Embed Size (px)
Citation preview

UNIVERSIDADE TECNOLÓGICA FEDERAL DO PARANÁ
DEPARTAMENTO ACADÊMICO DE CIÊNCIAS AGRÁRIAS
PROGRAMA DE PÓS-GRADUAÇÃO EM AGRONOMIA
AMANDA PACHECO CARDOSO MOURA
ESTRUTURA POPULACIONAL DE JABOTICABEIRAS NO
SUDOESTE DO PARANÁ
DISSERTAÇÃO
PATO BRANCO
2017

UNIVERSIDADE TECNOLÓGICA FEDERAL DO PARANÁ
DEPARTAMENTO ACADÊMICO DE CIÊNCIAS AGRÁRIAS
PROGRAMA DE PÓS-GRADUAÇÃO EM AGRONOMIA
AMANDA PACHECO CARDOSO MOURA
ESTRUTURA POPULACIONAL DE JABOTICABEIRAS NO
SUDOESTE DO PARANÁ
DISSERTAÇÃO
PATO BRANCO
2017

AMANDA PACHECO CARDOSO MOURA
ESTRUTURA POPULACIONAL DE JABOTICABEIRAS NO
SUDOESTE DO PARANÁ
Dissertação apresentada ao Programa dePós-Graduação em Agronomia daUniversidade Tecnológica Federal do Paraná,Câmpus Pato Branco, como requisito parcialà obtenção do título de Mestre em Agronomia- Área de Concentração: Produção Vegetal.
Orientador: Prof. Dr. Moeses Andrigo Danner
PATO BRANCO
2017

M929e Moura, Amanda Pacheco CardosoEstrutura populacional de jaboticabeiras no sudoeste do Paraná /
Amanda Pacheco Cardoso Moura. Pato Branco. UTFPR, – 2017
52 f. : il. ; 30 cm
Orientador: Prof. Dr. Moeses Andrigo DannerDissertação (Mestrado) - Universidade Tecnológica Federal do
Paraná. Programa de Pós-Graduação em Agronomia. Pato Branco, PR2017.
Bibliografia: f. 45 – 52.
1. Jaboticabeira. 2. Recursos Naturais. - Conservação. 3. Jaboticabeira– Regeneração (Biologia). I. Danner, Moeses Andrigo, orient. II. .Universidade Tecnológica Federal do Paraná. Programa de Pós-Graduação em Agronomia. III. Título.
CDD (22. ed.): 630
Ficha Catalográfica elaborada por:Suélem Belmudes Cardoso CRB9/1630Biblioteca da UTFPR Câmpus Pato Branco

Ministério da EducaçãoUniversidade Tecnológica Federal do Paraná
Câmpus Pato BrancoDiretoria de Pesquisa e Pós-Graduação
Programa de Pós-Graduação em Agronomia
TERMO DE APROVAÇÃO
Título da Dissertação n° 145
Estrutura Populacional de Jaboticabeiras no Sudoeste do Paraná
por
AMANDA PACHECO CARDOSO MOURA
Dissertação apresentada às treze horas e trinta minutos do dia vinte e três defevereiro de dois mil e dezessete, como requisito parcial para obtenção do título deMESTRE EM AGRONOMIA, Linha de Pesquisa – Sistema de Produção Vegetal,Programa de Pós-Graduação em Agronomia (Área de Concentração: Produçãovegetal) da Universidade Tecnológica Federal do Paraná, Câmpus Pato Branco. Ocandidato foi arguido pela Banca Examinadora composta pelos membros abaixodesignados. Após deliberação, a Banca Examinadora considerou o trabalhoAPROVADO.
Banca examinadora:
Prof. Dr. Moeses AndrigoDannerOrientadot
UTFPR
Prof. Dr. Idemir CitadinUTFPR/Pato Branco
Prof. Dr. Joel DonazzoloUTFPR/Dois Vizinhos
Prof. Dr. Jean Carlos BudkeURI/Erechim-RS
Prof. Dr. Moeses Andrigo DannerCoordenador do PPGA/UTFPR
A Folha de Aprovação assinada encontra-se na Coordenação do Programa.

Dedico este trabalho aos meus pais.

AGRADECIMENTOS
Agradeço aos meus pais Alene e Valdemir pelo apoio durante toda a
minha formação profissional. Mãe você sempre foi um exemplo para mim.
Ao meu orientador, Moeses Danner, pela confiança e oportunidade, o
qual me inspira como exemplo de profissional.
Aos integrantes do Grupo de Pesquisa em Fruteiras Nativas do
Sudoeste do Paraná que me auxiliaram na coleta de dados a campo. Anelise,
Demétrios, Everton, José Carlos, Gean, Heliatrissy e Vanessa, construímos mais
que um grupo de pesquisa, construímos uma grande família.
Aos proprietários dos remanescentes estudados, por cederem os locais
para a realização da pesquisa.
Ao Programa de Pós-Graduação em Agronomia da UTFPR-Pato
Branco pelo suporte oferecido.
Ao laboratório de análises do solo da UTFPR, Pato Branco.
À CAPES pela concessão de bolsa.
À todos que de alguma forma fizeram parte da minha caminhada até
aqui, meus sinceros agradecimentos.

RESUMO
MOURA, Amanda P.C. Estrutura populacional de jaboticabeiras no sudoeste doParaná. 52 f. Dissertação (Mestrado em Agronomia) – Programa de Pós-Graduaçãoem Agronomia (Área de Concentração: Produção vegetal), UniversidadeTecnológica Federal do Paraná. Pato Branco, 2017.
Resumo: No Paraná, foram identificados 14 fragmentos florestais de FlorestaOmbrófila Mista, onde ocorre a espécie Plinia cauliflora. A maioria dessaspopulações encontra-se em áreas de reserva legal e sofrem com a colheitaextrativista dos frutos (jaboticabas) e outras ações antrópicas. O presente trabalhoteve como objetivo determinar classes ontogenéticas, regeneração e distribuiçãoespacial em populações de jaboticabeira (Plinia cauliflora) no Sudoeste do Paraná.Foram determinados os estádios ontogenéticos por meio de amostragem aleatória(n=50). Foram delimitadas unidades amostrais em seis fragmentos localizados emquatro municípios do sudoeste do Paraná. Obteve-se diâmetro, altura ecoordenadas cartesianas (x,y). Os dados foram avaliados por meio de estatísticadescritiva. A distribuição espacial e a dependência espacial foram verificadasatravés da função O’ring uni e bivariada. Foram encontrados cinco estádiosontogenéticos para jaboticabeira: plântulas (h ≤ 0,10 m); juvenis (h >0,10 m até 1,70m); imaturo (altura >1,70 m até 5 m); virgem (h > 5 m, sem estruturas reprodutivas);reprodutivos (h > 5 m com estruturas reprodutivas). Os resultados mostram que hádiferença estrutural entre os remanescentes, acarretando no potencial deregeneração das populações. As populações de Clevelândia, Pato Branco I e IIapresentaram todas as classes ontogenéticas estabelecidas, um maior número deindivíduos nas classes que representam a regeneração (plântulas, juvenis eimaturos) e distribuição de altura e diâmetro do tipo J-invertido. O contrário foiobservado nas outras populações. Esses resultados estão atribuídos, principalmenteao histórico de uso das áreas e consequentemente nas maiores chances deexistência de possíveis animais dispersores. Esse fato poderá aumentar as chancesde germinação e estabelecimento de plântulas de jaboticabeira. A distribuiçãoespacial variou entre populações, apresentando distribuição aleatória na maioria dosestádios ontogenéticos, com exceção das populações de Clevelândia e ChopinzinhoII. Não houve dependência espacial entre pares de estádios ontogenéticos emnenhuma das populações. Esses resultados podem estar atribuídos à dispersão ououtros fatores, necessitando de mais estudos. Pode-se concluir que o histórico demanejo das áreas e a contínua ação antrópica em alguns locais parece ser um dosprincipais empecilhos na regeneração de indivíduos de jaboticabeira e consequenterecrutamento em classes de tamanho maiores.
Palavras-chave: Plinia cauliflora. FOM. Frutas Nativas. Dinâmica de Populações.

ABSTRACT
MOURA, Amanda P.C. Populational structure of jaboticabeiras in southwesternParaná. 52 f. Dissertation (Masters in Agronomy) - Graduate Program in Agronomy(Concentration Area: Crop), Federal University of Technology Paraná. Pato Branco,2017.
Abstract: In the Brazilian state of Paraná were identified 14 forest fragments ofMixed Ombrophilous Forest where species Plinia cauliflora occurs. Most of thosepopulations are in a legal reserve area and suffer from extractive harvesting of fruits(jaboticabas) and other anthropogenic actions. Our objective with this work was todetermine ontogenetic stages, regeneration and spatial distribution on populations ofjaboticabeira (Plinia cauliflora) in the Southwest of Paraná. We determine theontogenetic stages by random sampling (n=50). We set sample units (70x70m) in sixfragments located in four cities in the southwest of Paraná. On those fragments wecollected diameter, height and Cartesian coordinates (x, y). Collected data wereevaluated through descriptive statistics. Spatial distribution and spatial dependencewere verified through the univariate and bivariate O'ring function. We found fiveontogenetic stages for jaboticabeira: seedlings (h ≤ 0.10 m); juveniles (h> 0.10 m to1.70 m); immature (height> 1.70 m to 5 m); virgin (h> 5 m, without reproductivestructures); reproductive (h> 5 m with reproductive structures). Results showed thatthere are structural differences between the remnants which have on effect onpotential regeneration of the populations. The populations of Clevelândia, PatoBranco I and II have all ontogenetic classes established. They also, showed agreater individuals numbers of classes which represent regeneration (seedlings,juveniles and immature) as well J-inverted height and diameter distribution. On theother half of populations the opposite situation was noticed. We attributed theseresults basically by the use of areas historically and consequently greater chances ofpossible existence dispersing animals which increases chances of germination andestablishment of jaboticabeira seedlings. Spatial distribution varied betweenpopulations showing a random distribution on most ontogenetic stages exceptionallyfor Clevelândia and Chopinzinho II populations. There was no spatial dependencebetween ontogenetic pairs of stages in any population. These results may beattributed to dispersion or other factors which requires further studies. Weaccomplished that management history of the areas combined with, a continuousanthropic action in some places, seems to be one of the main obstacles in theregeneration of jaboticabeira individuals and consequent recruitment in larger sizeclasses.
Keywords: Plinia cauliflora. Mixed Ombrophilous Forest. Brazilian Fruits. PopulationDynamics.

LISTA DE ILUSTRAÇÕES
Figura 1 – Localização dos remanescentes estudados em quatro municípios do sudoeste do Paraná.UTFPR, Câmpus Pato Branco, 2017................................................................................19
Figura 2 – Estágios ontogenéticos de jaboticabeiras. A. Plântula. B. Juvenil. C. Imaturo. D. Virgem. E.Reprodutivo. F. Início de gemas floríferas. UTFPR, Câmpus Pato Branco, 2017.............22
Figura 3 – Histograma para a altura (m) de jaboticabeira (P. cauliflora) em cada local de estudo.UTFPR, Câmpus Pato Branco, 2017................................................................................27
Figura 4 – Histograma para a diâmetro (cm) de jaboticabeira (P. cauliflora) em cada local de estudo.UTFPR, Câmpus Pato Branco, 2017................................................................................29
Figura 5 – Padrão de distribuição espacial de P. cauliflora em estádios ontogenéticos em diferentesremanescentes e representação da distribuição dos indivíduos de acordo comcoordenadas cartesianas (x,y). UTFPR, Câmpus Pato Branco, 2017..............................31
Figura 6 – Padrão de distribuição espacial de P. cauliflora em estádios ontogenéticos em diferentesremanescentes e representação da distribuição dos indivíduos de acordo comcoordenadas cartesianas (x,y). UTFPR, Câmpus Pato Branco, 2017..............................32
Figura 7 – Independência espacial de P. cauliflora entre pares de estádios ontogenéticos emdiferentes remanescentes e representação da distribuição dos indivíduos de acordo comcoordenadas cartesianas (x,y). UTFPR, Câmpus Pato Branco, 2017..............................33
Figura 8 – Independência espacial de P. cauliflora entre pares de estádios ontogenéticos emClevelândia e representação da distribuição dos indivíduos de acordo com coordenadascartesianas (x,y). UTFPR, Câmpus Pato Branco, 2017....................................................34
Figura 9 – Independência espacial de P. cauliflora entre pares de estádios ontogenéticos em PatoBranco I e representação da distribuição dos indivíduos de acordo com coordenadascartesianas (x,y). UTFPR, Câmpus Pato Branco, 2017....................................................35
Figura 10 – Independência espacial de P. cauliflora entre pares de estádios ontogenéticos em PatoBranco II e representação da distribuição dos indivíduos de acordo com coordenadascartesianas (x,y). UTFPR, Câmpus Pato Branco, 2017....................................................36

LISTA DE TABELAS E QUADROS
Quadro 1 – Locais, coordenadas geográficas, elevação (m), área do fragmento (ha), número total dejaboticabeiras (P. cauliflora) adultas e características/histórico de manejo de seisremanescentes florestais localizado na região sudoeste do Paraná. UTFPR, Pato Branco,2017.................................................................................................................................. 20
Tabela 1 – Análise descritiva das variáveis diâmetro, número de ramos e altura de uma população dejaboticabeira (P. cauliflora) em diferentes classes ontogenéticas. UTFPR, Câmpus PatoBranco, 2017..................................................................................................................... 23
Tabela 2 – Densidade absoluta (ind/ha) de jaboticabeiras (P. cauliflora) em cada local de estudo deacordo com os estádios ontogenéticos: plântula, juvenil, imaturo, virgem e reprodutivo.UTFPR, Câmpus Pato Branco, 2017................................................................................24
Tabela 3 – Análise descritiva das variáveis altura e diâmetro do colo (dc) de jaboticabeiras (P.cauliflora) nos locais de estudos em diferentes estádios ontogenéticos. UTFPR, CâmpusPato Branco, 2017............................................................................................................25
Tabela 4 – Análise descritiva das variáveis altura e diâmetro a altura do peito (DAP) de jaboticabeiras(P. cauliflora) nos locais de estudo em diferentes estádios ontogenéticos. UTFPR,Câmpus Pato Branco, 2016..............................................................................................26

SUMÁRIO
1 INTRODUÇÃO.........................................................................................................11
2 REVISÃO BIBLIOGRÁFICA....................................................................................13
2.1 Jaboticabeira.........................................................................................................13
2.2 Floresta Ombrófila Mista.......................................................................................13
2.3 Padrão Espacial de Espécies Arbóreas................................................................14
2.4 Estádios Ontogenéticos de Espécies Arbóreas....................................................17
3 MATERIAL E MÉTODOS........................................................................................18
3.1 Classificação de jaboticabeira em Estádios Ontogenéticos.................................18
3.2 Distribuição Espacial e Regeneração de jaboticabeiras no Sudoeste do Paraná.....................................................................................................................................19
4 RESULTADOS.........................................................................................................22
4.1 Estádios Ontogenéticos da jaboticabeira..............................................................22
4.2 Distribuição Espacial e Regeneração...................................................................23
5 DISCUSSÃO............................................................................................................37
6 CONCLUSÕES........................................................................................................43
7 CONSIDERAÇÕES FINAIS.....................................................................................44
REFERÊNCIAS...........................................................................................................45

