Embed Size (px)
Citation preview

INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA (INPA)
PROGRAMA DE PÓS-GRADUAÇÃO EM GENÉTICA, CONSERVAÇÃO E
BIOLOGIA EVOLUTIVA (GCBEv)
Monandria e Poliandria como estratégia evolutiva no complexo de subespécies
de Melipona seminigra Friese, 1903 (Apidae, Meliponini) na Amazônia.
IZAURA BEZERRA FRANCINI
MANAUS-AM
2013

IZAURA BEZERRA FRANCINI
Monandria e Poliandria como estratégia evolutiva no complexo de subespécies
de Melipona seminigra Friese, 1903 (Apidae, Meliponini) na Amazônia.
ORIENTADOR (A): Dra. Gislene Almeida Carvalho-Zilse
CO - ORIENTADOR: Dr. Gustavo Campos Silva Kuhn
Tese apresentada ao Programa de Pós-
graduação do INPA como parte dos
requisitos para obtenção do título de
Doutor em Genética, Conservação e
Biologia Evolutiva.
MANAUS-AM
2013

iii
FICHA CATALOGRÁFICA
F817 Francini, Izaura Bezerra
Monandria e Poliandria como estratégia evolutiva no complexo
de subespécies de Melipona seminigra Friese, 1903 (Apidae,
Meliponini) na Amazônia / Izaura Bezerra Francini. --- Manaus :
[s.n.], 2014.
xvi, 93 f. : il. color.
Tese (Doutorado) --- INPA, Manaus, 2013.
Orientadora : Gislene Almeida Carvalho-Zilse.
Coorientador : Gustavo Campos Silva Kuhn.
Área de concentração : Genética, Conservação e Biologia
Evolutiva.
1. Abelha sem ferrão. 2. Melipona seminigra. 3. Eussocialidade.
I. Título.
CDD 595.799
Sinopse
A espécie politípica Melipona seminigra, com ampla distribuição na bacia Amazônica,
foi estudada molecularmente, usando-se DNA nuclear (locos microssatélites), para analisar
paternidade e parentesco. Adicionalmente, foi usado o gene mitocondrial citocromo c oxidase
subunidade 1 (COI) para identificar, molecularmente, seis subespécies do complexo M.
seminigra e feita a análise de segregação de sexos. A análise de segregação de sexos evidenciou
a produção de machos diploides, a análise de DNA nuclear indicou que o sistema de
acasalamento de M. seminigra é a poliandria; a análise do COI revelou alta distância genética
entre as subespécies, quando comparado com a distância genética entre espécies do mesmo
subgênero, indicando que se encontram separadas geograficamente por um longo tempo.
Palavras-chave: Melipona seminigra, poliandria, eussocialidade, Meliponini, paternidade,
parentesco, DNA microssatélites, DNA barcoding, macho diploide.

iv
Dedico esta Tese aos meus filhos Carlo Leopoldo B.
Francini (Leo) e Ronaldo Bastos Francini-Filho (Ro), e a
minha neta Naiara Alves Francini (Nai). O longo tempo que
passei sem vocês para realizar este trabalho, não tem volta.
Seja ele meu pedido de desculpas e a herança que lhes
deixo!

v
Acima de tudo o amor
(Coríntios 1-13)
Ainda que eu falasse línguas,
As dos homens e dos anjos,
Se eu não tivesse o amor,
Seria como sino ruidoso e estridente.
Ainda que eu tivesse o dom da profecia,
O conhecimento de todos os mistérios e de toda a ciência,
Ainda que eu tivesse toda a fé, a ponto de transportar montanhas,
Se eu não tivesse o amor, eu não seria nada.
Ainda que eu distribuísse todos os meus bens aos famintos,
Ainda que entregasse o meu corpo às chamas,
Se não tivesse o amor, nada disso me adiantaria.
O amor é paciente, o amor é prestativo, não é invejoso,
Não se ostenta e não se incha de orgulho.
Nada faz de inconveniente, não procura seu próprio interesse,
Não se irrita, não guarda rancor.
Não se alegra com a injustiça, mas se regozija com a verdade.
Tudo desculpa e tudo crê; tudo espera e tudo suporta.
O amor, jamais passará.
As profecias desaparecerão, as línguas cessarão e a ciência também desaparecerá,
Pois nosso conhecimento é limitado, limitada é também a nossa profecia,
Mas, quando vier A Perfeição, desaparecerá o que é limitado.
Quando eu era criança, falava como criança, pensava como criança,
Raciocinava como criança.
Depois me tornei adulto, deixei o que era próprio de criança.
Agora vemos como em espelho e de maneira confusa;
Mas depois vemos face a face.
Agora meu conhecimento é limitado, mas, depois conhecerei como sou conhecido.
Agora, portanto, permanecem três coisas: a fé, a esperança e o amor.
A maior dela, porém é o amor.

vi
Este trabalho foi possível devido:
Ao Programa de Genética Conservação de Biologia Evolutiva (GCBEv) do Instituto
Nacional de Pesquisas da Amazônia (INPA).
Ao Laboratório Temático de Biologia Molecular (LTBM), onde as análises
moleculares foram desenvolvidas.
Ao Grupo de Pesquisas em Abelhas (GPA), pela orientação e muito dos recursos
utilizados.
Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq).
Ao Instituto Chico Mendes de Conservação da Biodiversidade (ICMBio),
particularmente, às chefias do Parque Nacional de Anavilhanas e do Parque Nacional do Jaú,
pelo apoio logístico.

vii
Agradecimentos
Agradeço a todas as pessoas que, direta ou indiretamente, possibilitaram a realização
deste trabalho.
À minha orientadora, Dra. Gislene Almeida Carvalho-Zilse, por ter assumido à
responsabilidade da minha orientação e disponibilizado recursos de seus projetos para que
este trabalho fosse realizado. Muito obrigada por sua coragem, paciência e ensinamentos. Sei
que poucos assumiriam a responsabilidade de orientar um estudante de doutorado aos 64 anos
de idade, sem direito a bolsa de estudos, portanto, sem direito a qualquer outro recurso
financeiro fornecido por fomentos de pesquisa.
Ao meu co-orientador, Dr. Gustavo Campos Silva Kuhn, que mesmo à distância
sempre me atendeu, pronta e cordialmente, quando precisei.
Aos membros da banca de qualificação por sugestões e ensinamentos: Dra Adna
Cristina Barbosa de Sousa, Dr. Carlos Schneider e Dr. Cleiton Fantin Rezende.
Ao Dr. Carlos Gustavo Nunes-Silva, pela parceria nas publicações, pela bolsa de DTI,
nos dois primeiros anos do doutorado e por sua amizade, sua alegria contagiante.
À Dra. Jacqueline da Silva Batista, por ter permitido que me tornasse uma abelha-
pirada. Se há louros não sei, mas em havendo você tem parte neles.
À MSc. Kyara Formiga pelo apoio e compreensão, desde os primeiros momentos deste
trabalho, obrigada mesmo!
Ao Dr. Márcio de Oliveira, por todas as vezes que gentilmente gastou de seu tempo
para me ensinar e possibilitar minhas consultas à Coleção de Invertebrados do INPA.
Ao filho sobressalente e parceiro de trabalho, Antônio Saulo Cunha Machado, pelo
muito que gastou de seu tempo comigo, pelos muitos cuidados, pela paciência, pelos
ensinamentos, pelos bons momentos compartilhados, por todos os abraços.
Ao amigo João Marcos Capurucho por sua disposição em colaborar, ensinando,
discutindo, fazendo. Valeu João! Você é uma dessas pessoas que ajuda fazer a diferença.
Á todas as pessoas que me auxiliaram nas coletas em Rondônia. D. Nazaré Ravani,
pessoa maior em todos os sentidos; ao Sr. Plinio Vanzini, criador de abelhas sem ferrão, pelas
amostras, pelo apoio logístico e acolhimento; ao Sr. Josuel Ravani pelo enorme apoio
logístico, desde o primeiro momento, na chegada a Porto Velho; a meu amigo Luciano Costa,
atualmente na Alemanha e naqueles dias trabalhando no resgate de abelhas na área de
desmatamento da hidrelétrica de Sto Antônio.

viii
Ao meu amigo de fé, meu irmão camarada, Sr. Jose Almir de Souza Machado, por ter
me acompanhado em quase todo o trabalho de campo, principalmente no Parque Nacional do
Jau, em locais de difícil acesso e alto risco. Obrigada por me fazer sentir segura, por não
medir esforços em me ajudar, carregando mochilas e às vezes até a canoa. Obrigada por ter
me acompanhado e me conduzido basicamente por todo este longo e difícil período de minha
vida: mudanças, mau humor, lágrimas, muitas idas a médicos e dentistas, bancos, mercados,
idas e vindas ao INPA (LTBM e GPA) a qualquer hora e dia da semana. Sua religiosidade,
sua capacidade de conversar, sua bondade e sabedoria, sua paciência e amizade foram
fundamentais para que eu não deixasse este trabalho pelo caminho. Sou grata a Deus por sua
presença e ajuda.
Aos funcionários do ICMBio (Parque Nacional de Anavilhanas e Jaú) pelo apoio
logístico. Agradeço especialmente a Mariana Macedo Leitão, chefe do Parque Nacional do
Jaú, sua disponibilidade em ajudar foi fundamental.
Ao Sr. Eduardo Elísio pelo trabalho de campo e hospedagem no Parque Nacional do
Jaú; ao seu filho Eduardinho, menino querido, por alegrar os dias de Jaú: jacarés macetas,
borboletas, passeios de canoas, se a Sra. entrou na canoa, então vai conseguir sair!
Ao Sr. Jair Arruda e demais pessoas de sua família que tornaram possível as coletas
em Boa Vista do Ramos e Maués.
Aos amigos do LTBM: Gisele, Saulo, Paulinha, Adriel, João, Karen, Karoline, Paola,
Larissa, Elisama, Santiago, Jane Maria, vocês fizeram o mundo das moléculas ficar melhor.
Ao Grupo de Pesquisas em Abelhas, pelo objetivo que nos une e por ter contado com
todos sempre que precisei. Agradeço, especialmente, ao Diego pelas muitas ajudas.
Finalmente, mas não por último, agradeço a minha família porque aceitaram minha
ausência sem reclamar, sem nada cobrar. Não teria realizado este trabalho se não tivesse me
colocado, no dizer de Platão “ao abrigo dos muros”. Sou especialmente grata quando vocês
me dizem, se você está feliz tudo bem, se era isto que você queria estamos contentes por você.
Senti demais a falta de todos vocês, mas não posso lhes dizer que não faria tudo novamente,
se o tempo voltasse, errado ou certo, eu percorreria o mesmo caminho. Não pelos títulos ou
qualquer glória advinda, nem sei se são tão importantes assim, mas pela aventura ao mundo
do conhecimento. Viver no Amazonas estes longos sete anos estudando abelhas sem ferrão foi
um presente Divino que ousei aceitar. As crianças ficaram adolescentes lindos e eu não estava
lá, outras nasceram e nem as vi ainda. Obrigada a Deus porque me permitiu o tempo de ainda
poder estar com vocês.

ix
RESUMO
A abelha sem ferrão Melipona seminigra (Apidae, Meliponini) é uma espécie politípica com
ampla distribuição na bacia Amazônica. Tem papel chave na polinização de plantas nativas e
cultivadas, assim como na dispersão de sementes. É altamente eussocial, assim como outros
Meliponini e Apini, o que se caracteriza pela evolução do sistema de castas; divisão do
trabalho reprodutivo; sobreposição de gerações, rainha mãe e operárias no mesmo ninho;
cuidado cooperativo à prole; rainha (fêmea fértil) e operárias (fêmeas estéreis),
morfologicamente, diferenciadas. O sistema de acasalamento predito para as abelhas sem
ferrão é a monandria, comum à maioria dos Hymenoptera solitários e sociais. Nosso objetivo
foi estudar a paternidade e o parentesco em colônias naturais e manejadas de M. seminigra,
usando DNA microssatélite, e identificar molecularmente as diferentes subespécies, usando
DNA barcode. Além do que, objetivamos estudar o mecanismo de determnação do sexo,
através da análise de segregação de machos e fêmeas. Amostramos seis subespécies, quatro
delas descritas (M. s. abunensis, M. s. merrillae, M. s. pernigra e M. s. seminigra) e duas
delas não descritas (M. s. ssp1 e M. s. ssp2). Genotipamos 4-5 locos DNA microssatélite em
576 operárias, amostradas em 24 colônias, de três subespécies (M. s. abunensis, M. s.
merrillae e M. s. seminigra). Amplificamos um fragmento de 526 pares de bases do gene
COI, em 31 operárias das seis subespécies, amostradas em diferentes ninhos. Observamos
poliandria nas 24 colônias de M. seminigra estudadas. A frequência de paternidade variou de
3 a 11 (em média 7,416 ± 2,5) e a paternidade efetiva variou de 2 a 11,613 (em média 6,324 ±
2,655). Este nível de poliandria produziu um grau de parentesco que variou 0,298 a 0,5 (em
média 0,346 ± 0,051). Na maioria das colônias estudadas (21/24) os valores de skew (S) foram
próximos de zero. Em 3/24 das colônias observamos um relativo monopólio da paternidade
por alguns machos (S ˃ 0,5) e em 6/24 delas observamos skew negativo, interpretado como
zero. A distância genética entre as subespécies foi, em média, 1,8 ± 0,47 %, enquanto a
distância genética entre espécies (mesmo subgênero) foi, em média, 2,13 ± 0,5 %.
Observamos razão sexual 1:1 em 3/31 das colônias, o que evidencia a produção de macho
diplóide. Rainhas de M. seminigra são altamente poliândricas e o grau de parentesco
intracolonial é baixo; além do que, as diferentes subespécies encontram-se geograficamente
isoladas por longo tempo. Argumentamos que poliandria em M. seminigra é uma estratégia
evolutiva para minimizar os efeitos nocivos da produção de macho diplóide. O conhecimento
sobre o sistema de acasalamento de M. seminigra é essencial, no que diz respeito, ao manejo e
conservação desta espécie. No entanto, a implicação maior de nossos resultados refere-se à
evolução de poliandria em Hymenoptera sociais.

x
ABSTRACT
The stingless bee Melipona seminigra (Apidae, Meliponini) is a polytypic species widespread
in the Amazon basin. It has a key role in the pollination of native and cultivated plants as well
as in seed dispersal. It is highly eusocial as well as other Meliponini and Apini, characterized
for evolution of the caste system; division of reproductive labor; overlapping of generations,
queen mother and workers in the same nest; cooperative care of the offspring; queen (fertile
female) and workers (sterile females) morphologically differentiated. The mating system
predicted to stingless bees is monandria, common to most solitary and social Hymenoptera.
Our aim was to study paternity and relatedness in feral and managed colonies of M. seminigra
by using DNA microsatellite, and to identify molecularly the different subspecies by using
DNA barcoding. Moreover, we aimed to study sex determination mechanism through male
and female segregation. We sampled six subspecies, four of them described (M. s. abunensis,
M. s. merrillae, M. s. pernigra and M. s. seminigra) and two of them undescribed (M. s. ssp1
and M. s. ssp2). We genotyped 4-5 DNA microsatellite loci in 576 workers, sampled in 24
colonies, of three subspecies (M. s. abunensis, M. s. merrillae and M. s. seminigra). We
amplified a fragment of 526 base pairs of COI gene in 31 workers of the six subspecies,
sampled in different nests. We observed poliandry in the 24 colonies of M. seminigra studied.
Paternity frequency ranged from 3 to 11 (in average 7.416 ± 2.5) and effective paternity
ranging from 2 to 11.613(in average 6.324 ± 2.655). This level of polyandry produced a
degree of relatedness of 0.298 to 0.5 (in average 0.346 ± 0.051). In the most studied colonies
(21/24) skew (S) values were close to zero. In 3/24 colonies we observed a relative monopoly
of the paternity for some males (S ˃ 0.5) and in 6/24 of them we observed negative skew
interpreted as zero. The genetic distance inter subspecies was on average of 1.8 ± 0.47 %
while the genetic distance inter species (same subgenus) was on average of 2.13 ± 0.5 %. We
observed a 1:1 sex ratio in 3/31 of the colonies, which evidences diploid male production.
Queens of M. seminigra are highly polyandrous and intracolonial relatedness is low;
moreoever, the different subspecies are geographically isolated by a long time. We argue that
polyandry in M. seminigra is an evolutionary strategy to minimize the harmful effects of
diploid male production. The knowledge of M. seminigra mating system is essential as
regards to the management and conservation of this species. However, the greater implication
of our results refers to the evolution of polyandy in social Hymenoptera.

xi
Sumário
I. Introdução Geral......................................................................................................................1
I.1. Considerações sobre as abelhas................................................................................1
I.2. Sistema de acasalamento e determinação do sexo em Meliponini...........................3
I.3. Sobre Melipona seminigra........................................................................................6
I.4. Hipóteses.................................................................................................................10
I.5. Objetivos: geral e específicos.................................................................................11
II. Material e métodos, resultados e discussão…………......…..…………………….…...…..12
II.1. Capítulo 1. Produção de macho diplóide de duas abelhas Melipona (Hymenoptera:
Apidae) amazônicas. Psyche: Journal of Entomology, 1:1-7, 2012 ............................13
II.2. Capítulo 2. Nível incomum de poliandria em colônias selvagens e manejadas de
Melipona seminigra abunensis (Apidae: Meliponini)..................................................29
II.3. Capítulo 3. Paternidade e parentesco em colônias manejadas de Melipona seminigra
merrrillae (Apidae, Meliponini)……………………………………….……...…...…46
II.4. Capítulo 4. Análise de DNA microssatélites revela alta frequência de paternidade em
colônias manejadas da abelha sem ferrão Melipona seminigra seminigra...………....63
II. 5. Capítulo 5. DNA Barcoding do grupo Melipona seminigra revela que a distância
genética entre as subespécies é alta ……………………………..…………...…….....73
III. Conclusões Gerais...............................................................................................................85
IV. Perspectivas........................................................................................................................86
V. Referências...........................................................................................................................87
VI. Apêndice.............................................................................................................................93

xii
Lista de Figuras
I. Introdução
Figura 1: Diferença morfológica entre as castas de rainha e operária em Melipona
seminigra.....................................................................................................................................2
Figura 2: Melipona seminigra coletando pólen em flor de urucum (Bixa orellana). Corbícula
com pólen destacada por círculo em vermelho...........................................................................3
Figura 3: Distribuição das subespécies descritas para Melipona seminigra: Msa, M. s.
abunensis; Msm, M. s. merrillae; Msp, M. s. pernigra; Mss, M. s. seminigra (Camargo e
Pedro,
2013)...........................................................................................................................................7
Figura 4: Subespécies de Melipona seminigra: a, M. s. abunensis cf; b, M. s. merrillae cf; c,
M. s. pernigra cf; d, M. s. seminigra cf.......................................................................................7
Figura 5: Ninho de Melipona seminigra em tronco natural, coletado a 25 m de altura..............9
Figura 6: Entrada característica de ninho de Melipona (Michmelia): a, M. seminigra ssp2; b,
M. s. abunensis............................................................................................................................9
II.1. Capítulo 1
Figura 1: Melipona seminigra merrillae (a) mostrando a rainha, marcada no pronoto; e
operárias, mostrando a coloração característica do escutelo, nesta subespécie. Melipona
interrupta manaosensis (b) coloração característica de rainha e operária……...…………….19
Figura 2: Condições da colônia: (a) colônia em caixa padrão, com discos de crias grandes e
potes de pólen ao redor (b) colônia em tronco de árvore, com muitos discos de cria. A idade
do estágio de desenvolvimento aumenta de cima para baixo, evidenciado pela coloração mais
escura dos discos de cria em desenvolvimento inicial. Em (c) operárias de Melipona

xiii
seminigra merrillae alimentando-se em potes de mel; em (d) potes de provisão, pólen e mel,
da colônia em tronco de árvore, mostrada em (b), com muitas operárias alimentando-
se..………………………………………………………………….............................………19
Figura 3: Melipona interrupta manaosensis, comportamento das operárias em colônias com
razão sexual 1:1; operárias estão atacando um macho (a) e matando a rainha (b), na mesma
colônia, ao mesmo tempo……………..………………………………………………………21
II.5. Capítulo 5
Figura 1: Sítios de amostragem de Melipona seminigra, na bacia Amazônica. Estrêlas pretas,
M. s. abunensis cf (Porto Velho, RO); círculo branco, M. s. merrillae cf (GPA/INPA, Manaus,
AM); triângulo preto, M. s. ssp1 (Puraquequara, Manaus, AM); quadrado branco, M. s.
pernigra (Belterra, PA); círculo preto, M. s. seminigra cf (Boa Vista do Ramos e Maués,
AM); quadrado preto, M. s. ssp2 (Parque Nacional do Jaú, AM)…………………….………75
Figura 2: Análise de agrupamento das seis subespécies de Melipona seminigra (BAPS5: 100
réplicas, 200 referências individuais e 10 interações por indivíduo), mostrando seis grupos
bem delimitados: C1, M. s. abunensis; C2, M. s. merrillae; C3, M. s. ssp1; C4, M. s. pernigra;
C5, M. s. seminigra; C6, M. s. ssp2…………………...……………………………………...78
Figura 3: Árvore de similaridade, por Neighbor-Joining (NJ), para as subespécies de M.
seminigra e espécies de Melipona (Michmelia) (sequências baixadas do GenBank). Valores
de suporte (nos nós) foram obtidos por 1.000 réplicas de bootstrap. Como grupo externo foi
utilizado Melipona crinita (sequência baixada do GenBank). Barra, em negrito, 0,5 cm…....80

xiv
Lista de tabelas
II.1. Capítulo 1
Tabela 1: Segregação de sexos em Melipona seminigra merrillae (colônias MSM) e M.
interrupta manaosensis (colônias MIM)………………………………………………..…….20
Table 2: Variação da razão sexual em colônias de Melipona seminigra merrillae. Rainhas que
produziram machos diplóides (razão sexual 1:1 no primeiro disco de cria) não mantiveram
essa razão sexual no segundo e terceiro disco de cria ………………………..…..…….…….21
II.2. Capítulo 2
Tabela 1: Genótipos de rainhas (R) e machos (M) nas nove colônias, selvagens e manejadas,
de Melipona seminigra abunensis estudadas neste trabalho. Foram genotipadas 24 operárias
por colônia, em quatro locos microssatélites (Francini et al. 2009). Para análise dos dados
usamos o programa COLONY v. 2.0.4.3 (http://www.zsl.org/science/research-
projects/software) (Wang 2004)………………………..………..……………………………36
Tabela 2: Frequências alélicas dos locos microssatélites (MSM02, MSM09, MSM12 e
MSM13) (Francini et al. 2009) genotipados em 192 operárias de Melipona seminigra
abunensis, com a finalidade de avaliar paternidade e parentesco. Para o loco mais polimórfico
(MSM09) foram computados 16 alleles. O loco MSM13 foi monomórfico em quatro colônias
(1E, 1F, 1G e 1H). A frequência alélica foi obtida usando-se o programa COLONY v. 2.0.4.3
(Wang 2004). Col., colônia; N, número de operárias genotipadas…………....……………...37
Tabela 3: Paternidade e parentesco, em colônias selvagens e manejadas, de Melipona
seminigra abunensis, pela genotipagem de 216 operárias (24 por colônia) em quatro locos
microssatélites (MSM02, MSM09, MSM12 e MSM13) (Francini et al. 2009). N, tamanho da
amostra; kobs, paternidade observada; ^ke3, paternidade efetiva; S (skew), contribuição
paternal; r, coeficiente de parentesco; dp, erro de não detecção……….……………………..38

xv
II.3. Capítulo 3
Tabela 1: Genótipos de rainhas (R) e machos (M) nas oito colônias de Melipona seminigra
merrillae estudadas neste trabalho. Os genótipos parentais foram inferidos através dos
genótipos das operárias. Foram genotipadas 192 operárias (24/colônia), em cinco locos
microssatélites [51]. A frequência de paternidade foi obtida implementando 10 corridas no
programa COLONY v. 2.0.4.3 [54]. Um loco, MSM07 foi monomórfico em todas as colônias
e excluído das análises de paternidade...……………………….………….………...…….….52
Tabela 2: Frequências alélicas dos locos microssatélites genotipados nas 192 operárias de
Melipona seminigra merrillae. O número de alelos na população variou de 1 a 9 (em média
4,2 ± 2,7). Os locos MSM08 e MSM02 foram os mais polimórficos com 9 e 5 alelos,
respectivamente; o loco MSM07 foi monomórfico em todas as colônias. As frequências
alélicas foram atualizadas usando-se o programa COLONY v. 2.0.4.3 [54]……………...….53
Tabela 3: Frequência de paternidade e coeficiente de parentesco em colônias manejadas de
Melipona seminigra merrillae. A paternidade foi avaliada pela genotipagem de 192 pupas de
operárias, em cinco locos microssatélites (MSM02, MSM04, MSM07, MSM08 e MSM09)
[51]. A análise de paternidade foi feita usando-se o programa COLONY v. 2.0.4.3 [54]. N,
tamanho da amostra; kobs, paternidade observada; ke3, paternidade efetiva, calculada de acordo
com Nielsen et al. [56]; S, índice de skew , calculado segundo Pamilo e Crozier [59], ver
também Jaffé et al. [15]; r, coeficiente de parentesco, calculado conforme Pamilo [58]; dp,
erro de não detecção, calculado segundo Boomsma e Ratnieks [13]…………….………….54
Tabela 4: Erro de não amostragem (não detecção de linhagens paternas) nas oito colônias de
Melipona seminigra merrillae estudadas. Para linhagens paternas com 29,1% ou mais filhas, o
erro de não amostrgem foi zero (duas linhagens nas colônias 2Ba, 2Ea, 2Fa, 2Ha, 2Ka, 2La;
uma linhagem na colônia 2Pa). Para linhagens paternas representadas por 25%; 20,8%;
16,6%; 12,5%; 8,3% e 4,1% da paternidade, o erro de não amostragem foi, respectivamente,
de 0,001; 0,003; 0,012; 0,04; 0,124 e 0,366. Para linhagens paternas com 4,1% da paternidade
(duas linhagens em 2Ba; três linhagens em 2Ha), o erro de não amostragem foi muito alto. P,
linhagem paterna; pi, proporção de filhas; nse, erro de não amostragem…………..……..….55

xvi
II.4. Capítulo 4
Tabela 1: Paternidade de parentesco, em colônias manejadas de Melipona seminigra
seminigra, foram avaliadas pela genotipagem de quatro locos microssatélites (MSM02,
MSM09, MSM12 e MSM13) [25] em 168 operárias, amostradas em sete colônias. N,
tamanho da amostra; Kobs, paternidade observada; ^ke3, paternidade efetiva, calculada
conforme Nielsen et al. [31]; S, índice de skew, calculado de acordo com Pamilo e Crozier
[34] usando a equação descrita em Jaffé et al. [21]; r, coeficiente de parentesco, calculado
como descrito em Pamilo [33]; dp, erro de não detecção, calculado de acordo com Boomsma
e Ratnieks [6]. Para análise dos dados foi usado o programa COLONY v. 2.0.4.3
(http://www.zsl.org/science/research-projects/software)m [27]...............................................67
II.5. Capítulo 5
Tabela 1: Diversidade do COI, entre subespécies de Melipona seminigra e entre espécies de
Melipona (Michmelia). N, número de indivíduos sequenciados; S, número de sítios variáveis;
Hn, número de haplótipos observados; Hd, diversidade de haplótipos; Pi, diversidade de
nucleotídeos…………………………………………………………….……………………..78
Tabela 2: Distância genética entre seis subespécies de Melipona (Michmelia) seminigra (1-6)
e dez espécies de Melipona (subgênero Michmelia) (7-16). Abaixo a diagonal, estão descrito
os valores de divergência; acima o desvio padrão (SD). Em negrito, a distância genética entre
M. seminigra spp2 (amostrada no Parque Nacional do Jaú) M. seminigra atrofulva (sequência
baixada do GenBank), indicando ser a mesma subespécie………………………………...…79
VI. Apêndice: Sobre conclusões gerais
Tabela 1. Paternidade e parentesco em Melipona seminigra, estimativas feitas a partir da
genotipagem de 576 operárias. Foram usados 4-5 locos microssatélites, como descrito nos
capítulos II, III e IV. Foram amostradas 24 colônias, três subespécies: *, colônia manejada;
**, colônia selvagem. Os valores de skew (índice de contribuição de cada pai) evidenciam a
tendência a equalização da paternidade, na maioria das colônias.............................................93

1
I. Introdução Geral
I.1. Considerações sobre as abelhas
As abelhas pertencem ao maior grupo da ordem Hymenoptera, os Aculeata (formigas,
vespas e abelhas), insetos cujas fêmeas têm ferrão (modificação do ovipositor de grupos
ancestrais) (Michener, 2007). Constituem um grupo monofilético, isto é, têm ancestral
comum (Alexander, 1992; Melo, 1999). A monofilia das abelhas tem suporte em diversas
sinapomorfias do grupo, dentre as quais destacamos o abandono do comportamento carnívoro
ancestral, salvo exceções, usam pólen como fonte de proteína para a alimentação larval e de
adultos (Silveira et al., 2002; Michener, 2007). As abelhas atuais dependem das plantas com
flores, que lhes fornecem alimento (néctar, pólen, seiva), material para a construção dos
ninhos (resinas) e local para nidificação (principalmente ocos em troncos) (Roubik, 1995,
1989). Em troca tornaram-se os principais polinizadores da flora nativa nos ecossistemas
terrestres (Barth, 1985; Michener, 2007) e de plantas cultivadas (Heard, 1999).
Abelhas e outros insetos, em geral, exibem uma gama de comportamentos, que varia
do solitário ao eussocial avançado (Wilson, 1971). O termo eussocial (verdadeiramente
social) foi usado primeiramente por Batra (1966) para descrever o comportamento de
nidificação em abelhas (Halictinae). Michener (1969) formalizou o sistema social em abelhas
e considerou que três características básicas definem a eussocialidade nestas: (i) evolução de
castas e divisão de trabalho no ninho; (ii) sobreposição de gerações (mãe e prole adulta no
mesmo ninho); (iii) trabalho cooperativo nas células. Wilson (1971; 1975) estendeu a
terminologia de Michener para outros insetos sociais e formulou a definição de
eussocialidade, comumente usada na atualidade: comportamento pelo qual, alguns indivíduos
reduzem seu valor adaptativo, para cuidar da prole de seus parentes (Nowak et al., 2010).
Em abelhas, a eussocialidade evoluiu pelo menos onze vezes em linhagens de
Halictidae e Apidae, atingindo o ápice em três tribos de Apidae: Bombini, Apini e Meliponini
(Wilson, 1990). As abelhas das tribos Apini e Meliponini são as únicas que apresentam
comportamento eussocial avançado ou altamente eussocial (Crozier e Pamilo, 1996;
Michener, 2007). Isto significa que, em suas colônias observa-se cuidado contínuo e cuidado
cooperativo à prole, divisão de trabalho reprodutivo, sobreposição de pelo menos duas
gerações no mesmo ninho, diferenciação fisiológica e morfológica (Figura 1) da fêmea fértil
(rainha) em relação às fêmeas da casta trabalhadora (operárias), considerada estéril.

