para assegurar que todas as células são infectadas
simultaneamente e que ocorra sincronismo dos eventos da replicação de vírus (King &
Possee, 1992).
O rendimento final do produto para células infectadas na metade da fase exponencial
de crescimento é geralmente maior do que para células infectadas no final da fase exponencial
de crescimento para uma MOI até 10, então é melhor infectar células com baixa MOI na
metade da fase exponencial para obter ótimos rendimentos (O'Reilly; Miller; Luckow, 1994).
2.4.3. Passagem seriada
A passagem seriada de baculovírus em cultura de células de inseto pode resultar uma
variedade de mutações (Chakraborty & Reid, 1999). As mutações mais freqüentes são:
mutantes FP (Few Polyhedra) e partículas interferentes defectivas (DIPs) .
O mutante FP é caracterizado por um decréscimo na formação de poliedros e um
aumento na formação de vírus extracelular. Os poliedros produzidos praticamente não contêm
partículas virais possuindo baixa ou nenhuma infectividade. Outro tipo freqüente de alteração
genética é a produção de DIPs (Souza et al., 2003).
_______________________________________________________________________ 14Andréa Farias de Almeida, abril/2005

___________________________________________________________________________ Aspectos teóricos
Quando o vírus é submetido a longos períodos de ciclos múltiplos de infecção, a
habilidade de infectar as culturas de células diminui devido à formação dos DIPs.
A geração destes mutantes durante a passagem seriada é conhecida como efeito
passagem. Os virions encontrados no Autographa californica nucleopoliedrovirus (AcMNPV)
têm cerca de 44% de perda do seu genoma, incluindo os genes da poliedrina e da DNA
polimerase (Kool et al., 1991; Wickham et al., 1991). Estas partículas contêm apenas parte do
genoma viral, replicando mais rápido que o vírus padrão, conseqüentemente sua quantidade
aumenta rapidamente com a passagem seriada contínua.
A maneira de evitar a formação de DIPs no estoque de vírus é infectar a cultura de
células com baixa MOI, assim minimizando a probabilidade da entrada de DIPs na célula
(Wickham et al., 1991).
_______________________________________________________________________ 15Andréa Farias de Almeida, abril/2005

Capítulo 3
Estado da arte

___________________________________________________________________________ Estado da arte
Vários fatores contribuem para a replicação in vitro de baculovírus em cultivo de
células, tais como cinética de infecção do vírus, tempo de infecção, consumo de nutrientes e
efeito de passagem. Para uma produção in vitro de alta qualidade e biologicamente ativa é
preciso ter conhecimento da replicação do vírus.
A maioria das pesquisas em relação à replicação in vitro de baculovírus tem se
concentrado no estudo do Autographa californica nucleopoliedrovirus (AcMNPV) devido a
sua fácil propagação em cultura de células e a seu amplo espectro de hospedeiro (Volkman &
Summers, 1977; Lee & Miller, 1978; Weiss et al., 1981; Carstens, 1982; Fraser; Smith;
Summers, 1983; Carstens; Krebs; Gallerneault, 1986; Kumar & Miller, 1987; Beames &
Summers, 1988; 1989; 1990; Elam; Vail; Schreiber, 1990; Kool et al., 1991; Morales;
Moscardi; Gravena, 1993; Ayres et al., 1994; Harrison & Summers, 1995a; 1995b; Braunagel
et al., 1999; Pijlman et al., 2001; Rosas-Acosta; Braunagel; Summers, 2001; Spenger et al.,
2002; Bull; Godfray; O'Reilly, 2003).
No entanto, no que se refere aos aspectos de produção e estabilidade durante a
passagem seriada em cultura de células, os vírus Helicoverpa armigera nucleopoliedrovirus
(HaSNPV) (Chakraborty, 1997; Chen et al., 1997; Chakraborty et al., 1999; Chakraborty &
Reid, 1999; Chen et al., 1999; Chen et al., 2000; Lua & Reid, 2000; Chen, 2001; Chen et al.,
2001; Lua et al., 2002; Lua & Reid, 2003; Lua; Nielsen; Reid, 2003; Pedrini, 2003; Pedrini et
al., no prelo) e Lismantria dispar nucleopoliedrovirus (LdMNPV) (Slavicek, 1991; Lynn,
1994; Slavicek; Hayes-Plazolles; Kelly, 1995; Bischoff & Slavicek, 1997; Riegel & Slavicek,
1997; Slavicek et al., 1998; Bischoff & Slavicek, 1999; Slavicek; Popham; Riegel, 1999;
Popham; Bischoff; Slavicek, 2001; Slavicek; Hayes-Plazolles; Kelly, 2001; Slavicek &
Hayes-Plazolles, 2003) também estão sendo bastante explorados.
Apesar do grande sucesso do uso do baculovírus Anticarsia gemmatalis
nucleopoliedrovirus no controle de pragas da soja no Brasil (Moscardi & Corso, 1981;
Moscardi, 1989; 1999; Moscardi & Souza, 2002; Embrapa-Soja, 2005), este vírus ainda é
produzido utilizando-se a coleta de lagartas no campo. Em vista da exigência de padronização
de produção e demanda por grandes quantidades de biopesticida, há a necessidade de
desenvolvimento de tecnologias modernas de culturas celulares para a produção dos
baculovírus. Desta forma, estudos têm sido realizados com intuito de desenvolver o processo
de produção in vitro deste vírus (Claus et al., 1993; Castro et al., 1997; Castro; Souza;
Bilimoria, 1999; Maruniak et al., 1999).
Vários outros baculovírus já foram identificados no Brasil, tais como: Spodoptera
frugiperda nucleopoliedrovirus (SfMNPV) que infecta a lagarta do cartucho do milho
________________________________________________________________________ 17Andréa Farias de Almeida, abril/2005

___________________________________________________________________________ Estado da arte
(Valicente & Cruz, 1991; Pedrini; Wolff; Reid, 2004), Dione juno juno nucleopoliedrovirus
(DjjNPV) que infecta a lagarta-do-maracujazeiro (Boiça Júnior; Lara; Oliveira, 1999; Ribeiro
et al., 1997), Condylorrhiza vestigialis nucleopoliedrovirus (CvMNPV) que infecta a lagarta
do álamo (Castro et al., 2003), Bombix mori nucleopoliedrovírus (BmMNPV) que infecta o
bicho-da-seda da amoreira (Brancalhão; Souza; Soares, 2002), Lonomia obliqua
nucleopoliedrovirus que infecta a lagarta Lonomia obliqua (Wolff et al., 2002) e Erinnyis ello
granulovirus (EeGV) que infecta a lagarta da mandioca (Farias, 1991; Smitt, 1988)
Ao contrário do que acontece com o controle da lagarta da soja, onde cerca de 10% da
área cultivada com soja é tratada com baculovirus AgMNPV, o controle da lagarta do
cartucho do milho é realizado normalmente utilizando-se pesticidas químicos. Apesar de
existirem diversos parasitóides e predadores que são inimigos naturais da praga Spodoptera
frugiperda, mas são poucos os que realmente têm sido estudados no Brasil, visando o controle
biológico dessa praga (Cruz, 1995). Entretanto, o baculovírus SfMNPV é utilizado em escala
experimental nas culturas de milho para o controle da lagarta-do-cartucho, Spodoptera
frugiperda (J.E. Smith) (Embrapa, 2004). A eficiência do baculovírus SfMNPV no controle
desta praga foi demostrado por Valicente & Cruz (1991) em condições laboratoriais e por
Valicente & Costa (1995) utilizando sistemas de irrigação.
Além do Brasil, o vírus SfMNPV foi isolado e caracterizado em vários outros países
da América, tais como Argentina (Barretta; Rios; De Cap, 1998), Nicarágua (Simon et al.,
2004), Venezuela (Agudelo et al., 1983) e Estados Unidos (Loh; Hamm; Huang, 1981; Loh et
al., 1982; Maruniak; Brown; Knudson, 1984). Esses isolados, geralmente, apresentam
diferentes perfis de restrição enzimática e características biológicas distintas (Maruniak;
Brown; Knudson, 1984; Escribano et al., 1999; Simon et al., 2004; Barreto et al., 2005).
No estudo feito por Barreto et al. (2005) foi avaliado o efeito de 22 isolados do
baculovírus SfMNPV em lagartas Spodoptera frugiperda. Estes isolados foram coletados em
plantações de milho em diferentes regiões do Brasil. Os autores observaram que existiam
diferenças significativas entre todos os isolados, no entanto, todos foram eficientes em relação
à mortalidade das lagartas quando a concentração de poliedros era igual ou superior a 106
poliedros/ml. Os melhores resultados foram obtidos com isolados obtidos no estado do Paraná
(Isolado 3, Melissa, PR e Isolado 19, Sertaneja, PR).
Existem poucos estudos sobre a sua replicação in vitro, e não há dados sobre a
produção volumétrica e específica de poliedros deste baculovírus (Harrap, 1972; Knudson &
Tinsley, 1974; Knudson & Harrap, 1975; Knudson & Tinsley, 1978; Maruniak; Brown;
________________________________________________________________________ 18Andréa Farias de Almeida, abril/2005

___________________________________________________________________________ Estado da arte
Knudson, 1984; Mcintosh; Ignoffo; Andrews, 1985; Liu & Maruniak, 1995; Liu & Bilimoria,
1997; Levy; Maruniak; Maruniak, 2002; Pedrini; Wolff; Reid, 2004).
Os nucleopoliedrovírus de múltiplos capsídeos possuem um maior espectro de
hospedeiros do que os nucleopoliedrovírus de simples capsídeo. No entanto, no estudo
realizado por McIntosh et al. (1985), o vírus SfMNPV somente obteve um título viral
significativo para a célula homóloga Spodoptera frugiperda (IPLB-SF-21), dentre as cinco
células testadas (Trichoplusia ni, TN-CL1; Heliothis virescens, BCIRL-HV-AM1;
Helicoverpa zea, BCIRL-HA-AM3; e Helicoverpa armigera, BCIRL-HA-AM1).
Estudos microscópios da seqüência dos eventos morfogênicos da infecção com
SfMNPV em cultivo contínuo de células de Spodoptera frugiperda apresentaram a ordem da
seqüência da replicação deste vírus. Os autores observaram que a formação do poliedro foi
detectada em 18 h.p.i. e continuada até sua membrana ser formada em 48 h.p.i. Este estudo da
microscopia da seqüência dos eventos morfogênicos da infecção do SfMNPV em células de
Spodoptera frugiperda definiu a natureza e o tempo da morfogênese viral (Knudson &
Harrap, 1975).
A passagem seriada de baculovírus em linhagens de células pode resultar em uma
variedade de mutações ou populações de partículas virais defectivas predominantes nos
cultivos. A geração destes mutantes durante as passagens do cultivo de células, pode
comprometer o sistema in vitro para produção de baculovírus em larga escala. Características
morfogênicas aberrantes do vírus em passagens sucessivas do SfMNPV em cultura de células
foram registradas por Kundson & Harrap (1975). No trabalho realizado por Pedrini; Wolff;
Reid (2004), foi detectada uma rápida geração do fenótipo FP após somente duas passagens
do isolado SfMNPV em culturas de células Sf9. O isolado viral utilizado no trabalho realizado
por Pedrini; Wolff; Reid. (2004) foi coletado em Sete Lagoas em 1989 e denominado SL-
SfMNPV.
O efeito passagem não é meramente um produto do cultivo in vitro. Os mutantes FP
são selecionados mesmo in vivo, desde que a técnica de propagação utiliza VEC como inóculo
(Potter; Jaques; Faulkner, 1978).
As linhagens de células geralmente não são infectadas utilizando-se poliedros, pois os
virions não são liberados em pH neutro ou levemente ácidos do meio de cultura, portanto, não
disponíveis para infecção (Elam; Vail; Schreiber, 1990). Em contraste, é relativamente fácil
estabelecer infecções in vitro com isolados virais em culturas de células utilizando-se vírus
extracelular derivado da hemolinfa de inseto infectado ou sobrenadante de células
previamente infectadas (King & Possee, 1992). Em um estudo comparativo entre a
________________________________________________________________________ 19Andréa Farias de Almeida, abril/2005

___________________________________________________________________________ Estado da arte
infectividade de VEC e VDP, Volkman et al.(1976) estimaram que VEC é entre 1700 a 1900
vezes mais infeccioso do que o VDP. Assim, o uso de VEC é usualmente a primeira escolha
para a propagação viral em cultura de células (King & Possee, 1992).
É possível a utilização de vírus derivados de poliedro (VDP) como fonte de virions
infeciosos, porém estes devem ser liberados dos poliedros utilizando-se soluções alcalinas. No
entanto, quando poliedros são dissolvidos em soluções alcalinas antes de serem usados para
infectar linhagens de células de inseto, o nível de infecção é baixo (Vail; Romine; Vaughn,
1979; Elam; Vail; Schreiber, 1990). Porém, utilizando-se proteinases este processo pode ser
melhorado. McIntosh et al (1988) obtiveram melhores resultados de infectividade quando
utilizaram um tratamento com proteinase K para dissolver poliedros de Helicoverpa zea
nucleopoliedrovírus (HzSNPV) para infectar células homólogas. O mesmo resultado foi
obtido por Lynn (1994) utilizando tripsina na dissolução de poliedros de Lymantria dispar
nucleopoliedrovirus (LdMNPV).
O modo de ação da tripsina ainda não foi elucidado. Em um novo trabalho, Lynn
(2003) verificou que, apesar do tratamento com tripsina aumentar a infectividade dos
baculovírus, este aumento não acontece para todos os sistemas de célula e vírus. A
infectividade do baculovírus AcMNPV para a linhagem de células TN-368 aumentou 4750
vezes com o tratamento com tripsina. Para o vírus Anagrapha falcifera nucleopoliedrovirus
(AfMNPV), este aumento foi 77000 vezes. Por outro lado, a infectividade destes dois vírus
para a linhagem de célula IPLB-LdEp foi levemente reduzida.
________________________________________________________________________ 20Andréa Farias de Almeida, abril/2005

Capítulo 4
Metodologia experimental

___________________________________________________________________________Metodologia experimental
4.1. Lavagem das vidrarias
O cultivo de células animais exige uma série de cuidados no que se refere ao material
utilizado para a sua manipulação, por tratar-se de uma cultura extremamente sensível, deve-se
ter um cuidado excessivo quanto à presença de resíduos na vidraria, a fim de se evitar
qualquer tipo de contaminação indesejada.
Deixa-se toda a vidraria de molho, por pelo menos 24 horas, em uma solução de 20L
de água de torneira com 250mL de água sanitária (hipoclorito de sódio 2,5%) e 2L de
detergente (Detertec – Vetec). A partir daí, lava-se 5 vezes em água de torneira, 3 vezes em
água destilada e por fim, 2 vezes em água purificada a 18,3 MΩ-cm, esta lavagem é feita de
modo a certificar ausência de resíduos de detergente na vidraria. Seca-se esta vidraria
naturalmente em escorredor.
Colocam-se as pipetas em recipiente com 10L de água de torneira e 50mL de água
sanitária (hipoclorito de sódio a 2,5%) e 1L de detergente (Detertec – Vetec) deixando-as
nesta solução por pelo menos 24h. Após este período, lava-se 10 vezes em água de torneira, 5
vezes em água destilada e 5 vezes em água purificada a 18,3 MΩ-cm.
Para as pipetas contaminadas com vírus, coloca-se uma quantidade maior de água
sanitária (hipoclorito de sódio a 12%), 200mL/10 L de água. Após este procedimento, lava-se
10 vezes em água de torneira, 5 vezes em água destilada e 5 vezes em água purificada a 18,3
MΩ-cm para retirada de qualquer resíduo de detergente e hipoclorito existentes e deixando-se
secar naturalmente em seguida.
Esteriliza-se todo este material, antes do uso, em autoclave (Sercon) a uma
temperatura de 130°C e 1,5 atm de pressão, por 20 minutos.
4.2. Meio de cultura de células
O meio HyClone (HyQ SFX-InsectTM) suplementado com 5% e 2,5% SFB era
utilizado na manutenção das células e nos experimentos de infecção em cultivo de células em
suspensão.
________________________________________________________________________ 22Andréa Farias de Almeida, abril/2005

___________________________________________________________________________Metodologia experimental
4.2.1. Preparação do meio de cultura de células (HyClone)
Para o preparo de 2000 mL de meio de cultura (HyQ SFX-InsectTM – HyClone) é
necessário um volume inicial de 1500 mL de água purificada a 18,3 MΩ-cm (Easyure RF -
Barnstead) colocados em béquer de 3000mL (Ilmabor, Germany) e pesam-se 82,64g de meio
de cultura em balança analítica (Explorer-Ohaus, Suíça).
A partir deste preparo inicia-se a agitação em um agitador magnético (Tecnal –
TE089) e, após 2h de agitação, retira-se uma alíquota desta suspensão para a medir o pH em
potenciômetro (Digimed). Adicionam-se 0,7g de NaHCO3 (QEEL) e 3mL de glicerina P.A
(Cromato) e após a dissolução completa mede-se novamente o pH. Para ajustar o pH entre 6 –
6,2 adiciona-se gradativamente NaOH (Cromato) a 5M, logo depois deste ajuste completa-se
os 2000mL de meio.
Ao final de 5h de agitação mede-se o pH final e, após este procedimento, filtra-se esta
solução, em ambiente estéril, câmara de fluxo laminar (Purifier Class II Biosafety Cabinet –
Labconco), em sistema de filtração (Millipore) com membranas estéreis (0,45 e 0,22µm,
respectivamente). Este meio é armazenado em frascos estéreis (Scott) com tampa, mantendo-
os em temperatura ambiente, por 3 a 4 dias, para verificar se há alguma contaminação.
Comprovada a não contaminação suplementa-se o meio HyClone (HyQ SFX-InsectTM) com
5% e 2,5% de SFB e posteriormente, conservando-os em geladeira (4°C).
4.3. Linhagens de células
As células mais comumente utilizadas são linhagens derivadas de células epiteliais do
ovário da lagarta, Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae). As linhagens
de células utilizadas neste trabalho foram Sf9 e Sf21 (American Type Culture Collection,
MD) cedidas gentilmente pela Embrapa – Recursos Genéticos e Biotecnologia (Cenargen), já
adaptadas ao meio de cultura TNMFH suplementado com 10% de SFB e ao cultivo em
monocamada.
________________________________________________________________________ 23Andréa Farias de Almeida, abril/2005

