Embed Size (px)
Citation preview

Um
inho |
2013
Outubro de 2013
Diana Patrícia Andrade Vilas Boas
Avaliação dos mecanismos de interaçãoentre hospedeiro e fago em biofilmes
Escola de Engenharia
Dia
na P
atr
ícia
Andra
de V
ilas
Boas
Ava
liaçã
o d
os
meca
nis
mos
de inte
raçã
oentr
e h
osp
edeiro e
fago e
m b
iofilm
es


Outubro de 2013
Dissertação de Mestrado
em Bioengenharia
Trabalho efetuado sob a orientação da
Professora Doutora Joana Azeredo
Co-Orientador: Doutor Nuno Azevedo
Co-Orientador: Doutora Ana Nicolau
Escola de Engenharia
Diana Patrícia Andrade Vilas Boas
Avaliação dos mecanismos de interaçãoentre hospedeiro e fago em biofilmes

DE ACORDO COM A LEGISLAÇÃO EM VIGOR, NÃO É PERMITIDA A REPRODUÇÃODE QUALQUER PARTE DESTA TESE Universidade do Minho, ___/___/______
Assinatura: ________________________________________________

Agradecimentos


Agradecimentos
Agradecimentos
Inicialmente gostava de agradecer a todas as pessoas que direta ou
indiretamente me ajudaram em todo o meu processo de aprendizagem e sucesso
durante toda realização desta dissertação. Em particular tenho que agradecer:
À minha orientadora Joana Azeredo, pelo apoio científico, simpatia, confiança,
dedicação e disponibilidade manifestada ao longo do trabalho. Aos meus co-
orientadores Nuno Azevedo e Ana Nicolau pela disponibilidade e ajuda durante este
processo.
Estou grata a todos os meus colegas de trabalho do Laboratório de
Microbiologia Aplicada II da Universidade do Minho pelo excelente ambiente de
trabalho. Leonel, Ana, Sónia, Franklin, Luís e Célia pelo grande apoio, realçando a
minha grande amiga Graça.
Ao grupo de Biotecnologia de Bacteriófagos, em particular ao Sílvio pela sua
“simpatia”, ao Hugo, Iolanda e Ana Henriques pela ajuda prestada durante o
desenvolvimento do meu trabalho.
Ao grupo “ganir” pela amizade, alegria e partilha de ideias, entre eles os meus
amigos de longa data Débora e Rui. Elisabete, Elísia, Vanessa, Pedro e Alexandra
pela amizade e carinho. Carla pela amizade e ajuda durante a realização do meu
trabalho e ao Mário, um amigo “enfadonho” mas sempre disponível para me ajudar
em tudo que necessitei. Vocês são espetaculares e grupo como o nosso não há.
A toda a minha família (Pai, Mãe, Irmão e Bernardo), os mais profundos
agradecimentos pelo grande carinho, compreensão, amor e motivação para nunca
desistir dos meus objetivos.
Sou grata a todos os amigos que fiz em Braga ao longo deste processo em
especial às minhas amigas Helena e Teresa, vocês terão sempre um lugar especial. Ao
meu namorado Tiago pelo carinho, força e compreensão na reta final do meu
percurso.
E por fim, mas não menos importante às minhas “co-co-orientadoras” Carina e
Sanna por serem as melhores “co-co’s” que alguém poderia ter. Obrigado pela ajuda
em todo o trabalho desenvolvido, amizade e disponibilidade para tudo o que
necessitei. Sem vocês esta conquista não teria acontecido.


Sumário/Abstract


Sumário Pseudomonas aeruginosa e Acinetobacter baumannii são duas bactérias
gram-negativas patogénicas altamente problemáticas em infeções hospitalares muito devido à
sua capacidade de aderir a variadíssimas superfícies e formar biofilmes que contribuem
bastante para o aumento da resistência aos agentes antimicrobianos, o que consequentemente
potencia infeções graves em pacientes. Apesar da terapia fágica ter vindo a ser referida como
uma estratégia alternativa no controlo de biofilmes, ainda há poucos estudos sobre a interação
de fagos com as respetivas bactérias hospedeiras em biofilme mistos.
Este trabalho descreve o efeito de fagos em biofilmes simples de A. baumannii AB1 e
biofilmes mistos formados por A. baumannii AB1 e P. aeruginosa PAO1. Para este fim,
foram desenvolvidos métodos de diferenciação das duas espécies no biofilme recorrendo à
técnica de FISH (do inglês, Fluorescent in situ hibridization). Para uma deteção multiplex de
bactérias e fagos foram desenhadas “in silico” sondas com base num novo ácido nucleico LNA (do inglês Locked Nucleic Acid), contudo só a sonda específica para a bactéria
P. aeruginosa foi sintetizada e aplicada ao estudo. O método de LNA FISH para a
identificação desta bactéria foi otimizado e testes laboratoriais em amostras de subespécies de
Pseudomonas confirmaram os valores teóricos previstos de especificidade e sensibilidade.
Os ensaios de infeção fágica contra biofilme simples de A. baumannii, mostraram que
os fagos são eficientes, mesmo usando uma razão de 1 fago para 1 célula hospedeira. Com
efeito, verificou-se uma redução de aproximadamente 2 log de células viáveis em biofilmes
de 24 h após 6 h de contacto com o fago. Contudo a eficácia dos fagos mostrou-se
significativamente superior contra células do biofilme raspado o que indica que a matriz do
biofilme constitui uma barreira para a eficiente atuação do fago.
Os biofilmes mistos mostraram uma predominância de P. aeruginosa em relação a A.
baumannii, mesmo quando P.aeruginosa é adicionada após colonização inicial por
A.baumannii. A aplicação de um cocktail formado por fagos específicos contra estas duas
espécies presentes em biofilme misto revelou uma redução de 2 log de P. aeruginosa e de
aproximadamente 1,5 log de A. baumannii após 6 h. No entanto, após 24 h de infeção, foi
possível observar, em todos os biofilmes estudados, um aumento de células viáveis podendo
este facto estar relacionado com a aquisição de resistência das bactérias aos respetivos fagos.
A técnica de LNA FISH demostrou ser eficiente na discriminação clara das
populações de biofilme misto, sendo um complemento na deteção / compreensão na interação
dos fagos com os seus hospedeiros em estruturas complexas como os biofilmes.
Em suma, este trabalho demonstrou o valor potencial dos fagos no controlo de
biofilmes, tornando-se no entanto necessário progredir na presente investigação, para se
definirem as condições ótimas de atuação dos fagos e as razões da diminuição da eficácia dos
fagos quando em contacto com o biofilme por períodos superiores a 6 h.

Abstract Pseudomonas aeruginosa and Acinetobacter baumannii are two pathogenic
gram-negative bacteria that are highly prevalent in hospital infections due to their natural
ability to adhere to surfaces and form biofilms. Cells in biofilms are more tolerant to
antimicrobial agents and this enhances serious infections in patients. Phage therapy has been
reported as an alternative strategy for biofilm control but the interaction of phages with the
respective bacteria in mixed biofilm in poorly understood.
The work described herein focuses on the effect of phages in single species of A.
baumannii biofilms and in mixed biofilms formed by A. baumannii AB1 and P. aeruginosa
PAO1. To this end, methods were developed for the differentiation of the two species in
biofilm using the FISH (Fluorescent in situ hibridization) technique. For multiplex detection
of bacteria and phage probes based on a novel nucleic acid LNA (Locked Nucleic Acid) were
designed "in-silic", but only the probe specific for the bacteria P. aeruginosa was synthesized
and applied to the study. The LNA FISH method was optimized, and laboratory testing on
representative strains from the Pseudomonas genus subspecies confirmed the predicted
theoretical values of specificity and sensitivity.
Phage infection of A. baumannii single biofilms shows that phages were efficient
even when a ratio of 1 phage to 1 host cell is used. Indeed, there was a reduction of 2 log in
viable cells of 24 h biofilms, after 6 h of contact with the phage. Furthermore, the efficiency
of phage was significantly higher against biofilm scraped cells indicating that the biofilm
matrix is hindering the efficient performance of the phage.
The composition of mixed biofilms revealed a predominance of P. aeruginosa
compared to that of A. baumannii... The application of a phage cocktail resulted in a reduction
of 2 log of P. aeruginosa and 1,5 log of A. baumannii after 6 h. Nevertheless, after 24 hours
of infection it was observed the increase of viable cells in all biofilms studied. This result can
be explained by the acquisition of resistance of the bacterial hosts towards the respective
phages.
The LNA FISH technique demonstrated to be efficient in the clear discrimination of
mixed biofilm populations, being a complement the detection / understanding the interaction
of phages with their hosts in complex structures such as biofilms.
Briefly, this study demonstrated the potential use of phages for biofilm control,
however become necessary to progress in this research, to define the optimal conditions of
phage infections and the reasons for the decreased effectiveness of phages when in contact
with the biofilm for periods longer than 6 h.

Índice


Índice
Índice
Lista de figuras, tabelas e siglas…………………………………………………….xv
Estrutura da tese……………………………………………………………………xxv
Capítulo 1 | Introdução Geral
1.1 Pseudomonas aeruginosa e Acinetobacter baumannii................................ 1
1.1.1 Pseudomonas aeruginosa .................................................................................... 1
1.1.2 Acinetobacter baumannii ..................................................................................... 3
1.2 Teoria da formação de biofilmes ................................................................ 4
1.2.1 Biofilmes formados por P. aeruginosa e A. baumannii....................................... 6
1.3 Bacteriófagos ................................................................................................ 7
1.3.1 Classificação ........................................................................................................ 7
1.3.2 Ciclo de vida ........................................................................................................ 9
1.3.3 Interações fago-biofilme .................................................................................... 11
1.4 Hibridação fluorescente in situ ................................................................. 13
1.4.1 Aplicação de mímicos de DNA à técnica de FISH ............................................ 14
1.4.2 Aplicação da técnica de FISH em biofilme ....................................................... 15
1.5 Referências .................................................................................................. 16
Capítulo 2 | Desenvolvimento de sondas de LNA para aplicação na técnica de
FISH
2.1 Materiais e métodos ................................................................................... 27
2.1.1 Desenho e avaliação das sondas ........................................................................ 27
2.1.2 Otimização das condições de hibridação para LNA FISH ................................ 29

Índice
2.2 Resultados e discussão ............................................................................... 31
2.2.1 Desenho e avaliação das sondas ........................................................................ 31
2.2.2 Otimização das condições de hibridação para LNA FISH ................................ 39
2.2.3 Otimização das condições de hibridação para LNA FISH .............................. 393
2.3 Referências .................................................................................................. 42
Capítulo 3 | Caracterização da infeção fágica de biofilmes de Acinetobacter
baumannii
3.1 Materiais e métodos ................................................................................... 47
3.1.1 Estirpes bacterianas, fagos e condições de crescimento .................................... 47
3.1.2 Curva de calibração densidade ótica vs células viáveis ..................................... 47
3.1.3 Curva de crescimento de A. baumannii ............................................................. 48
3.1.4 Produção e concentração do fago phiIBB-Aba1 ................................................ 48
3.1.5 Determinação da concentração do fago ............................................................. 49
3.1.6 Curva de Crescimento do fago (OSGC) ............................................................ 49
3.1.7 Formação de biofilme ........................................................................................ 50
3.1.8 Quantificação da biomassa total ........................................................................ 50
3.1.9 Infeção Fágica de Biofilmes .............................................................................. 50
3.1.10 Análise estatística .............................................................................................. 51
3.1.11 Análise das proteínas estruturais do fago .......................................................... 51
3.1.12 Preparação de amostras de biofilmes e análise microscópica ............................ 52
3.2 Resultados e Discussão .............................................................................. 53
3.2.1 Curva de calibração D.O. vs células viáveis e Curva de crescimento de
A. baumannii ................................................................................................................... 53
3.2.1 Formação e infeção de biofilme......................................................................... 54
3.2.2 Análise das proteínas estruturais do fago .......................................................... 59
3.2.3 Curva de Crescimento do fago (OSGC) ............................................................ 61

Índice
3.2.4 Conclusão .......................................................................................................... 61
3.3 Referências .................................................................................................. 62
Capítulo 4 | Caracterização dos biofilmes mistos e avaliação da eficácia de fagos
específicos para A. baumannii e P. aeruginosa
4.1 Materiais e métodos ................................................................................... 67
4.1.1 Estirpe bacteriana, bacteriófagos e condições de cultura ................................... 67
4.1.2 Formação de biofilmes mistos ........................................................................... 68
4.1.3 Infeção Fágica de Biofilmes mistos ................................................................... 68
4.1.4 Análise estatística .............................................................................................. 68
4.1.5 Aplicação de hibridação in situ fluorescente (FISH) em biofilme misto ........... 68
4.2 Resultados e Discussão .............................................................................. 69
4.2.1 Formação de biofilmes mistos ........................................................................... 69
4.2.2 Infeção fágica de biofilmes mistos .................................................................... 70
4.2.3 Conclusão .......................................................................................................... 77
4.3 Referências .................................................................................................. 78
Capítulo 5 | Conclusão final e sugestões de trabalho futuro
Conclusão final e sugestões de trabalho futuro ....................................................... 83
Anexos ......................................................................................................................... 87


Lista de Figuras, Tabelas e Siglas


xix
Lista de Figuras
Capítulo 1 | Introdução Geral
Figura 1.1 - Representação esquemática das diferentes fases de desenvolvimento do
biofilme bacteriano. Na primeira fase (1), as células bacterianas aderem à superfície.
De seguida, na segunda fase (2), as células aderem umas às outras. Na fase seguinte
(3), a maturação é atingida e inicia-se a formação do biofilme. Uma segunda etapa de
maturação é atingida na quarta fase (4) momento da formação do biofilme quando se
apresenta totalmente desenvolvido. Na fase de dispersão (5), células individuais
(células escuras na figura) desprendem-se a partir das microcolônias. Adaptado de 47. 5
Figura 1.2 - Morfologia dos fagos. Adaptado de 68. ..................................................... 8
Figura 1.3 - Ciclo de vida lítico e lisogénico; (A) O fago reconhece um recetor
específico no hospedeiro e injeta do seu material genético via esse recetor; (B)
Circularização do material genético do fago; (C, D, E e F) Produção de fagos
descendentes; (G) Lise da parede celular do hospedeiro e consequentemente
libertação de uma centena de novos fagos produzidos; (H) Integração do material
genético do fago no genoma do hospedeiro (profago); (I) Divisão celular do
hospedeiro; (J) Divisão celular do hospedeiro originando células descendentes com
profago. Adaptado de 72. .............................................................................................. 10
Figura 1.4 - Modelo de infeção fágica em biofilme. Fase 1: Transporte dos fagos
através da matriz do biofilme e dos canais de água por difusão e/ou convecção; Fase:
Ligação dos fagos às células hospedeiras e progressão de ciclo lítico; Fase 3:
Libertação dos fagos descendentes por lise do hospedeiro e infeção das células
vizinhas, causando redução de biomassa; Fase 4: Desintegração de porções do
biofilme e progressão do ciclo lítico em células planctónicas. Adaptado de 63. .......... 11
Figura 1.5 - Estrutura química de LNA, DNA, RNA, PNA e 2’-O-metil RNA
respetivamente. Adaptado de 92. .................................................................................. 14

xx
Capítulo 2 | Desenvolvimento de sondas de LNA para aplicação na técnica de
FISH
Figura 2.1 - Alinhamento das sequências de rRNA das espécies em estudo com as
sondas Psaer (A) e ACA (B) no ClustralW2. .............................................................. 33
Figura 2.2 - Análise CoreGenes3.0 das sequências genómicas dos fagos de
Acinetobacter e Pseudomonas. .................................................................................... 34
Figura 2.3 - Imagens de microscopia de epifluorescência de P. aeruginosa PAO1
aplicando a sonda de LNA/2-O-metil Pser. São apresentadas amostra com uma
temperatura de hibridação de 55 ºC (A); 57 ºC (B) e 59 ºC (C). ................................. 39
Figura 2.4 - Imagens de microscopia de epifluorescência para as estripes
P. aeruginosa ATCC 10145, P. fluorescence PF7a e A. baumannii AB1 após
hibridação com a sonda Psaer a 59 ºC. Nas colunas estão representados os filtro de
microscopia (FITC e TRITC) e nas linhas as espécies utilizadas. A experiência foi
realizada simultaneamente, e as imagens foram obtidas com tempos de exposição
iguais. ........................................................................................................................... 41
Capítulo 3 | Caracterização da infeção fágica de biofilmes de Acinetobacter
baumannii
Figura 3.1 – Curva de crescimento bacteriano de A. baumannii AB1. As barras de
erros representam o desvio padrão em relação à média. .............................................. 53
Figura 3.2 - Infeção de biofilmes formados durante 24 h de A. baumannii AB1 e
infetadas durante 6 h e 24 h com o fago phiIBB-Aba1. A) Número de células viáveis
antes e após da infeção fágica; B) Quantificação da biomassa usando CV antes e após
da infeção fágica; C) Número de fagos (PFU) presentes nos biofilmes (aderidos) e o
número de fagos libertados durante a infeção fágica (suspensos). D) Imagens de
microscopia de epifluorescência de biofilme de 30 h em superfícies de poliestireno; E)
Imagens de microscopia de epifluorescência de biofilme infetado durante 6 h em
superfícies de vidro; F) Imagens de microscopia de epifluorescência de biofilme de 48

xxi
h em superfícies de poliestireno; G) Imagens de microscopia de epifluorescência de
biofilme infetado durante 24 h em superfícies de poliestireno. As barras de erros
representam o desvio padrão em relação à média. ** p < 0,01 e *** p < 0,001. ....... 55
Figura 3.3 - Infeção de biofilmes formados durante 12 h de A. baumannii AB1 com o
fago phiIBB-Aba1. A) Número de células viáveis antes e após da infeção fágica; B)
Quantificação da biomassa usando CV antes e após da infeção fágica. As barras de
erros representam o desvio padrão em relação à média. *** p < 0,01. ....................... 57
Figura 3.4 - Número de células viáveis (CFU) e fagos (PFU) após raspagem de
biofilme de A. baumannii AB1 formados durante 24 h e posteriormente infetadas com
o fago phiIBB-Aba1. A) Número de células viáveis antes e após infeção fágica; B)
Número de fagos presentes nos biofilmes suspensos. As barras de erros representam o
desvio padrão em relação à média. *** p < 0,01. ........................................................ 58
Figura 3.5 - Imagem de placas fágicas do fago phiIBB-Aba1. ................................... 59
Figura 3.6 - Análise por SDS-PAGE de proteínas estruturais de A. baumannii dos
fagos phiIBB-Aba2 e phiIBB-Aba1 respetivamente. .................................................. 60
Figura 3.7 - Curva One-Step-Growth (OSG) para fago phiIBB-Aba1. As barras de
erros representam o desvio padrão em relação à média. .............................................. 61
Capítulo 4 | Caracterização dos biofilmes mistos e avaliação da eficácia de fagos
específicos para A. baumannii e P. aeruginosa
Figura 4.1 - Número de células viáveis presentes de P. aeruginosa PAO1 e
A. baumannii AB1 em biofilme misto de 24 h. As barras de erros representam o
desvio padrão em relação à média. *** p < 0,01. ........................................................ 69
Figura 4.2 - Número de células viáveis (CFU), biomassa total e fagos (PFU) de
P. aeruginosa PAO1 e A. baumannii AB1 em biofilmes mistos de 24 h após infeção
com os fagos phiIBB-Aba1 e phiIBB-PAP1 separadamente ou em cocktail durante 6 h
e 24 h. A) Número de células viáveis (CFU) nos biofilmes mistos de após 6 h de
infeção; B) Número de células viáveis (CFU) nos biofilmes mistos de após 24 h de
infeção fágica; C) Quantificação da biomassa utilizando CV; D) Número de fagos

xxii
(PFU) de P. aeruginosa phiIBB-PAP1 libertados pelo biofilme (suspensos) e
presentes no biofilme (aderidos); E) Número de fagos (PFU) de A. baumannii
phiIBB-Aba1 libertados pelo biofilme (suspensos) e presentes no biofilme (aderidos).
As barras de erros representam o desvio padrão em relação à média. ** p < 0,01 e
*** p < 0,001. .............................................................................................................. 71
Figura 4.3 - Número de células viáveis e biomassa total após infeção de biofilme
misto de P. aeruginosa PAO1 e A. baumannii AB1 de 24 h com o fago phiIBB-PAP1
e phiIBB-PAA2 separadamente. A) Número de células viáveis (CFU); B)
Quantificação da biomassa utilizando CV. As barras de erros representam o desvio
padrão em relação à média. * p < 0,05. ....................................................................... 74
Figura 4.4 - Imagens de microscopia de epifluorescência de biofilme misto de
P. aeruginosa PAO1 e A. baumannii AB1 usando a sonda Pser e o corante DAPI.
A) biofilme misto de 30 h ( controlo da infeção de 6 h)em superfície de poliestireno;
B) biofilme misto de 24 h infetado durante 6 h com o fago de P. aeruginosa (phiIBB-
PAA2) em superfície de poliestireno, C) biofilme misto de 24 h infetado durante 6 h
com os fagos de A. baumannii (phiIBB-Aba1) e P. aeruginosa (phiIBB-PAA2) em
superfície de poliestireno; D) biofilme misto de 24 h infetado durante 6 h com o fago
de A. baumannii (phiIBB-Aba1) em superfície de poliestireno. ................................. 75
Figura 4.5 - Formação de biofilmes mistos durante 24 h. Foram testadas duas
condições, uma em que se adicionaram ambas as espécies em simultâneo e outra em
que se formou durante 12 h um biofilme de A. baumannii Ab1 e posteriormente
P. aeruginosa PAO1 até perfazer 24 h. A) Número de células viáveis (CFU); B)
Quantificação da biomassa usando CV. As barras de erros representam o desvio
padrão em relação à média. ** p < 0,01. ..................................................................... 76
Figura 4.6 - Imagens de microscopia de epifluorescência com utilização da sonda
fluorescente Pser e o corante DAPI. A) Imagens de microscopia de epifluorescência
de biofilmes mistos durante 24 h com aplicação de ambas as espécies em simultâneo
em superfície de poliestireno; B) Imagens de microscopia de epifluorescência de
biofilmes mistos durante 24 h com uma formação inicial durante 12 h de um biofilme
de A. baumannii AB1 e posteriormente a adição de P. aeruginosa PAO1 até perfazer
24 h em superfície de vidro. ......................................................................................... 77

xxiii
Lista de Tabelas
Capítulo 1 | Introdução Geral
Tabela 1.1 - Classificação dos bacteriófagos. Adaptado de 63 ...................................... 8
Capítulo 2 | Desenvolvimento de sondas de LNA para aplicação na técnica de
FISH
Tabela 2.1 - Sondas descritas na literatura para P. aeruginosa e A. baumannii,
juntamente com algumas das suas características mais importantes ........................... 31
Tabela 2.2 - Avaliação da sensibilidade (S) e especificidade (E) das sondas descritas
na literatura .................................................................................................................. 32
Tabela 2.3 - Sequências possíveis param a sonda dos fagos phiIBB-Aba1 e
phiIBB-Aba2 e os seus parâmetros termodinâmicos. A sonda do fago phiIBB-PAA2
foi também incluída para comparação ......................................................................... 36
Tabela 2.4 - Tm para as sondas de LNA/2-O-metil para as bactérias e fagos ............ 37
Tabela 2.5 - Valores dos parâmetros termodinâmicos das sondas. Energia livre de
Gibbs (ΔG), entropia (ΔS), entalpia (ΔH) e teor em GC (GC) .................................... 38
Tabela 2.6 - Fluorocromos associados às sondas ........................................................ 38
Tabela 2.7 - Resultados do teste de especificidade e sensibilidade da sonda de Pser 40

xxiv
Lista de siglas
ATCC American Type Culture Collection
cdDNA Ácido desoxirribonucleico de cadeia dupla
cdRNA Ácido ribonucleico de cadeia dupla
CFU Colony Forming Unit
CLSM Confocal Laser Scanning Microscopy
csDNA Ácido desoxirribonucleico de cadeia simples
csRNA Ácido ribonucleico de cadeia simples
D.O. Densidade Óptica
DAPI 4',6-diamidino-2-phenylindole, dihydrochloride
DNA Ácido desoxirribonucleico
EPS Substâncias Poliméricas Extracelulares (“Extracellular Polymeric Substances”)
ICTV Comité Internacional de Taxonomia de Vírus
LB Luria-Bertani broth
MOI Multiplicity of Infection
pb Pares de bases
PEG Polietilenoglicol
PFU Plaque Forming Units
PIA Pseudomona isolation agar
RNA Ácido ribonucleico
rpm Rotação por minuto
SDS-PAGE Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis
TSB Tryptic Soy Broth

Estrutura da tese


Estrutura da tese
xxvii
Estrutura da tese
A tese encontra-se organizada em 5 capítulos.
O primeiro capítulo apresenta uma breve introdução teórica sobre biofilmes
bacterianos dando realce aos que são formados pelas bactérias A. baumannii e P.
aeruginosa , sobre bacteriófagos e a sua utilização no controlo de biofilmes, sobre a
evolução da tecnologia de FISH na identificação de microrganismos e o surgimento
de moléculas de LNA como sondas moleculares.
O capítulo 2 descreve o desenvolvimento de novas sondas de LNA para a
deteção de A. baumannii e P. aeruginosa e para os seus fagos específicos (phiIBB-
Aba2 e phiIBB-PAA2). Descreve também os resultados dos testes de sensibilidade,
especificidade e otimizações de hibridações para a sonda da bactéria P. aeruginosa.
Os capítulos 3 e 4 descrevem a aplicação dos bacteriófagos específicos em
biofilmes simples de A. baumannii (capítulo 3) e em biofilme mistos de A. baumannii
e P. aeruginosa (capítulo 4), conjuntamente com a aplicação da técnica de LNA FISH
nas diferentes condições testadas.
No último capítulo são descritas as principais conclusões deste trabalho e
sugerem-se também alguns temas para trabalhos futuros no âmbito dos biofilmes,
fagos e uso de sondas de LNA.


