Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DA BAHIA INSTITUTO DE CIÊNCIAS DA SAÚDE
PROGRAMA DE PÓS-GRADUAÇÃO EM IMUNOLOGIA
ANA CRISTINA DE OLIVEIRA GONZALEZ
PROCESSOS IMUNOBIOLÓGICOS QUE REGULAM A BIOSSÍNTESE E O REMODELAMENTO DA MATRIZ
EXTRACELULAR NA CICATRIZAÇÃO DE FERIMENTOS CUTÂNEOS FOTOBIOMODULADOS (670nm) E NO
CONTEXTO TUMORAL.
Salvador, BA 2015

ANA CRISTINA DE OLIVEIRA GONZALEZ
PROCESSOS IMUNOBIOLÓGICOS QUE REGULAM A BIOSSÍNTESE E O REMODELAMENTO DA MATRIZ
EXTRACELULAR NA CICATRIZAÇÃO DE FERIMENTOS CUTÂNEOS FOTOBIOMODULADOS (670nm) E NO
CONTEXTO TUMORAL.
Tese apresentada ao Programa de Pós-graduação em Imunologia, da Universidade Federal da Bahia como requisito parcial para obtenção do título de Doutor em Imunologia. Orientador: Prof. Dr. Zilton de Araújo Andrade Co-orientadora: Profa Dra Alena Ribeiro Alves Peixoto Medrado
Salvador, BA 2015

Ficha Catalográfica elaborada pela Biblioteca do
Centro de Pesquisas Gonçalo Moniz / FIOCRUZ - Salvador - Bahia.
Gonzalez, Ana Cristina de Oliveira
G643p Processos imunobiológicos que regulam a biossíntese e o remodelamento da
matriz extracelular na cicatrização de ferimentos cutâneos fotobiomodulados
(670nm) e no contexto tumoral. / Ana Cristina de Oliveira Gonzalez. - 2015.
115 f. : il. ; 30 cm.
Orientador: Prof. Dr. Zilton de Araújo Andrade, Laboratório de Patologia
Experimental.
Tese (Doutorado em Imunologia) – Universidade Federal da Bahia, Instituto
de Ciências da Saúde, 2015.
1. Cicatrização de feridas. 2. Irradiação a laser de baixa potência. 3.
Imuno-histoquímica. 4. Proteínas Hedgehog. 5. Câncer bucal. I. Título.
CDU 616-003.9


Dedico este trabalho:
Aos meus pais (in memorian), por absolutamente tudo, dos mais simples conselhos
aos mais belos ensinamentos e por terem sempre acreditado em
mim.
Aos meus filhos Bruno, Ingrid e Henrique por me permitirem conhecer o amor incondicional,
pelo carinho e força em muitos momentos.
Ao esposo Almir, pelo companheirismo e apoio na busca de novos conhecimentos.

AGRADECIMENTOS
Ao Dr. Zilton Andrade, fonte de sabedoria e conhecimentos pela orientação e
colaboração neste trabalho, assim como pelos princípios éticos que regem sua
conduta pessoal e profissional.
Minha homenagem carinhosa à Alena Medrado pela competente e criteriosa
orientação, pela constante disposição em resolver dúvidas e incertezas. O
resultado desse processo é que acabamos fortalecendo mais ainda a sólida
amizade que carregaremos para sempre!
Ao Prof. Dr. Moysés Sadigursky, por ter plantado em mim as primeiras
sementes da pesquisa, seus ensinamentos fundamentais e amizade, a quem
terei gratidão eterna!
Sou muito grata à amiga Eli, pela indescritível solidariedade e afeto, que se
traduziram sempre em entusiasmada e valiosa parceria.
À Maíra e Tila, pelas sugestões e pelas inúmeras demonstrações práticas
sobre o ato de compartilhar.
À Mariana Lobão e Tamires, pelas inestimáveis colaborações neste estudo e
pelos momentos de alegria e companheirismo.
Ao Luciano, pela colaboração imprescindível na confecção das fotos deste
estudo e pela amizade.
À Isabela, pela atenção, carinho, por muitos conselhos e conversas
incentivadoras.
À Dinalva e Márcia, pelas palavras acolhedoras, sábias e a certeza de que o
trabalho seria concluído.
À Ariane, Rita, Fátima, Sr. Antônio, Miguel, Everton, Léo, Tiago Pereira, Iuri e
Tiago Landin pelo privilégio da amizade e estímulo para que eu conseguisse
completar este percurso.
Aos funcionários Lia e Presciano pela ajuda e carinho de sempre.
À Daniela Andrade, pelo apoio e colaboração na utilização do citômetro de
fluxo.
À Fiscina e Vânia, funcionárias da Biblioteca do CPqGM, pelo apoio
imprescindível dispensado.
Ao CPqGM pela oportunidade que me proporcionou.

Ao PPGIM (UFBA) pelo comprometimento na realização do curso de
doutorado, assim como os professores e funcionários. À Dilcea, secretária
deste departamento, por sua dedicação.
À banca examinadora pela disponibilidade em aceitar e compartilhar comigo
deste momento.
Finalmente, a todos aqueles que participaram direta ou indiretamente desta
conquista.

“Aqueles que passam por nós, não vão sós, não nos deixam sós. Deixam um pouco de si, levam um pouco de nós”
Antoine de Saint-Exupéry
“Ninguém pode voltar ao passado e fazer um novo começo, mas todos podem começar de novo e fazer um novo fim”
Chico Xavier

RESUMO
Os processos imunobiológicos que regulam a biossíntese e o remodelamento da matriz extracelular na cicatrização de ferimentos fotobiomodulados e no contexto tumoral foram contemplados no presente trabalho. Na primeira parte, foram descritas as fases da cicatrização integrantes do processo de reparo tecidual cutâneo, com ênfase no papel biológico desempenhado por diferentes células e suas moléculas sinalizadoras. Na segunda seção, foi realizado estudo experimental, duplo cego a fim de avaliar o efeito do laser de baixa potência na fase tardia da cicatrização, em especial em relação à angiogênese, células do sistema imune e a via de sinalização Hedgehog. Quarenta ratos foram alocados randomicamente em dois grupos experimentais, controle e tratado com laser diodo semicondutor de AsGaAl (670nm). Após indução de ferimento cutâneo padronizado, os espécimes foram eutanasiados em 14, 21, 28 e 35 dias. As secções teciduais foram submetidas à coloração com hematoxilina-eosina e à técnica de imuno-histoquímica com os anticorpos anti CD31, NG2, actina alfa de músculo liso, CD8, CD68, Ptch, Ihh e Gli-2. Os níveis séricos das citocinas IL-2, IL-4, IL-10 e IFNɣ foram avaliados através da técnica de citometria de fluxo. Todos os dados histomorfométricos foram submetidos à análise estatística e o nível de significância adotado foi p<0,05. Observou-se que a neoangiogênese persistiu, pois houve presença de CD31+(p=0,016), NG2+ e alfa actina de músculo liso (p=0,025) em ambos os grupos, sendo que no grupo tratado com laser ocorreu mais cedo. No grupo laser, aos 21 dias houve decréscimo de células CD68+ (p=0,032) e aumento de CD8+ (p=0,038). Os níveis sanguíneos de IL-2 e IL-10 foram indetectáveis em ambos os grupos experimentais, ao passo que o perfil de IL-4 e IFNɣ não foi significativo (p>0,05). Na fase do remodelamento, a sinalização da via Hedgehog pareceu estar ativada, haja vista a presença da expressão fenotípica de algumas proteínas desta via, embora, a expressão de Ihh e Ptch parece não ter sido fotobiomodulada, pois não houve diferenças estatisticamente significativas entre os grupos experimentais. A terceira parte do trabalho abordou o processo de transformação neoplásica através de um estudo comparativo de lesões pré-cancerizáveis de mucosa oral e carcinoma escamocelular, no qual foi investigada a presença de algumas proteínas da via Hedgehog em dezesseis casos humanos. Observou-se significativo aumento progressivo da expressão de todas as proteínas da via Hedgehog, tanto no epitélio quanto na lâmina própria, quando se comparou as secções de mucosa oral normal, displasia e carcinoma (p<0,05). Através da realização do primeiro estudo, conclui-se que o laser de baixa potência foi capaz de modular a fase do remodelamento da matriz extracelular durante a cicatrização experimental, uma vez que os tecidos fotobiomodulados apresentaram alterações mais precoces em relação ao grupo controle e os eventos que ocorrem no reparo tecidual parecem ser cíclicos e continuam se desenvolvendo no tecido, independente do aspecto clínico da lesão que evidencia o seu fechamento e completa cicatrização. Em relação ao estudo realizado em mucosa oral humana, observou-se que a ativação da via Hedgehog pareceu influenciar na transformação neoplásica, pois foi demonstrado aumento progressivo de células Hedgehog positivas nas lesões pré-cancerizáveis presentes no mesmo indivíduo.

Palavras-chave: Cicatrização de feridas. Laser de baixa potência. Imuno-histoquímica. Sinalização Hedgehog. Câncer bucal.

ABSTRACT
This study evaluated the immunological processes that regulate the biosynthesis and extracellular matrix remodeling in the context of photobiomodulated wound healing and tumors. Initially, the stages of the skin tissue repair process were described, with emphasis on the biological role played by different cells and their signaling molecules. Subsequently, a double-blind experimental study was conducted, in order to assess the effect of low power laser in the late stages of healing, with emphasis on angiogenesis, immune cells and Hedgehog signaling. Forty rats were allocated randomly in two groups; control and those treated with laser diode of GaAlAs (670 nm). After induction of standardized skin wounds, the specimens were euthanized at 14, 21, 28 and 35 days. Tissue sections were subjected to hematoxylin-eosin and immunohistochemical techniques with antibodies to CD31, NG2, smooth muscle alpha actin, CD8, CD68, Ptch, Gli-2 and Ihh. Serum levels of the cytokines IL-2, IL-4, IL-10 and IFNɣ were assessed by flow cytometry. All histomorphometric data were statistically analyzed and significance level was at p<0,05. We observed that neoangiogenesis persisted, because of the presence of CD31+ (p=0,016), NG2+ and smooth muscle alpha actin (p=0,025) in both groups. The laser-treated group responded earlier. By day 21, the laser-treated group had decreased CD68+ cells (p=0,032) and increased CD8+ (p=0,038).Blood levels of IL-2 and IL-10 were undectable in both experimental groups, whereas IL-4 and IFNɣ profile was not significant (p>0,05). At the remodeling stage, the Hedgehog signaling pathway seemed to be activated, given the presence of phenotypic expression of some of its proteins, although the expression of Ihh and Ptch did not seem to have been photobiomodulated because there were no statistically significant differences between the experimental groups. The third part of our study dealt with neoplastic transformation process, through a comparative study of pre-cancerous lesions of the oral mucosa and squamous cell carcinoma, in which we investigated the presence of some proteins of the Hedgehog pathway in sixteen human cases. There was a significant progressive increase in the expression of all proteins in the Hedgehog pathway, both in the epithelium and lamina propria, when comparing lesions of normal mucosa, dysplasia and carcinoma (p<0,05). We concluded that laser therapy was able to modulate the remodeling phase because laser treated tissues showed modifications earlier than control group and the events that occur in tissue repair appear to be cyclical and continue to remodel the tissue, independent of the clinical aspects of the lesion such as its closure and completed healing. Regarding the study of human oral mucosa, it was observed that Hedgehog pathway activation seems to influence the neoplastic transformation. Progressive increase of Hedgehog positive cells was observed in precancerous lesions present in the same individual. Keywords: Wound healing. Low level laser. Immunohistochemistry. Hedgehog signaling. Oral cancer.

LISTA DE ABREVIATURAS E SIGLAS AC (actinic cheilitis; queilite actínica)
AsGaAl (Arseneto de Gálio e Alumínio)
Bmp (Bone morphogenetic protein)
CBA (Cytometric Bead Array)
CEC (Carcinoma escamocelular)
CD4 (Linfócitos T CD4 (auxiliares)
CD8 (Linfócitos T CD8 (supressores/citotóxicos)
CEUA (Comitê de Ética no Uso de Animais)
EROs (Espécies reativas de oxigênio)
FGF (Fator Crescimento de Fibroblasto)
Gli (Fator de transcrição Gli - Glioma associated oncogene homolog)
Hh (Hedgehog)
Dhh (Desert Hedgehog)
HE (Hematoxilina e eosina)
Ihh (Indian Hedgehog)
IL (Interleucina)
IFNɣ (Interferon gama)
MCP-1 (Proteína 1 quimiotática para monócitos)
MIF (Fator inibição de macrófago)
PDGF (Fator de crescimento derivado de plaquetas)
Ptch (Patched)
rpm (rotação por minuto)
SCC (squamous cell carcinoma)
Shh (Sonic Hedgehog)
Smo (Smoothened)
TEM (Transição epitélio mesênquima)
TGF β (Fator de crescimento e transformação beta)
Th (T auxiliador ou T helper)
TLBP (Terapia laser de baixa potência)
TNF (Fator de necrose tumoral)
VEGF (Fator de crescimento endotelial vascular)
Wnt (Via de sinalização Wingless)

LISTA DE SÍMBOLOS cm2 (centímetro quadrado)
CO2 (gás carbônico)
0C (grau centígrado)
g (grama)
J (joule)
J/cm2 (Joule por centímetro quadrado)
mg/kg (miligrama por quilo)
ml (mililitro)
µm (micrômetro)
mM (milimolar)
mW (miliwatt)
nm (nanômetro)
NO (óxido nítrico)
PE (ficoeritrina)
pg/ml (picrogramas por mililitro)
® - (marca registrada)
W/cm2 (watt por centímetro quadrado)

LISTA DE FIGURAS Figura 1: Acúmulo de fibrina (asterisco) em meio à área de edema na
matriz extracelular (Microscopia eletrônica de transmissão. 4.500X).
25
Figura 2: Eletromicrografia de um leucócito com evidente degeneração
hidrópica em fase exsudativa do processo inflamatório (Microscopia
eletrônica de transmissão. 7.000X).
26
Figura 3: Eletromicrografia que ilustra pericito (seta) em íntimo contato com
a célula endotelial (asterisco), compartilhando a membrana basal desta
célula que constitui a parede do vaso sanguíneo (Microscopia eletrônica de
transmissão. 7.000X).
29
Figura 4: Eletromicrografia que apresenta visão de parte do citoplasma de
um fibroblasto que exibe hiperplasia de retículo endoplasmático e
mitocôndrias que ilustram intensa atividade de síntese. (Microscopia
eletrônica de transmissão. 12.000X).
32
Figura 5: Neoangiogênese evidenciada através da imunomarcação com
anticorpo anti alfa actina de músculo liso em pele de rato 3 dias após a
indução de ferimento cutâneo padronizado (Imuno-histoquímica. 400X).
33
Figura 6 - Área correspondente à fibroplasia que evidencia ausência de
anexos cutâneos na matriz extracelular e re-epitelização completada
epiderme (Hematoxilina-eosina. 100X).
34
Figura 7 – Eletromicrografia que exibe feixes colagênicos com diferentes
orientações na matriz extracelular que indicam o ínicio do processo de
fibroplasia a partir do sétimo dia após a realização de um ferimento
cutâneo.
36
Figura 8: Intensa celularidade na área do ferimento, grupo Controle (100x)
(A). Presença de células basofílicas na proximidade do revestimento
endotelial. Grupo tratado com laser (400x) (B).
63

Figura 9: (A) Expressão imuno-histoquímica de alfa-actina em células
perivasculares em tecido cutâneo de animal controle 14 dias, (B) Idem
para tecido submetido à fotobiomodulação.
63
Figura 10: Expressão imuno-histoquímica de Ptch (A), Ihh (B) e Gli-2 (C),
respectivamente, em células dispostas na área da fibroplasia do grupo
controle14 dias (100x), Idem para animal tratado com laser (D,E,F). Imuno-
histoquímica, anticorpo anti-Ihh, anti-Ptch e anti-Gli2.
65
Figura 11: Expressão de células CD31+ em grupo Controle 21 dias (A),
idem para grupo Laser 21 dias (B). Aumento 200x.
66
Figura 12: Expressão de alfa-actina de músculo liso (A) e NG2 (B),
respectivamente em secções teciduais obtidas de animal controle 21 dias.
Idem para animal do grupo tratado com laser (C,D).
67
Figura 13: Distribuição celular na matriz conjuntiva correspondente à área
de fibroplasia em animal controle (A) – 28 dias. Idem para animal tratado
com laser (B).
68
Figura 14: Expressão imuno-histoquímica de NG2 em células
perivasculares na área do ferimento em animal controle 28 dias (A). Idem
para o grupo Laser (B). Aumento 200x.
69
Figura 15: Expressão de células Gli-2 em grupo Controle 35 dias. Idem
para grupo laser. Aumento 200x.
70
Figure 16 (a-l). Hedgehog proteins expression in normal oral mucosa,
dysplasia and carcinoma, respectively (GLi 2 + cells, [a-c]; Shh+ cells [d-f];
SMO+ cells [g-i] and Ptch+ cells [j-l]). Immunohistochemistry, streptavidin-
biotin peroxidase (Anti Gli2, Anti Sonic, Anti SMO and Anti Patched
antibodies). 400X.
87
Figure 17 (a-h). Hedgehog proteins expression in lamina propria of normal
oral mucosa (Gli 2+ cells [a], SMO+ cells [c] and Shh+ cells [e]) and
immunostained area of Ptch [g] of displastic and neoplastic lesions (Gli 2+
89

cells [b], SMO+ cells [d] and Shh+ cells [f]) and the positive immunostained
area of Ptch [h]. Immunohistochemistry, streptavidin-biotin peroxidase (Anti
Gli2, Anti Sonic, Anti SMO and Anti Patched antibodies). 400X.

LISTA DE TABELAS
Tabela 1: Caracterização dos grupos e respectivos dias de eutanásia.
58
Tabela 2: Relação dos anticorpos primários monoclonais produzidos em
coelho.
59
Tabela 3: Relação dos anticorpos primários monoclonais produzidos em
camundongo.
60
Tabela 4: Expressão imuno-histoquímica de CD8, CD31, CD68, Alfa actina de
músculo liso e NG2.
64
Tabela 5: Expressão imuno-histoquímica de Gli-2, Ihh e Ptch.
65
Tabela 6: Dosagem das citocinas (pg/ml), através da técnica de Citometria de tria de
Fluxo.
67
Table 7: Positive cell area for Patched, Sonic Hedgehog, Gli-2 and
Smoothened immunostaining in normal oral mucosa epithelial tissue (control),
dysplasia and squamous cell carcinoma.
88
Table 8: Positive area for Patched, Smoothened, Gli- 2 and Sonic Hedgehog in
normal oral lamina propria (control group) and in underlying connective tissue
dysplasia and oral squamous cell carcinoma.
90
Table 9: Immunohistochemical expression of Patched, Smoothened, Gli-2 and
Sonic Hedgehog in normal oral mucosa cells (control) and groups with
different degrees of dysplasia and oral squamous cell carcinoma.
91

LISTA DE QUADROS
Quadro 1: Relação das citocinas utilizadas do kit Cytometric Bead
Array.
61
Box 2: Immunostaining pattern of Sonic Hedgehog, Patched, Gli-2 and
Smoothened.
85

SUMÁRIO
1
INTRODUÇÃO GERAL 20
2 REVISÃO DE LITERATURA: A cicatrização cutânea:
células, biomoléculas e laserterapia – uma revisão
narrativa.
22
2.1 Introdução 22
2.2
2.2.1
2.2.2
2.2.3
2.2.4
2.2.5
2.2.6
2.3
Revisão de Literatura
Fase Inflamatória
Fase Proliferativa
Fase de Remodelamento
Interação Epitélio-Mesênquima na cicatrização
Sinalização da via Hedgehog
Uso da laserterapia na cicatrização
Considerações finais
23
24
28
35
38
39
41
43
Referências 45
3 HIPÓTESE E OBJETIVOS 53
4 CAPÍTULO 1: Artigo Científico 1: Angiogênese,
participação de pericitos, do sistema imune e da via de
sinalização Hedgehog, no remodelamento de ferimentos
cutâneos submetidos à fotobiomodulação laser.
55
4.1 Introdução 55
4.2
4.2.1
4.2.2
4.2.3
4.2.4
2.2.5
4.2.6
Materiais e métodos
Grupos experimentais
Histopatologia
Imuno-histoquímica
Histomorfometria
Citometria de Fluxo
Análise estatística
57
57
58
59
60
60
61
4.3 Resultados 62
4.4 Discussão 70
4.5 Considerações finais 70
Referências
77
5 CAPÍTULO 2: Artigo Científico 2: Immunohistochemical
Evaluation of Hedgehog signaling in
Epithelial/Mesenchymal Interactions in Squamous Cell
Carcinoma Transformation: A Pilot Study.
80
5.1 Introduction 80
5.2
5.2.1
Materials and methods
Case Selection
80
82

5.2.2 Histopathology 82
5.2.3 Immunohistochemical Procedures 83
5.2.4 Histomorphometry 84
5.2.5 Statistical Analysis 85
5.3 Results 86
5.4
Discussion
References
91
95
6 CONCLUSÃO GERAL 99
REFERÊNCIAS BIBLIOGRÁFICAS
ANEXOS

20
1 INTRODUÇÃO GERAL
A cicatrização tem sido um tema, desde há muito tempo, descrito na
literatura, pois se trata de um processo biológico fundamental que engloba a
inflamação e geralmente evolui para o reparo dos ferimentos.
Tal processo compreende a restauração da integridade do tecido, pois
envolve a organização de células, sinais químicos e matriz extracelular.
Alterações essencialmente vasculares e proliferativas têm sido descritas e as
células que participam destas, exercem um papel importante na síntese e
remodelamento da matriz extracelular.
Com o decorrer do processo de maturação e remodelamento, a maioria
dos vasos, fibroblastos e células inflamatórias desaparece do local da lesão,
mediante processos de emigração, apoptose ou outros mecanismos de morte
celular.
Contudo, após a cicatrização, em especial a cutânea, quando a camada
epitelial é recomposta e o processo é tido como concluído, há indícios de
algumas modificações ainda possam ocorrer por um tempo variável e tais
alterações ainda não foram plenamente elucidadas (VELNAR et al., 2009).
Estas envolvem a ação de sinais bioquímicos que contribuem para a
remodelação, “limpeza” e posterior diferenciação em tecidos. Tais alterações
aqui têm sido estudadas como elementos participantes da diferenciação
morfogênica, tal como a sinalização Hedgehog que tivemos oportunidade de
investigar.
Em adição a este fato, o mecanismo dos efeitos sobre a cicatrização
tardia, no sistema imunológico induzido pelo laser de baixa potência, in vivo,
ainda não está bem compreendido, embora alguns estudos tenham fornecido
evidências das atividades imunomodulatórias desta fototerapia nas etapas
iniciais do reparo tecidual. De fato, há um vasto acervo de manuscritos que
descreve os efeitos anti-inflamatórios, analgésicos e proliferativos
desencadeados no tecido em razão da utilização do laser terapêutico até 14
dias, em média, após a indução de um ferimento (MEDRADO et al., 2003;
REIS et al., 2006).

21
O presente estudo foi realizado para verificar, por meio da análise
histológica, imuno-histoquímica e citométrica, o papel da terapia laser de baixa
potência (TLBP), mais precisamente com o laser de arseneto de gálio e
alumínio (AsGaAl) no processo de reparo tecidual tardio e sua relação com a
biossíntese de fibras colágenas, infiltrados celulares, pericitos e proteínas da
via de sinalização Hedgehog, promovendo a reorganização tecidual. Mediante
a análise histológica e histomorfométrica, este trabalho pretende avaliar o
comportamento imunológico de feridas cutâneas provocadas na região dorsal
de ratos, que foram submetidos à TLBP, em especial no tocante à
angiogênese, proliferação fibroblástica e pericítica e expressão de proteínas
sinalizadoras da via Hedgehog.
A segunda parte deste trabalho de pesquisa objetivou analisar, através
de métodos imuno-histoquímicos, a expressão das proteínas integrantes da via
Hedgehog, em queilite actínica (QA) e carcinoma escamocelular oral (CEC) e,
desta forma, avaliar o papel da interação epitélio-mesênquima na progressão
tumoral em um mesmo indivíduo.
A queilite actínica é considerada uma lesão cancerizável do lábio, com
forte ligação com a exposição solar crônica e costuma se estabelecer, em sua
maior parte, no lábio inferior. A relação entre o grau de displasia e a
probabilidade de progressão maligna para carcinoma escamocelular já está
bem estabelecida (GURGEL et al., 2014). Diversas vias de sinalização são
deflagradas e estimulam a transformação maligna do epitélio displásico assim
como alterações significativas da matriz extracelular. Atualmente, a relevante
interação epitélio-mesênquima tem sido demonstrada através de estudos que
realizaram abordagens imuno-histoquímicas para caracterizar uma importante
via de sinalização celular conhecida como Hedgehog.
A participação da via de sinalização já foi descrita em associação à
patogênese do carcinoma basocelular. Embora existam alguns trabalhos
científicos que estudaram sua relação com o CEC, são escassos os que
avaliaram a progressão da displasia para o referido carcinoma oral.
Deste modo, o segundo manuscrito apresentado aprofundou a sua análise
apenas nesta temática a fim de caracterizar comparativamente a expressão
das proteínas da via Hedgehog em lesões displásicas e neoplásicas.