___________________________________________________________________________Metodologia experimental
4.3.1. Adaptação e manutenção das células
As linhagens de células Sf9 e Sf21 (American Type Culture Collection, MD) foram
gradualmente adaptadas ao cultivo em suspensão e ao meio HyClone (HyQ SFX-InsectTM)
suplementado com diferentes concentrações de SFB, Figura A.1 (Anexo).
A adaptação inicial para o cultivo em suspensão foi realizada a partir das células
cultivadas em monocamada. Estas células foram transferidas dos frascos de cultura de tecidos
(Frascos-TC) para um Erlenmeyer de 125mL (Schott), quando atingiam 50% de confluência,
ou seja, ocupavam cerca de 50% de toda a superfície destinada para o crescimento celular. O
volume utilizado foi 20mL, com agitação inicial de 70 rpm e temperatura de 28°C, a agitação
em shaker (Certomat MO II – B. Braun) era aumentada gradualmente até atingir a agitação de
120 rpm.
Depois de adaptadas, as células foram mantidas em culturas de suspensão com 20mL
colocados em frascos Erlenmeyers de 125mL (Schott) agitados a 120 rpm, 28°C, em shaker
(Certomat MO II – B. Braun), e cultivadas por 3 a 4 dias, onde amostras eram coletadas para
contagem e diluídas a uma concentração de 1x106 células/mL com meio fresco TNMFH +
10% SFB. Esta diluição era realizada para as células não entrarem na fase estacionária e de
declínio, mantendo-as, assim, na fase de crescimento logarítmico.
Realizada a adaptação ao cultivo em suspensão, iniciou-se a adaptação ao meio de
cultura HyClone (HyQ SFX-InsectTM) em diferentes concentrações de SFB (5% e 2,5%,
respectivamente). Para esta adaptação, as células foram mantidas inicialmente em meio
combinado com TNMFH + 10% SFB e HyClone (HyQ SFX-InsectTM) (75%, 50% v/v,
respectivamente).
Realizava-se contagem diária das células e, após atingirem a concentração de 3x106
células viáveis/mL, em 72/96 horas de cultivo, eram diluídas a uma concentração de 1x106
células viáveis/mL e, ao atingirem a viabilidade igual ou superior a 90% indicava a adaptação
ao meio de cultura estabelecido.
Após a adaptação ao cultivo de suspensão e ao meio de cultura HyClone (HyQ SFX-
InsectTM), as células eram readaptadas ao meio HyClone (HyQ SFX-InsectTM) com diferentes
concentrações de SFB (5 e 2,5%, respectivamente). Os resultados da adaptação estão descritos
no item 5.1.
________________________________________________________________________ 24Andréa Farias de Almeida, abril/2005

___________________________________________________________________________Metodologia experimental
4.3.2. Contagem de células
A concentração celular era estimada utilizando um microscópio fase-contraste
(Olympus, Japão) e um hemocitômetro, modelo Neubauer (Bright-Line Hemacytometer,
Sigma), como mostrado na Figura 4.1. A contagem em triplicada era realizada em cada
amostra, conforme a Equação (1), a fim de obter um erro de 15%, com 200 células contadas
em ambos os lados do hemocitômetro (Nielsen; Smyth; Greenfield, 1991).
A concentração de células viáveis e a viabilidade eram determinadas pela contagem
em volume conhecido da suspensão previamente diluído, através da técnica de exclusão, pela
coloração com azul de tripan (Sigma-Aldrich) a uma concentração final de 0,1%. As células
que permitem a entrada do corante e tornam-se azuladas, não apresentam a membrana intacta,
e devem ser consideradas não-viáveis, enquanto que as células transparentes são consideradas
viáveis.
Figura 4.1. Câmara de Neubauer – hemocitômetro (Invitrogen, 2002).
A viabilidade é definida como a porcentagem de células viáveis em relação à
quantidade total de células viáveis e não viáveis, e a concentração celular é expressa em
células por mililitro (células/mL).
4.3.2.1. Cálculos da contagem
A concentração celular era calculada da seguinte forma:
________________________________________________________________________ 25Andréa Farias de Almeida, abril/2005

___________________________________________________________________________Metodologia experimental
1018 trohemacitôme do lados 2 de volume
4→
−×⋅
=⎥⎦⎤
⎢⎣⎡ DN
mLcélulasC células (1)
Onde: C: concentração de células
: número de células contadas célulasN
: fator de diluição D
4.4. Cinética de crescimento celular, consumo de substrato e produção de
poliedros
O crescimento celular foi determinado por contagem de células viáveis, como descrito
no item 4.3.2. O consumo de glicose foi determinado pelo método do ácido dinitro-salicílico
(DNS), que consiste na redução do ácido 3,5 dinitro-salicílico a ácido 3-amino-5-
nitrosalicílico ao mesmo tempo em que o grupo aldeído do açúcar é oxidado a grupo
carboxílico, com o desenvolvimento de uma coloração avermelhada, lida em
espectrofotômetro (Genesys, série 10) a 600nm. Esta leitura nos fornece a quantidade de
monossacarídeos existente no meio. A partir deste método, estimou-se a glicose consumida
nos experimentos utilizando-se uma curva padrão de glicose, Figura A.5 (Anexo). A
concentração de poliedros produzidos foi determinada pela contagem de poliedros, descrita no
item 4.7.
As velocidades específicas máximas de crescimento específico (µxmáx), de consumo de
substrato (µsmáx) e de produção de poliedros (µpmáx) foram determinadas pelos coeficientes
angulares das retas ajustadas para a variação do logaritmo neperiano das concentrações, com
o tempo. Ajustes feitos com auxílio do software Origin 6.0. (Anexo)
O tempo de duplicação celular (td) é o intervalo de tempo necessário para dobrar o
valor da concentração celular, este foi determinado conforme Equação 2:
dx t
Lnmáx
2=µ (2)
________________________________________________________________________ 26Andréa Farias de Almeida, abril/2005

___________________________________________________________________________Metodologia experimental
4.5. Vírus
O vírus de NPV da Spodoptera frugiperda (SfMNPV) isolado pelo Dr. Fernando
Hercos Valicente foi cedido pelo Centro Nacional de Pesquisa de Milho e Sorgo (CNPMS) da
Embrapa de Sete Lagoas (MG), e a hemolinfa, do Isolado 18, foi obtida no Laboratório de
Genética Molecular de Microorganismos e Invertebrados (LGM) - Núcleo de Controle
Biológico – Embrapa/Cenargen.
4.5.1. Obtenção da hemolinfa
As lagartas Spodoptera frugiperda, cedidas pela Embrapa/Cenargen, foram infectadas
através de dieta artificial contendo o inóculo viral do SfMNPV com concentração de
1x107poliedros/dieta. Após 4 dias pós-infecção (d.p.i), retirou-se a hemolinfa das larvas com
uma micro-seringa. Após a retirada, diluiu-se esta hemolinfa em um tubo (eppendorf) com
450 µL de meio de cultura sem soro (TNMFH) e 50µL de cisteína a 0,1 M. Em seguida,
filtrou-se com membrana de 0,45µm.
Utilizou-se 0,2 mL da hemolinfa para infectar 20 mL da cultura de células em
suspensão, com 1,0x106células/mL em meio HyClone (HyQ SFX-InsectTM) + 5% SFB, mas
esta infecção foi contaminada, portanto não utilizada nos experimentos posteriores.
4.5.2. Multiplicação viral in vivo do SfMNPV
A amplificação viral foi realizada a partir do Isolado 18 (macerado de lagartas do
campo infectadas) onde se utilizou cerca de 300 lagartas do tipo Spodoptera frugiperda do
terceiro para o quarto instar cedidas gentilmente pela Embrapa/Cenargen e mantidas em dieta
artificial.
A concentração do SfMNPV (Isolado 18) foi de 3,5x109 poliedros/mL, foram
utilizados 3µL deste isolado adicionando-se 27µL de água destilada estéril, correspondendo
30µL do inóculo viral em cada dieta (1x107poliedros/dieta).
As lagartas Spodoptera frugiperda foram colocadas individualmente (por serem
canibais) com suas respectivas dietas em copos plásticos descartáveis e vedados com tampas
________________________________________________________________________ 27Andréa Farias de Almeida, abril/2005

___________________________________________________________________________Metodologia experimental
de acrílico. Estas lagartas foram mantidas em ambiente com temperatura e umidade
controladas.
Depois de 9 d.p.i, as lagartas foram coletadas e maceradas em liquidificador por 3
minutos. O macerado foi filtrado através de uma camada de gaze com lã de vidro, para
remoção de partes da cutícula do inseto. Este filtrado foi centrifugado a 1000xg por 15
minutos, a 4°C, em rotor SS34, descartou-se o sobrenadante, coletou-se o pellet e este foi
ressuspenso em 1mL de dodecil sulfato de sódio (DSS) a 0,5% por inseto.
A suspensão foi centrifugada novamente a 1000xg por 15 minutos, a 4°C, em rotor
SS34, o sobrenadante foi descartado e o pellet formado foi novamente ressuspendido em igual
volume de NaCl 0,5M, a suspensão formada foi centrifugada, posteriormente, a 1200xg por
10 minutos, a 4°C, em rotor SS34, o novo pellet foi ressuspendido, e por fim, em água
destilada estéril (na proporção de 0,5 mL por inseto), sendo esta suspensão conservada a
temperatura de –20°C.
4.5.3. Multiplicação viral in vitro do SfMNPV
A multiplicação viral in vitro foi realizada a partir dos poliedros produzidos em cultura
de células. A infecção original para obtenção destes poliedros foi realizada por VDP do
Isolado 18, onde se utilizaram 300µL deste isolado, com concentração de
3,5x109poliedros/mL, para a liberação das partículas virais pelo método Lynn (1994), esta
suspensão de vírus foi utilizada como inóculo para infectar 20mL de suspensão de células a
uma concentração de 1x106células viáveis/mL, Figura A.2 (Anexo).
Após 10 d.p.i., obteve-se uma concentração de poliedros de 2,5x108 poliedros/mL. 10
mL da suspensão foram centrifugados a 100xg por 10 minutos, para separar células do meio
de cultura. O sobrenadante contendo os poliedros foi centrifugado a 1400xg por 30 minutos, o
pellet formado foi resuspendido com água destilada estéril e quantificado por contagem em
hemocitômetro.
________________________________________________________________________ 28Andréa Farias de Almeida, abril/2005

___________________________________________________________________________Metodologia experimental
4.6. Infecção in vitro com SfMNPV
Duas diferentes formas de vírus foram utilizadas neste estudo. O poliedro produzido
pela amplificação viral in vitro e o vírus extracelular produzido em cultivo de células em
suspensão.
4.6.1. Preparação do estoque de vírus
O estoque de vírus foi preparado segundo o método de Lynn (1994), 450µL de
poliedros produzidos, a uma concentração de 1x109poliedros/mL, foram utilizados para
liberação do VDP, e este foi o inóculo utilizado para infecção de 30 mL da suspensão de
células com o tempo de infecção (TOI) de 5x105 células viáveis/mL, Figura A.3 (Anexo).
4.6.2. Infecção com vírus derivado de poliedro (VDP)
Para infecção com VDP, 30mL do cultivo em suspensão de células Spodoptera
frugiperda eram infectados com 3mL de VDP com concentração final de células de 5,0x105
células viáveis/mL. Para obtenção do vírus extracelular a ser utilizado como inóculo nos
demais experimentos, a metade do volume da suspensão celular foi coletada com 72 h.p.i,
permanecendo os outros 50% da suspensão celular para a produção de poliedros.
Após 96 h.p.i, uma amostra de 1mL da suspensão das células infectadas era coletada e
centrifugada a 100xg por 10 min, o sobrenadante era retirado e utilizado como inóculo. O
pellet foi fixado com 3% de gluteraldeído (Sigma) em tampão fosfato, pH 7.2, para futura
análise em microscopia eletrônica de transmissão (MET). As concentrações e viabilidades das
células infectadas de todos os cultivos eram determinadas diariamente utilizando-se o método
de contagem, descrito no item 4.4. Ao final de 10 d.p.i, a produção de poliedros era
determinada também pela contagem em hemocitômetro, descrito no item 4.7. Todos estes
experimentos eram realizados em triplicata.
________________________________________________________________________ 29Andréa Farias de Almeida, abril/2005

___________________________________________________________________________Metodologia experimental
4.7. Liberação e quantificação de poliedros das células infectadas
Adicionava-se 100µL de uma solução de 1% de DSS (Cromato) em 100µL da
suspensão de células infectadas, esta mistura é mantida a 28°C por pelo menos uma hora para
proporcionar a total dissolução da membrana celular e consequentemente a liberação dos
poliedros das células, Figura A.4 (Anexo).
Diluia-se a suspensão, contendo os poliedros, com água purificada (18,3 MΩ-cm)
permitindo a contagem de 100 a 150 poliedros no quadrante central de cada lado do
hemocitômetro. Os poliedros eram contados com auxílio do microscópio de fase-contraste
800x (Olympus, Japão) após 10 minutos do carregamento, para fixação na superfície do
hemocitômetro (Bright-Line Hemacytometer, Sigma). A contagem era realizada em triplicata.
Calculava-se a produção volumétrica (poliedros/mL) e específica (poliedro/célula)
dividindo a concentração de poliedro pelo pico total da concentração celular do cultivo, após a
infecção.
A função de Boltzmann produz uma curva sigmoidal, representativa para a produção
de poliedros:
Valor inicial: A1=0 Valor final: A2=1 Ponto central: x0=0 Tempo constante: dx=1
4.8. Passagem seriada com SfMNPV
O vírus SfMNPV era sucessivamente replicado de 1 até 7 passagens utilizando a
linhagem de célula Sf9 em meio HyClone (HyQ SFX-InsectTM) suplementado com 5% de
SFB, na passagem seriada era realizada utilizando cultivo em suspensão em frascos
Erlenmeyers de 125 mL agitados a 120rpm e temperatura de 28°C.
A primeira passagem foi realizada utilizando como inóculo viral o VDP do SfMNPV
produzido in vitro. Este VDP foi liberado através de soluções alcalinas (Lynn, 1994) que
infectou uma suspensão com 30 mL de células na concentração inicial de 5x105 células
viáveis/mL. Após o estabelecimento da infecção, retirava-se metade do seu volume para ________________________________________________________________________ 30Andréa Farias de Almeida, abril/2005

___________________________________________________________________________Metodologia experimental
centrifugação (100xg, por 10 minutos), onde o sobrenadante (vírus extracelular) obtido era o
inóculo para passagem subseqüente.
Em cada passagem, a infecção era estabelecida utilizando 3 mL de VEC obtido da
passagem anterior (após 72/96h.p.i) e 27mL da suspensão, com uma concentração final de
células de 5x105células viáveis/mL. O VEC foi obtido por centrifugação (100xg, por 10
minutos) da suspensão infectada. As células não infectadas (controle) também possuíam uma
concentração final celular de 5x105 células viáveis/mL. Em cada passagem da cultura
infectada era retirada amostra após 72h.p.i. para análise de microscopia eletrônica de
transmissão (resultados não apresentados neste trabalho), contagens diárias eram realizadas
para quantificar a produção volumétrica de poliedros e após 10 d.p.i. era estimada a produção
específica máxima, dividindo o pico de produção volumétrica pelo pico de concentração das
células infectadas.
4.9. Bioensaios para teste de virulência
As amostras de poliedros produzidos in vitro, coletadas a 10 d.p.i, foram enviadas para
Embrapa Recursos Genéticos e Biotecnologia para realização de testes de virulência a fim de
verificar a bioatividade do biopesticida produzido. Os resultados não farão parte deste
documento e serão parte de publicações futuras.
________________________________________________________________________ 31Andréa Farias de Almeida, abril/2005

Capítulo 5
Resultados e discussão

___________________________________________________________________________ Resultados e discussão
5.1. Adaptação das células Sf9 e Sf21
As linhagens de células Sf9 e Sf21 foram adaptadas ao meio HyClone (HyQ SFX-
InsectTM) suplementado com diferentes concentrações de SFB, Figura 5.1.
A Figura 5.2 mostra as curvas de adaptação e manutenção das células Sf9 em meio
HyClone (HyQ SFX-InsectTM) suplementado com 5% e 2,5% SFB, respectivamente.
A adaptação ao meio HyClone (HyQ SFX-InsectTM) + 5% SFB foi realizada com uma
concentração celular inicial de 1x106 células viáveis/mL. Esta concentração foi utilizada
inicialmente, para garantir um bom número células na fase inicial deste processo, já que as
células não estavam adaptadas a este meio. As células eram diluídas em cada 72/96 horas de
cultivo (perfil zig-zag do gráfico). Como esperado, as concentrações celulares após 3 dias de
cultivo, são menores que as presentes após 4 dias. Após 24 passagens, esta concentração foi
reduzida a 5x105 células viáveis/mL, Figura 5.2A, concentração tal utilizada nos experimentos
de infecção celular. Dez passagens após a redução da concentração inicial, as células já
estavam adaptadas à nova condição estabelecida para a concentração inicial de cultivo.
Depois de 15 passagens, as viabilidades das células atingiram valores acima de 90% e, desta
forma, foram consideradas adaptadas. Além disso, houve um aumento na concentração obtida,
passando de aproximadamente 4x106 células viáveis/mL para valores superiores a 7x106
células viáveis/mL.
As células Sf9 em meio HyClone (HyQ SFX-InsectTM) + 5% SFB atingiram picos de
concentração celular de 7-8,5x106 células viáveis/mL entre as passagens 33 e 55,
apresentando um ótimo desempenho no crescimento celular, Figura 5.2A.
Para as células Sf9 em meio HyClone (HyQ SFX-InsectTM) + 2,5% SFB, a adaptação
foi realizada com uma concentração inicial de 5x105 células viáveis/mL, Figura 5.2B. Esta
concentração foi utilizada devido à adaptação prévia destas células ao mesmo meio
suplementado com 5% SFB.
Observa-se na Figura 5.2B que as células Sf9 cultivadas em meio com 2,5% de SFB
atingiram picos de concentração de 7-9,5x106 células viáveis/mL entre as passagens 11 - 25,
indicando um período bem menor de adaptação ao meio suplementado com 2,5% de SFB do
que ao meio suplementado com 5% de SFB. Um melhor desempenho quanto ao crescimento
celular foi destacado, comparando com as células desenvolvidas em meio HyClone (HyQ
SFX-InsectTM) + 5%SFB.
Alguns pontos próximos a 80% de viabilidade foram observados, mesmo após a
________________________________________________________________________ Andréa Farias de Almeida, abril/2005 33

