Embed Size (px)
Citation preview

Universidade Federal de Ouro Preto
Núcleo de Pesquisa em Ciências Biológicas
Nívia Carolina Nogueira de Paiva
Influência da Infecção por uma Cepa Policlonal do
Trypanosoma cruzi sobre a Resposta Imune no Coração e
Cólon Durante a Fase Aguda da Infecção Experimental de
Camundongos
Ouro Preto
2015

Nívia Carolina Nogueira de Paiva
Influência da Infecção por uma Cepa Policlonal do
Trypanosoma cruzi sobre a Resposta Imune no Coração e
Cólon Durante a Fase Aguda da Infecção Experimental de
Camundongos
Tese de Doutorado apresentada ao Programa de Pós-Graduação em Ciências Biológicas do Núcleo de Pesquisas em Ciências Biológicas da Universidade Federal de Ouro Preto, como requisito parcial para a obtenção do Título de Doutor em Ciências Biológicas, Área de concentração, Imunobiologia de Protozoários.
Orientadora: Profa Cláudia Martins Carneiro
Co-orientadora: Profª Paula Melo de Abreu Vieira
Ouro Preto 2015



Nogueira-Paiva, NC Colaboradores
i
Laboratório de Imunopatologia – NUPEB/UFOP
Dr. Alexandre Barbosa Reis
Dr. Bruno Mendes Roatt
Msc. Kátia da Silva Fonseca
Larissa Maris Rezende Oliveri
Prof. Washington Luiz Tafuri (in memorian)
Laboratório de Farmácia Clínica – CIPHARMA/UFOP
Dra. Vanja Maria Veloso
Laboratório de Doença de Chagas – NUPEB/UFOP
Dra. Marta de Lana
Msc. Maykon Tavares de Oliveira
Laboratório de Biomarcadores de Diagnóstico e Monitoração – Centro de Pesquisas
René Rachou/FIOCRUZ
Dra. Andréa Teixeira de Carvalho
Laboratório de Neurogenômica – Centro de Pesquisas René Rachou/FIOCRUZ
Dra. Karina Barbosa Queiroz

Nogueira-Paiva, NC Dedicatória
ii
Ao meu eterno e querido amigo, Carlos Henrique Gonçalves.

Nogueira-Paiva, NC Agradecimentos
iii
“(...) e que eu me lembre de Ti, com tranquilidade e determinação, mesmo quando for difícil
dizer que Te amo (...)” (Aleph, Paulo Coelho)
À Deus, obrigada pela presença e luz em todos os momentos.
À professora Cláudia Martins Carneiro, pela oportunidade e carinho. Por confiar no meu
trabalho ao longo de todos esses anos. Por guiar meus passos, compreender minhas limitações
e acima de tudo pela incrível orientação. Obrigada por mais essa etapa, minha eterna mestre.
À minha querida amiga, professora, co-orientadora, Paula, pela disponibilidade e
cumplicidade. Obrigada por fazer parte de cada capítulo de mais essa história. Eterna gratidão
da sua eterna aprendiz!!!!!
À professora Vanja, pela disponibilidade e generosidade durante a realização desse trabalho.
À minha filhinha científica, Larissa, cuja parceria ao longo desse trabalho foi imprescindível e
igualmente prazerosa.
Ao meu mais novo eterno colaborador, Maykon Tavares. Foi incrível conhecê-lo e trabalhar
ao seu lado. Obrigada!!!!
À Marta de Lana, por todo carinho de sempre...
À Dra Andréa Teixeira de Carvalho. Sua atenção, disponibilidade e grande experiência em
citometria de fluxo, contribuíram para o resultado final do nosso trabalho.
Ao professor Alexandre Reis, exemplo de paixão pelo que faz. Obrigada por inspirar, desde o
início, a minha caminhada acadêmica.
Ao Bruno, por toda a paciência de sempre e por ser tão generoso em compartilhar
conhecimento.
À Karina, pela realização da técnica de qPCR.

Nogueira-Paiva, NC Agradecimentos
iv
À Kátia, amiga de todos os momentos.
Aos demais colaboradores, obrigada pela contribuição, disponibilidade e atenção.
À Luciana, Renata Rezende, Fernando, Rory e Levi... Esse trabalho tem um pedacinho de
vocês... Obrigada!!!!
À minha querida Maria, pelo carinho e paciência durante os meus primeiros passos e sempre.
Obrigada pela eterna parceria!
Ao professor Evandro Machado, por seu auxílio na infecção in vitro.
Às minhas queridas amigas-irmãs, Gleise, Maria Cláudia, Aline, Débora, Lucilene, Nádia,
Jamille, Emília, Mariana. Obrigada por alegrarem esse caminho!!!!!
À minha nova filhinha científica, Karla.
Aos meus amigos e parceiros de trabalho, Rodrigo, Luísa Perin, Narjara, Lúcia, Thaís, Lívia,
Flávia, Beatriz, Henrique, João Felipe, João Veloso, Jéssica, Marcelo, Diogo, Sidney, Luísa
Couto, Élcio, Iago, George, Josefa, Irmázio, pela disponibilidade e carinho com que sempre
me ajudaram durante o desenvolvimento desse trabalho.
Aos colegas de trabalho, e acima de tudo, amigos do Centro de Ciência Animal-CCA/UFOP
pelo apoio, pela agradável companhia e pelo carinho e ética dedicados ao trabalho com
animais.
Aos saudosos amigos e companheiros que com brilhantismo estão trilhando seus caminhos,
Amanda, Juliana, Sheler, Tânia, Laser, Ludmilla, Mariana Trevisan, Wendel, Flávia
Bittencourt, Samuel, Micheline, Dayane, Lara, Edith, Ana Luiza, Caio, Caroline, Nathália,
Diogo, Jaqueline, Jerusa, João Lino...
Aos colegas da secretaria do Núcleo de Pesquisa em Ciências Biológicas, NUPEB/UFOP.
Obrigada!!!!

Nogueira-Paiva, NC Agradecimentos
v
Aos nossos guardiões, Maria, Celso, José Martins e Jorge. Obrigada pela amizade!
Às meninas que deixam nossa rotina mais agradável, Sônia e Cristiana.
À alguém muito especial, pela compaixão e solidariedade, a quem serei sempre grata.
Obrigada por devolver vida às cores e cores à minha vida!!!
À minhas irmãs Alquimistas, pela amizade e segurança. É muito bom saber que vocês estão
ao meu lado!!!!!
Ao meu pai, exemplo de perseverança. Obrigada por tornar meus sonhos possíveis!
À minha mãe, anjo e luz em todos os momentos. Sua “presença” acalma sempre meu coração.
Ao meu irmão “Dri”, maior razão da minha vida.
À minha tia Quitéria, simplesmente por tudo!!!
À minha família, por compreenderem minha ausência e nunca se esquecerem desta prima-
irmã, sobrinha, neta, e agora titia, que ama muito vocês!!!!!!!
Ao meu grande amor, Tiago, por ser o abraço que me mantém segura, o coração que me
mantém em paz. Seu amor faz com que cada dia renasça cheio de esperança.
À todos que contribuíram de forma direta ou indireta para que esse trabalho fosse realizado,
minha sincera gratidão!!!!!!

Nogueira-Paiva, NC Epígrafe
vi
“... O bom é ser inteligente e não entender. É uma benção estranha,
como ter loucura sem ser doida. É um desinteresse manso, é uma doçura
de burrice. Só que de vez em quando vem a inquietação: quero entender
um pouco. Não demais: mas pelo menos entender que não entendo.”
Clarice Lispector

Nogueira-Paiva, NC Resumo
vii
Fatores intrínsecos ao Trypanosoma cruzi e relacionados ao hospedeiro agem sobre a patogênese da doença de Chagas. Neste contexto, o polimorfismo genético do parasito parece ter um papel crítico no prognóstico da doença, e tem sido demonstrada uma associação entre o desenvolvimento das distintas formas clínicas e a distribuição geográfica de cepas do T. cruzi. Diante disso, esse projeto propôs avaliar a influência do polimorfismo biológico e molecular de subpopulações do T. cruzi sobre a resposta inflamatória no coração e cólon durante a fase aguda da infecção experimental. A primeira etapa consistiu na caracterização biológica (meio de cultura acelular e celular) e do perfil gênico (Low-Stringency Single Specific Primer-Polymerase Chain Reaction/LSSP-PCR) de subpopulações do parasito obtidas a partir de hemoculturas periódicas (3, 6 e 12 meses após a infecção-mai) em 24 camundongos experimentalmente infectados com 5000 formas tripomastigotas sanguíneas da cepa Berenice-78 (Be-78 parental) do T. cruzi. Foi possível identificar isolados com comportamento in vitro, assim como perfis de assinatura gênica, distintos do observado para a cepa Be-78 parental. Em alguns casos, os isolados mostraram certo grau de semelhança com a cepa Berenice-62 (Be-62), isolada da paciente Berenice, em fase crônica tardia, 16 anos antes do xenodiagnóstico que originou a cepa Be-78. Esses dados sugerem a presença de distintas subpopulações na cepa Be-78 e reafirmam a plasticidade do parasito frente à pressão seletiva exercida pela interação parasito-hospedeiro. Na segunda etapa, a infecção com um isolado selecionado na primeira etapa, que apresentou perfil distinto da cepa parental, foi comparada com a infecção com as cepas Be-78 parental e Be-62 em modelo murino com o objetivo de investigar a participação de diferentes subpopulações do parasito sobre o desenvolvimento do processo inflamatório durante os estágios iniciais da doença experimental. Nossos dados demonstraram um perfil cardiomiotrópico e retardo no controle da parasitemia na infecção pela cepa Be-78 parental, seguido por processo inflamatório intenso ao final da fase aguda, ao contrário da cepa Be-62 e do isolado Be-78is5-3mai, que por sua vez, apresentou-se preferencialmente parasitando o cólon, com parasitemia significativamente superior, porém, controlada precocemente, e processo inflamatório intenso ao longo da fase aguda. No coração, apesar de já observada no grupo Be-62 aos 14dai, no 28º, a densidade de macrófagos, linfócitos T CD4+ e CD8+ foi semelhante entre as cepas Be-62 e Be-78 parental, com sobreposição de células T CD8+ em relação às células T CD4+. O isolado Be-78is5-3mai apresentou miocardite, embora de menor intensidade, também apenas no 28ºdai, composta essencialmente por macrófagos e linfócitos T CD8+. De forma diferente ao observado no coração, o perfil de resposta imunológica no cólon foi cepa-dependente. Apenas o grupo Be-78 parental apresentou aumento precoce de células T CD4+, que foram substituídas por células T CD8+ e linfócitos B, no 14ºdai. Todos esses resultados são condizentes com o comportamento da parasitemia e do parasitismo tecidual nesse grupo. Embora com menor intensidade, o perfil imunofenotípico detectado nos animais infectados com o isolado Be-78is5-3mai foi próximo àquele demonstrado pelo grupo Be-62, e sobretudo, distinto da cepa Be-78 parental. Nas primeiras duas semanas de infecção, o processo inflamatório foi constituído principalmente por macrófagos e células T CD8+, com aumento expressivo de linfócitos T CD4+ apenas ao final da fase aguda. Estes resultados indicam que o isolado Be-78is5-3mai, consistente com os dados da avaliação in vitro, demonstrou durante a fase aguda da infecção de camundongos BALB/c, um comportamento intermediário entre a cepas de referência utilizadas, apresentando virulência (parasitemia e mortalidade) compatível com a cepa Be-78 parental, no entanto, perfil histotrópico compartilhado com a cepa Be-62, reforçando a hipótese de que a infecção a longo prazo com a cepa Be-78 foi capaz de selecionar uma subpopulação que compartilha características com a cepa Be-62, ambas originada da paciente Berenice, com 16 anos de intervalo entre as coletas. Palavras-chave: Trypanosoma cruzi, Polimorfismo, Resposta Imune

Nogueira-Paiva, NC Abstract
viii
Factors intrinsic to Trypanosoma cruzi and other host-related act in Chagas disease pathogenesis. In this context, the parasite genetic polymorphism seems to have a critical role in disease prognosis, and have been demonstrated an association between the distincts clinical forms development and geographic distribution of T. cruzi strains. Polyclonal strains are common in the natural infections, and may involve specific interactions parasite-organ. Therefore, this project proposed to evaluate the influence of T. cruzi subpopulations polymorphism on the inflammatory response in the heart and colon during the acute phase of the experimental infection. The first step consisted in biological (cellular and acellular growth media culture) and molecular (Low-Stringency Single-Specific Primer Polymerase Chain Reaction/LSSP-PCR) profile characterization of parasite subpopulations obtained from periodic blood cultures (3, 6, 12 months after the infection-mai) in 24 mice experimentally infected with 5000 blood trypomastigotes forms of Berenice-78 (Be-78 parental strain) T. cruzi strain. It was possible to identify isolates with in vitro behavior, as well as gene signature profile distincts from that observed for the Be- 78 parental strain. In some cases, the isolates showed some degree of resemblance with the Berenice-62 (Be- 62) strain, isolated from patient Berenice in Chagas disease chronic phase, 16 years before the xenodiagnostic that originated the Be- 78 strain. These data suggest the presence of distinct subpopulations in Be-78 strain and underline the plasticity of the parasite against the selective pressure exerted by the parasite-host interaction. In the second stage, infection with one isolate selected in the first step, presenting distinctive profile of the to parental strain, will be compared to infection with Be-78 parental and Be-62 strains in isogenic murine model, in order to investigate the participation of the different subpopulations on the inflammatory process during the early stages of experimental disease. Our data demonstrated a cardiomyotropic profile and a delay in parasitemia controlling in Be-78 parental infected mice, followed by severe inflammation at the end of the acute phase, unlike the Be-62 strain and Be-78is5-3mai isolate, which in turn is preferably parasitizing the colon, with significantly higher parasitaemia, however, early controlled, and intense inflammation during the acute phase. At heart, despite already observed in the Be-62 group to 14dai, on the 28th, the density of macrophages, CD4+ and CD8+ T cells were similar between the Be-62 and Be-78 parental strains, with T-cell overlap CD8+ over CD4+ T cells. The isolated Be-78is5-3mai presented myocarditis, although of lower intensity, also in 28ºdai essentially composed of macrophages and CD8+ T lymphocytes. Differently to that observed in the heart, the immune response profile in the colon was strain-dependent. As Be-78 parental group showed an early increase of CD4+ T cells, which were replaced by CD8+ T cells and B lymphocytes in 14ºdai. All these results are consistent with the parasitaemia and tissue parasitism behavior in that group. Although to a lesser extent, the immunophenotypic profile detected in the animals infected with the isolated Be-78is5-3mai was close to that shown by Be-62 group. In the first two weeks of infection, the inflammation was composed primarily of macrophages and CD8+ T cells, with significant increase of CD4+ T cells only at the end of the acute phase. These results indicate that the isolate Be-78is5-3mai, consistent with the in vitro evaluation, shown during the acute phase of infection of BALB/c mice, an intermediate behavior between the both reference strains used, with virulence (parasitemia and mortality) compatible with the Be-78 parental strain, however, shared histotropic profile with the Be-62 strain, reinforcing the hypothesis that long-term infection with the Be-78 strain was able to select a subpopulation that shares characteristics with the Be-62 strain, both originated from Berenice patient, 16-year interval between collections. keywords: Trypanosoma cruzi, Polymorphism, Immune Response

Nogueira-Paiva, NC Lista de Figuras
ix
Figura 1: Esquema representando as citocinas e as populações celulares envolvidas na geração da resposta protetora ou patogênica na doença de Chagas .......................................................................... 9
Figura 2: Fluxograma representando as atividades relacionadas à obtenção e identificação de isolados da cepa Be-78 do T. cruzi (ETAPA I) ................................................................................................... 28
Figura 3: Fluxograma representando as atividades relacionadas à avaliação in vivo (ETAPA II) ...... 32
Figura 4: Fluxograma representando a distribuição dos animais infectados com diferentes subpopulações do Trypanosoma cruzi utilizados para determinação da curva de parasitemia e da taxa de mortalidade ....................................................................................................................................... 33
Figura 5: Desenho esquemático representando a distribuição de camundongos BALB/c entre os tratamentos/grupos no ensaio por citometria de fluxo. ......................................................................... 38
Figura 6: Desenho esquemático representando as combinações de anticorpos marcados com fluorocromos utilizadas no ensaio de citometria de fluxo ..................................................................... 41
Figura 7: Representação da estratégia de análise da frequência de células inflamatórias com distintos fenótipos em suspensão de células do coração ou cólon de camundongos infectados com o isolado Be-78is5-3mai, ou com as cepas de referência Berenice-62 e Berenice-78 parental do Trypanosoma cruzi ............................................................................................................................................................... 42
Figura 8: Representação da estratégia de análise da frequência de macrófagos em suspensão de células do coração ou cólon de camundongos infectados com o isolado Be-78is5-3mai, ou com as cepas de referência Berenice-62 e Berenice-78 parental do Trypanosoma cruzi ................................ 43
Figura 9: Representação da estratégia de análise da frequência de linfócitos B em suspensão de células do coração ou cólon de camundongos infectados com o isolado Be-78is5-3mai, ou com as cepas de referência Berenice-62 e Berenice-78 parental do Trypanosoma cruzi ................................ 43
Figura 10: Representação da estratégia de análise da frequência de linfócitos T CD4+ ou CD8+, expressando IFN- em suspensão de células do coração ou cólon de camundongos infectados com o isolado Be-78is5-3mai, ou com as cepas de referência Berenice-62 e Berenice-78 parental do Trypanosoma cruzi ................................................................................................................................ 44
Figura 11: Cinética de crescimento de epimastigotas, cultivadas em meio LIT no período de 0-20 dias, de isolados do parasito obtidos a partir de hemoculturas aos 3, 6 ou 12 meses após a infecção de camundongos Swiss com 5x103 formas tripomastigotas sanguíneas da cepa Be-78 do Trypanosoma cruzi ....................................................................................................................................................... 51
Figura 12: Ensaio de infecção in vitro em culturas de células Vero usando as cepas Berenice-62, Berenice-78 parental e subpopulações/isolados da cepa Berenice-78 do Trypanosoma cruzi, obtidos em hemoculturas realizadas aos 3, 6 ou 12 meses após a infecção de camundongos Swiss com a cepa parental .................................................................................................................................................. 54
Figura 13: Ensaio de infecção in vitro em culturas de células Vero usando as cepas Berenice-62, Berenice-78 parental e subpopulações/isolados da cepa Berenice-78 do Trypanosoma cruzi, obtidos em hemoculturas realizadas aos 3, 6 ou 12 meses após a infecção de camundongos Swiss com a cepa parental .................................................................................................................................................. 55
Figura 14: Fotomicrografias representativas do ensaio de infecção in vitro em culturas de células Vero com as cepas Berenice-62, Berenice-78 parental e subpopulações/isolados da cepa Berenice-78 do Trypanosoma cruzi, obtidos em hemoculturas realizadas aos 3, 6 ou 12 meses após a infecção de camundongos Swiss com a cepa parental. ............................................................................................. 56

Nogueira-Paiva, NC Lista de Figuras
x
Figura 15: Assinaturas gênicas do fragmento de 330pb da região do minicírculo do kDNA reveladas em gel de poliacrilamida a 8% corado pela prata .................................................................................. 59
Figura 16: Fenograma construído a partir de UPGMA (Unweighted Pair Group Method using Arithmetic averages) resultante das análises dos perfis de assinatura gênica obtido por LSSP-PCR de isolados obtidos a partir de hemoculturas aos 3, 6 ou 12 meses após a infecção de camundongos Swiss com 5x103 formas tripomastigotas sanguíneas da cepa Be-78 do Trypanosoma cruzi em comparação à cepa Be-78 parental e à cepa Be-62. ..................................................................................................... 60
Figura 17: Curvas de parasitemia de camundongos BALB/c infectados com o isolado Be-78is5-3mai, ou com as cepas de referência Berenice-62 e Berenice-78 parental do Trypanosoma cruzi ................. 63
Figura 18: Taxa de sobrevida referente aos camundongos BALB/c infectados com o isolado Be-78is5-3mai, ou com as cepas de referência Berenice-62 e Berenice-78 parental do Trypanosoma cruzi ............................................................................................................................................................... 64
Figura 19: Avaliação da carga parasitária no coração e cólon de camundongos BALB/c necropsiados ao longo da fase aguda (7, 14 ou 28 dias após a infecção-dai) da infecção experimental com o isolado Be-78is5-3mai, ou com as cepas de referência Berenice-62 e Berenice-78 parental do Trypanosoma cruzi ....................................................................................................................................................... 65
Figura 20: Análise morfométrica do processo inflamatório no coração e cólon de camundongos BALB/c necropsiados ao longo da fase aguda (7, 14 ou 28 dias após a infecção-dai) da infecção experimental com o isolado Be-78is5-3mai, ou com as cepas de referência Berenice-62 e Berenice-78 parental do Trypanosoma cruzi ............................................................................................................. 66
Figura 21: Fotomicrografias representativas da análise semi-quantitativa do processo inflamatório em cortes histológicos do coração de camundongos BALB/c necropsiados ao longo da fase aguda (7, 14 ou 28 dias após a infecção-dai) da infecção experimental com o isolado Be-78is5-3mai, ou com as cepas de referência Berenice-62 e Berenice-78 parental do Trypanosoma cruzi .................................. 67
Figura 22: Fotomicrografias representativas da análise semi-quantitativa do processo inflamatório em cortes histológicos do cólon de camundongos BALB/c necropsiados ao longo da fase aguda (7, 14 ou 28 dias após a infecção-dai) da infecção experimental com o isolado Be-78is5-3mai, ou com as cepas de referência Berenice-62 e Berenice-78 parental do Trypanosoma cruzi. ........................................... 68
Figura 23: Análise imunofenotípica do percentual médio de macrófagos (CD45+ F4/80+) em relação às células CD45+ presentes no coração e cólon de camundongos BALB/c necropsiados ao longo da fase aguda (7, 14 ou 28 dias após a infecção-dai) da infecção experimental com o isolado Be-78is5-3mai, ou com as cepas de referência Berenice-62 e Berenice-78 parental do Trypanosoma cruzi ....... 70
Figura 24: Análise imunofenotípica do percentual médio de linfócitos B (CD45+ CD19+) em relação às células CD45+ presentes no coração e cólon de camundongos BALB/c necropsiados ao longo da fase aguda (7, 14 ou 28 dias após a infecção-dai) da infecção experimental com o isolado Be-78is5-3mai, ou com as cepas de referência Berenice-62 e Berenice-78 parental do Trypanosoma cruzi ....... 71
Figura 25: Análise imunofenotípica do percentual médio de Linfócitos T CD4+ (CD45+ CD3+ CD4+) em relação às células CD45+ presentes no coração e cólon de camundongos BALB/c necropsiados ao longo da fase aguda (7, 14 ou 28 dias após a infecção-dai) da infecção experimental com o isolado Be-78is5-3mai, ou com as cepas de referência Berenice-62 e Berenice-78 parental do Trypanosoma cruzi ............................................................................................................................................................... 72
Figura 26: Análise imunofenotípica do percentual médio de Linfócitos T CD8+ (CD45+ CD3+ CD8+) em relação às células CD45+ presentes no coração e cólon de camundongos BALB/c necropsiados ao longo da fase aguda (7, 14 ou 28 dias após a infecção-dai) da infecção experimental com o isolado Be-78is5-3mai, ou com as cepas de referência Berenice-62 e Berenice-78 parental do Trypanosoma cruzi ............................................................................................................................................................... 73

Nogueira-Paiva, NC Lista de Figuras
xi
Figura 27: Análise imunofenotípica e funcional do percentual médio de Linfócitos T CD4+ (CD45+ CD3+ CD4+) ou CD8+ (CD45+ CD3+ CD8+) expressando IFN- (IFN-+) em relação aos seus respectivos fenótipos auxiliar (CD4+) ou citotóxico (CD8+) presentes no coração e cólon de camundongos BALB/c necropsiados ao longo da fase aguda (7, 14 ou 28 dias após a infecção-dai) da infecção experimental com o isolado Be-78is5-3mai, ou com as cepas de referência Berenice-62 e Berenice-78 parental do Trypanosoma cruzi......................................................................................... 74
Figura 28: Representação esquemática dos resultados obtidos na ETAPA II, comparando o perfil da parasitemia, da carga parasitária e do processo inflamatório ao longo dos tempos avaliados apresentado por cada uma das diferentes subpopulações do T. cruzi utilizadas nesse estudo no coração e cólon de camundongos BALB/c ......................................................................................................... 77
Figura 29: Representação esquemática dos resultados obtidos na ETAPA II, comparando o perfil fenotípico do infiltrado inflamatório ao longo dos tempos avaliados apresentado por cada uma das diferentes subpopulações do T. cruzi utilizadas nesse estudo no coração e cólon de camundongos BALB/c ................................................................................................................................................. 78
Figura 30: Representação esquemática dos resultados obtidos na ETAPA II, comparando as diferentes subpopulações do T. cruzi, em relação à carga parasitária e ao processo inflamatório (intensidade e fenótipo) no coração de camundongos BALB/c necropsiados ao longo da fase aguda (7, 14 ou 28 dias após a infecção-dai) da infecção experimental com o isolado Be-78is5-3mai, ou com as cepas de referência Berenice-62 e Berenice-78 parental do Trypanosoma cruzi .............................................. 117
Figura 31: Representação esquemática dos resultados obtidos na ETAPA II, comparando as diferentes subpopulações do T. cruzi, em relação à carga parasitária e ao processo inflamatório (intensidade e fenótipo) no cólon de camundongos BALB/c necropsiados ao longo da fase aguda (7, 14 ou 28 dias após a infecção-dai) da infecção experimental com o isolado Be-78is5-3mai, ou com as cepas de referência Berenice-62 e Berenice-78 parental do Trypanosoma cruzi .............................................. 118

Nogueira-Paiva, NC Lista de Quadros
xii
Quadro 1: Classificação atual das cepas do Trypanosoma cruzi em Discrete Typing Units ............... 16
Quadro 2: Classificação das cepas Berenice-62 e Berenice-78 segundo diferentes critérios. ............. 20
Quadro 3: Especificações dos anticorpos utilizados no ensaio de citometria de fluxo. ....................... 40

Nogueira-Paiva, NC Lista de Tabelas
xiii
Tabela 1: Avaliação das hemoculturas realizadas aos 3, 6 e 12 meses após a infecção (mai) de camundongos Swiss com a cepa Berenice-78 do Trypanosoma cruzi. ................................................. 48
Tabela 2: Código dos isolados obtidos a partir de hemoculturas aos 3, 6 ou 12 meses após a infecção de camundongos Swiss com a cepa Berenice-78 do Trypanosoma cruzi. ............................................. 49
Tabela 3: Resumo dos resultados obtidos na ETAPA I, comparando todos os parâmetros avaliados entre os isolados e as cepas Be-78 parental e Be-62 do Trypanosoma cruzi ........................................ 61
Tabela 4: Determinação dos períodos pré-patente e patente, dia e pico máximo de parasitemia e área sob a curva nos animais infectados com as cepas Be-62, Be-78 ou isolado Be-78is5-3mai do Trypanosoma cruzi. ............................................................................................................................... 63

Nogueira-Paiva, NC Lista de Abreviaturas e Siglas
xiv
APC: Allophycocyanin (Aloficocianina)
APCs: Antigen-Presenting Cells (Células apresentadoras de antígenos)
AUC: Area Under the Curve (Área sob a curva)
Be-62: Cepa Berenice-62 do Trypanosoma cruzi
Be-78: Cepa Berenice-78 Trypanosoma cruzi
Be-78is1-12mai: isolado obtido por hemocultura do animal 1, 12 meses após a infecção
Be-78is15-12mai: isolado obtido por hemocultura do animal 15, 12 meses após a infecção
Be-78is15-6mai: isolado obtido por hemocultura do animal 15, 6 meses após a infecção
Be-78is1-6mai: isolado obtido por hemocultura do animal 1, 6 meses após a infecção
Be-78is21-3mai: isolado obtido por hemocultura do animal 21, 3 meses após a infecção
Be-78is2-6mai: isolado obtido por hemocultura do animal 2, 6 meses após a infecção
Be-78is5-3mai: isolado obtido por hemocultura do animal 5, 3 meses após a infecção
Be-78is5-6mai: isolado obtido por hemocultura do animal 5, 6 meses após a infecção
BFA: Brefeldin A (Brefeldina A)
BSA: Bovine Serum Albumin (Albumina Sérica Bovina)
CD: Cluster of differentiation (antígeno de membrana celular)
CEUA: Comitê de Ética no Uso de Animais
Cel: Controle negativo sem anticorpo
CO2: Dióxido de Carbono
COII: Cytochrome Oxidase II (Subunidade II da Citocromo Oxidase mitocondrial)
dai: Dias após a infecção
DCh: Doença de Chagas
DIA-NADH: Diaphorase-NADH
DMSO: Dimethyl sulfoxide (Dimetilsulfóxido)
DNA: Deoxyribonucleic Acid (Ácido Desoxirribonucléico)
dNTP: Deoxynucleotide Triphosphate (Desoxinuleotídeo Trifosfatado)
DPmáx: Dia do pico máximo de parasitemia
DTT: Dithiothreitol (Ditiotreitol)
DTU: Discret Typing Units
EDTA: Ácido etilendiminotetracetico sal dissódico
EGC: Enteric Glial Cells (Células Gliais Entéricas)
FITC: Fluorescein Isothiocyanate (Isotiocianato de fluoresceína)
FL: Fluorescência
FSC: Forward Scatter (dispersão para frente)
GFAP: Glial Fibrillary Acidic Protein (Proteína Acídica Fibrilar da Glia)
gp63: Glicoproteínas de superfície

Nogueira-Paiva, NC Lista de Abreviaturas e Siglas
xv
Gran A: Granzima A
gRNA: RNA guia
HBSS: Hank’s Buffered Salt Solution (Tampão de Hank)
HE: Hematoxilina-Eosina
HEPES: 2- [4- (2-hidroxietil) piperazin-1-il] etanossulfônico
HVR: Hipervariable Region (regiões hipervariáveis do minicírculo do kDNA)
ICC: Insuficiência Cardíaca Congestiva
IFN- Interferon-gamma
IgG: Imunoglobulina G
IL-10: Interleucina-10
IL-12: Interleucina-12
IL-4: Interleucina-4
iNOS: Inducible Nitric Oxide Synthase (Enzima óxido nítrico sintase induzível)
kDNA: Kinetoplastid DNA (DNA do cinetoplasto)
LIT: Liver Infusion Tryptose
LSSP-PCR: Low-Stringency Single Specific Primer-Polymerase Chain Reaction
mai: Meses após a infecção
MASP: Mucin Associated Surface Proteins (Proteínas de superfície associadas à mucinas)
MEC: Matriz extracelular
MHC: Major Histocompatibility Complex (Complexo de histocompatibilidade principal)
MOI: Multiplicity of infection
NI: Animais não-infectados
NK: Natural Killer Cells (Células Natural Killer)
pb: Pares de Base
PBS: Phosphate-Buffered Saline (Tampão Salina-Fosfato)
PBS-P: Phosphate-Buffered Saline-Permeabilization (Tampão Salina-Fosfato de permeabilização)
PBS-W: Phosphate-Buffered Saline-Wash (Tampão Salina-Fosfato de lavagem)
PCR: Reação em Cadeia da Polimerase
PE: R-phycoerythrin (Ficoeritrina)
PerCP: Peridinin-Chlorophyll-protein (Proteína clorofila peridinina)
PGM: Phosphoglucomutase
PM: Peso molecular
Pmáx: Pico máximo de parasitemia
PP: Período patente
PPP: Período pré-patente

Nogueira-Paiva, NC Lista de Abreviaturas e Siglas
xvi
qPCR: quantitative Polymerase Chain Reaction (Reação em Cadeia da Polimerase em tempo real ou
quantitativa)
RAPD: Random Amplified Polymorphic DNA (Amplificação aleatória do DNA polimórfico)
rDNA: DNA ribossomal
RFLP: Restriction Fragment-Length Polymorfism (Polimorfismo de tamanho de produtos de digestão
do kDNA)
RNA: Ribonucleic Acid (Ácido Ribonucléico)
RPMI: Roswell Park Memorial Institute medium
SFB: Soro Fetal Bovino
SMF: Sistema Mononuclear Fagocitário
SNA: Sistema Nervoso Autônomo
SNE: Sistema Nervoso Entérico
SSC: Side Scatter (dispersão lateral)
Tc: DTU Tc
TGI: Trato Gastrointestinal
Th: Linfócito T helper
TH: Tirosina Hidroxilase
TLR: TOLL Like Receptor (Receptor do tipo TOLL)
TNF-: Fator de Necrose Tumoral-alfa
TS: Trans-sialidases
UPGMA: Unweighted Pair Group Method using Arithmetic averages
Z: Zimodema

Nogueira-Paiva, NC Sumário
1. INTRODUÇÃO ................................................................................................................................. 1
1.1 Aspectos clínicos e epidemiológicos da doença de Chagas .......................................................... 2
1.2 Aspectos imunopatogênicos da doença de Chagas ....................................................................... 5
1.3 Polimorfismo e Plasticidade do Trypanosoma cruzi ................................................................... 10
1.4 As cepas “Berenice” do Trypanosoma cruzi ............................................................................... 18
2. JUSTIFICATIVA ............................................................................................................................ 22
3. OBJETIVOS .................................................................................................................................... 24
3.1 Objetivo Geral ............................................................................................................................. 25
3.2 Objetivos Específicos .................................................................................................................. 25
3.2.1 Caracterização in vitro e seleção de um isolado da cepa Berenice-78 do T. cruzi (ETAPA I) ....................................................................................................................................................... 25
3.2.2 Avaliação in vivo da influência de infecções experimentais por diferentes populações do T. cruzi derivadas da paciente Berenice (ETAPA II) ........................................................................ 25
4. MATERIAL E MÉTODOS ............................................................................................................ 26
4.1 Caracterização in vitro e seleção de isolados da cepa Be-78 do T. cruzi (ETAPA I) ................. 27
4.1.1 Curvas de Crescimento em meio de cultura acelular ........................................................... 28
4.1.2 Perfis de infectividade e desenvolvimento in vitro .............................................................. 28
4.1.3 Perfil de assinatura gênica .................................................................................................... 29
4.2 Avaliação in vivo da influência de infecções experimentais por diferentes populações do Trypanosoma cruzi originadas da paciente Berenice (ETAPA II) .................................................... 31
4.2.1 Infecção dos animais ............................................................................................................ 31
4.2.3 Parasitemia e sobrevida ........................................................................................................ 32
Abordagens 1 e 2 ........................................................................................................................... 34
4.2.4 Necropsia .............................................................................................................................. 34
4.2.5 Carga parasitária no coração e cólon .................................................................................... 34
4.2.6 Intensidade do processo inflamatório no coração e cólon .................................................... 36
Abordagem 3 ................................................................................................................................. 38
4.2.7 Necropsia .............................................................................................................................. 38
4.2.8 Perfil fenotípico do processo inflamatório no coração e cólon ............................................ 38
4.2.9 Estratégia de análise do perfil fenotípico do processo inflamatório no coração e cólon ...... 41
4.3 Análises estatísticas ..................................................................................................................... 45
5. RESULTADOS ................................................................................................................................ 46

Nogueira-Paiva, NC Sumário
ETAPA I............................................................................................................................................ 47
5.1 Hemoculturas .............................................................................................................................. 47
5.2 Curva de crescimento em meio de cultura acelular ..................................................................... 49
5.3 Perfis de infectividade e desenvolvimentos ................................................................................ 52
5.4 Perfil de assinatura gênica ........................................................................................................... 57
5.5 Representação esquemática do perfil e da caracterização gênica dos isolados da cepa Be-78 parental (ETAPA I) ........................................................................................................................... 61
ETAPA II .......................................................................................................................................... 62
5.6 Parasitemia e sobrevida ............................................................................................................... 62
5.7 Carga parasitária no coração e cólon durante a fase aguda da doença de Chagas experimental . 64
5.8 Processo inflamatório no coração e cólon durante a fase aguda da doença de Chagas experimental ...................................................................................................................................... 65
5.8.1 Intensidade do processo inflamatório ................................................................................... 65
5.8.2 Perfil fenotípico do processo inflamatório ........................................................................... 69
5.8.2.1 Macrófagos .................................................................................................................... 69
5.8.2.2 Linfócitos B ................................................................................................................... 70
5.8.2.3 Linfócitos T ................................................................................................................... 71
5.8.3 Perfil fenotípico e funcional dos Linfócitos T ...................................................................... 73
5.9 Representação esquemática do perfil da infecção no coração e cólon (ETAPA II) .................... 75
6. DISCUSSÃO .................................................................................................................................... 79
7. CONCLUSÃO ................................................................................................................................. 95
8. REFERÊNCIAS BIBLIOGRÁFICAS .......................................................................................... 97
9. ANEXOS ........................................................................................................................................ 114
9.1 Anexo 1: Protocolo nº 2011/30- CEUA/UFOP ......................................................................... 115
9.2 Anexo 2: Protocolo nº 2014/29- CEUA/UFOP ......................................................................... 116
9.3 Anexo 3: Resumo dos resultados obtidos na ETAPA II ........................................................... 117
9.4 Anexo 4: Manuscrito publicado no periódico PLOs One/ETAPA I ......................................... 119

1
1. INTRODUÇÃO

Nogueira-Paiva, NC Introdução
2
1.1 Aspectos clínicos e epidemiológicos da doença de Chagas
O Trypanosoma cruzi, agente etiológico da doença de Chagas (DCh), é um
protozoário hemoflagelado pertencente à ordem Kinetoplastida e família Tripanosomatidae
que infecta o homem e uma ampla variedade de mamíferos domésticos e silvestres. Os
hospedeiros invertebrados, vetores da DCh, são insetos hematófagos da subfamília
Triatominae, popularmente conhecidos como “barbeiros”. Durante seu repasto sanguíneo, o
triatomíneo elimina formas tripomastigotas metacíclicas que podem penetrar no hospedeiro
vertebrado através da pele ou mucosa adjacente, infectando principalmente células do sistema
mononuclear fagocitário (SMF). No interior dessas células, as formas tripomastigotas
metacíclicas se diferenciam em formas amastigotas, arredondadas e com flagelo involuído,
que dão início a uma série de replicações. Após vários ciclos de multiplicação as amastigotas
se diferenciam em tripomastigotas, que são liberadas na corrente sanguínea prontas para
infectar outras células hospedeiras e dar início a novas replicações, disseminando a infecção
para os diferentes tecidos e órgãos. O ciclo biológico do T. cruzi continua quando o
triatomíneo ingere as formas tripomastigotas sanguíneas durante hematofagia em mamíferos
infectados. No estômago do inseto, as formas tripomastigotas se transformam nas formas
epimastigotas (não-infectivas) e seguem para o intestino onde se reproduzem
extracelularmente por divisão binária simples. Uma vez que elas alcançam o reto, as
epimastigotas se diferenciam em formas tripomastigotas metacíclicas que podem ser
eliminadas nas fezes do triatomíneo durante o seu repasto sanguíneo reiniciando o ciclo
(Brener, 1987).
A via de transmissão vetorial, descrita acima, representa a principal e mais frequente
forma de transmissão, sendo o Triatoma infestans, o Rhodnius prolixus e o Triatoma
dimidiata os mais importantes vetores no ciclo doméstico da DCh (Zeledon e Rabinovich,
1981; Who, 2002). O T. infestans, embora atualmente considerado eliminado no Brasil (Dias,
1997), é historicamente associado às regiões endêmicas sub-amazônicas, enquanto as espécies
R. prolixus e T. dimidiata geralmente são encontradas ao norte da América do Sul e Central,
sendo que o último alcança o norte do México (Zeledon e Rabinovich, 1981; Who, 2002).
Contudo, a DCh ainda pode ser transmitida por mecanismos não-vetoriais através de
transfusão sanguínea, transplante de órgãos, transmissão vertical, acidentes de laboratório e
por via oral (Prata, 2001; Benchimol Barbosa, 2006; Bern et al., 2007).

Nogueira-Paiva, NC Introdução
3
Duas fases são clinicamente reconhecidas na DCh humana durante sua evolução
natural. Após a infecção pelo T. cruzi inicia-se a fase aguda, caracterizada por elevada
parasitemia, parasitismo tecidual e processo inflamatório intensos. Nesta fase iniciam-se as
lesões do sistema nervoso autônomo (SNA) que geralmente estão relacionadas a um infiltrado
inflamatório adjacente, associado ou não a ninhos do parasito (Lisboa, 1960; Andrade e
Andrade, 1966; Tafuri, 1971; Tafuri et al., 1971). A fase crônica inicia-se quando o sistema
imune do hospedeiro controla a multiplicação do parasito, ocorrendo simultaneamente à
redução do parasitismo tecidual e sanguíneo. Consequentemente, o processo inflamatório
tende a reduzir, porém a manutenção de uma resposta inflamatória ativa e progressiva está
associada às alterações graves da DCh e ao desenvolvimento da fase crônica determinada ou
aparente. Na forma determinada ou sintomática, manifestações clínicas mais comuns
envolvem o coração e o trato digestório, caracterizando as formas cardíaca ou digestiva
(megacólon ou megaesôfago), respectivamente, que podem ocorrer isoladas ou em associação
(Tafuri, 1971; Tafuri et al., 1971).
Indivíduos na fase crônica da infecção também podem apresentar a forma
indeterminada ou inaparente, considerada a mais frequente de todas as formas
anatomoclínicas da DCh (Andrade, 2005). O critério para o diagnóstico da forma
indeterminada, adotado pela Sociedade Brasileira de Medicina Tropical, prevê “indivíduos
assintomáticos, com evidências sorológicas e/ou parasitológicas da infecção pelo T. cruzi,
com eletrocardiograma e estudo radiológico contrastado do esôfago e cólon dentro dos limites
normais” (Andrade, 2005). Os indivíduos infectados com o T. cruzi podem permanecer
assintomáticos pelo resto de suas vidas, no entanto, aproximadamente 30% deles, após vários
anos de infecção, podem apresentar alterações morfofuncionais no coração e/ou trato
digestório e assim evoluírem para a forma determinada da DCh (Andrade et al., 1997).
Portanto, a forma inaparente ou indeterminada parece ter um complexo significado
anatomoclínico e epidemiológico e tem sido considerada por muitos autores apenas como
uma fase ou forma latente da DCh. A despeito da normalidade nos exames radiológicos e
eletrocardiográficos do tórax, lesões histológicas podem estar presentes nestes indivíduos e
serem indicativas da evolução para a forma aparente mesmo que em níveis leves (Macedo,
1980; Coura et al., 1985; Castro et al., 1994; Andrade et al., 1997; Macedo, 1999; Prata,
2001).
Um estudo longitudinal realizado na cidade de Mambaí/Goiás entre os anos de
1975/76 a 1986/91 revelou que 120 dos 190 pacientes com DCh apresentavam a forma

Nogueira-Paiva, NC Introdução
4
indeterminada no ano inicial do trabalho. Entretanto, 39 desses pacientes portadores da forma
indeterminada tiveram evolução progressiva em 1986/91 (Castro et al., 2001). Destes 39
indivíduos, 33 evoluíram para a forma digestiva, sendo que 23 pacientes desenvolveram
megaesôfago e 10 pacientes manifestaram sintomas do megacólon (Castro et al., 2001).
O substrato anatômico da forma inaparente é representado por lesões inflamatórias
microscópicas focais, associadas ou não à presença de parasitos que são raramente
demonstrados pelos métodos histológicos comuns (Andrade, 2005), mas têm sido revelados
por técnicas mais sensíveis, como a imuno-histoquímica e a PCR (Polymerase Chain
Reaction-Reação em Cadeia da Polimerase) (Higuchi et al., 1993; Jones et al., 1993; Vago et
al., 2000; Vago et al., 2003). A cardiomiopatia chagásica é caracterizada por alterações
eletrocardiográficas como bloqueio atrioventricular, extra-sístoles ventriculares, síndrome do
nódulo sinusal e taquicardia supraventricular (Gascon et al., 2007). As manifestações clínicas
incluem arritmias, tromboembolismo periférico, aneurisma de ponta e insuficiência cardíaca
congestiva, podendo ocorrer morte súbita (Dias et al., 1956). Em relação à forma digestiva,
embora lesões associadas ao intestino delgado (megaduodeno, megajejuno e/ou megaíleo)
possam ocorrer, o megaesôfago e o megacólon são as manifestações gastrointestinais mais
comuns da DCh, ocorrendo em áreas endêmicas numa taxa de 15-20% dos pacientes
chagásicos, associada ou não à forma cardíaca (Prata, 2001). No megaesôfago pode-se
observar relaxamento anormal do esfíncter esofageal e diminuição da peristalse
caracterizando uma alteração motora conhecida como acalasia com disfagia, regurgitação,
pirose e dor torácica, enquanto o megacólon está associado à constipação crônica, com
obstrução parcial a total de segmentos colônicos (em especial, no segmento retossigmóide) e
complicações como desnutrição e vólvulo (torção) sigmóide (De Rezende, 1984; De Oliveira
et al., 1998; Prata, 2001; Sanchez-Guillen Mdel et al., 2006).
No contexto epidemiológico, o número estimado de pessoas infectadas pelo T. cruzi,
segundo a Organização Mundial de Saúde, é de aproximadamente 10 milhões, distribuídas em
21 países da América Latina, onde a doença ainda é considerada endêmica com o predomínio
da transmissão vetorial, como Argentina, Chile, Bolívia, Venezuela e Colômbia, e em regiões
não-endêmicas como Canadá, Estados Unidos, Austrália, Japão, França, Itália, Espanha,
Portugal e Bélgica (Who, 2010a; b). Nestes últimos a incidência da DCh está relacionada a
mecanismos de transmissão não-vetoriais facilitados por fenômenos de mobilidade
populacional (Schmunis, 2007; Who, 2010a). No Brasil, um inquérito sorológico nacional
para DCh, realizado entre os anos 1975 e 1980, estimou uma soroprevalência de 4,22%,

Nogueira-Paiva, NC Introdução
5
todavia, com o controle da transmissão vetorial, a incidência da DCh no país caiu para 1,3%
no ano de 1995 (Akhavan, 2000; Bocanegra, 2008). As maiores prevalências foram
encontradas no Rio Grande do Sul (8,84%), Minas Gerais (8,83%), Goiás (7,40%), Sergipe
(5,97%) e Bahia (5,44%) (Camargo et al., 1984; Bocanegra, 2008).
A gravidade e prevalência das diferentes formas clínicas da DCh variam entre
diferentes regiões geográficas (Andrade, 1985), no entanto, a causa dessa heterogeneidade
ainda não é clara. Uma grande diversidade de fatores atua na patogênese dessa doença,
determinando maior ou menor intensidade do infiltrado inflamatório e da destruição tecidual.
Entre estes, alguns estão relacionados ao hospedeiro, como constituição genética, estado
imunológico, nutricional e idade, enquanto outros, são inerentes ao T. cruzi, ligados ao seu
polimorfismo biológico e molecular, associado à diferenças relacionadas ao histotropismo,
constituintes antigênicos, sensibilidade à drogas, entre outros.
1.2 Aspectos imunopatogênicos da doença de Chagas
Pacientes com acometimento cardíaco apresentam miocardite crônica difusa
progressiva, com consequente destruição das miofibras e substituição por tecido conjutivo,
culminando no desenvolvimento da insuficiência cardíaca congestiva (ICC), consistindo em
um quadro clínico grave da DCh (Andrade, 1985; Barros et al., 2001; Barros et al., 2003).
Achados histopatológicos tanto na doença humana quanto em modelos experimentais
demonstram a presença de processo inflamatório, com caráter fibrosante e degenerativo,
constituído predominantemente por células mononucleares, acompanhadas por uma fibrose
intersticial e áreas de degeneração e miocitólise (Andrade, 1985; De Lana et al., 1988; Tafuri
et al., 1988; Rossi, 1995; Caliari et al., 2002b; Barros et al., 2003; Dutra e Gollob, 2008;
Dutra et al., 2009). Os gânglios nervosos epicárdicos aparecem com número reduzido de
neurônios, com acometimento do sistema excito-condutor relacionado às alterações
eletrocardiográficas (Andrade et al., 1978; Andrade, 1985; Rossi e Bestetti, 1995).
Alterações em outros componentes da matriz extracelular (MEC), além do colágeno,
são observadas em experimentos in vitro e in vivo. A fibronectina, uma glicoproteína da
MEC, tem sido associada à interação parasito-célula hospedeira durante o processo de invasão
e a mecanismos de adesão e migração de células inflamatórias (Wirth e Kierszenbaum, 1984).
A laminina, principal constituinte da membrana basal, por sua vez, precisa ser degradada para
invasão do parasito e, dessa forma, sua participação nos fenômenos iniciais do processo