Capítulo 1
Introdução Geral


Introdução Geral
1
1 Introdução Geral
1.1 Pseudomonas aeruginosa e Acinetobacter
baumannii
1.1.1 Pseudomonas aeruginosa
P. aeruginosa é uma bactéria gram-negativa, aeróbia pertencente à família
Pseudomonadacea 1 que se apresenta sob a forma de bastonetes retos ou ligeiramente
curvos, isolados ou aos pares. O seu metabolismo é não-fermentativo, mas em certas
condições pode crescer na ausência de oxigénio caso possua nitrato disponível para ser
utilizado como aceitador de eletrões 2. P. aeruginosa apresenta poucas exigências para o
crescimento e grande versatilidade nutricional 3, tolerando variações das condições
físicas, resistência a altas concentrações de sais e pigmentos, anti-sépticos fracos e
muitos antibióticos comumente utilizados 4. Possui capacidade de crescer entre 5 ºC a
42°C, sendo a temperatura ideal 37°C. Tem um tamanho aproximado de 0,5 μm a
0,8 μm por 1,5 μm a 3,0 μm, um odor característico a fruta 5,4 e produz um pigmento
hidrossolúvel designado por piocianina que confere uma coloração esverdeada ao meio
de cultura 6. Este microrganismo é ubíquo, sendo encontrado no solo, água, vegetais e
matéria orgânica em decomposição e pode ser encontrado na natureza ligado a uma
superfície ou substrato (biofilme), como sob forma plantónica, movendo-se através de
flagelos polares 2.
P. aeruginosa é um patogénico oportunista, exibindo uma ampla variedade de
mecanismos de virulência, que podem originar diversas infeções 7. A infeção inicia-se
com a adesão bacteriana, seguindo-se a fixação e posteriormente a dispersão 8,9. As
fimbrias estendem-se a partir da superfície celular e promovem a adesão da bactéria aos
recetores presentes na superfície das células epiteliais do hospedeiro 8. Resíduos dos
recetores das células e a neuraminidade (que elimina o ácido siálico) facilitam a ligação
das fímbrias aos recetores 10,11. A expressão de exopolissacarídeo mucóide (ex. alginato)
funciona como mediador de aderência a superfícies epiteliais dos pulmões de pacientes
com fibrose cística e como mecanismos de defesa na formação de biofilmes 12–14.

Capítulo 1
2
Vários outros fatores extracelulares como a produção de elastases, protease
alcalina, fosfolipase C, exoenzima S, neuraminidase e exotoxinas facilitam o
rompimento da integridade epitelial do hospedeiro promovendo a invasão
bacteriana 15,16. Entre as proteases extracelulares, destacam-se a elastase e a protease
alcalina, que contribuem para a aderência e degradação de vários componentes do
sistema imune 17. A exotoxina A inibe a síntese protéica, efeitos citopáticos e interfere
nas funções celulares imunes do hospedeiro 18,19. A fosfolipase C tem a capacidade de
destruir a membrana citoplasmática e inativar as opsoninas. O lipolissacarídeo presente
na membrana externa da bactéria composto por lípido A, um núcleo oligosacarídeo
interno e externo ao qual se liga ao O-antigénio de banda A ou B permite a adesão
bacteriana mediando a entrada nas células eucarióticas 20,21.
Outra das propriedades importantes desta espécie é a capacidade de adesão a
superfícies e formação posterior de biofilmes que diminuem a ação de agentes
antimicrobianos. Na propagação da inflamação, há um grande risco de colonização de
outros locais devido à sua mobilidade 9. Para além disso, há um sistema de comunicação
entre as células denominado quorum sensing, em que pequenos compostos chamados
autoindutores são libertados pelas bactérias para o ambiente circundante e detetadas
pelas bactérias vizinhas que coordenam a expressão dos genes regulando a densidade
populacional 22.
A capacidade de P. aeruginosa de sobreviver em condições mínimas
nutricionais e a sua tolerância a uma variedade de condições físicas permite que este
organismo persista principalmente em ambientes hospitalares 23, sendo um importante
patogénico humano. Esta espécie aparece associada a infeções do trato respiratório,
fundamentalmente em doentes com fibrose cística, do trato urinário relacionadas com o
uso de cateteres urinários, da corrente sanguínea e dos locais que sofreram intervenção
cirúrgica (ex. locais de inserção de próteses ou que sofreram operação) 24. Pode assim
ser isolada a partir de uma variedade de fontes, como equipamentos de terapia
respiratória e cateteres e também, de pias, sabão, medicamentos e piscinas de
hidroterapia 25. Os pacientes com imunidade diminuta possuem um maior risco de
colonização por este microrganismo 26. Dados do Sistema Nacional de Vigilância
(EUA) de 1986 a 1998 indicam que P. aeruginosa é o quinto patogénico nosocomial
mais isolado, sendo responsável por 9 % de todas as infeções hospitalares nos Estados
Unidos 27,28. É também a segunda principal causa de pneumonia nosocomial (14 % a

Introdução Geral
3
16 %), a terceira causa mais comum de infeções do trato urinário (7 % a 11 %), o quarto
microrganismo mais isolado em infeções em local cirúrgico (8 %) e o sétimo
responsável por infeções na corrente sanguínea (2 % a 6 %) 23.
Fora do ambiente hospitalar, este microrganismo pode ser encontrado em
piscinas, banheiras de hidromassagens, soluções de lentes de contacto, humidificadores
de casa, solo e vegetais 23.
1.1.2 Acinetobacter baumannii
A. baumannii é uma bactéria gram-negativa pertencente à família
Moraxellaceae 29. É uma bactéria estritamente aeróbia, não-fermentadora, não-
fastidiosa, com motilidade, catalase positiva e oxidase-negativa 30. Durante a fase de
crescimento, a bactéria é um bacilo, com bastonetes até 2,5 μm de comprimento.
Durante a fase estacionária do crescimento bacteriano, as células tornam-se mais curtas
e arredondadas, assemelhando-se a pequenos cocos ao encolher para cerca de metade do
seu tamanho inicial e encontram-se em geral em pares ou em grupos densos de células
interligadas, possuindo as colónias uma cor esbranquiçada 30. Possui capacidade de
crescer a 37 ºC, apresentando requisitos mínimos nutricionais 31 e crescendo bem em
meios sólidos e em meios laboratoriais (ex. TSB, do inglês Tryptic Soy Broth) e
Luria-Bertani (LB)) 30,32. É um microrganismo ubíquo, de climas quentes e húmidos
que se adaptam a diferentes ambientes (água ou solo), encontrado frequentemente como
parasita em humanos ou em objetos inanimados 33,34.
A. baumannii é um agente patogénico oportunista e ao longo dos anos tem
demonstrado capacidade de adquirir resistências aos antibióticos 33,35. Apresenta vários
mecanismos de resistência como: enzimáticos, não enzimáticos e genéticos. Um dos
mecanismos de resistência enzimáticos está relacionado com a produção de enzimas no
meio intracelular da bactéria, nomeadamente as β-lactamases como as cefalosporinases,
que hidrolisam alterando a estrutura química dos antimicrobianos modificando assim a
sua atividade sobre o microrganismo 29. A permeabilidade da membrana externa é
alterada pela perda de proteínas (OMPs, do inglês Outer Membrane Proteins)
constituintes dos canais de porina no transporte de substâncias através da membrana
externa. A. baumannii tem poucas porinas em comparação com outras gram-negativas, o
que pode explicar, algumas das suas resistências à passagem dos antimicrobianos. Estas

Capítulo 1
4
alterações na membrana associadas à produção de β-lactamases conferem resistência
aos antibióticos β-lactâmicos 29. A alteração do local alvo aos antibióticos, como as
proteínas de ligação à penicilina (PBPs, do inglês Penicillin-Binding-Proteins), as
topoisomerases e as frações ribossómicas, provocam mutações nas células alvo
reduzindo a sua afinidade, diminuindo ou impedindo a ligação entre eles 29. A nível
genético, a resistência é conferida pela transferência horizontal através da troca de
plasmídeos, podendo ocorrer entre microrganismos da mesma espécie ou entre espécies
não correlacionadas. A. baumannii pode adquirir genes de resistência de outros
microrganismos ou desenvolver ao longo do tempo mutações, propiciando o
aparecimento de estirpes com capacidade de resistência antimicrobiana 34.
A. baumannii também possui a capacidade de formar biofilmes e permanecer em
ambientes propícios à sua colonização, como ambientes hospitalares 36. Acinetobacter é
um patogénico associado à pneumonia bacteriana, infeções do trato urinário, infeções de
feridas cirúrgicas e meningite. Os principais fatores de risco associados à colonização e
infeção de Acinetobacter são os procedimentos cirúrgicos, hospitalizações prévias e
procedimentos invasivos como ventilação mecânica, uso de cateter venoso central ou
cateteres urinários 37,38. A mortalidade hospitalar atribuída está compreendida entre 7,8
% a 23 %. Nas unidades de cuidados intensivos, este valor varia entre os 10 % - 43 % 38,39.
1.2 Teoria da formação de biofilmes
Atualmente, um biofilme é definido como uma associação de células
microbianas que formam uma pelicula multi-hidratada fixada a superfícies bióticas ou
abióticas, presa por uma complexa matriz extracelular de substâncias poliméricas
produzidas pelos microrganismos, fornecendo uma estratégia importante de
sobrevivência bacteriana 40,41. Na natureza, mais de 99 % dos microrganismos
(bactérias, algas, fungos e protozoários) formam biofilmes 42 e podem ser compostos
por uma população que se desenvolveu a partir de uma única espécie ou de uma
comunidade derivada de múltiplas espécies microbianas 43.
A primeira etapa de formação do biofilme inicia-se pela adesão dos
microrganismos à superfície. Quando as bactérias formam uma associação estável,
iniciam a divisão celular e formam micro-colonias que sintetizam a matriz polimérica

Introdução Geral
5
começando a desenvolver o biofilme 44,45. Uma vez estabelecido o biofilme, as
populações bacterianas a ele associadas podem desprender-se periodicamente,
colonizando novas superfícies e formando novos biofilmes em novos locais, causando
eventualmente infeções crónicas 46 (Figura 1.1).
Figura 1.1 - Representação esquemática das diferentes fases de desenvolvimento do biofilme
bacteriano. Na primeira fase (1), as células bacterianas aderem à superfície. De seguida, na segunda
fase (2), as células aderem umas às outras. Na fase seguinte (3), a maturação é atingida e inicia-se a
formação do biofilme. Uma segunda etapa de maturação é atingida na quarta fase (4) momento da
formação do biofilme quando se apresenta totalmente desenvolvido. Na fase de dispersão (5),
células individuais (células escuras na figura) desprendem-se a partir das microcolônias. Adaptado
de 47.
Ainda que cresçam mais lentamente que os microrganismos plantónicos, os
biofilmes são seguramente mais eficientes metabolicamente e conseguem utilizar de
uma forma mais extensiva todos os nutrientes existentes. Assim, pode-se considerar que
o potencial metabólico de um biofilme é bastante superior ao de uma comunidade
plantónica 48.
Devido a vários fatores, a composição dos biofilme varia entre os vários
microrganismos. Entre esses fatores destacam-se: o estado fisiológico dos
microrganismos, o ambiente físico e a superfície a que estão aderidos 43,49. Análises
estruturais têm demonstrado que 15 % do volume de biofilme é composto por células
incorporadas em 85 % de matriz extracelular 50. Esta matriz é apontada como
fundamental para a sobrevivência dos biofilmes 51. Grande parte dos compostos da
matriz são provenientes das secreções dos próprios organismos ou então resultantes de
restos celulares de microrganismos em decomposição, o que pode explicar a diferença
da composição dos biofilmes de organismo para organismo 51. Esta matriz é

Capítulo 1
6
essencialmente composta por: água (97 %), substâncias poliméricas extracelulares
contendo proteínas (incluindo enzimas) (1 % a 2 %), polissacarídeos (homo e
heteropolissacarídeos) (1 % a 2 %) e ácidos nucleicos (1 % a 2 %) 52. A matriz é um dos
grandes responsáveis pela resistência dos biofilmes pois age como barreira física,
protegendo as células para que não sejam arrastadas pelo fluxo de substâncias,
auxiliando a célula a resistir à diminuição e exaustão de nutrientes e de água, à presença
de biocidas e outros agentes antimicrobianos e a outras condições ambientais 44.
Na indústria, os biofilmes são responsáveis por graves perdas económicas
quando associados a problemas de corrosão, à perda de carga de fluidos em tubagens ou
a contaminações em produtos alimentares. No contexto clínico, os biofilmes contribuem
para a progressão de infeções associadas à colonização de dispositivos médicos
inseridos no corpo humano, mas também à colonização direta de tecidos 53.
1.2.1 Biofilmes formados por P. aeruginosa e A. baumannii
P. aeruginosa e A. baumannii são dois dos microrganismos comumente
encontrados em comunidades mistas de biofilmes em ambientes hospitalares. Ambas
são suscetíveis a antibióticos β-lactâmicos e a sua resistência a diversos antibióticos tem
crescido ao longo dos anos 54. Oliveira et al. 55.detetou em amostras do aspirado
traquial, 3,3 % e 20 % de A. baumannii e P. aeruginosa respetivamente num total de
30 pacientes internados nas unidades de terapia intensiva, concluindo que a presença
destes patogénicos serve como reservatório para microrganismos associados à
pneumonia nosocomial. Também Gil et al. 56 demostraram que A. baumannii e
P. aeruginosa estão entre as bactérias prevalentes envolvidas em infeções respiratórias
crónicas (45 %) e verificaram a existência de biofilmes (56 %) num total de 87 % dos
pacientes internados na unidade de terapia intensiva que necessitaram de ventilação
mecânica num período mínimo de 24 horas.
Storti 57demonstrou, num estudo com 118 pontas de cateteres intravenosos
centrais, que 34 estavam colonizadas. Dessas 34 pontas de cateteres colonizadas, foram
isolados 55 microrganismos, dos quais 10,9 % e 5,4 % eram A. baumannii e
P. aeruginosa, respetivamente. Também Storti et al. 58 num estudo anterior, tinha
observado que microrganismos isolados das superfícies de cateteres estavam
colonizadas com A. baumannii (14.3 %) e P. aeruginosa (21,4 %), e ainda que na

Introdução Geral
7
corrente sanguínea as amostras continham A. baumannii (18,2 %) e P. aeruginosa
(27,3 %), sugerindo que a presença de biofilmes nas superfícies dos cateteres podem
tornar-se num nicho de infeção na corrente sanguínea dos pacientes, causando graves
infeções bacterianas.
1.3 Bacteriófagos
Ao longo dos anos, os antibióticos foram a única forma de tratamento das
doenças infeciosas, limitando o crescimento de bactérias ou promovendo a sua
destruição. Contudo, devido à utilização inadequada destes medicamentos, as bactérias
adquiriram mecanismos de resistências contra os mesmos, sendo atualmente, segundo a
Organização Mundial de Saúde, uma das maiores ameaças à Saúde Pública. Devido a
esta problemática, ressurgiu um grande interesse pelos bacteriófagos (fagos), agentes
terapêuticos descobertos em 1915 pelo bacteriologista Inglês Frederick Twort e pelo
microbiologista franco-canadiano Félix d'Herelle 59.
Os fagos definem-se como vírus bacterianos que, afetam o metabolismo das
bactérias causando a sua morte 60,61. Sendo vírus, os fagos não possuem um
metabolismo próprio e para tal necessitam da maquinaria de biossíntese do respetivo
hospedeiro para que se possam replicar 62,63. Estima-se também que sejam os
organismos mais simples e abundantes na Terra, e que para cada célula bacteriana
existam dez partículas fágicas diferentes, 60 superando claramente as bactérias em todos
os ambientes estudados 64–66.
Para além de serem eficazes no combate a bactérias resistentes a antibióticos, os
fagos são também capazes de controlar de forma eficaz bactérias no estado séssil. Os
fagos apresentam uma toxicidade nula, uma vez que o seu principal constituinte são
ácidos nucleicos 67, apresentam-se assim como agentes terapêuticos muito promissores
no combate a biofilmes infeciosos 63.
1.3.1 Classificação
Há uma diversidade de fagos com formas, tamanhos, simetrias da cápside e
estruturas variadas. De acordo com o Comité Internacional de Taxonomia de Vírus
(ICTV, do inglês International Committee on Taxonomy of Viruses) os fagos podem ser
distinguidos com base na taxonomia e no tipo de ácidos nucleicos do seu genoma
(Tabela 1.1, Figura 1.2). Estima-se que 96 % de todos os fagos até hoje reportados

Capítulo 1
8
pertençam à ordem Caudovirales 63 que podem ser divididos em três famílias
(Podoviridae, Siphoviridae, Myoviridae) de acordo com as características da cauda.
Tabela 1.1 - Classificação dos bacteriófagos. Adaptado de 63
Família Características Ácido Nucleico
Corticoviridae Sem envelope; cápside icosaédrica dsDNA
Cystoviridae Com envelope; cápside icosaédrica dsRNA
Fuselloviridae Com envelope; pleomórfico; sem cápside dsDNA
Inoviridae Sem envelope; filamentoso ssDNA
Leviviridae Sem envelope; cápside icosaédrica ssRNA
Lipothrixviridae Com envelope; simetria helicoidal dsDNA
Microviridae Sem envelope; cápside icosaédrica ssDNA
Rudiviridae Sem envelope; simetria helicoidal dsDNA
Plasmaviridae Sem envelope; pleomórfico dsDNA
Podoviridae Sem envelope; cauda curta não contráctil dsDNA
Siphoviridae Sem envelope; cauda longa não contráctil dsDNA
Myoviridae Sem envelope; cauda contráctil dsDNA
Tectiviridae Sem envelope, cápside icosaédrica dsDNA
Figura 1.2 - Morfologia dos fagos. Adaptado de 68.

Introdução Geral
9
1.3.2 Ciclo de vida
No que diz respeito ao seu ciclo de vida, os bacteriófagos podem ser de dois
tipos: os fagos virulentos, que apresentam um ciclo de vida lítico e os fagos temperados,
que apresentam ciclo de vida lisogénico mas que, em determinadas condições, poderão
iniciar o ciclo lítico 69. A replicação fágica inicia-se por ligação do fago ao hospedeiro
através das suas fibras da cauda que se fixam a recetores específicos existentes na
superfície da bactéria (proteínas, carbohidratos, moléculas de lipopolissacarídeos e
flagelos) 60. A maioria dos fagos é altamente específica para os seus recetores e há
pouca ou nenhuma interação com recetores de estruturas ligeiramente diferentes 70. De
seguida, o seu material genético sob a forma de DNA (do inglês Deoxyribonucleic Acid)
ou RNA (do inglês Ribonucleic Acid) é introduzido no interior da célula. Os fagos
litícos (ex. tipo- T) são reproduzidos rapidamente, formando novas partículas virais no
interior do seu hospedeiro, terminando o ciclo de vida com a lise de bactérias mediada
pela produção das proteínas tardias (holina e endolisina), codificadas no genoma de
fagos de dsDNA. O número de novos fagos produzidos, depende do tipo de fago, da
espécie hospedeira e das condições de replicação, no entanto, estima-se que cada fago é
capaz de produzir cerca de 200 novos fagos por cada ciclo lítico 71. No ciclo de vida
lisogénico, o DNA viral, após atingir o citoplasma, integra-se no genoma bacteriano e
replica-se conjuntamente com este, passando para todas as células filhas durante a
divisão celular sem causar lise. Permanece assim num estado dormente como um
profago por períodos de tempo prolongados 60 e somente por ação de estímulos
externos, normalmente fatores de stress, poderá ocorrer o ciclo lítico 63 (Figura 1.3).

Capítulo 1
10
Figura 1.3 - Ciclo de vida lítico e lisogénico; (A) O fago reconhece um recetor específico no
hospedeiro e injeta do seu material genético via esse recetor; (B) Circularização do material
genético do fago; (C, D, E e F) Produção de fagos descendentes; (G) Lise da parede celular do
hospedeiro e consequentemente libertação de uma centena de novos fagos produzidos; (H)
Integração do material genético do fago no genoma do hospedeiro (profago); (I) Divisão celular do
hospedeiro; (J) Divisão celular do hospedeiro originando células descendentes com profago.
Adaptado de 72.
Kropinski, A. 73refere que os fagos temperados não devem ser utilizados na
terapia fágica, uma vez que podem integrar-se no genoma da bactéria e não causar a lise
celular, podendo mesmo potenciar a virulência das bactérias segundo um fenómeno
denominado “ conversão lisogénica”.
O maior interesse na terapia fágica assenta essencialmente na capacidade dos
fagos lizarem as células bacterianas, por esse facto, fagos filamentosos (ex. M13) não
terem interesse, uma vez que a expulsão do fago é realizada por mecanismos extrusão,
não causando a lise das células 74.

Introdução Geral
11
1.3.3 Interações fago-biofilme
A interação fago-biofilme é um processo complexo que depende de diversos
fatores. A estrutura e composição de um biofilme, bem como a fisiologia das células
que o compõem, limitam a infeção fágica 63.
Um dos maiores obstáculos na infeção fágica é a matriz do biofilme, uma vez
que é um reservatório de enzimas proteolíticas que conduzem à inativação do fago,
dificultando a sua passagem até às camadas mais profundas do biofilme 63.
Alguns fagos possuem a capacidade de produzir ou induzir a produção de
depolimerases que degradam polissacarídeos da matriz, facilitando a sua progressão no
biofilme através da matriz. Quando isto não acontece, os fagos penetram no interior do
biofilme através de canais de água que compõem a estrutura do biofilme 63.
Quando o fago é transportado através da matriz do biofilme e entra em contacto
com as bactérias, inicia uma ligação que se torna forte e irreversível, injetando o seu
material genético pela parede bacteriana e iniciando o seu ciclo lítico. Seguidamente os
fagos descendentes são libertados para o meio exterior 63 (Figura 1.4).
Figura 1.4 - Modelo de infeção fágica em biofilme. Fase 1: Transporte dos fagos através da matriz
do biofilme e dos canais de água por difusão e/ou convecção; Fase: Ligação dos fagos às células
hospedeiras e progressão de ciclo lítico; Fase 3: Libertação dos fagos descendentes por lise do
hospedeiro e infeção das células vizinhas, causando redução de biomassa; Fase 4: Desintegração de
porções do biofilme e progressão do ciclo lítico em células planctónicas. Adaptado de 63.

Capítulo 1
12
O estado fisiológico das células, também influencia a eficácia dos fagos uma vez
que células na fase exponencial entram mais rápida e eficazmente em lise do que as
células na fase estacionária. Como o biofilme é composto por células em diferentes
fases metabólicas e de crescimento, é expectável que a taxa de lise nos biofilmes seja
mais reduzida do que nas células plantónicas em fase exponencial 63. Outro fator
limitante é a presença de células mortas que faz diminuir o número de partículas virais
ativas 63.
Outro obstáculo na implementação da terapia fágica é o desenvolvimento de
resistência aos fagos por parte das bactérias 60,75. As bactérias podem tornar-se
resistentes aos fagos diminuindo ou eliminando a sensibilidade a estes, desenvolvendo
subpopulações com fenótipos estáveis e resistentes, com o intuito de se protegerem
contra a invasão de DNA estranho. Estes mecanismos podem ocorrer através de trocas
genética com outras bactérias, aquisição de plasmídeos, e conversão lisogénica 60,75.
Uma bactéria pode tornar-se resistente aos fagos também pela mutação ou perda de
recetores, mas esta questão pode não ser de todo benéfica para a bactéria, uma vez que o
recetor é um determinante de virulência, e a perda deste recetor irá diminuir
drasticamente a virulência das bactérias 60. Por vezes, após a injeção do genoma do fago
no hospedeiro pode também ocorrer a sua degradação pelas DNases bacterianas 60.
Finalmente, pode haver imunidade pela restrição-modificação das bactérias ou dos
plasmídeos, pois estes podem codificar enzimas de restrição que degradam o genoma do
fago 75.
Para tentar diminuir este fenómeno, tem sido sugerido a utilização de cocktails
fágicos de modo a alargar o espetro de ação, reduzindo o aparecimento de fenótipos
resistentes 76,77.
Se por um lado os fagos são bastante resistentes a fatores externos, por outro
lado, a viabilidade destes seres está comprometida quando presentes em condições
desfavoráveis. De entre os fatores que podem causar danos na estrutura do fago que
levam à inativação do mesmo, estão a temperatura, a salinidade e a acidez 78,79.

Introdução Geral
13
1.4 Hibridação fluorescente in situ
FISH (do inglês, Fluorescent in situ hibridization) é uma técnica molecular
utilizada na identificação de microrganismos 80. O método baseia-se na utilização de
sondas de ácidos nucleicos marcados com fluorescência que irão ligar-se
especificamente a determinadas regiões de RNA ribossomal (rRNA), devido ao seu
elevado número de cópias por célula 81. No caso das bactérias, o rRNA 16S e 23S são os
mais utilizados.
Dada a sua sensibilidade, especificidade e rapidez, a técnica de FISH tem sido
utilizada como uma ferramenta frequente tanto para o estudo da diversidade microbiana
em amostras ambientais, como também no diagnóstico de comunidades microbianas
que colonizam partes integrantes do tecido humano 82,83.
Este método assenta em quatro passos: fixação / permeabilização da amostra,
hibridação das sondas com as sequências alvo, lavagem para a remoção das sondas não
ligadas, e finalmente, observação das células fluorescentes ao microscópio de
fluorescência ou deteção por citometria de fluxo 82,83.
As sondas de ácidos nucleicos utilizadas na técnica de FISH têm geralmente
entre 15 a 30 nucleótidos de comprimento e estão ligados covalentemente a um
fluorocromo que confere assim fluorescência quando ocorre a hibridação. A hibridação
deve ocorrer numa câmara húmida, escura, e com uma temperatura geralmente
compreendida entre os 37 ºC e 60 ºC, por um período que pode variar entre os
30 minutos e várias horas 81.
A crescente necessidade de identificar simultaneamente várias espécies
microbianas, conduziu à utilização de fluorocromo diferentes que permitem discriminar
diferentes sondas e assim, distinguir diferentes populações. A utilização simultânea de
corantes não específicos (como o DAPI) permite ainda fazer a coloração simultânea da
população total, proporcionando uma combinação de cores que permite detetar
múltiplos alvos. Assim, a fluorescência dos diferentes fluorocromos deve ser claramente
distinta, de forma a criar um espectro único para cada espécie presente na população em
estudo 81,84.

