Embed Size (px)
Citation preview

Universidade Federal do Rio Grande do Norte Centro de Biociências - Departamento de Oceanografia e Limnologia
Programa de Pós Graduação em Bioecologia Aquática
DIVERSIDADE DE MACROALGAS MARINHAS BENTÔNICAS DOS RECIFES DE MARACAJAÚ, ÁREA DE PRESERVAÇÃO AMBIENTAL DOS RECIFES DE CORAIS,
RIO GRANDE DO NORTE, BRASIL.
Ingrid Balesteros Silva
NATAL, 2006

Ingrid Balesteros Silva
DIVERSIDADE DE MACROALGAS MARINHAS BENTÔNICAS DOS RECIFES DE MARACAJAÚ, ÁREA DE PRESERVAÇÃO AMBIENTAL DOS RECIFES DE CORAIS,
RIO GRANDE DO NORTE, BRASIL.
ORIENTADORA: Profa. Dra. Eliane Marinho-Soriano
Dissertação de Mestrado apresentada ao Programa de Pós-Graduação do Departamento de Oceanografia e Limnologia, Centro de Biociências, Universidade Federal do Rio Grande do Norte, como parte dos requisitos para obtenção do Título de Mestre em Bioecologia Aquática.
NATAL, 2006

Universidade Federal do Rio Grande do Norte Departamento de Oceanografia e Limnologia
Programa de Pós Graduação em Bioecologia Aquática
DIVERSIDADE DE MACROALGAS MARINHAS BENTÔNICAS DOS RECIFES DE MARACAJAÚ, ÁREA DE PRESERVAÇÃO AMBIENTAL DOS RECIFES DE CORAIS,
RIO GRANDE DO NORTE, BRASIL.
Ingrid Balesteros Silva
Esta dissertação, apresentada pela aluna Ingrid Balesteros Silva ao Programa de Pós Graduação em
Bioecologia Aquática do Departamento de Oceanografia e Limnologia da Universidade Federal do Rio
Grande do Norte, foi julgada adequada e aprovada pelos Membros da Banca Examinadora, na sua redação
final, para a conclusão do Curso e à obtenção do título de Mestre em Bioecologia Aquática.
MEMBROS DA BANCA EXAMINADORA
Profa. Dra. Eliane Marinho-Soriano (Orientadora)
Prof. Marcos Rogério Câmara (DOL/CB/UFRN)
Profa. Ivaneide Soares (CB/UFRN)
Profa. Dra. Mutue Toyota Fujii (Instituto de Botânica/SP)
Profa. Dra. Maria das Dores Mello (Universidade Potiguar)
Natal, 23 de junho de 2006.

IV
Aos meus queridos pais, Alcides e Vicentina, por tudo o que eles representam.

V
AGRADECIMENTOS
Meus sinceros agradecimentos a todos os que apoiaram na realização deste trabalho.
À Dra. Eliane Marinho-Soriano, por aceitar me orientar neste trabalho, pela confiança depositada, por sempre incentivar seus alunos;
À Dra. Mutuê Toyota Fujii, pelo uso de seu laboratório, por disponibilizar bibliografia e ajudar na identificação das algas, por apesar da distância estar sempre disposta a colaborar e por fazer parte da banca examinadora;
Ao Dr. Marcos Rogério Câmara, Dra. Ivaneide Soares e Dra. Maria das Dores Melo, por aceitar fazer parte da banca examinadora e pelas valorosas sugestões;
Ao Dr. Ricardo Amaral, coordenador do Projeto Corais de Maracajaú, por toda ajuda (imprescindível) e por toda a atenção;
Às operadoras de mergulho, Maracajaú Diver, Ma-Noa, Portal de Maracajaú e Corais de Maracajaú, por todo a ajuda logística;
Aos amigos Estevão Martins e Sandra Magalhães, pela ajuda em campo, por toda a hospitalidade, pelas fotografias e por toda a amizade;
Aos colegas Vanessa Benigno e Rafael (Japa), por toda a ajuda em campo;
Aos colegas Marcella Carneiro, Aline Martins, José Garcia Jr., Igor, pela ajuda nas análises estatísticas e por estarem sempre dispostos a ajudar;
À Dra. Silvia M.P.B. Guimarães, por disponibilizar sua bibliografia, por toda a atenção e hospitalidade;
Aos colegas do Laboratório de Macroalgas, em especial Rafaela, Dina, Marcella, Sdena, Danielton Leal e Carol, que estiveram sempre dipostos a ajudar, triando material, buscando água salgada na praia..., pela boa convivência e companheirismo;
À Diógina Barata, Valéria Cassano, Mariana Pacheco, Carolina Nanacoto, Aline Martins e Rita Manso, pela ajuda com identificação de material, auxilio no programa de captura de imagens, pela calorosa recepção e pelo companheirismo no laboratório da secção de Ficologia do Instituto de Botânica de São Paulo;
Ao Ricardo Garla e prof. Marcos Rogério pelas correções no abstract; e Euriel pela ajuda na apresentação;
Aos amigos Maria Isabel, Mônica Link e José Garcia, por toda a hospitalidade no momento de minha chegada à cidade, por toda a força e incentivo;
Aos funcionários do Departamento de Oceanografia e Limnologia que colaboraram com este trabalho, em especial Sr. Geraldo (in memorian) e Sr.Wellington, e às secretárias Cleide e Michelle, por me ajudarem em diversos momentos;
À CAPES, pela concessão da bolsa de mestrado, que me possibilitou realizar este estudo;
Aos meus queridos pais, Alcides e Vicentina, por tudo que fizeram. Sem a ajuda deles este estudo não seria possível.

VI
SUMÁRIO
RESUMO.................................................................................................................IX
ABSTRACT...............................................................................................................X
1. INTRODUÇÃO......................................................................................................1
2. ÁREA DE ESTUDO..............................................................................................6
3. MATERIAIS E MÉTODOS....................................................................................9
3.1. As estações de coleta.............................................................................9
3.2. Estratégia de amostragem .................................................................. 11
3.2.1. Determinação da biomassa ................................................... 11
3.2.2. Identificação do material......................................................... 12
3.3. Descritores analíticos........................................................................... 12
3.4. Descritores sintéticos simples. ............................................................ 13
3.4.1. Riqueza................................................................................... 13
3.4.2. Freqüência. ............................................................................ 13
3.4.3. Índice de diversidade de Shannon-Hill................................... 14
3.4.4. Índice de dominância de Simpson.......................................... 14
3.5. Descritores complexos: análise de similaridade.................................. 14
3.5.1. Análise de agrupamento em Modo Q (entre amostras).......... 15
3.6. Análises estatísticas............................................................................. 15
4. RESULTADOS.. .................................................................................................16
4.1. Composição biológica da comunidade estudada..................................16
4.2. Descritores sintéticos simples...............................................................19
4.2.1. Riqueza de espécies. .............................................................19
4.2.2. Índice de diversidade de Shannon-Hill.. .................................21
4.2.3. Índice de Dominância de Simpson..........................................22
4.3. Freqüência .......................................................................................... 23
4.4. Biomassa.. ........................................................................................... 25
4.5. Padrão de distribuição das espécies mais conspícuas........................ 27
4.6. Descritores complexos..........................................................................34
4.6.1. Análise de similaridade em Modo Q (entre amostras).............34

VII
5. DISCUSSÃO.......................................................................................................37
6. CONCLUSÃO ................................................................................................... 42
7. REFERÊNCIAS BIBLIOGRÁFICAS ................................................................. 44
ANEXO 1. Cronograma de coletas e níveis de marés............................................51
ANEXO 2. Sinopse dos táxons encontrados..........................................................52
ANEXO 3. Valores obtidos para riqueza, diversidade, dominância, biomassa úmida e seca (média e desvio padrão ±) e resultados de teste de significância (Teste-t)..............................................................................................................................54
ANEXO 4. Freqüência de ocorrência das espécies encontradas na área estudada, expressa em porcentagem (%) de presença nas amostragens mensais...................................................................................................................57

III
LISTA DE TABELAS
Tabela 1. Valores obtidos para os índices de riqueza, diversidade e dominância (média ± desvio padrão), e análise de variância (ANOVA) durante o período de estudo...........................................20
Tabela 2. Biomassa úmida e seca registrada na área estudada e resultados obtidos na análise de variância (ANOVA). ..........................................................................................................................25
LISTA DE FIGURAS
Figura 1. Recifes de Maracajaú..........................................................................................................6
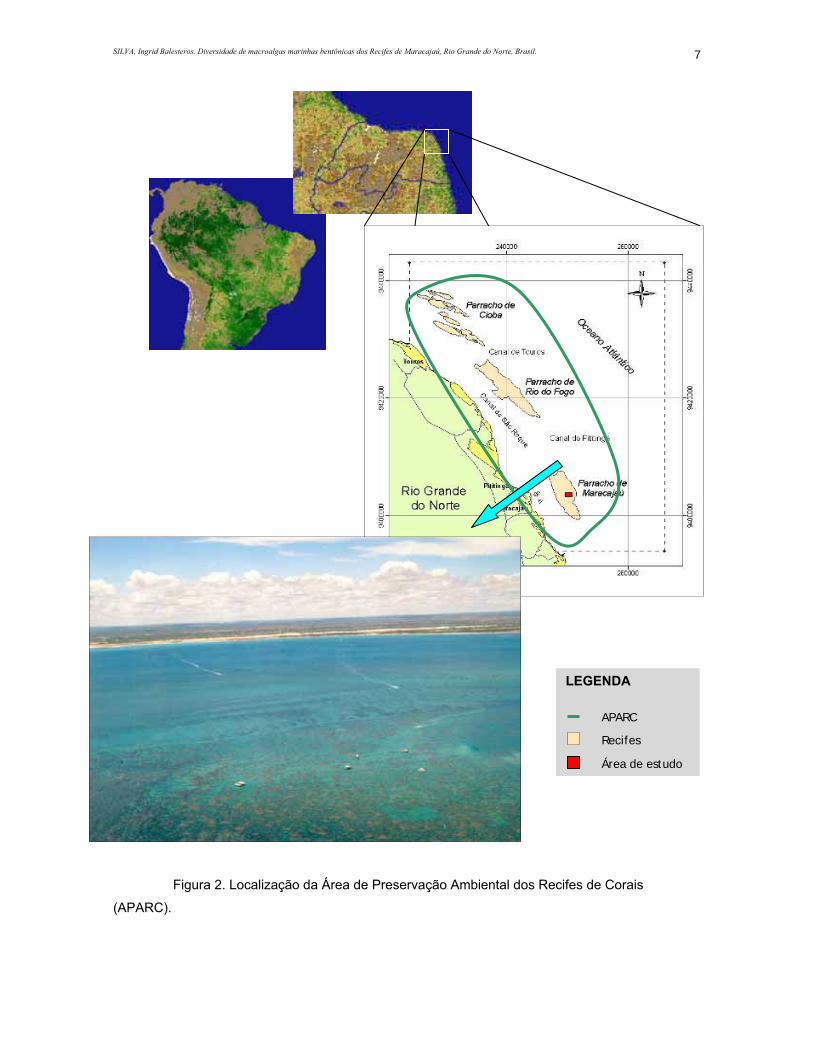
Figura 2. Localização da Área de Preservação Ambiental dos Recifes de Corais (APARC).............7
Figura 3. Imagens aéreas das estações de coleta............................................................................10
Figura 4. Representação gráfica do percentual de espécies encontradas para cada filo de macroalga..........................................................................................................................................16
Figura 5. Distribuição do número de espécies encontradas durante a realização do estudo entre as famílias de Rhodophyta....... .......................................... .................................................17
Figura 6. Distribuição (%) dos filos encontrados na área impactada (a) e na área não impactada (b)......................................................................................................................................................18
Figura 7. Número de espécies encontradas para cada família de macroalgas na área de estudo................................................................................ ...............................................................19
Figura 8. Índice de riqueza especifica registrado nas estações de coleta (± desvio padrão)............21
Figura 9. Diversidade de Shannon-Hill (médias mensais) baseada no efetivo das espécies encontradas na área estudada (± desvio padrão)..............................................................................21
Figura 10: Dominância de Simpson (médias mensais) baseada no efetivo das espécies encontradas na área de estudo (± desvio padrão)..................................................................................................22
Figura 11. Freqüência de ocorrência das espécies encontradas na área de estudo.........................24
Figura 12. Evolução da biomassa úmida (a) e seca (b) durante o período de estudo (média em
g.m2)....................................................................................................................................................26
Figura 13. Contribuição de biomassa úmida (%) das espécies mais conspícuas na área não impactada (a) e na área impactada (b).............................................................................................29
Figura 14. Dendograma de similaridade qualitativa entre amostras (modo Q).................................35
Figura 15. Aspecto geral da flora encontrada na área não impactada (a) e na área impactada (b)..41

IX
RESUMO
Apesar de toda a importância dos recifes para a humanidade, estes ambientes vêm
sendo ameaçados em todo o mundo. Diversos são os fatores que podem contribuir para a
degradação destes ambientes. Os recifes de Maracajaú, na Área de Preservação
Ambiental dos Recifes de Corais, localizada no litoral norte-oriental do Rio Grande do
Norte, vêm recebendo um número cada vez maior de turistas, o que está causando
preocupações quanto a integridade do local. Neste estudo, as comunidades de macroalgas
dos recifes de Maracajaú foram estudadas com a finalidade comparação entre duas
subáreas diferenciadas pela presença/ausência de turistas. Foram estabelecidas duas
estações de coleta: uma (I) onde as atividades de mergulho praticadas por turistas são
intensas e outra (II), onde estas práticas não ocorrem. Em cada estação foi estabelecida
transectos de 10 metros, onde foram distribuídos de forma aleatória 5 quadrados (625cm2).
O material coletado foi triado, identificado e teve a biomassa quantificada. A partir dos
dados de biomassa foram calculados os índices de riqueza, diversidade de Shannon-Hill e
dominância de Simpson. A similaridade entre as duas estações foi analisada pelo índice de
similaridade e distância de Bray-Curtis.
Foram coletadas 58 espécies de macroalgas no total, sendo 7 espécies de
Chlorophyta, 13 espécies de Phaeophyta e 38 espécies de Rhodophyta. Destas, 49
espécies foram encontradas na área não impactada e 42 na área impactada. Entre as
famílias encontradas, as mais representativas na área não impactada foram Dictyotaceae
e Corallinaceae, e na área impactada, Rhodomelaceae e Dictyotaceae.
A área não impactada apresentou maior biomassa e os maiores índices de riqueza
e diversidade. Na área impactada o índice de dominância foi maior, sendo C. racemosa a
espécie dominante nesta área.
O dendograma formado a partir do índice de similaridade apresentou-se dividido
em dois grandes grupos e um elemento isolado ao centro do dendograma, que
corresponde a uma amostra da área impactada. No primeiro agrupamento o predomínio foi
de amostras da área não impactada, onde as algas pardas foram mais conspícuas. Este
tipo de alga se torna dominante, geralmente, em recifes onde ocorreu a redução do
número de herbívoros. O segundo agrupamento foi composto por uma maioria de
amostras da área impactada, onde C. racemosa e associações de algas filamentosas e
calcárias eretas crescendo como tapetes foram encontradas cobrindo grandes extensões
nesta área. Estas associações compreenderam grupos de algas adaptadas a ambientes
sujeitos a perturbações freqüentes e, assim como C. racemosa, crescem rapidamente
sobre espaços recém desocupados nos recifes, não permitindo o estabelecimento de
outras espécies. Os resultados do presente estudo demonstraram que a atividade turística
é um fator impactante que têm causado alterações nas comunidades de macroalgas da
Área de Preservação Ambiental dos Recifes de Corais.

