Embed Size (px)
Citation preview

Bol. Mus. Para. Emílio Goeldi. Ciências Naturais, Belém, v. 2, n. 1, p. 155-182, jan-abr. 2007
155
Osteologia craniana de Cerylinae Osteologia craniana de Cerylinae Osteologia craniana de Cerylinae Osteologia craniana de Cerylinae Osteologia craniana de Cerylinae (Coraciiformes: Alcedinidae)(Coraciiformes: Alcedinidae)(Coraciiformes: Alcedinidae)(Coraciiformes: Alcedinidae)(Coraciiformes: Alcedinidae)Cranial osteology of Cerylinae Cranial osteology of Cerylinae Cranial osteology of Cerylinae Cranial osteology of Cerylinae Cranial osteology of Cerylinae (Coraciiformes: Alcedinidae)(Coraciiformes: Alcedinidae)(Coraciiformes: Alcedinidae)(Coraciiformes: Alcedinidae)(Coraciiformes: Alcedinidae)
Andrés Calonge-Méndez I
Elizabeth Höfling II
Resumo:Resumo:Resumo:Resumo:Resumo: A anatomia craniana de oito espécies de martins-pescadores da subfamília Cerylinae (Megaceryle torquata, M.lugubris, M. alcyon, Ceryle rudis, Chloroceryle amazona, C. americana, C. inda e C. aenea) foi comparada com a deoutros Alcedinidae e com representantes da maioria das famílias de Coraciiformes (Coraciidae, Leptosomidae,Bucerotidae, Upupidae, Todidae, Momotidae e Meropidae), a fim de verificar o padrão da região temporal e dabarra jugal. Esses dados foram comparados também com os disponíveis na literatura referentes aos Ramphastidae,Picidae, Galbulidae, Capitonidae e Cuculidae. A região temporal com duas fossas bem delimitadas, a barra jugalexpandida rostrolateralmente e o processo ótico do quadrado com três saliências articulares, entre outros, sãocaracteres comuns para Cerylinae. Quatro características permitiram separar os gêneros de Cerylinae: expansãodorsal do processo orbital do lacrimal (ausente em Chloroceryle [danificado em Ceryle rudis], presente em Megaceryle);côndilo caudal do quadrado (forma cotilar em Chloroceryle e Ceryle rudis, forma condilar em Megaceryle); cótilalateral e caudal da mandíbula (indistintas em Chloroceryle e Ceryle rudis, distintas em Megaceryle); e parede dorsalda fossa caudal da mandíbula (ausente em Chloroceryle e Ceryle rudis, ausente Megaceryle).
PPPPPalavras-chavesalavras-chavesalavras-chavesalavras-chavesalavras-chaves: Coraciiformes. Alcedinidae. Crânio. Osteologia. Sistemática.
AbstractAbstractAbstractAbstractAbstract: Skull osteology of eight species of Cerylinae Kingfishers (Megaceryle torquata, M. lugubris, M. Alcyon, Ceryle rudis,Chloroceryle amazona, C. americana, C. inda and C. aenea) were compared with other Alcedinidae andrepresentatives of most families of Coraciiformes (Coraciidae, Leptosomidae, Bucerotidae, Upupidae, Todidae,Momotidae and Meropidae) with the purpose of verifying the pattern of the temporal region and the jugal arch.These data were also compared with the information on the bibliography about the Ramphastidae, Picidae, Galbulidae,Capitonidae and Cuculidae. The temporal region with two well delimited fossas, the jugal arch expanded rostrolaterallyand quadrate with three saliences on the otical process, are common characters on the Cerylinae kingfishers. Fourcharacters were diagnostic to separate the genera of Cerylinae studied: the dorsal expansion of the orbital processof the lacrimal (absent in Chloroceryle [unavailable in Ceryle rudis], present in Megaceryle); caudal condyle ofquadrate (cotylar-shaped in Chloroceryle and Ceryle rudis, condylar-shape in Megaceryle); lateral and caudalcotyles of mandible (non distinct in Chloroceryle and Ceryle rudis, distinct in Megaceryle) and dorsal wall of thecaudal fossa of jaw (absent in Chloroceryle and Ceryle rudis, present in Megaceryle).
KKKKKeywordseywordseywordseywordseywords: Coraciiformes, Alcedinidae, Skull, Osteology, Systematic.
I Universidade de São Paulo. Instituto de Biociências. Departamento de Zoologia. São Paulo, Brasil ([email protected]).I I Universidade de São Paulo. Instituto de Biociências. Departamento de Zoologia. São Paulo, Brasil ([email protected]).

156
Osteologia craniana de Cerylinae (Coraciiformes: Alcedinidae)
INTRODUÇÃO
Os martins-pescadores (Alcedinidae) são um grupocosmopolita que habita, principalmente, regiõestropicais e subtropicais do Velho Mundo. Existemaproximadamente 90 espécies, sendo a regiãoNeotropical representada por apenas seis(HOWARD; MOORE, 1994).
São aves que, geralmente, apresentam corpo robusto,cabeça relativamente grande, pescoço curto, línguacurta, bico longo, robusto e pontiagudo. Possuemnarinas do tipo holorrino (GARROD, 1873), palatodesmognato, sendo o vômer e o processobasipterigóide ausentes. Os pés são bastante pequenos,impróprios para nadar (SICK, 1997), com sindactiliaavançada e os três artelhos anteriores unindo-sebasalmente e o III e o IV até a porção mediana.
Atualmente, reconhecem-se três grupos de martins-pescadores: Alcedininae, Daceloninae e Cerylinae(MILLER, 1912; MAURER; RAIKOW, 1981) ouAlcedinidae, Dacelonidae e Cerylidae (SIBLEY;AHLQUIST, 1990; FRY et al., 1992).
O relacionamento filogenético entre os Alcedinidaeé incerto. Miller (1912) acredita que os Alcedininaesão mais relacionados aos Daceloninae do que aosCerylinae. Maurer e Raikow (1981) citam osDaceloninae como basais em relação aos Alcedininaee Cerylinae. No entanto, Sibley e Ahlquist (1990)acreditam serem os Alcedinidae basais em relaçãoa Cerylidae e Dacelonidae, verificando-se, destamaneira, as três hipóteses possíveis derelacionamento filogenético para três táxons. Aquestão do relacionamento filogenético entre assubfamílias de Alcedinidae é, ainda, um assuntocontrovertido e de fundamental importância ao setentar elucidar a história evolutiva do grupo,principalmente sob os pontos de vista biogeográficoe da evolução do hábito alimentar.
Os Cerylinae distribuem-se em três gêneros(Megaceryle, Ceryle e Chloroceryle), comportandonove espécies (Ceryle rudis, Megaceryle maxima,
M. lugubris, M. alcyon, M. torquata, Chloroceryleamazona, C. americana, C. inda e C. aenea) e váriassubespécies (HOWARD; MOORE, 1994). Váriosautores reconhecem apenas Ceryle e Chloroceryle(SCHAUNSEE, 1982).
Segundo Miller (1912), os Cerylinae “constituem umgrupo bem definido separado das outras duas subfamíliaspor um excelente número de caracteres”. Estasassertivas são corroboradas por Stresemann eStresemann (1961), através do padrão de muda dasrêmiges primárias, por Maurer e Raikow (1981),através da musculatura dos membros posteriores, epor Sibley e Ahlquist (1990), através de hibridação deDNA. Estes últimos apresentam um cladograma paraas espécies de Cerylinae, onde Chloroceryle é o grupobasal e Ceryle rudis grupo-irmão dos Megaceryle.
Fry et al. (1992) acreditam que o relacionamentoentre as espécies de Chloroceryle seja de fácilinterpretação. Segundo estes autores, as quatroespécies de Chloroceryle são produto de especiaçãodicotômica: Chloroceryle amazona e Chloroceryleamericana seriam espécies-irmãs, assim comoChloroceryle inda e Chloroceryle aenea. Estashipóteses são corroboradas pelos hábitos de C. indae C. aenea, espécies de interior de mata, ao contráriode C. amazona e C. americana, que se associam aambientes aquáticos. Outra característica que apóiao relacionamento das espécies de Chloroceryle é acoloração das fêmeas de C. inda e C. aenea, queapresentam uma faixa esverdeada no peito.
Este trabalho objetiva descrever e comparar aosteologia craniana dos martins-pescadoresneotropicais, caracterizar os gêneros de Cerylinae combase na osteologia craniana, discutir e comparar opadrão da região temporal, do arco jugal e do quadrado.
MATERIAL E MÉTODOS
Foram analisados os crânios de 43 espécimes deaves, representando 41 espécies de sete famílias deCoraciiformes, além de um representante das

Bol. Mus. Para. Emílio Goeldi. Ciências Naturais, Belém, v. 2, n. 1, p. 155-182, jan-abr. 2007
157
famílias Trogonidae e Cuculidae (Apêndice 1). Destes41 crânios, analisaram-se 33 de Alcedinidae (martins-pescadores), amostrando-se as três subfamíliaspropostas por Miller (1912): Daceloninae (n=7),Alcedininae (n=2) e Cerylinae (n=24).
Um exemplar de Chloroceryle americana (AZ 511),fixado em formol 4% e conservado em álcool70% v. v., foi dissecado para se estabeleceradequadamente os limites da região temporal.
A descrição anatômica baseou-se em Chloroceryleamazona que, posteriormente, foi comparada comos outros martins-pescadores, produzindo umadescrição única. As outras espécies de Coraciiformes,como também Trogon melanurus (Trogonidae) eCoccyzus americanus (Cuculidae), foram analisadas,principalmente, com o intuito de comparar o padrãoda região temporal e da barra jugal, sendo tal análiseincluída na discussão.
O material ornitológico faz parte das coleções doMuseu Paraense Emílio Goeldi, Belém, PA, Brasil(MPEG); do Departamento de Zoologia, Institutode Biociências, Universidade de São Paulo, SãoPaulo, SP, Brasil (AZ); do United States NationalMuseum, Smithsonian Institution, Washington,USA (USNM); do Louisiana State UniversityMuseum of Zoology, Louisiana, USA (LSUMNS);e do Museu de História Natural de Taubaté, SP,Brasil (MHNT).
O material osteológico pertencente aoDepartamento de Zoologia do Instituto deBiociências da Universidade de São Paulo foipreparado com o auxílio de larvas de Dermestessp. (Coleoptera), segundo Alvarenga (1992). Omaterial oriundo do Museu Paraense Emílio Goeldi,do United States National Museum, SmithsonianInstitution, do Louisiana State University Museumof Zoology e do Museu de História Natural deTaubaté encontrava-se preparado.
Os crânios foram observados e descritos utilizando-seesteroscópio Wild M3 (9,6x - 60x), fotografados e,posteriormente, desenhados.
O quadrado foi desarticulado da caixa craniana,com o intuito de observar detalhes da articulaçãoentre o quadrado e a caixa craniana. Adesarticulação foi precedida de um banho em águapor, aproximadamente, 20 minutos.
Devido ao anquilosamento observado noscomponentes ósseos do crânio das aves, optou-sepor referir-se a eles como simplesmente regiões.
A nomenclatura util izada na descrição doscomponentes ósseos segue a Nomina AnatomicaAvium (BAUMEL et al., 1993), com exceção daregião temporal, que segue Zusi e Livezey (2000).
A nomenclatura das famílias segue Maurer e Raikow(1981) e a das espécies Howard e Moore (1994).
RESULTADOS
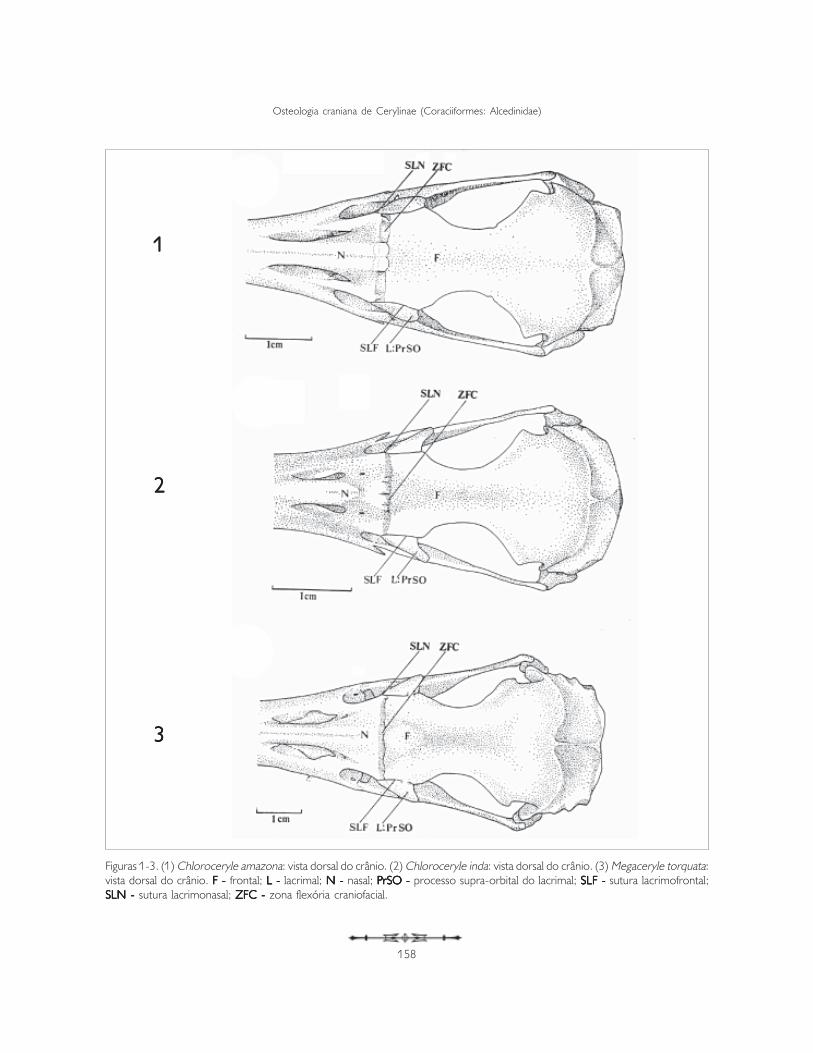
Ossa cranii: os frontale, os lacrimale, osOssa cranii: os frontale, os lacrimale, osOssa cranii: os frontale, os lacrimale, osOssa cranii: os frontale, os lacrimale, osOssa cranii: os frontale, os lacrimale, osectethmoidale, os mesethmoidaleectethmoidale, os mesethmoidaleectethmoidale, os mesethmoidaleectethmoidale, os mesethmoidaleectethmoidale, os mesethmoidaleA região do osso frontal (os frontale) (F) articula-serostralmente com a região do osso nasal (os nasale)(N) através da zona flexória craniofacial (zona flexoriacraniofacialis) (ZFC). Esta estrutura apresenta-secomo um sulco perpendicular ao eixo rostrocaudaldo crânio, bastante evidente e característico em todasas espécies analisadas (Figuras 1-3).
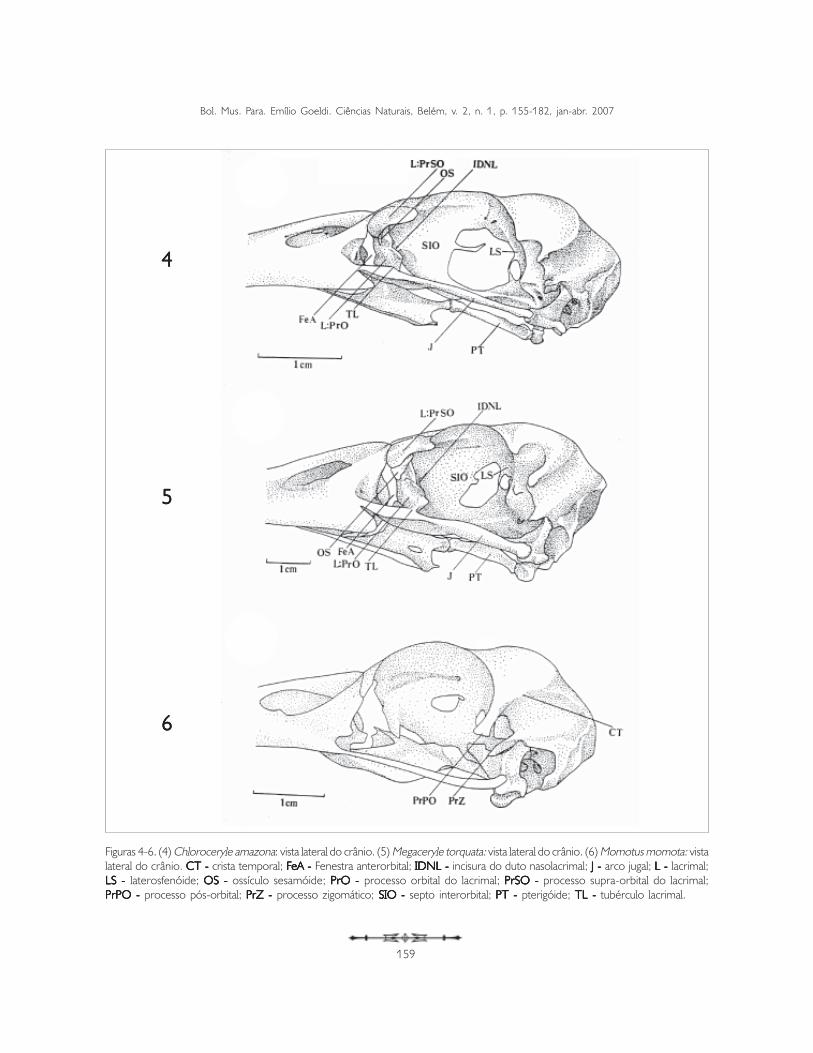
Os lacrimais (os lacrimale) (L) são um par de ossosorientados quase perpendicularmente ao eixorostrocaudal do crânio. Localizam-se entre a fenestraanterorbital (fenestra antorbitalis) (FeA) e a órbita,delimitando-as caudal e rostralmente, respectivamente(Figuras 4 e 5).
O lacrimal, na sua face lateral, apresenta umaconspícua depressão mediana por onde passa o dutonasolacrimal (ductus nasolacrimalis), sendodenominada incisura do duto nasolacrimal (incisuraductus nasolacrimalis) (IDNL) (Figuras 4 e 5). Estaestrutura delimita os processos supraorbital e orbitaldo lacrimal. O processo supraorbital (PrSO) localiza-sedorsalmente à incisura do duto nasolacrimal, em
158
Osteologia craniana de Cerylinae (Coraciiformes: Alcedinidae)
Figuras 1-3. (1) Chloroceryle amazona: vista dorsal do crânio. (2) Chloroceryle inda: vista dorsal do crânio. (3) Megaceryle torquata:vista dorsal do crânio. F - F - F - F - F - frontal; L - L - L - L - L - lacrimal; N - N - N - N - N - nasal; PPPPPrSO - rSO - rSO - rSO - rSO - processo supra-orbital do lacrimal; SLF - SLF - SLF - SLF - SLF - sutura lacrimofrontal;SLN - SLN - SLN - SLN - SLN - sutura lacrimonasal; ZFC - ZFC - ZFC - ZFC - ZFC - zona flexória craniofacial.
11111
22222
33333
Bol. Mus. Para. Emílio Goeldi. Ciências Naturais, Belém, v. 2, n. 1, p. 155-182, jan-abr. 2007
159
Figuras 4-6. (4) Chloroceryle amazona: vista lateral do crânio. (5) Megaceryle torquata: vista lateral do crânio. (6) Momotus momota: vistalateral do crânio. CT - CT - CT - CT - CT - crista temporal; FFFFFeA - eA - eA - eA - eA - Fenestra anterorbital; IDNL - IDNL - IDNL - IDNL - IDNL - incisura do duto nasolacrimal; J - J - J - J - J - arco jugal; L - L - L - L - L - lacrimal;LS - LS - LS - LS - LS - laterosfenóide; OS - OS - OS - OS - OS - ossículo sesamóide; PrO - PrO - PrO - PrO - PrO - processo orbital do lacrimal; PrSO - PrSO - PrSO - PrSO - PrSO - processo supra-orbital do lacrimal;PrPOPrPOPrPOPrPOPrPO - - - - - processo pós-orbital; PrZ - PrZ - PrZ - PrZ - PrZ - processo zigomático; SIO - SIO - SIO - SIO - SIO - septo interorbital; PT - PT - PT - PT - PT - pterigóide; TL - TL - TL - TL - TL - tubérculo lacrimal.
44444
55555
66666

