Embed Size (px)
DESCRIPTION
Impactos do incremento de água salgada na lagoa de Piratininga
Citation preview

UNIVERSIDADE FEDERAL FLUMINENSE INSTITUTO DE BIOLOGIA PROGRAMA DE PÓS-GRADUAÇÃO EM BIOLOGIA MARINHA
ICTIO E CARCINOFAUNA DO SISTEMA LAGUNAR PIRATININGA -ITAIPU,
NITERÓI, RJ - BRASIL, ANTES E DEPOIS DA ABERTURA DO CANAL DO TIBAU
LUANA PRESTRELO PALMEIRA
Dissertação de Mestrado apresentado ao Programa de Pós-Graduação em Biologia Marinha do Departamento de Biologia Marinha, Instituto de Biologia, Universidade Federal Fluminense.
Orientador: Dr. Cassiano Monteiro Neto
Niterói Fevereiro de 2012
Biologia Marinha
Pós-Graduação - u
ff

II
Agradecimentos
Agradeço,
Ao professor e orientador Cassiano, por todo apoio e aprendizado que
me proporcionou durante esses cinco anos que estive no laboratório, sempre
presente e estimulando a busca pelo conhecimento, sendo fundamental no
meu crescimento e amadurecimento profissional.
Aos meus pais, Eleonora e Guido, pelo apoio e compreensão durante
todos esses anos. Ao Felipe, companheiro de tanto tempo, pela paciência,
carinho e apoio durante essa jornada. A toda minha família pelo apoio.
Ás minhas amigas/irmãs de longa data, Maria, Marina e Pati, que
sempre me apoiaram e me ajudaram a superar todas as adversidades da vida.
A todos os meus amigos, Tomás, Rafael, Victor, Juliana, Mayumi, Ju, JuR,
SuRzy, Renatinha, por facilitarem os momentos difíceis, ajudarem a relaxar e
estarem sempre prontos para descontrair.
A todo o pessoal do ECOPESCA por deixar o trabalho mais fácil e
divertido, em especial à Nathália e Amanda pela grande ajuda nas saídas de
campo, viabilizando este trabalho.
Aos membros da banca por terem aceitado meu convite para avaliar e
enriquecer meu trabalho.
Ao CNPQ pela bolsa concedida, o que possibilitou um melhor
cumprimento deste trabalho.

III
ÍNDICE Página
LISTA DE TABELAS.........................................................................................IV
LISTA DE FIGURAS .........................................................................................VI
RESUMO ........................................................................................................VIII
ABSTRACT.......................................................................................................IX
1. INTRODUÇÃO ...............................................................................................1
2. OBJETIVOS ...................................................................................................7
2.1 Objetivo Geral.......................................................................................7
2.2 Objetivos Específicos ...........................................................................7
3. MATERIAIS E MÉTODOS..............................................................................8
3.1 Área de Estudo .....................................................................................8
3.2 Programa Amostral.............................................................................11
3.3 Análise de Dados................................................................................13
4. RESULTADOS .............................................................................................18
5. DISCUSSÃO ................................................................................................47
6. CONCLUSÃO...............................................................................................62
7. PERSPECTIVAS.......................................................................................... 63
8. REFERÊNCIAS BIBLIOGRÁFICAS .............................................................64

IV
LISTA DE TABELAS
Tabelas Página
1: Lista de abundância total das espécies de peixes e crustáceos (*) das lagoas
de Itaipu e Piratininga, coletados em 2010-11, por época do ano, em
ordem decrescente de abundância total. Linhas em branco = zero. Em
destaque, a classificação das frequências de ocorrência relativas (FO %)
de cada espécie, por lagoa, época do ano e total..................................... 21
2: Lista da biomassa (kg) das espécies de peixes e crustáceos (*) das lagoas
de Itaipu e Piratininga, por época do ano e totais, em ordem decrescente
de biomassa total. Linhas em branco = zero. ........................................... 22
3: Parâmetros ambientais de cada lagoa, por ponto e época do ano e suas
médias (MD ± desvio padrão). .................................................................. 23
4: Tabela comparativa das espécies presentes nas campanhas de 2006 e
2010-11, por lagoa, época do ano e totais, em ordem decrescente de
abundância total, somados os dois períodos. Linhas em branco = zero. Em
destaque, a classificação das frequências de ocorrência relativas (FO %)
de cada espécie, por período amostral, lagoa, época do ano e total. * -
espécies exclusivas de 2006. ** - espécies exclusivas de 2010-11.......... 27
5: Tabela comparativa da biomassa (em quilogramas) das espécies presentes
nas campanhas de 2006 e 2010-11, por lagoa, época do ano e totais, em
ordem decrescente de biomassa total somados os dois períodos. Linhas
em branco = zero. * - espécies exclusivas de 2006. ** - espécies exclusivas
de 2010-11................................................................................................ 30

V
6: Resultado das análises de variância (ANOVA) realizadas para cada
parâmetros ambiental nas lagoas de Itaipu e Piratininga por período,
estação do ano e sua interação. ............................................................... 33
7: Resultado da PERMANOVA comparando a influência dos períodos
amostrais (2006 vs 2010-11) na estruturação das comunidades.............. 35
8: Resultado da análise discriminante da lagoa de Itaipu, com os valores de
significância, F-valor e a quantidade de explicação (λ) de cada espécie na
separação dos períodos amostrais. * - espécies exclusivas de 2006. ** -
espécies exclusivas de 2010-11. .............................................................. 45
9: Resultado da análise discriminante da lagoa de Piratininga, com os valores
de significância, F-valor e a quantidade de explicação (λ) de cada espécie
na separação dos períodos amostrais. * - espécies exclusivas de 2006. ** -
espécies exclusivas de 2010-11. .............................................................. 46

VI
LISTA DE FIGURAS
Figuras Página
1: Mapa das lagoas costeiras do norte fluminense ............................................ 4
2: Mapa da área de estudo, especificando os pontos amostrais (Piratininga: P1
– P6, Itaipu: I1 – I6) no sistema lagunar. .................................................... 8
3: Porcentagem de lama e areia no sedimento de cada lagoa, por área e época
do ano....................................................................................................... 24
4: Curva cumulativa de espécies em função do número de amostras para as
lagoas de Itaipu e de Piratininga nos dois períodos. As linhas pontilhadas
representam o intervalo de confiança (95% para cada curva). ................. 26
5: Analise de componentes principais (PCA) para a abundância das lagoas de
Itaipu e Piratininga, nos períodos amostrais de 2006 e 2010-11. As elipses
correspondem a 10% da concentração dos dados. .................................. 35
6: “Loadings” (± desvio padrão) das espécies responsáveis pela distribuição
das amostras no plano do PCA. As espécies cujos desvios padrões não
cruzam o eixo X podem ser consideradas significativas para a distribuição
encontrada no PCA................................................................................... 36
7: Analise de correspondência canônica (CCA) para a lagoa de Itaipu nos dois
períodos amostrais. Os vetores representam as variáveis ambientais que
apresentaram uma relação significativa com a distribuição dos dados. ... 38
8: Analise de correspondência canônica (CCA) para a lagoa de Piratininga para
os dois períodos amostrais. Os vetores representam as variáveis
ambientais que apresentaram uma relação significativa com a distribuição
dos dados. ................................................................................................ 40

VII
9: Diagrama t-valor biplot da lagoa de Piratininga dos fatores ambientais que
apresentaram relação significativa com as espécies. Os círculos
representam a área significativa do teste. As espécies que aparecem
dentro desta área apresentam uma relação significativa com o fator
ambiental.. ................................................................................................ 41
10: Esquema ilustrando a porcentagem de variação de cada parâmetro
ambiental testado na partição de variância da lagoa de Itaipu. (A) dados
ambientais, (B) correlação entre os dados ambientais e o tempo, (C)
tempo, (D) porcentagem de variação que não é explicada por nenhum dos
parâmetros medidos. ................................................................................ 43
11: Esquema ilustrando a porcentagem de variação de cada parâmetro
ambientail testado na partição de variância da lagoa de Piratininga. (A)
dados ambientais, (B) correlação entre os dados ambientais e o tempo, (C)
tempo, (D) porcentagem de variação que não é explicada por nenhum dos
parâmetros medidos. ................................................................................ 43
12: Esquema ilustrando a porcentagem de variação de cada parâmetro
ambiental testado na partição de variância da lagoa de Piratininga. (A)
dados ambientais, sem a salinidade, (B) correlação entre os dados
ambientais e a salinidade, (C) salinidade isoladamente, (D) porcentagem
de variação que não é explicada por nenhum dos parâmetros testados. . 44

VIII
RESUMO
Estudos comparativos “antes vs depois” possibilitam a avaliação de ações de
manejo em ecossistemas. Este estudo analisou as modificações provocadas
pela abertura do canal canal do Tibau no sistema lagunar Piratininga-Itaipu
(Niterói, RJ). Dados abióticos e bióticos (peixes e crustáceos decápodes)
coletados antes (2006) e depois (2010-11) da abertura do canal foram
utilizados para comparação. As espécies foram amostradas com uso de tarrafa
nas estações seca e chuvosa. Foi observado uma diminuição na riqueza,
abundância e biomassa de espécies, com 22 espécies ocorrendo
exclusivamente em 2006, 10 em 2010-11 e 22 ocorrendo nos dois períodos. As
análises de Componentes Principais (PCA), Multivariada de Variância por
Permutação (PERMANOVA) e de Similaridade (ANOSIM) indicaram a
separação espacial entre as lagoas, que foram consideradas sub-sistemas
independentes. As análises de correspondência canônica (CCA) e partição de
variância, para a lagoa de Piratininga, indicaram significativa influência da
salinidade na alteração da fauna da lagoa, caracterizada pela diminuição na
abundância da espécie dulcícula dominante (Oreochromis niloticus) e um
aumento na ocorrência e abundância de espécies marinhas (Diapterus
rhombeus e Eucinostomus spp.). As maiores mudanças na composição
faunística observadas em Itaipu, aparentemente não estão diretamente
relacionadas com a abertura do canal do TIbau. O aumento na eutrofização na
lagoa de Itaipu provavelmente influenciou as mudanças observadas na biota,
provocando o desaparecimento de espécies incluindo Farfantepenaeus spp. e
Centropomus parallelus. O presente estudo comparativo só foi possível devido
ao conhecimento das características da comunidade de peixes e crustáceos
antes da abertura do Canal do Tibau. Este aspecto reforça a necessidade do
monitoramento contínuo destes ambientes costeiros, frequentemente sujeitos a
intervenções de manejo. Assim, o monitoramento deveria ter uma
continuidade, visando avaliar a evolução e reestruturação do sistema lagunar
Piratininga-Itaipu (Niterói, RJ)
Palavras-chaves: Estudos comparativos antes–depois (ECAD); lagoas costeiras; comunidade de peixes; impacto antrópico.

IX
ABSTRACT
Before-after contol Impact (BACI) studies enable the evaluation of management
actions in ecosystems. This study analyzed changes induced by the opening of
the Tibau Channel, in the Piratinnga-Itaipu lagoon system (Niterói, RJ). Abiotic
and biotic (fishes and decapod crustaceans) data collected before (2006) and
after (2010-11) the channel opening were used for comparison. Species were
sampled using castnets during the dry and rainy seasons. A decrease in
species richness, abundance and biomass was observed, with 22 species
occurring exclusively in 2006, 10 in 2010-11 and 22 occurring in both periods.
Principal Components Analysis (PCA), Permutational Multivariate Analysis of
Variance (PERMANOVA) and Simirarity Analysis (ANOSIM) indicated a spacial
segregation between lagoons that were then considered as independent
subsystems. Canonical Correspondence and variance patitioning analyses for
Piratininga lagoon indicated that salinity had a significant influence in the faunal
change, characterized by the decrease in abundance of the dominant
freshwater species (Oreochromis niloticus) and increase in the occurrence and
abundance of marine species (Diapterus rhombeus e Eucinostomus spp.). The
greatest changes observed in the faunal composition in Itaipu lagoon,
apparently were not directly related with the opening of the Tibau channel. The
increased eutrophication of Itaipu lagoon probably influenced the observed
biological changes inducing the disappearance of species including
Farfantepenaeus spp. and Centropomus parallelus. The present comparative
study was only possible due to our knowledge about the fish and shelfish
community features prior to the opening of the Tibau Channel. This aspect
further enhances the necessity of continuous monitoring of these coastal
ecosystems frequently subjected to management interventions. Therefore, the
monitoring should be continued, to evaluate the evolution and restructuring of
the Piratininga-Itaipu lagoon system (Niterói, RJ).
Keywords: Before-after control impacts (BACI); coastal lagoons; fish
community; anthropic impact.

1
1. INTRODUÇÃO
A região leste Fluminense do estado do Rio de Janeiro apresenta uma
série de 15 lagunas entre os municípios de Niterói e Cabo Frio (BAPTISTA &
FERNANDES 2009). Nos últimos 50 anos, com a construção da ponte Costa e
Silva (ponte Rio-Niterói), houve um crescimento intenso na região, movido pelo
desenvolvimento turístico, que teve como consequência um adensamento
urbano sem o devido planejamento de ocupação. Isso provocou a degradação
ambiental de diversos ecossistemas, incluindo as lagoas costeiras da região
(WASSERMAN et al. 1999).
Nesses ambientes, a entrada de nutrientes de origem continental
naturalmente já excede a quantidade extravasada para o mar, principalmente
em lagoas sufocadas, promovendo um processo de eutrofização natural
(SUZUKI et al. 1998). Em lagoas isentas de impactos antrópicos, esse aporte
de nutrientes induz à elevada produtividade, sustentando cadeias tróficas
diversificadas e baseadas tanto em detritos (MORENO & CASTRO 1998) como
na produção primária (LIN et al. 1999). Já em ambientes que sofrem pelo
adensamento populacional, o aporte de águas continentais geralmente
transporta grandes quantidades de esgotos domésticos, aumentando a entrada
de nutrientes e matéria orgânica, provocando a deterioração na qualidade da
água e dos sedimentos nas lagoas (BAPTISTA & FERNANDES 2009). Além
disso, a promoção de aterros para ocupação das margens lagunares reduz o
espelho d’água podendo bloquear, muitas vezes, as conexões da lagoa com o
mar (SAAD et al. 2002).

2
Lagoas costeiras são importantes ecossistemas e precisam ser
preservados por serem locais de berçário, alimentação e reprodução de muitas
espécies incluindo algumas importantes para a pesca (VALESINI et al. 1997;
VASCONCELOS et al. 2010). Estudos da comunidades de peixes destes
sistemas têm como objetivo aumentar o conhecimento e a compreensão
desses habitats, possibilitando uma gestão adequada da região (SCHIFINO et
al. 2004). Esteves et al. (2008), analisando lagoas costeiras tropicais,
ressaltaram a importância de estudos posteriores que mostrem como
mudanças na diversidade destas lagoas afetam a sua capacidade de realizar
os processos ecológicos vitais e fornecer serviços essenciais para a sociedade
local.
Visando recuperar ecossistemas lagunares impactados pela ação
humana, diversas intervenções dos órgãos gestores têm sido conduzidas
(WASSERMAN et al. 1999). A mais frequente é a abertura de canais artificiais,
perenizando conexões das lagoas com o mar visando aumentar a circulação e,
com isso, reduzir o tempo de residência das águas, melhorar a qualidade
ambiental, minimizar os efeitos das inundações das residências nas margens
das lagoas nos períodos de chuvas e cheias e proporcionar a entrada de
espécies marinhas importantes para a pesca (SUZUKI et al. 1998 e 2002; LIMA
et al. 2001). Assim, a abertura de canais artificiais é considerada uma ação
essencial para suavizar os efeitos da eutrofização nos sistemas lagunares
(SAAD et al. 2002).
Porém, mudanças no estado hídrico das lagoas, em função da abertura
ou fechamento de canais de comunicação com o mar, acarretam em alterações

3
imprevisíveis na composição e abundância da flora e fauna local, sendo estas
muitas vezes irreversíveis (WASSERMAN et al. 1999; MACEDO-SOARES et
al. 2007). A abertura do canal na lagoa de Saquarema (Figura 1), Barra Franca
de Saquarema em 2003, por exemplo, provocou um aumento na salinidade do
sistema, porém não melhorou a qualidade da água, provavelmente devido ao
grande aporte de efluentes domésticos, decorrente do crescimento regional da
população. Segundo Wasserman (2000) e Azevedo (2005), as previsões para o
futuro da lagoa de Saquarema não são favoráveis. A implementação dos
canais da Costa (lagoa de Maricá) e de Ponta Negra (lagoa de Guarapina)
também não provocaram resultados muito favoráveis para a melhoria ambiental
do sistema lagunar Maricá-Guarapina (Figura 1). Das quatro lagoas contidas no
sistema, apenas a lagoa de Guarapina passou a apresentar melhores
condições sanitárias e obteve um aumento na pesca do camarão e da tainha
(CRUZ et al. 1996). A abertura temporária da barra da lagoa de Iquipari em
São João da Barra (RJ), no ano de 1996, causou mudanças nas características
da água; áreas secas e diminuição do nível da água; perda de vegetação;
mortandade expressiva de peixes dulcícolas; e entrada de peixes marinhos
(LIMA et al. 2001; SUZUKI et al. 2002).
Já na lagoa de Imboassica, no norte fluminense (Figura 1), onde o canal
é aberto uma vez por ano, estudos feitos por Saad et al. (2002) e Macedo-
Soares et al. (2007) demonstraram que os períodos de abertura podem alterar
a estrutura da comunidade de peixes, aumentar a abundância e diversidade
das espécies e melhorar a pesca local, principalmente devido às variações nos
fatores físicos desses ambientes. Ainda, nas lagoas costeiras da região do

4
PARNA de Jurubatiba (2005), estudos feitos por Santos et al. (2007)
evidenciaram uma diminuição nos níveis de poluição, aumento da
balneabilidade e da abundância de peixes, principalmente de interesse
comercial no momento logo após a abertura de um canal, permitindo uma
ligação com o mar, porém não foram feitos novos estudos para determinar
mais alterações na estrutura da comunidade. Estes foram os únicos estudos
realizados nas lagoas da Região Leste Fluminense que demonstraram um
efeito benéfico da abertura de canais na comunidade de peixes.
Consequências negativas já foram observadas no complexo Piratininga-Itaipu
após a abertura do Canal de Camboatá em 1946 e do Canal de Itaipu em 1979,
porém não existe nenhum registro da ictio e carcinofauna antes dessas
modificações, tornando difíceis avaliações comparativas.
Figura 1: Mapa das lagoas costeiras do norte fluminense

5
Se por um lado os resultados das ações de mitigação são bastante
imprevisíveis, por outro observa-se uma carência de informações detalhadas,
não apenas da qualidade da água, mas também das características das
comunidades biológicas, tanto nos estágios que antecedem quanto nos que
sucedem às intervenções efetuadas nos sistemas. Segundo Agostinho et al.
(2005), o monitoramento de ações de manejo efetuadas pelo poder público é
muitas vezes inadequado ou ausente. Assim, a ausência de dados prévios que
possam eventualmente auxiliar na análise do desenvolvimento bio-ecológico de
um sistema modificado, impossibilita avaliar de forma correta, o impacto das
obras sobre o ambiente. Por outro lado, quando essas informações são
disponibilizadas, é possível comparar situações “antes” vs “depois” (Before-
After Control Impacts – BACI) buscando avaliar os resultados e propor medidas
que favoreçam a recuperação da produtividade natural do sistema.
Estudos BACI são normalmente realizados em áreas de conservação, e
raramente apresentam amostragens antes da instalação desta, normalmente
tendo estudos apenas com comparações espaciais entre locais dentro e fora
da área de preservação. Assim, um dos maiores problemas desse tipo de
estudo é exatamente o de amostragem, uma vez que é comum duas
populações se comportarem de maneira diferente em locais distintos
(UNDERWOOD 1994). Porém quando se tem a chance de saber a priori como
a comunidade de um local se comporta, ao identificar nela um posterior
distúrbio, esse problema está solucionado. Isso porque podemos então
comparar a comunidade antes e depois do acontecimento analisando o quanto
o distúrbio a afetou.