X
ABSTRACT
Despite the importance of coral reefs to humanity, these environments have been
threatened throughout the world. Several factors contribute to the degradation of these
ecosystems. The Maracajaú Reef Complex, in Rio Grande do Norte state is part of the
Coral Reefs Environment Preservation Area in northeastern Brazil. This area has been
receiving an increasing influx of tourism and the integrity of the local reefs is a matter of
concern. In this study, the reef macroalgae communities were studied and compared within
two areas distinguished by the presence or absence of tourism activities. Two sample sites
were chosen: the first one, where diving activities are intense; and the other, where these
practices do not occur. Samples were collected at both sites within a quadrate of 625 cm2
of area randomly thrown 5 times along a 10 meters transect line. Richness, Shannon-Hill
diversity and Simpson dominance indices were determined based on biomass data.
Similarity between sites was analyzed with Bray-Curtis similarity and distance index.
Fifty-eight macroalgae species were observed, including 7 Chlorophyta, 13
Phaeophyta and 38 Rhodophyta. In the non-disturbed site, 49 species were found, while at
the disturbed site, there were 42 species. Dictyotaceae and Corallinaceae were the most
representative families at the non-disturbed site, and Rhodomelaceae and Dictyotaceae at
the disturbed site. The non-disturbed site presented a higher biomass and the greatest
richness and diversity indices. In the disturbed site the dominance index was greater and
Caulerpa racemosa was the dominant species.
The dendogram based on similarity index showed two major clusters, and an
isolated element at the center that corresponds to a sample from the disturbed site. In the
first cluster, samples from the non-impacted site were predominant and fleshy brown algae
were more conspicuous. The second cluster was composed primarily of samples from the
impacted site, where C. racemosa and red filamentous and erect calcareous algae
associations (turf forming) were observed covering large extensions. These associations
are represented by groups of algae adapted to environments where disturbances are
frequent. They can grow rapidly on substrate where benthic community was removed and
do not allow the establishment of other species. The results of the present study show that
tourism activity is an impacting factor that has been causing shifts in macroalgae
communities in the Maracajaú Reef Complex.

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 1
1. INTRODUÇÃO
Os ecossistemas recifais são formações tipicamente tropicais, que
abrigam uma extraordinária fauna e flora, e junto às florestas tropicais, podem ser
considerados como os mais diversos ecossistemas do mundo (Connell, 1978).
A heterogeneidade do ambiente e a topografia irregular proporcionam aos
organismos diversos nichos passíveis de serem ocupados (Connell, 1978; Odum,
1988). Os organismos presentes nestes ambientes são o resultado da
especialização e coevolução, ao longo de bilhões de anos, culminado com a alta
diversidade e abundância biológica (Buddemeier, 1998). Os corais, uns dos
principais organismos formadores destes ambientes, apresentam uma faixa de
tolerância aos fatores ambientais muito estreita, fato que aliado à vulnerabilidade
das interações interespecíficas e às altas temperaturas comuns nestes ambientes
(que potencializa o efeito das substâncias tóxicas) fazem destes uns dos
ecossistemas mais frágeis do mundo (Pastorok & Bilyard, 1985). Tal diversidade e
abundância são fundamentais para a manutenção das comunidades ecológicas
dos próprios recifes e de outros ambientes marinhos que dependem de alguma
forma dos recifes (Hubbart, 1997). Este sistema é sustentado por um complexo
fluxo de energia, representado por cadeias tróficas de grande complexidade, as
quais apresentam grande variedade de espécies economicamente importantes,
seja para a indústria alimentícia, para a produção de fármacos e outros bens.
Estes ambientes também proporcionam áreas de recreação e lazer para a
humanidade. Além de toda a importância ecológica e sócio-econômica (Ferreira &
Maida, 2000; Cesar, 2000; Pollnac, 2000) estes ambientes proporcionam proteção
contra a erosão marinha ao longo da costa (Castro, 2002).
As algas são organismos de grande relevância nos ambientes recifais
brasileiros, estando presentes em diversas situações e desempenhando várias
funções. As macroalgas, além de serem base de diversas cadeias tróficas, servem
de abrigo, berçário e refúgio para várias espécies de invertebrados e pequenos
vertebrados. As algas calcárias, principalmente os gêneros que formam crostas,

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 2
fornecem resistência aos recifes porque preenchem os espaços vazios e
consolidam remanescentes de organismos mortos, contribuindo na sedimentação
destes ambientes (Figueiredo, 2000). Servem também como substrato para uma
grande variedade de organismos, inclusive outras macroalgas, sendo em muitos
recifes, dominantes sobre os corais em termos de abundância, e apresentam
fundamental importância no ciclo biogeoquímico, principalmente do carbono
(Figueiredo, 2000). Estes organismos absorvem o carbono do meio em forma de
carbonatos para sintetizar o CaCO3 que constituirá seus tecidos, e possivelmente
este carbonato que precipita no meio aquático se relaciona ao carbono
atmosférico. Oliveira (1996) e Buddemeier (1998) mencionaram sobre a
importância dos organismos que sintetizam carbonato de cálcio na manutenção
dos níveis de dióxido de carbono na atmosfera.
Apesar de toda a importância para a humanidade de manter a integridade
destes ambientes, os recifes vêm sofrendo fortes interferências antrópicas que
estão levando a degradação, muitas vezes de forma irreversível. Dentre as
atividades humanas que prejudicam os ecossistemas recifais, podemos citar a
pesca predatória e a explotação de seus recursos, que causa desequilíbrio nas
populações, como uma das atividades humanas que mais contribuem para a
destruição dos recifes de corais, bem como a poluição das águas costeiras
causada pelo desenvolvimento costeiro sem planejamento, as atividades agrícolas
(Leão, 1996) e de aqüicultura, a produção demasiada de lixo e esgoto, e a
destruição dos manguezais.
Há indícios de degradação em recifes de todo o mundo, evidenciados
principalmente pelas alterações nas estruturas das populações de corais e
macroalgas (Szmant, 2001). McCook et al (2001) revisaram as evidências e
mecanismos que envolvem as competições entre corais e macroalgas, e
concluíram que apesar da quantidade de sedimento disperso e da concentração
de nutrientes na coluna d’água serem fatores importantes para facilitar o
crescimento rápido de macroalgas, a herbivoria é o fator chave mediando os
efeitos das algas sobre os corais pois a habilidade da alga de competir pode
depender do acúmulo de biomassa suficiente para crescer sobre corais; poucos

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 3
táxons de algas podem realmente crescer sobre corais saudáveis e as
observações de sobreposições de algas em corais são freqüentemente resultado
de uma injúria ou morte do coral (McCook et al 2001). O fato é que estes fatores
que interferem na biota recifal podem estar, de certa forma, associados a
atividades antrópicas. Hughes (1994) relatou um drástico aumento na abundância
de macroalgas e diminuição na cobertura de corais nos recifes caribenhos que
vem ocorrendo desde os anos 80, após a mortalidade de Diadema antillarum e a
passagem de um furacão. Supostamente a sobrepesca na região teve relação
com estas alterações nas populações de algas e corais. Outros casos em que a
degradação de áreas recifais levou ao aumento das populações de macroalgas e
diminuição da cobertura de corais foram estudados por McCook (1999) na Grande
Barreira de Corais, por Lirman (2001) na Flórida, McClanahan & Muthiga (1998)
em Belize e Shulman & Robertson (1996) em San Blas, no Panamá.
Atividades de turismo e recreação em áreas de recifes também são causas
de preocupações em relação a conservação destes ambientes. O mergulho
recreativo é um segmento do mercado internacional de turismo que vem
crescendo e ganhando grande importância (Coutinho et al. 1993; Davis & Tisdell,
1996). O aumento da visitação a áreas recifais pode causar danos físicos à biota
através do pisoteamento, toque e ressuspensão de sedimentos provocada pelos
turistas pela má operação das nadadeiras (Bonilha, 2003), o que pode ocasionar a
morte de organismos, como os corais, por sufocamento. Há ainda influências
decorrentes do tráfego e fundeio de embarcações, e da extrapolação da
capacidade de suporte de cada sítio de mergulho – que, na maioria das unidades
de conservação ainda é desconhecida ou pouco conhecida (Amaral et al 2005).
Creed & Amado Filho (1999), através de experimentos entre as Ilhas Siriba e
Redonda, no Arquipélago de Abrolhos, observaram que cerca de 0,5% do campo
de gramíneas marinhas Halodule wrightii era danificado anualmente pelo
ancoramento de embarcações, e que a recuperação de uma área deteriorada por
apenas uma âncora demora cerca de nove meses.
O litoral do Rio Grande do Norte possui uma faixa costeira de praias
arenosas com aproximadamente 400 km de extensão, entremeadas pela

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 4
desembocadura de rios com a presença de manguezais, lagoas costeiras e
falésias, e apresenta cordões de recifes de arenito descontínuos ao longo da
costa. Esta área está compreendida na região fitogeográfica reconhecida por
Oliveira Filho (1977) como Zona Nordeste-oriental, com limites entre a costa oeste
do Ceará e o norte do Rio de Janeiro, caracterizada pela presença de águas
oligotróficas e abundância de substratos duros que abrigam a flora mais
diversificada do país. O litoral potiguar está dividido em dois setores ou zonas: o
Oriental que se estende da fronteira com o Estado da Paraíba até o município de
Maxaranguape e o Norte que abrange desde o Município de Touros até a divisa
com o Ceará (Moura 1992).
Os estudos sobre algas marinhas bentônicas do litoral potiguar foram
iniciados por Câmara Neto (1966) quando estudou várias espécies com
possibilidades de aproveitamento industrial. A seguir, foram referidas algumas
espécies quando se realizaram inventários para o litoral nordestino (Ferreira-
Correia & Pinheiro-Vieira 1969, Pinhero-Vieira & Ferreira Correia 1970). Câmara
Neto (1971) apresentou os primeiros levantamentos sobre a flora da região entre-
marés do Estado, complementado no segundo trabalho com dados quantitativos.
Diversas pesquisas sobre algas de importância econômica foram realizadas
(Durairatnam 1980, 1993; Alencar & Durairatnam 1993). Outras pesquisas sobre
levantamento de espécies e grupos taxonômicos foram feitas (Paes e Mello 1983,
Araújo 1983, Durairatnam & Nascimento 1983, Durairatnam et al. 1986, Moura e
Araújo 1990, Moura 1992, Marinho-Soriano, 1999) além de estudos sobre
sazonalidade de biomassa e de ficocolóides de espécies de interesse econômico
(Marinho-Sariano et al, 2001).
As formações recifais de Maracajaú, na Área de Preservação Ambiental dos
Recifes de Corais (APARC), no litoral norte-oriental do Rio Grande do Norte, são
visitadas por centenas de turistas (diariamente, na alta temporada). O aumento na
pressão de uso do local vem causando degradação na área e tem como principais
responsáveis a pesca predatória, a ocupação costeira e a exploração turística
desordenada em franco processo de expansão (Amaral et al. 2005).

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 5
O presente trabalho teve por objetivo contribuir para o conhecimento das
comunidades de macroalgas que crescem sobre os recifes de Maracajaú, e
detectar possíveis diferenças entre comunidades de áreas que sofrem maior
impacto da visitação turística e de áreas onde estas visitas não ocorrem.

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 6
2. ÁREA DE ESTUDO
O estudo foi realizado nos recifes de Maracajaú (Figura 1), situados no
litoral norte-oriental potiguar, no município de Maxaranguape, entre as
coordenadas 5º21’12” e 5º25’30” S e 35º14’30” e 35º17’12” W (Arantes, 2004).
Estes recifes estão inseridos na Área de Preservação Ambiental dos Recifes de
Corais, uma unidade de conservação criada em junho de 2001 que abrange a
faixa costeira e a plataforma rasa associada aos municípios de Maxaranguape,
Rio do Fogo e Touros (Marcelino & Amaral, 2003; Amaral et al, 2005) (Figura 2).
Figura 1. Recifes de Maracajaú. (Foto: Sandra Magalhães)
A região apresenta águas mornas (temperatura média de 28ºC) e límpidas
na maior parte do ano (Marcelino & Amaral, 2003). A salinidade (média de 36‰)
também não apresenta muitas variações ao longo do ano. Recobertos por rochas
biogênicas e organismos vivos, principalmente vermetídeos e algas calcárias
incrustantes (Laborel,1970 apud Castro, 2002), estes recifes apresentam-se
praticamente monoespecíficos em termos de coral, sendo Siderastrea stellata a
espécie dominante (Laborel, 1970 apud Castro, 2002; Arantes, 2004).
SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 7
LEGENDA
Recifes
APARC
Área de estudo
Figura 2. Localização da Área de Preservação Ambiental dos Recifes de Corais
(APARC).

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 8
O complexo recifal de Maracajaú é formado por diferentes tipos de
ambientes bentônicos, além dos corpos recifais, que são distinguidos pelo tipo de
fundo (substrato) e pelos organismos que ocorrem nestes locais. São eles: fundos
de rodólitos, as pradarias de fanerógamas e o fundo arenoso (Amaral et al. 2005).
Os corpos recifais apresentam-se na área como manchas de corais, arenito e
algas calcárias, os quais crescem a partir do fundo arenoso até cerca de três
metros de altura. A densidade com que estes corpos recifais estão dispostos na
área varia de alta, quando estes corpos apresentam muitas vezes fundidos uns
aos outros na porção sudeste da área, a baixa, quando estes corpos estão
dispersos espaçadamente sobre fundo arenoso, na porção mais a oeste (Amaral
et al. 2005). Estes ambientes apresentam grande importância para a extração de
recursos visando à subsistência e a mercantilização de pequena escala, por parte
de uma extensa comunidade extrativista regional de pescadores artesanais
(Bonilha, 2003).

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 9
3. MATERIAIS E MÉTODOS
3.1. As estações de coleta
Foi realizada uma visita preliminar ao local para a definição dos recifes a
serem estudados no dia 08/03/05. Foi levado em consideração como critério de
escolha a dimensão da formação recifal para que um transecto de dez metros
fosse devidamente posicionado. A riqueza aparente da estrutura recifal, a direção
das correntes, o tipo de fundo e a distância entre estações e base de apoio
flutuante também influenciaram na escolha do local.
Foram estabelecidas duas estações de coleta nos recifes de Maracajaú
(Figura 3). Uma estação corresponde a uma área que sofre intensa pressão
turística e foi, no presente trabalho, definida como área impactada. A outra
estação de coleta dista cerca de 800m da primeira e não é visitada por turistas
conforme parte das normas da APARC. Esta estação de coleta foi aqui definida
como área não impactada, por não sofrer impacto de turismo (Figura 3). A área
impactada compreendeu uma mancha de recife localizada exatamente ao lado da
estrutura flutuante utilizada como base para os visitantes, bem como para os
funcionários, da operadora de mergulho “Corais de Maracajaú” (Figura 3). Trata-se
de uma área onde as atividades de mergulho recreativo são intensas.
A área não impactada está localizada a aproximadamente 800 m da
primeira com a diferença que neste local o mergulho recreativo não é permitido
aos turistas. Nesta área foram observados alguns pescadores artesanais que
utilizam embarcações simples (sem motor) e com tripulação nunca superior a três
pessoas, mergulhando sem a utilização de equipamento de mergulho autônomo
para pescar lagostas e polvos.

