Embed Size (px)
Citation preview

INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA – INPA UNIVERSIDADE FEDERAL DO AMAZONAS – UFAM
Programa de Pós-Graduação em Biologia Tropical e Recursos Naturais - PPGBTRN
Efeitos bióticos e abióticos de ambientes alagáveis nas assembléias de aves aquáticas e piscívoras no Pantanal, Brasil
DALCI MAURICIO MIRANDA DE OLIVEIRA
Tese apresentada à Coordenação do Programa de Pós-Graduação em Biologia Tropical e Recursos Naturais, do convênio INPA/UFAM, como parte dos requisitos para obtenção do título de Doutor em Ciências Biológicas, área de concentração em Ecologia.
MANAUS-AM 2006

ii
INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA – INPA UNIVERSIDADE FEDERAL DO AMAZONAS – UFAM
Programa de Pós-Graduação em Biologia Tropical e Recursos Naturais - PPGBTRN
Efeitos bióticos e abióticos de ambientes alagáveis nas assembléias de aves aquáticas no Pantanal, Brasil
DALCI MAURICIO MIRANDA DE OLIVEIRA
ORIENTADOR: Dr. RENATO CINTRA SOARES
Tese apresentada à Coordenação do Programa de Pós-Graduação em Biologia Tropical e Recursos Naturais, do convênio INPA/UFAM, como parte dos requisitos para obtenção do título de Doutor em Ciências Biológicas, área de concentração em Ecologia.
Fonte Financiadora: CAPES (Bolsa PICD de Doutorado), Projeto Ecologia do Pantanal (PEP)
MANAUS-AM 2006

iii
Ficha catalográfica Oliveira, Dalci Mauricio Miranda de
Efeitos bióticos e abióticos de ambientes alagáveis nas assembléias de aves aquáticas e piscívoras no Pantanal, Brasil. / Dalci Mauricio Miranda de Oliveira. – Manaus, 2006.
198 p. : il. Tese (Doutorado) – INPA/UFAM, Manaus,2006.
1- Ecologia de aves 2- Aves piscívoras 3- Efeitos bióticos e abióticos 4 - Pantanal
Mato-grossense 5- Áreas alagáveis CDD19.ed. 598.045 Sinopse: Assembléias de aves aquáticas e piscívoras foram estudadas durante quatro anos no Pantanal Matogrossense. Foram realizados censos populacionais em transectos lineares em 1999 e 2000 ao longo da Estrada Parque Transpantaneira em Poconé-MT, e em 2002 e 2003 foram realizados censos das aves em 20 lagos ao longo da mesma rodovia. Foram determinadas a composição de espécies e a variação da abundância ao longo do ano e em anos subseqüentes. Foi verificado o papel de fatores abióticos (pluviosidade, temperatura do ar, insolação, tamanho e profundidade do lago, temperatura, transparência e pH da água) e bióticos (cobertura e riqueza de espécies de macrófitas aquáticas, e biomassa e riqueza de espécies de peixes) em prever a variação na riqueza e abundância e na composição de espécies de aves aquáticas e piscivoras no Pantanal. Palavras-chave: Ecologia de aves, aves piscívoras, efeitos bióticos e abióticos, áreas alagáveis, Pantanal mato-grossense

1
INTRODUÇÃO GERAL
Os fatores que determinam a estrutura e composição da comunidade de aves têm
recebido extensa e entusiástica atenção, tanto empírica quanto teórica (Kushlan et al., 1985;
Remsen Jr., 1985; Stiles, 1985; Willard, 1985; Wiens, 1989; Bierregaard, 1990; Robinson
& Terbough, 1990; Terbough et al.;1990; Loiselle & Blake, 1991; Cody, 1996; Cintra,
1997; Silva et al., 1997; Caziani et al., 2001).
A competição interespecifica por recursos limitados, entre outros, incluem também
as respostas espécie-especifica às condições ambientais, à predação, ao parasitismo, ao
comensalismo, e às interações mutualísticas, e os eventos históricos (Wiens, 1989). Apesar
da atenção que esses diferentes fatores têm recebido, a importância relativa dos mesmos é
ainda altamente controversa (Johnson, 1996), porque o efeito de nenhum deles ainda foi
satisfatoriamente demonstrado. Além disto, a maioria dos estudos está relacionada a aves
terrestres.
Aves aquáticas são aquelas associadas a ambientes aquáticos na maior parte de suas
atividades, para satisfazer suas necessidades energéticas de dieta e reprodução (Waterbirds
Conservation for the Americas, 2005). A riqueza de espécies e abundância de aves
aquáticas em determinados locais é um dos critérios utilizados pela Convenção Ramsar
para considerá-las como relevantes para conservação da biodiversidade no Planeta
(Ramsar, 2005).
Apesar destas aves serem alvo de muitos estudos há algumas décadas, ainda faltam
estudos de ecologia comportamental, e ecologia evolutiva necessários, para ampliar os
conhecimentos sobre a relação dessas aves com seu hábitat (Hafner, 1997). Segundo este

2
autor estes conhecimentos poderiam ser aplicados para conservação dessas aves e seus
ambientes. As aves aquáticas são muito sensíveis às mudanças do regime hidrológico
(Kushlan, 1976; Morales et al., 1981; Pailisson et al., 2002). A flutuação do nível da água
influencia a estrutura física do habitat, a disponibilidade e o acesso à fonte alimentar e o
aparecimento de local seguro para repouso e nidificação (González, 1996b, 1997; Bancroft
et al., 2002). Todas essas características são fatores determinantes na escolha do local pelas
aves aquáticas.
É provável que o primeiro estudo a levar em consideração a influência do nível dos
corpos de água, sobre a população de aves aquáticas, tenha sido realizado nos Everglades,
na Flórida, na década de 50 (Johnsgaard, 1956). Posteriormente, outras pesquisas
consideraram este fator como importante na estruturação da comunidade de aves aquáticas,
nos Everglades (Kushlan et al., 1975; Kushlan, 1976a b, 1986) e nos lhanos da Venezuela
(Pacheco et al., 1976, Kushlan et al., 1985). Estes autores concluíram que o aumento do
nível e duração da inundação, determina o incremento de aves aquáticas nestes locais, por
disporem de áreas mais extensas para exploração de recursos alimentares. Estudos
posteriores realizados em outras partes do mundo corroboraram este pensamento, como por
exemplo, nos lagos Bogoria, Naivasha, Elmenteria e Nakuru no Quênia (Owino et al.,
2001; Harper et al., 2003); na África do Sul (Velásquez, 1992); na Austrália (Taylor, 2004);
no lago Izunuma-Uchinuma no Japão (Shimada et al., 2000); no lago de Pozuelos na
Argentina (Masciatti, 2001) e na Ilha da Marchantaria no Brasil (Petermann, 1997).
No Brasil existem poucos estudos com aves aquáticas, sendo encontrado cerca de
253 publicações sobre o tema em uma revisão para os últimos 35 anos (Oliveira et al. in
prep.). A maioria dos estudos foi realizada com aves marinhas, principalmente as
migradoras intercontinentais, e os mesmos tratam de novos registros e expansão na

3
distribuição (Olmos, 2002; Ross & Piacentini, 2003), censos, anilhamento, monitoramento,
tamanho de populações (Azevedo-Júnior & Larrazábal, 1999; Larrazábal et al., 2002;
Telino-Júnior et al, 2003; Lyra-Neves et al., 2004), biologia reprodutiva (Olmos & Silva e
Silva, 2001; Branco, 2003; Martins & Dias, 2003; Fedrizzi et al., 2004) e comportamento
alimentar (Fonseca et al., 2001, Naves et al., 2002; Bugoni & Vooren, 2004).
Para as aves aquáticas continentais os trabalhos referem-se á biologia reprodutiva
(e.g Oliveira, 1997; Nascimento, 1990; Bouton & Bouton, 2001), comportamento alimentar
(Oliveira, 1997; Andrade et al., 2001) e o uso dessas aves como recurso turístico no
Pantanal (Bouton & Frederick, 2003). Alves & Pereira (1998), discutem o efeito da
precipitação anual sobre a riqueza e abundância da comunidade de aves aquáticas e
terrestres.
No Brasil ocorrem cerca de 1796 espécies de aves (Sick, 1997, Comitê Brasileiro de
Registros Ornitológicos http://www.ib.usp.br/cbro/2006); para o Pantanal mato-grossense
são registradas aproximadamente 463 espécies de aves (Tubelis & Tomas, 2003a). Estes
autores compararam a riqueza de espécies de aves das áreas úmidas do Pantanal, com as da
China (Vales do Yancheng, Dafeng e Poyang) e da África (Delta do Okavango, lagos
Naivasha e Bogoria no Quênia), e concluíram que o Pantanal é o local de maior diversidade
de aves das áreas inundáveis da Terra. Esses autores indicam que o conhecimento sobre as
aves do Pantanal ainda necessita de muitos trabalhos de coleta e informações biológicas
sistematizadas. Prova disto é que já foram registradas para o Pantanal de Barão de Melgaço
18 novas ocorrências (Antas & Palo Jr, 2003, Oliveira & Olivera in prep.), e para o
Pantanal de Poconé mais 24 novas ocorrências, elevando para 505 o número de espécies de
aves para o Pantanal (Pinho, 2005; Oliveira & Olivera in prep.).

4
Informações gerais sobre a avifauna do Pantanal, incluindo também as aves
aquáticas, são encontradas em Brown jr. (1986), Cintra & Yamashita (1990); Sick (1997);
Petermann (2000); Yabe & Marques (2001); Pearson & Beletsky (2002), Antas & Palo Jr
(2003), Tubelis & Thomas (2003ab), Figueira et al. (2006); Pinho (2005). Sobre frugívoria,
dispersão de sementes e a ornitofilia (Sazima et al., 2001; Araújo & Sazima, 2003; Galleti
& Guimarães, 2004); sobre biologia reprodutiva de psitacídeos (Pinho & Nogueira, 2000 e
2003).
Das 156 espécies de aves aquáticas que aparecem no Pantanal 32 se alimentam
principalmente de peixes (Cintra & Antas, 1996). Oliveira (1996) dividiu estas aves em três
grupos conforme a estratégia utilizada para forragear. O primeiro grupo é formado pelas
aves vadeadoras “wader”(20 espécies), que são aquelas que caminham nos ambientes
inundados para capturarem os peixes (entre elas os colhereiros, tuiuiú, cabeça-seca e
garças); o segundo, composto pelas aves que pescam em vôo, ocasião que podem percorrer
longas distancias “crusier”(4 espécies), são aquelas que sobrevoam ambientes aquáticos e
capturam suas presas com suas patas e garras (e.g a águia-pescadora e o gavião-belo); e o
terceiro, formado por aves que pescam mergulhando “diving” (8 espécies) sendo a presa
capturada com o bico. Neste grupo aparecem os biguás, biguatingas e os martins-
pescadores, gaivotas, trinta-réis (Figura 1, 2, 3 e 4).
Dados exclusivos para as aves aquáticas referem-se aos endoparasitas de ciconídeos
(Pinto et al., 2004), migração e deslocamentos (Antas,1994; Antas & Nascimento, 1996;
Nunes & Thomas, 2004), variabilidade genética de ciconídeos (Del Lama et al., 2002;
Rocha et al., 2004), recursos comunitários e conservação (da Silva et al., 2002; Bouton &
Frederick, 2003), comportamento alimentar (Oliveira & Cavalcanti, 2002; Magalhães,
1990ab; Andrade et al., 2001) e biologia reprodutiva (Yamashita & Valle, 1990;

5
Willis, 1995, Oliveira & Cavalcanti, 2002; Bouton & Bouton, 2001; Campos & Coutinho,
2004; Oliveira in prep.).
Segundo Hafner (1997) é de extrema importância e urgência, estudar a ecologia
alimentar das aves aquáticas, as interações com outras espécies e com o hábitat freqüentado
pelas mesmas durante a migração ou no período de repouso. Existe ainda segundo o mesmo
autor, a necessidade de maiores informações sobre as áreas de invernagem dessas aves,
considerado como um imenso campo de investigação, que poderá ser o fator-chave para
muitas questões relativas as dinâmicas populacionais de várias espécies, propiciando
condições para o manejo e conservação das mesmas.
As aves são essenciais quando se discute estratégias para a conservação da
biodiversidade no planeta (Alves & Silva, 2000; Birdlife International, 2004). Segundo
Alves & Silva (2000), as aves apresentam várias características que contribuem para isso
tais como: a) serem em sua grande maioria, diurnas e relativamente fáceis de observar,
facilitando a coleta de dados; b) sua sistemática e distribuição são relativamente bem
conhecidas quando comparadas com outros grupos animais e c) ocupam diferentes habitats,
sendo algumas espécies especialistas, reagindo facilmente às mudanças ambientais.
Considerando o exposto, esta pesquisa tem como objetivos analisar no Pantanal o
efeito da sazonalidade na composição da assembléia de aves aquáticas e piscívoras; 2)
Avaliar como as características físicas dos lagos (tamanho, profundidade e transparência)
influenciam a composição da assembléia de aves piscívoras; 3) Verificar como a cobertura
de macrófitas aquática nos lagos influencia a composição da assembléia de aves piscívoras;
4) Investigar se a variação na biomassa de peixes influencia na composição (riqueza e
abundância) da assembléia de aves piscívoras.

6
Figura 1. Duas aves vadeadoras no Pantanal: a) Maguari (Ardea cocoi) com um peixe-cachorro (Acestrorhynchus pantaneiro) no bico (Foto: Tom J. Ulrich); b) Socó-boi (Tigrisoma lineatum), com um Lobó (Hoplias malabaricus) no bico (Foto: Edson Endrigo/Avesfoto).
A
B

7
Figura 2. Duas aves mergulhadoras no Pantanal: a) Martim-pescador (Ceryle torquata) com uma Pacu-peva (Metynnis mola) no bico (Foto: Tom J. Ulrich); b) Biguatinga (Anhinga anhinga) com um pimelodídeo no bico (Foto: Marcos Bergamaso/Sematur/MT).
B
A

8
Figura 3. Diferentes maneiras de forragear das aves aquáticas no Pantanal. A) quando os lagos estão com o nível da água mais baixo as aves forrageam em seu leito e nas suas margens ainda úmidas. B) as aves vadeadoras e mergulhadoras ocupam diferentes profundidades nestes locais, repartindo os recursos. C) as aves que voam longas distâncias como as águias-pescadoras aparecem nos lugares mais profundos, junto com taiamãs, gaivotas e trinta-réis.
C
A
B

9
Figura 4. Aves aquáticas do Pantanal: mergulhadoras 1) Mergulhãozinho (Podilymbus podiceps ), 2) Biguá (Phalacrocorax brasilianus), 3) Biguatinga (Anhinga anhinga). Aves vadeadoras 4) Tuiuiú (Jabiru mycteria), 5) Tabuiaiá (Ciconia maguari) e 6) Cabeça-seca (Mycteria americana). Desenho de Rolf Grantsau para o Livro Aves aquáticas do Mato Grosso, História natural e Etnobiologia (Oliveira in prep.).

10
ESTRUTURA DA TESE
A tese está apresentada em 4 capítulos, com 3 seções. As 3 primeiras fornecem uma
introdução geral às aves aquáticas, uma introdução geral sobre o Pantanal e uma descrição
da área de estudo, e em seguida são apresentados quatro capítulos com a mesma estrutura.
Capítulo 1. Dinâmica e sazonalidade da assembléia de aves aquáticas e piscívoras no
Pantanal
Estudou-se a variação na dinâmica da assembléia de aves aquáticas e piscívoras, ao longo
do ano e entre dois anos consecutivos, relacionando essa variação com a pluviosidade
média mensal, temperatura média do ar e insolação total nestes períodos.
Capítulo 2. Efeitos bióticos e abióticos na composição da assembléia de aves piscívoras
Avaliou-se a variação na composição da assembléia de aves piscívoras nos lagos do
Pantanal em função da mudança nas variáveis físicas e biológicas dos lagos.
Capítulo 3. Efeito da cobertura de macrófitas aquáticas na assembléia de aves piscívoras
Analisou-se os efeitos da cobertura de macrófitas aquáticas dos lagos sobre a variação na
composição da assembléia de aves piscívoras.
Capítulo 4. Efeitos da variação da assembléia de peixes sobre a variação na assembléia de
aves piscívoras
Verificou-se os efeitos da variação na riqueza de espécies e biomassa de peixes na variação
da composição da assembléia de aves piscívoras durante dois anos consecutivos.

11
INTRODUÇÃO AO PANTANAL
2.1 Descrição e características gerais do Pantanal
O Pantanal mato-grossense ocupa uma área de aproximadamente 138.183Km², nos Estados
de Mato Grosso (compreendendo parte dos municípios de Poconé, Santo Antonio do
Leverger, Cáceres, Barão de Melgaço e Nossa Senhora do Livramento) e Mato Grosso do
Sul (Corumbá, Ladario, Rio Verde de Mato Grosso, Coxim, Miranda e Aquidauna) Da
Silva & Abdon (1998). Localiza-se na porção central da América do Sul, extremo Norte da
Bacia Platina (entre os paralelos 16º a 22º S e os meridianos 55º a 58ºW). Trata-se de um
dos maiores sistemas de áreas alagáveis continuas do continente, sendo formado pela
coalescência dos cursos da bacia do Alto Paraguai (da Silva, 1990). Segundo Westalake
(1963), sistemas alagáveis como o Pantanal, que recebem influxos anuais de água e
nutrientes, estão entre os mais produtivos, em função da estabilidade de pulso, representada
pela flutuação do nível da água (Margalef, 1968). (Figura 1).
Junk et al. (1989), consideraram o pulso de inundação o principal fator controlador
do sistema nas áreas alagáveis tropicais. Para estes autores as áreas alagáveis, ou Zona de
Transição Terrestre Aquática (Aquatic Terrestrial Transition Zone-ATTZ), representa uma
grande diversidade de hábitats periodicamente secos e inundados. Pode ser ocupada por
diversos tipos de vegetação terrestre e aquática, sendo também permeada por hábitats
permanentemente aquáticos, como os rios e lagos.
A flutuação anual do nível da água no Pantanal provoca uma série de
transformações na estrutura e funcionamento dos seus ecossistemas e influencia na
organização espacial de sua diversidade, através da variação do nível, duração e extensão

12
das enchentes. Contribui também influenciando a diversidade, a forma e duração de contato
das baías (lagos) com os rios ou a planície alagável, seu manejo, assim como a origem,
idade e fertilidade dos solos e sedimentos ai depositados (Da Silva, 2000). Em decorrência
destas condições, processos ecológicos como a produtividade, decomposição,
importação/exportação de nutrientes, variáveis limnológicas, a estrutura e a organização
espacial das comunidades biológicas variam no tempo (período de cheia e vazante) e no
espaço (da Silva, 1990, da Silva & Esteves, 1995, da Silva, 2000). Segundo Brown Jr.
(1986), as inundações sazonais influenciam fortemente a distribuição da fauna no Pantanal,
e o papel principal exercido por este sistema é o de corredor de dispersão e criadouro para
muitos animais. Em função da diversidade de habitat aquáticos que o Pantanal proporciona
e das características fisiológicas e comportamentais das aves aquáticas, estas encontram
neste sistema as condições propícias para sua alimentação e reprodução.
Brown Jr. (1986) relatou a presença de 657 espécies de aves para o Pantanal mato-
grossense incluíndo áreas periféricas como algumas serras, porém em uma revisão mais
crítica, baseada em coleções brasileiras e estrangeiras, além de observações de campo,
concluiu-se que a planície pantaneira tem cerca de 463 espécies (Tubelis & Tomas, 2003a),
das quais, segundo Cintra & Antas (1996), 156 dependem de alguma forma de ambientes
aquáticos; este grupo representa quase 34% das aves que ocorrem no Pantanal.
Das 32 espécies de aves piscívoras que ocorrem no Pantanal, 28 ocorrem
regularmente na Estrada Parque Transpantaneira, no Pantanal de Poconé. Estas espécies
têm sido bastante estudadas em outras regiões, com extensa literatura sobre a biologia das
mesmas podendo-se citar Coulter et al., (1991); Carboneras (1992ab), Elliott (1992),
Hancock et al., (1992); Martinez-Vilalta & Motis (1992); Matheu & del Hoyo (1992), Orta
(1992ab) Gochfeld & Burger (1996) e (Sick, 1997). Apesar disto, estudos ecológicos de

13
fatores ambientais e biológicos influenciando a estrutura e composição de comunidades de
aves aquática piscívoras na região neotropical são escassos (Remsen, 1991; Petermann,
1997; Cowx, 2003).
Figura 1. O Pantanal de Poconé, e as outras sub-divisões da planície pantaneira, segundo Adámoli (1981), imagem modificada de Nunes da Cunha & Junk, 2004. A Estrada Parque Transpantaneira, também é conhecida como MT-060.

14
2.2 Vegetação do Pantanal
Revisão sobre a vegetação herbáceo-arbórea do Pantanal mato-grossense pode ser
encontrada em Prance & Schaller (1982), Nunes da Cunha (1990), Guarim-Neto (1991,
1992), Lemes do Prado et al. (1992), Ponce & Nunes da Cunha (1993), Pott & Pott (1994,
1997, 1999 e 2000), Zeihoffer & Schessl (1999), Nunes da Cunha & Junk (1999, 2001) e
Damasceno-Junior et al.(2005).
Pott & Pott (1999), indicaram para o Pantanal Mato-grossense a ocorrência de 1647
espécies de plantas, distribuídas em 134 famílias e 693 gêneros, sendo o maior número de
espécies registrado para as famílias Poaceae e Leguminosae, 210 e 213 respectivamente.
Nunes da Cunha & Leitão-Filho (1996), identificaram 41 famílias com 138 espécies de
árvores no Pantanal de Poconé. Os mesmos autores dividiram, Pantanal, em relação à
vegetação arbórea em dois tipos básicos: Savana (Cerrado) e Florestas. As Savanas,
ocorrem nas áreas periféricas situadas na região norte do Pantanal de Poconé, tendo o rio
Pixaim como o limite Sul de sua distribuição. Abaixo deste rio, segundo os mesmos
autores, esta região estaria sob a influência maior dos períodos de inundação, quer pelo
tempo, quer pela altura da lamina de água. A ocorrência de vegetação de cerrado abaixo do
rio Pixaim, é esporádica e a riqueza florística das espécies arbóreas é menor segundo Nunes
da Cunha (1990).
Ainda conforme a mesma autora, nos cerrados do Pantanal de Poconé, algumas
espécies formam densos agrupamentos quase homogêneos, denominados localmente como
lixeiro (Curatella americana), carvoeiro (Callisthene fasciculata) e o paratudal (Tabebuia
caraiba). As Florestas da região de Poconé, segundo Nunes da Cunha (1990), apresentam-
se com três unidades fitofisionômicas: os capões; as florestas de inundação e as florestas de

15
cordilheiras. Os capões estão distribuídos nas proximidades dos atuais leitos dos principais
rios da região, como o Bento Gomes, o Pixaim, o Cassanges, o Paraguaizinho e o rio
Cuiabá.
As Florestas de Cordilheiras, também denominadas na região como Cordilheiras,
segundo Nunes da Cunha (1990), têm origem semelhante à dos capões. A diferença mais
marcante entre eles estaria na extensão, essas formações ocorrem em todo o Pantanal de
Poconé. Florestas inundáveis, ocorrem em áreas que ficam inundadas a maior parte do ano,
secando apenas nos meses de setembro a novembro.
No Pantanal mato-grossense ocorrem cerca de 242 espécies de macrófitas aquáticas
distribuídas em 106 gêneros e 54 famílias (Pott & Pott, 1997). Segundo estes autores,
estudos sobre a composição de plantas de lagoas e áreas inundadas no Pantanal ainda são
escassos. Uma síntese sobre a biologia, interações e sistemática das espécies de macrófitas
aquáticas do Pantanal é apresentado em Pott & Pott (2000).
Para a Estrada Parque Transpantaneira, em Poconé, foram listadas 25 espécies com
predominância dos aguapés: Eichhonia crassipes e E. azurea (da Silva & Pinto-Silva,
1989). Os corpos de água ao longo da EPT, como rios, corixos, vazantes, campos
inundáveis, lagos, lagoas, brejos e caixas de empréstimo, apresentam diferente composição
e distribuição dessas macrófitas ao longo do ano, sendo essa variação dependente do grupo
ecológico dessas comunidades, do nível da água, de fatores físico-químicos e estágio
sucessional, como demonstrado para as macrófitas aquáticas de acordo com da Silva &
Silva (1989). Essa cobertura varia nos ambientes lóticos e lênticos proporcionando
diferentes condições para que as aves tenham acesso às suas presas.
Os poucos estudos com macrófitas aquáticas em Poconé, tratam dos efeitos da
variação da inundação na dinâmica do crescimento e produção primária líquida de uma

16
espécie (Penha et al., 1998a, b), ou da estrutura de uma comunidade de macrófitas
aquáticas (Lemes do Prado et al. (1994).
Thomaz & Bini, (2003) fizeram uma revisão com análise crítica sobre os estudos
desenvolvidos no Brasil com macrófitas aquáticas, demonstrando a inexistência de estudos
com interações entre aves aquáticas e macrófitas.
2.3 A Fauna do Pantanal
Estudos gerais sobre a fauna do Pantanal Mato-grossense são encontrados em
Schaller (1983); Brown Jr (1986); Alho (1986); Cintra & Yamashita (1990); Dubs (1992);
Antas (1994) e Cintra & Antas (1996); PCBAP (1997); Heckman, (1998b); Britski et al.,
(1999); Pearson & Beletksky (2002); Antas & Palo Jr (2003); Tubelis & Tomas (2003 a e
b); Marques, et al., (2005) e Figueiras et al., (2006).
O conhecimento da flora e fauna pantaneira, notadamente do Pantanal de Poconé,
teve grande incremento nos últimos anos principalmente devido aos estudos realizados pela
Universidade Federal de Mato Grosso no referido Município, através do Projeto Ecologia
do Pantanal, convênio Brasil (UFMT/IB) Alemanha (Instituto Max-Planck para
Limnologia) a lista dos 122 trabalhos publicados ver em http://www.mpil-
ploen.mpg.de/mpils2d.htm. Os resultados desta cooperação cientifica esta apresentada em
Nunes da Cunha et al. (2003). Atualmente esses estudos continuam por meio dos
pesquisadores integrantes do Núcleo de Estudos Ecológicos do Pantanal - NEPA. Até o
momento 111 dissertações de mestrado foram defendidas no Programa de Pós-graduação
em Ecologia e Conservação da Biodiversidade da UFMT, destas, 57 foram realizadas no
Pantanal (lista e textos completos em www.ufmt.br/ib/ppgecb/dissertacoes.html).
Os invertebrados foram estudados por Heckman (1994a), ao propor novos conceitos
limnológicos para as regiões inundáveis nos trópicos. Estratégias de sobrevivência a

17
inundação de alguns artrópodos estão em (Adis et al., 2001). A estrutura e composição de
comunidades de artrópodos associados à copa de Vochysia divergens (Marques et al., 2001)
e Attalea phalerata (Santos et al., 2003, Batirola et al., 2004 e 2005) demonstram que a
técnica de nebulização de copas é uma ferramenta promissora para estudos desses
organismos no Pantanal; Callil & Junk (1999 e 2001) ao estudarem o papel de gastrópodes
e bivalves como organismos incorporadores de mercúrio, proveniente das atividades
garimpeiras na região concluíram que Anadontites trapesialis, Castalia ambigua e
Pomacea escalaris, seriam os melhores moluscos utilizados para um biomonitoramento. As
estratégias de sobrevivência dos moluscos ao pulso de inundação, foram avaliadas por
(Kretzchmar & Heckman, 1995).
O Pantanal Mato-grossense possui cerca de 263 espécies de peixes (Britisk et al.,
1999), mas, este número tende a aumentar com novas espécies que estão sendo descritas
para a região (Willick et al., 2000, Benine et al., 2004). Com relação aos aspectos da
biologia, que é ainda pouco conhecida, são encontrados para o Pantanal estudos sobre
táticas de forrageamento (Sazima & Caramaschi, 1989; Sazima & Machado, 1982;
Machado 1983, 2003) e estratégias defensivas contra predação (Sazima & Machado, 1989;
Sazima & Zamprogno, 1990 e Machado, 2003). Alguns poucos estudos tratam de fatores
bióticos (presença de predadores e macrófitas aquáticas) e abióticos (e.g pH, profundidade
e distância dos lagos aos rios) influenciando a riqueza e abundância de peixes nos lagos do
Pantanal (Mourão et. al., 1998; Catella, 1992; Pazin, 2000; Súarez et al., 2001 e 2004).
A importância da pesca no Pantanal bem como suas implicações, no turismo, na
vida dos ribeirinhos, e no conflito de uso com as aves aquáticas e na própria
sustentabilidade deste recurso são discutidos também (Ferraz de Lima, 1986; Coutinho et
al., 1994; Alho & Vieira, 1997; Da Silva, 2000; Oliveira & Nogueira, 2000).

18
Quanto aos répteis, Strussmann (1992 e 1997) e Strussmann & Sazima (1990, 1991
e 1993) apresentaram a composição, a história natural e a ecologia de uma comunidade de
serpentes, registrando a presença de 26 espécies para Poconé. Estudos sobre a ecologia do
jacaré-do-pantanal, referente à nidificação, cuidado parental encontra-se em Crawshaw &
Schaler (1980), Campos (1993), Cintra (1985; 1988) e Coutinho & Campos (1996); sobre o
manejo e efeito da caça ao jacaré informações são encontradas em Cintra (1989), Mourão et
al. (1996) e Coutinho et al. (1994).
A avifauna do Pantanal de Poconé foi inventariada por Cintra & Yamashita (1990),
que encontraram 317 espécies de aves, relatando o seu hábitat, abundância relativa e
sazonalidade. Oliveira & Olivera (no prelo) realizaram um levantamento na Fazenda
Ipiranga em Poconé é encontraram 352 espécies para o local, com algumas novas
ocorrências para o Estado de Mato Grosso. Yamashita & Valle (1990), mapearam e
monitoraram 13 ninhais, no município de Poconé, entre 1979 a 1984, ocasião na qual os
autores registraram o número de indivíduos, as espécies presentes, caracterizaram de
maneira geral os sítios de nidificação, os predadores e causas de possíveis declínios destes
locais para nidificação. Magalhães (1990a) estudou a biologia do Gavião-caramujeiro
(Rostrhamus sociabilis), relatando as presas principais dessa ave no Pantanal, os caramujos
(Pomacea caniculata e Pomacea scalaris). No Pantanal as aves mais estudadas até o
momento são o Tuiuiú (Jabiru mycteria), e a Arara-azul (Anodorhynchus hyacinthinus).
Estudos com uso de anilhas alares e rastreadas por satélite, bem como monitoramento
terrestre e aéreos de ninhos de tuiuiú foram realizados no Mato Grosso do Sul (Antas &
Nascimento, 1996; Campos & Coutinho, 2004), verificando que esta ave faz grandes
deslocamentos dentro do Pantanal, podendo voar em um só dia 400Km. Oliveira (1986),
avaliou as estratégias utilizadas pelo Tuiuiú (Jabiru mycteria) para capturar suas presas no

19
Pantanal de Poconé, além do comportamento alimentar dos seus filhotes, concluindo que o
tuiuiú é generalista quanto a carnívoria, e que o principal alimento dos seus filhotes é o
Muçum (Symbranchus marmoratus). Oliveira (1997) e Oliveira & Cavalcanti (2002),
pesquisaram a biologia reprodutiva do Tuiuiú como locais preferências de nidificação,
estabelecendo que no Pantanal de Poconé a Piúva (Tabebuia heptaphyla) é a espécie
vegetal preferida como local de nidificação, e o sucesso reprodutivo é o maior já registrado
para as áreas já estudadas nas Américas. Propostas para manejo e conservação desta ave no
Pantanal foram feitas por Oliveira (1997) e Oliveira & Cavalcanti (2002). Quanto à Arara-
azul do Pantanal Guedes (1993) no Mato Grosso do Sul e Pinho & Nogueira (2002) no
Mato Grosso estudaram a biologia reprodutiva desta mesma espécie, relatando a
necessidade e importância da conservação dos capões e cordilheiras, pois estes locais
fornecem seu principal alimento, frutos da palmeira Acuri (Sheelea phalerata) e árvores
para locais de ninho como o Manduvi (Sterculia striata e S. apétala). Jonhson et al. (1997)
estudaram a densidade de indivíduos jovens de Manduvi submetidos a diferentes tipos de
manejo, verificando que existe falha no estabelecimento de novos indivíduos em áreas
submetidas a queimadas e à presença do gado, o que pode comprometer a conservação da
Arara-azul em longo prazo. Entretanto, nenhuma destas pesquisas analisou a influência de
fatores ambientais e biológicos na estrutura e composição de aves piscívoras no Pantanal.
O número de espécies de mamíferos registrados para a planície pantaneira ainda é
muito controverso, pois depende do entendimento de cada autor sobre esta região. Assim,
encontram-se listas com 122 espécies (Fonseca et al., 1996); 89 espécies (PCBAP, 1997)
ou 93 espécies de mamíferos (Rodrigues et al., 2002). Novos elementos serão acrescidos
nesta lista, pois muitas regiões do Pantanal não foram ainda sistematicamente
inventariadas. Em relação a mastofauna do Pantanal de Poconé, em observações de campo

20
durante esta pesquisa, foram registradas 25 espécies de mamíferos, excluindo os morcegos
e pequenos roedores. Entre as espécies identificadas, algumas estão na categoria de animais
vulneráveis como a Onça-pintada (Panthera onça) (L)., o Tatu-canastra (Priodontes
giganteus) (Kerr), o Cervo-do-Pantanal (Blastocerus dichotomus) (Iliger), o Tamanduá-
bandeira (Mirmecophaga tetradactila)(L.), e a Ariranha (Pteronura brasiliensis) (Gmelin)
segundo o MMA (2003).
Nas últimas décadas o futuro do Pantanal, este Patrimônio Mundial da Humanidade,
está sendo bem discutido por entidades governamentais e não-governamentais, visando o
manejo e conservação deste ecossistema (Alho, 1986; Mittermeier et al., 1980; Alho et al.,
1988; Seidl & Moraes, 2000; da Silva, 2000; Harris et al., 2005). Várias são as causas que
contribuem para a alteração ambiental da região, dentre elas, as construções de estradas e
diques (Campos-Filho & Machado, 1999), hidrelétricas e obras hidráulicas (Ponce, 1995,
Wantzen et al., 1999; Girard et al., 2003). A agricultura intensiva, pastagens cultivadas e
efluentes urbanos também representam um drástico efeito no ambiente (Alho et al., 1988;
Ferreira, et al., 1994; Seidl et al., 2001). Algumas destas atividades estão concentradas no
planalto, causando contaminação do sistema por agrotóxicos e metais pesados como o
mercúrio, oriundo das atividades garimpeiras, que provocam danos à biota e ao homem
pantaneiro (Lacerda et al., 1990 e 1991; Ferreira, et al., 1994; Alho et al., 1997; Nogueira
et al., 1997; Guimarães et al., 1998 e 1999; da Silva, 2000; Hylander et al., 2000; Fonseca
et al., 2005). A introdução de espécies exóticas como o Búfalo (Bubalus bubalis) causam
também a destruição de locais de alimentação e nidificação de algumas espécies de aves
aquáticas (Oliveira obs. pess.).
Portanto, é de extrema urgência e importância a compreensão e entendimento deste
ecossistema para propor medidas visando a sua manutenção. As aves aquáticas são

21
excelentes bioindicadoras da qualidade ambiental (Fox et al., 1987; Browder et al., 2002;
Paillisson et al., 2002; Stapanian et al., 2004), por serem de fácil observação e possuírem
alta sensibilidade às alterações ambientais. Desta maneira este estudo propõe analisar os
fatores que influenciam a distribuição e abundância dessas aves visando subsidiar ações
conservacionistas.
3. Área de estudo:
O Município de Poconé está situado na região Norte do Pantanal e ocupa uma área
de 16.642 Km2. Cerca de 80% desta área está localizada no Pantanal, onde a criação de
gado é a principal atividade econômica (Nunes da Cunha & Junk ,1999). Os estudos foram
desenvolvidos na Estrada Parque Transpantaneira, que corta o município de Poconé
longitudinalmente, ligando esta cidade à localidade de Porto Jofre na margem direita do Rio
Cuiabá (Figura 1e 2).
Esta rodovia atravessa gradientes de inundação deste Pantanal por 145 km, sendo
utilizada como principal ponto de turismo. Os trabalhos foram concentrados nos primeiros
46km desta rodovia, incluindo desde o portão do IBAMA, até a margem direita do rio
Pixaim (Figura 1 e 2), pela facilidade logística para realização dos estudos e da existência
de muitos lagos neste trecho. Esta rodovia funciona atualmente como um dique, pois para
sua construção foi feito um aterro elevado. Sua implantação causou desequilíbrio ao
sistema de vazão e, conseqüentemente, impactos ambientais (Oliveira, 1997; Campos-Filho
& Machado, 1999), que ainda não foram avaliados em toda a sua extensão. O aterro da
rodovia foi construído com o solo de caixas de empréstimo laterais. A retirada do solo
formou grandes canais ao longo da mesma, servindo de reservatório de água para o gado no
período da seca. Com as cheias, esses reservatórios são povoados por muitos organismos

22
aquáticos (insetos, crustáceos, moluscos, anfíbios, répteis e peixes) que servem de alimento
para numerosas aves aquáticas. As aves encontram nestas caixas de empréstimo fontes
perenes para sua alimentação e da sua prole, atuando, portanto, como elementos
importantes no sucesso reprodutivo de aves aquáticas como o Tuiuiú (Jabiru mycteria)
(Oliveira, 1997; Oliveira & Cavalcanti, 2000). Sistema semelhante encontra nos lhanos
venezuelanos onde estudos das comunidades de aves aquáticas foram também realizados
(Pinowski et al., 1980; Morales et al., 1981, 1986; Pietro, 1988; González, 1996a,b, 1997).
O Pantanal, como as outras áreas alagáveis também apresenta sazonalidade quanto
ao nível da água, o chamado ciclo hidrológico. A variação anual de inundação das áreas
alagáveis foi denominado por Junk et al. (1989) de pulso de inundação. A região pantaneira
está submetida a ciclos plurianuais, caracterizados por revezamento de anos muitos
chuvosos (ciclos de enchentes) com anos relativamente secos (ciclo de secas) conforme
Ádamoli (1986) e Ponce (1995). Segundo Ab’Saber (1983), estes ciclos plurianuais estão
ligados a eventos regionais.
O Pantanal apresenta quatro fases distintas quanto ao regime hídrico, relacionados
com a variação anual do nível da água: a) fase da cheia (janeiro a março), ocorre o
transbordamento do dique marginal dos rios, as baías perdem os seus limites, ligando suas
águas entre si (da Silva, 1990); b) fase de vazante (abril a junho), os movimentos das águas
convergem na direção dos lagos (‘baías’)-sangradouros-rios; corta-se o fluxo entre os lagos
(‘baías’) e destes com os rios; c) fase de estiagem ou seca (julho a setembro), alguns corpos
de água secam, quando então os lagos(‘baías’) apresentam como corpos de água isolados
uns dos outros, conforme da Silva (1990); d) fase da enchente (outubro a dezembro),
caracterizada pelo começo das chuvas, momento em que os corpos de água começam a
receber os primeiros fluxos de água.

23
O Pantanal de Poconé, segundo Fonseca (2003), é drenado pelas bacias do rio Bento
Gomes, seus afluentes rios Claro, Novo, Paraguaizinho, Cassange e Alegre e os corixos
Nacional e Sararé, com a maioria desaguando em áreas de inundação permanente. O rio
Bento Gomes drena toda a região na direção geral Nordeste-Sudoeste, com inflexões para
Norte-Sul, Este-Oeste e Noroeste-Sudeste e praticamente sua bacia hidrográfica é a mais
extensa seccionando a Estrada Rodovia Transpantaneira e drenando suas adjacências.

24
Figura 2. Vista aérea parcial da Estrada Parque Transpantaneira no Pantanal de Poconé-MT, no alto no período da cheia (Foto: Arne Sucksdorf); em baixo na vazante do Rio Bento Gomes, quando os lagos começam a tomar forma definida (Foto: Dalci Oliveira).

