Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DE MATO GROSSO
FACULDADE DE ENGENHARIA FLORESTAL
Programa de Pós-Graduação em Ciências Florestais e Ambientais
DIVERSIDADE DE FORMIGAS (HYMENOPTERA:
FORMICIDAE) EM PLANTIOS DE TECA, PAU-DE-
BALSA E ÁREA NATIVA NO BIOMA CERRADO
SILVIO EDUARDO DE OLIVEIRA THOMAS
CUIABÁ – MT
2017

SILVIO EDUARDO DE OLIVEIRA THOMAS
DIVERSIDADE DE FORMIGAS (HYMENOPTERA:
FORMICIDAE) EM PLANTIOS DE TECA, PAU-DE-
BALSA E ÁREA NATIVA NO BIOMA CERRADO
Orientador: Prof. Dr. Alberto Dorval
Dissertação apresentada à Faculdade de
Engenharia Florestal da Universidade Federal
de Mato Grosso, como parte das exigências do
Curso de Pós-Graduação em Ciências
Florestais e Ambientais, para obtenção do título
de mestre.
CUIABÁ – MT
2017



À minha família, especialmente aos meus pais Renato Thomas e
Suzelei de Oliveira Mineiro.
DEDICO

AGRADECIMENTOS
- A Deus sobre todas as coisas;
- Aos meus pais pelo apoio, incentivo e colaboração;
- À CAPES, pela bolsa de pesquisa ofertada;
- À Faculdade de Engenharia Florestal, da Universidade Federal de Mato
Grosso juntamente com seus professores e técnicos, em especial ao Sr.
Manoel Lauro da Silva do Laboratório de Proteção Florestal, pela amizade
ajuda e aprendizado;
- Ao Dr. Alberto Dorval pela orientação, paciência, apoio e dedicação;
- Ao Dr. Otávio Peres Filho pela disponibilidade e apoio dado;
- Ao Dr. Jacques H. C. Delabie do Laboratório de Mirmecologia do
CEPLAC/CEPEC (Ilhéus – BA) pela gentil hospitalidade e pelas
identificações taxonômicas das formigas coletadas neste estudo;
- A Drª. Lilian Guimarães de Favare pela colaboração e auxilio prestado;
- Ao Dr. Marcelo Dias de Souza pelos ensinamentos e contribuições dadas
a este trabalho;
- A empresa Teca do Brasil por disponibilizar a área para instalação dessa
pesquisa e todo apoio dado;
- Ao Geison e demais funcionários da Teca do Brasil que sempre nos
receberam prestativos e atenciosos;

- Aos meus amigos que participaram direta e indiretamente no
desenvolvimento e conclusão deste trabalho, à Fabi, Thayza, Carol, Diego,
Marcelo, Mariana, Livio e Aquirya que me ajudaram e participaram nessas
inúmeras idas e vindas a campo;
- À Jeane, parceira de mestrado e de pesquisa, entramos juntos nesse
trabalho com as formigas;
- A Danielly e Tiago, pela grande parceria e amizade, sempre juntos nas
correrias do dia a dia do mestrado, em campo, em laboratório, em viagens,
sempre me ajudando quando precisava.

SUMÁRIO
RESUMO ................................................................................................. XII
ABSTRACT ............................................................................................ XIII
1. INTRODUÇÃO ....................................................................................... 1
2. REVISÃO DE LITERATURA ................................................................. 2 2.1. IMPORTÂNCIA ECOLÓGICA DA FAMÍLIA FORMICIDAE ................. 2 2.2. FATORES AMBIENTAIS QUE AFETAM A MIRMECOFAUNA ........... 4 2.3. EFEITOS DAS MONOCULTURAS FLORESTAIS NA
MIRMECOFAUNA ..................................................................................... 6 2.4. AMOSTRAGEM DE FORMIGAS EM AMBIENTES NATURAIS ......... 9
3. MATERIAL E MÉTODOS .................................................................. 11 3.1. ÁREA DE ESTUDO ........................................................................... 11 3.2. CARACTERÍSTICA DOS LOCAIS DE ESTUDO ............................... 12 3.2.1. Teca ............................................................................................... 12 3.2.2 Pau-de-balsa ................................................................................... 12 3.2.3 Área com vegetaçâo de cerrado ..................................................... 13 3.3. COLETA DE FORMIGAS .................................................................. 13 3.4. TRIAGEM E IDENTIFICAÇÃO DO MATERIAL ENTOMOLÓGICO ... 13 3.5. ANÁLISE DOS DADOS ..................................................................... 14 3.5.1. Análises Estatísticas ...................................................................... 14 3.5.2. Análise Faunística .......................................................................... 14
4. RESULTADOS E DISCUSSÃO ........................................................ 19 4.1. ANÁLISES QUANTITATIVA E QUALITATIVA .................................. 19 4.2. SAZONALIDADE NA DIVERSIDADE DE FORMIGAS ...................... 29 4.2. CURVA DE ACUMULAÇÃO DE ESPÉCIES DE FORMIGAS ............ 38 4.3. ANÁLISES ESTATÍSTICAS .............................................................. 39 4.4. ANÁLISE FAUNÍSTICA ..................................................................... 42 4.5. ANÁLISE DE SIMILARIDADE ........................................................... 53
5. CONCLUSÕES ................................................................................. 56
6. BIBLIOGRAFIA ................................................................................. 57
ANEXOS .................................................................................................. 69

LISTA DE TABELAS
Página
1.ESPÉCIES DE FORMIGAS AMOSTRADAS EM DIFERENTES
AMBIENTES FLORESTAIS NO MUNICÍPIO DE NOSSA SENHORA DO
LIVRAMENTO, ESTADO DE MATO GROSSO, 2015/2016. .................... 21 2. HÁBITO ALIMENTAR DE ESPÉCIES DE FORMIGAS AMOSTRADAS
EM PLANTIO DE TECA, PAU-DE-BALSA E CERRADO NO MUNICÍPIO DE
NOSSA SENHORA DO LIVRAMENTO-MT, 2015/2016. .......................... 22 3. HÁBITO ALIMENTAR DE ESPÉCIES DE FORMIGAS AMOSTRADAS
POR AMBIENTE NO MUNICIPIO DE NOSSA SENHORA DO
LIVRAMENTO-MT, 2015/2016. ................................................................ 23 4. HÁBITO ALIMENTAR DE ESPÉCIES DE FORMIGAS POR SUBFAMÍLIA
AMOSTRADAS NO MUNICIPIO DE NOSSA SENHORA DO
LIVRAMENTO-MT, 2015/2016. ................................................................ 24 5. RELAÇÃO DAS SUBFAMÍLIAS E ESPÉCIES EM PORCENTAGEM (%) E NÚMEROS DE INDIVÍDUOS (I) COLETADOS NOS TRÊS AMBIENTES
FLORESTAIS NO MUNICÍPIO DE NOSSA SENHORA DO LIVRAMENTO, ESTADO DE MATO GROSSO, 2015/2016 .............................................. 28 6. AÇÃO DAS SUBFAMÍLIAS, GÊNEROS E PERCENTAGENS (%) DE
ESPÉCIES (E) E DE NÚMEROS (I) DE FORMIGAS COLETADAS NO
PERÍODO DE SECA NOS TRÊS AMBIENTES FLORESTAIS NO
MUNICÍPIO DE NOSSA SENHORA DO LIVRAMENTO, ESTADO DE
MATO GROSSO, 2015/2016. .................................................................. 31 7. RELAÇÃO DAS SUBFAMÍLIAS, GÊNEROS E PERCENTAGENS (%) DE
ESPÉCIES (E) E DE NÚMEROS (I) DE FORMIGAS COLETADAS NO
PERÍODO DE CHUVA NOS TRÊS AMBIENTES FLORESTAIS NO
MUNICÍPIO DE NOSSA SENHORA DO LIVRAMENTO, ESTADO DE
MATO GROSSO, 2015/2016. .................................................................. 32 8. RELAÇÃO DAS SUBFAMÍLIAS E PORCENTAGEM DE NÚMEROS
INDIVÍDUOS (I) COLETADOS NO PERÍODO SECO NOS TRÊS
AMBIENTES FLORESTAIS NO MUNICÍPIO DE NOSSA SENHORA DO
LIVRAMENTO, ESTADO DE MATO GROSSO, 2015/2016. .................... 35 9. RELAÇÃO DAS SUBFAMÍLIAS E PORCENTAGEM DE NÚMEROS
INDIVÍDUOS (I) COLETADOS NO PERÍODO CHUVA NOS TRÊS
AMBIENTES FLORESTAIS NO MUNICÍPIO DE NOSSA SENHORA DO
LIVRAMENTO, ESTADO DE MATO GROSSO, 2015/2016. .................... 36 10. ANÁLISE DE VARIÂNCIA PARA NÚMERO DE INDIVÍDUOS
COLETADOS EM TRÊS DIFERENTES AMBIENTES FLORESTAIS NO
MUNICÍPIO DE NOSSA SENHORA DO LIVRAMENTO, ESTADO DE
MATO GROSSO, 2015/2016. .................................................................. 39

11. ANÁLISE DE VARIÂNCIA PARA NÚMERO DE ESPÉCIES
COLETADAS EM TRÊS DIFERENTES AMBIENTES FLORESTAIS NO
MUNICÍPIO DE NOSSA SENHORA DO LIVRAMENTO, ESTADO DE
MATO GROSSO, 2015/2016. .................................................................. 40 12. TESTE DE MÉDIAS PARA NÚMEROS DE ESPÉCIES COLETADAS
EM TRÊS AMBIENTES FLORESTAIS NO MUNICÍPIO DE NOSSA
SENHORA DO LIVRAMENTO, ESTADO DE MATO GROSSO, 2015/2016. ................................................................................................................. 40 13. TESTE DE MÉDIAS PARA NÚMEROS DE INDIVÍDUOS E DE
ESPÉCIES COLETADOS NO PERÍODO DE SECA EM TRÊS AMBIENTES
FLORESTAIS NO MUNICÍPIO DE NOSSA SENHORA DO LIVRAMENTO, ESTADO DE MATO GROSSO, 2015/2016. ............................................. 41 14. TESTE DE MÉDIAS PARA NÚMEROS DE INDIVÍDUOS E DE
ESPÉCIES COLETADOS NO PERÍODO DE CHUVA EM TRÊS
AMBIENTES FLORESTAIS NO MUNICÍPIO DE NOSSA SENHORA DO
LIVRAMENTO, ESTADO DE MATO GROSSO, 2015/2016. .................... 42 15. RELAÇÃO DAS ESPÉCIES DE FORMIGAS MAIS
REPRESENTATIVAS NA ANÁLISE FAUNÍSTIVA AMOSTRADAS COM
ARMADILHA PITFALL EM TALHÃO DE TECTONA GRANDIS NOS PERÍODOS
DE SECA, CHUVA E ANUAL, NO MUNICÍPIO DE NOSSA SENHORA DO
LIVRAMENTO, ESTADO DE MATO GROSSO, 2015/2016. .................... 44 16. RELAÇÃO DAS ESPÉCIES DE FORMIGAS MAIS
REPRESENTATIVAS NA ANÁLISE FAUNÍSTIVA AMOSTRADAS COM
ARMADILHA PITFALL EM TALHÃO DE OCHROMA PYRAMIDALE NOS
PERÍODOS DE SECA, CHUVA E ANUAL, NO MUNICÍPIO DE NOSSA
SENHORA DO LIVRAMENTO, ESTADO DE MATO GROSSO, 2015/2016. ................................................................................................................. 47 17. RELAÇÃO DAS ESPÉCIES DE FORMIGAS MAIS
REPRESENTATIVAS NA ANÁLISE FAUNÍSTICA AMOSTRADAS COM
ARMADILHA “PITFALL” EM VEGETAÇÃO DE CERRADO, NOS
PERÍODOS DE SECA, CHUVA E ANUAL, NO MUNICÍPIO DE NOSSA
SENHORA DO LIVRAMENTO, ESTADO DE MATO GROSSO, 2015/2016. ................................................................................................................. 49 18. ÍNDICE DE DIVERSIDADE DE ACORDO COM A SÉRIE DE NÚMEROS
DE HILL E EQUITATIVIDADE DAS ESPÉCIES DE FORMIGAS
AMOSTRADOS EM PLANTIOS DE TECA (TECTONA GRANDIS) E DE PAU-DE-BALSA (OCHROMA PYRAMIDALE) E EM VEGETAÇÃO DE CERRADO, NO
MUNICÍPIO DE NOSSA SENHORA DO LIVRAMENTO- MT, 2015/2016. 52 19. SIMILARIDADE ENTRE OS AMBIENTES, PARA OS PERIODOS
SECO, DE CHUVA E ANUAL, AMOSTRADOS NO MUNICÍPIO DE NOSSA
SENHORA DO LIVRAMENTO-MT, 2015/2016. ....................................... 54

LISTA DE FIGURAS
1. LOCALIZAÇÃO DA FAZENDA CAMPINA NO MUNICÍPIO DE NOSSA
SENHORA DO LIVRAMENTO, ESTADO DE MATO GROSSO. .............. 11 2. DISTRIBUIÇÃO DE INDIVÍDUOS COLETADOS EM AMBIENTES DE
PLANTIO DE TECA, PAU-DE-BALSA E ÁREA NATIVA DO CERRADO, NO
MUNICÍPIO DE NOSSA SENHORA DO LIVRAMENTO, MATO GROSSO, 2015/2016. ............................................................................................... 19 3. DISTRIBUIÇÃO DAS SUBFAMÍLIAS DE FORMICÍDEOS COLETADOS
EM AMBIENTES DE PLANTIO DE TECA, PAU-DE-BALSA E ÁREA
NATIVA DO CERRADO, NO MUNICÍPIO DE NOSSA SENHORA DO
LIVRAMENTO, MATO GROSSO, 2015/2016. ......................................... 20 4. CURVA DE ACUMULAÇÃO DE ESPÉCIES DE FORMIGAS, POR
NÚMERO DE COLETAS, EM PLANTIO DE TECA, PAU-DE-BALSA E EM
ÁREA NATIVA DO BIOMA CERRADO NO MUNICÍPIO DE NOSSA
SENHORA DO LIVRAMENTO, ESTADO DE MATO GROSSO, 2015/2016. ................................................................................................................. 38

xii
RESUMO
THOMAS, Silvio Eduardo de Oliveira. Diversidade de formigas (Hymenoptera: Formicidae) em plantios de teca, pau-de-balsa e área nativa no bioma Cerrado. 2017. Dissertação (Mestrado em Ciências Florestais e Ambientais) – Universidade Federal de Mato Grosso, Cuiabá-MT. Orientador: Prof. Dr. Alberto Dorval. Este trabalho teve por objetivo avaliar qualitativamente e quantitativamente as espécies de formigas (Hymenoptera) amostradas nos seguintes ambientes florestais: a) ambiente com vegetação de cerrado; b) ambiente com plantio de Tectona grandis; c) ambiente com plantio de Ochroma pyramidale. Dentro de cada ambiente foi instalado um transecto, com 20 armadilhas tipo pitfall, equidistantes 20 metros entre si. Foram realizadas coletas de julho de 2015 a junho de 2016. Os indivíduos coletados foram triados e identificados taxonômicamente ao nível de gênero e espécie. Foram realizados análise de variância (Teste de Tukey) para o número de indivíduos e de espécies amostradas e estudos faunísticos e de similaridade entre os ambientes. Pela análise de variância, há diferença significativa entre o número de espécies registradas por ambiente no decorrer do ano e no período de seca. As análises faunísticas possibilitaram concluir que Labidus coecus (Dorylinae) foi, quantitativamente, uma das espécies mais importantes nos três ambientes e Solenopsis sp. foi a espécie com maior potencial de adaptação aos ambientes; Crematogaster tenuicula e Camponotus blandus (Myrmicinae) foram a espécies mais representativas no cerrado; Labidus coecus e Solenopsis sp. foram as mais representativas para o plantio com pau-de-balsa e para o ambiente com plantio teca, Labidus coecus e Atta sexdens foram mais representativos. O índice de similaridade de Sorensen indicou alta similaridade na riqueza de formigas entre os ambientes estudados. Palavras-chave: Riqueza de espécies, formicídeos, mirmecofauna, monoculturas florestais.

xiii
ABSTRACT
THOMAS, Silvio Eduardo de Oliveira. Diversity of ants (Hymenoptera:
Formicidae) in teak, balsa-wood and native areas in the Cerrado
biome. 2017. Dissertation (Master of Forestry and Environmental Sciences)
- Federal University of Mato Grosso, Cuiabá, MT, Brazil. Adviser: Prof. Dr.
Alberto Dorval.
The objective of this work was to evaluate, qualitatively and quantitatively,
ant species (Hymenoptera) Sampled in the following forest environments:
a) Environment with cerrado vegetation; B) Environment with planting of
Tectona grandis; C) Environment with planting of Ochroma pyramidale.
Within each environment was installed a transect, with 20 pitfall traps,
equidistant 20 meters from each other. Collections were made from July
2015 to June 2016. The individuals collected were taxed and taxonomically
identified at the genus and species level. Variance analysis (Tukey Test)
was performed for the number of individuals and species sampled, faunistic
studies and similarity among the environments. By the analysis of variance,
there is a significant difference between the number of species recorded per
environment during the year and during the dry period. The faunal analysis
made it possible to conclude that Labidus coecus (Dorylinae) was
quantitatively one of the most important species in the three environments
and Solenopsis sp. Was the species with the greatest potential of adaptation
to the environments; Crematogaster tenuicula and Camponotus blandus
(Myrmicinae) were the most representative species in the cerrado
vegetation Labidus coecus and Solenopsis sp. Were the most
representative for the planting with balsa wood and for the environment with
teak planting, Labidus coecus and Atta sexdens were more representative.
Sorensen's similarity index indicated high similarity in ant richness among
the studied environments.
Key words: Species richness, formicides, ant fauna, monocultures

1
1. INTRODUÇÃO
No Mato Grosso, o bioma Cerrado vem sendo utilizado para a
implantação de florestas comerciais. Por serem monoculturas, esses
plantios simplificam o ambiente e geram alterações nos habitats naturais
como perda da biodiversidade local e predominância de algumas espécies
em detrimento de outras.
Dentre os animais presentes no cerrado, as formigas são o
grupo de insetos mais importante em termos de biomassa, número de
indivíduos e impacto ecológico (SILVA et al., 2004). Consideradas agentes
do meio, estão envolvidas em processos ambientais como aeração,
decomposição e ciclagem de nutrientes.
As formigas apresentam alta sensibilidade às variáveis
ambientais e capacidade de reagir rapidamente as mudanças do meio. Por
esses motivos, vem sendo amplamente utilizadas para avaliar o estado de
conservação/degradação dos ambientes. A biodiversidade das formigas
tem sido estudada com o objetivo de compreender as perturbações
ocasionadas pelas constantes simplificações dos ambientes naturais, como
é o caso das monoculturas. Apesar da proporção de florestas plantadas
existentes, os trabalhos sobre a resposta sucessional da fauna em áreas
de plantios florestais são escassos.
Considerando a importância das formigas nos ecossistemas e a
necessidade de ampliar os conhecimentos sobre a diversidade desse grupo
em ambientes alterados como em áreas de plantios florestais, este estudo
objetivou avaliar qualitativamente e quantitativamente as espécies de
formigas (Hymenoptera: Formicidae) que ocorram associadas às áreas de
plantios florestais de teca (Tectona grandis) e pau-de-balsa (Ochroma
pyramidale) e em área de cerrado, no município de Nossa Senhora do
Livramento – MT.

