Embed Size (px)
Citation preview

Inhibitors of the 5-lipoxygenase pathway activate pannexin1 channels inmacrophages via the thromboxane receptor
Hercules A. da Silva-Souza,1,2,3 Maria Nathália de Lira,1 Naman K. Patel,3 David C. Spray,3
Pedro Muanis Persechini,1,2 and Eliana Scemes3
1Instituto de Biofisica Carlos Chagas Filho, Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brazil; 2InstitutoNacional de Ciência e Tecnologia de Pesquisa Translacional em Saúde e Ambiente da Região Amazônica–INPeTAm,Rio de Janeiro, Brazil; and 3Dominick P. Purpura Department of Neuroscience, Albert Einstein College of Medicine,Bronx, New York
Submitted 21 March 2014; accepted in final form 25 July 2014
da Silva-Souza HA, de Lira MN, Patel NK, Spray DC, Per-sechini PM, Scemes E. Inhibitors of the 5-lipoxygenase pathwayactivate pannexin1 channels in macrophages via the thromboxanereceptor. Am J Physiol Cell Physiol 307: C571–C579, 2014. Firstpublished July 30, 2014; doi:10.1152/ajpcell.00087.2014.—A multi-tude of environmental signaling molecules influence monocyte andmacrophage innate and adaptive immune responses, including ATPand prostanoids. Interestingly, purinergic (P2) and eicosanoid receptorsignaling interact such that the activation of P2 receptors leads toprostanoid production, which can then interfere with P2Y-mediatedmacrophage migration. Recent studies suggest that blockade of 5-lipoxygenase (5-LOX) in macrophages can activate a permeationpathway involved in the influx of dye and the release of ATP. Here,we provide evidence that pannexin1 (Panx1) is a component of thispathway and present the intracellular signaling molecules linking thethromboxane (TP) receptor to Panx1-mediated dye influx and ATPrelease. Using pharmacological tools and transgenic mice deficient inPanx1, we show that two 5-LOX pathway inhibitors induce ATPrelease and influx of dye in a Panx1-dependent manner. Electrophys-iological recordings performed in wild-type and Panx1-deficient mac-rophages confirmed that these 5-LOX pathway inhibitors activatecurrents characteristic of Panx1 channels. We found that the mecha-nism by which Panx1 channels are activated under this conditioninvolves activation of the TP receptor that is mediated by the cAMP/PKA pathway. This is to our knowledge the first evidence for theinvolvement of Panx1 in the TP receptor signaling pathway. Futurestudies aimed to clarify the contribution of this TP-Panx1 signalingnetwork to macrophage immune responses are likely to be importantfor targeting inflammatory and autoimmune diseases.
ATP release; dye uptake; macrophages; pannexin; prostanoids
PURINES/PYRIMIDINES AND EICOSANOIDS are important modulatorsof monocyte function, inducing the production and release ofcytokines, migration, differentiation, and apoptosis. Interest-ingly, the nucleotide and eicosanoid signaling pathways inter-act. For instance, activation of P2 purinergic receptors inmacrophages has been reported to lead to prostanoid produc-tion through the generation of arachidonic acid (AA) (7, 10, 24,54). On the other hand, prostaglandin E2 (PGE2) has beenshown to interfere with P2Y-mediated macrophage migrationby inhibiting Ca2� mobilization and AKT (v-Akt murinethymoma viral oncogene homolog) phosphorylation inducedby P2Y receptors (53). Moreover, blockade of 5-LOX, the
enzyme that transforms AA into leukotrienes (12, 37, 38, 57),has been shown to decrease intracellular levels of ATP (2) andto attenuate ATP-induced macrophage cell death (7).
ATP is well known to induce membrane permeabilization inmacrophages through the activation of P2X7 receptors andpannexin1 (Panx1) channels (20, 24). Panx1, a member of thegap junction gene family, forms plasma membrane channelsthat are permeable to relatively large molecules (up to 1.5kDa), including ATP (3).
Interestingly, it was recently found that 5-LOX inhibitorsinduced the influx of cationic dyes in macrophages indepen-dently of P2X7 receptor activation (36, 46) and that such influxdid not occur after inhibiting the activity of cyclooxygenases(COX-1 and COX-2), enzymes that transform AA into prosta-noids (12, 37, 38, 57). Thus these recent studies raise thepossibility that the production of prostanoids resultant from theblockade of the 5-LOX pathway activates a permeation path-way with properties (influx of dye and ATP release) similar tothose described for Panx1. Therefore, the present study wasundertaken to identify and characterize the signaling pathwayinvolved in macrophage dye and ATP permeabilities inducedby blockers of the 5-LOX pathway. Using transgenic mice andpharmacological tools, we found that Panx1 is activated fol-lowing the addition of 5-LOX inhibitors through a mechanisminvolving cAMP production due to thromboxane (TP) receptoractivation. Moreover, our data showing that macrophages re-lease ATP in response to 5-LOX inhibitors have importantimplications for the understanding of the interplay betweenpurinergic and prostanoid signaling in the biology of mono-cytes.
MATERIAL AND METHODS
Materials
DMEM, fetal bovine serum, penicillin and streptomycin were ob-tained from Gibco; 1-[(4-chlorophenyl)methyl]-3-[(1,1-dimethylethyl)-thio]-�,�-dimethyl-5-(1-methylethyl)-1H-indole-2-propanoic acid (MK-886), {1S-[1�,2�(Z),3�(1E, 3S*),4�]}-7-{3-[3-hydroxy-4-(4-iodophe-noxy)-1-butenyl]-7-oxabicyclo [2.2.1]hept-2-yl}-5heptenoic acid(I-BOP), and {1S-[1�,2�(Z),3�,4�]}-7-[3-({2-[(phenylam) carbonyl]-hydrazine}methyl)-7-oxabicyclo[2.2.1]hept-2-yl]-5-heptenoic acid(SQ29548) were from Cayman Chemical. Thioglycolate medium wasfrom Difco; phosphate-buffered saline (PBS), EGTA, NaCl, MgCl2,CaCl2, KCl, CsCl, nordihydroguaiaretic acid (NDGA), HEPES, dimethylsulfoxide (DMSO), N6,2=-O-dibutyryladenosine 3=:5=-cyclic monophos-phate sodium (db-cAMP), �,4-dihydroxy-2-(6-hydroxy-1-heptenyl)-4-cyclopentanecrotonic acid �-lactone (brefeldin-A: BFA), 3�-hydroxy-11-oxoolean-12-en-30-oic acid 3-hemisuccinate (carbenoxolone diso-
Address for reprint requests and other correspondence: E. Scemes, Domin-ick P. Purpura Dept. of Neuroscience, Albert Einstein College of Medicine,1410, Pelham Parkway, Kennedy Center, Bronx, NY 10461 (e-mail:[email protected]).
Am J Physiol Cell Physiol 307: C571–C579, 2014.First published July 30, 2014; doi:10.1152/ajpcell.00087.2014.