22
2 REVISÃO DE LITERATURA: A CICATRIZAÇÃO CUTÂNEA: CÉLULAS,
BIOMOLÉCULAS E LASERTERAPIA – UMA REVISÃO NARRATIVA
2.1 Introdução
A cicatrização de feridas na pele é um processo fisiológico essencial que
consiste na colaboração de muitas linhagens celulares e seus produtos.¹ As
tentativas de restaurar a lesão induzida por uma agressão local começam
muito cedo na fase inflamatória. No final, resultam em reparo, que consiste na
substituição de estruturas especializadas pela deposição de colágeno, e
regeneração, que corresponde ao processo de proliferação celular e posterior
diferenciação a partir de células pré-existentes no tecido e/ou células tronco.²
Estes mecanismos não se excluem mutuamente, ou seja, após a lesão cutânea
no mesmo tecido, pode haver regeneração e reparo, a depender das linhagens
celulares comprometidas pela injúria.
Os processos de regeneração e reparo tecidual ocorrem após o
estabelecimento de uma lesão, quer esta seja de origem traumática ou
proveniente de uma determinada condição patológica. Uma lesão é criada por
todo estímulo que rompe a continuidade física de tecidos funcionais. Os
estímulos que causam lesões podem ser externos ou internos; podem ser
físicos, químicos, elétricos ou térmicos. As lesões podem resultar em danos em
organelas específicas ou nas células como um todo.¹
O reparo não é um simples processo linear no qual os fatores de
crescimento deflagram a proliferação celular e sim, uma integração de
alterações dinâmicas que envolvem mediadores solúveis, células do sangue,
produção de matriz extracelular e proliferação de células parenquimatosas. O
processo de cicatrização em pele, segundo Mitchel et al.³, é ilustrativo dos
princípios de reparo para a maioria dos tecidos.
Os eventos celulares e bioquímicos, no reparo das feridas, podem ser
divididos em fases, a saber, reação inflamatória, proliferação celular e síntese
dos elementos que compõem a matriz extracelular, e o período posterior

23
denominado como remodelamento.4 Essas fases não são mutuamente
excludentes e sim, sobrepostas no tempo3 (Gráfico 01).
Gráfico 1: Ilustração sequencial das fases envolvidas no reparo tecidual.
O objetivo desta revisão de literatura é destacar os processos biológicos
que estão envolvidos durante a cicatrização cutânea, com ênfase nas células,
fatores de crescimento e citocinas que participam no processo de reparo
tecidual.
2.2 Revisão da literatura
2.2.1 Fase inflamatória
Na resposta inflamatória vascular, os vasos lesados contraem e o
sangue extravasado coagula, contribuindo para a manutenção da sua
integridade. O coágulo consiste na agregação de trombócitos ou plaquetas

24
numa rede de fibrina mediante ação de alguns fatores através da ativação e
agregação destas células.5 A rede de fibrina, além de restabelecer a
homeostasia e formar uma barreira contra a invasão de micro-organismos,
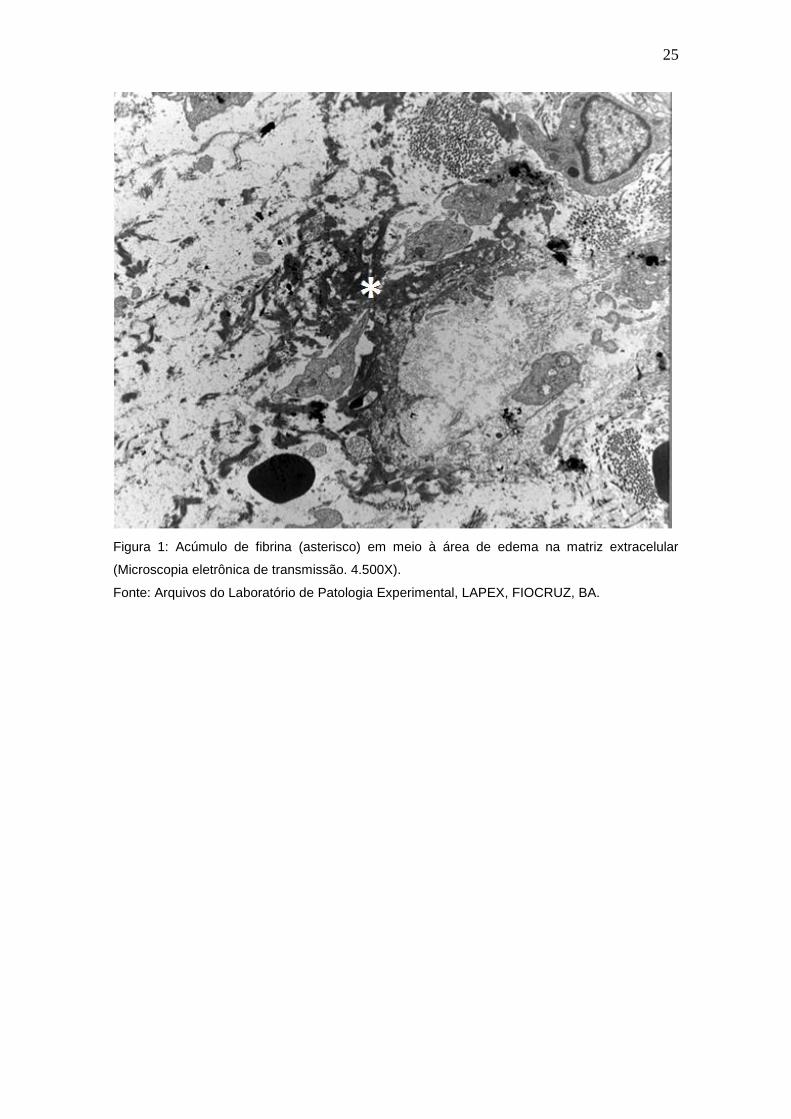
organiza a matriz provisória necessária para migração celular (Figura 1).
Assim, esta matriz temporária restaura a função da pele como uma barreira
protetora, mantendo a integridade do tecido¹. Possibilita ainda, a migração de
células para o microambiente da lesão, e o estímulo para a proliferação dos
fibroblastos.
A resposta celular da fase inflamatória é caracterizada pelo influxo de
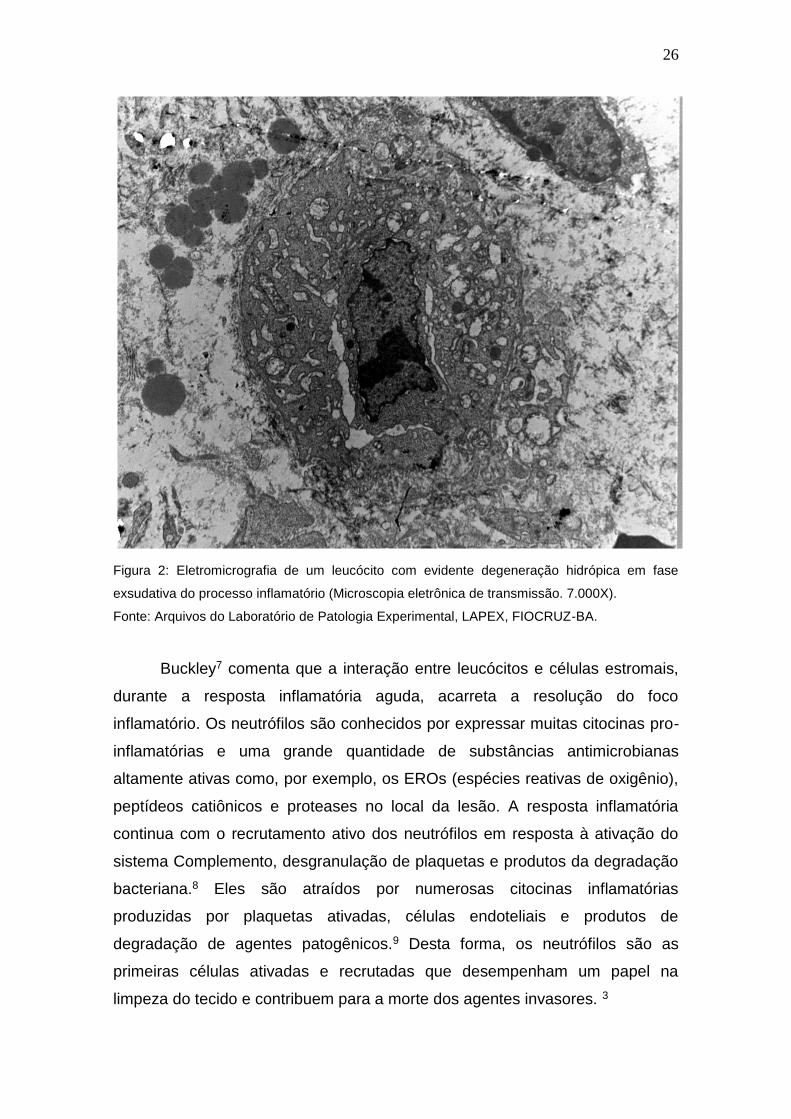
leucócitos na área da ferida (Figura 2). Tal resposta é muito rápida e coincide
com os sinais cardinais da inflamação, que são evidenciados através do edema
e eritema no local da lesão. Normalmente, ela se estabelece nas primeiras 24
horas e pode se estender até dois dias. Acontece também uma rápida ativação
de células imunes residentes no tecido a exemplo de mastócitos, células gama-
delta e células de Langerhans, que por sua vez secretam quimiocinas e
citocinas. A inflamação é uma resposta tecidual localizada, protetora,
desencadeada pela lesão, com destruição de tecido. As células inflamatórias
têm um importante papel durante a cicatrização das feridas e contribuem para
a liberação de enzimas lisossomais e espécies reativas de oxigênio, bem como
facilitam a limpeza de debris celulares diversos.6
25
Figura 1: Acúmulo de fibrina (asterisco) em meio à área de edema na matriz extracelular
(Microscopia eletrônica de transmissão. 4.500X).
Fonte: Arquivos do Laboratório de Patologia Experimental, LAPEX, FIOCRUZ, BA.
26
Figura 2: Eletromicrografia de um leucócito com evidente degeneração hidrópica em fase
exsudativa do processo inflamatório (Microscopia eletrônica de transmissão. 7.000X).
Fonte: Arquivos do Laboratório de Patologia Experimental, LAPEX, FIOCRUZ-BA.
Buckley7 comenta que a interação entre leucócitos e células estromais,
durante a resposta inflamatória aguda, acarreta a resolução do foco
inflamatório. Os neutrófilos são conhecidos por expressar muitas citocinas pro-
inflamatórias e uma grande quantidade de substâncias antimicrobianas
altamente ativas como, por exemplo, os EROs (espécies reativas de oxigênio),
peptídeos catiônicos e proteases no local da lesão. A resposta inflamatória
continua com o recrutamento ativo dos neutrófilos em resposta à ativação do
sistema Complemento, desgranulação de plaquetas e produtos da degradação
bacteriana.8 Eles são atraídos por numerosas citocinas inflamatórias
produzidas por plaquetas ativadas, células endoteliais e produtos de
degradação de agentes patogênicos.9 Desta forma, os neutrófilos são as
primeiras células ativadas e recrutadas que desempenham um papel na
limpeza do tecido e contribuem para a morte dos agentes invasores. 3

27
Poucas horas após a lesão, uma quantidade de neutrófilos transmigra
através das células endoteliais, presentes nas paredes dos capilares
sanguíneos, os quais são ativados por citocinas pró-inflamatórias como IL-1 β,
TNF α (fator de necrose tumoral alfa), e IFN-γ (interferon gama) no local da
lesão. Tais citocinas promovem a expressão de várias classes de moléculas de
adesão. As moléculas de adesão são determinantes para a diapedese dos
neutrófilos e entre elas, pode-se citar as selectinas e integrinas (CD11a/CD18
(LFA-1); CD11b/CD18 (MAC-1) e CD11c/CD18 (gp150, 95) e CD11d/CD18),²
que interagem com as já presentes na superfície da membrana das células
endoteliais. As referidas células também influenciam em muitos outros
aspectos do reparo, como na resolução do coágulo de fibrina e da matriz
extracelular, indução da angiogênese e reepitelização.¹
A partir de 48 horas após o estabelecimento da lesão, intensifica-se a
migração de monócitos de vasos vizinhos que também infiltram a área da lesão
e a partir do novo perfil de expressão gênica, diferenciam-se em macrófagos.
Estes, ativados através de sinais quimiotáticos, podem atuar como células
apresentadoras de antígenos e auxiliar os neutrófilos na fagocitose.²
Assim, além dos macrófagos residentes, a principal população de
macrófagos na lesão é recrutada do sangue em resposta aos produtos
quimiotáticos, a exemplo de fragmentos de proteína da matriz extracelular, TGF
β, MCP-1 (proteína 1 quimiotática para monócitos).10
Com base nos perfis de expressão gênica, os macrófagos podem ser
classificados como classicamente ativados (M1 pro-inflamatório) e
alternativamente ativados (M2 anti-inflamatório e pro-angiogênico).11 Os
macrófagos liberam fatores de crescimento como o PDGF e o VEGF que são
provavelmente necessários para iniciação e propagação de novo tecido na
área lesionada, pois animais com depleção de macrófagos apresentam defeito
no reparo das feridas, conferindo a estas células um papel central na transição
da fase exsudativa para a proliferativa do processo de reparo.12
Os macrófagos desempenham as funções de fagocitose de debris
musculares, produção e liberação de citocinas e fatores pro-angiogênicos,
inflamatórios e fibrogênicos, bem como de radicais livres.13 Ademais, os
macrófagos ao secretarem fatores quimiotáticos, atraem outras células
inflamatórias ao local da ferida. Produzem também prostaglandinas, que

28
funcionam como potentes vasodilatadores afetando a permeabilidade dos
microvasos. Juntos, tais fatores deflagram a ativação das células endoteliais.14
As referidas células, conforme Mendonça & Coutinho Netto15, também
produzem PDGF, TGF β, FGF e VEGF, que se destacam como as principais
citocinas capazes de estimular a formação do tecido de granulação.
2.2.2 Fase Proliferativa
O objetivo da fase proliferativa é diminuir a área de tecido lesionado por
contração e fibroplasia, estabelecendo uma barreira epitelial viável para a
atividade dos queratinócitos. Esta fase é responsável pelo fechamento da lesão
propriamente dita e compreende a angiogênese, a fibroplasia e a
reepitelização. Estes processos têm início no microambiente da lesão a partir
das primeiras 48 horas e podem se desenrolar até o 14º dia após o
estabelecimento da lesão.14
O remodelamento vascular induz mudanças no fluxo sanguíneo. A
angiogênese é um processo coordenado, envolvendo proliferação celular
endotelial, ruptura e rearrumação da membrana basal, migração e associação
em estruturas tubulares e recrutamento de células perivasculares. Há muito
tempo, a angiogênese tem sido descrita como essencial para diversas
condições fisiológicas e patológicas, tais como a embriogênese, cicatrização,
crescimento tumoral e metástase.16
A elaboração subsequente de vasos, conforme Gonçalves et al.17,
envolve produção de veias colaterais por meio de dois mecanismos:
brotamento e divisão celular. Os plexos vasculares resultantes são
remodelados para se diferenciar em grandes e pequenos vasos. O endotélio é
então preenchido pelas células acessórias e células musculares lisas. A nova
microvasculatura que se forma, viabiliza transporte de fluido, oxigênio,
nutrientes e de células imunocompetentes para o estroma.18
Além da participação ativa de células endoteliais e dos linfócitos neste
processo biológico, os pericitos constituem um grupo celular proveniente da
linhagem mesenquimal de células musculares lisas, descritos há muitas
29
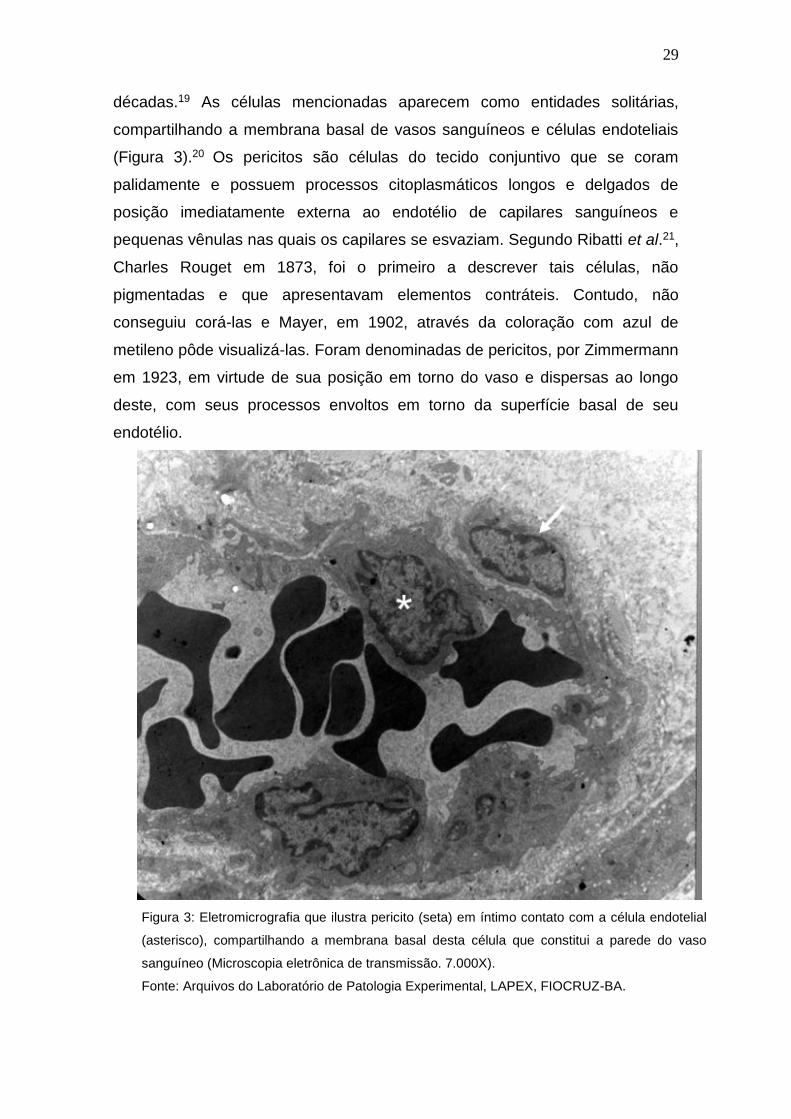
décadas.19 As células mencionadas aparecem como entidades solitárias,
compartilhando a membrana basal de vasos sanguíneos e células endoteliais
(Figura 3).20 Os pericitos são células do tecido conjuntivo que se coram
palidamente e possuem processos citoplasmáticos longos e delgados de
posição imediatamente externa ao endotélio de capilares sanguíneos e
pequenas vênulas nas quais os capilares se esvaziam. Segundo Ribatti et al.21,
Charles Rouget em 1873, foi o primeiro a descrever tais células, não
pigmentadas e que apresentavam elementos contráteis. Contudo, não
conseguiu corá-las e Mayer, em 1902, através da coloração com azul de
metileno pôde visualizá-las. Foram denominadas de pericitos, por Zimmermann
em 1923, em virtude de sua posição em torno do vaso e dispersas ao longo
deste, com seus processos envoltos em torno da superfície basal de seu
endotélio.
Figura 3: Eletromicrografia que ilustra pericito (seta) em íntimo contato com a célula endotelial
(asterisco), compartilhando a membrana basal desta célula que constitui a parede do vaso
sanguíneo (Microscopia eletrônica de transmissão. 7.000X).
Fonte: Arquivos do Laboratório de Patologia Experimental, LAPEX, FIOCRUZ-BA.

30
Através de longos prolongamentos citoplasmáticos que se estendem e
circundam o tubo endotelial, o pericito faz contatos focais com o endotélio por
meio de junções especializadas.22 Adicionalmente, tal célula influencia a
estabilidade do vaso mediante a deposição de matriz e/ou liberação e ativação
de sinais que promovem a diferenciação ou quiescência das células
endoteliais.23,24
Os pericitos são células murais dos microvasos sanguíneos, que estão
envolvidos na membrana basal, que é contínua com a membrana basal
endotelial. Alguns pericitos são provavelmente células da linhagem
mesenquimal ou progenitoras, os quais originam os adipócitos, cartilagem,
osso e músculo.22 Há evidências substanciais de que os pericitos retêm na vida
adulta uma potencialidade mesenquimal suficiente para dar origem não só a
fibroblastos como também a células musculares lisas.25 Estes elementos
celulares podem apresentar características de células pluripotentes,
constituindo-se em uma importante “fonte de reserva celular”. Embora a
plasticidade dos pericitos ainda não tenha sido amplamente estudada,
Farrington-Rock et al.26 relataram o seu potencial de diferenciação em
osteoblastos, condrócitos, fibroblastos, leiomiócitos e adipócitos. Esta
propriedade parece ser muito pertinente para o reparo tecidual, uma vez que
estas células poderiam contribuir sobremaneira para o repovoamento do tecido
cicatricial.
O tecido de granulação começa a se formar, aproximadamente quatro
dias após a lesão. Seu nome deriva da aparência granular do tecido recém-
formado, conferindo esta característica para o novo estroma. Segundo Calin et
al.27, o tecido de granulação é formado através dos seguintes mecanismos:
aumento da proliferação fibroblástica, biossíntese colagênica e elástica, a qual
forma uma rede tridimensional extracelular de tecido conjuntivo e produção de
fatores quimiotáticos e IFN-beta pelos fibroblastos. Fibroblastos e células
endoteliais expressam receptores de integrina, e através desses, invadem o
coágulo no espaço da lesão.28
Para que o processo de reparo tecidual seja compreendido, não se pode
deixar de mencionar algumas particularidades do sistema imune como a
participação de linfócitos B e mais especificamente a multifuncionalidade dos
linfócitos T. Morfologicamente, os linfócitos T se subdividem em populações

31
funcionais: CD4 (linfócitos T auxiliares) e CD8 (linfócitos T
supressores/citotóxicos). As células T CD4 são caracterizadas com base em
seus perfis de produção de citocinas, como a sub-população Th1, produtoras
de Il-2 e IFN gama, a Th2, que produz IL-4, IL-5 e IL-10 e a Th17, que se
caracteriza pela produção de IL-17.2 Outras subpopulações de Th foram
descobertas como: Th22, Th9 e Treg. As citocinas produzidas por estas
subpopulações desempenham papeis críticos principalmente na diferenciação
das células imunes.29
Quando ocorre uma lesão tecidual, o processo de reparo é modulado
pela atividade das células da resposta inflamatória, das células dos bordos da
lesão (queratinócitos) e pela variedade de citocinas e fatores de crescimento
que influenciam a migração, proliferação e a diferenciação celular local.30
Medrado et al.25 comentaram que a fibroplasia tem início na formação do
tecido de granulação, caracterizada pela proliferação de fibroblastos, principais
responsáveis pela deposição de nova matriz (Figura 4). O principal
componente de uma cicatriz de tecido conjuntivo maduro é o colágeno. Os
fibroblastos, produtores colagênicos, são recrutados da derme da margem da
ferida para sintetizar essa proteína. A formação da membrana basal intacta,
entre a epiderme e a derme, é essencial para o restabelecimento da
integridade e função. Durante a fase inicial do reparo, o colágeno tipo III é
predominante, sintetizado pelos fibroblastos no tecido de granulação. 31

32
Figura 4: Eletromicrografia que apresenta visão de parte do citoplasma de um fibroblasto que
exibe hiperplasia de retículo endoplasmático e mitocôndrias que ilustram intensa atividade de
síntese. (Microscopia eletrônica de transmissão. 12.000X).
F/onte: Arquivos do Laboratório Patologia Experimental, LAPEX, FIOCRUZ-BA.
Medrado et al.25 observaram que nesta fase, inicia-se o processo de
contração da ferida, desempenhada pelos fibroblastos, ricos em alfa actina de
músculo liso, sendo conhecidos como miofibroblastos. Tais células,
acumuladas nas margens das feridas, executam atividades contráteis e
contraem as bordas da lesão para o centro.27
A angiogênese ocorre na matriz extracelular do leito da ferida com a
migração e estimulação mitogênica das células endoteliais.15 Tal
neovascularização acompanha a fase fibroblástica, comentada acima. A boa
irrigação das bordas da ferida é essencial para a cicatrização, pois, permite o
aporte adequado de nutrientes e oxigênio32 e de células imunocompetentes
para o estroma (Figura 5).33