160
Osteologia craniana de Cerylinae (Coraciiformes: Alcedinidae)
contato com os ossos nasal e frontal. O processoorbital (PrO) localiza-se ventralmente à incisura edireciona-se ao arco jugal (Figuras 4, 5, 13 e 14).
O processo supra-orbital do lacrimal, na sua facearticular frontonasal, articula-se rostralmente como nasal através da sutura lacrimonasal (suturalacrimonasalis) (SLN) e caudalmente com o frontalatravés da sutura lacrimofrontal (suturalacrimofrontalis) (SLF: Figuras 1-3).
A face dorsal do processo supra-orbital – face articularfrontonasal – possui formato ligeiramente retangular,orienta-se obliquamente ao eixo rostrocaudal docrânio, com a porção caudal voltada para a órbita e aporção rostral curvada ventralmente, voltada para afenestra anterorbital (FeA: Figuras 4 e 5).
Na face rostrodorsolateral da face articularfrontonasal, logo acima da incisura do dutonasolacrimal, observa-se um pequeno osso triangular– ossículo sesamóide (OS: Figuras 4, 5, 19 e 20).Este ossículo recobre a incisura do duto nasolacrimal,porém, não se observa oclusão entre eles. Origina-se rostralmente na margem externa da face ventralda face articular frontonasal em Chloroceryleamazona, C. inda e C. americana, e medialmenteem Megaceryle torquata. Isto faz com que osossículos difiram na forma e na sutura entre oossículo e o corpo do lacrimal. Esta sutura apresenta-se reta em Chlorocery le amazona e C.americana, sendo curva em Megaceryle torquata,devido ao fato de acompanhar a borda arredondadacom a qual se articula (Figuras 4 e 5).
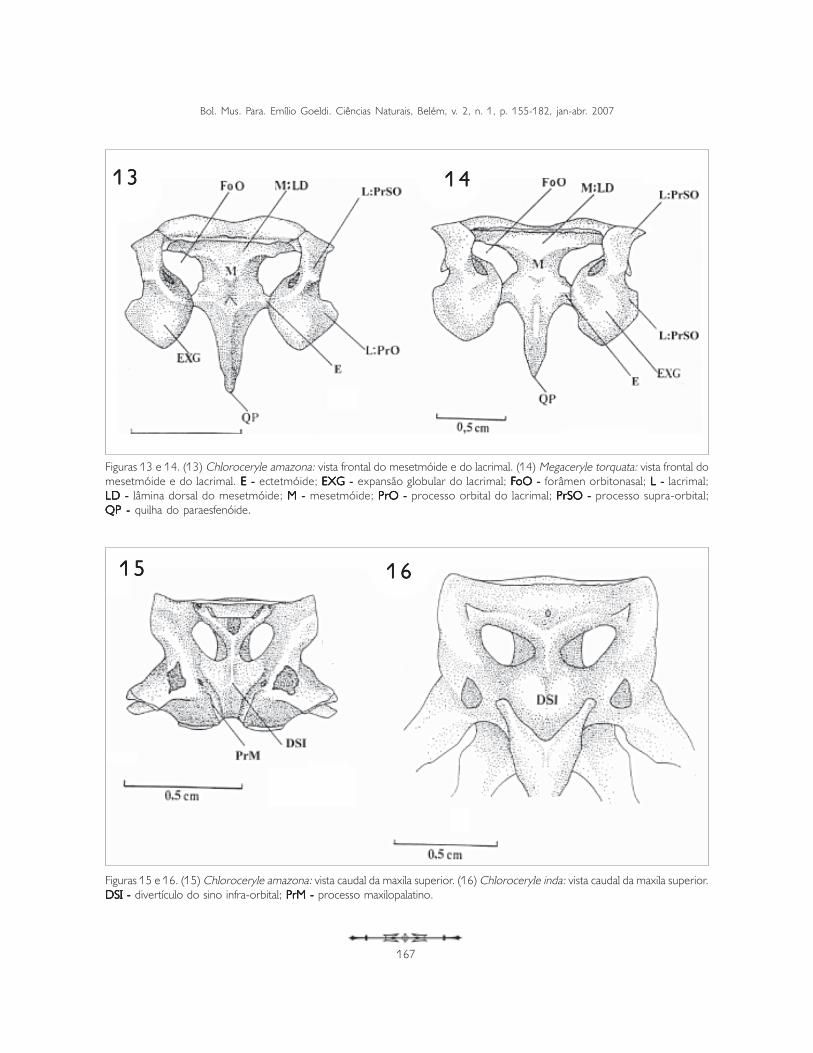
A parede medial do processo supra-orbital forma aparede lateral do forâmen orbitonasal (foramenorbitonasalis) (FoO: Figuras 13, 14, 19 e 20), queé amplo e conspícuo.
O processo orbital do lacrimal (PrO: Figuras 4, 5,13 e 14) localiza-se abaixo da incisura do dutonasolacrimal; sua face rostral delimita a parede caudalda fenestra anterorbital (fenestra antorbitalis) (FeA:Figuras 4 e 5) e sua face caudal forma a porção
rostroventral da órbita. A porção rostral do processoorbital do lacrimal apresenta-se pneumatizada ecom uma expansão globular (expansão processoorbital do lacrimal) (EXG: Figuras 13 e 14). Naparede medial deste processo observa-se umconspícuo entalhe (En), onde o ectetmóideencaixa-se. Isto é observado em Chloroceryle(danificado em Ceryle rudis) e não em Megaceryletorquata, M. Alcyon e M. lugubris. Ventralmente oprocesso orbital dispõe-se direcionado ao arco jugal,onde se apóia, porém, não se observa fusão entreeles (Figuras 4 e 5).
Os ectetmóides (os ectethemoidale) (E: Figuras 13,14, 19 e 20) originam-se lateralmente a partir daregião mediana do mesetimóide (M: Figuras 13 e14), como um delgado osso triangular em formaespicular, perpendicular ao eixo rostrocaudal dosepto interorbital (septum interorbitalis) (SIO: Figuras4-6), formando praticamente um ângulo de 90o
entre os dois (Figuras 13, 14, 19 e 20). Oectetmóide possui uma base larga que se afina atéalcançar o processo orbital do lacrimal, a partir doqual se torna distalmente pontiagudo (Figuras 19 e20). Em Megacery le torquata (Figura 20),M. Alcyon e M. lugubris pouco se observa aporção distal do ectetmóide, devido a umaconspícua expansão lateral do processo orbital dolacrimal (Exl), que recobre dorsalmente oectetmóide. Isto não é observado nas espécies dogênero Chloroceryle (Figuras 19 e 20).
Os mesetimóides (os mesethmoidale) (M)apresentam formato triangular, com o vérticedisposto ventralmente (Figuras 13 e 14); formam aparte óssea rostral do septo interorbital e,dorsalmente, mantém-se em contato com asuperfície ventral do frontal, através da lâminadorsal (lamina dorsalis) (LD: Figuras 13 e 14).Esta lâmina dispõe-se tranversalmente ao planosagital e distalmente direciona-se ao processosupra-orbital do lacrimal. O mesetimóide projeta-seventrocaudalmente e é limitado caudalmente pelaquilha do paraesfenóide (QP: Figuras 7, 8, 13 e 14).