___________________________________________________________________________ Resultados e discussão
adaptação. Estes declínios correspondem aos períodos onde houve falta de energia elétrica,
portanto, agitação e temperatura não poderiam ser controladas, afetando assim, a viabilidade
das células.
A adaptação das células Sf9 em meio HyClone (HyQ SFX-InsectTM) em ausência de
soro foi tentada, mas não houve sucesso. Este resultado foi inesperado, pois este meio
HyClone (HyQ SFX-InsectTM) foi desenvolvido para ser utilizado sem presença de soro.
Figura 5.1. Células Sf9 adaptadas ao cultivo em suspensão
________________________________________________________________________ Andréa Farias de Almeida, abril/2005 34

___________________________________________________________________________ Resultados e discussão
A
0 5 10 15 20 25 30 35 40 45 50 55 600
1
2
3
4
5
6
7
8
9
10
Concentração celular Viabilidade celular
Número de passagens
Con
cent
raçã
o ce
lula
r x 1
06 (cél
ulas
viá
veis
/mL)
0
10
20
30
40
50
60
70
80
90
100
Viabilidade (%)
B
0 5 10 15 20 25 30 35 400
1
2
3
4
5
6
7
8
9
10
Concentração celular Viabilidade celular
Número de passagens
Con
cent
raçã
o ce
lula
r x 1
06 (cél
ulas
viá
veis
/mL)
0
10
20
30
40
50
60
70
80
90
100
Viabilidade (%)
Figura 5.2. Adaptação e manutenção das células Sf9
A) Adaptação e manutenção das células Sf9 em meio HyClone + 5% de SFB. B) Adaptação e manutenção das células Sf9 em meio HyClone + 2,5% de SFB
________________________________________________________________________ Andréa Farias de Almeida, abril/2005 35

___________________________________________________________________________ Resultados e discussão
O mesmo método foi utilizado para a adaptação das células Sf21. Para esta célula, o
crescimento celular apresentou-se mais elevado quando comparado ao crescimento das
células Sf9.
A Figura 5.3 mostra as curvas de adaptação e manutenção das células Sf21 em meio
HyClone (HyQ SFX-InsectTM) suplementado com 5% e 2,5% de SFB, respectivamente.
A adaptação no meio HyClone (HyQ SFX-InsectTM) +5% SFB foi iniciada com uma
concentração de 1x106 células viáveis/mL, Figura 5.3A. Após 15 passagens, esta
concentração foi reduzida a 5x105 células viáveis/mL, como citado anteriormente,
concentração na qual foram realizados os experimentos de infecção. Cinco passagens após
esta diminuição de concentração inicial, as células já apresentavam uma viabilidade igual ou
superior a 90%, indicação de adaptação à nova condição estabelecida.
As células Sf21 cultivadas em meio HyClone (HyQ SFX-InsectTM) + 2,5% SFB,
Figura 5.3B, atingiram picos de concentração de 8-9,5x106 células viáveis/mL a partir da
quinta passagem, podendo atingir 1,1x107 células viáveis/mL, indicando também um período
bem menor de adaptação ao meio suplementado com 2,5% de SFB.
Apesar do bom rendimento no crescimento celular das células Sf21, escolheu-se para
os experimentos de infecção a linhagem Sf9, devido ao seu bom desempenho quanto à
produção de poliedros, que é o foco principal deste trabalho, resultados estes comentados no
item 5.3.
________________________________________________________________________ Andréa Farias de Almeida, abril/2005 36
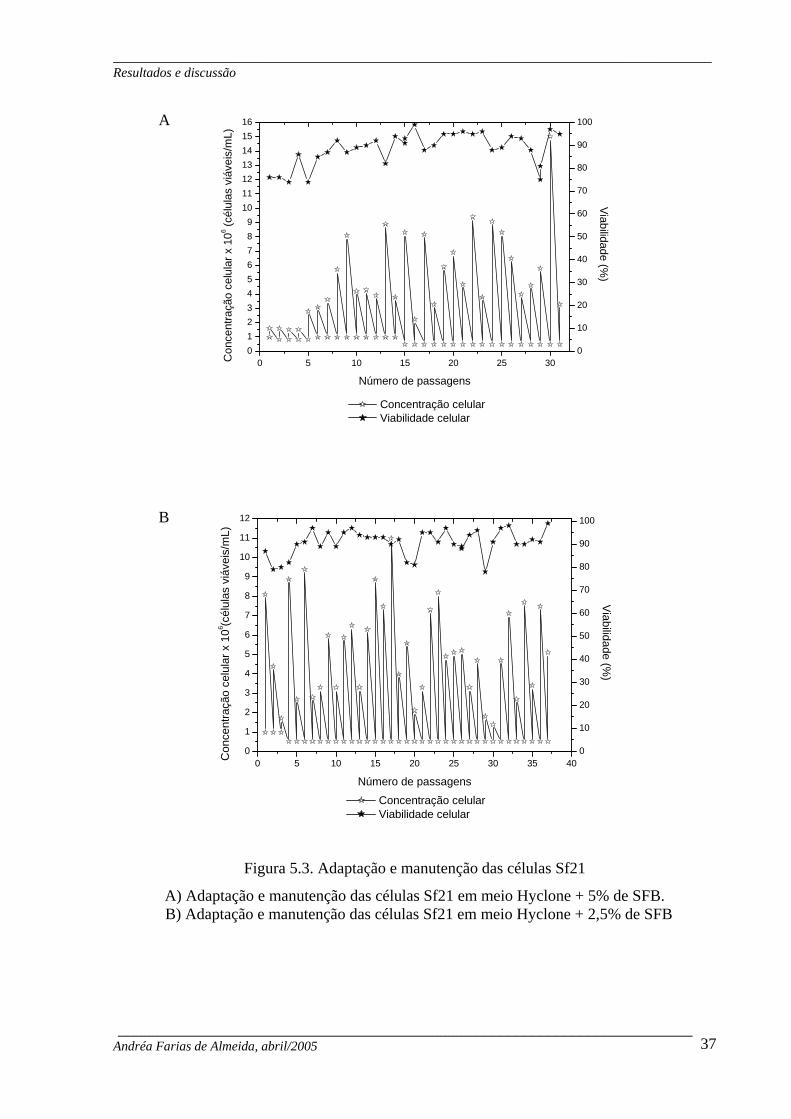
___________________________________________________________________________ Resultados e discussão
A
0 5 10 15 20 25 300123456789
10111213141516
Concentração celular Viabilidade celular
Número de passagens
Con
cent
raçã
o ce
lula
r x 1
06 (cél
ulas
viá
veis
/mL)
0
10
20
30
40
50
60
70
80
90
100
Viabilidade (%)
B
0 5 10 15 20 25 30 35 400
1
2
3
4
5
6
7
8
9
10
11
12
Concentração celular Viabilidade celular
Número de passagens
Con
cent
raçã
o ce
lula
r x 1
06 (cél
ulas
viá
veis
/mL)
0
10
20
30
40
50
60
70
80
90
100
Viabilidade (%)
Figura 5.3. Adaptação e manutenção das células Sf21
A) Adaptação e manutenção das células Sf21 em meio Hyclone + 5% de SFB. B) Adaptação e manutenção das células Sf21 em meio Hyclone + 2,5% de SFB
________________________________________________________________________ Andréa Farias de Almeida, abril/2005 37

___________________________________________________________________________ Resultados e discussão
5.2. Teste preliminar de infecção
A Figura 5.4 mostra a curva de crescimento celular do teste preliminar da infecção
com o VDP do Isolado 18 vírus SfMNPV em células Sf9, adaptadas ao cultivo em suspensão
e com concentração inicial de 1x106 células viáveis/mL.
Inicialmente a preocupação não foi a otimização do tempo de infecção (TOI), mas
verificar a viabilidade de se obter bons resultados de infecção.
Nas primeiras 48 h.p.i., o crescimento celular foi equivalente tanto para as células não
infectadas (controle) como para as infectadas. Após 48 h.p.i., houve uma diferenciação no
crescimento das células controle e infectadas, indicando assim o estabelecimento da infecção.
As células infectadas com VDP (SfMNPV) em meio HyClone (HyQ SFX-InsectTM)
suplementado com 5% SFB estão representadas na Figura 5.5, onde se pode observar que 100
% das células foram infectadas após 72 h.p.i. Após este período, os efeitos citopatológicos
característicos de infecção celular por baculovírus são evidentes, o aumento do volume das
células e a presença de poliedros em seu núcleo. Após 10 dias de cultivo, obteve-se uma
produção volumétrica de 2,5x108 poliedros/mL, correspondente a uma produção específica de
110 poliedros/célula. A Figura 5.6 mostra a produção de poliedros nas células Sf9 em meio
HyClone (HyQ SFX-InsectTM) + 5% SFB.
Diante destes resultados, constata-se uma potencial viabilidade de produção in vitro do
vírus SfMNPV em cultivo em suspensão, a partir do Isolado 18. No entanto, a quantidade
deste Isolado era insuficiente para a realização dos demais experimentos. Então, este Isolado
foi multiplicado in vivo para obtenção de mais inóculo viral.
A ampliação viral in vivo foi realizada na Embrapa/Cenargen, com a colaboração das
pesquisadoras Maria Elita B. de Castro e Zilda Maria B. Ribeiro, ambas do Laboratório de
Genética Molecular de Microorganismos e Invertebrados (LGM) - Núcleo de Controle
Biológico.
________________________________________________________________________ Andréa Farias de Almeida, abril/2005 38

___________________________________________________________________________ Resultados e discussão
0 24 48 72 96 120 144 168 192 216 2400
1
2
3
4
5
6
7
8
9
10
Concentração de células não infectadas Concentração de células infectadas Viabilidade das células não infectadas Viabilidade das células infectadas
Tempo (h)
Con
cent
raçã
o ce
lula
r x 1
06 (cél
ulas
viá
veis
/mL)
0
10
20
30
40
50
60
70
80
90
100
Viabilildade (%
)
Figura 5.4. Teste preliminar de infecção com VDP em células Sf9 (HyClone + 5%SFB)
Figura 5.5. Células Sf9 infectadas com Isolado SfMNPV a 72 h.p.i
________________________________________________________________________ Andréa Farias de Almeida, abril/2005 39

___________________________________________________________________________ Resultados e discussão
24 48 72 96 120 144 168 192 216 2400,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
4,0
Produção volumétrica de poliedros Produção específica de poliedros
Tempo (h)Prod
ução
vol
umét
ica
de P
olie
dros
x 1
08 (pol
iedr
os/m
L)
0
20
40
60
80
100
120
140
160
Produção específica de poliedros (poliedros/célula)
Figura 5.6. Produção de poliedros em células Sf9 (HyClone + 5% SFB)
Os poliedros produzidos in vivo foram utilizados como fonte de inóculo para os
demais experimentos, mas, os resultados não foram representativos, pois não foi obtida uma
produção significativa de poliedros.
A Figura 5.7 mostra um dos resultados obtidos da infecção com o VDP de poliedros
produzidos in vivo. Pode-se observar, Figura 5.7A, que houve crescimento tanto nas células
controle como nas células infectadas, estas atingindo uma concentração de 3,5x106 células
viáveis/mL e uma viabilidade de 90%, indicando que a infecção não foi bem sucedida.
A produção de poliedros, mostrada na Figura 5.7B, foi de 65 poliedros/célula, menor
que a obtida no teste preliminar já apresentado, cerca de 110 poliedros/célula.
O baixo desempenho desta infecção pode ser devido ao método de purificação de
poliedro utilizado, onde se empregou DSS na limpeza de cutículas de inseto existentes no
macerado. O DSS pode ter rompido o envelope dos poliedros e, conseqüentemente, a
partícula viral foi retirada do interior deste poliedro, prejudicando assim a infecção (Lua;
Nielsen; Reid, 2003).
Diante destes resultados, foram utilizados para os demais experimentos os poliedros
________________________________________________________________________ Andréa Farias de Almeida, abril/2005 40

___________________________________________________________________________ Resultados e discussão
produzidos in vitro do teste preliminar de infecção. O VDP utilizado foi obtido a partir do
poliedro através da liberação utilizando-se soluções alcalinas (Lynn, 1994).
________________________________________________________________________ Andréa Farias de Almeida, abril/2005 41

___________________________________________________________________________ Resultados e discussão
A
0 24 48 72 96 120 144 1680
1
2
3
4
5
6
7
8
9
10
Concentração de células não infectadas Concentração de células infectadas Viabilidade das células não infectadas Viabilidade das células infectadas
Tempo (h)
Con
cent
raçã
o ce
lula
r x 1
06 (Cél
ulas
viá
veis
/mL)
0
10
20
30
40
50
60
70
80
90
100
Viabilidade (%)
B
24 48 72 96 120 144 168 192 2160
50
100
150
200
250
300
350
400
450
Tempo (h)Prod
ução
vol
umét
rica
de p
olie
dros
x 1
06 (Pol
iedr
os/m
L)
0
10
20
30
40
50
60
70
80
Produção volumétrica de poliedros Produção específica de poliedros
Produção específica de poliedros (Poliedros/célula)
Figura 5.7. Infecção, em células Sf9, com VDP (SfMNPV) de poliedros multiplicados in vivo.
A) Curva de crescimento das células controle e infectadas com VDP (SfMNPV) B) Curva de produções volumétrica e específica de poliedros
________________________________________________________________________ Andréa Farias de Almeida, abril/2005 42

___________________________________________________________________________ Resultados e discussão
A Figura 5.8 mostra a curva de crescimento das células Sf9 infectadas e a produção de
poliedros, utilizando VDP proveniente do cultivo de células como fonte de inóculo para a
infecção.
Pode-se observar, Figura 5.8A, que as células cresceram ao redor de 2x106 células
viáveis/mL, em 72 h.p.i., sem diferenciação do crescimento das células controle. Então, para
evitar a limitação de nutrientes, a suspensão foi diluída a 1x106 células viáveis/mL. Em 96
h.p.i., a indicação da infecção viral foi observada através da diferenciação no crescimento das
células controle e infectadas, estabelecendo assim a infecção após 72 h.p.i.. Observou-se
ainda, que 100% das células foram infectadas. O vírus extracelular foi coletado, após estas 72
h.p.i., e utilizado como inóculo para o experimento seguinte.
A produção de poliedros está representada na Figura 5.8B. Após 10 dias de infecção,
observou-se uma produção volumétrica de 6x108 poliedros/mL, correspondente a uma
produção específica de 190 poliedros/célula. Como descrito na seção anterior, estes poliedros
produzidos in vitro foram utilizados com fonte de baculovírus para a primeira passagem,
através da infecção das células Sf9 com VDP (SfMNPV) liberado utilizando-se soluções
alcalinas (Lynn, 1994). É possível que, adotando-se a diluição aplicada neste experimento, a
infecção com VDP provenientes dos poliedros obtidos na amplicação in vivo, realizada na
Embrapa Cenargen, tenha um resultado semelhante. Porém, como a produção dos poliedros
neste experimento foi de aproximadamente 60% superior à obtida com o VDP do Isolado 18
proveniente de Sete Lagoas, tomou-se a decisão de considerar esta infecção como fonte de
vírus para as infecções subsequentes deste trabalho.
________________________________________________________________________ Andréa Farias de Almeida, abril/2005 43

___________________________________________________________________________ Resultados e discussão
0 24 48 72 96 120 144 168 192 216 2400
1
2
3
4
5
6
7
8
9
10
Concentração de células não infectadas Concentração de células infectadas Viabilidade das células não infectadas Viabilidade das células infectadas
Tempo (h)
Con
cent
raçã
o ce
lula
r x 1
06 (cél
ulas
viá
veis
/mL)
0
10
20
30
40
50
60
70
80
90
100
Viabilidade (%)
A
B
72 96 120 144 168 192 216 240 264 2883,5
4,0
4,5
5,0
5,5
6,0
6,5
7,0
7,5
Produção volumétrica de poliedros Produção específica de poliedros
Tempo (h)
Prod
ução
vol
umét
rica
de p
olie
dros
x 1
08 (pol
iedr
os/m
L)
100
125
150
175
200
225
Produção específica de poliedros (poliedros/célula)
Figura 5.8. Infecção, em células Sf9, com VDP (SfMNPV) de poliedros multiplicados in vitro
A) Curva de crescimento das células controle e infectadas com VDP (SfMNPV) B) Curva de produções volumétrica e específica de poliedros
________________________________________________________________________ Andréa Farias de Almeida, abril/2005 44

___________________________________________________________________________ Resultados e discussão
5.3. Produção de poliedros em células Sf9 e Sf21
As células Sf21 e Sf9 foram testadas com relação à produção de poliedros. Apesar das
células serem originadas da mesma linhagem, IPLB-SF-21 (Vaughn et al., 1977) diferentes
níveis de produção podem ser observados entre estes dois clones de célula (King & Possee,
1992; O'Reilly; Miller; Luckow, 1994)
A Figura 5.9 mostra as curvas de produção de poliedros das células Sf9 e Sf21
infectadas com o vírus extracelular (VEC) da segunda passagem da infecção com VDP.
As curvas foram ajustadas para obtenção da produção máxima de poliedros nas células
Sf9, Figura 5.9A, e Sf21, Figura 5.9B. Este ajuste foi realizado pelo software Origin 6.0,
utilizando a Equação de Boltzman, que melhor se ajustou aos pontos experimentais,
resultados apresentados na Tabela 5.1.
Os valores da produção de poliedros obtidos da infecção das células Sf9 foram 2 vezes
maiores para a produção volumétrica e 1,3 vezes para a produção específica quando
comparados aos valores obtidos da infecção com as células Sf21, conforme a Tabela 5.1.
Para confirmar este resultado foi realizada uma análise de variância (ANOVA),
Tabelas 5.2 e 5.3, onde se mostra que as médias são significativamente diferentes para
produção volumétrica e específica de poliedros, com nível de confiança de 95%.
Uma razão da diferenciação poderia ser o estado da célula, se ela está bem adaptada às
condições de operação utilizadas (O'Reilly; Miller; Luckow, 1994). As células Sf21, na
realidade, apresentaram melhores resultados de crescimento celular, demonstrando está bem
saudável e adaptada, mostrados no item 5.1. Apesar do crescimento celular das células Sf21
ter sido superior ao crescimento das células Sf9, conforme se mencionou anteriormente, o
objetivo deste trabalho é a produção de poliedros, assim a célula Sf9 foi escolhida para os
demais experimentos.
________________________________________________________________________ Andréa Farias de Almeida, abril/2005 45