25
RESUMO
As aves aquáticas e piscívoras do Pantanal, foram estudadas mensalmente durante
quatro anos, nos dois primeiros anos (1999-2000), em um transecto linear de 46km na
Estrada Parque Transpantaneira em Poconé-MT, e em 2001 e 2002 os estudos foram feitos
somente com aves piscívoras em 20 lagos marginais nesta mesma rodovia. Nos dois
primeiros anos (1999-2000), as aves foram observadas a olho nu e com auxilio de binóculo
(8x40). A riqueza da comunidade de aves aquáticas foi formada por 39 e 37 espécies,
enquanto a comunidade de aves piscívoras foi formada por 18 e 19 espécies de aves em
1999 e 2000 respectivamente, com abundância total de 6374 indivíduos no primeiro ano e
de 5107 no segundo. As espécies mais abundantes foram respectivamente em 1999 e 2000:
a Garça-branca grande (Ardea alba) (1115 e 728 indivíduos); o Gavião-caramujeiro
(Rostrhamus sociabilis) (861 e 880 indivíduos), o Carão (Aramus guaruna) (800 e 594
indivíduos) e a Garça branca pequena (Egretta thula) (490 e 417 indivíduos). Nos dois anos
a comunidade de aves piscívoras contribuiu com mais de 50% da abundância total de
indivíduos. A riqueza de espécies e abundância de aves aquáticas e piscívoras não
apresentaram diferenças significativa entre os anos. Porém, a composição quantitativa e
qualitativa da comunidade de aves piscívoras variou significativamente entre os meses. A
pluviosidade, e a temperatura do ar afetaram significativamente a abundância e a
composição quantitativa e qualitativa da comunidade de aves piscívoras sugerindo que
essas aves podem ser afetadas por variações climáticas no Pantanal. Nos dois últimos anos
(2002 e 2003), as aves piscívoras do Pantanal foram estudadas em 20 lagos marginais a
Estrada Parque Transpantaneira em Poconé, Mato Grosso. As aves também foram
observadas a olho nu ou com auxilio de binóculo (8X40) e Luneta (15x60), para esta

26
amostragem foram consideradas as aves que estavam dentro ou nas margens de cada lago.
Concomitante, com este censo, foram registradas as características limnológicas do lago
(tamanho, profundidade, transparência, oxigênio dissolvido e temperatura da água), a
riqueza de espécies de peixes e a cobertura e riqueza de espécies de macrófitas aquáticas. A
comunidade de peixes foi formada por 79 espécies distribuídas em 21 famílias, as mais
abundantes foram Characidae e Cichlidae, juntas contribuíram com 44.3% do total dos
peixes. As espécies de peixes mais freqüentes nos lagos foram Odontostilbe paraguayensis;
Odontostilbe calliura, Characidium laterallis, Aphyocarax rathbuni; Pyrrulina australis e
Hoplias malabaricus. A composição da comunidade macrófitas aquáticas foi formada por
26 espécies distribuídas em 21 famílias. As espécies de macrófitas mais freqüentes nos
lagos foram: Eichhornia azurea, Eichhornia crassipes e Pontederia cordata. A riqueza de
espécies de macrófitas variou na cheia entre 15 a 21 espécies e na seca entre 8 a 15 espécies
por lago. A maioria dos lagos apresentou cobertura de macrófitas entre 50 a 100%. A
riqueza da comunidade de aves piscívoras foi formada por 17 espécies, as aves mais
abundantes foram: a Garça-branca grande (Ardea alba) com 1221 e 447 indivíduos e a
Garça-branca-pequena (Egretta thula) com 428 e 258 indivíduos em 2002 e 2003
respectivamente. A abundância total das aves piscívoras foi de 2896 indivíduos em 2002 e
de 1415 em 2003 em todos os lagos. O lago do Bento Gomes foi o que apresentou a maior
abundância e riqueza de espécies de aves piscívoras nos dois anos de estudos. A riqueza de
espécies de aves piscívoras foi positivamente afetada pela composição quantitativa de
peixes na primeira dimensão. Entretanto a abundância das aves piscívoras não foi afetada
por nenhum dos fatores bióticos ou abióticos analisados. A riqueza de espécies e a
composição qualitativa de macrófitas aquáticas não influenciaram significativamente nem a
riqueza e abundância, nem a composição da comunidade de aves piscívoras no Pantanal. A

27
composição quantitativa da comunidade de aves piscívoras foi significadamente
influenciada pela composição quantitativa da comunidade de peixes, pela transparência da
água e pela profundidade do lago. A profundidade dos lagos também teve influencia na
composição qualitativa das aves piscívoras no Pantanal.
Palavras chaves: Aves aquáticas, Aves piscívoras, Peixes e Macrófitas, Fatores bióticos e
abióticos, Pantanal

28
ABSTRACT
I studied the aquatic birds of the Pantanal in the Estrada Parque Transpantaneira, Poconé,
Mato Grosso, Brazil. During two years (1999-2000), I quantified species richness and
abundance by observing individuals, using binoculars (8x40), along a 46-km long transect.
I recorded a total of 39 species and 6374 individuals in 1999 and 37 species and 5107
individuals in 2000. Of these, I recorded 18 species of piscivorous birds in1999 and 19
species in 2000. The most abundant species in 1999 and 2000 were as follows: the Great
Egret, Ardea alba (1115 and 728 individuals); the Snail Kite, Rostrhamus sociabilis (861
and 880 individuals), the Limpkin, Aramus guaruna (800 and 594 individuals), and the
Snowy Egret, Egretta thula (490 and 417 individuals). During both years of the study,
piscivorous birds accounted for more than 50% of overall abundance. Species richness and
abundance of the aquatic and piscivorous avian community remained the same during both
years of the study. However, the quantitative and qualitative composition of the piscivorous
avian community varied significantly among months. Rainfall and air temperature
significantly affected the abundance and quantitative and qualitative composition of the
piscivorous avian community, suggesting that they could be affected by climatic variations
in the Pantanal. I studied the piscivorous avian community of the Pantanal over two years
(2002 and 2003), in 20 marginal lakes in the Estrada Parque Transpantaneira, Poconé, Mato
Grosso, Brazil. I recorded all individuals within the lakes or along their margins using
binocular (8X40) and spotting scope (15x 60). Along with a census of birds, I also
quantified limnological characteristics of the lakes (size, depth, transparency, dissolved
oxygen content, and water temperature), coverage and species richness of aquatic
macrophytes, and fish species richness. The fish community of the lakes was composed of
79 species of fish from 21 families. Species from the families Characidae and Cichlidae

29
were the most abundant, accounting for 44.3% of all individuals recorded. The most
frequently encountered species were Odontostilbe paraguayensis, Odontostilbe calliura,
Characidium laterallis, Aphyocarax rathbuni, Pyrrulina australis, and Hoplias
malabaricus. Over all lakes, I found 26 species of aquatic macrophytes from 21 families.
The three most frequently encountered species were Eichhornia azurea, Eichhornia
crassipes, and Pontederia cordata. Macrophyte species richness per lake varied between 15
– 21 species in the wet season and 8 – 15 species in the dry season. The majority of lakes
had macrophyte coverages of 50 – 100%. Overall, I found 17 species of piscivorous birds
using the 20 study lakes; I recorded 2896 individuals in 2002 and 1415 in 2003. In 2002
and 2003, respectively, the two most common species I encountered on the lakes were the
Great Egret (Ardea alba), with 1221 and 447 individuals, and the Snowy Egret (Egretta
thula), with 428 and 258 individuals. Of the 20 lakes, lake Bento Gomes exhibited the
highest abundance and species richness of piscivorous birds during both years of the study.
Species richness of piscivorous birds is directly related to the quantitative composition. I
found no relation between piscivore abundance and any of the biotic or abiotic factors
measured in the study. The quantitative community composition of piscivorous birds was
significantly influenced by the quantitative community composition of fish, as well as by
lake transparency and depth. Lake also depth influenced the qualitative composition of
piscivorous birds in the Pantanal. The quantitative community composition of piscivorous
birds was significantly influenced by the quantitative community composition of fish, as
well as by lake transparency and depth. Lake also depth influenced the qualitative
composition of piscivorous birds in the Pantanal. I found no relationship between
macrophyte species richness and the abundance of piscivorous birds in the Pantanal.
Key words: Avian community, piscivorous birds, biotic and abiotic factors, Pantanal

30
Capítulo 1. A dinâmica e a sazonalidade da comunidade de aves aquáticas no
Pantanal
1. Introdução
O ciclo de vida das plantas e animais tende a se organizar temporalmente (Wolda,
1987). Isto ocorre entre espécies, ou entre diferentes populações de algumas espécies.
Apreciáveis diferenças acontecem em diferentes fases ao longo do ciclo de vida dos
organismos. De um modo geral a população de aves varia sazonalmente em abundância em
função de fatores bióticos e abióticos. A variação em abundância ao longo do tempo pode
ser resultado de movimentos locais dentro do habitat, ou entre habitats com topografia
semelhantes, ou entre habitats de diferentes altitudes (Martin & Karr, 1986; Stiles, 1980;
1988; e Loiselle & Blake, 1991; 1994).
Esses movimentos estão ligados à variação temporal e espacial da oferta alimentar,
especialmente frutos e néctar (Loiselle & Blake, 1991 e 1993; Levey & Stiles, 1992;
Stouffer & Bierregaard, 1996; Sazima et al., 1996). Também quando ocorre aumento na
abundância local de invertebrados como insetos aquáticos (Colwell, 1993) ou vertebrados
como peixes, de forma natural (Pinowski & Morales, 1981; González, 1996 e 1997; Wolter
& Pawlizki, 2003), ou em tanques de piscicultura (Carpentier et al., 2003; Gwiazda, 2003),
ou ainda através de descartes de pesca industrial de camarões e ou de peixes (Branco,
2001).
As variações na composição de espécies em assembléia de aves podem, ainda, estar
relacionadas com a migração (Sanaiotti & Cintra, 2001), estrutura da vegetação (Esler,
1992; Bancroft et al., 2002), efeito do fogo (Sannaiotti & Magnusson, 1995; Marini &

31
Cavalcanti, 1996; Cavalcanti & Alves, 1997; Epanchin et al., 2002; Cintra & Sanaiotti,
2006), efeito de barragem (Morales & Pacheco, 1986), variação na precipitação local
(Alves & Pereira, 1998), eventos climáticos fortuitos como o El Niño (Kalmbach et al.,
2001; Zavalaga et al., 2002.) ou mudanças climáticas globais (Boere & Taylor, 2004).
Algumas atividades humanas podem provocar perturbações locais e regionais na
comunidade de aves aquáticas, afetando a sua variação, tais como a caça, principalmente de
patos, marrecas e gansos (Madesen, 1988, Paillisson et al., 2002), e a movimentação
intensa de barcos nos rios e veículos nas estradas adjacentes aos corpos da água (Stolen,
2003). A visitação turística desordenada às colônias afeta drasticamente o sucesso
reprodutivo destas aves, levando em último caso ao abandono ou a extinção da área por elas
(Bouton & Frederick, 2003). A extração de guano na época reprodutiva (Malacalzá &
Bertellotti, 2001), a contaminação por petróleo, ingestão de material sintético, bem como a
morte das aves provocadas pelas atividades da pesca industrial (Kock, 2001; Petry et al.,
2002) são outras fonte de variação na comunidade local. Porém a maior fonte de impacto é
a transformação das áreas inundáveis em zonas de agropecuária e expansão das cidades,
neste caso o efeito na comunidade pode ser regional e global (Mitsch & Gosselink, 2000;
Ma et al., 2004)
Um dos fatores fundamentais para a manutenção local e regional da riqueza e
abundância das aves aquáticas em comunidades naturais nos trópicos é o ciclo hidrológico,
ou seja a variação sazonal quanto ao nível e duração da inundação (Kushlan, 1976, 1981;
Kushlan et al., 1975, 1985; Frederick & Bildstein, 1992; Gonzaléz, 1996b; Bancroft et al.,
2002). Junk et al., (1989), denominaram esta variação anual de inundação das áreas
alagáveis de pulso de inundação. O pulso de inundação no Pantanal, é um dos fatores
determinante na estrutura de populações e comunidades de vários organismos aquáticos e

32
terrestres. Como já foi verificado para a comunidade fitoplanctônica, em alguns lagos sul
mato-grossenses (Oliveira & Calheiros, 2000). Para as plantas terrestres (Nunes da Cunha
& Junk, 1999 e 2004) e para as aquáticas (Da Silva, 1990; Da Silva & Esteves; 1995;
Lemes do Prado et al., 1994; Penha et al., 1998a; b; Pott et al., 1999). Já os animais, como
os insetos aquáticos, moluscos, caranguejos (Kretzchmar & Heckman, 1995, Adis et al.,
2001), e até mesmo os vertebrados no Pantanal, como jacarés, capivaras, cervo-do-
Pantanal, respondem as alterações do nível da água na região tendo a sua riqueza e
abundância relacionada com o regime hídrico local (Alho & Rondon, 1987; Campos, 1993;
Coutinho & Campos, 1996; Tomas et al., 2001).
Entre as aves, algumas espécies ocorrem em vários habitats, mas as aquáticas
tendem a estarem associadas aos corpos de água e são muito abundantes ao longo da
Estrada Parque Transpantaneira. Esta abundância tende a variar temporariamente e entre os
lagos, entretanto, a causa, ainda não é totalmente conhecida, devido à ausência de estudos
avaliando nessa área os efeitos de fatores ambientais climáticos e ecológicos na
comunidade de aves.
No Pantanal, Cintra & Yamashita (1990) citam muitas espécies de aves de
ocorrência sazonal; como por exemplo, as garças os gaviões caramujeiros, mais abundantes
a partir do mês de junho, quando a oferta alimentar é mais abundante, sendo este também o
período reprodutivo das aves que formam ninhos coloniais (viveiros) como biguás, garças,
cabeça-seca, etc. Entretanto, esses autores não fizeram nenhuma análise para verificar se
componentes do clima ou fatores ecológicos afetaram as comunidades de aves no Pantanal.
Figueira et al.(2006) analisando os dados do trabalho de Cintra & Yamashita (1990)
verificaram que a maioria das espécies de aves residentes e sazonais são generalistas e
podem usar os mais variados hábitats. Eles encontraram que o número de espécies sazonais

33
permaneceu constante durante o ano em hábitats de florestas e aumentou em outros
habitats. Também encontraram que a abundância das espécies residentes parece estar
relacionada ao uso de múltiplos habitats por elas. Entretanto, nenhum componente
climático foi considerado em suas análises. Além disso não houve até agora um estudo
analítico só para aves de ambientes aquáticos no Pantanal.
O objetivo deste estudo foi verificar se a variação sazonal e anual na assembléia de
aves aquáticas e um sub-grupo delas as píscivoras no Pantanal está relacionada com fatores
ambientais como a pluviosidade, a insolação total e a temperatura do ar.
2. Material e Métodos
2.1 - Área de Estudo
O Município de Poconé, está situado na região Norte do Pantanal e ocupa uma área
de 16.642 Km2, cerca de 80% dela está localizada no Pantanal, delimitado entre as
coordenadas geográficas de latitudes 16º 16’ e 16º 48’ Sul e longitudes 56º 35’e 56º
58’Oeste, e onde a criação de gado é a principal atividade econômica (Nunes da Cunha &
Junk ,1999). O estudo foi desenvolvido na Estrada Parque Transpantaneira (EPT), que corta
o município de Poconé longitudinalmente, ligando esta cidade à localidade de Porto Jofre,
na margem direita do Rio Cuiabá.
Esta rodovia atravessa gradientes de inundação no Pantanal por 145 km, sendo
utilizada como principal ponto de turismo. Os trabalhos foram concentrados nos primeiros
46 km desta rodovia, ou seja do portão do IBAMA, até a margem direita do rio Pixaim,
pela facilidade logística para realização dos estudos e porque tem muitos lagos neste trecho.
Esta rodovia funciona atualmente como um dique, pois para sua construção foi feito um
aterro elevado. Sua implantação causou desequilíbrio ao sistema de vazão, e

34
conseqüentemente impactos ambientais (Oliveira, 1997, Campos-Filho & Machado, 1999),
que ainda não foram avaliados em toda a sua extensão. O aterro da rodovia foi construído
com o solo de caixas de empréstimo laterais. A retirada do solo formou ‘lagos’ ao longo da
mesma, servindo de reservatório de água para o gado no período da seca. Com as cheias,
esses reservatórios são povoados por muitos organismos aquáticos (insetos, crustáceos,
moluscos, anfíbios, répteis e peixes) que servem de alimento para numerosas aves
aquáticas. As aves encontram nestas caixas de empréstimo fontes perenes para sua
alimentação e da sua prole, atuando portanto como elementos importantes no sucesso
reprodutivo de várias espécies de aves aquáticas como o Tuiuiú Jabiru mycteria (Oliveira,
1997; Oliveira & Cavalcanti, 2000). Sistema semelhante encontra nos lhanos venezuelanos
onde estudos das comunidades de aves aquáticas foram também realizados (Pinowski et al.,
1980, 1981; Kushlan et al., 1985; Morales et al., 1987; Pietro, 1988; Frederick & Bildstein,
1992; González, 1996a,b, 1997).
2.1 - Desenho amostral
Durante dois anos (1999 e 2000) mensalmente de março a dezembro ao longo de um
transecto linear de 46 Km., a EPT foi percorrida de carro a uma velocidade de 20 Km/h. As
observações das aves ao longo do transecto foram realizadas das 06 ás 10h da manhã e das
14 ás 18hs, período de maior atividade das mesmas. O censo foi feito por pontos, com
paradas em vários pontos nos quais a rodovia era percorrida a pé. Para o censo visual, foi
utilizado um binóculo (8x40), e para cada espécie de ave observada foi anotado o número
de indivíduos, a idade (jovem, adulto) e o sexo (quando possível). Foram consideradas nos
censos todas as aves que estavam localizadas até cerca de 300 m de ambas as margens da
rodovia, dentro desta faixa estão concentradas a maioria das caixas de empréstimo

35
(‘lagos’artificiais) e dos lagos naturais. Tanto os lagos criados pela construção da Estrada,
que ocorreu a mais de trinta anos atrás, quanto os lagos naturais, foram usados para o
estudo porque ambos estão atuando como lagos funcionais pois são sujeitos ás mesmas
variações da dinâmica de inundação do Pantanal e visitação das aves. Como a maioria
desses lagos são perenes, oferecem uma boa oportunidade para estudar a dinâmica das
comunidades de aves durante o ano em um mesmo tipo de habitat aquático. Portanto, neste
estudo foi removido a priori o efeito de habitats nas comunidades de aves.
Os dados metereológicos índice de pluviosidade, insolação e temperatura média do ar para
os anos de 1999 e 2000 foram cedidos pela Estação de Metereología do Ministério da
Agricultura de Cuiabá. A lista sistemática das aves segue a adotada pelo Comitê Brasileiro
de Registro Ornitológico (CBRO, 2006).
2.3 – Análises dos dados
Para verificar se existia variação sazonal significativa na riqueza e abundância de aves
aquáticas e piscívoras no Pantanal, foram realizadas Análise de Variância de dois níveis
ANOVA (II). Um modelo usou como variável dependente a riqueza de espécies de aves e o
outro a abundância de aves ambas em relação ao nível I (meses, março a dezembro) e nível
II (anos, 1999 e 2000). As análises foram realizadas usando o Programa Systat (Wilkinson,
1997).
Para verificar a existência de variação sazonal na riqueza e abundância de aves
piscívoras no Pantanal, foram realizadas Análises de Variância de dois níveis ANOVA (II),
uma usando como variável dependente riqueza de espécies de aves e a outra usando
abundância de aves em relação estações climáticas (seca e chuvosa).

36
Para verificar se existia variação anual e sazonal significativa na composição da
assembléia de aves aquáticas e piscívoras no Pantanal, foi usada uma análise de ordenação
do tipo escalonamento multidimensional (MDS). Foi construída, com os dados brutos, uma
matriz multidimensional com as espécies de aves nas colunas (objetos) e os locais, lagos
(atributos) nas linhas. Nesta matriz foi aplicado o índice de Bray-Curtis para reduzir a sua
dimensionalidade e construir a matriz de dissimilaridade. Este índice tem sido o mais
recomendado para estudos de comunidades de aves (MacNally, 1994) e estudos ecológicos
em geral (Minchin, 1987; Cintra, 1997). Em seguida uma análise de ordenação do tipo
MDS foi feita sobre a matriz de dissimilaridade resultando em uma lista de scores para
duas dimensões que foram plotados no espaço bidimensional entre dois eixos (MDS1 e
MDS2). Duas ordenações foram feitas, usando valores de abundância das espécies (dados
quantitativos), os quais colocam mais peso nas espécies mais abundantes, e outra
ordenação usando presença e ausência (dados qualitativos), os quais dão mais peso para
espécies raras. Para verificar se alguns grupos de espécies são diferentes de outros nesta
bidimensionalidade, foi usada uma Análise de Variância Multivariada (MANOVA). Nesta
análise os escores de cada eixo (MDS1 e MDS2) foram usados como variáveis dependentes
no seguinte modelo: MDS1, MDS2 = constante + variável categórica. O robusto teste
inferencial de Pillai-Trace foi usado para testar a significância da MANOVA. As análises
de MDS foram feitas usando o programa PATN (Belbin, 1982) e aquelas para a MANOVA
e testes inferênciais no programa Systat (Wilkinson, 1997).
3 - Resultados
Em 1999, o mês mais chuvoso foi o de novembro (269,7mm) e o mais seco foi
agosto (0,0mm). A pluviosidade total anual foi 1318,7mm, a média mensal 109,89 mm. A
insolação média anual foi de 230,8 com valores máximos em janeiro (230,8) e mínimos em

37
setembro (153,3). A temperatura média anual foi de 26 ºC. O mês de outubro apresentou a
maior média mensal de temperatura do ar (28,7 ºC) enquanto a menor média foi em julho
(23,4 ºC).
Neste ano a composição da comunidade de aves piscívoras foi formada por 18
espécies, enquanto que a comunidade de aves aquáticas foi formada por 39 espécies
distribuídas em 13 famílias (Tabela 4). A abundância total neste ano foi de 6374 indivíduos,
as espécies mais abundantes foram: Ardea alba L. (1115 indivíduos); Rostrhamus
sociabilis (Vieillot) (861 indivíduos); Aramus guaruna L. (800 indivíduos); Mycteria
americana L. (580 indivíduos) e Egretta thula (Molina) com 490 indivíduos (Figura 1).
Neste ano as aves piscívoras contribuíram com 54,26% da comunidade total de aves
aquáticas.
Em 2000 a pluviosidade total anual foi de 1264 mm, e a média mensal 105,3mm. O
mês mais chuvoso foi o de março (307,4mm) e o mais seco foi junho (0,0mm). A insolação
média anual foi de 202,2 o valor máximo foi em maio (247,5) e fevereiro (138,2). A
temperatura média anual foi de 26,5 ºC a maior média mensal foi em outubro (28,3 ºC), e o
com a menor em julho (22,8 ºC).
Neste ano a comunidade de aves piscívoras foi composta por 19 espécies, e a
comunidade de aves aquáticas foi formada por 37 espécies distribuídas em 13 famílias
(Tabela 4). A abundância total neste ano foi de 5107 indivíduos, as espécies mais
abundantes foram: Rostrhamus sociabilis (880 indivíduos); Ardea alba (728 indivíduos);
Aramus guaruna (594 indivíduos); Egretta thula (417 indivíduos) e Jabiru mycteria
(Lichtenstein) com 320 indivíduos (Figura 2). As aves piscívoras compuseram 54,16% da
comunidade total.

38
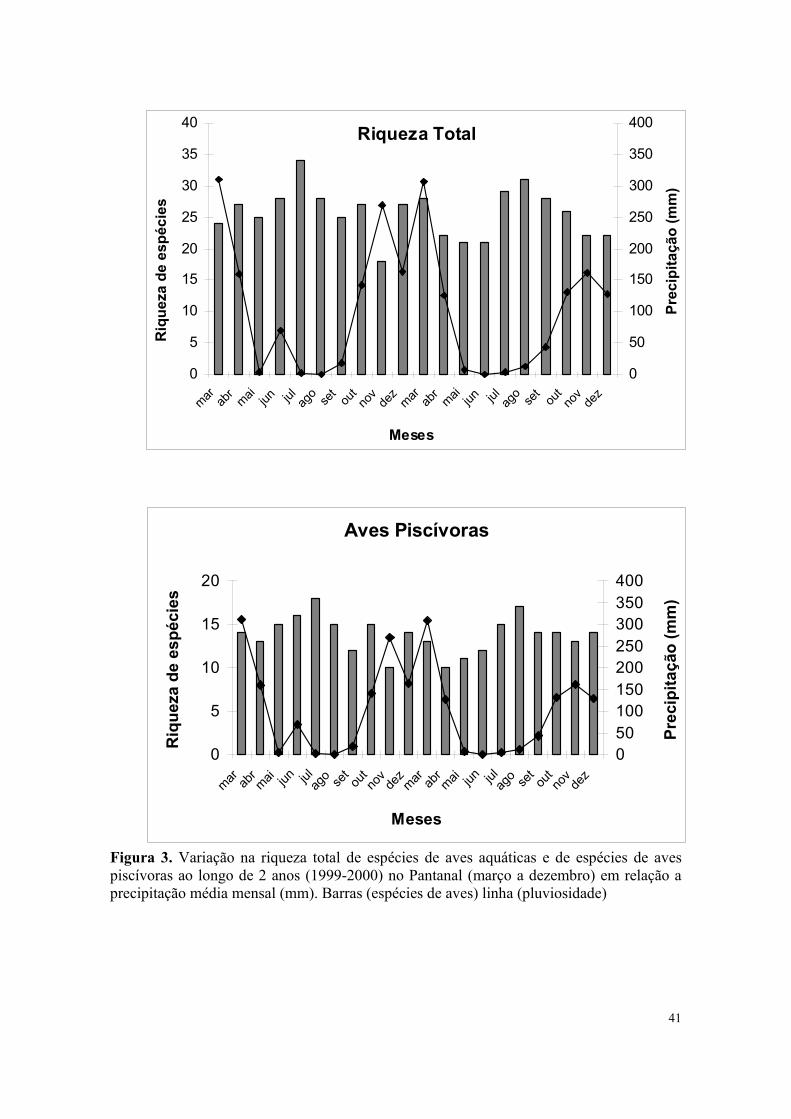
A riqueza total das aves associadas aos ambientes aquáticos e das piscívoras variou pouco
em função da quantidade de chuvas no Pantanal (Figura 3), entretanto a variação na
abundância das mesmas foi bem acentuada (Figura 4), mostrando a importância do nível
dos corpos da água estruturando a comunidade de aves aquáticas.

39
Figura 1. Estrutura da Comunidade total de aves aquáticas em 1999 no Pantanal de Poconé, MT. No eixo (x) o número total de indivíduos de cada espécie
0 200 400 600 800 1000 1200Ardea alba
Rostrhamus sociabilis
Aramus guarauna
Mycteria americana
Egretta thula
Bubulcus ibis
Jabiru mycteria
Jacana jacana
Ardea cocoi
Phalacrocorax brasilianus
Phimosus infuscatus
Polyborus plancus
Busarellus nigricollis
Coragyps atratus
Ceryle torquata
Tigrisoma lineatum
Pratalea ajaja
Nycticorax nycticorax
Cathartes aura
Theristicus caerulescens
Butorides striatus
Mesembrinibis cayennensis
Dendrocygna bicolor
Buteogallus meridionalis
Chloroceryle amazona
Buteogallus urubitinga
Cloroceryle americana
Dendrocygna viduata
Ciconia maguari
Dendrocygna autumnalis
Theristicus caudatus
Syrigma sibilatrix
Anhinga anhinga
Pilherodius pileatus
Chauna torquata
Cathartes burrovianus
Cairina moschata
Amazonetta brasiliensis
Milvago chimachima

40
Figura 2. Estrutura da Comunidade total de aves aquáticas em 2000 no Pantanal de Poconé, MT. No eixo (x) o número total de indivíduos de cada espécie
0 100 200 300 400 500 600 700 800Ardea alba
Aramus guarauna
Egretta thula
Jabiru mycteria
Mycteria americana
Phalacrocorax brasilianus
Jacana jacana
Ardea cocoi
Coragyps atratus
Caracara plancus
Busarellus nigricollis
Bubulcus ibis
Ciconia maguari
Phimosus infuscatus
Ceryle torquata
Tigrisoma lineatum
Butorides striatus
Buteogallus meridionalis
Cathartes aura
Theristicus caerulescens
Nycticorax nycticorax
Pratalea ajaja
Dendrocygna autumnalis
Mesembrinibis cayennensis
Chloroceryle amazona
Buteogallus urubitinga
Syrigma sibilatrix
Pilherodius pileatus
Chloroceryle americana
Dendrocygna viduata
Anhinga anhinga
Theristicus caudatus
Chauna torquata
Cairina moschata
Amazonetta brasiliensis
Egretta caerulea
41
Figura 3. Variação na riqueza total de espécies de aves aquáticas e de espécies de aves piscívoras ao longo de 2 anos (1999-2000) no Pantanal (março a dezembro) em relação a precipitação média mensal (mm). Barras (espécies de aves) linha (pluviosidade)
Riqueza Total
0
5
10
15
20
25
30
35
40
mar abr
mai jun jul ago se
tou
tno
vde
zmar ab
rmai jun jul ag
o set
out
nov
dez
Meses
Riq
ueza
de
espé
cies
0
50
100
150
200
250
300
350
400
Prec
ipita
ção
(mm
)
Aves Piscívoras
0
5
10
15
20
mar abrmai jun jul ag
o set
outno
vde
zmar ab
rmai jun jul ag
o set
outno
vde
z
Meses
Riq
ueza
de
espé
cies
050100150200250300350400
Prec
ipita
ção
(mm
)

42
Figura 4. Variação da abundância total de espécies de aves aquáticas e de espécies de aves piscívoras ao longo de 2 anos (1999-2000) no Pantanal (março a dezembro) em relação a precipitação média mensal (mm). Barras (espécies de aves) linha (pluviosidade)
Abundância Total
0
200
400
600
800
1000
1200
1400
mar abr mai jun jul ago set out nov dez mar abr mai jun jul ago set out nov dez
Meses
Abu
ndân
cia
0
50
100
150
200
250
300
350
400
Prec
ipita
ção
(mm
)
Aves Piscívoras
0
50
100
150
200
250
300
350
mar abr mai jun jul ago set out nov dez mar abr mai jun jul ago set out nov dez
Meses
Abu
ndân
cia
0
50
100
150
200
250
300
350
400
Prec
ipita
ção
(mm
)

43
3.1 - Variação na riqueza e abundância de aves aquáticas e piscívoras entre os anos de
1999-2000.
Para a comunidade total de aves aquáticas não houve diferença significativa na riqueza de
espécies entre os anos de 1999 e 2000 (ANOVA, F1, 18 = 0,641; p = 0,434), o mesmo
acontecendo para a abundância de aves aquáticas (ANOVA, F1, 18 = 0,955; p = 0,341)
(Figura 3). A comunidade de aves piscívoras tampouco apresentou diferença significativa
na riqueza de espécies entre os anos 1999 e 2000 (ANOVA, F1, 18 = 0,915; p = 0,352); o
mesmo acontecendo para a abundância de aves piscívoras (ANOVA, F1, 18 = 0,496; p =
0,490). Figura (4)
3.2 - Variação na Composição das Comunidades de aves aquáticas e piscívoras no
Pantanal entre os anos de 1999-2000.
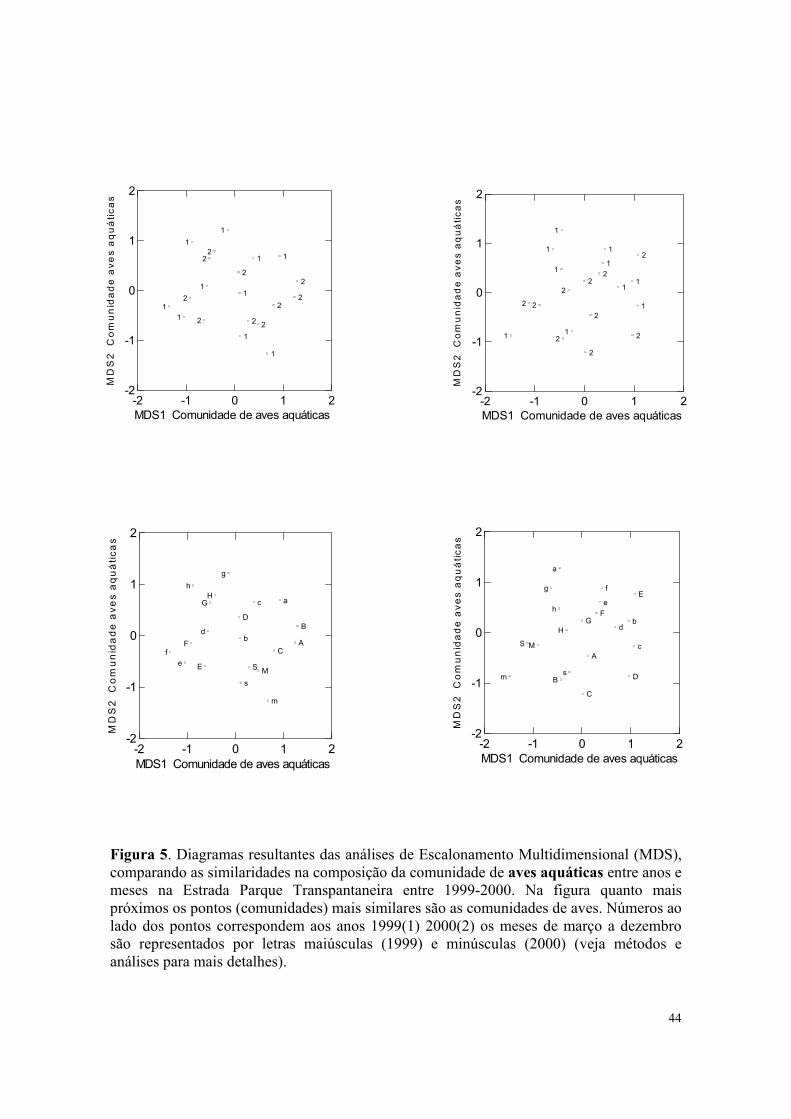
Os resultados das análises indicaram que houve mudança significativa na composição
quantitativa da comunidade de aves aquáticas entre os meses (MANOVA Pillai-Trace =
1;724; N = 20; p = 0,001); o mesmo foi verificado entre a composição qualitativa de aves
aquáticas entre os meses (Pillai-Trace = 1;324 N = 20; p = 0,047). Entretanto, a composição
quantitativa e qualitativa da comunidade de aves aquáticas não mudou significativamente
entre os anos (Quantitativa, Pillai-Trace = 0,057; N = 20; p = 0,610; Qualitativa, Pillai-
Trace = 0,134; N = 20; p = 0,294).
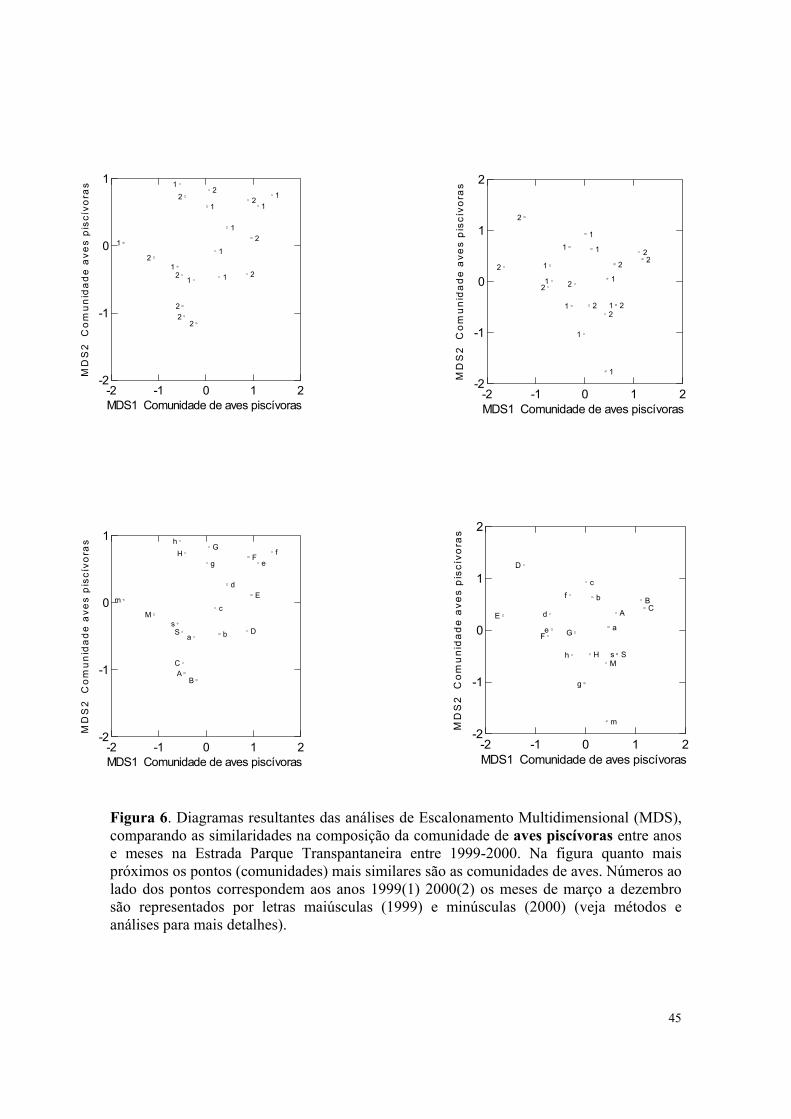
A composição quantitativa da comunidade de aves piscívoras mudou
significativamente entre os meses do ano (Pillai Trace = 1;789; N = 20 p = 0,001) , como
também verificado para a composição qualitativa da comunidade de aves piscívoras entre
os mesmos meses (Pillai-Trace = 1,601; N = 20 p = 0,001) (Figura 5 e 6).
44
Figura 5. Diagramas resultantes das análises de Escalonamento Multidimensional (MDS), comparando as similaridades na composição da comunidade de aves aquáticas entre anos e meses na Estrada Parque Transpantaneira entre 1999-2000. Na figura quanto mais próximos os pontos (comunidades) mais similares são as comunidades de aves. Números ao lado dos pontos correspondem aos anos 1999(1) 2000(2) os meses de março a dezembro são representados por letras maiúsculas (1999) e minúsculas (2000) (veja métodos e análises para mais detalhes).
-2 -1 0 1 2MDS1 Comunidade de aves aquáticas
-2
-1
0
1
2
MD
S2
C
om
un
ida
de
ave
s a
qu
átic
as
m
S M
g
h
a
B
H
s
G
E
c
D
bd
f
eF
A
C
-2 -1 0 1 2MDS1 Comunidade de aves aquáticas
-2
-1
0
1
2
MD
S2
C
om
un
ida
de
ave
s a
qu
áti c
as
fe
F
h
E
d
GH
g
DB
A
a
C
m
M
c
S
s
b
-2 -1 0 1 2MDS1 Comunidade de aves aquáticas
-2
-1
0
1
2
MD
S2
C
om
un
ida
de
ave
s a
qu
átic
as
1
2 2
1
1
1
2
2
1
2
2
1
2
11
1
12
2
2
-2 -1 0 1 2MDS1 Comunidade de aves aquáticas
-2
-1
0
1
2
MD
S2
C
om
un
ida
de
av e
s a
qu
át ic
as
11
2
1
2
1
22
1
22
2
1
2
1
2
1
2
1
1
45
Figura 6. Diagramas resultantes das análises de Escalonamento Multidimensional (MDS), comparando as similaridades na composição da comunidade de aves piscívoras entre anos e meses na Estrada Parque Transpantaneira entre 1999-2000. Na figura quanto mais próximos os pontos (comunidades) mais similares são as comunidades de aves. Números ao lado dos pontos correspondem aos anos 1999(1) 2000(2) os meses de março a dezembro são representados por letras maiúsculas (1999) e minúsculas (2000) (veja métodos e análises para mais detalhes).
-2 -1 0 1 2MDS1 Comunidade de aves piscívoras
-2
-1
0
1
MD
S2
C
om
un
ida
de
ave
s p
iscí
vora
s
1
21
1
2
22
2
1
2
11
2
2
2
1
1
1
2
1
-2 -1 0 1 2MDS1 Comunidade de aves piscívoras
-2
-1
0
1
2
MD
S2
C
om
un
ida
de
ave
s p
iscí
vora
s
2
2
2
1
1
1
1
2
1
1
22
21
2
1
1
2
1
2
-2 -1 0 1 2MDS1 Comunidade de aves piscívoras
-2
-1
0
1
MD
S2
C
om
un
ida
de
ave
s p
iscí
vora
s
m
Ms
h
S
CA
H
a
B
fe
E
F
D
d
b
c
G
g
-2 -1 0 1 2MDS1 Comunidade de aves piscívoras
-2
-1
0
1
2
MD
S2
C
om
un
ida
de
ave
s p
iscí
vor a
s
E
D
F
d
e
f
h
G
g
c
CB
Ss
A
a
m
M
b
H

46
Figura 7. Parciais da regressão múltipla linear da variação na abundância de aves aquáticas e piscívoras no Pantanal em relação a pluviosidade média, mensal (mm). As parciais correspondem aos resultados apresentados na Tabela 2. Alguns números nos eixos são negativos porque a regressão parcial representa os desvios dos resultados esperados se todas as outras variáveis são mantidas constantes nas suas médias observadas. Só estão apresentadas as figuras cujos resultados foram significativos
-100 0 100 200Pluviosidade (X parcial 1)
-500
0
500
1000
Ab
un
dâ
nci
a d
e a
ves
aq
uá
tica
s (Y
pa
rcia
l 1)
-200 -100 0 100 200Pluviosidade (X parcial 1)
-300
-200
-100
0
100
200
300
400
500
Ab
un
dâ
nci
a d
e a
ves
pis
cívo
ras
( Y p
arc
ial 1
)

47
Figura 8. Regressão linear simples da variação na abundância de aves aquáticas no Pantanal em relação a temperatura do ar (ºC). Os dois anos (1999-2000) estão incluídos na mesma figura. Os resultados correspondem a aqueles dos modelos apresentados na Tabela 2.
22 23 24 25 26 27 28 29Temperatura do ar
0
500
1000
1500
Ab
un
dâ
nci
a d
e a
v es
aq
uá
tica
s
22 23 24 25 26 27 28 29Temparatura do ar
0
100
200
300
400
500
600
700
800
900
1000
Ab
un
dâ
nci
a d
e a
ves
pis
cívo
ras

48
A correlação entre as variavéis das características físicas e biológicas dos lagos
Como algumas variavéis independentes (pluviosidade, insolação e temperatura do ar);
foram significativamente correlacionadas entre si (Tabela 1), elas foram usadas em modelos
estatisticos separados. As variáveis pluviosidade e insolação, foram usadas juntas no
mesmo modelo e a temperatura do ar em outro modelo.
3.3. Efeito das variavéis ambientais na riqueza e abundância de aves aquáticas e
piscívoras no Pantanal entre os anos de 1999-2000
A variação na riqueza de espécies aves aquáticas não foi afetada significativamente pela insolação total (Tabela 2) nem pela pluviosidade média mensal (Tabela 2) e tampouco pela temperatura do ar (Tabela 2). Entretanto, a abundância de aves aquáticas foi afetada significativamente pela pluviosidade média mensal (Tabela 2 e Figura 7) e pela temperatura média mensal do ar (Tabela 2, Figura 8). Contudo, a insolação não afetou significativamente a abundância de aves aquática (Tabela 2). A variação na riqueza de espécies aves piscívoras não foi afetada significativamente pela pluviosidade média mensal (Tabela 2) nem pela insolação total (Tabela 2) e nem também pela temperatura do ar (Tabela 2). Contudo a abundância de aves piscívoras foi afetada significativamente pela pluviosidade média mensal (Tabela 2, Figura 7) e pela temperatura média mensal do ar (Tabela 2, Figura 8). Todavia, a insolação não afetou significativamente a abundância de aves piscivoras (Tabela 2).
3.4- Efeito das variavéis ambientais na composição das aves aquáticas e piscívoras no Pantanal entre os anos de 1999-2000 Os resultados das análises indicaram que houve relação significativa entre a composição
quantitativa da comunidade de aves aquáticas e a pluviosidade (Tabela 3, Figura 9).
Entretanto, a insolação total e a temperatura do ar não afetaram significativamente a
composição quantitativa de aves aquáticas (Tabela 3).
A composição qualitativa da comunidade de aves aquáticas foi afetada significativamente
pela a temperatura média mensal do ar e pela pluviosidade média mensal (Tabela 3) e a
insolação mensal total não afetaram significativamente a composição qualitativa de aves
aquáticas (Tabela 3).

49
Os resultados das análises indicaram que houve relação significativa entre a composição
quantitativa da comunidade de aves piscivoras e a pluviosidade média mensal (Tabela 3,
Figura 10) e a temperatura média mensal do ar (Tabela 3, Figura 10). Entretanto, a
insolação mensal total não afetou significativamente a composição quantitativa de aves
piscívoras. (Tabela 3)
Os resultados das análises indicaram que houve relação significativa entre a composição
qualitativa da comunidade de aves piscivoras e a pluviosidade média mensal (Tabela 3,
Figura 10), e a temperatura média mensal do ar (Tabela 3, Figura 11). Entretanto, a
insolação mensal total não afetou significativamente a composição qualitativa de aves
piscívoras (Tabela 3).
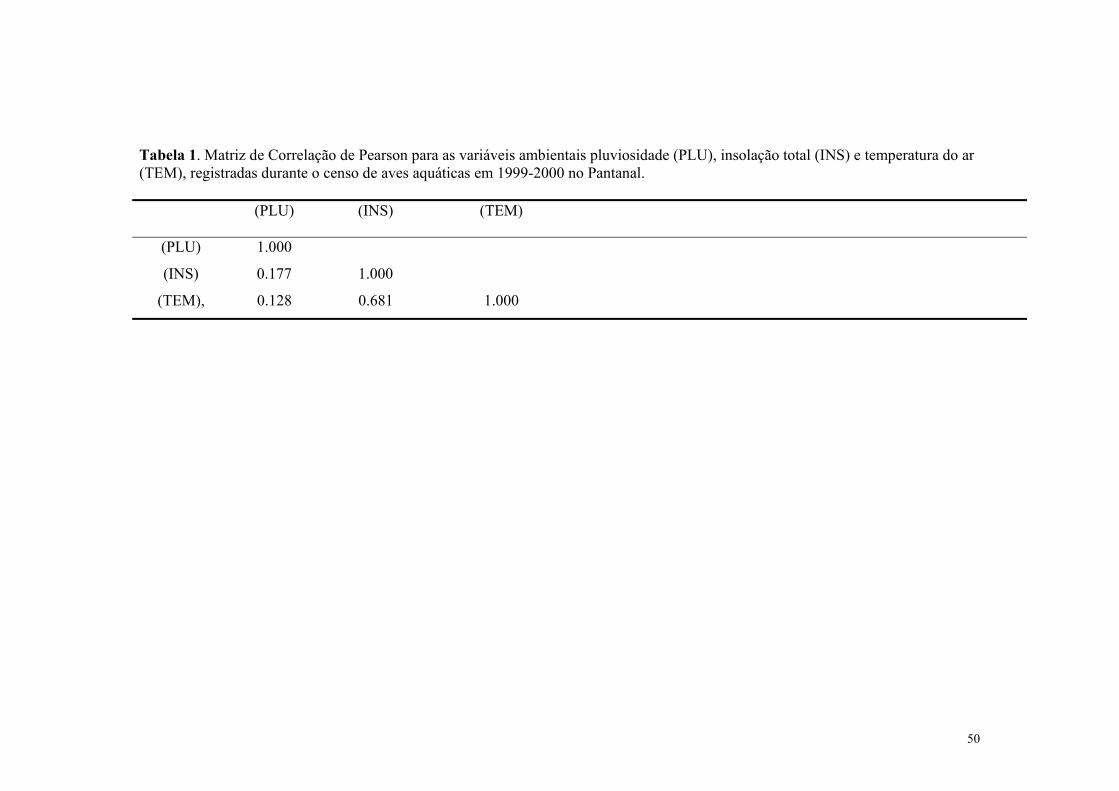
50
Tabela 1. Matriz de Correlação de Pearson para as variáveis ambientais pluviosidade (PLU), insolação total (INS) e temperatura do ar (TEM), registradas durante o censo de aves aquáticas em 1999-2000 no Pantanal.
(PLU) (INS) (TEM)
(PLU) 1.000
(INS) 0.177 1.000
(TEM), 0.128 0.681 1.000

51
Tabela 2. Resultados das análises de regressão múltipla linear dos efeitos das variavéis ambientais (Pluviosidade, Temperatura do ar e Insolação) na riqueza e abundância das aves aquáticas e piscívoras em 1999 e 2000 no Pantanal __________________________________________________________________________________________________________ Riqueza de aves Abundância de aves R 2 N T P R 2 N T P Aves aquáticas Pluviosidade 0.108 20 -1.423 0.173 0.384 20 -3.244 0.005 Temperatura do ar 0.098 20 -1.396 0.180 0.279 20 -2.638 0.017 Insolação total 0.108 20 -0.411 0.686 0.384 20 -0.321 0.752 Aves piscívoras Pluviosidade 0,157 20 -1.692 0,109 0.417 20 -3.485 0.003 Temperatura do ar 0.102 20 -1.430 0.170 0.203 20 -2.142 0.046 Insolação total 0.157 20 -0.243 0.811 0.417 20 -0.426 0.675

52
Tabela 3.. Resultados das análises de regressão múltipla multivariada dos efeitos da pluviosidade, temperatura do ar e insolação na composição quantitativa e qualitativa da comunidade de aves aquáticas e piscívoras em 1999 e 2000 no Pantanal ______________________________________________________________________________________________________________ Composição quantitativa da comunidade de aves Composição qualitativa da comunidade de aves Pillai-Trace N P Pillai-Trace N P Aves aquáticas Pluviosidade 0.413 20 0.014 0.311 20 0.051 Temperatura do ar 0.107 20 0.384 0,498 20 0.003 Insolação total 1.103 20 0.418 0.067 20 0.576 Aves piscívoras Pluviosidade 0.581 20 0.001 0.375 20 0.023 Temperatura do ar 0.510 20 0.002 0.527 20 0.002 Insolação total 0.077 20 0.525 0.058 20 0.618

53
Figura 9. Relação da variação na composição quantitativa (gráfico superior) e qualitativa (gráfico inferior) da comunidade de aves aquáticas no Pantanal em relação a pluviosidade média mensal (mm). Para simplificar somente a primeira dimensão (MDS1) da ordenação foi plotada contra a pluviosidade.
0 100 200 300 400Pluviosidade
-2
-1
0
1
2
MD
S 1
Com
unid
a de
de a
v es
aquá
ti cas
0 100 200 300 400Pluviosidade
-2
-1
0
1
2
MD
S 1
Com
unid
ade
de a
ves
aquá
tica s
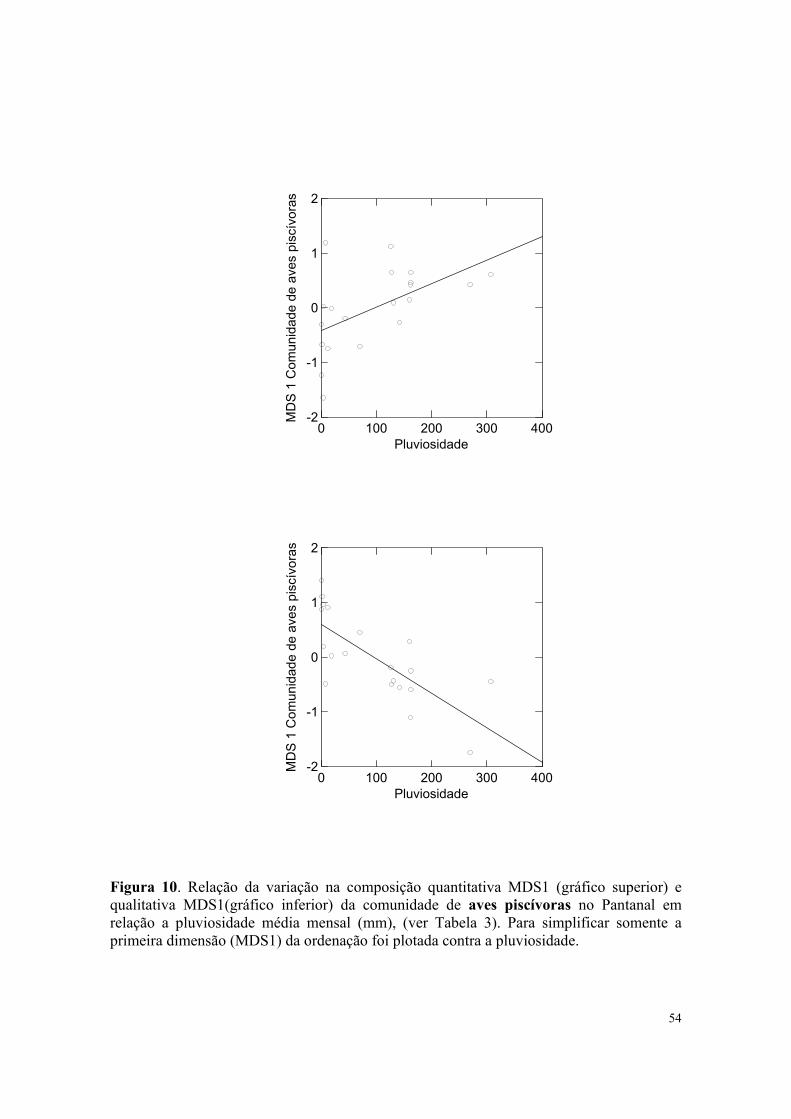
54
Figura 10. Relação da variação na composição quantitativa MDS1 (gráfico superior) e qualitativa MDS1(gráfico inferior) da comunidade de aves piscívoras no Pantanal em relação a pluviosidade média mensal (mm), (ver Tabela 3). Para simplificar somente a primeira dimensão (MDS1) da ordenação foi plotada contra a pluviosidade.
0 100 200 300 400Pluviosidade
-2
-1
0
1
2
MD
S 1
Com
unid
ade
de a
v es
pisc
ívor
as
0 100 200 300 400Pluviosidade
-2
-1
0
1
2
MD
S 1
Com
unid
ade
de a
ves
pisc
ívor
as

55
Figura 11. Relação da variação na composição qualitativa da comunidade de aves piscívoras no Pantanal (anos 1999-2000) em relação a temperatura do ar (ºC), (ver Tabela 3)
22 23 24 25 26 27 28 29Temperatura do ar
-2
-1
0
1
2
MD
S 1
Com
uni d
ade
d e a
ves
pisc
ívor
as

56
Figura 12. Distribuição das espécies de aves aquáticas em relação a média da pluviosidade mensal (mm). As barras mostram a abundância relativa dentro das espécies, mas não são comparadas entre as espécies, porque o número de indivíduos variou entre as espécies

57
4 – Discussão
4.1 Efeitos da pluviosidade, temperatura do ar e da insolação na riqueza e abundância de aves aquáticas e piscívoras no Pantanal.
Entre os dois anos de estudo (1999-2000) não ocorreram diferenças significativas na
riqueza de aves aquáticas e das piscívoras (Tabela 2). Neste bioma o pulso de inundação é a
principal força geradora dos padrões e processos ecológicos (Junk & Silva, 1999; Oliveira
& Calheiros, 2000), esta região esta submetida a ciclos plurianuais, caracterizados por
revezamento de anos muitos chuvosos, ciclos de enchentes; com anos relativamente secos,
ciclos de secas (Ádamoli 1986; Ponce,1995). Em decorrência das flutuações sazonais do
nível da água no Pantanal, o ciclo de vida das aves da região, estão ajustadas a este ciclo
hidrológico.
No Pantanal ocorrem 156 espécies de aves associadas aos ambientes aquáticos,
dessas muitas são sazonais, e poucas residentes (Cintra & Yamashita, 1990; Pinto, 2005). O
número de espécies sazonais permaneceu constante em ambientes de florestas e aumentou
em outros habitats, como nas áreas alagáveis conforme Figueiras et al., (2006). Na área de
estudo não ocorrem bandos de nenhuma espécie migratória intercontinental, como os
maçaricos. Outro grupo com muitas espécies são os anatídeos (patos, marrecos), dos quais
foram registradas somente 5 espécies no local. Das 32 espécies de aves piscívoras que
ocorrem no Pantanal, 28 espécies já foram registradas para o Pantanal de Poconé (Cintra &
Yamashita, 1990; Tubelis & Tomas, 2003a). Porém, neste local, não foram registradas aves
praieras continentais como o Taiamã (Rhynchops nigra) o Trinta-réis (Sterna superciliares)
ou espécies migratórias solitárias como por exemplo a Águia-pescadora (Pandion halietus).
Outras espécies de piscívoras não foram amostradas pelo método empregado, por causa dos
seus hábitos. Com exemplo deste caso pode ser citado o Arapapá (Cochlearius

58
cochlearius), ave noturna, solitária, que vive dentro das matas; embora o Socó-dorminhoco
(Nycticorax nycticorax), também seja de hábito noturno, o mesmo, vive em bandos e pode
ser observado nas primeiras horas da manhã e no crepúsculo, horário do censo populacional
nas margens da Transpantaneira; o Socó-beija-flor (Agamia agami) é raríssimo na região e
só foi avistado nas matas ciliares do Rio Pixaim, fora da área do censo. Estes, entre outros
aspectos, fazem com que a variação na riqueza de espécies de aves aquáticas e piscívoras
não seja significativa de ano para ano.
A abundância das aves aquáticas e das piscívoras (Figura 9) foi afetada
significativamente pela pluviosidade média mensal (Tabela 2). A abundância destas aves
aumentou com o decréscimo da pluviosidade. A diminuição das chuvas tem reflexo direto
no nível das áreas alagáveis da região. Com a retração das águas ocorre a concentração de
peixes, principalmente aqueles que resistem às baixas condições de oxigênio dissolvido na
água (Catella, 1982; Súarez, 2004; ver Capítulo 4). Estes peixes tornam-se presas fáceis
para este grupo de aves, formado em sua maioria por aves piscívoras, dentre estas as
pernaltas foram mais abundantes. Um dos fatores limitantes das aves pernaltas é o tamanho
das pernas (tarso-metatarso), que permitem ou não alcançar os locais mais profundos nos
lagos a procura de suas presas (Kushlan, 1976, 1986; Morales & Pacheco, 1986). Elas
aparecem no Pantanal em maior número na época da vazante e da seca, quando formam
grandes bandos. O mesmo é citado para os lhanos da Venezuela, onde a diminuição do
nível da água modifica o comportamento das aves aquáticas; as que antes caçavam
solitárias ou em pequenos grupos na cheia, na seca formam grandes bandos (Kushlan,
1986; González, 1996, 1997).

