Embed Size (px)
Citation preview

0
Alane de Medeiros Silva
PADRÃO DE DISTRIBUIÇÃO E CARACTERIZAÇÃO
MORFOLÓGICA DE FIBRAS SEROTONÉRGICAS NOS NÚCLEOS DA
LINHA MÉDIA/ INTRALAMINARES DO TÁLAMO DO MOCÓ
(Kerodon rupestris)
Dissertação apresentada à Universidade
Federal do Rio Grande do Norte, como pré-
requisito para obtenção do título de Mestre em
Psicobiologia.
NATAL-RN
2013

1
Alane de Medeiros Silva
PADRÃO DE DISTRIBUIÇÃO E CARACTERIZAÇÃO
MORFOLÓGICA DE FIBRAS SEROTONÉRGICAS NOS NÚCLEOS DA
LINHA MÉDIA/ INTRALAMINARES DO TÁLAMO DO MOCÓ
(Kerodon rupestris)
Dissertação apresentada à Universidade
Federal do Rio Grande do Norte, como pré-
requisito para obtenção do título de Mestre em
Psicobiologia.
Orientador: Prof. Dr. Expedito Silva do
Nascimento Júnior
NATAL-RN
2013


2
TÍTULO: PADRÃO DE DISTRIBUIÇÃO E CARACTERIZAÇÃO MORFOLÓGICA DE
FIBRAS SEROTONÉRGICAS NOS NÚCLEOS DA LINHA MÉDIA/INTRALAMINARES
DO TÁLAMO DE MOCÓ (Kerodon rupestris)
AUTOR: ALANE DE MEDEIROS SILVA
DATA DA DEFESA: 27/03/2013
BANCA EXAMINADORA:
Prof. Dr. Expedito Silva do Nascimento Júnior – Orientador
Universidade Federal do Rio Grande do Norte
Prof. Dr. Fausto Pierdoná Guzen
Universidade do Estado do Rio Grande do Norte
Prof. Dr. Judney Cley Cavalcante
Universidade Federal do Rio Grande do Norte

3
AGRADECIMENTOS
- Gostaria de agradecer primeiramente a Deus, que durante essa fase de minha vida me
permitiu entender mais e sentir mais sua presença, me possibilitando fortalecimento e
motivação, tornando-se meu eterno amigo e companheiro.
- Aos meus pais, Francimar e Maria do Rosário, que sempre deram forças em todas as
minhas decisões e se fazem presentes passo a passo de minha vida e dos meus irmãos, com
muito amor e carinho.
- Aos meus irmãos, Aline e Alisson pelo incentivo, amor, carinho e compreensão diante das
ausências.
- Aos meus amigos que se fizeram sempre presentes escutando minhas angústias e anseios
diante desse projeto de vida e me acompanharam me dando força e incentivo constante;
destaco especialmente todas as minhas amigas enfermeiras, o maior presente que o curso de
enfermagem poderia me dar; amigos da vida, amigos que adquiri no mestrado, especialmente
Clarissa, não só pelo apoio emocional, mas também pelas grandes contribuições
metodológicas do trabalho.
- Ao professor/orientador Expedito por todos os ensinamentos, contribuições e
especialmente paciência diante de meus anseios. Agradeço também pelo voto de confiança
quando me aceitou como orientanda.
- Aos professores Ruthnaldo e Judney, pois, através deles, meu interesse pela anatomia,
especialmente neuroanatomia, aflorou e incentivou minha busca pela docência. Judney
principalmente pela grande contribuição na minha inserção no LabNeuro.
- A toda equipe do Labneuro: Joacil e Kayo pela amizade, risadas constantes, como também
pela disposição em ajudar e colaborar; André e Janaína pelo companheirismo e também
diversas colaborações no decorrer desta etapa; Rovena pelo GRANDE apoio não só no
Labneuro como também através de telefonemas sempre disposta a ajudar e tirar dúvidas
frequentes; Rodolfo, Melquisedec, Paulo, karen e Nayra por todas as ajudas nos
experimentos e pela manutenção de um ambiente descontraído no laboratório e a Twyla que
além de companheira de turma tornou-se grande amiga e “compartilhadora” de angústias e
anseios.
- A Ronaldo por todo apoio, disponibilidade e contribuição metodológica.

4
- À Regina, pelo preparo das soluções, pela garantia da ordem do laboratório no tocante à
execução dos experimentos, como também pelos momentos de descontração.
- Aos professores Jeferson e Miriam Stela pela disponibilidade, esclarecimentos de dúvidas
e apoio.
- Ao Departamento de Morfologia – DMOR/UFRN pela garantia da infraestrutura
necessária a plena execução deste trabalho.
- Ao Ibama, pela autorização e licença para realização deste trabalho.
- Ao CNPq, FAPERN e a CAPES pelo apoio financeiro.

5
LISTA DE TABELAS E FIGURAS
Tabela 1: Divisão dos núcleos da linha média e intralaminares do tálamo do mocó em níveis
rostral, médio e caudal através das 16 imagens obtidas das secções coronais (S).
Tabela 2: DOR através dos níveis rostral, médio e caudal dos núcleos da linha
média/intralaminares do tálamo.
Figura 1: Grupamentos serotonérgicos B1 a B9.
Figura 2: Representação esquemática dos principais grupos nucleares do tálamo de humanos.
Figura 3: O mocó (Kerodon rupestris) em ambiente natural.
Figura 4: Mocó posicionado em aparelho estereotáxico e com alinhamento dorsoventral do
bregma e lambda.
Figura 5: Mocó posicionado em aparelho estereotáxico, após remoção da calota craniana e
consequente exposição do encéfalo.
Figura 6: Encéfalo do mocó em vistas dorsal (a) e ventral (b).
Figura 7: Reconstrução esquemática das secções coronais do encéfalo do mocó, ilustrando a
morfologia dos núcleos da linha média e intralaminares do tálamo, seguidos de
fotomicrografias submetidas ao método de coloração de Nissl no nível correspondente.
Figura 8: DOR de fibras 5-HT – IR nos núcleos da linha média do tálamo de mocós (n=4).
Figura 9: DOR de fibras 5-HT – IR nos núcleos intralaminares do tálamo de mocós (n=4).
Figura 10: DOR de fibras 5-HT – IR entre os núcleos da linha média e intralaminares do tálamo de
mocós (n=4)
Figura 11: Fotomicrografias em campo claro de secções coronais do encéfalo do mocó
submetidas à imunoistoquímica para 5-HT ao nível de aproximadamente 1,44 mm p.b.
Figura 12: Fotomicrografias em campo claro de secções do encéfalo do mocó submetidas à
imunoistoquímica para 5-HT ao nível de aproximadamente 2,34 mm p.b.
Figura 13: Fotomicrografias em campo claro de secções coronais do encéfalo do mocó
submetidas à imunoistoquímica para 5-HT ao nível de aproximadamente 3,24 mm p.b.
Figura 14: Fotomicrografias em campo claro de secção coronal do encéfalo do mocó
submetida à imunoistoquímica para 5-HT ao nível de aproximadamente 4,86 mm p.b.

6
ABREVIATURAS
3V - Terceiro ventrículo
5-HIAA - Ácido 5-hidroxiindolacético
5-HT - Serotonina
ABC - Complexo avidina-biotina peroxidase
AD - Núcleo anterodorsal do tálamo
AM - Núcleo anteromedial do tálamo
B9 - Núcleo supralemniscal
CL - Núcleo central lateral do tálamo
Cli - Núcleo linear caudal da rafe
CM - Núcleo central medial do tálamo
CMn - Núcleo central mediano do tálamo
DAB - Diaminobenzidina
DCAA - Descarboxilase dos aminoácidos aromáticos
DOR - Densidade óptica relativa
DR - Núcleo dorsal da rafe
f - Fórnix
fr - Fascículo retroflexo
GABA - Ácido gamaminobutírico
Hb - Núcleo habenular
IAD - Núcleo interanterodorsal do tálamo

7
IAM - Núcleo interanteromedial do tálamo
IMD - Núcleo intermediodorsal do tálamo
IR - Imunorreativo
LSD - Ácido-dietilamida-D-lisérgico
MAO - Monoamina oxidase
MD - Núcleo mediodorsal do tálamo
MnR - Núcleo mediano da rafe
mt - Trato mamilotalâmico
PaXi - Núcleo paraxifóide do tálamo
PC - Núcleo paracentral do tálamo
PF - Núcleo parafascicular do tálamo
PMnR - Núcleo paramediano da rafe
PnR - Núcleo pontino da rafe
PT - Núcleo paratenial do tálamo
PV - Núcleo paraventricular do tálamo
PVA - Núcleo paraventricular anterior do tálamo
PVM - Núcleo paraventricular médio do tálamo
PVP - Núcleo paraventricular posterior do tálamo
Re - Núcleo reuniens do tálamo
Rh - Núcleo rombóide do tálamo
RIP - Núcleo interpósito da rafe
RLi - Núcleo linear rostral da rafe

8
RMg - Núcleo magno da rafe
Rob - Núcleo obscuro da rafe
RPa - Núcleo pálido da rafe
sm - Estria medular do tálamo
Sub - Núcleo submédio do tálamo
VRe - Núcleo reuniens ventral
Xi - Núcleo xifóide do tálamo
ZI - Zona incerta

9
SUMÁRIO
RESUMO 10
ABSTRACT 11
1. INTRODUÇÃO 12
1.1 Sistema serotonérgico 12
1.2 Tálamo
1.2.1 Características gerais dos núcleos da linha média/ intralaminares
1.2.2 Núcleos da linha média do tálamo
1.2.3 Núcleos intralaminares do tálamo
16
18
19
21
1.3 Projeções serotonérgicas aos núcleos da linha média e intralaminares do
tálamo
23
1.4. Modelo experimental 24
2. JUSTIFICATIVA 26
3. OBJETIVOS 27
4. METODOLOGIA
4.1 Sujeitos
4.2 Procedimentos
4.2.1 Anestesia
4.2.2 Perfusão
4.2.3 Remoção do encéfalo
4.2.4 Microtomia
4.2.5 Imunoistoquímica
4.2.6 Obtenção e análise de imagens
4.2.7 Densidade óptica relativa
4.2.8 Análise estatística
28
28
29
29
29
29
31
31
32
33
34
5. RESULTADOS
5.1 Análise citoarquitetônica: Nissl
5.1.1 Núcleos da linha média
5.1.2 Núcleos intralaminares
5.2 Densidade óptica relativa
5.3 Caracterização morfológica das fibras
5.3.1 Núcleos da linha média
5.3.2 Núcleos intralaminares
35
35
35
36
42
45
46
46
6. DISCUSSÃO 51
7. CONCLUSÕES 58
8. PERSPECTIVAS 59
REFERÊNCIAS 61

10
RESUMO
O complexo nuclear da linha média/intralaminar forma um notável grupo de núcleos
da parte medial e dorsal do tálamo. Os núcleos da linha média, em ratos, são compreendidos
pelos núcleos paratenial (PT), paraventricular (PV), intermediodorsal (IMD), reuniens (Re) e
rombóide (Rh). Já os intralaminares compreendem os núcleos central medial (CM),
paracentral (PC), central lateral (CL) e parafascicular (PF). Tais núcleos apresentam densa
inervação serotonérgica oriunda do tronco encefálico, a partir, principalmente, do denominado
sistema de ativação ascendente. Esses núcleos, por sua vez, emitem projeções para diversas
áreas corticais e subcorticais, mais especificamente para áreas límbicas, o que propõe o
importante papel desse neurotransmissor na circuitaria límbica. O objetivo desse trabalho foi
caracterizar o padrão de distribuição e morfologia das fibras de serotonérgicas nos núcleos da
linha média e intralaminares do tálamo do mocó (Kerodon rupestris), um roedor típico da
região Nordeste. Para isso, foram utilizados 4 mocós adultos jovens. Após etapas de perfusão
e microtomia, foi realizada imunoistoquímica para serotonina (5-HT), técnica de Nissl e
posterior coleta e análise de imagens a fim de caracterizar a citoarquitetura desses núcleos,
bem como as fibras de 5-HT neles visualizadas. Foi realizada ainda uma análise de Densidade
Óptica Relativa (DOR) para semiquantificar a concentração de fibras de 5-HT nas áreas de
interesse. Sendo assim, observamos uma distribuição citoarquitetônica desses núcleos
semelhante ao observado em ratos. Em se tratando da distribuição de fibras, aquelas
imunorreativas a 5-HT apresentaram-se em maior concentração, conforme a DOR, nos
núcleos da linha média em relação aos intralaminares, sendo o Re o núcleo que apresenta
maior valor de pixels, seguido do PV, Rh, IMD e PT. Nos intralaminares o CL apresentou
maior valor de pixels, seguido dos núcleos CM, PC e PF. As fibras serotonérgicas foram
classificadas conforme número de varicosidades, bem como diâmetro axonal. Assim,
encontramos três tipos de fibras distribuídas através desse complexo nuclear: fibras rugosas,
granulares e semi-granulares. No PV predominaram fibras rugosas; PT predominaram fibras
granulares; IMD, CL e PF foram representados por fibras semi-granulares e Re, Rh, PC e CM
apresentaram fibras granulares e semi-granulares. A caracterização morfológica das fibras
serotonérgicas encontradas e as diferenças de densidades entre os núcleos permite sugerir
diferentes padrões de organização sináptica deste neurotransmissor além de confirmar seu
grande repertório funcional. Palavras-chave: Serotonina, Tálamo, Complexo de núcleos da
linha média/ intralaminares, Densidade óptica relativa, Mocó.

11
ABSTRACT
The midline/intralaminar nuclei form a remarkable group of nuclei of the medial and dorsal
thalamus. The midline nuclei, in rats, comprises the paratenial nuclei (PT), paraventricular
(PV), intermediodorsal (IMD), reuniens (Re) and rhomboid (Rh). The intralaminar nuclei
comprises the central medial (CM), paracentral (PC), central lateral (CL) and parafascicular
(PF). Such nuclei have dense serotonergic innervation originating from the brainstem,
especially from the so-called ascending activation system. These nuclei, in turn, send
projections to various cortical and subcortical areas, specifically to limbic areas, which
suggests the important role of this neurotransmitter in the limbic circuitry. The aim of this
study was to characterize the distribution pattern and morphology of serotonin fibers in the
nuclei of the midline and intralaminar thalamic of rocky cavy (Kerodon rupestris), a tipical
rodent from brazilizan northeast. To reach this aim we used four rock cavies adults. Following
the transcardially perfusion with paraformaldehyde and brain microtomy steps was performed
immunohistochemistry for serotonin (5-HT), Nissl technique and subsequent achievement and
image analysis to characterize the cytoarchitecture of these nuclei and the serotonergic fibers
visualized. An analysis was made of Relative Optical Density (ROD) to semi-quantify the
concentration of serotonin fibers in the areas of interest. Thus, we observed a
cytoarchitectonic arrangement of these nuclei similar to that found in rats. In case of fibers
distribution, those immunoreactive to 5-HT were presented in a higher concentration
according as ROD in the midline nuclei relative to intralaminar; Re being the core which has
a higher pixel value followed by the PV , Rh, IMD and PT. In intralaminar CL showed higher
pixels, followed by nuclei CM, PC and PF. The serotonergic fibers were classified as number
of varicosities and axon diameter, therefore find three types of fibers distributed through this
nuclear complex: fibers rugous, granular and semi-granular. In PV fibers predominated
rugous; in PT fibers predominated granular; IMD, CL and PF fibers were represented by
semi-granular and Re, Rh, PC and CM fibers showed granular and semi-granular.
Morphological characterization of serotonergic fibers and differences in density between the
nuclei may suggest different patterns of synaptic organization of this neurotransmitter beyond
confirming his large repertoire functional. Keywords: Serotonin, Thalamus, Midline /
intralaminar nuclei, Relative Optical Density, Rocy cavy.