2
2. REVISÃO DE LITERATURA
2.1. IMPORTÂNCIA ECOLÓGICA DA FAMÍLIA FORMICIDAE
As formigas são insetos pertencentes a família Formicidae, da
ordem Hymenoptera (HÖLLDOBLER e WILSON, 1990). Distribuídas em 21
subfamílias, 301 gêneros e 12.545 espécies descritas, considerando assim,
as formigas são um dos grupos de organismos dominantes do planeta,
tanto numérica quanto ecologicamente e, dentre os insetos, um dos táxons
mais importantes em termos de biomassa, representando mais de 15% da
biomassa animal em florestas tropicais (FITTKAU e KLINGE, 1973). Na
região Neotropical, são conhecidas 15 subfamílias, 122 gêneros e 3.100
espécies de formigas (FERNÁNDEZ, 2003).
Para Fernández e Sendoya (2004), o Brasil e a Colômbia são os
países da região Neotropical mais ricos em números de espécies de
formigas. No Brasil ocorre cerca de 2.000 espécies descritas, sendo que
destas, algumas dezenas são consideradas pragas (BUENO e CAMPOS-
FARINHA, 1999). São animais que vivem em diversos habitats, podendo
ser encontrado desde desertos a florestas úmidas (KASPARI, 2000).
As formigas constituem o maior grupo de insetos sociais,
apresentam uma organização colonial extremamente estruturada e são
provavelmente os mais bem sucedidos de todos os grupos de insetos
(BORROR e DELONG, 1969). Este sucesso pode ser atribuído,
principalmente, ao fato de terem sido o primeiro grupo predador social a
explorar o solo e a vegetação e, de se especializarem em grupos de
indivíduos executando a mesma tarefa (DIEHL – FLEIG, 1995).
Amplamente distribuídos geograficamente, as formigas são
encontradas desde regiões subpolares até o Equador e em todas as ilhas
oceânicas, exceto nos polos e nos mares, sendo mais abundante em locais
de clima tropical (WILSON, 1987).
Nos ambientes onde se encontram, estão envolvidas em
diversos processos ambientais como aeração, decomposição e ciclagem
de nutrientes (ANDERSEN, 1991). A família Formicidae está relacionada
com inúmeros processos importantes em ambientes terrestres, agindo

3
como herbívoros, polinizadores, dispersores de sementes e predadores.
Exercem relações mutualísticas com plantas e homópteras e contribuem
como biomassa alimentar para níveis tróficos superiores (FREITAS et al.,
2006).
Este grupo também apresenta função essencial na participação
da cadeia alimentar nos ambientes terrestres pois, estão entre os principais
predadores de insetos e pequenos invertebrados, atuando como agentes
de controle das populações de outros artrópodes, e também são herbívoros
fortemente destrutivos (HÖLLDOBLER e WILSON, 1990).
No solo, as formigas podem nidificar a ciclagem de nutrientes,
através do enriquecimento do solo, pela transferência de nutrientes para as
camadas mais superficiais durante a construção e relocação de ninhos
(MOUTINHO et al., 2003). Através das trilhas de forrageamento, podem
induzir a alteração do solo através de excreção, perda de alimento durante
translado e morte de indivíduos, que estende o impacto das formigas além
do perímetro da colônia (COSTA, 2007).
Os hábitos alimentares das formigas são os mais diversos
possíveis, havendo predomínio de forrageadoras oportunistas e
generalistas que se alimentam de secreções vegetais, sementes, e material
animal vivo ou morto (KASPARI, 2000). Outros grupos apresentam dieta
mais especializada, como o caso das formigas da subfamília Myrmicinae,
que cultivam fungos que crescem sobre um substrato composto por
material vegetal e/ou animal recolhido pelas operárias (WEBER, 1972).
Formigas das subfamílias Dolichoderinae e Formicinae possuem hábito
alimentar restrito a uma alimentação líquida que pode ser obtida através de
nectários extraflorais ou outras estruturas secretoras como interações com
alguns hemípteros que liberam uma excreção açucarada conhecida como
“honeydew” (DELABIE e FERNÁNDEZ, 2003). Muitos gêneros incluem
ainda formigas predadoras, que podem ser generalistas ou especializadas,
como por exemplo, em Cerapachys (Cerapachynae), Neivamyrmex
(Ecitoninae), Strumigenys (Myrmicinae) e Thaumatomyrmex (Ponerinae)
que se alimentam de grupos restrito de artrópodes (KASPARI, 2000).
No setor florestal, as formigas cortadeiras, do gênero Atta e
Acromyrmex constituem as formigas mais conhecidas por serem

4
consideradas pragas severas de culturas florestais e pastagens naturais ou
cultivadas (DELLA LUCIA et al., 2008). Embora os hymenópteros
apresentem esse grupo de insetos pragas, muitas espécies dessa ordem,
dos gêneros Solenopsis, Wasmannia e Pheidole são predadores e inimigos
naturais e representam um dos mais importantes fatores biológicos que
regulam o tamanho e a biomassa das populações de espécies
desfolhadoras (FREITAS et al., 2002).
As formigas apresentam capacidade de adaptação às condições
extremas do ambiente, ocupando locais intensamente degradados com
estreita condição de oxigênio, inundações periódicas, contaminação por
produtos químicos e exposição à radiação que, contribuem na sua ampla
distribuição geográfica (SILVESTRE, 2000). Assim, a presença ou a
ausência de determinadas espécies da mirmecofauna podem fornecer uma
indicação de várias características do ambiente. Desta forma, diversas
espécies da comunidade de formigas podem constituir um bom indicador
das condições ambientais (DIEHL-FLEIG, 1995).
Além de responderem ao estresse do meio, as formigas
apresentam características que são importantes nos estudos com objetivo
para compreender as perturbações ocasionadas pela simplificação dos
ecossistemas naturais, como alta diversidade, dominância numérica e de
biomassa em quase todos os habitats e facilidade na amostragem e
identificação (MAJER, 1996; ALONSO e AGOSTI, 2000). Os estudos
voltados ao levantamento de Formicidae são necessários, pois o atual
conhecimento taxonômico e biogeográfico para a maioria dos grupos de
organismos terrestres é incompleto, especialmente para os grupos
hiperdiversos, como as formigas (SILVA e BRANDÃO, 1999).
2.2. FATORES AMBIENTAIS QUE AFETAM A MIRMECOFAUNA
Embora facilmente encontradas em diversos ambientes, as
comunidades de formigas podem ser afetadas por diversos mecanismos,
como sensibilidade a inundações (MERTL et al., 2009), condições edáficas
da composição do solo, gradientes altitudinais e a variações em tolerância
térmica e umidade (ANGILLETTA et al., 2007).

5
Mudanças na complexidade da cobertura vegetal e,
consequentemente, no microclima têm grande influência na diversidade de
formigas (MORAES e BENSON, 1988). Há o aumento da densidade de
riqueza de espécies de formigas de acordo com o aumento da densidade
da vegetação local (RIBAS et al., 2003). Habitats complexos,
estruturalmente, permitem a instalação e sobrevivência de maior número
de espécies decorrentes da capacidade de suporte do meio onde são
disponibilizados maior variedade de recursos como alimentação e
esconderijo (LAWTON, 1983). Em levantamento da fauna de formigas em
áreas reabilitadas de mineração no Rio de Janeiro, Pereira et al. (2007)
observaram variação na composição de espécies de formicídeos, de
acordo com a variação da estrutura da vegetação.
Em fragmentação de florestas tropicais ocorre perda de
espécies por meio da intervenção do habitat natural. Esta modificação no
meio reduz do tamanho da população, inibe ou reduz a migração, promove
o efeito de borda alterando o microclima e a imigração de espécies exóticas
para as áreas desmatadas circundantes e, posteriormente, para o
fragmento, aumentando a competição por recursos (WINSTON, 1995;
TURNER, 1996).
Fagundes et al. (2009) defendem que a disponibilidade de
alimento é um dos principais fatores relacionados com a abundância de
formigas. A especificidade de habitat como a disponibilidade de alimento
específico, pode determinar a permanência de uma espécie no ambiente.
Em relação aos fatores abióticos, a umidade afeta de forma
direta (precipitação) e indireta (umidade do ar e do solo, evaporação e
disponibilidade de água) e pode influenciar a vida dos insetos de modo
geral, visto que, certas espécies têm preferências, como evitar excesso de
água ou sua escassez (LARA, 1992). Outros fatores como altitude, latitude
e condições climáticas, como temperatura e umidade, podem influenciar na
riqueza e composição de espécies de formigas (ALBUQUERQUE e DIEHL,
2009).

6
2.3. EFEITOS DAS MONOCULTURAS FLORESTAIS NA
MIRMECOFAUNA
A demanda mundial por celulose e por madeira certificada,
possibilidade da expansão de sistemas lavoura-pecuária-floresta e uso de
florestas para sequestro de carbono promovem o aumento da área dos
plantios florestais. Entre os biomas brasileiros utilizados para esses
plantios, encontra-se o Cerrado, no qual, as condições edafoclimáticas
(solo e clima) e fisiográficas são fatores favoráveis para o estabelecimento
e expansão dos plantios florestais nesse bioma (JUVENAL e MATTOS,
2002; DA SILVA, et al. 2010).
A simplificação do ambiente somado a fragmentação da floresta
nativa ocasionada pela implantação de monoculturas, acarretam em
mudanças que afetam de forma diferenciada os parâmetros demográficos
de mortalidade e natalidade de diferentes espécies (DA SILVA et al. 2010).
Para Viana e Pinheiro (1998), estas simplificações do ambiente promovem
alteração na abundância de polinizadores, dispersores, predadores e
patógenos. Além de mudanças microclimáticas, que atingem as bordas dos
fragmentos, alterando a taxa de mortalidade de árvores e influenciando
diretamente na diversidade de espécies do ambiente.
Pesquisas sobre a biodiversidade no cerrado são necessárias
para uma avaliação de impactos ambientais decorrentes das atividades
antrópicas, criação de unidades de conservação e para a adoção de
técnicas de manejo (FELFILI e FELFILI, 2001).
Mesmo com essa importância, o número de levantamentos de
formigas no Cerrado ainda continua insatisfatório (KEMPF, 1978; RIBAS et
al., 2003; SILVA et al., 2004). Este fato se torna um problema devido ao
alto grau de destruição a que esse habitat vem sendo submetido, causado
principalmente, pela expansão da agricultura, pecuária e de monocultura
como plantios florestais (KLINK e MOREIRA, 2002).
Há evidências que em áreas de plantações florestais ocorre
menor riqueza de espécies da mirmecofauna que em áreas de vegetação
nativa (MAJER, 1992). A influência dos plantios sobre populações de
insetos está relacionada com a diversidade e a concentração de recursos,

7
pois, ecossistemas complexos e mais persistentes suportam maior número
de espécies (LAWTON e STRONG JR., 1981; FREITAS et al., 2002).
Para Della-Lucia et al. (1982) a estrutura do dossel,
luminosidade e espaçamento entre plantas podem afetar a comunidade de
formicídeos. Deste modo, mudando a natureza do dossel, pode-se mudar
a espécie de formiga dominante mas, como um todo, há uma estabilidade
na diversidade.
As monoculturas florestais tendem a simplificar o ambiente
original pela modificação do estrato arbóreo, da vegetação arbustiva e
herbácea do sub-bosque, reduzindo a diversidade de substratos de
nidificação e alimentação e levando a uma redução na riqueza de espécies
(SOARES et al., 1998; SANTOS et al., 2006). As formigas dos gêneros
Atta e Acromyrmex são frequentementes relatadas como pragas de
monoculturas agrícolas e florestais pois, nesses ambientes, encontram
abundantes fonte de recursos alimentar e poucos inimigos naturais (ANJOS
et al., 1993).
Em florestas preservadas, a maior heterogeneidade e
quantidade de nichos alimentares e estruturais promove aumento da
variedade de espécies de insetos, geralmente com pequeno número
relativo de indivíduos. Nesses ambientes, a população de cada espécie é
controlada pelas diversas relações interespecíficas (LARA, 1992).
Em ambientes que sofreram intervenção, ocorre alteração na
comunidade de formigas ocasionadas pela mudança ecológica que
acontecem dentro da floresta e as influências físicas e bióticas como luz,
microclima e temperatura que agem sobre a região (CARVALHO e
VASCONCELOS, 1999). Os plantios florestais são continuamente
alterados criando micro-climas desfavoráveis para parasitoides e
predadores que controlam as populações (ALTIERI et al., 1993). Observa-
se nestes ambientes a presença de grandes populações e reduzido número
de espécies (LARA, 1992).
Diversos são os estudos objetivando comparar a biodiversidade
da mirmecofauna com diferentes ambientes como em plantios de eucalipto
e área nativa (SOARES et al., 1998; MARINHO et al., 2002; FREITAS et
al., 2002; MENTONE et al., 2011), com Pinus (SOUZA, 2006; BOSCARDIN

8
et al., 2014) e em fragmentos de florestas nativas no cerrado (SOARES et
al., 2006; ALVARENGA et al., 2007; CAMPOS et al., 2008; CANTARELLI
et al., 2015).
Entre os plantios florestais implantados no bioma do cerrado
mato-grossense, a teca (Tectona grandis L.f.) e pau-de-balsa (Ochroma
pyramidale Cav.ex Lam). Estimativas apontam que, atualmente são 64 mil
ha de teca plantado no estado. Plantios de eucalipto, que representam a
maior área das florestas plantadas no estado, ocupam 187 mil ha. Pau-de-
balsa é uma espécie que começa a ter um aumento significativo na sua
área plantada que representam 3 mil ha (FAMATO, 2013).
A teca, originária da Ásia, encontra-se em expansão nas regiões
centro-oeste e norte do Brasil. O principal produto desta espécie é a
madeira de alta qualidade, muito utilizada em móveis finos e na construção
naval. É uma espécie florestal de grande porte, podendo atingir alturas
entre 25 e 35m, de rápido crescimento. Tem grande procura no mercado
mundial, podendo alcançar preço até três vezes superior aos do mogno,
sendo utilizada na produção de móveis, esquadrias de alto padrão,
embarcações e decoração (MACEDO et al. 1999; PELISSARI, 2012;
SOUZA, 2011).
A espécie Ochroma pyramidale Urban, família Bombacaceae,
popularmente conhecida como Pau-de-balsa, é uma espécie arbórea que
atinge de 18 a 24 m de altura (LAMPRECHT,1990). Amplamente utilizada
em plantios mistos destinados à recomposição de áreas degradadas e de
preservação permanente, graças ao seu rápido crescimento e tolerância à
luminosidade (SANTOS et al., 2014). Sua madeira, apesar da baixa
densidade volumétrica (0,07-0,15 g.cm-3), possui elevada resistência
mecânica (VASQUEZ-YANES, 1974); sendo utilizado na confecção de
aeromodelos, revestimento de navios, balsas, embalagens especiais, bem
como sucedâneo da cortiça na fabricação de coletes salva-vidas
(LAMPRECHT, 1990).

9
2.4. AMOSTRAGEM DE FORMIGAS EM AMBIENTES NATURAIS
Entre as armadilhas utilizadas para a amostragem de formigas
de solo, destacam-se: pitfall, extrator de Winkler, iscas, aspirador e o funil
de Berlese (BESTELMEYER et al., 2000). As armadilhas de queda do tipo
pitfall constituem uma metodologia amplamente utilizada em estudos de
formicídeos (GARCÍA e LÓPEZ, 2001).
A armadilha do tipo pitfall é um dos métodos mais simples para
amostrar artrópodes de serapilheira (AQUINO et al., 2006). Entre as
vantagens desta armadilha, está o fato desta operar continuamente por
longos períodos e ser eficiente na coleta de espécies predadoras e
saprófagas (FARIA et al., 2001; MILHOMEM et al., 2003).
A armadilha pitfall consiste de qualquer equipamento
confeccionado para que, uma vez coletado, o animal não consiga mais sair
(ALMEIDA et al., 2003). Esse tipo de armadilha é destinado para animais
que vivem no solo, que caminham sobre o mesmo, porque não podem voar
ou porque passam uma fase da vida nele. Os diferentes tipos de solos,
cobertura vegetal e a escala temporal e regional, são fatores importantes
que determinam a composição e a riqueza dos artrópodes coletados
(PETILLON et al., 2006; LACHAT et al., 2006).
A armadilha apresenta uma estrutura simples, consiste, em
geral, de um recipiente de plástico enterrado ao nível de solo com líquido
para matar e conservar os animais capturados (AQUINO et al., 2006). O
volume da armadilha é de acordo com a solução utilizada, podendo ser
conservante ou isca atrativa. Se possível, utilizar recipientes que
contenham tampa para facilitar o transporte do campo até o laboratório
usando a mesma armadilha. O diâmetro da armadilha interfere na captura
dos espécimes. É recomendado o uso de armadilhas de mesmo tamanho
nos diferentes locais (PARR e CHOWN, 2001).
Para Sarmiento (2003), essas armadilhas devem ser
preenchidas com uma mistura contendo água, detergente líquido e álcool
etílico. Bestelmeyer et al. (2000) explicam que o detergente auxilia na
captura, pois quebra a tensão superficial, prevenindo a fuga dos insetos na
armadilha. Quanto à distribuição das armadilhas para captura de formigas,

10
alguns estudos apontam que não há diferença na forma de distribuir as
unidades amostrais (SARMIENTO, 2000). Segundo o autor, a distância
entre as armadilhas pode ser considerada um elemento importante, apesar
de alguns estudos sugerirem que não há variação significativa da
mirmecofauna dentro de uma floresta a distâncias de até 100 metros,
recomenda-se que as unidades amostrais devem ser distribuídas a cada
10 metros (AQUINO et al., 2006).
O sucesso da utilização da pitfall da-se pela adequada
montagem, com cuidado na perfuração do solo para não enterrar a
armadilha; quando enterrada deve simular o ambiente original como
espalhar uniformemente a serapilheira no entorno da armadilha; evitar
perturbação local após a instalação da armadilha; deve-se ter cuidado com
queda de material dentro das armadilhas, principalmente no momento da
colocação ao solo, mantendo o líquido limpo e facilitando a separação da
amostra (AQUINO et al., 2006; RÉ, 2007).

11
3. MATERIAL E MÉTODOS
3.1. ÁREA DE ESTUDO
A pesquisa foi conduzida de julho de 2015 a junho de 2016, na
Fazenda Campina, de propriedade da Empresa Teca do Brasil, localizada
próximo à Vila Pirizal, distrito do Município de Nossa Senhora de
Livramento, na região centro-sul do estado de Mato Grosso (16º 12’ 03”
latitude sul e 56º 22’ 44” longitude oeste). A propriedade está localizada na
Planície Alagável da Bacia do Alto Paraguai a 75 km do perímetro urbano
da cidade de Cuiabá, com acesso pela BR-070, seguida da MT-080. A
região caracteriza-se pelo bioma Cerrado com altitude média de 232 metros
acima do nível do mar (Figura 1).
FIGURA 1 - LOCALIZAÇÃO DA FAZENDA CAMPINA NO MUNICÍPIO DE NOSSA SENHORA DO LIVRAMENTO, ESTADO DE MATO GROSSO.
O clima da região, segundo o sistema de classificação climática
de Koppen, é do tipo AW – quente e úmido, com precipitação média anual
de 1.250 mm e temperatura média anual de 25,8ºC. Na região, dois
períodos climáticos distintos são observados durante o ano, sendo uma
seca e a outra chuvosa (REBELLATO e CUNHA, 2005).

12
O solo predominante nas áreas de estudo é classificado como
latossolo com bom teor de matéria orgânica, coloração vermelho escuro,
textura argilo-arenoso, boa capacidade de retenção de água, aeração e
drenagem (CALDEIRA e OLIVEIRA, 2008).
3.2. CARACTERÍSTICA DOS LOCAIS DE ESTUDO
As áreas de estudos situam-se dentro dos limites da fazenda
Campina e correspondiam a três ambientes, sendo um talhão de teca
(Tectona grandis), outro talhão de pau-de-balsa (Ochroma pyramidale) e
uma área de Cerrado.
A escolha das áreas de pesquisa foi em função das diferentes
composições vegetais dentro da propriedade. As áreas com os plantios de
teca e pau-de-balsa, antes de serem destinados para cultivo florestal, foram
utilizadas para o cultivo de pastagem para prática da pecuária.
3.2.1. Teca
O talhão de teca, utilizado no estudo, foi implantado no ano de
1999, em uma área de relevo plano e espaçamento inicial de 3m x 3m. Na
ocasião do plantio, os tratos silviculturais, foram o preparo do solo com
arado e grade niveladora; e controle mecânico de plantas infestantes com
roçadeira. Na ocasião do estudo, as áreas circunvizinhas ao plantio eram
constituídas por Área de Preservação Permanente (APP). Durante o
decorrer do experimento, não houve emprego de tratos culturais.
3.2.2. Pau-de-balsa
O plantio de pau-de-balsa ocorreu no ano de 2006 com
espaçamento de 4,0 m x 3,0 m, com o objetivo de reflorestar uma área de
pastagem próxima aos plantios de teca. Foram utilizados arado e grade
niveladora para o plantio e o uso de roçadeira para o controle de plantas
invasoras. Não foi realizado qualquer outro trato silvicultural na área.