33
Figura 5: Neoangiogênese evidenciada através da imunomarcação com anticorpo anti alfa
actina de músculo liso em pele de rato 3 dias após a indução de ferimento cutâneo
padronizado (Imuno-histoquímica. 400X).
Fonte: Arquivos do Laboratório de Patologia Experimental, LAPEX, FIOCRUZ-BA.
Paralelamente a todos os eventos supracitados, as células epiteliais de
revestimento, sob a ação de citocinas específicas, proliferam e migram a partir
das bordas do ferimento na tentativa de fechá-lo. Este processo é denominado
de reepitelização. A reepitelização de uma ferida por queratinócitos, é realizada
pela combinação da fase proliferativa e migração de células próximas à lesão.14
A migração dos queratinócitos ocorre na direção da pele remanescente da
lesão às extremidades desta.
Células epidérmicas dos folículos pilosos rapidamente removem o
coágulo e o estroma danificado. Li et al.14 comentaram que as células
germinativas epidérmicas do folículo piloso que originam o bulbo capilar,
servem de reservatório para os queratinócitos no processo de cicatrização.
Após aproximadamente doze horas do estabelecimento da lesão, há
desenvolvimento e alongamento de projeções tipo pseudópodo dos
queratinócitos, perda dos contatos matriz extracelular-célula e célula-célula,
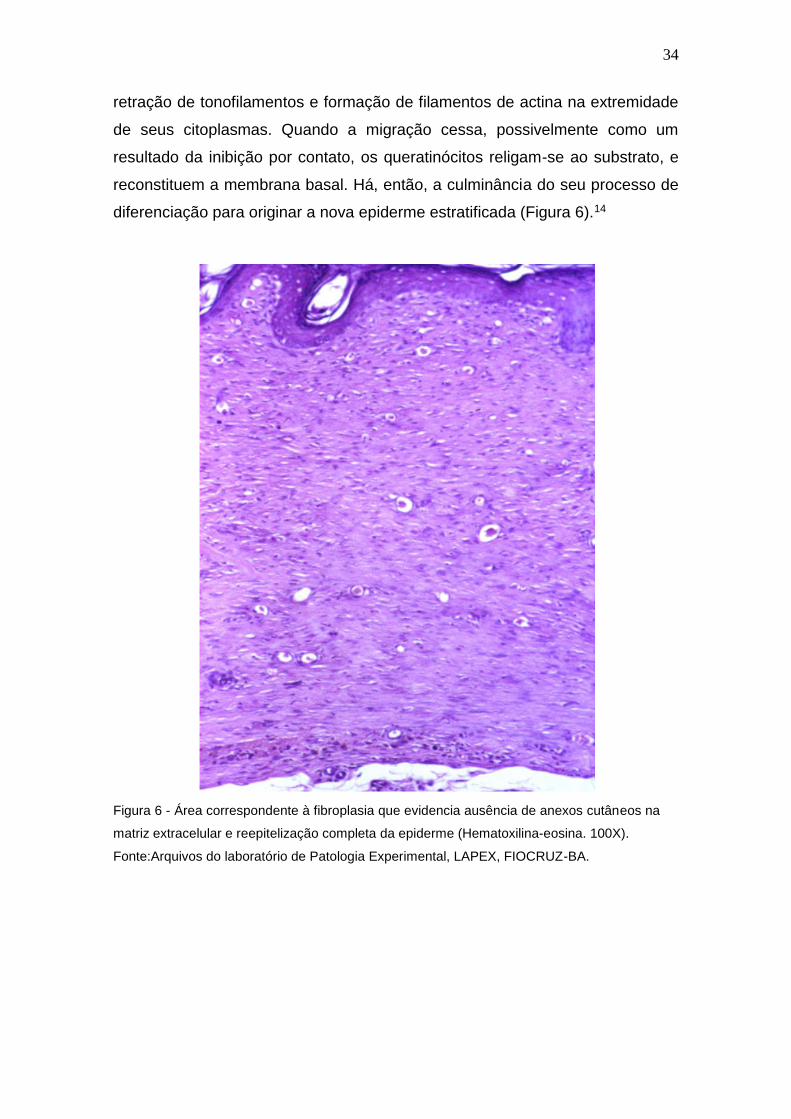
34
retração de tonofilamentos e formação de filamentos de actina na extremidade
de seus citoplasmas. Quando a migração cessa, possivelmente como um
resultado da inibição por contato, os queratinócitos religam-se ao substrato, e
reconstituem a membrana basal. Há, então, a culminância do seu processo de
diferenciação para originar a nova epiderme estratificada (Figura 6).14
Figura 6 - Área correspondente à fibroplasia que evidencia ausência de anexos cutâneos na
matriz extracelular e reepitelização completa da epiderme (Hematoxilina-eosina. 100X).
Fonte:Arquivos do laboratório de Patologia Experimental, LAPEX, FIOCRUZ-BA.

35
2.2.3 Fase de Remodelamento
A terceira fase da cicatrização consiste no remodelamento que começa
duas a três semanas após a lesão e pode durar um ano ou mais. O principal
objetivo da fase de remodelamento é alcançar a máxima força tênsil pela
reorganização, degradação e ressíntese da matriz extracelular. Nesta última
fase do processo de cicatrização de lesões, ocorre uma tentativa de
recuperação da estrutura tecidual normal e o tecido de granulação vai sendo
gradualmente remodelado, formando um tecido cicatricial, que é menos celular
e vascular³ e que exibe um aumento progressivo da concentração de fibras
colágenas (Figura 7). Este estágio é marcado pela maturação dos elementos
com profundas alterações na matriz extracelular e resolução da inflamação
inicial. Assim que a superfície da lesão é coberta por uma monocamada de
queratinócitos, sua migração epidérmica cessa e uma nova epiderme
estratificada com lâmina basal subjacente é restabelecida das margens da
ferida ao interior desta.5 Nesta etapa há deposição de matriz e subsequente
mudança em sua composição.14 Com o fechamento da ferida, o colágeno tipo
III sofre degradação e a síntese do colágeno tipo I aumenta. Ao longo do
remodelamento, há redução de ácido hialurônico e fibronectina, os quais são
degradados por células e metaloproteinases plasmáticas e, assim processa-se
concomitantemente a crescente expressão de colágeno do tipo I já citada.17
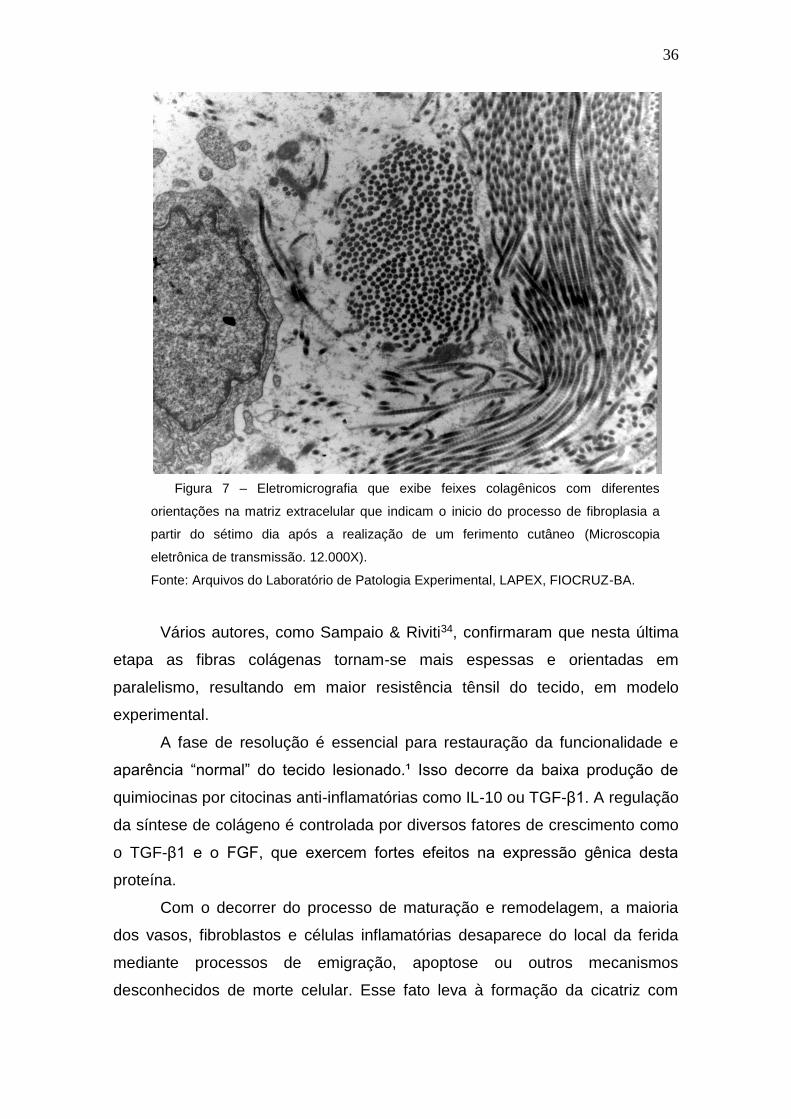
36
Figura 7 – Eletromicrografia que exibe feixes colagênicos com diferentes
orientações na matriz extracelular que indicam o inicio do processo de fibroplasia a
partir do sétimo dia após a realização de um ferimento cutâneo (Microscopia
eletrônica de transmissão. 12.000X).
Fonte: Arquivos do Laboratório de Patologia Experimental, LAPEX, FIOCRUZ-BA.
Vários autores, como Sampaio & Riviti34, confirmaram que nesta última
etapa as fibras colágenas tornam-se mais espessas e orientadas em
paralelismo, resultando em maior resistência tênsil do tecido, em modelo
experimental.
A fase de resolução é essencial para restauração da funcionalidade e
aparência “normal” do tecido lesionado.¹ Isso decorre da baixa produção de
quimiocinas por citocinas anti-inflamatórias como IL-10 ou TGF-β1. A regulação
da síntese de colágeno é controlada por diversos fatores de crescimento como
o TGF-β1 e o FGF, que exercem fortes efeitos na expressão gênica desta
proteína.
Com o decorrer do processo de maturação e remodelagem, a maioria
dos vasos, fibroblastos e células inflamatórias desaparece do local da ferida
mediante processos de emigração, apoptose ou outros mecanismos
desconhecidos de morte celular. Esse fato leva à formação da cicatriz com

37
reduzido número de células. Em fase mais tardia, os fibroblastos do tecido de
granulação alteram seu fenótipo, passando a expressar temporariamente
actina de músculo liso, recebendo a denominação especial de
miofibroblastos.6,27
Os miofibroblastos, segundo Calin et al.27, adquirem algumas
propriedades contráteis das células musculares lisas, aproximando as margens
do ferimento, sendo responsáveis pela contração deste. Desta forma, as
referidas células apresentam bandas bem desenvolvidas de microfilamentos
contráteis compostos de actina. São mantidas unidas através de junções
comunicantes e os seus filamentos citoplasmáticos de actina se ligam através
de receptores de integrinas às fibrilas de fibronectina e os colágenos I e III da
matriz extracelular.1,14 É digno de nota que os miofibroblastos são os principais
produtores de matriz extracelular nos processos de fibrose.31
De acordo com Midwood et al.35 e Badylak36, a matriz extracelular não é
um elemento estático e é capaz de desempenhar um papel relevante nesta
fase do reparo através da interação de seus constituintes estruturais e os
diferentes tipos celulares presentes no tecido. Tais constituintes estruturais
representados por proteínas como colágeno, fibronectina, fibrina, entre outros,
proveem sinais e desencadeiam atividades celulares específicas na área do
ferimento. A fibronectina, por exemplo, gera um arcabouço que viabiliza a
adesão e a migração celular. Outra glicoproteína adesiva, a vitronectina, pode
contribuir para a contração do tecido mediada pelo colágeno produzido pelos
fibroblastos. Em virtude da existência destes processos, o controle local das
interações células/matriz tem sido alvo de promissoras abordagens
terapêuticas.
Em todos esses processos citados, urge salientar que fatores exógenos
e endógenos podem modular tais eventos e influenciar o curso da cicatrização.
Em especial desordens sistêmicas como diabetes, imunossupressão, estase
venosa e aquelas provenientes de agentes externos como uso de
corticoterapia e tabagismo, podem dificultar o fechamento precoce da ferida.
Em adição a esses possíveis fatores complicadores, procede o
estabelecimento de cicatrizes hipertróficas e queloides.37

38
2.2.4 Interação Epitélio-Mesênquima na cicatrização
As células endoteliais sofrem transição epitélio-mesênquima (TEM) e
migram para os órgãos para se diferenciarem em seus componentes
mesenquimais incluindo fibroblastos, células musculares lisas dos vasos e mais
provavelmente também os pericitos.22
A pele assim como os intestinos, o fígado, os pulmões e os tecidos
glandulares apresenta células epiteliais e mesenquimais. As células epiteliais
estão firmemente aderidas umas as outras, formando camadas nas quais
exibem polaridade basoapical. As mesenquimais são não polarizadas, capazes
de movimentação como células individuais devido à perda de conexões
intercelulares.38
O processo biológico que ocorre na transição epitélio-mesênquima
possibilita a uma célula epitelial polarizada sofrer alterações moleculares,
adquirindo um fenótipo mesenquimal, com capacidade migratória através da
matriz extracelular, resistência à apoptose e aumento de produção dos
componentes da matriz.39
A expressão de genes regulatórios normais promotores e/ou inibidores
do crescimento celular que se expressam nas células presentes na matriz
extracelular ocorre na cicatrização, embora venha sendo estudada
primariamente no contexto tumoral. Liu et al. descreveram que a transição
epitélio-mesênquima pode ser regulada por microRNAs, a exemplo do miR-
221, assim como por outros oncogenes. Tome-Garcia et al. relataram que a
superexpressão do gene ras e do ERBB2 resultaram em um aumento da
mobilidade celular na matriz extracelular e no potencial metastático do câncer
de próstata. Embora estas alterações tenham sido descritas no microambiente
tumoral, tais resultados podem ser observados também na cicatrização.40,41
São conhecidos três tipos de transição epitélio-mesênquima. O tipo I
acontece quando os tecidos são construídos durante a embriogênese, a
exemplo dos fibroblastos dérmicos do tecido conjuntivo que fornecem sinais
determinantes para o posicionamento, tipos de pelos e outros apêndices
cutâneos que se diferenciarão na epiderme sobrejacente.5 A transição epitélio-
mesênquima também ocorre em tecidos adultos em reação ao remodelamento

39
e fibrose (tipo II).39 O processo metastático (tipo III) compreende células de
carcinoma que sofrem conversão fenotípica e adquirem mobilidade, usando o
programa de transição epitélio-mesênquima, que normalmente serve a
formação de fibroblastos adultos.42
A transição epitélio-mesênquima tipo II está associada à cicatrização do
tecido, regeneração e fibrose. Tal evento, ligado ao reparo, origina fibroblastos
e outras células relacionadas com objetivo de reconstruir tecidos seguidos de
traumas e inflamação. Esse tipo de transição epitélio-mesênquima, associado à
inflamação cessa assim que esta é atenuada.42
Através de análises moleculares, foi possível identificar a maioria dos
fatores morfogênicos que compreendem uma rede de sinais entre o epitélio e o
mesênquima, que incluem Bmp, Wnt (wingless), Notch/Delta e Hedgehog.43 Os
mecanismos de sinalização de Wnt e Notch estão criticamente envolvidos na
determinação da diferenciação terminal das células que participam do reparo
cutâneo. Shi et al.44 realizaram um estudo experimental no qual foi avaliada a
participação destas vias de sinalização durante a cicatrização. Os autores
observaram que o incremento na atividade de tais vias sinalizadoras promoveu
o fechamento das feridas cutâneas nos espécimes estudados e que a sua
inibição ou ativação pode afetar a proliferação de células tronco, assim como a
diferenciação e migração de queratinócitos e regeneração dos folículos pilosos.
Adicionalmente, segundo Sicklick et al.45, há hipóteses de que os
miofibroblastos possam produzir o ligante Hedgehog, proteína transmembranar
que controla também a construção tissular e o remodelamento, regulando a
viabilidade e atividade migratória de vários tipos celulares responsivos ao
referido ligante.
2.2.5 Sinalização da Via Hedgehog
Hedgehog (Hh) é uma família de moléculas sinalizadoras secretadas
que está envolvida em muitos processos, inclusive, agentes-chave na
padronização de numerosos tipos de tecihos.46 Compreende uma cascata de
proteínas que regula diversos processos biológicos como: desenvolvimento

40
embriológico, homeostase, tumorigênese e reparo de tecidos. Considerando-se
que o ligante Hedgehog pode ser regulador de angiogênese, sua sinalização
poderia influenciar o remodelamento tecidual.
O gene Hedgehog foi identificado em trabalhos genéticos sobre
segmentação de corpo em Drosophila melanogaster, a mosca da fruta. Os
pesquisadores alemães Nüsslein e Wieschaus,47 em 1980, estudando o
controle genético de embriões de Drosophila melanogaster, identificaram que a
perda de um gene causava projeções nestes. Tal fenótipo mutante adquiria
uma forma oval, com dentículos desorganizados, lembrando um ouriço-caixeiro
(tradução da língua portuguesa para hedgehog).
Os homólogos de Hh têm sido identificados em muitos invertebrados e
vertebrados e se mantêm conservados, desempenhando papeis similares vitais
no controle da padronização tissular, diferenciação e proliferação celular no
desenvolvimento embriológico e no controle do comportamento da célula
germinativa e homeostase no adulto.48 Essa via é extremamente essencial no
desenvolvimento, principalmente na formação dos membros e tubo neural.49
Muitos genes têm sido identificados para codificar componentes
citoplasmáticos da maquinaria de transdução de sinal Hh. Três homólogos,
identificados em vertebrados, do gene Hh foram designados pelos prefixos:
Sonic (Shh), Indian (Ihh) e Desert (Dhh).50,51
A via Hh pode ser iniciada, conforme Omeneti et al.52, por sinais
autócrinos, parácrinos ou endócrinos. Os ligantes Hh são sintetizados e sofrem
quebra autocatalítica para gerar um fragmento N-terminal, que é modificado
pela adição de colesterol e palmitato, antes de ser liberado para o espaço
extracelular.53
A ativação da via de sinalização Hedgehog é feita pela ligação de um
dos membros da família no receptor de doze passagens na membrana,
denominado Patched (Ptch). Este trabalha associado a um correceptor de sete
passagens na membrana, denominado Smoothened (Smo).54 Na ausência do
ligante Hedgehog, o receptor Ptch inibe o correceptor Smo, que fica em
vesículas na base do cílio primário e, portanto longe dos fatores de transcrição
Gli-1, 2 e 3 abundantes na superfície dessa organela.53 Isso faz com que Gli-3
seja transportado via microtúbulos até o proteassoma, onde é parcialmente
degradado originando um fragmento Gli-3 repressor que vai no núcleo e

41
reprime a transcrição dos genes alvo envolvidos na diferenciação,
sobrevivência e proliferação celular.
Em contrapartida, quando acontece a ligação de Hh ao Ptch, ocorre a
internalização desse receptor, ocorrendo uma mudança conformacional em sua
estrutura, liberando o correceptor Smo, que migra para o topo do cílio primário,
onde ativa a família Gli de fatores de transcrição. A ativação de Smo inibe o
processamento proteolítico dos referidos fatores, liberando-os para ativação
transcricional.43 Assim, Gli-1, 2 e 3 migram ao núcleo ativando a transcrição de
genes alvo desta via. 48
As desordens de desenvolvimento na sinalização Hedgehog podem ser
através da inativação ou superatividade. Teglund & Tofgard49 (2010)
descreveram que a ativação inapropriada da sinalização Hedgehog contribui
para vários cânceres e síndromes. Luo et al 55 evidenciaram que a atividade da
sinalização Sonic hedgehog (Shh) é necessária para a cicatrização normal e
que em camundongos com diabetes, tal via é impedida, sendo que a aplicação
exógena de Shh acelera a cicatrização por aumentar a função de NO (óxido
nítrico).
2.2.6 O uso da Laserterapia na Cicatrização
Ao longo dos anos, diversos estudos têm sido realizados para
compreender o processo de reparo tecidual, bem como os possíveis efeitos da
fotobiomodulação laser no processo de cicatrização das feridas.56
Dentre as várias técnicas e métodos, a laserterapia apresenta grande
eficácia no processo cicatricial e seu êxito se deve às respostas que induz nos
tecidos, como redução de edema e diminuição do processo inflamatório.17,57
O emprego do raio laser para estudar o processo de reparação dos
tecidos foi demonstrado pela primeira vez por Mester, em 1968, quem primeiro
empregou o termo terapia laser de baixa potência, que foi considerado um
bioestimulador, daí esse sinônimo para definir este tipo de laser.59 Mester
utilizou esta terminologia, pois esta terapia com laser acelerava o processo de

42
cicatrização. Na época, não se conhecia bem seu mecanismo de ação, mas,
observou-se excelente resultado nos tratamentos de úlceras abertas.
Em 1971, Mester60 e colaboradores observaram evidências de maior
acúmulo de fibras colágenas e vesículas intracitoplasmáticas eletrodensas nos
fibroblastos estimulados com laser. A partir daí, vários outros estudos foram
realizados e comprovaram o efeito benéfico do laser na cicatrização. Por
exemplo, Reddy61 após estudar a hipótese da fotobioestimulação pelo laser
induzir aceleração da cicatrização em ratos diabéticos, concluiu que o processo
de bioestimulação pela luz laser, acelerou a produção de colágeno.
Acrescentou ainda que promoveu uma estabilidade de todo o tecido conjuntivo
na cicatrização das referidas lesões.
Segundo Dederich & Bushick62, o efeito fotobiológico dos lasers de hélio-
neônio, arseneto-gálio e arseneto, gálio e alumínio, altera o comportamento
dos fibroblastos, regulando sua proliferação, maturação e locomoção. Muitos
estudos têm sido descritos e publicados sobre seus efeitos como: promoção de
cicatrização, alívio da dor, melhora de circulação local e da inflamação e ainda,
efeitos bactericidas63.
Além disso, o laser é capaz de aumentar a proliferação e ativação de
linfócitos, bem como elevar a capacidade fagocítica dos macrófagos, secreção
de fatores de crescimento de fibroblasto e intensificar a reabsorção de fibrina58.
No estudo de Gál et al., o tratamento com laser proporcionou efeitos
anti-inflamatórios, reduziu o dano e acelerou a regeneração da ferida em ratos
durante a fase inflamatória aguda57.
O mecanismo de TLBP na cicatrização, segundo Yasukawa et al.64 pode
induzir uma moderada reação inflamatória, traduzida pelo estímulo à secreção
do cortisol endógeno. Na cicatrização, a atividade de linfócitos por TLBP pode
fazê-los mais responsivos a mediadores presentes em tecidos lesionados.65 De
acordo com os autores, o referido hormônio, liberado pelas glândulas adrenais,
age como anti-inflamatório natural que estimula a angiogênese em volta da
lesão com subsequente aumento do fluxo sanguíneo, promovendo, então, a
cicatrização.
Apesar de existirem poucos trabalhos científicos que analisaram a
interação da fotobiomodulação laser com o pericito e células do sistema imune,
em um estudo, desenvolvido por Medrado et al.6, demonstrou-se um padrão de

43
marcação positiva para alfa actina de músculo liso em células que se
apresentavam em íntima justaposição às células endoteliais e dispersas em
meio ao tecido de granulação durante o processo cicatricial. Este dado foi
observado tanto nos grupos irradiados com laser de baixa potência quanto no
grupo controle.
É sabido que a alfa actina de músculo liso pode ser expressa por células
que exibem um aparato contrátil, a exemplo de pericitos e miofibroblastos.
Assim, Medrado et al.25 investigaram a análise ultraestrutural com foco no
estudo de miofibroblastos ratificando a sua presença, porém, não foi realizada
descrição detalhada dos aspectos morfológicos apresentados pelos pericitos.
É digno de nota, que muitos autores têm demonstrado que a
interferência do laser de baixa potência se faz tanto quantitativa como
qualitativamente66,67. Tem sido relatado aumento da proliferação celular e da
síntese de colágeno, assim como dos demais elementos que constituem a
matriz extracelular68. Todavia, as mudanças mais significativas ocorrem no
âmbito qualitativo, a exemplo da distribuição e orientação mais organizada das
fibras colágenas sintetizadas na área da lesão após a irradiação com laser.69,70
Ademais, outros autores documentaram um incremento da circulação
local após a irradiação com o laser.71,72 De fato, esta melhora circulatória pode
ser atribuída a dois fenômenos biológicos, a saber, vasodilatação e nova
proliferação vascular ou angiogênese.
2.3 Considerações finais
A cicatrização cutânea reproduz a maioria dos fenômenos biológicos que
caracterizam este processo em diferentes tecidos e se constitui um excelente
modelo de estudo para avaliar as múltiplas fases do reparo tecidual. Estas
etapas são finamente reguladas por moléculas sinalizadoras produzidas por
uma grande variedade de células presentes na matriz extracelular. Novos
estudos são necessários a fim de detalhar as possíveis interações epitélio-
mesênquima que contribuem para a histodiferenciação, em especial,
investigações que contemplem a influência de adjuvantes, a exemplo do laser

44
de baixa potência, sobre as vias sinalizadoras e processos biológicos
deflagrados pela lesão inicial.

