Embed Size (px)
Citation preview

Papel da Fosfolipase A2 cálcio independente nas vias endocítica e exocítica de Leishmania
amazonensis
A N N E C R I S T I N E S I L V A F E R N A N D E S
Dissertação apresentada ao Programa de Pós-Graduação em Ciências (Microbiologia) do Instituto de Microbiologia Paulo de Góes da Universidade Federal do Rio de Janeiro, como parte dos requisitos necessários à obtenção do título de Mestre em Ciências (Microbiologia).
Orientador (a): Thaïs Souto-Padrón
Rio de Janeiro Julho de
1

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

FICHA CATALOGRÁFICA
2
Fernandes, Anne Cristine Silva
O papel da fosfolipase A2 cálcio independente nas vias endocítica/exocítica de
Leishmania amazonensis/ Anne Cristine Silva Fernandes– Rio de Janeiro, Instituto de
Microbiologia Prof. Paulo de Góes, UFRJ, 2010.
xiv, 66f.
Dissertação (Mestrado em Ciências Biológicas) - UFRJ/ Instituto de Microbiologia Prof.
Paulo de Góes/ Programa de Pós-graduação em Ciências (Microbiologia), 2010.
Orientadora: Thaïs Souto-Padrón
Referências bibliográficas: f . 57 - 66
1. Leishmania amazonensis 2. Fosfolipase A2 cálcio independente 3. Bromoenol lactona
4. Via endocítica/exocítica 5. Microscopia eletrônica 6. Citometria de fluxo I. Souto-
Padrón, Thaïs. II. UFRJ/ Instituto de Microbiologia Prof. Paulo de Góes, mestrado em
ciências biológicas. Programa de Pós-graduação em Ciências (Microbiologia), 2010. III.
“O papel da fosfolipase A2 cálcio independente nas vias endocítica/exocítica de
Leishmania amazonensis”.
.

Anne Cristine Silva Fernandes
O papel da fosfolipase A2 cálcio independente nas vias endocítica/exocítica de Leishmania amazonensis
Rio de Janeiro, 15 de julho de 2010
(Thais Souto-Padron, sua titulação Institudo de Microbiologia prof Paulo de Góes)
(Antônio, sua titulação Institudo de Microbiologia prof Paulo de Góes)
(Rosiane, sua titulação e Institudo de Biofísica Carlos Chagas Filho)
(Técia, sua titulação e Institudo de Biofísica Carlos Chagas Filho)
(Elvira Maria Saraiva, sua titulação Institudo de Microbiologia prof Paulo de Góes)
3

O presente trabalho foi realizado no Laboratório de Biologia Celular e Ultraestrutura,
Departamento de Microbiologia Geral, Instituto de Microbiologia Prof. Paulo de Góes (IMPPG),
Centro de Ciências da Saúde (CCS), Universidade Federal do Rio de Janeiro, sob a orientação do(a)
Profª. Thaïs Souto-Padrón.
4

Num piscar de olhos lá estou eu, na seleção do mestrado e num minuto depois, lá estava eu, “passei!”. Sem pausa, chega agosto e começa a correria, experimento para um lado, disciplinas para o outro, mas arranja um espacinho, pois ainda tem os seminários e o treinamento didático. Não pisca, pois um ano passa e você tem que defender seu projeto. Corre, dá um jeitinho, pois ainda tem os congressos, até porque o mundo também quer ver o seu trabalho, mas atenção, não pare de ler e estudar nem por um segundo, pois em um segundo “n” novas publicações surgem e você não pode ficar para trás, até porque o mestrado é apenas o início. Que início longo, 2 anos. Pode parecer longo na teoria, mas na prática voa e quando você se dá conta já está na hora de escrever a dissertação. Corre, corre, seu tempo está acabando, não se esqueça: chore, se desespere, não durma, entre em pânico, praticamente suma, isso faz parte. Falta um mês, entrega para o revisor, 10 dias, 1 semana, entrega para a banca, a apresentação... espero que já esteja pronta. Noite anterior. Noite ou dia? Alguém dorme? Chega o grande dia, você defende. UFA acabou, mas o que predomina é a realização e o gostinho de quero mais, então começa a torcida para que venha o doutorado.
Anne Cristine Silva Fernandes
5

Esta tese é dedicada primeiramente a Deus,
Por ser minha esperança, meu refúgio e minha fortaleza. Obrigado por estar presente em todos os momentos da minha vida e por me fazer chegar até aqui, sem Ti não sou nada. Te amo!“Entrego a Ele o meu futuro, pois sei que estou seguro nas promessas do meu Deus”.
6

Esta tese é dedicada aos meus pais, Luiz e Emília.
Dedico a vocês todas as vitórias da minha vida, pois devo a vocês tudo o que sou até hoje, que fizeram destes 23 anos, muito mais que uma simples vida, me fazem viver um sonho do qual eu não quero acordar nunca. Amos vocês mais que tudo nessa vida.
7

AGRADECIMENTOS
Primeiramente gostaria de agradecer a Thaïs, por fazer desse meu sonho realidade, por todos os ensinamentos, que só aumentam minha admiração, por esclarecer TODAS as minhas dúvidas, por toda a dedicação mesmo em momentos difícies, nesses últimos meses admiro cada vez mais, sua
garra, coragem e determinação. Obrigada por tudo.
As Cristines da minha vida, Caroline (Ká), Emanuele (Nú) e Gabriele (Bi), obrigada por todo carinho, apoio, paparicação, brincadeiras, confidências, empréstimos de roupa (com certeza o Fundão ficou mais bonito com a ajudinha do armário de vocês). Obrigado por vocês serem as
melhores irmãs do mundo e por estarem sempre comigo. Amo vocês mais que tudo nessa vida, vocês são meus exemplos, quando “crescer” quero ser igual a vocês, sem vocês não teria chegado
até aqui.
Como te agradecer, toda a atenção, compreensão, paciência, força, carinho. Obrigado por nesses dois anos você ter sido mais que um namorado, e muitas vezes ter confiado mais em mim do que eu
mesma. Obrigado por toda digitação, help no inglês, almocinhos diários, obrigada por me aturar chorando, rindo, em pânico, desesperada, feliz, triste, irritada, sempre com toda a calma e com a famosa frase “calma, eu resolvo para você” e sempre deu certo, você sempre resolveu TUDO.
Te amooo muito Caiq!
As duas pessoas mais incríveis que nos fazem hoje ser oito, Fabio e Thiago.Fabitcho, obrigado por todo o design, aulas de photoshop, ajudinhas no inglês e dedinho para
relaxar. Said brother!!! Torto, obrigado pelos toques, com certeza minha voz está menos estridente agora pelos seus comentários “delicados”. Amo vocês.
A melhor família que uma pessoa pode ter, tios, tias, primos, primas e Avós, aqui ou no céu. Vocês são a base da minha vida, meu porto seguro. Amo muuuuito todos vocês
Elas, as de todo o dia, o dia todo. Camila, Moara, Roberta, Cyntia e Gabi. Agradeço a vocês por toda a diversão, besteiras, ajuda teórico pratica, companheirismo, puxão de orelha, ombro amigo.
Mesmo vocês me perguntando até hoje quem me trouxe, eu sei que vocês me amam e com certeza o laboratório não seria a mesma coisa sem mim, mas posso afirmar que sem vocês ao meu lado não
conseguiria ter chegado até aqui.
Luana, Natália, Marcela, Juliana, Elcia, Patrícia, Gabriela, Bizinha, Mizulene, Teka, Bia, Camilas e Clara. Obrigado por vocês me acharem a pessoa mais inteligente do mundo. E obrigado por toda a amizade de muuuito tempo, confidências, conselhos, que me fortaleceram para chegar até aqui.
A turma de farmácia UFRJ 2004/2, obrigado por serem mais que uma turma, serem uma segunda família, amo todos vocês, mas especialmente queria agradecer as farmacêuticas mais lindas desse mundo: Alicia, Letícia, Mariah e Taissa, obrigado por essa amizade que não tem fim. E como não
agradecer ao pessoal papo de pimba major, Stephan, Mariana, Said, Luzia, Kassia, Lidi, Geisa, Bruno. E claro ele, Fernandinho, obrigado por toda a amizade, conselhos e por ter aturado todos
meus surtos. Amo todos vocês.
A minhas companheiras de Recreio X Fundão: Lívia, Ana Clarissa, Bia e Rô, obrigado pelas conversas variadas que transformam 2h de trânsito em pura diversão.
A professora Elvira por toda a ajuda, ensinamento e revisão da dissertação. Muito obrigada.
8

Aos professores Norton, Zé Roberto, Rosângela, Ângela, Ulysses, Obrigada por toda a disponibilidade, sugestões e ensinamentos.
A todos meus professores da graduação, que me despertaram esse amor pela UFRJ e por este mundo acadêmico.
Deived, obrigado por aturar minhas ligações semanais e por toda Leishmania cedida. Carol, obrigada por todos os toques com os géis. Alexandre, obrigado pelas dosagens. Fernanda, obrigada
pelos toques de microscopia. Rafael, obrigada pela atividade da 3`nucleotidase.
Venício, obrigado por adoçar meus dias com suas balinhas e por toda a ajuda com as pirâmides e com os probleminhas no MET.
Obrigada a todos que contribuíram nessa fase tão importante da minha vida.
9

RESUMOAnne Cristine Silva Fernandes
PAPEL DA FOSFOLIPASE A2 CÁLCIO INDEPENDENTE NAS VIAS ENDOCÍTICA E EXOCÍTICA DE Leishmania amazonensis
Orientadora: Thaïs Souto-PadrónResumo da Dissertação de Mestrado submetida ao Programa de Pós-Graduação em Ciências (Microbiologia), Instituto de Microbiologia Prof. Paulo de Góes da Universidade Federal do Rio de Janeiro, como parte dos requisitos necessários para a obtenção do título de Mestre em Ciências Biológicas.
Compartimentos das vias endocítica e exocítica em protozoários da família Trypanosomatidae apresentam características estruturais e funcionais distintos daqueles descritos para os mesmo compartimentos em células de mamíferos. A dinâmica desses compartimentos é de importância fundamental em processos como a secreção, remoção e exposição de moléculas na superfície do parasita, assim como a modulação processo de adesão, de virulência e defesa contra o sistema imune do hospedeiro. Embora a estrutura e a composição de muitos compartimentos das vias endo / exocítica em Leishmania foram elucidadas, pouco se sabe sobre os mecanismos que controlam o reconhecimento e afusão de membranas. Várias proteínas entre elas algumas enzimas, como a fosfolipase A2 (PLA2), estão envolvidos no processo de fusão de membrana de compartimentos no citoplasma em diferentes células. Neste estudo, nós investigamos o efeito de Bromoenol lactona (BEL), um inibidor específico e irreversível da fosfolipase A2 cálcio independentes (iPLA2), na dinâmica e ultra-estrutura dos compartimentos das vias endo/exocítica em Leishmania amazonensis. Os parasitos foram cultivados por 72h em meio de Schneider, incubadas por 1h com 2,5 µM de BEL e analisados por microscopia eletrônica, imunofluorescência, citometria de fluxo (FACS), SDS-PAGE, immunoblotting, e ensaios bioquímicos. BEL promoveu mudanças significativas na estrutura de Golgi com o alargamento e a fragmentação de suas cisterna. BEL também alterou o aspecto e distribuição dos túbulos agregados e vesículas translúcidas na região anterior do corpo perto da bolsa flagelar, a estrutura e a dimensão do TMV e a distribuição intra-extracelular da atividade da fosfatase ácida. BEL inibiu a atividade endocítica de promastigotas. Parasitos tratados, incubados por 2hs na presença de BSA-TRITC, Transferrina-FITC e ConcanavalinaA-FITC e analisadas por citometria de fluxo mostraram cerca de 60 e 75% de inibição da atividade endocítica da fase fluida (BSA) e da endocitose mediada por receptor (transferrina) , respectivamente. Não houve mudança na endocitose de Concanavalina A. BEL também mudou a atividade exocítica dos parasitos. Observamos uma redução de aproximadamente 24% na atividade secretora detectado pela concentração de proteína total pelo método de Bradford. Análise do perfil de polipeptídeos secretado por SDS-PAGE mostrou uma redução de 48% na intensidade das bandas determinada por densitometria após o tratamento com BEL. A secreção de gp63 detectado por immunoblotting, e da fosfatase ácida, detectada por ensaio bioquímico da atividade da enzima, foram reduzidas em 38% e 43%, respectivamente. Os resultados sugerem que iPLA2 está envolvido no controle da fusão de diferentes compartimentos da via endocítica e exocítica em Leishmania.
Palavras-chave: 1. Leishmania amazonensis 2. Fosfolipase A2 cálcio independente 3. Bromoenol lactona 4. Via endocítica/exocítica 5. Microscopia eletrônica 6. Citometria de fluxo.
Rio de Janeiro Julho de 2010
10

ABSTRACT
Anne Cristine Silva FernandesROLE OF CALCIUM INDEPENDENT PHOSPHOLIPASE A2 IN THE PATHWAYS ENDOCYTIC
AND EXOCYTIC OF Leishmania amazonensis
Orientadora: Thaïs Souto-Padrón
Resumo da Dissertação de Mestrado submetida ao Programa de Pós-Graduação em Ciências (Microbiologia), Instituto de Microbiologia Prof. Paulo de Góes da Universidade Federal do Rio de Janeiro, como parte dos requisitos necessários para a obtenção do título de Mestre em Ciências Biológicas.
Compartments of the endocytic and exocytic pathways in protozoans of the Trypanosomatidae family present structural and functional characteristics distinct from those described for the same compartments in mammalian cells. The dynamics of these compartments is of fundamental importance in processes such as secretion, removal and exposure of molecules on the surface of the parasite thus modulating process like adhesion, virulence, and defense against the host immune system. Although the structure and composition of many compartments of the endo/exocytic pathway in Leishmania have been elucidated, very little is known about mechanisms controlling their recognition and fusion. Several proteins among them some enzymes, such as phospholipase A2 (PLA2), are involved in the process of fusion of membrane bound compartments in the cytoplasm in different cells. In this study, we investigated the effect of Bromoenol lactone (BEL), a specific and irreversible inhibitor of a calcium independent phospholipase A2 (iPLA2), in the dynamics and ultrastructure of the compartments of the endo/exocytic pathways in Leishmania amazonensis. Parasites were grown for 72h in Schneider medium, incubated for 1 h with 2.5 mM of BEL and analyzed by electron microscopy, immunofluorescence, flow cytometry (FACS), SDS-PAGE, immunoblotting, and biochemical assays. BEL promoted significant changes in Golgi structure with enlargement and fragmentation of its cisterna. BEL also changed the aspect and distribution of the clustered tubules and translucent vesicles in the anterior region of the body near the flagellar pocket, the structure and size of MVT and the intra-and extracellular distribution of the acid phosphatase activity. BEL inhibited the endocytic activity of promastigotes. Treated parasites incubated for 2hs in the presence of BSA-TRITC, Transferrin-TRITC and ConcanavalinaA-FITC and analyzed by flow cytometry showed approximately 60 and 75% inhibition of endocytic activity of fluid phase (BSA) and of receptor-mediated endocytosis (transferrin), respectively. There were no change in the endocytosis of Concanavalin A. BEL also changed the exocytic activity of the parasites. We observed a reduction of approximately 24% in secretory activity detected by total protein concentration by the Bradford method. Analysis by SDS-PAGE profiles of secreted polypeptides showed a 48% reduction in the intensity of the bands determined by densitometry after treatment with BEL. The secretion of gp63 detected by immunoblotting, and of acid phosphatase, detected by the biochemical detection of enzyme activity, were reduced by 38% and 43%, respectively. The results suggest that iPLA2 is involved in the control fusion of different compartments of the endocytic and exocytic pathway in Leishmania.
Palavras-chave: 1. Leishmania amazonensis 2. calcium-independent phospolipase A2 3. Bromoenol lactone 4. Endocytic and exocytic pathways 5. Electron microscopy 6. Flow cytometr
Rio de Janeiro Julho de 2010
11

ÍNDICE
1. INTRODUÇÃO..............................................................................................................................1
1.1. Tripanosomatídeos....................................................................................................................1
1.2. Leishmania e Leishmaniose......................................................................................................2
1.3. O papel da via secretória e endocítica na modificação da superfície celular...........................6
1.4. O controle de fusão de vesículas...........................................................................10
1.4.1 As proteínas Rab.........................................................................................................10
1.4.2. As PI3-cinases.............................................................................................................12
1.4.3. As fosfolipases.............................................................................................................13
1. 5 Inibidores da Fosfolipase A2 independente de cálcio (iPLA2)...............................................15
1.6. Tripanosomatídeos – o que sabemos sobre os compartimentos envolvidos em processos
endocíticos/exocíticos e dos mecanismos moleculares que os controlam..................................................16
1.6.1 Distribuição de compartimentos da via endocítica/exocítica em tripanosomatídeos....................17
1.6.1.1 Via endocítica/exocítica em T. brucei...........................................................................18
1.6. 1.2 Via endocítica/exocítica em T. cruzi............................................................................19
1.6. 1.3 Via endocítica/exocítica em Leishmania......................................................................20
1.6. 1.4 O controle da fusão de vesículas em Tripanosomatídeos.............................................22
1.7 Fosfatase ácida: atividade presente nas vias endocítica/exocítica de Leishmania................................23
2. OBJETIVOS................................................................................................................................25
2.1. Objetivo...................................................................................................................................25
2.2. Objetivos específicos..............................................................................................................25
3. MATERIAIS E MÉTODOS.......................................................................................................26
3.1. Parasitos..................................................................................................................................26
3.2. Materiais.................................................................................................................................26
3.3 Curva de Crescimento na presença de Bromoenol lactona (BEL)..........................................26
3.4. Tratamento com Bromoenol lactona (BEL) ...........................................................................26
3.4.1 Controle com DMSO.............................................................................................................26
3.5. Dosagem de proteína...............................................................................................................27
3.6. Endocitose...............................................................................................................................27
3.7. Ensaio de Viabilidade.............................................................................................................27
3.8. Eletroforese em gel de poliacrilamida sob condições desnaturantes (SDS-PAGE)...............27
12

3.9 Transferência das proteínas para membrana de nitrocelulose...............................................28
3.10. Immunoblotting...................................................................................................................28
3.11. Atividade fosfatase ácida .....................................................................................................29
3.12. Citometria de Fluxo..............................................................................................................29
3.14. Microscopia Eletrônica de Transmissão Convencional.......................................................30
3.15 Citoquimica da atividade da enzima fosfatase ácida.............................................................31
3.16. Análises estatísticas.............................................................................................................30
4. RESULTADOS..................................................................................32
4.1 Ação da BEL sobre o cerscimento de L. amazonensis............................................................33
4.2. Ensaios de Viabilidade...........................................................................................................33
4.3. Análise ultraestrutural...........................................................................................................33
4.4. Interferência da BEL na aquisição de moléculas pelo parasito............................................37
4.5. Efeito da BEL sobre a rota secretora e a secreção protéica.................................................42
5. DISCUSSÃO.........................................................................................50
5. CONCLUSÃO.........................................................................................56
6. REFERÊNCIAS BIBLIOGRÁFICAS.............................................57
13

LISTA DE ABREVIATURAS
LPG -lipofosfoglicano
Gp63 - glicoproteina de 63kDa
GPI - glicosilfosfatidilinositol
PPG - proteofosfoglicano
RE -retículo endoplasmático
CG -complexo de Golgi
LDL-lipoproteína de baixa densidade
Aps - proteínas adaptadoras
BF -bolsa flagelar
EI/ ET - endossoma inicial/tardio
TGN - trans-Golgi-Network
Rab -família de pequenas GTPase
SNARE -receptor de SNAPs
SNAPS -proteína solúvel de ligação ao NSF
NSF -proteína de fusão sensível a N-etilmaleimida
NEM -N-etilmaleimida
GTP/ GDP – trifosfato/difosfato de guanidina
ATP - trifosfato de adenosina
EEA1 -autoantígeno 1 de endossomo inicial
PL - fosfolipase
PLA -fosfolipase A
iPLA2-fosfolipase A2 cálcio independente
sPLA2 - fosfolipase A2 secretada
cPLA2 - fosfolipase A2 cácio dependente
VSG -glicoproteína variante de superfície
Tbclh - cadeia pesada de clatrina
MVB -corpos multivesiculares
TMV -tubulo mutivesiculares
BEL -bromoenol lactona
MET - microscopia eletrônica de transmissão
14

1. INTRODUÇÃO
Tripanosomatídeos tem sido modelo de estudos em nosso laboratório durante vários anos
devido a características, tanto estruturais pela presença de organelas típicas, como fisiológica pela
relação com os diferentes hospedeiros. Nesta dissertação em particular utilizamos a Leishmania
como modelo de estudo por motivos que serão mostrados a medida que desenvolvermos o tema.
1.1. Tripanosomatídeos
Os tripanosomatídeos compreendem um grupo de protozoários classificados em uma única
ordem, Kinetoplastida, que abriga duas famílias de protozoários, os Bodonidae e a
Trypanosomatidae, em função da presença de uma estrutura subcelular típica denominada
cinetoplasto, que é um complexo arranjo de fibrilas do DNA mitocondrial, denominado kDNA
(MEYER, 1968; Revisado por DE SOUZA, 2002a). O ciclo biológico dos tripanosomatídeos é
extremamente variado conforme a espécie e, pode depender de mais de um tipo de hospedeiro,
sendo classificado em monoxênicos e heteroxênicos.
Os tripanosomatídeos heteroxênicos compreendem um grande grupo de protozoários, alguns
dos quais causam importantes doenças em diversos mamíferos incluindo os humanos. Dentre os
tripanosomatídeos podemos citar o Trypanosoma brucei, agente causador da doença do sono no
continente Africano, o Trypanosoma cruzi, agente causador da doença de Chagas na América
Central e do Sul e Leishmania spp., representada por diferentes espécies causadoras de 4 diferentes
tipos de Leishmaniose nas regiões tropicais e subtropicais. Todos esses parasitos são transmitidos
por insetos vetores e ao invadir os hospedeiros vertebrados podem completar seu ciclo de vida no
espaço extracelular (parasitos extracelulares; Ex.: sangue - T. brucei) e uma gama de diferentes
tipos de células (parasitos intracelulares obrigatórios; Ex.: T. cruzi e Leishmania).
A presença dos parasitos em diferentes ambientes durante o ciclo de vida acarreta em
mudanças morfológicas e fisiológicas específicas (HOARE & WALLACE, 1996). As variações
morfológicas dão origem às diferentes formas de desenvolvimento. A definição das diferentes
formas do parasito é baseada na forma do corpo celular, na posição do cinetoplasto em relação ao
núcleo e na região do corpo do parasito a partir do qual o flagelo emerge. Além das alterações na
forma, a sobrevivência dos parasitos em diferentes ambientes extra e intracelulares também exige
mudanças moleculares, como por exemplo, a presença de moléculas de superfície específicas para
cada forma em desenvolvimento. Essas mudanças exigem a síntese e direcionamento para a
superfície através de um transporte intracelular de glicoproteinas, glicolipídeos, proteínas
transportadoras, enzimas de superfície e receptores.
15

1.2. Leishmania e Leishmanioses
Os parasitos do gênero Leishmania são protozoários dimórficos, apresentam uma forma
flagelada extracelular, presente no trato digestivo do inseto-vetor, denominada promastigota e uma
forma intracelular, sem flagelo livre, denominada amastigota, encontrada no interior dos
fagolisossomos de macrófagos do hospedeiro vertebrado. Esses parasitos são os agentes causadores
da leishmaniose, uma doença que atinge milhões de pessoas anualmente em todo mundo (CHAPPUIS
et al, 2007). Até o momento foram descritas mais de 20 espécies do gênero Leishmania que se
dividem em espécies do Velho e do Novo Mundo e que apresentam um amplo espectro de
manifestações clínicas que vão de lesões cutâneas superficiais (Leishmaniose cutânea algumas de
cura espontânea), a lesões cutâneas mais profundas que caracterizam a forma mucocutânea
(Leishmaniose mucocutânea ou cutâneo-mucosa), leões cutâneas disseminadas e não ulcerativas
(Leishmaniose cutâneo-difusa) e a forma visceral (Leishmaniose viceral) e fatal da doença
(SILVEIRA et al, 2004; VAN GRIENSVEN et al, 2010). Os agentes etiológicos, sua localização e formas
da doença podem ser observados no quadro abaixo
O ciclo de vida deste parasito (Fig.1) no inseto vetor se inicia quando formas amastigotas
são ingeridas pelo flebotomínio durante o repasto sanguíneo em um hospedeiro vertebrado
Forma da Doença
Espécies
Velho Mundo Novo Mundo
Leishmaniose Cutânea
L. majorL. aethiopicaL. tropicaL. infantun
Complexo mexicanaL. amazonensisL. garnhamiL. mexicanaL. pifanoiComplexo braziliensisL. braziliensisL. colombiensis L. guyanensisL. lainsoniL. panamensisL. peruviana
Leishmaniose Mucocutânea
(somente descrita no Novo Mundo)
L. braziliensisL. peruviana
Leishmaniose Cutâneo Difusa L. aethiopica L. amazonensisL. mexicanaL. pifanoi
Leishmaniose Visceral
L. archibaldiL. donovaniL. infantumL. tropica
L. chagasiL. amazonensis
16

infectado. A etapa seguinte ocorre no interior da matriz peritrófica, formada principalmente por
quitina secretada pelas células do epitélio intestinal, onde se dá a digestão do sangue e a
transformação das formas amastigotas em promastigotas procíclicas. Os promastigotas procíclicos
que possuem o corpo celular alongado e o comprimento do flagelo semelhante ao tamanho do
corpo, se multiplicam ativamente. Após o rompimento da matriz peritrófica, por ação de uma
quitinase secretada pelos parasitos (SCHLEIN et al., 1991), alguns procíclicos se ligam às células
epiteliais do intestino pela inserção de seus flagelos entre as microvilosidades através da interação
do (lipofosfoglicano) LPG presente na superfísie do parasito a moléculas de galectina da célula
hospedeira. Após a completa eliminação dos restos da digestão, os promastigotas migram para a
região anterior do trato digestivo alcançando a probóscide. À medida que esta migração acontece,
as formas promastigotas procíclicas sofrem um processo de diferenciação denominado
metaciclogênese, em que os promastigotas passam de um estágio não-infectante (procíclico), para
um estágio infectante (metacíclico).
Somente formas metacíclicas são inoculadas no hospedeiro vertebrado pelo vetor no
momento da picada. No vertebrado, essas formas se aderem às células do sistema fagocítico
mononuclear, e são fagocitadas. Nos macrófagos, as promastigotas metacíclicas são direcionadas e
permanecem em um ambiente inóspito, o fagolisossoma, onde se diferenciam em formas
amastigotas que sobrevivem e se multiplicam. Com a multiplicação das formas amastigotas as
células hospedeiras se rompem liberando essa forma no meio extracelular que então infectam novas
células, mantendo assim o ciclo no hospedeiro vertebrado.
A diferenciação entre metacíclicos e procíclicos é acompanhada tanto por mudanças
morfológicas, os metacíclicos possuem corpo celular pequeno e delgado e um longo flagelo, como
por mudanças moleculares reguladas, principalmente em moléculas de superfície dos
promastigotas. Uma das mudanças mais significativas descritas até o momento são as alterações no
padrão de glicosilação LPG. Outras diferenças são observadas no perfil de proteínas de superfície,
sendo a mais expressiva aquela relacionada ao aumento na expressão de uma glicoproteína de
63kDa (gp63), a principal protease de superfície dos promastigotas que é ancorada a membrana por
uma âncora de glicosilfosfatidilinositol (GPI). Em algumas espécies como a Leishmania
braziliensis, o aumento de expressão da gp63 está correlacionado ao aumento da virulência
(KWEIDER et al, 1987).
17
Amastigota intracelular