Nogueira-Paiva, NC Introdução
6
infeccioso tem sido avaliado (Moody et al., 2000). De forma interessante, foi observado um
aumento na expressão de laminina no coração de camundongos C3H/He infectados com a
cepa Colombiana do T. cruzi. Ao contrário do observado para fibronectina, em áreas
associadas à infiltrado inflamatório, não foi possível observar imunomarcação de laminina
(Marino et al., 2003). Assim pode-se sugerir que o remodelamento da matriz extracelular,
tanto na fase aguda quanto na fase crônica da DCh, em resposta ao parasito e seus antígenos,
está envolvido na disseminação do parasito, no estabelecimento da inflamação e dos
processos citotóxicos associados (Higuchi et al., 1999; Marino et al., 2003).
A forma clínica digestiva da DCh é caracterizada por alterações anatômicas como
dilatação em graus variáveis, aumento do lúmen, ausência de obstrução mecânica e
espessamento da parede, especialmente relacionada à hipertrofia muscular.
Microscopicamente é observado infiltrado inflamatório constituído predominantemente por
células mononucleares na muscular da mucosa, submucosa e na muscular própria; ganglionite
e periganglionite, especialmente circundando o plexo de Auerbach; ulceração da mucosa e
fibrose intermuscular intersticial (Tafuri, 1971; Tafuri et al., 1971; Adad et al., 1991; Caliari
et al., 1996). Lesões inflamatórias regressivas do sistema nervoso entérico (SNE) afetam
neurônios, células gliais e fibras nervosas e são consideradas elementos essenciais na
patogênese da desordem gastrointestinal (Köberle, 1957; Koberle e De Alcantara, 1960;
Tafuri e Raso, 1962; Andrade e Andrade, 1966; Tafuri e Brener, 1967a; Tafuri, 1970; 1971;
Tafuri et al., 1971; Köberle, 1974; Adad et al., 1991; Adad et al., 2001; Lages-Silva et al.,
2001; Da Silveira et al., 2005; Da Silveira et al., 2007a; Da Silveira et al., 2007b; Da Silveira
et al., 2008; Da Silveira et al., 2009; Nascimento et al., 2010; Nogueira-Paiva et al., 2014).
Iantorno et al. (2007), utilizando enolase como marcador pan-neuronal e S-100 como
marcador pan-glial, descreveram perda significativa de neurônios e células gliais em pacientes
com megacólon chagásico. Desnervação, avaliada pela marcação de áreas PGP9.5, também
foi observada em pacientes com megacólon chagásico em estudo realizado por Da Silveira et
al. (2007a). Além de lesões neuronais, alterações no número e fenótipo das células gliais
também são encontradas em pacientes com a forma digestiva (Da Silveira et al., 2007b;
Nascimento et al., 2010).
Os mecanismos imunopatogênicos relacionados ao desenvolvimento do amplo
espectro de formas clínicas da DCh ainda não são totalmente compreendidos. A escassez de
parasitos detectados pela microscopia óptica convencional nas lesões teciduais ou no sangue
durante a fase crônica suscitou dúvidas sobre seu papel patogênico direto e, juntamente com a

Nogueira-Paiva, NC Introdução
7
presença de auto-anticorpos, levantou a hipótese da autoimunidade na patogênese da DCh.
Contudo, técnicas mais sensíveis, como a imuno-histoquímica e a Reação em Cadeia da
Polimerase (PCR), demonstram uma forte correlação entre a presença do parasito e a
ocorrência de lesões teciduais reforçando assim, a participação do parasito na patogênese das
formas crônicas da doença (Higuchi et al., 1993; Jones et al., 1993; Vago et al., 1996a;
Andrade et al., 1999; Anez et al., 1999; Vago et al., 2000; Lages-Silva et al., 2001; Vago et
al., 2003). No entanto, as teorias de autoimunidade e de manutenção de um parasitismo
tecidual deixaram de ser mutuamente excludentes para se tornarem complementares (Soares
et al., 2001). A persistência do parasito a longo prazo parece ter um papel-chave para a
manutenção de um processo inflamatório ativo capaz de causar lesões histológicas (Tarleton e
Zhang, 1999; Lages-Silva et al., 2001; Andersson et al., 2003; Da Silveira et al., 2005). No
entanto, a habilidade de mimetismo antigênico do T. cruzi tem sido demonstrada em infecções
naturais ou experimentais, relacionando-se ao desenvolvimento de resposta imune contra
células próprias (Cunha-Neto e Kalil, 1995). Independente da origem dos antígenos que
modulam a resposta imunológica durante a infecção crônica, é sabido que o sistema imune do
hospedeiro, particularmente a resposta imune celular, tem um papel central no
desenvolvimento das manifestações clínicas (Andersson et al., 2003; Dutra et al., 2009).
Durante a infecção e o desenvolvimento da fase aguda da DCh, fatores genéticos,
moleculares e/ou biológicos relacionados ao parasito, em associação com o perfil imune dos
hospedeiros podem ter um papel preditivo importante sobre o prognóstico da doença. É
sabido que nos estágios iniciais da infecção, o desenvolvimento da resposta imune Th1 e a
expressão de citocinas pro-inflamatórias são importantes para o controle da replicação do
parasito. No entanto, a modulação dessa resposta é fundamental para prevenir a lesão tecidual
com consequentes manifestações clínicas (Dutra et al., 2009).
Os primeiros eventos da resposta imune inata iniciam-se quando células do sistema
mononuclear fagocitário infectadas passam a sintetizar as citocinas Interleucina-12 (IL-12) e
TNF(Fator de Necrose Tumoral-alpha) que agem ativando as células Natural Killer (NK) e
linfócitos T e, consequentemente, induzindo a expressão de IF (Interferon-gamma) e
intermediando o processo de recrutamento de células inflamatórias para o local da infecção.
Essa citocina, por sua vez, ativa os macrófagos, mediando a morte dos parasitos
intracelulares, através da expressão da enzima iNOS (óxido nítrico sintase induzível) e
produção de NO, controlando a replicação do parasito (Torrico et al., 1991; Vespa et al.,
1994; Graefe et al., 2003). A indução de uma resposta celular efetiva requer estimulação

Nogueira-Paiva, NC Introdução
8
apropriada via células apresentadoras de antígenos. Adicionalmente, a ativação das células T
pela IL-12 e a manutenção dos altos níveis de IFN- por essas células consiste na interface
entre a resposta imune inata e adaptativa. Células T CD4+ e CD8+ têm sido descritas como
essenciais ao desenvolvimento da resposta imune contra o parasito e distintos fenótipos dessas
células podem estar associadas aos distintos quadros clínicos da DCh (De Alencar et al.,
2009; Dutra et al., 2009).
Pacientes com a forma indeterminada da doença representam uma situação de
equilíbrio entre a infecção e a resposta inflamatória. Estudos têm demonstrado uma alta
frequência de células T CD4+, que não expressam a molécula co-estimuladora CD28, em
pacientes com a forma indeterminada da doença. Adicionalmente, estes pacientes
apresentaram altos níveis de expressão de IL-10, sugerindo que o fenótipo CD4+ CD28- possa
estar associado à capacidade de modulação da resposta inflamatória (Menezes et al., 2004;
Dutra et al., 2009).
O infiltrado inflamatório, presente principalmente na camada mucosa e adjacente aos
gânglios mientéricos, associado às lesões desenvolvidas em pacientes com megaesôfago ou
megacólon, é constituído predominantemente por macrófagos, eosinófilos, células NK,
mastócitos e células T CD4+ (D'avila Reis et al., 2001; Da Silveira et al., 2007b). Ao contrário
do observado na forma digestiva, o intenso processo inflamatório observado no miocárdio de
pacientes com a forma cardíaca da doença, é composto especialmente por células T CD8+
associados à expressão de citocinas pro-inflamatórias, como TNFe IFN- e, moléculas
citotóxicas, como perforina e granzima A (Higuchi, 1999). Camundongos nocautes para
linfócitos T CD8+ expressando perforina apresentaram menores níveis de lesões teciduais e
alterações cardíacas (Silverio et al., 2010). No entanto, pacientes com a doença cardíaca
também apresentam células T CD4+, sendo demonstrado que a deficiência dessas células leva
a uma redução global da resposta imunológica do hospedeiro e consequente aumento de
parasitismo tecidual, provavelmente porque linfócitos T CD4+ promovem a ativação de
macrófagos e a proliferação de linfócitos T CD8+ e B (Rottenberg et al., 1995).
De forma geral, a influência da resposta imune sobre as manifestações clínicas tem
sido relacionada a um desequilíbrio entre as repostas Th1 e Th2 (Figura 1), levando a
persistência do parasito, a exacerbação da resposta inflamatória e ao consequente dano
tecidual. Nesse contexto, o balanço entre as citocinas pro-inflamatórias e anti-inflamatórias
parece ser imprescindível para a modulação da resposta imune. O tratamento de camundongos
com anti-IFN-impediu o controle da replicação do parasito, enquanto o bloqueio de IL-10

Nogueira-Paiva, NC Introdução
9
ou IL-4 resultou em controle mais efetivo da infecção (Mccabe et al., 1991; Torrico et al.,
1991).
Figura 1: Esquema representando as citocinas e as populações celulares envolvidas na geração da resposta protetora ou patogênica na doença de Chagas. Durante a fase aguda, células da resposta imune inata e adaptativa trabalham para controlar a replicação do parasito. As células NK são as principais responsáveis pela ativação de monócitos/macrófagos através da síntese de IFN- nos estágios iniciais da fase aguda. Macrófagos e células dendríticas apresentam antígenos às células T, além de sintetizarem citocinas e proverem os sinais co-estimuladores para ativação dessas células. Células T CD4+ auxiliam na ativação de células T CD8+, que através de mecanismos citotóxicos, passam a ser as principais responsáveis pelo controle do parasitismo tecidual. A parasitemia patente é controlada pela ação do sistema imune e os indivíduos entram na fase crônica da doença. A interação inicial parasito-hospedeiro é a chave para o estabelecimento do controle efetivo da parasitemia patente e ao mesmo tempo é crucial na formação do microambiente de citocinas que podem orquestrar a subsequente diferenciação das subpopulações de linfócitos T efetores e reguladores. Diferentes perfis de resposta imune podem ser montados: resposta imune eficaz e controlada, associada à forma indeterminada, onde as lesões histológicas não são suficientes para causar alterações anatomoclínicas, ou ainda uma resposta contínua, que controla a replicação do parasito, no entanto, não é modulada e acaba sendo patogênica, causando as lesões crônicas das formas sintomáticas cardíaca e digestiva. A forma cardíaca está associada à uma sobreposição de linfócitos T CD8+, e citocinas como IFN- e TNF-. A forma digestiva está principalmente associada à maior frequência de linfócitos TCD4+, células NK, mastócitos e a citocinas como TNF-, IL-1 e IL-6. Pacientes com a forma indeterminada apresentam densidades inferiores de linfócitos T tanto CD4+ quanto CD8+, populações de células Treg e uma expressão elevada de IL-10. Fonte: Adaptado de (Dutra et al., 2009)
Resposta imune modulada/ausência de lesão
Resposta imune não modulada/presença de lesão
Forma Cardíaca
Forma Digestiva
Forma Indeterminada
Controle da parasitemia patente
Fatores determinantes: resposta imune, cepa do parasito, genética do hospedeiro
Lesão tecidual
Lesão tecidual

Nogueira-Paiva, NC Introdução
10
1.3 Polimorfismo e Plasticidade do Trypanosoma cruzi
O desenvolvimento de pesquisas com o intuito de reproduzir a DCh em modelos
experimentais, a partir do inóculo de subpopulações do parasito isoladas de hospedeiros
vertebrados ou invertebrados, mostraram a existência de uma variação intraespecífica do T.
cruzi com potenciais implicações fisiopatogênicas. Essas subpopulações receberam a
designação de cepas, referindo-se à amostra derivada de um "isolado", mantida em cativeiro
em reprodução contínua por passagens seriadas, em cultura ou animais de laboratório
(Lumsden, 1965).
A heterogeneidade intraespecífica do T. cruzi, observada pela primeira vez por Carlos
Chagas (1909), foi em seguida estudada e confirmada por Brener e Chiari (1963) que
demonstraram a ocorrência de três grupos morfologicamente distintos do parasito, de acordo
com a predominância de formas delgadas, largas e muito largas. Em 1969, Brener demonstrou
que as formas delgadas penetravam mais rapidamente nas células do hospedeiro e estas eram
mais susceptíveis aos mecanismos imunes, enquanto, as formas largas persistiam por um
tempo maior no sangue periférico, sendo, portanto, mais resistentes aos mecanismos
imunológicos do hospedeiro. Dessa maneira, as formas delgadas parecem mais adaptadas a
penetrarem nas células que as formas largas e muito largas, que tendem a acumular-se no
sangue (Brener, 1969). Corroborando com essa hipótese, as taxas de mortalidade foram
maiores nos animais infectados com as subpopulações constituídas predominantemente por
formas delgadas quando comparadas àquelas constituídas por formas largas ou muito largas,
onde muitos dos animais sobreviveram até a fase crônica (Brener, 1965), indicando que o
curso da infecção poderia depender da predominância relativa de uma forma sobre a outra.
Além dos aspectos morfológicos e sua influência sobre a resposta imune do
hospedeiro, a capacidade do T. cruzi de infectar vários tipos celulares garante a ele uma
adaptação bem sucedida e determina a manifestação de outro aspecto polimórfico, descrito
como cito/histotropismo (Andrade, 1985; Andrade et al., 1999; Vago et al., 2000; Andersson
et al., 2003; Macedo et al., 2004). A maioria das cepas apresenta preferência por células
musculares (Tarleton e Zhang, 1999) e dessa forma, os principais órgãos afetados são o
coração, o esôfago e o cólon, nos quais ocorrem as lesões mais graves que caracterizam as
formas cardíaca e/ou digestiva da doença. No entanto, estudos comprovam a presença de
parasitos no fígado, baço, músculo esquelético, pâncreas, intestino delgado, cérebro, bexiga,
tecido adiposo entre outros (Tafuri e Raso, 1962; Tafuri e Brener, 1967b; Melo e Brener,
1978; Saldanha et al., 2001; Meneghelli, 2004; Vieira et al., 2009; Ferreira et al., 2011) entre

Nogueira-Paiva, NC Introdução
11
outros órgãos. De acordo com a preferência tecidual é atribuído às diferentes cepas a
designação de reticulotrópicas ou macrofagotrópicas, para aqueles que apresentam
histotropismo para células do sistema mononuclear fagocitário (SMF) e miotrópicas, para
subpopulações com afinidade para células musculares (Melo e Brener, 1978).
Ao longo das últimas décadas do século XX, vários parâmetros morfológicos,
biológicos e/ou moleculares de diferentes “amostras” do T. cruzi foram investigados com o
objetivo de estabelecerem-se critérios de classificação que pudessem, além de indicar alguma
relação com aspectos clínicos e epidemiológicos da doença, criar bases satisfatórias para a
identificação das cepas (Oliveira, 1986). A observação de diferenças morfológicas e
comportamentais entre essas amostras, relacionadas à virulência e patogenicidade, levou à
proposta de distribuição das cepas em BIODEMAS, onde pertenciam ao Tipo I aquelas que
apresentavam predomínio de formas delgadas, rápida taxa de multiplicação, pico de
parasitemia e mortalidade máxima entre 7º-12º dia após a infecção (dai) e macrofagotropismo
na fase aguda (ex.: cepas Y e Peruana); ao Tipo II, cepas com predomínio de formas largas,
taxas de multiplicação menores, picos irregulares de parasitemia e taxa de mortalidade
máxima entre o 12º-20º dai e cardiomiotropismo (ex.: amostra 12SF); e, por último, ao Tipo
III, parasitos preferencialmente largos, com lenta taxa de multiplicação, picos parasitêmicos
altos e tardios, ocorrendo entre o 20º-30º dai, elevada patogenicidade e especial tropismo para
o músculo esquelético (ex.: Colombiana) (Andrade, 1974; Andrade e Magalhaes, 1996).
Além dos padrões biológicos, análises dos perfis de isoenzimas também foram
utilizados para distinguir subpopulações do T. cruzi. A avaliação das combinações entre
diferentes perfis isoenzimáticos permitiu a classificação das cepas em ZIMODEMAS, capaz
de identificar inicialmente no Brasil, três padrões isoenzimáticos, Z1, Z2 e Z3,
correlacionados aos ciclos silvestre (Z1, Z3) ou doméstico (Z2) com circulação independente
em determinada área endêmica e transmissão por diferentes espécies de vetores, sendo todos
capazes de causar a doença aguda em humanos, embora o zimodema 2 esteja mais
relacionado às manifestações clínicas da fase crônica (Miles et al., 1977; Miles et al., 1978;
Miles et al., 1980; Tibayrenc e Miles, 1983; Romanha e Brener, 1988). Carneiro et al. (1990)
submeteram isolados obtidos por hemocultura de pacientes em fase crônica à avaliação de
oito loci enzimáticos antes e após manipulação em condições de laboratório e identificaram
quatro grupos isoenzimáticos, ZA, ZB e ZC, além da possibilidade de alteração no perfil de
isoenzimas após sucessivas passagens em camundongos.

Nogueira-Paiva, NC Introdução
12
Embora diferenças relacionadas à genética, dieta e outros fatores, não sejam excluídas,
a associação entre a distribuição epidemiológica dos diferentes perfis isoenzimáticos e a
prevalência das manifestações clínicas foi observada durante a caracterização de isolados
originados de distintos hospedeiros na Venezuela, e nas regiões amazônica e central do Brasil.
Os zimodemas Z1 e Z3 foram identificados nas duas primeiras, onde prevalecem as formas
cardíaca e indeterminada, enquanto, na região central do Brasil, onde é comum o
desenvolvimento da forma digestiva, 98 dos 99 isolados de pacientes apresentaram perfil
compatível ao Z2 (Miles et al., 1981). Estudos convergindo as classificações em Biodemas e
Zimodemas confirmaram a distribuição epidemiológica dos zimodemas entre diferentes
regiões do Brasil, e demonstraram a equivalência entre o biodema Tipo I e o zimodema Z2b
(variante de Z2), biodema Tipo II e zimodema Z2 e, biodema Tipo III e zimodema Z1, sendo
o Tipo II/Z2, comumente encontrado parasitando cardiomiócitos e/ou células do sistema
nervoso entérico e predominante na região central do Brasil e o Tipo III/Z1, prevalente ao
norte do Brasil, e apresentando miotropismo (Andrade e Magalhaes, 1996). Sistemas
enzimáticos adicionais permitiram ainda a identificação de uma diversidade genética maior e
sendo proposta a existência de 43 genótipos, com quatro grupos considerados os genótipos
principais, genótipo 19, genótipo 20, genótipo 32, genótipo 39 (Tibayrenc et al., 1986;
Tibayrenc e Ayala, 1988). Análises posteriores sugeriram a existência de duas linhagens
maiores filogeneticamente heterogêneas, Linhagens 1 e 2, diferentes entre si em várias
propriedades biológicas (Tibayrenc, 1995).
Com o crescente interesse em correlacionar o polimorfismo do T. cruzi às variações
clinico-epidemiológicos da DCh, critérios de classificação baseados na caracterização
genética do DNA do parasito começaram a ser aplicadas aos estudos de campo e
experimentais. É sabido que o T. cruzi apresenta evolução predominantemente clonal, no
entanto, a possibilidade da ocorrência rara de recombinação sexual sugere a propagação de
genótipos clonais e o surgimento de linhagens do parasito que podem ser identificadas por
seus genótipos e/ou fenótipos distintos (Tibayrenc et al., 1986; De Freitas et al., 2006;
Zingales et al., 2009). O genoma do parasito é diploide e contém aproximadamente 23.000
genes, com cerca de 50% do genoma consistindo de sequências repetitivas, incluindo genes
codificadores de proteínas de superfície, como as trans-sialidases (TS), mucinas,
glicoproteínas de superfície (gp63) e proteínas de superfície associadas à mucinas (MASP)
(El-Sayed et al., 2005; Bartholomeu et al., 2009). Como outros flagelados da ordem
Kinetoplastida, o T. cruzi possui uma única mitocôndria que contém uma rede complexa de

Nogueira-Paiva, NC Introdução
13
DNA mitocondrial circular, conhecida como DNA do cinetoplasto (Kinetoplastid DNA-
kDNA), constituindo aproximadamente 15% do DNA total da célula (Baptista et al., 2006). O
kDNA é organizado em anéis concatenados entre si denominados maxicírculos e
minicírculos. Os maxicírculos, equivalentes ao DNA mitocondrial clássico, estão presentes
em algumas dezenas de cópias e contêm 20 genes codificadores de proteínas envolvidas no
processo de fosforilação oxidativa, importantes para transformação em formas epimastigota
no intestino do vetor, ou proteínas da via glicolítica, usadas pelo parasito no hospedeiro
mamífero (Westenberger et al., 2006). Cerca de 20.000-30.000 minicírculos são encontrados
em estrita relação com os maxicírculos e são formados por quatro regiões equidistantes, com
sequências nucleotídicas de aproximadamente 100pb (pares de base) quase idênticas,
constituindo os domínios ou regiões conservadas separadas entre si por quatro regiões
hipervariáveis (Hipervariable Region-HVR) de 280-320pb (Liu et al., 2005). A maioria dos
transcritos primários originados dos genes dos maxicírculos não podem ser diretamente
traduzidos por conterem “erros” e são submetidos a um processo de edição, conduzida por
RNAs guia (gRNA), codificados por genes presentes nos minicírculos, capaz de modificar sua
sequência nucleotídica (“RNA edition”) alterando assim a estrutura primária do RNA e
produzindo uma sequência funcional (Avila e Simpson, 1995; Westenberger et al., 2006).
Dessa forma, maxicírculos e minicírculos têm funções complementares que contribuem para
mecanismos responsáveis por mudanças no estágio do ciclo de vida e no metabolismo
energético em diferentes hospedeiros (Avila e Simpson, 1995).
Os domínios hipervariáveis do minicírculo apresentam uma grande diversidade entre
as linhagens do T. cruzi, e foram os primeiros alvos da caracterização do kDNA na busca de
marcadores genéticos do polimorfismo. A classificação em “ESQUIZODEMAS”, proposta
por Morel et al. (1980) é baseada no polimorfismo de tamanho de produtos de digestão do
kDNA (Restriction Fragment-Length Polymorfism-RFLP) e subpopulações agrupadas dentro
do mesmo esquizodema apresentam padrões similares de produtos de endonuclease de
restrição, sendo genotipicamente relacionadas. kDNA total obtidos de isolados classificados
em diferentes zimodemas foram submetidos à RFLP da região hipervariável do minicírculo e
os resultados demonstraram que as seis amostras pertencentes ao zimodema C apresentaram
um único perfil de esquizodema, enquanto três subpopulações pertencentes ao zimodema A
demonstraram três diferentes esquizodemas (Morel et al., 1980), o que indica uma
complementaridade entre zimodemas e esquizodemas, no entanto, evidencia maior capacidade
em detectar variabilidade para a técnica de RFLP em detrimento à caracterização de

Nogueira-Paiva, NC Introdução
14
isoenzimas. Avila et al. (1990) submeteram 56 amostras de T. cruzi isoladas de pacientes,
animais e insetos de diferentes regiões à RFLP do domínio variável do minicírculo a partir do
kDNA total ou de regiões de 330pb do minicírculo do kDNA previamente amplificadas
utilizando iniciadores específicos para a região conservada do minicírculo. Os padrões de
digestão obtidos da região variável do minicírculo amplificada apresentaram mais
informações que àquelas obtidas a partir do kDNA total, permitindo a classificação das cepas
em esquizodemas adicionais (Avila et al., 1990).
A amplificação aleatória do DNA polimórfico ou RAPD (Random Amplified
Polymorphic DNA) é baseada na utilização de iniciadores não-específicos que amplificam
aleatoriamente regiões do DNA do parasito, gerando perfis de RAPD que consistem de uma
complexa série de produtos de DNA de intensidade e tamanhos variados e potencialmente
capazes de fornecer informações geneticamente úteis devido à sua reprodutibilidade (Steindel
et al., 1993). A comparação entre “RAPDEMAS” e “ZIMODEMAS” de 32 isolados do T.
cruzi revelou que os perfis de RAPD de cepas pertencentes ao mesmo zimodema tenderam a
ser muito similares, enquanto expressivas mudanças no padrão de bandas produzidas foram
observadas quando um zimodema foi comparado a outro (Steindel et al., 1993), sugerindo
uma associação entre esses dois critérios de classificação.
Análises dos perfis isoenzimáticos e por RAPD sugeriram a redistribuição das cepas
da linhagem 2, em cinco subgrupos filogenéticos cada um compreendendo as cepas de
referência: T. cruzi IIa (CanIII cl1), T. cruzi IIb (Esmeraldo cl3), T. cruzi IIc (M5631 cl5), T.
cruzi IId (MN cl2) e T. cruzi IIe (CL Brener). Os dois primeiros grupos foram relacionados
principalmente ao ciclo silvestre, enquanto os demais grupos foram restritos ao ambiente
doméstico. A linhagem 1 (Z1), por sua vez, não apresentou perfis adicionais (Brisse et al.,
2000a). Baseado nesses dados, Brisse et al. (2000a; 2000b), propuseram a subdivisão do
táxon T. cruzi em seis Discrete Typing Units (DTUs: I, IIa, IIb, IIc, IId e IIe), com variações
geográficas e ecológicas.
Além do kDNA, sequências do DNA do parasito também foram submetidas à
genotipagem e o alinhamento comparativo de sequências do gene do mini-exon (ME) de
diferentes cepas do T. cruzi mostraram a presença de sequências altamente conservadas,
(exons e introns) e de regiões intergênicas divergentes, cuja análise por PCR permitiu a
identificação de dimorfismo entre as cepas de acordo com os produtos de amplificação. Cepas
pertencentes ao grupo 1, com produtos de 330pb foram diferentes das cepas designadas grupo
2, com produtos apresentando 350pb (Souto et al., 1996). A avaliação inicial do DNA

Nogueira-Paiva, NC Introdução
15
ribossomal (rDNA) entre diferentes tripanosomatídeos permitiu a amplificação de sequências
de 100pb ou 125pb para duas cepas do T. cruzi, Tulahuen e Y, respectivamente, sugerindo a
aplicação dessa técnica para avaliação do polimorfismo intraespecífico observado para esse
parasito. A caracterização da região 24S do gene do rRNA de isolados/cepas do T. cruzi
obtidos de diferentes hospedeiros permitiu a divisão e dois grupos, 1 e 2, que incluíram cepas
que apresentaram amplificação de fragmentos de 125 ou 100pb, respectivamente (Souto e
Zingales, 1993). A avaliação simultânea de diferentes marcadores moleculares, região 24S do
rDNA, região intergênica do mini-exon e RAPD, confirmaram posteriormente a existência
dos dois grupos discretos e um terceiro grupo apresentando ambos os produtos de PCR e
denominado grupo 1/2 (Souto et al., 1996).
Microssatélites são unidades repetitivas pequenas de 2-6 nucleotídeos presentes em
número variável, extremamente polimórficas e distribuídas por todo o genoma nuclear
(Polymorphic tandem repeats of 2-6 bases long simple motifs) amplamente utilizadas para
investigações filogenéticas e, ainda, determinação da natureza monoclonal ou policlonal de
diferentes cepas do T. cruzi (Oliveira et al., 1998; Oliveira et al., 1999; Macedo et al., 2001).
Dados obtidos por Oliveira et al. (1998; 1999), analisando cepas e clones do T. cruzi
utilizando oito marcadores de microssatélites, confirmaram o genoma diploide, a organização
clonal e o alto polimorfismo do parasito.
A classificação atual (Quadro 1) demonstra a classificação geral em Discrete Typing
Units (DTU) que identifica as cepas do T. cruzi em grupos geneticamente mais relacionados
baseada em características moleculares e biológicas: TcI, TcII, TcIII, TcIV, TcV e TcVI
(Zingales et al., 2009).
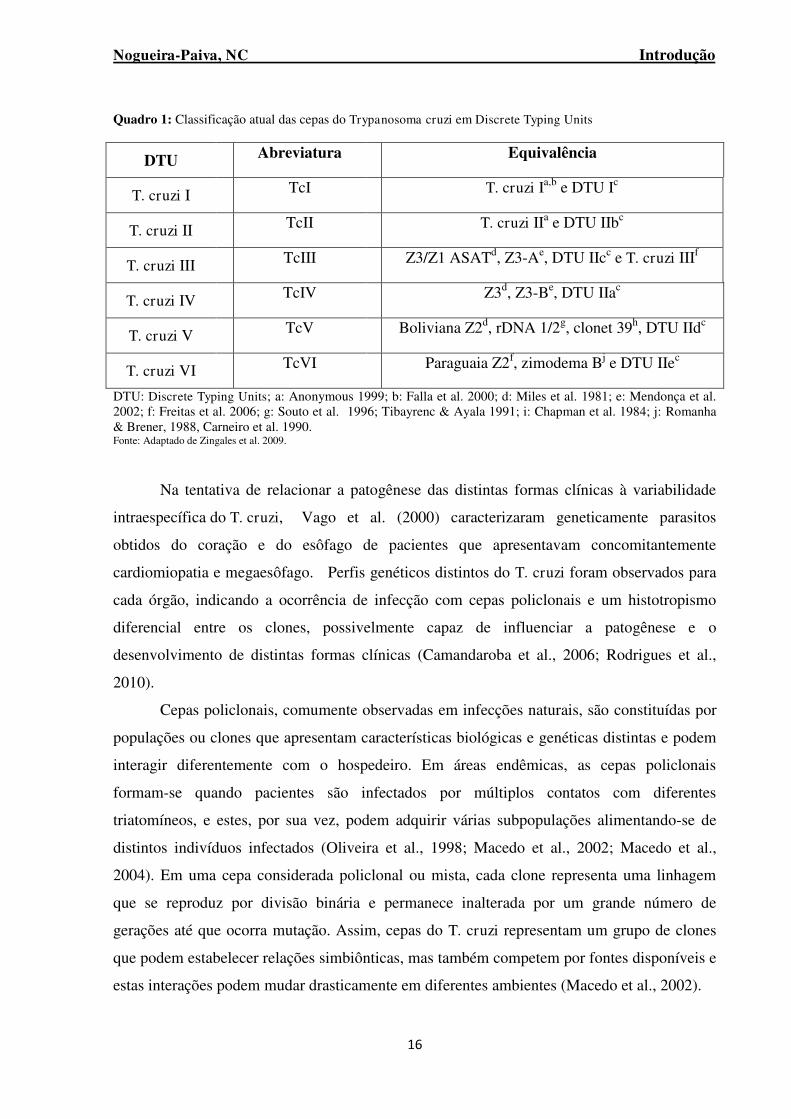
Nogueira-Paiva, NC Introdução
16
Quadro 1: Classificação atual das cepas do Trypanosoma cruzi em Discrete Typing Units
DTU Abreviatura Equivalência
T. cruzi I TcI T. cruzi Ia,b e DTU Ic
T. cruzi II TcII T. cruzi IIa e DTU IIbc
T. cruzi III TcIII Z3/Z1 ASATd, Z3-Ae, DTU IIcc e T. cruzi IIIf
T. cruzi IV TcIV Z3d, Z3-Be, DTU IIac
T. cruzi V TcV Boliviana Z2d, rDNA 1/2g, clonet 39h, DTU IIdc
T. cruzi VI TcVI Paraguaia Z2f, zimodema Bj e DTU IIec
DTU: Discrete Typing Units; a: Anonymous 1999; b: Falla et al. 2000; d: Miles et al. 1981; e: Mendonça et al. 2002; f: Freitas et al. 2006; g: Souto et al. 1996; Tibayrenc & Ayala 1991; i: Chapman et al. 1984; j: Romanha & Brener, 1988, Carneiro et al. 1990. Fonte: Adaptado de Zingales et al. 2009.
Na tentativa de relacionar a patogênese das distintas formas clínicas à variabilidade
intraespecífica do T. cruzi, Vago et al. (2000) caracterizaram geneticamente parasitos
obtidos do coração e do esôfago de pacientes que apresentavam concomitantemente
cardiomiopatia e megaesôfago. Perfis genéticos distintos do T. cruzi foram observados para
cada órgão, indicando a ocorrência de infecção com cepas policlonais e um histotropismo
diferencial entre os clones, possivelmente capaz de influenciar a patogênese e o
desenvolvimento de distintas formas clínicas (Camandaroba et al., 2006; Rodrigues et al.,
2010).
Cepas policlonais, comumente observadas em infecções naturais, são constituídas por
populações ou clones que apresentam características biológicas e genéticas distintas e podem
interagir diferentemente com o hospedeiro. Em áreas endêmicas, as cepas policlonais
formam-se quando pacientes são infectados por múltiplos contatos com diferentes
triatomíneos, e estes, por sua vez, podem adquirir várias subpopulações alimentando-se de
distintos indivíduos infectados (Oliveira et al., 1998; Macedo et al., 2002; Macedo et al.,
2004). Em uma cepa considerada policlonal ou mista, cada clone representa uma linhagem
que se reproduz por divisão binária e permanece inalterada por um grande número de
gerações até que ocorra mutação. Assim, cepas do T. cruzi representam um grupo de clones
que podem estabelecer relações simbiônticas, mas também competem por fontes disponíveis e
estas interações podem mudar drasticamente em diferentes ambientes (Macedo et al., 2002).

Nogueira-Paiva, NC Introdução
17
O modelo “histotrópico clonal” leva em consideração todos esses fatores e sugere uma
hipótese para a formação de cepas do T. cruzi policlonais e como isto pode afetar a
patogênese da DCh. Segundo esse modelo, algumas associações são seletivamente vantajosas
formando cepas estáveis, no entanto o background genético do hospedeiro pode determinar a
susceptibilidade à infecção e o desenvolvimento da doença. Dessa forma, quando mamíferos
são infectados, o repertório clonal pode determinar quais tecidos podem ser afetados. Como
nem todos os clones são capazes de estabelecer a infecção, clones que conseguem se
estabelecer provavelmente competem entre si e colonizam tecidos diferentes. Dessa forma, a
interação de distintos clones/isolados com tecidos específicos poderia ser um fator
determinante para o desenvolvimento das distintas formas clínicas (Macedo et al., 2002).
A classificação atual das subpopulações do T. cruzi em Discrete Typing Units (DTU)
coloca as cepas Be-62 e Be-78 como pertencente ao DTU TcII (Zingales et al., 2009), no
entanto, dados obtidos em nosso laboratório mostraram que, cepas pertencentes ao mesmo
DTU TcII, cepas Y e Be-78, foram capazes de alcançar e parasitar o esôfago e o cólon de cães
da raça Beagle e induzir processo inflamatório considerável nas camadas submucosa e
muscular e ainda, em regiões adjacentes ao gânglio mientérico. Em contrapartida, animais
infectados com a cepa Y controlaram o parasitismo e retornaram o sistema imune a
homeostase, enquanto animais infectados com a cepa Be-78 mantiveram o parasitismo ao
longo de 2 anos de infecção e apresentaram lesões de caráter aparentemente progressivo.
Embora sejam linhagens suficientemente relacionadas para serem classificados dentro do
mesmo DTU, TcII, essas duas cepas são morfologicamente distintas e a interação parasito-
hospedeiro pode interferir com o desenvolvimento da resposta imune do hospedeiro e
determinar o sucesso no controle do parasitismo tecidual e das lesões associadas (Nogueira-
Paiva et al., 2014).
Variações com importância epidemiológica nos padrões de distribuição geográfica das
diferentes linhagens do parasito têm sido observadas atualmente e sua associação com as
formas clínicas da DCh, embora ainda não clara (Florez et al., 2010), têm sido amplamente
investigadas. Na América Central e na região norte da América da Sul, há um predomínio de
subpopulações pertencentes ao grupo TcI, enquanto as outras linhagens (TcII-VI) são
encontradas predominantemente ao sul da América do Sul. Na região central da América do
Sul (Brasil e Bolívia) parece ocorrer uma sobreposição entre as linhagens do T. cruzi (HIGO
et al., 2004). Além da distribuição geográfica, o polimorfismo das subpopulações do T. cruzi
parece também ter relação com as diferentes manifestações anatomoclínicas. A forma

Nogueira-Paiva, NC Introdução
18
digestiva é principalmente atribuída ao TcII (Miles et al., 1981; Zingales et al., 1998; Virreira
et al., 2006; Freitas et al., 2009). Estudos epidemiológicos realizados na Colômbia
demonstraram, porém, que a cardiomiopatia parece não ser mais a única forma clínica da DCh
associada ao TcI, uma vez que foram descritas ocorrências da forma digestiva em pacientes
infectados com parasitos pertencentes ao TcI (Florez et al., 2010; Mantilla et al., 2010) e da
forma cardíaca associada ao TcII (Zafra et al., 2008). Na Bolívia, as subpopulações
pertencentes ao TcIId (atual TcV em Zingales et al. (2009) são as mais comumente
encontradas, seguidas pela linhagem TcI, e não parecem estar associadas a nenhuma forma
clínica específica, cujo desenvolvimento deve relacionar-se mais a fatores relacionados ao
hospedeiro (Del Puerto et al., 2010).
1.4 As cepas “Berenice” do Trypanosoma cruzi
A infecção com as cepas Berenice-62 (Be-62) e Berenice-78 (Be-78) do T. cruzi tem
sido estudada em modelos experimentais, tendo algumas de suas características bem definidas
como polimorfismo, patogenicidade, tropismo, resistência à quimioterapia, resposta imune
sistêmica e compartimentalizada (Brener, 1965; Lana e Chiari, 1986; Bahia et al., 2002;
Caliari et al., 2002a; Caliari et al., 2002b; Carneiro et al., 2007; Guedes et al., 2007; Veloso et
al., 2008; Vieira et al., 2009; Vieira et al., 2012; Nogueira-Paiva et al., 2014). Originalmente é
associada ao primeiro caso clínico humano descrito de DCh (Chagas, 1909) em uma paciente
residente em área endêmica para doença (município de Pirapora, MG/Brasil). Um
acompanhamento longitudinal das condições clínicas e parasitológicas dessa paciente
permitiu a observação de algumas peculiaridades, como por exemplo, a possibilidade de uma
infecção crônica com duração superior a 50 anos estar associada à ausência de manifestações
clínicas da doença.
Além da população isolada durante a fase aguda, cujo ciclo biológico foi reproduzido
em hospedeiro vertebrado e invertebrado (Chagas, 1909), duas outras amostras do parasito,
posteriormente denominadas Berenice-62 e Berenice-78, foram isoladas da mesma paciente
em momentos distintos, através de xenodiagnóstico utilizando T. infestans e Dipetalogaster
maximus, respectivamente (Salgado et al., 1962; Lana e Chiari, 1986). De forma interessante,
o isolado Be-78 apresenta comportamento biológico in vitro e aspectos relacionados à
virulência e patogenicidade distintos (Lana, 1981), sendo que duas hipóteses são consideradas
para explicar a alteração no perfil dessas subpopulações. Embora a paciente negue a

Nogueira-Paiva, NC Introdução
19
ocorrência de contato posterior com vetor, a possibilidade de reinfecção não é totalmente
descartada, uma vez que continuou residindo em área endêmica. No entanto, se não houve
reinfecção, as diferenças entre as duas amostras coletadas da mesma paciente com 16 anos de
intervalo sugerem a ocorrência de uma seleção de população do parasito através de
mecanismos imunológicos que atuam durante a fase crônica da doença (Lana, 1981).
A cepa Be-62 é predominantemente constituída por formas delgadas, apresenta pico de
parasitemia ao redor do sétimo dia e taxa de mortalidade de 100% aos 15 dias após a infecção
(Brener, 1965; Lana, 1981; Lana e Chiari, 1986), enquanto a cepa Be-78 é composta por
formas largas, apresenta pico de parasitemia mais tardio (15º dia) e taxa de sobrevida de
100% (Lana, 1981; Lana e Chiari, 1986; Carneiro et al., 2007; Guedes et al., 2007). Quando
introduzidas em meio de cultura acelular, a amostra Be-78 apresentou menor capacidade de
crescimento e altas taxas de diferenciação, enquanto o isolado Be-62 demonstrou, sob as
mesmas condições, grande capacidade de multiplicação e taxas médias de diferenciação
(Lana, 1981; Lana e Chiari, 1986).
A avaliação de cães infectados com as cepas Be-62 e Be-78 também demonstrou
aspectos particulares entre elas. Miocardite intensa tem sido constantemente associada à cepa
Be-78 durante a fase aguda da infecção, enquanto animais infectados com a cepa Be-62
demonstram um perfil variável, porém menos intenso, dependendo da forma infectiva e da via
de inoculação (Caliari et al., 1995; Bahia et al., 2002).
A análise de isoenzimas revelou que ambos os isolados pertenciam ao zimodema A
(Carneiro et al., 1990), no entanto, o perfil eletroforético permitiu a nítida diferenciação entre
as duas subpopulações (Lana, 1981; Lana e Chiari, 1986). É importante ressaltar que os perfis
de isoenzimas e de endonucleases de restrição não se modificaram ao longo de sucessivas
passagens sanguíneas e/ou em culturas acelulares ou em diferentes situações de isolamento
(Lana, 1981; Lana e Chiari, 1986). Comparando o tropismo tecidual, a cepa Be-78 é
amplamente observada parasitando células musculares (miotrópica) e a cepa Be-62, por sua
vez, tem histotropismo preferencial por células do sistema mononuclear fagocitário
(macrofagotrópica) (Lana e Chiari, 1986).
Embora ambos os isolados estejam atualmente classificados dentro do mesmo táxon
(Quadro 2), a caracterização genética das cepas Be-62 e Be-78 através da determinação dos
perfis de isoenzimas, endonucleases de restrição, e microssatélites reforçaram a existência de
importantes diferenças com potenciais implicações imunopatogênicas. O padrão
isoenzimático, baseado em 18 sistemas, confirmou a diferença em relação à PGM

Nogueira-Paiva, NC Introdução
20
(Phosphoglucomutase), já descrita por Lana e Chiari (1986), somando à DIA-NADH
(Diaphorase-NADH) (Cruz et al., 2006). Os dados obtidos por RAPD mostraram expressivas
diferenças em três dos nove iniciadores utilizados (Cruz et al., 2006). Adicionalmente, para
verificar se as duas amostras eram populações mono ou policlonais, as cepas parentais foram
submetidas ao processo de clonagem por diluição seriada e os quatro clones obtidos foram
comparativamente analisados pela técnica de microssatélite, utilizando sete loci, revelando
que ambas as cepas são populações provavelmente monoclonais, uma vez que cada cepa e
seus respectivos clones mostraram o mesmo perfil de amplificação com um ou dois picos em
todos os loci testados, que refletem homozigose ou heterozigose (Cruz et al., 2006). No
entanto, a presença de diferentes subpopulações clonais nas duas cepas avaliadas não é
descartada, uma vez que os métodos convencionais de clonagem não são capazes de
identificar todos os clones presentes em uma cepa podendo favorecer alguns indivíduos na
população original (Pinto et al., 1998) e o uso de marcadores moleculares mais sensíveis teria
maior chance de distingui-las (Cruz et al., 2006). Baseadas nas técnicas utilizadas neste
estudo, Cruz et al. (2006) concluem que as cepas Be-62 e Be-78, embora apresentem
propriedades biológicas e moleculares distintas, são filogeneticamente relacionadas e
consideradas pertencentes ao zimodema A (Romanha et al., 1979) ou Z2 (Miles et al., 1978),
Linhagem I (Souto et al., 1996) e, atualmente ao DTU TcII (Zingales et al., 2009).
Quadro 2: Classificação das cepas Berenice-62 e Berenice-78 segundo diferentes critérios.
Táxon Referência Bibliográfica
Genótipo 32 Tibayrenc & Ayala 1988
Zimodema A Romanha et al. 1979
Zimodema 2 Miles et al. 1978
Linhagem 1 Souto et al. 1996
T. cruzi II Anonymous 1999
Tc II Zingales et al. 2009
Em contrapartida, Veloso et al. (2005) avaliaram a influência da infecção crônica de
longa duração (2-7 anos) em hospedeiro vertebrado sobre as propriedades biológicas e
genéticas de quatro isolados obtidos de cães infectados com a cepa Be-78 do T. cruzi em
comparação à amostra parental, confirmando a heterogeneidade dessa cepa, sua plasticidade

Nogueira-Paiva, NC Introdução
21
frente a diferentes pressões de seleção e a importância das particularidades da interação
parasito-hospedeiro. Estas subpopulações, após isolamento, foram inoculadas em
camundongos e mantidas em passagens sanguíneas sucessivas sendo reisolados, por
hemocultura, após a primeira e 25ª passagem e submetidas à determinação das lesões
histológicas e caracterização dos perfis de isoenzimas e RAPD, utilizando cinco iniciadores
aleatórios. Os dados demonstraram diferenças significativas entre a cepa parental e seus
isolados em relação à parasitemia, virulência e patogenicidade em camundongos.
Adicionalmente, a avaliação molecular revelou perfis distintos de isoenzimas e RAPD entre
os diferentes isolados e ainda a presença de dois zimodemas dentro do mesmo isolado,
indicando a policlonalidade da cepa Be-78 parental. A capacidade de alguns isolados de
reverter os perfis genéticos, após a 25ª, passagem tornando-os semelhantes àqueles
observados para a cepa parental evidencia que a transição de uma subpopulação para outra é
gradual e que, eventualmente, uma pode sobrepor-se em relação à outra, dependendo da
resposta à pressão do ambiente (Veloso et al., 2005). Recentemente, outros isolados da cepa
Be-78, obtidos de diferentes passagens em camundongos, foram inoculados em cães da raça
Beagle. Seus perfis de RAPD, avaliados após hemocultura, revelaram a existência de dois
grupos, embora geneticamente relacionados, resultantes da possível influência de fatores do
hospedeiro, reforçando os dados de Veloso et al. (2005) (Veloso et al., 2012).
Valadares et al. (2012) submeteram as cepas Be-62 e Be-78 e dois isolados da cepa B-
78 a uma metodologia adaptada de Sorting celular (Fluorescence-Activated Cell Sorter-
FACS) e foram capazes de isolar uma única célula de cada uma dessas subpopulações do T.
cruzi para posterior genotipagem usando marcadores polimórficos para o genes da subunidade
II da Citocromo Oxidase mitocondrial (Cytochrome Oxidase II-COII) e nuclear (24S rDNA),
além da caracterização de quatro loci de microssatélites. Os resultados demonstraram que os
isolados da cepa Be-78 avaliados eram constituídos por três subpopulações, sendo uma
compatível com o perfil observado para Be-62, outra equivalente à Be-78 parental e uma
terceira apresentando um novo padrão, baseado nos quatro loci de microssatélites, ainda não
identificado (Valadares et al., 2012), reforçando a hipótese de que mudanças a nível biológico
e molecular podem ocorrer após passagens sucessivas ou infecção crônica de longa duração,
como resultado da pressão seletiva e do crescimento de subpopulações não previamente
detectadas mais constituintes da cepa original (Valadares et al., 2012; Veloso et al., 2012).