2
Figura 1: Diferença morfológica entre as castas de rainha e operária em Melipona seminigra.
A tribo Meliponini Lepeletier, tem distribuição tropical e subtropical em todo o
mundo, (http://www.nhm.ac.uk/research-curation/research/projects/bombus/pic_apini.html)
com a bacia Amazônica abrigando o maior número de subgêneros. Muitas espécies são
capazes de forragear e nidificar em terras dominadas pela atividade humana, mas, enquanto
grupo, os Meliponini estão relacionados ao ambiente florestado, onde se observa maior
abundância de indivíduos e riqueza de espécies (Brosi et al., 2008; Brosi, 2009), portanto,
dependem da manutenção das florestas. Têm papel relevante como polinizadores da flora
nativa dos trópicos (Michener, 2007) e de plantas cultivadas (Heard, 1999), além do potencial
para polinizar em casa de vegetação (Slaa et al., 2006).
Os Meliponini possuem o ferrão e estruturas associadas extremamente reduzidas,
razão pela qual foram denominadas “abelhas sem ferrão” (Kerr e Lello, 1962; Wille, 1983).
Duas outras sinapomorfias distinguem facilmente as abelhas da tribo Meliponini, redução e
fragilidade da venação das asas e a presença de penicillum (pincel de cerdas longas e rígidas,
localizado na margem apical externa da tíbia posterior) (Wille, 1983). A tribo Meliponini é
constituída por um total de 54 gêneros (33 são exclusivamente Neotropicais, sendo um
extinto) e 412 espécies (Camargo e Pedro, 2013). Os Meliponini, assim como, outros Apinae
1 1cm

3
(Apini, Bombini, Euglossini) (Michener, 2007) são abelhas corbiculadas, ou seja, armazenam
e transportam o pólen do campo para a colônia na corbícula (concavidade da tíbia posterior)
(Figura 2).
Figura 2: Melipona seminigra coletando pólen em flor de urucum (Bixa orellana). Corbícula com pólen,
destacada por círculo em vermelho.
O gênero Melipona Illiger tem distribuição exclusivamente Neotropical, ocorre do
México à Argentina, com maior diversificação na bacia amazônica (Michener, 2007). São
reconhecidas 74 espécies de Melipona (Camargo e Pedro, 2013), distribuídas em quatro
subgêneros: Melipona s. str.; Melikerria; Eomelipona e Michmelia (Moure, 1975; 1992;
Camargo e Pedro, 2013). As abelhas do gênero Melipona são de tamanho médio a grande (6 a
15 mm) (Silveira et al., 2002; Velthuis et al., 2003; Michener, 2007) e asas geralmente, não
ultrapassando o ápice do abdome.
I.2. Sistema de acasalamento e determinação do sexo em Meliponini
Colônias de Meliponini, assim como de outros Hymenoptera sociais, podem ser
administradas por uma rainha (monoginia) ou por duas ou mais rainhas (poliginia) (Kerr et
al., 1962; Kerr, 1969; Boomsma e Ratnieks, 1996; Strassmann, 2001). O predito para abelhas 11cm
1cm

4
e outras Hymenoptera (solitários e sociais) é a monoginia, considerada a condição ancestral
(Hughes et al. 2008). Casos de poliginia têm sido descritos para Meliponini, por exemplo,
Carvalho et al., (2011) observaram cinco rainhas em colônia de Melipona scutellaris e Alves
et al., (2011) observaram oito rainhas, por quatro meses, em uma colônia de Melipona
quadrifasciata.
Rainhas de Meliponini e de outros Hymenoptera (solitários e sociais) são comumente,
inseminadas por apenas um macho (monandria) (Kerr, 1969; Strassmann, 2001) enquanto a
inseminação da rainha por dois ou mais machos (poliandria) é considerada uma exceção. Mas,
poliandria foi bem documentada em três taxa altamente derivados de Hymenoptera sociais, os
gêneros Atta, Vespula e Apis (formiga, vespa e abelha, respectivamente) (Boomsma e
Ratnieks, 1996). Em abelhas, o gênero Apis evoluiu poliandria extrema (inseminação da
rainha por seis machos ou mais) (Boomsma e Ratnieks, 1996) com rainhas inseminadas por
até 100 machos (Wattanachaiyingcharoen et al., 2003).
De acordo com Hughes et al. (2008), monandria em Hymenoptera haplodiplóides é a
condição ancestral e a poliandria evoluiu como fenômeno secundário, após a eussocialidade
ter se tornado irreversível, isto é, após a evolução das castas.
Em Meliponini, a regra parece ser a monandria (Kerr et al., 1962; Peters et al., 1999;
Strassmann, 2001). Estudos prévios sobre paternidade e grau de parentesco em colônias de
Meliponini, usando marcadores moleculares microssatélites, observaram até seis machos
inseminando rainhas de Scaptotrigona postica (Paxton et al., 1999) e até três machos
inseminando rainhas de Melipona beecheii (Paxton et al., 2003), mas a paternidade efetiva
estimada foi de 1,4 e 1,1 respectivamente, o que não caracteriza poliandria (paternidade
efetiva ˃ 2). Dois outros estudos, Carvalho (2001) e Francini (2009) usando dados de
segregação, relatam inseminação de rainhas de Melipona scutellaris e Melipona seminigra
merrillae por dois e quatro machos, respectivamente. Mas, estes autores não estimaram a
paternidade efetiva nem grau de parentesco nas colônias estudadas. Portanto, podemos
considerar que a poliandria não foi descrita para Meliponini até o presente.
Em colônias monogínicas, se a rainha for inseminada por um único macho, suas filhas
(operárias) serão irmãs completas, compartilhando 75% dos seus genes por descendência
comum. Este elevado grau de parentesco é consequência do mecanismo haplodiploide de
determinação do sexo: as irmãs compartilham 100% dos genes do pai e 50% dos genes da
mãe. Cada operária recebe 1/2 de seus genes do pai e outra 1/2 da mãe, logo o coeficiente de
parentesco é dado por: r = (1 x 1/2) + (1/2 x 1/2) = 3/4 (Wilson, 1971). Este elevado grau de

5
parentesco foi considerado crucial para evolução da eussocialidade por seleção de parentesco
(Hamilton, 1964, 1970, 1972).
Mas, se a rainha for poliândrica, o grau de parentesco intracolonial diminui, por isto, a
poliandria foi interpretada como contrária às forças seletivas que mantém a eussocialidade.
Aparentemente, anula os benefícios da haplodiploidia, tornando-se uma evidência contrária ao
modelo de evolução da eussocialidade por seleção de parentesco (Hamilton, 1964; Wilson,
1971; Lin e Michener, 1972; Trivers e Hare, 1976). Adicionalmente, a poliandria aumenta o
custo da reprodução (maior gasto energético e maior risco de predação e contaminação)
(Keller e Reeve, 1994). Portanto, a evolução de poliandria em Hymenoptera eussociais, é uma
questão controversa. Atualmente, é bem aceito que o aumento da variabilidade genética, per
se vantajosa em muitas direções, é a explicação plausível (Getz et al.,1982; Crozier e Page,
1985; Hamilton et al., 1990; Schmid-Hempel, 1994; Page et al., 1995; Simmons, 2005;
Crozier e Fjerdingstad, 2001). Em espécies haplo-diplóides sob Determinação Complementar
do Sexo (CSD, Complementary Sex Determination) (Whiting, 1943), um sistema de
acasalamento poliândrico pode ter sido primordial contra os efeitos nocivos da produção de
machos diplóides (DMP, diploid male production) (Page, 1980; Page e Metcalf, 1982).
Em abelhas, assim como em outros Hymenoptera, o mecanismo de determinação do
sexo é a haplodiploidia do tipo partenogênese arrenótoca. Por este mecanismo, a fêmea fértil
(rainha) põe ovos não fertilizados, que se desenvolvem em indivíduos haplóides (n), que são
machos e ovos fertilizados que dão origem a indivíduos diplóides (2n), que são fêmeas
(Dzierzon, 1845; Crozier e Pamilo, 1996).
Desde o trabalho de Whiting (1943) o mecanismo CSD tem sido descrito para diversas
espécies haplo-diplóides. Nestes casos, a determinação do sexo dos indivíduos diplóides
depende da composição alélica do gene csd (complementary sex determiner) (Hasselmann et
al., 2008), em geral, altamente polimórfico. A homozigose do csd produz machos diplóides,
que são efetivamente estéreis (Mackensen, 1951; Hung et al., 1974; Adams et al., 1977; Kerr
1987; Crozier e Pamilo, 1996; Carvalho, 2001; Francini, 2009). Portanto, em colônias
monogínicas, se a rainha for inseminada por apenas um macho e compartilhar um alelo sexual
com o mesmo, a razão sexual de sua prole será 1:1, ou seja, 50% de todos os seus
descendentes serão machos diplóides, o que dizima a força operária (Carvalho, 2001).
Em Hymenoptera sociais sob CSD, a produção de machos diplóides (DMP, diploid
male production) varia de acordo com a espécie e com a população em estudo, podendo ser
extraordinariamente alta (Roubik et al.,1996). Em populações naturais, onde normalmente a
variabilidade genética é alta, a frequência de machos diplóides é inferior a 10% (Roubik,

6
1989). Populações com baixa variabilidade genética e/ou expostas ao endocruzamento
produzem machos diplóides com alta frequência (Kerr, 1987; Zayed, 2004). A produção de
machos diplóides pode representar um risco para a colônia e para população, dependendo do
tamanho da população e da frequência com que são produzidos (Kerr, 1987; Carvalho, 2001;
Zayed et al., 2003).
De acordo com estudos prévios, em abelhas, colônias monogínicas e monândricas, que
produzem machos diplóides, o esperado é que as operárias eliminem os irmãos diplóides e a
rainha mãe (Woyke, 1963a, 1963b; Kerr, 1987). Este comportamento não foi observado em
colônias de Melipona seminigra merrillae, que produziram macho diplóide (Francini et al.,
2012), ou seja, as operárias não foram vistas atacando os machos e a rainha não foi eliminada.
Nestas colônias, a razão sexual, observada nos diferentes discos de cria de uma mesma rainha,
variou de 1:1 (segregação característica da produção de machos diploides) a 0:1, indicando
que a rainha foi inseminada por mais e um macho. Portanto, as espécies que estão sob CSD
evoluíram diferentes estratégias para minimizar os efeitos nocivos da produção de machos
diplóides. Matar os machos diplóides e a rainha que os produz pode não ser a regra, em sendo,
pode haver exceções. Poliandria em espécies haplo-diplóides sob CSD pode ser uma
estratégia evolutiva para fazer frente aos efeitos nocivos da DMP.
I.3. Sobre Melipona seminigra
Melipona (Michmelia) seminigra Friese, 1903, conhecida como uruçu-boca-de-renda,
é uma espécie politípica com ampla distribuíção na bacia amazônica (Figura 3) (Schwarz,
1932; Moure e Kerr, 1950; Camargo e Pedro, 2013). Existe pelo menos sete subespécies,
quatro descritas (M. s. abunensis, M. s. merrillae, M. s. pernigra e M. s. seminigra) e três não
descritas (Camargo e Pedro, 2013). As subespécies descritas de M. seminigra têm distribuição
mutuamente exclusiva, havendo zonas de hibridação, por exemplo, entre M. s. merrillae e M.
s. seminigra e entre M. s. pernigra e M. s. seminigra. Para este trabalho amostramos seis
subespécies, as quatro descritas e duas outras que denominamos M. s. ssp1 (amostrada nas
proximidades do lago Puraquequara, Manaus, AM) e M. s. ssp2 (amostrada no Parque
Nacional do Jaú) (Figura 4).

7
Figura 3: Distribuição das subespécies descritas de Melipona seminigra: Msa, M. s. abunensis; Msm, M. s.
merrillae; Msp, M. s. pernigra; Mss, M. s. seminigra (Camargo e Pedro, 2013).
Figura 4: Subespécies de Melipona seminigra: a, M. s. abunensis cf; b, M. s. merrillae cf; c, M. s. pernigra cf;
d, M. s. seminigra cf; e, M. s. ssp1; f, M. s. ssp2.

8
M. seminigra nidifica em ocos de árvores (Figura 5), acima de quatro metros do solo
(Kerr et al. 1996), dependendo da localização do tronco, se em mata de terra firme ou
florestas alagáveis. Encontramos ninhos a cerca de 5 metros de altura em terra firme, em
Puraquequara, Manaus e ninhos a mais de 25m de altura, em florestas alagadas, no Parque
Nacional do Jaú, Amazonas, e no Rio Madeira, em Rondônia. Os seus ninhos foram
identificados pela entrada conhecida como “boca-de-renda” (Figura 6a, b).

9
Figura 5: Ninho de Melipona seminigra em tronco natural, coletado a 25 m de altura.
Figura 6: Entrada característica de ninho de Melipona (Michmelia): a, M. seminigra spp2; b, M. s. abunensis.
1111cm
a b
1cm 1111cm

10
M. seminigra tem importância como polinizador da flora nativa nos ecossistemas da
Amazônia e de plantas cultivadas (Absy e Kerr, 1977; Carvalho-Zilse, 2006) e na dispersão
de sementes (Bacelar-Lima et al., 2006; Nunez et al., 2008); é a principal espécie de abelha
sem ferrão criada na Amazônia Brasileira, como alternativa econômica (Cortopassi-Laurino et
al., 2006) e com o objetivo de preservação (Carvalho-Zilse e Nunes-Silva, 2012).
Estudamos o sistema de acasalamento em M. seminigra, usando DNA microssatélites;
fizemos a identificação molecular de seis subespécies, usando DNA barcoding e analisamos a
segregação de sexos, para verificar a produção de macho diplóide. Esse trabalho, justifica-se
por sua importância para o manejo e conservação desta espécie, principalmente, por suas
implicações, no que se refere a evolução de poliandria em Hymenoptera sociais, ao uso de
DNA barcoding para identificação de subespécies e a frequência macho diplóide no nível
colonial.
I.4. Hipóteses:
I.4.1. Sobre o sistema de acasalamento
H0: Rainhas de Melipona seminigra são monândricas
H1: Rainhas de Melipona seminigra são poliândricas
I.4.2. Sobre o mecanismo de determinação do sexo
H0: Melipona seminigra não está sob Determinação Complementar do Sexo
H1: Melipona seminigra está sob Determinação Complementar do Sexo
I.4.3. Sobre a identificação molecular
H0: DNA barcoding não delimita as subespécies de M. seminigra
H1: DNA barcoding delimita as subespécies de M. seminigra

11
I.5. Objetivos:
I.5.1. Geral
Estudar paternidade e parentesco no complexo de subespécies de Melipona
(Michmelia) seminigra Friese, 1903, em populações naturais e manejadas usando DNA
microssatélite; identificar molecularmente as diferentes subespécies, usando DNA barcoding;
estudar o mecanismo de determinação complementar do sexo, por análise de segregação.
I.5.2. Específicos
Determinar paternidade e parentesco no complexo de subespécies de M. seminigra
utilizando marcadores moleculares DNA microssatélites;
Estimar a paternidade efetiva e determinar o grau de parentesco intracolonial em M.
seminigra, nas populações estudadas;
Determinar o indíce de skew entre as linhagens paternas em cada colônia estudada e
sua relação com a paternidade efetiva e o grau de parentesco das filhas da rainha;
Delimitar as subespécies de M. seminigra usando DNA barcoding (COI);
Verificar a produção de macho diplóide em colônias de M. seminigra, com base na
razão sexual da prole da rainha.

12
II. Material e métodos, resultados e discussão
O material e os métodos utilizados para o estudo do sistema de acasalamento, análise
do mecanismo de determinação do sexo e identificação molecular das subespécies de
Melipona seminigra, assim como, os resultados observados e a discussão destes, são descritos
nos capítulos a seguir, como artigos científicos; Esses artigos estão escritos em português,
conforme regulamento do PPG do INPA, com as referências (texto e lista) formatadas de
acordo com o periódico onde foram publicados ou para o qual serão submetidos. Com relação
à paternidade e parentesco, optamos por manter a descrição por subespécies (Capítulos 2, 3 e
4), de acordo com muitos estudos realizados nas três últimas décadas, sobre paternidade e
parentesco em Apis mellifera.
Relação dos capítulos:
II.1. Capítulo 1: Produção de macho diplóide de duas abelhas Melipona (Hymenoptera:
Apidae) amazônicas. Artigo publicado no periódico Psyche: Journal of Entomology,
1:1-7, 2012.
II.2. Capítulo 2: Nível incomum de poliandria em colônias selvagens e manejadas de
Melipona seminigra abunensis (Apidae: Meliponini). Artigo para nova submissão à
Insectes Sociaux.
II.3. Capítulo 3: Paternidade e parentesco em colônias manejadas de Melipona seminigra
merrrillae, abelha sem ferrão produtora de mel. Artigo para submissão à ISRN
Entomology.
II.4. Capítulo 4: Análise de DNA microssatélites revela alta frequência de paternidade em
colônias manejadas da abelha sem ferrão amazônica Melipona seminigra seminigra.
Artigo para submissão ao Journal of Insects.
II. 5. Capítulo 5: DNA Barcoding do grupo Melipona seminigra revela que a distância
genética entre as subespécies é alta. Para nova submissão, no formato nota científica, à
Apidologie.

13
II.1. Capítulo 1: Produção de macho diplóide de duas abelhas Melipona
(Hymenoptera: Apidae) amazônicas.
Francini, I.B., Nunes Silva, C.G., Carvalho Zilse, G.A. 2012. Diploid male production of two
amazonian Melipona bees (Hymenoptera: Apidae). Psyche:Journal of Entomology, 1:1-7

14

15
II.1. Introdução
O mecanismo haplo-diplóide de determinação do sexo do tipo arrenotoquia, é uma
característica de insetos Hymenoptera (formigas, abelhas, vespas e sawflies) e é amplamente
distribuído em ordens de invertebrados. Evoluiu independentemnte pelo menos 17 vezes [1,
2]. Neste mecanismo de determinação do sexo, uma fêmea fértil (rainha) põe ovos fertilizados
e não fertilizados, que se desenvolvem em fêmeas diplóides e machos haplóides,
respectivamente [1, 2, 3, 4, 5, 6]. A diversidade de mecanismos de determinação do sexo que
os insetos evoluíram incluem heterogamia, haplodiploidia, perda de genoma paterno,
eliminação do cromossomo X e determinação complementar do sexo (CSD, complementary
sex determination) [5, 7, 8, 9]. Desde Whiting [9], machos diplóides têm sido descritos para
várias espécies arrenótocas [10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20]. Nestas espécies, a
produção de macho diplóide (DMP, diploid male production) depende da composição alélica
no gene csd (complementary sex determiner) [21, 22]. Sob CSD, animais que são hemizigotos
no loco csd tornam-se machos haplóides, enquanto os indivíduos diplóides podem se
desenvolver em fêmeas ou machos, quando eles são heterozigotos ou homozigotos,
respectivamente. A produção de descendentes diplóides numa razão sexual 1:1, ocorre entre
machos e fêmeas (rainha) que compartilham um alelo no loco csd [9, 16, 23, 24]. De acordo
com estudos prévios, quando machos diplóides são viáveis, eles são completamente estéreis.
Em muitas espécies, esses machos são assassinados pelas operárias na fase larval ou logo após
emergirem [10, 13, 25, 26, 27, 28, 29]. Se viáveis e férteis machos diplóides produzem
espermatozóides diplóides e leva a formação de fêmeas triplóides, o que poderia ser um “o
fim da linha” porque estas são estéreis [4, 30, 31]. O paradigm associado a carga genética da
DMP não foi confirmado em alguns vespídeos [6, 19, 32].
Em colônias de insetos sociais, o efeito negativo da DMP leva à perda de metade da
força operária por geração [16, 33, 34]. Portanto, estas espécies evoluíram alto polimorfismo
no loco csd para evitar o impacto de frequência elevada de machos diplóides estéreis [29].
Na vespa parasitóide Habrobracon hebetor (Braconidae) foram registrados 9-20 alelos
de determinação do sex [9, 35, 36]. Para Apis, Adams et al. [23] estimaram 18.9 alelos sexuais
e Tarpy et al. [37] relataram 8-27. Para abelhas sem ferrão, foram estimados 20 alelos sexuais
em Melipona compressipes fasciculata [13], 24 em Melipona scutellaris [16], 22 em
Melipona interrupta manaosensis (neste trabalho) e 16 em Melipona seminigra merrillae

16
(neste trabalho). Em populações nativas e introduzidas da formiga de fogo Solenopsis invicta,
Ross et al. [38] relataram 115-120 alelos de determinação do sexo.
Semelhante ao sistema de auto incompatibilidade em plantas, o alto polimorfismo do
gene csd é mantido por forte pressão de seleção [22, 23, 39, 40, 41, 42]. Se k é o número
efetivo de alelos sexuais em uma população panmítica, a probabilidade de um acasalamento
entre indíviduos que compartilham um alelo sexual idêntico é 2/k, e o número de indivíduos
diplóides machos, que é esperado, é igual a 1/k [23, 39, 40, 43]. Portanto, em populações
naturais, a frequência de machos diplóides, que é esperada, é baixa [40]; mas, havendo
endogamia, em geralmente, observa-se alterações.
Estudos moleculares em Apis mellifera mostraram a localização do gene csd, sinal
primário do desenvolvimento sexual, no cromossomo [44] e isolaram e identificaram esse
gene [24]. O gene csd ainda não foi mapeado ou isolado em abelhas sem ferrão, mas a
produção de machos diplóides tem sido documentada para algumas espécies de Melipona [11,
13, 16, 25].
Desde que o número de acasalamentos aumenta a variabilidade genética e, assim, o
número de alelos sexuais, as frequências de cruzamento das fêmeas são parâmetros
importantes em estudos de sistemas de acasalamento. As frequências de acasamentos das
rainhas em Hymenoptera solitários e sociais variam de exclusivamente monandria (rainha
acasala uma vez) [45, 46, 47] a poliandria extrema (rainha acasala mais de seis vezes) [45, 48,
49, 50, 51, 52].
A frequência de acasalamento em rainhas de abelhas também é variável. As abelhas
são em sua maioria solitárias, com rainhas acasalando uma vez, o que é suportado por estudos
químicos e ecológicos [53, 54]. Contudo, muitas abelhas solitárias acasalam multiplas vezes
[48]. Assim, são necessários mais estudos sobre o sistema de acasalamento das abelhas
solitárias [53]. Estudos sobre a frequência de acasalamento no gênero Bombus (bumble bees)
mostrou monandria para a maioria das espécies [55]. Nas abelhas sem ferrão estudadas até
agora, acasalamento único parece ser a regra [45, 56, 57]. Todavia, casos de acasalamento
com dois ou mais machos têm sido relatados [58, 16]. Apesar de rara em Hymenoptera
sociais, poliandria foi bem documentada em formigas (gênero Atta), vespas (genero Vespula)
e em abelhas eussociais avançadas (gênero Apis) [51]. O gênero Apis evoluiu para poliandria
extrema, com a frequência de acasalamento e a paternidade efetiva variável, entre espécies e
em alguns casos dentro da mesma espécie [37, 59, 60, 61]. Os níveis mais baixos de
poliandria foram registrados para Apis florea (rainhas acasalam com 5-14 machos [59]), e os

17
níveis mais elevados foram registrados para Apis dorsata (rainhas acasalam com 47-102
machos) [62].
Pesquisas sobre o mecanismo de determinação do sexo de M. s. merrillae e M. i.
manaosensis foram realizadas, e machos diplóides foram observados em ambas as espécies. A
diversidade genética foi calculada através da frequência de macho diplóide. O comportamento
das operárias em colônias produzindo machos diplóides (razão sexual 1:1) foi registrado
diariamente. O comportamento esperado para operárias de Melipona em colônias DMP, com
base em estudos prévios [11, 13, 16, 25], foi validado em M. i. manaosensis. Todavia, o
mesmo comportamento não foi observado para M. s. merrillae, de acordo com esse estudo.
II. 2. Material e Métodos
Foram produzidas trinta e uma colônias de M. s. merrillae (Figura 1a) e trinta de duas
colônias de M. i. manaosensis (Figura 1b). Isto foi realizado, pela reprodução de 63 colônias
em excelentes condições, do meliponário do Grupo de Pesquisas em Abelhas (GPA) do
Instituto Nacional de Pesquisas da Amazônia (INPA), Manaus, AM, Brasil, durante 2007 e
2008.
As colônias manejadas e reproduzidas estavam em boas condições quando
apresentavam grandes discos de cria em diferentes estágios de desenvolvimento, continham
provisões (potes de mel e potes de pólen circundando as células de cria) em grande
quantidade e possuiam uma população massiva de operárias adultas (Figura 2a-d) [63]. As
colônias originais, das quais as colônias adicionais foram derivadas, foram chamadas colônias
“mães”. As abelhas foram criadas em caixa padrão, o que facilitou a produção de colônia a
partir de uma colônia “mãe”. Como resultado desta produção, uma das colônias novas tornou-
se orfã (sem abelha rainha), levando alguns poucos dias, até uma nova rainha acasalar e se
estabelecer. As novas rainhas foram marcadas no pronoto com uma mancha branca de tinta
não tóxica (Figura 1a). Após a confirmação do acasalamento da nova rainha, identificado pelo
início da oviposição, a postura subsequente foi monitorada. Cada uma das 63 colônias nova
foi inspecionada e amostrada quarenta dias após o início da oviposição. Para verificar a razão
sexual, foram removidos das colônias fragmentos com 30 a 100 células do primeiro disco de
cria das novas rainhas e colocados em estufa, com temperatura controlada a 28 ºC, para
completar o desenvolvimento. Destes, foram amostrados, no estágio de pupa, 2.995
indivíduos de M. s. merrillae e 2.020 indivíduos de M. i. manaosensis. O número de pupas de
machos e de fêmeas foi quantificado para estimar a razão sexual.

18
A análise dos dados foi focalizada no acasalamento entre indíviduos que compartilham
alelos sexuais idênticos ou cruzamentos que produzem descendência com razão sexual 1:1. O
ajuste ao teste de χ2 foi feito usando o teste G de Willians [64]. O polimorfismo no loco de
determinação do sexo foi estimado pela equação de Laidlaw [65, 16] [n = 2M (N+1)/(H+1)],
onde n = número de alelos sexuais, N = número de colônias amostradas, H = número de
colônias que segregaram macho diplóide e M = número de machos que fertilizaram a rainha.
De acordo com a técnica descrita por Imai et al. [66, 67], em cada colônia com razão sexual
1:1, fizemos análise citogenética de 20-30 machos para confirmar a diploidia destes. Apenas
1-2% dos machos analisados citogenicamente não eram diplóides. Nestas colônias, o
comportamento das operárias foi observado diariamente e foto documentado. Nós também
amostramos aleatóriamente colônias com razão sexual diferente de 1:1 para fazer análise
citogenética dos machos.
II.3. Resultados
Os dados de segregação e da análise citogenética confirmaram a produção de macho
diplóide em ambas as espécies (Tabela 1). Três das trinta e uma colônias de M. s. merrillae
monitoradas apresentaram descendência com razão sexual 1:1, enquanto o mesmo foi
verificado em duas das trinta e duas colônias de M. i. manaosensis. Nestas colônias, a maioria
dos machos analisados citogeneticamente mostraram um número diplóide de 2n=18 em M. i.
manaosensis e 2n=22 em M. s. merrillae. Nós também observamos machos diplóides em
colônias com razão sexual diferente de 1:1. Assumindo monandria (rainha acasala uma vez),
que é o predito para rainhas de Meliponini [16, 63], estimamos 16 alelos de determinação do
sexo em M. s. merrillae e 22 em M. i. manaosensis.
Em M. i. manaosensis, o comportamento das operárias em colônias com razão sexual
1:1, que foi observado confirmou o que é previsto para o gênero Melipona [11, 13, 16]: as
operárias mataram os machos diplóides (Figura 3a) e a rainha mãe (Figura 3b) que os
produziu, assim que, os machos diplóides emergiram. Todavia, esse comportamento não foi
observado nas colônias de M. s. merrillae com razão sexual 1:1.
Para verificar se rainhas de M. s. merrillae mantinham a razão sexual em todos os
discos de cria, amostramos o segundo e o terceiro disco de crias em adição ao primeiro, em
colônias que tinham razão sexual 1:1 no primeiro disco. Observamos que em colônias de M. s.
merrillae com razão sexual 1:1 no primeiro disco, esta razão não foi mantida nos discos
subsequentes. Registramos um desvio nesta razão, no segundo e no terçeiro disco de crias,
tanto a favor das fêmeas quanto a favor dos machos (Tabela 2).

19
Figura 2: Condições da colônia: (a)
colônia em caixa padrão, com
discos de crias grandes e potes de
pólen ao redor (b) colônia em
tronco de árvore, com muitos
discos de cria. A idade do estágio
de desenvolvimento aumenta de
cima para baixo, evidenciado pela
coloração mais escura dos discos
de cria em desenvolvimento
inicial. Em (c) operárias de
Melipona seminigra merrillae
alimentando-se em potes de mel;
em (d) potes de provisão, pólen e
mel, da colônia em tronco de
árvore, mostrada em (b), com
muitas operárias alimentando-se.
Figura 1: Melipona seminigra
merrillae (a) mostrando a rainha,
marcada no pronoto; e operárias,
mostrando a coloração
característica do escutelo, nesta
subespécie. Melipona interrupta
manaosensis (b) coloração
característica de rainha e
operária.

20
Tabela 1: Segregação de sexos em Melipona seminigra merrillae (colônias MSM) e M. interrupta
manaosensis (colônias MIM)
Colônias
MSM ♂ ♀ Razão sexual Teste G
Colônias
MIM ♂ ♀ Razão sexual Teste G
01 06 89 0,06 S 01 00 74 0,00 S
02 00 74 0,00 S 02 00 36 0,00 S
03 07 55 0,11 S 03 00 63 0,00 S
04 09 78 0,10 S 04 00 31 0,00 S
05 00 60 0,00 S 05 00 53 0,00 S
06 00 63 0,00 S 06 00 69 0,00 S
07 14 55 0,20 S 07 00 81 0,00 S
08 00 69 0,00 S 08 08 32 0,20 S
09 06 82 0,07 S 09 05 78 0,06 S
10 13 21 0,38 S 10 39 56 0,41 NS
11 09 33 0,21 S 11 00 65 0,00 S
12 07 46 0,13 S 12 00 84 0,00 S
13 00 52 0,00 S 13 01 64 0,02 S
14 27 66 0,29 S 14 00 57 0,00 S
15 00 81 0,00 S 15 01 62 0,02 S
16 87 00 1,00 S 16 13 59 0,18 S
17 00 70 0,00 S 17 11 51 0,18 S
18 35 76 0,31 S 18 00 88 0,00 S
19 04 55 0,07 S 19 03 66 0,04 S
20 53 54 0,49 NS 20 00 78 0,00 S
21 03 80 0,04 S 21 00 46 0,00 S
22 16 43 0,27 S 22 03 57 0,05 S
23 16 84 0,16 S 23 00 53 0,00 S
24 86 13 0,87 S 24 00 58 0,00 S
25 03 56 0,05 S 25 00 64 0,00 S
26 22 61 0,26 S 26 00 51 0,00 S
27 39 54 0,42 NS 27 00 57 0,00 S
28 77 52 0,60 S 28 01 62 0,02 S
29 26 32 0,45 NS 29 00 48 0,00 S
30 3 97 0,03 S 30 00 63 0,00 S
31 12 48 0,20 S 31 00 79 0,00 S
32 21 29 0,42 NS
*Hipótese nula, razão sexual 1:1; teste G, valores críticos (G=3,841; DF=1; p=0,95 e α=0,05); S = significante;
NS = não significante; , colônias com razão sexual 1:1.