Figura 1: Representação esquemática do ciclo celular, mostrando os hospedeiros invertebrado e vertebrado e as duas formas evolutivas. (1) Formas promastigotas metacíclicas são fagocitadas por células do hospedeiro vertebrado; no interior do fagolisossomo da célula hospedeira se diferenciam em amastigotas; (2) as formas amastigoas se multiplicam intensamente no interior desse vacúolo; (3) após sucessivas divisões a célula hospedeira é rompida e as formas infectivas são liberadas no meio extracelular, podendo infectar novas células (4) ou serem ingeridas pelo inseto vetor (5); (6) no intestino do hospedeiro invertebrado as formas amastigotas se diferenciam em formas promastigotas procíclicas; (7) essas se multiplicam, realizam metaciclogênese e migram em direção a probóscide, as formas infectivas serão inoculadas no próximo repasto sanguíneo (8)as formas metacíclicas se aderem à superfície dos macrófagos e são fagocitadas por essas células reiniciando o ciclo de desenvolvimento do parasito. (adaptado de CHAPPUIS et al.,200
Macrófago
Promastigotas metacíclicas
Promastigotas procíclicas
Fagocitose Lise
Proliferação
Re-invasão
Repasto sanguíneo do inseto vetor
Proliferação no intestino médio
Adesão
Repasto Sanguíneo do inseto vetor
(1)
(5)
(4)
(3)
(2)
(8)
(7)
(6)
Amastigota
18
Fagolisossomo

No caso das Leishmanias, o principal alvo no hospedeiro vertebrado são os macrófagos.
Após a adesão à superfície dos macrófagos, as formas metacíclicas são interiorizadas através da
fagocitose, formando o fagossoma que só irá se fundir ao lisossoma após a transformação de forma
promastigota metacíclicas em amastigotas, uma vez que a primeira não resistiria à ação das enzimas
lisossomais. Este atraso na fusão do fagossomo ao lisossomo se deve à presença de actina na forma
filamentosa que recobre o vacúolo formado e que se desfaz após completa transformação do
parasito em amastigota (RASMUSSOM & DESCOTEAUX, 2004). A modulação da formação do
fagolisossomo em algumas espécies de Leishmania parece ocorrer devido à ação do LPG presente
na superfície do parasito. O LPG é transferido do parasito para a membrana dos macrófagos durante
a fagocitose e induz a ação da F-actina inibindo a maturação do fagossomo. As propriedades
biofísicas do LPG sugerem que ele pode ser intercalado na membrana celular do hospedeiro
(LODGE & DESCOTEAUX, 2005; WINBERG et al., 2009).
Dentre as várias funções que a superfície de parasitos apresenta, usando como exemplo a
Leishmania, estão o reconhecimento e adesão à célula hospedeira seguida da invasão e
sobrevivência no interior das mesmas. Um dos principais grupos de moléculas relacionados às
funções citadas acima estão os carboidratos que constituem o glicocálice ou “coat” celular. Nas
Leishmanias as cadeias de açúcar que constituem o glicocálice estão principalmente associadas a
proteínas transmembrana, ancoradas na membrana através de âncoras de GPI, ou a lipídios
formando os glicolipídios como o LPG, próteofosfoglicano (PPG) e outros (FERGUSON, 1999).
A principal glicoproteína de superfície de Leishmania é a gp63 (glicoproteina de 63kDa)
que é uma metaloprotease dependente de zinco ancorada à membrana através de uma âncora de
GPI. Algumas funções específicas foram identificadas nesta proteína. A gp63 pode clivar proteínas
do sistema complemento inativando-as como, por exemplo, na clivagem de C3b para C3bi
(BRITTINGHAM, et al., 1995). Este procedimento facilita a sobrevivência das formas promastigotas
no meio extracelular além de mediar o reconhecimento dos parasitos por macrófagos de mamíferos
através do receptor CR3 e CR1 (YAO et al.,2003), entretanto estudos com mutantes em gp63
mostraram que a atividade enzima não é necessária para a ligação a CR3, sugereindo que esta
molécula pode estar ligada diretamente na interação com a célula hospedeira. A gp63 foi também
reconhecida como sendo um fator de virulência (CHANG, et al., 1990) estando sua ação relacionada
com a proteção do parasito sobre a ação de enzimas lisossomais e da atividade citotóxica de
macrófagos (ALEXANDER, et al., 1999). Assim, gp63 pode desempenhar um papel crucial no
reconhecimento da Leishmania pelos macrófagos, na resistência a lise pelo sistema complemento e
na sobrevivência de formas amastigotas no interior de macrófagos (CORRADIN, et al., 2002; YAO et
al.,2003 ).
19

Outro importante glicoconjugado de superfície sintetizado pelo parasito é o LPG que
também é ancorado a membrana via âncora de GPI. É o glicoconjugado mais abundante na
superfície dos promastigotas, e tem um papel importante em mediar interações parasito-inseto e
parasito-célula hospedeira além de promover a resistência dos metacíclicos à lise mediada pelo
sistema complemento (PUENTES et al., 1990) e de modular a transdução de sinal em macrófagos
gerando resistência ao ataque oxidativo permitindo que o parasito possa estabelecer uma infecção
com sucesso. Esta cobertura proporciona também proteção da forma promastigota contra a ação de
hidrolases extracelulares (PUENTES et al., 1990). Formas promastigotas e amastigotas do parasito
apresentam ainda outra molécula secretada, os PPG que podem facilitar a transmissão de parasitos
aos flebotomínios durante a picada (NAHIRE et al., 2004). Os PPGs secretados podem degradar
componentes do sistema complemento em lesões em mamíferos, através da inativação de
precursores da cascata (PETERS et al., 1997) e também modulam o número e o tamanho dos vacúolos
parasitóforos assim como a produção de citocinas por macrófagos (PIANI et al., 1999).
Durante o processo de diferenciação de promastigotas não infectivos em promastigotas
metacíclicas, e também entre promastigota e amastigota e durante a interação parasito-célula
hospedeira as proteínas de superfície, LPG, gp63, PPG e moléculas secretadas podem variar em sua
composição e/ou quantidades expressas o que implica em alta síntese, trânsito intracelular mediado
pela fusão de compartimentos da via exocítica e finalmente a exposição/secreção dessas moléculas
pelo mecanismo de exocitose (MORGAN et al, 2002b). Alterações no processo de reconhecimento
entre compartimentos intracelulares do parasito podem interferir na exposição de moléculas
inviabilizando a sobrevivência destes nos seus hospedeiros.
A existência de diferentes rotas endocíticas e exocíticas de moléculas com diferentes
caracteristicas estruturais torna o modelo Leishmania particulamente atrativo para o estudo do
controle de suas vias endocítica e exocítica.
1.3. O papel da via secretória e endocítica na modificação da superfície celular
A breve explanação sobre o ciclo do parasito deixa claro o papel da sua superfície no
processo de reconhecimento da/pela célula hospedeira assim como na sobrevivência em ambientes
agressivos em ambos os hospedeiros. Modificações na superfície celular pressupõem mecanismos
de aporte e remoção de componentes através de mecanismos como a endocitose, exocitose e
“shedding”.
As células eucarióticas desenvolveram um complexo sistema de compartimentos
citoplasmáticos que permite a captação, processamento e secreção de macromoléculas. A rota
biossintética-secretora possibilita que a célula sintetize moléculas, que a elas sejam adicionados os
açúcares, que sejam ligadas a âncoras lipídicas, que sejam estocadas e finalmente expostas ao meio
20

extracelular ou secretadas por um processo denominado exocitose. Ao mesmo tempo as moléculas
endocitadas seguem trajetos distintos através de compartimentos citoplasmáticos que podem ser
compartilhados pelas vias exocíticas.
Em células eucarióticas o clássico caminho secretório envolve o reconhecimento de uma
sequencia sinal em proteínas recém sintetizadas para serem exportadas, proteínas exocitadas são
sintetizadas no retículo endoplasmático (RE) e se movem ao longo do caminho secretório através de
transporte vesicular. Em eucariotos superiores estas vesículas são revestidas por uma camada de
proteínas do complexo II (COPII) que promovem o direcionamento de RE ao Compartimento
Intermediário entre o Retículo e o Golgi (ERGIC) (LEE et al., 2004). COPII é composto por pequenas
GTPase (Sar1), pelos complexos de proteínas interior (Sec23-Sec24) e exterior (Sec13-Sec31). A
exportação de proteínas transmembranares do RE baseia-se no reconhecimento pelo domínio
citosólico Sec24 do revestimento da vesícula COPII desencadeando sinais para o transporte. Em
contrapartida, proteínas ancoradas a membrana por âncoras de GPI não conseguem vincular
diretamente a Sec24 e devem utilizar receptores de carga ou mecanismos alternativos para a
exportação do RE. Pouco se sabe sobre o mecanismo subjacente à exportação de proteínas
ancorada a GPI do RE em eucariontes superiores. Porém resultados recentes sugerem que a
exportação de proteínas ancorada às GPI a partir do RE depende de proteínas transmembrana
denominadas Emp24p (complexos protéicos que permitem o ciclo entre RE, ERGIC e Golgi)
(JENNE et al., 2002) e da participação de “lipid rafts” presentes na membrana do RE (BONNON et al.,
2010). As moléculas recém sintetizadas transportadas do RE ao complexo de Golgi (G) sofrem
alterações pós-transducionais sendo encaminhados posteriormente ao seu destino final (SCHARZ &
DOBBERSTEIN, 1996; LEE et al., 2004).
Outro tipo de secreção é baseada na fusão de corpos multivesiculares à membrana
plasmática com a liberação de exossomos (KELLER et al., 2006). A liberação de exossomos já foi
descrita em diferentes tipos celulares e em diferentes situações como, por exemplo, células
infectadas com bactérias ou vírus (BHATNAGAR et al., 2007; NGUYEN et al., 2003), células tumorais
(GRANER et al., 2009), em Caenorhabditis elegans (LIEGEOIS et al., 2006) e Cryptococcus neoformans
(RODRIGUES et al., 2008).
Outra forma de secreção considerada atualmente é a remoção de componentes da superfície
pelo mecanismo de shedding, que pode ser definido como a liberação de constituintes da superfície
celular na forma solúvel ou associados a vesículas sem afetar a viabilidade celular (TARABOLETTI et
al., 2002). A liberação de constituintes solúveis depende da atividade proteolítica que cliva
ectodomínios das proteínas transmembrana enquanto que as vesículas são liberadas a partir da
membrana plasmática (DELLA SBARBA & RODOVIDA 2002).
21

Já foram descritos 4 tipos de processos endocíticos baseados no tamanho do material a ser
endocitado e nas espécies moleculares envolvidas na formação do compartimento responsável pela
endocitose. São eles: a fagocitose, a macropinocitose, a pinocitose e a endocitose mediada por
receptor (Fig.2). Abordaremos apenas os dois últimos processos, pois, são os únicos descritos em
tripanosomatídeos.
O termo pinocitose também conhecido como endocitose de fase fluida ou endocitose não
adsortiva é atribuído à aquisição de fluidos e moléculas solúveis através da formação de pequenas
vesículas que brotam da membrana em direção a região mais central da célula. A pinocitose é um
processo constitutivo que permite não só a obtenção de nutrientes do meio extracelular como
representa um dos mecanismos responsáveis pela manutenção do volume celular. Vesículas de
pinocitose se fundem a compartimentos da via endocítica (vide a seguir) onde descarregam seu
conteúdo.
Outro processo que também pemite a aquisição de moléculas só que desta vez de maneira
específica, é a endocitose mediada por receptor. Este processo, que proporciona maior eficiência na
aquisição de moléculas, está normalmente associado à captação de moléculas essenciais e é
conservado nos mais diferentes organismos eucarióticos. A endocitose mediada por receptor requer
especificidade de interação, envolvendo reconhecimento receptor-ligante. Na região onde ocorrem
estas interações iniciam-se diversos processos de associação de proteínas à face citoplasmática da
membrana plasmática, que levam à formação de um coat visível ao microscópio eletrônico de
transmissão (ROTH & PORTER, 1964). As regiões especializadas da membrana plasmática onde se
inicia o processo de endocitose mediada por receptor são denominadas “coated pits”. No lado
citoplasmático a proteína majoritária das “coated pits” é a clatrina que possui 190kDa, e que
promove não só a agregação de determinados tipos de receptores como os receptores ligantes de
LDL (low density lipoprotein), manose-6-fosfato, transferrina, como também promove a curvatura
da membrana plasmática levando à formação de vesículas. A essa região também se associam
outras proteínas como as proteínas adaptadoras (APs) e proteínas transmembrana (ROBINSON, 1987).
Uma vez estando a clatrina ligada ao complexo adaptador na membrana alvo, entram em
ação pequenas GTPases denominadas dinaminas. Sua atividade causa o estrangulamento da “coated
pits” e a formação de vesículas revestidas ou “coated vesicles”. No citoplasma, as “coated vesicles”
perdem rapidamente o seu revestimento de clatrina, expondo proteínas necessárias à fusão da
vesícula com a membrana dos endosomas (ROBINSON, 1994). Uma vez formadas, as vesículas
perdem rapidamente seu revestimento e se fundem ao Endossoma Inicial (EI) (YOUNG, BARRAL &
ULRICH HARTL, 2003). O endossomo inicial é o sítio de distribuição das moléculas internalizadas.
Estas podem ser direcionadas de volta à superfície diretamente do EI ou através de segundo
compartimento denominado Endossoma de Reciclagem (ER). Além das vias de reciclagem as
22
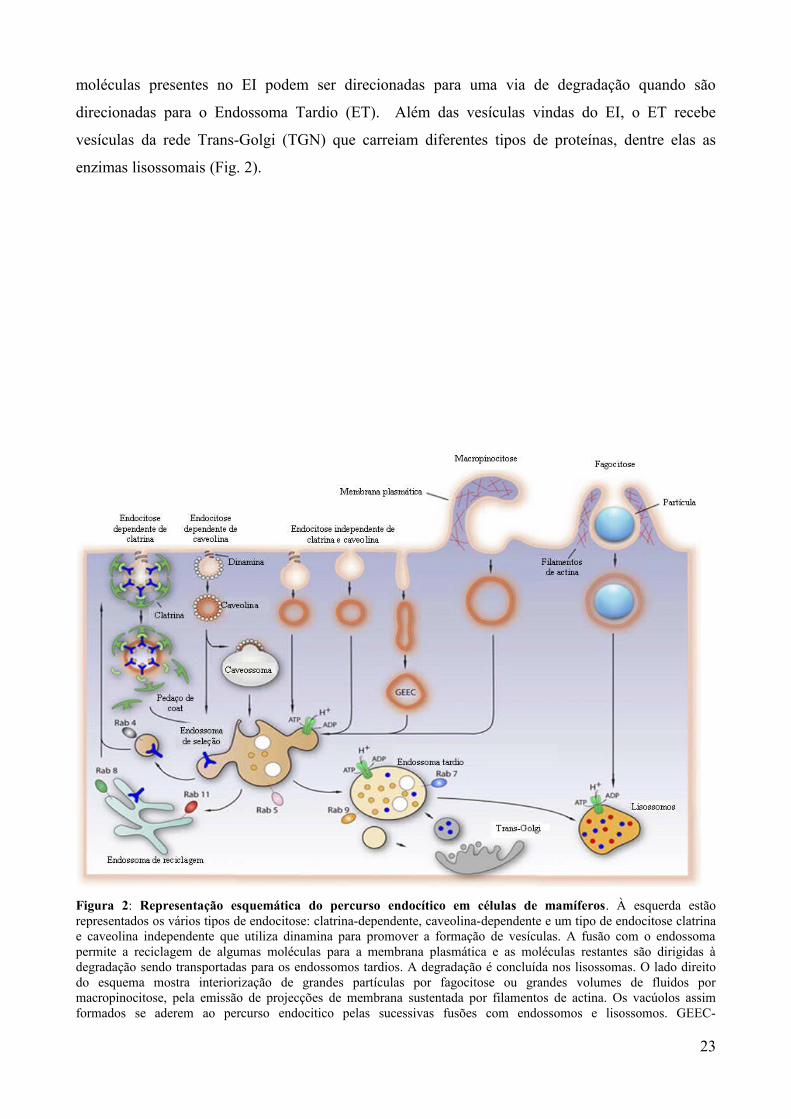
moléculas presentes no EI podem ser direcionadas para uma via de degradação quando são
direcionadas para o Endossoma Tardio (ET). Além das vesículas vindas do EI, o ET recebe
vesículas da rede Trans-Golgi (TGN) que carreiam diferentes tipos de proteínas, dentre elas as
enzimas lisossomais (Fig. 2).
Figura 2: Representação esquemática do percurso endocítico em células de mamíferos. À esquerda estão representados os vários tipos de endocitose: clatrina-dependente, caveolina-dependente e um tipo de endocitose clatrina e caveolina independente que utiliza dinamina para promover a formação de vesículas. A fusão com o endossoma permite a reciclagem de algumas moléculas para a membrana plasmática e as moléculas restantes são dirigidas à degradação sendo transportadas para os endossomos tardios. A degradação é concluída nos lisossomas. O lado direito do esquema mostra interiorização de grandes partículas por fagocitose ou grandes volumes de fluidos por macropinocitose, pela emissão de projecções de membrana sustentada por filamentos de actina. Os vacúolos assim formados se aderem ao percurso endocitico pelas sucessivas fusões com endossomos e lisossomos. GEEC-
23

Compartimento endossomal inicial enriquecido em proteínas ancoradas por GPI (DE SOUZA, SANT`ANNA. e CUNHA E SILVA, 2009).
A estação final da via endocítica é o lisossoma. Nele encontram-se todas as moléculas
destinadas à degradação. Os lisossomas possuem características comuns que são a grande variedade
de enzimas hidrolíticas, alta permeabilidade, um glicocálice com um alto ter de ácido neuramínico e
o pH em torno de 5,0 que é mantido por ATPases vácuolares (ESKELINEN, TANAKA & SAFTIG, 2003).
A transferência de carga entre o endossomo tardio e o lisossomo, é controversa. Bright e
colaboraores. (2005), mostraram que os dois compartimentos temporariamente se fundem, formando
uma organela híbrida, e posterior separação com re-estruturação do lisossomo.
Pouco tempo depois do descobrimento do revestimento de clatrina, Palade e Bruns (1968)
descreveram a presença de invaginações abundantemente distribuídas no endotélio relacionadas
com a passagem de material de uma extremidade a outra da célula através da formação de
caveossomos com fusão com os endossomas iniciais (PARTON E SIMONS, 2007). Rothberg e
colaboradores (1992) demonstraram que estas invaginações, denominadas cavéolas, possuíam um
revestimento granular, no lado citoplasmático da membrana, que se dispunha em forma de espiral.
Estes mesmos autores revelaram a presença de uma proteína de 28kDa, a caveolina, como sendo o
principal componente protéico deste revestimento. Existem evidências da presença de receptores
como o receptor para insulina (GOLDENBERG, SMITH & JARETT, 1987) e para toxina colérica e
tetânica que se concentram nas cavéolas, sugerindo um papel seletivo destas regiões. Demonstrou-
se que drogas que ligam colesterol alteram funcionalmente e morfologicamente as cavéolas,
sugerindo que estas organelas constituem um sítio de concentração e interiorização destes lipídeos
(PARTON E SIMONS, 2007). A endocitose mediada por cavéola também é conhecida como potocitose,
que é a aquisição de fluidos do meio extracelular através da região rica em caveolina.
1.4 - O controle de fusão de vesículas
A fusão de membranas é um evento chave no funcionamento de um organismo vivo. O
transporte de macromoléculas ao longo das vias endocítica e exocítica requer uma série de
eventos de fusão de membranas, que são coordenados por múltiplos fatores presentes tanto no
citosol quanto na membrana dos compartimentos. O processo de fusão envolve duas etapas, a
agregação de vesículas e a fusão propriamente dita. Já são conhecidas algumas proteínas que
participam deste processo algumas delas inclusive já descritas em tripanosomatídeos conforme
será comentado a seguir.
1.4.1. As proteínas Rab
24

As proteínas Rab constituem a maior família de pequenas GTPases monoméricas. Estas
proteínas estão envolvidas na regulação do tráfego de membranas, na fusão e no brotamento de
vesículas, assim como na reciclagem de membrana ao longo das vias endocítica e exocítica em
diferentes tipos celulares e na interação das vesículas com o citoesqueleto celular (ZERIAL &
MCBRIDE, 2001). Proteínas Rab atuam juntamente com moléculas efetoras e com componentes da
maquinaria SNARE (descrito mais adiante), para a iniciação do processo de fusão de membranas
dos compartimentos intracelulares. O princípio regulatório das proteínas Rab, assim como para
outras GTPases, está na habilidade em funcionar como um interruptor molecular, que oscila entre
sua conformação ligada ao GTP (forma ativa) ou ao GDP (forma inativa). Quando ligada ao GTP
esta proteína é endereçada para a membrana do compartimento onde atuará, já quando está ligada
ao GDP esta proteína encontra-se livre no citosol.
Cada proteína Rab apresenta uma distribuição intracelular específica, sendo assim estas
proteínas podem servir como importantes marcadores sub-celulares (ZERIAL & MCBRIDE, 2001).
Dentre elas, a Rab5 e a Rab4 encontram-se localizadas em domínios distintos de endossomas
iniciais. A Rab5 está intimamente envolvida no processo de fusão homotípica de endossomas
iniciais, enquanto que a Rab4 está envolvida na reciclagem de receptores para a membrana
plasmática. A Rab7 e a Rab9 estão localizadas nos endossomas tardios. Suas funções são as de
regular a fusão heterotípica de endossomas iniciais e tardios, e de vesículas provenientes do ET para
o CG, respectivamente. A Rab11 está localizada nos endossomas de reciclagem, atuando no
processo de reciclagem de receptores para a membrana plasmática (RODMAN & WANDINGER-NESS,
2000; TAKAI, SASAKI & MATOZAKI, 2001).
Além das proteínas Rab, um outro grupo de moléculas garante a especificidade aos eventos
de fusão de membranas intracelulares. A proteína de fusão sensível a N-etilmaleimida (NSF) que é
um fator importante para a realização de vários eventos de fusão intracelular. Esta molécula foi
originalmente identificada como um fator que iniciava a fusão de membrana do Golgi quando
tratadas com N-etilmaleimida (NEM) (BLOCK et al., 1988). NSF faz parte de uma família de
proteínas ligadoras de ATP, delineadas por uma ou mais regiões conservadas que correspondem aos
sítios de ligação para o ATP. Sendo assim, a hidrólise do ATP possui um papel muito importante na
fusão de vesícula com sua membrana alvo. NSF requer SNAPs (proteínas solúveis de ligação ao
NSF) para ligar-se às membranas (COLOMBO, 1996). NSF medeia a liberação de α-SNAP das
membranas dos endossomas numa etapa anterior a fusão dos compartimentos intracelulares.
A especificidade dos eventos de fusão intracelular se dá, pelo menos em parte, pelas
moléculas SNAREs (receptores de SNAPs), que possuem distribuição intracelular distinta. As
moléculas SNAREs mais relevantes são: Sinaptobrevina (v-SNARE), sintaxina 1 e SNAP-25 (t-
SNARE). Uma v-SNARE representa uma SNARE predominantemente associada com a vesícula 25
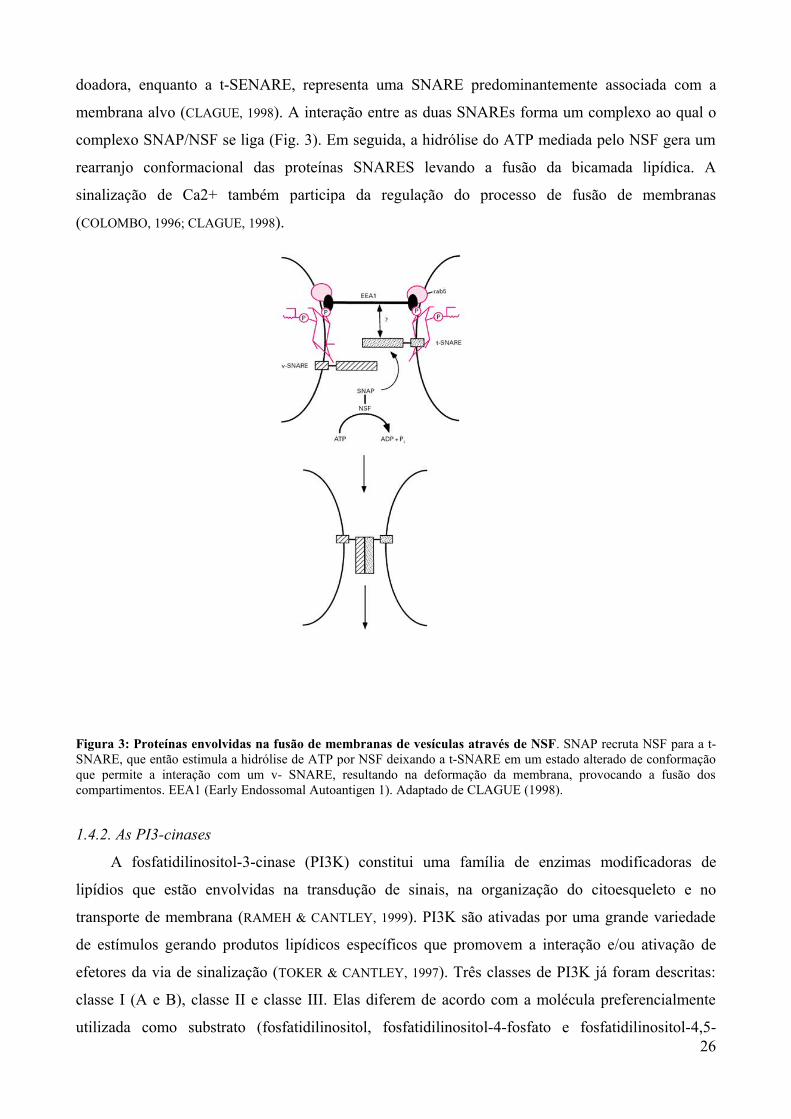
doadora, enquanto a t-SENARE, representa uma SNARE predominantemente associada com a
membrana alvo (CLAGUE, 1998). A interação entre as duas SNAREs forma um complexo ao qual o
complexo SNAP/NSF se liga (Fig. 3). Em seguida, a hidrólise do ATP mediada pelo NSF gera um
rearranjo conformacional das proteínas SNARES levando a fusão da bicamada lipídica. A
sinalização de Ca2+ também participa da regulação do processo de fusão de membranas
(COLOMBO, 1996; CLAGUE, 1998).
Figura 3: Proteínas envolvidas na fusão de membranas de vesículas através de NSF. SNAP recruta NSF para a t-SNARE, que então estimula a hidrólise de ATP por NSF deixando a t-SNARE em um estado alterado de conformação que permite a interação com um v- SNARE, resultando na deformação da membrana, provocando a fusão dos compartimentos. EEA1 (Early Endossomal Autoantigen 1). Adaptado de CLAGUE (1998).
1.4.2. As PI3-cinases
A fosfatidilinositol-3-cinase (PI3K) constitui uma família de enzimas modificadoras de
lipídios que estão envolvidas na transdução de sinais, na organização do citoesqueleto e no
transporte de membrana (RAMEH & CANTLEY, 1999). PI3K são ativadas por uma grande variedade
de estímulos gerando produtos lipídicos específicos que promovem a interação e/ou ativação de
efetores da via de sinalização (TOKER & CANTLEY, 1997). Três classes de PI3K já foram descritas:
classe I (A e B), classe II e classe III. Elas diferem de acordo com a molécula preferencialmente
utilizada como substrato (fosfatidilinositol, fosfatidilinositol-4-fosfato e fosfatidilinositol-4,5-26