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 10
I
II
Área de uso intensivo
Dunas
Estações de coleta
Cidades e vilas
LEGENDA
II Estação não impactada
I Estação impactada
Figura 3. Imagens aéreas das estações de coleta. (Fonte: Amaral et al, 2005)
A proximidade entre as duas estações de coleta minimiza o gradiente
ambiental que possa ocorrer de um local para o outro, fazendo com que o principal
fator que atua sobre as populações de macroalgas de maneira diferente nas duas
áreas seja as práticas de mergulho recreativo e todos os impactos relacionados
(toque e pisoteamento por parte dos visitantes, movimentação das embarcações,
óleo proveniente das embarcações e ressuspensão de sedimentos). As estações
de coleta foram definidas visando diferenciar uma da outra pela
presença/ausência de atividades de mergulho recreativo.

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 11
3.2. Estratégia de amostragem
A biomassa foi estimada a partir de amostragens mensais realizadas no
período de abril a setembro de 2005, perfazendo um total de seis meses. Foram
escolhidos os dias de maré baixa, baseado na tábua de marés do Departamento
de Hidrografia da Marinha do Brasil; as coletas foram realizadas em mergulho livre
com o uso de equipamento básico de mergulho (cronograma de coletas e níveis
de marés no ANEXO 1).
Para amostrar os organismos optou-se pelas amostragens aleatórias,
tendo-se como elemento amostral um quadrado de PVC de 25 cm de lado. Em
cada estação de coleta foi posicionado um transecto que consistiu em uma corda
de dez metros de comprimento marcada a cada dois metros com fita adesiva e
lastreada com fita de chumbo. Todos os meses esta corda era posicionada no
mesmo local no recife. Os quadrados foram dispostos ao longo dos dez metros de
corda de forma aleatória a cada amostragem, realizando-se 5 repetições em cada
estação amostrada. Todos os organismos ocorrentes dentro da unidade amostral
foram removidos manualmente com o auxílio de uma espátula de metal e
colocados dentro de sacolas de rede.
3.2.1. Determinação da biomassa
Após as coletas, os organismos foram mantidos em baldes com água do
mar até serem preservados. Em laboratório as amostras foram preservadas em
solução de formol a 4% preparada com água do mar. Posteriormente o material foi
triado macroscopicamente e a biomassa fresca de cada espécie foi quantificada.
Cada grupo de organismos de mesma espécie presente em cada elemento
amostral teve a biomassa úmida quantificada em balança eletrônica. Para as
espécies cujo valor de biomassa úmida foi menor que 5 g foi atribuído um valor
arbitrário (um) que significa que a espécie esteve presente na amostra. Espécies
com efetivo igual ou maior que 5 g tiveram o peso seco obtido após secar em
estufa a 60ºC até ter seu peso estabilizado, o que variou entre 24-48 horas,

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 12
dependendo da espécie. O material seco foi pesado em balança de 0,005 g de
precisão.
Após o processo de quantificação da biomassa presente em cada amostra,
obteve-se o total de biomassa presente nas cinco repetições e, a partir desta
soma determinou-se o percentual de contribuição de cada espécie nas
amostragens. A média das repetições foi extrapolada para uma área de 1m2 com
a finalidade de padronização.
3.2.2. Identificação do material
Os indivíduos sem táxon definido no momento da triagem permaneceram
fixados até serem identificados. A identificação de gêneros e espécies foi baseada
em bibliografia específica sobre floras (Taylor, 1960; Joly, 1957, 1965; Ugadim,
1974; Cordeiro-Marino, 1978; Yoneshigue, 1985; Horta, 2000) e em trabalhos de
revisão de grupos taxonômicos (Joly et al, 1969; Oliveira Filho, 1969; Santelices,
1978; Paes e Mello, 1983; Schneider, 1983; Guimarães, 1990; Guimarães &
Oliveira, 1996; Fernandes & Guimarães, 1998; Moura, 2000; Barreto e
Yoneshigue-Valentin, 2001; Guimarães et al, 2004; Barata, 2004; Bacci; 2005;
Fujii & Sentíes, 2005). A nomenclatura e o sistema de classificação utilizados
seguiu Wynne (1998). O material estudado encontra-se depositado na coleção do
Departtamento de Oceanografia e Limnologia da UFRN.
3.3. Descritores analíticos
No presente trabalho optou-se pela utilização do peso úmido e seco (plano
quantitativo) como descritor analítico. A partir dos dados de biomassa úmida das
espécies encontradas nas amostragens obteve-se os descritores de riqueza,
diversidade e dominância, bem como os dendogramas de similaridade entre
amostras. O uso deste descritor deveu-se ao fato do peso seco ser restrito às
espécies que apresentaram peso fresco igual ou superior a 5 g por elemento
amostral, se o peso seco fosse utilizado ao invés do peso fresco, muitas espécies
com peso inferior a 5 gramas simplesmente não seriam contabilizadas na

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 13
aplicação destes índices. Muitas vezes, espécies com baixos valores de biomassa
apresentaram-se muito freqüentes, e na maior parte dos casos suas associações
formaram expressivas comunidades nos recifes. Estas comunidades são formadas
por algas filamentosas e calcárias de hábito ereto, que apresentam rápido
desenvolvimento e crescem entrelaçadas umas às outras, formando densos
emaranhados. Este tipo de associação será designado no presente trabalho como
tapetes de algas, ou simplesmente tapetes, a exemplo de Coimbra (1998) que
aplicou a mesma terminologia referindo-se a estes tapetes como ‘unidades
taxonômicas operacionais’, as quais se comportam como uma entidade ecológica
única (como uma espécie desempenhando seu nicho, por exemplo), porém podem
ser formado por organismos de grupos evolucionalmente bastante distantes.
3.4. Descritores sintéticos simples
3.4.1. Riqueza
A riqueza expressa o número de espécies, sendo aqui adotada a riqueza
como a média entre o número total de espécies presentes em cada elemento
amostrado por mês. Este descritor diz pouco sobre a organização da comunidade
e está relacionado ao tamanho da amostra, podendo aumentar em função da área
(Rosso, 1990).
3.4.2. Freqüência
A freqüência de ocorrência expressa em termos percentuais a quantidade
de elementos amostrais em que certa espécie se faz presente numa amostragem.
A freqüência nos permite expressar dados qualitativos (presença/ausência)
num plano semiqualitativo, pois expressa em porcentagem a média de ocorrências
de certa espécie nas amostragens de determinado local. Isso nos possibilita
hierarquizar as variáveis, no entanto, sem permitir estabelecer uma distância entre
os níveis das categorias, uma vez que não traz informações acerca do efetivo (da
quantidade) com que cada espécie se apresentou na amostra (Rosso, 1990).

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 14
3.4.3. Índice de diversidade de Shannon-Hill
O índice de diversidade de Shannon-Hill é o índice de diversidade de
Shannon-Wiener (Shannon & Weaver, 1949) sob a forma exponencial de Hill
(1973). O índice baseia-se na composição efetiva de cada espécie, além da
riqueza específica, e se relaciona mais ao fator dominância que riqueza, uma vez
que está envolvendo as espécies ocorrentes e seus efetivos, no caso, a biomassa.
O índice é calculado pela fórmula:
H’= pi log2 pi
Onde pi é o efetivo (no caso o peso) da espécie i em relação à amostra, ou
seja, a proporção de peso de uma espécie na amostra.
3.4.4. Índice de dominância de Simpson
O índice de Simpson (1949) expressa o quanto uma comunidade está
sendo dominada ou não por uma, ou um pequeno grupo, de espécies. O produto
final da fórmula varia de 0 a 1, sendo igual a 0 quando a amostra é composta de
espécies que apresentam o mesmo efetivo, ou seja, a dominância é nula, e igual a
1 se todos os indivíduos da amostra forem da mesma espécie, representando
dominância total.
O índice de dominância de Simpson é calculado pela fórmula:
= pi2
Onde pi é o efetivo (no caso o peso) da espécie i em relação à amostra, ou
seja, a proporção de peso de uma espécie na amostra.
3.5. Descritores complexos: análise de similaridade
Optou-se neste estudo pela utilização da análise de similaridade entre
amostras (modo Q) com o objetivo de determinar agrupamentos formados de
acordo com a área de ocorrência. Desta forma poderemos detectar possíveis
diferenças entre a composição específica das duas áreas estudadas.

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 15
3.5.1. Análise de agrupamento em Modo Q (entre amostras)
O modo Q representa a comparação dos conjuntos de espécies presentes
em cada amostra, resultando em agrupamentos de amostras que tenham
espécies em comum. No estudo da similaridade em modo Q (entre amostras)
foram desconsideradas as espécies que foram coletadas fora dos limites
amostrais, ou seja, aquelas coletadas na amostragem preliminar, as quais não tem
determinação de área não impactada ou impactada.
Uma matriz de dados binários (0= ausência; 1=presença) foi gerada a partir
dos dados coletados nas duas estações de amostragens, no aplicativo PRIMER 5
e a análise de agrupamento (CLUSTER) foi aplicada em ligação completa e
utilizado o índice de Bray-Curtis para determinar as distâncias das similaridades
dos agrupamentos.
3.6. Análises estatísticas
As diferenças nos índices ecológicos (riqueza, diversidade e dominância) e
na biomassa entre as duas estações de coleta foram analisadas quanto a
significância pelo Teste-t para amostras independentes. A variação destes
descritores no período de estudo foi analisada empregando-se análise de
variância (one way ANOVA). Ambos os testes foram realizados através do
aplicativo Statistica 6.0, StatSoft®, 1998.

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 16
4. RESULTADOS
4.1. Composição biológica da comunidade estudada
Um total de 58 táxons foi observado, sendo 55 identificados ao nível de
espécie e 3 ao nível de gênero, das quais 10 estão sendo citadas pela primeira
vez para o litoral do estado, e 3 são novas citações para a região Nordeste
(ANEXO 2). No total foram observadas 7 espécies de Chlorophyta (12%),
representadas pelas famílias Ulvaceae (uma espécie), Siphonocladaceae (uma
espécie) e Caulerpaceae (5 espécies) ; 13 espécies de Phaeophyta (22%)
representadas pelas famílias Dictyotaceae (9 espécies) e Sargassaeae (4
espécies); e 38 espécies de Rhodophyta (66%) (Figura 4), distribuídas em 13
famílias representadas pela Figura 5.
12%
22%
66%
Chlorophyta Phaeophyta Rhodophyta
Figura 4. Representação gráfica do percentual de espécies encontradas para cada filo de
macroalga.
Das 58 espécies encontradas na área de estudo, 53 estiveram
presentes dentro dos limites amostrais, ou seja, nos limites da área de cada
elemento amostral, sendo categorizadas como ocorrência para área não
impactada e/ou impactada.

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 17
0 2 4 6 8 10
Acrochaetiaceae
Corallinaceae
Gelidiaceae
Gelidiellaceae
Bonnemaisoniaceae
Gigartinaceae
Hypneaceae
Solieriaceae
Halymeniaceae
Rhodymeniaceae
Lomentariaceae
Ceramiaceae
Rhodomelaceae
Número de espécies
Figura 5. Distribuição do número de espécies encontradas durante a realização do estudo
entre as famílias de Rhodophyta.
As outras 5 espécies (Enteromorpha sp, Bryopsis plumosa, Botryocladia
occidentalis, Gracilaria cervicornis e Polysiphonia subtilissima) foram coletadas
apenas na amostragem preliminar, considerando-se assim que estas estiveram
presentes na área de estudo, porém fora dos limites amostrais propostos para a
realização deste trabalho e portanto, tiveram sua presença registrada mas foram
desconsideradas em todas as análises comparativas entre estações de coleta.
Na área impactada foram encontradas 4 espécies de Chlorophyta, 8
espécies de Phaeophyta e 30 espécies de Rhodophyta, totalizando 42 espécies
de macroalgas. Na área não impactada, assim como na impactada, 4 espécies de
Chlorophyta foram encontradas. Quanto aos representantes de Phaeophyta e
Rhodophyta, a área não impactada mostrou-se mais rica, apresentando 13 e 32
espécies respectivamente, contabilizando um total de 49 espécies. A Figura 6
mostra estes valores em termos percentuais.
Tanto na área não impactada como na impactada, 13 famílias de rodofíceas
e 2 de feofíceas foram amostradas. Entre os representantes de clorofíceas, 2
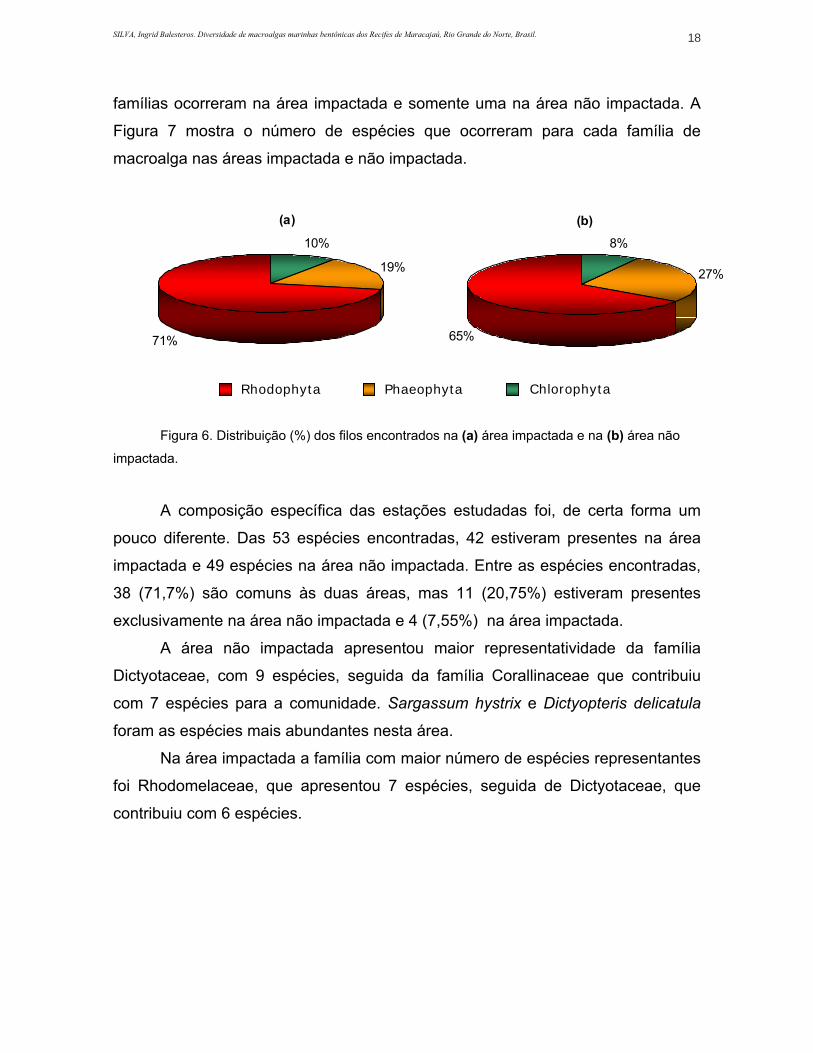
SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 18
famílias ocorreram na área impactada e somente uma na área não impactada. A
Figura 7 mostra o número de espécies que ocorreram para cada família de
macroalga nas áreas impactada e não impactada.
(a)
10%
19%
71%
(b)
8%
27%
65%
Chlorophyta Rhodophyta Phaeophyta
Figura 6. Distribuição (%) dos filos encontrados na (a) área impactada e na (b) área não
impactada.
A composição específica das estações estudadas foi, de certa forma um
pouco diferente. Das 53 espécies encontradas, 42 estiveram presentes na área
impactada e 49 espécies na área não impactada. Entre as espécies encontradas,
38 (71,7%) são comuns às duas áreas, mas 11 (20,75%) estiveram presentes
exclusivamente na área não impactada e 4 (7,55%) na área impactada.
A área não impactada apresentou maior representatividade da família
Dictyotaceae, com 9 espécies, seguida da família Corallinaceae que contribuiu
com 7 espécies para a comunidade. Sargassum hystrix e Dictyopteris delicatula
foram as espécies mais abundantes nesta área.
Na área impactada a família com maior número de espécies representantes
foi Rhodomelaceae, que apresentou 7 espécies, seguida de Dictyotaceae, que
contribuiu com 6 espécies.