___________________________________________________________________________ Resultados e discussão
72 96 120 144 168 192 2160
1
2
3
4
5
6
Produção volumétrica de poliedros Produção específica de poliedros
Tempo (h)
Prod
ução
vol
umét
rica
de p
olie
dros
x 1
08 (pol
iedr
os/m
L)
0
50
100
150
200
250
300
350
400
450 Produção específica de poliedros (poliedros/célula)
A
B
72 96 120 144 168 192 2160,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
Produção volumétrica de poliedros Produção específica de poliedros
Tempo (h)
Prod
ução
vol
umét
rica
de p
olie
dros
x 1
08 (pol
iedr
os/m
L)
0
50
100
150
200
250
300
350 Produção específica de poliedros (poliedros/célula)
Figura 5.9. Produção de poliedros com VEC (Passagem 2).
A) Produção de poliedros em células Sf9 (HyClone + 5%SFB) B) Produção de poliedros em células Sf21 (HyClone + 5%SFB)
________________________________________________________________________ Andréa Farias de Almeida, abril/2005 46

___________________________________________________________________________ Resultados e discussão
Tabela 5.1 – Resultados dos ajustes a curva de produção de poliedros (células Sf9 e Sf21)
Células Sf9
Produção Volumétrica de Poliedros Produção Específica de Poliedros
R2 = 0,99985 R2 = 0,99988 *A1 = -0,00022 ± 0,05845 *A1 = 1,22789 ± 4,0767 *A2 = 5,00658 ± 0,03415 *A2 = 399,63782 ± 2,36968 *X0 = 129,29468 ± 0,70899 *X0 = 129,0107 ± 0,61954 *dx = 20,45171± 0,70236 *dx = 20,06974 ± 0,60911
Células Sf21
Produção Volumétrica de Poliedros Produção Específica de Poliedros
R2 = 0,99596 R2 = 0,99181 *A1 = 0,0574 ± 0,15746 *A1 = 16,71736 ± 18,77961 *A2 = 2,50065 ± 0,06524 *A2 = 299,89875 ± 6,13494 *X0 = 118,38515 ± 3,40264 *X0 = 117,93496 ± 3,43472 *dx = 17,12589 ± 3,00047 *dx = 14,70963 ± 2,87298
*Parâmetros ajustados da Equação de Boltzmann,
Tabela 5.2. Resultado da ANOVA para a produção volumétrica de poliedros
Células Média Variância N
SF9 4,69852x108 8,13091x1014 3
SF21 2,42083x108 5,62994x1013 4
F = 247,71767 p= 1,88282 x10-5
As médias são significativamente diferentes,
com 95% de confiança
________________________________________________________________________ Andréa Farias de Almeida, abril/2005 47

___________________________________________________________________________ Resultados e discussão
Tabela 5.3. Resultado da ANOVA para a produção especifica de poliedros
Célula Média Variância N
SF9 376,23667 469,90243 3
SF21 300,89330 54,55563 3
F = 32,47134 p= 0,00469
As médias são significativamente diferentes,
com 95% de confiança.
________________________________________________________________________ Andréa Farias de Almeida, abril/2005 48

___________________________________________________________________________ Resultados e discussão
5.4. Produção de poliedros em diferentes concentrações de SFB
A Figura 5.10 mostra a produção de poliedros das células Sf9 infectadas, com o VEC
da terceira passagem, em meio HyClone (HyQ SFX-InsectTM) suplementado com 5% e 2,5%
de SFB, respectivamente.
Os resultados demonstraram, para a infecção de cultivos contendo 5% de SFB, Figura
5.10A, uma produção volumétrica de poliedros de 2,4x108 poliedros/mL. Não se observando
diferença significativa em relação à obtida com 2,5% de soro, Figura 5.10B. Pode-se destacar
que a produção volumétrica foi elevada quando comparada com outros sistemas: Lua & Reid,
(2000) com o sistema Helicoverpa armigera NPV em células Helicoverpa zea obtiveram uma
produção máxima de 1,1x108 poliedros/mL; Claus et al.,(1993) com o sistema Anticarsia
gemmatalis NPV em células Spodoptera frugiperda (SF21), atingiram uma produção de
3x107 poliedros/mL; Kariuki; McIntosh, Goodman, (2000) utilizando o sistema Plutella
xylostella MNPV em células Plutella xylostella obtiveram a produção de 2,8x106
poliedros/mL.
A produção específica obtida em meio contendo 5% de SFB foi de 397,79 (± 7,453)
poliedros/célula. Em meio contendo 2,5% de SFB, obteve-se uma produção específica de
264,72 (± 7,721) poliedros/célula, resultados apresentados na Tabela 5.4. Com base nestes
resultados, a produção específica de poliedros utilizando meio HyClone (HyQ SFX-InsectTM)
+ 5% SFB atingiu uma produtividade superior ao meio suplementado com 2,5% SFB, Tabela
5.4.
No trabalho de Claus et al.(1993), os autores também obtiveram uma produção de
poliedros superior quando utilizaram uma concentração maior de soro adicionado ao meio. Já
no estudo realizado por Chakraborty et al. (1999), os autores analisaram a produção de
poliedros em três diferentes concentrações de SFB e obtiveram resultados diferentes dos
observados neste trabalho. A produção específica de poliedros em meio suplementado com
10% de SFB foi menor do que as produções obtidas em meio com 1% de SFB e no meio
isento de soro. A produção volumétrica de poliedros para o meio isento soro foi superior
quando comparada com a produção volumétrica de poliedros em meio com a adição de soro.
No entanto, a produção volumétrica para 1% e 10% não apresentaram diferenças, atingindo
valores em torno de 4 x 107 poliedros/ml.
Pode-se concluir, que no caso do sistema usado por Chakraborty et al. (1999)
(HaSNPV em células Helicoverpa zea), a adição de soro, independente da concentração, inibe
________________________________________________________________________ Andréa Farias de Almeida, abril/2005 49

___________________________________________________________________________ Resultados e discussão
a produção de poliedros. A maioria dos autores considera a produção específica mais
importante do ponto de vista de produção, provavelmente devido ao fato das células
consumirem mais substrato e, em consequência, o custo de produção seria maior.
________________________________________________________________________ Andréa Farias de Almeida, abril/2005 50

___________________________________________________________________________ Resultados e discussão
A
48 72 96 120 144 1680
1
2
3
4
Produção volumétrica de poliedros Produção específica de poliedros
Tempo (h)
Prod
ução
vol
umét
rica
de p
olie
dros
x 1
08 (pol
iedr
os/m
L)
50
100
150
200
250
300
350
400
450 Produção específica de poliedros (poliedros/célula)
B
48 72 96 120 144 168 1920,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
4,0
Produção volumétrica de poliedros Produção específica de poliedros
Tempo (h)
Prod
ução
vol
umét
rica
de p
olie
dros
x 1
08 (pol
iedr
os/m
L)
0
50
100
150
200
250
300
350 Produção específica de poliedros (poliedros/célula)
Figura 5.10. Produção de poliedros com VEC (Passagem 3) em células Sf9
A) Produção de poliedros em meio HyClone + 5% SFB B) Produção de poliedros em meio HyClone + 2,5% SFB
________________________________________________________________________ Andréa Farias de Almeida, abril/2005 51

___________________________________________________________________________ Resultados e discussão
Tabela 5.4 – Resultados dos ajustes a curva de produção de poliedros (5% e 2,5%SFB)
HyClone + 5% SFB
Produção Volumétrica de Poliedros Produção Específica de Poliedros
R2 = 0.99753 R2 = 0.99724 *A1 = 0.32282 ± 0.14189 *A1 = 49.9056 ± 25.66869 *A2 = 2.43321 ± 0.0428 *A2 = 397.79439 ± 7.47534 *X0 = 76.20365 ± 2.50726 *X0 = 76.05112 ± 2.76678 *dx = 12.29002 ± 2.05993 *dx = 12.58274 ± 2.24645
HyClone + 2,5% SFB
Produção Volumétrica de Poliedros Produção Específica de Poliedros
R2 = 0.99527 R2 = 0.99495 *A1 = -0.06521 ± 0.19753 *A1 = -6.35608 ± 22.94554 *A2 = 2.29856 ± 0.06544 *A2 = 264.7257 ± 7.72114 *X0 = 89.79827 ± 4.27426 *X0 = 89.98243 ± 4.32043 *dx = 18.10855 ± 3.5258 *dx = 17.8751 ± 3.58084
*Parâmetros ajustados da Equação de Boltzmann:
Tabela 5.5. Resultado da ANOVA para a produção volumétrica de poliedros
Meio Média Variância N
2,5% SFB 2,40033x108 8,03333x1013 3
5% SFB 2,17750x108 1,79583x1014 4
F = 6,25019 p= 0,05449
As médias não são significativamente
diferentes, com 95% de confiança
Tabela 5.6. Resultado da ANOVA para a produção específica de poliedros
Meio Média Variância N
2,5% SFB 251,13 227,59593 3
5% SFB 392,47 240,11890 4
F = 147,22925 p= 6,71772x10-5
As médias são significativamente diferentes,
com 95% de confiança
________________________________________________________________________ Andréa Farias de Almeida, abril/2005 52

___________________________________________________________________________ Resultados e discussão
5.5. Cinética de crescimento das células Sf9 e Sf21
Os parâmetros cinéticos tais como: velocidade específica máxima de crescimento das
células não infectadas (µxmáx), velocidade específica máxima de crescimento das células
infectadas (µxmáx*), velocidade específica máxima de consumo de substrato das células não
infectadas (µsmáx), velocidade específica máxima de consumo de substrato das células
infectadas (µsmáx*) e velocidade específica máxima de produção de poliedros (µpmáx) foram
estimados com base nas infecções realizadas com as duas linhagens de células (Sf9 e Sf21) e
em diferentes concentrações de SFB (2,5 e 5%).
A Figura 5.11 apresenta a curva de crescimento das células Sf9 adaptadas ao meio
HyClone suplementado com 5% SFB e variação da concentração de substrato durante o
cultivo, tanto para as células controle, como para as infectadas. Percebe-se que no cultivo das
células controle, estas atingiram um pico de concentração celular de 7,4x106 (± 0,37) células
viáveis/mL e uma concentração final de substrato correspondente de 0,61 (±0,03054) g/L.
Para as células infectadas foi observada uma concentração praticamente constante de 8,5x105
(±0,0425) células viáveis/mL e, como esperado, uma superior concentração final de substrato
de 1,4 (±0,07163) g/L devido à menor quantidade de células viáveis para consumir todo o
substrato (glicose) disponível no meio de cultivo.
0 24 48 72 96 120 144
0
2
4
6
8
Concentração de células não infectadas Concentração de células infectadas Concentração de substrato - glicose das células não infectadas Concentração de substrato - glicose das células infectadas
Tempo (h)
Con
cent
raçã
o ce
lula
r x 1
06 (cél
ulas
viá
veis
/mL)
0,5
1,0
1,5
2,0
Concentração de substrato - glicose (g/L)
Figura 5.11. Curva de crescimento e consumo de glicose das células Sf9 (HyClone + 5%SFB)
________________________________________________________________________ Andréa Farias de Almeida, abril/2005 53

___________________________________________________________________________ Resultados e discussão
Tabela 5.7. Resultado da regressão linear para as células Sf9 controle (HyClone + 5% SFB)
Parâmetros Valor Erro R
µx (h-1) 0,03264 ±0,00353 0,98293
µs (h-1) -0,00747 ± 0,00176 0,9733
td (h) 21,23613
Tabela 5.8. Resultado da regressão linear para as células Sf9 infectadas (HyClone + 5% SFB)
Parâmetros Valor Erro R
µx* (h-1) -0,00544 ±0,00233 0,85525
µs* (h-1) 0,00036 ± 8,8527x10-8 1,0
µp (h-1) 0,02926 ±0,00317 0,99419
As Tabelas 5.7 e 5.8 apresentam os resultados das velocidades específicas máximas de
crescimento e de consumo de substrato das células não infectadas e infectadas, como também,
a velocidade específica máxima de produção de poliedros e o tempo de duplicação celular.
Os cálculos das velocidades específicas máximas de crescimento celular, consumo de
substrato e produção de poliedros estão expressos na regressão linear, utilizando o software
Origin 6.0, dos pontos experimentais obtidos da infecção das células Sf9 em meio HyClone
(HyQ SFX-InsectTM) + 5% SFB, Figuras A.10, A.11 e A.12, respectivamente (Anexo).
A Figura 5.12 apresenta os resultados obtidos com uma concentração de 2,5 % SFB.
Nota-se que o crescimento das células controle e infectadas e o consumo de substrato foram
equivalentes ao observado nas células cultivadas em meio contendo 5% de SFB. Atingiu-se
um pico de concentração celular de 6,8x106 (±0,34) células viáveis/mL e concentração final
de substrato de 0,62 (±0,03103) g/L. Para as células infectadas os valores foram 1,8x106
(±0,09) células viáveis/mL para concentração celular e 1,3 (±0,06365) g/L para a
concentração final de substrato.
________________________________________________________________________ Andréa Farias de Almeida, abril/2005 54

___________________________________________________________________________ Resultados e discussão
0 24 48 72 96 120 144
0
2
4
6
8
10
Concentração de células não infectadas Concentração de células infectadas Concentração de substrato - glicose das células não infectadas Concentração de substrato - glicose das células infectadas
Tempo (h)
Con
cent
raçã
o ce
lula
r x 1
06 (cél
ulas
viá
veis
/mL)
0,5
1,0
1,5
2,0
Concentração de substrato - glicose (g/L)
Figura 5.12. Curva de crescimento e consumo de glicose das células Sf9 (HyClone +
2,5%SFB)
Tabela 5.9. Resultado da regressão linear para as células Sf9 controle (HyClone + 2,5% SFB)
Parâmetros Valor Erro R
µx (h-1) 0,03229 ± 0,00444 0,98162
µs (h-1) -0,00632 ± 0,00164 0,96804
td (h) 21,46631
Tabela 5.10. Resultado da regressão linear para as células Sf9 infectadas (HyClone + 2,5%
SFB)
Parâmetros Valor Erro R
µx* (h-1) -0,00252 ± 0,00307 0,63407
µs* (h-1) -0,00186 ± 0,00153 0,77409
µp (h-1) 0,02405 ± 0,0022 0,99583
Os ajustes para os cálculos das velocidades específicas de crescimento, consumo de
________________________________________________________________________ Andréa Farias de Almeida, abril/2005 55

___________________________________________________________________________ Resultados e discussão
substrato, produção de poliedros para as células Sf9 (não infectadas e infectadas) cultivadas
em meio HyClone (HyQ SFX-InsectTM) com 2,5% de SFB estão apresentados nas Figuras
A.13, A.14 e A15 (Anexo).
Para a linhagem de célula (Sf21) foi observado, conforme a Figura 5.13, um pico de
concentração celular de 1x107 (±0,5) células viáveis/mL e concentração final de substrato
igual ao apresentado na célula Sf9, 0,64 (±0,03223)g/L. Para as células infectadas observou-
se uma concentração de 9,4x105 (±0,047) células viáveis/mL e uma concentração final de
substrato de 1,4 (±0,07078) g/L.
0 24 48 72 96 120 144
0
2
4
6
8
10
Concentração de células não infectadas Concentração de células infectadas Concentração de substrato - glicose das células não infectadas Concentração de substrato - glicose das células infectadas
Tempo (h)
Con
cent
raçã
o ce
lula
r x 1
06 (cél
ulas
viá
veis
/mL)
0,5
1,0
1,5
2,0
Concentração de substrato - glicose (g/L)
Figura 5.13. Curva de crescimento e concentração de glicose das células Sf21 em meio
HyClone + 5%SFB
Tabela 5.11. Resultado do ajuste linear para as células Sf21 controle (HyClone + 5% SFB)
Parâmetros Valor Erro R
µx (h-1) 0,03582 ±0,00316 0,99232
µs (h-1) -0,00496 ± 0,00262 0,80134
td (h) 19,35084
________________________________________________________________________ Andréa Farias de Almeida, abril/2005 56

___________________________________________________________________________ Resultados e discussão
Tabela 5.12. Resultado do ajuste linear para as células Sf21 infectadas (HyClone + 5% SFB)
Parâmetros Valor Erro R
µx* (h-1) -0,00329 ±0,00609 0,35677
µs* (h-1) -0,00107 ± 0,00294 0,2495
µp (h-1) 0,01962 ±0,00265 0,99097
Com base na regressão linear foram ajustados os parâmetros cinéticos para a infecção
das células Sf21 com o VEC da terceira passagem em meio HyClone (HyQ SFX-InsectTM) +
5% SFB, Figuras A.16, A.17 e A.18 (Anexo).
Os resultados apresentados nas Tabelas 5.8 e 5.11 mostraram que as velocidades
máximas específicas de crescimento celular são equivalentes, portanto, o crescimento e o
tempo de duplicação celular estão de acordo como o crescimento das células Sf9 e Sf21,
citado anteriormente no item 2.1.1, demonstrando a adaptação das células às condições de
cultivo.
Tabela 5.13. Parâmetros cinéticos para as células Sf9 e Sf21.
Células SFB
(%)
µxmáx
(h-1)
µsmáx
(h-1)
µpmáx
(h-1)
Sf9 5 0,03264 -0,00747 -
Sf9* 5 -0,00544 0,00036 0,02926
Sf9 2,5 0,03229 -0,00632 -
Sf9* 2,5 -0,00252 -0,00186 0,02405
Sf21 5 0,03582 -0,00496 -
Sf21* 5 -0,00329 -0,00107 0,01962 * Célula infectada
A velocidade específica máxima de crescimento das células Sf9 infectadas em meio
contendo 5% de soro obteve um valor superior aos apresentados pelas células Sf9 infectadas
em meio contendo 2,5% de soro e Sf21 infectadas em meio contendo 5% de soro. Nota-se que
os valores para as velocidades específicas máximas de crescimento das células infectadas são
negativos, pois as células não conseguem entrar na fase exponencial. A dinâmica de
crescimento é invertida, ao invés de produzir proteínas necessárias para o seu crescimento, as
células são utilizadas para expressar as proteínas necessárias para a replicação viral.
________________________________________________________________________ Andréa Farias de Almeida, abril/2005 57

___________________________________________________________________________ Resultados e discussão
A velocidade específica máxima de produção de poliedros (µpmáx) das células Sf9 e
Sf21 apresentou valores distintos, resultados que podem ser comparados pela Tabela 5.13. As
células Sf9 apresentaram uma velocidade máxima específica de produção de poliedros maior
do que as células Sf21, confirmando que as células Sf9 são melhores produtoras de poliedros
que a Sf21.
5.6. Passagem seriada com SfMNPV
A Figura 5.14 mostra as curvas de crescimento das células controle e infectadas, assim
como os dados de viabilidade destas células das passagens 1 a 7.
A primeira passagem já foi descrita anteriormente, item 5.2. Nas demais passagens, o
vírus extracelular foi coletado após 72/96 h.p.i., tempo no qual houve um decréscimo na
concentração celular, indicando uma alta MOI que é ideal para passagem subsequente.
Figura 5.14. Passagem seriada em células Sf9 (HyClone + 5%SFB)
A Figura 5.15 apresenta a produção específica de poliedros (poliedros/célula).Os
resultados mostram um pico de produção específica de 399 poliedros/célula, esta
produtividade é superior quando comparada a outros sistemas de produção in vitro de
baculovírus, tais como: Helicoverpa armigera SNPV em células de Helicoverpa zea– 222
________________________________________________________________________ Andréa Farias de Almeida, abril/2005 58