bisfosfato, respectivamente), assim como na sua regulação. PI3Ks são importantes componentes no
tráfego de receptores da via endocítica de mamíferos e no transporte de vesículas do CG para os
vacúolos de leveduras (RAMEH & CANTLEY, 1999). A ancoragem e fusão de EI, que são regulados
pela Rab5, requerem a atividade da PI3K, esta enzima regula o recrutamento para membrana do
EEA1 (Autoantígeno 1 do EI), uma molécula efetora da Rab5 e um componente central da
maquinaria de ancoragem e fusão de endossomas iniciais (Fig.3). Além da EEA1, a PI3K também
atua como uma molécula efetora da Rab5 (CHRISTOFORIDIS et al, 1999). A Rab5 estimula a PI3K,
que faz com que o fosfatidilinositol-3-fosfato forme um domínio localizado na membrana do
endossoma que irá atuar como um sítio de ligação para os componentes da maquinaria de fusão de
membrana. Isto também contribui para a ligação de proteínas que irão atuar no processo de
motilidade dos endossomas ao longo dos microtúbulos.
1.4.3. As fosfolipases
Fosfolipases são enzimas hidrolíticas que clivam fosfolipídios. A posição de clivagem da
cadeia do glicerol identifica a família de fosfolipases. As principais fosfolipases são as fosfolipases
A (PLA), C (PLC) e D (PLD) (Fig.4).
A fosfolipase A2 (PLA2) é uma super-família de enzimas constituída por muitos membros.
Estas enzimas clivam fosfolipídios de membrana (sn-2 ésteres de glicerofosfolipídios) gerando dois
produtos, lisofosfolipídios e ácidos graxos livres. Existem diferentes tipos de PLA2 que são
distinguidas pelo seu peso molecular, dependência de íons cálcio, mecanismo hidrolítico, e no caso
das espécies de baixa massa molecular, pelo número das pontes dissulfeto e modo de secreção. As
fosfolipases A2 podem ser subdivididas em dois principais tipos: as PLA2 secretadas (sPLA2) e as
PLA2 intracelulares. As PLA2 secretadas são enzimas de baixa massa molecular (14 kDa) que
27
Figura 4: Representação esquemática do local de hidrólise das diversas fosfolipases (PL) em fosfolipídios de membrana. Adaptado de Brown e colaboradores (2003).

requerem concentrações milimolares de Ca2+ para exercer sua atividade enzimática. São sintetizadas
por estímulos, encontradas no interior de grânulos citoplasmáticos e então secretadas
extracelularmente. O grupo das sPLA2 parece estar relacionado ao desenvolvimento de alguns
processos patológicos, iniciando a produção de numerosos metabólitos que irão atuar no processo
inflamatório (YEDGAR, LICHTENBERG & SCHNITZER, 2000). Por outro lado, muitos estudos têm
sugerido o papel em potencial das sPLA2 (de diferentes grupos) em vários eventos biológicos não
relacionados com a produção de mediadores lipídicos. Estes eventos incluem o metabolismo de
lipoproteínas envolvido na aterosclerose, na hidrólise de surfactantes no pulmão e também na
imunidade inata contra infecções bacterianas (KINI, 2005).
As PLA2 intracelulares incluem dois grupos de enzimas: as PLA2s citosólicas (cPLA2) que
requerem concentrações micromolares de Ca2+ e as PLA2 independentes de cálcio (iPLA2). As
cPLA2s estão livres no citosol, porém podem ser translocadas para as membranas dos
compartimentos intracelulares dependendo do estímulo sofrido pelas células (MURAKAMI, 2002). As
iPLA2 estão localizadas tanto no citoplasma como em frações de membrana (AKIBA & SATO, 2004).
As iPLA2s são enzimas de 85-88 kDa que existem em forma de agregados e que estão classificadas
em grupos ( VIA-1, VIA-2, VIB, VIC, VID VIE e VIF). O grupo VIA é o grupo das PLA2s
independentes de cálcio mais bem caracterizado. Elas possuem atividade de lisofosfolipase, são
capazes de hidrolisar ambas as cadeias pequena e longa de fosfolipídios e exibir atividade de PAF
(fator ativador de plaquetas), acetilhidrolase, além de exercer sua atividade de fosfolipase A2. Sua
principal função celular é mediar o remodelamento dos fosfolipídios (WINSTEAD, BALSINDE &
DENNIS, 2000). Existem evidências de que o processo de fusão esta diretamente ligado a mudanças
na bicamada lipídica, sugerindo que as propriedades fisiológica dos lipídeos desempenhem um
papel importante no processo de fusão (ROUX et al., 2005). As modificações lipídicas pela enzima
PLA2 e especialmente a iPLA2, estão envolvidas em vários eventos de tráfego intracelular de
membrana, como a formação de tubulos de membrana a partir do CG e endossomos, e eventos de
fusão de membrana nos caminhos endocíticos e exocíticos (DE FIGUEIREDO et al., 2000). As iPLA2
geram ácido graxos livres e lisofosfolipídeos que alteram a curvatura da membrana e as
propriedades intrínsecas de multicomponentes membranares além de modificarem a tensão nas
regiões de “lipid raft”, sendo assim considerada uma indutora de fissão e fusão de vesículas. As
iPLA2 participam também no processo de apoptose. Sua ativação pela caspase-3, leva à produção de
moléculas de lisofosfatidilcolina que são secretadas pelas células apoptóticas gerando sinais
quimioatrativos para que células fagocíticas profissionais as reconheçam e as englobem (LAUBER et
aL, 2003; MURAKAMI & KUDO, 2004). Outra função descrita para as iPLA2s é a de restaurar o estoque
de Ca+2 intracelular, permitindo o influxo destes íons em células com baixo estoque dos mesmos
(MURAKAMI & KUDO, 2004).
28

A participação da enzima iPLA2 nos processos de tráfego de membranas esta relacionada à
produção de moléculas sinalizadoras que poderão estimular indiretamente ou inibir uma via de
transdução de sinais. Através da produção de lisofosfolipídios e ácido araquidônico (duas moléculas
que afetam a estabilidade da membrana), a iPLA2 pode também ter um efeito direto na estrutura e
função da membrana, influenciando mais intimamente os eventos de fusão entre compartimentos
intracelulares, como no caso dos endossomos (BROWN, CHAMBERS & DOODY, 2003). Estudos
utilizando uma série de inibidores da PLA2, mostraram que a fagocitose mediada por anticorpos
assim como a adesão e o espalhamento de macrófagos são bloqueados por esses inibidores. Este
efeito foi revertido quando ácido aracdônico foi adicionado ao meio, o mesmo não acontecendo
quando da adição de lisofosfolipídeos. Desta forma foi sugerido que o ácido aracdônico é
necessário juntamente com outros fatores para a fusão de endossomos (MAYORGA et al., 1993).
1.5 Inibidores da Fosfolipase A2 independente de cálcio (iPLA2)
Nos últimos anos tem havido grande interesse no estudo do metabolismo de fosolipídeos de
células intactas. Tal interesse decorre sobre tudo da descoberta que fosfolipídeos de membrana
celular podem não só interferir na estrutura de membranas biológicas, mas também participar na
formação de segundos mensageiros fundamentais na sinalização celular.
A abordagem mais simples para avaliar a função de uma enzima é através da inibição da sua
atividade utilizando inibidores específicos. Existem diferentes compostos que agem como potentes
inibidores de iPLA2, e estes podem ser protótipos para o desenvolvimento de drogas mais seletivas.
Inibidores da iPLA2, como a bromoenol lactona (BEL) e anestésicos locais como a dibucaína,
quinacrina e procaína alteram processos como a fagocitose mediada por anticorpos assim como a
adesão e o espalhamento de macrófagos (LENNARTZ et al., 1997), a endocitose e exocitose em
fibroblastos, células musculares e mastócitos (MICHALIK et al., 2003; HAGIWARA & OZAWA, 1990;
FENSOME-GREEN et al., 2007) e a fusão de compartimentos da via endocítica e do Golgi (MAYORGA
et al.,1993). A bromoenol lactona (BEL) é um membro de uma família de compostos conhecida
como lactonas haloenol (Fig.5) que foram primeiramente descritos como substratos suicidas de
quimiotripsina e serina proteases (FENSOME-GREEN et al., 2007). Posteriormente, foi relatado que esta
droga é um potente inibidor específico de iPLA2, que apresenta uma especificidade 1000 vezes
maior para a inibição de iPLA2 em comparação com cPLA2 (FENSOME-GREEN et al., 2007). Devido a
esta seletividade, BEL, tem sido utilizada para identificar o papel de iPLA2 em células. Foi descrita
também a ação da BEL sobre uma outra enzima envolvida no metabolismo de fosfolipídeos, uma
fosfohidrolase citosólica dependente de magnésio, a PAP1 (FUENTES et al., 2003).
Estudos com inibidores de PLA2 mostraram que estes alteram morfologicamente e
funcionalmente estruturas envolvidas no processo endocítico. Foi mostrado que esses inibidores
29

causam uma separação em largos fragmentos do complexo de Golgi, sugerindo que a arquitetura
deste requer a atividade de PLA2 (DE FIGUEIREDO et al., 1998). Posteriormente, De Figueiredo e
colaboradores (2001), mostraram que estes antagonistas inibiam a reciclagem de receptores e
moléculas endocitadas para a membrana plasmática, se acumulando em regiões da via endocítica de
acordo com a concentração do inibidor utilizada e também impediam a formação de túbulos em
compartimentos endossomais. Estes resultados sugerem que a PLA2 desempenha um papel geral na
regulação ou medeia diretamente os eventos de trafego de membrana em células de mamíferos.
Em Tripanosomatídeos pouco se sabe sobre a ação de inibidores da iPLA2 na vias
endocítica/exocítica. Souto-Padrón e colaboradores (2006) mostraram que a dibucaína, anestésico
local que também age como inibidor específico de iPLA2, foi capaz de reduzir a taxa de endocitose
de albumina (BSA), a velocidade de sua degradação e também a quantidade de cisteína proteinase
encontrada nos reservossomos sugerindo uma interferência da dibucaína na fusão de diferentes
compartimentos da via endocítica da forma epimastigota do T. cruzi.
Figura 5.: Estrutura química da bromoenol lactona (BEL), inibidor seletivo e irreversível de iPLA2.
1.6. Tripanosomatídeos – o que sabemos sobre os compartimentos envolvidos em processos
endocíticos/exocíticos e dos mecanismos moleculares que os controlam.
Em tripanosomatídeos os processos endocítico e exocítico têm sido mais bem estudados em
T. brucei, T. cruzi e Leishmania. A atividade endocítica não foi ainda identificada em
tripanosomatídeos monoxênicos. O modelo de endocitose em tripanosomatídeos desperta grande
interesse, pois como já mostrado, apresenta diferenças morfológicas e funcionais nítidas em relação
aos modelos já bem estabelecidos de células de mamíferos além de apresentarem diferenças entre as
espécies analisadas (MCCONVILLE et al., 2002).
Leishmania, T. brucei e T. cruzi tem pelo menos uma forma de desenvolvimento com alta
atividade endocítica e exocítica que são as formas promastigota, tripomastigota e epimastigota,
respectivamente. O processo endocítico nestes protozoários está relacionado à aquisição de
nutrientes além da evasão da resposta imune e virulência. A bolsa flagelar se caracteriza por ser a
30

região do corpo desses protozoários desprovida de microtúbulos subpeliculares (Fig.6) cuja
presença e arranjo molecular com proteínas fibrosas, formando um arcabouço no lado
citoplasmático da membrana do corpo celular, impedem a fusão e brotamento de vesículas. O
processo endocítico e exocítico através da bolsa flagelar foi identificado em Leishmania, T. brucei
e T. cruzi. Em T. cruzi, no entanto, e principalmente na forma epimastigota, a endocitose se dá
preferencialmente através de uma estrutura denominada citóstomo que leva o material endocitado
através de uma invaginação tubular, a citofaringe, até regiões mais centrais do corpo do parasito
onde vesículas que brotam da citofaringe se fundem a outros compartimentos da via endocítica
(PORTO-CARREIRO et al., 2000)
1.6.1. Distribuição de compartimentos da via endocítica/exocítica em tripanosomatídeos
Embora em alguns aspectos do tráfego endocítico sejam semelhantes nos tripanosomatídeos
citados, existem diferenças nítidas na organização estrutural dos compartimentos das suas via
endo/exocítica.
31
GD
F
E
A B C

Figura 6: Representação esquemática das organelas que compõem as vias endocítica e exocítica em tripanosomatídeos. A) compartimentos da via endocítica de formas tripomastigotas sanguínea de T. brucei; B) em formas promastigotas de Leishmania; e C) em formas epimastigotas de T. cruzi; D) microscopia eletrônica de transmissão enfatizando a formação de vesículas a partir da membrana flagelar em formas tripomastigotas do T. brucei. As cabeças de seta indicam a formação de uma coated-pit; E) microscopia eletrônica de transmissão da região anterior de promastigotas de L. mexicana; F) vesículas contendo coat em brotamento a partir da mmbrana da bolsa flagelar de Leishmania; G) ensaio de pulso e caça tendo como marcador o complexo BSA-ouro coloidal mostrando a sua concentração nos reservossomos das formas epimastigotas do T. cruzi (seta).Ac.: acidocalcissoma ; L: lisossoma;m: tubulo multivesicular; FL: flagelo; FP: bolsa flagelar; bz: zona de brotamento; sc: revestimento de superfície e CCP: revestimento marcado com anticorpo anti-clatrina. Figura adaptada de MCCONVILLE et al, 2002, PORTO-CARREIRO et al., 2000; DE SOUZA, SANT`ANNA. & CUNHA E SILVA, 2009.
1.6.1.1. Via endocítica/exocítica em T. brucei
Em T. brucei, o processo endocítico é o mais bem conhecido e é o que apresenta maior
semelhança com os processos endocíticos descrito em células de mamíferos. Nas formas
tripomastigotas do T. brucei, a bolsa flagelar e os compartimentos das vias endocítica e exocítica,
incluindo os lisossomos e o CG, estão localizados na região posterior do corpo do parasito. Estudos
em T. brucei têm mostrado que o processo pode envolver a presença de receptores de superfície
(ex.: transferrina e LDL), a fusão das vesículas endocíticas, EI (endossoma inicial), ET (endossoma
tardio) e lisossomos.
Os compartimentos celulares que fazem parte da via endocítica de formas tripomastigotas do
T. brucei pode ser visualizadas na figura 6. Macromoléculas presentes no meio extracelular são
endocitadas através da região da bolsa flagelar de onde brotam vesículas que se dirigem à região
mais central do corpo do parasito. No caso do T. brucei, estas vesículas se fundem a uma rede
interconectada de túbulos e cisternas que compõem o EI. Vesículas que brotam deste
compartimento poderão se fundir ao endossoma de reciclagem ou então ao ET. A partir deste
último compartimento duas rotas de tráfego ainda podem ser seguidas pelas moléculas endocitadas:
1) a rota de reciclagem, onde vesículas carreadoras que brotam do ET transportarão o material
endocitado até o ER; ou 2) a rota de degradação, com a subsequente fusão do ET ao lisossoma, que
é o compartimento final da via endocítica deste parasito (OVERATH & ENGSTLER, 2004).
Como já foi visto, tripanosomatídeos podem possuir receptores para auxiliar o processo de
endocitose. O mecanismo de endocitose mediado por receptor em T. brucei foi primeiramente
descrito por Coppens e colaboradores (1987), neste estudo a transferrina foi utilizada como traçador,
mostrando que a captura desta era rápida, saturável, sensível à temperatura e ao tratamento com
tripsina e limitada a região da bolsa flagelar. O receptor de transferrina de T. brucei é um
heterodímero codificado pela associação de dois genes associados ao sítio de expressão (ESAG6 e
ESAG7) (STEVERDING et al., 1994). Este não apresenta homologia com o receptor de transferrina 32

homodimérico de células de mamíferos. Normalmente os genes ESAGs são co-transcritos
juntamente com o gene da glicoproteína variante de superfície (VSG), como um grande transcrito
policistrônico. Como existem aproximadamente 20 sítios de expressão do gene da VSG (cada um
com pelo menos uma cópia do ESAG6 e ESAG7), o T. brucei pode, teoricamente, fazer 20
receptores de transferrina diferentes, porém somente um sítio de expressão está ativo em um
determinado momento (CHAVES et al., 1999). No T. brucei após ser capturado, o complexo
transferrina-receptor é transportado através de um sistema vesículo-tubular que está presente entre a
região da bolsa flagelar e o núcleo até chegar a um compartimento que é positivo para TbRab5A,
uma GTPase homóloga a Rab5 de mamífero (PAL et al. 2002) onde a transferrina libera a molécula de
ferro contida em sua estrutura, sendo uma parte das moléculas de transferrina degradada e outra
parte reciclada juntamente com seu receptor para a membrana plasmática como sugeriu Steverding
(2000).
Outra molécula que serve como marcador de processos de endocitose é o LDL que é uma
importante fonte de esteróis para os hemoflagelados. Proteínas específicas ligadoras de LDL
parecem ser expressas em Leishmania. T. cruzi e T. brucei. A caracterização da afinidade do LDL
em T. brucei revelou sítios de ligação à lipoproteínas de alta e baixa afinidade, que são
provavelmente duas formas de um único receptor (COPPENS et al., 1988; PAL et al., 2002).
Através de numerosas análises ultraestruturais e bioquímicas têm-se proposto a existência
de endocitose mediada por clatrina em tripanosomatídeos. Em T. brucei, foi caracterizada a cadeia
pesada da molécula de clatrina (TbCLH) que está distribuída em várias estruturas, dentre elas, os
túbulos coletores associados à bolsa flagelar e também à rede trans-Golgi e em vesículas localizadas
na região posterior do corpo celular do parasita (MORGAN et al. 2002b). Evidências ultraestruturais
demonstraram VSGs sendo endocitadas por vesículas contendo revestimento de clatrina
(GRUNFELDER et al, 2003).
Recentemente, foi demonstrado que a depleção de actina interfere na via endocítica, porém
não afeta o transporte de proteínas secretadas na membrana plasmática, sugerindo que, pelo menos
em T. brucei, que os caminhos endocíticos e secretórios são independentes (NOLAN & GARCIA-
SALCEDO, 2008).
As taxas de endocitose e de exocitose são elevadas na forma sanguínea de T. brucei e são
dependentes do fluxo de vesículas do Golgi para os endossomos de reciclagem e para a superfície
ou além da célula pela região da bolsa flagelar.
Uma importante molécula de superfície em T. brucei é a VSG, sendo esta um bom exemplo
para descrevermos a rota biossintética-secretora em T. brucei. A biossíntese de VSG é iniciada no
RE, onde esta se liga com a âncora de GPI, sofre N-glicosilação e posteriormente é transportada
para o CG (BANGS et al., 1986; VIDUGIRIENE & MENON, 1994). A exocitose de VSG para a superfície
33

da célula se dá através da bolsa flagelar (GRUNFELDER et al., 2003). O transporte de moléculas do RE
a superfície celular é rápido e ocorre as custas da atividade de GTPases (GEIGER et al., 2010).
1.6.1.2. Via endocítica/exocítica em T. cruzi
Estudos em T.cruzi mostraram que este protozoário apresenta algumas peculiaridades no seu
percurso endocítico que o distingue de outras células.
A distribuição de organelas é bastante diferente da apresentada por T. brucei e Leishmania
(Fig.6). Em T. cruzi a forma epimastigotas é aquela que apresenta uma atividade endocítica mais
evidente. Além da existência de "duas portas de entrada", bolsa flagelar e citóstoma, que estão
situados na região anterior da célula, as moléculas endocitadas seguem através de vesículas e
compartimentos tubulares (endossomas iniciais) que se dispõem periféricamente, para a região
posterior do corpo do parasito de onde retornam para a região central para encontrar os
reservossomos (PORTO-CARREIRO et al., 2000). Os reservossomos são organelas típicas da forma
epimastigota do T. cruzi, classificados inicialmente como endossomas tardios, que apresentam
enzimas lisossomais no seu interior, pH em torno de 6,0, próton ATPases do tipo P e que acumulam
macromoléculas ingeridas pelo parasito através do processo endocítico (SOARES & DE SOUZA, 1988;
SOARES et al., 1992; VIEIRA et al., 2005). Mais recentemente, os reservossomas da forma epimastigota
assim como compartimentos semelhantes encontrados nas fomas amastigota e tripomastigota foram
classificados como compartimentos semelhantes a lisossomas (SANT`ANNA et al., 2008).
Alguns dados experimentais sugerem a existência de endocitose mediada por receptor em T.
cruzi, no entanto o receptor de transferrina assim como o receptor para LDL ainda não foram
isolados. A presença de proteínas ligadoras de transferrina foi descrita em amastigotas (LIMA &
VILLATA, 1990) em formas epimastigotas (PORTO-CARREIRO, 2004). Em epimastigotas, estudos de
imunocitoquímica localizaram as proteínas ligadoras de transferrina na citofaringe, no citóstoma e
na bolsa flagelar (PORTO-CARREIRO, 2004).
Vesículas revestidas de clatrina foram mostradas pela primeira vez brotando da região do CG
(SANT'ANNA et al., 2004). Posteriores análises comprovaram a presença de clatrina e adaptina em T.
cruzi (CORRÊA et al., 2007),além disso, a expressão de clathrina em T. cruzi foi demonstrada por
immunoblotting utilizando anticorpos policlonais contra a cadeia pesada de clatrina bovina.
TcClathrin foi localizada na região do complexo Golgi e da bolsa flagelar nestes parasitos (CORRÊA
et al., 2008).
Um estudo em T. cruzi utilizando compostos que estabilizam ou despolimerizam o
citoesqueleto (microtúbulos e microfilamentos) mostrou que em formas epimastigotas a utilização
dessas drogas acarretou em um decréscimo de 50% na quantidade de transferrina endocitada
34

quando da utilização de drogas que afetam microtúbulos, enquanto que para as drogas que afetam
microfilamentos a inibição da atividade endocítica foi ainda maior (PORTO-CARREIRO, 2004),
mostrando que de fato a participação do citoesqueleto é essencial no processo endocítico no T. cruzi.
Em T. cruzi, assim como descrito em T. brucei, a bolsa flagelar é o sítio de exocitose de moléculas
secretadas ou expostas na membrana plasmática (DE SOUZA, 1995).
1.6.1.3. Via endocítica/exocítica em Leishmania
Em Leishmania, o processo endocítico é bem estabelecido e apresenta estruturas distintas
daquela encontradas em eucariotos superiores. Assim como em T. brucei, a endocitose em
Leishmania esta relacionada com a aquisição de nutrientes e evasão do sistema imune, sendo um
mecanismo de sobrevivência durante a infecção pelo parasito (DE SOUZA et al., 2009). O caminho
endocítico em Leishmania apresenta compartimentos próprios como o túbulo multivesicular
(TMV), que abrange todo o corpo do parasito e que apresenta um caráter ácido semelhante ao
lisossoma de células de mamíferos (GHEDIN et al., 2001). O TVM é considerado a organela final da
via endocítica em Leishmania. (WEISE et al., 2000) (Fig.6).
Na forma promastigota as vesículas que brotam na região da membrana da bolsa flagelar se
fundem ao endossoma inicial, de onde irão brotar vesículas que transportarão o material endocitado
para MVB. Destes compartimentos brotam vesículas contendo o material destinado a degradação
que irão se fundir ao túbulo multivesicular, que corresponde ao sistema endo-lisossomal deste
parasito. Nas formas amastigotas de Leishmania o lisossoma forma um grande vacúolo eletron-
denso denominado megassomo (MCCONVILLE et al., 2002) que foi descrito pela primeira vez no
complexo Leishmania mexicana e mais recentemente em L. chagasi (ALBERIO et al., 2004) ( Fig.6)
O processo de endocitose mediada por receptor também foi identificado em Leishmania.
Voyiatzaky e Soteriadou (1992) purificaram e caracterizaram bioquimicamente o receptor para
transferrina das formas promastigotas de L.major e L. infantum como uma proteína monomérica
integral de membrana de 70kDa. Além do receptor para transferrina já foram também
caracterizados a presença de proteínas específicas ligadoras de LDL em amastigotas e
promastigotas de L. donovani (BASTIN et al., 1996) e uma proteína transmembrana de 46 KDa, como
o receptor para hemoglobina localizado na bolsa flagelar de formas promastigotas de L. donovani
(SINGH et al., 2003)
Em Leishmania, estudos de microscopia eletrônica de células congeladas por alta pressão e
submetidas a métodos de substituição a frio demonstraram a presença de “coated pits” e vesículas
revestidas que brotam da bolsa flagelar e da rede trans-Golgi (WEISE et al., 2000), sugerindo a
existência de um processo endocítico clatrina dependente. A análise genômica confirmou a
presença da cadeia pesada de clatrina (LmCHC) no genoma de Leishmania(DENNY et al., 2005).
35