45
Referências
1. Shaw T, Martin P. Wound repair at a glance. J Cell Sci. 2009;122(18):3209-
3213.
2. Eming S, Krieg T, Davidson J. Inflammation in wound repair: molecular and
cellular mechanisms. Journal Invest Dermatol 2007;127:514-525.
3. Mitchel RN, Kumar V,Abbas AK, Fausto N, Aster JC Robbins and Cotran:
Pathologic. Basis of disease . 8 ed, Philadelphia; 2012.
4. Nayac BS, Sandifor S, Maxiwell A. Evaluation of wound healing of ethanolic
extract of Morinda cetrifolia L leaf. Evid Based Complement Alternat Med.
2009;6(3):351-356.
5. Martin P. Wound Healing - Aiming for a perfect skin regeneration. Science
1997;276:75-81.
6. Medrado ARAP, Pugliese LS, Reis SRA, Andrade ZA. Influence of low level
laser therapy on wound healing and its biological action upon myofibroblasts.
Lasers Surg Med 2003;32:239-244.
7. Buckley C. Why does chronic inflammation persist: na unexpected role of
fibroblastos. Immunol Lett. 2011;138:12-14.
8. Gurtner GC, Werner S, Barrandon Y, Longaker M. Wound Repair and
Regeneration. Nature 2008;453:314-321.
9. Nunes PS, Albuquerque-Junior RLC, Cavalcante DRR, Dantas MDM,
Cardoso JC, Bezerra MS et al. Collagen-based films containing liposomes
loaded using acid as dressing for dermal burn healing. J Biomed Biotech
2011;2011:01-09.

46
10. Thuraisingam T, Xu YZ, Eadie K. MAPKAPK-2 signaling is critical for
cutaneous wound healing. J Invest Dermato 2010: 130:278-278.
11. Rodero MP, Khosrotehrani K. Skin wound healing modulation by
macrophages. Int J Clin Exp Pathol. 2010;3(7):643–653.
12. Singer A, Clark R. Cutaneous Wound Healing. N Engl J Med
1999;341(10):738-746.
13. Tidball JG. Inflammation process in muscle injury and repair. Am J Physiol
Regul Integr Comp Physiol 2005;288:345-353.
14. Li J, Chen J, Kirsner R. Pathophisiology of acute wound healing. Clin
Dermatol 2007;25:9-18.
15. Mendonça RJ, Coutinho-Netto J. Aspectos celulares da cicatrização. An
Bras Dermatol 2009;84(3):257-262.
16. Rosen LS. Clinical experience with angiogenesis signaling inhibitors:focus
on vascular endothelial growth factor (VEGF) blockers. Cancer Control 2002;
9(2):36-44.
17. Gonçalves RV, Souza NTA, Silva PH, Barbosa FS, Neves CA. Influência
do laser de arseneto de gálio-alumínio em feridas cutâneas de ratos. Fisoter
Mov 2010;23(3):381-388.
18. Carmeliet P. Angiogenesis in health and disease Nature Med
2003;9(6):653-660.
19. Crocker DJ, Murad TM, Geer JC. Role of the pericyte in wound healing an
ultrastructural study. Exp Mol Pathol 1970;13:51-65.
20. Alon R, Nourshargh S. Learning in motion: pericytes instruct migrating
innate leukocytes. Nature Immunol 2013;4(1):14-15.

47
21. Ribatti D, Nico B, Crivelatto E. The role of pericytes in Angiogenesis. Int J
Dev Biol 2011; 55: 261-268.
22. Armulik A, Genove G, Betsholtz C. Pericytes: Developmental, Physiological
and Pathological Perspectives, Problems and Promises. Dev Cell 2011;21:193-
215.
23. Xian X, Hakansson J, Stahlberg A, Lindblom P, Betsholtz C, Gerhardt H, et
al. Pericytes limit tumor cell metastasis. J Clin Invest 2006;116:642-651.
24. Takakura N. Role of hematopoietic lineage cells as accessory components
in blood vessel formation. Cancer Sci 2006;97:568-574.
25. Medrado AP, Costa T, Prado T, Reis SRA, Andrade ZA. Phenotype
characterization of pericytes during tissue repair following low-level laser
therapy. Photodermatol Photoimmunol Photomed 2010;26:192-197.
26. Farrington-Rock C, Crofts NJ, Doherty MJ, Ashton BA, Phil D, Griffi-Jones
C, Canfield AE. Chondrogenic and adipogenic potential of microvascular
pericytes. Circulation 2004;110:2226-2232.
27. Calin MA, Coman T, Calin MR. The effect of low level laser therapy on
surgical wound healing. Rom Rep in Phys 2010;62(3):617-627.
28. Tonnesen M, Feng X, Clark R. Angiogenesis in wound healing. JID Symp
Proc 2000;5(1):40-46.
29. Raphael I, Nalawade S, Eagar TN, Forsthuber TG T cells subsets and
theur signature cytokines in autoimmune and inflammatory diseases Cytokine
2015; 74: 5-17.
30. Mason DE, Mitchell KE, Li Y, Finley MR, Freeman LC. Molecular basis of
voltage-dependent potassium currents in porcine granulosa cells. Mol
Pharmacol. 2002 Jan;61(1):201-13.

48
31. Isaac C, Ladeira PRS, Rego FMP, Aldunate JCB, Ferreira MC. Processo de
cura das feridas: Cicatrização Fisiológica. Rev. Med. 2010; 89(3/4):125-131.
32. Tazima MFGS, Vicente YAMV, Moriya T. Biologia da Ferida e
Cicatrização Simpósio – Fundamentos em Clínica Cirúrgica 2008; 4(3): 259-
264.
33. Rutter DJ, Schlingemann RO, Westphal JR, Rietveld EJR, de Waal RMW.
Angiogenesis in wound healing and tumor metastasis. Behring Inst Mitt, 1993;
92:258-272.
34. Sampaio SA, Rivitti E. Dermatologia. 2a ed. São Paulo: Artes Médicas;
2001
35. Midwood KS, Williams LV, Schwarzbauer JE. Tissue repair and the
dynamics of the extracellular matrix. Int J iochem Cell Biol 2004;36:1031–1037.
36. Badylak SF The extracellular matrix as a scaffold for tissue reconstruction.
Cell Developm Biol 2002;13:377–383.
37. Fonseca MA, Almeida RR, Reis SRA, Medrado ARAP. Repercussão de
doenças sistêmicas no reparo tecidual. Rev Bahian Odont 2012;3(1):63-75.
38. Friedman SL. Mechanisms of hepatic fibrogenesis Gastroenterology.
2008;134:1655-1669.
39. Choi S, Diehl AM. Epithelial-to-Mesenchymal transitions in liver.
Hepatology. 2009;50:2007–2013.
40. Liu J, Cao J, Zhao X. miR-221 facilitates the TGFbeta1-induced epithelial-
mesenchymal transition in human bladder cancer cells by targeting STMN1.
BMC Urol 2015;28(15):1-9.

49
41. Tome-Garcia J, Ghazaryan S, Shu L, Wu L. ERBB2 increases metastatic
potentials specifically in androgen-insensitive prostate cancer cells. PLoS One
2014;9(6):1-12.
42. Zeisberg M, Neilson EG. Biomarkers of epithelial-mesenchymal transitions.
J Clin Invest 2009;119(6):1429-1437.
43. DeRouen MC, Oro AE. The primary cillium: a small yet mighty organelle. J
Invest Dermatol 2009; 129(2): 264-265.
44. Shi Y, Shu B, Yang R, Xu Y, Xing B, Liu J, Chen L, Qi S, Liu X, Wang P,
Tang J, Xie J. Wnt and Notch signaling pathway involved in wound healing by
targeting separately c-Myc and Hes1. Stem Cell Res Ther 2015;6(1):120.
45. Sicklick J, Li Y, Jayaraman A, Kanngai R, Qi Y, Vivekanadan P, Ludlow JW,
Owzar K, et al. Dysregulation of the Hedgehog pathway in human
hepatocarcinogenesis Carcinogenesis 2006;27(4):746-757.
46. Nybakken K, Perrimon N. Hedgehog signal transduction: recent findings.
Curr. Opin. Genet. Dev. 2002;2(5):503-511.
47. Nüsslein-Volhard, C, Wieschaus, E. Mutations affecting segment number
and polarity in Drosophila. Nature 1980;287:795-801.
48. King PJ, Guasti L, Laufer E. Hedgehog signalling in endocrine development
and disease. J Endocrinol. 2008;198(3):439-50.
49. Teglund S, Toftgard R. Hedgehog beyond medulloblastoma and basal cell
carcinoma Biochim. Biophys. Acta 2010;181–208.
50. Echelard Y, Epstein DJ, St-Jacques B, Shen L, Mohler J, McMahon JA, et
al. Sonic Hedgehog, a member of a family of putative signaling molecules, is
implicated in the regulation of CNS polarity. Cell 1993;75(7): 1417–1430.

50
51. Ingham PW, McMahon AP. Hedgehog signaling in animal development:
paradigms and principles. Genes & Dev. 2001;15:3059-3087.
52. Omenetti A, Choi S, Michelotti G, Diehl AM. Hedgehog signaling in the liver.
J. Hepatol 2011; 54(2):336-373.
53. Pepinsky RB, Zeng C, Wen D, Rayhorn P, Baker DP, Williams KP, Bixler
SA, et al. Identification of a palmitic acid-modified form of human Sonic
Hedgehog. J Biol Chem. 1998;273(22):14037-45.
54. Cohen Jr MM. Hedgehog signaling update. Am J Med Gen
2010;152(8):1875–1914.
55. Luo J-D, Hu T-P, Wang L, Chen M-S, Liu S-M, Chen AF. Sonic Hedgehog
improves wound healing via enhancing cutaneous nitric oxide function in
diabetes. Am J Physiol. Endocrinol Metab 2009;297:525-531.
56. Pereira ANP, Eduardo CP, Matson E,Marques MM Effect of low-power
laser irradiation on cell growth and procollagen synthesis of cultured fibroblasts
Lasers Surg Med, 2002; 31:307-313.
57. Gál,P; Vidinsky B, Toporcer T, Mokry M, Mozes S, Longauer F.
Histological assessement of the effect of laser irradiation on skin wound
healing in rats. Photomedicine and Laser Surgery, 2006; 24(4): 480-488.
58. Gonçalves RV, Souza, NTA, Silva PH; Barbosa FS, Neves CA. Influência
do laser arseneto de gálio-alumínio em feridas cutâneas de ratos. Fisioter Mov,
2010;23(3): 381-388.
59. Mester E, Lúdany G, Sellyei M, Szende B, Tota J. The stimulating effect of
low power laser rays on biological systems. Laser Rev 1968; 1:3.
60. Mester E, Spiry T, Szende B, Tota JG. Effect of laser rays on wound
healing. Am J Surg 1971; 122(4):532-535.

51
61. Reddy GK. Comparison of the photostimulatory effects of visible He-Ne and
infrared Ga-As lasers on healing impaired diabetic rat wounds. Lasers Surg
Med 2003; 33(5): 344-351.
62. Dederich DN, Bushick RD. Lasers in dentistry: separating science from
hype. Journal Am Dent Assoc 2004; 135(2): 204-212.
63. Prockt AP, Takahashi A, Pagnoncelli RM. Uso da terapia com laser de
baixa intensidade na cirurgia bucomaxilofacial. Rev Port Estomatol Ci Maxilofac
2008; 49(4): 247-255.
64. Yasukawa A, Hrui H, Koyama Y, Nagai M, Takakuda K. The effect of low
reactive-level laser therapy (LLLT) with helium-neon laser on operative wound
healing in a rat model. J Vet Med Sci 2007; 69(8): 799-806.
65. Vilela DDC, Chamusca FV, Andrade JCS, Vallve MLF, Gonzalez AC,
Andrade ZA, Medrado ARAP, Reis SRA. Influence of the HPA axis on the
inflammatory response in cutaneous wounds with the use of 670-nm laser
photobiomodulation. Journal Photochemistry Photobiology B:Biology 2012; 116:
114-120.
66. Fung DTC, NG GYF, Leung MCP, Tay DKC. Effects of a therapeutic laser
on the ultrastructural morphology of repairing medial collateral ligament on a rat
model. Lasers Surg Med 2003; 32:286-293.
67. Enwemeka CS; Parker JC, Dowdy DS, Harkness EE, Sanford LE, Woodruff
LD. The efficacy of low-power lasers in tissue repair and pain control. A meta-
analysis study. Photomedicine Laser Surg 2004; 22:323-329.
68. Pugliese LS, Medrado ARAP, Reis SRA, Andrade ZA. The influence of
therapy of laser of low level in biomodulation of collagen and elastic fibers.
Pesqui Odontol Bras 2003;17(4):307-313.

52
69. Garavello I, Baranauskas V, da Cruz-Hofling MA. The effects of low laser
irradiation on angiogenesis in injured rat tibiae. Histol Histopathol 2004; 19(1):
43-48.
70. Reis SRA, Medrado ARAP, Marchionni AMT, Figueira C, Fracassi LD, Knop
LAH. Effect of 670-nm laser therapy and dexamethasone on tissue repair: a
histological and ultrastructural study. Photomed Laser Surg 2008; 26:307-313.
71. Schindl A, Schindl L, Jurecka W, Honisgsmann H, Breier F. Increased
dermal angiogenesisafter low-intensity laser therapy for a chronic radiation ulcer
determined by a video measuring system. J acad Dermatol 1999; 40:481-484.
72. Medrado ARAP, Trindade E, Reis SRA. Andrade ZA Action of low-level
laser therapy on living fatty tissue of rats. Lasers Med Sci 2006; 21:19-23.

53
3 HIPÓTESES E OBJETIVOS
HIPÓTESES
O laser de baixa potência é capaz de imunomodular a expressão das
proteínas da via de sinalização Hedgehog, ativação e proliferação de
pericitos, células endoteliais e células imunocompetentes durante a
etapa tardia do reparo tecidual, em ferimentos cutâneos induzidos
experimentalmente em ratos.
A via de sinalização Hedgehog induz a transformação neoplásica de
lesões displásicas orais para carcinoma escamocelular.
OBJETIVOS
Geral
Identificar a expressão das proteínas da via de sinalização Hedgehog,
ativação e proliferação de pericitos, células endoteliais e células
imunocompetentes sob a ação do laser de arseneto de gálio e alumínio,
no leito do ferimento durante a fase tardia do reparo tecidual, em um
modelo experimental já desenvolvido e divulgado na literatura e no
contexto de lesões displásicas e neoplásicas orais.
Específicos
Analisar através da técnica de imuno-histoquímica, o perfil biológico dos
pericitos perivasculares, células endoteliais e células do sistema
imunológico durante a fase de remodelamento da matriz extracelular sob
a ação do laser de baixa potência.

54
Compreender a expressão de pericitos e células endoteliais na
angiogênese estimulada pela luz laser.
Identificar e avaliar, através da técnica de citometria de fluxo, o perfil de
citocinas representativas das subpopulações Th1 e Th2, a partir de
amostras sorológicas coletadas de espécimes representativas dos
diferentes grupos experimentais.
Identificar através de métodos imuno-histoquímicos a presença de
ligantes e fatores de transcrição da via de sinalização Hedgehog,
durante o reparo tecidual de ferimentos submetidos à laserterapia.
Caracterizar comparativamente a presença de ligantes e fatores de
transcrição da via de sinalização Hedgehog em lesões displásicas e
neoplásicas orais.

55
4 CAPÍTULO 1: Angiogênese, participação de pericitos, do sistema imune
e da via de sinalização Hedgehog, no remodelamento de ferimentos
cutâneos submetidos à fotobiomodulação laser.
4.1 Introdução
Muitas pesquisas têm demonstrado que a terapia com laser de baixa
potência (TLBP) acelera a fase aguda da inflamação, estimula os fenômenos
proliferativos, aumenta a contração de feridas cutâneas e acelera a
reepitelização (MEDRADO et al., 2008; REIS et al., 2008; FELICE et al., 2009;
GONÇALVES et al., 2014). Contudo, o mecanismo de reparo numa fase mais
tardia da cicatrização, sob o efeito primário da laserterapia, ainda não está bem
esclarecido e não se sabe ao certo por quanto tempo os benefícios oriundos da
fotobiomodulação poderiam se perpetuar nos tecidos irradiados.
Após as evidências clínicas do final da cicatrização, quando a camada
epitelial é recomposta e o processo é tido como concluído, há indícios de que
algumas modificações no tecido podem ocorrer por um tempo variável
(VELNAR et al., 2009). Nestes termos, procedem a perpetuação da
angiogênese e a permanência de células imunorreguladoras no leito do
ferimento (DELAVARY et al., 2011).
Até o momento, as descrições das ações desencadeadas pelo laser de
baixa potência concentraram-se nas fases iniciais do reparo tecidual. No
entanto, sabe-se que o estudo da presença ativa das células do sistema imune
e seu papel interativo no reparo, até mesmo na fase de remodelamento deste,
são relevantes para ampliar a compreensão das alterações morfológicas e
funcionais que se desenrolam durante a fase tardia da cicatrização, sob a
influência da laserterapia (PUGLIESE et al., 2003).
Medrado et al., (2008), descreveram alterações nos processos de
biossíntese de colágeno e proliferação de células alfa-actina positivas na etapa
tardia do processo de reparo cutâneo em feridas induzidas experimentalmente
em ratos, submetidos à luz laser de 670 nm. Os autores relataram que a
maturação das fibras colágenas e a angiogênese se perpetuaram até 60 dias

56
após a realização da cirurgia cutânea. Em virtude destas observações, foi
hipotetizado se a laserterapia poderia atuar como agente biomodulador sobre
os pericitos, células endoteliais e do sistema imune, assim como deflagrar,
ainda que numa fase mais avançada do reparo tecidual, a ativação de uma via
de sinalização até então descrita como Hedgehog.
As proteínas que integram esta via de sinalização foram identificadas por
estudos genéticos em Drosophila e se mantêm conservadas em vertebrados
(INGHAN et al., 2011). São conhecidos três ligantes da família Hedgehog:
Shh, Ihh e Dhh. Para que a sinalização ocorra, são necessárias também as
atividades de duas proteínas transmembranares: Ptch e Smo. Ptch é
considerado o receptor primário. A ativação de Smo, seu correceptor, é
indispensável para a transdução do sinal. Assim, este correceptor desloca-se
para o topo do cílio primário, o qual quando ativado, inicia uma série de
eventos intracelulares que resulta na estabilização de fatores de transcrição de
genes-alvo Hh, denominados de Gli, os quais induzem a transcrição de
proteínas essenciais para a morfogênese e histodiferenciação (BYRD &
GRABEL, 2004).
A via Hedgehog participa ativamente em muitos aspectos do
desenvolvimento embriológico, incluindo morfogênese, angiogênese e reparo
(BIELEFELD et al., 2013). Contudo, sua ação no reparo cutâneo, ainda é um
tópico pouco explorado em comparação ao papel desta via no desenvolvimento
da pele.
Mediante a realização de análises sorológicas, histológicas e
histomorfométricas, este trabalho objetivou caracterizar as células imunes e
endoteliais, miofibroblastos e pericitos, bem como a expressão de algumas
proteínas da via de sinalização Hedgehog no reparo tecidual tardio de feridas
cutâneas na região dorsal de ratos tratados com TLBP de 670nm.

57
4.2 Materiais e Métodos
4.2.1 Grupos Experimentais
Quarenta ratos machos da linhagem Wistar, Rattus norvegicus albinus,
com peso variando entre 200 a 300 gramas, foram cedidos pelo Centro de
Pesquisas Gonçalo Moniz, Fiocruz, Salvador-Bahia. Os animais foram
mantidos em sala com ar filtrado, sob condições controladas de temperatura,
umidade e iluminação e permaneceram em gaiolas individuais. Receberam
dieta comercial padrão e água ad libitum, ambas esterilizadas durante todo o
experimento. Os procedimentos foram conduzidos conforme a legislação do
Centro de Pesquisas Gonçalo Moniz, Fiocruz, sob aprovação do Comitê de
Ética no Uso de Animais (CEUA) com protocolo de número 010/11.
A anestesia dos espécimes foi realizada por via intraperitoneal com
associação de quetamina (50mg/Kg) e xilazina (10mg/kg), ministrando 0,2 mL
para cada 100 gramas de peso corpóreo. Após a anestesia, procedeu-se a
tricotomia e posterior assepsia da região dorsal (coxim adiposo), com iodóforo
(Bioquímica, Rio de Janeiro, Brasil).
Foi realizado um ferimento cutâneo através da remoção de um
fragmento de tecido padronizado, com um bisturi circular de oito milímetros de
diâmetro (Biopsy Punch-Stiefel, Germany). Não foi realizada nenhuma sutura
nos ferimentos para que a cicatrização ocorresse por segunda intenção.
Após a realização do ferimento cirúrgico, os animais foram mantidos sob
isolamento individual e diariamente inspecionados para análise do
sangramento, perda da crosta e formação de exsudado purulento.
Os espécimes foram randomicamente distribuídos em dois grupos
experimentais descritos a seguir.
Grupo Laser – Após o procedimento cirúrgico, foi realizada aplicação de
laser diodo semicondutor de AsGaAl, em emissão contínua (9mW, 670 nm,
0,031W/cm²), com área da ponta ativa do aparelho de 0,28 cm² (Laser VR-KC-
610 – Dentoflex, Brasil) e fluência da ordem de 4 J/cm2. Aplicou-se 1 J/cm2 nos

58
quatro pontos correspondentes aos vértices das linhas diametrais da área
circular. Tal aplicação foi realizada no dia do ferimento cirúrgico e no 3° e 5°
dias após este, até que se obtivesse uma dose total de 12 J/cm2.
Grupo Controle - Foi realizado um procedimento caracterizado pelo
contato da ponta ativa do laser de baixa potência com o tecido cruento, sem
haver emissão de luz. A referida ação aconteceu nos mesmos moldes descritos
para o grupo tratado com laser.
Foram determinados quatro períodos para a realização da eutanásia de
cada grupo experimental, listados na tabela 01.
Tabela 1: Caracterização dos grupos e respectivos dias de eutanásia.
Grupos Experimentais
Períodos de Eutanásia (dias)
14 21 28 35
Controle (n) 5 5 5 5
Laser 4 J/cm2 (n) 5 5 5 5
Nos respectivos tempos, os animais foram eutanasiados em câmara de
CO2. Constatado o óbito, foi retirada amostra de sangue da veia mesentérica,
que após retração do coágulo, foi submetida à centrifugação para obtenção do
soro. Os soros foram identificados e acondicionados em freezer -80°C.
4.2.2 Histopatologia
O tecido cutâneo da área lesionada foi removido e fixado em solução de
formalina tamponada a 4% com pH 7,4 por um período de vinte e quatro horas.
Seguiu-se a utilização de álcool a 70%, para continuação do processamento
histológico, até inclusão em blocos de parafina.
Os fragmentos teciduais incluídos em parafina foram cortados em
micrótomo, obtendo-se cortes com 4 m de espessura para a coloração com
Hematoxilina-eosina (HE), assim como para os testes imuno-histoquímicos.

59
4.2.3 Imuno-histoquímica
As lâminas foram previamente tratadas com adesivo à base de
organosilano (3-aminopropyltriethoxi-silano, SIGMA, St. Louis, MO, USA). Foi
realizada a padronização de todos os anticorpos utilizados no estudo, conforme
especificado nas tabelas 2 e 3. O método de imunomarcação utilizado foi o da
peroxidase.
Os cortes histológicos foram submetidos à desparafinização em xilol,
reidratados com álcool absoluto e depois com água à temperatura ambiente. A
recuperação antigênica foi realizada com Tampão Citrato 10 mM, pH 6,0 em
banho-maria a 96°C, com posterior bloqueio da peroxidase endógena,
utilizando o Dako Dual Enzyme Block (Dako, Carpinteria, CA, USA).
A eliminação das ligações inespecíficas foi realizada com Protein Block
Serum Free (Dako, Carpinteria, CA, USA) e assim, os cortes foram incubados a
4°C com os anticorpos primários, em câmara úmida por uma noite.
O anticorpo secundário foi o Polymer Dako Envision Peroxidase (Dako,
Carpinteria, CA, USA) incubado por 30 minutos, à temperatura ambiente. A
reação foi revelada através de DAB (Dako, Carpinteria, CA, USA) com uma
solução de peróxido de hidrogênio a 3%. As lâminas foram contracoradas com
hematoxilina de Harris e montadas com Bálsamo do Canadá.
Adotou-se uma graduação qualiquantitativa padrão para indicar a
intensidade da reação imuno-histoquímica, na qual zero foi considerado como
ausência de reatividade, + (discreto), ++ (moderado) e +++ (intenso).
Tabela 2: Relação dos anticorpos primários monoclonais produzidos em
coelho.
Isótipo igG Diluição Padrão Especific. Fonte Imunolocalização
Anti-CD31 1:1000 ++ SC-1506 S Cruz Biotech
Citoplasma
Anti Ptch1 1:1000 ++ NB200-118 Novus Biological
memb/citoplasma
Anti-Gli-2 1:1000 +++ GWB-CE7858
GenWay Biotech
Nuclear
Anti-Ihh 1:250 +++ AB 39634 Abcam Citoplasma

60
Tabela 3: Relação dos anticorpos primários monoclonais produzidos em
camundongo.
Isótipo igG Diluição Padrão Especific. Fonte Imunolocalização
Anti-NG2 1:200 ++ MAB 5384 Millipore
Citoplasma
Anti-Actina 1:200 +++ M0851 DakoCytomation
Citoplasma
Anti-CD8 1:10 +++ AR0801 Biosource
Citoplasma
Anti-CD68 1:200 ++ MCA341R AbD Serotec Citoplasma
4.2.4 Histomorfometria
As análises histológicas e imuno-histoquímicas, de todas as secções,
foram realizadas sem o conhecimento prévio dos grupos aos quais os animais
pertenciam. Na análise imuno-histoquímica, as áreas com padrão de marcação
positiva, para os anticorpos supracitados, foram mensuradas através de
morfometria e os resultados tabulados e submetidos à análise estatística.
A histomorfometria foi realizada através de imagens capturadas através
do software Motic Images Advanced 3.0. Foi estabelecida uma área padrão
para análise de todos os casos, a saber, 12288,000000 pixels. Foram
analisadas cinco áreas de cada secção, retiradas da área da lesão de cada
lâmina histológica, no aumento de 400 vezes.
A análise foi feita pelo programa Adobe Photoshop CS5 Extended,
Versão 12.0x32, Adobe Systems Incorporated, USA, que quantifica a
porcentagem de tecido corado. Em seguida, estes dados foram comparados
em relação à área total selecionada. Todas as análises foram feitas por um
examinador calibrado para evitar viés de tendenciosidade.
4.2.5 Citometria de Fluxo
Os níveis séricos de IL-2, IFN-ɣ, IL-4 e IL-10 foram avaliados pela citometria de
fluxo. Os soros dos ratos foram testados sem diluição (Tabela 4).