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 19
0 2 4 6 8 10
SiphonocladaceaeCaulerpaceaeDictyotaceae
SargassaceaeAcrochaetiaceae
CorallinaceaeGelidiaceae
GelidiellaceaeBonnemaisoniaceae
GigartinaceaeHypneaceaeSolieriaceae
HalymeniaceaeRhodymeniaceae
LomentariaceaeCeramiaceae
Rhodomelaceae
Número de espécies
Área impactada Área não impactada
Figura 7. Número de espécies encontradas para cada família de macroalgas na área de
estudo.
4.2. Descritores sintéticos simples
A Tabela 1 mostra os resultados obtidos durante o estudo para o índice de
riqueza, de diversidade de Shannon-Hill e de dominância de Simpson. Estes
valores não variaram significantemente ao longo do período analisado segundo
análise de variância (ANOVA) (Tabela 1).
4.2.1. Riqueza de espécies
A Figura 8 mostra os valores obtidos para riqueza, aqui representando a
média mensal do total de espécies coletadas em cada elemento amostral, na área
impactada e na área não impactada.

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 20
Tabela 1. Valores obtidos para os índices de riqueza, diversidade e dominância (média ±
desvio padrão), e análise de variância (ANOVA) durante o período de estudo.
ANOVA Mínimo-máximo
média ± desvio
padrão Fcal p
Área não impactada
Riqueza 5,4 - 14,2 10,24 ± 2,93 0,39 0,86
Diversidade 1,60 - 2,48 1,98 ± 0,31 1,13 0,38
Dominância 0,31 - 0,44 0,37 ± 0,06 0,28 0,93
Área impactada
Riqueza 6,8 - 11 8,7 ± 1,53 0,52 0,73
Diversidade 1,02 - 1,96 1,37 ± 0,33 0,54 0,75
Dominância 0,42 - 0,68 0,57 ± 0,09 0,33 0,90
Os índices mais altos de riqueza foram registrados no mês de maio, para a
área não impactada (14,2 ± 5,76) sendo Dictyotaceae (7 espécies) e Corallinaceae
(6 espécies) as famílias mais representativas neste mês. Na área impactada, o
índice de riqueza foi maior no mês de abril (9,6 ± 1,51). Neste mês, as famílias
Corallinaceae e Ceramiaceae foram as que apresentaram o maior número de
espécies (3 espécies cada família). Os meses em que a riqueza apresentou os
menores valores foram abril na área não impactada (5,4 ± 1,34) e junho na área
impactada (6,8 ± 5,67) (ANEXO 3).
As diferenças observadas para este índice não tiveram variação
significativa entre as duas estações de coleta (p>0,05), com exceção do mês de
abril (p<0,05), quando a riqueza amostrada na área não impactada apresentou-se
muito baixa em relação à área impactada e aos outros meses amostrados
(ANEXO 3).

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 21
0
5
10
15
20
25
abr/05 mai/05 jun/05 jul/05 ago/05 set/05
Índ
ice
de
riq
uez
a
Área não impactada Área impactada
Figura 8. Índice de riqueza especifica registrado nas estações de coleta (± desvio padrão).
4.2.2. Índice de diversidade de Shannon-Hill
Os índices apontaram uma maior diversidade na área não impactada,
durante todo o período do estudo (Figura 9). O maior valor de diversidade nesta
estação ocorreu no mês de maio (2,48 ± 1,04) e o menor foi registrado no mês de
abril (1,60 ± 0,27). Na área impactada, a diversidade teve valores mais altos em
julho (1,96 ± 1,28) e os mais baixos em setembro (1,02 ± 1,10) (ANEXO 3).
0
0,5
1
1,5
2
2,5
3
3,5
4
abr/05 mai/05 jun/05 jul/05 ago/05 set/05
Div
ersi
dad
e d
e S
han
no
n-H
ill
Área não impactada Área impactada
Figura 9. Diversidade de Shannon-Hill (médias mensais) baseada no efetivo das espécies
encontradas na área estudada (± desvio padrão).

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 22
Quando estes valores foram comparados (Teste t), somente o mês de maio
apresentou diferença significativa (p<0,05) entre os valores de diversidade das
duas estações (ANEXO 3). Neste mês a área não impactada apresentou alto
índice de diversidade, que obviamente se relaciona à riqueza que esteve elevada
neste mês, em relação às outras amostragens.
4.2.3. Índice de Dominância de Simpson
A dominância foi maior na área impactada (Figura 10), porém este índice,
assim como o de diversidade, não variou significativamente durante o estudo e
não diferiu significantemente entre as duas estações (ANEXO 3). Na estação de
coleta impactada a espécie dominante foi Caulerpa racemosa, a qual durante todo
o período de realização do trabalho apresentou altos valores de biomassa e
freqüência.
O mês em que a área impactada mostrou o maior índice de dominância foi
o mês de setembro, justamente o mês em que Caulerpa racemosa esteve mais
abundante.
0
0,2
0,4
0,6
0,8
1
1,2
abr/05 mai/05 jun/05 jul/05 ago/05 set/05
Do
min
ânci
a d
e S
imp
son
Área não impactada Área impactada
Figura 10: Dominância de Simpson (médias mensais) baseada no efetivo das espécies
encontradas na área de estudo (± desvio padrão).

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 23
Na área não impactada este índice se apresentou relativamente baixo,
sendo o mês em que a dominância esteve mais alta o mês de setembro (0,44 ±
0,28), sendo C. racemosa a espécie dominante. Nesta área a comunidade é
dominada por Caulerpa racemosa e Sargassum hystrix, além de Dictyopteris
delicatula. A diferença é que nesta área os valores de biomassa das espécies
dominantes não apresentam diferenças tão grandes em relação aos valores das
espécies menos abundantes, configurando assim uma comunidade menos
dominada e portanto, mais diversificada.
4.3. Freqüência
A Figura 11 mostra com que freqüência as espécies ocorreram nas
amostragens das duas estações de coleta ao longo do período de estudo, em
termos percentuais. Os valores mensais estão apresentados no ANEXO 4. Pode-
se observar que a espécie mais freqüente em ambas as estações foi Dictyopteris
delicatula, seguida de Sargussum hystrix na área não impactada e de Caulerpa
racemosa na área impactada.

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 24
0 20 40 60 80 100 120
Dictyopteris delicatula
Sargassum hytrix
Caulerpa racemosa
Jania adherens
Lobophora variegata
Amphiroa beauvoisii
Gelidiella acerosa
Gelidium coarctatum
Dictyota menstrualis
Halipt ilon subulatum
Laurencia f iliformis
Dictyota cervicornis
Bryothamniom triquetum
Dictyopteris just ii
Amphiroa anastomosans
Falkenbergia hillebrandii
Gelidiopsis planicaulis
Laurencia intricata
Cryptonemia crenulata
Spatoglossum schroederi
Jania sp2
Hypnea spinella
Sargassum vulgare
Pterocladiella caerulensces
Gelidiopsis variabilis
Jania sp1
Antithamnionella graef fei
Botryocladia pyriformis
Hypnea musciformis
Chondracanthus acicularis
Sargassum f ilipendula
Caulerpa cupressoides
Caulerpa mexicana
Jania pumila
Neosiphonia gorgoniae
Lomentaria corallicola
Dictyota mertensii
Dictyota ciliolata
Ceramium f laccidum
Ceramium vagans
Gelidiella ligulata
Chondria dangeardii
Caulerpella ambigua
Acrochaet ium f lexuosum
Dipterosiphonia dentrit ica
M erist iella gelidium
Dictyopteris polypodioides
Cryptonemia bengryi
Padina sanctae-crucis
0 20 40 60 80 100 120
Dictyopteris delicatula
Caulerpa racemosa
Hypnea spinella
Jania adherens
Gelidiella acerosa
Gelidiopsis planicaulis
Amphiroa beauvoisii
Pterocladiella caerulensces
Ceramium vagans
Gelidium coarctatum
Gelidiella ligulata
Amphiroa anastomosans
Laurencia intricata
Chondracanthus acicularis
Lomentaria corallicola
Ceramium f laccidum
Neosiphonia gorgoniae
Laurencia f iliformis
Ceramium dawsonii
Dictyota cervicornis
Caulerpella ambigua
Jania pumila
Ventricaria ventricosa
Sargassum f ilipendula
Sargassum vulgare
Dictyota menstrualis
Cryptonemia crenulata
Herposiphonia bipinnata
Chondria dangeardii
Ant ithamnionella graeffei
Halipt ilon subulatum
Hypnea musciformis
Gelidiopsis variabilis
Dictyota ciliolata
Dictyota mertensii
Lobophora variegata
Dipterosiphonia dentrit ica
Acrochaet ium f lexuosum
Falkenbergia hillebrandii
Herposiphonia secunda
Caulerpa mexicana
Dictyopteris just ii
M erist iella gelidium
Enteromorpha sp
Freqüência de ocorrência (%)
Área não impactada Área impactada
Figura 11. Freqüência de ocorrência das espécies encontradas na área de estudo.

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 25
4.4. Biomassa
A biomassa coletada foi, de forma geral, maior na área não impactada,
exceto em setembro quando a biomassa na área impactada esteve maior que na
área não impactada. Este descritor apresentou grande variação entre
subamostras, principalmente na área impactada, apresentando assim altos valores
de desvio padrão.
A média da biomassa úmida para a área não impactada foi de 2595,2 g.m-2
enquanto que na área impactada a média ficou em 2424,14 g.m-2 (Tabela 2). Este
descritor apresentou diferença significativa entre as duas áreas somente no mês
de junho (p<0,05) (ANEXO 3). Neste mês, as algas da ordem Dyctyotales foram
abundantes na área não impactada e os indivíduos de Caulerpa racemosa
estavam pequenos e pouco numerosos nas duas áreas.
A Figura 12 mostra a evolução da biomassa úmida e seca registrada ao
longo do período de realização do trabalho. Ao longo do período de amostragens
este descritor apresentou pouca variação, tanto na área não impactada (p=0,16)
como na impactada (p>0,05) (Tabela 2). Em termos de biomassa seca, a área não
impactada teve uma média de 237,78 g.m-2 enquanto que na área impactada a
média foi de 118,02 g.m-2 (Tabela 2).
Tabela 2. Biomassa úmida e seca registrada na área estudada e resultados obtidos na
análise de variância (ANOVA).
ANOVA Mínimo-máximo
(g/m2)
Média ± desvio padrão
(g/m2) F p
Área não impactada
Biomassa úmida 1475,20 - 3452,80 2595,20 ± 791,12 1,79 0,16
Biomassa seca 145,04 - 313,51 237,78 ± 65,63 1,78 1,60
Área impactada
Biomassa úmida 973,60 - 4323,2 2424,14 ± 1148,13 1,01 0,44
Biomassa seca 37,12 - 213,43 118,02 ± 59,14 1,31 0,30

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 26
(a)
0
2000
4000
6000
8000
10000
g.m
-2
(b)
0
200
400
600
800
abr/05 mai/05 jun/05 jul/05 ago/05 set/05
g.m
-2
Área impactada Área não impactada
Figura 12. Evolução da biomassa úmida (a) e seca (b) durante o período de estudo (média
em g.m-2).
Quanto aos valores obtidos em peso seco, a diferença entre as estações de
coleta foi significativa no mês de abril (p<0,05) e altamente significativa em junho
(p<0,001) (ANEXO 3).
O valor mínimo de biomassa úmida registrado na área não impactada foi
em maio, quando a espécie C. racemosa apresentou freqüência baixa e pouca
biomassa. No que se refere a biomassa seca, o mínimo registrado para esta
estação de coleta foi em julho, apesar do efetivo C. racemosa ter sido

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 27
considerável, neste mês Sargassum hystrix, L. variegata e espécies de Dyctyota
apresentaram-se pouco abundantes. Os valores máximos de biomassa úmida e
seca para esta estação de coleta foram em setembro e agosto, respectivamente.
Nesta área, a espécie mais representativa em termos de biomassa foi S. hystrix,
seguido por C. racemosa, Lobophora variegata e Dictyopteris delicatula. Embora
tenham contribuído com uma considerável biomassa, Cryptonemia crenulata,
Dictyopteris justii e Dictyota mertensii estiveram presentes somente em alguns
meses (Figura 13a), sendo que estas não apresentaram um padrão definido de
variação de acordo com as estações do ano, exceto Dictyota mertensii que foi
mais abundante a partir do mês de junho até o final do estudo (setembro). Outras
espécies foram representativas apenas pontualmente, como Gelidium coarctatum,
foram bastante expressivas no mês de junho, formando verdadeiros tapetes, e
Spatoglossum schroederii, que foi mais abundante no mês de julho.
A área impactada teve como espécie mais representativa C. racemosa, e
seus altos valores de abundância relativa fizeram com que os índices de
dominância para esta área ficassem altos. Este fato foi notável em campo durante
todo o período de realização do trabalho. Outra espécie que foi relativamente
abundante nesta área foi D. delicatula, a qual muitas vezes crescia associada a C.
racemosa, constituindo um estrato inferior nos bancos desta espécie. Gelidiella
acerosa, Gelidiopsis planicaulis, Hypnea spinella e algas calcárias pertencentes às
espécies Jania adhaerens e Amphiroa beauvoisii foram muito freqüentes e
representativas nas amostragens realizadas nesta área (Figura 11). Embora a
biomassa destas espécies tenha sido baixa, visto que se trata de espécies de
pequeno porte, a alta freqüência com que apareceram ao longo do período de
estudo mostra sua expressividade nesta área, sendo estas espécies as
descritoras da comunidade.
4.5. Padrão de distribuição das espécies mais conspícuas
Neste item será discutida a presença das espécies consideradas mais
representativas nas estações de amostragem ao longo do período estudado,

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 28
sendo estas espécies as descritoras da comunidade. No presente estudo, optou-
se por considerar como espécies mais conspícuas (ou representativas) aquelas
que obtiveram biomassa relativa igual ou maior que 1% em mais de uma
amostragem ou biomassa relativa igual ou maior que 2% em pelo menos um mês.
Como certas espécies que apresentam pouca biomassa apresentaram-se muito
freqüentes nas amostragens, a freqüência de ocorrência também foi utilizada
como critério para categorizar uma espécie como conspícua. Assim as espécies
com freqüência igual ou maior que 40% foram agrupadas nesta categoria.
A Figura 13 mostra a contribuição (em porcentagem) de cada uma das
espécies mais expressivas, conforme critério mencionado, para as duas áreas. Na
área não impactada 14 espécies foram consideradas conspícuas, o que
representou 28,57% do total das espécies encontradas. Caulerpa racemosa,
Sargassum hystrix e Dictyopteris delicatula foram as três espécies mais
representativas nesta área. Na área impactada 10 espécies (23,8% do total) foram
conspícuas. As espécies mais representativas nesta área foram C. racemosa e D.
delicatula, sendo que Hypnea spinella apresentou 80 ± 17,89% de freqüência
(Figura 11).
A espécie Caulerpa racemosa apresentou um padrão semelhante de
ocorrência nas duas estações de coleta ao longo do período estudado (Figura 13).
Os valores de biomassa proporcional registrados no início do estudo (mês de abril)
apresentaram uma redução progressiva até o mês de junho, quando foi registrado
a menor contribuição da espécie tanto para a área não impactada (18,8%) como
na área impactada (69,95%). Na área não impactada, a maior contribuição foi no
mês de setembro, quando a biomassa relativa foi 55,14%, enquanto que na área
impactada o valor máximo chegou a 89,65% no mês de agosto.
No mesmo mês de setembro quando C. racemosa foi mais abundante na
área não impactada, Dictyopteris delicatula apresentou a menor contribuição para
a área, representando apenas 4,63% da amostragem. Em julho, D. delicatula
representou 31,75% da amostragem, a maior proporção registrada para a espécie
na área não impactada.