Como em células de mamíferos também foi identificada em Leishmania uma banda de
43kDa referente a actina no extrato de L. amazonensis e L. donovani, uma vez que a
despolimerização desta poderia interferir no percursso endocítico (MORTARA, 1989,
SAHASRABUDDHE, BAJPAI & GUPTA, 2004) o citoesqueleto pode estar participando no processo de
endocitose, porém ainda não foi mostrado o envolvimento direto do filamento de actina auxiliando
no processo endocítico.
Especializados sistemas de secreção são usados por patógenos para exportar fatores de
virulência para a célula hospedeira, como mecanismos de defesa e como meio de interação entre
parasito-célula hospedeira (SILVERMAN et al., 2010). Leishmania secreta para o meio extracelular
enzimas envolvidas na interação parasita-célula hospedeira, na virulência e na defesa do sistema
imune como, fosfatase ácida (GOTTLIEB & DWYER, 1982), a maior protease de superfície (gp63)
(MCGWIRE et al.,2002) e uma metalopeptidase homóloga a gp63, que é localizada
predominantemente em estruturas vesiculares do caminho exocítico (CUERVO et al., 2008).
Em Leishmania foi mostrado a presença de proteínas secretadas por microvesículas do tipo
exossomos em L. donovani, L. mexicana, L. major e que estas vesículas poderiam auxiliar na
comunicação com macrófagos e neutrófilos (SILVERMAN et al., 2009, GEIGER et al., 2010).
1.6.1.4.Controle de fusão de vesículas em tripanosomatídeos
A endocitose e exocitose em alguns protozoários unicelulares também são reguladas pela
fusão de vesículas mediada por proteínas da família Rab. Homólogos da Rab4, Rab5, Rab7 e Rab11
já foram identificados em T. brucei, L. donovani e T. cruzi. Em T. brucei, TbRab5Ap e TbRab5Bp,
que são homólogas à Rab5 de células de mamífero e foram detectadas em compartimentos túbulo-
vesiculares, localizados entre a bolsa flagelar e o núcleo, tais compartimentos são similares aos
endossomos iniciais. TbRab4p é uma outra proteína descrita, que também pode ser detectada em
compartimentos pertencentes a via endocítica deste parasito (FIELD et al., 1998). A proteína
homóloga a Rab2- TbRab2p, foi localizada na região do retículo endoplasmático (FIELD et al., 1999),
enquanto que a proteína homóloga a Rab11 – TbRab11, esta presente nas vesículas endocíticas que
contêm materiais a serem reciclados (JEFFRIES, MORGAN & FIELD, 2001).
Embora ainda existam poucas informações sobre as proteínas Rab no T.cruzi, sabe-se que
este protozoário também expressa genes para estas proteínas (MENDONÇA et al., 1993). A proteína
Rab7, que em células de mamífero está associada com os endossomos tardios, possui um homólogo
que está presente nas três formas evolutivas deste parasito, porém sua localização intracelular ainda
é incerta (LEAL et al., 2000). A proteína homóloga a Rab11 foi também descrita nas formas
epimastigotas, localizando-se na região posterior do corpo celular em compartimentos muito
semelhantes aos reservossomos (MENDONÇA et al., 2000).
36

Foi determinado o envolvimento de proteína Rab no caminho endocítico de L. donovani,
mostrando que várias funcionalidades da fusão de membrana deste parasito são semelhantes a
eventos de fusão descritos em células de mamíferos. Um homólogo de Rab5 de mamífero foi
identificado em Leishmania (LmRab5). LmRab5 está localizada na região anterior do parasito,
responsavel pela fusão homotípica nos endossomas iniciais (SINGH et al., 2003) e foi mostrado que
Leishmania requer Rab7 para mediar a degradação de hemoglobina endocitada para auxiliar o
crescimento (PATEL et al., 2008)
Ainda não foi relatada a ação de PI3k auxiliando a fusão de vesículas em tripanosomatídeos,
porém não se pode descartar essa possibilidade, uma vez que já foi mostrada a presença de
homólogos da PI3K de levedura em T. cruzi e a superexpressão desta resulta em alterações
morfológicas e funcionais relacionadas com o tráfico vesicular (SCHOIJET et al., 2008). Desta forma,
PI3K poderia tambem agir na fusão de vesículas em tripanosomatídeos.
Já foram descritos em tripanosomatídeos diferentes tipos de fosfolipases ligadas ao
remodelamento da membrana celular e ao processo de invasão (BERTELLO et al., 2000; BRONIA et al.,
1999) porém seu papel vinculado diretamente ao tráfego intracelular ainda não foi elucidado em
protozoários.
1.7. Fosfatase ácida: atividade presente nas vias endocítica/exocítica de Leishmania.
Leishmania ssp., assim como outros patógenos possuem a capacidade de hidrolisar fosfatos
orgânicos e esta hidrolise é catalizada por fosfomonoesterases ácidas, as fosfatases ácidas, que
podem estar presente em organelas intracelulares, na superfície celular associadas a membrana ou
no meio extracelular na sua forma secretada (GOTTLIEB & DWYER, 1981).
Em 1981, Gottlied e Dwyer mostraram a presença da atividade da fosfatase ácida distribuida
pela superfície externa de promastigotas de L. donovani. Esta atividade também foi detectada em
vesículas intracelulares que são oriundas do CG e fundem-se com compartimentos endossomais e
então são transferidas ao longo da via endocítica até os lisossomas. A existência de altas
concentrações de fosfatase ácida em Leishmania foi e confirmada por Glew e colaboradores (1982),
que também mostraram que promastigotas contém baixo nível de outras hidrolases ácidas. A
fosfatase ácida associada à superfície foi posteriormente dividida em 3 grupos distintos de acordo
com o peso molecular, resistência a tartarato de sódio e especificidade de substrato (REMALEY et al.,
1985). Uma atividade extracelular foi detectada em meio de crescimento de promastigota de
Leishmania (GOTTLIEB & DWYER, 1981) sendo esta descrita com a principal proteína secretada por
algumas espécies (BATES & DWYER, 1987; BATES e tal., 1989, 1990). Esta enzima é sintetizada no RE,
37
D

exportada ao Golgi onde é glicosilada e provalvemente fosforilada estando pronta para ser
direcionada para região da bolsa flagelar para inserção na superfície celular ou secreção para o meio
extracelular, ou direcionadas ao lisossomo através de vias intracelulares (BATES et al,,1989; MOULAY
& ROBERT-GERO, 1995). Logo, a fosfatase ácida é uma enzima que está presente em compartimentos
na via endocítica e exocítica, sendo um bom parêmetro para avaliar modificações nestes sistemas.
Em Leishmania, a função das fosfatases parece estar relacionada com a sobrevivência do
parasito no interior da célula hospedeira (BARBIERI et al., 1990). Esta enzima inibe a produção de
ânions superóxidos por neutrófilos e macrófagos sendo a fosfatase ácida um importante fator de
virulência deste parasito (ELLIS et al., 1998; BATES & DWYER, 1987; KATAKURA & KOBAYASHI, 1988).
No presente estudo procuramos analisar o papel de iPLA2 na fusão de membranas de
compartimentos da via endo/exocítica de promastigotas de L. amazonensis. As diferenças
estruturais entre os compartimentos dessa via em Leishmania em relação a outros
Tripanosomatídeos e em relação a células de mamíferos assim como a possibilidade de mapear o
tráfego de diferentes moléculas como: glicolipídios, glicoproteínas, glicoproteínas ancoradas por
âncoras lipídicas, proteínas integrais de membrana e proteínas secretadas torna o modelo
Leishmania particularmente atraente. Tendo em vista que a fusão de compartimentos é primordial
para o transporte intracelular de moléculas e que estas rotas são essenciais para a sobrevivência e
virulência de parasitos, isto torna a via endo/exocítica em Tripanosomatídeos um potencial alvo
quimioterápico.
38

2- OBJETIVO
Analisar o papel da iPLA2 nos processos endocítico e exocítico em promastigota de
Leishmania amazonensis.
Objetivos específicos
- Analisar as mudanças estruturais provocadas pelo inibidor específico de iPLA2 sobre as diferentes
organelas das via endocíticas /exocíticas de Leishmania através da visualização por microscópio
eletrônico de transmissão (MET) e técnicas associadas.
- Analisar o efeito da BEL sobre o processo endocítico utilizando marcadores de endocitose de fase
fluida e endocitose mediada por receptor passivos de análise e quantificação por citometria de fluxo
e microscopia de fluorescência e MET.
- Analisar a interferência da BEL sobre a exocitose através da dosagem protéica e SDS-PAGE.
- Analisar o efeito da BEL na localização e secreção de moléculas do parasito como a gp63
(protease mais abundante, ancorada por âncora de GPI presente na membrana e no sobrenadante) e
a fosfatase ácida (apresenta pelo menos duas formas: uma ligada a membrana e outra secretada)
através da análise de atividade enzimática, técnicas citoquímicas, citometria de fluxo,
imunofluorescência e immunoblotting.
39

3- MATERIÁIS E MÉTODOS
3.1-Parasito
O modelo celular utilizado foi a forma promastigota de Leishmania amazonensis, cepa
WHOM/BR/75/Josefa (isolada de um caso de leishmaniose cutânea humana, cedida pelo DR.
Cuba-Cuba C.A, Universidade de Brasília), que foi mantida em meio Schneider (Schneider Insert
Medium,Sigma) suplementado com 10% de soro fetal bovino (SFB-Gibco BRL, Gaithersburg MD,
USA) a 26°C por 72horas.
3.2 – Materiais
Bromoenol lactona (BEL), diluída em DMSO formando a solução estoque a 10mM foi
armazenada a -20° e obtida da Sigma-Aldrich. Anticorpo policlonal anti-gp63 foi gentilmente
cedido pela Prof. Marta Branquinha (IMPPG-UFRJ) e o anticorpo monoclonal anti-α-tubulina foi
cedido pela professora Narcisa Cunha e Silva (IBCCF-UFRJ). Anticorpos e traçadores da via
endocítica fluorescentes foram adquiridos da Molecular Probes.
3.3-Curva de Crescimento na presença de Bromoenol lactona (BEL)
A curva de crescimento foi realizada em meio Scheneider com 10% de SFB. As culturas
foram incubadas a 26°C e o número de parasitos avaliados por contagem em câmara de Neubauer
com intervalo de 24 horas por 5 dias consecutivos. A curva de crescimento foi iniciada com 1 x 106
parasitos por ml de meio na ausência (controle) ou na presença de diferentes concentrações de
Bromoenol lactona (BEL).
3.4-Tratamento com Bromoenol lactona (BEL)
A partir da curva de crescimento, em todos os experimentos em que a BEL foi utilizada
ocorreu tratamento com a BEL na concentração final de 2,5µM por 1h a 26°C em meio RPMI sem
soro. A seguir, dependendo do experimento, a droga foi ou não mantida nos meios de incubação
posteriores.40

3.4.1-Controle com DMSO
Em todos os experimentos em que a BEL foi utilizada ocorreu tratamento somente na
presença de DMSO na concentração final de 2,5µM por 1h a 26°C em meio RPMI sem soro. Como
controle , uma vez que este é utilizado na solubilização do inibidor.
3.5-Dosagem de proteína
Formas promastigotas controle e tratadas com BEL por 1 hora, foram centrifugadas a 1500
× g por 8 minutos e lavadas uma vez em salina balanceada de Hank`s (HBSS) sem soro. Foi
avaliada a dosagem de proteínas totais referente a 1×108 parasitos para padronizar a quantidade de
proteínas por ml. Em seguida as células foram incubadas em HBSS sem soro em temperatura
ambiente por 3h. Após a incubação as células foram centrifugadas a 13.000 × g por 4 minutos o
sedimento descartado e as proteínas presentes no sobrenadante foram dosadas pelo método de
Bradford (1976), utilizando solução de albumina (BSA) a 1mg/ml para construção da curva-padrão.
A leitura foi realizada, após 10 min de contato dos sistemas reacionais com o corante, em leitor de
ELISA a 595 nm.
3.6-Endocitose
Parasitos controle e previamente tratados com BEL foram obtidos por centrifugação a 1.500
×g por 8 minutos e lavados 3 vez em PBS, pH 7,2, depois foram incubadas por 15mim em em PBS,
pH 7,2 a 26°C . Os promastigotas foram então submetidos a uma cinética de endocitose na presença
de marcadores fluorescentes: BSA-TRITC (tetramethyl Rhodamina Isothiocyanate) (30µg/ml),
Transferrina-AlexaFluor (25µg/ml) e ConcanavalinaA-AlexaFluor (20µg/ml). A incubação foi de 1
e 2h a 26°C, em meio RPMI sem soro na presença ou na ausência (controle) de BEL. Ao final, as
amostras foram lavadas 1× em PBS, pH 7,2, fixadas para citometria de fluxo ou microscopia de
fluorescência.
3.7-Ensaio de Viabilidade
Parasitos controle e tratados com BEL, que foram expostos a diferentes condições de
incubação, de uma hora e meia e três horas, em HBSS ou RPMI sem soro a 26°C tiveram sua
viabilidade analisada pela incorporação de Iodeto de Propídio (IP). A 5×106 parasitos foram
adicionados IP para uma concentração final de 10µg/ml deixando agir por 15 minutos a 26°C. A
marcação foi analisada por citometria de fluxo ou por microscopia de fluorescência.
41

3.8-Eletroforese em gel de poliacrilamida sob condições desnaturantes (SDS-PAGE)
Analisamos o perfil protéico total do parasito e das proteínas presentes no sobrenadante dos
parasitos controle e tratados com BEL. Após 1 hora de tratamento os parasitos foram lavados 1x em
HBSS sem soro. Os parasitos (1×108 parasitos/ml) controle e previamente tratados com BEL foram
então incubados por 3h em meio HBSS a 26ºC. Foram avaliadas as proteínas liberadas no
sobrenadante após 3 horas de incubação em meio HBSS sem soro em temperatura ambiente para
padronizar a quantidade de proteínas por ml. Após os diferentes tratamentos as células foram
lavadas 2× em PBS, pH 7,2 por 10 min, enquanto, que as proteínas presentes no sobrenadante
foram precipitadas com 100 % de etanol -20ºC (1:10, respectivamente). Após incubação por 10 min
a -20ºC, as proteínas foram centrifugadas 10.000 × g por 5 minutos a 4ºC. Em seguida, as proteínas
precipitadas foram lavadas com 90% de etanol a -20ºC e centrifugadas a 10.000 × g por 5 min 40C.
As proteínas precipitadas e os parasitos suspensos em PBS foram diluídos (1:1), respectivamente,
em tampão de amostra 4× (20% de glicerol, 20% de SDS, 0,02% de azul de bromo fenol e 0, 120M
Tris-HCl pH 6,8) contendo 5% de β-mercaptoetanol no volume final, e foram fervidos durante 5
min a 100°C e então centrifugados a 4.800 × g por 1 min. O perfil protéico foi determinado através
de um SDS-PAGE 12% (LAEMMLI, 1970), utilizando um padrão de peso molecular PageRulerTM
Plus Prestained Protein Ladder (Fermentas). A separação das proteínas ocorreu sob tampão de
corrida 1× (0,192 M de glicina, 0,1% de SDS, Tris-HCl 0,025 M, pH 8,8), em um sistema vertical
Mini Protean II (BioRad Laboratories, Richmond, Calif). A revelação das bandas do gel foi
realizada por coloração com Coomassie Blue R-250 (LAEMMLI, 1970) e por coloração de prata, pelo
método rápido SilverQuestTM. Fast Staining Protocolo, Invitrogen.
3.9-Transferência das proteínas para membrana de nitrocelulose -
Através do método de Towbim e colaboradores (1979), os polipeptídios presentes nos
extratos dos parasitos e do sobrenadante separados por SDS-PAGE foram transferidos
eletroforeticamente para membrana de nitrocelulose (Amersham Hybond™ ECL™ GE Healthcare
Bio-Science Corp., USA) em tampão de transferência (0,025 M de Tris, pH 8,3, 0,192 M de
Glicina e 20% de metanol), sob amperagem constante de 0,35 A0/cm por 90 mim e banho de gelo,
utilizando um sistema vertical Mini Trans-Blot Cell (BioRad Laboratories, Richmond, Calif). A
transferência das proteínas foi confirmada pela coloração da membrana com a solução vermelho de
Ponceau por 2 min.
3.10-Immunoblotting
42

A membrana de nitrocelulose contendo as proteínas transferidas foi incubada em solução de
bloqueio [10 mM de Tris-HCl, pH 7,5, 150 mM de NaCl (TBS); 5% leite em pó desnatado] por 1
hora á temperatura ambiente, com troca da solução bloqueio após 30 min, e sob constante agitação
em todas as etapas. Em seguida, a membrana de nitrocelulose foi lavada 3 × em TBS contendo
0,05% de Tween 20 (TBS-Tween) por 10 min e 1× com TBS por 5 min a temperatura ambiente. A
membrana de nitrocelulose foi então incubada com anticorpos em TBS contendo 3% BSA, 0,05%
NaN3 por 18h a 4°C. Após esse período, a membrana de nitrocelulose foi lavada 3 × com TBS-
Tween 20 por 10 min e 1 × em TBS por 5 min, a temperatura ambiente. Em seguida, a membrana
de nitrocelulose foi incubada com o anticorpo secundário acoplado a fosfatase alcalina (1:1000) em
TBS contendo 3% de BSA por 1 hora a temperatura ambiente. Após três ciclos de lavagem da
membrana de nitrocelulose com TBS-Tween 20 por 10 min e 1 × com TBS por 5 min, as proteínas
foram reveladas com uma solução de 0,03% NBT/ 0,02% BCIP (Fermentas Inc., Glen Burnie, MD,
U.S.A.) em tampão de fosfatase alcalina (100mM Tris-HCl pH 9,5, 150 mM NaCl, 5 mM MgCl2 e
0,05% Tween 20), a temperatura ambiente. Após o aparecimento das bandas proteicas sob a
membrana de nitrocelulose, a reação de revelação foi interrompida através de lavagens sucessivas
da mesma com água destilada.
3.11-Atividade fosfatase ácida
A atividade da fosfatase ácida foi medida pela concentração de p-nitrofenol no sobrenadante
e no extrato celular (p-NP, produto de hidrólise do p-nitrofenilfosfato (p-NPP). Células intactas e
tratadas com 2,5µM de BEL (1x108 parasitos/ml) foram incubadas por 1hora a temperatura
ambiente em 0,5ml de mistura de reação contendo, tampão HEPES-tris, pH5,4 e 25mM de p-NPP
como substrato, na presença ou ausência do inibidor específico, tartarato de sódio na concentração
de 10 mM. A reação foi iniciada pela adição de célula ou sobrenadante e interrompida pela adição
de 1,0ml de NaOH 1N. Controles, onde as células foram adicionadas após a interrupção da reação,
foram uitlizados como branco. Para determinar a concentração de p-NP liberado, os tubos foram
centrifugados a 1500 xg por 20 min e os sobrenadantes foram medidos espectofotometricamnete a
425nm. Nós também testamos a atividade da 3`nucleotidase utilizando com substrato o 3`AMP.
Neste caso, as atividades hidrolíticas foram analisadas espectofotometricamente a 425nm pela
medida de fosfato inorgânico liberado apartir deste substrato. Os valores apresentados representam
média de três experimentos independentes.
3.12-Citometria de Fluxo
Como marcadores utilizamos: (a) iodeto de propídeo (PI), se liga em ácidos nucleicos,
somente em células lisadas (GROOTEN et al., 1993); (b) anticorpo primário associados a anticorpo
43

secundário específico marcadores de antígenos de superfície; (C) marcadores do processo
endocítico, BSA-TRITC, ConA-AlexaFluor e transferrina-AlexaFluor.
Formas promastigotas foram analisadas seguindo o seguinte protocolo: os parasitos (1x106
céls/ml) foram incubados com os composto em meio RPMI sem soro a 26ºC por 1 e 2 horas, após
incubação (b) e (c) foram fixadas em formaldeído a 2% em PBS, pH 7,2 durante 10 minutos a 4ºC.
As amostras foram analisadas em citômetro de fluxo Becton Dickinson FACS Calibur,
100.000 eventos foram analisados. As fluorescências de Rodamina, AlexaFluor, PI e anticorpos
fluorescentes, foram captadas pelos seus respectivos filtros (FL-1: 530±30 nm, FL-1 e FL-2:585±26
e FL-3: acima de 670 nm para os dois últimos). Os dados foram analisados com o auxílio do
programa “Windows Multiple Document Interface Flow Citometry Application” (WinMDI).
3.13-Imunofluorescência
Células controle e tratadas com BEL foram colhidas por centrifugação a 1.500 x g por 8
minutos, lavadas em PBS. Após essa etapa os parasitos foram fixados com 3,7% de formalina em
PBS, pH 7,2 por 10 minutos a 4°C. Após o período de fixação, as amostras foram lavadas em PBS,
e aderidas em lamínulas previamente cobertas com poli-L-lisina (0,1 % em PBS) por 20 minutos.
As lamínulas contendo os parasitos foram lavadas com PBS, pH 7,2 e incubadas em solução de
cloreto de amônio a 50mM em PBS, pH 7,2, durante 30 minutos a temperatura ambiente. Em
seguida foi adicionado sobre as amostras uma solução de bloqueio contendo 0,2% de gelatina, 0,1%
de azida em PBS, pH 7,2 (PGN) por 30 minutos. Após essa etapa os parasitas foram incubados com
o anticorpo primário diluído em PGN (1:100) em temperatura ambiente por 1h. Após esta
incubação as lamínulas foram lavadas 3x em PBS, pH 7,2 e incubadas na presença do anticorpo
secundário conjugado ao marcador fluorescente AlexaFluor diluído (1:400) em PGN por 1h a
temperatura ambiente. Após a marcação as amostras foram lavadas extensivamente em PBS e
montadas sobre lâminas de vidro contendo solução de 0,2N de n-propil-galato diluído em glicerol
90% em PBS. As amostras foram analisadas ao microscópio de fluorescência ZEISS Axioplan.
3.14-Microscopia Eletrônica de Transmissão Convencional
Parasitos controle e cultivados por 1h na presença de BEL, foram fixados em 2,5% de
glutaraldeído em tampão cacodilato de sódio 0,1M, pH 7,2, 5 mM de cloreto de cálcio e 3,7% de
sacarose durante 1 hora a temperatura ambiente. Após fixação essas amostras foram lavadas 2 vezes
em PBS e pós-fixadas durante uma hora no escuro em solução de tetróxido de ósmio a 1% em
cacodilato de sódio 0,1M, pH 7,2, 0,8% de ferrocianeto de potássio e 5mM de cloreto de cálcio. As
amostras foram desidratadas em séries crescentes de acetona e incluídas em resina PolyBed. A
seguir a resina foi polimerizada durante 72 h a 60°C. Secções ultra-finas (60 nm) foram obtidas em
44

um ultramicrótomo Leica e contrastadas com acetato de uranila 5% durante 30 min em água e em
citrato de chumbo durante 5 mim (REYNOLDS, 1963). As secções foram observadas em microscópio
eletrônico de transmissão FEI Morgagni F-268.
3.15-Citoquimica da atividade da enzima fosfatase ácida
Células controle e tratadas de acordo com os protocolos dos experimentos foram
centrifugadas a 1.500 x g por 8 minutos e lavadas duas vezes em PBS e fixados em 1% de
glutaradeído em tampão cacodilato 0,1M, pH 7,2, e 5% de sacarose durante 10 mim a 4°C. Em
seguida as células foram lavadas duas vezes em tampão cacodilato 0,1M, pH 7,2, contendo 5% de
sacarose e posteriormente em tampão tris-maleato de sódio, pH 5,0, contendo 5% de sacarose. As
células foram então incubadas em meio contendo β-glicerofosfato de sódio 2mM, 5% de sacarose e
CeCl3 2mM em tampão Tris-maleato de sódio, pH 5,0, por 1h a 37oC com agitação na presença e
na ausência de 10mM de tartarato de sódio, inibidor da fosfatase ácida. Após o período de
incubação as células foram lavadas em solução de 5% de sacarose em tampão Tris-maleato de
sódio, pH 5,0, e em solução de 5% de sacarose em tampão cacodilato 0,1M, pH 7,2. As células
foram re-fixadas em solução 3% de glutaraldeido, e 3,7% de sacarose em tampão cacodilato 0.1 M,
durante 30 mim em temperatura ambiente. Após esta etapa as amostras foram lavadas em solução
de sacarose 3,7 % em tampão cacodilato 0,1 M, e pós-fixadas durante uma hora, em solução de
tetróxido de ósmio a 1% em cacodilato de sódio 0,1M, pH 7,2. A seguir foi feita desidratação em
série crescente de acetona, inclusão e obtenção de secções ultrafinas como descrito no item 3.13.
3.16- Análises estatísticas
Os resultados foram analisados estatisticamente pelo teste t de student usando o programa
Bio Estat 2.0. Os valores de P menores ou iguais a 0,05 foram considerados significativos.
45