13
3.2.3. Área com vegetaçâo de cerrado
A área com vegetação do cerrado está localizada próximo aos
plantios de teca e desde a implantação dos plantios, em 1999, não sofreu
intervenções significativas. A vegetação localiza-se sobre uma cordilheira,
um cordão arenoso livre de inundações anuais, e formada por um estrato
lenhoso arbóreo-arbustivo dominante, apresenta as características de
áreas de cerradão, como foi observado por Da Costa et al. (2010) após
realizar um levantamento florístico das áreas remanescentes de cerrado
em uma propriedade próximo a fazenda Campina. As áreas circunvizinhas
são plantios de teca e remanescentes de floresta nativa do bioma Cerrado.
3.3. COLETA DE FORMIGAS
A amostragem ocorreu de julho de 2015 a junho de 2016,
utilizando armadilhas de queda do tipo pitfall, composta de uma garrafa de
plástico transparente de 15 cm de altura e 2,5cm de diâmetro com
capacidade para 300 ml.
Em cada ambiente foram definidos transectos de 200 metros,
com espaçamento de 40 metros entre eles. Em cada transecto foram
distribuídas armadilhas em intervalos de 20 metros, totalizando 20
armadilhas por ambiente. Cada armadilha foi enterrada com suas bordas
ao nível do solo, preenchidas com solução conservante contendo água,
detergente neutro (utilizado para quebrar a tensão superficial do líquido) e
sal (AQUINO, 2006). As armadilhas permaneceram no campo por um
período de 15 dias a cada mês. Após este período, as garrafas foram
retiradas e o conteúdo coletado foi devidamente individualizado por
armadilha, sendo armazenado e etiquetado. As coletas foram realizadas
no período de 12 meses, totalizando 780 amostras.
3.4. TRIAGEM E IDENTIFICAÇÃO DO MATERIAL ENTOMOLÓGICO
O material coletado foi transportado para o Laboratório de
Proteção Florestal (LAPROFLOR), da Faculdade de Engenharia Florestal
(FENF), da Universidade Federal de Mato Grosso (UFMT) para triagem e

14
contagem das formigas. Após as triagens, os espécimes foram
encaminhado ao Prof. Dr. Jacques H. C. Delabie, do Laboratório de
Mirmecologia do CEPLAC-CEPEC, Ilhéus, Bahia – para a devida
identificação taxonômica.
3.5. ANÁLISE DOS DADOS
3.5.1. Análises Estatísticas
Foi realizado um delineamento em blocos casualizados com três
tratamentos e 20 repetições, sendo o número total de indivíduos por
armadilha transformado através do uso de , atendendo assim a
normalidade de erros e homogeneidade das variâncias.
O cálculo da Variância (ANOVA) das médias de ocorrência do
número de indivíduos por coletas e o número de espécies por área foram
testadas ao Teste de Tukey a 5% de probabilidade pelo programa
ASSISTAT 7.0.
3.5.2. Análise Faunística
Nos estudos faunísticos foram realizados cálculos de Constância,
Dominância, Abundância e Frequência através do programa ANAFAU
(MORAES et al., 2003). Foi calculado índices faunísticos para cada
comunidade.
Os índices utilizados foram os seguintes:
Índice de constância
Foi calculado utilizando-se dados mensais de coleta pela
fórmula de Dajóz (1974):
C (%) P
N
.100, em que
C: constância;
P: quantidade de coletas, em que a espécie foi capturada;

15
N: quantidade total de coletas efetuadas.
As espécies foram distribuídas em categorias, conforme
classificação de Bodenheimer (1955):
Espécies constantes (W): presentes em mais de 50% das coletas;
Espécies acessórias (Y): presentes de 25 a 50% das coletas;
Espécies acidentais (Z): presentes em menos de 25% das coletas.
Índice de Dominância
Foi obtido pelas equações propostas por Sakagami e Matsumura (1967):
Limite superior (LS)
n F
n n F
1 0
2 1 0
100.
., em que
n1: 2(K +1);
n2: 2(N - K + 1)
Limite inferior (LI) 100.
.1
012
01
Fnn
Fn , em que
n1: 2(N - K + 1);
n2: 2(K + 1).
N: número total de indivíduos capturados;
K: número de indivíduos de cada espécie;
Fo: valor obtido da tabela de distribuição de F ao nível de 5% de
probabilidade.
Serão consideradas espécies dominantes aquelas, cujo valor do
LI for maior que S, calculado para o valor de K = 0.
Índice de Frequência
Foi obtido calculando-se a percentagem de indivíduos de cada
espécie em relação ao total de indivíduos capturados em cada ambiente,
conforme Silveira Neto et al. (1976):

16
IF (%) = N
T 100em que,
IF: índice de frequência (%);
N: número de indivíduos de cada espécie coletada;
T: número total de indivíduos coletados
Índice de Abundância
Para o cálculo deste índice, foi utilizado uma medida de
dispersão (SILVEIRA NETO et al., 1976), determinando-se o intervalo de
confiança (IC) e o teste “t” a 5% e a 1% de probabilidade. Para se estimar
a abundância das espécies, foram consideradas as seguintes classes:
Rara (r): número de indivíduos menor do que o limite inferior do IC
a 1% de probabilidade;
Dispersa (e): número de indivíduos situado entre os limites
inferiores do IC a 5% e 1% de probabilidade;
Comum (c): número de indivíduos situado dentro do IC a 5% de
probabilidade;
Abundante (a): número de indivíduos situado entre os limites
superiores de IC a 5% e 1% de probabilidade;
Muito abundante (m): número de indivíduos maior do que o limite
superior do IC a 1% de probabilidade.
Índice de Riqueza
Foi calculado o índice de Riqueza de Margaleff que demonstra a
riqueza de espécies existentes dentro de cada área e expressa a
diversidade de espécies, ou seja, refere-se à abundância de uma
determinada área geográfica, região ou comunidade (RODRIGUES et al.,
2004).

17
Índice de Diversidade
Para cada comunidade foi calculado um índice de diversidade,
mostrando a relação existente entre o número de espécies e de indivíduos
e comparando-as entre si.
Conforme LUDWIG e REYNOLDS (1988), os números de
diversidade de Hill (N0, N1 e N2) são recomendados para medir a
diversidade de espécies, por serem mais interpretativos que outros índices
de diversidade e terem a vantagem de sua unidade de medida ser dada em
espécie. A equação utilizada foi a de Hill (1973), citado por LUDWIG e
REYNOLDS (1988):
NA = 1/(1-A), em que:
Pi: proporção de indivíduos da i-enésima espécie;
A: pode ser igual a 0, 1 e 2;
*A = 0 N0 S, sendo S: o número total de espécies.
*A = 1 N1= eH', sendo H' o índice de Shannon, que transformado
fornece o número de espécies abundantes:
H'= - ( pi ln pi), em que
*A2 N2 = 1/λ
O índice de Simpson (λ) dá mais peso a espécies muito
abundantes do que a espécies raras. Portanto, da sua transformação
obtém-se o número de espécies muito abundantes:
, em que
( )pii
S
1
i
S
1
S
i NN
nini
1 )1(
1

18
S: número total de espécies consideradas;
N: número total de indivíduos da população;
Ni: número de indivíduos em cada uma das i-ésimas espécies.
Índice de equitatividade
O índice de equitatividade (E) foi obtido por meio da relação entre
N2 e N1, conhecida como Razão de Hill modificada, conforme a fórmula:
E = (N2 - 1)/(N1 - 1), em que:
E= equitatividade;
N1= número 1 de diversidade de Hill;
N2= número 2 da diversidade de Hill.
Índice de Sorensen (Cs) (1984)
É um índice qualitativo e geralmente é empregado para estimar
a semelhança em termos de espécies entre amostras sucessivas coletadas
em diferentes ecossistemas ou ao longo de um gradiente ambiental
(WOLDA, 1981);
Cs = 2c ÷ a + b → onde:
a = número de espécies presentes na amostra a;
b = número de espécies presentes na amostra b;
c = número de espécies comuns ás amostras a e b.

19
4. RESULTADOS E DISCUSSÃO
4.1. ANÁLISES QUANTITATIVA E QUALITATIVA
Nos três ambientes foram coletados 60.506 indivíduos, sendo
25.027 (41,36%) no talhão de teca, 23.723 (39,21%) no planto com pau-
de-balsa e 11.756 (19,43%) na área de cerrado (Figura 2).
O levantamento quantitativo dos indivíduos coletados na
vegetação de cerrado foi o menor número em relação aos demais
ambientes. Isto pode ter ocorrido em função da competição existente na
área, proporcionada pela maior heterogeneidade de recursos disponíveis,
que possibilita a adaptação de diversas populações de espécies que
competem em equilíbrio e assim, afetando e reduzindo o número de
indivíduos coletados (FOWLER e DELABIE, 1995; RIBAS et al., 2003;
MARQUES e DEL-CLARO, 2006).
FIGURA 2 - DISTRIBUIÇÃO DE INDIVÍDUOS COLETADOS EM AMBIENTES DE PLANTIO DE TECA, PAU-DE-BALSA E ÁREA NATIVA DO CERRADO, NO MUNICÍPIO DE NOSSA SENHORA DO LIVRAMENTO, MATO GROSSO, 2015/2016.
No total foram registradas cinco subfamílias. Do número total de
espécimes coletados nos ambientes amostrados, 0,27% pertencem a

20
subfamília Dolichoderinae, 47,33% a subfamília Dorylinae, 4,35% a
subfamília Formicinae, 47,63% a subfamília Myrmicinae e 0,40% a
subfamília Ponerinae. A subfamília Myrmicinae foi a mais representativa
em número de espécies e indivíduos (Figura 3).
Esses resultados corroboram com os resultados de Souza
(2008) e Silva (2007) que observaram a predominância da subfamília
Myrmicinae com experimentos em diferentes ambientes florestais com o
mesmo método de coleta pois, Myrmicinae possui a maior riqueza e
distribuição em todos os ambientes no mundo (exceto no Ártico e Antártida)
(SHATTUCK, 1999).
FIGURA 3 - DISTRIBUIÇÃO DAS SUBFAMÍLIAS DE FORMICÍDEOS COLETADOS EM AMBIENTES DE PLANTIO DE TECA, PAU-DE-BALSA E ÁREA NATIVA DO CERRADO, NO MUNICÍPIO DE NOSSA SENHORA DO LIVRAMENTO, MATO GROSSO, 2015/2016.
Nos três ambientes amostrados, observou-se a ocorrência de 23
espécies de formigas na área com plantio de teca, 23 para no plantio de
pau-de-balsa e 24 na vegetação de cerrado. Foi constatado 20 espécies
comuns a todos os ambientes (Erro! Fonte de referência não e
ncontrada.). Em Dolichoderinae, a espécie Dorymyrmex pyramicus
ocorreu nos três ambientes observados, enquanto Tapinoma
melanocephalum ocorreu nos ambientes com pau-de-balsa e área nativa
(Tabela 1).

21
TABELA 1 - ESPÉCIES DE FORMIGAS AMOSTRADAS EM DIFERENTES AMBIENTES FLORESTAIS NO MUNICÍPIO DE NOSSA SENHORA DO LIVRAMENTO, ESTADO DE MATO GROSSO, 2015/2016.
Espécies/subfamília Ambientes* Hábito alimentar
Dolichoderinae
Dorymyrmex pyramicus A, B, C Onívora
Tapinoma melanocephalum B Onívora
Dorylinae
Labidus coecus A, B, C Predador
Labidus praedator A, B, C Predador
Nomamyrmex esenbeckii A, B, C Predador
Formicinae
Brachymyrmex heeri A, B, C Onívora Camponotus blandus A, B, C Onívora
Camponotus crassus A, B, C Onívora
Camponotus melanoticus A, B, C Onívora
Camponotus novogranadensis A, B, C Onívora
Myrmicinae
Pheidole araneoides A, B, C Onívora Acromyrmex subterraneus subterraneus
A,C Cultivadora de fungo
Atta sexdens A, B, C Cultivadora de fungo
Cephalotes atratus B, C Onívora
Crematogaster tenuicula A, B, C Onívora
Pheidole grupo fallax sp. 1 A, B, C Onívora
Pheidole grupo flavens sp. 2 A, B, C Onívora
Pheidole radoszkowskii A, C Onívora
Pheidole grupo tristis sp. 3 A, B Onívora
Solenopsis globularia A, B, C Onívora
Solenopsis sp. A, B, C Onívora
Sericomyrmex sp. 1 A, B, C Cultivadora de fungo
Strumigenys denticulata A, B, C Predador
Ponerinae
Odontomachus bauri A, B, C Predador
Neoponera commutata A, B, C Predador
Pseudoponera gilberti C Predador
*Ambientes: (A) teca; (B) pau-de-balsa; (C) cerrado

22
As espécies de Dorylinae, Labidus coecus, Labidus praedator e
Nomamyrmex esenbeckii foram comuns nos três ambientes, constituindo-
se na segunda subfamília com maior quantidade de indivíduos coletados
neste estudo. Formicinae ocorreu com as espécies, Brachymyrmex heeri,
Camponotus blandus, Camponotus crassus, Camponotus melanoticus e
Camponotus novogranadensis comuns aos três ambientes (Tabela 1).
Myrmicinae foi a mais diversificada em quantidade de espécies,
sendo Atta sexdens, Crematogaster tenuicula, formigas do gênero Pheidole
dos grupos Fallax e Flavens, Solenopsis sp., Solenopsis globularia,
Sericomyrmex sp. e Strumigenys denticulata, as quais foram coletadas nos
três ambientes de estudo, enquanto Acromyrmex subterraneus
subterraneus e Pheidole radoszkowskii ocorreram nos ambientes de plantio
de teca e em área nativa. Cephalotes atratus ocorreu em ambiente de pau-
de-balsa e na área nativa e Pheidole grupo tristis sp. 3 foi observada nos
ambientes com teca e pau-de-balsa (Tabela 1).
Em Ponerinae, Odontomachus bauri e Neoponera commutata
foram espécies comuns a todos os ambientes, enquanto Pseudoponera
gilberti foi encontrada apenas no ambiente de cerrado (Tabela 1). Entre as
espécies observadas, a maioria (16 espécies), são onívoras,
representando 61,54% do total de espécies amostradas. Sete espécies são
predadoras (26,92%) e três cultivadoras de fungo (11,54%). Esses valores
confirmam a variedade de hábitos alimentares que as formigas apresentam
e maior quantidade de espécies de formigas onívoras que não apresentam
especificidade alimentar e assim maior facilidade para adaptar em
diferentes ambientes (FAGUNDES et al., 2009) (Tabela 2).
TABELA 2 - HÁBITO ALIMENTAR DE ESPÉCIES DE FORMIGAS AMOSTRADAS EM PLANTIO DE TECA, PAU-DE-BALSA E CERRADO NO MUNICÍPIO DE NOSSA SENHORA DO LIVRAMENTO-MT, 2015/2016.
Total de espécies
Espécies Hábito alimentar Porcentagem (%)
16 Onívoras 61,54
7 Predadores 26,92
3 Cultivadora de fungo 11,54

23
No geral, os ambientes apresentaram a mesma proporção de
espécies por diferentes hábitos alimentares assim, os três ambientes
apresentaram suporte para o estabelecimento de espécies com
comportamentos distintos. Espécies onívoras foram representadas em
maior número nos três ambientes, corroborando com Baccaro et al. (2015)
que uma parcela significativa das espécies de formigas seja onívora. A
menor proporção de espécies predadoras e cultivadoras de fungos devem-
se a estas espécies serem mais específicas na busca por alimento e com
isso, mais exigentes com a oferta de alimento disponível (BACCARO et al.,
2015) (Tabela 3).
TABELA 3 - HÁBITO ALIMENTAR DE ESPÉCIES DE FORMIGAS AMOSTRADAS POR AMBIENTE NO MUNICIPIO DE NOSSA SENHORA DO LIVRAMENTO-MT, 2015/2016.
Ambiente Hábito alimentar Espécies Porcentagem (%)
Teca
Onívora 14 63,64
Predadora 6 22,73
Cultivadora de fungo 3 13,04
Pau-de-balsa
Onívora 15 61,9
Predadora 6 28,57
Cultivadora de fungo 2 9,52
Cerrado
Onívora 14 63,64
Predadora 7 27,27
Cultivadora de fungo 3 9,09
Em relação ao hábito alimentar, Dolichoderinae e Formicinae foram
representadas somente por espécies onívoras, que é característico da
maioria das espécies dessa subfamília. Em Dorylinae e Ponerinae
ocorreram somente espécies predadoras, sendo algumas espécies de
Dorylinae conhecidas popularmente como formigas-de-correição, enquanto
em Ponerinae ocorrem espécies predadoras generalistas até especialistas
extremas, sendo utilizadas em controle biológico. Myrmicinae é a
subfamília mais diversa de Formicidae, por isso a variedade de hábitos
alimentares das espécies amostradas neste trabalho (Tabela 4).
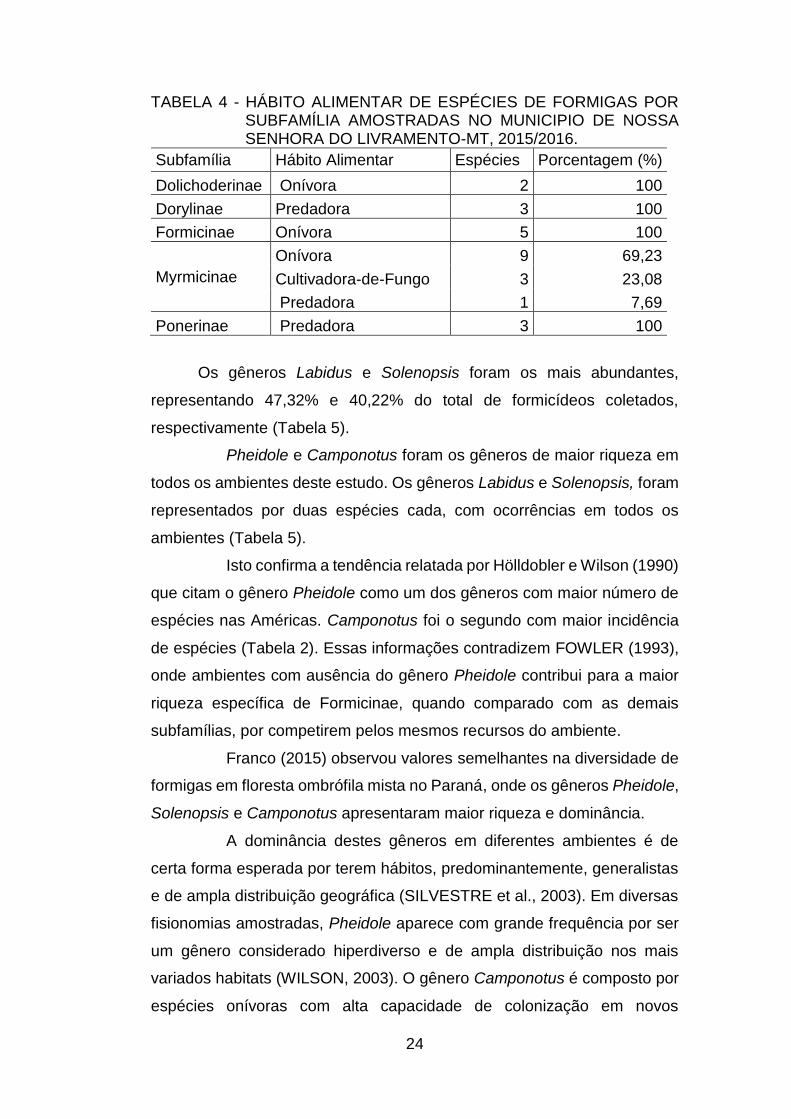
24
TABELA 4 - HÁBITO ALIMENTAR DE ESPÉCIES DE FORMIGAS POR SUBFAMÍLIA AMOSTRADAS NO MUNICIPIO DE NOSSA SENHORA DO LIVRAMENTO-MT, 2015/2016.
Subfamília Hábito Alimentar Espécies Porcentagem (%)
Dolichoderinae Onívora 2 100
Dorylinae Predadora 3 100
Formicinae Onívora 5 100
Myrmicinae
Onívora 9 69,23
Cultivadora-de-Fungo 3 23,08
Predadora 1 7,69
Ponerinae Predadora 3 100
Os gêneros Labidus e Solenopsis foram os mais abundantes,
representando 47,32% e 40,22% do total de formicídeos coletados,
respectivamente (Tabela 5).
Pheidole e Camponotus foram os gêneros de maior riqueza em
todos os ambientes deste estudo. Os gêneros Labidus e Solenopsis, foram
representados por duas espécies cada, com ocorrências em todos os
ambientes (Tabela 5).
Isto confirma a tendência relatada por Hölldobler e Wilson (1990)
que citam o gênero Pheidole como um dos gêneros com maior número de
espécies nas Américas. Camponotus foi o segundo com maior incidência
de espécies (Tabela 2). Essas informações contradizem FOWLER (1993),
onde ambientes com ausência do gênero Pheidole contribui para a maior
riqueza específica de Formicinae, quando comparado com as demais
subfamílias, por competirem pelos mesmos recursos do ambiente.
Franco (2015) observou valores semelhantes na diversidade de
formigas em floresta ombrófila mista no Paraná, onde os gêneros Pheidole,
Solenopsis e Camponotus apresentaram maior riqueza e dominância.
A dominância destes gêneros em diferentes ambientes é de
certa forma esperada por terem hábitos, predominantemente, generalistas
e de ampla distribuição geográfica (SILVESTRE et al., 2003). Em diversas
fisionomias amostradas, Pheidole aparece com grande frequência por ser
um gênero considerado hiperdiverso e de ampla distribuição nos mais
variados habitats (WILSON, 2003). O gênero Camponotus é composto por
espécies onívoras com alta capacidade de colonização em novos

25
ambientes (RAMOS et al., 2003). Algumas espécies do gênero Solenopsis
apresentam ninhos com diversas rainhas, o que oferece potencial para a
expansão de suas colônias e maior dominância no ambiente (KASPARI,
2003).
No ambiente com teca, a maior quantidade de gêneros e
espécies pertencem a subfamília Myrmicinae, representada por 16,57%
dos espécimes coletados neste ambiente. Dorylinae, com dois gêneros e
três espécies, correspondeu com 81,98% dos indivíduos coletados. As
subfamílias Formicinae e Ponerinae foram representadas por duas
espécies com 0,96% e 0,34% do total de indivíduos coletados,
respectivamente. A subfamília Dolichoderinae foi a menos representativa
com um gênero e uma espécie e com 0,14% de espécimes coletados
(Tabela 5).
No ambiente de pau-de-balsa, Myrmicinae ocorreu com maior
número em gêneros (sete) e espécies (11) e correspondeu a 73,95% dos
indivíduos coletados. Dolichoderinae, Dorylinae, Formicinae e Ponerinae
apresentam dois gêneros e, corresponderam em conjunto com 26,05% do
total de indivíduos coletados (Tabela 5).
No ambiente com vegetação de cerrado, Myrmicinae teve maior
representatividade, com 60,67% do total de indivíduos coletados,
distribuídos em oito gêneros e 12 espécies. Dorylinae e Formicinae
contribuíram em conjunto com 38,48% dos indivíduos coletados, enquanto
Dolichoderinae e Ponerinae foram responsáveis apenas por 0,85% dos
espécimes coletados, sendo as menos representativas em todos os
ambientes, sendo responsáveis por 0,27% e 0,40% do total de indivíduos
amostrados, respectivamente (Tabela 5).
Espécies de Ponerinae são encontradas de forma abundante na
serapilheira sendo mais diversas em ambientes arborizados, consideradas
grandes predadoras e generalistas, se alimentam de diversos artrópodes,
tendo preferencia cupins do gênero Syntermes spp. (BROWN JR., 2000;
DELABIE, et al., 2000; RÉ, 2007). A pequena quantidade de indivíduos
coletados desta subfamília pode ser explicada pela característica das
espécies serem dominantes em locais, onde o estresse e a perturbação
limitam a produtividade e diversidade de formigas (ANDERSEN, 2000).