Capítulo 1
14
1.4.1 Aplicação de mímicos de DNA à técnica de FISH
É conhecido que a natureza das sondas é um dos fatores que influencia o sucesso
da técnica de FISH, uma vez que a sua estabilidade e afinidade com as sequências
complementares influenciam o sucesso da hibridação 85. A técnica de FISH utiliza
tradicionalmente sondas de DNA 86, mas recentemente têm sido desenvolvidas novas
moléculas sintéticas, análogas ao DNA que permitem melhorar a performance do
método e encurtar substancialmente o procedimento de FISH 87,88.
Várias moléculas sintéticas têm surgido como alternativas promissoras na
aplicação da técnica de FISH. Alguns dos mímicos de DNA mais conhecidos incluem:
PNA (do inglês Peptide Nucleic Acid) 87, LNA 89, 2’-O-metil-(2’-OMe) RNA 90 e ácido
2’-desoxi-2´-fluoro-β-D-ribonucleico (2’-F RNA) 91 (Figura 1.5). O PNA tem sido o
mímico mais estudado e aplicado à técnica de FISH, existindo já uma panóplia de
sondas descritas para variadíssimos microrganismos 85.
Figura 1.5 - Estrutura química de LNA, DNA, RNA, PNA e 2’-O-metil RNA respetivamente.
Adaptado de 92.
O PNA foi sintetizado em 1991 por Nielson et al. 93 e possuía características de
hibridação superiores às sondas de oligonucleótidos de DNA então existentes. Estas
particularidades levaram à aplicação do PNA às técnicas de biologia molecular,
tornando os procedimentos de FISH mais robustos e rápidos.
Ao contrário do DNA que se baseia num “esqueleto” formado por pentoses e
grupos fosfato (carregados negativamente), a estrutura do PNA assenta num “esqueleto”
poliamida de carga neutra formado por unidades repetitivas de N-(2-aminoetil) glicina.
Esta estrutura confere-lhe uma maior estabilidade térmica (temperaturas de melting
(Tm) mais elevadas) 94 e permite que a hibridação seja realizada em concentrações
salinas mais baixas proporcionando um melhor acesso às sequências alvo.
Adicionalmente, devido à sua natureza sintética, estas moléculas são mais resistentes às

Introdução Geral
15
nucleases e protéases, e o seu caracter hidrofóbico permite que se difundam mais
facilmente na matriz de EPS (do inglês, Extracellular Polymeric Substances) dos
biofilmes 85.
Outros mímicos de DNA, apesar de pouco aplicados a FISH, apresentam
também um enorme potencial. Este é o caso do LNA. As molécula de LNA são
derivados do RNA que possuem o anel de ribose bloqueado numa conformação C3
'endo’, através de uma ligação extra entre o oxigénio na posição 2 e o carbono na
posição 4 da ribose (Figura 1.5) 95. A sua estrutura mais rígida aumenta a especificidade
de hibridação relativamente às sondas de DNA convencionais, o que permite diferenciar
melhor os alvos altamente homólogos 96.
Os nucleótidos de LNA também podem ser combinados com resíduos de DNA e
RNA (num processo comumente designada por síntese mista), o que permite um
controlo minucioso das temperaturas de hibridação e parâmetros termodinâmicos. Isto
reflete-se também num aumentando a especificidade de hibridação, e maior controlo no
desenho de abordagens multiplex 92,97,98.
Apesar de ainda pouco desenvolvida, já existem alguns estudos in vivo que
demostram a eficácia das sondas de LNA em aplicações terapêuticas 92,100. Existem
também alguns trabalhos, ainda que escassos, que aplicam estas sondas às técnicas de
FISH com bastante sucesso em ensaios in situ na expressão de miRNAs, na deteção
específica de células de leveduras, entre outros 100–103. Contudo, não são ainda
conhecidos estudos da técnica de LNA FISH em biofilmes.
1.4.2 Aplicação da técnica de FISH em biofilme
A técnica de FISH é uma ferramenta muito importante no estudo de biofilmes
uma vez que permite a compreensão e distribuição espacial das células. A sua aplicação
na caracterização in situ de populações em biofilme, tem facilitado a compreensão das
interações microbianas 104,105 contudo a estrutura do biofilme apresenta algumas
barreiras à aplicação de FISH.
A arquitetura dos biofilmes não permite uma análise quantitativa nem estrutural
de uma população abundante e complexa de microrganismos com recurso a técnicas
simples de microscopia. Contudo este problema pode ser ultrapassado com recurso à
utilização microscopia confocal de varrimento a laser (CLSM, do inglês Confocal Laser

Capítulo 1
16
Scanning Microscopy). CLSM é uma técnica capaz de produzir imagens nítidas e
precisas de amostras espessas 106, controlando a profundidade de campo. Permite
recolher séries de uma amostra em vários planos XY correspondentes a diferentes
profundidades, empilhando na vertical os planos e reconstruindo assim uma imagem
tridimensional da amostra 107.
Desta forma, a combinação de FISH e CLSM permite uma análise simultânea da
organização espacial de bactérias gram-negativas e gram-positivas, sem perturbar a
estrutura do biofilmes, fornecendo informações importantes sobre a organização e
interação dos microrganismos nestas estruturas 108.
1.5 Referências
1. Palleroni, N. J., Kunisawa, R., Contopoulou, R. & Doudoroff, M. Nucleic Acid Homologies in the Genus Pseudomonas. Int. J. Syst. Bacteriol. 23, 333–339 (1973).
2. Todar K. Pseudomonas aeruginosa . Disponivel em: www.textbookofbacteriology.net. Consultado em 2/07/2013.
3. Murray, P.R., Baron, E. J., Pfaller, M. A., Tenover, F.C. & Yolken, R. H. in Man. Clin. Microbiol. (O L Kiska, P. H. G.) Americait Society for Microbiology, 516–526 (2003).
4. Pollack, M. in Princ. Pract. Infect. Dis. (Mandell, G. L.) 2310–35 (2000).
5. Visca, P. et al. Metal regulation of siderophore synthesis in Pseudomonas aeruginosa and functional effects of siderophore-metal complexes. Appl. Environ. Microbiol. 58, 2886–93 (1992).
6. Merino, L. A. Pseudomonas aeruginosa: a bacterium with multiple personalities. Rev. Argent. Microbiol. 39, 143 (2007).
7. Pires, D., Sillankorva, S., Faustino, A. & Azeredo, J. Use of newly isolated phages for control of Pseudomonas aeruginosa PAO1 and ATCC 10145 biofilms. Res. Microbiol. 162, 798–806 (2011).
8. Hahn, H. P. The type-4 pilus is the major virulence-associated adhesin of Pseudomonas aeruginosa . Gene.Rev. 192, 99–108 (1997).
9. Van Alst, N. E., Picardo, K. F., Iglewski, B. H. & Haidaris, C. G. Nitrate sensing and metabolism modulate motility, biofilm formation, and virulence in Pseudomonas aeruginosa . Infect. Immun. 75, 3780–90 (2007).

Introdução Geral
17
10. Cacalano, G., Kays, M., Saiman, L. & Prince, A. Production of the Pseudomonas aeruginosa neuraminidase is increased under hyperosmolar conditions and is regulated by genes involved in alginate expression. J. Clin. Invest. 89, 1866–74 (1992).
11. Ghazaei, C., Ahmadi, M. & Jazani, N. H. Detection of Neuraminidase Activity in Pseudomonas aeruginosa PAO1. Iran. J. Basic Med. Sci. 13, 69–75 (2010).
12. Boyd, A. & Chakrabarty, A. M. Pseudomonas aeruginosa biofilms: role of the alginate exopolysaccharide. J. Ind. Microbiol. 15, 162–8 (1995).
13. Hassett, D. J. Anaerobic production of alginate by Pseudomonas aeruginosa: alginate restricts diffusion of oxygen. J. Bacteriol. 178, 7322–5 (1996).
14. Yorgey, P., Rahme, L. G., Tan, M. W. & Ausubel, F. M. The roles of mucD and alginate in the virulence of Pseudomonas aeruginosa in plants, nematodes and mice. Mol. Microbiol. 41, 1063–76 (2001).
15. Pollack, M. The Virulence of Pseudomonas aeruginosa . Clin. Infect. Dis. 6, 617–626 (1984).
16. Hamood, A. N., Griswold, J. A. & Duhan, C. M. Production of extracellular virulence factors by Pseudomonas aeruginosa isolates obtained from tracheal, urinary tract, and wound infections. J. Surg. Res. 61, 425–32 (1996).
17. Bardoel, B. W., van Kessel, K. P. M., van Strijp, J. A. G. & Milder, F. J. Inhibition of Pseudomonas aeruginosa virulence: characterization of the AprA-AprI interface and species selectivity. J. Mol. Biol. 415, 573–83 (2012).
18. Chieda, Y. et al. Virulence of an exotoxin A-deficient strain of Pseudomonas aeruginosa toward the silkworm, Bombyx mori. Microb. Pathog. 51, 407–14 (2011).
19. Pollack, M. The role of exotoxin A in Pseudomonas disease and immunity. Rev. Infect. Dis. 5, 979–84 (1983).
20. Cryz, S. J., Pitt, T. L., Fürer, E. & Germanier, R. Role of lipopolysaccharide in virulence of Pseudomonas aeruginosa . Infect. Immun. 44, 508–13 (1984).
21. Pier, G. B. Pseudomonas aeruginosa lipopolysaccharide: a major virulence factor, initiator of inflammation and target for effective immunity. Int J Med Microbiol. 297, 277–95 (2008).
22. Veesenmeyer, J. L., Hauser, A. R., Lisboa, T. & Rello, J. Pseudomonas aeruginosa virulence and therapy: evolving translational strategies. Crit. Care Med. 37, 1777–86 (2009).

Capítulo 1
18
23. Lister, P. D., Wolter, D. J. & Hanson, N. D. Antibacterial-resistant Pseudomonas aeruginosa: clinical impact and complex regulation of chromosomally encoded resistance mechanisms. Clin. Microbiol. Rev. 22, 582–610 (2009).
24. Lopes, S., Azevedo, N.F. & Pereira, M. O. in Biofilmes Na Saúde, no Ambient. na Indústria. Impacto dos biofilmes na fibrose cística. (Azevedo, N. F., Cerca, N.) Porto: Publindústria. 1, 45–57 (2012).
25. Pollack, M. Pseudomonas aeruginosa . In GL Mandell, R. Dolan, and JE Bennett (ed.). Princ. Pract. Infect. Dis. Churchill Livingstone, New York, NY 1820–2003 (1995).
26. Morrison, A. J. & Wenzel, R. P. Epidemiology of infections due to Pseudomonas aeruginosa . Rev. Infect. Dis. 6, 627–42 (1984).
27. National Nosocomial Infections Surveillance (NNIS) System report, data summary from October 1986-April 1998, issued June 1998. Am. J. Infect. Control 26, 522–33 (1998).
28. Emori, T. G. & Gaynes, R. P. An overview of nosocomial infections, including the role of the microbiology laboratory. Clin. Microbiol. Rev. 6, 428–42 (1993).
29. Gordon, N. C. & Wareham, D. W. Multidrug-resistant Acinetobacter baumannii: mechanisms of virulence and resistance. Int. J. Antimicrob. Agents 35, 219–26 (2010).
30. Peleg, A. Y., Seifert, H. & Paterson, D. L. Acinetobacter baumannii: emergence of a successful pathogen. Clin. Microbiol. Rev. 21, 538–82 (2008).
31. Warskow, A. L. & Juni, E. Nutritional requirements of Acinetobacter strains isolated from soil, water, and sewage. J. Bacteriol. 112, 1014–6 (1972).
32. Dorsey, C. W., Tomaras, A. P. & Actis, L. A. Genetic and Phenotypic Analysis of Acinetobacter baumannii Insertion Derivatives Generated with a Transposome System. Appl. Environ. Microbiol. 68, 6353-60 (2002).
33. Tomaras, A. P. Attachment to and biofilm formation on abiotic surfaces by Acinetobacter baumannii: involvement of a novel chaperone-usher pili assembly system. Microbiology 149, 3473–84 (2003).
34. Fournier, P. E. & Richet, H. The epidemiology and control of Acinetobacter baumannii in health care facilities. Clin. Infect. Dis. 42, 692–9 (2006).
35. Sanchez, C. J. et al. Biofilm formation by clinical isolates and the implications in chronic infections. BMC Infect. Dis. 13, 47 (2013).
36. Rodríguez-Baño, J. et al. Biofilm formation in Acinetobacter baumannii: associated features and clinical implications. Clin. Microbiol. Infect. 14, 276–8 (2008).

Introdução Geral
19
37. Fournier, P. E. & Richet, H. The epidemiology and control of Acinetobacter baumannii in health care facilities. Clin. Infect. Dis. 42, 692–9 (2006).
38. Falagas, M. E., Bliziotis, I. A. & Siempos, I. I. Attributable mortality of Acinetobacter baumannii infections in critically ill patients: a systematic review of matched cohort and case-control studies. Crit. Care 10, 48 (2006).
39. Fishbain, J. & Peleg, A. Y. Treatment of Acinetobacter infections. Clin. Infect. Dis. 51, 79–84 (2010).
40. Donlan, R. M. & Costerton, J. W. Biofilms: survival mechanisms of clinically relevant microorganisms. Clin. Microbiol. Rev. 15, 167–93 (2002).
41. Flemming, H.-C., Neu, T. R. & Wozniak, D. J. The EPS matrix: the “house of biofilm cells”. J. Bacteriol. 189, 7945–7 (2007).
42. Vu, B., Chen, M., Crawford, R. J. & Ivanova, E. P. Bacterial extracellular polysaccharides involved in biofilm formation. Molecules 14, 2535–54 (2009).
43. Davey, M. E. & O’toole, G. A. Microbial biofilms: from ecology to molecular genetics. Microbiol. Mol. Biol. Rev. 64, 847–67 (2000).
44. Boari, C. A. et al. Formação de biofilme em aço inoxidável por Aeromonas hydrophila e Staphylococcus aureus usando leite e diferentes condições de cultivo. Cienc.Tecnol.Aliment.2008, 886–95 (2009).
45. Donlan, R. M. Biofilm formation: a clinically relevant microbiological process. Clin. Infect. Dis. 33, 1387–92 (2001).
46. Costerton, J. W. Bacterial Biofilms: A Common Cause of Persistent Infections. Science (80-. ). 284, 1318–1322 (1999).
47. Stoodley, P., Sauer, K., Davies, D. G. & Costerton, J. W. Biofilms as complex differentiated communities. Annu. Microbiol. Rev. 56, 187–209 (2002).
48. Cerca, N., Jefferson, K. K. in Biofilmes Na Saúde, no Ambient. na Indústria. Que vanatgens têm os microrganismos em formar um biofilme? (Azevedo, N. F., Cerca, N.) Porto: Publindústria. 1, 11–16 (2012).
49. Stoodley, P., Dodds, I., Boyle, J. D. & Lappin-Scott, H. M. Influence of hydrodynamics and nutrients on biofilm structure. J. Appl. Microbiol. 85, 19–28 (1998).
50. Costerton, W. et al. The application of biofilm science to the study and control of chronic bacterial infections. J. Clin. Invest. 112, 1466–77 (2003).
51. Azevedo, N. F., Cerca, N. in Biofilmes Na Saúde, no Ambient. na Indústria. A matriz dos biofilmes o truque para Sobreviver nas mais hostis condições ambientais. (Azevedo, N. F., Cerca, N.) Porto: Publindústria. 1,23–26 (2012).

Capítulo 1
20
52. Sutherland, I. W. The biofilm matrix--an immobilized but dynamic microbial environment. Trends Microbiol. 9, 222–7 (2001).
53. Azevedo, N. F., Vieira & M. J. & Keevil, C. W. in Biofilmes Na Saúde, no Ambient. na Indústria. O impacto dos biofilmes na actividade humana (Azevedo, N F., Cerca, N.) Porto: Publindústria, 1, 17–21 (Publindústria, 2012).
54. Paterson, D. L. Serious Infections in the Intensive Care Unit: Pseudomonas aeruginosa and Acinetobacter baumannii. Clin. Infect. Dis. 43, S41–S42 (2006).
55. Oliveira, L., Carneiro, P., Fischer, R. & Tinoco, E. A Presença de Patógenos Respiratórios no Biofilme Bucal de Pacientes com Pneumonia Nosocomial. Rev. Bras. Ter. Intensiva 19, 428–33 (2007).
56. Gil, S. et al. Implications of endotracheal tube biofilm in ventilator-associated pneumonia response: a state of concept. Crit. Care 16, 93 (2012).
57. Storti, A. Colonização de cateteres venosos centrais por biofilme microbiano. Tese de mestrado. Univ. Estadual Paulista. Fac. Ciências Farmacêuticas. Câmpus Araraquara; 140 (2006).
58. Storti, A., Pizzolitto, A. & Pizzolitto, E. Detection of mixed microbial biofilms on central venous catheters removed from intensive care unit patients. Brazilian J. Microbiol. 36, 275–80 (2005).
59. Duckworth, D. H. “Who discovered bacteriophage?”. Bacteriol. Rev. 40, 793–802 (1976).
60. Skurnik, M. & Strauch, E. Phage therapy: facts and fiction. Int. J. Med. Microbiol. 296, 5–14 (2006).
61. Sulakvelidze, A., Alavidze, Z., Morris, J. Minireview Bacteriophage Therapy. J. Antimicrob. Agents Chemother. 45, 649–59 (2001).
62. Sulakvelidze, A. Safety by Nature: Potential Bacteriophage Applications. Microbe Mag. (2009).
63. Melo, L., Sillankorva, S. & Azeredo, J. in Biofilmes Na Saúde, no Ambient. na Indústria. Uso bacteriófagos para o Controlo de biofilmes. (Azevedo, N. & Cerca, N.) Porto: Publindústria. 1, 227–236 (2012).
64. Ashelford, K. E., Day, M. J. & Fry, J. C. Elevated Abundance of Bacteriophage Infecting Bacteria in Soil. Appl. Environ. Microbiol. 69, (2003).
65. Chibani-Chennoufi, S., Bruttin, A., Dillmann, M.-L. & Brüssow, H. Phage-host interaction: an ecological perspective. J. Bacteriol. 186, 3677–86 (2004).

Introdução Geral
21
66. Wommack, K. E. & Colwell, R. R. Virioplankton : Viruses in Aquatic Ecosystems Virioplankton : Viruses in Aquatic Ecosystems. Microbiol. Mol. Biol. Rev. 64, (2000).
67. Loc-Carrillo, C. & Abedon, S. T. Pros and cons of phage therapy. Bacteriophage 1, 111–14 (2011).
68. Maniloff, J. Bacteriophages. eLS (2012).
69. Thiel, K. Old dogma, new tricks-21st Century phage therapy. Nat. Biotechnol. 22, 31–6 (2004).
70. Sillankorva, S. Use of bacteriophages to control biofilms. Tese de doutoramento em Engenharia Biológica. Univ. do Minho Port. 7–8 (2008).
71. Carlton, R. M. Phage therapy: past history and future prospects. Arch. Immunol. Ther. Exp. (Warsz). 47, 267–74 (1999).
72. Materón, L. A. Disponivel em: http://faculty.utpa.edu/lmateron/4404/images/cycle1.jpg. Consultado em 1/09/2013.
73. Kropinski, A. Phage therapy – Everything old is new again. AMMI Canada Annu. Meet. Symp. 17, 297–306 (2006).
74. São-José, C., Nascimento, J., Parreira, R., Santos, M. . Release of progeny phages from infected cells. Bacteriophages Genet. Mol. Biol. 309–36 (2007).
75. Levin, B. R. & Bull, J. J. Population and evolutionary dynamics of phage therapy. Nat. Rev. Microbiol. 2, 166–73 (2004).
76. Sulakvelidze, A. and Kutter, E. in Bacteriophages Biol. Apl. (eds E. Kutter A. Sulakvelidze) 381–436 (2005).
77. Barbosa, C., Venail, P., Holguin, A. V & Vives, M. J. Co-Evolutionary Dynamics of the Bacteria Vibrio sp. CV1 and Phages V1G, V1P1, and V1P2: Implications for Phage Therapy. Microb. Ecol. 66, 897–905 (2013).
78. Jończyk, E., Klak, M., Międzybrodzki, R., Górski, A. The influence of external factors on bacteriophages,. 56, 191–200
79. Mara, D., Horan, N. The handbook of Water and Wastewater Microbiology. (2003).
80. Amann, R., Glöckner, F.-O. & Neef, A. Modern methods in subsurface microbiology: in situ identification of microorganisms with nucleic acid probes. FEMS Microbiol. Rev. 20, 191–200 (2006).

Capítulo 1
22
81. Bottari, B., Ercolini, D., Gatti, M. & Neviani, E. Application of FISH technology for microbiological analysis: current state and prospects. Appl. Microbiol. Biotechnol. 73, 485–94 (2006).
82. Amann, R., Fuchs, B. M. & Behrens, S. The identification of microorganisms by fluorescence in situ hybridisation. Environ. Biotechnol. 12, 231–36 (2001).
83. Moter, A. & Gobel, U. B. Fluorescence in situ hybridization ( FISH ) for direct visualization of microorganisms. J. Microbiol. Methods 41, 85–112 (2000).
84. Levsky, J. M. & Singer, R. H. Fluorescence in situ hybridization : past , present and future. J. Cell Sci. 116, 2833–38 (2003).
85. Cerqueira, L. et al. DNA mimics for the rapid identification of microorganisms by fluorescence in situ hybridization (FISH). Int. J. Mol. Sci. 9, 1944–60 (2008).
86. Lehtola, M. J., Torvinen, E., Miettinen, I. T. & Keevil, C. W. Fluorescence in situ hybridization using peptide nucleic acid probes for rapid detection of Mycobacterium avium subsp. avium and Mycobacterium avium subsp. paratuberculosis in potable-water biofilms. Appl. Environ. Microbiol. 72, 848–53 (2006).
87. Stender, H., Fiandaca, M., Hyldig-Nielsen, J. J. & Coull, J. PNA for rapid microbiology. J. Microbiol. Methods 48, 1–17 (2002).
88. Priya, N. G., Pandey, N. & Rajagopal, R. LNA probes substantially improve the detection of bacterial endosymbionts in whole mount of insects by fluorescent in-situ hybridization. BMC Microbiol. 12, 81 (2012).
89. Jepsen, J. S., Sørensen, M. D. & Wengel, J. Locked nucleic acid: a potent nucleic acid analog in therapeutics and biotechnology. Oligonucleotides 14, 130–46 (2004).
90. Cummins, L. L. et al. Characterization of fully 2’-modified oligoribonucleotide hetero- and homoduplex hybridization and nuclease sensitivity. Nucleic Acids Res. 23, 2019–24 (1995).
91. Pallan, P. S. et al. Unexpected origins of the enhanced pairing affinity of 2’-fluoro-modified RNA. Nucleic Acids Res. 39, 3482–95 (2011).
92. Kubota, K., Ohashi, A., Imachi, H. & Harada, H. Improved in situ hybridization efficiency with locked-nucleic-acid-incorporated DNA probes. Appl. Environ. Microbiol. 72, 5311–7 (2006).
93. Nielsen, P. E., Egholm, M., Berg, R. H. & Buchardt, O. Sequence-selective recognition of DNA by strand displacement with a thymine-substituted polyamide. Science 254, 1497–500 (1991).

Introdução Geral
23
94. Justé, A., Thomma, B. P. H. J. & Lievens, B. Recent advances in molecular techniques to study microbial communities in food-associated matrices and processes. Food Microbiol. 25, 745–61 (2008).
95. Amann, R. & Fuchs, B. M. Single-cell identification in microbial communities by improved fluorescence in situ hybridization techniques. Nat. Rev. Microbiol. 6, 339–48 (2008).
96. Sigma-Aldrich. Locked Nucleic Acids ® ( LNA ® ) FAQ.
97. McTigue, P. M., Peterson, R. J. & Kahn, J. D. Sequence-dependent thermodynamic parameters for locked nucleic acid (LNA)-DNA duplex formation. Biochemistry 43, 5388–405 (2004).
98. Koshkin, A. A. et al. LNA ( Locked Nucleic Acid ): An RNA Mimic Forming Exceedingly Stable LNA : LNA Duplexes. Am. Chem. Soc. 13252–53 (1998).
99. Kloosterman, W. P., Wienholds, E., de Bruijn, E., Kauppinen, S. & Plasterk, R. H. A. In situ detection of miRNAs in animal embryos using LNA-modified oligonucleotide probes. Nat. Methods 3, 27–9 (2006).
100. Nelson, P. T. et al. RAKE and LNA-ISH reveal microRNA expression and localization in archival human brain. RNA 12, 187–91 (2006).
101. Thomsen, R., Nielsen, P. S. & Jensen, T. H. Dramatically improved RNA in situ hybridization signals using LNA-modified probes. RNA 11, 1745–8 (2005).
102. Silahtaroglu, A. N., Tommerup, N. & Vissing, H. FISHing with locked nucleic acids (LNA): evaluation of different LNA/DNA mixmers. Mol. Cell. Probes 17, 165–9 (2003).
103. Kloosterman, W. P., Wienholds, E., de Bruijn, E., Kauppinen, S. & Plasterk, R. H. A. In situ detection of miRNAs in animal embryos using LNA-modified oligonucleotide probes. Nat. Methods 3, 27–9 (2006).
104. Almeida, C., Azevedo, N. F., Santos, S., Keevil, C. W. & Vieira, M. J. Discriminating multi-species populations in biofilms with peptide nucleic acid fluorescence in situ hybridization (PNA FISH). PLoS One 6, e14786 (2011).
105. Silva, S. et al. Candida glabrata and Candida albicans co-infection of an in vitro oral epithelium. J. Oral Pathol. Med. 40, 421–7 (2011).
106. Qvist, M. H. et al. Application of confocal laser scanning microscopy in characterization of chemical enhancers in drug-in-adhesive transdermal patches. AAPS PharmSci 4, E3 (2002).
107. Moghe, P. Principles of Confocal Laser scanning Microscopy. Rutgers Univ. (2012).