4- RESULTADOS
4.1- Ação da BEL sobre o crescimento de L. amazonensis
Para avaliar o efeito da BEL no crescimento das formas promastigotas e determinar a
melhor concentração a ser utilizada nos experimentos, parasitos foram crescidos durante 4 dias em
meio Schneider contendo 10% de soro fetal bovino, na presença de diferentes concentrações do
composto. Nas primeiras 24h as concentrações de 0,5; 1; 2,5 µM não observamos grandes
alterações no crescimento dos parasitos quando comparado á curva de crescimento dos parasitos
controle, porém nas 24h seguintes observamos uma inibição de 56% do crescimento na
concentração de 0,5 µM e uma inibição de aproximadamente 75% nas concentrações de 1 e 2,5 µM.
No decorrer da curva, o crescimento do parasito se iguala nas concentrações de 0,5; 1; 2,5 µM
reduzindo o mesmo em aproximadamente 25% em relação a curva controle (Fig. 7). Em todos os
tempos analisados os parasitos contabilizados na presença das diferentes concentrações de BEL se
apresentavam viáveis como os parasitos controle. Já as concentrações de 5 e 10 µM provocaram
uma inibição no crescimento durante os 5 dias de cultura. Foram testadas então as concentrações de
0,5; 1; 2,5µM sendo a concentração de 2,5µM escolhida para ser usada nos demais experimentos.
46

Figura 7: Efeito da BEL sobre a proliferação de formas promastigotas de L. amazonensis. Formas promastigotas de L. amazonensis foram cultivadas por até 96 horas na presença de 5 diferentes concentrações de BEL. A concentração de 2,5µM foi escolhida para o desenvolvimento do restante dos experimentos (n=3).
4.2- Ensaios de Viabilidade
Uma vez escolhida a concentração de BEL a ser usada verificamos a viabilidade dos
parasitos na presença e ausência do inibidor para nos certificarmos de que em todos os tempos de
incubação que foram usados nos demais experimentos, os parasitos se apresentam íntegros e viáveis
e que as alterações observadas ocorriam devido à ação da BEL e não por lise. Realizamos o ensaio
de viabilidade através da marcação com iodeto de propídeo (IP). Foram analisados os tempos de 1;2
e 3 horas e estes comparados com células controle na ausência da BEL (Fig. 8). O resultado por
citometria de fluxo da captação de IP foi plotado pela densidade de população. Células na ausência
e na presença de 2,5µM de BEL durante 1; 2 e 3 horas não mostraram uma intensidade de
fluorescência significativamente diferente do controle (Fig. 8), com um pequeno numero de células
marcadas. Células incubadas na presença de saponina foram utilizadas como controle positivo para
a marcação com IP e neste caso aproximadamente 82% das células apresentaram-se marcadas.
47

Figura 8: Análise quantitativa da viabilidade celular de células controle e submetidas à incubação na presença de BEL. Histograma de fluorescência associado com a marcação com IP em promastigotas de L. amazonensis. O eixos Y e X representam o número de eventos e a intensidade de fluorescência log (FL3), respectivamente. Os tempos de incubação analisados foram 1h (verde); 2h (amarelo); e 3h (azul), os dados foram normalizados por células lisadas com saponina (rosa) [10 µM, 15 min., 26oC] servindo de controle positivo para IP. Células controle (preto).
4.3-Análise ultraestrutural
Os efeitos do inibidor da iPLA2 sobre a ultraestrutura de L. amazonensis foram avaliados
por MET. Promastigotas controle apresentaram uma mitocôndria única e ramificada que se estende
a partir do cinetoplasto localizado próximo a bolsa flagelar (BF) (Fig. 9B). Há um grande número
de perfis de acidocalcissomos na região posterior da célula. O Golgi pode ser observado próximo ao
núcleo juntamente ao retículo endoplasmático (RE) (Fig.9C), além de estruturas semelhantes a
glicossomos (Fig. 9G). Sob a membrana plasmática, localizam-se microtúbulos subpeliculares
dispostos regularmente. Estes microtúbulos não são observados na membrana da BF (Fig. 9E, F) e
pode-se observar um elevado número de vesículas na bolsa. Na bolsa encontra-se a estrutura
paraxial, o axonema e um longo flagelo (Fig. 9F). Observamos a presença de diversas estruturas
túbulo-vesiculares (seta rosa nas Figs. 9B, C, D, H) contendo pequenas vesículas com tamanho e
estrutura variável (detalhe na Fig. 9A), estes se estendem ao longo do corpo do parasito. Autores
sugerem que tais estruturas sejam compartimentos originados do Golgi e podem ser considerados
funcionalmente equivalentes a endossomos tardios/lisossomos, que correspondem ao TMV,
também são encontrada vesículas contendo partículas de 45nm em células controle (seta azul na
Fig. 9I) (WEISE et al., 2000).
Após o tratamento de promastigotas com 2,5µM de BEL por 1h, observamos alterações em
estruturas envolvidas com o tráfego exocítico/endocítico. Células tratadas apresentaram um Golgi
fragmentado com cisternas alargadas principalmente na região Trans (Figs 10A e B). Também
observamos alteração nos TMV caracterizada por uma diminuição no número de vesículas internas
e com um aparente aumento do diâmetro das mesmas (Fig. 10A). Não se observou alterações na
estrutura da BF, porém foi observado em algumas micrografias uma redução brusca, próxima a
ausência do número de vesículas no seu interior (Figs 10B e D). A estrutura da região cis do Golgi
também apresentou um aspecto fragmentado (Fig. 10C).
Figura 9: Ultraestrutura das principais organelas das vias endocítica/exocítica de formas promastigotas de L. amazonensis controle – As micrografias mostram as principais organelas de promastigotas de L. amazonensis; (A) detalhe de um túbulo multivesicular (TMV) isolado mostrando as vesículas internas; (B) secção longitudinal mostrando a localização do núcleo, mitocôndria, cinetoplasto, acidocalcissomos (seta preta), várias secções do TMV (seta rosa); (C-D) secção longitudinal mostrando o Golgi e uma porção do TMV (seta rosa); (E-F) secções transversais da bolsa
48

flagelar mostrando seu lumen preenchido por vesículas de aproximadamente 50 nm de diâmetro (seta verde) e perfis de membrana (cabeça de seta em F) e o brotamento de vesículas a partir da membrana da bolsa flagelar (seta preta em F); (G) secção transversal mostrando organelas semelhantes a glicossomos; (H) vesículas com coat podem ser observadas na região Trans do Golgi (cabeça de seta); (I) D) e (H) estrutura de TMVe (I) secção longitudinal da região da bolsa flagelar mostrando os túbulos agregados (cabeça de seta), as vesículas contendo partículas de 45nm (seta azul) e uma vesícula translúcida (vT). Golgi -G; Bolsa flagelar – bf; mitocôndria – m; cinetoplasto-K; vesícula translúcida- VT e núcleo- N. Barras de aumento correspondem a 300nm.
49

50
VT

Figura 10: Ultraestrutura das principais organelas das vias endocítica/exocítica de formas promastigotas de L. amazonensis incubadas na presença de BEL – (A) células apresentando alteração no TMV caracterizada pela redução no número de vesículas internas (seta rosa) e um Golgi fragmentado (seta preta); (B e D) secção transversal da região da bolsa flagelar mostrando um aspecto ramificado dos túbulos agregados (seta preta em B e D), o alargamento das cisternas Trans do Golgi (seta verde) e a presença de uma vesícula translúcida (VT) que parece se fundir à membrana da bolsa flagelar; (C) o Golgi apresenta aspecto fragmentado na região de brotamento (seta vermelha) e na região do Trans-Golgi (seta azul). Golgi -G; Bolsa flagelar – bf e núcleo- N. Barras de aumento correspondem a 300nm.
4.4- Interferência da BEL na aquisição de moléculas pelo parasito
Para estudar o efeito da BEL no processo endocítico foi realizado um monitoramento da
endocitose de traçadores fluorescentes por citometria de fluxo e microscopia de fluorescência,
utilizando BSA (Fig.11) como marcador do processo endocítico de fase fluida, Transferrina
(Fig.13) como marcador de endocitose mediada por receptor e Concanavalina- A como marcador
dos compartimentos tubulares presentes no trêfego endocítico (Fig.12).
A intensidade de fluorescência das células controle após 2h de incubação na presença de
BSA fluorescente foi de aproximadamente 3 vezes a observada no tempo de 1 hora enquanto a
intensidade de marcação nas células tratadas com BEL sofreu um acréscimo de aproximadamente
60% (Tabela I, Fig.11A). Quando comparado ao controle a captação de BSA-TRITC por 2 h, na
presença de BEL apresentou uma redução de aproximadamente 66% em relação ao observado no
controle (Fig. 11A). Ao microscópio de fluorescência observamos que a incubação de
promastigotas por 2 h na presença de BSA-fluorescente resultou em aproximadamente 70% de
células marcadas enquanto na população de células tratadas com BEL a porcentagem de células
marcadas foi de aproximadamente 50%. Logo a BEL provocou tanto redução na porcentagem de
células marcadas (aproximadamente 25%) como na quantidade de marcador adquirido. A marcação
das células controle pode ser observada em vesículas na região posterior da célula, em
compartimentos próximos a bolsa flagelar e no interior da bolsa flagelar (Figs 11B e C). Após o
tratamento a localização da marcação não se apresentou muito diferente do controle, porém, em
algumas células os compartimentos marcados puderam ser observados na região entre o
cinetoplasto e o núcleo (Figs 11D e E).
A endocitose de transferrina seguiu os mesmos padrões apresentados pela endocitose de
BSA no que diz respeito à intensidade de marcação observada por citometria de fluxo. Após 2
horas de incubação a intensidade de fluorescência nas células controle foi de aproximadamente 3
vezes a observada no tempo de 1 hora enquanto que nas células tratadas com BEL e incubadas com
transferrina por 2 h, a intensidade de marcação foi apenas 25% maior do que aquela observada no
tempo de 1 hora de incubação. BEL provocou uma inibição de aproximadamente 70% na aquisição
de transferrina fluorescente em relação às células controle tendo como parâmetro o tempo de 2 51
B
C
D
VTCD

horas de incubação. Ao microscópio de fluorescência observamos que a incubação de
promastigotas por 2 h na presença de transferrina-fluorescente resultou em aproximadamente 87%
de células marcadas enquanto na população de células tratadas com BEL a porcentagem de células
marcadas foi de aproximadamente 19%. Observamos também que um padrão de marcação
significativamente diferente, onde células controle apresentam após 2 h, um grande número de
vesículas marcadas na região posterior das células (Figs 12B e E) enquanto nos promastigotas
tratados com BEL a marcação parece ser mais restrita a superfície celular (Figs 12F e G).
No ensaio com a ConA (Fig. 13A) observamos, nas células controle, uma alta intensidade
de marcação após 1 h de incubação que não foi absolutamente diferente daquela observada após 2
hs. A intensidade de marcação após tratamento com a BEL não apresentou diferença significativa
com a marcação do controle em nenhum dos tempos analisados (Fig 13A) e a diferença entre o
valor captado pelas células controle e tratadas com BEL entre 1 e 2 horas de incubação é
semelhante ficando abaixo de 10% (Tabela I). Ao microscópio de fluorescência observamos um
padrão de marcação bem distinto do apresentado pela BSA. A marcação com a ConA é vista em
aproximadamente 98% dos parasitos controle e dos tratados com BEL e se concentra na superfície
da célula e na bolsa flagelar com poucos compartimentos marcados no citoplasma (Figs 13B e E).
Há apenas uma sugestão de marcação em compartimentos tubulares no controle que precisa ser
averiguado por microscopia eletrônica de transmissão
TABELA I
Distribuição de marcadores da via endocítica em L. amazonensis em condições controle e na presença de BEL
Tratamen
to
Tempo BSA ConA Transferrina% cel1 Intens2 Local3 % cel1 Intens2 Local3 %
cel1
Intens2 Local3
Controle 1h - 4 - - 90 - 32hs 70 11 Região
posterior100 98 Superfície celular,
bolsa flagelar e regiões
vesiculares
87 9 Região posterior
do parasito
BEL 1h - 2,5 - - 91 - 2
2hs 54 4 Região posterior e próximo ao
núcleo
93 97 Superfície celular e bolsa flagelar
19 2,5 superfície celular
1% de células marcadas observadas por microscopia de fluorescência2Intensidade de marcação obtida por análise em citometria de fluxo3Localização da marcação observada por microscopia de fluorescência
52
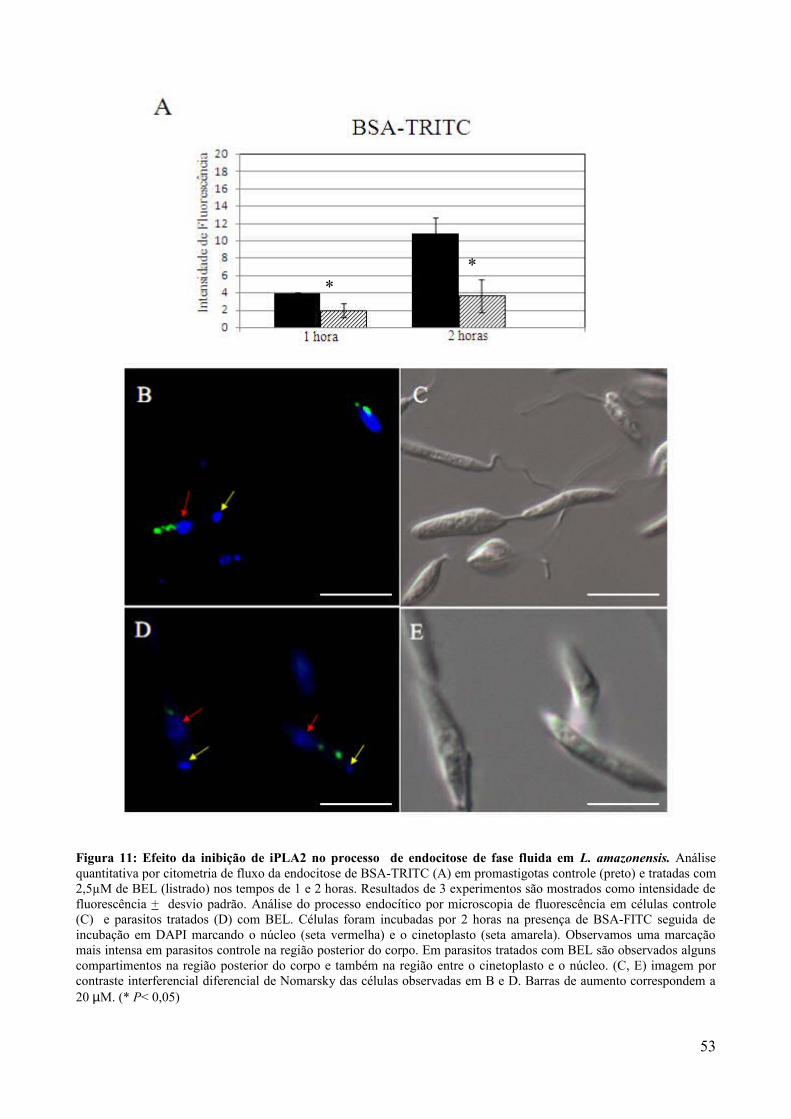
Figura 11: Efeito da inibição de iPLA2 no processo de endocitose de fase fluida em L. amazonensis. Análise quantitativa por citometria de fluxo da endocitose de BSA-TRITC (A) em promastigotas controle (preto) e tratadas com 2,5µM de BEL (listrado) nos tempos de 1 e 2 horas. Resultados de 3 experimentos são mostrados como intensidade de fluorescência + desvio padrão. Análise do processo endocítico por microscopia de fluorescência em células controle (C) e parasitos tratados (D) com BEL. Células foram incubadas por 2 horas na presença de BSA-FITC seguida de incubação em DAPI marcando o núcleo (seta vermelha) e o cinetoplasto (seta amarela). Observamos uma marcação mais intensa em parasitos controle na região posterior do corpo. Em parasitos tratados com BEL são observados alguns compartimentos na região posterior do corpo e também na região entre o cinetoplasto e o núcleo. (C, E) imagem por contraste interferencial diferencial de Nomarsky das células observadas em B e D. Barras de aumento correspondem a 20 µM. (* P< 0,05)
53
**

Figura 12: Efeito da inibição de iPLA2 no processo endocítico mediado por receptor em L. amazonensis. Análise quantitativa por citometria de fluxo da endocitose de transferrina-AlexaFluor 546 (A) em promastigotas controle (preto) e tratadas com 2,5µM de BEL (listrado) nos tempos de 1 e 2 horas. Resultados de 3 experimentos são mostrados como intensidade de fluorescência + desvio padrão. Análise do processo endocítico por microscopia de fluorescência em células controle (B,D) e tratadas (F) com BEL. Células foram incubadas por 2 horas na presença de transferrina-AlexaFluor 488 seguida de incubação em DAPI marcando o núcleo (seta vermelha) e o cinetoplasto (seta amarela). A marcação nas células controle está presente em compartimentos na região posterior do corpo celular. Após tratamento
54

com BEL a fluorescência é observada na superfície das células. (C, E, G) imagem por contraste interferencial diferencial de Nomarsky das células observadas em B, D e F. Barras de aumento correspondem a 20 µM. (* P< 0,05).
Figura 13: Efeito da inibição de iPLA2 no processo endocítico de ConA em L. amazonensis. Análise quantitativa por citometria de fluxo da endocitose de ConA-AlexaFluor 488 (A) em promastigotas controle (preto) e tratadas com 2,5µM de BEL (listrado) nos tempos de 1 e 2 horas. Resultados de 3 experimentos são mostrados como intensidade de fluorescência + desvio padrão. Análise do processo endocítico por microscopia de fluorescência em células controle (B) e parasitos tratados (D) com BEL. Células foram incubadas por 2 horas na presença de ConA-AlexaFluor 488 seguida de incubação em DAPI marcando o núcleo (seta vermelha) e o cinetoplasto (seta amarela). A marcação se apresentou de forma semelhante em parasitos controle e tratados com BEL, estando distribuída por toda a superfície celular e acentuadamente na região da bolsa flagelar (seta roxa). Em imagens das células controle podemos observar marcações no interior celular, possivelmente em compartimentos tubulares (seta branca); (C, E) imagem por contraste interferencial diferencial de Nomarsky das células observadas em B e D. Barras de aumento correspondem a 20 µM.
55
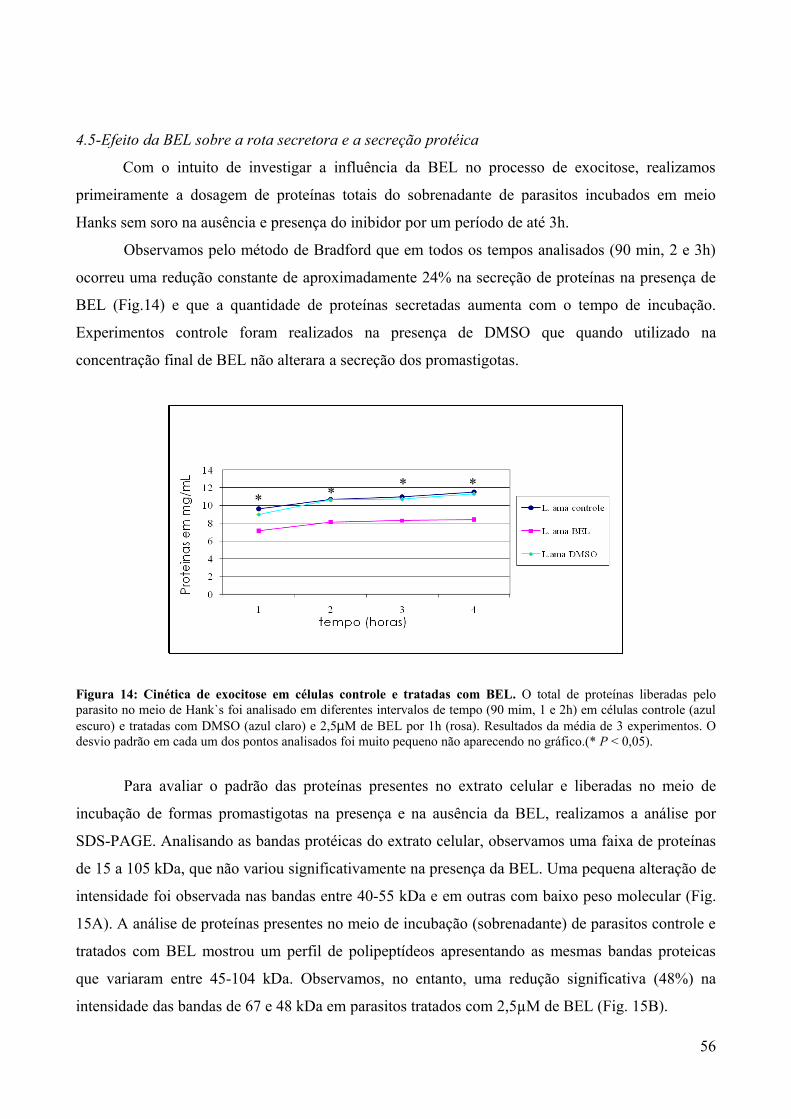
4.5-Efeito da BEL sobre a rota secretora e a secreção protéica
Com o intuito de investigar a influência da BEL no processo de exocitose, realizamos
primeiramente a dosagem de proteínas totais do sobrenadante de parasitos incubados em meio
Hanks sem soro na ausência e presença do inibidor por um período de até 3h.
Observamos pelo método de Bradford que em todos os tempos analisados (90 min, 2 e 3h)
ocorreu uma redução constante de aproximadamente 24% na secreção de proteínas na presença de
BEL (Fig.14) e que a quantidade de proteínas secretadas aumenta com o tempo de incubação.
Experimentos controle foram realizados na presença de DMSO que quando utilizado na
concentração final de BEL não alterara a secreção dos promastigotas.
Figura 14: Cinética de exocitose em células controle e tratadas com BEL. O total de proteínas liberadas pelo parasito no meio de Hank`s foi analisado em diferentes intervalos de tempo (90 mim, 1 e 2h) em células controle (azul escuro) e tratadas com DMSO (azul claro) e 2,5µM de BEL por 1h (rosa). Resultados da média de 3 experimentos. O desvio padrão em cada um dos pontos analisados foi muito pequeno não aparecendo no gráfico.(* P < 0,05).
Para avaliar o padrão das proteínas presentes no extrato celular e liberadas no meio de
incubação de formas promastigotas na presença e na ausência da BEL, realizamos a análise por
SDS-PAGE. Analisando as bandas protéicas do extrato celular, observamos uma faixa de proteínas
de 15 a 105 kDa, que não variou significativamente na presença da BEL. Uma pequena alteração de
intensidade foi observada nas bandas entre 40-55 kDa e em outras com baixo peso molecular (Fig.
15A). A análise de proteínas presentes no meio de incubação (sobrenadante) de parasitos controle e
tratados com BEL mostrou um perfil de polipeptídeos apresentando as mesmas bandas proteicas
que variaram entre 45-104 kDa. Observamos, no entanto, uma redução significativa (48%) na
intensidade das bandas de 67 e 48 kDa em parasitos tratados com 2,5µM de BEL (Fig. 15B).
* ** *
56