6
O sistema lagunar de Piratininga-Itaipu, quando em sua situação
primitiva, era submetido a três grandes fases hidrológicas: a) estagnação
mesoalina predominante na maior parte do ano; b) desequilíbrio de diluição e
cheias, causado pelas chuvas, culminando na abertura da barra para o mar,
mais frequentemente em Piratininga; c) com a barra aberta, as lagoas
passavam a se submeter ao regime das marés (BARROSO et al. 2000).
Em 2007 o Laboratório de Biologia do Nécton e Ecologia Pesqueira
conduziu um programa de monitoramento da íctio e carcinofauna das lagoas de
Piratininga e Itaipu, antevendo as possíveis mudanças que ocorreriam no
sistema após a abertura do novo canal. Nele foram coletados dados de ictio e
carcinofauna com diferentes petrechos, avaliando diferenças de captura e
relacionando-as com dados ambientais medidos in situ (Fortes 2007). Assim,
no caso deste complexo, temos uma situação privilegiada em que uma base de
dados anterior ao distúrbio foi concluída com êxito, permitindo assim o
desenvolvimento de um verdadeiro Experimento Comparativo Antes-Depois
(ECAD).

7
2. OBJETIVOS
2.1 Objetivo Geral
Verificar possíveis alterações na ictio e carcinofauna do complexo
lagunar de Piratininga-Itaipu, provocadas pelas alterações no regime de
circulação após a abertura do canal do Tibau.
2.2 Objetivos Específicos
• Descrever a íctio e carcinofauna das lagoas de Piratininga e Itaipu, com
relação aos parâmetros: biomassa, abundância e frequência de
ocorrência das espécies, em função das diferentes estações do ano
(estação seca - julho-agosto e estação chuvosa –janeiro-fevereiro ),.
• Comparar a ocorrência e a abundância da ictio e carcinofauna antes
(2006) e depois (2010-11) da abertura do canal do Tibau.
• Correlacionar as variações observadas na biota com a salinidade,
temperatura, pH, oxigênio dissolvido, transparência, profundidade,
granulometria e matéria orgânica de cada lagoa em cada período
amostral.
• Identificar as principais variáveis ambientais que influenciam as
modificações nas comunidades.

8
3. MATERIAIS E MÉTODOS
3.1 Área de Estudo
O sistema lagunar Piratininga-Itaipu está situado na região costeira do
município de Niterói, RJ (de 22°56’ S a 22°58’ S e 43°02’ W a 43°06’ W), a
cerca de 40 Km do centro do município do Rio de Janeiro. É formado por duas
lagoas, a Lagoa de Piratininga e a Lagoa de Itaipu, unidas apenas por um
canal com 2,4 Km de extensão, o Canal de Camboatá (Figura 2). O sistema
está localizado na região tropical, e apresentam taxas equilibradas de
evaporação e precipitação, sendo 1,3 m e 1,4 m, respectivamente
(KNOPPERS et al. 1991).
Figura 2: Mapa da área de estudo, especificando os pontos amostrais (Piratininga: P1 – P6, Itaipu: I1 – I6) no sistema lagunar.
A Lagoa de Piratininga apresenta uma área total de 3 Km2, com uma
bacia de drenagem de aproximadamente 23 Km² e profundidade média de 0,6
m (KNOPPERS et al. 1991; BARROSO et al. 2001). O Rio Jacaré representa o

9
seu aporte fluvial mais importante, seguido pelo Córrego do Arrozal. Já a lagoa
de Itaipu apresenta uma área de 2 Km2, uma bacia contribuinte de 22,5 Km² e
uma profundidade média de 1,0 m. Seu principal afluente é o Rio João Mendes
(KNOPPERS et al. 1991; BARROSO et al. 2001). Apesar das diferenças em
tamanho e profundidade, o volume das lagoas é o mesmo, correspondendo a
0,002 Km³ (KNOPPERS op cit.).
Por consistir em duas lagoas pouco ligadas ao mar, o sistema pode ser
considerado como sufocado segundo a classificação proposta por Kjerfve
(1986). Em relação a quantidade de nutrientes dissolvidos, clorofila a e o tempo
de residência das águas, Piratininga e Itaipu foram classificadas como
hipertrófica e mesotrófica, respectivamente (KNOPPERS et al. 1991).
Quando a lagoa de Itaipu era a única em contato direto com o mar a
amplitude média de maré era de 0,3 m, excedendo bastante a média da de
Piratininga de 0,01 m (KNOPPERS et al. 1991). Entre os anos de 2005-2006 o
tempo aproximado de residência da água na lagoa de Itaipu era de nove dias,
enquanto que na de Piratininga era de 83 dias. Atualmente, depois da abertura
do canal em Piratininga, não se tem registro da amplitude de maré, mas o
tempo de residência de Piratininga e Itaipu é de 39 e oito dias, respectivamente
(CERDA et al. no prelo).
Uma das primeiras grandes intervenções no sistema ocorreu em 1946,
com a abertura do canal de Camboatá, ligando as duas lagoas com o objetivo
de permitir o equilíbrio hidráulico entre elas, já que o espelho d’água de Itaipu
recebia maior aporte continental da bacia hidrográfica adjacente. Assim, o
excedente de água seria drenado para Piratininga, e depois seguiria para o mar

10
pela sua ligação natural. Porém, com a obra, o nível da água da lagoa de
Piratininga não mais atingiu o mínimo necessário para provocar a abertura da
barra (MACH & LONGO 1998; WASSERMAN et al. 1999), impedindo a
renovação das águas, essencial para a regulação do seu ciclo biológico
(BARROSO et al. 2000). Além disso, o canal passou a drenar a Lagoa de
Piratininga, sendo essa função intensificada com a abertura permanente da
barra de Itaipu promovida pelo empreendimento imobiliário da Veplan em 1979,
que constituiu o atual bairro de Camboinhas. Com isso, a renovação das águas
na lagoa de Itaipu passou a ser controlada pelo fluxo de marés, enquanto que
em Piratininga a dinâmica era regulada quase que exclusivamente pelo aporte
de água doce (KJERFVE 1994; BARROSO et al. 2000). Essas intervenções
provocaram a redução da lâmina d’água em ambas as lagoas (MARCOLINI &
CORREIA 1989; WASSERMAN et al. 1999), levando ao surgimento de áreas
marginais secas que, somadas aos frequentes aterros irregulares, levaram à
redução de seus perímetros (BARROSO et al. 2000).
Em 2005, A Superintendência Estadual de Rios e Lagoas – SERLA
iniciou o “Projeto de Renovação do Sistema Lagunar de Piratininga-Itaipu”,
objetivando a melhoria da qualidade ambiental do sistema lagunar, através de
uma série de intervenções que aumentassem o nível e altura do espelho
d’água, principalmente da Lagoa de Piratininga, favorecendo a renovação, a
circulação e a melhoria da qualidade das águas (TAVARES 2003). Em abril de
2008 foi concluida a abertura do canal de Tibau, em Piratininga, que consiste
em um túnel de 988 m de comprimento, 5 m de largura e 4,5 m de altura,
escavado no costão rochoso de Piratininga, ligando a lagoa de Piratininga ao

11
mar de forma permanente, permitindo a intrusão de água salgada com o
movimento das marés. Um sistema de comportas impede o retorno da água
pelo canal do Tibau, forçando sua saída pelo canal de Camboatá na outra
extremidade, garantindo a circulação e a manutenção de uma lâmina d’água
mínima adequada.
3.2 Programa Amostral
O programa amostral foi realizado com base no apresentado por Fortes
(2007), modificado de forma a não comprometer o estudo comparativo. Para
análise da ictio e carcinofauna somente os dados de tarrafa foram utilizados,
para uma comparação fiel dos dados. Foram realizadas quatro campanhas em
duas estações: duas durante o verão (estação chuvosa, nos meses de janeiro
e março de 2011) e duas durante o inverno (estação seca, nos meses de julho
e agosto de 2010).
Cada lagoa foi dividida em seis áreas de coleta (Figura 2), conforme
estudo anterior (FORTES 2007):
- Lagoa de Piratininga:
• P1: Área de maior interferência do canal de Camboatá, com influência
da Ilha do Modesto.
• P2: Área localizada na saída do rio Jacaré.
• P3: Área sem ocupação urbana. Com vegetação ciliar.
• P4: Área ao fundo da lagoa, no lado oposto ao canal de Camboatá e sob
maior influência do canal de Tibau, consequentemente com uma maior
influência marinha e urbana.

12
• P5: Área com influência urbana e vegetação ciliar predominantemente
de Typha dominguensis e Acrostichum sp.
• P6: Área de maior profundidade da lagoa, com a circulação sobre
influência do canal de Camboatá.
- Lagoa de Itaipu:
• I1: Área do canal de Itaipu, consequentemente, com maior influência
marinha.
• I2: Área com plantas de mangue e algas no sedimento.
• I3: Área sob influência do rio da Vala, com a presença de plantas de
mangue Laguncularia racemosa, Avicennia schaueriana e Rizophora
mangle.
• I4: Região oposta ao canal de Itaipu, porém mais próxima ao canal de
Camboatá; com influência do rio João Mendes.
• I5: Área com vegetação ciliar e sob influência do canal de Itaipu.
• I6: Área central da lagoa, região mais profunda e com bastante
influência das águas oriundas do canal de Itaipu.
As coletas da ictio e carcinofauna foram realizadas com a utilização de
duas tarrafas com tamanhos de malhas diferentes: 12 mm e 20 mm entre nós
adjacentes, ambas com perímetro de 12 braças e 32,2 m² de área. Ambos os
estudos utilizaram as mesmas redes e o mesmo tarrafeador, padronizando a
coleta de dados.
Em cada área foram escolhidos, de maneira aleatória, três pontos
amostrais para o lançamento das tarrafas. Em cada ponto foram realizados
quatro lances de tarrafa de malha 12 mm e quatro de malha 20 mm (formando

13
uma unidade amostral), totalizando 18 amostras (três pontos x seis áreas) por
campanha amostral em cada lagoa. Foram escolhidos, também de forma
aleatória, quatro pontos em cada área para a coleta dos seguintes parâmetros:
temperatura (°C) e salinidade (termo-salinômetro – YSI 30-25 FT),
transparência (em centímetros, com o disco de Sechii), pH (phmetro), oxigênio
dissolvido (mg/L, com oxímetro – WTW Oxi315i) e profundidade da água (cm).
Em três destes pontos foram coletadas amostras de sedimento para
caracterização granulométrica (aparelho CILAS 1026), e quantificação de
matéria orgânica (através do método de diferença de massa, CORDEIRO,
comunicação pessoal). Os valores médios de precipitação foram obtidos a
partir da Tabela de dados pluviométricos mensais da Fundação Instituto de
Geotécnica do Município do Rio de Janeiro (2011).
Os indivíduos coletados foram levados ao Laboratório de Biologia do
Nécton e Ecologia Pesqueira (ECOPESCA), fixados em formol 10% e
conservados em álcool 70%. Depois foram identificados (WILLIAMS 1974;
COELHO & RAMOS-PORTO 1992 para siris; COSTA et al.2003 para
camarões; FIGUEIREDO & MENEZES 1978, 1980; MENEZES & FIGUEIREDO
1980, 1985 para peixes), contados e pesados (Kg).
3.3 Análise de Dados
A compilação de uma lista de espécies de peixes e crustáceos que
ocorreram no complexo lagunar de Piratininga-Itaipu durante o período de
amostragem, forneceu uma caracterização atual da ictio e carcinofauna. Para
cada espécie foi computada a abundância (numero de indivíduos), frequência
de ocorrência e biomassa, por lagoa e por estação de coleta.

14
Os resultados obtidos foram comparados com os resultados anteriores
encontrados por Fortes (2007). A comparação considerou as distribuições das
amostras de cada lagoa, em cada período sazonal e os principais fatores
ambientais monitorados que contribuíram, em parte, para estas distribuições.
Os dados de abundância foram transformados em raiz quarta para diminuir o
efeito das espécies dominantes e foi utilizada a distância de Hellinger para que
não houvesse grandes distorções dos dados (RAO 1995). Para todas as
análises foi considerando uma significância de 95% (p = 0,05).
A análise granulométrica foi realizada com o auxílio do equipamento
CILAS 1026 com o método de análise de Blott & Pye (2001) no programa
GRADISTAT. A quantificação da matéria orgânica foi realizada de acordo com
o método de diferença de massa, antes e depois da queima do sedimento na
mufla a 550 °C durante 2 horas (CORDEIRO, comunicação pessoal).
Para os dados abióticos foi aplicada a análise de variância bifatorial
(two-way ANOVA) a qual testa se há diferença de cada fator entre os período
amostrados (2006 e 2010-11), entre as estações do ano (inverno e verão) e a
interação destes parâmetros. Para tal foi utilizado o programa STATISTICA 7
(STATSOFT 2005).
Para os dados bióticos foi realizada uma curva cumulativa de espécies,
visando esclarecer se o esforço de coleta foi suficiente para avaliar a riqueza
de cada lagoa nos dois períodos amostrais. Para tal foi utilizado o método de
reamostragem Mau Tao (COLWELL et al. 2004) com 10000 permutações,
através do programa EstimateS 8.2 (COLWELL 1994-2004).

15
Para verificar o grau de similaridade entre as diferentes dimensões
consideradas, foi utilizada a análise de similaridade (ANOSIM) realizada
através do aplicativo PRIMER 6 (CLARKE & GORLEY 2006). Esta análise
consiste num recurso estatístico que permite testar se há semelhanças
significativas entre dois ou mais grupos de dados multivariados (CLARKE
1993).
Os padrões de distribuição das amostras com relação às espécies foram
representados através de uma análise de componentes principais (“Principal
Components Analysis” – PCA), proposto por Hotelling (1933). Esta análise
consegue simplificar em poucas dimensões a variabilidade da dispersão dos
dados de uma matriz com muitos fatores, assim, a maior parte da variação
entre os dados fica retida ao primeiro eixo, sendo facilmente visualizada em um
gráfico bi-dimensional (LEGENDRE & LEGENDRE 1998). O valor atribuído a
cada espécie, relativo ao primeiro componente do gráfico de PCA (“loadings”)
foram representados em um segundo gráfico, com o objetivo de avaliar quais
foram as espécies mais importantes para a distribuição.
Visando identificar a influência do espaço, tempo, e da interação entre
estes fatores nas diferenças entre os períodos, foi realizada a análise de
variância multivariada com permutação (“Permutational Multivariate Analysis of
Variance” - PERMANOVA). Esta análise possibilita testar simultaneamente a
resposta de uma ou mais variáveis a um ou mais fatores, utilizando um
desenho amostral idêntico ao da ANOVA (ANDERSON 2005).
A análise de correspondência canônica (“Canonical Correspondence
Analysis” - CCA) foi utilizada para descrever as relações entre as variáveis

16
ambientais com cada amostra com a ictio e carcinofauna. A matriz de CCA
sintetiza, em um gráfico de eixos perpendiculares, a variação multidimensional
de um conjunto de parâmetros. Este método utiliza duas matrizes, uma
representando a abundância das amostras e a outra os fatores ambientais.
Ambas relacionadas com os diferentes períodos amostrais em cada lagoa.
Outro recurso utilizado deste programa foi o diagrama de t-valor biplot, o qual
realiza consecutivas análises de regressão par a par, visando estabelecer
relações entre os dados ambientais e cada uma das espécies (LEPS &
SMILAUER 2003). Deste modo, pode ser avaliado como as espécies do
conjunto de cada amostra respondem aos fatores ambientais monitorados. A
análise foi realizada usando o programa “CANOCO for Windows 4.5” (BRAAK
& SMILAUER 2002; LEPS & SMILAUER 2003).
Para revelar o quanto cada fator (dados ambientais e mudanças
temporais) influenciou a comunidade de cada lagoa foi realizada uma análise
de partição de variância, que revela a porcentagem de explicação de cada fator
na distribuição dos dados e a correlação entre estes fatores. Essa análise se
baseia no princípio que a variação ecológica pode ser dividida em fatores
(espacial, temporal, ambiental, suas correlações e desconhecidos) a serem
testados individualmente, possibilitando observar o quanto cada um deles
influencia na distribuição dos dados (BORCARD et al. 1992). A análise é
conduzida através de uma CCA, usando o programa “CANOCO for Windows
4.5” (BRAAK & SMILAUER 2002), e seguindo os quatro passos propostos por
Borcard et al. (1992).

17
Devido a sua alta sensibilidade a espécies raras, todas as análises
realizadas no “CANOCO for Windows 4.5” (BRAAK & SMILAUER 2002)
tiveram as espécies com frequência de ocorrência menor que 0,5 % retiradas
da análise.
Para identificar quais espécies foram responsáveis pela separação das
lagoas entre os períodos amostrais, foi realizada uma analise discriminante,
utilizando-se o programa CANOCO, onde a matriz solicitada de espécies foi
representada por uma matriz “dummy” dos períodos amostrais (2006 e 2010-
11) e a matriz requerida de dados ambientais foi a matriz original de
comunidade.