21
Figura 3: Melipona interrupta manaosensis, comportamento das operárias em colônias com razão sexual
1:1; operárias estão atacando um macho (a) e matando a rainha (b), na mesma colônia, ao mesmo tempo.
Tabela 2. Variação da razão sexual em colônias de Melipona seminigra merrillae. Rainhas que produziram
machos diplóides (razão sexual 1:1 no primeiro disco de cria) não mantiveram essa razão sexual no segundo
e terceiro discos de crias.
Colônia ♂ ♀ Razão sexual Trste G
20/D1 53 54 0,49 NS
20/D2 28 15 0,65 S
20/D3 19 99 0,16 S
27/D1 39 54 0,42 NS
27/D2 44 36 0,55 NS
27/D3 17 41 0,29 S
29/D1 26 32 0,45 NS
29/D2 33 65 0,34 S
29/D3 03 75 0,04 S
II. 4. Discussão
A frequência de macho diplóide para a mioria dos Hymenoptera estudados, é um
indicador da diversidade genética e de sua perda [18]. Este é um parâmetro que deve ser
destacado em abelhas sem ferrão, tanto no que se refere à questão da conservação, como para
o desenvolvimento da meliponicultura, como alternativa econômica [16, 42, 68]. A
viabilidade de machos diplóides foi descrita previamente para três espécies de Melipona. Em
todos os casos, as operárias mataram seus irmãos diplóides e a rainha mãe que os produziu
[11, 13, 16], conforme documentado neste trabalho para M. i. manaosensis (Figura 3). Não
*Hipétese nula, razão sexual 1:1; teste G, valores críticos (G=3,841; DF=1; p=0,95 and α=0,05); S = significante;
NS = não significante; D1, primeiro disco de cria; D2, segundo disco de cria; D3, terceiro disco de cria.

22
observamos as operárias de M. s. merrillae matando seus irmãos diplóides nem a rainha mãe,
em colônias que produziram machos diplóides. Portanto, em M. s. merrillae o comportamento
das operárias, em colônia que produzem macho diplóide, parece contradizer o que foi
observado previamente para Melipona [16].
Os dados indicaram que rainhas de M. s. merrillae acasalam com dois ou mais
machos. Adicionalmente, observamos machos diplóides em colônias com razão sexual
diferente de 1:1, corroborando os dados da tabela 1. A poliandria aumenta a diversidade
genética, o que é vantajoso em um sistema de determinação complementar do sexo [69]. Um
aumento da frequência de acasalamento leva a rainha a produzir macho diplóide em uma
frequência de 1/n da população, em condições de panmixia [70]. Portanto, poliandria pode
explicar a variação da razão sexual dos discos de cria da mesma rainha, observado aqui em M.
s. merrillae. Poliandria pode ser uma boa estratégia evoluída por M. s. merrillae para
minimizar o custo da DMP [11, 13, 16]. Este comportamento, é provavelmente único para
Melipona, todavia, mais estudos são necessários. Apesar das evidências observadas, o número
de alelos de determinação do sexo foi estimado sob o pressuposto que as rainhas são
monândricas, de acordo com a informação disponível para Melipona [63]. Considerando que
usamos a equação de Laidlaw, o número de alelos seria ainda maior havendo poliandria,
portanto, nossas estimativas são provavelmente baixas para M. s. merrillae.
Entre os problemas de conservação da flora nativa de abelhas, na America Latina,
existe a necessidade de informação básica em taxonomia, genética, ecologia, e biologia da
reprodução [70, 71]. Os resultados apresentados aqui representam uma contribuição ao
conhecimento básico necessário para manter a biodiversidade local associada com a
polinização por abelhas selvagens [68]. Este trabalho é um esforço para preencher a falta de
conhecimento indispensável para a conservação de abelhas nativas da região Neotropical,
especialmente da bacia Amazônica.
Agradecimentos
Os autores agradecem, ao Conselho Nacional de Desenvolvimento Científico e Tecnológico
CNPq), à Fundação de Amparo a Pesquisa do Estado do Amazonas (FAPEAM), à
Financiadora de Estudos e Projetos (FINEP) e ao Instituto Nacional de Pesquisas da
Amazônia (INPA) por subsídios financeiros.

23
Referências
[1] R. H. Crozier and P. Pamilo, Evolution of Social Insect Colonies Sex Alocation and Kin
Selection, Oxford, University Press Inc., New York, USA, 1996.
[2] B. K. Mable and S. P. Otto, “The evolution of life cycles with haploid and diploid
phases”, BioEssays, vol. 20, pp.453-462, 1998.
[3] J. Dzierzon: “On the development of bees”, Transl., Eichstadt, Bienenzeitung 1:113, 1845.
[4] R. H. Crozier, “Evolutionary Genetics of the Hymenoptera”, Annual Review of
Entomology, vol. 22, pp. 263-288, 1977.
[5] J. J. Bull, “An advantage for the evolution of male haploidy and systems with similar
genetic transmission” Heredity, vol. 43, no 3, pp. 361-381, 1979.
[6] J. K. Stahlhut and D. P. Cowan, “Single-locus complementary sex determination in the
inbreeding wasp Euodynerus foraminatus Saussure (Hymenoptera: Vespidae)”,
Heredity, vol. 92, pp. 189-196, 2004.
[7] J. J. Bull, Evolution of Sex Determining Mechanisms. Benjamin Cummings, Publishing
Company, Menlo Park, California, 1983.
[8] L. Sanchez and A. L. P. Perondini, “Sex determination in sciarid flies: a model for the
control of differential X-Chromosome elimination”, Journal of Theoretical Biology,
vol. 197, pp 247-259, 1999.
[9] P. W. Whiting, “Multiple alleles in complementary sex determination of Habrobracon”,
Genetics, vol. 28, pp 365–382, 1943.
[10] O Mackensen, “Viability and sex determination in the honey bee (Apis mellifera)”,
Genetics, vol. 36, pp. 500-509, 1951.
[11] C. A. Camargo, “Sex determination in bees. XI. Production of diploid males and sex
determination in Melipona quadrifasciata, Journal of Apicultural Research, vol. 18,
no 2, pp. 77–84, 1979.
[12] J.Woyke, Effect of sex allele homo-heterozygosity on honeybee colony populations and
on their honey production I. Favourable development condition and unrestricted
queens, Journal of Apicultural Research, vol. 19, no. 1, pp.51-63, 1979.
[13] W. E. Kerr “Sex determination in bees. XXI. Number of XO-heteroalleles in a natural
population of Melipona compressipes fasciculata, Insects Sociaux, vol. 34, pp. 274–
279, 1987.

24
[14] D. W. Roubik, L. A. Weigt, and M. A. Bonilla, “Population genetics, diploid males, and
limits to social evolution of euglossine bees”, Evolution, vol.50, no. 2, pp. 931-935,
1996.
[15] B. Polaczek, P. Neumann, B. Schricker, and R. F. A. Moritz, “A new, simple method for
rearing diploid drones in the honeybee (Apis mellifera L.)”, Apidologie, vol. 31, no. 4,
pp. 525-530, 2000.
[16] G. A. Carvalho “The Number of Sex Alleles (CSD) in a Bee Population and its Pratical
Importance (Hymenoptera: Apidae)”, Journal of Hymenoptera Research, vol. 10, no
1, pp. 10-15, 2001.
[17] A. Zayed and L. Packer, “High levels of diploid male production in a primitively
eusocial bee (Hymenoptera: Halictidae)”, Heredity, vol. 87, pp. 631-636, 2001.
[18] A. Zayed, D. W. Roubik, and P. Laurence, “Use of diploid male frequency data as an
indicator of pollinator decline”, Proceedings of Royal Society of London (S
uppl.):03SBL0243.S1-S4, 2003.
[19] A. E. Liebert, A. Sumana, and P. T. Starks, “Diploid males and their triploid offspring in
the paper wasp Polistes dominulus”, Biology Letters, vol. 1, no. 2, pp. 200-203, 2005.
[20] S. Armitage, J. Boomsma, and B. Baer, “Diploid male production in an leaf-cutting ant”,
Ecological Entomology, vol. 35, pp. 175-182, 2010.
[21] M. Beye, G. J. Hunt, R. E. Page, M. K. Fondrk, L. Grohmann, and R. F. A. Moritz
“Unusually high recombination rate detected in the sex locus region of the honey bee
(Apis mellifera)”, Genetics, vol. 153, pp. 1701-1708, 1999.
[22] M. Hasselmann, T. Gempe, M. Schiǿtt., C. G. Nunes-Silva, M. Otte, and M. Beye,
“Evidence for the evolutionary nascence of a novel sex determination pathway in
honeybees”, Nature, vol. 454 pp. 519-522, 2008.
[23] J. Adams, E. D. Rothman, W. E. Kerr, and Z. L. Paulino, “Estimation of the number of
sex alleles and queen matings from diploid male frequencies in a population of Apis
mellifera”, Genetics,vol. 86, pp. 583-596, 1977.
[24] M. Beye, M. Hasselmann, M. K. Fondrik, R. E. Page Jr., and S. W. Omholt, “The Gene
csd Is the Primary Signal for Sexual Development in the Honeybee and Encodes an
SR-Type Protein”, Cell, vol. 114, pp. 419-429, 2003.
[25] J. Woyke, “The hatchability of lethal eggs in a two allele fraternity of honeybee”,
Journal of Apicultural Research, vol. 1 pp. 6-13, 1962.
[26] J. Woyke, “What happens to diploid drone larvae in a honeybee colony”, Journal of
Apicultural Research, vol. 2, pp. 73–76, 1963.

25
[27] J. Woyke, “Rearing and viability of diploid drone larvae”, Journal of Apicultural
Research, vol. 2, pp. 77-84, 1963.
[28] R. Stouthamer, R. F. Luck, and J. H. Werren, “Genetics of sex determination and the
improvement of biological control using parasitoids”, Environmental Entomology, vol.
21, no.3, pp. 427-435, 1992.
[29] G. A. Carvalho, W. E. Kerr, and V. A. Nascimento, “Sex determination in bees. XXXIII.
Decrease of xo heteroalleles in a finite population of Melipona scutellaris (Apidae,
Meliponini)”, Brazilian Journal of Genetics, vol. 18, no. 1, pp. 13-16, 1995.
[30] T. Naito and H. Suzuki, “Sex Determination in the Sawfly, Athalia rosae ruficornis
(Hymenoptera): Occurrence of Triploid Males”, Journal of Heredity, vol. 82, pp. 101-
104, 1991.
[31] A. Zayed and L. Packer, “Complementary sex determination substantially increases
extinction proneness of haplodiploid populations, Proceedings of the National
Academy of Sciencies, vol. 102, no. 30 pp. 10742–10746, 2005.
[32] D. P. Cowan and J. K. Stahlhut, “Functionally reproductive diploid and haploid males in
an inbreeding hymenopteran with complementary sex determination” Proceedings of
National Academy of Sciences, vol. 28, no.101, pp. 10374–10379, 2004.
[33] K. G. Ross and D. J. C. Fletcher, “Diploid male production – a significant colony
mortality factor in the fire ant, Solenopsis invicta”, Behavioral Ecology and
Sociobiology, vol. 19, pp. 283-291, 1986.
[34] E. van Wilgenburg, G. Driessen, and L. W. Beukeboom, “Single locus complementary
sex determination in Hymenoptera: an “unintelligent” design? Frontiers in Zoology,
2006 - http://www.Frontiersinzoology.com/content/3/1/1 Acesso em 06/06/2007.
[35] G. E. Heimpel, M. F. Antolin, and M. R. Strand, “Diversity of sex-determining alleles in
Bracon hebetor”, Heredity, vol. 82, pp. 282-291, 1999.
[36] M. F. Antolin, P. J. Ode, G.E. Heimpel, R. B. O’Hara, and M.R. Strand, “Population
structure, mating system, and sex-determining allele diversity of the parasitoid wasp
Habrobracon hebetor”, Heredity, vol. 91, pp. 373-381, 2003.
[37] D. R. Tarpy, J. R. Caren, D. A. Delaney, D. Sammataro, J. Finley, G. M. Loper, and G.
DeGrandi-Hoffman, “Mating frequencies of Africanized honey bees in the south
western USA”, Journal of Apicultural Research and Bee World, vol. 49, pp. 302-310,
2010.

26
[38] K. G. Ross, E. L. Vargo, L. Kelller, and J. C. Trager, “Effect of a Founder Event on
Variation in the Genetic Sex-Determining System of the Fire Ant Solenopsis invicta”,
Genetics, vol. 135, pp.843-854, 1993.
[39] S. Yokoyama and M. Nei “Population dynamics of sex-determining alleles in honey bees
and self-incompatinility alleles in plants”, Genetics, vol. 91, pp. 609-626, 1979.
[40] R. E. Owen and L. Packer, “Estimation of the proportion of diploid males in populations
of Hymenoptera”, Heredity, vol. 72, pp. 219-227, 1994.
[41] D. Charlesworth, “Self-incompatibility: how to stay incompatible”, Current Biology,
vol.12, pp. 424-426, 2002.
[42] A. Zayed, “Bee genetics and conservation”, Apidologie, vol 40, pp. 237-262, 2009.
[43] J. M. Cook and R. H. Crozier, “Sex determination and population biology in the
Hymenoptera”, Trends in Ecology and Evolution, vol. 10 pp 281-286, 1995.
[44] M. Beye, R. F. A. Moritz, R. H. Crozier, and Y. C. Crozier, “ Mapping the Sex Locus of
the Honeybee (Apis mellifera)”, Naturwissenschaften, vol. 83, pp. 424-426, 1996.
[45] J. Strassmann, “The rarity of multiple mating by females in the Hymenoptera”, Insectes
Sociaux, vol. 48, pp. 01-13, 2001.
[46] K. A. Palmer and B. P. Oldroyd, “Evolution of multiple mating in the genus Apis”,
Apidologie, vol. 31, pp. 235-248, 2000.
[47] W. O. H. Hughes, B. P. Oldroyd, M. Beekmann, F. L. W. Ratnieks, “Ancestral
monogamy shows Kin Selection is key to evolution of eusociality”, Science, vol. 30,
pp. 1213-1216, 2008.
[48] W. E. Kerr, “Some aspects of the evolution of social bees (Apidae)”, Evolutionary
Biology, vol. 3, pp. 119-175, 1969.
[49] B. J. Cole, “Multiple Mating and the Evolution of Social Behavior in the Hymenoptera”,
Behavioral Ecology and Sociobiology, vol. 12, pp. 191-201, 1983.
[50] H. H. Laidlaw Jr. and R. E. Page Jr., “Polyandry in honey bees (Apis mellifera L.): sperm
utilization and intracolony genetic relationships”, Genetics, vol. 108, pp. 985-997,
1984.
[51] J. J. Boomsma and F. L. W. Ratnieks, “Paternity in eusocial Hymenoptera”,
Philosophical Transactions of the Royal Society B, vol. 351, pp. 947-975, 1996.
[52] B. P. Oldroyd, M. J. Clifton, S. Wongsiri, T. E. Rinderer, H. A. Sylvester, and R. H.
Crozier, “Polyandry in the genus Apis, particularly Apis andreniformis”, Behavioral
Ecology and Sociobiology, vol. 40, pp. 17-26, 1997.

27
[53] R. J. Paxton, “Male mating behavior and mating systems of bees: an overview”,
Apidologie, vol. 36, pp. 145-156, 2005.
[54] Y. Zimmermann, D. W. Roubik, J. J. G. Quezada-Euan, R. J. Paxton, and T. Eltz,
“Single mating in orchid bees (Euglossa, Apinae): implications for mate choice and
social evolution”, Insects Sociaux, vol. 56, pp. 241-249, 2009.
[55] R. Schmid-Hempel and P. Schmid-Hempel, “Female mating frequencies in Bombus spp.
From Central Europe”, Insectes Sociaux, vol. 47, pp.36-41, 2000.
[56] W. E. Kerr, R. Zucchi, J. T. Nakadaira, and J. E. Butolo, “Reproduction in the social bees
(Hymenoptera: Apidae)”, Journal of the New York Entomological Society, vol. 70, pp.
265-276, 1962.
[57] J. M. Peters, D.C. Queller, V. L. Imperatriz-Fonseca, D. W. Roubik, and J. E.
Strassmann, “Mate number, kin selection and social conflicts in stingless bees and
honeybees”, Proceedings of the Royal Society of London B, vol. 266, pp. 379-384,
1999.
[58] R. J. Paxton, “Not only single mating in stingless bees”, Naturwissenschaften, vol. 86,
pp. 143-146, 1999.
[59] B. P. Oldroyd, A. J. Smolenski, J-M. Cornuet, S. Wongsiri, A. Estoup, T. E. Rinderer, R.
H. Crozier, “Levels of polyandry and intracolonial genetic relationships in Apis
florea”, Behavioral Ecology and Sociobiology, vol 37, pp. 329-335, 1995.
[60] K. A. Palmer, B. P. P. Oldroyd, J. J. G. Quezada-Euán, R. J. Paxton, and W. de J. May-
Itza, “Paternity frequency and maternity of males in some stingless bee species”,
Molecular Ecology, vol. 11, pp. 2107-2113, 2002.
[61] R. F. A. Moritz, P. Kryger, G. Koeniger, N. Koeniger, A. Estoup, and S. Tinger, “High
degree of polyandry in Apis dorsata queens detected by DNA microsatellite
variability”, Behavioral Ecology and Sociobiology, vol. 37, pp. 357-363, 1995.
[62] W. Wattanachaiyingcharoen, B. P. Oldroyd, S. Wongsiri, K. Palmer, and J. Paar, “A
scientific note on the mating frequency of Apis dorsata”, Apidologie, vol. 34, pp. 85-
86, 2003.
[63] W. E. Kerr, G. A. Carvalho, and V. A. Nascimento, Uruçu bee: Biology, Handling and
Conservation, Foundation Acangaú, Paracatú, MG, Brazil, 1996 (in Portuguese).
[64] J. H. Zar, Biostatistical analysis, 4th edition, Prentice Hall, Upper Saddle River, NJ,
1999.

28
[65] H. H. Laidlaw, F. P. Gomes, and W. E. Kerr, “Estimation of the number of lethal alleles
in a panmitic population of Apis mellifera”, Genetics, vol. 41, no.2, pp. 179–188,
1956.
[66] H. T. Imai, R. W. Taylor, M. W. J. Crosland, and R. H. Crozier, “Modes of spontaneous
chromosomal mutation and karyotype evolution in ants with reference to the minimum
interaction hypothesis”, Japanese Journal of genetics, vol. 63, no. 2, pp. 159-185,
1988.
[67] I. B. Francini, M. C. Gross, C. G. Nunes-Silva, and G. A. Carvalho-Zilse “Cytogenetic
analysis of the Amazon stingless bee Melipona seminigra
merrillae reveals different chromosome number for the genus”, Scientia Agricola, vol.
68, no 5, pp. 592-593, 2011.
[68] M. Cortopassi-Laurino, V. L. Imperatriz-Fonseca, D. W. Roubik, A. Dollin, T. Heard, I.
Aguilar, G. C. Venturieri, C. Eardley, and P. Nogueira-Neto, “Global meliponiculture:
challenges and opportunities”, Apidologie, vol. 37, pp. 275-292, 2006.
[69] J. Loman, T. Madsen, and T. Hakansson,”Increased fitness from multiple mating, and
genetic heterogeneity: a model of a possible mechanism”, Oikos, vol. 52, pp.69-72,
1988.
[70] R. E.Page and R. A. Metcalf, “Multiple mating, sperm utilization and social evolution”,
American Naturalist, vol. 119, pp. 263-281, 1982.
[71] B. M. Freitas, V. L. Imperatriz-Fonseca, L. M. Medina, A. M. P. Kleinert, L. Galetto, G.
Nates-Parra, and J. J. G. Quezada-Euán,“Diversity, threats and conservation of native
bees in the Neotropics”, Apidologie, vol. 40, pp. 332-346, 2009.
[72] J. A. Foley et al. “Global Consequences of Land Use”, Science, vol. 309, pp.570-574,
2005.

29
II.2. Capítulo 2: Nível incomum de poliandria em colônias selvagens e
manejadas de Melipona seminigra abunensis (Apidae: Meliponini).
Francini, I.B.; Nunes-Silva, C.G.; Machado, A.S.C.; Batista, J.S.; Kuhn, G.C.S Carvalho-
Zilse, G.A. Unusual level of polyandry in feral and managed Melipona seminigra abunensis
colonies (Apidae: Meliponini). Insectes Sociaux (Artigo para ressumissão).
Abstract
Paternity and relatedness were studied in the Amazonian stingless bee, Melipona seminigra
abunensis, by genotyping four microsatellite loci in 216 workers from nine colonies, both
feral and managed, 24 individuals per colony. Sampling were carried out at Rondonia State,
Brazil; feral colonies sampled in areas under deforestation for hydroelectric power station and
managed colonies in meliponary, which was installed within a forested area. The observed
paternity frequency, Kobs, ranged from 6 to 11 (average 8.777 ± 1.715); the effective paternity,
^ke3, ranged from 5.779 to 11.613 (average 8.625 ± SD 1.910); the coefficient of relatedness
among nestmate workers, r, ranged from 0.293 to 0.336 (average 0.309 ± 0.014).The
measures of paternity skew, S, ranged from -0.163 to 0.203 (average 0.212 ± SD 0.1). Our
results evidence high paternity frequency in association with low paternity skew. This
maximizes colony genetic diversity and its benefits like the reduction of diploid male
production variation. We speculate about the key role of this unusual level of polyandry in
stingless bees, as an evolutionary strategy, in minimizing the dangerous effects of diploid
male production.
Keywords: paternity, high polyandry, relatedness, diploid male, paternity skew

30
II.2.1. Introdução
A história evolutiva das abelhas sem ferrão (Apidae, Meliponini) remonta há mais de
65 milhões de anos do presente (Michener e Grimaldi 1988; Engel 2000). Elas constituem um
grupo pantropical com proeminente abundância e riqueza de espécies nos Neotrópicos
(Michener 2007). Como grupo, as espécies de Meliponini estão relacionadas ao ambiente
florestado (Brosi 2009), desempenhando um papel fundamental na polinização da flora nativa
nas florestas tropicais (Roubik 1989; Michener 2007). Além do que, são polinizadores de
culturas (Heard 1999) e têm valor econômico como potenciais polinizadores em estufas (Slaa
et al. 2006). Abelhas sem ferrão são insetos eussociais avançados, os quais são definidos por:
evolução de castas, rainha e operária; divisão de trabalho reprodutivo; sobreposição de
gerações (mãe e prole adulta vivendo no mesmo ninho); cuidado cooperativo à prole; e
diferenciação morfológica das castas de rainha (fêmea fértil) e operárias (fêmeas estéreis)
(Michener 1969; Wilson 1971).
Rainhas de Meliponini geralmente acasalam com um único macho (monandria) (Kerr
1969; Strassmann 2001), assim como, ocorre na maioria dos outros Hymenoptera sociais
(Boomsma e Ratnieks 1996; Oldroyd et al. 1997; Oldroyd e Fewell 2007; Hughes et al. 2008).
Poliandria (acasalamentos múltiplos), definida por paternidade efetiva maior que 2, evoluiu
independentemente diversas vezes em differentes linhagens de Hymenoptera sociais
(formigas, abelhas e vespas) (Boomsma e Ratnieks 1996; Boomsma et al. 2009; Jaffé et al.
2012) e, é bem documentada nos gêneros Atta (formigas), Apis (abelhas) e Vespula (vespas)
(Boomsma e Ratnieks 1996). Rainhas de Apis são altamente poliândricas, sendo inseminadas
por até 100 machos (Wattanachaiyingcharoen et al. 2003). Em abelhas sem ferrão,
acasalamentos múltiplos pelas rainhas já foram relatado em estudos prévios (Carvalho 2001;
Strassmann 2001; Palmer et al. 2002; Paxton et al. 2003; Francini et al. 2012), no entanto, em
todos os casos, a paternidade efetiva foi baixa ( ˂ 2) e não caracterizou poliandria. Portanto,
neste cenário, pode-se dizer que poliandria não foi descrita para abelhas sem ferrão até o
presente.
À primeira vista, poliandria em Hymenoptera sociais parece ser um fenômeno
contrário às forças seletivas que mantêm a eussocialidade por seleção de parentesco
(Hamilton 1964, 1972), porque reduz o parentesco médio entre as operárias de um ninho
(Wilson 1971; Lin e Michener 1972; Trivers e Hare 1976). Como consequência do
mecanismo haplo-diplóide de determinação do sexo, em colônias dirigidas por uma rainha,

31
que acasalou com um macho, as operárias são irmãs completas e compartilham 75% de seus
genes por descendência comum. Irmãs completas compartilham 100% dos genes paternos e
50% dos genes maternos, e porque herdam 1/2 de seus genes da mãe e 1/2 do pai, o
coeficiente de parentesco é r = (1x 1/2) + (1/2 x 1/2) = 3/4 (Wilson 1971). O alto grau de
parentesco intracolonial tem sido considerado a pré-condição para evolução da eussocialidade
(Crozier e Pamilo 1996; Boomsma, 2007, 2009; Crozier 2008). Dadas as controversias,
poliandria em Hymenoptera sociais tem sido largamente discutida em pesquisas sobre seleção
de parentesco, por décadas (Page e Metcalf 1982; Boomsma e Ratnieks 1996; Boomsma
2009). Atualmente, a explicação mais plausível para a evolução de poliandria em
Hymenoptera sociais é o aumento da diversidade genética (Jaffé et al. 2012). Poliandria em
Hymenoptera sociais é, muito provavelmente, um fenômeno secundário, evoluiu após a
eussocialidade tornar-se irreversível devido à interdependência das castas (Hughes et al.
2008).
Melipona seminigra abunensis tem distribuição na Amazônia Brasileira (Estados do
Acre, Rondônia e Mato Grosso) e Bolívia (El Beni) (Schwarz 1932; Camargo e Pedro 2013),
em áreas impactadas por desmatamento frequente, por vezes, em uma escala grande e súbita
(por exemplo, desmatamento para a construção de hidrelétricas). O desmatamento representa
uma grande ameaça para esta e outras espécies de abelhas sem ferrão, que nidificam em ocos
de troncos e dependem de recursos fornecidos pelas plantas com flores (Roubik 1989;
Michener 2007; Carvalho-Zilse e Nunes-Silva, 2012). M. s. abunensis, assim como, outras
subespécies de M. seminigra é boa produtora de mel, razão pela qual, é cultivada na atividade
de meliponicultura (observação pessoal).
O sistema de acasalamento de M. s. abunensis foi pouco estudado até o presente,
assim como, o da maioria das espécies de abelhas sem ferrão, especialmente na Bacia
Amazônica, e marcadamente usando ferramentas moleculares. Com base em dados de
segregação de sexos descritos para a subespécie M. s. merrillae (Francini et al. 2012),
hipotetizamos que as rainhas de M. seminigra são poliândricas ao contrário do esperado para
abelhas sem ferrão (Strassmann 2001). O objetivo central deste trabalho foi verificar essa
hipótese para a subespécie M. s. abunensis, analisando paternidade e parentesco em colônias
selvagens e colônias manipuladas.

32
II.2.2. Material e Métodos
II.2.2.1. Amostragem
Foram amostradas um total de 216 operárias de Melipona seminigra abunensis de
nove colônias, manejadas (1A, 1B, 1C, 1D) e selvagens (1E, 1F, 1G, 1H e 1I) (Tabela 1). A
amostragem foi realizada em quatro diferentes populações, todas elas no Estado de Rondônia,
Brasil. Colônias selvagens; foram amostradas em ocos de troncos de árvores, três delas (1G,
1H e 1I) na área de desmatamento para a construção da usina hidréletrica de Sto. Antônio, no
Rio Madeira (margem esquerda) (coordenadas 8° 45' 48.1'' S 64° 05' 09.0'' W). Duas outras
(1E e 1F) foram amostradas próximo à barragem de Samuel no Rio Jamari (tributário do Rio
Madeira) em populações distantes 5 Km uma da outra (coordenadas 8° 55' 22.6'' S 63° 25'
17.4'' W e 8° 56' 55.4'' S 63° 23' 45.1'' W, respectivamente), também em áreas sob impacto de
desmatamento. As colônias manejadas foram amostradas no meliponário Sta. Marcelina (BR
364, Km 17, coordenadas 8° 47' 33.4'' S 63° 44' 21.4'' W), sendo originárias da área sob
desmatamento para contrução da hidrelétrica de Sto Antônio (coordenadas acima). Foram
coletadas vinte e quarto operárias, em cada entrada de ninho, fixadas em etanol a 96 % e
armazenadas em freezer a -20oC, para posterior extração de DNA.
II.2.2.2. Extração de DNA e genotipagem dos produtos de PCR
O DNA Genômico DNA foi extraído do tecido do tórax de cada operária, usando Kit
Promega, seguindo o protocolo do fabricante. A digestão do tecido foi feita em um mix
contendo 500 µL de lise de núcleo, 120 µL de EDTA 0,5M pH 8; adicionando-se 17,5 µL de
proteinase K a 10 mg/mL e incubado-se por uma noite à 55oC (com agitação suave). As PCRs
foram feitas em um volume total de 10 μL, usando-se: 10ng de DNA; Tampão10x com KCl;
primer forward 4μM marcado com cauda M13 (Schuelke 2000); primer reverse 4μM e
fluorecência (FAM, HEX ou NED). Foram amplificados quatro locos microssatélites
(MSM02, MSM09, MSM12, MSM13) (Francini et al. 2009) em 216 operárias (24 por
colônia). Os produtos de PCR foram multiplexados e analisados em sequenciador ABI 3130
xl (Applied Biosystems) usando-se ROX™ 500 como padrão de marcador de genotipagem. O
tamanho dos alelos microssatélites, em pares de bases (bp, base pairs), foi editado no
programa GENEMAPPER V. 4.0 (Applied Biosystems).

