Embed Size (px)
Citation preview

Programa de Pós-graduação em Tecnologias aplicadas a saúde
Daniela Oliveira Lima Magalhães
IDENTIFICAÇÃO E ANÁLISE DE REDES DE GENES ENVOLVIDOS NO CÂNCER
DE MAMA E CÉLULAS-TRONCO CANCEROSAS
Belo Horizonte, MG
2018

Daniela Oliveira Lima Magalhães
IDENTIFICAÇÃO E ANÁLISE DE REDES DE GENES ENVOLVIDOS NO CÂNCER
DE MAMA E CÉLULAS-TRONCO CANCEROSAS
Belo Horizonte, MG
Setembro, 2018
Dissertação apresentada ao Programa de
Mestrado em Tecnologias aplicadas a saúde
das Faculdades Promove, como requisito
parcial para obtenção do título de mestre.
Orientadora: Professora Dra. Ludmilla Regina
de Souza David

Daniela Oliveira Lima Magalhães
IDENTIFICAÇÃO E ANÁLISE DE REDES DE GENES ENVOLVIDOS NO CÂNCER
DE MAMA E CÉLULAS-TRONCO CANCEROSAS
Aprovada em ___/___/___ pela banca constituída dos seguintes professores:
_______________________________________ Profa. Dra. Carla Silvana de Oliveira e Silva
_________________________________________ Profa. Dra. Ludmilla Regina de Souza David
________________________________________ Profa. Dra. Rosangela Silqueira Hickson Rios
Belo Horizonte, de de 2018.
Faculdades Promove de Belo Horizonte Rua Aimorés 123 – 31.256-000 – Belo Horizonte, MG
www.faculdadepromove.com.br
Dissertação apresentada ao Programa de
Mestrado em Tecnologias aplicadas a
saúde das Faculdades Promove, como
requisito parcial para obtenção do título
de mestre.

Dedico este estudo a minha mãe,
guerreira, fonte inspiradora deste
trabalho. Na qual me inspirei em
aumentar cada minuto de sua vida.
Às minhas filhas, Isadora e Isabela, que
tanto sentiram a minha ausência.
Ao meu esposo, Ítalo, companheiro de
sempre.
Dedico também àquelas mulheres que já
receberam e, principalmente, às que
ainda irão receber o diagnóstico de
câncer de mama.
Desejo que este estudo possa contribuir
para a detecção precoce da doença e
para o início e a conclusão do tratamento
em um tempo mais curto possível.

AGRADECIMENTOS
- A Deus por me iluminar e guiar sempre para um bom caminhar.
- Meus queridos pais, Rosarinha e Lima, agradeço a vocês por terem me dado
condições e exemplos para eu ter um bom “alicerce de vida”.
- A minhas amadas filhas, Isadora e Isabela, minhas razões de viver. Minha principal
vontade de crescer, de ser melhor, sempre foi para poder dar exemplo a vocês, e
para que pudéssemos ter condições de um futuro melhor. Amo demais vocês!
- A meu amado esposo, Ítalo, pelo companheirismo, paciência e amizade.
- Minha orientadora e agora amiga, Ludmilla, agradeço-lhe pela oportunidade de tê-
la como orientadora. Ao seu lado aprendi novos conhecimentos científicos e de vida.
Obrigada pelo carinho, amizade e compreensão em cada momento de dificuldade.
- A enfermeira Franciele Ornelas e ao mestre em T.I. Diego Vinicius Castro pela
parceria e amizade.
- Muito obrigada ao meu sogro e minha sogra, Edmilson e Serva, pelo carinho e
cuidado com minha filha, Isadora, nas viagens mensais.
- Minhas amigas, agradeço por todos os elogios, convivência, risadas, conversas e
apoio... Amigas adoçam o viver e ao mesmo tempo nos ajudam a ver a vida quando
uma “névoa” cisma em cobrir nossos olhos.
- Meus irmãos (ãs) e cunhados (das), obrigada por sempre acreditarem em mim.

“A mente que se abre a uma nova ideia jamais retorna ao seu tamanho
original”. (EINSTEIN)

RESUMO
As células-tronco são células indiferenciadas presentes em tecidos embrionários e
extraembrionários. Estudos têm evidenciado a presença de uma população de
células com características de células-tronco em muitos tipos de neoplasias
malignas, entre elas o câncer de mama. As células-tronco cancerosas têm
contribuído com a progressão da doença e evolução para um quadro clínico de
metástase e pior sobrevida em pacientes diagnosticados com o câncer de mama.
Portanto, o objetivo do corrente estudo consistiu em analisar as redes de interações
gênicas das células-tronco cancerosas (CTC), células-tronco cancerosas no câncer
de mama (CTCM), câncer de mama (CM) e metástase de origem no câncer de
mama (MM). Para construir as redes de interações gênicas das condições estudas,
foram identificados os genes de cada condição na base de dados Genecards.
Marcadores de células-tronco cancerosas foram selecionados a partir de revisão da
literatura e analisados quanto à interação com outros genes em cada rede. Os
mapas de interações genômicas foram elaborados com o auxílio do software
STRING, e as análises estatísticas das interações foram realizadas no programa
SPSS® PASS Statistic 18. As redes CTC, CTCM, CM e MM apresentaram 386, 81,
396 e 160 genes, respectivamente. Em cada rede foram selecionados 8 genes
líderes, sendo eles os genes que apresentaram os maiores números de interações
dentro de cada rede: CTC (TP53, AKT1, CTNNB1, MYC, JUN, STAT3, EGFR e
NOTCH1), CTCM (STAT3, EGFR, CTNNB1, JUN, NFKB1, NOTCH1, FOS e ESR1),
CM (TP53, PIK3CA, AKT1, EGFR, MAPK1, EP300, SRC e CDK2) e MM (TP53,
EGFR, STAT3, JUN, HDAC1, ERBB2, VEGFA e HDAC2). Os grupos CTC e CTCM
apresentaram o maior número de genes líderes em comum. TP53, AKT1 e EGFR
foram genes líderes da rede de CTC que também se encontraram na rede de CM.
TP53, EGFR, STAT3 e JUN foram genes comuns aos grupos CTC e MM. EGFR foi
um dos genes líderes de todas as condições estudas. Dentre os marcadores
putativos de células-tronco cancerosas (WNT1, NOTCH1, NANOG, CD44 e
ALDHA1), apenas o CD44 foi encontrado nas 4 condições apresentadas neste
trabalho. NOTCH1 realizou interações nas redes de células-tronco (CTC e CTCM) e
CM, e foi o marcador que apresentou melhores combinações nas redes de células-
tronco cancerosas referentes à alta interação dentro da rede (alto WNL) e poucas
interações em redes globais (baixo TIS). NANOG, WNT1 e SOX2 apresentaram
interações apenas nas redes de células-tronco. ALDH1A1 apresentou-se apenas

nas redes CTC e CM, mas com apenas 1 interação em cada rede. Dessa forma,
percebe-se que o estudo dos mapas de interações gênicas revela diversos genes
relevantes para cada condição estudada, e genes que são importantes para os
processos relativos ao câncer de mama (CTCM, CM e MM), como os genes EGFR e
CD44. NOTCH1 foi o único marcador de CTC que também foi considerado um dos
genes líderes. Muitas outras interpretações podem ser realizadas a partir dos
resultados obtidos, destacando-se diferentes genes importantes para as condições
biológicas estudadas.
Palavras-chave: Células-tronco tumorais, carcinoma de mama, metástase,
biomarcadores.

ABSTRACT
Stem cells are undifferentiated cells found in embryonic and extra-embryonic tissues.
Studies have shown the presence of a population of cells with characteristics of stem
cells in many types of malignant neoplasms among them the breast cancer. Cancer
stem cells have contributed with disease progression and evolution to a clinical
picture of metastasis and worse survival in patients diagnosed with breast cancer.
Therefore, the objective of this study was to analyze gene interactions networks of
cancer stem cells (CSC), cancer stem cells in breast cancer (BCCSC), breast cancer
(BC) and source metastasis in breast cancer (BCM). To build networks of gene
interactions studying conditions, were identified the genes of each condition in the
Genecards. Cancer stem cells markers were selected from literature review and
analyzed regarding interaction with other genes in each network. The genomic
interactions maps were drawn up with the aid of software STRING, and the statistical
analysis of the interactions were performed in the program SPSS® PASS Statistic 18.
The CSC, BCCSC, BC and BCM networks, presented 386, 396, 81 and 160 genes,
respectively. In each network 8 genes were selected leaders with genes that showed
the greatest numbers of interactions with in each network: CSC (TP53, AKT1,
CTNNB1, MYC, JUN, STAT3, EGFR e NOTCH1), BCCSC (STAT3, EGFR, CTNNB1,
JUN, NFKB1, NOTCH1, FOS e ESR1), BC (TP53, PIK3CA, AKT1, EGFR, MAPK1,
EP300, SRC e CDK2) e BCM (TP53, EGFR, STAT3, JUN, HDAC1, ERBB2, VEGFA
e HDAC2). The CSC and BCCSC groups presented the largest number of leading
common genes. TP53, AKT1 and EGFR genes were leaders of the CSC network
which also met in BC. TP53, EGFR, STAT3, and JUN genes were common to the
CSC and BCM groups. EGFR was one of the leading genes of all conditions studied.
Among the putative markers of cancer stem cells (WNT1, NOTCH1, NANOG, CD44
e ALDHA1), only the CD44 was found on 4 conditions presented in this work.
NOTCH1 realized interactions in the stem cells networks (CSC and BCCSC), and
BC, and was the marker that presented bets combinations of cancer stem cells
networks, regarding the high interaction within the network (WNL) and few
interactions in global networks (low TIS). NANOG, WNT1 and SOX2 showed
interactions only in the stem cell networks. ALDH1A1 presented only in the CSC and
BC networks, but with only 1. Thus, the study of gene interaction maps reveals
several genes relevant to each condition studied, and genes that are important for

breast cancer processes (BCCSC, BC and BCM), such as EGFR and CD44 genes.
NOTCH1 was the only CsC marker that was considered one of the leading genes.
Many other interpretations can be made from the obtained results, highlighting
different genes important for the biological conditions studied.
Key-Words: Tumor stem cells, breast carcinoma, metastasis, biomarkers.

LISTA DE ILUSTRAÇÕES
FIGURA 1- Distribuição proporcional dos dez tipos de câncer mais incidentes
estimados para 2016 por sexo no Brasil, exceto pele não melanoma...................... 25
FIGURA 2 - Distribuição proporcional dos dez tipos de câncer mais incidentes
estimados para 2018 por sexo no Brasil, exceto pele não melanoma...................... 26
FIGURA 3 - Distribuição proporcional dos dez tipos de câncer mais incidentes
estimados para 2018 por sexo nos Estados Unidos da América, exceto pele não
melanoma.................................................................................................................. 26
FIGURA 4- Etapas de iniciação e promoção relativos ao processo da
carcinogênese........................................................................................................... 28
FIGURA 5- Bloqueio da progressão do ciclo celular desencadeado pela proteína
supressora de tumor p53 para a realização do reparo do dano no
DNA........................................................................................................................... 29
FIGURA 6- Visão esquemática do microambiente tumoral....................................... 30
FIGURA 7- Fontes de heterogeneidade no câncer................................................... 37
FIGURA 8- Autorrenovação e diferenciação de células-tronco normais e células-
tronco tumorais.......................................................................................................... 38
FIGURA 9- Os principais mecanismos de resistência terapêutica em células-tronco
de câncer de mama................................................................................................... 40
FIGURA 10- Mapa de interação dos 386 genes envolvidos na rede de células-tronco
cancerosas................................................................................................................ 52
FIGURA 11- Mapa de interação dos 81 genes envolvidos na rede das células-tronco
cancerosas do câncer de mama............................................................................... 53
FIGURA 12- Mapa de interação dos 396 genes envolvidos na rede do câncer de
mama......................................................................................................................... 54
FIGURA 13- Mapa de interação dos 160 genes potencialmente envolvidos na rede
de metástase de origem no câncer de mama........................................................... 55
FIGURA 14- Distribuição dos genes da rede de células-tronco cancerosas em
função das interações gênicas e análise de agrupamento....................................... 57
FIGURA 15- Distribuição dos genes da rede de células-tronco cancerosas da mama
em função das interações gênicas e análise de agrupamento................................. 57

FIGURA 16- Distribuição dos genes da rede de câncer de mama em função das
interações gênicas e análise de agrupamento.......................................................... 58
FIGURA 17- Distribuição dos genes da rede de metástase de mama em função das
interações gênicas e análise de agrupamento.......................................................... 58
FIGURA 18- Representação dos marcadores putativos de células-tronco
cancerosas................................................................................................................ 60
FIGURA 19- Número de interações realizadas por marcadores de células-
tronco......................................................................................................................... 62

LISTA DE TABELAS
TABELA 1- Genes líderes das condições estudadas................................................ 56
TABELA 2- Interações realizadas por marcadores de células-tronco cancerosas................................................................................................................ 63

LISTA DE ABREVIATURAS E SIGLAS
PI3K Fosfatidilinositol-4,5-bisfosfato 3-quinase
ABC Cassete de ligação a ATP
AKT1 Serina/treonina quinase 1
ALDH Enzima aldeído desidrogenase
ALDH1 Aldeído desidrogenase 1
ALDH1A1 Aldeído desidrogenase 1, membro A1
APC Polipose adenomatosa Coli
ATM Quinase serina/treonina ATM
CD133 Antígeno CD133 ou prominina 1
CD24 Molécula de adesão do grupo de diferenciação 24 ou antígeno
estável ao calor
CD44 Molécula de adesão do grupo de diferenciação 44, que pertence
ao grupo sanguíneo Indiano
CD49f Subunidade alfa da integrina 6
CD61 Subunidade beta da integrina 3
CDK2 Quinase dependente de ciclina 2
ChK Quinase de ponto de checagem;
CI Carcinoma localmente invasivo
CIS Carcinoma in situ
CM Câncer de mama
c-Myc Proto-oncongene MYC, fator de transcrição BHLH
CTC Células-tronco cancerosas
CTCM Células-tronco cancerosas do câncer de mama
CTNNB1 Beta catenina
DNA Ácido desoxirribonucleico
DP1 Fator de transcrição DP1
E2F Fator de transcrição da família E2
EGFR Receptor do fator de crescimento epidérmico
TEM Transição epitélio-mesenquimal
EP300 Proteína de Ligação E1A P300

ERBB2 Receptor de proteínas tirosina-quinase tipo 2
ESR1 Receptor de estrogênio 1
EUA Estados Unidos da América
FAK Proteína quinase de adesão focal
FN1 Fibronectina 1
FOS Proto-oncogene Fos, Subunidade Fator de Transcrição AP-1
G0 Fase de repouso por tempo indeterminado
G1 Primeira fase de intervalo na intérfase no ciclo celular
G2 Segunda fase de intervalo na intérfase do ciclo celular
GPI Glicosilfosfatidilinositol
HDAC1 Histona desacetilase 1
HDAC2 Histona desacetilase 2
HER-2 Receptor tipo 2 do fator de crescimento epidérmico
HIF1A Subunidade alfa do fator 1 induzível por hipóxia
HUGO Organização do Genoma Humano
IGF1R Receptor do fator de crescimento semelhante à insulina 1
INCA Instituto Nacional do Câncer
JAG1 Proteína Jagged-1
JUN Proto-Oncogene Jun, Subunidade Fator de Transcrição AP-1
KLF4 Fator de transcrição 4 da família Kruppel
Let-7 Precursor do microRNA de Let-7
LIF Citocina da família da interleucina 6
LIN28 Lin-28 homólogo
LIN28A Lin-28 homólogo A
MAPK1 Proteína quinase ativada por mitógeno 1
MDR1 Proteína de resistência a multidrogas 1;
MeSH Medical Subject Headings
MET Proto-oncogene MET, Receptor tirosina quinase
MiARNs Pequenos ácidos ribonucléicos
miR-452 MicroRNA 452
MM Metástase originada do câncer de mama
MMP2 Metaloproteinase da matriz 2
MMP9 Metaloproteinase da matriz 9
MTOR Alvo mecanicista da rapamicina quinase

MYC Proto-oncogene MYC, Fator de transcrição BHLH
NANOG Fator de transcrição homeobox Nanog
NFKB Subunidade Kappa B do Fator Nuclear
NFKB1 Subunidade Kappa B do Fator Nuclear 1
NICD Domínio intracelular de Notch (NICD).
Notch Via de sinalização Notch
NOTCH1 Proteína Notch associada à translocação TAN-1
Oct-4 Proteína de Ligação a Octómero 4
P21 Proteína Interativa com CDK 1
P53 Proteína Tumoral P53
P-gp Glicoproteína P
PIK3CA Subunidade catalítica de fosfatidilinositol-4,5-Bisfosfato 3-
Quinase Alfa
POU5F1 POU Classe 5 Homeobox 1
ROS Espécies reativas de oxigênio
SNAI2 Membro da família de fatores de transcrição dedo de zinco
SOX2 Sexo determinando região Y-Box 2
SPP1 Fosfoproteína Secreta 1
SRC Proto-oncogene SRC, tirosina quinase não receptora
SRR2 Reforço junsante de SOX2
STAT3 Transdutor de Sinais e Ativador da Transcrição 3
STRING Ferramenta de Pesquisa para Recuperação de Interações entre
Genes/Proteínas
TCF Fator de transcrição funcional
TGFβ Fator de crescimento transformador beta
TIS Pontuação de interações totais
TNM T=tumor, N= acometimento linfonodal e M= metástase à
distância
TP53 Proteína supressora de tumor P53
TWIST1 Fator de transcrição BHLH da Família Twist 1
UICC União Internacional Contra o Câncer
VEGF Fator de Crescimento Endotelial Vascular
VEGFA Fator de Crescimento Endotelial Vascular A
WNL Número ponderado de ligações

WNT Membro da família WNT
WNT1 Membro da família WNT 1

SUMÁRIO
1 INTRODUÇÃO ................................................................................................................................. 19
2 OBJETIVOS ..................................................................................................................................... 22
2.1 Objetivo geral: .......................................................................................................................... 22
2.2 Objetivos específicos: ............................................................................................................. 22
3 JUSTIFICATIVA .............................................................................................................................. 23
4 REFERENCIAL TEÓRICO ............................................................................................................ 24
4.1 Câncer ....................................................................................................................................... 24
4.2 Câncer de mama ..................................................................................................................... 32
4.3 Células-tronco cancerosas ..................................................................................................... 35
4.4 Células-tronco cancerosas em câncer de mama ............................................................... 40
4.5 Marcadores putativos de células-tronco cancerosas ......................................................... 43
4.6 Bioinformática ........................................................................................................................... 45
5 METODOLOGIA .............................................................................................................................. 48
5.1 Caracterização do estudo ....................................................................................................... 48
5.2 Identificação dos genes .......................................................................................................... 48
5.3 Construção e análise do mapa de interações genômico ................................................. 48
5.4 Definição dos genes marcadores putativos de células-tronco cancerosas .................... 50
5.5 Análises das interações realizadas pelos genes marcadores putativos de CTC .......... 50
6 RESULTADOS ................................................................................................................................ 51
7 DISCUSSÃO .................................................................................................................................... 64
8 CONCLUSÃO .................................................................................................................................. 73
REFERÊNCIAS .................................................................................................................................. 74

19
1 INTRODUÇÃO
As células-tronco são células indiferenciadas ou com pequeno percentual de
diferenciação presentes em tecidos embrionários e extraembrionários. Podem
permanecer em estado quiescente até a fase adulta ou diferenciar-se em diversos
tecidos a partir da expressão de determinados genes. Pesquisas têm demonstrado
resultados benéficos na utilização dessas células para o tratamento de doenças, de
modo que as células-tronco representam as potenciais terapias para o tratamento de
muitas doenças no futuro (SOUZA et al., 2003; YANG et al., 2016; LEON, et al.
2015). Os constantes estudos com células-tronco resultaram na identificação de
células-tronco cancerosas (CTC) em um conjunto de tumores humanos, o que
sugere que estas células também possuem potencial para desencadearem o
desenvolvimento de cânceres (YANG et al., 2016; KONRAD et al., 2017).
[...] apenas um pequeno número de células da massa tumoral é capaz de proliferar e formar novos tumores quando enxertado em animais imunodeficientes, como demonstrado no câncer de mama, próstata e leucemia. As células com essas habilidades são denominadas de células-tronco tumorais e compartilham diversas características com sua contraparte normal (SOUZA, 2006, p. 379).
As células-tronco tumorais foram inicialmente identificadas na leucemia
mielóide aguda humana por Bonnet e Dick (1997). Posteriormente, foram descritas
em tumores sólidos, próstata, cérebro, ovário, cólon, pele, pulmão, e, em particular,
no câncer de mama (PICCIRILLO e VESCOVI, 2007). Os estudos com essas células
têm indicado que tumores mais agressivos possuem uma maior quantidade de
células-tronco cancerosas em sua população celular (HANAHAN e WEINBERG,
2011).
As CTCs apresentam capacidade de auto-renovação, diferenciação em pelo
menos uma linhagem específica, heterogeneidade, quiescência, atividade de
telomerase ativa, resistência à apoptose e consequente longevidade, além da
habilidade de migrar e favorecer o processo de metástase (SOUZA, 2006). Essas
células são ainda resistentes aos tratamentos e aumentam em número após
pequenas doses de radiação fracionada, caracterizando um processo de
repopulação acelerada das células tumorais observada durante intervalos de tempo