18
4. RESULTADOS
Foram realizadas 144 amostras no período 2010-11. Foram capturados
1180 indivíduos da ictiofauna pertencentes a 17 famílias e 28 espécies e 147
indivíduos da carcinofauna pertencentes a 2 famílias e 4 espécies (Tabela 1).
Foi capturada uma biomassa total de 38,3 Kg, sendo apenas 2,03 Kg
pertencentes a carcinofauna (Tabela 2).
No total, o complexo lagunar apresentou maiores riqueza de espécies e
abundância de indivíduos no verão. A lagoa de Piratininga seguiu este mesmo
padrão, porém a de Itaipu apresentou uma maior abundância de indivíduos no
inverno, muito embora a maior biomassa tenha ocorrido no verão. Apesar
disso, as diferenças entre as estações do ano, de um modo geral, não foram
muito acentuadas.
As cinco espécies mais abundantes para ambas as lagoas foram
Diapterus rhombeus, Callinectes danae (Crustacea, Portunidae), Eucinostomus
argenteus, Elops saurus, e Eucinostomus gula, respectivamente, que juntas
somaram 75,36 % do total de indivíduos capturados. D. rhombeus foi a espécie
mais abundante e representou sozinha 50,94% do total, porém foi muito mais
representativa em Itaipu do que em Piratininga (Tabela 1). Em Itaipu, as cinco
espécies mais abundantes foram D. rhombeus, C. danae, E. saurus, E. gula e
Sardinella brasiliensis, respectivamente, somando 84,78% do total de
indivíduos capturados. Em Piratininga, as espécies mais abundantes foram E.
argenteus, C. danae, Brevoortia pectinata, E. saurus e E. gula,

19
respectivamente, com um total de 65,40% dos indivíduos capturados na lagoa
(Tabela 1).
As cinco espécies mais frequentes foram também as mais abundantes
(Tabela 1), porém em ordem diferente. D. rhombeus também foi a mais
frequente aparecendo em 48,6% nas amostras (sendo 88,90% só em Itaipu e
apenas 8,3% em Piratininga), seguida por E. saurus (aparecendo em 29,2%
das amostras), C. danae (em 25,0% da amostras), E. gula (em 24,3%) e E.
argenteus (em 20,8%). Na lagoa de Itaipu as espécies mais frequentes também
foram, na mesma ordem, as mais abundantes com exceção de E. gula que não
apresentou uma ocorrência tão alta e Litopenaeus schmitti (Crustacea,
Penaeidae) que apareceu como a quinta espécie mais frequente. Já em
Piratininga as cinco espécies mais frequentes foram E. argenteus, E. gula, E.
saurus, L. schmitti e Brevoortia aurea, respectivamente.
As cinco espécies com maior representatividade em biomassa foram D.
rhombeus, E. saurus, Mugil liza, Mugil curema e Oreochromis niloticus,
respectivamente, com 69,80% do total de biomassa capturada. Em Itaipu as
cinco espécies com maior biomassa foram D. rhombeus, E. saurus, M. liza, S.
brasiliensis e M. curema representando 82,96% da biomassa total. Em
Piratininga as cinco espécies foram E. saurus, O. niloticus, Micropogonias
furnieri, B. aurea e M. curema, respectivamente, com 68,99% da biomassa total
da lagoa (Tabela 2).
Com relação aos parâmetros ambientais, as médias de temperatura
variaram ao longo do ano apresentando maiores médias no verão (30,1°C em
Itaipu e 29,8°C em Piratininga) e menores no inverno (23,8°C em Itaipu e

20
24,6°C em Piratininga), para as duas lagoas. A média de salinidade não
apresentou uma variação muito grande por época do ano dentro da mesma
lagoa, porém entre as lagoas foram encontrados valores bastante diferentes,
variando de 30,5 (verão) a 31,2 (inverno) em Itaipu e de 23,8 (inverno) a 24,4
(verão) em Piratininga. Os valores médios de pH, oxigênio dissolvido e
transparência não apresentaram um padrão muito claro entre as épocas do ano
nem entre as lagoas. Já os níveis de matéria orgânica no sedimento em
Piratininga foram, em média, mais altos (19,4 no inverno e 17,9 no verão) que
em Itaipu (12,8 no inverno e 11,4 no verão), independente da época do ano. A
taxa média de precipitação nos meses de verão foi maior do que nos meses de
inverno, justificando a separação entre as estações seca (inverno) e chuvosa
(verão) (Tabela 3).
A proporção de areia e lama em Itaipu variou pouco com as estações do
ano, mas pode ser observado um predomínio de lama no inverno e areia no
verão. Já em Piratininga não houve essa mudança de padrão granulométrico
entre as estações do ano, sendo a lama predominante o ano todo (Figura 3).

21
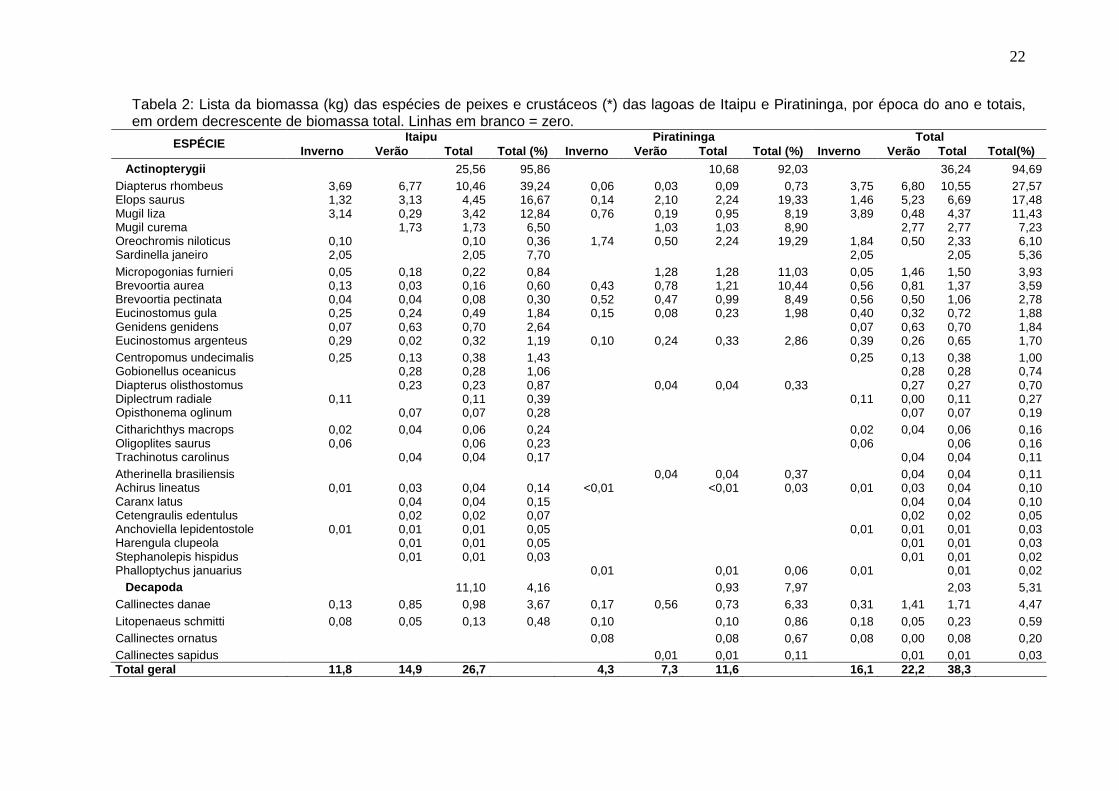
Tabela 1: Lista de abundância total das espécies de peixes e crustáceos (*) das lagoas de Itaipu e Piratininga, coletados em 2010-11, por época do ano, em ordem decrescente de abundância total. Linhas em branco = zero. Em destaque, a classificação das frequências de ocorrência relativas (FO %) de cada espécie, por lagoa, época do ano e total.
Itaipu Piratininga Total Espécies Inverno Verão Total Total (%) Inverno Verão Total T otal (%) Inverno Verão Total Total (%)
Actinopterygii 951 91,62 193 66,78 1180 86,21 Diapterus rhombeus 337 327 664 63,97 6 6 12 4,15 343 333 676 50,94 Eucinostomus argenteus 31 2 33 3,18 14 34 48 16,61 45 36 81 6,10 Elops saurus 21 38 59 5,68 4 3 34 11,76 25 68 66 4,97 Eucinostomus gula 27 16 43 4,14 18 1 28 9,69 45 26 62 4,67 Brevoortia aurea 3 3 6 0,58 16 24 4 1,38 19 27 46 3,47 Sardinella janeiro 43 43 4,14 43 43 3,24 Brevoortia pectinata 2 3 5 0,48 21 14 35 12,11 23 17 40 3,01 Mugil liza 13 2 15 1,45 5 1 6 2,08 18 3 21 1,58 Mugil curema 1 11 12 1,16 5 5 1,73 1 16 17 1,28 Gobionellus oceanicus 11 11 1,06 11 11 0,83 Micropogonias furnieri 1 4 5 0,48 6 6 2,08 1 1 11 0,83 Oreochromis niloticus 1 1 0,10 6 3 9 3,11 7 3 10 0,75 Diapterus olisthostomus 8 8 0,77 1 1 0,35 9 9 0,68 Anchoviella lepidentostole 5 3 8 0,77 5 3 8 0,60 Genidens genidens 2 6 8 0,77 2 6 8 0,60 Achirus lineatus 2 4 6 0,58 1 1 0,35 3 4 7 0,53 Centropomus undecimalis 3 2 5 0,48 3 2 5 0,38 Citharichthys macrops 2 3 5 0,48 2 3 5 0,38 Atherinella brasiliensis 3 3 1,04 3 3 0,23 Caranx latus 3 3 0,29 3 3 0,23 Oligoplites saurus 3 3 0,29 3 3 0,23 Cetengraulis edentulus 2 2 0,19 2 2 0,15 Trachinotus carolinus 2 2 0,19 2 2 0,15 Diplectrum radiale 1 1 0,10 1 1 0,08 Harengula clupeola 1 1 0,10 1 1 0,08 Opisthonema oglinum 1 1 0,10 1 1 0,08 Phalloptychus januarius 1 1 0,35 1 1 0,08 Stephanolepis hispidus 1 1 0,10 1 1 0,08 Decapoda 87 8,38 60 20,76 147 11,08 Callinectes danae 13 58 71 6,84 3 41 44 15,22 16 99 115 8,67 Litopenaeus schmitti 12 4 16 1,54 14 14 4,84 26 4 30 2,26 Callinectes ornatus 1 1 0,35 1 1 0,08 Callinectes sapidus 1 1 0,35 1 1 0,08 Total de famílias 14 16 18 10 11 12 16 16 19 Total de espécies 19 24 28 13 14 18 21 27 32 Total de indivíduos 523 515 1038 110 179 289 633 694 1327 Total de amostras 36 36 72 36 36 72 72 72 144
FO a acima de 40 % FO entre 20% e 40% FO entre 10% e 20% FO Menor que 10 %
22
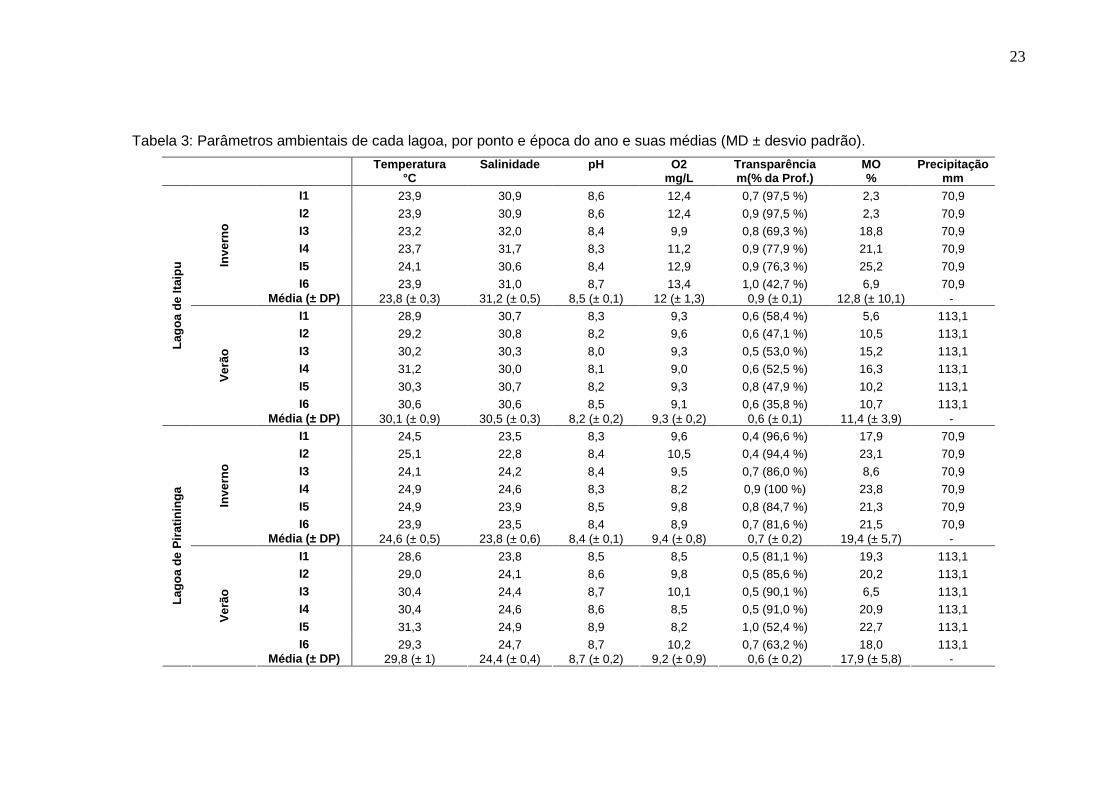
Tabela 2: Lista da biomassa (kg) das espécies de peixes e crustáceos (*) das lagoas de Itaipu e Piratininga, por época do ano e totais, em ordem decrescente de biomassa total. Linhas em branco = zero.
Itaipu Piratininga Total ESPÉCIE Inverno Verão Total Total (%) Inverno Verão Total Total (%) Inverno Verão Total T otal(%)
Actinopterygii 25,56 95,86 10,68 92,03 36,24 94,69 Diapterus rhombeus 3,69 6,77 10,46 39,24 0,06 0,03 0,09 0,73 3,75 6,80 10,55 27,57 Elops saurus 1,32 3,13 4,45 16,67 0,14 2,10 2,24 19,33 1,46 5,23 6,69 17,48 Mugil liza 3,14 0,29 3,42 12,84 0,76 0,19 0,95 8,19 3,89 0,48 4,37 11,43 Mugil curema 1,73 1,73 6,50 1,03 1,03 8,90 2,77 2,77 7,23 Oreochromis niloticus 0,10 0,10 0,36 1,74 0,50 2,24 19,29 1,84 0,50 2,33 6,10 Sardinella janeiro 2,05 2,05 7,70 2,05 2,05 5,36 Micropogonias furnieri 0,05 0,18 0,22 0,84 1,28 1,28 11,03 0,05 1,46 1,50 3,93 Brevoortia aurea 0,13 0,03 0,16 0,60 0,43 0,78 1,21 10,44 0,56 0,81 1,37 3,59 Brevoortia pectinata 0,04 0,04 0,08 0,30 0,52 0,47 0,99 8,49 0,56 0,50 1,06 2,78 Eucinostomus gula 0,25 0,24 0,49 1,84 0,15 0,08 0,23 1,98 0,40 0,32 0,72 1,88 Genidens genidens 0,07 0,63 0,70 2,64 0,07 0,63 0,70 1,84 Eucinostomus argenteus 0,29 0,02 0,32 1,19 0,10 0,24 0,33 2,86 0,39 0,26 0,65 1,70 Centropomus undecimalis 0,25 0,13 0,38 1,43 0,25 0,13 0,38 1,00 Gobionellus oceanicus 0,28 0,28 1,06 0,28 0,28 0,74 Diapterus olisthostomus 0,23 0,23 0,87 0,04 0,04 0,33 0,27 0,27 0,70 Diplectrum radiale 0,11 0,11 0,39 0,11 0,00 0,11 0,27 Opisthonema oglinum 0,07 0,07 0,28 0,07 0,07 0,19 Citharichthys macrops 0,02 0,04 0,06 0,24 0,02 0,04 0,06 0,16 Oligoplites saurus 0,06 0,06 0,23 0,06 0,06 0,16 Trachinotus carolinus 0,04 0,04 0,17 0,04 0,04 0,11 Atherinella brasiliensis 0,04 0,04 0,37 0,04 0,04 0,11 Achirus lineatus 0,01 0,03 0,04 0,14 <0,01 <0,01 0,03 0,01 0,03 0,04 0,10 Caranx latus 0,04 0,04 0,15 0,04 0,04 0,10 Cetengraulis edentulus 0,02 0,02 0,07 0,02 0,02 0,05 Anchoviella lepidentostole 0,01 0,01 0,01 0,05 0,01 0,01 0,01 0,03 Harengula clupeola 0,01 0,01 0,05 0,01 0,01 0,03 Stephanolepis hispidus 0,01 0,01 0,03 0,01 0,01 0,02 Phalloptychus januarius 0,01 0,01 0,06 0,01 0,01 0,02 Decapoda 11,10 4,16 0,93 7,97 2,03 5,31 Callinectes danae 0,13 0,85 0,98 3,67 0,17 0,56 0,73 6,33 0,31 1,41 1,71 4,47 Litopenaeus schmitti 0,08 0,05 0,13 0,48 0,10 0,10 0,86 0,18 0,05 0,23 0,59 Callinectes ornatus 0,08 0,08 0,67 0,08 0,00 0,08 0,20 Callinectes sapidus 0,01 0,01 0,11 0,01 0,01 0,03 Total geral 11,8 14,9 26,7 4,3 7,3 11,6 16,1 22,2 38,3
23
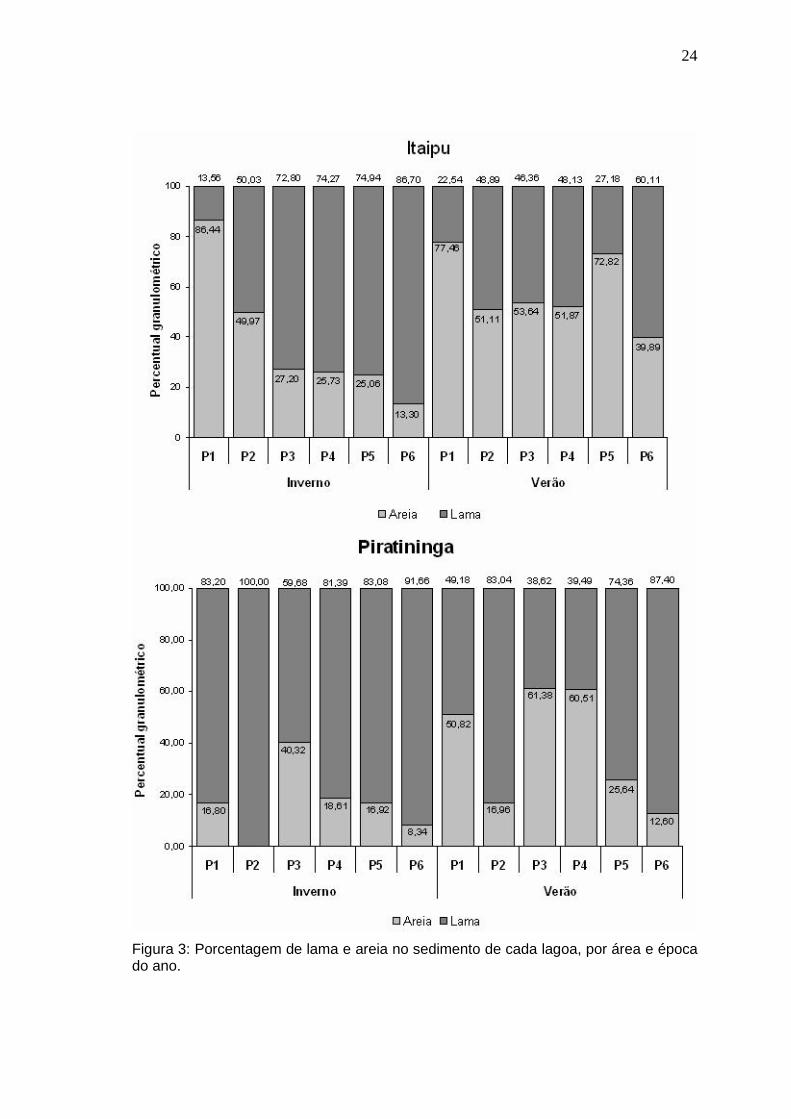
Tabela 3: Parâmetros ambientais de cada lagoa, por ponto e época do ano e suas médias (MD ± desvio padrão).
Temperatura
°C Salinidade
pH
O2
mg/L Transparência m(% da Prof.)
MO %
Precipitação mm
I1 23,9 30,9 8,6 12,4 0,7 (97,5 %) 2,3 70,9
I2 23,9 30,9 8,6 12,4 0,9 (97,5 %) 2,3 70,9 I3 23,2 32,0 8,4 9,9 0,8 (69,3 %) 18,8 70,9 I4 23,7 31,7 8,3 11,2 0,9 (77,9 %) 21,1 70,9
I5 24,1 30,6 8,4 12,9 0,9 (76,3 %) 25,2 70,9 I6 23,9 31,0 8,7 13,4 1,0 (42,7 %) 6,9 70,9
Inve
rno
Média (± DP) 23,8 (± 0,3) 31,2 (± 0,5) 8,5 (± 0,1) 12 (± 1,3) 0, 9 (± 0,1) 12,8 (± 10,1) - I1 28,9 30,7 8,3 9,3 0,6 (58,4 %) 5,6 113,1 I2 29,2 30,8 8,2 9,6 0,6 (47,1 %) 10,5 113,1
I3 30,2 30,3 8,0 9,3 0,5 (53,0 %) 15,2 113,1 I4 31,2 30,0 8,1 9,0 0,6 (52,5 %) 16,3 113,1
I5 30,3 30,7 8,2 9,3 0,8 (47,9 %) 10,2 113,1 I6 30,6 30,6 8,5 9,1 0,6 (35,8 %) 10,7 113,1
Lago
a de
Itai
pu
Ver
ão
Média (± DP) 30,1 (± 0,9) 30,5 (± 0,3) 8,2 (± 0,2) 9,3 (± 0,2) 0 ,6 (± 0,1) 11,4 (± 3,9) - I1 24,5 23,5 8,3 9,6 0,4 (96,6 %) 17,9 70,9 I2 25,1 22,8 8,4 10,5 0,4 (94,4 %) 23,1 70,9
I3 24,1 24,2 8,4 9,5 0,7 (86,0 %) 8,6 70,9 I4 24,9 24,6 8,3 8,2 0,9 (100 %) 23,8 70,9
I5 24,9 23,9 8,5 9,8 0,8 (84,7 %) 21,3 70,9 I6 23,9 23,5 8,4 8,9 0,7 (81,6 %) 21,5 70,9
Inve
rno
Média (± DP) 24,6 (± 0,5) 23,8 (± 0,6) 8,4 (± 0,1) 9,4 (± 0,8) 0 ,7 (± 0,2) 19,4 (± 5,7) -
I1 28,6 23,8 8,5 8,5 0,5 (81,1 %) 19,3 113,1 I2 29,0 24,1 8,6 9,8 0,5 (85,6 %) 20,2 113,1
I3 30,4 24,4 8,7 10,1 0,5 (90,1 %) 6,5 113,1 I4 30,4 24,6 8,6 8,5 0,5 (91,0 %) 20,9 113,1
I5 31,3 24,9 8,9 8,2 1,0 (52,4 %) 22,7 113,1 I6 29,3 24,7 8,7 10,2 0,7 (63,2 %) 18,0 113,1
Lago
a de
Pira
tinin
ga
Ver
ão
Média (± DP) 29,8 (± 1) 24,4 (± 0,4) 8,7 (± 0,2) 9,2 (± 0,9) 0,6 (± 0,2) 17,9 (± 5,8) -
24
Figura 3: Porcentagem de lama e areia no sedimento de cada lagoa, por área e época do ano.

