Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DE PERNAMBUCO
CENTRO DE CIÊNCIAS BIOLÓGICAS
PROGRAMA DE PÓS-GRADUAÇÃO EM BIOLOGIA ANIMAL
ARTHUR VINICIUS DE OLIVEIRA MARROCOS DE MELO
DISSERTAÇÃO DE MESTRADO
RECRUTAMENTO E SUCESSÃO DE ANIMAIS SÉSSEIS E SEDENTÁRIOS EM SUBSTRATO
ARTIFICIAL SUBMERSO NA ÁREA PORTUÁRIA DO RECIFE – PE, COM ENFOQUE PARA A
BIOINVASÃO
RECIFE
2012

ARTHUR VINICIUS DE OLIVEIRA MARROCOS DE MELO
RECRUTAMENTO E SUCESSÃO DE ANIMAIS SÉSSEIS E SEDENTÁRIOS EM SUBSTRATO
ARTIFICIAL SUBMERSO NA ÁREA PORTUÁRIA DO RECIFE – PE, COM ENFOQUE PARA A
BIOINVASÃO
RECIFE
2012
Dissertação apresentada ao Programa de Pós-
Graduação em Biologia Animal da Universidade Federal de Pernambuco como parte dos requisitos para obtenção do título de Mestre em Biologia
Animal.
ORIENTADOR: RALF SCHWAMBORN CO-ORIENTADOR: CRISTIANE MARIA ROCHA FARRAPEIRA


ARTHUR VINICIUS DE OLIVEIRA MARROCOS DE MELO
RECRUTAMENTO E SUCESSÃO DE ANIMAIS SÉSSEIS E SEDENTÁRIOS EM SUBSTRATO
ARTIFICIAL SUBMERSO NA ÁREA PORTUÁRIA DO RECIFE – PE, COM ENFOQUE PARA A
BIOINVASÃO
Dissertação defendida e Aprovada em 31 de agosto de 2012.
BANCA EXAMINADORA: TITULARES
Dra. Maria Eduarda de Lacerda Larrazábal
Depto. de Zoologia - UFPE
Dra. Andréa Karla Pereira da Silva
Depto. de Biologia - UPE
Dra. Fernanda Maria Duarte do Amaral
Depto. de Biologia - UFRPE
SUPLENTES:
Dr. Múcio Luiz Banja Fernandes
Depto. de Biologia - UPE
Dr. André Morgado Esteves
Depto. de Zoologia - UFPE

“Os ventos que às vezes tiram
algo que amamos, são os
mesmos que trazem algo que
aprendemos a amar...
Por isso não devemos chorar
pelo que nos foi tirado e sim,
aprender a amar o que nos foi
dado. Pois tudo aquilo que é
realmente nosso, nunca se vai
para sempre...” Bob Marley

AGRADECIMENTOS
À minha mãe Cleide Oliveira, por todos os momentos e auxílios que me foram
dados, mesmo na minha distância e frieza em alguns momentos.
À Cristiane Farrapeira, pelo companheirismo e auxílio, como sempre digo é
mais que uma co-orientadora, ex-orientadora, é uma amiga, a minha mãe científica;
minha eterna admiração por tudo.
Ao Professor Dr. Ralf Schwamborn, por ter aceitado ser meu orientador junto ao
PPGBA, acreditando no trabalho a ser desenvolvido e por todas as conversas.
Às Drªs Fernanda Amaral, Andréa Karla e Maria Eduarda por aceitarem
participar da banca e contribuírem para conclusão dessa etapa.
Aos colegas Lis Stegmann, David Oliveira, Vithor Macêdo, Gustavo Valença,
Yuri Valença e Roberta Salgado pelos cafés e conversas sobre os conteúdos aprendidos
em sala.
A Edson Andrade por tudo que me fez aprender nesse período, me fez ver a vida
por outros lados, por outros ângulos, pela força na elaboração da parte prática, as idas
semanais ao porto, as triagens no laboratório, as discussões nos locais mais
improváveis... Pelo carinho, companheirismo e por deixar tudo mais tranquilo e depois
mais complicado e difícil de ser feito.
A Gabriel Oliveira, meu filho, só posso pedir desculpas pela ausência e
agradecer por todo o carinho que me deu nesses meses.
À Nathália Paiva, pela ajuda nas idas ao porto, e pelas conversas nas horas que
eu tava meio preocupado com tudo e com nada.
À Mônica Mota, por toda a compreensão e ouvidos abertos para ouvir meus
choros e desesperos, por saber me ouvir e dizer o que eu precisava ouvir em cada
momento, por me fazer rir quando eu só queria chorar, pelo companheirismo e amizade
sincera e verdadeira.
A José Divard Oliveira Filho e à administração do Porto do Recife, por
permitirem o desenvolvimento desse trabalho na bacia portuária e por todas as
informações passadas.
Aos professores do PPGBA por toda contribuição e auxilio desenvolvido,
especialmente ao Prof. Andre Esteves, pelos auxílios nas horas pertinentes.
À Ana Fraga, por nos atender de forma clara e objetiva na secretaria do PPGBA.
À CAPES pela concessão da bolsa que auxiliou na realização desse trabalho.
A todos os que contribuíram para a realização deste trabalho, de forma direta ou
indireta. Obrigado.

SUMÁRIO
PÁGINA
RESUMO 7
ABSTRACT 9
INTRODUÇÃO 10
REFERÊNCIAS 17
MANUSCRITO A SER SUBMETIDO PARA PUBLICAÇÃO 24
RESUMO 24
ABSTRACT 25
INTRODUÇÃO 26
MATERIAL E MÉTODOS 28
RESULTADOS 32
DISCUSSÃO 41
CONCLUSÕES 46
REFERÊNCIAS BIBLIOGRÁFICAS 47
CONSIDERAÇÕES FINAIS 54
REGRAS DO PERIÓDICO IHERINGIA, SÉRIE ZOOLOGIA 55

RESUMO
O Porto do Recife recebe uma grande quantidade de navios de vários tipos e de muitas
partes do mundo, podendo trazer espécies exóticas incrustadas no casco ou em água de
lastro. O objetivo do presente estudo foi registrar e analisar os padrões de recrutamento
e a dinâmica de sucessão de espécies bentônicas sésseis, assim como investigar uma
possivel introdução e estabelecimento de espécies bentônicas exóticas e criptogênicas
em uma área portuária com grande fluxo de navios. Utilizaram-se placas de metal
galvanizado para estudo do recrutamento e sucessão de animais sésseis e sedentários em
três estações no porto, com três réplicas cada, totalizando nove pontos, durante seis
meses (outubro de 2010 a março de 2011), considerado como período seco. As placas
de recrutamento foram retiradas a cada 30 dias durante seis meses, as placas de sucessão
foram retiradas após 60 dias e depois de sucessivos 30 dias até completarem 180 dias de
submersão. As principais espécies de acordo com a área de cobertura foram Mytella
charruana (72%) e Amphibalanus improvisus (22%) e espaços vazios (7,5%). Não
houve diferença significativa quando comparadas as três estações no porto do Recife,
sendo encontradas diferenças quando comparadas as áreas de cobertura das espécies
mais abundantes no recrutamento. Foram elas: Clytia gracilis, Obelia dichotoma, M.
charruana e A. improvisus. Na comparação entre as estações de acordo com a área de
cobertura das espécies mais abundantes na sucessão houve diferenças significativas
apenas para M. charruana e A. improvisus. Observou-se o estabelecimento das espécies
exóticas Haliplanella lineata (Cnidaria) e Sinelobus stanfordi (Arthropoda). Mytella
charruana foi considerada a espécie clímax na sucessão ecológica, já que foi a única
espécie encontrada em grande quantidade e ocupando todas as placas. Observou-se a

maior riqueza das espécies exóticas e criptogênicas em detrimento das espécies nativas,
tanto no recrutamento quanto na sucessão.
Palavras chaves: Animais Bentônicos, predação, espécies exóticas, sucessão ecológica.

ABSTRACT
The Port of Recife receives a large number of vessels of various types and from many
parts of the world, and can bring exotic species encrusted hull or in ballast water. The
aim of this study was to record and analyze the patterns of recruitment and succession
dynamics of sessile benthic species, as well as investigating a possible introduction and
establishment of exotic and cryptogenic benthic species in a port area with great flow
vessels. We used galvanized metal plates to study the recruitment and succession of
sessile and sedentary animals at three stations in the harbor, with three replicas each,
totaling nine points, six months (October 2010 to March 2012), considered the dry . The
plates recruitment were taken every 30 days for six months, the plates were removed
succession after 60 days and after successive 30 days up to 180 days of submersion. The
main species according to coverage area were Mytella charruana (72%) and
Amphibalanus improvisus (22%) and voids (7.5%). There was no significant difference
when comparing the three stations in the Port of Recife, differences were found when
comparing the coverage areas of most abundant species in recruitment. They were:
Clytia gracilis, Obelia dichotoma, M. charruana and A. improvisus. Comparing the
stations according to the coverage area of the most abundant species in succession
significant differences only for M. charruana and A. improvisus. We observed the
establishment of exotic species Haliplanella lineata (Cnidaria) and Sinelobus stanfordi
(Arthropoda). Mytella charruana was considered the climax species in ecological
succession, since it was the only species found in large quantities and occupying all
boards. We observed the greatest wealth of exotic and cryptogenic to the detriment of
native species, both in recruiting and succession.
Keywords: benthic animals, predation, alien species, ecological succession.

INTRODUÇÃO
No ambiente aquático, qualquer objeto duro, natural ou artificial, é objeto de
uma bioincrustação (biofouling ou simplesmente fouling), ou seja, desenvolvimento de
uma comunidade bentônica constituída por organismos sésseis, algas e animais, que se
fixam em substratos naturais ou artificiais submersos. A eles, juntam-se outros
invertebrados bentônicos sedentários de movimentação esporádica ou lenta, ou de
hábito tubícola, escavador e perfurador (WHOI, 1952; BAKER et al., 2004).
O processo de ocupação do substrato por esses organismos é conhecida como
recrutamento, que consiste de colonização de uma superfície sólida, viva ou morta,
(WAHL, 1989). O processo de colonização de habitats é assegurado pela liberação de
propágulos (esporos e larvas), da maioria dos vegetais e animais bentônicos que
constituem o principal meio de ampliação da distribuição geográfica (dispersão) natural,
sobretudo para as espécies que possuem larvas com desenvolvimento prolongado no
plâncton (alguns dias a algumas semanas) (MILEIKOVSKY, 1974; MARIANI et al.,
2006). Na transição entre a fase de dispersão pelágica e existência bentônica do adulto,
os propágulos necessitam primeiro encontrar um substrato e, seguindo sua chegada a
esse, eles necessitam determinar a adequação do mesmo para as necessidades do adulto,
usando uma variedade de pistas, as quais podem incluir o contorno da superfície, o tipo
de substrato, sua natureza química e presença de um filme microbiano (ABELSON &
DENNY, 1997).
Para Keough & Downes (1982), a colonização envolve três fases:
desenvolvimento, exploração do habitat e estabelecimento. Absalão (1993) define
recrutamento como o processo de assentamento das larvas dos organismos, que, no

caso, daqueles com larva planctônica, implica na transformação de seu hábito de vida
para o bentônico, bem como a sobrevivência até o momento do recenseamento. O
recrutamento, de maneira geral é definido como a adição de novos indivíduos para
populações ou como um estágio sucessivo do ciclo de vida de uma população.
Menge & Sutherland (1976); Keough & Downes (1982); Done (1982) e Keough,
(1988) destacam que o recrutamento de organismos bentônicos marinhos é definido
como o número de novos indivíduos que assentam e sobrevivem no substrato, sendo
considerado um indicador dos padrões de suprimento de larvas, das diferenças na
escolha ativa da larva pelo substrato e das taxas de mortalidade pós-assentamento. Este
processo é muito importante para a compreensão da extensão de um fenômeno
ecológico, para a estrutura da comunidade (CALEY et al., 1996).
As fases iniciais da colonização de organismos incrustantes nos substratos
submersos na água são compostas predominantemente por bactérias, protozoários,
diatomáceas e algas (WHOI, 1952; PERSONNE, 1968). Logo após a imersão dos
substratos as bactérias iniciam sua fixação atingindo um número apreciável em poucas
horas (WHOI, 1952). A presença de diatomáceas, algas e protozoários recrutam mais
lentamente, atingindo o seu desenvolvimento máximo depois de alguns dias ou semanas
(HEULEKIAN & CROSBY, 1956; SKERMAN, 1956; O׳NEILL & WILCOX, 1971;
RELINI, 2003).
São conhecidos dois fatores primários que influenciam a estrutura de uma
comunidade incrustante no seu estágio inicial de desenvolvimento e na direção
subsequente da sucessão (LEVINTON, 1982). O primeiro são os eventos anteriores ao
assentamento, como dispersão (SCHELTEMA, 1986) e a probabilidade de colonização
de um habitat (GAINES & ROUGHARDEN, 1985) e, o segundo, os processos de
seleção pós-assentamento (SAMMARCO, 1980).