22
2. JUSTIFICATIVA

Nogueira-Paiva, NC Justificativa
23
Variações dos padrões de distribuição geográfica das diferentes linhagens ou
subpopulações do T. cruzi associadas ao amplo espectro clínico da DCh têm apontado o
polimorfismo intraespecífico do T. cruzi como um dos fatores importantes para o prognóstico
da doença. Essas distintas subpopulações presentes no inóculo durante a infecção natural,
segundo o modelo histotrópico clonal, podem apresentar características intrínsecas com
repercussão sobre o estabelecimento da infecção e colonização dos tecidos (Macedo et al.,
2002). Embora estudos genéticos possam comprovar a heterogeneidade intraespecífica do
parasito, avaliações do comportamento biológico e da interação parasito-hospedeiro,
poderiam esclarecer a importância das diferentes cepas, na determinação das manifestações
clínico-epidemiológicas da doença (Andrade e Magalhaes, 1996). Assim sendo, a
compreensão a respeito da participação de diferentes subpopulações do parasito no processo
patogênico, em relação à modulação da resposta imune e ao desenvolvimento das lesões
histológicas durante a fase aguda, poderá contribuir não só para elucidação dos fenômenos
relacionados aos danos teciduais, como será de fundamental importância para o
esclarecimento da evolução da doença, além de auxiliar no estabelecimento de estratégias
terapêuticas eficazes para o tratamento e prevenção da fibrose cardíaca e da lesão entérica na
infecção chagásica.

24
3. OBJETIVOS

Nogueira-Paiva, NC Objetivos
25
3.1 Objetivo Geral
Avaliar a influência da infecção por uma cepa policlonal do T. cruzi sobre a resposta
imune e o desenvolvimento das lesões histológicas no coração e cólon durante a fase aguda da
DCh experimental.
3.2 Objetivos Específicos
3.2.1 Caracterização in vitro e seleção de um isolado da cepa Berenice-78 do T. cruzi
(ETAPA I)
Identificar e selecionar um isolado da cepa Berenice-78 do T. cruzi com características
biológicas e/ou moleculares distintas da cepa parental.
3.2.2 Avaliação in vivo da influência de infecções experimentais por diferentes
populações do T. cruzi derivadas da paciente Berenice (ETAPA II)
Comparar os perfis das curvas de parasitemia;
Quantificar o parasitismo no coração e cólon;
Avaliar o processo inflamatório total no coração e cólon;
Fenotipar o perfil de células mononucleares no coração e cólon;
Analisar as células T CD4+ e CD8+ em relação à expressão de IFN- no coração e cólon.

26
4. MATERIAL E MÉTODOS

Nogueira-Paiva, NC Material e Métodos
27
4.1 Caracterização in vitro e seleção de isolados da cepa Be-78 do T. cruzi (ETAPA I)
Vinte e quatro camundongos Swiss com 30 dias de idade foram inoculados com 5 x
103 formas tripomastigotas sanguíneas da cepa Be-78 por via intraperitoneal e mantidos no
Centro de Ciência Animal - CCA/UFOP com ração e água ad libitum durante 12 meses após
a infecção (mai). A confirmação da infecção e a curva de parasitemia, segundo metodologia
de Brener (1962), foram determinadas em exame de sangue a fresco. Os animais foram
submetidos à coleta de sangue para hemocultura aos três, seis e 12 mai (Figura 2). Em
ambiente estéril, 200l de sangue colhido do plexo orbital de cada camundongo foram
adicionados a um tubo plástico cônico estéril de 15mL (Falcon®, Becton Dickinson, EUA)
(1 tubo/animal), previamente identificados, contendo LIT suplementado com 10% de soro
fetal bovino inativado (LIT-10%SFB, Liver Infusion Tryptose-10% Soro Fetal Bovino) e
mantidos em estufa biológica refrigerada BOD (FANEM® modelo 347) à temperatura de
28°C ± 1°C, para avaliação aos 30, 60 e 90 dias após coleta.
A partir dos tubos positivos na hemocultura, foi realizada uma passagem em meio de
cultura acelular (LIT-10%SFB). Todos os tubos foram incubados a 28°C e, após um período
de 7 dias, foram centrifugados a 2200×g a temperatura ambiente durante 10min, seguido da
remoção do sobrenadante para um tubo plástico cônico de 15mĿ e acréscimo de 3mL de
LIT. Esse procedimento foi realizado com a finalidade de limpar a cultura. Posteriormente, a
cultura foi transferida para erlenmeyer, sendo examinada periodicamente para a adição de
LIT (1:3) em média a cada sete dias, para o crescimento e multiplicação do T. cruzi em fase
logarítmica até a concentração de 1 × 1010 epimastigotas/mĿ (adaptado de D’Ávila et al.,
2009). Na tentativa de evitar seleção de subpopulações do parasito por prolongada
manutenção em meio artificial, todos os cultivos permaneceram aproximadamente 40 dias
em meio LIT quando os parasitos foram criopreservados em nitrogênio líquido, ou lavados
três vezes com tampão PBS 10% (Phosphate-Buffered Saline) estéril, pH 7,2, por
centrifugação a 3400 × g a 4ºC durante 30min para obtenção de sedimento de parasitos
(massa úmida) que foi estocado a -80ºC até o momento da extração do DNA. Além disso,
simultaneamente, amostras dessas culturas foram submetidas à determinação do
comportamento em meio de cultura acelular (LIT) ou celular (células Vero) (Figura 2).
Todos os procedimentos estão sendo realizados de acordo com os princípios éticos
preconizados pelo COBEA (Colégio Brasileiro de Experimentação Animal), tendo sido
aprovados pelo Comitê de Ética no Uso de Animais da Universidade Federal de Ouro Preto-
CEUA/UFOP (Anexo 1: Protocolo nº 2011/30).

Nogueira-Paiva, NC Material e Métodos
28
Figura 2: Fluxograma representando as atividades relacionadas à obtenção e identificação de isolados da cepa Be-78 do T. cruzi (ETAPA I). Be-62 ( ): animais infectados com a cepa Be-62; Be-78 isolado ( ): animais infectados com o isolado Be-78is5-3mai; Be-78 parental ( ): animais infectados com a cepa Berenice-78. do T. cruzi; mai: meses após e infecção, LIT: Liver Infusion Tryptose.
4.1.1 Curvas de Crescimento em meio de cultura acelular
O comportamento das curvas de crescimento in vitro dos isolados em meio de cultura
acelular LIT-10% SFB (Liver Infusion Tryptose-10% Soro Fetal Bovino) foi determinado a
partir de um inóculo padrão de 1 x 107 parasitos/mL em um volume final de 3mL. Para isso,
o volume utilizado no inóculo referente a cada cultura foi determinado com auxílio da
câmara de Neubauer, e as culturas, em triplicata, foram mantidas em estufa biológica
refrigerada BOD (FANEM® modelo 347) à temperatura de 28°C ± 1°C durante o período de
avaliação. O crescimento numérico foi acompanhado diariamente através de contagem em
câmara de Neubauer, em aumento de 400x ao longo de 20 dias.
4.1.2 Perfis de infectividade e desenvolvimento in vitro
Be-78is
Be-62
Expansão das culturas em meio LIT
Comportamento em cultura celular
(Células Vero)
Assinatura gênica(LSSP-PCR)
Comportamento em cultura acelular
(Meio LIT)
Hemoculturas Positivas
Be-78
1 2 3 4 5 6 7 8 9 10 11 24...
Hemoculturas
3, 6 e 12 MAI
24 camundongos SwissETAPA I

Nogueira-Paiva, NC Material e Métodos
29
Previamente ao ensaio de infecção in vitro, amostras de cultura de epimastigotas dos
parasitos foram incubadas em meio Grace (Grace’s Insect Medium, Sigma), e mantidas
durante 15 dias, para indução de metaciclogênese. O perfil de infecção e desenvolvimento in
vitro dos isolados da cepa Be-78 foi comparado aos perfis de crescimento da cepa Be-78
parental e da cepa Be-62, em culturas de células Vero (células renais extraídas de
Cercopithecus aethiops) submetidas à infecção e avaliadas após 24, 48 e 72 horas, em
triplicatas, segundo adaptações da metodologia utilizada por Andrade et al. (2010). Após 24
horas de cultura, monocamadas semiconfluentes de células Vero (104), fixadas em lamínulas,
foram incubadas em estufa a 5% de CO2 e à 37ºC, durante 18 horas (overnight), com formas
tripomastigotas metacíclicas de culturas axênicas das diferentes subpopulações do T. cruzi
(105), a uma MOI (multiplicity of infection) de 10. Após lavagem com tampão PBS 10% para
eliminação de parasitos extracelulares, as culturas foram mantidas nas mesmas condições de
incubação anteriores até o momento da coleta das células (24, 48 e 72 horas após o inóculo).
Em seguida, as culturas foram fixadas em metanol, coradas pelo Panótico Rápido e as taxas
de infecção e desenvolvimento ou multiplicação foram analisadas por microscopia óptica em
aumento de 1000x. A taxa de infecção foi obtida como número de células infectadas em
relação a 100 células contadas, enquanto a taxa de desenvolvimento foi avaliada como
número de parasitos em relação ao número de células infectadas.
4.1.3 Perfil de assinatura gênica
A avaliação do polimorfismo gênico entre os diferentes isolados e a cepa parental
Berenice-78, através da LSSP-PCR, consistiu na obtenção, amplificação e purificação do
DNA template acompanhadas por uma segunda amplificação utilizando um único primer sob
condições de baixa estringência (Pena et al., 1994). A amplificação específica do fragmento
de 330pb (pares de base) do kDNA do T. cruzi foi realizada conforme protocolo de (Gomes
et al., 1998). Os produtos amplificados foram revelados em gel de agarose 1,5% (1,0%
agarose + 0,5% agarose Low Melting Point/Invitrogen) e corados por brometo de etídio. Os
fragmentos de 330pb, visualizados pela radiação ultravioleta, foram removidos do gel com o
auxílio de um pipetador de plástico. Após aquecimento em banho-maria, esses fragmentos
foram homogeneizados, diluídos em água Milli-Q estéril na proporção de 1:10 e utilizados
em um segundo passo de amplificação. Para as reações de LSSP-PCR as amostras de DNA
foram re-amplificadas utilizando um único iniciador específico (correspondente ao iniciador
S35), denominado S35G* (5’-AAA TAA TGT ACG GGG GAG AT-3’). A mistura da

Nogueira-Paiva, NC Material e Métodos
30
reação totalizou um volume final de 10 μL, contendo 6,38 μL de água milli-Q estéril; 2,0 μL
de tampão de amostra; 0,2 μL de cada desoxinucleotídeo (dNTPs: dATP, dCTP, dGTP,
dTTP – Sigma, St. Louis, MO, EUA); 0,1 μL do iniciador S35G* (450μM) e 0,32 μL de Taq
DNA polimerase (5U/L - Go Taq- Promega). Nessa mistura foi acrescentado 1μL do DNA
diluído na proporção 1:10. A amplificação ocorreu nas seguintes condições: uma etapa
inicial de desnaturação do DNA a 94°C por 5min, anelamento a 30º por 1 minuto, extensão a
72º por 1 minuto, seguida de 40 ciclos de amplificação constituídos de um passo de
desnaturação a 94ºC por 1min, um de anelamento a 30°C por 1min, finalizado com uma
etapa de extensão a 72ºC por 10min.
Os produtos dessa segunda amplificação foram revelados, em duplicata, em gel de
poliacrilamida 8%, corados por prata e submetidos à análise do perfil do compartilhamento
de bandas e construção do fenograma para identificação de polimorfismos.

Nogueira-Paiva, NC Material e Métodos
31
4.2 Avaliação in vivo da influência de infecções experimentais por diferentes populações
do Trypanosoma cruzi originadas da paciente Berenice (ETAPA II)
A avaliação in vivo foi dividida em três abordagens (Figura 3), nas quais foram
empregados 330 camundongos BALB/c jovens (5-8 semanas de idade), fêmeas, nascidos no
Setor de Criação do Centro de Ciência Animal-CCA da Universidade Federal de Ouro Preto.
Os animais, distribuídos em quatro grupos experimentais, não infectado (NI), infectado com
a cepa Berenice-78 do T. cruzi parental (Be-78 parental), infectado com um isolado da cepa
Be-78 (Be-78isolado) e infectado com a cepa Be-62 (Be-62), foram necropsiados aos 7, 14 e
28 dias após a infecção (dai). As abordagens 1 e 2 envolveram 90 animais distribuídos entre
os diferentes grupos experimentais (Be-78, n=22; Be-78is, n=22; Be-62, n=34; e NI, n=12),
sendo quantificados o parasitismo tecidual (qPCR) e o processo inflamatório total (HE). Dez
animais de cada grupo infectado foram submetidos à avaliação da curva de parasitemia e
taxa de mortalidade. A Abordagem 3 incluiu os demais 240 camundongos BALB/c
distribuídos entre os diferentes grupos experimentais (Be-78, n=48; Be-78is, n=48; Be-62,
n=96; e NI, n=48). Nessa abordagem, as amostras do coração e cólon, coletadas durante a
necropsia, foram submetidas à avaliação imunofenotípica das células inflamatórias por
citometria de fluxo (macrófagos, linfócitos B, linfócitos T CD4+ e CD8+). As três abordagens
foram realizadas simultaneamente. Todos os procedimentos foram realizados de acordo com
os princípios éticos preconizados pelo COBEA (Colégio Brasileiro de Experimentação
Animal), tendo sido aprovados pelo Comitê de Ética no Uso de Animais da Universidade
Federal de Ouro Preto-CEUA/UFOP (Anexo 2: Protocolo nº 2014/29).
4.2.1 Infecção dos animais
Duzentos e setenta camundongos BALB/c com idade variando entre 5-8 semanas
foram distribuídos em quatro grupos experimentais: infectados com a cepa Be-78 parental
(Be-78, n=70), infectados com um isolado da cepa Be-78 (Be-78is, n=70), infectados com a
cepa Be-62 (Be-62, n=130) e não-infectados (NI, n=60). O grupo infectado com a cepa Be-
62 foi realizado em número maior considerando os altos índices de mortalidade descritos na
fase aguda da infecção.Os animais infectados foram inoculados, por via intraperitoneal, com
5 x 103 formas tripomastigotas sanguíneas obtidas após descongelamento das amostras
utilizadas na ETAPA I. A infecção foi confirmada por observação de tripomastigotas

Nogueira-Paiva, NC Material e Métodos
32
sanguíneos, em exame de sangue a fresco, e os animais mantidos no Centro de Ciência
Animal-CCA/UFOP com ração e água ad libitum durante o tempo de avaliação (28 dias).
Figura 3: Fluxograma representando as atividades relacionadas à avaliação in vivo (ETAPA II) proposta neste trabalho. Os 330 animais foram distribuídos em três abordagens. Abordagem 1: quantificação do parasitismo tecidual; Abordagem 2: avaliação do processo inflamatório; Abordagem 3: análise imunofenotípica e funcional das células inflamatórias. Todas as avaliações foram realizadas em amostras do coração e cólon coletadas durante as necropsias aos 7, 14 e 28 dias após a infecção. NI ( ): grupo de animais não-infectados; Be-62 ( ): animais infectados com a cepa Be-62; Be-78 isolado ( ): animais infectados com o isolado Be-78is5-3mai; Be-78 parental ( ): animais infectados com a cepa Berenice-78. do T. cruzi; dai: Dias após a infecção; qPCR: Polymerase Chain Reaction quantitativa.
4.2.3 Parasitemia e sobrevida
Para determinação da curva de parasitemia, 10 camundongos, pertencentes a cada um
dos grupos infectados, foram avaliados diariamente, a partir do 3o dai até a negativação do
exame, por cinco dias consecutivos, segundo a metodologia de Brener (1962) (Figura 4).
Cinco µL de sangue da veia caudal foram analisados diariamente ao microscópio óptico
(aumento de 400x) e a curva de parasitemia plotada, para cada grupo experimental,
empregando a média diária da parasitemia detectada nos animais. Esta foi expressa em
número de tripomastigotas sanguíneos por 0,1mL de sangue. O período pré-patente (PPP),
NI
(n=12)
NI
(n=20)
NI
(n=48)
ETAPA I
I
330 camundongos BALB/c
Abordagens 1 e 2
Abordagens 3
Necropsia (7, 14 e 28 DAI)
Coração e Cólon
Be-78
(n=20)
Be-78 parental
(n=12+10)
Be-78 parental
(n=48)
Be-62
(n=24+10)
Be-78cl2
(n=20)
Be-62
(n=96)
Be-78 isolado
(n=12+10)
Be-78cl1
(n=20)
Be-78 isolado
(n=48)
Fenotipagem de Leucócitos
Mononucleares
Abordagem 3
Citometria de Fluxo
Parasitismo Tecidual(qPCR)
Abordagem 1
PCR Microscopia Óptica
Abordagem 2
Infiltrado Inflamatório
ParasitemiaMortalidade

Nogueira-Paiva, NC Material e Métodos
33
período patente (PP), pico máximo de parasitemia (Pmáx), dia do pico máximo de
parasitemia (DPmáx) também foram definidos.
Período pré-patente (PPP)
O PPP corresponde ao período compreendido entre a inoculação e o primeiro dia de
detecção do parasito no sangue periférico do animal.
Período patente (PP)
O PP corresponde ao período compreendido entre o primeiro dia de exame a fresco
positivo até a completa negativação da parasitemia e foi expresso em dias.
Pico máximo de parasitemia (Pmáx)
O Pmáx corresponde ao máximo de parasitemia e foi expresso em número de
tripomastigotas sanguíneos/0,1 mL de sangue.
Dia do pico máximo de parasitemia (DPmáx)
O DPmáx corresponde ao dia de ocorrência do máximo de parasitemia e foi expresso
em dias.
Além da parasitemia, a taxa de sobrevida foi avaliada nestes mesmos animais,
diariamente, até 42 dias após a infecção sendo a sobrevida registrada e expressa em
porcentagem cumulativa.
Figura 4: Fluxograma representando a distribuição dos animais infectados com diferentes subpopulações do Trypanosoma cruzi utilizados para determinação da curva de parasitemia e da taxa sobrevida. Be-62 ( ): grupo de animais infectados com a cepa Be-62; Be-78 isolado ( ): grupo de animais infectados com o isolado Be-78is5-3mai obtido a partir da cepa Be-78 parental, selecionado na ETAPA I; Be-78 parental ( ): animais infectados com a cepa parental Be-78.
30 camundongos BALB/c
Parasitemia e taxa de sobrevida(diariamente)
Be-78 isolado
(n=10)
Be-62
(n=10)
Be-78 parental
(n=10)

Nogueira-Paiva, NC Material e Métodos
34
Abordagens 1 e 2
4.2.4 Necropsia
Para a realização das atividades propostas nas Abordagens 1 e 2, quatro animais de
cada grupo foram eutanasiados, por deslocamento cervical, aos 7, 14 e 28 dai. Durante a
necropsia, amostras de coração in totum e cólon (região retossigmóide) foram coletadas
sendo um fragmento de cada órgão submerso em nitrogênio líquido e posteriormente
armazenado em freezer -80ºC para quantificação do parasitismo tecidual (quantitative-PCR),
enquanto um segundo fragmento foi fixado em solução Metanol 80%-Dimetilsulfóxido
(DMSO) 20% para posterior avaliação, do processo inflamatório (Figura 3).
4.2.5 Carga parasitária no coração e cólon
O parasitismo no coração e cólon foi quantificado por Real-Time PCR (Cummings e
Tarleton, 2003). Essa técnica foi dividida em duas etapas, sendo a primeira constituída pela
extração de DNA e a segunda pela amplificação específica do kDNA do T. cruzi.
Cerca de 20 mg de coração e cólon foram utilizados para obtenção do DNA genômico,
utilizando o kit Wizard Genomic® DNA Purification (Promega, São Paulo, Brasil), seguindo
as recomendações do protocolo do fabricante. Os fragmentos armazenados em freezer -80ºC
foram transferidos para tubos de microcentrífuga de 1,5mL e, para aperfeiçoar o processo de
digestão, foram previamente trituradas com tesoura estéril em 0,25 mL de tampão de lise (Tris
HCl 50mM pH 7,5; EDTA 1mM; 1% N-lauril sarcosina). O processo de digestão foi realizado
com a incubação das amostras em 0,025 mL de Proteinase K (25 mg/ mL) (Sigma, São Paulo,
Brasil) a 37°C por 16 horas. Em seguida, foram adicionados 0,6 mL de Solução de Lise
Nuclear (Nuclear Lysis Solution), e as amostras foram mantidas a 65° C por 15 minutos.
Posteriormente, foram adicionados 0,003 mL de Solução de RNase (RNase Solution) e as
amostras incubadas a 37° C por 15 minutos. Depois de 5 minutos em temperatura ambiente,
as amostras receberam 0,2 mL de Solução de Precipitação Protéica (Protein Precipitations
Solution) e foram vortequizadas vigorosamente. Após 5 minutos no gelo, foram centrifugadas,
a 12.000 x g por 5 minutos, e então, a fase aquosa foi recuperada e recebeu 0,6 mL de
isopropanol gelado (1:1). Os tubos foram invertidos 3x e centrifugados (12.000 x g por 5
minutos). Após centrifugação, o sobrenadante foi descartado e adicionou-se 0,6 mL de etanol
70% v/v gelado (preparado com água livre de DNAse). Depois de centrifugados (12.000 x g

Nogueira-Paiva, NC Material e Métodos
35
por 5 minutos), o sobrenadante foi descartado e os tubos ficaram abertos no fluxo laminar
para secar os resíduos de álcool. O DNA foi hidratado em 0,05 ml de Solução de Re-
hidratação de DNA (DNA Rehidratation Solution) e incubado a 37° C por 2 horas. O controle
de qualidade do DNA foi realizado a partir da quantificação e avaliação do grau de pureza do
mesmo pelo aparelho NanoDrop® (GE Healthcare, São Paulo, Brasil).
Para a determinação da carga parasitária foi utilizada a técnica de PCR em tempo real.
As reações foram realizadas pelo kit SYBR® Green PCR Master Mix (Applied Biosystems,
São Paulo, Brasil) em placas de 96 poços (MicroAmp® Optical 96 Well Reaction Plate –
Applied Biosystems, São Paulo, Brasil) e seladas com adesivo óptico (MicroAmp® Optical
Adhesive Film – Applied Biosystems, São Paulo, Brasil) ao final do procedimento.
Foram pipetados 3 μL de DNA (30 ng) em duplicata e 7 μL de um mix de reação
contendo 2 μL dos iniciadores (na concentração de 2,5 μM) e 5 μL de SYBR® Green Master
Mix, totalizando um volume de reação, em cada poço, de 10 μL. A reação de PCR foi
conduzida conforme programação contida no aparelho ABI 7500 Applied Biosystems; os
valores de baseline foram ajustados para 3-15 ciclos e o threshold foi ajustado à região
associada ao crescimento exponencial do produto da PCR (default).
As análises foram feitas pelo método da quantificação absoluta. O método da
quantificação absoluta baseia-se na comparação com um padrão cujo número de moléculas é
conhecido, e utiliza da equação da reta gerada pela diluição seriada da curva padrão (y=ax+b)
para quantificar a quantidade de DNA do parasita na amostra (x). Os iniciadores para o DNA
de T. cruzi foram desenhados para o gene TCZ (F 5' GCTCTTGCCCACAMGGGTGC 3 ', em
que M = A ou C, e R 5' CCAAGCAGCGGATAGTTCAGG 3'; amplicon de 195 pb) (Moser
et al., 1989; Cummings e Tarleton, 2003).
A curva padrão foi representada por um gráfico de regressão linear semi-log do valor
de Cq (eixo Y) em comparação ao log da quantidade inicial do ácido nucléico (eixo X). Foi
utilizado o DNA de epimastigotas da cepa Y do T. cruzi, na quantidade inicial de 600 pg, em
7 diluições seriadas de 1:10. O slope (inclinação) da curva padrão foi usado para estimar a
eficiência de amplificação (~100%) e o coeficiente de determinação (R2) estabelecido como
adequado foi de 0,98.
Em cada placa foi realizada uma curva padrão para determinar a quantidade de DNA
do parasita nas amostras. Separadamente, reações contendo 50 ng de DNA genômico, 0,5M
de iniciadores específicos para Fator de Necrose Tumoral-TNF murino TNF-5241 F 5’-
TCCCTCTCATCAGTTCTATGGCCCA-3’ e R 5’

Nogueira-Paiva, NC Material e Métodos
36
AGCAAGCATCTATGCACTTAGACCCC (desenhado por Vector NTI), 10l SYBR™
Green, e água livre de nucleases para um volume final total de 20L, foram utilizadas como
controle endógeno. Além disso, cada reação no termociclador contou com dois controles
negativos que consistiram de uma reação sem adicionar DNA e outra reação com adição
apenas de DNA de tecido não-infectado. Como os resultados apresentados estavam em escala
logarítmica de DNA, foi feita uma conversão da quantidade de DNA encontrada para número
de cópias do parasita. Para tal, considerou-se que um genoma de T. cruzi possui,
aproximadamente, 0,33 pg de DNA (Lanar et al., 1981; Gonzalez et al., 1984; Souza et al.,
2011), efetuando a conversão e expressando os dados na forma de parasitas por mg de tecido.
Os valores corrigidos para cada grupo experimental foram apresentados como média e desvio
padrão. A curva de eficiência foi determinada usando o seguinte cálculo (Stordeur et al.,
2002):
Eficiência (E) = 10(−1/slope)
4.2.6 Intensidade do processo inflamatório no coração e cólon
As amostras de coração e cólon, fixadas em Metanol/DMSO, foram processadas
rotineiramente, incluídas em parafina, e submetidas à microtomia para obtenção de cortes
histológicos com 4m de espessura.
Para avaliação do infiltrado inflamatório, os cortes foram corados pela técnica de
Hematoxilina-Eosina. Após desparafinização em dois banhos de xilol, os cortes foram
hidratados em concentrações decrescentes de álcool (100, 90, 80 e 70%) e então lavados em
água corrente. Em seguida, foram corados pela Hematoxilina, por 10 minutos e lavados em
água corrente, para retirada do excesso do corante. Os cortes foram diferenciados em álcool
acidulado (100 mL de álcool absoluto e 10 gotas de ácido clorídrico) e novamente lavados em
água corrente para evitar acidificação excessiva. Posteriormente, foram corados pela Eosina,
por 30 segundos. Após a última lavagem em água corrente, foram desidratados, em 2 banhos
de álcool absoluto e levados à estufa a 56ºC para secagem e montados com lamínula e
Entellan.
A avaliação semi-quantitativa foi realizada nos tecidos sendo atribuídas as seguintes
classificações: processo inflamatório ausente, discreto, moderado e intenso.

Nogueira-Paiva, NC Material e Métodos
37
Para quantificação do processo inflamatório os núcleos celulares foram quantificados
separadamente na submucosa e muscular, sendo 20 imagens (campos) aleatórias em cada uma
das camadas, totalizando uma área percorrida igual a 1,5 x 106 m2. As imagens foram
visualizadas pela objetiva de 40x e digitalizadas através da microcâmera Leica DFC340FX
associada ao microscópio Leica DM5000B e todas as imagens foram analisadas com o auxílio
do software de análise e processamento de imagem Leica QwinV3 no Laboratório
Multiusuários do Núcleo de Pesquisas em Ciências Biológicas da Universidade Federal de
Ouro Preto. O processo inflamatório foi determinado pela diferença significativa (p<0,05)
entre o número de núcleos celulares presentes nos animais infectados pelo T. cruzi e aquele
observado nos animais não-infectados ± desvio padrão (Caliari, 1997; Maltos et al., 2004;
Ferreira Junior et al., 2015). Os resultados foram demonstrados como média do número de
núcleos detectados nas 20 imagens em uma área de um frame (imagem/janela) igual a
75.183,8m2.
Todas as análises das células inflamatórias foram realizadas apenas nas camadas
musculares (miocárdio e camadas musculares interna e externa do cólon).

Nogueira-Paiva, NC Material e Métodos
38
Abordagem 3
4.2.7 Necropsia
Para a obtenção do número de células necessário à avaliação imunofenotípica por
citometria de fluxo, proposta na Abordagem 3, cada tratamento, referente à subpopulação
infectante do T. cruzi e os diferentes tempos das necropsias, foi composto por 16 animais,
onde pools independentes de macerado de coração ou cólon de quatro camundongos
constituíram um único elemento dentro do tratamento (Figura 5). Dessa forma, 16 animais de
cada grupo foram eutanasiados, por deslocamento cervical, aos 7, 14 e 28 dias após a
infecção sendo procedida a coleta dos órgãos, posteriormente submetidos a protocolos para a
obtenção de suspensões celulares.
Figura 5: Desenho esquemático representando a distribuição de camundongos BALB/c entre os tratamentos/grupos no ensaio por citometria de fluxo.
4.2.8 Perfil fenotípico do processo inflamatório no coração e cólon
As suspensões de células do coração e cólon obtidas em pool (4 animais em cada
pool), conforme observado anteriormente, foram preparadas de acordo com o método
descrito por Cuervo et al. (2011) e Figueiredo et al. (2011). Em ambiente estéril, após a
Elemento 1 Elemento 2 Elemento 3 Elemento 4
Tratamento (ex.: grupo Be-78)
Elem
ento
1
Elem
ento
2
Elem
ento
3
Elem
ento
4
Elem
ento
1
Elem
ento
2
Elem
ento
3
Elem
ento
4
Coração Cólon

Nogueira-Paiva, NC Material e Métodos
39
eutanásia e coleta dos órgãos em meio completo RPMI (5% Soro Fetal Bovino-SFB), as
amostras foram cortadas em fragmentos de 1mm, lavadas em tampão Hank’s Buffered Salt
Solution-HBSS + 5% de SFB + 25mM de HEPES e incubadas em dois banhos de 30 minutos
em solução de pré-digestão em Dithiothreitol-DTT a 1mM à 37º C sob agitação (120rpm).
Após lavagem em tampão HBSS + 5% de SFB + 25mM de HEPES foi procedida a digestão
em duas incubações de 60 minutos cada em solução de colagenase a 0,01% (Collagenase
type II from Clostridium histolyticum) sob agitação (120rpm) e à 37ºC. As suspensões
celulares foram centrifugadas a 1800 rpm por 10 minutos, ressupendidas em 10mL de
solução HBSS + 5% de SFB + 25mM de HEPES e filtradas em um primeiro sistema de
filtros com malha de 100m. Após centrifugação sob as mesmas condições anteriores, as
suspensões celulares foram ressuspendidas em 10mL de solução HBSS + 25mM de HEPES
+ 2mM EDTA e novamente filtradas em um segundo sistema utilizando malha de 40m para
retirada dos debris celulares. Após nova centrifugação, as suspensões celulares
resuspendidas em meio completo RPMI (5% de SFB) foram submetidas à quantificação em
câmara de Neubauer. Alíquotas de 100l de células cardíacas e entéricas (1 x 105células/mL)
foram incubadas na presença de meio de cultura RPMI (5% de SFB) por 12 horas em estufa
de CO2 com 5% de umidade a 37ºC. Os tubos foram retirados da estufa e foi adicionada
Brefeldina A - BFA (10 µg/ml, Sigma) para paralisação da liberação de citocinas mantendo-
as no interior do complexo de Golgi. Após 4 horas de incubação em BFA, as culturas foram
centrifugadas e foram coletados 1mL de sobrenadante de cultura para avaliação de citocinas.
As culturas foram então tratadas com 200μl EDTA (Sigma), numa concentração final de
20mM, e incubadas por 10 minutos à temperatura ambiente. Este procedimento bloqueia o
processo de ativação posterior das células e garante a obtenção de resultados padronizados.
Posteriormente ao tratamento com EDTA, as células foram lavadas com 3 mL de tampão de
lavagem – PBS-W (0,015M de PBS 1X, 0,5% albumina sérica bovina – BSA e 0,1% de
azida sódica), por centrifugação a 1200 rpm durante 7 minutos a 18ºC. Após a última
lavagem, as células foram ressuspendidas em 2 mL de PBS-W. Em seguida, as amostras
foram incubadas com anticorpos anti-moléculas de superfície (CD45, CD3, CD4, CD8,
CD19 e F4/80 – Figura 6 e Quadro 3) por 30 minutos ao abrigo da luz. As especificações dos
anticorpos, fluorocromos, assim como a titulação padronizada são mostrados no Quadro 3.
Após a etapa da identificação das populações celulares, procedeu-se à lise dos eritrócitos e à
fixação dos leucócitos pelo tratamento com 2mL de solução de lise por 10 minutos à
temperatura ambiente e ao abrigo da luz. Após a fixação, as suspensões de leucócitos foram

Nogueira-Paiva, NC Material e Métodos
40
centrifugadas a 1200 rpm durante 7 minutos a 18ºC, os sobrenadantes descartados e as
células permeabilizadas com 2mL de solução permeabilizante – PBS-P (PBS-W e 0,5% de
saponina – Sigma), por 10 minutos à temperatura ambiente e ao abrigo da luz. Após a
permeabilização, as suspensões de leucócitos foram centrifugadas a 1200 rpm durante 7
minutos, o sobrenadante descartado e as células lavadas com 3mL de PBS-W. Após a
ressuspensão das células em PBS-W, procedeu-se à marcação intracitoplasmática em placas
de 96 poços e fundo em “U” (IFN-). Para isso, alíquotas de 30μL das suspensões celulares
previamente marcadas com as combinações de anticorpos anti-superfície CD45CD3CD4 ou
CD45CD3CD8 foram incubadas por 30 minutos à temperatura ambiente, ao abrigo da luz na
presença de 20μL da suspensão de anticorpos anti-citocinas conjugados com o fluorocromo
PE (anti-IFN-– Figura 6 e Quadro 3) e previamente diluídos a 1:50 em PBS-P estéril com
10% de soro normal de Rato. Após a incubação, as células foram lavadas com 100μl de PBS-
P e, em seguida, com 200μl de PBS-W. As preparações celulares foram então fixadas em
200μl de solução fixadora e estocadas a 4ºC ao abrigo da luz até a sua leitura no citômetro de
fluxo.
Quadro 3: Especificações dos anticorpos utilizados no ensaio de citometria de fluxo.
Anticorpos (Marca) Fluorocromo
Especificações/Isotipo Clone Titulação
Fenótipo alvo
CD45 (eBioscience)
APC Rat anti-mouse/IgG2b Clone: 30F11
1:10 Pan-
leucocitário
F4/80 (eBioscience)
FITC Rat anti-mouse/IgG2a Clone: BM8
1:10 Macrófagos
CD19 (eBioscience)
FITC Rat anti-mouse/IgG2a Clone: eBIO1D3
1:10 Linfócitos B
CD3 (Biolegend)
FITC Rat anti-mouse/IgG2b Clone: 17A2
1:10 Linfócitos T
CD4 (Biolegend)
PerCp Rat anti-mouse/IgG2a Clone: RM4-5
1:10 Células CD4+
CD8 (Biolegend)
PerCp Rat anti-mouse/IgG2b Clone: 53-6.7
1:10 Células CD8+
IFN- (Serotec)
PE Rat anti-mouse/IgG2b Clone: XMG1.2
1:10 Células
produtoras de IFN-
CD: cluster of differentiation; APC: Aloficocianina; FITC: Isotiocianato de Fluoresceína; PercP: Proteína Clorofila Peridinina; PE: Ficoeritrina; IgG: Imunoglobulina G.

Nogueira-Paiva, NC Material e Métodos
41
Figura 6: Desenho esquemático representando as combinações de anticorpos marcados com fluorocromos utilizadas no ensaio de citometria de fluxo. As cores representam os espectros de emissão dos fluorocromos.
4.2.9 Estratégia de análise do perfil fenotípico do processo inflamatório no coração e cólon
A aquisição de eventos foi realizada no citômetro de fluxo FACScalibur - Becton
Dickinson, utilizando o programa CELLQuestTMPro. As células das suspensões celulares de
coração (20.000 eventos) e cólon (100.000 eventos) foram adquiridas em escala linear e
inicialmente identificadas pelos parâmetros tamanho da célula (dispersão para frente [FSC]) e
complexidade interna (dispersão lateral [SSC]). A seleção das populações de células
leucocitárias foi baseada na utilização de marcadores de linhagem e análise multiparamétrica
com combinações de regiões em gráficos bidimensionais de distribuição pontual
(Pseudocolor “Large Dot”).
A análise dos dados foi realizada utilizando o software de análise de dados de
citometria de fluxo FlowJo X. Os resultados foram expressos como frequência (%) das
populações de macrófagos, de linfócitos B e das subpopulações de linfócitos T CD4+ e CD8+
em relação às células com fenótipo CD45+. Além disso, foi analisado o perfil funcional
linfócitos T CD4+ e CD8+ em relação à expressão de IFN-, também determinado como
frequência em relação às células com fenótipo CD45+.
A Figura 7 representa a distribuição da população, em gráficos bidimensionais de
distribuição pontual com as diferentes combinações, nas suspensões celulares de coração e
cólon em tubo controle “Cel” (onde não foram adicionados anticorpos). Os controles
negativos foram utilizados para determinar a localização do gates utilizados para identificar
as populações positivas para cada fenótipo, sendo os limites aplicados como cut-off, as
regiões que abrangiam população negativa inferior a 1%.
CD
45-A
PC
CD
4-P
erc
P
CD
45-A
PC
IFN
g-P
E
IFN
g-P
E
CD
8-P
erc
P
Cel
Co
ntr
ole
d
e Is
oti
po
CD
3-F
TIC
CD
3-F
TIC
CD
45-A
PC
CD
19
CD
45-A
PC
F4/8
0-F
ITC

Nogueira-Paiva, NC Material e Métodos
42
Figura 7: Representação da estratégia de análise da frequência de células inflamatórias com distintos fenótipos em suspensão de células do coração ou cólon de camundongos infectados com o isolado Be-78is5-3mai, ou com as cepas de referência Berenice-62 e Berenice-78 parental do Trypanosoma cruzi: (A) Gráfico de distribuição pontual Tamanho (FSC) versus Granulosidade (SSC) demonstrando o padrão de distribuição da população nas suspensões celulares de coração e cólon (B) Gráfico de distribuição pontual FL4 versus Granulosidade (SSC) no tubo controle demonstrando ausência de fluorescência; (C) Gráfico de distribuição pontual FL1 (CD3 FITC) versus SSC no tubo controle demonstrando ausência de fluorescência; (D) Gráfico de distribuição pontual FSC versus FL3 no tubo controle demonstrando ausência de fluorescência; E) Gráfico de distribuição pontual FSC versus FL2 no tubo controle demonstrando ausência de fluorescência.
A subpopulação de leucócitos foi identificada aplicando os parâmetros expressão
característica do antígeno pan-leucocitário CD45 (SSC / FL4) e SSC, onde a região
inicialmente selecionada foi constituída das populações positivas “low” e “high” para o
marcador CD45 e granulosidade inferior a 500 (Figura 8A, 9A e 10A). A seleção dessa
subpopulação foi comum à análise de todos os fenótipos avaliados (Macrófagos, Linfócitos B,
Linfócitos T CD4+IFN-+, Linfócitos T Linfócitos T CD8+IFN-+).
Para determinação da frequência de macrófagos (Figura 8B) e linfócitos B (Figura
9B), as subpopulações de interesse foram selecionadas a partir da análise combinada F4/80
FITC / SSC (FL1 / SSC) ou CD19 FITC / SSC (FL1 / SSC), respectivamente.
CORAÇÃO
CÓLON
A) B) C) D) E)
A) B) C) D) E)
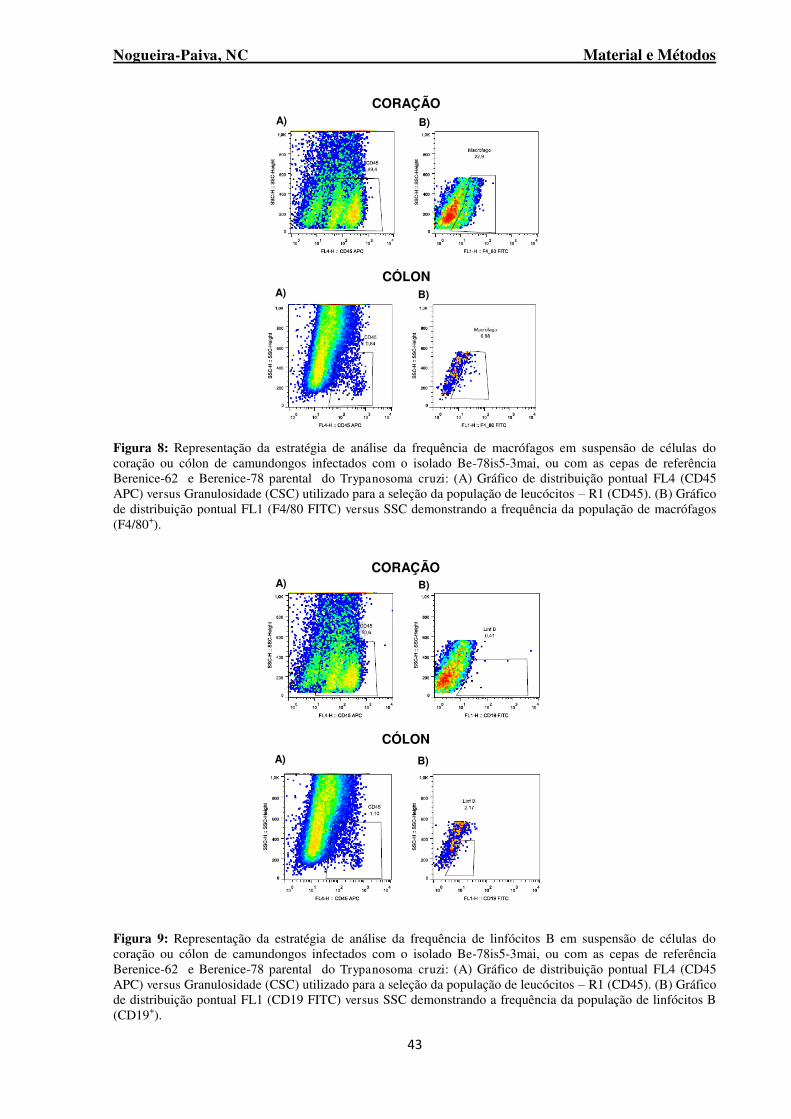
Nogueira-Paiva, NC Material e Métodos
43
Figura 8: Representação da estratégia de análise da frequência de macrófagos em suspensão de células do coração ou cólon de camundongos infectados com o isolado Be-78is5-3mai, ou com as cepas de referência Berenice-62 e Berenice-78 parental do Trypanosoma cruzi: (A) Gráfico de distribuição pontual FL4 (CD45 APC) versus Granulosidade (CSC) utilizado para a seleção da população de leucócitos – R1 (CD45). (B) Gráfico de distribuição pontual FL1 (F4/80 FITC) versus SSC demonstrando a frequência da população de macrófagos (F4/80+).
Figura 9: Representação da estratégia de análise da frequência de linfócitos B em suspensão de células do coração ou cólon de camundongos infectados com o isolado Be-78is5-3mai, ou com as cepas de referência Berenice-62 e Berenice-78 parental do Trypanosoma cruzi: (A) Gráfico de distribuição pontual FL4 (CD45 APC) versus Granulosidade (CSC) utilizado para a seleção da população de leucócitos – R1 (CD45). (B) Gráfico de distribuição pontual FL1 (CD19 FITC) versus SSC demonstrando a frequência da população de linfócitos B (CD19+).
A) B)
CORAÇÃO
CÓLON A) B)
A) B)
CORAÇÃO
CÓLON
A) B)
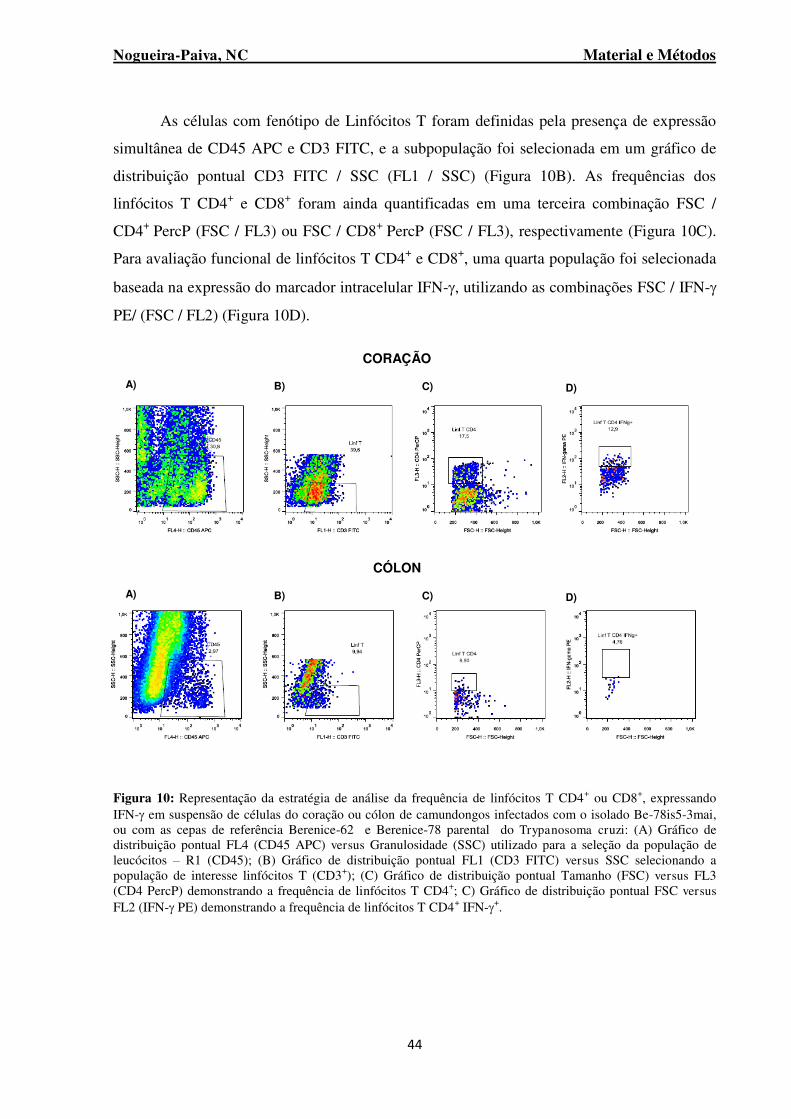
Nogueira-Paiva, NC Material e Métodos
44
As células com fenótipo de Linfócitos T foram definidas pela presença de expressão
simultânea de CD45 APC e CD3 FITC, e a subpopulação foi selecionada em um gráfico de
distribuição pontual CD3 FITC / SSC (FL1 / SSC) (Figura 10B). As frequências dos
linfócitos T CD4+ e CD8+ foram ainda quantificadas em uma terceira combinação FSC /
CD4+ PercP (FSC / FL3) ou FSC / CD8+ PercP (FSC / FL3), respectivamente (Figura 10C).
Para avaliação funcional de linfócitos T CD4+ e CD8+, uma quarta população foi selecionada
baseada na expressão do marcador intracelular IFN-, utilizando as combinações FSC / IFN-
PE/ (FSC / FL2) (Figura 10D).
Figura 10: Representação da estratégia de análise da frequência de linfócitos T CD4+ ou CD8+, expressando IFN- em suspensão de células do coração ou cólon de camundongos infectados com o isolado Be-78is5-3mai, ou com as cepas de referência Berenice-62 e Berenice-78 parental do Trypanosoma cruzi: (A) Gráfico de distribuição pontual FL4 (CD45 APC) versus Granulosidade (SSC) utilizado para a seleção da população de leucócitos – R1 (CD45); (B) Gráfico de distribuição pontual FL1 (CD3 FITC) versus SSC selecionando a população de interesse linfócitos T (CD3+); (C) Gráfico de distribuição pontual Tamanho (FSC) versus FL3 (CD4 PercP) demonstrando a frequência de linfócitos T CD4+; C) Gráfico de distribuição pontual FSC versus FL2 (IFN- PE) demonstrando a frequência de linfócitos T CD4+ IFN-+.
A) B) C) D)
A) B) C) D)
CORAÇÃO
CÓLON

Nogueira-Paiva, NC Material e Métodos
45
4.3 Análises estatísticas
As análises estatísticas foram realizadas com o apoio instrumental do software
GraphPad Prism 4.03 (Prism Software, Irvine, CA, USA). Em todos os casos, as diferenças
foram consideradas significativas quando valores de p foram inferiores a 0,05 (p<0,05).
Todos os dados foram submetidos à testes de normalidade, e, com exceção da análise
da AUC para os dados de parasitemia, todos os demais dados demonstraram-se não
paramétricos e testes adequados foram utilizados para determinar as diferenças estatísticas.
Para avaliação dos perfis de crescimento em meio de cultura axênico, foi considerada
a análise da área sob a curva (area under the curve-AUC) através do teste de análise de
variância Kruskal-Wallis, para dados não paramétricos. Quando as alterações foram
significativas, o pós-teste de Dunns foi realizado para determinar as diferenças específicas
entre as medianas referentes às diferentes subpopulações do T. cruzi em comparação às cepas
de referência usadas neste trabalho, Be-62 e Be-78 parental.
Em relação ao ensaio de infecção in vitro em culturas de células Vero, considerando as
subpopulações do parasito utilizadas na infecção e os tempos de incubação avaliados, as
diferenças significativas foram determinadas usando análise de variância, ANOVA two-way.
Diferenças significativas entre as médias referentes às diferentes subpopulações do T. cruzi
em comparação às cepas de referência, Be-62 e Be-78 parental, foram determinadas pelo pós-
teste de Bonferroni.
A área sob a curva (AUC) utilizada para comparar o perfil de parasitemia entre os
grupos infectados, cujos dados foram considerados paramétricos, foi submetida à análise
estatística utilizando o teste de análise de variância ANOVA One-way, e pós-teste de Tukey.
Para comparação da carga parasitária, da intensidade e do perfil imunofenotípico das
células inflamatórias no coração e cólon entre os diferentes grupos experimentais,
considerando tanto as subpopulações do T. cruzi quanto os diferentes tempos avaliados, foi
utilizado o teste de análise de variância ANOVA Two-way. Quando as alterações foram
significativas, o pós-teste de Bonferroni foi realizado para determinar as diferenças
específicas entre as médias referentes às diferentes subpopulações do T. cruzi.

