Embed Size (px)
Citation preview

1
DIVERSIDADE E DISTRIBUIÇÃO DOS CETÁCEOS NA
COSTA CENTRO-NORTE DO ESTADO DO RIO DE
JANEIRO, BRASIL.
HELIO KINAST CRUZ SECCO
UNIVERSIDADE ESTADUAL DO NORTE FLUMINENSE DARCY RIBEIRO
CAMPOS DOS GOYTACAZES - RJ
JANEIRO DE 2011

2
DIVERSIDADE E DISTRIBUIÇÃO DOS CETÁCEOS NA
COSTA CENTRO-NORTE DO ESTADO DO RIO DE
JANEIRO, BRASIL.
HELIO KINAST CRUZ SECCO
Orientador: Salvatore Siciliano
Supervisor Acadêmico: Alvaro Ramon Coelho Ovalle
Campos dos Goytacazes, RJ
Janeiro de 2011
Monografia apresentada ao Centro de Biociências
e Biotecnologia da Universidade Estadual do Norte
Fluminense, como parte das exigências para a
obtenção do título de Bacharel em Ciências
Biológicas.

3
DIVERSIDADE E DISTRIBUIÇÃO DOS CETÁCEOS NA
COSTA CENTRO-NORTE DO ESTADO DO RIO DE
JANEIRO, BRASIL.
HELIO KINAST CRUZ SECCO
Monografia aprovada em 20 de janeiro de 2011.
Comissão Avaliadora:
_________________________________________________________________
Dr. Salvatore Siciliano (ENSP / FIOCRUZ)
_________________________________________________________________
Dr. Álvaro Ramon Coelho Ovalle (LCA / UENF)
_________________________________________________________________
Dr. Ana Paula Madeira Di Beneditto (LCA / UENF)
_________________________________________________________________
Msc.Bruno Pereira Masi (LCA / UENF)
Monografia apresentada ao Centro de Biociências
e Biotecnologia da Universidade Estadual do Norte
Fluminense, como parte das exigências para a
obtenção do título de Bacharel em Ciências
Biológicas, ênfase em Ciências Ambientais.

4
“O bom marinheiro se
prova nos dias de
tempestade”

5
AGRADECIMENTOS
Agradeço inicialmente a pessoa que mais admiro na vida e que tenho o
orgulho de ser filho e carregar o mesmo nome. Pai, obrigado por tudo, sua
confiança e apoio incondicionais foram minha motivação para batalhar pelos
meus objetivos em todos os momentos da minha vida.
À minha família por todo o incentivo ao longo de todos esses anos,
principalmente minha Mãe, Mônica Kinast e minha Tia Avó, Lucia Bustamante.
Ao meu supervisor acadêmico Professor Álvaro Ramon Ovalle, por ter sido
um exemplo para mim desde que entrei nesta Universidade, por ter proporcionado
minhas primeiras oportunidades acadêmicas e por ter sido mais do que um
orientador em diversas situações ao longo da minha formação. Obrigado pela
confiança e amizade durante esses quatro anos.
Ao orientador Salvatore Siciliano, a quem sempre serei grato por ter me
concedido a oportunidade que impulsionou minha vida acadêmica profissional.
Obrigado pela orientação, pela atenção despendida nos momentos em que
precisei, pelo conhecimento passado e pela confiança depositada em mim.
Espero ter correspondido à altura de sua expectativa.
Agradeço também a todos os professores da UENF que de alguma forma
contribuíram efetivamente para a minha formação profissional, principalmente à
Professora Ana Paula Di Beneditto, pelas críticas e conselhos que sempre me
despertaram novas perspectivas, e à Professora Marina Suzuki, minha primeira
orientadora, sempre atenciosa e disposta a me auxiliar.
Ao meu companheiro de trabalho, conselheiro e grande amigo, Jailson
Fulgencio de Moura, pelas conversas científicas extremamente relevantes para o
meu desenvolvimento acadêmico e pelo exemplo de garra que sempre me
inspirou.
Ao amigo Renato Aguiar da Silva, por toda a atenção, paciência e tempo
gastos na produção dos mapas deste trabalho.
Aos colegas Bruno Rennó e Luciano Lima, pela convivência semanal
durante o ano de 2009, ano este em que eu iniciava minha experiência em
estudos sobre a fauna marinha da Bacia de Campos e tive a oportunidade de
aprender muito com as experiências passadas pelos dois.

6
Agradeço enormemente aos meus amigos e colegas de faculdade por
todos esses anos de convívio, se não fossem vocês, eu provavelmente não
estaria aqui até o final, nós fizemos uma verdadeira família na UENF.
Agradeço principalmente a Henrique Carneiro, Ricardo Alves, Felipe
Araújo, Marcello Carvalho, Douglas Braga, Hudson Lemos, Natiele Gentil e Alice
Guiotti, pela imensa amizade que com toda certeza levarei pro resto da minha
vida.
À Priscilla Carla dos Santos Costa, pelo extremo companheirismo durante
o ano de 2010, sua enorme paciência em ter ouvido muitas das minhas
“alucinações científicas” e pelo seu carinho ininterrupto.
Aos amigos doutorandos Bruno Masi, Marcos Franco, Daniela Sampaio e
Frederico Teixeira, pelos conselhos e pelas conversas sobre assuntos
considerados chatos pela maioria, mas que foram prazerosamente fundamentais
na minha rotina.
Ao CENPES-PETROBRAS pelo apoio através do Projeto Habitat:
Heterogeneidade Ambiental da Bacia de Campos, no período de 2009 a 2010.
Por último agradeço à Universidade Estadual do Norte Fluminese Darcy
Ribeiro por tudo que eu vivenciei aqui dentro e por ter me proporcionado tamanho
crescimento pessoal e profissional. Hoje posso dizer que aprendi a gostar deste
lugar como nunca havia imaginado. Tenho orgulho de me formar nesta instituição,
criada sobre o paradigma de ser “Universidade do Terceiro Milênio”, e carregar
em suas origens não só o nome, mas também os princípios de Darcy Ribeiro.

7
SUMÁRIO
Página
LISTA DE FIGURAS ....................................................................................................... 08
LISTA DE TABELAS ....................................................................................................... 14
RESUMO ......................................................................................................................... 16
ABSTRACT ..................................................................................................................... 17
1.INTRODUÇÃO ............................................................................................................. 18
1.1 Considerações gerais sobre os cetáceos ........................................................... 18
1.2 Dificuldade do estudo com cetáceos ................................................................... 21
1.3 Os encalhes como ferramenta de estudo com cetáceos ..................................... 21
1.4 Consideração sobre o estudo de cetáceos no Brasil e sua viabilidade................ 23
2.OBJETIVOS ................................................................................................................. 24
2.1 Objetivo geral ..................................................................................................... 24
2.2 Objetivos específicos .......................................................................................... 24
3.JUSTIFICATIVA ........................................................................................................... 25
4.MATERIAS E MÉTODOS ............................................................................................. 26
4.1 Descrição da área de estudo .............................................................................. 26
4.2 Acesso aos dados .............................................................................................. 28
4.3 Coleta dos dados ................................................................................................ 28
4.4 Protocolo de encalhe padrão .............................................................................. 29
4.5 Análise dos dados .............................................................................................. 31
5.RESULTADOS E DISCUSSÃO .................................................................................... 32
5.1 Resultados gerais ............................................................................................... 32
5.1.1 Representatividade dos encalhes .................................................................... 32
5.1.2 Variação temporal dos encalhes ...................................................................... 34
5.1.3 Proporção dos encalhes de todas as espécies por estações ........................... 37
5.1.4 Proporção dos encalhes de misticetos por estações........................................ 38
5.1.5 Proporção dos encalhes de odontocetos por estações .................................... 39
5.1.6 Distribuição geral dos encalhes ....................................................................... 41
5.2 Resultados específicos ....................................................................................... 43
5.2.1 Análise por espécie.......................................................................................... 43
6.CONCLUSÕES .......................................................................................................... 102
7.REFERÊNCIAS BIBLIOGRÁFICAS ........................................................................... 104

8
LISTA DE FIGURAS
Pág.
Figura 1: Imagem ilustrativa de um cetáceo misticeto (Eubalaena
australis)........................................................................................................ 20
Figura 2: Imagem ilustrativa de um cetáceo odontoceto (Orcinus orca)...... 20
Figura 3: Mapa da área de estudo................................................................ 27
Figura 4: Imagem ilustrativa biometria externa............................................. 30
Figura 5: Variação temporal dos encalhes de cetáceos entre 1993 e 2010. 34
Figura 6: Correlação entre a série temporal de encalhes e comportamento
dos eventos de El Niño e La Niña entre os anos de 1995 e
2010............................................................................................................... 36
Figura 7: Imagem de satélite da costa centro-norte fluminense. As setas
amarelas representam a ação do vento nordeste e a direção em que a
água é empurrada, devido ao efeito de Coriolis. As setas brancas
representam o vento sudoeste e a direção em que a água é empurrada,
devido ao efeito de Coriolis............................................................................ 41
Figura 8: Mapa com todos os encalhes de cetáceos na costa centro-norte
do Estado do Rio de Janeiro entre 1993 e 2010............................................ 42
Figura 9: Espécime de baleia-jubarte encalhado.......................................... 43
Figura 10: Série temporal dos encalhes de Baleia-jubarte na costa centro-
norte do Estado do Rio de Janeiro, Brasil, entre fevereiro de 1993 e junho
de 2010........................................................................................................... 44
Figura 11: Distribuição dos encalhes de baleia-jubarte pelas localidades
da área de estudo.......................................................................................... 44

9
Figura 12: Distribuição dos encalhes de baleia-jubarte por estação do
ano.................................................................................................................
44
Figura 13: Espécime de baleia-de-Bryde encalhado.................................... 46
Figura 14: Série temporal dos encalhes de baleia-de-Bryde na costa
centro-norte do Estado do Rio de Janeiro, Brasil, entre fevereiro de 1993 e
junho de 2010................................................................................................
47
Figura 15: Distribuição dos encalhes de baleia-de-Bryde pelas localidades
da área de estudo.......................................................................................... 47
Figura 16: Distribuição dos encalhes de baleia-de-Bryde por estação do
ano................................................................................................................. 47
Figura 17: Espécime de baleia minke-anã encalhado.................................. 49
Figura 18: Série temporal dos encalhes de baleia minke-anã na costa
centro-norte do Estado do Rio de Janeiro, Brasil, entre fevereiro de 1993 e
junho de 2010................................................................................................ 50
Figura 19: Distribuição dos encalhes de baleia minke-anã pelas
localidades da área de estudo....................................................................... 50
Figura 20: Distribuição dos encalhes baleia minke-anã por estação do
ano................................................................................................................. 50
Figura 21: Espécime de baleia minke-antártica encalhado vivo................... 52
Figura 22: Série temporal dos encalhes de baleia minke-antártica na costa
centro-norte do Estado do Rio de Janeiro, Brasil, entre fevereiro de 1993 e
junho de 2010................................................................................................. 52
Figura 23: Espécime de baleia franca-do-sul encalhado.............................. 54

10
Figura 24: Série temporal dos encalhes de baleia franca-do-sul na costa
centro-norte do Estado do Rio de Janeiro, Brasil, entre fevereiro de 1993 e
junho de 2010.................................................................................................
55
Figura 25: Distribuição dos encalhes de baleia franca-do-sul pelas
localidades da área de estudo....................................................................... 55
Figura 26: Distribuição dos encalhes de baleia franca-do-sul por estação
do ano............................................................................................................ 55
Figura 27: Mapa da costa centro-norte fluminense com a distribuição dos
encalhes de baleia-jubarte e baleia franca-do-sul......................................... 57
Figura 28: Mapa da costa centro-norte fluminense com a distribuição dos
encalhes de baleia minke-anã, baleia minke-antártica e baleia-de-Bryde.... 58
Figura 29: Espécime de boto-cinza encalhado............................................. 59
Figura 30: Série temporal dos encalhes de boto-cinza na costa centro-
norte do Estado do Rio de Janeiro entre fevereiro de 1993 e junho de
2010............................................................................................................... 60
Figura 31: Distribuição dos encalhes de boto-cinza pelas localidades da
área de estudo............................................................................................... 60
Figura 32: Distribuição dos encalhes de boto-cinza por estação do ano..... 60
Figura 33: Mapa da costa centro-norte fluminense com a distribuição dos
encalhes de boto-cinza.................................................................................. 62
Figura 34: Espécime de golfinho-nariz-de-garrafa encalhado...................... 63
Figura 35: Série temporal dos encalhes de golfinho-nariz-de-garrafa na
costa centro-norte do Estado do Rio de Janeiro, Brasil, entre fevereiro de
1993 e junho de 2010..................................................................................
64

11
Figura 36: Distribuição dos encalhes de golfinho-nariz-de-garrafa pelas
localidades da área de estudo....................................................................... 64
Figura 37: Distribuição dos encalhes de golfinho-nariz-de-garrafa por
estação do ano............................................................................................... 64
Figura 38: Mapa da costa centro-norte fluminense com a distribuição dos
encalhes de golfinho-nariz de-garrafa........................................................... 66
Figura 39: Espécime de golfinho-de-dentes-rugosos encalhado................. 67
Figura 40: Série temporal dos encalhes do golfinho-de-dentes-rugosos na
costa centro-norte do Estado do Rio de Janeiro entre fevereiro de 1993 e
junho de 2010................................................................................................ 68
Figura 41: Distribuição dos encalhes de golfinho-de-dentes-rugosos pelas
localidades da área de estudo....................................................................... 68
Figura 42: Distribuição dos encalhes de golfinho-de-dentes-rugosos por
estação do ano.............................................................................................. 68
Figura 43: Mapa da costa centro-norte fluminense com a distribuição dos
encalhes de golfinho-de-dentes-rugosos....................................................... 70
Figura 44: Espécime de golfinho-pintado-do-Atlântico encalhado............... 71
Figura 45: Distribuição dos encalhes de golfinho-pintado-do-Atlântico
pelas localidades da área de estudo............................................................. 72
Figura 46: Distribuição dos encalhes de golfinho-pintado-do-Atlântico
pelas localidades da área de estudo............................................................. 72
Figura 47: Distribuição dos encalhes de golfinho-pintado-do-Atlântico por
estação do ano.............................................................................................. 72
Figura 48: Golfinho-pintado-pantropical em vida livre.................................. 74

12
Figura 49: Série temporal de encalhes de golfinho-pintado-pantropical na
costa centro-norte do Estado do Rio de Janeiro, entre fevereiro de 1993 e
junho de 2010................................................................................................ 74
Figura 50: Espécime de golfinho-de-Clymene encalhado........................... 76
Figura 51: Série temporal de encalhes de golfinho-de-Clymene na costa
centro-norte do Estado do Rio de Janeiro, entre fevereiro de 1993 e junho
de 2010.......................................................................................................... 76
Figura 52: Especime de golfinho-de-Fraser encalhado................................ 78
Figura 53: Série temporal de encalhes de golfinho-de-Fraser na costa
centro-norte do Estado do Rio de Janeiro, entre fevereiro de 1993 e junho
de 2010.......................................................................................................... 78
Figura 54: Mapa da costa centro-norte fluminense com a distribuição dos
encalhes de golfinhos do gênero Stenella e golfinho-de-Fraser................... 80
Figura 55: Espécime de golfinho-comum encalhado................................... 81
Figura 56: Série temporal dos encalhes de golfinho-comum costa centro-
norte do Estado do Rio de Janeiro, Brasil, entre fevereiro de 1993 e junho
de 2010.......................................................................................................... 82
Figura 57: Distribuição dos encalhes de golfinho-comum pelas localidades
da área de estudo......................................................................................... 82
Figura 58: Distribuição dos encalhes de golfinho-comum por estação do
ano................................................................................................................ 82
Figura 59: Mapa da costa centro-norte fluminense com a distribuição dos
encalhes de golfinho-comum......................................................................... 84
Figura 60: Grupo de orcas avistado em Arraial do Cabo............................. 85

13
Figura 61: Série temporal dos encalhes de orca na costa centro-norte do
Estado do Rio de Janeiro, Brasil, entre fevereiro de 1993 e junho de 2010..
85
Figura 62: Orca-pigméia em vida livre.......................................................... 87
Figura 63: Série temporal dos encalhes de orca-pigméia na costa centro-
norte do Estado do Rio de Janeiro, Brasil, entre fevereiro de 1993 e junho
de 2010.......................................................................................................... 87
Figura 64: Baleia-piloto-de-peitorais-curtas em vida livre............................. 89
Figura 65: Série temporal dos encalhes de baleia-piloto-de-peitorais-
curtas na costa centro-norte do Estado do Rio de Janeiro, Brasil, entre
fevereiro de 1993 e junho de 2010................................................................ 89
Figura 66: Mapa da costa centro-norte fluminense com a distribuição dos
encalhes de Baleia-piloto-de-peitorais-curtas, orca e orca
pigméia........................................................................................................... 91
Figura 67: Espécime de cachalote encalhado.............................................. 92
Figura 68: Série temporal dos encalhes de cachalote na costa centro-
norte do Estado do Rio de Janeiro, Brasil, entre fevereiro de 1993 e junho
de 2010.......................................................................................................... 92
Figura 69: Espécime de cachalote-pigmeu encalhado................................. 94
Figura 70: Série temporal dos encalhes de cachalote-pigmeu na costa
centro-norte do Estado do Rio de Janeiro, Brasil, entre fevereiro de 1993 e
junho de 2010................................................................................................ 94
Figura 71: Cachalote-anão em vida livre...................................................... 96
Figura 72: Série temporal dos encalhes de cachalote-anã na costa centro-
norte do Estado do Rio de Janeiro, Brasil, entre fevereiro de 1993 e junho
de 2010.......................................................................................................... 96