33
II.2.2.3. Análise dos dados
II.2.2.3.1. Paternidade e parentesco
Todas as colônias de M. s. abunensis estudadas neste trabalho eram monogínicas, o
que ocorre, geralmente, para a maioria das espécies de Melipona (Kerr 1969). Além disto, nós
assumimos, a priori, paternidade única (monandria) para M. s. abunensis, por ser o sistema de
acasalamento ancestral de Hymenoptera sociais (Hughes et al. 2008). Linhagens paternas,
genótipos parentais e frequência alélica, foram estimados implementando 10 corridas no
programa COLONY v. 2.0.4.3 (http://www.zsl.org/science/research-projects/software) (Wang
2004). Esse programa utiliza dados de genótipos multi-locos para implementar métodos
probabilísticos de genealogia completa e inferior, ao mesmo tempo, paternidade e parentesco
(Wang e Santure 2009). Adicionalmente, o COLONY implementa uma abordagem par a par,
e usando o mesmo modelo de erro de genotipagem, pode acomodar e estimar erros de
genotipagem (alelos dropouts e outros erros, tais como mutações) de cada indivíduo por loco.
Alelos dropouts resultam de um erro comum na genotipagem de microssatélites, em
organismos diplóides, por falha do processo de PCR (Pompanon et al. 2005). Esse erro
produz homozigotos espúrios ou perda de dados, porque uma ou ambas as cópias alélicas em
um loco não amplicam.
A paternidade efetiva (^ke3) foi calculada de acordo com Nielsen et al. (2003) usando
a equação a seguir (ver Tarpy et al. 2004):
Onde, N = tamanho da amostra; k = número observado de pais (famílias de irmãs completas);
pi = frequência relativa de filhas do iésimo pai.
O coeficiente de parentesco (r) dentro de cada colônia foi computado por Pamilo (1993) por:

34
II.2.2.3.2. Contribuição paternal (skew)
O skew ou partilha desigual dos benefícios derivados da vida de grupo, é medido,
geralmente, no contexto da criação cooperativa. Os índices de skew comparam entre grupos se
um ou poucos indivíduos reprodutivos produzem toda a prole, ou seja, calculam a
contribuição de cada pai com a prole de uma fêmea poliândrica. Calculamos os valores de
skew pelo número efetivo (S) (Pamilo e Crozier 1996), pela equação descrita em Jaffé et al.
(2012): S = (kobs - ^ke3) ∕ (kobs - 1). Este índice tem sido usado em estudos recentes sobre
insetos sociais. Comumente, os valores de skew variam de 0 a 1, mas, valores negativos
podem ser observados, e indicam que o tamanho da amostra é inadequado para calcular
paternidade efetiva e o índice skew, mas também podem ser interpretados como skew igual a
zero (Jaffé et al. 2012).
II.2.2.3.3. Erro de não detecção e erro de não amostragem
Erro de não detecção (dp) é a probabilidade de dois machos tendo alelos idênticos em
todos os locos não seja detectado, porque as operárias, por chance possuem genótipos
idênticos, apesar de serem filhas de pais diferentes. Esse tipo de erro depende dos locos
marcadores (número e polimorfismo) usados para a análise da amostra. Assumindo o
equilíbrio de Hardy-Weinberg, calculamos a probabilidade de filhas com paternidade dual de
acordo com Boomsma e Ratnieks (1996), usando a equação descrita em Paxton et al. (2003):
Onde: n = número de alelos; s= número de locos; qi = frequência do iésimo alelo de n alelos, no
loco jésimo de s locos.
O erro de não amostragem (pais não detectados) foi calculado como (1-p)n, onde p é a
proporção de filhas observadas em uma linhagem paterna e n é o número amostral (Boomsma
e Ratnieks 1996). De acordo com Oldroyd et al. (1995) a genotipagem de 24 operárias fornece
uma boa chance de amostrar todas as linhagens paternas. Para n = 24, usado neste trabalho, o
erro de não amostragem foi zero para linhagens paternas constituídas por 29,16% ou mais
filhas de uma rainha. Para linhagens paternas representadas por valores menores, o erro de
não amostragem aumenta com a diminuição da contribuição paterna, tornando-se máximo
(0,366) para linhagens representadas por 4,16% das filhas.

35
II.2.3. Resultados
Encontramos alto nível de poliandria e baixo nível de skew nas nove colônias
estudadas, que é o esperado para Hymenoptera sociais (Jaffé et al. 2012). O baixo parentesco
observado em colônias de M. s. abunensis é consistente com evolução de poliandria após o
estabelecimento da eussocialidade (Hughes et al. 2008).
Os quatro locos microssatélites (MSM02, MSM09, MSM12, MSM13) amplificados
nas 216 operárias de M. s. abunensis, de nove colônias, selvagens e manejadas, discriminaram
os genótipos parentais com sucesso na maioria dos casos (Tabela 1), exceto o loco MSM13
que foi monomórfico nas quatro colônias selvagens (1E, 1F, 1G e 1H). O número de alelos
por loco, variou de 4 a 16 (4 alelos nos locos MSM02 e MSM13; 5 alelos no loco MSM12 e
16 no loco MSM09); as frequências alélicas estão descritas na Tabela 2.
A frequência de paternidade observada (kobs) em M. s. abunensis, nas colônias
analisadas variou de 6 a 11 (média aritmética 8,777 ± 1,715). A paternidade efetiva (^ke3)
variou de 5,779 a 11,613 (média harmônica 8,625 ± SD 1,910). O parentesco observado (r)
foi em média 0,309 ± 0,014 (colônia 1A, r=0,303; colônia 1B, r=0,298; colônia 1C, r=0,293;
colônia 1D, r=0,313; colônia 1E, r=0,336; colônia 1F, r=0,329; colônia 1G, r=0,306; colônia
1H, r=0,300; colônia 1I, r=0,307). O número efetivo de skew (S) variou de -0,163 a 0,203 (S =
0, se todos os pais têm igual contribuição; S = 1, quando um único macho é pai de toda a
prole). O erro de não detecção variou de 0,080 a 0,359 (em média 0,213 ± SD 0,1) (Tabela 3).
Não foram observados erros de genotipagem, por exemplo, alelos dropouts e mutações. O
erro de não amostragem (não detecção de machos) variou de zero a 0,366 para linhagens
paternas representadas por 29,16% e 4,16% da paternidade, respectivamente. Do total de 79
linhagens paternas, 22 foram representadas por 4.16% da paternidade (uma linhagem/colônia
1A; duas linhagens/colônia 1B; quatro linhagens/colônia 1C; duas linhagens/colônia 1D; duas
linhagens /colônia 1F; cinco linhagens/colônia 1G; quatro linhagens/colônia 1H; duas
linhagens/colônia 1I).
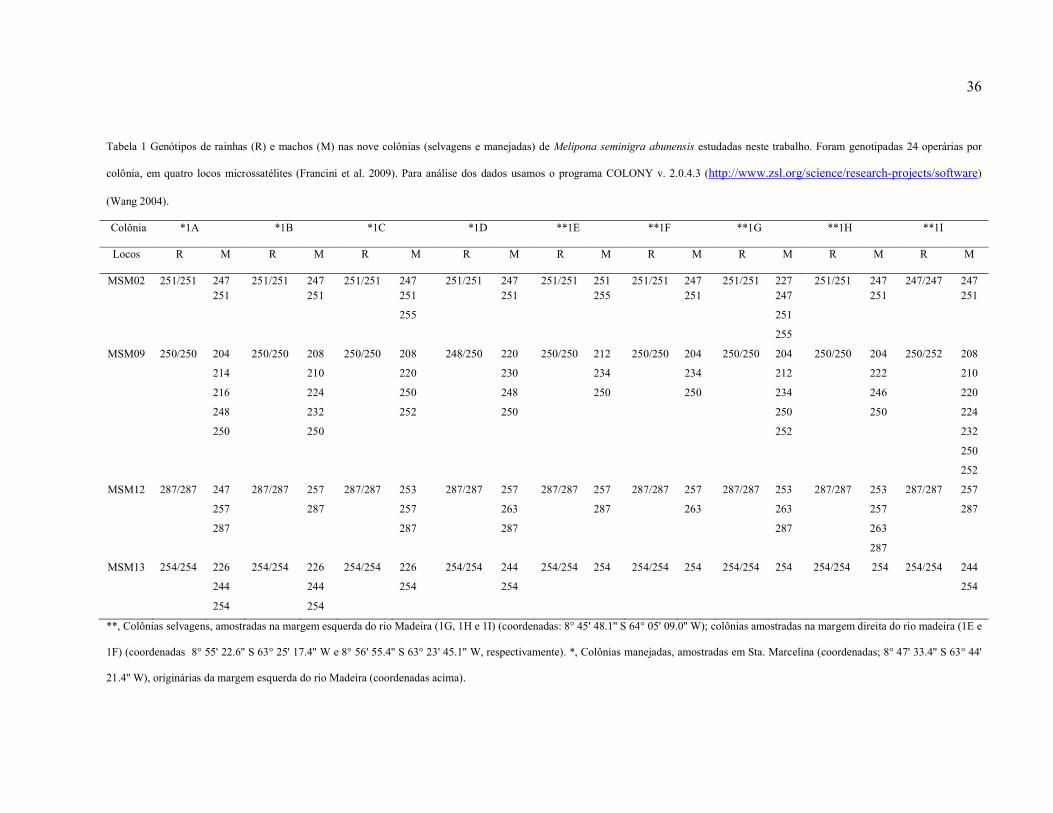
36
Tabela 1 Genótipos de rainhas (R) e machos (M) nas nove colônias (selvagens e manejadas) de Melipona seminigra abunensis estudadas neste trabalho. Foram genotipadas 24 operárias por
colônia, em quatro locos microssatélites (Francini et al. 2009). Para análise dos dados usamos o programa COLONY v. 2.0.4.3 (http://www.zsl.org/science/research-projects/software)
(Wang 2004).
Colônia *1A *1B *1C *1D **1E **1F **1G **1H **1I
Locos R M R M R M R M R M R M R M R M R M
MSM02 251/251 247 251/251 247 251/251 247 251/251 247 251/251 251 251/251 247 251/251 227 251/251 247 247/247 247
251 251 251 251 255 251 247 251 251
255 251
255
MSM09 250/250 204 250/250 208 250/250 208 248/250 220 250/250 212 250/250 204 250/250 204 250/250 204 250/252 208
214 210 220 230 234 234 212 222 210
216 224 250 248 250 250 234 246 220
248 232 252 250 250 250 224
250 250 252 232
250
252
MSM12 287/287 247 287/287 257 287/287 253 287/287 257 287/287 257 287/287 257 287/287 253 287/287 253 287/287 257
257 287 257 263 287 263 263 257 287
287 287 287 287 263
287
MSM13 254/254 226 254/254 226 254/254 226 254/254 244 254/254 254 254/254 254 254/254 254 254/254 254 254/254 244
244 244 254 254 254
254 254
**, Colônias selvagens, amostradas na margem esquerda do rio Madeira (1G, 1H e 1I) (coordenadas: 8° 45' 48.1'' S 64° 05' 09.0'' W); colônias amostradas na margem direita do rio madeira (1E e
1F) (coordenadas 8° 55' 22.6'' S 63° 25' 17.4'' W e 8° 56' 55.4'' S 63° 23' 45.1'' W, respectivamente). *, Colônias manejadas, amostradas em Sta. Marcelina (coordenadas; 8° 47' 33.4'' S 63° 44'
21.4'' W), originárias da margem esquerda do rio Madeira (coordenadas acima).
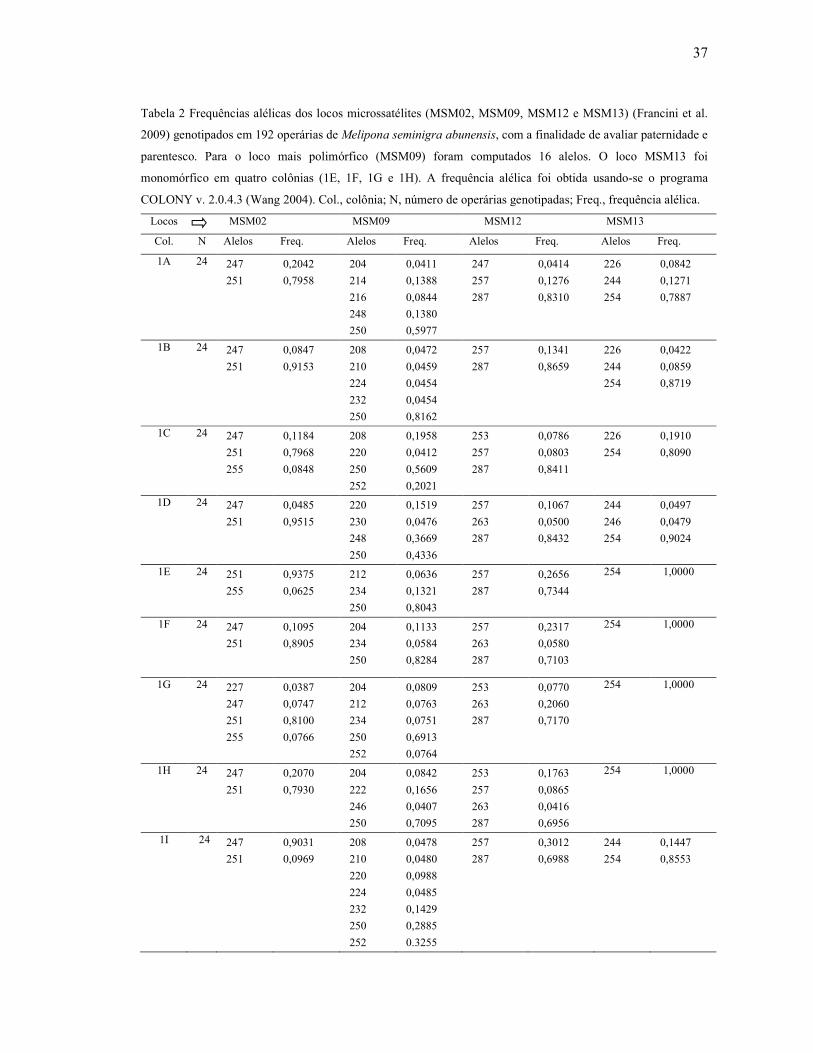
37
Tabela 2 Frequências alélicas dos locos microssatélites (MSM02, MSM09, MSM12 e MSM13) (Francini et al.
2009) genotipados em 192 operárias de Melipona seminigra abunensis, com a finalidade de avaliar paternidade e
parentesco. Para o loco mais polimórfico (MSM09) foram computados 16 alelos. O loco MSM13 foi
monomórfico em quatro colônias (1E, 1F, 1G e 1H). A frequência alélica foi obtida usando-se o programa
COLONY v. 2.0.4.3 (Wang 2004). Col., colônia; N, número de operárias genotipadas; Freq., frequência alélica.
Locos MSM02 MSM09 MSM12 MSM13
Col. N Alelos Freq. Alelos Freq. Alelos Freq. Alelos Freq.
1A 24 247
251
0,2042
0,7958
204
214
216
248
250
0,0411
0,1388
0,0844
0,1380
0,5977
247
257
287
0,0414
0,1276
0,8310
226
244
254
0,0842
0,1271
0,7887
1B 24 247
251
0,0847
0,9153
208
210
224
232
250
0,0472
0,0459
0,0454
0,0454
0,8162
257
287
0,1341
0,8659
226
244
254
0,0422
0,0859
0,8719
1C 24 247
251
255
0,1184
0,7968
0,0848
208
220
250
252
0,1958
0,0412
0,5609
0,2021
253
257
287
0,0786
0,0803
0,8411
226
254
0,1910
0,8090
1D 24 247
251
0,0485
0,9515
220
230
248
250
0,1519
0,0476
0,3669
0,4336
257
263
287
0,1067
0,0500
0,8432
244
246
254
0,0497
0,0479
0,9024
1E 24 251
255
0,9375
0,0625
212
234
250
0,0636
0,1321
0,8043
257
287
0,2656
0,7344
254 1,0000
1F 24 247
251
0,1095
0,8905
204
234
250
0,1133
0,0584
0,8284
257
263
287
0,2317
0,0580
0,7103
254 1,0000
1G 24 227
247
251
255
0,0387
0,0747
0,8100
0,0766
204
212
234
250
252
0,0809
0,0763
0,0751
0,6913
0,0764
253
263
287
0,0770
0,2060
0,7170
254 1,0000
1H 24 247
251
0,2070
0,7930
204
222
246
250
0,0842
0,1656
0,0407
0,7095
253
257
263
287
0,1763
0,0865
0,0416
0,6956
254 1,0000
1I 24 247
251
0,9031
0,0969
208
210
220
224
232
250
252
0,0478
0,0480
0,0988
0,0485
0,1429
0,2885
0.3255
257
287
0,3012
0,6988
244
254
0,1447
0,8553
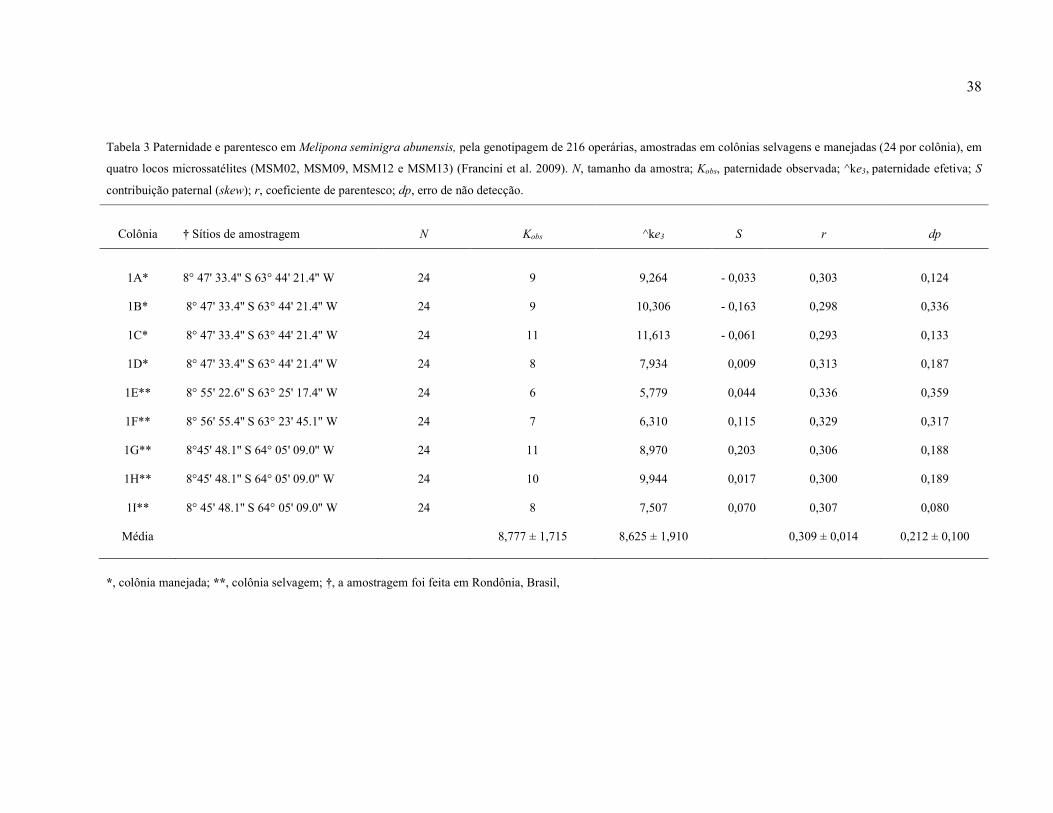
38
Tabela 3 Paternidade e parentesco em Melipona seminigra abunensis, pela genotipagem de 216 operárias, amostradas em colônias selvagens e manejadas (24 por colônia), em
quatro locos microssatélites (MSM02, MSM09, MSM12 e MSM13) (Francini et al. 2009). N, tamanho da amostra; Kobs, paternidade observada; ^ke3, paternidade efetiva; S
contribuição paternal (skew); r, coeficiente de parentesco; dp, erro de não detecção.
Colônia
† Sítios de amostragem
N
Kobs
^ke3
S
r
dp
1A* 8° 47' 33.4'' S 63° 44' 21.4'' W 24 9 9,264 - 0,033 0,303 0,124
1B* 8° 47' 33.4'' S 63° 44' 21.4'' W 24 9 10,306 - 0,163 0,298 0,336
1C* 8° 47' 33.4'' S 63° 44' 21.4'' W 24 11 11,613 - 0,061 0,293 0,133
1D* 8° 47' 33.4'' S 63° 44' 21.4'' W 24 8 7,934 0,009 0,313 0,187
1E** 8° 55' 22.6'' S 63° 25' 17.4'' W 24 6 5,779 0,044 0,336 0,359
1F** 8° 56' 55.4'' S 63° 23' 45.1'' W 24 7 6,310 0,115 0,329 0,317
1G** 8°45' 48.1'' S 64° 05' 09.0'' W 24 11 8,970 0,203 0,306 0,188
1H** 8°45' 48.1'' S 64° 05' 09.0'' W 24 10 9,944 0,017 0,300 0,189
1I** 8° 45' 48.1'' S 64° 05' 09.0'' W 24 8 7,507 0,070 0,307 0,080
Média 8,777 ± 1,715 8,625 ± 1,910 0,309 ± 0,014 0,212 ± 0,100
*, colônia manejada; **, colônia selvagem; †, a amostragem foi feita em Rondônia, Brasil,

39
II.2.4. Discussão
Estudos de paternidade em abelhas sem ferrão têm mostrado, geralmente, o esperado
em Hymenoptera sociais, monandria (Strassmann 2001; Oldroyd e Fewell 2007), que é
sistema de acasalamento ancestral (Boomsma 2007; Hughes et al. 2008). Estudos prévios
reportaram paternidade múltipla para abelhas sem ferrão, mas em todos os casos a paternidade
efetiva observada foi muito próxima de um (Strassmann 2001). Por outro lado, monandria em
abelhas sem ferrão, em princípio, estaria assegurada porque de acordo com estudos prévios,
os machos colocam um tampão pós-acasalamento (mating plug) nas fêmeas, o que,
teoricamente, evita um próximo acasalamento (Imperatriz-Fonseca e Zucchi, 1995;
Strassmann 2001; Boomsma e Ratnieks 1996). No entanto, observamos múltiplos
acasalamentos para rainhas de M. s. abunensis. De acordo com Campos e Melo (1990), o
reacasalamento da rainha fisogástrica pode ocorrer para abelhas sem ferrão, enquanto a
colônia está sendo manejada, mas, isto não foi observado nas colônias que estudamos.
Para as nove colônias de M. s. abunensis analisadas neste trabalho, foi observado um
alto nível de poliandria, o que contradiz o esperado para Hymenoptera sociais e, potanto, para
abelhas sem ferrão em geral (Kerr 1969; Strassmann 2001). Rainhas acasalaram pelo menos
6-11 vezes e a paternidade efetiva foi consistente com poliandria extrema (^ke3 ˃ 6) (Oldroyd
e Fewell 2007) para a maioria das colônias (8/9) (Tabela 3). Este nível de poliandria, reduziu
o grau de parentesco (r) para menos da metade do esperado nas colônias com uma única
rainha, que acasalou com um macho (r = 0,75). Nas colônias de M. s. abunensis estudadas
neste trabalho, as operárias compartilharam, em media, apenas 0,309 ± 0,143 de seus genes
por herança comum (Tabela 3). O baixo parentesco observado, teoricamente, reduz os ganhos
dos benefícios oriundos do fitness inclusivo, em colônias de insetos sociais (Hamilton 1964),
não suportando, assim, a evolução da eussocialidade por seleção de parentesco (Wilson 1971;
Lin e Michener 1972; Trivers e Hare 1976).
Por outro lado, com os baixos valores de skew de paternidade que foram observados
em colônias de M. s. abunensis (Tabela 3), espera-se maximizar os ganhos da diversidade
genética, que é o predito para Hymenoptera sociais com níveis altos de poliandria (Jaffé et al.
2012). Grupos de baixo parentesco foram provavelmente favorecidos por ganhos extras de
benefícios da diversidade genética (Nonacs e Kapheim 2007; Nonacs 2011). Assim sendo,
poliadria evoluiu, independentemente, diversas vezes em Hymenoptera sociais (Boomsma e
Ratnieks 1996; Boomsma et al. 2009).

40
O aumento da diversidade genética, oriunda da poliandria, reduz a variância genética
entre colônias, minimizando os efeitos negativos da produção de macho diplóide (DMP)
(Page 1980). Em colônias monogínicas e monândricas de insetos sociais, a DMP leva à perda
de metade da força operária por geração (Crozier e Pamilo 1996; Carvalho 2001; Wilgenburg
et al. 2006; Francini et al. 2012) porque machos diplóides (DM, diploid male) estéreis são
produzidos em uma razão sexual igual a 1:1. Poliandria leva as rainhas a produzirem macho
diplóide em uma frequência de 1/n da população, sob a condição de panmixia (Page 1980,
Page e Metcalf 1982, Crozier e Page 1985). Portanto, rainhas poliândricas ajustam a
freqüência de macho diplóide da colônia com a da população.
Machos diplóides foram descritos para uma subespécie de M. seminigra (Francini et al
2012) e observados para outras subespécies, incluindo M. s. abunensis (dados não
publicados). O predito para Melipona, é que as operárias matem os machos e a rainha mãe,
em colônias que produzem machos diplóides (Ratnieks 1990; Page e Kerr 1990), o que não
foi observado por Francini et al. (2012) para M. seminigra merrillae. Mas, esse
comportamento precisa ser estudado melhor.
Dado o número de colônias amostradas (9), o número de locos genotipados (quatro), o
polimorfismo dos locos (1-7 alelos/colônia) (Tabela 2) e o tamanho da amostra (24 operárias
por colônia), nossos resultados estão, muito provavelmente, subestimados. Como descrito
acima, os erros de não detecção e de não amostragem foram muito altos. Portanto, a
genotipagem de maior número de locos, de locos mais polimórficos e a utilização de um
número amostral maior, pode surpreender. Nossas estimativas de paternidade e parentesco
para M. s. abunensis são conservativas, ainda assim, são relevantes, representam fato novo em
abelhas sem ferrão.
Poliandria em M. s. abunensis é, provavelmente, um fenômeno secundário, evoluiu
após o estabelecimento da eussocialidade, que é o predito para outros Hymenoptera sociais
(Hughes et al. 2008). No entanto, outras características, além do sistema de acasalamento,
devem ser invocadas para explicar evolução da eusocialidade (Nowak et al. 2010).
Adicionalmente, as prováveis perdas oriundas do baixo grau de parentesco, entre as famílias
de irmãs incompletas, em colônias de Hymenoptera sociais, podem ser menores que os
ganhos advindos de maior diversidade genética. Isto pode ser especialmente verdadeiro para
M. s. abunensis, cujas rainhas são altamente poliândricas e tendem à equalizar a paternidade
(skew próximo de zero).

41
Especulamos, que o nível incomum de poliandria em abelhas sem ferrão observado
para M. s. abunensis, em associação com baixos valores de skew, é uma estratégia evolutiva-
chave para minimizar os efeitos nocivos da DMP.
Agradecimentos
Os autores agradecem ao Conselho Nacional de Desenvolvimento Científico e
Tecnológico (CNPq), à Financiadora de Estudos e Projetos (FINEP), e ao Instituto Nacional
de Pesquisas da Amazônia (INPA) por apoio financeiro. À Fundação de Amparo a Pesquisa
do Estado do Amazonas (FAPEAM) por fornecer bolsa de estudos para Antonio Saulo Cunha
Machado.

42
Referências
Boomsma JJ (2007) Kin selection versus sexual selection: why the ends do not meet. Curr
Biol 17:R673–R683
Boomsma JJ (2009) Lifetime Monogamy and the evolution of eusociality. Phil Trans R Soc
Lond B 364:3191-3207
Boomsma JJ, Ratnieks FLW (1996) Paternity in eusocial Hymenoptera. Phil Trans R Soc
Lond B 351:947-975
Boomsma JJ, Kronauer DJC, Pederson JS (2009) The evolution of social insect mating
systems. Pp.3-25 in Gadau J and Fewell J eds. Organization of insect societies–from
genome to sociocomplexity. Harvard Univ Press, Cambridge, Massachusetts
Brosi BJ (2009) The complex responses of social stingless bees (Apidae: Meliponini) to
tropical deforestation. For Ecol Manag. 258:1830-1837
Camargo JMF, Pedro SEM (2013) Meliponini Lepeletier, 1836. In: Moure JS, Urban D, Melo
GAR (Orgs). Catalogue of Bees (Hymenoptera, Apoidea) in the Neotropical Region.
Online version available at http://www.moure.cria.org.br/catalogue. Accessed 22 Jul
2013
Campos, LAO and GAR Melo (1990) Physogastric-queen mating in Melipona quadrifasciata
Lep. (Hymenoptera, Apidae) Rev Bras Genet 13:491-500
Carvalho GA (2001) The number of sex alleles (CSD) in a bee population and its practical
importance (Hymenoptera: Apidae). Jour Hym Res 10:10-15
Carvalho-Zilse, G.A. and Nunes-Silva, C.G. (2012). Threats to the Stingless Bees in the
Brazilian Amazon: How to Deal with Scarce Biological Data and an Increasing Rate
of Destruction. In: Florio, RM (Ed), Bees: Biology, Threats and Colonies. Nova
Science Publishers, Chapter 4:147-168
Crozier RH, Page RE (1985) On being the right size: male contributions and multiple mating
in social Hymenoptera. Behav Ecol Sociobiol 18:105-115
Crozier RH (2008) Advanced eusociality, kin selection and male haploidy. Aust J Entomol
47:2–8
Crozier RH, Pamilo P (1996) Evolution of social insect colonies, sex allocation and kin
selection. Oxford University Press

43
Engel MS (2000) A new interpretation of the Oldest Fossil bee (Hymenoptera: Apidae). Am
Mus Nov 3296:1-11
Francini IB, Sforça DA, Sousa ACB, Campos T, Zucchi MI, Souza AP, Nunes-Silva CG,
Carvalho-Zilse GA (2009) Microsatellite loci for an endemic stingless bee Melipona
seminigra merrillae (Apidae, Meliponini) from Amazon. Cons Gen Res1:487-49
Francini IB, Nunes-Silva CG, Carvalho-Zilse GA (2012) Diploid male production of two
amazonian Melipona bees (Hymenoptera: Apidae). Psyche: J Entomol 1:1-7
Hamilton WD (1964) The genetic evolution of social behavior. J Theor Biol 7:1-52
Hamilton WD (1972) Altruism and related phenomena, mainly in social insects. Ann Rev
Ecol Syst 3:193-232
Heard TA (1999) The role of stingless bees in crop pollination. Ann Rev Entomol 44:183–
206
Hughes WOH, Oldroyd BP, Beekmann M, Ratnieks FLW (2008) Ancestral monogamy shows
kin selection is key to evolution of eusociality. Science 30:1213-1216
Imperatriz-Fonseca VL, Zucchi R (1995) Virgin queens in stingless bee (Apidae,
Meliponinae) colonies: a review. Apidologie 26:231-244
Jaffe R, Garcia-Gonzalez F, den Boer SPA, Simmons LW (2012) Patterns of paternity skew
among polyandrous social insects: what can they tell us about the potential for sexual
selection? Evolution 66:3778-3788.
Jones OR;Wang J (2010) COLONY: a program for parentage and sibship inference from
multilocus genotype data. Mol Ecol Res 10:551-555
Kerr WE (1969) Some aspects of the evolution of the social bees. Evol Biol 3:119-175
Lin N, Michener CD (1972) Evolution of sociality in insects. Quart Rev Biol 47:131-159
Michener CD (1969) Comparative social behavior of bees. Ann Rev Entomol 14:299-342
Michener CD (2007) The Bees of the World. Johns Hopkins, University Press, Baltimore
Michener CD, Grimaldi DA (1988) The oldest fossil bee: Apoid history evolutionary stasis,
and antiquity of social behavior. Proc Natl Acad Sci 85:6424-26
Nielsen R, Tarpy DR, Reeve HK (2003) Estimating effective paternity number in social
insects and the effective number of alleles in a population. Mol Ecol 12:3157-3164.
Nonacs P, Kaphein KM (2007) Social heterosis and the maintenance of genetic diversity. J
Evol Biol 20:2253-2265
Nonacs P (2011) Monogamy and high relatedness do not preferentially favor the evolution of
cooperation. BMC Evol Biol 11:58
Nowak MA, Tarnita CE, Wilson EO (2010) The evolution of sociality. Nature, 466:10571062