Capítulo 1
24
108. Thurnheer, T., Gmür, R. & Guggenheim, B. Multiplex FISH analysis of a six-species bacterial biofilm. J. Microbiol. Methods 56, 37–47 (2004).

Capítulo 2
Desenvolvimento de sondas de LNA para aplicação na técnica de FISH


Desenvolvimento de sondas de LNA para aplicação na técnica de FISH
27
2 Desenvolvimento de sondas de LNA para aplicação na técnica de FISH
Este capítulo tem como principal objetivo a descrição do desenvolvimento de
sondas de LNA para as bactérias Pseudomonas aeruginosa e Acinetobacter baumannii e
respetivos fagos. Estas sondas serão posteriormente utilizadas na análise microscópica
dos biofilmes simples e mistos formados pelas bactérias anteriormente mencionadas.
Para além da avaliação em termos da diferenciação e distribuição das espécies, as
sondas específicas para a deteção dos fagos, permitirão, em trabalhos futuros, estudar a
infeção fágica neste tipo de estruturas.
Neste sentido, foi efetuado inicialmente o desenho e/ou avaliação teórica do
desempenho das diferentes sondas com recurso a ferramentas bioinformáticas. Alguns
parâmetros termodinâmicos foram também determinados. No âmbito do trabalho
desenvolvido, e uma vez que a aplicação de moléculas de LNA é muito recente em
FISH, apenas uma das sondas desenhadas foi sintetizada e testada laboratorialmente
para verificar o seu funcionamento, como prova de conceito.
2.1 Materiais e métodos
2.1.1 Desenho e avaliação das sondas
Avaliação teórica das sondas de P. aeruginosa e A. baumannii
Usando a base de dados probebase (http://www.microbial-
ecology.net/probebase/) foram identificadas sondas já descritas na literatura para
A. baumannii e P. aeruginosa .
Para a determinação da sensibilidade e especificidade das sondas que hibridam
na subunidade menor do rRNA (16S) foi usado o programa Probe Match
(http://www.microbial-ecology.net/probebase/default.asp?mode=probeset) e o browser
da base de dados de RNA ribossomal RDP II (do inglês Ribosomal Database Project)
(http://rdp.cme.msu.edu/hierarchy/hb_intro.jsp). Uma vez que a base de dados RDP II
não comtempla sequências para a subunidade maior (23S), para as sondas que hibridam
na região 23S usou-se o programa ProbeCheck (http://131.130.66.200/cgi-
bin/probecheck/content.pl?id=home) disponível no site ARB Silva.

Capítulo 2
28
A especificidade foi calculada através da fórmula nSs / (TnS) × 100, onde nSs
representa o número de não- A. baumannii que não reagem com a sonda e TnS é o total
de não-A. baumannii presentes na base de dados. A sensibilidade foi calculada como
Ss / (TSs) × 100, onde Ss representa o número de A. baumannii detetadas pela sonda e
TSs é o número total de estirpes A. baumannii presentes na base de dados 1. Os mesmos
cálculos foram efetuados para a bactéria P. aeruginosa , substituindo A. baumannii por
P. aeruginosa .
Desenho e avaliação das sondas dos fagos
Foi usada a base de dados do National Center for Biotechnology Information
(NCBI) (http://www.ncbi.nlm.nih.gov/genomes/GenomesGroup.cgi?taxid=35237) para
recolher um número significante de genomas completos de fagos de A. baumannii e
P. aeruginosa . Seguidamente foi utilizado o programa CoreGenes3.0
(http://binf.gmu.edu:8080/CoreGenes3.0/) que realiza uma análise BLAST fornecendo
uma tabela de genes conservados entre os genomas analisados 2.
As sequências dos genes selecionados pelo CoreGene 3.0 foram alinhadas e
comparadas pelo programa ClustralW2 (http://www.ebi.ac.uk/Tools/msa/clustalw2/).
Dada a inexistência de regiões conservadas nos genes analisados, o genoma dos fagos
foram sequenciados e as sondas foram desenhadas para o mRNA (RNA mensageiro)
que codifica para a proteína que constituiu maioritariamente a cápside do fago (major
head protein).
Adaptação para LNA
As sondas para as bactérias e para os seus respetivos fagos foram modificadas
para sondas de LNA/2-O-metil, de acordo com o descrito em Søe et al.3. Com recurso
ao programa RNA Chemistry Laboratory
(http://rnachemlab.ibch.poznan.pl/calculator2.php) foram calculadas as temperaturas de
fusão (Tm, do inglês melting temperature) e os restantes parâmetros termodinâmico
para as diferentes sequências. Foram selecionas sequências com temperaturas de
melting semelhantes e com valores de energia livre de ligação (∆G) baixos 4.

Desenvolvimento de sondas de LNA para aplicação na técnica de FISH
29
2.1.2 Otimização das condições de hibridação para LNA FISH
Estirpes bacterianas e condições de cultura
As estirpes bacterianas utilizadas estão listadas na Tabela 2.7. Todas as estirpes
foram cultivadas em meio nutritivo Luria-Bertani (LB) (nzytech, Portugal) com
1,2 % (p/v) de ágar (Liofilchem, Itália). As bactérias foram crescidas a 37 ºC durante
16 h - 20 h, sendo repicadas a cada 24 h.
Hibridação in situ fluorescente (FISH)
Após a seleção das diferentes sondas, a sonda para a bactéria P. aeruginosa foi
sintetizada por uma empresa especializada (Ribotask, Dinamarca). Todos os
procedimentos da preparação e utilização das sondas foram realizados em condições de
baixa luminosidade com o intuito de evitar a ação negativa que a luz provoca na
integridade dos fluorocromos.
A alíquota original da sonda foi preparada à concentração de 100 µM em água
ultrapura estéril. Após a preparação da alíquota original, foram preparadas alíquotas de
stock a uma concentração de 4 μM, também em água ultrapura, alíquotas de uso foram
preparadas à concentração de 200 μM em solução de hibridação (20 mM Tris-HCl
(pH 7,2) (Biorad, Portugal), 0,01 % SDS (p/v) (Biorad), 900 mM NaCl (Panreac,
Espanha) e 30 % formamida (v/v) (Sigma, Portugal)).
Para aplicação da técnica FISH, foi utilizado o protocolo descrito em
Almeida et al. 5 com algumas alterações.
Na otimização do processo de hibridação, foram testadas diferentes temperaturas
de hibridação (55 ºC, 57 ºC e 59 ºC). Na análise experimental da especificidade e
sensibilidade da sonda, foram testadas várias estirpes listadas na Tabela 2.7.
Uma ansa de cultura fresca foi dissolvida em 1 ml de PBS. Foi colocado 20 µl
do inóculo anterior num poço da uma lâmina de microscopia adequada ao uso de
fluorescência (revestimento com epoxy, Marienfeld, Alemanha). Posteriormente a
lâmina foi colocada numa incubadora com temperatura entre 50 ºC a 60 ºC até secar o
inóculo.
De seguida, foram adicionados 30 µl de paraformaldeido a 4 % (v/v) (Sigma)
durante 10 min para promover a fixação celular. Seguidamente, o excesso foi removido

Capítulo 2
30
utilizando papel absorvente. Foram adicionados 30 µl de etanol 50 % (v/v) durante
10 min e depois, o excesso foi novamente removido.
Após a etapa de fixação, foram aplicados 20 µl da alíquota de uso da sonda. Foi
incluído para cada amostra um controlo negativo, sujeito às mesmas condições mas sem
adição de sonda na solução de hibridação. Cada poço foi coberto com lamela para
garantir que toda a amostra fique em contacto com a sonda. As lâminas foram colocadas
em placas de petri envolvidas em folha de alumínio, contendo papel adsorvente húmido,
e foram a incubar durante 1 h. A solução de lavagem (20 mM Tris-HCl (pH 7,2),
0,01 % SDS (v/v) e 900 mM NaCl) foi colocada em coplin-jars na estufa, junto com as
lâminas, para pré-aquecer.
No final da hibridação, as lamelas foram retiradas e as lâminas mergulhadas em
solução de lavagem durante 30 min para retirar o excesso de sondas 6. Após a lavagem,
as lâminas foram retiradas e secas durante 5 min na incubadora. No final, as lâminas
foram guardadas nas caixas de petri protegidas da luz e armazenadas a 4 ºC até à sua
observação.
Visualização microscópica
Os resultados da hibridação in situ foram analisados por microscopia de
epifluorescência com aplicação de uma gota de óleo de imersão não fluorescente
(Merck, EUA). O microscópio de epifluorescência Olympus BX51 (Olympus Portugal
SA, Porto, Portugal) possui 3 conjuntos de filtros (DAPI - 365-370/421, FITC - 470-
490/516 e TRITC - 530-550/591), objetivas a seco (10 ×, 20 ×, 40 × de ampliação) e de
imersão (60 ×, 100 × de ampliação), e uma câmara a cores para aquisição de imagem
(Olympus DP71). O software Cell-B (Olympus) foi utilizado para captação e tratamento
das imagens. Para a sonda da bactéria P. aeruginosa, o fluorocromo utilizado foi o
FAM que emite fluorescência na zona do verde utilizando-se por isso o filtro para FITC.

Desenvolvimento de sondas de LNA para aplicação na técnica de FISH
31
2.2 Resultados e discussão
2.2.1 Desenho e avaliação das sondas
Sondas para as bactérias P. aeruginosa e A. baumannii
Neste estudo foram avaliadas sondas que detetam especificamente as bactérias
A. baumannii e P. aeruginosa . Através da base de dados probebase foram pesquisadas
as sondas já descritas para estes dois microrganismos patogénicos. Foram identificadas
4 sondas, uma sonda de DNA para a região 16S do rRNA de A. baumannii e três sondas
para P. aeruginosa, das quais duas são de DNA (para as regiões 16S e 23S) e uma outra
de PNA para a região16S (Tabela 2.1).
Tabela 2.1 - Sondas descritas na literatura para P. aeruginosa e A. baumannii, juntamente com
algumas das suas características mais importantes
Microrganismo Sonda Sequência (5´-3´) Subunidade
rRNA Sonda Posição Referência
P. aeruginosa PseaerA GGTAACCGTCCCCCTTGC 16S DNA 1449 Hogardt et al.7
P. aeruginosa PseaerB TCTCGGCCTTGAAACCCC 23S DNA 1499 Hogardt et al.7
P. aeruginosa Psaer AACTTGCTGAACCAC 16S PNA 583 Malic et al.8
A. baumannii ACA ATCCTCTCCCATACTCTA 16S DNA 651 Wagner et al.9
Na avaliação teórica, as sequências das sondas de 16S foram comparadas, na
base de dados de rRNA 16S (RDP II), com um total de 200 062 sequências com mais de
1200 pares de bases (pb) e consideradas de boa qualidade. A sequência da sonda de 23S
de P. aeruginosa foi comparada, na base de dados de rRNA 23S (23S LSU, ARB
Silva), com um total de 28 681 sequências com mais de 1900 pb e consideradas
igualmente de boa qualidade.
De entre os parâmetros mais importantes no desempenho de uma sonda, os
valores da sensibilidade e especificidade são os mais informativos 6. A sensibilidade de
uma sonda mede a capacidade desta identificar corretamente a presença da bactéria
numa amostra, sendo inversamente proporcional à probabilidade de ocorrência de falsos
negativos. A especificidade mede a capacidade do teste excluir corretamente as
amostras não portadoras das bactérias para as quais foi concebida, sendo por isso
indicativa da probabilidade de ocorrência de falsos positivos.

Capítulo 2
32
Na Tabela 2.2 estão representados os valores de sensibilidade e especificidade
relativos a cada sonda, determinados com descrito no ponto 2.1.1.
Tabela 2.2 - Avaliação da sensibilidade (S) e especificidade (E) das sondas descritas na literatura
Microrganismo Sonda S (%) E (%)
P. aeruginosa PseaerA 48 99
P. aeruginosa PseaerB 64 99,7
P. aeruginosa Psaer 86 99
A. baumannii ACA 91 99
Com base nos valores apresentados pela Tabela 2.2, a sonda para P. aeruginosa
que apresentou valores mais satisfatórios foi a sonda Psaer com uma sensibilidade de
86 % e uma especificidade de 99 %. Quanto maior for o valor de sensibilidade, melhor
será o desempenho da sonda durante a hibridação.
Uma vez que a única sonda estudada de A. baumannii apresentou valores de
sensibilidade e especificidade satisfatórios (91 % e 99 %, respetivamente), esta foi
considerada uma sonda de boa qualidade.
Para além de assegurar uma elevada especificidade e sensibilidade das sondas, é
importante verificar se poderá ocorrer hibridação cruzada de cada uma das sondas com
a outra espécie a utilizar nos ensaios de biofilme misto. Além disso, é desejável que
exista mais que um mismatch (um nucleótico não emparelhado) entre a sonda e a
sequência não alvo, de forma a proporcional uma excelente descriminação das espécies.
Por este motivo, após a escolha das sondas, procedeu-se à verificação de analogia entre
as estirpes e as sondas (Figura 2.1), através do programa ClustralW2
(http://www.ebi.ac.uk/Tools/msa/clustalw2/), que permite o alinhamento e análise
múltipla de sequências conforme a sua semelhança.

Desenvolvimento de sondas de LNA para aplicação na técnica de FISH
33
Figura 2.1 - Alinhamento das sequências de rRNA das espécies em estudo com as sondas Psaer (A) e ACA (B) no ClustralW2.
Através da Figura 2.1, é possível verificar que a sonda de Acinetobacter não
apresentava complementaridade com a sequência de rRNA 16S de P. aeruginosa e vice-
versa. Na verdade, cada uma das sondas possui 3 mismatches para a sequência não-alvo,
o que indicia um elevado poder de discriminação sem necessidade de condições de
restringência extremas na etapa de hibridação.
Sondas para os fagos de P. aeruginosa e A. baumannii
A primeira abordagem no desenho das sondas para fagos, passou por tentar
encontrar regiões altamente conservadas nos diferentes genomas depositados nas bases
de dados. Esta abordagem iria permitir a utilização das sondas num grupo alargado de
fagos.
Na análise da base de dados do NCBI, foram identificados 49 genomas de fagos
de P. aeruginosa e 5 de A. baumannii. Dos 49 genomas de fagos de P. aeruginosa
foram escolhidos 5 completos e anotados, pertencentes à família Siphoviridae, família à
qual pertence o fago que será utilizado neste estudo. Foram também selecionados os
únicos 5 fagos de A. baumannii. Este conjunto de sequências foi depois utilizado para
identificar regiões genómicas altamente conservadas com recurso ao software
CoreGenes3.0. Após a introdução dos números de acesso dos fagos no programa foram
obtidos os genes das regiões mais conservadas (Figura 2.3).
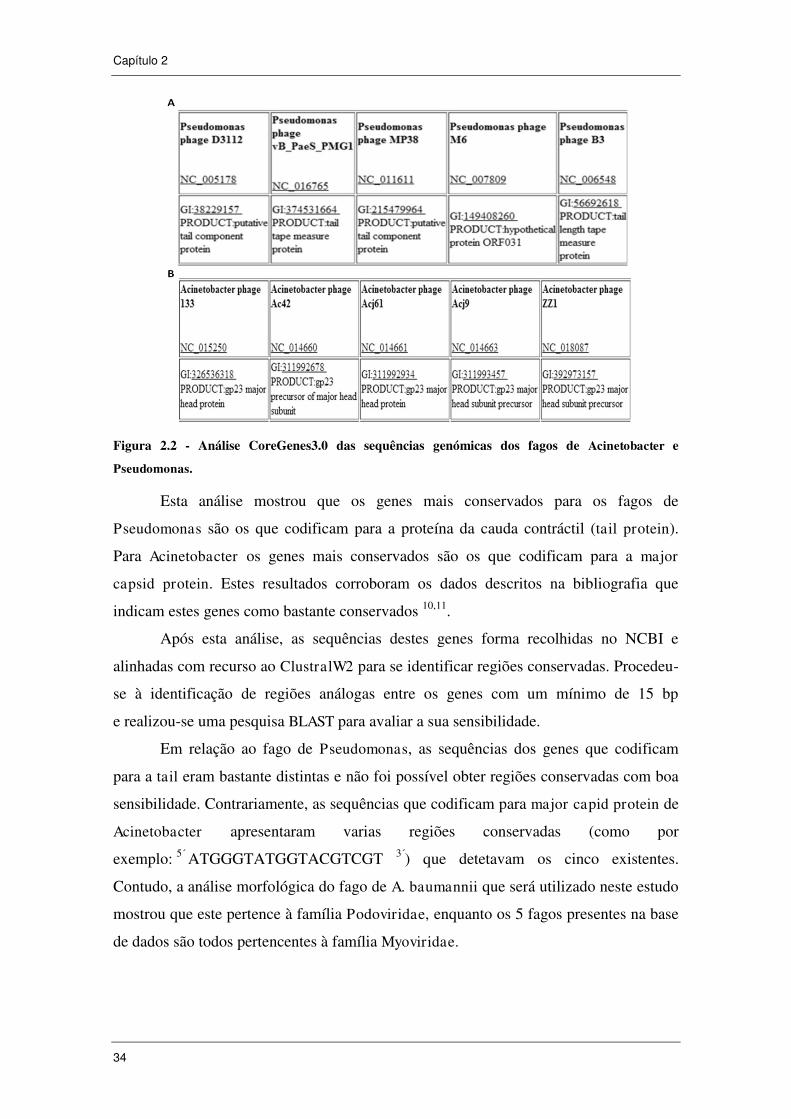
Capítulo 2
34
Figura 2.2 - Análise CoreGenes3.0 das sequências genómicas dos fagos de Acinetobacter e
Pseudomonas.
Esta análise mostrou que os genes mais conservados para os fagos de
Pseudomonas são os que codificam para a proteína da cauda contráctil (tail protein).
Para Acinetobacter os genes mais conservados são os que codificam para a major
capsid protein. Estes resultados corroboram os dados descritos na bibliografia que
indicam estes genes como bastante conservados 10,11.
Após esta análise, as sequências destes genes forma recolhidas no NCBI e
alinhadas com recurso ao ClustralW2 para se identificar regiões conservadas. Procedeu-
se à identificação de regiões análogas entre os genes com um mínimo de 15 bp
e realizou-se uma pesquisa BLAST para avaliar a sua sensibilidade.
Em relação ao fago de Pseudomonas, as sequências dos genes que codificam
para a tail eram bastante distintas e não foi possível obter regiões conservadas com boa
sensibilidade. Contrariamente, as sequências que codificam para major capid protein de
Acinetobacter apresentaram varias regiões conservadas (como por
exemplo: 5´ ATGGGTATGGTACGTCGT 3´) que detetavam os cinco existentes.
Contudo, a análise morfológica do fago de A. baumannii que será utilizado neste estudo
mostrou que este pertence à família Podoviridae, enquanto os 5 fagos presentes na base
de dados são todos pertencentes à família Myoviridae.

Desenvolvimento de sondas de LNA para aplicação na técnica de FISH
35
Assim, a única estratégia possível no desenho das sondas passou por sequenciar
total ou parcialmente os genomas dos fagos em estudo, phiIBB-PAP1 (fago de
Pseudomonas) e phiIBB-Aba1 (fago de Acinetobacter), de forma a desenhar sondas
específicas para as suas sequências. Assim, optou-se por sequenciar o gene que codifica
para a major capsid protein, uma vez que, tal como mencionado anteriormente, está
descrita como muito conservada 10,11. Contudo, para o fago de Pseudomonas utilizado
neste estudo, phiIBB-PAP1, não foi possível obter a sequência que codifica a major
capsid protein. Portanto, um fago muito semelhante em termos de morfologia e espectro
de ação (fago phiIBB-PAA2) foi selecionado para o desenho da sonda e foi depois
usado nos ensaios de microscopia.
Após a obtenção das sequências dos genes da major capsid protein dos fagos
phiIBB-Aba1 (Acinetobacter) e phiIBB-PAA2 (Pseudomonas), foram realizadas
análises com recurso ao BLAST. Apesar de não ser necessário procurar regiões
conservadas, uma vez que a sequência exata do gene é conhecida, esta análise permitiu
escolher uma região que se mantenha noutros fagos de Acinetobacter ou Pseudomonas,
tentando assim obter uma maior abrangência para as sondas desenvolvidas.
Desta análise, foi possível verificar que o gene da major capsid protein do fago
phiIBB-PAA2 apresentava regiões análogas com os fagos LUZ24 e o PaP3, também
específicos para Pseudomonas. Os genes foram alinhados no ClustralW e escolhidas
sequências conservadas com 15 bp. Com uma nova análise BLAST destas sequências
verificou-se que uma das sequências permitia detetar 7 sequências de fagos para
Pseudomonas (AACAACGAGAAGTCC). Esta sequência foi então escolhida para o
desenho da sonda, e denominada de PGPseLO.
Para o fago de Acinetobacter (phiIBB-Aba1) a sequência do gene da
major capsid protein foi comparada com a sequência de um fago muito semelhante
(phiIBB-Aba2 – fago do grupo de investigação e não depositado na base de dados) com
recurso ao ClustralW e selecionadas regiões conservadas nos dois fagos.
Após uma análise BLAST das regiões selecionadas, foram obtidas 6 sequências
que detetavam somente os dois fagos em estudo. Para selecionar a sonda com as
melhores características, foram calculadas as temperaturas de melting e energias livres
de ligação (∆G) para duplexes DNA / DNA usando as metodologias de cálculo
empregadas nos estudos de SantaLucia et al. 12 (Tabela 2.3). Apesar das sondas
selecionadas serem ainda alvo de adaptação na LNA na próxima secção, e portanto
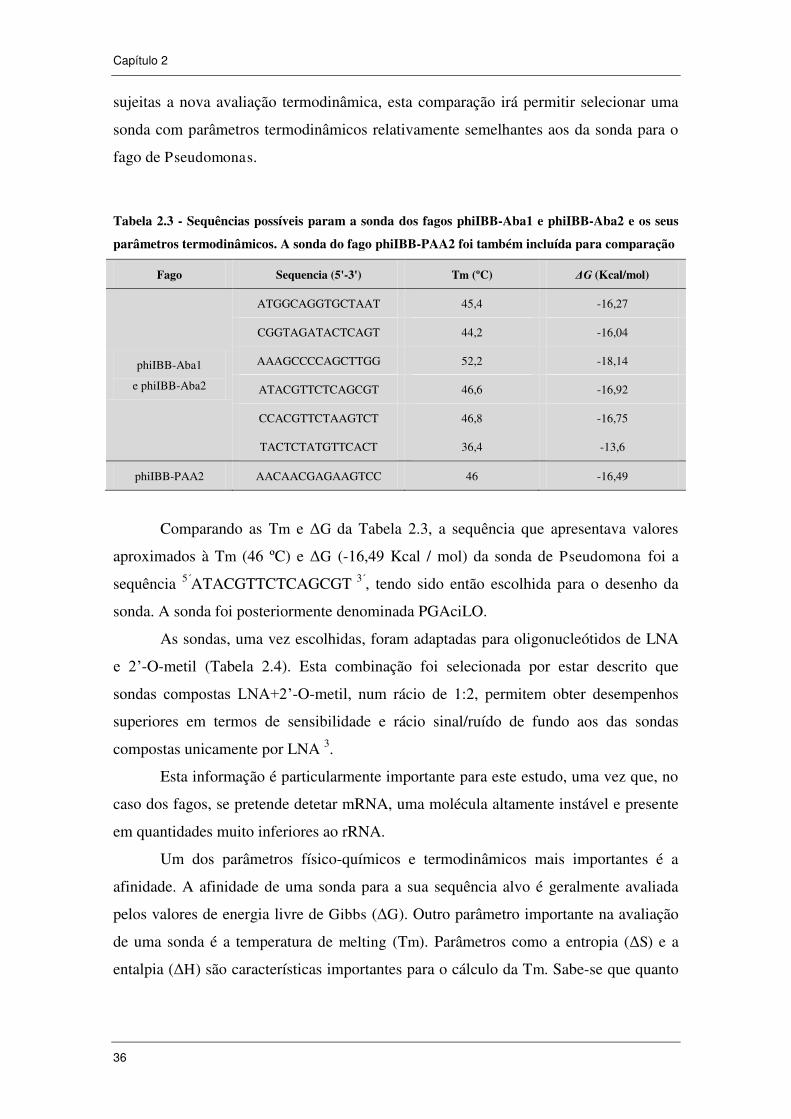
Capítulo 2
36
sujeitas a nova avaliação termodinâmica, esta comparação irá permitir selecionar uma
sonda com parâmetros termodinâmicos relativamente semelhantes aos da sonda para o
fago de Pseudomonas.
Tabela 2.3 - Sequências possíveis param a sonda dos fagos phiIBB-Aba1 e phiIBB-Aba2 e os seus
parâmetros termodinâmicos. A sonda do fago phiIBB-PAA2 foi também incluída para comparação
Fago Sequencia (5'-3') Tm (ºC) ΔG (Kcal/mol)
phiIBB-Aba1
e phiIBB-Aba2
ATGGCAGGTGCTAAT 45,4 -16,27
CGGTAGATACTCAGT 44,2 -16,04
AAAGCCCCAGCTTGG 52,2 -18,14
ATACGTTCTCAGCGT 46,6 -16,92
CCACGTTCTAAGTCT 46,8 -16,75
TACTCTATGTTCACT 36,4 -13,6
phiIBB-PAA2 AACAACGAGAAGTCC 46 -16,49
Comparando as Tm e ΔG da Tabela 2.3, a sequência que apresentava valores
aproximados à Tm (46 ºC) e ΔG (-16,49 Kcal / mol) da sonda de Pseudomona foi a
sequência 5´ATACGTTCTCAGCGT 3´, tendo sido então escolhida para o desenho da
sonda. A sonda foi posteriormente denominada PGAciLO.
As sondas, uma vez escolhidas, foram adaptadas para oligonucleótidos de LNA
e 2’-O-metil (Tabela 2.4). Esta combinação foi selecionada por estar descrito que
sondas compostas LNA+2’-O-metil, num rácio de 1:2, permitem obter desempenhos
superiores em termos de sensibilidade e rácio sinal/ruído de fundo aos das sondas
compostas unicamente por LNA 3.
Esta informação é particularmente importante para este estudo, uma vez que, no
caso dos fagos, se pretende detetar mRNA, uma molécula altamente instável e presente
em quantidades muito inferiores ao rRNA.
Um dos parâmetros físico-químicos e termodinâmicos mais importantes é a
afinidade. A afinidade de uma sonda para a sua sequência alvo é geralmente avaliada
pelos valores de energia livre de Gibbs (ΔG). Outro parâmetro importante na avaliação
de uma sonda é a temperatura de melting (Tm). Parâmetros como a entropia (ΔS) e a
entalpia (ΔH) são características importantes para o cálculo da Tm. Sabe-se que quanto