Figura 15: Análise por SDS-PAGE do extrato total (A) e do sobrenadante (B) de promastigotas controle e tratadas com BEL 2,5µM por 1h. Um painel similar de bandas foi encontrado no extrato celular e no sobrenadante na presença e na ausência de BEL. Notar a redução acentuada na intensidade das bandas do sobrenadante total entre os parasitos controle e tratados. Resultados representaivos de 3 experimentos.
Na tentativa de identificar as proteínas liberadas no sobrenadante utilizamos a técnica de
“Western blotting” e o anticorpo anti-gp63, anticorpo policlonal. A gp63 é a proteína mais
abundante da superfície de Leishmania. Ela é transportada através de vesículas para a superfície da
célula e incorporada a “lipd rafts” quando da sua chegada à membrana plasmática (DENNY et
al.,2001). Esta proteína também apresenta uma forma solúvel encontrada no sobrenadante que é
resultado de um processo de autoproteólise (MCGUIRE et al.,2002).
Os resultados obtidos com a técnica de “Western blotting” mostraram uma redução de
aproximadamente 38% na intensidade da banda de maior peso molecular relativa à gp63 e uma
maior diminuição, próximo da inibição na banda de menor peso molecular resultante do processo
de degradação observada no sobrenadante de parasitos tratados com BEL (Fig. 16A). Para avaliar
se a BEL era capaz de interferir na síntese protéica analisamos a presença de tubulina, proteína de
citoesqueleto e não secretada pelo parasito, utilizando um anticorpo monoclonal anti-α-tubulina
(Fig. 16B). Não foram observadas variações no perfil das bandas entre células controle e tratadas
com BEL. Ao analisar a expressão de gp63 por fluorescência notamos que a BEL não causa
alterações significativas na intensidade da marcação total de gp63 observada por citometria de fluxo
(Fig. 16C), ao microscópio de fluorescência não observamos alteração na distribuição desta
molécula entre parasitos na ausência (Figs 16D e E) ou presença de BEL (Figs 16F e G).
57
72KDa
250KDa
130KDa
95KDa
55KDa
36KDa
17KDa
Sobrenadante totalExtrato total
controlecontrole tratado tratado
28KDa
A B
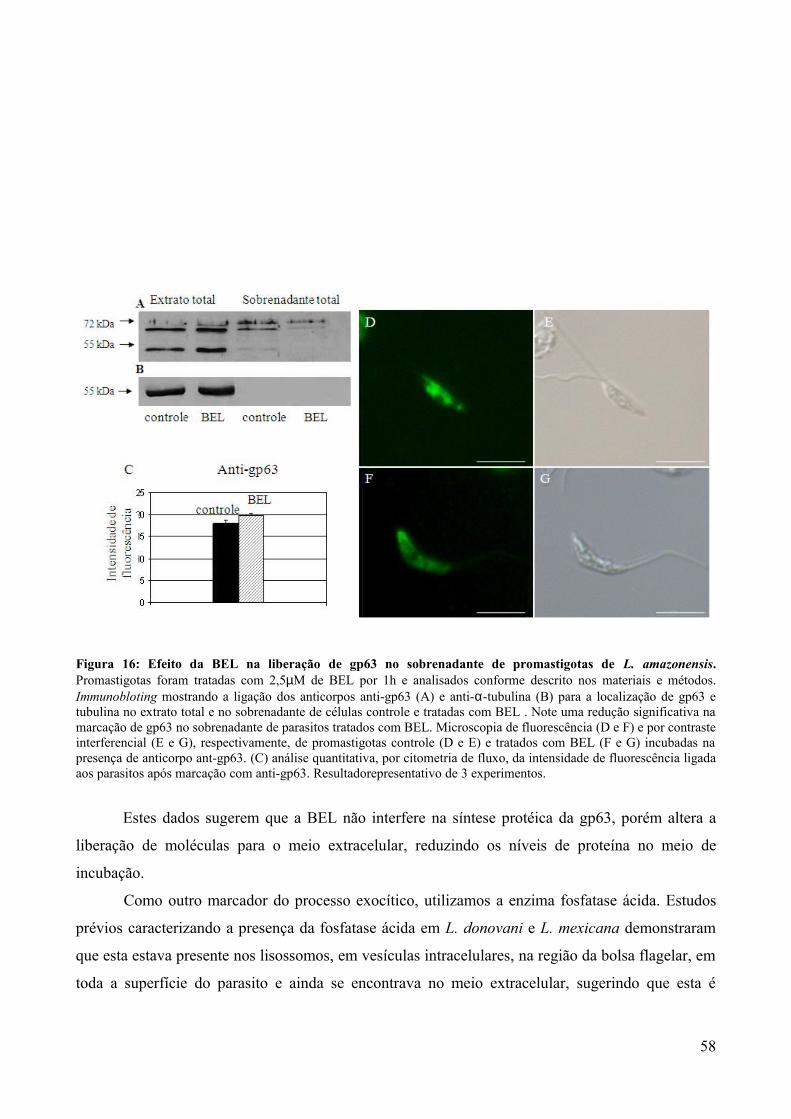
Figura 16: Efeito da BEL na liberação de gp63 no sobrenadante de promastigotas de L. amazonensis. Promastigotas foram tratadas com 2,5µM de BEL por 1h e analisados conforme descrito nos materiais e métodos. Immunobloting mostrando a ligação dos anticorpos anti-gp63 (A) e anti-α-tubulina (B) para a localização de gp63 e tubulina no extrato total e no sobrenadante de células controle e tratadas com BEL . Note uma redução significativa na marcação de gp63 no sobrenadante de parasitos tratados com BEL. Microscopia de fluorescência (D e F) e por contraste interferencial (E e G), respectivamente, de promastigotas controle (D e E) e tratados com BEL (F e G) incubadas na presença de anticorpo ant-gp63. (C) análise quantitativa, por citometria de fluxo, da intensidade de fluorescência ligada aos parasitos após marcação com anti-gp63. Resultadorepresentativo de 3 experimentos.
Estes dados sugerem que a BEL não interfere na síntese protéica da gp63, porém altera a
liberação de moléculas para o meio extracelular, reduzindo os níveis de proteína no meio de
incubação.
Como outro marcador do processo exocítico, utilizamos a enzima fosfatase ácida. Estudos
prévios caracterizando a presença da fosfatase ácida em L. donovani e L. mexicana demonstraram
que esta estava presente nos lisossomos, em vesículas intracelulares, na região da bolsa flagelar, em
toda a superfície do parasito e ainda se encontrava no meio extracelular, sugerindo que esta é
58
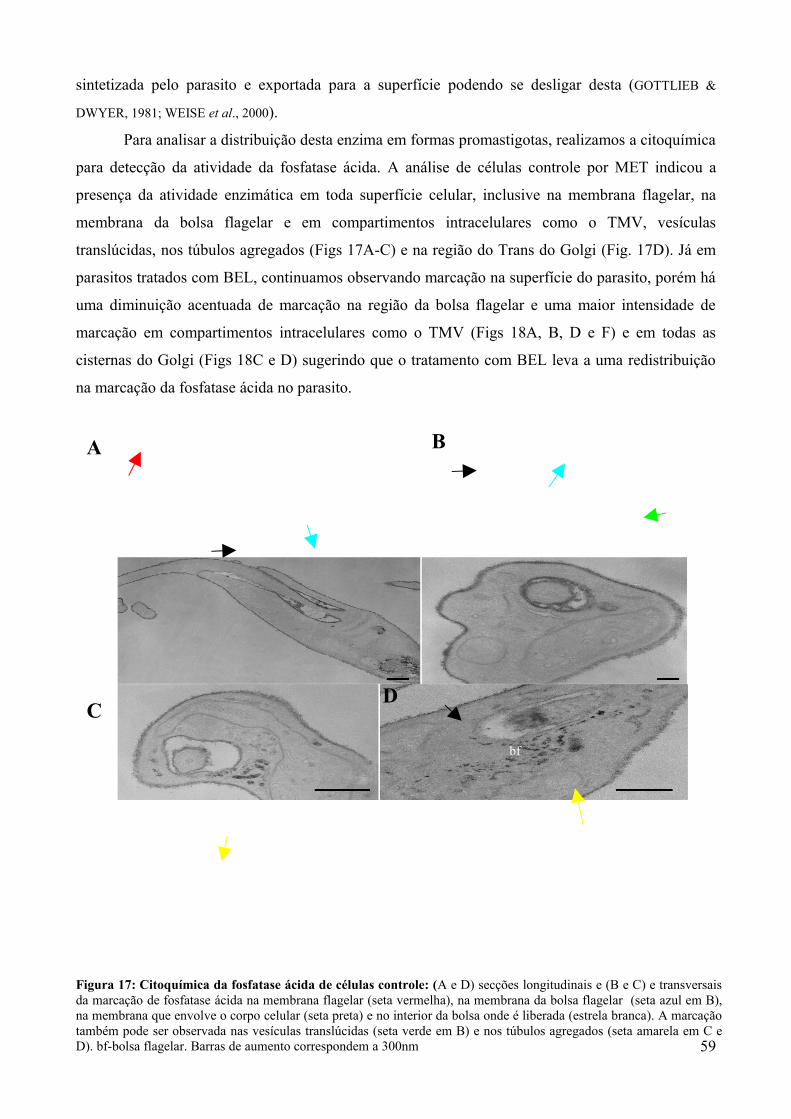
sintetizada pelo parasito e exportada para a superfície podendo se desligar desta (GOTTLIEB &
DWYER, 1981; WEISE et al., 2000).
Para analisar a distribuição desta enzima em formas promastigotas, realizamos a citoquímica
para detecção da atividade da fosfatase ácida. A análise de células controle por MET indicou a
presença da atividade enzimática em toda superfície celular, inclusive na membrana flagelar, na
membrana da bolsa flagelar e em compartimentos intracelulares como o TMV, vesículas
translúcidas, nos túbulos agregados (Figs 17A-C) e na região do Trans do Golgi (Fig. 17D). Já em
parasitos tratados com BEL, continuamos observando marcação na superfície do parasito, porém há
uma diminuição acentuada de marcação na região da bolsa flagelar e uma maior intensidade de
marcação em compartimentos intracelulares como o TMV (Figs 18A, B, D e F) e em todas as
cisternas do Golgi (Figs 18C e D) sugerindo que o tratamento com BEL leva a uma redistribuição
na marcação da fosfatase ácida no parasito.
Figura 17: Citoquímica da fosfatase ácida de células controle A-corte longitudinal B e C corte transversal observe a marcação acentuada na superfície e na região da bolsa flagelar e D-vesículas marcadas no caminho do Golgi a região da bolsa flagelar.
59
Figura 17: Citoquímica da fosfatase ácida de células controle: (A e D) secções longitudinais e (B e C) e transversais da marcação de fosfatase ácida na membrana flagelar (seta vermelha), na membrana da bolsa flagelar (seta azul em B), na membrana que envolve o corpo celular (seta preta) e no interior da bolsa onde é liberada (estrela branca). A marcação também pode ser observada nas vesículas translúcidas (seta verde em B) e nos túbulos agregados (seta amarela em C e D). bf-bolsa flagelar. Barras de aumento correspondem a 300nm
A B
CD
bf
bf
bf
bf
G
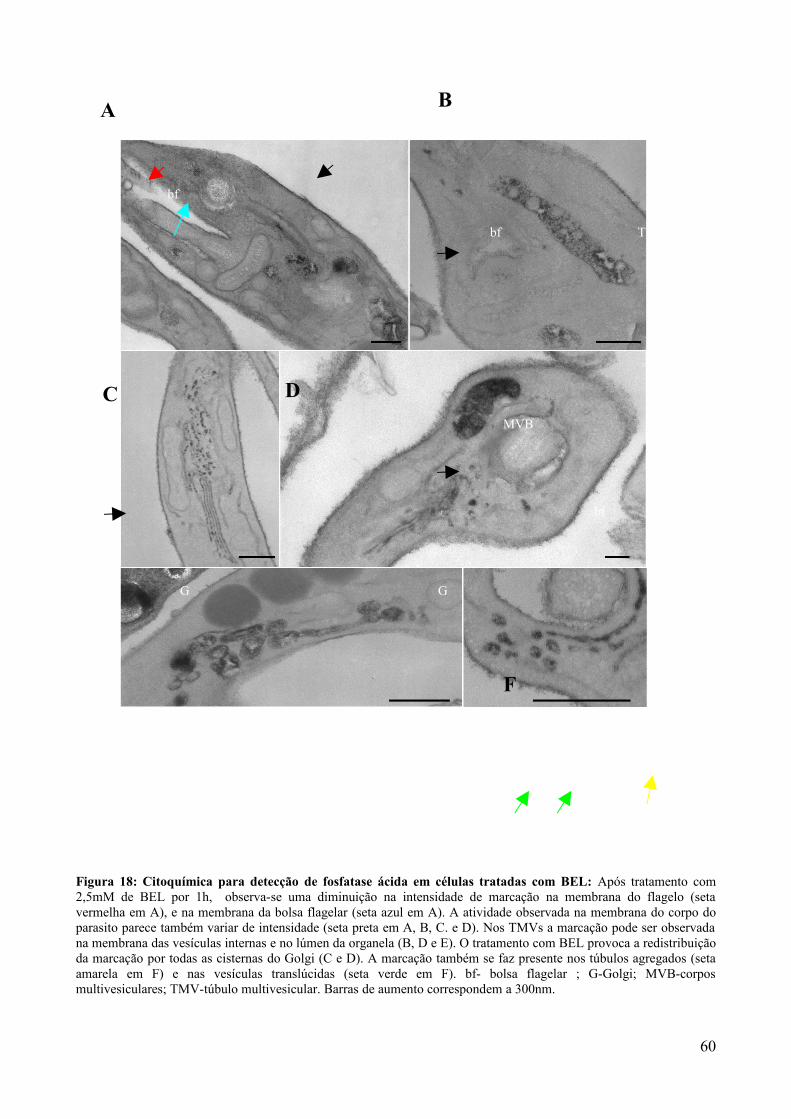
Figura 18: Citoquímica para detecção de fosfatase ácida em células tratadas com BEL: Após tratamento com 2,5mM de BEL por 1h, observa-se uma diminuição na intensidade de marcação na membrana do flagelo (seta vermelha em A), e na membrana da bolsa flagelar (seta azul em A). A atividade observada na membrana do corpo do parasito parece também variar de intensidade (seta preta em A, B, C. e D). Nos TMVs a marcação pode ser observada na membrana das vesículas internas e no lúmen da organela (B, D e E). O tratamento com BEL provoca a redistribuição da marcação por todas as cisternas do Golgi (C e D). A marcação também se faz presente nos túbulos agregados (seta amarela em F) e nas vesículas translúcidas (seta verde em F). bf- bolsa flagelar ; G-Golgi; MVB-corpos multivesiculares; TMV-túbulo multivesicular. Barras de aumento correspondem a 300nm.
60
A B
C D
E F
bf
bf
bf
bf
GG
MVB
TMV
TMV

Além da análise citoquímica realizamos estudos bioquímicos para a caracterização da
atividade de fosfatase ácida das formas promastigotas (Fig. 19). Como comentado anteriormente a
atividade de fosfatase ácida em Leishmania apresenta características próprias como a localização
(ligada à membrana ou secretada), à especificidade em relação ao substrato (GOTTLIEB E DWYER,
1982) e em relação a diferentes inibidores (ALMEIDA-AMARAL et al., 2006). Observamos que a
atividade fosfatase ácida sensível a tartarato é majoritária tanto no extrato celular como no meio
extracelular e que a atividade tartarato resistente é mais expressiva no extrato celular. Quando
comparamos a atividade das células controle com as células tratadas com BEL observamos uma
redução de 43% (10,1 nM/ml) na atividade fosfatásica sensível ao tartarato observada no meio de
incubação (Fig. 19, comparar barras brancas com pontos pretos ligadas por linha tracejada
vermelha). Coincidentemente observamos um aumento na atividade fosfatásica do extrato celular
de células incubadas com BEL de 10,2 nM/ml (Fig. 19, comparar barras listradas ligadas por linha
tracejada vermelha) indicando que a inibição da iPLA2 leva a uma diminuição na secreção da
fosfatase ácida e consequentemente um acúmulo no parasito tratado. Na figura 19B observamos a
atividade da enzima 3`nucleotidase como controle de síntese protéica e como marcador de
integridade de membrana plasmática, uma vez que somente está presente na membrana plasmática
não sendo secretada para o meio extracelular.
A seguir fomos analisar a distribuição da fosfatase ácida por citoquímica ultraestrutural na
presença do inibidor tartarato de sódio. Em células tratadas com tartarato observamos uma inibição
significativa da atividade enzimática em toda superfície celular e em compartimentos tubulares
intracelulares. Pequena atividade pode ser observada próxima a bolsa flagelar e na região do Golgi
(Figs. 20B e C). Em parasitos tratados com BEL e tartarato, observamos total inibição de marcação
na superfície do parasito e na região da bolsa flagelar e uma menor intensidade de marcação em
compartimentos intracelulares próximos a bolsa flagelar, nos túbulos agregados (Figs. 20D e F).
61

Figura 19: (A) Análise quantitativa da atividade da fosfatase ácida total (coluna preta), sensível (coluna listratada) e resistente (coluna branca) a tartarato de sódio observada no extrato celular de células controle e tratadas com 2,5µM de BEL por 1h. Observar também a atividade de fosfatase ácida secretada total (coluna cinza) sensível (coluna pontilhada) e resistente (coluna azul) ao tartarato de células controle e tratadas com BEL. (B) Análise quantitativa da atividade da enzima 3`nucleotidase, marcador de membrana plasmática, nas células controle e após tratamento com BEL. Não foi observada diferença significativa nos níveis desta atividade. Resultados representativos de 3 experimentos. (* p<0,05)
62
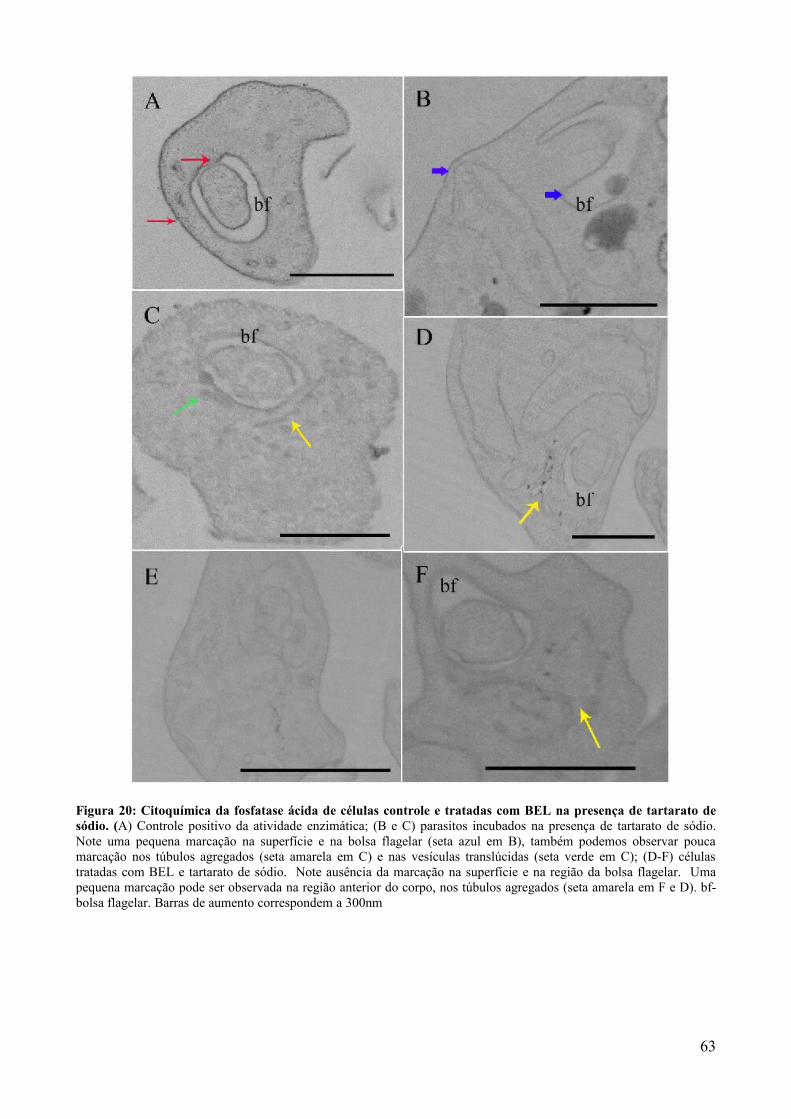
Figura 20: Citoquímica da fosfatase ácida de células controle e tratadas com BEL na presença de tartarato de sódio. (A) Controle positivo da atividade enzimática; (B e C) parasitos incubados na presença de tartarato de sódio. Note uma pequena marcação na superfície e na bolsa flagelar (seta azul em B), também podemos observar pouca marcação nos túbulos agregados (seta amarela em C) e nas vesículas translúcidas (seta verde em C); (D-F) células tratadas com BEL e tartarato de sódio. Note ausência da marcação na superfície e na região da bolsa flagelar. Uma pequena marcação pode ser observada na região anterior do corpo, nos túbulos agregados (seta amarela em F e D). bf-bolsa flagelar. Barras de aumento correspondem a 300nm
63

5- DISCUSSÃO
Os estudos com fosfolipases A2 em mamíferos descrevem seu importante papel no processo
inflamatório. Porém outro papel vem sendo sugerido para esta enzima: o do seu envolvimento na
fusão de membranas dos compartimentos envolvidos nas rotas endo/exocíticas de diferentes tipos
celulares (MAYORGA et al., 1993; DE FIGUEIREDO, 2000). A hidrólise de glicerofosfolipídios na
membrana pelas PLA2s gera lisofosfolipídios, que assumem a forma de cones invertidos
provocando a formação de curvaturas nas membranas e favorecendo a formação de túbulos em
membranas (BROWN, CHAMBERS, DOODY, 2003; STAVENA, ANGELOVA & KOUMANOV, 2004;
TANAKA et AL., 2004; NAKANO et al., 2009), e também ácidos graxos livres como o ácido aracdônico
(AA) cuja participação nos eventos de fusão foi também demonstrada (MEERS et al., 1988).
Diferentes grupos mostraram que a atividade de PLA2 está relacionada diretamente com os
processos de endocitose e exocitose em diferentes modelos celulares (LENNARTZ ET al., 1997;
TAKUMA & ICHIDA, 1997; DARIOS, CONNEL, DAVLETOV, 2007).
Já foram descritos em T. cruzi diferentes tipos de fosfolipases ligadas ao remodelamento de
membrana celular e ao processo de invasão (BERTELLO et al., 2000; BRONIA et al., 1999). Outros
patógenos também usam a fosfolipase no processo infeccioso, como: Toxoplasma gondii,
Cryptosporidium parvum e Entamoeba histolytica (PASSERO et al.,2008;VARGAS-VILLARREAE et al.,
1998). A análise genômica e a atividade enzimática em Leishmania indicaram a presença da enzima
PLA2 em promastigotas de L. amazonensis (PASSERO et al., 2008).
Pouco ainda se sabe sobre a dinâmica do processo endocítico/exocítico em
tripanosomatídeos, portanto demonstrar o envolvimento do iPLA2 na modificação da membrana
plasmática e seu envolvimento na fusão de vesículas no processo de aquisição e secreção de
moléculas em tripanosomatídeos poderia ajudar a entender o mecanismo de transferência de
material vesicular nestes processos, uma vez que estas vias são essenciais para a nutrição, defesa e
virulência do parasito, sendo assim, possíveis alvos quimioterápicos.
A abordagem mais simples para avaliar a função de uma enzima se dá através da inibição de
sua atividade. Anestésicos locais, como a dibucaína e a quinacrina foram algumas das primeiras
drogas utilizadas como inibidores da atividade de iPLA2 em células de mamíferos resultando na
inibição da fusão entre compartimentos da via endo/exocítica com reduções significativas nos
níveis de proteínas endocitadas e excretadas, respectivamente (TOLLESHAUG, BERG & HOLT, 1982;
HAGIWARA & OZAWA, 1990; LENNARTZ et al., 1997; DE FIGUEIREDO et al., 1998). Como anestésicos
locais podem potencialmente interagir com diferentes tipos de proteínas dentre elas receptores de
membrana, enzimas e proteínas do citoesqueleto (MAYORGA et al., 1993; HOLMANN et al., 2001;
MONDAL & CHAKRABARTI, 2002) além de provocarem efeitos diretos na bicamada lipídica, novos
64

compostos inibidores da atividade de PLA2 foram desenvolvidos apresentando uma alta
especificidade para diferentes tipos de fosfolipases. Dentre os compostos mais específicos para a
inibição da iPLA2 está a bromoenol lactona que foi escolhida neste estudo como ferramenta para
investigar a participação desta enzima na fusão de compartimentos da via endo/exocítica de
Leishmania (LENNARTZ et al., 1997; MURAKAME & KUDO, 2004).
Mais recentemente foi observado que a BEL além de inibir a iPLA2, pode inibir uma
fosfohidrolase Mg2+dependente cuja atividade está associada a mudanças no metabolismo de
fosfolipídios podendo promover a morte celular (BAUSINDE & DENNIS, 1996; FUENTES et al., 2003).
Por este motivo, primeiramente realizamos a curva de crescimento para mapear a ação da BEL em
promastigotas de L. amazonensis em função da concentração e tempo de tratamento. Alcançamos
uma concentração ideal do inibidor de 2,5 µM para a realização dos experimentos que é
intermediária àquelas utilizadas em diferentes modelos de células de mamíferos (ACKERMANN,
CONDE-FRIEBOES & DENNIS, 1995; DE FIGUEIREDO et al., 1998, 2001). Os testes de viabilidade
mostraram que nas doses e tempos de incubação utilizados nos nossos diferentes experimentos a
porcentagem de morte celular não foi significativa.
A primeira questão levantada sobre a ação da BEL foi se ela alteraria a morfologia e
localização de organelas das vias endocítica e secretória de tripanosomatídeos. Estudos prévios em
células de mamíferos utilizando diferentes inibidores de iPLA2 mostraram a formação de “blebs” e
fragmentação do complexo de Golgi (DE FIGUEIREDO et al., 1998) e que esta ação é específica, uma
vez que não são observadas em lisossomos e compartimentos pré-lisossomais (DE FIGUEIREDO,
2000). Com base nestes dados, analisamos o efeito da BEL na ultra-estrutura da L. amazonensis por
MET. As alterações observadas no Golgi dos promastigotas, e que podem ser descritas como
fragmentação, são bastante semelhantes àquelas observadas em células de mamíferos e que são
atribuídas a uma alteração no tráfego de vesículas entre o retículo endoplasmático e o Golgi
(MAYORGA et al., 1993; DE FIGUEIREDO et al., 1998). Outras alterações que observamos nos parasitos e
que também foram descritas em outros modelos celulares foram a do alargamento das cisternas na
região do Trans-Golgi e do aumento do diâmetro dos diferentes compartimentos localizados na
região anterior do corpo do parasito entre o núcleo e a bolsa flagelar. Nesta região onde na verdade
se dá toda a movimentação de moléculas a serem endocitadas e exocitadas notamos um aparente
aumento no número de vesículas provavelmente derivadas dos túbulos agregados e das vesículas
translúcidas. A alteração do TMV, considerado um compartimento endossomal/lisossomal foi
também caracterizada pelo aumento aparente do diâmetro dos túbulos e das vesículas internas. Em
outros modelos celulares o aumento do diâmetro das vesículas indica alteração no tráfego (inibição
ou retardo no processo de fusão de compartimentos) que leva a uma inibição tanto do processo de
endocitose como de exocitose (MAYORGA et al., 1993; LENNARTZ et al., 1997; DE FIGUEIREDO et al.,
65