Bol. Mus. Para. Emílio Goeldi. Ciências Naturais, Belém, v. 2, n. 1, p. 155-182, jan-abr. 2007
161
Figuras 7 e 8. (7) Chloroceryle amazona: vista lateral do crânio, sem o osso quadrado. (8) Chloroceryle inda: vista lateral do crânio,sem o osso quadrado. CFT - CFT - CFT - CFT - CFT - crista da fossa temporal; CLCLCLCLCLT - T - T - T - T - crista do laterosfenóide; CNS - CNS - CNS - CNS - CNS - crista nucal sagital; CNT - CNT - CNT - CNT - CNT - crista nucaltranversa; CoO - CoO - CoO - CoO - CoO - cótila ótica; Co1 - Co1 - Co1 - Co1 - Co1 - porção rostral da cótila esquamosal; Co2 - Co2 - Co2 - Co2 - Co2 - porção caudal da cótila esquamosal; CT - CT - CT - CT - CT - cristatemporal; DP - DP - DP - DP - DP - depressão do paraesfenóide; FST - FST - FST - FST - FST - fossa subtemporal; FT - FT - FT - FT - FT - fossa temporal; FRM - FRM - FRM - FRM - FRM - fossa rostral do músculo adutormandibular externo rostral (MAE); LS - LS - LS - LS - LS - laterosfenóide; MAE - MAE - MAE - MAE - MAE - meato acústico externo; OT - OT - OT - OT - OT - óstio timpânico; OTOTOTOTOTA - A - A - A - A - óstio canaldo tubo auditivo; PPPPPrLP - rLP - rLP - rLP - rLP - processo lateral do paraesfenóide; P P P P PrLS - rLS - rLS - rLS - rLS - processo ventral do laterosfenóide; PPPPPrPrPrPrPrPA - A - A - A - A - processoparaoccipital; PrPO - PrPO - PrPO - PrPO - PrPO - processo pós-orbital; PrSM - PrSM - PrSM - PrSM - PrSM - processo suprameático; PrZ - PrZ - PrZ - PrZ - PrZ - processo zigomático; QP - QP - QP - QP - QP - quilha doparaesfenóide; RC - RC - RC - RC - RC - recesso da columela; RTD - RTD - RTD - RTD - RTD - recesso timpânico dorsal; TFTFTFTFTF - tubérculo frontal.
77777
88888

162
Osteologia craniana de Cerylinae (Coraciiformes: Alcedinidae)
O septo interorbital (SIO) é uma delgada placa ósseasagital que tem origem na porção caudal domesetimóide e estende-se pela porção ventral do frontal,não atingindo a parede caudal das órbitas (Figuras 4 e 5).
Entre a extremidade rostral do mesetimóide e a bordacaudal do septo nasal ocorre um espaço denominadohiato craniofacial (hiatus craniofacialis septi).
A fenestra anterorbital (fenestra antorbitalis) (FeA) éum espaço limitado rostralmente pelos processosnasal do maxilar e maxilar do nasal (BAUMEL et al.,1993), ventralmente pelo processo jugal do maxilar,dorsalmente pelo nasal e pelo processo supra-orbital
do lacrimal e, caudalmente, pela parede rostral doprocesso orbital do lacrimal (Figuras 4 e 5).
Os parasphenoidale, os basisphenoidale,Os parasphenoidale, os basisphenoidale,Os parasphenoidale, os basisphenoidale,Os parasphenoidale, os basisphenoidale,Os parasphenoidale, os basisphenoidale,os laterosphenoidale, os squamosumos laterosphenoidale, os squamosumos laterosphenoidale, os squamosumos laterosphenoidale, os squamosumos laterosphenoidale, os squamosumOs basisfenóides não são distinguíveis devido à suatotal fusão rostral com a lâmina do paraesfenóide(lamina parasphenoidalis) (LP: Figuras 9 e 10) e caudalcom os basioccipitais (os basioccipitale).
No osso paraesfenóide, duas regiões sãoprontamente discerníveis nas espécies analisadas:lâmina do paraesfenóide (LP) e rostro paraesfenóide(rostrum parasphenoidale) (RP: Figuras 9 e 10).
Figuras 9 e 10. (9) Chloroceryle amazona: vista ventral do crânio. (10) Megaceryle torquata: vista ventral do crânio. CL - CL - CL - CL - CL - crista lateral;COC - COC - COC - COC - COC - côndilo occipital; CV - CV - CV - CV - CV - crista ventral; FFFFFoMA -oMA -oMA -oMA -oMA - forâmen magno; FP - FP - FP - FP - FP - fossa parabasal; FS - FS - FS - FS - FS - fossa sub-condilar; FV - FV - FV - FV - FV - fossaventral; J - J - J - J - J - arco jugal; LP - LP - LP - LP - LP - lâmina do paraesfenóide; PPPPPoC - oC - oC - oC - oC - porção coanal; P P P P Pol - ol - ol - ol - ol - porção lateral; P P P P PrLP - rLP - rLP - rLP - rLP - processo lateral doparaesfenóide; PrM - PrM - PrM - PrM - PrM - processo maxilopalatino; PT - PT - PT - PT - PT - pterogóide; RP - RP - RP - RP - RP - rostro do paraesfenóide; TB - TB - TB - TB - TB - tubérculo basilar; ZFJ - ZFJ - ZFJ - ZFJ - ZFJ - zonaflexória do arco jugal; ZFP - ZFP - ZFP - ZFP - ZFP - zona flexória palatina.
99999
1010101010

Bol. Mus. Para. Emílio Goeldi. Ciências Naturais, Belém, v. 2, n. 1, p. 155-182, jan-abr. 2007
163
A lâmina do paraesfenóide é limitada caudalmentepelos tubérculos basilares (tuberculum basilare) (TB)e fossa subcondilar (fossa subcondilaris) (FS) dosossos basioccipitais (Figuras 9 e 10). Tal lâminaapresenta formato triangular, com o vértice dispostorostralmente e localiza-se ventralmente ao rostroparaesfenóide.
A base da lâmina do paraesfenóide estende-selateralmente, formando o processo lateral doparaesfenóide (processus lateralis parasphenoidalis)(PrLP). Este processo é conspícuo e assume formatriangular com o vértice disposto em direção ventral(Figuras 7-10). O processo lateral do paraesfenóidedireciona-se caudodorsalmente encontrando oprocesso paroccipital (PrPA: Figuras 7 e 8), ondeforma a parede caudal do meato acústico externo(meatus acusticus externus) (MAE: Figura 7). Omeato é limitado dorsalmente pela fossasubtemporal (fossa subtemporalis) (FST) e peloprocesso suprameático (processum suprameaticus)(PrSM) do esquamosal (Figuras 7 e 8).
O processo lateral do paraesfenóide seguecaudodorsalmente em direção à cótila ótica doesquamosal (cotyla quadratica otici) (CoO:Figura 7), formando a parede rostral do meatoacústico externo, o qual se limita dorsalmente pelacótila supracitada (Figura 7). Tal parede apresenta-se expandida lateralmente, formando um meatobastante profundo (Figura 7). Ventrocaudalmenteeste processo limita-se pelo processo paroccipitaldo exoccipital (PrPA: Figuras 7 e 8).
No interior do meato acústico externo observam-setrês pequenos orifícios, sendo dois situadosdorsalmente e um ventralmente. Entre os primeiros,o óstio timpânico (ostium tympanicum) tem posiçãorostral (OT) e o óstio canal do tubo auditivo (ostiumcanalis tubae auditivae) tem posição caudal (OTA).O orifício situado ventralmente é o recesso dacolumela (recessus columellae) (RC: Figura 7).
Disposto a partir do óstio timpânico e lateralmenteno rostro paraesfenóide, dispõe-se o tubo auditivo.
Seus ramos direito e esquerdo confluemrostralmente em direção à quilha do paraesfenóide.
O rostro paraesfenóide (RP) apresenta um formatotriangular com o vértice disposto rostralmente(Figuras 9 e 10) e limita-se caudalmente pela lâminado paraesfenóide (LP) e pela parede rostral do meatoacústico externo (Figuras 9 e 10). Dorsalmentelimita-se pelo laterosfenóide (LS: Figuras 4 e 8) eventralmente pela lâmina do paraesfenóide. Talrostro apresenta uma expansão dorsal mediana deesboço triangular, localizada logo abaixo do fontículointerorbital (fonticulis interorbitalis) e uma placa ósseaventral, entre este e a lâmina do paraesfenóide(Figuras 7 e 8). Rostralmente à placa óssea ventralobserva-se uma acentuada depressão rostral dorostro paraesfenóide (DP), seguida da acentuadaquilha do paraesfenóide (QP), que repousa sobre aarticulação entre o pterigóide e o palatino (articulatiopterygopalatina), limitando rostralmente o rostroparaesfenóide (Figuras 7 e 8).
Os laterosfenóides (os laterosphenoidale) (LS)limitam-se medialmente com o septo interorbital(SIO), dorsalmente com o frontal e ventralmentecom o rostro paraesfenóide. Ventrolateralmenteobserva-se a área muscular áspera, de formatoglobular, onde medialmente forma-se um pequenoprocesso (PrLS: processo do laterosfenóide).
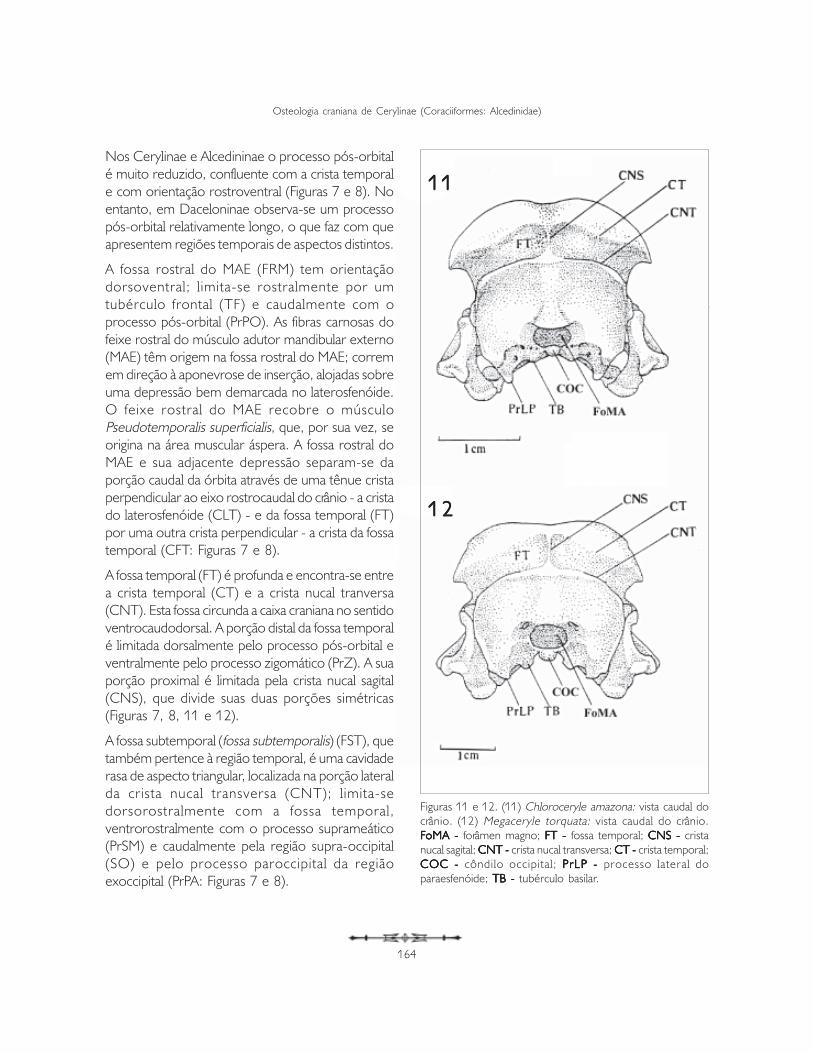
A região temporal (regio temporalis) correspondeàs fossas e impressões decorrentes da origem docomplexo muscular adutor mandibular externo(ZUSI; LIVEZEY, 2000). A porção mais conspícuadesta região é a fossa temporal (fossa temporalis),que, por definição, se situa caudalmente ao processopós-orbital (processum postorbitalis) e dorsal aomeato acústico externo (BAUMEL et al., 1993),sendo seu principal componente o osso esquamosal.
Os Cerylinae apresentam duas fossas na regiãotemporal: a fossa rostral do músculo adutormandibular externo rostral (feixe pós-orbital Burton1984), denominada a partir daqui como fossa rostraldo MAE (FRM) e fossa temporal (FT).
164
Osteologia craniana de Cerylinae (Coraciiformes: Alcedinidae)
Nos Cerylinae e Alcedininae o processo pós-orbitalé muito reduzido, confluente com a crista temporale com orientação rostroventral (Figuras 7 e 8). Noentanto, em Daceloninae observa-se um processopós-orbital relativamente longo, o que faz com queapresentem regiões temporais de aspectos distintos.
A fossa rostral do MAE (FRM) tem orientaçãodorsoventral; l imita-se rostralmente por umtubérculo frontal (TF) e caudalmente com oprocesso pós-orbital (PrPO). As fibras carnosas dofeixe rostral do músculo adutor mandibular externo(MAE) têm origem na fossa rostral do MAE; corremem direção à aponevrose de inserção, alojadas sobreuma depressão bem demarcada no laterosfenóide.O feixe rostral do MAE recobre o músculoPseudotemporalis superficialis, que, por sua vez, seorigina na área muscular áspera. A fossa rostral doMAE e sua adjacente depressão separam-se daporção caudal da órbita através de uma tênue cristaperpendicular ao eixo rostrocaudal do crânio - a cristado laterosfenóide (CLT) - e da fossa temporal (FT)por uma outra crista perpendicular - a crista da fossatemporal (CFT: Figuras 7 e 8).
A fossa temporal (FT) é profunda e encontra-se entrea crista temporal (CT) e a crista nucal tranversa(CNT). Esta fossa circunda a caixa craniana no sentidoventrocaudodorsal. A porção distal da fossa temporalé limitada dorsalmente pelo processo pós-orbital eventralmente pelo processo zigomático (PrZ). A suaporção proximal é limitada pela crista nucal sagital(CNS), que divide suas duas porções simétricas(Figuras 7, 8, 11 e 12).
A fossa subtemporal (fossa subtemporalis) (FST), quetambém pertence à região temporal, é uma cavidaderasa de aspecto triangular, localizada na porção lateralda crista nucal transversa (CNT); l imita-sedorsorostralmente com a fossa temporal,ventrorostralmente com o processo suprameático(PrSM) e caudalmente pela região supra-occipital(SO) e pelo processo paroccipital da regiãoexoccipital (PrPA: Figuras 7 e 8).
Figuras 11 e 12. (11) Chloroceryle amazona: vista caudal docrânio. (12) Megaceryle torquata: vista caudal do crânio.FFFFFoMAoMAoMAoMAoMA - - - - - forâmen magno; FT - FT - FT - FT - FT - fossa temporal; CNS - CNS - CNS - CNS - CNS - cristanucal sagital; CNT - CNT - CNT - CNT - CNT - crista nucal transversa; CT - CT - CT - CT - CT - crista temporal;COC - COC - COC - COC - COC - côndilo occipital; PrLP - PrLP - PrLP - PrLP - PrLP - processo lateral doparaesfenóide; TB - TB - TB - TB - TB - tubérculo basilar.
1111111111
1212121212