Desenvolvimento de sondas de LNA para aplicação na técnica de FISH
37
maior a Tm maior será a estabilidade térmica e afinidade da sonda 13. Assim, quanto
mais estável for a sonda de LNA, mais tolerante será às altas temperaturas.
Uma vez que o estudo assenta num estudo multiplex, é essencial que as sondas
apresentem Tm próximas. Assim, a primeira etapa foi calcular a Tm pelo programa
RNA Chemistry Laboratory (http://rnachemlab.ibch.poznan.pl/calculator2.php) e
verificou-se que a sonda ACA apresentava valores bastante elevados quando comparada
com as restantes sondas, assim a sonda ACA foi reduzida para 15 nucleótidos com a
finalidade de aproximar a Tm às restantes sondas, retirando os nucleótidos das
extremidades (Tabela 2.4). Esta redução do número de bases não comprometeu os
valores de sensibilidade e especificidade.
Tabela 2.4 - Tm para as sondas de LNA/2-O-metil para as bactérias e fagos
Microrganismo Sonda Sequência (5´-3´) Tm (ºC)
A. baumannii ACA lTmCmClTmCmTlCmCmClAmTmAlCmTmC 122,9
P. aeruginosa Psaer lAmAmClTmTmGlCmTmGlAmAmClCmAmC 113,9
Fago de A. baumannii
(phiIBB-Aba1) PGAciLO lAmTmAlCmGmTlTmCmTlCmAmGlCmGmT 112,3
Fago de P. aeruginosa
(phiIBB-PAA2) PGPseLO lAmAmClAmAmClGmAmGlAmAmGlTmCmC 111
Após esta análise, foram calculados os restantes parâmetros físico-químicos
utilizando o mesmo programa de cálculo (Tabela 2.5).
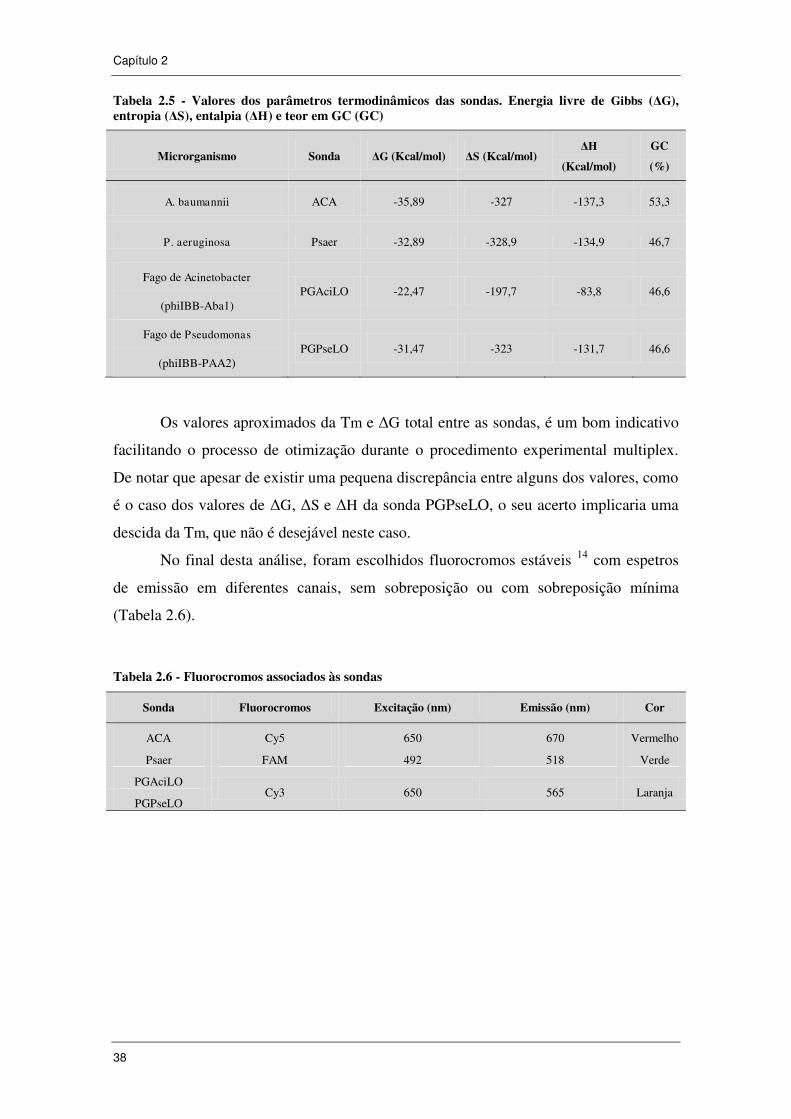
Capítulo 2
38
Tabela 2.5 - Valores dos parâmetros termodinâmicos das sondas. Energia livre de Gibbs (ΔG), entropia (ΔS), entalpia (ΔH) e teor em GC (GC)
Microrganismo Sonda ΔG (Kcal/mol) ΔS (Kcal/mol) ΔH
(Kcal/mol)
GC
(%)
A. baumannii ACA -35,89 -327 -137,3 53,3
P. aeruginosa Psaer -32,89 -328,9 -134,9 46,7
Fago de Acinetobacter
(phiIBB-Aba1) PGAciLO -22,47 -197,7 -83,8 46,6
Fago de Pseudomonas
(phiIBB-PAA2) PGPseLO -31,47 -323 -131,7 46,6
Os valores aproximados da Tm e ΔG total entre as sondas, é um bom indicativo
facilitando o processo de otimização durante o procedimento experimental multiplex.
De notar que apesar de existir uma pequena discrepância entre alguns dos valores, como
é o caso dos valores de ΔG, ΔS e ΔH da sonda PGPseLO, o seu acerto implicaria uma
descida da Tm, que não é desejável neste caso.
No final desta análise, foram escolhidos fluorocromos estáveis 14 com espetros
de emissão em diferentes canais, sem sobreposição ou com sobreposição mínima
(Tabela 2.6).
Tabela 2.6 - Fluorocromos associados às sondas
Sonda Fluorocromos Excitação (nm) Emissão (nm) Cor
ACA Cy5 650 670 Vermelho
Psaer FAM 492 518 Verde
PGAciLO Cy3 650 565 Laranja
PGPseLO

Desenvolvimento de sondas de LNA para aplicação na técnica de FISH
39
2.2.2 Otimização das condições de hibridação para LNA FISH
Para garantir uma boa aplicação das sondas na técnica de FISH, as condições de
hibridação e de lavagem devem estar entre os valores de temperatura, pH, força iónica e
formamida bem definidos, de modo a garantir que a sonda tenha acesso e hibridize com
sucesso a sequência alvo 6.
Quando cumpridos todos os requisitos teóricos no desenho de sondas, a variação
da temperatura é geralmente suficiente para maximizar o sinal de fluorescência obtido.
Assim, a otimização das condições de hibridação começou com o teste de temperaturas
de hibridação entre 55 ºC e 59 ºC (Figura 2.3).
Figura 2.3 - Imagens de microscopia de epifluorescência de P. aeruginosa PAO1 aplicando a sonda
de LNA/2-O-metil Pser. São apresentadas amostra com uma temperatura de hibridação de
55 ºC (A); 57 ºC (B) e 59 ºC (C).
Após análise microscópica, verificou-se que a temperatura que permitiu obter
um sinal mais forte e, simultaneamente, com menos ruído foi 59 ºC, sendo a
temperatura de hibridação aplicada nos trabalhos posteriores. Uma vez que o método de
hibridação foi otimizado, a especificidade e sensibilidade da sonda Pser foram testadas.
Procedeu-se assim, à aplicação da técnica de FISH em 12 estirpes de Pseudomonas,
3 espécies pertencentes à família Pseudomonadaceae e uma estirpe de Acinetobacter
(família Moraxellaceae). Na Tabela 2.7 estão representados os resultados do teste de
especificidade e sensibilidade da sonda Pser. Enquanto que na Figura 2.4 é possível
visualizar imagens de microscopia, usando os filtros verde e vermelho para algumas das
estirpes, alvo e não alvo, de forma a garantir que o sinal obtido é de facto o resultado da
hibridação bem-sucedida da sonda e não da autofluorescência, como da possível
interferência com as restantes sondas posteriormente aplicadas em multiplex.

Capítulo 2
40
Tabela 2.7 - Resultados do teste de especificidade e sensibilidade da sonda de Pser
Microrganismo Resultado
LNA FISH
P. aeruginosa PAO1 +
P. aeruginosa ATCC 10145 +
P. aeruginosa ATCC Wbpl +
P. aeruginosa ATCC M1 +
P. aeruginosa ATCC M3 +
P. aeruginosa ATCC M4 +
P. aeruginosa ATCC M5 +
P. aeruginosa ATCC M6 +
P. aeruginosa W2XKO +
P. aeruginosa PMKW +
Pseudomonas fluorescence (PF7a) -
Pseudomonas putida -
A. baumannii Aba1 -
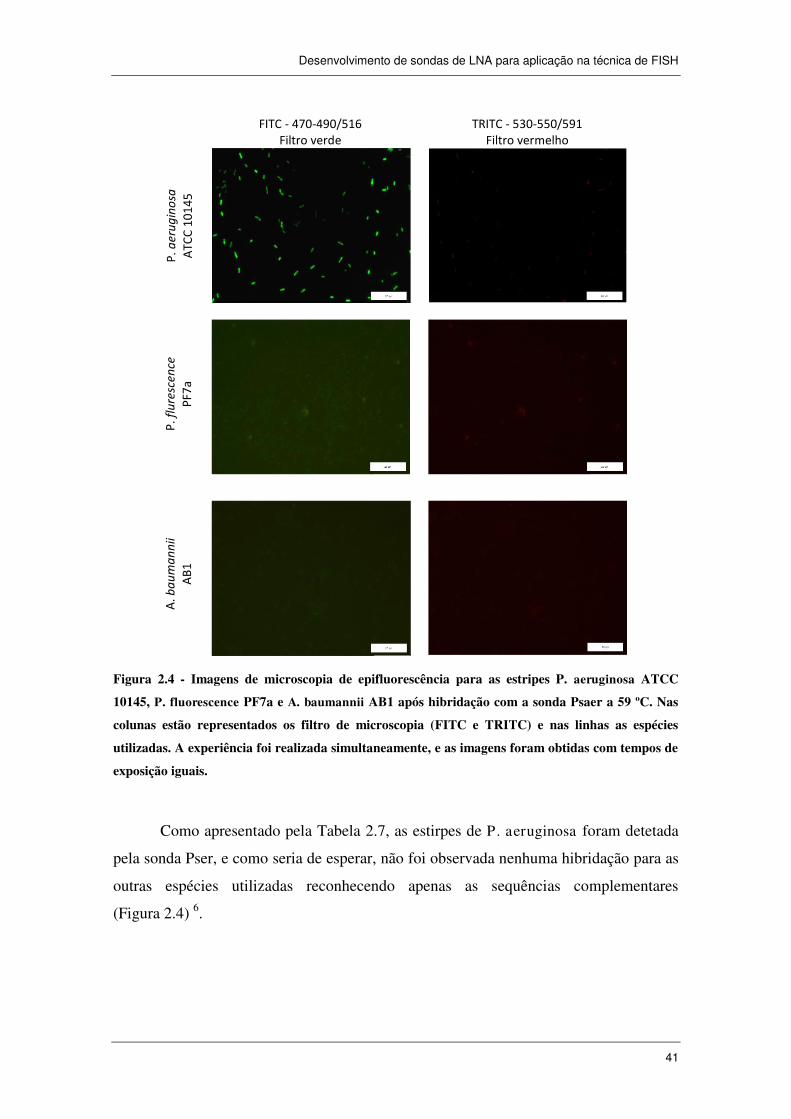
Desenvolvimento de sondas de LNA para aplicação na técnica de FISH
41
FITC - 470-490/516
Filtro verde
TRITC - 530-550/591
Filtro vermelho
P.
ae
rug
ino
sa
AT
CC
10
14
5
P.
flu
resc
en
ce
PF
7a
A.
ba
um
an
nii
AB
1
Figura 2.4 - Imagens de microscopia de epifluorescência para as estripes P. aeruginosa ATCC
10145, P. fluorescence PF7a e A. baumannii AB1 após hibridação com a sonda Psaer a 59 ºC. Nas
colunas estão representados os filtro de microscopia (FITC e TRITC) e nas linhas as espécies
utilizadas. A experiência foi realizada simultaneamente, e as imagens foram obtidas com tempos de
exposição iguais.
Como apresentado pela Tabela 2.7, as estirpes de P. aeruginosa foram detetada
pela sonda Pser, e como seria de esperar, não foi observada nenhuma hibridação para as
outras espécies utilizadas reconhecendo apenas as sequências complementares
(Figura 2.4) 6.

Capítulo 2
42
2.2.3 Conclusão
Em suma é possível concluir que o processo de FISH utilizando a sonda de LNA
é um método sensível e especifico para a deteção da bactéria P.aeruginosa apresentando
uma temperatura ótima de hibridação de 59 ºC.
2.3 Referências
1. Almeida, C., Azevedo, N. F., Fernandes, R. M., Keevil, C. W. & Vieira, M. J. Fluorescence in situ hybridization method using a peptide nucleic acid probe for identification of Salmonella spp. in a broad spectrum of samples. Appl. Environ. Microbiol. 76, 4476–85 (2010).
2. Mahadevan, P., King, J. F. & Seto, D. CGUG: in silico proteome and genome parsing tool for the determination of “core” and unique genes in the analysis of genomes up to ca. 1.9 Mb. BMC Res. Notes 2, 168 (2009).
3. Søe, M. J., Møller, T., Dufva, M. & Holmstrøm, K. A sensitive alternative for microRNA in situ hybridizations using probes of 2’-O-methyl RNA + LNA. J. Histochem. Cytochem. 59, 661–72 (2011).
4. Igloi, G. L. Variability in the stability of DNA-peptide nucleic acid (PNA) single-base mismatched duplexes: real-time hybridization during affinity electrophoresis in PNA-containing gels. Proc. Natl. Acad. Sci. U. S. A. 95, 8562–7 (1998).
5. Almeida, C., Azevedo, N.F., Fernandes, Iversen, C., Fanning, S., R.M, Keevil, C.W., Vieira, J. V. Development and Amplication of a Novel Pepetide Nucleic Acid Probe for the Specific Detection of Cronobacter Genomospecies (Enterobacter sakasakii) in Powdered Infant Formula. Appl. Environ. Microbiol. 75, 2925–2930 (2009).
6. Cerqueira, L. et al. DNA mimics for the rapid identification of microorganisms by fluorescence in situ hybridization (FISH). Int. J. Mol. Sci. 9, 1944–60 (2008).
7. Hogardt, M. et al. Specific and rapid detection by fluorescent in situ hybridization of bacteria in clinical samples obtained from cystic fibrosis patients. J. Clin. Microbiol. 38, 818–25 (2000).
8. Malic, S. et al. Detection and identification of specific bacteria in wound biofilms using peptide nucleic acid fluorescent in situ hybridization (PNA FISH). Microbiology 155, 2603–11 (2009).
9. Wagner, M. et al. Development of an rRNA-targeted oligonucleotide probe specific for the genus Acinetobacter and its application for in situ monitoring in activated sludge. Appl. Environ. Microbiol. 60, 792–800 (1994).

Desenvolvimento de sondas de LNA para aplicação na técnica de FISH
43
10. Hambly, E. et al. A conserved genetic module that encodes the major virion components in both the coliphage T4 and the marine cyanophage S-PM2. Proc. Natl. Acad. Sci. U. S. A. 98, 11411–6 (2001).
11. Xu, J., Hendrix, R. W. & Duda, R. L. Conserved translational frameshift in dsDNA bacteriophage tail assembly genes. Mol. Cell 16, 11–21 (2004).
12. SantaLucia, J., Allawi, H. T. & Seneviratne, P. A. Improved nearest-neighbor parameters for predicting DNA duplex stability. Biochemistry 35, 3555–62 (1996).
13. Armitage, B. A. The impact of nucleic acid secondary structure on PNA hybridization. Drug Discov. Today 8, 222–8 (2003).
14. Gu, Q., Sivanandam, T. M. & Kim, C. A. Signal stability of Cy3 and Cy5 on antibody microarrays. Proteome Sci. 4, 21 (2006).


Capítulo 3
Caracterização da infeção fágica de biofilmes de Acinetobacter baumannii


Caracterização da infeção fágica de biofilmes de Acinetobacter baumannii
47
3 Caracterização da infeção fágica de biofilmes de Acinetobacter baumannii As infeções causadas por A. baumannii têm aumentado drasticamente nos últimos
anos muito devido à sua capacidade de formar biofilmes e à crescente resistência
deste microrganismo aos antimicrobianos convencionais. O uso de fagos como
alternativa preventiva e terapêutica tem sido descrito como uma solução promissora 1.
No trabalho descrito neste capítulo caracterizou-se a formação de biofilmes de
A. baumannii AB1 e avaliou-se a eficácia do fago philBB-Aba1 nos biofilmes
formados.
Nesta secção, encontram-se também descritas algumas das características não só
do fago acima mencionado mas também do fago phiIBB-Aba2 específico para o
hospedeiro A. baumannii AB2.
3.1 Materiais e métodos
3.1.1 Estirpes bacterianas, fagos e condições de crescimento
Neste estudo, utilizou-se a bactéria A. baumannii (isolado clínico AB1),
cultivada em meio nutritivo Luria-Bertani (LB) (nzytech, Portugal) em placas de petri
com meio sólido de LB (1,2 % (p/v) de agar (Liofilchem, Itália)), preparados de
acordo com as instruções do fabricante e devidamente esterilizados no autoclave a
120 ºC durante 15 min.
Os bacteriófagos utilizados neste estudo foram o philBB-Aba1 e o philBB-
Aba2, previamente isolados, pertencentes à coleção de fagos do grupo de
Biotecnologia de Bacteriófagos da Universidade do Minho.
3.1.2 Curva de calibração densidade ótica vs células viáveis
Uma vez que em todos os processos de infeção fágica é determinante saber
exatamente o número de células viáveis para estimar a multiplicidade de infeção
(do inglês Multiplicity of Infection (MOI) (razão entre o número de fagos / número de
células)) foi necessário realizar uma curva de calibração entre a Densidade Ótica
(D.O.) e o número de células viáveis existentes em biofilme. Para tal, uma suspensão
bacteriana foi diluída de forma a obterem-se suspensões celulares com D.O.620nm
compreendidas num intervalo de 0,1 a 1,4. Cada suspensão foi diluída e após

Capítulo 3
48
plaqueamento as placas de petri foram incubadas a 37 ºC para posterior determinação
do número de células viáveis existentes (CFU, do inglês Colony Forming Unit)
através da equação:
3.1.3 Curva de crescimento de A. baumannii
Para a curva de crescimento de A. baumannii (AB1), inoculou-se num matraz
de 100 ml, 50 ml de meio LB com um pré-inóculo crescido durante 16 h de forma a
perfazer uma D.O.620nm inicial de 0,1. A suspensão foi incubada (Biosan, Letónia) a
37 ºC com agitação (120 rpm) e foram retiradas amostras de 2 em 2 h durante um
período de 24 h. Para cada amostra efetuaram-se diluições seriadas (1:10) para a
determinação do número de células viáveis.
3.1.4 Produção e concentração do fago phiIBB-Aba1
A produção de fago foi efetuada de acordo com a técnica de dupla camada 2
usando placas de petri com meio sólido de LB às quais foram adicionados 200 µl de
suspensão bacteriana (crescida durante 16 h) e 3 ml de LB com 6 % (p/v) de agar
(top agar, 45 ºC). Deixou-se solidificar a camada de agar e de seguida adicionaram-se
uma gota (5 µl a 10 µl) de fago philBB-Aba1 que, com a ajuda de uma tira de papel
estéril, foi distribuída uniformemente pela superfície do top agar, de forma a garantir a
máxima cobertura (máximo de replicação) pelos fagos. Este procedimento foi repetido
em várias placas e incubadas durante 12 h a 16 h a 37 ºC. Seguidamente adicionou-se
a cada placa 3 ml a 5 ml de SM Buffer (5,8 gl-1 de NaCl (Panreac, Espanha), 2,0 gl-1
de MgSO4.7H2O (Panreac, Espanha) e 5 % do preparado 1M Tris-HCl (pH de 7,5)
(Biorad, Portugal)) e incubou-se durante 16 h a 4 ºC sob lenta agitação (80 rpm), para
permitir a passagem para a fase líquida para posterior concentração e purificação.
Resumidamente, após a recolha do SM Buffer contendo fagos, para matrazes
adicionou-se NaCl (0.584 g / 10 ml de solução) e após uma agitação lenta para
dissolução do NaCl incubou-se o matraz a 4 ºC (sob agitação lenta). Posteriormente,
centrifugou-se (Sigma, Portugal) a solução a 9500 × g, 10 min, 4 ºC, verteu-se o
sobrenadante para um novo matraz e adicionou-se o polietilenoglicol (PEG 8000,

Caracterização da infeção fágica de biofilmes de Acinetobacter baumannii
49
Fisher) a 10 % (p/v) 3. A solução foi incubada durante 16 h (4 ºC a 100 rpm) e
centrifugada (9500 × g, 10 min, 4 ºC). O pellet contendo os fagos foi resuspendido
num volume de 5-6 ml de SM Buffer e a esta solução adicionou-se clorofórmio (25 %
(v/v), Sigma), a solução foi bem homogeneizada no vortex durante 30 s. De seguida
procedeu-se à centrifugação (3000 × g, 15 min, 4 ºC), recolheu-se e filtrou-se
(0,2 µm) a fase aquosa superior contendo fago e este foi guardado a 4 ºC até à sua
utilização.
3.1.5 Determinação da concentração do fago
Para a determinação do número de placas fágicas, foi utilizado o método da
microgota 4, e para tal realizaram-se diluições sucessivas de 1:10 em microplacas de
96 poços (Orange Scientific, Bélgica) (20 µl de suspensão fágica em 180 µl de
tampão SM Buffer). Foram adicionados 20 µl de cada diluição do fago a 20 µl
suspensão bacteriana crescida durante 16 h, seguida de incubação de 10 min (37 ºC,
120 rpm). Após a incubação 20 µl de cada diluição foram plaqueadas em placas de
LB agar de 1,2 % (p/v) e as gotas deixadas secar antes de as placas de petri serem
colocadas, invertidas, na incubadora (16 h, 37 ºC). Posteriormente determinou-se o
número de fagos (tendo em conta a diluição que continha um número de placas
fágicas compreendida entre 3 a 30) segundo a equação:
3.1.6 Curva de Crescimento do fago (OSGC)
As curvas de crescimento do fago phiIBB-Aba1 (OSGC, do inglês One Step
Growth Curve) foram realizadas como descrito por Rahman et al. 5 mas com algumas
modificações. Resumidamente, culturas bacterianas na fase exponencial
(D.O.620nm: 0,5), foram centrifugadas 10 ml (7000 × g, 5 min, 4 ºC) e ressuspendidas
em 5 ml de meio LB. A esta suspensão foram adicionados 5 ml de solução de fago até
atingir uma MOI de 1 e deixou-se o fago adsorver ao hospedeiro durante 5 min a
37 ºC (120 rpm). A suspensão foi centrifugada (7000 × g, 5 min, 4ªC) e o pellet
ressuspenso em 10 ml de meio LB. A suspensão, contendo apenas os fagos
adsorvidos, foi incubada a 37 ºC (120 rpm) e foram retiradas 2 amostras a

Capítulo 3
50
determinados tempos durante a duração da experiência (1 h) para determinação do
número de fagos (3.1.5).
3.1.7 Formação de biofilme
Os biofilmes de A. baumannii foram formados no fundo de microplacas de 24
poços (Orange Scientific, Bélgica) contendo 1 ml de meio LB e 10 µl de suspensão
bacteriana a uma D.O.620nm de 1. As microplacas foram incubadas a 37 ºC, sob
agitação (120 rpm), durante diferentes tempos, tendo-se feito uma mudança de meio
após 24 h de formação de biofilme. Antes do plaqueamento das amostras, os poços
foram lavados duas vezes com NaCl (0,9 %) (v/v) para remover todas as células
plantónicas, de seguida foram raspados (com a ajuda de uma ponta amarela) para
desprender os biofilmes e foram efetuadas diluições seriadas (1:10) com NaCl em
microplacas de 96 poços. Gotas de 20 µl de cada diluição foram plaqueadas em placas
de petri contendo LB agar de 1,2 % (p/v). A determinação do número de células
viáveis existentes nos biofilmes foi realizada tal como descrita anteriormente (3.1.2) e
a biomassa total no biofilme quantificada pelo método de violeta cristal 6 (ver ponto
3.1.8), sendo que foram realizados 3 ensaios independentes.
3.1.8 Quantificação da biomassa total
Após a lavagem do biofilme (2 ×) com solução salina (0,9 % NaCl (v/v)),
adicionou-se 1ml de metanol (100 %, Fisher EUA) e deixou-se a placa durante 15
min. Após este período, o metanol foi retirado e a placa de 24 poços deixada a secar.
Seguidamente, adicionou-se 1ml de violeta cristal (1 % (v/v), Merck, EUA) a cada
poço e deixou-se a placa à temperatura ambiente durante 5 min. O cristal violeta foi
removido, os poços lavados com água e finalmente adicionou-se 1ml de ácido acético
(33 % (v/v), Fisher). Foi retirado um volume de 250 µl de cada poço que foi
transferido para uma microplaca de 96 poços e a absorvância lida num leitor de
ELISA (Bio-Tek, EUA) a 570 nm. Para cada experiência, foram realizados três
ensaios independentes.
3.1.9 Infeção Fágica de Biofilmes
Os biofilmes de A. baumannii, foram formados como descrito em 3.1.7, sob
condições dinâmicas (120 rpm, 37 ºC). Após o crescimento durante 12 h e/ou 24 h, o