1998). Todas as observações feitas no nosso trabalho necessitam de confirmação por estudos
morfométricos e reconstrução tridimensional além de estudos de pulso e caça com marcadores
fluorescentes ou complexados a ouro coloidal para avaliação da cinética dos processos de fusão e
técnicas imunocitoquímicas para identificação dos diferentes compartimentos.
A pergunta que se seguiu foi se a BEL estaria inibindo a endocitose de macromoléculas em
promastigotas.
O processo endocítico em eucariotos é de grande importância para a captação de nutrientes,
ajuste do volume celular e para a modulação da sinalização celular. Lipídeos são importantes
reguladores de diversas etapas do processo endocítico (SORKIN & VON ZASTROW, 2002).
Em Leishmania a captação de moléculas ocorre pela bolsa flagelar. Já foi descrito a
endocitose de fase fluida e a endocitose mediada por receptor, sendo caracterizados receptores de
transferrina (VOYIATZAKY & SOTERIADOU et al.,1992), LDL (BASTIN et al., 1996) e hemoglobina
(SINGH et al., 2003). Analisamos, por citometria de fluxo e fluorescência, a endocitose de
macromoléculas marcadoras dos dois processos endocíticos citados acima em promastigotas
incubadas com BEL uma vez que já existem dados na literatura mostrando que inibidores da iPLA2
como a dibucaína inibem o processo endocítico de transferrina em células musculares de rato
(HAGIWARA & OZAWA, 1990) de BSA em T. cruzi (SOUTO-PADRÓN et al., 2006) por diferentes
inibidores de PLA2. A endocitose de BSA e transferrina por promastigotas de L. amazonensis
apresentou características bem distintas entre si. O tratamento com BEL provocou uma inibição de
aproximadamente 64% na endocitose de BSA, que foi bem semelhante à inibição observada em
epimastigotas de T. cruzi incubados na presença de dibucaína (SOUTO-PADRÓN et al., 2006). O padrão
da marcação com a BSA é intracelular nas células controle e tratadas. A diminuição na intensidade
de marcação mostra diminuição na quantidade adquirida pelo parasito.
A endocitose de transferrina é relativamente mais afetada pelo tratamento com BEL. A
inibição foi de 72% e o padrão de marcação dos parasitos tratados é bem diferente daquele
observado nos parasitos controle. Nas células controle a presença de transferrina é intracelular
(vesículas observadas na região posterior do corpo dos promastigotas) enquanto nos parasitos
tratados a marcação permanece na superfície sugerindo que há realmente uma inibição do processo
endocítico.
Estudos prévios sugerem um possível envolvimento de domínios de membrana ricos em
colesterol (lipid rafts) na captação de transferrina em epimastigotas de T. cruzi (PORTO-CARREIRO et
al.,2000). Estas são regiões de membrana resistentes a detergentes que têm sido demonstrados em T.
brucei, T. cruzi, L. major e L. (Vianna) braziliensis (NOLAN et al., 2000; DENNY et al., 2005;
YONCYAMA et al., 2006). Estes autores demonstram uma redução dramática na captação de
transferrina em parasitos submetidos a tratamento com filipina e metil-β-ciclodextrina (MbCD),
66

estes compostos afetam o dinamismo de membrana em microdomínios ricos em colesterol. Já foi
mostrado que a atividade de PLA2 na célula é importante para o disparo, desenvolvimento e
finalização do processo de formação de “vesículas transportadoras de rafts” derivadas tanto da
membrana plasmática como de organelas e que a interferência neste mecanismo poderia
comprometer o tráfego intracelular de vesículas (STANEVA, ANGELOVA & KOUMANOV, 2004). A
ação da PLA2 sobre a movimentação das rafts poderia explicar tanto a diminuição na quantidade de
transferrina fluorescente ligada ao parasito (menor quantidade de receptores voltando para ou se
incorporando à superfície) como o padrão difuso de marcação mostrando uma dispersão dos
receptores já presentes na superfície por todo o corpo do parasito. Outro aspecto importante é que a
inibição simples dos processos de fusão entre compartimentos, independentemente da contribuição
das lipid rafts, podem atrasar o processo de aquisição de transferrina ou qualquer outra molécula. A
célula se encontraria em um estado de paralisia.
A análise da marcação com ConA não mostrou diferença significativa na intensidade de
fluorescência medida por citometria de fluxo entre células controle e tratadas ao contrário do
observado em células CHO (SCHWARZ, HARPER & JULIANO, 1982). Selecionamos uma concentração
não aglutinante (20µg/ml) mediante experimentos realizados em outros tripanosomatídeos e
observação do comportamento dos promastigotas após incubação com lectina fluorescente quando
não observamos grumos de células. Ao microscópio de fluorescência as células controle se
destacam das células tratadas com BEL apenas por uma pequena marcação que pode ser observada
no interior celular próximo ao núcleo, região correspondente ao TMV, já que a marcação de
superfície e da bolsa flagelar parecem bem semelhantes entre controle e tratado. Uma melhor
análise desta marcação será feita utilizando a lectina complexada a particulas de ouro coloidal e
análise ao microscópio eletrônico de transmissão. Chamamos, no entanto, a atenção para o fato de
que a marcação de ConA em L. mexicana, diferentemente do observado em T. brucei é
caracterizada pela marcação extracelular (BRICKMAN, COOK & BALBER, 1995).
Uma vez que a BEL está diminuindo a internalização de moléculas no processo endocítico e
este pode compartilhar compartimentos citoplasmáticos com a via exocítica e ambas as vias
parecem compartilhar compartimentos intracelulares semelhantes aos endossomos de células de
mamíferos o passo seguinte foi verificar se a iPLA2 também estaria relacionada com a fusão de
membranas no tráfego exocítico. Fomos então analisar a ação da BEL sobre a secreção de
moléculas em promastigotas de L. amazonensis uma vez que já foi demonstrado que inibidores da
iPLA2 podem inibir a exocitose em mastócitos (FENSOME-GEEN et al., 2007).
Observamos que o tratamento com BEL reduz em aproximadamente 50 % a liberação de
proteínas no meio extracelular. Surpreendentemente, não observamos o acúmulo destas proteínas
não secretadas no gel do extrato celular em promastigotas tratados. Se em células tratadas com BEL
67

menos proteínas eram encontradas no meio de incubação e estatisticamente não se observava um
aumento no número de proteínas no extrato celular de parasitos tratados, será que a BEL estaria
interferindo na síntese de proteínas pelo parasito? Estudos prévios já demonstraram que o ácido
araquidônico e seus metabólitos podem interferir na proliferação de diferentes tipos celulares,
incluindo células endoteliais (HERBERT et al., 2005). Mediante esta informação o nosso passo
seguinte foi analisar se BEL interferiria na síntese protéica de Leishmania amazonensis o que não
aconteceu (Fig. 16B). Então, como explicar a mesma intensidade, em relação ao controle,
apresentada por algumas bandas no extrato celular de promastigotas com BEL. Na verdade em
algumas delas observamos até mesmo um pequeno decréscimo (Fig. 15).
A superfície celular de todo tripanosomatídeo, incluindo espécies de Leishmania é
constituída predominantemente por moléculas ancoradas na membrana por âncoras de
glicosilfosfatidilinositol (GPI). GPI é sintetizado a partir de fosfatidilinositóis convencionais (PIs)
que contém uma cadeia de ácido graxo insaturado na posição sn-2, enquanto que os GPIs maduros,
ancorados à proteína, têm diacilglicerol com ácido graxo de cadeia longa na posição sn-2 (SIPOS et
al., 1997). Após a fixação do GPI à proteína, o grupo acila ligado ao resíduo inositol da âncora de
GPI é eliminado, sendo este um processo necessário para o controle de qualidade de GPI ancorado
a proteína e para seu transporte eficiente do RE para o Golgi e posteriormente à superfície celular
(TANAKA et al., 2004; FUJITA et al., 2006). Em células de mamíferos e em leveduras, o sítio de
remodelação da âncora é o Golgi (TASHIMA et al., 2006) ou o RE e o Golgi, respectivamente (SIPOS et
al., 1997). O mecanismo molecular que evolve a remodelação lipídica não é bem compreendido,
porém dados sugerem que uma PLA2 poderia estar envolvida na remoção de uma cadeia acila na
posição sn-2 das âncoras de GPI durante a remodelação da âncora em leveduras e células de
mamíferos (FUJITA et al., 2006).
A partir dos dados mostrados acima, podemos sugerir que uma vez que em parasitos
tratados com BEL não estaria ocorrendo remodelação lipídica da âncora de GPI, comprometendo,
assim a exportação das moléculas recém-sintetizadas e promovendo o acúmulo destas no RE. Estes
processos levariam à degradação das mesmas explicando então a menor intensidade nas bandas do
extrato celular dos parasitos tratados com BEL em relação ao observado no controle. Ainda são
necessários mais estudos para confirmar esta hipótese.
O estudo de proteínas secretadas tem contribuído para um melhor entendimento dos
mecanismos envolvidos na rota secretora de parasitos e outros tipos celulares (WEISE et al., 2000).
Para confirmarmos a ação da iPLA2 no processo exocitico em L. amazonensis utilizamos como
marcadores duas moléculas específicas do parasito, gp63 e fosfatase ácida, que são direcionadas
para a membrana plasmática podendo também ser encontradas no meio extracelular. A gp63 que é
uma das proteínas predominantemente expressa na superfície de Leishmania (membrana celular e
68

membrana da bolsa flagelar) é ancorada por ancora de GPI e associada à “lipid rafts” no RE ou
compartimentos muito iniciais da via exocítica (WEISE et al., 2000; DENNY, FIELD & SMITH, 2001;
DENNY et al., 2004). Observamos que o tratamento com BEL reduziu a quantidade de gp63 liberada
no meio extracelular mas não modificou, pelo menos nas imagens obtidas por imunofluorescência,
a localização da gp63 na superfície da célula que normalmente se encontra distribuída
homogeneamente por toda a superfície do parasito (WEISE et al., 2000). A quantificação desta
marcação utilizando o anticorpo anti-gp63 será efetuada em estudos de imunocitoquímica
utilizando cortes ultrafinos, quando a distribuição da proteína poderá ser também avaliada nos
diferentes compartimentos intracelulares.
Atualmente já esta bem estabelecida que as vesículas transportadoras contendo enzimas
hidrolíticas destinadas aos lisossomos são oriundas do Golgi e fundem-se com endossomas precoce
e então são transferidas ao longo da via endocítica até os lisossomos (GRUENBERG, 2003) ou
transferidas pela via exocítica a superfície celular e ao meio extracelular. A fosfatase ácida é uma
enzima que está presente em endossomos, lisossomos, Golgi e membrana plasmática de
Leishmania sendo um bom parâmetro bioquímico e citoquímico para avaliar modificações nestes
sistemas. A inibição de iPLA2 não alterou a atividade total de fosfatase ácida nos promastigotas. Ela
altera a distribuição da atividade tartarato sensível (atividade de fosfatase ácida secretada) entre
extrato celular e meio de incubação mostrando uma inibição na secreção desta enzima. A
observação da marcação citoquímica ao MET das células tratadas com BEL mostrou um aumento
do número e/ou volume de compartimentos intracelulares marcados e a diminuição da atividade
fosfatásica no meio extracelular principalmente aquela liberada no interior da bolsa flagelar. Para
avaliar as modificações estruturais associadas à detecção da atividade enzimática serão necessários
estudos de reconstrução tridimensional por microscopia eletrônica de transmissão e o uso de
substratos e inibidores específicos para as diversas atividades de fosfatase ácida existentes em
Leishmania (GOTTLIEB & DWYER, 1982; ALMEIDA-AMARAL et al., 2006).
69

6- CONCLUSÕES
- Inibição da atividade de iPLA2 parece não interferir na disposição das organelas de promastigotas,
no entanto provocou alterações no formato de organelas como Golgi, túbulos agregados e vesículas
translúcidas.
- BEL interferiu significativamente no processo endocítico de BSA (endocitose de fase fluida) e de
transferrina (endocitose mediada por receptor). No caso da endocitose de transferrina notamos
também uma redistribuição dos receptores pela superfície dos promastigotas. A marcação em
compartimentos tubulares intracelulares pela ConcanavalinaA foi ausente em parasitos tratados com
BEL, porem a intensidade de fluorescência não foi alterada pela análise por citometria de fluxo.
-A quantidade de gp63 liberada no meio de incubação foi reduzida com o tratamento com BEL. A
falta de acúmulo da proteína no extrato celular sugere que a BEL pode estar interferindo no
remodelamento da âncora de GPI.
- BEL inibiu a secreção de fosfatase ácida no meio extracelular provocando um acúmulo no interior
do parasito em organelas como o TMV.
- iPLA2 parece estar envolvida na regulação dos processos endocítico e exocíticos de promastigotas de L. amazonensis, sendo seu efeito mais pronunciado no processo exocítico e na endocitose mediada por receptor.
70

7- REFERÊNCIAS BIBLIOGRÁFICAS
ACKERMANN EJ, CONDE-FRIEBOES K, DENNIS EA. 1995. Inhibition of macrophage Ca(2+)-independent phospholipase A2 by bromoenol lactone and trifluoromethyl ketones. J Biol Chem. 270:445-450.
AKIBA S & SATO T. 2004. Cellular function of calcium-independent phospholipase A2. Biol Pharm Bull. 27 :1174-1178.
ALBERIO SO, DIAS SS, FARIA FP, MORTARA RA, BARBIÉRI CL, FREYMÜLLER HAAPALAINEN E. 2004. Ultrastructural and cytochemical identification of megasome in Leishmania ( Leishmania ) chagasi . Parasitol Res. 92: 246-254.
ALEXANDER J, SATOSKAR AR, RUSSELL DG. 1999. Leishmania species: models of intracellular parasitism. J Cell Sci. 18:2993-3002.
AMLABU E, NOK AJ, SALLAU AB. 2009. Purification and biochemical characterization of lysosomal acid phosphatases (EC 3.1.3.2) from blood stream forns, Trypanosoma brucei brucei. Parasitol Int. 58: 238-242.
ANTOINE JC, JOUANNE C, RYTER A, BENICHOU JC. 1988. Leishmania amazonensis: acidic organelles in amastigotes. Exp Parasitol. 67:287-300.
ANTOINE JC, LANG T, PRINA E, COURRET N, HELLIO R. 1999. H-2M molecules, like MHC class II molecules, are targeted to parasitophorous vacuoles of Leishmania-infected macrophages and internalized by amastigotes of L. amazonensis and L. mexicana. J Cell Sci. 112:2559–2570.
BALSINDE J & DENNIS EA. 1996. Bromoenol lactone inhibits magnesium-dependent phosphatidate phosphohydrolase and blocks triacylglycerol biosynthesis in mouse P388D1 macrophages. J Biol Chem. 271:31937-31941.
BANGS JD, ANDREWS NW, HART GW, ENGLUND PT. 1986. Posttranslational modification and intracellular transport of a trypanosome variant surface glycoprotein. J Cell Biol. 103:255-263.
BARBIERI CL, DOINE AL, FREYMULLER F. 1990. Lysosomal depletion in macrophages from spleen and foot lesions Leishmania-infected hamster. Exp. Parasitol. 71: 218-228
BASTIN P, STEPHAN A, RAPER J, SAINT-REMY JM, OPPERDOES FR, COURTOY PJ. 1996. An M(r) 145,000 low-density lipoprotein (LDL)-binding protein is conserved throughout the Kinetoplastida order. Mol Biochem Parasitol. 76:43-56
BATES PA & DWYER DM. 1987. Biosynthesis and secretion of acid phosphatase by Leishmania donovani promastigotas. Mol Biochem Parasitol 26: 289-296.
BATES PA, HERMES I, DWYER M. 1990. Golgi-mediated pos-translation processing of secretory acid phosphatase by Leishmania donovani promastigotas. Mol Biochem Parasitol 39: 247-55.
BATES PA, HERMS I, DWYER M. 1989: Leishmania donovani: Immunochemical localization and secretory mechanism of soluble acid phosphatase. Exp Parasitol 68: 335-346
BERTELLO LE, ALVES MJ, COLLI W, DE LEDERKREMER RM. 2000. Evidence for phospholipases from Trypanosoma cruzi active on phosphatidylinositol and inositolphosphoceramide. Biochem J. 345:77-84.
BHATNAGAR S, SHINNAGAWA K, CASTELLINO FJ, SCHOREY JS. 2007. Exosomes released from macrophages infected with intracellular pathogens stimulate a pro-inflammatory response in vitro and in vivo. Blood 110: 3234-3244.
BLOCK MR, GLICK BS, WILCOX CA, WIELAND FT, ROTHMAN JE. 1988. Purification of an N-ethylmaleimide-sensitive protein catalyzing vesicular transport. Proc Natl Acad Sci U S A. 85:7852-7856.
BONNON C, WENDELER MW, PACCAUD JP, HAURI HP. 2010. Selective export of human GPI-anchored proteins from the endoplasmic reticulum. J Cell Sci. 123:1705-1715.
71

BRADFORD M. 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72:248-254.
BRIGHT NA, GRATIAN MJ, LUZIO JP. 2005. Endocytic delivery to lysosomes mediated by concurrent fusion and kissing events in living cells. Curr Biol. 15:360–365.
BRITTINGHAM A, MORRISON CJ, MCMASTER WR, MCGWIRE BS, CHANG KP, MOSSER DM. 1995. Role of the Leishmania surface protease gp63 in complement fixation, cell adhesion, and resistance to complement-mediated lysis. J Immunol. 155:102-111.
BRONIA DH, PEREIRA BM, LUJÁN HD, FRETES RE, FERNÁNDEZ A, PAGLINI PA. 1999. Ganglioside treatment of acute Trypanosoma cruzi infection in mice promotes long-term survival and parasitological cure. Ann Trop Med Parasitol. 93:341-350.
BROWN TE, WRIGHT JW, SPETH RC. 2003. Phospholipase C inhibitor U73122 attenuates the dipsogenic response induced by angiotensin II. Proc West Pharmacol Soc. 46: 61-63.
BROWN WJ, CHAMBERS K, DOODY A. 2003. Phospholipase A2 (PLA2) enzymes in membrane trafficking: mediators of membrane shape and function. Traffic 4: 214-221.
BROWN WJ. 2001. Inhibition of transferring recycling and Endosome Tubulation by Phospholipase A2 Antagonists. J. Biol. Chem. 276: 47361-47370.
CHANG KP, CHAUDHURI G, FONG D. 1990. Molecular determinants of Leishmania virulence. Annu Rev Microbiol. 44: 499-529.
CHAPPUIS F, SUNDAR S, HAILU A, GHALIB H, RIJAI S, PEELING RW,ALVAR J, BOELAERT M 2007. Visceral Leishmania: what are the needs for diagnosis treatment and control? Nat Rev Microbiol. 5: 873-882.
CHAVES I, RUDENKO G, DIRKS-MULDER A, CROSS M, BORST P. 1999. Control of variant surface glycoprotein gene-expression sites in Trypanosoma brucei . EMBO J. 18:4846-4855.
CHEN J, RAUCH CA, WHITE JH, ENGLUND PT, COZZARELLI NR. 1995. The topology of kinetoplast. Cell 80: 61-69.
CHISTOFORIDIS S, MIACZYNSKA M, ASHMAN K, WILM M, ZHAO L, YIP SC, WATERFIELD MD, BACKER JM, ZERIAL M. 1999. Phosphatidylinosital-3-OH kinases are Rab are Rab5 effectors. Nat. Cell Biol. 1: 249-252.
CLAGUE MJ. 1998. Molecular aspects of the endocytic pathway. Biochem. J. 336: 271-282.
COLOMBO MI. 1996. Role for NSF on vesicular transport: insights from in vitro endosome fusion. Bio Cell 20:317-323.
COPPENS I, BALDHUIN P, OPPERDOES F, COURTOY PJ. 1988. Receptor for the host low lipoproteins on the hemoflagellate Trypanosoma brucei: purification and involvement in the growth of the parasite. Proc. Natl. Acad. Sci. USA 85: 6753-6757.
COPPENS I, OPPERDOES FR, COURTOY PJ, BAUDHUIN P. 1987. Receptor-mediated endocytosis in the bloodstream form of Trypanosoma brucei. J. Protozool. 34: 465-473.
CORRADIN S, RANSIJN A, CORRADIN G, BOUVIER J, DELGADO MB, FERNANDEZ-CARNEADO J, MOTTRAM JC, VERGÈRES G, MAUËL J. 2002. Novel peptide inhibitors of Leishmania gp63 based on the cleavage site of MARCKS (myristoylated alanine-rich C kinase substrate)-related protein. Biochem J. 367:761-769.
CORRÊA JR., ATELLA GC, BATISTA MM, SOARES MJ. 2008. Transferrin uptake in Trypanosoma cruzi is impaired by interference on cytostome associated cytoskeleton elements and stability of membrane cholesterol, but not by obstruction of clathrin-dependent endocytosis. Exp Parasitol. 119: 58–66.
72

CORRÊA JR, ATELLA GC, MENNA-BARRETO RS, SOARES MJ. 2007. Clathrin in Trypanosoma cruzi: in silico gene identification, isolation, and localization of protein expression sites. J Eukaryot Microbiol. 54: 297-302.
CREUTZ CE, MOSS S, EDWARDSON JM, HIDE I, GOMPERTS B. 1992. Differential recognition of secretory vesicles by annexins. European Molecular Biology Organization Course "Advanced Techniques for Studying Secretion". Biochem Biophys Res Commun. 184:347-352.
CUERVO P, SANTOS AL, ALVES CR, MENEZES GC, SILVA BA, BRITTO C. 2008: Cellular localization and expression of gp63 homologous metalloproteases in Leishmania (Viannia) braziliensis strains. Acta Trop 106: 143-148.
DE FIGUEIREDO P, DRECKTRAH D, KATZENELLENBOGEN JA, STRANG M, BROWN WJ. 1998. Evidence that phospholipase A2 activity is required for Golgi complex and Trans-Golgi network membrane tubulation. Proc Natl Acad Sci U S A. 95:8642-8647.
DE FIGUEIREDO P, DRECKTRAH D, POLIZOTTO RS, COLE NB, LIPPINCOTT-SCHWARTZ J, BROWN WJ, 2000. Phospholipase A2 antagonists inhibit constitutive retrograde membrane traffic to the endoplasmic reticulum. Traffic. 1:504-511.
DE FIGUEIREDO P, DOODY A, POLIZOTTO RS, DRECKTRAH D, WOOD S, BANTA M, STRANG MS, BROWN WJ. 2001. Inhibition of transferrin recycling and endosome tubulation by phospholipase A2 antagonists. J Biol Chem. 276:47361-70.
DENNY PW, MORGAN GW, FIELD MC, SMITH DF. 2005. Leishmania major : clathrin and adaptin complexes of an intra-cellular parasite. Exp Parasitol. 109:33-37.
DE SOUZA W, SANT`ANNA C, CUNHA E SILVA NL. 2009. Electron microscopy and cytochemistry analyses of the endocytic pathway of phatogenic protozoa. Prog. Histochem. Cytochem. 44:67-124.
DE SOUZA, 1995. Structural organization of the cell surface of Trypanososmatidae. Micron 26: 405-430.
DE SOUZA W. 2002a. Basic cell biology of Trypanosoma cruzi. Curr. Pharm. Des. 8: 269-283.
DE SOUZA W. 2002b. Special organelles of some patogenic protozoa. Parasitol. Res. 88: 1013-1025.
DELLA SBARBA P, RODOVIDA E. 2002. Transmodulation of cell surface regulatory molecules via ectodomain shedding. Biol. Chem. 383: 69-83.
DENNY PW, FIELD MC, SMITH DF. 2001. GPI-anchored proteins and glycoconjugates segregate into lipid rafts in Kinetoplastida. FEBS Lett.491:148-53
DOMIN J & WATERFIELD MD. 1997. Using structure to define the function of phosphoinositide 3-kinase family members. FEBS Lett. 410:91-95.
ELLIS SL, SHAKARIAN AM, DWYER DM. 1998: Leishmania amastigotes synthesize conserved secretory acid phosphatase during human infection. Exp Parasitol 89: 161-168.
ESKELINEN EL TANAKA Y, SAFTIG P. 2003. At the acidic edge: emerging functions for lysosomal membrane proteins. Trens Cell Biol. 13:137-145.
FENSOME-GREEN A, STANNARD N, LI M, BOLSOVER S, COCKCROFT S. 2007. Bromoenol lactone, an inhibitor of Group V1A calcium-independent phospholipase A2 inhibits antigen-stimulated mast cell exocytosis without blocking Ca2+ influx. Cell Calcium. 41:145-153.
FERGUSON MA. 1999. The structure, biosynthesis and functions of glycosylphosphatidylinositol anchors, and the contributions of trypanosome research. J Cell Sci.112: 2799-2809.
FIELD H, FARJAH M, PAL A, GULL K, FIELD MC. 1998. Complexity of trypanosomatid endocytosis pathways revealed by Rab4 and Rab5 isoforms in Trypanosome brucei. J. Biol. Chem. 273: 32102-32110.
73