___________________________________________________________________________ Resultados e discussão
poliedros/célula (Lua et al., 2002); Trichoplusia ni NPV em células Trichoplusia ni – 70
poliedros/célula (Potter; Faulkner; Mackinnon, 1976) e Lymantria dispar MNPV em células
Lymantria díspar – 57 poliedros/célula (Slavicek et al., 1996).
Durante a passagem do vírus em cultivo de células, várias alterações podem ocorrer no
genoma viral e acumular em cada passagem (Chakraborty & Reid, 1999). Uma destas
alterações é o acúmulo de mutantes FP (Few Polyhedra). A característica mais marcante do
fenótipo FP é o rápido declínio na produção de poliedros (Pedrini et al., no prelo)
No sistema utilizado neste estudo, foi observado o efeito de passagem devido à
redução de 399 a 109 no número de poliedros produzidos durante sete passagens do SfMNPV
(Isolado 18) em células Sf9. Mesmo assim, apresentou uma elevada produtividade em relação
aos sistemas citados anteriormente, onde apresentaram as seguintes reduções: Helicoverpa
armigera SNPV em células de Helicoverpa zea– 222 a 2 poliedros/célula em seis passagens
(Lua et al., 2002) e Lymantria dispar MNPV em células Lymantria díspar – 57 a 35
poliedros/célula em cinco passagens (Slavicek et al., 1996).
Além da performance em relação à produtividade, o SfMNPV (Isolado18) parece
apresentar uma maior estabilidade. No trabalho realizado por Pedrini; Wolff; Reid, (2004) foi
utilizado um isolado de SfMNPV (SfMNPV – SL) onde mostrou uma rápida geração de
mutantes FP. Após somente duas passagens em cultura de células, os autores observaram que
a maioria das células já demonstrava o fenótipo FP. Como foi citado anteriormente, e apesar
dos autores não apresentarem dados de produtividade, a geração de mutantes FP é um
indicativo da diminuição desta produtividade. Já para o Isolado 18 esta redução foi observada
a partir da quarta passagem. Isto pode ser explicado pela variação genotípica existente entre
isolados de um mesmo baculovírus que é causado pela recombinação destes vírus. A
recombinação é um importante mecanismo no aumento da variabilidade genotípica do vírus
no campo (Pereira et al., 2002; Ribeiro; Pavan; Muotri, 1997).
Observa-se ainda, Figura 5.15, um aumento na produção específica de poliedros (190
para 400 poliedros/célula) entre a primeira e a segunda passagens e uma redução (400 para
250 poliedros/célula) entre a terceira e quarta passagens do baculovirus SfMNPV. O aumento
foi devido à forma viral utilizada na infecção em cultivo de células. Para a primeira passagem
utilizava-se o VDP do baculovirus SfMNPV, enquanto a segunda passagem utilizava-se o
VEC retirado da primeira passagem, confirmando que o virus extracelular é mais eficiente em
cultivo de células do que o virus derivado de poliedros. A redução observada a partir da
quarta passagem foi devido à formação de mutantes FP ou DIPs.
________________________________________________________________________ Andréa Farias de Almeida, abril/2005 59

___________________________________________________________________________ Resultados e discussão
1 2 3 4 5 6 70
50
100
150
200
250
300
350
400
450
500
Polie
dros
/cél
ula
Número de passagens Produção específica de poliedros
Figura 5.15. Produção específica de poliedros em células Sf9 (HyClone + 5%SFB)
Para obter-se uma boa produção de virus é necessário uma concentração celular de
1x107células/mL como descrito por Greenfield et al. (1999), concentrações superiores a esta,
o custo de substrato para produzir as células se torna excessivo.
Considerando a concentração celular de 1x107 células/mL e a produção de 400
poliedros/célula obtidos na terceira passagem do virus SfMNPV, Figura 5.15, obtêm-se 4x109
poliedros/mL, ou seja, 4x1012 poliedros por litro de suspensão. Para pulverizar 1 hectare são
necessários 8,75x1011 poliedros de SfMNPV (Valicente & Cruz, 1991), ou seja, 0,22 litros de
baculovírus por hectare de cultura de milho. Portanto, com um litro de baculovirus pode-se
tratar 4 hectares de área cultivada.
O custo de produção do baculovírus SfMNPV não se pode estimar pois o mesmo foi
utilizado somente experimentalmente no campo. Mas, admitindo as mesmas condições de
produção que foram utilizadas para o HaSNPV (Greenfield et al., 1999), este produto pode ser
produzido por US$15-20/ha.
________________________________________________________________________ Andréa Farias de Almeida, abril/2005 60

Capítulo 6
Conclusões

___________________________________________________________________________ Conclusões
As linhagens Sf9 e Sf21 adaptaram-se ao meio HyClone (HyQ SFX-InsectTM)
suplementado com 5 e 2,5% de SFB em 50 passagens. A adaptação ao meio HyClone (HyQ
SFX-InsectTM) sem suplementação de SFB não foi bem sucedida, pois, estas linhagens não
conseguiram atingir a fase exponencial de crescimento celular.
A infecção preliminar realizada constatou uma potencial viabilidade na produção in
vitro do vírus SfMNPV em cultivo submerso a partir do Isolado 18. Diante deste resultado, a
realização da comparação de produção de poliedros em células Sf9 e Sf21 foi oportuna para o
andamento dos demais experimentos.
As células Sf9 produziram 2 vezes mais poliedros por mL de suspensão e 1,3 vezes
mais poliedros/célula do que as células Sf21. As células Sf9 foram escolhidas para os
experimentos de infecção celular, conseqüentemente, para a produção in vitro dos vírus
SfMNPV.
A comparação da produção de poliedros de SfMNPV em meio HyClone (HyQ SFX-
InsectTM) com concentrações de 5 e 2,5% SFB obteve uma resposta contrária à maioria dos
sistemas de produção in vitro de baculovírus estudados. Neste estudo, a produção de poliedros
foi maior em cultivos com maior concentração de SFB, ou seja, o cultivo onde se utilizou 5%
de SFB obteve-se uma produção de 397 poliedros/célula e 2,5x108 poliedros/mL enquanto
que no cultivo com 2,5% de SFB obteve-se 264 poliedros/célula e 2,3x108 poliedros/mL.
Alguns parâmetros cinéticos foram estimados para os cultivos com as células Sf9 e
Sf21 contendo 5 e 2,5% de SFB no meio. As velocidades específicas máximas de crescimento
celular para as duas linhagens de células cultivadas foram equivalentes: µxmáx = 0,03264 h-1
para Sf9 em 5% SFB, µxmáx = 0,03229 h-1 para Sf9 em 2,5% SFB e µxmáx = 0,03582 h-1 para
Sf21. Porém, apresentaram diferentes velocidades especificas máximas de produção de
poliedros: µpmáx = 0,02926h-1 para Sf9 em 5% SFB, µpmáx = 0,02405h-1 para Sf9 em 2,5% SFB
e µpmáx = 0,01962 h-1 para Sf21. O que justifica a melhor produção de poliedros pela linhagem
Sf9.
No sistema utilizado neste estudo, observou-se, através da passagem seriada do
SfMNPV, uma elevada produtividade em relação a outros sistemas de produção in vitro de
baculovírus, mesmo após sete passagens sucessivas. Verificada esta produtividade, é
conveniente otimizar o processo de produção in vitro deste baculovírus.
________________________________________________________________________ 62Andréa Farias de Almeida, abril/2005

Capítulo 7
Referências bibliográficas

___________________________________________________________________________ Referências bibliográficas
AGUDELO, F.; ROMANO, M.; WASSINK, H.; UCATEGUI, H. C. Una poliedrosis de Spodoptera frugiperda en Venezuela. Turrialba, v.33, p.219-222. 1983. AHRENS, C. H.; RUSSELL, R. L.; FUNK, C. J.; EVANS, J. T.; HARWOOD, S. H.; ROHRMANN, G. F. The sequence of the Orgyia pseudotsugata multinucleocapsid nuclear polyhedrosis virus genome. Journal of Virology, v.229, n.2, p.381-399. 1997. AYRES, M. D.; HOWARD, S. C.; KUZIO, J.; LOPEZ-FERBER, M.; POSSEE, R. D. The complete DNA sequence of Autographa californica nuclear polyhedrosis virus. Journal of Virology, v.202, n.2, p.586-605. 1994. BARRETO, M. R.; GUIMARÃES, C.T; TEIXEIRA, F. F.; PAIVA, E.; VALICENTE, F.H. Effect of baculovirus spodoptera isolates in Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) larvae and their characterization by RAPD. Neotropical Entomology, v.34, n.1, p.067-075. 2005. BARRETTA, M. F.; RIOS, M. L.; DE CAP, A. S. Caracterization of a nuclear polyhedrosis virus of Spodoptera frugiperda from Argentina. Journal of Invertebrate Pathology, v.71, p.280-282. 1998. BATISTA, F.R.X.; PEREIRA, C.A.; MENDONÇA, R.Z.; MORAES, A.M. Avaliação de suplementos na formulação de meios de cultura para células de inseto visando à substituição do soro fetal bovino. In: Simpósio Nacional de Bioprocessos,T146p. Florianópolis. 2003. BEAMES, B.; SUMMERS, M. D. Comparisons of host cell DNA insertions and altered transcription at the site of insertions in few polyhedra bacilovirus mutants. Journal of Virology, v.162, n.1, p.206-220. 1988. BEAMES, B.; SUMMERS, M. D. Location and nucleotide sequence of the 25K protein missing from baculovirus few polyhedra (FP) mutants. Journal of Virology, v.168, n.2, p.344-353. 1989. BEAMES, B.; SUMMERS, M. D. Sequence comparison of cellular and viral copies of host cell DNA insertions found in Autographa californica nuclear polyhedrosis virus. Journal of Virology, v.174, n.2, p.354-363. 1990. BISCHOFF, D. S.; SLAVICEK, J. M. Molecular analysis of an enhancin gene in the Lymantria dispar nuclear polyhedrosis virus. Journal of Virology, v.71, n.11, p.8133-8140. 1997. BISCHOFF, D. S.; SLAVICEK, J. M. Phenotypic and genetic analysis of Lymantria dispar nucleopolyhedrovirus few polyhedra mutants: mutations in the 25K FP gene may be caused by DNA replication errors. Journal of Virology, v.71, n.2, p.1097-1106. 1997. BISCHOFF, D. S.; SLAVICEK, J. M. Impact of deletion of the Lymantria dispar nucleopolyhedrovirus PEP gene on viral potency: Expression of the green fluorescent protein prevents larval liquefaction. Biological Control, v.14, n.1, p.51-59. 1999. BOIÇA JÚNIOR, A. L.; LARA, F. M.; OLIVEIRA, J. C. Flutuação populacional de Dione juno juno (Cramer, 1779) (Lepidoptera: Nymphalidae) em maracujazeiros (Passiflora spp.), métodos de amostragem e resistência de genótipos. Scientia agricola, v.56, n.2. 1999.
________________________________________________________________________ 64Andréa Farias de Almeida, abril/2005

___________________________________________________________________________ Referências bibliográficas
BRANCALHÃO, R.M.C.; SOUZA, V. B. C.; SOARES, M.A.M. Infecção causada por nucleopolyhedrovirus nas células gordurosas de Bombyx mori L., 1758 (Lepidoptera: Bombycidae). Arq. Instit. Biol., v.69, n.2, p.57-63. 2002. BRAUNAGEL, S. C.; BURKS, J. K.; ROSAS-ACOSTA, G.; HARRISON, R. L.; MA, H.; SUMMERS, M. D. Mutations within the Autographa californica nucleopolyhedrovirus FP25K gene decrease the accumulation of ODV-E66 and alter its intranuclear transport. Journal of Virology, v.73, n.10, p.8559-8570. 1999. BULACH, D. M.; KUMAR, A. C.; ZAIA, A.; LIANG, B.; TRIBE, D.E. Group II nucleopolyhedrovirus subgroups revealed by phylogenetic analysis of polyhedrin and DNA polymerase gene sequences. Journal of Invertebrate Pathology, v.73, n.1, p.59-73. 1999. BULL, J. C.; GODFRAY, H. C. J.; O'REILLY, D. R. A few-polyhedra mutant and wild-type nucleopolyhedrovirus remain as a stable polymorphism during serial coinfection in Trichoplusia ni. Appl. Environ. Microbiol., v.69, n.4, p.2052-2057. 2003. CARSTENS, E. B. Mapping the mutation site of an Autographa californica nuclear polyhedrosis virus polyhedron morphology mutant. Journal of Virology, v.43, p.809-818. 1982. CARSTENS, E. B.; KREBS, A.; GALLERNEAULT, C. E. Identification of an amino acid essential to the normal assembly of Autographa californica nuclear polyhedrosis virus polyhedra. Journal of Virology, v.58, p.684-688. 1986. CASTRO, M. E. B.; RIBEIRO, Z. M. A.; SOUZA, M. L.; SOUSA, N.J.; MOSCARDI, F. Identificação do baculovírus da lagarta-do-álamo Condylorrhiza vestigialis (Guenée, 1854)(Lepidoptera Pyralidae). Comunicado Técnico - Embrapa Recursos Genéticos e Biotecnologia, v.87, p.1-3. 2003. CASTRO, M. E. B.; SOUZA, M. L.; ARAUJO, S.; BILIMORIA, S. L. Replication of Anticarsia gemmatalis nuclear polyhedrosis virus in four lepidopteran cell lines. Journal of Invertebrate Pathology, v.69, n.1, p.40-45. 1997. CASTRO, M. E. B.; SOUZA, M. L.; BILIMORIA, S. L. Host-specific transcription of nucleopolyhedrovirus gene homologues in productive and abortive Anticarsia gemmatalis MNPV infections. Arch Virol, v.144, n.6, p.1111-1121. 1999. CASTRO, M. E. B.; SOUZA, M. L.; SIHKER, W.; RODRIGUES, J. C. M.; RIBEIRO, B. M. Biologia molecular de baculovírus e seu uso no controle biológico de pragas no Brasil. Pesquisa Agropecuária Brasileira, v.34, n.10, p.1733-1761. 1999. CHAKRABORTY, S. In vitro production studies with a wild-type Helicoverpa Baculovirus. 1997. 276f., University of Queensland, [St. Lucia, Qld.]. CHAKRABORTY, S.; MONSOUR, C.; TEAKLE, R.; REID, S. Yield, biological activity, and field performance of a wild-type Helicoverpa nucleopolyhedrovirus produced in H. zea cell cultures. Journal of Invertebrate Pathology, v.73, n.2, p.199-205. 1999.
________________________________________________________________________ 65Andréa Farias de Almeida, abril/2005

___________________________________________________________________________ Referências bibliográficas
CHAKRABORTY, S.; REID, S. Serial passage of a Helicoverpa armigera nucleopolyhedrovirus in Helicoverpa zea cell cultures. Journal of Invertebrate Pathology, v.73, n.3, p.303-308. 1999. CHAN, L. C.; GREENFIELD, P. F.; REID, S. Optimising fed-batch production of recombinant proteins using the baculovirus expression vector system. Biotechnol Bioeng, v.59, n.2, p.178-188. 1998. CHAN, L.C.L. Optimization of recombinant protein production in the baculovirus expression vector system using batch, fed-batch and perfusion cultures. 1998. 1 v.f. [St. Lucia, Qld.],. CHEN, X. Genomics and genetic engineering of Helicoverpa armigera nucleopolyhedrovirus. 2001. 154f. (PhD thesis). Wageningen University, Netherlands. CHEN, X.; HU, Z.; JEHLE, J. A.; ZHANG, Y.; VLAK, J. M. Analysis of the ecdysteroid UDP-glucosyltransferase gene of Heliothis armigera single-nucleocapsid baculovirus. Virus Genes, v.15, p.219-225. 1997. CHEN, X.; IJKEL, W.F. J.; DOMINY, C.; ZANOTTO, P. M. A. ; HASHIMOTO, Y.; FAKTOR, O.; HAYAKAWA, T. ; WANG, C.H.; PREKUMAR, A.; MATHAVAN, S.; KRELL, P J.; HUA, Z.; VLAK, J. M. Identification, sequence analysis and phylogeny of the lef-2 gene of Helicoverpa armigera single-nucleocapsid baculovirus. Virus Research, v.65, n.1, p.21-32. 1999. CHEN, X.; SUN, X.; HU, Z.; LI, M.; O'REILLY, D. R.; ZUIDEMA, D.; VLAK, J. M. Genetic engineering of Helicoverpa armigera single-nucleocapsid nucleopolyhedrovirus as an improved pesticide. Journal of Invertebrate Pathology, v.76, n.2, p.140-146. 2000. CHEN, X.; WF, I. JKEL; TARCHINI, R.; SUN, X.; SANDBRINK, H.; WANG, H.; PETERS, S.; ZUIDEMA, D.; LANKHORST, R. K.; VLAK, J. M.; HU, Z. The sequence of the Helicoverpa armigera single nucleocapsid nucleopolyhedrovirus genome. Journal General Virology, v.82, n.Pt 1, p.241-257. 2001. CLAUS, J. D.; REMONDETTO, G. E.; GUERRERO, S. A.; DEMONTE, A. M.; MURGUIA, M.; MARCIPAR, A. J. Anticarsia gemmatalis nuclear polyhedrosis virus replication in serum- free and serum-reduced insect cell cultures. Journal of Biotechnology, v.31, n.1, p.1-15. 1993. CRUZ, I. A lagarta-do-cartucho na cultura do milho Circular Técnica n° 21 1995. DREWS, M.; DOVERSKOG, M.; ÖHMAN, L.; CHAPMAN, B.E.; JACOBSSON, U.; KUCHEL, P.W.; HÄGGSTRÖM, L. Pathways of glutamine metabolism in Spodoptera frugiperda (Sf9) insect cells: evidence for the presence of the nitrogen assimilation system, and a metabolic switch by 1H/15N NMR. Journal of Biotechnology, v.78, p.23 -37. 2000. ELAM, P.; VAIL, P. V.; SCHREIBER, F. Infectivity of Autographa californica nuclear polyhedrosis virus extracted with digestive fluids of Heliothis zea, Estigmene acrea, and carbonate solutions. Journal of Invertebrate Pathology, v.55, p.278-283. 1990. EMBRAPA. Controle biológico da lagarta-do-cartucho do milho. http://www.embrapa.br/linhas_acao/temas/controle_bio/lagarta.htm. 20.01. 2004.
________________________________________________________________________ 66Andréa Farias de Almeida, abril/2005