Thorson (1946) sugeriu que muitas larvas pelágicas têm a capacidade de
retardar a sua fixação até o encontro com o substrato adequado, hipótese essa
confirmada por Bayne (1965). Paranaguá (1972) apontou a alta taxa de mortalidade no
estágio larval da Mytella falcata em aquário, por causa da ausência de um substrato para
o seu assentamento.
Fatores abióticos, como a presença de adultos, predadores e competidores por
espaço (CONNEL, 1975; GROSBERG, 1982); e fatores abióticos como luminosidade,
temperatura, salinidade, rugosidade do substrato e sedimentação têm sido observados
influenciando os padrões no processo de recrutamento, podendo induzir ou inibir o
assentamento, características químicas no substrato também podem exercer ação
atrativa para os animais bentônicos no processo de recrutamento e também na sucessão
(SAMMARCO, 1980; CARLETON & SAMMARCO, 1987; HOGDSON, 1990;
MAIDA et al., 1994).
Sucessão é um processo ordenado e direcional, resultante de modificações do
ambiente físico pelos organismos presentes, este resultado é gerado por diferenças no
comportamento evolutivo entre espécies mais oportunistas e menos oportunistas
(ABSALÃO 1993). Odum (1985) define sucessão como uma modificação direcional
observada na composição da comunidade, ao longo do tempo. Ao longo do processo de
recrutamento e sucessão, a dinâmica de uma comunidade vai definindo seus
componentes e neste processo alguns grupos de organismos expressam características
próprias muito importantes. Para Brower & Zar (1984), o processo de sucessão e uma
reposição progressiva de uma comunidade por outra envolve, não somente uma
mudança na composição de espécies, mas também alterações na biomassa e nas
características do ambiente. Sendo que a análise de um processo de sucessão primária,

utilizando um substrato limpo, pode mostrar importantes aspectos estruturais da
comunidade ao longo do tempo.
Pinto & Coelho (2000) destacam que a modificação na comunidade pode estar
ligada a distúrbios externos e perturbações causadas pelo homem. Distúrbios bióticos e
abióticos podem alterar a estrutura da comunidade marinha, alterando padrões de
sucessão ecológica formando um conjunto de manchas que, em conjunto com o
recrutamento aleatório, podem formar comunidades com características de mosaicos
(SAUER-MACHADO et al., 1992).
Características físicas ou as variáveis biológicas, tais como: a competição e a
predação explicam geralmente diferenças espaciais e mudanças temporais nas faixas dos
recifes, que são dominadas por diferentes espécies. A presença de determinadas
espécies, sua abundância relativa, qualidade e quantidade de larvas no plâncton são
fatores que influenciam na estrutura das comunidades (APOLINÁRIO, 1999).
A dinâmica da sucessão em comunidades pode ainda ser influenciada por
interações entre o recrutamento de espécies e por características das aberturas criadas
por distúrbios. Essas aberturas criadas dentro das comunidades sésseis em substrato
emergente podem ser colonizadas pelas larvas e propágulos que vêm da coluna de água,
e pela migração lateral da espécie vizinha, incluindo o crescimento vegetativo, de
macroalgas e espécies coloniais (TANAKA & MAGALHÃES, 2002).
A área de estudo foi o Porto do Recife localizado na cidade do Recife, capital do
estado de Pernambuco nas margens das desembocaduras dos rios Capibaribe e
Beberibe, localiza-se entre as coordenadas geográficas 8º 03' de latitude sul e 34º 51' de
longitude oeste, limitados a leste pelo Oceano Atlântico, ao norte por uma linha leste-
oeste a quatro quilômetros ao norte do Farol do Picão (VILLAR, 1992).

De acordo com Ottmann & Coutinho (1963) e Mabessone & Coutinho (1970),
podem-se distinguir as seguintes zonas, em relação aos processos de sedimentação do
Porto do Recife: uma composta por sedimentos de areia grossa e que forma a Coroa dos
Passarinhos (Zona do Pina), outra composta por lama fina (Zona do Porto) e a terceira
que é a zona de transição ou central. Sendo a Zona do Pina composta por areia de
quartzo e fragmentos de organismos. A Zona do Porto é constituída por sedimento fino
e de coloração escura de origem fluvial. Na Zona central encontramos uma lama rígida,
estratificada e cinza escuro com a superfície oxidada de origem fluvial marinha.
Em um determinado ambiente, as espécies podem ser consideradas como nativas,
criptogênicas ou exóticas, relativas aos seus aspectos de origem, distribuição e
comportamento bioecológico em um determinado local (ENO et al., 1997; BAKER et al.,
2004; CLARKE et al., 2003; FARRAPEIRA et al., 2011). O enquadramento de uma
espécie nestas categorias requer, necessariamente, o conhecimento prévio da biota do local
em questão, da biologia e biogeografia da espécie-alvo e das possíveis rotas de introdução
(CHAPMAN & CARLTON, 1991).
Assim, consideram-se espécies nativas (indígenas), aquelas que ocorrem
naturalmente em um ambiente, sendo um membro da comunidade da região, cuja presença
geralmente tem registro paleontológico. As espécies exóticas (introduzidas, não-indígenas,
alienígenas) são aquelas observadas em uma área fora de seu limite natural historicamente
conhecido de distribuição, como resultado de dispersão acidental ou transferência
intencional por atividades humanas. A designação de espécies criptogênicas foi criada por
Carlton (1996) e revisada pelo mesmo autor (CARLTON, 2009), para contemplar aquelas
que possuem ampla distribuição geográfica em baías, portos e estuários do mundo, podendo
até ter registros paleontológicos em várias regiões do mundo, incluindo, portanto, as
espécies cuja origem não é facilmente demonstrada a partir de uma data provável.

De acordo com Crooks & Soulé (1999), a partir do momento em que uma
espécie exótica ou criptogênica é introduzida em um determinado local, só um pequeno
número de organismos sobreviverá e se estabelecerá no novo ambiente e uma porção
ainda menor, provocará impactos negativos. Além de serem necessárias muitas
inoculações de espécimes no novo ambiente para que a introdução das espécies seja
bem sucedida, há que ser considerada a condição fisiológica do propágulo na sua
chegada à região receptora, que depende, por sua vez, do organismo (tamanho, ciclo de
vida e/ou idade), do vetor usado para a sua transferência e das condições ambientais dos
locais (alimento, temperatura, estação do ano, hora do dia e outras variáveis) (RUIZ et
al., 1997; CHAPIN III et al., 1998; SMITH et al., 1999).
Se a espécie introduzida ocupar o mesmo nicho ecológico ou usar o mesmo
recurso ambiental das espécies nativas residentes, esses organismos, livres de seus
predadores, parasitas e competidores naturais, podem competir com os nativos por
alimento e/ou espaço dividindo nichos, atingindo altas densidades populacionais,
acarretar em um decréscimo de biodiversidade e se dispersar de maneira rápida e
agressiva para além das proximidades dos focos de introdução, quando passam a ser
denominados de (bio)invasores (RUIZ et al., 1997; 2000; ORENSANZ et al., 2002;
BAX et al., 2001; GODWIN, 2003; TILMAN, 2004; STREFTARIS et al., 2005).
O processo de bioinvasão é definido, portanto, como a chegada, o
estabelecimento, a proliferação e a expansão de distribuição geográfica pela dispersão
local e regional de uma espécie exótica ou criptogênica em um local que não é o seu
habitat natural, afetando as comunidades nativas (GROSHOLZ & RUIZ 1995;
ABRAMS, 1996; JOHNSON & PADILLA, 1996; REUSCH & WILLIAMS, 1998; MACK
et al., 2000; OCCHIPINTI-AMBROGI & GALIL, 2004). É importante ressaltar que toda a
trajetória geográfica de introdução e invasão de uma espécie em uma nova localidade, desde

seus estágios iniciais até os subsequentes, caracterizando a bioinvasão, é influenciada pela
combinação de condições ecológicas requeridas pelo invasor e a dinâmica de sua dispersão
(JOHNSON & PADILLA, 1996).

REFERÊNCIAS
ABELSON, A.; DENNY, M. 1997. Settlement of marine organisms in flow. Annual
Review in Ecology and Systematics, Palo Alto, v. 28, p. 317–39.
ABSALÃO, R. S. 1993. Colonização primária e sequência de substituições, em
substratos artificial, na comunidade epibêntica da Baia da Guanabara, RJ - Brasil.
Tese: Doutorado. Instituto de Biociências, Universidade de São Paulo. 220pp.
ABRAMS, P.A. 1996.Evolution and the consequences of species introductions and
deletions. Ecology, Tempe, v. 77, p. 1321-1328.
APOLINÁRIO, M. 1999.Temporal variations in community structure in and around
intertidal barnacle Chthamalus challengeri Hoek patches on a plebby shore in Japan.
Revista Brasileira de Biologia, v. 59, n. 1, p. 4-53.
BAKER, P.; BAKER, S. M.; FAJANS, J. 2004. Nonindigenous marine species in the greater
Tampa Bay ecossystem. Final Report. Department of Fisheries and Aquatic Sciences,
University of Florida, pp. 131
BAX, N.; CARLTON, J.T.; MATHEWS-AMOS, A.; HAEDRICH, R.L.; HOWARTH,
F.G.; PURCELL, J.E; RIESE, A.; GRAY, A. 2001. The control of biological invasions
in the world’s ocean. Conservation Biology, Boston, v. 15, n. 5, p. 234-1246.
BAYNE, B.L. 1965. Growth and the delay of metamorphosis of the larva of Mytilus
edulis (L.). Ophelia, Helsinki, v. 2, n. 1, p. 1-47.
BROWER, J.E.; ZAR, J.H. 1984. Field and laboratory methods for general ecology.
2.ed. Dubuque: W. C. Brown Company, 226p.

CALEY, M.J.; CARR, M.H.; HIXON, M.A.; HUGHES, T.P.; JONES, G.P.; MENGE,
B. A. 1996. Recruitment and the local dynamics of open marine populations. Annual
Review of Ecology and Systematics, Palo Alto, v. 27, p. 477-500.
CARLTON, J.T. 1996. Biological invasions and cryptogenic species. Ecology, Tempe,
v. 77, p. 1653-1655.
CARLTON, J.T. 2009. Deep invasion ecology and the assembly of communities in
historical time. In: Rilov, G.; Crooks, J.A. (eds.). Biological invasions in marine
ecosystems. Ecological Studies 204. Berlin: Springer-Verlag, p. 13-56.
CARLETON, J.C.; SAMMARCO, P.W. 1987. Effects of substrate irregularity on
success of coral settlement: Quantification by comparative geomorphological
techniques. Bulletin of Marine Science, Miami, v. 40, n. 1, p. 85-98.
CHAPIN III, F.S.; SALA, O.E.; BURKE, I.C.; GRIME, J.P.; HOOPER, D.U.;
LAURENROTH, W.K.; LOMBARD, A.; MOONEY, H.A.; MOISER, A.R.; NAEEM,
S.; PACALA, S.W.; ROY, J.; STEFFEN, W.L.; TILMAN, D. 1998. Ecosystem
consequences of changing biodiversity; experimental evidence and a research agenda
for the future. Bioscience, Washington, v. 48, p. 45-52.
CHAPMAN, J.W.; CARLTON, J.T. 1991 .A test of criteria for introduced species: the
global invasion by the isopod Synidotea laevidorsalis (Miers, 1881). Journal of
Crustacean Biology, Woods Hole, v. 11, p. 386-400.
CLARKE, C.; HILLIARD, R. ; JUNQUEIRA, A.O.R. ; NETO, A.C.L.; POLGLAZE,
J.; RAAYMAKERS, S. 2003. Ballast water risk assessment, Port of Sepetiba, Federal
Republic of Brazil, December 2003: final report. London: GloBallast Monograph Series
No.14. IMO, 131p.

CONNELL, J.H. 1975. Some mechanisms producing structure in natural communities:
a model and some evidence from field experiments. In: Cody M.L.; Diamond, J.M.
(eds.) Ecology and evolution of communities. Cambridge: Belknap Press, pp. 460-490.
CROOKS, J.A.; SOULÉ, M.E. 1999. Lag times in population explosions of invasive
species: causes and implications. In: Sandlund, O.T.; Schei, P.J.; Viken, A. (eds.).
Invasive species and biodiversity management. Dordrecht: Kluwer Academic
Publishers, p. 103-125.
DONE, T. J. 1982. Patterns in the distribution of coral communities across the central
Great Barrier Reef. Coral reefs, 1: 95-107.
ENO, N.C.; CLARK, R.A.; SANDERSON, W.G. (Eds.) 1997. Non-native marine
species in British waters: a review and directory. Peterborough: Joint Nature
Conservation Committee, 136 p.
FARRAPEIRA, C.M.R. 2011. Invertebrados macrobentônicos detectados na costa
brasileira transportados por resíduos flutuantes sólidos abiogênicos. Revista de Gestão
Costeira Integrada, Lisboa, v. 12, n. 1, p. 1-28,
GAINES, S. S. B.; ROUGHGARDEN, J. 1985. Larval settlement rate: A leading
determinant of structure in an ecological community of the marine intertidal zone.
Proceedings of the National Academy of Sciences of the United States of America,
Washington, v. 82, p. 3707-3711,
GODWIN, L.S. 2003. Hull fouling of maritime vessels as a pathway for marine species
invasions to the Hawaiian Islands. Biofouling, Chur, v. 19 (Supplement), p. 123-131,
GROSBERG, R. K. 1982. Competitive ability influences habitat choice in marine
invertebrates. Nature, London, v. 290, p. 270-702,

GROSHOLZ, E.D.; RUIZ, G.M. 1995. Spread and potential impact of the recently
introduced European green crab, Carcinus maenas, in central California. Marine
Biology, Berlin, v. 122, n. 2, p. 239-247,
JOHNSON, L.E.; PADILLA, D.K. 1996. Geographic spread of exotic species:
ecological lessons and opportunities from the invasion of the zebra mussel Dreissena
polymorpha. Biological Conservation, Essex, v. 78, p. 23-33,
HEULEKIAN, H.; CROSBY, E. S. 1956. Slime formation in polluted waters. III-
Nature and composition of slimes. Sewage Industrial Wastes, v. 28, n. 2, p. 206-210,
HOGDSON, G. 1990. Sediment and the settlement of larvae of the reef coral
Pocillopora daminicornis. Coral Reefs. 9: 41-43.
KEOUGH, M. J. 1988. Benthic populations: is recruitment limiting or just fashionable?
In: Internacional Coral Reef Symposium, 6, Townsville, 1988. Proceedings...
Townsville, 1: 141-148.
KEOUGH, M.J.; DOWNES, B.J. 1982. Recruitment of marine invertebrates: The roles
of active larval choice and early mortality. Oecologia, Berlin, v. 54, p. 348-352, 1982.
LEVINTON, J. S. Marine ecology. Prentice hall inc. Englewoods Cliffs New Jersey.
USA.
MABESSONE, J. M. COUTINHO, P. N. 1970. Litoral and shalow marine geology of
norheastern Brasil. Trabalhos do Instituto Oceanográfico. Universidade Federal de
Pernambuco, Recife, v. 12. p. 1-214.
MACK, R.N.; SIMBERLOFF, D.; LONSDALE, W.M.; EVANS, H.; CLOUT, M.;
BAZZAZ, F.A. 2000. Biotic invasions: causes, epidemiology, global consequences, and
control. Ecological Applications, Tempe, v. 10, n. 3, p. 689-710.