59
Em um único local na área estudada foi observado no mês de julho uma grande
agregação de aves aquáticas formadas entre outras por 745 garça-branca grande; 230 tuiuis
e 350 garça-branca-pequena.
As aves aquáticas formam diferentes guildas como piscívoras, malacófagas, insetívoras,
herbívoras, etc. Para obterem seus alimentos cada uma destas guildas utilizam diferentes
técnicas. Assim, na época das cheias, quando o nível da água esta mais alto, ocorre
predominância de aves mergulhadoras, como o Biguá (Phalacrocorax brasilianus) e
Biguatinga (Anhinga anhinga). Também é neste momento que a população do Gavião-
caramujeiro (Rosthramus sociabilis) aumenta no Pantanal, com o incremento de muitos
jovens oriundos provavelmente do Sul do Brasil. Neste período estes gaviões capturam os
caramujos e caranguejos, suas principais presas no Pantanal, na superfície dos corpos de
água. Ao longo dos meses, com o escoamento das águas, as aves mergulhadoras já
começam a enfrentar dificuldades para procurar suas presas, por causa da diminuição do
nível da água, aumento da turbidez, aumento da cobertura de macrófitas aquáticas e
também pela competição com as aves vadeadoras. Então, realizam deslocamentos no
Pantanal ou fora dele à procura de locais mais propícios.
Também a temperatura média do ar (Tabela 2), teve influencia significativa tanto na
abundância das aves aquáticas quanto das piscívoras (Figura 10). Branco (2002), ao estudar
as flutuações sazonais na abundância de Biguá (Phalacrocorax olivaceus) em Santa
Catarina, verificou que ocorreram variações significativas na abundância desta ave entre os
meses do ano, entretanto, entre os cinco anos estudados tal fato não aconteceu. Entretanto, a
literatura cita para o Gaivotão (Larus dominicanus), na mesma região, que além das
variações significativas na abundância entre os meses, também ocorreram variações entre
os anos (Branco & Ebert, 2002). Os autores atribuíram essas variações entre os anos, a uma

60
maior abundância dessas aves durante um dos anos de estudo. E atribuíram este fato ao
ciclo de vida desta ave, as oscilações diárias no estuário e ao período de atividade da frota
pesqueira. Nos lagos do Pantanal ocorre situação similar, o número de indivíduos de
algumas espécies, como por exemplo a Garça-branca grande (Ardea alba), no primeiro ano
foi registrado 1115 exemplares no segundo ano somente 728 indivíduos. A abundância na
composição de espécies das aves piscívoras no local está fortemente influenciada pela
presença das aves que nidificam em colônias ou viveiros, como as garças (Ardea alba,
Egretta thula), o Cabeça-seca (Mycteria americana) e o Biguá (Phalacrocorax
brasilianus). Alguns destes viveiros situam-se a menos de 15km da rodovia, servindo de
fonte segura de alimento para essas espécies, contribuindo assim para o sucesso reprodutivo
dessas aves coloniais, da mesma forma como foi mostrado nos Everglades (Kushlan et
al.,1975; Frederick & Collopy, 1989) e nos Lhanos da Venezuela (González, 1999). O
constante movimento das várias espécies de aves indo do ninhal para as proximidades da
estrada deve contribuir para que o número de espécies e a abundância de indivíduos se
mantenham similares nas comunidades, apesar das comunidades mudarem na composição.
Estudos realizados em lagos da América do Norte mostraram que estas aves podem realizar
longos deslocamentos desde o ninhal até a fonte de alimentação, por exemplo o Cabeça-
seca (Mycteria americana) foi observado forrageando até 130km da colônia. Entretanto,
apesar desses valores extremos, a distância média de deslocamento observado ficou em
torno de 24km para o conjunto de espécies de aves que nidificaram neste local (Custer &
Osborn, 1978). Forragear próximo ao local de reprodução aumenta o sucesso reprodutivo
tanto de aves que nidificam em colônias como das que fazem ninhos solitários. Dentre
outros fatores o que contribui para esse sucesso é o fornecimento de farta alimentação aos
filhotes, o que faz com que eles cresçam mais rápido, e também a presença dos pais no

61
ninho, que inibe a predação (Frederick & Callopy, 1988; Oliveira, 1997; González, 1999).
Estes locais de nidificação podem ser usados por vários anos, o que para as aves colônias é
denominado tenacidade ao ninhal ‘colony site tenacity,’. A tenacidade também ocorre para
algumas espécies de aves aquáticas que nidificam solitariamente (Burger, 1984, Oliveira,
1997). Esta fidelidade ao local de alimentação e a tenacidade à área de nidificação
poderiam explicar a grande similaridade na composição da comunidade de aves aquáticas e
piscívoras entre estes dois anos deste estudo.
Uma outra explicação para esta aparente “estabilidade” na composição é que os lagos
maiores da área estudada são perenes e podem manter comunidade de aves durante a
maioria dos meses do ano. As espécies raras no local, como Trinta-réis (Sterna
superciliares) são sazonais, enquanto que as espécies menos abundantes como os martins-
pescadores são residentes, contribuindo assim para a manutenção da estabilidade da
comunidade entre os anos.
Diferentes espécies de aves aquáticas, vivem em diferentes habitats. Algumas
espécies utilizam vários ambientes, porém algumas possuem um habitat favorito
dependendo da sua morfologia, da fisiologia, da táticas de forrageamento, e do
comportamento alimentar (Meyerriecks, 1962, Kushland, 1978 e 1981; Thomas, 1985;
Willard, 1985; Frederick & Bildstein, 1999; Zeffer et al., 2003). As aves piscívoras
utilizam diferentes táticas de forrageamento, para obterem suas presas. Assim, na época das
cheias, quando o nível da águas esta mais alto, ocorre predominância de aves
mergulhadoras, como o Biguá e Biguatinga. Neste momento a transparência da água nos
lagos é maior, as macrófitas aquáticas estão mais dispersas pela superfície do lago,
facilitando a detecção dos peixes por estas aves, que são visualmente orientadas. Ao longo
dos meses, com o escoamento das águas, as aves mergulhadoras já começam a enfrentar

62
dificuldades para procurar suas presas, por causa da diminuição na profundidade dos
ambientes aquáticos, aumento da turbidez, aumento da cobertura de macrófitas aquáticas e
também enfrentam a competição com as aves vadeadoras. Então estas realizam
deslocamento dentro e fora do Pantanal acompanhando o fluxo da água. Na vazante
predominam as aves piscívoras vadeadoras ou pernaltas. A diminuição do volume de água
faz com que as aves aquáticas tenham sua riqueza e abundância mudadas acompanhando a
diferença da flutuação do nível da água e disponibilidade de presas no Pantanal. Situação
semelhante é observada nos lhanos da Venezuela. Assim as aves aquáticas aproveitam a
farta alimentação que está disponível em toda a planície pantaneira na época da vazante.
Portanto, o esperado é que ocorra uma diferença na composição qualitativa e quantitativa
das aves aquáticas e piscívoras ao longo dos meses, como os resultados deste estudo
sugerem.
Esta região parece não ser propicia para os patos e marrecos, que formam grandes
bandos em locais relativamente próximos, entre 35 a 80Km, como nos campos inundáveis
da estrada de Porto Cercado e Campo do Jofre onde podem ser observados bandos mistos
de mais de 1000 indivíduos de Marreca caneleira (Dendrocygna bicolor) e Irerê
(Dendrocigna viduata). Durante os dois anos de amostragem nos lagos só foram
observados poucos exemplares dessas aves.
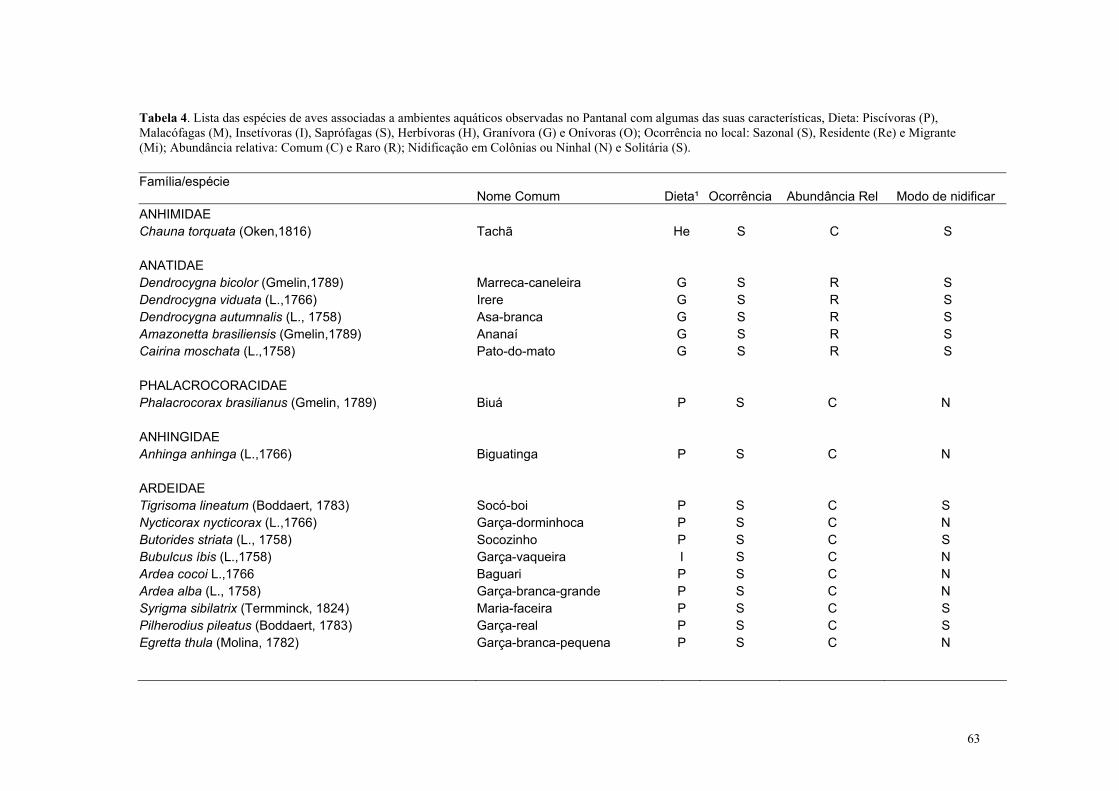
63
Tabela 4. Lista das espécies de aves associadas a ambientes aquáticos observadas no Pantanal com algumas das suas características, Dieta: Piscívoras (P), Malacófagas (M), Insetívoras (I), Saprófagas (S), Herbívoras (H), Granívora (G) e Onívoras (O); Ocorrência no local: Sazonal (S), Residente (Re) e Migrante (Mi); Abundância relativa: Comum (C) e Raro (R); Nidificação em Colônias ou Ninhal (N) e Solitária (S). Família/espécie Nome Comum Dieta¹ Ocorrência Abundância Rel Modo de nidificar ANHIMIDAE Chauna torquata (Oken,1816) Tachã He S C S ANATIDAE Dendrocygna bicolor (Gmelin,1789) Marreca-caneleira G S R S Dendrocygna viduata (L.,1766) Irere G S R S Dendrocygna autumnalis (L., 1758) Asa-branca G S R S Amazonetta brasiliensis (Gmelin,1789) Ananaí G S R S Cairina moschata (L.,1758) Pato-do-mato G S R S PHALACROCORACIDAE Phalacrocorax brasilianus (Gmelin, 1789) Biuá P S C N ANHINGIDAE Anhinga anhinga (L.,1766) Biguatinga P S C N ARDEIDAE Tigrisoma lineatum (Boddaert, 1783) Socó-boi P S C S Nycticorax nycticorax (L.,1766) Garça-dorminhoca P S C N Butorides striata (L., 1758) Socozinho P S C S Bubulcus íbis (L.,1758) Garça-vaqueira I S C N Ardea cocoi L.,1766 Baguari P S C N Ardea alba (L., 1758) Garça-branca-grande P S C N Syrigma sibilatrix (Termminck, 1824) Maria-faceira P S C S Pilherodius pileatus (Boddaert, 1783) Garça-real P S C S Egretta thula (Molina, 1782) Garça-branca-pequena P S C N

64
Tabela 4 (Continuação) Família/espécie Nome Comum Dieta¹ Ocorrência Abundância Rel Modo de nidificar¹ Egretta caerulea (L., 1758) Garça-azul P S R N* THRESKIORNITHIDAE Mesembrinibis cayennensis (Gmelin, 1789) Corocoró O S C S Theristicus caerulescens (Vieillot, 1817) Curicaca-cinza O S C S Theristicus caudatus (Boddaert, 1783) Curicaca O S C S Phimosus infuscatus (Lichtenstein, 1823) Frango-d'água O S C S Pratalea ajaja (L., 1758) Colhereiro P S R N CICONIIDAE Ciconia maguari (Gmelin, 1789) Tabuiaiá P S R S Jabiru mycteria (Lichtenstein, 1819) Tuiuiú P S C S Mycteria americana (L.,1758) Cabeça-seca P S C N CATHARTIDAE Cathartes aura (L.,1758) Urubu-caçador Sa Re C S Cathartes burrovianus Cassin, 1845 Urubutinga Sa Re C S Coragyps atratus (Bechstein, 1793) Urubu-preto Sa Re C S ACCIPITRIDAE Rostrhamus sociabilis (Vieillot, 1817) Gavião-caramujeiro M S C N* Rupornis magnirostris (Gmelin, 1788) Gavião-carijó C S C S Busarellus nigricollis (Latham, 1790) Gavião-belo P S C S Buteogallus meridionalis (Latham,1790) Gavião-fumaça C S C S Buteogallus urubitinga (Gmelin, 1788) Gavião-preto C S C S Geranospiza caerulescens (Vieillot,1817) Gavião-pernilongo C S C S

65
Tabela 4. (Continuação) Família/espécie Nome Comum Dieta¹ Ocorrência Abundância Rel Modo de nidificar FALCONIDAE Caracara plancus (Miller, 1777) Caracará O Re C S ARAMIDAE Aramus guarauna (L.,1766) Carão M S C S RALLIDAE Aramides cajanea (Statius Muller, 1776) Três-potes O S C S EURYPYGIDAE Eurypyga helias (Pallas,1781) Pavãozinho O S C S JACANIDAE Jacana jacana (L.1766) Cafezinho O S C S CHARADRIIDAE Vanellus cayanus (Latham, 1790) Batuira-de-esporão S C S SCOLOPACIDAE Tringa solitaria Wilson, 1813 Maçarico-solitario I Mi-VN R S RECURVIROSTIDAE Himantopus melanurus Vieillot, 1817 Pernilongo I S R S LARIDAE Sternula superciliaris (Vieillot, 1819) Trinta-reis-anão P S R N
66
Tabela 4. (Continuação) Família/espécie Nome Comum Dieta¹ Ocorrência Abundância Rel Modo de nidificar ALCEDINIDAE Ceryle torquatus (L.,1766) Martim-pescador-grande P Re C S Chloroceryle amazona (Latham, 1790) Martim-pescador-verde P Re C S Chloroceryle americana (Gmelin, 1788) Martim-pescador-pequeno P Re C S *N = Aves que nidificam em colônias, porém não há registro da nidificação delas para o Pantanal; Dieta¹ e nidificação¹ Carboneras (1992ab); Elliott (1992); Gochfeld & Buger ,1996; Hancock et al., 1992; Martinez-Vilalta & Motis ,1992; Sick, 1997.

67
Capítulo 2. Efeitos das características bióticas e abióticas dos ambientes alagáveis na
composição da assembléia de aves piscívoras
1. Introdução
Padrões hidrológicos afetam muitos aspectos da ecologia das aves piscívoras em vários
ecossistemas, como por exemplo nos Everglades, Lhanos, Amazônia e Pantanal (Bancroft
et al., 2002; González, 1996a b, 1997; Oliveira, 1997; Petermann, 1997; Cintra et al., no
prelo). Das variáveis hidrológicas tais como profundidade, oxigênio dissolvido, salinidade,
temperatura e transparência; a profundidade dos corpos da água é a que tem um papel
fundamental na determinação dos locais de forrageamento dessas aves nas áreas alagáveis
(Kushlan, 1976; Chow-Fraser et al., 1998; Bancroft et al., 2002).
As aves piscívoras respondem de diferentes maneiras às variações sazonais do nível
da água desses ambientes, pois utilizam diferentes táticas para capturar suas presas. Essas
diferenças estão relacionadas com a sua anatomia (principalmente tamanho do tarso-
metatarso), fisiologia e comportamento (Meyerriecks, 1962, Kushland, 1978 e 1981;
Thomas, 1985; Willard, 1985; Frederick & Bildstein, 1999; Zeffer et al. 2003). Estudos
realizados nos Everglades e nos Llanos venezuelanos demonstraram que existe correlação
positiva entre a profundidade do nível da água e a presença de determinadas espécies de
aves aquáticas nestes locais (Kushlan et al., 1975; Kushlan, 1976; 1986; Dreitz et al. 2004).
Similar situação foi verificada para o Pantanal (ver Capítulo 1). O acesso a presas e
abundância das mesmas, é um fator determinante no sucesso reprodutivo dessas aves,

68
notadamente das que fazem ninhos em colônias (Davis & Graham, 1991; Wagner, 1997;
Collis et al., 2002; Dreitz et al., 2004).
A disponibilidade de recursos alimentares é um dos fundamentos da ecologia de
comunidades para explicar a riqueza de espécies e abundância de aves em diferentes
regiões (Chow-Fraser et al., 1998; Elmberg et al., 2003). Assim, os peixes como o principal
recurso alimentar para as aves piscívoras, têm sido estudados em muitas regiões. Esses
trabalhos demonstraram a existência de correlação positiva entre a abundância deste
recurso e a abundância de aves (Wagner, 1997; Paskowski &Tonn, 2000). Também a
competição por alimento entre aves e peixes pode afetar a distribuição e abundância delas
nos lagos (Wagner & Hansson, 1998). Muitos destes peixes vivem associados a macrófitas
aquáticas, à procura de alimento, abrigo ou refúgio contra predadores em ambientes
naturais (Junk, 1973; Araujo-Lima et al., 1986; Soares et al., 1986; Meschiatti et al., 2000;
Pazin, 2000; Sanches-Botero et al., 2001; Petry et al., 2003), ou artificiais como represas de
hidrelétricas (Casatti et al., 2003; Pelecice et al., 2005). O efeito dessa associação na
população de peixes tem sido longamente negligenciado pelos pesquisadores (Miranda &
Hodges, 2000). Entretanto, o sucesso de captura dessas presas pelas aves piscívoras é
influenciado também pelo tipo e tamanho da cobertura que estas plantas oferecem nos
ambientes aquáticos (Esler, 1990 e 1992; Allouche & Gaudin, 2001). Assim, estudar
interações entre aves, peixes, macrófitas e as outras variavéis ambientais é importante para
entender o funcionamento do ecossistema pantaneiro.
No Pantanal ocorrem 32 espécies de aves aquáticas, que alimentam-se
primariamente de peixes (Magalhães, 1990b; Cintra & Antas, 1996; Oliveira, 1986; 1997).
Estas aves formam grupos de vadeadoras, pescadoras de superfície e mergulhadoras de
fundo (Oliveira, 1996). Elas utilizam diferentes estratégias para capturarem suas presas, daí

69
a abundância e a riqueza estarem também relacionadas com o pulso de inundação destes
locais (Kushlan, et al., 1985; Frederick & Bildstein, 1999)
O objetivo geral deste estudo foi investigar como as características bióticas e
abióticas dos lagos influenciam na variação da composição da assembléia de aves
piscívoras no Pantanal.
2. Material e Métodos
2.1. Desenho amostral
As aves foram estudadas durante dois anos (2002 e 2003), mensalmente de maio a
setembro, em 20 lagos marginais ao longo de 46km da Estrada Parque Transpantaneira, em
Poconé (Figura 1). Estes lagos foram previamente sorteados utilizando uma tabela de
números aleatórios (Rohf & Sokal, 1969). Para as observações das aves a rodovia que
margeia os lagos foi percorrida a pé. O censo visual foi feito a olho nu ou com auxílio de
binóculo (8x40) e as observações foram realizadas das 06 ás 10hs da manhã e das 14 ás
18hs. Foram consideradas nos censos todas as aves que estavam nas margens ou dentro dos
lagos. Esporadicamente o comportamento destas aves foi filmado com câmera digital
(700x) para identificação dos peixes consumidos por elas. Foram também coletados
regurgitos dessas aves (Figura 2).
Os lagos deste local, em geral têm a forma retangular, no mês de maio eles foram
medidos com trena de 50m. Para medir mensalmente o recuo das águas, foram colocadas
duas estacas de madeira uma em cada margem, de modo que ao medir a distância da estaca
até a margem no mês subseqüente, foi possível estimar a área ocupada pelas águas do lago.
A profundidade dos lagos foi medida sempre a 1m da margem do lago, com uma corda
graduada de 10 em 10 cm, com um peso amarrado na extremidade, e a transparência com

70
um disco de Secchi de 20 cm de diâmetro; o pH, oxigênio dissolvido e a temperatura da
água foram medidos com Phmetro e oxímetro digitais. O censo populacional das aves, a
tomada das medidas limnológicas a amostragem das macrófitas aquáticas (ver Métodos
Capítulo 3), e a coleta de peixes (ver Métodos Capítulo 4) foram realizadas
concomitantemente.

71
Figura 1. Mapa da distribuição dos pontos de observação das aves nos lagos amostrados ao longo da Estrada Parque Transpantaneira em 2002 e 2003
Baia Rio Bento Gomes
Rio Pi i
1
2
3
5 4
6
7 8
9
1110
12
14 15
16
13
18
17
19
20

72
Figura 2. Confirmação da dieta através do hábito alimentar e do regurgito de aves piscívoras no Pantanal: a) regurgito do Tuiuiú (Jabiru mycteria), peixes como Lobó (Hoplias malabaricus), Tuvira (Hypopomus sp), Piaus (Leporinus sp) Acarás (Aequidens plagiozonatus). b) regurgito da Curicaca-cinza (Theristicus caerulescens), peixes como Lobó; Camboatá (Callichthys callichthys) e um Acará. Fotos: Dalci Oliveira.

73
2.2. Análise dos dados
Neste capítulo e no capítulo 3 e 4 a unidade amostral, foram 20 lagos, e não mais o
transecto linear de 46 km como no capítulo 1. Como estes lagos possuem tamanhos (área)
diferentes, tanto a riqueza de espécies como a abundância ou biomassa de indivíduos de
cada lago foi dividida pela área do lago para padronizar as amostras. Também para este
capítulo e os seguintes às análises foram feitas somente para as aves piscívoras. Para
verificar como as características físicas e biológicas dos ambientes alagáveis influenciam
na variação da composição da assembléia de aves piscívoras foram usadas duas Análises de
regressão linear múltipla nas quais as variáveis dependentes foram para a primeira a riqueza
de espécies de aves, e para a segunda a abundância de aves. As variáveis independentes
para cada uma das três análises foram à profundidade e o tamanho do lago, a transparência
da água, a cobertura de macrófitas aquáticas do lago, e a composição qualitativa e
quantitativa da comunidade de peixes. A composição da assembléia de aves, uma outra
variável usada, foi gerada a partir das análises de ordenação do tipo MDS. Os scores MDS1
e MDS2 resultantes da ordenação foram usados como variavéis dependentes, em 4 modelos
de regressão multi multivariada usando as variavéis abióticas acima e em grupos separados
mas ora usando nos modelos incluindo a composição qualitativa de peixes e ora usando a
composição qualitativa da comunidade de peixes (usando os escores dos dois primeiros
vetores resultantes das análises de MDS feita para a comunidade de peixes). As análises de
ordenação foram feitas usando o programa PATN (Belbin, 1982), seguindo o mesmo
protocolo do capítulo 1. As regressões lineares múltiplas foram feitas com o programa
Systat (Wilkinson, 1997).

74
3. Resultados Em 2002 a variável limnológica pH variou na cheia entre 7,39 a 6,32 e na seca entre 9,3 a
6,69. A temperatura média da água do lago apresentou variação na cheia entre 29ºC a 26ºC
e na seca entre 31ºC a 24ºC, a porcentagem de oxigênio dissolvido na água oscilou na cheia
entre 75% a 42% e na seca entre 30% a 8%. A profundidade máxima dos lagos na cheia foi
de 162 cm e a mínima na seca foi 25cm, a transparência máxima da água mudou na cheia
de 116 cm., para a mínima na seca de 9cm. O maior lago na cheia tinha uma área de 1,8
Km² e o menor com 0,01 Km²
Em 2003 a variável limnológica pH variou na cheia entre 9,30 a 6,42 e na seca o
mesmo teve variação entre 8,09 a 5,58. A temperatura da água do lago apresentou variação
na cheia entre 29ºC a 22ºC e na seca entre 32ºC a 20ºC, a porcentagem de oxigênio
dissolvido na água oscilou na cheia entre 78% a 25% e na seca entre a 35% a 1%. A
profundidade máxima na cheia foi 152 cm e a mínima na seca foi de14 cm, a transparência
máxima da água mudou de 84cm na cheia, para a mínima de 4cm nos lagos que não
secaram completamente. Neste ano 6 lagos estavam completamente secos mês de setembro.
A comunidade de aves piscívoras em 2002 foi formada por 17 espécies. As cinco
espécies mais abundantes foram a Garça-branca-grande (Ardea alba) (1121), Garça-branca-
pequena (Egretta thula) (428);Tuiuiú (Jabiru mycteria) (337) Cabeça-seca (Mycteria
americana) (200) e o Socó-dorminhoco (Nycticorax nycticorax) com 119 indivíduos
(Figura 3).
A comunidade de aves piscívoras em 2003 foi formada também por 17 espécies. As
cinco espécies mais abundantes foram a Garça-branca-grande (Ardea alba) (447), Garça-
branca-pequena (Egretta thula) (258); Socó-dorminhoco (Nycticorax nycticorax) (187),
Biguá (Phalacrocorax brasilianus) (107) e o Cabeça-seca (Mycteria americana) com 92

75
indivíduos (Figura 4). Em 2002 o conjunto das aves pernaltas formou 91.43% da
assembléia de aves piscívoras, enquanto que em 2003 elas tiveram 97.56% de participação
na composição das mesmas.
A correlação entre as variavéis das características físicas e biológicas dos lagos
Em 2002 e 2003, antes de realizar as análises de regressão, as variavéis independentes
foram avaliadas quanto a colinearidade usando matriz de correlação de Pearson e também
por simulação, usando correlação múltipla na qual cada variável independente foi analisada
como dependente contra todas as demais como independentes, e verificados os valores de
tolerância em cada modelo. Somente valores de tolerância acima de 0.1, foram
considerados (Wilkinson, 1997).
Em 2002, as variavéis (transparência da água e profundidade do lago; tamanho do
lago e biomassa de peixes e riqueza de peixes foram significativamente correlacionadas
entre si, portanto, elas foram usadas em modelos separados. A variável transparência da
água foi usada junta no mesmo modelo e as outras como biomassa de peixe e transparência
da água em outro modelo (Tabela 1).
Em 2003 algumas variavéis independentes (tamanho e profundidade do lago;
transparência da água e profundidade do lago; e biomassa e riqueza de peixes) foram
significadamente correlacionadas entre si. Elas foram usadas em modelos separadas. A
variável tamanho do lago, cobertura de macrófitas aquáticas foram usadas juntas no mesmo
modelo e as outras e transparência da água em outro modelo (Tabela 2).

76
3.1-Efeitos bióticos e abióticos dos lagos e da composição da comunidade de peixes na
riqueza de espécies de aves piscívoras
Em 2002, a variação na riqueza de espécies de aves piscívoras foi afetada
significativamente pela composição quantitativa da comunidade dos peixes, considerando
apenas a primeira dimensão resultante da análise de ordenação (MDS1), (Tabela 3; Figura
5). Porém, o tamanho do lago, a profundidade do lago, e a cobertura de macrófitas
aquáticas, bem como a composição quantitativa da comunidade dos peixes considerando a
segunda dimensão (MDS2), não afetaram significativamente a riqueza de espécies de aves
piscívoras (Tabela 3).
Ainda neste ano a variação na riqueza de espécies de aves piscívoras não foi afetada
significativamente pelo tamanho e profundidade do lago, nem pela cobertura de macrófitas
aquáticas, e tampouco pela composição qualitativa da comunidade dos peixes (na primeira
e segunda dimensão resultante da análise de ordenação (MDS1) (MDS2), não afetaram
significativamente a riqueza de espécies de aves piscívoras (Tabela 3).
Neste mesmo ano (2002), a variação na riqueza de espécies de aves piscívoras foi
afetada significativamente pela composição quantitativa da comunidade dos peixes
considerando apenas a primeira dimensão resultante da análise de ordenação (MDS1)
(Figura 6). Contudo, a transparência da água e a composição quantitativa da comunidade
dos peixes, considerando a segunda dimensão (MDS2), não afetaram significativamente a
riqueza de espécies de aves piscívoras (Tabela 3).
Ainda em 2002, a variação na riqueza de espécies de aves piscívoras não foi afetada
significativamente pela transparência da água. Também a composição qualitativa da
comunidade dos peixes, considerando a primeira dimensão resultante da análise de

77
ordenação (MDS1), e nem pelo segundo vetor (MDS2), afetaram significativamente a
riqueza de espécies de aves piscívoras (Tabela 3).
Em 2003, a variação na riqueza de espécies de aves piscívoras não foi afetada pelo
tamanho do lago, nem pela cobertura de macrófitas aquáticas nem pela transparência da
água, tampouco pelo pH. Também a composição quantitativa da comunidade dos peixes
(considerando apenas a primeira dimensão resultante da análise de ordenação MDS1), e a
composição quantitativa da comunidade dos peixes (considerando a segunda dimensão
MDS2) não afetaram significativamente a riqueza de espécies de aves piscívoras (Tabela
4).
Ainda neste ano (2003), a variação na riqueza de espécies de aves piscívoras não foi
afetada significativamente pelo tamanho do lago e profundidade do lago, nem pela
cobertura de macrófitas aquáticas, transparência e pH da água do lago, e tampouco pela
composição qualitativa da comunidade dos peixes na primeira dimensão resultante da
análise de ordenação (MDS1) e nem pelo segundo vetor (MDS2) não afetaram
significativamente a riqueza de espécies de aves piscívoras (Tabela 4). Neste mesmo ano, a
variação na riqueza de espécies de aves piscívoras não foi afetada significativamente pela
% de oxigênio dissolvido na água, nem pela profundidade e nem pela temperatura da água.
A composição quantitativa da comunidade dos peixes considerando apenas a primeira
dimensão resultante da análise de ordenação (MDS1) e a composição quantitativa da
comunidade dos peixes considerando a segunda dimensão (MDS2) não afetaram
significativamente a riqueza de espécies de aves piscívoras (Tabela 4).
Ainda em 2003, a variação na riqueza de espécies de aves piscívoras não foi afetada
significativamente pela % de oxigênio dissolvido na água, nem pela profundidade do lago e
nem pela temperatura da água. Também a composição qualitativa da comunidade dos

78
peixes na considerando a primeira dimensão resultante da analise de ordenação (MDS1) e
nem pelo segundo vetor (MDS2) afetaram significativamente a riqueza de espécies de aves
piscívoras (Tabela 4).
3.2-Efeitos bióticos e abióticos dos lagos e da composição da comunidade de peixes na
abundância de aves piscívoras
Em 2002, a variação na abundância de aves piscivoras não foi afetada significativamente
pelo tamanho nem pela profundidade do lago e nem pela cobertura de macrófitas aquáticas.
Também a composição quantitativa da comunidade dos peixes na primeira dimensão
resultante da análise de ordenação (MDS1) e nem pelo segundo vetor (MDS2). Não
afetaram significativamente abundância de aves piscívoras (Tabela 3).
Ainda em 2002, a variação na abundância de aves piscívoras não foi afetada
significativamente pelo tamanho do nem pela profundidade do lago nem pela cobertura de
macrófitas aquáticas. Também a composição qualitativa da comunidade dos peixes na
primeira dimensão resultante da analise de ordenação (MDS1) e o segundo vetor (MDS2).
Não afetaram significativamente abundância de aves piscívoras (Tabela 3).
Ainda em 2002, a variação na abundância de aves piscívoras não foi afetada
significativamente pela transparência da água e nem pela composição qualitativa da
comunidade dos peixes na primeira dimensão resultante da análise de ordenação (MDS1) e
nem pelo segundo vetor (MDS2) (Tabela 3).
Em 2003, a variação na abundância de aves piscivoras não foi afetada
significativamente pelo tamanho do lago, pela cobertura de macrófitas aquáticas, nem pela
transparência e tampouco pelo pH da água. Também a composição quantitativa da
comunidade de peixes na primeira dimensão resultante da analise de ordenação (MDS1) e

79
nem pelo segundo vetor (MDS2). Não afetaram significativamente abundância de aves
piscívoras (Tabela 4).
Ainda em 2003, a variação na abundância de aves piscívoras não foi afetada
significativamente pelo % oxigênio dissolvido, nem pela profundidade do lago e nem pela
temperatura da água. A composição quantitativa da comunidade dos peixes na primeira
dimensão resultante da analise de ordenação (MDS1) e o segundo vetor (MDS2). Não
afetaram significativamente abundância de aves piscívoras. (Tabela 4).
Ainda em 2003, A variação na abundância de aves piscívoras não foi afetada
significativamente pelo oxigênio dissolvido, nem pela profundidade e nem pela temperatura
da água. A composição qualitativa da comunidade dos peixes na primeira dimensão
resultante da analise de ordenação (MDS1) e nem pelo segundo vetor (MDS2), não
afetaram significativamente a abundância de aves piscívoras (Tabela 4).
3.3 Efeitos bióticos e abióticos dos lagos e da composição da comunidade de peixes na
abundância de aves piscívoras
Em 2002 os resultados das análises indicaram que houve relação significativa entre
a profundidade do lago, e a composição quantitativa e qualitativa da comunidade de aves
piscívoras, nos modelo 1 e 2 (Tabela 5). Ainda neste ano a composição quantitativa da
comunidade de peixes considerando a primeira dimensão (MDS1QTP) teve efeito
significativo na composição quantitativa da comunidade de aves piscívoras. As demais
variavéis analisadas não afetaram significativamente a composição quantitativa e
qualitativa das aves piscívoras (Tabela 5).

80
Em 2003 os resultados das análises indicaram que não houve relação significativa
entre as variavéis bióticas e abióticas e composição quantitativa e qualitativa de peixes na
composição quantitativa e qualitativa de aves piscívoras nos modelos analisados (Tabela 6).
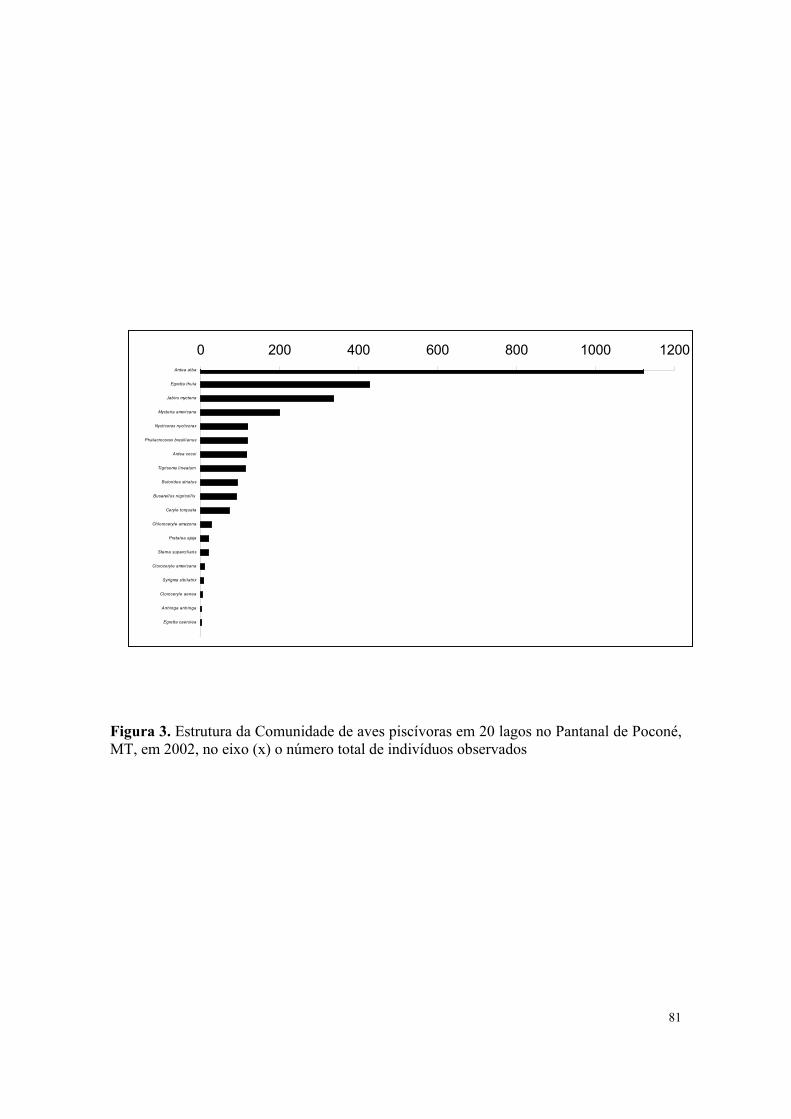
81
Figura 3. Estrutura da Comunidade de aves piscívoras em 20 lagos no Pantanal de Poconé, MT, em 2002, no eixo (x) o número total de indivíduos observados
0 200 400 600 800 1000 1200Ardea alba
Egretta thula
Jabiru mycteria
Mycteria americana
Nycticorax nycticorax
Phalacrocorax brasil ianus
Ardea cocoi
Tigrisoma lineatum
Butorides striatus
Busarellus nigricoll is
Ceryle torquata
Chloroceryle amazona
Pratalea ajaja
Sterna supercil iaris
Cloroceryle americana
Syrigma sibilatrix
Cloroceryle aenea
Anhinga anhinga
Egretta caerulea
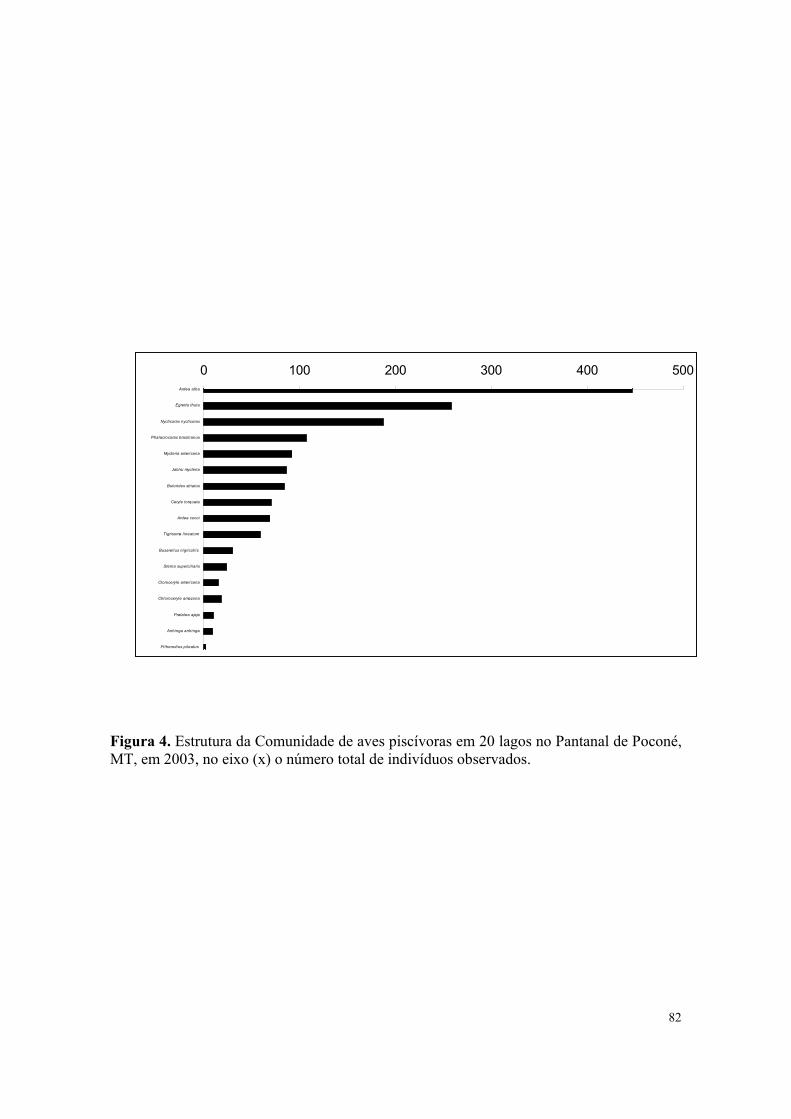
82
Figura 4. Estrutura da Comunidade de aves piscívoras em 20 lagos no Pantanal de Poconé, MT, em 2003, no eixo (x) o número total de indivíduos observados.
0 100 200 300 400 500Ardea alba
Egretta thula
Nycticorax nycticorax
Phalacrocorax brasil ianus
Mycteria americana
Jabiru mycteria
Butorides striatus
Ceryle torquata
Ardea cocoi
Tigrisoma lineatum
Busarellus nigricoll is
Sterna supercil iaris
Cloroceryle americana
Chloroceryle amazona
Pratalea ajaja
Anhinga anhinga
Pilherodius pileatus