0363-6143/14 Copyright © 2014 the American Physiological Societyhttp://www.ajpcell.org C571

dium salt: CBX), and firefly luciferase (EC 1.13.12.7) were purchasedfrom Sigma-Aldrich; YO-PRO1 was from Life Technologies; N-[2-(methylamino)ethyl] isoquinoline-5-sulfonamide dihydrochloride (H-8.2HCl) was from Santa Cruz Biotechnology; Enliten ATP assay systemwas from Promega; BCA reagents were from Thermo Scientific; Super-script VILO cDNA Synthesis Kit and oligofectamine reagent werepurchased from Invitrogen.
Most reagents were prepared as �1,000 stock solutions either inDMSO (MK-886, NDGA, BFA, CBX, SQ29548, and YO-PRO1) orethanol (I-BOP) or Milli-Q water (db-cAMP), and aliquots were keptfrozen until use. NDGA was prepared fresh on the day of theexperiment.
Animals
Male adult (2– 4 mo old) wild-type and Panx1-deficient[Panx1tm1a(KOMP)Wtsi] mice on the C57Bl/6 background were usedin this study. Panx1-deficient mice originally purchased from KOMP(Knockout Mouse Project) were bred and maintained in the Institutefor Animal Studies at Albert Einstein College of Medicine. Aspreviously described (15), these Panx1-deficient mice are hypomorph,not complete knockouts at the gene level; these mice are here referredas Panx1-hypo. All experiments were preapproved by the InstitutionalAnimal Care and Use Committee.
Cell Cultures
Macrophage cultures. Macrophages were obtained from the intra-peritoneal cavity of mice collected 4 days after thioglycolate injection,as previously described (8). In brief, cells collected in PBS wereplated for 1 h on glass-bottomed dishes (MatTek), on coverslips, or in24-well plates containing DMEM, supplemented with 10% FCS and1% of penicillin/streptomycin at 37°C in an incubator with humidifiedatmosphere containing 5% CO2. Nonadherent cells were washed outand the macrophage cultures maintained for at least 2 days beforeexperimentation. All experiments were performed at room tempera-ture (RT).
J774 mouse macrophage cell line. Parental and Panx1-deficientJ774 cells were obtained and maintained as previously described (20).Briefly, J774 cells were cultured in 60-mm dishes containing DMEM,supplemented with 10% FCS, and 1% penicillin/streptomycin andmaintained in a humidified chamber with 5% CO2 at 37°C. Beforeexperimentation, cells were trypsinized and transferred to glass-bottomed dishes.
Dye influx
The influx of the dye YO-PRO1 (Invitrogen) was performed aspreviously described (19, 52). Briefly, J774 (parental and Panx1deficient) and peritoneal macrophages (WT and Panx1-hypo) platedon glass-bottomed dishes were bathed in a basal salt solution (BSS inmM: 145 NaCl, 5 KCl, 1 MgCl2, 1 CaCl2, and 10 Na-HEPES, pH 7.4)containing the cell-impermeant dye YO-PRO1 (5 �M). After a 5-minmeasurement of basal fluorescence intensity, cells were exposed to the5-LOX inhibitor NDGA (50 �M), the thromboxane mimetic I-BOP(50 �M), or to db-cAMP (500 �M) and fluorescence intensity wasmeasured for 10 min. For blocking experiments, cells were preincu-bated for 30 min with the TP receptor antagonist SQ29548 (10 �M)or the PKA inhibitor H-8 (50 �M), before the addition of NDGA,I-BOP, or db-cAMP. All compounds were dissolved in BSS contain-ing YO-PRO1 (5 �M), and experiments were performed at RT.YO-PRO1 fluorescence intensity obtained from regions of interestplaced randomly on cells was captured using an Orca-ER charge-coupled device camera (Hamamatsu) attached to an inverted micro-scope (Eclipse 2000; Nikon), equipped with a �20 dry objective and488/512 nm excitation/emission filter set and Metafluor softwareversion 7.1 (Molecular Devices). Dye influx was reported as the ratioof YO-PRO1 fluorescence intensity normalized to basal values (F/F0).
Electrophysiology
Whole cell voltage-clamp recordings were performed at RT onWT and Panx1-hypo peritoneal macrophages plated on coverslipsand bathed in BSS (see above). Heat-polished micropipettes (5–10M) were filled with internal solution (in mM: 130 CsCl, 10EGTA, 10 HEPES, and 0.5 CaCl2). Cell membrane potential washeld at 60 mV after forming high-resistance seals (1–10 G),and current amplitudes were evaluated using a ramp (60 to �80mV, 5-s duration, 20 trials) followed by step pulse (60 to �140mV, 10-mV steps, 800-ms duration) protocols. Cells were stimu-lated with NDGA (5 �M), MK-886 (5 �M), I-BOP (5–50 �M),and db-cAMP (50 �M), and a voltage-ramp protocol was repeat-edly applied until maximal current amplitude was attained (4 min:NDGA, MK-886, I-BOP; 10 min: db-cAMP). Current amplitudeswere then measured from voltage-step pulses. For experimentsusing blockers, cells were incubated for 30 min with SQ29548 (10�M) or H-8 (50 �M) before the addition of the tested compounds.Data were acquired with an Axopatch-1C amplifier and digitizedusing an Axon Instruments Digitizer (Molecular Devices). Clam-pex and Clampfit software (version 10.2) were used for recordingand analysis, respectively.
Measurements of ATP
The amount of ATP released from WT and Panx1-hypo peritonealmacrophages was measured as previously described (50, 51). Cellsplated in 24-well plates were washed twice with BSS (in mM: 145NaCl, 5 KCl, 1 MgCl2, 1 CaCl2, and 10 Na-HEPES, pH 7.4). Cellswere then incubated for 1 min at RT with 50 �M NDGA or I-BOP inthe absence or presence (30-min preincubation) of CBX (50 �M),BFA (10 �M), SQ-29548 (10 �M), and H-8 (50 �M). Aliquots ofbathing solutions were collected before and after cell stimulation andkept at 20°C until analysis that was performed using the luciferin/luciferase assay (Enliten ATP Assay System; Promega) and a Turnerluminometer. The concentrations of ATP released from cells wereobtained from standard curves and normalized to the total amount ofprotein measured using the BCA reagents (Thermo Scientific).
Quantitative RT-PCR
WT and Panx1-hypo peritoneal macrophages were used to quantifythe levels of Panx1 transcripts, as previously described (15). TotalRNA was extracted using the plus mini kits (Qiagen) according to themanufacturer’s protocol. Complementary DNA was synthesized from1 �g/10 �l of RNA, using a Superscript VILO cDNA Synthesis Kit(Invitrogen). Primers used were Panx1 (F: AGCCAGAGAGTG-GAGTTCAAAGA; R: CATTAGCAGGACGGATTCAGAA) and18S ribosomal RNA (F: CACGGCCGGTACAGTGAAAC; R: AGAGG-AGCGAGCGACCAAA). 18S was used as housekeeping gene fornormalization. Real-time RT-PCR was performed using SYBRGreen PCR Master Mix with 7300 Fast Real-Time PCR system(Applied Biosystems). Reaction mixtures were denatured at 95°Cfor 10 min, followed by 40 PCR cycles. Each cycle consisted of thefollowing three steps: 94°C for 15 s, 57°C for 15 s, and 72°C for1 min. Each sample was normalized against internal controls (18Sribosomal); the relative values for target abundance were extrap-olated from standard curves generated from the reference standard.