25
Ao comparar o conjunto de dados obtidos nos dois períodos, foi possível
observar que a comunidade de peixes e crustáceos das lagoas mudou
bastante. De uma maneira geral, pode ser observado uma diminuição no
número de espécies, famílias e indivíduos. Das 44 espécies encontradas em
2006, 22 foram exclusivas. Já dentre as 32 espécies amostradas em 2010-11,
10 foram exclusivas, restando 22 espécies comuns aos dois períodos (Tabela
4). A curva cumulativa de espécies indicou que as amostragens foram
suficientes para a detecção de uma parcela significativa da fauna do local
(Figura 4).
Das cinco espécies mais capturadas, a espécie mais abundante em
ambos os períodos foi D. rhombeus, alterando apenas o número absoluto de
indivíduos, e sendo sempre mais representativa em Itaipu que em Piratininga.
Já o total de capturas das demais espécies variou bastante entre os anos: a
segunda espécie mais abundante em 2006 foi O. niloticus, seguida por
Farfantepenaeus brasiliensis (Crustacea, Penaeidae), L. schmitti e E. saurus,
respectivamente; enquanto que em 2010-11 as outras quatro espécies mais
representativas foram C. danae, E. saurus, E. argenteus e E. gula,
respectivamente. Apesar desta diferença entre os períodos, as cinco espécies
mais frequentes se mantiveram praticamente as mesmas, com exceção de O.
niloticus, cuja frequência de ocorrência e abundância em 2010-11 foram menor
(Tabela 4).

26
Figura 4: Curva cumulativa de espécies em função do número de amostras para as lagoas de Itaipu e de Piratininga nos dois períodos. As linhas pontilhadas representam o intervalo de confiança (95% para cada curva).

27
Tabela 4: Tabela comparativa das espécies presentes nas campanhas de 2006 e 2010-11, por lagoa, época do ano e totais, em ordem decrescente de abundância total, somados os dois períodos. Linhas em branco = zero. Em destaque, a classificação das frequências de ocorrência relativas (FO %) de cada espécie, por período amostral, lagoa, época do ano e total. * - espécies exclusivas de 2006. ** - espécies exclusivas de 2010-11.
Lagoa de Itaipu Lagoa de Piratininga 2006 2010 2006 2010 Espécie
Inverno Verão Total Inverno Verão Total Inverno Verão Total Inverno Verão Total
Total 2006
Total 2010
Total geral
Actinopterygii 1387 951 566 1953 1180 3133 Diapterus rhombeus 246 665 911 337 327 664 5 2 7 6 6 12 918 676 1594 Oreochromis niloticus 11 11 1 1 209 58 267 6 3 9 278 10 288 Elops saurus 11 10 21 21 38 59 6 117 123 4 30 34 144 93 237 Eucinostomus argenteus 32 68 100 31 2 33 15 1 16 14 34 48 116 81 197 Eucinostomus gula 27 45 72 27 16 43 5 5 18 10 28 77 71 148 Mugil liza 7 25 32 13 2 15 16 21 37 5 1 6 69 21 90 Brevoortia aurea 2 1 3 3 3 6 40 40 16 24 40 43 46 89 Mugil curema 7 26 33 1 11 12 3 1 4 5 5 37 17 54 Sardinella janeiro** 43 43 43 43 Gobionellus oceanicus 8 22 30 11 11 30 11 41 Brevoortia pectinata** 2 3 5 21 14 35 40 40 Cetengraulis edentulus 35 2 37 2 2 37 2 39 Atherinella brasiliensis 7 7 16 9 25 3 3 32 3 35 Diapterus olisthostomus 10 15 25 8 8 1 1 25 9 34 Citharichthys spilopterus* 23 7 30 30 30 Centropomus undecimalis 1 1 3 2 5 23 23 24 5 29 Micropogonias furnieri 11 11 1 4 5 7 7 6 6 18 11 29 Harengula clupeola 19 19 1 1 19 1 20 Centropomus parallelus* 3 4 7 10 10 17 17 Achirus lineatus 5 1 6 2 4 6 1 1 6 7 13 Anchoviella lepidentostole** 5 3 8 8 8 Genidens genidens** 2 6 8 8 8 Anchoa tricolor* 5 5 5 5 Caranx latus 2 2 3 3 2 3 5 Citharichthys macrops** 2 3 5 5 5 Symphurus plagusia* 4 4 4 4

28
Oligoplites saurus** 3 3 3 3 Opisthonema oglinum 2 2 1 1 2 1 3 Prionotus punctatus* 3 3 3 3 Chilomycterus spinosus* 2 2 2 2 Eucinostomus melanopterus* 1 1 2 2 2 Synodus foetens* 2 2 2 2 Trachinotus carolinus** 2 2 2 2 Anchovia clupeoides* 1 1 1 1 Archosargus rhomboidalis* 1 1 1 1 Chloroscombrus chrysurus* 1 1 1 1 Diplectrum formosum* 1 1 1 1 Diplectrum radiale** 1 1 1 1 Haemulon plumierii* 1 1 1 1 Microgobius meeki* 1 1 1 1 Phalloptychus januaris** 1 1 1 1 Selene vomer* 1 1 1 1 Sphoeroides greeleyi* 1 1 1 1 Sphoeroides testudines* 1 1 1 1 Stephanolipis hispidus** 1 1 1 1 Stephanolipis setifer* 1 1 1 1 Trachinotus sp.* 1 1 1 1
Decapoda 391 87 307 60 698 147 845 Callinectes danae 57 9 66 13 58 71 65 65 3 41 44 131 115 246 Farfantepenaeus brasiliensis* 87 87 109 109 196 196 Litopenaeus schmitti 124 3 127 12 4 16 26 26 14 14 153 30 183 Callinectes ornatus 65 65 43 43 1 1 108 1 109 Farfantepenaeus paulensis* 46 46 36 36 82 82 Callinectes bocourti* 17 3 20 20 20 Callinectes sapidus 8 8 1 1 8 1 9 TOTAL DE ESPÉCIES 29 27 40 19 24 28 19 10 21 13 14 18 44 32 54 TOTAL DE FAMÍLIAS 18 18 24 14 16 18 11 7 12 10 11 12 24 19 26 TOTAL DE INDIVÍDUOS 828 950 1778 523 515 1038 628 245 873 110 179 289 2651 1327 3978 TOTAL DE AMOSTRAS 36 36 72 36 36 72 36 36 72 36 36 72 144 144 288

29
Diapterus rhombeus foi a espécie mais abundante nos dois períodos na
lagoa de Itaipu. Em 2006 as quatro espécies que se seguiram foram L. schmitti,
E. argenteus, F. brasiliensis e E. gula, respectivamente. Já em 2010-11 as
quatro espécies foram C. danae, E. saurus, E. gula e S. brasiliensis,
respectivamente. Em Piratininga, as cinco espécies mais abundantes nas
amostragens de 2006 foram O. niloticus, E. saurus, F. brasiliensis, C. danae e
Callinectes ornatus, respectivamente. Já em 2010-11 foram E. argenteus, C.
danae, B. aurea, B. pectinata e E. saurus, respectivamente (Tabela 4).
A riqueza de espécies e a abundância de indivíduos, quer seja em
número total de indivíduos ou em biomassa total capturada, foi bem menor na
campanha de 2010-11 do que na campanha anterior. No total, a espécie com
maior biomassa foi O. niloticus, com maior captura em 2006 (35,7% do total).
Se considerarmos apenas a campanha de 2010-11, a espécie com maior
captura em peso foi D. rhombeus, enquanto que O. niloticus passou a
representar a quinta espécie com maior biomassa. Além disso, em ambos os
anos esta espécie foi mais capturada em Piratininga, com um total de 28 Kg em
2006 e 2,24 Kg em 2010-11 somente nesta lagoa (Tabela 5).
As outras quatro espécies mais representativas em peso total foram E.
saurus, D. rhombeus, M. liza e M. curema, respectivamente, e as demais
espécies apresentaram uma biomassa total menor que 0,5 Kg (Tabela 5).

30
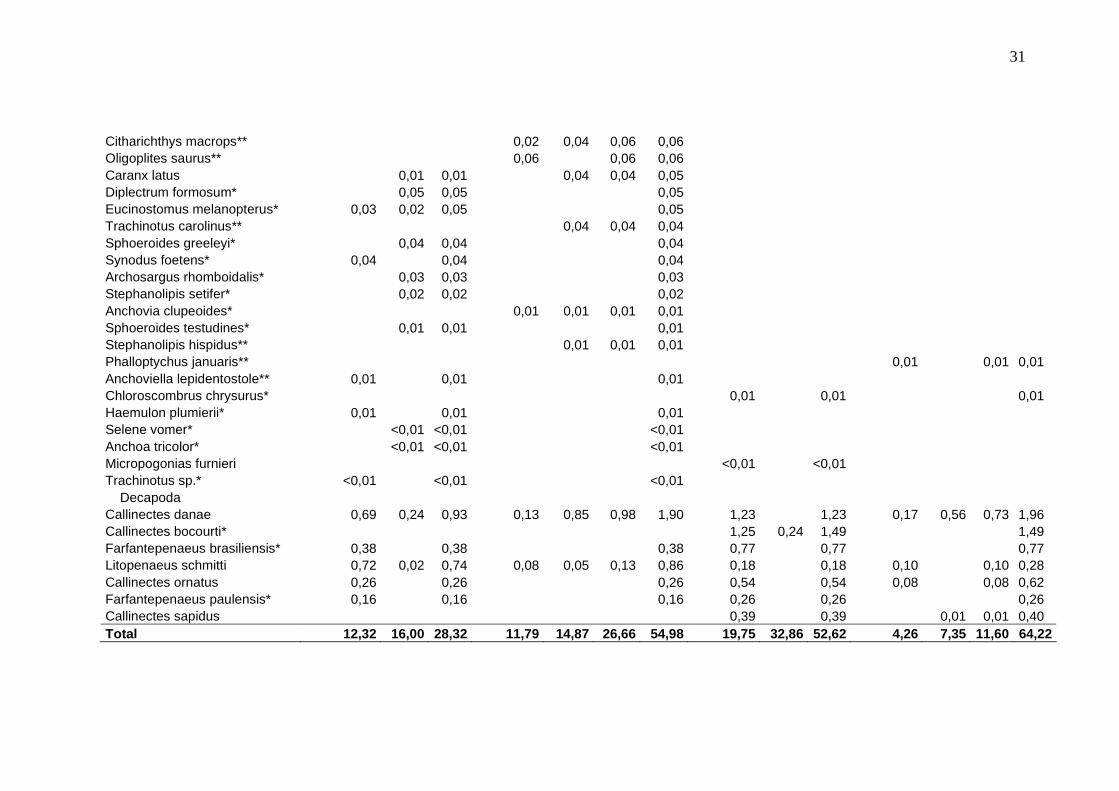
Tabela 5: Tabela comparativa da biomassa (em quilogramas) das espécies presentes nas campanhas de 2006 e 2010-11, por lagoa, época do ano e totais, em ordem decrescente de biomassa total somados os dois períodos. Linhas em branco = zero. * - espécies exclusivas de 2006. ** - espécies exclusivas de 2010-11.
Lagoa de Itaipu Lagoa de Piratininga 2006 2010-11 2006 2010-11 Espécie
Inverno Verão Total Inverno Verão Total Total
Inverno Verão Total Inverno Verão Total Total
Actinopterygii Oreochromis niloticus 0,86 0,86 0,10 0,10 0,95 12,15 15,86 28,01 1,74 0,50 2,24 30,25 Elops saurus 0,77 0,83 1,60 1,32 3,13 4,45 6,04 0,25 12,16 12,41 0,14 2,10 2,24 14,65 Diapterus rhombeus 2,68 6,26 8,93 3,69 6,77 10,46 19,40 0,01 0,05 0,06 0,06 0,03 0,09 0,15 Mugil liza 2,04 3,74 5,78 3,08 0,29 3,36 9,14 1,38 2,09 3,48 0,76 0,19 0,95 4,43 Mugil curema 0,31 1,87 2,19 0,06 1,73 1,79 3,98 0,04 0,04 0,08 1,03 1,03 1,11 Centropomus undecimalis 0,15 0,15 0,25 0,13 0,38 0,53 1,91 1,91 1,91 Microgobius meeki* 0,60 0,60 0,05 0,18 0,22 0,83 0,23 0,23 1,28 1,28 1,51 Sardinella janeiro** 2,05 2,05 2,05 Brevoortia aurea 0,02 0,02 0,04 0,13 0,03 0,16 0,21 0,53 0,53 0,43 0,78 1,21 1,74 Cetengraulis edentulus 1,43 0,06 1,49 0,02 0,02 1,51 Eucinostomus gula 0,28 0,32 0,59 0,25 0,24 0,49 1,08 0,03 0,03 0,15 0,08 0,23 0,26 Eucinostomus argenteus 0,19 0,34 0,53 0,29 0,02 0,32 0,85 0,11 0,01 0,12 0,10 0,24 0,33 0,45 Brevoortia pectinata** 0,04 0,04 0,08 0,08 0,52 0,47 0,99 0,99 Gobionellus oceanicus 0,17 0,51 0,68 0,28 0,28 0,97 Diapterus olisthostomus 0,20 0,30 0,50 0,23 0,23 0,73 0,04 0,04 0,04 Genidens genidens** 0,07 0,63 0,70 0,70 Atherinella brasiliensis 0,09 0,09 0,09 0,40 0,14 0,54 0,04 0,04 0,58 Centropomus parallelus* 0,17 0,05 0,22 0,22 0,38 0,38 0,38 Citharichthys spilopterus* 0,31 0,19 0,50 0,50 Harengula clupeola 0,37 0,37 0,01 0,01 0,38 Prionotus punctatus* 0,14 0,14 0,14 Opisthonema oglinum 0,06 0,06 0,07 0,07 0,13 Chilomycterus spinosus* 0,13 0,13 0,13 Symphurus plagusia* 0,12 0,12 0,12 Diplectrum radiale** 0,11 0,11 0,11 Achirus lineatus 0,02 0,01 0,03 0,01 0,03 0,04 0,07 <0,01 <0,01 <0,01
31
Citharichthys macrops** 0,02 0,04 0,06 0,06 Oligoplites saurus** 0,06 0,06 0,06 Caranx latus 0,01 0,01 0,04 0,04 0,05 Diplectrum formosum* 0,05 0,05 0,05 Eucinostomus melanopterus* 0,03 0,02 0,05 0,05 Trachinotus carolinus** 0,04 0,04 0,04 Sphoeroides greeleyi* 0,04 0,04 0,04 Synodus foetens* 0,04 0,04 0,04 Archosargus rhomboidalis* 0,03 0,03 0,03 Stephanolipis setifer* 0,02 0,02 0,02 Anchovia clupeoides* 0,01 0,01 0,01 0,01 Sphoeroides testudines* 0,01 0,01 0,01 Stephanolipis hispidus** 0,01 0,01 0,01 Phalloptychus januaris** 0,01 0,01 0,01 Anchoviella lepidentostole** 0,01 0,01 0,01 Chloroscombrus chrysurus* 0,01 0,01 0,01 Haemulon plumierii* 0,01 0,01 0,01 Selene vomer* <0,01 <0,01 <0,01 Anchoa tricolor* <0,01 <0,01 <0,01 Micropogonias furnieri <0,01 <0,01 Trachinotus sp.* <0,01 <0,01 <0,01 Decapoda Callinectes danae 0,69 0,24 0,93 0,13 0,85 0,98 1,90 1,23 1,23 0,17 0,56 0,73 1,96 Callinectes bocourti* 1,25 0,24 1,49 1,49 Farfantepenaeus brasiliensis* 0,38 0,38 0,38 0,77 0,77 0,77 Litopenaeus schmitti 0,72 0,02 0,74 0,08 0,05 0,13 0,86 0,18 0,18 0,10 0,10 0,28 Callinectes ornatus 0,26 0,26 0,26 0,54 0,54 0,08 0,08 0,62 Farfantepenaeus paulensis* 0,16 0,16 0,16 0,26 0,26 0,26 Callinectes sapidus 0,39 0,39 0,01 0,01 0,40 Total 12,32 16,00 28,32 11,79 14,87 26,66 54,98 19,75 32,86 52,62 4,26 7,35 11,60 64,22