Bol. Mus. Para. Emílio Goeldi. Ciências Naturais, Belém, v. 2, n. 1, p. 155-182, jan-abr. 2007
165
processo zigomático e a porção rostral da mesmacótila; seu limite caudal é estabelecido com oprocesso suprameático (PrSM), ventralmente coma cótila ótica (CoO) e ventrocaudalmente com orecesso timpânico dorsal (recessum tympanicusdorsalis) (RTD: Figuras 7 e 8).
A cótila ótica (CoO) é a mais ventral entre as três eapresenta orientação lateral. Limita-se dorsalmentecom a porção caudal da cótila esquamosal,ventralmente com a porção dorsal do meatoacústico externo e caudalmente com o recessotimpânico dorsal (RTD: Figuras 7 e 8).
Os basioccipitale, os exoccipitale, osOs basioccipitale, os exoccipitale, osOs basioccipitale, os exoccipitale, osOs basioccipitale, os exoccipitale, osOs basioccipitale, os exoccipitale, ossupraoccipitalesupraoccipitalesupraoccipitalesupraoccipitalesupraoccipitaleO conjunto destas três regiões forma a parede caudalque delimita a caixa cranaina, transversalmente aoplano sagital. Ela é limitada dorsalmente pela cristanucal tranversal e ventralmente pelos tubérculosbasilares (TB) e pela fossa parabasal (FP: Figuras 9,10, 11 e 12).
No adulto não é possível estabelecer os limitesdestas três regiões devido ao seu fusionamento,porém, a procedência de suas estruturas maisconspícuas, tais como o côndilo occipital (condylusoccipitalis) (COC), o tubérculo basilar (TB), a fossaparabasal (FP), o processo paroccipital (PrPO) e acrista nucal transversa (CNT), é apresentada porBaumel et al. (1993) e adotada no presente trabalho.
A crista nucal tranversal limita dorsalmente as trêsregiões (basioccipital, exoccipital e supra-occipital),além de formar o limite caudal da fossa temporal einterromper-se lateralmente por meio da fossasubtemporal (FST: Figuras 7 e 8).
Caudalmente, estas três regiões limitam-se pelospouco conspícuos tubérculos basilares (TB), fossasubcondilar (FS) e fossas parabasais (FP: Figuras 9 e 10).Dorsalmente às estruturas supracitadas, observa-seo côndilo occipital (COC), que, por sua vez, limitaventralmente o forâmen magno (foramen magnum)(FoMa: Figuras 9). Laterorostralmente ao forâmen
O esquamosal sempre está associado à fossatemporal e à região que recebe o processo óticodo quadrado (Figuras 7 e 8). Porém, no adulto éimpossível determinar sua abrangência e estabelecerseus l imites, principalmente pelo grandedesenvolvimento da fossa temporal, que recebecontribuições de ossos adjacentes.
A cótila esquamosal (CoS), o processo zigomático(PrZ) e o processo suprameático (PrSM) são asprincipais e mais conspícuas estruturas doesquamosal (Figuras 7 e 8). A cótila ótica (CoO)também está associada à região esquamosal, mas éoriginária embriologicamente do ósso ótico (ossaotica), principalmente do proótico (os prooticum)(BAUMEL et al., 1993).
O processo zigomático (PrZ) limita ventrorostralmentea fossa temporal e apresenta-se como uma pequenaparede achatada com orientação oblíqua ao eixorostrocaudal do crânio (Figuras 7 e 8).
O processo suprameático (PrSM) forma a partecaudal da cótila esquamosal e contribui para o limitedorsal do meato acústico externo (BAUMEL et al.,1993) (Figuras 7 e 8).
O quadrado articula-se com o crânio através de duascótilas: cótila esquamosal (CoS) e ótica (CoO). Caberessaltar que a cótila esquamosal apresenta-se bemdesenvolvida e dividida em duas porções, observando-se, portanto, três cavidades cotilares para receber oprocesso ótico do quadrado. A primeira cavidadecotilar (Co1: porção rostral da cótila esquamosal)recebe a porção rostral do capítulo esquamosal(CaS1). Esta primeira cavidade cotilar apresenta-seorientada caudalmente e limita-se dorsalmente coma fossa temporal (FT), rostralmente com o processozigomático (PrZ), caudalmente com o processosuprameático (PrSM) e ventralmente com a porçãocaudal da cótila esquamosal (CoS2: Figuras 7 e 8).
A porção caudal da cótila esquamosal (CoS2) é amais desenvolvida entre as três cavidades cotilarese apresenta formato de meia-lua. Orienta-seventralmente e limita-se rostralmente com o

166
Osteologia craniana de Cerylinae (Coraciiformes: Alcedinidae)
leve orientação lateral e em forma de asas, formandoas paredes dorsolaterais da fossa coanal (Figura 23).
Dorsocaudalmente encontram-se as cristasdorsolaterais (CDL), que não são fusionadas e poristo formam uma fossa dorsocaudal (FPP: fossapalatoparaesfenoidal) por onde a quilha do rostroparaesfenóide desliza no momento da abertura dasmaxilas (Figura 23).
A fossa coanal orienta-se ventrocaudalmente e écircundada pelas cristas ventrais (CV), que se fusionamcaudalmente, formando o processo do pterigóide(processum pterygoideus), que conecta o palatinocom o pterigóide (PT: Figuras 9 e 10).
Ventralmente observa-se a fossa ventral (fossaventralis) (FV), limitada pelas crista ventrais (CV) ecrista laterais (CL), sendo que estas últimas, nasespécies de Chloroceryle, ao contrário doobservado em Megaceryle torquata, são poucoconspícuas, percebendo-se apenas uma tênue linha(Figuras 9 e 10). Esta fossa está relacionada àmusculatura do pterigóide (BAUMEL et al., 1993).
Os ângulos caudolaterais (angulus caudolateralis) (ACL),representados pela intersecção entre as margens caudaise laterais das porções laterais, são conspícuos epossuem uma projeção óssea, com orientação caudalem forma de espinho (Figuras 9, 10 e 23).
O processo maxilopalatino (processummaxillopalatinus) (PrM) do maxilar apresenta-secomo a principal estrutura para a definição do tipode palato desmognato (Figuras 9, 10 e 15). Os doisprocessos unem-se rostroventralmente na linhamediocaudal do rostro maxilar e formam a porçãorostral da fossa coanal do palatino, a qual separacaudalmente ambos os processos (Figura 15);caudalmente cada processo possui duas porçõesseparadas e expandidas em forma de asa.Dorsalmente ao processo observa-se o divertículodo sino infra-orbital (DSI), que se apresenta comouma evaginação da cavidade nasal (Figuras 15 e 16).Tal divertículo tem formato ligeiramente triangular,com o vértice ventral (Figura 15).
magno observam-se acentuadas fossas parabasais(FP), que, possivelmente, contribuem, por meio dacrista fossa parabasal, para o bem desenvolvidoprocesso lateral do paraesfenóide (Figuras 9, 10,11 e 12).
O processo paroccipital (PrPA) do exoocipital é bemdesenvolvido e também recebe contribuições doosso ótico (BAUMEL et al., 1993). Tal processoforma a parede caudal do meato acústico externo elimita caudalmente a fossa subtemporal (Figuras 4,5, 7 e 8).
Os palatinum, os pterygoideum, os jugaleOs palatinum, os pterygoideum, os jugaleOs palatinum, os pterygoideum, os jugaleOs palatinum, os pterygoideum, os jugaleOs palatinum, os pterygoideum, os jugaleOs palatinos (os palatinum) (PA: Figura 23) fundem-serostromedialmente aos maxilares (os maxillare)através dos processos maxilares dos palatinos(processum maxillaris) (PrMP), com os quais formama zona flexória palatina (zona flexoria palatina)(ZFP: Figuras 9 e 10). Rostralmente a esta zona, oscomponentes que formam a maxila superiorcoalescem, fechando-a ventralmente. Estaconformação, acrescida da fusão mediana dosprocessos maxilopalatinos (PrM), caracteriza osAlcedinidae como desmognatos (HUXLEY, 1867);não se observa o vômer (Figuras 9 e 10).
Os palatinos caudalmente encontram-se com ospterigóides (os pterigoideum) (PT) e o rostroparaesfenóide através das face articular do pterigóide(facies articularis pterygoidea) da porção coanal (parschoanalis) (PoC) e face articular do paraesfenóide(facies articularis parasphenoidalis) da porção lateral(pars lateralis) (BAUMEL et al., 1993) (Figura 9,10 e 23).
Duas partes principais são identificáveis nos palatinos:a porção coanal (pars choanalis) (PoC), associada àfossa coanal (fossa choanalis) (FC); e a porção lateral(pars lateralis) (PoL), geralmente lateral à precedentee relacionada com a musculatura do pterigóide(BAUMEL et al., 1993) (Figuras 9, 10 e 23).
As lamelas coanais (lamellas choanalis) (LC)projetam-se dorsalmente (Figura 23), apresentam
Bol. Mus. Para. Emílio Goeldi. Ciências Naturais, Belém, v. 2, n. 1, p. 155-182, jan-abr. 2007
167
Figuras 13 e 14. (13) Chloroceryle amazona: vista frontal do mesetmóide e do lacrimal. (14) Megaceryle torquata: vista frontal domesetmóide e do lacrimal. E - E - E - E - E - ectetmóide; EXG - EXG - EXG - EXG - EXG - expansão globular do lacrimal; FFFFFoO - oO - oO - oO - oO - forâmen orbitonasal; L - L - L - L - L - lacrimal;LD - LD - LD - LD - LD - lâmina dorsal do mesetmóide; M - M - M - M - M - mesetmóide; PrO - PrO - PrO - PrO - PrO - processo orbital do lacrimal; PrSO - PrSO - PrSO - PrSO - PrSO - processo supra-orbital;QP -QP -QP -QP -QP - quilha do paraesfenóide.
Figuras 15 e 16. (15) Chloroceryle amazona: vista caudal da maxila superior. (16) Chloroceryle inda: vista caudal da maxila superior.DSI - DSI - DSI - DSI - DSI - divertículo do sino infra-orbital; PrM - PrM - PrM - PrM - PrM - processo maxilopalatino.
1313131313 1414141414
16161616161515151515

168
Osteologia craniana de Cerylinae (Coraciiformes: Alcedinidae)
Os pterigóides (PT) são barras delgadas, que searticulam rostralmente com os palatinos através daface articular palatina (facies articularis palatina) ecaudalmente com os ossos quadrados através daface articular do quadrado (facies articularisquadratica).
O arco jugal (formado pelo processo jugal domaxilar, jugal e quadradojugal) (J) é uma barra queliga o quadrado ao maxilar. Caudalmente articula-secom o quadrado, através do conspícuo côndilo doquadrado (condylus quadraticus) (Figuras 4-6, 9 e10). Rostralmente este arco encontra-se com oprocesso jugal do maxilar (processus jugalis de osmaxillare), onde se observa a zona flexória do jugal(zona flexoria arcus jugalis) (ZFJ: Figuras 9 e 10).
O arco jugal, nos Cerylinae e Alcedininae, a partirde sua porção mediana, apresenta uma expansãorostrolateral, formando uma placa achatadadorsoventralmente, cerca de três vezes mais largado que a porção caudal do jugal (J: Figuras 9 e 10),
A porção rostral do jugal apresenta-se ventralmentecôncava e, dorsalmente na margem externa,observa-se o conspícuo tubérculo do lacrimal(tuberculum lacrimale) (TL: Figuras 4 e 5).Exatamente na margem oposta ao tubérculo dolacrimal existe uma pequena incisura. Rostralmenteao tubérculo lacrimal nota-se uma pequena fossa,extremamente acentuada em Megaceryle torquata.
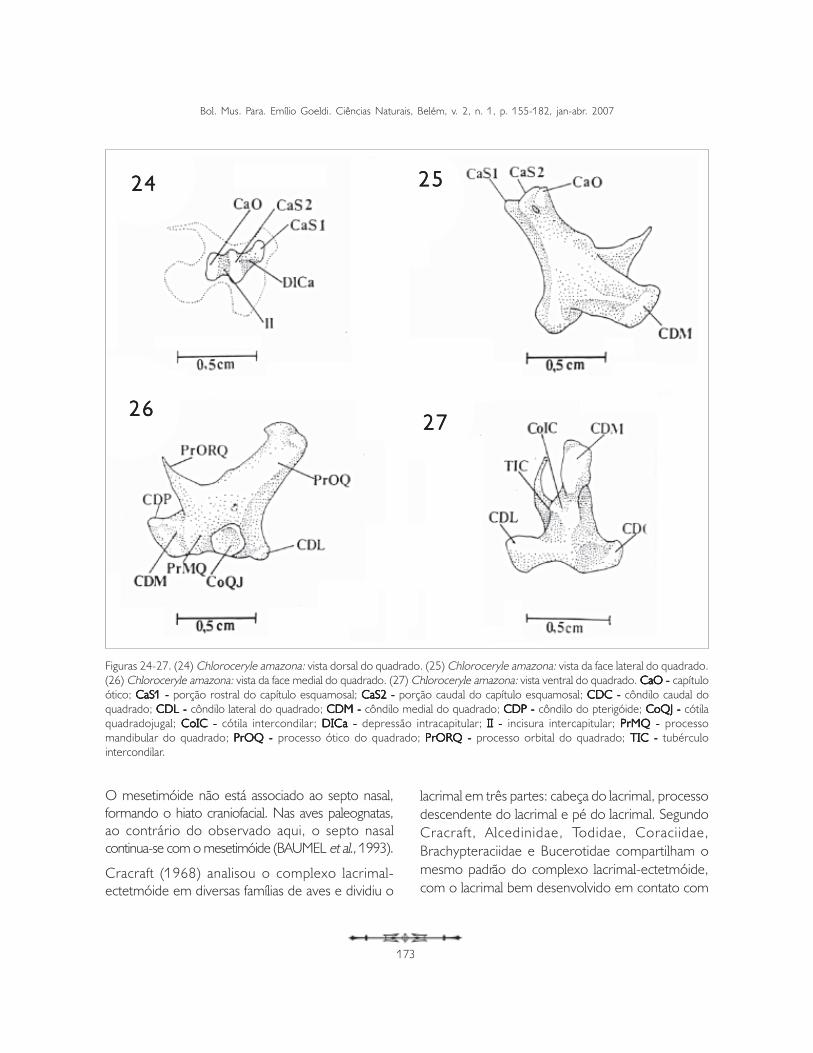
Os quadratumOs quadratumOs quadratumOs quadratumOs quadratumTrês principais processos são distinguíveis no corpodeste osso (corpus ossis quadrati ): processo ótico(processum oticum) (PrOQ), processo orbital(processum orbitalis) (PrORQ) e processomandibular (processum mandibularis) (PrMQ:Figuras 24-27).
O quadrado articula-se com o crânio através doprocesso ótico, por meio dos capítulos esquamosal(capitulum squamosum) (CaS) e ótico (capitulumoticum) (CaO: Figuras 24-26). Estes articulam-serespectivamente às cótilas esquamosal (cotyla
quadratica squamosi ) e ótica (CoO) (cotylaquadratica otici ) (CoS: Figuras 7 e 8).
O capítulo esquamosal é o maior entre os doiscapítulos e possui uma expansão rostral que seprolonga através da borda do processo ótico doquadrado até sua extremidade rostral. Destamaneira, forma-se uma nova saliência, que se articulacom a porção rostral da cótila esquamosal (Figuras24-26).
A porção rostral do capítulo esquamosal (CaS1)localiza-se na extremidade rostral do processo óticodo quadrado, sendo menos arredondada que aporção caudal. Ela se posiciona rostralmente emrelação à porção caudal do capítulo esquamosal(CaS2) e do capítulo ótico (CaO: Figuras 24 e 26),separando-se da porção caudal do capítuloesquamosal por meio de uma conspícua depressão(DICa: depressão intracapitular). Porém, taldepressão não é completa devido ao capítuloesquamosal prolongar-se rostralmente através daface medial do processo ótico do quadrado,resultando na segunda saliência articular do capítuloesquamosal (CaS1: Figuras 24 e 26). Cabe ressaltarque esta conformação poderia ser considerada comoduas estruturas distintas e, assim, lhes seriamatribuídos nomes distintos. No entanto, optou-sepor descrever uma estrutura única com duasporções.
A porção caudal do capítulo esquamosal (CaS2) é amaior entre as três saliências articulares e localiza-seentre a porção rostral do capítulo esquamosal(CaS1) e capítulo ótico (CaO: Figuras 24 e 26).
O capítulo ótico é menor que o esquamosal,situando-se caudalmente em relação às duas porçõesdeste último capítulo. Separa-se do capítuloadjacente pela incisura intercapitular (incisuraintercapitularis) (II: Figuras 24 e 26).
O processo orbital (processum orbitalis) (PrORQ)projeta-se rostralmente na forma de um fino espinhoe situa-se na porção rostral do quadrado, entre os