12
1. INTRODUÇÃO
O tálamo é caracterizado como o principal regulador do fluxo de informações
direcionadas ao córtex cerebral através de suas projeções tálamo-corticais. Os núcleos
talâmicos são modulados a partir de um sistema de ativação ascendente originário do tronco
encefálico (Jones, 2007). Tal sistema utiliza neurotransmissores específicos como
noradrenalina, acetilcolina, ácido gamaminobutírico (GABA), histamina e serotonina (5-HT)
(McCormick, 1992). A 5-HT, produzida predominantemente no tronco encefálico, apresenta
forte influência nos núcleos talâmicos (McCormick, 1992; Monckton e McCormick, 2001;
Jones, 2007).
Regiões específicas do sistema serotonérgico de ratos apresentaram maior aporte de
fibras ascendentes para núcleos talâmicos, como núcleos da linha média e intralaminares do
tálamo (Cropper et al., 1984; Vertes, 1991; Vertes et al., 2010).
Nessa perspectiva, o trabalho em questão aborda o padrão de distribuição e
caracterização morfológica de fibras serotonérgicas nos núcleos da linha média e
intralaminares do tálamo do mocó, um roedor típico do semiárido do nordeste brasileiro.
1.1 Sistema serotonérgico
A 5-HT é um neurotransmissor pertencente à classe das aminas biogênicas, mais
especificamente monoaminas, categorizada como indolamina (Halbach e Dermietzel, 2006).
Possui funções tanto de neurotransmissor, como de hormônio nas porções central e periférica
do sistema nervoso (Julius, 1991).
Em 1947, essa substância foi descrita como um fator sanguíneo que promovia
vasoconstrição (Rapport, 1949). Na década de 50, foi isolada e sua estrutura molecular
elucidada, apresentando características químicas semelhantes ao ácido-dietilamida-D-lisérgico
(LSD), sendo posteriormente classificada como um neurotransmissor (Hamlin e Fisher, 1951;
Halbach e Dermietzel, 2006).
Esse neurotransmissor é produzido a partir de um aminoácido essencial, o L-
triptofano, o qual é convertido em 5-hidroxitriptofano, por ação da primeira enzima dessa
cadeia, a triptofano-hidroxilase; posteriormente, 5-hidroxitriptofano é descarboxilado, pela
segunda enzima envolvida, a descarboxilase dos aminoácidos aromáticos (DCAA),
resultando, portanto, em 5-hidroxitriptamina ou 5-HT. Nesse sentido, o triptofano circulante

13
do organismo é um fator limitante para a produção de 5-HT (Jacobs e Azmitia, 1992; Kandel
et al., 2000b; Halbach e Dermietzel, 2006).
Após liberada, a 5-HT atuará em seus receptores e então poderá ser recaptada ou
degradada. Assim, seu principal metabólito é o ácido 5-hidroxiindolacético (5-HIAA), o qual
se forma a partir da ação da monoamina oxidase (MAO), que promove uma desaminação na
5-HT, seguida pela ação da aldeído desidrogenase (Halbach e Dermietzel, 2006).
Estudos iniciais como os de Ramón y Cajal (1911) já apresentavam descrições de
células aglomeradas na linha média do tronco encefálico, sendo caracterizadas como
multipolares e de projeções indefinidas. Posteriormente, a partir de estudos citoarquitetônicos,
aglomerados celulares, denominados núcleos da rafe, foram descritos em gatos como um
grupo de 8 núcleos ao longo da linha média, organizados do sentido rostral para o caudal em:
núcleos lineares rostral e caudal da rafe, dorsal da rafe, central superior (ou mediano) da rafe,
pontino da rafe, magno da rafe, pálido da rafe e obscuro da rafe (Taber et al., 1960).
Uma descrição posterior evidenciou a presença de um sistema neuronal
monoaminérgico na região do tronco encefálico de ratos revelados pela técnica de
fluorescência induzida por formaldeído (Falck et al., 1962; Dahlström e Fuxe, 1964; 1965);
esses grupamentos foram denominados de B1 a B9 do sentido caudal para o rostral
(Dahlstrom e Fuxe, 1964; 1965) (Figura 1).
Figura 1: Grupamentos serotonérgicos B1 a B9, segundo Dahlstrom e Fuxe (1964).

14
A partir do advento da técnica de imunoistoquímica, esta classificação foi integrada a
nomenclatura citoarquitetônica do sistema da rafe (Steinbush et al., 1978; Steinbush, 1981;
Törk, 1985). Muito embora os principais grupos celulares do sistema serotonérgico sigam as
divisões citoarquitetônicas dos núcleos da rafe, essa superposição não é exata, considerando
que numerosos neurônios imunorreativos a 5-HT (5-HT-IR) estão presentes em outros setores
da formação reticular, além dos limites dos núcleos da rafe; assim como alguns neurônios não
serotonérgicos estão presentes nos núcleos da rafe (Törk, 1985; Harding et al., 2004).
A anatomia destes grupos foi revisada em muitas espécies, incluindo rato (Parent et
al., 1981; Lidov e Molliver, 1982; Takeuchi et al., 1982; Törk, 1985; Paxinos e Watson,
2007), coelho (Bjarkam et al., 1997), gato (Takeuchi et al., 1982; Jacobs et al., 1984),
humanos (Törk, 1990) e macacos do Novo e Velho Mundo (Felten et al., 1974; Azmitia e
Gannon, 1983; Felten e Sladek, 1983; Hornung e Fritschy, 1988). A classificação mais
recente dos núcleos da rafe (grupamentos serotonérgicos) no cérebro do rato, conforme
Paxinos e Watson (2007) indica a existência de 10 grupamentos nucleares da rafe, em
sequência rostro-caudal: no mesencéfalo, os núcleos linear rostral da rafe (RLi),
frequentemente referido como dopaminérgico e não serotonérgico, linear caudal da rafe (Cli),
dorsal da rafe (DR), mediano da rafe (MnR) e paramediano da rafe (PMnR). Na ponte, os
núcleos pontino da rafe (PnR) e interpósito da rafe (RIP). Por último, no bulbo, os núcleos
magno da rafe (RMg), pálido da rafe (RPa) e obscuro da rafe (Rob) (Paxinos e Watson, 2007).
No mocó (Kerodon rupestris), a imunorreatividade à 5-HT nestes núcleos apresentou-
se similar ao que tem sido previamente descrito em outros mamíferos, sendo relatado, numa
direção rostro-caudal os núcleos: RLi, Cli, DR, MnR, PMnR, supralemnsical ou B9, situado
imerso no lemnisco medial, PnR, RIP, RMg, RPa e Rob; além de grupamentos ventrolateral
rostral e ventrolateral caudal da região bulbar. De forma geral os núcleos da rafe
serotonérgicos do mocó se assemelham aos núcleos do mesmo nome encontrados em outros
roedores já estudados, com exceção do RLi, o qual não foi mencionado como sendo núcleo
serotonérgico em estudos em outras espécies de roedores (Soares et al., 2012).
Estudos farmacológicos e fisiológicos têm mostrado que muitas ações da 5-HT são
mediadas por diversos subtipos de receptores (Richardson e Engel, 1986). Desse modo, sabe-
se que estes podem ser classificados conforme três constituintes: um transportador, um canal
iônico e a proteína G acoplada ao canal iônico. No entanto, a grande maioria dos receptores
serotonérgicos pertence à subfamília de receptores acoplados a proteína G (Halbach e

15
Dermietzel, 2006). Tais receptores foram inicialmente classificados em dois subtipos: M e D
(Gaddum e Picarelli, 1957; Osman e Ammar, 1975), porém, a partir do advento de estudos
bioquímicos e moleculares foi possível caracterizar 7 subtipos de receptores de 5-HT (5-HT1
a 5-HT7), sendo os mais frequentemente citados: 5-HT1A, 5-HT1B, 5-HT1D, 5-HT1E, 5-HT1F,
5-HT2A, 5-HT2B, 5-HT2C, 5-HT3, 5-HT4, 5-HT5, 5-HT6 e 5-HT7 (Halbach e Dermietzel, 2006).
Em ratos, as fibras serotonérgicas possuem características específicas, como o calibre
fino, não-mielinização na maioria das fibras e elevada colateralização; além disso sua
localização concentra-se preferencialmente no plano mediano (Jacobs e Azmitia, 1992;
(Kosofsky e Molliver, 1987). As fibras serotonérgicas de espécies de escala evolutiva de
maior complexidade, como primatas, apresentam-se mielinizadas em sua maioria, com poucas
ramificações e com algumas fibras localizadas mais lateralmente (lateralização) (Hornung e
Fritschy, 1988). Isso parece indicar especializações evolutivas do sistema serotonérgico
(Jacobs e Azmitia, 1992).
Os núcleos da rafe recebem aferentes, bem como emitem eferências para praticamente
todas as regiões do SNC, no entanto a densidade dessas projeções apresenta variações (Törk,
1985; 1990; Jacobs e Azmitia, 1992; Harding et al., 2004).
Tais características hodológicas dos núcleos da rafe, bem como a existência de
diversos tipos de receptores para 5-HT ajudam a justificar a variedade funcional desse
sistema. Por exemplo, estudos relatam que a 5-HT tem sido associada ao controle do humor
(Montgomery, 1995; Lowry et al., 2008), comportamento alimentar (Takase et al., 2000;
Takase e Nogueira, 2008), função locomotora (Noga et al., 2009), regulação do ciclo sono-
vigília (Jacobs e Azmitia, 1992; Carlson, 2002; Monti, 2011; Franco-Pérez et al., 2012),
comportamento sexual, termorregulação, processamento sensorial nociceptivo, alerta e
atenção (Jacobs e Azmitia, 1992; Sakai e Crochet., 2001), memória e aprendizagem (Vertes et
al., 1999), regulação circadiana, entre outras (Jacobs e Azmitia, 1992; Carlson, 2002). Além
disso, estudos elucidaram que alterações funcionais desse sistema podem favorecer processos
neuropatológicos, como distúrbios de sono e ansiedade, depressão e distúrbios
neurodegenerativos como as doenças de Parkinson (Politis e Loane, 2011) e Alzheimer
(Michelsen et al., 2008).
Foi observado que a maioria das fibras serotonérgicas ascendentes se origina nos
núcleos DR e MnR (Vertes, 1991; Vertes et al., 1999); por exemplo, em ratos,
aproximadamente 50% dos neurônios serotonérgicos são encontrados no DR (Azmitia e

16
Segal, 1978; Jacobs e Azmitia, 1992). Técnicas utilizando traçadores permitiram a observação
de robustas projeções eferentes do DR e MnR ao tálamo (Vertes, 1991; Vertes et al., 1999;
Vertes e Linley, 2007;2008). O principal alvo dessas fibras são núcleos anteriormente
denominados “não-específicos” do tálamo, como o núcleo anterior do tálamo e partes do
“tálamo visual” (Vertes, 1991; Morin e Meyer-Bernstein 1999; Vertes et al., 1999; Vertes et
al., 2010). Além disso, os núcleos da linha média e núcleos intralaminares do tálamo de ratos
(considerados não-específicos) apresentaram elevada densidade de fibras serotonérgicas
quando analisadas em imunoistoquímica para transportador de 5-HT (Vertes et al., 2010),
bem como para a própria 5-HT (Cropper et al., 1984).
1.2 Tálamo
O tálamo é parte integrante do diencéfalo, assim como o epitálamo, hipotálamo e
subtálamo (Parent, 1996). Praticamente todas as informações sensoriais atravessam os núcleos
do tálamo. Tais informações ascendem através das radiações tálamo-corticais e se projetam
para áreas corticais. A informação que atravessa o tálamo geralmente sofre modificação, com
isso, muitos impulsos são alterados antes de ascenderem ao córtex, sofrendo, portanto,
modulação (Parent, 1996; Jones, 2007).
Além do sistema sensorial, o tálamo também integra informações inerentes aos
sistemas límbico e motor, através de conexões com os núcleos da base, cerebelo e estruturas
límbicas (Bentivoglio et al., 1991; Freedman e Cassel, 1991; Burk e Mair, 2001; Glimcher e
Lau, 2005). O sistema límbico é composto por diversas estruturas corticais e subcorticais que
estão intimamente interconectadas, como córtex pre-frontal, córtex cingulado, córtex
entorrinal, hipocampo, núcleo accumbens, núcleo pálido ventral e hipotálamo anterior
(Swanson e Petrovich, 1998; Heimer, 2003); está especialmente relacionado à geração e
expressão de estados motivacionais e emocionais. Em humanos, disfunções desse sistema
podem gerar diversos transtornos como esquizofrenia, depressão, abuso de drogas, entre
outros (Morgane et al., 2005). Estudos afirmam também a participação do tálamo em circuitos
de processamento de memória (Aggleton e Brown, 1999; Aggleton et al., 2011; Hembrook e
Mair, 2011).
O tálamo é constituído, na maioria dos mamíferos, por duas grandes massas ovoides
de tecido nervoso, situando-se na porção laterodorsal do diencéfalo, acima do sulco
hipotalâmico. Sua superfície dorsal é coberta por uma lâmina de substância branca, o extrato

17
zonal do tálamo, que ao se estender a sua face lateral, denomina-se lâmina medular externa.
Ao penetrar e percorrer longitudinalmente o tálamo, o extrato zonal passa a constituir a
lâmina medular interna, a qual representa um ponto de referência fundamental para a divisão
do tálamo em grupos nucleares (Parent, 1996; Jones, 2007). Tais grupos nucleares apresentam
variações interespecíficas evidentes, como por exemplo, entre roedores, principalmente ratos
e camundongos, em comparação a outros mamíferos. A diferença chave, com exceção do
tamanho, é o grau de distinção entre os vários núcleos (Jones, 2007).
Figura 2: Representação esquemática dos principais grupos nucleares do tálamo de humanos. Fonte:
(Kandel et al., 2000a)
De maneira geral, o tálamo pode ser dividido em quatro porções, a partir da lâmina
medular interna, sendo um grupo nuclear anterior, no polo anterior, um grupo nuclear medial,
localizado medialmente à lâmina medular interna, grupo nuclear ventrolateral e um grupo
nuclear posterior, ambos localizados lateralmente à lâmina medular interna. Há ainda um
grupamento de neurônios localizados intrinsicamente à lâmina medular interna, os quais são
denominados intralaminares (Figura 2) (Kandel et al., 2000a). Tendo como base as regiões de
saída do telencéfalo, o tálamo pode ser subdividido em: núcleos de retransmissão sensorial
específicos, especialmente os da via somatossensorial (tálamo sensorial); núcleos

18
relacionados aos núcleos da base (tálamo motor) e grupamentos nuclear anterior, mediodorsal
(MD) e linha média/intralaminar (tálamo límbico) (Csillag e Montagnese, 2005). O complexo
nuclear linha média/ intralaminares compõe uma porção significante do tálamo, e
supostamente, apresenta influência global no funcionamento cortical (Van der Werf et al.,
2002).
1.2.1 Características gerais dos núcleos da linha média/ intralaminares
O complexo nuclear da linha média/intralaminares forma um notável grupo de núcleos
da parte medial e dorsal do tálamo. Os da linha média constituem uma faixa estreita de
pequenos núcleos que estão distribuídos medialmente no tálamo da porção dorsal à ventral. Já
os núcleos intralaminares estão localizados lateralmente ao MD e embebidos na lâmina
medular interna, o que caracteriza sua denominação (Parent, 1996; Groenewegen e Witter,
2004; Jones, 2007).
Com base nos estudos eletrofisiológicos de Dempsey e Morison (1943) esses núcleos
foram considerados como parte integrante do sistema talâmico não específico. Isso
aconteceu, pois, ao serem estimulados eletricamente, os núcleos retransmissores ou
“específicos” do tálamo emitiam respostas de baixa frequência; enquanto se a estimulação
ocorresse nos núcleos talâmicos “não específicos”, aconteciam alterações de longa latência e
amplamente difusas na atividade cortical. Além disso, essa denominação de “não específicos”
foi suportada pelo fato de o complexo de núcleos da linha média/intralaminares receber
extensas aferências da formação reticular da ponte e mesencéfalo (Hallanger et al., 1987;
Cornwall e Phillipson, 1988; Newman e Ginsberg, 1994), bem como emitirem eferências para
alvos corticais de maneira difusa (Scheibel e Scheibel, 1967; Jones e Leavitt, 1974;), gerando
o efeito chamado recrutamento cortical (Moruzzi e Magoun, 1949; Jasper, 1960).
Posteriormente, estudos elucidaram que esses núcleos também apresentam projeções
específicas, as quais se dirigem, especialmente, para o córtex pré-frontal (Berendse e
Groenewegen, 1991; Groenewegen e Berendse, 1994; Van der Werf et al., 2002; Vertes,
2006); e que a estimulação elétrica de núcleos individuais da linha média produz efeitos
seletivos em seus alvos corticais, em vez de respostas espalhadas por todo o córtex
(Dolleman-Van der Weel et al., 1997; Bertram e Zhang, 1999; Kung e Shyu, 2002; Zhang e
Bertram, 2002; Viana di Prisco e Vertes, 2006).
O complexo de núcleos da linha média/ intralaminares possui papel associado a
diversas modalidades sensoriais, especialmente relacionadas à regulação da atenção/vigília

19
(Edelstyn et al., 2012). Por exemplo, estudos utilizando tomografia por emissão de pósitrons
em humanos afirmaram a importância funcional desses núcleos na vigilância auditiva (Paus et
al., 1997). Além disso, podem estar relacionados à conscientização de informação visual
(Hunsperger e Roman, 1976; Schlag e Schlag-Ray, 1984), nociceptiva e tátil, sendo estas
últimas especialmente ligadas aos aspectos emocionais (Hautvast et al.,1997; Johannsen et al.,
1997; Rainville et al., 1997).
Esse complexo nuclear é especialmente conhecido por receber projeções oriundas da
formação reticular do tronco encefálico e assim integrar o sistema de ativação ascendente, o
qual emite projeções difusas para todo o córtex cerebral. Sendo assim, estão diretamente
associados à regulação da atividade cerebral, especialmente funções sensoriais, cognitivas e
motoras (Bentivoglio et al., 1991; Van der Werf et al., 2002). Estudos anteriores em roedores
e primatas relataram que as células desse complexo nuclear apresentam imunorreatividade a
determinadas substâncias, como GABA (Miller e Ferrendeli, 1990), glutamato (Bokor et al.,
2002), proteínas ligantes de cálcio, principalmente calretinina e parvoalbumina (Jones e
Hendry, 1989; Arai et al., 1994), entre outros, que podem estar participando ativamente de
diversas circuitarias relacionadas ao comportamento, como a circuitaria límbica, por exemplo.
1.2.2 Núcleos da linha média do tálamo
A linha média do tálamo é constituída pelos núcleos: paraventricular (PV), paratenial
(PT), intermediodorsal (IMD), reuniens (Re) e rombóide (Rh). No rato, esses núcleos
constituem cerca de um terço do tálamo (Parent, 1996; Van der Werf et al., 2002;
Groenewegen e Witter, 2004).
O PV está localizado medianamente no tálamo de rato, abrangendo toda sua extensão
rostro-caudal do complexo linha média/intralaminares. Localiza-se medial e dorsalmente aos
núcleos mediodorsais e limita-se ventralmente com o terceiro ventrículo (3V), podendo ser
dividido numa porção anterior (PVA), média (PVM) e outra posterior (PVP). Utilizando
como referencial o atlas do rato (Paxinos e Watson, 2007), o PVa inicia-se imediatamente
após o forame interventricular, ocupando toda extensão dorsoventral do tálamo, dorsalmente
ao hipotálamo anterior, medialmente ao núcleo PT do tálamo, seguido da estria medular do
tálamo. Estende-se posteriormente, mantendo associação com o núcleo paratenial e a estria
medular. A transição da parte anterior para o nível médio do PV ocorre no nível em que o
núcleo das habênulas medial aparece medialmente à estria medular. Sua margem dorsal está