14
Figura 73: Mapa da costa centro-norte fluminense com a distribuição dos
encalhes de cachalote, cachalote-pigmeu e cachalote-anão........................ 98
Figura 74: Espécime de toninha encalhado................................................. 99
Figura 75: Série temporal dos encalhes de toninha na costa centro-norte
do Estado do Rio de Janeiro, Brasil, entre fevereiro de 1993 e junho de
2010................................................................................................................
100
Figura 76: Distribuição dos encalhes de toninha pelas localidades da área
de estudo........................................................................................................ 100
Figura 77: Distribuição dos encalhes de toninha por estação do ano........... 100
Figura 78: Mapa da costa centro-norte fluminense com a distribuição dos
encalhes de toninha....................................................................................... 102
LISTA DE TABELAS
Tabela 1: Esforço de monitoramento GEMM-Lagos..................................... 38
Tabela 2: Freqüência relativa das espécies registradas na área de estudo 32
Tabela 3: Número de encalhes (N) e porcentagem representativa (%) por estação
do ano para todos os cetáceos...................................................................... 37
Tabela 4: Significância das diferenças entre as proporções de cetáceos
encalhados na costa centro-norte do Estado do Rio de Janeiro, considerando o
período chuvoso e período seco. Para p < 0,05, a diferença entre as proporções é
estatísticamente significativa........................................................................ 37
Tabela 5: Significância das diferenças entre as proporções de cetáceos
encalhados na costa centro-norte do Estado do Rio de Janeiro, considerando o

15
período de ressurgência e período sem ressurgência. Para p < 0,05, a diferença
entre as proporções é estatísticamente significativa................................... 37
Tabela 6: Número de encalhes (N) e porcentagem representativa (%) por estação
do ano para os cetáceos misticetos ........................................................... 38
Tabela 7: Significância das diferenças entre as proporções de misticetos
encalhados na costa centro-norte do Estado do Rio de Janeiro, considerando o
período chuvoso e período seco. Para p < 0,05, a diferença entre as proporções é
estatísticamente significativa............................................................................ 38
Tabela 8: Significância das diferenças entre as proporções de misticetos
encalhados na costa centro-norte do Estado do Rio de Janeiro, considerando o
período de ressurgência e período sem ressurgência. Para p < 0,05, a diferença
entre as proporções é estatísticamente significativa................................. 38
Tabela 9: Número de encalhes (N) e porcentagem representativa (%) por estação
do ano para os cetáceos odontocetos...................................................... 39
Tabela 10: Significância das diferenças entre as proporções de odontocetos
encalhados na costa centro-norte do Estado do Rio de Janeiro, considerando o
período chuvoso e período seco. Para p < 0,05, a diferença entre as proporções é
estatísticamente significativa............................................................................ 39
Tabela 11: Significância das diferenças entre as proporções de odontocetos
encalhados na costa centro-norte do Estado do Rio de Janeiro, considerando o
período de ressurgência e período sem ressurgência. Para p < 0,05, a diferença
entre as proporções é estatísticamente significativa................................. 39

16
RESUMO
Historicamente, as pesquisas com cetáceos foram conduzidas a partir do que se
podia extrair de informação de carcaças, tanto de animais encalhados em praias
quanto de animais mortos nas mãos de baleeiros ou pescadores. Os encalhes
podem gerar informações valiosas quanto à distribuição e abundância relativa,
status populacional, morfologia, doenças e história natural dos cetáceos, desde
que se faça o melhor uso possível dessas oportunidades, gerando séries
amostrais de longo prazo. A costa centro-norte do Estado do Rio de Janeiro
(Saquarema 22o55'S, 42o30'W - São Francisco do Itabapoana 21°25'S-41°00'W) é
uma importante área de uso por parte de diversas espécies de cetáceos, incluindo
espécies migratórias e residentes. O objetivo do presente estudo foi revisar e
atualizar a informação sobre a diversidade e distribuição de cetáceos encalhados
na costa centro-norte do Estado do Rio de Janeiro, Brasil, entre os anos de 1993
e 2010. Ao todo foram registrados 210 espécimes de cetáceos encalhados na
área de estudo, representando 20 espécies, sendo cinco pertencentes à
subordem Mysticeti e 15 da subordem Odontoceti. Os maiores valores de
freqüência relativa (FR) foram observados para Sotalia guianensis e Megaptera
novaeangliae, 0,424 e 0,110, respectivamente. A maioria dos encalhes foram
registrados nos meses de inverno devido à intensificação do vento sudoeste. A
península de Arraial do Cabo e a foz do Rio Paraíba do Sul são consideradas
áreas de concentração de encalhes de cetáceos na área de estudo. Tanto Sotalia
guianensis quanto Pontoporia blainvillei estão associadas à área de influência da
foz do Rio Paraíba do Sul. Os registros de Pontoporia blainvillei são mais
escassos devido ao pequeno porte da espécie, o que diminui a probabilidade do
encalhe. Os encalhes das espécies de hábitos oceânicos-costeiros (Tursiops
truncatus, Steno bredanensis, Stenella frontalis e Dephinus sp.) refletem
fortemente a influência do fenômeno da ressurgência de Cabo Frio e o
estreitamento da plataforma no seu padrão de ocorrência. Entre os cetáceos
misticetos, Megaptera novaeangliae e Balaenoptera edeni são as espécies mais
representativas entre os encalhes. Encalhes de Feresa attenuata e Stenella
clymene constituem registros inéditos para a costa do Estado do Rio de Janeiro.

17
ABSTRACT
Historically, cetaceans researchs were conducted from information about
carcasses of stranded and accidentally captured animals. Long-term sampling
series of stranding cetaceans events can produce valuable information on
distribution and abundance relative, population status, morphology, diseases and
natural history. The north-central coast of Rio de Janeiro State (Saquarema
22o55'S, 42o30'W - São Francisco do Itabapoana 21°25'S, 41°00'W) is an
important use area by several cetaceans species, including migratory and resident
species. The aim of this study was to review and update the information about
diversity and distribution of cetaceans stranded along the north-central coast of
Rio de Janeiro State, Brazil, between 1993 and 2010. There were reported 210
cetaceans stranded in study area, representing 20 species, five of suborder
Misticety and 15 of suborder Odontoceti. The highest values of relative frequency
were observed for Sotalia guianensis and Pontoporia blainvillei, 0,424 and 0,110,
respectively. Most strandings were recorded in winter months due to the
intensification of southwest wind. The peninsula of Arraial do Cabo and the mouth
of the Paraiba do Sul river are considered “hotspots” of strandings in the study
area. Both Sotalia guianensis and Pontoporia blainvillei are associated with the
influence area of the mouth of Paraiba do Sul river. The Pontoporia blainvillei
records are scarce due to the small size of this specie, wich decreases the
probability of stranding. The narrowing of the platform and the Cabo Frio upwelling
phenomenon has a strong influence on the stranding pattern occurrence of
species of oceanic-coastal habits (Tursiops truncates, Steno bredanensis, Stenella
frontalis and Delphinus sp.). Among baleen whales, Megaptera novaeangliae and
Balaenoptera edeni are the most representative species. Strandings of Feresa
attenuata and Stenella clymene are new records for the coast of the State of Rio
de Janeiro.

18
1. INTRODUÇÃO
1.1 - Considerações gerais sobre os cetáceos
Os cetáceos compreendem uma ordem de mamíferos que apresentam
adaptações à vida exclusivamente aquática. Esta ordem de mamíferos evoluiu de
ancestrais terrestres há cerca de 60 milhões de anos atrás quando passaram a
explorar de maneira mais efetiva recursos oriundos do meio aquático.
Durante sua evolução, os cetáceos desenvolveram adaptações fisiológicas
anatômicas únicas, tais como: corpo alongado e fusiforme, aberturas nasais na
parte superior da cabeça, membros anteriores achatados em forma de
nadadeiras, estruturas desenvolvidas a partir de tecidos não ósseos como as
nadadeiras dorsal e caudal, perda dos membros posteriores e cintura pélvica,
internalização dos órgãos genitais, glândulas mamárias e aparato auditivo, pele
extremamente lisa, camada de gordura subcutânea espessa e ausência ou
escassez de pêlos (Jefferson et al. 1993).
Considerados mamíferos de alto valor adaptativo, alto nível de inteligência e
mobilidade, os cetáceos, apresentam considerável habilidade em explorar uma
grande diversidade de habitats (Nowak, 2003). Cada espécie de cetáceo tem seu
padrão de distribuição particular de acordo com seus comportamentos,
necessidades ecológicas e características ambientais (Jefferson et al. 1993).
Atualmente existem pelo menos 88 espécies viventes, as quais se dividem em
dois grupos: subordem Mysticeti e subordem Odontoceti. Os cetáceos que
apresentam cerdas bucais e aberturas respiratórias separadas constituem os
misticetos (Figura 1), enquanto que os odontocetos apresentam dentes e apenas
uma abertura respiratória (Figura 2).
Os misticetos são conhecidos como rorquais ou “grandes baleias”,
representados por quatro famílias e 16 espécies viventes. De acordo com a
espécie, a expectativa de vida varia entre 50 e 80 anos, a maturidade sexual entre
quatro e 10 anos e apresentam período de gestação com cerca de 10 a 12 meses
(Lockyer, 1984). Esses animais se adaptaram a ingerir pequenos organismos

19
presentes no ambiente marinho como estratégia de alimentação. As grandes
baleias privilegiam recursos alimentares aparentemente de baixo valor energético,
porém, de alta disponibilidade e densidade na coluna d’água, como por exemplo;
pequenos crustáceos componentes do zooplâncton e pequenos peixes pelágicos.
Dessa forma, as cerdas bucais funcionam como um “filtro”, separando as
pequenas presas do grande volume de água engolfado. Geralmente, as áreas de
alimentação se separam por milhares de quilômetros das áreas de reprodução,
obrigando algumas espécies a realizarem, anualmente, extensas migrações.
Algumas populações de misticetos são altamente migratórias, apresentando
padrão de movimento sazonal, entre zonas produtivas de alta latitude durante o
período de alimentação e zonas de baixa latitude durante o período reprodutivo
(Stevick et al. 2002).
Os odontocetos compõem a subordem com maior riqueza de espécies, com
ampla variação morfológica, representada principalmente pelos golfinhos ou
botos. Dependendo da espécie, a expectativa de vida pode variar entre 20 e um
pouco mais de 60 anos, a maturidade sexual é atingida com cerca de dois a 10
anos e o período de gestação pode durar entre 10 e 16 meses (Perrin & Reilly,
1984). Quanto à alimentação, presas de maior tamanho, incluindo crustáceos,
cefalópodes, peixes e outros itens alimentares mais energéticos, constituem seus
principais recursos. Dentre suas características mais peculiares, os cetáceos
odontocetos apresentam sistema de ecolocalização, responsável por promover a
detecção e reconhecimento do ambiente por meio da emissão de ondas sonoras
de alta frequência, as quais interagem com o ambiente ao redor e geram um eco
de resposta. Utilizando-se desse sistema, os odontocetos são capazes de
capturar suas presas com apreciável êxito. As espécies desta subordem não
promovem migrações em larga escala, especialmente aquelas que vivem em
latitudes tropicais e temperadas. Os cetáceos odontocetos apresentam uma
radiação geográfica extremamente extensiva, podendo estar entre os grupos de
mamíferos mais bem distribuídos pelo mundo (Nowak, 2003).

20
Figura 1: Imagem ilustrativa de um cetáceo misticeto (Eubalaena australis).
[Obtido de www.killerwhalemuseum.com.au/Images/right.gif]
Figura 2: Imagem ilustrativa de um cetáceo odontoceto (Orcinus orca).
[Obtido de www.horta.uac.pt/species/newSite/Cetacea]
Os cetáceos são considerados componentes vitais da biodiversidade aquática
marinha e fluvial. Sua importância ecológica no ecossistema se relaciona, entre
outros aspectos, com a manutenção do equilíbrio da estrutura trófica (Katona &
Whitehead, 1988; Parsons, 1992). Além disso, esses mamíferos aquáticos se
apresentam como potenciais bioindicadores da qualidade ambiental (Norse, 1993;
Marine Mammal Commission, 1999; Van Bressem et al., 2003).

21
1.2 - Dificuldades de estudo com cetáceos
O estudo de cetáceos apresenta uma série de adversidades práticas.
Primeiramente, a dificuldade de acesso ao habitat utilizado por estes muitas
vezes impossibilita a realização de diversos modelos de estudos aplicados a
outros animais. Além disso, o tamanho do objeto de estudo (baleias e golfinhos),
normalmente, dificulta o manejo do animal vivo. A coleta de informações da
maioria das espécies de cetáceos tende a ser pontual, em função da raridade das
mesmas em águas próximas da costa e da dificuldade de acesso aliada a falta de
infra-estrutura adequada (Santos, 1999). Sendo assim, muitas informações sobre
cetáceos provêm de avistagens, exemplares encalhados em praias ou capturados
acidentalmente em redes de pesca. No entanto, o número amostral destes
eventos normalmente é reduzido, tornando-se necessário um longo período de
estudos para se obter dados confiáveis sobre a biologia e ecologia dos cetáceos.
1.3 - Os encalhes como ferramenta de estudo de cetáceos
Historicamente, as pesquisas com cetáceos foram conduzidas a partir do que
se podia extrair de informação de carcaças, tanto de animais encalhados em
praias quanto de animais mortos nas mãos de baleeiros ou pescadores. Segundo
Samuels & Tyack (2000), era necessário “fazer o melhor de uma situação ruim”,
ou seja, aproveitar o máximo possível do que uma carcaça pode oferecer.
Descrições originais de várias espécies foram baseadas exclusivamente em
carcaças encontradas em praias (Hofman, 1991), esses materiais muitas vezes
são os únicos disponíveis sobre certas espécies consideradas raras (Odell, 1991).
Eventos de mortalidade de cetáceos representam oportunidades ímpares para
incrementar o conhecimento sobre sua história de vida e subsidiar a implantação
de medidas de conservação eficientes e adequadas a realidade de cada região
(Norman et al., 2004). Nos últimos 40 anos, as informações geradas a partir de
cetáceos encalhados passaram a representar fontes de pesquisa e gerar
subsídios adicionais para a elaboração de planos de conservação e de manejo de
espécies em países do hemisfério norte (Geraci & Lounsbury, 1993).

22
Programas de coleta e monitoramento esporádicos ou sistematizados
passaram a se estruturar em diversos locais do mundo devido a crescente
notificação e divulgação dos encalhes de cetáceos em praias e o recente
aumento do número de pesquisadores especialistas nessa área.
Atualmente, sabe-se que os principais fatores geradores de encalhes podem
estar relacionados com as condições oceanográficas e topográficas locais
(Brabyn & McLean, 1992), como rápidas variações de marés associadas a
regiões costeiras com formações de bancos de areia; perseguição de presas e
fuga de predadores em águas rasas (Nores & Perez, 1988; Casinos & Vericad,
1976); doenças virais (Duignan, et al., 1995; Dhermain et al., 2002); infecções
parasitárias pronunciadas (Ridgway & Dailey, 1972); contaminação por toxinas
geradas por marés vermelhas ou compostos organoclorados (Geraci et al., 1989);
falhas no direcionamento de navegação pela possível sensibilidade dos cetáceos
em relação aos contornos magnéticos da Terra (Klinowska, 1986; Klinowska
1991) e outros impactos de natureza antrópica como capturas acidentais (Perrin
et al., 1994; Read et al., 2003) e colisões com embarcações (Jensen & Silber,
2003).
Os encalhes podem gerar informações valiosas quanto à distribuição e
abundância relativa (Berrow, 2001), “status” populacional, morfologia, doenças e
história natural dos cetáceos, desde que se faça o melhor uso possível dessas
oportunidades, gerando séries amostrais de longo prazo (Geraci & St. Aubin,
1979; Reynolds III & Odell, 1991).
Neste contexto, apesar de todas as limitações existentes, os encalhes são
uma excelente fonte de informações para o estudo de cetáceos, apresentando-se
como uma ótima oportunidade de se gerar subsídios para o melhor conhecimento
das espécies.

23
1.4 - Consideração sobre os estudos de cetáceos no Brasil e sua viabilidade
As informações pertinentes a espécies de cetáceos em águas brasileiras
ainda são incipientes, se comparadas com o conhecimento acumulado em países
como Estados Unidos, Canadá e outros do continente europeu (Santos, 1999).
Os trabalhos envolvendo tomada sistemática de dados no Brasil vêm sendo
desenvolvidos desde a década de 70, quando os eventos de encalhes de
cetáceos passaram a ser reportados com maior frequência (Carvalho, 1975;
Gianuca & Castello, 1976). Durante a década de 80, os primeiros censos de
espécies encalhadas ao longo de determinadas áreas da costa brasileira foram
publicados (Bittencourt, 1984; Geise & Borobia, 1987; Ximenez et al., 1987).
O cenário nacional da pesquisa relacionada à fauna silvestre ainda não é
considerado condizente com a biodiversidade e extensão territorial, incluindo a
zona costeira brasileira. Nesse sentido, a dedicação adicional e o rigor
metodológico por parte dos interessados em estudos sobre a fauna selvagem,
principalmente marinha, passam a ser essenciais ao desenvolvimento de
pesquisas dessa natureza (Di Beneditto & Siciliano 2004).
Em contrapartida à falta de oportunidade, estrutura e financiamento de
estudos com cetáceos, no Brasil, os registros de encalhes se tornaram a principal
forma de estudo com esse grupo em particular. Em algumas regiões do país,
grupos de estudos e pesquisas com cetáceos vêm se empenhando para gerar
extensos bancos de dados ao longo das últimas décadas.