MAIDA, M.; COLL, J. C.; WILSON, E. O. 1994. Sheddeng new ligh on sclerectinian
coral recruitment. Journal of Experimental. Marine Biology and. Ecology. 180. 180 –
202.
MARIANI, S.; URIZ, M.-J.; TURON, X.; ALCOVERRO, T. 2006. Dispersal strategies
in sponge larvae: integrating the life history of larvae and the hydrologic component.
Oecologia, Berlin, v. 149, n. 1, p. 174-184.
MENGE, B. A.; SUTHERLAND, J. P. 1976. Species diversity gradients: synthesis of
the roles of predation, competition, and temporal heterogeneity. Am Nat 110:351–369
MILEIKOVSKY, S.A. 1974. Types of larval development in marine bottom
invertebrates: an integrated ecological scheme. Thalassia Jugoslavica, Zagreb, v. 10, n.
1/2, p. 171-180.
OCCHIPINTI-AMBROGI, A.; GALIL, B.S. 2004. A uniform terminology on
bioinvasions: a chimera or an operative tool? Marine Pollution Bulletin, Oxford, v. 49,
p. 688-694,
ODUM. E. P. 1985. Ecología. Rio de Janeiro: Interamericana, 434 pp.
ORENSANZ, J.M.; SCHWINDT, E.; PASTORINO, G.; BORTOLUS, A.; CASAS, G.;
DARRIGRAN, G.; ELÍAS, R.; GAPPA, J.J.L.; OBENAT, S.; PASCUAL, M.;
PENCHASZADEH, P.; PIRIZ, M.L.; SCARABINO, F.; SPIVAK, E.D.;
VALLARINO, E. A. 2002. No longer the pristine confines of the world ocean: a survey
of exotic marine species in the southwestern Atlantic. Biological Invasions, New York,
v. 4, p. 115-143.
O ' NEILL, T. B.; WILCOX, G. L. 1971. The formation of a “primary film” on the
materials submerged in the sea at Port Hueneme, California. Pacific Science, Honolulu,
v. 25, p. 1-12.

OTTMANN, F. ; COUTINHO, P. N. 1963. Études sedimentologiques dans le Port de
Recife (Brasil). Cahiers Oceanographiques, n. 15, p. 161-169.
PARANAGUÁ, M. N. 1972. Distribuição, ecologia e desenvolvimento de Mytella
charruana nos estados do Rio de Janeiro e Guanabara, RJ – Brasil. Tese de Doutorado.
Instituto de Biociências e Instituto de Pesquisa da Marinha do R.J, Universidade Federal
de Pernambuco. p.64.
PERSONNE, G. 1968. Ecologie des infusoires dans les salissures de substrats immerges
dans un port de mer. I- Le film primaire et le recouvremert primaire. Protistologiaca, v.
4, n. 2, p 187-194.
PINTO-COELHO, R. M. 2000. Fundamentos em ecologia. Porto Alegre: Artes Médicas
Sul.
RELINI, G. 2003. Il biofouling. Parte prima. Il macrofouling. Biologia Marina
Mediterranea, s.l., v. 10, p. 285-326.
REUSCH, T.B.H.; WILLIAMS, S. L. 1998. Variable responses of native eelgrass
Zostera marina to a non-indigenous bivalve Musculista senhousia. Oecologia, Berlin, v.
113, p. 428-441.
RUIZ, G.M.; CARLTON, J.T.; GROSHOLZ, E.D.; HINES. A.H. 1997. Global
invasions of marine and estuarine habitats by non-indigenous species: mechanisms,
extent, and consequences. American Zoologist, Thousand Oaks, v. 37, n. 6, p. 619-630.
RUIZ, G.M.; FOFONOFF, P.; CARLTON, J.T.; WONHAM, M.J.; HINES, A.H. 2000.
Invasion of coastal marine communities in North America: apparent patterns, processes,
and biases. Annual Review in Ecology and Systematics, Palo Alto, v. 31, p. 481-531.

SAMMARCO, P. W. 1980. Diadema and its relationship to coral spat mortality:
grazing, competition and biological disturbance. Journal of Experimental Marine
Biology and Ecology, Amsterdan, v. 45, p. 245-272.
SAUER-MACHADO, K. R. S.; CHAPMAN, A. R. O.; COUTINHO, R. 1992. Patch
structure in a tropical rocky shore community in Brazil: a mosaic of sucessional states?
Ophelia, v. 35, p. 187 – 195.
SCHELTEMA, R. S. 1986. .On dispersal and planktonic larvae of benthic invertebrates.
An eclectic overview and summary problems. Bulletin of Marine Science, Coral Gables,
v. 39, p. 290-322.
SKERMAN, T. M. 1956. The nature and development of primary films on surfaces
submerged in the ses. N. Z. JLSCI. Technol., B. 38(1): 44-57.
SMITH, L. D.; WONHAM, M. J.; MCCANN, L. D.; RUIZ, G. M.; HINES, A. H.;
CARLTON, J. T. 1999. Invasion pressure to a ballast-flooded estuary and an
assessment of inoculant survival. Biological Invasions, New York, v. 1, n. 1, p. 67-87,
STREFTARIS, N.; ZENETOS, A.; PAPATHANASSIOU, E. 2005. Globalization in
marine ecosystems: the story of non-indigenous marine species across European seas.
Oceanography and Marine Biology: An Annual Review, Aberdeen, v. 43, p. 419-453,
TANAKA, M. O.; MAGALHÃES, C. A. 2002. Edge effects and succession dynamics
in Brachidontes mussel beds. Marine Ecology Progress Series, v. 237, p. 151-158,
TILMAN, D. 2004. Niche tradeoffs, neutrality, and community structure: a stochastic
theory of resource competition, invasion, and community assembly. Proceedings of the
National Academy of Sciences of the United States of America, Washington, v. 101, n.
30, p. 10854-10861.

THORSON, G. 1946. Reproduction and larval development of danish marine botton
invertebrates. Medd. Fisk. Og. Havunders., ser. Plankton, 4: 1-523.
VILLAR, G. J. V. 1992. Porto do Recife – Uma Abordagem Estrutural. Monografia,
Universidade Católica de Pernambuco – PE.
WAHL, M. 1989. Epibiosis. I. Fouling and antifouling: some basic aspects. Marine
Ecology Progress.Series. 58: 189.
WHOI. 1952. Marine fouling and its prevention. Woods Hole Oceanographic
Institution, Annapolis, 388 p.

RECRUTAMENTO E SUCESSÃO DE ANIMAIS SÉSSEIS E SEDENTÁRIOS
EM SUBSTRATO ARTIFICIAL SUBMERSO NA ÁREA PORTUÁRIA DO
RECIFE – PE, COM ENFOQUE PARA A BIOINVASÃO.
ARTHUR V. O. M. MELO 1; RALF
SCHWAMBORN,
1; CRISTIANE M. R.
FARRAPEIRA 2
1. Depto. de Zoologia, Universidade Federal de Pernambuco (UFPE). Recife, Brasil.
*E-mail – [email protected]
2. Depto. de Biologia, Universidade Federal Rural de Pernambuco (UFRPE). Recife,
Brasil.
RESUMO
O Porto do Recife recebe uma grande quantidade de navios de vários tipos e de muitas
partes do mundo, podendo trazer espécies exóticas incrustadas no casco ou em água de
lastro. O objetivo do presente estudo foi registrar e analisar os padrões de recrutamento
e a dinâmica de sucessão de espécies bentônicas sésseis, assim como investigar uma
possivel introdução e estabelecimento de espécies bentônicas exóticas e criptogênicas
em uma área portuária com grande fluxo de navios. Utilizaram-se placas de metal
galvanizado para estudo do recrutamento e sucessão de animais sésseis e sedentários em
três estações no porto, com três réplicas cada, totalizando nove pontos, durante seis
meses (outubro de 2010 a março de 2011), considerado como período seco. As placas
de recrutamento foram retiradas a cada 30 dias durante seis meses, as placas de sucessão
foram retiradas após 60 dias e depois de sucessivos 30 dias até completarem 180 dias de
submersão. As principais espécies de acordo com a área de cobertura foram Mytella
charruana (72%) e Amphibalanus improvisus (22%) e espaços vazios (7,5%). Não
houve diferença significativa quando comparadas as três estações no porto do Recife,
sendo encontradas diferenças quando comparadas as áreas de cobertura das espécies
mais abundantes no recrutamento. Foram elas: Clytia gracilis, Obelia dichotoma, M.
charruana e A. improvisus. Na comparação entre as estações de acordo com a área de
cobertura das espécies mais abundantes na sucessão houve diferenças significativas

apenas para M. charruana e A. improvisus. Observou-se o estabelecimento das espécies
exóticas Haliplanella lineata (Cnidaria) e Sinelobus stanfordi (Arthropoda). Mytella
charruana foi considerada a espécie clímax na sucessão ecológica, já que foi a única
espécie encontrada em grande quantidade e ocupando todas as placas. Observou-se a
maior riqueza das espécies exóticas e criptogênicas em detrimento das espécies nativas,
tanto no recrutamento quanto na sucessão.
Palavras chaves: Animais Bentônicos, predação, espécies exóticas, sucessão ecológica.
ABSTRACT
The Port of Recife receives a large number of vessels of various types and from many
parts of the world, and can bring exotic species encrusted hull or in ballast water. The
aim of this study was to record and analyze the patterns of recruitment and succession
dynamics of sessile benthic species, as well as investigating a possible introduction and
establishment of exotic and cryptogenic benthic species in a port area with great flow
vessels. We used galvanized metal plates to study the recruitment and succession of
sessile and sedentary animals at three stations in the harbor, with three replicas each,
totaling nine points, six months (October 2010 to March 2011), considered the dry . The
plates recruitment were taken every 30 days for six months, the plates were removed
succession after 60 days and after successive 30 days up to 180 days of submersion. The
main species according to coverage area were Mytella charruana (72%) and
Amphibalanus improvisus (22%) and voids (7.5%). There was no significant difference
when comparing the three stations in the Port of Recife, differences were found when
comparing the coverage areas of most abundant species in recruitment. They were:
Clytia gracilis, Obelia dichotoma, M. charruana and A. improvisus. Comparing the
stations according to the coverage area of the most abundant species in succession

significant differences only for M. charruana and A. improvisus. We observed the
establishment of exotic species Haliplanella lineata (Cnidaria) and Sinelobus stanfordi
(Arthropoda). Mytella charruana was considered the climax species in ecological
succession, since it was the only species found in large quantities and occupying all
boards. We observed the greatest wealth of exotic and cryptogenic to the detriment of
native species, both in recruiting and succession.
Keywords: benthic animals, predation, alien species, ecological succession.
INTRODUÇÃO
O recrutamento é a ocupação inicial de um substrato por organismos incrustantes
“fouling” (TOMMASI et al., 1972; BAKER et al., 2004), que sobrevivem durante certo
período de tempo após o assentamento (RODRIGUEZ et al., 1993; BOOTH & BROSNAM,
1995). Em uma placa experimental submersa, desenvolvem-se microrganismos após
alguns minutos, formando um biofilme ou micro-incrustação (CALLOW & CALLOW,
2002). Logo após esta fase, larvas de vários organismos bentônicos começam a recrutar
(WHOI, 1952; OTSUKA & DAUER, 1982), ocorrendo sucessão ecológica (ODUM, 1988).
De acordo com SUTHERLAND (1974) quatro semanas seria um período suficiente
para estudar o recrutamento sobre substratos submersos, Nery et al. (2008) destacou que
cinco meses são suficiente para descrever a sucessão ecológica na Bacia Portuária do
Recife.
A sucessão ecológica é um fenômeno natural que envolve mudanças nos
ecossistemas ao longo do tempo, como consequência da colonização de um substrato, e
das interações biológicas estruturadoras das comunidades (SAUER-MACHADO et al.,
1992). O padrão e a velocidade da sucessão são determinados pelo ambiente (ODUM,
1988), e resultam em um incremento de informação no ecossistema (MARGALEF, 1968).