61
Foi utilizado o kit CBA (Cytometric Bead Array) Rat Soluble Protein Flex
Set System, da BD Biosciences e todo o procedimento foi realizado conforme
instruções do fabricante (San Jose, CA, USA). O kit possui esferas de captura
específicas para cada citocina, essas são ligadas a um anticorpo específico
com intensidades de fluorescência distintas, e um anticorpo de detecção
conjugado à ficoeritrina (PE).
Os anticorpos evidenciaram um sinal fluorescente proporcional com a
quantidade de citocinas capturadas. Para a realização da dosagem, as
amostras de soro foram incubadas com as esferas e o PE, por três horas a
4°C. Após a incubação foi realizada a lavagem (centrifugação refrigerada, 1500
rpm, 10min). Depois da calibração do citômetro de fluxo e ajustes dos seus
canais/filtros, assim como a aquisição da curva padrão (fornecida pelo kit), foi
realizado o procedimento de leitura no Citômetro de Fluxo FACS ARRAY (BD
Biosciences, USA).
A análise foi realizada pelo Software FCAP Array (BD Biosciences
Pharmingen,San Diego, CA, USA) considerando uma curva padrão com
valores conhecidos (pg/mL). Na leitura, os dados foram importados para
planilha do software Excel para processamento das análises.
Quadro 1: Relação das citocinas utilizadas do kit Cytometric Bead Array.
Proteína Especificação Fonte
Rat IL-2
51-9007072
BD Biosciences - CBA
Rat IL-4
51- 9004113
BD Biosciences – CBA
Rat IL-10
51-9004112
BD Biosciences – CBA
Rat IFNγ 51-9004111 BD Biosciences – CBA
BD BiosciencePharmingen, San Diego, CA, USA
4.2.6 Análise estatística
O banco de dados foi criado no Excel 2003 Microsoft® e analisado no
software R (versão 3.1.3), onde foi realizada uma análise descritiva (mediana e

62
quartis) com a finalidade de identificar as características gerais e específicas da
amostra estudada.
Foi adotado o teste de Shapiro-Wilk para a verificação da normalidade
da distribuição dos dados. De acordo com esta análise, as varáveis
paramétricas foram analisadas pelo teste T de Student e para as não
paramétricas, utilizou-se o teste de Mann-Whitney para amostras
independentes.
Para identificar as diferenças significativas nos níveis de CD-31, NG-2,
Ihh, Ptch e Gli-2, com o decorrer do tempo de estudo, foi utilizado o teste
paramétrico ANOVA para medidas repetidas ou o não paramétrico de
Friedman, seguidos quando necessário, pelos testes a posteriori de Bonferroni
e de Dunn, respectivamente. O nível de significância estabelecido, para este
estudo, foi de 5% (p <0,05).
4.3 Resultados
14 dias
No período de 14 dias, os diferentes grupos experimentais apresentaram
derme com intensa celularidade na área correspondente à fibroplasia. Nesta
região, não foram visualizados anexos cutâneos, mas apenas células
fusiformes e de morfologia romboidal com intensa basofilia e presença de
capilares sanguíneos. No grupo submetido à fotobiomodulação com laser de
baixa potência, havia ainda moderada vascularização e alguns vasos
sanguíneos exibiam células basofílicas que proliferavam para a matriz
extracelular a partir da parede do endotélio vascular (Figura 8). Estas células
apresentaram também positividade para imunomarcação com os anticorpos
anti-alfa actina de músculo liso e anti-NG2.

63
Figura 8: intensa celularidade na área do ferimento, grupo Controle (100x) (A). e presença de
céls. basofílicas na proximidade do revestimento endotelial. Grupo tratado com laser (400x)
(B).
Na análise morfométrica realizada a partir da técnica imuno-
histoquímica, foi observada a presença de células monomorfonucleares, CD8+
e CD68+, em ambos os grupos experimentais, sem diferenças estatisticamente
significativas (p> 0,05). Houve um maior número de células CD31+ no grupo
controle em relação ao tratado (p> 0,05). Células alfa actina de músculo liso e
NG2+ foram mais numerosas no grupo controle, e esta diferença foi
estatisticamente significativa apenas para células alfa actina de músculo liso
(p< 0,001) (Figura 9); (Tabela 4).
Figura 9: (A) Expressão imuno-histoquímica de alfa-actina em células perivasculares
em tecido cutâneo de animal controle -14 dias, (B) Idem para tecido submetido à
fotobiomodulação.
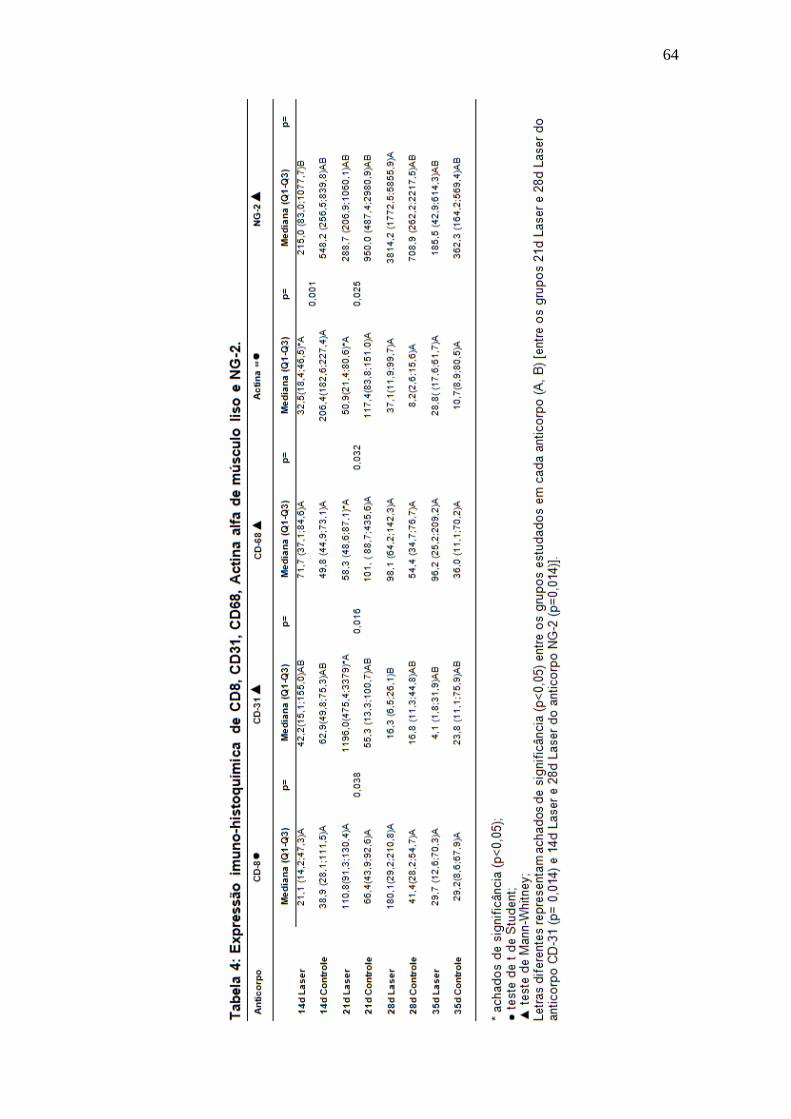
64
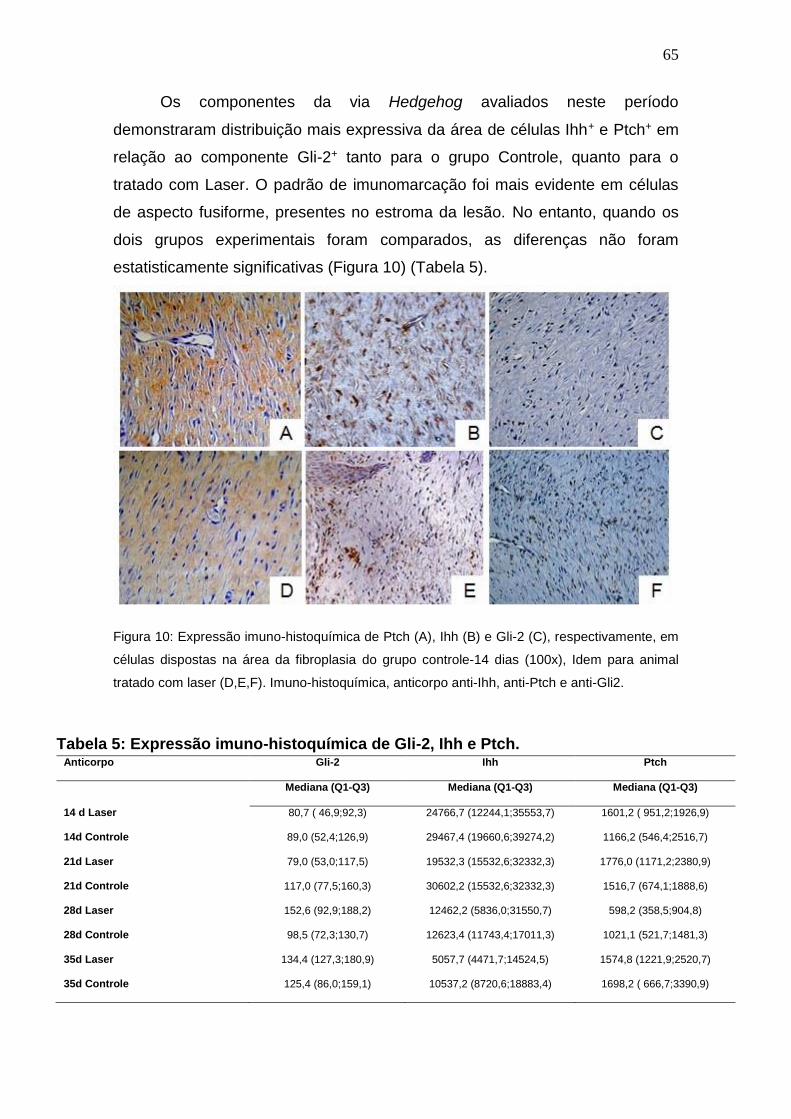
65
Os componentes da via Hedgehog avaliados neste período
demonstraram distribuição mais expressiva da área de células Ihh+ e Ptch+ em
relação ao componente Gli-2+ tanto para o grupo Controle, quanto para o
tratado com Laser. O padrão de imunomarcação foi mais evidente em células
de aspecto fusiforme, presentes no estroma da lesão. No entanto, quando os
dois grupos experimentais foram comparados, as diferenças não foram
estatisticamente significativas (Figura 10) (Tabela 5).
Figura 10: Expressão imuno-histoquímica de Ptch (A), Ihh (B) e Gli-2 (C), respectivamente, em
células dispostas na área da fibroplasia do grupo controle-14 dias (100x), Idem para animal
tratado com laser (D,E,F). Imuno-histoquímica, anticorpo anti-Ihh, anti-Ptch e anti-Gli2.
Tabela 5: Expressão imuno-histoquímica de Gli-2, Ihh e Ptch. Anticorpo Gli-2 Ihh Ptch
Mediana (Q1-Q3) Mediana (Q1-Q3) Mediana (Q1-Q3)
14 d Laser 80,7 ( 46,9;92,3) 24766,7 (12244,1;35553,7) 1601,2 ( 951,2;1926,9)
14d Controle 89,0 (52,4;126,9) 29467,4 (19660,6;39274,2) 1166,2 (546,4;2516,7)
21d Laser 79,0 (53,0;117,5) 19532,3 (15532,6;32332,3) 1776,0 (1171,2;2380,9)
21d Controle 117,0 (77,5;160,3) 30602,2 (15532,6;32332,3) 1516,7 (674,1;1888,6)
28d Laser 152,6 (92,9;188,2) 12462,2 (5836,0;31550,7) 598,2 (358,5;904,8)
28d Controle 98,5 (72,3;130,7) 12623,4 (11743,4;17011,3) 1021,1 (521,7;1481,3)
35d Laser 134,4 (127,3;180,9) 5057,7 (4471,7;14524,5) 1574,8 (1221,9;2520,7)
35d Controle 125,4 (86,0;159,1) 10537,2 (8720,6;18883,4) 1698,2 ( 666,7;3390,9)
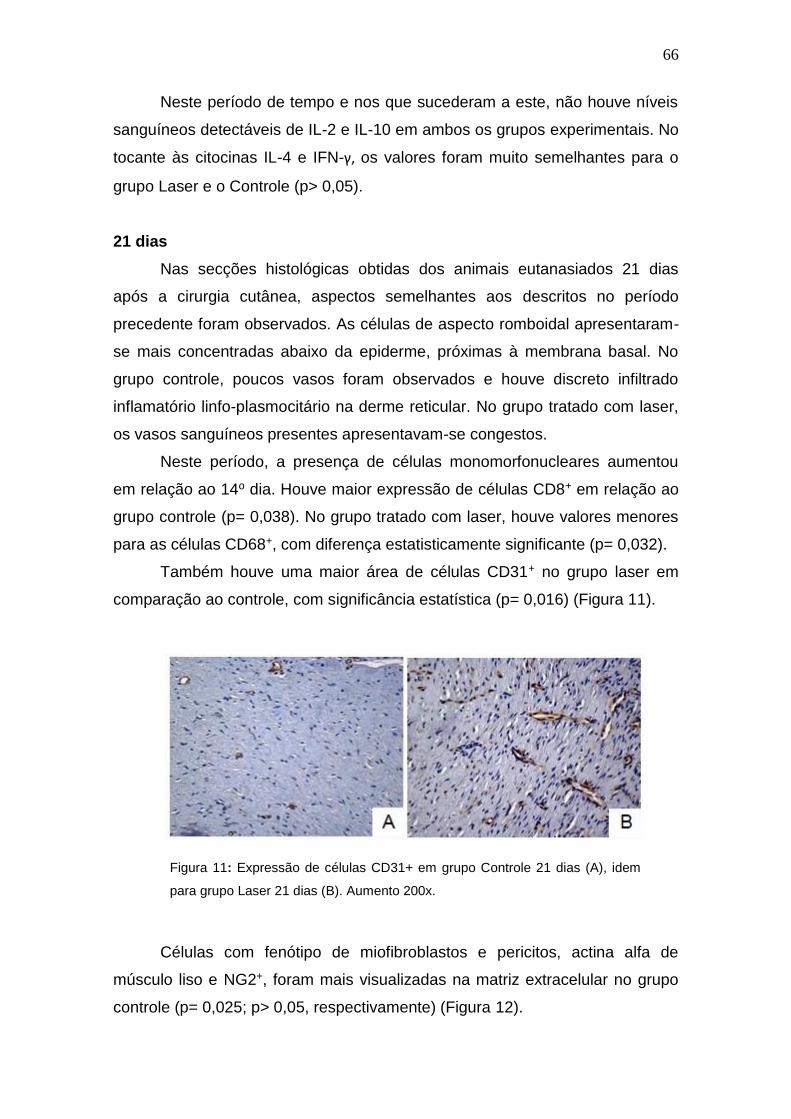
66
Neste período de tempo e nos que sucederam a este, não houve níveis
sanguíneos detectáveis de IL-2 e IL-10 em ambos os grupos experimentais. No
tocante às citocinas IL-4 e IFN-γ, os valores foram muito semelhantes para o
grupo Laser e o Controle (p> 0,05).
21 dias
Nas secções histológicas obtidas dos animais eutanasiados 21 dias
após a cirurgia cutânea, aspectos semelhantes aos descritos no período
precedente foram observados. As células de aspecto romboidal apresentaram-
se mais concentradas abaixo da epiderme, próximas à membrana basal. No
grupo controle, poucos vasos foram observados e houve discreto infiltrado
inflamatório linfo-plasmocitário na derme reticular. No grupo tratado com laser,
os vasos sanguíneos presentes apresentavam-se congestos.
Neste período, a presença de células monomorfonucleares aumentou
em relação ao 14o dia. Houve maior expressão de células CD8+ em relação ao
grupo controle (p= 0,038). No grupo tratado com laser, houve valores menores
para as células CD68+, com diferença estatisticamente significante (p= 0,032).
Também houve uma maior área de células CD31+ no grupo laser em
comparação ao controle, com significância estatística (p= 0,016) (Figura 11).
Figura 11: Expressão de células CD31+ em grupo Controle 21 dias (A), idem
para grupo Laser 21 dias (B). Aumento 200x.
Células com fenótipo de miofibroblastos e pericitos, actina alfa de
músculo liso e NG2+, foram mais visualizadas na matriz extracelular no grupo
controle (p= 0,025; p> 0,05, respectivamente) (Figura 12).
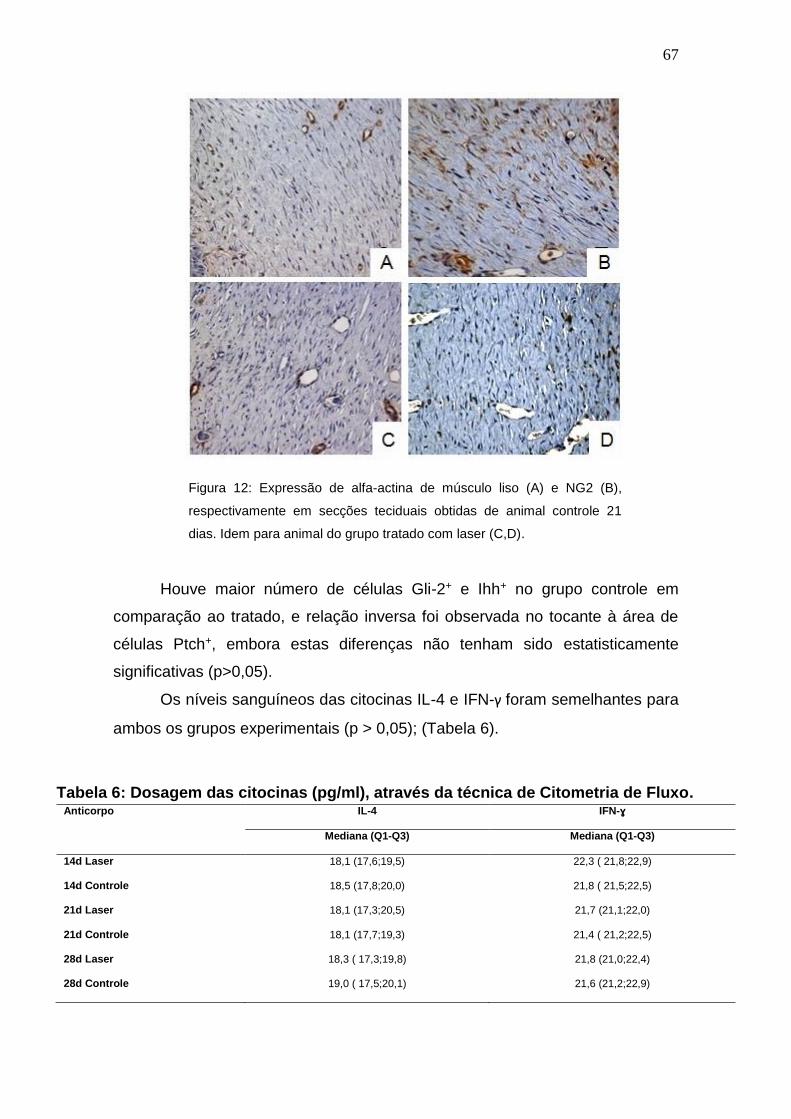
67
Figura 12: Expressão de alfa-actina de músculo liso (A) e NG2 (B),
respectivamente em secções teciduais obtidas de animal controle 21
dias. Idem para animal do grupo tratado com laser (C,D).
Houve maior número de células Gli-2+ e Ihh+ no grupo controle em
comparação ao tratado, e relação inversa foi observada no tocante à área de
células Ptch+, embora estas diferenças não tenham sido estatisticamente
significativas (p>0,05).
Os níveis sanguíneos das citocinas IL-4 e IFN-γ foram semelhantes para
ambos os grupos experimentais (p > 0,05); (Tabela 6).
Tabela 6: Dosagem das citocinas (pg/ml), através da técnica de Citometria de Fluxo. Anticorpo IL-4 IFN-ɣ
Mediana (Q1-Q3) Mediana (Q1-Q3)
14d Laser 18,1 (17,6;19,5) 22,3 ( 21,8;22,9)
14d Controle 18,5 (17,8;20,0) 21,8 ( 21,5;22,5)
21d Laser 18,1 (17,3;20,5) 21,7 (21,1;22,0)
21d Controle 18,1 (17,7;19,3) 21,4 ( 21,2;22,5)
28d Laser 18,3 ( 17,3;19,8) 21,8 (21,0;22,4)
28d Controle 19,0 ( 17,5;20,1) 21,6 (21,2;22,9)

68
35d Laser 17,4 (17,1;17,7) 21,3 (21,1;21,5)
35d Controle 17,9 ( 17,3;18,9) 21,6 (21,3;22,2)
28 dias
Neste período do estudo, houve diminuição da celularidade na derme
em ambos os grupos experimentais, embora tenha sido observada uma
expressiva concentração de células basofílicas de aspecto ora fusiforme, ora
romboidal, na área subepitelial no grupo tratado com Laser (Figura 13). Poucos
vasos foram visualizados nas secções teciduais e estes, quando presentes,
concentravam-se mais na derme reticular.
Figura 13: Distribuição celular na matriz conjuntiva correspondente à área de fibroplasia em
animal controle (A) – 28 dias. Idem para animal tratado com laser (B).
Embora a área correspondente às células inflamatórias
monomorfonucleares tenha sido superior no grupo tratado com laser quando
este foi comparado ao controle, esta diferença não foi significativa (p>0,05).
As medianas relativas às células CD31+ foram semelhantes para ambos
os grupos experimentais. No entanto, no grupo submetido à fotobiomodulação,
o número de células CD31+ foi significativamente menor em comparação ao
período de tempo anterior (p=0,014).
A expressão de células NG2+ foi maior neste período, quando
comparado ao 14odia (p=0,014); (Figura 14). No entanto, quando os grupos
experimentais foram comparados entre si, neste ponto específico do sacrifício,
as diferenças observadas não foram estatisticamente significativas, nem em

69
relação à presença de células actina alfa de músculo liso e NG2+, nem para as
células Gli-2+, Ihh+ e Ptch+ (p> 0,05).
Figura 14: Expressão imuno-histoquímica de NG2 em células perivasculares na
área do ferimento em animal controle 28 dias (A). Idem para o grupo Laser (B).
Aumento 200x.
Embora, o grupo Controle tenha evidenciado uma discreta elevação dos
níveis sanguíneos de IL-4 em relação ao grupo Laser, esta diferença não foi
estatisticamente significativa (Tabela 6).
35 dias
Secções teciduais obtidas de animais tratados e controles evidenciaram
extensa área de fibroplasia e ausência de anexos cutâneos. Nestes espécimes
pode-se observar redução de células CD8+ e CD68+ em comparação ao
período de 28 dias (p>0,05).
O valor correspondente à mediana de células CD31+ no grupo tratado
com laser foi menor do que aqueles observados no grupo controle nesta fase
da cicatrização, embora sem significância estatística (p> 0,05). No tocante à
presença de células actina alfa de músculo liso, também não houve diferença
estatisticamente significativa entre os grupos, sendo o maior valor descrito para
o grupo laser neste período. Relação inversa foi observada para as células
NG2+, embora sem significância estatística (p>0,05).
A área de células Gli2+ foi maior no grupo submetido à laserterapia,
(Figura 15); ao passo que a área correspondente às células Ihh+ e Ptch+
predominaram no grupo controle, embora esta diferença não tenha sido
estatisticamente significativa (p> 0,05). A expressão de células Gli-2+ foi maior