24
2. OBJETIVOS
2.1 - Objetivo geral
Revisar e atualizar a informação sobre diversidade e distribuição de cetáceos
encalhados na costa centro-norte do Estado do Rio de Janeiro, Brasil, entre
1993 e 2010.
2.2 - Objetivos específicos
Analisar a distribuição espacial e temporal dos encalhes das espécies de
cetáceos que ocorrem na região.
Contribuir para a atualização da lista de espécies de cetáceos com ocorrência
comprovada para a Bacia de Campos.

25
3. JUSTIFICATIVA
A costa centro-norte do Estado do Rio de Janeiro é uma importante área de
uso por parte de diversas espécies de cetáceos, incluindo espécies migratórias e
residentes (Siciliano et al. 2006).
Deve se ressaltar que a região Sudeste do Brasil possui fortes
adensamentos populacionais por toda sua linha de costa, onde a população
associada a esta área promove uma degradação crônica do ambiente,
acarretando em distúrbios na dinâmica marinha costeira.
A intensa atividade pesqueira é tida como uma característica marcante da
região centro-norte fluminense, a qual representa sérios problemas em relação à
exploração e perda de biodiversidade marinha, principalmente para as espécies
de cetáceos que sofrem pressão de captura acidental.
A região abriga a Bacia de Campos, principal fonte produtora de petróleo
no Brasil, contribuindo com mais de 80% da produção total do país. Em função
das atividades petrolíferas, o tráfego de embarcações de médio e grande porte
intensificou-se na região, passando a representar maior ameaça à fauna marinha
local (Di Beneditto & Ramos 2001).
Além disso, a exploração e produção de petróleo, apesar de licenciadas e
monitoradas, representam potenciais riscos ambientais, tais como vazamentos e
derrames de substâncias derivadas de hidrocarbonetos do petróleo,
extremamente danosas a saúde do ecossistema marinho, as quais requerem
atenção contínua.
Diante deste cenário, o monitoramento da fauna marinha na costa centro-
norte fluminense mostra grande relevância, uma vez que a mesma está exposta a
diversas atividades antrópicas que interferem diretamente no funcionamento
ecossistêmico marinho. Os cetáceos, além de serem considerados bioindicadores
da qualidade ambiental, são componentes essenciais ao equilíbrio da
estruturação trófica marinha, logo, justificam o esforço de estudo.

26
4. MATERIAIS E MÉTODOS
4.1 – Descrição da área de estudo
A área de estudo faz parte da costa centro-norte do Estado do Rio de Janeiro,
entre as localidades de Saquarema (22o55'S, 42o30'W) e São Francisco de
Itabapoana (21°25'S, 41°00'W), englobando grande parte da Bacia de Campos
(Figura 3).
A área é caracterizada por uma plataforma continental que se alarga no
sentido norte-sul até a altura de Cabo Frio. O talude continental é interrompido por
cânions submarinos, como os que ocorrem ao largo da porção norte do Estado do
Rio de Janeiro, situando-se a cerca de 100 metros de profundidade e entre 70 e
110 quilômetros da linha da costa. Próximo da região de Cabo Frio ocorre uma
drástica modificação no perfil da plataforma, de modo que a distância entre o
talude e a costa diminui consideravelmente, e em torno de nove quilômetros da
praia já se verifica profundidades em torno de 100 metros (Siciliano et al. 2006).
A região da plataforma continental entre Campos e o Rio de Janeiro apresenta
mistura de massas d’água oriundas da corrente do Brasil, corrente Costeira e
Água Central do Atlântico Sul (ACAS). A corrente do Brasil é caracterizada por
águas oligotróficas de temperatura mais elevada. A corrente Costeira contribui
com as massas d’água junto à costa, recebendo importantes descargas de água
doce, devido à ação das desembocaduras de rios de pequeno e médio porte,
destacando-se o Rio Paraíba do Sul. A ACAS, por sua vez, se encontra entre 200
e 400 metros de profundidade, caracterizada por apresentar menor salinidade,
baixa temperatura e altas concentrações de nutrientes (Matsuura, 1995).
Os ventos predominantes na região da Bacia de Campos são os de
nordeste durante a maior parte do ano, considerados importantes mecanismos na
geração de correntes nesta plataforma. Durante o verão, quando uma termoclina
sazonal se estabelece, os ventos NE/E entre 21o e 23o S se intensificam e
controlam o evento de ressurgência, resultando no afloramento de ACAS na
plataforma interna (Valentin, 1988). Devido a esse fenômeno, a região sul da área

27
de estudo é caracterizada por elevada produtividade primária, diversidade e
abundância de organismos aquáticos (Valentin, 2001).
Ao sul da área de estudo, a ressurgência de Cabo Frio é o principal agente
controlador das condições oceanográficas costeiras, enquanto que ao norte do
Cabo de São Tomé (Campos dos Goytacazes), o rio Paraíba do Sul possui maior
influência. O rio Paraíba do Sul aumenta drásticamente sua vazão durante o
verão (Pezzini, 2006), o que faz com que sua área de influência na região costeira
próxima a sua desembocadura aumente. Além disso, a maior vazão interfere
diretamente na disponibilidade de nutrientes que chegam até o oceano (Kruger et
al. 2006), alterando assim a produtividade biológica e o equilíbrio trófico do
ambiente costeiro.
Figura 3: Mapa da área de estudo.

28
4.2 – Acesso aos dados
Os resultados presentes neste trabalho foram gerados a partir da revisão
de informações referentes aos registros de encalhes de cetáceos para a costa
centro-norte fluminense, contidas no banco de dados do Projeto Cetáceos, da
Fundação Brasileira para a Conservação da Natureza (FBCN), entre fevereiro de
1993 e agosto de 2001, e do Grupo de Estudos de Mamíferos Marinhos da
Região dos Lagos (GEMM-Lagos), da Escola Nacional de Saúde
Pública/FIOCRUZ, entre março de 1999 e junho de 2010. Adicionalmente, foi
revisada a literatura especializada sobre encalhes de cetáceos na costa centro-
norte fluminense.
4.3 – Coleta dos dados
O Projeto Cetáceos atuava na área compreendida entre Macaé (22º22'S,
41º46'W) e São Francisco de Itabapoana (21°25'S, 41°00'W), monitorando o
desembarque pesqueiro dos principais portos da região norte fluminense, com a
finalidade de recolher carcaças de cetáceos capturados acidentalmente em redes
de pesca artesanal. Apesar de não realizar monitoramentos de praia, encalhes na
região eram eventualmente relatados à equipe do Projeto Cetáceos, que por sua
vez confirmavam o registro in situ.
O GEMM-Lagos monitora o trecho compreendido entre Saquarema
(22o55'S, 42o30'W) e Quissamã (22o06'S, 41o28'W), totalizando aproximadamente
235 quilômetros de linha de costa (Tabela 1).
Tabela 1: Esforço de monitoramento.
Período Área Periodicidade Grupo de Pesquisa
1999 - 2003 Saquarema – Arraial do Cabo 15 dias GEMM-LAGOS
2003 - 2009 Saquarema - Quissamã 30 dias GEMM-LAGOS
2010 Saquarema – Arraial do Cabo 30 dias GEMM-LAGOS
2010 Cabo Frio - Quissamã 7 dias GEMM-LAGOS

29
Oportunamente, também foram registrados encalhes a partir de
informações reportadas através do programa de “Disque-Carcaça” difundido em
toda costa centro-norte fluminense, contando com o apoio de membros de
comunidades locais, colônias de pesca, turistas, corpo de bombeiros e secretarias
municipais de meio ambiente.
Ressalta-se o fato de que o esforço amostral para a coleta dos dados
utilizados neste trabalho não foi sistemático, tendo em vista as dificuldades em
aplicar esforço contínuo ao longo de toda a área estudada.
4.3 – Protocolo de encalhe padrão
Como mencionado anteriormente, eventos de mortalidade de cetáceos
representam excelentes oportunidades de se obter diversas informações
normalmente inacessíveis. Para tanto, a padronização dos métodos de
investigação é essencial, permitindo que as informações e amostras relevantes
não sejam desconsideradas ou perdidas. Os dados devem ser confiáveis, a ponto
de permitir comparação de informações oriundas de diversas fontes e para que o
potencial intercâmbio entre os pesquisadores seja ampliado. Dessa forma, a
qualidade das informações obtidas sobre carcaças coletadas a partir de encalhes,
terá seu valor científico de acordo com a acuidade em que as atividades no
campo e laboratório são conduzidas (Di Beneditto & Siciliano, 2004).
Registro
Cada carcaça recebeu um número/código de tombo, caracterizando um
registro único, o qual acompanhou todas as amostras referentes à mesma.
Sempre que possível foram obtidas fotografias da carcaça e examinaram-se
detalhes do corpo, e possíveis ferimentos ou marcas. Os dados de registro
continham: data, local, coordenadas geográficas, coletores, dados relacionados a
marcas de interação com atividade pesqueira, material osteológico coletado e
tipos de amostras coletadas.

30
Identificação
A identificação da espécie foi realizada pela observação dos caracteres
morfológicos, métricos, merísticos e de coloração, seguindo literatura específica
(Bastida et al., 2007, Jefferson et al., 1993, Leatherwood & Reeves 1983, Pinedo
et al., 1992 e Wada et al., 2003).
Avaliação e caracterização dos espécimes
O estado de decomposição da carcaça foi avaliado de acordo com Geraci
& Lounsbury (1993). A determinação do sexo foi realizada seguindo
características morfológicas externas ou, de acordo com o estado de
decomposição da carcaça, observando o aparelho reprodutor. A biometria externa
foi realizada segundo Norris (1961), a partir de medidas em projeção retilínea
(Figura 4). Quando houve impossibilidade de efetuar a biometria como um todo,
se registrou apenas o comprimento total da carcaça. As contagens do número de
dentes, cerdas bucais e sulcos ventrais, foram feitas nos casos em que o estágio
de decomposição das carcaças ainda era moderado.
Figura 4: Imagem ilustrativa biometria externa
[Obtido de DiBeneditto & Siciliano 2004.]

31
4.4 – Análise dos dados
Apenas as informações referentes à data e local dos encalhes de cada
espécie foram utilizadas nas análises deste trabalho.
De acordo com as informações coletadas no campo, os dados foram
digitalizados em planilhas do programa Microsoft Excel v.12.0, servindo como
base para a formulação dos gráficos.
As frequências relativas (FR) foram determinadas através do cálculo da
razão entre o número de exemplares de cada espécie e o número total de
exemplares recolhidos encalhados. Além disso, foram analisadas as proporções
de cetáceos encalhados entre o período com ressurgência e sem ressurgência,
entre período chuvoso e período seco e entre as estações do ano. O grau de
significância das diferenças entre as proporções de cetáceos encalhados no
período com ressurgência / chuvoso e o período sem ressurgência / seco foi
testado estatisticamente através do programa Statistica for Windows vs. 5.5.
Visando uma análise espacial adequada da distribuição dos encalhes na
área de estudo, todos os encalhes foram georeferenciados e plotados no mapa da
costa centro-norte do Estado do Rio de Janeiro, utilizando ferramenta SIG
(Sistema de Informações Geográficas) ArcGis 9.3.

32
5. RESULTADOS E DISCUSSÃO
5.1 – Resultados Gerais
5.1.1 – Representatividade dos encalhes
Ao todo foram registrados 210 espécimes de cetáceos encalhados na costa
centro-norte do Estado do Rio de Janeiro entre fevereiro de 1993 e junho de
2010. No total foram identificadas 20 espécies de cetáceos, sendo cinco
pertencentes à subordem Mysticeti e 15 da subordem Odontoceti (Tabela 2).
.
Tabela 2: Freqüência relativa das espécies registradas na área de estudo.
Família Espécie Nome comum FR
Balaenopteridae
Megaptera novaeangliae Baleia-jubarte 0,110
Balaenoptera edeni Baleia-de-Bryde 0,048
Balaenoptera acutorostrata Baleia-minke-anã 0,029
Balaenoptera bonaerensis Baleia-minke-antártica 0,005
Balaenidae
Eubalaena australis Baleia-franca-do-sul 0,029
Delphinidae
Sotalia guianensis Boto-cinza 0,424
Tursiops truncatus Golfinho-nariz-de-garrafa 0,072
Steno bredanensis Golfinho-de-dentes-rugosos 0,057
Stenella frontalis
Stenella attenuata
Stenella clymene
Delphinus sp.
Lagenodelphis hosei
Orcinus orca
Golfinho-pintado-do-Atlântico
Golfinho-pintado-Pantropical
Golfinho-de-Clymene
Golfinho comum
Golfinho-de-Fraser
Baleia orca
0,048
0,009
0,005
0,038
0,009
0,009

33
Pontoporiidae
Kogiidae
Physeteridae
Feresa attenuata
Globicephala macrorhynchus
Pontoporia blainvillei
Kogia beviceps
Kogia sima
Physeter macrocephalus
Orca-pigméia
Baleia-piloto-de-peitorais-longas
Toninha
Cachalote-pigmeu
Cachalote-anão
Cachalote
0,005
0,009
0,062
0,014
0,009
0,009
Os maiores valores de freqüência relativa (FR) foram observados para
Sotalia guianensis e Megaptera novaeangliae, 0,424 e 0,110, respectivamente.
No caso de Sotalia guianensis, é uma espécie de hábitos exclusivamente
costeiros, normalmente associada à desembocadura de rios e regiões estuarinas
(Flores & Silva 2009), como por exemplo, na foz do Rio Paraíba do Sul, logo, a
probabilidade da carcaça vir a encalhar aumenta à medida que os indivíduos
vivem próximos da costa. Para Megaptera novaeangliae, o grande número de
registros de encalhes se justifica uma vez que esta espécie utiliza a costa centro-
norte fluminense como parte de sua rota migratória durante o inverno e a
primavera (Siciliano et al. 2006), migrando de áreas antárticas e sub-antárticas
em direção a áreas tropicais e sub-tropicais para se reproduzir (Dawbin, 1966).
Em alguns locais, as baleias desta espécie passam bem próximo da linha de
costa, como no caso da península de Arraial do Cabo, onde a plataforma
continental é estreita e a profundidade é maior, aumentando assim as chances
das carcaças de indivíduos que não resistiram à migração chegarem até a praia.

34
5.1.2 – Variação temporal dos encalhes
A influência do esforço aplicado
Esta série temporal (Figura 5) é o testemunho da história recente de pesquisa
com mamíferos marinhos na costa centro-norte do Estado do Rio de Janeiro.
Entre os anos de 1993 e 1999, o único grupo de pesquisa com mamíferos
marinhos que atuava na região era o do Projeto Cetáceos, o qual se dedicava
prioritariamente em pesquisas com animais capturados acidentalmente em redes
de pesca no norte fluminsense e não realizavam monitoramento de praia na
região. A partir de 1999, o GEMM-Lagos começa a atuar na região, monitorando o
litoral entre Saquarema e Arraial do Cabo. De 1999 a 2001, com os dois grupos
de pesquisa atuando na costa centro-norte fluminense, há um aumento do
número de encalhes registrados. Em 2002, o Projeto Cetáceos encerra suas
atividades, e o GEMM-Lagos passa a ser o único grupo de pesquisa atuando na
área, o que resulta em uma diminuição temporária na quantidade de encalhes
registrados. A partir de 2003 o GEMM-Lagos extende sua área de monitoramento
até Quissamã, passando a monitorar aproximadamente 235 quilômetros de linha
de costa. Conforme o esperado, este aumento do esforço amostral aplicado
reflete em um pico de encalhes (n=26) no ano de 2004. Entre janeiro e junho de
2010, os monitoramentos de praia passam a ser semanais entre Cabo Frio e
Quissamã, e entre Arraial do Cabo e Saquarema continuam sendo executados
mensalmente, justificando, mais uma vez, um incremento da quantidade de
encalhes registrados (n=21).
A influência de condições climáticas
A variação de condições climáticas e acontecimentos de fenômenos como o
El Niño, podem influenciar na mortalidade e dinâmica dos encalhes de cetáceos,
uma vez que interferem nas condições oceanográficas.
Apesar do pouco conhecimento em relação a real influência do El Niño sobre a
fauna marinha como um todo, eventos pretéritos deste fenômeno tiveram
consequências devastadoras na produtividade marinha da costa oeste da

35
América do Sul, além de afetar o clima de diversas regiões do mundo (Bastida et
al. 2007).
O El Ninõ consiste em um desequilíbrio térmico no Pacífico gerando a
aparição de águas superficiais mais quentes que o habitual na costa Pacífica da
América do Sul (Bastida et al. 2007). Normalmente, os ventos alísios na região
equatorial do Pacífico predominam no sentido leste-oeste, transportando águas
quentes para o extremo Ocidental do Pacífico (Austrália e Indonésia), onde nível
do mar se torna aproximadamente 50 centímetros mais elevado (Ricklefs, 2010).
Esta dinâmica permite que a Corrente de Humboldt (composta por águas de
ressurgência ricas em nutrientes) suba até o norte da costa Pacífica da América
do Sul. O El Niño acontece quando os ventos alísios se debilitam, permitindo que
a massa d’água quente chege até a costa Pacífica da América do Sul. Uma faixa
superficial de água quente, pobre em nutrientes se forma, impedindo o
afloramento de águas profundas e comprometendo a produtividade biológica local
(Bastida et al. 2007).
Os efeitos climáticos e oceanográficos do El Niño se estendem para muitas
outras partes do mundo, afetando ecossistemas em áreas distantes. Em algumas
regiões ocorrem anomalias na temperatura da superfície do mar e drásticas
mudanças nas taxas de precipitação, podendo gerar tempestades contínuas ou
períodos de seca severa (Ricklefs, 2010).
O El Niño acontece em média entre intervalos de três a sete anos,
normalmente intercalados com eventos de La Ninã, que é o fenômeno inverso
(Bastida et al. 2007). Durante os períodos de La Niña, os ventos alísios são
fortalecidos, acentuando as correntes de ressurgência e de superfície no oceano
Pacífico (Bastida et al. 2007). A La Niña é caracterizada por chuvas pesadas em
muitas regiões dos trópicos, secas na regiões temperadas do norte, e aumento da
atividade de furacões no Oceano Atlântico Norte (Ricklefs, 2010). Períodos de La
Niña rigorosa causam a intensificação de frentes frias em regiões tropicais, o que
deve ser considerado como um fator favorável ao aumento de encalhes de
cetáceos uma vez que carcaças à deriva estarão sujeitas a ação dos ventos.