___________________________________________________________________________ Referências bibliográficas
EMBRAPA-SOJA. Controle biológico: Baculovirus anticarsia. http://www.cnpso.embrapa.br/html/baculov.htm. 2005. ESCRIBANO, A.; WILLIAMS, T.; GOULSON, D.; CAVE, R. D.; CHAPMAN, J. W.; CABALLERO, P. Selection of a nucleopolyhedrovirus for control of Spodoptera frugiperda (Lepidoptera: Noctuidae): structural, genetic, and biological comparison of four isolates from the Americas. Journal Economical Entomology, v.92, n.5, p.1079-1085. 1999. FARIAS, A. R. N. Controle biológico do mandarová da mandioca com baculovirus erinnyis. Embrapa - CNPMF, v.7. 1991. FRASER, M. J.; SMITH, G. E.; SUMMERS, M. D. Acquisition of host cell DNA sequences by baculovirus: relationship between host DNA insertion and FP mutants of Autographa californica and Galleria mellonella nuclear polyhedrosis viruses. Journal of Virology, v.47, p.287-300. 1983. GERK, A. O.; KITAJIMA, E. W.; SOUZA, M. L. Identificação e caracterização de isolado brasileiro do vírus de poliedrose nuclear da lagarta do cartucho do milho. An. Soc. Entomol. Brasil, v.26, n.3, p.507-515. 1997. GOMI, S; MAJIMA, K; MAEDA, S. Sequence analysis of the genome of Bombyx mori nucleopolyhedrovirus. Journal General Virology, v.80, n.5, p.1323-1337. 1999. GUIA RURAL. Briga limpa contra a lagarta do cartucho. São Paulo. 1990 GREENFIELD, P.F.; REID, S.;WEISS, S.; CHRISTIAN, P.; SCHOLZ, B. Baculoviruses as biological control agents: research, production and commercial issues. In: The Asia-Pacific Biochemical Engineering Conference (APBioChEC99). Thailand. 1999 HARRAP, K. A. The structure of nuclear polyhedrosis viruses. III Virus assembly. Virology, v.50, n.1, p.133-139. 1972. HARRISON, R. L.; SUMMERS, M. D. Biosynthesis and Localization of the Autographa californica nuclear polyhedrosis virus 25K gene product. Journal of Virology, v.208, n.1, p.279-288. 1995a. HARRISON, R. L.; SUMMERS, M. D. Mutations in the Autographa californica multinucleocapsid nuclear polyhedrosis virus 25 kDa protein gene result in reduced virion occlusion, altered intranuclear envelopment and enhanced virus production. Journal of General Virology, v.76, n.Pt 6, p.1451-1459. 1995b. HAYAKAWA, T.; KO, R.; OKANO, K.; SEONG, S.I.; GOTO, C.; MAEDA, S. Sequence analysis of the Xestia c-nigrum granulovirus genome. Journal of Virology, v.262, n.2, p.277-297. 1999. IJKEL, W. F. J.; VAN STRIEN, E. A.; HELDENS, J. G. M.; BROER, R.; ZUIDEMA, D.; GOLDBACH, R. W.; VLAK, J. M. Sequence and organization of the Spodoptera exigua multicapsid nucleopolyhedrovirus genome. Journal General Virology, v.80, n.12, p.3289-3304. 1999.
________________________________________________________________________ 67Andréa Farias de Almeida, abril/2005

___________________________________________________________________________ Referências bibliográficas
INVITROGEN. Growth and maintenance of insect cell lines - A manual insect cell lines. http://www.invitrogen.com. 03.03.2004. KALMAKOFF; WARD. Baculoviruses. http://microbes.otago.ac.nz/. 26.01. 2004. KARIUKI, C.W.; McINTOSH, A.H.; GOODMAN, C.L. In vitro host range studies with a new baculovirus isolate from the Diamondback Moth Plutella xylostella (L.) (Plutellidae: Lepidoptera). In Vitro Cell. Dev. Biol - Animal, v.36, p.271 - 276. 2000. KING, L. A.; POSSEE, R. D. The baculovirus expression system : a laboratory guide London ; New York: Chapman & Hall, 1992. p. xiv, 229 p. KNUDSON, D. L.; HARRAP, K. A. Replication of nuclear polyhedrosis virus in a continuous cell culture of Spodoptera frugiperda: microscopy study of the sequence of events of the virus infection. Journal of Virology, v.17, n.1, p.254-268. 1975. KNUDSON, D. L.; TINSLEY, T. W. Replication of a nuclear polyhedrosis virus in a continuous cell line of Spodoptera frugiperda: purification, assay of infectivity, and growth characteristics of the virus. Journal of Virology, v.14, p.934-944. 1974. KNUDSON, D. L.; TINSLEY, T. W. Replication of a nuclear polyhedrosis virus in a continuous cell line of Spodoptera frugiperda: partial characterization of the viral DNA, comparative DNA-DNA hybridization, and patterns of DNA synthesis. Journal of Virology, v.87, n.1, p.42-57. 1978. KOOL, M.; VONCKEN, J. W.; VAN LIER, F. L.; TRAMPER, J.; VLAK, J. M. Detection and analysis of Autographa californica nuclear polyhedrosis virus mutants with defective interfering properties. Journal of Virology, v.183, n.2, p.739-746. 1991. KUMAR, S.; MILLER, L. K. Effects of serial passage of Autographa californica nuclear polyhedrosis virus in cell culture. Virus Research, v.7, n.4, p.335-349. 1987. KUZIO, J.; PEARSON, M. N.; HARWOOD, S. H.; FUNK, C. J.; EVANS, J. T.; SLAVICEK, J. M.; ROHRMANN, G. F. Sequence and analysis of the genome of a baculovirus pathogenic for Lymantria dispar. Journal of Virology, v.253, n.1, p.17-34. 1999. LEE, H. H.; MILLER, L. K. Isolation of genotypic variants of Autographa californica nuclear polyhedrosis virus. Journal of Virology, v.27, n.3, p.754-767. 1978. LEVY, H. C; MARUNIAK, A. G.; MARUNIAK, J. E. Strain identification of Spodoptera frugiperda (Lepidoptera: Noctuidae) insects and cell line: PCR-RFLP of cytochrome oxidase C subunit I gene. Florida Entomologist, v.85, n.1, p.186-190. 2002. LIU, H.-S.; BILIMORIA, S. L. Regulation of infected-cell-specific protein synthesis in SF IPLB-21 cells productively infected with Spodoptera frugiperda multicapsid nuclear polyhedrosis virus. Intervirology, v.40, p.50-54. 1997. LIU, J. C.; MARUNIAK, J. E. Nucleotide sequence and transcriptional analysis of the gp41 gene of Spodoptera frugiperda nuclear polyhedrosis virus. Journal of General Virology, v.76, n.Pt 6, p.1443-1450. 1995.
________________________________________________________________________ 68Andréa Farias de Almeida, abril/2005

___________________________________________________________________________ Referências bibliográficas
LOH, L. C.; HAMM, J. J.; HUANG, E. S. Spodoptera-frugiperda nuclear polyhedrosis-virus genome - physical maps for restriction endonucleases Bamhi and Hindiii. Journal of Virology, v.38, n.3, p.922-931. 1981. LOH, L. C.; HAMM, J. J.; KAWANISHI, C.; HUANG, E. S. Analysis of the Spodoptera-frugiperda nuclear polyhedrosis-virus genome by restriction endonucleases and electron-microscopy. Journal of Virology, v.44, n.2, p.747-751. 1982. LUA, L. H. L.; PEDRINI, M. R. S.; REID, S.; ROBERTSON, A.; TRIBE, D.E. Phenotypic and genotypic analysis of Helicoverpa armigera nucleopolyhedrovirus serially passaged in cell culture. Journal General Virology, v.83, n.4, p.945-955. 2002. LUA, L. H.; REID, S. Virus morphogenesis of Helicoverpa armigera nucleopolyhedrovirus in Helicoverpa zea serum-free suspension culture. Journal of General Virology, v.81, n.10, p.2531-2543. 2000. LUA, L. H.; REID, S. Effect of time of harvest of budded virus on the selection of baculovirus FP mutants in cell culture. Biotechnology Progress, v.19, n.1, p.238-242. 2003. LUA, L.H. L.; NIELSEN, L. K.; REID, S. Sensitivity of Helicoverpa armigera nucleopolyhedrovirus polyhedra to sodium dodecyl sulfate. Biological Control, v.26, n.1, p.57-67. 2003. LYNN, D. E. Enhanced infectivity of occluded virions of Gypsy Moth nuclear polyhedrosis virus for cell cultures. Journal of Invertebrate Pathology, v.63, p.268-274. 1994. LYNN, D. E. Development and characterization of insect cell lines. Cytotechnology, v.20, p.3 - 11. 1996. MARTIGNONI, M.E. Baculovirus: an attractive biological alternative. In: W. Y. Garner; J. Harvey (Ed.). Chemical and biological controls in forestry. Washington, D.C.: American Chemical Society, 1984. p.55-67. MARUNIAK, J. E.; BROWN, S. E.; KNUDSON, D. L. Physical maps of SfMNPV baculovirus DNA and its genomic variants. Journal of Virology, v.136, n.1, p.221-234. 1984. MARUNIAK, J. E.; GARCIA-MARUNIAK, A.; SOUZA, M. L.; ZANOTTO, P. M.; MOSCARDI, F. Physical maps and virulence of Anticarsia gemmatalis nucleopolyhedrovirus genomic variants. Arch Virol, v.144, n.10, p.1991-2006. 1999. McINTOSH, A. H.; IGNOFFO, C. M.; ANDREWS, P. L. In vitro host range of five baculoviruses in lepidopteran cell lines. Intervirology, v.23, n.3, p.150-156. 1985. McINTOSH, A. H.; IGNOFFO, C. M.; CARLO, M. Effect of larval extract and protease K on the in vitro infectivity of Heliothis zea nonoccluded and alkali-liberated virions. Applied Entomology and Zoology, v.23, p.175-180. 1988. MORALES, L.; MOSCARDI, F.; GRAVENA, S. Potential of a baculovirus of Autographa-californica (Speyer) for the control of chreysodeixis-includens (Walker) and Anticarsia-gemmatalis Hubner (Lep, Noctuidae). Pesquisa Agropecuaria Brasileira, v.28, n.2, p.237-243. 1993.
________________________________________________________________________ 69Andréa Farias de Almeida, abril/2005

___________________________________________________________________________ Referências bibliográficas
MOSCARDI, F. Use of viruses for pest-control in brazil - the case of the nuclear polyhedrosis-virus of the soybean caterpillar, Anticarsia-gemmatalis. Memorias Do Instituto Oswaldo Cruz, v.84, p.51-56. 1989. MOSCARDI, F. Assessment of the application of baculoviruses for control of Lepidoptera. Annual Review of Entomology, v.44, p.257-289. 1999. MOSCARDI, F.; CORSO, I. C. Ação de baculovirus anticarsia sobre a lagarta da soja (Anticarsia gemmatalis) e outros Lepidópteros. In: Anais do Seminário Nacional de Pesquisa de Soja,. 1981, Londrina, Brasil EMBRAPA-CNPSO, v.2, p. 52-61. MOSCARDI, F.; SOUZA, M. L. Baculovírus para o controle de pragas: Panacéia ou realidade? Biotecnologia Ciência & Desenvolvimento, v.24, p.22-29. 2002. NIELSEN, L. K.; SMYTH, G. K.; GREENFIELD, P. F. Hemocytometer cell count distributions: implications of non-poisson behaviour. Biotechnology Progress, v.7, p.560-563. 1991. O'REILLY, D.R.; MILLER, L.K.; LUCKOW, V.A. Baculovirus expression vectors: a laboratory manual New York: Oxford University Press, 1994. p. PEARSON, M. N.; GROTEN, C.; ROHRMANN, G. F. Identification of the Lymantria dispar nucleopolyhedrovirus envelope fusion protein provides evidence for a phylogenetic division of the Baculoviridae. Journal of Virology, v.74, n.13, p.6126-6131. 2000. PEDRINI, M. R. S. Characterization of few polyhedra mutants of Helicoverpa armigera nucleopolyhedrovirus cultived in vitro. 2003. 136f. (PhD). Chemical Engineering Department, The University of Queensland, Brisbane -Australia. PEDRINI, M. R. S.; CHAN, L. C. L.; NIELSEN, L. K.; REID, S. In vitro production of Helicoverpa armigera single-nucleocapsid nucleopolyhedrovirus. Brazilian Archives of Biology and Technology. no prelo. PEDRINI, M. R. S.; WOLFF, J. L. C.; REID, S. Fast accumulation of few polyhedra mutants during passage of a Spodoptera frugiperda multicapsid nucleopolyhedrovirus (Baculoviridae) in Sf9 cell cultures. Annals of Applied Biology, v.145, n.1, p.107-112. 2004. PEREIRA, E.R.; SIQUEIRA, C.B.; MOSCARDI, F.; SOUZA, M. L. Comparação de isolados temporais do nucleopoliedrovirus de Anticarsia gemmatalis por análise de restrição do DNA com as enzimas Eco RI e Pst I. In: VII Encontro do Talento Estudantil da Embrapa Recursos Genéticos e Biotecnologia. 2002, Brasília - DF. PIJLMAN, G. P.; BORN, E. V.D.; MARTENS, D.E.; VLAK, J. M. Autographa californica baculoviruses with large genomic deletions are rapidly generated in infected insect cells. Journal of Virology, v.283, n.1, p.132-138. 2001. POPHAM, H. J. R.; BISCHOFF, D. S.; SLAVICEK, J. M. Both Lymantria dispar nucleopolyhedrovirus Enhancin genes contribute to viral potency. Journal of Virology, v.75, n.18, p.8639-8648. 2001.
________________________________________________________________________ 70Andréa Farias de Almeida, abril/2005

___________________________________________________________________________ Referências bibliográficas
POTTER, K. N.; JAQUES, R. P.; FAULKNER, P. Modification of Trichoplusia ni nuclear polyhedrosis virus passage in vivo. Intervirology, v.9, p.76-85. 1978. POTTER, K.N.; FAULKNER, P.; MACKINNON, E.A. Strain selection during serial passage of Trichoplusia ni nuclear polyhedrosis virus. Journal of Virology, v.18, n.3, p.1040-1050. 1976. RIBEIRO, B. M.; ZANOTTO, P. M. A.; McDOWELL, S.; SOUZA, M. L.; KITAJIMA, E. W. Characterization of a baculovirus infecting the passion fruit caterpilar Dione Juno Juno. Biocell, v.21, n.1, p.71 - 82. 1997. RIBEIRO, H.C.T.; PAVAN, O.H.O.; MUOTRI, A.R. Comparative susceptibility of two different hosts to genotypic variants of the Anticarsia gemmatalis nuclear polyhedrosis virus. Entomologia Experimentalis et Applicata.v.83, n.2, p.233-237.1997 RIEGEL, C. I.; SLAVICEK, J. M. Characterization of the replication cycle of the Lymantria dispar nuclear polyhedrosis virus. Virus Res, v.51, n.1, p.9-17. 1997. ROHRMANN, G. F. Nucleopolyhedrovirus. In: A. Granoff; R. G. Webster (Ed.). Encyclopedia of Virology. New York: Academic Press, v.1, 1999. p.146-152. ROSAS-ACOSTA, G.; BRAUNAGEL, S. C.; SUMMERS, M.D. Effects of deletion and overexpression of the Autographa californica nuclear polyhedrosis virus FP25K gene on synthesis of two occlusion-derived virus envelopeproteins and their transport into virus-induced intranuclear embranes. Journal of Virology, v.75, n.22, p.10829-10842. 2001. SCHLAEGER, E. J. Medium design for insect cell culture. Cytotechnology, v.20, p.57-70. 1996. SCHMITT, A.T. Using baculovirus erinnyis in the biological control of cassava hornworm.Cassava Newsletter, v.12, n.1, p.1-4. 1988 SCHMID, G. Insect cell cultivation: growth and kinectics. Cytotechnology, v.20, p.43 - 56. 1996. SIMON, O.; WILLIAMS, T.; LOPEZ-FERBER, M.; CABALLERO, P. Genetic structure of a Spodoptera frugiperda nucleopolyhedrovirus population: high prevalence of deletion genotypes. Applied and Environmental Microbiology, v.70, n.9, p.5579-5588. 2004. SLAVICEK, J. M. Temporal analysis and spatial mapping of Lymantria dispar nuclear polyhedrosis virus transcripts and in vitro translation polypeptides. Virus Res, v.20, n.3, p.223-236. 1991. SLAVICEK, J. M.; HAYES-PLAZOLLES, N; KELLY, M.E. Identification of a Lymantria dispar nucleopolyhedrovirus isolate that does not accumulate few-polyhedra mutants during extended serial passage in cell culture. Biological Control, v.22, n.2, p.159-168. 2001. SLAVICEK, J. M.; HAYES-PLAZOLLES, N. The Lymantria dispar nucleopolyhedrovirus contains the capsid-associated p24 protein gene. Virus Genes, v.26, n.1, p.15-18. 2003.
________________________________________________________________________ 71Andréa Farias de Almeida, abril/2005