26
Schmidt et al. (2005) afirmam que a maioria das espécies de
formigas neotropicais são generalistas. No entanto, existem grupos
especializados que vivem em associação com plantas, como algumas
espécies de Dolichoderinae, onde vivem, principalmente em plantas do
gênero Cecropia, atuando em sua defesa contra a ocorrência de herbivoria
e restringindo a diversidade de ambientes onde as espécies dessa
subfamília são dominantes. A subfamília Myrmicinae foi a mais
diversificada em número de espécies, entretanto, as espécies do gênero
Solenopsis foram quantitativamente, as mais representativas dentro desta
subfamília. Nos ambientes com pau-de-balsa e vegetação de cerrado, o
gênero Solenopsis foi o mais representativo com 70,72% e 44,99% do total
de indivíduos amostrados para ambos os ambientes, respectivamente.
L. coecus foi a espécie com maior representatividade em
quantidade de indivíduos coletados nos ambientes de teca e pau-de-balsa,
ocorrendo com 27.544 indivíduos, o que representou 45,52% do total
coletado, contribuindo para que Dorylinae seja a segunda mais abundante
(Tabela 5).
A maior representatividade de L. coecus ocorreu no ambiente
com plantio de teca, onde representou 81,47% dos indivíduos amostrados.
Nos ambientes com pau-de-balsa e vegetação de cerrado, a espécie ficou
entre as mais importantes em números de indivíduos com 19,46% e
21,60% do total coletados, respectivamente (Tabela 5).
O ambiente com plantio de pau-de-balsa apresentou a menor
complexidade estrutural dentre os ambientes amostrados, e isto em função
da idade do plantio e pela ausência de sub-bosques. O que pode justificar
a dominância das espécies do gênero Solenopsis neste ambiente, por
serem formigas onívoras (BUENO e CAMPOS-FARINHA, 1999), e estão
entre as mais agressivas na utilização de recursos, podendo suportar
ambientes com restrição de alimento (MARINHO et al., 2002). A menor
população de Solenopsis nos demais ambientes pode ser justificada pelo
aumento da população de espécies da subfamília Dorylinae que são
predadoras e possuem dieta especializada em formigas, incluindo deste
gênero (POWELL e BACKER, 2008).

27
A maior representatividade de L. coecus em plantio de teca é
justificada pela simplificação do ambiente. Esta espécie é considerada
como formigas de correição, predadora com alta tolerância ecológica e
ampla distribuição (WETTERER e SNELLING, 2015). Essas formigas são
conhecidas pelo seu forrageamento agressivo e pelo grande número de
indivíduos forrageando dentro de uma determinada área. Não constroem
ninhos permanentes, suas colônias se movem constantemente em uma
síndrome conhecida por “comportamento legionário” (BRADY, 2003). As
formigas do gênero Labidus emigram de forma irregular, frequentemente
formando ninhos em um mesmo ponto durante vários meses
(RETTENMEYER, 1963).
As formigas de correição, representadas neste trabalho pelos
gêneros Labidus e Nomamyrmex foram amostradas nos três ambientes.
Rettenmeyer et al. (1983) defende que a maior diversidade desse tipo de
formigas se encontram em florestas úmidas ou pluviais. As atividades
podem ser subterrâneas e/ou superficiais ao solo, podendo ser de dia ou a
noite, intensificando em dias nublados e em períodos com sol ameno, como
no amanhecer. Esse comportamento é característico dessas espécies que
apresentam sensibilidade ao sol direto. O cerrado, que apresenta um
período de seca a cada ano e estrutura de savana, gera pouca sombra no
solo durante o dia ao longo de todo o ano, apresentando uma comunidade
de formigas de correição ativa e diversa, porém menos aparente por
apresentarem baixíssima atividade sobre a superfície na maior parte do dia
(POWELL e BACKER, 2008).
As fomigas de correição são predadoras de diversos artrópodes,
incluindo outros tipos de formigas. Nomamyrmex esenbeckii é a única
formiga de correição que é um predador confirmado de formigas
cortadeiras do gênero Atta, atacando com sucesso colônias maduras
enormes que possuem soldados grandes (RETTENMEYER et al. 1983;
SWARTZ, 1998; SÁNCHEZ-PEÑA e MUELLER 2002; POWELL e CLARK,
2004). As formigas do gênero Labidus predam sobre formigas e outros
artropodes como aranhas, larvas de formigas e lepidópteros adultos ou
jovens (POWELL e BACKER, 2008).

28
TABELA 5 - RELAÇÃO DAS SUBFAMÍLIAS E ESPÉCIES EM PORCENTAGEM (%) E NÚMEROS DE INDIVÍDUOS (I) COLETADOS NOS TRÊS AMBIENTES FLORESTAIS NO MUNICÍPIO DE NOSSA SENHORA DO LIVRAMENTO, ESTADO DE MATO GROSSO, 2015/2016
Subfamília/ Espécie AMBIENTES TOTAL TECA PAU-DE-BALSA CERRADO
Indivíduos % Indivíduos % Indivíduos % Indivíduos %
Dolichoderinae
Dorymyrmex pyramicus 36 0,14 100 0,42 28 0,24 164 0,27
Tapinoma melanocephalum − − 1 0,00 − − 1 0,00
Dorylinae
Labidus coecus 20.389 81,47 4.616 19,46 2.539 21,60 27.544 45,52
Labidus praedator 127 0,51 498 2,10 461 3,92 1086 1,79
Nomamyrmex esenbeckii 1 0,00 5 0,02 5 0,04 11 0,02
Formicinae 0,00
Brachymyrmex heeri 26 0,10 14 0,06 18 0,15 58 0,10
Camponotus blandus 56 0,22 122 0,51 977 8,31 1.155 1,91
Camponotus crassus 12 0,05 152 0,64 26 0,22 190 0,31
Camponotus melanoticus 122 0,49 434 1,83 134 1,14 690 1,14
Camponotus novogranadensis 26 0,10 150 0,63 365 3,10 541 0,89
Myrmicinae
Pheidole araneoides 65 0,26 176 0,74 170 1,45 411 0,68
Acromyrmex subterraneus subterraneus 227 0,91 − − 1 0,01 228 0,38
Atta sexdens 871 3,48 17 0,07 1 0,01 889 1,47
Cephalotes atratus − − 1 0,00 9 0,08 10 0,02
Crematogaster tenuicula 2 0,01 3 0,01 1.447 12,31 1.452 2,40
Pheidole grupo fallax sp. 1 615 2,46 481 2,03 147 1,25 1.243 2,05
Pheidole grupo flavens sp. 2 3 0,01 56 0,24 7 0,06 66 0,11
Pheidole radoszkowskii 4 0,02 − − 21 0,18 25 0,04
Pheidole grupo tristis sp. 3 61 0,24 14 0,06 − − 75 0,12
Solenopsis sp. 2.233 8,92 16.753 70,62 5.280 44,91 24.266 40,11
Solenopsis globularia 38 0,15 23 0,10 11 0,09 72 0,12
Sericomyrmex sp. 1 14 0,06 2 0,01 29 0,25 45 0,07
Strumigenys denticulata 14 0,06 18 0,08 8 0,07 40 0,07
Ponerinae
Neoponera commutata 53 0,21 16 0,07 42 0,36 111 0,18
Odontomachus bauri 32 0,13 71 0,30 12 0,10 115 0,19
Pseudoponera gilberti − − − − 18 0,15 18 0,03
TOTAL 25.027 100 23.723 100 11.756 100 60.506 100

29
4.2. SAZONALIDADE NA DIVERSIDADE DE FORMIGAS
A sazonalidade (seca e chuva) no período de coletas interferiu
na composição da diversidade e na quantidade de formigas coletadas. No
período de seca foram coletados 21.244 indivíduos, enquanto no período
chuvoso foram coletados 39.262 indivíduos (Tabelas 6 e 7).
Quantitativamente, observou-se diferenças entre os três
ambientes. No plantio de teca, ocorreu aumento no número de indivíduos
coletados, no período da seca foram coletados 5.984 e nos meses de chuva
foram 19.043 indivíduos. O mesmo fato foi observado na área com o plantio
de pau-de-balsa, onde nos meses de seca foram coletados 8.812
espécimes, enquanto no período de chuva coletaram-se 14.911 indivíduos
(Tabelas 6 e 7).
Na área com vegetação de cerrado, o maior número de
indivíduos coletados foi no período da seca, quando foram amostrados
6.448 indivíduos, enquanto no período chuvoso foram coletados 5.308
indivíduos (Tabelas 6 e 7).
A diversidade de espécies também alterou com a sazonalidade.
No período de seca, as espécies presentes foram 20 em plantio de teca,
21 para pau-de-balsa e 23 para cerrado, enquanto no período chuvoso, as
quantidades de espécies foram de 19 em plantio de teca, 18 em plantio de
pau-de-balsa e 19 na vegetação de cerrado (Tabelas 6 e 7).
Em relação aos períodos estudados, seca e chuva, as formigas
dos gêneros Cephalotes e Sericomyrmex só ocorreram na seca e o
primeiro no cerrado e e o segundo em todos os ambientes. No período
chuvoso não foi observado a ocorrência de espécies desses gêneros, visto
que todos os registros ocorreram no período de seca (Tabela 6 e 7).
Wink et al. (2005) registraram ocorrência maior de formigas
Sericomyrmex em eucaliptais e na vegetação nativa, devido,
provavelmente, a sua facilidade de adaptação em ambientes que ocorre
substituição da vegetação. Enquanto que Cephalotes pusillus e
Thaumatomyrmex foram registradas tanto na vegetação nativa e na área
de eucalipto.

30
No ambiente com teca, as formigas do gênero Nomamyrmex
ocorreu somente no período de chuva, enquanto Crematogaster e
Sericomyrmex só ocorreram no período de seca. No plantio com pau-de-
balsa, a única espécie de formigas do gênero Atta só ocorreu no período
de chuva. Crematogaster, Sericomyrmex, Strumigenys e Tapinoma
ocorreram apenas no período da seca. Na vegetação de cerrado,
Nomamyrmex só ocorreu nos meses de chuva, Acromyrmex, Atta,
Cephalotes, Sericomyrmex ocorreram apenas no período seco (Tabelas 6
e 7).
Pode-se observar nas Tabelas 5 e 6 que todas as espécies de
ocorrência no período de chuva, foram comuns no período de seca, exceto
T. melanocephalum, C. atratus e Sericomyrmex sp. 1 que só ocorreram no
primeiro período.

31
TABELA 6 - AÇÃO DAS SUBFAMÍLIAS, GÊNEROS E PERCENTAGENS (%) DE ESPÉCIES (E) E DE NÚMEROS (I) DE FORMIGAS COLETADAS NO PERÍODO DE SECA NOS TRÊS AMBIENTES FLORESTAIS NO MUNICÍPIO DE NOSSA SENHORA DO LIVRAMENTO, ESTADO DE MATO GROSSO, 2015/2016.
Subfamília/Gênero/Espécie PERÍODO: SECO TOTAL
AMBIENTES
TECA PAU-DE-BALSA CERRADO
E % I % E % I % E % I % I %
Dolichoderinae
Dorymyrmex 1 5 1 0,02 1 5 13 0,15 1 4,35 10 0,15 24 0,11
Tapinoma - - - - 1 5 1 0,01 - - - - 1 0,00
Dorylinae
Labidus 2 10 3.743 62,55 2 10 775 8,88 2 8,70 194 2,99 4.712 22,22
Nomamyrmex - - - - 1 5 55 0,63 - - - 55 0,26
Formicinae
Brachymyrmex 1 5 26 0,43 1 5 40 0,46 1 4,35 5 0,08 71 0,33
Camponotus 4 20 120 2,01 4 20 277 3,17 4 17,39 1.263 19,44 1660 7,83
Myrmicinae
Pheidole 3 15 86 1,44 3 15 493 5,65 4 17,39 209 3,22 788 3,72
Acromyrmex 1 5 15 0,25 1 5 13 0,15 1 4,35 1 0,02 29 0,14
Atta 1 5 255 4,26 - - - - 1 4,35 1 0,02 256 1,21
Cephalotes - - - - - - - 1 4,35 9 0,14 9 0,04
Crematogaster 1 5 2 0,03 1 5 1 0,01 1 4,35 1.096 16,87 1099 5,18
Solenopsis 2 10 1.689 28,23 2 10 7.010 80,32 2 8,70 3.638 55,99 12.337 58,17
Sericomyrmex 1 5 14 0,23 1 5 14 0,16 1 4,35 29 0,45 57 0,27
Strumigenys 1 5 6 0,10 1 5 2 0,02 1 4,35 5 0,08 13 0,06
Ponerinae
Neoponera 1 5 20 0,33 1 5 10 0,11 1 4,35 26 0,40 56 0,26
Odontomachus 1 5 7 0,12 1 5 23 0,26 1 4,35 9 0,14 39 0,18
Pseudoponera - - - 1 5 1 0,01 1 4,35 3 0,05 4 0,02
TOTAL 20 100 5.984 100 21 100 8.728 100 23 100 6.498 100 21.244 100

32
TABELA 7 - RELAÇÃO DAS SUBFAMÍLIAS, GÊNEROS E PERCENTAGENS (%) DE ESPÉCIES (E) E DE NÚMEROS (I) DE FORMIGAS COLETADAS NO PERÍODO DE CHUVA NOS TRÊS AMBIENTES FLORESTAIS NO MUNICÍPIO DE NOSSA SENHORA DO LIVRAMENTO, ESTADO DE MATO GROSSO, 2015/2016.
PERÍODO: CHUVA
Subfamília/Gênero/Espécie
AMBIENTES
TOTAL TECA PAU-DE-BALSA CERRADO
E % I % E % I % E % I % I %
Dolichoderinae
Dorymyrmex 1 4,76 35 0,17 1 5,6 163 1,09 1 5,3 18 0,34 216 0,53
Tapinoma - - - - - - - - - - - - 0,00
Dorylinae
Labidus 2 9,52 18.280 88,36 2 11,1 3.864 25,91 2 10,5 2.806 52,86 24.950 60,99
Nomamyrmex 1 4,76 1 0,00 1 5,6 1 0,01 1 5,3 5 0,09 7 0,02
Formicinae
Brachymyrmex 1 4,76 1 0,00 1 5,6 60 0,40 1 5,3 13 0,24 74 0,18
Camponotus 4 19,05 105 0,51 4 22,2 437 2,93 4 21,1 239 4,50 781 1,91
Myrmicinae
Pheidole 5 23,81 671 3,24 4 22,2 566 3,80 3 15,8 136 2,56 1.373 3,36
Acromyrmex 1 4,76 218 1,05 1 5,6 5 0,03 - - - - 223 0,55
Atta 1 4,76 705 3,41 1 5,6 17 0,11 - - - - 722 1,77
Cephalotes - - - - - - - - - - - - -
Crematogaster - - - - - - - - 1 5,3 401 7,55 401 0,98
Solenopsis 2 9,52 588 2,84 1 5,6 9.744 65,35 2 10,5 1.653 31,14 11.985 29,30
Sericomyrmex - - - - - - - - - - - - -
Strumigenys 1 4,76 11 0,05 - - - - 1 5,3 3 0,06 14 0,03
Ponerinae
Neoponera 1 4,76 42 0,20 1 5,6 6 0,04 1 5,3 16 0,30 64 0,16
Odontomachus 1 4,76 30 0,15 1 5,6 48 0,32 1 5,3 3 0,06 81 0,20
Pseudoponera - - - - - - - - 1 5,3 15 0,28 15 0,04
TOTAL 19 100 19.043 100 18 100 14.911 100 19 100 5.308 100 39.262 100

33
L. coecus e Solenopsis sp. foram as espécies mais abundantes,
com 21,66% e 57,85% dos espécimes coletados no período de seca,
respectivamente e 58,44% e 30,51% dos indivíduos coletados no período
chuvoso, respectivamente. (Tabelas 8 e 9).
No ambiente com teca, no período de seca, as espécies mais
abundantes foram L. coecus, A. sexdens e Solenopsis sp., enquanto no
período da chuva, L. coecus, P. grupo tristis sp. 3, A. sexdens e P. grupo
fallax sp. 1. foram as mais representativas. No ambiente com plantio de
pau-de-balsa, as espécies mais abundantes no período da seca foram
Solenopsis sp., L. coecus. L. praedator e P. grupo fallax sp. 1. e no período
de chuva, Solenopsis sp., L. coecus e P. grupo fallax sp.1,
quantitativamente, foram as mais expressivas. Na vegetação de cerrado,
as espécies, L. coecus, Solenopsis sp., Crematogaster tenuicula e L.
praedator foram as mais importantes no período da chuva. No período da
seca, P. grupo flavens sp. 2, C. tenuicula e Solenopsis sp., foram as
espécies mais expressivas em números de indivíduos coletados (Tabelas
8 e 9).
Santos et al. (2007) registraram resultados semelhantes para a
riqueza de formigas em fragmento de floresta amazônica, sendo espécies
do gênero Pheidole, Solenopsis e Crematogaster como as mais
abundantes nos períodos muito chuvosos e menos chuvosos, durante o
ano.
Essas variações temporais dos espécimes de formigas no
decorrer do ano podem ser em função da oferta de alimento no ambiente.
A disponibilidade de alimentação é defendida como um dos principais
fatores relacionados à abundância de formigas. Comportamentos
específicos no ambiente como hábito de armazenar alimentos,
especificidades de habitat podem determinar a permanência de uma
espécie durante e após as variações das condições climáticas
(FAGUNDES et al., 2009).
No período de seca, as espécies do gênero Solenopsis
representam 58,17% do total de indivíduos coletados, com as quantidades
mais expressivas coletas na vegetação de cerrado e no plantio de pau-de-
balsa com 56,42% e 79,66% dos espécimes coletados respectivamente.

34
Em plantio de teca, o gênero Labidus foi, quantitativamente, a mais
importante, correspondendo com 62,55% dos indivíduos coletados,
seguido de Solenopsis com 28,23% do total de indivíduos coletados
(Tabela 8 e 9).