70
neste período quando comparado aos períodos de 14 e 21 dias, apenas para o
grupo submetido ao tratamento com laser (p> 0,05).
Figura 15: Expressão de células Gli-2 em grupo Controle 35 dias. Idem para grupo
laser. Aumento 200x.
O grupo Controle evidenciou uma diminuição das citocinas IL-4 e IFN-γ
em relação aos períodos de tempo anteriores, mas os níveis séricos das
referidas citocinas ainda mantiveram-se maiores quando comparados ao grupo
Laser, embora estas diferenças não tenham sido estatisticamente significativas
(Tabela 6).
4.4 Discussão
Há evidências crescentes de que as alterações celulares, bioquímicas e
moleculares que se desenvolvem precocemente em um tecido submetido à
uma nova agressão, muitas vezes se perpetuam por mais tempo durante o
reparo tecidual (VELNAR et al., 2009). O aspecto clínico final de um ferimento
pode não corresponder à ocorrência simultânea de fenômenos biológicos
complexos que se desenrolam no contexto cicatricial. Sendo assim, a presente
investigação objetivou caracterizar a população de células imunocompetentes,
assim como miofibroblastos, pericitos e quaisquer outros tipos celulares que
evidenciassem, através da técnica de imuno-histoquímica, a expressão
fenotípica de algumas proteínas integrantes da via de sinalização Hedgehog,
em fases mais avançadas do reparo tecidual. Em adição a este aspecto, foi

71
investigado se os efeitos primários desencadeados pelo laser de baixa potência
poderiam de alguma forma influenciar o curso dos eventos biológicos com o
avançar do reparo.
O modelo experimental de cicatrização cutânea em ratos tem sido muito
utilizado ao longo dos anos para investigar as diferentes etapas do reparo (GÁL
et al., 2006). Em especial, a ação de possíveis fatores endógenos e exógenos
considerados como agentes biomodulatórios, a exemplo do laser de baixa
potência (ROCHA et al., 2006; GONÇALVES et al., 2010; MEDRADO et al.,
2003, 2008, 2010). Em um estudo desenvolvido por Medrado et al., (2003),
estes autores descreveram a biomodulação da laserterapia de 670 nm na pele
de ratos e observaram um significativo incremento da transdiferenciação de
miofibroblastos durante a evolução da cicatrização. No entanto, neste estudo
os autores investigaram apenas as fases iniciais do reparo e estenderam a sua
análise até o 140 dia. Posteriormente, em 2008, os mesmos autores avaliaram
a cicatrização cutânea agravada pela ação do dióxido de silício até sessenta
dias após a indução do ferimento cutâneo padronizado. Embora não houvesse
diferenças estatisticamente significativas, entre os grupos experimentais nos
períodos mais avançados do reparo, correspondentes a 30 e 60 dias, houve
algumas alterações discretas a exemplo da angiogênese e presença de
miofibroblastos, fato este que instigou a realização do presente estudo.
Nesta pesquisa, optou-se por iniciar a análise do período cicatricial, a
partir do 140 dia após a realização da cirurgia cutânea. Neste período,
observou-se que a área correspondente às células CD8+ apresentava-se
menor no grupo tratado com laser, em comparação ao grupo controle. Tal fato
pode ser respaldado por resultados semelhantes descritos em inúmeros
estudos experimentais e clínicos, os quais ratificam a potente ação anti-
inflamatória do laser de baixa potência, que tem início nos períodos iniciais do
reparo e podem se estender até o período de tempo citado (CATÃO, 2004;
ROCHA JR et al., 2006).
A cicatrização é uma sequência de eventos celulares e bioquímicos
reguladas para restaurar a injúria tecidual (PARK & BARBOUL, 2004). Há uma
ampla evidência de trabalhos in vivo e in vitro de que os linfócitos T CD8+
participam da cicatrização e exercem efeitos regulatórios (BARBOUL et al.,
1988). Boyce et al., (2000) demonstraram o aumento de células T CD8

72
supressoras em fases mais avançadas da cicatrização, sugerindo o
envolvimento destes linfócitos neste processo, provavelmente mediado pela
liberação de citocinas/fatores de crescimento, modulando o reparo, através do
papel regulador das citocinas do perfil CD 4 Th-1. De fato, no grupo submetido
à laserterapia houve um aumento crescente da expressão de células CD8+ até
o período de 28 dias, sendo que esta diferença só foi estatisticamente
significante no período de 21 dias, quando este grupo foi comparado ao
Controle.
No tocante à expressão de células CD68+, observou-se que esta foi
sempre maior no grupo laser em quase todos os tempos de estudo. Isso está
de acordo com trabalhos no qual o laser de baixa potência promove o estímulo
da atividade macrofágica (SOUZA et al., 2004). Somente no período de 21 dias
o grupo laser apresentou diminuição e esta diferença foi estatisticamente
significativa em comparação ao grupo controle (p <0,005). Este decréscimo na
expressão de células CD 68+ e aumento de células T CD 8+, neste mesmo
período, pode ser explicado pela ação regulatória dos linfócitos T CD8+ através
da produção de moléculas sinalizadoras que afetam tanto macrófagos e
fibroblastos como MIF, IFN ɣ, TGF e outros. Sabe-se que o macrófago é capaz
de produzir simultaneamente mediadores inflamatórios e pró-fibróticos
(DELAVARY et al., 2011). Este segundo grupo de mediadores químicos
poderia determinar a fibroplasia mais rápida no grupo laser (MEDRADO et al.,
2008). Esta aparente modulação sugere que a permanência de tais células no
microambiente da cicatriz fibrosa pode estar relacionada à ação de uma
possível via sinalizadora estimulatória para o aumento na biossíntese do
colágeno.
A expressão de alfa actina de músculo liso, biomodulada pelo laser de
baixa potência tem sido investigada na literatura (HOURELD et al., 2014).
Embora esta proteína indique o processo de transdiferenciação do fibroblasto
para miofibroblasto, outros tipos celulares podem expressar tal miofilamento.
Medrado e colaboradores (2010) sugeriram que os miofibroblastos presentes
em secções teciduais de animais tratados com laser, poderiam ser
provenientes de células perivasculares (pericitos) ou de fibroblastos pré-
existentes na matriz extracelular, uma vez que estas células compartilharam a
expressão de NG2, actina alfa de músculo liso e desmina ao longo da

73
cicatrização. No presente estudo, houve maior expressão de células NG2+ no
grupo laser em relação ao grupo controle apenas no período de 28 dias,
embora sem significância estatística. Isto sugere que o processo de contração
máxima da ferida neste tempo, pode estar relacionado com o aumento
significativo de vasos neoformados no período precedente, observado nas
secções teciduais de animais tratados com laser.
Os papeis desempenhados por células actina alfa de músculo liso
positivas e NG2+, no leito do ferimento, abrangem um amplo espectro de
processos biológicos, como por exemplo, a produção de colágeno, o
repovoamento do tecido e as suas respectivas funções essencialmente
estruturais (CHERNG et al., 2008). Em estudo prévio, desenvolvido por nosso
grupo de pesquisa, (Medrado et al., 2003), observou-se um aspecto
semelhante com expressão de células alfa actina de músculo liso positivas,
maior no grupo laser. No entanto, a diferença estatística só se fez presente no
70 dia após a realização do ferimento, uma vez que por volta do 140 dia houve
um decréscimo da expressão deste miofilamento. Frente a estes achados e
aos do presente estudo, tais oscilações do fenótipo celular podem sugerir que a
contração da ferida parece ser um fenômeno cíclico, que sob o efeito da
fotobiomodulação costuma se apresentar mais precocemente. Em adição a
esta hipótese, procede ao fato de que o estado imunológico exerce um efeito
determinante na atividade de certos elementos celulares, a exemplo dos
miofibroblastos e pericitos (PARK & BARBOUL, 2004).
A angiogênese, ou seja, a formação de novos capilares a partir dos pré-
existentes, é um componente essencial para a cicatrização devido ao aumento
da demanda de nutrientes (DELAVARY et al., 2011). Segundo Andrade (2013),
a angiogênese é um processo biológico que exerce um impacto positivo no
remodelamento de feridas. Sendo assim, utilizou-se na presente pesquisa, o
anticorpo CD31 para avaliar a participação das células endoteliais na evolução
da cicatrização fotobiomodulada. Observou-se que a máxima neoangiogênese
aconteceu apenas no período de 21 dias no grupo de animais irradiados com
laser, quando comparado ao controle (p< 0,05). A permanência de vasos no
microambiente da lesão, neste período, pode ter contribuído não só para
garantir o maior aporte de oxigênio e nutrientes, mas, também para o
repovoamento celular neste local (TAZIMA et al., 2008).

74
Vários estudos têm demonstrado que pericitos presentes nos capilares
proliferados podem se destacar da parede do capilar e se acumular nos tecidos
lesionados onde se diferenciam em miofibroblastos e passam a depositar
matriz extracelular (LEE et al., 2007). Em especial, a maior quantidade de
células actina alfa de músculo liso e NG2+ no período imediatamente posterior,
ou seja, 28 dias, pode indicar que os miofibroblastos e pericitos foram
provenientes da parede dos vasos neoformados. Sendo assim, a
neoangiogênese seria responsável também por fornecer uma “fonte de reserva
celular” para o tecido em formação. É digno de nota que este mesmo processo
ocorreu no grupo controle, mas com pelo menos uma semana de atraso.
Tem sido documentada na literatura a via de sinalização Hedgehog, que
está envolvida em muitos aspectos do desenvolvimento embriológico, incluindo
morfogênese, angiogênese e reparo (BIELEFELD et al., 2013). E sua ação no
reparo, ainda é um tópico pouco explorado, comparando-se ao papel desta via
no desenvolvimento da pele.
Em relação à imunomarcação de Ptch, sua presença foi mais expressiva
no grupo laser nos períodos de 14 e 21 dias, embora sem significância
estatística, evidenciando a possibilidade de ligação com alguma proteína da via
na histodiferenciação da pele. Este achado é biologicamente relevante e pode
indicar que as células, sob a ação da laserterapia, aumentaram o número de
receptores em preparação para a possível ativação da via Hedgehog. Contudo,
embora esse dado possa estar relacionado com a maior expressão de Gli-2
nos tempos de 28 e 35 dias de animais tratados com laser, a expressão de Ihh,
neste grupo experimental, parece não ter sido influenciada. De fato, a
expressão de tal anticorpo foi sempre maior no grupo controle, embora sem
significância estatística. De acordo com Athar et al., (2006), a proteína
transdutora Shh tem sido demonstrada como a mais atuante no processo de
cicatrização cutânea. Neste caso, o laser de baixa potência poderia ter inibido a
expressão de Ihh e ativado Shh ou outra proteína da via Hedgehog, que não foi
objeto de análise neste estudo.
Foi observado que a área correspondente às células que expressaram o
fator de transcrição Gli-2 esteve aumentada no grupo tratado com laser nos
períodos de 28 e 35 dias, em relação ao grupo controle. Desta forma, sugere-
se que a via Hh possa ter promovido a proliferação de miofibroblastos, haja

75
vista a maior expressão de actina alfa de músculo liso nos mesmos períodos,
ou seja, 28 e 35 dias, sob a ação do laser de baixa potência. Isso é apoiado no
estudo de Sicklick et al., (2006). Tais autores descreveram que os
miofibroblastos podem produzir o ligante Hedgehog (Hh), o qual controla a
construção tissular e o remodelamento. Tal ligante pode também regular a
viabilidade e a atividade migratória de vários tipos celulares estimulados pela
sinalização.
Investigações adicionais que avaliem a expressão de outras proteínas
transdutoras de sinais, no modelo de cicatrização cutânea sob a ação do laser
de baixa potência, serão necessárias a fim de caracterizar o impacto da
fotobiomodulação sobre o reparo do tecido cutâneo.
Embora o microambiente da área do ferimento apresentasse diferentes
perfis de populações celulares ao longo do tempo, nos grupos experimentais
estudados, no presente estudo não se conseguiu demonstrar diferenças
significativas nos níveis sanguíneos de IL-2, IL-4, IL-10 e IFN-ɣ. A ação
sistêmica da laserterapia tem sido objeto de discussão na literatura. Vilela et
al., (2012) demonstraram a atividade sistêmica do laser de baixa potência e
relataram que a resposta ao laser de 670 nm não dependeu exclusivamente do
eixo hipotálamo-pituitária-adrenal e assim, sugeriram que outros tecidos extra-
adrenais poderiam influenciar a resposta imune a este tipo de fototerapia. No
entanto, estes autores e alguns outros avaliaram a ação sistêmica do laser em
períodos de tempo diferentes dos da presente investigação (RODRIGO et al.,
2009; MAYER et al, 2013). De fato, sua análise se concentrou nas fases iniciais
do reparo tecidual. A aparente discrepância dos nossos resultados pode indicar
que a ação sistêmica do laser exerce o seu maior impacto no início da
cicatrização, ao passo que na etapa correspondente ao remodelamento, esta
parece não se manter mais tão ativa, uma vez que o nível das citocinas não foi
significativo, em nenhum dos períodos estudados. Além disso, os níveis
indetectáveis das citocinas IL-2 e IL-10 podem ser justificados em virtude do
grande espaço de tempo decorrido entre a última sessão da laserterapia e os
períodos de eutanásia do presente estudo.
Contudo, ainda assim, os fenômenos biológicos que ocorreram no leito
do ferimento demonstraram a influência do laser localmente no tecido, haja

76
vista a presença de células que expressaram diferentes perfis na área sujeita à
remodelação.
4.5 Considerações Finais
Os achados do presente estudo sugerem que o laser de baixa potência
foi capaz de modular a fase de remodelamento da matriz extracelular durante a
cicatrização experimental, uma vez que os tecidos fotobiomodulados
apresentaram alterações mais precoces em relação ao grupo controle.
Embora as ações primárias da laserterapia sejam mais marcantes no
período inicial do reparo, elas podem se perpetuar ao longo do tempo no
microambiente da lesão, mesmo que estas sejam discretas ou até mesmo
imperceptíveis do ponto de vista clínico.

77
REFERÊNCIAS
Andrade, Z A Double and paradoxical role for angiogenesis Rev Patol Trop 2013; 42 (3): 259-264. Athar, M; Tang, X; Lee, J L; Kopelovich, L; Kim, A L Hedgehog signalling in skin development and cancer Experimental Dermatology 2006; 15: 667-677. Barboul, A; Breslin, R J; Woodyard, J P; Wasserkrug, H L;Efron, G The effect of in vivo T helper and T supressor Lymphocyte depletion on wound healing Annals of Surgery 1989; 209(4): 479- 483. Bielefeld, K A; Amini-Nik, S; Alman, B A Cutaneous wound healing: recruiting developmental pathways for regeneration Cellular and Molecular Life Sciences 2013; 70: 2059-2081. Byrd, N & Grabel, L Hedgehog signalling in murine vasculogenesis and angiogenesis Trends Cardiovascular Medicine 2004; 14: 308-313. Boyce, D E; Jones, W D; Ruge, F; Harding, K G; Moore, K The role of lymphocytes in human dermal wound healing British Journal of Dermatology 2000; 143: 59-65. Catao, M H C V Os benefícios do laser de baixa intensidade na clínica odontológica e na estomatologia Revista Bras. de Patol. Oral 2004: 3: 214-8. Cherng, S; Young, J; Ma, H Alpha-smooth muscle actin The Journal of American Science 2008; 4(4): 7-9. Delavary, B M; Veer, W M; Egmond, M; Niessem, F B; Beelen, R H Macrophages in skin injury and repair Immunobiology 2011; 216: 753-762. Felice, T; Pinheiro, A; Menchik, E; Silva, A; Souza, L; Caires, C; Abel, A; Bartemeyer, C; Oliveira, J; Assis, T; Silva, L; Lopes, T; Felippe, L; Pinheiro A utilização do laser de baixa potência na Ccicatrização de feridas Interbio 2009; 3 (2): 42-52. Gál, P; Vidinsky, B; toporcer, T; Mokry, M; Mozes, S; Longauer, F Histological assessement of the effect of laser irradiation on skin wound healing in rats Photomedicine and Laser Surgery 2006: 24(4): 480-488. . Gonçalves, R V; Souza, N T A; Silva, P H: Barbosa, F S; Neves, C A Influência do laser de arseneto de gálio-alumínio em feridas cutâneas de ratos Fisoter Mov 2010; 23 (3): 381-388. Houreld, N N; Ayuk, S M; Abrahamse, H Expression of genes in normal fibroblasts cells (WS1) in response to irradiation at 660 nm Journal of Photochemistry and Photobiology B: Biology 2014; 130: 146-152.

78
Inghan, P W; Nakano, Y; Seger, C Mechanisms and functions of Hedgehog signaling across the metazoan Nature Reviews Genetcs 2011; 12: 393-406 Lee, J S; Semela, D; Iredale, J; Shah, V H Sinusoidal remodeling and angiogenesis: a new function for the liver-specific pericyte? Journal of Hepatology 2007; 45: 817-823. Medrado, A R A P; Pugliese, L S; Reis, S R A; Andrade, Z A. Influence of low level laser therapy on wound healing and its biological action upon myofibroblasts. Lasers in Surgery and Medicine 2003; 32: 239-244. Medrado, A R A P; Trindade, E; Reis, S R A; Andrade, Z A. Action of low-level laser therapy on living fatty tissue of rats. Lasers Med Sci 2006; 21: 19-23. Medrado, A P; Soares, A P ; Santos, E T; Reis, S R A; Andrade, Z A Influence of laser photobiomodulation upon connective tissue remodeling during wound healing Journal of Photochemistry and Photobiology B: Biology 2008; 92(3): 144-152. Medrado, A P; Costa, T; Prado, T; Reis, S R A; Andrade, Z A. Phenotype characterization of pericytes during tissue repair following low-level laser therapy. Photodermatology, Photoimmunology & Photomedicine 2010; 26: 192-197. Mendonça, R J & Coutinho-Netto, J Aspectos celulares da cicatrização An Bras Dermatol 2009; 84 (3): 257-262. Park, J E & Barbul, A Understanding the role of immune regulation in wound healing The American Journal of Surgery 2004; 187: 11S-16S. Pereira, A N P; Eduardo, C P; Matson, E; Marques, M M. Effect of Low-Power Laser irradiation on cell growth and procolagen synthesis of cultured fibroblasts. Lasers Surg Med 2002; 31:263-267. Pugliese, L S; Medrado, A P; Reis, S R A; Andrade, Z A. The influence of low-level laser therapy on biomodulation of collagen and elastic fibers. Pesquisa Odontológica Brasileira 2003; 17(4): 307-313. Raitz, R O papel da matriz extracellular ma cicatrização de feridas bucais Revista Inst Ciênc Saúde 2008; 26 (3): 351-356. Reis, S R A; Medrado, A P; Marchionni, A M T; Figueira, C; Fracassi, L D; Knop, L A H. Effect of low level laser therapy and dexametasone on tissue repair: a histologic and histologic and ultrastructural study. Lasers Surg Med 2008. Rocha Jr, A M; Oliveira, R G; Farias, R E; Andrade, L C F; Aarestrup, F M Modulação da proliferação fibroblástica e da resposta inflamatória pela terapia a laser de baixa intensidadeno processo de reparo tecidual Anais Brasileiros de Dermatologia 2006; 81(2): 150-156.

79
Rodrigo, S M; Cunha, A; Pozza, D H; Blaya, D S; Moraes, J F; Weber, J B B; Oliveira, M G Analysis of the systemic effect of red and infrared laser therapy on wound repair Photomedicine and Laser Surgery 2009; 27(6): 929-935 Ribatti, D; Nico, B: Crivelatto, E The role of pericytes in Angiogenesis Int J Dev Biol 2011; 55: 261-268 Rutter, D J; Schlingemann, R O; Westphal, J R; Rietveld E J R; de Waal, R M W. Angiogenesis in wound healing and tumor metastasis. Behring Inst Mitt 1993; 92: 258-272. Souza, N H C; Ferrari, R A M; Silva, D F T; Nunes, F D; Bussadori, S K; Fernandes, K P S Effect of low-level laser therapy on the modulation of mitochondrial activity of macrophages Brazilian Journal of Physical Therapy 2014; 18(40: 308-314. Tazima, M F G S; Vicente, Y A M V: Moriya, T Biologia da Ferida e Cicatrização Simpósio – Fundamentos em Clínica Cirúrgica 2008; 4(3): 259-264. Tidball, J G Inflammation process in muscle injury and repair American Journal of Physiology Regulation Integrative and Comparative Physiology 2005; 288: 345-353. Velnar, T; Bailley, T; Smrkolj, V The wound healing process: an overview of the cellular and molecular mechanisms The Journal of International Medical Research 2009; 37:1528-1542. Vilela, D D C; Chamusca, F V; Andrade, J C S; Vallve, M L F; Gonzalez, A C; Andrade, Z A; Medrado, A R A P; Reis, S R A Influence of HPA axis on the inflammatory response in cutaneous wounds with the use of 670-nm laser photobiomodulation Journal of photochemistry and Photobiology B: Biology 2012; 116: 114-120.