20
no tratamento radioterápico. São também resistentes à quimioterapia e possuem
capacidade de acentuar certas formas de dormência tumoral, cujas células
cancerosas latentes persistem por anos ou décadas após remoção cirúrgica ou
radio/quimioterapia e são capazes de regenerar o tumor após o final da terapia
(HANAHAN e WEINBERG, 2011).
O câncer de mama é considerado o segundo tipo de câncer mais comum
entre as mulheres e responsável por 25% de casos novos a cada ano (INCA, 2016).
A presença de uma pequena população de células-tronco cancerosas e
heterogeneidade tumoral tem sido observado neste tipo de neoplasia (LEON, et al.
2015). As células deste tipo de câncer possuem um fenótipo de superfície
CD44+/CD24-, que apresentam propriedades iniciadoras de tumor com
características de pluripotência e capacidade invasiva. A presença desse fenótipo de
superfície sugere uma transição de um fenótipo epitelial (CD44-/CD24+) para um
fenótipo mesenquimal (CD44+/CD24-), o que capacita a célula a mover-se do tumor
primário para um sítio metastático (MANI et al., 2008). A disseminação das células
tumorais da mama se inicia precocemente e normalmente acontece através das vias
linfáticas. Os implantes secundários são frequentemente encontrados em vértebras
e arcos costais, caracterizando a metástase óssea (SLEEMAN e THIELE, 2009).
Nesse tipo de tumor, a disseminação metastática é a principal causa da baixa
sobrevida dos pacientes (FREUDENBERG, 2009; RUITERKAMP, 2011).
A partir dessas evidências, metodologias que contribuam para a
compreensão da biologia do câncer de mama se fazem necessárias, e a
identificação de biomarcadores que auxiliem na eliminação das células-tronco
cancerosas da mama são fundamentais (CALIFF, 2018). Estudos de bioinformática
têm recebido bastante atenção no cenário mundial das ciências da saúde na
interpretação e na compreensão de grandes volumes de dados. Em câncer de
mama, estudos in silico identificaram 390 novos pares de fármacos anticâncer
capazes de exibir efeitos sinérgicos antitumorais, muitos deles estão em fase de
testes in vitro (JAERGER et al., 2017). Em outros estudos, ferramentas
computacionais são utilizadas para a identificação de padrões de mutação em
grandes estudos de genômica de câncer e para a realização de simulações
moleculares e construção de estruturas proteicas (VERLI, 2014; HOU et al., 2016).
Dessa forma, estudos que visam contribuir com conhecimentos acerca da biologia
tumoral, especialmente referente aos processos de proliferação e progressão dos

21
tumores associados às características das CTCs, são de fundamental importância
para futuras estratégias terapêuticas e representam uma abordagem promissora no
tratamento do câncer de mama.
Desta forma, a presente pesquisa teve o intuito de analisar as redes de
interação gênica de quatro condições, células-tronco cancerosas (CTC), células-
tronco cancerosas do câncer de mama (CTCM), câncer de mama (CM) e a
metástase originada do câncer de mama (MM). Além disso, objetivou-se identificar
as interações realizadas por marcadores de células-tronco cancerosas com os
genes das redes em estudo.

22
2 OBJETIVOS
2.1 Objetivo geral:
Avaliar as redes de interações gênicas das células-tronco cancerosas,
células-tronco cancerosas no câncer de mama, câncer de mama e da metástase de
origem do câncer de mama.
2.2 Objetivos específicos:
- Construir o mapa de interação genômico das células-tronco cancerosas,
células-tronco cancerosas no câncer de mama, câncer de mama e da
metástase de origem do câncer de mama;
- Identificar os genes líderes de cada condição estudada;
- Avaliar as interações realizadas por genes marcadores de células-tronco
cancerosas em cada condição.
- Realizar um estudo comparativo dos genes líderes e genes marcadores de
células-tronco cancerosas nas condições estudadas.

23
3 JUSTIFICATIVA
Estimativas apontam que o câncer apresenta elevada incidência em todo o
mundo e que até 2030 cerca de 21,4 milhões de novos casos serão diagnosticados
no Brasil, e o número de óbitos será de 13,2 milhões de pessoas (BRASIL, 2015).
Especificamente para o câncer de mama, que é o segundo tipo de câncer mais
frequente entre as mulheres, foram estimados que 57.960 novos casos seriam
registrados para o biênio 2016-2017 no Brasil (INCA, 2016). A presença de uma
população de CTCs em cânceres de mama representa um importante fator na
promoção de resistência clínica, permitindo a evolução para um tumor mais
agressivo ao longo do tempo. Nesse cenário, as disseminações metastáticas
contribuem para uma alta taxa de mortalidade e pior prognóstico dos pacientes
(VELASCO-VELÁZQUEZ et al., 2013).
Diante disso, percebeu-se a necessidade de identificar os genes que mais
interagem nas células-tronco do câncer de mama, e comparar com os genes
evidenciados na população de células-tronco cancerosas. Além disso, é relevante
para a compreensão da biologia do câncer de mama, conhecer os genes mais
importantes para o câncer de mama e sua condição metastática, além de verificar
como os genes mais importantes da condição CTCs interagem nas condições de
câncer de mama e metástase originada do câncer de mama. E ainda, a
compreensão das interações realizadas por genes marcadores de células-tronco
cancerosas nas redes de CTCs, CTCs da mama, câncer de mama e metástase
originada do câncer de mama pode assessorar estudos futuros sobre tais genes nas
condições patológicas descritas.

24
4 REFERENCIAL TEÓRICO
4.1 Câncer
A palavra câncer foi utilizada pela primeira vez por Hipócrates, que é
considerado o pai da medicina, e viveu entre 460 e 377 a.C. (BRASIL, 2011).
“Câncer” veio do grego karkínos, que significa caranguejo, e como definição é
compreendido como um grupo de mais de 100 doenças com crescimento celular
desordenado e com capacidade de se infiltrar em órgãos e tecidos vizinhos (INCA,
2012). Pesquisas demonstram relatos de câncer em múmias do Egito há mais de
3000 anos a.C. (BRASIL, 2011).
O câncer é considerado uma doença insidiosa e a segunda causa de morte
global, estando atrás somente das doenças cardiovasculares. Trata-se de um
problema de saúde pública, diante da sua relevância no âmbito epidemiológico,
social e econômico (INCA, 2012; BRENNER, 2016). Segundo pesquisas realizadas
em 2015, foram evidenciados 17,5 milhões de novos casos de câncer no mundo e
8,7 milhões de pessoas morreram com esta doença (FITZMAURICE, 2017). As
causas dessa doença associam-se à exposição a fatores externos ao hospedeiro,
como tabaco, micro-organismos infecciosos e uma dieta não saudável, e por fatores
intrínsecos, como mutações genéticas, hormônios e condições do sistema
imunológico (AMERICAN CANCER SOCIETY, 2015).
Nas últimas décadas vem ocorrendo no Brasil um fenômeno chamado de
transição epidemiológica. Tal fenômeno diz respeito a três mudanças específicas:
aumento da morbimortalidade pelas doenças e agravos não transmissíveis e pelas
causas externas; deslocamento da carga de morbimortalidade dos grupos mais
jovens para grupos mais idosos; e predomínio da morbidade em função da
mortalidade. O câncer está entre as doenças responsáveis por essas mudanças do
perfil epidemiológico do país (BRASIL, 2011). Pesquisas apontam que até 2030
cerca de 21,4 milhões de novos casos serão diagnosticados no Brasil, e o número
de óbitos será de 13,2 milhões de pessoas (BRASIL, 2015).
O câncer de mama é o segundo tipo de câncer mais recorrente nas regiões
Sul, Sudeste, Centro-Oeste e Nordeste do Brasil e nos EUA (INCA, 2013; DE

25
SANTIS et al., 2017). Estudo desenvolvido nos EUA afirma que aproximadamente
252.710 novos casos de câncer de mama invasivo e cerca de 63.410 casos de
carcinoma in situ foram estimados para 2017 (DE SANTIS et al., 2017).
No Brasil, segundo o Instituto Nacional do Câncer (INCA), foram
diagnosticados 57.120 novos casos de câncer no ano de 2014 (INCA, 2014). Em
2016, foram estimados 57.960 novos casos (INCA, 2016). Os cânceres mais
incidentes em 2016 foram os cânceres de próstata, pulmão, cólon e reto e estômago
para o sexo masculino; e os cânceres de mama, colo do útero, cólon e reto e
glândula tireoide para o sexo feminino (Figura 1).
Figura 1- Distribuição proporcional dos dez tipos de câncer mais incidentes
estimados para 2016 por sexo no Brasil, exceto pele não melanoma.
Fonte: INCA Estimativa 2012 - Incidência de câncer no Brasil.
Para 2018 são estimados 600 mil novos casos de câncer no Brasil, sendo o
câncer de próstata o mais incidente no sexo masculino (61.200 casos), seguido dos
cânceres em traqueia, brônquio e pulmão com 17.330, cólon e reto com 16.660 e
estômago com 12.920 diagnósticos. Já no sexo feminino, os cânceres de mama,
cólon e reto, e colo do útero lideram em incidência, com 59.700, 18.980 e 16.370
novos casos, respectivamente (Figura 2) (INCA, 2017).

26
Figura 2 - Distribuição proporcional dos dez tipos de câncer mais incidentes
estimados para 2018 por sexo no Brasil, exceto pele não melanoma.
Fonte: INCA Estimativa 2016 - Incidência de câncer no Brasil.
Nos EUA, estima-se que em 2018, 856.370 indivíduos do sexo masculino
terão algum tipo de câncer, sendo que 19% deles apresentarão câncer de próstata,
14% câncer de pulmão e brônquios e 9% câncer colorretal. Nas mulheres, o câncer
de mama será responsável por 30% dos casos, seguido por pulmão e brônquios
(13%) e colorretal (7%), conforme representado na figura 3 (SIEGEL et al., 2018).
Figura 3 - Distribuição proporcional dos dez tipos de câncer mais incidentes
estimados para 2018 por sexo nos Estados Unidos da América, exceto pele não
melanoma.
Fonte: Estimativa 2018 – A Cancer Journal for Clinicians. Inglês/Português: Males/Machos;
Prostate/Próstata; Lung & bronchus/Pulmão e Brônquios; Colon & rectum/Cólon e reto; Urinary
bladder/Bexiga urinária; Melanoma of the skin/Melanoma; Kidney & renal pélvis/Rim e Pelve renal;
Non-Hodgkin lymphoma/Linfoma não-Hodgkin; Oral cavity & pharynx/Cavidade oral e faringe;
Leukemia/Leucemia; Liver & intrahepatic bile duct/Fígado e ducto biliar intra-hepático;

27
Females/Fêmeas; Breast/Mama; Colon & rectum/Cólon e reto; Uterine corpus/Colo uterino;
Thyroid/Tireoide; Melanoma of the skin/Melanoma; Non-Hodgkin lymphoma/Linfoma não-Hodgkin;
Pancreas/Pâncreas; Leukemia/Leucemia; Kidney & renal pélvis/Rim e Pelve renal.
O estilo de vida da população pode definir o tipo de câncer mais prevalente
de acordo com cada região, além de apresentar diferenças de acordo com o
desenvolvimento do país, sendo que alguns deles são mutáveis e evitados por meio
da conscientização da sociedade (PEREIRA, 2016). Pesquisadores têm buscado
identificar mutações genéticas relativas ao processo da carcinogênese desde o
sequenciamento do genoma humano, para assim tentarem promover a prevenção e
cura dessa doença. As alterações genéticas hereditárias decorrentes de mutações
na linhagem germinativa são responsáveis por cerca de 15% dos casos. Em
contrapartida, 85% dos tumores decorrem de alterações genéticas que surgem de
maneira aleatória como consequência da interação do homem com o meio em que
vive (GONÇALVES et al., 2010; INCA, 2014).
Os principais fatores de risco e associados ao câncer são o uso de tabaco,
alimentação inadequada, inatividade física, obesidade, consumo de álcool, agentes
infecciosos, radiação, poluição ambiental, comportamento sexual, nível
socioeconômico, idade, raça e gênero (INCA, 2017). Além destes, a história familiar
destaca-se como importante fator de risco (CGHFBC, 2001).
As neoplasias que dão origem ao câncer, em sua grande maioria, surgem
quando uma única célula exposta a agentes carcinogênicos sofre alterações
genéticas ou epigenéticas que influenciam no seu mecanismo de proliferação e
apoptose (Figura 4). Após esse processo, inicia-se a fase de promoção, em que as
células sofrem mitoses consecutivas, formando vários clones celulares. O terceiro
estágio, a progressão, caracteriza-se pelo descontrole da multiplicação celular e a
célula possui autonomia para se proliferar. Na fase de conversão, as células tornam-
se invasivas (INCA, 2008).

28
Figura 4: Etapas de iniciação e promoção relativos ao processo da carcinogênese.
Fonte: INCA, 2008.
As células neoplásicas possuem seis características que as difere das
células normais: autossuficiência em sinais proliferativos, insensibilidade aos sinais
antiproliferativos, evasão por apoptose e potencial replicativo ilimitado. O
descontrole da proliferação celular ocorre por meio da ativação de oncogenes e a
repressão de genes supressores de tumor (WICHA, 2006; GOUVEA et al., 2014).
Mutações em proto-oncogenes, que correspondem a um grupo de genes que
controlam a proliferação e diferenciação celular, fazem com que eles se tornem
oncogenes, favorecendo assim a proliferação celular descontrolada (WARD, 2002).
Os genes supressores de tumor são perdidos ou inativados em diversos tipos de
cânceres. Os genes supressores de tumor mais comuns são os genes TP53 e Rb.
Quando ativados, a célula interrompe a divisão celular na fase G1 por intermédio da
proteína p21, permitindo o reparo do DNA danificado antes que o fator de
transcrição E2F ative a síntese de novos filamentos de DNA (Figura 5) (WARD,
2002).

29
Figura 5. Bloqueio da progressão do ciclo celular desencadeado pela proteína
supressora de tumor p53 para a realização do reparo do dano no DNA.
Fonte: WARD, 2002. Painel à esquerda: ciclo celular prosseguindo normalmente sob a vigilância
passiva de p53, cujos níveis se mantém baixos. Painel à direita: níveis aumentados de p53 em função
de um dano no DNA e consequente ativação de p21 para impedir a ligação da ciclina B com a
quinase CDK2. Dessa forma, a proteína Rb não será fosforilada, impedindo assim a liberação do fator
de transcrição formado pelo complexo E2F-DP1, que não poderá ativar os genes essenciais para a
progressão na fase de síntese (fase S). G0: fase de repouso do ciclo celular; G1 e G2: fases de
intervalo no ciclo celular.
Estudos demostram que o microambiente tumoral é também fundamental no
processo da carcinogênese, contribuindo com a progressão, agressividade e
disseminação metastática. O microambiente tumoral é um ambiente organizado e
dinâmico, onde células cancerosas e matriz extracelular interagem para favorecer a
migração das células tumorais. O aumento do volume tumoral é marcado pelo
surgimento de uma área em hipóxia, a qual torna-se o principal desencadeador da
angiogênese. Essa última caracteriza-se pela proliferação de células endoteliais,
necrose de células tumorais, permeabilidade de vasos sanguíneos, degradação da
matriz extracelular e formação de novos vasos. Nesse ambiente, células
inflamatórias, que surgem na tentativa de conter o tumor, possuem um importante
papel no fornecimento de fatores de crescimento mutagênicos para as células
cancerígenas, contribuindo para a angiogênese e resistência à morte celular (Figura
6) (TATE e AGHI, 2009; GORKEM, 2015).

30
Devido à angiogênese, formam-se vasos sanguíneos imaturos que
contribuem para o aumento da hipóxia. Estudos têm demonstrado também que os
vasos sanguíneos anormais criam um nicho vascular que abriga células-tronco
tumorais, as quais correspondem a uma pequena fração de células capazes de
originarem um novo tumor e conferir resistência às terapias (TATE e AGHI, 2009;
GORKEM, 2015).
Figura 6: Visão esquemática do microambiente tumoral.
Fonte: GORKEM, 2015. Inglês/Português: Extracellular matrix proteins/Proteínas da matriz
extracelular; Cancer cell/Células cancerosas; Cancer stem cell/Células-tronco cancerosas;
Hypoxic/Hipóxia; Apoptotic cancer cell/Célula cancerosa apoptótica; Cancer associated
fibrosblasts/Fibroblastos associados ao câncer; Tumor-associated macrophages/Macrófagos
associados ao tumor; T-cells/Células T; Natural killer cells/Células assassinas naturais; B-cell/Células
B; Pericyte/Pericitos.
A formação de novos vasos, sendo muitos deles considerados imaturos,
contribui para a ocorrência do fenômeno metastático. A metástase consiste no
processo de deslocamento de células cancerígenas de um determinado local para
outro, encontrando possíveis locais para formar um novo tumor. O deslocamento
das células tumorais pode ser feito através da circulação sanguínea, linfática e por
cavidades do corpo, como peritônio, pleura ou espaços subaracnóideos (SLEEMAN
e THIELE, 2009). Das células que entram em circulação, cerca de 0,01% chega a
desenvolver novos tumores. A grande maioria das células metastáticas são

31
eliminadas por apoptose ou através da ação lítica de células citotóxicas. As células
sobreviventes a esse processo se estabelecem no novo microambiente, podendo
desenvolver novos tumores ou permanecer em repouso (CHAMBERS et al., 2002).
Trata-se da principal causa de morbidade e mortalidade por câncer, chegando a
cerca de 90% das mortes, talvez por ser o obstáculo mais difícil e incompreendido
quando se trata de tumores malignos (CHAFFER e WEINBERG, 2011; BACAC e
STAMENKOVIC, 2008).
Está cada vez mais claro que as células tumorais dependem do seu microambiente para estabelecer as metástases e que dependem das células do estroma do tecido do hospedeiro para sua progressão, tais como a diversidade de atividade proteolítica que elas próprias podem não possuir (BACAC; STAMENKOVIC, 2008, p.222,223. Tradução do autor)**
Estima-se que cerca de 10% das pacientes são diagnosticados com
metástase (FREUDENBERG, 2009). Os subtipos específicos de câncer são
caracterizados por padrões diferentes de metástase, com impactos diversos sobre a
sobrevida do paciente (GERRATANA, 2015). Por isso, os métodos e incentivos
públicos para o diagnóstico precoce do câncer e as melhorias no tratamento
sistêmico têm contribuído para a redução da mortalidade (BRASIL, 2011; WANG et
al., 2017).
As modalidades de tratamento para o câncer são cirurgia, radioterapia,
quimioterapia, terapia hormonal, terapia de imunossupressão e terapia direcionadas
a moléculas específicas (AMERICAN CANCER SOCIETY, 2015).
A radioterapia pode ser utilizada como modalidade pré-operatória, que visa
reduzir o tamanho do tumor para facilitar o procedimento cirúrgico, como
radioterapia pós-operatória ou pós-quimioterapia (radioterapia profilática) e como
radioterapia paliativa, que objetiva o tratamento local do tumor primário ou de
metástases (BRASIL, 2011).
A quimioterapia utiliza de quimioterápicos administrados em intervalos
regulares com a finalidade de tornar os tumores ressecáveis ou melhorar o
prognóstico, e visando a cura após tratamento cirúrgico, para o tratamento de
** It is increasingly clear that tumor cells depend upon their microenvironment to metastasize and that they even rely on host
tissue stromal cells to provide functions, such as a diversity of proteolytic activity, that they themselves may lack (BACAC;
STAMENKOVIC, 2008, p.222,223)