111 INTRODUÇÃO
A jaboticabeira (Plinia sp.) é uma espécie de ocorrência natural do
Brasil (MATTOS, 1983). Na região sudoeste do Paraná, foram identificados 14
fragmentos florestais de Floresta Ombrófila Mista, onde ocorre a espécie de
jaboticabeira Plinia cauliflora (Mart.) Kausel(DANNER et al., 2010). A maioria das
populações naturais de jaboticabeira do sudoeste do Paraná encontra-se em áreas
de reserva legal, normalmente são fragmentos circundados por lavouras, sofrem
com a colheita extrativista dos frutos (jaboticabas) e outras ações antrópicas (ex.
pastejo de gado, roçadas, corte de madeira). Uma vez que não há registro na
literatura de outra região do país com tantas populações naturais de jabuticabeira
ainda existentes e devido a espécie ser endêmica do Brasil, estudos de ecologia
básica ainda inexistentes são extremamente importantes para a definição de
estratégias de conservação in situ e também ex situ da espécie.
Para iniciar e desenvolver estratégias de manejo visando à
conservação e uso da espécie é imprescindível o entendimento biológico das
mesmas (PALUDO et al., 2011; NAZARENO; REIS, 2014). Uma forma de analisar a
ecologia de populações se dá mediante estudo do padrão de distribuição espacial
das espécies (HAASE, 1995; CONDIT et al. 2000; CAPRETZ et al., 2012). Aliado a
isso, deve-se considerar a distribuição espacial em diferentes estágios de vida
(SEIDLER; PLOTKIN, 2006; DALMASO et al., 2013; ZHU et al., 2013; MIRANDA et
al. 2016), pois, é uma forma de mensurar a influência da dispersão de sementes na
população (CAUGHLIN et al., 2014), sendo um conhecimento indispensável para
entender o funcionamento da população (HIGUCHI et al. 2015).
Ações antrópicas, em populações podem influenciar sua estrutura,
diminuindo o número de indivíduos adultos e consequentemente fornecedores de
sementes. Além disso, diminuem o número de habitats e o número de animais
dispersores efetivos, mudam processos de polinização e consequentemente afetam
a regeneração e o recrutamento de indivíduos para classes etárias mais velhas
(FAHRIG, 2003; SOUZA-MARTINS, 2004; LAURENCE; VASCONCELOS, 2009;
ABREU et al., 2014; GUARINO et al., 2014).
Apesar desta importância, não há estudos de distribuição espacial e

12regeneração em jaboticabeira, tampouco do efeito antrópico em populações da
espécie. Estudo mais próximo a espécie foi desenvolvido por Gomes et al. (2016),
os autores pesquisaram a estrutura demográfica e o padrão espacial para três
espécies da família Myrtaceae: Campomanesia xanthocarpa, Myrciaria delicatula e
Myrcianthes gigantea.
Levando em consideração o que foi exposto, o intuito do Grupo de
Pesquisa em Fruteiras Nativas da UTFPR Câmpus Pato Branco é realizar estudos
sobre aspectos da ecologia de jaboticabeiras (Plinia cauliflora) em populações
(remanescentes florestais) no Sudoeste do Paraná, visto que não há relatos de
estudos destes parâmetros em outros locais do Brasil, no qual a espécie é
endêmica. Aliado a isso, existe a importância socioeconômica que a espécie
desempenha na região, auxiliando na renda dos produtores e também empregando
famílias carentes, pois, essas complementam sua renda por meio da colheita
extrativista e venda dos frutos (jaboticabas) nas rodovias regionais.
O presente trabalho teve como objetivo determinar classes
ontogenéticas, regeneração e distribuição espacial em populações de jaboticabeira
(Plinia cauliflora) no Sudoeste do Paraná. Especificamente, visou-se elucidar as
seguintes questões: Existem diferenças na regeneração natural em fragmentos com
históricos de manejo distintos? Há recrutamento de classes intermediárias nas
populações de jaboticabeira no Sudoeste do Paraná? Quais são as classes
ontogenéticas que a espécie apresenta? O padrão de distribuição espacial das
plântulas de jaboticabeira é agregado e se modifica para aleatório nas classes
ontogenéticas mais avançadas? Há dependência espacial entre as classes
ontogenéticas descritas?

132 REVISÃO BIBLIOGRÁFICA
2.1 Jaboticabeira
No Brasil a jaboticabeira (Plinia sp.) ocorre naturalmente no
Centro/Sul/Sudeste, principalmente no Bioma Mata Atlântica. Também pode ser
encontrada no Paraguai e Argentina. São conhecidas no total, nove espécies,
algumas já classificadas como espécie em extinção. De ocorrência natural no Brasil
temos: Plinia trunciflora (O.Berg) Kausel; Plinia cauliflora (Mart.) Kausel e Plinia
jaboticaba (Vell.) Kausel (MATTOS, 1983). É uma espécie da família Myrtaceae e
suas principais características morfológicas são: ramos terminais glabros,
achatados; folhas adultas glabras, opacas e com base aguda; frutos no formato de
baga globosa, lisa, negra, de 2,2 a 2,8 cm de comprimento e 2,2 a 2,9 cm de
diâmetro (LORENZI, 2009). Os principais grupos polinizadores são abelhas da
família Meliponinae e Halictidae (GRESSLER et al. 2006), destacando-se as abelhas
africanizadas (Apis mellifera) (MALERBO-SOUZA et al., 2004). Os dispersores de
sementes mais comuns dessa espécie são aves e macacos (GRESSLER et al.
2006). Os frutos de jaboticabeira (jaboticabas) apresentam grande potencial de
comercialização, pois são muito apreciados para consumo in natura e também para
fabricação de geleia, bebidas fermentadas, vinagre e licores. Além disso, a
jaboticaba apresenta alto teor de antocianinas e flavonoides na casca, o que pode
ser aproveitado pela indústria farmacêutica e alimentícia com elevado potencial
como alimento funcional (DANNER et al., 2011a).
2.2 Floresta Ombrófila Mista
A floresta ombrófila mista (FOM) conhecida como mata de araucária,
ocorre principalmente no planalto meridional e a ocorrência de gêneros como
Drymis, Podocarpus e da espécie Araucaria angustifolia caracterizam esta floresta.
Em sua formação original, sua área abrangia desde o sul dos estados de São Paulo,
Rio de Janeiro e Minas Gerais e nos estados do Paraná, Santa Catarina, Rio Grande
do Sul (VELOSO et al., 1991). Atualmente, a FOM também pode ser denominada

14como Floresta mista lati-acicufoliada, a qual apresenta uma única espécie
acicufoliada (Araucaria angustifolia) que representa mais de 50% do dossel da
floresta, com altura média de 30 m. O restante das espécies que compõe o estrato
arbóreo é latifoliada, exceto Podocarpus lambertii (OLIVEIRA-FILHO, 200;
EISENLOHR; OLIVEIRA-FILHO, 2014).
No estado do Paraná a FOM era a formação de maior abrangência
territorial, ocupando cerca de 7.387.000 de hectares, o que correspondia a 37% da
superfície do estado (MAACK, 1981). No último inventário realizado no Paraná,
verificou-se que fragmentos florestais mais conservados (estágio avançado de
sucessão) representavam apenas 0,8% da área original da FOM (CASTELLA;
BRITEZ, 2004). O intenso desmatamento para cultivo agrícola e a utilização da
madeira de araucária e de espécies da família Lauraceae no século XX foram os
responsáveis pela redução de área da floresta (GUERRA et al., 2002).
Uma maneira de incentivar a conservação da floresta se dá pelo
manejo sustentável de espécies com potencial não madeireiro. Em áreas muito
alteradas, como as florestas de araucária no estado do Paraná, essa é uma das
alternativas mais viáveis. Em áreas de FOM, a riqueza de espécies frutíferas da
família Myrtaceae e espécies com potencial medicinal é muito grande (SANQUETTA
et al., 2010). Além da parte econômica e ambiental, o uso da floresta proporciona o
aumento do conhecimento ecológico das espécies pela população e melhora a
relação do homem com a natureza, consequentemente, isso aumentará o nível de
conscientização da população no que se refere à preservação das florestas
(ZUCHIWSCKI et al., 2010).
2.3 Padrão Espacial de Espécies Arbóreas
Para traçar metodologias que visem o manejo sustentável é muito
importante ter como informação dados sobre a ecologia das populações, e o
conhecimento sobre o padrão espacial das espécies é uma das ferramentas mais
utilizadas. Estudos deste âmbito fornecerão subsídios para entender as
características espaciais e temporais e relacioná-las a processos ecológicos de
competição, mortalidade, reprodução e dispersão. Isso possibilitará compreender a

15forma de distribuição de determinada população (HAASE, 1995; CAPRETZ et al.,
2012), definir estratégias de recuperação de áreas degradadas (GIEHL et al., 2007),
além de poder relacionar como a espécie utiliza os recursos acessíveis no meio
ambiente (CONDIT et al., 2000).
Os padrões espaciais possíveis são três: aleatório, agregado ou regular
(MATTEUCI; COLMA, 1984). Quando o padrão espacial resultar em aleatório,
indicará que qualquer indivíduo pode ocupar qualquer ponto no espaço,
independentemente da posição do outro. Um padrão espacial que resulta em
uniforme ou regular indicará tendência do indivíduo em evitar outros indivíduos. E o
padrão agregado indicará que o indivíduo tende a ser atraído a locais específicos,
resultando em indivíduos mais próximos do que o esperado (RIPLEY, 1977).
Normalmente presume-se que o padrão espacial aleatório pode
resultar da homogeneidade ambiental ou então devido à espécie apresentar um
comportamento não seletivo. O padrão uniforme pode ser resultado da ocorrência da
espécie em locais com recursos limitados ou com competição intraespecífica e o
padrão agregado poderá ser resultado de dispersão de sementes próximas à planta
matriz ou resultado da ocorrência da espécie em nichos ecológicos específicos
(WIEGAND; MOLONEY, 2014).
O resultado de um padrão espacial ser agregado, regular ou aleatório
depende do histórico de vida da planta e da escala em que o padrão é observado
(GETZIN et al., 2006). Esses padrões ocorrerão devido a fatores de ordem biótica
(fenologia, síndromes de dispersão, competição, dinâmica da vegetação), fatores
históricos (perturbação antrópica) ou fatores ambientais (fertilidade do solo,
luminosidade e relevo) (McINTIRE; FARJADO, 2009; ABREU et al., 2014). Fatores
ambientais são frequentemente relacionados como causador de padrões espaciais,
porém, onde esses não são limitantes, fatores bióticos definirão os padrões
espaciais (GIEHL et al., 2007).
Em florestas tropicais e subtropicais, a maioria das populações
apresentam padrões agregados, enquanto que, em florestas de clima temperado o
padrão espacial mais frequente é aleatório. O padrão uniforme é uma forma muito
rara (ARMESTO et al., 1986). É muito comum encontrar variação nos padrões
espaciais, conforme ocorrem mudanças nos estágios de vida das espécies

16(PALUDO et al., 2009; ZHANG et al., 2015; MIRANDA et al., 2016). Um exemplo
disso é a espécie Ocotea odorifera, a qual apresentou um padrão agregado para
indivíduos regenerantes e um padrão aleatório em indivíduos adultos na FLONA de
Irati – PR, mostrando a possível influência da competição entre os indivíduos
regenerantes (DALMASO et al., 2013).
Existem várias maneiras de estimar esses padrões, desde a aplicação
estatística de variância entre amostras até a aplicação da geoestatística (PERRY et
al., 2002). Com a geoestatística se analisará os padrões espaciais por meio de
padrões de pontos, os quais consistem na localização mapeada de determinados
organismos em uma área (WIEGAND et al., 2013). Assume-se então que os objetos
ecológicos em estudo podem ser aproximados por pontos e não têm nenhum
tamanho ou forma (WIEGAND; MOLONEY, 2014). Um dos métodos mais utilizado e
recomendado para análise espacial do padrão de pontos é a análise de segunda
ordem, permitindo a dispersão dos pontos como uma função da escala (DALE et al.,
2001).
Dentre as funções de análise de padrão de pontos de segunda ordem,
destacam-se as funções K de Ripley (RIPLEY, 1977) e O’ring (WIEGAND;
MOLONEY, 2004), sendo que ambas podem ser encontradas na forma uni e
bivariada. A função univariada determina os padrões espaciais, enquanto que a
análise bivariada explica a relação de dependência entre a distribuição de dois
grupos distintos, resultando em atração ou repulsão entre os grupos, por exemplo
entre adultos e juvenis (RIPLEY, 1977).
A função L (K de Ripley modificada), dada por ,
determina o padrão espacial mediante círculos de raios centricos (r) para cada
indivíduo amostrado e dentro desses raios, o número de plantas é computado
(BESAG, 1977). A função O’ring baseia-se na função L e utiliza anéis de raios em
vez de círculos concêntricos, proporcionando uma avaliação melhor do padrão por
meio de escalas. A utilização dos anéis ao invés dos círculos proporciona o
isolamento de classes de distância específicas, uma vez que a função K de Ripley é
é cumulativa e confunde efeitos de distâncias maiores e menores (WIEGAND;
MOLONEY, 2014).
Em análises de distribuição espacial é necessário testar modelos nulos

17para verificação dos padrões (aleatório, agregado ou regular) (SCALON et al., 2012).
Para função univariada, a hipótese nula testada é a de Completa Aleatoriedade
Espacial (CSR) e sua significância é geralmente avaliada por comparação com os
dados observados em envelopes de Monte Carlo (WIEGAND; MOLONEY, 2004).
Essa hipótese determina que um número aleatório de indivíduos siga uma
distribuição uniforme (LAW et al., 2009). A função bivariada é calculada entre dois
grupos, utilizando a hipótese nula de Completa Independência Espacial e indicará a
independência entre os grupos testados, por simulações toroidais (WIEGAND;
MOLONEY, 2004).
2.4 Estádios Ontogenéticos de Espécies Arbóreas
Como forma de auxiliar na padronização em estudos de estrutura,
dinâmica e distribuição espacial de populações, é possível realizar a definição de
estádios ontogenéticos (SMIRNOVA et al., 2002; MIRANDA-MELO et al., 2007;
PORTELA et al., 2011). O desenvolvimento de organismos passa por fases que vão
do nascimento a morte, constituindo a idade cronológica. Dentro dessas fases há
mudanças morfológicas, anatômicas, fisiológicas ou bioquímicas, por isso, um
indivíduo não deve ser caracterizado apenas pela sua idade, mas também, por
critérios que consigam indicar o seu status de desenvolvimento (GATSUK et al.,
1980). Portanto, considera-se que há dois processos relacionados a isso: temporais
e espaciais, uma vez que deve-se levar em consideração a idade cronológica da
planta, bem como sucessivas mudanças estruturais (SMIRNOVA et al., 2002).
Essas mudanças são caracterizadas pela ontogenia, e a partir dela, é
possível criar uma classificação dos indivíduos denominada de “estado etário”, mais
conhecido como estádios ontogenéticos. Os estádios estão distribuídos em
diferentes períodos ontogenéticos (latente, pré-reprodutivo, reprodutivo e pós
reprodutivo) totalizando dez estádios (GATSUK et al., 1980). A partir de estudos
como este, pode-se tirar conclusões mais precisas acerca das condições de
populações em seus ambientes naturais e também comparar com mais segurança o
comportamento de populações sob diferentes pressões ambientais e antrópicas
(PORTELA et al, 2011).