Num determinado ambiente as espécies podem ser consideradas nativas,
criptogênicas ou exóticas (ENO et al., 1977). CARLTON (2009) definiu como nativas as
espécies ocorrentes naturalmente em um ambiente, com registro paleontológico; como
exóticas, as observadas fora de seu limite de distribuição historicamente conhecido,
resultado de dispersão acidental ou transferência intencional por atividades humanas; e
como criptogênicas, aquelas cuja origem não é facilmente demonstrada e possuem
ampla distribuição geográfica em baías, portos e estuários do mundo.
Nenhum habitat está imune à introdução de espécies exóticas, mas comumente
aquelas que se tornam invasoras ocorrem em portos e marinas e circunvizinhanças
(GOULLETQUER et al., 2002; COHEN et al., 2005; RAMADAN et al., 2006; FARRAPEIRA et
al., 2009; ROCHA et al., 2012). Estudos de colonização de invertebrados permitem
conhecer a fauna local, assim como as mudanças ocorrentes na comunidade ao longo do
tempo (CARVALHO & UIEDA, 2004). As atividades de navegação constituem um dos
principais meios de introdução de espécies em novas áreas (LEWIS et al., 2003), fato que
vem aumentando devido ao aumento do comércio internacional (FOFONOFF et al., 2003;
CARLTON, 2009; FARRAPEIRA et al., 2011).
Estudos com placas de recrutamento têm sido realizados com o intuito de
observar os processos de colonização e sucessão ecológica, e a introdução de espécies
exóticas, em várias partes do mundo (COE, 1932; PECH et al., 2002; RAMADAN et al.,
2006; FREESTONE et al., 2009). No Brasil, vários autores demonstraram a técnica como
bom instrumento de gestão e diagnóstico ambiental, ao definir composição faunística e
introdução de exóticas (MAYER-PINTO et al., 2000, SILVA et al., 2001, BREVES-RAMOS
et al., 2005, FERNANDES et al., 2006, NEVES et al., 2007, XAVIER et al., 2008, NERY et
al., 2008, CANGUSSU et al., 2010).

No Porto de Recife, por receber muitas embarcações de outros países (Farrapeira
et al., 2007), faz-se necessário entender os processos de recrutamento e sucessão dos
animais bentônicos, e monitorar a introdução de espécies exóticas no litoral
pernambucano. Assim, o objetivo do presente estudo foi investigar uma possível
introdução e estabelecimento de espécies bentônicas exóticas e/ou criptogênicas em
uma área portuária com grande fluxo de navios. Buscou-se registrar e analisar os
padrões de recrutamento e a dinâmica de sucessão de espécies bentônicas sésseis.
MATERIAIS E MÉTODOS
O trabalho foi realizado no Porto do Recife (8º03' S e 34º51' W) no Estado
de Pernambuco. A bacia portuária é formada pela desembocadura do rio Capibaribe e
dos rios componentes da Bacia do Pina (rios Tejipió, Jiquiá, Pina e Jordão
(FARRAPEIRA, 2006). O porto apresenta grande impacto causado pelo número de
atracamento de embarcações (exportação, importação e transatlânticos), recebe uma
média anual de 491 atracações de navios de (FARRAPEIRA et al., 2007).
O local, de acordo com FARRAPEIRA et al. (2010), funciona também como um
reservatório de descarga de indústria e esgoto doméstico, com coliformes e bactérias
termo-tolerantes presentes na água. O ambiente é considerado como um complexo
estuarino–fluvial, com grande aporte de água doce (CPRH, 2001). A salinidade varia
com o aumento da pluviometria no local, de cinco a 22, durante as marés baixas nos
meses chuvosos, quando ocorre um forte aporte de água doce, enquanto que na estação
seca alcança valores de 32, chegando ao máximo de 37 em preamares (SOMERFIELD et
al., 2003; NERY et al., 2008; FARRAPEIRA et al., 2009). Por conta da grande
sedimentação que ocorre na área, FERNANDES et al. (2006) observaram que a
transparência da água atinge apenas uma profundidade média de 0,66cm.

A área apresenta um clima do tipo As’ denominado Tropical Quente Úmido,
segundo a classificação de Köppen, com uma média anual de temperatura de 25°C e da
água variando entre 24 a 32°C, e uma precipitação média de 1.763 mm, dos quais, 80%
caem durante o período de março a agosto (inverno) (SOMERFIELD et al., 2003).
CARACTERIZAÇÃO DO EXPERIMENTO
O período de estudo foi de 30 de setembro de 2010 a 27 de março de 2011,
totalizando seis meses. O paredão de atracagem do porto foi dividido em três pontos:
estação I (Terminal Açucareiro), II (Terminal Graneleiro) e III (Cais inativo) (Fig. 1). A
estação I é o cais de atracagem dos navios açucareiros e sofre maior influência marinha
e dos estuários dos rios Capibaribe e Beberibe. A estação II localiza-se na região
mediana da Bacia Portuária, sendo a área de maior atividade de atracagem de navios de
grãos e transatlânticos. A estação III é conhecida como Marco Zero e fica na parte mais
interna do porto, onde não ocorreram atividades portuárias durante o período do
experimento. Em cada estação três equipamentos foram instalados de forma
equidistante, a 100 metros um do outro, como réplicas nas suas respectivas estações.
Figura 1. Vista aérea da Bacia Portuária de Recife- PE, sinalizando as áreas de instalação dos
experimentos. Estação I, local de atracagem dos navios açucareiros. Estação II, local de atracagem de
navios graneleiros. Estação III, sem atividades portuárias. Fonte: Google earth.

Foram realizados dois experimentos para retratar a dinâmica de recrutamento e
sucessão dos animais sésseis. Para estudar o recrutamento era removida uma placa de
cada equipamento, a cada 30 dias, para análise do recrutamento ocorrido neste período
(SUTHERLAND, 1974). Após a remoção, outra placa limpa era instalada em substituição,
a fim de que esta representasse o recrutamento do mês subsequente. Este procedimento
foi repetido até o fim do período de estudo. Após 60 dias, uma placa de cada
equipamento foi removida a fim de representar a sucessão ocorrida no período e,
passados trinta dias, mais uma placa era removida, representando a sucessão ocorrida
em 90 dias, e assim sucessivamente até os 180 dias de submersão. Esse tempo foi
baseado nas observações de NERY et al. (2008), que verificaram que o desenvolvimento
da comunidade bentônica em placas na área estudada atinge seu clímax em cinco meses.
Cada equipamento era formado por uma base de vergalhão de ferro e continham
seis placas de metal galvanizado com 20 cm de comprimento por 10 cm de largura e 0,5
cm de espessura,distando 20 cm entre si, totalizando 54 placas. A estrutura foi mantida
presa às defensas do porto, a um metro do menor nível de maré, durante a baixa mar
(SILVA et al. 2001; NERY et al., 2008).
Os corpos de provas (placas) removidos foram acondicionados em sacos
plásticos etiquetados, contendo água do local e posteriormente fixadas em
formolaldeído a 4%, para ulterior triagem e identificação. Para a leitura das placas,
utilizou-se uma placa de leitura com 10 cm x 8 cm, contendo 80 quadrículas e
interseções de 1 x 1 cm. A leitura das placas foi feita pelo método de contagem dos
organismos presentes no ponto de interseção das quadrículas, considerando que este
método é um dos mais eficientes quando comparado com outros (SILVA et al., 2001;
MACEDO et al., 2006; NERY et al., 2008).

A área da placa que não se enquadrou na rede de quadrículas foi observada para
identificação taxonômica dos invertebrados, sendo considerada como fauna associada,
para melhor caracterização da biota, seguindo NERY et al. (2008).
Nos corpos de prova (placas) de sucessão foram ainda observadas a
sobreposição de espécies, para identificação de basibiontes e epibiontes, na acepção de
WAHL (1989), observando-se também os que estavam vivos ou mortos, assim como a
presença de predadores naturais. Foram considerados basibiontes, aqueles indivíduos
que recrutaram na placa, e epibiontes, os que recrutaram sobre estes (MAYER-PINTO et
al., 2000). Em cada mês foram contados os estratos de organismos, calculado o tamanho
médio dos epibiontes e a medidas das espessuras das placas.
Foram calculadas as áreas de cobertura e as frequências relativas das espécies
em todos os meses, tanto no recrutamento mensal, quanto na sucessão. Segundo NERY
et al. (2008), as espécies foram classificadas quanto à frequência, em: muito frequente
(>70%), frequente (30% a 70%), pouco frequente (10% a 30%) e raras (<10%).
A classificação das espécies quanto ao status de distribuição geográfica e de
origem seguiu a definição de FARRAPEIRA et al. (2011), considerando espécies nativas
aquelas com uma longa presença na área geográfica estudada; as exóticas, as que foram
trazidas de outras regiões, por apresentarem descontinuidade geográfica grande entre a
região onde foram originalmente descritas como nativas e a área onde foram
encontradas na costa brasileira, e criptogênicas, aquelas em que não havia nenhum tipo
de informação sobre sua localidade de origem ou que não foram classificadas como
nativas ou exóticas de nenhuma região específica.
As abundâncias de cada estação foram comparadas usando a ANOVA Kruskal-Wallis,
usando o software BioEstat 5.0. Foi gerada uma matriz de similaridade de Bray-Curtis sobre o
log (x+1) da abundância dos animais em recrutamento e a sucessão mais recente das

abundâncias das populações mais abundantes em cada equipamento. A partir desta matriz foi
feita uma ANOSIM (CLARKE & WARWICK, 2001) usando o software Primer 6.0. Os dados
pluviométricos foram obtidos do Instituto Nacional de Meteorologia – Inmet.
RESULTADOS
Os experimentos foram eficientes por registrarem as dinâmicas de recrutamento
e sucessão nas condições ambientais do Porto do Recife. Espécies exóticas,
criptogênicas e nativas foram registradas, sobrevivendo aos impactos locais.
A preciptação variou de 23,5 a 329,5 mm (Fig. 2), destacando-se que no período
de dezembro de 2010 a fevereiro de 2011 houve mais chuvas do que a média histórica
registrada para o período.
0
50
100
150
200
250
300
350
Outubro Novembro Dezembro Janeiro Fevereiro Março
2010 - 2011
Plu
vio
sid
ad
e (
mm
)
Media mensal Media histórica
Fig 2. Pluviosidade média mensal do Porto do Recife, Pernambuco, durante os meses de outubro de 2010
a março de 2011. INMET.
Experimentos de Recrutamento
Nos corpos de provas (placas) foram encontrados 30 táxons pertencentes a seis
filos animais, dentre os quais foram identificadas 16 espécies, os demais, pertencentes
ao Filo Polychaeta, identificados no nível de Família (Tab. 1). Foram perdidas três
placas, uma no mês de fevereiro (estação I), duas no mês de março (estação III).
Com relação à fauna associada, além das espécies já mencionadas (Tab. I),
foram encontrados o mexilhão sedentário Mytilopsis leucophaeta Conrad, 1831 e a
fauna vágil: planária Distylochus martae Marcus, 1947, gastrópodos Melanoides

tuberculatus Müller, 1774, Nassarius vibex Say, 1822, Rissoina cancelata Philippi,
1847, poliquetas das famílias Ampharetidae, Cirratulidae, Dorvilleidae, Eunicidae,
Hesionidae, Nereididae, Pholoididae, Phyllodocidae, Polynoidae, Syllidae e
Terebellidae e pantópodo Anoplodactylus californicus Hall, 1912.
Foram encontradas 11 espécies não nativas do Brasil (cinco exóticas e cinco
criptogênicas) e seis nativas. Foram mais frequentemente e recrutadoras iniciais as
espécies criptogênicas: Clytia gracilis Sars, 1850, Obelia dichotoma Linnaeus, 1758,
Amphibalanus improvisus Darwin, 1854, A. amphitrite Darwin, 1854 e Bowerbankia
gracilis Leidy, 1855, porém sem regularidade mensal na frequência. Dentre as espécies
exóticas duas merecem destaque pelas estratégias de ocupação espacial, em interstícios
de cirrípedes e interior de testas vazias destes animais: anêmona Haliplanella lineata
Verrill, 1869 e tanaidaceo Sinelobus stanfordi Richardson 1901. A espécie nativa
Mytella charruana d´Orbigny, 1842 recrutou todos os meses, ocupando, na maioria das
vezes, no sentido periferia das placas para o centro. As ostras Crasssostrea rhizophorae
Guilding, 1828 recrutaram nos meses de janeiro a março de 2011 sendo encontrados
apenas indivíduos jovens.
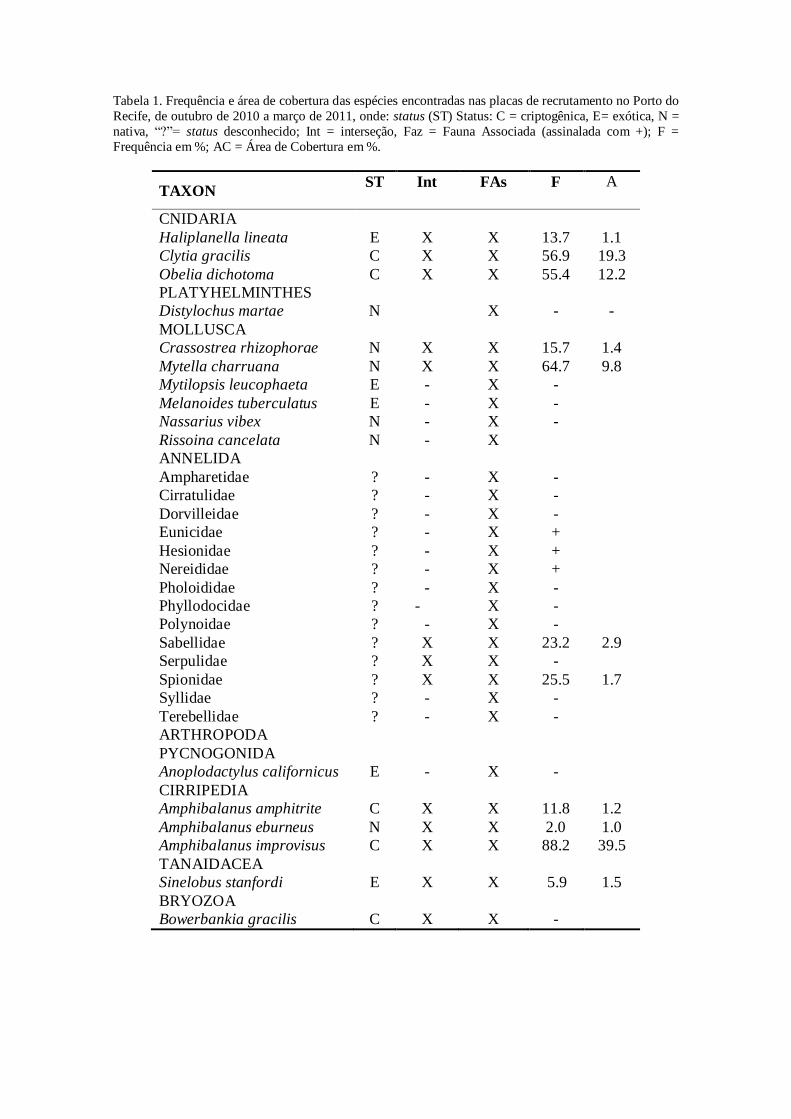
Tabela 1. Frequência e área de cobertura das espécies encontradas nas placas de recrutamento no Porto do
Recife, de outubro de 2010 a março de 2011, onde: status (ST) Status: C = criptogênica, E= exótica, N =
nativa, “?”= status desconhecido; Int = interseção, Faz = Fauna Associada (assinalada com +); F =
Frequência em %; AC = Área de Cobertura em %.
TAXON ST Int FAs F A
CNIDARIA
Haliplanella lineata E X X 13.7 1.1
Clytia gracilis C X X 56.9 19.3
Obelia dichotoma C X X 55.4 12.2
PLATYHELMINTHES
Distylochus martae N X - -
MOLLUSCA
Crassostrea rhizophorae N X X 15.7 1.4
Mytella charruana N X X 64.7 9.8
Mytilopsis leucophaeta E - X -
Melanoides tuberculatus E - X -
Nassarius vibex N - X -
Rissoina cancelata N - X
ANNELIDA
Ampharetidae ? - X -
Cirratulidae ? - X -
Dorvilleidae ? - X -
Eunicidae ? - X +
Hesionidae ? - X +
Nereididae ? - X +
Pholoididae ? - X -
Phyllodocidae ? - X -
Polynoidae ? - X -
Sabellidae ? X X 23.2 2.9
Serpulidae ? X X -
Spionidae ? X X 25.5 1.7
Syllidae ? - X -
Terebellidae ? - X -
ARTHROPODA
PYCNOGONIDA
Anoplodactylus californicus E - X -
CIRRIPEDIA
Amphibalanus amphitrite C X X 11.8 1.2
Amphibalanus eburneus N X X 2.0 1.0
Amphibalanus improvisus C X X 88.2 39.5
TANAIDACEA
Sinelobus stanfordi E X X 5.9 1.5
BRYOZOA
Bowerbankia gracilis C X X -