32
neoplasias hematopoiéticas de evolução crônica ou ainda para a paliação de sinais
e sintomas que comprometem a capacidade funcional do paciente (BRASIL, 2011).
Terapia hormonal é a classe terapêutica com maior eficiência no tratamento
dos tumores receptores hormonais de estrogênio e progesterona positivos (KALIKS,
2016). As células cancerígenas podem responder de diversas formas ao tratamento
(LIEDKE, 2006).
O tratamento do câncer teve uma grande evolução nas últimas décadas, e
juntamente com o avanço no diagnóstico precoce colabora com o aumento nos anos
de sobrevida. A taxa de sobrevida dobrou nos últimos 40 anos, porém com
diferenças entre os países. Na Inglaterra, metade das pessoas diagnosticadas com
câncer sobrevive por 10 anos ou mais, e as taxas de sobrevivência são maiores nas
mulheres que nos homens (CRUK, 2011). Na Finlândia a taxa de sobrevivência após
5 anos de diagnóstico aumentou em 5% entre os anos de 2000 e 2014 (CANCER
SOCIETY OF FINLAND, 2016).
A sobrevida em cinco anos dos cânceres de cólon, reto e mama aumentou de
forma constante nos países mais desenvolvidos (ALLEMANI et al., 2015). Para
indivíduos que foram diagnosticados entre 2005 e 2009, a sobrevida para câncer de
cólon e reto atingiu 60% ou mais em 22 países ao redor do mundo; para o câncer de
mama, a sobrevida em 5 anos aumentou para 85% ou mais em 17 países do mundo
(ALLEMANI et al., 2015).
As melhorias na sobrevida do câncer nos últimos 15 anos têm acontecido
sobretudo devido a melhores investimentos na saúde, monitoramento público dos
hospitais quanto ao tempo de espera e melhoras nos tratamentos em diversos
países no mundo (WEGWARTH, 2015). A meta da Organização Mundial da Saúde é
de uma redução de 25% nas mortes por câncer outras doenças não transmissíveis
em pessoas com idade entre 30-69 anos até 2025 (WON H., 2012).
4.2 Câncer de mama
O câncer de mama é uma neoplasia heterogênea causada pelo acúmulo
progressivo de aberrações genéticas e epigenéticas, tais como mutações,
amplificações cromossômicas, deleções, rearranjos, translocações e duplicações
(WOODS, 2007). É uma doença complexa, com apresentações esporádica e

33
familiar, como a maioria dos canceres, e se classifica como a neoplasia que mais
acomete a população feminina e a maior causa de morte por câncer no mundo entre
as mulheres (INCA, 2016).
O câncer mamário é o segundo tipo de câncer mais recorrente nas regiões
Sul, Sudeste, Centro-Oeste e Nordeste do Brasil, bem como EUA (INCA, 2013; DE
SANTIS et al., 2017). Estudo desenvolvido nos EUA afirmou que aproximadamente
252.710 novos casos de câncer de mama invasivo e cerca de 63.410 novos casos
de carcinoma in situ foram estimados para 2017 (DE SANTIS et al., 2017). No Brasil,
foram diagnosticados 57.120 novos casos de câncer de mama no ano de 2014
(INCA, 2014), e foram estimados 57.960 novos casos para 2016 (INCA, 2016).
Estimativas brasileiras para o ano de 2018 apontam cerca de 59.700 novos
casos de câncer de mama no Brasil. Provavelmente a região sudeste será a mais
acometida, com 30.880 novos diagnósticos, sendo que São Paulo terá o maior
número de mulheres com câncer de mama, chegando a 16.340 casos, superior aos
índices para toda a região Sul, que tem estimativa de 11.030 novos cânceres (INCA,
2017). Em virtude do acima exposto, esta condição destaca-se como um dos
principais problemas de saúde pública, além de ser a principal causa de morte por
câncer em alguns países que estão em desenvolvimento (SIEGEL, 2012; OMS,
2013).
O câncer de mama pode iniciar-se a partir de lesões hiperplásicas, cujo
epitélio apresenta alterações genéticas e na sua estrutura, porém ainda não
possuem atipias citológicas. Quando as hiperplasias chegam a apresentar atipias
celulares, ocorre a oclusão dos ductos resultando no carcinoma intraductal in situ
(CIS). Neste estágio, as atipias são mais evidentes e há um aumento significativo da
atividade mitótica epitelial e maiores alterações genéticas. Assim, o processo
progride chegando ao carcinoma localmente invasivo (CI), no qual ocorre o
rompimento da membrana basal e extravasamento das células epiteliais para o
estroma circundante, o que predispõem à disseminação metastática para vários
órgãos, como pulmões, ossos e fígado (TLSTY et al., 2004).
De acordo com a classificação chamada de estadiamento clínico TNM
(T=tumor, N= acometimento linfonodal e M= metástase a distância), preconizada
pela União Internacional Contra o Câncer (UICC), e mais aceito, o câncer de mama
também pode ser classificado em: Tx: tumor não pode ser avaliado; T0: não há
evidência de tumor primário; Tis - carcinoma intraductal in situ/ carcinoma

34
intraductal; Tis - carcinoma intralobular in situ/carcinoma intralobular; Tis (Paget) –
doença de Paget do mamilo sem tumor na mama, ou T1 – tumor ≤ 2 cm em sua
maior dimensão, T1mic (microinvasão) - ≤ 0,1 cm, T1a - >0,1 ≤0,5 cm, T1b - >0,5 ≤
1,0cm, T1c - >1,0 ≤2,0cm, T2 - tumor >2cm ≤5cm em sua maior dimensão, T3 -
tumor >5cm em sua maior dimensão, T4 - tumor de qualquer tamanho, com
extensão direita à parede torácica e/ou à pele, T4a - extensão à parede torácica
(costelas, músculos intercostais e serrátil anterior), T4b - edema (inclusive tipo casca
de laranja), ulceração da pele ou nódulos cutâneos satélites confinados à mesma
mama, T4c - associação de T4a e T4b, T4d - carcinoma inflamatório (BRASIL, 2011;
BRASIL, 2018).
Além dos sistemas de classificação de tumores tradicionais, existem também
os biomarcadores moleculares que auxiliam o diagnóstico e a conduta terapêutica
empregada, como os receptores hormonais de estrogênio e progesterona e o
receptor de crescimento epidérmico humano-2 (HER2) (WEIGEL e DOWSETT,
2010; BRASIL, 2018).
Os receptores de estrogênio e progesterona são os fatores preditivos mais
utilizados para a escolha do tratamento hormonal, sendo que o receptor de
estrogênio tem sido o indicador prognóstico mais estudado. Tal fato se deve à
relação positiva entre a presença do receptor de estrogênio e melhorias na
sobrevida do paciente (ONCOGUIA, 2017; BRASIL, 2018). No tumor receptor
hormonal negativo, a terapia hormonal é menos benéfica e os mesmos tendem a
crescer mais rápido. No câncer de mama triplo negativo, como não há receptor de
estrogênio, progesterona e da proteína HER2, estes crescem e se disseminam mais
rapidamente, e nesse caso o tratamento quimioterápico é a opção mais indicada
(ONCOGUIA, 2017; BRASIL, 2018).
Segundo Weigelt et al. (2005), o óbito dos portadores de tumor de mama é
consequência da disseminação metastática, e não do desenvolvimento do tumor
primário. A disseminação metastática ocorre nos estágios iniciais da doença
(FREUDENBERG, 2009; RUITERKAMP, 2011). Devido a isso, cerca de 80% dos
pacientes com câncer de mama recebem terapia adjuvante. No entanto, 40% destes
pacientes apresentam recidiva e morte em decorrência das metástases (WEIGELT
et al., 2005). Cerca de 36% dos pacientes com câncer de mama metastático morrem
no primeiro ano de acompanhamento médico (ORDING, 2016). Os principais alvos
de metástases do câncer de mama são os ossos, pulmões, fígado e cérebro.

35
Indivíduos que apresentam metástase no cérebro têm o maior risco de mortalidade,
chegando a 62% no primeiro ano de diagnóstico, enquanto os acometidos nos ossos
e pulmões têm 32% de chance de óbito no mesmo período (ORDING, 2016).
Os tumores que são receptores hormonais positivos, como o câncer de
mama, geralmente têm metástases nos ossos. Porém, esse tipo de câncer pode
apresentar metástase em qualquer órgão do corpo, o que dificulta o tratamento e a
sobrevida do paciente (HO, 2015; LOBBEZOO, 2015; ORDING, 2017).
Há aproximadamente uma década, o tratamento do câncer de mama era
conduzido após evidências da morfologia do tumor, e a abordagem cirúrgica
consistia em procedimentos mutilantes de retirada da mama com uma grande
margem de segurança (PACHNICKI et al., 2012; TIEZZI, 2007). Atualmente,
adotam-se procedimentos menos mutilantes, uma vez que a elevada margem de
segurança demonstrou exercer pouca influência no prognóstico. Assim, operações
conservadoras associadas ao tratamento radioterápico tornaram-se eficientes no
controle da doença local (TIEZZI, 2007). Pacientes com câncer mamário não
metastático geralmente utilizam terapias pré-operatórias, o que proporciona a
redução do tamanho do tumor primário (AMOROSO et al., 2015).
No tratamento realizado com terapias hormonais, as drogas utilizadas atuam
de forma a bloquear a ação do estrogênio, e retardam a progressão do tumor, sendo
a primeira escolha para pacientes com câncer de mama em estágio avançado e que
apresente receptores. As drogas mais utilizadas no tratamento hormonal do câncer
de mama são estrogênios, antiestrogênio, progestagênios e androgênios hormonais
(INSTITUTO ONCOGUIA, 2014).
Já a quimioterapia adjuvante é um dos tratamentos que aumenta a sobrevida
do paciente e reduz a taxa de recorrência, com aumento de 2 a 3% da sobrevida
global em dez anos em pacientes que já passaram pela menopausa e que tenham
tumores receptores hormonais positivos. Antraciclinas e taxanos são alguns dos
quimioterápicos utilizados (TESTA e MANO, 2011).
4.3 Células-tronco cancerosas
As células-tronco são definidas como células que possuem a capacidade de
se perpetuarem através da autorrenovação e de se diferenciarem em células

36
maduras de um tecido particular (REYA et al., 2001). Elas são classificadas em
células-tronco embrionárias, presentes apenas nos primeiros estágios do
desenvolvimento e são capazes de originar todos os tipos celulares, e em células-
tronco adultas, células multipotentes, capazes de se diferenciarem em múltiplas
linhagens com expressão de marcadores de superfície específicos do tecido de
origem. As células-tronco adultas podem ser divididas em células-tronco
hematopoiéticas, que dão origem às células sanguíneas, e em células-tronco não
hematopoiéticas (células-tronco mesenquimais), encontradas em todo o corpo,
incluindo medula óssea, cérebro, sangue periférico, vasos sanguíneos, músculo
esquelético, pele, polpa dental, coração, intestino, fígado, epitélio do ovário e do
testículo (ISSCR, 2018).
Muitos tumores contêm células do câncer fenotipicamente e funcionalmente
heterogêneas (MAGEE et al., 2012). Para se explicar a teoria da heterogeneidade
nos tumores, tem-se apresentado dois modelos específicos. No modelo da evolução
clonal, qualquer célula com capacidade proliferativa que sofra mutações genéticas
ou epigenéticas pode levar à formação de novos tumores. No modelo das células-
tronco cancerosas (CTC), apenas uma pequena população de células dentro do
tumor é capaz de proliferar e formar novos tumores. Fatores ambientais e a
diferenciação das células-tronco cancerosas fornecem mecanismos para gerar
heterogeneidade fenotípica e funcional (Figura 7) (REYA et al., 2001; VISVADER e
LINDEMAN, 2008; MAGEE et al., 2012).

37
Figura 7. Fontes de heterogeneidade no câncer
Fonte: MAGEE, PISKOUNOVA et al., 2012. (A) Fatores genéticos ou mudanças epigenéticas; (B)
Fatores do microambiente tumoral; (C) Modelo de células-tronco tumorais, com a hierarquia celular.
Contribuição dos fatores A, B e C para o aumento dessa heterogeneidade (D).
Há cerca de 150 anos observou-se semelhanças entre as células-tronco
embrionárias e adultas com as células cancerosas (WICHA et al., 2006). 82 anos
após essas especulações, pesquisadores confirmaram a existência de células-
tronco cancerosas, mostrando que uma célula tumoral, à semelhança de uma célula-
tronco, foi capaz de gerar células descendentes e favorecer o crescimento de um
novo tumor (MAKINO, 1956; BRUCE, VAN DER GAAG, 1963; KLEINSMITH,
PIERCE, 1964). Autores como Hamburger e Salmon (1977) confirmam a hipótese de
que alguns cânceres podem conter pequenas subpopulações de células idênticas às
células-tronco normais. Bonnet e Dick (1997) identificaram células-tronco
cancerosas em leucemia mielóide aguda humana ao observarem que um pequeno
grupo de células tumorais foi capaz de se replicar facilmente in vitro, enquanto o
mesmo não ocorreu com as demais células leucêmicas. Posteriormente, as células-
tronco foram descritas em diversos tipos tumorais, como próstata, cérebro, ovário,
cólon, pele, pulmão, e, em particular no câncer de mama (PICCIRILLO e VESCOVI,
2007).

38
Atualmente o surgimento, progressão e recidiva de tumores, a ocorrência de
metástases e falhas em terapias têm sido relacionadas às funções orquestradas
pelas células-tronco cancerosas (CTC). Trata-se de uma subpopulação de células
dentro do tumor que apresentam um metabolismo único, autofágico, invasivo e
propriedades de quimiorresistência que lhes permitem renovar continuamente e
resistir às terapias oncológicas (VALLE et al., 2018). Acredita-se que as CTC
possuem a capacidade de auto-renovação e tendem a acumular alterações
genéticas ao longo do tempo, evadindo-se dos mecanismos de controle do
microambiente e, com isso, contribuindo para a evolução tumoral (SHIPITSIN e
POLYAK, 2008) (Figura 8).
Figura 8. Autorrenovação e diferenciação de células-tronco normais e células-tronco
tumorais.
Fonte: JORDAN et al., 2006. A seta curva indica a habilidade de autorrenovação das células-tronco
normal e tumoral. O potencial de multidiferenciação é indicado pelas setas retas contínuas. A seta
pontilhada indica o surgimento das células-tronco tumorais a partir das células progenitoras. As
células-tronco tumorais surgem a partir de mutações nas células-tronco normais ou nas células
progenitoras, que crescem e se diferenciam para produzir o tumor primário.
As CTC também podem se diferenciarem em células maduras de um tecido
particular, assim como possuir longas durações nos ciclos celulares (REYA et al.,
2001). Tais características proporciona a essas células a capacidade de produção
de todos os tipos celulares de um tumor, o que confere a heterogeneidade tumoral
(CAMPBELL e POLYAK, 2007). Possuem ainda, capacidade de quiescência,

39
permanecendo-se em estágio G0, e tornam-se ativadas apenas quando recebem
sinais apropriados do seu nicho. Apresentam também atividade de telomerase ativa,
impedindo que os telômeros se encurtem e consequentemente parem de reproduzir,
e elevado nível de proteínas anti-apoptóticas, como os membros da família Bcl-2 e
inibidores da apoptose, o que confere a elas resistência à apoptose e longevidade
(SOUZA, 2006; WICHA et al., 2006).
Além disso, as CTC possuem habilidade de induzir a angiogênese tumoral,
migração e metástase. A resistência das células-tronco tumorais ao tratamento
quimioterápico é baseada na hipótese de que estas células reúnem características
fundamentais ao fenômeno de resistência, incluindo a capacidade em reparar danos
no DNA, a expressão aumentada de proteínas que realizam o efluxo de agentes
quimioterápicos para fora da célula (transportadores ABC) e a baixa taxa de divisão
celular, fazendo com que não respondam à maioria dos agentes quimioterápicos que
têm como alvo as células em divisão (Figura 9) (DEAN, 2005). Segundo Bozorgi,
Khazaei (2015), as células-tronco cancerosas possuem altos níveis de proteínas
anti-apoptóticas e da enzima aldeído desidrogenase (ALDH). A ALDH, encontra-se
diretamente envolvida, na radiorresistência das CTC por meio da remoção direta de
radicais de oxigênio (ROS) que são produzidos no meio intracelular e pela produção
indireta de nicotinamida adenina dinucleotídeo (fosfato), um composto antioxidante
(considerados aceptores de ROS - ROS scavenging). O paradigma que a resistência
da droga origina no fenótipo de células-tronco pode estimular novas estratégias para
o desenvolvimento de terapias anti-câncer (DEAN, 2005).
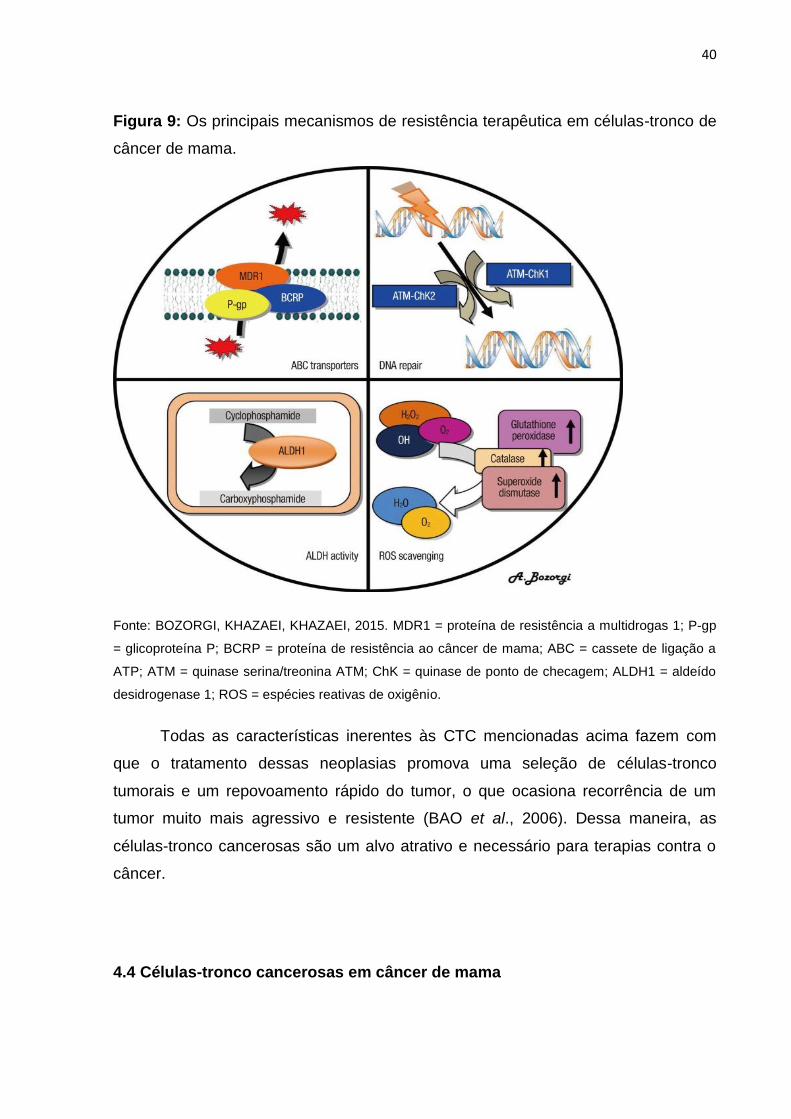
40
Figura 9: Os principais mecanismos de resistência terapêutica em células-tronco de
câncer de mama.
Fonte: BOZORGI, KHAZAEI, KHAZAEI, 2015. MDR1 = proteína de resistência a multidrogas 1; P-gp
= glicoproteína P; BCRP = proteína de resistência ao câncer de mama; ABC = cassete de ligação a
ATP; ATM = quinase serina/treonina ATM; ChK = quinase de ponto de checagem; ALDH1 = aldeído
desidrogenase 1; ROS = espécies reativas de oxigênio.
Todas as características inerentes às CTC mencionadas acima fazem com
que o tratamento dessas neoplasias promova uma seleção de células-tronco
tumorais e um repovoamento rápido do tumor, o que ocasiona recorrência de um
tumor muito mais agressivo e resistente (BAO et al., 2006). Dessa maneira, as
células-tronco cancerosas são um alvo atrativo e necessário para terapias contra o
câncer.
4.4 Células-tronco cancerosas em câncer de mama