36
Apesar de estarmos levando em consideração o alcance de eventos
climáticos como El Niño e La Niña na dinâmica dos encalhes, não se identificou
nenhum padrão de resposta quando correlacionamos a variação dos encalhes
com períodos de incidência desses eventos (Figura 6). A série temporal do
presente estudo não permite tirar maiores conclusões, entretanto ressalta-se que
a continuação do monitoramento de encalhes é essencial para o maior
conhecimento acerca deste assunto.
Figura 5: Variação dos encalhes de cetáceos entre fevereiro de 1993 e junho de 2010.
Figura 6: Correlação entre a série temporal de encalhes e o comportamento dos eventos de El
Niño e La Niña entre os anos de 1995 e 2010 (Adaptado de
http://www.esrl.noaa.gov/psd/people/klaus.wolter/MEI).

37
5.1.3 – Proporção dos encalhes de todas as espécies por estações
Tabela 3: Número de encalhes (N) e porcentagem representativa (%) por estação do ano.
Tabela 4: Significância das diferenças entre as proporções de cetáceos encalhados na costa
centro-norte do Estado do Rio de Janeiro, considerando o período chuvoso e período seco. Para p
< 0,05, a diferença entre as proporções é estatísticamente significativa.
Tabela 5: Significância das diferenças entre as proporções de cetáceos encalhados na costa
centro-norte do Estado do Rio de Janeiro, considerando o período de ressurgência e período sem
ressurgência. Para p < 0,05, a diferença entre as proporções é estatísticamente significativa.
Período N encalhes Porcentagem (%)
Com Ressurgência (setembro – fevereiro)
100
47,6%
Sem Ressurgência (março – agosto) 110 52,4%
Significância - p = 0,4916
Estação N encalhes Porcentagem (%)
Verão (dez,jan,fev) 42 20%
Outono (mar.abr,mai) 39 18,60%
Inverno (jun,jul,ago) 72 34,30%
Primavera (set,out,nov) 57 27,10%
Período N encalhes Porcentagem (%)
Chuvoso (outubro – março)
91
43,3%
Seco (abril – setembro) 119 56,7%
Significância - p = 0,0556

38
5.1.4 – Proporção dos encalhes de misticetos por estações
Tabela 6: Número de encalhes (N) e porcentagem representativa (%) por estação do ano.
Tabela 7: Significância das diferenças entre as proporções de misticetos encalhados na costa
centro-norte do Estado do Rio de Janeiro, considerando o período chuvoso e período seco. Para p
< 0,05, a diferença entre as proporções é estatísticamente significativa.
Tabela 8: Significância das diferenças entre as proporções de misticetos encalhados na costa
centro-norte do Estado do Rio de Janeiro, considerando o período de ressurgência e período sem
ressurgência. Para p < 0,05, a diferença entre as proporções é estatísticamente significativa.
Período N encalhes Porcentagem (%)
Com Ressurgência (setembro – fevereiro)
31
67,4%
Sem Ressurgência (março – agosto) 15 32,6%
Significância - p = 0,0309
Estação N encalhes Porcentagem (%)
Verão (dez,jan,fev) 10 21,7%
Outono (mar.abr,mai) 3 6,5%
Inverno (jun,jul,ago) 12 26,1%
Primavera (set,out,nov) 21 45,7%
Período N encalhes Porcentagem (%)
Chuvoso (outubro – março)
18
40%
Seco (abril – setembro) 28 60%
Significância - p = 0,1876

39
5.1.5 – Proporção dos encalhes de odontocetos por estações
Tabela 9: Número de encalhes (N) e porcentagem representativa (%) por estação do ano.
Tabela 10: Significância das diferenças entre as proporções de odontocetos encalhados na costa
centro-norte do Estado do Rio de Janeiro, considerando o período chuvoso e período seco. Para p
< 0,05, a diferença entre as proporções é estatísticamente significativa.
Tabela 11: Significância das diferenças entre as proporções odontocetos encalhados na costa
centro-norte do Estado do Rio de Janeiro, considerando o período de ressurgência e período sem
ressurgência. Para p < 0,05, a diferença entre as proporções é estatísticamente significativa.
Período N encalhes Porcentagem (%)
Com Ressurgência (setembro – fevereiro)
69
42%
Sem Ressurgência (março – agosto) 95 58%
Significância - p = 0,0465
Estação N encalhes Porcentagem (%)
Verão (dez,jan,fev) 32 19,5%
Outono (mar.abr,mai) 36 22%
Inverno (jun,jul,ago) 60 36,5%
Primavera (set,out,nov) 36 22%
Período N encalhes Porcentagem (%)
Chuvoso (outubro – março)
73
45,5%
Seco (abril – setembro) 91 55,5%
Significância - p = 0,2048

40
Apesar da maioria das análises apontarem para uma maior frequência de
encalhes nos meses de inverno (Tabelas 3 e 9), período seco (Tabelas 4,7 e 10)
e sem ressurgência (Tabela 5 e 11), apenas duas delas foram estatísticamente
significativas.
A diferença nas proporções de misticetos encalhados no período com
ressurgência e sem ressurgência é estatísticamente significativa (p < 0,05) ao
demonstrar maior número de encalhes no período com ressurgência (Tabela 8).
Os misticetos apresentam tendência inversa aos outros resultados devido ao
padrão de ocorrência distinto da espécie mais representativa do grupo. A baleia-
jubarte utiliza a costa centro-norte fluminense como parte de sua rota migratória
durante o inverno e a primavera (Siciliano et al. 2006), o que faz com que a
maioria dos seus encalhes sejam registrados durante a primavera, início do
período com ressurgência na região. No caso dos odontocetos, a diferença entre
as proporções de indivíduos encalhados no período com ressurgência e sem
ressurgência é estatísticamente significante (p < 0,05) ao demonstrar maior
número de encalhes no período sem ressurgência (Tabela 11).
Durante o verão, o vento nordeste ganha força, tornando-se o agente
responsável pela intensificação do processo de ressurgência. A medida em que
ele afasta a massa d’água superficial próxima à costa, possibilita o afloramento de
ACAS, que por sua vez impulsiona toda a produtividade biológica local (Valentin,
2001). Porém, quando ocorre esse deslocamento de massa d’água, as chances
das carcaças de cetáceos à deriva chegarem até a praia diminuem
consideravelmente (Figura 7), uma vez que as mesmas serão afastadas da costa.
Durante o inverno, o oposto acontece quando o vento sudoeste passa a ser
predominante na região (subsidência) (Gonzalez-Rodriguez et al. 1992). A ação
do vento sudoeste empurra a água superficial na direção do continente, causando
o “empilhamento” da água na costa, e consequentemente as chances das
carcaças à deriva encalharem aumentam (Figura 7), o que explica a tendência
observada nos resutados das análises, principalmente para os odontocetos.

41
Figura 7: Imagem de satélite da costa centro-norte fluminense. Setas amarelas: vento nordeste e
a direção em que a água superficial é empurrada devido ao efeito de Coriolis / Setas brancas:
vento sudoeste e a direção em que a água superficial é empurrada devido ao efeito de Coriolis.
5.1.6 – Distribuição geral dos encalhes
De uma maneira geral, a maioria dos encalhes concentram-se nas
penínsulas de Arraial do Cabo e Armação dos Búzios, além da área costeira onde
o rio Paraíba do Sul desemboca. A espécie mais abundante na área de estudo
(Sotalia guianensis) esta predominantemente associada à região estuarina da foz
do Rio Paraíba do Sul (Di Beneditto & Ramos 2004), contribuindo assim com um
grande número de encalhes nesta área. No caso das penínsulas, a incidência de
ventos na região é determinante para que haja este maior número de encalhes.
Carcaças à deriva ao sul da área de estudo podem ter a península de Arraial do
Cabo como um anteparo ao norte, assim como carcaças oriundas do norte da
área de estudo podem ter a península de Armação dos Búzios como um anteparo
ao sul. Além disso, a região de Arraial do Cabo e Cabo Frio apresenta plataforma
continental estreita, onde a linha isobatimétrica dos 100 metros está localizada
extremamente próxima da costa. Esta característica propicia que espécies
tipicamente oceânicas se aproximem da costa (De Moura et al. 2010),
contribuindo para um maior número contribuindo de encalhes também.

42
.
Figura 8: Mapa com todos os encalhes de cetáceos na costa centro-norte do Estado do Rio de
Janeiro entre 1993 e 2010.

43
5.2 – Resultados Específicos
5.2.1 – Análise por espécie
Misticetos
Família Balaenopteridae
Baleia-jubarte (Megaptera novaeangliae):
Esta espécie totalizou 23 encalhes (Figura 10), estando distribuídos ao
longo de toda a área de estudo (Figura 11 e 27). A maioria dos encalhes foram
registrados na primavera e no inverno (Figura 12). Indivíduos jovens e adultos (de
ambos os sexos), além de um filhote foram constatados. O comprimento total dos
espécimes variou entre 4,63 e 17 metros.
Figura 9: Espécime de baleia-jubarte encalhado [GEMM 227 – 04/10/2010 – Praia Seca/Araruama].

44
Figura 10: Série temporal dos encalhes de baleia-jubarte na costa centro-norte do Estado do Rio
de Janeiro, Brasil, entre fevereiro de 1993 e junho de 2010.
Figura 11: Distribuição dos encalhes de baleia-jubarte pelas localidades da área de estudo
Figura 12: Distribuição dos encalhes de baleia-jubarte por estação do ano.

45
A baleia-jubarte ocorre em todos os oceanos do planeta e apresenta
padrões migratórios sazonais semelhantes às demais espécies de misticetos
(Jefferson et al. 1993).
No litoral brasileiro são abundantes na região do Arquipélago de Abrolhos,
Bahia (Siciliano, 1997), o qual é um reconhecido sítio reprodutivo desta população
que se desloca das Ilhas Geórgia do Sul/Sandwich do Sul em direção a costa
nordeste do Brasil.
Apesar da baleia-jubarte se manter predominantemente afastada do
continente durante sua migração, na região próxima de Arraial do Cabo (onde a
plataforma continental é estreita e a profundidade é maior) é comum avistar
indivíduos desta espécie passando bem próximos da linha de costa entre os
meses de junho e dezembro (Siciliano et al. 2006). A maior proporção de
encalhes constatada neste mesmo período está de acordo com o esperado. O
fato de haver registros de encalhe de indivíduos recém-nascidos, jovens e adultos
(de ambos os sexos), corrobora com a existência de um corredor migratório da
espécie na Bacia de Campos (Siciliano et al. 2006).
Apesar do crescente conhecimento sobre a distribuição desta população de
baleia-jubarte (Zerbini et al. 2004), outros aspectos sobre a ocorrência desta
espécie no litoral do Brasil necessitam de mais informações principalmente em
relação às rotas migratórias que a espécie utiliza quando chega e/ou retorna para
áreas de reprodução.

46
Baleia-de-Bryde (Balaenoptera edeni):
Ao todo, 10 registros de encalhe de Baleia-de-Bryde foram contabilizados
(Figura 14), estando distribuídos entre Quissamã e Arraial do Cabo (Figura 15 e
28). Dois indivíduos foram identificados como machos e os outros oito não
puderam ser identificados quanto ao sexo devido ao estágio de decomposição
avançado. Foram constatados encalhes em todas as estações do ano, porém não
houve predomínio em nenhuma delas (Figura 16). O comprimento total dos
espécimes encalhados variou entre 7 e 14 metros.
Figura 13: Espécime de baleia-de-Bryde encalhado [GEMM 157 – 11/10/2008 – Praia do
Afonso/Arraial do Cabo].

47
Figura 14: Série temporal dos encalhes de baleia-de-Bryde na costa centro-norte do Estado do Rio
de Janeiro, Brasil, entre fevereiro de 1993 e junho de 2010.
Figura 15: Distribuição dos encalhes de baleia-de-Bryde pelas localidades da área de estudo.
Figura 16: Distribuição dos encalhes de baleia-de-Bryde por estação do ano.

48
A baleia-de-Bryde é o único balenopterídeo que habita exclusivamente
águas tropicais e temperadas acima de 20 oC e raramente excede sua distribuição
além dos 35 o de latitude norte e sul. Esta espécie tende a se distribuir pelas
zonas restritas de alta produtividade tropicais e subtropicais (Bastida et al. 2007).
No Brasil existem encalhes registrados desde a costa do Maranhão até o
Rio Grande do Sul, porém, a região Sudeste é a região com maior quantidade de
avistagens registradas (Siciliano et al. 2006).
Na Bacia de Campos e na região Sudeste de uma forma geral, esta
espécie é freqüentemente avistada em águas próximas da costa (Siciliano et al.
2004). Os encalhes registrados em todas as estações do ano, além das
avistagens freqüentes, sugerem que esta espécie possa ter uma população
residente na Bacia de Campos. O encalhe de um indivíduo de sete metros de
comprimento total, corrobora esta hipótese, uma vez que um filhote só atinge este
tamanho quando se aproxima dos seis meses de vida, idade esta em que passa a
não ser mais dependente do leite materno. Além disso, avistagens de pares mãe
e filhote nas proximidades de Armação dos Búzios e Arraial do Cabo já foram
comprovadas (Siciliano et al. 2006).
De acordo com seu padrão local de distribuição, esta espécie pode ser
considerada componente da comunidade costeira de cetáceos da Bacia de
Campos.

49
Baleia minke-anã (Balaenoptera acutorostrata):
Seis encalhes foram registrados (Figura 18) entre as localidades de
Quissamã e Saquarema (Figura 19 e 28). Apenas um indivíduo pode ter seu sexo
identificado (macho). Houve registros para três estações do ano, exceto no
outono (Figura 20). O comprimento total dos espécimes encalhados variou entre
3,13 e 4,5 metros.
Figura 17: Espécime de baleia minke-anã encalhado [GEMM 200 – 31/07/2010 – Praia do
Abricó/Rio das Ostras].

50
Figura 18: Série temporal dos encalhes de baleia minke-anã na costa centro-norte do Estado do
Rio de Janeiro, Brasil, entre fevereiro de 1993 e junho de 2010.
Figura 19: Distribuição dos encalhes de baleia minke-anã pelas localidades da área de estudo
Figura 20: Distribuição dos encalhes de baleia minke-anã por estação do ano.

51
Esta espécie é restrita ao hemisfério sul, onde ocorre em águas tropicais,
sub-tropicais, antárticas e sub-antárticas. Aparentemente apresentam um padrão
migratório característico, habitando águas frias no verão e águas mais quentes
durante o inverno e a primavera.
A baleia-minke-anã ocorre no oeste do Oceano Atlântico Sul em latitudes
maiores que 7oS (Zerbini et al. 1996). Na costa brasileira é o balenopterídeo com
maior número de encalhes registrados, desde o litoral do Rio Grande do Sul até a
costa da Paraiba (Siciliano et al. 2006).
Entretanto, a maior porcentagem dos encalhes estão concentrados nas
regiões Sul e Sudeste, estando de acordo com a hipótese de que esta espécie é
mais comum em médias latitudes do Oceano Atlântico Sul Ocidental (Siciliano et
al. 2006). Acredita-se que a área entre o sudeste do Brasil e o norte da Argentina
seja área de permanência de imaturos e mães acompanhadas de seus filhotes,
enquanto indivíduos reprodutivamente ativos podem utilizar baixas latitudes
exclusivamente como área de acasalamento (Siciliano et al. 2006).
Os resultados encontrados neste estudo corroboram esta hipótese, uma
vez que todos os exemplares encalhados desta espécie eram jovens com menos
de 4,5 metros de comprimento total. Alguns estudos demonstram que a espécie é
relativamente comum sobre a plataforma continental (Best, 1985; Zerbini et al.
1997). No Brasil, aproximadamente 80% do total de encalhes de baleia-minke
correspondem a indivíduos da forma anã, sugerindo que sua distribuição seja
predominantemente costeira (Siciliano et al. 2006).
Adicionalmente, a maioria das avistagens confirmadas de Balaenoptera
acutorostrata em águas brasileiras, provém de áreas situadas entre a costa e o
limite de quebra da plataforma continental, onde normalmente a batimetria gira
em torno dos 200 metros. Em Arraial do Cabo, durante os meses de verão é
comum avistar indivíduos de baleia minke-anã a partir de observações por ponto-
fixo (Hassel et al. 2003).