Bol. Mus. Para. Emílio Goeldi. Ciências Naturais, Belém, v. 2, n. 1, p. 155-182, jan-abr. 2007
169
côndilos medial (condylus medialis) (CDM) e lateral(condylus lateralis) (CDL: Figuras 25 e 27).
O processo mandibular (processum mandibularis)(PrMQ) é o mais dorsal entre os três e articula-secom a mandíbula através de três côndilos de formatosirregulares: côndilo medial (condylus medialis)(CDM), côndilo lateral (condylus lateralis) (CDL)e côndilo caudal (condylus caudalis) (CDC). Ocôndilo medial é o mais ventral e os outros doisencontram-se no mesmo plano (Figuras 25 e 27).
O côndilo medial (CDM: Figuras 25-27) é o maisdesenvolvido entre os três e apresenta-seligeiramente arredondado. Além de articular-se coma cótila medial (CoM) da mandíbula, articula-serostralmente com o pterigóide, através do côndilodo pterigóide (condylus pterygoideus), que seapresenta dorsal em relação ao côndilo medial(Figura 25).
Entre os côndilos medial e lateral nota-se umconspícuo tubérculo intercondilar (TIC: Figura 27);a presença deste processo na parede que separa oscôndilos faz com que se formem duas cavidadescotilares. A formação cotilar ventral (CoIC: cótilaintercondilar) articula-se com o tubérculo intercotilarda mandíbula (Figura 27).
O côndilo lateral localiza-se na face lateral doprocesso mandibular do quadrado, onde se observauma conspícua e profunda cótila quadradojugal(cotyla quadratojugalis) (CoQJ), que recebe a barrajugal (Figura 25). Este côndilo articula-se com amandíbula através da cótila lateral (CoL) da fossaarticular quadrática (fossa articularis quadratica) damandíbula.
O côndilo caudal (condylus caudalis) (CDC)encontra-se adjacente à porção caudal do côndilolateral e é o menos desenvolvido entre os três(Figuras 17, 18 e 27), situando-se no mesmo planodo côndilo lateral (Figuras 17, 18 e 27). Em Cerylerudis e nas espécies de Chloroceryle o côndilopossui natureza cotilar, com a face ventral formando
uma superfície ventral côncava, ou seja, cotilar, quese articula com a mandíbula através do processocaudal cotilar da mandíbula de natureza condilar(Figura 17).
Em Megaceryle torquata, M. alcyon e M. lugubrisnão se observa esta condição. O côndilo caudalrealmente possui natureza condilar, articulando-secom a cótila caudal da mandíbula (Figura 18).
Ossa mandibulaeOssa mandibulaeOssa mandibulaeOssa mandibulaeOssa mandibulaeA mandíbula apresenta formato pontiagudo e trêspartes principais são prontamente discerníveis:porção sinfisial (pars symphysialis) (PSI), porçãointermediária (pars intermedia) (PI) e porção caudal(pars caudalis) (PC: Figura 31).
A porção sinfisial (PSI) é a porção mais rostral, comambos os ramos mandibulares unidos. A partir desua separação, até o ângulo mandibular (angulusmandibulae) (AM), situa-se a porção intermediária(PI), seguida da porção caudal (PC) (Figura 31).
A porção sinfisial é achatada dorsoventralmente e asua metade caudal torna-se gradativamente achatadalateralmente. A partir do ângulo mandibular, emdireção caudal, é achatada lateralmente em suatotalidade. A extremidade caudal, que se articula como quadrado, por meio da fossa articular do quadrado,apresenta formato irregular.
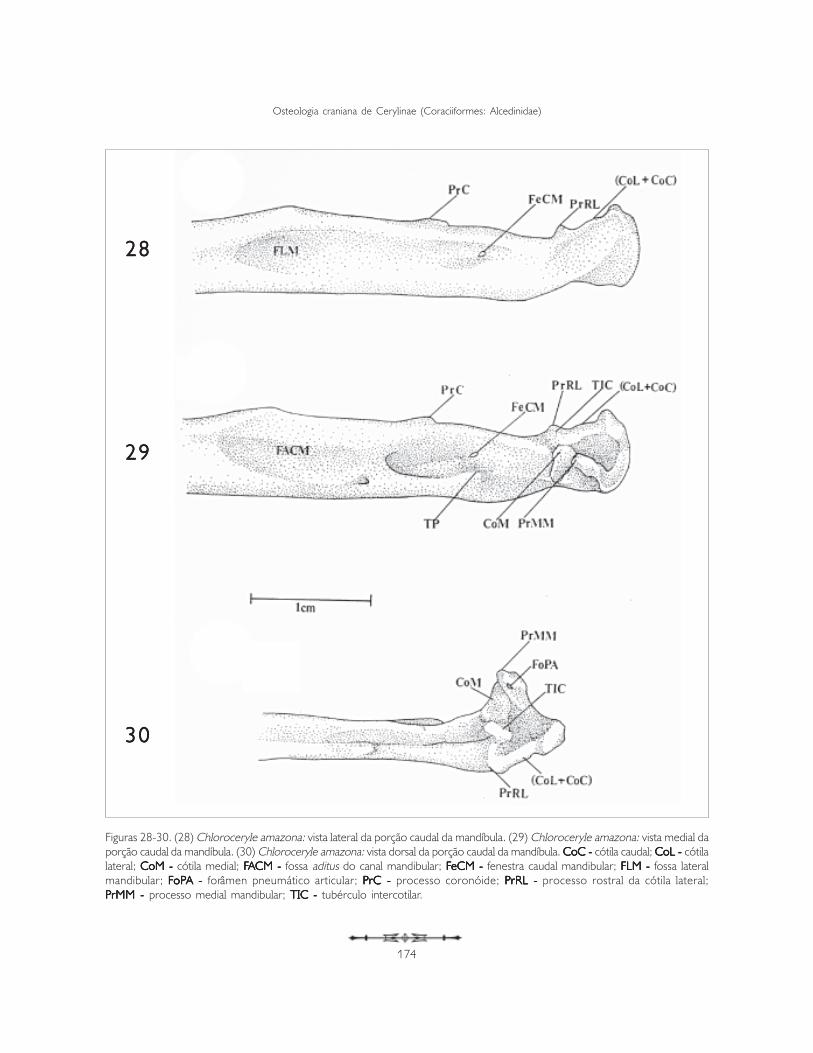
A porção caudal limita-se rostralmente pelo ângulomandibular e caudalmente pela fossa caudal (fossacaudalis) (FOC: Figura 21 e 22). Esta porção exibemaior complexidade estrutural por servir como elode ligação entre a mandíbula e a caixa craniana, atravésdo quadrado, e sustentar musculatura, aponevrosese tendões relacionados com a movimentação damandíbula (Figuras 28-30, 31-34).
Esta região pode ser dividida em porção rostral ecaudal. A primeira estende-se do ângulo mandibularaté a porção rostral da fossa articular do quadrado,enquanto a porção caudal é a própria fossa articulardo quadrado (Figuras 29, 30, 31 e 33).

170
Osteologia craniana de Cerylinae (Coraciiformes: Alcedinidae)
A porção rostral apresenta fossas em ambas as faces(lateral e medial): fossa aditus do canal mandibular(fossa aditus canalis mandibulae) (FACM) e fossalateral mandibular (fossa lateralis mandibulae) (FLM:Figuras 28, 29, 32 e 34).
Na face medial nota-se a fossa aditus do canalmandibular, que é longa e se estende desde o ângulocaudomedial até a porção rostral da fossa articulardo quadrado. Observam-se duas fenestras: fenestrarostral mandibular (fenestra rostralis mandibulae)(não representada) e fenestra caudal mandibular(fenestra caudalis mandibulae) (FeCM: Figuras 28,29, 32 e 34).
A fenestra caudal mandibular (FeCM) situa-se entreo processo coronóide (processum coronoideus)(PrC) e o tubérculo pseudotemporal (tuberculumpseudotemporale) (TP: Figuras 28, 29, 32 e 34).
Na face dorsal da extremidade rostral da porçãocaudal observa-se o conspícuo processo coronóide(PrC), que se situa entre o ângulo caudomedial e aporção caudal da fossa articular do quadrado (Figuras28, 29, 32-34).
Na porção medioventral da fossa aditus do canalmandibular, e exatamente abaixo da fenestra caudalmandibular, observa-se o conspícuo tubérculopseudotemporal (TP: Figuras 29, 32 e 33).
A fossa articular do quadrado recebe os côndilos doquadrado através das cótilas da fossa articular doquadrado (cotylae fossae articularis).
As cótilas medial, lateral e caudal (cotyla medialis,cotyla lateralis e cotyla caudalis) (CoM, CoL, e CoC)recebem, respectivamente, os côndilos medial,lateral e caudal do quadrado.
As cót i las la tera l (CoL) e cauda l (CoC)encontram-se na face lateral da fossa articulardo quadrado (Figuras 28-30, 32-34). Orientam-serostrocaudalmente e são adjacentes, ocupando omesmo plano, ao contrário da cótila medial, emforma de meia-lua, que não se posiciona no mesmo
plano que as outras, situando-se dorsalmente aambas (Figuras 28-30, 32-34).
A cótila lateral limita-se rostralmente por umpequeno processo (processo rostral da cótila lateral)(PrRL: Figuras 17, 18, 28-30, 32 e 34). EmMegaceryle torquata, M. alcyon e M. lugubris estacótila limita-se caudalmente por um pequenoprocesso (PrMIC: processo medial intercotilar), quetambém marca o início da cótila caudal (Figuras 18,32 e 34), que, por sua vez, se limita caudalmentepor outro pequeno processo (processo caudal dacótila caudal) (PrCC: Figuras 17, 18 e 34).
O processo medial intercotilar é ausente em Cerylerudis e nas espécies de Chloroceryle. Esta condiçãofaz com que, em Ceryle rudis e em Chloroceryle,ambas as cótilas (lateral e caudal) confundam seuslimites, aparentando existir apenas uma grande e únicacótila (Figuras 17, 28-30). No entanto, Megaceryletorquata, M. alcyon e M. lugubris possuem as duascótilas bem definidas (Figuras 18, 32-34).
Em Ceryle rudis e nas espécies de Chloroceryle, oprocesso caudal da cótila caudal (PrCC) assume papelimportante na articulação quadrado mandibulardevido à ausência do processo medial intercotilar.Tal ausência faz com que a articulação entre as cótilasmandibulares (exetuando a medial) e os côndilosdo quadrado (exetuando o medial) apresentem-setotalmente distintas nos gêneros em questão (Figuras17 e 18).
Em Megaceryle torquata, M. alcyon e M. lugubrisnota-se a articulação entre as duas cótilasmandibulares e os dois côndilos do quadrado(excetuando a cótila e o côndilo medial) (Figura 18).No entanto, as espécies de Chloroceryleapresentam uma condição contrária, no que serefere à cótila e ao côndilo caudal. Esta articulaçãoocorre na forma de uma saliência condilar namandíbula e uma cavidade cotilar do quadrado(Figura 17), o que se deve à ausência do processomedial intercotilar, formando, aparentemente, umaúnica grande cótila (cótila lateral mais a caudal) e,

Bol. Mus. Para. Emílio Goeldi. Ciências Naturais, Belém, v. 2, n. 1, p. 155-182, jan-abr. 2007
171
principalmente, pela condição do côndilo caudal doquadrado, que forma uma aba côncava (portanto,de natureza cotilar), que recobre e articula-se como processo caudal da cótila caudal (portanto, denatureza condilar) (Figuras 17 e 18).
A porção caudal da fossa articular do quadrado temformato de V e nela observa-se a fossa caudal (fossacaudalis) (FOC: Figuras 21 e 22), que estárelacionada ao músculo depressor mandibular. Nas
espécies de Chloroceryle e Ceryle rudis esta fossaé rasa e estreita. No entanto, Megaceryle torquata,M. alcyon e M. lugubris possuem a fossa mais ampla,devido à formação de uma parede dorsal (PaFC:parede dorsal da fossa caudal), que praticamentefecha a porção caudal do ramo mandibular,descaracterizando o formato de V supracitado, paraassumir uma conformação triangular com o vérticesituado ventralmente (Figura 22).
Figuras 17-22. (17) Chloroceryle amazona: vista lateral da articulação entre o quadrado e a mandíbula. (18) Megaceryle torquata: vistalateral da articulação entre o quadrado e a mandíbula. (19) Chloroceryle amazona: vista caudal do lacrimal e do ectetmóide.(20) Megaceryle torquata: vista caudal do lacrimal e do ectetmóide. (21) Chloroceryle amazona: vista caudal da mandíbula. (22) Megaceryletorquata: vista caudal da mandíbula. CDC - CDC - CDC - CDC - CDC - côndilo caudal do quadrado; CDL - CDL - CDL - CDL - CDL - côndilo lateral do quadrado; CoC - CoC - CoC - CoC - CoC - cótila caudalda mandíbula; CoL - CoL - CoL - CoL - CoL - cótila lateral da mandíbula; E - E - E - E - E - ectetmóide; EXL - EXL - EXL - EXL - EXL - expansão do processo orbital; FFFFFoO - oO - oO - oO - oO - forâmen orbitonasal;FOC - FOC - FOC - FOC - FOC - fossa caudal da mandíbula; L - L - L - L - L - lacrimal; PPPPPAFC - AFC - AFC - AFC - AFC - parede dorsal da fossa caudal; PPPPPrCC - rCC - rCC - rCC - rCC - processo caudal da cótila caudal damandíbula; PrRL - PrRL - PrRL - PrRL - PrRL - processo rostral da cótila lateral da mandíbula; PrMIC - PrMIC - PrMIC - PrMIC - PrMIC - processo medial intercotilar; OS - OS - OS - OS - OS - ossículo sesamóide.
1717171717 1818181818 1919191919
2020202020 2121212121 2222222222