20
em contato com as células ependimárias do 3V em toda sua extensão antero-posterior. A
transição do nível médio para a parte posterior ocorre com o aparecimento do fasciculo
retroflexo. O PVp se estende mantendo contato com o núcleo médio-dorsal e desaparecendo
lateralmente à linha média do tálamo coincidindo com um nível em que o núcleo pré-
comissural se torna evidente.
Estudos com traçadores relatam uma forte aferência aminérgica no PV, sendo
histaminérgica originada no núcleo tuberomamilar (Panula et al., 1989), dopaminérgica da
área tegmentar ventral e região retrorubral (Takada et al., 1990), noradrenérgica do locus
coeruleus e do núcleo do trato solitário e serotonérgica originada especialmente do DR
(Otake e Nakamura, 1995; Otake e Ruggiero, 1995; Krout et al., 2002). O PV também recebe
aferências do núcleo parabraquial, colículo superior e do núcleo supramamilar (Vertes, 1992;
Bester et al., 1999; Krout et al., 2001; Li et al., 2012). Além disso, são relatadas conexões
recíprocas com o núcleo supraquiasmático e com o núcleo dorsomedial do hipotálamo,
regiões fundamentais na regulação dos ritmos circadianos (Cornwall e Phillipson, 1988; Chen
e Su, 1990; Moga et al., 1995; Vertes e Hoover, 2008; Li et al., 2012). Estudos com
traçadores em macaco (Callithrix jacchus) relataram ainda projeções aferentes provenientes
da retina ao PV (Cavalcante et al., 2005). Em se tratando de suas projeções eferentes, o PV
direciona-se ao estriado e ao córtex cerebral, destacando-se os córtices pré-límbico e
infralímbico, à amígdala e ao accumbens, bem como algumas fibras ao córtex cingulado
anterior e córtex orbital medial (Van der Werf et al., 2002).
O PT é um núcleo estreito alongado localizado na metade anterior do tálamo dorsal,
lateralmente ao PVA. Na sua porção posterior funde-se com o núcleo MD do tálamo (Jones,
2007). Recebe aferências do córtex infralímbico, do subículo ventral, da metade rostral do
claustro (Chen e Su, 1990), e fraca aferência do DR e MnR, locus coeruleus, parabraquial e
núcleo do trato solitário (Bobillier et al., 1979; Cornwall e Phillipson, 1988; Chen e Su, 1990;
Newman e Ginsberg, 1994). Suas projeções eferentes dirigem-se, em geral, ao estriado ou a
áreas do córtex pré-frontal (Van der Werf et al., 2002).
O IMD está localizado entre o núcleo MD esquerdo e direito. Este núcleo não é
reconhecido em todas as espécies, sendo descrito com mais detalhes em ratos (Jones, 2007).
Com base em estudos de traçadores, foi relatado que o IMD recebe aferências do núcleo
supramamilar em ratos (Vertes, 1992); e em gatos foram observadas aferências originadas do
córtex límbico anterior (Kaitz e Robertson, 1981). Com relação às eferências, o núcleo

21
também se projeta especialmente ao estriado e ao córtex pré-frontal medial (Van der Werf et
al., 2002).
O Re está localizado no terço anterior do tálamo de rato. Anteriormente é dividido em
componentes direito e esquerdo pelo 3V. Caudalmente esses componentes se unem e tornam-
se uma massa de células situada imediatamente dorsal 3V (Jones, 2007). Na sua borda inferior
o Re é margeado pelo núcleo reuniens ventral (VRe) em ambos os lados. O Re recebe
projeções do córtex pré-frontal medial, de outros núcleos talâmicos como o núcleo reticular e
núcleo geniculado lateral, bem como regiões do tronco encefálico, como a área tegmentar
ventral, o colículo superior, o núcleo parabraquial e os núcleos DR e MnR (Herkenham, 1978;
Bobillier et al., 1979; Vertes, 1992; Vertes, 2002). Suas projeções eferentes direcionam-se aos
córtices pré-limbicos, infralímbicos, cingulado anterior e hipocampo, além das projeções ao
estriado ventral (Su e Bentivoglio, 1990; Van der Werf et al., 2002; Hoover e Vertes, 2011).
O Rh está localizado abaixo da lâmina medular interna (Berendse e Groenewegen,
1991). Rostralmente é confluente com o núcleo anteromedial e possui duas extensões laterais
em forma de asa (Groeneweger e Witter, 2004). Suas aferências têm sido relatadas oriundas
do tronco encefálico, incluindo os núcleos da rafe, especificamente o MnR; além das
formações reticulares medular, pontina e mesencefálica (Bobillier et al., 1979; Krout et al.,
2002) e do núcleo supramamilar (Vertes, 1992). Semelhantemente ao núcleo Re, suas
projeções eferentes direcionam-se especialmente aos córtices pré-límbico, infralímbico e
cingulado anterior, além das projeções ao estriado ventral (Van der Werf et al., 2002).
1.2.3 Núcleos intralaminares do tálamo
Possuem um grupo rostral, formado pelos núcleos central medial (CM), paracentral
(PC) e central lateral (CL). O grupo caudal é composto do núcleo parafascicular (PF) e central
mediano (CMn). Este último está presente apenas em alguns mamíferos, de modo que em
ratos o grupo posterior é composto apenas pelo PF (Groenewegen e Witter, 2004; Jones,
2007).
O CM é uma estrutura ímpar situada através da linha média, sendo contínuo com o PC
em ambos os lados. Facilmente localizado por apresentar forte coloração quando coradas pelo
Nissl e células achatadas, claramente distintas dos núcleos da linha média (Van der Werf et
al., 2002; Groenewegen e Witter, 2004; Jones, 2007). A maioria das suas aferências são
originadas de estruturas subcorticais como a formação reticular (Peschanski e Besson, 1984),

22
núcleo supramamilar (Vertes, 1992), alguns núcleos cerebelares, como o denteado, o fastigial
e o interpósito posterior (Haroian et al., 1981) e também o colículo superior (Yamasaki et al.,
1986). Suas eferências direcionam-se, de maneira geral, ao estriado, mais especificamente sua
porção ventral e a amígdala (Van der Werf et al., 2002).
O PC corresponde a uma fina lâmina de células contínua com o CM medialmente e
lateralmente com o CL. O PC é difícil de distinguir do CL, no entanto as células daquele
apresentam-se mais achatadas (Van der Werf, 2002). Em quase toda a extensão esse núcleo
repousa entre o MD e o núcleo ventral lateral (Jones, 2007). Suas aferências são em maioria
originadas de estruturas subcorticais, podendo-se citar o colículo superior (Yamasaki et al.,
1986), o núcleo supramamilar e a formação reticular do tronco encefálico (Peschanski e
Besson, 1984; Vertes e Martin, 1988; Vertes, 1991). Suas fibras eferentes direcionam-se
especialmente para os córtices parietal, cingulado anterior, visual e para o estriado (Van der
Werf, 2002).
O CL é bem semelhante ao PC em termos de coloração e formato de células, sendo
localizado posteriormente e dorsalmente ao mesmo. O CL apresenta-se em forma de Y na
região frontal de diversas espécies. No entanto, muitas vezes, o CL e o PC são considerados
uma única estrutura (Van der Werf, 2002; Jones, 2007). O CL apresenta uma densa aferência
do tronco encefálico (Peschanski e Besson, 1984; Yamasaki et al., 1986; Hallenger et al.,
1987). A formação reticular medular, pontina e mesencefálica, especialmente os núcleos DR e
MnR, parte reticulada da substância negra, núcleo supramamilar e colículo superior são áreas
que emitem projeções ao CL. Suas projeções eferentes direcionam-se mais abundantemente
ao córtex cingulado e ao estriado (Van der Werf, 2002).
O PF representa um componente caudal do complexo intralaminar. Destaca-se por sua
coloração escura, especialmente na sua porção lateral. Ventralmente, o PF repousa quase
diretamente sobre o mesencéfalo e dorsalmente é margeado pelo núcleo CL e complexo
habenular. Como aferências pode-se destacar aquelas oriundas da formação reticular medular,
pontina e mesencefálica, principalmente DR e de alguns outros núcleos do tronco encefálico,
como o núcleo vestibular, parabraquial, tegmental, ambíguo e núcleo do trato solitário (Van
der Werf, 2002). O PF emite fibras direcionadas aos córtices frontal e parietal, e em menor
quantidade aos córtices cingulado, parahipocampal e pré-límbicos. Além disso, destacam-se
suas projeções ao estriado (Van der Werf, 2002).

23
1.3 Projeções serotonérgicas aos núcleos da linha média e intralaminares do
tálamo
Estudos utilizando imunoistoquímica para 5-HT em ratos (Sprague-Dawley) (Cropper
et al., 1984) e em macacos (Saimiri sciureus) (Lavoie e Parent, 1991) descreveram os padrões
de distribuição desse neurotransmissor no tálamo dos respectivos animais. Cropper et al.
(1984) observaram uma densa concentração de fibras serotonérgicas nos núcleos da linha
média e intralaminares do tálamo. Em macacos, Lavoie e Parent (1991) demonstraram
similaridade desse padrão de distribuição. Vertes et al. (2010) observaram, em ratos, um
padrão de distribuição de fibras serotonérgicas similar a Cropper et al. (1984) com algumas
particularidades que podem estar relacionadas aos diferentes marcadores utilizados.
Pesquisas utilizando técnica de traçadores relataram que as fibras serotonérgicas
ascendentes aos núcleos da linha média e intralaminares originam-se principalmente dos
núcleos DR e MnR (Vertes, 1991; Vertes et al., 1999).
Krout et al. (2002) analisaram as projeções do tronco encefálico aos núcleos da linha
média e intralaminares do tálamo de rato utilizando a técnica de injeção de traçador
retrógrado; assim, observaram intensas projeções de fibras serotonérgicas do DR para todos
os núcleos intralaminares e moderada projeção do MnR aos núcleos CL, CM e PF; o núcleo
PF ainda apresentou projeções oriundas do RPa; e a injeção no PC resultou em marcações em
diversas áreas do tronco encefálico, como a área tegmental dorsoventral e a porção reticulada
da substância negra, porém poucas projeções oriundas do DR. Tal relato difere do que foi
observado por Vertes et al. (1999), o qual afirmou robusta projeção do DR ao PC. Nos
núcleos da linha média, o padrão de marcação foi semelhante, de modo que houve intenso
registro de fibras localizadas no DR e MnR. Também foram visualizadas marcações no RPa
após injeções de traçadores retrógrados no PV (Krout et al., 2002). Esse padrão de projeções
foi semelhante ao que foi observado por Vertes (1991) utilizando traçador anterógrado no DR.
Nesse sentido, os núcleos da linha média e intralaminares recebem diversas projeções
serotonérgicas (Krout et al., 2002; Van der Werf et al., 2002; McKenna e Vertes, 2004) e
projetam para regiões corticais e subcorticais em geral, mas especialmente para áreas
límbicas, o que supõe o importante papel na circuitaria límbica (Berendse e Groenewegen,
1991; Vertes, 2006).
Tendo em vista a elevada densidade de projeções serotonérgicas recebidas pelos
núcleos da linha média e intralaminares do tálamo de alguns animais, como roedores e

24
primatas, o presente estudo analisará a diferença de densidade e as características
morfológicas dessas fibras num roedor típico do Nordeste brasileiro, cujos hábitos
comportamentais e características morfofuncionais se diferenciam dos demais roedores.
1.4 Modelo experimental
O mocó é um roedor cuja distribuição predomina nos estados do Nordeste e norte de
Minas Gerais, o qual habita preferencialmente a caatinga do semi-árido do Nordeste
Brasileiro (Cabrera, 1961; Lacher Jr., 1981).
Pode ser classificado taxonomicamente como membro da superfamília Cavioidea,
família Caviidae, subfamília Caviinae, gênero Kerodon, juntamente com os gêneros Cavia,
Galea e Microcavia (Moojen, 1952; Cabrera, 1961; Lacher Jr, 1981). A este grupo pertencem
pequenos animais já reconhecidamente domésticos, como porquinho da índia (Cavia
porcellus), os quais vivem em pequenas colônias feitas em buracos na terra ou usam
cavidades nas bordas das rochas. Possuem hábitos gregários e diurnos (Crandall, 1964).
A família Caviinae apresenta animais adaptáveis a diversos tipos de ambiente,
podendo ser arborícolas, rupícolas e terrícolas; costumam ser herbívoros em consequência de
sua dentição ser desprovida de caninos (Moojen, 1952).
Estudos de morfologia (Silva Neto, 2000) e de biologia molecular (Rowe e Honeycutt,
2002) afirmaram relação do gênero Kerodon com a família Hydrochaeridae, à qual também
pertence a capivara (Hydrochaeris).
O mocó é um animal que habita locais rochosos com diversas fendas onde se abrigam
dos predadores e passam boa parte do tempo. São facilmente adaptáveis às condições
ecológicas locais como calor, escassez de água e de alimentos, especialmente nos períodos de
grandes secas do semi-árido nordestino. Extraem sua alimentação de galhos de árvores e
rochas, principalmente por serem excelentes saltadores e escaladores. Tal alimentação é
constituída de cascas de árvores e gramíneas em geral, sendo as árvores mais procuradas o
mufumbo (Cobretum leprosum), faveleira (Cnidoscolus phyllacanthus) e a parreira brava
(Cissampelos pareira) (Carvalho, 1969; Lacher Jr, 1981; Mendes, 1985; 1987). Em cativeiro
aceitam bem frutas (maçã, banana, melão, melancia, manga) e raízes.
Apresenta coloração cinza clara com pelos pretos e amarelos ou esbranquiçados na
região dorsal, castanho-ferruginoso na região caudal, um pouco acastanhada nas patas e
branco na região cervical. Suas patas possuem coxins calosos pouco excedidos pelas unhas

25
rígidas que lhes proporcionam a habilidade de saltar e escalar (Moojen, 1952). Seu olfato e
audição são bastante aguçados, detectando seu predador a longas distâncias (Carvalho, 1969).
Atingem a fase adulta aos 200 dias, podendo alcançar até 50 cm de comprimento e 1 quilo de
peso corporal (Moojen, 1952; Carvalho, 1969; Roberts et al., 1984).
A reprodução ocorre durante todo o ano, com exceção do período que vai de abril a
junho. Dentre os cavinnaes, o Kerodon possui a gestação mais longa (65±1,34 dias), a menor
média de tamanho da ninhada (1,28±0,09), o mais baixo peso de filhotes e o mais longo
tempo de maturidade sexual (Roberts et al., 1984). Apesar das poucas crias por parto, o curto
período gestacional garante uma elevada produção de crias durante o ano (Lacher Jr, 1981).
Com relação ao seu padrão de atividade locomotora, tem sido relatado que nos dias
mais escuros o mocó sai para se alimentar pela manhã e a tarde, enquanto nos dias mais
claros sua atividade se concentra no período noturno (Carvalho, 1969). Estudos realizados por
Sousa e Menezes (2006) elucidaram que embora o mocó apresente atividade ao longo de 24
horas, os períodos de transição de fases claro-escuro são os de maior concentração de
atividade, sugerindo, portanto, comportamento predominantemente crepuscular.
Nesse sentido, em consequência das singularidades morfofuncionais e hábitos
comportamentais dos mocós em relação a outros animais da ordem Rodentia é que este
animal vem sendo adotado como modelo para estudos neuroanatômicos do Laboratório de
Neuroanatomia da Universidade Federal do Rio Grande do Norte – UFRN.
Figura 3: O mocó (Kerodon rupestris) em ambiente natural. Fonte: LabNeuro.

26
2. JUSTIFICATIVA
A revisão apresentada reflete a importância evidente de novas pesquisas acerca dos
componentes do sistema nervoso do animal em questão; além disso, também se faz
justificativa do presente estudo a relevância do papel modulatório da 5-HT como
neurotransmissor nos núcleos da linha média e intralaminares do tálamo influenciando em
diversos aspectos comportamentais.
Nesse sentido, apesar da existência de trabalhos relatando a presença de terminais
serotonérgicos na linha média e núcleos intralaminares do tálamo de primatas e roedores, não
existe descrição quanto ao padrão de densidade relativa ou uma abordagem qualitativa das
fibras descritas. Desta forma, esse estudo visa contribuir na mudança na forma de análise de
tais fibras, acrescentando uma qualificação diferencial no padrão de distribuição entre os
diversos núcleos estudados. Adicionalmente, o estudo permitirá uma análise filogenética no
padrão de distribuição das fibras serotonérgicas no tálamo de roedores e, ao mesmo tempo,
contribuirá para consolidar o mocó como modelo experimental regional.

27
3. OBJETIVOS
Geral
Caracterizar o padrão de distribuição e a morfologia das fibras de 5-HT nos núcleos da
linha média e intralaminares do tálamo do mocó
Específicos
Delimitar a citoarquitetura desses núcleos utilizando o método de coloração de Nissl.
Verificar, a partir da análise de densidade óptica relativa (DOR), as diferenças nos
padrões de densidade de fibras 5-HT-IR distribuídas no complexo de núcleos da linha
média/intralaminares do tálamo de mocó.
Caracterizar morfologicamente as fibras serotonérgicas em cada um dos núcleos.