46
5. RESULTADOS

Nogueira-Paiva, NC Resultados
47
ETAPA I 5.1 Hemoculturas
Para investigar potenciais diferenças em relação à virulência e patogenicidade de
diferentes subpopulações presentes no inóculo de uma cepa policlonal/mista do T. cruzi,
comum na infecção natural, este trabalho propôs obter, caracterizar e selecionar isolados com
comportamento, in vitro, distinto da cepa Be-78 parental e posteriormente, avaliar, in vivo, a
resposta inflamatória no coração e cólon ao longo da fase aguda da DCh experimental.
Camundongos Swiss infectados com 5x103 tripomastigotas sanguíneas, via
intraperitoneal, da cepa Be-78 do T. cruzi foram submetidos à coleta de sangue para
hemoculturas aos 3, 6 e 12 mai e as populações do parasito obtidas foram mantidas em
crescimento exponencial em meio LIT suplementado com 10% de SFB. A Tabela 1
representa os resultados das hemoculturas realizadas nos diferentes tempos de coleta. Embora
nem todos os animais tenham apresentado hemocultura positiva, foi possível isolar parasitos
em todos os tempos de coleta, ao longo de um ano de infecção, provavelmente devido às
diferenças na interação parasito-hospedeiro que permitiram ou não a manutenção de
parasitemia patente. Reforçando essa hipótese, alguns animais apresentaram positividade em
mais de um tempo (animais 1, 2, 5 e 15), entretanto, outros não foram positivos em nenhuma
das hemoculturas. Ao longo dos 12 meses de avaliação, houve perda de sete animais. Os
isolados obtidos nos diferentes tempos receberam um código de identificação demonstrado na
Tabela 2.

Nogueira-Paiva, NC Resultados
48
Tabela 1: Avaliação das hemoculturas realizadas aos 3, 6 e 12 meses após a infecção (mai) de camundongos Swiss com a cepa Berenice-78 do Trypanosoma cruzi.
Animais Hemoculturas
3MAI 6MAI 12MAI
1 - + +
2 - + +*
3 - - -
4 - - +*
5 + + -
6 - - -
7 - - -
8 - - -
9 - - -
10 - - -
11 - - -
12 - - -
13 - - +*
14 - - na
15 - + +
16 - na na
17 - - -
18 - na na
19 - - na
20 - - na
21 + na na
22 - - -
23 - na na
24 - - -
(na): não avaliado; (+) hemocultura positiva; (-) hemocultura negativa; (*) crescimento insuficiente em meio LIT.

Nogueira-Paiva, NC Resultados
49
Tabela 2: Código dos isolados obtidos a partir de hemoculturas aos 3, 6 ou 12 meses após a infecção de camundongos Swiss com a cepa Berenice-78 do Trypanosoma cruzi.
Hemocultura Animal Código
3mai 5 Be-78is5-3mai
21 Be-78is21-3mai
6mai
1 Be-78is1-6mai
2 Be-78is2-6mai
5 Be-78is5-6mai
15 Be-78is15-6mai
12mai 1 Be-78is1-12mai
15 Be-78is15-12mai
Be-78: Berenice-78; is: isolado; mai: meses após a infecção;
5.2 Curva de crescimento em meio de cultura acelular
Um dos parâmetros utilizados para identificar potenciais diferenças entre os isolados
obtidos foi a determinação do perfil de crescimento de formas epimastigotas cultivadas em
meio de cultura acelular, durante 20 dias após o inóculo de 107 parasitos/mL de cultura de
cada isolado, obtido nas hemoculturas realizadas aos 3, 6 ou 12 mai com a cepa Be-78 (cepa
Be-78 parental) do T. cruzi (Figura 11). Os perfis de crescimento dos isolados estão
apresentados em comparação à amostra da cepa Be-78 utilizada no inóculo (parental) e à cepa
Be-62.
Foi possível observar diferença significativa na área sob a curva (Area Under the
Curve-AUC) entre as duas cepas utilizadas como referência, com a cepa Be-78 (5,3x108)
parental apresentando maior número de parasitos que a cepa Be-62 (3,7x108). Além disso, os
isolados Be-78is15-6mai (4,2x108) e Be-78is1-12mai (4,2x108) apresentaram redução
significativa da AUC em relação à cepa Be-78 utilizada no inóculo (5,3x108), indicando que
essas duas subpopulações apresentam capacidade reduzida de crescimento em meio de cultura
em relação à cepa parental. O crescimento em meio de cultura simula o comportamento das
formas epimastigotas no intestino do hospedeiro invertebrado e, sendo assim, esses dados

Nogueira-Paiva, NC Resultados
50
podem refletir em uma habilidade reduzida de multiplicação dessas subpopulações, com
potenciais implicações sobre o sucesso da infecção vetorial (Figura 11).
Além da diferença na AUC, foi confirmado o comportamento diferenciado entre as
cepas Be-62 e Be-78 em culturas axênicas, evidenciando um crescimento com pico superior e
precoce e rápida entrada em fase de declínio, para a cepa Be-62, em comparação com a cepa
Be-78 (Figura 11).
Ao comparar as diferentes etapas de crescimento (Fase Lag, Log, Estacionária e de
Declínio) entre as subpopulações do parasito, independente da AUC, foi verificado que alguns
isolados, aqueles coletados 3mai (Be-78is5-3mai e Be-78is21-3mai), apresentaram perfil da
curva intermediário entre as cepa Be-62 e Be-78 parental (Figura 11), enquanto os demais
mostraram curvas mais próximas àquela exibida pela cepa Be-78 parental. Este dado
evidencia a presença, na população constituinte da cepa Be-78, de pelo menos uma
subpopulação que compartilha características biológicas com a cepa Be-62.
Considerando os distintos perfis de crescimento das formas epimastigotas observados
para as subpopulações avaliadas, a próxima pergunta desse trabalho refere-se ao
comportamento, em meio celular, para compreender como determinadas populações do
parasito comportam-se sob diferentes pressões ambientais.

Nogueira-Paiva, NC Resultados
51
Figura 11: Cinética de crescimento de epimastigotas, cultivadas em meio LIT no período de 0-20 dias, de isolados do parasito obtidos a partir de hemoculturas aos 3, 6 ou 12 meses após a infecção de camundongos Swiss com 5x103 formas tripomastigotas sanguíneas da cepa Be-78 do Trypanosoma cruzi. O eixo Y representa o número médio de parasitos x 107/mL quantificados em triplicata e o eixo X representa os dias de cultivo. As curvas de crescimento dos isolados são apresentadas em comparação às curvas da cepa utilizada no inóculo (Berenice-78) e da cepa Berenice-62. Be-62; Be-78 utilizada no inóculo (Be-78 parental); Be-78is5-3mai; Be-78is21-3mai; Be-78is1-6mai; Be-78is2-6mai; Be-78is5-6mai; Be-78is15-6mai; Be-78is1-12mai; Be-78is15-12mai. a: diferença significativa na Área sob a Curva (AUC) em relação à cepa Be-62; b: diferença significativa na AUC em relação à cepa parental Be-78. Os dados representam as medianas de triplicatas.
0 1 2 3 4 5 6 7 8 9 1011121314151617181920
2.0×10 07
4.0×10 07
6.0×10 07
8.0×10 07
1.0×10 08
0 1 2 3 4 5 6 7 8 9 1011121314151617181920
2.0×10 07
4.0×10 07
6.0×10 07
8.0×10 07
1.0×10 08
0 1 2 3 4 5 6 7 8 9 1011121314151617181920
2.0×10 07
4.0×10 07
6.0×10 07
8.0×10 07
1.0×10 08
0 1 2 3 4 5 6 7 8 9 1011121314151617181920
2.0×10 07
4.0×10 07
6.0×10 07
8.0×10 07
1.0×10 08
0 1 2 3 4 5 6 7 8 9 1011121314151617181920
2.0×10 07
4.0×10 07
6.0×10 07
8.0×10 07
1.0×10 08
0 1 2 3 4 5 6 7 8 9 1011121314151617181920
2.0×10 07
4.0×10 07
6.0×10 07
8.0×10 07
1.0×10 08
0 1 2 3 4 5 6 7 8 9 1011121314151617181920
2.0×10 07
4.0×10 07
6.0×10 07
8.0×10 07
1.0×10 08
1 2 3 4 5 6 7 8 9 101112131415161718192021220.0
2.0×10 07
4.0×10 07
6.0×10 07
8.0×10 07
1.0×10 08
Nú
mer
o d
e p
aras
itos
x10
7 /m
L d
e cu
ltura
Dias de cultivo
2
6
10
0
Be-78is5-3mai Be-78is21-3mai
Be-78is1-6mai Be-78is2-6mai
Be-78is5-6mai Be-78is15-6mai
Be-78is1-12mai Be-78is15-12mai
4
8
2
6
10
0
4
8
2
6
10
0
4
8
2
6
10
0
4
8
a
b
a
a
b

Nogueira-Paiva, NC Resultados
52
5.3 Perfis de infectividade e desenvolvimento in vitro
Para avaliar o comportamento em meio celular, os isolados foram submetidos à
avaliação dos perfis de infectividade e multiplicação em cultura de células Vero após 24, 48 e
72 horas de incubação.
A Figura 12 representa as taxas de infecção e desenvolvimento in vitro ao longo das
72 horas avaliadas. Todas as subpopulações foram observadas parasitando células Vero após
24 horas de incubação. De forma geral, as taxas de infectividade duplicaram a partir de 48
horas após o inóculo, correspondendo ao período de multiplicação do parasito no interior da
célula hospedeira. Em contrapartida, o isolado Be-78is2-6mai apresentou perfis de
infectividade e desenvolvimento intracelular três vezes superior às demais subpopulações já
no tempo de 24 horas, mantendo-se estável nos demais tempos avaliados. Entre 48 e 72 horas
após a incubação, as taxas de infectividade e multiplicação das subpopulações avaliadas
permaneceram estáveis, com exceção do aumento na taxa de desenvolvimento intracelular
observado para a cepa parental, sugerindo um comportamento mais lento em cultura de
células. Todas as outras subpopulações apresentaram perfil ascendente em relação ao número
de células infectadas, variando as taxas de multiplicação ao longo do tempo.
As Figuras 13 e 14 representam as taxas de infecção e desenvolvimento in vitro das
diferentes subpopulações avaliadas em comparação às cepas utilizadas como referência neste
trabalho, cepa Be-78 parental e Be-62, em cada tempo avaliado. A cepa Be-62 e os isolados
Be-78is5-3mai e Be-78is21-3mai foram capazes de parasitar mais células em relação à cepa
Be-78, 48 horas após o inóculo, no entanto, o número de amastigotas por célula infectada foi
semelhante entre as duas cepas de referência, refletindo, nesse momento, habilidade infectiva
inferior para a cepa parental. Após 72h de incubação, embora não haja diferença entre o
número de células infectadas, a taxa de multiplicação das formas amastigotas da cepa Be-78
foi superior à observada para a cepa Be-62 (p<0,001) e para as subpopulações isoladas aos
3mai (p<0,05). Esses dados mostram que a cepa Be-62 apresenta aumento precoce da
infectividade, em contrapartida, à maior capacidade replicativa in vitro da cepa Be-78 e à um
perfil de infectividade e desenvolvimento intracelular mais lento. Além disso, o
comportamento em cultura celular descrito para os isolados Be-78is5-3mai e Be-78is21-3mai
sugerem, de forma aditiva à curva de crescimento, mais uma semelhança entre esses isolados
e a cepa Be-62.
O isolado Be-78is2-6mai apresentou maior taxa de infectividade que as cepas Be-62
(p<0,001) e Be-78 (p<0,001) já no tempo de 24h, mantendo-se estável ao longo das 72 horas

Nogueira-Paiva, NC Resultados
53
avaliadas. O número de amastigotas foi maior em comparação às duas cepas de referência
(Be-62 e Be-78) nos tempos de 24 e 48h e, embora a contagem de formas intracelulares por
célula infectada não tenha sido diferente nesses dois tempos, o maior número de células
infectadas observado para este isolado permite que este seja considerado como portador de
um perfil de infectividade rápido e eficiente. Setenta e duas horas após o inóculo, o número de
amastigotas por célula infectada do isolado Be-78is2-6mai tornou-se inferior ao observado
para Be-78. Essa redução reflete o aumento da taxa de multiplicação das formas intracelulares
da cepa Be-78 observada neste tempo, no entanto, a taxa de infecção superior mantida pelo
isolado Be-78is2-6mai, reafirma o sucesso dessa subpopulação em desenvolver-se e invadir
novas células do tecido hospedeiro. Este comportamento parece ser resultado de uma seleção
exercida pela pressão do sistema imune do hospedeiro vertebrado (camundongo da linhagem
Swiss), capaz de favorecer a prevalência de uma subpopulação mais infectiva.
Os demais isolados não apresentaram alteração significativa no perfil de infectividade
em comparação às cepas Be-78 parental e Be-62, e ainda, mantiveram o número de
amastigotas por célula infectada inferior a estas cepas em todos os tempos avaliados, sendo
considerados isolados com baixas taxas de replicação. Uma exceção, entretanto, é o aumento
na taxa de desenvolvimento intracelular, observada 48 horas após o inóculo, para o isolado
Be-78is15-12mai em comparação à Be-78 parental e Be-62, demonstrando um pico de
proliferação precoce em relação à Be-78 parental e uma maior capacidade replicativa
comparada à Be-62. É importante ressaltar que esse isolado apresenta uma curva de
crescimento em meio acelular significativamente menor que a observada para a cepa Be-78
parental, ressaltando as especificidades comportamentais frente a diferentes pressões
ambientais.

Nogueira-Paiva, NC Resultados
54
0
5
10
15
0
200
400
600
0
20
40
60
80
100
Amostras/Tempo de cultura (horas)
Nú
mer
o d
e cé
lula
s in
fect
adas
/100
cé
lula
s
Nú
mer
o d
e am
asti
go
tas
Nú
mer
o d
e am
asti
go
tas/
Cél
ula
s in
fect
adas
A)
B)
C)
*
*
**
** * * *
*
*
#
*#
24 48 72 24 48 72 24 48 72 24 48 72 24 48 72 24 48 72 24 48 72 24 48 72 24 48 72 24 48 72
*
*
*#
*#
Figura 12: Ensaio de infecção in vitro em culturas de células Vero usando as cepas Berenice-62 ( ), Berenice-78 (parental, ) e subpopulações/isolados da cepa Berenice-78 do Trypanosoma cruzi, obtidos em hemoculturas realizadas aos 3, 6 ou 12 meses após a infecção de camundongos Swiss com a cepa parental (Be-78is5-3mai ; Be-78is21-3mai ; Be-78is1-6mai ; Be-78is2-6mai ; Be-78is5-6mai ; Be-78is15-6mai ; Be-78is1-12mai ; Be-78is15-12mai ). Após 18 horas de exposição ao parasito, as células foram lavadas para eliminar parasitos extracelulares e mantidas em meio até a coleta, fixação e coloração 24 (barra sólida, ), 48 (barra tracejada, ) ou 72 horas (barra pontilhada, ) após o inóculo. A) Número de células infectadas em 100 células contadas; B) Número de amastigotas intracelulares em 100 células contadas; C) Número de parasitos intracelulares por célula infectada. *: diferença significativa em relação à cultura de 24horas; #: diferença significativa em relação à cultura de 48horas. Os dados representam as médias de triplicatas ± erro padrão.

Nogueira-Paiva, NC Resultados
55
Amostras/Tempo de cultura (horas)
0
20
40
60
80
100
0
20
40
60
80
100
0
20
40
60
80
100
0
200
400
600
0
200
400
600
0
200
400
600
0
5
10
15
0
5
10
15
0
5
10
15
Nú
mer
o d
e am
asti
go
tas
Nú
mer
o d
e am
asti
go
tas/
Cél
ula
s in
fect
adas
24h 48h 72h
a, b
b b b
ba, b
a, b
b
a, b
a, b
bb
b b b b b
a, b
a, b
b bb
b b bb
b b
a aa
a
a
a
a
A)
B)
C)
Nú
mer
o d
e cé
lula
s in
fect
adas
/100
cé
lula
s
Figura 13: Ensaio de infecção in vitro em culturas de células Vero usando as cepas Berenice-62 ( ), Berenice-78 (parental, ) e subpopulações/isolados da cepa Berenice-78 do Trypanosoma cruzi, obtidos em hemoculturas realizadas aos 3, 6 ou 12 meses após a infecção de camundongos Swiss com a cepa parental (Be-78is5-3mai ; Be-78is21-3mai ; Be-78is1-6mai ; Be-78is2-6mai ; Be-78is5-6mai ; Be-78is15-6mai ; Be-78is1-12mai ; Be-78is15-12mai ). Após 18 horas de exposição ao parasito, as células foram lavadas para eliminar parasitos extracelulares e mantidas em meio até a coleta, fixação e coloração 24, 48 ou 72 horas após o inóculo. A) Número de células infectadas em 100 células contadas; B) Número de amastigotas intracelulares em 100 células contadas; C) Número de parasitos intracelulares por célula infectada. a: diferença significativa em relação à cepa Be-62; b: diferença significativa em relação à cepa parental Be-78. Os dados representam as médias de triplicatas ± erro padrão.
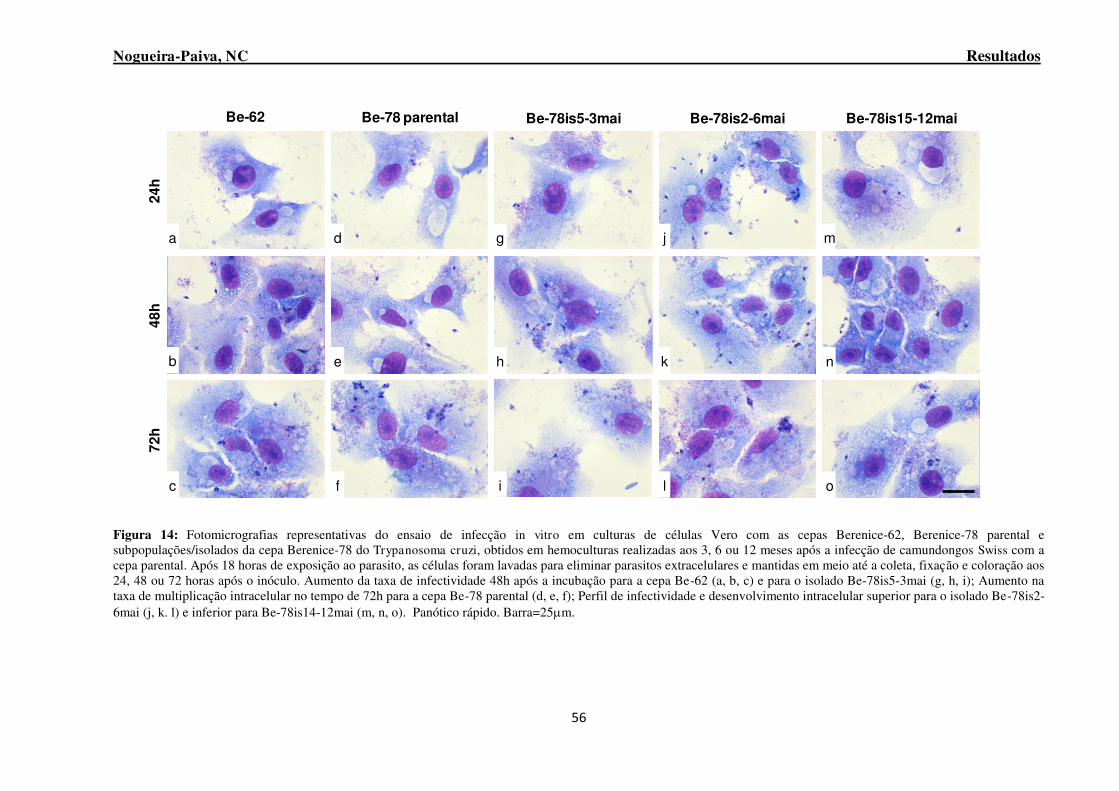
Nogueira-Paiva, NC Resultados
56
Figura 14: Fotomicrografias representativas do ensaio de infecção in vitro em culturas de células Vero com as cepas Berenice-62, Berenice-78 parental e subpopulações/isolados da cepa Berenice-78 do Trypanosoma cruzi, obtidos em hemoculturas realizadas aos 3, 6 ou 12 meses após a infecção de camundongos Swiss com a cepa parental. Após 18 horas de exposição ao parasito, as células foram lavadas para eliminar parasitos extracelulares e mantidas em meio até a coleta, fixação e coloração aos 24, 48 ou 72 horas após o inóculo. Aumento da taxa de infectividade 48h após a incubação para a cepa Be-62 (a, b, c) e para o isolado Be-78is5-3mai (g, h, i); Aumento na taxa de multiplicação intracelular no tempo de 72h para a cepa Be-78 parental (d, e, f); Perfil de infectividade e desenvolvimento intracelular superior para o isolado Be-78is2-6mai (j, k. l) e inferior para Be-78is14-12mai (m, n, o). Panótico rápido. Barra=25m.
Be-62 Be-78 parental Be-78is5-3mai Be-78is2-6mai Be-78is15-12mai
24h
48h
72h
a d g j m
b e h k n
c f i l o

Nogueira-Paiva, NC Resultados
57
5.4 Perfil de assinatura gênica
Para determinar se a pressão ambiental, exercida pelo sistema imune do hospedeiro
vertebrado durante a infecção crônica, foi capaz de selecionar isolados, não apenas com perfis
biológicos específicos, mas também geneticamente distintos, as subpopulações avaliadas
neste estudo foram submetidas à caracterização molecular por LSSP-PCR. A vantagem da
utilização dessa técnica refere-se à possibilidade de comparar os perfis gênicos dos isolados
obtidos na ETAPA I, aos perfis gênicos das subpopulações encontradas, parasitando os
diferentes órgãos, na infecção in vivo pela cepa Be-78 parental. Essa avaliação, juntamente
com a caracterização das lesões histológicas, permitirá esclarecer um pouco mais os aspectos
propostos pelo modelo histotrópico clonal, comparando possíveis especificidades
comportamentais, relacionadas ao histotropismo e à patogenicidade.
Foi possível confirmar os dados da literatura em relação à diferença genética entre as
amostras do parasito isoladas da paciente Berenice, com 16 anos de intervalo entre as coletas,
Be-62 e Be-78, adicionando padrões distintos de assinatura gênica, aos já descritos diferentes
perfis de isoenzimas, RAPD e microssatélites (Figura 15). Em relação às subpopulações da
cepa Be-78, obtidas por hemocultura em diferentes tempos durante a fase crônica, pode-se
observar que o isolado Be-78is5-3mai apresentou um perfil com bandas comuns tanto à cepa
Be-78 parental quanto à cepa Be-62. Essa banda, localizada próxima à região de 100pb,
inaparente no perfil gênico da cepa parental, foi visível no padrão de bandas da cepa Be-62
(Figura 15). Embora menos expressiva, essa banda também foi aparente nos isolados Be-
78is1-6mai e Be-78is5-6mai, revelando alguma similaridade entre essas amostras e a/as
subpopulações constituintes da cepa Be-62, no entanto, a presença de um perfil variado entre
as duplicatas pode ser explicada pelo fato de que os isolados não foram obtidos por
metodologia de clonagem e portanto podem ser formados por mais de uma subpopulação
(Figura 15).
De forma interessante, o isolados Be-78is5-3mai, que apresentou uma banda comum à
cepa Be-62 (Figura 15), mostrou comportamento em meios de cultura mais próximo à cepa
Be-62 do que o observado para a cepa Be-78 parental. Em contrapartida, o isolado Be-78is2-
6mai, que apresentou taxas de multiplicação e desenvolvimento in vitro significativamente
superiores à cepa Be-78 parental, foi geneticamente similar, pela técnica de LSSP-PCR, à
amostra do inóculo parental (Figuras 12, 13, 14 e 15), mostrando que mudanças no
comportamento biológico não necessariamente refletem variações genéticas.

Nogueira-Paiva, NC Resultados
58
Oito bandas obtidas na análise do perfil de assinatura gênica por LSSP-PCR foram
usadas para construção do fenograma a partir de UPGMA (Unweighted Pair Group Method
using Arithmetic averages). A escolha das bandas foi baseada em sua resolução,
reprodutibilidade entre as duplicatas e intensidade. O fenograma mostrou dois grupos
distintos (Figura 16), o grupo I, incluindo a cepa Be-78 parental e os isolados obtidos, em
diferentes tempos, ao longo da fase crônica experimental (Be-78is5-3mai, Be-78is21-3mai,
Be-78is1-6mai, Be-78is2-6mai, Be-78is5-6mai, Be-78is15-6mai, Be-78is1-12mai e Be-
78is15-12mai) e, o grupo II, compreendendo a cepa Be-62. No grupo I, as subpopulações da
cepa Be-78 subdividiram-se em dois grupos, sendo o isolado Be-78is5-3mai, o único a
distanciar-se da cepa parental. Os demais isolados formaram um grupo correlacionado
geneticamente com a cepa parental (Figura 16).
A análise do perfil de LSSP-PCR demonstrou a ocorrência de quatro bandas
compartilhadas entre a amostra Be-78 parental e o isolado Be-78is5-3mai, enquanto o número
de bandas compartilhadas entre a cepa Be-62 e o isolado Be-78is5-3mai foi igual a três. De
forma interessante, a cepa Be-78 parental compartilhou apenas 2 bandas com a cepa Be-62.
Estes dados indicam que o isolado Be-78is5-3mai é geneticamente mais próximo à cepa Be-
78 parental que à cepa Be-62, entretanto, a distância ente este isolado e a cepa Be-62 é menor
que a distância genética entre a cepa Be-78 parental e a cepa Be-62.
A próxima etapa do trabalho propõe investigar, in vivo, a influência do polimorfismo
observado para a cepa Be-78, utilizando a amostra parental e um isolado com variação no
perfil biológico e/ou molecular. De acordo com os resultados apresentados, os isolados Be-
78is5-3mai, Be-78is2-6mai e Be-78is15-12mai representam diferenças em nível biológico
e/ou molecular em relação à cepa parental, mostrando-se como potenciais candidatos a
avaliação de possíveis especificidade no comportamento in vivo (Tabela 3).

Nogueira-Paiva, NC _____________________________ _________ _Resultados
59
Figura 15: Assinaturas gênicas do fragmento de 330pb da região do minicírculo do kDNA reveladas em gel de poliacrilamida a 8% corado pela prata. Peso molecular (PM) de 100pb na canaleta 1 e demais canaletas referentes aos perfis gênicos dos isolados obtidos a partir de hemoculturas aos 3, 6 ou 12 meses após a infecção de camundongos Swiss com 5x103 formas tripomastigotas sanguíneas da cepa Be-78 do Trypanosoma cruzi em comparação à cepa Be-78 parental e à cepa Be-62. Be-62 (2-3); Be-78 parental (4-5); Be-78is5-3mai (6-7); Be-78is21-3mai (8-9); Be-78is1-6mai (10-11); Be-78is2-6mai (12-13); Be-78is5-6mai (14-15); Be-78is15-6mai (16-17); Be-78is1-12mai (18-19); Be-78is15-12mai (20-21); Cabeças de setas indicam as bandas, compartilhadas entre o isolado Be-78is5-3mai e as cepas de referência, Be-62 e Be-78 parental.
Be-
78is
53m
ai
Be-
78is
213m
ai
Be-
78is
16m
ai
Be-
78is
26m
ai
Be-
78is
56m
ai
Be-
78is
156m
ai
Be-
78is
112
mai
Be-
78is
1512
mai
PM
Be-
62
Be-
78
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21
100pb
200pb
300pb
400pb500pb
1000pb

Nogueira-Paiva, NC ___________ _______Resultados
60
Be-78is1-12mai
Be-78is15-12mai
Be-78is5-6mai
Be-78is15-6mai
Be-78
Be-78is21-3mai
Be-78is1-6mai
Be-78is2-6mai
Be-78is5-3mai
Be-62
Figura 16: Fenograma construído a partir de UPGMA (Unweighted Pair Group Method using Arithmetic averages) resultante das análises dos perfis de assinatura gênica obtido por LSSP-PCR de isolados obtidos a partir de hemoculturas aos 3 (Be-78is5-3mai; Be-78is21-3mai) , 6 (Be-78is1-6mai; Be-78is2-6mai; Be-78is5-6mai; Be-78is15-6mai) ou 12 (Be-78is1-12mai; Be-78is15-12mai) meses após a infecção de camundongos Swiss com 5x103 formas tripomastigotas sanguíneas da cepa Be-78 do Trypanosoma cruzi em comparação à cepa Be-78 parental e à cepa Be-62.

Nogueira-Paiva, NC ___________ _______Resultados
61
5.5 Representação esquemática do perfil in vitro e da caracterização gênica dos isolados
da cepa Be-78 parental (ETAPA I)
Foi possível identificar isolados com comportamento em cultura acelular ou celular, e
perfis de assinatura gênica, distintos do observado para a cepa Be-78 parental. Em alguns
casos, os isolados mostraram certo grau de semelhança com a cepa Be-62, isolada da paciente
Berenice, em fase crônica tardia, 16 anos antes do xenodiagnóstico que originou a cepa Be-
78.
Tabela 3: Resumo dos resultados obtidos na ETAPA I, comparando todos os parâmetros avaliados entre os isolados e as cepas Be-78 parental e Be-62 do Trypanosoma cruzi. pb: pares de base.
Subpopulação do T. cruzi
Curva de Crescimento
(meio de cultura acelular)
Infectividade e desenvolvimento intracelular
(meio de cultura celular)
Perfil LSSP-PCR
Be-62 precoce/rápido infectividade precoce (48h)
menor taxa de multiplicação
Be-78 parental tardio/lento infectividade tardia (72)
maior taxa de multiplicação
Be-78is5-3mai precoce/rápido infectividade precoce (48h)
menor taxa de multiplicação
Be-78is21-3mai precoce/rápido infectividade precoce (48h)
taxa de multiplicação intermediária
Be-78is1-6mai tardio/lento infectividade precoce (48)
menor taxa de multiplicação
Be-78is2-6mai tardio/lento infectividade precoce (24h)
maior taxa de multiplicação
Be-78is5-6mai tardio/lento infectividade baixa
menor taxa de multiplicação
Be-78is15-6mai menos replicativo infectividade baixa
menor taxa de multiplicação
Be-78is1-12mai tardio/lento infectividade baixa
menor taxa de multiplicação
Be-78is15-12mai menos replicativo infectividade baixa
menor taxa de multiplicação
100pb
200pb
100pb
200pb
100pb
200pb
100pb
200pb
100pb
200pb
100pb
200pb
100pb
200pb
100pb
200pb
100pb
200pb
100pb
200pb

Nogueira-Paiva, NC ___________ _______Resultados
62
ETAPA II
5.6 Parasitemia e sobrevida
Para avaliar se o comportamento in vitro do isolado selecionado na ETAPA I (Be-
78is5-3mai) seria reproduzido in vivo em comparação com as cepas de referência utilizadas
nesse trabalho, Be-62 e Be-78 parental do T. cruzi, camundongos BALB/c foram infectados
individualmente com essas subpopulações do T. cruzi e acompanhados ao longo da fase aguda
da infecção.
As curvas de parasitemia estão representadas na Figura 17, e os dados referentes ao
período pré-patente, período patente, dia e pico máximo de parasitemia estão demonstrados na
Tabela 4. Foi possível observar um perfil diferenciado entre as três subpopulações estudadas
já em relação à curva de parasitemia. Não houve diferença significativa no período pré-
patente referente aos diferentes grupos infectados, sendo observados parasitos no sangue
periférico cerca de 4 a 5 dias após o inóculo. A cepa Be-62 apresentou PP de 25 dias enquanto
animais infectados com a cepa Be-78 parental ou com o isolado Be-78is apresentaram
parasitos circulantes por 49 ou 35 de avaliação, respectivamente. O dia do pico de parasitemia
foi precoce (9 dai) e o número de parasitos observados nesse dia foi significativamente
superior nos animais infectados com a cepa Be-62 em relação aos demais grupos infectados.
Ainda em relação ao pico de parasitemia, na curva referente ao isolado Be-78is foi observado
pico máximo no 15º dai, ocorrendo com quatro dias de antecedência em relação do DPmáx
para a cepa Be-78 parental (19º dai). Embora precoce, o número máximo de parasitos
demonstrado pelo grupo Be-78is foi significativamente inferior ao observado para o grupo
infectado com a cepa parental (Tabela 4). A média da área sob a curva de parasitemia (AUC)
dos animais infectados com a cepa Be-62 (3,1 x106) foi significativamente superior àquela
observada para animais infectados com a cepa Be-78 parental (2,1 x106) ou mesmo com o
isolado Be-78is (0,71 x106) (Figura 17). Comparando a infecção com a cepa parental (Be-78
parental) à infecção com o isolado (Be-78 isolado), pôde-se notar que o isolado apresentou
AUC significativamente inferior (Figura 17).

Nogueira-Paiva, NC ___________ _______Resultados
63
Tabela 4: Determinação dos períodos pré-patente e patente, dia e pico máximo de parasitemia e área sob a curva nos animais infectados com as cepas Be-62, Be-78 ou isolado Be-78is5-3mai do Trypanosoma cruzi.
PPP PP DPmáx Pmáx AUC
Be-62 4 25 9 1.151.455 ±
424.241
3.089.682 ±
674.745
Be-78 isolado 5 49 15 75.000 ±
66.957 a, b
715.650 ±
377.948 a, b
Be-78
parental 5 38 19
173.455 ±
54.149 a
2.136.545 ±
464.028 a
a: diferença significativa em relação à Be-62; b: diferença significativa em relação à Be-78; PPP (Período pré-patente), PP (Período Patente), DPmáx (Dia do Pico máximo de parasitemia) expressos em dias; Pmáx (Pico máximo de parasitemia) expresso em número de tripomastigotas sanguíneos/0,1mL de sangue
Figura 17: Curvas de parasitemia de camundongos BALB/c infectados com o isolado Be-78is5-3mai ( ), ou com as cepas de referência Berenice-62 ( ) e Berenice-78 parental ( ) do Trypanosoma cruzi. Cada curva representa a média de 10 animais. a: diferença significativa na Área sob a Curva (AUC) em relação à cepa Be-62; b: diferença significativa na AUC em relação à cepa parental Be-78.
As taxas de sobrevida dos animais infectados com as cepas Be-62 e Be-78 do T. cruzi
estão apresentadas na Figura 18. Os animais infectados com a cepa Be-62 apresentaram
mortalidade a partir do 16º dai, sendo que, ao final de 28 dias, a taxa de mortalidade alcançou
90%. Animais infectados com o isolado ou com a cepa Be-78 parental apresentaram taxa de
sobrevida de 100% até o final da avaliação.
0 5 10 15 20 25 30 35 40 45 50 55 600
50000
100000
150000
200000
250000
300000
350000
400000600000.0
1200000.0
Dias após a infecção
Tri
po
mas
tig
ota
s/0,
1mL
de
san
gu
e
a
a, b

Nogueira-Paiva, NC ___________ _______Resultados
64
Figura 18: Taxa de sobrevida referente aos camundongos BALB/c infectados com o isolado Be-78is5-3mai ( ), ou com as cepas de referência Berenice-62 ( ) e Berenice-78 parental ( ) do Trypanosoma cruzi. Cada curva representa a avaliação de 10 animais ao longo de 28 dias após a infecção.
5.7 Carga parasitária no coração e cólon durante a fase aguda da doença de Chagas experimental
As subpopulações do T. cruzi avaliadas comportaram-se de forma específica em
relação à carga parasitária, que foi determinada pela técnica de PCR em tempo real. A
intensidade do parasitismo no cólon foi expressivamente superior ao observado no coração
para todas as subpopulações do T. cruzi e em todos os tempos avaliados (Figura 19). No
coração, animais infectados com a cepa Be-78 parental apresentaram parasitismo
significativamente superior aos demais grupos infectados, nos tempos de 14 e 28dai (Figura
19). Embora animais infectados com a cepa Be-78 parental tenham apresentado, no cólon,
densidade parasitária superior àquela observada para o coração, os animais infectados com a
cepa Be-62 ou com o isolado Be-78is-3mai demonstraram quantidade significativamente
superior de parasitos de tecido quando comparados ao grupo Be-78 parental. A cepa Be-62
mostrou-se ainda mais eficiente em parasitar o cólon que o isolado Be-78is5-3mai (Figura
19).
So
bre
vid
a (%
)
Dias após a infecção
0 5 10 15 20 25 30 35 40 45 50 55 600
50
100
150

Nogueira-Paiva, NC ___________ _______Resultados
65
Figura 19: Avaliação da carga parasitária no coração e cólon de camundongos BALB/c necropsiados ao longo da fase aguda (7, 14 ou 28 dias após a infecção-dai) da infecção experimental com o isolado Be-78is5-3mai, ou com as cepas de referência Berenice-62 e Berenice-78 parental do Trypanosoma cruzi. Os dados estão representados como Média ± Erro Padrão. Grupos com n=4. Be-62 ( ): grupo de animais infectados com a cepa Be-62; Be-78 isolado ( ): grupo de animais infectados com o isolado Be-78is5-3mai; Be-78 parental (
): grupo de animais infectados com a cepa Berenice-78. As linhas conectoras indicam diferenças significativas entre os grupos experimentais em um determinado tempo de avaliação; *: diferença significativa em relação à 7dai; #: diferença significativa em relação à 14dai.
5.8 Processo inflamatório no coração e cólon durante a fase aguda da doença de Chagas experimental
5.8.1 Intensidade do processo inflamatório
Para determinar se as distintas subpopulações do T. cruzi apresentaram especificidades
em relação à capacidade de provocar resposta inflamatória no coração e cólon dos animais
infectados, amostras desses órgãos, obtidas durante a necropsia, foram submetidas à avaliação
da intensidade do processo inflamatório em cortes histológicos corados pela técnica
Hematoxilina-Eosina.
No coração, foi possível detectar infiltrado inflamatório significativo nos animais
infectados com a cepa Be-62, 14 dias após a infecção (dai), sendo o processo inflamatório
mantido com intensidade semelhante no 28ºdai. Demonstrando um comportamento mais
tardio, animais infectados com a cepa Be-78 parental ou com o isolado Be-78is5-3mai
apresentaram infiltrado inflamatório significativo apenas aos 28dai em comparação aos
animais não-infectados. No entanto, nos animais pertencentes ao grupo Be-78is5-3mai, o
número de células inflamatórias foi significativamente inferior em relação aos animais dos
grupos Be-62 e Be-78parental no tempo 28dai (Figura 20 e 21). Embora nenhuma diferença
0
300
600
900
50000
100000
150000
10000000200000003000000040000000
0
10
20
30
40
50
300
600
900
Nú
mer
o d
e p
aras
ito
s/m
g d
e te
cid
o
Grupos experimentais /Tempo (dias após a infecção)
CORAÇÃO CÓLON
*
7 14 28 7 14 28
*
* #* #
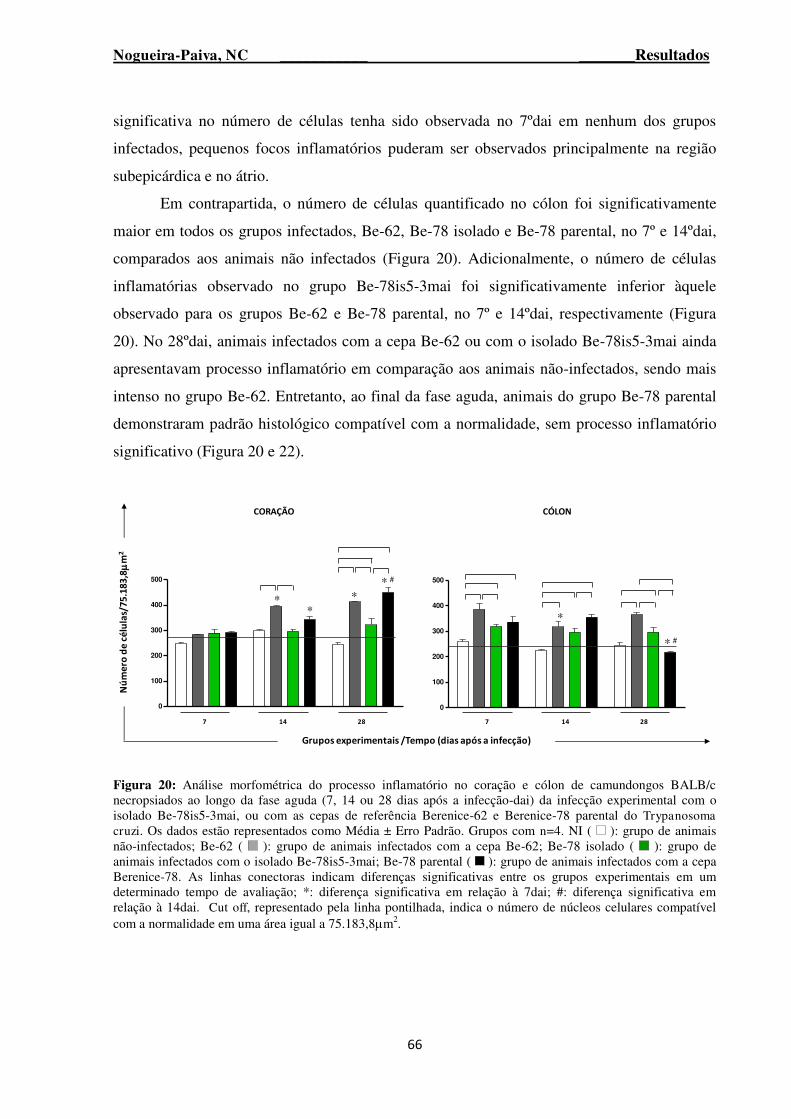
Nogueira-Paiva, NC ___________ _______Resultados
66
significativa no número de células tenha sido observada no 7ºdai em nenhum dos grupos
infectados, pequenos focos inflamatórios puderam ser observados principalmente na região
subepicárdica e no átrio.
Em contrapartida, o número de células quantificado no cólon foi significativamente
maior em todos os grupos infectados, Be-62, Be-78 isolado e Be-78 parental, no 7º e 14ºdai,
comparados aos animais não infectados (Figura 20). Adicionalmente, o número de células
inflamatórias observado no grupo Be-78is5-3mai foi significativamente inferior àquele
observado para os grupos Be-62 e Be-78 parental, no 7º e 14ºdai, respectivamente (Figura
20). No 28ºdai, animais infectados com a cepa Be-62 ou com o isolado Be-78is5-3mai ainda
apresentavam processo inflamatório em comparação aos animais não-infectados, sendo mais
intenso no grupo Be-62. Entretanto, ao final da fase aguda, animais do grupo Be-78 parental
demonstraram padrão histológico compatível com a normalidade, sem processo inflamatório
significativo (Figura 20 e 22).
Figura 20: Análise morfométrica do processo inflamatório no coração e cólon de camundongos BALB/c necropsiados ao longo da fase aguda (7, 14 ou 28 dias após a infecção-dai) da infecção experimental com o isolado Be-78is5-3mai, ou com as cepas de referência Berenice-62 e Berenice-78 parental do Trypanosoma cruzi. Os dados estão representados como Média ± Erro Padrão. Grupos com n=4. NI ( ): grupo de animais não-infectados; Be-62 ( ): grupo de animais infectados com a cepa Be-62; Be-78 isolado ( ): grupo de animais infectados com o isolado Be-78is5-3mai; Be-78 parental ( ): grupo de animais infectados com a cepa Berenice-78. As linhas conectoras indicam diferenças significativas entre os grupos experimentais em um determinado tempo de avaliação; *: diferença significativa em relação à 7dai; #: diferença significativa em relação à 14dai. Cut off, representado pela linha pontilhada, indica o número de núcleos celulares compatível com a normalidade em uma área igual a 75.183,8m2.
0
100
200
300
400
500
0
100
200
300
400
500
Nú
mer
o d
e cé
lula
s/75
.183
,8m
2
Grupos experimentais /Tempo (dias após a infecção)
CORAÇÃO CÓLON
7 14 28 7 14 28
**
* #
* #
*
*

Nogueira-Paiva, NC ___________ _______Resultados
67
Figura 21: Fotomicrografias representativas da análise semi-quantitativa do processo inflamatório em cortes histológicos do coração de camundongos BALB/c necropsiados ao longo da fase aguda (7, 14 ou 28 dias após a infecção-dai) da infecção experimental com o isolado Be-78is5-3mai, ou com as cepas de referência Berenice-62 e Berenice-78 parental do Trypanosoma cruzi. Aspecto histológico normal em animais não-infectados ao longo da fase aguda (a, b, c); presença de infiltrado inflamatório discreto em animais infectados com a cepa Be-62 no 7ºdai (d), evoluindo para moderado a intenso aos 14 (e) e 28dai (f); infiltrado inflamatório discreto em animais infectados com o isolado Be-78is5-3mai no 7ºdai (g) e 14ºdai (h), evoluindo para discreto a moderado aos 28dai (i); infiltrado inflamatório discreto em animais infectados com a cepas Be-78 parental no 7ºdai (j), evoluindo para moderado aos 14 (k) e intenso aos 28dai (l). NI: grupo de animais não-infectados; Be-62: grupo de animais infectados com a cepa Be-62; Be-78 isolado: grupo de animais infectados com o isolado Be-78is5-3mai; Be-78 parental: grupo de animais infectados com a cepa Berenice-78. Hematoxilina-Eosina. Barra= 50m.
Be-62 Be-78 parentalBe-78is5-3mai
7dai
14d
ai28
dai
NI
a d g j
b e h k
f i lc