52
Baleia minke-antártica (Balaenoptera bonaerensis):
Registrou-se apenas um encalhe de baleia minke-antártica na costa centro-
norte fluminense entre 1993 e 2010 (Figura 22). Este encalhe ocorreu em agosto
de 2004, na Praia dos Ossos, Armação dos Búzios (Figura 28), quando um
indivíduo encalhou ainda vivo.
Figura 21: Espécime de baleia minke-antártica encalhado vivo [GEMM 062 – 01/08/2004 – Praia
dos Ossos/Armação dos Búzios]
Figura 22: Série temporal dos encalhes de baleia minke antártica na costa centro-norte do Estado
do Rio de Janeiro, Brasil, entre fevereiro de 1993 e junho de 2010.

53
A espécie Balaenoptera bonaerensis possui distribuição circumpolar no
hemisfério sul. No verão se encontra em águas da Antártida e durante o inverno e
a primavera ocorre em águas oceânicas tropicais e sub-tropicais de todos os
oceanos do hemisfério sul (Bastida et al. 2007).
No Brasil, a espécie ocorre em áreas profundas sobre ou além do talude
continental, desde o Rio Grande do Sul até o litoral do Nordeste, sendo este
último uma provável área de reprodução desta espécie (IWC 1991).
Na Bacia de Campos, a baleia minke-antártica pode ser observada durante
sua migração, entre julho e setembro, especialmente no talude continental, entre
300 e 3.000 metros de profundidade (Siciliano et al. 2006).
O único encalhe de Balaenoptera bonaerensis registrado neste estudo
ocorreu em agosto, o que está de acordo com o período migratório conhecido
para a espécie.
O fato de haver um número elevado de encalhes de filhotes e juvenis no
litoral sul do Brasil, Uruguai e norte da Argentina, aliado à falta de avistagens de
fêmeas com filhotes na região nordeste, levanta a suspeita de que fêmeas
grávidas e os nascimentos possam estar ocorrendo em médias latitudes no
Oceano Atlântico Sul Ocidental (Zerbini et al, 1997).

54
Família Balaenidae
Baleia-franca-do-sul (Eubalaena australis):
Foram registrados seis encalhes de baleia-franca-do-sul (Figura 24) entre São
Francisco do Itabapoana e Saquarema (Figura 25). Cinco espécimes não
puderam ser identificados quanto ao sexo devido ao estágio de decomposição
avançado, e um foi identificado como macho. Os encalhes foram predominantes
no segundo semestre dos anos (Figura 26) e o comprimento total dos espécimes
variou entre 7,8 e 15 metros.
Figura 23: Espécime de baleia-franca-do-sul encalhado [GEMM 051 – 17/01/2004 - Praia de Itaúna/Saquarema]

55
Figura 24: Série temporal dos encalhes de baleia-franca-do-sul na costa centro-norte do Estado do
Rio de Janeiro, Brasil, entre fevereiro de 1993 e junho de 2010.
Figura 25: Distribuição dos encalhes de baleia-franca-do-sul pelas localidades da área de estudo.
Figura 26: Distribuição dos encalhes de baleia-franca-do-sul por estação do ano.

56
A baleia-franca-do-sul é a única espécie da família Balaenidae presente no
hemisfério sul, ocorrendo entre 15oS e 65oS (Bastida et al. 2007).
Durante o verão as baleias desta espécie se concentram em águas próximas
da Convergência Antártica, onde podem se alimentar em grande quantidade em
virtude da alta produtividade local. Durante o inverno e a primavera elas realizam
extensos movimentos migratórios até as áreas de reprodução e cria, situadas em
regiões tropicais e sub-tropicais (Bastida et al. 2007). A Península de Valdez, na
Argentina, e a costa sul do Estado de Santa Catarina são os principais sítios
reprodutivos da espécie no Oceano Atlântico Sul Ocidental (Bastida et al. 2007).
No Brasil, as baleias-franca-do-sul podem ser avistadas entre os meses de
julho e novembro desde o litoral sul da Bahia até o litoral do Rio Grande do Sul
(Siciliano et al. 2006).
Na Bacia de Campos, grupos de baleia-franca-do-sul podem ser observados
próximos da costa de Arraial do Cabo (Siciliano et al. 2006), porém encalhes já
foram registrados tanto na Região dos Lagos quanto no extremo norte fluminense,
adjacente ao Estado do Espírito Santo (Figura 27). Dos seis encalhes registrados
na área de estudo, cinco aconteceram entre agosto e setembro e apenas um foi
registrado fora do período esperado.

57
Figura 27: Mapa da costa centro-norte fluminense com a distribuição dos encalhes de baleia-
jubarte e baleia-franca-do-sul.

58
Figura 28: Mapa da costa centro-norte fluminense com a distribuição dos encalhes de baleia
minke-anã, baleia minke-antártica e baleia-de-Bryde.

59
Odontocetos
Família Delphinidae
Boto-cinza (Sotalia guianensis):
Esta espécie foi representada por 89 encalhes (Figura 30), com uma maior
concentração de encalhes ao norte de Armação dos Búzios (Figura 31 e 33).
Todas as estações do ano tiveram encalhe desta espécie, porém a maioria dos
encalhes ocorreu no inverno e na primavera, ou seja, no segundo semestre dos
anos (Tabela 32). Foram identificados 29 indivíduos machos, 20 fêmeas e 40 com
sexo não identificado devido ao estágio de decomposição. Foram confirmados 20
casos de interação negativa com artefatos de pesca, de acordo com marcas
externas no corpo dos espécimes. O comprimento total variou entre 0,70 e 2,12
metros (um dos maiores espécimes já registrados pela literatura).
Figura 29: Espécime de boto-cinza encalhado [GEMM 175 – 29/01/2010 - Praia do Parque
Nacional da Jurubatiba/Macaé]

60
Figura 30: Série temporal dos encalhes de boto-cinza na costa centro-norte do Estado do Rio de
Janeiro entre fevereiro de 1993 e junho de 2010.
Figura 31: Distribuição dos encalhes de boto-cinza pelas localidades da área de estudo
Figura 32: Distribuição dos encalhes de boto-cinza por estação do ano.

61
O delfinideo Sotalia guianensis é uma espécie tipicamente costeira, que se
encontra em toda a costa atlântica tropical e subtropical da América do Sul e
Central, desde Honduras até Florianópolis, Brasil, onde águas temperadas ao sul
determinam o limite de sua distribuição (Flores & Silva 2009). É considerada uma
espécie costeira e associada a baías e regiões estuarinas. Acredita-se que esteja
representada por várias populações ao longo da costa da América do Sul (Flores
& Silva 2009).
O boto-cinza é o cetáceo com maior abundância na costa centro-norte
fluminense, o que justifica a maior freqüência entre os encalhes de todas as
espécies na área de estudo. Esta população está fortemente associada à
desembocadura do Rio Paraíba do Sul, o que é comprovado pela clara diminuição
dos encalhes na direção sul da área de estudo.
A maior quantidade de encalhes durante os meses de inverno e primavera
ocorre devido à maior quantidade de frentes frias originadas pela incidência do
vento sudoeste (predominante no inverno), trazendo carcaças à deriva até a
praia. O maior número de encalhes a partir de 2002 pode ser explicado pelo
aumento do esforço do GEMM-Lagos na região e pelo término do monitoramento
das capturas acidentais por parte do Projeto Cetáceos, acarretando em um maior
número de carcaças lançadas ao mar pelos pescadores, os quais anteriormente
levavam os animais capturados acidentalmente até os pesquisadores.
Aproximadamente 22,5% dos espécimes encalhados apresentavam marcas
indicativas de interação negativa com artefatos de pesca, esta proporção é menor
do que o observado em estudos pretéritos (De Moura et al. 2009).
Como a potencial área de distribuição desta espécie se dá até a isóbata de 30
metros (Di Beneditto 2003), os indivíduos desta população ficam expostos à ação
de redes de pesca que operam na região entre cinco e 67 metros de profundidade
(Di Beneditto 2003). As redes de pesca podem causar sérios impactos na
comunidade de golfinhos, principalmente nas espécies costeiras, como no caso
do boto-cinza.

62
Figura 33: Mapa da costa centro-norte fluminense com a distribuição dos encalhes de boto-
cinza.

63
Golfinho-nariz-de-garrafa (Tursiops truncatus):
Esta espécie foi representada por 15 encalhes (Figura 35), distribuídos por
grande parte da área de estudo, porém com uma predominância de encalhes na
Região dos Lagos (Figura 36 e 38). Entre os encalhes, foram identificados quatro
indivíduos machos, duas fêmeas e nove não tiveram o sexo identificado devido ao
estágio de decomposição avançado. Encalhes foram registrados em três estações
do ano (verão, outono e inverno), com maior incidência nos meses de inverno
(Figura 37). O comprimento total dos espécimes encalhados variou entre 1,06 e
3,35 metros.
Figura 34: Espécime de golfinho-nariz-de-garrafa encalhado [GEMM 081 – 26/06.2005 – Praia de
Manguinhos/Búzios]

64
Figura 35: Série temporal dos encalhes de golfinho-nariz-de-garrafa na costa centro-norte do
Estado do Rio de Janeiro, Brasil, entre fevereiro de 1993 e junho de 2010.
Figura 36: Distribuição dos encalhes de golfinho-nariz-de-garrafa pelas localidades da área de
estudo.
Figura 37: Distribuição dos encalhes de golfinho-nariz-de-garrafa por estação do ano.

65
Esta espécie ocupa todos os mares tropicais e temperados, distribuindo-se
principalmente em zonas costeiras, inclusive lagoas e estuários (Bastida et al.
2007). Nos últimos anos populações oceânicas foram descobertas em frente à
costa norte-americana do Oceano Pacífico e no Oceano Atlântico em frente à
costa do Brasil (Bastida et al. 2007). É considerada uma espécie de grande
plasticidade ambiental, pois suporta diferentes condições de temperatura,
salinidade e turbidez, além de locais com alto nível de contaminação (Bastida et
al. 2007).
No Brasil, o golfinho-nariz-de-garrafa é largamente distribuído pela costa,
onde se identifica populações residentes em regiões estuarinas, lagoas costeiras
e desembocaduras de rios no Rio Grande do Sul e Santa Catarina (Castello &
Pinedo 1977; Simões-Lopes et al. 1998), além de populações oceânicas na região
nordeste, incluindo o Arquipélago de São Pedro e São Paulo (Moreno et al. 2009).
Campanhas oceanográficas na Bacia de Campos, realizadas entre os anos de
2003 e 2005, demonstraram que a espécie é a mais freqüente em áreas
afastadas da costa (Siciliano et al. 2006).
A distribuição dos encalhes de Tursiops truncatus neste estudo reflete uma
tendência de encalhes nas praias da Região dos Lagos (Figura 38), onde
normalmente a água é mais fria, salobra, e existem locais de maior profundidade,
o que poderia apontar uma preferência desta população por águas de
características oceânicas. Os encalhes em pelo menos três estações do ano
(Figura 37) demonstram que a espécie é comum na região.

66
Figura 38: Mapa da costa centro-norte fluminense com a distribuição dos encalhes de golfinho-
nariz de-garrafa.

67
Golfinho-de-dentes-rugosos (Steno bredanensis):
Esta espécie foi representada por 12 encalhes (Figura 40) na área, entre
São João da Barra e Saquarema, porém com uma maior proporção de encalhes
entre Arraial do Cabo e Cabo Frio (Figura 41 e 43). Dos 12 espécimes
encalhados, quatro eram machos, um era fêmea e sete não puderam ser sexados
devido ao estágio de decomposição avançado. Registrou-se encalhes em três
estações do ano, exceto na primavera (Figura 42). O comprimento total dos
espécimes variou entre 2,40 e 2,74 metros.
Figura 39: Espécime de golfinho-de-dentes-rugosos encalhado [GEMM080 – 24/06/2005 – Praia
de Tucuns/Búzios]

68
Figura 40: Série temporal dos encalhes do golfinho-de-dentes-rugosos na costa centro-norte do
Estado do Rio de Janeiro entre fevereiro de 1993 e junho de 2010.
Figura 41: Distribuição dos encalhes de golfinho-de-dentes-rugosos pelas localidades da área de
estudo.
Figura 42: Distribuição dos encalhes de golfinho-de-dentes-rugosos por estação do ano.

69
O golfinho-de-dentes-rugosos é encontrado em águas tropicais e
temperadas quentes de todo o mundo (Bastida et al. 2007). Habita
predominantemente águas oceânicas profundas, com raras exceções, como no
caso da costa brasileira (Bastida et al. 2007).
No Brasil esta espécie pode ser encontrada não apenas em regiões
pelágicas, mas também em águas de menor profundidade, próximas de praias,
ilhas costeiras, baías e recifes de corais (Bastida et al. 2007).
Na Bacia de Campos, apesar de existir registros de encalhe desde a foz do
Rio Paraíba do Sul até Saquarema (Figura 41), existe uma área específica em
que grupos de golfinho-de-dentes-rugosos são constantemente avistados. Esta
área está localizada entre a Prainha em Arraial do Cabo e a Praia do Forte em
Cabo Frio, onde inclusive, barcos de passeio turístico exploram o local, como rota
de navegação para as pessoas observarem os golfinhos. Os encalhes registrados
neste estudo se concentram na mesma área (Figura 43) em que os animais são
constantemente avistados, o que pode ser mais um indício da existência de uma
pequena população residente nesta região. Apesar dos encalhes serem
exclusivamente de adultos, indivíduos de ambos os sexos já foram constatados.

70
Figura 43: Mapa da costa centro-norte fluminense com a distribuição dos encalhes de golfinho-de-
dentes-rugosos.

71
Golfinho-pintado-do-Atlântico (Stenella frontalis):
Esta espécie foi representada por 10 encalhes (Figura 45) entre Armação
dos Búzios e Saquarema (Figura 46). Entre os 10 espécimes encalhados, quatro
eram machos, quatro fêmeas e dois não puderam ter o sexo identificado devido
ao estágio de decomposição avançado. Houve um predomínio de encalhes
durante o inverno (60%) (Figura 47). Os espécimes tiveram o comprimento total
variando entre 1,58 e 2,08 metros.
Figura 44: Espécime de golfinho-pintado-do-Atlântico encalhado [GEMM 046 – 16/10/2003 – Praia
de Geribá/Armação dos Búzios].

72
Figura 45: Série temporal dos encalhes de golfinho-pintado-do-Atlântico na costa centro-norte do
Estado do Rio de Janeiro, Brasil, entre fevereiro de 1993 e junho de 2010.
Figura 46: Distribuição dos encalhes de golfinho-pintado-do-Atlântico pelas localidades da área.
Figura 47: Distribuição dos encalhes de golfinho-pintado-do-Atlântico por estação do ano.

73
O golfinho-pintado-do-Atlântico é uma espécie endêmica do Oceano
Atlântico, distribuindo-se em águas tropicais, subtropicais e temperadas quentes.
É uma das espécies mais comumente avistadas sobre a plataforma continental,
podendo ser encontrada em águas costeiras com menos de 20 metros de
profundidade e raramente encontrada em áreas de talude (Jefferson & Schiro
1997).
Os registros de encalhes e avistagens desta espécie para a costa brasileira
parecem indicar uma descontinuidade na sua distribuição, formando
aparentemente duas populações separadas por um hiato entre o norte do Espírito
Santo e sul da Paraíba (Moreno et al. 2005). A princípio, Stenella frontalis é a
espécie do gênero Stenella mais abundante das regiões Sul e Sudeste do Brasil
(Moreno et al. 2005), ocorrendo predominantemente sobre a plataforma
continental, entre 100 e 200 metros de profundidade (Zerbini et al 2004).
Na região Sul, os encalhes de golfinho-pintado-do-Atlântico acontecem
predominantemente nos meses mais quentes, quando a área está sob maior
influência da Corrente do Brasil (Moreno et al. 2005), deixando a água em média
6oC mais quente que nos meses de inverno. O presente estudo demonstra uma
tendência inversa para região Sudeste, onde 80% dos encalhes na costa centro-
norte do Estado do Rio de Janeiro ocorreram no inverno, e nenhum encalhe foi
registrado durante o verão.
A distribuição dos encalhes está restrita ao sul da área de estudo (entre
Armação dos Búzios e Saquarema) (Figura 54), onde águas de ressurgência são
predominantes. Apesar de haver registros de capturas acidentais desta espécie
no norte do Estado do Rio de Janeiro (Di Beneditto 2003), aparentemente ela está
mais concentrada nas proximidades da Região dos Lagos. Moreno et al. (2005)
apontam uma preferência desta espécie por áreas influenciadas por correntes
quentes e por áreas de ressurgência.

74
Golfinho-pintado-pantropical (Stenella attenuata):
Foram registrados apenas dois encalhes desta espécie na área de estudo
(Figura 49). Os dois espécimes eram machos, um deles encalhou em Arraial do
Cabo e tinha 2 metros de comprimento total, enquanto o outro tinha 1,64 metros e
seu encalhe foi registrado em Araruama. Os dois encalhes ocorreram no mês de
junho (inverno) nos anos de 2001 e 2006.
Figura 48: Golfinho-pintado-pantropical em vida livre.
[Obtido de www.flickr.com/photos/chrismorgan/2318309082]
Figura 49: Série temporal de encalhes de golfinho-pintado-pantropical na costa centro-norte do
Estado do Rio de Janeiro, entre fevereiro de 1993 e junho de 2010.