32
Considerando as diferenças nos valores totais de biomassa entre as
lagoas, em 2006, a lagoa de Piratininga apresentou uma biomassa total maior
que Itaipu, enquanto que em 2010-11 esta situação se inverteu. Na verdade a
inversão desta relação se deu principalmente pela queda em biomassa da
lagoa de Piratininga, uma vez que a biomassa total de Itaipu se manteve
semelhante (Tabela 5).
A Tabela 6 mostra o resultado da ANOVA bifatorial realizada com os
dados ambientais para cada lagoa. Em Itaipu, a temperatura, pH, oxigênio
dissolvido e profundidade apresentaram diferença significativa tanto entre os
períodos como entre as estações do ano e sua interação. Enquanto que a
granulometria só apresentou diferença significativa entre os períodos, a
transparência entre as estações e os demais fatores não foram
significativamente diferentes.
Em Piratininga, a salinidade, pH e granulometria apresentaram diferença
significativa entre os períodos, as estações do ano e suas interações. A
temperatura só apresentou diferença significativa entre estações, o oxigênio
dissolvido e a profundidade só entre os períodos e a transparência somente
entre a interação entre os dois fatores.
As taxas de precipitação foram iguais para ambas as lagoas,
apresentando uma diferença significativa entre períodos, estações e suas
interações.

33
Tabela 6: Resultado das análises de variância (ANOVA) realizadas para cada parâmetros ambiental nas lagoas de Itaipu e Piratininga por período, estação do ano e sua interação.
Período Estação Período X Estação
r² F p F p F p
Temperatura 0,73 4,36 0,04 104,19 0,00* 10,65 0,00 Salinidade 0,20 0,37 0,55 9,01 0,00 1,96 0,17 pH 0,73 30,24 0,00* 30,03 0,00* 57,58 0,00* O2 (mg.L-1) 0,73 96,12 0,00* 6,31 0,02 15,27 0,00 Transparência (%) 0,18 0,70 0,41 5,90 0,02 3,23 0,08 Profundidade 0,13 5,74 0,02 28,90 0,00* 28,90 0,00* Matéria orgânica (%) 0,07 0,01 0,92 3,28 0,08 0,01 0,92 Granulometria 0,52 41,67 0,00* 2,64 0,11 2,64 0,11
Lgoa
de
Itaip
u
Precipitação 0,70 4,57 0,04 78,00 0,00* 20,91 0,00*
Período Estação Período X Estação
r² F p F p F p
Temperatura 0,75 0,22 0,64 130,99 0,00* 2,10 0,15 Salinidade 0,92 320,69 0,00* 91,47 0,00* 87,33 0,00* pH 0,53 5,62 0,02 27,48 0,00* 17,42 0,00 O2 (mg.L-1) 0,20 10,66 0,00 0,00 0,97 0,18 0,68 Transparência (%) 0,32 2,08 0,16 1,12 0,30 17,91 0,00 Profundidade 0,11 4,66 0,04 0,31 0,58 0,31 0,58 Matéria orgânica (%) 0,03 0,86 0,36 0,22 0,64 0,07 0,79 Granulometria 0,35 14,84 0,00 4,24 0,05 4,24 0,05
Lago
a de
Pira
tinin
ga
Precipitação 0,70 4,57 0,04 78,00 0,00* 20,91 0,00* * p < 0,0001
A análise de componentes principais (PCA), usando os dados de
abundância de espécies, mostrou uma separação entre as lagoas de Itaipu e
Piratininga, independente do período amostral, com os dois primeiros eixos
correspondendo a 22,1 % da variação dos dados (Figura 5). Os resultados da
PERMANOVA reforçam a distribuição espacial do PCA (Tabela 7), indicando
que, independente do período amostral, as lagoas diferem entre si em relação
a íctio e carcinofauna, podendo ser tratadas como dois sub-sistemas
independentes. Assim, as análises subsequentes consideram as duas lagoas
como componentes distintos, permitindo a observação das mudanças em cada
uma, separadamente.

34
De acordo com os “loadings” da análise de PCA, as espécies que
influenciaram significativamente na distribuição das amostras foram: A.
brasiliensis, B. aurea, B. pectinata, Callinectes bocourti (Crustácea,
Portunidae), Centropomus undecimalis, Diapterus auratus, D. rhombeus, E.
gula, Gobionellus oceanicus, M. liza e O. niloticus (Figura 6).
A análise de similaridade (ANOSIM) mostrou diferença significativa, com
um R global igual a 0,26 (p = 0,0001) para as duas lagoas corroborando a
hipótese de que ambas lagoas podem ser analisadas independentemente.

35
Eixo 1 = 12,6 %
Eix
o 2
= 9
,5 %
Itaipu 2010-11
Itaipu 2006
Piratininga 2010-11
Piratininga 2006
Figura 5: Analise de componentes principais (PCA) para a abundância das lagoas de Itaipu e Piratininga, nos períodos amostrais de 2006 e 2010-11. As elipses correspondem a 10% da concentração dos dados.
Tabela 7: Resultado da PERMANOVA comparando a influência dos períodos amostrais (2006 vs 2010-11) na estruturação das comunidades.
Fator F p r²
Espaço 5,665 0,0001 0,49
Tempo 12,266 0,0001 0,1
Espaço x Tempo 3,72 0,0001 0,02

36
Figura 6: “Loadings” (± desvio padrão) das espécies responsáveis pela distribuição das amostras no plano do PCA. As espécies cujos desvios padrões não cruzam o eixo X podem ser consideradas significativas para a distribuição encontrada no PCA.
A análise de correspondência canônica (CCA) para a lagoa de Itaipu
revelou uma maior separação entre os períodos amostrais na época de
inverno, enquanto que as amostras do verão não apresentaram distinção no
espaço canônico. Os dois primeiros eixos, juntos, explicaram 14,1 % da
variação dos dados. A precipitação apresentou uma relação positiva com as
amostras do verão, assim como a quantidade de areia no sedimento. A
transparência, profundidade e porcentagem de lama no sedimento
apresentaram relações negativas com este período amostral (Figura 7). A
salinidade não apresentou uma diferença significativa entre os períodos,
indicando que a abertura do canal do Tibau não teve grande influência nas
variações da salinidade na lagoa de Itaipu.

37
Em relação às amostras do inverno, o oxigênio dissolvido esteve
positivamente relacionado com as amostras de 2010-11, assim como a
profundidade e o pH, sendo estes, provavelmente os responsáveis pela
separação entre os períodos, principalmente o oxigênio, que apresentou uma
relação negativa com as amostras do inverno de 2006. Já a transparência e a
porcentagem de lama no sedimento estiveram positivamente relacionadas com
as amostras do inverno de 2006.
Segundo os resultados obtidos pelo diagrama t-valor biplot, nenhuma
delas apresentou relação significativa com os fatores ambientais. Porém, de
acordo com a CCA, foi possível observar uma tendência de agrupamento das
espécies H. clupeola, M. furnieri, Trachinotus carolinus, M. curema e Caranx
latus, com as amostras do verão nos dois períodos. Além disso, S. brasiliensis,
Oligoplites saurus e B. pectinata apresentaram uma maior relação com as
amostras do inverno de 2010-11, enquanto O. niloticus, C. ornatus,
Farfantepenaeus paulensis (Crustacea, Penaeidae), F. brasiliensis, Symphurus
plagusia, Prionotus punctatus e C. spilopterus com as do inverno de 2006.
Destas, a única que também esteve presente nas amostras de 2010-11 foi O.
niloticus, porém em quantidade bem menor (Figura 7).

38
Figura 7: Analise de correspondência canônica (CCA) para a lagoa de Itaipu nos dois períodos amostrais. Os vetores representam as variáveis ambientais que apresentaram uma relação significativa com a distribuição dos dados.
Em Piratininga, os dois primeiros eixos da CCA corresponderam a 19,3
% da variação dos dados. O primeiro eixo explicou a maior parte da variação
(11,8 %), sendo que a salinidade foi o fator que apresentou maior relação na
estruturação dos dois grupos de amostras formados, devido ao posicionamento
e tamanho do vetor que representa esta variável no espaço canônico. Em
relação as estações do ano, em 2010-11 não foi observado um padrão claro de
separação, ao contrário das amostras de 2006, as quais tiveram a

39
profundidade e precipitação separando-as, estando a primeira relacionada com
as amostras de inverno e a segunda com as de verão (Figura 8).
As espécies mais relacionadas com as amostras de 2010-11 foram B.
pectinata, E. gula, E. argenteus, C. danae e M. furnieri. Já, L. schmitti, C.
sapidus, C. ornatus, F. brasiliensis e F. paulensis estiveram relacionados com
as amostras do inverno e C. parallelus e C. undecimalis com as do verão de
2006. Dois grupos de espécies estiveram presentes em ambos períodos,
contribuindo pouco para a separação das amostras no espaço canônico. Ainda
assim, D. rhombeus e M. curema foram mais importantes para as amostras de
2010-11 e M. liza, A. brasiliensis e O. niloticus, para as de 2006 (Figura 8).
Em Piratininga, de acordo com o diagrama t-valor biplot, três fatores
ambientais apresentaram uma relação significativa com as espécies
capturadas. A precipitação e a transparência apresentaram uma relação
negativa significativa com as espécies de camarão (F. brasiliensis e F.
Paulensis), as quais tiveram seus vetores representados dentro do círculo de
relação negativa ao fator ambiental (Figuras 9a e b). A salinidade foi o fator
ambiental mais significativamente relacionado com as espécies (positivamente
com E. gula, E. argenteus, e B. pectinata, e negativamente com C. bocourti, F.
brasiliensis, F. paulensis, C. undecimalis e C. parallelus), evidenciando, mais
uma vez, sua importância na estruturação da comunidade em Piratininga
(Figura 9c).

40
Figura 8: Analise de correspondência canônica (CCA) para a lagoa de Piratininga para os dois períodos amostrais. Os vetores representam as variáveis ambientais que apresentaram uma relação significativa com a distribuição dos dados.

41
Figura 9: Diagrama t-valor biplot da lagoa de Piratininga dos fatores ambientais que apresentaram relação significativa com as espécies. Os círculos representam a área significativa do teste. As espécies que aparecem dentro desta área apresentam uma relação significativa com o fator ambiental..

42
De acordo com a análise de partição de variância foi observado que
para a lagoa de Itaipu, 28,89% da variação da fauna de peixes e crustáceos
coletados ocorreu devido aos fatores ambientais monitorados e 3,22% devido a
diferenças temporais (Figura 10). O resultado desta análise para a lagoa de
Piratininga mostrou que 34,03% da variação da fauna foi devido aos fatores
ambientais, apenas 3,50% devido as diferenças temporais entre as amostras e
6,64% devido a colinearidade entre os fatores. Um total de 50,84% da variação
não pode ser explicada (Figura 11).
A partição de variância entre os dados ambientais e a salinidade
separadamente mostrou que 8,79% da variação na comunidade de peixes e
crustáceos na lagoa de Piratininga foi devido à salinidade (Figura 12). Esse
resultado corrobora o padrão encontrado no CCA onde a salinidade esteve
positivamente correlacionada às amostras de 2010-11.

43
Figura 10: Esquema ilustrando a porcentagem de variação de cada parâmetro ambiental testado na partição de variância da lagoa de Itaipu. (A) dados ambientais, (B) correlação entre os dados ambientais e o tempo, (C) tempo, (D) porcentagem de variação que não é explicada por nenhum dos parâmetros medidos.
Figura 11: Esquema ilustrando a porcentagem de variação de cada parâmetro ambientail testado na partição de variância da lagoa de Piratininga. (A) dados ambientais, (B) correlação entre os dados ambientais e o tempo, (C) tempo, (D) porcentagem de variação que não é explicada por nenhum dos parâmetros medidos.

44
Figura 12: Esquema ilustrando a porcentagem de variação de cada parâmetro ambiental testado na partição de variância da lagoa de Piratininga. (A) dados ambientais, sem a salinidade, (B) correlação entre os dados ambientais e a salinidade, (C) salinidade isoladamente, (D) porcentagem de variação que não é explicada por nenhum dos parâmetros testados.
Em Itaipu, as espécies que foram significativas para separação dos dois
períodos foram: Citharichthys spilopterus, D. auratus, F. paulensis, D.
rhombeus, M. liza, Genidens genidens e Anchoviella lepidentostole (Tabela 8).
Destas, C. spilopterus e F. paulensis foram exclusivas de 2006 e A.
lepidentostole e G. genidens foram exclusivas de 2010-11. As demais espécies
apresentaram maior abundancia em 2006, mas estiveram presentes nos dois
períodos amostrais.
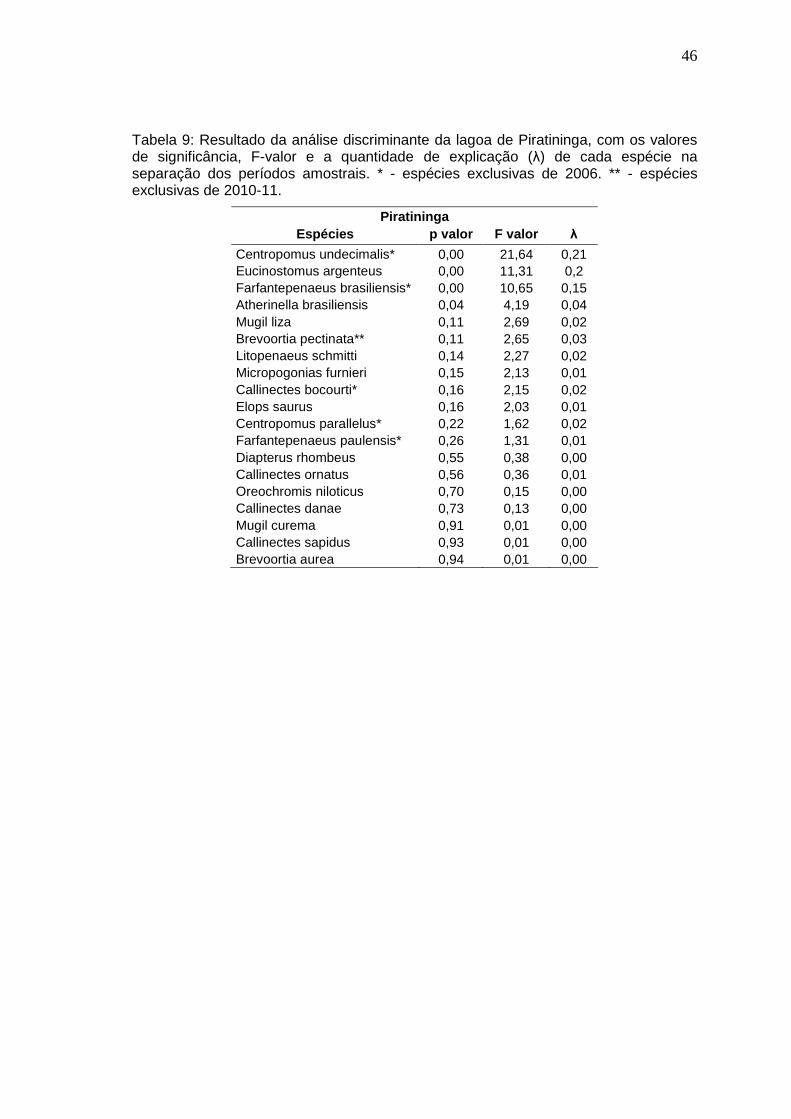
A análise discriminante revelou que as espécies que contribuíram
significativamente para a distinção da fauna entre os anos de 2006 e 2010-11
na lagoa de Piratininga foram: C. undecimalis, E. argenteus, F. brasiliensis e A.
brasiliensis (Tabela 9). Centropomus undecimalis e F. brasiliensis só foram
capturados em 2006 na lagoa de Piratininga. Atherinella brasiliensis e E.