183 MATERIAL E MÉTODOS
3.1 Classificação de jaboticabeira em Estádios Ontogenéticos
O trabalho foi realizado em um fragmento de Floresta Ombrófila Mista
(26°26'17" S; 52°19'20" W; 963 m de altitude) localizado no município de Clevelândia
(PR). A classificação climática de acordo com Köppen é do tipo Cfb (subtropical
úmido, sem estação seca, com verão ameno), apresentando invernos severos
(temperatura média inferior a 18 ºC e geadas frequentes) e verão com temperaturas
inferiores a 22 ºC. A precipitação média anual varia entre 1900 a 2200 mm
(ALVARES et al., 2013).
O local de estudo é mantido como reserva legal da propriedade,
possuindo área total de 12,3 hectares e 930 jaboticabeiras adultas (DANNER et al.,
2010). Atualmente, o fragmento é circundado por lavouras com culturas anuais
(trigo, soja e milho).
Foram amostrados de forma aleatória 50 indivíduos para cada estádio
ontogenético, os quais foram definidos com base em Gatsuk et al. (1980) para
espécies arbóreas. Foram escolhidas para o trabalho as fases ocorrentes no período
pré-reprodutivo (plântula, juvenil, imaturo e virgem) e reprodutivo. Neste estudo, não
foram amostrados os períodos latentes (semente) e pós-reprodução (sub senil e
senil), devido a não estarem presentes na amostragem a campo. Foram definidos
cinco estádios ontogenéticos: plântulas, juvenis, imaturos, virgens e reprodutivos.
Foram coletadas medidas quantitativas e qualitativas importantes para
auxiliar na diferenciação dos estádios ontogenéticos. As medidas quantitativas
foram: altura, circunferência a altura do peito, diâmetro do colo, número de ramos,
altura de inserção dos primeiros ramos e área de copa. As medidas qualitativas
foram: coloração das folhas, início da descamação do caule, presença de gemas
floríferas e mortalidade de ramos. As medidas de diâmetro do colo, número de
ramos e altura foram analisadas por meio de estatística descritiva (média ± erro
padrão da média), pois foram as principais variáveis para a definição dos estádios
ontogenéticos. O intervalo de altura entre as classes ontogenéticas foi definido
previamente, mediante uma pré-amostragem na população.

193.2 Distribuição Espacial e Regeneração de jaboticabeiras no Sudoeste do Paraná
O estudo foi realizado em seis fragmentos de Floresta Ombrófila Mista,
localizados em quatro municípios da região sudoeste do Paraná: Chopinzinho,
Clevelândia, Coronel Vivida e Pato Branco (Figura 1).
Os remanescentes foram escolhidos conforme informações contidas
em Danner et al. (2010) e foram coletadas e observadas informações para a
caracterização e diferenciação dos locais, principalmente segundo histórico de
manejo (Quadro 1). Para caracterizar os locais conforme o histórico de manejo foi
aplicado um questionário semiestruturado composto por 42 questões aos
proprietários. Antes de iniciar a entrevista, os entrevistados eram informados sobre o
objetivo da pesquisa e assinavam o Termo de Consentimento Livre Esclarecido
(TCLE). Este questionário foi submetido e aprovado pelo Comitê de Ética em
Pesquisa da UTFPR, em julho de 2014.
Figura 1 – Localização dos remanescentes estudados em quatro municípios do sudoeste do Paraná.UTFPR, Câmpus Pato Branco, 2017.
20Para a coleta dos dados das jaboticabeiras em todos os fragmentos foi
demarcada uma unidade amostral (U.A), parcela, de 70x70 m, subdividida em 49
subparcelas de 10x10 m. Dentro de cada U.A, procurou-se identificar todos os
indivíduos de jaboticabeira (Plinia cauliflora), os quais foram numerados, localizados
por coordenadas cartesianas (x, y) e mensurados por altura e diâmetro. Os
indivíduos de jaboticabeira amostrados foram divididos em cinco classes
ontogenéticas: 1) plântulas (indivíduos com altura ≤ 0,10 m); 2) juvenis (indivíduos
com altura > 0,10 e ≤ 1,70 m); 3) imaturos (indivíduos com altura >1,70 m e ≥ 5,0 m);
4) virgens (indivíduos com altura > 5 m, sem presença de estruturas reprodutivas); 5)
reprodutivos (indivíduos com altura > 5 m, com presença de estruturas reprodutivas).
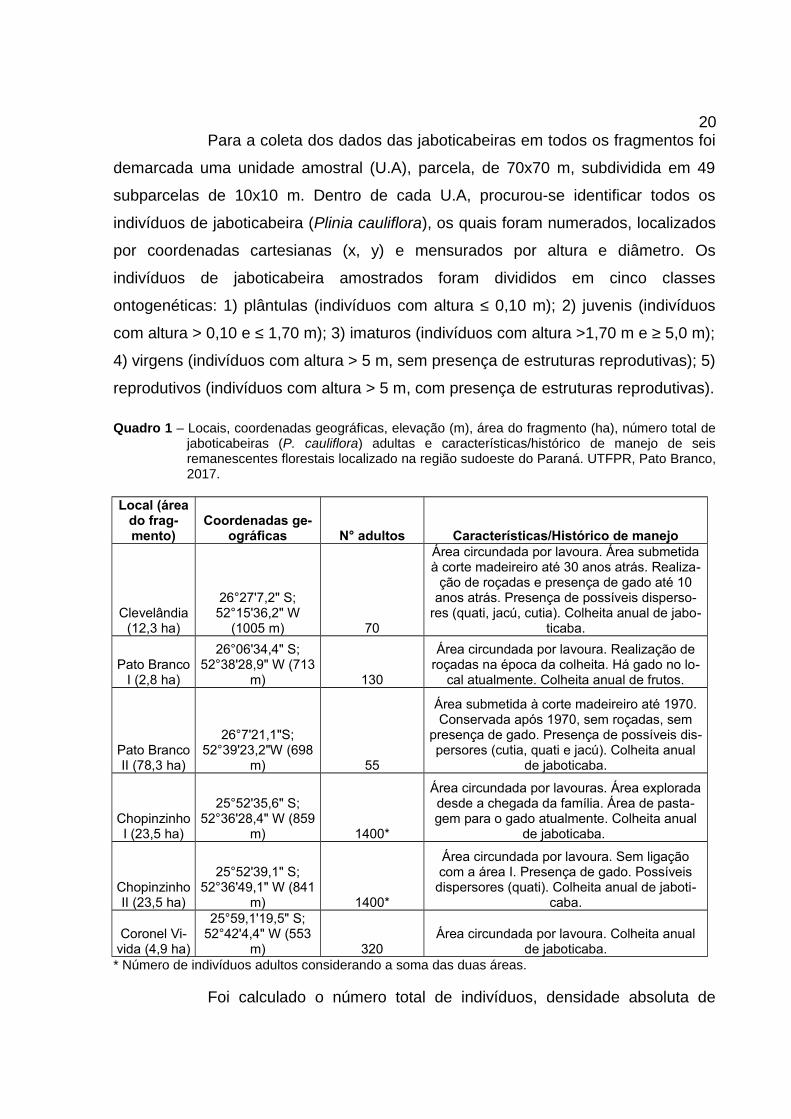
Quadro 1 – Locais, coordenadas geográficas, elevação (m), área do fragmento (ha), número total dejaboticabeiras (P. cauliflora) adultas e características/histórico de manejo de seisremanescentes florestais localizado na região sudoeste do Paraná. UTFPR, Pato Branco,2017.
Local (áreado frag-mento)
Coordenadas ge-ográficas N° adultos Características/Histórico de manejo
Clevelândia(12,3 ha)
26°27'7,2" S;52°15'36,2" W
(1005 m) 70
Área circundada por lavoura. Área submetidaà corte madeireiro até 30 anos atrás. Realiza-
ção de roçadas e presença de gado até 10anos atrás. Presença de possíveis disperso-
res (quati, jacú, cutia). Colheita anual de jabo-ticaba.
Pato BrancoI (2,8 ha)
26°06'34,4" S;52°38'28,9" W (713
m) 130
Área circundada por lavoura. Realização deroçadas na época da colheita. Há gado no lo-
cal atualmente. Colheita anual de frutos.
Pato BrancoII (78,3 ha)
26°7'21,1"S;52°39'23,2"W (698
m) 55
Área submetida à corte madeireiro até 1970.Conservada após 1970, sem roçadas, sem
presença de gado. Presença de possíveis dis-persores (cutia, quati e jacú). Colheita anual
de jaboticaba.
ChopinzinhoI (23,5 ha)
25°52'35,6" S;52°36'28,4" W (859
m) 1400*
Área circundada por lavouras. Área exploradadesde a chegada da família. Área de pasta-gem para o gado atualmente. Colheita anual
de jaboticaba.
ChopinzinhoII (23,5 ha)
25°52'39,1" S;52°36'49,1" W (841
m) 1400*
Área circundada por lavoura. Sem ligaçãocom a área I. Presença de gado. Possíveis
dispersores (quati). Colheita anual de jaboti-caba.
Coronel Vi-vida (4,9 ha)
25°59,1'19,5" S;52°42'4,4" W (553
m) 320Área circundada por lavoura. Colheita anual
de jaboticaba.* Número de indivíduos adultos considerando a soma das duas áreas.
Foi calculado o número total de indivíduos, densidade absoluta de

21indivíduos (ind/ha-1) e média de altura e diâmetro para cada local em cada classe
ontogenética. Após isso, foi calculada a distribuição de frequência para diâmetro em
cada local estudado. Essa distribuição é representada por gráficos de histograma e
serve para verificar se a população segue o padrão de distribuição J-invertido
(LEAK, 1965). Complementar ao histograma foi calculado o coeficiente de assimetria
por meio da fórmula corrigida: (JOANES; GILL, 1998); o intervalo de
confiança a 95% de probabilidade (IC95%): , em que SES é o
erro padrão (standard error) do coeficiente de assimetria, calculado por:
. (BULMER, 1979) e a significância de G1 pelo teste proposto
por Cramer (1997): .
A avaliação da distribuição espacial de cada estádio ontogenético em
cada local foi realizada por meio da função O’ring univariada (WIEGAND;
MOLONEY, 2004). O anel empregado para o cálculo do K foi de 1 metro e a
distância máxima empregada foi de 35 m (correspondente à metade do lado da
parcela). Para testar a completa aleatoriedade espacial, realizou-se o cálculo do
intervalo de confiança mediante 999 simulações de Monte Carlo. Também foi
examinada a associação espacial entre as classes ontogenéticas de cada local, com
o uso da função O’ring bivariada (WIEGAND; MOLONEY, 2004). Para a análise da
distribuição espacial e independência espacial, não foram utilizados todos os
estádios ontogenéticos amostrados, aqueles que possuíam número de indivíduos
menor que cinco não foram analisados. Para o teste da completa independência
espacial, realizou-se o cálculo do intervalo de confiança com 999 simulações
toroidais. Para testes com 95% de confiança, 500 simulações são suficientes para a
construção dos intervalos de confiança, entretanto, quanto maior o número de
simulações melhor o valor p estimado (BAROT et al., 1999). Os índices de
distribuição espacial foram calculados pelo programa computacional ‘Programita’
(WIEGAND; MOLONEY, 2014).