Nogueira-Paiva, NC ___________ _______Resultados
68
Figura 22: Fotomicrografias representativas da análise semi-quantitativa do processo inflamatório em cortes histológicos do cólon de camundongos BALB/c necropsiados ao longo da fase aguda (7, 14 ou 28 dias após a infecção-dai) da infecção experimental com o isolado Be-78is5-3mai, ou com as cepas de referência Berenice-62 e Berenice-78 parental do Trypanosoma cruzi. Aspecto histológico normal em animais não-infectados ao longo da fase aguda (a, b, c); presença de infiltrado inflamatório focal de caráter discreto a moderado em animais infectados com a cepa Be-62 no 7ºdai (d), 14dai (e) e moderado a intenso no 28ºdai (f); infiltrado inflamatório discreto em animais infectados com o isolado Be-78is5-3mai no 7ºdai (g), 14ºdai (h) e 28ºdai (i); infiltrado inflamatório discreto em animais infectados com a cepas Be-78 parental no 7º (j) e 14ºdai (k), e aspecto compatível com a normalidade aos 28dai (l). NI: grupo de animais não-infectados; Be-62: grupo de animais infectados com a cepa Be-62; Be-78 isolado: grupo de animais infectados com o isolado Be-78is5-3mai; Be-78 parental: grupo de animais infectados com a cepa Berenice-78. Hematoxilina-Eosina. Barra= 50m.
Be-62 Be-78 parentalBe-78is5-3mai
7dai
14d
ai28
dai
NI
a d g j
b e h k
c f i l

Nogueira-Paiva, NC _ ___Resultados
69
5.8.2 Perfil fenotípico do processo inflamatório
Além da intensidade do processo inflamatório, a participação das distintas
subpopulações do T. cruzi na indução da resposta imune inata e adaptativa durante a fase
aguda da DCh experimental, foi avaliada através da determinação fenotípica e funcional do
infiltrado inflamatório, utilizando a técnica de imunofenotipagem por citometria de fluxo em
suspensão de células de coração e cólon.
5.8.2.1 Macrófagos
Aumento significativo no percentual de macrófagos foi observado no tecido muscular
cardíaco dos animais infectados com a cepa Be-62 nos tempos 14 e 28dai, quando comparado
aos demais grupos experimentais. Aos 14dai, animais do grupo infectado com o isolado Be-
78is5-3mai começaram a demonstrar percentual superior dessas células em relação aos
animais não-infectados, sustentando essa elevação no 28ºdai. A cepa Be-78 parental, por sua
vez, foi capaz de induzir aumento no percentual de macrófagos apenas no 28ºdai (Figura 23).
A análise ao longo do tempo no coração demonstrou ainda que, para o grupo Be-62,
no 28ºdai, embora ainda elevado, o percentual de macrófagos foi inferior em relação ao
observado 14dai.
No cólon, animais pertencentes ao grupo Be-62 e Be-78 isolado mostraram percentual
superior de macrófagos no 7º e 14ºdai em comparação com os grupos não-infectado e
infectado com a cepa Be-78 parental. O aumento observado nos animais infectados com a
cepa Be-62 aos 14dai foi superior àquele observado para o grupo Be-78 isolado. Entretanto,
este grupo, ao contrário do grupo Be-62, elevou a taxa de macrófagos no 28ºdai, mantendo-a
superior em todos os tempos avaliados. Animais do grupo Be-78 parental não apresentaram
alteração na densidade dos macrófagos ao longo da fase aguda (Figura 23).
A avaliação longitudinal no cólon revelou um perfil diferenciado entre as três
subpopulações estudadas, no qual, Be-62 retorna os níveis de macrófagos à normalidade ao
final da fase aguda, Be-78 isolado permanece com níveis elevados em todos os tempos e, Be-
78 parental, por sua vez, mantêm taxas compatíveis com a normalidade ao longo dos tempos
avaliados (Figura 23).
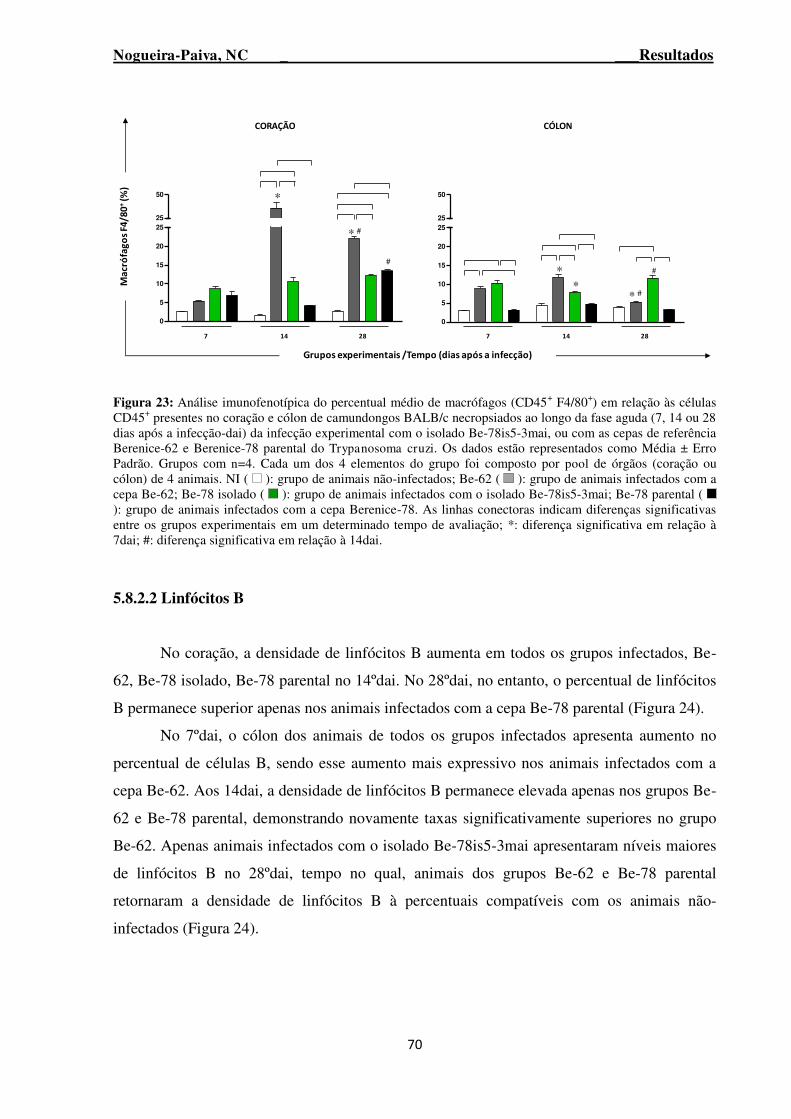
Nogueira-Paiva, NC _ ___Resultados
70
Figura 23: Análise imunofenotípica do percentual médio de macrófagos (CD45+ F4/80+) em relação às células CD45+ presentes no coração e cólon de camundongos BALB/c necropsiados ao longo da fase aguda (7, 14 ou 28 dias após a infecção-dai) da infecção experimental com o isolado Be-78is5-3mai, ou com as cepas de referência Berenice-62 e Berenice-78 parental do Trypanosoma cruzi. Os dados estão representados como Média ± Erro Padrão. Grupos com n=4. Cada um dos 4 elementos do grupo foi composto por pool de órgãos (coração ou cólon) de 4 animais. NI ( ): grupo de animais não-infectados; Be-62 ( ): grupo de animais infectados com a cepa Be-62; Be-78 isolado ( ): grupo de animais infectados com o isolado Be-78is5-3mai; Be-78 parental (
): grupo de animais infectados com a cepa Berenice-78. As linhas conectoras indicam diferenças significativas entre os grupos experimentais em um determinado tempo de avaliação; *: diferença significativa em relação à 7dai; #: diferença significativa em relação à 14dai.
5.8.2.2 Linfócitos B
No coração, a densidade de linfócitos B aumenta em todos os grupos infectados, Be-
62, Be-78 isolado, Be-78 parental no 14ºdai. No 28ºdai, no entanto, o percentual de linfócitos
B permanece superior apenas nos animais infectados com a cepa Be-78 parental (Figura 24).
No 7ºdai, o cólon dos animais de todos os grupos infectados apresenta aumento no
percentual de células B, sendo esse aumento mais expressivo nos animais infectados com a
cepa Be-62. Aos 14dai, a densidade de linfócitos B permanece elevada apenas nos grupos Be-
62 e Be-78 parental, demonstrando novamente taxas significativamente superiores no grupo
Be-62. Apenas animais infectados com o isolado Be-78is5-3mai apresentaram níveis maiores
de linfócitos B no 28ºdai, tempo no qual, animais dos grupos Be-62 e Be-78 parental
retornaram a densidade de linfócitos B à percentuais compatíveis com os animais não-
infectados (Figura 24).
0
5
10
15
20
2525
50
0
5
10
15
20
2525
50
Ma
cró
fago
s F4
/80+
(%)
Grupos experimentais /Tempo (dias após a infecção)
7 14 28 7 14 28
CORAÇÃO CÓLON
*
*#
* #*
#
* #

Nogueira-Paiva, NC _ ___Resultados
71
Figura 24: Análise imunofenotípica do percentual médio de linfócitos B (CD45+ CD19+) em relação às células CD45+ presentes no coração e cólon de camundongos BALB/c necropsiados ao longo da fase aguda (7, 14 ou 28 dias após a infecção-dai) da infecção experimental com o isolado Be-78is5-3mai, ou com as cepas de referência Berenice-62 e Berenice-78 parental do Trypanosoma cruzi. Os dados estão representados como Média ± Erro Padrão. Grupos com n=4. Cada um dos 4 elementos do grupo foi composto por pool de órgãos (coração ou cólon) de 4 animais. NI ( ): grupo de animais não-infectados; Be-62 ( ): grupo de animais infectados com a cepa Be-62; Be-78 isolado ( ): grupo de animais infectados com o isolado Be-78is5-3mai; Be-78 parental (
): grupo de animais infectados com a cepa Berenice-78. As linhas conectoras indicam diferenças significativas entre os grupos experimentais em um determinado tempo de avaliação; *: diferença significativa em relação à 7dai; #: diferença significativa em relação à 14dai.
5.8.2.3 Linfócitos T
a) Linfócitos T CD4+
No coração dos animais infectados com a cepa Be-62, pôde-se observar aumento
significativo no percentual de linfócitos T CD4+ ao longo de toda a fase aguda e, em relação a
todos os demais grupos, não-infectado, Be-78 isolado e Be-78 parental, sendo mais
expressivo no 14ºdai. A cepa Be-78 parental induziu elevação nos índices de células T CD4+
apenas aos 28dai. Contudo, não foi possível demonstrar alteração no percentual dessas células
nos animais infectados com o isolado Be-78is5-3mai em nenhum dos tempos avaliados
(Figura 25).
O perfil de células T CD4+ demonstrado no cólon foi, no entanto, o oposto do
observado no coração, onde o percentual dessas células foi superior no início da fase aguda
(7dai) apenas nos animais pertencentes ao grupo Be-78 parental em relação aos grupos NI,
Be-62 e Be-78 isolado. Os animais infectados com a cepa Be-62 ou com o isolado Be-78is5-
3mai apresentaram aumento nas taxas de linfócitos T CD4+ somente no 28ºdai (Figura 25).
0
1
2
3
4
5
6
0
1
2
3
4
5
6
Lin
fóci
tos
B C
D1
9+ (%
)
Grupos experimentais /Tempo (dias após a infecção)
7 14 28 7 14 28
CORAÇÃO CÓLON
*
* #
#
*
*
*
#

Nogueira-Paiva, NC _ ___Resultados
72
Figura 25: Análise imunofenotípica do percentual médio de Linfócitos T CD4+ (CD45+ CD3+ CD4+) em relação às células CD45+ presentes no coração e cólon de camundongos BALB/c necropsiados ao longo da fase aguda (7, 14 ou 28 dias após a infecção-dai) da infecção experimental com o isolado Be-78is5-3mai, ou com as cepas de referência Berenice-62 e Berenice-78 parental do Trypanosoma cruzi. Os dados estão representados como Média ± Erro Padrão. Grupos com n=4. Cada um dos 4 elementos do grupo foi composto por pool de órgãos (coração ou cólon) de 4 animais. NI ( ): grupo de animais não-infectados; Be-62 ( ): grupo de animais infectados com a cepa Be-62; Be-78 isolado ( ): grupo de animais infectados com o isolado Be-78is5-3mai; Be-78 parental ( ): grupo de animais infectados com a cepa Berenice-78. As linhas conectoras indicam diferenças significativas entre os grupos experimentais em um determinado tempo de avaliação; *: diferença significativa em relação à 7dai; #: diferença significativa em relação à 14dai.
b) Linfócitos T CD8+
De forma semelhante ao demonstrado para as células T CD4+ no coração dos animais
infectados com a cepa Be-62, pôde-se observar aumento significativo no percentual de
linfócitos T CD8+ ao longo de toda a fase aguda em relação aos grupos não-infectado e Be-78
isolado. Entretanto, a elevação destas células foi gradual e mostrou-se significativamente
superior no tempo de 28dai em comparação aos demais tempos avaliados. Animais infectados
com a cepa Be-78 parental ou com o isolado Be-78is5-3mai induziram elevação nos índices
de células T CD8+ apenas aos 28dai (Figura 26).
No cólon, animais infectados com o isolado Be-78is5-3mai apresentaram aumento no
percentual de linfócitos T CD8+, de forma gradual, em todos os tempos avaliados ao longo da
fase aguda. Os animais dos grupos Be-62 demonstraram elevação na densidade dessas células
aos 14 e 28dai, enquanto nos animais pertencentes ao grupo Be-78 parental, o aumento na
taxa de células CD8+ pode ser percebido, e discretamente, apenas no 14ºdai, sendo
significativamente inferior ao aumento observado nos demais grupos infectados (Figura 26).
0
5
10
15
2020
30
0
5
10
15
2020
30
Lin
fóci
tos
T C
D4
+ (%
)
Grupos experimentais /Tempo (dias após a infecção)
7 14 28 7 14 28
CORAÇÃO CÓLON
*
* #
#
* #

Nogueira-Paiva, NC _ ___Resultados
73
Figura 26: Análise imunofenotípica do percentual médio de Linfócitos T CD8+ (CD45+ CD3+ CD8+) em relação às células CD45+ presentes no coração e cólon de camundongos BALB/c necropsiados ao longo da fase aguda (7, 14 ou 28 dias após a infecção-dai) da infecção experimental com o isolado Be-78is5-3mai, ou com as cepas de referência Berenice-62 e Berenice-78 parental do Trypanosoma cruzi. Os dados estão representados como Média ± Erro Padrão. Grupos com n=4. Cada um dos 4 elementos do grupo foi composto por pool de órgãos (coração ou cólon) de 4 animais. NI ( ): grupo de animais não-infectados; Be-62 ( ): grupo de animais infectados com a cepa Be-62; Be-78 isolado ( ): grupo de animais infectados com o isolado Be-78is5-3mai; Be-78 parental ( ): grupo de animais infectados com a cepa Berenice-78. As linhas conectoras indicam diferenças significativas entre os grupos experimentais em um determinado tempo de avaliação; *: diferença significativa em relação à 7dai; #: diferença significativa em relação à 14dai.
5.8.3 Perfil fenotípico e funcional dos Linfócitos T
A frequência da expressão de IFN- nas células T é demonstrada na Figura 25 como
frequência de linfócitos T CD4+ IFN-+ ou linfócitos T CD8+ IFN-+ no coração e cólon.
Os animais do grupo Be-78 apresentaram níveis significativamente superiores de
células T CD4+ IFN-+ e CD8+ IFN-+, no coração, ao longo da fase aguda quando
comparados aos demais grupos experimentais, com produção mais elevada no 28ºdai.
Adicionalmente, uma discreta redução foi observada no 14ºdai, nesse grupo, em relação a
ambos os perfis. A cepa Be-62, por sua vez, foi capaz de induzir aumento na densidade de
linfócitos T CD4+ IFN-+ e CD8+ IFN-+ apenas no 28ºdai, e significativamente inferior ao
observado no grupo Be-78 parental, enquanto animais infectados com o isolado Be-78is5-
3mai não apresentaram alteração na expressão dessa citocina por nenhum dos fenótipos de
células T avaliados (Figura 27).
No cólon, padrão semelhante de frequência de linfócitos T CD4+ IFN-+ foi
demonstrado para o grupo Be-78 parental, no entanto o pico de expressão dessa citocina
ocorreu no 7ºdai. Linfócitos T CD8+ IFN-+ foram detectados em níveis superiores aos
0
5
10
15
2020
30
0
5
10
15
2020
30
Lin
fóci
tos
T C
D8
+ (%
)
7 14 28 7 14 28
CORAÇÃO CÓLON
Grupos experimentais /Tempo (dias após a infecção)
*
* #
#
* #* #
* #*
**
* #

Nogueira-Paiva, NC _ ___Resultados
74
animais não-infectados apenas no 28ºdai para o grupo infectado com a cepa Be-78 parental.
Ao contrário do observado no coração, animais do grupo Be-78 isolado apresentaram
aumento significativo de linfócitos T CD4+ IFN-+ e CD8+ IFN-+ em relação aos animais
infectados ao final da fase aguda. Em contrapartida, somente aos 28dai, a cepa Be-62 foi
capaz de estimular a síntese dessa citocina apenas por células T CD8+ (Figura 27).
Além disso, foi possível demonstrar uma redução na expressão de IFN- tanto por
linfócitos TCD4+ quanto por T CD8+ ao longo dos 28 dias de avaliação nos animais não
infectados (Figura 27).
Figura 27: Análise imunofenotípica e funcional do percentual médio de Linfócitos T CD4+ (CD45+ CD3+ CD4+) ou CD8+ (CD45+ CD3+ CD8+) expressando IFN- (IFN-+) em relação aos seus respectivos fenótipos auxiliar (CD4+) ou citotóxico (CD8+) presentes no coração e cólon de camundongos BALB/c necropsiados ao longo da fase aguda (7, 14 ou 28 dias após a infecção-dai) da infecção experimental com o isolado Be-78is5-3mai, ou com as cepas de referência Berenice-62 e Berenice-78 parental do Trypanosoma cruzi. Os dados estão representados como Média ± Erro Padrão. Grupos com n=4. Cada um dos 4 elementos do grupo foi composto por pool de órgãos (coração ou cólon) de 4 animais. NI ( ): grupo de animais não-infectados; Be-62 ( ): grupo de animais infectados com a cepa Be-62; Be-78 isolado ( ): grupo de animais infectados com o isolado Be-78is5-3mai; Be-78 parental ( ): grupo de animais infectados com a cepa Berenice-78. As linhas conectoras indicam diferenças significativas entre os grupos experimentais em um determinado tempo de avaliação; as letras indicam diferença significativa dentro de um mesmo grupo experimental ao longo dos tempos avaliados (a: diferença significativa em relação à 7dai; b: diferença significativa em relação à 14dai).
0
1
2
3
4
5
66
2020
30
0
1
2
3
4
5
66
2020
30
0
1
2
3
4
5
66
2020
30
0
1
2
3
4
5
66
2020
30
Lin
fóci
tos
T C
D8
+IF
N+
(%)
Lin
fóci
tos
T C
D4
+IF
N+
(%)
Grupos experimentais /Tempo (dias após a infecção)
CORAÇÃO CÓLON
7 14 28 7 14 28
* #
* #
*
*
#* #
* #
**
#
* #
* #
* ** #

Nogueira-Paiva, NC _ ___Resultados
75
5.9 Representação esquemática do perfil da infecção no coração e cólon (ETAPA II)
A Figura 28 representa esquematicamente o comportamento da infecção, com as
distintas subpopulações do T. cruzi utilizadas nesse trabalho, comparando a parasitemia, a
carga parasitária e o processo inflamatório ao longo da fase aguda.
Observa-se o perfil cardiomiotrópico e o retardo no controle da parasitemia na
infecção pela cepa Be-78 parental, seguido por processo inflamatório intenso ao final da fase
aguda.
A cepa Be-62, por sua vez, apresentou-se preferencialmente parasitando o cólon, com
parasitemia superior no início da fase aguda, no entanto, capaz de controlá-la precocemente.
Apesar da redução do parasitismo cardíaco, o processo inflamatório mostrou-se intenso nos
dois órgãos avaliados no 28ºdai para essa cepa.
O isolado Be-78is5-3mai, consistente com os dados da avaliação in vitro,
demonstraram durante a fase aguda da infecção de camundongos BALB/c, um
comportamento intermediário entre a cepas de referência utilizadas, apresentando virulência
(parasitemia e mortalidade) compatível com a cepa Be-78 parental, no entanto, perfil
histotrópico compartilhado com a cepa Be-62.
Um resumo do perfil imunofenotípico apresentado pelos diferentes grupos
experimentais está esquematizado na Figura 29. No coração, apesar de já observada no grupo
Be-62 aos 14dai, no 28º, a densidade de macrófagos, linfócitos T CD4+ e CD8+ é semelhante
entre as cepas Be-62 e Be-78 parental, com sobreposição de células T CD8+ em relação às
células T CD4+. O isolado Be-78is5-3mai apresentou resposta inflamatória, embora de menor
intensidade, também apenas no 28ºdai, composta essencialmente por macrófagos e linfócitos
T CD8+.
De forma contrária ao observado no coração, o perfil de resposta imunológica no
cólon foi cepa-dependente. Apenas o grupo Be-78 parental apresentou aumento precoce de
células T CD4+, no 7ºdai, que foram substituídas por células T CD8+ e linfócitos B, no 14ºdai.
No 28ºdai, não foi observado processo inflamatório no cólon. Todos esses resultados são
condizentes com o comportamento da parasitemia e do parasitismo tecidual nesse grupo.
Embora com menor intensidade, o perfil imunofenotípico detectado nos animais infectados
com o isolado Be-78is5-3mai foi próximo àquele demonstrado pelo grupo Be-62, e sobretudo,
distinto da cepa Be-78 parental. Nas primeiras duas semanas de infecção, o processo

Nogueira-Paiva, NC _ ___Resultados
76
inflamatório foi constituído principalmente por macrófagos e células T CD8+, com aumento
expressivo de linfócitos T CD4+ apenas ao final da fase aguda.
.

Nogueira-Paiva, NC ___________ ____ _Resultados
77
Figura 28: Representação esquemática dos resultados obtidos na ETAPA II, comparando o perfil da parasitemia, da carga parasitária e do processo inflamatório ao longo dos tempos avaliados apresentado por cada uma das diferentes subpopulações do T. cruzi utilizadas nesse estudo no coração e cólon de camundongos BALB/c. Os camundongos foram necropsiados durante da fase aguda (7, 14 ou 28 dias após a infecção-dai) da infecção experimental com o isolado Be-78is5-3mai, ou com as cepas de referência Berenice-62 e Berenice-78 parental do Trypanosoma cruzi. Cada dado (parasitemia, carga parasitária e número de células) foi separadamente transformado em percentual e os valores plotados na figura apenas para comparar o comportamento entre os grupos experimentais, sem considerar a intensidade em relação a cada variável analisada. Be-62: grupo de animais infectados com a cepa Be-62; Be-78 isolado: grupo de animais infectados com o isolado Be-78is5-3mai; Be-78 parental: grupo de animais infectados com a cepa Berenice-78; dai: dias após a infecção. ( ): parasitemia; ( ): carga parasitária (qPCR); ( ): processo inflamatório (HE).
7 14 28DAI
7 14 28DAI
7 14 28DAI
7 14 28DAI
7 14 28DAI
7 14 28DAI
Be-
78 is
ola
do
Be-
62B
e-78
par
enta
lCORAÇÃO CÓLON
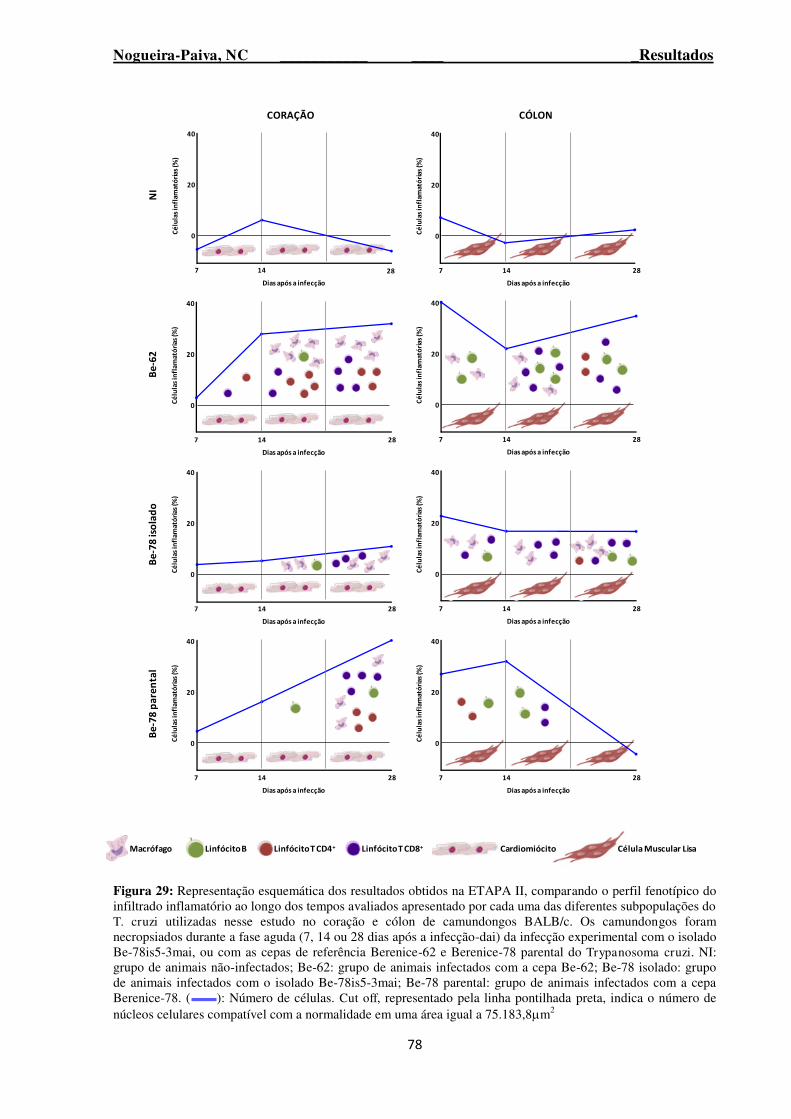
Nogueira-Paiva, NC ___________ ____ _Resultados
78
Figura 29: Representação esquemática dos resultados obtidos na ETAPA II, comparando o perfil fenotípico do infiltrado inflamatório ao longo dos tempos avaliados apresentado por cada uma das diferentes subpopulações do T. cruzi utilizadas nesse estudo no coração e cólon de camundongos BALB/c. Os camundongos foram necropsiados durante a fase aguda (7, 14 ou 28 dias após a infecção-dai) da infecção experimental com o isolado Be-78is5-3mai, ou com as cepas de referência Berenice-62 e Berenice-78 parental do Trypanosoma cruzi. NI: grupo de animais não-infectados; Be-62: grupo de animais infectados com a cepa Be-62; Be-78 isolado: grupo de animais infectados com o isolado Be-78is5-3mai; Be-78 parental: grupo de animais infectados com a cepa Berenice-78. ( ): Número de células. Cut off, representado pela linha pontilhada preta, indica o número de núcleos celulares compatível com a normalidade em uma área igual a 75.183,8m2
Be-
78 is
ola
do
Be-
62
CORAÇÃO CÓLON
NI
Dias após a infecção
7 14 28
40
20
0
Cé
lula
s in
flam
ató
rias
(%)
7 14 28
40
20
0
Cé
lula
s in
flam
ató
rias
(%)
Dias após a infecção
Be-
78
pa
ren
tal
7 14 28
40
20
0
Cé
lula
s in
flam
ató
rias
(%)
Dias após a infecção
7 14 28
40
20
0
Cé
lula
s in
flam
ató
rias
(%)
Dias após a infecção
7 14 28
40
20
0
Cé
lula
s in
flam
ató
rias
(%)
Dias após a infecção
7 14 28
40
20
0
Cé
lula
s in
flam
ató
rias
(%)
Dias após a infecção
7 14 28
40
20
0
Cé
lula
s in
flam
ató
rias
(%)
Dias após a infecção
7 14 28
40
20
0
Cé
lula
s in
flam
ató
rias
(%)
Dias após a infecção
Macrófago Linfócito T CD8+Linfócito T CD4+Linfócito B Cardiomiócito Célula Muscular Lisa

79
6. DISCUSSÃO

Nogueira-Paiva, NC Discussão
80
De acordo com a classificação atual das subpopulações do T. cruzi em Discrete Typing
Units (DTU), ambas as cepas de referência utilizadas nesse trabalho, Be-62 e Be-78,
pertencem a DTU Tc II (Zingales et al., 2009; Zingales et al., 2012). Embora essas duas
linhagens sejam suficientemente relacionadas para serem classificadas dentro do mesmo
DTU, elas apresentam peculiaridades já descritas na literatura que incluem, morfologia,
comportamento biológico e composição isoenzimática (Brener, 1965; Lana, 1981; Lana e
Chiari, 1986). A cepa Be-62 é composta, predominantemente, por formas delgadas, apresenta
pico de parasitemia em torno do sétimo dia e taxa de mortalidade de 100% em 15 dias após a
infecção em camundongos (Brener, 1965; Lana, 1981; Lana e Chiari, 1986), enquanto a cepa
Be-78 é composta por formas largas, apresenta parasitemia com pico mais tardio (dia 15) e
uma taxa de sobrevida de 100% (Lana, 1981; Lana e Chiari, 1986; Carneiro et al., 2007;
Guedes et al., 2007). Quando introduzida em meio de cultura acelular, a cepa Be-78
apresentou menor capacidade de crescimento e altos índices de diferenciação, enquanto que,
nas mesmas condições a cepa Be-62 mostrou uma alta capacidade de multiplicação e taxas
médias de diferenciação (Lana, 1981; Lana e Chiari, 1986).
Além disso, dados obtidos em nosso laboratório mostraram que outras linhagens do T.
cruzi pertencentes ao mesmo DTU (Tc II), as cepas Y e Be-78, comportaram-se drasticamente
diferente em relação à patogenicidade nos principais órgãos afetados pela DCh, coração e
trato digestório, em infecção experimental de cães da raça Beagle (Veloso et al., 2008;
Nogueira-Paiva et al., 2014). No trato digestório, animais infectados com a cepa Y mostram
controle do parasitismo e homeostasia da resposta imunológica ao final da fase aguda,
enquanto animais infectados com a cepa Be-78 mantiveram parasitismo dois anos após a
infecção e apresentaram lesões de caráter aparentemente progressivo (Nogueira-Paiva et al.,
2014). As subpopulações presentes em cepas policlonais podem interagir de forma específica
com o hospedeiro, o que pode interferir com o desenvolvimento da resposta imune e
determinar o sucesso em controlar o parasitismo tecidual e as lesões associadas.
Têm sido descrito na literatura que após a infecção com uma cepa policlonal do T.
cruzi, a interação parasito-hospedeiro pode conduzir à ausência de parasitemia e/ou
parasitismo tecidual em alguns indivíduos, enquanto outros com hemoculturas positivas
podem diferir em relação ao desenvolvimento da resposta imunopatogênica, com aspectos
particulares de histotropismo, lesões histológicas, manifestações clínicas e prognóstico da
DCh (Silva et al., 2013). Um dos fatores investigados como potencial responsável é a genética
relacionada tanto ao hospedeiro quanto ao parasito (Andrade, 1990; Vago et al., 1996b; Vago

Nogueira-Paiva, NC Discussão
81
et al., 2000; Macedo et al., 2002; Silva et al., 2013). Variações, com importância
epidemiológica, dos padrões de distribuição geográfica das diferentes linhagens ou
subpopulações e do amplo espectro clínico da DCh têm apontando o polimorfismo
intraespecífico do T. cruzi como um dos fatores importantes para o prognóstico da doença.
Essas distintas subpopulações presentes no inóculo durante a infecção natural, segundo o
modelo histotrópico clonal, podem apresentar características intrínsecas com repercussão
sobre o estabelecimento da infecção e colonização dos tecidos (Macedo et al., 2002). Embora
estudos genéticos possam comprovar a heterogeneidade intraespecífica do parasito, avaliações
do comportamento biológico e da interação parasito-hospedeiro, poderiam esclarecer a
importância das diferentes cepas, na determinação das manifestações clínico-epidemiológicas
da doença (Andrade e Magalhaes, 1996). Assim sendo, a participação de diferentes
subpopulações do parasito no processo imunopatogênico durante a fase aguda da DCh
experimental foi investigada nesse estudo, dividido em duas etapas.
Na primeira etapa, a fim de testar a capacidade de um indivíduo em selecionar uma
subpopulação a partir de uma cepa policlonal do T. cruzi, 24 camundongos Swiss foram
infectados com cepa Be-78 e amostras de sangue coletadas por hemoculturas foram avaliadas
em três momentos após a infecção, durante a fase crônica recente (3 meses após a infecção-
mai), intermediária (6mai) e tardia (12mai). Os parasitos obtidos nas hemoculturas positivas
foram caracterizados in vitro quanto à cinética de crescimento em meio acelular, à
infectividade e desenvolvimento intracelular em meio de células Vero e ao perfil de assinatura
gênica por LSSP-PCR. A linhagem heterogênea de camundongos Swiss foi selecionada para o
presente estudo, em vez de uma população isogênica, devido à sua variabilidade genética
condizente com o perfil da população de pacientes com DCh.
Nossos resultados demonstraram que as assinaturas gênicas das cepas Be-62 e Be-78
parental, obtidas por LSSP-PCR, foram diferentes, confirmando a heterogeneidade genética
dessas duas cepas isoladas da mesma paciente com um intervalo de 16 anos. Nossos dados
complementam àqueles anteriormente descritos na literatura, usando caracterização dos perfis
isoenzimáticos ou caracterização de microssatélites (Lana, 1981; Lana e Chiari, 1986; De
Lana et al., 1996; Cruz et al., 2006; Valadares et al., 2012). As amostras das cepas Be-62 e
Be-78 têm sido mantidas em nosso laboratório, ao longo dos últimos 30 anos, em passagens
alternadas entre camundongos e cultura axênica, com intervalos onde as mesmas são
criopreservadas. Nós gostaríamos de ressaltar que a alternância entre a manutenção, in vivo ou
in vitro, não tem alterado a virulência, a patogenicidade ou o perfil molecular dessas cepas, o

Nogueira-Paiva, NC Discussão
82
que tem sido constantemente demonstrado em diversos trabalhos do nosso grupo (Lana e
Chiari, 1986; Veloso et al., 2005; Carneiro et al., 2007; Veloso et al., 2008; Vieira et al.,
2009; Valadares et al., 2012; Vieira et al., 2012).
O perfil de kDNA de um dos isolados, Be-78is5-3mai, compartilhou bandas
específicas com ambas as cepas de referência, Be-62 e Be-78 parental (Figura 15 e 16). Além
disso, esse mesmo isolado apresentou comportamento em meio de cultura acelular (meio LIT)
ou celular (células Vero) mais semelhante ao perfil da cepa Be-62 que àquela do grupo Be-78
(Tabela 3). Em contrapartida, o isolado Be-78is2-6mai, que mostrou taxas de infectividade e
desenvolvimento intracelular cerca de 3 vezes superior à cepa Be-78 parental, foi similar em
relação à assinatura gênica obtida por LSSP-PCR, demonstrando que mudanças no
comportamento biológico não necessariamente refletem variações genéticas (Tabela 3).
O isolado Be-78is5-3mai, que mais se distanciou geneticamente da cepa parental, foi
coletado do animal 5 aos 3 meses após a infecção-mai. Outro isolado foi originado desse
mesmo animal aos 6mai, Be-78is5-6mai. Embora esse isolado também tenha apresentado, em
seu perfil gênico, uma banda discreta próxima à região de 100pb similar à cepa Be-62, suas
propriedades biológicas in vitro foram similares à cepa Be-78 parental. Esta observação
sugere um processo de reversibilidade da modulação genética ao longo do tempo durante a
infecção crônica por T. cruzi, em que diferentes subpopulações podem ser favorecidas em um
determinado momento em detrimento de outras (Veloso et al., 2005; Veloso et al., 2012).
A manutenção dos isolados em meio de cultura foi realizada durante pouco tempo a
fim de minimizar sua influência sobre o processo de seleção. Assim, consideramos que a
pressão exercida pelo sistema imune do hospedeiro vertebrado (camundongo da linhagem
Swiss) seja a variável principal responsável por favorecer o processo de seleção, observado
nesse trabalho para o isolado Be-78is5-3mai. No entanto, é importante considerar a
possibilidade de que a hemocultura e a manutenção dos isolados in vitro, realizados
previamente à análise fenotípica e molecular, possam ter exercido alguma influência adicional
à seleção, alterando a organização de suas subpopulações podendo favorecer a expansão ou a
redução de determinados clones presentes na amostra parental (Macedo & Pena, 1998), mas
não a constituição de subclones da amostra obtida por hemocultura.
Além da possibilidade de que o isolado Be-78is5-3mai seja uma subpopulação
selecionada dentro da população policlonal da cepa Be-78 parental, o comportamento em
meios de cultura e o perfil gênico intermediários, observados para esse isolado, em relação às
cepas de referência, Be-78 parental e Be-62, também podem ser atribuídos à uma associação

Nogueira-Paiva, NC Discussão
83
entre subclones, com participação de dois ou mais subclones, determinando um perfil
biológico e molecular misto. A caracterização por microssatélites da cepa Be-78 realizada por
Valadares et al. (2012), de forma semelhante ao observado nesse estudo, confirmou o status
multiclonal da cepa Be-78, e sugeriu a presença de uma subpopulação, dentro da amostra de
Be-78, semelhante à Be-62.
Nossos dados confirmam a natureza policlonal ou mista da cepa Be-78, já relatada na
literatura (Veloso et al., 2005; Valadares et al., 2012; Veloso et al., 2012), e reforça a hipótese
de que a pressão seletiva exercida pelo hospedeiro durante a fase crônica pode favorecer o
desenvolvimento de uma ou mais subpopulação(s) sobre a(s) outra(s). Este fenômeno pode
estar relacionado com a plasticidade do T. cruzi e não menos importante, à interação parasito-
hospedeiro, uma vez que todos os animais receberam o mesmo inóculo e responderam à
infecção selecionando subpopulações distintas.
Três isolados obtidos após a infecção de camundongos Swiss com a cepa Be-78, Be-
78is5-3mai, Be-78is2-6mai e Be-78is15-12mai, mostraram diferenças nos parâmetros
biológicos e/ou moleculares avaliados em comparação com a cepa Be-78 parental (Tabela 3),
no entanto, um deles em especial, o isolado Be-78is5-3mai, parece apresentar características
peculiares que o colocam como principal candidato para investigação da influência do
polimorfismo genético, dentro de uma cepa policlonal, sobre a imunopatogênese da DCh.
Dessa forma, na segunda etapa do nosso trabalho, camundongos isogênicos, da linhagem
BALB/c, foram infectados com um dos isolados selecionados na primeira etapa, Be-78is5-
3mai e com as cepas de referência Be-78 parental e Be-62 com o objetivo de investigar a
participação de diferentes subpopulações do parasito no processo patogênico, no que diz
respeito à modulação da resposta imune e ao desenvolvimento das lesões histológicas durante
a fase aguda na tentativa de esclarecer aspectos propostos pelo modelo histotrópico clonal,
comparando possíveis especificidades comportamentais, relacionadas ao histotropismo e à
patogenicidade.
Foi possível confirmar o comportamento distinto da curva de parasitemia e taxa de
sobrevida entre as cepas de referência utilizadas nesse trabalho, Be-62 e Be-78 parental.
Animais infectados com a cepa Be-62 apresentaram pico de parasitemia precoce e
significativamente superior, em relação aos animais do grupo Be-78 parental. Além disso, a
mortalidade alcançou índice de 92% no grupo Be-62 no 28ºdai, em detrimento à 0% nos
demais grupos. Nossos dados corroboram com aqueles descritos por Lana e Chiari (1986),
que observaram padrão semelhante para essas cepas em camundongos de linhagens distintas,

Nogueira-Paiva, NC Discussão
84
CH3 isogênicos ou albinos, tanto para a parasitemia quanto para a taxa de sobrevida. Embora
com PPP, Pmáx e DPmáx distintos, Bahia et al. (2002) também descreveram comportamento
tardio da cepa Be-78 em comparação com a cepa Be-62 em cães infectados com
tripomastigotas sanguíneas pela via intraperitoneal.
Concordando com os dados de Camandaroba et al. (2006), que demonstraram o
mesmo padrão de parasitemia para cinco dos sete clones da cepa Colombiana analisados, os
animais infectados com o isolado Be-78is5-3mai apresentaram PPP, PP, Pmáx e DPmáx
semelhantes à cepa Be-78 parental. No entanto, houve uma redução significativa na área sob a
curva, sugerindo infectividade inferior para o isolado Be-78is5-3mai quando comparado à
cepa Be-78 parental. Campos e Andrade (1996) e Camandaroba et al. (2001), ainda que
utilizando método de isolamento distinto, também detectaram alterações nas curvas
parasitêmicas de clones das cepas Colombiana e 21 SF, respectivamente, onde alguns dos
clones apresentaram menor virulência, com menores taxas de parasitemia e mortalidade, em
relação à cepa parental. Em nosso estudo, a baixa infectividade pode ser uma característica
inerente ao isolado ou pode ainda estar relacionada ao número reduzido de passagens em
culturas ou camundongos, que foi necessário neste trabalho para evitar variáveis adicionais ao
processo de seleção. A ascensão lenta e gradual da parasitemia de subpopulações
recentemente isoladas tem sido descrita na literatura por outros autores (Lana e Chiari, 1986;
Camandaroba et al., 2001). No entanto, na maioria das vezes, não estão associadas à
alterações no tropismo e imunogenicidade.
De forma semelhante aos resultados apresentados neste trabalho, diferentes autores
observaram taxa de sobrevida igual a 100% para animais infectados com a cepa Be-78, em
contrapartida a 100% de mortalidade para camundongos infectados com a cepa Be-62
(Brener, 1965; Lana e Chiari, 1986). Os animais infectados com o isolado Be-78is5-3mai
apresentaram sobrevida compatível com a cepa Be-78 parental, o que também foi observado
por outros autores para clones obtidos da cepa Colombiana e 21 SF, sugerindo estabilidade da
cepa parental e indicando homogeneidade clonal de suas subpopulações (Campos e Andrade,
1996; Camandaroba et al., 2001; Camandaroba et al., 2006).
Cepas do T. cruzi representam um equilíbrio entre as subpopulações, sejam elas
homogêneas ou heterogêneas, onde ocorre a predominância de um clone principal, que é
responsável por um padrão de comportamento da cepa parental e de seus clones (Engel et al.,
1982; Gomes et al., 1991; Campos e Andrade, 1996; Camandaroba et al., 2001; Camandaroba
et al., 2006). Entretanto, segundo o modelo histotrópico clonal, nem todos os clones são

Nogueira-Paiva, NC Discussão
85
capazes de estabelecer a infecção, e aqueles que conseguem ainda podem competir entre si e
colonizar tecidos distintos (Macedo e Pena, 1998; Macedo et al., 2002). A capacidade de
diferentes subpopulações em interagir com tecidos específicos poderia ser um fator
determinante para o desenvolvimento do amplo espectro anatomoclínico da DCh. Para testar
essas duas hipóteses, onde o isolado Be-78is5-3mai represente um clone principal
apresentando virulência e patogenicidade compatíveis com a cepa de origem ou ainda
apresente-se como uma subpopulação que interaja de forma específica com os tecidos do
hospedeiro, a infecção experimental com o isolado foi ainda investigada em relação ao
histotropismo e à intensidade e fenótipo da resposta inflamatória nos principais órgãos
acometidos pela DCh.
A cepa Be-62 inicia precocemente a infecção de células no tecido cardíaco já aos 7dai,
mas esse parasitismo é reduzido ao final da fase aguda (28dai). Ao contrário, a cepa Be-78
parental apresenta aumento da carga parasitária cardíaca aos 14dai, mantendo-a elevada até o
28ºdai. Esse comportamento está diretamente relacionado ao perfil da curva de parasitemia
dessas duas cepas, onde se pode demonstrar compatibilidade entre os dias onde são
observados índices parasitêmicos e parasitismo tecidual maiores. Somado a isso, também
nota-se a queda do parasitemia em consequência ao controle da replicação do parasito pelo
sistema imune do hospedeiro, com redução do parasitismo tecidual.
Inversão no histotropismo entre as cepas Be-62 e Be-78 parental foi observada no
cólon, onde se pode perceber que a cepa Be-62 aumenta gradualmente o parasitismo tecidual
nesse órgão ao longo da fase aguda, alcançando pico no 28ºdai, enquanto a cepa Be-78 tem
sucesso na infecção já no 7ºdai, com flutuações até mostrar-se significativamente inferior em
relação aos demais grupos infectados no 28ºdai. Diferente do coração, o perfil da carga
parasitária no cólon parece estar relacionado ao momento onde ocorre o controle da
multiplicação do parasito pela resposta imunológica do hospedeiro. Dados obtidos por Lewis
et al. (2015) têm identificado o trato gastrointestinal como principal nicho onde o parasito
permanece, evadindo do sistema imune, durante a infecção crônica. Portanto, nossos dados
apontam uma associação entre o comportamento mais rápido da cepa Be-62, no que se refere
à infectividade, taxa de multiplicação e controle precoce da parasitemia (25dai) e o intenso
parasitismo no cólon observado no 28ºdai. Da mesma forma, o perfil tardio da cepa Be-78
parental parece ser o responsável pela manutenção de uma carga parasitária inferior à
observada para cepa Be-62 até o final da fase aguda. Uma vez que o controle da parasitemia
nos animais infectados com a cepa parental ocorreu apenas no 38ºdai, a avaliação da carga

Nogueira-Paiva, NC Discussão
86
parasitária no cólon desses animais em tempos mais tardios poderia revelar se a cepa Be-78
apresentaria histotropismo preferencial para o coração ou se, de forma semelhante às demais
subpopulações investigadas, evadiria o sistema imune “escondendo-se” no TGI na fase
crônica da infecção.
Dados obtidos em nosso laboratório no modelo cão, comparando achados
histopatológicos no coração, esôfago e cólon, revelaram uma preferência por órgãos do TGI
para a cepa Be-78, sustentando a participação de aspectos genéticos do hospedeiro sobre
aspectos patogênicos das distintas formas clínicas da DCh (Veloso et al., 2008; Nogueira-
Paiva et al., 2014). Corroborando com esses dados, a análise das respostas de diferentes
linhagens de camundongos, C57BL/6 e Swiss versus BALB/c e DBA-2, à infecção com uma
mistura das cepas JG e Coll.7G2 (Andrade et al., 2002) mostraram que, para os camundongos
BALB/c e DBA-2, o mesmo padrão de distribuição tecidual foi observado, com
predominância da cepa JG no coração e Col1.7G2 no reto e demais tecidos. No entanto,
camundongos C57BL/6 e Swiss apresentaram maior parasitismo cardíaco em relação à cepa
Col1.7G2, enquanto que, a cepa JG parasitou preferencialmente o reto e demais tecidos.
Dessa forma, a distinta distribuição tecidual observada para BALB/c e DBA-2 versus
C57BL/6 e Swiss foi associada às diferenças entre os perfis genéticos destes animais (Andrade
et al., 2002).
De forma interessante, o isolado Be-78is5-3mai apresentou, tanto no coração quanto
no cólon, comportamento mais próximo à cepa Be-62, com parasitismo reduzido no coração e
elevado no cólon no 28ºdai. Esses dados também reforçam a associação entre o perfil da curva
de parasitemia e o escape do parasito para o TGI, uma vez que, o DPmáx foi anterior ao
observado para a cepa Be-78 parental e, embora parasitos no sangue tenham sido observados
durante 45 dias, os índices parasitêmicos mostravam-se bem baixos a partir do 20ºdai. Essa
alteração no histotropismo demonstrada do isolado em comparação à cepa parental,
contrariam os resultados obtidos por Campos et al. (1996) e Camandaroba et al. (2001; 2006),
cujos clones das cepas Colombiana e 21 SF, respectivamente, apresentaram histotropismo
semelhante às suas cepas parentais. Essas diferenças podem estar relacionadas ao método
utilizado no isolamento das subpopulações, obtendo subpopulações monoclonais, além das
hemoculturas terem sido realizadas na fase aguda, onde ainda não é possível observar
claramente a ação do sistema imune no processo de seleção.
Estudos realizados por Andrade et al. (1999) envolvendo duas diferentes populações
do T. cruzi, JG e Col1.7G2, originalmente isoladas de pacientes com formas clínicas distintas