45
argenteus estiveram presentes nos dois períodos, porém a primeira ocorreu em
maior quantidade em 2006, enquanto que a segunda em 2010-11.
Tabela 8: Resultado da análise discriminante da lagoa de Itaipu, com os valores de significância, F-valor e a quantidade de explicação (λ) de cada espécie na separação dos períodos amostrais. * - espécies exclusivas de 2006. ** - espécies exclusivas de 2010-11.
Itaipu
Espécies p valor F valor λ
Citharichthys spilopterus* 0,00 35,58 0,44 Diapterus auratus 0,01 7,27 0,07 Farfantepenaeus paulensis* 0,02 5,57 0,06 Diapterus rhombeus 0,02 6,15 0,04 Mugil liza 0,03 4,89 0,04 Genidens genidens** 0,03 4,84 0,03 Anchoviella lepidentostole** 0,04 4,56 0,03 Centropomus parallelus* 0,05 3,88 0,04 Micropogonias furnieri 0,05 4,03 0,03 Brevoortia pectinata** 0,07 3,51 0,02 Oreochromis niloticus 0,08 3,17 0,01 Elops saurus 0,08 3,16 0,02 Atherinella brasiliensis 0,09 3,06 0,01 Synodus foetens* 0,09 3,07 0,02 Eucinostomus gula 0,10 2,91 0,01 Caranx latus 0,11 2,72 0,01 Farfantepenaeus brasiliensis* 0,13 2,47 0,01 Litopenaeus schmitti 0,13 2,39 0,01 Mugil curema 0,14 2,29 0,01 Achirus lineatus 0,16 2,14 0 Brevoortia aurea 0,17 2,13 0 Citharichthys macrops** 0,21 1,62 0,01 Eucinostomus argenteus 0,23 1,56 0,01 Prionotus punctatus* 0,27 1,26 0,01 Anchoa tricolor* 0,31 1,1 0,01 Cetengraulis edentulus 0,35 0,92 0 Harengula clupeola 0,38 0,83 0 Oligoplites saurus** 0,43 0,66 0 Symphurus plagusia* 0,44 0,63 0 Eucinostomus melanopterus* 0,47 0,55 0 Callinectes ornatus 0,48 0,52 0 Centropomus undecimalis 0,51 0,45 0,01 Sardinella brasiliensis** 0,51 0,45 0 Opisthonema oglinum 0,52 0,42 0 Chilomycterus spinosus* 0,53 0,39 0 Callinectes danae 0,85 0,04 0 Gobionellus oceanicus 0,88 0,03 0
46
Tabela 9: Resultado da análise discriminante da lagoa de Piratininga, com os valores de significância, F-valor e a quantidade de explicação (λ) de cada espécie na separação dos períodos amostrais. * - espécies exclusivas de 2006. ** - espécies exclusivas de 2010-11.
Piratininga Espécies p valor F valor λ
Centropomus undecimalis* 0,00 21,64 0,21 Eucinostomus argenteus 0,00 11,31 0,2 Farfantepenaeus brasiliensis* 0,00 10,65 0,15 Atherinella brasiliensis 0,04 4,19 0,04 Mugil liza 0,11 2,69 0,02 Brevoortia pectinata** 0,11 2,65 0,03 Litopenaeus schmitti 0,14 2,27 0,02 Micropogonias furnieri 0,15 2,13 0,01 Callinectes bocourti* 0,16 2,15 0,02 Elops saurus 0,16 2,03 0,01 Centropomus parallelus* 0,22 1,62 0,02 Farfantepenaeus paulensis* 0,26 1,31 0,01 Diapterus rhombeus 0,55 0,38 0,00 Callinectes ornatus 0,56 0,36 0,01 Oreochromis niloticus 0,70 0,15 0,00 Callinectes danae 0,73 0,13 0,00 Mugil curema 0,91 0,01 0,00 Callinectes sapidus 0,93 0,01 0,00 Brevoortia aurea 0,94 0,01 0,00

47
5. DISCUSSÃO
Diversos estudos descrevendo a ictio e carcinofauna de lagoas e
ambientes estuarinos buscaram avaliar possíveis relações entre a fauna e
determinados fatores abióticos, tais como oxigênio dissolvido, pH, taxas de
precipitação, turbidez, maré, temperatura e principalmente salinidade da água
(e.x. MONTEIRO-NETO et al. 1990; CASTILLO-RIVERA et al. 2010;
CONTENTE et al. 2011a; FRANÇA et al. 2011). Normalmente estes estudos
são realizados avaliando (a) um gradiente longitudinal dos fatores ao longo de
um estuário (ex. LOEBMANN et al. 2008; SANCHEZ-BOTERO et al. 2008;
PLAVAN et al. 2010; VENDEL et al. 2010; CONTENTE et al. 2011a; FRANÇA
et al. 2011); (b) mudanças sazonais em lagoas ou estuários com ligações
intermitentes com o mar (YOUNG et al. 1997; GRIFFITHS 2001; YOUNG &
POTTER 2002; FERREIRA & FREIRE 2009); ou (c) a comunidade após a
abertura de um canal, sem ter um estudo anterior (WASSERMAN 2000; SAAD
et al. 2002). Até o momento, nenhum trabalho comparou as características da
íctio e carcinofauna e condições pré e pós uma intervenção, como a abertura
do canal do Tibau no sistema lagunar Piratininga Itaipu.
A riqueza de espécies do complexo lagunar Piratininga-Itaipu foi similar
a de outros estudos feitos em lagoas costeiras tropicais apresentando em torno
de 30 espécies (GRIFFITHS 2001; SCHIFINO et al. 2004; GARCIA et al. 2007;
MACEDO-SOARES et al. 2007; LOEBMANN et al. 2008; SANCHEZ-BOTERO
et al. 2008). A comunidade de peixes e crustáceos das lagoas apresentou
semelhanças com algumas comunidades estudadas em lagoas costeiras
brasileiras, principalmente do Rio de Janeiro, com representantes da família

48
Gerreidae e espécies como M. liza, M. curema, B. aurea, B. pectinata, G.
genidens, A. brasiliensis, F. brasiliensis, L. schmitti e Callinectes spp. bastante
frequentes, estando fortemente associadas a estes ambientes (ANDREATA et
al. 1989; 1997; ALBERTONI 1998; BARROSO et al. 2000; ARAÚJO &
AZEVEDO 2001; FERREIRA & FREIRE 2009).
Diferenças encontradas na ictio e carcinofauna entre o presente estudo
e os anteriormente citados podem ser devido ao uso da arte de pesca
escolhida. Contudo, estudos demonstraram que não há grande diferença de
captura entre os diferentes amostradores em lagoas costeiras (ANDREATA et
al. 1989; MONTEIRO-NETO et al. 1990; FORTES 2007). Andreata et al.
(1989), na lagoa de Marapendi, observaram que a tarrafa e a pesca de arrasto
apresentaram uma captura semelhante em relação a composição de espécies,
com a maioria das espécies em comum, e sugeriram que o arrasto é mais rico
em espécies e amostra melhor as margens enquanto a tarrafa é mais eficiente
para as áreas mais profundas. Entretanto, Monteiro-Neto et al. (1990) sugerem
que o arrasto apresenta fortes limitações práticas em ambientes lamosos,
como é o caso do complexo lagunar estudado. Além disso a comparação de
diversas artes de pesca no complexo lagunar Piratininga-Itaipu feita por Fortes
(2007), destacou que a tarrafa foi o amostrador que capturou mais espécies
exclusivas. Com base nisso e considerando que o presente estudo tem fins
comparativos, a utilização apenas da tarrafa se mostrou satisfatória,
principalmente por ela apresentar outras vantagens como o fácil manuseio,
diminuindo o esforço amostral e a possibilidade do cálculo da área amostrada,
podendo-se estimar a densidade de cada espécie.

49
A maioria das espécies exclusivas de 2010-11 apareceram em Itaipu e
não em Piratininga, com exceção de Phalloptychus januarius (com apenas um
indivíduo) e de B. pectinata. Além disso, duas espécies, A. lineatus e D.
auratus (ambas com apenas um indivíduo), que antes estavam presentes
apenas em Itaipu, em 2010-11 foram registradas também em Piratininga.
A ausência de B. pectinata em 2006 pode estar relacionada a problemas
de identificação, uma vez que sua separação taxonômica de B. aurea é bem
difícil devido a similaridades entre as espécies. Assim, a relação positiva de B.
pectinata com a salinidade pode não representar uma relação direta com este
fator. Por outro lado a ocorrência de P. januarius em 2010-11 pode ter sido
ocasional, uma vez que a tarrafa não é o melhor amostrador para as margens
da lagoa, onde essa espécie normalmente é encontrada. Segundo Fortes
(2007), esta espécie esteve presente em Piratininga, mas foi capturada com
outro petrecho, assim como D. auratus. Já o aparecimento de A. lineatus, que
antes só estava presente em Itaipu, pode indicar que o aumento da salinidade
e da profundidade em Piratininga, facilitou o trânsito desta espécie de uma
lagoa para a outra, uma vez que ela já esteve associada a regiões mais salinas
(ANDREATA et al. 1989; BARLETTA et al. 2005) e a um gradiente de
profundidade (ALLEN & BALTZ 1997).
Todas as novas ocorrências em Itaipu (S. brasiliensis, Anchoviella
lepidentostole, Oligoplites saurus, Citharichthys macrops, Trachinotus
carolinus, Diplectrum radiale, Stephanolepis hispidus e G. genidens), com
exceção de C. macrops, são espécies registradas para a região costeira de
Itaipu, reforçando a idéia de que a fauna presente em estuários e lagoas

50
apresenta grande conectividade com as áreas rasas marinhas adjacentes
(MONTEIRO-NETO et al. 2008). Além disso, três destas espécies, S.
brasiliensis, A. lepidentostole e D. radiale, foram capturadas com outro
petrecho na lagoa de Itaipu em estudos anteriores (FORTES 2007).
Diapterus rhombeus e E. argenteus, espécies mais abundantes tanto em
Itaipu quanto em Piratininga, são frequentemente encontradas em maior
abundância nas regiões mais baixas de estuários, onde a salinidade é maior
(ANDREATA et al. 1989; CONTENTE et al. 2011a), sendo muitas vezes
classificadas como espécies visitantes-marinhas (GARCIA & VIEIRA 2001;
GARCIA et al. 2001). GARCIA et al. (2001) observaram que tais espécies tem
sua abundância reduzida com a diminuição da salinidade. Os resultados
encontrados neste trabalho corroboram a hipótese de que ambas espécies
preferem áreas mais salinas, já que o aumento da salinidade após a abertura
do canal do Tibau, resultou no aumento na abundância das duas, sugerindo
que, em águas lagunares mais salinas, elas encontrariam um ambiente
apropriado para o seu desenvolvimento.
Diapterus rhombeus é bastante comum em estuários e lagoas (ARAÚJO
et al. 1998; ARAÚJO & AZEVEDO 2001; CASTILLO-RIVERA et al. 2005),
normalmente estando entre as espécies mais abundantes sendo, inclusive,
importante para a pesca em diversas lagoas do leste fluminense (BARROSO et
al. 2000). Por outro lado E. argenteus é menos frequente, sendo superada em
número por outras espécies. Assim, sua abundância elevada neste trabalho
pode indicar que a degradação do complexo lagunar e o aumento da salinidade
em Piratininga desfavoreceram espécies que normalmente tem vantagem

51
competitiva sobre ela, possibilitando a sua prevalência. Inclusive, de acordo
com as análises realizadas, a salinidade foi importante para o aumento das
duas espécies de Eucinostomus presentes em Piratininga.
Das 22 espécies exclusivas em 2006, as mais representativas (com
abundância superior a 5 indivíduos) foram: F. brasiliensis, F. paulensis, C.
parallelus, C. spilopterus e C. bocourti.
Perez-Castaneda & Defeo (2001) observaram em uma lagoa no México,
a preferência de F. brasiliensis por regiões mais salinas, indicando que a média
de salinidade ótima para esta espécie situa-se entre 15 e 35‰ (BRITO et al.
2000). Outros estudos (ex. ALBERTONI 1998; ALBERTONI et al. 2003)
observaram também que a abertura esporádica da conexão entre lagoas e o
mar adjacente aumenta o recrutamento de pós-larvas de camarões F.
brasiliensis e F. paulensis, renovando os estoques nesses ambientes. Assim,
era de se esperar um padrão de acréscimo destas espécies no complexo
Piratininga-Itaipu após a abertura do canal. Todavia, o padrão encontrado foi o
oposto e com estas espécies apresentando uma relação negativa com a
salinidade.
Uma vez que F. brasiliensis e F. paulensis também se relacionaram
negativamente com a taxa de precipitação e transparência da água em
Piratininga, estes fatores poderiam ser julgados importantes para a não
ocorrência destas espécies. No entanto, elas também não foram capturadas
em Itaipu, onde não houve diferença significativa de salinidade e nenhuma
relação direta delas com os dados ambientais. Assim, o padrão encontrado
deve estar relacionado a outros fatores como: o aumento da pesca, já que

52
estas espécies são importantes recursos pesqueiros (ALBERTONI 1998;
BARROSO et al. 2000); diminuição da vegetação (observação dos pescadores
e pessoal, quando comparada à descrição dos habitats feita por FORTES
2007), visto que esta aumenta a complexidade de habitats, provocando um
crescimento na riqueza e diversidade do ambiente (ARAÚJO & AZEVEDO
2001; MARTINO & ABLE 2003) e, mais especificamente, na abundância
dessas duas espécies (GARCIA et al. 1996; PEREZ-CASTANEDA & DEFEO
2001), além de ser um agregador de sedimento fino, fator relacionado com as
espécies em Itaipu.
O desaparecimento de C. parallelus em 2010-11 pode estar relacionado
ao aumento no aporte de contaminantes domésticos, uma vez que esta
espécie é sensível a poluentes surfactantes, principalmente em altas
salinidades, podendo inclusive ser um indicador deste tipo de contaminante em
ambientes estuarinos (ROCHA et al. 2007). Além disso, esta espécie
normalmente é encontrada associada a regiões mais altas de estuários e
lagoas, onde a salinidade é menor (ANDREATA et al. 1989; CONTENTE et al.
2011a). Como C. parallelus foi capturada apenas no período de 2006, nas duas
lagoas, podemos inferir que o aumento da eutrofização do sistema (CERDA
2012), reduzindo a qualidade da água, também pode ter influenciado as
alterações encontradas na ictiofauna das lagoas (MACEDO & SIPAÚBA-
TAVARES 2010). Ainda, o aumento da salinidade em Piratininga, o qual
apresentou uma relação negativa com esta espécie, provavelmente agravou
ainda mais a sensibilidade dela, impossibilitando sua sobrevivência no
complexo lagunar.

53
Já a ausência de C. spilopterus pode estar relacionada com o aumento
da concentração de oxigênio dissolvido e de substrato mais grosso, uma vez
que Allen & Baltz (1997) sugeriram que C. spilopterus estaria mais associado a
regiões com menores concentrações de oxigênio dissolvido e substratos mais
finos, indicando que estes fatores podem ter sido importantes na diminuição da
densidade desta espécie, acarretando sua ausência nas capturas no período
de 2010-11.
O único Portunidae exclusivo de 2006, C. bocourti, foi capturado apenas
em Piratininga e, neste trabalho, apresentou uma relação negativa com a
salinidade. Embora considerada uma espécie pouco abundante em outros
estudos (MONTEIRO-NETO et al. 2000), a total ausência dela no período de
2010-11 e sua relação significativa com a salinidade indicam que esta espécie
pode ser a mais sensível a alterações neste fator. O aumento da salinidade na
lagoa também pode ter influenciado a diminuição da abundância de C. sapidus,
espécie mais relacionada a áreas menos salinas de estuários (Monteiro-Neto et
al. 2000). A diminuição da riqueza e abundancia da família Portunidae pode ser
reflexo também da diminuição da vegetação das lagoas e o aumento de C.
danae em Itaipu pode ter ocorrido devido a sua melhor adaptação em áreas
com menos vegetação. Segundo Garcia et al. (1996) C. sapidus é mais
abundante em áreas com mais vegetação na lagoa dos patos, enquanto que C.
danae não apresenta relação direta com a quantidade de vegetação, indicando
que a ocorrência desta espécie independe deste fator.
Atherinella brasiliensis é uma espécie estuarino-residente (FROTA &
CARAMASCHI 1998; GARCIA & VIEIRA 2001) que tolera longos períodos de

54
fechamento de lagoas com o mar (SERGIPENSE & VIEIRA 1999; CONTENTE
et al. 2011b), porém no primeiro momento de abertura sua população sofre
uma emigração para áreas marinhas adjacentes à lagoa (FROTA &
CARAMASCHI 1998). Esta espécie era considerada abundante e frequente na
lagoa de Piratininga (SERGIPENSE & VIEIRA 1999), porém desde o primeiro
período amostral ela apresentou uma queda em abundância, que se mostrou
ainda mais evidente neste trabalho. Possivelmente este último acontecimento
está ligado à abertura do canal, já que ela foi apontada como uma das
espécies responsáveis pela diferenciação entre os dois períodos.
A perda em biomassa na lagoa de Piratininga foi consequência da
diminuição nas capturas de O. niloticus. O raro contato com o mar, provocado
pela ausência de um canal de ligação direta durante um longo período,
ocasionou uma mudança de estado na lagoa de Piratininga, com o
desenvolvimento de uma comunidade dominada por espécies dulcícolas
(FROTA & CARAMASCHI 1998), tendo como espécie dominante O. niloticus. A
maioria das espécies dulcícolas encontradas em lagoas e estuários não são
residentes e, por serem mais sensíveis a mudanças na salinidade, ficam
restritas às áreas mais altas de estuários, perto dos rios e onde a amplitude de
salinidade é menor (FROTA & CARAMASCHI 1998; MARTINO & ABLE 2003).
Oreochromis niloticus, espécie exótica utilizada em aquicultura, é uma das
poucas espécies de água doce que pode ser aclimatada a ambientes
estuarinos e lagunares. Porém um aumento na salinidade, como o que ocorreu
na lagoa de Piratininga, pode ter provocado tanto um aumento na mortalidade
quanto a emigração da espécie para áreas mais altas dos rios, conforme