28
4. METODOLOGIA
4.1 Sujeitos:
Foram utilizados quatros mocós adultos jovens machos obtidos através de captura no
município de Serra Negra do Norte (RN), mediante autorização do IBAMA (Instituto
Brasileiro do Meio Ambiente e Recursos Naturais Renováveis, SISBIO no. 21440-1). O
processo de captura se realizou a partir do uso de armadilhas e/ou contenção química
(ketamina + xilazina) a fim de reduzir o estresse causado ao animal.
Após a captura, os animais foram alojados no Centro de Biociências, UFRN, em um
recinto de alvenaria e tela de arame, medindo 3,00 x 2,00 x 2,50 m, onde ficaram expostos a
luminosidade, temperatura e umidade naturais, com água e comida ad libitum. Os animais
permaneceram nesses recintos por um período gradativo conforme os mesmos foram sendo
utilizados, já que o contingente de animais não pode ser processado em um único dia. No dia
anterior ao experimento os animais foram transferidos para gaiolas medindo 0,90 x 0,60 x
0,75m.
Os procedimentos foram realizados no laboratório de Neuranatomia, departamento de
Morfologia, UFRN, vinculado ao Programa de pós-graduação em Psicobiologia. Os devidos
cuidados foram tomados com o objetivo de evitar dor e sofrimento aos animais seguindo
estritamente às normas estabelecidas pelo National Research Council of the National
Academy publicadas no livro “Guidelines for the Care and Use of Mammals in Neuroscience
and Behavioral Research”. Uma versão em formato pdf está disponível gratuitamente no site
da Sociedade de Neurociência e Comportamento (SBNeC) – http://www.sbnec.gov.br/links.
É importante ressaltar que parte dos animais utilizados nesse projeto foram os mesmos
de outro projeto em andamento no laboratório, previamente aprovado pelo Comitê de Ética no
Uso de Animais (CEUA), voltado para a caracterização anatômica e neuroquímica da retina
do mocó e sua relação direta com o Sistema de Temporização Circadiana (Protocolo
015/2009). Os experimentos do projeto foram aprovados pelo comitê de ética em uso animal
(CEUA – UFRN nº 033/2012).

29
4.2 Procedimentos:
4.2.1 Anestesia
Os animais receberam como medicação pré-anestésica cloridrato de tramadol e xilazina,
ambos na dose de 5 mg/Kg por via intramuscular. O tramadol é um opióide necessário para
potencializar o efeito da analgesia adequada ao procedimento, já a xilazina é um relaxante
muscular. Após 10 minutos, os animais foram induzidos e mantidos em anestesia inalatória
com isoflurano e oxigênio 100% administrado através de máscara, até atingir o plano
anestésico, ou seja, o terceiro plano do terceiro estágio de Guedel (Massone, 2008).
4.2.2 Perfusão
Ao atingir o plano anestésico, cada animal foi submetido à perfusão transcardíaca, que
compreende os seguintes passos:
1. Posicionamento do animal em decúbito dorsal sobre tela de arame sob ponto de água.
2. Toracotomia, com incisão de pele, músculos e arco costal, sendo estes removidos em
bloco, para exposição do coração.
3. Cardiopunção no ventrículo esquerdo, utilizando uma agulha de 17 mm x 1,5 mm, a qual
é direcionada para o cone arterioso, seguindo-se uma incisão no átrio direito. A agulha é
conectada a uma bomba peristáltica (Cole-Parmer), passando-se 300 ml de solução salina
a 0,9% em tampão fostato 0,1M, pH 7,4 com heparina (Parinex, Hipolabor, 2ml/1000 ml
de solução salina), seguida de 700ml de solução de paraformaldeido 4% em tampão
fosfato 0,1M, pH 7.4, passando-se metade desta solução a um fluxo de 6,0 ml por minuto
e a outra metade a 3,0 ml por minuto, durando todo o procedimento em média de 30
minutos.
4.2.3 Remoção do encéfalo
Após concluída a perfusão, os animais foram posicionados no aparelho estereotáxico
para roedores. Depois de fazer uma incisão longitudinal na pele e rebatê-la lateralmente, foi
feita a limpeza da superfície óssea, facilitando a visualização do bregma e do lambda, os quais
foram nivelados na mesma altura dorsoventral, ajustando-se a barra dos incisivos,
padronizando assim o plano de corte coronal para todos os animais. Após anotação das
coordenadas do bregma e do lambda, o osso da calota craniana foi removido com o uso de

30
broca e trocater, expondo-se o encéfalo. Ainda no aparelho estereotáxico, o encéfalo é
seccionado em 3 blocos, através de 2 secções coronais: uma no nível do bregma e outra no
nível do lambda. Os encéfalos foram retirados delicadamente para evitar danos, preservando
os olhos e nervos ópticos para o outro projeto envolvido, e em seguida fotografados. Logo
após, os três blocos foram armazenados em uma solução de sacarose a 30% em tampão
fosfato 0,1M, pH 7,4, a 4 ºC, até serem submetidos a microtomia.
Figura 4: Mocó posicionado em aparelho estereotáxico e com alinhamento dorsoventral do bregma e
lambda.
Figura 5: Mocó posicionado em aparelho estereotáxico, após remoção da calota craniana e
consequente exposição do encéfalo. Realização dos cortes coronais em nível de bregma e lambda para
formação dos blocos padronizados.

31
4.2.4 Microtomia
Após a etapa de pós-fixação e imersão em tampão sacarose 30%, os encéfalos foram
submetidos à microtomia cuja espessura dos cortes foi padronizada em 30 µm.
Os encéfalos foram congelados por gelo seco e seccionados em micrótomo de
deslizamento, obtendo-se secções coronais. Estas foram coletadas em um meio líquido de
tampão fostato 0,1M, pH 7,4, distribuídas sequencialmente em seis compartimentos, de
maneira cíclica e sequenciada, de modo a manter a distância entre uma secção e a outra
imediatamente seguinte de um mesmo compartimento de aproximadamente 180 µm.
Os cortes de um compartimento foram imediatamente montados em lâminas de vidro
gelatinizadas e submetidas à coloração pelo método de Nissl para permitir uma melhor
demarcação das estruturas. Os cortes dos demais compartimentos foram transferidos para
solução anticongelante e conservados a -20 ºC para utilização posterior em procedimentos de
imunoistoquímica.
4.2.5 Imunoistoquímica
Antes da técnica de imunoistoquímica, foi necessário realizar um pré-tratamento dos
cortes, na perspectiva de atenuar ou abolir a presença de artefatos. O pré-tratamento inclui
uma etapa de lavagem em tampão fosfato 0,1M, numa frequência de seis vezes em cinco
minutos; incubação em peróxido de hidrogênio (H2O2) a 0,3% em tampão fosfato 0,1M, pH
7,4 por vinte minutos, com a finalidade de abolir artefatos causados pela liberação de
peroxidases endógenas e posteriormente novas lavagens em tampão fosfato 0,1M, pH 7,4.
Em seguida foi processada a incubação dos cortes em anticorpo primário, uma solução
formada pelo anticorpo anti-5-HT obtido em coelho (Sigma) em diluição de 1:5.000,
acrescido de soro normal de cabra a 2% em Triton X-100 a 0,4% permanecendo em
incubação por 12 a 24 horas em rotor com rotação lenta. Ao fim deste período os cortes
foram colocados em contato com o anticorpo secundário anti-coelho obtido em cabra (Vector
Labs) diluído a 1:200 no mesmo veículo anterior, por 90 minutos à temperatura ambiente, sob
agitação lenta, em rotor. Posteriormente, os cortes foram imersos em solução do complexo
avidina-biotina-HRP (Protocolo ABC, Kit elite da Vector) (2% da solução total) em Triton X-
100 a 0,4%, contendo NaCl por 90 minutos.
Para visualização da reação, as secções foram postas em meio contendo H2O2 como
substrato e a 3,3’,4,4’tetrahidrocloreto-diaminobenzidina (DAB), utilizada como cromógeno.
A H2O2 foi oferecida indiretamente, colocando-se na solução glicose oxidase e D-glicose,

32
provocando uma reação em que a primeira agindo sobre a segunda libera H2O2 (Itoh et al.,
1979). Esta reação dura em torno de 15 minutos e após esta, os cortes foram submetidos a
mais seis lavagens em tampão fosfato 0,1 M em agitador orbital.
Os cortes foram montados em lâminas gelatinizadas, as quais após secarem a
temperatura ambiente, foram imersas em solução de tetróxido de ósmio a 0,05% para
intensificar a reação. Após as etapas de desidratação, em baterias de álcool de graduação
crescente até o álcool absoluto, e diafanização em xilol, foram montadas as lamínulas. As
demais séries serão utilizadas para repetição de imunoistoquímica para 5-HT, ou utilizadas em
outros projetos.
Como controle, algumas secções serão submetidas ao protocolo específico de
imunoistoquímica, com a omissão do anticorpo primário.
4.2.6 Obtenção e análise de imagens
As secções dos núcleos da linha média e intralaminares do tálamo coradas pelo
método de Nissl e/ou submetidas à imunoistoquímica para 5-HT foram examinadas ao
microscópio óptico (Olympus BX41) em campo claro e então selecionadas. A área dos
núcleos considerada para efeito de quantificação de fibras 5-HT - IR foi delimitada com base
na secção adjacente correspondente corada pelo Nissl. Imagens digitais foram obtidas usando
uma videocâmara digital (Nikon DXM1200) acoplada ao microscópio, ajustado com a
objetiva de aumento 2x para a DOR, 2x para a montagem dos esquemas e 100x para
visualização detalhada de fibras. Nas imagens correspondentes à caracterização morfológica
de fibras, realizamos uma classificação com base no diâmetro axonal, bem como número de
varicosidades de tais fibras.
Os resultados foram documentados em fotomicrografias corrigidas para brilho e
contraste com auxílio do software Adobe photoshop CS2®
e esquemas construídos a partir das
mesmas com auxílio do software Adobe Illustrator CS5®
e tomando como base o atlas de
cérebro de rato (Paxinos e Watson, 2007).
Elas foram comparadas e classificadas conforme número de varicosidades e espessura
do diâmetro axonal. Foram observadas preliminarmente quanto a aspectos relativos de sua
microestrutura, como espessura, comprimento, varicosidades e eixo de orientação no interior
dos núcleos de interesse do estudo.

33
4.2.7 Densidade óptica relativa
Para análise de DOR, as imagens foram obtidas num mesmo momento, estando
submetidas a uma mesma intensidade de luz e sem modificação de brilho e contraste, já que
os valores de pixels encontrados em cada imagem correspondem a uma intensidade maior ou
menor de marcação da substância analisada no tecido correspondente. Para cada animal foram
obtidas 16 imagens que representaram as áreas de interesse desde o nível rostral ao caudal. A
Tabela 1 retrata a divisão dos núcleos em rostral, médio e caudal através dessas 16 imagens.
Usando o programa Image J (versão 1.46i, NIH) campos com dimensões de 0,8 x 0,6
mm foram feitos nos núcleos de interesse e medido a intensidade da imunomarcação. Como
os núcleos apresentam tamanhos ou formatos diferentes ao longo dos níveis, houve variações
no número de medições por núcleos. Assim, no PV, IMD, Re foram obtidos 2 campos de
medição em todos os níveis, enquanto que no PT e Rh apenas um campo foi selecionado em
cada imagem em todos os níveis. Em se tratando dos núcleos intralaminares, no CM, PC e CL
apenas um campo foi medido; no núcleo PF, em seus níveis mais rostrais, foi obtido apenas
um campo e nos níveis mais caudais, três campos. Para cada imagem foi feito um campo de
igual tamanho em uma área controle, com pouca ou quase nenhuma marcação de 5-HT no
tecido, o qual serviu para a normalização dos valores em todos os núcleos, uma vez que as
imunoistoquímicas foram realizadas em momentos diferentes e assim poderiam apresentar
backgrounds diferenciados. Naqueles núcleos em que foi feita mais de uma captura, uma
média foi realizada e então o valor médio de pixels obtido em todos os núcleos foram
subtraídos do valor de pixels da área controle do mesmo tecido.
A partir deste método foi possível semiquantificar a concentração de fibras de 5-HT
nas áreas de interesse com a correspondência dos valores de DOR observados.

34
Tabela 1: Divisão dos núcleos da linha média e intralaminares do tálamo do mocó em níveis rostral,
médio e caudal através das 16 imagens obtidas das secções coronais (S).
4.2.8 Análise estatística
Para analisar se houve diferença estatística nos valores de DOR entre os núcleos da linha
média e entre os núcleos intralaminares foi utilizada uma ANOVA de uma via, seguida do
teste de Tukey. A diferença estatística entre a região dos núcleos da linha média e a região dos
intralaminares foi analisada através do test t pareado a partir da média dos valores de DOR
dos núcleos que compoem essas regiões para cada animal. O nível de significância foi
definido como p < 0,05. Os valores foram analisados a partir das médias geradas e estão
representados pela média ± o erro padrão médio (EPM).
TABELA METODOLÓGICA
Núcleo Rostral Médio Caudal
Núcleos da
linha média
PV S1 – S6 S7 – S11 S12 – S16
PT S1 – S2 S3 S4
Re S1 – S4 S5 – S9 S10 – S13
Rh S5 – S7 S8 – S9 S10 – S13
IMD S8 – S9 S10 – S11 S12 – S13
Núcleos
intralaminares
CM S2 – S7 S8 – S10 S11 – S13
PC S3 – S7 S8 – S10 S11 – S13
CL S8 – S9 S10 – S11 S12 – S13
PF S14 S15 S16

35
5. RESULTADOS
O comprimento rostro-caudal do encéfalo, da extremidade anterior do bulbo olfatório
ao limite bulbo-espinal, foi em média 3,72 cm (Fig. 6).
Figura 6: Encéfalo do mocó em vistas dorsal (a) e ventral (b). Barra: 0,54 cm.
5.1 Análise Citoarquitetônica: Nissl
5.1.1 Núcleos da linha média
Nas secções coronais analisadas o PV do mocó foi visualizado como um núcleo
distinto, com forte afinidade pelo corante tionina, com formato regularmente triangular
(Figura 7A-N). Estando localizado na porção dorsal da linha média do tálamo, é o primeiro
núcleo a surgir em seu pólo rostral. Situa-se dorsalmente ao hipotálamo anterior e
medialmente ao PT. Pode ser dividido em três porções: anterior (PVA), média (PVM),
marcada pelo surgimento do núcleo MD, e posterior (PVP), evidenciada pela presença do
fascículo retroflexo (Figura 7A-N).
O PT é o segundo núcleo a aparecer rostralmente e está localizado entre o PVA e a
estria medular do tálamo. Apresenta-se na região dorsal da linha média, latero-inferiormente
ao PVA, fundindo-se com o núcleo MD ao longo de seu comprimento rostro-caudal (Figura
7A-H).
O Re está localizado na região anterior, na porção mais ventral talâmica, sendo
imediatamente dorsal ao 3V. Em seus níveis mais rostrais inicia como duas massas ovóides
que se unem ao longo do comprimento rostro-caudal. Na sua borda inferior, assim como em
ratos, o Re é margeado pelo VRe em ambos os hemisférios (Figura 7C-M).

36
O Rh do mocó evidenciou-se ventralmente ao núcleo anteromedial do tálamo e dorsal
ao núcleo Re. É possível observá-lo na porção média do tálamo (Figura 7H-K).
No mocó, o IMD é um núcleo retangular que aparece entre os núcleos MD direito e
esquerdo sendo evidenciado mais claramente nas secções coronais que apresentam o PVM. O
IMD estende-se caudalmente até o nível do fascículo retroflexo ao longo do eixo rostro-
caudal (Figura 7I-M).
5.1.2 Núcleos Intralaminares
No polo anterior o CM encontra-se ventralmente ao PVA e é contínuo ao núcleo PC.
(Figura 7E-M). O PC do mocó repousa anteriormente sob o núcleo PC e medialmente sob o
núcleo MD, sendo contínuo ao CL rostromedialmente. Devido à relativa semelhança em
relação a formato e compactação celular, o CM, PC e CL são dificilmente distinguíveis pelo
método de Nissl através das secções coronais (Figura 7E-M).
O CL está localizado lateralmente e ventralmente ao núcleo MD, estendendo-se
dorsalmente até o nível da estria medular do tálamo (Figura 7J-N).
Por último, o PF é o mais caudal dos núcleos intralaminares. Este núcleo é margeado
pelo CL e apresenta-se medial ao grupo nuclear talâmico posterior na maior parte de sua
extensão rostro-caudal, envolvendo o fascículo retroflexo (Figura 7L-O).
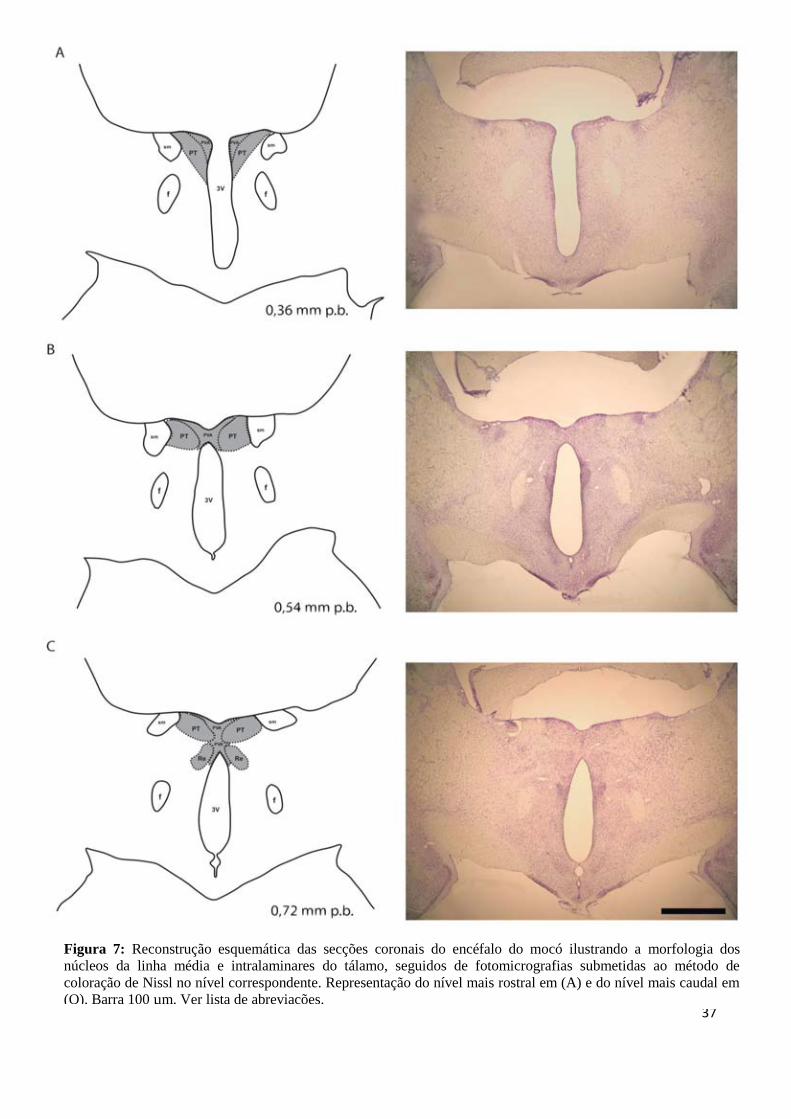
37
Figura 7: Reconstrução esquemática das secções coronais do encéfalo do mocó ilustrando a morfologia dos
núcleos da linha média e intralaminares do tálamo, seguidos de fotomicrografias submetidas ao método de
coloração de Nissl no nível correspondente. Representação do nível mais rostral em (A) e do nível mais caudal em
(O). Barra 100 µm. Ver lista de abreviações.