Data Analysis
Statistical significance was determined using ANOVA followed byNewman-Keuls multiple comparison test or unpaired t-test. A P value�0.05 was considered significant. Numbers of animals used areindicated in parentheses.
C572 TP RECEPTORS ACTIVATE PANX1 CHANNELS
AJP-Cell Physiol • doi:10.1152/ajpcell.00087.2014 • www.ajpcell.org

RESULTS
Membrane Permeabilization Induced by Inhibitors of the5-LOX Pathway Involves Panx1 Channels
Recently, it was reported that blockers of the 5-LOX path-way resulted in the uptake of fluorescent molecules in macro-phage through an unknown, P2X7 receptor-independent mech-anism (36, 46). The possible involvement of Panx1 in thephenomenon of NDGA-induced membrane permeabilizationwas here investigated using the J774 macrophage cell line andperitoneal macrophages in which Panx1 expression was ma-nipulated by shRNA knockdown and transgenesis, respec-tively. As shown in Fig. 1, A and B, no influx of YO-PRO1 wasrecorded from cells deficient in Panx1 when stimulated withNDGA (50 �M). Further confirmation that NDGA activatesPanx1 channels was obtained by means of electrophysiologicalrecordings on peritoneal macrophages derived from WT andPanx1-hypo mice (Fig. 1, C–F). Bath application of 5 �MNDGA to macrophages induced a time-dependent increase inoutward currents that stabilized around 4 min after drug appli-
cation (Fig. 1C). Four minutes after NDGA application, theconductance measured at �80 mV doubled, increasing from1.64 � 0.06 nS at control condition to 3.40 � 0.03 nS (Fig. 1C,inset). After this stabilization period, cells were subjected to avoltage-step protocol (Fig. 1D) and the amplitudes of NDGA-induced currents were measured in WT and Panx1-hypo mac-rophages. As shown in Fig. 1E, 4 min after NDGA application,current amplitudes recorded from WT macrophages were sig-nificantly larger compared with untreated cells, while inPanx1-hypo macrophages, NDGA did not induce changes incurrents. The Panx1 channel blocker CBX (5 �M) preventedthe NDGA-induced 1.8-fold increase in outward currents inWT macrophages (control: 183.8 � 51.2 pA; NDGA: 328.3 �97.8 pA; CBX � NDGA: 184.8 � 48.1 pA; n 3). Similarresults were obtained when using the more specific 5-LOXinhibitor MK-886 (Fig. 1F). At 5 �M, MK-886 inducedoutward currents (388.4 � 52.9 pA; n 3) with amplitudessimilar to those generated by 5 �M NDGA (461.2 � 30.3 pA;n 4); no changes in current amplitudes were recorded fromPanx1-hypo macrophages exposed to MK-886 (Fig. 1F).
Fig. 1. Pannexin 1 (Panx1) channel is activated follow-ing inhibition of the 5-lipoxygenase (5-LOX) pathway.A and B: dye influx. Bar histograms of the means � SEvalues of YO-PRO1 fluorescence intensity (F/F0) in amacrophage cell line J774 (A) and peritoneal macro-phages (B) measured before (control) and 10 min afterbath application of nordihydroguaiaretic acid (NDGA;50 �M). Black and white bars in A are for parental and3 clones of Panx1 shRNA treated J774 cells, respec-tively. Black and white bars in B are for wild-type(WT) and Panx1-deficient (Panx1-hypo) macrophages,respectively; n 4 independent experiments for J774;n 10 and 7 mice for WT and Panx1-hypo, respec-tively. C–F: electrophysiological recordings. C: meanvalues of currents amplitudes recorded from WT mac-rophages (n 3) before (0 min) and after 1- to 4-minapplication of NDGA (5 �M). Inset: time course of themeans � SE conductances recorded before and afterNDGA application to cells subjected to a voltage-rampprotocol (60 to � 80 mV; 5 s). D: examples ofcurrents recorded from WT macrophages before (top)and after 4-min NDGA (5 �M) application recordedusing a voltage-step protocol (60 to �140 mV;10-mV steps for 800 ms) illustrating the increase inoutward current. E: current-voltage (I–V) relation ob-tained from WT and Panx1-hypo macrophages after4-min exposure to NDGA. Each point in the I–V curverepresents the mean current value recorded at a partic-ular membrane potential ranging from 60 to �100mV obtained from 3 experiments as illustrated in D. F:bar histograms of the means � SE values of Panx1current amplitudes induced by NDGA (5 �M) andMK-886 (MK: 5 �M) in WT (black bars) and Panx1-hypo (white bars) macrophages. Note that the morespecific 5-LOX inhibitor MK-886 induced Panx1 cur-rents with similar amplitudes to those induced byNDGA. In Panx1-deficient macrophages, no currentswere evoked by the two 5-LOX inhibitors. *P �0.05, **P � 0.01, ***P � 0.001, ANOVA followed byNewman-Keuls multiple comparison test; n 3–4mice per genotype.
C573TP RECEPTORS ACTIVATE PANX1 CHANNELS
AJP-Cell Physiol • doi:10.1152/ajpcell.00087.2014 • www.ajpcell.org

Inhibitors of the 5-LOX Pathway Induce ATP Release fromPeritoneal Macrophages
Previous studies indicated that inhibitors of the 5-LOXpathway reduced intracellular levels of ATP and caused therelease of cytosolic ATP from peritoneal macrophages (2, 46).
To gain insight into the mechanisms involved in ATP release,C57Bl/6 peritoneal macrophages were treated for 30 min withthe Panx1 channel blocker CBX (50 �M) or with the vesiculartransport blocker BFA (10 �M) before NDGA (50 �M) appli-cation. As shown in Fig. 2A above, CBX and BFA significantly
Fig. 2. The 5-LOX pathway inhibitor NDGA induces ATP release from peritoneal macrophages. A: dot plot with bar histograms of the means � SE values ofATP released from C57Bl/6 peritoneal macrophages (n 4 mice per group) before (control) and after 1 min stimulation with NDGA (50 �M) in the absenceand presence of brefeldin-A (BFA: 10 �M) and carbenoxolone (CBX: 50 �M). Blockers were applied 30 min before NDGA. ***P � 0.001, control vs. NDGA;###P � 0.001, NDGA vs. blockers, ANOVA followed by Newman-Keuls multiple comparison test. B: bar histograms of the means � SE values of ATP releasedfrom WT (black bar) and Panx1-hypo (white bar) macrophages before (control) and after 1-min stimulation with NDGA (50 �M). ***P � 0.001, control vs.NDGA, ANOVA followed by Newman-Keuls multiple comparison test. C: bar histograms of the means � SE levels of Panx1 mRNA in WT (black bars) andPanx1-hypo (white bars) macrophages showing 70% reduced transcript in the transgenic mice, consistent with their hypomorph phenotype (14). ***P � 0.001,unpaired t-test. Numbers of animals used are in parentheses.