Não houve diferença significativa entre as abundâncias das estações, indicando
que o recrutamento na Bacia Portuária do Recife é bem parecido em todos as estações.
A Anosim feita entre as estações a partir das espécies mais abundantes indicou que não
existe diferença significativa na composição das espécies entre as estações. Já na análise
entre as espécies por mês, foram encontradas diferenças significantes, A. improvisus
entre os meses 1 e 6; 2 e 3; 2 e 4; 2 e 5; 2 e 6, os hidróides entre os meses 1 e 3; 2 e 3; 2
e 4, os espaços vazios entre os meses 3 e 4 (Tab. II).
Tabela II. Anosim das espécies mais abundantes entre os meses de outubro de 2010 a março de 2011 no
Porto do Recife - PE. Onde (R) = Recrutamento, (S) = sucessão, (-) = sem registro.
Espécie H(R) p(R) H(S) p(S)
Clytia gracilis 23,18 0,01 - -
Obelia dichotoma 23,14 0,01 - -
Mytella charruana 7,16 0,02 10,79 0,03
Polychaeta - - 7,53 0,11 Amphibalanus improvisus 27,46 0,01 12,18 0,01
Espaço vazio 13,67 0,02 1,54 0,82
Verificou-se uma maior presença de A. improvisus, O. dichotoma, C. gracilis e
M. charruana nas estações I e II; na Estação II, registrou-se também a ocorrência de A.
amphitrite e espaços vazios. Placas de todos os meses amostrados findaram seu período
de exposição com espaços vazios, sobretudo nas estações II e III.
Durante os meses de recrutamento a espécie A. improvisus foi a que mais
recrutou nos dois primeiros meses, tendo O. dichotoma e C. gracilis aumentado sua área
de cobertura principalmente por causa do alto índice pluviométrico de dezembro a
fevereiro de 2011. O mesmo aconteceu com os espaços vazios e o recrutamento de M.
charruana, que também teve sua população influenciada pelas chuvas (Fig. 3).

0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Out Nov Dez Jan Fev Mar
Meses
Áre
a d
e C
ob
ertu
ra
Ai Hi My vz Outros
Fig. 3. Comparação mês a mês das espécies mais frequentes na dinâmica do recrutamento durante os
meses de outubro de 2010 a março de 2011 no Porto do Recife - PE. Ai = Amphibalanus improvisus; Hi =
hidróides (Obelia dichotoma, Clytia gracilis); My = Mytella charruana; Vz = Vazio; Outros =
Haliplanella lineata, Crassostrea rhizophorae, A. amphitrite, A. eburneus, polychaetas, Sinelobus
stanfordi, Bowerbankia gracilis.
Nas placas de recrutamento foi observada a mortalidade de diversos espécimes
de cirrípedes, sobretudo nas placas onde havia planárias Distylochus martae. No mês de
novembro, dos 552 indivíduos de A. improvisus fixados, 123 estavam mortos, enquanto
se encontraram 43 planárias. Fato semelhante foi observado em relação às colônias de
hidróides C. gracilis e O. dichotoma e a presença de Anoplodactylus californicus. Nas
placas em que este conjunto apareceu havia muitos espaços vazios e colônias predadas.
Experimento de sucessão
Nas placas de sucessão totalizaram-se treze espécies, pertencentes a sete filos
(Tab. III). Foram perdidas 10 placas, sendo uma no mês de novembro (Estação I), duas
em dezembro (estações I e III), duas em fevereiro (estações II e III,) e cinco placas no
mês de março (uma na Estação I, e duas em cada uma das estações II e III).
Foram registradas quatro espécies nativas (D. martae, C. rhizophorae, M.
charruana e A. eburneus), três exóticas (H. lineata, Mytilopsis leucophaeta e
Melanoides tuberculatus) e cinco criptogênicas (C. gracilis, O. dichotoma, A.
amphitrite e A. improvisus, Bowerbankia gracilis). Foi mantida a proporção de espécies

exóticas/criptogênicas versus nativas. Com o passar do tempo observou-se o aumento
numérico de espécimes de: M. charruana, de A. improvisus e dos espaços vazios com o
aumento das chuvas (Fig. 4).
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Nov Dez Jan Fev Mar
Meses
Áre
a d
e C
ob
ert
ura
Ai Hi My vz Outros
Fig. 4. Comparação mês a mês das espéceis mais frequentes na sucessão durante os meses de novembro
de 2010 a março de 2011. Ai = Amphibalanus improvisus; Hi = hidróides (Obelia dichotoma, Clytia
gracilis); My = Mytella charruana; Vz = Vazio; Outros = Haliplanella lineata, poliquetas, A. amphitrite,
A. eburneus, Sinelobus stanfordi, Bowerbankia gracilis.
Os primeiros indivíduos a recrutarem e servirem como basibiontes iniciais foram
os cirrípedes A. improvisus, que criam condições adequadas para o recrutamento de M.
charruana. Quando os hidróides foram os primeiros basibiontes não houve
recrutamento de A. improvisus, com isso, os bivalves recrutaram das margens das
placas, migrando posteriomente para o centro.

Tabela III. Frequência e área de cobertura das espécies encontradas nas placas de sucessão no Porto do
Recife de outubro de 2010 a março de 2011, onde: status (ST) Status: C= criptogênica, E= exótica, N=
nativa, “?”= status desconhecido; Int= interseção, FAs= Fauna Associada (assinalada com +); FT=
Frequência Total em %; AC= Área de Cobertura em %.
TÁXON ST Int FAs FT AC
CNIDARIA
Haliplanella lineata E X 31.4 8.2
Clytia gracilis C X X 10.9 5.8
Obelia dichotoma C X X 11.4 5.8
PLAYHELMINTHES
Distylochus martae N - X - 1.6
MOLLUSCA
Crassostrea rhizophorae N - - - -
Mytella charruana N X X 97.1 59.2
Mytilopsis leucophaeta E - - - -
Melanoides tuberculatus E - X - 1.4
ANNELIDA
Eunicidae ? - X - 1.2
Hesionidae ? - X - 1.4
Nereididae ? - X - 2.0
Pholoididae ? - X - 1.2
Polynoidae ? - X - 1.2
Sabellidae ? - X - 1.4
Serpulidae ? X - 51.2 5.0
Spionidae ? X X 54.3 5.0
Syllidae ? - X - -
Terebellidae ? - X - -
ARTHROPODA
PYCNOGONIDA
Anoplodactylus californicus E - X + -
CIRRIPEDIA
Amphibalanus amphitrite C X X 5.7 1.8
Amphibalanus eburneus N X X 22.9 1.8
Amphibalanus improvisus C X X 62.9 18.8
BRYOZOA
Bowerbankia gracilis C - X + -

No processo de sucessão quando comparadas as abundâncias das espécies mais
frequentes como basibiontes e epibiontes (A. improvisus, H. lineata, C. gracilis, O.
dichotoma, M. charruana e espaços vazios), houve diferença significante apenas para os
hidróides O. dichotoma (p = 0,03) e C. gracilis (p = 0,03). Quando comparadas as áreas
de cobertura em cada mês por espécie A. improvisus e M. charruana tiveram diferenças
significantes, A. improvisus diferindo entre os meses 2 e 3, M. charruana teve diferença
significante, mas não diferiu em nenhum mês específico (Tab. III).
Após 30 dias do recrutamento (placas de 60 dias), a espécie mais frequente
como basibionte foi A. improvisus (59% de frequência) e, secundariamente, M.
charruana (28%). O cirrípede foi também o principal epibionte (e.g. 1.454 indivíduos
na estação I), seguido de M. charuana (454), A. amphitrite (32) e A. eburneus (18),
bem como hidróides O. dichotoma (12), a anêmona H. lineata (21) e os poliquetas
Sabellidae, Serpulidae e Spionidae foram também encontrados sobre os basibiontes.
Nesse período, as placas apresentaram uma espessura média de 2,5 cm, possuindo dois
estratos, o primeiro composto por A. improvisus, o segundo com M. charruana ainda
juvenis (média de 1,9 cm). Nesse período, houve uma média de 267 indivíduos de M.
charruana por placa.
Passados mais 30 dias (placas de 90 dias), a comunidade apresentou como
basibionte e também epibionte principal M. charruana, (muito frequente, 89% e até
1.211 espécimes). Outros indivíduos epibiontes foram: A. improvisus e H. lineata. A
espessura média encontrada no período foi de 4,5 cm, com 2 estratos. Constatou-se
aumento na espessura, mas não aumento dos estratos, resultado do aumento no tamanho
dos indivíduos, sendo os estratos compostos pelas mesmas espécies do período anterior,
tendo os mexilhões M. charruana um tamanho médio de 2,8 cm.

No período de 120 dias de imersão das placas, M. charruana continuou muito
frequente (76%), as espécies consideradas pouco frequentes foram: A. improvisus
(18%), C. gracilis e O. dichotoma (14%), H. lineata (13%), as classificadas como raras:
os poliquetas Nereididae, Syllidae e Terebellidae (0.2%). Foi observado um aumento de
espaços vazios nas placas, quando comparado com o período anterior, possivelmente
reflexo das intensas chuvas do período. Foram contados 598 espécimes de A.
improvisus e 256 de M. charruana, todos mortos, nas áreas adjacentes à placa de leitura,
assim como foi registrada a presença de inúmeros recém-recrutas de A. improvisus,
demonstrando uma colonização recente, semelhante ao registrado nas placas de
recrutamento.
A fauna epibêntica foi composta por apenas cinco espécies, sendo mais
frequente M. charruana (932 indivíduos), seguida de: A. improvisus (282), H. lineata
(134), D. martae (43), O. dichotoma (15), C. gracilis (5). Houve um aumento na
espessura média das placas (média de 5,9 cm) e também um aumento no número de
estratos (três), sendo o primeiro composto por A. improvisus e os dois superiores com
M. charruana. Os individuos do segundo estrato possuíam uma média de 3,1 cm de
tamanho e no terceiro estrato 1,5cm.
No quinto mês de sucessão (placas de 150 dias) houve uma diminuição na área
de ocorrência de M. charruana (62%), considerada apenas frequente. Neste período
houve grande precipitação pluviométrica. Todos os outros animais foram considerados
raros, os espaços vazios também diminuíram (58%). Os epibiontes encontrados foram
A. improvisus (384 indivíduos), M. charruana (377), H. lineata (90), O. dichotoma (3),
C. gracilis (2) e B. gracilis (6). A espessura média das placas foi de 6,1 cm, três
estratos. O tamanho médio de M. charruana foi de 4,3cm e 2,1 cm no último estrato.