41
As células-tronco cancerosas em câncer de mama têm sido amplamente
estudadas desde a década de 90. Elas têm apresentado importante influência na
agressividade do tumor e na resistência aos tratamentos em função da capacidade
de auto-renovação e a existência de canais de íons que facilitam o bombeamento de
medicamentos para o meio extracelular (YARAK e OKAMOTO, 2010).
Marcadores de superfície são usados para o isolamento e identificação de
células-tronco cancerosas no câncer de mama, como as proteínas
transmembrânicas com fenótipo CD44+CD24-, identificadas pela primeira vez em
2003 por Al-hajj et al. (2003). Desde então, esse fenótipo tem sido utilizado como um
fenótipo de confiança para o isolamento de células-tronco cancerosas da mama
(BOZORGI et al., 2015).
A glicoproteína de superfície celular CD44 é um receptor específico para o
ácido hialurônico, colágeno, fibronectina e sulfato de condroitina, considerados
componentes importantes da matriz extracelular. Regula a adesão célula-célula e
interação célula-matriz, bem como a migração celular e também a ligação à citocina
osteopontina (BOZORGI et al., 2015 e ORIAN-ROUSSEAU, 2010). Formas variantes
de CD44, em particular CD44v6, possuem expressão tecido-específica e estão
associadas a tumores mais agressivos e pior prognóstico, incluindo o câncer de
mama (AFIFY et al., 2009).
Já a glicoproteína CD24, quando expressa em níveis baixos, CD24-, aumenta
a capacidade de crescimento e metástase tumoral (BOZORGI et al., 2015). Trata-se
de uma proteína mucinosa glicosilada pesada, ligada à membrana celular via âncora
de GPI (Glicosilfosfatidilinositol). CD24 é o ligante de P-seletina, um transportador do
tipo ABC superexpresso em células resistentes a multidrogas e expresso em
receptores de adesão de células endoteliais ativadas e plaquetas, indicando
importante papel no processo de quimiorresistência e metástase (AIGNER et al.,
1998; KIM et al., 2007).Está envolvida na regulação da proliferação celular e na
interação célula-célula, encontra-se expressa em pré-linfócitos B normais, assim
como em vários cânceres hematológicos e em tumores sólidos (LIM, 2005).
Outro importante marcador de células-tronco do câncer da mama é a
atividade da enzima aldeído desidrogenase (ALDH), uma proteína que provém de
uma família de enzimas presentes no citoplasma responsáveis por catalisar a
oxidação de aldeídos intracelulares, além de oxidar o retinol no ácido retinóico
durante a diferenciação de células-tronco rudimentares. Algumas isoformas da

42
família ALDHA1 têm sido implicadas na progressão e resistência a terapias
(THOMAS et al., 2016). Outros marcadores utilizados para identificar as células-
tronco cancerosas na mama são CD133, CD49f e CD61 (BOZORGI et al., 2015;
SUN et al., 2015).
Algumas vias de sinalização celular são fundamentais para o
desenvolvimento do câncer de mama. As CTC tipicamente demonstram ativação
persistente de um ou mais mecanismos de transdução de sinal envolvidos no
desenvolvimento e homeostase tecidual, incluindo as vias de sinalização Notch, Wnt
e Hedgehog. Alterações nessas vias estão relacionadas com a resistência às
terapias e à proliferação descontrolada de células-tronco cancerosas durante e após
o tratamento (SUN et al., 2015). Em mama, as vias Notch e Hedgehog alteradas
resultam em um fenótipo de células-tronco cancerosas. Os receptores
transmembrana Notch (Notch 1 a Notch 4) são primariamente envolvidos na
comunicação entre células, e apresentam papeis importantes na dormência e
recorrência do câncer de mama. Notch1 tem sido ativado em pacientes
adenocarcinoma negativo para o receptor de estrógeno (GU et al., 2012;
ABRAVANEL et al., 2015). As proteínas da sinalização Hedgehod estão envolvidas
com o desenvolvimento e reparo de tecidos normais embrionários e transição das
células do epitélio para o mesênquima. A cascata de sinalização Wnt promove a
ativação do fator de transcrição β-catenina, a regulação dos níveis de cálcio
intracelular e o citoesqueleto, e desempenha um papel fundamental na auto-
renovação das células-tronco e na preservação de um estado indiferenciado
(BOZORGI et al., 2015).
Os fatores de transcrição também interferem na progressão e resistência do
câncer de mama. SOX2 e NANOG são fatores de transcrição que atuam como
reguladores da pluripotência, além de manter o estado indiferenciado das células.
NANOG é capaz de promover a pluripotência independente da fosforilação de
STAT3 ativada por LIF. Ele também regula 14 dos 22 genes alvos de STAT3
envolvidos na manutenção do estágio indiferenciado. NANOG também regula o ciclo
celular por ligação ao promotor da ciclina D1. Indução de NANOG causa regulação
aumentada de CD133 e ALDH1 (KAUFHOLD et al., 2016). CD133 é uma
glicoproteína transmembrana codificada pelo gene PROM1, está localizado
preferencialmente em protuberâncias e microvilosidades da membrana plasmática,
sendo relacionada com a organização das membranas, metabolismo da glicose e

43
modulação do citoesqueleto. CD133 é o antígeno de superfície celular mais
frequentemente utilizado para detectar e isolar CTCs de tumores sólidos, entre eles
o câncer de mama (GLUMAC e LEBEAU 2018). 43% dos carcinomas basais de
mama possuem expressão aumentada de SOX2 (BOZORGI et al., 2015). Ativação
de SOX2 promove o aumento da proliferação via indução de uma das proteínas
reguladoras do ciclo celular, a ciclina D3 (KAUFHOLD et al., 2016). Wang et al.
(2017) mencionaram em seu estudo que a superexpressão de SOX2 pode aumentar
a proliferação e a invasão de células de câncer de mama e promover o processo da
transição epitélio-mesenquimal (TEM). SOX2 pode ser um biomarcador preditivo do
prognóstico do câncer de mama, e um potencial alvo terapêutico.
4.5 Marcadores putativos de células-tronco cancerosas
Em estudos das células-tronco cancerosas tem-se identificado genes
biomarcadores preditivos para o mau prognóstico em vários carcinomas (YOU,
2018). Sabe-se que detectar células-tronco cancerosas no sangue é importante para
monitorar pacientes com câncer, afim de se identificar as metástases em estágio
inicial e personalização da terapia empregada (MANSOORI et al., 2017).
As vias de sinalização geralmente não atuam sozinhas, interagem com outras
formando uma rede biológica complexa e coordenada, contribuindo para a
diversidade celular das células-tronco durante a embriogênese e a homeostase dos
tecidos. Essa interação pode desempenhar papeis essenciais no câncer e na
biologia da células-tronco cancerosas. A exemplo, observa-se que, ao passo que a
proteína Mel-18, considerada reguladora negativa da autorrenovação da células-
tronco cancerosas do câncer de mama, ao ser inibida, ativa a sinalização da via
Wnt, que, por sua vez aumenta a expressão do gene alvo WNT JAGGED-1, levando
à ativação da via Notch e consequente autorrenovação das células-tronco
cancerosas. Assim como a regulação positiva do gene JAGGED-1, induz a
sinalização da via Notch, elevando a atividade do oncogene β-catenina das
sinalizações de Wnt e Notch em células de câncer colorretal (WON et al., 2012;
KOURY et al., 2017).
Marcadores como SOX2 e NANOG são responsáveis pela regulação da
pluripotência e auto-renovação de células-tronco e por isso desempenham papéis

44
fundamentais na carcinogênese em vários tumores malignos, como de pulmão,
mama e endométrio. O marcador SOX2 está associado positivamente com a
ocorrência de metástases em linfonodos (SHAN et al., 2012; YOU, 2018). O
aumento da expressão do marcador NANOG demonstrou também maior resistência
a terapêuticos, maior metástase e invasão tumoral, tornando o NANOG um
biomarcador de células-tronco cancerosas, fundamental para a auto-renovação e
disseminação de tais células através da via de sinalização do fator de crescimento
IGF1R (SHAN et al., 2012).
Sabe-se que a via de sinalização Notch quando sofre mutações promove a
tumorigênese, inibição da apoptose, além do comprometimento da divisão e
diferenciação celular (SHEN et al., 2016). Shen et al., (2016) retratam que NOTCH
não apenas regula a expressão de células cancerosas, como também facilita a
ativação de células tumorais associadas a macrófagos mediado por TGFβ. Shen et
al., (2016) afirmam ainda que os subtipos de câncer de mama, Câncer de mama
básico (BLBC) e o Câncer de mama Triplo Negativo, são ativados por NOTCH e
altamente infiltrados por macrófagos.
Koury et al., (2017), afirmam que a ativação anormal da via de sinalização
Notch desempenha um papel fundamental nas células-tronco cancerosas no câncer
de mama, pâncreas e glioblastoma. Mencionaram ainda que a inibição da proteína
fascina (proteína agregadora da actina) reduziu o fenótipo de CTCs através da
regulação negativa de genes pluripotentes de células-tronco cancerosas, como
OCT4, NANOG, SOX2 e KLF4, e com isso as células apresentaram menor potencial
para a formação de colônias e tumoresferas. Em contrapartida, a ativação da via de
sinalização de Notch apresentou maior tumorigênese.
Células do câncer de próstata submetidas à transição epitélio mesenquimal
(TEM) exibiram características de células-tronco, caracterizadas pelo aumento da
expressão de Notch 1, além de outros genes pluripotentes como SOX2, NANOG,
OCT4 e LIN28, indicando que a via de sinalização Notch está intimamente envolvida
na TEM (KOURY et al., 2017).
Em pesquisa realizada por Kim et al., (2017), demonstrou-se que níveis
elevados da expressão dos genes relacionados aos marcadores de células-tronco
cancerosas ALDH1 e NOTCH, principalmente o NOTCH3, estavam associados a
células-tronco cancerosas e com o Carcinoma Seroso de Ovário quimiorresistentes
e células de câncer de ovário resistentes ao paclitaxel. Além disso, a elevada

45
expressão do gene NOTCH3 foi associado ao mau prognóstico, ou seja, estágio
avançado, linfonodos e metástases à distância.
A desregulação da sinalização Wnt está ligada à tumorigênese de muitos
tipos de câncer, assim como a resistência aos medicamentos utilizados nas terapias
oncológicas (GENECARDS, 2018; YANG et al, 2017). A via de sinalização Wnt está
envolvida no processo da transição epitélio-mesenquimal (TEM), processo biológico
que permite que a célula epitelial sofra modificações bioquímicas, moleculares e
morfológicos, resultando na aquisição de um fenótipo de células mesenquimais, com
capacidade de migração, invasão, resistência e apoptose. TEM é considerado um
dos passos iniciais na cascata de invasão e metástase tumoral (BEKAII-SAAB,
2017).
A desregulação da via Wnt também está associada à mutação dos genes
APC e β-catenina, visto que a inativação da função da proteína APC resulta da
ativação da sinalização Wnt e fosforilação e degradação de β-catenina. Tais
mutações foram descritas em vários cânceres, incluindo 10% dos cânceres
colorretais. Alterações na via de sinalização Wnt também interrompe o crescimento e
a diferenciação de células-tronco (KOURY et al., 2017).
Identificado pela primeira vez em 2003, o marcador de superfície de
células-tronco cancerosas CD44 é uma glicoproteína de superfície celular e um
receptor específico para o ácido hialurônico. Sua interação com a osteopontina,
proteína de ligação codificada pelo gene SPP1, leva à progressão tumoral, além de
contribuir para a progressão, adesão, movimentação e migração, levando à
angiogênese e metástase tumoral (BOZORGI et al., 2015; KIN, 2018). Kim et al.
(2017) indicaram que as células com fenótipo CD44+/CD24- possuíam maior
resistência à irradiação e superior capacidade de reparar danos ao DNA quando
comparadas com outras células.
4.6 Bioinformática
A bioinformática tem auxiliado pesquisadores e tem sido uma das áreas mais
conspícua na ciência moderna. Esta área do conhecimento envolve aspectos
multidisciplinares e disponibiliza um conjunto variado de ferramentas para
armazenar, processar, analisar e avaliar dados genéticos, bioquímicos e de biologia
molecular, através de experimentos in silico, por meio de simulações

46
computacionais. Além disso, tem sido utilizada em diversas áreas, como a
construção de banco de dados, mineração de dados, análises de sequências,
identificação e análise de genes, relacionamento entre genes e proteínas. A
bioinformática também tem auxiliado os estudos laboratoriais ao prever a
conformação tridimensional das proteínas, construir árvores filogenéticas, modelos
evolutivos, construir bibliotecas genômicas, estudar as funções biológicas, desenho
de drogas, entre muitas outras aplicações (NOTARI, 2012; PEREIRA, 2016).
Uma das áreas pertencente ao estudo da bioinformática é a biologia de
sistemas, campo que investiga as interações entre os componentes de um sistema
biológico e auxilia a compreender como acontecem as interações entre os genes. As
redes biológicas são construídas a partir da teoria dos grafos. Os grafos são
representados por conjuntos de nós ou vértices, conectados entre si por conectores
ou arestas. Os nós podem representar genes, proteínas, indivíduos, enquanto que
as arestas representam a relação entre cada par de nós. Esta representação gráfica
é denominada rede (MOREIRA, 2015; VERLI, 2014).
O estudo de redes de genes ou proteínas depende da integração das
informações disponíveis em bases de dados. A ferramenta GeneCards é um banco
de dados integrado de fácil utilização, que fornece informações genômicas,
proteômicas, transcriptômicas, genética, clínica e funcional sobre todos os genes
humanos anotados e previstos. Dados sobre as funções celulares e o envolvimento
com doenças são também fornecidos por essa base de dados (GENECARDS,
2018). A base de dados Search Tool For The Retrieval Of Interacting Genes/Proteins
(STRING – Ferramenta de Pesquisa para Recuperação de Interações entre
Genes/Proteínas) é uma base de dados de interações entre proteínas ou genes que
incluem associações físicas diretas, indiretas e interações funcionais, que resultem
de previsões computacionais, a partir da transferência de conhecimentos entre
organismos. Interações obtidas de outros bancos de dados interligados, conhecidos
como banco de dados primários, também estão presentes no STRING
(SZKLARCZYK, 2017). Essas interações ajudam a descrever as funções de um
gene. O conhecimento sobre os parceiros na interação é um requisito importante
para entender melhor a função de um gene (FRANCESCHINI et al., 2015).
A base de dados STRING abrange atualmente 9.643.763 proteínas de 2.031
organismos. Todas as associações do STRING são fornecidas com uma pontuação
de confiança probabilística. Essas pontuações são escaladas entre zero e um. Elas

47
indicam a probabilidade estimada de que uma dada interação seja biologicamente
significativa, específica e reprodutível, tendo em vista a evidência de suporte, ou
seja, os canais de evidência. O resultado é uma rede de associações previstas para
um determinado grupo de gene. Os nós da rede são os genes e as arestas
representam as associações funcionais previstas. Uma aresta pode ser
representada com até sete linhas de cores diferentes, sendo que estas linhas
indicam a existência dos tipos de provas utilizadas na previsão das associações
(SZKLARCZYK, 2017).
Os canais de evidência do STRING são o canal de experiências (a evidência
vem de experimentos reais no laboratório), o canal de banco de dados (utiliza dados
que foram afirmados por um curador perito humano), o canal de mineração de textos
(baseado em menções de nomes de proteínas em todos os resumos da PubMed e
em uma coleção interna de mais de três milhões de artigos de texto completo),
o canal de coexpressão (dados de expressão gênica provenientes de uma variedade
de experiências de expressão), canal de vizinhança (canal de predição baseados em
genoma), canal de fusão (os pares de proteínas recebem uma pontuação de
associação quando há pelo menos um organismo onde seus respectivos ortológos
se fundem em um único gene codificador de proteínas) e o canal de co-ocorrência
(avalia a distribuição filogenética de ortólogos de todas as proteínas em um
determinado organismo) (SZKLARCZYK, 2017).
Estudos de bioinformática envolvendo redes de interação gênica em células-
tronco cancerosas do câncer de mama podem assessorar a compreensão a respeito
da biologia dessa condição, e servir de base para estudos celulares e moleculares
futuros.

48
5 METODOLOGIA
5.1 Caracterização do estudo
O presente estudo caracteriza-se como uma pesquisa exploratória de
natureza computacional.
5.2 Identificação dos genes
A identificação dos genes envolvidos com as condições estudadas foi
realizada através de pesquisas na base de dados de genética humana GeneCards®,
utilizando a nomenclatura oficial para genes humanos Human Genoma Organzation
(HUGO). Os descritores adotados para cada condição foram definidos de acordo
com a biblioteca de descritores de assuntos médicos Medical Subject Headings
(MeSH). Como critério de inclusão para a condição “Cancer stem cells”, o termo
selecionado a partir dos descritores do MeSH foi aquele que apresentou o maior
número de genes na rede. Para a condição “Breast cancer”, o termo selecionado foi
o sinônimo que referenciava à condição maligna da mama. Não havia termos válidos
para as condições de “metastasis of breast cancer” e “Breast cancer stem cells”. O
descritor utilizado para a identificação dos genes relativos às células-tronco
cancerosas foi "cancer stem cells". Para o estudo das células-tronco cancerosas no
câncer de mama utilizou-se o descritor "breast cancer stem cells". Os descritores
para câncer de mama e metástase originada do câncer de mama foram “breast
cancer” e "metastasis of breast cancer", respectivamente. Com exceção da condição
câncer de mama, nas demais condições em estudo foram utilizados todos os genes
relacionados ao homo sapiens fornecido pela base de dados genética humana
GeneCards®. No câncer de mama, foram identificados 8.610 genes relacionados a
essa condição, sendo selecionados para o corrente estudo 400 genes com as
pontuações (scores) mais elevadas. Todos os genes estudados possuíram um nível
de confiança acima de 0,9 fornecido pela base GeneCards®.
5.3 Construção e análise do mapa de interações genômico

49
Após a identificação dos genes, foi construído um mapa genômico de cada
condição estudada com o auxílio do software STRING (Search Tool for the Retrieval
of Interacting Genes/Proteins) versão 10.0. Somente as interações gênicas com grau
de confiança acima de 0,9 foram utilizadas nas análises. Foram utilizadas todas as
fontes de interações ativas (mineração de texto, experimentos, base de dados, co-
expressão, vizinhança, fusão de genes e co-ocorrência). No mapa genômico, os nós
das redes representam os genes enquanto as arestas representam as associações
proteína-proteína. Uma aresta pode ser representada por até oito linhas de cores
diferentes, sendo que tais linhas indicam a existência dos tipos de prova utilizadas
nas associações, sejam elas por interações conhecidas ou previstas (SZKLARCZYK
et al., 2017).
A análise do mapeamento da rede de genes na base de dados STRING
resultou em um arquivo Text Output contendo as pontuações para todas as
associações realizadas por cada gene em cada rede em estudo. Assim, tais
pontuações foram tabuladas no editor de planilhas Excel 2013 e multiplicados por
1000. Posteriormente foi realizada a soma da pontuação de interação de cada gene,
com o intuito de encontrar a pontuação única denominada Weighted Number of
Links (WNL), número ponderado de ligações (POSWAR et al., 2015). O WNL refere-
se ao cálculo da soma das associações de cada gene dentro da rede a que pertence
(PEREIRA, 2016). Na presente pesquisa, também foi calculada a pontuação de
interação global dos genes denominada Significant Global Connectivity (TIS),
conectividade global significativa. Os valores de TIS foram estabelecidos somando-
se as associações de cada gene a partir dos valores obtidos do arquivo de
interações globais Protein Network Data gerado pelo software STRING. Ele refere-se
ao cálculo dos valores das associações de cada gene em todas as redes disponíveis
na base de dados do software STRING (PEREIRA, 2016). A razão entre WNL/TIS
representa os genes mais influentes dentro da rede em estudo. Genes que não
realizaram interações são chamados de genes órfãos (POSWAR et al., 2015;
PEREIRA, 2016; SANTOS et al., 2016).
Os genes de cada mapa genômico foram agrupados em função dos seus
valores de WNL por meio da análise de agrupamento realizada pelo algorítmo K-
means no programa de estatística SPSS® PASS Statistic 18.0. Para avaliar as
diferenças entre os grupos gerados pela análise de K-means, foi realizada a análise
de variância (ANOVA) e pós-teste de Tukey Kramer. A significância estatística foi

50
fixada em um P valor < 0,001. Os genes que apresentaram maiores valores de WNL
foram considerados genes líderes (SANTOS et al., 2016).
Gráficos da relação do WNL/TIS e os resultados do agrupamento através do
algoritmo K-means foram construídos no programa SPSS® PASS Statistic 18.0.
5.4 Definição dos genes marcadores putativos de células-tronco cancerosas
Os marcadores putativos de células-tronco cancerosas foram definidos com
base em pesquisas realizadas em artigos científicos publicados na biblioteca
Medical Literature Analysis and Retrieval System Online, MEDLINE, através da base
de dados online de referências de ciências da vida, PubMed. O critério de inclusão
para os genes marcadores de células-tronco cancerosas nesse estudo foi estar
entre os genes mais comumente utilizados nos artigos científicos como marcadores
putativos e estar presente na rede de células-tronco cancerosas do corrente estudo.
Portanto, os genes marcadores putativos de células-tronco cancerosas estudados
foram WNT1, NOTCH1, NANOG, SOX2, CD44 e ALDH1A1.
5.5 Análises das interações realizadas pelos genes marcadores putativos de
CTC
O número de interações realizadas por cada gene marcador de células-tronco
cancerosas foi realizada em cada condição biológica estudada. O gráfico
representativo dessa condição foi construído no programa GraphPad Prisma® 5.0.
Os genes que interagem com os marcadores de células-tronco cancerosas WNT1,
NOTCH1, NANOG, SOX2, CD44 e ALDH1A1 também foram definidos para cada
condição estudada.