Fig. 3. Activation of Panx1 via thromboxane receptor. A: cartoon illustrating a putative signaling pathway leading to the activation of Panx1 channels followingNDGA stimulation involving thromboxane (TP) receptors used as a model for experiments designed in parts B–E. AA, arachidonic acid; COX, cyclooxygenase;PGH2, prostaglandin H2; TXA2, thromboxane A2; AC, adenylyl cyclase. B: means � SE values of the amount of ATP released from macrophages before(control) and after 1-min stimulation with NDGA (5 �M) and with the TXA2 mimetic (I-BOP: 50 �M) in the absence and presence (30-min preincubation) ofthe TP receptor antagonist SQ-29548 (10 �M). **P � 0.01, ***P � 0.001, ANOVA followed by Newman-Keuls test; n 3 mice. C: bar histogram of themeans � SE values of YO-PRO1 fluorescence intensity (F/F0) measured from WT (black bars) and Panx1-hypo (white bars) macrophages before (control) and10 min after bath application of NDGA (5 �M) and I-BOP (50 �M) in the absence and presence of SQ-29548 (10 �M). ***P � 0.001, control vs. NDGA;###P � 0.001, NDGA vs. NDGA � blockers, ANOVA followed by Newman-Keuls test. Numbers of mice are in parentheses. D: means � SE. values of Panx1current amplitudes induced by I-BOP (5–50 �M) recorded from wild-type macrophages. **P � 0.01, ***P � 0.001, ANOVA followed by Newman-Keuls test;n 3- 4 mice. E: inhibition of Panx1 currents induced by NDGA (5 �M), MK-886 (5 �M), and I-BOP (10 �M) by the TP receptor antagonist SQ-29548 (10�M); n 2–3 mice.
C574 TP RECEPTORS ACTIVATE PANX1 CHANNELS
AJP-Cell Physiol • doi:10.1152/ajpcell.00087.2014 • www.ajpcell.org
reduced the amount of ATP released by NDGA from 1.75 �0.10 to 1.15 � 0.12 and 1.06 � 0.14 nM·�g1·�l protein1,respectively. Although reduced, the latter values were signifi-cantly higher than the amount of ATP released under control,untreated condition (0.49 � 0.10 nM·�g1·�l protein1). Co-application of CBX and BFA reduced the amount of ATPreleased by NDGA to levels (0.71 � 0.09 nM·�g1·�l pro-tein1) that were not significantly different from control, un-treated conditions. As a further test for the participation ofPanx1 in response to NDGA application, we used peritonealmacrophages derived from Panx1-deficient mice. As expectedfrom the CBX experiments, the release of ATP induced byNDGA was prevented in Panx1-hypo mice (Fig. 2, B and C).Together, these data suggest that ATP release involves twopathways, one being vesicular and the other through ionchannels mainly formed by Panx1.
Activation of Panx1 via TP Receptors
Given that our results showed that NDGA and MK-886affected Panx1 channels in a similar way, we speculated thattheir effects could be mediated by AA metabolites other thanleukotrienes, in particular those derived from COX activity,such as thromboxane (Fig. 3A above). To test this possibility,we assessed Panx1 activity by measuring the amount of ATPrelease and influx of YO-PRO1, as well as the amplitudes ofPanx1 currents in WT and Panx1-hypo macrophages treated
with TP receptor agonist and antagonist (Fig. 3). As indicatedin Fig. 3B, NDGA-induced ATP release was prevented by a30-min preincubation with 10 �M of the TP receptor antago-nist SQ-29548. SQ-29548 also prevented ATP release inducedby the TP receptor agonist I-BOP (50 �M). The influx ofYO-PRO1 induced by NDGA (1.19 � 0.02; n 10) and I-BOP(1.25 � 0.04; n 3) was significantly reduced to 0.97 � 0.01 and1.11 � 0.02, respectively, when WT macrophages were pre-treated with the TP receptor antagonist (Fig. 3C). No influx ofdye was recorded from Panx1-hypo macrophages treated withNDGA or I-BOP (Fig. 3C). Electrophysiological recordingsprovided further support for the involvement of TP receptor inPanx1 channel activation following 5-LOX inhibition. Panx1current amplitudes were significantly increased when cellswere stimulated with I-BOP (�10 �M; Fig. 3D). The foldincrease in Panx1 currents induced by I-BOP (1.46 � 0.07;n 3) was not significantly different from that induced byMK-866 (1.63 � 0.22; n 3) and by NDGA (1.56 � 0.10;n 4; P � 0.05 ANOVA; compare Fig. 3D and 2E). The TPreceptor antagonist SQ-29548 totally prevented Panx1 cur-rents induced by NDGA, MK-886, and I-BOP (Fig. 3E).Because percent inhibition was estimated relative to controlvalues (before the addition of NDGA), inhibition by �100%likely represents the presence of variable numbers of activePanx1 channels under control conditions. Together, theseresults indicate that the mechanism linking the blockade of
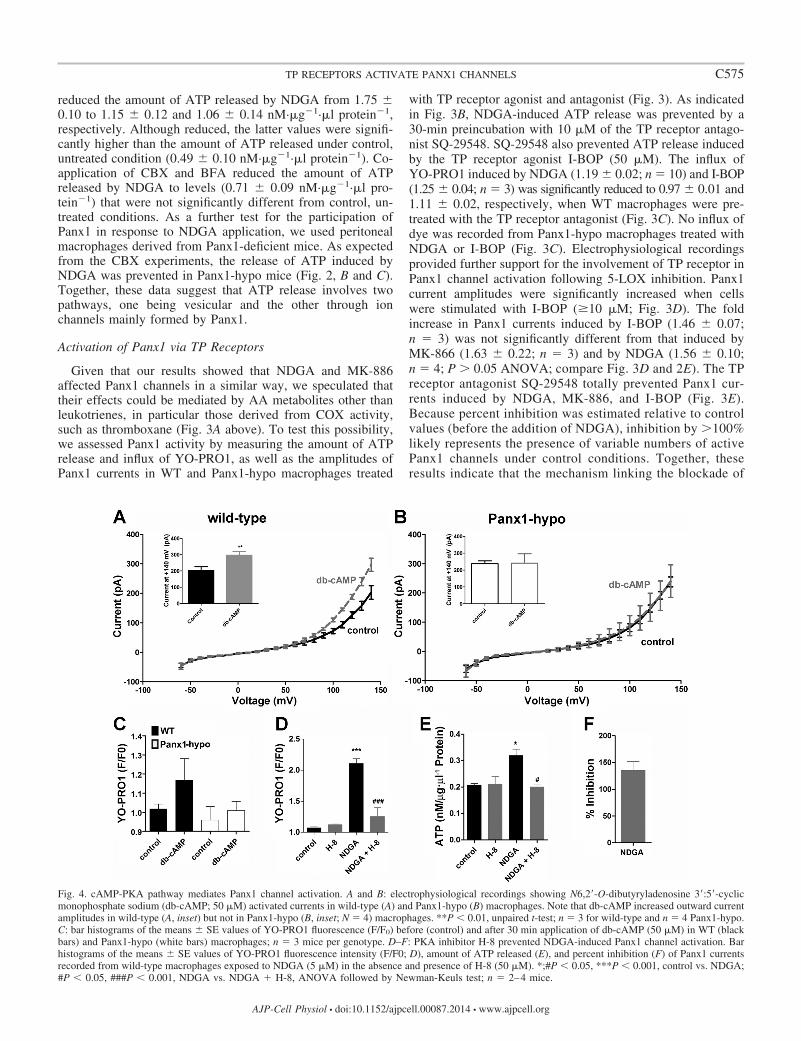
Fig. 4. cAMP-PKA pathway mediates Panx1 channel activation. A and B: electrophysiological recordings showing N6,2=-O-dibutyryladenosine 3=:5=-cyclicmonophosphate sodium (db-cAMP; 50 �M) activated currents in wild-type (A) and Panx1-hypo (B) macrophages. Note that db-cAMP increased outward currentamplitudes in wild-type (A, inset) but not in Panx1-hypo (B, inset; N 4) macrophages. **P � 0.01, unpaired t-test; n 3 for wild-type and n 4 Panx1-hypo.C: bar histograms of the means � SE values of YO-PRO1 fluorescence (F/F0) before (control) and after 30 min application of db-cAMP (50 �M) in WT (blackbars) and Panx1-hypo (white bars) macrophages; n 3 mice per genotype. D–F: PKA inhibitor H-8 prevented NDGA-induced Panx1 channel activation. Barhistograms of the means � SE values of YO-PRO1 fluorescence intensity (F/F0; D), amount of ATP released (E), and percent inhibition (F) of Panx1 currentsrecorded from wild-type macrophages exposed to NDGA (5 �M) in the absence and presence of H-8 (50 �M). *;#P � 0.05, ***P � 0.001, control vs. NDGA;#P � 0.05, ###P � 0.001, NDGA vs. NDGA � H-8, ANOVA followed by Newman-Keuls test; n 2–4 mice.