83
Tabela 1. Matriz de Correlação de Pearson para as variavéis abióticas: tamanho do lago (TAM), profundidade do lago(PRO), transparência da água (TRA),
oxigênio dissolvido (OXI), pH da água (PHA), temperatura da água (TEA) e biótica cobertura de macrófitas aquáticas (CMA) registradas em 20 lagos no
Pantanal em 2002.
(TAM) (m) (PRO) (m) (TRA) (m) (OXI) (%) (PHA) (TEA) (ºC) (CMA) (%)
(TAM) (m) 1.000
(PRO) (m) 0.074 1.000
(TRA) (m) 0.085 0.827** 1.000
(OXI) (%) 0.054 0.106 0.076 1.000
(PHA) -0.108 0.353 0.208 0.172 1.000
(TEA) (ºC) 0.218 0.085 -0.266 0.071 -0.026 1.000
(CMA) (%) 0,054 -0.073 0.093 -0.438 -0.210 -0.137 1.000
* Asterísticos correspondem aos valores de “p” resultantes da matriz de probabilidade de Bonferroni, usada para avaliar se as correlações entre as variáveis na matriz de correlação de Person são significativas. *=p<0,05; **=p<0,001
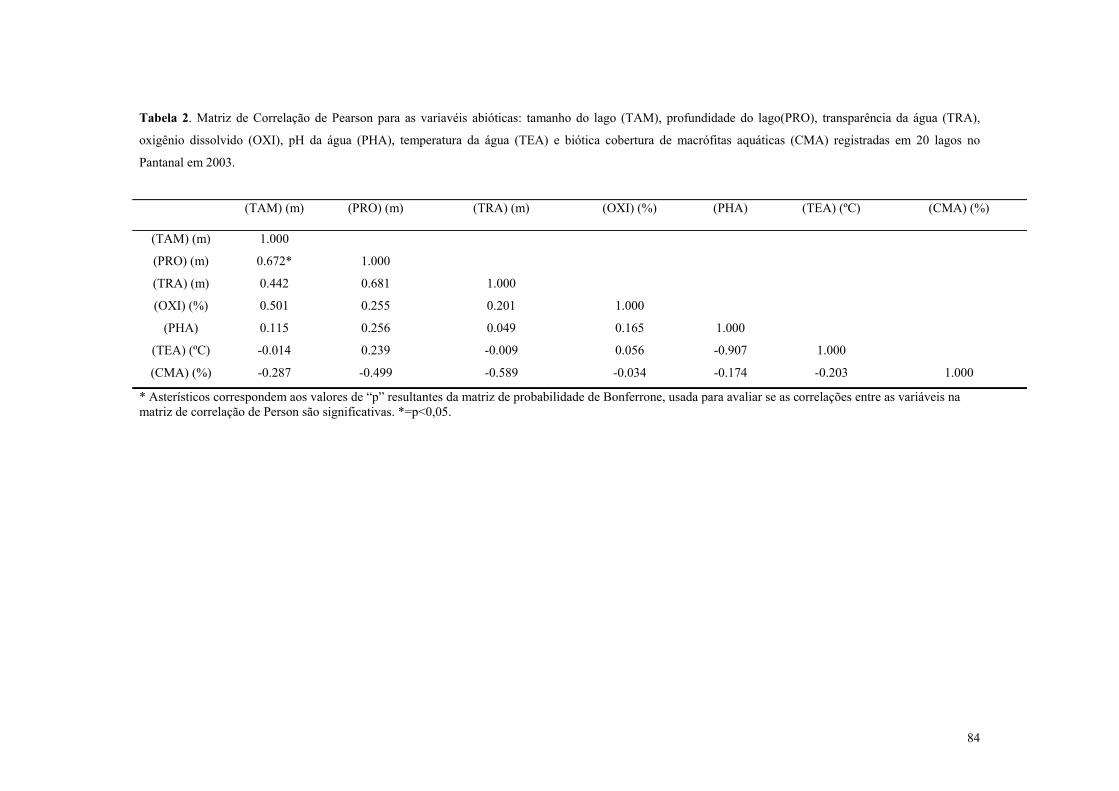
84
Tabela 2. Matriz de Correlação de Pearson para as variavéis abióticas: tamanho do lago (TAM), profundidade do lago(PRO), transparência da água (TRA),
oxigênio dissolvido (OXI), pH da água (PHA), temperatura da água (TEA) e biótica cobertura de macrófitas aquáticas (CMA) registradas em 20 lagos no
Pantanal em 2003.
(TAM) (m) (PRO) (m) (TRA) (m) (OXI) (%) (PHA) (TEA) (ºC) (CMA) (%)
(TAM) (m) 1.000
(PRO) (m) 0.672* 1.000
(TRA) (m) 0.442 0.681 1.000
(OXI) (%) 0.501 0.255 0.201 1.000
(PHA) 0.115 0.256 0.049 0.165 1.000
(TEA) (ºC) -0.014 0.239 -0.009 0.056 -0.907 1.000
(CMA) (%) -0.287 -0.499 -0.589 -0.034 -0.174 -0.203 1.000
* Asterísticos correspondem aos valores de “p” resultantes da matriz de probabilidade de Bonferrone, usada para avaliar se as correlações entre as variáveis na matriz de correlação de Person são significativas. *=p<0,05.
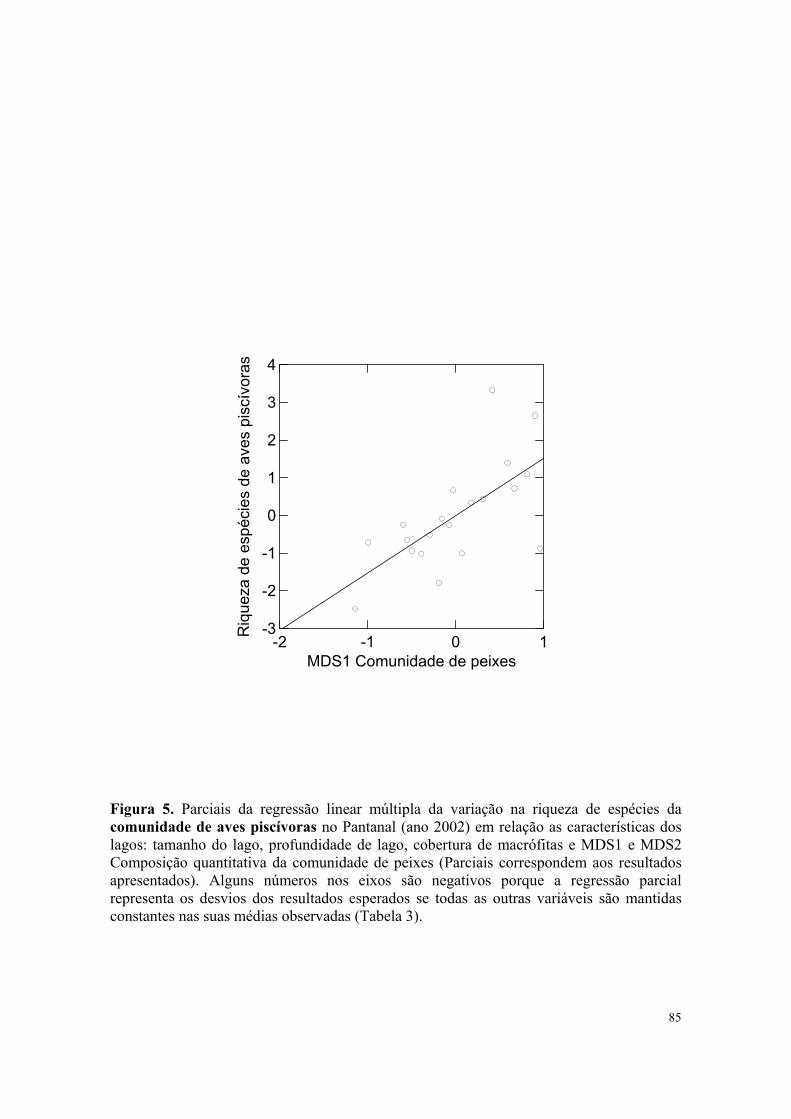
85
Figura 5. Parciais da regressão linear múltipla da variação na riqueza de espécies da comunidade de aves piscívoras no Pantanal (ano 2002) em relação as características dos lagos: tamanho do lago, profundidade de lago, cobertura de macrófitas e MDS1 e MDS2 Composição quantitativa da comunidade de peixes (Parciais correspondem aos resultados apresentados). Alguns números nos eixos são negativos porque a regressão parcial representa os desvios dos resultados esperados se todas as outras variáveis são mantidas constantes nas suas médias observadas (Tabela 3).
-2 -1 0 1MDS1 Comunidade de peixes
-3
-2
-1
0
1
2
3
4
Riq
ueza
de
esp é
cies
de
ave s
pis
cívo
ras
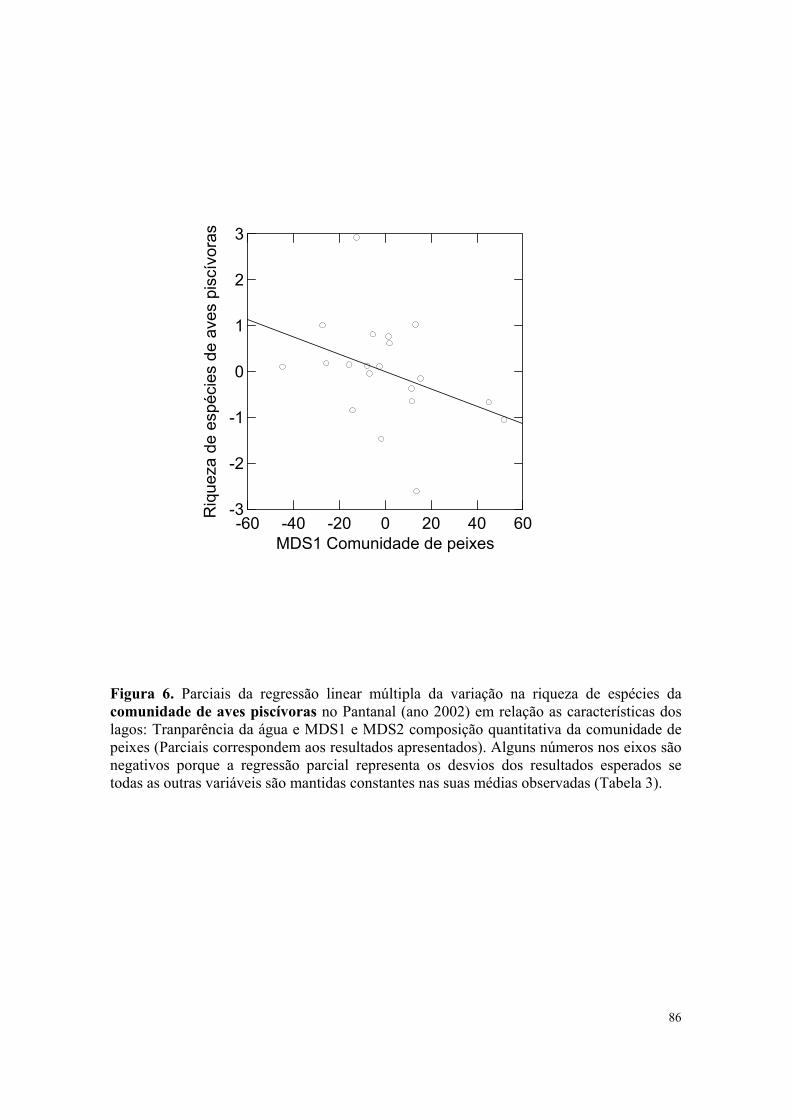
86
Figura 6. Parciais da regressão linear múltipla da variação na riqueza de espécies da comunidade de aves piscívoras no Pantanal (ano 2002) em relação as características dos lagos: Tranparência da água e MDS1 e MDS2 composição quantitativa da comunidade de peixes (Parciais correspondem aos resultados apresentados). Alguns números nos eixos são negativos porque a regressão parcial representa os desvios dos resultados esperados se todas as outras variáveis são mantidas constantes nas suas médias observadas (Tabela 3).
-60 -40 -20 0 20 40 60MDS1 Comunidade de peixes
-3
-2
-1
0
1
2
3R
ique
za d
e es
p éci
es d
e av
e s p
isc í
vora
s

87
Figura 7. Análise de escalonamento multidimensional (MDS), para as comunidades de aves piscívoras em 20 lagos no Pantanal de Poconé em 2002. Os números na figura são referente aos lagos (ver apêndice) na figura quanto mais próximos os números (lagos) mais similar é a composição da comunidades de aves piscívoras nestes lagos.
-2 -1 0 1 2MDS - axis 1
-2
-1
0
1
2
MD
S -
axis
2
1
16
9
8
14
123
17
11
13
5
4
18
6
7
15
2
10
20
19

88
Figura 8. Análise de escalonamento multidimensional (MDS), para as comunidades de
aves piscívoras em 20 lagos no Pantanal de Poconé em 2003. Os números na figura são
referentes aos lagos (ver apêndice) na figura quanto mais próximos os números (lagos)
mais similar é a composição das comunidades de aves piscívoras nestes lagos.
-2 -1 0 1 2MDS - axis 1
-2
-1
0
1
2
MD
S -
axis
2
3
2
15
4
207
6
16
12
5
19
10
13
8
11
9
1718
14
1
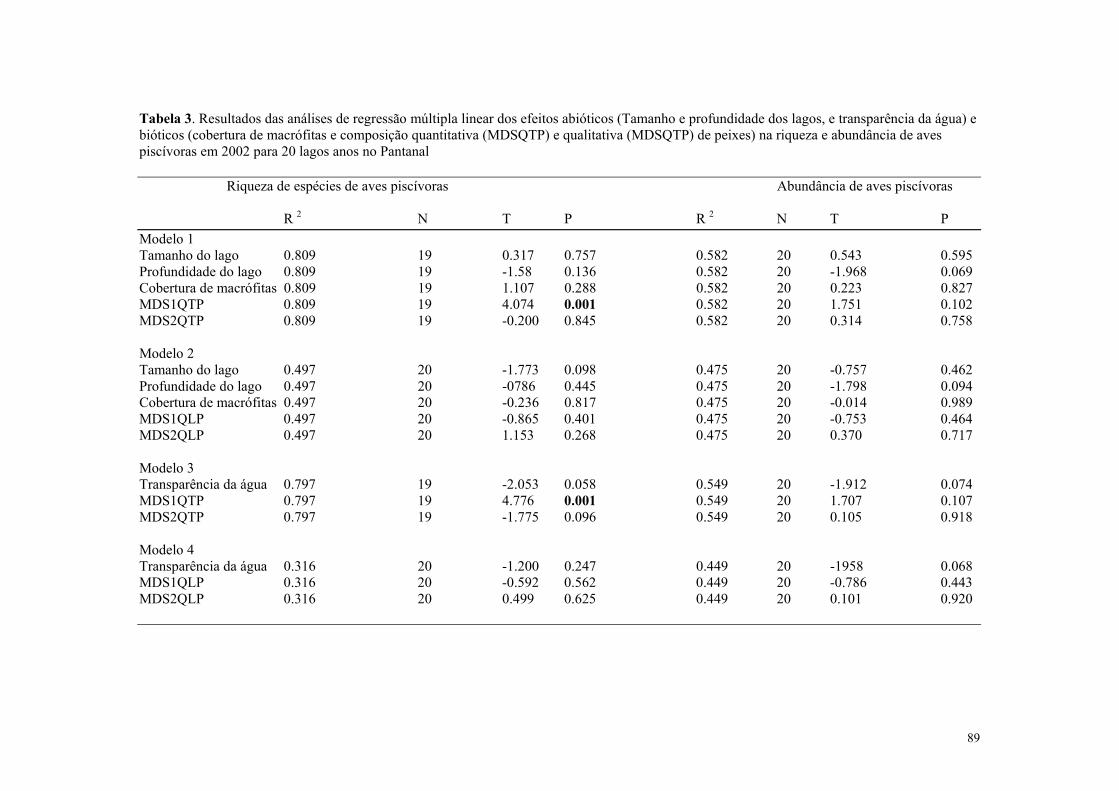
89
Tabela 3. Resultados das análises de regressão múltipla linear dos efeitos abióticos (Tamanho e profundidade dos lagos, e transparência da água) e bióticos (cobertura de macrófitas e composição quantitativa (MDSQTP) e qualitativa (MDSQTP) de peixes) na riqueza e abundância de aves piscívoras em 2002 para 20 lagos anos no Pantanal Riqueza de espécies de aves piscívoras Abundância de aves piscívoras R 2 N T P R 2 N T P Modelo 1 Tamanho do lago 0.809 19 0.317 0.757 0.582 20 0.543 0.595 Profundidade do lago 0.809 19 -1.58 0.136 0.582 20 -1.968 0.069 Cobertura de macrófitas 0.809 19 1.107 0.288 0.582 20 0.223 0.827 MDS1QTP 0.809 19 4.074 0.001 0.582 20 1.751 0.102 MDS2QTP 0.809 19 -0.200 0.845 0.582 20 0.314 0.758 Modelo 2 Tamanho do lago 0.497 20 -1.773 0.098 0.475 20 -0.757 0.462 Profundidade do lago 0.497 20 -0786 0.445 0.475 20 -1.798 0.094 Cobertura de macrófitas 0.497 20 -0.236 0.817 0.475 20 -0.014 0.989 MDS1QLP 0.497 20 -0.865 0.401 0.475 20 -0.753 0.464 MDS2QLP 0.497 20 1.153 0.268 0.475 20 0.370 0.717 Modelo 3 Transparência da água 0.797 19 -2.053 0.058 0.549 20 -1.912 0.074 MDS1QTP 0.797 19 4.776 0.001 0.549 20 1.707 0.107 MDS2QTP 0.797 19 -1.775 0.096 0.549 20 0.105 0.918 Modelo 4 Transparência da água 0.316 20 -1.200 0.247 0.449 20 -1958 0.068 MDS1QLP 0.316 20 -0.592 0.562 0.449 20 -0.786 0.443 MDS2QLP 0.316 20 0.499 0.625 0.449 20 0.101 0.920

90
Tabela 4.. Resultados das análises de regressão múltipla linear dos efeitos abióticos (Tamanho e profundidade dos lagos, e transparência da água) e bióticos (cobertura de macrófitas e composição quantitativa (MDSQTP) e qualitativa (MDSQLP) de peixes) na riqueza e abundância de aves piscívoras em 2003 para 20 lagos anos no Pantanal Riqueza de espécies de aves piscívoras Abundância de aves piscívoras R 2 N T P R 2 N T P Modelo 1 Tamanho do lago 0.468 20 -0.885 0.392 0.563 20 0.012 0.991 Transparência da água 0.468 20 -0.734 0.476 0.563 20 -1.162 0.266 Cobertura de macrófitas 0.468 20 0.468 0.721 0.563 20 0.222 0.827 pH da água 0.468 20 0.572 0.577 0.563 20 1.396 0.186 MDS1QTP 0.468 20 -0.341 0.739 0.563 20 0.329 0.747 MDS2QTP 0.468 20 0.062 0.952 0.563 20 0.623 0.544 Modelo 2 Tamanho do lago 0.600 20 -1.424 0.178 0.618 20 -0.548 0.593 Transparência da água 0.600 20 -0.540 0.598 0.618 20 -1.493 0.159 Cobertura de macrófitas 0.600 20 0.687 0.504 0.618 20 0.098 0.924 pH da água 0.600 20 1.101 0.291 0.618 20 1.672 0.118 MDS1QLP 0.600 20 -0.724 0.482 0.618 20 -0.520 0.612 MDS2QLP 0.600 20 -1.450 0.171 0.618 20 -1.192 0.255 Modelo 3 Oxigênio dissolvido 0.440 20 0.543 0.595 0.471 20 -0.016 0.987 Profundidade 0.440 20 -1.632 0.125 0.471 20 -1.025 0.323 Temperatura da água 0.440 20 0.845 0.412 0.471 20 1.213 0.245 MDS1QTP 0.440 20 -0.194 0.849 0.471 20 0.266 0.794 MDS2QTP 0.440 20 -0.493 0.629 0.471 20 0.261 0.798 Modelo 4 Oxigênio dissolvido 0.554 20 0.887 0.390 0.544 20 -0.099 0.923 Profundidade 0.554 20 -2.144 0.050 0.544 20 -1.843 0.087 Temperatura da água 0.554 20 0.046 0.964 0.544 20 1.074 0.301 MDS1QLP 0.554 20 -1.315 0.210 0.544 20 -0.513 0.616 MDS2QLP 0.554 20 -0.152 0.882 0.544 20 0.750 0.466

91
Tabela 5. Resultados dos efeitos das características físicas e biológicas dos lagos e da composição quantitativa (MDSQTP) e qualitativa (MDSQLP) da comunidade de peixes na composição quantitativa e qualitativa de espécies de aves piscivoras em 2002 no Pantanal (usando modelo de regressão múltipla multivariada) Composição quantitativa da comunidade de aves piscívoras Composição qualitativa da comunidade de aves piscívoras Pillai-Trace N P Pillai-Trace N P Modelo 1 Tamanho do lago 0.170 20 0.479 0.219 20 0.200 Profundidade do lago 0.470 20 0.016 0.451 20 0.020 Cobert. Macrófitas 0.108 20 0.477 0.195 20 0.244 MDS1QTP 0.563 20 0.005 0.054 20 0.698 MDS2QTP 0.070 20 0.622 0.127 20 0.413 Modelo 2 Tamanho do lago 0.166 20 0.308 0.150 20 0.348 Profundidade do lago 0.533 20 0.007 0.436 20 0.024 Cobert. Macrófitas 0.101 20 0.500 0.250 20 0.154 MDS1QLP 0.126 20 0.416 0.055 20 0.694 MDS2QLP 0.169 20 0.301 0.091 20 0.538

92
Tabela 6. Resultados dos efeitos das características físicas e biológicas dos lagos e da composição da comunidade de peixes na composição de espécies de aves piscívoras em 2003 no Pantanal (usando modelo de regressão múltipla multivariada) Composição quantitativa da comunidade de aves piscívoras Composição qualitativa da comunidade de aves piscívoras Pillai-Trace N P Pillai-Trace N P Modelo 1 Tamanho do lago 0.226 20 0.216 0.056 20 0.707 Tranparência da água 0.321 20 0.098 0.069 20 0.650 Cobert. Macrófitas 0.020 20 0.888 0.055 20 0.712 pH da água 0.098 20 0.538 0.139 20 0.408 MDS1QTP 0.142 20 0.398 0.029 20 0.837 MDS2QTP 0.251 20 0.176 0.260 20 0.164 Modelo 2 Tamanho do lago 0.275 20 0.146 0.026 20 0.855 Tranparência da água 0.321 20 0.098 0.064 20 0.672 Cobert. Macrófitas 0.036 20 0.804 0.062 20 0.679 pH da água 0.234 20 0.201 0.092 20 0.559 MDS1QLP 0.022 20 0.874 0.022 20 0.875 MDS2QLP 0.360 20 0.069 0.013 20 0.926 Modelo 3 Oxigênio dissolvido 0.040 20 0.769 0.003 20 0.982 Profundidade do lago 0.121 20 0.432 0.134 20 0.393 Temperatura da água 0.077 20 0.594 0.069 20 0.627 MDS1QTP 0.074 20 0.607 0.026 20 0.842 MDS2QTP 0.112 20 0.463 0.193 20 0.248 Modelo 4 Oxigênio dissolvido 0.042 20 0.758 0.132 20 0.401 Profundidade do lago 0.360 20 0.055 0.070 20 0.534 Temperatura da água 0.037 20 0.782 0.200 20 0.196 MDS1QLP 0.061 20 0.664 0.198 20 0.256 MDS2QLP 0.154 20 0.336 0.135 20 0.407

93
4. Discussão Nas áreas alagáveis a profundidade dos corpos da água é o fator preponderante na
estruturação da composição das aves aquáticas (Kushlan et al., 1975 e 1985; González,
1996ab, 1997; Oliveira, 1997; Petermann, 1997; Bancroft et al., 2002; Cintra et al. no
prelo).
A riqueza de espécies de aves piscívoras nos lagos do Pantanal em 2002 e 2003 foi
composta por 19 espécies mudando apenas a composição das espécies. É esperado que esta
composição não tenha grande variação de ano para ano, porque as características físicas dos
lagos não oferecem condições para que outras aves piscívoras possam procurar suas presas
nestes locais. Estes lagos não possuem margens arenosas que poderiam formar praias
possibilitando a nidificação de por exemplo do Trinta-réis (Sterna superciliaris), do Taiamã
(Rynchops nigra); outros possuem uma declividade abrupta, que causa dificuldade para
aves com pernas curtas procurarem suas presas; outros não são suficientes grandes e
profundos para possibilitar a pesca pela Águia-pescadora (Pandion haliaetus) por exemplo.
A Garça-azul (Egretta caerulea) é raríssima na região apenas 2 indivíduos foram avistados
nos lagos nos dois anos de observação. O Tabuiaiá (Ciconia maguari) não foi observado
forrageando dentro dos lagos, apenas nos campos inundáveis. Desse modo a riqueza de
espécies das aves piscívoras tendem a ser constante, além do mais esta comunidade foi
formada predominantemente por aves pernaltas e mergulhadoras que fazem ninhos
coloniais em regiões próximas. Paszkowski & Tonn (2000), estudando lagos no Canadá,
encontraram três padrões, para a distribuição das aves aquáticas um para lagos pequenos e
rasos encontraram entre 3 a 8 espécies de aves, outro para lagos grandes e profundos,
encontraram um pouco mais, entre 11 a 16 espécies, incluindo grandes aves piscívoras
cruzadoras ou pescadoras de longa distância como águias -pescadoras, e em lagos grandes e

94
rasos encontraram o maior número de espécies entre 15 a 23, incluindo várias espécies de
anatídeos. Os lagos do Pantanal são rasos, com menos de 2m de profundidade. Nestes
locais, a riqueza de espécies de aves piscívoras seguiu o padrão verificado para estes lagos
das zonas temperadas. Nos lagos menores ocorreram entre 4 a 6 espécies, enquanto que no
maior lago 15 espécies se fizeram presentes ao longo do ano. A abundância variou
mensalmente, entretanto, a profundidade não teve efeito significativo nesta, sendo muito
provável que isto tenha acontecido porque mais de 90% dos indivíduos encontrados foram
de aves pernaltas de grande porte que podem forragear desde as margens úmidas dos lagos
até uma profundidade de 40cm, e também porque um único lago que é permanente manteve
uma abundância de 47.65% das aves piscívoras em 2002 e 27.56% em 2003. Outros fatores
abióticos analisados como tamanho do lago não tiveram efeito na riqueza das aves
piscívoras, provavelmente por causa da padronização das amostras. Alguns estudos
mostraram uma variação na riqueza de aves aquáticas e piscívoras em função do aumento
da área dos lagos (Paszkowski & Tonn, 2000). A cobertura de macrófitas aquáticas também
não teve influencia significativa na riqueza de espécies das aves piscívoras, é provável que
isto esteja relacionado com o tipo biológico predominante das macrófitas dos lagos que foi
o flutuante. Estudos realizados tanto em lagos da Europa, América do Norte, como também
em lagos andinos da América do Sul, mostraram uma correlação positiva da riqueza e
abundância das aves aquáticas com a presença de macrófitas submersas (Esler, 1990 e
1992; Paszkowski & Tonn, 2000; Caziani, 2001).
Todavia, a riqueza de aves piscívoras foi significativamente afetada pela
composição quantitativa da comunidade de peixes (Tabela 3, Figura 5), a abundância e
disponibilidade de peixes para as aves piscívoras é um dos fatores fundamentais para o
sucesso reprodutivo destas aves, principalmente das que fazem ninhos em colônias (Erwin,

95
1983; Frederick & Callopy, 1989; González, 1996; Collis et al., 2001). No Pantanal um
grande ninhal de 15.000 aves consome por temporada reprodutiva cerca de 150T de peixes,
consumo estimado com base na fórmula de Nilsson & Nilsson (1976) apud Petermann
(1997), para o ninhal do Corutuba, localizado no Pantanal de Barão de Melgaço (Obs.
pess). Embora o resultado aqui obtido da abundância da composição da comunidade de
peixes que foi baseada na biomassa destes e não na abundância de cada espécie, possa
sugerir que as aves não teriam preferência por determinadas espécies (ver capítulo 4);
outros trabalhos indicam que existe preferência por tamanho da presa, e a dieta pode mudar
em função da idade dos filhotes (Thomas, 1985; Oliveira, 1997; Collis et al., 2001; Da
Silva et al., 2002).
A composição quantitativa e qualitativa da comunidade de aves piscívoras em 2002
foram afetada significativamente pela profundidade dos lagos (Tabela 5), embora a maioria
da comunidade das aves piscívoras tenha sido formada por grandes aves vadeadoras ou
pernaltas. Ainda assim, quando se analisa a comunidade na sua totalidade são encontradas
também aves vadeadoras de pernas curtas como o Socozinho (Butorides striatus), a Garça-
branca pequena (Egretta thula) e o Socó-dorminhoco (Nycticorax nycticorax).
Principalmente estas duas últimas espécies, que formam grandes bandos no Pantanal, têm
as suas distribuições relacionadas com a baixa profundidade desses corpos de água. O
contrário ocorre com as aves mergulhadoras que necessitam de um mínimo de
profundidade para realizar a sua pesca. Desse modo a composição muda em função da
profundidade e teve efeito significativo na composição destas aves no Pantanal, ao longo do
período deste estudo.
Também a composição quantitativa da comunidade de peixes na primeira dimensão
teve um efeito altamente significativo na composição quantitativa da comunidade de aves

96
piscívoras em 2002 (Tabela 5). É esperado que quanto mais peixes houver no ambiente,
mais aves piscívoras ocorram no local. Isto também foi verificado para alguns grupos de
aves piscívoras, que tiveram sua riqueza correlacionada com a abundância de peixes.
A transparência da água dos lagos afetou a composição quantitativa da comunidade
de aves piscívoras, indicando que com a mudança de transparência da água ocorre uma
variação na composição da comunidade de aves. As aves mergulhadoras são as mais
prejudicadas com o aumento da turbidez da água, que ocorre normalmente com a
diminuição da profundidade dos lagos. Nessas ocasiões, as aves vadeadoras ou pernaltas
adentram em grande número nestes lagos revolvendo o sedimento e diminuindo a
transparência da água. No entanto, estas aves, notadamente as que formam grandes bandos
na época da vazante no Pantanal, como o Cabeça-seca (Mycteria americana) o Tuiuiú
(Jabiru mycteria) e as garças (Ardea alba e Egretta thula) usam para forragear tanto a
tática visual e táctil como ambas simultaneamente, dependendo da qualidade da água
(Thomas, 1985).

97
Capítulo 3. Efeitos da comunidade de macrófitas aquáticas na assembléia de aves piscívoras no Pantanal.
Introdução:
As macrófitas aquáticas constituem elemento estrutural característico em áreas alagáveis
(Esteves, 1998). A presença da vegetação em corpos de água é um dos principais fatores
ligados à estruturação do habitat ocupado pelas comunidades de animais aquáticos (Junk &
Robertson,1997; Agostinho et al., 2003).
Macrófitas aquáticas são importantes na cadeia trófica de ecossistemas aquáticos,
porque fornecem abrigo, locais de desova e substrato de alimentação a peixes (Junk, 1973;
Araujo-Lima et al., 1986; Soares et al., 1986; Pazin, 2000; Sanches-Botero et al., 2001;
Agostinho et al., 2003; Cassiati et al. 2003; Petry et al., 2003; Pelecice et al., 2005). A
insetos aquáticos (Vieira & Adis, 1993; Nunes & Adis, 1994; Junk & Robertson, 1997;
Amorin & Adis, 1995 e 1996; Marco Jr. et al., 2001, Gimenez, 2002), aranhas, crustáceos e
moluscos (Raizer & Amaral, 2001; Mansur & Hebling, 2002), itens muito importantes na
dieta das aves aquáticas. Além disso, elas próprias são consumidas por algumas espécies de
peixes e aves, que afeta a abundância e riqueza de espécies dessas plantas (Santos, 1981 e
1982; Anderson & Nilsson, 1999; Perrow et al., 1997; Marklund et al., 2002).
As macrófitas aquáticas são importantes sítios de nidificação para muitas espécies
de aves aquáticas, como o Cafezinho (Jacana jacana) (L.) que constrói seu ninho sobre
Salvinia auriculata Aubl. e Ludwigia elegans (Cambess.) Hara, a Viuvinha (Arundinicola
leucocephala) (L.), faz seu ninho nos talos de Echinodorus spp. ou Thalia geniculata L.
(Pott & Pott, 2000, Oliveira presente estudo). Essas plantas também fornecem material

98
verde para confecção e forro dos ninhos de várias espécies de aves aquáticas (Nores &
Yzurieta, 1980; Oliveira, 1997; Oliveira obs pess). Esse material verde pode ter um papel
anti-parasitário, mas, para uma afirmação mais categórica, ainda são necessários estudos
experimentais mais acurados (Rodgers Jr, 1988).
No Pantanal Mato-grossense, ocorrem mais de 242 espécies de macrófitas aquáticas
distribuídas em 106 gêneros e 54 famílias (Pott & Pott, 1997). Para a Estrada Parque
Transpantaneira (EPT) foram listadas 25 espécies com predominância dos aguapés
Eichhonia crassipes (Martius) Solms-Laubach e Eichhonia azurea (Sw.) Kunth (Da Silva
& Pinto-Silva, 1989). Os corpos de água ao longo da EPT, como rios, corixos, vazantes,
campos inundáveis, lagos, lagoas, brejos e caixas de empréstimo, apresentam diferente
composição e distribuição dessas plantas ao longo do ano. Esta variação depende do grupo
ecológico dessas comunidades, do nível da água, de fatores físico-químicos e do estágio
sucessional, como demonstrado por Da Silva & Silva (1989). Essa cobertura varia nos
ambientes lóticos e lênticos proporcionando diferentes condições para que as aves tenham
acesso às suas presas. Apesar da grande importância ecológica das macrófitas aquáticas
estudos sobre a composição desse grupo de plantas em lagoas e áreas inundadas no
Pantanal ainda são escassos (Pott & Pott, 1997).
Estudos ecológicos analisando a interações entre aves e plantas têm dado ênfase ao
efeito de uma ou mais espécie de plantas sobre populações ou comunidades de aves (e.g
Stiles, 1978; Terborg et al., 1990; Loiselle & Blake, 1994; Poulin et al., 1992; Cintra, 1997;
Robinson & Terborg, 1997; Ortiz-Pulido, 2000; Sazima et al., 2001). A maioria destes foi
realizada em ambientes de florestas, cerrados (savanas) e campos, portanto não
considerando comunidades de ambientes aquáticos.

99
A maior parte dos trabalhos mostrando as interações entre aves e macrófitas
aquáticas foram realizados principalmente nas zonas temperadas. Esses trabalhos
mostraram uma forte correlação entre a presença de determinadas espécies de macrófitas
submersas e abundância de aves aquáticas, principalmente gansos, cisnes e patos (Elmberg
et al., 2002; Bos et al., 2004; Thórarinsson & Einarsson, 2004). Ainda nestas regiões,
estudos demonstraram o importante papel desempenhado pelas aves aquáticas na dispersão
de sementes de macrófitas (Figuerola & Green, 2002) e no controle populacional das
mesmas através do pastejo (Anderson & Nilsson, 1999; Perrow et al., 1997; Marklund et
al., 2002).
Raros são os trabalhos que relacionam a comunidade de macrófitas aquáticas e a
comunidade de aves na região Neotropical. Caziani et al. (2001), encontraram uma forte
correlação positiva entre a abundância de aves aquáticas e a presença de macrófitas
aquáticas submersas em lagos de Altiplano na Argentina. Não foi encontrado nenhum
trabalho com este enfoque para o Brasil, e para o Pantanal, este é o primeiro com esta
abordagem (Esteves, 1999; Thomaz & Bini, 2003; Oliveira et al. in prep.).
O objetivo deste estudo foi verificar como a composição da comunidade de
macrófitas aquáticas afeta a composição e estrutura da comunidade de aves aquáticas no
Pantanal.
Material e Método
1) Desenho Amostral
Este trabalho foi realizado no mesmo período em que foram realizados os censos
populacionais das aves aquáticas (conforme capítulo II). A área de cobertura das macrófitas
aquáticas nos lagos foi estimada e dividida em quatro categorias: menos de 25% de

100
cobertura, de 25 a 50%, de 50 a 75% e de 75 a 100%. A identificação das macrófitas
aquáticas foi efetuada por comparação, com o manual de identificação de Pott & Pott
(2002). A lista dos nomes científicos das macrófitas aquáticas e suas formas de vida estão
de acordo com Pott & Pott (2002).
2) Análise dos dados
Análises de regressão linear múltipla foram usadas para avaliar (em dois modelos
separados), os efeitos das características físicas dos lagos (fatores abióticos) na riqueza e
cobertura de macrófitas aquáticas (variavéis dependentes).
Análises de regressão linear múltipla também foram usadas para avaliar (em dois
modelos separados) os efeitos das características físicas dos lagos, e da composição
qualitativa da comunidade de macrófitas aquáticas (variável independente) na riqueza e
abundância de aves piscívoras (variavéis dependentes)
Para verificar o efeito da composição de espécies de macrófitas na composição de
espécies de aves piscívoras foi usada uma análise de regressão multivariada com o seguinte
modelo: composição da comunidade de aves piscívoras (MDS1, MDS2) = constante +
cobertura de macrófitas + composição da comunidade de macrófitas aquáticas (MDS1,
MDS2).
3. Resultados
A comunidade de macrófitas aquáticas foi formada por 26 espécies distribuídas em 21
famílias (Tabela 6). As espécies de macrófitas mais freqüentes nos lagos foram Eichhornia
azurea (80%), Eichhornia crassipes (75%) e Pontederia cordata presente em 73% dos
lagos. A riqueza de espécies de macrófitas variou na cheia entre 15 a 21 espécies e na seca

101
entre 8 a 15 espécies por lago. Na Estrada Parque Transpantaneira foi observado a presença
de formas emergentes (E), anfíbias (A), flutuantes fixas (FF), flutuantes livres (FL),
submersas fixas (SF) e submersas livres (SL). Na cheia, a espécie predominante foi
Pontederia cordata, enquanto que nos períodos de vazante e seca, predominaram
Eichhonia azurea, E. crassipes nos ambientes lóticos, enquanto que nos lênticos Pistia
stratiotes L., foi a espécie a recobrir totalmente as superfícies dos corpos de água. Quanto
ao hábito das macrófitas o mais abundante foi o emergente com 10 espécies (nº 2, 6, 7, 9,
10, 11, 15, 16, 20, 26) seguido das flutuantes fixas com 8 espécies (nº 4, 14, 17, 18, 20, 21,
22, 25). As plantas submersas foram as mais raras, uma flutuante Ludwigia inclinata (L.F)
P.H. Raven, e uma livre Ultricalia poconensis Fromm-Trinta (Figura 1). Várias interações
diretas e indiretas entre as macrófitas aquáticas e as aves e outros organismos puderam ser
observadas; alguns exemplos são apresentados (Figura 2).
A comunidade de aves piscívoras foi formada por 17 espécies, as cinco espécies
mais abundantes foram a Garça-branca-grande (Ardea alba)L., com 447 indivíduos
(31.59%), a Garça-branca-pequena (Egretta thula) (Molina), com 258 (18.23%); o Socó-
dorminhoco (Nycticorax nycticorax) L., com 187; (13.21%), o Biguá (Phalacrocorax
brasilianus) (Gmelin) com 107 (7.56%) e o Cabeça-seca (Mycteria americana) (L.), com
92 indivíduos representando 6.50% das aves piscívoras. Neste ano as aves pernaltas ou
vadeadoras tiveram 97.56% de participação na composição da assembléia de aves
piscívoras.

102
A correlação entre as variavéis das características físicas e biológicas dos lagos
Em 2003, antes de realizar as análises de regressão, as variavéis independentes foram
avaliadas quanto a colinearidade usando matriz de correlação de Pearson e também por
simulação usando correlação múltipla na qual cada variável independente foi analisada
como dependente contra todas as demais como independentes, e verificados os valores de
tolerância em cada modelo, somente valores de tolerância acima de 0.1, forma considerados
(Wilkinson, 1997). Como algumas variavéis independentes (tamanho e profundidade do
lago; transparência, temperatura e pH da água); foram significativamente correlacionadas
entre si, elas foram usadas em modelos de regressão separados. As variáveis tamanho do
lago, transparência e pH da água, foram usadas em um mesmo modelo e Profundidade do
lago ,% de oxigênio dissolvido e temperatura da água em outro modelo (Tabela 1).
3.1 - Efeitos das características físicas e biológicas dos lagos na riqueza de espécies de
macrófitas aquáticas.
Em 2003, A variação na riqueza de espécies macrófitas aquáticas não foi afetada
significativamente pelo tamanho dos lagos e nem pelo Ph da água. Tampouco a
transparência da água afetou significativamente a variação na riqueza de espécies de
macrófitas aquáticas (Tabela 2). Neste mesmo ano, a variação na riqueza de espécies de
macrófitas aquáticas foi afetada significativamente pela temperatura da água do lago,
porém, a % de oxigênio dissolvido na água e nem a profundidade influenciaram
significativamente na riqueza de macrófitas (Tabela 2).

103
3.2 - Efeitos das características físicas e biológicas dos lagos na composição da
comunidade de macrófitas aquáticas.
Em 2003 não houve relação significativa entre a composição qualitativa da comunidade de
macrófitas aquáticas e transparência da água do lago nem com o tamanho do lago e
tampouco com o Ph da água não afetaram significativamente a composição qualitativa de
macrófitas aquáticas. Ainda neste ano a composição qualitativa da comunidade de
macrófitas aquáticas não foi afetada significativamente pela profundidade do lago nem pela
% de oxigênio dissolvido na água dos lagos e nem pela a temperatura da água (Tabela 3).
3.3 - Efeitos das características físicas e biológicas dos lagos e da composição da
comunidade de macrófitas aquáticas na riqueza de espécies de aves piscívoras
Em 2003, a variação na riqueza de espécies de aves piscívoras não foi afetada
significativamente pelo tamanho do lago nem pela transparência da água. A composição
qualitativa da comunidade das macrófitas aquáticas na primeira dimensão resultante da
análise de ordenação MDS1 e o segundo vetor resultante da composição qualitativa da
comunidade das macrófitas aquáticas MDS2 não afetaram significativamente a riqueza de
espécies de aves piscívoras (Tabela 4).
Neste ano a variação na riqueza de espécies de aves piscívoras não foi afetada
significativamente pela profundidade e nem pela temperatura da água do lago. A
composição qualitativa da comunidade das macrófitas aquáticas na primeira dimensão
resultante da analise de ordenação - MDS1 e o segundo vetor da mesma ordenação MDS2
não afetaram significativamente a riqueza de espécies de aves piscívoras (Tabela 4).

104
3.4 - Efeitos das características físicas e biológicas dos lagos e da composição
qualitativa da comunidade de macrófitas aquáticas na abundância de aves piscívoras
Em 2003, a variação na abundância de aves piscívoras não foi afetada significativamente
pelo tamanho do lago, nem pela transparência da água. A composição qualitativa da
comunidade das macrófitas aquáticas (na primeira dimensão resultante da análise de
ordenação - MDS1) e o segundo vetor resultante da composição qualitativa da comunidade
das macrófitas aquáticas - MDS2, não afetaram significativamente a abundância de aves
piscívoras (Tabela 4).
Neste ano a variação na abundância de aves piscívoras não foi afetada significativamente
pela profundidade, nem pela temperatura da água do lago. A composição qualitativa da
comunidade das macrófitas aquáticas (na primeira dimensão resultante da análise de
ordenação - MDS1) e segundo vetor da mesma ordenação MDS2). Também não afetaram
significativamente a riqueza de espécies de aves piscívoras (Tabela 4).
3.5 - Efeitos das características físicas e biológicas dos lagos e da composição
qualitativa da comunidade de macrófitas aquáticas na composição quantitativa da
comunidade de aves piscívoras
Em 2003 os resultados das análises indicaram que não houve uma relação significativa
entre a composição quantitativa da comunidade de aves piscívoras, o tamanho do lago e a
sua transparência. A composição qualitativa da comunidade das macrófitas aquáticas (na
primeira dimensão resultante da análise de ordenação - MDS1) e o segundo vetor da mesma
ordenação MDS2), não afetaram significativamente a composição quantitativa de aves
piscívoras (Tabela 5). Ainda neste ano os resultados das análises indicaram que não houve
relação significativa entre a composição quantitativa da comunidade de aves piscívoras e a

105
profundidade, e a temperatura da água do lago, nem tampouco com a composição
qualitativa de macrófitas aquáticas MDS1 e nem com a composição qualitativa de
macrófitas aquáticas MDS2 (Tabela 5).
3.6 - Efeitos das características físicas e biológicas dos lagos e da composição da
qualitativa da comunidade de macrófitas aquáticas na composição qualitativa da
comunidade de aves piscívoras
Em 2003 os resultados das análises indicaram que não houve uma relação significativa
entre a composição qualitativa da comunidade de aves piscivoras e o tamanho do lago e a
transparência da água do lago e o mesmo ocorreu com a composição qualitativa da
comunidade das macrófitas aquáticas na primeira dimensão resultante da analise de
ordenação - MDS1e nem com o segundo vetor da mesma ordenação MDS2 não afetaram
significativamente a composição qualitativa de aves piscívoras (Tabela 5). Neste ano os
resultados das análises indicaram que não houve relação significativa entre a composição
qualitativa da comunidade de aves piscívoras e a profundidade e temperatura da água do
lago, nem tampouco com a composição qualitativa de macrófitas aquáticas na primeira
dimensão MDS1 e nem na segunda dimensão MDS2 (Tabela 5).