51
6 RESULTADOS
Identificação e análise dos genes associados às condições biológicas estudadas
A pesquisa através da base de dados genética GeneCards® permitiu a
identificação de um total de 1116 genes para todas condições estudadas, sendo
que, 1023 genes foram inseridos no estudo por estarem presentes em Homo
sapiens. Na rede de células-tronco cancerosas (CTC) e células-tronco cancerosas
do câncer de mama (CTCM) foram estudados 386 genes e 81 genes,
respectivamente. Na rede do câncer de mama (CM) e na metástase originada do
câncer de mama (MM) foram identificados 396 e 160 genes, respectivamente. Os
mapas das interações gênicas de cada condição podem ser visualizados nas figuras
10 a 13. Os genes que mais interagem apresentam maior número de ligações.
Dentre os 1023 genes inseridos no estudo, 178 (17,4%) genes não
realizaram nenhuma interação gênica dentro da rede, sendo por esse motivo
considerados genes órfãos. A porcentagem de genes órfãos em cada rede foi de
14% em CTC (55 genes), 21% em CTCM (17 genes), 15% em CM (60 genes) e 29%
em MM (46 genes).

52
Figura 10: Mapa de interação dos 386 genes envolvidos na rede de células-tronco
cancerosas.
Fonte: Dados da pesquisa. Construído com nível de confiança > 0,9. Os genes considerados órfãos
são aqueles que não apresentam interação com outros genes da rede. A associação vermelha indica
fusão de genes; verde: genes vizinhos; azul escuro: co-ocorrência de genes; roxa: homologia
proteica; rosa: relação determinada experimentalmente; amarela: relação determinada por estudos de
mineração de textos; azul claro: relação proveniente de um banco de dados com curadoria; preta: co-
expressão gênica.

53
Figura 11: Mapa de interação dos 81 genes envolvidos na rede das células-tronco cancerosas do câncer de mama.
Fonte: Dados da pesquisa. Construído com nível de confiança > 0,9. Os genes considerados órfãos
são aqueles que não apresentam interação com outros genes da rede. A associação vermelha indica
fusão de genes; verde: genes vizinhos; azul escuro: co-ocorrência de genes; roxa: homologia
proteica; rosa: relação determinada experimentalmente; amarela: relação determinada por estudos de
mineração de textos; azul claro: relação proveniente de um banco de dados com curadoria; preta: co-
expressão gênica. Fonte: Dados da pesquisa.

54
Figura 12: Mapa de interação dos 396 genes envolvidos na rede do câncer de
mama.
Fonte: Dados da pesquisa. Construído com nível de confiança > 0,9. Os genes considerados órfãos
são aqueles que não apresentam interação com outros genes da rede. A associação vermelha indica
fusão de genes; verde: genes vizinhos; azul escuro: co-ocorrência de genes; roxa: homologia
proteica; rosa: relação determinada experimentalmente; amarela: relação determinada por estudos de
mineração de textos; azul claro: relação proveniente de um banco de dados com curadoria; preta: co-
expressão gênica.

55
Figura 13: Mapa de interação dos 160 genes potencialmente envolvidos na rede de
metástase de origem no câncer de mama.
Fonte: Dados da pesquisa. Construído com nível de confiança > 0,9. Os genes considerados órfãos
são aqueles que não apresentam interação com outros genes da rede. A associação vermelha indica
fusão de genes; verde: genes vizinhos; azul escuro: co-ocorrência de genes; roxa: homologia
proteica; rosa: relação determinada experimentalmente; amarela: relação determinada por estudos de
mineração de textos; azul claro: relação proveniente de um banco de dados com curadoria; preta: co-
expressão gênica.
A análise do número ponderado de interações dentro da rede (WNL) em
função das interações realizadas na rede global (TIS) permitiu a distribuição dos
genes por grupo e a identificação dos genes considerados líderes de cada condição.
As figuras 14 a, 15 a, 16 a e 17 a apresentam as distribuições de todos os genes em
função dos valores de WNL e TIS. Percebe-se em tais figuras que todos os genes
líderes se encontram acima da linha de tendência de regressão (R2 linear). Em cada

56
rede estudada, foram considerados genes líderes os 8 genes com maiores valores
de WNL. A tabela 1 apresenta os genes líderes de cada condição estudada.
O gene TP53 apresentou maior valor de WNL nas redes de células-tronco
cancerosas (CTC), metástase originada do câncer de mama (MM) e na rede do
câncer de mama (CM). Em CTC e MM, o gene TP53 foi o único gene do grupo A
(grupo portador do maior WNL), enquanto na rede de CM o grupo A foi composto
pelos genes TP53 e PIK3CA. Já na rede das células-tronco cancerosas do câncer
de mama, os genes STAT3, EGFR, CTNNB1 e JUN foram evidenciados no grupo A.
A comparação dos genes líderes em cada condição estudada demonstra que
EGFR foi um gene líder presente em todas as condições estudadas. TP53, STAT3 e
JUN também se destacaram porque foram evidenciados em 3 condições. Os grupos
de CTC e CTCM apresentaram o maior número de genes líderes em comum
(STAT3, EGFR, CTNNB1, JUN e NOTCH1). NFKB1, FOS e ESR1 foram genes
líderes da rede de CTCM que diferiram dos líderes da rede de CTC. TP53, AKT1 e
EGFR foram genes líderes da rede de CTC que também se encontraram na rede de
CM. TP53, EGFR, STAT3 e JUN foram genes comuns aos grupos CTC e MM.
Tabela 1: Genes líderes das condições estudadas
Células-tronco cancerosas
Células-tronco cancerosas do câncer de mama
Câncer de Mama Metástase originada do câncer de mama
Genes
Grupos Genes Grupos Genes Grupos Genes Grupos
TP53 AKT1
CTNNB1 MYC JUN
STAT3 EGFR
NOTCH1
A B B B C C D D
STAT3 EGFR
CTNNB1 JUN
NFKB1 NOTCH1
FOS ESR1
A A A A B B B C
TP53 PIK3CA AKT1 EGFR
MAPK1 EP300 SRC CDK2
A A B C C C C C
TP53 EGFR STAT3
JUN HDAC1 ERBB2 VEGFA HDAC2
A B C C C D D D
Fonte: Dados da pesquisa.
As análises dos dados relacionadas ao agrupamento e distribuição de genes
por grupos estão representadas nas figuras 5b, 6b, 7b e 8b. A análise preliminar de
k-means revelou os genes pertencentes a cada grupo nas condições em estudo,
sendo 14 grupos para a rede de células-tronco cancerosas e para a rede do câncer
de mama, 6 grupos para células-tronco cancerosas do câncer de mama e 9 grupos

57
para a rede de metástase originada do câncer de mama. A partir da análise Anova
One Way, identificou-se que houve diferença estatisticamente significante nos
valores de WNL dos grupos de cada condição estudada, todos os p valores foram p<
0,001.
Figura 14: Distribuição dos genes da rede de células-tronco cancerosas em função
das interações gênicas e análise de agrupamento.
Fonte: Dados da pesquisa. Representação dos genes em função dos valores de WNL (número ponderado de ligações) e TIS (Conectividade Global Significativa) (a). Distribuição dos genes por análise de agrupamento k-means (b).
Figura 15: Distribuição dos genes da rede de células-tronco cancerosas da mama
em função das interações gênicas e análise de agrupamento.
Fonte: Dados da pesquisa. Representação dos genes em função dos valores de WNL (número
ponderado de ligações) e TIS (Conectividade Global Significativa) (a). Distribuição dos genes por
análise de agrupamento k-means (b).

58
Figura 16: Distribuição dos genes da rede de câncer de mama em função das
interações gênicas e análise de agrupamento.
Fonte: Dados da pesquisa. Representação dos genes em função dos valores de WNL (número ponderado de ligações) e TIS (Conectividade Global Significativa) (a). Distribuição dos genes por análise de agrupamento k-means (b).
Figura 17: Distribuição dos genes da rede de metástase de mama em função das
interações gênicas e análise de agrupamento.
Fonte: Dados da pesquisa. Representação dos genes em função dos valores de WNL (número
ponderado de ligações) e TIS (Conectividade Global Significativa) (a). Distribuição dos genes por
análise de agrupamento k-means (b).

59
Análise de marcadores putativos de células-tronco cancerosas nas redes de CTC,
CTCM, CM e MM
Os marcadores putativos de células-tronco cancerosas WNT1, NOTCH1,
NANOG, SOX2, CD44 e ALDH1A1 foram obtidos a partir de informações da
literatura. Todos estão presentes na rede de células-tronco cancerosas e percebeu-
se na figura 18 a que tais genes possuem distintos valores de WNL e TIS. Destacou-
se também na figura 18, a presença dos marcadores de células-tronco cancerosas
nas condições CTCM (Figura 18 b), CM (Figura 18 c) e MM (Figura 18 d). O único
marcador presente em todas as quatro condições é o CD44, cujo valor de WNL foi
abaixo da linha de tendência regressiva (R2 Linear) em CTC, CTCM e CM. Na rede
de metástase do câncer de mama, o marcador CD44 possui uma maior interação
com a rede de metástase (WNL acima da linha de tendência regressiva) e menor
interação com outras redes (baixo TIS). O marcador NOTCH1 foi encontrado nas
redes de CTC, CTCM e CM, estando acima da linha de tendência regressiva em
todas as redes (Figuras 18 a, 18 b, 18 c). Esse marcador foi o marcador de células-
tronco cancerosas com melhores combinações de alto WNL e baixo TIS nas redes
de células-tronco cancerosas (CTC e CTCM). Já o marcador WNT1 foi encontrado
nas redes de CTC e CTCM (Figuras 18 a e 18 b), em ambas acima da linha de
tendência regressiva. Dentre os marcadores em estudo, NANOG foi o que
apresentou o maior valor de TIS, e apresentou-se nas redes de CTC e CTCM
(Figuras 18 a, 18 b). SOX2 também foi evidenciado apenas nas redes de CTC e
CTCM apresentando valores de WNL acima da linha de tendência regressiva e
baixo valor de TIS (Figuras 18 a, 18 b), enquanto que o marcador ALDH1A1 foi
encontrado nas redes de CTC e CM apresentando os menores valores de WNL
quando comparado aos demais marcadores (Figuras 18 a, 18 c).
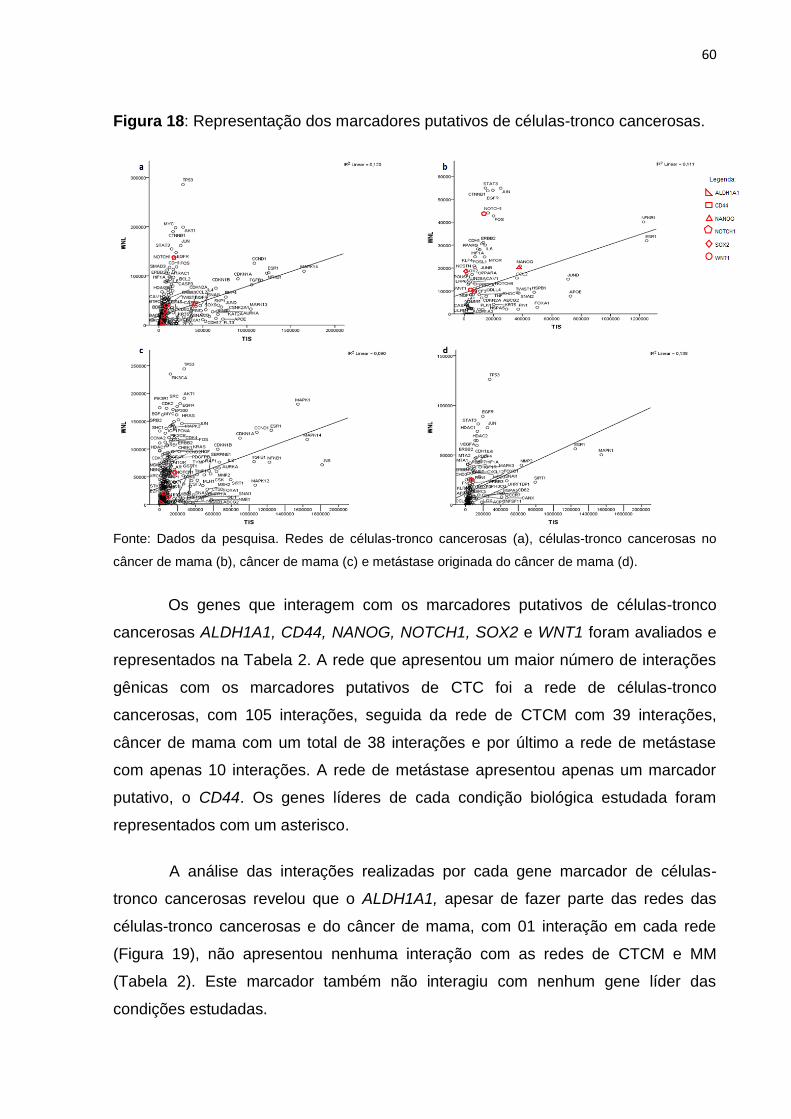
60
Figura 18: Representação dos marcadores putativos de células-tronco cancerosas.
Fonte: Dados da pesquisa. Redes de células-tronco cancerosas (a), células-tronco cancerosas no
câncer de mama (b), câncer de mama (c) e metástase originada do câncer de mama (d).
Os genes que interagem com os marcadores putativos de células-tronco
cancerosas ALDH1A1, CD44, NANOG, NOTCH1, SOX2 e WNT1 foram avaliados e
representados na Tabela 2. A rede que apresentou um maior número de interações
gênicas com os marcadores putativos de CTC foi a rede de células-tronco
cancerosas, com 105 interações, seguida da rede de CTCM com 39 interações,
câncer de mama com um total de 38 interações e por último a rede de metástase
com apenas 10 interações. A rede de metástase apresentou apenas um marcador
putativo, o CD44. Os genes líderes de cada condição biológica estudada foram
representados com um asterisco.
A análise das interações realizadas por cada gene marcador de células-
tronco cancerosas revelou que o ALDH1A1, apesar de fazer parte das redes das
células-tronco cancerosas e do câncer de mama, com 01 interação em cada rede
(Figura 19), não apresentou nenhuma interação com as redes de CTCM e MM
(Tabela 2). Este marcador também não interagiu com nenhum gene líder das
condições estudadas.

61
O gene CD44 foi o único marcador selecionado que apresentou interação
com as quatro condições estudadas. Na rede de CTC ele realizou 11 interações, em
CTCM 04, no CM 09 interações e na rede de MM 10 interações (Figura 19). Além
disso, esse marcador realizou interações com genes líderes em todas as redes,
EGFR, MYC, STAT3 e TP53 (CTC), EGFR e STAT3 (CTCM), TP53 (CM) e EGFR,
ERBB2, STAT3 e TP53 na (MM). Também foram evidenciadas interações com
genes relacionados à carcinogênese, como o gene supressor de tumor TP53, o fator
de transcrição e oncogene STAT3, os receptores de fator de crescimento epidérmico
(EGFR e ERBB2), bem como com genes que participam do processo de metástase,
como FN1, MMP2 e MMP9.
O marcador NANOG realizou interações apenas nas redes de CTC (13
interações) e CTCM (08 interações), sendo que na primeira ele interagiu com quatro
genes líderes (CTNNB1, STAT3, TP53 e NOTCH1) e na rede de CTCM com três
genes líderes (NOTCH1, STAT3 e CTNNB1). NANOG interagiu com os genes KLF4,
LIN28A, MTOR, POU5F1, SOX2 em ambas as redes CTC e CTCM.
Já o gene marcador de células-tronco cancerosas NOTCH1 foi detectado
nas redes de CTC, CTCM e CM. Apesar de não ter realizado interações com a rede
de metástase do câncer de mama, foi o marcador que fez mais interações, um total
de 94, sendo que 54 interações foram realizadas na rede de células-tronco
cancerosas, 17 na rede de CTCM e 23 interações em CM (Figura 19). Este
marcador interagiu com os genes CTNNB1 e HIF1A nas três condições CTC, CTCM
e no CM. Apresentou interações com os genes líderes AKT1, CTNNB1, STAT3,
JUN, TP53, EGFR e MYC na rede de CTC, CTNNB1, STAT3, JUN, EGFR, FOS e
NFKB1 na rede de CTCM e os líderes AKT1 e EGFR no CM. NOTCH1 e CD44
foram os marcadores de células-tronco cancerosas que mais fizeram interações com
os genes líderes (Tabela 2).
O marcador SOX2 realizou 13 interações na rede de CTC e 06 interações na
rede CTCM. Em ambas as redes, SOX2 interagiu com apenas um gene líder, o
STAT3 (tabela 2). KLF4, LIN28A, NANOG, POU5F1 e TWIST1 interagiram com
SOX2 nas redes CTC e CTCM.

62
O marcador WNT1 apresentou interações com as redes de CTC (13
interações), CTCM (04 interações) e CM (05 interações). Nas três condições, ele
realizou interação com o gene CTNNB1, sendo que nas redes de CTC e CTCM este
gene aparece como gene líder.
Figura 19:Número de interações realizadas por marcadores de células-tronco.
CSC CSCM CM MM0
20
40
60ALDH1A1
CD44
NANOG
NOTCH1
SOX2
WNT1
Condições
Nú
mero
de i
nte
raçõ
es
Fonte: Dados da pesquisa. Células-tronco cancerosas (CTC); Células-tronco do câncer de mama (CTCM); Câncer de mama (CM) e Metástase originada do câncer de mama (MM).

63
Tabela 2: Interações realizadas por marcadores de células-tronco cancerosas.
Fonte: Dados da pesquisa. Células-tronco Cancerosas (CTCs); Células-tronco cancerosas do câncer
de mama (CTCM); Câncer de Mama (CM) e Metástase originada do câncer de mama (MM). Genes
em destaque são os considerados líderes na pesquisa. *Genes líderes.
ALDH1A1
CTC CYP2C9
CTCM
- Câncer de mama
CYP1A1
Metástase
-
CD44
CTC CDH1, EGFR*, ERBB2, HLA-A,
ICAM1, IQGAP1, MMP9, MYC*,
SPP1, STAT3*, TP53*
CTCM
CDH1, EGFR*, ERBB2,
STAT3*
Câncer de mama
ERBB2, FN1, HMMR,
MMP2, MMP9, PTPRC,
SPP1, TP53*, TRIM25
Metástase
B2M, CDH1,
EGFR*,
ERBB2*,
FN1, MMP2,
MMP9, SPP1,
STAT3*,
TP53*
NANOG
CTC CDX2, CTNNB1*, FGF2, GATA6,
KLF4, LIN28A, MTOR,
NOTCH1*, POU5F1, SOX2,
STAT3*, TDGF1, TP53*
CTCM
CTNNB1*, KLF4, LIN28A,
MTOR, NOTCH1*,
POU5F1, SOX2, STAT3*
Câncer de mama
- Metástase
-
NOTCH1
CTC
ADAM17, AKT1*, APC, ATOH1,
BCL2, BMP4, CCND1, CDH1,
CDKN1A, CTBP1, CTNNB1*,
DLL1, DLL4, E2F1, E2F3,
EGFR*, ERBB2, ERBB3, EZH2,
FBXW7, FOS, FOSL1, GLI1,
HDAC3, HEY1, HIF1A, IGF1R,
IKZF1, IL4, JAG1, JUN*, KLF4,
MMP9, MYC*, NANOG, NCSTN,
NFKB1, NOTCH3, NOTCH4,
NUMB, POGLUT1, POSTN,
PTEN, SKP1, SKP2, SMAD3,
SNAI1, SNAI2, SOX9, STAT3*,
TCF3, TFDP1, TP53*, WNT5A
CTCM
CDH1, CTNNB1*, DLL4,
EGFR*, ERBB2, FOS*,
FOSL1, HIF1A, JUN*,
KLF4, NANOG, NCSTN,
NFKB1*, NOTCH4, SNAI2,
STAT3*, TCF3
Câncer de mama
AKT1*, APC, CCND1,
CDKN1A, CTNNB1,
E2F1, E2F3, EGFR*,
EP300, ERBB2,
FBXW7, FOS, GATA3,
HDAC1, HDAC4,
HDAC6, HIF1A, JUN,
MYC, NOTCH4,
PSEN2, RBBP8, SKP2
Metástase
-
SOX2
CTC ATOH1, CDX1, CDX2, FGF2,
KLF4, LIN28A, NANOG,
POU5F1, POU5F1B, PROM1,
STAT3*, TDGF1, TWIST1
CTCM
KLF4, LIN28A, NANOG,
POU5F1, STAT3*, TWIST1
Câncer de mama
- Metástase
-
WNT1
CTC
BMP4, CCND1, CTNNB1*,
HDAC3, LRP6, NOTCH2,
PPARG, PPARGC1A, RAC1,
SHH, WIF1, WNT5A, WNT5B
CTCM
CTNNB1*, LRP6, PPARG,
WIF1
Câncer de mama
CTNNB1, GSK3B,
RAC1, WNT10B,
WNT5A
Metástase
-