224 RESULTADOS
4.1 Estádios Ontogenéticos da jaboticabeira
As plântulas de jaboticabeira caracterizam-se por indivíduos com até
0,10 m, com poucas ramificações (0 a 4) e ausência de escamação do caule. Os
juvenis caracterizam-se por jaboticabeiras com altura >0,10 m até 1,70 m, com
elevado número de ramos (2 a 86) e escamação do caule (Figura 2, Tabela 1).
Figura 2 – Estágios ontogenéticos de jaboticabeiras. A. Plântula. B. Juvenil. C. Imaturo. D. Virgem. E.Reprodutivo. F. Início de gemas floríferas. UTFPR, Câmpus Pato Branco, 2017.
Para classificação do estádio imaturo levou-se em consideração a
possibilidade de medição do CAP, altura (>1,70 m até 5,0 m) e presença de caule
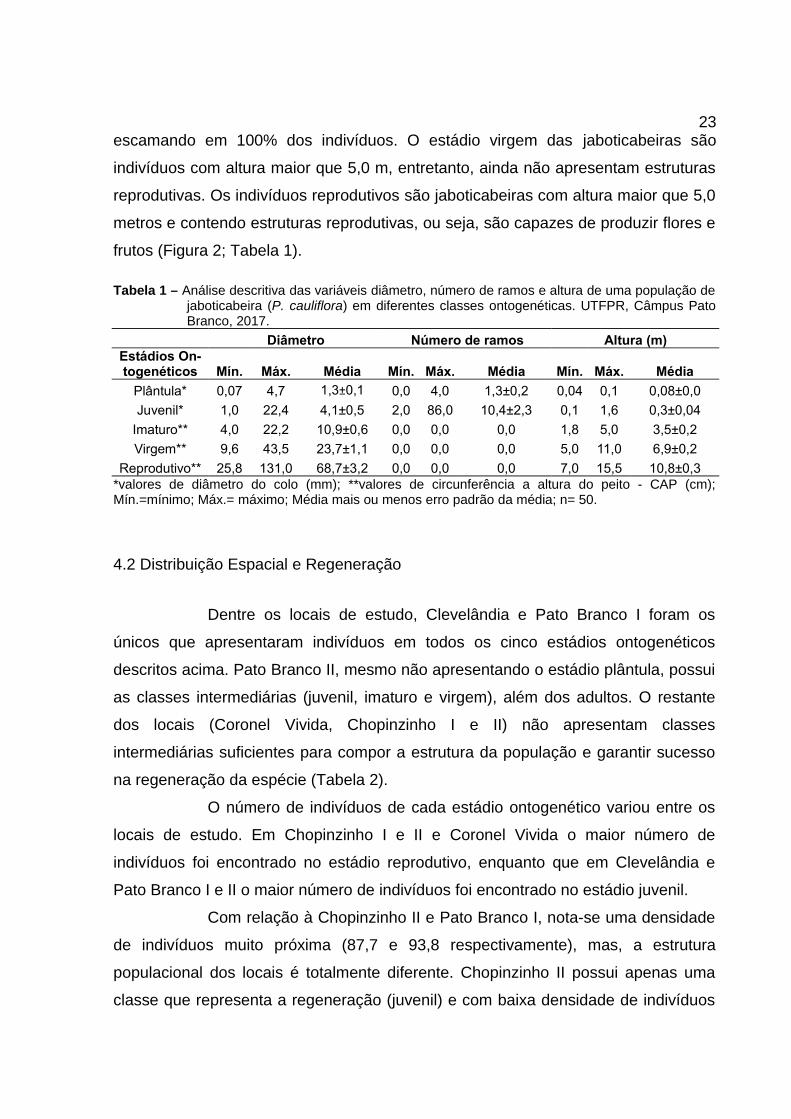
23escamando em 100% dos indivíduos. O estádio virgem das jaboticabeiras são
indivíduos com altura maior que 5,0 m, entretanto, ainda não apresentam estruturas
reprodutivas. Os indivíduos reprodutivos são jaboticabeiras com altura maior que 5,0
metros e contendo estruturas reprodutivas, ou seja, são capazes de produzir flores e
frutos (Figura 2; Tabela 1).
Tabela 1 – Análise descritiva das variáveis diâmetro, número de ramos e altura de uma população dejaboticabeira (P. cauliflora) em diferentes classes ontogenéticas. UTFPR, Câmpus PatoBranco, 2017.
Diâmetro Número de ramos Altura (m)Estádios On-togenéticos Mín. Máx. Média Mín. Máx. Média Mín. Máx. Média
Plântula* 0,07 4,7 1,3±0,1 0,0 4,0 1,3±0,2 0,04 0,1 0,08±0,0
Juvenil* 1,0 22,4 4,1±0,5 2,0 86,0 10,4±2,3 0,1 1,6 0,3±0,04
Imaturo** 4,0 22,2 10,9±0,6 0,0 0,0 0,0 1,8 5,0 3,5±0,2
Virgem** 9,6 43,5 23,7±1,1 0,0 0,0 0,0 5,0 11,0 6,9±0,2
Reprodutivo** 25,8 131,0 68,7±3,2 0,0 0,0 0,0 7,0 15,5 10,8±0,3*valores de diâmetro do colo (mm); **valores de circunferência a altura do peito - CAP (cm);Mín.=mínimo; Máx.= máximo; Média mais ou menos erro padrão da média; n= 50.
4.2 Distribuição Espacial e Regeneração
Dentre os locais de estudo, Clevelândia e Pato Branco I foram os
únicos que apresentaram indivíduos em todos os cinco estádios ontogenéticos
descritos acima. Pato Branco II, mesmo não apresentando o estádio plântula, possui
as classes intermediárias (juvenil, imaturo e virgem), além dos adultos. O restante
dos locais (Coronel Vivida, Chopinzinho I e II) não apresentam classes
intermediárias suficientes para compor a estrutura da população e garantir sucesso
na regeneração da espécie (Tabela 2).
O número de indivíduos de cada estádio ontogenético variou entre os
locais de estudo. Em Chopinzinho I e II e Coronel Vivida o maior número de
indivíduos foi encontrado no estádio reprodutivo, enquanto que em Clevelândia e
Pato Branco I e II o maior número de indivíduos foi encontrado no estádio juvenil.
Com relação à Chopinzinho II e Pato Branco I, nota-se uma densidade
de indivíduos muito próxima (87,7 e 93,8 respectivamente), mas, a estrutura
populacional dos locais é totalmente diferente. Chopinzinho II possui apenas uma
classe que representa a regeneração (juvenil) e com baixa densidade de indivíduos
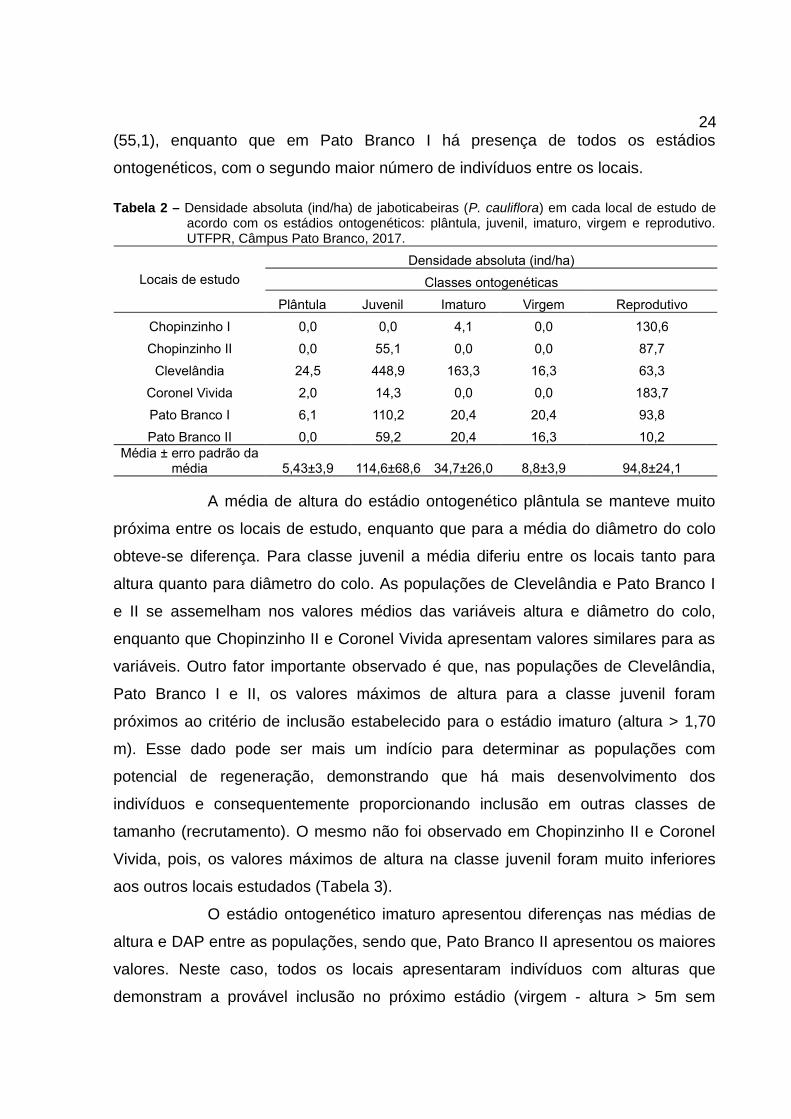
24(55,1), enquanto que em Pato Branco I há presença de todos os estádios
ontogenéticos, com o segundo maior número de indivíduos entre os locais.
Tabela 2 – Densidade absoluta (ind/ha) de jaboticabeiras (P. cauliflora) em cada local de estudo deacordo com os estádios ontogenéticos: plântula, juvenil, imaturo, virgem e reprodutivo.UTFPR, Câmpus Pato Branco, 2017.
Locais de estudo
Densidade absoluta (ind/ha)
Classes ontogenéticas
Plântula Juvenil Imaturo Virgem Reprodutivo
Chopinzinho I 0,0 0,0 4,1 0,0 130,6
Chopinzinho II 0,0 55,1 0,0 0,0 87,7
Clevelândia 24,5 448,9 163,3 16,3 63,3
Coronel Vivida 2,0 14,3 0,0 0,0 183,7
Pato Branco I 6,1 110,2 20,4 20,4 93,8
Pato Branco II 0,0 59,2 20,4 16,3 10,2Média ± erro padrão da
média 5,43±3,9 114,6±68,6 34,7±26,0 8,8±3,9 94,8±24,1
A média de altura do estádio ontogenético plântula se manteve muito
próxima entre os locais de estudo, enquanto que para a média do diâmetro do colo
obteve-se diferença. Para classe juvenil a média diferiu entre os locais tanto para
altura quanto para diâmetro do colo. As populações de Clevelândia e Pato Branco I
e II se assemelham nos valores médios das variáveis altura e diâmetro do colo,
enquanto que Chopinzinho II e Coronel Vivida apresentam valores similares para as
variáveis. Outro fator importante observado é que, nas populações de Clevelândia,
Pato Branco I e II, os valores máximos de altura para a classe juvenil foram
próximos ao critério de inclusão estabelecido para o estádio imaturo (altura > 1,70
m). Esse dado pode ser mais um indício para determinar as populações com
potencial de regeneração, demonstrando que há mais desenvolvimento dos
indivíduos e consequentemente proporcionando inclusão em outras classes de
tamanho (recrutamento). O mesmo não foi observado em Chopinzinho II e Coronel
Vivida, pois, os valores máximos de altura na classe juvenil foram muito inferiores
aos outros locais estudados (Tabela 3).
O estádio ontogenético imaturo apresentou diferenças nas médias de
altura e DAP entre as populações, sendo que, Pato Branco II apresentou os maiores
valores. Neste caso, todos os locais apresentaram indivíduos com alturas que
demonstram a provável inclusão no próximo estádio (virgem - altura > 5m sem
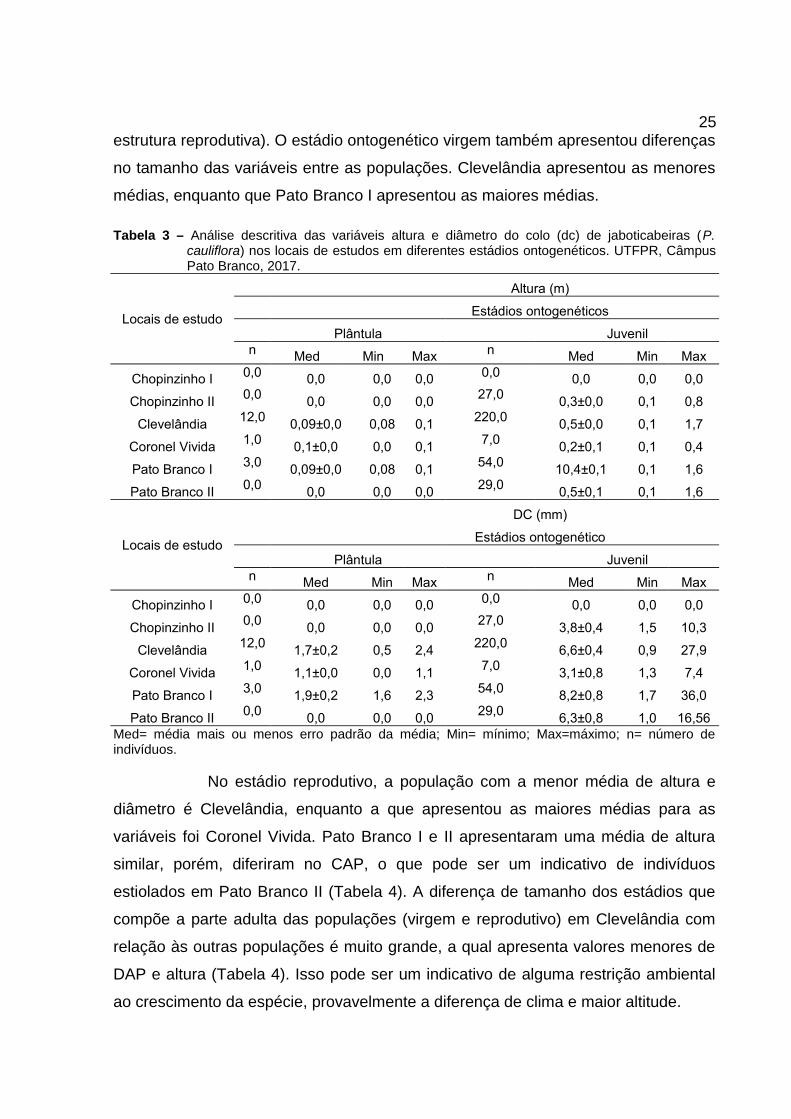
25estrutura reprodutiva). O estádio ontogenético virgem também apresentou diferenças
no tamanho das variáveis entre as populações. Clevelândia apresentou as menores
médias, enquanto que Pato Branco I apresentou as maiores médias.
Tabela 3 – Análise descritiva das variáveis altura e diâmetro do colo (dc) de jaboticabeiras (P.cauliflora) nos locais de estudos em diferentes estádios ontogenéticos. UTFPR, CâmpusPato Branco, 2017.
Locais de estudo
Altura (m)
Estádios ontogenéticos
Plântula Juveniln Med Min Max n Med Min Max
Chopinzinho I 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Chopinzinho II 0,0 0,0 0,0 0,0 27,0 0,3±0,0 0,1 0,8
Clevelândia 12,0 0,09±0,0 0,08 0,1 220,0 0,5±0,0 0,1 1,7
Coronel Vivida 1,0 0,1±0,0 0,0 0,1 7,0 0,2±0,1 0,1 0,4
Pato Branco I 3,0 0,09±0,0 0,08 0,1 54,0 10,4±0,1 0,1 1,6
Pato Branco II 0,0 0,0 0,0 0,0 29,0 0,5±0,1 0,1 1,6
Locais de estudo
DC (mm)
Estádios ontogenético
Plântula Juveniln Med Min Max n Med Min Max
Chopinzinho I 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Chopinzinho II 0,0 0,0 0,0 0,0 27,0 3,8±0,4 1,5 10,3
Clevelândia 12,0 1,7±0,2 0,5 2,4 220,0 6,6±0,4 0,9 27,9
Coronel Vivida 1,0 1,1±0,0 0,0 1,1 7,0 3,1±0,8 1,3 7,4
Pato Branco I 3,0 1,9±0,2 1,6 2,3 54,0 8,2±0,8 1,7 36,0
Pato Branco II 0,0 0,0 0,0 0,0 29,0 6,3±0,8 1,0 16,56Med= média mais ou menos erro padrão da média; Min= mínimo; Max=máximo; n= número deindivíduos.
No estádio reprodutivo, a população com a menor média de altura e
diâmetro é Clevelândia, enquanto a que apresentou as maiores médias para as
variáveis foi Coronel Vivida. Pato Branco I e II apresentaram uma média de altura
similar, porém, diferiram no CAP, o que pode ser um indicativo de indivíduos
estiolados em Pato Branco II (Tabela 4). A diferença de tamanho dos estádios que
compõe a parte adulta das populações (virgem e reprodutivo) em Clevelândia com
relação às outras populações é muito grande, a qual apresenta valores menores de
DAP e altura (Tabela 4). Isso pode ser um indicativo de alguma restrição ambiental
ao crescimento da espécie, provavelmente a diferença de clima e maior altitude.
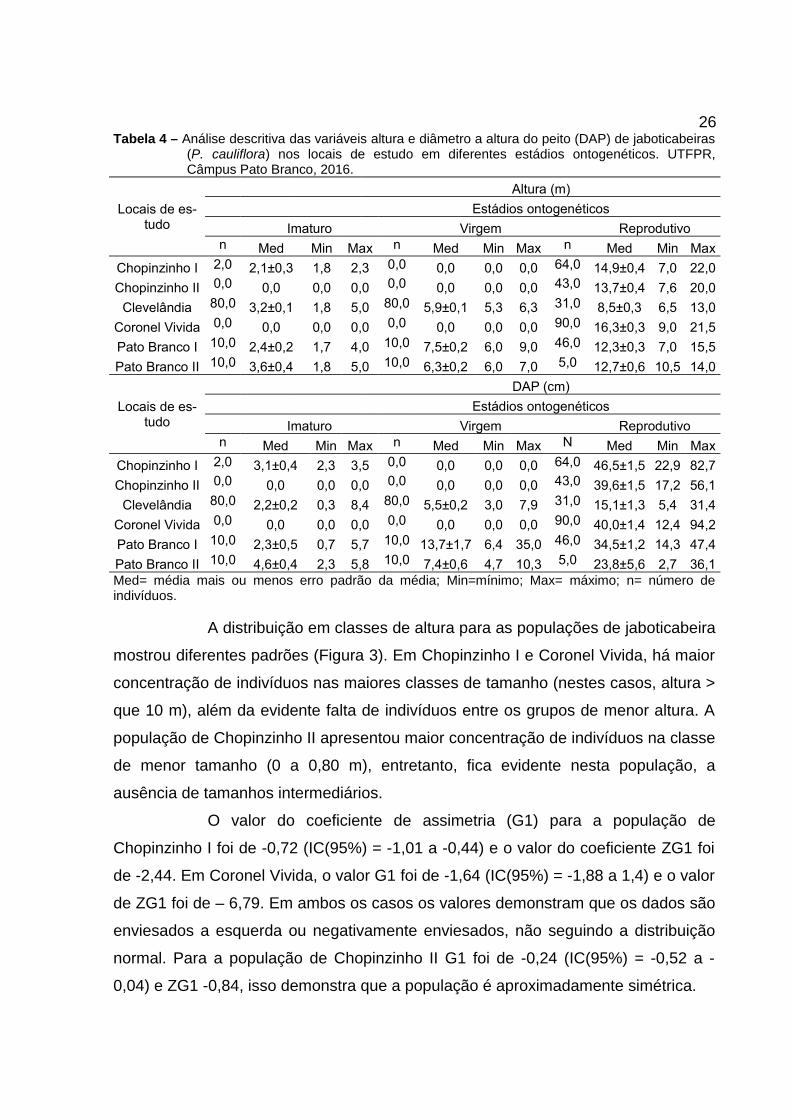
26Tabela 4 – Análise descritiva das variáveis altura e diâmetro a altura do peito (DAP) de jaboticabeiras
(P. cauliflora) nos locais de estudo em diferentes estádios ontogenéticos. UTFPR,Câmpus Pato Branco, 2016.
Locais de es-tudo
Altura (m)
Estádios ontogenéticos
Imaturo Virgem Reprodutivon Med Min Max n Med Min Max n Med Min Max
Chopinzinho I 2,0 2,1±0,3 1,8 2,3 0,0 0,0 0,0 0,0 64,0 14,9±0,4 7,0 22,0
Chopinzinho II 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 43,0 13,7±0,4 7,6 20,0
Clevelândia 80,0 3,2±0,1 1,8 5,0 80,0 5,9±0,1 5,3 6,3 31,0 8,5±0,3 6,5 13,0
Coronel Vivida 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 90,0 16,3±0,3 9,0 21,5
Pato Branco I 10,0 2,4±0,2 1,7 4,0 10,0 7,5±0,2 6,0 9,0 46,0 12,3±0,3 7,0 15,5
Pato Branco II 10,0 3,6±0,4 1,8 5,0 10,0 6,3±0,2 6,0 7,0 5,0 12,7±0,6 10,5 14,0
Locais de es-tudo
DAP (cm)
Estádios ontogenéticos
Imaturo Virgem Reprodutivon Med Min Max n Med Min Max N Med Min Max
Chopinzinho I 2,0 3,1±0,4 2,3 3,5 0,0 0,0 0,0 0,0 64,0 46,5±1,5 22,9 82,7
Chopinzinho II 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 43,0 39,6±1,5 17,2 56,1
Clevelândia 80,0 2,2±0,2 0,3 8,4 80,0 5,5±0,2 3,0 7,9 31,0 15,1±1,3 5,4 31,4
Coronel Vivida 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 90,0 40,0±1,4 12,4 94,2
Pato Branco I 10,0 2,3±0,5 0,7 5,7 10,0 13,7±1,7 6,4 35,0 46,0 34,5±1,2 14,3 47,4
Pato Branco II 10,0 4,6±0,4 2,3 5,8 10,0 7,4±0,6 4,7 10,3 5,0 23,8±5,6 2,7 36,1Med= média mais ou menos erro padrão da média; Min=mínimo; Max= máximo; n= número deindivíduos.
A distribuição em classes de altura para as populações de jaboticabeira
mostrou diferentes padrões (Figura 3). Em Chopinzinho I e Coronel Vivida, há maior
concentração de indivíduos nas maiores classes de tamanho (nestes casos, altura >
que 10 m), além da evidente falta de indivíduos entre os grupos de menor altura. A
população de Chopinzinho II apresentou maior concentração de indivíduos na classe
de menor tamanho (0 a 0,80 m), entretanto, fica evidente nesta população, a
ausência de tamanhos intermediários.
O valor do coeficiente de assimetria (G1) para a população de
Chopinzinho I foi de -0,72 (IC(95%) = -1,01 a -0,44) e o valor do coeficiente ZG1 foi
de -2,44. Em Coronel Vivida, o valor G1 foi de -1,64 (IC(95%) = -1,88 a 1,4) e o valor
de ZG1 foi de – 6,79. Em ambos os casos os valores demonstram que os dados são
enviesados a esquerda ou negativamente enviesados, não seguindo a distribuição
normal. Para a população de Chopinzinho II G1 foi de -0,24 (IC(95%) = -0,52 a -
0,04) e ZG1 -0,84, isso demonstra que a população é aproximadamente simétrica.