106
Tabela 1. Matriz de Correlação de Pearson para as variavéis abióticas: tamanho do lago (TAM), profundidade do lago(PRO),
transparência da água (TRA), oxigênio dissolvido (OXI), pH da água (PHA) e temperatura da água (TEA), registradas em 20 lagos no
Pantanal em 2003.
(TAM) (m) (PRO) (m) (TRA) (m) (OXI) (%) (PHA) (TEA) (ºC)
(TAM) (m) 1.000
(PRO) (m) 0.067* 1.000
(TRA) (m) 0.442 0.681* 1.000
(OXI) (%) 0.501 0.255 0.201 1.000
(PHA) 0.115 0.256 0.049 0.165 1.000
(TEA) (ºC) -0.014 0.239 -0.009 -0.034 -0.907** 1.000
* Asterísticos correspondem aos valores de “p” resultantes da matriz de probabilidade de Bonferroni, usada para avaliar se as correlações entre as variáveis na matriz de correlação de Pearson são significativas. *=p<0,05; **=p<0,001
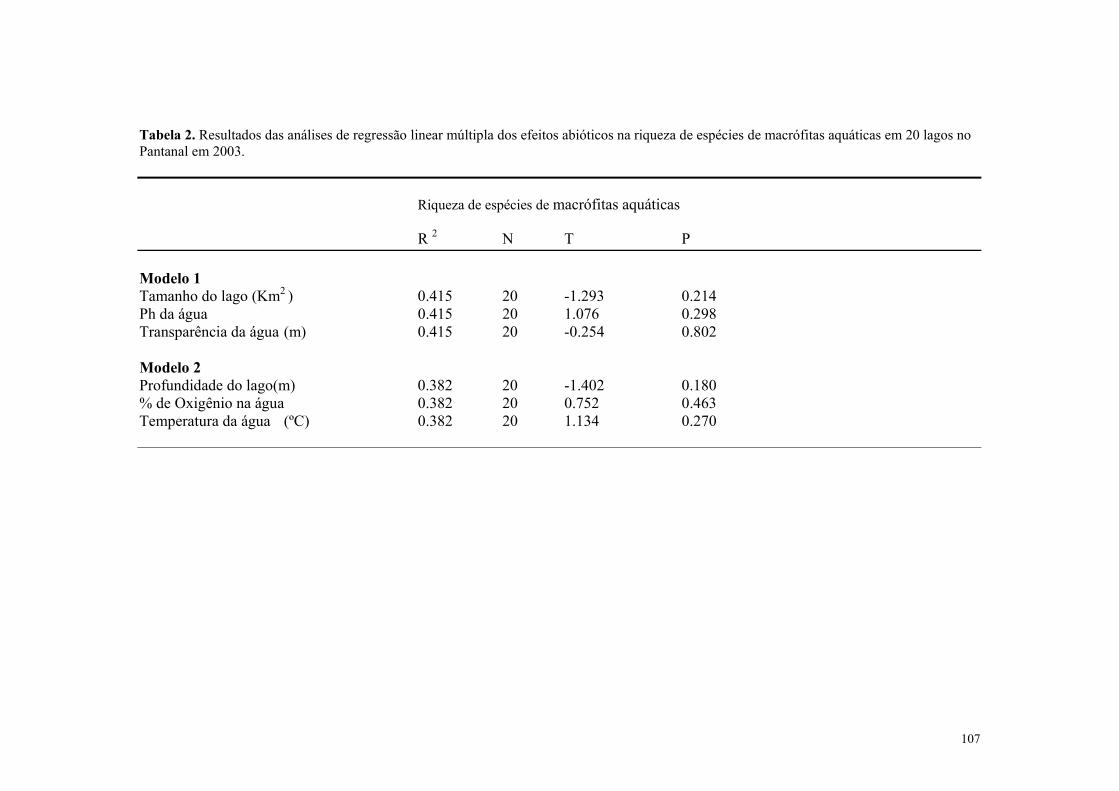
107
Tabela 2. Resultados das análises de regressão linear múltipla dos efeitos abióticos na riqueza de espécies de macrófitas aquáticas em 20 lagos no Pantanal em 2003. Riqueza de espécies de macrófitas aquáticas R 2 N T P Modelo 1 Tamanho do lago (Km2 ) 0.415 20 -1.293 0.214 Ph da água 0.415 20 1.076 0.298 Transparência da água (m) 0.415 20 -0.254 0.802 Modelo 2 Profundidade do lago(m) 0.382 20 -1.402 0.180 % de Oxigênio na água 0.382 20 0.752 0.463 Temperatura da água (ºC) 0.382 20 1.134 0.270

108
Tabela 3. Resultados das análises de regressão múltipla multivariada dos efeitos das características físicas e biológicas dos lagos na composição qualitativa das macrófitas aquáticas em 2003 para 20 lagos no Pantanal. __________________________________________________________________________________________________________ Composição qualitativa da comunidade de macrófitas aquáticas Pillai-Trace N P Modelo 1 Tamanho do lago 0.169 20 0.249 Ph da água 0.234 20 0.136 Transparência da água 0.019 20 0.869 Modelo 2 Profundidade do lago 0.138 20 0.329 % de Oxigênio na água 0.072 20 0.570 Temperatura da água 0.380 20 0.028
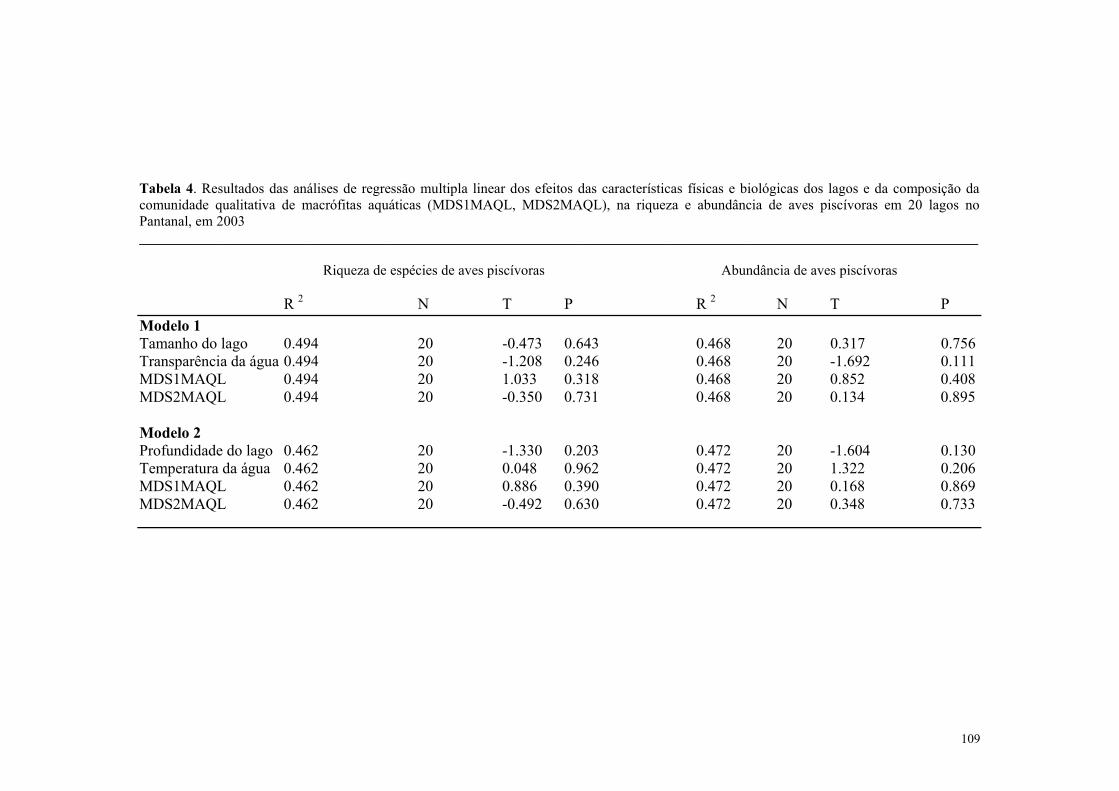
109
Tabela 4. Resultados das análises de regressão multipla linear dos efeitos das características físicas e biológicas dos lagos e da composição da comunidade qualitativa de macrófitas aquáticas (MDS1MAQL, MDS2MAQL), na riqueza e abundância de aves piscívoras em 20 lagos no Pantanal, em 2003 __________________________________________________________________________________________________________ Riqueza de espécies de aves piscívoras Abundância de aves piscívoras R 2 N T P R 2 N T P Modelo 1 Tamanho do lago 0.494 20 -0.473 0.643 0.468 20 0.317 0.756 Transparência da água 0.494 20 -1.208 0.246 0.468 20 -1.692 0.111 MDS1MAQL 0.494 20 1.033 0.318 0.468 20 0.852 0.408 MDS2MAQL 0.494 20 -0.350 0.731 0.468 20 0.134 0.895 Modelo 2 Profundidade do lago 0.462 20 -1.330 0.203 0.472 20 -1.604 0.130 Temperatura da água 0.462 20 0.048 0.962 0.472 20 1.322 0.206 MDS1MAQL 0.462 20 0.886 0.390 0.472 20 0.168 0.869 MDS2MAQL 0.462 20 -0.492 0.630 0.472 20 0.348 0.733
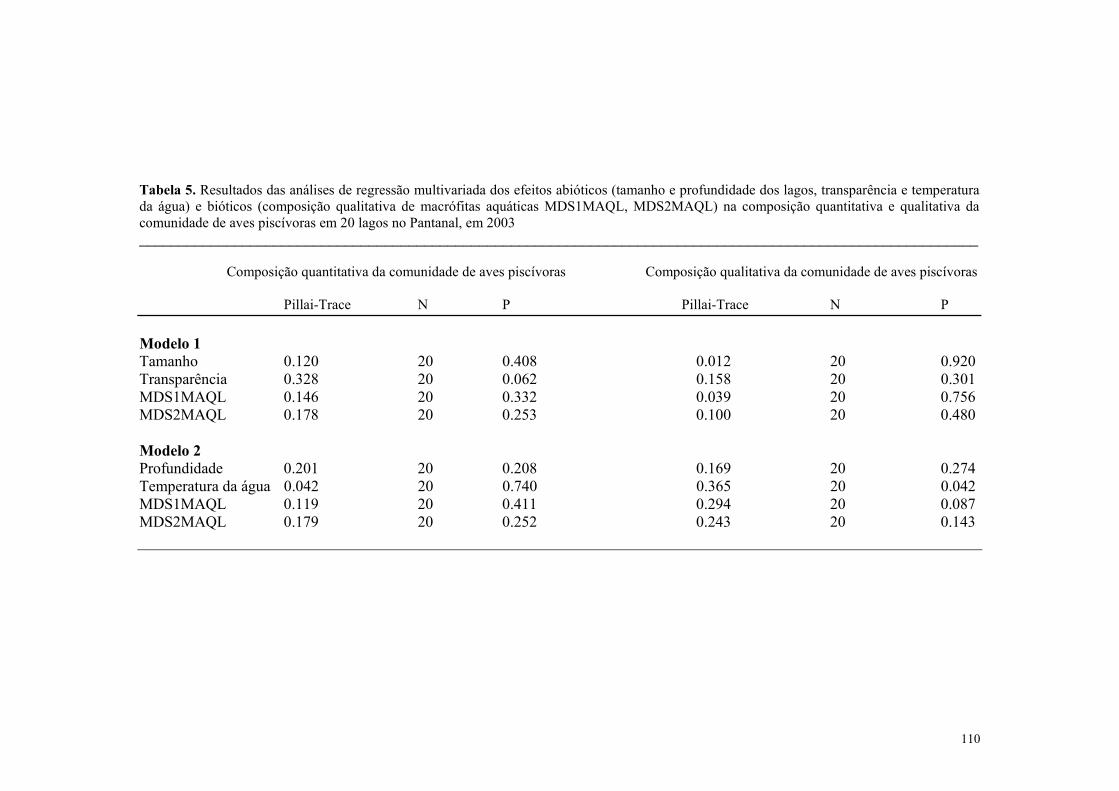
110
Tabela 5. Resultados das análises de regressão multivariada dos efeitos abióticos (tamanho e profundidade dos lagos, transparência e temperatura da água) e bióticos (composição qualitativa de macrófitas aquáticas MDS1MAQL, MDS2MAQL) na composição quantitativa e qualitativa da comunidade de aves piscívoras em 20 lagos no Pantanal, em 2003 __________________________________________________________________________________________________________ Composição quantitativa da comunidade de aves piscívoras Composição qualitativa da comunidade de aves piscívoras Pillai-Trace N P Pillai-Trace N P Modelo 1 Tamanho 0.120 20 0.408 0.012 20 0.920 Transparência 0.328 20 0.062 0.158 20 0.301 MDS1MAQL 0.146 20 0.332 0.039 20 0.756 MDS2MAQL 0.178 20 0.253 0.100 20 0.480 Modelo 2 Profundidade 0.201 20 0.208 0.169 20 0.274 Temperatura da água 0.042 20 0.740 0.365 20 0.042 MDS1MAQL 0.119 20 0.411 0.294 20 0.087 MDS2MAQL 0.179 20 0.252 0.243 20 0.143

111
4- Discussão
A composição da comunidade macrófitas aquáticas nos lagos do Pantanal de Poconé foi
formada por 26 espécies distribuídas em 21 famílias. A riqueza de espécies de macrófitas
aquáticas nas áreas alagáveis depende do grupo ecológico dessas comunidades, do nível da
água, de fatores físico-químicos e do estágio sucessional da planta (Da Silva & Silva, 1989;
Esteves, 1998; Pott & Pott, 2000). Das características analisadas como tamanho e
profundidade do lago, pH, oxigênio dissolvido (%), transparência e temperatura da água do
lago, nenhuma delas teve efeito significativo na riqueza de espécies de macrófitas aquáticas
(Tabela 2).
A distribuição de macrófitas aquáticas no Pantanal é heterogênea, havendo espécies
exclusivas de certos habitats, como por exemplo, Eichhornia diversifolia (Vahl) Urb.
encontrada somente em solos argilosos férteis no inicio da planície de inundação de
Cáceres, enquanto outras são generalistas (Pott & Pott, 2000). Isto porque as macrófitas
aquáticas apresentam grande capacidade de adaptação e grande amplitude ecológica, além
disso o grupo é capaz de suportar longos períodos de seca. (Esteves, 1998). Também
contribui para distribuição das macrófitas nos lagos a turbidez da água e o vento,
favorecendo o crescimento de diferentes grupos ecológicos (Esteves, 1998; Scheffer, 1999).
Na Amazônia as macrófitas submersas, raramente ocorrem nas várzeas e igapós,
porque as condições de luz são desfavoráveis (Junk & Piedade, 1997). Situação similar
deve ter ocorrido nos lagos do Pantanal, onde foram registradas apenas duas espécies de
macrófitas do tipo submersa, Ludwigia inclinata (L.F) P.H.Raven e Ultricalia poconensis
Fromm-Trinta (Tabela 6), pois a maioria dos lagos estudados tiveram mais de 50% de sua
superfície coberta por outras macrófitas, reduzindo assim a penetração direta de luz, e
provavelmente os nutrientes e sedimentos em suspensão nestes locais talvez tenham

112
contribuído para isto. É provável que em lagos onde espécies que formam tapetes não
sejam dominantes, ocorra uma diversidade maior de macrófitas, mas isto ainda não foi
estudado no Pantanal. Esses lagos do Pantanal, tal como acontece em reservatórios da
Amazônia, apresentam baixa turbulência da água e um aumento da concentração de
nutrientes, tendo como conseqüência devido à ausência de espécies competidoras e
predadoras, espécies como Eichhonia crassipes, Pistia stratiotes e Salvinia auriculata,
apresentam alta taxa de crescimento formando grandes tapetes nestes lagos (Esteves, 1998).
No Pantanal ocorrem as ‘Salinas’ denominação local, para lagoas alcalinas, nestas
lagoas o pH varia entre 3 a 4. Geralmente estas lagoas não apresentam macrófitas, mas,
apenas algas (Pott & Pott). Em reservatórios na Amazônia a diversidade de macrófitas foi
positiva e fortemente influenciada pelo pH e concentração iônica elevados (Junk & Melo
Nunes, 1987). Nos lagos do Pantanal o pH teve uma variação média de 6.7 provavelmente
não tendo nenhum efeito sobre o estabelecimento de macrófitas nestes ambientes.
Na Amazônia corpos de água que apresentam água com turbidez elevada, possuem
conseqüentemente maior condutividade e pH próximo ao neutro. Nestes ambientes as
comunidades de macrófitas aquáticas são abundantes e produzem maior biomassa (Junk
1979; Junk & Howard-Williams, 1984; Junk 1993). Já nos corpos que possuem coloração
preta, vinda de bacias de drenagem pobres em nutrientes, apresentam menor diversidade de
plantas aquáticas e menor biomassa (Junk & Howard-Williams, 1984). Nos lagos ao longo
da Estrada Parque Transpantaneira, funcionam na cheia como corpos de água branca, e na
seca água preta. Os mecanismos que explicam os padrões identificados ainda necessitam de
elucidação para comunidades de macrófitas aquáticas (Thomaz & Bini, 2003).
O conhecimento das interações entre as plantas aquáticas e os tipos de ambientes
bem como com outros organismos, é muito importante para o manejo é conservação dos

113
recursos naturais. Também essa riqueza de espécies e abundância pode ser influenciada
pela presença de peixes pastadores como a carpa Ctnepharyngodon idella Val.(Dall
Armellina et al., 1999), de aves aquáticas herbívoras como patos, gansos (Marklund et al.,
2002), de mamíferos herbívoros, como por exemplo do Manati (Trichecus manatus L.) na
Florida e do Peixe-boi (Trichechus inunguis Natterer) na Amazônia, da Capivara
(Hydrochoeris hydrochaeris) e do Cervo do Pantanal (Blastocerus dichotomus) no Pantanal
e na Amazônia (Junk & Silva, 1997; Junk & Piedade, 2000; Hauxwell et al., 2004; Silva,
2004). Porém, o efeito da herbívoria na distribuição de espécies de macrófitas no Pantanal
ainda não foi estudado.
As análises dos efeitos da composição e cobertura da comunidade de macrófitas
aquáticas não mostrou nenhuma influencia significativa na riqueza de espécies, na
abundância nem tampouco na composição da comunidade de aves piscívoras no Pantanal
(Tabela 4 e 5). A cobertura de macrófitas tem um efeito indireto sobre a riqueza e
abundância destas aves. Isto porque, dependendo do tipo biológico destas plantas, elas
podem oferecer melhor refúgio e alimento para vários organismos, incluindo os peixes,
principal presas das mesmas (ver Capítulo 4). Em um lago na Escócia foi verificado à
influência positiva da associação de peixes com algas (Fucus versicolosus e Ascophyllum
nodosum), mostrando que a Garça-real-européia (Ardea cinerea) prefere forragear nos
locais onde predominam estas algas, ocorrendo um aumento na abundância destas aves e
melhorando o seu sucesso reprodutivo (Cars & Elstron, 2003). Nos Everglades um estudo
com quatro espécies de aves vadeadoras, mostrou que a abundância delas estão
relacionadas com as comunidades de plantas terrestres e aquáticas, e esta relação é alterada
com a variação do nível da água (Bancroft et al., 2002). Estes autores sugerem que neste
padrão de distribuição, existem outros fatores interagindo, como por exemplo a

114
disponibilidade de presas, o que necessitaria ser melhor estudado. Em lagos andinos na
Argentina a riqueza de espécies de aves aquáticas foi altamente significativa com a
presença de macrófitas aquáticas submersas (principalmente flamingos, patos, gansos,
gaivotas). Trabalhos realizados em lagos temperados da Europa e da América do Norte têm
mostrado uma correlação positiva entre a presença de macrófitas e algas submersas
influenciando a riqueza e abundância de aves aquáticas (Cars & Elstron, 2003). Esses
estudos sugerem um padrão entre a presença de macrófitas submersas e aves aquáticas, no
entanto, no Pantanal com seus lagos muito rasos, e com poucas espécies de macrófitas
submersas,outros fatores devem ser investigados.

115
Figura 1. Espécies de macrófitas aquáticas dos lagos do Pantanal: a) Forma submersa fixa Ludwigia inclinata e sua flor (b); c) Forma submersa livre Utricalia poconensis, d) Forma emergente e anfíbia Justicia sp, e) Forma flutuante fixa Nymphoides crayana f) Forma flutuante fixa Hydroclerys parvifolia. Fotos: Dalci Oliveira

116
Figura 2. Algumas interações entre macrófitas aquáticas e outros organismos nos lagos do Pantanal. a) Ninho de Cafezinho (Jacana jacana) sobre Pistia statiotes e em b) sobre Ludwigia sedoides, c) Ninho de beija-flor-de-bico-curvo (Polytmus gauinumbi) no talo de Ipomea, d) Caranguejo (Dilocarcinus pagei) sobre caule de Eichhornia, e) desova de caramujo (Pomacea caniculata) sobre Pontederia lanceolata, f) Filhotes de jacaré (Caiman yacare), refugiando entre bancos de Pistia stratiotes. Fotos de Dalci Oliveira
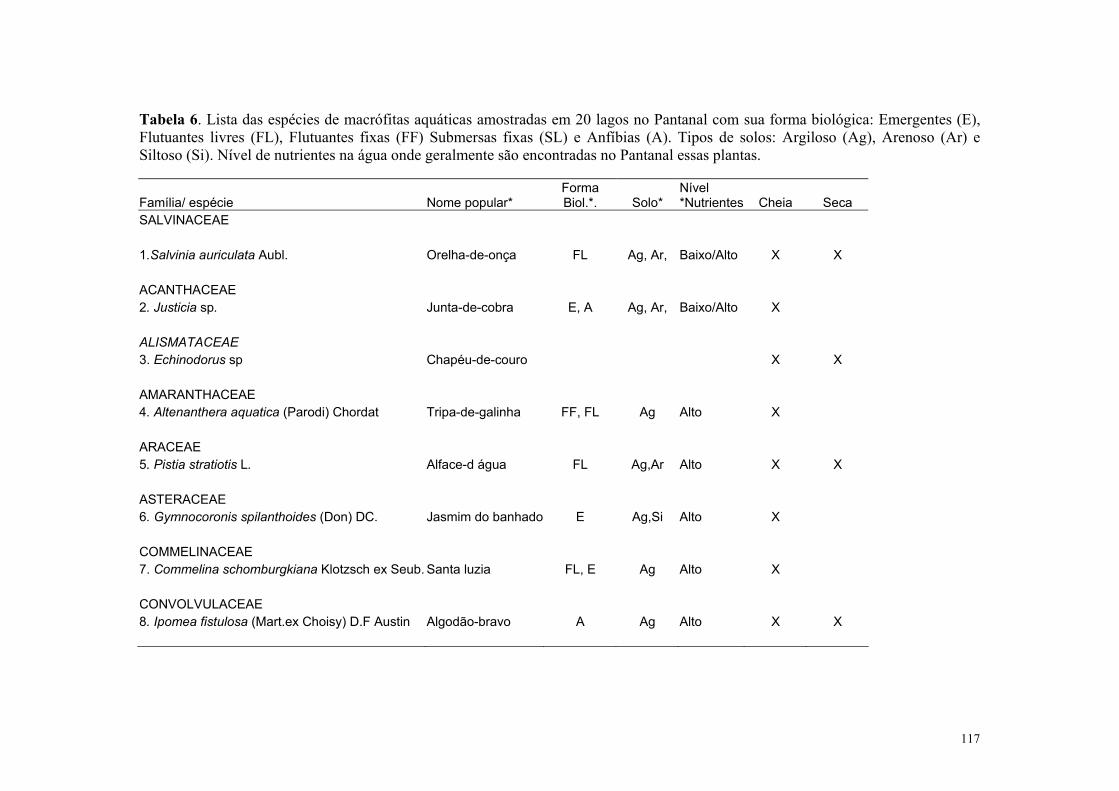
117
Tabela 6. Lista das espécies de macrófitas aquáticas amostradas em 20 lagos no Pantanal com sua forma biológica: Emergentes (E), Flutuantes livres (FL), Flutuantes fixas (FF) Submersas fixas (SL) e Anfíbias (A). Tipos de solos: Argiloso (Ag), Arenoso (Ar) e Siltoso (Si). Nível de nutrientes na água onde geralmente são encontradas no Pantanal essas plantas.
Família/ espécie Nome popular* Forma Biol.*. Solo*
Nível *Nutrientes Cheia Seca
SALVINACEAE 1.Salvinia auriculata Aubl. Orelha-de-onça FL Ag, Ar, Baixo/Alto X X ACANTHACEAE 2. Justicia sp. Junta-de-cobra E, A Ag, Ar, Baixo/Alto X ALISMATACEAE 3. Echinodorus sp Chapéu-de-couro X X AMARANTHACEAE 4. Altenanthera aquatica (Parodi) Chordat Tripa-de-galinha FF, FL Ag Alto X ARACEAE 5. Pistia stratiotis L. Alface-d água FL Ag,Ar Alto X X ASTERACEAE 6. Gymnocoronis spilanthoides (Don) DC. Jasmim do banhado E Ag,Si Alto X COMMELINACEAE 7. Commelina schomburgkiana Klotzsch ex Seub. Santa luzia FL, E Ag Alto X CONVOLVULACEAE 8. Ipomea fistulosa (Mart.ex Choisy) D.F Austin Algodão-bravo A Ag Alto X X
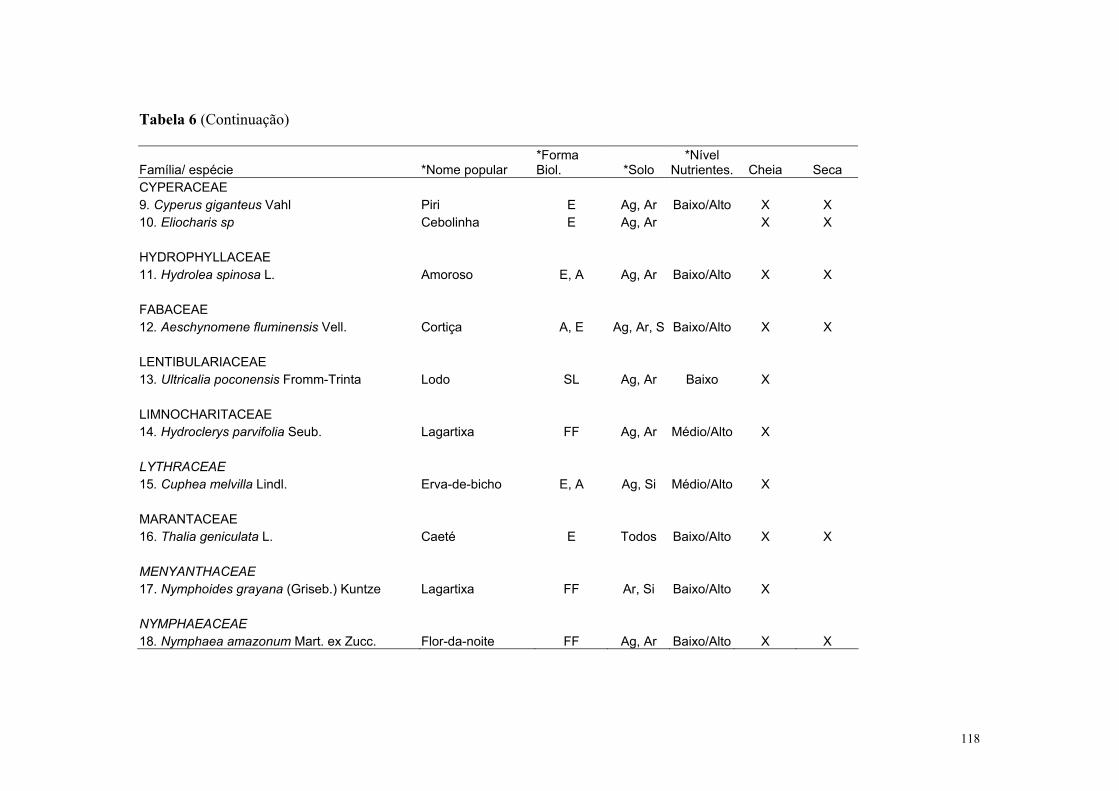
118
Tabela 6 (Continuação)
Família/ espécie *Nome popular *Forma Biol. *Solo
*Nível Nutrientes. Cheia Seca
CYPERACEAE 9. Cyperus giganteus Vahl Piri E Ag, Ar Baixo/Alto X X 10. Eliocharis sp Cebolinha E Ag, Ar X X HYDROPHYLLACEAE 11. Hydrolea spinosa L. Amoroso E, A Ag, Ar Baixo/Alto X X FABACEAE 12. Aeschynomene fluminensis Vell. Cortiça A, E Ag, Ar, S Baixo/Alto X X LENTIBULARIACEAE 13. Ultricalia poconensis Fromm-Trinta Lodo SL Ag, Ar Baixo X LIMNOCHARITACEAE 14. Hydroclerys parvifolia Seub. Lagartixa FF Ag, Ar Médio/Alto X LYTHRACEAE 15. Cuphea melvilla Lindl. Erva-de-bicho E, A Ag, Si Médio/Alto X MARANTACEAE 16. Thalia geniculata L. Caeté E Todos Baixo/Alto X X MENYANTHACEAE 17. Nymphoides grayana (Griseb.) Kuntze Lagartixa FF Ar, Si Baixo/Alto X NYMPHAEACEAE 18. Nymphaea amazonum Mart. ex Zucc. Flor-da-noite FF Ag, Ar Baixo/Alto X X
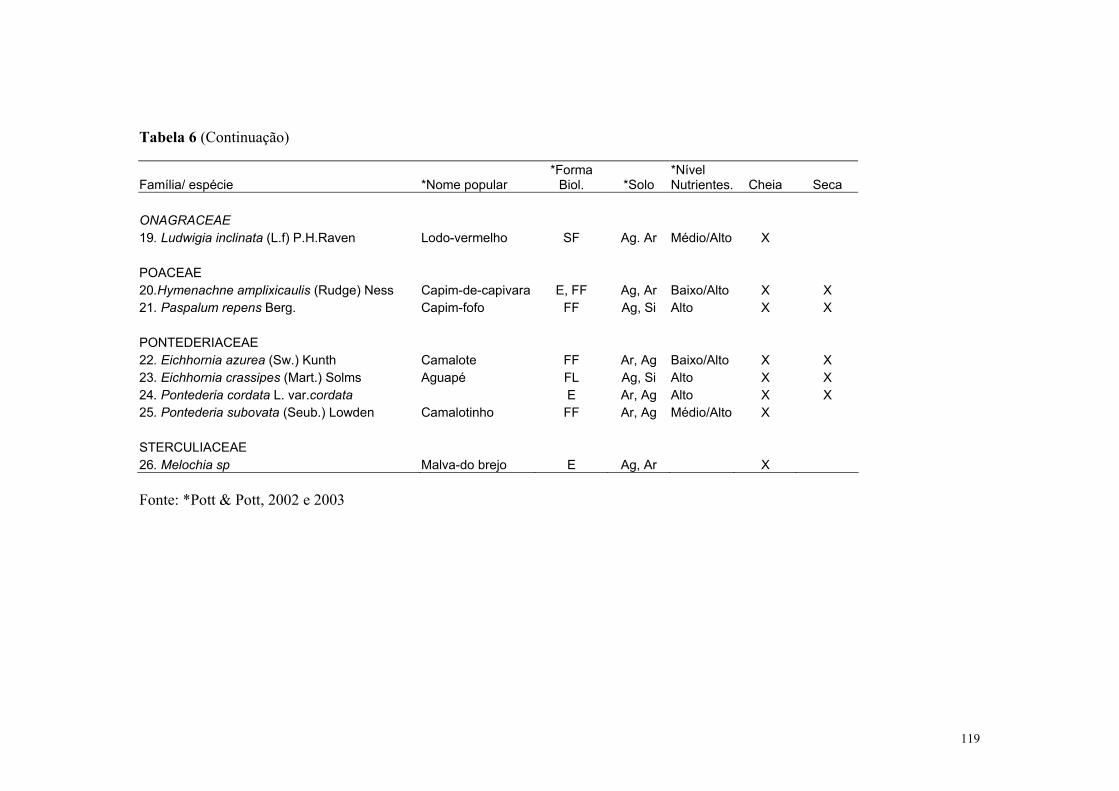
119
Tabela 6 (Continuação)
Família/ espécie *Nome popular *Forma
Biol. *Solo *Nível Nutrientes. Cheia Seca
ONAGRACEAE 19. Ludwigia inclinata (L.f) P.H.Raven Lodo-vermelho SF Ag. Ar Médio/Alto X POACEAE 20.Hymenachne amplixicaulis (Rudge) Ness Capim-de-capivara E, FF Ag, Ar Baixo/Alto X X 21. Paspalum repens Berg. Capim-fofo FF Ag, Si Alto X X PONTEDERIACEAE 22. Eichhornia azurea (Sw.) Kunth Camalote FF Ar, Ag Baixo/Alto X X 23. Eichhornia crassipes (Mart.) Solms Aguapé FL Ag, Si Alto X X 24. Pontederia cordata L. var.cordata E Ar, Ag Alto X X 25. Pontederia subovata (Seub.) Lowden Camalotinho FF Ar, Ag Médio/Alto X STERCULIACEAE 26. Melochia sp Malva-do brejo E Ag, Ar X Fonte: *Pott & Pott, 2002 e 2003

120
Capítulo 4. Efeito da variação da assembléia de peixes sobre a variação na assembléia
de aves piscívoras
1. Introdução
Uma das principais metas em ecologia de comunidades é tentar entender quais são os
mecanismos e processos responsáveis por padrões de variações espaço-temporais. O
número de espécies que ocorrem em um dado local é regulado por um balanço entre
processos de larga escala e de pequena escala espacial. Processos de larga escala,
especiação e dispersão, geralmente determinam o pool de espécies disponíveis para ocorrer
regionalmente, e processos de pequena escala (recurso alimentar, local de nidificação,
competição, perturbações ambientais e predação) geralmente limitam o número de espécies
que presentemente ocorrem em um dado local (Ricklefs, 1987; González, 1997; Englund &
Krupa, 2000; Collis et al., 2002; Bryan et al., 2003; Súarez et al., 2004).
A variação espaço-temporal na distribuição dos organismos é uma característica em
todos os sistemas ambientais. Em regiões temperadas e tropicais, um volume crescente de
estudos demonstrou que existe algum grau de variação em características estruturais e
funcionais de comunidades de aves e peixes (Junk et al., 1983; Kushlan et al., 1985;
Remsen Jr, 1985; 1991; Stiles, 1985; Saint-Paul & Soares, 1987; Loiselle & Blake, 1991;
1992; 1994; Merona & Bettencout, 1993, Lowe-McConnel, 1999; Hoeinghaus et al., 2003,
2004; Petry et al., 2003; Súarez et al., 2001 e 2004).
Em lagos da região temperada, um estudo mostrou uma significativa concordância
entre assembléias de aves piscívoras e peixes. Ambas são fortemente afetadas por alguns
fatores ambientais, principalmente o tamanho, a profundidade, a produtividade e o

121
isolamento geográfico do lago, como também pelas interações diretas entre aves e peixes,
como predação e competição (Pazkowski & Tonn, 2000).
Pelo menos 156 espécies de aves aquáticas ocorrem no Pantanal (Cintra & Antas,
1996), e 32 se alimentam primariamente de peixes (Oliveira, 1986 e 1997; Magalhães,
1990; Cintra & Antas, 1996).
Entre as aves piscívoras são encontradas espécies que utilizam diferentes estratégias
para capturar suas presas, as quais estão diretamente relacionadas com o pulso de
inundação (Kushlan et al., 1975; Kushlan et al., 1985). Nos períodos de enchente (outubro
a dezembro) e cheia (janeiro a março) o aumento da área inundada favorece a ictiofauna
que pode explorar diversos habitats. A retração das águas limita o espaço, o que atinge o
máximo de importância na seca (julho a agosto), quando então os lagos ficam isolados na
paisagem. Para compreender a dinâmica da fauna desse sistema aquático é fundamental
investigar os fatores bióticos e abióticos que afetam a distribuição e abundância de peixes e
a composição da assembléia de aves piscívoras.
Estudos realizados com comunidades de peixes em lagos de várzea da Amazônia
Central, identificaram deslocamentos sazonais (Junk et al., 1983) e circadianos (Saint-Paul
& Soares, 1987), relacionados com a variação do oxigênio, mostrando a importância desse
fator abiótico, determinante na distribuição de peixes. Com relação a reprodução é
conhecida à relação entre a desova e a mudança no nível de água, como também a
importância das várzeas da Amazônia Central como locais de dispersão de larvas
(Goulding, 1980; Araujo-Lima, 1984; Junk, 1984; Araujo-Lima & Oliveira, 1988). Por
outro lado, estes fatores poderiam também influenciar a distribuição de peixes nestes locais.
Estudos preliminares indicam que situação similar ocorre no Pantanal. Mourão et al.
(1988), relacionaram fatores limnológicos com a ictiofauna ao estudarem baías (lagos)

122
salinas no Pantanal de Nhecolândia, e verificaram que o pH elevado é um fator limitante
para os peixes da região.
Os primeiros estudos com comunidade de peixes no Pantanal, relacionados com
estrutura trófica (Resende & Palmeira, 1999; Resende, 2000), demonstraram que existe
uma variação de abundância dos mesmos entre os diferentes ambientes. Trabalhos indicam
que há forte variação espaço-temporal nestas comunidades (Junk et al., 1983; Saint-Paul &
Soares, 1987; Cox-Fernandes, 1997; Sánchez-Botero et al., 2001). Entretanto, ainda pouco
se sabe sobre os fatores responsáveis por tais variações.
Somente recentemente foram realizados os primeiros trabalhos no Pantanal
relacionando fatores abióticos (profundidade, transparência, distância dos lagos ao rio e
tempo de isolamento dos lagos) e bióticos (cobertura de macrófitas abundância de
predadores) atuando em conjunto na estruturação da comunidade de peixes nos lagos
(Súarez et al., 2001 e 2004).
Estudos verificando os efeitos das interações entre comunidade de peixes e
comunidades de aves na região neotropical são escassos (Gawlik, 2002; Steinmetz et al.,
2003; Mendonça et al., 2004). Portanto, mais estudos que enfoquem os fatores que
determinam a abundância e riqueza de aves são necessários para entender como as
comunidades de aves e peixes são estruturadas ajudando a entender o funcionamento do
ecossistema do Pantanal e propondo medidas para a sua conservação.
O objetivo deste estudo foi avaliar se variações espaço-temporais na comunidade de
aves piscívoras estão relacionadas com as características bióticas e abióticas dos lagos e
com as variações na comunidade de peixes.

123
2. Material e Métodos
2.1 Desenho amostral
Nos mesmos lagos utilizados para observação das aves, além da medição das
variáveis limnologicas e verificação da proporção de cobertura e riqueza de espécies de
macrófitas aquáticas (conforme Capítulo 2 e 3), também foi efetuado o registro das espécies
de peixes. Para a coleta e determinação da ictiofauna nos corpos da água selecionados
foram efetuadas coletas com armadilhas feitas de telas de nylon tipo mosquiteiro em
armação de alumínio fosco de 2 m x 1,5 m (Figura 1). Este peneirão manejado por duas
pessoas que adentravam o lago, foi lançado dez vezes, aleatoriamente dentro do lago, a
soma das dez coletas foram consideradas como uma amostra daquele lago. Esta
amostragem foi realizada concomitantemente com o censo populacional das aves nos 20
lagos. Estas coletas foram realizadas em dois anos consecutivos uma vez na cheia e outra
na seca.
As espécies de peixes coletadas, foram preparadas em solução de formol a 10%,
transportadas para laboratório, lavadas em água corrente e transferidas para álcool 70%.
Após a triagem e identificação os peixes foram pesados em balança analítica com 0,001g de
precisão, acondicionados em sacos plásticos e depositados em tambores plásticos. A
identificação dos peixes foi efetuada através de comparação com espécimes depositadas na
coleção de Vertebrados da UFMT, ou com o manual de identificação (Britski et al., 1999),
ou ainda e com a colaboração de especialistas. As espécies testemunho foram depositadas
na mesma coleção. Para as observações das aves a rodovia que margeia os lagos, foi
percorrida a pé, o censo visual foi feito a olho nu ou com auxilio de binóculo (8x40), as
observações foram realizadas das 06 ás 10h da manhã e das 14 ás 18h. Foram consideradas
nos censos populacionais todas as aves que estavam nas margens ou dentro dos lagos.

124
Esporadicamente o comportamento destas aves foi filmado com câmera digital (700x) para
identificação dos peixes consumidos por elas e para ajudar na confirmação da dieta também
foram coletadas regurgitos dessas aves (para mais detalhes ver Capítulo 2).
2.2 Análise dos dados
Para verificar se a riqueza de espécies e abundância de aves são influenciadas pela riqueza
de espécies de peixes e a biomassa de peixes foi usada em análises de regressão linear
múltipla disponíveis no programa Systat (Wilkinson, 1997). Para verificar responder se a
variação na composição da assembléia de aves piscívoras é influenciada pela composição
qualitativa e qualitativa da comunidade de peixes (riqueza de espécies de peixes e a
biomassa de peixes) os dados de riqueza de espécies de peixes foram usados para construir
uma matriz qualitativa de espécie por local amostrado (lago), e uma matriz quantitativa de
biomassa de peixe por local, a partir das quais foram criadas duas matrizes de similaridade
usando o índice de Bray-Curtis. Este índice tem sido o mais recomendado para estudos de
comunidades de aves (MacNally, 1994) e estudos ecológicos em geral (Minchin, 1987;
Cintra, 1997).
Nesta matriz foi usada uma análise de ordenação do tipo escalonamento multidimensional
(MDS). Os valores dos dois vetores principais de análise de ordenação para os peixes foram
usados como variável independente em modelos de regressão com as variáveis
dependentes, riqueza de espécies de aves, abundância de aves, através de uma análise de
regressão linear simples, para verificar se a relação à composição da comunidade de peixes
tem relação significativa com a riqueza de espécies de aves, e a abundância delas nos
ambientes alagáveis no Pantanal.

125
Os escores MDS1, e MDS2 resultantes das análises de ordenação para a comunidade de
peixes também foram usados como variáveis dependentes em uma Análise de Variância
Multivariada (MANOVA) tendo os locais (lagos) como variáveis categóricas no modelo. O
teste de Pillai-Trace, por ser mais robusto, foi usado para testar a significância da
MANOVA e verificar se as assembléias de peixes dos lagos diferem significativamente
entre si na sua composição. Para as aves piscívoras foram construídas matrizes qualitativas
(presença/ausência) e quantitativa (abundância) de espécies por local (lagos). Antes de
construir a matriz a abundância de indivíduos de cada espécie foi dividida pelo tamanho do
lago para padronizar as amostras. A matriz quantitativa foi construída com a densidade de
espécies de aves piscívoras (colunas) por locais ou lagos (linhas). O índice de Bray-Curtis
aplicado em dados de presença e ausência, é conhecido como medida de distância de
Sorensen (Legendre & Legendre, 1998). Nessas matrizes também foram usadas analises de
MDS, os dois vetores MDS1, MDS2 foram usados como variáveis dependentes em
modelos de regressão múltipla multivariada para verificar como a composição qualitativa e
quantitativa da comunidade de aves piscívoras (MDS1, MDS2 aves), estava sendo afetada
pela transparência da água e a composição da comunidade de peixes (MDS1, MDS2
peixes). As análises de MDS foram feitas usando o programa PATN (Belbin, 1982) e a
MANOVA e o teste inferêncial de Pillai-Trace no programa Systat (Wilkinson, 1997).
3. Resultados
3.1. Comunidade de peixes em 2002 e 2003
Nos dois anos de coleta 2002 e 2003 a comunidade de peixes foi formada por 79
espécies de peixes distribuídas em 21 famílias (Tabela 1). As famílias mais abundantes
foram Characidae e Cichlidae, juntas contribuíram com 44.3% do total de peixes coletados.