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 29
(a)
0%
20%
40%
60%
80%
100%
abr/05 mai/05 jun/05 jul/05 ago/05 set/05
Caulerpa racemosa Dictyopteris delicatula Distyopteris justii
Dictyota mertensii Lobophora variegata Sargassum hystrix
Sargassum vulgare Bryothamnion triquetum Cryptonemia crenulata
Gelidiella acerosa L filiformis Gelidium coarctactum
OUTRAS
0%
20%
40%
60%
80%
100%
abr/05 mai/05 jun/05 jul/05 ago/05 set/05
Caulerpa racemosa Ventricaria verrucosa Hypnea spinella
Dictyopteris delicatula Sargassum vulgare Gelidiella acerosa
L filiformis
(b)
OUTRAS
Figura 13. Contribuição de biomassa úmida (%) das espécies mais conspícuas na área não
impactada (a) e na área impactada (b).

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 30
Na área impactada, D. delicatula também foi mais abundante com relação
às outras espécies no mês de setembro, representando 3,4% da amostragem. A
maior contribuição da espécie nesta área aconteceu no mês de junho, sendo
16,87% do total.
Sargassum hystrix foi uma espécie muito representativa na área não
impactada, porém esta espécie não foi amostrada na área impactada. S. hystrix
apresentou os maiores valores de biomassa relativa no mês de junho, assim como
D. delicatula, representando 48,75% da amostragem. Em abril esta espécie
apresentou sua menor contribuição para a comunidade, apenas 5,36% da
biomassa relativa.
As outras espécies que foram conspícuas na área não impactada foram
Lobophora variegata, Cryptonemia crenulata, Gelidiella acerosa, Dictyopteris
Justii, Dictyota mertensii, Gelidium coarctatum, Laurencia filiformis, Bryothamnion
triquetum e Sargassum vulgare, pelo critério da biomassa, e Jania adhaerens e
Amphiroa beauvoisi pela alta freqüência. Estas espécies apresentaram-se
ausentes em algumas amostragens (Figura 13a), porém quando presentes
apresentaram valores de biomassa expressivos o bastante para se enquadrarem
entre as espécies mais conspícuas.
Na área impactada, além de C. racemosa e D. delicatula, as outras
espécies consideradas conspícuas foram Hypnea spinella, G. acerosa, L. filiformis,
Ventricaria ventricosa e S. vulgare, pela biomassa registrada, e J. adhaerens, A.
beauvoisii e Gelidiopsis planicaulis pela freqüência. Destas, J. adhaerens, A.
beauvoisii e Gelidiopsis planicaulis estiveram presentes em todas as amostragens,
ou seja, em todos os meses, na área não impactada, e as outras espécies
estiveram ausentes em certos períodos (Figura 13b).

Prancha 1
SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 31
Ventricaria ventricosa
Dictyopteris justii Lobophora variegataLobophora variegata
Dictyota mertensii
Dictyopteris delicatula Caulerpa racemosa

Prancha 2
SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 32
Sargassum hystrix
Jania adhaerens
Gelidiella acerosaSargassum vulgare
Gelidium coarctatum
Amphiroa beauvoisii

Prancha 3
SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 33
Hypnea spinella
Cryptonemia crenulataGelidiopsis planicaulis
Bryothamnion triquetum Laurencia filiformis

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 34
4.6. Descritores complexos
4.6.1. Análise de similaridade em Modo Q (entre amostras)
A Figura 14 apresenta o dendograma gerado a partir da análise de
similaridade em modo Q (entre amostras).
Formaram-se dois grandes grupos de amostras (G1 e G2), e um elemento
isolado ao centro do gráfico (G3). Os dois grandes grupos foram ainda divididos
em subgrupos (G1a e b, que corresponde ao Grupo 1, e G2a e b formados a partir
de G2).
No primeiro grupo (G1), o predomínio foi de amostras da área não
impactada. O agrupamento G1a apresentou 18 elementos amostrais, destes
apenas dois foram da área impactada. Estes elementos tiveram em comum
Caulerpa racemosa, Dictyopteris delicatula e Gelidium coarctatum, e associações
de algas calcárias, além de Sargassum hystrix para amostras da área não
impactada. As amostras da área impactada que estiveram unidas neste subgrupo
apresentaram Dictyota menstrualis e D. cervicornis como variáveis comuns aos
outros elementos e agrupou estas amostras ao subgrupo G1a. O subgrupo G1b
apresentou 17 elementos, dos quais 6 são da área impactada e os outros nove da
área não impactada. Estes elementos tiveram em comum Caulerpa racemosa,
Dictyopteris delicatula e não apresentaram espécies de algas calcárias em
comum. G1b apresentou ainda agrupamentos de elementos que tiveram
Sargassum vulgare, Lobophora variegata, Gelidiella acerosa e/ou Laurencia
filiformis como variáveis comuns.
O subgrupo G2 foi formado por 7 elementos, todos da área impactada,
unidos pela presença de Dictyopteris delicatula, Pterocladiella caerulensces,
Hypnea spinella e Gelidium coarctatum, e pela ausência de Caulerpa racemosa.
Estes elementos foram representados por áreas ocupadas principalmente por
espécies que crescem emaranhadas, formando extensos tapetes sobre os recifes.

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 35
Jun_
4PJu
l_5P
Jun_
3PJu
n_2P
Jun_
5PS
et_3
PJu
l_1P
Jul_
2PA
go_1
IJu
l_4I
Set
_5P
Ago
_1P
Ago
_5P
Jul_
3PM
ai_1
PA
go_2
PA
go_3
PS
et_1
PA
br_4
PA
br_5
PA
go_4
PJu
n_1P
Set
_2P
Jul_
4PJu
n_3I
Jun_
4IS
et_3
IJu
n_2I
Jul_
5IA
br_1
PA
br_2
PA
br_3
PS
et_4
PM
ai_5
PS
et_5
IS
et_2
IA
go_3
IJu
l_1I
Jul_
3IJu
n_1I
Ago
_4I
Abr
_5I
Ago
_5I
Mai
_3P
Abr
_4I
Jul_
2IJu
n_5I
Mai
_4P
Set
_1I
Mai
_3I
Set
_4I
Mai
_5I
Mai
_2I
Ago
_2I
Abr
_2I
Mai
_2P
Mai
_4I
Mai
_1I
Abr
_1I
Abr
_3I
100
80
60
40
20
0
Sim
ilari
ty
SIMILARIDADE modoQ
G1 G3 G2
Figura 14. Dendograma de similaridade qualitativa entre amostras (modo Q).
No G2b ocorreram amostras de dois tipos: amostras em que C. racemosa
esteve ausente e as amostras que tinham C. racemosa. No primeiro agrupamento
de G2b, em que C. racemosa esteve ausente, as amostras foram unidas pela
presença de Gelidiella acerosa, Lomentaria corallicola, Gelidiopsis planicaulis,
Pterocladiella caerulensces e suas epífitas, Ceramium dawsonii, C. vagans e
Neosiphonia gorgoniae. Este agrupamento apresentou 6 elementos amostrais,
sendo que dois são da área não impactada e os outros quatro da área impactada.
Os elementos da área não impactada deste agrupamento não apresentaram
espécies de Dictyotales e possivelmente por isso estiveram agrupados aos demais
elementos deste grupo. No segundo agrupamento gerado em G2b, todos os
elementos tiveram a presença de Caulerpa racemosa e foi composto por 11
amostras das quais apenas uma foi da área não impactada. Estes elementos

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 36
também tiveram em comum Dictypteris delicatula. Hypnea spinella, Jania
adhaerens, Gelidiopsis planicaulis, Gelidiella acerosa e Ceramium dawsonii. A
amostra da área não impactada presente neste agrupamento foi composta
também de Sargassum hystrix, Dictyota cervicornis e Lobophora variegata.
O elemento isolado ao centro do dendograma (G3) corresponde a uma
subamostra coletada no mês de setembro na área impactada, composta
exclusivamente de G. acerosa.
O primeiro grande grupo discutido (G1) foi formado sobretudo por
elementos nos quais espécies de Dictyotales estiveram presentes. Este grupo foi
representado principalmente por elementos da área não impactada, sendo que
todos os elementos da área impactada que se apresentaram neste agrupamento
tiveram a presença de espécies de Dictyotales (como Lobophora variegata,
Dictyota mertensii e D. cervicornis) que supostamente foram as variáveis decisivas
na geração do agrupamento. No segundo grupo (G2) ocorreram um número maior
de amostras da área impactada, as quais estiveram unidas pela presença de
associações de Gelidiopsis planicaulis com algas calcárias ou de Pterocladiella
caerulensces e ceramiales epífitas, além da presença de Caulerpa racemosa e de
Dictyopteris delicatula. O dendograma apresentou-se bastante fragmentado,
refletindo a heterogeneidade dos povoamentos encontrados nos recifes.

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 37
5. DISCUSSÃO
No complexo recifal de Maracajaú, as duas áreas escolhidas para a
realização deste estudo apresentaram diferenças na composição específica, na
freqüência de ocorrência das espécies, na biomassa e nos índices de diversidade
e de dominância obtidos. Outro resultado que mostrou diferenças entre os dois
locais foi o dendograma gerado a partir da análise de similaridade entre as duas
estações de amostragem. Este se apresentou dividido em dois grandes
agrupamentos. No primeiro grupo formado, o predomínio foi de amostras da área
não impactada, e no segundo grupo houve maior representatividade de amostras
da área impactada, indicando que estas áreas possuem características distintas
com relação a composição florística (Figura 15).
A área impactada mostrou-se dominada por populações de Caulerpa
racemosa, que muitas vezes ocorreram associadas a Dictyopteris delicatula.
Outros tipos de associações que foram muito comuns nesta estação de coleta
foram as de algas que formam tapetes. Estas espécies formadoras de tapetes que
cobriram extensas áreas na estação impactada, apesar de não terem apresentado
biomassa expressiva, mostraram-se bastante freqüentes durante todo o período
de estudo, mostrando assim sua importância (Figura 15b). Foram verificados três
tipos de associações, categorizados de acordo com a composição biológica. A
primeira associação foi composta por algas calcárias articuladas (Jania
adhaerens, Amphiroa beauvoisii, Amphiroa anastomosans) formando extensos
bancos junto com algas corticadas (Hypnea spinella, Laurencia intricata,
Gelidiopsis planicaulis, Lomentaria corallicola e Chondria dangaerdii) e
filamentosas (Ceramium dawsonii, C. flaccidum, Neosiphonia gorgoniae). O
segundo tipo foi formado por Pterocladiella caerulensces, G. planicaulis e suas
epífitas (principalmente Ceramium dawsonii, C. vagans e Neosiphonia gorgoniae).
E o terceiro tipo foi composto de Gelidiella ligulata e epífitas (principalmente C.
dawsonii). Estas associações, em geral, envolveram algas que se adaptam

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 38
facilmente a ambientes sujeitos a freqüentes perturbações (Kendrick, 1991;
Semple, 1997; Soltan et al. 2001).
Na área impactada freqüentemente ocorre a ressuspensão dos sedimentos
de fundo, causada pela movimentação das embarcações e pelo mau uso das
nadadeiras por parte de muitos turistas. Estes fatores contribuem para a remoção
da cobertura biológica em certos locais e favorecem a colonização dos espaços
desocupados por estas associações de algas. Outras espécies que se mostraram
expressivas na área em questão em determinados períodos do estudo foram
Sargassum vulgare no primeiro mês, e Laurencia filiformis nos dois últimos meses
amostrados. As outras espécies que ocorreram nas amostragens desta área foram
muito pouco expressivas em termos de biomassa, com relação à área não
impactada. As famílias mais representativas nesta área (impactada) foram
Rhodomelaceae, Dictyotaceae e Corallinaceae, sendo que nesta área os
indivíduos representantes de Dictyotaceae não apresentaram biomassa
expressiva, exceto Dictyopteris delicatula, que crescia freqüentemente associada
a Caulerpa racemosa, e Dictyota cervicornis e D. ciliolata, que estiveram
presentes apenas no mês de setembro. Assim, o domínio nesta área foi da alga
verde Caulerpa racemosa.
Caulerpa racemosa é uma espécie de alga verde amplamente distribuída
nos mares tropicais e foi dominante em todas as amostragens realizadas na
estação impactada. Esta espécie é reconhecida por sua habilidade de invadir
áreas e por sua habilidade em competir com outras espécies (Piazzi et al. 2001;
Verlaque et al. 2004). No Mar Mediterrâneo, a invasão de Caulerpa racemosa em
certas áreas diminuiu a riqueza, a diversidade e o percentual de cobertura de
espécies nativas (Piazzi et al, 2001). O rápido crescimento dos estolões possibilita
aos indivíduos uma rápida e ampla disseminação, facilitando o crescimento desta
espécie sobre outras espécies de macroalgas (Piazzi et al. 1999).
Na área não impactada, Sargassum hystrix foi a espécie mais
representativa, seguida de Caulerpa racemosa, Cryptonemia crenulata,
Dictyopteris justii, Lobophora variegata, Dictyopteris delicatula e Gelidium
coarctatum (Figura 15a). Os extensos bancos de associações em tapetes