27
Figura 3 – Histograma para a altura (m) de jaboticabeira (P. cauliflora) em cada local de estudo.UTFPR, Câmpus Pato Branco, 2017.
Chopinzinho I Chopinzinho II
Clevelândia Coronel Vivida
Pato Branco I Pato Branco II

28Com relação às populações de Clevelândia, Pato Branco I e II, há
maior concentração de indivíduos nas menores classes de altura e um menor
número de indivíduos nas maiores classes de altura, neste caso, as populações
seguem o padrão J-invertido. Para a população de Clevelândia o valor de G1 foi de
1,87 (IC(95%) = 1,40 a 1,88). O valor de ZG1 foi 14,46, valor alto e significativo. Para
a população de Pato Branco I o valor de G1 foi de 0,44 (IC(95%) = 0,23 a 0,65) e o
valor ZG1 foi 2,08. A população de Pato Branco II o valor de G1 foi de 1,48 (IC(95%)
= 1,17 a 1,79) e o valor ZG1 foi 4,71. Neste caso, as três populações supracitadas
são enviesadas à direita e positivamente, demonstrando que as populações não
seguem distribuição normal.
Encontrou-se diferenças na distribuição em classes de diâmetro para
as populações de jaboticabeira (Figura 4). Em Chopinzinho I e Coronel Vivida, há
maior concentração de indivíduos na classe de tamanho 36 a 48 cm, não seguindo
um padrão de distribuição J invertido. As populações de Chopinzinho II e Pato
Branco II apresentaram maior concentração de indivíduos na classe de menor
tamanho, entretanto, houve ausência de indivíduos em algumas classes.
O valor do coeficiente de assimetria (G1) para a população de
Chopinzinho I foi de -1,03 (IC(95%) = -0,95 a -1,11) e o valor do coeficiente ZG1 foi
de 12,9. Em Chopinzinho II, o valor G1 foi de -1,10 (IC(95%) = -0,5 a -1,7) e o valor
de ZG1 foi de – 3,69. Em ambos os casos os valores sugerem que as populações
não seguem distribuição normal. Para a população de Coronel Vivida o valor de G1
foi 0,41 (IC(95%) = 0,07 a 0,24) e ZG1 1,72, demonstrando que a população é
aproximadamente simétrica.
Com relação às populações de Clevelândia, Pato Branco I e II, há
maior concentração de indivíduos nas menores classes de diâmetro e um menor
número de indivíduos nas maiores classes de altura, neste caso, as populações
seguem o padrão J-invertido. Para a população de Clevelândia o valor de G1 foi de
1,00 (IC(95%) = -0,70 a 1,26) e o valor de ZG1 foi 7,69. Para a população de Pato
Branco I o valor de G1 foi de 1,03 (IC(95%) = -0,59 a 1,47) e o valor ZG1 foi 4,68. A
população de Pato Branco II o valor de G1 foi de 7,4 (IC(95%) = 6,74 a 8,06) e o
valor ZG1 foi 22,42. Neste caso, as três populações supracitadas são enviesadas à
direita e positivamente, não seguindo distribuição normal.
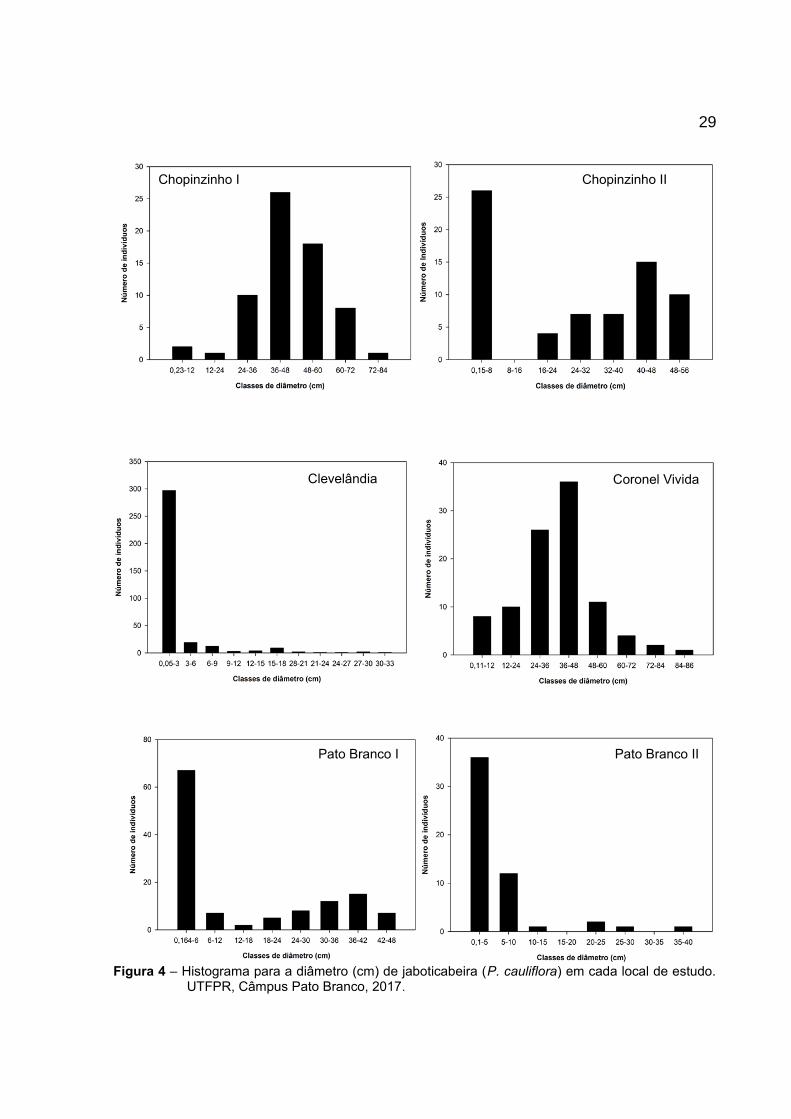
29
Figura 4 – Histograma para a diâmetro (cm) de jaboticabeira (P. cauliflora) em cada local de estudo.UTFPR, Câmpus Pato Branco, 2017.
Chopinzinho II
Clevelândia Coronel Vivida
Chopinzinho I
Pato Branco I Pato Branco II