126
No ano de 2002 a composição da comunidade peixes foi formada por 65 espécies
distribuídas em 21 famílias. A riqueza de espécies de peixes variou nos lagos na cheia entre
13 a 42 espécies, e na seca entre 7 a 25 espécies de peixes. As espécies de peixes mais
freqüentes nos lagos foram Odontostilbe paraguayensis Eigenmann & Kennedy, 1903,
presente em todos os lagos na cheia, e na seca esteve presente em 16 deles; Characidium
laterallis (Boulerger, 1895) esteve presente em 95% dos lagos, na cheia em 19 e na seca em
12; Aphyocarax rathbuni (Eigenmann, 1907) foi registrado em 90% dos lagos; na cheia
ocorreu em 18 e na seca em 14 lagos; Pyrrulina australis Eigenmann & Kennedy, 1903,
esteve presente também em 90% dos lagos, na cheia em 18 e na seca em 14; e Hoplias
malabaricus (Block, 1794) também em 90% dos lagos, na cheia em 18 e na seca em 10
lagos (Figura 1).
Em 2003 a composição da comunidade peixes foi formada por 67 espécies
distribuídas em 18 famílias. A riqueza de espécies de peixes variou nos lagos na cheia entre
(9 a 27 espécies) e na seca (5 a 19 espécies). As espécies de peixes mais freqüentes nos
lagos foram as piquiras Odontostilbe calliura (Boulerger, 1900) presente em todos os lagos
na cheia e ela ocorreu na seca apenas em 13 lagos; Odontostilbe paraguayensis também foi
registrada em todos os lagos; na cheia em 19 e na seca em 13; Aphyocarax rathbuni esteve
presente em 95% dos lagos; na cheia em 19 e na seca em 8; Pyrrulina australis esteve
também em 95% dos lagos na cheia em 16 e na seca em 7; e Corydoras hastatus
Eigenmann & Eigenmann, 1888 em 90% dos lagos, na cheia em 17 e na seca em 11 lagos.
A composição da comunidade de peixes mostrou um melhor padrão de similaridade
entre os lagos para a comunidade qualitativa de peixes considerando o primeiro eixo
(Figura 2 e 3).
3.2 Comunidade de Aves piscívoras em 2002 e 2003

127
A comunidade de aves piscívoras foi formada por 17 espécies em 2002 (Tabela 2).
As cinco espécies mais abundantes foram a Garça-branca-grande (Ardea alba)L., com 1121
indivíduos representando 42.76% do total das aves piscívoras, a Garça-branca-pequena
(Egretta thula) (Molina) com 428 indivíduos (16.32%); o Tuiuiú (Jabiru mycteria)
(Lichtenstein) com 337 (12.85%), o Cabeça-seca (Mycteria americana) L., com 200
(7.63%) e o Socó-dorminhoco (Nycticorax nycticorax) (L.), com 119 (4.54%).
Em 2003, a comunidade de aves piscívoras foi formada por 17 espécies (Tabela 2).
As cinco espécies mais abundantes foram a Garça-branca-grande (Ardea alba) com 447;
representando 31.59% do total das aves piscívoras; a Garça-branca-pequena (Egretta thula)
(258; 18.23%); o Socó-dorminhoco (Nycticorax nycticorax) (187; 13.21%), o Biguá
(Phalacrocorax brasilianus) (Gmelin), (107; 7.56%) e o Cabeça-seca (Mycteria
americana) (92; 6.50%).
Em 2002 o conjunto das aves pernaltas formou 91.43% da assembléia de aves
piscívoras, enquanto que em 2003 elas tiveram 97.56% de participação na composição das
mesmas.
A composição das aves piscívoras em 2002 mostrou similaridade entre os lagos
principalmente na composição qualitativa separando em um eixo os lagos com 3 a 5
espécies de aves e em outro eixo lagos cuja riqueza de espécies variou entre 8 a 15 espécies
de aves piscívoras (Figura 4). Em 2003 este padrão não foi tão evidente (Figura 5).
A correlação entre as variavéis das características bióticas e abióticas dos lagos em
2002 e 2003
Em 2002, antes de realizar as análises de regressão, as variavéis independentes
foram avaliadas quanto a colinearidade usando matriz de correlação de Pearson e também

128
por simulação usando correlação múltipla na qual cada variável independente foi analisada
como dependente contra todas as demais como independentes, e verificados os valores de
tolerância em cada modelo, somente valores de tolerância acima de 0.1 forma considerados
(Wilkinson, 1997). Como algumas variavéis independentes estavam significativamente
correlacionadas como por exemplo tamanho e profundidade do lago; transparência da água
e temperatura da água; oxigênio dissolvido na água (%) e cobertura de macrófitas aquáticas
(Tabela 3), elas foram usadas em dois modelos estatísticos separados. As variáveis
temperatura da água e tamanho e profundidade do lago, % de oxigênio dissolvido e foram
usadas juntas no modelo 1 e as outras como transparência da água e pH da água e cobertura
de macrófitas no modelo 2 de regressão múltipla.
Em 2003, Como algumas variavéis independentes (tamanho e profundidade do lago;
transparência da água e profundidade do lago; e biomassa e riqueza de peixes) foram
significativamente correlacionadas entre si, (Tabela 4), elas foram usadas em modelos
separados. A variável tamanho do lago, cobertura de macrófitas aquáticas e pH e
transparência da água foram usadas juntas no mesmo modelo e as outras como
Profundidade do lago, % de oxigênio dissolvido e temperatura da água em outro modelo.
3.1.1 - Efeitos das características bióticas e abióticas dos lagos na riqueza de espécies de peixes
Em 2002 e 2003 a riqueza de espécies de peixes não foi afetada significativamente
por nenhuma das características bióticas e abióticas dos lagos (Tabela 3 e 4).
3.1.2 - Efeitos das características bióticas e abióticas dos lagos na biomassa de peixes
Em 2002 e 2003 a biomassa de peixes não foi afetada significativamente por
nenhuma das características bióticas e abióticas dos lagos (Tabela 3 e 4).
3.1.3 - Efeitos das características bióticas e abióticas dos lagos na composição da comunidade de peixes

129
Em 2002 os resultados das análises indicaram que houve relação significativa entre
a composição quantitativa e qualitativa da comunidade de peixes e cobertura de macrófitas
aquáticas no modelo 1 (Tabela 5). Entretanto, nenhuma das outras características bióticas e
abióticas dos lagos afetaram significativamente a composição quantitativa e qualitativa da
comunidade de peixes (Tabela 5).
Em 2003 os resultados das análises indicaram que houve relação significativa entre
a composição quantitativa da comunidade de peixes e a profundidade dos lagos no modelo
1 (Tabela 6). Também neste mesmo modelo ocorreu relação significativa entre a
composição qualitativa da comunidade de peixes e a % de oxigênio dissolvido na água e
temperatura da água (Tabela 6). As demais características bióticas e abióticas dos lagos não
afetaram significativamente a composição quantitativa e qualitativa de peixes (Tabela 6).
3.2.1 - Efeito da transparência da água dos lagos e da composição da comunidade de peixes na composição da comunidade de aves piscívoras
Em 2002 os resultados das análises indicaram que houve relação significativa entre
a transparência da água e a composição quantitativa da comunidade de aves piscívoras,
(modelo 2). A composição quantitativa da comunidade de aves piscívoras também foi
afetada pela composição quantitativa da comunidade de peixes na primeira dimensão
(MDS1) (Tabela 9). Contudo no modelo1 a transparência da água não afetou
significativamente a composição quantitativa e qualitativa das aves piscívoras (Tabela 9).
Ainda, neste ano houve relação significativa entre a composição quantitativa da
comunidade de peixes na primeira dimensão (MDS1) e a composição quantitativa de aves
piscívoras no modelo 1 (Tabela 9). A composição qualitativa de aves piscívoras não foi
afetada significativamente pela composição qualitativa da comunidade de peixes na
primeira e segunda dimensão (MDS1 e MDS2 ;Tabela 9).
Em 2003 houve relação significativa entre a transparência da água e a composição
quantitativa da comunidade de aves piscívoras nos modelos 1 e 2 (Tabela 10). Entretanto, a
composição quantitativa e qualitativa de aves piscívoras não foi afetada significativamente
pela composição quantitativa e qualitativa de peixes (Tabela 10).
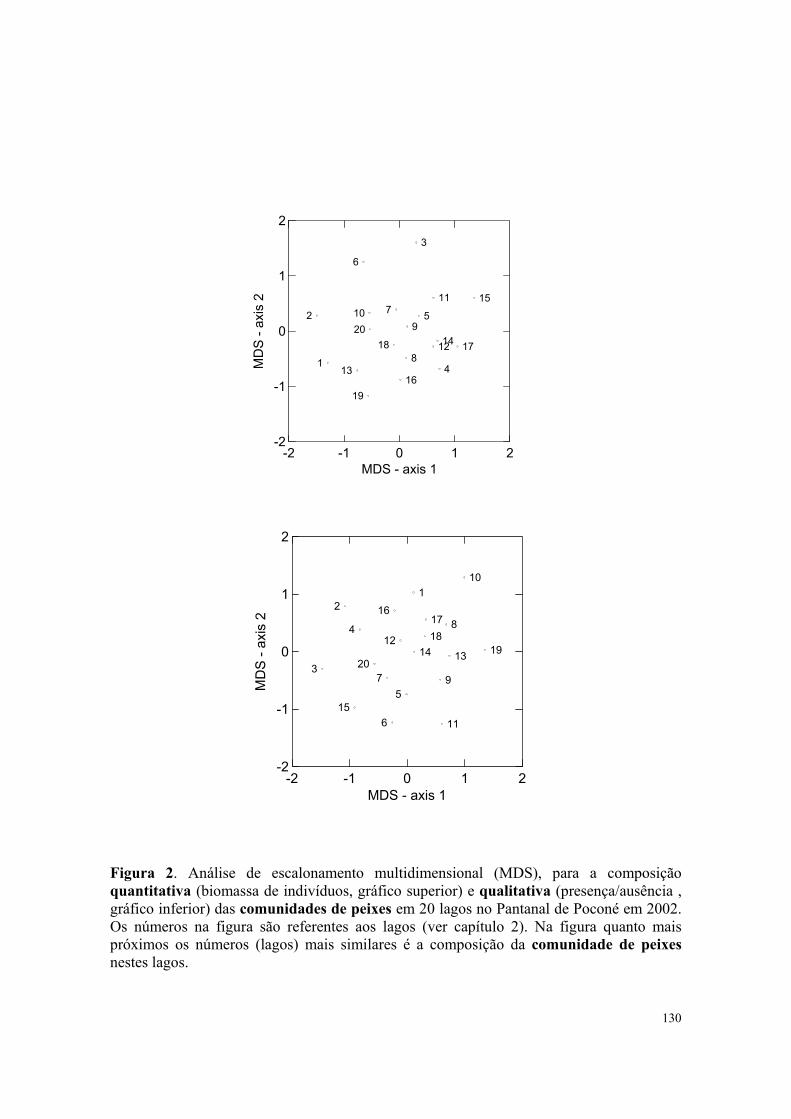
130
Figura 2. Análise de escalonamento multidimensional (MDS), para a composição quantitativa (biomassa de indivíduos, gráfico superior) e qualitativa (presença/ausência , gráfico inferior) das comunidades de peixes em 20 lagos no Pantanal de Poconé em 2002. Os números na figura são referentes aos lagos (ver capítulo 2). Na figura quanto mais próximos os números (lagos) mais similares é a composição da comunidade de peixes nestes lagos.
-2 -1 0 1 2MDS - axis 1
-2
-1
0
1
2
MD
S -
axis
2
2
113
6
19
1020
18
7
16
15
17
4
14
11
12
5
3
9
8
-2 -1 0 1 2MDS - axis 1
-2
-1
0
1
2
MD
S -
a xis
2
3
2
15
4
207
6
16
12
5
19
10
13
8
11
9
1718
14
1

131
Figura 3. Análise de escalonamento multidimensional (MDS), para a composição quantitativa (biomassa de indivíduos, gráfico superior) e qualitativa (presença/ausência , gráfico inferior) das comunidades de peixes em 20 lagos no Pantanal de Poconé em 2003. Os números na figura são referentes aos lagos (ver capítulo 2). Na figura quanto mais próximos os números (lagos) mais similares é a composição da comunidade de peixes nestes lagos.
-2 -1 0 1 2MDS - axis - 1
-2
-1
0
1
2
MD
S -
a xis
- 2
1
7
26
4
18
15
3
9
17
11
16
1410
5
1920
13
12
8
-1 0 1 2MDS - axis - 1
-2
-1
0
1
2
MD
S -
a xis
- 2
18
9
14
15
8
10
20
1319
12
1
7
4
16
5
63
2
11
17

132
Figura 4. Análise de escalonamento multidimensional (MDS), para a composição quantitativa (abundância de indivíduos, gráfico superior) e qualitativa (presença/ausência , gráfico inferior) das comunidades de aves piscívoras em 20 lagos no Pantanal de Poconé em 2002. Os números na figura são referentes aos lagos (ver capítulo 2). Na figura quanto mais próximos os números (lagos), mais similar é a composição da comunidade de aves
-2 -1 0 1 2MDS - axis 1
-2
-1
0
1
MD
S - a
xis
2
14
4
3
17
11
16
10
12
5
7
18
6
20
19
8
9
15
2
13
1
-1 0 1 2MDS - axis 1
-2
-1
0
1
2
MD
S -
axis
2
19
13
2
43
8
10
1112
16
1517
9
18
6
20
7
5
1
14
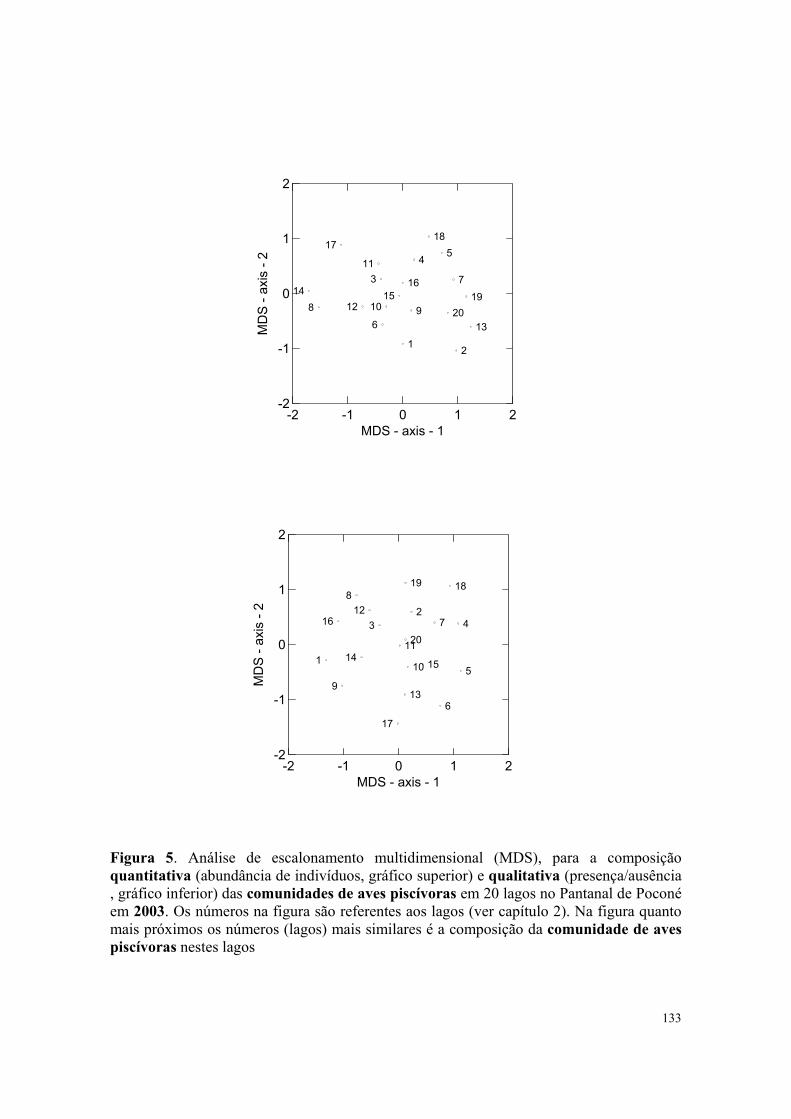
133
Figura 5. Análise de escalonamento multidimensional (MDS), para a composição quantitativa (abundância de indivíduos, gráfico superior) e qualitativa (presença/ausência , gráfico inferior) das comunidades de aves piscívoras em 20 lagos no Pantanal de Poconé em 2003. Os números na figura são referentes aos lagos (ver capítulo 2). Na figura quanto mais próximos os números (lagos) mais similares é a composição da comunidade de aves piscívoras nestes lagos
-2 -1 0 1 2MDS - axis - 1
-2
-1
0
1
2
MD
S -
axi s
- 2
148
17
12
113
6
1015
1
13
19
2
7
20
518
4
9
16
-2 -1 0 1 2MDS - axis - 1
-2
-1
0
1
2
MD
S -
a xis
- 2
1
16
9
8
14
123
17
11
13
5
4
18
6
7
15
2
10
20
19

134
Tabela 3. Matriz de Correlação de Pearson para as variavéis bióticas e abióticas: tamanho do lago (TAM), profundidade do lago (PRO),
transparência da água (TRA), cobertura de macrófitas aquáticas (CMA), biomassa de peixes (BPE) e riqueza de espécies de peixe
(RPE), registradas em 20 lagos no Pantanal em 2002.
(TAM) (m) (PRO) (m) (TRA) (m) (CMA) (%) (BPE) (Kg)
(RPE) (m)
(TAM) (m) 1.000
(PRO) (m) 0.074 1.000
(TRA) (m) 0.085 0.827** 1.000
(CMA) (%) 0.054 -0.073 0.093 1.000
(BPE) (m) 0.790* 0.199 0.197 -0.098 1.000
(RPE) (m) 0.129 0.187 0.258 -0.026 0.591 1.000
* Asterísticos correspondem aos valores de “p” resultantes da matriz de probabilidade de Bonferroni, usada para avaliar se as correlações entre as variáveis na matriz de correlação de Pearson são significativas. *=p<0,05; **=p<0,001
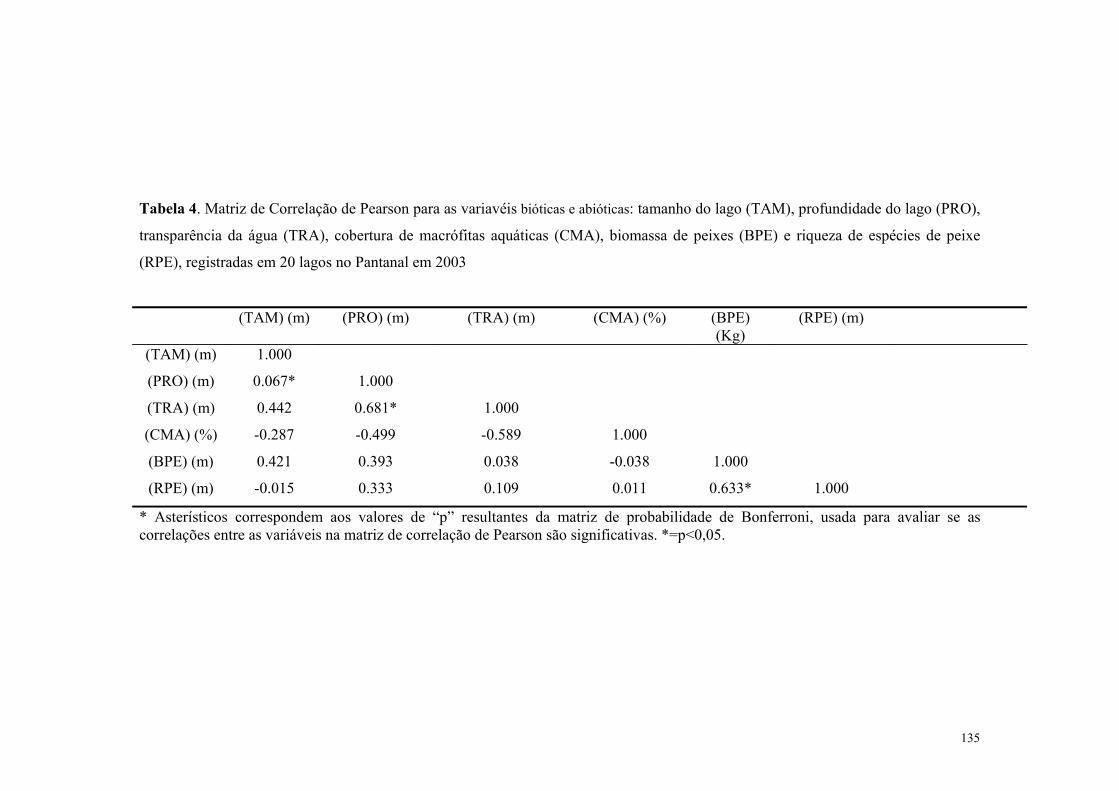
135
Tabela 4. Matriz de Correlação de Pearson para as variavéis bióticas e abióticas: tamanho do lago (TAM), profundidade do lago (PRO),
transparência da água (TRA), cobertura de macrófitas aquáticas (CMA), biomassa de peixes (BPE) e riqueza de espécies de peixe
(RPE), registradas em 20 lagos no Pantanal em 2003
(TAM) (m) (PRO) (m) (TRA) (m) (CMA) (%) (BPE) (Kg)
(RPE) (m)
(TAM) (m) 1.000
(PRO) (m) 0.067* 1.000
(TRA) (m) 0.442 0.681* 1.000
(CMA) (%) -0.287 -0.499 -0.589 1.000
(BPE) (m) 0.421 0.393 0.038 -0.038 1.000
(RPE) (m) -0.015 0.333 0.109 0.011 0.633* 1.000
* Asterísticos correspondem aos valores de “p” resultantes da matriz de probabilidade de Bonferroni, usada para avaliar se as correlações entre as variáveis na matriz de correlação de Pearson são significativas. *=p<0,05.
136
Tabela 5. Resultados das análises de regressão múltipla linear dos efeitos das variavéis bióticas e abióticas na riqueza e biomassa de peixes em 2002 para os 20 lagos no Pantanal Riqueza de Peixes Biomassa de Peixes R 2 N T P R 2 N T P Modelo 1 Tamanho 0.375 20 -1.497 0.155 0.357 19 -1.248 0.233 Profundidade 0.375 20 0.253 0.804 0.357 19 -0.422 0.679 Oxigênio dissolvido 0.375 20 0.433 0.671 0.357 19 -0.065 0.949 Temperatura da água 0.375 20 0.438 0.668 0.357 19 0.728 0.470 Modelo 2 Transparência 0.349 20 -0.678 0.507 0.364 20 0.189 0.852 pH da água 0.349 20 1.187 0.253 0.364 20 0.342 0.737 Cobertura macrófitas 0.349 20 -0.460 0.652 0.364 20 -1.396 0.182 __________________________________________________________________________________________________________________________________

137
Tabela 6. Resultados das análises de regressão múltipla linear dos efeitos das variavéis bióticas e abióticas na riqueza e biomassa de peixes em 2003 para os 20 lagos no Pantanal Riqueza de Peixes Biomassa de Peixes R 2 N T P R 2 N T P Modelo 1 Tamanho 0.441 20 -1.022 0.323 0.420 20 -0.694 0.499 Transparência 0.441 20 0.304 0.766 0.420 20 -0.033 0.974 pH da água 0.441 20 1.262 0.226 0.420 20 1.356 0.195 Cobertura macrófitas 0.441 20 0.972 0.346 0.420 20 0.750 0.465 Modelo 2 Profundidade 0.304 20 -0.769 0.453 0.345 20 -0.721 0.481 Oxigênio dissolvido 0.304 20 0.291 0.775 0.345 20 -0.021 0.984 Temperatura da água 0.304 20 1.183 0.254 0.345 20 1.396 0.182 __________________________________________________________________________________________________________________________________
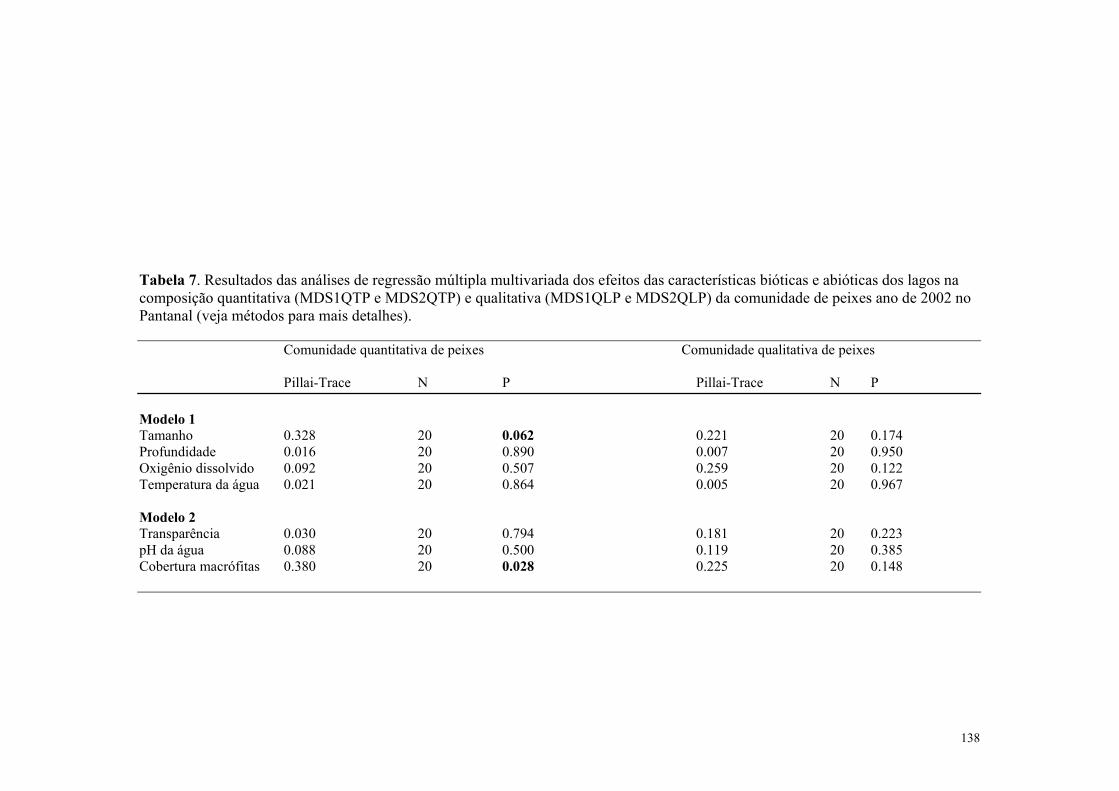
138
Tabela 7. Resultados das análises de regressão múltipla multivariada dos efeitos das características bióticas e abióticas dos lagos na composição quantitativa (MDS1QTP e MDS2QTP) e qualitativa (MDS1QLP e MDS2QLP) da comunidade de peixes ano de 2002 no Pantanal (veja métodos para mais detalhes). Comunidade quantitativa de peixes Comunidade qualitativa de peixes Pillai-Trace N P Pillai-Trace N P Modelo 1 Tamanho 0.328 20 0.062 0.221 20 0.174 Profundidade 0.016 20 0.890 0.007 20 0.950 Oxigênio dissolvido 0.092 20 0.507 0.259 20 0.122 Temperatura da água 0.021 20 0.864 0.005 20 0.967 Modelo 2 Transparência 0.030 20 0.794 0.181 20 0.223 pH da água 0.088 20 0.500 0.119 20 0.385 Cobertura macrófitas 0.380 20 0.028 0.225 20 0.148

139
Tabela 8. Resultados das análises de regressão múltipla multivariada dos efeitos das características bióticas e abióticas dos lagos na composição quantitativa (MDS1QTP, MDS2QTP) e qualitativa (MDS1QLP, MDS2QLP) da comunidade de peixes no ano de 2003 no Pantanal (veja métodos para mais detalhes). Comunidade quantitativa de peixes Comunidade qualitativa de peixes Pillai-Trace N P Pillai-Trace N P Modelo 1 Tamanho 0.029 20 0.813 0.200 20 0.209 Transparência 0.236 20 0.152 0.041 20 0.745 pH da água 0.003 20 0.976 0.309 20 0.076 Cobertura macrófitas 0.014 20 0.907 0.053 20 0.684 Modelo 2 Profundidade 0.370 20 0.031 0.044 20 0.715 Oxigênio dissolvido 0.063 20 0.615 0.382 20 0.027 Temperatura da água 0.158 20 0.275 0.588 20 0.001
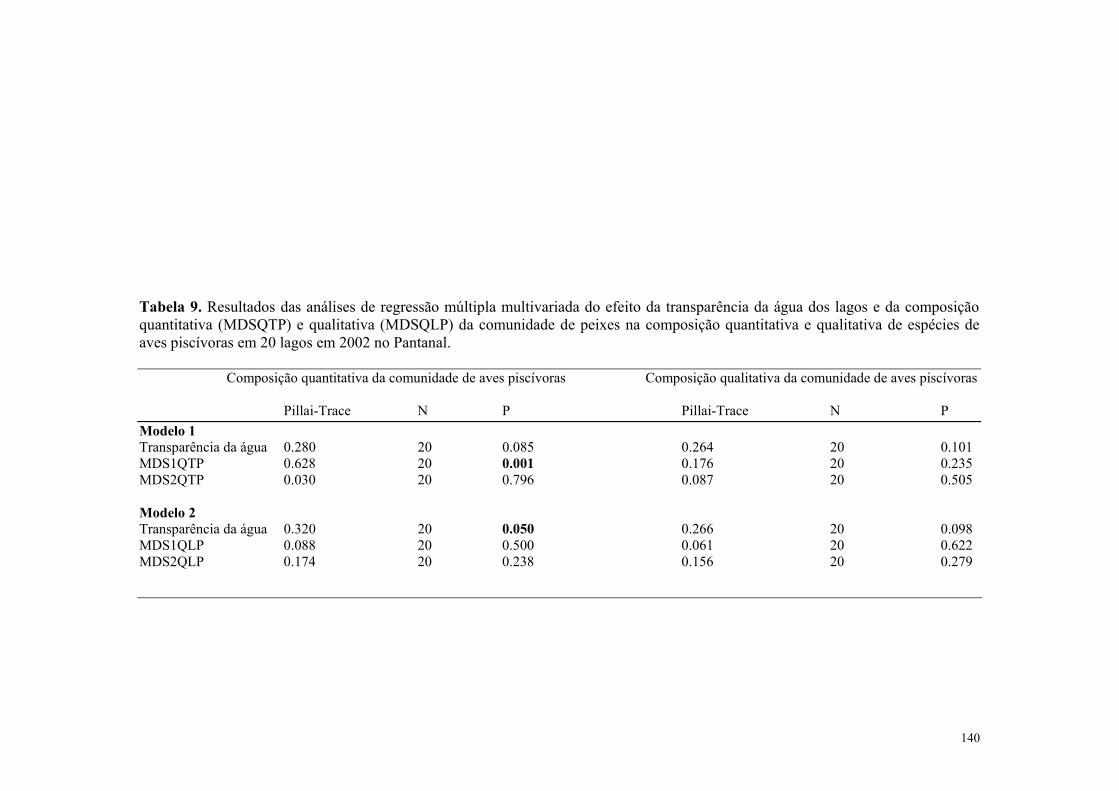
140
Tabela 9. Resultados das análises de regressão múltipla multivariada do efeito da transparência da água dos lagos e da composição quantitativa (MDSQTP) e qualitativa (MDSQLP) da comunidade de peixes na composição quantitativa e qualitativa de espécies de aves piscívoras em 20 lagos em 2002 no Pantanal. Composição quantitativa da comunidade de aves piscívoras Composição qualitativa da comunidade de aves piscívoras Pillai-Trace N P Pillai-Trace N P Modelo 1 Transparência da água 0.280 20 0.085 0.264 20 0.101 MDS1QTP 0.628 20 0.001 0.176 20 0.235 MDS2QTP 0.030 20 0.796 0.087 20 0.505 Modelo 2 Transparência da água 0.320 20 0.050 0.266 20 0.098 MDS1QLP 0.088 20 0.500 0.061 20 0.622 MDS2QLP 0.174 20 0.238 0.156 20 0.279
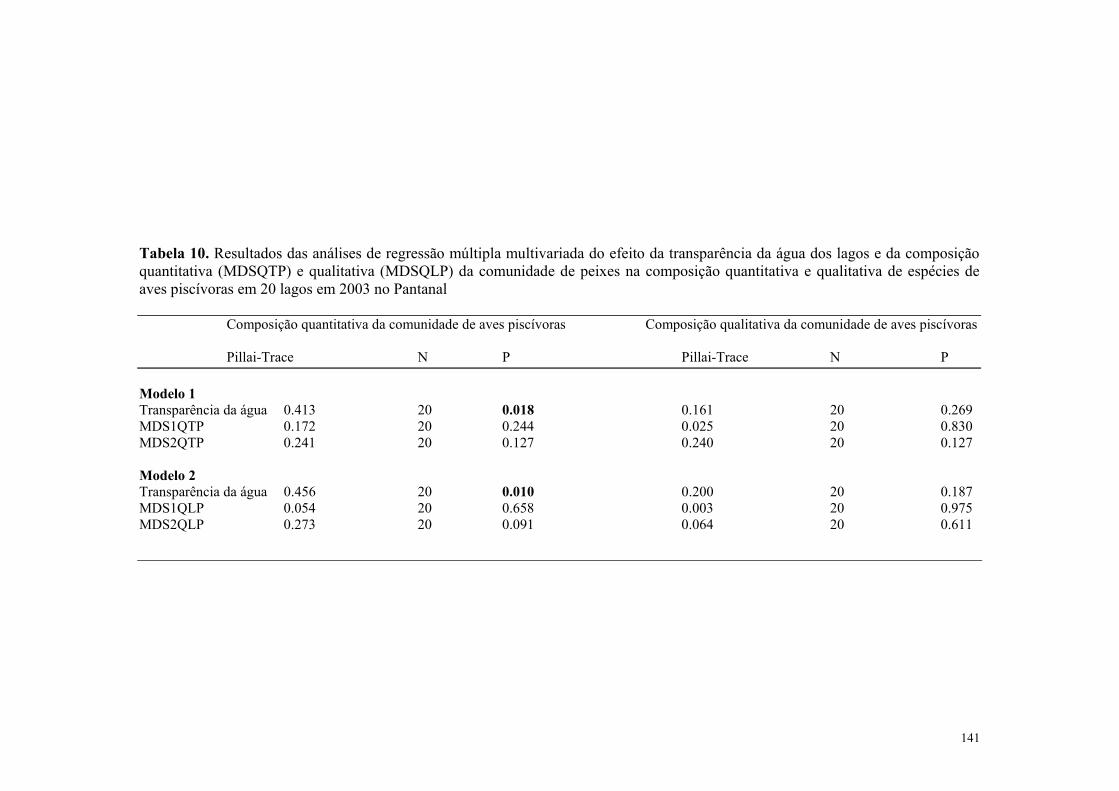
141
Tabela 10. Resultados das análises de regressão múltipla multivariada do efeito da transparência da água dos lagos e da composição quantitativa (MDSQTP) e qualitativa (MDSQLP) da comunidade de peixes na composição quantitativa e qualitativa de espécies de aves piscívoras em 20 lagos em 2003 no Pantanal Composição quantitativa da comunidade de aves piscívoras Composição qualitativa da comunidade de aves piscívoras Pillai-Trace N P Pillai-Trace N P Modelo 1 Transparência da água 0.413 20 0.018 0.161 20 0.269 MDS1QTP 0.172 20 0.244 0.025 20 0.830 MDS2QTP 0.241 20 0.127 0.240 20 0.127 Modelo 2 Transparência da água 0.456 20 0.010 0.200 20 0.187 MDS1QLP 0.054 20 0.658 0.003 20 0.975 MDS2QLP 0.273 20 0.091 0.064 20 0.611

142
4. Discussão
A Ictiofauna dos rios da América do Sul é caracterizada pela dominância de
Characiformes sobre Siluriformes (Lowe-McConnell, 1989). Contudo, em lagos e represas
essa dominância é intensificada (Agostinho et al., 1993; Súarez et al., 2001, Machado
2003). Essa dominância pode ser atribuída à capacidade dos pequenos characiformes em
explorar o oxigênio na camada superficial da lâmina da água (Súarez et al., 2001), o que até
agora foi confirmado pelo presente estudo.
A riqueza de espécies de peixes encontrada nos lagos de planície de inundação da
Amazônia é a mais alta das Américas, variando entre 132 no lago Camaleão (Junk et al.,
1983) a 155 no lago do Rei ambos próximos da cidade de Manaus (Mérona & Bittencout,
1993). Nos lagos de inundação do Pantanal, a riqueza de espécie de peixe é bem menor; no
Pantanal de Nhecolandia foram encontrados de 7 a 26 espécies por lago e a riqueza total foi
de 49 espécies (Súarez et al., 2004). Entretanto, no presente estudo realizado nos lagos do
Pantanal de Poconé o número de espécies de peixes encontrado variou entre 9 a 41. A
riqueza total foi de 79 espécies de peixes sendo a maior já registrada para a região para um
mesmo conjunto de lagos. Diversidade semelhante é encontrada para os lagos de inundação
dos Lhanos venezuelanos onde foram registradas entre 26 a 60 espécies de peixes (Lowe-
McConnell, 1989; Hoeinghaus et al., 2003).
Nestas áreas inundáveis dos trópicos variáveis abióticas como pH, oxigênio,
turbidez, transparência e profundidade afetam a distribuição de peixes (Junk, et al., 1983,
Mourão et al., 1988; Rodrigues & Lewis, 1997; Tejerina-Garro et al., 1998, Súarez et al.,
2001). Porém, à influência da heterogeneidade estrutural dos habitats e variáveis bióticas
atuando nas assembléias de peixes ainda é pouco conhecida (Petry et al., 2003, Súarez et
al., 2004).

143
Um dos principais fatores ligados à estruturação do hábitat ocupados pelas
comunidades animais aquáticos é a presença de macrófitas aquáticas (Meschiatti et al.,
2000; Agostinho et al., 2003, Petry et al., 2003). Outra é a abundância de predadores como
aves piscívoras, peixes carnívoros, mamíferos, etc (Englund & Krupa, 2000; Súarez et al.,
2004). Conseqüentemente, estes fatores atuando em conjunto podem afetar a riqueza e
abundância de peixes em determinado hábitat, todavia, no presente estudo algumas
variáveis abióticas (e.g tamanho do lago, profundidade, oxigênio dissolvido, transparência,
pH e temperatura da água dos lagos); e biótica como cobertura de macrófitas aquáticas não
tiveram influencia significativa na variação da riqueza e biomassa de peixes no local.
Catella (1992), sugere que as espécies que sobraram nos lagos são representantes
individuais que não retornaram, ou não tiveram oportunidade de retornar ao rio antes do
isolamento, ou espécies que possuem características que permitem sobreviverem nestes
ambientes que sofrem hipóxia e dissecação, refugiando-se em macrófitas ou enterrando-se
no solo, como acontece também nos lagos inundáveis da Venezuela e da Amazônia
brasileira (Allisson-Machado, 1990, Junk et al., 1983). De fato 80% das espécies de peixes
coletados nestes lagos vivem associados a bancos de macrófitas aquáticas no Pantanal
como por exemplo Markiana nigripinnis, Gymnocorymbus ternetzi, Odontostilbe
paraguaensis e Eignmannia trilineata etc.(Pazin, 2000, Willink et al., 2000) (Tabela 1;
Figura 6). Outros apresentam alta tolerância a hipóxia. Muitos peixes sul-americanos,
possuem adaptações para conviver com baixos níveis de oxigênio, estas adaptações incluem
o uso de camadas superficiais oxigenadas para respiração branquial, isto é auxiliado por
projeções vascularizadas dos lábios. Em várias espécies, órgãos respiratórios são derivados
do trato alimentar: cavidade bucal, em Hypopomus spp. (com respiração aérea obrigatória)
e Synbranchus marmoratus (com respiração aérea facultativa) que ainda pode viver

144
enterrado no solo; o estômago cheio de ar, em alguns bagres loricariideos (e.g Loricaria,
Rhineloricaria); o intestino em alguns Calictídeos (e.g Callicthys, Hoplosternum
Corydoras e Brochis) Figura 8. A bexiga natatória altamente vascularizada, é usada por
Hoplerythrynus unitaeniatus por exemplo (Junk et al., 1983, Almeida-Val & Farias, 1996;
Val, 1996, Lowe McConnel, 1999). O uso do estômago ou intestino parece ser uma
especialidade sul-americana, talvez relacionada com alimentação reduzida nos períodos de
seca, quando as adaptações respiratórias são mais necessárias (Lowe McConnel, 1999).
Provavelmente, a retenção destes peixes nos lagos, explicaria a relação significativa entre a
composição qualitativa da comunidade de peixes e o oxigênio dissolvido na água do lago
em 2003 (ver Tabela 7), pois as espécies que ficaram nestes lagos foram na sua grande
maioria as que tinham algum mecanismo para tolerar a hipóxia.
A riqueza e abundância de peixes, nos lagos tropicais estão diretamente relacionadas
com o tamanho do lago (Amarasinghe & Welcomme, 2002). O que também aconteceu no
Pantanal, quando são analisados somente os dados brutos da amostragem, nos maiores
lagos a riqueza de espécies variou entre 27 a 12, enquanto que nos menores teve uma
variação entre 8 a 5 espécies de peixes. Entretanto, como foi feita uma padronização das
amostras, dividindo a riqueza pela área de cada lago, as análises realizadas não mostraram
efeito significativo com este parâmetro.
No Pantanal a comunidade de peixes nos habitats sazonalmente inundados é
regulada pelo tempo de isolamento dos lagos, distancia deles ao rio, abundância de
predadores e não por eventos imprevisíveis, é o que sugere os resultados obtidos no
Pantanal de Nhecolandia (Súarez et al., 2004). Estes autores indicam também que os lagos
do Pantanal não é um bom modelo para predizer riqueza e regularidade de comunidade de
peixes. Isto porque os grandes lagos são geralmente rasos e desconectados dos rios. O que

145
talvez explique os resultados não significativos (para tamanho do lago, transparência, pH e
temperatura da água Tabela 7 e 8), aqui obtidos, a maioria dos lagos do Pantanal de
Poconé, não tem conectividade com os rios na maior parte do ano.
Contudo, a influencia positiva da cobertura de macrófitas na diversidade de peixes
foi considerada por Tonn & Magnusson (1982) e Pihl et al., (1994) como resultado da
diminuição da pressão biótica (competição, predação e parasitismo) e aumento da
heterogeneidade do habitat. Os resultados aqui obtidos (Tabela 7) concordam com os estes
obtidos para as zonas temperadas, e discordam com os resultados de Súarez et al., (2004).
Estes autores, argumentam que esta situação aconteceu no Pantanal porque os lagos com
ampla cobertura de macrófitas são isolados dos rios por longos períodos, aumentando assim
as condições de stress para a sobrevivência dos peixes. Contudo a amostragem que eles
fizeram foi durante um único mês, neste trabalho a amostragem foi realizada em meses
diferentes e em dois anos consecutivos, é provável que amostragem em meses de cheia e
seca tenha influenciado estes resultados, mostrando a significativa importância da cobertura
de macrófitas na composição qualitativa e quantitativa da comunidade de peixes (Tabela 7).
Súarez et al., (2004) mostraram a importância da cobertura de macrófitas para a
determinação da abundância relativa de muitas espécies representativas, indicando a
influencia da cobertura dessas plantas na dinâmica populacional de algumas espécies de
peixes. A importância da cobertura de macrófitas aquáticas para a distribuição dos peixes,
está mais relacionada com o tipo biológico da macrófita do que com a sua diversidade
(Petry et al., 2003, Pelicice et al., 2005). Assim, a abundância de algumas espécies de
macrófitas ou o conjunto delas é mais importante para o refúgio e pastejo dos peixes que
outras. O que explicaria a relação significativa entre a composição qualitativa e quantitativa
da comunidade de peixes em 2002 e cobertura de macrófitas aquáticas. Os tapetes de

146
macrófitas foram formados por duas espécies predominantes Eichhornia azurea e Pistia
stratiotes estas são do tipo flutuantes, isto ocorreu em lagos rasos. Estudos realizados nas
zonas temperadas têm mostrado uma forte correlação positiva entre a presença de algumas
espécies de macrófitas submersas e algumas aves aquáticas principalmente herbívoras e
piscívoras (Maklund et al., 2002), o que também ocorreu para lagos andinos na América do
Sul (Caziani et al., 2003).
Nos lagos do Pantanal, a composição quantitativa da comunidade de aves piscívoras
foi fortemente influenciada pela composição quantitativa (biomassa) da comunidade de
peixes considerando a primeira dimensão.
A disponibilidade de recursos alimentares é um dos fundamentos da ecologia de
comunidades para explicar a riqueza de espécies e abundância de aves em diferentes
regiões (Chow-Fraser et al., 1998; Elmberg et al., 2003). Trabalhos realizados em várias
regiões com interações aves piscívoras e peixes, demonstraram a existência de correlação
positiva entre a abundância deste recurso e a abundância de aves (Wagner, 1997;
Paskowski &Tonn, 2000). Os resultados aqui obtidos sugerem que este padrão também
acontece no Pantanal, embora outros trabalhos tenham usado abundância de peixes, neste
foi usado biomassa de cada espécie de peixe, o que poderia sugerir estas aves não teriam
preferência por determinada espécie de peixes e sim a pela disponibilidade e facilidade em
captura-los. Mas isto merece mais estudos, pois, outros trabalhos indicam que algumas
espécies de aves piscívoras têm uma preferência por determinados tamanhos e espécies de
peixes (Thomas, 1985, Willard, 1985; Oliveira, 1997; Da Silva et al., 2002)
A transparência da água dos lagos afetou a composição quantitativa da comunidade
de aves piscívoras, isto indica que com a mudança de transparência da água ocorre uma
variação na composição da comunidade destas aves. As aves mergulhadoras são as mais

147
prejudicadas com o aumento da turbidez da água, isto acontece de modo geral com a
diminuição da profundidade dos lagos. A turbidez da água, tem sido utilizada por
piscicultores europeus, como medida preventiva para diminuir a predação por parte destas
aves nos tanques de piscicultura (Cezilly, 1992), quando as aves vadeadoras ou pernaltas
adentram em grande número nestes lagos, revolvendo o sedimento e diminuindo a
transparência da água. No entanto, estas aves, que neste período do ano formam grandes
bandos no Pantanal, como o Cabeça-seca (Mycteria americana) o Tuiuiú (Jabiru mycteria)
e as garças (Ardea alba e Egretta thula) usam igualmente tática visual e táctil para
forragear ou usam ambas simultaneamente dependendo da qualidade da água (Thomas,
1985).