38
Figura 7: Continuação.
Figura 7: Continuação.

39
Figura 7: Continuação.

40
Figura 7: Continuação.

41
Figura 7: Continuação.
Figura 7: Continuação

42
5.2 Densidade óptica relativa
A análise dos valores de DOR foi feita para fibras 5-HT-IR observando um contínuo
de 16 cortes coronais correspondentes aos núcleos da linha média/ intralaminares. A ANOVA
de uma via revelou diferença significativa na DOR entre os núcleos da linha média do tálamo
[F(4,19) = 6,301, p=0,004]. A figura 8 sugere uma maior DOR no núcleo Re, seguido do PV,
Rh, IMD e PT. Entretanto, a partir do teste de correção de Tukey, apenas foi observada
diferença significativa de DOR do PV [p=0,022] e Re [p=0,003] em relação ao PT.
Em se tratando dos núcleos intralaminares, a ANOVA de uma via também revelou
diferença significativa para DOR [F(3,15) = 8,506, p=0,003], com uma DOR maior no CL,
seguido do CM, PC e PF (Figura 9). O teste de Tukey evidenciou uma DOR
significativamente maior apenas do CL em relação ao CM [p=0,038], ao PC [p=0,005] e ao
PF [p=0,004] (Figura 9).
A partir do teste t pareado mostramos que houve DOR estatisticamente maior dos
núcleos da linha média em relação aos intralaminares [t(3)=7,553, p=0,005] (Figura 10).
A tabela 2 evidencia a DOR dos núcleos da linha média/intralaminares do tálamo de
mocó através dos níveis rostral, médio e caudal, sendo representada a partir da média ± desvio
padrão (DP). Conforme observação da distribuição de DOR dos níveis rostral ao caudal é
possível inferir que há certa homogeneidade na distribuição das fibras serotonérgicas
rostrocaudalmente, no entanto, algumas particularidades podem ser destacadas: entre os
núcleos da linha média, o PV apresentou uma predominância quantitativa de fibras
serotonérgicas nos níveis médios e caudais, o Re nos níveis médios, o Rh nos níveis rostrais e
médios, o PT mostrou-se homogêneo por toda extensão rostro-caudal e o IMD predominou
nos níveis rostrais, porém com elevada variação nos demais níveis entre os animais estudados.
Já entre os intralaminares, no CM observa-se uma maior DOR nos níveis médios e caudais,
CL e PF nos níveis rostrais e o PC mostrou-se homogêneo (Tabela 2).

43
Figura 8: DOR de fibras 5-HT – IR nos núcleos da linha média do tálamo de mocós (n=4). A
ANOVA de uma via revelou diferença significativa entre os núcleos analisado. Valores são expressos
como média e EPM. *p<0,05 e **p<0,005 (Teste post-hoc de Tukey).
Figura 9: DOR de fibras 5-HT – IR nos núcleos intralaminares do tálamo de mocós (n=4). A ANOVA
de uma via revelou diferença significativa entre os núcleos analisados. Valores são expressos como
média e EPM. *p<0,05 e **p<0,005 (Teste post-hoc de Tukey).

44
Figura 10: DOR de fibras 5-HT – IR entre os núcleos da linha média e intralaminares do tálamo de
mocós (n=4). O teste t pareado revelou DOR estatisticamente maior dos núcleos da linha média em
relação aos intralaminares. Valores são expressos como média e EPM. *p<0,05.
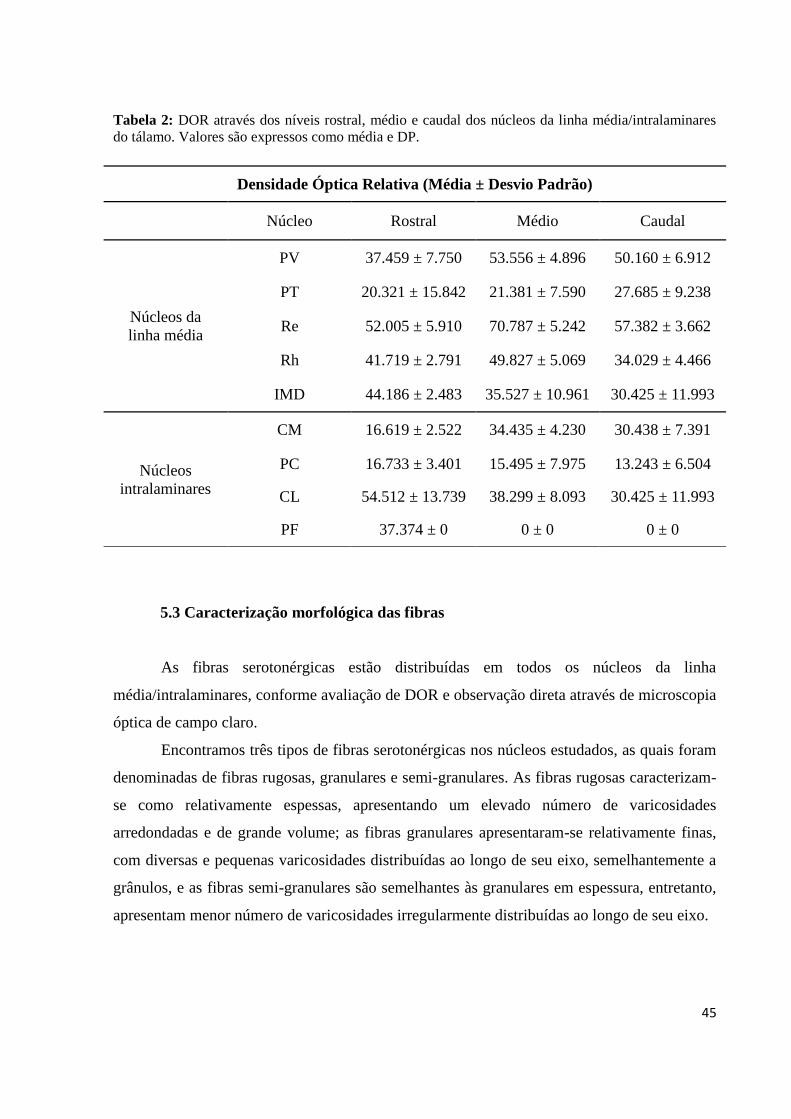
45
Tabela 2: DOR através dos níveis rostral, médio e caudal dos núcleos da linha média/intralaminares
do tálamo. Valores são expressos como média e DP.
5.3 Caracterização morfológica das fibras
As fibras serotonérgicas estão distribuídas em todos os núcleos da linha
média/intralaminares, conforme avaliação de DOR e observação direta através de microscopia
óptica de campo claro.
Encontramos três tipos de fibras serotonérgicas nos núcleos estudados, as quais foram
denominadas de fibras rugosas, granulares e semi-granulares. As fibras rugosas caracterizam-
se como relativamente espessas, apresentando um elevado número de varicosidades
arredondadas e de grande volume; as fibras granulares apresentaram-se relativamente finas,
com diversas e pequenas varicosidades distribuídas ao longo de seu eixo, semelhantemente a
grânulos, e as fibras semi-granulares são semelhantes às granulares em espessura, entretanto,
apresentam menor número de varicosidades irregularmente distribuídas ao longo de seu eixo.
Densidade Óptica Relativa (Média ± Desvio Padrão)
Núcleo Rostral Médio Caudal
Núcleos da
linha média
PV 37.459 ± 7.750 53.556 ± 4.896 50.160 ± 6.912
PT 20.321 ± 15.842 21.381 ± 7.590 27.685 ± 9.238
Re 52.005 ± 5.910 70.787 ± 5.242 57.382 ± 3.662
Rh 41.719 ± 2.791 49.827 ± 5.069 34.029 ± 4.466
IMD 44.186 ± 2.483 35.527 ± 10.961 30.425 ± 11.993
Núcleos
intralaminares
CM 16.619 ± 2.522 34.435 ± 4.230 30.438 ± 7.391
PC 16.733 ± 3.401 15.495 ± 7.975 13.243 ± 6.504
CL 54.512 ± 13.739 38.299 ± 8.093 30.425 ± 11.993
PF 37.374 ± 0 0 ± 0 0 ± 0

46
5.3.1 Núcleos da linha Média
O PV está densamente preenchido por fibras e terminais serotonérgicos, conforme
DOR. As fibras encontradas nesse núcleo foram predominantemente do tipo rugosa, com suas
varicosidades se apresentando conspícuas, variando de 1 a 3 botões por fibra. Tais fibras
apresentam orientação predominantemente dorsoventral, sendo relativamente curtas (Figura
11B).
O PT foi o núcleo com menor marcação de fibras serotonérgicas, conforme
demonstrado pela DOR. A maioria das fibras encontradas nesse núcleo foi do tipo granular,
apresentando comprimento relativamente maior em relação àquelas observadas no PV. A
orientação das fibras no interior do PT é semelhante à observada no PV, ou seja, dorsoventral
(Figura 11C).
O Re é o mais povoado núcleo da linha média em relação a fibras 5-HT-IR, conforme
análise de DOR. Neste núcleo observamos fibras do tipo granular e semi-granular. Tais fibras
apresentam seu maior eixo direcionado dorsoventralmente e uma menor parte com orientação
horizontal; em geral, são relativamente curtas. Uma característica observada nas fibras
serotonérgicas no Re é uma ampla ramificação (Figura 11D).
O Rh é povoado por terminais serotonérgicos com DOR próxima ao observado no PV
e IMD. Suas fibras também se caracterizaram especialmente do tipo granular e semi-granular,
estando orientadas predominantemente no eixo horizontal (Figura 12C).
A análise do IMD permitiu identificar presença de fibras serotonérgicas do tipo semi-
granular. A orientação de tais fibras também se caracterizou dorsoventral, apresentando maior
comprimento relativo (Figura 12B).
5.3.2 Núcleos Intralaminares
A análise qualitativa das fibras serotonérgicas nos núcleos intralaminares do tálamo do
mocó revela fibras com predominância de orientação dorsoventral no PC e PF; no CM e CL a
orientação das fibras é predominantemente horizontal. Nos núcleos PC e CM foram
encontradas fibras granulares e semi-granulares (Figura 13); no CL e no PF sobressaíram
fibras do tipo semi-granulares (Figuras 13D e 14). Aparentemente não existem muitas
ramificações, entretanto, uma característica das fibras em todos os núcleos intralaminares do
mocó é o grande comprimento quando comparadas àquelas observadas nos núcleos da linha
média.

47
.

48

49

50

51
6. DISCUSSÃO
No presente estudo, mostramos as características morfológicas dos núcleos da linha
média e intralaminares do tálamo do mocó, a partir da construção de esquemas baseados em
secções coradas pelo método de Nissl. Além disso, através da utilização da técnica de
imunoistoquímica foi possível caracterizar o padrão de distribuição dos terminais
serotonérgicos em tais núcleos, bem como descrever os aspectos morfológicos associados às
fibras examinadas. Verificamos, também, as diferenças relacionadas à densidade de fibras
serotonérgicas entre os núcleos da linha média e intralaminares através da técnica de DOR. Os
resultados revelaram que a organização deste complexo de núcleos é, em parte, similar ao que
tem sido previamente descrito em outros mamíferos.
Em comparação à organização citoarquitetônica do rato (Paxinos e Watson, 2007),
também identificamos, no mocó, os núcleos da linha média, do sentido rostral ao caudal:
núcleos PV, PT, Re, Rh e IMD; e os intralaminares, no mesmo sentido: núcleos CM, PC, CL
e PF.
A distribuição de fibras serotonérgicas através de núcleos talâmicos foi observada por
Vertes et al. (2010) em ratos (Sprague-Dawley) a partir de imunoistoquímica para
transportador de 5-HT. Visualmente os autores identificaram os núcleos da linha média e
intralaminares do tálamo de rato como densamente marcados. Dentre os núcleos da linha
média, classificaram o PV, Rh, Re e IMD como intensamente marcados, enquanto o PT como
moderadamente marcado. Dentre os núcleos intralaminares, classificaram o CM e CL como
intensamente marcados, enquanto que o PC e PF moderadamente marcados.
Cropper et al. (1984) também descreveram o padrão de distribuição de 5-HT em ratos
albinos a partir da técnica de imunocitoquímica indireta usando um anticorpo contra um
conjugado 5-HT-hemocianina em animais tratados com L-triptofano e MAO. Os autores
identificaram os núcleos da linha média e intralaminares com elevada densidade de marcação,
destacando-se, dentre os da linha média, os núcleos PV, Rh e Re; poucas fibras foram
observadas no PT. Dentre os intralaminares relatou o CM como levemente marcado.
Em primatas, a distribuição de 5-HT foi descrita no tálamo de macacos-de-cheiro
(Saimiri sciureus) a partir de imunoistoquímica contra a própria 5-HT. Dentre os núcleos que
apresentaram maior densidade de inervação destacam-se os da linha média/intralaminares do
tálamo; entre os da linha média ressaltam-se os núcleos PV, Re e o núcleo central, localizado

52
entre o PV e Re, região comparável ao núcleo IMD em ratos ou, conforme o nível observado,
ao Rh. Dentre os intralaminares destaca-se o núcleo CM como intensamente marcado (Lavoie
e Parent, 1991). Em outra espécie de primata, (Macaca Mulata), a distribuição de fibras
serotonérgicas no PV é visivelmente maior em relação aos demais núcleos da linha média e
intralaminares (Hsu e Price, 2009).
Em hamsters sírios (Mesocricetus auratus), a partir de imunoistoquímica para 5-HT,
foi observada densa marcação nos núcleos da linha média e intralaminares do tálamo.
Especificamente, foi identificado que os núcleos PV e Re apresentaram-se intensamente
marcados, seguidos do PT e IMD, este último considerado moderadamente marcado. Dentre
os intralaminares, o CM foi considerado densamente marcado e o CL fracamente marcado;
não houve relato do PC (Morin e Meyer-Bernstein, 1999).
Diferentemente dos autores supracitados, analisamos a distribuição de fibras
serotonérgicas através dos núcleos da linha média/intralaminares a partir de uma técnica de
DOR, o que permite a obtenção de dados numéricos na semi-quantificação dessas fibras. Até
o presente, nenhum estudo havia empregado métodos para semi-quantificação de terminais
serotonérgicos nos núcleos da linha média e intralaminares. Tais estudos têm como base a
simples análise visual feita através de microscopia óptica. Considerando que a análise de
DOR equivale à marcação do neurotransmissor analisado, identificamos, de maneira geral,
uma intensidade de marcação de fibras serotonérgicas estatisticamente maior nos núcleos da
linha média em relação aos intralaminares, esse dado corrobora com estudos realizados em
ratos (Cropper, et al.,1984; Vertes, et al., 2010), primatas (Lavoie e Parent, 1991) e hamsters
Morin e Meyer-Bernstein, 1999). Dentre os da linha média, nesta ordem, os núcleos Re, PV,
Rh e IMD apresentaram maior intensidade de marcação. O PT foi o núcleo que apresentou
menor marcação, semelhante ao observado em ratos e primatas (Lavoie e Parent, 1991; Vertes
et al., 2010), diferentemente do observado em hamster que apresenta uma densa inervação
serotonérgica nesse núcleo (Morin e Meyer-Bernstein, 1999).
Dentre os intralaminares foi identificada, através da DOR e análise estatística, intensa
marcação no CL em relação aos núcleos CM, PC e PF que apresentaram baixa densidade de
fibras 5-HT-IR. Tal padrão de distribuição contrasta com os resultados descritos em hamster
(Morin e Meyer-Bernstein, 1999) e macaco (Lavoie e Parent, 1991) em que uma maior
quantidade de fibras é encontrada no CM. No entanto, Vertes et al. (2010) destacaram o CL
como intensamente marcado em ratos, corroborando com os resultados encontrados no mocó.