80
5 CAPÍTULO 2: ARTIGO CIENTÍFICO 2: Immunohistochemical Evaluation
of Hedgehog signaling in Epithelial/Mesenchymal Interactions in
Squamous Cell Carcinoma Transformation: A Pilot Study
5.1 Introduction
Actinic cheilitis (AC) is a premalignant lesion of the lip attributed to
chronic sun exposure that occurs most often in the lower lip (1,2).Clinically, AC
may have different characteristics, for example: keratosis; crust formation and
cracking; erythema and atrophy (3). In histopathological examinations, atypical
cells are found, having altered nucleus-cytoplasm ratio, hyperchromatic nuclei,
cellular and nuclear pleomorphism, large numbers of irregular mitoses, islands
of possibly invasive malignant epithelial cells and keratin pearls (4).
Although all of the above features can be related to AC, the loss of
sharpness of the red edge of the lip and the infiltrative aspect is clinically
correlated with neoplastic transformation (4).The role of existing inflammatory
components in AC is a recurrent subject of study in the literature, because they
can act as immunomodulators in the process of malignant transformation, from
its beginning to the final development of tumours (1,5).
The relationship between the degree of dysplasia and the probability of
malignant transformation of lesions is well established. According to some
authors, the presence of inflammatory components is related to greater
disorganisation of keratinocytes in epithelial dysplasia (5).
In previous studies, the clinical aspects of oedema and erythema were
seen in AC cases that progressed to squamous cell carcinoma (SCC) (1,5).
There are reports that the only histological characteristics associated with the
degree of epithelial atypia and the presence of invasive spinous carcinoma cells
is the intensity of the inflammatory process observed in AC (1).
SCC is defined as a malignant neoplasm arising from squamous
epithelial cells that originate in the stratified epithelium of the oral mucosa. It
accounts for 90% of malignant lesions of the stomatognathic system and is the
most common type of oral cancer. In SCC, several signalling pathways are

81
triggered and stimulate malignant transformation of the dysplastic epithelium, as
well as significant changes in the extracellular matrix. Recently, the relevant
epithelial/mesenchymal interaction has been demonstrated by
immunohistochemical studies to involve an important cellular pathway known as
Hedgehog signaling. The participation of this signaling pathway has previously
been described in association with the pathogenesis of basal cell carcinoma (6,
7). Although there are some scientific studies regarding its relationship with
SCC (8, 9), there are few studies that evaluate the progress from dysplasia to
squamous cell carcinoma in relation to Hedgehog signalling. As a matter of fact,
several proteins in the Hedgehog signaling pathway have been studied as
prognostic biomarkers for oral squamous cell carcinoma (10).
Proteins that belong to the Hedgehog (Hh) signaling pathway are active
in the embryogenesis of insects and vertebrates (11). In humans, the Hedgehog
signaling pathway is quiescent in adulthood, but its activation has been
described in several neoplastic processes (12,13).The proteins involved are
Sonic Hedgehog (Shh), Indian Hedgehog (Ihh) and Desert Hedgehog (Dhh) and
they act as secreted ligands for the transmembrane receptor proteins Patched 1
(Ptch1) and Patched 2 (Ptch2). The three morphogens are homologous and
have the same function: to induce Hh signal transduction, varying in the way
they bind to the receptor and their signaling potential. This process controls the
activity of a membrane signal transducer, named Smoothened (Smo). Once
stimulated, the Smoothened protein begins an intracellular cascade of
transcription modulated by the glycoproteins, Gli-1 Gli-2 and Gli-3, which have
nuclear effects.
The Gli-1 protein acts as a positive transcriptional factor and Gli-3 as a
negative factor, while Gli-2, depending on its post-transcriptional modification,
can act positively or negatively (13). Sonic Hedgehog (Shh) is the most highly
expressed homologue of Hh in vertebrates, participating in the formation of
various organs of the plate and neural tube, brain, axial skeleton, teeth and hair.
Epithelial tissue has great influence in the maintenance of stem cells. The
mutation of the protein Patched (Ptch) is the cause of Gorlin-Goltz syndrome,
which results in various tissue abnormalities including basal cell carcinoma (14).
Given the involvement of Hedgehog signaling pathway components in
physiological and pathological processes, this study aims to analyse the

82
expression of the proteins of the Hedgehog signaling pathway in actinic cheilitis
and squamous cell carcinoma using immunohistochemistry, and thus evaluate
the role of epithelium/mesenchyme interactions in tumour transformation.
5.2 Materials and methods
The project was submitted to the Human Research Ethics Committee
and was approved with protocol number 495.219.
5.2.1 Case Selection
Sixteen AC cases with associated lip SCC were selected from lower-lip
biopsies of the oral pathology laboratory archives of a private higher education
institution. The anatomical and pathological diagnoses had been reviewed and
confirmed by two experienced pathologists using slides stained with
hematoxylin-eosin (HE) and analysed morphologically by light microscopy. As a
control group, five cases of normal tissue resections mucocele were used,
which were chosen at random in order to observe the immunohistochemical
pattern of the antibodies in normal lip epithelium and lamina propria.
5.2.2 Histopathology
For the analysis of AC lesions, the histological criteria for diagnosis of
epithelial dysplasia defined by the World Health Organization were used (WHO,
1978). Each section was examined for the presence or absence of: irregular
stratification; epithelial hyperplasia of basal cells; reticular drop-shaped
processes; loss of polarity of basal cells; increased nucleus/cytoplasm ratio;

83
nuclear polymorphism; increase in size of nucleoli; nuclear hyperchromatism;
reduced intercellular adhesion and suprabasal mitosis and dyskeratosis. To
measure the degree of dysplasia, the standards suggested by Banoczy and
Csiba (1976) were used: a) epithelium with no atypia; b) light epithelial
dysplasia where two features are present; c) moderate epithelial dysplasia
characteristics where four or more features are present; d) severe epithelial
dysplasia, where five or more features are present.
The histological evaluations of squamous cell carcinomas were
performed according to the malignancy grading system developed by the WHO
(WHO, 2005), that classifies tumours as poorly, moderately or well-
differentiated. There were no poorly-differentiated tumours found in the study.
Well-differentiated tumours presented a tissue architecture similar to a normal
squamous epithelium standard, while moderately-differentiated lesions
presented a certain degree of cellular pleomorphism and mitotic activity, with
little keratinisation.
5.2.3 Immunohistochemical Procedures
The material was embedded in paraffin, sectioned at a thickness of four
microns and mounted on glass slides which had been previously prepared by
treatment with 3-aminopropyl triethoxysilane (Sigma Aldrich, St. Louis, Missouri,
USA).
Peroxidase was employed as the immunohistochemical method.
Monoclonal primary antibodies used were: anti-Ptch (1:1000) (Novus Biological,
USA), anti-Gli-2 (1:2000) (Gen way Biothec Inc, San Diego, California, USA),
anti-Shh (1:6000) (Epitomics, Burlingame, California, USA) and anti-Smo
(1:100) (Epitomics, Burlingame, California, USA). The slides underwent
deparaffinization followed by sequential baths of xylene, alcohol (absolute, 95%,
70%, 50%, 30%) and distilled water. Antigen retrieval was performed in 10mM
citrate buffer, pH 6.0 in a water bath at 96°C, with subsequent blocking of
endogenous peroxidase with Duo Enzyme Block (DAKO, Corporation, USA)
and non-specific binding with Serum Free Protein Block (DAKO, Corporation,

84
USA). Slides were then incubated at 4°C overnight with primary antibody.
Secondary antibody was the Polymer Dako Envision Peroxidase (Dako
Corporation, USA) and the developing was performed using DAB with
3% hydrogen peroxide. The slides were counterstained with Harris
haematoxylin and mounted with cover slips and Canada balsam for
morphological and quantitative analysis. Mucocele cuts, removed by previous
surgical resection, were used as positive controls.
5.2.4 Histomorphometry
Images captured from tissue sections were submitted to
immunohistochemical analysis for Sonic Hedgehog, Patched, Gli-2 and
Smoothened, performed using the Advanced Motic Images 3.0 software. A
standard area of analysis of 13107.200000 pixels was established. Fifteen
standard images of the established dimensions were captured and the area
corresponding to the immunostaining pattern of each antibody studied was
measured using Adobe Photoshop CS5 Extended software, Version 12.0x32
(Adobe Systems Incorporated, USA). These data were compared to the total
area selected. All analyses were performed using an examiner calibrated to
avoid bias. The immunostaining pattern of each antibody is described in Box 01.
The immunohistochemical scoring system used to determine staining positivity
was based in the degree of cells staining and was considered as absent,
discreet, moderate and intense.
85
Box 2: Immunostaining pattern of Sonic Hedgehog, Patched, Gli-2
and Smoothened.
Antibody Intensity of staining Locations of expression
Sonic Moderate Cytoplasmic
Patched Intense Extracellular matrix and
epithelial cell cytoplasm
Gli-2 Intense Nuclei
Smoothened Discreet Cytoplasmic
5.2 .5 Statistical Analysis
The database was created in Microsoft Excel® 2010 (version
14.0.7132.5000), Microsoft® Office Professional Plus 2010, and analysed using
R software (version 3.1.1). A descriptive analysis was performed
(absolute/relative frequency, mean, median and standard deviation and
quartiles).The distribution of the data was tested for normality with the Shapiro–
Wilk test. According to this analysis, the variables of Smo and Gli-2 in epithelial
tissue were considered non-parametric, as were Ptch and Shh in the lamina
propria tissue. Methods for analysis of nonparametric variables were therefore
applied.
For comparisons among the control, dysplasia and carcinoma
(epithelium) samples, the areas labelled by Ptch and Shh antibodies were
submitted to ANOVA followed by the Bonferronipost-hoc test. For Smo and Gli-
2 antibodies, we used the Kruskal–Wallis test followed by Dunn’s test. To
analyse Smo and Gli-2 expression, Student’s t-test was used; for Ptch and Shh,
the Mann–Whitney U test was used. The established significance level for this
work was p<0.05.

86
5.3 Results
Tissue sections stained with haematoxylin-eosin from dysplasia (AC) and
carcinoma were classified according to the histological grade of their
abnormalities. It was observed that of the total dysplasias, four (25%) were
classified as light, nine (56.3%) as moderate and three (18.7%) as severe.
Among dysplasic histological changes, atypical epithelial stratification was the
most common, followed by nuclear hyperchromatism, presence of reticular drop
processes and basal layer hyperplasia. Regarding the classification of
carcinomas, 11 (69.3%) were well-differentiated and five (30.7%) moderately-
differentiated. There were no cases of poorly-differentiated carcinoma among
the samples.
The immunohistochemical staining pattern was more intense in tissue
sections subjected to treatment with anti Gli-2, which labelled the nuclei of
dysplasic and neoplasic epithelial cells (Figure 16). Other antibodies exhibited
cytoplasmic staining in the same cells. In the lamina propria nuclear Gli-2
expression and cytoplasmic expression of Shh and Smo was observed.

87
Figure 16 (a-l). Hedgehog proteins expression in normal oral mucosa, dysplasia
and carcinoma, respectively (GLi 2 + cells, [a-c]; Shh+ cells [d-f]; SMO+ cells [g-i]
and Ptch+ cells [j-l]). Immunohistochemistry, streptavidin-biotin peroxidase (Anti
Gli2, Anti Sonic, Anti SMO and Anti Patched antibodies). 400X.
Only endothelial cells of blood vessels dispersed in the matrix were
positive for the expression of both Smo and Shh. Anti-Ptch antibody stained the
extracellular matrix.
Table 7 shows the number of cells that had a positive staining pattern for
all antibodies used. A progressive and significant increase in Gli-2-positive (Gli-
2+), Smoothened-positive (Smo+), Patched-positive (Ptch+) and Sonic
Hedgehog-positive (Shh+) cell areas was observed in dysplasia and carcinoma
tissue sections compared with the normal mucosal oral epithelium (p<0.05). In
particular, Gli-2+ and Smo+ areas were significantly increased in carcinoma

88
cells compared with normal epithelial cells (p = 0.011 and p = 0.001,
respectively) (Figure 17). Ptch+ cell expression, was significantly different
between the control group and carcinoma (p = 0.032) and between the
dysplasia group and carcinoma (p = 0.017).
Table 7: Positive cell area for Patched, Sonic Hedgehog, Gli-2 and Smoothened
immunostaining in normal oral mucosa epithelial tissue (control), dysplasia and squamous cell
carcinoma.
Ptch+ ♦
Shh+ ♦
Gli-2 +►
Smo+►
Groups Median
D.
Standard (DP)
Average
D.
Standard (DP)
Median
Quartiles
(Q3-Q5) Median
Quartiles
(Q3-Q5)
ORAL
MUCOSA
126.1 B
109
167.6 B
103.7
66 B
26.9
- 240.5
2.6 B
1.6
- 4.3
DYSPLASIA 201.8 B 186.4
380.5 AB 273.1
491.7 AB 311.2
- 681.4
120.1 AB 25.8
- 385.5
CARCINOMA 469.1 A 318.2 1007.4 A 597.4 642.5 A 247.6
- 681.6
419.9 A 209.3
- 1107.4
♦1-way ANOVA and Bonferroni correction.
► Kruskal–Wallis and Dunn.
Different letters represent findings of significance (p <0.05) between the groups in each
antibody (A, B).
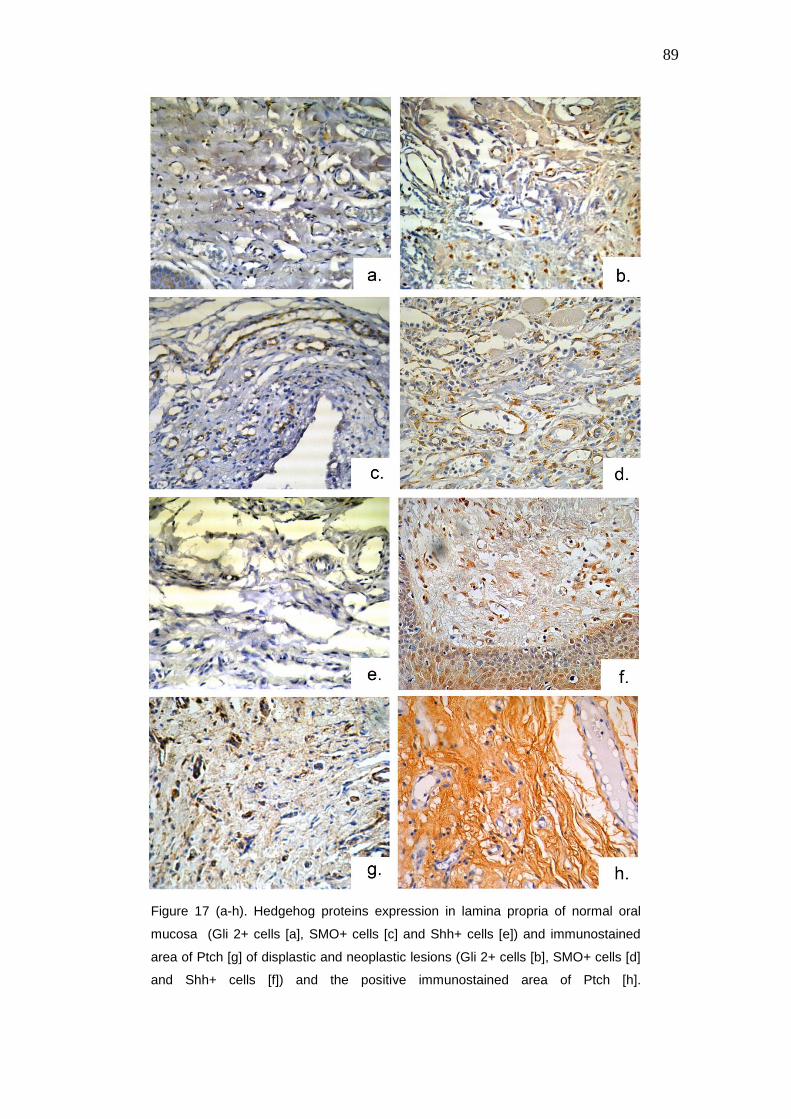
89
Figure 17 (a-h). Hedgehog proteins expression in lamina propria of normal oral
mucosa (Gli 2+ cells [a], SMO+ cells [c] and Shh+ cells [e]) and immunostained
area of Ptch [g] of displastic and neoplastic lesions (Gli 2+ cells [b], SMO+ cells [d]
and Shh+ cells [f]) and the positive immunostained area of Ptch [h].

90
Immunohistochemistry, streptavidin-biotin peroxidase (Anti Gli2, Anti Sonic, Anti
SMO and Anti Patched antibodies). 400X.
Similar changes were found for the Shh+ cells in these study groups (p =
0.002 and p = 0.001, respectively) (Figure 17).
Immunohistochemical expression of the proteins in this study was further
assessed in the lamina propria (Table 8). Although an increase in the area
corresponding to the expression of all of the proteins was observed, this
difference was statistically significant only for Smo+, Ptch+ and Shh+ (p = 0.05,
p <0.0001 and p = 0.001 respectively) (Figure 17).
Table 8: Positive area for Patched, Smoothened, Gli- 2 and Sonic Hedgehog in normal oral lamina propria (control group) and in underlying connective tissue dysplasia and oral squamous cell carcinoma.
●Ptch+
Shh+●
Gli-2+▲
Smo+▲
Groups Median
Quartiles
(Q3-Q5) Median
Quartiles
(Q3-Q5) Average
D.
Standard
(DP)
Average
D.
Standard
(DP)
ORAL
MUCOSA
52.2 B
31.05
- 164
5.6 B
7.1
51.5 A
15.85
- 117.8
106.4 B
57.71
- 234.55
Dysplasia / CARCINOMA
101.7 A 29.27
- 159.27
114.3 A 90.4
200 A 111.87
- 368.05
186.4 A 144.22
- 268.55
● Mann–Whitney U test;
▲ Student's t-test;
Different letters represent significant findings (p <0.05) between groups in each antibody studied
(A, B).
Table 9 shows the area of cells that exhibited positive staining for the
proteins examined in the study. A progressive increase in Hedgehog signalling
pathway expression was found in neoplastic lesions compared with the control
group (p <0.05). Although similar observations were made for the dysplasia
group, this increase was not statistically significant (p> 0.05). There was a
statistically significant difference between the light and moderate dysplasia
groups compared with the different degrees of carcinoma, in particular from light
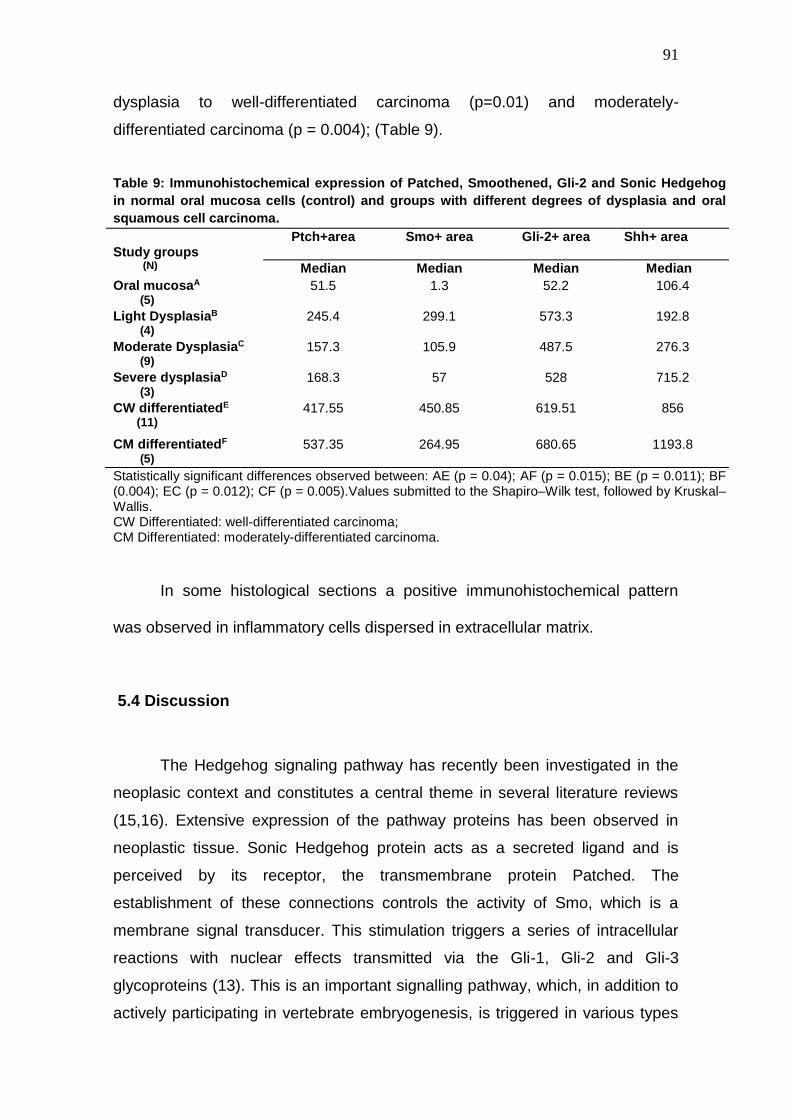
91
dysplasia to well-differentiated carcinoma (p=0.01) and moderately-
differentiated carcinoma (p = 0.004); (Table 9).
Table 9: Immunohistochemical expression of Patched, Smoothened, Gli-2 and Sonic Hedgehog
in normal oral mucosa cells (control) and groups with different degrees of dysplasia and oral
squamous cell carcinoma. Study groups (N)
Ptch+area Smo+ area Gli-2+ area Shh+ area
Median Median Median Median Oral mucosaA (5)
51.5 1.3 52.2 106.4
Light DysplasiaB (4)
245.4 299.1 573.3 192.8
Moderate DysplasiaC (9)
157.3 105.9 487.5 276.3
Severe dysplasiaD (3)
168.3 57 528 715.2
CW differentiatedE (11)
417.55 450.85 619.51 856
CM differentiatedF (5)
537.35 264.95 680.65 1193.8
Statistically significant differences observed between: AE (p = 0.04); AF (p = 0.015); BE (p = 0.011); BF (0.004); EC (p = 0.012); CF (p = 0.005).Values submitted to the Shapiro–Wilk test, followed by Kruskal–Wallis. CW Differentiated: well-differentiated carcinoma; CM Differentiated: moderately-differentiated carcinoma.
In some histological sections a positive immunohistochemical pattern
was observed in inflammatory cells dispersed in extracellular matrix.
5.4 Discussion
The Hedgehog signaling pathway has recently been investigated in the
neoplasic context and constitutes a central theme in several literature reviews
(15,16). Extensive expression of the pathway proteins has been observed in
neoplastic tissue. Sonic Hedgehog protein acts as a secreted ligand and is
perceived by its receptor, the transmembrane protein Patched. The
establishment of these connections controls the activity of Smo, which is a
membrane signal transducer. This stimulation triggers a series of intracellular
reactions with nuclear effects transmitted via the Gli-1, Gli-2 and Gli-3
glycoproteins (13). This is an important signalling pathway, which, in addition to
actively participating in vertebrate embryogenesis, is triggered in various types

92
of neoplasms such as those present in the lung (17), epithelium, gastric mucosa
(18), and oesophagus among others (19). In immunohistochemical and PCR
analyses, a relationship was found between the expression of Shh pathway
proteins and the prognosis for cancer in patients with non-small cell lung
cancer, and it was established that Shh has an important role in the
transformation of these tumours (17). The transformation of lesions to malignant
neoplasms was not analysed by these authors, but it was one of the goals of
the present study. As a progressive increase in the expression of all protein
components of the Hedgehog pathway in AC and SCC was observed, it can be
suggested that they do participate in the transformation of epithelial dysplasia to
squamous cell carcinoma of the lip.
According to authors who have analysed Gli-2 function in osteosarcoma
pathogenesis, some drugs like arsenic trioxide, vismodegib and GANT61, were
able to inhibit the transcriptional activity of Gli-2 as well as migration and
metastasis of osteosarcoma cells, confirming that Gli-2 regulates the
transformation of this specific type of tumour (20). As a gradual increase of Gli-2
was seen in the cases studied in the present work, it is possible that this protein
also has some participation in tumour transformation of lip SCC. Zang et al
have demonstrated that the expression of Gli-2 is strongly associated with a
poor clinical outcome in oral squamous cell carcinoma patients, and their data
suggested that a subset of these patients might benefit from anti-HH/GLI
therapies (21).
In dentistry, the Shh protein has been reported as an important mediator
of epithelial/mesenchymal interactions in odontogenesis (11, 22) and in the
pathogenesis of odontogenic tumours — keratocysts and ameloblastomas (6,
23,24). In the present study, the values corresponding to Shh protein
expression were statistically significant in both epithelium and lamina propria
ofdifferent studied groups (control, dysplasia and carcinoma). In particular,
endothelial Gli-2 and Shh positive cells were observed, which may suggest
vascular endothelium involvement in neoplastic transformation.
In the present investigation, positive immunostaining for all proteins of
the Hedgehog family was found. Although the most obvious expression was
observed in dysplastic and neoplastic cells located in the epithelium, the
underlying stroma also showed a positive staining pattern, in particular in the

93
matrix-dispersed endothelial capillaries. The Hedgehog signalling pathway has
been shown to have an active role in the inflammatory process (23). These
findings are in agreement with the present study, because the inflammation was
present in epithelial dysplasia and carcinoma extracellular matrix and some
inflammatory cells were positive for immunohistochemical staining.
It has been reported that the Hh pathway produces morphological or
migratory changes in endothelial cells and interferes with angiogenesis by
increasing the expression of pro inflammatory cytokines, such as VEGF (25).
Hh signalling has also been linked to tumorigenesis of SCC of the oral cavity
and pharynx (26).PCR analysis of the Hh pathway in oral carcinoma and
odontogenic tumours (ameloblastomas and keratocysts) showed
overexpression of Shh, Smo, and Ptch genes. These results were similar to our
findings in the present study (8, 24). The above studies did not observe the
expression of Hh pathway proteins in normal oral mucosa (control). In the
present study protein expression was observed even in normal oral mucosa,
which can be explained by the great potential of cell renewal and repair in this
tissue. The involvement of the Hh pathway has also been reported in tissue
repair and healthy adult tissues (26, 27). The pathological process of tumor
transformation has not been analysed yet in relation to the Hh pathway in
actinic cheilitis. In this study, Ptch and Shh proteins were shown to have a
progressive and statistically significant expression between normal oral
mucosa, dysplasic and neosplastic cells, while Smo and Gli-2 protein
expression was significantly increased between normal oral tissue and
carcinoma cells.
Inhibition of the Hedgehog pathway has shown positive results when
applied in cancer treatment. The inhibition of Gli-2 proved to be effective in
osteosarcoma (20) and gastric cancer (18) as well as in non-surgical treatment
of basal cell carcinomas (28). The findings of the present work, in conjunction
with previously published data, can open the way for clinical studies analyzing
the performance of Hh pathway inhibitors in the treatment of squamous lip
carcinoma.
The results of this study strongly suggest that the Hedgehog signalling
pathway influences neoplastic transformation and that both - epithelium and
connective tissue participate in the process. In view of these findings, a medical

94
approach with inibitors for such Hedgehog proteins would be very useful for
dysplastic and neoplastic lesions treatment.

95
REFERENCES
1. Paiva MAF, Soares MSM, Figueiredo CRLV, Luna AH, Oliveira VEN, et al.
Associação entre displasia e inflamação em queilite actínica. Bras Patol Med
Lab 2012;6(48):455-8.
2. Abreu MAMM, Silva OMP, DRN Pimentel, Hirata CHW, Weckx LLM, Alchorne
MMA, Michalany NS et al. Actinic cheilitis adjacent to squamous carcinoma of
the lips as an indicator of prognosis. Rev Bras Otorrinolaringol 2006;72(6):767-
71.
3. Marques KP, Lorenço VS, Silva LFF, Sotto MN, Carneiro PC. Actinic lesions in
fishermen’s lower lip: clinical, cytopathological and histopathologic analysis.
CLINICS 2010;65(4):363-7.
4. Markopoulos A, Albanidou-Farmaki E &Kayavis I.Actinic cheilitis: clinical and
pathologic characteristics. Oral dis. 2004;10:212–6.
5. Gomes APN, Johann JE, Lovato GG, Ferreira AM. Comparative Analysis of the
Mast Cell Density in Normal Oral Mucosa, Actinic Cheilitis and Lip Squamous
Cell Carcinoma. Braz Dent J 2008;19(3):186-189.
6. Zedan W, Robinson A, Markham AF & High AS. Expression of the Sonic
Hedgehog receptor ‘PATCHED’. J Pathol 2001;194:473–477.
7. Hoff DD, Lo Russo PM, Rudin CM, Reddy JC, Launch RL, Tibes R, et al.
Inhibition of the Hedgehog Pathway in advanced Basal-Cell Carcinoma. N Engl
J Med. 2009;361(9):1164-72.
8. Buim MEC, Gurgel CAS, Ramos EAG, Lourenço SV & Soares FA. Activation of
sonic Hedgehog signaling in oral squamous cell carcinomas: a preliminary
study. Human Pathology 2011;42:1484–90.