Nogueira-Paiva, NC Discussão
87
da DCh, também mostraram alteração no padrão de histotropismo. A cepa JG, isolada de um
paciente com megaesôfago, causou parasitismo cardíaco em camundongos infectados por ela
isoladamente ou em associação com a cepa Col1.7G2. Independente da infecção monoclonal
ou mista, para os autores, esses dados reforçam o modelo histotrópico clonal. Na infecção
mista pode ter havido uma interação entre as duas cepas que foi capaz de alterar a colonização
tecidual, enquanto, a inversão de histotropismo observada na infecção apenas com a cepa JG
confirma o envolvimento de características do hospedeiro.
Adicionalmente à complexidade da interação parasito-hospedeiro, a resposta imune
dirigida contra moléculas antigênicas, independente da fonte desses antígenos estar direta ou
indiretamente associada ao parasito (próprio parasito ou fenômenos de autoimunidade
induzidos pela infecção), tem um papel central sobre mecanismo patogênico da DCh (Dutra et
al., 2009). A ativação do sistema imune nos estágios iniciais da infecção é imprescindível para
o controle da parasitemia e do parasitismo tecidual, no entanto, a capacidade de controlar essa
resposta ao longo da infecção crônica, através da indução de mecanismos imunomodulatórios
parece ter papel central na manutenção de um equilíbrio favorável ao hospedeiro com bom
prognóstico da doença (Dutra et al., 2009).
Miocardite significativa foi notada no coração dos animais infectados com a cepa Be-
62 duas semanas antes do observado para os demais grupos infectados. Além disso, 28dai, o
isolado Be-78is5-3mai induziu uma resposta inflamatória inferior comparada à cepa parental e
à cepa Be-62. Resposta inflamatória exuberante ao final da fase aguda tem sido descrita por
vários autores em diferentes modelos experimentais para a cepa Be-78 (Bahia et al., 2002;
Caliari et al., 2002a; Vieira et al., 2009; Vieira et al., 2012) e Be-62 (Bahia et al., 2002).
No cólon, o comportamento da isolado foi mais próximo ao detectado para a cepa Be-
62, com um processo inflamatório sustentado, embora inferior, até o final da fase aguda
(28dai), em detrimento da resolução observada para o grupo de animais infectados com a cepa
Be-78 parental nesse tempo. De forma geral, uma associação pode ser vista entre o
parasitismo tecidual e a resposta imune no cólon, com histotropismo acompanhado de
processo inflamatório predominante para a cepa Be-62 e para o isolado Be-78is5-3mai.
Em relação ao coração, nossos dados estão em concordância com aqueles obtidos por
diferentes autores avaliando a infecção com isolados/clones e suas respectivas cepas
parentais, onde o processo inflamatório total apresentou caráter semelhante (Camandaroba et
al., 2001; Camandaroba et al., 2006). Entretanto, para o cólon, nossos resultados
demonstraram uma especificidade para o isolado Be-78is5-3mai no padrão de inflamação, que

Nogueira-Paiva, NC Discussão
88
concorda com o parasitismo tecidual dando a este características intermediárias entre a cepa
parental e a cepa Be-62. A possibilidade de o isolado compartilhar padrões comportamentais
com a cepa Be-62 é reforçada no estudo realizado por Valadares et al. (2012), no qual, uma
subpopulação semelhante à Be-62 foi detectada dentro da população policlonal da cepa Be-78
pela técnica de microssatélite. Achados histopatológicos após infecção monoclonal com duas
subpopulações do T. cruzi, cepa JG e o clone CL-Brener, revelaram que, durante a fase aguda,
o clone CL-Brener foi detectado parasitando diferentes tecidos dos camundongos infectados,
enquanto a cepa JG mostrou-se preferencialmente cardiomiotrópica (Franco et al., 2003).
Adicionalmente, na fase crônica da infecção somente a cepa JG foi encontrada no coração,
confirmando as especificidades dessas duas subpopulações em relação ao histotropismo.
Na infecção pelo T. cruzi, monócitos e macrófagos são reconhecidamente a primeira
barreira contra a disseminação do parasito pelo organismo, seja na doença humana ou
experimental. Os parasitos circulantes são fagocitados por monócitos, que uma vez ativados
por citocinas pró-inflamatórias (IFN) produzidas por células da resposta imune inata (Células
NK), representam o mecanismo inicial de resistência do hospedeiro (Fabrino et al., 2011). Os
parasitos que conseguem evadir a ação do sistema complemento e da atividade microbicida
dependente de NO de monócitos/macrófagos, conseguem se desenvolver e replicar dentro das
células parasitadas (Ouaissi e Ouaissi, 2005). Nos tecidos, o rompimento das células
parasitadas libera moléculas antigênicas que induzem a expressão de quimiocinas e citocinas
pro-inflamatórias, estimulando a mobilização de células inflamatórias para o local da infecção
promovendo a montagem de uma resposta imune celular eficiente, Th1, que resulte no
controle da replicação do parasito (Dosreis et al., 2005). O desenvolvimento da resposta
celular Th2 está associado ao fracasso do hospedeiro em controlar a disseminação da
infecção. Nesses tecidos, macrófagos infectados agem como células apresentadoras de
antígenos (APCs) produzindo citocinas, como IL-12, IFN-e TNF- e ainda providenciam os
sinais co-estimuladores necessários à ativação das células T, evidenciando sua importância na
interação entre a resposta imune inata e adaptativa (Gazzinelli et al., 1992; Van Voorhis,
1992; Vespa et al., 1994; Silva et al., 1995; Aliberti et al., 1996; Rodrigues et al., 2000;
Michailowsky et al., 2001; Ropert et al., 2002; Vieira et al., 2009).
Em concordância com dados da literatura (Melo e Machado, 1998; 2001; Arnaiz et al.,
2002; Paiva et al., 2009), um elevado percentual de macrófagos foi observado no coração dos
animais infectados, aumento esse que foi consistente com as curvas de parasitemia, sendo
mais intenso e precoce nos animais infectados com a cepa Be-62 (14dai), gradual e discreto

Nogueira-Paiva, NC Discussão
89
nos animais infectados com o isolado, e tardio, porém, moderado, no grupo Be-78 parental
(28dai). A depleção de macrófagos em ratos Holtzman, por pré-tratamento com sílica (1 dia
antes da infecção), infectados com a cepa Y, demonstrou aumento da parasitemia e
parasitismo cardíaco durante a fase aguda em relação aos animais apenas infectados,
apontando a importância dessas células no controle da replicação do parasito nas fases iniciais
da infecção (Melo, 1999; Melo e Machado, 2001).
Apesar de mais discreto, o aumento no percentual de macrófagos também foi
demonstrado no cólon dos animais infectados, 7 e 14dai, com exceção do grupo Be-78
parental. Interessantemente, o isolado Be-78is5-3mai foi a única subpopulação estudada a
manter a densidade de macrófagos elevada até o final da fase aguda, o que pode estar
relacionado à manutenção da parasitemia durante 49dai e ao parasitismo elevado no cólon.
A participação de linfócitos B no processo inflamatório foi discreta, aproximadamente
1,5 e 2,5-5% dos leucócitos totais CD45+, no coração e no cólon respectivamente. No
coração, foi possível observar aumento em todos os grupos infectados no 14ºdai, no entanto,
foi sustentado no 28ºdai, apenas nos animais infectados com a cepa Be-78 parental. Sete dias
após a infecção, todos os animais infectados apresentaram elevação na densidade de linfócitos
B no cólon. Nos grupos Be-62 e Be-78 parental, houve um aumento de cerca de 30% no
14ºdai em relação ao observado no 7ºdai, no entanto, os índices dessas células retornou a
normalidade no 28ºdai. Ao final da fase aguda, taxa elevada de linfócitos B foi observada no
cólon apenas nos animais infectados com o isolado Be-67is5-3mai.
O envolvimento de linfócitos B e plasmócitos nas lesões teciduais que ocorrem
durante a fase aguda nos principais órgãos envolvidos na DCh tem sido escassamente
examinado em modelos experimentais. Cardillo et al. (2007) demonstraram o papel crítico de
linfócitos B para a resistência do hospedeiro nos eventos iniciais da infecção pelo T. cruzi.
Camundongos C57BL/6 nocautes para linfócitos B, infectados com 500 formas
tripomastigotas sanguíneas da cepa Talahuen, foram mais susceptíveis à infecção,
apresentando maiores níveis parasitêmicos e redução na síntese de citocinas pró-inflamatórias
(IFN- e IL-12), em relação aos animais controle células B-competentes. Adicionalmente, em
detrimento dos animais imunocompetentes, que demonstraram aumento de células T CD4+ e
CD8+ no baço, camundongos nocautes não alteraram a frequência de células T CD4+ e ainda
reduziram pela metade o número de linfócitos T CD8+ em relação aos animais não infectados
(Cardillo et al., 2007). O processo inflamatório no músculo esquelético também foi reduzido
nos animais deficientes para célula B, sendo observada uma inversão na razão CD4+/CD8+

Nogueira-Paiva, NC Discussão
90
(sobreposição de CD4+), em relação aos animais imunocompetentes (sobreposição de CD8+)
(Cardillo et al., 2007). A associação entre a frequência de linfócitos B e a carga parasitária
também pode ser observada no nosso estudo, onde animais infectados com a cepa Be-78 e
com o isolado Be-78is5-3mai mantiveram, no coração e no cólon, respectivamente, níveis
mais altos de células B condizentes com a manutenção do parasitismo tecidual.
O papel protetor da resposta imune celular Th1, mediada por linfócitos T e IFN- na
infecção por T. cruzi tem sido descrito na literatura tanto em modelos experimentais quanto na
DCh humana (Gazzinelli et al., 1992; Munoz-Fernandez et al., 1992; Cardillo et al., 2007;
Sales et al., 2008). A polarização da resposta imunológica para o eixo Th1 é associada a um
perfil de controle da replicação do parasito nos eventos iniciais da infecção. Por outro lado, a
modulação do processo inflamatório, relacionada à produção de citocinas como IL-10, IL-17,
TGF-, por macrófagos e linfócitos T reguladores (Treg CD4+ CD25+ Foxp3+) é
imprescindível para prevenir uma resposta imune exacerbada com consequentes danos
teciduais secundários, sendo assim associada ao melhor prognóstico da doença (Mariano et
al., 2008; Sanoja et al., 2013).
Densidade elevada de células T CD4+ entre o infiltrado de células inflamatórias
mononucleares no tecido cardíaco são achados frequentes durante a fase aguda da infecção
experimental pelo T. cruzi (Mariano et al., 2008; Sales et al., 2008; Sanoja et al., 2013). Essas
células atuam na resistência à infecção através da produção de IFN- responsável por
estimular a síntese e NO por macrófagos e ainda, auxiliando na ativação e expansão de
linfócitos T CD8+ citotóxicos, sendo ambos os mecanismos importantes para o controle da
replicação intracelular do parasito (Brener e Gazzinelli, 1997).
A infecção com a cepa Be-62 foi capaz de manter níveis superiores de linfócitos T
CD4+ ao longo de toda a fase aguda no coração, alcançando maior frequência no 14ºdai, ao
contrário do observado para animais do grupo Be-78 parental, que apresentaram elevação na
densidade dessas células somente no 28ºdai. Aumento na frequência de células T CD4+ no
coração de camundongos C57BL/6 e BALB/c infectados com baixo (50 TS) ou alto (200 TS)
inóculo da cepa Y, no 14ºdai, mantido até o 22ºdai (menores níveis), também foi observado
por outros autores (Sanoja et al., 2013). Embora ainda elevado em relação aos animais não-
infectados, a redução na densidade dessas células no 22ºdai observada por Sanoja et al. (2013)
foi compatível com a redução da parasitemia, o que foi consistente com nossos dados que
demonstraram esse mesmo perfil aos 28dai condizente com o controle da parasitemia no
grupo Be-62. Pode-se sugerir que o comportamento da cepa Be-78 parental também parece

Nogueira-Paiva, NC Discussão
91
acompanhar o perfil descrito por Sanoja et al. (2013), com elevação do percentual de
linfócitos T CD4+ no 28º, próximo ao pico de parasitemia. A associação entre linfócitos T
CD4+ e controle da parasitemia durante a fase aguda também foi demonstrada por Russo et al.
(1988) em camundongos tratados com anticorpos anti-CD4 e infectados com a cepa CL.
Segundo os autores, após inativação das células T helper, miocardite reduzida, e aumento
significativo no número de parasitos no sangue, bem como no coração, foram detectadas nos
animais infectados e tratados em relação aos infectados não tratados.
Linfócitos T CD8+ são essenciais para o controle da infecção quando o parasito
encontra-se no estágio de amastigota intracelular (Rottenberg et al., 1993). Células T CD8+
reconhecem antígenos específicos derivados de proteínas citosólicas ligados à moléculas do
MHC de classe I em qualquer célula nucleada infectada (Tarleton, 2015). Após os estímulos
apropriados, em resposta ao reconhecimento de antígenos do T. cruzi, os linfócitos citolíticos
ativados podem agir sintetizando IFN- ou provocando a morte das células infectadas por
mecanismos que envolvem proteínas como perforina e granzimas A ou B (Martin e Tarleton,
2004).
Todos os animais infectados apresentaram aumento na frequência de linfócitos T
CD8+ no coração e cólon durante a fase aguda da infecção, entretanto, a cepa Be-78 parental e
o isolado Be-785-3mai, mostraram um atraso nessa reposta em relação à cepa Be-62. De
forma interessante, apenas camundongos infectados com a cepa Be-62 e com o isolado Be-
785-3mai, mantiveram, no cólon, elevação na densidade dessas células no 28ºdai. A
associação positiva entre o parasitismo tecidual e a resposta imune celular associada às células
T CD8+ parece evidente em nossos dados. De modo geral, ao final da fase aguda, observa-se
uma relação entre a densidade de células T CD8+ e maior carga parasitária no coração e no
cólon, para o grupo Be-78 parental e para os grupos Be-62 e Be-78 isolado, respectivamente.
Significante aumento de células T CD8+ também foi observado no coração de ratos Wistar
Furth (WF) infectados com a cepa Y, entre o 3º e 18ºdai, quando os animais sucumbiram à
infecção (Sato et al., 1992).
Cães infectados com a cepa Be-78 e necropsiados durante a fase aguda (33dai), de
forma similar ao observado em nosso estudo, apresentaram elevação na frequência de células
T CD4+ e CD8+, com sobreposição do fenótipo CD8+ em relação ao fenótipo CD4+ (Caliari et
al., 2002a). Interessantemente, animais infectados com essa cepa e mantidos ao longo da fase
crônica tardia apresentaram densidades similares em relação aos dois fenótipo de células T
estudados, enquanto animais infectados com a cepa Be-62, necropsiados também na fase

Nogueira-Paiva, NC Discussão
92
crônica tardia, demonstraram uma inversão na proporção CD4+/CD8+, com níveis superiores
de linfócitos T CD4+ (Caliari et al., 2002a). Embora nossas análises tenham sido realizadas
em modelo murino, os resultados obtidos durante a fase aguda da infecção, mostram um perfil
particular para a cepa Be-62, com um aumento precoce (14ºdai) e expressivamente superior
de células T CD4+, entretanto, ao final da fase aguda, ocorre sobreposição do fenótipo CD8+.
As especificidades do comportamento da cepa Be-62 devem-se não somente ao momento
investigado ao longo da infecção (fase aguda ou fase crônica), mas também à fatores
associados ao hospedeiro (camundongo ou cão). Avaliações ao longo da fase crônica no
modelo murino, poderão responder se ambas as cepas apresentarão comportamento
compatível com o observado em cães, além de mostrar a influência do polimorfismo da cepa
Be-78 nos animais infectados com o isolado Be-78is5-3mai sobre o fenótipo do infiltrado
inflamatório, bem como sobre o histotropismo.
Para investigar o efeito do polimorfismo do T. cruzi sobre a ativação da resposta
imune durante a fase aguda da infecção, a habilidade de linfócitos T CD4+ ou CD8+ em
produzir IFN- também foi avaliada em camundongos infectados com as diferentes
subpopulações do parasito.
A cepa Be-78 foi capaz de estimular ao longo de toda a fase aguda, no coração e
cólon, a síntese de IFN- tanto por linfócitos T CD4+ quanto por linfócitos T CD8+, sendo a
expressão maior observada nesse último fenótipo. De forma contrária, animais infectados com
a cepa Be-62 apenas apresentaram aumento na síntese de IFN- no 28ºdai. Interessantemente,
as células T presentes no infiltrado inflamatório induzido pelo isolado Be-78is5-3mai foram
capazes de sintetizar IFN- somente no cólon e de forma tardia (28dai). A maior frequência
de linfócitos T CD8+ IFN-+ no 28ºdai para o grupo Be-78 isolado concorda com o aumento
na frequência do fenótipo CD8+ no processo inflamatório do cólon. Entretanto, não houve
aumento significativo de células T CD4+ no cólon desses animais, e a maior densidade de
linfócitos T CD4+IFN-+ em relação às células CD45+ está relacionada ao fato de que mais de
65% das células T helper estão sintetizando essa citocina, em detrimento aos 20% observados
para o grupo não-infectado (dados não mostrados).
Nossos resultados sugerem que a participação de IFN- sobre o controle do
parasitismo tecidual parece ocorrer de forma distinta no coração no cólon, além de ter um
fator cepa-dependente. No coração dos animais infectados com a cepa Be-62 e com o isolado
Be-78is5-3mai uma associação entre o aumento da expressão dessa citocina e o controle da
carga parasitária pode ser observada no 28ºdai. No entanto, no cólon, o atraso na produção de

Nogueira-Paiva, NC Discussão
93
IFN- pelos animais pertencentes a estes dois grupos pode estar associado à manutenção da
elevada carga parasitária. Em contrapartida, parece não haver relação entre a produção dessa
citocina e o parasitismo cardíaco nos animais do grupo Be-78 parental, uma vez que níveis
elevados dessa citocina ao longo de toda a fase aguda não foram capazes de controlar a
replicação do parasito. A avaliação de outras citocinas pró-inflamatórias envolvidas nos
eventos iniciais da infecção (IL-12, TNF- bem como de citocinas imunomoduladoras (IL-
10, TGF-) pode ser a chave para explicar o comportamento distinto da cepa Be-78 parental
com o seu isolado (Be-78is5-3mai) e com a cepa Be-62.
Michailowsky et al. (2001) demonstraram a importância do INF- na resistência de
camundongos BALB/c e C57BL6 à infecção com a cepa Colombiana do T. cruzi.
Camundongos nocautes para IFN- foram altamente susceptíveis à infecção, apresentando
elevação da parasitemia, maior número de ninhos de amastigotas no coração e sistema
nervoso central e 100% de mortalidade aos 20dai (Michailowsky et al., 2001). Esses dados
são consistentes com àqueles obtidos em nosso estudo para a cepa Be-62, onde o aumento
discreto e atrasado de IFN- associado à alta parasitemia e à elevada carga parasitária parecem
ter um papel importante na mortalidade (100%) observada 28dai. Por outro lado, embora
também tenha sido observado para o grupo Be-78 isolado, o perfil atrasado de produção dessa
citocina neste caso está associado a uma baixa virulência compatível com à observada para a
cepa Be-78 parental, reforçando que o isolado Be-78is5-3mai apresenta características
intermediárias entre a cepa Be-62 e a cepa Be-78 parental, no que diz respeito ao
comportamento in vitro, ao tropismo tecidual e à montagem da resposta imune nos eventos
iniciais da infecção.
Em suma, nossos resultados mostraram um perfil cardiomiotrópico e retardo no
controle da parasitemia na infecção pela cepa Be-78 parental, seguido por processo
inflamatório intenso ao final da fase aguda, ao contrário da cepa Be-62, que por sua vez,
apresentou-se preferencialmente parasitando o cólon, com parasitemia significativamente
superior, no entanto, controlada precocemente, e processo inflamatório intenso nos dois
órgãos avaliados ao longo da fase aguda. O isolado Be-78is5-3mai, consistente com os dados
da avaliação in vitro, demonstrou durante a fase aguda da infecção de camundongos BALB/c,
um comportamento intermediário entre a cepas de referência utilizadas, apresentando
virulência (parasitemia e mortalidade) compatível com a cepa Be-78 parental, no entanto,
perfil histotrópico compartilhado com a cepa Be-62. O perfil da resposta imunológica

Nogueira-Paiva, NC Discussão
94
apresentado pelos animais infectados com o isolado foi, de forma geral, consistente com
àquele observado para a cepa Be-62, sobretudo no cólon.
Estudos abordando aspectos imunopatogênicos, tanto em modelos experimentais de
infecção quanto em pacientes portadores da DCh, indicam que a situação ideal para ambos
parasito e hospedeiro ocorre quando há um balanço entre a resposta imune efetiva capaz de
manter o parasitismo em níveis baixos e a modulação desta, prevenindo seus efeitos
citotóxicos (Dutra et al., 2009). O efeito direto da resposta imunológica desenvolvida durante
a fase aguda da infecção sobre lesões crônicas, assim como a influência do polimorfismo
biológico e molecular do T. cruzi, associado também à fatores intrínsecos do hospedeiro
suscitam a necessidade de estudos complementares, no que diz respeito à modulação da
resposta imune e ao desenvolvimento das lesões histológicas durante a fase crônica,
contribuindo não só para elucidação dos fenômenos relacionados aos danos teciduais, como
sendo de fundamental importância para o esclarecimento da evolução da doença, além de
auxiliar no estabelecimento de estratégias terapêuticas eficazes para o tratamento e prevenção
da fibrose cardíaca e da lesão entérica na infecção chagásica.
Nesse sentido, a avaliação de camundongos BALB/c infectados com as cepas Be-62,
Be-78 parental e com o isolado Be-78is5-3mai está sendo conduzida por nosso grupo de
pesquisa, com o objetivo de elucidar melhor a influência da infecção por uma cepa policlonal
sobre as lesões cardíaca e digestiva ao longo da DCh experimental.

95
7. CONCLUSÃO

Nogueira-Paiva, NC Conclusão
96
Nossos dados reforçam a hipótese de que a infecção a longo prazo com a cepa Be-78 foi
capaz de selecionar uma subpopulação que compartilha características com a cepa Be-62.
Adicionalmente, confirmamos o modelo histotrópico clonal, indicando heterogeneidade
clonal capaz de influenciar de forma distinta os eventos iniciais da infecção, com possíveis
repercussões para o curso crônico da doença de Chagas.
A subpopulação Be-78is5-3mai, isolada de camundongos Swiss infectados com a cepa
policlonal Be-78 do Trypanosoma cruzi, demonstrou comportamento in vitro e perfil
gênico intermediários entre as cepas de referência, Be-78 parental e Be-62.
Durante a fase aguda da infecção de camundongos, o isolado Be-78is5-3mai manteve esse
mesmo comportamento, entre a cepas de referência utilizadas, apresentando virulência
(parasitemia e mortalidade) compatível com a cepa Be-78 parental, no entanto, perfil
histotrópico e resposta imune compartilhados com a cepa Be-62.

97
8. REFERÊNCIAS BIBLIOGRÁFICAS

Nogueira-Paiva, NC Referências Bibliográficas
98
ADAD, S. J.; ANDRADE, D. C.; LOPES, E. R.; CHAPADEIRO, E. [Pathological anatomy of chagasic megaesophagus]. Rev Inst Med Trop Sao Paulo, v. 33, p. 443-50, 1991. ADAD, S. J.; CANCADO, C. G.; ETCHEBEHERE, R. M.; TEIXEIRA, V. P.; GOMES, U. A.; CHAPADEIRO, E.; LOPES, E. R. Neuron count reevaluation in the myenteric plexus of chagasic megacolon after morphometric neuron analysis. Virchows Arch, v. 438, p. 254-8, 2001. AKHAVAN, D. Análise de custo efetivo do programa de controle da doença de chagas no Brasil. 2000. ALIBERTI, J. C.; CARDOSO, M. A.; MARTINS, G. A.; GAZZINELLI, R. T.; VIEIRA, L. Q.; SILVA, J. S. Interleukin-12 mediates resistance to Trypanosoma cruzi in mice and is produced by murine macrophages in response to live trypomastigotes. Infect Immun, v. 64, p. 1961-7, 1996. ANDERSSON, J.; ORN, A.; SUNNEMARK, D. Chronic murine Chagas' disease: the impact of host and parasite genotypes. Immunol Lett, v. 86, p. 207-12, 2003. ANDRADE, L. O.; GALVAO, L. M.; MEIRELLES MDE, N.; CHIARI, E.; PENA, S. D.; MACEDO, A. M. Differential tissue tropism of Trypanosoma cruzi strains: an in vitro study. Mem Inst Oswaldo Cruz, v. 105, p. 834-7, 2010. ANDRADE, L. O.; MACHADO, C. R.; CHIARI, E.; PENA, S. D.; MACEDO, A. M. Differential tissue distribution of diverse clones of Trypanosoma cruzi in infected mice. Mol Biochem Parasitol, v. 100, p. 163-72, 1999. ANDRADE, L. O.; MACHADO, C. R.; CHIARI, E.; PENA, S. D.; MACEDO, A. M. Trypanosoma cruzi: role of host genetic background in the differential tissue distribution of parasite clonal populations. Exp Parasitol, v. 100, p. 269-75, 2002. ANDRADE, S. G. Caracterização de cepas de Trypanosoma cruzi isolados no Recôncavo Baiano. Revista de Patologia Tropical, v. 3, p. 65-121, 1974. ANDRADE, S. G. Influence of Trypanosoma cruzi strain on the pathogenesis of chronic myocardiopathy in mice. Mem Inst Oswaldo Cruz, v. 85, p. 17-27, 1990. ANDRADE, S. G.; ANDRADE, Z. A. [Chagas' disease and neuron changes in Auerbach's plexus. (Experimental study in mice)]. Rev Inst Med Trop Sao Paulo, v. 8, p. 219-24, 1966. ANDRADE, S. G.; MAGALHAES, J. B. Biodemes and zymodemes of Trypanosoma cruzi strains: correlations with clinical data and experimental pathology. Rev Soc Bras Med Trop, v. 30, p. 27-35, 1996. ANDRADE, Z. A. [The pathology of Chagas disease in man]. Ann Soc Belg Med Trop, v. 65 Suppl 1, p. 15-30, 1985. ANDRADE, Z. A. A forma indeterminada da doença de Chagas em tempos de controle do Triatoma infestans. Revista de Patologia Tropical, v. 34, p. 105-111, 2005. ANDRADE, Z. A.; ANDRADE, S. G.; OLIVEIRA, G. B.; ALONSO, D. R. Histopathology of the conducting tissue of the heart in Chagas' myocarditis. Am Heart J, v. 95, p. 316-24, 1978. ANDRADE, Z. A.; ANDRADE, S. G.; SADIGURSKY, M.; WENTHOLD, R. J., JR.; HILBERT, S. L.; FERRANS, V. J. The indeterminate phase of Chagas' disease: ultrastructural

Nogueira-Paiva, NC Referências Bibliográficas
99
characterization of cardiac changes in the canine model. Am J Trop Med Hyg, v. 57, p. 328-36, 1997. ANEZ, N.; CARRASCO, H.; PARADA, H.; CRISANTE, G.; ROJAS, A.; FUENMAYOR, C.; GONZALEZ, N.; PERCOCO, G.; BORGES, R.; GUEVARA, P.; RAMIREZ, J. L. Myocardial parasite persistence in chronic chagasic patients. Am J Trop Med Hyg, v. 60, p. 726-32, 1999. ARNAIZ, M. R.; FICHERA, L. E.; POSTAN, M. Cardiac myocyte hypertrophy and proliferating cell nuclear antigen expression in Wistar rats infected with Trypanosoma cruzi. J Parasitol, v. 88, p. 919-25, 2002. AVILA, H.; GONCALVES, A. M.; NEHME, N. S.; MOREL, C. M.; SIMPSON, L. Schizodeme analysis of Trypanosoma cruzi stocks from South and Central America by analysis of PCR-amplified minicircle variable region sequences. Mol Biochem Parasitol, v. 42, p. 175-87, 1990. AVILA, H. A.; SIMPSON, L. Organization and complexity of minicircle-encoded guide RNAs in Trypanosoma cruzi. RNA, v. 1, p. 939-47, 1995. BAHIA, M. T.; TAFURI, W. L.; CALIARI, M. V.; VELOSO, V. M.; CARNEIRO, C. M.; COELHO, G. L.; LANA, M. Comparison of Trypanosoma cruzi infection in dogs inoculated with blood or metacyclic trypomastigotes of Berenice-62 and Berenice-78 strains via intraperitoneal and conjunctival routes. Rev Soc Bras Med Trop, v. 35, p. 339-45, 2002. BAPTISTA, C. S.; VENCIO, R. Z.; ABDALA, S.; CARRANZA, J. C.; WESTENBERGER, S. J.; SILVA, M. N.; PEREIRA, C. A.; GALVAO, L. M.; GONTIJO, E. D.; CHIARI, E.; STURM, N. R.; ZINGALES, B. Differential transcription profiles in Trypanosoma cruzi associated with clinical forms of Chagas disease: Maxicircle NADH dehydrogenase subunit 7 gene truncation in asymptomatic patient isolates. Mol Biochem Parasitol, v. 150, p. 236-48, 2006. BARROS, M. V.; RIBEIRO, A. L.; MACHADO, F. S.; ROCHA, M. O. Doppler tissue imaging to assess systolic function in Chagas' disease. Arq Bras Cardiol, v. 80, p. 36-40, 31-5, 2003. BARROS, M. V.; ROCHA, M. O.; RIBEIRO, A. L.; MACHADO, F. S. Doppler tissue imaging to evaluate early myocardium damage in patients with undetermined form of Chagas' disease and normal echocardiogram. Echocardiography, v. 18, p. 131-6, 2001. BARTHOLOMEU, D. C.; CERQUEIRA, G. C.; LEAO, A. C.; DAROCHA, W. D.; PAIS, F. S.; MACEDO, C.; DJIKENG, A.; TEIXEIRA, S. M.; EL-SAYED, N. M. Genomic organization and expression profile of the mucin-associated surface protein (masp) family of the human pathogen Trypanosoma cruzi. Nucleic Acids Res, v. 37, p. 3407-17, 2009. BENCHIMOL BARBOSA, P. R. The oral transmission of Chagas' disease: an acute form of infection responsible for regional outbreaks. Int J Cardiol, v. 112, p. 132-3, 2006. BERN, C.; MONTGOMERY, S. P.; HERWALDT, B. L.; RASSI, A., JR.; MARIN-NETO, J. A.; DANTAS, R. O.; MAGUIRE, J. H.; ACQUATELLA, H.; MORILLO, C.; KIRCHHOFF, L. V.; GILMAN, R. H.; REYES, P. A.; SALVATELLA, R.; MOORE, A. C. Evaluation and treatment of chagas disease in the United States: a systematic review. JAMA, v. 298, p. 2171-81, 2007. BOCANEGRA, J. M. C. Estudo clínico parasitológico de pacientes com doença de Chagas, de Mambaí-GO, compreendendo um período de 30 anos. 2008. 138 (PhD). Universidade de Brasília BRENER, Z. Therapeutic activity and criterion of cure on mice experimentally infected with Trypanosoma cruzi. Rev Inst Med Trop Sao Paulo, v. 4, p. 389-96, 1962.

Nogueira-Paiva, NC Referências Bibliográficas
100
BRENER, Z. (Comparative Studies of Different Strains of Trypanosoma Cruzi.). Ann Trop Med Parasitol, v. 59, p. 19-26, 1965. BRENER, Z. The behavior of slender and stout forms of Trypanosoma cruzi in the blood-stream of normal and immune mice. Ann Trop Med Parasitol, v. 63, p. 215-20, 1969. BRENER, Z. Pathogenesis and immunopathology of chronic Chagas’Disease. Memórias do Instituto Oswaldo Cruz, v. 82, p. 205-213, 1987. BRENER, Z.; CHIARI, E. [Morphological Variations Observed in Different Strains of Trypanosoma Cruzi]. Rev Inst Med Trop Sao Paulo, v. 5, p. 220-4, 1963. BRENER, Z.; GAZZINELLI, R. T. Immunological control of Trypanosoma cruzi infection and pathogenesis of Chagas' disease. Int Arch Allergy Immunol, v. 114, p. 103-10, 1997. BRISSE, S.; BARNABE, C.; TIBAYRENC, M. Identification of six Trypanosoma cruzi phylogenetic lineages by random amplified polymorphic DNA and multilocus enzyme electrophoresis. Int J Parasitol, v. 30, p. 35-44, 2000a. BRISSE, S.; DUJARDIN, J. C.; TIBAYRENC, M. Identification of six Trypanosoma cruzi lineages by sequence-characterised amplified region markers. Mol Biochem Parasitol, v. 111, p. 95-105, 2000b. CALIARI, E. R.; CALIARI, M. V.; DE LANA, M.; TAFURI, W. L. [Quantitative and qualitative studies of the Auerbach and Meissner plexuses of the esophagus in dogs inoculated with Trypanosoma cruzi]. Rev Soc Bras Med Trop, v. 29, p. 17-20, 1996. CALIARI, M. V. Pricípios da morfometria digital: KS300 para iniciantes. Editora UFMG, Belo Horizonte, 149p, 1997. CALIARI, M. V.; DE LANA, M.; CAJA, R. A.; CARNEIRO, C. M.; BAHIA, M. T.; SANTOS, C. A.; MAGALHAES, G. A.; SAMPAIO, I. B.; TAFURI, W. L. Immunohistochemical studies in acute and chronic canine chagasic cardiomyopathy. Virchows Arch, v. 441, p. 69-76, 2002a. CALIARI, M. V.; DE LANA, M.; CALIARI, E. R.; TAFURI, W. L. Cardiac plexus of dogs experimentally infected with Trypanosoma cruzi: inflammatory lesions and quantitative studies. Rev Soc Bras Med Trop, v. 28, p. 13-7, 1995. CALIARI, M. V.; DO PILAR MACHADO, R.; DE LANA, M.; CAJA, R. A.; CARNEIRO, C. M.; BAHIA, M. T.; DOS SANTOS, C. A.; MAGALHAES, G. A.; SAMPAIO, I. B.; TAFURI, W. L. Quantitative analysis of cardiac lesions in chronic canine chagasic cardiomyopathy. Rev Inst Med Trop Sao Paulo, v. 44, p. 273-8, 2002b. CAMANDAROBA, E.; THE, T. S.; PESSINA, D. H.; ANDRADE, S. G. Trypanosoma cruzi: clones isolated from the Colombian strain, reproduce the parental strain characteristics, with ubiquitous histotropism. Int J Exp Pathol, v. 87, p. 209-17, 2006. CAMANDAROBA, E. L.; CAMPOS, R. F.; MAGALHAES, J. B.; ANDRADE, S. G. Clonal structure of Trypanosoma cruzi Colombian strain (biodeme Type III): biological, isoenzymic and histopathological analysis of seven isolated clones. Rev Soc Bras Med Trop, v. 34, p. 151-7, 2001.

Nogueira-Paiva, NC Referências Bibliográficas
101
CAMARGO, M. E.; DA SILVA, G. R.; DE CASTILHO, E. A.; SILVEIRA, A. C. [Serological survey of the prevalence of Chagas' infection in Brazil, 1975/1980]. Rev Inst Med Trop Sao Paulo, v. 26, p. 192-204, 1984. CAMPOS, R. M.; ANDRADE, S. G. Characterization of subpopulations (clones and subclones) of the 21 SF strain of Trypanosoma cruzi after long lasting maintenance in the laboratory. Mem Inst Oswaldo Cruz, v. 91, p. 795-800, 1996. CARDILLO, F.; POSTOL, E.; NIHEI, J.; AROEIRA, L. S.; NOMIZO, A.; MENGEL, J. B cells modulate T cells so as to favour T helper type 1 and CD8+ T-cell responses in the acute phase of Trypanosoma cruzi infection. Immunology, v. 122, p. 584-95, 2007. CARNEIRO, C. M.; MARTINS-FILHO, O. A.; REIS, A. B.; VELOSO, V. M.; ARAUJO, F. M.; BAHIA, M. T.; DE LANA, M.; MACHADO-COELHO, G. L.; GAZZINELLI, G.; CORREA-OLIVEIRA, R.; TAFURI, W. L. Differential impact of metacyclic and blood trypomastigotes on parasitological, serological and phenotypic features triggered during acute Trypanosoma cruzi infection in dogs. Acta Trop, v. 101, p. 120-9, 2007. CARNEIRO, M.; CHIARI, E.; GONCALVES, A. M.; DA SILVA PEREIRA, A. A.; MOREL, C. M.; ROMANHA, A. J. Changes in the isoenzyme and kinetoplast DNA patterns of Trypanosoma cruzi strains induced by maintenance in mice. Acta Trop, v. 47, p. 35-45, 1990. CASTRO, C.; MACEDO, V.; REZENDE, J. M.; PRATA, A. [Longitudinal radiologic study of the esophagus, in an endemic area of Chagas disease, in a period of 13 years]. Rev Soc Bras Med Trop, v. 27, p. 227-33, 1994. CASTRO, C.; PRATA, A.; MACEDO, V. [A 13-year clinical study on 190 chronic chagasic patients from Mambai, Goias, Brazil]. Rev Soc Bras Med Trop, v. 34, p. 309-18, 2001. CHAGAS, C. Nova tripanozomiase humana. Estudos sobre a morfolojia e o ciclo evolutivo do Schizotrypanum cruzi n. gen., n. sp., ajente etiolójico de nova entidade mórbida do homem. Memórias do Instituto Oswaldo Cruz, v. 1, p. 159-218, 1909. COURA, J. R.; DE ABREU, L. L.; PEREIRA, J. B.; WILLCOX, H. P. [Morbidity in Chagas' disease. IV. Longitudinal study of 10 years in Pains and Iguatama, Minas Gerais, Brazil]. Mem Inst Oswaldo Cruz, v. 80, p. 73-80, 1985. CRUZ, R. E.; MACEDO, A. M.; BARNABE, C.; FREITAS, J. M.; CHIARI, E.; VELOSO, V. M.; CARNEIRO, C. M.; BAHIA, M. T.; TAFURI, W. L.; LANA, M. Further genetic characterization of the two Trypanosoma cruzi Berenice strains (Be-62 and Be-78) isolated from the first human case of Chagas disease (Chagas, 1909). Acta Trop, v. 97, p. 239-46, 2006. CUERVO, H.; GUERRERO, N. A.; CARBAJOSA, S.; BESCHIN, A.; DE BAETSELIER, P.; GIRONES, N.; FRESNO, M. Myeloid-derived suppressor cells infiltrate the heart in acute Trypanosoma cruzi infection. J Immunol, v. 187, p. 2656-65, 2011. CUMMINGS, K. L.; TARLETON, R. L. Rapid quantitation of Trypanosoma cruzi in host tissue by real-time PCR. Mol Biochem Parasitol, v. 129, p. 53-9, 2003. CUNHA-NETO, E.; KALIL, J. Autoimmunity in Chagas' heart disease. Sao Paulo Med J , v. 113, p. 757-66, 1995. D'AVILA, D. A.; MACEDO, A. M.; VALADARES, H. M.; GONTIJO, E. D.; DE CASTRO, A. M.; MACHADO, C. R.; CHIARI, E.; GALVAO, L. M. Probing population dynamics of Trypanosoma

Nogueira-Paiva, NC Referências Bibliográficas
102
cruzi during progression of the chronic phase in chagasic patients. J Clin Microbiol, v. 47, p. 1718-25, 2009. D'AVILA REIS, D.; LEMOS, E. M.; SILVA, G. C.; ADAD, S. J.; MCCURLEY, T.; CORREA-OLIVEIRA, R.; MACHADO, C. R. Phenotypic characterization of the inflammatory cells in chagasic megaoesophagus. Trans R Soc Trop Med Hyg, v. 95, p. 177-8, 2001. DA SILVEIRA, A. B.; ADAD, S. J.; CORREA-OLIVEIRA, R.; FURNESS, J. B.; D'AVILA REIS, D. Morphometric study of eosinophils, mast cells, macrophages and fibrosis in the colon of chronic chagasic patients with and without megacolon. Parasitology, v. 134, p. 789-96, 2007a. DA SILVEIRA, A. B.; ARANTES, R. M.; VAGO, A. R.; LEMOS, E. M.; ADAD, S. J.; CORREA-OLIVEIRA, R.; D'AVILA REIS, D. Comparative study of the presence of Trypanosoma cruzi kDNA, inflammation and denervation in chagasic patients with and without megaesophagus. Parasitology, v. 131, p. 627-34, 2005. DA SILVEIRA, A. B.; FREITAS, M. A.; DE OLIVEIRA, E. C.; NETO, S. G.; LUQUETTI, A. O.; FURNESS, J. B.; CORREA-OLIVEIRA, R.; D'AVILA REIS, D. Neuronal plasticity of the enteric nervous system is correlated with chagasic megacolon development. Parasitology, v. 135, p. 1337-42, 2008. DA SILVEIRA, A. B.; FREITAS, M. A.; DE OLIVEIRA, E. C.; NETO, S. G.; LUQUETTI, A. O.; FURNESS, J. B.; CORREA-OLIVEIRA, R.; REIS, D. Glial fibrillary acidic protein and S-100 colocalization in the enteroglial cells in dilated and nondilated portions of colon from chagasic patients. Hum Pathol, v. 40, p. 244-51, 2009. DA SILVEIRA, A. B.; LEMOS, E. M.; ADAD, S. J.; CORREA-OLIVEIRA, R.; FURNESS, J. B.; D'AVILA REIS, D. Megacolon in Chagas disease: a study of inflammatory cells, enteric nerves, and glial cells. Hum Pathol, v. 38, p. 1256-64, 2007b. DE ALENCAR, B. C.; PERSECHINI, P. M.; HAOLLA, F. A.; DE OLIVEIRA, G.; SILVERIO, J. C.; LANNES-VIEIRA, J.; MACHADO, A. V.; GAZZINELLI, R. T.; BRUNA-ROMERO, O.; RODRIGUES, M. M. Perforin and gamma interferon expression are required for CD4+ and CD8+ T-cell-dependent protective immunity against a human parasite, Trypanosoma cruzi, elicited by heterologous plasmid DNA prime-recombinant adenovirus 5 boost vaccination. Infect Immun, v. 77, p. 4383-95, 2009. DE FREITAS, J. M.; AUGUSTO-PINTO, L.; PIMENTA, J. R.; BASTOS-RODRIGUES, L.; GONCALVES, V. F.; TEIXEIRA, S. M.; CHIARI, E.; JUNQUEIRA, A. C.; FERNANDES, O.; MACEDO, A. M.; MACHADO, C. R.; PENA, S. D. Ancestral genomes, sex, and the population structure of Trypanosoma cruzi. PLoS Pathog, v. 2, p. e24, 2006. DE LANA, M.; CHIARI, C. A.; CHIARI, E.; MOREL, C. M.; GONCALVES, A. M.; ROMANHA, A. J. Characterization of two isolates of Trypanosoma cruzi obtained from the patient Berenice, the first human case of Chagas' disease described by Carlos Chagas in 1909. Parasitol Res, v. 82, p. 257-60, 1996. DE LANA, M.; TAFURI, W. L.; CALIARI, M. V.; BAMBIRRA, E. A.; CHIARI CDE, A.; LEITE, V. H.; BARBOSA, A. J.; TOLEDO, M. J.; CHIARI, E. [Chronic fibrotic cardiac stage of experimental trypanosomiasis cruzi in dogs]. Rev Soc Bras Med Trop, v. 21, p. 113-21, 1988. DE OLIVEIRA, R. B.; TRONCON, L. E.; DANTAS, R. O.; MENGHELLI, U. G. Gastrointestinal manifestations of Chagas' disease. Am J Gastroenterol, v. 93, p. 884-9, 1998.

Nogueira-Paiva, NC Referências Bibliográficas
103
DE REZENDE, J. M. The digestive tract in Chagas'disease. Memórias do Instituto Oswaldo Cruz, v. 79, p. 97-106, 1984. DEL PUERTO, R.; NISHIZAWA, J. E.; KIKUCHI, M.; IIHOSHI, N.; ROCA, Y.; AVILAS, C.; GIANELLA, A.; LORA, J.; VELARDE, F. U.; RENJEL, L. A.; MIURA, S.; HIGO, H.; KOMIYA, N.; MAEMURA, K.; HIRAYAMA, K. Lineage analysis of circulating Trypanosoma cruzi parasites and their association with clinical forms of Chagas disease in Bolivia. PLoS Negl Trop Dis, v. 4, p. e687, 2010. DIAS, E.; LARANJA, F. S.; MIRANDA, A.; NOBREGA, G. Chagas' disease; a clinical, epidemiologic, and pathologic study. Circulation, v. 14, p. 1035-60, 1956. DIAS, J. C. P., COURA, J. R. Clínica e terapêutica da doença de Chagas: uma abordagem prática para o clínico geral [online]. 16, 1997. DOSREIS, G. A.; FREIRE-DE-LIMA, C. G.; NUNES, M. P.; LOPES, M. F. The importance of aberrant T-cell responses in Chagas disease. Trends Parasitol, v. 21, p. 237-43, 2005. DUTRA, W. O.; GOLLOB, K. J. Current concepts in immunoregulation and pathology of human Chagas disease. Curr Opin Infect Dis, v. 21, p. 287-92, 2008. DUTRA, W. O.; MENEZES, C. A.; VILLANI, F. N.; DA COSTA, G. C.; DA SILVEIRA, A. B.; REIS, D.; GOLLOB, K. J. Cellular and genetic mechanisms involved in the generation of protective and pathogenic immune responses in human Chagas disease. Mem Inst Oswaldo Cruz, v. 104 Suppl 1, p. 208-18, 2009. EL-SAYED, N. M.; MYLER, P. J.; BARTHOLOMEU, D. C.; NILSSON, D.; AGGARWAL, G.; TRAN, A. N.; GHEDIN, E.; WORTHEY, E. A.; DELCHER, A. L.; BLANDIN, G.; WESTENBERGER, S. J.; CALER, E.; CERQUEIRA, G. C.; BRANCHE, C.; HAAS, B.; ANUPAMA, A.; ARNER, E.; ASLUND, L.; ATTIPOE, P.; BONTEMPI, E.; BRINGAUD, F.; BURTON, P.; CADAG, E.; CAMPBELL, D. A.; CARRINGTON, M.; CRABTREE, J.; DARBAN, H.; DA SILVEIRA, J. F.; DE JONG, P.; EDWARDS, K.; ENGLUND, P. T.; FAZELINA, G.; FELDBLYUM, T.; FERELLA, M.; FRASCH, A. C.; GULL, K.; HORN, D.; HOU, L.; HUANG, Y.; KINDLUND, E.; KLINGBEIL, M.; KLUGE, S.; KOO, H.; LACERDA, D.; LEVIN, M. J.; LORENZI, H.; LOUIE, T.; MACHADO, C. R.; MCCULLOCH, R.; MCKENNA, A.; MIZUNO, Y.; MOTTRAM, J. C.; NELSON, S.; OCHAYA, S.; OSOEGAWA, K.; PAI, G.; PARSONS, M.; PENTONY, M.; PETTERSSON, U.; POP, M.; RAMIREZ, J. L.; RINTA, J.; ROBERTSON, L.; SALZBERG, S. L.; SANCHEZ, D. O.; SEYLER, A.; SHARMA, R.; SHETTY, J.; SIMPSON, A. J.; SISK, E.; TAMMI, M. T.; TARLETON, R.; TEIXEIRA, S.; VAN AKEN, S.; VOGT, C.; WARD, P. N.; WICKSTEAD, B.; WORTMAN, J.; WHITE, O.; FRASER, C. M.; STUART, K. D.; ANDERSSON, B. The genome sequence of Trypanosoma cruzi, etiologic agent of Chagas disease. Science, v. 309, p. 409-15, 2005. ENGEL, J. C.; DVORAK, J. A.; SEGURA, E. L.; CRANE, M. S. Trypanosoma cruzi: biological characterization of 19 clones derived from two chronic chagasic patients. I. Growth kinetics in liquid medium. J Protozool, v. 29, p. 555-60, 1982. FABRINO, D. L.; LEON, L. L.; GENESTRA, M.; PARREIRA, G. G.; MELO, R. C. Rat models to investigate host macrophage defense against Trypanosoma cruzi. J Innate Immun, v. 3, p. 71-82, 2011. FERREIRA, A. V.; SEGATTO, M.; MENEZES, Z.; MACEDO, A. M.; GELAPE, C.; DE OLIVEIRA ANDRADE, L.; NAGAJYOTHI, F.; SCHERER, P. E.; TEIXEIRA, M. M.; TANOWITZ, H. B.