30Houve diferença na distribuição espacial dos indivíduos entre os
estádios ontogenéticos (plântula, juvenil, imaturos, virgens e reprodutivos). É
possível observar que os indivíduos reprodutivos nas populações de Chopinzinho II
e Clevelândia apresentaram distribuição aleatória (Figura 5).
Na população de Clevelândia, o estádio plântula apresentou
distribuição aleatória e em Coronel Vivida, foi observado que o único estádio que
representa a regeneração (juvenil) também apresentou distribuição aleatória (Figura
5).
A distribuição aleatória encontrada para as populações supracitadas
ocorre devido os valores estimados estarem dentro do intervalo de confiança
construído por meio de simulações de Monte Carlo.
A maioria dos estádios que representam a regeneração apresentaram
distribuição agregada nas populações de Chopinzinho II (juvenis) e Clevelândia
(juvenis e imaturo), com exceção do estádio plântula já citado. A distribuição
agregada é caracteriza devido os valores estimados se encontrarem acima dos
intervalos de confiança construídos (Figura 5).
A agregação em Chopinzinho II ocorreu em 16 m para indivíduos
adultos e até aproximadamente 8 m de distância para indivíduos juvenis. Em
Clevelândia a agregação variou conforme o estádio ontogenético, aumentando a
agregação do estádio juvenil (agregação até aproximadamente 2 m) até imaturo
(agregação até 8 m) e virgem (agregação até aproximadamente 10 m) (Figura 5).
Com relação ao estádio juvenil da população de Clevelândia, nota-se
que a agregação ocorreu dos 10 aos 12 metros novamente (Figura 5).
Nas populações Coronel Vivida (estádio reprodutivo), Pato Branco I e
Pato Branco II nenhum dos estádios ontogenéticos apresentou grau de agregação,
sendo todos eles distribuídos de forma aleatória, pois os valores estimados
apresenta-se dentro dos limites do intervalo de confiança (Figura 6).
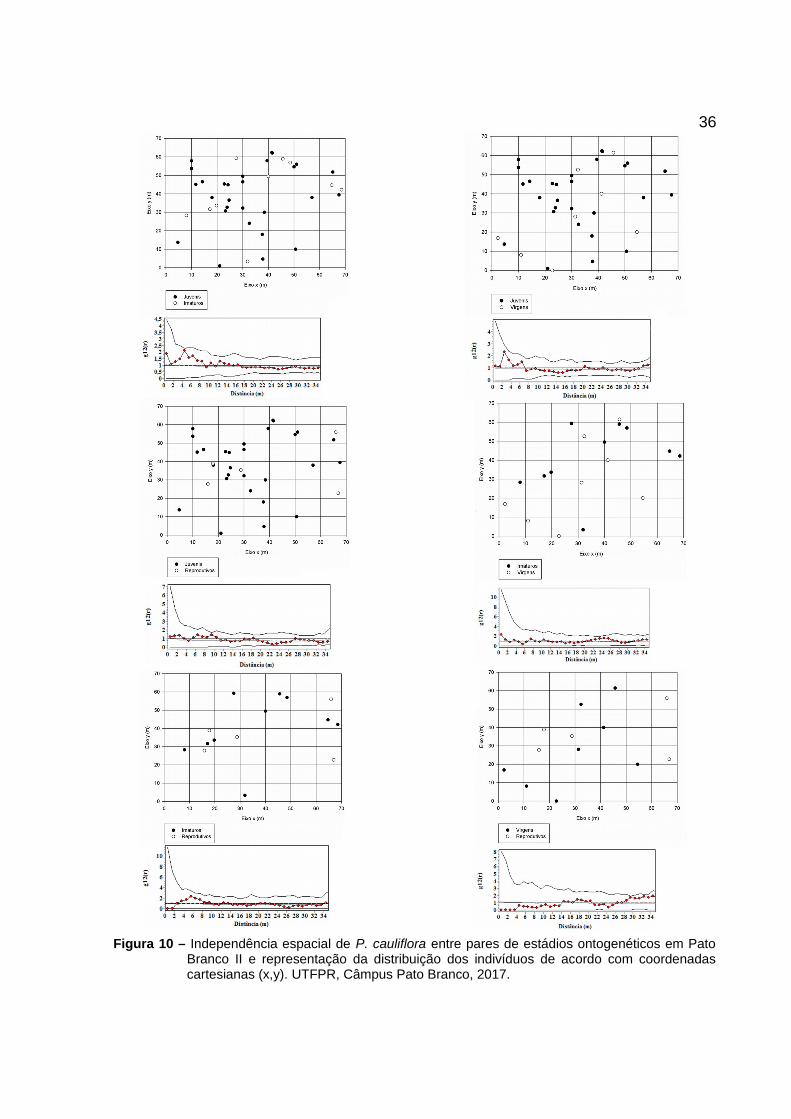
A análise da independência espacial demonstrou que em nenhuma das
populações estudadas houve associação espacial entre os pares de estádios
ontogenéticos estudados, uma vez que, os valores se encontram dentro dos limites
do intervalo de confiança (Figuras 7, 8, 9 e 10).
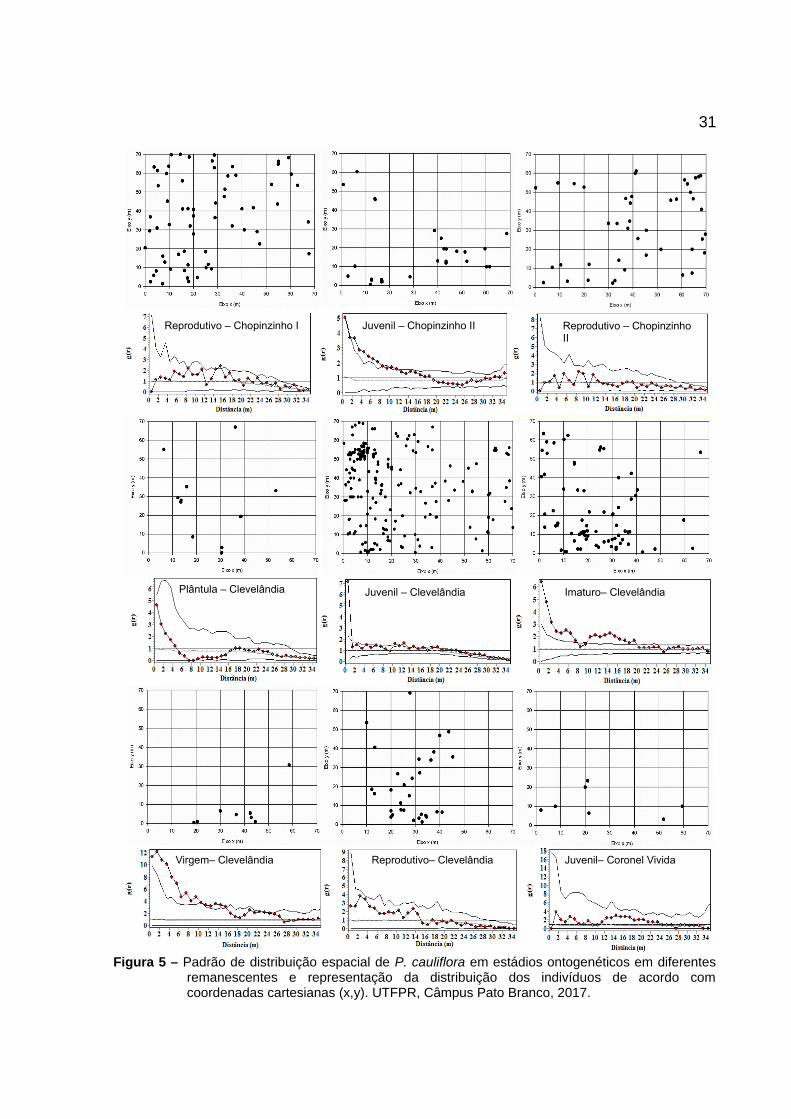
31
Figura 5 – Padrão de distribuição espacial de P. cauliflora em estádios ontogenéticos em diferentesremanescentes e representação da distribuição dos indivíduos de acordo comcoordenadas cartesianas (x,y). UTFPR, Câmpus Pato Branco, 2017.
Reprodutivo – Chopinzinho I Juvenil – Chopinzinho II Reprodutivo – Chopinzinho II
Plântula – Clevelândia Juvenil – Clevelândia Imaturo– Clevelândia
Virgem– Clevelândia Reprodutivo– Clevelândia Juvenil– Coronel Vivida
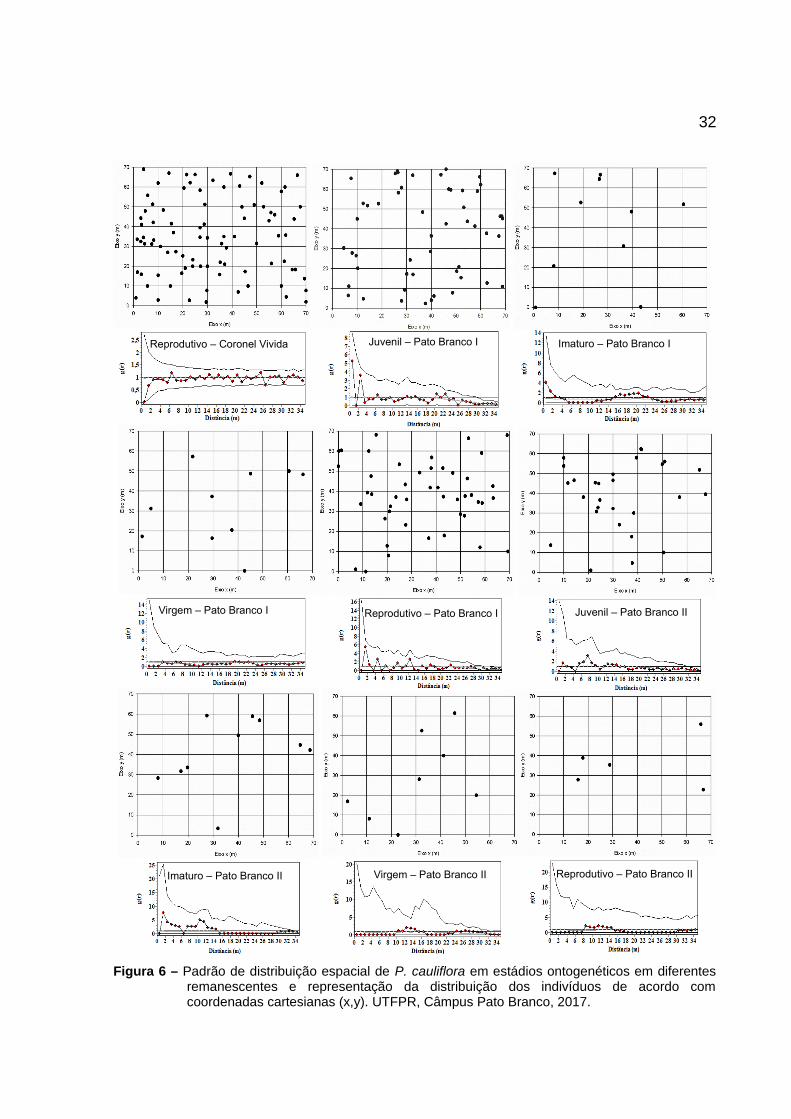
32
Figura 6 – Padrão de distribuição espacial de P. cauliflora em estádios ontogenéticos em diferentesremanescentes e representação da distribuição dos indivíduos de acordo comcoordenadas cartesianas (x,y). UTFPR, Câmpus Pato Branco, 2017.
Reprodutivo – Coronel Vivida Juvenil – Pato Branco I Imaturo – Pato Branco I
Virgem – Pato Branco I Reprodutivo – Pato Branco I Juvenil – Pato Branco II
Imaturo – Pato Branco II Virgem – Pato Branco II Reprodutivo – Pato Branco II

33
Figura 7 – Independência espacial de P. cauliflora entre pares de estádios ontogenéticos emdiferentes remanescentes e representação da distribuição dos indivíduos de acordo comcoordenadas cartesianas (x,y). UTFPR, Câmpus Pato Branco, 2017.

34
Figura 8 – Independência espacial de P. cauliflora entre pares de estádios ontogenéticos emClevelândia e representação da distribuição dos indivíduos de acordo com coordenadascartesianas (x,y). UTFPR, Câmpus Pato Branco, 2017.

35
Figura 9 – Independência espacial de P. cauliflora entre pares de estádios ontogenéticos em PatoBranco I e representação da distribuição dos indivíduos de acordo com coordenadascartesianas (x,y). UTFPR, Câmpus Pato Branco, 2017.
36
Figura 10 – Independência espacial de P. cauliflora entre pares de estádios ontogenéticos em PatoBranco II e representação da distribuição dos indivíduos de acordo com coordenadascartesianas (x,y). UTFPR, Câmpus Pato Branco, 2017.

375 DISCUSSÃO
Na literatura consultada, não existem estudos sobre a estrutura
populacional de jaboticabeiras (Plinia cauliflora) e sua regeneração em áreas
naturais. Os resultados mostram que é possível classificar a espécie em diferentes
estádios ontogenéticos. Esta classificação ocorreu por meio de uma adaptação da
escala proposta por Gatsuk et al. (1980), devido as diferenças entre a jaboticabeira e
a espécie estuda pelos autores.
Além disso, demonstram que existem diferenças estruturais nas
populações e que há interferência no processo de recrutamento de indivíduos para
classes ontogenéticas intermediárias em três das seis populações estudadas,
provavelmente, devido a ação antrópica que ainda existem nesses locais. Em todas
as populações o estádio ontogenético reprodutivo apresentou distribuição aleatória,
enquanto que, os outros estádios apresentaram diferenças entre os locais de
estudo. Não foi relatada dependência espacial entre os estádios ontogenéticos das
populações. Por meio desses resultados foi possível obter uma caracterização geral
da estrutura populacional de cada remanescente estudado.
Não foi possível identificar se as plântulas de jaboticabeira ainda
utilizam reservas nutricionais das sementes, conceito de Gatsuk et al. (1980) para
plântulas. Portanto, a utilização de outros parâmetros para classificação foi
essencial. É possível notar que o número de ramos, a altura e a escamação do
caule foram as principais variáveis para diferenciar os estádios plântulas e juvenis. A
média de ramos do estádio plântula é muito inferior ao estádio juvenil (Tabela 2) e a
escamação não ocorreu em indivíduos classificados com plântulas.
A diferenciação entre os estádios juvenis e imaturas se deu pela altura
e a possibilidade de medição do CAP, uma vez que, plantas maiores que 1,70 m, há
a possibilidade de medição da variável. E a diferenciação entre virgens e
reprodutivas se deu pela presença ou ausência de estruturas reprodutivas. Isso
porque, as estruturas morfológicas que as plantas desses dois últimos estádios
apresentam são muito semelhantes. É possível notar por meio dos valores máximos
e mínimos tanto para a variável CAP como para a variável altura que existem um
intervalo onde os valores se intercalam (Tabela 1).

38A definição dos estádios ontogenéticos baseia-se nas classes criadas
por Gatsuk et al. (1980). Os autores utilizaram para a espécie arbórea Fraxinus
excelsior características quantitativas e qualitativas, porém, é muito importante que
haja o aparecimento de estruturas que não existiam antes, para que as classes
possam ser separadas. No texto o autor classifica os estádios ontogenéticos em
quatro grupos: latente -semente; pré-reprodutivo - plântulas, juvenis, imaturos e
virgens; reprodutivo - jovem, maduro e velho; pós-reprodutivos – subsenil e senil.
Em jaboticabeiras não foi possível determinar todos esses estádios, sendo utilizados
os grupos pré-reprodutivo e reprodutivo, este último sem subdivisões, uma vez que,
não foram encontradas diferenças estruturais suficientes para estas separações.
É comum ocorrer adaptações na utilização dos estágios ontogenéticos
para cada espécie, pois, muitas apresentam grandes variações morfológicas
(PORTELLA et al., 2011). Para a espécie Psychotria tenuinervis, foram identificados
apenas quatro estádios: plântulas, juvenis, imaturos e adultos (fase vegetativa e
reprodutiva) (ROSSETO et al., 2013), enquanto que, para a palmeira Attalea humilis,
os estádios definidos foram cinco: plântulas, juvenis, imaturos, virgem e reprodutivos
(SOUZA et al., 2000), os mesmos definidos no presente estudo.
Na literatura é possível encontrar trabalhos que definem os estádios
ontogenéticos de algumas formas de plantas, como palmeiras (SOUZA et al., 2000;
MIRANDA-MELO et al., 2007; BERNARCCI et al., 2008; PORTELA et al., 2011),
arbustos (ROSSETO et al., 2013) e árvores (SILVEIRA et al., 2012). Porém, ainda é
notável a escassez de estudos com este objetivo e para jaboticabeiras não foram
encontrados relatos.
Com relação à estrutura populacional, fica evidente a diferença entre
as populações. Clevelândia, Pato Branco I e II, apresentam praticamente todas as
classes ontogenéticas, um maior número de indivíduos na regeneração, e um
padrão de distribuição de altura do tipo J-invertido, o que não ocorreu nos outros
locais. Os fatores ecológicos que afetam o recrutamento de novas plântulas pode
ser explicada pela insuficiência de frutos/sementes (limitação de fonte) ou qualidade
de dispersão (limitação de dispersão) (JORDANO et al., 2006), competição, ataque
por inimigos naturais e requisitos ambientais específicos para crescimento e
sobrevivência (CLARK; CLARK, 1987; GRAU, 2000; BECKMAN; ROGERS, 2013).

39A presença de animais frugívoros de grande porte é importante, pois,
esses possuem a capacidade de dispersar sementes a longas distâncias, o que
aumenta a possibilidade de desenvolvimento da espécie (JORDANO et al., 2006),
diminuindo a taxa de predação por herbívoros em plântulas (JANZEL, 1970;
JANZEL, 1980) e a competição por recursos entre as plântulas (COLUSSI;
PRESTES, 2011). Ainda não existem relatos de dispersores efetivos da
jaboticabeira, mas em Myrciaria trunciflora as aves atuam como agentes
dispersores, mas com baixa eficiência, pois a maioria consome os frutos diretamente
no tronco (COLUSSI; PRESTES, 2011). Também há registro de consumo de
jaboticaba pelo cachorro-do-mato ou graxaim (Dusicyon thous), pois foram
encontradas sementes nas fezes, as quais não apresentaram germinação (MOTTA-
JUNIOR et al., 1994).
A presença de possíveis animais consumidores de jaboticabas em
Clevelândia e Pato Branco I e (Quadro 1) provavelmente são os principais aspectos
que favoreceram a regeneração nestes locais. O maior grau de conservação auxilia
na presença de possíveis animais consumidores dos frutos da espécie, os quais são
fundamentais para a germinação da espécie, auxiliando na retirada da polpa e
aumentando a chance de estabelecimento. Nos locais de estudo não foram feitos
estudos sobre os animais dispersores da espécie, apenas relatos de possíveis
consumidores. Em uma população de Bertholletia excelsa, com aproximadamente
93% de frutos colhidos, densidades razoáveis de plantas juvenis foram encontradas.
Dos frutos deixados, muitos foram consumidos por cutias (Dasyprocta spp.),
principais dispersores da espécie (ZUIDEMA; BOOT, 2002).
Nos locais com o menor número de indivíduos reprodutivos
(Clevelândia e Pato Branco II), obteve-se o maior número de indivíduos nos estádios
ontogenéticos que representam a regeneração. Em Chopinzinho I e II e Coronel
Vivida, temos um grande número de indivíduos reprodutivos (fornecedores de
sementes), entretanto, um baixo número de plantas que compõe a regeneração
(Tabela 2). Esperava-se que, em locais com maior número de indivíduos
reprodutivos houvesse maior número de indivíduos regenerantes devido ao maior
número de sementes produzidas. Neste caso podemos inferir que a ação antrópica
pode ser o fator principal, o que influenciaria na dispersão das sementes e no grau