55
observado por FROTA & CARAMASCHI (1998) em outras regiões. Pescadores
de Piratininga também identificaram esta redução da espécie nas suas
capturas (Senhor Manoel, comunicação pessoal). Tal fato, pode elucidar a
perda em abundância e biomassa da espécie entre os períodos amostrais em
Piratininga, reforçando a mudança significativa na estrutura da ictio e
carcinofauna nesta lagoa.
As principais consequências imediatas da abertura de canais sobre a
comunidade de peixes normalmente são: a mortalidade (pelo aumento na
salinidade) e emigração de espécies dulcículas; a possibilidade de imigração
de espécies marinhas (FROTA & CARAMASCHI 1998). A abertura do canal de
Tibau, aparentemente não favoreceu a entrada de espécies marinhas na lagoa,
já que não houve o aumento de tais registros no período de 2010-11 em
Piratininga onde o canal foi aberto. Isso pode ter acontecido pelo fato do canal
não ter ligação direta com a zona de arrebentação, por ser submerso,
dificultando a entrada de juvenis que normalmente estão nessas áreas
aguardando o momento certo para adentrarem as lagoas (BELL et al. 2001).
Além disso, a lagoa ficou fechada por muitos anos impossibilitando que
espécies marinhas que entram nas lagoas para reprodução sincronizassem
seus ciclos reprodutivos com a abertura desta (HARRISON & WHITFIELD
1995; FROTA & CARAMASCHI 1998).
Alterações nas condições de lagoas e estuários após ligações com o
mar, como foi visto neste estudo, são bem documentadas na literatura (ex.
GRIFFITHS 2001; YOUNG & POTTER 2002; BONETTI et al. 2005;), inclusive
no Brasil (e.x. ARAÚJO & AZEVEDO 2001; LOEBMANN et al. 2008;

56
FERREIRA & FREIRE 2009) e no estado do Rio de Janeiro (ex. SANTOS et al.
2007; SANCHEZ-BOTERO et al. 2008). Frota & Caramaschi (1998), na lagoa
de Imboacica, verificaram pouca atividade pesqueira no período em que a
barra esteve fechada, mas com a sua abertura houve um crescimento da pesca
devido à entrada de espécies marinhas. Lima et al. (2001) observaram um
aumento da pesca de espécies dulcícolas durante a abertura da barra,
atribuída a sua maior vulnerabilidade devido às modificações hídricas
provocadas pelo canal.
Mudanças nas características ambientais são apontadas como sendo
fatores capazes de limitar a distribuição de algumas espécies, favorecer outras
e facilitar o acesso de espécies costeiras a esses ambientes (SAAD et al. 2002;
MACEDO-SOARES et al. 2007). Na lagoa de Piratininga, assim como em
outros estudos (FROTA & CARAMASCHI 1998; LIMA et al. 2001, ALBERTONI
et al. 2003, MACEDO-SOARES et al. 2007; SANTOS et al. 2007), estes
processos ficaram evidenciados pelo aumento na riqueza de peixes e
crustáceos de origem marinha e diminuição de espécies dulcículas,
principalmente de O. niloticus. Dentre os fatores monitorados neste trabalho, a
salinidade foi a variável determinante nas mudanças faunísticas, corroborando
a idéia de que ela é a grande responsável pelas variações espaciais e
temporais na fauna de ecossistemas lagunares e estuarinos (MONTEIRO-
NETO et al. 1990; SUZUKI et al. 1998; LIMA et al. 2001; MARTINO & ABLE
2003; AKIN et al. 2005; BARLETTA et al. 2005; POMBO et al. 2005;
CASTILLO-RIVERA et al. 2010; CONTENTE et al. 2011a).

57
Estudos propõem que diferenças de salinidade podem promover fortes
respostas biológicas espécie-específicas uma vez que, apesar de muitos
peixes serem capazes de aguentar diferentes níveis de salinidade, existem
níveis ótimos para cada espécie (AKIM et al. 2005; CONTENTE et al. 2011a).
Por isso, pode-se dizer que a abertura de estuários e lagoas altera as
condições normais do sistema, interferindo na sua estrutura hidrológica (CRUZ
et al. 1996; SUZUKI et al. 1998, 2002; BONETTI et al. 2005, WEBSTER 2010),
planctônica (BRANCO et al. 2007; SANTANGELO et al. 2007) e,
consequentemente, na composição e recrutamento de peixes (POLLARD 1994;
YOUNG & POTTER 2002; POMBO et al. 2005) e crustáceos (ALBERTONI
1998; ALBERTONI et al. 2003; FERREIRA & FREIRE 2009), eventos que
também puderam ser observados neste trabalho.
O presente estudo observou uma alteração na estrutura da comunidade
de peixes e crustáceos do complexo lagunar de Piratininga-Itaipu decorrente
das alterações no estado das lagoas, especialmente da elevação da salinidade,
decorrente principalmente da abertura do canal de Tibau. Resultado diferente
foi encontrado por Griffiths (2001), que observou que a ictiofauna de uma lagoa
na Austrália se mostrou estável e resistente às mudanças físico-químicas do
ambiente provocadas pela sua ligação esporádica com o mar. Todavia, ele
relacionou esta estabilidade à baixa mudança na salinidade, mantendo-se
numa faixa intermediária de tolerância para espécies estuarinas e marinhas. No
caso deste estudo a elevação da salinidade, principalmente na lagoa de
Piratininga, foi muito grande, provocando as diferenças na ictio e carcinofauna
observadas.

58
Aparentemente, a abertura do canal de Tibau não se mostrou totalmente
benéfica, pelo menos na escala de tempo analisada neste estudo. As
alterações observadas levaram a uma diminuição na diversidade, abundância e
biomassa da ictio e carcinofauna das lagoas, e segundo os pescadores locais,
provocou mortandade de peixes e uma drástica diminuição da pesca local
(Senhor Manoel, comunicação pessoal). Resultados opostos foram
encontrados em outros estudos, os quais sugerem que a ligação de lagoas e
estuários com o mar pode ser benéfica para as comunidades de peixes, se o
ecossistema apresentar uma resistência aos distúrbios gerados por ela (SAAD
et al. 2002; MACEDO-SOARES et al. 2007). Por exemplo, em seus estudos em
lagoas costeiras com ligação esporádica com o mar, Young et al (1997) e Saad
et al. (2002) perceberam uma renovação dos indivíduos sempre que o canal é
aberto, prevalecendo espécies marinhas devido ao aumento da salinidade, mas
com o tempo a comunidade conseguiu se equilibrar novamente. Ainda, de
acordo com Sanchez-Botero et al. (2008) e Young & Potter (2002), a falta de
ligação com o mar por um longo período de tempo constitui um distúrbio para
lagoas costeiras e estuários, diminuindo a estabilidade e riqueza de espécies
no sistema e por consequência o número de espécies marinhas.
Estudos sugerem que processos esporádicos de abertura de canais
representam uma perturbação de pequena duração, possibilitando a posterior
reestruturação da comunidade, enquanto que distúrbios de longa duração
podem não ter o mesmo resultado (LIMA et al. 2001; SAAD et al. 2002).
Segundo Frota & Caramaschi (1998), se a abertura do canal é feita entre curtos
intervalos de tempo, a comunidade não tem tempo de se estabilizar devido a

59
frequente emigração e imigração de determinadas espécies. No caso da lagoa
de Piratininga, a intervenção do canal do Tibau fez com que a lagoa, que antes
não apresentava nenhuma ligação direta com o mar, passasse a estar
constantemente aberta, gerando um grande distúrbio e induzindo uma queda
da diversidade no momento imediatamente após a abertura do canal. Esse
distúrbio de longa duração pode provocar outros tipos de alterações nas
comunidades, como a mudança nas espécies dominantes, podendo alterar
todo o funcionamento e dinâmica do sistema.
A relação entre o aumento no grau de distúrbio do ambiente e a
diminuição na diversidade de peixes também foi visto por Pollard (1994) ao
comparar três lagoas com diferentes regimes de ligação com o mar. Seu
estudo indicou que a única que apresentava ligação permanente com o mar era
a mais rica e com a ictiofauna mais distinta, enquanto que as outras (com
abertura intermitente) eram mais parecidas entre si e estariam empobrecidas,
justamente devido ao distúrbio gerado nos momentos de ligação. Ainda, Araújo
et al. (1998) encontraram uma alta riqueza na Baia de Sepetiba e atribuíram à
relativa estabilidade dos fatores ambientais na região. Com base nisso
podemos inferir que o complexo lagunar Piratininga-Itaipu se encontra num
estágio transicional, razão pela qual ainda não pode ser observado um
equilíbrio no sistema. O esperado é que, com o tempo, suas características
ambientais se estabilizem e sua comunidade consiga se reestruturar, mesmo
que as espécies chaves não sejam as mesmas de antes. Por isso, é
indispensável o monitoramento futuro que avalie o grau de recuperação das
lagoas.

60
Com base nos dados observados sustentamos a idéia de que a
tolerância de organismos em um gradiente abiótico é o que parece determinar
a estrutura de uma comunidade (MARTINO & ABLE 2003; AKIN et al. 2005),
com algumas espécies optando por locais com características ambientais
especificas (POMBO et al. 2005). Isso também foi discutido por Andreata et al.
(1989), que observou que a distribuição espacial de peixes dividiu a lagoa de
Marapendi em dois grupos influenciados principalmente pela salinidade: um
constituído pela área do canal com salinidade mais elevada, e o outro
compreendendo as demais localidades da lagoa.
Em Piratininga os componentes abióticos foram responsáveis por quase
35% da variação nos dados de fauna e a salinidade sozinha por quase 9%,
enquanto em Itaipu eles foram responsáveis por quase 30% desta variação.
Porém, eles não foram os únicos influenciando a mudança na ictio e
carcinofauna das lagoas, ainda restando em torno de 50% a ser explicada por
outros fatores. Em muitos casos, extinções locais e alterações abruptas da
estrutura das comunidades ocorrem como resultado de mudanças no tempo de
retenção e qualidade da água, aumento da poluição e eutrofização do sistema
(AGOSTINHO et al. 2005). Sendo assim, esta parcela não explicada pode ser
atribuída também a fatores biológicos não medidos como predação,
competição e o comportamento de algumas espécies (MARTINO & ABLE
2003), ou mesmo pela mudança na vegetação local.
Padrões temporais não ligados às características ambientais
modificados pela abertura do canal, provavelmente tiveram pouca influência na
mudança da comunidade em Piratininga. Em outras lagoas e estuários a

61
distribuição de peixes se mostrou dependente de variações bióticas e abióticas,
com as mudanças espaciais e temporais sendo um reflexo de fatores
ambientais (WHITFIELD 1999; SANCHEZ-BOTERO et al. 2008; CONTENTE et
al. 2011a). Martino & Able (2003) observaram que a comunidade estudada
durante três anos se manteve praticamente estável especialmente pelo fato de
não ter havido grandes alterações nos padrões abióticos.
Como pode ser observado neste trabalho, a construção do canal de
Tibau causou consequências ao complexo lagunar Piratininga-Itaipu, muitas
delas diferentes às registradas em estudos similares. Mesmo quando trabalhos
são feitos na mesma região, podem gerar resultados diferentes devido as
constantes mudanças inerentes aos ecossistemas. Por exemplo, Sanchez-
Botero et al. (2008) realizaram um trabalho semelhante a dois anteriores na
lagoa de Cabíunas e encontraram resultados bastante distintos com relação a
comunidade de peixes. Por essa razão, projetos de monitoramento visando
verificar o desenvolvimento das lagoas são bastante importantes, já que a
preservação destes ambientes depende de estudos específicos e constantes
em cada sistema (SCHIFINO et al. 2004). Isso porque uma mesma intervenção
pode gerar consequências distintas em diferentes ecossistemas devido
principalmente: (1) às diferenças no tamanho de suas bacias de drenagem, (2)
às variações na evolução natural dos ambientes e (3) às diferentes formas de
uso antrópico em cada um deles (ESTEVES 1998; LIMA et al. 2001).

62
6. CONCLUSÃO
Alterações na ictio e carcinofauna do complexo lagunar Piratininga-Itaipu
foram registradas como consequência da abertura do canal de Tibau. Dentre
elas, ficou evidente a diminuição na riqueza e abundância de espécies nas
duas lagoas e a diminuição da biomassa em Piratininga.
A elevação da salinidade na lagoa de Piratininga, provocada diretamente
pela sua ligação com o mar através do canal do Tibau, ocasionou a mudança
na ictio e carcinofauna, promovendo uma troca de dominância de espécies,
levando ao predomínio de espécies marinhas e prejudicando a principal
espécie de água doce que antes dominava a lagoa. Ainda assim, este estudo
sugere que ictio e carcinofauna da lagoa se encontram em um estágio de
transição, onde as espécies dominantes sejam de origem marinha e não de
água doce.
Já em Itaipu, apesar de ter ocorrido uma supressão de algumas
espécies e surgimento de outras, não foi possível identificar o principal fator
responsável pela modificação da comunidade. Processos de eutrofização local
assim como a própria ligação direta com o mar, podem ter sido mais
importantes na estruturação da comunidade de peixes e crustáceos em Itaipu,
do que os possíveis efeitos da abertura do canal do Tibau neste sistema.

63
7. PERSPECTIVAS
É necessário um acompanhamento constante da fauna da região para
monitorar as mudanças a longo prazo no complexo lagunar, para que se possa
acompanhar a evolução do sistema. Além disso, houve um assoreamento no
canal de Tibau que fechou novamente a ligação da lagoa de Piratininga com o
mar. Por esses motivos é fundamental que haja novos estudos acompanhando
a evolução da comunidade do Complexo lagunar Piratininga-Itaipu, avaliando
as consequências deste novo fechamento na comunidade e visando antecipar
o que novas alterações podem gerar, para que medidas de manejo adequadas
sejam realizadas.

64
8. REFERÊNCIAS BIBLIOGRÁFICAS
Agostinho AA, Thomaz SM & Gomes LC (2005) Conservação da
biodiversidade em áreas continentais do Brasil. Megadiversidade, 1(1):
70-78.
Akin S, Buhan E, Winemiller WO & Yilmaz H (2005) Fish assemblage structure
of Koycegiz Lagoone - Estuary, Turkey: Spatial and temporal distribution
patterns in relation to environmental variation. Estuarine, Coastal and
Shelf Science, 64: 671-684.
Albertoni EF (1998) Ocorrência de camarões Peneídeos e Paleomonídeos nas
lagoas Imboassica, Cabiúnas, Comprida e Carapebus. In: Esteves, SA
(ed) Ecologia de lagoas costeiras do Parque Nacional da Restinga de
Jurubatiba e do Município de Macaé (RJ). NUPEM/UFRJ, Rio de
Janeiro, p. 351-358.
Albertoni EF, Palma-Silva C & Esteves FA (2003) Crescimento e fator de
condição na fase juvenil de Farfantepenaeus brasiliensis (Latreille) e F.
paulensis (Perez-Farfante) (Crustácea, Decapoda, Penaeidae) em uma
lagoa costeira tropical do Rio de Janeiro, Brasil. Revista Brasileira de
Zoologia, 20(3): 409-418.
Allen RL & Baltz DM (1997) Distribution and microhabitat use by flatfishes in a
Louisiana estuary. Environmental Biology of Fishes, 50: 85–103.
Anderson MJ (2005) PERMANOVA: a FORTRAN computer program for
permutational multivariate analysis of variance. Departamento de
estatística, Universidade de Auckland, Nova Zelândia, 24 p.

65
Andreata JV, Marca AG, Soares CL & Santos RS (1997) Distribuição mensal
dos peixes mais representativos da lagoa Rodrigo de Freitas, Rio de
Janeiro, Brasil. Revista Brasileira de Zoologia, 14(1): 121-134.
Andreata JV, Saad AM & Barbieri LRR (1989) Associação e distribuição das
espécies de peixes na laguna de Marapendi, Rio de Janeiro, no período
de marçode 1985 a fevereiro de 1987. Memórias do Instituto Oswaldo
Cruz, 84(4): 45-51.
Araújo FG & Azevedo MCC (2001). Assemblages of southeast-south brazilian
coastal systems based on the distribution of fishes. Estuarine, Coastal
end Shelf Science, 52: 729-738.
Araujo FG, Cruz-Filho AG, Azevedo MCC & Santos ACA (1998) Estrutura da
comunidade de peixes demersais da Baía de Sepetiba, RJ. Revista
Brasileira de Biologia, 58(3): 417-430.
Azevedo FBB (2005) Modelagem da capacidade de suporte da laguna de
Saquarema – RJ após a abertura de uma conexão permanente com o
mar. Dissertação de mestrado. Programa de Pós-Graduação em Ciência
Ambiental, Universidade Federal Fluminense, Niterói, 104 p.
Baptista JV & Fernandes VF (2009) Alterações ambientais em decorrência do
processo de urbanização acelerada na bacia hidrográfica do Rio Jacaré,
Niterói – RJ. II seminário de recursos hídricos da bacia Hidrográfica do
Paraíba do Sul: recuperação de áreas degradadas, serviços ambientais
e sustentabilidade. Taubaté, SP. 9-11 de Dezembro, IPABHi, 537-544.

66
Barletta M, Barletta-Bergan A, Saint-Paul U & Hubold G (2005) The role of
salinity in structuring the fish assemblages in a tropical estuary. Journal
of Fish Biology, 66: 45-72.
Barroso LV, Abdo OE & Xavier-da-Silva J (2001) Geoprocessamento aplicado
aos sistemas lagunares de Niterói, Maricá e Saquarema, região costeira
leste-fluminense (RJ). X Simpósio Brasileiro de Sensoriamento Remoto.
Foz do Iguaçu, PR. 21-26 de Abril, INPE, 915-919.
Barroso LV, Medina RS, Moreira-Turcq TF & Bernardes MC (2000) Aspectos
ambientais e atividade de pesca em lagoas costeiras fluminenses.
IBAMA, Brasília, 49p.
Bell KNI, Cowley PD & Whitfield AK (2001) Seasonality in frequency of marine
access to an intermittently open estuary: implications for recruitment
strategies. Estuarine, Coastal and Shelf Science, 52: 327–337.
Blott SJ & Pye K (2001) GRADISTAT: a grain size distribution and statistics
package for the analysis of unconsolidated sediments. Earth Surface
Processes and Landforms, 26: 1237-1248.
Bonetti C, Bonetti J & Beltrame E (2005) Mudanças nas características
composicionais das águas da lagoa de Ibiraquera (SC) em resposta a
dinâmica de abertura e fechamento de sua desembocadura. Brazilian
Journal of Aquatic Sciences and Technology, 9(2): 39-47.
Borcard D, Legendre P & Drapeau P (1992) Partialling out the spacial
component of ecological vatiation. Ecology, 73(3): 1045-1055.