75
O golfinho-pintado-pantropical se distribui em todas as zonas oceânicas
equatoriais e subtropicais, entre os 400 N e 400 S (Bastida et al. 2007). As
populações mais abundantes se concentram em águas tropicais de baixa latitude
(Bastida et al. 2007).
No Brasil, existem registros desde o nordeste até o Rio de Janeiro, que
parece ser o limite sul de distribuição da espécie no Oceano Atlântico Sul
Ocidental (Siciliano et al. 2006).
Habita águas profundas (Siciliano et al. 2006), normalmente além do
talude, o que justifica o pequeno número de encalhes desta espécie. Na Bacia de
Campos, esta espécie pode ser avistada próxima a plataformas de petróleo, em
águas de 500 a 1.000 metros de profundidade (Siciliano et al. 2006). Apenas dois
encalhes foram registrados até o presente momento na costa centro-norte
fluminense.

76
Golfinho-de-clymene (Stenella clymene):
Entre 1993 e 2010 foi registrado apenas um encalhe desta espécie na
costa centro-norte do Estado do Rio de Janeiro (Figura 51 e 54). Esse encalhe
ocorreu em abril de 2010 e se tratava de um macho adulto de 2,02 metros de
comprimento total.
Figura 50: Espécime de golfinho-de-Clymene encalhado [GEMM 189 – 16/04/2010 – Praia de
Tatagiba/São Francisco do Itabapoana].
Figura 51: Série temporal de encalhes de golfinho-de-Clymene na costa centro-norte do Estado do
Rio de Janeiro, entre fevereiro de 1993 e junho de 2010.

77
Esta espécie de golfinho é uma das menos conhecidas do gênero Stenella
(Fertl et al. 2003). Sua distribuição está restrita a águas tropicais e temperadas
quentes do Oceano Atlântico, habitando predominantemente águas oceânicas
(Bastida et al. 2007).
No Brasil, a maioria dos encalhes são registrados na região Nordeste, além
de escassos registros para a região Sul (Moreno et al. 2005).
Até recentemente não existiam registros de encalhes nem de avistagens
confirmadas desta espécie para a Bacia de Campos, mas sua ocorrência era
considerada provável (Siciliano et al. 2006). O presente trabalho apresenta o
primeiro registro de encalhe desta espécie para a região Sudeste do Brasil.
Este encalhe foi registrado em abril de 2010, no extremo norte do Estado
do Rio de Janeiro, em São Francisco do Itabapoana (Figura 54). Tratava-se de
um espécime macho adulto com comprimento total de 2,02 metros, o maior já
registrado para a espécie. A identificação da espécie foi confirmada a partir da
examinação da morfologia e medidas do crânio.

78
Golfinho-de-Fraser (Lagenodelphis hosei):
No presente estudo, foram registrados dois encalhes desta espécie (Figura
53), um em Armação dos Búzios e o outro em Arraial do Cabo (Figura 54). Os
dois encalhes aconteceram no mês de setembro, nos anos de 1999 e 2004. Um
dos espécimes estava em avançado estágio de decomposição, não sendo
possível identificar o sexo e o comprimento total, o outro foi identificado como um
macho de 2,2 metros de comprimento.
Figura 52: Espécime de golfinho-de-Fraser encalhado. [Arquivo NEMA - RS].
Figura 53: Série temporal de encalhes de golfinho-de-Fraser na costa centro-norte do Estado do
Rio de Janeiro, entre fevereiro de 1993 e junho de 2010.

79
O golfinho-de-Fraser está distribuído por todos os oceanos do mundo, em
águas temperadas e tropicais entre 300N e 300S (Bastida et al. 2007). É
considerada uma espécie freqüente em área oceânicas do Oceano Pacífico,
Índico e Atlântico, geralmente em águas com mais de 1.000 metros de
profundidade (Bastida et al. 2007).
Avistagens costeiras só são possíveis em regiões onde o talude se
encontra muito próximo da costa, como por exemplo, na Indonésia (Bastida et al.
2007).
A espécie foi registrada pela primeira vez na costa brasileira em 1997,
quando ocorreu uma série de encalhes de animais vivos no Rio Grande do Sul e
no Rio de Janeiro (Moreno et al. 2003.). Depois desse evento, foram registrados
encalhes esporádicos ao longo da costa, como os apresentados neste trabalho.
Além do pequeno número de encalhes, não existem registros publicados
de avistagens desta espécie em vida livre para o Brasil, o que sugere que
Lagenodelphis hosei possa ser uma espécie relativamente rara em águas
brasileiras.

80
Figura 54: Mapa da costa centro-norte fluminense com a distribuição dos encalhes de golfinhos do
gênero Stenella e golfinho-de-Fraser.

81
Golfinho-comum (Delphinus sp.):
Neste levantamento, foram registrados oito encalhes de golfinho-comum
(Figura 56) e todos estavam distribuídos na porção mais ao sul da área de estudo,
entre Cabo Frio e Saquarema (Figura 57). Um dos espécimes foi identificado
como uma fêmea, enquanto os outros não tiveram o sexo determinado em
decorrência do avançado estágio de decomposição. Foram registrados encalhes
em todas as estações do ano (Figura 58). O comprimento total dos espécimes
variou entre 1,68 e 1,86 metros.
Figura 55: Espécime de golfinho-comum encalhado. [GEMM 221 – 19/09/2010 – Praia de Monte
Alto/Arraial do Cabo].
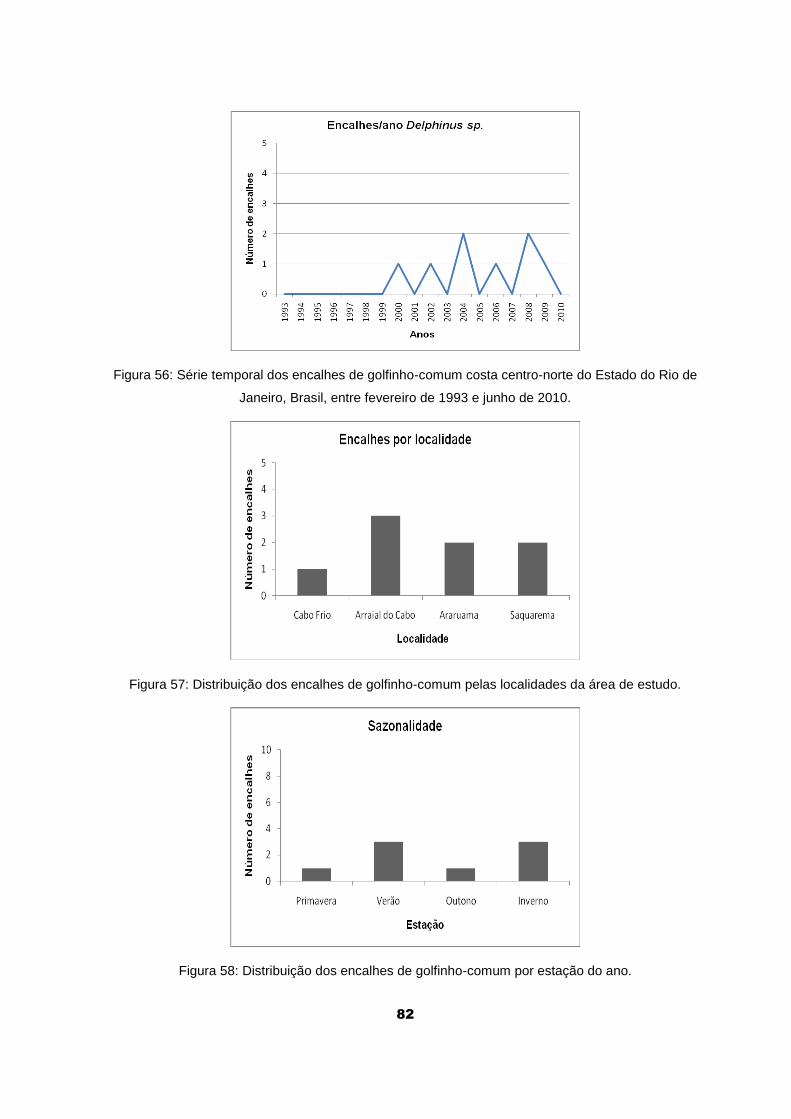
82
Figura 56: Série temporal dos encalhes de golfinho-comum costa centro-norte do Estado do Rio de
Janeiro, Brasil, entre fevereiro de 1993 e junho de 2010.
Figura 57: Distribuição dos encalhes de golfinho-comum pelas localidades da área de estudo.
Figura 58: Distribuição dos encalhes de golfinho-comum por estação do ano.

83
O gênero Delphinus é atualmente dividido em duas espécies, golfinho
comum-de-bico-longo (Delphinus capensis) e o golfinho comum-de-bico-curto
(Delphinus delphis). Há pouco tempo atrás, acreditava-se que as duas espécies
apresentavam ecótipos próprios distintos, com características morfológicas que os
diferenciavam basicamente, como por exemplo: padrão de coloração, número de
dentes e razão rostral (Bastida et al. 2007, Heyning e Perrin 1994).
Porém, os recentes estudos têm demonstrado que a taxonomia aplicada
para estas espécies não é adequada (Tavares et al. 2010). Ambas as espécies se
distribuem pelos dois hemisférios, em águas tropicais e subtropicais (Bastida et al.
2007).
Apesar de Delphinus capensis ocorrer prodominantemente em áreas águas
costeiras e Delphinus delphis estar presente em diversas regiões pelágicas, no
Atlântico Ocidental Sudoeste as espécies são simpátricas e apresentam três
estoques distintos (Tavares et al. 2010). Segundo Tavares et al. (2010) existem
dois padrões diferentes de habitats utilizados por Delphinus sp. no Brasil. Na
região Sudeste, as avistagens desses indivíduos são restritas a área entre 18 e
70 metros de profundidade, enquanto que entre a região Sul do Brasil e a
Argentina Central a grande maioria das avistagens são registradas em locais de
maior profundidade, entre 71 e 1435 metros. Um dos estoques identificados está
associado à ressurgência de Cabo Frio, o que é confirmado pelos resultados
apresentados no presente estudo.
Os encalhes de Delphinus sp. na costa centro-norte fluminense se
restringem a área entre Cabo Frio e Saquarema (Figura 59), refletindo assim a
associação deste estoque com o processo de ressurgência da região. Os
encalhes desta espécie podem ser registrados ao longo de todas as estações do
ano, assim como avistagens nas proximidades de Arraial do Cabo (Siciliano et al.
2006).

84
Figura 59: Mapa da costa centro-norte fluminense com a distribuição dos encalhes de golfinho-
comum.

85
Orca (Orcinus orca):
Esta espécie foi representada por dois encalhes na área de estudo (Figura
63), um em Arraial do Cabo e o outro em Cabo Frio (Figura 66). O primeiro era
uma fêmea de 4,69 metros de comprimento total e o segundo se tratava de um
macho juvenil com 3,5 metros de comprimento total.
Figura 62: Grupo de orcas avistado em Arraial do Cabo [Arquivo GEMM-Lagos].
Figura 63: Série temporal dos encalhes de orca na costa centro-norte do Estado do Rio de
Janeiro, Brasil, entre fevereiro de 1993 e junho de 2010.

86
A orca é uma das espécies de cetáceo mais bem estudadas no mundo. É a
espécie de maior distribuição geográfica, presentes em todos os oceanos.
Embora sejam cosmopolitas, as orcas são mais abundantes em áreas costeiras
temperadas frias de alta produtividade (Bastida et al. 2007).
No oceano Atlântico Sul Ocidental, as orcas habitam preferencialmente
águas temperadas, desde a Argentina até o Sudeste do Brasil (Bastida et al.
2007).
No Brasil, as orcas têm sido registradas tanto em águas afastadas da costa
desde a região Nordeste até o Rio Grande do Sul (Ott & Danilewicz 1997; Secchi
& Vaske Jr 1998), quanto em águas costeiras nas regiões Sul e Sudeste (Siciliano
et al. 1999).
Nas penínsulas de Búzios e Arraial do Cabo é possível avistar
eventualmente orcas passando próximo dos costões, onde inclusive, orcas
acompanhadas de filhotes já foram fotografadas (Figura 62).
Aparentemente, as orcas não são abundantes na região, o que é refletido
pelo pequeno número de encalhes desta espécie.

87
Orca-pigméia (Feresa attenuata):
Apenas um encalhe de orca-pigméia foi registrado na área de estudo
desde 1993 (Figura 65). Este encalhe foi registrado em junho de 2007
(Saquarema) e representa o único encalhe conhecido de Feresa attenuata para a
costa do Estado do Rio de Janeiro.
Figura 64: Orca-pigméia em vida livre.
[Obtido de http://www.saveyourlogo.org/en/encyclopedia/species/8551]
Figura 65: Série temporal dos encalhes de orca-pigméia na costa centro-norte do Estado do Rio
de Janeiro, Brasil, entre fevereiro de 1993 e junho de 2010.

88
A orca-pigméia é um golfinho oceânico que habita águas tropicais e sub-
tropicais, entre 400 N e 350 S, sendo encontrado no Oceano Pacífico, Índico e
Atlântico (Bastida et al. 2007).
Apesar do constante aumento do esforço em pesquisas de campo com
mamíferos marinhos, pouco se sabe sobre a distribuição, biologia e status de
conservação desta espécie. A maior parte da informação conhecida a respeito
desta espécie provém de eventos de encalhe ou captura acidental.
O encalhe registrado neste estudo foi previamente descrito por De Moura
et al. (2010), de modo a inferir que a região próxima do encalhe (Figura 66)
apresenta um conjunto de características particulares (plataforma continental
estreita e abundância de recursos alimentares associados ao processo de
ressurgência), as quais oferecem condições ideais para que várias espécies
tipicamente oceânicas se aproximem da costa.

89
Baleia-piloto-de-peitorais-curtas (Globicephala macrorhynchus):
Foram registrados dois encalhes desta espécie (Figura 61), ambos em
Arraial do Cabo (Figura 66). Os dois registros aconteceram no inverno dos anos
de 2000 e 2004. Um dos espécimes era um macho de 4,15 metros de
comprimento total, enquanto o outro não pode ter o sexo e o comprimento
identificados devido ao avançado estágio de decomposição.
Figura 60: Baleia-piloto-de-peitorais-curtas em vida livre.
[Obtido de www.animal.discovery.com/tvblood-dolphins/dolphins10-targeted-dolphins-04.html]
Figura 61: Série temporal dos encalhes de baleia-piloto-de-peitorais-curtas na costa centro-norte
do Estado do Rio de Janeiro, Brasil, entre fevereiro de 1993 e junho de 2010.

90
A baleia-piloto-de-peitorais-curtas habita águas tropicais e temperadas de
todos os oceanos entre 500N e 450S (Bastida et al. 2007).
No Atlântico Sul Ocidental, onde a espécie tem seu limite de distribuição na
latitude de 250S, o primeiro registro da baleia- piloto de peitorais-curtas só
aconteceu em 1986, quando um crânio foi encontrado na Ilha Comprida, costa do
Estado de São Paulo (Schmiegelow & Paiva-Filho 1989).
A outra espécie do mesmo gênero (Globicephala melas) passa a ser
predominante em águas mais frias ao sul de São Paulo, onde já não existem
registros reconhecidos de Globicephala macrorhynchos.
Os encalhes conhecidos desta espécie para a costa centro-norte do Estado
do Rio de Janeiro estão apresentados neste trabalho. Assim como a orca e a
orca-pigméia, esta espécie parece ser atraída pelo o ambiente de ressurgênica
(onde os encalhes foram registrados), devido à abundância de recursos
alimentares e ao perfil batimétrico peculiar.

91
Figura 66: Mapa da costa centro-norte fluminense com a distribuição dos encalhes de baleia-
piloto-de-peitorais-curtas, orca e orca-pigméia.

92
Família Physeteridae
Cachalote (Physeter macrocephalus):
Esta espécie teve apenas dois encalhes registrados na área de estudo
(Figura 68). Um macho de 12 metros de comprimento total encalhou em março de
1999 em Arraial do Cabo, e um neonato de 3,26 metros encalhou em 2005 em
Rio das Ostras (Figura 67).
Figura 67: Espécime de cachalote encalhado. [GEMM 092 – 07/11/2005 - Praia do Centro/Rio das
Ostras].
Figura 68: Série temporal dos encalhes de cachalote na costa centro-norte do Estado do Rio de
Janeiro, Brasil, entre fevereiro de 1993 e junho de 2010.

93
Os cachalotes representam os maiores cetáceos odontocetos viventes.
Apresentam distribuição cosmopolita, sendo encontrados do Ártico até a Antártida
(Bastida et al. 2007). Em geral, as fêmeas e machos jovens permanecem entre
40oN e 400S e os machos adultos migram para águas polares no verão (Bastida et
al. 2007).
Os cachalotes habitam predominantemente regiões oceânicas de grande
profundidade e as áreas de maior concentração estão localizadas em águas
oceânicas tropicais e subtropicais de alta produtividade, muitas das vezes onde
existem canyons submarinos onde eles encontram lulas como alimento (Bastida
et al. 2007).
No Brasil, existem registros de cachalotes desde o Rio Grande do Sul até o
Ceará (Clarke et al. 1980; Ramos et al. 2001). Os encalhes de um adulto e de um
neonato na área de estudo comprovam que a Bacia de Campos é área de
ocorrência da espécie.

94
Família Kogiidae
Cachalote-pigmeu (Kogia breviceps):
Foram registrados três encalhes desta espécie (Figura 70 e 73), em Macaé
(Figura 69), Arraial do Cabo e Saquarema. No ano de 2005 ocorreu dois encalhes
desta espécie, um durante o mês de setembro e o outro em dezembro. Um dos
espécimes era uma fêmea, enquanto os outros não tiveram o sexo identificado.
Figura 69: Espécime de cachalote-pigmeu encalhado [GEMM 192 – 19/05/2010 – Praia do Parque
Nacional de Jurubatiba/Macaé].
Figura 70: Série temporal dos encalhes de cachalote-pigmeu na costa centro-norte do Estado do
Rio de Janeiro, Brasil, entre fevereiro de 1993 e junho de 2010.