44
Oldroyd BP, Fewell JH (2007) Genetic diversity promotes homeostasis in insect colonies.
Ecol and Evol 22: 408-412
Oldroyd BP, Smolenski AJ, Cornuet JM, Wongsiri S, Estoup A, Rinderer TE, Crozier RH
(1995) Levels of polyandry and intracolonial genetic relationships in Apis florea.
Behav Ecol Sociobiol 37:329-335
Oldroyd BP, Clifton MJ, Wongsiri S, Rinderer TE, Sylvester HA, Crozier RH (1997)
Polyandry in the genus Apis, particularly Apis andreniformis. Behav Ecol Sociobiol
40:17-26
Page RE Jr (1980) The evolution of multiple mating behavior by honey bee queens (Apis
mellifera). Genetics 96:263–273
Page RE Jr, Kerr WE (1990) The evolution of monandry and queen replacement in Melipona
(Hymenoptera: Apidae). Rev Bras de Gen 13: 209–229
Page RE, Metcalf RA (1982) Multiple mating, sperm utilization and social evolution. Am.
Nat.119:263-281
Palmer KA, Oldroyd BP, Quezada-Euán, Paxton RJ, May-Itza W (2002) Paternity frequency
and maternity of males in some stingless bee species. Mol. Ecol. 11:2107-2113
Pamilo P (1993) Polyandry and allele frequency differences between the sexes in the ant
Formica aquilonia. Heredity 70:472-480
Pamilo P, Crozier RH (1996) Reproductive skew simplified. Oikos 75:533-535.
Paxton RJ, Bego LR, Shah MM, Mateus S (2003) Low mating frequency of queens in
stingless bee Scaptotrigona postica and worker maternity of males. Behav Ecol
Sociobiol 53:174–181
Pompanon F, Bonin A, Bellemain E, Taberlet P (2005) Genotyping errors: causes,
consequences and solutions. Nat Rev Gen 6: 847-859.
Ratnieks FLW (1990) The evolution of polyandry by queens in social Hymenoptera: the
significance of the timing of removal of diploid males. Behav Ecol Sociobiol 26:343–
348
Roubik DW (1989) Ecology and Natural History of Tropical Bees. Cambridge, University
Press
Schuelke M (2000) An economic method for the fluorescent labeling of PCR fragments. Nat
Biotech 18: 233-234
Schwarz HF (1932) The genus Melipona. Bull Am Mus Nat Hist 63:231-480
Slaa EJ, Chaves LAS, Malgodi-Braga KS, Hofstede FE (2006) Stingless bees in applied
pollination: practice and perspectives. Apidologie 37:293-315

45
Strassmann J (2001) The rarity of multiple mating by females in the Hymenoptera. Insectes
Soc 48:01-13
Tarpy DR, Nielsen R, Nielsen DI (2004) A scientific note on the revised estimates of effective
paternity frequency in Apis. Insectes Soc 51:203-204
Trivers RL, Hare H (1976) Haplodiploidy and the evolution of social insects. Science
191:294-263
Wang J (2004) Sibship reconstruction from genetic data with typing errors. Genetics
166:1963-1979
Wang J (2012) Computationaly efficient sibship and parentage assignment from multilocus
marker data. Genetics 191:183-194
Wang J, Santure AW (2009) Parentage and sibship inference from multi-locus genotype data
under polygamy. Genetics 181:1579-1594
Wilgenburg van E, Driessen G, Beukeboom LW (2006) Single locus complementary sex
determination in Hymenoptera: an unintelligent design? Front Zool 3:1-15
Wilson EO (1971) The Insects Societies. Cambridge, Harvard University Press
Wattanachaiyingcharoen W, Oldroyd BP, Wongsiri S, Palmer K, Paar J (2003) A scientific
note on the mating frequency of Apis dorsata. Apidologie 34:85-86

46
II.3. Capítulo 3: Paternidade e parentesco em colônias manejadas de Melipona
seminigra merrillae (Apidae, Meliponini).
Francini, I.B.; Machado, A.S.C.; Batista, J.S.; Zucchi, M.I.; Nunes-Silva, C.G; Carvalho-
Zilse, G.A. Paternity and relatedness in managed colonies of Melipona seminigra merrillae
(Apidae, Meliponini). ISRN Entomology (submissão a ser feita).
Abstract
Mating system in Melipona seminigra merrillae managed colonies was investigated by using
five microsatellite DNA loci. Genotypes were obtained for 192 workers from eight colonies
sampled at Manaus, Amazonas, Brazil. Paternity frequency (kobs) ranged from 3 to 7 (on
average 4.5 ± 1.309); the effective paternity number (^ke3) ranged from 2.496 to 6.938 (on
average 4.161 ± 1.376); the relatedness coefficient (r) ranged from 0.322 to 0.450 (on average
0.380 ± 0.039); paternity skew (S) in 6/8 colonies was close to zero. Polyandry lowers genetic
relatedness among worker patrilines (r ˂ 0.5), thus diminishing within-colony conflict and
worker inclusive fitness while it increases the queen’ fitness. The negative correlation
between polyandry and paternity skew herein observed for M. s. merrillae, is expected to
maximize genetic diversity benefits. We suggest that polyandrous queens of M. s. merrillae
reduce the proportion of diploid males (DMP) within their diploid offspring and ensure the
benefit of worker force. Within-colony DMP minimization may be particularly advantageous
for managed populations. Polyandry in stingless bee is a novelty and has implication refers to
the current understanding of the evolution of sociality in social Hymenoptera.
Keywords: parentage, polyandry, relatedness, microsatellite, Meliponini.

47
II.3.1. Introdução
Abelhas sem ferrão (Meliponini) têm distribuição nas florestas tropicais e subtropicais
em todo o mundo [1] e uma maior diversidade na bacia amazônica. São reconhecidos 54
gêneros e 412 espécies de abelha sem ferrão [2]. Abelhas da tribo Meliponini desempenham
um papel essencial na polinização da flora nativa [1,3] e de plantas cultivadas [4]. Algumas
espécies vivem em ambiente com distúrbio antropogênico [5], mas, a maioria depende das
florestas. Abelhas sem ferrão, principalmente espécies do gênero Melipona, foram
domesticadas há muito tempo, por povos nativos da América Central e do Sul, para obtenção
de mel [6]. Melipona, gênero com distribuição exclusivamente Neotropical (do México à
Argentina), tem o maior número de espécies [2].
Colônias de abelhas sem ferrão são, geralmente, monogínicas (uma rainha por colônia)
[7, 8], mas colônias com mais de uma rainha (poliginia) foram observadas [9, 10]. Quanto ao
sistema de acasalamento, monandria (rainha inseminada por um macho) parece ser a regra em
Meliponini [8], é o comum em Hymenoptera solitários e sociais [8, 11, 12]. No entanto,
poliandria evoluiu, independente, várias vezes, em diferentes linhagens de Hymenoptera
sociais [13, 14, 15]. Em abelhas, o gênero Apis evoluiu poliandria extrema, com a menor
freqüência de paternidade (rainha inseminada por cinco machos) descrita para Apis florea [16]
e a maior freqüência de paternidade registrada para Apis dorsata (cerca de 100 machos
inseminam as rainhas) [17]. Para abelhas sem ferrão, alguns estudos anteriores relataram
múltiplos acasalamentos, mas em todos os casos a paternidade efetiva observada foi próxima
de um, o que não caracteriza poliandria [8], exceto para M. seminigra abunensis [dados não
publicados].
O alto grau de parentesco das operárias (r = 0,75), em colônias monogínicas e
monândricas de insetos sociais, é considerado essencial para evolução da eussocialidade por
seleção de parentesco [18, 19, 20]. Neste sentido, a monandria é primordial, ou seja, é pré-
condição para a eussocialidade [21, 22]. O comportamento de ajuda, em princípio, é
favorecido pela seleção natural, quando o parentesco é maior do que a relação custo/benefício
(r > c/b) [18]. Se as irmãs compartilham 75% de seus genes por descendência comum, então,
cuidar da prole de um parente ou de sua própria prole, é igualmente valioso [23].
Em insetos eussociais, rainhas poliândricas reduzem o parentesco intracolonial [14], o
que, à primeira vista, contradiz o modelo evolução do comportamento social por seleção de
parentesco [18, 19, 20]. Atualmente, considera-se que a diversidade genética é a explicação

48
mais plausível para evolução de poliandria em Hymenoptera sociais [14, 15, 25, 26, 27, 28,
29, 30, 31, 32, 33, 34, 35].
Em Hymenoptera, solitários e sociais, a haplodiploidia é o mecanismo de
determinação do sexo prevalente [11], ou seja, fêmeas férteis (rainhas) põem ovos fertilizados
e não fertilizados, que se desenvolvem em fêmeas (indivíduos diplóides) e machos
(indivíduos haplóides), respectivamente. Mas, desde Whiting [36], o mecanismo de
Determinação Complementar do Sexo (CSD, Complementary Sex Determination) tem sido
descrito para muitas espécies de haplo-diplóides [37, 38, 39, 40]. Sob CSD, os diplóides
heterozigotos para o gene csd (csd, complementary sex determiner) são fêmeas e os diplóides
homozigotos são machos [39], que são anômalos e estéreis [11]. O gene csd é altamente
polimórfico (por exemplo: csd1, csd2, csd3,...,csdn), o que minimiza a chance de homozigose e,
assim, de produção de machos diplóides (DM, diploid males). A homozigose do csd ocorre
quando a rainha compartilha um alelo sexual, com pelo menos um de seus parceiros sexuais
(por exemplo, a rainha é csd1csd2 e o macho é csd1 ou csd2). Se a rainha é monândrica e
compartilha um alelo csd com o seu parceiro sexual, então 50% de todos os seus descendentes
serão machos diplóides [11], o que dizima a força operária e pode causar a morte da colônia
[37, 38, 40]. Nestes casos, espera-se que as operárias matem os machos diplóides e também a
rainha mãe que os produziu [40, 41, 42, 43, 40].
M. s. merrillae tem sua distribuição limitada ao estado do Amazonas, Brasil [2],
ocorrendo de Manaus para o Norte, seguindo o Rio Negro, e ao longo do Rio Branco. É um
polinizador essencial da flora nativa em ecossistemas da Amazônia Central e de plantas
cultivadas [44], além de ser um dispersor de sementes [45, 46]. É cultivada pelos povos do
Amazonas para extração de mel e como medida de conservação [47]. Com base no observado
em estudo prévio [40] e no descrito para M. s. abunensis [dados não publicados], formulamos
a hipótese que o sistema de acasalamento de M. s. merrillae é poliandria. Nós testamos a
hipótese de poliandria, em população manejada, de M. s. merrillae, usando marcadores
moleculares microssatélites para analisar paternidade e parentesco.
II.3.2. Material e Métodos
II.3.2.1. Material biológico
Para esse trabalho foram amostradas 192 pupas de operárias de M. s. merrillae, em
oito colônias manejadas, 24 indivíduos por colônia. Optamos por amostrar as operárias no
estágio de pupa como recomendado por Palmer et al. [49] e, assim, eliminando a possibilidade

49
de deriva das operárias entre colônias. A amostragem foi feita no meliponário do Instituto
Nacional de Pesquisas da Amazônia (INPA), Manaus, AM, Brail. Nas colônias estudadas, as
rainhas foram marcadas logo após iniciarem a primeira postura, assegurando que não foram
substituídas até a data da amostragem (40-45 dias após início da postura). As colônias foram
coberta com um vidro para permitir observação frequente sem serem abertas, desta forma,
eliminando a possibilidade de reacasalamento da rainha fisogástrica, relatado para outras
espécies [48]. As vinte e quarto pupas de operárias foram amostradas aleatoriamente, no
primeiro disco de cria, fixadas em etanol 96 %, estocadas em freezer a -20oC para extração de
DNA, posteriormente.
II.3.2.2. Análises de DNA
O DNA genômico foi extraído de tecido do tórax usando-se kit Promega, seguindo o
protocolo do fabricante. As reações de polimerase em cadeia (PCRs) foram feitas em um
volume total de 10 μL, usando: 10 ng de DNA, Tampão 10x com KCl; primer forward 4µM
marcado com cauda M13 [50], primer reverse 4µM e fluorescência 4µM (FAM, HEX ou
NED). As 192 operárias de M. s. merrillae foram genotipadas em cinco locos microssatélites
(MSM02, MSM04, MSM07, MSM08 e MSM09) [51]. Os produtos de PCRs foram
multiplexados, levando-se em conta o tamanho dos locos microssatélites e a fluorescência
usada. A genotipagem foi feita em sequenciador ABI 3130 xl (Applied Biosystems), usando-se
ROX™ 500 como marcador de genotipagem. O tamanho dos alelos foi determinado em pares
de bases (bp, base pairs) e editado usando-se o programa GENEMAPPER v. 4.0 (Applied
Biosystems).
II.3.2.3. Análise de dados
II.3.2.3.1. Paternidade e parentesco
Os genótipos dos pais foram inferidos a partir dos genótipos das operárias. Os alelos
maternos, em cada loco, são os alelos comuns a todas as operárias ou um dos alelos mais
frequentes, para rainhas homozigotas e heterozigotas, respectivamente [52]. A probabilidade
de deriva de operárias entre as colônias foi eliminada, uma vez que foram amostradas no
estágio de pupa. A frequência de paternidade (número de famílias de irmãs completas), os
genótipos parentais, as frequências alélicas e erros de genotipagem (alelos dropouts e outros
erros, por exemplo, mutações) [53] foram estimados implementando 10 corridas no programa
COLONY v. 2.0.4.3 [54, 55] disponível em http://www.zsl.org/science/research-
projects/software.

50
Assumindo que cada macho contribuiu igualmente com a paternidade, a paternidade
efetiva (^ke3) foi calculada por Nielsen et al. [56], usando a equação descrita em Tarpy et al.
[57]:
O coeficiente de parentesco (r) foi calculado de acordo com Pamilo [58]:
Onde, N = operárias amostradas por colônia; k = número de pais observados; pi = frequência
relativa de operárias filhas do iésimo pai.
II.3.2.3.2. Contribuição paternal (skew)
Os indices de skew, ou seja, o quanto cada pai contribui com a prole da rainha, foram
calculado de acordo com o proposto por Pamilo e Crozier [59], usando a equação descrita em
Jaffé et al. [15]: S = (Kobs - ^ke3) ∕ (Kobs - 1). Os valores de skew são medidos de zero a um
(skew = 0, indica equalização da paternidade entre os prováveis pais; skew = 1, indica que a
paternidade foi monopolizada por um dos prováveis pais). Valores negativos de skew
(paternidade efetiva > paternidade observada) podem ser interpretados como N amostral
inadequado para cálculo de paternidade efetiva e índice de skew [15] ou como skew igual a
zero.
II.3.2.3.3. Erro de não detecção e erro de não amostrgem
O erro de não detecção, dp, foi calculado de acordo com Boomsma e Ratnieks [13] e
usando a equação descrita em Paxton et al. [60]. Erro de não detecção é a probabilidade de
dois machos tendo alelos idênticos em todos os loci não serem detectados, porque duas
operárias possuem genótipos idênticos, por acaso, apesar de terem pais diferentes. Assumindo
o equilíbrio de Hardy-Weinberg, calculamos a probabilidade de filhas com paternidade dual
como:
Onde n é o número de alelos; s, é o número de locos; e qi, é a frequência alélica do iésimo alelo,
no jésimo loco.

51
O erro de não amostragem é outro erro comum nas estimativas de frequência de
paternidade. É a probabilidade que uma linhagem paterna não seja detectada devido ao
tamanho da amostra. Amostras pequenas, geralmente, resultam em perda de linhagens
paternas. Calculamos o erro de não amostragem de acordo com Boomsma e Ratnieks [13]
como (1-pi)n, onde pi é a proporção de operárias na iésima linhagem paterna e n é o tamanho da
amostra.
II.3.3. Resultados
Os locos microssatélites, que utilizamos para análise de paternidade e parentesco em
M. s. merrillae, discriminaram os genótipos parentais de modo satisfatório. Observamos
poliandria e baixos valores de skew em todas as colônias estudadas (Tabela 1). O
polimorfismo alélico na população estudada variou de 1 a 9 (em média 4,2 ± 2,7). Os locos
MSM08 e MSM02 foram os mais polimórficos, com 9 e 5 alelos, respectivamente. Os locos
MSM04 e MSM09 foram igualmente polimórficos, com 3 alelos cada um (Tabela 2). Um
loco, MSM07, foi monomórfico para todas as colônias; o loco MSM02 foi monomórfico em
cinco colônias (2Ba, 2Fa, 2Ka, 2La, 2Pa), e o loco MSM04, em duas colônias (2Ba e 2Ka).
Os locos monomórficos foram excluídos da análise dos dados em todas as colônias. O alelo
166 (colônia 2Ba, loco MSM09) teve a menor frequência (2,08%), sendo, provavelmente, um
alelo raro (Tabela 2).
Nas colônias de M. s. merrillae estudadas, a paternidade observada (kobs) variou de 3 a
7 (média aritmética 4,5 ± 1,309), a paternidade efetiva (^ke3) variou de 2,496 a 6,938 (média
harmônica 4,161 ± 1,376), o coeficiente de parentesco (r) variou de 0,322 a 0,450 (em média
0,380 ± 0,039) e os valores de skew (S) variaram de -0,187 a 0,50 (Tabela 3). Em duas
colônias (2Ba e 2Ha) observamos que alguns machos tiveram maior contribuição com a
paternidade. Na colônia 2Ba, a rainha acasalou com quatro machos e dois deles contribuíram
com 91,8% da paternidade (45,8% cada), os dois outros contribuíram apenas com 8,2 % da
paternidade (4,1% cada). A rainha da colônia 2Ha, acasalou com sete machos, três deles
contribuíram com 79,0% da paternidade (dois contribuíram com 29,1% cada; um com
20,8%); os outros quarto machos contibuíram com 21% da paternidade (um contribuiu com
8,3% e três contribuíram com 4,1% cada). Nas demais colônias a paternidade foi igualmente
compartilhada (interpretamos os valores negativos de skew como zero). Não foram
observados erros de genotipagem. O erro de não detecção variou de 0,046 a 0,416 (em média
0,210 ± 0,141) (Tabela 3) e o erro de não amostragem de zero a 0,365 (Tabela 4).
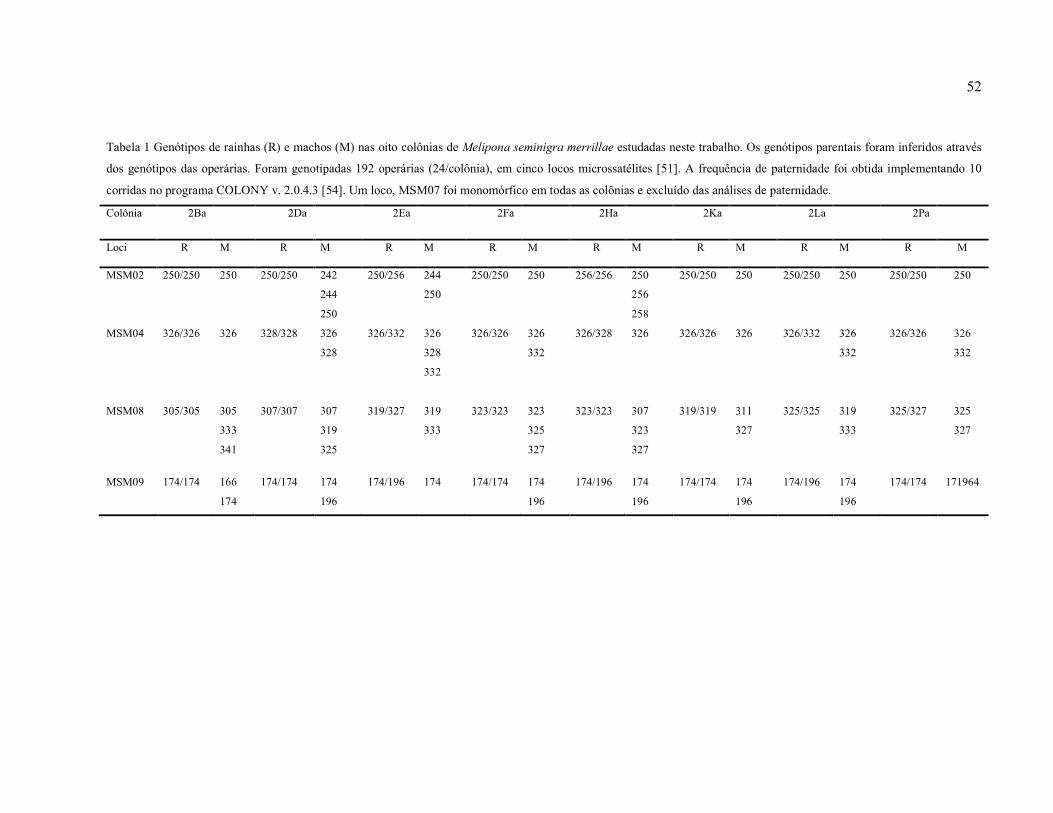
52
Tabela 1 Genótipos de rainhas (R) e machos (M) nas oito colônias de Melipona seminigra merrillae estudadas neste trabalho. Os genótipos parentais foram inferidos através
dos genótipos das operárias. Foram genotipadas 192 operárias (24/colônia), em cinco locos microssatélites [51]. A frequência de paternidade foi obtida implementando 10
corridas no programa COLONY v. 2.0.4.3 [54]. Um loco, MSM07 foi monomórfico em todas as colônias e excluído das análises de paternidade.
Colônia 2Ba 2Da 2Ea 2Fa 2Ha 2Ka 2La 2Pa
Loci R M R M R M R M R M R M R M R M
MSM02 250/250 250 250/250 242 250/256 244 250/250 250 256/256 250 250/250 250 250/250 250 250/250 250
244 250 256
250 258
MSM04 326/326 326 328/328 326 326/332 326 326/326 326 326/328 326 326/326 326 326/332 326 326/326 326
328 328 332 332 332
332
MSM08 305/305 305 307/307 307 319/327 319 323/323 323 323/323 307 319/319 311 325/325 319 325/327 325
333 319 333 325 323 327 333 327
341 325 327 327
MSM09 174/174 166 174/174 174 174/196 174 174/174 174 174/196 174 174/174 174 174/196 174 174/174 171964
174 196 196 196 196 196

53
Tabela 2 Frequências alélicas dos locos microssatélites genotipados nas 192 operárias de Melipona seminigra merrillae. O número de alelos na população variou de 1 a 9 (em
média 4,2 ± 2,7). Os locos MSM08 e MSM02 foram os mais polimóricos com 9 e 5 alelos, respectivamente; o loco MSM07 foi monomórfico em todas as colônias. As
frequências alélicas foram atualisadas usando-se o programa COLONY v. 2.0.4.3 [54].
Locos MSM02 MSM04 MSM07 MSM08 MSM09
Colônia Amostra Alelo Frequência Alelo Frequência Alelo Frequência Alelo Frequência Alelo Frequência
2Ba 24 250 1,0000 326 1,0000 247 1,0000 305 0,6574 166 0,0994
--- --- --- --- --- --- 333 0,1141 174 0,9006
--- --- --- --- --- --- 341 0,2285 --- ---
2Da 24 242 0,1492 326 0,2893 247 1,0000 307 0,5403 174 0,9457
244 0,1363 328 0,7107 --- --- 319 0,2176 196 0,0543
250 0,7145 --- --- --- --- 325 0,2420 --- ---
2Ea 24 244 0,2060 326 0,2806 247 1,0000 319 0,5554 174 0,8250
250 0,6185 328 0,4250 --- --- 327 0,1670 196 0,1750
256 0,1755 332 0,9440 --- --- 333 0,2775 --- ---
2Fa 24 250 1,0000 326 0,5000 247 1,0000 323 0,5641 174 0,8745
--- --- 332 0,5000 --- --- 325 0,1354 196 0,1255
--- --- --- --- --- --- 327 0,3005 --- ---
2Ha 24 250 0,1940 326 0,7232 247 1,0000 307 0,2015 174 0,7641
256 0,6716 328 0,2768 --- --- 323 0,6058 196 0,2359
258 0,1344 --- --- --- --- 327 0,1927 --- ---
2Ka 24 250 1,0000 326 1,0000 247 1,0000 311 0,2220 174 0,7929
--- --- --- --- --- --- 319 0,5632 196 0,2071
--- --- --- --- --- --- 327 0,2148 --- ---
2La 24 250 1,0000 326 0,7274 247 1,0000 319 0,2183 174 0,7272
--- --- 332 0,2726 --- --- 325 0,5666 196 0,2728
--- --- --- --- 333 0,2152 --- ---
2Pa 250 1,0000 326 0,9087 247 1,0000 325 0,7275 174 0,9046
--- --- 332 0,0913 --- --- 327 0,2725 196 0,0954
Número de alelos 5 3 1 9 3

54
Tabela 3 Frequência de paternidade e coeficiente de parentesco em colônias manejadas de Melipona seminigra merrillae. A paternidade foi avaliada pela genotipagem de 192
pupas de operárias, em quatro locos microssatélites (MSM02, MSM04, MSM07, MSM08 e MSM09) [51]. A análise de paternidade foi feita usando-se o programa COLONY
v. 2.0.4.3 [54]. N, tamanho da amostra; kobs, paternidade observada; ^ke3, paternidade efetiva, calculada de acordo com Nielsen et al. [56]; S, índice de skew calculado segundo
Pamilo e Crozier [59], ver também Jaffé et al. [15]; r, coeficiente de parentesco, calculado conforme Pamilo [58]; dp, erro de não detecção, calculado segundo Boomsma e
Ratnieks [13].
Colônia N kobs ^ke3 S r dp
2Ba 24 4 2,496 0,501 0,450 0,408
2Da 24 6 6,938 -0,187* 0,322 0,115
2Ea 24 4 3,745 0,085 0,383 0,046
2Fa 24 3 2,948 0,026 0,419 0,166
2Ha 24 7 5,137 0,310 0,347 0,086
2Ka 24 4 4,201 -0,067* 0,369 0,277
2La 24 4 4,075 -0,025* 0,372 0,168
2Pa 24 4 3,745 0,085 0,383 0,416
Média 4,500 ± 1,309 4,161 ± 1,376 0,380 ± 0,039 0,210 ± 0,141
*, valores negativos de skew (paternidade efetiva ˃ paternidade observada), podem ser interpretados como resultado de número amostral inadequado para cálculo de skew ou
como skew zero.

55
Tabela 4 Erro de não amostragem (não detecção de linhagens paternas) nas oito colônias de Melipona seminigra merrillae estudadas. Para linhagens paternas com 29,1% ou
mais filhas, o erro de não amostrgem foi zero (duas linhagens nas colônias 2Ba, 2Ea, 2Fa, 2Ha, 2Ka, 2La; uma linhagem na colônia 2Pa). Para linhagens paternas
representadas por 25%; 20,8%; 16,6%; 12,5%; 8,3% e 4,1% da paternidade, o erro de não amostragem foi, respectivamente, de 0,001; 0,003; 0,012; 0,04; 0,124 e 0,366. Para
linhagens paternas com 4,1% da paternidade (duas linhagens em 2Ba; três linhagens em 2Ha), o erro de não amostragem foi muito alto. P, linhagem paterna; pi, proporção de
filhas; nse, erro de não amostragem.
P1 P2 P3 P4 P5 P6 P7
Colônia pi nse pi nse pi nse pi nse pi nse pi nse pi nse
2Ba 0,041 0,365 0,458 0,000 0,458 0,000 0,041 0,365 --- --- --- --- --- ---
2Da 0,250 0,001 0,166 0,012 0,125 0,040 0,125 0,040 0,166 0,012 0,166 0,012
2Ea 0,250 0,001 0,291 0,000 0,083 0,124 0,375 0,000 --- --- --- --- --- ---
2Fa 0,208 0,003 0,333 0,000 0,458 0,000 --- --- --- --- --- --- --- ---
2Ha 0,291 0,000 0,208 0,003 0,041 0,365 0,291 0,000 0,041 0,365 0,041 0,365 0,083 0,124
2Ka 0,166 0,012 0,333 0,000 0,208 0,003 0,291 0,000 --- --- --- --- --- ---
2La 0,333 0,000 0,250 0,001 0,125 0,040 0,291 0,000 --- --- --- --- --- ---
2Pa 0,291 0,016 0,375 0,000 0,083 0,124 0,250 0,001 --- --- --- --- --- ---

56
II.3.4. Discussão
Nosssos resultados foram consistentes com poliandria em colônias manejadas de M. s.
merrillae (Tabela 3). Para o nível de poliandria observado (rainhas inseminadas por 3 a 7
machos), em associação com baixos valores de skew, epera-se que ocorra a maximização dos
ganhos da diversidade genética nestas colônias, o que é predito para Hymenoptera sociais
[15]. O relativo monopólio da paternidade, por alguns machos, em duas colônias (2Ba, S =
0,501; 2Ha, S = 0,310), provavelmente, explica-se pelo pequeno número amostral usado.
O grau de parentesco, descrito acima, mostra que em todas as colônias de M. s.
merrillae estudadas, as operárias compartilharam menos de 50% dos seus genes por
descendência comum. Estes resultados, contradizem o esperado para abelhas sem ferrão e
para Hymenoptera sociais haplo-diplóides, alto grau de parentesco intracolonial (r = 0,75) em
colônias monogínicas (uma rainha) e monândricas (rainha inseminada por um macho) [11,61].
O baixo grau de parentesco em colônias de insetos sociais, teoricamente reduz os ganhos dos
benefícios do fitness inclusivo [18]. Assim, a poliandria é considerado uma força contrária à
evolução da eussocialidade por seleção de parentesco [18, 19, 61], porque o alto grau de
parentesco é a pré-condição para evolução da eussocialidade [12]. Todavia, grupos de baixo
grau de parentesco são, provavelmente, favorecidos pelos ganhos extras de benefícios da
diversidade genética [62, 63].
O número de operárias que genotipamos por colônia (N = 24) foi determinado com
base em Oldroyd et al. [16]. Esses autores consideram que a genotipagem de 24 filhas,
fornece uma boa chance de amostrar todas as linhagens paternas. No entanto, nosso erro de
não amostragem foi alto (Tabela 4). Para linhagens paternas compreendendo 29,1% da prole
da rainha ou mais, o erro de não amostragem foi zero; para linhagens paternas que compõem
menos de 29,1% da prole da rainha, o erro de não amostragem aumenta na medida em que a
contribuição paternal diminui, sendo máximo para linhagens paternas representadas por 4,1%
da paternidade. Portanto, a frequência de paternidade observada para M. s. merrillae neste
trabalho está, provavelmente, subestimada.
Poliandria aumenta a diversidade genética e reduz a variação na produção de machos
diplóides (DMP, diploid male production), consequência do mecanismo de Determinação
Complementar do Sexo (CSD, complementary sex determination) [11, 64]. Em outras
palavras, poliandria leva a rainha a ajustar a frequência de macho diplóide intracolonial com a
frequência de macho diplóide da população que é 1/n, em condições de panmixia [24, 26].