148
Figura 1. Coleta e triagem dos peixes a) peneirão; b) triagem. Algumas das espécies de peixes mais abundantes nos lagos no Pantanal:, c) Camboatázinho (Corydoras hastatus); d) Douradinho (Aphyocarax rathbuni); e) Pirrhulina australis. Fotos: Dalci Oliveira

149
1
Figura 6. Algumas espécies de peixes que vivem associados a macrófitas aquáticas nos lagos no Pantanal: a) Tetra-preto (Gymnocorymbus ternetzi); b) Pacu-peva (Metynis mola); c) Mato-grosso (Hypssobrycon equens); d) Sauá (Markiana nigripinnis); e) Cará (Chaetobranchopis australis); f) Acaraçu (Astronotus ocellatus). Fotos: Dalci Oliveira

150
Figura 7. Algumas espécies de peixes que resistem a hipóxia nos lagos do Pantanal: a) Camboatazinho (Brochis splendens); b) Jeju (Hopleterinus unitaeniatus); c) Camboatá (Hoplosternum sp); d) Lobo (Hoplias malabaricus); e) e f) Tuviras (Eigenmannia spp). Fotos: Dalci Oliveira

151
Tabela 1. Lista das espécies de peixes coletados nos lagos do Pantanal nos anos de 2002 e 2003 com algumas características como, Amplitude de Comprimento Total em cm (CT); Atividade alimentar (AA): diurna(D), noturno(N) e Crepuscular(C), e associação com macrófitas aquáticas (AMA).
2002 2003 Família/espécie Nome comum CT* AA* AMA**Cheia Seca Cheia Seca CHARACIFORMES CHARACIDAE Triportheus nematurus ( Kner, 1860 ) Sardinha 9.0-12.0 D X x x x X Tetragonopterus argenteus (Curvier, 1817) Sauá 11 D x X Markiana nigripinnis (Perugia, 1991) Lambari-campo 10.4 D X x x x X Bryconamericus sp 5.7 x x X Gymnocorymbus ternetzi (Boulerger, 1895) Tetra-preto 3.3-7.0 C X x x x X Moenkhausia sanctaefilomenae (Steidachner, 1907) Lambari 7 X x x x X Moenkhausia dichroura (Kner, 1858) Lambari 10 X x x x X Astyanax bimaculatus (Linnaeus, 1758) Lambari 15 X x x x X Hemigrammus ulreyi (Boulerger, 1859) Lambari 1.7-1.9 D X x x x X Phenacogaster tegatus (Eigenmann, 1911) Lambari 2.1-3.5 C x x x X Psellogrammus kennedyi (Eigenmann, 1903) Lambari 5.5 x x x X Hyphessobrycon eques (Steidanchner, 1882) Mato-grosso 2.0-3.0 D X x x x X Prionobrama paraguayensis (Eigenmann, 1914) 5 X x x x Aphyocharax rathbuni Eigenmann, 1907 Douradinho 2.6 X x x x X Odontostilbe calliura (Boulerger, 1900) Piquira 2.0-3.6 D X x X Cyanopotamus kincaidi (Schultz, 1950) Saicanga 4.5-15.0 N x x x X Roeboides bonariensis Steidanchner, 1879 Saicanga 20 N x x x X Roeboides prognathus Boulerger, 1895 Saicanga 12 N x X Charax leticiae Lucena, 1987 Saicanga 10 Acestrorhynchus pantaneiro Menezes, 1992 Peixe-cachorro 9.0-18.0 D X x x Poptella paraguayensis (Eigenmann, 1907) Saia-branca 3.1-4.5 D X x x x X Metynnis mola Eigenmann & Kennedy, 1903 Pacu-peva 4.0-6.5 D X x x x X Serrasalmus spilopleura Kener,1860 Piranha 3.5-9.5 D X x x x
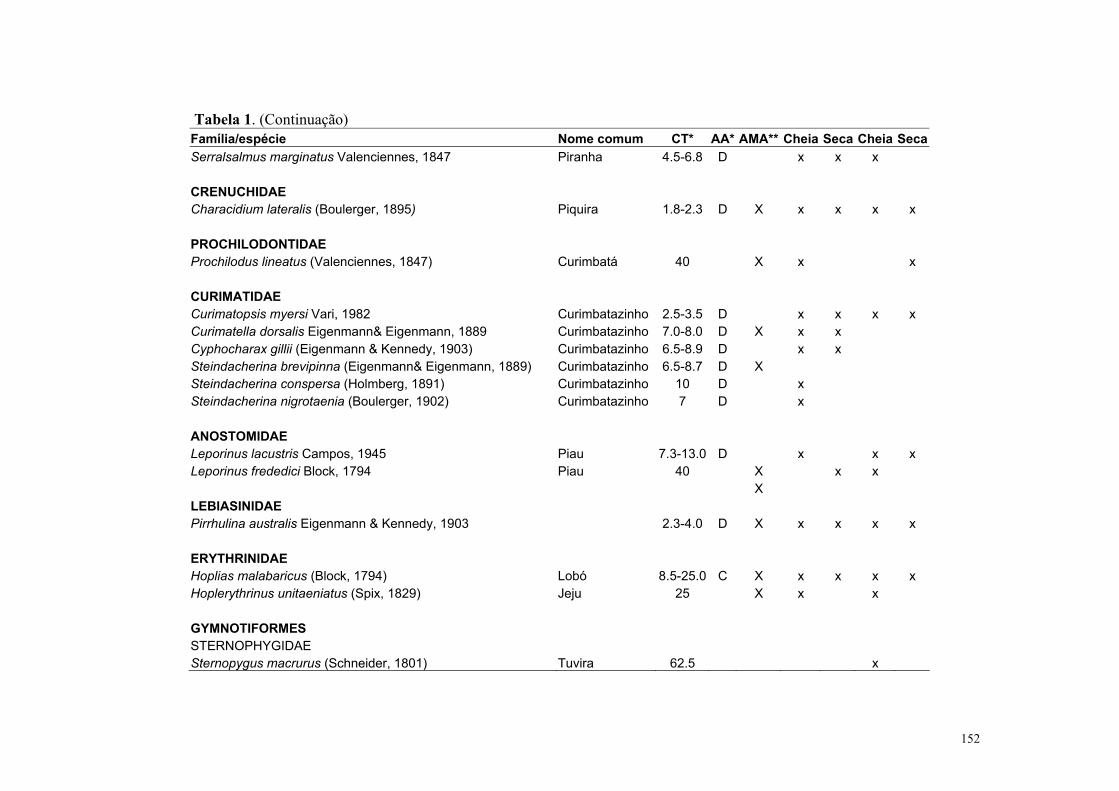
152
Tabela 1. (Continuação) Família/espécie Nome comum CT* AA* AMA** Cheia Seca Cheia SecaSerralsalmus marginatus Valenciennes, 1847 Piranha 4.5-6.8 D x x x CRENUCHIDAE Characidium lateralis (Boulerger, 1895) Piquira 1.8-2.3 D X x x x x PROCHILODONTIDAE Prochilodus lineatus (Valenciennes, 1847) Curimbatá 40 X x x CURIMATIDAE Curimatopsis myersi Vari, 1982 Curimbatazinho 2.5-3.5 D x x x x Curimatella dorsalis Eigenmann& Eigenmann, 1889 Curimbatazinho 7.0-8.0 D X x x Cyphocharax gillii (Eigenmann & Kennedy, 1903) Curimbatazinho 6.5-8.9 D x x Steindacherina brevipinna (Eigenmann& Eigenmann, 1889) Curimbatazinho 6.5-8.7 D X Steindacherina conspersa (Holmberg, 1891) Curimbatazinho 10 D x Steindacherina nigrotaenia (Boulerger, 1902) Curimbatazinho 7 D x ANOSTOMIDAE Leporinus lacustris Campos, 1945 Piau 7.3-13.0 D x x x Leporinus frededici Block, 1794 Piau 40 X x x X LEBIASINIDAE Pirrhulina australis Eigenmann & Kennedy, 1903 2.3-4.0 D X x x x x ERYTHRINIDAE Hoplias malabaricus (Block, 1794) Lobó 8.5-25.0 C X x x x x Hoplerythrinus unitaeniatus (Spix, 1829) Jeju 25 X x x GYMNOTIFORMES STERNOPHYGIDAE Sternopygus macrurus (Schneider, 1801) Tuvira 62.5 x

153
Tabela 1. (Continuação) Família/espécie Nome comum CT* AA* AMA** Cheia Seca Cheia SecaEigenmannia trilineata Lopez & Castello, 1966 Tuvira 21.2 x x x x Eigenmannia virescens (Valenciennes, 1847) Tuvira 7.5-16.5 N Eigenmannia sp Tuvira x x HYPOPOMIDAE Hypopomus spB Tuvira 8.0-12.0 N x x x x
SILURIFORMES TRICHOMYCTERIDAE Trichomycterus amazonicus Steindachner, 1883 2.3-35.0 N X x x x x PIMELODIDAE Rhamdia sp 9.5-15.3 N x Pimelodella gracilis (Valenciennes, 1840) Chum-chum 4.2-7.9 N x x Pimelodella mucosa Eigenmann & Ward, 1907 Chum-chum 13.5 x x x AUCHENIPTERIDAE Entomocorus benjamini Eigenmann, 1917 3.5-5.9 N X x x Trachelyopterus coriaceus Valenciennes, 1840 10 x x x Parauchenipterus striatulus (Steindachner, 1876) Jauzinho 9.0-16.0 N x x Paurachenipterus galeatus (Linnaeus, 1766) Jauzinho 20 x x x DORADIDAE Gen nov. 70 X x x Anadoras weddelii (Castelnau, 1855) x x Platydoras armatulus (Valenciennes, 1840) Roque-roque 20 x x x SCOLAPLACIDAE Scoloplax empousa Schaefer, Weitzman & Britski, 1989 1.3-1.65 N X x
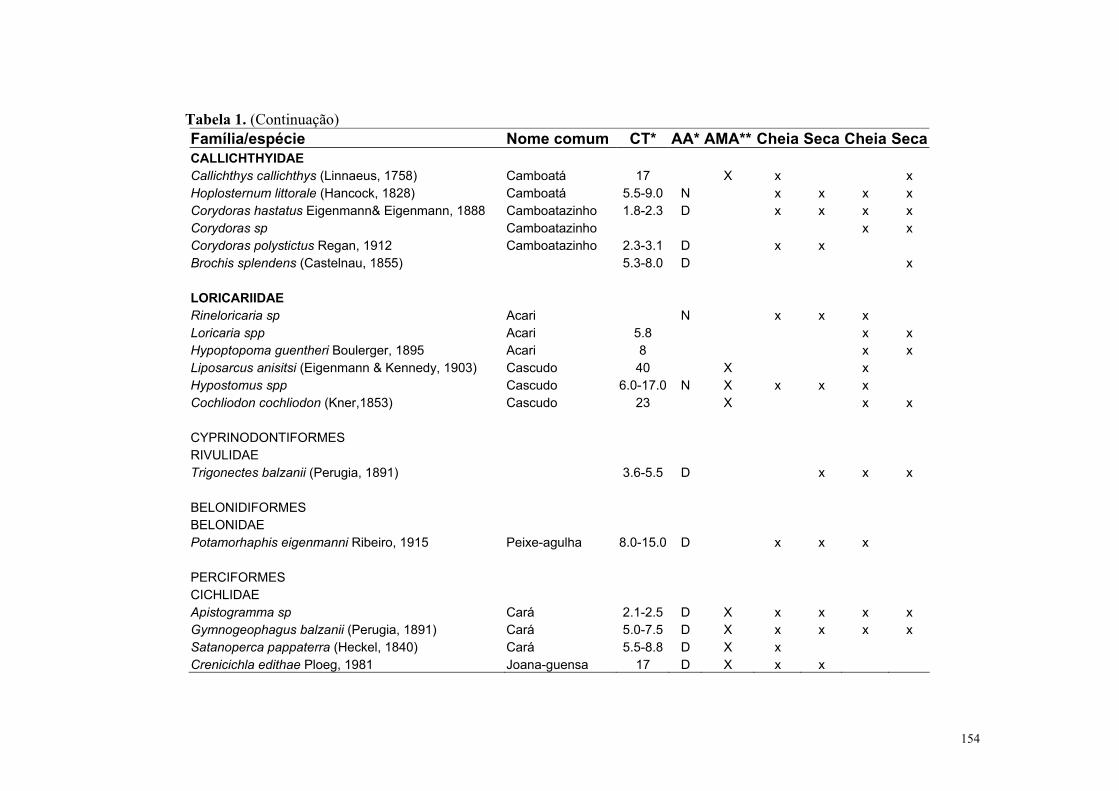
154
Tabela 1. (Continuação) Família/espécie Nome comum CT* AA* AMA** Cheia Seca Cheia SecaCALLICHTHYIDAE Callichthys callichthys (Linnaeus, 1758) Camboatá 17 X x x Hoplosternum littorale (Hancock, 1828) Camboatá 5.5-9.0 N x x x x Corydoras hastatus Eigenmann& Eigenmann, 1888 Camboatazinho 1.8-2.3 D x x x x Corydoras sp Camboatazinho x x Corydoras polystictus Regan, 1912 Camboatazinho 2.3-3.1 D x x Brochis splendens (Castelnau, 1855) 5.3-8.0 D x LORICARIIDAE Rineloricaria sp Acari N x x x Loricaria spp Acari 5.8 x x Hypoptopoma guentheri Boulerger, 1895 Acari 8 x x Liposarcus anisitsi (Eigenmann & Kennedy, 1903) Cascudo 40 X x Hypostomus spp Cascudo 6.0-17.0 N X x x x Cochliodon cochliodon (Kner,1853) Cascudo 23 X x x CYPRINODONTIFORMES RIVULIDAE Trigonectes balzanii (Perugia, 1891) 3.6-5.5 D x x x BELONIDIFORMES BELONIDAE Potamorhaphis eigenmanni Ribeiro, 1915 Peixe-agulha 8.0-15.0 D x x x PERCIFORMES CICHLIDAE Apistogramma sp Cará 2.1-2.5 D X x x x x Gymnogeophagus balzanii (Perugia, 1891) Cará 5.0-7.5 D X x x x x Satanoperca pappaterra (Heckel, 1840) Cará 5.5-8.8 D X x Crenicichla edithae Ploeg, 1981 Joana-guensa 17 D X x x

155
Tabela 1. Continuação) Família/espécie Nome comum CT* AA* AMA** Cheia Seca Cheia SecaCrenicichla vittata Heckel, 1840 Joana-guensa 10.3-16.8 D X x x x Bujurquina vittata (Heckel, 1840) Cará 4.0-5.9 D X x x x Aequidens plagiozonatus Kullander, 1984 Cará 10.3 D X x x x x Laetacara dorsigera (Heckel, 1840) Cará 2.9-4.0 D X x x x Astronotus ocellatus (Cuvier,1829) Acará-açu 13.0-19.0 D x x x x Mesonauta festivus (Heckel, 1840) Cará 4.6-7.9 D X x x x x Cichlasoma dimerus (Heckel, 1840) Cará 5.7-8.3 D X x x x SYNBRANCHYFORMES SYNBRANCHYDAE Synbranchus marmoratus Block, 1795 Mussum 5.5-21.0 N X x x x x PLEURONECTIFORMES ARCHIRIDAE Catathyridium jenynsii (Gunther, 1862) Solha 2-21.0 D x
*Machado, 2003; ** Pazin, 2000 e Willik et al., 2000.

156
Tabela 2. Lista das espécies de aves piscívoras observadas em 20 lagos no Pantanal em 2002 e 2003, com algumas das suas características: ocorrência no local: Sazonal (S), Residente (Re) e Migrante (Mi); modo de forragear: Vadeadoras ou pernaltas (V), Mergulhadoras (M), buscadoras ou cruzadoras (C); Modo de nidificar: em Colônias ou Ninhal (N) e Solitária (S).
Família/espécie Nome Comum Dieta¹ Ocorrência
Modo de forragear Modo de nidificar
PHALACROCORACIDAE Phalacrocorax brasilianus (Gmelin, 1789) Biuá P S M N ANHINGIDAE Anhinga anhinga (L.,1766) Biguatinga P S M N ARDEIDAE Tigrisoma lineatum (Boddaert, 1783) Socó-boi P S V S Nycticorax nycticorax (L.,1766) Garça-dorminhoca P S V N Butorides striatus (L., 1758) Socozinho P S V S Ardea cocoi L.,1766 Baguari P S V N Ardea alba (L., 1758) Garça-branca-grande P S V N Pilherodius pileatus (Boddaert, 1783) Garça-real P S V S Egretta thula (Molina, 1782) Garça-branca-pequena P S V N Egretta caerulea (L., 1758) Garça-azul P S V N* THRESKIORNITHIDAE Pratalea ajaja (L., 1758) Colhereiro P S V N CICONIIDAE Mycteria americana (L.,1758) Cabeça-seca P S V N Ciconia maguari (Gmelin, 1789) Tabuiaiá P S V S Jabiru mycteria (Lichtenstein, 1819) Tuiuiú P S V S
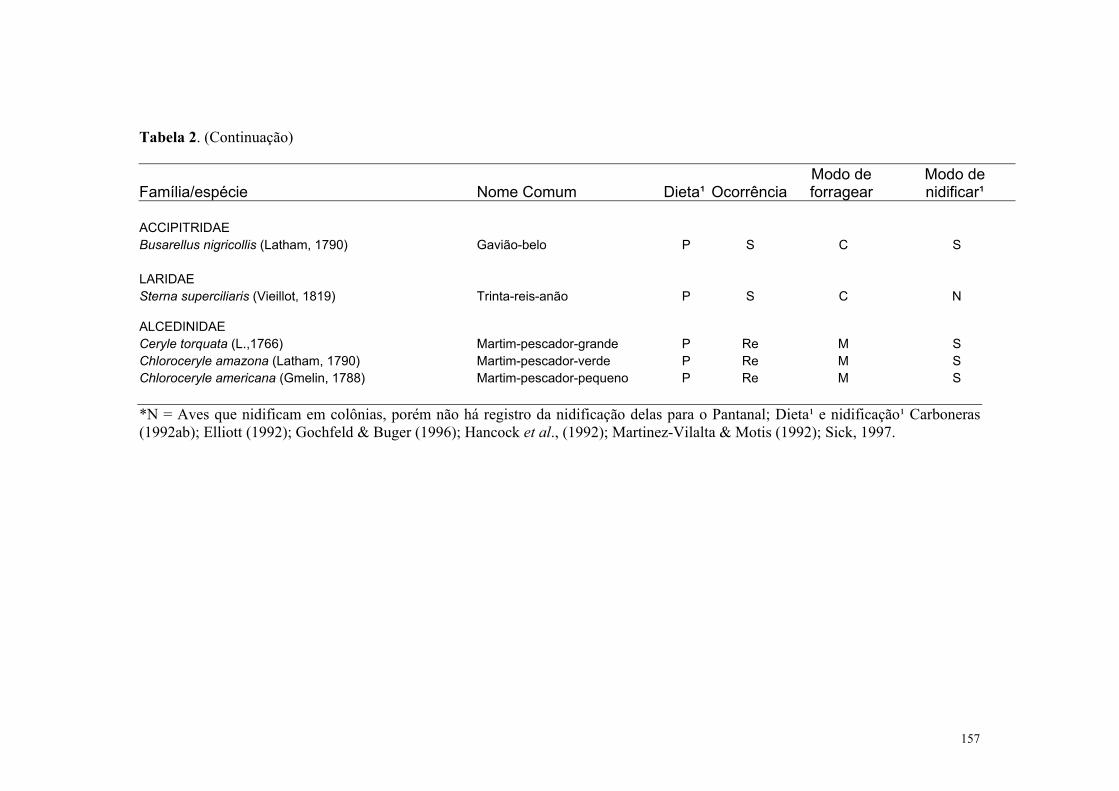
157
Tabela 2. (Continuação) Família/espécie Nome Comum Dieta¹ Ocorrência
Modo de forragear
Modo de nidificar¹
ACCIPITRIDAE Busarellus nigricollis (Latham, 1790) Gavião-belo P S C S LARIDAE Sterna superciliaris (Vieillot, 1819) Trinta-reis-anão P S C N ALCEDINIDAE Ceryle torquata (L.,1766) Martim-pescador-grande P Re M S Chloroceryle amazona (Latham, 1790) Martim-pescador-verde P Re M S Chloroceryle americana (Gmelin, 1788) Martim-pescador-pequeno P Re M S *N = Aves que nidificam em colônias, porém não há registro da nidificação delas para o Pantanal; Dieta¹ e nidificação¹ Carboneras (1992ab); Elliott (1992); Gochfeld & Buger (1996); Hancock et al., (1992); Martinez-Vilalta & Motis (1992); Sick, 1997.

158
Conclusões e considerações finais
Os resultados das análises obtidas no presente estudo permitem concluir que, de um modo
geral, fatores bióticos e abióticos podem determinar mudanças significativas na composição
da comunidade das aves aquáticas e piscívoras no Pantanal.
A composição (qualitativa e quantitativa) da comunidade de aves aquáticas e
piscívoras mudam sazonalmente entre os meses no Pantanal, e esse padrão pode não
ocorrer entre intervalos curtos de anos, porém, podendo ocorrer á longo prazo.
A composição quantitativa da comunidade das aves aquáticas e piscívoras é
significativamente influenciada pela variação anual da pluviosidade no Pantanal; com a
diminuição das chuvas aumenta a variação na comunidade de aves aquáticas e piscívoras.
A composição qualitativa da comunidade das aves piscívoras é significativamente
influenciada pela variação da temperatura do ar no Pantanal; nos meses mais quentes do
ano, aumenta a variação nesta comunidade de aves.
A composição (qualitativa e quantitativa) da comunidade de aves piscívoras no
Pantanal é afetada significativamente por fatores abióticos como profundidade; quanto mais
rasos ficam os lagos a diversidade e abundância das aves aumenta. A transparência da água
dos lagos tem efeito similar, com o aumento da turbidez aumenta a riqueza e abundância
das aves piscívoras. A composição da comunidade de peixes é o único fator biótico a ter
efeito significativo na mudança da comunidade de aves piscívoras no Pantanal, quanto mais
peixes mais diversa e abundante é a comunidade de aves piscívoras.

159
A composição (qualitativa e quantitativa) da comunidade de aves piscívoras no
Pantanal não é direta e significativamente afetada por fatores abióticos como tamanho do
lago, % de oxigênio dissolvido na água, temperatura e pH da água dos lagos, e biótico
como cobertura de macrófitas aquáticas. Porém esses fatores devem influir indiretamente
na composição da comunidade de peixes, que tem influencia direta na composição de aves
piscívoras, entretanto são necessários mais estudos de longo prazo destas interações
testando este e outros modelos para tentar entender que fatores determinam a dinâmica das
comunidades de aves aquáticas e piscívoras no Pantanal.

160
Bibliografia Citada
Ab’Sáber, A. 1988. O Pantanal Mato-grossense e a teoria dos refúgios. Rev. Brasil. Geogr.,
1(2): 9-57
Almeida-Val, V.M.F; Farias, I.P. 1996. Respiration in fish of the Amazon: Metabolic
adjustments to chonic hypoxia 20. in: p257-271 Val, A.L; Almeida-Val, V.M.F.;
Randall, D.J.(eds.). Physiology and Biochemistry of the fishes of Amazon. INPA,
Manaus.
Adamoli, J. 1981. O Pantanal e suas relações fitogeográficas com os cerrados. in: Anais do
XXXII Cong. Nac. Botânica. Teresina p.109-119
Adis, J.; Marques, M.I.; Wantzen, K.M. 2001. First observations on the survival strategies
of terricolous arthropods in the northern Pantanal wetland of Brazil. Andrias, 15: 127-
128
Agostinho, A.A; Gomes, L.C.; Julio Jr., H.F. 2003. Relações entre macrófitas aquáticas e
fauna de peixes. p.261-279. in Thomas, S.M.; Bini, L.M. (eds.) 2003. Ecologia e
Manejo de macrófitas aquáticas. Eduem, Maringá.
Alho, C.J.R. 1986. Manejo de fauna silvestre. in: Anais do I Simpósio sobre recursos
naturais e sócio-econômicos do Pantanal. Corumbá/Brasília. Embrapa-DDT. p.183-
197.
Alho, C.J.R.; Lacher, T.E.; Gonçalves, H.C. 1988. Environmental degradation in the
Pantanal Ecosystem. BioScience, 38(3): 164-171
Alho, C.J.R.; Rondon, N.L. 1987. 1987. Habitat, populations densities, and social structure
of capybaras (Hydrochaeris hydrochaeris, Rodentia) in the Pantanal, Brasil. Rev.
bras. Zool. 4(2): 71-88.

161
Alho, C.J.R.; Viera, L. 1997. Fish and wildlife resouses in the Pantanal of Brazil and
potential disturbances from the of environmental contaminants. Environm. Toxicol.
Chemistry., 16(1):71-74.
Alves, M.A.S.; Pereira, E.F. 1998. Richness, abundance and seasonality of birds species in
lagoon of urban area (Lagoa Rodrigo de Freitas) of Rio de Janeiro, Brazil.
Ararajuba., 6(2): 110-116
Alves, M.S.A.; Silva, J.M.C. 2000. A Ornitologia no Brasil: Desenvolvimento, Tendências
atuais e Perspectivas. p. 327-344 in: Alves, M.S.A.; Silva, J.M.C.; Sluys, M.V.;
Bergallo, H.G.; Rocha, C.F.D. (orgs.). A ornitologia no Brasil, pesquisa atual e
perspectivas. Eduerj, Rio de Janeiro.
Allouche, S.; Gaudin, P. 2001. Effects of avian predation threat, water flow and cover on
growth and habitat use by chub, Leuciscus cephalus, in an experimental stream. Oikos,
94: 481-492
Amarasinghe, U.S.; Welcomme, R.L. 2002. An analysis of fish species richness in natural
lakes. Environm. Biol. Fish.,65: 327-339
Amorim, M.A; Adis, J. 1996. Consumo de alimentos por gafanhoto neotropical, Stenacris
fissicauda fissicauda (BRUNER, 1908) (Orthoptera: Acrididae) da várzea
Amazônica. Acta Amazonica, 24(3/4): 289-302
Andersson, G.; Nilsson, L. 1999. Autumn waterfowl abundance in Lake Ringsjön, 1968-
1996. Hydrobiologia, 404-41-51
Andrade, M.A.; Leite, E.B.; Carvalho, C.E. 2001. Predação de jovem do jacaré-do-pantanal
(Caiman yacare) pelo gavião-padre (Busarellus nigricollis) no Pantanal Sul Mato-
grossense, Brasil: um registro fotográfico. Tangara, 1(2): 88-89

162
Antas, P.T.Z. 1994. Migration and other movements among the lower Paraná River valley
wetlands, Argetina, and the south Brazil/Pantanal. Bird Conservation International,
4:181-190
Antas, P.T.Z.; Nascimento, I.L.S. 1996. Sob os céus do Pantanal: biologia e conservação
do tuiuiú . São Paulo. Empresa das Artes. 169p. il.
Antas, P.T.Z., Palo Jr., H. 2003. Aves do Pantanal, guia da RPPN Sesc Pantanal. Rio de
Janeiro
Araújo, A.C.; Sazima, M. 2003. The assemblages of flowers visited by hummerbirds in the
capões of Southern Pantanal, Mato Grosso do Sul. Brazil. Flora, 198: 427-435
Araújo-Lima, C.A.R.M.; Portugal, L.P.S.; Ferreira, E.G. 1986. Fish-macrophyte
relationship in the Anavilhanas Archipelago, a black water system in the Central
Amazon. J. Fish Biol., 29: 1-11
Azevedo Júnior, S.M.; Larrazabal, M.E. 1999. Captura e anilhamento de Calidris pusilla
(Scolapacidae) na costa de Pernambuco. Ararajuba, 7(2): 63-69
Bancroft, G.T.; Gawlik, D.E.; Rutchey, K. 2002. Distribution of wading birds to vegetation
and water depths in the Northern Everglades of Florida, USA. Waterbirds, 25(3): 265-
391
Batirola, L.D.; Marques, M.I.; Adis, J.; Brescovit, A.D. 2004. Aspectos ecológicos da
comunidade de Araneae (Arthopoda, Arachnidae) em copas de palmeiras Attalea
phalerata Mart.(Aracaceae) no Pantanal de Poconé, Mato Grosso, Brasil. Rev. Brasil.
Entomologia, 48(3):421-430
Batirola, L.D.; Marques, M.I.; Adis, J.; Delabie, J.H. 2005. Composição da comunidade de
Formicidae (Insecta, Hymenoptera) em copas de Attalea phalerata Mart.(Aracaceae) no
Pantanal de Poconé, Mato Grosso, Brasil. Rev. Brasil. Entomologia, 49(1):107-117

163
Belbin, L. 1992. PATN Pattern Analysis Package. CSIRO, Canberra, Australia
Benine, R.C., Castro, R.M.C.; Sabino, J. 2004. Moenkhausia bonita: A new small characin
fish from the Rio Paraguay basin, Southwestern Brazil (Characiformes: Characidae).
Copeia, 1:68-73
Bennetts, R.E.; Kitchens, W.M. 2000. Factors influencing movement probabilities of a
nomadic food specialist: proximate foraging benefits or ultimate gains from
exploration? Oikos, 91: 459-467
Bierregaard Jr., R. O. 1990. Species composition and trofic organization of the understory
birds community in a Central Amazonian terra firme forest. in: Gentry, A.H (ed.)
Four Neotropical Rainforests. Yale Univ. Press. New Haven
BirdLife International, 2004. Threatened birds of the world. Cambridge/BirdLife
International.
Boere, G.C.; Taylor, D. 2004. Global and regional governmental policy and treaties as tools
towards the mitigation of the effect of climate change on waterbirds. Ibis, 146
(Suppl.1):111-119
Bos, D.; van de Koppel, J.; Weissing, F.J. 2004. Dark-billied Brent geese agrégate to COPE
with increased levels of primary production. Oikos, 107: 485-496
Bouton, S.; Bouton, T. 2001. Interspecific interactions and nest building behavior in Buff-
necked Ibis in the Brazilian Pantanal. Waterbirds, 24 (2): 294-295
Bouton, S.; Frederick, P. 2003. Stakeholders`perceptions of a wading birds colony as a
community resource in the Brasilian Pantanal. Conserv. Biol., 17 (1): 297-306.
Branco, J.O. 2001. Descartes da pesca do camarão sete-barbas como fonte de alimento para
aves marinhas. Rev. Brasil. Zool., 18(1): 293-300

164
Branco, J.O. 2002. Flutuações sazonais na abundância de Phalacrocorax brasilianus
(Gmelin) no estuário do Saco da Fazenda, Itajaí, Santa Catarina, Brasil. Rev.Brasil.
Zool., 19(4): 1057-1062
Branco, J.O. 2003. Reprodução de Sterna hirundinacea Lesson e S. eurygnatha Sauders
(Aves, Laridae), no litoral de Santa Catarina, Brasil. Rev. Brasil. Biol., 20 (4): 655-659
Branco, J.O.; Ebert, L.A. 2002. Estrutura populacional de Larus dominicanus Lichtenstein,
1823 no estuário do Saco da Fazenda, Itajaí, Santa Catarina, Brasil. Ararajuba, 10 (1):
Britski, H.A.; Silimon, K.Z.S.; Lopes, B.S. 1999. Peixes do Pantanal. Manual de
identificação. Embrapa/Cpac. Brasilia/Corumbá.
Browder, S.F.; Johnson, D.H.; Ball, I.J. 2002. Assemblages of feeding birds as indicators of
grassland condition. Ecological Indicators, 2: 257-270
Brown Jr., K. S. 1986. Zoogeografia da região do Pantanal Mato-grossense in: I Simpósio
sobre recursos naturais e sócio-econômicos do Pantanal. Anais... Corumbá/Brasília.
Embrapa-DDT. 137-178.
Bryan, J.C; Miller, S.J.; Yates, C.S; Minno, M. 2003. Variation in size and location of
wading birds colonies in the upper St. Johns River Basin, Florida, USA. Waterbirds, 26
(2): 239-251
Bugoni, M.; Vooren, C.M. 2004. Feeding ecology of the Common Tern Sterna hirundo in a
wintering area in southern Brazil. Ibis, 146: 438-453
Burger, J. 1984. Colony stability in Least Terns. Condor, 86(1): 61-67
Callil, C.; Jung, W.J. 1999. Concentração e incorporação de mercúrio por moluscos
bivalves Anadontites trapesialis (LAMARCK, 1819) e Castalia ambigua
(LAMARCK, 1819) no Pantanal de Poconé-MT, Brasil. Biociências, 7(1): 3-28

165
Callil, C.; Jung, W.J. 2001. Aquatic gastropods as mercury indicators in the Pantanal of
Poconé region (Mato Grosso, Brazil). Water, Air and Soil Pollution, 319: 319-330
Campos Filho, L.V.; Machado, M.F.R. 1999. Quando a modernidade não é adequada a
todos os lugares: O caso dos diques no Pantanal de Poconé-MT. in: II Simpósio sobre
recursos naturais e sócio-econômicos do Pantanal. Anais... Corumbá: Embrapa
Pantanal. p 487-495
Campos, Z.M.S. 1993. Effect of habitat on survival of eggs and sex ratio of hatchlings of
Caiman crocodilus yacare in the Pantanal, Brazil. J. Herpetol., 27(2): 127-132
Campos, Z.M.S.; Coutinho, M.E. 2004. Levantamento aéreo de ninhos de tuiuiú, Jabiru
mycteria, no Pantanal Sul. Bol. Pesq. Desenvol., 56: 1-14. Disponível em
www.cpap.embrapa.br/publicações/online/BP56. acessado em 7/12/04
Carboneras, C. 1992a. Family Anhimidae p.528-535 in: Del Hoyo, J.; Elliott, A.; Sargatal,
J. (eds.). 1992. Handbook of the Birds of the World.-Ostrich to Ducks. Lynx edicions.
Barcelona
Carboneras, C. 1992b. Family Anatidae p.536-628 in: Del Hoyo, J.; Elliott, A.; Sargatal, J.
(eds.). 1992. Handbook of the Birds of the World.-Ostrich to DucksI. Lynx edicions.
Barcelona
Carpentier, A.; Paillisson, J.M.; Marion, L. 2003. Assessing the interation between
cormorants and fisheries. P187-195 in: Cowx, I.G. (ed.). Interactions between fish
and birds: Implications for management. Blackwell Publishing Company. Oxford.
376p.
Cars, D.N.; Elstron, D.A 2003. Patterns of association between algae, fishes and Grey
Herons Ardea cinerea in the rocky littoral zone of a Scottish sea loch. Estuarine
Coastal and Shelf Science, 58: 265-277

166
Casiatti, L.; Mendes, H.F.; Ferreira, K.M. 2003. Aquatic macrophytes as feeding site for
small fishes in the reservoir, Paranapanema River, Southeastern Brazil. Braz. J. Biol.,
63(2): 213-222.
Catella, A C. 1992. Estrutura da comunidade e alimentares dos peixes da Baia do Onça,
uma lagoa do Pantanal do Rio Aquidauana-MS. Dissertação de Mestrado. Unicamp.
Campinas, São Paulo.215p.
Cavalcanti, R.B.; Alves, M.A.S. 1997. Effects of fire on savanna birds in Central Brazil
savanna. Orn. Neotrop., 8(1): 85-87
Caziani, S.M.; Derlindati, E.J.; Tálamo, A.; Sureda, A.L.; Trucco, C.E.; Nicolossi, G. 2001.
Waterbird richeness in altiplano wetlands of northwestern Argentina. Waterbirds,
24(1): 103-117
Cintra, R. 1985. Nascimento de filhotes de Caiman yacare (Daudin, 1802) (Crocodylia,
Alligatoridae) em condições seminaturais no Pantanal Mato-grossense. Pap. Avulsos
Zool. São Paulo 36: 91-101
Cintra, R. 1988. Nesting Ecology of the Paraguayan Caiman (Caiman yacare) in the
Brasilian Pantanal. J. Herpetol,. 22 (2): 219-222
Cintra, R. 1989. A comparasion of sizes of caiman in hunted and non hunted areas in
Brazilian Pantanal. Proc. Working Meeting Crocodile Specialist Group., 8: 125-127
Cintra, R. 1997. Spatial distribution and foraging tactics of Tyrant Flycatchers in two
habitats in the Brasilian Amazon. Stud. Neotrop. Fauna Environm., 32: 17-27
Cintra, R.; Antas, P.T.Z. 1996. Distribuição geográfica, história natural e conservação das
espécies de aves da região do Pantanal no Brasil. in: Resumos do II Simpósio sobre
Recursos Naturais e Sócio-Econômicos do Pantanal. Corumbá. Embrapa-SPI 200p.

167
Cintra, R.; Santos, P.M.R.; Leite, C.B. no prelo. Composition and structure of the lacustrine
birds communities of seasonally flooded wetlands of western Brazilian Amazonia.
Cintra, R.; Sanaiotti, T.; Cohn-Haft, M. 2005. Spatial distribution and habitat of the
Anavilhanas Archipelago birds community in the brazilian Amazon. Biodiversity
Conserv. 1-23
Cintra, R.; Sanaiotti, T. 2005. Fire effects on the composition of birds community in na
Amazonian savanna (Brazil). Braz. J. Biol., 65 (4): 29-41
Cintra, R.; Yamashita, C. 1990. Habitats, abundância e ocorrência das espécies de aves no
Pantanal de Poconé, Mato Grosso, Brasil. Papeis Avulsos Zool. São Paulo, 37(1): 1-
21.
Chow-Fraser, P.; Lougheed, V; Le Thiec, V.; Crosbie, B.; Simser, L.; Lord, J. 1998. Long-
term response of the biotic community to fluctuating water levels and changes in
water quality in Cootes Paradise marsh, a degraded coastal wetland of Lake Ontario.
Wetlands Ecology Management, 6: 19-42
Cody, M.L. 1996. Bird communities in the Central Rocky Mountains. p.291-342. in: Cody,
M.L.; Smallwood, J.A.(eds.). Long-Term Studies of Vertebrate Communities.
Academic Press, San Diego
Collis, K.; Roby, D.D.; Craig, D.P; Adamany, S.; Adkins, J.Y.; Lyons, D.E. 2002. Colony
size and diet composition of piscivorous waterbirds on lower Columbia Rivers:
Implications for losses of juvenile salmonids to avian predation. Transactions of the
American Fisheries Society, 131: 537-550
Colwell, M.A. 1993. Shorebird community patterns in a seasonally dynamic estuary.
Condor, 95: 104-114

168
CBRO, 2005. Lista das aves do Brasil. Versão 22/03/2005. disponível em <
http://www.ib.usp.br/crbo>. Acesso em 22/03/2005.
Cox-Fernandes, C. 1997. Lateral migration of fishes in Amazon floodplains. Ecol.
Freswater Fish, 6: 36-44
Coulter, M.C. et al. 1991. A bibliography of Storks, ibises and Spoonbills. Aiken, Savannah
River Ecology Laboratory, Aiken, 191 p.
Coutinho, M.; Campos, Z. 1996. Effects of habitat and seasonality on densities of caiman in
southern Pantanal, Brazil. J. Trop. Ecol., 12: 741-747
Coutinho, M.; Mourão, G.M.; Silva, M.P. ; Campos, Z. 1994. The sustainable use of natural
resources and the conservation of the Pantanal wetland, Brazil. Acta Limnol. Bras., 5:
165-176
Cowx, I.G. (ed.). Interactions between fish and birds: Implications for management.
Blackwell Publishing Company. Oxford. 376p.
Crawshaw, P.G.; Schaller, G.B. 1980. Nesting of Paraguayan caiman, Caiman yacare, in
Brazil. Pap. Avulsos Zool. São Paulo 33: 283-292
Custer, T.; Orborn, R. 1978. Feeding habitats use by colonially breeding herons, egrets, and
ibises in North Carolina. Auk, 95: 733-743
Dall Armellina, A.A.; Bezic, C.R.; Gajardo, O.A. 1999. Submerged macrophyte control
with herbivorous fish in irrigation channels of semiarid Argentina. Hydrobiologia,
415: 265-269
Dasmaceno-Junior, G.A.; Semir, J; Santos, F.A.M.; Leitão-Filho, H.F. 2005. Structure,
distribution of species and inundation in riparian forest of Rio Paraguay, Pantanal,
Brazil. Flora, 200:109-135
Da Silva, C.J. 1990. Influência da variação do nível d’água sobre a estrutura e

169
funcionamento de uma área alagável do Pantanal Mato-grossense (Pantanal de
Barão de Melgaço, Município de Santo Antônio do Leverger e Barão de Melgaço,
MT), Tese de Doutorado, UFScar, São Carlos, São Paulo
Da Silva, C.J. 2000. Ecological basis for the management of the Pantanal-upper paraguay
river basin. p 97-117 in: Smits, A.J.M. et. al., New approaches to river management.
Backhuys Publishers, Leiden
Da Silva, C.J.; Esteves, F. 1995. Dinâmica das caracterizações limnológicas das baias porto
de fora e acurizal (Pantanal de Mato Grosso) em função da variação do nível da água.
Oecologia Brasiliensis, 1: 47-60
Da Silva, C.J; Pinto-Silva, V. 1989. Macrófitas aquáticas e condições fisico-químicas dos
alagados, rios e corixos ao longo da Estrada Parque Transpantaneira. Rev. Brasil.
Biol., 49(3): 691-697.
Da Silva, C.J.; Abdo, M.S.A.; Oliveira, D.M.M.; Girardi, P. 2002. Caracterização do ninhal
Corutuba, Pantanal de Barão de Melgaço, MT. in: Dantas, M. (Coord.), 3º Simpósio
sobre Recursos Naturais e Sócio-econômicos do Pantanal. Disponível em
http://www.cpap.embrapa.br/agencia/congresso/Bioticos/Silva acessado em
7/12/2004
Da Silva, J.S.V.; Abdon, M.M. 1998. Delimitação do Pantanal Brasileiro e suas sub-
regiões. Pesquisa Agropecuária Brasileira, 33: 1703-1711
Da Silva, V.M.F. 2004. O peixe-boi da Amazônia Trichechus inunguis (Sirenia:
Trichechidae). p283-290 in: Cintra, R. (Cood.) História Natural, Ecologia e
Conservação de algumas espécies de plantas e animais da Amazônia. Manaus,
EDUA/INPA/FAPEAM.

170
Davis, W.J.; Graham, D.J. 1991. The influence of food on reproductive strategies in a
monogamous kingfisher (Chloroceryle amazona). Auk, 108: 780-789
Del Lama, S.; Lopes, I.F.; Del Lama, M.A. 2002. Genetic variability and level of
differentiation among Brazilian Pantanal Wood Stork populations. Biochemical
Genetics. 40(3/4): 87-99
Dreitz, V.J.; Kitchens, W.M; DeAngelis, D.L. 2004. Effects of natal departure and water
level on survival of juvenile Snail Kites (Rostrhamus sociabilis) in Florida. Auk, 121(3):
894-903
Dubs, B. 1992. Birds of Southwestern Brazil. Catalogue and Guide to the Pantanal of Mato
Grosso and its Border Areas. Betrona. Betrona-Verlag. 164p.il.
Elliott, A. 1992. Family Ciconiidae p. 436-465 in: Del Hoyo, J.; Elliott, A.; Sargatal, J.
(eds.). 1992. Handbook of the Birds of the World.-Ostrich to Ducks. Lynx edicions.
Barcelona
Elmberg, J.; Nummi, P.; Pöysä, H.; Sjöberg, K. 2003. Breending success of sympatric
dabbling ducks in relation to population density and food resources. Oikos, 100: 333-
341
Englund, G.; Krupa, J.J. 2000. Habitat use by crayfish in stream pools: influence of
predators, depth and body size. Freshwater Biology, 43: 75-83
Epanchin, P.N.; Heath, J.A.; Frederick, P.C. 2002. Effects of fire on foraging and breeding
wading birds in the Everglades. Wilson Bull., 114(1): 139-141
Erwin, R.M. 1983. Feeding habitats of nesting wading birds: spatial use and social
influences. Auk, 100:960-970
Esler, D. 1990. Avian community response to Hydrilla invasion. Wilson Bull., 102(3):427-
440

171
Esler, D. 1992. Habitat use by piscivorous birds on a power plant cooling reservoir. J. Field
Ornith., 63(3): 241-392
Esteves, F.A. 1998. Fundamentos de Limnologia. Interciências, Rio de Janeiro, 602p. 2.ed.
Fedrizzi, C.E.; Azevedo Júnior, S.M.; Larrazábal, M.E.L. 2004. Boby mass and acquisition
of breending plumage of wintering Calidris pusilla (Linnaeus) Aves, Scolapacidae) in
the coast of Pernambuco, north-eastern Brazil. Rev. Brasil. Zool., 21(2): 249-252
Ferraz de Lima, J.A. 1986. A pesca no Pantanal de Mato Grosso. Rio Cuiabá: importância
dos peixes migradores. Acta Amazonica, 16: 87-94
Fereira, C.J.A.; Soriano, B.M.A.; Galdino, S.; Hamilton, S.K. 1994. Anthropogenic factors
affecting waters of the Pantanal wetland and associated rivers in the upper Paraguay
River basin of Brazil. Acta Linmol. Brasil., 5: 135-148
Figueira, J.E.C.; Cintra, R.; Viana, R.;Yamashita, C. 2005. Spatial and temporal patterns of
birds species diversity in the Pantanal of Mato Grosso, Brazil: implications for
conservations. Braz. J. Biol.,
Figuerola, J.; Green, A.J. 2002. Dispersal of aquatic organisms by waterbirds: a review of
past research and priorities for future studies. Freshwater Biology, 47: 483-494
Finney, S.K.; Pearce-Higgins, J.W.; Yalden, D.W. 2005. The effect of recreational
disturbance on an upland breending bird, the golden plover Pluvialis apricaria. Biol.
Conserv., 121: 53-63
Fonseca, E.M. 2003. Uma caracterização geoambiental da Rodovia Transpantaneira e
suas relações com a biodiversidade do Pantanal Mato-grossense no Estado de Mato
Grosso. Dissertação de Mestrado. IB/UFMT, Cuiabá, MT. 110p.

172
Fonseca, F.R.D.; Malm, O.; Waldemarin, H.F. 2005. Mercury levels in tissues of Giant
otters (Pteronura brasiliensis) from the Rio Negro, Pantanal, Brazil. Environm.
Reserch, 98:368-371
Fonseca, G.A.B. et al. 1996. Lista anotada dos mamíferos do Brasil. Conserv. Biol., 4: 1-38
Fonseca, V.S.S.; Petry, M.V.; Jost, A.H. 2001. Diet of the Magellanic Penguin on the coast
of Rio Grande do Sul, Brazil. Waterbirds, 24(2): 290-293
Fox, G.A.; Weseloh, D.V. 1987. Colonial Waterbirds as Bio-indicators of environmental
contamination in the Great Lakes. p209-216 in: Diamomd, A.W.; Filion, F.L.(eds.)
The Value of Birds. ICBP Tecnical Publication 6. Cambridge
Frederick, P.; Bildstein, K.L. 1992. Foraging ecology of seven species of neotropical ibises
(Threskiornithidae) during the dry season in the llanos of Venezuela. Wilson Bull.,
104(1):1-21
Frederick, P.;Collopy, M.W., 1989. Nesting success of five ciconiiforms species in relation
to water conditions in the Florida Everglades. Auk, 106: 625-634
Galeti, M.; Guimarães Jr., P.R. 2004. Seed dispersal of Attalea phalerata (Palmae) by
Crested Caracara (Caracara plancus) in the Pantanal and a reveiw of frugivory by
raptors. Ararajuba, 13(1) 133-135
Gawlik, D.E. 2002. The effects of prey availability on the numerical response of wading
birds. Ecol. Monogr., 73 (3): 329-346
Gimenes, M. 2002. Interactions between bees and Ludwigia elegans (Camb.) Hara
(Onagraceae) flowers at different altitudes in São Paulo, Brazil. Rev.Brasil. Zool.,19(3):
681-689

173
Girard, P.; Da Silva, C.J.; Abdo, M. 2003. River-groundwater interactions in the Brazilian
Pantanal. The case of the Cuiabá River. J. Hydrology, 283: 57-66
Gochfeld, M.; Burger, J. 1996. Family Sternidae, p. 624-667. in: Del Hoyo, J.; Elliott, A.;
Sargatal, J. (eds.). 1992. Handbook of the Birds of the World.-Hoatzin to Auks 3.
Lynx edicions
González, J.A. 1996a. Breeding Biology of the Jabiru in the Southern Llanos of Venezuela.
Wilson Bull., 108(3): 524-534
González, J.A. 1996b. Densidad y dinamica espaco-temporal de las poblaciones de
Cigüeñas (Ciconiidae) en los llanos inundables de Venezuela. Condor, 99: 671-680
González, J.A. 1997. Seasonal variation in the foraging ecology of the Wood Stork in the
Southern Llanos of Venezuela. Condor, 99:671-680
Goulding, M. 1980. The fish and the forest. University California Press. Los Angeles. 280p.
Guarim-Neto, G. 1991. Plantas do Brasil-Angiospermas do Estado de Mato Grosso-
Pantanal. Acta.bot.bras., 5(1): 25-47.
Guarim-Neto, G. 1992. Biodiversidade do ecossistema pantaneiro: a vegetação do Pantanal.
Anais II Congr. Nac. Essências Nativas, 106-110.
Guedes, N.M.R. 1993. Biologia reprodutiva da arara azul (Anodorhynchus hyacinthinus)
no Pantanal MS, Brasil. Dissertação de Mestrado, ESALQ. Piracicaba, São Paulo.
122pp.
Guimarães, J.R.D.; Mieli, M.; Malm, O.;Brito, E.M.S. 1998. Hg methylation in sediments
and floating meadows of a tropical lake in the Pantanal floodplain, Brazil. Sci. total
Environ., 213: 165-175
Guimarães, J.R.D.; Mieli, M.;Hylander, L.D.; Castro e Silva, E.; Roulet, M.; Maero, J.B.N.;
Lemos, R.A. 2000. Mercury net methylations in five tropical flood plain regions of

174
Brazil: high in the root zone of floating macrophyte mast but low in surface sedments
and flooded soils. Sci. total Environm., 261: 99-107
Gwiazda, R. 2003. Changes in the piscivorous birds commmunity at a Polish submontane
resevoir between 1990-2000 in relation to water level. p 211-220 in: Cowx, I.G. (ed.).
Interactions between fish and birds: Implications for management. Blackwell
Publishing Company. Oxford.
Hafner, H. 1977. Ecology of Wading Birds. Colonial Waterbirds, 20(1): 115-120
Hancock, J.A.; Kushlan, J.; Kahl, M.P. 1992. Storks, ibises and Spoonbills of the world.
London Academic Press. London. 385 p. il.
Harper, D.M. et al. 2003. Aquatic biodiversity and saline lakes: Lake Bogoria National
Reserve, Kenya. Hydrobiologia, 500: 259-276
Harris, M.; Tomas, W.; Mourão, G.; Da Silva, C.J.; Guimarães, E.; Sonoda, F.; Fachim, E.
2005. Safeguarding the Pantanal wetlands: Treats and conservation iniciatives.
Conserv. Biol., 19(3):714-720
Heckman, C.W. 1998b. The Pantanal of Poconé. Biota and Ecology in the Northern
section of the World´s largest pristine Wetland. Dordrecht. Kluwer Academic
Publishers. 622p.
Hoeinghaus, D.J.; Layman, C.A.; Arrington, D.A.; Winemiller, K.O. 2003. Spatiotemporal
variation in fish assemblage structure in tropical floodplain creeks. Environm. Biol.
fish.; 67: 379-387
Hoeinghaus, D.J.; Winemiller, K.O.; Taphorn, D. C.; 2004. Compositional changes in fish
assemblages along the Andean piedmont- Lhanos floodplain gradient of río Portuguesa,
Venezuela. Neotropical Ichtyology, 2(2): 85-92

175
Howard-Williams, C.; Junk, W. 1977. The chemical composition of central Amazoniam
aquatic macrophytes with special reference to their role in the ecosystem. Archiv. Für
Hydrobiologie. 79: 446-464
Hylander, L.D.; Pinto, F.N.; Guimarães, J.R.D.; Meili, M.; Oliveira, L.J.; Castro e Silva, E.
2000. Fish mercury concentration in the Alto Pantanal Brazil: influence of season and
water parameters. Sci. Total Environm., 261: 9-20
Johnsgaard, P.A. 1956. Effects of water fluctuation and vegetation change on birds
populations, particularly waterfowl. Ecology, 37: 689-701
Johnson, D.H. 1996. Waterfowl communities in the northern plains. pp 391-418. in: Cody,
M.L.; Smallwood, J.A. (eds.). Long-Term Studies of Vertebrate Communities.
Academic Press. San Diego
Johnson, M.A.; Tomas, W.M.; Guedes, N.M.R. 1997. On the Hyacinth macaw’s nesting
tree: density of young adult trees under three different manegement conditions in the
Pantanal wetland, Brazil. Ararajuba, 5(2): 185-188
Junk, W.J. 1973. Investigations on the ecology and production-biology of the “floating
meadows” (Paspalo-Echinochloetum) on the middle Amazon. Amazoniana, 4(1): 9-
102
Junk, W.J. 1984. Ecology of the varzea, floodplain of Amazon white water rivers. pp215-
244 in: Sioli, H. (ed.) The Amazon-Limnology and landscape ecology of a mighy
tropical river and its basin. Junk.W. Dordrecht
Junk, W.J.; Bayley, P.B.; Sparks, R.E. 1989. The flood pulse concept in river-floodplain
systems, in: Dodge, D.P. (ed.) Proceedings of the international large river
symposium. Can. Spec. Publ. Fish. Aquat. Sci., 106: 110-127
Junk, W.J.; Da Silva, C.J. 1995. Neotropical floodplains: A comparison between the

176
Pantanal of Mato Grosso and the large Amazonian river floodplains. pp195-217 in:
Tundisi et al. (eds.) Limnology in Brazil, ABC/SBL, Rio de Janeiro.
Junk, W.J.; Da Silva, C.J. 1999. O Conceito do Pulso de inundação e suas implicações para
o Pantanal de Mato Grosso. in: II Simpósio sobre recursos naturais e sócio-
econômicos do Pantanal. Anais... Corumbá: Embrapa Pantanal. pp 17-28
Junk, W.J.;Howard-Willians, C. 1984. Ecology of aquatic macrophytes in Amazonia.
pp269-293 in: Sioli, H. (ed.) The Amazon, limnology and landscape of a mighty
tropical river and its basin.
Junk, W.J.; Nunes da Cunha, C. 2005. Pantanal: a large South American wetland at a
crossroads. Ecological Engineering, 24:391-401
Junk, W.J.;Mello Nunes, J.A.S. 1987. Impactos ecológicos das represas hidroelétricas na
bacia amazônica brasileira. Tubinger Geogr. Stud., 95: 367-385
Junk, W.J.; Piedade, M.T.F. 1997. Plant life in the floodplain with special reference to
herbaceous plants. Pp147-185 in: Junk, W.J (ed.). The Central Amazon Floodplain:
Ecology of a pulsing system, Springer-Verlag. Berlin/Heidelberg.
Junk, W.J.; Piedade, M.T.F. 2000. Natural grasslands and herbaceus plants in the amazon
floodplain and their use. pp 269-290 in: Junk, W.J et al.(eds.). The Central Amazon
Floodplain: Actual use and options for a sustainable management. Backhuys
Publishers. Leiden, The Netherlands.
Junk, W.J.; Robertson, B.A. 1997. Aquatic Invertebrates. pp 279-298 in: Junk, W.J (ed.).
The Central Amazon Floodplain: Ecology of a pulsing system, Springer-Verlag.
Berlin/Heidelberg.