172
Osteologia craniana de Cerylinae (Coraciiformes: Alcedinidae)
O parietal, no entanto, não possui um conjunto deestruturas características, sendo a crista nucaltransversa a única estrutura listada para este osso(BAUMEL et al., 1993). Porém, devido ao grandedesenvolvimento da fossa temporal e a localizaçãoda crista nucal transversa, é plausível supor que oparietal também contribua para a supracitada fossa.
Barnikol (1952, ap. Burton 1984) descreve o crâniodo tipo streckschädel, com o forâmen magno situadoventralmente no crânio, os pterigóides longos erepousando no mesmo plano dos palatinos e rostroparaesfenoidal. Este autor acredita que talconformação de crânio seria uma forma arcaica,relacionada à piscivoria. Nos martins-pescadoresanalisados esta condição encontra-se bastanteevidente e Burton (1984) afirma que nos martins-pescadores piscívoros (Cerylinae e Alcedininae) estacondição é mais marcada do que nos martins-pescadores florestais (Daceloninae).
As narinas são tipicamente do tipo holorrino(GARROD, 1873), que, juntamente com uma zonaflexória craniofacial bem desenvolvida e a presençado hiato craniofacial (hiatus craniofacialis septi ),proporcionam um eficiente movimento deprocinese. A condição holorrina, geralmente, estáassociada ao procinetismo das aves (BOCK, 1964;ZUSI, 1984). O hiato craniofacial é importante parao cinetismo craniano e está presente em todas asaves modernas procinéticas, como também emmuitas neognatas rincocinéticas (BÜHLER, 1985).
Huxley (1867), ao classificar os tipos de palato, defineos desmognatos como apresentando fusão medialdos processos maxilopalatinos, totalmente emconcordância com este trabalho. A condição de palatodesmognato parece ter surgido independentementeem diferentes grupos de aves, já que vários gruposnão estreitamente relacionados (Ardeidae, Anatidae,Alcedinidae) possuem a mesma condição de palato.O processo maxilopalatino, segundo Baumel et al.(1993), apresenta considerável variação emdecorrência da sua associação com o divertículomaxilar do sino infra-orbital.
O processo medial mandibular (processum medialismandibulae) (PrMM) é conspícuo e mostra-seorientado dorsalmente (Figuras 29, 30, 32 e 33).Na sua face medial encontra-se o forâmenpneumático articular (foramen pneumaticumarticularis) (FoPA: Figuras 30 e 33).
A cótila medial (CoM) é a mais desenvolvida entreas três cótilas, tem forma de meia-lua e orienta-seperpendicularmente ao eixo rostrocaudal do crânio(Figuras 29, 30, 32 e 33). Esta cótila, na sua facemedial, limita-se com o processo medial mandibular(PrMM) e na sua face lateral apresenta o tubérculointercotilar (TIC: Figuras 29, 30, 32 e 33), que searticula com a cótila intercondilar (CoIC: Figura 27)do quadrado.
DISCUSSÃO
Dois ossos foram suprimidos da descrição e dascorrespondentes figuras: o esquamosal e o parietal.O esquamosal não foi citado nominalmente porquesuas principais estruturas estão descritas erepresentadas (processo zigomático, processosuprameático e cótila do esquamosal).
Figura 23. Chloroceryle amazona: vista dorsal do palatino.ACL - ACL - ACL - ACL - ACL - ângulo caudolateral; CDL - CDL - CDL - CDL - CDL - crista dorsolaterais; FPP -FPP -FPP -FPP -FPP - fossapalato-paraesfenoidal; LC - LC - LC - LC - LC - lamelas coanais.
2323232323
Bol. Mus. Para. Emílio Goeldi. Ciências Naturais, Belém, v. 2, n. 1, p. 155-182, jan-abr. 2007
173
O mesetimóide não está associado ao septo nasal,formando o hiato craniofacial. Nas aves paleognatas,ao contrário do observado aqui, o septo nasalcontinua-se com o mesetimóide (BAUMEL et al., 1993).
Cracraft (1968) analisou o complexo lacrimal-ectetmóide em diversas famílias de aves e dividiu o
lacrimal em três partes: cabeça do lacrimal, processodescendente do lacrimal e pé do lacrimal. SegundoCracraft, Alcedinidae, Todidae, Coraciidae,Brachypteraciidae e Bucerotidae compartilham omesmo padrão do complexo lacrimal-ectetmóide,com o lacrimal bem desenvolvido em contato com
Figuras 24-27. (24) Chloroceryle amazona: vista dorsal do quadrado. (25) Chloroceryle amazona: vista da face lateral do quadrado.(26) Chloroceryle amazona: vista da face medial do quadrado. (27) Chloroceryle amazona: vista ventral do quadrado. CaO - CaO - CaO - CaO - CaO - capítuloótico; CaS1 - CaS1 - CaS1 - CaS1 - CaS1 - porção rostral do capítulo esquamosal; CaS2 - CaS2 - CaS2 - CaS2 - CaS2 - porção caudal do capítulo esquamosal; CDC - CDC - CDC - CDC - CDC - côndilo caudal doquadrado; CDL - CDL - CDL - CDL - CDL - côndilo lateral do quadrado; CDM - CDM - CDM - CDM - CDM - côndilo medial do quadrado; CDP - CDP - CDP - CDP - CDP - côndilo do pterigóide; CoQJ - CoQJ - CoQJ - CoQJ - CoQJ - cótilaquadradojugal; CoIC - CoIC - CoIC - CoIC - CoIC - cótila intercondilar; DICa - DICa - DICa - DICa - DICa - depressão intracapitular; II - II - II - II - II - incisura intercapitular; PrMQ - PrMQ - PrMQ - PrMQ - PrMQ - processomandibular do quadrado; PrOQ - PrOQ - PrOQ - PrOQ - PrOQ - processo ótico do quadrado; PrORQ - PrORQ - PrORQ - PrORQ - PrORQ - processo orbital do quadrado; TIC - TIC - TIC - TIC - TIC - tubérculointercondilar.
2424242424
26262626262727272727
2525252525
174
Osteologia craniana de Cerylinae (Coraciiformes: Alcedinidae)
Figuras 28-30. (28) Chloroceryle amazona: vista lateral da porção caudal da mandíbula. (29) Chloroceryle amazona: vista medial daporção caudal da mandíbula. (30) Chloroceryle amazona: vista dorsal da porção caudal da mandíbula. CoC - CoC - CoC - CoC - CoC - cótila caudal; CoL -CoL -CoL -CoL -CoL - cótilalateral; CoM - CoM - CoM - CoM - CoM - cótila medial; FFFFFAAAAACM - CM - CM - CM - CM - fossa aditus do canal mandibular; FFFFFeCM - eCM - eCM - eCM - eCM - fenestra caudal mandibular; FLM - FLM - FLM - FLM - FLM - fossa lateralmandibular; FFFFFoPoPoPoPoPA - A - A - A - A - forâmen pneumático articular; P P P P PrC - rC - rC - rC - rC - processo coronóide; PPPPPrRL - rRL - rRL - rRL - rRL - processo rostral da cótila lateral;PPPPPrMM -rMM -rMM -rMM -rMM - processo medial mandibular; TIC - TIC - TIC - TIC - TIC - tubérculo intercotilar.
2828282828
2929292929
3030303030

Bol. Mus. Para. Emílio Goeldi. Ciências Naturais, Belém, v. 2, n. 1, p. 155-182, jan-abr. 2007
175
Figuras 31-34. (31) Megaceryle torquata: vista dorsal esquemática da mandíbula. (32) Megaceryle torquata: vista medial da porçãocaudal da mandíbula. (33) Megaceryle torquata: vista dorsal da porção caudal da mandíbula. (34) Megaceryle torquata: vista lateral daporção caudal da mandíbula. CoC - CoC - CoC - CoC - CoC - cótila caudal; CoL - CoL - CoL - CoL - CoL - cótila lateral; CoM - CoM - CoM - CoM - CoM - cótila medial; FFFFFAAAAACM - CM - CM - CM - CM - fossa aditus do canalmandibular; F F F F FeCM - eCM - eCM - eCM - eCM - fenestra caudal mandibular; FFFFFoPoPoPoPoPA - A - A - A - A - forâmen pneumático articular; PC - PC - PC - PC - PC - porção caudal; PI - PI - PI - PI - PI - porçãointermediária; PSI - PSI - PSI - PSI - PSI - porção sinfisial; PrC - PrC - PrC - PrC - PrC - processo coronóide; PrCC - PrCC - PrCC - PrCC - PrCC - processo caudal da cótila caudal; PrMIC - PrMIC - PrMIC - PrMIC - PrMIC - processomedial intercotilar; PrMM - PrMM - PrMM - PrMM - PrMM - processo medial mandibular; PrRL - PrRL - PrRL - PrRL - PrRL - processo rostral da cótila lateral; TIC - TIC - TIC - TIC - TIC - tubérculo intercotilar;TPTPTPTPTP - - - - - tubérculo pseudotemporal.
3131313131
3232323232
3434343434
3333333333

176
Osteologia craniana de Cerylinae (Coraciiformes: Alcedinidae)
a zona flexória craniofacial e o ectetmóide delgadocom forma de ‘espora’. Também descreve, semmaiores detalhes, um lacrimal com um entalhemedial para receber o ectetmóide e a ocorrênciaocasional de ossos sesamóides na sua parede lateral.
Nos Alcedinidae, como na maioria das aves, olacrimal articula-se com os ossos frontal e nasalatravés de suturas que permitem movimentação docomplexo lacrimal-ectetmóide em relação à maxilasuperior, o que auxilia na procinese (ZUSI, 1962;CRACRAFT, 1968; BÜHLER, 1985). O lacrimalpode articular-se somente com o frontal(Psittacidae), apenas com o nasal (Treskiornithidae)(Cracraft 1968) ou, ainda, fusionar-se ao ectetmóide(Ramphastidae), delimitando rostralmente a órbita(HÖFLING; GASC, 1984; HÖFLING, 1995).
Nos Galbulidae, Donatelli (1992) verificou que olacrimal articula-se ao frontal e ao nasal, enquantonos Picidae (DONATELLI, 1996) encontra-sefusionado ao frontal e à maxila superior.
Em Cracidae, exceto em Mitu tuberosa, o nasal eo frontal encontram-se fundidos, “formando umaúnica sutura nasofrontolacrimal” (MARCELIANO,1996). Ophistocomus hoazin, Tauraco hartlaubi,Tauraco erythrolophus e Tauraco leucotisapresentam as suturas nasolacrimal e frontolacrimal,como também os cuculídeos Coccyzus americanus,Coccyzus melacoryphus, Piaya cayana, Crotophagaani e Guira guira.
As únicas variações observadas na forma dos processossupra-orbital e orbital dos martins-pescadores foi apresença de uma apófise na porção caudal da facearticular frontonasal em Dacelo novaguineae e apresença da expansão dorsal do processo orbitalrecobrindo o ectetmóide nos Megaceryle. Umaapófise no lacrimal é reportada por Höfling (1995)para a maioria dos Ramphastidae e Capitonidae daregião Neotropical. Na porção rostral do processoorbital do lacrimal, a expansão globular confere umaspecto bulboso, tal qual como reportado por Gennip(1986) para Strigidae e Tytonidae.
Miller (1912) aponta diferenças quanto ao local decontato do ectetmóide e o processo orbital dolacrimal. Segundo este autor, em Cerylinae, istoocorre próximo ao limite dorsal do processo orbitaldo lacrimal; em Alcedininae, na porção mediana; eem Daceloninae, o contato ocorre ventral à porçãomediana. Este autor também aponta diferençasentre Megaceryle e Chloroceryle, com oectetmóide formando a porção ventral de umagrande vacuidade em Megaceryle e uma pequenavacuidade em Chloroceryle. O ectetmóide nosmartins-pescadores apresenta formato de espinhoe é totalmente independente do lacrimal, nãoapresentando problemas quanto à sua individualização,como observado em outros grupos de aves(Ramphastidae). Cabe ressaltar que em Picidae oectetmóide ancora-se na barra jugal e em Galbulidaee Bucconidae observa-se uma delgada barra ósseacom origem dorsomedial no lacrimal, que se fundeà barra jugal (DONATELLI, 1992, 1996;HÖFLING, 1995; LADEIRA, 1997).
O ossículo sesamóide do lacrimal é bastante frágil,podendo ser facilmente perdido durante apreparação. Este ossículo não foi observado em algunsAlcedinidae, porém, isto deve ser, provavelmente,decorrente de sua fragilidade. Cracraft (1968), semmaiores detalhes, reporta ossos sesamóides naparede lateral do lacrimal em Alcedinidae.
No processo maxilar do palatino observa-se umaconspícua zona de flexão denominada zona flexóriapalatina, que é bem marcada nos martins-pescadorese, junto com a zona flexória do arco jugal e zonaflexória craniofacial, devem exercer papel importantena procinese relacionada à dieta piscívora.
O arco jugal, na sua porção rostral, possui umagrande expansão rostrolateral, que faz com que abarra duplique-se em largura. Esta expansão éobservada em todos os Cerylinae e Alcedininae, masnão nos Daceloninae; segundo Miller (1912), estaestrutura está relacionada à piscivoria eprovavelmente estaria presente em Ceryle rudis, oque foi confirmado no presente trabalho.