53
Na análise referente ao contínuo rostro-caudal, revelamos homogeneidade na
distribuição de fibras serotonérgicas através dos núcleos da linha média/intralaminares do
tálamo do mocó, semelhante ao observado por Vertes et al. (2010). No entanto, algumas
particularidades podem ser destacadas: mostramos que o núcleo PV apresentou uma
predominância quantitativa de fibras serotonérgicas nos níveis médios e caudais, o Re nos
níveis médios, o Rh nos níveis rostrais e médios, o PT mostrou-se homogêneo por toda
extensão rostro-caudal e o IMD predominou nos níveis rostrais. Vertes et al. (2010),
diferentemente dos nossos resultados, mostraram, em ratos, que havia maior presença de
fibras serotonérgicas em níveis mais rostrais do PV, Re e PT em detrimento dos níveis médios
e caudais e que o IMD não apresentava variação na distribuição de fibras entre os níveis
avaliados. Entre os intralaminares, no CM foi revelada uma maior DOR nos níveis médios e
caudais, CL e PF nos níveis rostrais e o PC mostrou-se homogêneo (Tabela 2).
Contrariamente aos dados apresentados por Vertes et al. (2010) que descrevem a inexistência
de diferenças significativas da distribuição de fibras serotonérgicas entre os núcleos
intralaminares do tálamo rato ao longo do eixo rostro-caudal.
Após análise dos resultados referentes à distribuição de fibras 5-HT-IR, em
comparação aos dados apresentados na literatura, podemos constatar que a distribuição das
fibras serotonérgicas através dos núcleos deste complexo nuclear talâmico, no mocó, é similar
à encontrada em outras espécies de roedores, assim como em alguns primatas, com algumas
variações. Por exemplo, nenhum artigo cita uma maior presença de terminais serotonérgicos
no Re em relação ao PV ou aos demais núcleos da linha média. Tais diferenças descritivas
podem estar associadas a não utilização de método dequantificação de fibras em trabalhos
prévios.
Os núcleos da linha média/intralaminares do tálamo podem ser classificados como
integrantes do tálamo límbico em consequência de emitirem projeções para diversas estruturas
relacionadas a funções cognitivas/afetivas. Van der Werf et al. (2002) analisaram diversos
casos de injeção com traçadores anterógrados nos núcleos da linha média e intralaminares do
tálamo de 128 ratos Wistar, e assim propuseram a organização desse complexo nuclear em
quatro grupamentos conforme suas eferências. Um grupo dorsal, composto pelo PV, PT e
IMD, cujas saídas direcionam-se especialmente para o estriado ventral, destacando-se a parte
medial do núcleo accumbens, córtices pré e infralímbico e a amígdala, podendo estar,
portanto, relacionado a funções víscero-límbicas. Um grupo lateral, composto pelo PC, CL e a

54
porção anterior do CM, cujas fibras direcionam-se principalmente ao estriado dorsal e córtex
cingulado, podendo estar associado a funções cognitivas. Um grupo ventral, cujos integrantes
são o Re, Rh e a parte posterior do CM, projetando poucas fibras ao estriado e substanciais
fibras às camadas superficiais e profundas da maioria das áreas corticais, além de emitir
projeções a regiões não límbicas, como córtex gustatório, visceral, insular, auditivo e motor.
O Re e provavelmente o Rh emitem projeções também ao hipocampo; sendo assim, o grupo
lateral pode estar relacionado ao processamento sensorial multimodal; e o grupo posterior
composto do PF, cujas eferências direcionam-se especialmente ao estriado, mas também ao
córtex motor e sensorial, podendo estar, portanto, envolvido em funções motoras límbicas.
Pesquisas utilizando técnica de traçadores relataram forte entrada serotonérgica nos
núcleos da linha média e intralaminares do tálamo, as quais se originam principalmente dos
núcleos DR e MnR (Vertes, 1991; Vertes et al., 1999). Sendo assim, pressupõe-se que esse
neurotransmissor possui influência direta na modulação desses núcleos, estando relacionado
provavelmente ao controle de funções límbicas.
Muitos pesquisadores têm concentrado seus esforços em entender o papel da 5-HT no
tálamo, especificamente em núcleos envolvidos com o processamento da informação visual
(Curtis e Davis 1962; Rogawski e Aghajanian 1980; Yoshida et al., 1984; Marks et al., 1987;
Kayama et al., 1989; Monckton e McCormick, 2001). Todos estes estudos sugerem uma ação
inibitória da 5-HT sobre o núcleo geniculado lateral dorsal do tálamo. Outros estudos
intracelulares in vivo também apontam ação inibitória direta da 5-HT em neurônios tálamo-
corticais devido ao aumento da condutância ao potássio (McCormick e Pape 1990; Lee e
McCormick, 1996). Em contraste a sua ação direta nos neurônios tálamo-corticais, a 5-HT
aplicada no núcleo reticular talâmico resulta em forte excitação nos neurônios GABAérgicos
desse núcleo (McCormick e Wang 1991; Sanchez-Vives et al., 1996), sugerindo um
mecanismo de ação indireta da 5-HT que pode inibir a entrada tálamo-cortical e,
consequentemente, alterar o estado comportamental dos animais. Em um estudo in vivo
utilizando o furão (Mustela putorius furo) (Monckton e McCormick, 2001), foi observado que
a administração local de 5-HT promove uma hiperpolarização dos neurônios registrados
eletrofisiologicamente nos núcleos CL e CM. O mesmo estudo sugere que os efeitos
hiperpolarizantes da 5-HT nestes núcleos sejam mediados pelo receptor 5HT-1A devido ao
fato da administração de antagonista desse receptor ter abolido tal resposta. Outros estudos
propõem uma ação excitatória dos núcleos da linha média do tálamo sobre o prosencéfalo

55
límbico (Van der Werf et al., 2002; Vertes, 2006); com isso, fibras serotonérgicas podem,
possivelmente, representar uma entrada excitatória aos núcleos da linha média ou,
alternativamente, um portão que pode bloquear ou permitir o fluxo de informações desses
núcleos a outras regiões límbicas (Vertes et al., 2006). Estudos mostraram, por exemplo, que
as projeções do DR, relatadas em sua maioria serotonérgicas, ao PV exercem efeitos no
estado de vigília; o PV é reciprocamente conectado com o núcleo supraquiasmático (Moga et
al., 1995; Novak et al., 2000; Peng e Bentivoglio 2004; Li e Kirouac 2008; Vertes e Hoover,
2008) e com o núcleo dorsomedial do hipotálamo (Vertes e Hoover, 2008), área crítica no
controle do ciclo sono e vigília. Além disso, as células do PV expressam elevado nível de c-
fos quando em estado de vigília (Peng et al., 1995; Novak et al., 2000).
A referida dualidade funcional da 5-HT no tálamo está amplamente associada aos
variados tipos de receptores presentes nos diversos campos receptivos destes axônios nesta
região. Além disso, tais diferenças funcionais podem ainda estar relacionadas às variações de
quantidade de fibras entre os núcleos, bem como aos diferentes padrões morfológicos das
mesmas distribuídas nos núcleos da linha média e intralaminares. Este trabalho é o primeiro a
descrever, com maior riqueza de detalhes, o padrão de distribuição de fibras serotonérgicas de
forma quantitativa, levando em consideração as diferenças morfológicas, como espessura,
comprimento e número de varicosidades das referidas fibras especificamente nos núcleos da
linha média e intralaminares do tálamo de um roedor. Em geral, as fibras que trazem
informações sensoriais específicas para os diversos núcleos talâmicos apresentam uma maior
quantidade de terminais, indicando um caráter controlador sobre os neurônios tálamo-
corticais. As fibras serotonérgicas, em particular, atuam promovendo mudanças na
condutância da membrana celular, alterando o limiar de excitação das células ou ativando
sistemas de segundo-mensageiro intracelular, sendo geralmente descritas como fibras
moduladoras devido a grande variabilidade de receptores distribuídos no tálamo (Pompeiano
et al., 1994; Kia et al., 1996; Mengod et al., 1996; Lopez-Gimenez et al., 1998, 2001; Chapin
e Andrade 2001a,b).
Nossos resultados mostram que as fibras observadas no complexo de núcleos da linha
média/intralaminares podem ser classificadas em três tipos: fibras rugosas, granulares e semi-
granulares. Em um estudo utilizando injeção de traçador anterógrado nos núcleos DR e MnR
de rato (Kosofsky e Molliver, 1987) foram visualizadas fibras morfologicamente distintas no
córtex cerebral. O estudo mostrou que as fibras originárias do núcleo MnR eram

56
relativamente espessas e com varicosidades de grande diâmetro, as quais foram chamadas de
fibras do tipo M. A análise comparativa permite sugerir que as fibras rugosas observadas no
mocó, devido às semelhanças morfológicas observadas com as fibras M descritas no rato
também podem ter origem no MnR. Analogamente, as fibras com origem no DR foram
classificadas como fibras do tipo D, sendo estas semelhantes morfologicamente às fibras
granulares descritas no PT, Re, Rh, PC e CM no mocó. Entretanto, não existe analogia entre
as fibras semi-granulares observadas no mocó e às fibras descritas no estudo de traçador
anterógrado realizado no rato (Kosofsky e Molliver, 1987).
Em um trabalho realizado em coelho (Bjarkam et al., 2005), as fibras serotonérgicas
que se projetam para as áreas retrohipocampais também foram caracterizadas
morfologicamente. Os autores demonstraram a presença de fibras serotonérgicas com axônios
de pequeno diâmetro e varicosidades pequenas, as quais foram classificadas como do tipo
"finas". Essas fibras são semelhantes morfologicamente às fibras do tipo granulares
observadas em nosso estudo. Outro tipo de fibra observada no coelho apresenta axônios de
pequeno diâmetro e grandes varicosidades que foram chamadas de fibras "frisadas",
semelhantes às fibras do tipo rugosa de nosso estudo. Um terceiro tipo de fibra foi descrita
como do tipo "tronco", com axônios retos, de grande diâmetro e sem nenhuma varicosidade.
De maneira geral, da mesma forma que o estudo em ratos (Kosofsky e Molliver, 1987), foram
observadas semelhanças com as fibras do tipo rugosas e granulares, no entanto, nenhuma fibra
vista em coelhos assemelha-se as do tipo semi-granulares. Assim, é possível inferir a presença
de diferentes padrões de organização sináptica, consequentemente, diferentes mecanismos
funcionais da 5-HT nos núcleos da linha média e intralaminares do tálamo do mocó em
consequência dessa variabilidade morfológica.
Os dados relativos à distribuição de fibras serotonérgicas no tálamo descritos na
literatura, em sua maioria, são baseados em observações diretas através de microscopia óptica.
Em nenhum deles foram empregados métodos adequados para quantificar as fibras
serotonérgicas em nível talâmico. Assim, a partir de tal quantificação, bem como das variadas
características morfológicas das fibras serotonérgicas encontradas, é possível sugerir
diferentes influências modulatórias desse neurotransmissor, concordando com o seu grande
repertório funcional exercido em nível talâmico para controlar a atividade cortical. As funções
serotonérgicas estão especialmente associadas aos seus receptores; no tálamo de ratos e
primatas, diversos receptores de 5-HT foram encontrados distribuídos heterogeneamente. Na

57
maioria das espécies, o principal tipo de receptor observado a nível talâmico é o subtipo 5-
HT1 (Glennon, 1990). Em primatas não-humanos, altos níveis do subtipo 5-HT1 foram
observados nos núcleos da linha média, bem como núcleo PF (Stuart, et. al., 1986). No tálamo
de humanos, especialmente o receptor 5-HT1A tem sido identificado (Pazos et al., 1987a).
Receptores 5-HT1A podem estar relacionados a uma série de funções como comportamento
sexual (Larsson e Ahlenius, 1999), termoregulação (Goodwin, et al., 1985) e comportamento
alimentar, além disso, pessoas que apresentaram doenças como depressão ou Azheimer
revelaram alteração na distribuição desses receptores (Meltzer, 1990; Nordberg, 1992). Um
pequeno número de receptores do tipo 5HT2 foi visualizado no tálamo, exceto no núcleo PF
onde uma intermediária concentração desse receptor pode ser mostrada em ratos e humanos
(Pazos et al., 1985, 1987b). Os níveis de receptores 5HT3 foram considerados baixos nos
núcleos talâmicos (Kilpatrick et al., 1987; Waeber et al., 1988). Receptores 5HT7 foram
identificados em elevada concentração nos núcleos da linha média e intralaminares do tálamo
(Gustafson et al., 1996; Vizuete et al., 1997); a funcionalidade desse receptor, identificado
recentemente, ainda está sendo mais aprofundada, no entanto sabe-se que gera importante
modulação da atividade neuronal, podendo, portanto, ser importante em estratégias
terapêuticas para doenças psiquiátricas (Goaillard e Vicent, 2002). Estudo em um paciente
com lesão talâmica, especialmente, nos núcleos CL, PV e CM revelou deficiência em funções
de atenção e de execução de atividades (Edelstyn et al., 2012). Nesse sentido, é possível
sugerir que as projeções serotonérgicas variadas em quantificação e caracterização
morfológica podem exercer nos núcleos da linha média e intralaminares diversas funções,
especialmente associadas à cognição.
Nessa perspectiva, comparando nossos resultados com a literatura analisada, é visto
que a ordem Rodentia, apesar das variações de fenótipo, história de vida e tamanho cerebral
não reflete grandes alterações da organização nuclear do complexo da linha
média/intralaminares do tálamo, bem como da distribuição serotonérgica nesses núcleos,
reforçando a sugestão de que o sistema serotoninérgico é um sistema de neurotransmissores
antigo e evolutivamente bem conservado (Parent, 1981). Com isso, o presente trabalho
fornece importantes contribuições para compreender a filogenia do sistema serotonérgico.

58
7. CONCLUSÕES
Os resultados do presente trabalho nos permitem chegar às seguintes conclusões com
relação aos núcleos da linha média e intralaminares do tálamo do mocó e às fibras
serotonérgicas neles encontradas:
1. A técnica de Nissl foi eficiente no que tange a delimitação e caracterização
citoarquitetônica dos núcleos da linha média e intralaminares do tálamo do
mocó, assim como a imunoistoquímica contra 5-HT para marcação de fibras
serotononérgicas presentes nesses núcleos.
2. O complexo nuclear linha média/intralaminares do tálamo do mocó evidenciou-
se semelhante ao observado em outros roedores sendo os da linha média
compostos pelos núcleos PV, PT, Re, Rh e IMD e os intralaminares pelos
núcleos CM, PC, CL e PF.
3. A imunorreatividade contra 5-HT mostrou-se presente em fibras/terminais por
toda a região de núcleos da linha média/intralaminares com variação através dos
núcleos. A partir da DOR visualizamos maior marcação de fibras nos núcleos da
linha média em detrimento dos intralaminares. Dentre os da linha média a
marcação seguiu esta ordem: Re, PV, Rh, IMD e PT; nos intralaminares: CL,
CM, PC e PF.
4. As fibras serotonérgicas estão diferencialmente distribuídas ao longo do eixo
rostro-caudal em alguns núcleos da linha média/intralaminares.
5. As fibras serotonérgicas apresentam diferentes padrões morfológicos entre os
diversos núcleos estudados.

59
8. PERSPECTIVAS
O mocó (Kerodon rupestris) vem sendo utilizado como modelo experimental no
Programa de pós graduação em Psicobiologia da UFRN desde 2003 e, no Laboratório de
Neuranatomia, desde 2004. Assim, estudos já foram desenvolvidos com este animal
envolvendo ritmos circadianos e projeções retininianas, por exemplo. Tais estudos geraram,
portanto, produtos como teses de doutorado, dissertações de mestrado e trabalhos de
conclusão de curso, além de publicações em revistas científicas de indexação internacional.
Atualmente, encontra-se em andamento uma pesquisa de doutorado cujo objetivo consiste em
abordar aspectos anatômicos do olho e neuroquímicos da retina do animal, relacionando-os
com o sistema de temporização circadiana.
Em se tratando de pesquisa que envolva mapeamento de centros cerebrais desta
espécie, esta pesquisa é a terceira que trabalha nesta perspectiva, tendo sido recentemente
concluído um trabalho de mestrado que fez a descrição dos núcleos serotonérgicos da rafe e
outro de núcleos dopaminérgicos do mesencéfalo deste animal. No entanto, o nosso é o
primeiro que trabalha com distribuição e caracterização morfológica de fibras serotonérgicas
em núcleos talâmicos de roedores, e mais especificamente, no mocó.
Sendo assim, este trabalho mostrou a similaridade existente entre os núcleos da linha
média e intralaminares do tálamo de mocó e de outros animais (roedores ou não) estudados.
Entretanto, variações na distribuição e caracterização de fibras serotonérgicas através dos
núcleos em questão foram observadas em relação aos demais animais estudados. Isto permite
sugerir diferentes padrões de organização sináptica, consequentemente, mecanismos
funcionais variados da 5-HT em nível talâmico no controle da atividade cortical.
Nesse sentido, nossa pesquisa lançou a perspectiva de aprofundamento de estudos, a
fim de esclarecer ainda mais algumas divergências existentes, assim como algumas lacunas
que surgem no decorrer dos estudos. Com isso, os mecanismos funcionais e a hodologia
devem ser estudados a fim de compreender a fundo as interrelações entre os
neurotransmissores e estruturas anatômicas das varias espécies estudadas.
Assim, faz-se relevante a continuidade de pesquisas no tocante ao sistema
serotonérgico desse animal, bem como caracterização citoarquitetônica das estruturas que o
compõem. Seguem abaixo algumas possibilidades:

60
Caracterização neuroquímica dos núcleos da linha média e intralaminares do
tálamo;
Estudos de morfometria desses núcleos
Estudos hodológicos do sistema serotonérgico envolvendo núcleos da linha
média/intralaminares
Estudo de caráter funcional que busque evidenciar e correlacionar com
respostas biológicas/comportamentais à distribuição e caracterização
morfológica de fibras serotonérgicas
Estudos filogenéticos e ontogenéticos
Contribuir com o “Projeto Atlas” do mocó

61
REFERÊNCIAS1
Aggleton, J.P., Brown, M.W., 1999. Episodic memory, amnesia, and the hippocampal –
anterior thalamic axis. Behav. Brain Sci. 22, 425-489.
Aggleton, J.P., Dumont, J.R., Warburton, E.C., 2011. Unraveling the contributions of the
diencephalon to recognition memory. Learn. Mem. 18, 384-400.
Arai, R., Jacobowitz, D.M., Deura, S., 1994. Distribution of calretinin, calbindin-D28k, and
parvalbumin in the rat thalamus. Brain Res. Bull. 33, 595-614.
Azmitia, E.C., Gannon, P.J., 1983. The ultrastructural localization of serotonin
immunoreactivity in myelinated and unmyelinated axons within the medial forebrain bundle
of rat and monkey. J. Neurosci. 3, 2083-2090.
Azmitia, E.C., Segal, M., 1978. An autoradiographic analysis of the differential ascending
projections of the dorsal and median raphe nuclei in the rat. J. Comp. Neurol. 179, 641-667.
Bentivoglio, M., Balercia, G., Kruger, L., 1991. The specificity of the nonspecific thalamus:
the midline nuclei. Prog. Brain Res. 87, 53-89.
Berendse, H.W., Groenewegen, H.J., 1991. Restricted cortical termination fields of the
midline and intralaminar thalamic nuclei in the rat. Neuroscience. 42, 73-102.
Bertram, E.H., Zhang, D.X., 1999. Thalamic excitation of hippocampal CA1 neurons: a
comparison with the effects of CA3 stimulation. Neuroscience. 92, 15-26.
Bester, H., Bourgeais, L., Villanueva, L., Besson, J.M., Bernard, J.F., 1999. Differential
projections to the intralaminar and gustatory thalamus from the parabrachial area: a PHA-L
study in the rat. J. Comp. Neurol. 405, 421-429.
Bjarkam, C.R, Sorensen, J.C., 2005. Distribuition and morphology of serotonin-
immunoreactive axons in the retrohippocampal areas of the New Zealand white rabbit. Anat.
Embriol. 210, 199-207.
Bjarkam, C.R., Sorensen, J.C., Geneser, F.A., 1997. Distribution and morphology of
serotonin-immunoreactive neurons in the brainstem of the New Zealand white rabbit. J.
Comp. Neurol. 380, 507-519.
Bobillier, P., Seguin, S., Degueurce, A., Lewis, B.D., Pujol, J.F., 1979. The efferent
connections of the nucleus raphe centralis superior in the rat as revelead by autoradiography.
Brain Res. 166, 1-8.
Bokor, H., Csaki, A., Kocsis, K., Kiss, J., 2002. Cellular architecture of the nucleus reuniens
thalami and its putative aspartatergic/glutamatergic projection to the hippocampus and medial
septum in the rat. Eur. J. Neurosci. 16, 1227-1239.
1 De acordo com as normas da revista Journal of Chemical Neuroanatomy.

