Embed Size (px)
Citation preview

CA
DHp
α2
α1
α2*
α1*
CA*
Membrana
90º
CA
DHp
α2
α1 nucleótido
CA*
α2*
α1* nucleótido
Patricia Casino y Alberto Marina
Departamento de Genómica y Proteómica, Laboratorio de Cristalografía de Macromoléculas Instituto de Biomedicina de Valencia (CSIC), Ciberer. ([email protected])
Histidina quinasa: Una proteína que se autofosforila, fosfotransfiere y defosforila
INTRODUCCIÓN Los sistemas de dos componentes son el mecanismo principal de transducción de señal en microorganismos. Un sistema paradigmático se compone de dos proteínas: una histidina quinasa homodimérica (HK) y un regulador de la respuesta (RR). El mecanismo de transducción de señal implica al menos tres reacciones (ver esquema): 1) Autofosforilación: el estímulo reconocido por el dominio extracelular sensor de la HK, modula la trans-fosforilación en un residuo de His conservado en el dominio de dimerización (DHp) de una subunidad por parte del dominio de unión de ATP (CA) de la otra subunidad en el dímero. 2) Fosfotransferencia: este grupo fosforilo es transferido desde la His a un residuo de Asp del dominio aceptor del correspondiente RR. 3) Defosforilación: el RR pierde el grupo fosforilo espontáneamente o mediado por la HK. El estado de fosforilación del dominio aceptor en el RR regula la actividad de su dominio efector, que es habitualmente un factor de transcripción. Con la intención de conocer el mecanismo de estas reacciones a nivel molecular hemos resuelto la estructura tridimensional del complejo formado por el dominio citoplasmático completo de la HK HK853 (HK853CP) de Thermotoga maritima en complejo su RR RR468. La comparación de dicha estructura con la de HK853CP en solitario ha permitido deducir valiosa información funcional y estructural. Por otra parte, la realización de ensayos bioquímicos nos han permitido dilucidar el mecanismo de la reacción de autofosforilación.
Dominio aceptor
Dominio efector
Dominio sensor
Dominio citoplasmático
Dominio trans-
membrana
Señal ATPHis
PAsp
P
HK RR
Autofosforilación
Fosfotransferencia
Defosforilación P
A
B
C
D
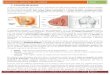
HK853CP es homodimérica y está formada por dos subunidades A (azul) y B (cian) RR468 contiene un sólo dominio correspondiente al aceptor. Moléculas C y D
CENTRO ACTIVO DEL COMPLEJO FORMADO ENTRE UNA HK Y SU RR Al observar la estructura del centro activo del complejo HK853CP:(RR468)2 advertimos dos cosas: 1) la presencia de un sulfato (que emula a un fosforilo) a una distancia de enlace entre la His260 catalítica de HK853 y el
Asp53 catalítico de RR468. Dichas distancias permiten proponer un mecanismo disociativo para la reacción de fosfotransferencia además de sugerir que la estructura del centro activo se corresponde con la reacción de defosforilación.
2) la presencia de una molécula de ADP en el sitio de unión de nucleótido cercana a la His260 aceptora dentro de una misma subunidad, ha permitido proponer un mecanismo de cis-autofosforilación (En semitransparente se muestra el posible mecanismo de autofosforilación entre confórmero (II) de la His260 y el fosfato γ del ATP).
► HK853CP
► HK853CP:RR468
► (HK853CP)2
►
HK853CP~P
►
HK853CP:RR468~P
►
(HK853CP)2~P
1’ 30’ 1’ 30’ 1’ 30’ K105A RRwt D53A
HK
HK +
DMS
HK+RR +
DMS SDS-PAGE
+ [γ-32P] ATP SDS-PAGE
EN HK853CP, LA CONFORMACIÓN DE AUTOFORILACIÓN Y DE FOSFOTRANSFERENCIA SE ENCUENTRAN MUY PRÓXIMAS
Ensayos de entrecruzamiento entre HK853CP y RR468 tanto silvestre como mutante (D53A y K105A) utilizando el entrecruzador DMS, han demostrado que el complejo HK853CP:RR468-D53A puede autofosforilarse estando éste entrecruzado. También prueba que el complejo HK853CP:RR468 silvestre puede fosfotransferir y defosforilar estando entrecruzado. Esto prueba que las conformaciones de fosfotransferencia y de defosforilación observadas en la estructura del complejo están muy cercanas a la de cis-autofosforilación que fue deducida de esta misma estructura.
MOVIMIENTOS QUE PERMITEN ADQUIRIR LOS DIFERENTES CAMBIOS CONFORMACIONALES
La superposición de las estructuras de HK853CP en solitario (morado) y HK853CP en complejo (azul) nos permite observar cambios conformacionales en los dos dominios citoplasmáticos DHp (haz de cuatro hélices formado por α1 (contiene la His260 fosforilable) y α2 que dimerizan α1* y α2*) y CA (dominio globular unido al DHp por α2 y que une el nucleótido) de cada subunidad. Estos cambios debe ser similares a los inducidos por la señal a la hora de controlar las diferentes reacciones del sistema.
La conexión entre las porciones citoplasmática y extracelular (donde la señal llega) se realiza por la hélice α1 del DHp. Al comparar ambas estructuras de HK853CP observamos que la parte de α1 más cercana a la membrana rota ~20 º obligando a rotar a α2 ~10 º a la vez que ambas hélices se desplazan ~4 Å. Dichos movimientos van disminuyendo hasta casi anularse en la Pro265 de α1, residuo que actuaría como bisagra. El dominio CA se asienta en α1 y α2, por lo que dichos movimientos afectan a la superficie de interacción entre DHp y CA provocando una rotación de CA de ~37º acercando la His aceptora a dicho dominio. De esta forma, la disposición relativa de los dominios DHp y CA, modulada por la señal, controlaría la autofosforilación y la fosfotransferencia.