FIELD H, SHERWIN T, SMITH AC, GULL K, CROFT SL, FIELD MC. 1999. TbRab2p, a marker of the endocytosis reticulum of Trypanosoma brucei, localizes to the ERGIC in mammalian cells. J. Cell Sci. 112: 147-156.
FUENTES L, PÉREZ B, NIETO ML, BALSINDE, BALBOA MA. 2003. Bromoenol lactone promotes cell death by a mechanism involving phosphatidate phosphohydrolase-1 rather than calcium-independent phospholipase A2. J Biol Chem. 278: 44683-44690.
FUJITA M, UMEMURA M, YOKO-O T, JIGAMI Y. 2006. PER1 is required for GPI-phospholipase A2 activity and involved in lipid remodeling of GPI-anchored proteins Mol Biol Cell.17:5253-5264.
GAULLIER, JM, SIMONSEN A, D’ARRIGO A, BREMNES B, STENMARK H, AASLAND R. 1998. FYVE fingers bind PtdIns(3)p. Nature 394: 432-433.
GEIGER A, HIRTZ C, BÉCUE T, BELLARD E, CENTENO D, GARGANI D, ROSSIGNOL M, CUNY G, PELTIER JB,2010. Exocytosis and protein secretion in Trypanosoma . BMC Microbiol. 10:20.
GHEDIN E, DEBRABANT A, ENGEL JC, DWYER DM. 2001. Secretory and endocytic pathways converge in a dynamic endosomal system in a primitive protozoan. Traffic. 2:175–188.
GLEW RH, CZUCZMAN MS, DIVEN WF, BERENS RL, POPE MT, KATSOULIS DE. 1982. Partial purification and characterization of particulate acid phosphatase of Leishmania donovani promastigotes. Comp Biochem Physiol B. 72:581-590.
GOLDENBERG RI, SMITH RM, JARETT L. 1987. Insulin and α2− macroglobulin-methilamine undergo endocytosis by different mechanisms in rat adipocytes. I. Comparison of cell surface events. J Cell Physiol. 133: 203-212.
GOTTLIEB M, DWYER DM. 1981: Leishamania donovani: surface membrane acid phosphatase activity of promastigotas. Exp Parasitol 52: 117-128.
GOTTLIEB M & DWYER DM 1982: Identification and partial characterization of an extracellular acid phosphatase activity of Leishmania donovani promastigotas. Mol Cel Biol. 2: 76-81.
GRANER MW, ALZATE O, DECHKOVSKAIA AM, KEENE JD, SAMPSON JH, MITCHELL DA, BIGNER DD. 2009. Proteomic and immunologic analyses of brain tumor exosomes. FASEB J 23: 1541-1557.
GROOTEN J, GOOSSENS V, VANHAESEBROECK B, FIERS W. 1993. Cell membrane permeabilization and cellular collapse, followed by loss of dehydrogenase activity: early events in tumour necrosis factor-induced cytotoxicity. Cytokine.5:546-55.
GRUENBERG J. 2003. Lipids in endocytic membrane transport and sorting. Curr Opin Cell Biol. 15: 382-388.
GRUNFELDER CG, ENGSTLER M, WEISE F, SCHWARZ H, STIERHOF YD, MORGAN GW, et al. 2003. Endocytosis of a glycosylphosphatidylinositol-anchored protein via clathrin-coated vesicles, sorting by default in endosomes, and exocytosis via RAB11-positive carriers. Mol Biol Cell. 14: 2029–2040.
HAGIWARA, Y. & OZAWA, E. 1990. Suppresion of transferring internalization in myogenic L6 cells by dibucaine. Biochim. Biophys. Acta 1051: 237-241.
HERBERT SP, PONNAMBALAM S, WALKER JH. 2005. Cytosolic phospholipase A2-alpha mediates endothelial cell proliferation and is inactivated by association with the Golgi apparatus. Mol Biol Cell. 16: 3800-3809.
HOARE CA & WALLACE FG. 1996. Developmental stages of trypanosomatid flagellates: a new terminology. Nature. 212: 1385-1386.
JEFFRIES TR, MORGAN GW, FIELD MC. 2001. A developmentally regulated Rab11 homologue in Trypanosoma brucei is involved in recycling processes. J Cell Sci. 114: 2617-2626.
JENNE N, FREY K, BRUGGER B, WIELAND FT. 2002. Oligomeric state and stoichiometry of p24 proteins in the early secretory pathway. J Biol Chem. 277: 46504-46511.
74

KAPETANOVICH L, BAUGHMAN C, LEE TH, 2005. Nm23H2 facilitates coat protein complex II assembly and endoplasmic reticulum export in mammalian cells. Mol Biol Cell. 16: 835-848.
KATAKURA K & KOBAYASHI A. 1988. Acid phosphatase activity of virulent and avirulent clones of Leishmania donovani promastigotas. Infect Immun. 56: 2856-2860.
KELLER S, SANDERSON MP, STOECK A, ALTEVOGT P. 2006. Exosomes from biogenesis and secretion to biological function. Immunology Letters 107: 102-108.
KINI RM. 2005. Structure-function relationship and mechanism ofanticoagulant phospholipase A2 enzyme from snake venoms. Toxicon. 45: 1147-61.
KWEIDER M, LEMESRE JL, DARCY F, KUSNIERZ JP, CAPRON A, SANTORO F. 1987. Infectivity of Leishmania braziliensis promastigotes is dependent on the increasing expression of a 65,000-dalton surface antigen. J Immunol. 138: 299-305.
LAEMMLI UK. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227: 680-685.
LANDFEAR SM, IGNATUSHCHENKO M. 2001. The flagellum and flagellar pocket of trypanosomatids. Mol Biochem Parasitol. 115: 1–17.
LAUBER K, BOHN E, KROBE SM, XIAO Y J, BLUMENTHAL SG, LINDEMANN RK, MARINI P, WIEDIG C, ZOBYWALSKI A, BAKSH S, XU Y, AUTENRIETH IB, SCHULZE-OSTHOFF K, BELKA C, STUHLER G, WESSELBORG S. 2003. Apoptotic cells induce migration of phagocytes via caspase-3-mediated release of a lipid attraction signal. Cell 113: 717-730.
LEAL ST, ARARIPE JR, URMÉNYI TP, CROSS GA, RONDINELLI E. 2000. Trypanosoma cruzi : cloning and characterization of a RAB7 gene. Exp Parasitol. 96: 23-31.
LEE MC, MILLER EA, GOLBERG J, ORCI L, SCHEKMAN R. 2004. Bi-directional protein transport between the ER and Golgi. Annu Rev Cell Dev Biol. 20: 87-123.
LENNARTZ MR, YUEN A, MASI SM, RUSSELL DG, BUTTLE KF, SMITH JJ. 1997. Phospholipase A2 inhibition results in sequestration of plasma membrane into electronlucent vesiscles during IgG-mediated phagocytosis. J Cell Sci. 110: 2041-2052.
LIEGEOIS S, BENEDETTO A, GARNIER JM, SCHWAB Y, LABOUESSE M. 2006. The vo-ATPase mediates apical secreton of exosomes containing hedgehog-related proteins in Caenorhabditis elegans. J. Cell Biol. 173: 949-961.
LIMA MT & VILLATA F. 1990. Trypanosoma cruzi receptor for human trasferrin and their role. Mol Biochem Parasitol. 38: 245-252.
LODGE R & DESCOTEAUX A. 2005. Modulation of phagolysosome biogenesis by the lipophosphoglycan of Leishmania . Clin Immunol. 114: 256-265.
MAYORGA LS, COLOMBO MI, LENNARTZ M, BROWN EJ, RAHMAN KH, WEISS R, LENNON PJ, STAHL PD. 1993. Inhibition of endosome fusion by phospholipase A2 (PLA2) inhibitors points to a role for PLA2 in endocytosis. Proc Natl Acad Sci USA. 90:10255-10259.
MCCONVILLE MJ, MULLIN KA, ILGOUTZ SC, TEASDALE RD. 2002. Secretory pathway of trypanosomatid parasites. Microbiol. Mol. Biol. Rev. 66 : 122-154.
MCGWIRE BS, O`CONNEL WA, CHANG KP, ENGMAN DM. 2002. Extracellular release of the glycosylphospfatidylinositol (GPI)-linked Leishmania surface metalloprotease, gp63, is independent of GPI phospholipolysis: implication for parasite virulence. J Biol Chem. 277: 8802-8809.
MEERS P, HONG K, PAPAHADJOPOULOS D. 1988. Free fatty acid enhancement of cation-induced fusion of liposomes: synergism with synexin and other promoters of vesicle aggregation. Biochemistry. 27: 6784-6794.
75

MENDONÇA SM, CAMPOS CB, GUEIROS-FILHO FJ, LOPES UG. 1993. Identification of GTPase genes in the protozoa parasites Trypanosoma cruzi and Leishmania amazonensis . Biol Res. 26: 3-9.
MENDONÇA SM, NEPOMUCENO DA SILVA JL, CUNHA E SILVA N, DE SUZA W, LOPES UG. 2000. Characterization of a Rab11 homologue in Trypanosoma cruzi. Gene 243: 179-185.
MEYER H. 1968. The fine structure of the flagellum and kinetoplast-chondriome of the Trypanosoma (Schizotrypanum) cruzi in tissue culture. J. Protozool. 15: 614-621.
MICHALIK M, PIERZCHALSKA M, PABIANCZYK-KULKA A, KOROHODA W. 2003. Procaine-induced enhancement of fluid-phase endocytosis and inhibition of exocytosis in human skin fibroblasts. Eur J Pharmacol. 475: 1-10.
MORGAN A, BURGOYNE RD. 1990. Relationship between arachidonic acid release and Ca2(+)-dependent exocytosis in digitonin-permeabilized bovine adrenal chromaffin cells. Biochem. J. 271:571-4. (thais a referência esta errada, esta aqui é o artigo em que tirei a informação, amanha posso te explicar minha ideia)
MORGAN GW, HALL BS, DENNY PW, CARRINGTON M, FIELD MC. 2002a. The kinetoplastida endocytic apparatus. Part I: a dynamic system for nutrition and evasion of host defences. Trends Parasitol. 18: 491-496.
MORGAN GW, HALL BS, DENNY PW, FIELD MC, CARRINGTON M. 2002b. The endocytic apparatus of the kinetoplastida. Part II: machinery and components of the system. Trends Parasitol. 18: 540-546.
MORTARA RA. 1989. Studies on trypanosomatid actin. I. Immunochemical and biochemical identification. J Protozool. 36: 8-13.
MOULAY L, ROBERT-GERO M. 1995. Leishmania donovani: enhanced expression of soluble acid phosphatase in the presence of sinefungin, an antiparasitic agent. Exp Parasitol. 80: 8-14.
MURAKAMI M. 2004. Diversity of Phospholipase A2 Enzymes. Hot Topics in Phospholipase A2 Field. Biol Pharm Bull. 27:1179-82.
MURAKAMI M. & KUDO I. 2004. Secretory phospholipase A2. Biol. Pharm. Bull. 27: 1158-1164.
NGUYEN DG, BOOTH A, GOULD SJ, HILDRETH JEK. 2003. Evidence that HIV budding in primary macrophages occurs through the exososome release pathway. J. Biol. Chem. 278: 52347-52354.
NAKANO T, INOUE I, SHINOZAKI R, MATSUI M, AKATSUKA T, TAKAHASHI S, TANAKA K, AKITA M, SEO M, HOKARI S, KATAYAMA S, KOMODA T. 2009. A possible role of lysophospholipids produced by calcium-independent phospholipase A(2) in membrane-raft budding and fission Biochim Biophys Acta 788: 2222-2228.
NOLAN DP, GARCIA-SALCEDO JA. 2008. Loss of actin does not affect export of newly synthesized proteins to the surface of Trypanosoma brucei. Mol Biochem Parasitol. 157:233–235.
OVERATH P & ENGSTLER M. 2004. Endocytosis, membrane recycling and sorting of GPI-anchored proteins: Trypanosoma brucei as model system. Mol. Microbiol. 53: 735-744.
PAL A, HALL BS, NESBETH DN, FIELD HI, FIELD MC. 2002. Differential endocytic functions of Trypanosoma brucei Rab5 isoforms reveal a glycosylphosphatidylinositol-specific endosomal pathway. J Biol Chem. 277:9529–9539.
PALADE GE, BRUNS RR.1986. Structural modulations of plasmalemmal vesicles. J Cell Biol. 37:633-649.
PARTON RG, SIMONS K. 2007. The multiple faces of caveolae. Nat Rev Mol Cell Biol. 8:185–194.
PASSERO LF, LAURENTI MD, TOMOKANE TY, CORBETT CE, TOYAMA MH. 2008. The effect of phospholipase A2 from Crotalus durissus collilineatus on Leishmania (Leishmania) amazonensis infection. Parasitol Res.102: 1025-1033.
PATEL N, SINGH SB, BASU SK, MUKHOPADHYAY A. 2008. Leishmania requires Rab7-mediated degradation of endocytosed hemoglobin for their growth. Proc Natl Acad Sci U S A. 105: 3980-3985.
76

PETERS C, KAWAKAMI M, KAUL M, ILG T, OVERATH P, AEBISCHER T. 1997. Secreted proteophosphoglycan of Leishmania mexicana amastigotes activates complement by triggering the mannan binding lectin pathway. Eur J Immunol. 27: 2666-2672.
PIANI A, ILG T, ELEFANTY AG, CURTIS J, HANDMAN E. 1999. Leishmania major proteophosphoglycan is expressed by amastigotes and has an immunomodulatory effect on macrophage function. Microbes Infect. 1: 589-599.
PIMENTA PF, TURCO SJ, MCCONVILLE MJ, LAWYER PG, PERKINS PV, SACKS DL. 1992. Stage-specific adhesion of Leishmania promastigotes to the sandfly midgut. Science. 256: 1812-1815.
PORTO-CARREIRO I, ATTIAS M, MIRANDA K, DE SOUZA W, CUNHA-E-SILVA N. 2000. Trypanosoma cruzi epimastigote endocytic pathway: cargo enters the cytostome and passes through a nearly endosomal network before storage in reservosomes. Eur J Cell Biol. 79: 858–869.
PORTO-CARREIRO I. 2004. Contribuição pelo estudo da via endocítica das formas epimastigotas de Trypanosoma cruzi. IBCCF-UFRJ.
PUENTES, S M., DA SILVA R P, SACKS DL, HAMMER CH, JOINER KA. 1990. Serum resistance of metacyclic stage Leishmania major promastigotes is due to release of C5b-9. J Immunol. 145: 4311-4316.
PUPKIS MF, TETLEYL, COOMBSGH. 1986. Leishmania mexicana: amastigote hydrolases in unusual lysosomes. Exp Parasitol, 62: 29–39.
RAMALEY AT, DAS S, CAMPBELL PI, LAROCCA GM, POPE MT, GLEW RH. 1985. Characterization of Leishmania donovani acid phosphatase. Biol Chem. 25: 880-886.
RAMEH LE & CANTLEY LC. 1999. The role of phosphoinositide 3-kinase lipid products in cell fuction. J. Biol. Chem. 247: 8347-8350.
RASMUSSON B & DESCOTEAUX A. 2004. Contribution of electron and confocal microscopy in the study of Leishmania -macrophage interactions. Microsc Microanal. 10: 656-661.
REYNOLDS RW. 1963. The use of lead Citrate at high pH as an electron opaque stain in electron microscopy. J. Cell Biol. 17:208.
ROBINSON MS. 1987. 100-KD coated vesicle proteins: Molecular heterogeneity and intracellular distribution studied with monoclonal antibodies. J Cell Biol. 104: 887-895.
ROBINSON MS. 1994. The role of clathrin, adaptors and dynamin in endocytosis. Curr. Oppin. Cell Biol. 6: 538-544.
RODMAN SJ & WANDINGER-NESS A. 2000. Rab GTPases coordinate endocytosis. J Cell Sci. 113: 183-192.
RODRIGUES ML, NAKAYASU ES, OLIVEIRA DL, NIMRICHTER L, NOSANCHUK JD, ALMEIDA LC, CASADEVALL A. 2008. Extracellular vesicles produced by Cryptococcus neoformans contain protein components associated with virulence. Eukaryotic Cell 7: 58-67.
ROTH TF & PORTER KR. 1964. Yolk protein uptake in the oocyte of mosquito Aedes aegypti. J Cell Biol. 20: 313-332.
ROTHBERG KG, HEUSER JE, DONZELL WC, TING YS, GLENNEY JR, ANDERSON RG. 1992. Caveolin, a protein component of caveolae membrane coats. Cell 68: 673-682
ROUX S, COLASANTE C, SAINT CLOMENT C, BARBIER J, CURIE T, GIRARD E, MOLGÓ J, BRÛLET P. 2005. Internalization of a GFP-tetanus toxin C-terminal fragment fusion protein at mature mouse neuromuscular junctions. Mol Cell Neurosci. 30: 572-582.
RUSSELL DG, WILHELM H. 1986. The envolvement of the major surface glyprotein (gp63) of leishmania promastigotes in attachment to macrophages. J Immunol. 136: 2613-20
77

SAHASRABUDDHE AA, BAJPAI VK, GUPTA CM. 2004. A novel form of actin in Leishmania: molecular characterisation, subcellular localisation and association with subpellicular microtubules. Mol Biochem Parasitol. 134: 105–114.
SALVATORE J, TURCO E , DESCOTEAUX . 1992. The lipophosphoglycan of Leishmania parasites, Annual Review of Microbiology. 46 : 65-92.
SANT'ANNA C, DE SOUZA W, CUNHA-E-SILVA N. 2004. Biogenesis of the reservosomes of trypanosoma cruzi. Microsc Microanal.10:637-46.
SANT'ANNA C, PARUSSINI F, LOURENÇO D, DE SOUZA W, CAZZULO JJ, CUNHA-E-SILVA NL. 2008. All Trypanosoma cruzi developmental forms present lysosome-related organelles. Histochem Cell Biol. 130:1187-1198.
SANTOS AL, SOUTO-PADRÓN T, ALVIANO CS, LOPES AH, SOARES RM, MEYER-FERNANDES JR. 2002. Secreted phosphatase activity induced by dimethyl sulfoxide in Herpetomonas samuelpessoai. Arch Biochem Biophys.405:191-198.
SCHARZ G, DOBBERSTEIN B. 1996. Common principles of protein translocation across membranes. Science 271: 1519-1526.
SCHLEIN Y, JACOBSON RL, SHLOMAI J. 1991. Chitinase secreted by Leishmania functions in the sandfly vector. Proc Biol Sci. 245: 121-126.
SCHNITZER JE, OH P, PINNEY E, ALLARD J. 1994. Filipin-sensitive caveolae mediated transport in endothelium: reduced transcytosis scavenger endocytosis, and capillary permeability of selected macromolecules. J Cell Biol. 127: 1217-1232.
SCHOIJET AC, MIRANDA K, GIRARD-DIAS W, DE SOUZA W, FLAWIÁ MM, TORRES HN, DOCAMPO R, ALONSO GD. 2008. A Trypanosoma cruzi phosphatidylinositol 3-kinase (TcVps34) is involved in osmoregulation and receptor-mediated endocytosis. J Biol Chem. 283: 31541-31550.
SCHWARZ MA, HARPER PA, JULIANO RL. 1982. Interactions of lectins with CHO cell surface membranes. II. Differential effects of local anesthetics on endocytosis of Con A and WGA binding sites. J Cell Physiol. 111:264-274
SEED JR, BYRAM J, GAM AA. 1967. Characterization and localization of acid phosphatase activity of trypanosome gambiense. J Protozool. 14: 117-125.
SILVEIRA FT, LAINSON R, CORBETT CE. 2004. Clinical and immunopathological spectrum of American cutaneous leishmaniasis with special reference to the disease in Amazonian Brazil: a review. Mem Inst Oswaldo Cruz. 99:239-51
SILVERMAN JM, CLOS J, OLIVEIRA CC, SHIRVANI O, RANG Y, WANG C, FOSTER LJ, REINER N. 2010. An exosome-based secretion pathway is responsible for protein export from Leishmania and communication with macrophages. J. Cell Sci.123: 842-852. (THAIS, O ANO DESTE ARTIGO QUE ESTAVA ERRADO)
SIMONSEN AC. 2008. Activation of phospholipase A2 by ternary model membranes. Biophys J. 94: 3966-3975.
SINGH SB, TANDON R, KRISHNAMURTHY G, VIKRAM R, SHARMA N, BASU SK, MUKHOPADHYAY A. 2003. Rab5-mediated endosome-endosome fusion regulates hemoglobin endocytosis in Leishmania donovani. EMBO J. 22: 5712-5722.
SIPOS G, REGGIORI F, VIONNET C, CONZELMANN A. 1997. Alternative lipid remodelling pathways for glycosylphosphatidylinositol membrane anchors in Saccharomyces cerevisiae. EMBO J. 16: 3494-3505.
SOARES MJ, DE SOUZA W. 1988. Cytoplasmic organelles of trypanosomatids: a cytochemical and stereological study. J Submicrosc Cytol Pathol. 20: 349-361.
SOARES MJ, SOUTO-PADRÓN T, DE SOUZA W. 1992. Identification of a large pre-lysosomal compartment in the pathogenic protozoon Trypanosoma cruzi . J Cell Sci. 102: 157-167.
78

SORKIN A, VON ZASTROW M. 2002. Signal transduction and endocytosis: close encounters of many kinds. Nat Rev Mol Cell Biol. 3: 600-614.
SOUTO-PADRÓN T, LIMA AP, RIBEIRO RO. 2006. Effects of dibucaine on the endocytic/exocytic pathways in Trypanosoma cruzi . Parasitol Res. 99: 317-320.
STEVERDING D. 2000. The transferring receptor of Trypanosoma brucei. Parasitol Int. 48: 191-198.
STEVERDING D, STIERHOF YD, CHAUDHRI M, LIGTENBERG M, SCHELL D, BECK-SICKINGER AG, OVERATH P. 1994. ESAG 6 and 7 products of Trypanosoma brucei form a transferring binding protein complex. Eur. J Cell Biol. 64: 78-87.
STIERHOF YD, BATES PA, JACOBSON RL, ROGERS ME, SCHLEIN Y, HANDMAN E, ILG T. 1999. Filamentous proteophosphoglycan secreted by Leishmania promastigotes forms gel-like three-dimensional networks that obstruct the digestive tract of infected sandfly vectors. Eur J Cell Biol. 78: 675-689.
SUBRAMANYA S, MENSA-WILMOT K. 2010. Diacylglycerol-stimulated endocytosis of transferrin in trypanosomatids is dependent on tyrosine kinase activity. PLoS One.1: 8538.
TAKAI Y, SASAKI T, MATOZAKI T. 2001. Small GTP-binding proteins. Physiol. Rev. 81: 153-208.
TANAKA S, MAEDA Y, TASHIMA Y, KINOSHITA T. 2004. Inositol deacylation of glycosylphosphatidylinositol-anchored proteins is mediated by mammalian PGAP1 and yeast Bst1p. J Biol Chem. 279: 14256-14263.
TARABOLETTI G, D'ASCENZO S, BORSOTTI P, GIAVAZZI R, PAVAN A, DOLO V. 2002. Shedding of the matrix metalloproteinases MMP-2, MMP-9, and MT1-MMP as membrane vesicle-associated components by endothelial cells. Am J Pathol. 160: 673-680.
TASHIMA Y, TAGUCHI R, MURATA C, ASHIDA H, KINOSHITA T, MAEDA Y. 2006. PGAP2 is essential for correct processing and stable expression of GPI-anchored proteins. Mol Biol Cell. 17: 1410-1420.
TOKER A, CANTLEY LC. 1997. Signaling through the lipid products of phosphoinositide-3-OH kinase. Nature. 387: 673-676.
TOLLESHAUG H, BERG T, HOLTE K. 1982. Effects of local anesthetics and related compounds on the endocytosis and catabolism of asialo-glycoproteins in isolated hepatocytes. Biochim Biophys Acta. 714: 114-121.
TOWBIN H, STAEHELIN T, GORDON J. 1979. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proc. Natl. Acad. Sci., 76: 4350-4354.
TURCO SJ. 1992. The lipophosphoglycan of Leishmania . Subcell Biochem, 18: 73-97.
UEDA-NAKAMURA T, ATTIAS M, DE SOUZA W. 2001. Megasome biogenesis in Leishmania amazonensis: a morphometric and cytochemical study. Parasitol Res. 87: 89–97.
VAN GRIENSVEN J, BALASEGARAM M, MEHEUS F, ALVAR J, LYNEN L, BOELAERT M. 2010. Combination therapy for visceral leishmaniasis. Lancet Infect Dis.10:184-94.
VARGAS-VILLARREAL J, OLVERA-RODRÍGUEZ A, MATA-CÁRDENAS BD, MARTÍNEZ-ROGRÍGUEZ HG, SAID-FERNÁNDEZ S, ALAGÓN-CANO A. 1998. Isolation of an Entamoeba histolytica intracellular alkaline phospholipase A2. Parasitol. 84: 310-314 VIDUGIRIENE J, MENON AK. 1994. The GPI anchor of cell-surface proteins is synthesized on the cytoplasmic face of the endoplasmic reticulum. J Cell Biol. 127: 333-341.
79

VIEIRA M, ROHLOFF P, LUO S, CUNHA-E-SILVA NL, DE SOUZA W, DOCAMPO R. 2005. Role for a P-type H+-ATPase in the acidification of the endocytic pathway of Trypanosoma cruzi. Biochem J.392:467-74.
VITETTA ES, UHR JW. 1972. Cell surface immunoglobulin. V. Release from murine splenic lymphocytes. J Exp Med. 136: 676-696.
VOYIATZAKY CS, SOTERIADOU KP. 1992. Identification and isolation of the Leishmania transferring receptor. J Biol Chem. 267: 9112-9117.
WEISE F, STIERHOF YD, KUHN C, WIESE M, OVERATH P. 2000. Distribution of GPI-anchored proteins in the protozoan parasite Leishmania, based on an improved ultrastructural description using high-pressure frozen cells. J Cell Sci. 113: 4587-4603.
WILSON ME, LEWIS TS, MILLER MA, MCCORMICK ML, BRITIGAN BE. 2002. Leishmania chagasi: uptake of iron bound to lactoferrin or transferrin requires an iron reductase. Exp Parasitol. 100: 196–207.
WINBERG ME, HOLM A, SÄRNDAHL E, VINET AF, DESCOTEAUX A, MAGNUSSON KE, RASMUSSON B, LERM M. 2008. Calcium-independent phospholipase A2: structure and function. Biochim Biophys Acta. 1488: 28-39.
WINBERG ME, HOLM A, SÄRNDAHL E, VINET AF, DESCOTEAUX A, MAGNUSSON KE, RASMUSSON B, LERM M. 2009. Leishmania donovani lipophosphoglycan inhibits phagosomal maturation via action on membrane rafts. Microbes Infect. 11: 215-222.
WINSTEAD MV, BALSINDE J, DENNIS EA. 2000. Calcium-independent phospholipase A(2): structure and function. Biochim Biophys Acta. 1488: 28-39.
YAO C, DONELSON JE, WILSON ME. 2003. The major surface protease (MSP or GP63) of Leishmania sp.Biosynthesis, regulation of expression, and function. Mol. Biochem. Parasitol. 132: 1-6
YEDGARS, LICHTENBERG D, SCHNITZER E. 2000. Inhibition of phospholipase A2 as a therapeutic target. Biochim Biophys Acta. 1488: 182-187.
YOUNG JC, BARRAL JM, ULRICH HARTL F. 2003. More than folding: localized functions of cytosolic chaperones. Trens Biochem. Sci. 28: 541-547.
ZERIAL M & MCBRIDE H. 2001. Rab proteins as membrane organizers. Nat Rev Mol Cell Biol. 2: 107-117.
80
Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas
Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo





