62
Burk, J.A., Mair, R.G., 2001. Effects of intralaminar thalamic lesions on sensory attention and
motor intention in the rat: a comparison with lesions involving frontal cortex and
hippocampus. Behav. Brain Res. 123, 49-63.
Cabrera, A., 1961. Catalogo de los mamíferos de América del Sur. II. (Sirenia,
Perrissodactyla, Lagomorpha, Rodentia, Cetacea). Revista del Museo Argentino de Ciencias
Naturales “Bernardino Rivadavia”, Ciencias Zoológicas. 4, 309-732.
Carlson, N.R., 2002. Fisiologia do comportamento. Editora Manole, São Paulo. 124-125.
Carvalho J.M.C., 1969. Notas de viagem de um zoólogo à região das caatingas e áreas
limítrofes. Fortaleza, Imprensa Universitária do Ceará.
Cavalcante, J.S., Costa, M.S.M.O., Santee, U.R., Britto, L.R.G., 2005. Retinal projections to
the midline and intralaminar thalamic nuclei in the common marmoset (Callithrix jacchus).
Brain Res. 1043, 42-47.
Chapin, E.M., Andrade, R.A., 2001a. 5HT(7) receptor mediated depolarization in the
anterodorsal thalamus. I. Pharmacological characterization. J. Pharmacol. Exp. Ther. 297,
395-402.
Chapin, E.M., Andrade, R. A., 2001b. 5HT(7) receptor mediated depolarization in the
anterodorsal thalamus. II. Involvement of the hyperpolarization-activated current Ih J.
Pharmacol. Exp. Ther. 297, 402-409.
Chen, S., Su., H.S., 1990. Afferent connections of the thalamic paraventricular and paratenial
nuclei in the rat – A retrograde tracing study with iontophoretic application of fluoro-gold.
Brain Res. 5222, 1-6.
Cornwall, J., Phillipson, O.T., 1988. Afferent projections to the thalamus dorsal of rat as
shown by retrograde lectin transport. II. The midline nuclei. Brain Res. Bull. 21, 147-161.
Crandall, L.S., 1964. Family caviidae, In: Manangement of the wild mammals captivity.
Chicago: University of Chicago Press. 247-250.
Cropper, E.C., Eisenman, J.S., Azmitia, E.C., 1984. An immunocytochemical study of the
serotoninergic innervation of the thalamus of the rat. J. Comp. Neurol. 224, 38-50.
Csillag, A., Montagnese, C.M., 2005. Thalamotelencephalic organization in birds. Brain Res.
Bull. 66, 303-310.
Curtis, D.R., Davis, R., 1962. Pharmacologic studies upon neurons of the lateral geniculate
nucleus of the cat. Brit. J. Pharmacol. 18, 21-246.
Dahlstrom, A., Fuxe, K., 1964. Evidence for the existence of monoamine-containing neurons
in the central nervous system. I. Demonstration of monoamines in the cell bodies of brain
stem neurons. Acta Physiol. Scand. 62 (suppl. 232), 1-55.
Dahlstrom, A., Fuxe, K., 1965. Evidence for the existence of monoamine-containing neurons
in the central nervous system. II. Experimentally induced changes intraneuronal amine levels
of bulbo-spinal neuron systems. Acta Physiol. Scand. 64 (suppl. 247), 1-36.

63
Dempsey, E.W., Morison, R.S., 1943. The electrical activity of a thalamo-cortical relay
system. Am. J. Physiol. 138, 283-296.
Dolleman-Van Der Weel, M.J., Lopes da Silva, F.H., Witter, M.P., 1997. Nucleus reuniens
thalami modulates activity in hippocampal field CA1 through excitatory and inhibitory
mechanisms. J. Neurosci. 17, 5640-5650.
Edelstyn, N.M.J., Mayes, a R., Ellis, S.J., 2012. Damage to the dorsomedial thalamic nucleus,
central lateral intralaminar thalamic nucleus, and midline thalamic nuclei on the right-side
impair executive function and attention under conditions of high demand but not low demand.
Neurocase. 37-41.
Falck, B., Hillarp, N.A., Thieme, G., Torp, A., 1962. Fluorescence of catecholamines and
related compounds condensed with formaldehyde. J. Histochem. Cytochem. 10, 348-354.
Felten, D.L., Laties, A.M., Carpenter, M.B., 1974. Monoamine-containing cell bodies in the
squirrel monkey brain. Am. J. Anat. 139, 153-166.
Felten, D.L., Sladek, J.R., 1983. Monoamine distribution in primate brain. V. Monoamine
distribution in primate brain. V. Monoaminergic nuclei: anatomy, pathways and local
organization. Brain Res. Bull. 10, 171-284.
Franco-Pérez, J., Ballesteros-Zebadúa, P., Custodio, V., Paz, C., 2012. Major
neurotransmiters involved in the regulation of sleep-wake cycle. Rev. Invest. Clin. 64, 182-
191.
Freedman, L.J., Cassel, M.D., 1991. Tralamic afferents of the rat infralimbic and lateral
agranular cortices. Brain Res. Bull. 26, 957-64.
Gaddum, J.H., Picarelll, Z.P., 1957. Two kinds of tryptamine receptor. Brit. J. Pharmacol. 12,
232.
Glennon, R.A., 1990. Serotonin receptors: Clinical implications. Neurosci. Biobehav. Rev.
14, 35-47.
Glimcher, P.W., Lau, B., 2005. Rethinking the thalamus. Nature Neurosci. 8, 983-984.
Goaillard, J.M., Vicent, P., 2002. Serotonin suppresses the slow afterhyperpolarization in rat
intralaminar and midline thalamic neurones by activating.. Physiol. 541, 453-465.
Goodwin, G.M., Souza, R.J., Green, A.R., 1985. The pharmacology of the hypothermic
response in mice to 8-hydroxy-2-(di-n-pro-p ylamino) tetralin (8-OH-DPAT). A model of
presynaptic 5-HT1 function, Neuropharmacology. 24, 1187-1194.
Groenewegen, H.J., Berendse, H.W., 1994. The specificity of the ‘nonspecific’ midline and
intralaminar thalamic nuclei. Trends Neurosci. 17, 52-57.
Groenewegen, H.J., Witter, M.P., 2004. Thalamus, in: Paxinos, G. (Ed.), The Rat Nervous
System, 3rd
ed., Elsevier Academic Press, New York., 407-453.

64
Gustafson, E.L., Durkin, M.M., Bard, J.A., Zgombick, J., Branchek, T.A., 1996. A receptor
autoradiographic and in situ hybridization analysis of the distribution of the 5-HT7 receptor in
rat brain. Brit. J. Pharmacol. 117, 657-666.
Halbach, O.V.B.U., Dermietzel, R., 2006. Serotonin (5-Hydroxytryptamine). In:
Neurotransmitters and Neuromodulators: Handbook of receptors and Biological Effects.
Weinheim: Wiley-VCH., 132-142.
Hallanger, A.E., Levey, A.I., Lee, H.J., Rye, D.B., Wainer, B.H., 1987. The origins of
cholinergic and other subcortical afferents to the thalamus in the rat. J.Comp. Neurol. 162,
105-124.
Hamlin, K.E., Fischer, F.E.,1951. The synthesis of 5-hydroxytryptamine. Journal of J. Am.
Chem. Soc. 73, 5007-5008.
Harding, A., Paxinos, G., Halliday, G., 2004. The serotonin and tachykinin systems, in:
Paxinos G. (Ed.), The Rat Nervous System, 3rd ed., Elsevier Academic Press, New York.,
1205-1256.
Haroian, A.J., Massopust, L.C., Young, P.A., 1981. Cerebellotralamic projections in the rat:
an autoradiographic and degeneration study. J. Comp. Neurol. 197, 217-236.
Hautvast, R.W., Ter Horst, G.J., De Jong, B.M., De Jongste, M.J., Blanksma, P.K., Paans,
A.M. Korf, J., 1997. Relative changes in regional cerebral blood flow during spinal cord
stimulation in patients with refractory angina pectoris. Eur. J. Neurosci. 9, 1178-1183.
Heimer, L., 2003. A new anatomical framework for neuropsychiatric disorders and drug
abuse. Am. J. Psych. 160, 1726-1739.
Hembrook, J.R., Mair, R.G., 2011. Lesions of reuniens and rhomboid thalamic nuclei impair
radial maze win-shift performance. Hippocampus. 21, 815-26.
Herkenham, M., 1978. The connections of the nucleus reuniens thalami: Evidence for a direct
thalamo – hippocampal pathway in the rat. J. Comp. Neurol. 177, 589-610.
Hoover, W.B., Vertes, R.P., 2012. Collateral projections from nucleus reuniens of thalamus to
hippocampus and medial prefrontal cortex in the rat: a single and double retrograde
fluorescent labeling study. Brain Struct. Funct. 217, 191-209.
Hornung, J.P., Fritschy, J.M., 1988. Serotonergic system in the brainstem of the marmoset: a
combined immunocytochemical and three-dimensional reconstruction study. J. Comp. Neurol.
270, 471-487.
Hsu, D.T., Price, J.L., 2009. Paraventricular thalamic nucleus: subcortical connections and
innervation by serotonin, orexin, and corticotropin-releasing hormone in macaque monkeys. J.
Comp. Neurol. 512, 825-48.
Hunsperger, R.W., Roman, D., 1976. The integrative role of the intralaminar system of the
thalamus in visual orientation and perception in the cat. Exp. Brain Res. 25, 231-246.

65
Itoh, K., Konish, A., Nomura, S., Mizuno, N., Nakamura, Y., Sugimoto, T., 1979. Application
of coupled oxidation reaction to electron microscopic demonstration of horseradish
peroxidase: cobalt-glucose oxidase method. Brain Res. 175, 341-346.
Jacobs, B.L., Azmitia, E.C., 1992. Structure and function of the brain serotonin system.
Physiol. Rev. 72, 165-229.
Jacobs, B.L., Gannon, P.J., Azmitia, E.C., 1984. Atlas of serotonergic cell bodies in the cat
brainstem: an immunocytochemical analysis. Brain Res. Bull. 13, 1-31.
Jasper, H.H., 1960. Unspecific thalamocortical relations, in: J. Field, H.W. Magoun, V.E. Hall
(Eds.), Handbook of Physiology, Am. Physiol. Soc. 2, 1307-1321.
Johannsen, P., Jakobsen, J., Bruhn, P., Hansen, S.B., Gee, A., Stodkilde-Jorgensen, H.,
Gjedde, A., 1997. Cortical sites of sustained and divided attention in normal elderly humans,
Neuroimage. 6, 145-155.
Jones, E.G., 2007. The Thalamus, Second Edition (NewYork, NewYork: Cambridge
University Press).
Jones, E.G., Hendry, S.H.C., 1989. Differential calcium binding protein immunoreactivity
distinguishes different classes of relay neurons in monkey thalamic nuclei. Eur. J. Neurosci.
1, 222-246.
Jones, E.G., Leavitt, R.Y., 1974. Retrograde axonal transport and demonstration of non-
specific projections to the cerebral cortex and striatum from thalamic intralaminar nuclei in
the rat, cat e monkey. J. Comp. Neurol. 154, 349-378.
Julius, D., 1991. Molecular biology of serotonin receptors. Annual Review Neuroscience. 14,
335-60.
Kaitz, S.S., Robertson, R.T., 1981. Thalamic connections with limbic cortex. II.
Corticothalamic projections. J. Comp. Neurol. 195, 527-545.
Kandel, E.R., Schwartz, J.H., Jessell, T.M., 2000a. The Functional Organization of Perception
and Movement. In: Kandel E.R., Schwartz, J.H., Jessell, T.M., Principles of neural science. 4ª
ed. McGraw-Hill, New York, 337-349.
Kandel, E.R., Schwartz, J.H., Jessell, T.M., 2000b. Disorders of mood: depression, mania and
anxiety disorders. In: Kandel E.R., Schwartz, J.H., Jessell, T.M., Principles of neural science.
4ª ed. McGraw-Hill, New York, 1216-1219.
Kayama, Y., Shimada, S., Hishikawa, Y., Ogawa, T., 1989. Effects of stimulating the dorsal
raphe nucleus of the rat on neuronal activity in the dorsal lateralgeniculate nucleus. Brain Res.
489, 1-11.
Kia, H.K., Miquel, M.C., Brisorgueil, M.J., Daval, G., Riad, M., El Mestikawy, S., Hamon,
M., Verge, D., 1996. Immunocytochemical localization of serotonin1A receptors in the rat
central nervous system. J. Comp. Neurol. 365, 289-305.
Kilpatrick, G.J., Jones, B.J., Tyers, M.B., 1987. Identification and distribution of 5-HT3
receptors in rat brain using radioligand binding. Brain Res. Bull. 13,l-31.

66
Kosofsky, B.E., Molliver, M.E., 1987. The serotoninergic innervation of cerebral cortex:
different classes of axon terminals arise from dorsal and median raphe nuclei. Synapse. 1,
153-268.
Krieg, W.J.S., 1944. The medial region of the thalamus of the albino rat. J. Comp. Neurol. 80,
381-415.
Krout, K.E., Loewy, A.D., Westby, G.W., Redgrave, P., 2001. Superior colliculus projections
to midline and intralaminar thalamic nuclei of the rat. J. Comp. Neurol. 431,198-216.
Krout, K.E., Belzer, R.E., Loewy, A.D., 2002. Brainstem projections to midline and
intralaminar thalamic nuclei of the rat. J. Comp. Neurol. 448, 53-101.
Kung, J.C., Shyu, B.C., 2002. Potentiation of local field potentials in the anterior cingulate
cortex evoked by the stimulation of the medial thalamic nuclei in rats. Brain Res. 953, 37-44.
Lacher Jr., T.E., 1981. The comparative social behavior os Kerodon rupestris and Galea
spixii and the evolution of behavior in the caviidea. Bull. Carnegie Museum Nat. 17, 1-71.
Larsson, K., Ahlenius, S., 1999. Brain and sexual behavior, Ann. NY Acad. Sci. 877, 292–
308.
Lavoie, B., Parent, A., 1991. Serotoninergic innervation of the thalamus in the primate: an
immunohistochemical study. J. Comp. Neurol. 312, 1-18.
Lee, K., McCormick, D.A., 1996. Abolition of spindle oscillations by seotonin and
norepinephrine in the ferret lateral geniculate and perigeniculate nucleus in vitro. Neuron. 17,
309-321.
Li, S., Kirouac, G.J., 2008. Projections from the paraventricular nucleus of the thalamus to the
forebrain, with special emphasis on the extended amygdala. J. Comp. Neurol. 506, 263-287.
Li, S., Kirouac, G.J., 2012. Sources of inputs to the anterior and posterior aspects of the
paraventricular nucleus of the thalamus. Brain Struct. Funct. 217, 257-73.
Lidov, H.G.W., Molliver, M.E., 1982. Immunohistochemical study of the development of
serotonergic neurons in the rat CNS. Brain Res. Bull. 9, 559-604.
Lopez-Gimenez, J.F., Vilaro, M.T., Palacios, J.M., Mengod, G., 1998. [3H]MDL 100,907
labels 5-HT2A serotonin receptors selectively in primate brain. Neuropharmacology. 37,
1147-1158.
Lopez-Gimenez, J.F., Vilaro, M.T., Palacios, J.M., Mengod, G., 2001. Mapping of 5-HT2A
receptors and their mRNA in monkey brain: [3H]MDL100,907 autoradiography and in situ
hybridization studies. J. Comp. Neurol. 429, 571-589.
Lowry, C.A., Hale, M.W., Evans, A.K., Heerkens, J., Staub, D.R., Gasser, P.J.,
Shekhar, A., 2008. Serotonergic systems, anxiety and affective disorder. Stress,
neurotrans-mitters, and hormones. Ann. N.Y. Acad. Sci. 1148, 86-94.