57
Argumentamos que poliandria em M. s. merrillae tem um papel chave em minimizar
os efeitos perigosos da DMP [40], principalmente, em populações manejadas, onde a DMP
pode ser favorecida devido a maior chance de endogamia.
Agradecimento
Agradecemos ao Conselho Nacional de Desenvolvimento Científico e Tecnológico
(CNPq), à Financiadora de Estudos e Projetos (FINEP) e Instituto Nacional de Pesquisas da
Amazônia (INPA) pelo suporte financeiro. Á Fundação de Amparo à Pesquisa do Estado do
Amazonas (FAPEAM) pelo suporte financeiro e pela bolsa de estudos de Antonio Saulo
Cunha Machado.
Referências
[1] C. D. Michener, The Bees of the World, Johns Hopkins University Press, Baltimore, 2007.
[2] J. M. F, Camargo, and S. E. M. Pedro, “Meliponini Lepeletier, 1836. In: J. S. Moure, D.
Urban, G. A. R. Melo, Orgs. Catalogue of Bees (Hymenoptera, Apoidea) in the
Neotropical Region.” Online version: http://www.moure.cria.org.br/catalogue,
accessed 22 October 2013.
[3] D. W. Roubik, Ecology and Natural History of Tropical Bees, Cambridge University
Press, 1989.
[4] T. A. Heard, “The role of stingless bees in crop pollination,” Annual Review of
Entomology, vol. 44, pp. 183–206, 1999.
[5] B. J. Brosi, G. C. Daily, T. M. Shih, F. Oviedo, and G. Duran, “The effects of forest
fragmentation on bee communities in tropical countryside,” Journal of Applied
Ecology, vol. 45 pp. 773–78, 2008.
[6] W. E. Kerr; G. A. Carvalho, and V. A. Nascimento, Uruçu bee: Biology, Handling and
Conservation, Foundation Acangaú, Paracatu, M. G., Brazil, 1996 (in Portuguese).
[7] W. E. Kerr, “Some aspects of the evolution of the social bees,” Evolutionary Biology, vol.
3, pp. 119-175, 1969.
[8] J. Strassmann, “The rarity of multiple mating by females in the Hymenoptera,” Insectes
Sociaux, vol. 48, pp. 01-13, 2001.
[9] C. A. L Carvalho, B. A. Souza, C. S. Dias, R. M. O Alves, A. F. L. Melo, and G. A.
Carvalho-Zilse, “Five egg-laying queens in a single colony of brazilian stingless bees
(Melipona scutellaris Latreille),” Acta Amazonica, vol. 41, pp. 123-126, 2011.

58
[10] D. A. Alves, C. Menezes, V. L. Imperatrix-Fonseca; and T. Wenseleers, “First Discovery
of a rare polygyne colony in the stingless bee Melipona quadrifasciata (Apidae,
Meliponini),” Apidologie, vol. 42 pp. 211-213, 2011.
[11] R. H. Crozier, and P. Pamilo, Evolution of social insect colonies. Sex allocation and kin
selection, Oxford University Press, Oxford, 1996.
[12] W. O. H. Hughes, B. P. Oldroyd, M. Beekmann, and F. L. W. Ratnieks, “Ancestral
monogamy shows kin selection is key to evolution of eusociality,” Science, vol. 30, pp.
1213-1216, 2008.
[13] J. J. Boomsma, and F. L. W. Ratnieks, “Paternity in eusocial Hymenoptera,”
Philosophical Transactions of the Royal Society of London B, vol. 351, pp. 947-975,
1996.
[14] J. J. Boomsma, D. J. C. Kronauer, and J. S. Pederson, The evolution of social insect
mating systems, pp.3-25 in J. Gadau and J. Fewell eds. Organization of insect
societies–from genome to sociocomplexity, Harvard University Press, Cambridge,
Massachusetts, 2009.
[15] R. Jaffe, F. Garcia-Gonzalez, S. P. A. den Boer, and L. W. Simmons, “Patterns of
paternity skew among polyandrous social insects: what can they tell us about the
potential for sexual selection?,” Evolution, vol. 66, pp. 3778-3788, 2012.
[16] B. P. Oldroyd, A. J. Smolenski, J. M. Cornuet, S. Wongsiri, A. Estoup, T. E. Rinderer,
and R. H. Crozier, “Levels of polyandry and intracolonial genetic relationships in Apis
florea,” Behavioral Ecology and Sociobiology, vol. 37, pp. 329-335, 1995.
[17] W. Wattanachaiyingcharoen, B. P. Oldroyd, S. Wongsiri, K. Palmer, and J. Paar, “A
scientific note on the mating frequency of Apis dorsata,” Apidologie, vol. 34 pp. 85-
86, 2003.
[18] W. D. Hamilton, “The genetic evolution of social behavior,” Journal of Theoretical
Biology, vol. 7 pp. 1-52, 1964.
[19] W. D. Hamilton, “Altruism and related phenomena, mainly in social insects,” Annual
Review of Ecology and Systematic, vol. 3 pp. 193-232, 1972.
[20] W. D. Hamilton, “Kinship, recognition, disease, and intelligence: constraints of social
evolution. In Animal societies: theories and facts (eds. Y. Ito, J. L. Brown, J.
Kikkawa),” pp. 81-102, Tokyo: Japan Science Society Press, 1987.
[21] J. J. Boomsma “Kin selection versus sexual selection: Why the ends do not meet,”
Current Biology, vol. 17, R673-R683, 2007.

59
[22] J. J. Boomsma “Lifetime monogamy and the evolution of eusociality,” Philosophical
Transactions of the Royal Society of London B, vol. 364 pp.3191-3207, 2009.
[23] P. Nonacs, “Monogamy and high relatedness do not preferentially favor the evolution of
cooperation,” BCM Evolutionary Biology (http://www.biomedcentral.com/1471-
2148/11/58), 2011.
[24] R. E. Page, and R. A. Metcalf, “Multiple mating, sperm utilization and social evolution,”
American Naturalist, vol. 119, pp. 263-281, 1982.
[25] D. R. Tarpy, and R. E. Page Jr., “The curious promiscuity of queen honey bees (Apis
mellifera): volutionary and behavioral mechanisms,” Annales Zoologica Fennici, vol.
38, pp. 255–265, 2001.
[26] R. H. Crozier, and R. E. Page, “On being the right size: male contributions and multiple
mating in social Hymenoptera,” Behavioral Ecology and Sociobiology, vol. 18, pp.
105-115, 1985.
[27] J. Loman, T. Madsen, and T. Hakansson “Increased fitness from multiple mating, and
genetic heterogeneity: a model of a possible mechanism”, Oikos, vol. 52, pp. 69-72,
1988.
[28] P. W. Sherman, T. D. Seely, and H. K. Reeve, “Parasites, pathogens and polyandry in
social hymenoptera”, American Naturalist, vol. 131, pp. 602-610, 1988.
[29] W. D. Hamilton, R. Axelrod, and R. Tanese, “Sexual reproduction as an adaptation to
resist parasites (a review),” Proceedings of the National Academy of Sciences USA,
vol. 87, pp. 3566-3573, 1990.
[30] R. H. Crozier, and E. J. Fjerdingstad, “Polyandry in social Hymenoptera: disunity in
diversity?” Annales Zoologica Fennici, vol. 38 pp. 267-285, 2001.
[31] W. H. O. Hughes, and J. J. Boomsma, “Does genetic diversity hinder parasite evolution
in social insect colonies?,” Journal of Evolutionary Biology, vol. 19, pp. 32-143, 2006.
[32] H. R. Mattila, and T. D. Seeley “Genetic diversity in honey bee colonies enhances
productivity and fitness,” Science, vol. 317, pp. 362-364, 2007.
[33] B. P. Oldroyd, and J. H. Fewell, “Genetic diversity promotes homeostasis in insect
colonies”, Ecology and Evolution, vol. 22, pp. 408-412, 2007.
[34] T. D. Seeley, and D. R. Tarpy, “Queen promiscuity lowers disease within honeybee
colonies,” Proceeding of the Royal Society of London B Biological Sciences, vol. 274,
pp. 67-72, 2007.

60
[35] C. R. Smith, A. L. Toth, A. V. Suarez, and G. E. Robinson, “Genetic and genomic
analyses of the division of labour in insect societies,” Nature Reviews Genetics, vol. 9,
735-748, 2008.
[36] P. W. Whiting, “Multiple alleles in complementary sex determination of Habrobracon,”
Genetics, vol. 28 pp. 365-382, 1943.
[37] G. A. Carvalho, “The number of sex alleles (CSD) in a bee population and its practical
importance (Hymenoptera: Apidae),” Journal of Hymenoptera Research, vol. 10, pp.
10-15, 2001.
[38] E. van Wilgenburg, G. Driessen, and L. W. Beukeboom, “Single locus complementary
sex determination in Hymenoptera: an unintelligent design?,” Frontiers in Zoology,
vol. 3 pp. 1-15, 2006.
[39] M. Hasselmann, T. Gempe, M. Schiøtt, C. G. Nunes-Silva, M. Otte, and M. Beye,
“Evidence for the evolutionary nascence of a novel sex determination pathway in
honeybees,” Nature, vol. 454 -7203, pp. 519–522, 2008.
[40] I. B. Francini, C. G. Nunes-Silva, and G. A. Carvalho-Zilse, “Diploid male production of
two amazonian Melipona bees (Hymenoptera: Apidae),” Psyche: a Journal of
Entomology, vol. 2012, pp. 7, 2012.
[41] J. Woyke, “What happens to diploid drone larvae in a honey bee colony,” Journal of
Apicultural Research, 2: 73-76, 1963a
[42] J. Woyke, “Rearing and viability of diploid drone larvae,” Journal of Apicultural
Research, vol. 2 pp. 77-84, 1963b.
[43] W. E. Kerr, “Sex determination in bees. XXI. Number of XO-heteroalleles in a natural
population of Melipona compressipes fasciculata,” Insectes Sociaux, vol. 34, pp. 274-
279, 1987.
[44] M. L. Absy, and W. E. Kerr, “Some plants visited by workers of Melipona seminigra
merrillae to obtain pollen, at Manaus”, Acta Amazonica, vol. 7, pp. 309-315 (in
Portuguese), 1977.
[45] C. G. Bacelar-Lima, D. C. B. Freire, A. Coletto-Silva, K. B Costa, J. P. B. Laray, H. C.
Vilas-Boas, and G. A. Carvalho-Zilse, “Melitocoria of Zygia racemosa (Ducke)
Barneby and Grimes by Melipona seminigra merrillae Cockerell, 1919 and Melipona
compressipes manaosensis Schwarz, 1932 (Hymenoptera, Meliponina) in Central
Amazon, Brazil,” Acta Amazonica, vol. 36, pp. 343-348, 2006.

61
[46] C. V. Nunez, M. L. Oliveira, R. D. Lima, I. E. C. Diaz, E. Sargentini Jr., O. L. Pereira Jr.,
and L. M. Araújo, “Chemical analyses confirm a rare case of seed dispersal by bees,”
Apidologie, vol. 39, pp. 618–626, 2008.
[47] G. A. Carvalho-Zilse, and C. G. Nunes-Silva, Threats to the Stingless Bees in the
Brazilian Amazon: How to Deal with Scarce Biological Data and an Increasing Rate
of Destruction. In: R. M. Florio (eds.) Bees: Biology, Threats and Colonies, Nova
Science Publishers, Chapter 4, pp. 147-168, 2012.
[48] L. A. O. Campos, and G. A. R. Melo, “Physogastric-queen mating in Melipona
quadrifasciata Lep. (Hymenoptera, Apidae) Brazil,” Journal of Genetics, vol. 13 pp.
491-500, 1990.
[49] K. A. Palmer, B. P. Oldroyd, J. J. G. Quezada-Euan, R. J. Paxton, and W. de J. May-Itza,
“Paternity frequency and maternity of males in some stingless bee species,” Molecular
Ecology, vol. 11, pp. 2107-2113, 2002.
[50] M. Schuelke, “An economic method for the fluorescent labeling of PCR fragments,”
Nature Biotechnology, vol. 18 pp. 233-234, 2000.
[51] I. B. Francini, D. A. Sforça, A. C. B. Sousa, T. Campos, M. I. Zucchi, A. P. Souza, C. G.
Nunes-Silva, and G. A. Carvalho-Zilse, “Microsatellite loci for an endemic stingless
bee Melipona seminigra merrillae (Apidae, Meliponini) from Amazon,” Conservation
Genetics Resources, vol. 1 pp. 487-49, 2009.
[52] B. P. Oldroyd, M. J. Clifton, S. Wongsiri, T. E. Rinderer, H. A. Sylvester, and R. H.
Crozier, “Polyandry in the genus Apis, particularly Apis andreniformis,” Behavioral
Ecology and Sociobiology, vol. 40, pp. 17-26, 1997.
[53] F. Pompanon, A. Bonin, E. Bellemain, and P. Taberlet, “Genotyping errors: causes,
consequences and solutions,” Nature Reviews Genetics, vol. 6, pp. 847-859, 2005.
[54] J. Wang, “Sibship reconstruction from genetic data with typing errors,” Genetics, vol.
166, pp.1963-1979, 2004.
[55] O. R. Jones, and J. Wang “COLONY: a program for parentage and sibship inference
from multilocus genotype data”, Molecular Ecology Resources, vol. 10 pp. 551-555,
2010.
[56] R. Nielsen, D. R. Tarpy, and H. K. Reeve, “Estimating effective paternity number in
social insects and the effective number of alleles in a population,” Molecular Ecology,
vol. 12, pp. 3157-3164, 2003.
[57] D. R. Tarpy, R. Nielsen, and D. I. Nielsen, “A scientific note on the revised estimates of
effective paternity frequency in Apis,” Insectes Sociaux, vol. 51, pp. 203-204, 2004.

62
[58] P. Pamilo, “Polyandry and allele frequency differences between the sexes in the ant
Formica aquilonia,” Heredity, vol. 70, 472-480, 1993.
[59] P. Pamilo, and R. H. Crozier, “Reproductive skew simplified,” Oikos, vol. 75, pp. 533-
535, 1996.
[60] R. J. Paxton, L. R. Bego, M. M. Shah, and S. Mateus, “Low mating frequency of queens
in stingless bee Scaptotrigona postica and worker maternity of males,” Behavioral
Ecology and Sociobiology, vol. 53, pp. 174–181, 2003.
[61] E. O. Wilson, The Insects Societies, Cambridge, Harvard University Press, 1971.
[62] P. Nonacs, and K. M. Kaphein, “Social heterosis and the maintenance of genetic
diversity,” Journal of Evolutionary Biology, vol. 20, pp. 2253-2265, 2007.
[63] P. Nonacs, “Monogamy and high relatedness do not preferentially favor the evolution of
cooperation,” BMC Evolutionary Biology, 11:58, 2011.
[64] D. R. Tarpy, and R. E. Page Jr., “Sex determination and the evolution of polyandry in
honey bees (Apis mellifera),” Behavioral Ecology and Sociobiology, vol. 52 pp. 143-
150, 2002.

63
II.4. Capítulo 4: Análise de DNA microssatélites revela alta frequência de
paternidade em colônias manejadas da abelha sem ferrão Melipona seminigra
seminigra.
Francini, I.B.; Nunes-Silva, C.G; Machado, A.S.C; Batista, J.S.; Machado, A.S.C.; Carvalho-
Zilse, G.A. DNA microsatellite analysis reveals high paternity frequency for the stingless bee
Melipona seminigra seminigra in managed colonies. Journal of Insects (submissão a ser feita)
Abstract
Four DNA microsatellite loci with a total of 28 alleles were amplified in 168 workers from
seven colonies of Melipona seminigra seminigra to assess paternity. The loci polymorphism,
mainly MSM09 with 16 alleles provided sufficient genetic variability to assign workers into
distinct paternal lineage. Paternity frequency ranged from 7 - 10 (average 9 ± SD 1) and the
effective paternity ranged from 2.000 - 9.446 (average 5.837 ± SD 2.341). This level of
polyandry produced within-colony relatedness (r) of 0.302 - 0.363 (average 0.355 ± SD
0.066). Paternity skew (S) ranges from 0.059 - 0.833 (average 0.43 ± SD 0.262). Thereby, we
observed polyandry in all colonies analized. In agreement with the current opnion we think
that mating multiple by M. s. seminigra queens may be explained by increased genetic
diversity benefits. We regard that polyandry in M. s. seminigra diminishes the harmful effects
of diploid male.
Keywords: paternity, polyandry, relatedness, diploid male, paternity skew

64
II.4.1. Introducão
As abelhas da tribo Meliponini são um grupo altamente diverso na bacia amazônica
[1], são abelhas altamente eusociais (rainhas e operárias diferenciadas morfologicamente),
vivem em colônias perenes com algumas dúzias a 100.000 operárias ou mais, são conhecidas
como “abelhas sem ferrrão” poque possuem o ferrão e estruturas associadas como órgãos
vestigiais [2]. Salvo exceções, usam pólen como fonte de proteínas e nectar como fonte de
carboidratos, portanto, dependem das plantas com flores, da mesma forma que estas
dependem de polinização das abelhas [3, 4].
Considerando o sistema de acasalamento, o esperado é que rainhas de Meliponini
sejam inseminadas por apenas um macho (monandria) [5], que é o comum para a maioria dos
Hymenoptera solitários e sociais [6, 7, 8]. Todavia, múltiplos acasalamentos (poliandria) têm
sido amplamente documentados para rainhas de Hymenoptera sociais [6, 9]. Em abelhas sem
ferrão, poucos estudos relataram rainhas sendo inseminadas por mais de um macho, em todos
os casos a paternidade efetiva foi sempre inferior a dois [5, 10, 11, 12, 13].
Devido aos prováveis benefícios do fitness inclusivo, o alto grau de parentesco
intracolonial foi considerado primordial para a evolução da eussocialidade, por seleção de
parentesco [14, 15, 16, 17]. Poliandria em Hymenoptera sociais é uma questão polêmica e
fascinante [6, 18, 19], tem sido considerada um fenômeno secundário, que evoluiu após o
estabelecimento e irreversibilidade da eussocialidade, quando as castas de operária e rainha
tornaram-se interdependentes [8]. Os benefícios da diversidade genética têm sido invocados,
frequentemente, para explicar evolução de poliandria em Hymenoptera sociais [7, 20, 21].
Melipona seminigra seminigra tem distribuição nos Estados do Amazonas e Pará,
Brasil [1]. É criada pelos povos locais, principalmente, para obtenção de mel [observação
pessoal]. Objetivamos verificar a hipótese de poliandria em colônias manejadas de M. s.
seminigra, o que foi observado para duas outras subespecies: M. s. abunensis e M. s. merrillae
[dados não publicados, capítulos 2 e 3]. Para isto, analisamos o sistema de acasalamento, em
sete colônias, usando marcadores moleculares microssatélites.
II.4.2. Material e Métodos
II.4.2.1. Amostragem
Foram amostradas 168 operárias de M. s. seminigra em sete colônias manejadas. A
amostragem foi feita em meliponários rurais, nos municípios de Boa Vista do Ramos e

65
Maués, Amazonas. As operárias foram coletadas na entrada de cada ninho, fixadas em etanol
a 96 % e armazenadas em freezer à -20oC. Posteriormente, foi extraído o DNA.
II.4.2.2. Análise molecular
O DNA foi extraído do tecido do tórax das 168 operárias de M. s. seminigra. As
extrações foram feitas utilizando kit Promega de acordo com o protocolo do fabricante.
Foram amplificados quatro locos microssatélites (MSM02, MSM09, MSM12, MSM13) [25],
via reação de polimerase em cadeia (PCR, polymerase chain reaction). As PCRs foram feitas
em um volume total de 10 μL, usando 10ng de DNA, tampão 10x com KCl; primer forward
4μM marcado com cauda M13 [24]; primer reverse 4μM e fluorescência 4μM (FAM, HEX
ou NED). A análise dos produtos de PCR foi feita em sequenciador ABI 3130 xl (Applied
Biosystems). Como marcador de genotipagem foi usado ROX™ 500. O tamanho dos alelos,
em pares de bases (bp, base pairs) foi estimado e editado usando-se o software
GENEMAPPER v. 4.0 (Applied Biosystems).
II.4.2.3. Análises de dados
II.4.2.3.1 Paternidade, parentesco e valores de skew
A paternidade em M. s. seminigra foi estimada a partir dos genótipos das operárias,
assumindo, a priori, monoginia e monandria, nas colônias estudadas [26]. Para determinar os
genótipos parentais, a frequência alélica, o número de famílias de irmãs completas (mesmo
pai e mesma mãe) e erros de genotipagem, foram implementadas 10 corridas no programa
COLONY v. 2.0.4.3 (http://www.zsl.org/science/research-projects/software) [27, 28, 29].
A paternidade efetiva (^ke3) foi calculada por Nielsen et al [31] pela equação [32]:
Onde, N é o número de operárais geneotipadas por colônia (24); k é a paterniade observada
(número de famílias de irmãs completas); pi, é a frequência relativa de filhas (operárias) do
iésimo pai. Para o cálculo do coeficiente de parentesco intracolonial (r) foi utilizada a equação
descrita em Pamilo [33]:
O skew, contribuição de cada pai com a prole da rainha foi calculado de acordo com
Pamilo e Crozier [34] pela equação descrita em Jaffé et al. [21]: S = (Kobs - ^ke3) ∕ (Kobs - 1).

66
II.4.2.3.2. Erro de não detecção e erro de não amostragem
O erro de não detecção (dp) é probabilidade de dois machos com alelos idênticos em
todos os locos, não serem detectados, porque as operárias possusem genótipos idênticos,
embora sejam filhas de pais diferentes. Assumindo-se o equilíbrio de Hardy-Weinberg, a
probabilidade de paternidade dual foi calculada de acordo com Boomsma e Ratnieks [6] pela
equação descrita em Paxton et al. [12]:
Sendo, n = número de alelos; s= número de locos; qi = frequência alélica do iésimo alelo no
loco jésimo de s locos.
O erro de não amostrgem (não detecção de uma linhagem paterna devido ao tamanho
da amostra) foi calculado de acordo com Boomsma e Ratnieks [6] por (1-p)n, onde p é a
proporção de filhas observadas em uma linhagem paterna e n o número amostral.
2.4.3. Resultados
Os quatro locos microssatélites empregados para genotipar as 168 operárias de M. s.
seminigra apresentaram variabilidade genética suficiente para distinguir diferentes linhagens
paternas. Registramos um total de 28 alelos, sendo o loco MSM09 o mais polimórfico com 16
alelos. O loco MSM02 foi monomórfico para a colônia 4F e o loco MSM13 monomórfico
para as colônias 4F e 4G. Os locos monomórficos foram excluídos das análises.
A paternidade observada (Kobs) nas colônias de M. s. seminigra estudadas neste
trabalho, variou de 7 a 10 (média aritmética 9 ± SD 1) e a paternidade efetiva (^ke3) variou de
2 a 9,446 (média harmônica 5,837 ± SD 2,341). O grau de parentesco observado (r) variou de
0,302 a 0,5 (média 0,355 ± SD 0,066) e o skew (S) variou de 0,059 a 0,833. O erro de não
detecção variou de 0,076 a 0,36 (em média 0,211 ± SD 0,103) (Tabela 1). O erro de não
amostragem foi de 0,000000003 para linhagens constituíndo 33,3% da paternidade; para
linhagens representando menos de 33,3% da paternidade o erro de não amostragem aumentou
na proporção que a paternidade diminui (para linhagens paternas que representaram 25%,
16,6%, 12,5%, 8,3% e 4,1% da paternidade, os erros de não amostragem foram,
respectivamente, 0,001, 0,003, 0,012, 0,04, 0,124 e 0,366). Erros de genotipagem (alelos
dropouts e mutações) não foram observados.
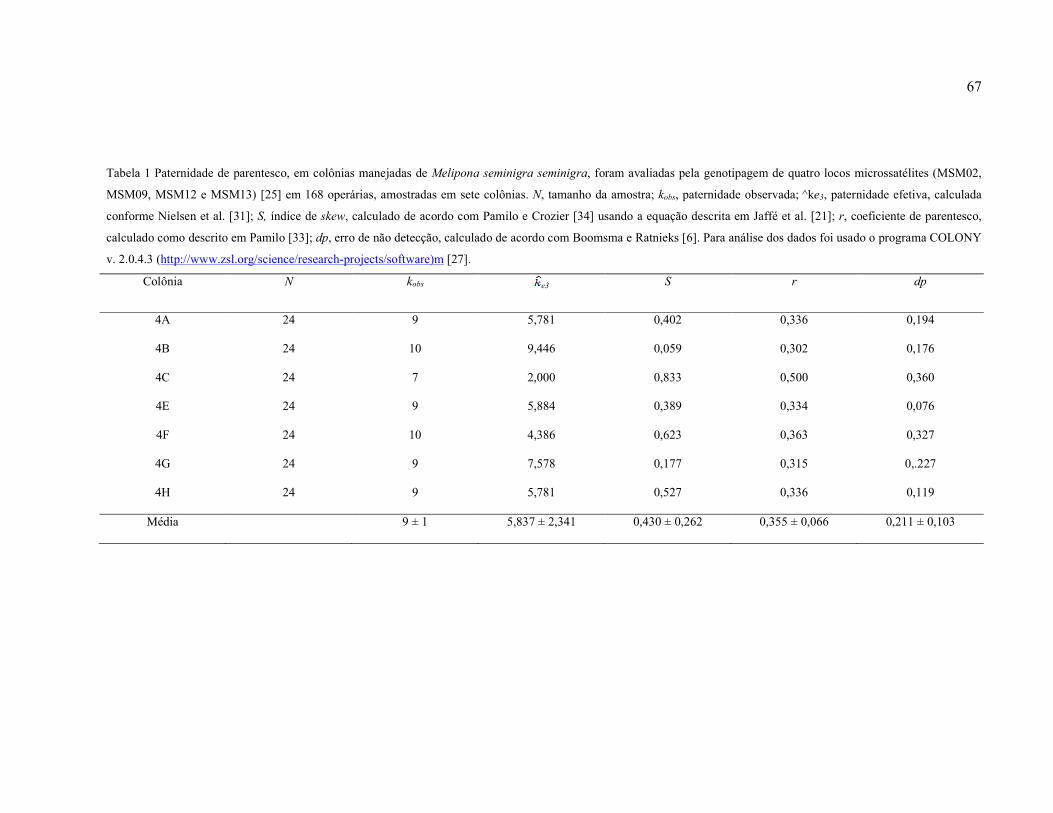
67
Tabela 1 Paternidade de parentesco, em colônias manejadas de Melipona seminigra seminigra, foram avaliadas pela genotipagem de quatro locos microssatélites (MSM02,
MSM09, MSM12 e MSM13) [25] em 168 operárias, amostradas em sete colônias. N, tamanho da amostra; kobs, paternidade observada; ^ke3, paternidade efetiva, calculada
conforme Nielsen et al. [31]; S, índice de skew, calculado de acordo com Pamilo e Crozier [34] usando a equação descrita em Jaffé et al. [21]; r, coeficiente de parentesco,
calculado como descrito em Pamilo [33]; dp, erro de não detecção, calculado de acordo com Boomsma e Ratnieks [6]. Para análise dos dados foi usado o programa COLONY
v. 2.0.4.3 (http://www.zsl.org/science/research-projects/software)m [27].
Colônia N kobs e3 S r dp
4A 24 9 5,781 0,402 0,336 0,194
4B 24 10 9,446 0,059 0,302 0,176
4C 24 7 2,000 0,833 0,500 0,360
4E 24 9 5,884 0,389 0,334 0,076
4F 24 10 4,386 0,623 0,363 0,327
4G 24 9 7,578 0,177 0,315 0,.227
4H 24 9 5,781 0,527 0,336 0,119
Média 9 ± 1 5,837 ± 2,341 0,430 ± 0,262 0,355 ± 0,066 0,211 ± 0,103

68
2.4.4. Discussão
Nossos resultados, sobre paternidade e parentesco em colônias manejadas de M. s.
seminigra, são consistentes com poliandria (as rainhas são inseminadas, em média, por nove
machos). Ao contrário do observado para outras espécies de Meliponini em estudos prévios
[5], a paternidade efetiva registrada para a maioria dos casos (6/7 das colônias) indica um alto
nível de poliandria (Tabela 1).
Em 3/7 das colônias alguns machos tiveram maior contribuição com a prole da rainha,
conforme evidenciado por valores altos de skew (S ˃ 0,5). Mas, em 4/7 das colônias
observamos que os valores de skew são baixos (S ˂ 0,5), que é o esperado para rainhas de
Hymenoptera sociais altamente poliândricas [21]. Supomos que os valores altos de skew,
verificados para 3/7 das colônias, podem ser explicados pelo tamanho da amostra que usamos
(N=24). À luz desta suposição, sugerimos que o alto nível de poliandria observado para
rainhas de M. s. seminigra está associado a baixos níveis de skew, ou seja, as rainhas tendem a
equalizar a paternidade e, assim, maximizam a diversidade genética e seus benefícios.
Nas colônias de M. s. seminigra estudadas neste trabalho, as operárias
compartilharam, em media, 0,355 ± SD 0,066 de seus genes, por descendência comum
(Tabela 1). Este nível de parentesco resulta do grau de poliandria observado e, aparentemente,
minimiza os ganhos do fitness inclusivo, o que, em princípio, contradiz o modelo de evolução
da eussocialidade por seleção de parentesco [14, 35, 36]. Poliandria foi bem documentada em
Hymenoptera eussociais [6, 9] e tem sido explicada pelos ganhos extras de benefícios de uma
maior diversidade genética [7, 21, 37, 38].
Em M. s. seminigra, poliandria, pode ser uma estratégia evolutiva para minimizar os
efeitos deletérios da produção de machos diplóides (DMP, diploid male production) [39]. Em
colônias monogínicas com rainhas monândricas, é esperado que a DMP dizime a força
operária [10, 13, 15, 40] porque machos diplóides (DM) são produzidos em uma razão sexual
igual a 1:1 [13]. Rainhas poliândricas, podem ajustar a produção de macho diplóide da
colônia com a frequência de macho diplóide da população (1/n dos indivíduos diplóides) [18,
39, 41, 42]. Isto é particularmente vantajoso em populações manejadas de meliponários, como
a população de M. s. seminigra, que estudamos, onde a DMP é favorecida pelo alto grau de
parentesco das colônias.
Nossas estimativas sobre paternidade e parentesco em M. s. seminigra são
conservativas, conforme evidenciado pelos erros de não detecção e erros de não amostragem,

69
que foram altos. Ainda assim, os nossos resultados são relevantes, validando a hipótese de
poliandria no grupo de subespécies de M. seminigra, o que tem implicações no que se refere à
sua criação, manejo e conservação. Pensando na evolução da eussocialidade por seleção de
parentesco, outros fatores, além do sistema de acasalamento, devem ser considerados [43, 44].
Agradecimentos
Os autores agradecem ao Conselho Nacional de Desenvolvimento Científico e Tecnológico
(CNPq), á Financiadora de Estudos e Projetos (FINEP), ao Instituto Nacional de Pesquisas da
Amazônia (INPA) e à Fundação de Amparo à Pesquisa do Estado do Amazonas (FAPEAM)
pelo suporte financeiro.
Referências
[1] J. M. F, Camargo, and S. R. M. Pedro, “Meliponini Lepeletier, 1836. In: J. S. Moure, D.
Urban, G. A. R. Melo, Orgs. Catalogue of Bees (Hymenoptera, Apoidea) in the
Neotropical Region.” Online version: http://www.moure.cria.org.br/catalogue,
accessed 22 October 2013.
[2] C. D. Michener, The Bees of the World, Johns Hopkins University Press, Baltimore, 2007
[3] D. W. Roubik, Ecology and Natural History of Tropical Bees, Cambridge University
Press, 1989.
[4] T. A. Heard, “The role of stingless bees in crop pollination,” Annual Review of
Entomology, vol. 44, pp. 183–206, 1999.
[5] J. Strassmann, “The rarity of multiple mating by females in the Hymenoptera,” Insectes
Sociaux, vol. 48, pp. 01-13, 2001.
[6] J. J. Boomsma, and F. L. W. Ratnieks, “Paternity in eusocial Hymenoptera,”
Philosophical Transactions of the Royal Society of London B, vol. 351, pp. 947-975,
1996.
[7] B. P. Oldroyd, and J. H. Fewell, “Genetic diversity promotes homeostasis in insect
colonies”, Ecology and Evolution, vol. 22, pp. 408-412, 2007.
[8] W. O. H. Hughes, B. P. Oldroyd, M. Beekmann, and F. L. W. Ratnieks, “Ancestral
monogamy shows kin selection is key to evolution of eusociality,” Science, vol. 30, pp.
1213-1216, 2008.
[9] J. J. Boomsma, D. J. C. Kronauer, and J. S. Pederson, The evolution of social insect
mating systems, pp.3-25 in J. Gadau and J. Fewell eds. Organization of insect

70
societies–from genome to sociocomplexity, Harvard University Press, Cambridge,
Massachusetts, 2009.
[10] G. A. Carvalho, “The number of sex alleles (CSD) in a bee population and its practical
importance (Hymenoptera: Apidae),” Journal of Hymenoptera Research, vol. 10, pp.
10-15, 2001.
[11] K. A. Palmer, B. P. Oldroyd, J. J. G. Quezada-Euan, R. J. Paxton, and W. de J. May-Itza,
“Paternity frequency and maternity of males in some stingless bee species,” Molecular
Ecology, vol. 11, pp. 2107-2113, 2002.
[12] R. J. Paxton, L. R. Bego, M. M. Shah, and S. Mateus, “Low mating frequency of queens
in stingless bee Scaptotrigona postica and worker maternity of males,” Behavioral
Ecology and Sociobiology, vol. 53, pp. 174–181, 2003
[13 I. B. Francini, C. G. Nunes-Silva, and G. A. Carvalho-Zilse, “Diploid male production of
two amazonian Melipona bees (Hymenoptera: Apidae),” Psyche: a Journal of
Entomology, vol. 2012, pp. 7, 2012.
[14] W. D. Hamilton, “The genetic evolution of social behavior,” Journal of Theoretical
Biology, vol. 7 pp. 1-52, 1964.
[15] R. H. Crozier, and P. Pamilo, Evolution of social insect colonies. Sex allocation and kin
selection, Oxford University Press, Oxford, 1996.
[16] J. J. Boomsma “Kin selection versus sexual selection: Why the ends do not meet,”
Current Biology, vol. 17, R673-R683, 2007.
[17] Crozier RH (2008) Advanced eusociality, kin selection and male haploidy. Aust J
Entomol 47:2–8
[18] R. E. Page, and R. A. Metcalf, “Multiple mating, sperm utilization and social evolution,”
American Naturalist, vol. 119, pp. 263-281, 1982.
[19] J. J. Boomsma “Lifetime monogamy and the evolution of eusociality,” Philosophical
Transactions of the Royal Society of London B, vol. 364 pp.3191-3207, 2009.
[20] C. R. Smith, A. L. Toth, A. V. Suarez, and G. E. Robinson, “Genetic and genomic
analyses of the division of labour in insect societies,” Nature Reviews Genetics, vol. 9,
735-748, 2008.
[21] R. Jaffe, F. Garcia-Gonzalez, S. P. A. den Boer, and L. W. Simmons, “Patterns of
paternity skew among polyandrous social insects: what can they tell us about the
potential for sexual selection?,” Evolution, vol. 66, pp. 3778-3788, 2012.