35
TABELA 8 - RELAÇÃO DAS SUBFAMÍLIAS E PORCENTAGEM DE NÚMEROS INDIVÍDUOS (I) COLETADOS NO PERÍODO SECO NOS TRÊS AMBIENTES FLORESTAIS NO MUNICÍPIO DE NOSSA SENHORA DO LIVRAMENTO, ESTADO DE MATO GROSSO, 2015/2016.
Subfamília/ Espécie
AMBIENTES TOTAL TECA PAU-DE-BALSA CERRADO
Indivíduos % Indivíduos % Indivíduos % Indivíduos %
Dolichoderinae
Dorymyrmex pyramicus 1 0,02 40 0,45 10 0,16 51 0,24
Tapinoma melanocephalum - - 1 0,01 - - 1 0,00
Dorylinae
Labidus coecus 3.716 62,10 754 8,54 131 2,03 4.601 21,66
Labidus praedator 27 0,45 321 3,64 63 0,98 411 1,93
Nomamyrmex esenbeckii - - 1 0,01 - - 1 0,00
Formicinae
Brachymyrmex heeri 26 0,43 14 0,16 5 0,08 45 0,21
Camponotus blandus 50 0,84 86 0,97 836 12,97 972 4,58
Camponotus crassus 4 0,07 46 0,52 24 0,37 74 0,35
Camponotus melanoticus 62 1,04 143 1,62 106 1,64 311 1,46
Camponotus novogranadensis 4 0,07 87 0,99 297 4,61 388 1,83
Myrmicinae
Acromyrmex subterraneus subterraneus 15 0,25 - - 1 0,02 16 0,08
Atta sexdens 255 4,26 - - 1 0,02 256 1,21
Cephalotes atratus - - 1 0,01 9 0,14 10 0,05
Crematogaster tenuicula 2 0,03 3 0,03 1.046 16,22 1051 4,95
Pheidole araneoides 41 0,69 13 0,15 148 2,30 202 0,95
Pheidole grupo fallax sp.1 38 0,64 169 1,92 39 0,60 246 1,16
Pheidole grupo flavens sp.2 - - 55 0,62 1 0,02 56 0,26
Pheidole radoszkowskii - - - - 21 0,33 21 0,10
Pheidole grupo tristis sp. 3 7 0,12 - - 7 0,03
Solenopsis sp. 1.651 27,59 7.009 79,42 3.629 56,28 12.289 57,85
Solenopsis globularia 38 0,64 21 0,24 9 0,14 68 0,32
Sericomyrmex sp.1 14 0,23 2 0,02 29 0,45 45 0,21
Strumigenys denticulata 6 0,10 13 0,15 5 0,08 24 0,11
Ponerinae
Neoponera commutata 20 0,33 10 0,11 26 0,40 56 0,26
Odontomachus bauri 7 0,12 23 0,26 9 0,14 39 0,18
Pseudoponera gilberti - - - - 3 0,05 3 0,01
TOTAL 5.984 100 8.812 100 6.448 100 21.244 100

36
TABELA 9 - RELAÇÃO DAS SUBFAMÍLIAS E PORCENTAGEM DE NÚMEROS INDIVÍDUOS (I) COLETADOS NO PERÍODO CHUVA NOS TRÊS AMBIENTES FLORESTAIS NO MUNICÍPIO DE NOSSA SENHORA DO LIVRAMENTO, ESTADO DE MATO GROSSO, 2015/2016.
Subfamília/ Espécie
AMBIENTES TOTAL TECA PAU-DE-BALSA CERRADO
Indivíduos % Indivíduos % Indivíduos % Indivíduos %
Dolichoderinae
Dorymyrmex pyramicus 35 0,18 60 0,40 18 0,34 113 0,29
Tapinoma melanocephalum - - - - - - - -
Dorylinae
Labidus coecus 16.673 87,55 3.862 25,90 2.408 45,37 22.943 58,44
Labidus praedator 100 0,53 177 1,19 398 7,50 675 1,72
Nomamyrmex esenbeckii 1 0,01 4 0,03 5 0,09 10 0,03
Formicinae
Brachymyrmex heeri - - - - 13 0,24 13 0,03
Camponotus blandus 6 0,03 36 0,24 141 2,66 183 0,47
Camponotus crassus 8 0,04 106 0,71 2 0,04 116 0,30
Camponotus melanoticus 60 0,32 291 1,95 28 0,53 379 0,97
Camponotus novogranadensis 22 0,12 63 0,42 68 1,28 153 0,39
Myrmicinae
Acromyrmex subterraneus subterraneus 212 1,11 - - - - 212 0,54
Atta sexdens 616 3,23 17 0,11 - - 633 1,61
Cephalotes atratus - - - - - - - -
Crematogaster tenuicula - - - - 401 7,55 401 1,02
Pheidole araneoides 24 0,13 163 1,09 22 0,41 209 0,53
Pheidole grupo fallax sp.1 577 3,03 312 2,09 108 2,03 997 2,54
Pheidole grupo flavens sp.2 3 0,02 1 0,01 6 0,11 10 0,03
Pheidole radoszkowskii 4 0,02 - - 4 0,01
Pheidole grupo tristis sp.3 54 0,28 14 0,09 68 0,17
Solenopsis sp. 582 3,06 9.744 65,35 1.651 31,10 11.977 30,51
Solenopsis globularia - - 2 0,01 2 0,04 4 0,01
Sericomyrmex sp. 1 - - - - - -
Strumigenys denticulata 8 0,04 5 0,03 3 0,06 16 0,04
Ponerinae
Neoponera commutata 33 0,17 6 0,04 16 0,30 55 0,14
Odontomachus bauri 25 0,13 48 0,32 3 0,06 76 0,19
Pseudoponera gilberti - - - - 15 0,28 15 0,04
TOTAL 19.043 100 14.911 100 5.308 100 39.262 100

37
No período chuvoso, Solenopsis foi o segundo gênero mais
expressivo em número de indivíduos coletados na área com pau-de-balsa
e vegetação de cerrado com 65,36% e 31,14% dos espécimes coletados,
respectivamente. No plantio de teca, este gênero foi o terceiro em
importância com 3,06% do total coletado (Tabelas 8 e 9).
A espécie L. coecus foi a mais abundante em todos os
ambientes estudados. No período de seca, foi a segunda espécie mais
importante em número de indivíduos com 21,66% do total coletado e a mais
abundante no plantio de teca, com 62,10% do total coletado. No período
chuvoso foi a espécie mais abundante com 58,44% do total de indivíduos
coletados e a mais expressiva na vegetação de cerrado e na área com
plantio de teca, com 87,55% e 45,37% do total coletado de indivíduos,
respectivamente (Tabelas 8 e 9).
A predominância da espécie L. coecus no ambiente com plantio
de teca durante o ano, demonstra a sua adaptação a esse monocultivo. A
sua incidência em todos os ambientes amostrados é uma característica das
espécies do gênero Labidus, com alta tolerância ecológica e ampla
distribuição e hábitos subterrâneos (BOLTON, 1990).
A redução da população de formigas no período da seca nos
ambientes de florestas plantadas deve-se a menor diversidade de
substratos de nidificação e alimentação em relação à vegetação nativa
neste período. A população de formigas no ambiente com teca teve maior
influência das mudanças sazonais, porque a teca é uma espécie heliófita
caducifólia, com queda de folhas no período de menor precipitação
pluviométrica (CHAVES, 2013), o que em Mato Grosso corresponde ao
período entre o fim do mês de junho e início de setembro. A queda das
folhas da teca na seca, diminui a disponibilidade de alimentos e de abrigo
afetando sensivelmente a densidade populacional de várias espécies de
formigas neste período.

38
4.3. CURVA DE ACUMULAÇÃO DE ESPÉCIES DE FORMIGAS
Na área do Cerrado e plantio de pau-de-balsa houve
estabilização do acúmulo de riqueza na nona coleta. A curva do coletor
para o ambiente com teca apresenta tendência de acúmulo crescente com
o aumento do esforço amostral. A não estabilização da curva de coleta,
para a comunidade de formigas é comum e pode estar ligada com a
distribuição agregada e a raridade das espécies nos ambientes amostrados
(SANTOS et al., 2006) (Figura 4).
FIGURA 4 - CURVA DE ACUMULAÇÃO DE ESPÉCIES DE FORMIGAS, POR NÚMERO DE COLETAS, EM PLANTIO DE TECA, PAU-DE-BALSA E EM ÁREA NATIVA DO BIOMA CERRADO NO MUNICÍPIO DE NOSSA SENHORA DO LIVRAMENTO, ESTADO DE MATO GROSSO, 2015/2016.
A curva de acumulação é uma forma de esquematizar a riqueza
observada e avaliar a suficiência amostral do ambiente. A riqueza das
espécies de formigas pode estar correlacionada com a complexidade
estrutural do ambiente pois, níveis elevados de perturbação em um
ambiente resulta na diminuição de espécies (SANTOS et al., 2006).
No ambiente com teca, a preferencia de algumas espécies em
forragear pode gerar faunas adaptativas a explorar diferentes tipos de
recursos, diminuindo a competição e aumentando o número de espécies
que podem co-existir em um mesmo local (TOBIN, 1997).

39
4.4. ANÁLISES ESTATÍSTICAS
Na análise de variância, concluiu-se que não ocorreu diferença
significativa entre os ambientes amostrados para a quantidades de
indivíduos coletados (Tabela 10). Este resultado indica que, cada ambiente
disponibiliza recursos suficientes para o desenvolvimento de suas
populações de acordo com as exigências de nidificação, forrageamento e
reprodução. Assim, a população de cada espécie se encontra adaptada ao
seu ambiente, em plena condição de colonização.
TABELA 10 - ANÁLISE DE VARIÂNCIA PARA NÚMERO DE INDIVÍDUOS COLETADOS EM TRÊS DIFERENTES AMBIENTES FLORESTAIS NO MUNICÍPIO DE NOSSA SENHORA DO LIVRAMENTO, ESTADO DE MATO GROSSO, 2015/2016.
Fontes de Variação G.L. S. Q. Q. M. F calculado
Blocos 19 2677,87187 140,94062 0,7826 ns
Ambientes 2 772,13767 386,06884 2,1436 ns
Resíduos 38 6843,88892 180,10234
Total 59 10293,89847
Coeficiente de Variação (CV): 60,14%
Na análise de variância para o número de espécies de formigas
foi observada diferença significativa entre cerrado e os plantios florestais.
(Tabela 11 e 12).

40
TABELA 11 - ANÁLISE DE VARIÂNCIA PARA NÚMERO DE ESPÉCIES COLETADAS EM TRÊS DIFERENTES AMBIENTES FLORESTAIS NO MUNICÍPIO DE NOSSA SENHORA DO LIVRAMENTO, ESTADO DE MATO GROSSO, 2015/2016.
Fontes de Variação G.L. S. Q. Q. M. F calculado
Blocos 19 76,316 4,0166 0,9622 ns
Ambientes 2 30,700 15,3500 3,6770 *
Resíduos 38 158,633 4,1745
Total 59 265,650 (n.s.): Não significativo (p>=0.5). Coeficiente de Variação (CV): 21,17% *: significativo ao nível de 5% de probabilidade (0,01 = < p < 0,05)
TABELA 12 - TESTE DE MÉDIAS PARA NÚMEROS DE ESPÉCIES
COLETADAS EM TRÊS AMBIENTES FLORESTAIS NO MUNICÍPIO DE NOSSA SENHORA DO LIVRAMENTO, ESTADO DE MATO GROSSO, 2015/2016.
AMBIENTES MÉDIAS
TECA 8,800 b PAU-DE-BALSA 9,60 ab CERRADO 10,55 a *Médias seguidas pela mesma letra não diferem estatisticamente entre si pelo Teste de Tukey ao nível de 5% de probabilidade
Este resultado foi semelhante ao trabalho feito por Soares et al.
(1998), que encontraram uma comunidade de formigas com maior número
de espécies em uma mata nativa do que em plantios de eucalipto. Para
esses autores, a diversidade de substratos de nidificação e alimentação é
menor nas monoculturas do que em vegetação de floresta nativa. Essas
características levam a uma redução na riqueza de espécies.
A vegetação de cerrado apresentou maior número de espécies
de formigas em comparação com os plantios florestais. Isto reforça o fato
de que áreas de vegetação nativa apresentam maior diversidade de
espécies em comparação com os plantios florestais que são ambientes
simplificados (OLIVEIRA et al., 1995).
No período de seca, observa-se na análise de variância que
houve diferença significativa entre os ambientes para a quantidade de
espécies e indivíduos de formigas amostradas (Tabela 13). Nos plantios
florestais, no período da seca, houve menor riqueza de espécies, em
comparação com o período de chuva. Nestes ambientes, no período de
seca, há menor oferta de recursos para nidificação. Assim, ocorre

41
competição entre as espécies por recursos, diminuindo a população
excedente e possibilitando a adaptação de populações de espécies
oportunistas que tendem a entrar em equilíbrio. Assim, o número de
indivíduos coletados no período de seca foi reduzido.
Já a ocorrência de maior número de espécies no período seco,
na vegetação de cerrado, pode ser em função do ambiente apresentar
maior equilíbrio, possibilitando que, maior quantidade de espécies usem os
recursos disponíveis de forma igualitária, sem dominância e de forma a
manter neste ambiente, maior riqueza de espécies de formigas.
Outro fator a ser considerado é que no período seco, ocorre o
acúmulo de serapilheira no solo, já que neste período, a vegetação do
cerrado perde grande parte do seu dossel. A serapilheira acumulada
fornece microclima favorável às espécies hipogéicas, equilibra a
temperatura do solo e propicia meios de nidificação e forrageamento para
as espécies de formigas que circulam no solo (ROCHA, 2012).
TABELA 13 - TESTE DE MÉDIAS PARA NÚMEROS DE INDIVÍDUOS E DE ESPÉCIES COLETADOS NO PERÍODO DE SECA EM TRÊS AMBIENTES FLORESTAIS NO MUNICÍPIO DE NOSSA SENHORA DO LIVRAMENTO, ESTADO DE MATO GROSSO, 2015/2016.
Variáveis Ambientes Estatísticas
Teca Pau-de-
balsa Cerrado CV(%) Pα DMS
Indivíduos 8,874 b 14,695ab 14,988
a 61,25 0,065** 6,066
Espécies 4,70 b 6,65 a 7,80 a 27,19 0,866** 1,337 ** Significativo a 5% de probabilidade de erro Médias seguidas de mesma letra nas linhas não diferem estatisticamente entre si pelo Teste de Tukey, ao nível de 5% de probabilidade de erro
Observa-se na análise de variância, que no período de chuva
não há diferença significativa entre os ambientes neste período em relação
as médias populacionais e espécies (Tabela 14).

42
TABELA 14 - TESTE DE MÉDIAS PARA NÚMEROS DE INDIVÍDUOS E DE ESPÉCIES COLETADOS NO PERÍODO DE CHUVA EM TRÊS AMBIENTES FLORESTAIS NO MUNICÍPIO DE NOSSA SENHORA DO LIVRAMENTO, ESTADO DE MATO GROSSO, 2015/2016.
Variáveis
Ambientes Estatísticas
Teca Pau-de-
balsa Cerrado CV(%) Pα DMS
Indivíduos 14,09 22,151 13,423 76,56 0.869NS 9,7668
Espécies 5,75 6,70 5,80 32,28 0,263NS 1,513 NS : Não significativo
4.5. ANÁLISE FAUNÍSTICA
Na análise anual pelo método de Laroca e Mielke (1975)
ocorreram no ambiente com teca, quatro espécies superdominantes,
quatorze dominantes e quatro não dominante. Enquanto que pelo método
de Sakagami e Laroca (1967) quatro espécies foram superdominantes,
sete dominantes e onze não dominantes. Quanto a abundância, quatro
espécies foram superabundantes, três muito abundantes, nove comuns e
sete raras. Com relação a frequência, quatro super frequentes, três muito
frequentes, nove frequentes e sete pouco frequentes. Quanto a constância,
doze espécies foram constantes, quatro acessórias e sete acidentais
(Anexo l).
Quanto a sazonalidade, ocorreram no período de seca, pelo
método de Laroca e Mielke (1975), três espécies superdominantes,
quatorze espécies dominantes e três não dominantes. Pelo método de
Sakagami e Laroca (1967), ocorreram três superdominantes, sete
dominantes e dez não dominantes. Quanto a abundância, três espécies
superabundantes, cinco muito abundantes cinco comuns e sete espécies
raras. Com relação a frequência, três espécies foram super frequentes,
cinco espécies muito frequentes, cinco espécies frequentes e seis espécies
pouco frequentes. Quanto à constância, nove espécies foram constantes,
oito acessórias e três acidentais (Anexo I).
No período de chuva, pelo método de Laroca e Mielke (1975)
ocorreram quatro espécies superdominantes, doze espécies dominantes e

43
três não dominantes, enquanto pelo método de Sakagami e Laroca (1967),
quatro espécies foram superdominantes, quatro dominantes e onze não
dominante. Em relação à abundância, quatro espécies foram
superabundantes, três muito abundantes, sete comuns, quatro dispersas e
uma rara. Em relação a frequência, quatro espécies foram super
frequentes, duas muito frequentes, sete frequentes e seis pouco
frequentes. Quanto à constância, 11 espécies foram constantes, três
acessórias e cinco acidentais (Anexo I).
As espécies L. coecus e A. sexdens foram as mais
representativas do ambiente com teca, ocorrendo como superdominantes,
superabundantes, super frequentes e constantes durante o ano. No período
de seca, também foram as mais importantes, ocorrendo como
superabundantes, super frequentes e acessórias (Tabela 15).
Neste ambiente, Camponotus blandus sofreu uma maior
influência dos períodos de chuva e seca, ocorrendo como muito frequente,
muito abundante e dominante no período de seca e pouco frequente,
acessória e não dominante no período de chuva (Tabela 15).

44
TABELA 15 - RELAÇÃO DAS ESPÉCIES DE FORMIGAS MAIS REPRESENTATIVAS NA ANÁLISE FAUNÍSTIVA AMOSTRADAS COM ARMADILHA PITFALL EM TALHÃO DE Tectona grandis NOS PERÍODOS DE SECA, CHUVA E ANUAL, NO MUNICÍPIO DE NOSSA SENHORA DO LIVRAMENTO, ESTADO DE MATO GROSSO, 2015/2016.
Subfamília/espécie
Anual
F(%) A C D
D1 D2
Dorylinae
Labidus coecus sf sa w sd sd
Myrmicinae
Atta sexdens sf sa w sd sd
Subfamília/espécie
Seca
F(%) A C D
D1 D2
Dorylinae
Labidus coecus sf sa y sd sd
Myrmicinae
Atta sexdens sf sa y sd sd
Subfamília/espécie
Chuva
F(%) A C D
D1 D2
Dorylinae
Labidus coecus sf sa w sd sd
Myrmicinae
Atta sexdens sf sa w sd sd
Programa ANAFAU: (1) Método de Laroca e Mielke (1975); (2) método de Sakagami e Laroca (1967). D: Dominância - (sd) superdominante; (d) dominante; (nd) não dominante. A: Abundância - (sa): superabundante; (ma) muito abundante; (a) abundante: (c) comum; (d) dispersa; (r) rara. F: Frequência: (sf) super frequente; (mf) muito frequente; (pf) pouco frequente. C: Constância - (w) constante; (y) acessória; (z) acidental
Na análise anual no ambiente com pau-de-balsa ocorreram, pelo
método de Laroca e Mielke (1975), duas espécies superdominates, quinze
dominantes e seis não dominantes. Enquanto que, pelo método de
Sakagami e Laroca (1967), ocorreram duas espécies superdominantes,
sete dominantes e doze não dominantes. Em relação à abundância, duas
espécies foram superabundantes, três muito abundantes, seis comuns,
uma dispersa e dez raras. Quanto à frequência, duas espécies super
frequentes, três muito frequentes, sete frequentes e 11 pouco frequentes.