Caracterização da infeção fágica de biofilmes de Acinetobacter baumannii
51
biofilme foi lavado tal como descrito no ponto 3.1.7 com NaCl 0,9 % (v/v) e
posteriormente adicionaram-se 900 µl de meio fresco de LB e 100 µl de fago de
forma a ter uma MOI de 1 sendo que no controlo a concentração de fago foi
substituída por 100 µl de SM Buffer. Após a adição de fago, as placas de 24 poços
foram a incubar (37 ºC, 120 rpm) e foram recolhidas amostras após 6 h de infeção em
biofilmes de 12 h e após 6 h e 24 h de infeção em biofilmes de 24 h. Para cada ensaio
foi efetuado um controlo. Foi também analisado o efeito do fago em biofilmes com a
matriz danificada e para tal, biofilmes de 24 h foram sujeitos a raspagem e posterior
infeção fágica.
As bactérias viáveis e os fagos presentes foram determinados pela contagem
de células viáveis e fagos tal como descrito no ponto 3.1.2 e 3.1.5, sendo realizados
3 ensaios independentes.
3.1.10 Análise estatística
Os resultados são apresentados como a média ± desvio padrão de pelo menos
duas repetições, utilizando o software Microsoft Office Excel como ferramenta de
análise.
Os resultados mais relevantes foram analisados pelo teste ANOVA aplicando o
modelo estatístico Tukey que compara várias condições em simultâneo. Uma
diferença foi considerada estatisticamente significativa quando p < 0,05. As análises
estatísticas foram realizadas utilizando o software GraphPad (San Diego, EUA).
3.1.11 Análise das proteínas estruturais do fago
As proteínas estruturais do fago foram analisadas usando a técnica de
SDS-PAGE 7, com algumas alterações. Para a preparação das proteínas do fago, 1ml
de fago concentrado foi precipitado por adição de acetona (4 % (v/v), Sigma) gelada.
As amostras foram centrifugadas (1600 × g, 20 min, 4 ºC), o sobrenadante foi
descartado e o pellet foi deixado a secar até toda a acetona estar evaporada 8.
Adicionou-se tampão de aplicação (2 ×) (Tris-HCl 125 mM (pH 6,8) (v/v), glicerol
(20 % (p/v), Biorad), β- mercaptoetanol (1 % (v/v), Sigma), azul de bromofenol
(0,01 % (p/v), Biorad) e SDS (4 % (p/v), Biorad)) às amostras e estas foram aquecidas
durante 5 min num banho a 100 ºC para desnaturar as proteínas e posteriormente
colocadas em gelo para arrefecer. Adicionaram-se 15 µl da amostra a cada poço de

Capítulo 3
52
géis de gradiente de policrilamida (Mini Precast Polyacrylamide Gel, Advansta,
California), tendo a eletroforese decorrido em 1× TGS (25 mM Tris), 192 mM glicina
(Sigma), e 91 % (p/v) SDS com pH 8,3), a 80 V durante 6 h a 7 h.
Após a eletroforese, os géis foram colocados em solução de fixação (etanol
(50 % (v/v) (Sigma)), ácido acético (12 % (v/v)); formaldeído (0,05 % (v/v) (Sigma))
e colocados sob agitação lenta durante aproximadamente 2 h. A solução foi descartada
e os géis foram colocados por duas vezes imersos em solução de lavagem (etanol
20 % (v/v)) durante 20 min. A solução foi removida e adicionou-se solução de
sensibilização (tiossulfato de sódio 0,02 % (p/v), Sigma) durante 2 min. A solução de
sensibilização foi retirada e foi adicionada água ultra pura (2 × 1 min). De seguida os
géis foram colocados numa solução de nitrato de prata 0,2 % (p/v), Sigma) e 0,076 %
(v/v) formaldeído durante 30 min. O nitrato de prata foi removido e os géis lavados
duas vezes com água ultra pura durante 10 seg. Posteriormente aos géis foi adicionada
a solução de revelação (0,05 % (v/v) formaldeído), carbonato de sódio (6 % (p/v),
Panreac)) e tiossulfato de sódio 0,0004 % (p/v)) até os géis revelarem a cor desejada.
Finalmente a reação foi parada com solução de paragem (ácido acético 12 % (v/v))
durante 1 min. O peso molecular das proteínas foi estimado com base na sequência
dos seus aminoácidos, utilizando os marcadores Kaleidoscope Prestained Standards
(Bio-Rad) e Low Molecular Weight Protein Marker (nzytech).
3.1.12 Preparação de amostras de biofilmes e análise
microscópica
As amostras de biofilme utilizadas para a análise microscópica foram
realizadas em superfícies de polistireno e de vidro sob as condições de crescimento,
infeção fágica e lavagem anteriormente descritas. Na formação de biofilmes em vidro,
superfícies de vidro com 2,5 × 2,5 cm2 foram introduzidos em microplacas de 6 poços
(Orange Scientific, Bélgica) seguindo o mesmo protocolo utilizado na superfície de
polistireno. O crescimento do biofilme em polistireno corresponde à sua formação no
fundo dos poços das microplacas.
Após a formação do biofilme ou infeção com fagos, procedeu-se à fixação das
amostras adicionando-se 1 ml de metanol (100 %) durante 20 min. Seguidamente foi
retirado o metanol e após estar totalmente seco, adicionaram-se 100 µl de
paraformaldeido a 4 % (v/v) (Sigma), solução essa que permaneceu em contacto
durante 10 min e no final foi removido e foram adicionados 100 µl de etanol 50 %
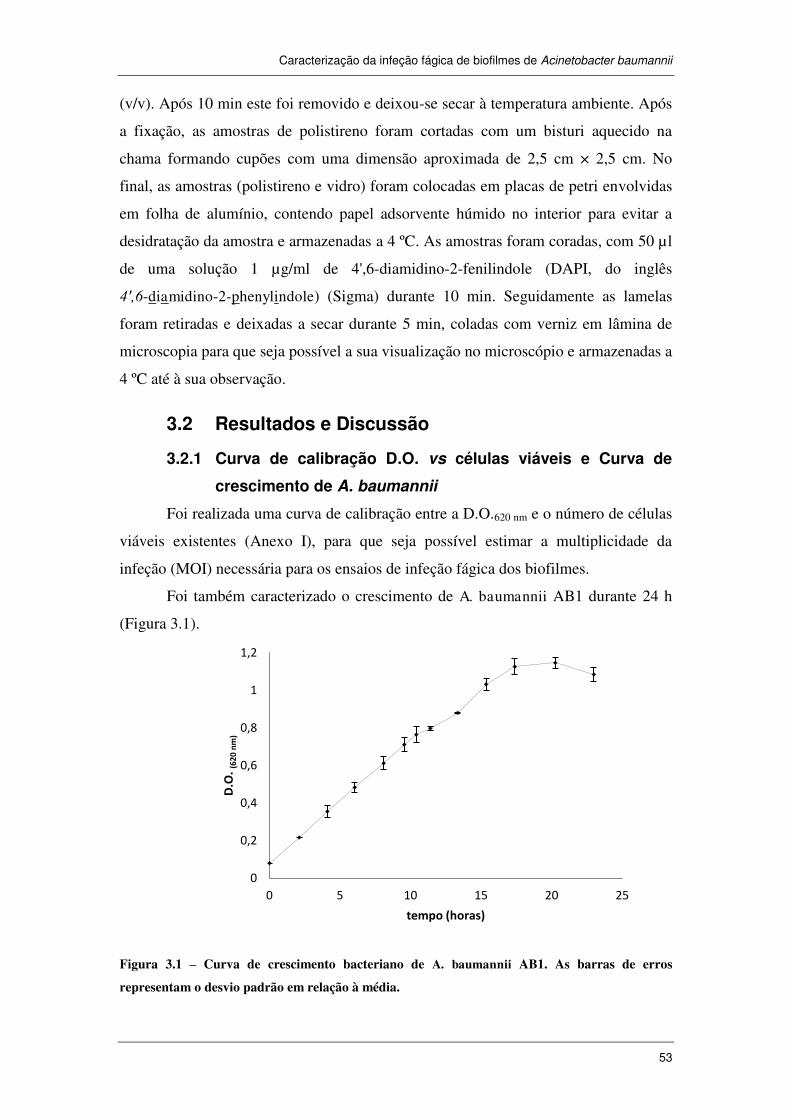
Caracterização da infeção fágica de biofilmes de Acinetobacter baumannii
53
(v/v). Após 10 min este foi removido e deixou-se secar à temperatura ambiente. Após
a fixação, as amostras de polistireno foram cortadas com um bisturi aquecido na
chama formando cupões com uma dimensão aproximada de 2,5 cm × 2,5 cm. No
final, as amostras (polistireno e vidro) foram colocadas em placas de petri envolvidas
em folha de alumínio, contendo papel adsorvente húmido no interior para evitar a
desidratação da amostra e armazenadas a 4 ºC. As amostras foram coradas, com 50 µl
de uma solução 1 µg/ml de 4',6-diamidino-2-fenilindole (DAPI, do inglês
4′,6-diamidino-2-phenylindole) (Sigma) durante 10 min. Seguidamente as lamelas
foram retiradas e deixadas a secar durante 5 min, coladas com verniz em lâmina de
microscopia para que seja possível a sua visualização no microscópio e armazenadas a
4 ºC até à sua observação.
3.2 Resultados e Discussão
3.2.1 Curva de calibração D.O. vs células viáveis e Curva de
crescimento de A. baumannii
Foi realizada uma curva de calibração entre a D.O.620 nm e o número de células
viáveis existentes (Anexo I), para que seja possível estimar a multiplicidade da
infeção (MOI) necessária para os ensaios de infeção fágica dos biofilmes.
Foi também caracterizado o crescimento de A. baumannii AB1 durante 24 h
(Figura 3.1).
Figura 3.1 – Curva de crescimento bacteriano de A. baumannii AB1. As barras de erros
representam o desvio padrão em relação à média.
0
0,2
0,4
0,6
0,8
1
1,2
0 5 10 15 20 25
D.O
. (62
0 n
m)
tempo (horas)

Capítulo 3
54
De acordo com os resultados obtidos a taxa específica de crescimento (µ) é de
0,14 h-1. A fase de crescimento exponencial dura em média 17 h, apresentando uma
fase estacionária já às 20 h.
3.2.1 Formação e infeção de biofilme
Biofilmes de A. baumannii AB1 foram formados em microplacas de
polistireno com e sem infeção. A caracterização dos respetivos biofilmes foi feita com
base na contagem de unidades formadoras de colónias (CFU) ( Figura 3.2 A) e
quantificação de biomassa pela colocação de violeta de cristal (CV) ( Figura 3.2 B).
Apesar desta estratégia de formação de biofilme em microplaca não ser a ideal pois
não mimetiza condições reais, esta é uma forma rápida, simples e funcional de
avaliação de diversos parâmetros durante a formação de biofilme.
Uma vez caracterizados os biofilmes (tempo 0 que corresponde a um biofilme
de 24 h sem infeção da Figura 3.2), o fago de A. baumannii (phiIBB-Aba1) foi
aplicado tendo a infeção prosseguido durante 6 h e 24 h (MOI de 1) sendo neste caso
também contabilizados os números de fagos presentes (PFU’s) ( Figura 3.2 C) e
foram também realizados ensaios de microscopia de epifluorescência, dos biofilmes
com e sem infeção corados com DAPI ( Figura 3.2 D a G).
0
0,1
0,2
0,3
0,4
0,5
0,6
0h 6h 24h
DO
(5
70
nm
)
Tempo de infeção
B
sem infeção com infeção
* * *
1,E+00
1,E+02
1,E+04
1,E+06
1,E+08
1,E+10
0h 6h 24h
log(
CFU
/ cm
2 )
Tempo de infeção
A
sem infeção com infeção
* * * 10
8
6
4
2
0

Caracterização da infeção fágica de biofilmes de Acinetobacter baumannii
55
Figura 3.2 - Infeção de biofilmes formados durante 24 h de A. baumannii AB1 e infetadas
durante 6 h e 24 h com o fago phiIBB-Aba1. A) Número de células viáveis antes e após da infeção
fágica; B) Quantificação da biomassa usando CV antes e após da infeção fágica; C) Número de
fagos (PFU) presentes nos biofilmes (aderidos) e o número de fagos libertados durante a infeção
fágica (suspensos). D) Imagens de microscopia de epifluorescência de biofilme de 30 h em
superfícies de poliestireno; E) Imagens de microscopia de epifluorescência de biofilme infetado
durante 6 h em superfícies de vidro; F) Imagens de microscopia de epifluorescência de biofilme
de 48 h em superfícies de poliestireno; G) Imagens de microscopia de epifluorescência de biofilme
infetado durante 24 h em superfícies de poliestireno. As barras de erros representam o desvio
padrão em relação à média. ** p < 0,01 e *** p < 0,001.
1,E+00
1,E+02
1,E+04
1,E+06
1,E+08
1,E+10
0h 6h 24h
log
(PFU
/ m
l)
Tempo de infeção
C
fagos suspensos fagos aderidos
* * * * 10
8
6
4
2
0

Capítulo 3
56
Os controlos de A. baumannii AB1 realizados mostram que não há grandes
variações tanto no número de células viáveis como na quantidade de biomassa total (
Figura 3.2 A e B). O fago testado nos biofilmes formados durante 24 h causou uma
redução de aproximadamente 2 log de células viáveis após 6 h (p <0,0001) de infeção,
no entanto, verificou-se que após uma exposição mais prolongada do biofilme com o
fago, existia um aumento da quantidade de células viáveis no biofilme para valores
idênticos ao número de células iniciais (t 0), o que poderá indicar o aparecimento de
fenótipos resistentes ao fago, uma conclusão já evidenciada em vários trabalhos 9–11.
Em biofilmes mais maduros (infeção durante 24 h) é normal que o fago atue de uma
forma mais lenta, uma vez que há uma predominância de células na fase estacionária,
atrasando assim o processo de infeção e de lise celular que se caracteriza por uma
atividade reduzida dos fagos.
As imagens de microscopia de epifluorescência efetuadas após 6 h de
exposição do fago ao biofilme ( Figura 3.2 E) evidenciam uma redução considerável
de células viáveis sobre o substrato, contudo as imagens microscópicas da infeção
fágica decorridas durante 24 h ( Figura 3.2 G) não apoiam os dados de células viáveis
e biomassa (gráficos da Figura 3.2A e B). Este fato pode dever-se aos processos de
tratamento das amostras para análise em microscopia. A fixação da amostra e o
processo de lavagem são métodos agressivos para o biofilme, podendo proporcionar
uma libertação da células e consequentemente redução de biofilme.
Em relação ao número de fagos, há uma maior concentração na fase plantónica
do que aderida tanto ao biofilme como à superfície do poço. Numa análise ao número
de fagos aderidos verifica-se um número estatisticamente significativo de fagos às 6 h
(p < 0,001) mas que se mantem idêntico às 24 h. Contudo, observa-se que a
concentração de fagos após a infeção é superior à inicial indicando que o fago
reproduziu-se eficientemente. O fato de haver menos fagos aderidos do que suspensos
poderá indicar que à medida que os fagos reproduzidos são libertados estes passam
maioritariamente para a fase planctónica e provavelmente só aqueles que se ligam ao
hospedeiro permanecem imersos no biofilme 12. Assumindo que as bactérias
adquiriram resistência aos fagos, este fator pode explicar o facto do número de fagos
não ter aumentado após 24 h de infeção, possivelmente devido à alteração dos
recetores específicos à superfície do hospedeiro aos quais o fago se ligava 13.
Para avaliar a influência que pode ter a idade dos biofilmes no desempenho do
fago e eventualmente compreender o mecanismo que leva à redução da atividade dos

Caracterização da infeção fágica de biofilmes de Acinetobacter baumannii
57
fagos em biofilmes de 24 h, foram realizados os mesmos ensaios mas em biofilmes
com metade do período de formação (12 h) com uma infeção de 6 h (Figura 3.3).
Figura 3.3 - Infeção de biofilmes formados durante 12 h de A. baumannii AB1 com o fago
phiIBB-Aba1. A) Número de células viáveis antes e após da infeção fágica; B) Quantificação da
biomassa usando CV antes e após da infeção fágica. As barras de erros representam o desvio
padrão em relação à média. *** representam p < 0,01.
Comparando os biofilmes de 12 h e 24 h ( Figura 3.2 A e B e Figura 3.3) não
se observou uma diferença significativa e portanto, os restantes ensaios foram todos
executados tendo em conta um período de formação de biofilmes de 24 h. Este
resultado permitiu concluir que um biofilme com uma idade inferior a 24 h não
influencia o desempenho do fago, havendo outros fatores que o possam condicionar a
sua atividade, tal como a presença da matriz.
Com efeito, vários autores referem que a matriz é um dos principais fatores
que impede a atuação de fagos em biofilmes, devido às limitações difusionais por ela
impostas e à capacidade de reterem o fago por ligações químicas mais ou menos
fortes 1,14. Assim, para analisar se a eficácia dos fagos está a ser prejudicada pela
presença da matriz, realizou-se um estudo com biofilmes formados durante 24 h mas
com raspagem do biofilme antes da infeção. Esta raspagem danifica de alguma forma
a matriz do biofilme, dispersando os componentes da matriz e desta forma, permitindo
uma maior exposição das células do biofilme ao meio envolvente, uma vez que as
células ou aglomerados de células passem para uma faze planctónica (Figura 3.4).
1,E+00
1,E+02
1,E+04
1,E+06
1,E+08
1,E+10
0h 6h
log(
CFU
/ cm
)2
Tempo de infeção
A
sem infeção com infeção
* * * 10
8
6
4
2
0
0
0,1
0,2
0,3
0,4
0,5
0,6
0h 6h
DO
(5
70
nm
)
Tempo de infeção
B
sem infeção com infeção
* * *

Capítulo 3
58
Figura 3.4 - Número de células viáveis (CFU) e fagos (PFU) após raspagem de biofilme de A.
baumannii AB1 formados durante 24 h e posteriormente infetadas com o fago phiIBB-Aba1. A)
Número de células viáveis antes e após infeção fágica; B) Número de fagos presentes nos
biofilmes suspensos. As barras de erros representam o desvio padrão em relação à média.
*** p < 0,01.
Este estudo demonstra que há uma significativa redução do número de células
viáveis às 6 h (p < 0,01) comparativamente com biofilmes não raspados; no entanto,
mais uma vez se observa um aumento de células viáveis após 24 h. Esta redução às
6 h pode ser explicada pela facilidade do contacto dos fagos com as bactérias ou
aglomerados na fase planctónica aumentando a probabilidade de um fago infetar uma
bactéria.
Apesar de as placas fágicas do fago phiIBB-Aba1 mostrarem um halo em
torno das mesmas (Figura 3.5), indicativo da presença de depolimerase associada ao
fago, não é muito funcional após uma exposição prolongada do fago (infeção de
24 h). Esta observação contraria o reportado por Hughes et al. 15, que observou que as
depolimerases são eficazes na remoção de grandes quantidades de matriz, uma vez
que degradam as substâncias poliméricas extracelulares que facilita o acesso do fago a
superfícies mais profundas do biofilme.
1,E+00
1,E+02
1,E+04
1,E+06
1,E+08
1,E+10
0h 6h 24h
log(
CFU
/ cm
2 )
Tempo de infeção
A
sem infeção com infeção
* * * 10
8
6
4
2
0
1,E+00
1,E+02
1,E+04
1,E+06
1,E+08
1,E+10
0h 6h 24h
log(
PFU
/ m
l)
Tempo de infeção
B
fagos suspensos
10
8
6
4
2
0
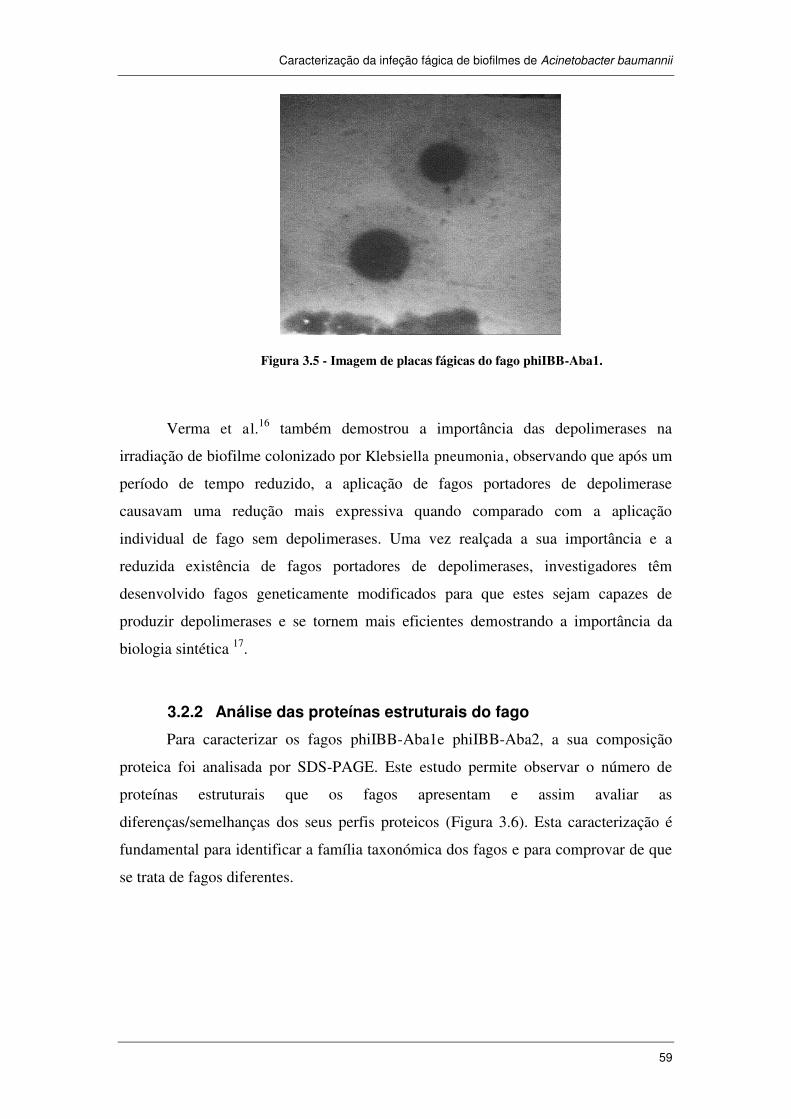
Caracterização da infeção fágica de biofilmes de Acinetobacter baumannii
59
Figura 3.5 - Imagem de placas fágicas do fago phiIBB-Aba1.
Verma et al.16 também demostrou a importância das depolimerases na
irradiação de biofilme colonizado por Klebsiella pneumonia , observando que após um
período de tempo reduzido, a aplicação de fagos portadores de depolimerase
causavam uma redução mais expressiva quando comparado com a aplicação
individual de fago sem depolimerases. Uma vez realçada a sua importância e a
reduzida existência de fagos portadores de depolimerases, investigadores têm
desenvolvido fagos geneticamente modificados para que estes sejam capazes de
produzir depolimerases e se tornem mais eficientes demostrando a importância da
biologia sintética 17.
3.2.2 Análise das proteínas estruturais do fago
Para caracterizar os fagos phiIBB-Aba1e phiIBB-Aba2, a sua composição
proteica foi analisada por SDS-PAGE. Este estudo permite observar o número de
proteínas estruturais que os fagos apresentam e assim avaliar as
diferenças/semelhanças dos seus perfis proteicos (Figura 3.6). Esta caracterização é
fundamental para identificar a família taxonómica dos fagos e para comprovar de que
se trata de fagos diferentes.