40de extrativismo dos frutos. Em todos os casos, necessita-se de mais estudos para
confirmação dessas hipóteses. Sabe-se que o estabelecimento de mudas e
consequentemente a regeneração de adultos pode ser extremamente sensível a
pequenas mudanças na produção de sementes e nas taxas de mortalidade entre
mudas. Além disso, o extrativismo poderá aumentar a taxa de predação sobre os
frutos remanescentes, o que também diminuirá o número de indivíduos ao longo das
gerações (HALL; BAWA, 1993).
Nas populações que possuem além da colheita de jaboticabas, outra
atividade antrópica (nos três casos, presença de bovinos), existem duas situações:
1º. Chopinzinho I e II: não apresenta todas as classes ontogenéticas e baixo número
de regenerantes; Pato Branco I: presença de todas as classes ontogenéticas e
número considerável de indivíduos regenerantes. É evidente que o pastejo por
bovinos em área natural afeta a estrutura populacional das espécies. Populações de
Butia eriospatha são severamente afetas por pastoreio: em locais alterados, foram
identificados apenas dois estágios ontogenéticos: adulto e plântulas; no local onde
não havia pastoreio foram detectados seis estágios: plântulas, infantil, juvenis,
imatura, reprodutivas e adultos. Isso ocorre principalmente pela herbivoria causada
pelo gado (NAZARENO; REIS, 2014). Outro ponto pode estar na degradação e
compactação do solo, afetando diretamente a germinação de sementes e o
desenvolvimento de plântulas. A diferença estrutural entre essas populações pode
ser explicada pela intensidade de pastejo. Em populações de Araucaria angustifolia,
a intensidade de pastejo influenciou a densidade de indivíduos nos três primeiros
estágios de vida (plântulas, infantil e jovem), os quais foram encontrados em maior
número nos locais com menor intensidade de pastejo (SAMPAIO; GUARINO, 2007).
Mediante a relação entre presença ou ausência de cada estádio
ontogenético proposto neste estudo e a densidade de indivíduos, podemos separar
os locais em dois grupos: Grupo I (Chopinzinho I e II e Coronel Vivida) – locais com
baixo potencial de regeneração; Grupo II (Clevelândia, Pato Branco I e II) – locais
com potencial de regeneração. Isso porque, nos locais com potencial de
regeneração, temos o recrutamento de novos indivíduos entre os estádios
ontogenéticos propostos. O recrutamento é o ingresso de indivíduos em uma
determinada população, e normalmente é quantificada por indivíduos que alcançam

41um limite de tamanho. Esse processo pode ser influenciado pelo estabelecimento,
crescimento e mortalidade dos indivíduos (LEXEROD; EID, 2005). Em áreas
naturais manejadas, o estudo do ingresso de indivíduos (regeneração e
recrutamento) é um dado essencial para trabalho em longo prazo e para prever a
produção da população (ROSSI et al., 2007).
É importante levar em consideração as características das sementes
de jaboticabeira para o entendimento da regeneração da espécie. As sementes são
recalcitrantes, perdendo totalmente a viabilidade com umidade abaixo de 30%
(VALIO; FERREIRA, 1992), indicativo de que a espécie se regenera por meio de
banco de plântulas (PAMMENTER; BERJAK, 2000). Além disso, a viabilidade das
sementes em atmosfera natural é de no máximo cinco dias (DANNER et al., 2011b).
Com a retirada da polpa, ocorre um aumento expressivo na taxa de germinação de
sementes de jaboticabeira (ROSSA et al., 2010). Estes fatores juntos mostram que a
germinação e consequente formação de plântulas em um ambiente natural (floresta)
podem ser facilitadas por meio do consumo e dispersão das sementes. A dispersão
promoveria a retirada da polpa, a provável deposição em local adequado para
germinação e evitaria a exposição da semente em ambiente natural por muitos dias,
auxiliando na formação do banco de plântulas.
A função O’ring univariada, mostrou que a maioria das populações
apresentou distribuição aleatória nos diferentes estádios ontogenéticos. Esperava-se
um padrão agregado nas menores classes de tamanho, o que ocorreu apenas nas
populações de Clevelândia e Chopinzinho II, a uma distância pequena (2 e 8 m
respectivamente). Padrões agregados podem ser reflexos do tipo de dispersão das
sementes (barocoria ou zoocoria) (SEIDLER; PLOTKIN, 2006; ABREU et al., 2014),
da capacidade de dispersão ou de fatores como disponibilidade de recursos e
condições microclimáticas (CAPRETZ et al., 2012). Em alguns locais de estudo
(Quadro 1), há relatos da ocorrência de cutias, serelepes e quatis, os quais se
alimentam dos frutos e podem ser dispersores. O comportamento destes animais
consiste em estocar as sementes próximas à planta matriz, contribuindo para a
agregação das plântulas (OLMOS et al., 1997). Além da zoocoria, a jaboticabeira
também é dispersa por meio da barocoria. Comumente, encontram-se padrões
agregados em classes menores e padrões aleatórios em classes maiores

42(CAPRETZ et al., 2012). Em Clevelândia, os estádios intermediários (imaturos e
virgens), também apresentaram agregação, o que pode ser reflexo da falta de
competição entre indivíduos ou ausência de predação de plântulas, contrariando o
modelo de mortalidade proposto por Janzel & Connell (STERNER et al., 1986).
Um padrão aleatório representa a chance de todos os organismos
ocuparem qualquer ponto no espaço (SILVA et al., 2009). O que deve ser levado em
consideração é o baixo número de indivíduos de algumas populações,
principalmente nos estádios ontogenéticos que representam a regeneração
(plântulas, juvenis e imaturos). Este fator dificulta a geração de um padrão por meio
da estatística O’Ring.
A análise de dependência espacial mostrou que não há relação entre
os estádios ontogenéticos nas populações estudadas, o que significa que ocorrem
de forma independente. Padrões de dependência espacial podem ser explicados
pela preferência da espécie por um nicho específico, por exemplo, associações
negativas (repulsão), mostram a procura por habitats diferentes, que proporcionarão
um melhor desenvolvimento do indivíduo (ZHANG et al., 2010). Neste caso, a
espécie não necessita de um nicho específico.
Acredita-se que os resultados encontrados são oriundos de três fatores
principais: histórico de manejo das áreas; falta de dispersores devido à antropização
dos locais; semente recalcitrante. Os resultados encontrados mostram um panorama
da situação dos remanescentes e os possíveis causadores desses fatores,
entretanto, mais estudos deverão ser realizados, principalmente, o que tange a
quantificação da produção e exploração dos frutos e a quantificação da intensidade
de pastejo nos locais.

436 CONCLUSÕES
A ação antrópica existente em Chopinzinho I e II e Coronel Vivida
parece ser um dos principais empecilhos na regeneração de indivíduos de
jaboticabeira e consequente recrutamento em classes de tamanho maiores.
A espécie pode ser classificada em cinco estádios ontogenéticos:
plântula, juvenil, imaturo, virgem e adulto.
A distribuição da maioria dos indivíduos se mostrou aleatória.
Não houve dependência espacial entre pares de classes ontogenéticas
estudadas.

447 CONSIDERAÇÕES FINAIS
A estrutura populacional e a distribuição espacial são ferramentas
muito úteis, pois, nos dão um diagnóstico da situação de determinada espécie ou
comunidade. Podemos inferir sobre quais são as melhores metodologias de
conservação ou definir estratégias que visem à conservação por meio do uso dos
produtos que a floresta pode nos oferecer. No caso das jaboticabeiras, é nítida a
importância econômica e social que a espécie exerce na região sudoeste do Paraná.
Portanto, ações que englobem a comercialização e a conservação devem ser
traçadas em conjunto.
Os resultados desse trabalho levantaram hipóteses e com isso a
possibilidade de novos estudos para afirmar com mais precisão os mecanismos
responsáveis pelos resultados. Estudos estão sendo planejados pelo Grupo UTF-
Nativas, tais como: estudo da dinâmica populacional de jaboticabeira, estudos da
fenologia em diferentes condições climáticas e estudos para determinação da idade
das populações. Acreditamos que esses resultados, aliados a estudos paralelos de
fluxo gênico e comercialização dos frutos, podem nos guiar a uma metodologia de
conservação aliada ao uso das populações.
De uma maneira geral, a dispersão por animais é muito importante
para o estabelecimento de jaboticabeiras. Seria muito importante a conservação dos
remanescentes para atração desses animais, bem como, a formação de corredores
ecológicos. Entretanto, essa pode ser uma realidade distante para nossa região, a
qual, os remanescentes se encontram em maioria isolados e circundados por
lavouras ou criação de gados. Nos locais com menor regeneração e falta de classes
intermediárias entre juvenis e adultos, sugere-se um plantio de enriquecimento.
Neste caso, a coleta de frutos e formação de muda em viveiro e posterior plantio,
poderia ser uma opção mais rápida de intervenção visando à conservação.

45REFERÊNCIAS
ABREU, Jadson C.; GUEDES, Marcelino C.; GUEDES, Ana Claudia L.; BATISTA,Edmilson M. Estrutura e distribuição espacial de andirobeiras (Carapa spp.) emfloresta de várzea do estuário amazônico. Ciência Florestal, v. 24, n. 4, p. 1009 –1019, 2014.
ALVARES, Clayton A.; STAPE, José L.; SENTELHAS, Paulo C.; GONÇALVES, JoséL. M.; SPAROVEK, Gerd. Köppen’s climate classification map for Brazil.Meteorologische Zeitschrift, v. 22, n. 6, p. 711 – 728, 2013.
ARAUJO, Maristela M.; LONGHI, Sólon J.; BARROS, Paulo L. C.; BRENA, Doádi A.Caracterização da chuva de sementes, banco de sementes do solo e banco deplântulas em Floresta Estacional Decidual ripária Cachoeira do Sul, RS, Brasil.Scientia Forestalis, n. 66, p. 128 – 141, 2004.
ARMESTO, J. J.; MITCHELL, J. D.; VILLAGRAN, C. A. comparison of spatialpatterns of trees in some tropical and temperate forests. Biotropica, v. 18, n. 1, p. 1– 11, 1986.
BAROT, Sebastien; GIGNOUX, Jacques; MENAUT, Jean Claude. Demography of asavanna palm tree: predictions from comprehensive spatial pattern analyses.Ecology, v. 80, n. 6, p. 1987 – 2005, 1999.
BECKMAN, Noelle G.; ROGERS, Haldre S. Consequences of seed dispersal forplant recruitment in tropical forests: interactions within the seed scape. Biotropica, v.45, n. 6, p. 666 – 681, 2013.
BERNACCI, Luís C.; MARTINS, Fernando R.; SANTOS, Flávio A. M. Estrutura deestádios ontogenéticos em população nativa de palmeira Syagrus romanzoffiana(Cham.) Glassman (Arecaceae). Acta Botanica Brasilica, v. 22, n. 1, p. 119 – 130,2008.
BESAG, Julian E. Comments on Ripley's paper. Journal of the Royal StatisticalSociety, vol. B 39, n. 2, p. 193 – 195, 1977.
BULMER, Michael G. Principles of Statistics. New York: Dover Publications, 1979.252 p.
CAPRETZ, Robson L.; BATISTA, João L. F.; SOTOMAYOR, Jaime F. M.; CUNHA,Camila R.; NICOLETTI, Marcos F.; RODRIGUES, Ricardo R. Padrão espacial dequatro formações florestais do estado de São Paulo, através da função K de Ripley.Ciência Florestal, v. 22, n. 3, p. 551 – 565, 2012.

46CASTELLA, Paulo R.; BRITEZ, Ricardo M. A. (Orgs.). Floresta com Araucária noParaná: conservação e diagnóstico dos remanescentes florestais. Brasília: Ministériodo Meio Ambiente, 2004. 233 p.
CAUGHLIN, Trevor T.; FERGUSON, Jake M.; LICHSTEIN, Jeremy W.; ZUIDEMA,Pieter A.; BUNYAVEJCHEWIN, Sarayudh; LEVEY, D.J. Loss of animal seeddispersal increases extinction risk in a tropical tree species due to pervasive negativedensity dependence across life stages. Proceeding B, v. 282, p. 2014 – 2095, 2014.
CLARK, Deborah A.; CLARK, David B. Análisis de la regeneración de árboles deldosel en bosque muy húmedo tropical: aspectos teóricos y práticos. Revista deBiologia Tropical, v. 35, n. 1, p. 41 – 54, 1987.
COLUSSI, Juliano; PRESTES, Nêmora P. Frugivoria realizada por aves emMyrciaria trunciflora (Mart) O. Berg (Myrtaceae), Eugenia uniflora L. (Myrtaceae) eIlex paraguariensis St. Hil. no norte do estado do Rio Grande do Sul. RevistaBrasileira de Ornitologia, v. 19, n. 1, p. 48 – 55, 2011.
CONDIT, Richard; ASHTON, Peter S.; BAKER, Patrick; BUNYAVEJCHEWIN,Sarayudh; GUNATILLEKE, Savithri; GUNATILLIKE, Nimal; HUBBELL, Stephen P.;FOSTER, Robin B.; ITOH, Akira; LaFRANKIE, James V.; LEE, Hua S.; LOSOS,Elizabeth; MANOKARAN, N.; SUKUMAR, R.; YAMAKURA, Takuo. Spatial patternsin the distribution of tropical tree species. Science, v. 288, p. 1414 – 1418, 2000.
CRAMER, Duncan. Basic Statistics for Social Research: Step-by-stepCalculations and Computer Techniques Using Minitab. New York: Routledge, 1997.401 p.
DALE, M.R.T; POWELL, R.D. A new method for characterizing point pattern in plantecology. Journal of Vegetation Science, v. 12, p. 597 – 608, 2001.
DALMASO, Cilmar A.; INOUE, Mario T.; OLIVEIRA FILHO, Paulo C.; MARCELINO,Vânia R. Padrões espaciais na regeneração de Ocotea odorifera na floresta nacionalde Irati, PR. Floresta, v. 43, n. 2, p. 301 – 312, 2013.
DANNER, Moeses A.; CITADIN, Idemir; SASSO, Simone A. Z.; TOMAZONI, Julio C.Diagnóstico ecogeográfico da ocorrência de jabuticabeiras nativas no Sudoeste doParaná. Revista Brasileira de Fruticultura, v. 32, n. 3, p. 746 – 753, 2010.
DANNER, Moeses A.; CITADIN, Idemir; SASSO, Simone A. Z.; SCARIOT, Silvia;BENIN, Giovane. Genetic dissimilarity among jabuticaba trees native toSouthwestern Paraná, Brazil. Revista Brasileira de Fruticultura, v. 33, n. 2, p. 517– 525, 2011a.
DANNER, Moses A.; CITADIN, Idemir; SASSO, Simone A. Z; AMBROSIO, Rodrigo;