Passados 180 dias de sucessão, houve um aumento do percentual de área de
cobertura de M. charruana (70%) e de A. improvisus (7%). Os espaços vazios passaram
a ser pouco frequentes (11%). Como epibiontes principais, mantiveram-se as espécies A.
improvisus (118 indivíduos) e M. charruana (117), C. rhizophorae (6), Mytilopsis
leucophaeta (6), O. dichotoma (5), C. gracilis (2), A. eburneus (2) e poliquetas
Serpulidae e Spionidae. Houve aumento na espessura média das placas (6,7 cm) e o
aumento de um estrato, sendo composta por: A. improvisus no primeiro estrato
(indivíduos mortos) e M. charruana nos demais, sendo que, no segundo estrato, os
indivíduos apresentaram tamanho médio de 5,9 cm, no terceiro 4,6 cm e no quarto 2,1
cm.
DISCUSSÃO
O presente estudo encontrou no recrutamento 16 espécies, sendo dessas 11
espécies não nativas do Brasil, as mais frequentes foram as criptogênicas C. gracilis e
A. improvisus e a exótica O. dichotoma, demonstrando o sucesso dessas espécies nesse
processo. Na sucessão foram encontradas 13 espécies, três exóticas e cinco
criptogênicas, tendo A. improvisus um aumento na área de cobertura e a M. charruana
com sua área diminuída, ligada às altas taxas pluviométricas no período.
Dispersão dos organismos
CARRIKER (1992) afirmou que a dispersão geográfica bem sucedida de uma
espécie é produto de interação entre propriedades fisiológicas do organismo e da
qualidade ambiental. Para PETERSON et al. (2003) embora os fatores bióticos sejam
importantes em tornar o ambiente mais suscetível à invasão, os fatores abióticos servem
como "filtro" para as introduções e limitam o estabelecimento de espécies exóticas ou
criptogênicas a condições ambientais similares às das suas regiões originais.

Os vetores de dispersão apontados como responsáveis pela introdução
inadvertida de muitos organismos bentônicos nas águas costeiras de todos os
oceanos em regiões litorâneas são, principalmente, a bioincrustação de animais nos
cascos das embarcações ou na água de lastro (SKERMAN, 1960; WILLIAMS et al.,
1988; RUIZ et al., 2000a; 2000b; GOLLASCH, 2002; FOFONOFF et al., 2003; COUTTS
& TAYLOR, 2004; FARRAPEIRA et al., 2007; DAVIDSON et al., 2008; FARRAPEIRA et
al., 2010; 2011), como ocorreu na área de estudo com H. lineata, O. dichotoma, M.
leucophaeta e B. gracilis (FARRAPEIRA et al. 2011).
Eventos abióticos
O período estudado é classificado como verão, pelo comportamento climático
histórico (SOMERFIELD et al., 2003), mas se apresentou particularmente chuvoso nos
meses de dezembro de 2010 a fevereiro de 2011. Assim, a semelhança encontrada tanto
no recrutamento quanto na sucessão para todos os meses indica que não houve
influência da pluviosidade e a consequente diminuição de salinidade local sobre os
animais. MUNIZ et al. (2005) verificaram que a circulação das correntes na bacia
portuária causam a mistura completa das águas e deixa o ambiente homogêneo, o que
também dá suporte aos resultados. Todos os táxons identificados são reconhecidamente
estuarinos ou, pelo menos, eurihalinos (FARRAPEIRA et al., 2011).
Segundo GLASBY e CONNELL (2001), as larvas planctônicas de
invertebrados marinhos bentônicos tendem a se fixar tão logo encontrem um
substrato natural ou artificial adequado, que facilite sua colonização. Porém,
segundo KENNISH (1994), os impactos ambientais associados a um processo de
intensa movimentação de sedimentos podem provocar efeitos tanto no habitat
quanto nos organismos, alterando a qualidade da água, relocando os sedimentos,
nesta dinâmica, aumentando a mortalidade dos organismos devido à ressuspensão

do sedimento de fundo e afetando, sobretudo os animais filtradores. Essas
condições foram relevantes para a dinâmica ecológica observada na área em
algumas estações, sobretudo na Estação II (Terminal Graneleiro), que,
coincidentemente era a estação de atracagem dos navios transatlânticos.
Para XAVIER et al. (2008) um distúrbio físico no ambiente abre novos espaços
para colonização e pode interferir nos processos de competição por uma determinada
fonte, sendo uma causa importante da heterogeneidade da estrutura e dinâmica natural
da comunidade, não apenas eliminando uma parte da cobertura da comunidade,
aumentando a quantidade de nutrientes disponíveis, e a morte dos indivíduos. A
ressuspensão de sedimentos motivada, tanto pela atracagem de navios transatlânticos
(Estação II), quanto pelo aporte maior de sedimentos nos meses mais chuvosos afetou
igualmente a dinâmica observada tanto no recrutamento quanto na sucessão.
Em janeiro de 2011 apenas seis espécies foram observadas fixadas nas placas,
sendo considerado um dos meses como menor número de espécies; essa diminuição da
diversidade estaria relacionado ao alto índice pluviométrico no período (139,10 mm), o
que fugia ao padrão de chuvas esperado para o mês. Para KROHLING & ZALMON (2008)
as chuvas trazem mais material particulado em suspensão e sedimentação inorgânica, o
que prejudica a colonização. Porém, favorece a aparição de espécies oportunistas,
conforme destacado por FLYNN & VALERIO-BERARDO (2012), que relataram a rápida
colononização de substrato artifical pelos hidróides O. dichotoma e C gracilis, sendo
essas espécies consideradas as mais abudantes no período onde ocorrem altos índices de
chuvas.
Dinâmica do Recrutamento
O fato de terem sido contabilizados 30 táxons nas placas de recrutamento
mostra, entretanto, que a área estudada, apresenta um aumento no número de espécies

no infralitoral quando comparado com NERY et al. (2008). Por outro lado, apesar da
bacia portuária ser qualificada como um ambiente poluído, o número de taxa excede o
encontrado por BREVES-RAMOS et al. (2005), em placas experimentais na Baía de
Guanabara, Rio de Janeiro, e se equipara ao encontrado por EMARA & BELAL (2004), no
Canal de Suez, ambientes igualmente poluídos.
Nesse estudo, o cirrípede criptogênico A. improvisus foi a única espécie
considerada mais frequente, em todos os meses pesquisados. CANGUSSU et al. (2010)
destacaram a sua grande quantidade em placas experimentais, classificando-o como
uma das primeiras espécies a recrutarem. Esta espécie compreende mais de 90% da
população bentônica fixada em superfícies artificiais (WEISS, 1947), sendo já relatada
como principal recrutadora em placas experimentais em diversas regiões (MAYER-
PINTO et al., 2000; BERNTSSON & JONSSON, 2003; SLUYS et al., 2005). FARRAPEIRA et
al. (2007) destacam que esta espécie foi a única presente em quase todas as
embarcações que acostavam no Porto do Recife. FARRAPEIRA et al. (2009) a relataram
na faixa inferior do mediolitoral fixada diretamente em substratos naturais e artificiais e
sobre outros organismos na Baía de Suape, Pernambuco, e este padrão de fixação foi
observado por FARRAPEIRA (2006) e FARRAPEIRA et al. (2009) em toda a área estuarina
do Recife, sendo dominante na faixa inferior dos médio e infralitoral, semelhantemente
ao que ocorreu neste trabalho. A alta porcentagem de animais mortos pode ser explicada
pela presença da planária D. martae. As planárias são reconhecidamente predadores de
cirrípedes (HURLEY, 1976; SLUYS et al., 2005; LEE et al., 2006).
As demais espécies de cirrípedes encontradas, a criptogênica A. amphitrite e a
nativa A. eburneus não estiveram bem representadas numericamente nas placas
estudadas. Amphibalanus amphitrite é uma das espécies mais estudadas do “fouling”
(EDMONDSON & INGRAM, 1939; GHOBASHY & EL-KOMY, 1981; MAYER-PINTO &

JUNQUEIRA, 2003; ZVYAGINTSEV, 2003; FLOERL et al., 2004; RAMADAN et al., 2006;
FREESTONE et al., 2009). Na área de estudo, porém, esta espécie é encontrada
preferencialmente na zona média do mediolitoral (FARRAPEIRA, 2006), não contemplada
neste estudo. Embora A. eburneus também seja igualmente encontrada em placas
(GHOBASHY & EL-KOMY, 1981; MAYER-PINTO et al., 2000; RAMADAN et al., 2006), ela
é incapaz de se fixar em superfícies artificiais de metal (WEISS, 1947), apesar de ser
uma das espécies mais adaptadas e resistentes a águas poluídas (RELINI, 2003). Na área
de estudo, FARRAPEIRA (2006) e FARRAPEIRA et al. (2009), encontrou-a apenas nas
áreas inferiores do mediolitoral e infralitoral, epibiôntica em conchas de bivalves.
Nas placas de recrutamento duas espécies de hidróides: C. gracilis e O.
dichotoma, foram registradas, com frequência variável em relação aos meses de estudo.
A presença do hidróide criptogênico O. dichotoma em placas experimentais tem sido
relatada com frequência (NANDAKUMAR, 1995; MAYER-PINTO et al., 2000; RELINI et al.,
2000; BREVES-RAMOS et al., 2005; FLYNN & VALERIO-BERARDO, 2012). Esta espécie é
reportada como tolerante em locais com índices elevados de eutrofização e baixa
salinidade (BREVES-RAMOS et al., 2005). Já o exótico C. gracilis, ocasionalmente
observado em placas (MIGOTTO, 1996; CANGUSSU et al., 2010; FLYNN & VALERIO-
BERARDO, 2012), é comumente mencionado como epizóica em vários invertebrados
bentônicos (MIGOTTO, 1996), fato também observado durante este experimento. Foram
classificadas como muito frequentes, nas placas de recrutamento, especialmente nos
meses chuvosos, e, posteriormente, apenas como epibiontes na sucessão, corroborando
as observações de MIGOTTO (1996) e MIGOTTO et al. (2001).
Sucesão de animais sésseis e sedentários
Classificada como muito frequente no experimento de sucessão, o mexilhão
nativo M. charruana forma densas agregações na faixa inferior do mediolitoral e

infralitoral (LEONEL & SILVA, 1988), fato também observado por FARRAPEIRA (2006) e
FARRAPEIRA et al. (2009), na área em estudo. Embora WHOI (1952) classifique os
mexilhões, genericamente, como colonizadores iniciais de substratos artificiais
recentemente submersos, este fato não foi confirmado nesse estudo. Definitivamente,
esta espécie constituíu o clímax da comumidade bentônica da Bacia Portuária do Recife.
MARGALEF (1974) defeniu que o clímax de uma comunidade é o ponto final da sucessão
das espécies. Estes dados se contrapõem aos observado por NERY et al. (2008), que
destacaram o mexilhão Mytilopsis leucophaeta como muito abundante para o local.
Outro importante destaque a ser feito é sobre as espécies móveis que se abrigam
em nichos produzidos pelos animais sésseis e sedentários pioneiros; estes animais
também integram o “fouling” (WHOI, 1952; WONHAM et al., 2000; ELDREDGE &
CARLTON, 2002; RAILKIN, 2004; HEWITT et al. 2009; MINCHIN et al., 2009;
FARRAPEIRA et al., 2011). As espécies exóticas H. lineata e M. leucophaeta foram
encontradas em cascos de diversas embarcações que acostavam no porto estudado
(FARRAPEIRA et al., 2007), e já foram reportadas como estabelecidos nos substratos
artificiais do mesmo (FARRAPEIRA et al , 2009). No presente estudo, o estabelecimento
das espécies na área se confirma, como fauna associada e epibionte. O pantópodo A.
californicus e o tanaidáceo S. stanfordi foram encontrados no mediolitoral do paredão
do porto, colônias do briozoário criptogênico B. gracilis também já foram encontradas
recobrindo espécies de mediolitoral na área de estudo, mas sua frequência em placas de
recrutamento e sucessão foram incipientes (FARRAPEIRA, dados não publicados).
CONCLUSÕES
Foram encontradas mais espécies exóticas e criptogênicas do que nativas
tanto no preocesso de recrutamento quanto no de sucessão, o estabelecimento das
espécies exóticas Haliplanella lineata e Sinelobus stanford e o aumento das chuvas

aumentaram o recrutamento dos hidróides Clytia gracilis e Obelia dichotoma, a
presença das espécies Dystilocus martae e Anoplodactylus californicus controlaram
a presença de Amphibalanus improvisus e dos hidróides C. gracilis e O. dichotoma
respectivamente. A. eburneus foi encontrada em maior quantidade nas placas de
sucessão, as espécies exóticas Mytilopsis leucophaeta e Melanoides tuberculatus só
foram encontradas em placas de sucessão. A biota portuária não diferiu nos três
pontos demonstrando uma uniformidade da fauna para o local.
AGRADECIMENTOS
À Fundação CAPES, pela bolsa do mestrado e à administração do Porto do
Recife S.A., pela autorização para o desenvolvimento do trabalho na área.
REFERÊNCIAS BIBLIOGRÁFICAS
BAKER, P.; BAKER, S. M. & FAJANS, J. 2004. Nonindigenous marine species in the
greater. TAMPA BAY ECOSYSTEM. TAMPA BAY ESTUARY PROGRAM TECH. PUBL., N. 02-
04
BERNTSSON, K. M. & JONSSON, P. R. 2003. Temporal and spatial patterns in recruitment
and succession of a temperate marine fouling assemblage: a comparison of static panels
and boat hulls during the boating season. Biofouling 19(3): 187–195.
BOOTH, D. J. & BROSNAN, D. M. 1995. The role of recruitment dynamics in rocky shore
and coral reef fi sh communities. Advances in Ecological Research 26: 309-385.
BREVES-RAMOS, A.; LAVRADO, H. P.; JUNQUEIRA, A. O. R. & SILVA, S. H. G. 2005.
Succession in rocky intertidal benthic communities in areas with different pollution
levels at Guanabara Bay (RJ-Brazil). Brazilian Archives of Biology and Technology
48(6): 951-965.
CALLOW, M. E. & CALLOW, J. A. 2002. Marine biofouling sticky problem. Biologist
49(1): 1-5.
CANGUSSU, L. C.; KREMER, L. P.; ROCHA, R. M.; PITOMBO, F. B.; HEYSE, H. &
BORNANCIN, E. C. 2010. Substrate type as a selective tool against colonization by non-
native sessile invertebrates. Brazilian Journal of Oceanography 58(3): 219-231.
CARLTON, J. T. 2009. Deep invasion ecology and the assembly of communities in
historical time. In: RILOV, G. & CROOKS, J.A. eds. Biological invasions in marine
ecosystems. Ecological Studies 204. Berlin, Springer-Verlag. p. 13–56.
CARRIKER, M.R. 1992. Introductions and transfers of molluscs: risk considerations and
implications. Journal of Shellfish Research 11: 507-510.