___________________________________________________________________________ Referências bibliográficas
SLAVICEK, J. M.; HAYES-PLAZOLLES, N.; KELLY, M. E. Rapid formation of few polyhedra mutants of Lymantria-Dispar multinucleocapsid nuclear polyhedrosis-virus during serial passage in cell-culture. Biological Control, v.5, n.2, p.251-261. 1995. SLAVICEK, J. M.; MERCER, M. J.; POHLMAN, D.; KELLY, M. E.; BISCHOFF, D. S. Identification of a novel Lymantria dispar nucleopolyhedrovirus mutant that exhibits abnormal polyhedron formation and virion occlusion. Journal of Invertebrate Pathology, v.72, n.1, p.28-37. 1998. SLAVICEK, J. M.; POPHAM, H. J. R.; RIEGEL, C. I. Deletion of the Lymantria dispar multicapsid nucleopolyhedrovirus ecdysteroid UDP-glucosyl transferase gene enhances viral killing speed in the last instar of the gypsy moth. Biological Control, v.16, n.1, p.91-103. 1999. SLAVICEK, J.M.; MERCER, M.J.; KELLY, M.E.; HAYES-PLAZOLLES, N. Isolation of a baculovirus variante that exhibits enhanced polyhedra production stability during serial passage in cell culture. Journal of Invertebrate Pathology, v.67, p.153-160. 1996. SOUZA, M. L.; CASTRO, M.E.B.; SIHKER, W.; DALMOLIN, C.C.; PEDRINI, M. R. S.; SILVA, F.R. Caracterização do gene 25K FP de Anticarsia gemmatalis nucleopoliedrovirus. Boletim de Pesquisa e Desenvolvimento. Brasilia: 16 p. 2003. SOUZA, M.L.; CASTRO, M. E. B.; SIHKER, W.; RIBEIRO, Z. M. A.; MOSCARDI, F. Caracterização de baculovírus utilizados no controle de pragas: técnicas de caracterização de vírus de insetos. Biotecnologia Ciência & Desenvolvimento, v.24. 2002. SPENGER, A.; GRABHERR, R.; TOLLNER, L.; KATINGER, H.; ERNST, W. Altering the surface properties of baculovirus Autographa californica NPV by insertional mutagenesis of the envelope protein gp64. Eur J Biochem, v.269, n.18, p.4458-4467. 2002. TONSO, A. Monitoramento e operação de cultivos de células animais em sistemas de perfusão. 2000. 109f. (Doutoramento). Engenharia Química, Escola Politécnica da Universidade de São Paulo, São Paulo. VAIL, P. V.; ROMINE, C. L.; VAUGHN, J. L. Infectivity of nuclear polyhedrosis virus extract with digestive juices. Journal of Invertebrate Pathology, v.33, p.328-330. 1979. VALICENTE, F.H; COSTA, E. F. Controle da lagarta do cartucho Spodoptera frugiperda (J. E. Smith) com baculovirus spodoptera, aplicado via água de irrigação.In: Anais da Sociedade de Entomológica Brasileira, v.24, p.61-67. 1995. VALICENTE, F.H.; CRUZ, I. Controle biológico da lagarta-do-cartucho, Spodoptera frugiperda, com o baculovírus. Circular Técnica n° 15 1991. VALICENTE, F.H.; PEIXOTO, M.J.; PAIVA, E.; KITAJIMA, E. W. Identificação e purificação de um vírus de poliedrose nuclear da lagarta-de-cartucho Spodoptera frugiperda. In: Anais da Sociedade de Entomológica Brasileira. 1989, v.18, p. 71 - 82. VAUGHN, J. L.; GOODWIN, R. H.; TOMPKINS, G. J.; MCCAWLEY, P. The establishment of two cell lines from the insect Spodoptera frugiperda (Lepidoptera; Noctuidae). In Vitro, v.13, n.4, p.213-217. 1977.
________________________________________________________________________ 72Andréa Farias de Almeida, abril/2005

___________________________________________________________________________ Referências bibliográficas
VOLKMAN, L. E.; SUMMERS, M. D. Autographa californica nuclear polyhedrosis virus: comparative infectivity of the occluded, alkali-liberated, and nonoccluded forms. Journal of Invertebrate Pathology, v.30, p.102-103. 1977. VOLKMAN, L. E.; SUMMERS, M. D.; HSIEH, C. H. Occluded and nonoccluded nuclear polyhedrosis virus grown in Trichoplusia ni: comparative neutralization comparative infectivity, and in vitro growth studies. Journal of Virology, v.19, n.3, p.820–832. 1976. WEISS, S. A.; SMITH, G. C.; KALTER, S. S.; VAUGHN, J. L.; DOUGHERTY, E. Improved replication of Autographa californica nuclear polyhedrosis virus in roller bottles: characterization of the progeny virus. Intervirology, v.15, n.4, p.213-222. 1981. WEISS, S. A.; THOMAS, D. W.; DUNLOP, B. F.; GEORGIS, R.; VAIL, P.; HOFFMAN, D. In vitro production of viral pesticides: key elements. In: Biopesticides: opportunities for Australian industry. 1994, Brisbane, Australia June 9-10. p. 137. WICKHAM, T. J.; DAVIS, T.; GRANADOS, R. R.; HAMMER, D. A.; SHULER, M. L.; WOOD, H. A. Baculovirus defective interfering particles are responsible for variations in recombinant protein production as a function of multiplicity of infection. Biotechnology Letters, v.13, p.483-488. 1991. WOLFF, J. L.; MORAES, R. H.; KITAJIMA, E.; LEAL, E. S.; ZANOTTO, P. M. Identification and characterization of a baculovirus from Lonomia obliqua (Lepidoptera: Saturniidae). Journal of Invertebrate Pathology, v.79, n.3, p.137-145. 2002. YOSHIFUMI, H.; TOHRU, H.; YASUMASA, U.; TOMONORI, F.; YOSHITAKA, S.; TSUGUO, M. Sequence analysis of the Plutella xylostella granulovirus genome. Journal of Virology, v.275, n.2, p.358-372. 2000. ZANOTTO, P. M.; KESSING, B. D.; MARUNIAK, J. E. Phylogenetic interrelationships among baculoviruses: evolutionary rate and host associations. Journal of Invertebrate Pathology, v.62, n.2, p.147-164. 1993.
________________________________________________________________________ 73Andréa Farias de Almeida, abril/2005

Anexos

__________________________________________________________________________________________ Anexos
Figura A.1. Fluxograma para a adaptação das células ao cultivo em suspensão.
Figura A.2. Fluxograma para infecção preliminar com SfMNPV.
_______________________________________________________________________________________ 75Andréa Farias de Almeida, abril/2005

__________________________________________________________________________________________ Anexos
Figura A.3. Fluxograma para infecção com VDP do SfMNPV.
Figura A.4. Fluxograma para a contagem de poliedros.
_______________________________________________________________________________________ 76Andréa Farias de Almeida, abril/2005

__________________________________________________________________________________________ Anexos
Tabela A.1 – Pontos experimentais da infecção das células Sf9 com a 2ª passagem SfMNPV
(HyClone + 5% SFB)
Infecção em
triplicata
Pico de concentração das células infectadas x 106
(células/mL)
Produção volumétrica de poliedros x 108
(poliedros/mL)
Produção específica de poliedros (poliedros/célula)
1 1,3 (± 0,195)* 3,6 (± 0,756) ** 276,9 (± 58,149) **
2 1,1 (± 0,165)* 5,4 (± 1,134) ** 490,9 (± 103,1) **
3 1,3 (± 0,195)* 5,8 (± 1,218) ** 446,2 (± 93,7) **
média 1,2 (± 0,180)* 4,9 (± 1,029) ** 404,7 (± 60,7) **
*Erro relativo ± 15% **Erro relativo ± 21%
Tabela A.2 – Pontos experimentais da infecção das células Sf21 com a 2ª passagem SfMNPV
(HyClone + 5% SFB)
Infecção em
triplicata
Pico de concentração das células infectadas x 106
(células/mL)
Produção volumétrica de poliedros x 108
(poliedros/mL)
Produção específica de poliedros (poliedros/célula)
1 1,0 (± 0,150)* 2,6 (± 0,546) ** 260 (± 54,6) **
2 0,75 (± 0,1125)* 2,6 (± 0,546) ** 347 (± 52,0) **
3 0,7 (± 0,105)* 2,1 (± 0,441) ** 300 (± 63,0) **
média 0,82 (± 0,1225)* 2,4 (± 0,365) ** 292,2 (± 61,5) **
*Erro relativo ± 15% **Erro relativo ± 21%
Tabela A.3 – Pontos experimentais da infecção das células Sf9 com a 3ª passagem SfMNPV
(HyClone + 5% SFB)
Infecção em
triplicata
Pico de concentração das células infectadas x 106
(células/mL)
Produção volumétrica de poliedros x 108
(poliedros/mL)
Produção específica de poliedros (poliedros/célula)
1 0,61 (± 0,0915)* 2,6 (± 0,546) ** 426,2 (± 89,5) **
2 0,56 (±0,084)* 2,4 (± 0,504) ** 428,6 (± 90,0) **
3 0,66 (± 0,099)* 2,4 (± 0,504) ** 363,6 (± 76,4) **
média 0,61 (± 0,0915)* 2,5 (± 0,518) ** 409,8 (± 86,1) **
*Erro relativo ± 15% **Erro relativo ± 21%
_______________________________________________________________________________________ 77Andréa Farias de Almeida, abril/2005

__________________________________________________________________________________________ Anexos
Tabela A.4 – Pontos experimentais da infecção das células Sf9 com a 3ª passagem SfMNPV
(HyClone + 2,5% SFB)
Infecção em
triplicata
Pico de concentração das células infectadas x 106
(células/mL)
Produção volumétrica de poliedros x 108
(poliedros/mL)
Produção específica de poliedros (poliedros/célula)
1 0,91 (± 0,1365)* 2,3 (± 0,483) ** 252,7 (± 53,1) **
2 0,85 (± 0,1275)* 2,4 (± 0,504) ** 282,4 (± 59,3) **
3 0,85 (± 0,1275)* 2,1 (± 0,441) ** 247,1 (± 51,9) **
média 0,87 (± 0,1305)* 2,3 (± 0,476) ** 264,4 (± 55,5) **
*Erro relativo ± 15% **Erro relativo ± 21%
Tabela A.5 – Pontos experimentais da infecção das células Sf9 com a 4ª passagem SfMNPV
(HyClone + 5% SFB)
Infecção em
duplicata
Pico de concentração das células infectadas x 106
(células/mL)
Produção volumétrica de poliedros x 108
(poliedros/mL)
Produção específica de poliedros (poliedros/célula)
1 0,75 (± 0,1125) * 2,1 (± 0,441) ** 280 (± 58,8) **
2 0,81 (± 0,1215) * 2,0 (± 0,420) ** 246,9 (± 51,9) **
média 0,78 (± 0,1700) * 2,05 (± 0,4305) ** 262,8 (± 55,2) **
*Erro relativo ± 15% **Erro relativo ± 21%
Tabela A.6 – Pontos experimentais da infecção das células Sf9 com a 5ª passagem SfMNPV
(HyClone + 5% SFB)
Infecção em
duplicata
Pico de concentração das células infectadas x 106
(células/mL)
Produção volumétrica de poliedros x 108
(poliedros/mL)
Produção específica de poliedros (poliedros/célula)
1 1,5 (± 0,225) * 2,5 (± 0,525) ** 166,7 (± 35,0) **
2 1,4 (± 0,21) * 2,5 (± 0,525) ** 178,6 (± 37,5) **
média 1,45 (± 0,2175) * 2,5 (± 0,525) ** 172,4 (± 36,2) **
*Erro relativo ± 15% **Erro relativo ± 21%
_______________________________________________________________________________________ 78Andréa Farias de Almeida, abril/2005

__________________________________________________________________________________________ Anexos
Tabela A.7 – Pontos experimentais da infecção das células Sf9 com a 6ª passagem SfMNPV
(HyClone + 5% SFB)
Infecção em
duplicata
Pico de concentração das células infectadas x 106
(células/mL)
Produção volumétrica de poliedros x 108
(poliedros/mL)
Produção específica de poliedros (poliedros/célula)
1 0,79 (± 0,1185) * 1,4 (± 0,294) ** 177,2 (± 37,21) **
2 0,72 (± 0,108) * 1,5 (± 0,315) ** 208,3 (± 43,75) **
média 0,76 (± 0,11325) * 1,45 (± 0,3045) ** 190,8 (± 40,07) **
*Erro relativo ± 15% **Erro relativo ± 21%
Tabela A.8 – Pontos experimentais da infecção das células Sf9 com a 7ª passagem SfMNPV
(HyClone + 5% SFB)
Infecção em
duplicata
Pico de concentração das células infectadas x 106
(células/mL)
Produção volumétrica de poliedros x 108
(poliedros/mL)
Produção específica de poliedros (poliedros/célula)
1 1,8 (± 0,27)* 1,3 (± 0,273)** 72,22 (± 15,17) **
2 1,6 (± 0,24)* 1,2 (± 0,252) ** 75 (± 15,75) **
média 1,7 (± 0,255)* 1,25 (± 0,2625) ** 73,53 (± 15,4) **
*Erro relativo ± 15% **Erro relativo ± 21%
Tabela A.9 – Pontos experimentais para a construção da curva de calibração da glicose
ABS 1
ABS 2
ABS (Med)
Conc (mg/mL)
0,019 0,02 0,0195 0,1 0,073 0,072 0,0725 0,2 0,123 0,127 0,1250 0,3 0,179 0,168 0,1735 0,4 0,233 0,244 0,2385 0,5 0,267 0,266 0,2665 0,6 0,325 0,328 0,3265 0,7 0,377 0,363 0,3700 0,8 0,428 0,414 0,4210 0,9 0,463 0,416 0,4395 1
_______________________________________________________________________________________ 79Andréa Farias de Almeida, abril/2005

__________________________________________________________________________________________ Anexos
0,0 0,1 0,2 0,3 0,4 0,50,0
0,2
0,4
0,6
0,8
1,0
Curva de Calibração da Glicose
Abs
orbâ
ncia
(AB
S)
Concentração de Glicose (mg/mL)
Figura A.5 – Curva de Calibração da Glicose
Regressão Linear : Y = A + B * X Parâmetro Valor Erro ------------------------------------------------------------ A 0,04263 0,01539 B 2,06877 0,05464 ------------------------------------------------------------ R = 0,99722
_______________________________________________________________________________________ 80Andréa Farias de Almeida, abril/2005

__________________________________________________________________________________________ Anexos
Tabela A.10 – Pontos experimentais para o cálculo dos parâmetros cinéticos das células Sf9
(HyClone + 5% SFB)
tempo (h)
Abs 1
Abs 2
Abs média
Glicose g/L diluição C. glic
g/L 0 0,409 0,319 0,364 0,155344 10 1,55343524 0,395 0,394 0,395 0,170087 10 1,70086648 0,32 0,312 0,316 0,132141 10 1,32141372 0,292 0,285 0,289 0,118848 10 1,18848496 0,241 0,236 0,239 0,094679 10 0,946794120 0,233 0,234 0,234 0,092263 10 0,922626C
élul
as C
ontro
le
144 0,168 0,17 0,169 0,061085 10 0,610846
tempo (h)
Abs 1
Abs 2
Abs média
Glicose g/L diluição C. glic
g/L 0 0,336 0,339 0,338 0,142534 10 1,42534024 0,458 0,388 0,423 0,183863 10 1,83862948 0,316 0,335 0,326 0,136733 10 1,36733472 0,395 0,347 0,371 0,158727 10 1,58727296 0,361 0,332 0,347 0,146884 10 1,468844120 0,357 0,366 0,362 0,154135 10 1,541351C
élul
as In
fect
adas
144 0,347 0,331 0,339 0,143259 10 1,432590
Tabela A.11 – Pontos experimentais para o cálculo dos parâmetros cinéticos das células Sf9
(HyClone + 2,5% SFB)
tempo (h)
Abs 1
Abs 2
Abs média
Glicose g/L diluição C. glic
g/L 0 0,342 0,342 0,342 0,144709 10 1,44709224 0,399 0,416 0,408 0,176371 10 1,76370548 0,339 0,332 0,336 0,141567 10 1,41567272 0,315 0,309 0,312 0,130208 10 1,30207896 0,245 0,25 0,248 0,09903 10 0,990299120 0,235 0,241 0,238 0,094438 10 0,944378C
élul
as c
ontro
le
144 0,171 0,171 0,171 0,062051 10 0,620514
tempo (h)
Abs 1
Abs 2
Abs média
Glicose g/L diluição C. glic
g/L 0 0,336 0,347 0,342 0,144467 10 1,44467524 0,394 0,392 0,393 0,169362 10 1,69361548 0,292 0,326 0,309 0,128758 10 1,28757772 0,348 0,378 0,363 0,15486 10 1,54860196 0,327 0,323 0,325 0,136492 10 1,364917120 0,332 0,339 0,336 0,141567 10 1,415672C
élul
as In
fect
adas
144 0,304 0,308 0,306 0,127308 10 1,273075
_______________________________________________________________________________________ 81Andréa Farias de Almeida, abril/2005

__________________________________________________________________________________________ Anexos
Tabela A.12 – Pontos experimentais para o cálculo dos parâmetros cinéticos das células Sf21
(HyClone + 5% SFB)
tempo (h)
Abs 1
Abs 2
Abs média
Glicose g/L diluição C. glic
g/L 0 0,35 0,338 0,344 0,145676 10 1,45675924 0,389 0,397 0,393 0,169362 10 1,69361548 0,274 0,324 0,299 0,123924 10 1,23923972 0,349 0,33 0,340 0,143501 10 1,43500796 0,261 0,273 0,267 0,108456 10 1,084557120 0,227 0,231 0,229 0,090087 10 0,900873C
élul
as C
ontro
le
144 0,173 0,179 0,176 0,064468 10 0,644683
tempo (h)
Abs 1
Abs 2
Abs média
Glicose g/L diluição C. glic
g/L 0 0,344 0,36 0,352 0,149543 10 1,49543024 0,38 0,395 0,388 0,166703 10 1,66702948 0,268 0,33 0,299 0,123924 10 1,23923972 0,369 0,374 0,372 0,158969 10 1,58968996 0,332 0,336 0,334 0,140842 10 1,408421120 0,299 0,363 0,331 0,139392 10 1,393920C
élul
as In
fect
adas
144 0,349 0,323 0,336 0,141809 10 1,418089
0 24 48 72 96 120 144 168 192 2160
1
2
3
4
5
6
7
8
9
10
Concentração das células não infectadas Concentração das células infectadas Viabilidade das células não infectadas Viabilidade das células infectadas
Tempo (h)
Cél
uas
viáve
is x
106 (c
élul
as/m
L)
0
10
20
30
40
50
60
70
80
90
100
Curva de Crescimento Celular - Sf9 - P2 (HyClone + 5% SFB)
Viabilidade (%)
Figura A.6. Curva de crescimento das células Sf9 infectadas com VEC – P2 (SfMNPV)
_______________________________________________________________________________________ 82Andréa Farias de Almeida, abril/2005