71
[22] I. B. Francini, C. G. Nunes-Silva, and G. A. Carvalho-Zilse, “Diploid male production of
two amazonian Melipona bees (Hymenoptera: Apidae),” Psyche: a Journal of
Entomology, vol. 2012, pp. 7, 2012.
[23] B. P. Oldroyd, A. J. Smolenski, J. M. Cornuet, S. Wongsiri, A. Estoup, T. E. Rinderer,
and R. H. Crozier, “Levels of polyandry and intracolonial genetic relationships in Apis
florea,” Behavioral Ecology and Sociobiology, vol. 37, pp. 329-335, 1995.
[24] M. Schuelke, “An economic method for the fluorescent labeling of PCR fragments,”
Nature Biotechnology, vol. 18 pp. 233-234, 2000.
[25] I. B. Francini, D. A. Sforça, A. C. B. Sousa, T. Campos, M. I. Zucchi, A. P. Souza, C. G.
Nunes-Silva, and G. A. Carvalho-Zilse, “Microsatellite loci for an endemic stingless
bee Melipona seminigra merrillae (Apidae, Meliponini) from Amazon,” Conservation
Genetics Resources, vol. 1 pp. 487-49, 2009.
[26] W. E. Kerr, “Some aspects of the evolution of the social bees,” Evolutionary Biology,
vol. 3, pp. 119-175, 1969.
[27] J. Wang, “Sibship reconstruction from genetic data with typing errors,” Genetics, vol.
166, pp.1963-1979, 2004.
[28] J. Wang, A. W. Santure, “Parentage and sibship inference from multi-locus genotype
data under polygamy”, Genetics, vol. 181, pp. 1579-1594, 2009.
[29] J. Wang, “Computationally efficient sibship and parentage assignment from multilocus
marker data”, Genetics, vol. 191, pp. 183-194, 2012.
[30] O. R. Jones, and J. Wang “COLONY: a program for parentage and sibship inference
from multilocus genotype data”, Molecular Ecology Resources, vol. 10 pp. 551-555,
2010.
[31] R. Nielsen, D. R. Tarpy, and H. K. Reeve, “Estimating effective paternity number in
social insects and the effective number of alleles in a population,” Molecular Ecology,
vol. 12, pp. 3157-3164, 2003.
[32] D. R. Tarpy, R. Nielsen, and D. I. Nielsen, “A scientific note on the revised estimates of
effective paternity frequency in Apis,” Insectes Sociaux, vol. 51, pp. 203-204, 2004.
[33] P. Pamilo, “Polyandry and allele frequency differences between the sexes in the ant
Formica aquilonia.” Heredity, vol. 70, 472-480, 1993.
[34] P. Pamilo, and R. H. Crozier, “Reproductive skew simplified,” Oikos, vol. 75, pp. 533-
535, 1996.
[35] E. O. Wilson, The Insects Societies, Cambridge, Harvard University Press, 1971.

72
[36] R. L. Trivers, and H. Hare, “Haplodiploidy and the evolution of social insects”, Science,
vol. 191, pp. 294-263, 1976.
[37] R. H. Crozier, and E. J. Fjerdingstad, “Polyandry in social Hymenoptera: disunity in
diversity?” Annales Zoologica Fennici, vol. 38 pp. 267-285, 2001.
[38] R. Jaffé, D. J. Kronauer, F. B. Kraus, J. J. Boomsma, R. F. A. Moritz, “Worker caste
determination in the army ant Ection burchellii.” Biology Letters, vol. 3, pp. 513-516,
2007.
[39] R. E. Page Jr., “The evolution of multiple mating behavior by honey bee queens (Apis
mellifera),” Genetics, vol. 96, pp. 263–273, 1980.
[40] E. van Wilgenburg, G. Driessen, and L. W. Beukeboom, “Single locus complementary
sex determination in Hymenoptera: an unintelligent design?,” Frontiers in Zoology,
vol. 3 pp. 1-15, 2006.
[41] A. Zayed, L. Packer, “Complementary sex determination substantially increases
extinction proneness of haplodiploid populations,” Proceedings of the National
Academy of Sciences, vol. 30, pp.10742–10746, 2005.
[42] R. H. Crozier, and R. E. Page, “On being the right size: male contributions and multiple
mating in social Hymenoptera,” Behavioral Ecology and Sociobiology, vol. 18, pp.
105-115, 1985.
[43] M. A. Nowak, C. E. Tarnita, and E. O. Wilson, “The evolution of sociality,” Nature, vol.
466 (7310), pp. 1057-1062, 2010.
[44] P. Nonacs, “Monogamy and high relatedness do not preferentially favor the evolution of
cooperation,” BMC Evolutionary Biology, 11:58, 2011

73
II. 5. Capítulo 5: DNA Barcoding do grupo Melipona seminigra revela que a
distância genética entre as subespécies é alta.
Francini, I.B.; Capurucho, J.M.; Machado, A.S.C.; Nunes-Silva, C.G.; Batista, J.S.; Costa, L.;
Carvalho-Zilse, G. A. DNA Barcoding of the Melipona seminigra group reveals high genetic
distances among them. Apidologie (nota científica)
Abstract
The stingless bee Melipona (Michmelia) seminigra, a polytypic species widely distributed in
Amazon Basin, is a honey-producer and essential pollinator of native flora and crop. We
sequenced partially the cytochrome c oxidase subunit 1 (COI) gene in six subspecies of M.
seminigra (M. seminigra abunensis, M. seminigra merrillae, M. seminigra pernigra, M.
seminigra seminigra, M. seminigra ssp1 and M. seminigra ssp2) and added public data
(sequences of ten other Melipona species), then we carried out data analyses. We aimed to test
COI to resolve the Melipona seminigra complex boundaries and compare it with interspecific
genetic divergence values. The genetic distance observed inter subspecies ranges from 0.4 to
2.7% (average 1.80 % ± 0.47) and inter species from 0.2 to 2.9% (average 2.13 % ± 0.5). The
average haplotype diversity was of 0.8770 ± 0.0140 and the average nucleotide diversity of
0.0166 ± 0.0004. Results showed well delimited clusters for all of the subspecies, which
indicates that the mtDNA fragment herein used has barcode property to identify Melipona
seminigra subspecies. The accurate identification of M. seminigra subspecies is essential for
its adequate handling and its conservation. Our study also suggests that a more comprehensive
study of the speciation of M. seminigra in the Amazon basin might be worthwile.
Keywords: DNA barcode/ Stingless bees/ mtDNA/ COI diversity/ eusociality

74
II.5.1. Introducão
As abelhas sem ferrão (Apidae, Meliponini) são abelhas eussociais avançadas, têm
distribuição tropical e subtropical em todo o mundo, com maior diversificação nos
Neotrópicos, marcadamente na bacia Amazônica (Roubik 1989; Michener 2007). O gênero
com maior riqueza de espécies é o gênero Melipona, abelhas geralmente robustas; com
tamanho médio (pelo menos 7 mm); possuem o ápice das asas, geralmente, não ultrapassando
a extremidade distal do metassoma (Silveira et al. 2002). As abelhas sem ferrão têm papel
significativo como polinizadores da flora nativa (Roubik 1989) e plantas cultivadas (Heard
1999; Slaa et al. 2006). São abelhas que vivem em colônias perenes que se caracterizam por:
sobreposição de gerações, mãe e filhas no mesmo ninho; divisão do trabalho reprodutivo;
cuidado cooperativo para com a prole; castas (rainha e operária) morfologicamente distintas
(Wilson 1971; Crozier e Pamilo 1996; Michener 2007). O tamanho da colônia varia de
algumas dezenas (menos de 10) a centenas de milhares de operária (Michener 2007). Muitas
espécies nidificam em ocos de troncos, outras constroem ninhos expostos e atados a troncos
de árvores; existem espécies que nidificam abaixo da superfície do solo e espécies que vivem
associadas à colônias de outros insetos sociais (Roubik 1989).
Melipona seminigra Friese é uma espécie politípica com ampla distribuíção na bacia
Amazônica, seguindo os grandes rios e seus tributários (Camargo e Pedro 2013). Existem sete
subespécies de M. seminigra, quatro delas descritas (M. s. abunensis, M. s. merrillae, M. s.
pernigra e M. s. seminigra) e três que não foram descritas até o presente (Camargo and Pedro
2013). É considerada boa produtora de mel (1,87 a 5,00 kg de mel/colônia/ano) e tem baixa
agressividade, por isto, é a abelha sem ferrão mais cultivada pelos povos da Amazônia
brasileira, como alternativa econômica e como medida de conservação (Carvalho-Zilse e
Nunes-Silva 2012). M. seminigra pode ser identificada pelos seguintes caracteres
morfológicos: bandas abdominais ausentes ou vestigiais; os dois primeiros segmentos
abdominais glabros no disco, a partir desses para a extremidade distal, lisos e brilhantes;
extremidades laterais do abdome acumulando cerdas grossas a partir dos segmentos 3-5, até
mesmo nos dois primeiros; pilosidade curta escassa ou quase nula; coloração principalmente
laranja a marron. Note-se que a subespécie M. s. pernigra é preta com o primeiro segmento
abdominal amarelado.
O nosso objetivo foi a delimitação das subespécies de M. seminigra usando o gene
citocromo c oxidase subunidade 1 (COI) (Hebert 2003a, 2003b). Formulamos a hipótese que
estas subespécies encontram-se geograficamente separadas por um longo tempo, de forma

75
que, a diversidade do DNA barcoding entre elas é suficiente para identificá-las
molecularmente.
II.5.2. Material e Métodos
II.5.2.1. Dados de amostragem de abelhas
Foram amostrados trinta e um ninhos de Melipona (Michmelia) seminigra (sete ninhos
de M. s. abunensis cf; três ninhos de M. s. merrillae cf; quatro ninhos de M. s. pernigra; sete
ninhos de M. s. seminigra cf; três ninhos de M. s. ssp1 e sete ninhos de M. s. ssp2), uma
operária por ninho. Todas as amostras foram coletadas pelos autores. Ninhos selvagens foram
amostrados acima de quatro metros do solo, a maioria deles a cerca de vinte e cinco metros de
altura. Os locais da amostragem do complexo de subespécies de M. seminigra estão descritos
na Figura 1. As subespécies analisadas foram identificadas morfologicamente, por
comparação com espécimes identificados por especialistas e depositdos na Coleção
Entomológica do Instituto Nacional de Pesquisas da Amazônia (INPA), Manaus, Amazonas,
Brasil. Todas as amostras foram preservadas em etanol 96% e armazenadas em freezer a -
20ºC.
Figura 1. Sítios de amostragem de Melipona seminigra, na bacia Amazônica. Estrêlas pretas, M. s.
abunensis cf (Porto Velho, RO); círculo branco, M. s. merrillae cf (GPA/INPA, Manaus, AM);
triângulo preto, M. s. ssp1 (Puraquequara, Manaus, AM); quadrado branco, M. s. pernigra (Belterra,
PA); círculo preto, M. s. seminigra cf (Boa Vista do Ramos e Maués, AM); quadrado preto, M. s. ssp2
(Parque Nacional do Jaú, AM)

76
II.5.2.2. Análise de DNA e sequenciamento
O DNA genômico foi extraído de tecido do tórax de cada operária, usando-se kit de
purificação de DNA genômico (Promega), baseado no protocolo do fabricante. O DNA foi
quantificado em gel de agarose a 1%. Um fragmento do gene citocromo c oxidase subunidade
1 (COI), foi amplificado nas trinta e uma operárias de M. seminigra, usando-se o par de
primers modificado de Simon et al. (1994) CI-F 1632 (5’TGTCAAATTTATAAT3’) e CI-R-
2191 (5’GGTAAAATTAAAATATAAACTTC3’) (Kambhampati e Smith 1995). As reações
de polimerase em cadeia (PCRs, polymerase chain reactions) foram feitas em um volume
total de 25μL, usando 2,0μL de DNA a 50ng, 2,5μL de tampão10x, 2,0μL MgCl2 a 50mM,
5.0μL de DNTPs a 1μM, 2μM de primer forward, 2μM de primer reverse, 0.2μL (5U/μL) de
polimerase Taq DNA e 8,3μL de água deionizada. A ciclagem das PCRs foi feita com
desnaturação inicial por dois minutos a 95°C, seguido por 30 ciclos de 30 s a 95°C, 1 min a
45°C, 1 min a 72°C e 10 min a 72°C de extensão final. Os produtos de PCR foram
visualizados em gel de agarose a 1% e purificados usando-se polietileno glicol 8000/etanol
(Paithankar e Prasad 1991). O sequenciamento foi realizado em ambos os sentidos, usando
sequenciador ABI 3130 xl (Applied Biosystems) baseado no protocolo do fabricante. Para o
sequenciamento usamos o protocolo de ciclagem de Platt et al. (2007).
II.5.2.3. Análise de dados: avaliação das sequências e delimitação de grupos
Sequências parciais de consenso do COI foram geradas usando o programa SeqScape
2.7 (Applied Biosystems). Após eliminação das regiões terminais, obtivemos uma matriz final
de subespécies com 526 bp (bp, base pairs). Então, adicionamos sequências públicas
(baixadas do GenBank) de espécies congenéricas do mesmo subgênero (M. fasciata, M. illota,
M. fulva, M. lateralis, M. fuscopilosa, M. sp MP70, M. sp. grupo eburnea, M. costaricaensis,
M. scutellaris, M. seminigra atrofulva) (Ramírez et al. 2010). O alinhamento das sequências
foi feito utilizando o ClustalW (Thompson et al. 1994) no BioEdit (Hall 1999) e comparado
pelo método Neighbor-Joining (NJ), corrigido por Kimura-2-Parâmetros (K2P) (Kimura
1980) implementado no MEGA 5 (Tamura et al. 2011). Consideramos duas matrizes, uma de
subespécies mais o grupo externo, outra formada com as seis subespécies mais as espécies e o
grupo externo. Como grupo externo, usamos M. crinita (sequência baixada do GenBank). A
reconstrução filogenética foi feita por máxima parcimônia (MP) usando o PAUP 4.0
(Swofford 2002) e máxima verossimilhança (ML) usando o RA x ML (Online) (Stamatakis et

77
al. 2008). O modelo de evolução molecular selecionado foi o GTR+I+G usando o programa
MODELTEST versão 3.7 (Posada e Crandall 1998) de acordo com o Critério de Informação
Akaike (AIC, Akaike Information Criterion). Para uma topologia de árvore com o menor
número de mudanças de caráteres, usamos uma busca heurística com bissecção e reconexão
de árvore (TBR, Tree Bissection - Reconection). Os valores de suporte nos nós foram obtidos
com 1.000 réplicas de bootstrap. Nós testamos a existência de mistura entre as subespécies,
usando uma análise de agrupamento implementada no BAPS5 (BAPS, Bayesian Analysis of
Population Structure) (Corander e Tang 2007; Corander et al. 2008) e a matriz de
subespécies. Primeiro, rodamos uma análise de mistura, que identificou o número de clusters
do grupo M. seminigra, para nosso conjunto de dados. Esta análise foi executada várias vezes
para confirmar a convergência dos resultados. Então, usamos os resultados da mistura para
executar uma análise de mistura adicional (admixture analysis), usando 100 réplicas, 200
referências individuais e 10 interações por indivíduo. Para executar os cálculos das distâncias
genéticas entre as subespécies usamos MEGA5. Para avaliar a riqueza de haplótipos, a
diversidade nucleotídica e a variabilidade genética, usamos o programa DNAsp v 5.0
(Librado e Rozas 2009).
II.5.3. Resultados
As sequências foram depositadas no GenBank sob os números de acesso KF113947 -
KF114020. Na matriz total, com 526 bp, do complexo de subespécies de Melipona seminigra,
encontramos 28 sítios variáveis e 30 mutações informativas. Não observamos inserções nem
deleções em nossas sequências, indicando ausência de pseudogenes (Gaziev e Shaikhaev
2010), nem stop códons. Observamos um total de 13 haplótipos, resultando em uma
diversidade haplotípica de 0,8770 ± 0,0140 e uma diversidade de nucleotídica de 0,0166 ±
0,0004 (Tabela 1). Encontramos seis clusters bem delimitados para as subespécies de
Melipona seminigra analisadas, evidenciado pela análise de agrupamento por admixture
(Figura 2) e análise de similaridade por Neighbor-Joining (NJ) (Figura 3).
Os valores de distância genética observados entre as espécies de Melipona
(Michmelia) e entre as subespécies do grupo M. seminigra estão descritos nas Tabelas 1 e 2.
Entre subespécies a distância genética variou de 0,4 a 2,7% (em média 1,8 % ± SD 0,47) e
entre espécies de Melipona do mesmo subgênero, a divergência foi de 0,2 a 2,9% (em média
2,13% ± SD 0,50). Para cálculo de distância genética (Tabela 1) e análise de agrupamento
(Figura 2), usamos as sequências específicas da Tabela 2. Algumas espécies foram
substituídas na análise de similaridade (Figura 3), por recomendação de revisores.
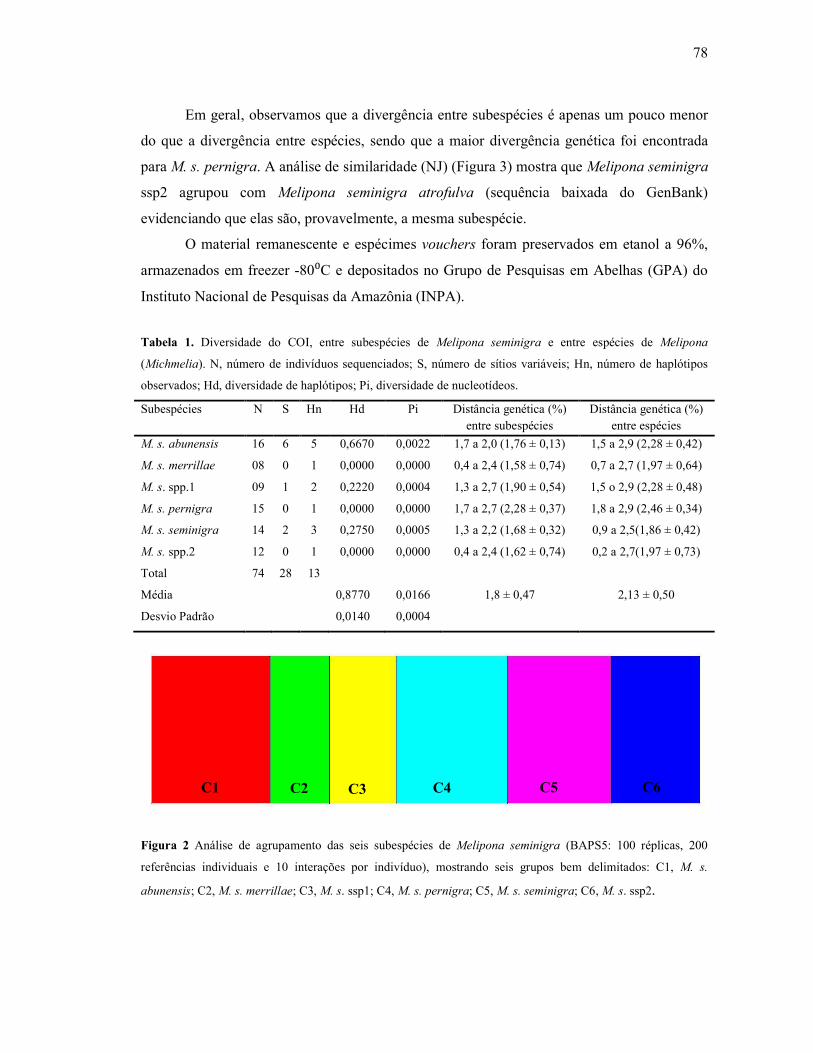
78
Em geral, observamos que a divergência entre subespécies é apenas um pouco menor
do que a divergência entre espécies, sendo que a maior divergência genética foi encontrada
para M. s. pernigra. A análise de similaridade (NJ) (Figura 3) mostra que Melipona seminigra
ssp2 agrupou com Melipona seminigra atrofulva (sequência baixada do GenBank)
evidenciando que elas são, provavelmente, a mesma subespécie.
O material remanescente e espécimes vouchers foram preservados em etanol a 96%,
armazenados em freezer -80⁰C e depositados no Grupo de Pesquisas em Abelhas (GPA) do
Instituto Nacional de Pesquisas da Amazônia (INPA).
Tabela 1. Diversidade do COI, entre subespécies de Melipona seminigra e entre espécies de Melipona
(Michmelia). N, número de indivíduos sequenciados; S, número de sítios variáveis; Hn, número de haplótipos
observados; Hd, diversidade de haplótipos; Pi, diversidade de nucleotídeos.
Subespécies N S Hn Hd Pi Distância genética (%)
entre subespécies
Distância genética (%)
entre espécies
M. s. abunensis 16 6 5 0,6670 0,0022 1,7 a 2,0 (1,76 ± 0,13) 1,5 a 2,9 (2,28 ± 0,42)
M. s. merrillae 08 0 1 0,0000 0,0000 0,4 a 2,4 (1,58 ± 0,74) 0,7 a 2,7 (1,97 ± 0,64)
M. s. spp.1 09 1 2 0,2220 0,0004 1,3 a 2,7 (1,90 ± 0,54) 1,5 o 2,9 (2,28 ± 0,48)
M. s. pernigra 15 0 1 0,0000 0,0000 1,7 a 2,7 (2,28 ± 0,37) 1,8 a 2,9 (2,46 ± 0,34)
M. s. seminigra 14 2 3 0,2750 0,0005 1,3 a 2,2 (1,68 ± 0,32) 0,9 a 2,5(1,86 ± 0,42)
M. s. spp.2 12 0 1 0,0000 0,0000 0,4 a 2,4 (1,62 ± 0,74) 0,2 a 2,7(1,97 ± 0,73)
Total 74 28 13
Média
Desvio Padrão
0,8770
0,0140
0,0166
0,0004
1,8 ± 0,47 2,13 ± 0,50
Figura 2 Análise de agrupamento das seis subespécies de Melipona seminigra (BAPS5: 100 réplicas, 200
referências individuais e 10 interações por indivíduo), mostrando seis grupos bem delimitados: C1, M. s.
abunensis; C2, M. s. merrillae; C3, M. s. ssp1; C4, M. s. pernigra; C5, M. s. seminigra; C6, M. s. ssp2.
C1 C5 C6 C4 C3 C2
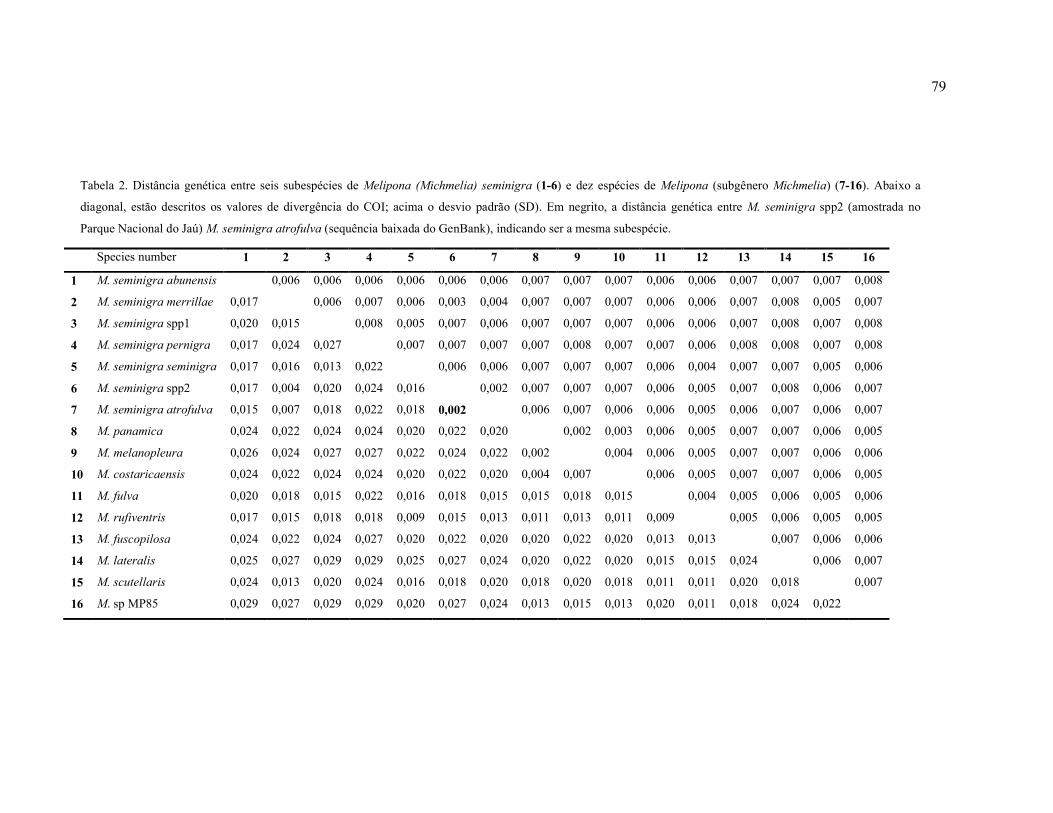
79
Species number 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
1 M. seminigra abunensis 0,006 0,006 0,006 0,006 0,006 0,006 0,007 0,007 0,007 0,006 0,006 0,007 0,007 0,007 0,008
2 M. seminigra merrillae 0,017 0,006 0,007 0,006 0,003 0,004 0,007 0,007 0,007 0,006 0,006 0,007 0,008 0,005 0,007
3 M. seminigra spp1 0,020 0,015 0,008 0,005 0,007 0,006 0,007 0,007 0,007 0,006 0,006 0,007 0,008 0,007 0,008
4 M. seminigra pernigra 0,017 0,024 0,027
0,007 0,007 0,007 0,007 0,008 0,007 0,007 0,006 0,008 0,008 0,007 0,008
5 M. seminigra seminigra 0,017 0,016 0,013 0,022 0,006 0,006 0,007 0,007 0,007 0,006 0,004 0,007 0,007 0,005 0,006
6 M. seminigra spp2 0,017 0,004 0,020 0,024 0,016
0,002 0,007 0,007 0,007 0,006 0,005 0,007 0,008 0,006 0,007
7 M. seminigra atrofulva 0,015 0,007 0,018 0,022 0,018 0,002
0,006 0,007 0,006 0,006 0,005 0,006 0,007 0,006 0,007
8 M. panamica 0,024 0,022 0,024 0,024 0,020 0,022 0,020 0,002 0,003 0,006 0,005 0,007 0,007 0,006 0,005
9 M. melanopleura 0,026 0,024 0,027 0,027 0,022 0,024 0,022 0,002
0,004 0,006 0,005 0,007 0,007 0,006 0,006
10 M. costaricaensis 0,024 0,022 0,024 0,024 0,020 0,022 0,020 0,004 0,007 0,006 0,005 0,007 0,007 0,006 0,005
11 M. fulva 0,020 0,018 0,015 0,022 0,016 0,018 0,015 0,015 0,018 0,015 0,004 0,005 0,006 0,005 0,006
12 M. rufiventris 0,017 0,015 0,018 0,018 0,009 0,015 0,013 0,011 0,013 0,011 0,009
0,005 0,006 0,005 0,005
13 M. fuscopilosa 0,024 0,022 0,024 0,027 0,020 0,022 0,020 0,020 0,022 0,020 0,013 0,013 0,007 0,006 0,006
14 M. lateralis 0,025 0,027 0,029 0,029 0,025 0,027 0,024 0,020 0,022 0,020 0,015 0,015 0,024
0,006 0,007
15 M. scutellaris 0,024 0,013 0,020 0,024 0,016 0,018 0,020 0,018 0,020 0,018 0,011 0,011 0,020 0,018
0,007
16 M. sp MP85 0,029 0,027 0,029 0,029 0,020 0,027 0,024 0,013 0,015 0,013 0,020 0,011 0,018 0,024 0,022
Tabela 2. Distância genética entre seis subespécies de Melipona (Michmelia) seminigra (1-6) e dez espécies de Melipona (subgênero Michmelia) (7-16). Abaixo a
diagonal, estão descritos os valores de divergência do COI; acima o desvio padrão (SD). Em negrito, a distância genética entre M. seminigra spp2 (amostrada no
Parque Nacional do Jaú) M. seminigra atrofulva (sequência baixada do GenBank), indicando ser a mesma subespécie.