67
Braak CJF ter & Šmilauer P (2002) CANOCO Reference Manual and
CanoDraw for Windows User's Guide: Software for Canonical
Community Ordination (version 4.5). Microcomputer Power, Ithaca, NY,
USA. 500 p.
Branco CWC, Kozlowsky-Suzuki B & Esteves F A (2007) Environmental
changes and zooplankton temporal and spatial variation in a disturbed
brazilian coastal lagoon. Brazilian Journal of Biology, 67(2): 251-262.
Brito R, Chimal ME & Rosas C (2000) Effect of salinity in survival, growth, and
osmotic capacity of early juveniles of Farfantepenaeus brasiliensis
(decapoda: Penaeidae). Journal of Experimental Marine Biology and
Ecology, 244: 253-263.
Castillho-Rivera M, Zárate R & Ortiz S (2005) Diel and seasonal variation of
abundance, richness and dominant species of fishes, in an estuarine soft
bottom environmental. Hidrobiológica, 15(2): 227-238.
Castillo-Rivera M, Zárate-Hernández R, Ortiz-Burgos S & Zavala-Hurtado J
(2010) Diel and seasonal variability in the fish community structure of a
mud-bottom estuarine habitat in the Gulf of Mexico. Marine Egology, 31:
633-642.
Cerda M, Barboza CDN, Carvalho CNS, Jandre KA & Marques Jr NA (2011)
Nutrient budgets in the Itaipu-Piratininga Lagoon system (Southeastern
Brazil): effects of sea-exchange management. Lajar no prelo.
Clarke KR & Gorley RN (2006) PRIMER v6: User Manual/Tutorial. PRIMER-E,
Plymouth, UK, 193 p.

68
Clarke KR (1993) Non-parametric multivariate analysis of changes in
community structure. Australian Journal of Ecology, 18: 117-143.
Coelho PA & Ramos-Porto M (1992) Sinopse dos crustáceos decápodos
brasileiros (Portunidae). Revista Brasileira de Zoologia, 9 (3/4): 291-298.
Colwell RK (1994–2004) EstimateS: statistical estimation of species richness
and shared species from samples.
^http://viceroy.eeb.uconn.edu/estimates&. [Persistent URL:
^http://purl.oclc.org/estimates&.]
Colwell RK Mao CX & Chang J (2004) Interpolating, extrapolating, and
comparing incidence-based species accumulation curves. Ecology, 85:
2717-2727.
Contente RF, Stefanoni MF & Spach HL (2011a) Fish assemblage structure in
an estuary of the Atlantic Forest biodiversity hotspot (Southern Brazil).
Ichthyological Research, 58: 38-50.
Contente RF, Stefanoni MF & Spach HL (2011b) Feeding ecology of the
Brazilian silverside Atherinella brasiliensis (Atherinopsidae) in a sub-
tropical estuarine ecosystem. Journal of the Marine Biological
Association of the United Kingdom, 91(6): 1197-1205.
Costa RC, Fransozo A, Melo GAS & Freire FAM (2003) Chave ilustrada para
identificação dos camarões dendrobranchiata do litoral norte do estado
de São Paulo, Brasil. Biota Neotropica, 3(1): 1-12.
Cruz CBM, Carvalho-Junior W, Barros RS, Argento MSF & Mayr LM (1996)
Impactos ambientais no sistema lagunar de Maricá-Guarapina. VIII

69
Simpósio Brasileiro de Sensoriamento Remoto. Salvador, BA. 14-19 de
Abril, INPE, 137-141.
Esteves FA (1998) Lagoa de Imboacica: impactos antrópicos, propostas
mitigadoras e sua importância para a pesquisa ecológica. In: Esteves, F.
A. (ed). Ecologia das lagoas costeiras do Parque Nacional da Restinga
de Jurubatiba e do Município de Macaé (RJ). UFRJ, NUPEM. Rio de
Janeiro, p. 401-429.
Esteves FA, Caliman A, Santagelo JM, Guariento RD, Farjalla VF & Bozelli RL
(2008) Neotropical coastal lagoons: an appraisal of their biodiversity,
functioning, threats and conservation management. Brazilian Journal of
Biology, 68 (4): 967-981.
Ferreira NC & Freire AS (2009) Spatio-temporal variation of the pink shrimp
Farfantepenaeus paulensis (Crustacea, Decapoda, Penaeidae)
associated to the seasonal overture of the sandbar in a subtropical
lagoon. Iheringia, Série Zoológica, 99(4): 390-396.
Figueiredo JL & Menezes NA (1978) Manual de peixes Marinhos do Sudeste do
Brasil: II teleostei (1). Museu de Zoologia, USP, São Paulo. 110 p.
Figueiredo JL & Menezes NA (1980) Manual de peixes Marinhos do Sudeste do
Brasil: III teleostei (2). Museu de Zoologia, USP, São Paulo. 90 p.
Fortes WLS (2007) Estrutura da comunidade de peixes e crustáceos do
sistema lagunar Piratiniga-Itaipu, RJ. Dissertação de Mestrado.
Programa de Pós-Graduação em Biologia Marinha, Universidade
Federal Fluminense, Niterói, 84 p.

70
França S, Costa MJ & Cabral HN (2011) Inter- and intra-estuarine fish
assemblage variability patterns along the Portuguese coast. Estuarine,
Coastal and Shelf Science, 91: 262-271.
Frota LO & Caramaschi EP (1998) Abertura da barra da lagoa de Imboassica e
seus efeitos sobre a fauna de peixes. In: Esteves SA (ed) Ecologia de
lagoas costeiras do Parque Nacional da Restinga de Jurubatiba e do
Município de Macaé (RJ). NUPEM/UFRJ. Rio de Janeiro, p. 327-350.
Fundação Instituto de Geotécnica do Município do Rio de Janeiro (2011).
http://www2.rio.rj.gov.br/georio/site/alerta/dados/acumuladas.asp? Data
de consulta: 24/08/2011.
Garcia AM & Vieira JP (2001) O aumento da diversidade de peixes no estuário
da Lagoa dos Patos durante o episódio el niño 1997-1998. Atlântica, Rio
Grande, 23: 85-96.
Garcia AM, Hoeinghaus DJ, Vieira JP & Winemiller KO (2007) Isotopic variation
of fishes in freshwater and estuarine zones of a large subtropical coastal
lagoon. Estuarine, Coastal and Shelf Science, 73: 399-408.
Garcia AM, Vieira JP & Winemiller KO (2001) Dynamics of the shallow-water
fish assemblage of the Patos Lagoon estuary (Brazil) during cold and
warm ENSO episodes. Journal of Fish Biology, 59: 1218-1238.
Garcia AM, Vieira JP, Bemvenuti CE & Geraldi RM (1996) Abundância e
diversidade da assembléia de crustáceos decápodos dentro e fora de
uma pradaria de Rupia maritima L. no estuário da Lagoa dos Patos (RS-
Brasil). Nauplius, Rio Grande, 4: 113-128.

71
Gladstone W, Hacking N & Owen V (2006) Effects of artificial openings of
intermittently opening estuaries on macroinvertebrate assemblages of
the entrance barrier. Estuarine, Coastal and Shelf Science, 67: 708-720.
Griffiths SP (2001) Factors influencing fish composition in an australian
intermittently open estuary. is stability salinity-dependent?. Estuarine,
Coastal and Shelf Science, 52: 739-751.
Harrison TD & Whitfield AK (1995) Fish community structure in three
temporarily open/closed estuaries on the Natal coast. Ichthyological
Bulletin, 64: 1-80.
Hotelling H (1933) Analysis of a complex of statistical variables into principal
components. The Journal Of Educational Psycholog, 24: 417-441.
Kjerfve B (1986) Comparative oceanography of coastal lagoons. In: Wolfe DA
(ed). Estuarine Variability. Academic Press, New York, p. 63-81.
Kjerfve B (1994) Coastal lagoons. In: Kjerfev B (ed) Coastal Lagoons
Processes. Elsevier, Amsterdan. 1-8 p.
Knoppers B, Kjerfve B & Carmouze JP (1991) Trophic state and water turn-over
time in six choked coastal lagoons in Brazil. Biogeochemistry, 14: 149-
166.
Legendre P & Legendre L (1998) Numerical Ecology. 2 ed. Elsevier Science B.
V., Amsterdam, 853 p.
Leps J & Smilauer P (2003) Multivariate analysis of ecological data using
CANOCO. Canbridge university press, New York. 269 p.

72
Lima NRW, Bizerril CRSF, Caniçali MR, Suzuki MS & Assumpção J (2001)
Atividade de pesca durante a abertura da barra da lagoa de Iquipari, São
João da Barra, RJ. Boletim do Instituto de Pesca, 27(2): 191–200.
Lin HJ, Shao KT, Kuo SR, Hsieh HL, Wong SL, Chen IM, Lo WT & Hung JJ
(1999) A trophic model of a sandy barrier lagoon at Chiku in
southwestern Taiwan. Estuarine, Coastal and Shelf Science, 48: 575-
588.
Loebmann D, Vieira JP, Bemvenuti MA, Astarloa JMD, Cousseau MB & Figeroa
D (2008) Composição e abundância da ictiofauna de duas lagunas
costeiras da América do Sul Austral: lagoa do Peixe (31ºS; 51ºW), Brasil
e laguna Mar Chiquita (37ºS; 57ºW), Argentina. Neotropical Biology and
Conservation, 3(1): 28-33.
Macedo CF & Sipaúba-Tavares LH (2010) Eutrofização e qualidade da água na
piscicultura: consequências e recomendações. Boletim do Instituto de
Pesca, 36(2):149-163.
Macedo-Soares PHM, Câmara EM & Caramaschi EP (2007) Efeito da abertura
de barra de uma lagoa costeira sobre a comunidade de peixes no norte
do estado do Rio de Janeiro. VIII Congresso de Ecologia do Brasil,
Caxambu, MG. 23- 28 de Setembro, SEB, 1-2.
Mach JSO & Longo OC (1998) Considerações sobre a gestão ambiental -
impacto da construção civil: um estudo de caso do sistema lagunar
Piratininga-Itaipu. 18° Encontro Nacional de Engenharia de Produção.
21-25 de Setembro, Niterói, UFF, 8.

73
Marcolini S & Correa KSS (1989) A Laguna de Piratininga, Município de Niterói.
Prefeitura Distrital da Região Oceânica. Niterói, 4p.
Martino EJ & Able KW (2003) Fish assemblages across the marine to low
salinity transition zone of a temperate estuary. Estuarine, Coastal and
Shelf Science, 56: 969-987.
Menezes NA & Figueiredo JL (1980) Manual de peixes Marinhos do Sudeste do
Brasil: IV teleostei (3). Museu de Zoologia, USP, São Paulo. 96 p.
Menezes NA & Figueiredo JL (1985) Manual de peixes Marinhos do Sudeste do
Brasil: V teleostei (4). Museu de Zoologia, USP, São Paulo. 105 p.
Monteiro-Neto C, Blacher C, Laurent AAS, Snizek FN, Canozzi MB & Tabajara
LLCA (1990) Estrutura da comunidade de peixes em águas rasas na
região de Laguna, Santa Catarina, Brasil. Atlântica, Rio Grande, 12(2):
53-69.
Monteiro-Neto C, Laurent, AAS, Blacher C, Tabajara LLAA & Cannozi MMB
(2000) Distribuição de juvenis de siris do gênero Callinectes no sistema
estuarino-lagunar de Laguna, Santa Catarina, Brasil. Arquivos de
Ciências do Mar, 33: 57-64.
Monteiro-Neto C, Tubino RA, Moraes LES, Mendonça-Neto JP, Esteves GV &
Fortes WL (2008) Associações de peixes na região costeira de Itaipu,
Niterói, RJ. Iheringia, Série Zoológica, 98(1): 50-59.
Moreno T & Castro JJ (1998) Trophic structure of the Maspalomas Lagoon
(Gran Canaria, Canary Islands), a regenerated ecosystem of brackish
water. Boletim do Museu Municipal do Funchal, 5: 245-261.

74
Perez-Castaneda R & Defeo O (2001) Population variability of four sympatric
penaeid shrimps (farfantepenaeus spp.) in a tropical coastal lagoon of
mexico. Estuarine, Coastal and Shelf Science, 52: 631-641.
Plavan AA, Passadore C & Gimenez L (2010) Fish assemblage in a temperate
estuary on the uruguayan coast: seasonal variation and environmental
influence. Brazilian Journal of Oceanography, 58(4): 299-314.
Pollard DA (1994) A comparison of fish assemblages and fisheries in
intermittently open and permanently open coastal lagoons on the south
coast of New South Wales, South-Eastern Australia. Estuaries, 17(3):
631-646.
Pombo L, Elliott M & Rebelo JE (2005) Environmental influences on fish
assemblage distribution of an estuarine coastal lagoon, Ria de Aveiro
(Portugal). Scientia Marina, 69(1): 143-159.
Ramos LA & Vieira JP (2001) Composição específica e abundância de peixes
de zonas rasas dos cinco estuários do Rio Grande do Sul, Brasil.
Boletim do Instituto de Pesca, 27(1): 109-121.
Rao CR (1995) A review of canonical coordinates and an alternative to
correspondence analysis using Hellinger distance. Questiió, 19(1, 2, 3):
23-63.
Rocha AJS, Ngan VGPV, Passos MJACR & Furia RR (2007) Effects of anionic
surfactant and salinity on the bioenergetics of juveniles of Centropomus
parallelus (Poey). Ecotoxicology and Environmental Safety, 68: 397-404.

75
Saad AM, Beaumord AC & Caramaschi EP (2002) Effects of artificial canal
openings on fish community structure of Imboassica coastal lagoon, Rio
deJaneiro, Brazil. Journal of Coastal Research, 36: 634-639.
Sanchez-Botero JI, Caramaschi EP & Garcez DS (2008) Spatiotemporal
variation in fish assemblage in a coastal lagoon without direct contact
with the sea (Southeastern Brazil). Journal of Coastal Research, 24(4):
225-238.
Santangelo JM, Rocha AM, Bozelli RL, Carneiro LS & Esteves FA (2007)
Zooplankton responses to sandbar opening in a tropical eutrophic
coastal lagoon. Estuarine, Coastal and Shelf Science, 71: 657-668.
Santos MC, Ferreira MIP, Pinheiro MRC & Boldrini JC (2007) Aberturas
artificiais de barras das lagoas costeiras - a experiência do Parque
Nacional da Restinga de Jurubatiba. V congresso Brasileiro de
Conservação. Foz do Iguaçu, PR. 17-21 de Junho, 1-11.
Schifino LC, Fialho CB & Verani JR (2004) Fish community composition,
seasonality and abundance in Fortaleza Lagoon, Cidreira. Brazilian
Archives of Biology and Technology, 47(5): 755-763.
Sergipense S & Vieira AC (1999) Aspectos sazonais de ocorrência e tamanho
de Xenomelaniris brasiliensis (Quoy e Gaimard, 1824) (Teleostei,
Atherinidae) na laguna de Piratininga, Niterói, RJ. Oecologia Brasiliensis,
7: 291-304.
Statsoft, Inc. (2005) Statistica (data analysis software system), version 7,1.
www.statsoft.com

76
Suzuki MS, Figueiredo RO, Castro SC, Silva CF, Pereira EA, Silva JA & Aragon
GT (2002) Sand bar opening in a coastal lagoon (Iquipari) in the northern
region of Rio de Janeiro state: hydrological and hydrochemical changes.
Brazilian Journal of Biology, 62(1): 51-62.
Suzuki MS, Ovalle ARC & Pereira EA (1998) Effects of sand bar openings on
some limnological variables in a hypertrophic tropical coastal lagoon of
Brazil. Hydrobiologia, 368: 111-122.
Tavares GVL (2003) O controle externo da gestão ambiental: competência e
prática nos tribunais de contas. 2º Encontro Nacional de Auditoria em
Obras Públicas, São Paulo, 28p.
Underwood AJ (1994) On beyond baci: sampling designs that might reliably
detect environmental disturbances. Ecological Applications, 4(1): 3-15.
Valesini FJ, Potter IC, Platell ME & Hyndes GA (1997) Ichthyofaunas of a
temperate estuary and adjacent marine embayment. Implications
regarding choice of nursery area and influence of environmental
changes. Marine Biology, 128: 317-328.
Vasconcelos RP, Reis-Santos P, Maia A, Fonseca V, França S, Wouters N,
Costa MJ & Cabral HN (2010) Nursery use patterns of commercially
important marine fish species in estuarine systems along the Portuguese
coast. Estuarine, Coastal and Shelf Science, 86: 613-624.
Vendel AL, Bouchereau J & Chaves PT (2010) Environmental and subtidal fish
assemblage relationships in two different brazilian coastal estuaries.
Brazilian Archives of Biology and Technology, 53(6): 1393-1406.

77
Wasserman JC (2000) Relatório de impacto ambiental da Barra Franca na
Lagoa de Saquarema – RJ. Rio de Janeiro, 214 p.
Wasserman JC, Cunha LC, Carneiro ME & Knopper B (1999) The impact of a
canal lock upon the water balance and the trophic state of Piratininga
lagoon, state of Rio de Janeiro, Brazil. Environmental Geochemistry of
Coastal Lagoon Systems, 6: 169-177.
Webster IT (2010) The hydrodynamics and salinity regime of a coastal lagoon -
The Coorong, Australia - seasonal to multi-decadal timescales.
Estuarine, Coastal and Shelf Science, 90: 264-274.
Whitfield AK (1999) Ichthyofaunal assemblages in estuaries: a South African
case study. Reviews in Fish Biology and Fisheries, 9: 151–186.
Williams AB (1974) Te swimming crabs of the genus Cllinectes (decapoda:
Portunidae). Fishery Bulletin, 72(3): 685-798.
Young GC & Potter IC (2002) Influence of exceptionally high salinities, marked
variations in freshwater discharge and opening of estuary mouth on the
characteristics of the ichthyofauna of a normally-closed estuary.
Estuarine, Coastal and Shelf Science, 55: 223-246.
Young GC, Potter IC, Hyndes GA & Lestang S (1997) The ichthyofauna of an
intermittently open estuary: implications of bar breaching and low
salinities on faunal composition. Estuarine, Coastal and Shelf Science,
45: 53-68.





