__________________________________________________________________________________________ Anexos
0 24 48 72 96 120 144 168 192 2160
2
4
6
8
10
12
Curva de Crescimento Celular - Sf21 - P2 (HyClone + 5% SFB)
Concentração das células não infectadas Concentração das células infectadas Viabilidade das células não infectadas Viabilidade das células infectadas
Tempo (h)
Cél
ulas
viá
veis
x 1
06 /mL
30
40
50
60
70
80
90
100
Viabilidade (%)
Figura A.7. Curva de crescimento das células Sf21 infectadas com VEC – P2 (SfMNPV)
0 24 48 72 96 120 144 1680
1
2
3
4
5
6
7
8
9
10
Curva de Crescimento Celular - Sf9 - P3 (HyClone + 2,5% SFB)
Concentração das células não infectadas Concentração das células infectadas Viabilidade das células não infectadas Viabilidade das células infectadas
Tempo (h)
Cél
ulas
viá
veis
x 1
06 /mL
0
10
20
30
40
50
60
70
80
90
100
Viabilidade (%)
Figura A.8. Curva de crescimento das células Sf9 infectadas com VEC – P3 (SfMNPV) –
(HyClone + 2,5%SFB)
_______________________________________________________________________________________ 83Andréa Farias de Almeida, abril/2005

__________________________________________________________________________________________ Anexos
0 24 48 72 96 120 1440
1
2
3
4
5
6
7
8
9
10
Curva de Crescimento Celular - Sf9 - P3 (HyClone + 5% SFB)
Concentração das células não infectadas Concentração das células infectadas Viabilidade das células não infectadas Viabilidade das células infectadas
Tempo (h)
Cél
ulas
viá
veis
x 1
06 /mL
0
10
20
30
40
50
60
70
80
90
100
Viabilidade (%)
Figura A.9. Curva de crescimento das células Sf9 infectadas com VEC – P3 (SfMNPV) –
(HyClone + 5% SFB)
_______________________________________________________________________________________ 84Andréa Farias de Almeida, abril/2005

__________________________________________________________________________________________ Anexos
Tabela A.6 - Ajustes para a curva de produção específica de poliedros em células Sf21 (HyClone + 5% SFB)
72 28,66118 75,25424 31,46461 78,50847 34,88198 81,76271 39,02545 85,01695 44,0167 88,27119 49,98231 91,52542 57,0461 94,77966 65,31818 98,0339 74,88062 101,2881 85,77068 104,5424 97,96351 107,7966 111,3577 111,0509 125,7676 114,3051 140,9261 117,5593 156,5003 120,8136 172,1183 124,0678 187,4043 127,322 202,0134 130,5763 215,6617 133,8305 228,1439 137,0848 239,3395 140,339 249,2071 143,5932 257,7712 146,8475 265,1049 150,1017 271,3133 153,3559 276,5182 156,6102 280,8463 159,8644 284,421 163,1186 287,3568 166,3729 289,7568
_______________________________________________________________________________________ 85
169,6271 291,7115 172,8814 293,2985 176,1356 294,5839 179,3898 295,6228 182,6441 296,4611 185,8983 297,1368 189,1525 297,6806 192,4068 298,1181 195,661 298,4697 198,9153 298,7522 202,1695 298,979 205,4237 299,1611 208,678 299,3072 211,9322 299,4244 215,1864 299,5184 218,4407 299,5938 221,6949 299,6543 224,9492 299,7028 228,2034 299,7416 231,4576 299,7728 234,7119 299,7978 237,9661 299,8178 241,2203 299,8339 244,4746 299,8468 247,7288 299,8571 250,9831 299,8654 254,2373 299,872 257,4915 299,8773 260,7458 299,8816
264 299,885
Andréa Farias de Almeida, abril/2005

__________________________________________________________________________________________ Anexos
Tabela A.7 - Ajustes para a curva de produção volumétrica de poliedros em células Sf21 (HyClone + 5% SFB)
72 0,21004 74,44068 0,23176 76,88136 0,25629 79,32203 0,28393 81,76271 0,31496 84,20339 0,3497 86,64407 0,3884 89,08475 0,43134 91,52542 0,47874 93,9661 0,53077 96,40678 0,58754 98,84746 0,64908 101,2881 0,7153 103,7288 0,78602 106,1695 0,86092 108,6102 0,93956 111,0509 1,02137 113,4915 1,10567 115,9322 1,19169 118,3729 1,27859 120,8136 1,36549 123,2542 1,45153 125,6949 1,53585 128,1356 1,61769 130,5763 1,69636 133,017 1,7713 135,4576 1,84206 137,8983 1,90833 140,339 1,96991 142,7797 2,02674
______________________________________________________________________________________ 86
145,2203 2,07881 147,661 2,12626 150,1017 2,16924 152,5424 2,20799 154,9831 2,24276 157,4237 2,27383 159,8644 2,3015 162,3051 2,32607 164,7458 2,3478 167,1864 2,36699 169,6271 2,3839 172,0678 2,39876 174,5085 2,4118 176,9492 2,42323 179,3898 2,43323 181,8305 2,44197 184,2712 2,4496 186,7119 2,45626 189,1525 2,46206 191,5932 2,46712 194,0339 2,47152 196,4746 2,47535 198,9153 2,47868 201,3559 2,48157 203,7966 2,48409 206,2373 2,48628 208,678 2,48818 211,1186 2,48983 213,5593 2,49126
216 2,4925
Andréa Farias de Almeida, abril/2005

__________________________________________________________________________________________ Anexos
Tabela A.8 - Ajustes para a curva de produção específica de poliedros em células Sf9 (HyClone + 5% SFB)
72 23,20697 74,44068 25,87334 76,88136 28,83946 79,32203 32,13303 81,76271 35,78286 84,20339 39,81846 86,64407 44,26962 89,08475 49,16579 91,52542 54,53538 93,9661 60,40486 96,40678 66,79781 98,84746 73,73379 101,2881 81,22712 103,7288 89,28562 106,1695 97,90939 108,6102 107,0896 111,0509 116,8077 113,4915 127,0342 115,9322 137,7288 118,3729 148,8403 120,8136 160,3069 123,2542 172,0575 125,6949 184,0132 128,1356 196,0892 130,5763 208,1973 133,017 220,2481 135,4576 232,1542 137,8983 243,8322 140,339 255,2053 142,7797 266,2048
______________________________________________________________________________________ 87
145,2203 276,7718 147,661 286,8582 150,1017 296,4267 152,5424 305,4511 154,9831 313,9155 157,4237 321,8139 159,8644 329,1486 162,3051 335,9294 164,7458 342,1722 167,1864 347,8978 169,6271 353,1308 172,0678 357,8982 174,5085 362,2289 176,9492 366,1524 179,3898 369,6986 181,8305 372,8967 184,2712 375,7753 186,7119 378,3618 189,1525 380,6821 191,5932 382,7607 194,0339 384,6204 196,4746 386,2824 198,9153 387,7661 201,3559 389,0895 203,7966 390,2689 206,2373 391,3193 208,678 392,2542 211,1186 393,0858 213,5593 393,8251
216 394,482
Andréa Farias de Almeida, abril/2005

__________________________________________________________________________________________ Anexos
Tabela A.9 - Ajustes para a curva de produção volumétrica de poliedros em células Sf9 (HyClone + 5% SFB)
72 0,28639 74,44068 0,3204 76,88136 0,35813 79,32203 0,39992 81,76271 0,44611 84,20339 0,49707 86,64407 0,55313 89,08475 0,61466 91,52542 0,68198 93,9661 0,7554 96,40678 0,83519 98,84746 0,92159 101,2881 1,01474 103,7288 1,11474 106,1695 1,22158 108,6102 1,33514 111,0509 1,45519 113,4915 1,58139 115,9322 1,71326 118,3729 1,85018 120,8136 1,99142 123,2542 2,13615 125,6949 2,28343 128,1356 2,43226 130,5763 2,58159 133,017 2,73036 135,4576 2,87754 137,8983 3,02211 140,339 3,16316 142,7797 3,29984
145,2203 3,43143 147,661 3,55733
150,1017 3,67707 152,5424 3,7903 154,9831 3,8968 157,4237 3,99645 159,8644 4,08926 162,3051 4,17532 164,7458 4,25479 167,1864 4,3279 169,6271 4,39492 172,0678 4,45616 174,5085 4,51196 176,9492 4,56267 179,3898 4,60864 181,8305 4,65022 184,2712 4,68775 186,7119 4,72157 189,1525 4,752 191,5932 4,77934 194,0339 4,80387 196,4746 4,82585 198,9153 4,84553 201,3559 4,86312 203,7966 4,87885 206,2373 4,89289 208,678 4,90542
211,1186 4,9166 213,5593 4,92656
216 4,93543
______________________________________________________________________________________ 88Andréa Farias de Almeida, abril/2005

__________________________________________________________________________________________ Anexos
Tabela A.10 - Ajustes para a curva de produção específica de poliedros em células Sf9 (HyClone + 5% SFB)
48 83,70201 50,0339 88,96552 52,0678 94,93125 54,10169 101,6555 56,13559 109,1873 58,16949 117,565 60,20339 126,811 62,23729 136,9282 64,27119 147,8951 66,30508 159,6626 68,33898 172,1519 70,37288 185,2547 72,40678 198,8349 74,44068 212,7338 76,47458 226,7767 78,50847 240,7815 80,54237 254,5682 82,57627 267,9677 84,61017 280,83 86,64407 293,0303 88,67797 304,4722 90,71186 315,0896 92,74576 324,8452 94,77966 333,7281 96,81356 341,7498 98,84746 348,94 100,8814 355,3419 102,9153 361,0081 104,9492 365,9968 106,9831 370,3686
109,017 374,1842 111,0509 377,5026 113,0848 380,3796 115,1186 382,8673 117,1525 385,0134 119,1864 386,861 121,2203 388,4491 123,2542 389,8119 125,2881 390,98 127,322 391,98 129,3559 392,8355 131,3898 393,5666 133,4237 394,1911 135,4576 394,7242 137,4915 395,179 139,5254 395,5668 141,5593 395,8975 143,5932 396,1793 145,6271 396,4194 147,661 396,6239 149,6949 396,7981 151,7288 396,9465 153,7627 397,0728 155,7966 397,1803 157,8305 397,2718 159,8644 397,3497 161,8983 397,416 163,9322 397,4724 165,9661 397,5204
168 397,5613
_______________________________________________________________________________________ 89Andréa Farias de Almeida, abril/2005

__________________________________________________________________________________________ Anexos
Tabela A.11 - Ajustes para a curva de produção volumétrica de poliedros em células Sf9 (HyClone + 5% SFB)
48 0,51603 50,0339 0,5471 52,0678 0,5825 54,10169 0,6226 56,13559 0,66773 58,16949 0,71818 60,20339 0,77412 62,23729 0,83561 64,27119 0,90254 66,30508 0,97465 68,33898 1,05146 70,37288 1,1323 72,40678 1,2163 74,44068 1,30246 76,47458 1,38964 78,50847 1,47667 80,54237 1,56236 82,57627 1,64561 84,61017 1,72545 86,64407 1,80107 88,67797 1,87184 90,71186 1,93734 92,74576 1,99735 94,77966 2,05182 96,81356 2,10083 98,84746 2,14459 100,8814 2,1834 102,9153 2,21761 104,9492 2,2476 106,9831 2,27377
_______________________________________________________________________________________ 90
109,017 2,29651 111,0509 2,31621 113,0848 2,33321 115,1186 2,34784 117,1525 2,36041 119,1864 2,37119 121,2203 2,38041 123,2542 2,38829 125,2881 2,39502 127,322 2,40075 129,3559 2,40564 131,3898 2,4098 133,4237 2,41333 135,4576 2,41634 137,4915 2,4189 139,5254 2,42107 141,5593 2,42291 143,5932 2,42447 145,6271 2,4258 147,661 2,42693 149,6949 2,42788 151,7288 2,42869 153,7627 2,42938 155,7966 2,42996 157,8305 2,43046 159,8644 2,43088 161,8983 2,43123 163,9322 2,43153 165,9661 2,43179
168 2,432
Andréa Farias de Almeida, abril/2005

__________________________________________________________________________________________ Anexos
Tabela A.12 - Ajustes para a curva de produção específica de poliedros em células Sf9 (HyClone + 2,5% SFB)
48 17,27487 50,44068 20,391 52,88136 23,86784 55,32203 27,73359 57,76271 32,01501 60,20339 36,73637 62,64407 41,91815 65,08475 47,57564 67,52542 53,7174 69,9661 60,34374 72,40678 67,44535 74,84746 75,00209 77,28814 82,9822 79,72881 91,34197 82,16949 100,0261 84,61017 108,9687 87,05085 118,0951 89,49153 127,3238 91,9322 136,5697 94,37288 145,7473 96,81356 154,7732 99,25424 163,5698 101,6949 172,0674 104,1356 180,2061 106,5763 187,9376 109,017 195,2254 111,4576 202,0446 113,8983 208,3818 116,339 214,2335 118,7797 219,6051
______________________________________________________________________________________ 91
121,2203 224,5095 123,661 228,9652 126,1017 232,995 128,5424 236,625 130,9831 239,8828 133,4237 242,797 135,8644 245,3961 138,3051 247,7083 140,7458 249,7603 143,1864 251,5777 145,6271 253,1843 148,0678 254,6023 150,5085 255,8521 152,9492 256,9521 155,3898 257,9193 157,8305 258,7689 160,2712 259,5146 162,7119 260,1685 165,1525 260,7416 167,5932 261,2435 170,0339 261,683 172,4746 262,0675 174,9153 262,4039 177,3559 262,698 179,7966 262,9551 182,2373 263,1798 184,678 263,3761 187,1186 263,5476 189,5593 263,6974
192 263,8282
Andréa Farias de Almeida, abril/2005

__________________________________________________________________________________________ Anexos
Tabela A.13 - Ajustes para a curva de produção volumétrica de poliedros em células Sf9 (HyClone + 2,5% SFB)
48 0,14858 50,44068 0,17628 52,88136 0,20711 55,32203 0,24131 57,76271 0,27909 60,20339 0,32066 62,64407 0,36617 65,08475 0,41575 67,52542 0,46945 69,9661 0,52726 72,40678 0,58908 74,84746 0,65473 77,28814 0,72393 79,72881 0,79629 82,16949 0,87134 84,61017 0,94852 87,05085 1,02719 89,49153 1,10666 91,9322 1,18623 94,37288 1,26517 96,81356 1,34279 99,25424 1,41843 101,6949 1,49151 104,1356 1,56155 106,5763 1,62812 109,017 1,69093 111,4576 1,74976 113,8983 1,80449 116,339 1,8551 118,7797 1,90163
______________________________________________________________________________________ 92
121,2203 1,94417 123,661 1,98289 126,1017 2,01797 128,5424 2,04962 130,9831 2,07808 133,4237 2,10359 135,8644 2,12638 138,3051 2,1467 140,7458 2,16476 143,1864 2,18079 145,6271 2,19499 148,0678 2,20755 150,5085 2,21863 152,9492 2,22841 155,3898 2,23703 157,8305 2,24461 160,2712 2,25127 162,7119 2,25713 165,1525 2,26227 167,5932 2,26679 170,0339 2,27075 172,4746 2,27422 174,9153 2,27726 177,3559 2,27992 179,7966 2,28226 182,2373 2,2843 184,678 2,28609 187,1186 2,28765 189,5593 2,28902
192 2,29022
Andréa Farias de Almeida, abril/2005

__________________________________________________________________________________________ Anexos
0 20 40 60 80 10012,5
13,0
13,5
14,0
14,5
15,0
15,5
16,0
Ln (c
once
ntra
ção
celu
lar)
Tempo (h) Ln (concentração de células não infectadas) Ln (concentração de células infectadas)
Figura A.10. Ln (concentração celular) versus tempo (HyClone+5%SFB)
0 24 48 72 96
0,15
0,20
0,25
0,30
0,35
0,40
0,45
0,50
0,55
Ln (c
once
ntra
ção
de s
ubst
rato
)
Tempo (h) Ln (concentração de substrato - células não infectadas) Ln (concentração de substrato - células infectadas)
Figura A.11. Ln (concentração de substrato) versus tempo (HyClone + 5% SFB)
______________________________________________________________________________________ 93Andréa Farias de Almeida, abril/2005

__________________________________________________________________________________________ Anexos
48 72 96 12017,6
17,8
18,0
18,2
18,4
18,6
18,8
19,0
19,2
Ln(C
once
ntra
ção
de p
olie
dros
)
Tempo (h)
Figura A.12. Ln (concentração de poliedros) versus tempo (HyClone+5%SFB)
24 48 72 96 120
13,5
14,0
14,5
15,0
15,5
16,0
Ln (c
once
ntra
ção
celu
lar)
Tempo (h)
Ln (concentração de células não infectadas) Ln (concentração de células infectadas)
Figura A.13 Ln (concentração celular) versus tempo (HyClone + 2,5%SFB)
______________________________________________________________________________________ 94Andréa Farias de Almeida, abril/2005

__________________________________________________________________________________________ Anexos
0 24 48 72 96
0,25
0,30
0,35
0,40
0,45
0,50
0,55
0,60
Ln (c
once
ntra
ção
de s
ubst
rato
)
Tempo (h)
Ln (concentração de substrato - células não infectadas) Ln (concentração de substrato - células infectadas)
Figura A.14. Ln (concentração de substrato) versus tempo (HyClone + 2,5%SFB)
72 96 12017,0
17,5
18,0
18,5
19,0
19,5
Ln(C
once
ntra
ção
de p
olie
dros
)
Tempo (h)
Figura A.15. Ln (concentração de poliedros) versus tempo (HyClone + 2,5%SFB)
______________________________________________________________________________________ 95Andréa Farias de Almeida, abril/2005

__________________________________________________________________________________________ Anexos
0 24 48 72 96 12013,0
13,5
14,0
14,5
15,0
15,5
16,0
Ln (c
once
ntra
ção
celu
lar)
Tempo (h) Ln (concentração de células não infectadas) Ln (concentração de células infectadas)
Figura A.16. Ln (concentração celular) versus tempo (HyClone + 5%SFB)
24 48 72 96 1200,0
0,1
0,2
0,3
0,4
0,5
0,6
Ln (c
once
ntra
ção
de s
ubst
rato
)
Tempo (h) Ln (concentração de substrato - células não infectadas) Ln (concentração de substrato - células infectadas)
Figura A.17. Ln (concentração de substrato) versus tempo (HyClone + 5%SFB)
______________________________________________________________________________________ 96Andréa Farias de Almeida, abril/2005

__________________________________________________________________________________________ Anexos
72 96 120 14418,0
18,5
19,0
19,5
Ln(C
once
ntra
ção
de p
olie
dros
)
Tempo (h)
Figura A.18. Ln (concentração de poliedros) versus tempo (HyClone + 5%SFB)
______________________________________________________________________________________ 97Andréa Farias de Almeida, abril/2005