Nogueira-Paiva, NC Referências Bibliográficas
104
Evidence for Trypanosoma cruzi in adipose tissue in human chronic Chagas disease. Microbes Infect, v. 13, p. 1002-5, 2011. FERREIRA JUNIOR, M.; BATISTA, S. A.; VIDIGAL, P. V.; CORDEIRO, A. A.; OLIVEIRA, F. M.; PRATA, L. O.; DINIZ, A. E.; BARRAL, C. M.; BARBUTO, R. C.; GOMES, A. D.; ARAUJO, I. D.; QUEIROZ, D. M.; CALIARI, M. V. Infection with CagA-positive Helicobacter pylori strain containing three EPIYA C phosphorylation sites is associated with more severe gastric lesions in experimentally infected Mongolian gerbils (Meriones unguiculatus). Eur J Histochem, v. 59, p. 2489, 2015. FIGUEIREDO, M. M.; BARBOSA, V. S.; AMORIM, I. F. G.; DEOTTI, B.; MICHALICK, M. S. M.; FARIA, A. C.; TAFURI, W. L. Obtaining cells from colon of dog with leishmaniasis for flow cytometric analysis. Protocol Exchange, p. 1-13, 2011. FLOREZ, O.; ESPER, J.; HIGUERA, S.; BARRAZA, M. F.; CABRERA, H. B.; MANTILLA, J. C.; RUGELES, C. I. Chagasic megacolon associated with Trypanosoma cruzi I in a Colombian patient. Parasitol Res, v. 107, p. 439-42, 2010. FRANCO, D. J.; VAGO, A. R.; CHIARI, E.; MEIRA, F. C.; GALVAO, L. M.; MACHADO, C. R. Trypanosoma cruzi: mixture of two populations can modify virulence and tissue tropism in rat. Exp Parasitol, v. 104, p. 54-61, 2003. FREITAS, J. M.; ANDRADE, L. O.; PIRES, S. F.; LIMA, R.; CHIARI, E.; SANTOS, R. R.; SOARES, M.; MACHADO, C. R.; FRANCO, G. R.; PENA, S. D.; MACEDO, A. M. The MHC gene region of murine hosts influences the differential tissue tropism of infecting Trypanosoma cruzi strains. PLoS One, v. 4, p. e5113, 2009. GASCON, J.; ALBAJAR, P.; CANAS, E.; FLORES, M.; GOMEZ I PRAT, J.; HERRERA, R. N.; LAFUENTE, C. A.; LUCIARDI, H. L.; MONCAYO, A.; MOLINA, L.; MUNOZ, J.; PUENTE, S.; SANZ, G.; TREVINO, B.; SERGIO-SALLES, X. [Diagnosis, management and treatment of chronic Chagas' heart disease in areas where Trypanosoma cruzi infection is not endemic]. Rev Esp Cardiol, v. 60, p. 285-93, 2007. GAZZINELLI, R. T.; OSWALD, I. P.; HIENY, S.; JAMES, S. L.; SHER, A. The microbicidal activity of interferon-gamma-treated macrophages against Trypanosoma cruzi involves an L-arginine-dependent, nitrogen oxide-mediated mechanism inhibitable by interleukin-10 and transforming growth factor-beta. Eur J Immunol, v. 22, p. 2501-6, 1992. GOMES, M. L.; MACEDO, A. M.; VAGO, A. R.; PENA, S. D.; GALVAO, L. M.; CHIARI, E. Trypanosoma cruzi: optimization of polymerase chain reaction for detection in human blood. Exp Parasitol, v. 88, p. 28-33, 1998. GOMES, M. L.; ROMANHA, A. J.; GONCALVES, A. M.; CHIARI, E. Stability of isoenzyme and kinetoplast DNA (k-DNA) patterns in successively cloned Trypanosoma cruzi populations. Mem Inst Oswaldo Cruz, v. 86, p. 379-85, 1991. GONZALEZ, A.; PREDIGER, E.; HUECAS, M. E.; NOGUEIRA, N.; LIZARDI, P. M. Minichromosomal repetitive DNA in Trypanosoma cruzi: its use in a high-sensitivity parasite detection assay. Proc Natl Acad Sci U S A, v. 81, p. 3356-60, 1984. GRAEFE, S. E.; JACOBS, T.; GAWORSKI, I.; KLAUENBERG, U.; STEEG, C.; FLEISCHER, B. Interleukin-12 but not interleukin-18 is required for immunity to Trypanosoma cruzi in mice. Microbes Infect, v. 5, p. 833-9, 2003.

Nogueira-Paiva, NC Referências Bibliográficas
105
GUEDES, P. M.; VELOSO, V. M.; CALIARI, M. V.; CARNEIRO, C. M.; SOUZA, S. M.; DE LANA, M.; CHIARI, E.; BAHIA, M. T.; GALVAO, L. M. Trypanosoma cruzi high infectivity in vitro is related to cardiac lesions during long-term infection in Beagle dogs. Mem Inst Oswaldo Cruz, v. 102, p. 141-7, 2007. HIGUCHI, M. D. L. Human chronic chagasic cardiopathy: participation of parasite antigens, subsets of lymphocytes, cytokines and microvascular abnormalities. Mem Inst Oswaldo Cruz, v. 94 Suppl 1, p. 263-7, 1999. HIGUCHI, M. L.; DE BRITO, T.; MARTINS REIS, M.; BARBOSA, A.; BELLOTTI, G.; PEREIRA-BARRETO, A. C.; PILEGGI, F. Correlation between Trypanosoma cruzi parasitism and myocardial inflammatory infiltrate in human chronic chagasic myocarditis: Light microscopy and immunohistochemical findings. Cardiovasc Pathol, v. 2, p. 101-6, 1993. HIGUCHI, M. L.; FUKASAWA, S.; DE BRITO, T.; PARZIANELLO, L. C.; BELLOTTI, G.; RAMIRES, J. A. Different microcirculatory and interstitial matrix patterns in idiopathic dilated cardiomyopathy and Chagas' disease: a three dimensional confocal microscopy study. Heart, v. 82, p. 279-85, 1999. IANTORNO, G.; BASSOTTI, G.; KOGAN, Z.; LUMI, C. M.; CABANNE, A. M.; FISOGNI, S.; VARRICA, L. M.; BILDER, C. R.; MUNOZ, J. P.; LISERRE, B.; MORELLI, A.; VILLANACCI, V. The enteric nervous system in chagasic and idiopathic megacolon. Am J Surg Pathol, v. 31, p. 460-8, 2007. JONES, E. M.; COLLEY, D. G.; TOSTES, S.; LOPES, E. R.; VNENCAK-JONES, C. L.; MCCURLEY, T. L. Amplification of a Trypanosoma cruzi DNA sequence from inflammatory lesions in human chagasic cardiomyopathy. Am J Trop Med Hyg, v. 48, p. 348-57, 1993. KÖBERLE, F. Patogenia da moléstia de Chagas. Estudos dos órgãos musculares ocos. Revista Goiana de Medicina, v. 3, p. 155-180, 1957. KÖBERLE, F. Pathogenesis of Chagas disease. Ciba Foundation Symposium. 20: 137-158 p. 1974. KOBERLE, F.; DE ALCANTARA, F. [Mechanism of destruction of the neurons of the peripheral nervous system in Changas' disease]. Hospital (Rio J), v. 57, p. 1057-62, 1960. LAGES-SILVA, E.; CREMA, E.; RAMIREZ, L. E.; MACEDO, A. M.; PENA, S. D.; CHIARI, E. Relationship between Trypanosoma cruzi and human chagasic megaesophagus: blood and tissue parasitism. Am J Trop Med Hyg, v. 65, p. 435-41, 2001. LANA, M. Caracterização do Trypanosoma cruzi, cepa Berenice, isolada da mesma paciente em diferentes períodos. 1981. 91 (PhD). Universidade Federal de Minas Gerais LANA, M.; CHIARI, C. A. [Comparative biological characterization of Berenice and Berenice-78 strains of Trypanosoma cruzi isolated from the same patient at different times]. Mem Inst Oswaldo Cruz, v. 81, p. 247-53, 1986. LANAR, D. E.; LEVY, L. S.; MANNING, J. E. Complexity and content of the DNA and RNA in Trypanosoma cruzi. Mol Biochem Parasitol, v. 3, p. 327-41, 1981. LEWIS, M. D.; FRANCISCO, A. F.; TAYLOR, M. C.; KELLY, J. M. A new experimental model for assessing drug efficacy against Trypanosoma cruzi infection based on highly sensitive in vivo imaging. J Biomol Screen, v. 20, p. 36-43, 2015.

Nogueira-Paiva, NC Referências Bibliográficas
106
LISBOA, A. C. Sobre a forma congenita da doença de Chagas. Estudo anatomopatológico de 6 casos. Revista do Instituto de Medicina Tropical de São Paulo, v. 2, p. 319-334, 1960. LIU, B.; LIU, Y.; MOTYKA, S. A.; AGBO, E. E.; ENGLUND, P. T. Fellowship of the rings: the replication of kinetoplast DNA. Trends Parasitol, v. 21, p. 363-9, 2005. LUMSDEN, W. H. Biological aspects of trypanosomiasis research. Adv Parasitol, v. 3, p. 1-57, 1965. MACEDO, A. M.; MACHADO, C. R.; OLIVEIRA, R. P.; PENA, S. D. Trypanosoma cruzi: genetic structure of populations and relevance of genetic variability to the pathogenesis of chagas disease. Mem Inst Oswaldo Cruz, v. 99, p. 1-12, 2004. MACEDO, A. M.; OLIVEIRA, R. P.; PENA, S. D. Chagas disease: role of parasite genetic variation in pathogenesis. Expert Rev Mol Med, v. 4, p. 1-16, 2002. MACEDO, A. M.; PENA, S. D. Genetic Variability of Trypanosoma cruzi:Implications for the Pathogenesis of Chagas Disease. Parasitol Today, v. 14, p. 119-24, 1998. MACEDO, A. M.; PIMENTA, J. R.; AGUIAR, R. S.; MELO, A. I.; CHIARI, E.; ZINGALES, B.; PENA, S. D.; OLIVEIRA, R. P. Usefulness of microsatellite typing in population genetic studies of Trypanosoma cruzi. Mem Inst Oswaldo Cruz, v. 96, p. 407-13, 2001. MACEDO, V. Forma indeterminada da Doença de Chagas. Brazilian Journal of Medical and Biological Research, v. 38, p. 34-40, 1980. MACEDO, V. Indeterminate form of Chagas disease. Mem Inst Oswaldo Cruz, v. 94 Suppl 1, p. 311-6, 1999. MALTOS, K. L.; MENEZES, G. B.; CALIARI, M. V.; ROCHA, O. A.; SANTOS, J. M.; ALVES, D. L.; DUARTE, I. D.; FRANCISCHI, J. N. Vascular and cellular responses to pro-inflammatory stimuli in rat dental pulp. Arch Oral Biol, v. 49, p. 443-50, 2004. MANTILLA, J. C.; ZAFRA, G. A.; MACEDO, A. M.; GONZALEZ, C. I. Mixed infection of Trypanosoma cruzi I and II in a Colombian cardiomyopathic patient. Hum Pathol, v. 41, p. 610-3, 2010. MARIANO, F. S.; GUTIERREZ, F. R.; PAVANELLI, W. R.; MILANEZI, C. M.; CAVASSANI, K. A.; MOREIRA, A. P.; FERREIRA, B. R.; CUNHA, F. Q.; CARDOSO, C. R.; SILVA, J. S. The involvement of CD4+CD25+ T cells in the acute phase of Trypanosoma cruzi infection. Microbes Infect, v. 10, p. 825-33, 2008. MARINO, A. P.; SILVA, A. A.; PINHO, R. T.; LANNES-VIEIRA, J. Trypanosoma cruzi infection: a continuous invader-host cell cross talk with participation of extracellular matrix and adhesion and chemoattractant molecules. Braz J Med Biol Res, v. 36, p. 1121-33, 2003. MARTIN, D.; TARLETON, R. Generation, specificity, and function of CD8+ T cells in Trypanosoma cruzi infection. Immunol Rev, v. 201, p. 304-17, 2004. MCCABE, R. E.; MEAGHER, S. G.; MULLINS, B. T. Endogenous interferon-gamma, macrophage activation, and murine host defense against acute infection with Trypanosoma cruzi. J Infect Dis, v. 163, p. 912-5, 1991.

Nogueira-Paiva, NC Referências Bibliográficas
107
MELO, R. C. Depletion of immune effector cells induces myocardial damage in the acute experimental Trypanosoma cruzi infection: ultrastructural study in rats. Tissue Cell, v. 31, p. 281-90, 1999. MELO, R. C.; BRENER, Z. Tissue tropism of different Trypanosoma cruzi strains. J Parasitol, v. 64, p. 475-82, 1978. MELO, R. C.; MACHADO, C. R. Depletion of radiosensitive leukocytes exacerbates the heart sympathetic denervation and parasitism in experimental Chagas' disease in rats. J Neuroimmunol, v. 84, p. 151-7, 1998. MELO, R. C.; MACHADO, C. R. Trypanosoma cruzi: peripheral blood monocytes and heart macrophages in the resistance to acute experimental infection in rats. Exp Parasitol, v. 97, p. 15-23, 2001. MENEGHELLI, U. G. Chagasic enteropathy. Rev Soc Bras Med Trop, v. 37, p. 252-60, 2004. MENEZES, C. A.; ROCHA, M. O.; SOUZA, P. E.; CHAVES, A. C.; GOLLOB, K. J.; DUTRA, W. O. Phenotypic and functional characteristics of CD28+ and CD28- cells from chagasic patients: distinct repertoire and cytokine expression. Clin Exp Immunol, v. 137, p. 129-38, 2004. MICHAILOWSKY, V.; SILVA, N. M.; ROCHA, C. D.; VIEIRA, L. Q.; LANNES-VIEIRA, J.; GAZZINELLI, R. T. Pivotal role of interleukin-12 and interferon-gamma axis in controlling tissue parasitism and inflammation in the heart and central nervous system during Trypanosoma cruzi infection. Am J Pathol, v. 159, p. 1723-33, 2001. MILES, M. A.; CEDILLOS, R. A.; POVOA, M. M.; DE SOUZA, A. A.; PRATA, A.; MACEDO, V. Do radically dissimilar Trypanosoma cruzi strains (zymodemes) cause Venezuelan and Brazilian forms of Chagas' disease? Lancet, v. 1, p. 1338-40, 1981. MILES, M. A.; LANHAM, S. M.; DE SOUZA, A. A.; POVOA, M. Further enzymic characters of Trypanosoma cruzi and their evaluation for strain identification. Trans R Soc Trop Med Hyg, v. 74, p. 221-37, 1980. MILES, M. A.; SOUZA, A.; POVOA, M.; SHAW, J. J.; LAINSON, R.; TOYE, P. J. Isozymic heterogeneity of Trypanosoma cruzi in the first autochthonous patients with Chagas' disease in Amazonian Brazil. Nature, v. 272, p. 819-21, 1978. MILES, M. A.; TOYE, P. J.; OSWALD, S. C.; GODFREY, D. G. The identification by isoenzyme patterns of two distinct strain-groups of Trypanosoma cruzi, circulating independently in a rural area of Brazil. Trans R Soc Trop Med Hyg, v. 71, p. 217-25, 1977. MOODY, T. N.; OCHIENG, J.; VILLALTA, F. Novel mechanism that Trypanosoma cruzi uses to adhere to the extracellular matrix mediated by human galectin-3. FEBS Lett, v. 470, p. 305-8, 2000. MOREL, C.; CHIARI, E.; CAMARGO, E. P.; MATTEI, D. M.; ROMANHA, A. J.; SIMPSON, L. Strains and clones of Trypanosoma cruzi can be characterized by pattern of restriction endonuclease products of kinetoplast DNA minicircles. Proc Natl Acad Sci U S A, v. 77, p. 6810-4, 1980. MOSER, D. R.; KIRCHHOFF, L. V.; DONELSON, J. E. Detection of Trypanosoma cruzi by DNA amplification using the polymerase chain reaction. J Clin Microbiol, v. 27, p. 1477-82, 1989.

Nogueira-Paiva, NC Referências Bibliográficas
108
MUNOZ-FERNANDEZ, M. A.; FERNANDEZ, M. A.; FRESNO, M. Synergism between tumor necrosis factor-alpha and interferon-gamma on macrophage activation for the killing of intracellular Trypanosoma cruzi through a nitric oxide-dependent mechanism. Eur J Immunol, v. 22, p. 301-7, 1992. NASCIMENTO, R. D.; DE SOUZA LISBOA, A.; FUJIWARA, R. T.; DE FREITAS, M. A.; ADAD, S. J.; OLIVEIRA, R. C.; D'AVILA REIS, D.; DA SILVEIRA, A. B. Characterization of enteroglial cells and denervation process in chagasic patients with and without megaesophagus. Hum Pathol, v. 41, p. 528-34, 2010. NOGUEIRA-PAIVA, N. C.; FONSECA KDA, S.; VIEIRA, P. M.; DINIZ, L. F.; CALDAS, I. S.; MOURA, S. A.; VELOSO, V. M.; GUEDES, P. M.; TAFURI, W. L.; BAHIA, M. T.; CARNEIRO, C. M. Myenteric plexus is differentially affected by infection with distinct Trypanosoma cruzi strains in Beagle dogs. Mem Inst Oswaldo Cruz, v. 109, p. 51-60, 2014. NOGUEIRA-PAIVA, N. C.; VIEIRA, P. M.; OLIVERI, L. M.; FONSECA KDA, S.; POUND-LANA, G.; DE OLIVEIRA, M. T.; DE LANA, M.; VELOSO, V. M.; REIS, A. B.; TAFURI, W. L.; CARNEIRO, C. M. Host-Parasite Interactions in Chagas Disease: Genetically Unidentical Isolates of a Single Trypanosoma cruzi Strain Identified In Vitro via LSSP-PCR. PLoS One, v. 10, p. e0137788, 2015. OLIVEIRA, A. M. Revisão da Taxonomia e classificação do Trypanosoma cruzi - importância da genética. Rev Pat Trop, v. 15, p. 13-16, 1986. OLIVEIRA, R. P.; BROUDE, N. E.; MACEDO, A. M.; CANTOR, C. R.; SMITH, C. L.; PENA, S. D. Probing the genetic population structure of Trypanosoma cruzi with polymorphic microsatellites. Proc Natl Acad Sci U S A, v. 95, p. 3776-80, 1998. OLIVEIRA, R. P.; MELO, A. I.; MACEDO, A. M.; CHIARI, E.; PENA, S. D. The population structure of Trypanosoma cruzi: expanded analysis of 54 strains using eight polymorphic CA-repeat microsatellites. Mem Inst Oswaldo Cruz, v. 94 Suppl 1, p. 65-70, 1999. OUAISSI, A.; OUAISSI, M. Molecular basis of Trypanosoma cruzi and Leishmania interaction with their host(s): exploitation of immune and defense mechanisms by the parasite leading to persistence and chronicity, features reminiscent of immune system evasion strategies in cancer diseases. Arch Immunol Ther Exp (Warsz), v. 53, p. 102-14, 2005. PAIVA, C. N.; FIGUEIREDO, R. T.; KROLL-PALHARES, K.; SILVA, A. A.; SILVERIO, J. C.; GIBALDI, D.; PYRRHO ADOS, S.; BENJAMIM, C. F.; LANNES-VIEIRA, J.; BOZZA, M. T. CCL2/MCP-1 controls parasite burden, cell infiltration, and mononuclear activation during acute Trypanosoma cruzi infection. J Leukoc Biol, v. 86, p. 1239-46, 2009. PENA, S. D.; BARRETO, G.; VAGO, A. R.; DE MARCO, L.; REINACH, F. C.; DIAS NETO, E.; SIMPSON, A. J. Sequence-specific "gene signatures" can be obtained by PCR with single specific primers at low stringency. Proc Natl Acad Sci U S A, v. 91, p. 1946-9, 1994. PINTO, A. S.; DE LANA, M.; BASTRENTA, B.; BARNABE, C.; QUESNEY, V.; NOEL, S.; TIBAYRENC, M. Compared vectorial transmissibility of pure and mixed clonal genotypes of Trypanosoma cruzi in Triatoma infestans. Parasitol Res, v. 84, p. 348-53, 1998. PRATA, A. Clinical and epidemiological aspects of Chagas disease. Lancet Infect Dis, v. 1, p. 92-100, 2001.

Nogueira-Paiva, NC Referências Bibliográficas
109
RODRIGUES, C. M.; VALADARES, H. M.; FRANCISCO, A. F.; ARANTES, J. M.; CAMPOS, C. F.; TEIXEIRA-CARVALHO, A.; MARTINS-FILHO, O. A.; ARAUJO, M. S.; ARANTES, R. M.; CHIARI, E.; FRANCO, G. R.; MACHADO, C. R.; PENA, S. D.; FARIA, A. M.; MACEDO, A. M. Coinfection with different Trypanosoma cruzi strains interferes with the host immune response to infection. PLoS Negl Trop Dis, v. 4, p. e846, 2010. RODRIGUES, M. M.; RIBEIRAO, M.; BOSCARDIN, S. B. CD4 Th1 but not Th2 clones efficiently activate macrophages to eliminate Trypanosoma cruzi through a nitric oxide dependent mechanism. Immunol Lett, v. 73, p. 43-50, 2000. ROMANHA, A. J.; BRENER, Z. Trypanosoma cruzi isoenzyme pattern as an epidemiological tool. Mem Inst Oswaldo Cruz, v. 83 Suppl 1, p. 383-6, 1988. ROMANHA, A. J.; DA SILVA PEREIRA, A. A.; CHIARI, E.; KILGOUR, V. Isoenzyme patterns of cultured Trypanosoma cruzi: changes after prolonged subculture. Comp Biochem Physiol B, v. 62, p. 139-42, 1979. ROPERT, C.; FERREIRA, L. R.; CAMPOS, M. A.; PROCOPIO, D. O.; TRAVASSOS, L. R.; FERGUSON, M. A.; REIS, L. F.; TEIXEIRA, M. M.; ALMEIDA, I. C.; GAZZINELLI, R. T. Macrophage signaling by glycosylphosphatidylinositol-anchored mucin-like glycoproteins derived from Trypanosoma cruzi trypomastigotes. Microbes Infect, v. 4, p. 1015-25, 2002. ROSSI, M. A. Pathogenesis of chronic Chagas' myocarditis. Sao Paulo Med J , v. 113, p. 750-6, 1995. ROSSI, M. A.; BESTETTI, R. B. The challenge of chagasic cardiomyopathy. The pathologic roles of autonomic abnormalities, autoimmune mechanisms and microvascular changes, and therapeutic implications. Cardiology, v. 86, p. 1-7, 1995. ROTTENBERG, M. E.; BAKHIET, M.; OLSSON, T.; KRISTENSSON, K.; MAK, T.; WIGZELL, H.; ORN, A. Differential susceptibilities of mice genomically deleted of CD4 and CD8 to infections with Trypanosoma cruzi or Trypanosoma brucei. Infect Immun, v. 61, p. 5129-33, 1993. ROTTENBERG, M. E.; RIARTE, A.; SPORRONG, L.; ALTCHEH, J.; PETRAY, P.; RUIZ, A. M.; WIGZELL, H.; ORN, A. Outcome of infection with different strains of Trypanosoma cruzi in mice lacking CD4 and/or CD8. Immunol Lett, v. 45, p. 53-60, 1995. RUSSO, M.; STAROBINAS, N.; MINOPRIO, P.; COUTINHO, A.; HONTEBEYRIE-JOSKOWICZ, M. Parasitic load increases and myocardial inflammation decreases in Trypanosoma cruzi-infected mice after inactivation of helper T cells. Ann Inst Pasteur Immunol, v. 139, p. 225-36, 1988. SALDANHA, J. C.; DOS SANTOS, V. M.; DOS REIS, M. A.; DA CUNHA, D. F.; ANTUNES TEIXEIRA, V. P. Morphologic and morphometric evaluation of pancreatic islets in chronic Chagas' disease. Rev Hosp Clin Fac Med Sao Paulo, v. 56, p. 131-8, 2001. SALES, P. A., JR.; GOLGHER, D.; OLIVEIRA, R. V.; VIEIRA, V.; ARANTES, R. M.; LANNES-VIEIRA, J.; GAZZINELLI, R. T. The regulatory CD4+CD25+ T cells have a limited role on pathogenesis of infection with Trypanosoma cruzi. Microbes Infect, v. 10, p. 680-8, 2008. SALGADO, J. A.; GARCEZ, P. N.; DE, O. C.; GALIZZI, J. [Current clinical review of the 1st described human case of Chagas' disease]. Rev Inst Med Trop Sao Paulo, v. 4, p. 330-7, 1962.

Nogueira-Paiva, NC Referências Bibliográficas
110
SANCHEZ-GUILLEN MDEL, C.; LOPEZ-COLOMBO, A.; ORDONEZ-TOQUERO, G.; GOMEZ-ALBINO, I.; RAMOS-JIMENEZ, J.; TORRES-RASGADO, E.; SALGADO-ROSAS, H.; ROMERO-DIAZ, M.; PULIDO-PEREZ, P.; PEREZ-FUENTES, R. Clinical forms of Trypanosoma cruzi infected individuals in the chronic phase of Chagas disease in Puebla, Mexico. Mem Inst Oswaldo Cruz, v. 101, p. 733-40, 2006. SANOJA, C.; CARBAJOSA, S.; FRESNO, M.; GIRONES, N. Analysis of the dynamics of infiltrating CD4(+) T cell subsets in the heart during experimental Trypanosoma cruzi infection. PLoS One, v. 8, p. e65820, 2013. SATO, M. N.; YAMASHIRO-KANASHIRO, E. H.; TANJI, M. M.; KANENO, R.; HIGUCHI, M. L.; DUARTE, A. J. CD8+ cells and natural cytotoxic activity among spleen, blood, and heart lymphocytes during the acute phase of Trypanosoma cruzi infection in rats. Infect Immun, v. 60, p. 1024-30, 1992. SCHMUNIS, G. A. Epidemiology of Chagas disease in non-endemic countries: the role of international migration. Mem Inst Oswaldo Cruz, v. 102 Suppl 1, p. 75-85, 2007. SILVA, J. C.; ASSIS, G. F.; OLIVEIRA, M. T.; VALADARES, H. M.; VALLE, I. F.; PAIVA, N. C.; MARTINS, H. R.; LANA, M. Molecular and biological characterization of Trypanosoma cruzi strains isolated from children from Jequitinhonha Valley, State of Minas Gerais, Brazil. Rev Soc Bras Med Trop, v. 46, p. 433-40, 2013. SILVA, J. S.; VESPA, G. N.; CARDOSO, M. A.; ALIBERTI, J. C.; CUNHA, F. Q. Tumor necrosis factor alpha mediates resistance to Trypanosoma cruzi infection in mice by inducing nitric oxide production in infected gamma interferon-activated macrophages. Infect Immun, v. 63, p. 4862-7, 1995. SILVERIO, J. C.; DE-OLIVEIRA-PINTO, L. M.; DA SILVA, A. A.; DE OLIVEIRA, G. M.; LANNES-VIEIRA, J. Perforin-expressing cytotoxic cells contribute to chronic cardiomyopathy in Trypanosoma cruzi infection. Int J Exp Pathol, v. 91, p. 72-86, 2010. SOARES, M. B.; PONTES-DE-CARVALHO, L.; RIBEIRO-DOS-SANTOS, R. The pathogenesis of Chagas' disease: when autoimmune and parasite-specific immune responses meet. An Acad Bras Cienc, v. 73, p. 547-59, 2001. SOUTO, R. P.; FERNANDES, O.; MACEDO, A. M.; CAMPBELL, D. A.; ZINGALES, B. DNA markers define two major phylogenetic lineages of Trypanosoma cruzi. Mol Biochem Parasitol, v. 83, p. 141-52, 1996. SOUTO, R. P.; ZINGALES, B. Sensitive detection and strain classification of Trypanosoma cruzi by amplification of a ribosomal RNA sequence. Mol Biochem Parasitol, v. 62, p. 45-52, 1993. SOUZA, R. T.; LIMA, F. M.; BARROS, R. M.; CORTEZ, D. R.; SANTOS, M. F.; CORDERO, E. M.; RUIZ, J. C.; GOLDENBERG, S.; TEIXEIRA, M. M.; DA SILVEIRA, J. F. Genome size, karyotype polymorphism and chromosomal evolution in Trypanosoma cruzi. PLoS One, v. 6, p. e23042, 2011. STEINDEL, M.; DIAS NETO, E.; DE MENEZES, C. L.; ROMANHA, A. J.; SIMPSON, A. J. Random amplified polymorphic DNA analysis of Trypanosoma cruzi strains. Mol Biochem Parasitol, v. 60, p. 71-9, 1993.

Nogueira-Paiva, NC Referências Bibliográficas
111
STORDEUR, P.; POULIN, L. F.; CRACIUN, L.; ZHOU, L.; SCHANDENE, L.; DE LAVAREILLE, A.; GORIELY, S.; GOLDMAN, M. Cytokine mRNA quantification by real-time PCR. J Immunol Methods, v. 259, p. 55-64, 2002. TAFURI, W. L. Pathogenesis of lesions of the autonomic nervous system of the mouse in experimental acute Chagas' disease. Light and electron microscope studies. Am J Trop Med Hyg, v. 19, p. 405-17, 1970. TAFURI, W. L. Light and electron microscope studies of the autonomic nervous system in experimental and human American trypanosomiasis. Virchows Arch A Pathol Pathol Anat, v. 354, p. 136-49, 1971. TAFURI, W. L.; BRENER, Z. [Injuries of Meissner and Auerback plexus of albino mice intestines in the chornic phase of experimental trypanosomiasis cruzi]. Rev Inst Med Trop Sao Paulo, v. 9, p. 149-54, 1967a. TAFURI, W. L.; BRENER, Z. Lesões do sistema nervoso autônomo do camundongo albino na tripanosomiase cruzi experimental, na fase aguda. Hospital, v. 69, p. 371-383, 1967b. TAFURI, W. L.; DE LANA, M.; CHIARI, E.; CALIARI, M. V.; BAMBIRRA, E. A.; RIOS-LEITE, V. H.; BARBOSA, A. J. [Dogs as experimental models for the study of the natural course of Chagas disease]. Rev Soc Bras Med Trop, v. 21, p. 77, 1988. TAFURI, W. L.; MARIA, T. A.; LOPES, E. R. [Myenteric plexus lesions in the esophagus, jejunum and colon of chronic chagasic patients. Electron microscopy study]. Rev Inst Med Trop Sao Paulo, v. 13, p. 76-91, 1971. TAFURI, W. L.; RASO, P. [Lesions of the autonomic nervous system in albino mice with trypanosomiasis]. Hospital (Rio J), v. 62, p. 1325-42, 1962. TARLETON, R. L. CD8+ T cells in Trypanosoma cruzi infection. Semin Immunopathol, v. 37, p. 233-8, 2015. TARLETON, R. L.; ZHANG, L. Chagas disease etiology: autoimmunity or parasite persistence? Parasitol Today, v. 15, p. 94-9, 1999. TIBAYRENC, M. Population genetics of parasitic protozoa and other microorganisms. Adv Parasitol, v. 36, p. 47-115, 1995. TIBAYRENC, M.; AYALA, F. J. Isozyme variability in Trypanosoma cruzi, the agent of Chagas’disease: genetical, taxonomical, and epidemiological significance. Evolution, v. 42, p. 277-292, 1988. TIBAYRENC, M.; HOFFMANN, A.; POCH, O.; ECHALAR, L.; LE PONT, F.; LEMESRE, J. L.; DESJEUX, P.; AYALA, F. J. Additional data on Trypanosoma cruzi isozymic strains encountered in Bolivian domestic transmission cycles. Trans R Soc Trop Med Hyg, v. 80, p. 442-7, 1986. TIBAYRENC, M.; MILES, M. A. A genetic comparison between Brazilian and Bolivian zymodemes of Trypanosoma cruzi. Trans R Soc Trop Med Hyg, v. 77, p. 76-83, 1983. TORRICO, F.; HEREMANS, H.; RIVERA, M. T.; VAN MARCK, E.; BILLIAU, A.; CARLIER, Y. Endogenous IFN-gamma is required for resistance to acute Trypanosoma cruzi infection in mice. J Immunol, v. 146, p. 3626-32, 1991.

Nogueira-Paiva, NC Referências Bibliográficas
112
VAGO, A. R.; ANDRADE, L. O.; LEITE, A. A.; D'AVILA REIS, D.; MACEDO, A. M.; ADAD, S. J.; TOSTES, S., JR.; MOREIRA, M. C.; FILHO, G. B.; PENA, S. D. Genetic characterization of Trypanosoma cruzi directly from tissues of patients with chronic Chagas disease: differential distribution of genetic types into diverse organs. Am J Pathol, v. 156, p. 1805-9, 2000. VAGO, A. R.; MACEDO, A. M.; ADAD, S. J.; REIS, D. D.; CORREA-OLIVEIRA, R. PCR detection of Trypanosoma cruzi DNA in oesophageal tissues of patients with chronic digestive Chagas' disease. Lancet, v. 348, p. 891-2, 1996a. VAGO, A. R.; MACEDO, A. M.; OLIVEIRA, R. P.; ANDRADE, L. O.; CHIARI, E.; GALVAO, L. M.; REIS, D.; PEREIRA, M. E.; SIMPSON, A. J.; TOSTES, S.; PENA, S. D. Kinetoplast DNA signatures of Trypanosoma cruzi strains obtained directly from infected tissues. Am J Pathol, v. 149, p. 2153-9, 1996b. VAGO, A. R.; SILVA, D. M.; ADAD, S. J.; CORREA-OLIVEIRA, R.; D'AVILA REIS, D. Chronic Chagas disease: presence of parasite DNA in the oesophagus of patients without megaoesophagus. Trans R Soc Trop Med Hyg, v. 97, p. 308-9, 2003. VALADARES, H. M.; PIMENTA, J. R.; SEGATTO, M.; VELOSO, V. M.; GOMES, M. L.; CHIARI, E.; GOLLOB, K. J.; BAHIA, M. T.; DE LANA, M.; FRANCO, G. R.; MACHADO, C. R.; PENA, S. D.; MACEDO, A. M. Unequivocal identification of subpopulations in putative multiclonal Trypanosoma cruzi strains by FACs single cell sorting and genotyping. PLoS Negl Trop Dis, v. 6, p. e1722, 2012. VAN VOORHIS, W. C. Coculture of human peripheral blood mononuclear cells with Trypanosoma cruzi leads to proliferation of lymphocytes and cytokine production. J Immunol, v. 148, p. 239-48, 1992. VELOSO, V. M.; GUEDES, P. M.; ANDRADE, I. M.; CALDAS, I. S.; MARTINS, H. R.; CARNEIRO, C. M.; MACHADO-COELHO, G. L.; DE LANA, M.; GALVAO, L. M.; BAHIA, M. T.; CHIARI, E. Trypanosoma cruzi: blood parasitism kinetics and their correlation with heart parasitism intensity during long-term infection of Beagle dogs. Mem Inst Oswaldo Cruz, v. 103, p. 528-34, 2008. VELOSO, V. M.; GUEDES, P. M.; DE LANA, M.; MARTINS, H. R.; CARNEIRO, C. M.; DA CAMARA, A. C.; D'AVILA, D. A.; CALDAS, I. S.; GALVAO, L. M.; CHIARI, E.; BAHIA, M. T. Genetic modulation in Be-78 and Y Trypanosoma cruzi strains after long-term infection in Beagle dogs revealed by molecular markers. Infect Genet Evol, v. 12, p. 1128-35, 2012. VELOSO, V. M.; ROMANHA, A. J.; LANA, M.; MURTA, S. M.; CARNEIRO, C. M.; ALVES, C. F.; BORGES, E. C.; TAFURI, W. L.; MACHADO-COELHO, G. L.; CHIARI, E.; BAHIA, M. T. Influence of the long-term Trypanosoma cruzi infection in vertebrate host on the genetic and biological diversity of the parasite. Parasitol Res, v. 96, p. 382-9, 2005. VESPA, G. N.; CUNHA, F. Q.; SILVA, J. S. Nitric oxide is involved in control of Trypanosoma cruzi-induced parasitemia and directly kills the parasite in vitro. Infect Immun, v. 62, p. 5177-82, 1994. VIEIRA, P. M.; FRANCISCO, A. F.; DE SOUZA, S. M.; MALAQUIAS, L. C.; REIS, A. B.; GIUNCHETTI, R. C.; VELOSO, V. M.; DE LANA, M.; TAFURI, W. L.; CARNEIRO, C. M. Trypanosoma cruzi: Serum levels of nitric oxide and expression of inducible nitric oxide synthase in myocardium and spleen of dogs in the acute stage of infection with metacyclic or blood trypomastigotes. Exp Parasitol, v. 121, p. 76-82, 2009.

Nogueira-Paiva, NC Referências Bibliográficas
113
VIEIRA, P. M.; FRANCISCO, A. F.; MACHADO, E. M.; NOGUEIRA, N. C.; FONSECA KDA, S.; REIS, A. B.; TEIXEIRA-CARVALHO, A.; MARTINS-FILHO, O. A.; TAFURI, W. L.; CARNEIRO, C. M. Different infective forms trigger distinct immune response in experimental Chagas disease. PLoS One, v. 7, p. e32912, 2012. VIRREIRA, M.; SERRANO, G.; MALDONADO, L.; SVOBODA, M. Trypanosoma cruzi: typing of genotype (sub)lineages in megacolon samples from bolivian patients. Acta Trop, v. 100, p. 252-5, 2006. WESTENBERGER, S. J.; CERQUEIRA, G. C.; EL-SAYED, N. M.; ZINGALES, B.; CAMPBELL, D. A.; STURM, N. R. Trypanosoma cruzi mitochondrial maxicircles display species- and strain-specific variation and a conserved element in the non-coding region. BMC Genomics, v. 7, p. 60, 2006. WHO. Expert Committee. Control of Chagas disease. Brasília, Brasil. WHO technical report series 905. 2002. WHO. Chagas disease control and prevention in Europe. Report of a WHO Informal Consultation. 2010a. WHO. First WHO Report on Neglected Tropical Diseases: working to overcome the global impact of neglected tropical diseases. 2010b. WIRTH, J. J.; KIERSZENBAUM, F. Fibronectin enhances macrophage association with invasive forms of Trypanosoma cruzi. J Immunol, v. 133, p. 460-4, 1984. ZAFRA, G.; MANTILLA, J. C.; VALADARES, H. M.; MACEDO, A. M.; GONZALEZ, C. I. Evidence of Trypanosoma cruzi II infection in Colombian chagasic patients. Parasitol Res, v. 103, p. 731-4, 2008. ZELEDON, R.; RABINOVICH, J. E. Chagas' disease: an ecological appraisal with special emphasis on its insect vectors. Annu Rev Entomol, v. 26, p. 101-33, 1981. ZINGALES, B.; ANDRADE, S. G.; BRIONES, M. R.; CAMPBELL, D. A.; CHIARI, E.; FERNANDES, O.; GUHL, F.; LAGES-SILVA, E.; MACEDO, A. M.; MACHADO, C. R.; MILES, M. A.; ROMANHA, A. J.; STURM, N. R.; TIBAYRENC, M.; SCHIJMAN, A. G.; SECOND SATELLITE, M. A new consensus for Trypanosoma cruzi intraspecific nomenclature: second revision meeting recommends TcI to TcVI. Mem Inst Oswaldo Cruz, v. 104, p. 1051-4, 2009. ZINGALES, B.; MILES, M. A.; CAMPBELL, D. A.; TIBAYRENC, M.; MACEDO, A. M.; TEIXEIRA, M. M.; SCHIJMAN, A. G.; LLEWELLYN, M. S.; LAGES-SILVA, E.; MACHADO, C. R.; ANDRADE, S. G.; STURM, N. R. The revised Trypanosoma cruzi subspecific nomenclature: rationale, epidemiological relevance and research applications. Infect Genet Evol, v. 12, p. 240-53, 2012. ZINGALES, B.; SOUTO, R. P.; MANGIA, R. H.; LISBOA, C. V.; CAMPBELL, D. A.; COURA, J. R.; JANSEN, A.; FERNANDES, O. Molecular epidemiology of American trypanosomiasis in Brazil based on dimorphisms of rRNA and mini-exon gene sequences. Int J Parasitol, v. 28, p. 105-12, 1998.

114
9. ANEXOS

Nogueira-Paiva, NC Anexos
115
9.1 Anexo 1: Protocolo nº 2011/30- CEUA/UFOP

Nogueira-Paiva, NC Anexos
116
9.2 Anexo 2: Protocolo nº 2014/29- CEUA/UFOP

Nogueira-Paiva, NC Anexos
117
9.3 Anexo 3: Resumo dos resultados obtidos na ETAPA II
CORAÇÃO
Tempo Grupos Carga
Parasitária Inflamação
Total Macrófago Linfócito B
Linf T CD4+
Linf T CD8+
Linf T CD4+IFN-+
Linf T CD8+IFN-+
7dai
NI x Be-62 = = =
NI x Be-78 isolado ~ = = = = = = =
NI x Be-78 parental ~ = = = = =
Be-62 x Be-78 isolado = = = = =
Be-62 x Be-78 parental = = =
Be-78 isolado x Be-78 parental = = = = = =
14dai
NI x Be-62 = =
NI x Be-78 isolado = = = = =
NI x Be-78 parental = = = = =
Be-62 x Be-78 isolado = = = =
Be-62 x Be-78 parental = = =
Be-78 isolado x Be-78 parental = = = = = =
28dai
NI x Be-62 =
NI x Be-78 isolado = = = =
NI x Be-78 parental
Be-62 x Be-78 isolado = =
Be-62 x Be-78 parental = = = =
Be-78 isolado x Be-78 parental = = =
Figura 30: Representação esquemática dos resultados obtidos na ETAPA II, comparando as diferentes subpopulações do T. cruzi, em relação à carga parasitária e ao processo inflamatório (intensidade e fenótipo) no coração de camundongos BALB/c necropsiados ao longo da fase aguda (7, 14 ou 28 dias após a infecção-dai) da infecção experimental com o isolado Be-78is5-3mai, ou com as cepas de referência Berenice-62 e Berenice-78 parental do Trypanosoma cruzi. NI: grupo de animais não-infectados; Be-62: grupo de animais infectados com a cepa Be-62; Be-78 isolado: grupo de animais infectados com o isolado Be-78is5-3mai; Be-78 parental: grupo de animais infectados com a cepa Berenice-78. (=): ausência de diferença estatística; (~): ausência de diferença estatística, no entanto, alguns parasitos presentes; ( ): e; ( ): estatisticamente superior para o grupo Be-62; ( ): estatisticamente superior para o grupo Be-78 isolado; ( ): estatisticamente superior para o grupo Be-78 parental.
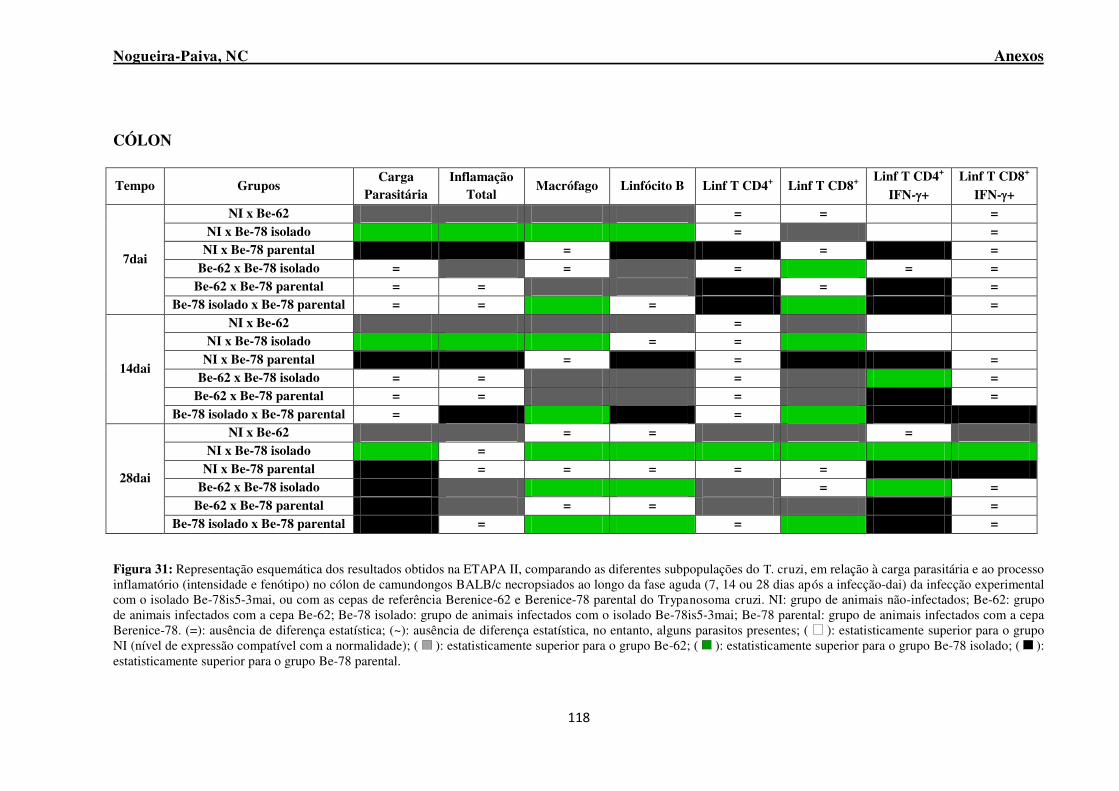
Nogueira-Paiva, NC Anexos
118
CÓLON
Tempo Grupos Carga
Parasitária Inflamação
Total Macrófago Linfócito B Linf T CD4+ Linf T CD8+
Linf T CD4+ IFN-+
Linf T CD8+ IFN-+
7dai
NI x Be-62 = = = NI x Be-78 isolado = =
NI x Be-78 parental = = = Be-62 x Be-78 isolado = = = = =
Be-62 x Be-78 parental = = = = Be-78 isolado x Be-78 parental = = = =
14dai
NI x Be-62 = NI x Be-78 isolado = =
NI x Be-78 parental = = = Be-62 x Be-78 isolado = = = =
Be-62 x Be-78 parental = = = = Be-78 isolado x Be-78 parental = =
28dai
NI x Be-62 = = = NI x Be-78 isolado =
NI x Be-78 parental = = = = = Be-62 x Be-78 isolado = =
Be-62 x Be-78 parental = = = Be-78 isolado x Be-78 parental = = =
Figura 31: Representação esquemática dos resultados obtidos na ETAPA II, comparando as diferentes subpopulações do T. cruzi, em relação à carga parasitária e ao processo inflamatório (intensidade e fenótipo) no cólon de camundongos BALB/c necropsiados ao longo da fase aguda (7, 14 ou 28 dias após a infecção-dai) da infecção experimental com o isolado Be-78is5-3mai, ou com as cepas de referência Berenice-62 e Berenice-78 parental do Trypanosoma cruzi. NI: grupo de animais não-infectados; Be-62: grupo de animais infectados com a cepa Be-62; Be-78 isolado: grupo de animais infectados com o isolado Be-78is5-3mai; Be-78 parental: grupo de animais infectados com a cepa Berenice-78. (=): ausência de diferença estatística; (~): ausência de diferença estatística, no entanto, alguns parasitos presentes; ( ): estatisticamente superior para o grupo NI (nível de expressão compatível com a normalidade); ( ): estatisticamente superior para o grupo Be-62; ( ): estatisticamente superior para o grupo Be-78 isolado; ( ): estatisticamente superior para o grupo Be-78 parental.