80
Figura 3. Árvore de similaridade, por Neighbor-Joining (NJ), para as subespécies de M. seminigra e espécies de
Melipona (Michmelia) (sequências baixadas do GenBank). Valores de suporte (nos nós) foram obtidos por 1.000
réplicas de bootstrap. Como grupo externo foi utilizado Melipona crinita (sequência baixada do GenBank).
Barra, em negrito, 0,5 cm.
5A2 5C1 5B1 5D1 5F5 5H3 5E1
EU163138 M. s. atrofulva 2Ca1 2Ka1 2Pa1 1H1
1 G3 1E1 1B1 1F1 1C1 1D1
2U11 2V1 2T2
4F1 4A4 4H1 4B1 4G1
4D1 4E1
EU163121 M. costaricaensis EU163159 M. sp grupo eburnea
EU163136 M. fuscopilosa EU163152 M. scutellaris
EU163125 M. fulva EU163144 M. lateralis
JX869617M. fasciata EU163148 Melipona sp MP70
EU163167 Melipona illota 3Bb1 3Ab1 3Ca1 3Aa25 3Cb1 3Ba1 3Da2
EU163133 Melipona crinita
99
99
38
98
7 9
4 6
62 79
60
65
79
67
41
30
18
26
58
14 25
21 25
20
M. s. pernigra
M. s. seminigra
M. s. ssp1
M. s. abunensis
M. s. merrillae
M. s. ssp2

81
II.5.4. Discussão e conclusões
O gene mitocôndrial citocromo c oxidase subunidade 1 (COI) foi usado com sucesso
na delimitação das subespécies de Melipona seminigra. As análises de agrupamento, por
admixture (Corander e Tang 2007; Corander et al. 2008) (Figura 2) e de similaridade, por NJ
corrigido por K2P (Kimura 1980) (Figura 3) mostraram que as subespécies analisadas
formam seis grupos perfeitamente distintos. O gene COI tem se mostrado uma ferramenta
eficiente para a identificação de espécies em vários grupos animais (Hebert et al. 2003a, b;
Hajibabaei et al. 2006, 2007; Waugh 2007), com o limiar de divergência genética entre
espécies variando, de acordo com o nível taxonômico analisado e o grupo animal (Avise e
Aquadro 1982). Para Lepidoptera, Hajibabaei et al. (2006) observaram uma divergência entre
espécies de 3%, exceto para espécies congenéricas, que mostraram uma divergência genética
de 0,6 a 2,0% (evidenciando origem recente). Garnery et al. (1992) observaram uma média de
1,3% de divergência para linhagens de Apis mellifera, evidência de isolamento geográfico por
longo tempo. Para abelhas sem ferrão do gênero Scaptotrigona foi observado uma média de
2,79% e 0,7% de divergência genética interespecífica e intraespecífica, respectivamente
(Hurtado-Burillo et al. 2013).
No grupo de subespécies de Melipona seminigra, encontramos uma divergência
genética entre as subespécies variando de 0,4 % (entre M. s. merrillae e M. s. ssp2) a 2,7 %
(entre M. s. pernigra e M. s. ssp1) (Tabela 1). A % de divergência genética entre subespécies
foi, em média, 1,80 ± 0,47 SD e entre espécies do mesmo subgênero, foi, em média, 2,13 % ±
0,50 SD. Comparando a divergência do COI entre as subespécies de M. seminigra com a
divergência de espécies congenéricas (mesmo subgênero) podemos inferir que as subespécies
de M. seminigra encontram-se isoladas por um longo tempo.
Nossos resultados indicam que M. seminigra atrofulva (Ramírez et al. 2010) e M.
seminigra ssp2 são o mesmo taxon (Figura 3). Destacamos também o valor de divergência
(0,2 %) observado entre M. melanopleura e M. panamica (o mesmo entre M. s. atrofulva e M.
seminigra ssp2) e o valor de divergência (0,7 %) observado entre Melipona melanopleura e
Melipona costaricaensis (Tabela 2), que de acordo com Camargo e Pedro (2013) são
sinônimos.
Acreditamos que outras subespécies de M. seminigra, não amostradas até agora,
podem ser encontradas na Bacia Amazônica, se um esforço amostral maior for feito.
Considerando a importância de M. seminigra como polinizador e produtor de mel, aspectos de

82
sua biologia e comportamento merecem ser vistos com cuidado. M. seminigra, muito
provavelmente, tem status derivado em relação à outras espécies de Melipona (Francini et al.
2011), de modo que, estudos sobre a sua história evolutiva pode nos ensinar sobre manejo
adequado e a preservação de seu habitat.
Agradecimentos
Agradecemos ao Dr. Hanno Schäfer (Technical University of Munich) por sugestões e
correções que melhoraram nosso manuscrito. Pelo suporte financeiro, agradecemos ao
Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), à Fundação de
Amparo a Pesquisa do Estado do Amazonas (FAPEAM), e ao Instituto Nacional de Pesquisas
da Amazônia (INPA).
Referências
Avise, J.C. Aquadro, C.F. (1982) A comparative summary of genetic distances in the
vertebrates: patterns and correlations. Evol. Biol. 15, 151-185
Camargo, J.M.F., Pedro, S.R.M. (2013) Meliponini Lepeletier, 1836. In: Moure J.S., Urban,
D., Melo, G.A.R. (Orgs). Catalogue of Bees (Hymenoptera, Apoidea) in the
Neotropical Region (http://www.moure.cria.org.br/catalogue acessed in february
3/2013)
Carvalho-Zilse, G.A., Nunes-Silva, C.G. (2012). Threats to the Stingless Bees in the Brazilian
Amazon: How to Deal with Scarce Biological Data and an Increasing Rate of
Destruction. In: Florio, R. M (Ed.). Bees: Biology, Threats and Colonies. Nova
Science Publishers, Chapter 4, pp.147-168
Corander, J., Tang, J. (2007). Bayesian analysis of population structure based on
linked molecular information. Math Biosci 205, 19-31
Corander. J., Marttinen, P., Sirén, J., Tang, J. (2008). Enhanced Bayesian modelling in
BAPS software for learning genetic structures of populations. BMC Bioinformatics 9, 539
Crozier, R.H., Pamilo, P. (1996) Evolution of social insect colonies. Sex allocation and kin
Selection, Oxford University Press, Oxford
Francini, I.B., Gross, M.C.; Nunes-Silva, C.G. Carvalho-Zilse,G.A. (2011) Cytogenetic
analysis of the Amazon stingless bee Melipona seminigra merrillae reveals different
chromosome number for the genus. Sci. Agric. 68(5), 592-593
Garnery, L., Cornuet, J.M., Solignac, M. (1992) Evolutionary history of the honey bee Apis
mellifera inferred from mitochondrial DNA analysis. Mol. Ecol. 1, 145-154

83
Gaziev, A.I., Skaikhaev, G.O. (2010) Nuclear mitochondrial pseudogenes. Mol. Biol. 44, 358-
368
Hajibabaei, M., Janzen, D.H., Burns, J.M., Hallwachs, W., Herbert, P.D.N. (2006) DNA
barcodes distinguish species of tropical Lepidoptera. Proc. Natl. Acad. Sci. U.S.A.
103(4), 968-971
Hajibabaei, M., Singer, G.A.C., Hebert, P.D.N., Hickey, D.A. (2007) DNA barcoding: how it
complements taxonomy, molecular phylogenetics and population genetics. Trends
Genet. 23(4), 167-172
Hall, T.A. (1999) BioEdit: a user-Friendly biological sequence alignment editor and analysis
program for Windows 95/98/NT
Heard, A. (1999) The role of stingless bees in crop pollination. Ann. Rev. Entomol. 44, 183-
206
Hebert, P.D.N., Cywinska, A., Ball, S.L., deWaard, J.R. (2003a) Biological identification
through DNA barcodes. Proc. R. Soc. Lond. Ser. B, 270, 313-322
Hebert, P.D.N., Ratnasingham S., deWaard, J.R. (2003b) Barcoding animal life: cytochrome c
oxidase subunit 1 divergences among closely related species. Proc. R. Soc. Lond. Ser.
B, 270, S96-S99
Hurtado-Burillo, M., Ruiz, C., May-Itzá, W., Quezada-Eúan, J.J G., De la Rúa, P. (2013)
Barcoding stingless bees: genetic diversity of the economically important genus
Scaptotrigona in Mesoamerica. Apidologie 44(1) 1-10
Kambhampati, S., Smith, P.T. (1995) PCR primers for the amplification of four insect
mitochondrial gene fragments. Ins. Mol. Biol. 4, 233-236
Kimura, M. (1980) A simple method for estimating evolutionary rates of base substitutions
through the comparative studies of sequence evolution. J. Mol. Evol. 16, 111-120
Librado, P., Rozas, J. (2009) DnaSP v5: A software for comprehensive analysis of DNA
polymorphism data. Bioinformatics 25, 1451–1452.
Michener, C.D. (2007) The Bees of the World, Johns Hopkins University Press, Baltimore
Paithankar, K.R., Prasad, K.S.N., (1991) Precipitation of DNA by polyethylene glycol and
ethanol. Nucleic Acids Res. 19(6), 1346
Platt, A.R., Woodhall, R.W., George Jr., A.L. (2007) Improved DNA sequencing quality and
efficiency using an optimized fast cycle sequencing protocol. Biotechniques 43, 58-62
Posada, D., Crandall, K.A. (1998). MODELTEST: Testing the model of DNA substitution.
Bioinformatics 14, 817–818

84
Ramírez, S.R., Nieh, J.C., Quental, T.B., Roubik, D.W., Imperatriz-Fonseca, V.L., Pierce, N.
E. (2010) A molecular phylogeny of the stingless bee genus Melipona (Hymenoptera:
Apidae). Mol. Phylogenet. Evol. 56, 519–525
Roubik, D.W. (1989) Ecology and Natural History of Tropical Bees. Cambridge University
Press, USA
Simon, C., Frati, F., Beckenbach, A., Crespi, B.,Liu, H., Flook, P. (1994) Evolution,
weighting and Phylogenetic utility of mitochondrial gene sequences and a compilation
of conserved polymerase chain reaction primers. Ann. Entomol. Soc. Am. 87, 651-701
Silveira, F.A., Melo, G.A.R., Almeida, E.A. (2002) Brazilian Bees: Systematic and
Identification (in Portuguese). Araucaria Foundation, BH, MG, Brazil
Slaa, E.J., Chaves, L.A.S., Malgodi-Braga, K.S., Hofstede, F.E. (2006) Stingless bees in
applied pollination: practice and perspectives. Apidologie, 37, 293-315
Stamatakis, A., Hoover, P., Rougemont, J. (2008) A Rapid Bootstrap Algorithm for the
RAxML Web-Servers Syst. Biol. 75(5), 758-771
Swofford, D.L. (2002) PAUP* Phylogenetic Analysis Using Parsimony and other methods.
Sinauer Associates, Sunderland, Massachusetts
Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei, M., Kumar, S. (2011) MEGA5:
molecular evolutionary genetics analysis using maximum likelihood, evolutionary
distance, and maximum parsimony methods. Mol. Biol. Evol. 28, 2731-2739
Thompson, J.D., Higgins, D.G., Gibson, T.J. (1994) CLUSTAL W: improving the sensitivity
of progressive multiple sequence alignment through sequence weighting, position-
specific gap penalties and weight matrix choice. Nucleic Acids Res. 22, 4673-4680
Waugh, J. (2007) DNA barcoding in animal species, progress, potencial and pitfalls.
Bioessays, 29, 188-197
Wilson, E.O. (1971) The Insects Societies. Cambridge, Harvard University Press

85
Conclusões Gerais
- A partir da análise de DNA microssatélites em 576 operárias de Melipona seminigra,
amostradas em colônias de três subespécies (M. s. abunensis, M. s. merrillae e M. s.
seminigra) em populações naturais e manejadas, concluimos que o sistema de acasalamento
de M. seminigra é poliandria. A frequência de paternidade (3 a 11 acasalamentos) e o número
de paternidade efetiva (2 a 11,613) evidenciam que rainhas de M. seminigra são inseminadas
por dois ou mais machos e, frequentemente, por mais de seis machos (poliandria extrema),
tanto em colônias naturais como em colônias manejadas, conforme descrito nos capítulos II,
III e IV e na tabela de resultados gerais (Tabela 1, Apêndice).
- O baixo grau de parentesco intracolonial observado em colônias de M. seminigra (as
operárias compartilharam, em média, 34,6 % de seus genes por descendência comum) (Tabela
1) corrobora o esperado para Hymenoptera sociais haplo-diplóides com sistema de
acasalamento poliândrico, é fato novo em abelhas sem ferrão (Apidae, Meliponini).
- Poliandria em M. seminigra, assim como em outros Hymenoptera sociais, explica-se porque
os benefícios da diversidade genética, que certamente são maiores do que os custos de
múltiplos acasalamentos.
- Os valores de skew (S < 0,5) (Tabela 1, Apêndice) indicam que na maioria das colônias a
rainha tende a equalizar a paternidade, maximizando os ganhos da diversidade genética.
Exceto, em três colônias de M. s. seminigra, onde alguns machos monopolizaram a
paternidade (S ˃ 0,5), o que pode ser explicado pelo N amostral usado.
- O aumento da diversidade genética em colônia de M. seminigra, resultante de múltiplos
acasalamentos da rainha, aumenta o fitness da colônia em várias direções, argumentamos a
favor da hipótese de ser uma estratégia evolutiva para minimizar os efeitos nocivos da
produção de macho diplóide, descritos para M. seminigra em estudo prévio (Capítulo I).
- A análise de DNA barcoding das seis subespécies de M. seminigra amostradas neste
trabalho, revelou que a distância genética entre as subespécies é alta, indicando que estão
separadas geograficamente por longo tempo. As subespécies foram perfeitamente delimitadas
pelo COI, o que também é fato novo, uma vez que, o DNA barcoding tem sido usado com

86
sucesso na identificação de espécies e taxons superiores e não de subespécies, porque
diversidade do COI entre subespécies é, em princípio, baixa e insuficiente para delimitá-las.
- O conhecimento produzido ao longo deste trabalho representa uma contribuição significativa
a respeito do sistema de acasalamento de M. seminigra, é essencial, no que diz respeito ao
manejo e conservação desta espécie. No entanto, a implicação maior de nossos resultados
refere-se à evolução de poliandria em Hymenoptera sociais.
Perspectivas:
- Os resultados das análises moleculares (DNA microssatélite e DNA barcoding) indicam que
M. seminigra é um livro aberto a muitos novos estudos, tais como: comportamento das
operárias em colônias que produzem macho diplóide; novos estudos sobre o sistema de
acasalamento; relação do sistema de acasalamento com a produtividade e tamanho das
colônia; citogenética do grupo de subespécies; taxonomia e filogeografia.

87
Referências
Absy, M.L.; Kerr, W.E. 1977. Some plants visited by workers of Melipona seminigra
merrillae to obtain pollen, at Manaus. Acta Amazonica, 7: 309-315 (in Portuguese)
Adams, J.; Rothman, E.D.; Kerr, W.E.; Paulino, Z.L. 1977. Estimation of the number of sex
alleles and queen matings from diploid male frequencies in a population of Apis
mellifera. Genetics, 86: 583-596.
Alexander, B.A. 1992. An exploratory analysis of cladistic relationships within the
superfamily Apoidea, with special reference to sphecid wasps. Journal of
Hymenoptera Research 1: 25-61
Alves, D.A.; Menezes, C.; Imperatrix-Fonseca, V.L.; Wenseleers, T. 2011. First Discovery o
a rare polygyne colony in the stingless bee Melipona quadrifasciata (Apidae,
Meliponini). Apidologie, 42: 211-213.
Bacelar-Lima, C.G.; Freire, D.C.B.; Coletto-Silva, A.; Costa, K.B.; Laray, J.P.B.; Vilas-Boas,
H.C.; Carvalho-Zilse, G.A. 2006. Melitocoria de Zygia racemosa (Ducke) Barneby &
Grimes por Melipona seminigra merrillae Cockerell, 1919 y Melipona compressipes
manaosensis Schwarz, 1932 (Hymenoptera, Meliponina) en la Amazonía Central,
Brasil. Acta Amazonica, 36(3): 343-348.
Barth, F.G. 1985. Insects and flowers. Princenton University Press, Princeton, New Jersey,
USA, 297pp.
Batra, S.W.T. 1966. Nests and social behavior of halictine bees of India. Indian Jounal of
Entomology, 28:375-393.
Boomsma, J.J.; Ratnieks, F.L.W. 1996. Paternity in eusocial Hymenoptera. Philosophical
Trasantions of the Royal Society of London, séries B, 351(1342): 947-975.
Brosi, B.J. 2009. The complex responses of social stingless bees (Apidae: Meliponini) to
tropical deforestation. Forest Ecology and Management 258: 1830-1837.
Brosi, B.J.; Daily, G.C.; Shih, T.M.; Oviedo, F.; Durán, G. 2008. The effects of forest
fragmentation on bee communities in tropical countryside. Journal of Applied
Ecology, 45:773–783.
Camargo, J.M.F.; Pedro, S.R.M. 2013. Meliponini Lepeletier, 1836. In: Moure, J. S.; Urban,
D.; Melo, G.A.R. (Orgs). Catalogue of Bees (Hymenoptera, Apoidea) in the
Neotropical Region. (http://www.moure.cria.org.br/catalogue). Acesso 02/09/2013.
Carvalho, G.A. 2001. The Number of Sex Alleles (CSD) in a Bee 3 Population and its Pratical
Importance (Hymenoptera: Apidae). Journal of Hymenoptera Research, 10(1): 10-15.

88
Carvalho, C.A.L.; Souza, B.A.; Dias, C.S.; Alves, R.M.O.; Melo, A.F.L.; Carvalho-Zilse,
G.A. 2011. Five egg-laying queens in a single colony of brazilian stingless bees
(Melipona scutellaris Latreille). Acta Amazonica, 41(1):123-126.
Carvalho-Zilse, G.A. 2006. Meliponicultura na Amazônia. Anais do VII Encontro sobre
Abelhas (Ribeirão Preto-SP): em CD ROM.
Carvalho-Zilse, G.A.; Nunes-Silva, C.G. 2012. Threats to the Stingless Bees in the Brazilian
Amazon: How to Deal with Scarce Biological Data and an Increasing Rate of
Destruction. In: Florio, R.M (Ed.). Bees: Biology, Threats and Colonies. Nova Science
Publishers, Chapter 4,147-168pp.
Cortopassi-Laurino M.; Imperatriz-Fonseca V.L.; Roubik, D.W.; Dollin, A.; Heard, T.
Aguilar, I.; Venturieri, G.C.; Eardley, C.; Nogueira-Neto, P. 2006. Global
meliponiculture: challenges and opportunities. Apidologie, 37: 275-292.
Crozier, R.H.; Page, R.E. 1985. On being the rigth size: male contributions and multiple
mating in social Hymenoptera. Behavioral Ecology and Sociobiology, 18: 105-115.
Crozier, R. H. e Pamilo, P. 1996. Evolution of Social Insect Colonies Sex Alocation and Kin
Selection. Oxford University Press Inc., New York, USA, 306 pp.
Crozier, R.H.; Fjerdingstad, E.J. 2001. Polyandry in social Hymenoptera: disunity in
diversity? Annales Zoologici Fennici, 38: 267-285.
Dzierzon, J. 1845. On the development of bees. Transl. Eichstadt. Bienenzeitung 1:113.
Francini, I.B. 2009. Variabilidade Genética do loco csd em populações de cativeiro de
Melipona interrupta manaosensis Schwarz, 1932 e Melipona seminigra merrillae
Cockerell, 1919 (Apidae, Meliponini) na Amazônia. Dissertação de Mestrado, Instituto
Nacional de Pesquisas da Amazônia (INPA), Manaus, Amazonas. 93pp.
Francini, I.B., Nunes-Silva, C.G., Carvalho-Zilse, G.A. 2012. Diploid male production of two
amazonian Melipona bees (Hymenoptera: Apidae). Psyche: Journal of Entomology,1:
1-7.
Getz, W.M.; Brückner D.; Parisian, T.R. 1982. Kin structure and the swarming behavior of
the honey bee Apis mellifera. Behavioral Ecology and Sociobiology, 10: 265-270.
Hamilton, W.D. 1964. The genetical evolution of social behavior, I e II. Jounal of Theoretical
Biology, 7: 1-52.
Hamilton, W.D. 1970. Selfish and behavior in an evolutionary model. Nature, 228: 1218-
1220.
Hamilton, W.D. 1972. Altruism and related phenomena, mainly in social insects. Annual
Review of Ecology and Systematics, 3: 193-232.

89
Hamilton, W.D.; Axelrod, R.; Tanese, R. 1990. Sexual reproduction as an adaptation to resist
parasites (A review). Proceedings of the National Academy of Science USA, 87: 3566-
3573.
Hasselmann M.; Gempe, T.; Schiott, M.; Nunes-Silva, C.G.; Otte, M.; Beye, M. 2008.
Evidence for the evolutionary nascence of a novel sex determination pathway in
honeybees. Nature, 454: 519-522.
Heard, T.A. 1999. The role of stingless bees in crop pollination. Annual Review of
Entomology, 44: 183-206.
Hughes, W.O.H.; Oldroyid, B.P.; Beekman, M.; Ratnieks, F.L.W. 2008. Ancestral
Monogamy Shows Kin Selection Is Key to the Evolution of Eusociality. Science, 320:
1213-1216.
Hung, A.C.F.; Vinson, S. B.; Summerlin, J. W. 1974. Male sterility in the red imported fire
ant, Solenopsis invicta. Annals of the Entomological Society of America, 67: 909-912.
Keeler, L.; Reeve, H. 1994. Genetic variability, Queen number, and polyandry in social
Hymenoptera. Evolution 48: 694-704.
Kerr, W.E. 1969. Some aspects of the evolution of the social bees. Evolution Biology, 3: 119-
175.
Kerr, W.E. 1987. Sex determination in bees. XXI. Number of XO-heteroalleles in a natural
population of Melipona compressipes fasciculata. Insects Sociaux, 34: 274–279.
Kerr, W.E.; Lello, E. 1962. Sting glands in stingless bees a vestigial character (Hymenoptera:
Apoidea). Journal of the New York Entomological Society, 70: 190-214.
Kerr, W.E.; Zucchi, R.; Nakadaiara, J.T.; Butolo, J.E. 1962. Reproduction in the social bees
Kerr, W.E.; Carvalho, G.A.; Nascimento, V. A. 1996. Abelha Uruçu: Biologia, Manejo e
Conservação. Fundação Acangaú, Paracatu, MG. 144 pp.
Lin, N.; Michener, C.D. 1972. Evolution of sociality in insects. Quarterly Review of Biology,
47: 131-159.
Melo, G.A.R. 1999. Phygenetic relationships and classification of the major lineages of
Apoidea (Hymenoptera), with emphasis on the crabronid wasps. Scientific Papers of
the Natural History Museum of the University of Kansas, 14: 1-55.
Mackensen, O. 1951. Viability and sex determination in the honey bee (Apis mellifera).
Genetics, 36: 500-509.
Michener, C.D. 1969. Comparative social behavior of bees. Annual Rev. Entomol., 14:299-
342.
Michener, C.D. 2007. The Bees of the World. Johns Hopkins University Press, Baltimore.

90
Moure, J.S. 1975. Notas sobre as espécies de Melipona descritas por Lepeletier em 1836
(Hymenoptera-Apidae). Revista. Brasileira de Biologia, 35 (4): 615-623
Moure, J.S.1992. Melikerria e Eomelipona, dois subgêneros novos em Melipona Illiger, 1806,
In: Anais do Encontro Brasileiro sobre Biologia de Abelhas e outros Insetos Sociais.
Naturalia, edição especial: 32-38.
Moure, J.S CMF; Kerr, W. E. 1950. Sugestões para a modificação da sistemática do gênero
Melipona (Hymenoptera, Apoidea). Dusenia, 1(2): 105-131.
Nowak, M.A.; Tarnita, C.E.; Wilson, E.O. 2010. The evolution of eusociality. Nature,
466(7310): 1057-1062
Nunez, C.V.; Oliveira, M.L.; Lima, R.D.; Diaz, I.E.C.; Sargentini Jr, E.;Pereira Jr, O.L.;
Araújo, L.M. 2008. Chemical analyses confirm a rare case of seed dispersal by bees.
Apidologie, 39: 618–626.
Page, R.E. 1980. The evolution of multiple mating behavior by honey bee queens (Apis
mellifera L). Genetics, 96: 263-273.
Page, R.E.; Metcalf, R.A. 1982. Multiple mating, sperm utilization and social evolution.
American Naturalist, 119: 263-281.
Page Jr, R.E.; Robinson, G.E.; Fondrk, M.K.; Nasr, M.E. 1995. Effects of worker genotypic
diversity on honey bee colony and bahavior (Apis mellifera L.). Behavioral Ecology
and Sociobiology, 36: 387-396.
Paxton, R.; Weibschuh, N.; Engels, W.; Hartfelder, K.; Quezada-Euan, J.G. 1999. Not only
single mating in stingless bees. Naturwissenschaften, 86: 143–146.
Paxton, R.J.; Bego, L.R.; Shah, M.M.; Mateus, S. 2003. Low mating frequency of queens in
stingless bee Scaptotrigona postica and worker maternity of males. Behavioral
Ecology and Sociobiology, 53: 174 – 181.
Peters, J.M.; Queller, D.C.; Imperatriz-Fonseca, V.L.; Roubik, D.W. 1999. Mate number, kin
selection and social conflicts in stingless bees and honeybees. Proceedings of Royal
Society of London, 266: 379-384.
Roubik, D.W. 1995. Pollination for cultivated plants in the tropics. FAO, Agricultural
Bulletin, 118: 150-154.
Roubik, D.W. 1989. Ecology and Natural History of Tropical Bees. Cambridge University
Press, USA. 514 pp.
Roubik, D.W.; Weight, L.A.; Bonilla, M.A. 1996. Population genetics, diploid males, and
limits to social evolution of euglossine bees. Evolution, 50:931-935.

91
Schmid-Hempel, P. 1994. Infection and colony viability in social insects. Philosophical
Trasantions of the Royal Society of London, B 346: 313-320.
Schwarz, H.F. 1932. The Genus Melipona: type Genus of the Meliponinae or Stingless Bees.
Bulletin of American Museum of Natural History, LXIII (IV): 231-460.
Silveira, F.A.; Melo, G.A.; Almeida, E.A.B. 2002. Abelhas brasileiras: sistemática e
identificação. Ed. MMA e Fundação Araucária, Belo Horizonte, MG, 253 pp.
Simmons, L.W. 2005. The evolution of polyandry: Sperm Competition, Sperm Selection, and
Offspring viability. Annual Review of Ecology, Evolution, and Systematics, 36: 125-
146.
Slaa, E.J.; Chaves, L.A.S.; Malgodi-Braga, K.S.; Hofstede, F.E. 2006. Stingless bees in
applied pollination: practice and perspectives. Apidologie, 37:293-315.
Strassmann, J. 2001 The rarity of multiplemating by females in the social hymenoptera,
Insectes Sociaux, 48(1): 1-13.
Trivers, R.L.; Hare, H. 1976. Haplodiploidy and the evolution of social insects. Science, 191:
294-263.
Velthuis, H.H.W.; Cortopassi-Laurino, M.; Z. Pereboom, Z.; Imperatriz-Fonseca, V. L. 2003.
The conservative egg of the genus Melipona and its consequences for speciation. In:
Melo, G.A.R. e Alves-dos-Santos, I. (eds). Apoidea Neotropica: Homenagem aos 90
anos de Jesus Santiago Moure. Editora UNESC, 320 pp.
Wattanachaiyinhcharoen, W.; Oldroyd, B.P.; Wongsiri, S.; Palmer, K.; Paar, S. 2003. A
scientific note on the mating frequency of Apis dorsata Fabricius. Apidologie, 34:85-
86.
Whiting, P.W. 1943. Multiple alleles in complementary sex determination of Habrobracon.
Genetics, 28: 365-382.
Wille, A. 1983. Biology of Stingles Bees. Annual Review of Entomology, 28:41-64
Wilson, E.O. 1971. The Insects Societies. Cambridge, Harvard University Press, 548 pp.
Wilson, E.O. 1975. Sociobiology. Cambridge, Massachusetts:Belknap
Wilson, E.O. 1990. Excellence in Ecology. Ecology Institute, Oldendorf/Luhe. Federal
Republic of Germany, 104 pp.
Woyke, J. 1963a What happens to diploid drone larvae in a honey bee colony. Journal of
Apicultural Research, 2: 73-76
Woyke, J. 1963b Rearing and viability of diploid drone larvae. Journal of Apicultural
Research, 2: 77-84, 1963b.

92
Zayed, A. 2004. Effective population size in Hymenoptera with complementary sex
determination. Heredity, 93: 627-630.
Zayed, A.; Roubik, D. W.; Laurence, P. 2003. Use of diploid male frequency data as an
indicator of pollinator decline. Proceedings of Royal Society of London (Suppl.):
03SBL0243.S1-S4.

93
Apêndice – Sobre conclusões gerais
Tabela 1. Paternidade e parentesco em Melipona seminigra, estimativas feitas a partir da genotipagem de 576 operárias. Foram genotipados 4-5 locos microssatélites, como
descrito nos capítulos II, III e IV. Foram amostradas 24 colônias, três subespécies: *, colônia manejada; **, colônia selvagem. Os valores de skew (contribuição de cada pai
com a prole da rainha) evidenciam a tendência a equalização da paternidade, na maioria das colônias.
Subespécie Colônia Operárias genotipadas Paternidade observada Paternidade efetiva skew Grau de parentesco
Melipona seminigra abunensis
1A* 24 09 9,264 - 0,033 0,303
1B* 24 09 10,306 - 0,163 0,298
1C* 24 11 11,613 - 0,061 0,293
1D* 24 08 7,934 0,009 0,313
1E** 24 06 5,779 0,044 0,336
1F** 24 07 6,310 0,115 0,329
1G** 24 11 8,970 0,203 0,306
1H** 24 10 9,944 0,017 0,300
1I** 24 08 7,507 0,070 0,307
M. s. merrillae
2Ba* 24 04 2,496 0,501 0,450
2Da* 24 06 6,938 -0,187 0,322
2Ea* 24 04 3,745 0,085 0,383
2Fa* 24 03 2,948 0,026 0,419
2Ha* 24 07 5,137 0,310 0,347
2Ka* 24 04 4,201 -0,067 0,369
2La* 24 04 4,075 -0,025 0,372
2Pa* 24 04 3,745 0,085 0,383
M. s. seminigra
4A* 24 09 5,781 0,402 0,336
4B* 24 10 9,446 0,059 0,302
4C* 24 07 2,000 0,833 0,500
4E* 24 09 5,884 0,389 0,334
4F* 24 10 4,386 0,623 0,363
4G* 24 09 7,578 0,177 0,315
4H* 24 09 5,781 0,527 0,336
Média Desvio padrão
7,416
2,500 6,324 2,655
0,176 0,028
0,346 0,051





