CARVALHO, E. M. & UIEDA, V. S. Colonização por macroinvertebrados bentônicos em
substrato artificial e natural em um riacho da serra de Itatinga, São Paulo, Brasil.
Revista Brasileira de Zoologia, 21(2) 287-293. 2004.
CLARKE, K. R. & WARWICK, R. M. 2001. Change in marine communities: an
approach to statistical analysis and interpretation. Bournemouth, Bourne Press. 128
p.
COE WR. 1932. Sexual phases in the American oyster (Ostrea virginica). Biol
Bull. 63:419–441.
COHEN, A. N.; HARRIS, L.; BINGHAM, B. L.; CARLTON, J.; CHAPMAN, J.; LAMBERT, C. C.;
LAMBERT, G.; LJUBENKOV, J. C.; MURRAY, S. N.; RAO, L.C.; REARDON, K. &
SCHWINDT, E. 2005. Rapid assessment survey for exotic organisms in southern
California bays and harbors, and abundance in port and non-port areas. Biological
Invasions 7(6): 995-1002.
COUTTS, A. D. M. & TAYLOR, M. D. 2004. A preliminary investigation of biosecurity
risks associated with biofouling on merchant vessels in New Zealand. New Zealand
Journal of Marine and Freshwater Research 38(2): 215-229.
CPRH. 2001. Relatório de monitoramento de bacias hidrográficas do Estado de
Pernambuco. Recife: Companhia Pernambucana de Controle de Poluição Ambiental e
de Administração dos Recursos Hídricos- CPRH. 110 p.
DAVIDSON, I. C.; MCCANN, L. D.; FOFONOFF, P. W.; SYTSMA, M. D. & RUIZ, G. M.
2008. The potential for hull-mediated species transfers by obsolete ships on their final
voyages. Diversity and Distributions 14(3): 518-529.
EDMONDSON, C.H. & INGRAM, W.M. 1939. Fouling organisms in Hawaii. Occasional
Papers of the Bernice Pauahi Bishop Museum of Polynesian Ethnology and
Natural History (14): 251-300.
ELDREDGE, L. G. & CARLTON, J. T. 2002. Hawaiian marine bioinvasions: A preliminary
assessment. Pacific Science 56(2): 211-212.
EMARA, A. M. & BELAL, A. A. 2004. Marine fouling in Suez Canal, Egypt. Egyptian
Journal of Aquatic Research 30(A): 189-206.
ENO, N.C.; CLARK, R.A. & SANDERSON, W.G. (Eds.) 1997. Non-native marine species
in British waters: a review and directory. Peterborough: Joint Nature Conservation Committee, 136 p.
FARRAPEIRA, C. M. R. 2006. Barnacles (Cirripedia Balanomorpha) of the estuarine
region of Recife, Pernambuco, Brazil. Tropical Oceanography 34(2): 100-119.
FARRAPEIRA, C. M. R.; MELO, A. V. O. M.; BARBOSA, D. F. & SILVA, K. M. E. 2007.
Ship hull fouling in the Port of Recife, Pernambuco. Brazilian Journal of
Oceanography 55(3): 207-221.
FARRAPEIRA, C. M. R.; RAMOS, C. A. C.; BARBOSA, D. F.; MELO, A. V. O. M.; PINTO, S.
L.; VERÇOSA, M. M.; OLIVEIRA, D. A. S. & FRANCISCO, J. A. 2009. Zonación vertical de

la macrofauna de sustratos sólidos del estuario del Río Massangana, Bahía de Suape –
Pernambuco, Brasil. Biota Neotropica 9(1): 1-12.
FARRAPEIRA, C. M. R.; MENDES, E. S.; DOURADO, J. & GUIMARÃES, J. 2010. Coliforms
accumulation in Amphibalanus amphitrite (Darwin, 1854) (Cirripedia) and its use as
organic pollution bioindicator in the estuarine area of Recife, Pernambuco, Brazil.
Brazilian Journal of Biology 70(2): 301-309.
FARRAPEIRA, C. M. R.; TENÓRIO, D. O. & AMARAL, F. M. D. 2011. Vessel biofouling as
an inadvertent vector of benthic invertebrates occurring in Brazil. Marine Pollution
Bulletin 62(3): 1538-1544.
FERNANDES, M. L. B.; PAIVA, R. J C.; GONÇALVES, K. L. F.; SOARES, A. M. C.; SILVA,
M. L.; SILVA. A. K. P. S. & CHAVES, A. C. 2006.Viabilidade do suururu Mytella
charruana (Mollusca: Bivalvia) como indicadora em estudos de monitoramento
ambiental na região portuária do Recife – PE. Lumen 14(1): 101-103.
FLOERL, O.; POOL, T. K.; INGLIS, G. J. 2004. Positive interactions between
nonindigenous species facilitate transport by human vectors. Ecological Applications
14(6): 1724-1736.
FLYNN, M. N. & VALÉRIO-BERARDO, M. T. 2012. Avaliação da toxicidade in situ através
do recrutamento de comunidade incrustante em painéis artificiais em terminal da
Petrobrás, Canal de São Sebastião, São Paulo. Revista Intertox de Toxicologia, Risco
Ambiental e Sociedade 5(1): 103-114.
FOFONOFF, P. F.; STEVES, B.; MILLER, W. A.; RUIZ, G. & CARLTON, J. 2003. In ships or
on ships? Unravelling the relative contribution of ballast tanks vs. hull fouling to ship-
mediated invasions of North America by marine species. In: Third International
Conference on Marine Bioinvasions. La Jolla, s. ed. p. 1- 42.
FREESTONE, A. L.; OSMAN, R. W. & WHITLATCH, R. B. 2009. Latitudinal gradients in
recruitment and community dynamics in marine epifaunal communities: Implications
for invasion success. In: LANG, M. A.; MACINTYRE, I. G.; RÜTZLER, K. eds.
Proceedings of the Smithsonian Marine Science Symposium. Washington, D.C.,
Smithsonian Contributions to the Marine Sciences. n. 38. pp. 247-258.
GHOBASHY, A. F. A. & EL-KOMY, M. M. 1981. Fouling in the southern region of the
Suez Canal. Hydrobiological Bulletin 14(3): 179-185.
GLASBY, T. M. & CONNEL, S. D. 2001. Orientation and position of a substratum have
large effects on epibiotic assemblages. Marine Ecology Progress Series 214: 127-135.
GOLLASCH, S. 2002. The importance of ship hull fouling as a vector of species
introductions into the North Sea. Biofouling 18(2): 105-121.
GOULLETQUER. P.; BACHELET, G.; SAURIAU, P. G. & NOEL, P. 2002 Open Atlantic coast
of Europe a century of introduced species into French waters. In: LEPPÄKOSKI, E.
GOLLASCH , S.; OLENIN, S. (eds) Invasive aquatic species of Europe: distribution,
impacts and management. Kluwer, Dordrecht, pp 276–290

HEWITT, C. L.; GOLLASCH, S. & MINCHIN, D. 2009. The vessel as a vector- biofouling,
ballast water and sediments. In: RILOV, G.; CROOKS, J. A. eds. Biological invasions in
marine ecosystems. Ecological Studies 204. Berlin, Springer-Verlag. p. 117-131.
HURLEY, A. C. 1976. The polycladid flatworm Stylochus tripartitus Hyman as a
barnacle predator. Crustaceana 31(1): 110-111.
KENNISH, M. J. 1994. Practical handbook of marine science. New York, John Wiley
& Sons, Inc. 562 p.
KROHLING, W. & ZALMON, I. R. 2008. Epibenthic colonization on an artificial reef in a
stressed environment off the north coast of the Rio de Janeiro State, Brazil. Brazilian
Archives of Biology and Technology 51(1): 213-221.
LEE, K.-M.; BEAL, M. A. & JOHNSTON, E.L. 2006. A new predatory flatworm
(Platyhelminthes, Polycladida) from Botany Bay, New South Wales, Australia. Journal
of Natural History 39(47): 3987-3995.
LEONEL, R. M. V. & SILVA, I. N. 1988. Estudo da sobrevivência e da capacidade de
isolamento de Mytella guyanensis (Mollusca: Bivalvia) em diferentes salinidades.
Revista Nordestina de Biologia 6: 35-41.
LEWIS, P. N.; HEWITT, C. L.; RIDDLE, M. & MCMINN, A. 2003. Marine introductions in
the Southern Ocean: an unrecognised hazard to biodiversity. Marine Pollution Bulletin
46(2): 213-223.
MACEDO, I. M.; MASI, B. P. & ZALMON, I. R. 2006. Comparison of rocky intertidal
community sampling methods at the northern coast of Rio de Janeiro State, Brazil.
Brazilian Journal of Oceanography 54(2/3): 147-154.
MARGALEF, R. 1968. Perspectives in ecological theory. University of Chicago Press,
Chicago. 111 pp.
MARGALEF, R. 1974. El ecossistema en el tempo In: Margalef, R. Ecología. Omega:
Barcelona. pp.737-788.
MAYER-PINTO, M. & JUNQUEIRA, A. O. R. 2003. Effects of organic pollution on the
initial development of fouling communities in a tropical bay, Brazil. Marine Pollution
Bulletin 46(11): 1495-1503.
MAYER-PINTO, M.; VIANA, M. S.; LAVRADO, H. P.; SILVA, T. A. & SILVA, S. H. G. 2000.
Epibiosis on barnacles at Angra dos Reis. RJ: eutrophication effects. Nauplius 8(1): 55-
61
MIGOTTO, A. E. 1996. Benthic shallow-water hydroids (Cnidaria, Hydrozoa) of the
coast of São Sebastião, Brazil, including a checklist of Brazilian hydroids. Zoologische
Verhandelingen 306: 1-125.
MIGOTTO, A. E.; MARQUES, A. C. & FLYNN, M. N. 2001. Seasonal recruitment of
hydroids (Cnidaria) on experimental panels in the São Sebastião Channel, southeastern
Brazil. Bulletin of Marine Science 68(2): 287-298.

MINCHIN, D.; GOLLASCH, S.; COHEN, A. N.; HEWITT, C. L. & OLENIN, S. 2009.
Characterizing vectors of marine invasion. In: RILOV, G.; CROOKS, J. A. eds. Biological
invasions in marine ecosystems. Ecological Studies 204. Berlin, Springer-Verlag. p.
109-116.
MUNIZ, P.; VENTURINI, N.; PIRES-VANIN, A. M. S.; TOMMASI, L. R. & BORJA, A. 2005.
Testing the applicability of Marine Index (AMBI) to assessing the ecological quality of
soft-bottom benthic communities, in the South America Region. Marine Pollution
Bulletin 50: 524-637.
NANDAKUMAR, K. 1995. Competitive interactions among sessile organisms in Tomioka
Bay, south Japan: importance of light conditions on the panel surface. Marine Biology
121(4): 713-719
NERY, P. P. C. F.; LEITÃO, S. N.; FERNANDES, M. L. B.; SILVA, A. K. P. & CHAVES, A. C.
2008. Recrutamento e sucessão ecológica da macrofauna incrustante em substratos no
Porto do Recife - PE, BRASIL. Revista Brasileira de Engenharia de Pesca 3(1): 51-
61.
NEVES, C. S.; ROCHA, R. M.; PITOMBO, F. B. & ROPER, J. J. 2007. Use of artificial
substrata by introduced and cryptogenic marine species in Paranaguá Bay, southern
Brazil. Biofouling 23(5): 319-30.
ODUM, E. P. 1988. Ecologia. Rio de Janeiro: Editora Guanabara.
OTSUKA, C. M. & DAUER, D. M. 1982. Fouling community dynamics in Lynnhaven
Bay, Virginia. Estuaries 5(1): 10-22.
PECH, D.; ARDISSON, P.-L. & BOURGET, E. 2002. Settlement of a tropical marine
epibenthic assemblage on artificial panels: Influence of substratum heterogeneity and
complexity scales. Estuarine, Coastal and Shelf Science 55: 743–750.
PETERSON, A. T.; PAPES, M. 7 KLUZA, D. A. Predicting the potential invasive distribuitions
of four Plant species in North American. Weed Science, V. 51, P. 863 - 868. 2003.
RAILKIN, A. I. 2004. Marine biofouling: colonization process and defenses. New
York, CRC Press. 303 p.
RAMADAN, SH. E.; KHEIRALLAH, A. M. & ABDEL-SALAM, KH. M. 2006. Marine fouling
community in the Eastern harbour of Alexandria, Egypt compared with four decades of
previous studies. Mediterranean Marine Science 7(2): 19-29.
RELINI, G.; RELINI, M. & MONTANARI, M. 2000. An offshore buoy as a small artificial
island and a fish-aggregating device (FAD) in the Mediterranean. Hydrobiologia
440(1-3): 65-80.
RELINI, G. 2003. II biofouling. Parte prima. Il macrofouling. Biologia Marina
Mediterranea 10: 285-326,
ROCHA, R. M.; BONNET, N. Y. K.; BAPTISTA, M. S. & BELTRAMIN, F. S. 2012.
Introduced and native Phlebobranch and Stolidobranch solitary ascidians (Tunicata:
Ascidiacea) around Salvador, Bahia, Brazil. Zoologia 29(1): 39-53.