Capítulo 3
60
Figura 3.6 - Análise por SDS-PAGE de proteínas estruturais de A. baumannii dos fagos
phiIBB-Aba2 e phiIBB-Aba1 respetivamente.
É possível observar a presença de 7 bandas no fago phiIBB-Aba2 sendo que as
mais evidentes são as que apresentam um tamanho aproximado de 37 kDa e inferior à
última banda do marcador (> 7,6 kDa) para o fago phiIBB-Aba2, enquanto que para o
fago phiIBB-Aba1 a mais evidente é a banda de 37 kDa. As duas bandas mais
intensas poderão corresponder à major capsid protein que são sempre as mais visíveis
em géis de SDS-PAGE 18,19. Tanto o fago philBB-Aba1 como o philBB-Aba2
pertencem ambos à família Podoviridae 6 20 o que realmente se confirma nos géis
havendo um elevado grau de homologia.
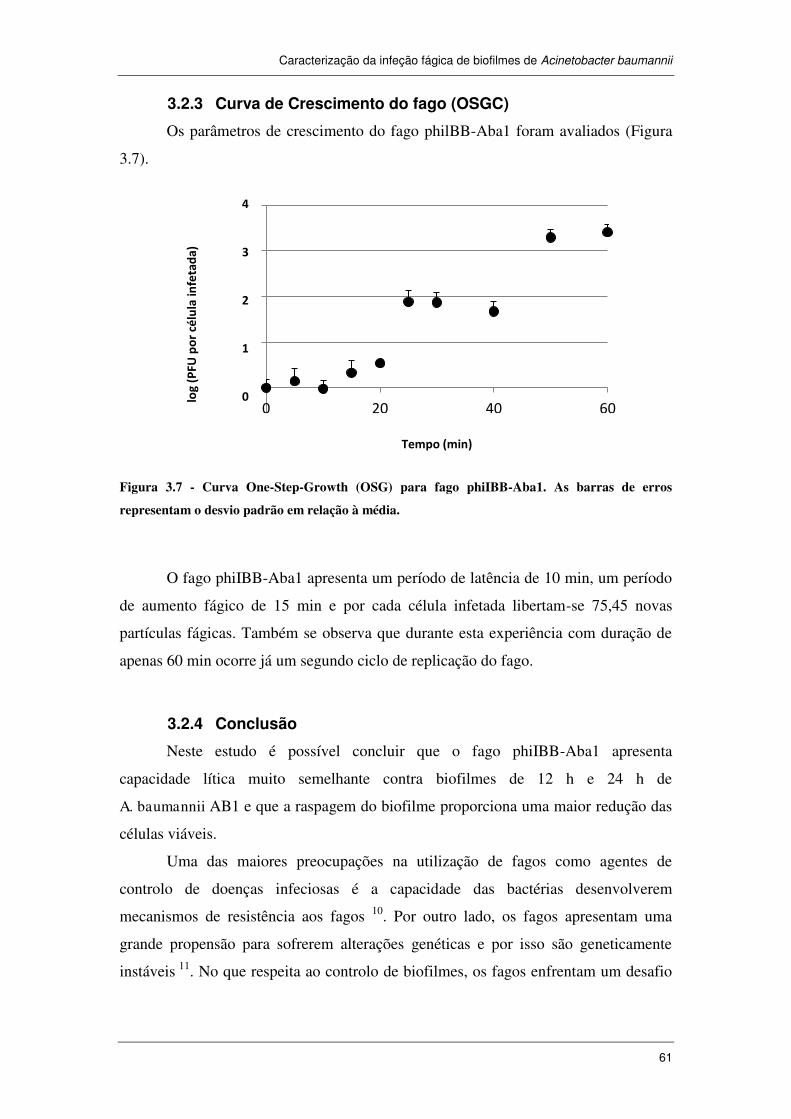
Caracterização da infeção fágica de biofilmes de Acinetobacter baumannii
61
3.2.3 Curva de Crescimento do fago (OSGC)
Os parâmetros de crescimento do fago philBB-Aba1 foram avaliados (Figura
3.7).
Figura 3.7 - Curva One-Step-Growth (OSG) para fago phiIBB-Aba1. As barras de erros
representam o desvio padrão em relação à média.
O fago phiIBB-Aba1 apresenta um período de latência de 10 min, um período
de aumento fágico de 15 min e por cada célula infetada libertam-se 75,45 novas
partículas fágicas. Também se observa que durante esta experiência com duração de
apenas 60 min ocorre já um segundo ciclo de replicação do fago.
3.2.4 Conclusão
Neste estudo é possível concluir que o fago phiIBB-Aba1 apresenta
capacidade lítica muito semelhante contra biofilmes de 12 h e 24 h de
A. baumannii AB1 e que a raspagem do biofilme proporciona uma maior redução das
células viáveis.
Uma das maiores preocupações na utilização de fagos como agentes de
controlo de doenças infeciosas é a capacidade das bactérias desenvolverem
mecanismos de resistência aos fagos 10. Por outro lado, os fagos apresentam uma
grande propensão para sofrerem alterações genéticas e por isso são geneticamente
instáveis 11. No que respeita ao controlo de biofilmes, os fagos enfrentam um desafio
1,E-01
1,E+00
1,E+01
1,E+02
1,E+03
1,E+04
0 20 40 60
log
(PFU
po
r cé
lula
infe
tad
a)
Tempo (min)
4
3
2
1
0

Capítulo 3
62
acrescido devido à presença da matriz polimérica que poderá dificultar o acesso dos
fagos as células 21. Neste estudo é percetível que a aplicação de fagos por curtos
períodos de tempo apresenta uma maior eficiência na redução de biofilmes. Visto que
após 24 h de infeção não foi verificada qualquer redução nem aumento do biofilme
quando comparado com o seu controlo, coloca-se assim a hipótese de que uma
exposição prolongada do fago permita às bactérias desenvolver fenótipos resistentes
aos fagos, levando a uma proliferação das mesmas em biofilme. Também,
Pires et al. 22, verificou um aumento da quantidade de células após 24 h de tratamento
com o fago phiIBB-PAP21 sugerindo a possibilidade de resistência bacteriana em
relação ao fago.
A. baumannii apresenta-se muitas vezes em comunidades mistas, associado a
outras bactérias como é o caso da bactéria P. aeruginosa 23, também formadora de
biofilme. Neste sentido torna-se relevante o estudo da eficácia de cocktails fágicos no
controlo de biofilmes mistos como será demostrado no capítulo seguinte.
3.3 Referências
1. Melo, L., Sillankorva, S. & Azeredo, J. in Biofilmes Na Saúde, no Ambient. na Indústria. Uso bacteriófagos para o Control. biofilmes. (Nuno F. Azevedo, N. C.) Porto: Publindústria. 227–236 (2012).
2. Adams, M. H. Bacteriophages. Interscience, New York, N.Y.(1959).
3. Sambrook, J. & Russell, D. Molecular Cloning A Laboratory manual 3rd ed. (2001).
4. Mazzocco, A., Waddell, T.E., Lingohr, E. & Johnson, R. P. in Bacteriophages Methods Protoc. Isolation, Characterization and Interactions, 81–8 (2008).
5. Rahman, M., Kim, S., Kim, S. M., Seol, S. Y. & Kim, J. Characterization of induced Staphylococcus aureus bacteriophage SAP-26 and its anti-biofilm activity with rifampicin. Biofouling. 27, 1087–93 (2011).
6. Pires, D., Sillankorva, S., Faustino, A. & Azeredo, J. Use of newly isolated phages for control of Pseudomonas aeruginosa PAO1 and ATCC 10145 biofilms. Res. Microbiol. 162, 798–806 (2011).
7. Schägger, H. & von Jagow, G. Tricine-sodium dodecyl sulfate-polyacrylamide gel electrophoresis for the separation of proteins in the range from 1 to 100 kDa. Anal. Biochem. 166, 368–79 (1987).

Caracterização da infeção fágica de biofilmes de Acinetobacter baumannii
63
8. O’Flaherty, S. et al. Genome of Staphylococcal Phage K: a New Lineage of Myoviridae Infecting Gram-Positive Bacteria with a Low G+C Content. J. Bacteriol. 186, 2862–2871 (2004).
9. Soothill, J. S. Treatment of experimental infections of mice with bacteriophages. J. Med. Microbiol. 37, 258–61 (1992).
10. Buckling, A. & Rainey, P. B. Antagonistic coevolution between a bacterium and a bacteriophage. Proc. Biol. Sci. 269, 931–6 (2002).
11. Balakshina, V. V, Kulakov, L. A. & Boronin, A. M. Dynamics of the interaction between Pseudomonas aeruginosa bacteriophage phimF81 and host bacteria. Mikrobiologiia. 61, 1051–5 (1992).
12. Sillankorva, S., Neubauer, P. & Azeredo, J. Phage control of dual species biofilms of Pseudomonas fluorescens and Staphylococcus lentus. Biofouling. 26, 567–75 (2010).
13. Sillankorva, S., Oliveira, R., Vieira, M. J. & Azeredo, J. Real-time quantification of Pseudomonas fluorescens cell removal from glass surfaces due to bacteriophage varphiS1 application. J. Appl. Microbiol. 105, 196–202 (2008).
14. Hanlon, G. W., Denyer, S. P., Olliff, C. J. & Ibrahim, L. J. Reduction in exopolysaccharide viscosity as an aid to bacteriophage penetration through Pseudomonas aeruginosa biofilms. Appl. Environ. Microbiol. 67, 2746–53 (2001).
15. Hughes, K. A., Sutherland, I. W. & Jones, M. V. Biofilm susceptibility to bacteriophage attack: the role of phage-borne polysaccharide depolymerase. Microbiology. 144, 3039–47 (1998).
16. Verma, V., Harjai, K. & Chhibber, S. Structural changes induced by a lytic bacteriophage make ciprofloxacin effective against older biofilm of Klebsiella pneumoniae. Biofouling. 26, 729–37 (2010).
17. Lu, T. K. & Collins, J. J. Dispersing biofilms with engineered enzymatic bacteriophage. Proc. Natl. Acad. Sci. U. S. A. 104, 11197–202 (2007).
18. Lee, C.-N. et al. Lytic myophage Abp53 encodes several proteins similar to those encoded by host Acinetobacter baumannii and phage phiKO2. Appl. Environ. Microbiol. 77, 6755–62 (2011).
19. Lavigne, R. et al. The structural proteome of Pseudomonas aeruginosa bacteriophage phiKMV. Microbiology. 152, 529–34 (2006).
20. Lin, N.-T., Chiou, P.-Y., Chang, K.-C., Chen, L.-K. & Lai, M.-J. Isolation and characterization of phi AB2: a novel bacteriophage of Acinetobacter baumannii. Res. Microbiol. 161, 308–14 (2010).

Capítulo 3
64
21. Briandet, R. et al. Fluorescence correlation spectroscopy to study diffusion and reaction of bacteriophages inside biofilms. Appl. Environ. Microbiol. 74, 2135–43 (2008).
22. Pires, D. P., Sillankorva, S. & Azeredo, J. The use of bacteriophages for P. aeruginosa biofilm control. Port. chapter IEEE EMBS 1–4 (2011).
23. Navon-Venezia, S., Ben-Ami, R. & Carmeli, Y. Update on Pseudomonas aeruginosa and Acinetobacter baumannii infections in the healthcare setting. Curr. Opin. Infect. Dis. 18, 306–13 (2005).

Capítulo 4
Caracterização dos biofilmes mistos e avaliação da eficácia de fagos específicos para A. baumannii e P. aeruginosa


Caracterização dos biofilmes mistos e avaliação da eficácia de fagos específicos para A. baumannii e P. aeruginosa
67
4 Caracterização dos biofilmes mistos e avaliação da eficácia de fagos específicos para A. baumannii e P. aeruginosa
Vários estudos demonstram que as bactérias P. aeruginosa e A. baumannii são
frequentemente isoladas de feridas crónicas, ambientes hospitalares, etc 1–4. Contudo,
não há qualquer estudo de biofilmes mistos de P. aeruginosa e A. baumannii e muito
menos de ensaios de infeção com fagos, uma terapia alternativa aos atuais agentes
antimicrobianos 5.
Seguindo a mesma estrutura da secção anterior, nesta secção encontram-se
descritos os principais resultados sobre a avaliação do desempenho do fago de
Acinetobacter (philBB-Aba1) e dos fagos de Pseudomonas (philBB-PAP1 ou philBB-
PAA2) e também de um cocktail constituído por fagos para ambas as espécies
(philBB-Aba1 e philBB-PAP1) presentes nos biofilmes mistos.
4.1 Materiais e métodos
4.1.1 Estirpe bacteriana, bacteriófagos e condições de cultura
Neste estudo, utilizou-se a bactéria A. baumannii (isolado clínico AB1) e
P. aeruginosa (PAO1). As estirpes foram cultivadas em meio nutritivo Luria-Bertani
(LB) (nzytech, Portugal) LB ou em placas de petri com meio sólido de LB com 1,2 %
(p/v) de agar (Liofilchem, Itália), preparados de acordo com as instruções do fabricante
e devidamente esterilizados no autoclave a 120 ºC durante 15 min. Para as contagens do
número de células viáveis e de fagos de P. aeruginosa utilizou-se o meio seletivo
Pseudomona Isolation Agar (PIA, Sigma, Portugal) enquanto que no caso de
A. baumannii utilizou-se meio LB contendo o antibiótico ceftazidime (Sigma, Portugal)
(concentração inicial de 5 × 10-4 g.ml-1) de forma a inibir o crescimento de
P. aeruginosa .
Nos vários ensaios de biofilmes mistos os fagos utilizados foram: philBB-Aba1,
philBB-PAP1 e philBB-PAA2 que se encontram depositados na coleção de fagos do
Grupo de Biotecnologia de Bacteriófagos da Universidade do Minho.

Capítulo 4
68
4.1.2 Formação de biofilmes mistos
Os biofilmes mistos foram formados em microplacas de 24 poços (Orange
Scientific, Bélgica) contendo 990 µl de meio LB e 10 µl de cada suspensão bacteriana
crescida durante 16 h (37 ºC, sob agitação (120 rpm)) e diluída de forma a ter uma
D.O.620nm de 1 durante tempos variáveis, com mudança de meio após 24 h. Foram
também formados biofilmes mistos com adição de apenas A. baumannii AB1 durante as
primeiras 12 h (37 ºC, 120 rpm), seguida de duas lavagens e adição de meio fresco LB e
10 µl de P. aeruginosa PAO1 ficando a incubar durante 12 h nas mesmas condições.
A determinação do número de células em biofilme e quantificação de biomassa
foram realizadas como descrito no ponto 3.1.2 e 3.1.8 respetivamente.
4.1.3 Infeção Fágica de Biofilmes mistos
Os biofilmes mistos foram sujeitos a dois tipos de infeção fágica: infeção com
apenas um fago (phiBB-Aba1, phiBB-PAP1 ou phiBB-AA2) e infeção com um cocktail
contendo dois fago (phiBB-Aba1 e phiBB-PAP1). A metodologia de infeção fágica foi
realizada como descrita no ponto 3.1.9 no entanto aquando da aplicação do cocktail de
fagos, adicionou-se 50 µl de cada fago de forma a garantir uma MOI inicial no poço
de 1 para cada fago-hospedeiro.
4.1.4 Análise estatística
Esta análise foi efetuada como descrito no ponto 3.1.10 do capítulo anterior.
4.1.5 Aplicação de hibridação in situ fluorescente (FISH) em
biofilme misto
Para aplicação da técnica de FISH em estruturas de biofilme misto, o
procedimento aplicado foi semelhante ao descrito no ponto 2.1.2. Após a formação de
biofilmes mistos descritos anteriormente, foram realizadas as etapas de fixação descritas
no ponto 3.1.9. Seguidamente, foram aplicados 50 µl de alíquota de uso da sonda em
cada cupão. As amostras foram cobertas com lamelas e incubadas durante 1 h a 59 ºC.
De seguida as amostras foram mergulhadas em solução de lavagem durante 30 min. No
final as amostras ficaram em repouso até secarem. A composição das soluções foi a
descrita no ponto 2.1.2.

Caracterização dos biofilmes mistos e avaliação da eficácia de fagos específicos para A. baumannii e P. aeruginosa
69
No final, foi feita uma coloração da população total do biofilme com DAPI
como descrito no ponto 3.1.12.
4.2 Resultados e Discussão
4.2.1 Formação de biofilmes mistos
As bactérias P. aeruginosa PAO1 e A. baumannii AB1 foram usadas para
produzir biofilmes mistos. Foram formados biofilmes de 24 h que consequentemente
foram infetados durante 6 h e 24 h. As condições de crescimento de P. aeruginosa
foram realizadas tendo em conta os estudos realizados por Pires et al 5. Os ensaios de
formação e infeção fágica dos biofilmes mistos e as respetivas caracterizações dos
biofilmes foram realizados tal como nos ensaios com biofilmes de A. baumannii
descritos anteriormente (Capítulo 3).
Na primeira fase da caracterização de biofilmes mistos, quantificou-se o
crescimento bacteriano individualizado de P. aeruginosa e A. baumannii nos biofilmes
de 24 h (Figura 4.1).
Figura 4.1 - Número de células viáveis presentes de P. aeruginosa PAO1 e A. baumannii AB1 em
biofilme misto de 24 h. As barras de erros representam o desvio padrão em relação à média.
*** p < 0,01.
1,E+00
1,E+01
1,E+02
1,E+03
1,E+04
1,E+05
1,E+06
1,E+07
1,E+08
1,E+09
P.aeruginosa PAO1 A.baumannii AB1
log
(CFU
/ cm
2 )
* * *
9
8
7
6
5
4
3
2
1
0
P.aeruginosa PAO 1 A. baumannii AB1

Capítulo 4
70
Os resultados revelam que a proliferação de A. baumannii AB1 foi claramente
inibida (p < 0,01) pela presença de P. aeruginosa PAO1 (aproximadamente 5 log)
comparativamente aos ensaios mono-espécie de A. baumannii apresentados no
capítulo 3. Por outro lado, verificou-se também que ao contrário dos estudos de
Pires et al. 5, em que as placas de 24 poços com biofilmes de P. aeruginosa formados
durante 24 h apresentavam 6 × 10 8 CFU/cm2, verificou-se um decréscimo de 2 log de
células cultiváveis por parte desta bactéria nos biofilmes crescidos na presença de
A. baumannii AB1. Estudos anteriormente realizados em comunidades mistas,
compostas por duas espécies, revelam também comportamentos competitivos ou
cooperativos entre as espécies que compõem o biofilme 6,7.
Também alguns estudos de biofilmes mistos têm demostrado a inibição de
diferentes microrganismos quando crescidos conjuntamente com P. aeruginosa 8–11.
Cerqueira et al. 8 demostrou que P .aeruginosa persiste melhor dentro do biofilme
quando cresce juntamente com a bactéria E. coli. Pires et al. 11 verificou também que P.
aeruginosa inibe eficazmente a proliferação de biofilmes de C.albicans após 24 h e 48 h
de formação de biofilme misto quando comparada com os biofilmes isolados de
C. albicans.
4.2.2 Infeção fágica de biofilmes mistos
Uma vez estabelecidas as condições para a formação de biofilmes mistos, foram
realizados estudos da sua interação com os fagos para analisar o comportamento de
A. baumannii AB1 e P. aeruginosa PAO1. A infeção fágica foi realizada usando apenas
uma infeção simples com os fagos phiIBB-Aba1 ou phiIBB-PAP1 e em cocktail
composto por ambos. Os tempos de infeção decorreram durante 6 h e 24 h tendo-se
usado uma MOI de 1. Contabilizou-se o número total de células viáveis pelo método
CFU (Figuras 4.2 A e B), a biomassa total (Figura 4.2 C) e a replicação fágica pelo
número de fagos (PFU) aderidos ao biofilme e na fase planctónica (Figura 4.2 D e E).
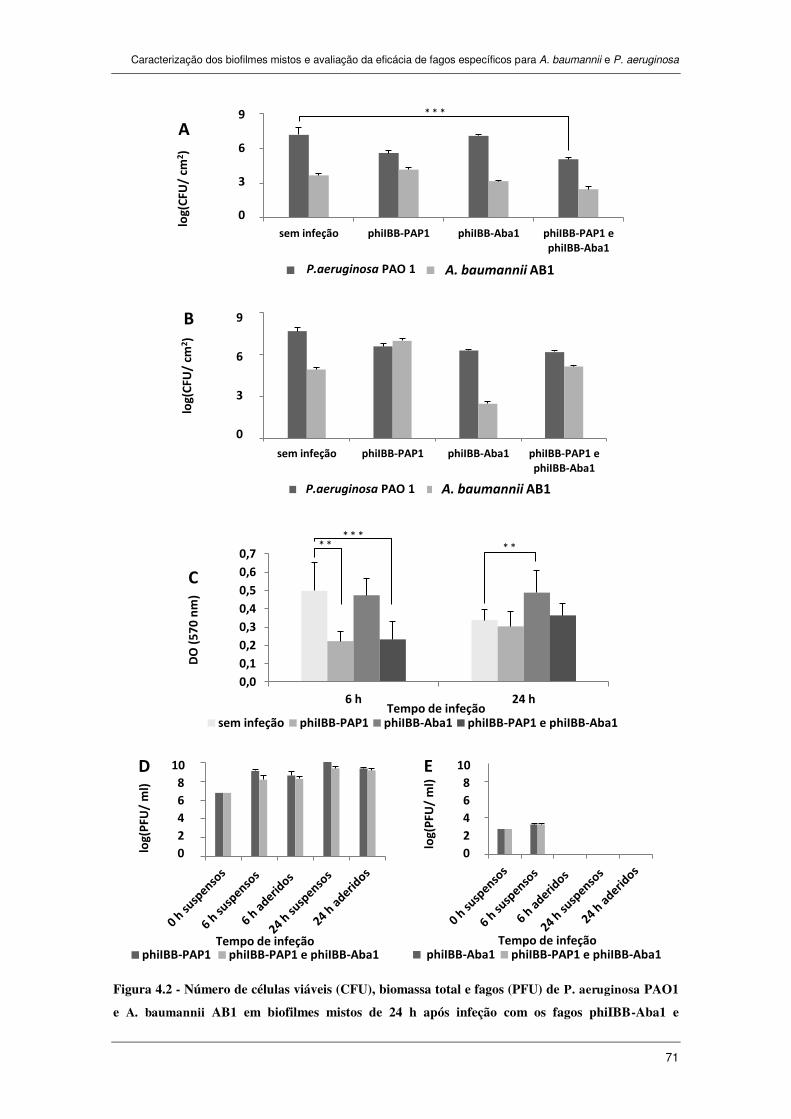
Caracterização dos biofilmes mistos e avaliação da eficácia de fagos específicos para A. baumannii e P. aeruginosa
71
Figura 4.2 - Número de células viáveis (CFU), biomassa total e fagos (PFU) de P. aeruginosa PAO1
e A. baumannii AB1 em biofilmes mistos de 24 h após infeção com os fagos phiIBB-Aba1 e
1,E+00
1,E+03
1,E+06
1,E+09
sem infeção phiIBB-PAP1 phiIBB-Aba1 phiIBB-PAP1 e phiIBB-Aba1
log(
CFU
/ cm
2 )
A
P.aeruginosa PAO A.baumannii AB1
* * *
1
9
6
3
0
P.aeruginosa PAO 1 A. baumannii AB1
1,E+00
1,E+03
1,E+06
1,E+09
sem infeção phiIBB-PAP1 phiIBB-Aba1 phiIBB-PAP1 e phiIBB-Aba1
log(
CFU
/ cm
2 )
B
P.aeruginosa PAO A.baumannii AB1 1
9
6
3
0
P.aeruginosa PAO 1 A. baumannii AB1
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
6 h 24 h
DO
(5
70
nm
)
Tempo de infeção
C
sem infeção phiIBB-PAP1 phiIBB-Aba1 phiIBB-PAP1 e phiIBB-Aba1
* * * * *
* *
1,E+00
1,E+02
1,E+04
1,E+06
1,E+08
1,E+10
log(
PFU
/ m
l)
Tempo de infeção
D
phiIBB-PAP1 phiIBB-PAP1 e phiIBB-Aba1
10
8
6
4
2
0
1,E+00
1,E+04
1,E+08
log(
PFU
/ m
l)
Tempo de infeção
E
phiIBB-Aba1 phiIBB-PAP1 e phiIBB-Aba1
10
8
6
4
2
0

Capítulo 4
72
phiIBB-PAP1 separadamente ou em cocktail durante 6 h e 24 h. A) Número de células viáveis
(CFU) nos biofilmes mistos de após 6 h de infeção; B) Número de células viáveis (CFU) nos
biofilmes mistos de após 24 h de infeção fágica; C) Quantificação da biomassa utilizando CV; D)
Número de fagos (PFU) de P. aeruginosa phiIBB-PAP1 libertados pelo biofilme (suspensos) e
presentes no biofilme (aderidos); E) Número de fagos (PFU) de A. baumannii phiIBB-Aba1
libertados pelo biofilme (suspensos) e presentes no biofilme (aderidos). As barras de erros
representam o desvio padrão em relação à média. ** p < 0,01 e *** p < 0,001.
Em biofilmes infetados durante 6 h (Figura 4.2 A), os resultados revelam que
após infeção individual com o fago de P. aeruginosa phiIBB-PAP1, ocorreu uma
diminuição de células de P. aeruginosa de aproximadamente 2 log, e um aumento
mínimo de A. baumannii. A diminuição de P. aeruginosa era esperada, uma vez que o
fago é específico para este microrganismo.
Na infeção individual com o fago de A. baumannii phiIBB-Aba1 após 6 h
(Figura 4.2 A), a quantidade de células viáveis de P. aeruginosa permaneceu
semelhante às presentes nos controlos, demonstrando que a redução de A. baumannii
causada pelo seu fago não facilitou a propagação de P. aeruginosa . Por outro lado, o
fago não se mostrou muito eficaz no controlo de A. baumannii. Possivelmente, o
reduzido número de células viáveis de A. baumannii nos biofilmes mistos e a presença
de matriz polimérica produzida pela P. aeruginosa 12. reduziram a probabilidade de
contacto entre o fago e o seu hospedeiro.
A infeção individual com o fago phiIBB-PAP1 durante 24 h (Figura 4.2 B),
apresenta uma redução de 1 log de células de P. aeruginosa e um grande aumento de
A. baumannii, conseguindo aproximar-se ou mesmo ser ligeiramente superior aos
níveis populacionais de P. aeruginosa . Na infeção individual do fago phiIBB-Aba1, o
número de células de A. baumannii manteve-se nos níveis iniciais da infeção, o que
demonstra que a infeção fágica do fago phiIBB-Aba1 não permitiu a proliferação de
A. Baumannii; contudo a P. aeruginosa não beneficiou com esta ação, visto que os
seus valores são um pouco inferiores aos verificados em biofilme de 24 h de formação
e 24 h de infeção. Este facto pode estar relacionado com a libertação, para a fase
planctónica, das bactérias não suscetíveis ao fago, como verificado nos estudos de
Sillankorva et al. 13.
Após estes primeiros ensaios com infeção individual dos fagos, seguiu-se a
aplicação, durante 6 h, de um cocktail contendo os dois fagos específicos das duas