Bol. Mus. Para. Emílio Goeldi. Ciências Naturais, Belém, v. 2, n. 1, p. 155-182, jan-abr. 2007
177
A zona flexória do arco jugal é bem marcada econseqüentemente também apresenta-se expandidalateralmente, o que aumenta consideravelmente aárea de f lexão do arco jugal. Levando emconsideração que o bico dos martins-pescadoresestá sujeito a uma grande pressão, quando elemergulha na água, a partir de um poleiro alto,parece plausível supor que esta condição seja umaadaptação para este tipo de piscivoria. Esta hipótesetem a favor o fato dos Daceloninae, de hábitosflorestais, não apresentarem nenhuma expansão nojugal, enquanto os piscívoros Cerylinae e Alcedininaepossuem tal expansão.
A região temporal nas aves está relacionada com oconjunto de impressões (impressio) e fossasdecorrentes da origem do músculo adutormandibular externo e seus distintos feixes (ZUSI;LIVEZEY, 2000). Nos martins-pescadores analisados,com exceção dos Daceloninae, a região temporalapresenta duas fossas bem delimitadas: fossa rostraldo MAE e a fossa temporal. No entanto, só a fossatemporal é observada em todas as famíliasexaminadas de Coraciiformes, como exemplificadopor Momotus momota (Figura 6). Este padrão deregião temporal é bastante comum em aves(Ramphastidae, Bucconidae, Galbulidae), com oprocesso pós-orbital longo e confluenteventralmente com a crista temporal, formando olimite rostral da fossa temporal. Interpreta-se queeste padrão de região temporal (Cerylinae eAlcedininae) deve-se à presença de um feixeadicional do músculo adutor mandibular externo(feixe pós-orbital) (BURTON, 1984), que devidoao seu grande desenvolvimento forma uma fossacomo local de origem.
A fossa rostral do MAE aloja um feixe do músculoadutor mandibular externo rostral. Este padrão deregião temporal poderia ser questionado quanto àinclusão ou não da fossa rostral do MAE na regiãotemporal, mas a origem deste feixe muscular justificaa sua inclusão. Mais ainda, observou-se por meiode dissecção de material fixado que o feixe que se
origina na fossa rostral do MAE percorre a regiãodo laterosfenóide e insere-se por meio deaponevrose àquela do feixe principal. Por sua vez,esta aponevrose (aponevrose 1) (BURTON, 1984)insere-se por um tendão achatado no processocoronóide da mandíbula.
Os Daceloninae, em vista lateral, apresentam umaregião temporal com uma única fossa (fossatemporal), cujo processo pós-orbital é relativamentelongo e confluente ventralmente com a cristatemporal. No entanto, na parede caudal da órbitanota-se uma conspícua fossa associada à áreamuscular áspera, com o processo ventral dolaterosfenóide extremamente pronunciado emforma de espinho. Esta fossa pode ser homóloga àfossa rostral do MAE.
Os martins-pescadores possuem um processo pós-orbital extremamente curto, ao contrário doobservado em Galbulidae, Bucconidae, Coraciidaee Leptosomidae, entre outros. O processo pós-orbital é o local de origem do ligamento pós-orbitale, segundo Zusi (1967), este ligamento surgiu noinício da evolução das aves modernas, relacionadoà coordenação do movimento da mandíbula. Esteligamento está ausente nos Alcedininae e presentenos Daceloninae e Cerylinae (BURTON, 1984).
Um padrão muito semelhante de fossa temporal éencontrado em Merops apiaster. Nesta espécie aregião temporal apresenta uma única fossa (fossatemporal), extremamente semelhante à fossatemporal dos martins-pescadores, por ser bastanteprofunda e circundar o crânio no mesmo sentidodaquela dos Alcedinidae. Esta conformação de fossatemporal concorda com a hipótese de Maurer eRaikow (1984), de que a família Meropidae seriagrupo-irmão de Alcedinidae.
Shufeldt (1884) não cita a região temporal com duasfossas. No entanto, cita a fossa rostral do MAE, como“parede do alisfenóide” e uma bem desenvolvidafossa temporal, dividida medianamente por umaconspícua linha (crista nucal sagital).

178
Osteologia craniana de Cerylinae (Coraciiformes: Alcedinidae)
Verheyen (1955) cita fossas temporais longas eprofundas para os Alcedinidae e também umprocesso pós-orbital “bicéfalo”, consistindo em dois“lobos vestigiais” separados por um “encurvamento”da borda da órbita.
Jollie (1957) cita sete centros de ossificação para oosso paraesfenóide, constituindo uma estruturamerecedora de estudos ontogenéticos, para verificarse este número varia em diferentes grupos de aves.
Ao analisar um exemplar previamente imerso emágua, verifica-se que a quilha do paraesfenóide deslizasobre a fossa palatoparaesfenoidal do palatino. Aquilha do paraesfenóide é discutida quanto ao seuaspecto funcional por Burton (1978), que acreditater surgido independentemente em várias linhagensde Alcedinidae, principalmente nos piscívoros maisespecializados.
O processo lateral do paraesfenóide, bastantepronunciado nos Alcedinidae, articula-se com oprocesso medial da mandíbula, provavelmente coma finalidade de prevenir a desarticulação entre a caixacraniana e a mandíbula (BOCK, 1960). Este termoé sinônimo de processo lateral basitemporal,proposto por Bock (1960), funcionando como umasegunda articulação entre o crânio e a mandíbula,ao formar uma articulação entre esta última e oparaesfenóide (articulatio mandibulosphenoidalis). Érelevante citar que pela extensão do processo lateraldo paraesfenóide, é possível que este recebacontribuições de ossos adjacentes, tais como oprocesso paroccipital e a crista da fossa parabasal doexoccipital, e não somente da lâmina doparaesfenóide, sendo importante a análise deindivíduos jovens suturados para corrobar taisassertivas. Höfling e Gasc (1984) não observaramesta estrutura nos Ramphastidae.
Walker (1888) analisa a forma do quadrado em váriasfamílias de aves e, basicamente, aponta paraAlcedinidae as mesmas divisões observadas nestetrabalho. O quadrado mostra-se pouco significativoem termos de padrão filogenético, devido ao fato
de grupos não estreitamente aparentadosapresentarem o mesmo padrão de quadrado, comoRamphastos e Upupa ou Alcedo e Psittacinae.
O capítulo esquamosal é o mais desenvolvido e podeser divido em duas porções: rostral e caudal. Estadivisão parece ser válida, não somente pela grandeextensão do capítulo, mas também pela efetivaformação de uma segunda saliência articular naporção rostral do capítulo esquamosal. Optou-sepor não nomear um novo capítulo (porção rostraldo capítulo esquamosal) devido à porção rostral doprocesso ótico do quadrado sofrer umestrangulamento no seu bordo lateral, porém, semse interromper e continuar-se através da face medialdo capítulo esquamosal, formando na sua porçãorostral uma nova saliência articular. Cabe ressaltarque, ao analisar as cótilas, verifica-se a diferenciaçãode uma cavidade articular adicional (porção rostralda cótila esquamosal) para receber a porção rostraldo capítulo esquamosal. Esta situação podeincentivar a reconhecer três capítulos e cótilasdist intas. No entanto, antes de um estudoontogenético, prefere-se individualizar somenteduas estruturas distintas.
Uma hipótese para a formação do capítuloesquamosal dividido em duas porções consiste emsupor que este capítulo sofreu uma expansão rostralatravés da face medial do processo ótico doquadrado, formando rostralmente a terceira saliênciaarticular. Walker (1888) cita para os martins-pescadores um capítulo externo (=esquamosal)escavado externamente, sem mencionar qualquersaliência articular adicional. Esta descrição sugere quea condição do capítulo esquamosal é derivada deum capítulo (grande) que sofreu uma escavação noseu bordo lateral, formando a terceira saliênciaarticular rostral, e não decorrente de uma expansãorostral por parte do mesmo.
Walker (1888) divide a face de articulação doquadrado com a mandíbula em duas partesprincipais: superfície articular anterior e superfíciearticular posterior, esta última ainda dividida em

Bol. Mus. Para. Emílio Goeldi. Ciências Naturais, Belém, v. 2, n. 1, p. 155-182, jan-abr. 2007
179
anterior e posterior. Dorsalmente à porção caudaldo côndilo medial, na parede que separa este côndilodo lateral, observa-se uma pequena proeminênciadenominada tubérculo intercondilar. Esta estruturafoi descrita para os Alcedinidae como “trócleaacessória”. A presença deste tubérculo forma umacavidade entre os côndilos medial e lateral, que searticula com o tubérculo intercotilar da mandíbula.
Os côndilos lateral e caudal são adjacentes eencontram-se no mesmo plano. Chloroceryle,Alcedo atthis e Ceyx erithacus apresentam o côndilolateral em forma de aba, formando uma superfíciecotilar ventral que se articula com o processo caudalda cótila caudal. No entanto, Megaceryle torquata,M. alcyon, M. lugubris e os Daceloninae possuem acondição padrão para aves, com os dois côndilosrealmente de natureza condilar (Figura 18).
A parte sinfisial da mandíbula corresponde aaproximadamente 50% do comprimento total damandíbula. Em Ramphastidae esta parte éextremamente menor do que os ramos separados(HÖFLING, 1995) e em Galbulidae apresenta-semaior (DONATELLI, 1992).
A região de articulação com o quadrado é a maisheterogênea morfologicamente. A cótila medial é amais desenvolvida entre as três cótilas mandibularese apresenta lateralmente o tubérculo intercotilar, quese articula com a cavidade intercondilar do quadrado.Esta mesma estrutura é descrita por Höfling (1995)para Capitonidae, com o tubérculo articulando-secom a face lateral do côndilo medial do quadrado.
Em algumas aves as cótilas lateral e caudal emergemem uma única superfície articular (Larus,Chloroceryle, Alcedo) e em outras aparecem duassuperfícies articulares distintas (Ardea, Morus, Sula,Megaceryle, Dacelo) (BAUMEL et al., 1993).
Os martins-pescadores possuem ambas condições.No entanto, independentemente da condiçãoencontrada, a cótila lateral limita-se rostralmente peloprocesso rostral da cótila lateral e a cótila caudal
limita-se pelo processo caudal da cótila caudal.Chloroceryle, Alcedo atthis e Ceyx erithacus possuemuma única cótila que recebe o côndilo lateral doquadrado, enquanto o côndilo caudal apresentaformato de aba e cobre o processo caudal da cótilacaudal. Cabe ressaltar que nesta última articulaçãoforma-se uma condição contrária ao esperado, já quehá ligação entre uma superfície cotilar do quadrado euma superfície condilar na mandíbula, sendo o padrãogeral exatamente contrário.
Megaceryle torquata, M. alcyon, M. lugubris e osDaceloninae possuem as duas cótilas (lateral ecaudal) distintas, devido à presença do processomedial intercotilar, que as separa medianamente.A articulação entre a mandíbula e o quadrado segueo padrão normal, com superfícies condilares porparte do quadrado e superfícies cotilares por parteda mandíbula.
O processo medial mandibular é bastantedesenvolvido em todas as espécies e articula-secom o processo medial do paraesfenóide. Estaarticulação (mandibuloesfenoidal) foi descritaoriginalmente por Bock (1960) e estariarelacionada à prevenção da desarticulação do crâniocom a mandíbula.
A fossa caudal esta relacionada à inserção do músculodepressor mandibular (BAUMEL et al., 1993) emostra significativa variação nas espécies analisadas.Ela é estreita em Chloroceryle, Ceryle rudis, Alcedoatthis e Ceyx erithacus, enquanto em Megaceryletorquata, M. alcyon, M. lugubris e nos Daceloninaeocorre uma ampla parede dorsal associada a estafossa, tal qual como observado nos Galbulidae porDonatelli (1992).
CONCLUSÕES
Os resultados contidos no presente trabalhofornecem informações da osteologia craniana dosmartins-pescadores neotropicais e pretendemsomar-se a outros trabalhos na mesma linha de

180
Osteologia craniana de Cerylinae (Coraciiformes: Alcedinidae)
pesquisa (HÖFLING; GASC, 1984; HÖFLING, 1995;DONATELLI, 1992, 1996, 1997; MARCELIANO,1996; LADEIRA, 1997), que tratam de grupospossivelmente aparentados (Piciformes ePasseriformes). Dois principais aspectos podemsurgir futuramente com a somatória e sistematizaçãodestes trabalhos: análises funcionais comparativas esubsídios morfológicos para uma análise filogenéticados grupos em questão.
As estruturas ósseas analisadas apresentam-sebastante homogêneas nos seus limites intragenéricos.No entanto, a osteologia craniana revelou-sebastante eficiente para separar os gêneros deCerylinae. Todos os caracteres que separam os doisgêneros não apresentam variação significativa parapoder estabelecer algum tipo de relação entre asespécies de Chloroceryle. Estas estruturas, nasespécies de Chloroceryle, variaram no tamanho,decorrente da grande disparidaridade de tamanhoentre seus membros, porém não se detecta variaçãoefetiva na forma.
Quatro caracteres consistentes separam os gênerosem questão: expansão dorsal do lacrimal sobre oectetmóide, ausente em Chloroceryle (danificadoem Ceryle rudis) e presente em Megaceryletorquata; côndilo caudal do quadrado, cotilar emChloroceryle e Ceryle rudis e condilar emMegaceryle torquata; cótilas lateral e caudal damandíbula, indistintas em Chloroceryle e Cerylerudis e distintas em Megaceryle torquata; fossacaudal da mandíbula, parede dorsal ausente emChloroceryle e Ceryle rudis e presente emMegaceryle torquata.
Com base nestes caracteres é possível afìrmar queos gêneros Chloroceryle e Ceryle são maissemelhantes entre si do que com Megaceryle.Entretanto, estes dados são inconclusivos para tentarestabelecer as relações filogenéticas entre osgêneros, assim como para validar ou invalidar ogênero Ceryle. Para tais finalidades, há a necessidadede analisar um número mais representativo de
Alcedinidae, tal qual de outros Coraciiformes e,também, se possível, fazer uma análise ontogenéticapara elucidar possíveis homoplasias.
AGRADECIMENTOS
Ao Dr. Herculano M. F. Alvarenga (MHNT), quegentilmente forneceu material e sempre esteve atentoa sanar dúvidas referentes à ornitologia. A SérgioPosso, pelas valiosas sugestões. A France M. Pereira,pelas ilustrações deste trabalho. A Alexandre Aleixo,que se empenhou para a obtenção de material. AoDr. J. V. Remsen (LSUMNS), ao Dr. S. Olson (USMN)e à Dra. Maria Luiza Videira Marceliano (MPEG), peloempréstimo do material. À Coordenação deAperfeicoamento de Pessoal de Nível Superior(CAPES), pela bolsa de mestrado de A. C. M., e aoConselho Nacional de Pesquisas (CNPq), pela bolsade Produtividade em Pesquisa de E. H.
REFERÊCIAS
ALVARENGA, H. Coleções osteológicas: perspectivas para aornitologia no Brasil. Bol. Mus. PBol. Mus. PBol. Mus. PBol. Mus. PBol. Mus. Para. Emilio Goeldiara. Emilio Goeldiara. Emilio Goeldiara. Emilio Goeldiara. Emilio Goeldi. Belém,v.8, n.1, p. 247-257, 1992.
BAUMEL, J. J. ; KING, A. S.; BREAZILE, J. E.; EVANS, H. E.; BERGE,J. C. van den. (Eds.) Handbook of avian anatomyHandbook of avian anatomyHandbook of avian anatomyHandbook of avian anatomyHandbook of avian anatomy: nominaanatomica avium. 2. ed. Cambridge: Nuttall Ornithological Club,1993. 779 p.
BOCK, W. J. Secondary articulation of avian mandible. Auk,Auk,Auk,Auk,Auk,Washington, v.77, n.1, p.19-55, 1960.
BOCK, W. J. Kinetics of the avian skull. J. Morphol.,J. Morphol.,J. Morphol.,J. Morphol.,J. Morphol., Philadelphia,v.114, n.1, p.1-52, 1964.
BÜHLER, P. 1985. Functional anatomy of the avian jaw apparatus.In: KING, A. S.; Mclelland, J. (eds). FormFormFormFormForm and function in birdsand function in birdsand function in birdsand function in birdsand function in birds.London: Academic Press., 1985. v.2. p. 439-465.
BURTON, P. J. K. The basisphenoid notch of kingfishers. Bull. Brit.Bull. Brit.Bull. Brit.Bull. Brit.Bull. Brit.Orn. Cl.Orn. Cl.Orn. Cl.Orn. Cl.Orn. Cl., London, v.98, n.2, p. 68-74, 1978.
BURTON, P. J. K. Anatomy and evolution of the feeding apparatus inavian orders Coraciiformes and Piciformes. Bull. Brit. Mus. Nat.Bull. Brit. Mus. Nat.Bull. Brit. Mus. Nat.Bull. Brit. Mus. Nat.Bull. Brit. Mus. Nat.Hist. ZoolHist. ZoolHist. ZoolHist. ZoolHist. Zool. London, n.47, p.331-443, 1984.
CRACRAFT, J. The lacrimal-ecthetmoid bone complex in birds: asingle character analysis. Am. Midl. NatAm. Midl. NatAm. Midl. NatAm. Midl. NatAm. Midl. Nat. New York, v.80, n.2,p.316-59, 1968.
DONATELLI, R. L. Cranial osteology and myology of the jawapparatus in the Galbulidae (Aves, Piciformes). Arq. ZoolArq. ZoolArq. ZoolArq. ZoolArq. Zool., SãoPaulo, v.33, n.1, p.1-32, 1992.