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 39
observados na área impactada não foram tão expressivos nesta área, além de
apresentarem duas espécies de Jania que não foram observadas na área
impactada.
Os efetivos das espécies que compunham as amostragens da área
impactada (onde C. racemosa apresentou grande representatividade em todos os
períodos do trabalho) eram muito distantes, fazendo com que a dominância nesta
área fosse maior e a diversidade menor que na área não impactada. Na área não
impactada, as espécies apresentaram efetivos mais próximos, configurando um
cenário de baixa dominância por parte de poucas espécies, como foi o caso da
área impactada.
A riqueza específica esteve maior na área não impactada somente nos
meses de maio e agosto. No entanto, os índices de diversidade sempre foram
maiores nesta estação, mesmo em abril quando somente 12 espécies foram
encontradas. Isso aconteceu porque as espécies mais freqüentes na área
impactada foram as que ocorreram associadas formando tapetes de algas,
representadas por indivíduos de pequeno porte e pouca biomassa, e estas
associações foram, conforme mencionado, bastante expressivas. Os resultados
sugerem que a riqueza da área impactada se deve às espécies associadas que
formaram extensos tapetes nesta área e às suas epífitas.
A biomassa fresca apresentou valores muito próximos para as duas áreas
estudadas, e esta proximidade dos valores obtidos se deve a grande quantidade
de água nos tecidos de Caulerpa racemosa, espécie predominante na área
impactada. Quando observamos os valores obtidos após a secagem do material, a
diferença entre as duas áreas se torna significante pois a maior parte do material
amostrado na área não impactada era de macroalgas pardas, que apresentam
menor proporção de água nos tecidos quando comparadas a C. racemosa.
Se por um lado a área impactada apresentou uma flora dominada por
Caulerpa racemosa e por extensas coberturas de tapetes de algas, a flora da área
não impactada apresentou maior representatividade de Sargassum hystrix,
Dictyopteris delicatula, Dictyota mertensii e Lobophora variegata. Estudos
anteriores descreveram comunidades recifais onde este tipo de alga parda se

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 40
torna dominante, ocupando a posição de domínio que os corais exerciam nestas
comunidades (Hughes, 1994; McClanahan et al, 1999; Szmant, 2001). As
macroalgas se tornam, em geral, abundantes em comunidades recifais sob
condições de redução nas populações de herbívoros (Scheibling, 1986; Hughes,
1994; Shulman & Robertson, 1996; McCook, 1999; Lirman, 2001; McCooK et al,
2001), ou em ambientes eutrofizados (Semple, 1997; Smith et al, 2001).
Smith et al (2001) constataram em experimentos que locais onde houve a
redução de herbívoros as macroalgas eretas (fleshy) apresentaram-se mais
abundantes e onde houve aumento de nutrientes as populações de algas calcárias
aumentaram. Este estudo mostrou que a ação de herbívoros exerce controle
sobre as populações de formas eretas de macroalgas, pois onde há
enriquecimento nutricional e herbívoros presentes as macroalgas não
apresentaram alterações nos padrões de crescimento populacional, ao passo que
onde os herbívoros estiveram ausentes, as macroalgas aumentaram a taxa de
crescimento, passando a competir com as algas calcárias e corais. Em condições
de redução na herbivoria, macroalgas podem aumentar sua densidade
populacional e assim, algumas espécies chegam a crescer sobre os tecidos de
corais, causando-lhes a morte (Jompa & McCook, 2000; McCook et al, 2001).
Desta forma, as relações populacionais entre algas e corais assumem importância
crucial no monitoramento de áreas recifais expostas a este tipo de ameaça
(Jompa & McCook, 2000; McCook et al, 2000, 2001). O aumento na densidade de
certas macroalgas formadoras de um estrato superior entre organismos bênticos
em recifes pode ainda inibir o assentamento de larvas de corais (Maypa et al,
2000).

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 41
a
b
Figura 15. Aspecto geral da flora encontrada na área não impactada (a) e na área
impactada (b). (Note a quantidade de C. racemosa na área impactada; a seta indica os tapetes de
algas)

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 42
6. CONCLUSÃO
Os resultados mostraram que as comunidades de macroalgas observadas
nas duas áreas amostradas são diferentes. Estas diferenças foram evidenciadas
pela composição biológica, pela biomassa e pelos índices ecológicos aplicados
neste estudo. Na área não impactada foi possível observar o predomínio de
espécies de algas pardas (Fucales e Dictyotales). Os outros grupos de algas que
ocorreram na flora deste local apresentaram valores de biomassa que não foram
muito menores que os valores obtidos para as espécies dominantes (Sargassum
hystrix e Dictyopteris delicatula), tornando esta área mais diversificada em relação
à outra área de estudo. Por outro lado, na área impactada o domínio de Caulerpa
racemosa sobre as outras espécies da comunidade foi muito maior. Nesta área, as
macroalgas se apresentaram pouco desenvolvidas, com exceção de C. racemosa
com o predomínio foi de espécies de crescimento rápido, como Hypnea spinella,
Lomentaria corallicola, Ceramiales e coralináceas articuladas. A cobertura vegetal
apresenta sinais de constantes renovações e Caulerpa racemosa pode assumir
um comportamento invasor, se aproveitando dos espaços abertos no substrato
recifal, crescendo por meio de seus estolões e tomando conta de grandes
extensões.
De fato, se a área impactada não estivesse sendo influenciada pelo turismo,
que foi o único fator operando de maneira diferente nas duas áreas, possivelmente
esta área apresentaria uma flora mais desenvolvida e diversificada, semelhante a
encontrada na área de estudo não impactada. Assim, a intensa pressão turística
sobre a Área de Preservação Ambiental dos Recifes de Corais pode ser um fator
que está influenciando a comunidade de forma negativa, uma vez que ficou
constatada a redução de diversidade de macroalgas e o aumento da dominância
por parte de C. racemosa. As conseqüências das intensificações nas atividades
turísticas podem abranger a redução da diversidade recifal como um todo.
Vale salientar, no entanto, que embora foram observados sinais de
degradação, várias espécies foram registradas para o local, das quais 10 foram

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 43
citadas pela primeira vez para a costa do Rio Grande do Norte e 3 para o litoral
nordestino. Isso nos mostra a necessidade da realização de novos estudos com a
finalidade de conhecer a diversidade da flora regional.

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 44
7. REFERÊNCIAS BIBLIOGRÁFICAS
Alencar, J.D. & Durairatnam, M. 1993. Estudos sobre liberação de esporos em Gracilaria sjoestedtii (Kylin) Dawson (Rhodophyta-Gigartinales). Resumos da XVII Reunião Nordestina de Botânica, Teresina, Soc. Bras. de Botânica do Brasil. p.115.
Amaral, R.F., Feitoza, B. M., Attayde, J. L., Batista, D. S., Mendes, L. F., Silva, I. B., Soriano, E. M., 2005. Diagnóstico Ambiental Preliminar da Área de Uso Turístico Intensivo (AUTI) no Parracho de Maracajaú. IDEMA-RN. Relatório Interno. 128p.
Arantes, S. V. 2004. Descrição do recobrimento e ocorrência de macroinvertebrados bentônicos nos Parrachos de Maracajaú, município de Maxaranguape – RN. Monografia de Bacharel em Ciências Biológicas. UFRN. 74p.
Araújo, M.S.V. 1983. Clorofíceas e Feofíceas do Rio Grande do Norte. Dissertação de Mestrado. Recife. UFRPE.
Bacci, D. 2005. Estudos taxonômicos do gênero Chondria (Ceramiales, Rhodophyta) no litoral dos estados de São Paulo e Espírito Santo, Brasil. Instituto de Botânica do Estado de São Paulo. Dissertação de mestrado. 114p.
Barata, D. 2004. Clorofíceas marinhas do Estado do Espírito Santo. Dissertação de mestrado. Instituto do Botânica do Estado de São Paulo.
Barreto, M.B.B. & Yoneshigue-Valentin, Y. 2001. Aspectos morfológicos do gênero
Ceramium Roth (Ceramiaceae – Rhodophyta) no Estado do Rio de Janeiro. Instituto
de Botânica de São Paulo. Hoenea, 28(1): 77-110.
Bonilha, L.E.C. 2003. Considerações quanto ao uso atual, conservação e situação sócio-ambiental no ecossistema recifal/coralino dos “parrachos” do Rio Grande do Norte - (Municípios de Maxaranguape a Rio do Fogo). Relatório GEREX - IBAMA / RN. 19p.
Buddemeier, 1998. Coral reefs and global change: adaptation, acclimation, or extinction? Environmental Conservation 25 (2): 172-174.
Câmara Neto, C. 1966. Algumas algas de provável aproveitameno industrial para o Rio Grande do Norte. Bolm. Inst. Biol. Mar. Univ. Fed. RN 3:53.
Câmara Neto, C. 1971. Primeira contribuição ao inventário das algas marinhas bentônicas do litoral do Rio Grande do Norte. Separata do Bolm. Do Inst. De Biol. Mar. Da UFRN. Natal 5:137-154.
Castro, C.B. 2002. Recifes de corais. Relatório do workshop para Avaliação e Ações prioritárias para conservação da biodiversidade das Zonas Costeira e Marinha. Porto Seguro. Fund. Bio-Rio; SECTAM; IDEMA; Soc. Nordestina de Ecologia; Sec. Meio Ambiente do Est. de SP.; FEPAM. Ministério do Meio Ambiente.

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 45
Cesar, H.S.J. 2000. the economic value of the invaluable Indonesian Coral Reefs. Proc. 9th Int. Coral Reef Sym. p 185.
Creed, J. C. & Amado Filho, G. M. 1999. Disturbance and recovery of the macroflora of a seagrass (Halodule wrightii Ascherson) meadow in the Abrolhos Marine National Park, Brazil: an experimental evaluation of anchor damage. J. Exp. Mar. Biol. Ecol. 235: 285-306.
Coimbra, C.S. 1998. Caracterização ecológica das comunidades bênticas da região entre-marés no costão da Praia do Itaguá, Ubatuba, São Paulo. Dissertação (mestrado). Inst. Biociências da USP. São Paulo.
Connell, J.H. 1978. Diversity in tropical rainforests and coral reefs. Science 199: 1302-1309.
Cordeiro-Marino, M. 1978. Rodofíceas Bentônicas Marinhas do Estado de Santa Catarina. Rickia – Arquivos de Botânica do Estado de São Paulo, 7:243.
Coutinho, R.; R.C. Villaça; C.A. Magalhães; M.A. Guimaraens; M. Apolinário & G. Muricy 1993. Influência antrópica nos ecossistemas coralinos da região de Abrolhos, Bahia, Brasil. Acta Biol. Leopoldensia, 15(1): 133-144.
Davis, D. & Tisdell, C. 1996. Economic managment of recreational scuba diving and the environmemnt. Journal of Environment Managment 48: 229-248.
Durairatnam, M. 1980. Studies on the agar producing seaweeds and their distribution in Northeast Brasil. Ciência e Culrura, 32 (10): 1358-1372.
Durairatnam, M. 1989. Explotation and managment of seaweed resources in Northeast Brazil. J. Mar. Biol. Ass. Índia, 31 (1&2): 117-121.
Duraraitnam, M. Studies on the preparation of Carragenan from Gigartina insignis J. Agardh. Resumos da VI Reunião Brasileira de Ficologia, Tramandaí, RS. p. 159.
Durairatnam, M. & Nascimento, H.C. 1983. Algas marinhas bentônicas da Praia de Maracajaú (Estado do Rio Grande do Norte). Resumos do XXXIV Congres. Nac. de Botânica. Porto Alegre.
Duraraitnam, M., Nascimento, H.C., Silva, E.M. 1986. algas marinhas ocorrentes na praia do Pipa e Baía Formosa no Estado do Rio Grande do Norte. Resumos do XXXVII Congr. Nac. Bot. Ouro Preto. Soc. Bras. Bot. p. 128.
Ferreira, B.P. & Maida, M. 2000. Relationship between artisanal fisheries and tourism in brazilian coastal reefs. Proc. 9th Coral Reef Sym. p.185.
Ferreira- Correia, M.M. & Pinheiro-Vieira, F. 1969. Estudos taxonômicos sobre o gênero Caulerpa Lamouroux no Nordeste Brasileiro (Chlorophyta: Caulerpaceae. Arq. Ciên. Mar. 9(2): 147-161.
Figueiredo, M. A. de O. 2000. Recifes de corais ou recifes de algas? Ciência Hoje 166 (28): 74-75.

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 46
Fujii, M.T. & Santíes, A.G. 2005. Taxonomia do Complexo Laurencia (Rhodomelaceae – Rhodophyta) do Brasil, com ênfase nas espécies dos estados de São Paulo e do Espírito Santo. Monografías ficológicas (2): 69-135.
Guimarães, S.M.P.B. 1990. Rodofíceas marinhas bentônicas do Estado do Espírito Santo: Ordem Cryptonemiales. Tese de Doutorado. Instituto de Biociências da Univ. São Paulo. 275pp.
Guimarães, S.M.P.B. & Oliveira, E.C. 1996. Taxonomy of flattened Solieriaceae (Rhodophyta) in Brazil: Agardhiella and Meristiella. J. Phycol. 32: 656-668.
Guimarães, S.M.P.B.; Fujii, M.T., Pupo, D. & Yokoya, N.S. 2004. Reavaliação das caracteristicas morfológicas e suas implicações taxonômicas no gênero Polysiphonia sensu lato (Ceramiales, Rhodophyta) do litoral dos Estados de São Paulo e Espírito Santo, Brasil. Rev. Bras. Bot. 27: 163-183.
Hawkins, J.P. & C.P. Roberts 1992. Effects of recreational SCUBA diving on fore-reef slope communities of coral reefs. Biological Conservation 26: 171-178.
Hill, M.O. 1973. Diversity and eveness; an unifying notation and its consequences. Ecology, 54 (2): 427-432.
Hubbard, D. K. 1997. Reefs as dynamic systems. In: Birkeland, C. (ed.). Life and death of coral reefs. New, york, Chapman & Hall. Pp. 43-67.
Hughes, T.P. 1994. Catastrophes, phase shifts, and large-scale degradation of a Caribbean Coral Reef. Science 265: 1547-1551.
Horta, P.A. 2000. Macroalgas do infralitoral do sul e sudeste do Brasil: Taxonomia e Biogeografia. Tese de doutorado, Universidade de São Paulo, São Paulo, 301 p.
Joly, A.B. 1957. Contribuição ao conhecimento da flora ficológica marinha da Baía de Santos e arredores. Bolm. Fac. Filos. Ciências Letrs., Univ. S. Paulo. Botânica, (14) 393 p.
Joly, A.B.; Ugadim, Y; Diaz, G.J.J.; Yamaguishi-Tomita, N; Cordeiro-Marino, M. & Yoneshigue-Braga, Y. 1969. Additions to the marine flora of Brazil – X. Rickia 4: 61-74.
Jompa, J. & McCook, L.J. 2000. Variable effects of macroalgae on hard corals. Proc. 9th
Int. Coral Reef Sym. p. 123.
Kendrick, G.A. 1991. Recruitment of coralline crusts and filamentous turf algae in the Galapagos archipelago: effect of simulated scour, erosion and accretion. J. Exp. Mar. Biol. Ecol. 147: 47-63.
Leão, Z.M.A.N. 1996. The coral reefs of Bahia: morphology, distribution and the major environmental impacts. An. Acad. bras. Ci. 68 (3): 339-452.
Lirman, D. 2001. Competition between macroalgae and corals: efects of herbivore exclusion and increased algal biomass on coral survivorship and growth. Coral Reefs 19: 392-399.