64
7 DISCUSSÃO
Após a descoberta de que uma população de células do tumor de mama,
conhecidas como células-tronco, apresenta importante papel na iniciação e
disseminação do tumor, uma série de pesquisas têm sido realizadas para identificar
as cascatas de sinalizações e as proteínas envolvidas com a manutenção das
características de células-tronco (MANI et al., 2008; DITTMER, 2018). Na busca por
novas terapias e alvos que possam identificar essas células na massa tumoral e
eliminá-las sem o comprometimento do bem estar do paciente, a identificação de
candidatos a biomarcadores torna-se um importante ponto de partida (CORRÒ e
MOCH, 2018). Assim, propusemos no corrente estudo avaliar por abordagem
computacional as redes de interações gênicas das condições células-tronco
cancerosas, células-tronco cancerosas do câncer de mama, câncer de mama e
metástase originada do câncer de mama, assim como fazer um paralelo entre genes
importantes para essas condições.
O corrente estudo revelou que a rede de células-tronco cancerosas apresenta
386 genes envolvidos com essa condição, sendo 8 deles (TP53 (Proteína
supressora de tumor p53), AKT1 (AKT Serina/Treonina Quinase 1 ) CTNNB1 (Beta
Catenina 1), MYC (Proto-Oncogene MYC, Fator de Transcrição BHLH), JUN (Jun
Proto-Oncogene, Subunidade Fator de Transcrição AP-1), STAT3 (Transdutor de
Sinais e Ativador da Transcrição 3), EGFR (Receptor do Fator de Crescimento
Epidérmico), NOTCH1 (Proteína Notch associada à translocação TAN-1))
considerados líderes em função das suas elevadas interações dentro da rede. O
gene TP53 se destacou como o maior gene líder dessa rede e também foi
considerado líder nas condições de câncer de mama e metástase originada do
câncer de mama. Conhecido como um guardião do genoma, o gene TP53 codifica
uma proteína chamada p53, que faz parte de uma família de proteínas com atividade
supressora de tumor (MAXIMOV e MAXIMOV, 2008). Em condições normais, essa
proteína induz o bloqueio do ciclo celular, senescência e apoptose em resposta aos
eventos de estresse celular, como hipóxia e danos no DNA (PERRI et al., 2016).
Mutações no gene TP53 evidenciadas no câncer de mama têm contribuído para a
sua progressão. Em células-tronco cancerosas, evidências indicam que a proteína
p53 está diretamente relacionada com processos de auto renovação, diferenciação,

65
quiescência e resistência a fármacos. Apesar de não ter sido considerado um gene
líder das células-tronco cancerosas da mama, a proteína p53 regula a pluripotência
celular por promover, por exemplo, bloqueio da expressão de Nanog e da proteína
quinase de adesão focal (FAK) (PRABHU et al., 2012). Na sinalização cruzada entre
a proteína quinase de adesão focal (FAK), p53, Mdm2 e Nanog, FAK fosforila Mdm2
para facilitar a ubiquitinação e degradação de p53, o que tem sido implicado na
proliferação de CTC, mobilidade, invasão e diferenciação. Em contrapartida, a
ativação de p53 inibe Nanog e FAK, favorecendo a manutenção das características
das células-tronco (OLIVOS e MAYO, 2016). Quando Nanog ativa FAK, e FAK
estável fosforila Nanog, esta sinalização cruzada favorece a mobilidade e invasão
das células-tronco cancerosas e tem um papel significante na metástase tumoral
(GOLUBOVSKAYA, 2013). Em tumores de mama, mutações em p53 foram
correlacionadas com expressão elevada de FAK (GOLUBOVSKAYA, 2013). A
desregulação de p53 tem sido descrita em vários estudos como um fator para o
favorecimento da migração celular, transição epitélio-mesenquimal e metástase.
Receptores de adesão celular, integrinas, ativação de fatores de crescimento, como
EGFR, entre outros, são componentes que também contribuem para a promoção de
metástases com a proteína p53 mutada (ARJONEN et al., 2014; CHANDER et al.,
2014).
No câncer de mama também foram evidenciados 8 genes líderes (TP53,
PIK3CA (Subunidade Catalítica de Fosfatidilinositol-4,5-Bisfosfato 3-Quinase Alfa),
AKT1, EGFR, MAPK1 (Proteína Quinase Ativada por Mitógeno 1), EP300 (Proteína
de Ligação E1A P300), SRC (Proto-oncogene SRC, tirosina quinase não receptora),
e CDK2 (Quinase Dependente de Ciclina 2) e 396 genes envolvidos com essa
condição. Os genes líderes desse estudo, com exceção do gene MAPK1, podem ser
considerados candidatos a drogas em função de suas elevadas interações nas
redes em estudo e baixa interação nas redes globais. MAPK1, diferente dos demais
líderes, apresentou alta interação na rede de CM e nas redes globais (elevado TIS).
O gene PIK3CA juntamente com o gene TP53 foram os genes que apresentaram as
maiores quantidades de interações na rede, sendo por isso evidenciados no grupo
A. Este gene é fundamental na via de sinalização fosfatidilinositol 3-quinase (PI3K),
com implicações em diversos eventos celulares, tais como proliferação,
diferenciação e migração. Em câncer de mama, cerca de 27% de mutações são

66
detectadas em PIK3CA (MANGONE et al., 2012). Evidências sugerem que
mutações em TP53 e PIK3CA são frequentes no câncer de mama e podem ser
considerados fatores chaves para a progressão desse tipo de câncer. PIK3CA e
TP53 são os genes mais comumente mutados em cânceres de mama positivos para
o receptor de estrógeno e negativos para esse receptor, respectivamente (KIM et al.,
2017). Estudos relatam associação entre a ocorrência de mutações do éxon 20 do
gene PIK3CA com a presença de mutações do gene TP53 (MANGONE et al., 2012;
KOTOULA et al., 2016). Em função disso, inibidores de PI3K já estão sendo
utilizados no tratamento de pacientes com câncer de mama apresentando mutações
em PI3KCA na triagem clínica (KIM et al., 2017).
Os genes STAT3, EGFR, CTNNB1, JUN, NFKB1 (Subunidade Kappa B do
Fator Nuclear 1), NOTCH1, FOS (Proto-Oncogene Fos, Subunidade Fator de
Transcrição AP-1) e ESR1 (Receptor de estrogênio 1) foram identificados como
líderes na rede de interação gênica das células-tronco cancerosas do câncer de
mama, sendo que STAT3, EGFR, CTNNB1 e JUN estão presentes no grupo A desta
condição com valores elevados de WNL e baixos de TIS. O gene STAT3 é um fator
de transcrição intrínseco na promoção da tumorigênese, atuando na invasão e
crescimento de tumores, sendo ativado pela fosforilação do resíduo de tirosina por
pela proteína JAK (LIU et al., 2016). Em nosso estudo, STAT3 foi considerado um
dos genes líderes em CTC, CTCM e MM. Thakuret et al. (2015) relataram que a
inibição da via de sinalização STAT3 compromete a manutenção das células-tronco
do câncer de mama no microambiente tumoral. O segundo líder desse grupo foi o
gene EGFR. A superexpressão ou ativação do gene EGFR está diretamente
relacionado a um prognóstico ruim em vários tipos de tumores sólidos, incluindo o
câncer de mama (KANG et al., 2017). Manupati et al. (2017) sugeriram que em
função da alta expressão de EGFR nas células-tronco cancerosas do câncer de
mama em comparação com as demais células cancerosas da mama, este gene
pode ser um bom alvo terapêutico. E ainda, a atenuação da via de sinalização EGFR
levou à reversão da TEM em CTC de mama, juntamente com uma maior capacidade
de resposta a quimioterápicos. Em nosso estudo, que não tem como proposta
avaliar a intensidade da expressão gênica, mas a importância dele na rede em
termos de número de interações realizadas, percebe-se uma importância do EGFR
como gene líder em ambas as redes. CTNNB1 também foi um gene evidenciado no
grupo A em CTCM. Ele é um membro chave na via de sinalização Wnt/β-catenina,

67
que está associada ao desenvolvimento e progressão do câncer e na manutenção
das CTCs. Esse gene encontra-se também envolvido na manutenção de
marcadores putativos de CTCs, como é o caso do CD44 e CD133 (MOON et al.,
2014). O gene JUN também se apresentou no grupo A da rede de células-tronco do
câncer de mama, sendo considerado líder em CTC, CTCM e MM. O proto-oncogene
Jun e a proteína β-catenina regulam a transcrição de NANOG, conferindo habilidade
de autorenovação às CTCs (IBRAHIM et al., 2012).
Conforme esperado, a rede de células-tronco cancerosas e a rede de
células-tronco cancerosas do câncer de mama apresentaram o maior número de
genes líderes em comum (STAT3, EGFR, CTNNB1, JUN e NOTCH1). As proteínas
sintetizadas por esses genes têm em comum a participação nos mecanismos que
regulam a pluripotência celular, modulando as atividades de proliferação,
diferenciação e apoptose (KIDDER et al., 2008; SOLOZOBOVA e BATTLER, 2011;
TANABE et al., 2016; AI et al., 2017). EGFR tem a capacidade de acelerar o
processo de proliferação das células-tronco pluripotentes e atrasar sua senescência,
atuando também em interação com outros genes, entre eles o gene STAT3 (AI et al.,
2017). A proteína beta-catenina 1 interage com moléculas de sinalização, como por
exemplo Wnt e Notch, culminando na transcrição de genes relacionados à
proliferação celular (TANABE et al., 2016).
A análise comparativa dos genes líderes das condições de CTC e CM
revelou que AKT1 e EGFR são fundamentais para ambas as condições. A proteína
Serina/Treonina Quinase 1 (AKT1) juntamente com PI3K são necessários para a
manutenção das características de células-tronco em CM, e participam de eventos
celulares como proliferação, sobrevivência, crescimento e angiogênese (GARGINI et
al., 2015). Elevada incidência de mutações no gene AKT1 relaciona-se à redução da
sobrevida das CTC, redução na geração de células CD44+/CD24-, perda do fenótipo
de células-tronco mesenquimais e recuperação de marcadores epitelais. Segundo
Gargini et al. (2015), a identificação de PIK3 e AKT são prováveis relevantes alvos
terapêuticos. De maneira similar, EGFR contribui com a expressão de marcadores
de auto-renovação, ativação da proliferação, invasão e resistência das células-
tronco aos tratamentos quimioterápicos (ABHOLD et al., 2012; HAN et al., 2013).
EGFR encontra-se superexpresso em 15-20% de todos os carcinomas mamários e
em 50-70% de câncer de mama triplo negativo, e sua alta expressão associa-se a

68
um maior volume tumoral, ocorrência de metástases e pior sobrevida (KOZLOVA et
al., 2017).
A comparação entre os genes líderes das CTC e MM revelou que EGFR,
STAT3 e JUN são relevantes para o perfil de CTCs assim como para as células
metastáticas originadas do câncer de mama. Conforme relatado anteriormente,
EGFR atua ativando vários outros genes, inclusive STAT3, atuando sobretudo no
aumento da proliferação e disseminação das células tumorais. C-jun, um proto-
oncogene superexpresso em CM, ativa fatores de transcrição e também contribui
para a auto renovação das CTCs, formação de angiogênese e disseminação
metastática, o que justifica o elevado número de interações realizadas por esses
genes em ambas as redes.
Nos últimos anos, esforços consideráveis foram investidos na detecção e
caracterização de marcadores de células-tronco cancerosas. O resultado é que há
um número cada vez maior de tais moléculas marcadoras, inclusive que diferem
entre tipos tumorais. A maioria dos marcadores já descritos são proteínas. Alguns
são lipídeos e carboidratos (KARSTEN e GOLETZ, 2013). WNT1, NOTCH1,
NANOG, SOX2, CD44 e ALDH1A1 foram os genes marcadores de células-tronco
cancerosas observados em nosso estudo.
O marcador putativo de células-tronco cancerosas NOTCH1 foi o único
marcador identificado como gene líder das CTCs, e inclusive das CTCM. Isso
demonstra que esse gene e, consequentemente, sua proteína, são importantes para
a identificação fenotípica bem como a manutenção das características típicas de
uma célula-tronco cancerosa. NOTCH1 foi também o marcador putativo de CTCs
que mais realizou interações nas redes do corrente estudo, um total de 94
interações. Ele somente não fez parte da rede de metástase. Nas redes de CTC e
CTCM foram registradas interações com 54 e 17 genes, respectivamente. Em CTC,
NOTCH1 interage com todos os genes líderes. Em CTCM, ele só não interagiu com
o gene líder ESR1. Na rede do CM, NOTCH1 interagiu com 23 genes, dentre eles
dois líderes (AKT1 e EGFR). Interações com CTNNB1 e HIF1A foram observadas
em todas as três redes.
A regulação de NOTCH1 na via Wnt está relacionada à sua ação sobre a
translocação de β-catenina para o núcleo, e consequente ativação da transcrição de

69
diversos genes (ISHIGURO et al., 2017). Conforme mencionado anteriormente, β-
catenina ativa a sinalização Wnt e, consequentemente, estimula a proliferação
celular. De maneira similar, HIF1A é uma subunidade de um fator de transcrição,
HIF1, que regula a resposta celular em condições de hipóxia. A hipóxia lidera o
recrutamento de HIF1A nos promotores responsivos a NOTCH1, e eleva a
expressão de genes regulados por NOTCH1. Segundo Gustafsson et al., (2005), a
hipóxia bloqueia a diferenciação de células progenitoras através da regulação de
Nocht1. Esses dados reforçam a importância de NOTCH1 como um gene necessário
à manutenção das células-tronco cancerosas. Estudos relatam a participação da
proteína Notch1 nos eventos relativos à proliferação, invasão e quimioresistência em
cânceres, entre eles o câncer de mama (VENKATESH et al., 2018) (BOSE, 2017;
BRZOZOWA-ZASADA et al., 2017; HARBUZARIU et al., 2016). A alta de expressão
de NOTCH1 tem sido correlacionada com menor diferenciação do câncer de mama,
com 80% das hiperplasias epiteliais do tipo usual, 67% dos carcinomas ductais in
situ, 89% dos carcinomas ductais invasivos e 57% dos carcinomas lobulares
invasivos, além de ser fundamental na transição epitélio mesenquimal. A ativação do
gene JAG1 por NOTCH1, evidenciada na rede de células-tronco cancerosas,
contribui diretamente para a indução da TEM e disseminação metastática ao ativar
NF-κB e seus genes alvo a jusante, as metaloproteinases da matriz MMP2, MMP9 e
o fator de crescimento VEGF (Li et al., 2014; BRZOZOWA-ZASADA et al., 2017;
RIZZO et al., 2008; SHAO et al., 2015).
Outro biomarcador que se destacou no presente estudo foi o gene CD44,
em especial porque foi o único marcador que realizou interações em todas as redes
em estudo. CD44 realizou 34 interações nas quatros redes, sendo 11 delas na rede
de CTC, 4 na rede de CTCM, e 9 em CM. Foi o único marcador que esteve presente
na rede de metástase originada do câncer de mama (MM), com 10 interações. Em
todas as redes o marcador CD44 apresentou interação com genes líderes, como o
gene supressor de tumor TP53, o fator de transcrição e oncogene STAT3, os
receptores de fator de crescimento epidérmico (EGFR e ERBB2), bem como com
genes que participam do processo de metástase, como FN1, MMP2 e MMP9. Nas
redes de CTCM, CM e MM, o marcador CD44 fez interações com os genes líderes
EGFR, ERBB2, STAT3 e TP53, o que reforça o envolvimento desse gene nos
processos de desenvolvimento e disseminação das células tumorais da mama
(OWCZAREK et al., 2017; WINCEWICZ et al., 2011). Por se tratar de uma

70
glicoproteína de superfície celular e um receptor específico para o ácido hialurônico,
sua ligação ao ácido hialurônico favorece a proliferação, migração e invasão de
células neoplásicas da mama (ESMAEILI et al., 2018). Foi também o marcador que
apresentou o menor valor de TIS, fazendo com que seja um importante candidato à
inativação pela sua baixa relevância em outras redes.
O marcador WNT1 realizou interações nas redes CTC, CTCM e CM. Em
todas interagiu com o gene CTNNB1, um dos genes líderes das CTC e CTCM. É
uma proteína de sinalização pertencente à família de ligantes WNT, implicada em
diversos fenômenos relativos à embriogênese, diferenciação e migração celular
(JANDA et al., 2012; ANSHULA et al., 2017). Ligantes de WNT ativam moléculas de
sinalização que estimulam a transcrição genética e acúmulo de β-catenina no
núcleo. β-catenina associa-se ao fator de transcrição funcional (TCF), um fator de
transcrição que interfere na transativação de genes envolvidos com a progressão do
tumor, invasão e consequente metástase. Estudos demonstram que muitos ligantes
de WNT promovem a transição epitélio-mesenquimal e a atividade de CTC em
vários tipos de tumores, e em especial o aumento na expressão de WNT1 promove
um enriquecimento da população de CTC (ALOK et al., 2017; MUZIO, 2001; ZHANG
et al., 2017). A superexpressão de Let-7 inibe a sinalização WNT1, sendo por esse
motivo considerado um bom indicador da intensidade de células-tronco cancerosas
(JIN et al., 2016).
O marcador NANOG, fator de transcrição envolvido com a auto-renovação
de células-tronco embrionárias indiferenciadas, tem sido estudado como um dos
genes mais expressos heterogeneamente em células-tronco cancerosas e em
células-tronco induzidas. NANOG desempenha importante papel na manutenção da
auto renovação celular e da pluripotência. Presente na rede de células-tronco
cancerosas e células-tronco cancerosas da mama, NANOG foi o marcador que
apresentou o maior valor de TIS, o que sugere que possui uma maior interação com
outras redes. Subpopulações de células-tronco com baixa expressão de Nanog são
mais propensas à diferenciação do que as que apresentam alta expressão (YU et
al., 2018; GLAUCHE et al., 2010).
Estudos tem mostrado que o marcador NANOG realiza interações com
importantes genes específicos de célula-tronco, como o SOX2. Essa interação
também foi evidenciada em nossa rede de células-tronco cancerosas (MITSUI et al.,
2003; YU et al., 2018). Além disso, ele realizou interações com quatro genes líderes

71
na rede de células-tronco cancerosas, CTNNB1, NOTCH1, STAT3 e TP53, e com
três genes líderes na rede de células-tronco cancerosas na mama, CTNNB1,
NOTCH1 e STAT3. NANOG interagiu ainda com os genes KLF4, LIN28A, MTOR,
POU5F1, SOX2 nas duas condições de CTC e CTCM. De maneira similar, Sox2
também realizou interações apenas nas redes de CTC e CTCM, e interagiu com
KLF4, LIN28A, POU5F1 e TWIST1, que também foram ligantes de NANOG.
KLF4, Kruppel Like Factor 4, é um membro da família KLF de fatores de
transcrição, envolvido na regulação da proliferação, diferenciação, apoptose e
reprogramação de células somáticas, exercendo papel fundamental na indução de
células-tronco pluripotentes (TAKAHASHI et al., 2007). Yori et al. 2011
demonstraram que o KLF4 inibiu a invasão e a metástase distal do câncer de mama
pela supressão da transição epitelial-mesenquimatosa, enquanto que Foster et. al.,
2005 apontaram que a expressão de KLF4 foi aumentada em 70% das amostras de
tecido de câncer de mama. É possível que KLF4 funcione ora como um oncogene,
ora como supressor de tumor, dependendo do tipo histológico e microambiente
(SONG et al., 2017; MOON et al., 2011). LIN28A, proteína de ligação de RNA que
regula a auto renovação de células-tronco, assim como POU5F1, também conhecido
como Oct-4, são considerados marcadores de células-tronco embrionárias humanas
indiferenciadas (TSIALIHAS e ROMER, 2015; CAI et al., 2016; MAHERALI et al.,
2007). Em estudo, Shen et al. (2017) identificou que Lin28A pode ativar o receptor
de andrógenos (AR) por meio da regulação de c-myc e promover o crescimento do
tumor mamário ER-/Her2+ através da proliferação e invasão de células de câncer de
mama. Sendo assim, LIN28A pode ser um alvo interessante para o tratamento do
câncer de mama ER-/Her2+. Kearney et al. (2010) identificaram baixa expressão de
LIN28A em câncer de mama e malignidades humanas avançadas. Cai et al. 2016
encontraram que a elevada expressão de POU5F1/Oct-4 em tecidos do câncer de
mama estão associados à metástase com linfonodos sentinelas e à distância. A
interação molecular entre Nanog, Sox2 e Oct-4 está diretamente ligada à
pluripotencia e regulação de células estaminais mesenquimais (GAGLIARDI et al.,
2013; NIU et al., 2011; MOHAMMADIAN e NEGAHDARI, 2017). SOX2 também
interagiu com oncogene TWIST1, presente na rede de CTC e CTCM. TWIST1 tem
sido expresso em vários carcinomas metastáticos, relacionando-se aos mecanismos
de angiogênese, extravasamento e instabilidade cromossômica, além de proteção
contra apoptose (PUISIEUX et al., 2006). Tais características sugerem que TWIST1

72
seja responsável pela manutenção de células-tronco cancerosas e consequente
resistência à quimioterapia (ROBERTS et al., 2016). Wu e Yang (2011) identificaram
que TWIST1 é expresso em condições de hipóxia, associando-se à resistência à
quimioterapia, já que células hipóxicas respondem menos a drogas quimioterápicas.
Relatos da literatura apontam também que as populações tumorais
enriquecidas para a atividade ALDH1, considerado também como um marcador de
células-tronco, mostraram ter maior mobilidade, potencial de invasão e metástase
do que a maior parte da população de células de câncer (LI et al., 2017). A enzima
ALDH1 catalisa a oxidação intracelular de aldeídos, em especial a oxidação de
retinol a ácido retinóico, o que é essencial para a diferenciação inicial das células-
tronco, e em contrapartida tem sido apontado como a causa da resistência dessas
células a drogas (CHUTE et al., 2006; ROUDI et al., 2015). A combinação entre a
alta razão CD44/CD24 e ALDH1+ é uma forma de se caracterizar células-tronco
cancerosas da mama (LI et al., 2017). Apesar de ser um marcador usual para CTC
na literatura, ALDH1A1 foi encontrado nas redes de CTC e CM apresentando os
menores valores de WNL quando comparado aos demais marcadores. Foi também
o marcador que menos realizou interações nas redes estudadas, interagindo apenas
com CYP2C9 em CTC e CYP1A1 em CM. Mesmo não tem sido evidenciadas muitas
interações nas redes estudadas, o gene ALDH1A1, assim como outros marcadores
de células-tronco avaliados nesse trabalho, são comprovadamente importantes no
estudo e identificação das células-tronco cancerosas, bem como são importantes
marcadores da evolução da neoplasia maligna. Estudos posteriores podem ser
realizados com outros genes relevantes para as CTC, em especial as CTCM, e que
não são considerados genes líderes.