45
Quanto a constância, 12 espécies constantes, três acessórias e oito
acidentais (Anexo II)
Ao analisar o período da seca, pelo método de Laroca e Mielke
(1975) observa-se que quatro espécies foram superdominantes, onze
dominantes e seis não dominantes. Pelo método de Sakagami e Laroca
(1967), quatro espécies superdominantes, quatro dominantes e treze não
dominantes. Em relação à abundância, três espécies foram
superabundantes, duas muito abundantes, sete comuns, quatro dispersas
e cinco raras. Em relação a frequência, três espécies super frequentes,
duas muito frequentes, seis frequentes e nove pouco frequentes. Quanto a
constância, treze espécies foram constantes, quatro acessórias e quatro
raras (Anexo II).
No período de chuva, pelo método de Laroca e Mielke (1975)
três espécies foram superdominantes, onze dominantes e quatro não
dominantes. Pelo método de Sakagami e Laroca (1967), três espécies
superdominantes, quatro dominantes e onze não dominantes. Quanto à
abundância, três espécies ocorreram como superabundantes, três muito
abundantes, cinco comuns, uma dispersa e seis raras. Quanto à
frequência, duas espécies foram super frequentes, quatro muito frequentes,
três frequente e oito pouco frequentes. Quanto a constância, doze espécies
foram constantes, cinco acessórias e um acidental (Anexo II).
Neste ambiente, L. coecus e Solenopsis sp. foram as espécies
mais representativas, ocorrendo como superdominantes,
superabundantes, super frequentes e constantes tanto nos períodos de
chuva, seca e anual. A espécie P. grupo Fallax sp.1 ocorreu como
superdominante, superabundante, super frequente e constante nos
períodos de chuva e seca. Na amostragem anual, essa espécie ocorreu
como dominante, superabundantes, super frequente e constante (Tabela
16).
Na área com pau-de-balsa, P. araneoides foi a espécie que
apresentou a maior variação em relação aos períodos analisados, pois
enquanto no período seco ocorreu como pouco frequente, dispersa e
dominante, no período de chuva ocorreu como muito frequente, muito

46
abundante e dominante, mostrando uma clara preferência pelos meses
mais úmidos.
Na análise anual no ambiente com vegetação de cerrado
ocorreram, pelo método de Laroca Mielke e (1975), quatro espécies
superdominantes, dezessete dominantes e três não dominantes. Pelo
método de Sakagami e Laroca (1967) quatro foram superdominantes, cinco
dominantes e quinze não dominantes. Para a abundância, quatro espécies
foram superabundantes, três muito abundantes, duas abundantes, quatro
comuns, oito dispersas e duas raras. Quanto à frequência, quatro espécies
foram super frequentes, cinco muito frequentes, cinco frequentes e dez
pouco frequentes. Quanto a constância, sete espécies foram constantes,
nove acessórias e oito acidentais (Anexo III).
No período de seca, pelo método de Laroca e Mielke (1975), três
espécies superdominantes, quatorze dominantes e cinco não dominantes.
Pelo método de Sakagami e Laroca (1967): três espécies
superdominantes, seis dominantes e quatorze não dominantes. Para a
abundância, temos três espécies superabundante, cinco muito abundante,
seis comuns, sete dispersas e duas raras. A frequência foi de três espécies
super frequente, quatro muito frequente, cinco frequente e nove pouco
frequente. Em relação a constância, doze espécies constantes e oito
acessórias (Anexo III).

47
TABELA 16 - RELAÇÃO DAS ESPÉCIES DE FORMIGAS MAIS REPRESENTATIVAS NA ANÁLISE FAUNÍSTIVA AMOSTRADAS COM ARMADILHA PITFALL EM TALHÃO DE Ochroma pyramidale NOS PERÍODOS DE SECA, CHUVA E ANUAL, NO MUNICÍPIO DE NOSSA SENHORA DO LIVRAMENTO, ESTADO DE MATO GROSSO, 2015/2016.
Subfamília/espécie
Anual
F(%) A C D
D1 D2
Dorylinae
Labidus coecus sf as y sd sd
Myrmicinae
Solenopsis sp sf as w sd sd
Crematogaster tenuicula
sf as w sd sd
Subfamília/espécie
Seca
F(%) A C D
D1 D2
Dorylinae
Labidus coecus mf ma y d d
Myrmicinae
Solenopsis sp sf as w sd sd
Crematogaster tenuicula
sf as w sd sd
Subfamília/espécie
Chuva
F(%) A C D
D1 D2
Dorylinae
Labidus coecus sf as w sd sd
Myrmicinae
Solenopsis sp. sf as w sd sd
Pheidole grupo Fallax sp. 1
mf ma y d d
Programa ANAFAU: (1) Método de Laroca e Mielke (1975); (2) método de Sakagami e
Laroca (1967). D: Dominância - (sd) superdominante; (d) dominante; (nd) não dominante.
A: Abundância - (sa): superabundante; (ma) muito abundante; (a) abundante: (c) comum;
(d) dispersa; (r) rara. F: Frequência: (sf) super frequente; (mf) muito frequente; (pf) pouco
frequente. C: Constância - (w) constante; (y) acessória; (z) acidental
No período da chuva, pelo método de Laroca e Mielke (1975):
quatro espécies superdominantes, oito dominantes e cinco não
dominantes. Pelo método de Sakagami e Laroca (1967): quatro espécies

48
superdominantes, três dominantes e doze não dominantes. Quanto à
abundância, quatro espécies superabundantes, três muito abundantes,
seis comuns e seis dispersas. Quanto à frequência, quatro espécies foram
super frequentes, três muito frequentes, cinco frequentes e seis pouco
frequentes. Em relação a constância, nove espécies foram constantes,
quatro acessórias e seis acidentais (Anexo III).
C. tenuicula e Solenopsis sp. foram as espécies mais
representativas neste ambiente, sendo superdominantes,
superabundantes, super frequentes e constantes para os períodos chuvoso
e seco. L. coecus ocorreu como super frequente, superabundante,
superdominante e constante no período chuvoso. Na coleta anual, esta
espécie ocorreu como super frequente, superabundante, superdominante
e acessória (Tabela 17).

49
TABELA 17 - RELAÇÃO DAS ESPÉCIES DE FORMIGAS MAIS REPRESENTATIVAS NA ANÁLISE FAUNÍSTICA AMOSTRADAS COM ARMADILHA “PITFALL” EM VEGETAÇÃO DE CERRADO, NOS PERÍODOS DE SECA, CHUVA E ANUAL, NO MUNICÍPIO DE NOSSA SENHORA DO LIVRAMENTO, ESTADO DE MATO GROSSO, 2015/2016.
Subfamília/espécie
Anual
F(%) A C D
D1 D2
Dorylinae
Labidus coecus sf as y sd sd
Myrmicinae
Solenopsis sp sf as w sd sd
Crematogaster tenuicula
sf as w sd sd
Subfamília/espécie
Seca
F(%) A C D
D1 D2
Dorylinae
Labidus coecus mf ma y d d
Myrmicinae
Solenopsis sp sf as w sd sd
Crematogaster tenuicula
sf as w sd sd
Subfamília/espécie
Chuva
F(%) A C D
D1 D2
Dorylinae
Labidus coecus sf as w sd sd
Myrmicinae
Solenopsis sp. sf as w sd sd Programa ANAFAU: (1) Método de Laroca e Mielke (1975); (2) método de Sakagami e Laroca (1967). D: Dominância - (sd) superdominante; (d) dominante; (nd) não dominante. A: Abundância - (sa): superabundante; (ma) muito abundante; (a) abundante: (c) comum; (d) dispersa; (r) rara. F: Frequência: (sf) super frequente; (mf) muito frequente; (pf) pouco frequente. C: Constância - (w) constante; (y) acessória; (z) acidental
De acordo com os resultados das análises faunísticas,
Solenopsis sp. ocorreu com os melhores índices em todos os ambientes
estudados, demonstrando adaptabilidade nesses ambientes. As espécies
do gênero Solenopsis são onívoras e apresentam agressividade na
utilização de recursos e um grande potencial de adaptabilidade a novos
ambientes e quase sempre com uma grande densidade populacional.

50
Considerando os resultados dos índices faunísticos, a espécie
A. sexdens foi superdominante, super frequente, constante e
superabundante no plantio de teca, dominante em plantio de pau-de-balsa
e não-dominante na área com vegetação nativa, mostrando que ambientes
mais preservados e estáveis, essa espécie tende a ocorrer com baixas
densidades populacionais. Mesmo presente em diversos ambientes, A.
sexdens é uma espécie adaptada para viver em ambientes mais abertos
(Delabie et al., 2009). Nessas condições, pode ocorrer dominância de
espécies vegetais mais palatáveis para essas formigas. Wilson (1987)
concluiu que a quantidade de alimento é importante para o estabelecimento
e desenvolviemnto das colônias dessa espécie.
Nos ambientes estudados, C. tenuicula apresentou potencial
para bioindicador de ambiente com vegetação inalterada, embora tenha
sido amostrada em todos os ambientes, na área nativa esta espécie
ocorreu como super frequente, superdominante, superabundante e
constante. Nas áreas de plantios florestais essa espécie ocorre como não
frequente e não dominante para ambos os plantios. Corassa et al. (2015)
amostraram essa espécie apenas em área de vegetação nativa
comparando com áreas agrícolas e com plantios de eucalipto.
Pseudoponera gilberti também pode ser utilizada como
bioindicador para regiões recuperadas do bioma Cerrado. Esta espécie foi
amostrada apenas no ambiente de área nativa, sendo frequente, constante
e dominante no período de chuva. Levantamento feito por Mackay e
Mackay (2010), esta espécie é encontrada em floresta tropical preservada,
com árvores de médio porte e terra firme que não sofre alagamento.
Utilizando o número de espécies amostradas nas coletas de
formigas, foram obtidos os índices de diversidade para comparar e
diferenciar os três ambientes estudados.
No ambiente com teca, ocorreram total de 23 espécies, sendo
três abundantes e duas muito abundantes. No período da seca, 23
espécies foram registradas para este ambiente, três abundantes e duas
muito abundantes. No período de chuva, ocorreram dezenove espécies e
destas, uma foi abundante e uma muito abundante. Os valores para os
índices de Simpson e Shannon indicam a maior dominância das espécies

51
presente devido a menor diversidade entre os ambientes amostrados.
Neste ambiente, o período de seca apresentou maior riqueza e maior
diversidade e, consequente, menor dominância das espécies (Tabela 18).
Os valores para o índice de riqueza de Margalef, para o total de
coletas no plantio com teca foi o menor entre os estudados, indicando que,
este é o ambiente mais alterado devido a simplificação causada pelo
plantio. Neste ambiente, que é homogêneo, há menor oferta de recursos
alimentares, restringindo o número de espécies raras e aumentando o
número de espécies mais comuns com grande número de indivíduos. O
valor da equitatividade, que expressa uma distribuição mediana dos
indivíduos das espécies amostradas, pouco se alterou entre os períodos de
chuva e seca, demonstrando uniformidade na abundância de espécies no
decorrer do ano (Tabela 18).
No ambiente com plantio de pau-de-balsa, ocorreram 23
espécies, sendo duas abundantes e duas muito abundantes. No período
de seca, o número de espécies foram 21, destas, duas foram abundantes
e uma muito abundante. No período de chuva, ocorreram dezoito espécies
sendo duas abundantes e duas muito abundantes. Os índices de Simpson
e de Shannon demonstraram menor dominância das espécies e maior
diversidade em relação ao plantio de teca para o total de amostras durante
o ano. No período de chuva, ocorreu uma maior diversidade de espécies,
enquanto na seca, ocorreu uma maior dominância. Nos três ambientes
estudados, o período de seca, no plantio de pau-de-balsa, foi onde ocorreu
a menor riqueza (Tabela 18).
Com o maior número de espécies amostradas entre os
ambientes estudados, na vegetação de cerrado apresentou 24 espécies
amostradas, cinco ocorreram como abundantes e três muito abundantes,
enquanto no período seco, dentre as 23 espécies amostradas, quatro foram
abundantes e duas muito abundantes. No período chuvoso, foram
registradas 19 espécies, sendo quatro abundantes e três muito
abundantes. Com base valores dos índices de Shannon e Simpson, pode-
se afirmar que este ambiente apresenta maior diversidade e menor
dominância entre as espécies, quando comparado com os demais

52
ambientes estudados, mostrando menor diferença entre os índices de
diversidade e dominância nos períodos de chuva e seca.
TABELA 18 - ÍNDICE DE DIVERSIDADE DE ACORDO COM A SÉRIE DE NÚMEROS DE HILL E EQUITATIVIDADE DAS ESPÉCIES DE FORMIGAS AMOSTRADOS EM PLANTIOS DE TECA (Tectona grandis) E DE PAU-DE-BALSA (Ochroma pyramidale) E EM VEGETAÇÃO DE CERRADO, NO MUNICÍPIO DE NOSSA SENHORA DO LIVRAMENTO- MT, 2015/2016.
Índice de diversidade
Períodos
Teca Pau-de-balsa Cerrado
Seco Chuva Anual Seco Chuva Anual Seco Chuva Anual
N0 20 19 23 21 18 23 23 19 24
N1 3,01 1,84 2,22 2,45 2,78 2,76 4,37 4,43 5,38
N2 2,15 1,29 1,48 1,55 2,01 1,85 2,75 3,17 3,65
H’ 1,10 0,61 0,80 0,89 1,02 1,01 1,47 1,48 1,68
E 0,71 0,70 0,66 0,63 0,72 0,67 0,62 0,71 0,67
β 0,46 0,76 0,67 0,64 0,49 0,53 0,36 0,31 0,27
α 2,18 1,82 2,17 2,20 1,76 2,18 2,50 2,09 2,45
N0 = número total de espécies; N1 = número de espécies abundantes; N2 = Número de espécies muito abundantes; H’= Índice de Shannon; E=equitatividade entre as espécies; β= Índice de Simpson; α = Índice de Riqueza de Margalef.
Estes resultados são características de ambientes com uma
maior complexidade estrutural, com maior riqueza de espécies e com
distribuição homogênea dos recursos ambientais que favorecem a
ocupação das formigas nos nichos presentes no ambiente. A maior
proximidade da diversidade do plantio de pau-de-balsa com a área de
cerrado é devido a baixa intervenção antrópica neste plantio. Apesar de ser
um monocultivo, desde a sua implantação, não foi realizado nenhum
tratamento silvicultural como desbaste ou poda dos ramos das árvores.
Resultados apresentados por Fleck (2011) comparando a diversidade de
formigas de áreas nativas com plantios de eucalipto com 28 anos de
implantação e pouca intervenção, a área com eucalipto apresentou
resultados semelhantes a áreas nativas. Corassa et al. (2015) encontraram
menor número de espécies de formigas em ambiente com plantios
agrícolas por causa das constantes intervenções e menor complexidade
estrutural desses plantios e defendem que no ambiente, a riqueza de

53
espécies apresenta comportamento descrescente ao aumento das
intervenções antrópicas.
O índice de equitatividade variou pouco entre os ambientes. Os
resultados foram 0,66, 0,67 e 0,67 para plantio de teca, pau-de-balsa e
cerrado, respectivamente, na análise anual. Para avaliar a
heterogeneidade, a equitabilidade leva em consideração o grau de
uniformidade na abundância de espécies (MAGURRAN, 2011). Os valores
observados nos três ambientes sugerem distribuição não uniforme da
abundância, ocorrendo dominância de algumas espécies visto que, o índice
de equitabilidade varia de zero a um (MAGURRAN, 1988). Desta forma, o
grau de uniformidade da distribuição das espécies de formigas nos três
ambientes é similar. Isso pode ser explicado pela proximidade entre os
ambientes e pelas suas características, que são ambientes florestais, com
porte arbóreo, dossel formado, apresentam sub-bosque e grande
quantidade de serapilheira, que oferta recursos essenciais para o
estabelecimento de uma variedade de espécies (FLECK, 2011).
4.6. ANÁLISE DE SIMILARIDADE
Na Tabela 19 estão contidos os resultados encontrados para a
similaridade entre os diferentes ambientes, estimada pelo índice de
Sorensen. Com os valores obtidos, observou-se maior similaridade entre
os ambientes com vegetação de cerrado e a área com plantio de teca. Nos
períodos de seca e chuva, a maior similaridade ocorreu entre com e pau-
de-balsa e vegetação de cerrado. Por isso, com esses resultados, concluiu-
se elevada semelhança entre as comunidades de formigas nos ambientes
por serem considerados altos valores de similaridade, uma vez que, valores
de similaridade de Sorensen acima de 0,5 são considerados altos (IBGE,
1990).

54
TABELA 19 - SIMILARIDADE ENTRE OS AMBIENTES, PARA OS PERIODOS SECO, DE CHUVA E ANUAL, AMOSTRADOS NO MUNICÍPIO DE NOSSA SENHORA DO LIVRAMENTO-MT, 2015/2016.
ANUAL SECA CHUVA
Ambientes Pau-de-Balsa
Cerrado Pau-de-
balsa Cerrado
Pau-de-Balsa
Cerrado
Teca 0,93 0,95 0,87 0,85 0,88 0,84 Pau-de-Balsa
- 0,89 - 0,90 - 0,89
Os resultados se aproximam com os valores de similaridade de
Sorensen observados por outros autores na fauna de formigas quando
comparada com diferentes ambientes. Tavares (1996) verificou uma
similaridade média de 0,707 na fauna de formigas de cerrado “strictu sensu”
e plantio de Eucalyptus cloziana. Souza (2008) avaliando interações entre
formigas e diásporos de espécies vegetais, encontrou em suas análises,
similaridade de 0,78 entre dois fragmentos de mata atlântica na Reserva
Biológica do Tinguá-RJ. Pereira et al. (2007), utilizando formigas como
ferramenta de monitoramento da recuperação de uma área antropizada por
mineração, constaram maior similaridade (0,405) entre área nativa com
área de plantios de espécies florestais nativas.
A similaridade da comunidade de formigas pode estar
relacionada com as características deste grupo de insetos como a riqueza
de espécies, hábitos generalistas e altas distribuição e abundância local
(SILVA e BRANDÃO, 1999). Os altos valores de similaridade encontrados
atribuem-se à proximidade entre os ambientes amostrados e a ocorrência
de um grande número de espécies de formigas com capacidade para
colonizar ambientes naturais e antrópicos desde que esses ambientes
estejam próximos (TAVARES, 1996).
A maior similaridade entre plantio teca e vegetação de cerrado, pode
ter ocorrido em função das condições similares entre esses ambientes que
acarretam em diversidade de formiga semelhantes. Entre os períodos
sazonais, observou-se maior similaridade entre as áreas com vegetação de
cerrado e pau-de-balsa nos períodos de chuva e seca. Este fato deve ter
ocorrido em função de uma maior dominância de espécies do gênero
Solenopsis, Camponotus e Pheidole que foram comuns nestes ambientes.

55
As espécies destes gêneros possuem potencial e capacidade de colonizar
ambientes naturais e antropizados, desde que os mesmos se encontrem
próximos (BRANDÃO, 1999).
A menor similaridade para o plantio de teca nos períodos
sazonais, deve-se, ao fato de que, espécies como C. atratus e P. grupo
flavens sp. 2, no período de seca, foram encontradas somente no plantio
de pau-de-balsa e na área de cerrado enquanto que P. grupo tristis sp.3 foi
amostrada somente no plantio de teca. No período chuvoso, S. globularia
foi amostrada em comum no plantio de pau-de-balsa e no ambiente
cerrado, enquanto que A. s. subterraneus e P. radoszkowskii foram
espécies encontradas apenas no plantio de teca.

56
5. CONCLUSÕES
Os plantios florestais de teca e pau-de-balsa caracterizam-se
como um importante reservatório de espécies de Formicidae;
- Os períodos de chuva e a seca influenciam a composição e
riqueza de formigas nos ambientes estudados;
- As diferentes complexidades estruturais dos ambientes afeta a
diversidade das formigas;
A proximidade dos ambientes favorece a maior similaridade
de riqueza de formigas;
O plantio de teca favorece a maior abundância de Atta
sexdens e de Acromyrmex subterraneus subterraneus em qualquer época
do ano;
Os índices de diversidade e dominância evidenciam maior
equilíbrio entre as espécies de formigas na vegetação de cerrado;
O índice de similaridade das populações de formigas sugere
que os três ambientes estudados apresentam as mesmas condições
ambientais para a ocorrência das espécies de formigas.

57
6. BIBLIOGRAFIA
ALBUQUERQUE, E.Z.; E. DIEHL. Análise faunística das formigas epígeas (Hymenoptera, Formicidae) em campo nativo no Planalto das Araucárias, Rio Grande do Sul. Revista Brasileira de Entomologia, v. 53, p. 398–403, 2009.
ALMEIDA, L. M.; RIBEIRO-COSTA, C. S. R.; MARINONI, L. Manual de coleta, conservação, montagem e identificação de insetos. Ribeirão Preto: Holos, 88 p. (Série Manuais Práticos em Biologia). 2003.
ALONSO, L. E.; AGOSTI, D. Biodiversity studies, monitoring, and ants: an overview. In: Agosti, D.; Majer, J. D.; Alonso, L. E. & Schultz, T. R. (Eds). Ants standart methods for measuring and monitoring biodiversity. Smithsonian Institution Press, Washington, 01-08p, 2000. Disponível em <http://antbase.org/ants/publications/20333/20333.pdf>. Acesso em 30/04/2016.
ALTIERI, M. A.; CURE, J.R.; GARCIA, M.A. The role and enhancement of parasitic Hymenoptera biodiversity in agroecosystems. In: LASALLE, J.; GAULD, I.D. (Ed.) Hymenoptera and biodiversity. London: CAB International, p.257-275, 1993.
ALVARENGA P. E. F.; FREITAS R. F.; ALGUSTO S. C. Diversidade de Euglossini (Hymenoptera, Apidae) em areas de cerrado do Triangulo Mineiro. Biosci, v. 23. p. 30-37, 2007.
ANDERSEN, A. N. A global ecology of rainforest ants: functional groups in relation to environmental stress and disturbance. Ants: Standard methods for measuring and monitoring biodiversity, Washington Smithsoniam Institution Press, v. 3, p.25-34, 2000.
ANDERSEN, A.N. Responses of ground-foraging ant communities to three experimental fire regimes in a savanna forest of tropical Australia. Biotropica, v. 23, n. 4, p. 575-585, 1991.
ANGILLETTA, M.J. et al. Urban physiology: City ants possess high heat tolerance. PLOS ONE, v. 2, n. 2, p. 258, 2007.
ANJOS, N.; MOREIRA, D. D. O.; DELLA LUCIA, T. M. C. Manejo integrado de formigas cortadeiras em reflorestamentos. As formigas cortadeiras. Viçosa: UFV, p. 212-241, 1993.
AQUINO, A. M.; AGUIAR-MENEZES, E. L.; QUEIROZ, J.M. Recomendações para coleta de artrópodes terrestres por armadilhas de queda ("pitfall-traps"). Seropédica, Embrapa Agrobiologia, p. 8, 2006
AQUINO, A. Manual para coleta de macrofauna do solo. Seropédica: Seropédica: Embrapa Agrobiologia. Série Documentos, n. 130, p. 24, 2001.