C575TP RECEPTORS ACTIVATE PANX1 CHANNELS
AJP-Cell Physiol • doi:10.1152/ajpcell.00087.2014 • www.ajpcell.org

the 5-LOX pathway by NDGA and MK-886 to the activationof Panx1 channels involves the production of prostanoids,one candidate being thromboxane A2 (TXA2) acting on TPreceptors.
cAMP/PKA Pathway Mediates Panx1 Activation
To gain insight into the possible intracellular signalingcascades involved in the activation of Panx1 via TP receptorstimulation, we evaluated whether cAMP and PKA were in-volved. This hypothesis was explored because macrophageshave been reported to express a TP receptor that is linked toadenylyl cyclase-Gs-coupled protein (28, 29, 48). As shown inFig. 4, A and B above, db-cAMP (50 �M) significantly increasedPanx1 currents (1.46 � 0.11 fold; n 4) and YO-PRO1 uptake(1.17 � 0.11 fold; n 4) in WT but not in Panx1-hypomacrophages. To test for the participation of PKA in theactivation of Panx1 mediated by TP receptors, we pretreatedthe cells with the PKA inhibitor H-8 before the addition ofNDGA. We used the 5-LOX inhibitor instead of the TPreceptor agonist (I-BOP) to further test the hypothesis linkingprostanoid production following 5-LOX inhibition and Panx1activation via the TP receptor. As expected, the PKA inhibitorH-8 prevented NDGA-induced YO-PRO1 uptake, ATP re-lease, and the augmentation of Panx1 currents (Fig. 4, D–F).Thus these data support the hypothesis that activation of Panx1channels in response to NDGA application involves the cAMP/PKA signaling cascade resultant from TP receptor stimulation,as illustrated in Fig. 5.
DISCUSSION
Monocytes and macrophages play a central role in complexinnate and adaptive immune responses, and their functions areinfluenced by a multitude of environmental signaling mole-
cules, including ATP and prostanoids (2, 7, 13, 53). It isbecoming apparent that prostanoid and purinergic receptorsignaling networks interact to modulate macrophage immuneresponses. For instance, large amounts of prostanoids areproduced by macrophages in response to ATP stimulation;released prostanoids then act in an autocrine/paracrine fashionto regulate immune responses (1, 9, 23). On the other hand,interference with the eicosanoid biosynthesis pathway, as inthe case of compounds that block the 5-LOX, has been shownto prevent P2X7 receptor-mediated apoptosis and to decreasecytosolic ATP levels (2, 7).
Interestingly, inhibitors of the 5-LOX pathway were re-ported to induce membrane permeabilization and ATP releasefrom macrophages through a yet not identified mechanism (36,46). We here show for the first time that Panx1 channels areactivated following the blockade of 5-LOX pathway and thatthese channels are involved, at least in part, in the release ofATP and influx of dyes. The intracellular signaling pathwaysleading to Panx1 activation likely involve a PKA-mediatedposttranslational modification resulting from the production ofcAMP following activation of TXA2 (TP) receptors. TheCOOH terminus of Panx1 has one threonine and six serineconsensus sites for phosphorylation (35); Panx1 phosphoryla-tion increases upon contraction of skeletal muscles (41). How-ever, there is no report demonstrating that a particular kinase(PKC, PKA) phosphorylates Panx1 at specific residues. Furtherstudies are necessary to evaluate whether or not PKA directlyphosphorylates Panx1 channels leading to its opening as re-ported here.
Our data showing that blockade of 5-LOX pathway byNDGA- and MK886-induced influx of dye and ATP release ina Panx1-dependent manner are corroborated by our resultsshowing that these inhibitors induced outward currents in WTbut not in Panx1-deficient peritoneal macrophages. AlthoughPanx1 channel seems to provide a conduit for YO-PRO1uptake following 5-LOX pathway blockade, additional mech-anisms for the release of ATP from macrophages are likely tobe in place. For instance, LPS-treated monocytes and NDGA/MK-866 treated macrophages release ATP mainly via exocy-tosis (11, 42, 46). In contrast, it has been recently shown thatlipoxin-A4, a nonclassical eicosanoid, induces ATP releasefrom airway epithelial cells mainly via Panx1 channels with aminor contribution of secretory vesicles (16). Our presentstudy showing that the Panx1 channel blocker (CBX) and thevesicular transporter blocker (BFA) partially reduced theamount of ATP released and that coapplication of both block-ers produced additive effects (Fig. 2A) indicates that twodistinct and parallel pathways are involved in the NDGA-induced ATP release from macrophages. Although future stud-ies are required to elucidate the relative contribution of each ofthese two pathways to macrophage immune responses, weinvestigated here the mechanism by which Panx1 is activatedfollowing blockade of 5-LOX pathway.
Panx1 channels are known to be activated basically by twomeans, via an intracellular signaling cascade and by the directaction of the stimulant on the channel itself. In the first case,activation of membrane (P2 and N-methyl-D-aspartate) recep-tors opens Panx1 channels through a mechanism involvingelevation of intracellular Ca2� (25) and/or Src-tyrosine kinase(20, 55). The alternative mode of Panx1 channel activationinvolves the direct action of a “stimulant” on the channel itself,
Fig. 5. Cartoon summarizing the pathway by which Panx1 channel is activatedby prostanoids. Blockade of 5-LOX pathway by NDGA or MK-866 leads to ashunt of the phospholipase A2 (PLA2) metabolite AA towards the productionof prostanoids (PGH2 and TXA2) via COX activity. TP receptor agonist(I-BOP, a TXA2 mimetic) is shown acting on TP receptor leading to theproduction of AC, an enzyme converting ATP into cAMP. Panx1 channelactivation, indicated by the influx of YO-PRO1 and release of ATP, ismodulated by protein kinase A (PKA) activity. Blockers of TP receptors(SQ-29548), PKA activity (H-8), Panx1 channels (CBX), and vesicular trans-port (BFA) are illustrated.