73
8 CONCLUSÃO
O presente estudo relatou, por meio de ferramentas de bioinformática, as
redes de interações gênicas evidenciadas nas condições de células-tronco
cancerosas, células-tronco cancerosas do câncer de mama, câncer de mama e na
metástase originada do câncer de mama.
Além disso, os resultados obtidos permitem concluir que:
1- Os mapas de interação gênica das redes de células-tronco cancerosas,
células-tronco cancerosas de mama, câncer de mama e metástase de mama
revelaram 386, 81, 396 e 160 genes, respectivamente.
2- Os genes TP53, AKT1, CTNNB1, MYC, JUN, STAT3, EGFR e NOTCH1
foram os genes que mais interagiram na rede de células-tronco cancerosas,
sendo por isso considerados os líderes dessa condição.
3- Os genes STAT3, EGFR, CTNNB1, JUN, NFKB1, NOTCH1, FOS e ESR1
foram os genes líderes da rede de células-tronco cancerosas do câncer de
mama.
4- Os genes TP53, PIK3CA, AKT1, EGFR, MAPK1, EP300, SRC e CDK2 foram
os genes líderes da rede do câncer de mama.
5- Os genes TP53, EGFR, STAT3, JUN, HDAC1, ERBB2, VEGFA e HDAC2
foram os genes líderes da rede de metástase de originada da mama.
6- As redes de CTC e CTCM apresentaram maior número de genes líderes em
comum. EGFR foi gene líder em todas as condições estudadas. TP53, STAT3
e JUN também foram genes líderes que se destacaram porque estiveram
presentes como líderes em três condições estudadas.
7- Todos marcadores de CTCs, WNT1, NOTCH1, NANOG, SOX2, CD44 e
ALDH1A1, apresentaram-se no grupo de CTC, destacando-se o CD44 que
realizou interações em todas as elas.
8- NOTCH1 realizou interações nas redes de células-tronco (CTC e CTCM) e
CM. NANOG, WNT1 e SOX2 apresentaram interações apenas nas redes de
células-tronco cancerosas. ALDH1A1 apresentou-se apenas nas redes CTC e
CM, mas com apenas 1 interação em cada rede.
9- NOTCH1 é considerado um marcador de células-tronco cancerosas e foi
também considerado gene líder dessa condição.

74
REFERÊNCIAS
ABHOLD EL, et al. EGFR kinase promotes acquisition of stem cell-like properties: a potential therapeutic target in head and neck squamous cell carcinoma stem cells. PLoS One, v. 7, n. 2, 27 fev., 2012.
ABRAVANEL, Daniel L. et al. Notch Promotes Recurrence of Dormant Tumor Cells Following HER2/neu-Targeted Therapy. The Journal of Clinical Investigation, v. 125, p 2484–2496,11 mai., 2015.
AFIFY A, PURNELL P, NGUYEN L. Role of CD44s and CD44v6 on human breast cancer cell adhesion, migration, and invasion. Exp Mol Pathol, v. 86, p 95–100, abr., 2009.
AI G, et al. Epidermal growth factor promotes proliferation and maintains multipotency of continuous cultured adipose stem cells via activating STAT signal pathway in vitro. Medicine (Baltimore), v. 96, jul., 2017.
AIGNER, S, et al. CD24 mediates rolling of breast carcinoma cells on P-selectin. FASEB J; v. 12, p 1241–1251, 12 set.,1998.
ALLEMANI, C., et al. Global surveillance of cancer survivak 1995-2009: analysis of individual data for 25 676 887 patients from 279 population-based registries in 67 countries (CONCORD-2). The lancet, v. 385, p. 977-1010, 14 mar., 2015.
ALOK, ANSHULA. et al. Wnt proteins synergize to activate β-catenin signaling. Journal of Cell Science, v. 130, p. 1532-1544, 13 mar., 2017.
AMERICAN CANCER SOCIETY. Global Cancer Facts & Figures 3rd edition. Atlanta: American Cancer Society; 2015.
AMOROSO V, et al. International expert consensus on primary systemic therapy in the management of early breast cancer: highlights of the fifth symposium on primary systemic therapy in the management of operable breast cancer, Cremona, Italy

75
(2013). JNCI Monographs, v 2015, p 90–6, 01 mai., 2015.
ARAÚJO, N. D. et. al. A era da bioinformática: seu potencial e suas implicações para as ciências da saúde. Estudos de Biologia: Ambiente e Diversidade, v.30, n.70, p.143-148, 2008.
ARJONEN, Antti. Mutant p53–associated myosin-X upregulation promotes breast cancer invasion and metastasis. The Journal of Clinical Investigation, v.124 n. 3, p. 1069–1082, 3 fev., 2014.
BACAC, M. e STAMENKOVIC, I. Metastatic Cancer Cell. Annual Review of Pathology Mechanisms of Disease, v. 3, n. 2, p. 221-247, 28 fev., 2008.
BAO, S. et al. Glioma stem cells promote radioresistance by preferential activation of the DNA damage response. Nature International Weekly Journal of Science, v. 444, n. 7120, p.756-760, 07 dez., 2006.
BEDNARZ-KNOLL, NATALIA, et al. Potential involvement of Jagged1 in metastatic progression of human breast carcinomas. Clin Chem, v. 62, n. 2, p. 378-386, jan. 2016.
BEKAII‐SAAB, T e EL‐RAYES, B. Identifying and targeting cancer stem cells in the treatment of gastric cancer. Cancer, v. 123, p 1303–1312. 24 Jan. 2017.
BONNET, D. e DICK, J. E. Human acute myeloid leukemia is organized as a hierarchy that originates from a primitive hematopoietic cell. Nature Medicine, v. 3, n.7 p. 730-737, jul., 1997.
BOSE, BIPASHA; SEM, UTSAV; SHENOY, SUDHEER. Breast cancer stem cell therapeutics, multiple strategies versus using engineered mesenchymal stem cells with notch inhibitory properties: Possibilities and Perspectives. J. Cell. Biochem, v.119, p.141-149, 2018.
BOZORGI, A, et al. New Findings on Breast Cancer Stem Cells: A Review. Journal of Breast Cancer, v. 18, n. 4, p. 303-312, 23 dez., 2015.

76
BRASIL. ABC do câncer: Abordagens para o controle do câncer. Instituto Nacional do Câncer INCA – Rio de Janeiro, 2011.
BRASIL. Ministério da Saúde. Secretaria de Atenção à Saúde Departamento de Atenção Básica. Controle dos Cânceres do Colo do Útero e da Mama. 2014.
BRASIL. Ministério da Saúde (BR). Instituto Nacional do Câncer. Estimativa 2016: incidência de câncer no Brasil. Brasília: Ministério da Saúde; 2015.
BRASIL. Instituto Nacional de Câncer José Alencar Gomes da Silva. Coordenação de Prevenção e Vigilância. Estimativa 2018: incidência de câncer no Brasil. Rio de janeiro, RJ, 2017.
BRASIL. Ministério da Saúde. Instituto Nacional De Câncer. O que é o câncer? Rio de Janeiro, RJ. 2 p. 2018.
BRENNER DR, et al. Breast cancer survival among young women: a review of the role of modifiable lifestyle factors. Cancer Causes Control. v. 27, p 459-472. abr., 2016.
BRUCE, W, et al. A quantitative assay for the number of murine lymphoma cells capable of proliferation in vivo. Nature, v. 199, p 79–80, 1963.
BRZOZOWA-ZASADA, MARLENA. et al. Notch and its oncogenic activity in human malignancies. Eur Surg, v. 49, p 199 -209, 2017.
CAI, SHOULIANG, et al. POU5F1/Oct-4 Expression in Breast Cancer Tissue Is Significantly Associated with Non-Sentinel Lymph Node Metastasis. BMC Cancer,
v.16, p 175, 01 mar., 2016.
CAMPBELL, L. L. & POLYAK, K. Breast tumor heterogeneity: cancer stem cells or clonal evolution? Cell Cycle, v. 6, p 2332–2338, 02 out., 2007.
CANCER SOCIETY OF FINLAND: Taxas de sobrevivência após o câncer na Finlândia. Disponível em: < https://www.cancersociety.fi/publications/reports/cancer-in-finland-2016/the-survival-of-cancer-patients/ > Acesso em 25 de agosto de 2018.

77
CGHFBC. Collaborative Group on Hormonal Factors in Breast Cancer (CGHFBC). Familial breast cancer: collaborative reanalysis of individual data from 52 epidemiological studies including 58,209 women with breast cancer and 101,986 women without the disease. The Lancet, v. 358, p 1389 - 1399, 27 out., 2001.
CHAFFER CL e WEINBERG RA. A perspective on cancer cell metastasis. Science,
v. 331, issue 6024, p 1559–1564, 25 mar., 2011.
CHAMBERS, A.F, et al. Dissemination and growth of cancer cells in metastatic sites. Nature Reviews Cancer, v. 2, p 563–572, 01 ago., 2002.
CHANDER, H, et al. Toca-1 is suppressed by p53 to limit breast cancer cell invasion and tumor metastasis. Breast Cancer Research, v. 16, p 3413, 30 dez., 2014.
CHUTE, J. P. et al. Inhibition of aldehyde dehydrogenase and retinoid signaling
induces the expansion of human hematopoietic stem cells. P Natl Acad Sci USA.
v.103, p 11707–11712, 01 ago., 2006.
CORRÒ, CLAUDIA e MOCH, HOLGER. Biomarker Discovery for Renal Cancer Stem Cells. The Journal of Pathology: Clinical Research, v. 4, n. 1, p 3–18, 18 dez., 2018.
CRUK: Taxas de sobrevivência após câncer na Inglaterra. Disponível em: <https://www.cancerresearchuk.org/> Acesso em 20 de agosto. DEAN, M, et al. Tumour stem cells and drug resistance. Nature Reviews Cancer, v. 5, n. 4, p. 275-285, 01 abr., 2005.
DE ROBERTIS, Edward M; HIB, José. Biologia Celular e Molecular. 16. Ed. Rio de Janeiro: Editora Guanabara Koogan, 2014.
DE SANTIS. Carol E., MPH et al., Breast Cancer Statistics, 2017, Racial Disparity in Mortality by State. CA: A Cancer Journal for Clinicians. v. 67, n. 6, p 439 – 448, 2017.
DICK, John. et al. Cancer Stem Cells—Perspectives on Current Status and Future Directions: AACR Workshop on Cancer Stem Cells. Cancer Research, v. 66, out., 2006.

78
E.M. SANTOS, et al. Obesity-related genes and oral cancer: a bioinformatics approach and systematic review. Journal of Applied Bioinformatics & Computacional Biology. v. 5, n. 1, 31 abr., 2016.
ESKIIZMIR, Görkem. Tumor Microenvironment in Head and Neck Squamous Cell Carcinomas. Turkish Archives of Otorhinolaryngology. v. 53, n.3 p. 120–127, 2015.
ESMAEILI, Rezvan, et al. Unique CD44 intronic SNP is associated with tumor grade in breast cancer: a case control study and in silico analysis. Cancer Cell Int. v. 18, p 28, 23 fev., 2018.
FITZMAURICE C. Global, Regional, and National Cancer Incidence, Mortality, Years of Life Lost, Years Lived With Disability, and DisabilityAdjusted Life-years for 32 Cancer Groups, 1990 to 2015 A Systematic Analysis for the Global Burden of Disease Study. JAMA Oncology. v. 3, Issue 4, p 524-548, 01 abr., 2017.
FOSTER K Wade, et al. Induction of KLF4 in basal keratinocytes blocks the proliferation-differentiation switch and initiates squamous epithelial dysplasia. Oncogene. v. 24, p. 1491–1500, 24 fev., 2005.
FREUDENBERG JA, et al. The role of HER2 in early breast cancer metastasis and the origins of resistance to HER2- targeted therapies. Exp Mol Pathol. v. 87, p 1-11, ago., 2009.
GAGLIARDI, ALESSIA, et al. A Direct Physical Interaction between Nanog and Sox2 Regulates Embryonic Stem Cell Self-Renewal. The EMBO Journal, v. 32, n. 16, p. 2231–2247, 14 ago., 2013.
GARGINI R, et al. Cancer stem cell-like phenotype and survival are coordinately regulated by Akt/FoxO/Bim pathway. Stem Cells. v. 33, p 646–660, 19 nov., 2015.
GENECARDS. The human gene database. www.genecards.org. Stelzer G. et al. The
GeneCards Suite: From Gene Data Mining to Disease Genome Sequence Analysis.
2018
GERRATANA L, et al. Pattern of metastasis and outcome in patients with breast cancer. Clinical Experimental Metastasis. v. 32, p 125–133, 29 jan., 2015.

79
GLAUCHE, INGMAR, et al. Nanog variability and pluripotency regulation of embryonic stem cells - insights from a mathematical model analysis. PLoS One. v. 5, n. 6, 21 jun., 2010.
GLUMAC, PM e LeBeau, AM. The roleof CD133 in cancer: a concise review. Clinical and Translational Medicine v. 7, p18, 09 jul., 2018.
GOLUBOVSKAYA, VITA M. FAK and Nanog Cross Talk with p53 in Cancer Stem Cells. Anti-cancer agents in medicinal chemistry. v. 13 n.4, p. 576–580, 01 mai., 2013.
GONÇALVES, LEILA LUÍZA CONCEIÇÃO, et al. Fatores de risco para câncer de mama em mulheres assistidas em ambulatório de oncologia. Rev. enferm. UERJ, Rio de Janeiro, v.18, n.3, p. 468-472, 2010.
GOUVEA, M, et al. Análise matemática de um modelo para crescimento de células-tronco cancerígenas em tumores. ForSci.: r. cient. IFMG campus Formiga, Formiga, v. 2, n. 1, p. 96-112, jan./jun. 2014.
GU J. W, et al. Notch signals in the endothelium and cancer "stem-like" cells: opportunities for cancer therapy. Vascular Cell. V. 4, 09 abr., 2012.
GUNSALUS, KEARNEY T.W, et al. Induction of the RNA Regulator LIN28A Is Required for the Growth and Pathogenesis of RESTless Breast Tumors. Cancer Res, v. 72 n. 13 p. 3207–3216, 2012.
GUSTAFSSON M.V., et al. Hypoxia requires notch signaling to maintain the
undifferentiated cell state. Developmental Cell. V. 9, p. 617-628, 01 nov., 2005.
HAMBURGER, A. W. & SALMON, S. E. Primary bioassay of human tumor stem cells. Science. v. 197, p 461–463, 1977.
HAN, Woody, et al . STAT1 gene expression is enhanced by nuclear EGFR and HER2 via cooperation with STAT3. Carcinogenese Molecular. v. 52, n.12, p 959-969, 12 jun., 2012.

80
HANAHAN, D. e WEINBERG, R.A. The hallmarks of cancer: The next generation. Cell. v.144, n.5, p 646-674, 2011.
HARBUZARIU, ADRIANA. et al. Leptin-Notch signaling axis is involved in pancreatic cancer progression. Oncotarget, v. 8, n. 5, p. 7740- 7752, 31 jan., 2017.
HOU, J. P. et al. A new correlation clustering method for cancer mutation analysis. Genome analysis. Bioinformatics. v. 32, n. 24, p. 3717-3728, 15 dez., 2016.
HO VK, et al. Survival of breast cancer patients with synchronous or metachronous central nervous system metastases. European Journal of Cancer. v. 51, p 2508–2516, nov., 2015.
INCA - Instituto Nacional de Câncer. Ações de enfermagem no controle do câncer: uma proposta de integração ensino-serviço. Fisiopatologia do câncer. Rio de Janeiro 3. ed. rev. atual. ampl. – Rio de Janeiro, 2008
INSTITUTO NACIONAL DO CÂNCER JOSÉ ALENCAR GOMES DA SILVA (INCA). Coordenação Geral de Ações Estratégicas. Coordenação de Educação ABC do câncer: abordagens básicas para o controle do câncer. 2ª ed. rev. e atual. Rio de Janeiro, 2012.
INSTITUTO NACIONAL DE CÂNCER JOSÉ DE ALENCAR GOMES DA SILVA (INCA). Atlas de mortalidade por câncer. Rio de Janeiro, 2013.
INSTITUTO NACIONAL DE CÂNCER JOSÉ DE ALENCAR GOMES DA SILVA (INCA). Tipos de câncer: mama. Rio de Janeiro, 2014.
INSTITUTO NACIONAL DE CÂNCER-INCA. Estimativa 2016: Incidência de Câncer no Brasil. Rio de Janeiro, 2016.
INSTITUTO NACIONAL DE CÂNCER JOSÉ ALENCAR GOMES DA SILVA (INCA). Estimativa 2018: incidência de câncer no Brasil. Coordenação de Prevenção e Vigilância. Rio de Janeiro, 2017.
INSTITUTO ONCOGUIA. Tratamentos para o câncer de mama. 2014. Disponível em: http://www.oncoguia.org.br/conteudo/tratamentos/15/12/. Acesso em: 20 de fevereiro de 2018.

81
INTERNATIONAL SOCIETY FOR STEM CELL RESEARCH (ISSCR). Stem Cell Facts. Deerfield, Florida, 2011.
JAEGER, S. et al. Quantification of pathway crosstalk reveals novel synergistic drug combinations for breast cancer. Cancer Research, v.77, n.2, p. 459-469, jan., 2017.
JANDA, CLAUDIA Y, et al. Structural Basis of Wnt Recognition by Frizzled. Science, v. 337, n. 6090, p. 59-64, 06 jul., 2012.
JIN, BIN, et al. Let-7 Inhibits Self-Renewal of Hepatocellular Cancer Stem-like Cells through Regulating the Epithelial-Mesenchymal Transition and the Wnt Signaling Pathway. BMC Cancer. v.16, p 863, 08 nov., 2016.
JORDAN, C.T, et al. Mechanisms of the disease: Cancer Stem Cells. The England Journal of Medicine, v. 355, p 1253-1261, 09 nov., 2006. KALIKS, R. A. Avanços em oncologia para o não oncologista. Hospital Israelita Albert Einstein, v. 14, jun, 2016.
KANG, HYE-MIN, et al. Phytosphingosine exhibits an anti-epithelial–mesenchymal transition function by the inhibition of EGFR signaling in human breast cancer cells. Oncotarget. v. 8, n. 44, p. 77794–77808, 2017.
KAUFHOLD, SAMANTHA, et al. Yin Yang 1 Is Associated with Cancer Stem Cell Transcription Factors (SOX2, OCT4, BMI1) and Clinical Implication. Journal of Experimental & Clinical Cancer Research. v. 35, n. 84, 25 mai., 2016.
KIDDER, BENJAMIN L, et al. Stat3 and c-Myc Genome-Wide Promoter Occupancy in Embryonic Stem Cells. PLoS ONE, v.3, n. 12, 11 dez., 2008.
KIM, M. J, et al. Correlation of ALDH1 and Notch3 Expression: Clinical implication in Ovarian Carcinomas. Journal of Cancer. v. 8, n. 16, p 3331–3342. 2017.
KIM, M, et al . VEGFA links self-renewal and metastasis by inducing Sox2 to repress miR-452, driving Slug. Oncogene. v. 36, n. 36, p 5199–5211, 07 set., 2017.