177
Junk, W.J.; Soares, M.G.; Carvalho, F.M. 1983. Distribution of fish species in a lake of the
Amazon river floodplain near Manaus (Lago Camaleão), with special reference to
extreme oxygen conditions. Amazoniana, 7(4): 397-431
Junk, W.J.; Soares, M.G.; Saint-Paul, U. 1997. The fish. pp385-408 in: Junk, W.J (ed.). The
Central Amazon Floodplain: Ecology of a pulsing system, Springer-Verlag.
Berlin/Heidelberg.
Kalmbach, E.; Ramsay, S.C.; Wendelln, H.; Becker, P. 2001. A study of Neotropic
Cormorants in Central Chile: possible effects of El Niño. Waterbirds, 24(3): 345-351
Kock, K.H. 2001. The direct influence of fishing and fishery-related activities on non-target
species in the Southern Ocean with particular emphasis on longline fishing and its
impact on albatrosses and petrels- a review. Fish Biology and Fisheries, 11: 31-56.
Kretzschmar, A.U.; Heckman, C.W. 1995. Estratégias de sobrevivência das espécies de
Ampullariidae durante mudanças das condições ambientais extremas do ciclo sazonal
sob o clima tropical úmido-seco. Acta Limnol. Brasil., 7: 60-66
Kushlan, J.A. 1976. Wading bird predation in a seasonally fluctuating pond. Auk, 93: 464-
476
Kushlan, J.A. 1978. Feeding ecology of wading birds. Nat. Audubon Soc. Res. Rep., 7: 247-
297.
Kushlan, J.A. 1981. Resource use strategies of wading birds. Wilson Bull., 93(2): 145-163
Kushlan, J.A. 1986. Response of wading birds to seasonally fluctuating water levels:
strategies and their limits. Colonial Waterbirds, 92(3): 155-162
Kushlan, J.A.; Ogden, J.C.; Higer, A.L. 1975. Relation of water level and fish availability
to Wood Stork reproduction in the Southern Everglades, Florida. U. S. National Park
Service, 75: 1-31

178
Kushlan, J.A.; Morales, L.G.; Frohring, P.C. 1985. Foraging niche relations of wading
birds in tropical savannas. Ornithological Monograph, 36: 663-682.
Lacerda, L. D. et. al. 1990. Mercury dispersal in water, sediments and aquatic biota of gold
mining tailing deposit drainage in Poconé, Brazil. Water Air Soil Pollut., 55: 283-
294.
Lacerda, L. D. et al. 1991. Mercury distribution in sediment profiles from lakes in the high
Pantanal, Mato Grosso State, Brazil. Biogeochemistry, 14(2): 91-98
Larrazábal, M.E.; Azevedo-Júnior, S.M.; Pena, O. 2002. Monitoramento de aves limícolas
na Salina Diamante Branco, Galinhos, Rio Grande do Norte, Brasil. Rev. Bras. Zool.
19(4): 1081-1089
Legendre, P. & Legendre, L. 1998. Numerical Ecology. Elsivier, Amesterdan. 853 pp
Lemes do Prado, A.; Heckman, C.; Martins, F.R. 1994. The Seasonal succession of biotic
communities in wetlands of the tropical wet-and-dry climatic zone: II. The Aquatic
macrophyte vegetation in the Pantanal of Mato Grosso, Brazil. Int. Revue. Ges.
Hydrobiol., 79(4): 569-589.
Levey, D.J.; Stiles, F.G. 1992. Evolutionary precursors of long-distance migration: resouse
availability and movement patterns in neotropical landbirds. Amer. Nat. 140: 447-476
Lima, A.L; Magnusson, W.E; Williams, D.G. 2000. Differences in diet among frogs and
lizards coexisting in subtropical forest of Australia. J. Herpetol., 34(1): 40-46
Loiselle, B.A.; Blake, J.G. 1991. Temporal variation in Birds and fruits along an
elevational gradient in Costa Rica. Ecology, 72(1): 180-193
Loiselle, B.A.; Blake, J.G. 1993. Spatial distribution of understory fruit-eating birds and
fruiting plants in a neotropical lowland wet forest. Vegetatio, 197/108: 177-189

179
Loiselle, B.A.; Blake, J.G. 1994. Annual variation in birds and plants of a tropical second-
growth woodland. Condor, 96: 368-380
Lowe-McConnell, R.H. 1999. Estudos Ecológicos de Comunidades de Peixes Tropicais.
Tradução Vazzoler, A.E.A; Agostinho, A.A; Cunnhinghan, P. São Paulo. Edusp.354p.
Lyra-Neves, R.M.; Azevedo-Júnior, S.M.; Telino-Júnior, W.R. 2004. Monitoramento do
maçarico-branco, Calidris alba (Pallas) (Aves, Scolapacidae), através de
recuperações de anilhas coloridas, na Coroa do Avião, Igarassu, Pernambuco, Brasil.
Rev. Brasil. Zool. 21(2): 319-324
Ma, Z.; Li, B.; Zhao, B.; Jing, K.; Tang, S.; Chen, J. 2004. Are artificial wetlands for
waterbirds? -A case study on Chongming Island, China. Biodiversity and Conservation,
13: 333-350.
Machado-Allison, A. 1990. Ecologia de los peces de las areas inundables de los llanos de
Venezuela. Interciencia, 15(6): 411-423
Machado, F.A. 1983. Comportamento e hábitos alimentares de quatro espécies de
Cichlidae (Teleostei) no Pantanal Mato-grossense, Dissertação de Mestrado,
Unicamp. Campinas, São Paulo. 80pp.
Machado, F.A. 2003. História natural de peixes do Pantanal: com destaque em hábitos
alimentares e defesa contra predadores. Tese de Doutorado, Unicamp. Campinas, São
Paulo. 96pp.
Machado, F.A.; Sazima, I. 1982. Comportamento alimentar do peixe hematófago
Branchioica bertonii (Siluriformes, Trichomycteridae). Ciência e Cultura, 35(3):
344-348
Mac Nally. R.C. 1994. On Charactering foraging versatility by using birds. Oikos, 69:95-
106

180
Magalhães, C.A. 1990a. Hábitos alimentares e estratégias de forrageamento de Rostrhamus
sociabilis no Pantanal de Mato Grosso, Brasil. Ararajuba, 1: 95-98
Magalhães, C.A. 1990b. Comportamento alimentar de Busarellus nigricollis no Pantanal de
Mato Grosso, Brasil. Ararajuba, 1: 119-120
Magnusson, W.E.; Lima, O.P.; Reis, F.Q.; Higuchi, N.; Ramos, J.F. 1999. Logging activity
and tree regeneration in Amazonian forest. Forest Ecol. Maneg., 113: 67-74
Malacalza, V.E.; Bertellotti, M. 2001. Cambios poblacionales de los cormoranes
(Phalacrocorax) in Punta Lobería, Patagonia, Argentina. Ornitología Neotropical,
12:83-86
Mansur, C.B.; Hebling, N.J. 2002. Análise comparativa entre fecundidade de Dilocarcinus
pagei Stimpson e Sylviocarcinus australis Magalhães; Turkay (Crustácea, Decapoda,
Trichodactylidae) no Pantanal do Rio Paraguai, Porto Murtinho, Mato Grosso do Sul.
Rev. Bras. Zool., 19(3): 797-805
Marco Jr., P.; Araújo, M.A.R.; Barcelos, M.K.; Santos, M.B.L. 2001. Aquatic invertebrates
associated with the Water-Hyacinth (Eichhornia crassipes) in an eutrophic reservoir
in tropical Brazil. Stud. Neotrop. Fauna Environm., 36(1): 73-80
Marini, M.Â.; Cavalcanti, R.B. 1996. Influência do fogo na avifauna de sub-bosque de uma
mata de galeria do Brasil Central. Rev. Brasil. Biol., 56(4): 749-754
Margalef, R. 1968. Ecología. Omega. Barcelona. xxv+951p
Marklund, O.; Stansten, H.; Hansson, L.; Blindow, I. 2002. Effects of waterfowl and fish
on submerged vegetation and macroinvertebrates. Freshwater Biology, 47: 2049-2059
Marques, M.I.; Adis, J.; Nunes da Cunha, C.; Santos, G.B. 2001. Arthropod biodiversity in
the canopy of Vochysia divergens (Vochysiaceae), a forest dominant in the Brasilian
Pantanal. Studies Neotrop. Fauna Environ., 36:205-210

181
Marques, O.A.V.; Eterovic, A.; Strussmann, C.; Sazima, I. 2005. Serpentes do Pantanal-
Guia ilustrado. Ribeirão Preto. Holos. 184p. il.
Martins, F.C.; Dias, M.M. 2003. Cuidado parental de Sula Leucogaster (Boddaert) (Aves,
Pelecaniformes, Sulidae) nas illhas dos Currais, Paraná, Brasil. Rev. Brasil. Biol. 20(4):
583-589
Martínez-Vilalta, A.; Motis, 1992. Family Ardeidae. p376-429 in: Del Hoyo, J.; Elliott, A.;
Sargatal, J. (eds). 1992. Handbook of the Birds of the World.-Ostrich to Ducks. Lynx
edicions. Barcelona
Martin, T.E.; Karr, J.R. 1986. Temporal dynamics of neotropical birds with special
reference to frugivores in second-growth woods. Wilson Bull., 98: 3-41
Mascitti, V. 2001. Habitat changes in Laguna de Pozuelos, Jujuy, Argentina: Implications
for South American Flamingo populations. Waterbirds, 24(1): 16-21
Matheu, E.; Del Hoyo, J. 1992. Family Threskiornithidae. p472-507 in: Del Hoyo, J.;
Elliott, A.; Sargatal, J. (eds.). 1992. Handbook of the Birds of the World.-Ostrich to
Ducks. Lynx edicions. Barcelona
Mendonça, L.B; Gimenes, M.R.; Anjos, L. 2004. Interactions between birds and other
organisms in the Upper Paraná River floodplain, Brazil. Pp-215-219 in: Agostinho,
A.A. et al.(eds.) 2004. Structure and functioning of the Paraná River and its floodplain:
LTER-Site 6(PELD- sitio-6). Maringá: EDUEM
Mérona, B.; Bittencout, 1993. Les peuplements de poissoins du “Lago do rei” , um lac
d’inondation d’Amazonie Centrale: description générale. Amazoniana, 12: 415-441
Meschiatti, A.J.; Arcifa, M.S.; Fenerich-Verani, N. 2000. Fish communities associated with
macrophytes in Brazilian floodplain lakes. Environ. Biol., 58: 133-143
Meyerriecks, A J. 1962. Diversity typifies heron feeding. Nat. Hist.,71: 16-57

182
Ministério do Meio Ambiente, 2004. Lista Nacional das espécies da Fauna Brasileira
Ameaçada de Extinção. Disponível em < http://www.mma.gov.br > Acesso em
30/11/2004
Minchin, P.R. 1987. An evalution of the relative robstness of techniques of ordination.
Vegetatio, 69: 89-107
Miranda, L.E.; Hodges, K.B. 2000. Role of aquatic vegetation coverage on hypoxia and
sunfish abundance in bays of a eutrophic reservoir. Hydrobiologia, 427: 51-57
Mitsch, J.W.; Gosselink, J.G. 2000. Wetlands. John Wiley; Sons. New York. 3ed. 920p.
Mittermeier, R.A; Gusmão-Camara, I; Jorge-Paduá, M.T.; Blanck, J. 1990. Conservation in
the Pantanal of Brazil. Oryx, 24: 103-112
Morales, G.; Pacheco, J. 1986. Effects of Diking of a venezuelan savanna on avian habitat,
species diversity, energy flow, and mineral flow though wading birds. Colonial
Waterbirds, 9(2): 236-242.
Morales, G.; Pinowski, J.; Pacheco, J.; Madriz, M.; Gomez, F. 1981. Densidades
poblacionales, flujo de energía y hábitos alimentarios de las aves ictiófagas de los
módulos de Apure, Venezuela. Acta Biol. Venez., 11(2):1-45
Mourão, G.M.; Campos, Z.; Coutinho, M.; Abercrombie, C. 1996. Size structure of ilegally
harvested and surviving caiman Caiman crocodilus yacare in Pantanal, Brazil. Biol.
Conserv., 75: 261-265
Mourão, G.M.; Ishii, T.H.; Campos, Z. 1988. Alguns fatores limnológicos relacionados
com a ictiofauna de baías e salinas do Pantanal de Nhecolândia, Mato Grosso do Sul,
Brasil. Acta Limnol. Bras., 2: 181-198.
Naves, L.C.; Brusque, L.F.; Vooren, C.M. 2002. Feeding ecology of Sula leucogaster,
Anous stolidus and Anous minutus at Saint Peter and Saint Paul’s Rocks, Brasil.

183
Ararajuba, 10 (1): 21-30
Nascimento, J.L.X. 1990. Reprodução de Agamia agami na Usina Hidrelétrica Balbina,
Amazonas, Brasil. Ararajuba, 1: 79-83
Nogueira, F.M.B.; Castro, E.; Silva, E.C.; Junk, W.J. 1997. Mercury from gold mining in
the Pantanal of Poconé (Mato Grosso, Brazil). Int. J. Environm. Healt Research., 7:
181-191
Nores, M.; Yzurieta, D. 1980. Aves de ambientes acuáticos de Cordoba y centro de
Argentina. Secr. Agricultura. y Ganaderia. Córdoba. 236p
Norris, K.; Atkinson, P.W.; Gill, J.A. 2004. Climate change and coastal waterbirds
populations-past declines and future impacts. Ibis, 146(Suppl.) 82-89
Nunes, A.L.; Adis, J. 1994. Comportamento populacional de Tucayaca gracilis (Giglio-Tos
1897) (Orthopetera-Acrididae) frente a oscilação do nível d’água na várzea da
Amazônia Central. Bol. Mus. Para. Emilio Goeldi. Sér. Zool., 10(2): 211-223
Nunes da Cunha, C. 1990. Estudo florístico e fitossociológico das principais formações
arbóreas do Pantanal de Poconé, MT., Dissertação de Mestrado, Unicamp.
Campinas, São Paulo. 102pp
Nunes da Cunha, C.; Junk, W.J. 1999. Composição florística de capões e cordilheiras:
localização das espécies lenhosas quanto ao gradiente de inundação no Pantanal de
Poconé-MT, Brasil in: II Simpósio sobre recursos naturais e sócio-econômicos do
Pantanal. Anais... Corumbá: Embrapa Pantanal. pp.387-405
Nunes da Cunha, C.; Junk, W.J. 2001. Distribution of wood plant communities along the
flood gradient in the Pantanal of Poconé, Mato Grosso, Brazil. J. Ecol. Environ. Sci.
27: 63-70

184
Nunes da Cunha, C.; Junk, W.J. 2004. Year-to-year changes in water level drive the
invasion of Vochysia divergens in Pantanal grassland. Appl. Veg. Sci., 7: 103-110
Nunes da Cunha, C.; Leitão-Filho, H.F. 1996. Composição florística arbórea do Pantanal
de Poconé-MT: Lista das espécies. in: Anais... do II Simpósio sobre Recursos
Naturais e Sócio-econômicos do Pantanal. Corumbá
Nunes da Cunha, C.; Wantzen, K.M.; Junk, W. 2003. The Pantanal Ecology Project:
Challenges and Progress of a Brazilian-German scientific collaboration (O Projeto
Ecológico Pantanal: Desafios e resultados em uma colaboração científica Brasil-
Alemanha). 56-67
Nunes, A P.; Tomas, W.M. 2004. Aves migratórias ocorrentes no Pantanal: caracterização e
conservação. Série Documentos, Embrapa Pantanal, 62: 1-25
Oliveira, D.M.M. 1986. Estratégias de caça do Tuiuiú Jabiru mycteria (Ciconiidae-Aves)
no Pantanal Matogrossense, Monografia de especialização, UFMT/IB, Cuiabá, Mato
Grosso. 40pp.
Oliveira, D.M.M. 1996. Aves Aquáticas do Pantanal Mato-Grossense in: Da Silva, C.J.
Conhecendo Pantanal -textos populares, Cuiabá, Projeto Ecologia do Gran-Pantanal.
2:1-8
Oliveira, D.M.M. 1997. Sucesso Reprodutivo e Conservação de tuiuiú Jabiru mycteria
(Aves: Ciconiidae) no Pantanal de Poconé, Mato Grosso, Dissertação de Mestrado,
UFMT/IB, Cuiabá, Mato Grosso. 68pp.
Oliveira, D.M.M in prep. Aves aquáticas de Mato Grosso, História natural e Etnobiologia.
Oliveira, D.M.M.; Cavalcanti, R.B. 2002. Sítios de nidificação de tuiuiú Jabiru mycteria
(Aves: Ciconiidae) no Pantanal de Poconé, Mato Grosso. in: Dantas, M. (Coord.) 3º
Simpósio sobre Recursos Naturais e Sócio-econômicos do Pantanal. Disponível em

185
http://www.cpap.embrapa.br/agencia/congresso/Bioticos/Oliveira-023.pdf acessado
em 7/12/2004
Oliveira, D.M.M.; Olivera, B.A.C. in prep. Aves da Fazenda Ipiranga, Poconé-MT.
Oliveira, D.M.M., et al. in prep. 1., Aves aquáticas no Brasil: uma revisão das pesquisas do
passado e prioridades para estudos futuros.
Oliveira, M.D.; Calheiros, D.F. 2000. Flood pulse influence on phytoplanckton
communities of the south Pantanal floodplain, Brazil. Hydrobiologia, 427-101-112
Oliveira, R.D.; Nogueira, F.M.B. 2000. Characterization of the fishes and subsistence
fishing in the Pantanal of Mato Grosso, Brazil. Rev. Brasil. Biol., 60 (3): 435-445
Olivera, B.A.C. 1999. Valor recreativo da Rodovia Transpantaneira: Uso turístico e
conservação no Pantanal Mato-Grossense. Dissertação de Mestrado. IB/UFMT,
Cuiabá, MT. 102p
Olmos, F. 2002. First record of Northern Royal Albatross (Diomedea sanfordi) in Brazil.
Ararajuba, 10 (2): 271-272
Olmos, F.; Silva e Silva, R. 2001. Breending biology and nest site characteristics of Scarlet
Ibis in southeastern Brazil. Waterbirds, 24(1): 58-67
Orta, J. 1992a. Family Phalacrocoracidae. p.326-353 in: Del Hoyo, J.; Elliott, A.; Sargatal,
J. (eds). 1992. Handbook of the Birds of the World.-Ostrich to Ducks. Lynx edicions,
Barcelona
Orta, J. 1992b. Family Anhingidae. p.354-3 in Del Hoyo, J.; Elliott, A.; Sargatal, J. (eds.).
1992. Handbook of the Birds of the World.-Ostrich to Ducks. Lynx edicions.
Barcelona
Ortiz-Pulido, R. 2000. Abundance of frugivorous birds and richness of fruit resource: is
there a temporal relationship? Caldasia, 22(1): 93-107

186
Owino, A.O.; Oyugi, J.O.; Nasirwa, O.O.; Bennun, L.A. 2001. Patterns of variation in
waterbirds numbers on four Rift Valley lakes in Kenya, 1991-1999. Hydrobiologia,
458: 45-53
Pacheco J.; Naranjo, C.J.; Madriz, M.; Morales, L.G.; Gomez, F. 1976. Relaciones entre los
niveles de inundación y la avifauna em los módulos de Apure. Acta Cient. Venezuela,
27:71-81
Paillisson, J.M.; Reeber, S.; Marion, L. 2002. Bird assemblages as bio-indicators of water
regime management and hunting disturbance in natural wet grasslands. Biological
Conservation, 106: 115-127
Paszkowski, C.A.; Tonn, W.M. 2000. Community concordance between the fish and
aquatic birds of lakes in northern Alberta, Canada: the relative importance of
environmental and biotic factors. Freshwater Biology, 43: 421-437
Pazin, V.F.V.2000. Alimentação de ciclídeos que ocorrem associados à macrófitas
marginais na lagoa do coqueiro, Pantanal de Poconé, Mato Grosso. Monografia de
conclusão de curso. UFMT/IB, Cuiabá, MT 68p.
PCBAP, 1997. Plano de Conservação da Bacia do Alto Paraguai. PNMA. Brasília. 3vol.
400p. il.
Pearson, D.L.; Beletsky, L. 2002. Brazil Amazon and Pantanal. The ecotravellers’wildlife
guide. Academic Press, San Diego. 476p
Penha, J.M.F.; Da Silva, C.J.; Bianchini-Júnior, I. 1998a. Impacto da Variação do Nível de
água no ciclo de vida da Macrófita aquática Pontederia cordata var. ovalis (Mart.)
Solms, em área alagável do Pantanal Mato-Grossense. Brazilian J. Ecol., 02: 30-35

187
Penha, J.M.F.; Da Silva, C.J.; Bianchini-Júnior, I. 1998b. Análise do Crescimento da
macrófita aquática Pontederia lanceolata em área alagável do Pantanal Mato-
Grossense, Brasil. Rev. Brasil. Biol., 5(2): 287-300
Pelicice, F.M.; Agostinho, A.A.; Thomaz, S.M. 2005. Fish assemblages associated with
Egeria in a tropical reservoir: investigating the effects of plant biomass and diel
period. Acta Oecologica, 27:9-16
Perrow, M.R.;Schutten, J.H.; Howes, J.R.; Holzer, T.; Madgwick, F.J.; Jowitt, A.J.D. 1997.
Interactions between coot (Fulica atra) and submerged macrophytes: the role of birds in
the restoraction process. Hydrobiologia, 342/343: 241-255.
Perrow, M.R; Jowitt, A.J.D; Stansfield, J.H.; Phillips, G.L. 1999. The practical importance
of the interactions between fish, zooplankton and macrophytes in lake restoration.
Hydrobiologia, 395/396: 199-210.
Petermann, P. 1997. The Birds. p 419-452 in: Junk, W.J (ed.). The Central Amazon
Floodplain: Ecology of a pulsing system.Springer-Verlag. Berlin Heidelberg
Petermann, P. 2000. Indicator value of avian communities in gallery forests in the region of
the Pantanal, Mato Grosso, Brazil: Preliminary results. Proceedings of the German-
Brazilian Workshop. Hamburgo
Petry, M.V.; Fonseca, V.S.S. 2002. Effects of human activities in the marine environment
on seabirds along the coast of Rio Grande do Sul, Brazil. Ornitologia Neotropical,
13:137-142
Petry, P.; Bayley, B.; Markle, D.F. 2003. Relationships between fish assemblages,
macrophytes and environmental in the Amazon River floodplain. J.Fish Biology, 63:
547-579

188
Piedade, M.T.F. 1993. Biologia e ecologia de Echinochloa polystachya (H.B.K.) Hitchcock
(Graminae=Poaceae) capim semi-aquático da várzea amazônica. Acta Limnol. Brasil.,
6: 173-185
Pietro, E.A. 1988. La Comunidade de Ibises (Threskiornithidae) en los Lhanos de
Venezuela. Mem. Soc. Cient. Nat. de La Salle, 48(130): 59-75
Pihl, L.; Wennhage, H.; Nilsson, S. 1994. Fish assemblage structure in relation to
macrophyte and filamentous epiphytes in shallow non-tidal and soft-bottom habitats.
Environm. Biol. Fish., 39: 271-288
Pinho, J.B.; Nogueira, F.M.B. 2000. Mostra da retirada de Psitacídeos em cativeiro na
Cidade de Cuiabá e Pantanal, MT, no período de 1995-1997. Ararajuba, 8:51-53
Pinho, J.B.; Nogueira, F.M.B. 2003. Hyacinth Macaw (Anadorhynchus hyacinthinus)
reproduction in the Northern Pantanal, Mato Grosso, Brazil. Ornitologia Neotropical,
14: 29-38.
Pinho, J.B. 2005. Riqueza de espécies, padrões de migração e biologia reprodutiva de aves
em quatro ambientes florestais do Pantanal de Poconé, MT. Tese de Doutorado,
UFMG, Belo Horizonte, Minas Gerais. 185pp
Pinowski, J.; Morales, G. 1981. Aspectos ecologicos de las aves de los modulos de Apure.
Bol. Soc. Venez. Cienc. Nat.,35 (139): 67-78
Pinowski, J. et al. 1980. Estimation of the food consumption of fish-eating birds in the
seasonally-flooded savannas (llanos) of Alto Apure, Venezuela. Bull. Acad. Pol. Sci.
Ser. Sci. Biol., 28: 163-170
Pinto, R.M.; Barros, L.A.; Tortelly, L.; Teixeira, R.F.; Gomes, D.C. 2004. Prevalence and
pathology of ciconiiformes birds from the Brazilian swamplands. J. Helminthology,
78: 1-6

189
Ponce, V.M. 1995. Impacto Hidrológico e Ambiental da Hidrovia Paraná-Paraguai no
Pantanal Matogrossense. Um Estudo de Referência.San Diego State University, San
Diego. 134p. il.
Ponce, V.M.; Nunes da Cunha, C. 1993. Vegetated earthmounds in tropical savannas of
Central Brazil: a synthesis. J. Biogeography, 20: 219-225
Pott, A.; Pott, V.J. 1994. Plantas do Pantanal. Corumbá. Embrapa, 320p.il.
Pott, A.; Pott, V.J. 1999. Flora do Pantanal-listagem atual de Fanerógamas. in: II Simpósio
sobre recursos naturais e sócio-econômicos do Pantanal. Anais... Corumbá: Embrapa
Pantanal. pp 297-325
Pott, V.J.; Pott, A. 1997. Checklist das macrófitas aquáticas do Pantanal, Brasil. Acta Bot.
Brasil., 11(12): 215-227.
Pott, V.J.; Pott, A. 2000. Plantas aquáticas do Pantanal. Embrapa. Brasília. 404p. il.
Pott, V.J.; Pott, A. 2003. Dinâmica da vegetação aquática do Pantanal. Pp 145-162. in
Thomas, S.M.; Bini, L.M. (eds.) 2003. Ecologia e Manejo de macrófitas aquáticas.
EdUEM. Maringá
Poulin, B.; Lefebvre, G.; McNeil, R. 1992. Tropical avian phenology in relation to
abundance and exploitation of food resources. Ecology, 73(6): 2295-2309.
Prance, G T.; Schaller, G.B. 1982. Preliminary study of some vegetation types of the
Pantanal, Mato Grosso, Brazil. Brittonia, 34:228-251.
Raizer, J.; Amaral, M.E. 2001. Does the structural complexity of aquatic macrophytes
explain the diversity of associated spider assemblages? J. Arachnology, 29: 227-237
Remsen Jr., J.V. 1985. Community organization and ecology of birds of high elevation
humid forest of the Bolivian Andes. Ornithological Monographs, 36: 733-756
Remsen Jr., J.V. 1991. Community ecology of neotropical Kingfischers. Univ. California

190
Publications, Berkley 124: 1-115.
Resende, E.K. 2000. Trophic stucture of fish assemblages in the lower Miranda River,
Pantanal, Mato Grosso do Sul State, Brazil. Rev. Brasil. Biol., 60(3): 389-403
Resende, E.K.; Palmeira, S.S. 1999. Estrutura e dinâmica das comunidades de peixes da
planície inundável do Rio Miranda, Pantanal de Mato Grosso do Sul. in: II Simpósio
sobre recursos naturais e sócio-econômicos do Pantanal. Anais... Corumbá: Embrapa
Pantanal pp 249-280
Ricklefs, R.E. 1987. Ecology. Chiron Press. New York. 996p
Robinson, S.K. 1997. Birds of a peruvian oxbow lake: populations, resources, predation,
and social behavior. Ornithological Monographs, 48: 613-639.
Robinson, S.K; Terborgh, J. 1990. Birds communities of the Cocha Cashu biological
station in amazonian Peru. pp.199-216. in: Gentry, A.H (ed.) Four Neotropical
Rainforests. Yale Univ. Press. New Haven
Robinson, S.K.; Terborg, J. 1997. Bird community dynamics along primary successional
gradients of an amazonian whitewater river. Ornithological Monographs, 48: 641-672.
Rocha, C.D; Del Lama, S.; Regitano, L.C.A. 2004. Lack of genetic structuring among
tropical brazilian Wood Stork populations and low genetic differentiation from North
American populations. Biotropica, 36(2): 248-258
Rodrigues, F.H.G.; Medri, I.M.; Tomas, W.M.; Mourão, G.M. 2002. Revisão do
conhecimento sobre ocorrência e distribuição de mamíferos do Pantanal.
Rodrigues, M.A.; Lewis, W.M. 1997. Structure of fish assemblages along environmental
gradients in floodplain lakes of Orinogo River. Ecol. Monogr., 67: 109-128
Rohf, J.F.; Sokal, R.E. 1969. Statistical Tables. W.H. Freeman Company, New York. 219p

191
Ross, A.L.; Piacentini, V.Q. 2003. Revisão dos registros sul-brasileiros do gênero
Phoebetria Reichenbach, 1853 e primeiro registro documentado de Phoebetria
palpeblata (Forster, 1785) (Procellariiformes: Diomedeidae) para Santa Catarina.
Ararajuba, 11(2): 223-225
Safina, C.; Burger, J. 1983. Effects of human disturbance on reproductive success in the
Black Skimmer. Condor, 85: 164-171
Saint-Paul, U. Zuanon, J.; Villacorta, M.A; Garcia, M.; Fabré, N.N.; Berger, U.; Junk, W.J.
2000. Fish communities in central Amazonian white-and blackwater floodplains. Env.
Biol. Fish., 57: 235-250
Saint-Paul, U.; Soares, M.G. 1987. Diurnal distribution and behavioral response of fishes to
extreme hypoxia in an Amazon floodplain lake. Environm. Biol. Fishes, 20 (2): 91-
104
Sanaiotti, T.M.; Cintra, R. 2001. Breending and migranting Birds in an Amazonian
Savanna. Stud. Neotrop. Fauna Environm., 36(1): 23-32
Sanaiotti, T.M.; Magnusson, W.E. 1995. Effects of annual fire on the production of fleshy
fruits eaten by birds in Brazilian Amazonian savanna. J. Trop. Ecol., 11: 53-65
Sanchez-Botero, J.I.; Araújo-Lima, C. 2001. As macrófitas aquáticas como berçário para a
ictiofauna da várzea do Rio Amazonas. Acta Amazonica, 31(3):437-447
Santos, G.B.; Marques, M.I.; Adis,J.; De Musis, C.R. 2003. Artrópodos associados a copa
de Attalea phalerata Mart. (Aracaceae), na região do Pantanal de Poconé, Mato Grosso,
Brasil. Rev. Brasil. Entom. 47: 211-224
Santos, G.M. 1981. Estudos de alimentação e hábitos alimentares de Schizodon fasciatus
Agassiz, 1829, Rhytiodus microlepis Kner, 1859 e Rhytiodus argenteofuscus Kner,

192
1859, do lago Januacá-AM. (Osteichthyes, Characoidei, Anastomidae). Acta
Amazonica, 11(2): 267-283
Santos, G.M. 1982. Caracterização, hábitos alimentares e reprodutivos de quatro espécies
de ‘aracus’ e considerações ecológicas sobre o grupo no lago Januacá-AM.
(Osteichthyes, Characoidei, Anastomidae). Acta Amazonica, 12(4): 713-739
Sazima, I.; Caramaschi, E.P. 1989. Comportamento alimentar de duas espécies de
Curimata, sintópicas no Pantanal de Mato Grosso (Osteichthyes, Characiformes).
Rev. Brasil. Biol., 49(2): 325-333
Sazima, I.; Machado, F.A. 1982. Hábitos e comportamento de Roeboides prognatus, um
peixe lepidófago (Osteichthyes, Characoidei). Bol. Zool. Univ. S. Paulo, 7:37-56
Sazima, I.; Machado, F.A. 1989. Melhor no seco que na água: uma tática defensiva do
peixe Laetacara dorsigera (Ciclidae). Ciência e Cultura, 41(10): 1014-1016
Sazima, I.; Zamprogno, C. 1984. Use of water hyacinths as shelter, foraging place and
transport by yuong piranhas Serrasalmus spilopleura. Environm. Biol. Fishes, 12(3):
237-240
Sazima, I., Buzato, S.; Sazima, M. 1996. An assemblage of hummingbird-pollinited flowers
in a montane forest in southeastern Brazil. Bot. Acta. 109: 149-160
Sazima, M.; Vogel, S.; Lemes do Prado, A.; Oliveira, D.M.M.; Franz, G.; Sazima, I. 2001.
The sweet jelly of Combretum lanceolatum flowers (Combretaceae): a cornucopia
resource for bird pollinators in the Pantanal, western Brazil. Plant Syst. Evol. 227: 195-
208
Schaller, G.B.1983. Mammals and their biomas on a Brasilian ranch. Arq. Zool. São Paulo,
31: 1-36

193
Scheffer, M. 1999. The effect of aquatic vegetation on turbidity; how important are the
filter feeders? Hydrobiologia, 408/409: 307-316
Seidl, A.F.; Moraes, A.S. 2000. Grobal valuation of ecosydtem services: application to the
Pantanal da Nhecolandia, Brazil. Ecol. Economics, 33: 1-6
Seidl, A.F.; Silva, J.S.V.; Moraes, A.S. 2001. Cattle ranching and deflorestation in the
Brazilian Pantanal. Ecol. Economics, 36: 413-425
Shimada, T.; Bowman, A.; Ishida, M. 2000. Effects of flooding on a wetland birds
community. Ecological Research, 15:229-235
Sick, H. 1997. Ornitologia Brasileira, uma Introdução. Editora Nova Fronteira. Rio de
Janeiro. 912p.il.
Silva, J.M.C.; Oren, D.C.; Roma, J.C.; Henriques, L.M.P. 1997. Composition and
distribution patterns of avifauna of an Amazonian upland savanna, Amapá, Brazil.
Ornithological Monographs, 38. 743-762
Soares, M.G.M. 1986. The tropic status of fish fauna in lago Camaleão and macrophyte
dominated floodplain lake in the middle Amazon. Amazoniana, 9(4): 511-526
Stapanian, M.A.; Waite, T.A.; Krzys, G; Mack, J.J.; Micacchion, M. 2004. Rapid
assessment indicator of wetland integrity as an unintended predictor of avian diversity.
Hydrobiologia, 520: 119-126
Steinmetz, J; Kohler, S.L.; Solur, D.A 2003. Birds are overlooked top predators in aquatic
food webs. Ecology, 84(5): 1324-1328.
Stiles, F.G. 1980. The annual cycle in a tropical wet forest hummingbird community. Ibis,
122: 322-343
Stiles, F.G. 1988. Altitudinal movements of birds on the Caribbean slope of Costa Rica:
implications for conservation. pp 243-258 in: Almeda, F.; Plinge, C.M. (eds.).

194
Tropical Rainforest: Diversity and Conservation. California Academy Sciences,
Memoirs 12.
Stolen, E.D. 2003. The effects of vehicle passage on foraging behavior of wading birds.
Waterbirds, 26(4):429-436
Stouffer, P.C.; Bierregaard, R. O. 1996. Forest fragmentation and seasonal patterns of
hummingbird abundance in Amazonian Brazil. Ararajuba, 4: 9-14
Strüssman, C. 1992. Serpentes do Pantanal de Poconé, Mato Grosso: composição
faunística história natural e ecologia comparada. Dissertação de Mestrado.
Unicamp, Campinas. São Paulo.
Strüssman, C. 1997. Hábitos alimentares da sucuri-amarela Eunectes notaeus Cope, 1862
no Pantanal Mato Grosso. Biociências, 5:35-52
Strüssman, C.; Sazima, I. 1990. Esquadrinhar com a cauda: uma tática de caça da serpente
Hydrodynastes gigas no Pantanal, Mato Grosso. Mem. Inst. Butantan, 52:57-61
Strüssman, C.; Sazima, I. 1991. Predation on avian eggs by the boid snake, Eunectes
notaeus. Herpt. Review, 22:118-120
Strüssman, C.; Sazima, I. 1993.The snake assemblage of Poconé, western Brazil: faunal
composition and ecological summary. Stud.Neotrop.Fauna Environ.,28:157-168
Súarez, Y.R; Petrere Jr, M.; Catella, A.C. 2001. Factors determining the structure of fish
communities in Pantanal lagoons (MS, Brazil). Fisheries Management and Ecology,
8: 173-186
Súarez, Y.R; Petrere Jr, M.; Catella, A.C. 2004. Factors regulating diversity and abundance
of fish communities in Pantanal lagoons, Brazil. Fisheries Management and Ecology,
11:45-50

195
Taylor, I.R. 2004. Foraging ecology of the Black-fronted Plover on Saline Lagoons in
Australia: The importance of receding water levels. Waterbirds, 27(3): 270-276
Tejerina-Garro, F.L.; Fortín, R.; Rodríguez, M.A. 1998. Fish community structure in
relation to environmental variation in floodplain lakes of the Araguaia River, Amazon
Basin. Environm. Biol. Fish., 51: 399-410.
Telino-Júnior, W. R., Azevedo-Júnior, S. M. & Neves, R. M. L. 2003. Censo de aves
migratórias (Charadriidae, Scolapacidae e Laridae) na Coroa do Avião, Igarassu,
Pernambuco, Brasil. Rev. Brasil. Zool., 20(3): 451-456
Terborgh, J.; Robinson, S.K.; Parker III, T.A.; Munn, C.A.; Pierpont, N. 1990. Structure
and organization of an Amazonian forest bird community. Ecol. Monogr., 60(2): 213-
238.
Thomas, B.T. 1985. Coexistence and behavior differences among the three western
hemisfhere Storks. pp931-931.in: Foster, M.A.(ed.). Neotropical Ornithhology. Allen
Press, Kansas.
Thomaz, S.M; Bini, L.M. 2003, Análise crítica dos estudos sobre macrófitas aquáticas
desenvolvidos no Brasil. Pp19-38. in Thomas, S. M.; Bini, L. M. (eds.) 2003.
Ecologia e Manejo de macrófitas aquáticas. Maringá. EDUEM.
Thórarinsson, T.L.; Einarsson, A. 2004. Dispersal of the horned grebe Podiceps auritus (L.)
(Aves) on Lake Myvatan, Iceland , in late Summer. Aquatic Ecology, 38: 309-315
Tomas, W.M; Salis, S.M; Silva, M.P.; Mourão, G.M. 2001. Marsh Deer (Blastoceros
dichotomus) distribution as a function of floods in the Pantanal wetland, Brazil. Stud.
Neotrop. Fauna Environm., 36(1): 9-13.
Tonn, W.M.; Magnusson, J.J. 1982. Patterns in the species composition and richness of fish
assemblages in nortern Wisconsin lakes. Ecology, 63: 1149-1116

196
Tubelis, D.P.; Tomas, W.M. 2003a. Bird species of the Pantanal wetland, Brazil.
Ararajuba, 11(1): 5-37.
Tubelis, D.P.; Tomas, W.M. 2003b.The contributions of museum collections and of records
not involving collections to the knowledge of bird species composition of the Pantanal,
Brazil. Ararajuba, 11(2): 207-214.
Val, A.L. 1996. Surviving low oxygen levels: Lessons from fishes of the Amazon. 6 p 59-
73 in: Val, A.L; Almeida-Val, V.M.F.; Randall, D.J.(eds.). Physiology and
Biochemistry of the fishes of Amazon. INPA, Manaus.
Velásques, C.R. 1992. Managing artificial saltpans as a waterbird habitat: species’response
to water level manipulation. Colonial Waterbirds, 15(1): 43-55
Vieira, M.F.; Adis, J. 1992. Abundância e biomassa de Paulinia acuminata (DE GEER,
1773) (Orthoptera: Pauliniidae) em um lago de várzea da Amazônia Central.
Amazoniana, 12(2): 337-352
Wagner, B.M.A. 1997. Influence of fish on the breending of the Red-necked Grebe
Podiceps grisegena (Boddaert, 1783). Hydrobiologia, 344: 57-63
Wagner, B.M.A.; Hansson, L. 1998. Food competition and niche separation between fish
and the Red-necked Grebe Podiceps grisegena (Boddaert, 1783). Hydrobiologia, 368:
75-81
Wantzen, K.M.; Da Silva, C.J.; Figueirdo, D.M.; Mogliácio,M.C. 1999. Recent impacts of
navigation on the upper Paraguay. Rev. Bol. Ecol. 6: 173-182
Westalake, D.F. 1963. Comparisons of plants productivity. Biol. Rev., 38: 385-425
Wiens, J.A. 1989. The Ecology of Birds Communities. Cambridge Univ. Press. Cambridge.
2V.

197
Willink, P.W. et al. 2000. Fishes from the rios Negro, Negrinho, Taboco, Aquidauna,
Taquari and Miranda, Pantanal, Brazil: diversity, distribution, critical habitats, and
value, p.63-81. in: Willink, P.W. et al.(eds.). RAP Bull. Biol. Assessment, 18
Willis, E.O. 1995. Black versus White waterbirds colonies (Aves) in the Bolivian-Brazilian
Pantanal. Iheringia. Ser. Zool., 78: 95-97
Wilkinson, L. 1991. SYSTAT The system for statistics. Inc. Evanston, Illinois.
Willard, D.E. 1985. Comparative feeding ecology of twenty-two tropical piscivores.
Ornithological Monographs, 36. 788-797
Wolda, H. 1987. Seasonality and the community. pp 69-95. in: Gee, J. H. R.; Giller, P.S.
(eds.). Organization of Communities, past and present. British Ecological Society ;
Blackwell. London
Wolter, C.; Pawlizki, R. 2003. Seazonal and Spatial variation in cormorant predation in a
lowland floodplain river. p 178-186 in: Cowx, I.G. (ed.). Interactions between fish
and birds: Implications for management. Blackwell Publishing Company. Oxford.
376p.
Yabe, R.; Marques, E.J. 2001. Deslocamentos de aves entre capões no Pantanal Mato-
grossense e sua relação com a dieta. Pp 103-123 in: Albuquerque, J.L.B. (eds.).
Ornitologia e conservação: da Ciência às estratégias. Tubarão, Unisul
Yamashita, C.; Valle, M.P. 1990. Sobre Ninhais de Aves no Pantanal do Município de
Poconé, Mato Grosso, Brasil. Vida Silvestre Neotropical, 2(2): 59-63
Zar, J.H. 1984. Biostatistical Analyses. Prentice-Hall. New Jersey 718p.
Zavalaga, C.B.; Frere, E.; Gandini, P. 2002. Status of the Red-legged Cormorant in Peru:
GAT factors affect distribution and numbers? Waterbirds, 25(1): 8-15

198
Zeffer, A.; Johansson, C.; Marmebro, A. 2003. Functional correlation between habitat use
and leg morphology in birds (Aves). Biol. J. Linnean Society, 79: 461-484
Zeihoffer, P.; Schessl, M. 1999. Relationship between vegatation and environmental
conditions in the northern Pantanal of Mato Grosso, Brazil. J. Biogeogr., 27: 159-168





