95
O cachalote-pigmeu se distribui por todos os oceanos tropicais,
subtropicais e temperados do mundo, habitando predominantemente águas
oceânicas profundas além da margem da plataforma continental (Caldwell and
Caldwell 1989; Lucas and Hooker 2000; Bastida et al. 2007).
Segundo Duguy (1994), o cachalote pigmeu pode realizar migração
costeira-oceânica sazonalmente, o que explica a sazonalidade dos encalhes em
algumas regiões do mundo.
Aparentemente registros de encalhe de Kogia breviceps e Kogia sima são
igualmente escassos ao longo da costa Atlântica da América do Sul (Muñoz-
Hincapié et al. 1998). Os registros dessa espécie na área de estudo não permitem
maiores conclusões.

96
Cachalote-anão (Kogia sima):
Esta espécie foi representada por dois encalhes (Figura 72). Um encalhe
ao norte da área de estudo, em Campos dos Goytacazes (2 metros de
comprimento total) e um em Cabo Frio (Figura 73).
Figura 71: Cachalote-anão em vida livre
[Obtido de www.cms.intreportssmall_cetaceans/dataK_simaK_sima.htm]
Figura 72: Série temporal dos encalhes de cachalote-anão na costa centro-norte do Estado do Rio
de Janeiro, Brasil, entre fevereiro de 1993 e junho de 2010.

97
O cachalote-anão é uma espécie oceânica que se distribui por todos os
oceanos tropicais, subtropicais e temperados do mundo (Bastida et al. 2007).
Aparentemente, Kogia sima tem uma potencial preferência por águas
ligeiramente mais quentes, próximo da plataforma continental, e não tende a
realizar migrações de maior escala (Caldwell and Caldwell 1989; Duguy 1994;
Lucas and Hooker 2000).
Na América do Sul existem registros de sua presença em todos os países
costeiros do Oceano Atlântico e do Oceano Pacífico (Bastida et al. 2007), equanto
que no Brasil, existem registros de encalhes desde o Rio Grande do Sul até o
Ceará (Pinedo, M. C. 1987; Meireles et al. 2009).
A maior parte da informação das espécies do gênero Kogia, é oriunda de
eventos de encalhe, os quais são responsáveis por propiciar a informação básica
necessária ao conhecimento sobre a biologia, distribuição e ecologia destas
espécies incomuns. Apesar da preferência por águas mais afastadas do
continente, ambas as espécies possuem registros relativamente freqüentes de
animais encalhados ainda vivos e principalmente de encalhes de fêmeas grávidas
e fêmeas com filhotes.

98
Figura 73: Mapa da costa centro-norte fluminense com a distribuição dos encalhes de cachalote,
cachalote-pigmeu e cachalote-anão.

99
Família Pontoporiidae
Toninha (Pontoporia blainvillei):
Foram registrados 13 encalhes desta espécie (Figura 75), distribuídos entre
São Francisco do Itabapoana e Barra de São João (Figura 76). Mais de 50% dos
encalhes se concentraram nas proximidades da foz do Rio Paraíba do Sul.
Registrou-se encalhes em todas as estações do ano, com uma maior incidência
no verão e no outono (Figura 77). Foram identificados três espécimes machos,
cinco fêmeas e em cinco exemplares foi inviável fazer a sexagem devido ao
avançado estágio de decomposição. Em três espécimes foi comprovada a
interação negativa com artefatos de pesca de acordo com marcas indicativas no
corpo. Os espécimes encalhados variaram entre 0,68 e 1,31 metros de
comprimento total.
Figura 74: Espécime de toninha encalhado [GEMM 199 – 18/07/2010 – Praia de
Fleicheiras/Quissamã].

100
Figura 75: Série temporal dos encalhes de toninha na costa centro-norte do Estado do Rio de
Janeiro, Brasil, entre fevereiro de 1993 e junho de 2010.
Figura 76: Distribuição dos encalhes de toninha pelas localidades da área de estudo.
Figura 77: Distribuição dos encalhes de toninha por estação do ano.

101
A toninha é um golfinho de pequeno porte endêmico da costa
atlântica da América do Sul. Esta espécie ocorre entre Itaúnas, Espírito Santo
(Siciliano, 1994) até o Golfo San Matias, na Província de Chubut na Argentina
(Crespo et al. 1998). Seu hábitat preferencial inclui regiões estuarinas e costeiras,
com menos de 50 metros de profundidade, entretanto, a maior parte dos registros
se dá em águas mais rasas, de até 30 metros de profundidade (Di Beneditto &
Ramos, 2001). A preferência por águas rasas próximas da costa faz com que a
toninha seja vulnerável às atividades humanas (Danilewicz et al. 2009).
A distribuição da toninha não é contínua ao longo da costa
sulamericana devido à presença de dois hiatos ao longo de sua área de
ocorrência (Siciliano et al. 2002). Acredita-se que os hiatos limitam a distribuição
da toninha devido a três fatores principais: temperatura da água, salinidade e
turbidez.
Com base em uma revisão das informações bioecológicas disponíveis
sobre a espécie, foi proposto a criação de áreas de manejo para a toninha (Secchi
et al. 2003). O Espírito Santo e o norte do Estado do Rio de Janeiro foram
denominados como Área de Manejo I (FMA I).
Os 13 espécimes registrados neste trabalho pertencem ao estoque de
FMA I, porém ainda faltam maiores informações sobre a estruturação
populacional deste estoque. Os encalhes apontam para uma concentração de
encalhes de toninhas na região próxima a desembocadura do Rio Paraíba do Sul
(Figura 78), o que poderia indicar uma população associada a esta região.
Em relação aos espécimes que apresentavam marcas de interação
negativa com artefatos de pesca, o número não reflete a real mortalidade de
toninha na região. Segundo De Moura et al. (2009), o pequeno tamanho do corpo
associado a fatores oceanográficos da região, tais como a dinâmica das correntes
e os perfis/relevos das praias, podem fazer com que as carcaças à deriva não
cheguem a encalhar na costa. Estudos pretéritos mostram que a toninha é a
espécie mais vulnerável à captura acidental em redes de pesca na região (Netto &
Di Beneditto 2008), e de acordo com Di Beneditto (2003), estima-se que as redes
de espera matam acidentalmente 110 toninhas por ano na região, o que causaria
um grande impacto e colocaria em risco a viabilidade desta população.

102
Figura 78: Mapa da costa centro-norte fluminense com a distribuição dos encalhes de toninha.

103
6 – CONCLUSÕES
A costa centro-norte do Estado do Rio de Janeiro é uma área de forte
incidência de encalhes, o que comprova a ocorrência de cetáceos na
região.
A península de Arraial do Cabo e a região próxima a foz do Rio Paraíba do
Sul são as áreas de maior concentração de encalhes de cetáceos na costa.
Encalhes de cetáceos na área de estudo ocorrem predominantemente
durante o inverno devido à ação de frentes frias originadas pelo vento
sudoeste.
O boto-cinza é a espécie com maior freqüência relativa de encalhes devido
ao fato de ser a espécie mais abundante na área de estudo e apresentar
hábitos exclusivamente costeiros. Sua distribuição está associada
principalmente à área de influência da desembocadura do Rio Paraíba do
Sul.
De uma forma geral, os encalhes das espécies de hábitos oceânicos-
costeiros (golfinho-nariz-de-garrafa, golfinho-de-dentes-rugosos, golfinho-
pintado-do-Atlântico e golfinho-comum) refletem fortemente a influência do
fenômeno da ressurgência de Cabo Frio e do estreitamento da plataforma
continental no seu padrão de ocorrência.
Os encalhes de toninha refletem uma forte associação com a foz do Rio
Paraíba do Sul, assim como o boto-cinza, porém, por ser um golfinho de
pequeno porte, dificilmente as carcaças chegam a encalhar e quando
encalham se decompõem rapidamente, diminuindo as chances do registro.
Entre os cetáceos misticetos, a baleia-jubarte e a baleia-de-Bryde são as
espécies com maior freqüência relativa entre os encalhes. A baleia-jubarte
utiliza a costa centro-norte do Estado do Rio de Janeiro como parte de sua
rota migratória, enquanto que os encalhes de baleia-de-Bryde sugerem que
a espécie possui uma população residente na Bacia de Campos.

104
As espécies que apresentaram poucos registros de encalhes são
predominantemente oceânicas, logo, as carcaças raramente se aproximam
da costa.
Os encalhes registrados de orca pigméia e golfinho-de-Clymene
comprovam a ocorrência dessas espécies para a Bacia de Campos e
representam os primeiros registros para a costa do Estado do Rio de
Janeiro.
Este trabalho demonstra a importância da formação de séries temporais de
longo prazo para a melhor interpretação dos parâmetros de ocorrência das
espécies.

105
6. REFERÊNCIAS BIBLIOGRÁFICAS
BASTIDA, R., RODRÍGUEZ, D., SECCHI, E., DA SILVA, V. 2007. Mamiferos
Acuaticos de Sudamerica y Antartida, Vázquez Mazzini Editores, Buenos Aires
v.1, 368p.
BEST, P. B. 1985. External characthers of southern minke whales and the
existence of a diminutive form. Scientific Reports of the Whales Research Institute
36:1-33.
BRABYN, M. W. & MCLEAN, I. G. 1992. Oceanography and coastal topography of
herd-strandings sites for whales in New Zealand. J. Mammal., 73(3): 469-476.
BERROW, S. 2001. Biological diversity of cetaceans (whales, dolphins and
porpoises) in Irish waters. In: J. D. Nunn (ed.) Marine biodiversity in Ireland and
adjacent waters. Belfast: Ulster Museum, pp. 115-120.
BITTENCOURT, M. L. 1984. Contribuições para a identificação de cetáceos na
costa e águas interiores do Brasil. Arquivos de Biologia e Tecnologia do Paraná,
27: 529-547.
CALDWELL, D. K. & CALDWELL, M. C. 1989. Pygmy Sperm Whale Kogia
breviceps (Blainvillei, 1838); Dwarf Sperm Whale Kogia simus (Owen, 1866). In:
Ridgway, S. H., Harrison, S. R. (Eds) Handbook of Marine Mammals Vol.4: River
Dolphins and the Larger Toothed Whales. Academic Press, London, pp. 235-260.
CARVALHO, C. T. 1975. Ocorrências de mamíferos marinhos no Brasil. Boletim
Técnico IF, São Paulo, v.16, p.13-32.
CASINOS, A. & VERICAD, J. R. 1976. The cetaceans of the Spanish coasts: a
survey. Mammalia, 40: 267-269.
CASTELLO, H. P. & PINEDO, M. C. 1977. Botos na Lagoa dos Patos. Natureza
em Revista 2:46-49.

106
CLARKE, M. R., MCLEOD, N., CASTELLO, H. P. & PINEDO, M. C. 1980.
Cephalopod remains from the stomach of sperm whale stranded at Rio Grande do
Sul in Brazil. Marine Biology, 59:235-239.
CRESPO, E. A., HARRIS, G. & GONZÁLES, R. 1998. Group size and distribution
range of the franciscana, Pontoporia blainvillei. Marine Mammal Science 14:845-
849.
DANILEWICZ, D., SECCHI, E. R., OTT, P. H., MORENO, I. B., BASSOI, M. &
BORGES-MARTINS, M. 2009. Habitat use patterns of franciscana dolphins
(Pontoporia blainvillei) off southern Brazil in relation to water depth. Journal of the
Marine Biological Association of the United Kingdom, 89: 943-949
DAWBIN , W. H. 1966. The seasonal migratory cycle of humpback whales . In
“Whales, Dolphins and Porpoises” ( K. S. Norris , ed. ) , pp. 145 – 170 . University
of California Press , Berkeley, CA .
DHERMAIN, F., SOULIER, L. & BOMPAR, J. M. 2002. Natural mortality factors
affecting cetaceans in the Mediterranean Sea. Cetaceans of the Mediterranean
and Black Seas: state of knowledge and conservation strategies. A report to the
ACCOBAMS Secretariat, Monaco, Section 15, 14pp.
DE MOURA, J. F., RODRIGUES, E. D. S., SHOLL, T. G. C. & SICILIANO,S. 2009.
Franciscana dolphin (Pontoporia blainvillei) on the north-east coast of Rio de
Janeiro State, Brazil, recorded during a long term monitoring programme. Marine
Biodiversity Records, v.2:e66 published online. Disponível em
http://www.mba.ac.uk/jmba/pdf/6332.pdf.
DE MOURA, J. F., SHOLL, T. G. C., RODRIGUES, E. D. S., HACON, S. &
SICILIANO, S. 2009. Marine tucuxi dolphin (Sotalia guianensis) and its interaction
with passive gill-net fisheries along the northern coast of the Rio de Janeiro State,
Brazil. Marine Biodiversity Records, v.2:e82 published online. Disponível em
http://www.mba.ac.uk/jmba/pdf/6327.pdf.
DE MOURA, J. F., DI DARIO, B. P. S., LIMA, M. L. & SICILIANO, S. 2010. A
stranded pygmy killer whale on the coast of Rio de Janeiro State, Brazil. Marine

107
Biodiversity Records, v.3:e11 published on line. Disponível em
http://www.mba.ac.uk/jmba/jmba2biodiversityrecords.php.
DI BENEDITTO, A. P. M. & RAMOS, R. 2001. Biology and the conservation of the
franciscana (Pontoporia blainvillei) in the north of Rio de Janeiro, Brazil. Journal of
Cetacean Research and Management, 2(3):185-192.
DI BENEDITTO, A. P. M. & RAMOS, R. 2001. Os cetáceos da Bacia de Campos.
Revista Ciência Hoje, v.29, n.171: 66-70.
DI BENEDITTO, A. P. M. & RAMOS, R. M. A. 2001. Biologia e Conservação de
Pequenos Cetáceos no Norte do Estado do Rio de Janeiro. Série Ciências
Ambientais do Laboratório de Ciências Ambientais, Centro de Biociências e
Biotecnologia, Universidade Estadual do Norte Fluminense, Campos dos
Goytacazes, v.1, 94p.
DI BENEDITTO, A. P. M. 2003. Interaction between gillnet fisheries and small
cetaceans in northern Rio de Janeiro, Brazil: 2001-2002. Latin American Journal
of Aquatic Mammals, 2: 79-86.
DI BENEDITTO, A. P. M. & SICILIANO, S. 2004. Guia para estudo de cetáceos:
Métodos para coleta de amostras em carcaças. Guia de estudos em ciências
ambientais, Campos dos Goytacazes, v.2, 50p.
DI BENEDITTO, A. P. M. & RAMOS, R. M. A. 2004. Biology of the marine tucuxi
dolphin (Sotalia fluviatilis) in south-eastern Brazil. Journal of the Marine Biological
Association of the United Kingdon, 84:1245-1250.
DUGUY, R. 1994. Kogia breviceps (Blainvillei, 1838) - Zwergpottwal In:
Niethammer, J., Krapp, F. (Eds) Handbuch der Saugetiere Europas. Band 6:
Meeressauger. Teil 1B: Wale unbd Delphine 2. Aula-Verlag, Wies-baden,
Germany, pp. 598-614.
DUIGNAN, P. J., HOUSE. C., GERACI, J. R., EARLY, G., COPLAND, H. G.,
WALSH, M. T., BOSSART, G. D., CRAY, C., SADOVE, S., ST.AUBIN, D. J. &

108
MOORE, M. 1995. Morbillivirus infection in two species of pilot whales
(Globicephala sp.) from western Atlantic. Marine Mammal Science,11(2): 150-162.
FERTL, D., JEFFERSON, T. A., MORENO, I. B., ZERBINI, A. N. & MULLIN, K. D.
2003. Distribution of Clymene dolphin Stenella clymene. Mammal Review 33(3):
253-271.
FLORES P. A. C. AND DA SILVA, V. M. F. Tucuxi and Guiana Dolphin Sotalia
fluviatilis and S. guianensis. In: Perrin, W. F., Wursig, B. and Thewissen, J. G. M.
(Ed.) Encyclopedia of Marine Mammals. 2nd ed. San Diego: Academic Press,
2009, p.1188-1191.
GEISE, L. & BOROBIA, M. 1987. New Brazilian records of Kogia, Pontoporia,
Grampus, and Sotalia (Cetacea, Physeteridea, Platanistidea, and Delphinidae).
Journal of Mammalogy, 68: 873-875.
GERACI, J. R., ANDERSON, D. M., TIMPERI, R. J., ST.AUBIN, D.J., EARLY, G.
A., PRESCOTT, J. H. & MAYO, C. A. 1989. Humpback whales (Megaptera
novaeangliae) fatally poisoned by dinoflagellate toxin. Canadian Journal of
Fisheries and Aquatic Sciences, 46:1895-1898.
GERACI, J. R. & ST. AUBIN, D. J. 1979. Biology of marine mammals: Insights
through strandings. National Technical Information Service, Springfield, VA.
343pp.
GERACI, J. R. & LOUNSBURY, V. J. 1993. Marine mammals ashore: A Field
guide for strandings. Texas A&M University Sea Grant College Program,
Galveston, TX. 305pp.
GIANUCA, N. M. & CASTELLO, H. P. 1976. First Record of the southern
bottlenose whale, Hyperodon planifrons from Brazil. Scientific Reports os the
Whales Research Institute, n.28, p.119-126, 1976.
GONZALEZ-RODRIGUEZ, E., VALENTIN, J. L., LACERDA ANDRE, D. & JACOB,
S. A. 1992. Upwelling and downwelling at Cabo Frio (Brazil): comparison of
biomass and primary production responses. Journal of Plankton Research 14(2):
289-306.