82
KIM, H, et al. Isolation of CD24 (high) and CD24 (low/-) cells from MCF-7: CD24 expression is positively related with proliferation, adhesion and invasion in MCF-7. Cancer Lett, v. 258, P 98–108, 08 dez. 2007.
KIN, YUN GYOUNG, et al. Breast cancer stem cells in HER2-negative breast cancer cells contribute to HER2-mediated radioresistance and molecular subtype conversion: clinical implications for serum HER2 in recurrent HER2-negative breast cancer. Oncotarget. v. 9, n. 5, dez., 2018.
KLEINSMITH, L. J. & PIERCE, G. B., JR. Multipotentiality of single embryonal carcinoma cells. Cancer Res. v. 24, p1544–1551, out., 1964.
KONRAD, C. V, et al. "The role of cancer stem cells in tumor heterogeneity and resistance to therapy." Canadian Journal Physiology Pharmacology. v. 95, n. 1, p 1-15, 03 set. 2017.
KOURY, JEFFREY, et al. Targeting Signaling Pathways in Cancer Stem Cells for Cancer Treatment. Stem Cells International. v. 2017, mar., 2017.
KOZLOVA, NINA, et al. Hypoxia-inducible factor prolyl hydroxylase 2 (PHD2) is a direct regulator of epidermal growth factor receptor (EGFR) signaling in breast cancer. Oncotarget. v.8 n. 6, p. 9885–9898, 2017.
KURUVILLA, JES G, et al. Krüppel-like Factor 4 Modulates Development of BMI1+ Intestinal Stem Cell-Derived Lineage Following γ-Radiation-Induced Gut Injury in Mice. Stem Cell Reports. v. 6, n. 6, p.815-824, 2016.
LEON, G, et al. Cancer Stem Cells in Drug Resistant Lung Cancer: Targeting Cell Surface Markers and Signaling Pathways. Pharmacology and Therapeutics, v. 158, p.71-90, 2016.
LIEDKE, P.E.R. Hormonioterapia Adjuvante em Câncer de Mama. Revista Brasileira Oncologia Clínica. v. 3 ,n. 8, p 23-27, Mai/Ago., 2006.
LI, LI, et al. Notch-1 signaling promotes the malignant features of human breast cancer through NF-κB activation. PLoS ONE, v. 9, n. 4, 23 abr., 2014.
LIM, S. C. CD24 and human carcinoma: tumor biological aspects. Biomed

83
Pharmacother. v. 59, p 351–354, 2005.
LI, WENZHE, et al., Unraveling the roles of CD44/CD24 and ALDH1 as cancer
stem cell markers in tumorigenesis and metastasis. Scientific Reports. v. 7, 23 out., 2017.
LI, XI XI, et al. Overexpression of CTNNB1: Clinical implication in Chinese de novo acute myeloid leukemia. Pathology - Research and Practice. v. 214, n.3, 2018.
LIU, YONGHONG, et al. Hepatitis B virus X protein stabilizes amplified in breast cancer 1 protein and cooperates with it to promote human hepatocellular carcinoma cell invasiveness. Hepatology. v.56, n.3, p 1015-1024, 2012.
LIU, YONGLEI, et al. EMMPRIN Down-regulating miR-106a/b Modifies Breast Cancer Stem-like Cell Properties via Interaction with Fibroblasts Through STAT3 and HIF-1α. Scientific Reports. v. 6, 21 jun., 2016.
LO MUZIO, L, et al. A possible role for the WNT-1 pathway in oral carcinogenesis. Crit Rev Oral Biol Med. v. 12, n. 2, p 152- 165, 2001.
LOBBEZOO DJ, et al. Prognosis of metastatic breast cancer: are there differences between patients with de novo and recurrent metastatic breast cancer? Br J Cancer. v. 112, p 1445 - 1451, 2015.
MAGEE, JEFFREY A, et al. Cancer Stem Cells: Impact, Heterogeneity, and Uncertainty. Cancer cell. v.21, n.3, p 283-296, 2012.
MAHERALI, NIMET, et al. Directly Reprogrammed Fibroblasts Show Global Epigenetic Remodeling and Widespread Tissue Contribution. Cell Stem Cell. v. 1, n.1, p. 55 – 70, 2007.
MAKINO, S. Further evidence favoring the concept of the stem cell in ascites tumors of rats. Ann. NY Acad. Sci. v. 63, p 818–830, 1956.
MANGONE, Flavia R, et al. PIK3CA Exon 20 Mutations Are Associated with Poor Prognosis in Breast Cancer Patients. Clinics. v.67, n. 11, p 1285–1290, 2012.

84
MANI, S.A. et al. The epithelial-mesenchymal transition generates cells with properties of stem cells. Cell. v. 133, n.4, p 704-715, 2008.
MANSOORI, Maryam, et al. Circulating cancer stem cell markers in breast carcinomas: a systematic review protocol. Systematic Reviews. v. 6, n. 1, p 1, 2017.
MANUPATI, Kanakaraju, et al. Inhibiting epidermal growth factor receptor signalling potentiates mesenchymal-epithelial transition of breast cancer stem cells and their responsiveness to anticancer drugs. The FEBS Journal. v. 284, p. 1830-1854, 2017.
MITSUI, Kaoru, et al. The Homeoprotein Nanog Is Required for Maintenance of Pluripotency in Mouse Epiblast and ES Cells. Cell. v. 112, n. 5, p 631-642, 11 abr., 2003.
MOHAMMADIAN, Farideh e NEGAHDARI, Babak. Isolation and characterization of mesenchymal stem cells and its antitumor application on ovarian cancer cell line. Artificial Cells, Nanomedicine, and Biotechnology. v. 3, p 1-10, 2017. MOHAMMED, Maryam K, et al. Wnt/β-catenin signaling plays an ever-expanding role in stem cell self-renewal, tumorigenesis and cancer chemoresistance. Genes diseases. v. 3, n. 1, p 11-41, 2016.
MOON, Jong-Seok, et al. Krüppel-like factor 4 (KLF4) activates the transcription of the gene for the platelet isoform of phosphofructokinase (PFKP) in breast cancer. J Biol Chem. v. 286, p 23808–23816, 2011.
MOREIRA, L.M. Ciências Genômicas: fundamentos e aplicações. Ribeirão Preto: Sociedade brasileira de genética. Cubo, 403p, 2015.
NISHIDA, Naoshi. et al. Characteristic patterns of altered DNA methylation predict emergence of human hepatocellular carcinoma. Hepatology. v. 56, n. 3, p 994 – 1003 , 2012.
NIU, Chao-Shi. et al. Expression of NANOG in human gliomas and its relationship with undifferentiated glioma cells. Oncology Reports. v. 26, p. 593-601, 2011.
NOTARI, D.L. Desenvolvimento de workflows científicos para a geração e análise de diferentes redes de interatomas. 2012.181f. Tese (Doutorado em biotecnologia), Universidade de Caxias do Sul, 2012.

85
NOWELL, Peter C.; JENSEN, Julie; GARDNER, Frank. Two complex translocations in chronic granulocytic leukemia involving chromosomes 22, 9 and a third chromosome. Humangenetik. v. 30, n.1, p. 13-21, 1975.
ORDING AG, et al. Site of metastasis and breast cancer mortality: a Danish nationwide registry-based cohort study. Clin Exp Metastasis.34:93-101, 2017.
ORGANIZAÇÃO MUNDIAL DA SAÚDE (OMS). GLOBOCAN 2012 v.1.0. Cancer Incidence and Mortality Worldwide, 2013. Disponível em: < http://globocan.iarc.fr. > Acesso em 24 de janeiro de 2018.
ORIAN-ROUSSEAU, et al.CD44 is required for two consecutive steps in HGF/c-Met signaling. Genes Dev. v. 16, p 3074–3086, 2002.
OWCZAREK, Witold, et al. The incidence and management of cutaneous adverse events of the epidermal growth factor receptor inhibitors. Postepy Dermatol Alergol. v.34, n. 5, p 418-428, 2017. PACHNICKI JPA, Czeczko NG, Tuon F, Cavalcanti TS, Malafaia AB, Tuleski AM. Avaliação imunoistoquímica dos receptores de estrogênio e progesterona no câncer de mama, pré e pós-quimioterapia neoadjuvante. Rev Col Bras Cir. V. 39, n 2, p 86-92, 2012.
PEREIRA, Bruna de Cássia Sales, et al. Conhecimento sobre o câncer de mama em usuárias do serviço público. Rev Inst Ciências Saúde. v. 26, n.1, p.10-5, 2008. PEREIRA, D.V.C. Identificação e análise da rede de genes envolvidos no câncer de pele de melanoma através da mineração de texto. 2016. 128f. Dissertação (Mestrado em Tecnologia da Informação Aplicada à Biologia Computacional) - Faculdades Infórium de Tecnologia. Belo Horizonte, 2016.
PIACENTINI, Amanda Bernardini. Novas perspectivas para a Biologia do Câncer:Compreendendo as metástases. 2012. 70 f. Trabalho de conclusão de curso (bacharel em ciências biológicas) Instituto de Biociências, Universidade Estadual Paulista, Rio Claro, 2012.

86
PICCIRILLO, S.G. M; VESCOVI, A.L. Brain tumour stem cells: possibilities of new therapeutic strategies. Journal Expert Opinion on Biological Therapy. v. 7, n. 8, p 1129- 1135, 2007. POSWAR, Fabiano de Oliveira, et al. Bioinformatics, Interaction Network Analysis, and Neural Networks to Characterize Gene Expression of Radicular Cystand Periapical Granuloma. Journal of Endodontics, V 41, Issue 6, p 877–883, Jun., 2015. PRABHU, Varun V. Therapeutic targeting of the p53 pathway in cancer stem cells. Expert OpiniononTherapeuticTargets. v. 16, n.12, p 1161–1174, 2012.
PUISIEUX, A, et al. A twist for survival and cancer progression. British Journal of Cancer. v. 94, p 13–17, 2006.
QUIN, Xing. et al. Cancer-associated Fibroblast-derived IL-6 Promotes Head and Neck Cancer Progression via the Osteopontin-NF-kappa B Signaling Pathway. Theranostics. v.8, n.4, 2018.
REYA, T. et al. Stem cells, cancer, and cancer stem cells. Nature International Weekly Journal of Science. v. 414, n. 6859, p105-111, 2001.
RIGGIO, Marina, et al. AKT1 and AKT2 isoforms play distinct roles during breast cancer progression through the regulation of specific downstream proteins. Scientific Reports. v. 7, 13 mar., 2017.
RIZZO, Paola, et al. Cross-talk between notch and estrogen receptor in breast cancer suggests novel therapeutic approaches. Cancer Res. v. 68 n. 13, p 5226–5235, 2008.
ROBERTs, Cai M, et al. TWIST1 Drives Cisplatin Resistance and Cell Survival in an Ovarian Cancer Model, via Upregulation of GAS6, L1CAM, and Akt Signalling. Scientific Reports. v. 6, 23 nov., 2016.
ROUDI R. et al., Differential Expression of Cancer Stem Cell Markers ALDH1 and
CD133 in Various Lung Cancer Subtypes. Cancer Investigation. v. 33, n.7, p 294 –
302, 05 jun, 2015.

87
RUDOLPH, Marion, et al. AKT1E17K mutation profiling in breast cancer: prevalence, concurrent oncogenic alterations, and blood-based detection. BMC Cancer. v. 16, 11 ago., 2016.
SHAN, Juanjuan, et al. Nanog regulates self-renewal of cancer stem cells through the insulin-like growth factor pathway in human hepatocellular carcinoma. Hepatology. v.56, n.3, p 1004-1014, 2012.
RUITERKAMP J, et al. Improved survival of patients with primary distant metastatic breast cancer in the period of 1995–2008. A nationwide population-based study in the Netherlands. Breast Cancer Res Treat. v.128, p 495-503, 2011.
SHAO, Shan, et al. Notch1 signaling regulates the epithelial-mesenchymal transition and invasion of breast cancer in a Slug-dependent manner. Mol Cancer. v. 15, 03 fev., 2015.
SHEN, Honghong, et al. Lin28A activates androgen receptor via regulation of c-myc and promotes malignancy of ER−/Her2+ breast cancer. Oncotarget. v. 7, n. 37, p 60407–60418, 2016.
SCHMIDT, C, et al. Acetyl-lupeolic acid inhibits Akt signaling and induces apoptosis in chemoresistant prostate cancer cells in vitro and in vivo. Oncotarget. v. 8, n. 33, p 55147–55161, 2017.
SHIPITSIN, M. e POLYAK, K. The cancer stem cell hypothesis: in search of definitions, markers, and relevance. Laboratory Investigation; a Journal of Technical Methods and Pathology. V. 88, n. 5, p 459–463, 2008.
SIEGEL R, Naishadham D e Jemal A. Cancer statistics, 2012. CA Cancer J Clin. v. 62, n. 1, p 10-29, 2012.
SIEGEL RL e Miller KD e Jemal A. Cancer Statistics, 2018. CA Cancer J clin. v. 68, n.1, p 7-30, 2018.
SLEEMAN, J.P e THIELE, W. Tumor metastasis and the lymphatic vasculature. International Journal of Cancer. v. 125, n. 12, p 2747-2756, 2009.

88
SOLOZOBOVA, Valeriya e BLATTNER, Christine. p53 in stem cells. World Journal of Biological Chemistry. v. 2, n. 9, p 202–214, 2011.
SONG, Xiang, et al. Expression of Krüppel-like factor 4 in breast cancer tissues and its effects on the proliferation of breast cancer MDA-MB-231 cells. Exp Ther Med. v. 13, n 5, p 2463-2467, 2017.
SOUZA, V. F. et al. Células-tronco: uma breve revisão. Revista de Ciências Médicas e Biológicas. v. 2, n. 2, p 251-256,2003. SOUZA, L.T. A persistência dos cânceres e as Células-tronco tumorais. Revista da Associação Médica Brasileira, São Paulo, v.52, n. 6, 2006.
SUN, Z, et al. Special AT-rich sequence-binding protein-1 participates in the maintenance of breast cancer stem cells through regulation of the Notch signaling pathway and expression of Snail1 and Twist1. Molecular Medicine Reports. v.11, n 5, p 3235–3242, 2015.
SUN, H, et al. Let-7 miRNAs sensitize breast cancer stem cells to radiation-induced repression through inhibition of the cyclin D1/Akt1/Wnt1 signaling pathway. Molecular Medicine Reports. v.14, p 3285-3292, 2016.
SZKLARCZYK. The STRING database in 2017: quality-controlled protein–protein association networks, made broadly accessible Nucleic Acids Research. v. 45, p 362 – 368, 04 Jan., 2017.
SZKLARCZYK, D, et al. STRING v10: protein–protein interaction networks, integrated over the tree of life. Nucleic Acids Research. v. 43, p 447– 452, 2015.
TAKAHASHI, Kazutoshi. et al. Induction of Pluripotent Stem Cells from Adult Human Fibroblasts by Defined Factors. Cell, v. 131, n. 5, p 861 – 872, 2007.
TANABE, Shihori, et al. Gene expression and pathway analysis of CTNNB1 in cancer and stem cells. World Journal of Stem Cells. v. 8, n.11, p 384–395, 2016.

89
TANG, Yanjuan, et al. Coexpression of hyperactivated AKT1 with additional genes activated in leukemia drives hematopoietic progenitor cells to cell cycle block and apoptosis. ISEH - Experimental Hematology. v. 43, n 7, p 554–564, jul., 2015.
TATE, Matthew C, et al. Biology of Angiogenesis and Invasion in Glioma. Neurotherapeutics. v. 6, n. 3, p 447–457, 2009.
TESTA, Laura e MANO, Max S. Quimioterapia em câncer de mama. Rev Bras Mastologia. v.20, n.4, p 190-198, 2011.
THAKUR, Ravi, et al. Inhibition of STAT3, FAK and Src mediated signaling reduces cancer stem cell load, tumorigenic potential and metastasis in breast cancer. Scientific Reports. v. 5, 14 mai., 2015.
THOMAS A, Margaret Lois, et al. Citral reduces breast tumor growth by inhibiting the cancer stem cell marker ALDH1A3. Molecular Oncology. v.10, n.9, p 1485 -1496, 2016.
TIEZZI, Daniel Guimarães. Cirurgia conservadora no câncer de mama. Rev. Bras. Ginecol. Obstet. Rio de Janeiro. v. 29, n. 8, p 428-434, 2007.
TSIALIKAS, Jennifer; ROMER-SEIBERT, Jennifer. LIN28: roles and regulation in development and beyond. Development. v.142, n.14, p 2397 – 2404, 2015.
TLSTY, T. D. et al. Genetic and Epigenetic Changes in Mammary Epithelial Cells May Mimic Early Events in Carcinogenesis. Journal of Mammary Gland Biology and Neoplasia. v. 9, n. 3, p 263-274, 2004.
VALLE, S, et al . The Ever-Evolving Concept of the Cancer Stem Cell in Pancreatic Cancer. Cancers, v. 10, n. 2, p 33, 2018. VERLI, H. Bioinformática da Biologia à Flexibilidade Molecular. 1. ed. São Paulo: SBBq, 282p, 2014.
VISVADER, et al. Cancer stem cells in solid tumours: accumulating evidence and unresolved questions. Nat. Rev. Cancer. v. 8, p 755-768, 2008.

90
WANG, Jiankui, et al. Expression of C-myc and β-catenin and their correlation in triple negative breast cancer. Minerva med. v.108, n. 6, p 513 – 517, 2017.
WARD, Laura Sterian. Entendendo o Processo Molecular da Tumorigênese. Arq Bras Endocrinol Metab. v. 46, n. 4, p 351-360, 2002.
WEGWARTH, Odette. Cancer Survival: the CONCORD-2 study. The lancet. v. 386, n. 9992, p 429, 2015.
WEIGEL MT, Dowsett M. Current and emerging biomarkers in breast cancer: prognosis and prediction. Endocr Relat Cancer. v. 17, n.4, p 245 – 262, 2010.
WEIGELT, B, et al. Breast cancer metastasis: markers and models. Nature reviews Cancer. v. 5, n. 8, p 591-602, 2005. WINCEWICZ A, et al. STAT3 and apoptosis regulators: Bak and Bcl-xL in endometrioid adenocarcinomas of different estrogen receptor-α immunoprofile.Gynecological Endocrinilogy Journal. v. 27, n.8, p 536 – 540, 04 jul., 2011.
WON H.-Y, et al., “Loss of Mel-18 enhances breast cancer stem cell activity and tumorigenicity through activating Notch signaling mediated by the Wnt/TCF pathway,” The FASEB Journal. v. 26, n. 12, p 5002–5013, 2012.
WOODS. Woods LD, et al., The genomic landscape of human breast and colorectal cancers. Science. v. 318, p 1108-1113, 2007.
WU, Kou-Juey e YANG ,Muh-Hwa . Epithelial-mesenchymal transition and cancer stemness: the Twist1-Bmi1 connection. Bioscience Reports. v. 31, n. 6, p 449–55, 2011.
WICHA MS, Liu S, Dontu G. Cancer stem cells: An old idea - A paradigm shift. Cancer Res. v. 66, p 1883-1890, 2006.
YANG, Shi-Wei, et al. ATG4A Promotes Tumor Metastasis by Inducing the

91
Epithelial-Mesenchymal Transition and Stem-like Properties in Gastric Cells. Oncotarget. v. 7, n.26, p 39279–39292, 2016. YARAK, Samira e OKAMOTO, Oswaldo Keith. Células-tronco derivadas de tecido adiposo humano: desafios atuais e perspectivas clínicas. An. Bras. Dermatol. v. 85, n. 5, p 647-656, 2010.
YORI,Jennifer L, et al. Krüppel-like factor 4 inhibits tumorigenic progression and metastasis in a mouse model of breast cancer. Neoplasia. v.13, n. 7, p 601–610, 2011.
YOU, Liuping, et al. Correlation of Cancer Stem-Cell Markers OCT4, SOX2, and NANOG with Clinicopathological Features and Prognosis in Operative Patients with Rectal Cancer. Yonsei Medical Journal. v. 59, n. 1, p. 32-42, 2017.
YU, peijia. et al. Nanog induced intermediate state in regulating stem cell differentiation and reprogramming. BMC Systems Biology. v. 12, n.1, 2018.
ZARDAWAI, Sarah J, et al. High Notch1 protein expression is an early event in breast cancer development and is associated with the HER2 molecular subtype. Histopathology. v. 56, n. 3, p 286–296.2010.
ZHANG, Ying. et al. Regulation of p53 Level by UBE4B in Breast Cancer. PLoS ONE. v. 9 n. 2, 26 fev. 2014.





