C576 TP RECEPTORS ACTIVATE PANX1 CHANNELS
AJP-Cell Physiol • doi:10.1152/ajpcell.00087.2014 • www.ajpcell.org

as is the case of elevated extracellular K�, which binds to oneof the extracellular loops of the protein (21, 47).
Inspection of Panx1 current development following NDGAor MK-886 application revealed that a moderately long timecourse is necessary for the full development of Panx1 currents(�4 min); this suggests that the action of the drugs is notdirectly on the channel, as would be expected from off-targeteffects reported for many 5-LOX inhibitors (27) but likelyinvolves AA intermediate molecules, such as those generatedby COX activity. The possibility that blockade of LOX activitycould lead to a metabolic shunt in a similar but oppositedirection than the arachidonate metabolic shunt reported tooccur from COX to LOX (24, 26) is supported by our datashowing that TP receptor stimulation leads to Panx1 channelactivation. In this regard, we found that blockade of the TPreceptor with SQ29458 prevents NDGA-induced dye influx,ATP release, and Panx1 currents. Further support for thehypothesis that blockade of 5-LOX shunts AA metabolitestowards the generation of prostanoids are our data showing thatthe TXA2 mimetic I-BOP had similar effects to those producedby NDGA and MK-866.
Although direct measurements of TXA2 levels generatedfollowing the blockade of 5-LOX are necessary, our proposedmodel for the synthesis of TXA2 in the absence of increasedPLA2-mediated AA production is based on reports indicatingthat thromboxane synthase is abundantly expressed in activatedmacrophages [see Tone et al. (52)] and that these cells, differ-ently from resting macrophages, convert AA mainly intoTXA2, which is released faster than PGE2 (4, 30).
Thus our data indicate that activation of Panx1 following theblockade of the 5-LOX pathway is likely mediated by TXA2acting on the G protein-coupled TP receptor. Differently fromhumans where TP receptors consist of two isoforms (TP� andTP�: 17, 18, 40), mice express only one isoform that is 76%identical to the human TP� (31, 33). As would be expectedfrom a TP� isoform that elevates cytosolic levels of cAMP(17), we show that Panx1 channels in peritoneal macrophagescan be activated by db-cAMP and that blockade of the cAMP-dependent protein kinase (PKA) prevents NDGA-inducedPanx1 currents, dye influx, and ATP release.
Interestingly, however, the effects db-cAMP on Panx1 chan-nels were less pronounced than those induced by I-BOP,suggesting the requirement of additional inputs from prostacy-clin receptors and/or from the TP receptor itself but viaGq/PLC signaling pathway. The involvement of Gq signalingarm of TP receptors seems unlikely given that NDGA-inducedmembrane permeabilization in peritoneal macrophages is in-dependent of intracellular calcium levels (46). It remains pos-sible that the heterodimerization of prostacyclin receptors andTP receptors is necessary to enhance cAMP levels in responseto TP� stimulation, as shown for human TP receptors (56).
Prostanoids and their receptors control several cellular pro-cesses, including acute inflammation and innate and adaptiveimmunity. They thus represent a complex system of immuno-regulation, with varied and distinct outcomes dependent onreceptor types and subtypes. Although monocytes/macro-phages are enriched in thromboxane synthase and release largeamounts of TXA2 upon stimulation (6, 30, 32, 52), the functionof TXA2-TP receptor signaling is not as well defined as it is thecase for other immune cells. For instance, dendritic and T-cellinteractions are inhibited by TXA2, and therefore, TXA2
negatively modulates acquired immunity (22). Nevertheless, itis possible that by activating Panx1 channels TXA2-TP recep-tor signaling contributes positively to macrophage immuneresponses.
Several lines of evidence support the notion that Panx1 is akey component of immune responses. Panx1 is ubiquitouslyexpressed and forms plasma membrane channels permeable tolarge molecules (reviewed in Refs. 5, 44, 45, 49). Not onlydoes Panx1 provide sites for the release of ATP (3, 19, 39, 51),an important immune modulator, but it is also a component ofthe inflammasome leading to caspase-1 activation and theprocessing of IL-1� (34, 47). Moreover, a recent studyproposed pannexin/innexin as being the gatekeepers of mi-croglia response to injury by showing that the dose-depen-dent inhibition of AA was correlated with pannexin/innexinactivity and microglia migration towards the site of lesion(43). In this situation, blockade of Panx1 channels by highlevels of AA would prevent inflammasome activation, whilelower levels of AA due to enzymatic catabolism wouldovercome the inhibition of Panx1 channels and promote aninflammatory response (43).
Many processes involved in immune responses are regulatedby a cross talk between different intracellular signaling net-works including the prostanoid (TP), Toll-like (TL), purinergic(P2), and cytokine receptors. Here we provide the first evi-dence for the involvement of Panx1 in the TP receptor signal-ing network. Future studies aimed at clarifying the contributionof this TP-Panx1 signaling network to macrophage immuneresponses are likely to be important for targeting inflammatoryand autoimmune diseases.
ACKNOWLEDGMENTS
We thank Adriana Mello for technical assistance.
GRANTS
H. A. da Silva-Souza was supported by fellowships from the ConselhoNacional de Desenvolvimento Científico e Tecnológico (CNPq; DoutoradoSanduíche-ECsF 246349/2012-3) and a Fogarty Training Grant (NHITW007129). P. M. Persechini is supported by Conselho Nacional de Desen-volvimento Científico e Tecnológico (CNPq), Fundação Carlos Chagas Filhode Amparo a Pesquisa do Estado do Rio de Janeiro (FAPERJ), and InstitutoNacional para Pesquisa Translacional em Saúde e Ambiente na RegiãoAmazônica (INCT-INPeTAm/CNPq/MCT).
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the author(s).
AUTHOR CONTRIBUTIONS
Author contributions: H.A.d.S.-S., P.M.P., and E.S. conception and designof research; H.A.d.S.-S., M.N.d.L., N.K.P., D.C.S., and E.S. performed exper-iments; H.A.d.S.-S., M.N.d.L., N.K.P., and E.S. analyzed data; H.A.d.S.-S.,D.C.S., and E.S. interpreted results of experiments; H.A.d.S.-S. and E.S.prepared figures; H.A.d.S.-S. and E.S. drafted manuscript; H.A.d.S.-S., D.C.S.,and E.S. edited and revised manuscript; H.A.d.S.-S., M.N.d.L., N.K.P., D.C.S.,P.M.P., and E.S. approved final version of manuscript.
REFERENCES
1. Atarashi K, Nishimura J, Shima T, Umesaki Y, Yamamoto M, OnoueM, Yagita H, Ishii N, Evans R, Honda K, Takeda K. ATP drives laminapropria T(H)17 cell differentiation. Nature 455: 808–812, 2008.