109
HASSEL, L. B. 2003. Monitoramento de pequenos cetáceos na costa de Arraial
do Cabo, Rio de Janeiro, com ênfase na ocorrência do golfinho-comum-de-bico-
longo (Delphinus capensis). Trabalho de conclusão de curso. Universidade
Federal do Rio de Janeiro, Brasil.
HASSEL, L. B., VENTUROTTI, A., MAGALHÃES, F. A., CUENCA, S., SICILIANO,
S. & MARQUES, F. F. C. 2003. Summer sightings of dwarf minke whales
(Balaenoptera acutorostrata) off eastern coast of Rio de Janeiro State, Brazil.
Latin American Journal of Aquatic Mammals, 2(1):47-50.
HOFMAN, R. J. 1991. History, goals, and achievements of the regional marine
mammal stranding networks in the United States. In: J. E. Reynolds III & D. K.
Odell (Eds) Marine Mammal Strandings in the United States. NOAA Tchenical
Report NMFS 98. 157pp.
IBAMA Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais
Renováveis. 2001. Mamíferos Aquáticos do Brasil. Plano de ação, 2.ed. Brasília:
Ed. Ministério do Meio Ambiente, 96p.
INTERNATIONAL WHALING COMMISSION 1991. Report of the Sub-Committee
on Southern Hemisphere minke whales. Report of the International Whaling
Commission 41:113-131.
JEFFERSON, T. A., LEATHERWOOD, S. & WEBBER, M. P. 1993. Marine
Mammals of the world. FAO Species Identification Guide, UNEP-FAO, Rome,
320pp.
JEFFERSON, T. A., SCHIRO, A. J. 1997. Distribution of cetaceans in the offshore
Gulf of Mexico. Mammal Review, 27:27-50.
LUCAS, Z. N. & HOOKER, S. K. 2000. Cetacean Strandings on Sable Island,
Nova Scotia, 1970-1998. Canadian Field-Naturalist, 114: 45-61.
JENSEN, A. S. & SILBER, G. K. 2003. Large whale ship strike database. NOAA
Technical Memorandum NMFSOPR. 37pp.

110
KATONA, S., WHITEHEAD, H. 1988. Are cetacea ecologically important ?
Oceanography and Marine Biology: An Annual Review 26: 553-568.
KLINOWSKA, M. 1986. The cetacean magnetic sense – evidence from strandings.
In: M. M. Bryden & R. J. Harrison (Eds) Research on Dolphins. Oxford University
Press, U. K. 428pp.
KLINOWSKA, M. 1991. Cetacean live stranding dates relate to geomagnetic
disturbances. Aquatic Mammals, 11: 109-119.
KRUGER, G. C. T., CARVALHO, C. E. V. & SUZUKI, M. S. 2006. Dissolved
nutrient, chlorophyll a and DOC dynamic under distinct riverine discharges and
tidal cycles regimes at the Paraiba do Sul river estuary, RJ. Brazil. Journal of
Coastal Research 39: 724-730.
LEATHERWOOD, S. & REEVES, R. R. 1983. The Sierra Club Handbook of
Whales and Dolphins. Sierra Club Books, San Francisco, 302pp.
LOCKYER, C. 1984. Review of baleen whale (Mysticeti) reproduction and
implications for management. In: W. F. Perrin, R. L. Brownell Jr and D. P
DeMaster (Eds) Reproduction in whales, dolphins and porpoises. Int.Whal.
Commn, Cambridge, U.K., special issue 6. 495pp.
MARINE MAMMALS COMMISSION. 1999. Marine Mammals and persistent
ocean contaminants. O’Shea, T. J.; Reeves, R. R. & Long, A. K. (Eds). In:
Proceedings of the Marine Mammals Commission Workshop, Keystone, 150pp.
MATSUURA, Y. 1995. Exploração Pesqueira. In: Os ecossistemas brasileiros e os
principais macrovetores de desenvolvimento: subsídios ao planejamento da
gestão ambiental. Secretaria de Coordenação dos Assuntos de Meio Ambiente.
PNMA-MMA, Brasília, pp.1-15.
MEIRELLES, A. C. O., MONTEIRO-NETO, C., MARTINS, A. M. A., COSTA, A. F.,
BARROS, H. M. D. R. & ALVES, M. D. O. 2009. Cetacean strandings on the coast
of Ceará, north-eastern Brazil (1992 – 2005). Journal of Marine Biological
Association of U. K. 89(5):1083-1090.

111
MORENO, I. B., DANILEWICZ, D., BORGES-MARTINS, M., OTT, P. H., CAON,
G. & OLIVEIRA, R. L. 2003. Fraser`s dolphin (Lagenodelphis hosei Fraser, 1956)
in southern Brazil. Latin American Journal of Aquatic Mammals. 2(1):39-46.
MORENO, I. B., ZERBINI, A. N., DANILEWICZ, D., SANTOS, M. C. O., SIMÕES-
LOPES, P. C., LAÍLSON-BRITO, J. & AZEVEDO, A. 2005. Distribution and habitat
characteristics of dolphins of the genus Stenella (Cetacea: Delphinidae) in the
southwest Atlantic Ocean. Marine Ecology Progress Series, 300:229-240.
MUÑOZ-HINCAPIÉ, M. F., MORA-PINTO, D. M., PALACIOS, D. M., SECCHI, E.
R. & MIGNUCCI-GIANNONI, A. A. 1998. First osteological record of the dwarf
sperm whale in Colombia, with notes on the zoogeography of Kogia in South
America. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y
Naturales, 22(84):433-444.
NETTO, R. F. & DI BENEDITTO, A. P. M. 2008. Interactions between fisheries
and cetaceans in Espírito Santo State, southeastern Brazil. Revista Brasileira de
Zoociências, 10:55-63.
NOREZ, C. & PEREZ, C. 1988. Multiple strandings of Stenella coeruleoalba and
Globicephala macrorhyncus on the coast of Spain. European Research on
Cetaceans, 2: 25-26.
NORMAN, S. A., BOWLBY, C. E., BRANCATO, M. S., CALAMBOKIDIS, J.,
DUFFIELD, D., GEARIN, P. J., GORNALL, T. A., GOSHO, M. E., HANSON, B.,
JEFRIES, S. J., LAGERQUIST, B., LAMBOURN, D.M., MATE, B., NORBER, B.,
OSBORNE, R. W., RASH, J. A., RIEMER, S. & SCORDING, J. 2004. Cetacean
strandings in Oregon and Washington between 1930 and 2002. Journal of
Cetacean Research and Management, 6:87-99.
NORRIS, K. 1961. Standardized methods for measuring and recording data on the
smaller cetaceans. Journal of Mammalogy, 42(4): 471-6.
NORSE, E. A. 1993. Global marine biological diversity: a strategy for building
conservation into decision making. Washington: Island Press. 383pp.

112
NOWAK, R. M. 2003. Walker’s Marine Mammals of the World. The Johns Hopkins
University Press, Baltimore, 264 pp.
ODELL, D. K. 1991. A review of the southeastern United States marine mammal
stranding network: 1978-1987. In: J. E. Reynolds III & D. K. Odell (Eds) Marine
Mammal Strandings in the United States. NOAA Tchenical Report NMFS 98.
157pp.
OTT, P. H. & DANILEWICZ, D. 1997. Presence of franciscana dolphins
(Pontoporia blainvillei) in the stomach of a killer whale (Orcinus orca) stranded in
southern Brazil. Mammalia, 62(4):605-609.
PARSONS, T. R. 1992. The removal of marine predators by fisheries and the
impacts of thophic structure. Marine Pollution Bulletin, 24(1-4): 51-53.
PERRIN, W. F., DONOVAN, G. P. & BARLOW, J. (Eds) 1994. Gillnets and
cetaceans. Int. Whal. Comm, Cambridge, U.K., special issue 15. 629pp.
PERRIN, W. F. & REILLY, S. B. 1984. Reproductive parameters of dolphins and
small whales of the family Delphinidae. In: W. F. Perrin, R. L. Brownell Jr & D. P.
DeMaster (Eds) Reproduction in whales, dolphins and porpoises. Int. Whal.
Commn, Cambridge, U.K., special issue 6. 495pp.
PEZZINI, A. 2006. Caracterização da variação sazonal do transporte de materiais
dissolvidos na saída da Bacia do Rio Paraíba do Sul, Campos dos Goytacazes,
RJ. Trabalho de Monografia. Universidade Estadual do Norte Fluminense Darcy
Ribeiro. 56pp.
PINEDO, M. C. 1987. First record of dwarf sperm whale from southwest Atlantic,
with reference to osteology, food habitats and reproduction. Scientific Reports
Whales Research Institute Tokyo, 38:171-186.
RAMOS, R., SICILIANO, S., BOROBIA, M., ZERBINI, A. N., PIZZORNO, J. L.,
FRAGOSO, A. B. L., BRITO JR, J. L., AZEVEDO, A. F., SIMÕES-LOPES, P. C. &
SANTOS, M. C. O. 2001. A note on strandings and age of sperm whales on the
Brazilian Coast. Journal of Cetacean Research and Management, 3(3):321-327.

113
READ, A. J., DRINKER, P. & NORTHRIDGE, S. 2003. By-catches of marine
mammals in US fisheries and a first attempt to estimate the magnitude of global
marine mammal by-catch. Cambridge, U.K: International Whaling Commission.
Scientific Committee Document SC/55/BC5.
REYNOLDS III, J. E. & ODELL, D. K. 1991. (Eds) Marine Mammal Strandings in
the United States, NOAA Technical Report, NMFS 98. 157pp.
RICKLEFS, R. E. 2010. A Economia da Natureza. Rio de Janeiro: Guanabara
Koogan 6ª Ed.,p.63-66.
RIDGWAY, S. H. & DAILEY, M. D. 1972. Cerebral and cerebellar involvement of
trematode parasites in dolphins and their possible role in stranding. J. Wild. Dis.,
8: 33-43.
SAMUELS, A. & TYACK, P. 2000. Flukeprints: a history of studying cetacean
societies. In: J. Mann, R. C. Connor, P. L. Tyack and H. Whitehead (Eds),
Cetacean Societes: Field Studies of Dolphins and Whales, 9-44. University of
Chicago Press, Chicago.
SANTOS, M. C. 1999. Novas informações sobre cetáceos no litoral sul de São
Paulo e norte do Paraná com base em estudos sobre encalhes e na aplicação da
técnica de foto-identificação individual de Sotalia fluviatilis (Cetacea, Delphinidae).
Dissertação de mestrado. Universidade de São Paulo, Brasil.
SCHMIEGELOW, J. M. M. 1990. Estudo sobre cetáceos odontocetes encontrados
em praias da região entre Iguapé (SP) e Baía de Paranaguá (PR) com especial
referência a Sotalia guianensis (Gervais, 1853) (Delphinidae). Dissertação de
Mestrado. Universidade de São Paulo, Brasil.
SECCHI, E. R. & VASKE JR, T. 1998. Killer whale (Orcinus orca) sightings and
depredation on tuna and sword-fish longline catches in southern Brazil. Aquatic
Mammals 21(2):105-108.
SECCHI, E. R., DANILEWICZ, D. & OTT, P. H. 2003. Applyng the
phylogeographic concept to identify franciscana dolphin stocks: implications to

114
meet management objectives. Journal of Cetacean Research and Management. 5
(1): 61-68.
SICILIANO, S. 1994. Review of small cetaceans and fishery interactions n coastal
Waters of Brazil. Reporto of International Whaling Commission. Special Issue,
15:241-245.
SICILIANO, S. 1997. Características da população de baleias-jubarte (Megaptera
novaeangliae) da costa brasileira, com especial referência aos Bancos de
Abrolhos. Dissertação de Mestrado. Universidade Federal Rural do Rio de
Janeiro, Brasil.
SICILIANO, S., LAÍLSON-BRITO, J. & AZEVEDO, A. 1999. Seasonal ocurrence of
killer whales (Orcinus orca) in waters of Rio de Janeiro, Brazil. Mammalian Biology
64(4):251-255.
SICILIANO, S., DI BENEDITTO, A. P. M. & RAMOS, R. M. A. 2002. A toninha,
Pontoporia blainvillei (Gervais & d’Orbigny, 1844) (Mammalia, Cetacea,
Pontoporiidae), nos Estados do Rio de Janeiro e Espírito Santo, costa sudeste do
Brasil: caracterização dos habitats e fatores de isolamento das populações.
Boletim do Museu Nacional (Zoologia), 476:1-15.
SICILIANO, S., SANTOS, M. C. O., VICENTE, A. F. C., ALVARENGA, F. S.,
ZAMPIROLLI, E., LAÍLSON-BRITO, J., AZEVEDO, A. F. & PIZZORNO, J. L. A.
2004. Strandings and feeding records of Bryde’s whales (Balaenoptera edeni) in
south-eastern Brazil. Journal of the Marine Biological Association of the United
Kingdon. 84:857-859.
SICILIANO, S., MORENO, I. B., DEMARI, E., ALVES, V. C. 2006. Baleias, botos e
golfinhos na Bacia de Campos, Série Guias de Campo: fauna marinha da Bacia
de Campos, Rio de Janeiro: ENSP/FIOCRUZ, 99p.
SIMÕES-LOPES, P. C., FABIÁN, M. E. & MENEGUETI, J. O. 1998. Dolphin
interactions with the mullet artisanal fishing on Southern Brazil: a quantitative and
qualitative approach. Revista Brasileira de Zoologia 15:709-726.

115
STEVICK, P. T., MCCONNELL, B. J. & HAMMOND, P. S. 2002. Patterns of
movement. In A. R. Hoezel (ed.), Marine Mammal Biology: An Evolutionary
approach, 185-216. Blackwell Science, Oxford.
TAVARES, M., MORENO, I. B., SICILIANO, S., RODRÍGUEZ, D., SANTOS, M. C.
O., LAÍLSON-BRITO, J. & FABIÁN, M. E. 2010. Biogeography of common
dolphins (genus Delphinus) in the Southwestern Atlantic Ocean. Mammal Review,
1:40-64.
VALENTIN, J. L. 1988. A dinâmica do plâncton na ressurgência de Cabo Frio –
RJ. Memórias do III Encontro Brasileiro de Plâncton. Cuiabá – Brasil. pp.25-35.
VALENTIN, J. L. 2001. The Cabo Frio Upwelling System, Brazil. In: U. Seeliger, B.
Kjerfve (eds) Ecological Studies: Coastal Marine Ecosystems of Latin America, vol
144. Springer-Verlag, Berlin Heidelberg, pp.97-105.
VAN BRESSEM, M. F., WAEREBEEK, K. V., RAGA, J. A., GASPAR, R., DI
BENEDITTO, A. P., RAMOS, R., SIEBERT, U. 2003. Tattoo diseases of
odontocetes as a potential indicator of a degrading or stressful enviromment: a
preliminary report. Internacional Whaling Commission, Berlin, 6pp.
MORENO, I. B., OTT. P. H., TAVARES, M., OLIVEIRA, L. R., DANILEWICZ, D.,
SICILIANO, S. & BONNATO, S. L. 2009. Os cetáceos com ênfase no golfinho-
nariz-de-garrafa, Tursiops truncatus (Montagu, 1821). In: Viana, D. L., Hazin, F. H.
V., De Souza, M. A. C. O Arquipélago de São Pedro e São Paulo: 10 anos de
Estação Científica. Brasília, DF: SECIRM.
XIMENEZ, A., SIMÒES-LOPES, P. C. & PRADERI, R. 1987. Notas sobre
mamíferos de Santa Catarina e Rio Grande do Sul (Pinnipedia-Cetacea). In
Abstracts of the II Reunião de Trabalhos de Especialistas em Mamíferos
Aquáticos da América do Sul, Rio de Janeiro, Brazil, pp. 100-103.
ZERBINI, A. N., SECCHI, E. R., SICILIANO, S. & SIMÕES-LOPES, P. C. 1996.
The dwarf form of the minke whale, Balaenoptera acurostrata (Lacépède, 1804) in
Brazil. Report of the International Whaling Commission 46:333-340.

116
ZERBINI, A. N., SECCHI, E. R., SICILIANO, S. & SIMÕES-LOPES, P. C. 1997.
Review of the ocurrence and distribution of whales of the genus Balaenoptera
along the Brazilian coast. Report of the International Whaling Commission 47:407-
417.
ZERBINI, A. N., SECCHI, E. R., BASSOI, M., DALLA-ROSA, L., HIGA, A., DE
SOUSA, L., MORENO, I. B., MOLLER, L. M. & CAON, G. 2004. Distribuição e
abundância relativa de cetáceos na Zona Econômica Exclusiva na Região
Sudeste-Sul do Brasil. Série Documentos Revizee – Score Sul. Instituto
Oceanográfico, Universidade de São Paulo, 40p.









![A sociedade subaquática do Boto-Cinza · 1 0 [Entrevista] sobre o comportamento de cetáceos. 12 O som na vida dos cetáceos. 14 A sociedade subaquática do Boto-Cinza. 19 Golfinhos-rotadores:](https://img.document.onl/doc/110x75/5f54d08725c3e16a50094f8f/a-sociedade-subaqutica-do-boto-cinza-1-0-entrevista-sobre-o-comportamento-de.jpg)


