58
BACCARO, F. B. et al. Guia para os gêneros de formigas do Brasil. Manaus: Editora INPA, 2015.
BESTELMEYER, B. T. et al. Fiel techiniques for the study of ground-dwelling ants: an overview, description, and evaluation. In: AGOSTI, D. et al. (Ed.). Ants: standard methods for measuring and monitoring biodiversity. Washington and London: Smithsonian Institution Press, cap. 9, p. 122-144, 2000.
BODENHEIMER, F. S. Precis Décologie Animale. Paris: Payout, p. 315, 1955.
BOLTON, B. Army ants reassessed: the phylogeny and classification of the doryline section (Hymenoptera, Formicidae). Journal of Natural History. v. 2, n. 24, p. 1339-1364, 1990.
BORROR, D. J.; DELONG, D. M. Introdução ao estudo dos insetos. São Paulo: Blucher/USP/EDUSP, 1969.
BOSCARDIN, J. et al. Efeito do fogo sobre a riqueza de formigas (Hymenoptera: Formicidae) associadas à Pinus elliottii Engelm. no sul do Brasil. Ciência Florestal, v. 24, n. 4, p. 1031-1040, 2014.
BRADY, S.G. Evolution of the army ant syndrome: the origin and long-term evolutionary stasis of a complex of behavioral and reproductive adaptations. Proceedings of the National Academy of Sciences, v. 100, n. 11, p. 6575-6579, 2003.
BRANDÃO, C. R. F. Família Formicidae. Biodiversidade do Estado de São Paulo, Brasil: síntese do conhecimento ao final do século XX, v. 5, p. 213-223, 1999.
BROWN JR, WILLIAM L. Diversity of ants. Ants: standard methods for measuring and monitoring biodiversity, p. 45-79, 2000.
BUENO, O. C.; CAMPOS-FARINHA, A. E. de C. As formigas domésticas. Insetos e outros invasores de residências. Piracicaba: FEALQ, p. 135-180, 1999.
CALDEIRA, S. F.; OLIVEIRA, D. L. C. Desbaste seletivo em povoamentos de Tectona grandis com diferentes idades. Acta Amazonica, v. 38, n. 2, p. 223-228, 2008.
CAMPOS, R. I. et al. Estratificação vertical de formigas em Cerrado strictu sensu no Parque Estadual da Serra de Caldas Novas, Goiás, Brasil. Iheringia, Série Zoologia, v. 98, n. 3, p. 311-316, 2008.
CANTARELLI, E. B. et al. Diversidade de formigas (Hymenoptera: Formicidae) da serrapilheira em diferentes sistemas de uso do solo. Ciência Florestal, v. 25, n.3, p.607-616, 2015.

59
CARVALHO, K. S.; VASCONCELOS, H. L. Forest fragmentation in central Amazonia and its effects on litter-dwellings ants. Biological Conservation, v. 91, n.2, p. 151-157, 1999.
CARVALHO, M. S. Manual de reflorestamento. Belém, PA. Sagrada Família, 2006. 115p.
CHAVES, A. G. S. Modelagem do crescimento e da redução de Tectona grandis L. f. até a idade de rotação. 2013. Dissertação. (Mestrado em Ciências Florestais e Ambientais) - Universidade Federal de Mato Grosso; Cuiabá.
CORASSA, J. D. N. et al. Biodiversidade da mirmecofauna em diferentes usos do solo no Norte Mato-Grossense. Comunicata Scientiae, v. 6, n. 2, 2015.
COSTA, C.B. Avaliação da consistência e evolução da reabilitação de áreas mineradoras nos aluniões do Rio Jequitinhonha com base nos condicionantes geológicos e na fauna de formigas. Dissertação (Mestrado) - Universidade federal de Ouro Preto. Escola Minas. 91f. 2007.
DA COSTA, C. P.; DA CUNHA, C. N.; COSTA, S. C. Caracterização da flora e estrutura do estrato arbustivo-arbóreo de um cerrado no Pantanal de Poconé, MT/Characterization of the flora and structure of the shrub-tree layer in a cerrado area in Pantanal do Poconé, MT. Biota Neotropica, v. 10, n. 3, p. 61, 2010.
DA SILVA, L. G. et al. Atributos físicos, químicos e biológicos de um Latossolo de cerrado em plantio de espécies florestais. Pesquisa agropecuária brasileira, v. 44, n.6, p. 613-620, 2010.
DAJÓZ, R. Tratado de ecologia. Madrid, Mundi, 487p, 1974.
DELABIE, J. H. C.; FERNÁNDEZ, F. Relaciones entre hormigas y“homopteros” (Hemiptera: Sternorrhyncha y Auchenorryncha). In: FERNÁNDEZ, F. (Ed.). Introducción a lashormigas de laregión Neotropical. Bogotá: Instituto Humboldt, p. 181-200, 2003.
DELABIE, J. H. C.; FRESNEAU, D.; PEZON, A. Notes on the Ecology of Thaumatomyrmex spp. (Hymenoptera: Formicidae: Ponerinae) in. Sociobiology, v. 36, n. 3, 2000.
DELABIE, Jacques HC et al. Ants as biological indicators of Wayana Amerindian land use in French Guiana. Comptes rendus biologies, v. 332, n. 7, p. 673-684, 2009.
DELLA LUCIA, T. M. C.; MARINHO, C. G. S.; RIBEIRO, M. M. R. Perspectivas no manejo de formigas cortadeiras. Insetos Sociais: da biologia à aplicação, p. 369-380, 2008.

60
DELLA-LUCIA, T.M.C. et al. Ordenação de comunidades de Formicidae em quatro agroecossistemas em Viçosa, Minas Gerais. Experientiae, v.28. n. 6, p. 67-94, 1982.
DIEHL-FLEIG, E. Formigas: Organização Social e Ecologia Comportamental. São Leopoldo: Ed. UNISINOS, p. 168, 1995.
FAGUNDES, R. et al. Efeito das mudanças climáticas sazonais no forrageio de Formigas em uma área de mata estacional semidecidual Montana. In: Anais IX Congresso de Ecologia do Brasil, São Lourenço, MG. 2009.
FARIA, F. S. B. et al. Uso da armadilha “pit-fall” na avaliação dos efeitos da fragmentação florestal em comunidades de formigas epígeas da mata atlântica. Anais do XV Encontro de Mirmecologia., v. 15, p. 365-368, 2001.
FEDERAÇÃO DA AGRICULTURA E PECUÁRIA DO ESTADO DE MATO GROSSO - FAMATO. Diagnóstico de florestas plantadas do Estado de Mato Grosso. Cuiabá: IMEA, 106 p. 2013.
FELFILI, M.; C.; FELFILI, J. M. Diversidade alfa e beta no cerrado sensu stricto da Chapada Pratinha, Brasil. Acta bot. bras, v. 15, n. 2, p. 243-254, 2001.
FERNÁNDEZ, F. Introducción a las hormigas de la región Neotropical. Bogotá: Instituto de investigación de recursos biológicos Alexander von Humboldt, 2003.
FERNÁNDEZ, F.; SENDOYA, S. Synonymic list of Neotropical ants (Hymenoptera: Formicidae). Biota Colombiana, Bogotá, v.5, n. 1, p. 3-105, jun. 2004.
FIGUEIREDO, E. O.; OLIVEIRA, A. D.; SCOLFORO, J. R. S. Análise econômica de povoamentos não desbastados de Tectona grandis L.f., na microrregião do baixo rio Acre. Cerne, Lavras, v. 11, n. 4, p. 342-353, 2005.
FITTKAU, E. J.; KLINGE, H. On biomass and trophic structure of the central Amazonian rain forest ecosystem. Biotropica, p. 2-14, 1973.
FLECK, M. D. Composição e diversidade de formigas (Hymenoptera: Formicidae) da serrapilheira em diferentes sistemas de uso do solo. 2011. Monografia (Graduação em Engenharia Florestal) - Universidade Federal De Santa Maria, Santa Maria - RS.
FONSECA, R.C.; DIEHL, E. Riqueza de formigas (Hymenoptera, Formicidae) epigéicas em povoamentos de Eucalyptus spp. (Myrtaceae) de diferentes idades no Rio Grande do Sul, Brasil. Revista Brasileira de Entomologia, v.48, n.1, p. 95-100, 2004.
FOWLER, H. G. Relative representation of Pheidole (Hymenoptera: Formicidae) in local ground ant assemblages of the Americas. Anales de Biologia. Seccion Biologia Animal (Espana), 1993.

61
FOWLER, H. G.; DELABIE, J. H. Resource partitioning among epigaeic and hypogaeic ants (Hymenoptera: Formicidae) of a Brazilian cocoa plantation. Ecología Austral, v. 5, p. 117-124, 1995.
FRANCO, W. Levantamento da fauna de formigas (Hymenoptera: Formicidae) do Parque Estadual de Vila Velha, 2015. 42 f. Monografia (Graduação em Ciências Biológicas) - Universidade Federal do Paraná, Curitiba - PR.
FREITAS, A. V. L.; FRANCINI, R. B.; BROWN JR, K. S. Insetos como bioindicadores ambientais. Métodos de estudos em biologia da conservação e manejo da vida silvestre, p. 127-151, 2006.
FREITAS, F. A. et al. Similaridade e abundância de Hymenoptera inimigos naturais em plantio de eucalipto e em área de vegetação nativa. Floresta e Ambiente, v. 9, p. 145-152, 2002.
GARCÍA, G. L.; LÓPEZ J. R. Muestreos de hormigas con trampas de caída: Tasa de captura diferencial según las especies. In: Boletin de la Asociacion Espanola de Entomologia. Salamanca. Resumo tec... Espanha: Asociacion Espanola de Entomologia, 2001, p. 43-51.
HÖLLDOBLER, B, WILSON, E. O. The ants. Cambridge: The Belknap Press of Harvard University Press, 732 p. 1990.
INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA Projeto Biogeografía do Bioma do Cerrado. Brasília: SEPLAN/FUB, 1990.
JUVENAL, T.L.; MATTOS, R.L.G. O setor florestal no Brasil e a importância do reflorestamento. BNDES Setorial, n.16, p.3-30, 2002.
KASPARI, M. 2003. Introducción a la ecologia de las hormigas, p. 97– 112. In: F. 34 Fernández (ed.). Introducción a las hormigas de la región Neotropical. Resumo tec... Bogotá, Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, 424 p. 2003,
KASPARI, M. Do imported fire ants impact canopy arthropods? Evidence from simple arboreal pitfall traps. The Southwestern Naturalist, p. 118-122, 2000.
KEMPF, W. W. A preliminary zoogeographical analysis of a regional ant fauna in Latin America.Studia Entomol, v. 20, p. 43-62, 1978.
KLINK, C. A.; MOREIRA, A. G. Past and current human occupation, and land use. The cerrados of Brazil: ecology and natural history of a neotropical savanna, p. 69-88, 2002.
KNOECHELMANN, C. M.; MORAIS, H. C. Visitas de formigas (Hymenoptera, Formicidae) a nectários extra-florais de Stryphnodendron adstringens (Mart.) Cov. (Fabaceae, Mimosoideae) em uma área de cerrado freqüentemente queimado. Revista Brasileira de Zoociências, v. 10, n. 1, 2009.

62
LACHAT, T. et al. Arthropod diversity in Lama forest reserve (South Benin), a mosaic of natural, degraded and plantation forests. Biodiversity and Caonservation, London, v. 15, n. 1, p. 3-23, 2006.
LAMPRECHT, H. Silvicultura nos trópicos: ecossistemas florestais e respectivas espécies arbóreas: possibilidade e métodos de aproveitamento sustentado. Eschborn: Dt Ges. Fur Techn. Zusammenarbeit (GTZ) GmbH, 343 p. 1990.
LARA, F.M. Princípios de Entomologia. São Paulo, Brasil: Ícone, 331p., 1992.
LAROCA, S.; MIELKE, O. H. H. Ensaio sobre ecologia de comunidade em Sphingidae na Serra do Mar, Paraná – Br, (Lepidoptera). Revista Brasileira de Biologia, Rio de Janeiro, v. 35, n. l, p. 1-19, 1975.
LAWTON, J. H. Plant architecture and the diversity of phytophagous insects. Annual review of entomology, v. 28, n. 1, p. 23-39, 1983.
LAWTON, J. H.; STRONG JR, D. R. Community patterns and competition in folivorous insects. The American Naturalist, v. 118, n. 3, p. 317-338, 1981.
LOUZADA, J. N.C.; SANCHES, N. M.; SCHILINDWEIN, M. N. Bioindicadores de qualidade e de impactos ambientais da atividade agropecuária. Informe Agropecuário, v. 21, n. 202, p. 57-71, 2000.
LUDWIG, J. A.; REYNOLDS, J. F. Statistical ecology: a primer on methods and computing. New York: John Wiley & Sons, p. 337, 1988.
MACEDO, L. P. M. Diversidade de formigas edáficas (Hymenoptera: Formicidae) em fragmentos da Mata Atlântica do Estado de São Paulo. 2004. 113 f. Tese (Doutorado em Entomologia) - Escola Superior de Agricultura Luiz de Queiroz. Universidade de São Paulo, Piracicaba - SP.
MACEDO, R. L. G.; BOTELHO, S. A.; SCOLFORO, J. R. Considerações preliminares sobre o estabelecimento da Tectona Grandis L. f. (TECA), introduzida na região noroeste do Estado de Minas Gerais. Simpósio Internacional sobre Ecossistemas Florestais, n. 5, p. 4, 1999
MACKAY W. P.; MACKAY E. The systematics and biology of the New World ants of the genus Pachycondyla (Hymenoptera: Formicidae). New York: Edwin Mellen Press, Lewiston, 2010.
MAGURRAN, A. E. Ecological Diversity and Its Measurement. Ecological diversity and its measurement. Princeton Univ. Press, Princeton, NJ, p. 167 1988.
MAGURRAN, A. E. Medindo a diversidade biológica. Curitiba: Editora da UFPR, v. 261, 2011.

63
MAJER J. D. Ants: bio-indicators of minesite rehabilitation, landuse, and land conservation. Environmental Management, v. 7, p. 375-85, 1983.
MAJER, J. D. Ant recolonization of rehabilitated bauxite mines at Trombetas, Pará, Brazil. Journal of Tropical Ecology, v. 12, n. 02, p. 257-273, 1996.
MAJER, J.D. Ant recolonization of rehabilitated bauxite mines of Poços de Caldas, Brasil. J. Appl. Ecol. v. 8, p. 97-108, 1992.
MARINHO, C. G. et al. Diversidade de Formigas (Hymenoptera: Formicidae) da Serapilheira em Eucaliptais (Myrtaceae) e Área de Cerrado de Minas Gerais. Neotropical Entomology, v. 31, n.2. 2002.
MARQUES, G. D. V.; DEL-CLARO, K. The ant fauna in a Cerrado area: the influence of vegetation structure and seasonality (Hymenoptera: Formicidae). Sociobiology, v. 47, n. 1, p. 235-25, 2006.
MENTONE, T. D. O. et al. Composição da fauna de formigas (Hymenoptera: Formicidae) de serapilheira em florestas semidecídua e de Eucalyptus spp., na região sudeste do Brasil. Biota Neotropica, p. 237-246, 2011.
MERTL, A.L; WILKIE, K.T.R.; TRANIELLO, J.F.A. Impact of flooding on the species richness, density and composition of amazonian litter-nesting Ants. Biotropica, v. 41, n. 5, p. 633–641, 2009.
MILHOMEM, M. S.; MELLO, F. Z.V.; DINIZ, I. R. Ténica de coleta de besouro copronecrófagos no cerrado. Pesquisa Agropecuária Brasileira, Brasília, DF, V. 38, n. 11, p. 1249-1256, 2003.
MORAES, H. C.; BENSON, W. W. Recolonização de vegetação de cerrado após queimadas por formigas arborícolas. Revista Brasileira de Biologia., v. 48, p. 459-466, 1988.
MORAES, R. B. C. et al. Software para análise faunística-ANAFAU. In: SIMPÓSIO DE CONTROLE BIOLÓGICO, 8., São Pedro, 2003. Resumos... São Pedro: s.ed., 2003. p.195.
MOUTINHO, P.; NEPSTAD, D.C.; DAVIDSON, E.D. Influence of leaf-cutting ant nests on secondary forest growth and soil properties. Amazonia. Ecology. v.84, p. 1265-1276, 2003.
OLIVEIRA, P.S.; KLITZKE C.; VIEIRA E. The ant fauna associated with the extrafloral nectaries of Ouratea hexasperma (Ochnaceae) in an area of cerrado vegetation in Central Brazil. Entomologist’s Monthly Magazine, 131, 77– 82, 1995.
PARR, C. L.; CHOWN, S. L. Inventory and bioindicator sampling: Testing pitfall and winkler methods wih ants in a South African savanna. Journal of Insect Conservation, Dordrecht, v. 5, p, 27-36, 2001.

64
PELISSARI, A. L. Silvicultura de precisão aplicada ao desenvolvimento de Tectona grandis l.f. na região sul do estado de Mato Grosso. 2012. 78f. Dissertação (Mestrado em Ciências Florestais e Ambientais) -Universidade Federal de Mato Grosso, Faculdade de Engenharia Florestal, Cuiabá - MT.
PEREIRA, M. P.et al. Influência da heterogeneidade da serapilheira sobre as formigas que nidificam em galhos mortos em floresta nativa e plantio de eucalipto. Neotrop Biol Conserv, v. 2, p. 161-164, 2007.
PETILLON, L.; CANARD, A.; YSNEL, F. Spiders as indicators of microbabitat changes after a grass invasion in salt-marshes: synthetic results from a case study in the Mont-Saint-Michel Bay. Cahiers de Biologie Marine, Paris, v. 47, n. 1, p. 11-18, 2006.
POWELL, S.; BAKER, B. Os grandes predadores dos neotrópicos: Comportamento, dieta e impacto das formigas de correição (ecitoninae). pp. 18-37, 2008. In: Vilela, E.F. et al. (eds.). Insetos sociais da biologia à aplicação. Viçosa, Universidade Federal de Viçosa. 442p., 2008.
POWELL, S.; CLARK, E. Combat between large derived societies: A subterranean army ant established as a predator of mature leaf-cutting ant colonies. Insectes Sociaux, v. 51, p. 342 – 351, 2004.
RAMOS, L. D. S. et al. Comunidades de formigas (Hymenoptera: Formicidae) de serapilheira em áreas de cerrado “stricto sensu” em Minas Gerais. Lundiana, v. 4, n. 2, p. 95-102, 2003.
RÉ, T.M. O uso de formigas como bioindicadores no monitoramento ambiental de revegetação de áreas mineradas. 2007. 244 f. Tese (Doutorado em Engenharia) - Escola Politécnica da Universidade de São Paulo, São Paulo - SP.
REBELLATO, L.; CUNHA, C. N. Efeito do “fluxo sazonal mínimo da inundação” sobre a composição e estrutura de um campo inundável no Pantanal de Poconé, MT, Brasil. Acta Botanica Brasilica, v. 19, n. 4, p. 789-799, 2005.
RETTENMEYER, C. W. et al. Comparative foraging by neotropical army ants. 1983. In: JAISSON, P. Social insects in the tropics. Social insects in the tropics, Paris. v. 2, p. 59-73., 1983.
RIBAS, C. R. et al. Mesa redonda formigas podem ser utilizadas como bioindicadoras de recuperação após impactos ambientais? Biológico, São Paulo, v. 69, n. 2, p. 57-60, 2007.
RIBAS, C. R. et al. Tree heterogeneity, resource availability, and larger scale processes regulating arboreal ant species richness. Austral Ecology, v. 28, n. 3, p. 305-314, 2003.
ROCHA, W. O. Estudos da mirmecofauna aplicada na avaliação de áreas de garimpo de diamantes no município de Poxoréu, MT. 2012.