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 47
Marcelino, A. & Amaral, R. 2003. Área de proteção ambiental estadual dos Recifes de Corais. In: Prates, A.P.L. (ed.). Atlas dos recifes de corais nas Unidades de Conservação Brasileiras. Brasília: MMA/SBF. 180p.
Marinho-Soriano, E. 1999. Species composition of seaweeds in Búzios Beach, Rio Grande do Norte, Brazil. Seaweed Res. Utiln, 21(1 & 2): 9-13.
Marinho-Soriano, E.; Silva, T.S.F.; Moreira, W.S.C. 2001. Seasonal variation in the biomass and agar yield from Gracilaria cervicornis and Hydropuntia cornea from Brazil. Bioresource Technology 77: 115-120.
Maypa, A.P.; Raymundo, L.J.H.; Calumpong, H.P. & de Leon, R.O. 2000. Algae-coral interactions: mediation of coral settlement, early survival and growth by macroalgae in central Philippines. Proc. 9th Int. Coral Reef Sym. p. 124
McCook, L.J. 1999. Macroalgae, nutrients and phase shifts on coral reefs: scientific issues and management consequences for the Great Barrier Reef. Coral Reefs 18 : 357-367.
McCook, L.J.; Jompa, J.; Diaz-Pulido, G.; Cumming, R.; Mundy, C. & Price, I. 2000. Competition between coral and benthic algae on inshore coral reefs of the Great Barrier Reef. Proc. 9th Int. Coral Reef Sym. p. 124.
McCook, L.J.; Jompa, J. & Diaz-Pulido, G. 2001. Competition between coral and algae: a rewiew of evidence and mechanisms. Coral Reefs 19: 400-417.
McClanahan,T.R.; Aronson, R.B.; Precht, W.F. & Muthiga, N.A. 1999. Fleshy algae dominate remote coral reefs of Belize. Coral Reefs 18: 61-62.
McClanahan, T. & Muthiga, N. 1998. An ecological shift in a remote coral atoll of Belize over 25 years. Environmental Conservation 25(2): 122-130.
Moura, C.W.N. 1992. Coralináceas com genículo (Corallinales, Rhodophyta) do litoral oriental do estado do Rio Grande do Norte – Brasil. Dessertação (mestrado). Univ. Fed. Rural de Pernambuco. Recife. 254 p
Moura, C.W.N. 2000. Coralináceas com genículo do litoral do Brasil. Tese de Doutorado. Instituto de Bociências da Univ. São Paulo.
Moura, C.W.N. & Araújo, M.S.V.B. 1990. Adições ao inventário das algas marinhas bentônicas do Estado do Rio Grande do Norte (1). Resumos da XIV Reunião Nordestina de Botânica, Recife. p. 31.
Nordemar, I. 2000. Degradation of fringing coral reefs in the Gulf of Thailand. 9th Int. Coral Reef Sym. p. 371.
Odum, E.P. Ecologia. Ed. Guanabara. Rio de Janeiro. 434pp.
Oliveira, E.C. 1996. Is there a relation among the global warming, the missing carbon and the calcareous algae? An. Acad. bras.Ci. 68 (1): 17-21

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 48
Oliveira Filho E.C. 1969. Ceramiales do sul do Estado do Espírito Santo (Brasil). Boletim da Faculdade de Filosofia, Ciências e Letras da Universidade de São Paulo, série Botânica 26: 1-277.
Oliveira Filho, E.C. 1977. Algas marinhas bentônicas do Brasil. (tese) Universidade de São Paulo. 407 p.
Paes e Mello, L.B. 1983. Estudos taxonômicos sobre a Família Rhodomelaceae (Rhodophyta – Ceramiales) no litoral oriental do Estado do Rio Grande do Norte. Dissertação de Mestrado. Recife. UFRPE.
Pastorok, R.A. & Bilyard, G.R. 1985. Effect of sewage pollution on coral-reef communities. Mar. Ecol. Prog. Ser. 21: 175-189.
Piazzi, L., Pardi, G. & Cinelli, F. 1999. Algal vertical zonation and seazonal dynamic along a subtidal cliff on Gorgonia Island (Tuscan Archipelago, Italy). Plant Biosyst 133 (1):3-13.
Piazzi, L.; Ceccherelli, G. & Cinelli, F. 2001. Threat to macroalgal diversity: effects of the introduced green alga Caulerpa racemosa in the Mediterranean. Mar. Ecol. Prog. Ser. 210: 149-159.
Pinheiro-Vieira, F. & Ferreira-Correia, M.M., 1970b. Quarta contribuição ao inventário das algas marinha bentônicas do nordeste brasileiro. Arq. Cienc. Mar. Fortaleza 6(2): 131-134.
Pollnac, R.B. 2000. Assessment of socioeconomic factors influencing the success of marine sactuaries in the Philippines. Proc. 9th Int. Coral Reef Sym. p. 187.
Rosso, S. 1990. Estrutura de comunidades intermareais de substrato consolidado das proximidades da baía de Santos (SP, Brasil): uma abordagem descritiva enfatizando aspectos metodológicos. 216pp Tese (doutorado). Universidade de São Paulo, São Paulo.
Santelices, B. 1978. The morphological variation of Pterocladia caerulesces (Gelidiales - Rhodophyta) in Hawaii. Phycologia 17: 53-59.
Scheffer, M.; Carpenter, S.; Foley, J.A.; Folkeß, C. & Walkerk, B. 2001. Catastrophic shifts in ecosystems. Nature 413: 591-596.
Scheibling, R. 1986. Increased macroalgal abundance following mass mortalties of sea urchins (Strongylocentrotus droebachiensis) along the Atlantic coast of Nova Scotia. Oecologia 68:186-189
Schneider, C.W. 1983. The red algal genus Audouinella Bory (Nemaniales-Acrochaetiaceae) from North Carolina. Smithsonian Contr. Mar. Sci. 22 III 25pp.
Semple, S., 1997. Algal growth on two sections of a fringing coral reef subject to different levels of eutrophication in Reunion Island. Oceanologica Acta 20(6): 851-861.
Shannon, C.E. & Weaver, W. 1949. The mathematical theory of communication. Urbana, The University of Illinois Press. 125p.

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 49
Shulman, M.J. & Robertson, D.R. 1996. Changes in the coral reefs of San Blas, Caribean Panama: 1983 to 1990. Coral Reefs 15: 231-236.
Simpson, E.H. 1949. Measurement of diversity. Nature 163: 688.
Smith, J.E.; Smith, C. & Hunter, C. 2001. An exprimental analysis of the effects of herbivory and nutrient enrichment on benthic community dynamics on a Hawaiian reef. Coral Reefs (19): 332-342.
Soltan, D.; Verlaque, M.; Boudouresque, C.F. & Francour, P. 2001. Changes in macroalgal commuties in the vicinity of a Mediterranean sewage outfall after the setting up of a treatment plant. Mar. Poll. Bull. 42 (1): 59-70.
Szmant, A.M. 2001. Why are coral reefs world-wide becoming overgrown by algae? ‘Algae, algae everywhere, and nowhere a bite to eat!’ Coral Reefs 19: 199-302.
Taylor, W.R. 1960. Marine algae of the eastern tropical and subtropical coasts of the America. University of Michigan Press, Ann Arbor, 870 p.
Ugadim, Y. 1974. Algas marinhas bentônicas do Litoral Sul do Estado de São Paulo e Litoral do Estado do Paraná. III. Divisão Rhodophyta (1): Goniotrichales, Bangiales, Nemalionales e Gelidiales. Bol. Bot. Univ. São Paulo. 2: 93-137.
Verlaque, M.; Durand, C.; Huisman, J.M.; Boudouresque, C.F. & Le Parco, Y. 2003. On the identity and origin of the Mediterranean invasive Caulerpa racemosa (Caulerpales, Chlorophyta). Eur. J. Phycol. 38: 325-339.
Walker, D.I. & G.A. Kendrick 1998. Threats to macroalgal diversity: Marine habitat destruction and fragmentation, pollution and introduced species. Bot. Mar., 41: 105-112.
Williams, S.L.; Breda, V.A.; Anderson, T.W. & Nyden, B.B. 1985. Growth and sediment disturbance of Caulerpa spp. (Chlorophyta) in a submarine canyon. Mar. Ecol. Prog. Ser. 21: 275-281.
Wynne, M.J. 1998. A checklist of benthic marine algae of the tropical and subtropical western Atlantic: first revision. Nova Hedwigia, 116: 1-155.
Yoneshigue Y. 1985. Taxonomie et ecologie des algues marines dans la region de Cabo Frio (Rio de Janeiro, Brésil) These de Docteur d'Etat- Sciences, Faculté des Sciences de Luminy, Univerisité d'' Áix-Marseille II, France, 466 p.

Anexos
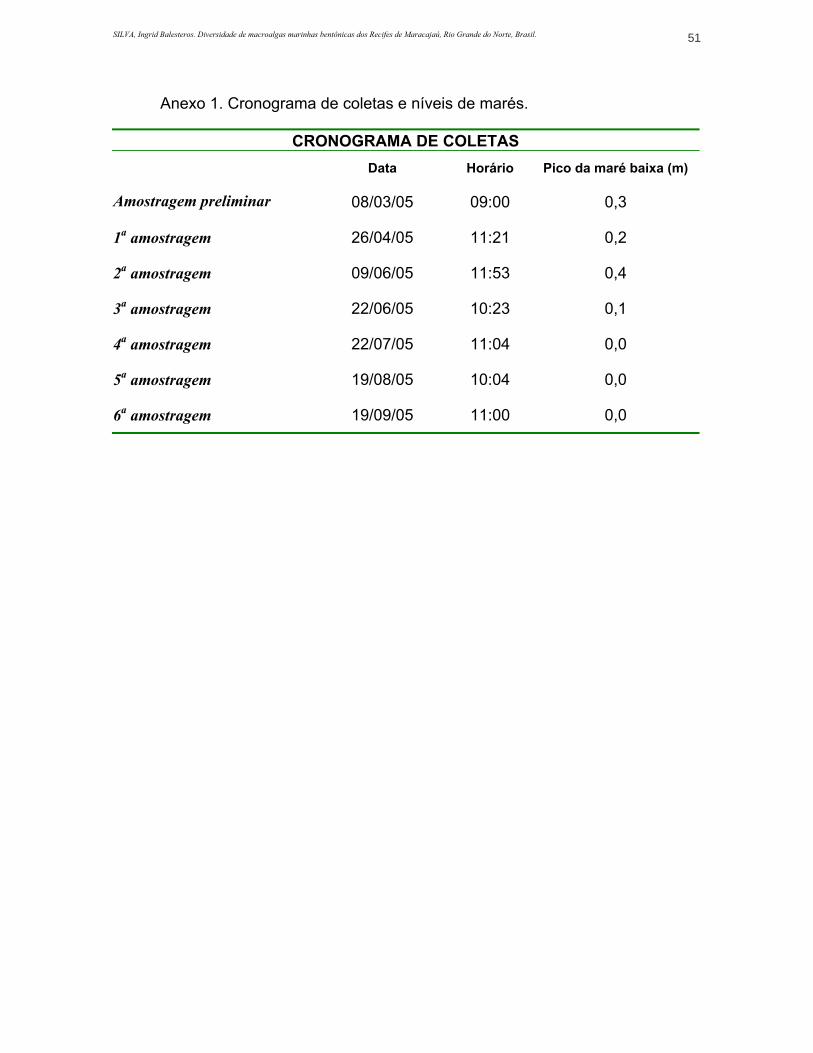
SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 51
Anexo 1. Cronograma de coletas e níveis de marés.
CRONOGRAMA DE COLETAS
Data Horário Pico da maré baixa (m)
Amostragem preliminar 08/03/05 09:00 0,3
1a amostragem 26/04/05 11:21 0,2
2a amostragem 09/06/05 11:53 0,4
3a amostragem 22/06/05 10:23 0,1
4a amostragem 22/07/05 11:04 0,0
5a amostragem 19/08/05 10:04 0,0
6a amostragem 19/09/05 11:00 0,0
SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 52
Anexo 2. Sinopse dos táxons encontrados.
CHLOROPHYTA Ulvophyceae
Ulvales
Ulvaceae
Enteromorpha sp
Cladophorales
Siphonocladaceae
Ventricaria ventricosa (J. Agardh) J.L. Olsen & J.A. West
Bryopsidales
Bryopsidaceae
Bryopsis plumosa (Hudson) C. Agardh
Caulerpaceae
Caulerpa cupressoides (H. West in Vahl) C. Agardh
Caulerpa mexicana Sond. ex Kütz.
Caulerpa racemosa (Forsskål) J. Agardh
Caulerpella ambigua (Okamura) Prud´homne & Lokhorst*
PHAEOPHYTA
Phaeophyceae
Dictyotales
Dictyotaceae
Dictyopteris delicatula J.L. Lamour.
Dictyopteris justii J.L. Lamour.
Dictyopteris polypodioides (DC in Lam & DC) Lamour.*
Dictyota cervicornis Kütz.
Dictyota ciliolata Sond. ex. Kütz.
Dictyota menstrualis (Hoyt) Schnetter, Hörnig & Weber-Bukert
Dictyota mertensii (Mart.) Kütz.
Lobophora variegata (J.V. Lamour.) Womersley ex. E.C. Oliveira
Padina sactae-crucis Børgesen*
Spatoglossum schoerederi (C. Agardh) Kützing
Fucales
Sargassaceae
Sargassum filipendula C. Agardh
Sargassum hystrix J. Agardh
Sargassum vulgare C. Agardh
RHODOPHYTA
Florideophycidae
Acrochaetiales
Acrochaetiaceae
Acrochaetium flexuosum Vickers
Corallinales
Corallinaceae
Amphiroa anastomosans Weber Bosse
Amphiroa beauvoisii J.V. Lamour.
Haliptilon subulatum (J. Ellis & Sol.) H.W. Johans.
Jania adhaerens J.V. Lamour.
Jania pumila J.V. Lamour.* Jania sp 1
Jania sp 2
Gelidiales
Gelidiaceae