HK853~P
90º
HK853 + RR~P HK853 + RR~P
TM2 TM2*
α1 α1* α1 α1*
HK853 libre HK853 complejo
α2 α2*
LuxQ
HtrII
HAMP Rotación 26º
Rotación de TM2 de 20-30º
Dimerización
90º
Rotación α1 (20º) y CA (~37º) CA CA CA CA 90º
37º
α2 α5
α4
α3
CA
DHp
P265
α1
α2
α1 α1*
α2*
Superposición de HK853CP en solitario y HK853CP en complejo
Visión de ambas proteínas superpuestas desde la membrana hacia el citoplasma
A
B
C Flechas blancas son indicadoras de desplazamientos y las flechas en curva indicadoras de rotación
A) Visión de los movimientos de las hélices que conforman el DHp (α1 y α2 de una subunidad y α1* y α2* de la subunidad contraria) en ambas estructuras HK853CP
B) La rotación de α1 se anula a la altura de la Pro265. Los movimientos de α1 y α2 provocan una rotación del CA (se muestran las hélices α3, α4 y α5) de ~37 º.
C) Esquema superior mostrando el modelo de transducción de señal desde la porción extracelular hacia la intracelular. D) Esquema circular mostrando el cambio conformacional adoptado por HK853CP para unir RR468 (amarillo) inducido por la transducción de la señal.
Porción extracelular
Región transmembrana
Porción intracelular
D
HATP
HATP
HATP
HATP
HATPHATP
HATP
HATP
HATPHATP
HATP
HATP
1
H HATP ATP
H
2
HATP
H
7
ATP ATP
H
8
ATP
H
3
HATP
H
9
ATP
4
HATP ATP
5
HATP
6
HATP
CIS y TRANS
TRANS
CIS
1
H HATP ATP
H HATP ATP
H
2
HATP
H
7
ATP ATP
H
8
ATP
H
3
HATP
H
9
ATP
4
HATP ATP
5
HATP
6
HATP
CIS y TRANS
TRANS
CIS
1 2 3 4 5 6 7 8 9 FL CP
FL
CP
FL/FL Heterodímero FL/CP
CP/CP
► ►
►
►
►
►
►
►
►
►
FL
CP
FL/FL
CP/CP
A) Nativo-PAGE
B) Nativo-PAGE + [γ-32P] ATP
C) SDS-PAGE + [γ-32P] ATP
D) SDS-PAGE + [γ-32P] ATP
De la banda correspondiente al
heterodímero
FL/FL CP/CP
FL: HK853 completa (Dominio periplásmico + Dominio citoplasmático completo) CD: Dominio citoplasmático completo de HK853 Cómo se puede observar en la columna 8, el heterodímero formado en A) aparece fosforilado en B) y tanto en el gel SDS C) como en el D) sólo aparece una banda fosforilada asignable a la especie FL, que es la única posible fosforilable según se observa en el esquema E). Esto prueba la fosforilación en CIS.
LA REACCIÓN DE AUTOFOSFORILACIÓN EN HK853 SE REALIZA DENTRO DE UNA MISMA SUBUNIDAD (MODO CIS) AL CONTRARIO QUE EN LAS HKs MODELO NtrB Y EnvZ,
DONDE SE REALIZA ENTRE SUBUNIDADES (MODO TRANS) Ensayos bioquímicos realizados con varias combinaciones de HK853 completa (HK853FL) y HK853CP tanto silvestres como mutantes (sin His aceptora o sin capacidad de unir ATP) de forma que formáramos heterodímeros FL/CP, han demostrado que la reacción de autofosforilación en esta HK se realiza de modo CIS, dentro de una misma subunidad, apoyando los datos estructurales.
Financiado por las ayudas BIO2005-00153 del Ministerio de Educación y Ciencia. Patricia Casino es contratada Ciberer.
Controles
Heterodímeros
Heterodímeros
Heterodímeros
Fosforilación?
Fosforilación?
Fosforilación?
E) Esquema mostrando los diferentes heterodímeros FL/CP resultado de mezclar HK853 bien FL o CP ya sean silvestres o mutantes. Los círculos con raya indican ausencia de la His aceptora o incapacidad de unión del ATP
Heterodímero FL/CP
FL
CP
E) Esquema gráfico
MODELO DE TRANSDUCCIÓN DE
SEÑAL
D53
D9
D10 T83
K105
H260
ATP
SO4
ATP-lid
6.0 Å
Mg
(II)
3.4 Å
2.4 Å D53
D9
D10
T83
K105
H260
SO4 Mg
1) El centro catalítico del complejo está formado por todos los residuos conservados del dominio aceptor del RR así como por la His aceptora de la HK. La posición adoptada por los residuos del RR (D9, D10, D53, T83 y K105) añadida a la presencia del sulfato y el Mg sugiere que estamos observando la conformación fosforilada de RR468.
2) Si modelamos dentro de una misma subunidad otra conformación para la His aceptora (II), así como el fosfato γ en el ADP (ambos en semitransparente), vemos que la distancia entre ambos es de 6.0 Å. Distancia que sugiere una autofosforilación dentro de una misma subunidad, es decir, en modo CIS.