65
53 f. Dissertação (Mestrado em Ciências Florestais e Ambientais) -Universidade Federal de Mato Grosso, Faculdade de Engenharia Florestal, Cuiabá - MT.
RODRIGUES, S. T. et al. Composição florística e abundância de pteridófitas em três ambientes da bacia do rio Guamá, Belém, Pará, Brasil. Acta Amazonica, Manaus, vol. 34, n. 1, p. 35-42, 2004.
SAKAGAMI, S. F.; LAROCA, S.; MOURE, J. S. Wild Bee Biocoenotics in São Jose dos Pinhais (PR), South. Zoology, v. 16, n. 2, p. 253-291, 1967.
SAKAGAMI, S. F.; MATSUMARA, T. Relative abundance, phenology and flower preference of andrenid bees in Sapporo, north Japan (Hymenoptera, Apoidea) Japanese Journal of Ecology, v. 17, n. 6, p. 237-50, 1967.
SÁNCHEZ-PEÑA, S. R.; MUELLER, U. G. A nocturnal raid of Nomamyrmex army ants on Atta leafcutting ants in Tamaulipas, Mexico. Southwestern Entomologist, v. 27, p. 221-223, 2002.
SANTOS M. P. et al. Fauna de formigas como ferramenta para monitoramento de área de mineração reabilitada na Ilha da Madeira, Itaguaí, RJ. Ciência Florestal, v. 17, n. 3, p. 197-204, 2007.
SANTOS, M. S. et al. Riqueza de formigas (Hymenoptera, Formicidae) da serapilheira em fragmentos de floresta semidecídua da Mata Atlântica na região do Alto do Rio Grande, MG, Brasil. Iheringia, Série Zoologia, v. 96, n. 1, p. 95-101, 2006.
SANTOS, S. R. et al. A riqueza das formigas relacionada aos períodos sazonais em Caxiuanã durante os anos de 2006 e 2007. Revista Brasileira de Meteorologia, v. 27, n. 3, 2012.
SANTOS, U. F. et al. Níveis de sombreamento na produção de mudas de Pau-de-balsa (Ochroma pyramidale). Bioscience Journal, Uberlândia, v. 30, n. 1, p. 129-136, 2014.
SANTOS, W. C. Análise de características dendrométricas e fatores edáficos no crescimento e produção de um povoamento de Araucaria angustifolia (Bert.) O. Ktze. 2006. 124 f. Dissertação (Mestrado em Engenharia Florestal) - Universidade Federal do Paraná, Curitiba-PR.
SARMIENTO, C. Comparacion de três clases de transecto para captura de hormigas (Hymenoptera: Formicidade) em dos formaciones vegetales. Caldasia, Bogotá, v. 22, n. 2, p. 317-326, 2000.
SARMIENTO, C. Metodologias de captura y studio de las hormigas. In: PALACIO, E. E.; FERNÁNDEZ, F. (Ed.) Introduction a las hormigas de las región Neotropical. Bogotá: Instituto de Investigación de Recursos Biológicos Alexander Von Humbolt, Cap. 12, p. 201-210. 2003.

66
SCHMIDT, K.; CORBETTA, R.; CAMARGO, J. A. Formigas (Hymenoptera: Formicidae) da Ilha João da Cunha, SC: composição e diversidade. Biotemas, v. 18, n. 1, p. 57 - 71, 2005.
SHATTUCK, S. O. Australian Ants: Their Biology and Identification. Series: Monographs on invertebrate taxonomy. v. 3. Collingwood Victoria, CSIRO Publishing. 226 p. 1999.
SILVA, R. R. da; BRANDÃO, C. R. F. Formigas (Hymenoptera: Formicidae) como indicadores da qualidade ambiental e da biodiversidade de outros invertebrados terrestres. Biotemas, v. 12, n. 2, p. 55-73, 1999.
SILVA, R. R.; BRANDÃO, C. R. F.; SILVESTRE, R. Similarity between cerrado localities in central and southeastern Brazil based on the dry season bait visitors ant fauna. Studies on Neotropical Fauna and Environment, v. 39, n. 3, p. 191-199, 2004.
SILVA, R. S.; BRANDÃO, C. R. F. Formigas (Hymenoptera: Formicidae) como indicadores de qualidade ambiental e da biodiversidade de outros invertebrados terrestres. Biotemas, Florianópolis, v. 12, Ed. 2, pg 75-100, 1999.
SILVA, T. G. M. estrutura e dinâmica de comunidade de formigas (Hymenoptera: formicidae) de serapilheira em eucaliptais tratados com herbicida na região de Mata Atlântica. 2007, Lavras: UFLA, 62p.: il
SILVEIRA NETO, S. et al. Manual de ecologia dos insetos. Piracicaba: Ceres, 419p. 1976.
SILVESTRE, R. C.; BRANDÃO C. R. F.; SILVA, R. R. Grupos funcionales de hormigas: el caso de los gremios del Cerrado, p.113-148. In F. Fernández (ed.). Introducción a las hormigas de la región neotropical. Bogotá: Instituto de Investigación de Recursos Biológicos Alexander Von Humbolt, p. 398.2003.
SILVESTRE, R. Estrutura de comunidades de formigas do cerrado. 2000. 216f. Tese de Doutorado. Tese (Doutorado em Ciências-Entomologia) - Faculdade de Filosofia, Ciências e Letras, USP, Ribeirão Preto - SP.
SOARES, N. S. et al. Levantamento da diversidade de formigas (Hymenoptera: Formicidae) na região urbana de Uberlândia, MG. Neotropical Entomology, v. 35, n. 3, p. 324-328, 2006.
SOARES, S. de M.; MARINHO, C. G. S.; DELLA LUCIA, T. M. C. Riqueza de espécies de formigas edáficas em plantação de eucalipto e em mata secundária nativa. Revista brasileira de Zoologia, v. 15, n. 4, p. 889-898, 1998.
SOARES, S. M.; MARINHO, C. G. S.; DELLA LUCIA, T. M. C. Diversidade de invertebrados edáficos em áreas de eucalipto e mata secundária. Acta Biol. Leopoldensia, v. 19, p. 157-164, 1998.

67
SORENSEN, T. A method of establishing groups of equal amplitude in plant sociology based on similarity of species and its application to analyses of the vegetation on Danish commons. Biol. Skr., v. 5, p. 1-34, 1948.
SOUZA, C. M. G. de. Densidade ótima de estradas para povoamentos de teca (Tectona grandis l.f.) no estado de Mato Grosso. 2011. 42f. Dissertação (Mestrado em Ciências Florestais e Ambientais) -Universidade Federal de Mato Grosso, Faculdade de Engenharia Florestal, Cuiabá - MT.
SOUZA, G. O. Interações entre formigas e diásporos de espécies vegetais da Mata Atlântica na Reserva Biológica do Tinguá, Nova Iguaçu, RJ, Brasil. 2008. 43 f. Dissertação (Mestrado em Biologia Animal) - Universidade Federal do Rio de Janeiro, Soropédica - RJ.
SOUZA, K. K. F. Diversidade de formigas (Hymenoptera: Formicidae) epigéicas em áreas de plantios de Pinus sp., Mata Nativa. 2006. 93 f. Dissertação (Mestrado em Engenharia Florestal) - Universidade Federal do Paraná, Curitiba-PR.
SWARTZ, M. B. Predation on an Atta cephalotes colony by an army ant, Nomamyrmex esenbeckii. Biotropica, v. 30, p. 682-684, 1998.
TAVARES, A.A. Diversidade de formigas não desfolhadoras (Hymenoptera: Formicidae) em áreas de cerrado "stricto sensu"e Eucalyptus cloeziana F. Muell. 1996. 57f. Dissertação (Mestrado em Engenharia Florestal) – Universidade Federal de Lavras, Lavras - MG.
TOBIN, J. Competition and coexistence of ants in a small patch of rainforest canopy in Peruvian Amazonia. Journal of New York Entomological Society, v. 105, n. 1, p.105-112, 1997.
TURNER, I. M. Species loss in fragments of tropical rain forest: a review of the evidence. Journal of applied Ecology, v.33, p. 200-219, 1996.
VALENTIM, C. L. et al. Formigas como bioindicadoras de impacto ambiental causado por arsênio. Biológico, XVIII Simpósio de Mirmecologia, p. 297-300, 2007.
VASCONCELOS, H. L. Diversidade de formigas cortadeiras no Brasil. In: XV Encontro de Mirmecologia, 2001, Londrina. Anais do XV Encontro de Mirmecologia. Londrina: Editora do Instituto Agronômico do Paraná (IAPAR), p. 53 – 54. 2001.
VASCONCELOS, H. L.; DELABIE, J. H. C. Ground ant communities from central Amazonia forest fragments. Sampling Ground-Dwelling Ants: Case Studies from the World’s Rain Forests, v. 18, p. 59-70, 2000.
VASQUEZ-YANES, C. Studies on the germination of seeds of Ochroma lagopus Sw. Turrialba, San José, v. 24, n. 2 p. 176-179, 1974.
VIANA, V. M.; PINHEIRO, L. A. F. V. Conservação da biodiversidade em fragmentos florestais. Série técnica IPEF, v. 12, n. 32, p. 25-42, 1998.

68
WEBER, N. A. Gardening ants: the attines (Memoirs of the American Philosophical Society, vol. 92). The American Philosophical Society, Independence Square Philadelphia, p. 146, 1972.
WETTERER J; SNELLING G. Geographic distribution of Labidus coecus (Latr.) (Hymenoptera, Formicidae), a subterranean army ant. Journal of Hymenoptera Research 44, p. 31-38, 2015.
WILSON, E. O. Causes of ecological sucess: the case of the ants. Journal of Animal Ecology, v. 56, p. 1-9, 1987.
WILSON, E. O. La hiperdiversidad como fenomeno real: el caso de Pheidole. p. 363–370. In: FERNÁNDEZ, F. (ed.). Introducción a las hormigas de la región Neotropical. Bogotá: Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, p. 424, 2003.
WINK, C. et al. Insetos edáficos como indicadores da qualidade ambiental= Soilborne insects as indicators of environmental quality. Revista de Ciências Agroveterinárias, v. 4, n. 1, p. 60-71, 2005.
WINSTON, M. R. Co-occurrence of morphologically similar species of stream fishes. American Naturalist, v. 145, p. 527–545, 1995.
WOLDA, H. Similarity indices, sample size and diversity. Oecologia, v. 50, n. 3, p. 296-302, 1981.

69
ANEXOS

70
ANEXO l - FREQUÊNCIA E ÍNDICES DE ABUNDÂNCIA (A), CONSTÂNCIA (C) E DE DOMINÂNCIA (D) DAS ESPÉCIES DE FORMIGAS AMOSTRADAS COM ARMADILHA PITFALL EM TALHÃO DE Tectona grandis NOS PERÍODOS DE SECA, CHUVA E ANUAL, NO MUNICÍPIO DE NOSSA SENHORA DO LIVRAMENTO, ESTADO DE MATO GROSSO, 2015/2016.
SUBFAMÍLIA/ESPÉCIE TECA (Tectona grandis)
PERÍODOS
SECA CHUVA ANUAL
F A C D F A C D F A C D
1 2 1 2 1 2
Dolichoderinae
Dorymyrmex pyramicus pf r z nd nd f C y d nd F c z d nd
Dorylinae
Labidus coecus sf sa y sd sd sf sa w sd sd Sf sa w sd sd
Labidus praedator f c y d d mf ma y d d Mf ma y d d
Nomamyrmex esenbeckii pf R z nd nd Pf r z nd nd
Formicinae
Brachymyrmex heeri f c w d d - - - - - F c w d nd
Camponotus blandus mf ma w d d pf D y d nd F c y d d
Camponotus crassus pf r z nd nd pf D z d nd Pf r z d nd
Camponotus melanoticus mf ma w d d f C w d d Mf ma w d d
Camponotus novogranadensis pf r y d nd f C w d nd F c y d nd
Myrmicinae
Pheidole araneoides mf ma w d d f C w d nd F c w d d
Acromyrmex subterraneus subterraneus f c y d nd mf ma w d d Mf ma w d d
Atta sexdens sf sa y sd sd sf sa w sd sd Sf sa w sd sd
Crematogaster tenuicula pf r z nd nd - - - - - Pf r z nd nd
Pheidole grupo Fallax sp. 1 mf ma w d d sf sa w sd sd Sf sa w sd sd
Pheidole grupo Flavens sp.2 - - - - - pf ma z nd nd Pf r z nd nd
Pheidole radoszkowskii - - - - - pf D z nd nd Pf r z nd nd
Pheidole grupo Tristis sp.3 pf r y d nd f C z d d F c z d d
“Continua...”
71
“ANEXO l, Cont.”
Teca SUBFAMÍLIA/ESPÉCIE PERÍODOS
SECA CHUVA ANUAL
F A C D F A C D F A C D
1 2 1 2 1 2
Myrmicinae
Strumigenys denticulata pf r y d nd Pf d w d nd Pf r w d nd
Ponerinae
Neoponera commutata f c w d nd F c w d nd F c w d d
Odontomachus bauri pf r y d nd F c W d nd F c w d nd Programa ANAFAU: (1) Método de Laroca e Mielke (1975); (2) método de Sakagami e Laroca (1967). D: Dominância - (sd) super dominante; (d) dominante; (nd) não dominante. A: Abundância - (sa): super abundante; (ma) muito abundante; (a) abundante: (c) comum; (d) dispersa; (r) rara. F: Frequência: (sf) super frequente; (mf) muito frequente; (pf) pouco frequente. C: Constância - (w) constante; (y) acessória; (z) acidental

72
ANEXO ll - FREQUÊNCIA E ÍNDICES DE ABUNDÂNCIA (A), CONSTÂNCIA (C) E DE DOMINÂNCIA (D) DAS ESPÉCIES DE FORMIGAS AMOSTRADAS COM ARMADILHA PITFALL EM TALHÃO DE Ochroma pyramidale NOS PERÍODOS DE SECA, CHUVA E ANUAL, NO MUNICÍPIO DE NOSSA SENHORA DO LIVRAMENTO, ESTADO DE MATO GROSSO, 2015/2016.
PAU-DE-BALSA
SUBFAMÍLIA/ESPÉCIE PERÍODOS
SECA CHUVA ANUAL
F A C D F A C D F A C D
1 2 1 2 1 2
Dolichoderinae
Dorymyrmex pyramicus f c W d nd f c w d nd f c W d nd
Tapinoma melanocephalum pf r Z nd nd - - - - - pf r Z nd nd
Dorylinae
Labidus coecus sf sa W sd sd sf sa w sd sd sf sa W sd sd
Labidus praedator mf ma Y d d mf ma w d d mf ma Y d d
Nomamyrmex esenbeckii pf r Z nd nd pf r y nd nd pf r Z nd nd
Formicinae
Brachymyrmex heeri pf d W d nd - - - - - pf r Y nd nd
Camponotus blandus f c W d d f c w d nd f c W d d
Camponotus crassus f c W d nd f c w d d f c W d d
Camponotus melanoticus mf ma W d d mf ma w d d mf ma W d d
Camponotus novogranadensis f c Y d d pf c w d nd f c W d d
Myrmicinae
Pheidole araneoides pf d W d nd mf ma w d d f c W d d
Atta sexdens pf d y d nd pf r Z d nd
Cephalotes atratus pf r Z nd nd - - - - - pf r Z nd nd
Crematogaster tenuicula pf r Z nd nd - - - - - pf r Z nd nd
Pheidole grupo Fallax sp.1 sf sa W sd sd mf sa w sd sd mf ma W d d
Pheidole grupo Flavens sp. 2 f c Y nd nd pf r z nd nd f c Z d nd
Pheidole grupo Tristis sp.3 - - - - - pf r y d nd pf r Z d nd
Solenopsis sp. sf sa W sd sd sf sa w sd sd sf sa W sd sd
“Continua...”

73
“ANEXO ll, Cont.”
PAU-DE-BALSA SUBFAMÍLIA/ESPÉCIE PERÍODOS
SECA CHUVA ANUAL
F A C D F A C D F A C D
1 2 1 2 1 2
Myrmicinae
Solenopsis globularia f c W d nd pf r y nd nd pf d w d nd
Sericomyrmex sp.1 pf r Y nd nd - - - - - pf r z nd nd
Strumigenys denticulata pf d W d nd pf r w nd nd pf r w d nd
Ponerinae
Neoponera commutata pf d W d nd pf r y d nd pf r y d nd
Odontomachus bauri f c W sd sd f c w d nd f c w d nd Programa ANAFAU: (1) Método de Laroca e Mielke (1975); (2) método de Sakagami e Laroca (1967). D: Dominância - (sd) super dominante; (d) dominante; (nd) não dominante. A: Abundância - (sa): super abundante; (ma) muito abundante; (a) abundante: (c) comum; (d) dispersa; (r) rara. F: Frequência: (sf) super frequente; (mf) muito frequente; (pf) pouco frequente. C: Constância - (w) constante; (y) acessória; (z) acidental

74
ANEXO lll – FREQUÊNCIA (F) E ÍNDICES DE ABUNDÂNCIA (A), CONSTÂNCIA (C) E DE DOMINÂNCIA (D) DAS ESPÉCIES DE FORMIGAS AMOSTRADAS COM ARMADILHA PITFALL EM VEGETAÇÃO DE CERRADO, NOS PERÍODOS DE SECA, CHUVA E ANUAL, NO MUNICÍPIO DE NOSSA SENHORA DO LIVRAMENTO, ESTADO DE MATO GROSSO 2015/2016.
SUBFAMÍLIA/ESPÉCIES VEGETAÇÃO DE CERRADO
PERÍODOS
SECA CHUVA ANUAL
F A C D F A C D F A C D
1 2 1 2 1 2
Dolichoderinae
Dorymyrmex pyramicus pf d W d nd f c y d nd f c y d nd
Dorylinae
Labidus coecus mf ma Y d d sf sa w sd sd sf sa y sd sd
Labidus praedator f c Y d d sf sa w sd sd mf ma y d d
Nomamyrmex esenbeckii pf d z nd nd pf d z nd nd
Formicinae
Brachymyrmex heeri pf d Y nd nd f c z d nd pf d z d nd
Camponotus blandus sf sa W sd sd mf ma w d d sf sa w sd sd
Camponotus crassus f c Z nd nd pf d z nd nd f c z d nd
Camponotus melanoticus mf ma W d d f c w d nd mf a w d d
Camponotus novogranadensis mf ma Y d d mf ma w d d mf ma y d d
Myrmicinae
Pheidole araneoides mf ma W d d f c w d nd mf ma w d d
Acromyrmex subterraneus subterraneus pf d W d nd pf r z nd nd
Atta sexdens pf ma W d d pf r z nd nd
Cephalotes atratus pf d W d nd pf d y d nd
Crematogaster tenuicula sf sa W sd sd sf sa w sd sd sf sa w sd sd
Pheidole grupo Fallax sp. 1 f c Y d nd mf ma y d d mf a y d d
Pheidole grupo Flavens sp. 2 pf r Z nd nd pf d z d nd pf d z d nd
Pheidole radoszkowskii f c Z d nd f c z d nd
Solenopsis sp. sf sa W sd sd sf sa w sd sd sf sa w sd sd
Solenopsis globularia pf d Y d nd pf d z nd nd pf d y d nd
“Continua...”

75
“ANEXO lll, Cont.”
SUBFAMÍLIA/ESPÉCIES VEGETAÇÃO DE CERRADO
PERÍODOS
SECA CHUVA ANUAL
F A C D F A C D F A C D
1 2 1 2 1 2
Myrmicinae
Sericomyrmex sp. 1 f c w d nd f c y D nd
Strumigenys denticulata pf d y nd nd pf d y nd nd pf d y D nd
Ponerinae
Neoponera commutata f c w d nd f c w d nd f c w D nd
Odontomachus bauri pf d w d nd pf d y nd nd pf d w D nd
Pseudoponera gilberti pf r y nd nd f c z d nd pf d z D nd PROGRAMA ANAFAU: (1) Método de Laroca e Mielke (1975); (2) método de Sakagami e Laroca (1967). D: Dominância - (sd) super dominante; (d) dominante; (nd) não dominante. A: Abundância - (sa) super abundante; (ma) muito abundante; (a) abundante; (c) comum; (d) dispersa; (r) rara. F: Frequência: (sf) super frequente; (mf) muito frequente; (f) frequente; (pf) pouco frequente. C: Constância - (w) constante; (y) acessória; (z) acidental





