67
Magalhães, C.P., Freitas, M.F.L., Nogueira, M.I., Campina, R.C.F., Marks, G.A.,
Speciale, S.G., Cobbey, K., Roffwarg, H.P.,1987. Serotonergic inhibition of the dorsal lateral
geniculate nucleus. Brain Res. 18, 76-84.
Massone, F., 2008. Anestesiologia Veterinária: Farmacologia e Técnicas, 1ª Ed. Guanabara
Koogan, Rio de Janeiro, 571p.
McCormick, B.Y.D.A., Wang, Z., 1991. Serotonin and noradenalin e excite GABAergic
neuronesof the guinea-pig and cat nucleus reticularis thalami. J. Physiol. 442, 235-255.
McCormick, D.A., 1992. Neurotransmitter actions in the thalamus and cerebral cortex and
their role in neuromodulation of thalamocortical activity. Prog. Neurobiol. 39, 337-388.
McCormick, D.A., Pape, H.C., 1990. Noradrenergic and serotonergic modulation of a
hyperpolarization-activated cation current in thalamic relay neurons. J. Physiol. 431, 319-342.
McKenna, J. T., Vertes, R. P., 2004. Afferent projections to nucleus reuniens of the thalamus.
J. Comp. Neurol. 480, 114-142.
Meltzer, H.Y., 1990. Role of serotonin in depression, Ann. NY Acad. Sci. 600, 486–500.
Mendes, B.V., 1985. Alternativas tecnológicas para a agropecuária do semi-árido. Nobel, São
Paulo, 171 p.
Mendes, B.V., 1987. Plantas e animais para o nordeste, 1ª Ed. Globo, Rio de Janeiro, 167 p.
Mengod, G., Vilaro, M.T., Raurich, A., Lopez-Gimenez, J.F., Cortes, R., Palacios, J.M., 1996.
5-HT receptors in mammalian brain: receptor autoradiography and in situ hybridization
studies of new ligands and newly identified receptors. Histochem. J. 28, 747-758.
Michelsen, K.A., Prockaerts, J., Steinbusch, H.W.M., 2008. The dorsal raphe nucleus and
serotonin: implications for neuroplasticity linked to major depression and Alzheimer’s
disease. Prog. Brain Res. 172, 233-264.
Miller, J.W., Ferrendelli, J.A., 1990. Characterization of gabaergic seizure regulation in the
midline thalamus. Neuropharmacology. 29, 649-655.
Moga, M.M., Weis, R.P., Moore, R.Y.M., 1995. Efferent projections os the paraventricular
thalamic nucleus in the rat. J. Comp. Neurol. 359, 221-238.
Monckton, J.E., McCormick, D.A., 2001. Neuromodulatory Role of Serotonin in the Ferret
Thalamus. J. Neurophysiol. 87, 2124-2136.
Montgomery, S., 1995. Serotonin, sertraline and depression. J. Psychopharmacol. 9,179-184
Monti, J.M., 2011. Serotonin control of sleep-wake behavior. Sleep Med. Rev. 15, 269-281.
Moojen, J., 1952. Os roedores do Brasil. Rio de Janeiro: Mininstério da Educação e Saúde/
Instituto Nacional do Livro. 214 p. (biblioteca científica brasileira. Série A - II).

68
Morin, L.P., Meyer-Bernstein, E.L., 1999. The ascending serotonergic system in the hamster:
comparison with projections of the dorsal and median raphe nuclei. Neuroscience. 91, 81-105.
Morgane, P.J., Galler, J.R., Mokler, D.J., 2005. A review of systems and networks of the
limbic forebrain/limbic midbrain. Prog. Neurobiol. 75, 143-160.
Moruzzi, G., Magoun, H.W., 1949. Brain stem reticular formation and activation of the EEG.
Electroencephalogr. Clin. Neurophysiol. 1, 455-473.
Newman, D.B., Ginsberg, C.Y., 1994. Brainstem reticular nuclei that project to the thalamus
in rats: A retrograde tracer study. Brain Behav. Evol. 44, 1-39.
Noga, B.R., Johnson, D.M.G., Riesgo, M.I., Pinzon, A., 2009. Locomotor-Activated Neurons
of the Cat. I. Serotonergic Innervation and Co-Localization of 5-HT7, 5-HT2A, and 5-HT1A
Receptors in the Thoraco-Lumbar Spinal Cord. J. Neurophysiol. 102, 1560-1576.
Nordberg, A., 1992. Neuroreceptor changes in Alzheimer disease, Cerebrovasc. Brain Metab.
Rev. 4, 303-328.
Novak, C.M., Harris, J.A., Smale, L., Nunez, A.A., 2000. Suprachiasmatic nucleus
projections to the paraventricular thalamic nucleus in nocturnal rats ( Rattus norvegicus) and
diurnal nile grass rats (Arviacanthis niloticus). Brain Res. 874, 147-157.
Osman, F.H., Ammar, E.M., 1975. 5-hydroxytryptamine receptors in uterine smooth
muscle. Jpn. J. Pharmacol. 25, 631-637.
Otake, K., Nakamura, Y., 1995. Sites of origin of corticotropin-releasing factor-like
immunoreactive projection fibers to the paraventricular thalamic nucleus in the rat, Neurosci.
Lett. 201, 84-86.
Otake, K., Ruggiero, D.A., 1995 Monoamines and nitric oxide are employed by afferents
engaged in midline thalamic regulation. J. Neurosci. 15, 1891-1911.
Panula, P., Pirvola, U., Auvinem, S., Airaksinen, M. S., 1989. Histamine – imunorreactive
nerve fibers in the rat brain. Neuroscience. 28, 586-610.
Parent, A., 1981. Comparative anatomy of the serotoninergic systems. J. de Physiologie 77,
147-156.
Parent, A. 1996. Human neuroanatomy. 9 ed. Canada: Willians & Wilkins.
Parent, A., Descarries, L., Beaudet, A., 1981. Organization of ascending serotonin systems in
the adult rat brain. A radioautographic study after intraventricular administration of [3H]5-
HT. Neuroscience. 6, 115-138.
Paus, T., Zatorre, R.J., Hofle, N., Caramanos, Z., Gotman, J., Petrides, M., Evans, A.C., 1997.
Evans, Time-related changes in neural systems underlying attention and arousal during the
performance of an auditory vigilance task. J. Cogn. Neurosci. 9, 392-408.
Paxinos, G., Watson, C., 2007. The rat brain in stereotaxic coordinates. 2ª Ed. Academic
Press, San Diego.

69
Pazos, A., Cartes, R., Palacios, J.M., 1985. Quantitative autoradiographic mapping of
serotonin receptors in the rat brain. 11. Serotonin-2 recep- tors. Brain Res. 346, 231-249.
Pazos, A., Probst, A., Palacios, J.M., 1987a. Serotonin receptors in the human brain. 111.
Autoradiographic mapping of serotonin-1 receptors. Neuroscience. 21, 97-122.
Pazos, A., Probst, A., Palacios, J.M., 1987b. Serotonin receptors in the human brain. IV.
Autoradiographic mapping of serotonin-2 receptors. Neuroscience. 21,123-139.
Peng, Z.C., Bentivoglio, M., 2004. The thalamic paraventricular nucleus relays information
from the suprachiasmatic nucleus to the amygdala: a combined anterograde and retrograde
tracing study in the rat at the light and electron microscopic levels. J. Neurocytol. 33, 101-
116.
Peng, Z.C., Grassi-Zucconi, G., Bentivoglio, M., 1995. Fos-related protein expression in the
midline paraventricular nucleus of the rat thalamus: basal oscillation and relationship with
limbic efferents. Exp Brain Res. 104, 21-29.
Peschanski, M., Besson, J.M., 1984. A spino-reticulo-thalamic pathway in the rat: an
anatomical study with reference to pain transmission. Neuroscience. 12, 165-178.
Politis, M., Loane, C., 2011. Serotonergic dysfunction in Parkinson’s disease and its relevanc
to disability. Scientif. World J. 11, 1726-1734.
Pompeiano, M., Palacios, J.M., Mengod, G., 1994. Distribution of serotonin 5-HT2 receptor
family mRNAs: comparison between 5-HT2A and 5-HT2C receptors. Mol. Brain Res. 23,
163-178.
Rainville, P., Duncan, G.H., Price, D.D., Carrier, B., Bushnell, M.C., 1997. Pain affect
encoded in human anterior cingulate but not somatosensory cortex. Science. 277, 968-971.
Ramón y Cajal, S., 1911. Histologie du Systeme Nerveux de L’homme et des Vertebres. Vol
II, Paris.
Rapport, M.M., 1949. Serum vasoconstrictor (Serotonin). V. The presence of creatinine in the
complex. A proposed structure of the vasoconstrictor principle. J. Biol. Chem.180, 961-969.
Richardson, B.P., Engel, G., 1986. The pharmacology and function of 5-HT receptors. Trends
Neurosci. 9, 424-428.
Roberts, M., Malini, A.K.E., Deal, M., 1984.The reprodutive biology of the rock cavy,
Kerodon rupestris, in captivity: A study of reproductive adaptation in a trophic specialist.
Mammalia. 48, 253-266.
Rogawski, M.A., Aghajanian, G.K., 1980. Norepinephrine and serotonin: opposite effects on
the activity of lateral geniculate neurons evoked by optic pathway stimulation. Exp. Biol. 69,
678-694.
Rowe, D.L., Honeycutt, R.L., 2002. Phylogenetic relationships, ecological correlates, and
molecular evolution within the Cavioidea (Mammalia, Rodentia). Mol. Biol. Evol. 19, 263-
277.

70
Sakai, K., Crochet, S., 2001. Differentiation of presumed serotonergic dorsal raphe neurons
in relation to behavior and wake-sleep states. Neuroscience. 104, 1141-1155.
Sanchez-Vives, M.V., Bal, T., Kim, U., Von Krosigk, M., McCormick, D.A., 1996. Are the
interlaminar zones of the ferret LGNd actually part of the perigeniculate nucleus? J. Neurosci.
16, 5923-5941.
Scheibel, M.E., Scheibel, A.B., 1967. Structural organization of nonspecific thalamic nuclei
and their projection toward cortex. Brain Res. 6, 60-94.
Schlag, J., Schlag-Rey, M., 1984. Visuomotor functions of central thalamus in monkey. II.
Unit activity related to visual events, targeting, and fixation. J. Neurophysiol. 51, 1175-1195.
Silva Neto, E.J., 2000. Morphology of the regions ethmoidalis and orbitotemporalis in Galea
musteloides Meyen 1832 and Kerodon rupestris (Wied-Neuwied 1820) (Rodentia: Caviidae)
with comments on the phylogenetic systematics of the Caviidae. J. Zool. Syst. Evol. Res. 38,
219-229.
Soares, J.G., Cavalcanti, J.R.L.P, Oliveira, F.G., Pontes, A.L.B., Sousa, T.B., Freitas, L.M.,
Cavalcante, J.S., Nascimento Júnior, E.S., Cavalcante, J.C., Costa, M.S.M.O., 2012. Nuclear
organization of the serotonergic system in the brain of rock cavy (Kerodon rupestris). J.
Chem. Neuroanat. 43, 112-119.
Sousa, R.A., Menezes, A.A.L., 2006. Circadian rhythms of motor activity of the Brazilian
rock cavy (Kerodon rupestris) under artificial photoperiod, Biol. Rhythm. Res. 37, 443-450.
Steinbusch, H.W., 1981. Distribution of serotonin-immunoreactivity in the central nervous
system of the rat-cell bodies and terminals. Neuroscience. 6, 557-618.
Steinbusch, H.W.M., Verhofstad, A.A., Joosten, H.W., 1978. Localization of serotonin in the
central nervous system by immunohistochemistry: description of a specific and sensitive
technique and some applications. Neuroscience. 3, 811-819.
Stuart, A.M., Mitchell, I.J., Slater, P., Unwin, H.L.P., Crossman, A.R., 1986. A semi-
quantitative atlas of 5-hydrolcytryptamine-1 receptors in the primate brain. Neuroscience. 18,
619-639.
Su, H. S., Bentivoglio, M., 1990. Thalamic midline cell populations projecting to the nucleus
accumbens, amygdala, and hippocampus in the rat, J. Comp. Neurol. 297, 582-593.
Swanson, L.W., Petrovich, G.D., 1998. What is the amygdala? Trends Neurosci. 21, 323-331.
Taber, E., Brodal, A., Walberg, F., 1960. The raphe nuclei of the brain stem in the cat. I.
Normal topography and cytoarchitecture and general discussion. J. Comp. Neurol. 114, 161-
187.
Takada, M., Campbell, K.J., Moriizumi, T., Hattori, T., 1990. On the origin of the
dopaminergic innervation of the paraventricular thalamic nucleus. Neurosci. Lett. 115, 33-36.
Takase, L. F., Barone, J. R., Nogueira, M. I., 2000. Involvement of the caudal raphe nuclei in
the feeding behavior of rats. Braz. J. Med. Biol. Res. 33, 223-228.

71
Takase, L.F., Nogueira, M.I., 2008. Patterns of fos activation in rat raphe nuclei during
feeding behavior. Brain Res. 1200, 10-18.
Takeuchi, Y., Kimura, H., Sano, Y., 1982. Immunohistochemical demonstration of the
distribution of serotonin neurons in the brainstem of the rat and the cat. Cell Tissue Res. 224,
247-267.
Törk, I., 1985. Raphe nuclei and serotonin containing systems, in: Paxinos, G. (Ed.), The rat
nervous system. Ed Academic Press Australia, Sidney. 43-78.
Törk, I., 1990. Anatomy of the serotonergic system. Ann. N. Y. Acad. Sci. 600, 9-35.
Van der Werf, Y.D., Witter, M.P., Groenewegen, H.J., 2002. The intralaminar and midline
nuclei of the thalamus. Anatomical and functional evidence for participation in processes of
arousal and awareness. Brain Res. Rev. 39, 107-140.
Vertes, R.P., 1991. A PHA-L analysis of ascending projections of the dorsal raphe nucleus in
the rat. J. Comp. Neurol. 313, 643-668.
Vertes, R.P., 1992. PHA-L analysis of projections from the supramammillary nucleus in the
rat, J. Comp. Neurol. 326, 595-622.
Vertes, R.P., 2002. Analysis of projections from the medial prefrontal cortex to the thalamus
in the rat, with emphasis on nucleus reuniens. J. Comp. Neurol. 442, 163-187.
Vertes, R.P., 2006. Interactions among the medial prefrontal cortex, hippocampus and midline
thalamus in emotional and cognitive processing in the rat. Neuroscience. 142, 1-20.
Vertes, R.P., Fortin, W.J., Crane, A. M., 1999. Projections of the median raphe nucleus in the
rat. J. Comp. Neurol. 407,555-58.
Vertes, R.P., Hoover, W.B., 2008. Projections of the paraventricular and paratenial nuclei of
the dorsal midline thalamus in the rat. J. Comp. Neurol. 508, 212-237.
Vertes, R.P., Hoover W.B., Valle, A.C., Sherman A., Rodriguez J.J., 2006. Efferent
projections of reuniens and rhomboid nuclei of the thala-mus in the rat. J. Comp. Neurol. 499,
768-796.
Vertes, R.P., Linley, S.B., 2007. Comparison of projections of the dorsal and median raphe
nuclei, with some functional considerations. International Congress Series. 1304, 98-120.
Vertes, R. P., Linley, S. B., 2008. Efferent and afferent connections of the dorsal and median
raphe nuclei in the rat. In: Monti, J.M., Pandi-Perumal, S.R., Jacobs, B.L., Nutt, D.J. (eds)
Serotonin and sleep: molecular, functional and clinical aspects. Birkhauser, Basel,
Switzerland. 69-102.
Vertes, R.P., Linley, S.B., Hoover, W.B., 2010. Pattern of distribution of serotonergic fibers
to the thalamus, Brain Struct. Funct. 215, 1-28.
Vertes, R.P., Martin, G.F., 1988. Autoradiographic analysis of ascending projections from the
pontine and mesencephalic reticular formation and the median raphe nucleus in the rat. J.
Comp. Neurol. 175, 511-541.

72
Viana di Prisco, G., Vertes, R.P., 2006. Excitatory actions of the ventral midline thalamus
(rhomboid/reuniens) on the medial prefrontal cortex in the rat. Synapse. 60, 45-55.
Vizuete, M.L., Venero, J.L., Traiffort, E., Vargas, C., Machado, A., Cano, J., 1997.
Expression of 5-HT7 receptor mRNA in rat brain during postnatal development. Neurosci.
Lett. 227, 53–56.
Waeber, C., Dixon, K., Hoyer, D., Palacios, J.M., 1988. Localization by autoradiography of
neuronal 5-HT3 receptors in the mouse CNS. Eur. J. Pharmacol. 15, 351-352.
Yamasaki, D.S.G., Krauthamer, G.M., Rhoades, R.W., 1986. Superior collicular projection to
intralaminar thalamus in rat. Brain Res. 378, 223-233.
Yoshida, M., Sasa, M., Takaori, S., 1984. Serotonin-mediated inhibition from dorsal raphe
nucleus of neurons in dorsal lateral geniculate and thalamicreticular nuclei. Brain Res. 290,
95-105.
Zhang, D.X., Bertram, E.H., 2002. Midline thalamic region: widespread excitatory input to
the entorhinal cortex and amygdala. J. Neurosc. 22, 3277-3284.





![Serotonina Medicina [Modo de Compatibilidade] · 14/11/2013 1 serotonina profa. sharlene lopes pereira universidade federal de juiz de fora instituto de ciÊncias biolÓgicas departamento](https://img.document.onl/doc/110x75/5bdbc9ae09d3f263278c8a5e/serotonina-medicina-modo-de-compatibilidade-14112013-1-serotonina-profa.jpg)