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 53
Gelidium coarctatum Kütz.
Pterocladiella caerulensces (Kütz.) Santel & Hommers.
Gelidiellaceae
Gelidiella acerosa (Forssk) Feldman & Hamel
Gelidiella ligulata Dawson*Bonnemaisoniales
Bonnemaisoniaceae
Asparagopsis taxiformis (Delile) Trevis [fase esporofítica]
Gigartinales
Gigartinaceae
Chondracanthus acicularis (Roth) Frederich
Hypneaceae
Hypnea musciformis (Wulfen in Jacqu.) J.V. Lamour.
Hypnea spinella (C. Agardh) Kützing
Solieriaceae
Meristiella gelidium (J. Agardh) D.P. Cheney & P.W. Gabrielson
Halymeniales
Halymeniaceae
Cryptonemia bengryi W.R. Taylor
Cryptonemia crenulata (J. Agardh) J. Agardh
Gracilariales
Gracilariaceae
Gracilaria cervicornis (Turner) J. Agardh
Rhodymeniales
Lomentariaceae
Lomentaria corallicola Børgesen**
Rhodymeniaceae
Botryocladia occidentalis (Børgesen) Kylin
Botryocladia pyriformis (Børgesen) Kylin
Gelidiopsis planicaulis (W.R. Taylor) W.R. Taylor
Gelidiopsis variabilis (Grev. ex J. Agardh) F. Schmitz
Ceramiales
Ceramiaceae
Antithamnionella graeffei (Grunow) Athanas.*
Ceramium dawsonii A.B. Joly
Ceramium flaccidum (Kütz.) Ardiss.
Ceramium vagans P.C. Silva*
Rhodomelaceae
Bryothamniom triquetrum (S.G. Gmel.) M. Howe
Chondria dangaerdii (J. Agardh) Cheney & Gabrielson**
Dipterosiphonia dentritica (C. Agardh) F. Schmitz in Engler & Prantl*
Herposiphonia bipinnata M. Howe* Herposiphonia secunda f tenella (C. Agardh) M.J. Wynne*
Laurencia filiformis (C. Agardh) Mont.
Laurencia intricita J.V. Lamour.
Neosiphonia gorgoniae (Harvey) S.M Guim. & M.T. Fujii**
Polysiphonia subtilissima Mont.*
* Primeira referência para o estado.
** Primeira referência para o Nordeste
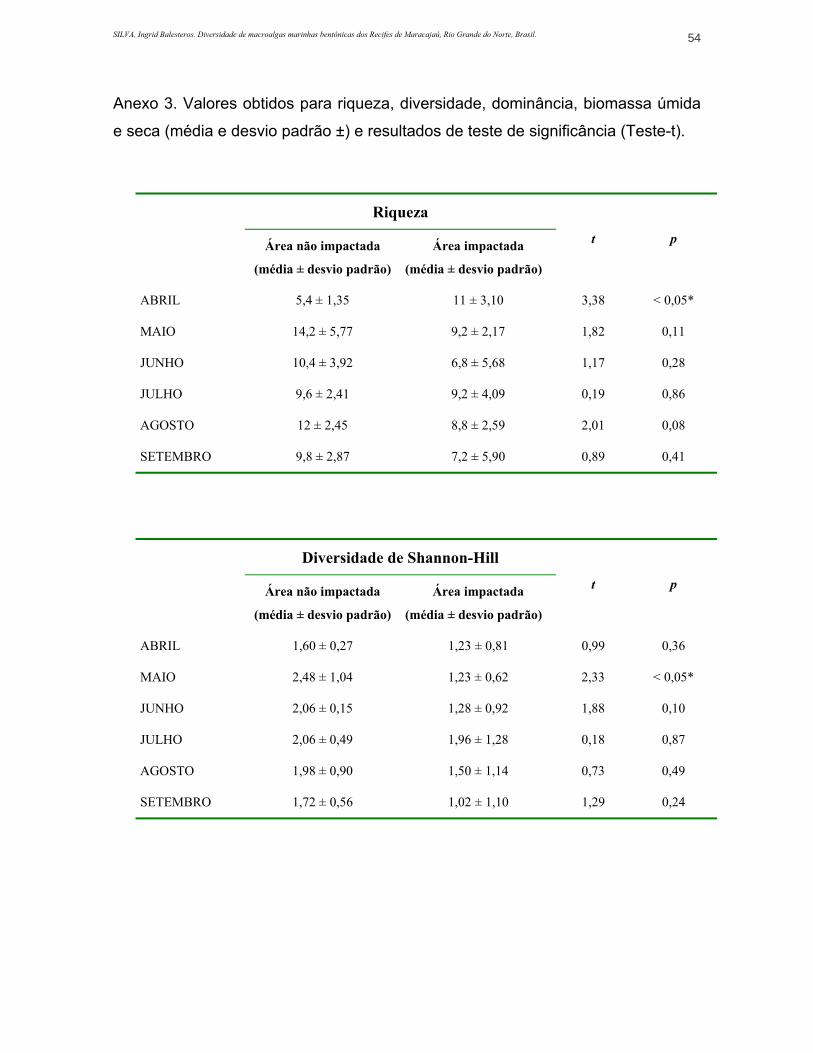
SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 54
Anexo 3. Valores obtidos para riqueza, diversidade, dominância, biomassa úmida
e seca (média e desvio padrão ±) e resultados de teste de significância (Teste-t).
Riqueza
Área não impactada
(média ± desvio padrão)
Área impactada
(média ± desvio padrão)
t p
ABRIL 5,4 ± 1,35 11 ± 3,10 3,38 < 0,05*
MAIO 14,2 ± 5,77 9,2 ± 2,17 1,82 0,11
JUNHO 10,4 ± 3,92 6,8 ± 5,68 1,17 0,28
JULHO 9,6 ± 2,41 9,2 ± 4,09 0,19 0,86
AGOSTO 12 ± 2,45 8,8 ± 2,59 2,01 0,08
SETEMBRO 9,8 ± 2,87 7,2 ± 5,90 0,89 0,41
Diversidade de Shannon-Hill
Área não impactada
(média ± desvio padrão)
Área impactada
(média ± desvio padrão)
t p
ABRIL 1,60 ± 0,27 1,23 ± 0,81 0,99 0,36
MAIO 2,48 ± 1,04 1,23 ± 0,62 2,33 < 0,05*
JUNHO 2,06 ± 0,15 1,28 ± 0,92 1,88 0,10
JULHO 2,06 ± 0,49 1,96 ± 1,28 0,18 0,87
AGOSTO 1,98 ± 0,90 1,50 ± 1,14 0,73 0,49
SETEMBRO 1,72 ± 0,56 1,02 ± 1,10 1,29 0,24
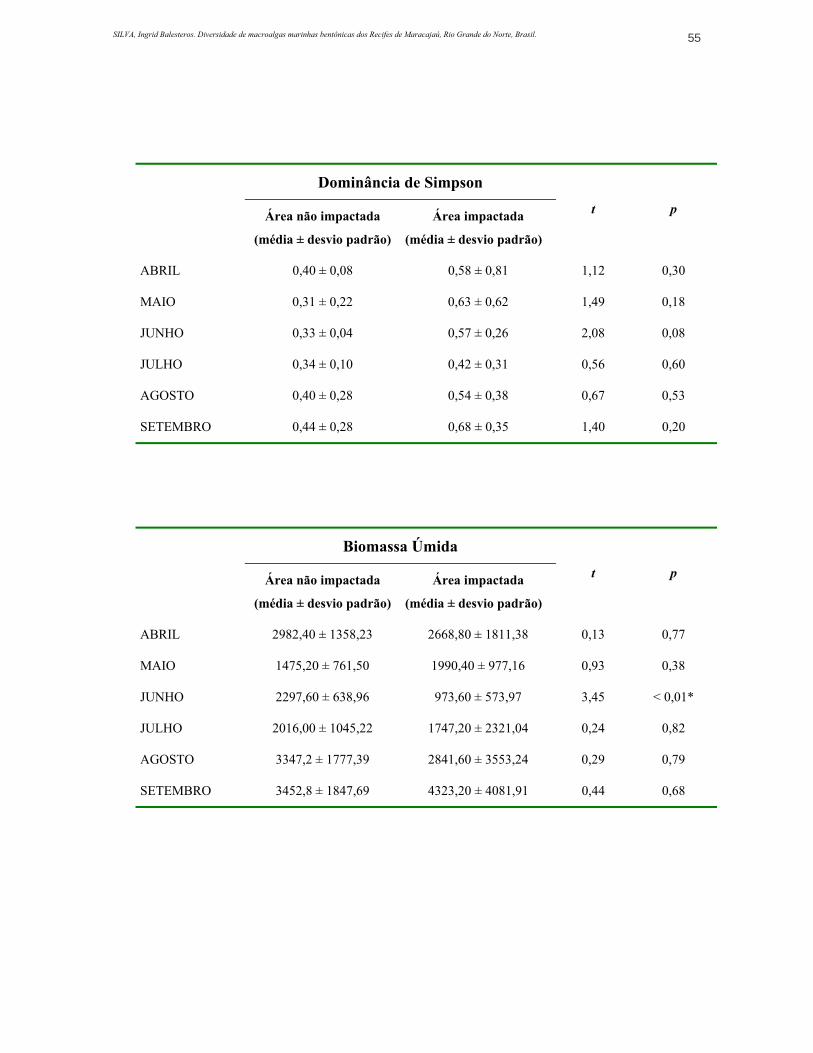
SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 55
Dominância de Simpson
Área não impactada
(média ± desvio padrão)
Área impactada
(média ± desvio padrão)
t p
ABRIL 0,40 ± 0,08 0,58 ± 0,81 1,12 0,30
MAIO 0,31 ± 0,22 0,63 ± 0,62 1,49 0,18
JUNHO 0,33 ± 0,04 0,57 ± 0,26 2,08 0,08
JULHO 0,34 ± 0,10 0,42 ± 0,31 0,56 0,60
AGOSTO 0,40 ± 0,28 0,54 ± 0,38 0,67 0,53
SETEMBRO 0,44 ± 0,28 0,68 ± 0,35 1,40 0,20
Biomassa Úmida
Área não impactada
(média ± desvio padrão)
Área impactada
(média ± desvio padrão)
t p
ABRIL 2982,40 ± 1358,23 2668,80 ± 1811,38 0,13 0,77
MAIO 1475,20 ± 761,50 1990,40 ± 977,16 0,93 0,38
JUNHO 2297,60 ± 638,96 973,60 ± 573,97 3,45 < 0,01*
JULHO 2016,00 ± 1045,22 1747,20 ± 2321,04 0,24 0,82
AGOSTO 3347,2 ± 1777,39 2841,60 ± 3553,24 0,29 0,79
SETEMBRO 3452,8 ± 1847,69 4323,20 ± 4081,91 0,44 0,68
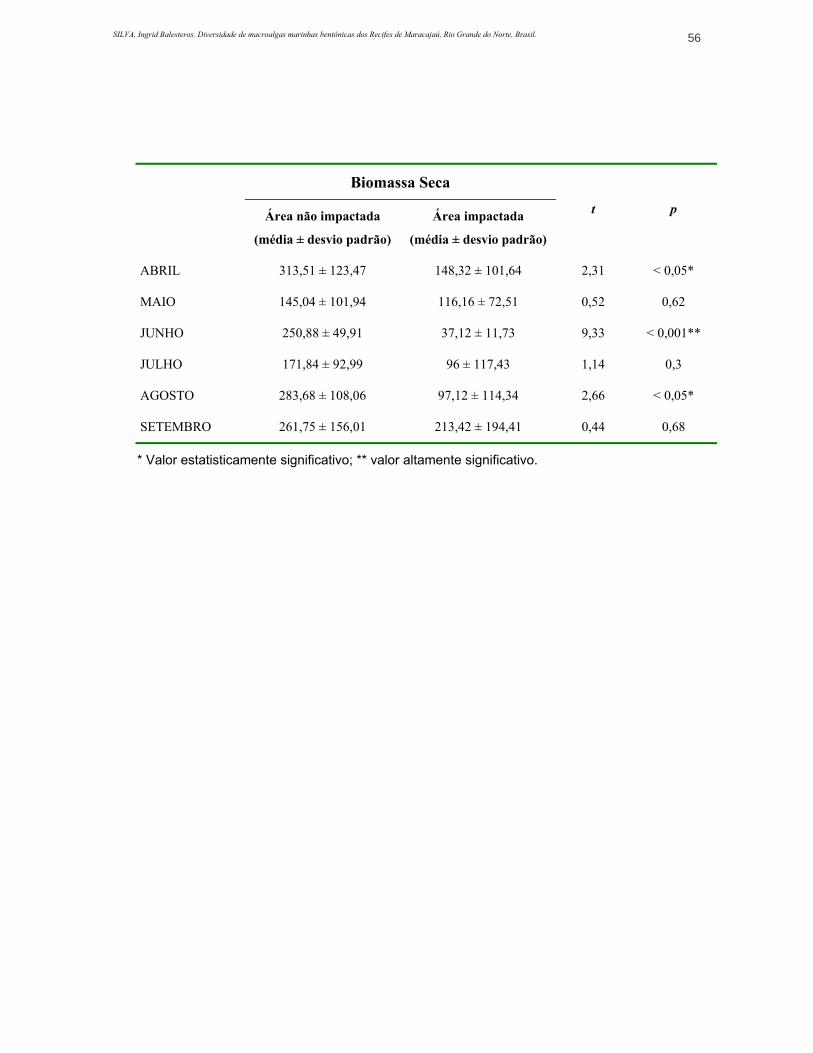
SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 56
Biomassa Seca
Área não impactada
(média ± desvio padrão)
Área impactada
(média ± desvio padrão)
t p
ABRIL 313,51 ± 123,47 148,32 ± 101,64 2,31 < 0,05*
MAIO 145,04 ± 101,94 116,16 ± 72,51 0,52 0,62
JUNHO 250,88 ± 49,91 37,12 ± 11,73 9,33 < 0,001**
JULHO 171,84 ± 92,99 96 ± 117,43 1,14 0,3
AGOSTO 283,68 ± 108,06 97,12 ± 114,34 2,66 < 0,05*
SETEMBRO 261,75 ± 156,01 213,42 ± 194,41 0,44 0,68
* Valor estatisticamente significativo; ** valor altamente significativo.

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 57
Anexo 4. Freqüência de ocorrência das espécies encontradas na área estudada, expressa em porcentagem (%) de presença nas amostragens mensais.
Área não impactada Área impactada ABR MAI JUN JUL AGO SET ABR MAI JUN JUL AGO SET
Caulerpa racemosa 80 40 80 80 40 80 100 100 100 100 60 80Caulerpa cupressoides 20 20Caulerpa mexicana 20 20 20Ventricaria ventricosa 20 20 20Enteromorpha sp 20Caulerpella ambigua 20 20 20 20 20Dictyopteris delicatula 100 80 100 100 100 100 100 100 100 80 100 80Dictyopteris justii 20 20 20 20 80 20Dictyopteris polypodioides 20Dictyota ciliolata 20 20Dictyota cervicornis 20 40 20 40 40 20 20 20 20Dictyota menstrualis 20 40 40 60 60 20 20Dictyota mertensii 20 20Lobophora variegata 60 40 100 60 60 20Padina sanctae-crucis 20Spatoglossum schroederi 20 20 60 20Sargassum hytrix 40 80 100 80 60 80Sargassum filipendula 20 20 20 20Sargassum vulgare 80 20 20 20Ceramium dawsonii 20 20 20 20Ceramium flaccidum 20 40 20 20 20Ceramium vagans 20 60 40 20 20 20Neosiphonia gorgoniae 40 20 40 20 20Bryothamniom triquetum 20 40 20 60 20Botryocladia pyriformis 20 40Laurencia filiformis 40 80 80 20 20 60Laurencia intricata 40 60 40 40 40 20 20Jania adherens 20 80 80 60 60 100 80 80 20 80 60 40Jania pumila 40 40 20 20Jania sp1 20 20 20Jania sp2 40 20 20 20Amphiroa anastomosans 60 40 20 20 20 20 60 20 20Amphiroa beauvoisii 80 60 60 40 40 80 40 20 60 40 20Haliptilon subulatum 20 60 40 60 40 20 20Gelidium coarctatum 80 60 60 80 20 20 20 80 20Gelidiella acerosa 40 60 20 80 80 80 40 20 40 60 40Gelidiella ligulata 20 40 20 20 20 20 20Gelidiopsis planicaulis 60 40 20 20 60 80 40 20 40 20Gelidiopsis variabilis 20 20 20 20Hypnea musciformis 20 20 40Hypnea spinella 60 40 80 100 60 100 80 60Cryptonemia crenulata 40 40 20 20 20 20Cryptonemia bengryi 20Pterocladiella caerulensces 60 20 20 40 60 80 20Chondracanthus acicularis 20 20 20 40 20 40

SILVA, Ingrid Balesteros. Diversidade de macroalgas marinhas bentônicas dos Recifes de Maracajaú, Rio Grande do Norte, Brasil. 58
Meristiella gelidium 20 20Lomentaria corallicola 40 60 20 20 20Herposiphonia secunda 20Herposiphonia bipinnata 20 20Chondria dangeardii 20 20 20Dipterosiphonia dentritica 20 20Antithamnionella graeffei 20 20 20 20 20Acrochaetium flexuosum 20 20Asparogopsis taxiformis (esporofito) 20 40 60 20 20





