RODRIGUEZ, S. R.; OJEDA, F. P. & INESTROSA, N. C. 1993. Settlement of benthic marine
invertebrates. Marine Ecology Progress Series 97: 193-207.
RUIZ, G. M.; FOFONOFF, P.; CARLTON, J. T.; WONHAM, M.J. & HINES, A.H. 2000a.
Invasion of coastal marine communities in North America: apparent patterns, processes,
and biases. Annual Review in Ecology and Systematics 31: 481-531.
RUIZ, G. M.; RAWLINGS, T. K.; DOBBS, F. C.; DRAKE, L. A.; MULLADY, T.; HUQ, A. &
COLWELL, R. R. 2000b. Global spread of microorganisms by ships. Ballast water
discharged from vessels harbours a cocktail of potential pathogens. Nature 408: 49-50.
SAUER-MACHADO, K.R.S., CHAPMAN, A.R.O. & COUTINHO, R. 1992. Patch structure in
a tropical rocky shore community in Brazil: a mosaic of sucessional states? Ophelia 35:
187- 195.
SILVA, A. K. P.; MAYAL, E. M.; MELLO, R. L. S. & FERNANDES, M. L. B. 2001. Estudo
preliminar da bioincrustação sobre substratos naturais, como indicador de impacto na
região do Complexo Portuário de Suape, PE. Tropical Oceanography 29(2): 139-146.
SKERMAN, T. M. 1960. Ship-fouling in New Zealand Waters: a survey of marine fouling
organisms from vessels of the coastal and overseas trades. New Zealand Journal of
Science 3(4): 620-648.
SLUYS, R.; FAUBEL, A.; RAJAGOPAL, S. & VAN DER VELDE, G. 2005. A new and alien
species of ‘‘oyster leech’’ (Platyhelminthes, Polycladida, Stylochidae) from the
brackish North Sea Canal, The Netherlands. Helgoland Marine Research 59(4): 310-
314.
SOMERFIELD, P. J., FONSÊCA-GENEVOIS, V. G., RODRIGUES, A. C. L., CASTRO, F. J. V. &
SANTOS, G. A. P. 2003. Factors affecting meiofaunal community structure in the Pina
Basin, an urbanized embayment on the coast of Pernambuco, Brazil. Journal of the
Marine Biological Association of the United Kingdom 83(6): 1209-1213.
SUTHERLAND, J. P. 1974. Multiple stable points in natural communities. American
Naturalist 108(964): 859-873.
TOMMASI, L. R.; BAUER, L. A. F.; LARA, G. A. 1972. Controle do desenvolvimento
de incrustações biológicas em tubulações de Fundações de ponte marítima, em
sistema de concretagem submersa "prepakt". Boletim Instituto Oceanográfico. ,
21:117-128
XAVIER, E. A.; GAMA, B. A. P.; PORTO, T. F.; ANTUNES, B. L. & PEREIRA, R. C. 2008.
Effects of disturbance area on fouling communities from a tropical environment:
Guanabara Bay, Rio de Janeiro, Brazil. Brazilian Journal of Oceanography 56(2): 73-
84
WAHL, M. 1989. Marine epibiosis. I. Fouling and antifouling: some basic aspects.
Marine Ecology Progress Series 58: 175-189.
WEISS, C. M. 1947. The comparative tolerances of some fouling organisms to cooper
and mercury. The Biological Bulletin 93(1): 56-63.

WHOI. 1952. Marine fouling and its prevention. Maryland, Woods Hole
Oceanographic Institute- WHOI. 388 p.
WILLIAMS, R. J.; GRIFFITHS, F. B.; VAN DER WAL, E. J. & KELLY, J. 1988. Cargo vessel
ballast water as a vector for the transport of nonindigenous marine species. Estuarine,
Coastal and Shelf Science 26(4): 409-420.
WONHAM, M. J.; CARLTON, J. T.; RUIZ, G. M. & SMITH, L. D. 2000. Fish and ships:
relating dispersal frequency to success in biological invasions. Marine Biology 136(6):
1111-1121.
ZVYAGINTSEV, A. Y. 2003.Introduction of species into the Northwestern Sea of Japan
and the problem of marine fouling. Russian Journal of Marine Biology 29(1): S10–
S21.

CONSIDERAÇÕES FINAIS
Foram encontradas 16 espécies no recrutamento sendo que dessas 11 espécies
não nativas do Brasil, dessas mais frequentes os hidróides Clytia gracilis, Obelia
dichotoma, o cirrípede Amphibalanus improvisus. Na sucessão, foram 12 espécies,
sendo três exóticas e cinco criptogênicas, tendo A. improvisus um aumento na sua área
de cobertura e Mytella charruana uma diminuição durante os meses de novembro de
2010 a janeiro de 2011. A espécie que serviu como basibionte determinou a forma como
ocorreu a sucessão nas placas de metal, caso fosse A. improvisus como basibionte as M.
charruana logo se fixavam matando-os, caso fossem os hidroides, os bivalves se
fixavam primeiro nas bordas das placas para depois migrarem para o centro.
Destacamos o estabelecimento na bacia portuária de Recife das espécies exóticas
Haliplanella lineata, Sinelobus stanfordi, Mytilopsis leucophaeta e Melanoides
tuberculatus ambas as espécies foram encontradas entre os bivalves nas placas de
sucessão, durante os meses de estudo. O aglomeramento formado pela M. charruana e o
agregamento de sedimento da área cria nichos ecológicos para as famílias de poliquetas
encontradas no estudo.
A M. charruana foi à única espécie que esteve presente em todos os meses
recrutando nas placas, as chuvas favoreceram o aumento no recrutamento dos hidroides.
A espécie de cirrípide A. amphitrite só foi encontrada nas placas de recrutamento
e a espécie A. eburneus nas placas de sucessão, nos interstícios das conchas de M.
charruana.

A presença das espécies Dystilocus martae e Anoplodactylus californicus
controlaram por predação a presença de A. improvisus e dos hidróides (C. gracillis e O.
dichotoma).
O uso de substrato artificial é uma técnica boa para estudar o recrutamento e
sucessão de espécies sésseis e sedentárias em bacias portuárias. Constatou-se que não
houve diferença significativa entre a biota portuária quando comparadas as estações, já
quando comparadas as taxas de cobertura das espécies por mês podemos constatar
diferenças principalmente nas espécies C. gracilis, O .dichotoma, M. charruana e A.
improvisus no processo de recrutamento, já na sucessão tivemos: M. charruana e A.
improvisus.
Sugerimos um melhor monitoramento da bacia portuária, bem como a utilização
de técnicas para controlar a dispersão de espécies exóticas para área a fim de proteger a
fauna nativa.

REGRAS DA REVISTA ILHERINGIA, SÉRIE ZOOLOGIA SCOPO E
POLÍTICA
O periódico Iheringia, Série Zoologia, editado pelo Museu de Ciências Naturais da
Fundação Zoobotânica do Rio Grande do Sul, destina-se a publicar trabalhos
completos originais em Zoologia, com ênfase em taxonomia e sistemática,
morfologia, história natural e ecologia de comunidades ou populações de espécies da
fauna Neotropical recente. Notas científicas não serão aceitas para publicação. Em
princípio, não serão aceitas listas faunísticas, sem contribuição taxonômica, ou que
não sejam o resultado de estudos de ecologia ou história natural de comunidades,
bem como chaves para identificação de grupos de táxons definidos por limites
políticos. Para evitar transtornos aos autores, em caso de dúvidas quanto à adequação
ao escopo da revista, recomendamos que a Comissão Editorial seja previamente
consultada. Também não serão aceitos artigos com enfoque principal em Agronomia,
Veterinária, Zootecnia ou outras áreas que envolvam zoologia aplicada. Manuscritos
submetidos fora das normas da revista serão devolvidos aos autores antes de serem
avaliados pela Comissão Editorial e Corpo de Consultores.
Forma e preparação de manuscritos
1. Submeter o manuscrito eletronicamente através do
site: http://submission.scielo.br/index.php/isz.
2. Os manuscritos serão analisados por, no mínimo, dois consultores. A aprovação do
trabalho, pela Comissão Editorial, será baseada no conteúdo científico, respaldado
pelos pareceres dos consultores e no atendimento às normas. Alterações substanciais
poderão ser solicitadas aos autores, mediante a devolução dos arquivos originais

acompanhados das sugestões.
3. O teor científico do trabalho é de responsabilidade dos autores, assim como a
correção gramatical.
4. O manuscrito, redigido em português, inglês ou espanhol, deve ser configurado em
papel A4, em fonte “Times New Roman”com no máximo 30 páginas numeradas
(incluindo as figuras) e o espaçamento duplo entre linhas. Manuscritos maiores
poderão ser negociados com a Comissão Editorial.
5. Os trabalhos devem conter os tópicos: título; nomes dos autores (nome e
sobrenome por extenso e demais preferencialmente abreviados); endereço completo
dos autores, com e-mail para contato; abstract e keywords (máximo 5) em inglês;
resumo e palavras-chave (máximo 5) em português ou espanhol; introdução; material
e métodos; resultados; discussão; agradecimentos e referências bibliográficas. As
palavras-chave não deverão sobrepor com aquelas presentes no título.
6. Não usar notas de rodapé.
7. Para os nomes genéricos e específicos usar itálico e, ao serem citados pela primeira
vez no texto, incluir o nome do autor e o ano em que foram descritos. Expressões
latinas também devem estar grafadas em itálico.
8. Citar as instituições depositárias dos espécimes que fundamentaram a pesquisa,
preferencialmente com tradição e infraestrutura para manter coleções científicas e
com políticas de curadoria definidas.
9. Citações de referências bibliográficas no texto devem ser feitas em Versalete (caixa
alta reduzida) usando alguma das seguintes formas: Bertchinger & Thomé (1987),
(Bryant, 1915; Bertchinger & Thomé, 1987), Holme et al. (1988).

10. Dispor as referências bibliográficas em ordem alfabética e cronológica, com os
autores em Versalete (caixa alta reduzida). Apresentar a relação completa de autores
(não abreviar a citação dos autores com "et al.") e o nome dos periódicos por extenso.
Alinhar à margem esquerda com deslocamento de 0,6 cm. Não serão aceitas citações
de resumos e trabalhos não publicados.
Exemplos:
Bertchinger, R. B. E. & Thomé, J. W. 1987. Contribuição à caracterização
de Phyllocaulis soleiformis (Orbigny, 1835) (Gastropoda, Veronicellidae). Revista
Brasileira de Zoologia 4(3):215-223.
Bryant, J. P. 1915. Woody plant-mammals interactions. In: ROSENTHAL, G. A. &
BEREMBAUM, M. R. eds. Herbivores: their interactions with secondary plants
metabolites. San Diego, Academic. v.2, p.344-365..
Holme, N. A.; Barnes, M. H. G.; Iwerson, C. W. R.; Lutken, B. M. & Mcintyre, A. D.
1988. Methods for the study of marine mammals. Oxford, Blackwell Scientific.
527p.
Platnick, N. I. 2002. The world spider catalog, version 3.0. American Museum of
Natural History. Disponível em:
<http://research.amnh.org/entomology/spiders/catalog81-87/index.html>. Acesso em:
10.05.2002.
11. As ilustrações (desenhos, fotografias, gráficos e mapas) são tratadas como figuras,
numeradas com algarismos arábicos sequenciais e dispostas adotando o critério de
rigorosa economia de espaço e considerando a área útil da página (16,5 x 24 cm) e da
coluna (8 x 24 cm). A Comissão Editorial reserva-se o direito de efetuar alterações na

montagem das pranchas ou solicitar nova disposição aos autores. As legendas devem
ser autoexplicativas. Ilustrações a cores implicam em custos a cargo dos autores. As
figuras devem ser encaminhadas apenas em meio digital de alta qualidade (ver
item 16).
12. As tabelas devem permitir um ajuste para uma (8 cm) ou duas colunas (16,5 cm)
de largura, ser numeradas com algarismos romanos e apresentar título conciso e
autoexplicativo.
13. Figuras e tabelas não devem ser inseridas, somente indicadas no corpo do texto.
14. A listagem do material examinado deve dispor as localidades de Norte a Sul e de
Oeste a Leste e as siglas das instituições compostas preferencialmente de até 4 letras,
segundo o modelo abaixo:
VENEZUELA, Sucre: San Antonio del Golfe, (Rio Claro, 5o57'N 74
o51'W, 430m) 5
♀, 8.VI.1942, S. Karpinski col. (MNHN 2547). PANAMÁ, Chiriquí: Bugaba
(Volcán de Chiriquí), 3 ♂, 3 ♀, 24.VI.1901, Champion col. (BMNH 1091).
BRASIL, Goiás: Jataí (Fazenda Aceiro), 3 ♂, 15.XI.1915, C. Bueno col.
(MZSP); Paraná: Curitiba, ♀, 10.XII.1925, F. Silveira col. (MNRJ); Rio Grande do
Sul: São Francisco de Paula (Fazenda Kraeff, Mata com Araucária, 28o30'S 52
o29'W,
915m), 5 ♂, 17.XI.1943, S. Carvalho col. (MCNZ 2147).
15. Recomenda-se que os autores consultem um artigo recentemente publicado na
Iheringia Série Zoologia para verificar os detalhes de formatação.
16. Enviar o arquivo de texto em Microsoft Word (*.doc) ou em formato "Rich Text"
(*.rtf). Para as imagens utilizar arquivos Bitmap TIFF (*.tif) e resolução mínima de
300 dpi (fotos) ou 600 dpi (desenhos em linhas). Enviar as imagens nos arquivos

digitais independentes (não inseridas em arquivos do MS Word, MS Power Point e
outros), nomeados de forma autoexplicativa (e. g. figura01.tif). Gráficos e tabelas
devem ser inseridos em arquivos separados (Microsoft Excel para gráficos e
Microsoft Word ou Excel para tabelas). Para arquivos vetoriais utilizar formato Corel
Draw (*.cdr).
17. Para cada autor será fornecido um exemplar da revista. Os artigos também estarão
na página do Scientific Electronic Library Online, SciELO/Brasil, disponível
em www.scielo.br/isz.





