Bol. Mus. Para. Emílio Goeldi. Ciências Naturais, Belém, v. 2, n. 1, p. 155-182, jan-abr. 2007
181
DONATELLI, R. L The jaw apparatus of the Neotopical andAfrotropical woodpeckers (Aves, Piciformes). Arq. ZoolArq. ZoolArq. ZoolArq. ZoolArq. Zool., São Paulo,v.33, n.1, p.1-70, 1996.
DONATELLI, R. L. Osteologia e miologia cranianas deDendrocolaptidae (Passeri formes, Tyranni) 1. GênerosGlyphorhyncus, Dendrocincla, Xiphorhynchus e Dendrocolaptes.Ararajuba,Ararajuba,Ararajuba,Ararajuba,Ararajuba, Rio de Janeiro, v.5, n.1, p. 19-37, 1997.
FRY, C, H.; FRY, K.; HARRIS, A. Kingf ishers, bee-eaters &Kingf ishers, bee-eaters &Kingf ishers, bee-eaters &Kingf ishers, bee-eaters &Kingf ishers, bee-eaters &rollersrollersrollersrollersrollers. New Jersey: Princeton Univ. Press, 1992. 324p.
GARROD, A. H. On the value in classification of a peculiarity in theanterior margin of the nasal bones of certain birds. Proc. Zool. Proc. Zool. Proc. Zool. Proc. Zool. Proc. Zool.Soc. LSoc. LSoc. LSoc. LSoc. Londonondonondonondonondon, London, v.30, n. 3, p.33-38, 1873.
GENNIP, E. M. S. J. van. The osteology, arthrology and myology ofthe jaw apparatus of the Pigeon (Columba livia L). Neth. J. Zool.Neth. J. Zool.Neth. J. Zool.Neth. J. Zool.Neth. J. Zool.,Wageningen, v.36, n.1, p.1-46, 1986.
HÖFLING, E.; GASC, J. P. Biomécanique du crâne et du bec chezRamphastos (Aves, Ramphastidae). 1 Description des élémentsanatomiques. GegenbaursGegenbaursGegenbaursGegenbaursGegenbaurs Morphol. JahrbMorphol. JahrbMorphol. JahrbMorphol. JahrbMorphol. Jahrb., Leipzig, v.130,n..1, p.125-147, 1984.
HÖFLING, E. Anatomia do crAnatomia do crAnatomia do crAnatomia do crAnatomia do crânio e da cintura escapularânio e da cintura escapularânio e da cintura escapularânio e da cintura escapularânio e da cintura escapulardos Ramphast idae (Aves: Pic i formes) e de alguns gruposdos Ramphast idae (Aves: Pic i formes) e de alguns gruposdos Ramphast idae (Aves: Pic i formes) e de alguns gruposdos Ramphast idae (Aves: Pic i formes) e de alguns gruposdos Ramphast idae (Aves: Pic i formes) e de alguns grupospróximos, com impl icações s istemáticaspróximos, com impl icações s istemáticaspróximos, com impl icações s istemáticaspróximos, com impl icações s istemáticaspróximos, com impl icações s istemáticas. Tese (Livre-Docência). Universidade de São Paulo,1995
HOWARD, R.; MOORE, A. A complete checklist of the birdsA complete checklist of the birdsA complete checklist of the birdsA complete checklist of the birdsA complete checklist of the birdsof the worldof the worldof the worldof the worldof the world. 2. ed. London: Academic Press, 1994. 630 p.
HUXLEY, T. H. On the classification of birds, and on the taxonomyvalues of the modification of certain of the cranial bones observablein that class. Proc. Zool. Soc. Lond.Proc. Zool. Soc. Lond.Proc. Zool. Soc. Lond.Proc. Zool. Soc. Lond.Proc. Zool. Soc. Lond., London, n.27, p.415-472, 1867.
JOLLIE, M. The head skeleton of the chicken and remarks on theanatomy of this region in other birds. J. MorpholJ. MorpholJ. MorpholJ. MorpholJ. Morphol., Philadelphia,v.100, n.4, p.389-436, 1957.
LADEIRA, L. M. C. E. B. Osteologia craniana de BucconidaeOsteologia craniana de BucconidaeOsteologia craniana de BucconidaeOsteologia craniana de BucconidaeOsteologia craniana de Bucconidae(Aves: Piciformes)(Aves: Piciformes)(Aves: Piciformes)(Aves: Piciformes)(Aves: Piciformes). Dissertação (Mestrado). Universidade deSão Paulo,.1997.
MARCELIANO, M. L. V. Estudo osteológico e miológico deOphistocomus hoazin (Müller, 1776) (Aves: Ophistocomidae)comparado com algumas espécies de Cracidae, Musophagidae eCuculidae. Bol. Mus. PBol. Mus. PBol. Mus. PBol. Mus. PBol. Mus. Para. Emíl io Goeldi. sér ie Zoolara. Emíl io Goeldi. sér ie Zoolara. Emíl io Goeldi. sér ie Zoolara. Emíl io Goeldi. sér ie Zoolara. Emíl io Goeldi. sér ie Zool.,Belém, v.12, n.2, p.95-246, 1996.
MAURER, R. D.; RAIKOW, R. J. Appendicular myology, phylogeny,and classification of the avian order Coraciiformes (includingTrogoniformes). Ann. Carnegie Mus. PAnn. Carnegie Mus. PAnn. Carnegie Mus. PAnn. Carnegie Mus. PAnn. Carnegie Mus. Pittsburgittsburgittsburgittsburgittsburg, v.50, n.18,p.417-34, 1981.
MILLER, W. DeW. A revision of the classification of the kingfishers.Bull. AmerBull. AmerBull. AmerBull. AmerBull. Amer. Mus. Nat. Hist. Mus. Nat. Hist. Mus. Nat. Hist. Mus. Nat. Hist. Mus. Nat. Hist., New York, v.22, n.31 p.239-311, 1912.
POSSO, S. R. Osteologia craniana dos Cucul i formes doOsteologia craniana dos Cucul i formes doOsteologia craniana dos Cucul i formes doOsteologia craniana dos Cucul i formes doOsteologia craniana dos Cucul i formes doNovo Mundo (Aves) com impl icações s istemáticasNovo Mundo (Aves) com impl icações s istemáticasNovo Mundo (Aves) com impl icações s istemáticasNovo Mundo (Aves) com impl icações s istemáticasNovo Mundo (Aves) com impl icações s istemáticas.Dissertação (Mestrado). Universidade Estadual Júlio de MesquitaFilho,1999
SCHAUNSEE, R. M. Meyer de A guide to the birds of SouthA guide to the birds of SouthA guide to the birds of SouthA guide to the birds of SouthA guide to the birds of SouthAmericaAmericaAmericaAmericaAmerica. [S. l.]: Intercollegiate Press, 1982. 498 p.
SHUFELDT, R. W. Osteology of Ceryle alcyon. J. Anat. PhysiolJ. Anat. PhysiolJ. Anat. PhysiolJ. Anat. PhysiolJ. Anat. Physiol.,London, n.18, p. 279-94, 1884.
SIBLEY, C. G.; AHLQUIST, J. E. Phylogeny and classif icationPhylogeny and classif icationPhylogeny and classif icationPhylogeny and classif icationPhylogeny and classif icationof birds.of birds.of birds.of birds.of birds. New Haven: Yale Univ. Press, 1990. 1111 p.
SICK, H. Ornitologia brasileiraOrnitologia brasileiraOrnitologia brasileiraOrnitologia brasileiraOrnitologia brasileira. Rio de Janeiro: Nova Fronteira,1997. 862 p.
STRESEMANN, V.; STRESEMANN, E.Die handschwingen-mauserder eisvögel (Alcedinidae). J.OrnitholJ.OrnitholJ.OrnitholJ.OrnitholJ.Ornithol., Leipzig, v.102 , n.4, p.439-55, 1961.
VERHEYEN, R. Analyse du potentiel morphologique et considérationssur la systématique des Coraciiformes (Wetmore, 1934). Bull.Bull.Bull.Bull.Bull.Inst. R. Sci. NaturInst. R. Sci. NaturInst. R. Sci. NaturInst. R. Sci. NaturInst. R. Sci. Natur. Belg.. Belg.. Belg.. Belg.. Belg., Bruxelles, v. 31, n. 92, p. 1-16, 1-19, 1-16, 1955.
WALKER, M. On the form of the quadrate bone in birds. Zool.Zool.Zool.Zool.Zool.UnivUnivUnivUnivUniv. Coll. Dundee.. Coll. Dundee.. Coll. Dundee.. Coll. Dundee.. Coll. Dundee. p.1-8, 1888.
ZUSI, R. L. Structural adaptat ion of the head and neck inStructural adaptat ion of the head and neck inStructural adaptat ion of the head and neck inStructural adaptat ion of the head and neck inStructural adaptat ion of the head and neck inthe black-skimmerthe black-skimmerthe black-skimmerthe black-skimmerthe black-skimmer RhyncopsRhyncopsRhyncopsRhyncopsRhyncops nigranigranigranigranigra. Cambridge: Nuttal lOrnithological Club, 1962. 101p. (Publication n. 3).
ZUSI, R. L. The role of the depressor mandibulae muscle in kinesis ofthe avian skull. PPPPProc. U. S. Nat. Musroc. U. S. Nat. Musroc. U. S. Nat. Musroc. U. S. Nat. Musroc. U. S. Nat. Mus., Washington, v.123,n.3607, p.1-28, 1967.
ZUSI, R. L. A functional and evolutionary analysis of rhynchokinesisin birds. Smithson. Contrib. Zool.Smithson. Contrib. Zool.Smithson. Contrib. Zool.Smithson. Contrib. Zool.Smithson. Contrib. Zool., Washington, n.395, p.1-40, 1984.
ZUSI, R. L.; LIVEZEY, B. C. Homology and phylogenetic implicationsof some enigmatic cranial features in Galliform ans Anseriform birds.Ann. Carnegie Mus.Ann. Carnegie Mus.Ann. Carnegie Mus.Ann. Carnegie Mus.Ann. Carnegie Mus., Pittsburg, v.69, n.3, p.157-193, 2000.
Recebido: 25/02/2000Aprovado: 17/01/2002

182
Osteologia craniana de Cerylinae (Coraciiformes: Alcedinidae)
Apêndice. Material examinado
CoraciidaeCoraciidaeCoraciidaeCoraciidaeCoraciidae (n=1): Eurystomus orientalis (MHNT:133); BucerotidaeBucerotidaeBucerotidaeBucerotidaeBucerotidae (n=2): Aceros undulatus(MHNT: 138), Tockus erythrorhyncus (MHNT:137); UpupidaeUpupidaeUpupidaeUpupidaeUpupidae (n=1): Upupa epops (MHNT:135); TTTTTodidaeodidaeodidaeodidaeodidae (n=1): Todus subulatus (MHNT:129); MomotidaeMomotidaeMomotidaeMomotidaeMomotidae (n=2): Baryphthengusruficapillus (MHNT: 1429), Momotus momota (AZ:65); MeropidaeMeropidaeMeropidaeMeropidaeMeropidae (n=1): Merops apiaster (MHNT:131); AlcedinidaeAlcedinidaeAlcedinidaeAlcedinidaeAlcedinidae (n=33); DaceloninaeDaceloninaeDaceloninaeDaceloninaeDaceloninae (n=7):Halcyon cyanoventris (LSUMNS: 131369),Halcyon sancta (LSUMNS: 105061), Halcyonfunebris (USNM: 557210), Halcyon cinnamomina(LSUMNS: 87038), Dacelo novaeguineae (n=2)(LSUMNS: 154937; MHNT: 126), Tanysiptera
galatea (USNM: 557213); AlcedininaeAlcedininaeAlcedininaeAlcedininaeAlcedininae (n=2):Alcedo atthis (MHNT: 128), Ceyx erithacus (USNM:559832); CerylinaeCerylinaeCerylinaeCerylinaeCerylinae (n=25): Megaceryle torquata(n=4) (MHNT: 127, 378, 1500; MPEG: 1047),Megaceryle alcyon (LSUMNS: 139939), Megacerylelugubris guttulata (USNM: 318238), Ceryle rudisleucomelanura (USNM: 343520), Chloroceryleamazona (n=4) (MHNT: 306, s.no, 701; AZ: 66),Chloroceryle americana (n=6) (MHNT: 611, 628,767; AZ: 62, 63, 511); Chloroceryle inda (n=4)(MHNT: 1371; AZ: 61, 64; MPEG 881),Chloroceryle aenea (n=4) (MHNT: 609, 1416;MPEG: 2773, 3742); TTTTTrogonidaerogonidaerogonidaerogonidaerogonidae (n=1) Trogonmelanurus (MHNT:410); CuculidaeCuculidaeCuculidaeCuculidaeCuculidae (n=1)Coccyzus americanus (AZ: 55).




























![Osteologia [Montalvo]](https://img.document.onl/doc/110x75/5596bd151a28aba02d8b468d/osteologia-montalvo.jpg)