47WAGNER JUNIOR, Américo. Armazenamento a vácuo prolonga viabilidade desementes de jabuticabeira. Revista Brasileira de Fruticultura, v. 33, n. 1, p. 246 –252, 2011b.
EISENLOHR, Pedro V.; OLIVEIRA-FILHO, Ary T. Tree species composition in areasof Atlantic Forest in southeastern Brazil is consistent with a new system forclassifying the vegetation of South America. Acta Botanica Brasilica, v. 28, n. 2, p.227 – 233, 2014.
FAHRIG, Lenore. Effects of habitat fragmentation on biodiversity. Annual Review ofEcology, Evolution and Systematics, v. 34, p. 487 – 515, 2003.
GATSUK, L.E.; SMIRNOVA, O.V.; VORONTZOVA, L.I.; ZAUGOLNOVA, L.B.;ZHUKOVA, L.A. Age states of plants of various growth forms: a review. Journal ofEcology, v. 68, p. 675 – 696, 1980.
GETZIN, Stephan; DEAN, Charmaine; FANGLIANG, He; TROFYNOW, John A.;WIEGAND, Kerstin; WIEGAND, Thorsten. Spatial patterns and competition of treespecies in a Douglas-fir chronosequence on Vancouver Island. Ecography, v. 29, p.671 – 682, 2006.
GIEHL, Eduardo L. H.; BUDKE, Jean C.; ATHAYDE, Eduardo A. Distribuiçãoespacial de espécies arbóreas em uma floresta estacional em Santa Maria, sul doBrasil. Pesquisas Botânicas, n. 58, p. 215 – 226, 2007.
GOMES, Juliano P.; FERREIRA, Paula I.; DACOREGIO, Helen M.; FRANÇA,Caroline S. S.; HIGUCHI, Pedro; MANTOVANI, Adelar; BORTULUZZI, Roseli L. C.Demographics and spatial pattern on three populations of Myrtaceae in theOmbrophilous Mixed Forest. Ciência Rural, v. 46, n. 9, p. 1579 – 1584, 2016.
GRAU, Hector R. Regeneration patterns of Cedrela lilloi (Meliaceae) in northwesternArgentina subtropical montane forests. Journal of Tropical Ecology, v. 16, p. 227 –242, 2000.
GRESSLER, Eliana; PIZO, Marco A.; MORELATTO, Patrícia L. C. Polinização edispersão de sementes em Myrtaceae do Brasil. Revista Brasileira de Botânica, v.29, n. 4, p. 509 – 530, 2006.
GUARINO, Ernestino S. G.; GESSNER, Camila M.; WADT, Lúcia H. O.; FONSECA,Fernanda L.; RAPOSO, Andréa. Estrutura etária e espacial de uma populaçãonatural de Carapa guianensis Aubl. (Meliaceae) na Amazônia Sul Ocidental.Scientia Forestalis, v. 42, n. 101, p. 91 – 99, 2014.
GUERRA, Miguel P.; SILVEIRA, Vanildo; REIS, Maurício S.; SCHNEIDER, Lineu.Exploração, manejo e conservação da araucária (Araucaria angustifolia). In:

48SIMÕES, Luciana L.; LINO, Clayton F. (Ed.). Sustentável Mata Atlântica: aexploração de seus recursos florestais. São Paulo: Editora SENAC, 2002. p. 85 –101.
HAASE, Peter. Spatial pattern analysis in ecology based on Ripley´s K-function:Introduction and methods of edge correction. Journal of Vegetation Science, v. 6,p. 575 – 582, 1995.
HALL, Pamela; BAWA, Kamaljit. Methods to assess the impact of extraction of non-timber tropical forest products on plant populations. Economic Botany, v. 47, n. 3, p.234 –247, 1993.
HIGUCHI, Pedro; SILVA, Ana C.; BUZZI JUNIOR, Fernando; NEGRINII, Marcelo;FERREIRA, Tiago S.; SOUZA, Sheila T.; SANTOS, Kristiana F.; VEFAGO, ManoelaB. Fatores determinantes da regeneração natural em um fragmento de floresta comaraucária no planalto catarinense. Scientia Forestalis, v. 43, n. 106, p. 251 – 259,2015.
JANZEN, Daniel H. Ecologia vegetal nos trópicos. São Paulo: EPU: Ed. daUniversidade de São Paulo. 1980. p. 55 – 59.
JANZEN, Daniel H. Herbivores and the number of tree species in tropical forests.The American Naturalist, v. 104, n. 940, p. 501 – 528, 1970.
JOANES, D.N.; GILL, C.A. Comparing measures of sample skewness and kurtosis.The Statistician, v. 47, n. 1, p. 183 – 189, 1998.
JORDANO, Pedro, GALETTI, Mauro; PIZO, Marco A; SILVA, Wesley R. LigandoFrugivoria e Dispersão de sementes à biologia da conservação. In: DUARTE, CarlosF.; BERGALLO, Helena G.; DOS SANTOS, Maria A. (Eds.). Biologia daConservação: essências. São Paulo: Editorial Rima. 2006. p. 1 – 26.
LAURENCE, Willian F; VASCONCELOS, Heraldo L. Consequências ecológicas dafragmentação florestal na Amazônia. Oecologia Brasiliensis, v. 13, n. 3, p. 434 –451, 2009.
LAW, Richard; ILLIAN, Janine; BURSLEM, David F. R. P.; GRATZER, Georg;GUNATILLEKE, C.V.S; GUNATILLEKE, I.A.U.N. Ecological information from spatialpatterns of plants: insights from point process theory. Journal of Ecology, v. 97, p.616 – 628, 2009.
LEAK, Willian B. The J-shaped probability distribution. Forest Science, v. 11, n. 4, p.405 – 409, 1965.
LEXEROD, Niels; EID, Tron. Recruitment models for Norway spruce, Scots pine,

49birch and other broadleaves in young growth forests in Norway. Silva Fennica, v. 39,n. 3, p. 391 – 406, 2005.
MAACK, Reinhard. Geografia física do estado do Paraná. 2 ed. Rio de Janeiro:José Olympio, 1981.
MALERBO-SOUZA, Darclet T.; NOGUEIRA-COUTO, Regina H.; TOLEDO, VagnerA. A. Abelhas visitantes nas flores da jabuticabeira (Myrciaria cauliflora Berg) eprodução de frutos. Acta Scientiarum – Animal Sciences, v. 26, n. 1, p. 1 – 4,2004.
MATTEUCI, Silvia D.; COLMA, Aída Metodología para el estudio de lavegetacion. Washington: OEA, 1982. 168 p.
MATTOS, João R. Frutíferas nativas do Brasil: jaboticabeiras. Porto Alegre: Nobel,1983. 92 p.
McINTIRE, Eliot J. B.; FAJARDO, Alex. Beyond description: the active and effectiveway to infer processes from spatial patterns. Ecology, v. 90, p. 46 – 56, 2009.
MIRANDA-MELO, Aneliza A.; MARTINS, Fernando R.; SANTOS, Flávio A. M. S.Estrutura populacional de Xylopia aromatica (Lam.) Mart. e de Roupala montanaAubl. em fragmentos de cerrado no Estado de São Paulo. Revista Brasileira deBotânica, v. 30, n. 3, p. 50 – 507, 2007.
MIRANDA, Alejandro; HERNANDEZ, Jaime H.; BUSTAMANTE, Ramiro.; DÍAZ,Matias E.; GONZALEZ, Alberto L.; ALTAMIRANO, Adison. Regeneración natural ypatrones de distribución espacial de la palma chilena Jubaea chilensis (Molina)Baillon en los bosques mediterrâneos de Chile central. Gayanna Botanica, v. 73, n.1, p. 54 – 63, 2016.
MOTTA-JUNIOR, José C.; LOMBARDI, Julio A.; TALAMONI, Sônia A. Notes oncrab-eating fox (Dusicyon thous) seed dispersal and food habits in southeasternBrazil. Mammalia, v. 58, n. 1, p. 156 – 159, 1994.
NAZARENO, Alison G.; REIS, Maurício S. At risk of population decline? Anecological and genetic approach to the threatened palm species Butia eriospatha(Arecaceae) of Southern Brazil. Journal of Heredity, v. 105, n. 1, p. 120 – 129,2014.
OLIVEIRA-FILHO, Ary T. Classificação das fitofisionomias da América do Sulcisadina tropical e subtropical: proposta de um novo sistema – prático e flexível – ouuma injeção a mais caos? Rodriguésia, v. 60, n. 2, p. 237 – 258, 2009.
OLMOS, Fabio.; PARDINI, Renata; BOULHOSA, Ricardo L. P.; BÜRGL, Roberto;

50MORSELLO, Carla. Do tapirs steal food from palm seed predators or give them alift? Biotropica, v. 31, n. 2, p. 375 – 379, 1999.
PALUDO, Giovani F.; MANTOVANI, Adelar; KLAUBERG, Carine; REIS, Maurício S.Estrutura demográfica e padrão espacial de uma população natural de Araucariaangustifolia (Bertol.) Kuntze (Araucariaceae), na reserva genética florestal deCaçador, estado de Santa Catarina. Revista Árvore, v. 33, n. 6, p. 1109 – 1121,2009.
PALUDO, Giovani F.; MANTOVANI, Adelar; REIS, Maurício S. Regeneração de umapopulação natural de Araucaria angustifolia (Araucariaceae). Revista Árvore, v. 35,n. 5, p. 1107 – 1119, 2011.
PAMMENTER, Norman W.; BERJAK, Patrícia. Aspects of recalcitrant seedphysiology. Revista Brasileira de Fisiologia Vegetal, v. 12, p. 56 – 69, 2000.
PERRY, J.N.; LIEBHOLD, A.M.; ROSENBERG, M.S.; DUNGAN, J.; MIRITI, M.;JAKOMULSKA, A.; CITRON-POUSTY, S. Illustrations and guidelines for selectingstatistical methods for quantifying spatial pattern in ecological data. Ecography, v.25, p. 578 – 600, 2002.
PORTELA, Rita C. Q.; SANTOS, Flávio A. M. Caracterização dos estádiosontogenéticos de três espécies de palmeiras: uma proposta de padronização paraestudos de dinâmica populacional. Revista Brasileira de Botânica, v. 34, n. 4, p.523 – 535, 2011.
RIPLEY, Brian D. Modeling spatial patterns. Journal of the Royal StatisticalSociety, v. 39, n. 2, p. 172 – 212, 1977.
ROSSA, Uberson B.; TRICHES, Gilmar P.; GROSSI, Fernando; NOGUEIRA,Antônio C.; REISSMANN, Carlos B.; RAMOS, Michelle R. Germinação de sementese qualidade de mudas de Plinia trunciflora (jabuticabeira) em função de diferentestratamentos pré-germinativos. Floresta, v. 40, n. 2, p. 371 – 378, 2010.
ROSSETO, Vanessa; CARNEIRO, Magda S.; RAMOS, Flávio N.; SANTOS, FlávioA. M. Ontogeny, allometry, and architecture of Psychotria tenuinervis (Rubiaceae).Acta Botanica Brasilica, v. 27, n. 4, p. 730 – 736, 2013.
ROSSI, Luiz M. B.; KOEHLER, Henrique S.; ARCE, Júlio E.; SANQUETTA, CarlosR. Modelagem de recrutamento em florestas. Florestas, v. 37, n. 3, p. 453 – 467,2007.
SAMPAIO; Maurício B.; GUARINO, Ernestino S. G. Efeitos do pastoreio de bovinosna estrutura populacional de plantas em fragmentos de Floresta Ombrófila Mista.Revista Árvore, v. 31, n. 6, p. 1035 – 1046, 2007.

51SANQUETTA, Carlos R.; FERNANDES, Lucila A. V.; MIRANDA, Dirceu L. C.;MOGNON, Francelo. Inventário de plantas fornecedoras de produtos nãomadeireiros da floresta ombrófila mista no estado do Paraná. Scientia Agraria, v.11, n. 5, p. 359 – 369, 2010.
SCALON, João D.; OLIVEIRA, Carlos A. P.; MELLO, José M. Análise espacial de umfragmento florestal baseado no mosaico de dirichlet. Revista Árvore, v. 36, n. 4, 733– 740, 2012.
SEIDLER, Tristam G.; PLOTKIN, Joshua B. Seed dispersal and spatial pattern intropical trees. Plos Biology, v. 4, n. 11, p. 2132 – 2137, 2006.
SILVA, Kátia E.; MARTINS, Sebastião V.; SANTOS, Nerilson T.; RIBEIRO, Carlos A.A. S. Padrões espaciais de espécies arbóreas tropicais. In: MARTINS, Sebastião V.(Ed). Ecologia de florestas tropicais do Brasil. Viçosa: UFV. 2009. p. 216 – 244.
SILVEIRA, Andréa P.; MARTINS, Fernando R; ARAÚJO, Francisca S. Are treeontogenetic structure and allometric relationship independent of vegetation formationtype? A case study with Cordia oncocalyx in the Brazilian caatinga. ActaOecologica, v. 43, p. 126 – 133, 2012.
SMIRNOVA, O.V.; PALENOVA, M.M.; KOMAROV, A.S. Ontogeny of different lifeforms of plants and specific features of age and spatial structure of their populations.Russian Journal of Development Biology, v. 33, n. 1, p. 1 – 10, 2002.
SOUZA, Alexandre F.; MARTINS, Fernando R. Population structure and dynamics ofa neotropical palm in fire-impacted fragments of the Brazilian Atlantic forest.Biodiversity Conservations, v. 13, p. 1611 – 1632, 2004.
SOUZA, Alexandre F.; MARTINS, Fernando R.; MATOS, Dalva M. S. Detectingontogenetic stages of the palm Attalea humilis in fragments of the Brazilian Atlanticforest. Canadian Journal of Botany, v. 78, p. 1227 – 1237, 2000.
STERNER, Robert W.; RIBIC, Christine A.; SCHATZ, George E. Testing for lifehistorical changes in spatial patterns of four tropical trees species. Journal ofEcology, v. 74, p. 621 – 633, 1986.
VALIO; Ivany F. M.; FERREIRA, Zenith L. Germination of seeds of Myrciariacauliflora (Mart) Berg. (Myrthaceae). Revista Brasileira de Fisiologia, v. 4, n. 2, p.95 – 98, 1992.
VELOSO, Henrique P.; RANGEL FILHO, Antonio L. R.; LIMA, Jorge C. A.Classificação da vegetação brasileira, adaptada a um sistema universal.Instituto Brasileiro de Geografia e Estatística: Rio de Janeiro, 1991.

52WIEGAND, Throsten; HE, Fangliang; HUBBELL, Stephen P. A systematiccomparison of summary characteristics for quantifying point patterns in ecology.Ecography, v. 36, p. 92 – 103, 2013.
WIEGAND, Throsten; MOLONEY, Kirk A. Rings, circles and null-modell for pointpattern analysis in ecology. Oikos, v. 104, n. 209 – 229, 2004.
WIEGAND, Throsten; MOLONEY, Kirk A. Handbook of Spatial Point-PatternAnalysis in Ecology. Boca Raton: Chapman and Hall/CRC Press, 2014. 550 p.
ZHANG, Jiachen; ZHANG, Yuqing; FAN, Dongqing; QIN, Shugao; JIA, Xin; WU, Bin;CHEN, Dong; GAO, Hao; ZHU, Linfeng. Effects of sand dune stabilization on thespatial pattern of Artemisia ordosica population in mu us desert, northwest China.PlosOne, v. 10, n. 6, p. e0129728, 2015.
ZHANG, Zhong-Hua, HU, Gang, ZHU, Jie-Dong, LUO, Dong-Hui, NI, Jian. Spatialpatterns and interspecific associations of dominant tree species in two old-growthkarst forests, SW China. Ecological Research, v. 25 p. 1151 – 1160, 2010.
ZHU, Yan; GETZIN, Stephan; WIEGAND, Thorsten; REN, Haibao.; MA, Keping. Therelative importance of Janzen-Connell effects in influencing the spatial patterns at theGutianshan Subtropical Forest. PlosOne, v. 8, n. 9, p. 1 – 8, 2013.
ZUCHIWSCHI, Elaine; FANTINI, Alfredo C.; ALVES, Antonio C.; PERONI, Nivaldo.Limitações ao uso de espécies florestais nativas pode contribuir com a erosão doconhecimento ecológico tradicional e local de agricultores familiares. Acta BotanicaBrasilica, v. 24, n. 1, p. 270 – 22, 2010.
ZUIDEMA, Pieter A.; BOOT, René G. A. Demography of the Brazil nut tree(Bertholletia excelsa) in the Bolivian Amazon: impact of seed extraction onrecruitment and population dynamics. Journal of Tropical Ecology, v. 18, p. 1 – 31,2002.





