2. Ballatori N, Wang W. Nordihydroguaiaretic acid depletes ATP andinhibits a swelling-activated, ATP-sensitive taurine channel. Am J PhysiolCell Physiol 272: C1429–C1436, 1997.
3. Bao L, Locovei S, Dahl G. Pannexin membrane channels are mechano-sensitive conduits for ATP. FEBS Lett 572: 65–68, 2004.
C577TP RECEPTORS ACTIVATE PANX1 CHANNELS
AJP-Cell Physiol • doi:10.1152/ajpcell.00087.2014 • www.ajpcell.org

4. Bezugla Y, Kolada A, Kamionka S, Bernard B, Scheibe R, Dieter P.COX-1 and COX-2 contribute differentially to the LPS-induced release ofPGE2 and TxA2 in liver macrophages. Prostaglandins Other Lipid Mediat79: 93–100, 2006.
5. Bond SR, Naus CC. The pannexins: past and present. Front Physiol 5: 58,2014.
6. Brune K, Glatt M, Kälin H, Peskar BA. Pharmacological control ofprostaglandin and thromboxane release from macrophages. Nature 274:261–263, 1978.
7. Costa-Junior HM, Mendes AN, Davis GH, da Cruz CM, Ventura AL,Serezani CH, Faccioli LH, Nomizo A, Freire-de-Lima CG, BisaggioRda C, Persechini PM. ATP-induced apoptosis involves a Ca2�-inde-pendent phospholipase A2 and 5-lipoxygenase in macrophages. Prosta-glandins Other Lipid Mediat 88: 51–61, 2009.
8. Coutinho-Silva R, Persechini PM. P2Z purinoceptor-associated poresinduced by extracellular ATP in macrophages and J774 cells. Am J PhysiolCell Physiol 273: C1793–C1800, 1997.
9. Di Virgilio F, Chiozzi P, Ferrari D, Falzoni S, Sanz JM, Morelli A,Torboli M, Bolognesi G, Baricordi OR. Nucleotide receptors: an emerg-ing family of regulatory molecules in blood cells. Blood 97: 587–600,2001.
10. El Ouaaliti M, Seil M, Dehaye JP. Activation of calcium-insensitivephospholipase A2 (iPLA2) by P2X7 receptors in murine peritoneal mac-rophages. Prostaglandins Other Lipid Mediat 99: 116–123, 2012.
11. Fujiwara T, Takami N, Misumi Y, Ikehara Y. Nordihydroguaiareticacid blocks protein transport in the secretory pathway causing redistribu-tion of Golgi proteins into the endoplasmic reticulum. J Biol Chem 273:3068–3075, 1998.
12. Gaddi A, Cicero AF, Pedro EJ. Clinical perspectives of anti-inflamma-tory therapy in the elderly: the lipoxigenase (LOX)/cycloxigenase (COX)inhibition concept. Arch Gerontol Geriatr 38: 201–212, 2004.
13. Gordon S. Alternative activation of macrophages. Nat Rev Immunol 3:3–35, 2003.
14. Hamad AM, Sutcliffe AM, Knox AJ. Aspirin-induced asthma: clinicalaspects, pathogenesis and management. Drugs 64: 417–2432, 2004.
15. Hanstein R, Negoro H, Patel NK, Charollais A, Meda P, Spray DC,Suadicani SO, Scemes E. Promises and pitfalls of a Pannexin1 transgenicmouse line. Front Pharmacol 4: 61, 2013.
16. Higgins G, Buchanan P, Perriere M, Al-Alawi M, Costello R, VerriereV, McNally P, Harvey BJ, Urbach V. Activation of P2RY11 and ATPrelease by LXA4 restores the airway surface liquid layer and epithelialrepair in cystic fibrosis. Am J Respir Cell Mol Biol 51: 178–190, 2014.
17. Hirata M, Hayashi Y, Ushikubi F, Yokota Y, Kageyama R, NakanishiS, Narumiya S. Cloning and expression of cDNA for a human throm-boxane A2 receptor. Nature 349: 617–620, 1991.
18. Hirata T, Ushikubi F, Kakizuka A, Okuma M, Narumiya S. Twothromboxane A2 receptor isoforms in human platelets. Opposite couplingto adenylyl cyclase with different sensitivity to Arg60 to Leu mutation. JClin Invest 97 949–956, 1996.
19. Iglesias R, Dahl G, Qiu F, Spray DC, Scemes E. Pannexin 1: themolecular substrate of astrocyte ’’hemichannels’’. J Neurosci 29: 7092–7097, 2009.
20. Iglesias R, Locovei S, Roque A, Alberto AP, Dahl G, Spray DC,Scemes E. P2X7 receptor-Pannexin1 complex: pharmacology and signal-ing. Am J Physiol Cell Physiol 295: C752–C760, 2008.
21. Jackson DG, Wang J, Keane RW, Scemes E, Dahl G. ATP andpotassium ions: a deadly combination for astrocytes. Sci Rep 4: 4576,2014.
22. Kabashima K, Murata T, Tanaka H, Matsuoka T, Sakata D, YoshidaN, Katagiri K, Kinashi T, Tanaka T, Miyasaka M, Nagai H, UshikubiF, Narumiya S. Thromboxane A2 modulates interaction of dendritic cellsand T cells and regulates acquired immunity. Nat Immunol 4: 694–701,2003.
23. Kihara Y, Gupta S, Maurya MR, Armando A, Shah I, QuehenbergerO, Glass CK, Dennis EA, Subramaniam S. Modeling of eicosanoidfluxes reveals functional coupling between cyclooxygenases and terminalsynthases. Biophys J 106: 966–75, 2014.
24. Lin WW, Lee YT. Pyrimidinoceptor-mediated activation of phospho-lipase C and phospholipase A2 in RAW 264.7 macrophages. Br J Phar-macol 119: 261–268, 1996.
25. Locovei S, Wang J, Dahl G. Activation of pannexin 1 channels by ATPthrough P2Y receptors and by cytoplasmic calcium. FEBS Lett 580:239–244, 2006.
26. Maxis K, Delalandre A, Martel-Pelletier J, Pelletier JP, Duval N,Lajeunesse D. The shunt from the cyclooxygenase to lipoxygenasepathway in human osteoarthritic subchondral osteoblasts is linked with avariable expression of the 5-lipoxygenase-activating protein. Arthritis ResTher 8: R181, 2006.
27. Meirer K, Steinhilber D, Proschak E. Inhibitors of the arachidonic acidcascade: interfering with multiple pathways. Basic Clin Pharmacol Toxi-col 114: 83–91, 2014.
28. Meja KK, Barnes PJ, Giembycz MA. Characterization of the prostanoidreceptor(s) on human blood monocytes at which prostaglandin E2 inhibitslipopolysaccharide-induced tumor necrosis factor-alpha generation. Br JPharmacol 122: 149–157, 1997.
29. Mogi C, Tobo M, Tomura H, Murata N, He XD, Sato K, Kimura T,Ishizuka T, Sasaki T, Sato T, Kihara Y, Ishii S, Harada A, OkajimaF. Involvement of proton-sensing TDAG8 in extracellular acidification-induced inhibition of proinflammatory cytokine production in peritonealmacrophages. J Immunol 182: 3243–3251, 2009.