Caracterização dos biofilmes mistos e avaliação da eficácia de fagos específicos para A. baumannii e P. aeruginosa
73
bactérias presentes nos biofilmes mistos (Figura 4.2 A). Foi evidente que a aplicação
do cocktail foi mais eficaz comparada com a aplicação individual dos fagos, reduzindo
até 2 log de células de P. aeruginosa (significativo (p < 0,001)) e aproximadamente
1,5 log de A. baumannii após 6 h (não significativo (p > 0,05)) . Possivelmente, a
presença do fago phiIBB-PAP1 de P. aeruginosa reduziu o número populacional,
permitindo desta forma um maior contacto dos fagos phiIBB-Aba1 com o hospedeiro.
Outro fator pode estar ligado à capacidade do fago phiIBB-Aba1 excretar enzimas que
degradam as substâncias poliméricas extracelulares da matriz dos biofilmes,
facilitando o acesso de ambos os fagos a superfícies mais profundas do biofilme.
A aplicação do cocktail fágico durante 24 h (Figura 4.2 B) não apresentou as
mesmas vantagens quando comparadas na aplicação de biofilmes infetados durante
6 h. A redução da população de P. aeruginosa foi semelhante à aplicação individual
do fago phiIBB-PAP1 (p < 0,001); no entanto, a redução de células viáveis de
A. baumannii não foi tão significativa comparada com a aplicação individual do fago
phiIBB-Aba1 e observou-se um aumento de células em comparação com os biofilmes
no início da infeção (biofilme de 24 h). Assim, conclui-se que a aplicação de cocktail
fágico num período superior a 24 h não é uma forma eficaz de controlar biofilmes
mistos. Estes resultados podem estar relacionados, como referidos anteriormente no
capítulo anterior, com a resistência que as bactérias adquirem.
Estudo realizado por Pires et al.11, já referido anteriormente, demostrou que em
biofilme misto de P.aeruginosa e C.albicans, o número destas aumenta após a infeção
por fagos de P.aeruginosa , sugerindo que as células sobreviventes de P.aeruginosa
após o ataque do fago adquiriram alterações no seu fenótipo e exibiram uma
diminuição da capacidade de inibir o crescimento de biofilme de C.albicans.
Em geral, o fago phiIBB-PAP1 foi capaz de se reproduzir em todas as
condições, observando-se valores superiores de PFU’s após as 24 h de infeção
(Figura 4.2 D). A reprodução do fago phiBB-Aba1 foi muito reduzida e não se
conseguiram contabilizar fagos aderidos aos biofilmes após 24 h de infeção
(Figura 4.2 E). Este resultado deve-se à baixa concentração de A. baumannii existente
nos biofilmes mistos obtidos em que P. aeruginosa claramente predominava. A baixa
multiplicação celular e a dificuldade de acesso às bactérias alvo, inevitavelmente reduz
a reprodução fágica.
Como referido no capítulo anterior, para a análise estrutural do biofilme foram
desenhadas diferentes sondas para a bactérias e fagos, mas não foi possível obter a
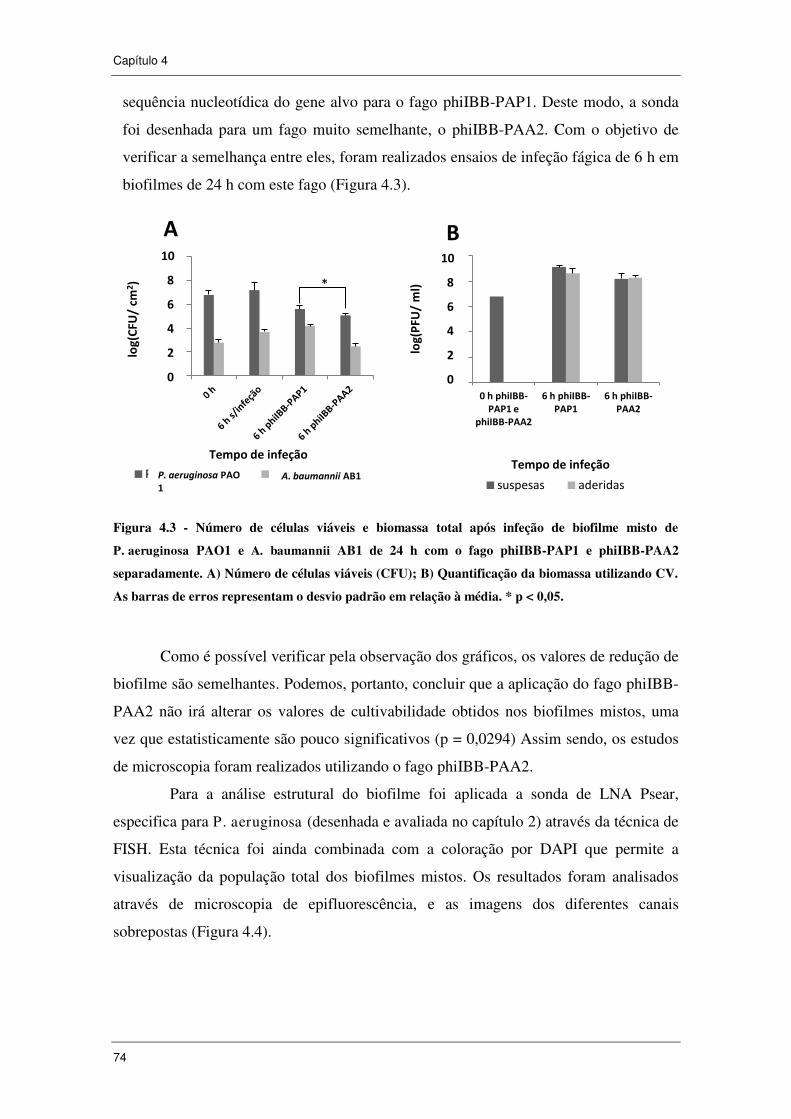
Capítulo 4
74
sequência nucleotídica do gene alvo para o fago phiIBB-PAP1. Deste modo, a sonda
foi desenhada para um fago muito semelhante, o phiIBB-PAA2. Com o objetivo de
verificar a semelhança entre eles, foram realizados ensaios de infeção fágica de 6 h em
biofilmes de 24 h com este fago (Figura 4.3).
Figura 4.3 - Número de células viáveis e biomassa total após infeção de biofilme misto de
P. aeruginosa PAO1 e A. baumannii AB1 de 24 h com o fago phiIBB-PAP1 e phiIBB-PAA2
separadamente. A) Número de células viáveis (CFU); B) Quantificação da biomassa utilizando CV.
As barras de erros representam o desvio padrão em relação à média. * p < 0,05.
Como é possível verificar pela observação dos gráficos, os valores de redução de
biofilme são semelhantes. Podemos, portanto, concluir que a aplicação do fago phiIBB-
PAA2 não irá alterar os valores de cultivabilidade obtidos nos biofilmes mistos, uma
vez que estatisticamente são pouco significativos (p = 0,0294) Assim sendo, os estudos
de microscopia foram realizados utilizando o fago phiIBB-PAA2.
Para a análise estrutural do biofilme foi aplicada a sonda de LNA Psear,
especifica para P. aeruginosa (desenhada e avaliada no capítulo 2) através da técnica de
FISH. Esta técnica foi ainda combinada com a coloração por DAPI que permite a
visualização da população total dos biofilmes mistos. Os resultados foram analisados
através de microscopia de epifluorescência, e as imagens dos diferentes canais
sobrepostas (Figura 4.4).
1,E+00
1,E+02
1,E+04
1,E+06
1,E+08
1,E+10
log(
CFU
/ cm
2 )
Tempo de infeção
A
P.aeruginosa PAO A.baumannii AB1
*
1 P. aeruginosa PAO 1
A. baumannii AB1
10
8
6
4
2
0
1,E+00
1,E+02
1,E+04
1,E+06
1,E+08
1,E+10
0 h phiIBB-PAP1 e
phiIBB-PAA2
6 h phiIBB-PAP1
6 h phiIBB-PAA2
log(
PFU
/ m
l)
Tempo de infeção
B
suspesas aderidas
10
8
6
4
2
0

Caracterização dos biofilmes mistos e avaliação da eficácia de fagos específicos para A. baumannii e P. aeruginosa
75
Figura 4.4 - Imagens de microscopia de epifluorescência de biofilme misto de P. aeruginosa PAO1 e
A. baumannii AB1 usando a sonda Pser e o corante DAPI. A) biofilme misto de 30 h ( controlo da
infeção de 6 h)em superfície de poliestireno; B) biofilme misto de 24 h infetado durante 6 h com o
fago de P. aeruginosa (phiIBB-PAA2) em superfície de poliestireno, C) biofilme misto de 24 h
infetado durante 6 h com os fagos de A. baumannii (phiIBB-Aba1) e P. aeruginosa (phiIBB-PAA2)
em superfície de poliestireno; D) biofilme misto de 24 h infetado durante 6 h com o fago de
A. baumannii (phiIBB-Aba1) em superfície de poliestireno.
As imagens de epifluorescência (Figura 4.4) corroboram os resultados
anteriormente verificados (Figura 4.2 A e B). Da análise da Figura 4.4 A pode concluir-
se que que em biofilme misto de 30 h (controlo da infeção de 6 h) o número de células
de A. baumannii (corado com a cor azul, DAPI), é muito escasso quando comparado
número de células de P. aeruginosa . Verifica-se também uma redução muito
significativa do número de P. aeruginosa , e, consequentemente, uma exposição mais
visível de células de A. baumannii, após a infeção com fago phIBB-PAA2 por um
período de 6 h (Figura 4.4 B). Já na Figura 4.4 D, que apresenta a infeção de 6 h pelo
fago phiIBB-Aba1, apesar de não se verificar uma redução muito significativa na
população de A. baumannii, é notória uma redução significativa do biofilme total. Como

Capítulo 4
76
esperado, a aplicação do cocktail fágico durante 6 h, foi a mais eficiente verificando-se
uma redução do número de células (Figura 4.4 C).
Na tentativa da se encontrar uma explicação para a baixa colonização por parte
da A. baumannii, procedeu-se a um ensaio de biofilme misto com aplicação das
bactérias em diferentes tempos. Assim, foi formado um biofilme de 12 h de
A. baumannii, de forma a permitir uma pré-colonização por esta bactéria, e
posteriormente, foi adicionada a P. aeruginosa (Figura 4.5). Biofilmes mistos de 24 h
destas duas espécies, foram utilizados como controlo.
Figura 4.5 - Formação de biofilmes mistos durante 24 h. Foram testadas duas condições, uma em
que se adicionaram ambas as espécies em simultâneo e outra em que se formou durante 12 h um
biofilme de A. baumannii Ab1 e posteriormente P. aeruginosa PAO1 até perfazer 24 h. A) Número
de células viáveis (CFU); B) Quantificação da biomassa usando CV. As barras de erros
representam o desvio padrão em relação à média. ** p < 0,01.
Analisando os gráficos, é possível verificar uma maior presença (> 2 log) de
A. baumannii (p < 0,001) quando ocorre uma adição posterior de P. aeruginosa .
Contudo observa-se também um ligeiro aumento da população de P. aeruginosa o que
demonstra que esta estratégia beneficiou de forma eficaz a colonização dos biofilmes
mistos.
1,E+00
1,E+02
1,E+04
1,E+06
1,E+08
1,E+10
24 h 12/24 h
log(
CFU
/ cm
2 )
Tempo de formação de biofilme
A
P.aeruginosa PAO A.baumannii AB1
* * *
1
10
8
6
4
2
0
P. aeruginosa PAO 1 A. baumannii AB1
0,0
0,1
0,2
0,3
0,4
0,5
0,6
24 h 12/ 24 h
DO
(5
70
nm
)
Tempo de formação do biofilme
B

Caracterização dos biofilmes mistos e avaliação da eficácia de fagos específicos para A. baumannii e P. aeruginosa
77
Figura 4.6 - Imagens de microscopia de epifluorescência com utilização da sonda fluorescente Pser
e o corante DAPI. A) Imagens de microscopia de epifluorescência de biofilmes mistos durante 24 h
com aplicação de ambas as espécies em simultâneo em superfície de poliestireno; B) Imagens de
microscopia de epifluorescência de biofilmes mistos durante 24 h com uma formação inicial
durante 12 h de um biofilme de A. baumannii AB1 e posteriormente a adição de P. aeruginosa
PAO1 até perfazer 24 h em superfície de vidro.
A análise das imagens de microscopia (Figura 4.6 A e B), evidenciaram os
resultados anteriormente obtidos (Figura 4.5). É possível verificar um aumento de
A. baumannii (Figura 4.6 B), contudo é notório que P.aeruginosa continua a
predominar no biofilme misto, mesmo quando coloniza “à posteriori”.
4.2.3 Conclusão
Em suma, verificou-se que a bactéria A. baumannii apresenta um crescimento
em biofilme muito diferente quando co-habita o mesmo substrato com P. aeruginosa ,
apresentando uma contagem de células viáveis muito inferior em comparação com o
crescimento individual. Este trabalho experimental indica também que um único fago
aplicado a um biofilme misto de duas espécies pode atingir eficazmente o hospedeiro e
reduzir o número de células do biofilme. Além disso, a aplicação de um cocktail fágico
composto pelos fagos específicos para cada espécie bacteriana do biofilme diminui o
número de células num curto período de infeção (6 h) mais eficientemente quando
comparado com a aplicação individual dos fagos.
Estudos têm demostrado que a aplicação de um cocktail fágico apresenta
melhores resultados na erradicação de biofilme quando comparados com a aplicação

Capítulo 4
78
individual 14,15,sugerindo que os que os cocktails de fagos pode ser utilizados para
diminuir a resistência das bactérias 16,17.
Kelly et al.18 verificou também uma remoção de biofilme de Staphylococus
aureus Xen 29 muito satisfatória durante um tratamento de 48 h e 72 h com a aplicação
de um cocktail fágico do fago K com seis dos seus derivados.
No final deste estudo, fica demostrado que a presença de um outro
microrganismo durante o crescimento de biofilme misto de duas espécies pode afetar
negativamente o seu crescimento como também o posterior tratamento fágico. No
entanto, com uma boa seleção de diferentes fagos para cada hospedeiro e otimização das
condições da sua aplicação, deverá ser possível utilizar eficientemente os fagos no
controlo de biofilmes simples e mistos.
4.3 Referências
1. Oliveira, L., Carneiro, P., Fischer, R. & Tinoco, E. A Presença de Patógenos Respiratórios no Biofilme Bucal de Pacientes com Pneumonia Nosocomial. Rev. Bras. Ter. Intensiva 19, 428–33 (2007).
2. Gil, S. et al. Implications of endotracheal tube biofilm in ventilator-associated pneumonia response: a state of concept. Crit. Care 16, 93 (2012).
3. Storti, A. Colonização de cateteres venosos centrais por biofilme microbiano. Tese de mestrado. Univ. Estadual Pauista. Fac. Ciências Farmacêuticas. Câmpus Araraquara; 140 (2006).
4. Storti, A., Pizzolitto, A. & Pizzolitto, E. Detection of mixed microbial biofilms on central venous catheters removed from intensive care unit patients. Brazilian J. Microbiol. 36, 275–80 (2005).
5. Pires, D., Sillankorva, S., Faustino, A. & Azeredo, J. Use of newly isolated phages for control of Pseudomonas aeruginosa PAO1 and ATCC 10145 biofilms. Res. Microbiol. 162, 798–806 (2011).
6. Hansen, S. K., Rainey, P. B., Haagensen, J. A. J. & Molin, S. Evolution of species interactions in a biofilm community. Nature 445, 533–6 (2007).
7. Adam, B., Baillie, G. S. & Douglas, L. J. Mixed species biofilms of Candida albicans and Staphylococcus epidermidis. J. Med. Microbiol. 51, 344–9 (2002).

Caracterização dos biofilmes mistos e avaliação da eficácia de fagos específicos para A. baumannii e P. aeruginosa
79
8. Cerqueira, L., Oliveira, J. A., Nicolau, A., Azevedo, N. F. & Vieira, M. J. Biofilm formation with mixed cultures of Pseudomonas aeruginosa/Escherichia coli on silicone using artificial urine to mimic urinary catheters. Biofouling 29, 829–40 (2013).
9. Purschke, F. G., Hiller, E., Trick, I. & Rupp, S. Flexible survival strategies of Pseudomonas aeruginosa in biofilms result in increased fitness compared with Candida albicans. Mol. Cell. Proteomics 11, 1652–69 (2012).
10. Kuznetsova, M. V, Maslennikova, I. L., Karpunina, T. I., Nesterova, L. Y. & Demakov, V. A. Interactions of Pseudomonas aeruginosa in predominant biofilm or planktonic forms of existence in mixed culture with Escherichia coli in vitro. Can. J. Microbiol. 59, 604–10 (2013).
11. Pires, D. P. et al. Evaluation of the ability of C. albicans to form biofilm in the presence of phage-resistant phenotypes of P. aeruginosa . Biofouling (2013).
12. Hanlon, G. W., Denyer, S. P., Olliff, C. J. & Ibrahim, L. J. Reduction in exopolysaccharide viscosity as an aid to bacteriophage penetration through Pseudomonas aeruginosa biofilms. Appl. Environ. Microbiol. 67, 2746–53 (2001).
13. Sillankorva, S., Neubauer, P. & Azeredo, J. Phage control of dual species biofilms of Pseudomonas fluorescens and Staphylococcus lentus. Biofouling 26, 567–75 (2010).
14. Gu, J. et al. A method for generation phage cocktail with great therapeutic potential. PLoS One 7, e31698 (2012).
15. Chan, B. K. & Abedon, S. T. Phage therapy pharmacology phage cocktails. Adv. Appl. Microbiol. 78, 1–23 (2012).
16. Sulakvelidze, A. & Kutter, E. Bacteriophage therapy in humans, em Bacteriophages: Biology and Application (eds E. Kutter A. Sulakvelidze, CRC Press,. Boca Raton, FL, 381–436 (2005).
17. Barbosa, C., Venail, P., Holguin, A. V & Vives, M. J. Co-Evolutionary Dynamics of the Bacteria Vibrio sp. CV1 and Phages V1G, V1P1, and V1P2: Implications for Phage Therapy. Microb. Ecol. 66, 897–905 (2013).
18. Kelly, D., McAuliffe, O., Ross, R. P. & Coffey, A. Prevention of Staphylococcus aureus biofilm formation and reduction in established biofilm density using a combination of phage K and modified derivatives. Lett. Appl. Microbiol. 54, 286–91 (2012).


Capítulo 5
Conclusão final e sugestões de trabalho futuro


Conclusão final e sugestões de trabalho futuro
83
5 Conclusão final e sugestões de trabalho futuro
Este trabalho teve como objetivo o estudo dos mecanismos de interação entre
hospedeiro e fago em biofilmes simples de A. baumannii e mistos de A. baumannii e
P. aeruginosa . Para tal utilizaram-se dois fagos recentemente isolados pelo grupo de
Biotecnologia de Bacteriófagos do CEB (phiIBB-Aba1 e phiIBB-PAP1) que foram
caracterizados no âmbito do trabalho descrito nesta dissertação.
Para a caracterização da interacção fago hospedeiro em biofilme misto foi
necessário o desenvolvimento de sondas de LNA específicas para as espécies
bacterianas e fagos que permitiriam a visualização da estrutura e organização espacial
do biofilme. As sondas foram desenhadas e otimizadas “in silico”, contudo apenas a
sonda para a bactéria P. aeruginosa foi sintetizada. Isto deveu-se não só ao elevado
custo das moléculas de LNA, mas também ao considerável esforço experimental
requerido na otimização de procedimentos multiplex. Foi possível conceber as 4
sondas com parâmetros termodinâmicos semelhantes, tendo sido ainda possível
otimizar as condições de hibridização da sonda de LNA para P. aeruginosa, sonda
esta que posteriormente foi utilizada nos ensaios de microscopia apresentados nos
diferentes capítulos.
A interação dos fagos com os respetivos hospedeiros foi estudada em
biofilmes simples ou mistos (ambos aderidos à superfície das microplacas) e até
mesmo em biofilmes com a matriz danificada. Os resultados das experiências com
biofilmes simples de A. baumannii sujeitos à infeção fágica mostraram que se deu
uma redução significativa de células viáveis presentes nos biofilmes após curtos
tempos de tratamento. Esta redução tornou-se menos expressiva para tempos de
infeção mais longos. Apesar disso, foi ainda possível averiguar que a redução de
células do biofilme era mais significativa quando a matriz era danificada, o que realça
o papel da matriz na resistência dos biofilmes à infeção por fagos.
Em biofilmes mistos de A. baumannii e P. aeruginosa verificou-se que a
aplicação combinada dos fagos específicos de P. aeruginosa e de A. baumannii foi
mais eficiente em relação à sua aplicação individual, resultando numa redução ainda
mais significativa das células das diferentes espécies presentes no biofilme misto.
Contudo, mais uma vez, a redução foi mais pronunciada nas primeiras horas de

Capítulo 5
84
infeção fágica (6 h de infeção), podendo a aparente recuperação do biofilme ser
atribuída ao aparecimento de fenótipos resistentes aos fagos. Outro facto que poderá
estar na origem deste comportamento é o estado metabólico do hospedeiro. Na
realidade, é bem sabido que um biofilme é constituído por células em diferentes
estados fisiológicos, predominado, em biofilmes maduros, células em fase
estacionária, e que a taxa de lise celular resultante da infeção fágica em bactérias perto
da fase estacionária é muito reduzida. No entanto, parece claro que existem outros
fatores envolvidos na formação de biofilme que afetaram a eficácia dos fagos após
longos períodos de exposição que deverão ser estudados com mais pormenor.
Os resultados de contagem de células viáveis foram suportados pelas análises
de microscopia de epifluorescência utilizando a sonda de LNA desenhada e a técnica
de FISH. Os ensaios de FISH utilizando esta sonda de LNA mostraram claramente
uma boa capacidade de distinguir e localizar bactérias diferentes, em populações
mistas (com ou sem infeção fágica), com recurso à utilização simultânea de um
corante genérico como o DAPI.
Trabalhos futuros devem focar essencialmente nos fenómenos que estarão a
reduzir a eficácia da infeção fágica em biofilme. Assim, sugere-se que sejam
abordados os seguintes pontos:
- Desenvolvimento da aplicação multiplex das 4 sondas de LNA, já
desenvolvidas neste estudo, para identificar as populações do biofilme que estão
efetivamente a ser alvo da ação fágica. Para tal, a técnica de FISH deverá ser
combinada com microscopia confocal de forma a permitir uma avaliação espacial
tridimensional;
- Combinação de fagos, individuais ou em cocktail, acoplados a agentes
químicos, como detergentes, desinfetantes e antibióticos; que poderá resultar numa
melhor eficácia de erradicação de biofilmes em ambientes diversos, tais como na área
clínica em procedimentos cirúrgicos, procedimentos invasivos com ventilação
mecânica, uso de cateter venoso central ou cateteres urinários;
- Manipulação genética para construir fagos específicos para o reconhecimento
dos hospedeiros. Para tal, deverão ser alterados os seus genes responsáveis pela sua
especificidade;
- Aplicação de depolimerases, combinadas com fagos e / ou antibióticos,
poderá também ser uma boa abordagem em biofilme. Estas proteínas, capazes de

Conclusão final e sugestões de trabalho futuro
85
degradar a matriz do biofilme, deverão proporcionar uma redução significativa na
formação de biofilmes e também, causar alterações estruturais na matriz;
- Estudo detalhado dos mecanismos de resistência / susceptibilidade das
células de biofilme expostas aos fagos por longos períodos de tempo;
- Estudo de biofilmes mistos com mais de duas espécies, tendo em conta que,
em biofilmes reais, verificam-se comunidades ainda mais complexas;
- Uma outra estratégia com potencial poderá envolver a utilização de
endolisinas (enzimas líticas que degradam peptidoglicano das bactérias) que
possivelmente serão capazes de, sozinhas ou juntamente com os fagos, causar uma
redução mais significativa nas populações bacterianas do biofilme.


Anexos

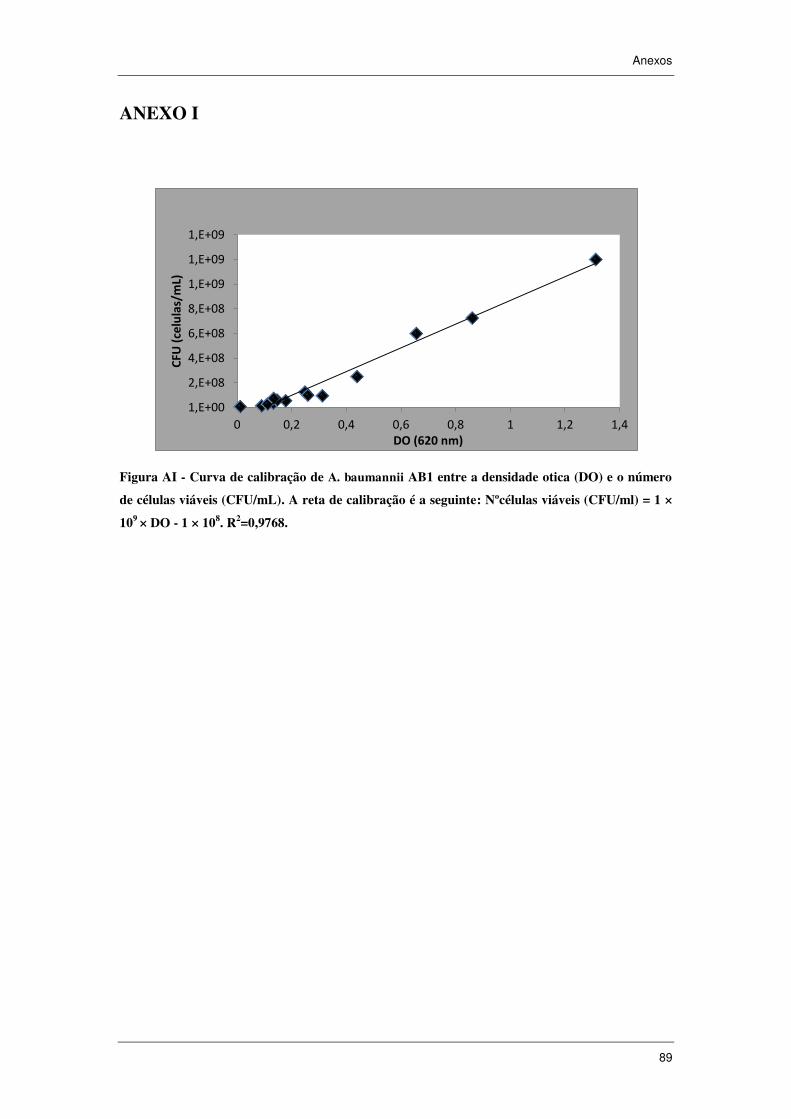
Anexos
89
ANEXO I
Figura AI - Curva de calibração de A. baumannii AB1 entre a densidade otica (DO) e o número
de células viáveis (CFU/mL). A reta de calibração é a seguinte: Nºcélulas viáveis (CFU/ml) = 1 ×
109 × DO - 1 × 108. R2=0,9768.
1,E+00
2,E+08
4,E+08
6,E+08
8,E+08
1,E+09
1,E+09
1,E+09
0 0,2 0,4 0,6 0,8 1 1,2 1,4
CFU
(ce
lula
s/m
L)
DO (620 nm)

















![Aldeias, vilas e_cidades[1]](https://img.document.onl/doc/110x75/559af5721a28ab2d2e8b45c2/aldeias-vilas-ecidades1.jpg)