Nogueira-Paiva, NC Anexos
119
9.4 Anexo 4: Manuscrito publicado no periódico PLOs One/ETAPA I
NOGUEIRA-PAIVA, N. C.; VIEIRA, P. M.; OLIVERI, L. M.; FONSECA KDA, S.; POUND-LANA, G.; DE OLIVEIRA, M. T.; DE LANA, M.; VELOSO, V. M.; REIS, A. B.; TAFURI, W. L.; CARNEIRO, C. M. Host-Parasite Interactions in Chagas Disease: Genetically Unidentical Isolates of a Single Trypanosoma cruzi Strain Identified In Vitro via LSSP-PCR. PLoS One, v. 10, p. e0137788, 2015.

RESEARCH ARTICLE
Host-Parasite Interactions in Chagas Disease:
Genetically Unidentical Isolates of a Single
Trypanosoma cruzi Strain Identified In Vitro
via LSSP-PCR
Nívia Carolina Nogueira-Paiva1, Paula Melo de Abreu Vieira1,3, Larissa Maris
Rezende Oliveri1, Kátia da Silva Fonseca1, Gwenaelle Pound-Lana1, Maykon Tavares de
Oliveira2, Marta de Lana2,5, Vanja Maria Veloso4, Alexandre Barbosa Reis1,5, Washington
Luiz Tafuri1†, Cláudia Martins Carneiro1,5*
1 Laboratório de Imunopatologia, Núcleo de Pesquisas em Ciências Biológicas, Instituto de Ciências Exatase Biológicas, Universidade Federal de Ouro Preto, Ouro Preto, MG, Brazil, 2 Laboratório de Doença deChagas, Núcleo de Pesquisas em Ciências Biológicas, Instituto de Ciências Exatas e Biológicas,Universidade Federal de Ouro Preto,Ouro Preto, MG, Brazil, 3 Departamento de Ciências Biológicas,Núcleo de Pesquisas em Ciências Biológicas, Instituto de Ciências Exatas e Biológicas, UniversidadeFederal de Ouro Preto, Ouro Preto, MG, Brazil, 4 Departamento de Farmácia, Escola de Farmácia,Universidade Federal de Ouro Preto, Ouro Preto, MG, Brazil, 5 Departamento de Análises Clínicas, Escolade Farmácia, Universidade Federal de Ouro Preto,Ouro Preto, MG, Brazil
†Deceased.* [email protected]
Abstract
The present study aims at establishing whether the diversity in pathogenesis within a genet-
ically diverse host population infected with a single polyclonal strain of Trypanosoma cruzi
is due to selection of specific subpopulations within the strain. For this purpose we infected
Swiss mice, a genetically diverse population, with the polyclonal strain of Trypanosoma
cruzi Berenice-78 and characterized via LSSP-PCR the kinetoplast DNA of subpopulations
isolated from blood samples collected from the animals at various times after inoculation (3,
6 and 12 months after inoculation). We examined the biological behavior of the isolates in
acellular medium and in vitro profiles of infectivity in Vero cell medium. We compared the
characteristics of the isolates with the inoculating strain and with another strain, Berenice
62, isolated from the same patient 16 years earlier. We found that one of the isolates had
intermediate behavior in comparison with Berenice-78 and Berenice-62 and a significantly
different genetic profile by LSSP-PCR in comparison with the inoculating strain. We hereby
demonstrate that genetically distinct Trypanosoma cruzi isolates may be obtained upon
experimental murine infection with a single polyclonal Trypanosoma cruzi strain.
PLOSONE | DOI:10.1371/journal.pone.0137788 September 11, 2015 1 / 14
OPEN ACCESS
Citation: Nogueira-Paiva NC, Vieira PMdA, Oliveri
LMR, Fonseca KdS, Pound-Lana G, de Oliveira MT,
et al. (2015) Host-Parasite Interactions in Chagas
Disease: Genetically Unidentical Isolates of a Single
Trypanosoma cruzi Strain Identified In Vitro via
LSSP-PCR. PLoS ONE 10(9): e0137788.
doi:10.1371/journal.pone.0137788
Editor: Alex Córdoba-Aguilar, Universidad Nacional
Autonoma de Mexico, MEXICO
Received: May 19, 2015
Accepted: August 21, 2015
Published: September 11, 2015
Copyright: © 2015 Nogueira-Paiva et al. This is an
open access article distributed under the terms of the
Creative Commons Attribution License, which permits
unrestricted use, distribution, and reproduction in any
medium, provided the original author and source are
credited.
Data Availability Statement: All relevant data are
within the paper and its Supporting Information file.
Funding: Funding was provided by the Universidade
Federal de Ouro Preto (UFOP), Fundação de
Amparo a Pesquisa do Estado de Minas Gerais
(FAPEMIG), Conselho Nacional de Desenvolvimento
Científico e Tecnológico (CNPq), Coordenação de
Aperfeiçoamento de Pessoal de Nível Superior
(CAPES) and research fellowships from CNPq
(Carneiro CM, Reis AB, Lana M, Oliveri LMR),
CAPES (Fonseca KS, Nogueira-Paiva NC) and

Introduction
More than a hundred years after the first human clinical case of Chagas disease was described,
the origin of the broad clinical spectrum of the disease, ranging from asymptomatic to cardiac
and gastro-enteric manifestations, has yet to be elucidated. The polymorphism of Trypanosoma
cruzi (T. cruzi), the etiologic agent of Chagas disease, was demonstrated via both biochemical
and genetic assays [1]. However, attempts to correlate the pathogenicity, tissue-tropism or
drug-susceptibility [2] to specific strains or distinct typing units (DTU) of T. cruzi have failed.
Polyclonal strains are commonly found in natural infections. They consist of a combination
of subpopulations of clones with distinct biological and genetic characteristics. According to
the clonal-histotropic model [3] some clonal associations are selectively advantageous resulting
in stable strains. The host genetic background may however determine its susceptibility
towards infection and towards the development of the disease [4, 5]. When mammals are
infected, the clonal repertory may determine which tissues will be affected [6]. Given that not
all clones are able to trigger infection, the clones that are able to establish themselves probably
compete among each other and colonize distinct tissues. Therefore the interaction between
each clone and a specific tissue may be a determining factor in the development of distinct clin-
ical forms of Chagas disease [3].
The Berenice strain was isolated from the first described human case of Chagas disease [7]
from a patient living in an endemic area for the disease (município de Pirapora, MG/Brasil). In
addition to the T. cruzi subpopulation isolated during the acute phase [7], another two parasite
samples, currently named Berenice 62 (Be-62) and Berenice-78 (Be-78) strains, were isolated
from this same patient with a 16-year interval between collections via xenodiagnostic using
Triatoma infestans and Dipetalogaster maximus, respectively [8, 9]. Interestingly both isolates
present different in vitro biological behavior, virulence and pathogenicity [10]. Two hypotheses
have been put forward to explain the alteration in profile of these strains. Even though the
patient denies any contact with the vector after contracting the disease, the possibility of re-
infection cannot be completely discarded given that she continued living in an endemic area.
However, should there have been no re-infection, the differences between the two isolates sug-
gest selection of one or more subpopulations from the initial strain via immunologic mecha-
nisms that take place during the chronic phase of the disease [10]. This hypothesis is supported
by the demonstrated polyclonality of the Berenice strain [11] but remains to be elucidated.
While genetic studies can demonstrate the intraspecific heterogeneity of the parasite, prob-
ing the biological behavior and host-parasite interactions help clarify the importance of differ-
ent strains and different subpopulations within a strain in determining the clinical and
epidemiologic manifestations of the disease [12]. In this respect, Vago et al. [13] demonstrated
that T. cruzi with distinct kinetoplast DNA (kDNA) signatures were detected in different tis-
sues in human cases of Chagas disease. Veloso and coworkers [14] studied the host-parasite
interaction in experimental infections in dogs with polyclonal T. cruzi strains and found dis-
tinct T. cruzi populations in each host, which composition could vary along the course of long-
term infection [14]. Rodrigues and coworkers [5] observed that the immune response of
human patients in the case of co-infection with two different T. cruzi strains does not follow
the pattern of the immune response observed in the case of infection with either of the two
strains studied. All the above-cited studies of natural and experimental co-infections have
pointed out the need to consider clonal interactions in understanding the pathology.
Due to possible adaptive clonal selection, in vitro and in vivo studies on the genetic variabil-
ity of T. cruzi in infected animals have long suffered from experimental limitations related to
culture and maintenance of the parasite [15, 16, 17]. Many of these limitations were overcome
recently with the introduction of the experimental technique low-stringency single specific
Trypanosoma cruzi and Parasite Selection
PLOS ONE | DOI:10.1371/journal.pone.0137788 September 11, 2015 2 / 14
FAPEMIG (Pound-Lana G, Oliveira MT). The funders
had no role in study design, data collection and
analysis, decision to publish, or preparation of the
manuscript.
Competing Interests: The authors have declared
that no competing interests exist.

primer polymerase chain-reaction (LSSP-PCR), a PCR-based technique that enables DNA pro-
filing directly from an infected tissue [18]. The technique is designed in such a way that minute
alterations in the genome, such as a single-base alteration can be detected [19]. As such it is a
very sensitive technique capable of differentiating between subpopulations within a given DTU
[20].
Although there is supporting evidence for the validity of the clonal-histotropic model, it
remains to be confirmed whether a single subpopulation should display tropism for a specific
organ or cell component. In an attempt to validate such hypothesis we designed the presently
reported study, where the Swiss mice host serves as a “filter” to isolate a clonal subpopulation
with distinct genetic characteristics from the infecting strain.
Animals, Materials and Methods
All procedures and experimental protocols were conducted in accordance with the directives
issued by the Brazilian College of Animal Experimentation (COBEA) and approved by the Eth-
ics Committee in Animal Research at the Universidade Federal de Ouro Preto (UFOP), Minas
Gerais, Brazil (protocol number 2011/30, the arrive guidelines checklist in S1 File). Prior to the
study, the animals were dewormed and vaccinated against several infectious diseases.
2.1 T. cruzi strainsBe-62 [8] and Be-78 [9] strains were maintained by successive passages in mice. Be-62 and Be-
78 belong to DTU TcII [21].
2.2 Animals and experimental infectionTwenty-four one-month-old Swiss mice from the Center for Animal Science CCA/UFOP,
weighed 23.5–24.5g, were inoculated intraperitoneally with 5.0 × 103 bloodstream trypomasti-
gotes of the Be-78 T. cruzi strain and maintained at the following environmental conditions: 12
h day/night cycle, temperature 22 ± 2°C, food and water ad libitum for 12 months after infec-
tion (MAI). The infection and curve of parasitemia were confirmed via examination of fresh
blood samples according to the methodology of [22]. The anesthetized mice (ketamine 60 mg/
kg and xylazine 7.5 mg/kg, [23]) underwent blood sampling for hemoculture at three, six and
twelve MAI, which corresponds to the chronic phase of the disease. Under sterile conditions,
200 μl of blood collected from the orbital plexus of each mouse were added to 15 ml sterile
tubes (Falcon, Becton Dickinson, USA) containing liver infusion tryptose (LIT) medium sup-
plemented with 10% inactivated fetal bovine serum (LIT-10% FCS, Liver Infusion Tryptose-
10% Fetal Bovine Serum) and stored in a refrigerated B.O.D incubator at 28°C ± 1°C for assess-
ment at 30, 60 and 90 days after collection. During the course of the experiment the animals
were monitored daily and not show signs of pain or distress. The criteria used for evaluating
their health and welfare were their weight, appetite, water consumption, skin and fur condition,
in comparison with a non-inoculated control group. At the end of the study, mice were eutha-
nized in a CO2 chamber and all efforts were made to minimize suffering.
Positive hemocultures were maintained in exponential growth in LIT medium-10% FCS,
for a short period of time to minimize parasite selection, under the same conditions
(28°C ± 1°C) [24] to determine the behavior in acellular culture medium (LIT) or cellular cul-
ture medium (Vero cells) and also for obtaining a sufficient number of epimastigotes for
molecular characterization. Samples of these cultures were subjected to evaluation of in vitro
growth kinetics of epimastigotes, incubated in Grace (Grace's Insect Medium, Sigma) to induce
metacyclogenesis (in vitro infection assay) or washed and harvested for LSSP-PCR. For this
purpose the cultures were transferred to 50 ml tubes (Falcon, Becton Dickinson, USA) and
Trypanosoma cruzi and Parasite Selection
PLOS ONE | DOI:10.1371/journal.pone.0137788 September 11, 2015 3 / 14

centrifuged at 3.400g for 30 min at 4°C. The supernatant was discarded and the pellet sus-
pended in 10 ml of sterile 10% PBS (Phosphate-Buffered Saline) at pH 7.2. The cells were
washed and centrifuged 3 times (3.400g for 10 min, 4°C) in 10% PBS. The epimastigotes
obtained were stored in a freezer at -70°C.
2.3 Growth kinetics in acellular culture mediumIn vitro growth kinetic curves of the isolates in acellular culture medium LIT-10% SFB were
obtained from a standard inoculum of 1 x 106 parasites / ml in a final volume 3 ml. For this
purpose the volume used in the inoculum for each culture was determined with a Neubauer
chamber, and the cultures were kept in triplicate in a refrigerated B.O.D incubator at
28°C ± 1°C during the evaluation period. Cell count was carried out daily in a Neubauer cham-
ber under 400x magnification over a period of 20 days [25].
2.4 Profile of infectivity and in vitro developmentThe in vitro profile of infectivity and growth of the isolates of Be- 78 strain were compared to
those of the parental Be-78 strain and of Be-62 strain in Vero cell cultures subjected to infection
and assessed after 24, 48 and 72h, in triplicate, according to a methodology adapted from
Andrade et al. [26]. Briefly, after 24h of culture, semiconfluent monolayers of Vero cells (104)
fixed on glass coverslips were incubated at 5% CO2 and 37°C for 18h (overnight) with axenic
cultures metacyclic trypomastigotes, to a MOI (multiplicity of infection) of 10, of different T.
cruzi subpopulations (105), previously submitted to metacyclogenesis in Grace medium. After
washing with PBS to remove extracellular parasites, cultures were kept under the same incuba-
tion conditions until the time of collection of the cells (24, 48 and 72h after inoculation). Subse-
quently, the cultures were fixed in methanol and stained with Fast Panoptic and the ratios of
infection and intracellular development or proliferation were examined by optical microscopy
under 1000x magnification. The infectivity ratio was calculated as the number of infected cells
per 100 cells counted, while the intracellular development ratio was calculated as the ratio of
the number of parasites counted over the number of infected cells.
2.5 Gene Signature by LSSP-PCRThe evaluation of gene polymorphism between different isolates and the Be-78 parental strain
by LSSP-PCR consisted in obtaining, purifying and amplifying the template DNA followed by
a second amplification using a single primer under conditions of low stringency [19]. The spe-
cific amplification of the 330bp (base pairs) fragment of the kDNA of T. cruzi was performed
according to the method of Gomes et al. [27]. The amplified products were revealed on 1.5%
agarose gel (1.0% agarose + 0.5% LowMelting Point agarose / Invitrogen) and stained with
ethidium bromide. The 330bp fragment, visualized under UV radiation, was removed from the
gel using a plastic pipettor. After heating in a thermostated water bath, these fragments were
homogenized, diluted in sterile Milli-Q water at a ratio of 1:10 (v/v) and used in a second
amplification step. For the LSSP-PCR reactions the DNA samples were re-amplified using a
single specific primer (corresponding to the primer S35), termed S35G� (5’-AAA TAA TGT
ACG GGG GAG AT-3’). The reaction mixture reached a final volume of 10 μL containing
6.38 μl of sterile milli-Q water; 2.0 μl of sample buffer; 0.2 μl of each deoxynucleotide (dNTP:
dATP, dCTP, dGTP, dTTP—Sigma, St. Louis, MO, USA); 0.1 μl of primer S35G � (450μM)
and 0.32 μl of Taq DNA polymerase (5U / μl—Go Taq Promega). To this mixture was added
1 μL of DNA diluted at 1:10 (v/v). The amplification was carried out under the following condi-
tions: initial DNA denaturation step at 94°C for 5 min, annealing at 30°C for 1 min, extension
at 72°C for 1 min, followed by 40 amplification cycles consisting of a denaturation step at 94°C
Trypanosoma cruzi and Parasite Selection
PLOS ONE | DOI:10.1371/journal.pone.0137788 September 11, 2015 4 / 14

for 1 min, annealing at 30°C for 1 min, ending with an extension step at 72°C for 10 min. The
products of this second amplification were revealed in duplicate on 8% polyacrylamide gel,
stained with silver and analyzed for band sharing.
2.6 Statistical analysesFor the evaluation of growth profiles in axenic growth medium the area-under-the-curve
(AUC) was considered via Kruskal-Wallis one-way analysis of variance for non-parametric
data followed by Dunn´s post-hoc test for comparison with the reference Be-78 and Be-62
strains. The in vitro infection test with Vero cells were compared using Anova two-way analysis
of variance followed by Bonferroni post-hoc test for comparison with the reference Be-78 and
Be-62 strains. In all cases, differences were considered statistically significant at p values lower
than 0.05.
Results
3.1 HemoculturesOut of the twenty-four mice infected with the Be-78 T. cruzi strain and subjected to blood col-
lection seven presented positive hemocultures at one sampling time at least (Table 1). Although
not all animals were found positive for hemocultures, it was possible to isolate parasites at all
three sampling times, over a year of infection.
3.2 Growth kinetics in acellular culture mediumGrowth curves were recorded over 20 days after inoculation of 106 parasites / ml of culture for
each isolate. We observed a significant difference in the area under the curve (AUC) between
the two strains used as reference. The Be-78 parental strain (5,3x108) presented a higher repli-
cation ratio than the Be-62 strain (3,7x108). In addition to the AUC, the differences in behavior
in the distinct phases of the growth epimastigote curve was confirmed between these strains in
axenic culture showing a higher and earlier growth with early entry into the decline phase for
the Be-62 strain, compared to the strain Be-78 (Fig 1), as described by other authors [9, 10].
Furthermore, a significant reduction in the AUC was identified in two isolates, Be-78is15-6mai
(4,2x108) and Be-78is1-12mai (4,2x108) in comparison with the Be-78 strain used in the inocu-
lum (5,3 x108), indicating that the former have reduced ability to grow in culture medium com-
pared to the parental strain. In comparing the different stages of growth (lag, log, stationary
and decline phases), regardless of the AUC, among the various isolates, it was found that both
Table 1. Hemocultures. Repartition of the positive hemocultures from blood samples collected frommiceinfected with Berenice-78 T. cruzi strains at three, six and twelve months after infection.
Hemoculture Animal Identification
3MAI 5 Be-78is5-3mai
21 Be-78is21-3mai
6MAI 1 Be-78is1-6mai
2 Be-78is2-6mai
5 Be-78is5-6mai
15 Be-78is15-6mai
12MAI 1 Be-78is1-12mai
15 Be-78is15-12mai
Be-78: Berenice-78; is: isolate; MAI/mai: months after infection.
doi:10.1371/journal.pone.0137788.t001
Trypanosoma cruzi and Parasite Selection
PLOS ONE | DOI:10.1371/journal.pone.0137788 September 11, 2015 5 / 14

isolates collected at three MAI (Be-78is5-3mai and Be-78is21-3mai) presented profiles similar
to the Be-62 strain (Fig 1). This data indicates the presence in the population of strain Be-78 of
at least one subpopulation that shares features similar to strain Be-62.
3.3 Infectivity and in vitro developmentIn order to evaluate the behavior of each parasite isolate in cell medium, the profiles of infectiv-
ity and development in Vero cell were determined at 24, 48 and 72h of incubation (Figs 2 and
3). All subpopulations successfully infected Vero cells after 24h. For most samples the
Fig 1. Growth kinetics in acellular culture medium.Growth kinetics of epimastigotes in LIT medium fromdays 0 to 20 of the parasites isolates obtained from hemoculture at 3, 6 or 12 months after infection of Swissmice with 5000 blood trypomastigotes from Be-78 Trypanosoma cruzi strain. The Y-axis represents thenumber of parasites triplicate median x 107/ml and the X axis represents the days of culture. The isolates’growth curves (hatched line) are presented in comparison with the curves of the strain used in the inoculum(Berenice-78 parental: continous black lineBe-78) and of Berenice-62 strain (continous grey line Be-62).
doi:10.1371/journal.pone.0137788.g001
Trypanosoma cruzi and Parasite Selection
PLOS ONE | DOI:10.1371/journal.pone.0137788 September 11, 2015 6 / 14

infectivity ratios doubled at 48h after inoculation, corresponding to the period of multiplica-
tion of the parasite inside the host cell. One isolate obtained at 6MAI, however, (Be-78is2-
6mai) presented ratios of infectivity and development threefold higher than the other isolates
within 24h of incubation only, remaining stable until the end of the experiment. Between 48
Fig 2. Infectivity and in vitro development. In vitro infection assay in Vero cell cultures using Berenice-62(grey square), Berenice-78 (parental, black square) strains and subpopulations/isolates of Be-78 strain,obtained from hemocultures at 3, 6 or 12 months after infection of Swiss mice with 5000 bloodtrypomastigotes from Be-78 Trypanosoma cruzi strain. After 18 exposure hours to the parasite, the cells werewashed to remove extracellular parasites and maintained in medium until collection, fixation and staining 24(solid bar), 48 (hatched bar) ou 72 (dotted bar) hours after inoculum. A) number of infected cells in 100counted cells; B) number of intracellular amastigotes in 100 counted cells; C) number of intracellularparasites per infected cell. *: Significant difference compared to the 24 hours of culture; #: Significantdifference compared to the 48 hours of culture; a: significant difference compared to the strain Be-62; b:significant difference compared to the Be-78 parental strain. The data represent the mean oftriplicates ± standard error.
doi:10.1371/journal.pone.0137788.g002
Fig 3. Infectivity and in vitro development illustration.Representative photomicrographs of in vitro assayinfection in Vero cell cultures with Berenice-62 and Berenice-78 parental strains and subpopulations/isolatesof Be-78 strain, obtained from hemocultures at 3, 6 or 12 months after infection of Swiss mice with 5000 bloodtrypomastigotes from Be-78 Trypanosoma cruzi strain. After 18 hours exposure to the parasite, the cells werewashed to remove extracellular parasites and maintained in medium until collection, fixed and stained at 24,48 or 72 hours after inoculation. Increasing the rate of infectivity (asterisks) 48h after incubation for Be-62strain (a, b, c) strain and for Be 78is5-3mai isolate (g, h, i); Increased rate of intracellular multiplication(asterisks) in 72h time for Be-78 parental strain (d, e, f); Infectivity and intracellular development profilesupper (asterisks) for isolate Be-78is2-6mai (j, k. G), and lower for Be-78is14-12mai (m, n, o). Fast Panotic.Bar = 25μm.
doi:10.1371/journal.pone.0137788.g003
Trypanosoma cruzi and Parasite Selection
PLOS ONE | DOI:10.1371/journal.pone.0137788 September 11, 2015 7 / 14

and 72h after incubation the ratios of infectivity and development of all samples remained sta-
ble, except for the ratio of development of the parental strain, lower in the early times, which
continued increasing with time, suggesting a slow behavior in cell culture of the latter. All
other subpopulations presented an increase in the ratio of infectivity with time and variable
ratio of development along the experiment.
Comparison of the in vitro ratios of infectivity and intracellular development of the different
isolates with the strains used as references in the present study (Be-62 and Be-78) is presented
in Fig 2. 48 h after inoculation Be-62 strain and the isolates Be-78is5-3mai and Be-78is21-3mai
were found to have parasitized more cells than Be-78 strain.
At 72h after infection, although there was no difference in the infectivity ratios, the ratio of
development of amastigotes from Be-78 was higher than that of Be-62 (p<0,001) and of the
isolates obtained at three MAI (p<0,05). This data demonstrates that strain Be-62 presents an
early increase in infectivity, whereas Be-78 shows a higher ability for in vitro development and
a slower profile of infectivity. The behaviors of isolates Be-78is5-3mai and Be-78is21-3mai
were similar to that of strain Be-62, which is in line with observations from the growth in acel-
lular medium assay.
Isolate Be-78is2-6mai showed a higher infectivity ratio than both strains Be-62 (p<0,001)
and Be-78 (p<0,001) already at 24h after inoculation, and remained stable for the rest of the
experiment. At 24 and 48h the number of amastigotes was higher than for Be-62 and Be-78.
Although the number of intracellular amastigotes per infected cell remained the same between
24 and 48h, the higher number of infected cells for this isolate indicates a fast and efficient
infectivity profile. 72 h after inoculation the number of infected cells for isolate Be-78is2-6mai
became lower than that of Be-78, which is due to a higher intracellular development ratio of
Be-78. Nonetheless, the higher infectivity ratio maintained by isolate Be-78is2-6mai reaffirms
the success of this particular isolate in invading new cells of the host tissue. This behavior may
be a result of selection by the vertebrate host immune system (Swiss mouse), which in the case
of this particular animal promoted a more virulent subpopulation.
The other isolates presented no significant change in infectivity profiles compared to those
of Be-62 and Be-78 strains, even though their ratios of development were lower. One exception,
however, is the increase in ratio of development, observed 48h after inoculation, for isolate Be-
78is15-12mai compared to Be-78 and Be-62, showing an early proliferation peak in compari-
son with Be-78 and greater replicative capacity compared to Be-62. This isolate showed a
growth curve in acellular medium significantly lower than that observed for the Be-78 parental
strain, highlighting the variable behavior of each strain under different environmental
pressures.
3.4 kDNA signature by LSSP-PCRMany of the bands in the multiband patterns by LSSP-PCR of the reference Be-78 and Be-62
strains were composed of bands shared by both reference strains. One band, however, in the
100bp region, is absent from the profile of strain Be-78 and visible in the Be-62 band profile
(Fig 4). Isolate Be-78is5-3mai presented a profile with bands shared with each of the strains,
the parental strain Be-78 and strain Be-62, including the band slightly below 100bp specific to
Be-62. This band was also visible, although with a lesser intensity, in the profiles of two other
isolates (Be-78is1-6mai and Be-78is5-6mai), indicating some similarities between the subpopu-
lation(s) present in the isolates and those present in strain Be-62. A slight variability in the
band profiles between duplicates may be due to the fact that the isolates were not obtained via
cloning and therefore may be composed of more than one subpopulation.
Trypanosoma cruzi and Parasite Selection
PLOS ONE | DOI:10.1371/journal.pone.0137788 September 11, 2015 8 / 14

Eight bands obtained in the gene signature profile analysis for LSSP-PCR were used to build
the phenogram from UPGMA (Unweighted Pair Group Method using Arithmetic Averages).
The choice of the bands was based on its resolution, reproducibility between duplicates and
intensity. The phenogram showed two distinct groups (Fig 5), group I, including the Be-78
parental strain and the isolates obtained at different times along the experimental chronic
phase (Be-78is5-3mai, Be-78is21-3mai, Be-78is1-6mai, Be-78is2-6mai, Be-78is5-6mai, Be-
78is15-6mai, Be-Be-78is1-12mai and 78is15-12mai) and group II, comprising the Be-62 strain.
In group I, the Be-78 strain of subpopulations divided into two groups, with isolated Be-78is5-
3mai, being the one who distanced itself from parental strain. The remaining isolates formed a
group more genetically correlated with the parental strain (Fig 5).
Isolate Be-78is5-3mai shared four bands in its LSSP-PCR profile with that of Be-78, versus
three with Be-62. Strain Be-78 shared only 2 bands with strain Be-62. This data indicates that
Fig 4. kDNA signature by LSSP-PCR.Gene signatures of 330bp fragment of the kDNAminicircle regionrevealed in 8% polyacrylamide gel and silver stained. 100bp molecular weight (MW) in channel one and otherchannels regarding the isolates genetic profiles obtained from hemocultures at 3, 6 or 12 months afterinfection of Swiss mice with 5000 blood trypomastigotes from Be-78 Trypanosoma cruzi strain compared tothe Be-78 parental and Be-62 strains. Be-62 (2–3); Be-78 parental (4–5); Be-78is5-3mai (6–7); Be-78is21-3mai (8–9); Be-78is1-6mai (10–11); Be-78is2-6mai (12–13); Be-78is5-6mai (14–15); Be-78is15-6mai (16–17); Be-78is1-12mai (18–19); Be-78is15-12mai (20–21). Arrow heads indicate the bands shared between theisolate Be-78is5-3mai and the reference strains, Be-78 parental and Be-62.
doi:10.1371/journal.pone.0137788.g004
Fig 5. UPGMA. Phenogram constructed from UPGMA (Unweighted Pair Group Method using Arithmeticaverages) resulting from the gene signature profile analysis obtained by LSSP-PCR of isolates obtained fromhemocultures at 3 (Be-78is5-3mai, Be-78is21-3mai), 6 (Be-78is1-6mai; Be-78is2-6mai; Be-78is5-6mai;78is15-6mai-Be) and 12 (Be-78is1-12mai; Be-78is15-12mai) months after infection of Swiss mice with 5000blood trypomastigotes from Be-78 Trypanosoma cruzi strain in comparison with the Be-78 parental and Be-62 strains.
doi:10.1371/journal.pone.0137788.g005
Trypanosoma cruzi and Parasite Selection
PLOS ONE | DOI:10.1371/journal.pone.0137788 September 11, 2015 9 / 14

the isolate Be-78is5-3mai is genetically closer to strain Be-78 than to strain Be-62, however,
this genetic distance between the isolate and Be-62 strain is less than the distance between
strain Be-78 and strain Be-62 (Fig 5).
Discussion
According to the current classification of T. cruzi subpopulations in Discrete Typing Units
(DTU) both Be-62 and Be-78 strains belong to DTU TcII [21]. Although these two strains were
sufficiently related to be classified within the same DTU they are morphologically distinct. The
Be-62 strain is predominantly composed of slender forms, presents peak parasitemia around the
seventh day and a mortality rate of 100% at 15 days after infection in mice [9, 10, 28], while the
Be-78 strain is composed of large shapes, presents peak parasitemia later (day 15) and a survival
rate of 100% [9, 10, 29, 30]. When introduced in acellular culture medium, Be-78 showed lower
growth capacity and high rates of differentiation, while under the same conditions Be-62
showed a high capacity of multiplication and average rates of differentiation [9, 10].
Furthermore, data obtained in our laboratory showed that strains belonging to the same
DTU, namely TcII, Y and Be-78 strains, had drastically different pathogenicities in experimen-
tal Beagle dog infection. Animals infected with the Y strain showed controlled parasitism and
their immune system returned to homeostasis, whereas animals infected with the Be-78 strain
maintained parasitism over two years after infection and showed lesions of apparently progres-
sive character [31]. The interaction of each subpopulation within the strain with the host may
interfere with the development of the host immune response and determine success in control-
ling tissue parasitism and associated lesions.
In order to test the ability of an individual to select a clonal subpopulation from a polyclonal
strain of T. cruzi we infected 24 Swiss mice with strain Be-78 and tested blood samples at three
different times after infection. The parasites from positive hemocultures were characterized in
vitro in terms of growth kinetics in acellular medium, profile of infection and development in
Vero cell medium and kDNA gene-signature by LSSP-PCR. Swiss mice were selected for the
present study, rather than a clonal population of mice, because of their genetic variability. It
was already observed that upon infection with a polyclonal strain of T. cruzi the host-parasite
interaction may lead to the absence of parasitemia in some individuals, whereas individuals
with positive hemocultures may differ in terms of pathologic response, in particular tissue-tro-
pism [4], most likely due to genetic factors.
We found that the gene signatures of Be-62 and Be-78 obtained by LSSP-PCR were differ-
ent, confirming the genetic heterogeneity of the two strains isolated from the same patient with
a 16 years interval. Our data complements previous literature reports using isoenzyme profiles,
RAPD and microsatellites [9, 10, 11, 32]. The Be-62 and Be-78 strain samples have been main-
tained in our laboratory for the past 30 years by alternating passages in mice and axenic cul-
ture, and including periods of cryopreservation. It is worth mentioning that, in spite of
alternating between in vivo and in vitromaintenance, no changes in the virulence, pathogenic-
ity or molecular profile have been observed in comparison with the original samples, as dem-
onstrated in various studies published by our group [9, 11, 14, 17, 29; 33; 34].
The kDNA profile of one of the isolates, Be-78is5-3mai, was closer to Be-62 and Be-78
strains (Fig 3). Interestingly, isolate Be-78is5-3mai, showed a behavior in Vero cell culture
medium closer to that of the Be-62 strain. In contrast, isolate Be-78is2-6mai, which showed in
vitro ratios of infectivity and development threefold higher than the Be-78 parental strain was
genetically similar in terms of gene signature by LSSP-PCR, showing that changes in the bio-
logical behavior do not necessarily reflect genetic variations. Growth in culture medium simu-
lates the behavior of epimastigotes in the gut of an invertebrate host and, therefore, a low ratio
Trypanosoma cruzi and Parasite Selection
PLOS ONE | DOI:10.1371/journal.pone.0137788 September 11, 2015 10 / 14

of infectivity or development may reflect a reduced ability for multiplication of these subpopu-
lations, with potential implications on the success of vector infection.
The isolate that stood out the most in terms of genetic distance to the parental strain, Be-
78is5-3mai, was collected from animal 1 at 3MAI. Another isolate was obtained from the same
animal three months later at 6MAI, Be-78is5-6mai. Although the latter also displayed in its
LSSP-PCR profile the characteristic band slightly below 100bp similar to Be-62, yet with a
lower intensity, its biological properties in vitro were similar to the parental strain Be-78. This
observation is a hint on the reversibility of the genetic modulation over time in the case of
chronic infection with T. cruzi, where a subpopulation may be favored at a certain time and
another one take over later on.
Even though maintenance of the isolates in culture medium was limited in time, it is impor-
tant to consider the possible influence of hemoculture and in vitromaintenance prior to phe-
notypic and molecular characterization, which could add up to clonal selection exerted by the
immune system of the vertebrate host (Swiss mice). Hence, hemoculture and in vitromainte-
nance may favor one or more subpopulation over others resulting in the expansion or reduc-
tion of selected clones that were present in the original sample [35]. However, this process not
allow the appearance of subclones that were absent from the original sample. Therefore it is
more likely that the selective process, which resulted in isolate Be-78is5-3mai was a conse-
quence of the pressure exerted by the immune system of the host rather than a consequence of
clonal selection during in vitromaintenance.
Microsatellite characterization of the Be-78 strain by Valadares et al. [11] confirmed its mul-
ticlonality, and suggested the presence of a subpopulation with similar characteristics to those
of strain Be-62. Therefore, isolate Be-78is5-3mai may have originated either from selection of a
subpopulation within the polyclonal parental strain Be-78, or due to the association of sub-
clones, with the participation of one or more subclones, resulting in mixed biological and
molecular profiles. The latter is supported by the isolate´s intermediate behavior in culture
medium and genetic profile in comparison with the reference strains, Be-78 and Be-62.
The present data confirms the polyclonal or mixed nature of strain Be-78 already reported
in the literature [11, 14, 17] and reinforces the hypothesis that the selective pressure exerted by
the host throughout the chronic phase may favor the development of one or more subpopula-
tion(s) over the other(s). This phenomenon may be related to the plasticity of T. cruzi and not
less importantly, to the parasite-host interaction, since all outbred animals received the same
inoculum and responded to infection in selecting distinct subpopulations.
In conclusion, three isolates were obtained upon infection of Swiss mice with Be-78, namely
Be-78is5-3mai, Be-78is2-6mai and Be-78is15-12mai, which showed differences in biological
and/or molecular parameters compared to the parental strain. As such, they are good candi-
dates for probing in vivo the influence of clonal subpopulations in the pathogenesis of the
disease.
Conclusion
T. cruzi isolates from Swiss mice infected with the same polyclonal parental strain Be-78 char-
acterized in vitro were found to have a kDNA gene signature significantly distinct from the
parental strain. The present work confirms the hypothesis that the maintenance of a T. cruzi
population in outbred mice results in the selection of genetically distinct subpopulations.
Subsequent in vivo studies are under way to evaluate the pathogenicity and histotropy of a
specific isolate in comparison with that of the parental strain in experimental murine infection.
Trypanosoma cruzi and Parasite Selection
PLOS ONE | DOI:10.1371/journal.pone.0137788 September 11, 2015 11 / 14

Supporting Information
S1 File. The ARRIVE Guidelines Checklist. Report of in vivo experiments in animal research.
(PDF)
Acknowledgments
We thank the Centro de Ciência Animal-CCA/UFOP for the availability and ethic work to ani-
mals and Renata Rezende Rocha Oliveira for help with the in vitro infection analyzes.
Author Contributions
Conceived and designed the experiments: CMC NCNP PMAVWLT. Performed the experi-
ments: NCNP LMROMTO KSF PMAV. Analyzed the data: NCNP LMRO PMAV VMV. Con-
tributed reagents/materials/analysis tools: ML ABR CMC PMAV. Wrote the paper: GPL
NCNP CMC PMAV.
References1. Macedo AM, Machado CR, Oliveira RP, Pena SD. Trypanosoma cruzi: Genetic structure of populations
and relevance of genetic variability to the pathogenesis of chagas disease. Mem Inst Oswaldo Cruz.2004; 99: 1–12.
2. Villarreal D, Barnabe C, Sereno D, Tibayrenc M. Lack of correlation between in vitro susceptibility tobenznidazole and phylogenetic diversity of Trypanosoma cruzi, the agent of chagas disease. Exp Para-sitol. 2004; 108: 24–31. PMID: 15491545
3. Macedo AM, Oliveira RP, Pena SD. Chagas disease: Role of parasite genetic variation in pathogene-sis. Exp Rev Mol Med. 2002; 4: 1–16.
4. Andrade LO, Machado CR, Chiari E, Pena SD, Macedo AM. Trypanosoma cruzi: Role of host geneticbackground in the differential tissue distribution of parasite clonal populations. Exp Parasitol. 2002;100: 269–275. PMID: 12128054
5. Rodrigues CM, Silva Valadares HM, Francisco AF, Arantes JM, Campos CF, Teixeira-Carvalho A,et al. Coinfection with different Trypanosoma cruzi strains interferes with the host immune response toinfection. PLoS Negl Trop Dis. 2010; 4: e846. doi: 10.1371/journal.pntd.0000846 PMID: 20967289
6. Andrade LO, Machado CR, Chiari E, Pena SD, Macedo AM. Differential tissue distribution of diverseclones of Trypanosoma cruzi in infected mice. Mol Biochem Parasitol. 1999; 100: 163–172. PMID:10391378
7. Chagas C. Nova trypanosomiase humana. Estudos sobre a morphologia e o cyclo evolutivo do Schizo-
trypanum cruzi n. gen., n. sp., agente etiológico de nova entidade mórbida do homem. Mem InstOswaldo Cruz. 1909; 1: 159–218.
8. Salgado JA, Garcez PN, Oliveira CA, Galizzi J. Revisão clínica atual do primeiro caso humano descritode Doença de Chagas. Rev Inst Med Trop São Paulo. 1962; 4: 330–337.
9. Lana M, Chiari CA. Comparative biological characterization of Berenice and Berenice-78 strains of Try-panosoma cruzi isolated from the same patient at different times. Mem Inst Oswaldo Cruz. 1986; 81:247–253. PMID: 3106753
10. Lana M. Caracterização do Trypanosoma cruzi, cepa Berenice, isolada da mesma paciente em dife-rentes períodos. PhD thesis. Universidade Federal de Minas Gerais. 1981. 91 pp.
11. Valadares HM, Pimenta JR, Segatto M, Veloso VM, Gomes ML, Chiari E, et al. Unequivocal identifica-tion of subpopulations in putative multiclonal Trypanosoma cruzi strains by facs single cell sorting andgenotyping. PLoS Negl Trop Dis. 2012; 6: e1722. doi: 10.1371/journal.pntd.0001722 PMID: 22802979
12. Andrade SG, Magalhães JB. Biodemes and zymodemes of Trypanosoma cruzi strains: correlationswith clinical data and experimental pathology. Rev Soc Bras Med Trop. 1997; 30: 27–35.
13. Vago AR, Andrade LO, Leite AA, D’avila RD, Macedo AM, Adad SJ, et al. Genetic characterization ofTrypanosoma cruzi directly from tissues of patients with chronic Chagas disease: differential distributionof genetic types into diverse organs. Am J Pathol. 2000; 156: 1805–1809. PMID: 10793092
14. Veloso VM, Guedes PM, Lana M, Martins HR, Carneiro CM, Camara AC, et al. Genetic modulation inBe-78 and Y Trypanosoma cruzi strains after long-term infection in beagle dogs revealed by molecularmarkers. Infect Genet Evol. 2012; 12: 1128–1135. doi: 10.1016/j.meegid.2012.03.019 PMID:22554652
Trypanosoma cruzi and Parasite Selection
PLOS ONE | DOI:10.1371/journal.pone.0137788 September 11, 2015 12 / 14

15. Carneiro M. Changes in the isoenzyme and kinetoplast DNA patterns of T. cruzi strains induced bymaintenance in mice. Acta Trop. 1990; 47: 35–45. PMID: 1967507
16. Romanha AJ, Dasilvapereira AA, Chiari E, Kilgour V. Isoenzyme patterns of cultured Trypanosoma
cruzi—changes after prolonged subculture. Comp Biochem Physiol. 1979; 62: 139–142.
17. Veloso VM, Romanha AJ, Lana M, Murta SM, Carneiro CM, Alves CF, et al. Influence of the long-termTrypanosoma cruzi infection in vertebrate host on the genetic and biological diversity of the parasite.Parasitol Res. 2005; 96: 382–389. PMID: 15940522
18. Vago AR, Macedo AM, Adad SJ, Reis DA, Corrêa-Oliveira R. PCR detection of Trypanosoma cruzi
DNA in esophageal tissues of patients with chronic Chagas disease. Lancet. 1996; 348: 891–892.PMID: 8826826
19. Pena SD, Barreto G, Vago AR, Demarco L, Reinach FC, Neto ED, et al. Sequence-specific gene signa-tures can be obtained by PCR with single specific primers at low stringency. Proc Natl Acad Sci U S A.1994; 91: 1946–1949. PMID: 8127912
20. Brito CM, Lima MM, Sarquis O, Pires MQ, Coutinho CF, Duarte R, et al. Genetic polymorphism in Try-
panosoma cruzi I isolated from brazilian northeast triatomines revealed by low-stringency single spe-cific primer-polymerase chain reaction. Parasitol Res. 2008; 103: 1111–1117. doi: 10.1007/s00436-008-1102-5 PMID: 18633644
21. Zingales B, Andrade SG, Briones MR, Campbell DA, Chiari E, Fernandes O, et al. A new consensus forTrypanosoma cruzi intraspecific nomenclature: second revision meeting recommends TcI to TcIV.Mem Inst Oswaldo Cruz. 2009; 104: 1051–1054. PMID: 20027478
22. Brener Z. Therapeutic activity and criterion of cure in mice experimentally infected with Trypanosoma
cruzi. Rev Inst Med Trop São Paulo. 1962; 4: 389–396. PMID: 14015230
23. Oliveira LT, Garcia GM, Kanoa EK, Tedesco AC, Mosqueira VCF. HPLC-FLDmethods to quantifychloroaluminum phthalocyanine in nanoparticles, plasma and tissue: application in pharmacokineticand biodistribution studies. J Pharm Biomed Anal. 2011; 56: 70–77. doi: 10.1016/j.jpba.2011.04.016PMID: 21596512
24. D’Ávila DA, Macedo AM, Valadares HMS, Gontijo ED, Castro AM, Machado CR, et al. Probing Popula-tion Dynamics of Trypanosoma cruzi during Progression of the Chronic Phase in Chagasic Patients. JClin Microbiol. 2009; 47: 1718–1725. doi: 10.1128/JCM.01658-08 PMID: 19357212
25. Rimoldi A, Alves R. T., Ambrósio D. L., Fernandes MZT, Martinez I, Araújo RF, et al. Morphological, bio-logical and molecular characterization of three strains of Trypanosoma cruzi Chagas, 1909 (Kinetoplas-tida, Trypanosomatidae) isolated from Triatoma sordida (Stal) 1859 (Hemiptera, Reduviidae) and adomestic cat. Par. 2012; 139: 37–44.
26. Andrade LO, Galvão LM, Meirelles MN, Chiari E, Pena SD, Macedo AM. Differential tissue tropism ofTrypanosoma cruzi strains: An in vitro study. Mem Inst Oswaldo Cruz. 2010; 105: 834–837. PMID:20945002
27. Gomes ML, Macedo AM, Vago AR, Pena SD, Galvão LM, Chiari E. Trypanosoma cruzi: Optimization ofpolymerase chain reaction for detection in human blood. Exp Parasitol. 1998; 88: 28–33. PMID:9571142
28. Brener Z. Comparative studies of different strains of Trypanosoma cruzi. Ann Trop Med Parasitol. 1965;59: 19–26. PMID: 14297351
29. Carneiro CM, Martins-Filho OA, Reis AB, Veloso VM, Aráujo FM, Bahia MT, et al. Differential impact ofmetacyclic and blood trypomastigotes on parasitological, serological and phenotypic features triggeredduring acute Trypanosoma cruzi infection in dogs. Acta Trop. 2007; 101: 120–129. PMID: 17296162
30. Guedes PM, Veloso VM, Caliari MV, Carneiro CM, Souza SM, Lana M, et al. Trypanosoma cruzi highinfectivity in vitro is related to cardiac lesions during long-term infection in Beagle dogs. Mem InstOswaldo Cruz. 2007; 102: 141–147. PMID: 17426876
31. Nogueira-Paiva NC, Fonseca KS, Vieira PM, Diniz LF, Caldas IS, Moura SA, et al. Myenteric plexus isdifferentially affected by infection with distinct Trypanosoma cruzi strains in beagle dogs. Mem InstOswaldo Cruz. 2013; 109: 51–60. doi: 10.1590/0074-0276130216 PMID: 24271001
32. Cruz RE, Macedo AM, Barnabe C, Freitas JM, Chiari E, Veloso VM, et al. Further genetic characteriza-tion of the two Trypanosoma cruzi Berenice strains (Be-62 and Be-78) isolated from the first humancase of Chagas disease (Chagas, 1909). Acta Trop. 2006; 97: 239–246. PMID: 16343412
33. Vieira PMA, Francisco AF, Souza SM, Malaquias LCC, Reis AB, Giunchetti RC, et al. Trypanosoma
cruzi: Serum levels of nitric oxide and expression of inducible nitric oxide synthase in myocardium andspleen of dogs in the acute stage of infection with metacyclic or blood trypomastigotes. Exp Parasitol.2009; 121: 76–82. doi: 10.1016/j.exppara.2008.09.023 PMID: 18983995
Trypanosoma cruzi and Parasite Selection
PLOS ONE | DOI:10.1371/journal.pone.0137788 September 11, 2015 13 / 14

34. Vieira PMA, Francisco AF, Machado EMM, Nogueira C, Fonseca KS, Reis AB, et al. Different InfectiveForms Trigger Distinct Immune Response in Experimental Chagas Disease. PLos One. 2012; 7:32912–32922.
35. Macedo AM, Pena SDJ. Genetic variability of Trypanosoma cruzi: implications for the pathogenesis ofChagas disease. Parasitol Today. 1998; 14: 119–124. PMID: 17040719
Trypanosoma cruzi and Parasite Selection
PLOS ONE | DOI:10.1371/journal.pone.0137788 September 11, 2015 14 / 14





![Referências Bibliográficas - Freelite...Internacionais [1]. Teste policlonal Freelite® "A introdução do teste de cadeia leve livre em soro revolucionou nossa capacidade de avaliar](https://img.document.onl/doc/110x75/6128608b711e0a12691c7225/referncias-bibliogrficas-freelite-internacionais-1-teste-policlonal.jpg)