30. Murota S, Kawamura M, Morita I. Transformation of arachidonic acidinto thromboxane B2 by the homogenates of activated macrophages.Biochim Biophys Acta 528: 507–511, 1978.
31. Nakahata N. Thromboxane A2: physiology/pathophysiology, cellularsignal transduction and pharmacology. Pharmacol Ther 118: 18 –35,2008.
32. Namba T, Sugimoto Y, Hirata M, Hayashi Y, Honda A, Watabe A,Negishi M, Ichikawa A, Narumiya S. Mouse thromboxane A2 receptor:cDNA cloning, expression and Northern blot analysis. Biochem BiophysRes Commun 184: 1197–1203, 1992.
33. Narumiya S, Sugimoto Y, Ushikubi F. Prostanoid receptors: structures,properties, functions. Physiol Rev 79: 1193–1226, 1999.
34. Pelegrin P, Surprenant A. The P2X7 receptor-pannexin connection todye uptake and IL-1beta release. Purinergic Signal 5: 129–137, 2009.
35. Penuela S, Bhalla R, Gong XQ, Cowan KN, Celetti SJ, Cowan BJ, BaiD, Shao Q, Laird DW. Pannexin 1 and pannexin 3 are glycoproteins thatexhibit many distinct characteristics from the connexin family of gapjunction proteins. J Cell Sci 120: 3772–3783, 2007.
36. Persechini PM, Silva-Souza HA, Silva-Vasconcelos JS, Schachter J. Acation transport mechanism activated by inhibitors of the 5-lipoxigenase-arachidonic acid dependent pathway in macrophages. 14th InternationalCongress of Immunology, PP-047–22, 2010.
37. Peters-Golden M, Canetti C, Mancuso P, Coffey MJ. Leukotrienes:underappreciated mediators of innate immune responses. J Immunol 174:89–94, 2005.
38. Peters-Golden M, Henderson WR Jr. The role of leukotrienes in allergicrhinitis. Ann Allergy Asthma Immunol 94: 609–618, 2005.
39. Qiu F, Wang J, Spray DC, Scemes E, Dahl G. Two non-vesicular ATPrelease pathways in the mouse erythrocyte membrane. FEBS Lett 585:3430–3435, 2011.
40. Raychowdhury MK, Yukawa M, Collins LJ, McGrail SH, Kent KC,Ware JA. Alternative splicing produces a divergent cytoplasmic tail in thehuman endothelial thromboxane A2 receptor. J Biol Chem 269: 19256–19261, 1994.
41. Riquelme MA, Cea LA, Vega JL, Boric MP, Monyer H, Bennett MV,Frank M, Willecke K, Sáez JC. The ATP required for potentiation ofskeletal muscle contraction is released via pannexin hemichannels. Neu-ropharmacology 75: 594–603, 2013.
42. Sakaki H, Tsukimoto M, Harada H, Moriyama Y, Kojima S. Auto-crine regulation of macrophage activation via exocytosis of ATP andactivation of P2Y11 receptor. PLoS One 8: e59778, 2013.
43. Samuels SE, Lipitz JB, Wang J, Dahl G, Muller KJ. Arachidonic acidcloses innexin/pannexin channels and thereby inhibits microglia cellmovement to a nerve injury. Dev Neurobiol 73: 621–631, 2013.
44. Scemes E, Spray DC, Meda P. Connexins, pannexins, innexins: novelroles of “hemi-channels”. Pflügers Arch 457: 1207–1226, 2009.
45. Scemes E, Suadicani SO, Dahl G, Spray DC. Connexin and pannexinmediated cell-cell communication. Neuron Glia Biol 3: 199–208, 2007.
46. Silva-Souza HA, de Lira MN, Costa-Junior HM, da Cruz CM, Vas-concellos JS, Mendes AN, Reis GP, Alvarez CL, Faccioli LH, SerezaniCH, Schachter J, Persechini PM. Inhibitors of the 5-lipoxygenasearachidonic acid pathway induce ATP release and ATP-dependent organiccation transport in macrophages. Biochim Biophys Acta 1838: 1967–1977,2014.
47. Silverman WR, de Rivero Vaccari JP, Locovei S, Qiu F, Carlsson SK,Scemes E, Keane RW, Dahl G. The pannexin 1 channel activates the
C578 TP RECEPTORS ACTIVATE PANX1 CHANNELS
AJP-Cell Physiol • doi:10.1152/ajpcell.00087.2014 • www.ajpcell.org

inflammasome in neurons and astrocytes. J Biol Chem 284: 18143–18151,2009.
48. Simmons TR, Cook JA, Moore JN, Halushka PV. Thromboxane A2receptors in equine monocytes: identification of a new subclass of TXA2receptors. J Leukoc Biol 53: 173–178, 1993.
49. Sosinsky GE, Boassa D, Dermietzel R, Duffy HS, Laird DW,MacVicar B, Naus CC, Penuela S, Scemes E, Spray DC, ThompsonRJ, Zhao HB, Dahl G. Pannexin channels are not gap junction hemi-channels. Channels (Austin) 5: 193–197, 2011.
50. Suadicani SO, Brosnan CF, Scemes E. P2X7 receptors mediate ATPrelease and amplification of astrocytic intercellular Ca2� signaling. JNeurosci 26: 1378–1385, 2006.
51. Suadicani SO, Iglesias R, Wang J, Dahl G, Spray DC, Scemes E. ATPsignaling is deficient in cultured Pannexin1-null mouse astrocytes. Glia60: 1106–1116, 2012.
52. Tone Y, Miyata A, Hara S, Yukawa S, Tanabe T. Abundant expressionof thromboxane synthase in rat macrophages. FEBS Lett 340: 241–244,1994.
53. Través PG, Pimentel-Santillana M, Carrasquero LM, Pérez-Sen R,Delicado EG, Luque A, Izquierdo M, Martín-Sanz P, Miras-PortugalMT, Boscá L. Selective impairment of P2Y signaling by prostaglandin E2in macrophages: implications for Ca2�-dependent responses. J Immunol190: 4226–4235, 2013.
54. Ulmann L, Hirbec H, Rassendren F. P2X4 receptors mediate PGE2release by tissue-resident macrophages and initiate inflammatory pain.EMBO J 29: 2290–300, 2010.
55. Weilinger NL, Tang PL, Thompson RJ. Anoxia-induced NMDA recep-tor activation opens pannexin channels via Src family kinases. J Neurosci32: 12579–12588, 2012.
56. Wilson SJ, Roche AM, Kostetskaia E, Smyth EM. Dimerization of thehuman receptors for prostacyclin and thromboxane facilitates thrombox-ane receptor-mediated cAMP generation. J Biol Chem 279: 53036–53047,2004.
57. Zeldin DC. Epoxygenase pathways of arachidonic acid metabolism. J BiolChem 276: 36059–36062, 2001.
C579TP RECEPTORS ACTIVATE PANX1 CHANNELS
AJP-Cell Physiol • doi:10.1152/ajpcell.00087.2014 • www.ajpcell.org





