96
9. Fan HX, Wang S, Zhao H, Liu N, Chen D, Sun M et al. Sonic Hedgehog
signaling may promote invasion and metastasis of oral squamous cell
carcinoma by activating MMP-9 and E-cadherin expression. Med Oncol 2014;
31(41): 1-8.
10. Yan M, Wang L, Zuo H, Zhang Z, Chen W, Mao L, et al. HH/GLI signaling as a
new therapeutic target for patients with oral squamous cell carcinoma. Oral
Oncol. 2011;47(6):504-9.
11. Nüsslein-Volhard C, Wieschaus E. Mutations affecting segment and polarity in
Drosophila. Nature. 1980; 287 (10): 795-801.
12. Hu X, Zhang S, Chen G, Lin C, Huang Z, Chen Y et al. Expression of SHH
signaling molecules in the developing human primary dentition. BMC
Developmental Biol. 2013;13(11);1-8.
13. Dahmane N, Lee J, Robins P, Heller P &Altaba AR. Activation of the
transcription factor Gli1 and the Sonic Hedgehog signaling pathway in skin
tumours. Nature 1997;389(23):878-81.
14. Gailani MR, Bale SJ, Leffell DJ, DiGiovanna JJ, Peck GL, et al. Developmental
defects in Gorlin Syndrome related to a putative tumor suppressor gene on
chromosome 9. Cell 1992; 3(69):11-17.
15. Altaba AR, Sánchez P &Dahmane N. Gli and Hedgehog in cancer: tumours,
embryos and stem cells. Nature Rev. 2002; (2):361-72.
16. Sekulic A, Mangolda AR, Northfeltb DW &LoRussoc PM. Advanced basal cell
carcinoma of the skin: targeting the Hedgehog pathway. Curr Opin Oncol 2013;
25:218–223.

97
17. Hwang J, Kang MH, Yoo YA, Quan YH, Kim HK, Oh SC. The effects of sonic
Hedgehog signaling pathway components on non-small-cell lung cancer
transformation and clinical outcome. World J Surg Oncol 2014, 12:268.
18. Zeng C, Wang Y, Lu Q, Chen J, Zhang J, Liu T et al. SPOP suppresses
tumorigenesis by regulating Hedgehog/Gli2 signaling pathway in gastric cancer.
J Exp Clin Cancer Res 2014; (75)1-12.
19. Isohata N, Aoyagi K, Mabuchi T, Daiko H, Fukaya M, Ohta H et al. Hedgehog
and epithelial-mesenchymal transition signaling in normal and malignant
epithelial cells of the esophagus. Int J Cancer 2009;125:1212-1221.
20. Kitamoto HN, Nagata M, Nagano S, Kitamoto S, Ishidou Y, Yamamoto T et al.
GLI2 is a novel therapeutic target for metastasis of osteosarcoma. Int. J. Cancer
2014;00, 00–00.
21. Wang YF, Chang CJ, Lin CP, Chang SY, Chu PY, Tai SK, et al. Expression oh
Hedgehog signaling molecules as a prognostic indicator of oral squamous cell
carcinoma. Head Neck. 2012; 34(11):1556-61.
22. Dassule HR, Lewis P, Bei M, Maas R & McMahon AP. Sonic Hedgehog
regulates growth and morphogenesis of the tooth. Development
2000;127:4775-85.
23. Zhang L, Sun ZL, Chen XM & Chen Z. Immunohistochemical expression of
SHH, PTC, SMO and GLI1 in glandular odontogenic cysts and dentigerous
cysts. Oral Dis. 2010;16:818–22.
24. Gurgel CAS, Buim MEC, Carvalho KC, Sales CBS,Reis MG, Souza RO et al.
Transcriptional profiles of SHH pathway genes in keratocystic odontogenic
tumor and ameloblastoma. J Oral Pathol Med 2014;43:619–626.

98
25. Pinter M, Prager G, Sieghart W, Dienes HP, Schmid M, Trauner M et al.
Hedgehog inhibition reduces angiogenesis by downregulation of tumoral VEGF-
A expression in hepatocellular carcinoma. United European Gastroenterol J
2013;265-75.
26. Schneider S, Thurnher D, Kloimstein P, Leitner V, Petzelbauer P, Pammer J.
Expression of the sonic Hedgehog pathway in squamous cell carcinoma of the
skin and the mucosa of the head and neck. Inc. Head Neck 2011;33:244–250.
27. Le H, Kleinerman R, Lerman OZ, Gurtner GC, Warren SM, Levine JP et al.
Hedgehog signaling is essential for normal wound healing. Wound Rep Reg
2008;16:768–773.
28. Maier T, Kulichova D, Ruzicka T &Berking C. Noninvasive monitoring of basal
cell carcinomas treated with systemic Hedgehog inhibitors: Pseudocysts as a
sign of tumor regression. J Am Acad Dermatol 2014;4(71):725-30.

99
6 CONCLUSÃO GERAL
A revisão de literatura que compôs o capítulo 1 do presente trabalho nos
permitiu ratificar que o reparo tecidual é um processo biológico finamente
regulado por células e moléculas sinalizadoras, entre as quais, destacam-se as
proteínas da via Hedgehog. No entanto, há escassez de estudos que abordam
a participação desta via sob a influência da laserterapia.
A técnica de imuno-histoquímica contribuiu para a visualização da
expressão fenotípica das proteínas estudadas na fase tardia da cicatrização
fotobiomodulada.
Observou-se que no modelo experimental de cicatrização cutânea,
houve uma diminuição inicial de células T CD8+ no grupo submetido ao laser
com posterior aumento destas até o período de 28 dias, o que sugere a sua
ação imunoreguladora neste processo biológico, em especial no tocante à sua
inter-relação com as células T CD68+.
Na fase de remodelamento do reparo tecidual houve continuidade do
processo de neoangiogênese confirmado através da presença de células
CD31+, actina alfa de músculo liso e NG2+ no microambiente da área de
fibroplasia, tanto para os animais controle quanto os tratados com laser.
Contudo, no grupo irradiado, este evento ocorreu mais precocemente.
Os níveis sanguíneos de IL-2 e IL-10 foram indetectáveis em ambos os
grupos experimentais e não houve diferenças estatisticamente significativas no
perfil de expressão de IL-4 e IFN ɣ entre os grupos de estudo. Tal fato sugere
que na etapa tardia da cicatrização, o efeito sistêmico desencadeado pelo
TLBP seja atenuado e que as alterações locais sejam mais evidentes e se
perpetuem por mais tempo a partir da dosimetria inicial empregada no tecido.
Os eventos que ocorrem no reparo tecidual parecem ser cíclicos e
continuam se desenvolvendo no tecido, independente do aspecto clínico da
lesão que evidencia o seu fechamento e completa cicatrização.
Houve indícios da ativação da via de sinalização Hedgehog na etapa de
remodelamento da matriz, haja vista a presença de células Ptch +, Ihh+ e Gli-2+.
Porém, a expressão de Ihh e Ptch parece não ter sido fotobiomodulada, uma

100
vez que não houve diferenças estatisticamente significativas entre os grupos
experimentais.
Em relação à ativação da via de sinalização Hedgehog no estudo
realizado em mucosa oral humana, observou-se que esta pareceu influenciar
positivamente a transformação neoplásica, uma vez que houve aumento
progressivo de células Hedgehog positivas nas lesões pré-cancerizáveis
presentes no mesmo indivíduo. Tanto o epitélio quanto a matriz conjuntiva
participaram deste processo.
Os resultados preliminares do presente estudo experimental, referentes
à via de sinalização Hedgehog, fortalecem a necessidade da realização de
investigações que contemplem os períodos iniciais do processo de cicatrização
cutânea.

REFERÊNCIAS
Abreu MAMM, Silva OMP, DRN Pimentel, Hirata CHW, Weckx LLM, Alchorne MMA, Michalany NS et al. Actinic cheilitis adjacent to squamous carcinoma of the lips as an indicator of prognosis. Rev Bras Otorrinolaringol 2006;72(6):767-71. African Journal of Child and Adolescent Psychiatry, 7: 75–88. Alon R, Nourshargh S. Learning in motion: pericytes instruct migrating innate
leukocytes. Nature Immunol 2013;4(1):14-15.
Altaba AR, Sánchez P &Dahmane N. Gli and Hedgehog in cancer: tumours, embryos and stem cells. Nature Rev. 2002; (2):361-72. Andrade, Z A Double and paradoxical role for angiogenesis Rev Patol Trop
2013; 42 (3): 259-264.
Armulik A, Genove G, Betsholtz C. Pericytes: Developmental, Physiological and
Pathological Perspectives, Problems and Promises. Dev Cell 2011;21:193-215.
Athar, M; Tang, X; Lee, J L; Kopelovich, L; Kim, A L Hedgehog signalling in skin
development and cancer Experimental Dermatology 2006; 15: 667-677.
Badylak SF The extracellular matrix as a scaffold for tissue reconstruction. Cell
Developm Biol 2002;13:377–383.
Barboul, A; Breslin, R J; Woodyard, J P; Wasserkrug, H L;Efron, G The effect of
in vivo T helper and T supressor Lymphocyte depletion on wound healing
Annals of Surgery 1989; 209(4): 479- 483.
Bielefeld, K A; Amini-Nik, S; Alman, B A Cutaneous wound healing: recruiting
developmental pathways for regeneration Cellular and Molecular Life
Sciences 2013; 70: 2059-2081.
Boyce, D E; Jones, W D; Ruge, F; Harding, K G; Moore, K The role of
lymphocytes in human dermal wound healing British Journal of Dermatology
2000; 143: 59-65.
Buckley C. Why does chronic inflammation persist: na unexpected role of
fibroblastos. Immunol Lett. 2011;138:12-14.
Buim MEC, Gurgel CAS, Ramos EAG, Lourenço SV & Soares FA. Activation of sonic Hedgehog signaling in oral squamous cell carcinomas: a preliminary study. Human Pathology 2011;42:1484–90.

Byrd, N & Grabel, L Hedgehog signalling in murine vasculogenesis and
angiogenesis Trends Cardiovascular Medicine 2004; 14: 308-313.
Calin MA, Coman T, Calin MR. The effect of low level laser therapy on surgical
wound healing. Rom Rep in Phys 2010;62(3):617-627.
Carmeliet P. Angiogenesis in health and disease Nature Med 2003;9(6):653-
660.
Catao, M H C V Os benefícios do laser de baixa intensidade na clínica
odontológica e na estomatologia Revista Bras. de Patol. Oral 2004: 3: 214-8.
Cherng, S; Young, J; Ma, H Alpha-smooth muscle actin The Journal of
American Science 2008; 4(4): 7-9.
Choi S, Diehl AM. Epithelial-to-Mesenchymal transitions in liver.
Hepatology. 2009;50:2007–2013.
Cohen Jr MM. Hedgehog signaling update. Am J Med Gen 2010;152(8):1875–
1914.
Crocker DJ, Murad TM, Geer JC. Role of the pericyte in wound healing an
ultrastructural study. Exp Mol Pathol 1970;13:51-65.
Dahmane N, Lee J, Robins P, Heller P &Altaba AR. Activation of the transcription factor Gli1 and the Sonic Hedgehog signaling pathway in skin tumours. Nature 1997;389(23):878-81. Dassule HR, Lewis P, Bei M, Maas R & McMahon AP. Sonic Hedgehog regulates growth and morphogenesis of the tooth. Development 2000;127:4775-85. Dederich DN, Bushick RD. Lasers in dentistry: separating science from hype.
Journal Am Dent Assoc 2004; 135(2): 204-212.
Delavary, B M; Veer, W M; Egmond, M; Niessem, F B; Beelen, R H
Macrophages in skin injury and repair Immunobiology 2011; 216: 753-762.
DeRouen MC, Oro AE. The primary cillium: a small yet mighty organelle. J
Invest Dermatol 2009; 129(2): 264-265.
Echelard Y, Epstein DJ, St-Jacques B, Shen L, Mohler J, McMahon JA, et al.
Sonic Hedgehog, a member of a family of putative signaling molecules, is
implicated in the regulation of CNS polarity. Cell 1993;75(7): 1417–1430.

Eming S, Krieg T, Davidson J. Inflammation in wound repair: molecular and
cellular mechanisms. Journal Invest Dermatol 2007;127:514-525.
Enwemeka CS; Parker JC, Dowdy DS, Harkness EE, Sanford LE, Woodruff LD.
The efficacy of low-power lasers in tissue repair and pain control. A meta-
analysis study. Photomedicine Laser Surg 2004; 22:323-329.
Fan HX, Wang S, Zhao H, Liu N, Chen D, Sun M et al. Sonic Hedgehog signaling may promote invasion and metastasis of oral squamous cell carcinoma by activating MMP-9 and E-cadherin expression. Med Oncol 2014; 31(41): 1-8. Farrington-Rock C, Crofts NJ, Doherty MJ, Ashton BA, Phil D, Griffi-Jones C,
Canfield AE. Chondrogenic and adipogenic potential of microvascular pericytes.
Circulation 2004;110:2226-2232.
Felice, T; Pinheiro, A; Menchik, E; Silva, A; Souza, L; Caires, C; Abel, A;
Bartemeyer, C; Oliveira, J; Assis, T; Silva, L; Lopes, T; Felippe, L; Pinheiro A
utilização do laser de baixa potência na Cicatrização de feridas Interbio 2009;
3 (2): 42-52.
Fonseca MA, Almeida RR, Reis SRA, Medrado ARAP. Repercussão de
doenças sistêmicas no reparo tecidual. Rev Bahian Odont 2012;3(1):63-75.
Friedman SL. Mechanisms of hepatic fibrogenesis Gastroenterology
2008;134:1655-1669.
Fung DTC, NG GYF, Leung MCP, Tay DKC. Effects of a therapeutic laser on
the ultrastructural morphology of repairing medial collateral ligament on a rat
model. Lasers Surg Med 2003; 32:286-293.
Gailani MR, Bale SJ, Leffell DJ, DiGiovanna JJ, Peck GL, et al. Developmental defects in Gorlin Syndrome related to a putative tumor suppressor gene on chromosome 9. Cell 1992; 3(69):11-17. Gál, P; Vidinsky, B; toporcer, T; Mokry, M; Mozes, S; Longauer, F Histological
assessement of the effect of laser irradiation on skin wound healing in rats
Photomedicine and Laser Surgery 2006: 24(4): 480-488.
Garavello I, Baranauskas V, da Cruz-Hofling MA. The effects of low laser
irradiation on angiogenesis in injured rat tibiae. Histol Histopathol 2004; 19(1):
43-48.

Gomes APN, Johann JE, Lovato GG, Ferreira AM. Comparative Analysis of the Mast Cell Density in Normal Oral Mucosa, Actinic Cheilitis and Lip Squamous Cell Carcinoma. Braz Dent J 2008;19(3):186-189. Gonçalves RV, Souza NTA, Silva PH, Barbosa FS, Neves CA. Influência do
laser de arseneto de gálio-alumínio em feridas cutâneas de ratos. Fisoter Mov
2010;23(3):381-388.
Gurgel CAS, Buim MEC, Carvalho KC, Sales CBS,Reis MG, Souza RO et al. Transcriptional profiles of SHH pathway genes in keratocystic odontogenic tumor and ameloblastoma. J Oral Pathol Med 2014;43:619–626. Gurtner GC, Werner S, Barrandon Y, Longaker M. Wound Repair and
Regeneration. Nature 2008;453:314-321.
Histological assessement of the effect of laser irradiation on skin wound
healing in rats. Photomedicine and Laser Surgery. 2006; 24(4): 480-488.
Hoff DD, Lo Russo PM, Rudin CM, Reddy JC, Launch RL, Tibes R, et al. Inhibition of the Hedgehog Pathway in advanced Basal-Cell Carcinoma. N Engl J Med. 2009;361(9):1164-72. Houreld, N N; Ayuk, S M; Abrahamse, H Expression of genes in normal
fibroblasts cells (WS1) in response to irradiation at 660 nm Journal of
Photochemistry and Photobiology B: Biology 2014; 130: 146-152.
Hu X, Zhang S, Chen G, Lin C, Huang Z, Chen Y et al. Expression of SHH signaling molecules in the developing human primary dentition. BMC Developmental Biol. 2013;13(11);1-8. Hwang J, Kang MH, Yoo YA, Quan YH, Kim HK, Oh SC. The effects of sonic Hedgehog signaling pathway components on non-small-cell lung cancer transformation and clinical outcome. World J Surg Oncol 2014, 12:268. Ingham PW, McMahon AP. Hedgehog signaling in animal development:
paradigms and principles. Genes & Dev. 2001;15:3059-3087.
Inghan, P W; Nakano, Y; Seger, C Mechanisms and functions of Hedgehog
signaling across the metazoan Nature Reviews Genetcs 2011; 12: 393-406.
Isaac C, Ladeira PRS, Rego FMP, Aldunate JCB, Ferreira MC. Processo de
cura das feridas: Cicatrização Fisiológica. Rev. Med. 2010; 89(3/4):125-131.
Isohata N, Aoyagi K, Mabuchi T, Daiko H, Fukaya M, Ohta H et al. Hedgehog and epithelial-mesenchymal transition signaling in normal and malignant epithelial cells of the esophagus. Int J Cancer 2009;125:1212-1221.

King PJ, Guasti L, Laufer E. Hedgehog signalling in endocrine development and
disease. J Endocrinol.2008;198(3):439-50.
Kitamoto HN, Nagata M, Nagano S, Kitamoto S, Ishidou Y, Yamamoto T et al. GLI2 is a novel therapeutic target for metastasis of osteosarcoma. Int. J. Cancer 2014;00, 00–00. Le H, Kleinerman R, Lerman OZ, Gurtner GC, Warren SM, Levine JP et al. Hedgehog signaling is essential for normal wound healing. Wound Rep Reg 2008;16:768–773. Lee, J S; Semela, D; Iredale, J; Shah, V H Sinusoidal remodeling and
angiogenesis: a new function for the liver-specific pericyte? Journal of
Hepatology 2007; 45: 817-823.
Li J, Chen J, Kirsner R. Pathophisiology of acute wound healing. Clin Dermatol
2007;25:9-18.
Liu J, Cao J, Zhao X. miR-221 facilitates the TGFbeta1-induced epithelial-
mesenchymal transition in human bladder cancer cells by targeting STMN1.
BMC Urol 2015;28(15):1-9.
Luo J-D, Hu T-P, Wang L, Chen M-S, Liu S-M, Chen AF. Sonic Hedgehog
improves wound healing via enhancing cutaneous nitric oxide function in
diabetes. Am J Physiol. Endocrinol Metab 2009;297:525-531.
Maier T, Kulichova D, Ruzicka T &Berking C. Noninvasive monitoring of basal cell carcinomas treated with systemic Hedgehog inhibitors: Pseudocysts as a sign of tumor regression. J Am Acad Dermatol 2014;4(71):725-30. Markopoulos A, Albanidou-Farmaki E &Kayavis I.Actinic cheilitis: clinical and pathologic characteristics. Oral dis. 2004;10:212–6. Marques KP, Lorenço VS, Silva LFF, Sotto MN, Carneiro PC. Actinic lesions in fishermen’s lower lip: clinical, cytopathological and histopathologic analysis. CLINICS 2010;65(4):363-7. Martin P. Wound Healing - Aiming for a perfect skin regeneration. Science
1997;276:75-81.
Mason DE, Mitchell KE, Li Y, Finley MR, Freeman LC. Molecular basis of
voltage-dependent potassium currents in porcine granulosa cells. Mol
Pharmacol. 2002 Jan;61(1):201-13.
Medrado AP, Costa T, Prado T, Reis SRA, Andrade ZA. Phenotype
characterization of pericytes during tissue repair following low-level laser

therapy. Photodermatol Photoimmunol Photomed 2010;26:192-197.
Medrado ARAP, Pugliese LS, Reis SRA, Andrade ZA. Influence of low level
laser therapy on wound healing and its biological action upon myofibroblasts.
Lasers Surg Med 2003;32:239-244.
Medrado ARAP, Trindade E, Reis SRA. Andrade ZA Action of low-level laser
therapy on living fatty tissue of rats. Lasers Med Sci 2006; 21:19-23.
Medrado, A P; Soares, A P ; Santos, E T; Reis, S R A; Andrade, Z A Influence
of laser photobiomodulation upon connective tissue remodeling during wound
healing Journal of Photochemistry and Photobiology B: Biology 2008;
92(3): 144-152.
Medrado, A R A P; Trindade, E; Reis, S R A; Andrade, Z A. Action of low-level
laser therapy on living fatty tissue of rats. Lasers Med Sci 2006; 21: 19-23.
Mendonça RJ, Coutinho-Netto J. Aspectos celulares da cicatrização. An Bras
Dermatol 2009;84(3):257-262.
Mester E, Lúdany G, Sellyei M, Szende B, Tota J. The stimulating effect of low
power laser rays on biological systems. Laser Rev 1968; 1:3.
Mester E, Spiry T, Szende B, Tota JG. Effect of laser rays on wound healing.
Am J Surg 1971; 122(4):532-535.
Midwood KS, Williams LV, Schwarzbauer JE. Tissue repair and the dynamics of
the extracellular matrix. Int J iochem Cell Biol 2004;36:1031–1037.
Mitchel RN, Kumar V,Abbas AK, Fausto N, Aster JC Robbins and Cotran:
Pathologic. Basis of disease . 8 ed, Philadelphia; 2012.
Nayac BS, Sandifor S, Maxiwell A. Evaluation of wound healing of ethanolic
extract of Morinda cetrifolia L leaf. Evid Based Complement Alternat Med.
2009;6(3):351-356.
Nunes PS, Albuquerque-Junior RLC, Cavalcante DRR, Dantas MDM, Cardoso
JC, Bezerra MS et al. Collagen-based films containing liposomes loaded using
acid as dressing for dermal burn healing. J Biomed Biotech 2011;2011:01-09.
Nüsslein-Volhard C, Wieschaus E. Mutations affecting segment and polarity in Drosophila. Nature. 1980; 287 (10): 795-801.

Nybakken K, Perrimon N. Hedgehog signal transduction: recent findings. Curr.
Opin. Genet. Dev. 2002;2(5):503-511.
Omenetti A, Choi S, Michelotti G, Diehl AM. Hedgehog signaling in the liver. J.
Hepatol 2011; 54(2):336-373.
Paiva MAF, Soares MSM, Figueiredo CRLV, Luna AH, Oliveira VEN, et al. Associação entre displasia e inflamação em queilite actínica. Bras Patol Med Lab 2012;6(48):455-8. Park, J E & Barbul, A Understanding the role of immune regulation in wound
healing The American Journal of Surgery 2004; 187: 11S-16S.
Pepinsky RB, Zeng C, Wen D, Rayhorn P, Baker DP, Williams KP, Bixler SA, et
al. Identification of a palmitic acid-modified form of human Sonic Hedgehog. J
Biol Chem. 1998;273(22):14037-45.
Pereira, A N P; Eduardo, C P; Matson, E; Marques, M M. Effect of Low-Power
Laser irradiation on cell growth and procolagen synthesis of cultured fibroblasts.
Lasers Surg Med 2002; 31:263-267.
Pinter M, Prager G, Sieghart W, Dienes HP, Schmid M, Trauner M et al. Hedgehog inhibition reduces angiogenesis by downregulation of tumoral VEGF-A expression in hepatocellular carcinoma. United European Gastroenterol J 2013;265-75. Prockt AP, Takahashi A, Pagnoncelli RM. Uso da terapia com laser de baixa
intensidade na cirurgia bucomaxilofacial. Rev Port Estomatol Ci Maxilofac
2008; 49(4): 247-255.
Pugliese LS, Medrado ARAP, Reis SRA, Andrade ZA. The influence of therapy
of laser of low level in biomodulation of collagen and elastic fibers. Pesqui
Odontol Bras 2003;17(4):307-313.
Raitz, R O papel da matriz extracellular ma cicatrização de feridas bucais
Revista Inst Ciênc Saúde 2008; 26 (3): 351-356.
Raphael I, Nalawade S, Eagar TN, Forsthuber TG T cells subsets and theur
signature cytokines in autoimmune and inflammatory diseases. Cytokine 2015;
74: 5-17.
Reddy GK. Comparison of the photostimulatory effects of visible He-Ne and
infrared Ga-As lasers on healing impaired diabetic rat wounds. Lasers Surg
Med 2003; 33(5): 344-351.

Reis SRA, Medrado ARAP, Marchionni AMT, Figueira C, Fracassi LD, Knop
LAH. Effect of 670-nm laser therapy and dexamethasone on tissue repair: a
histological and ultrastructural study. Photomed Laser Surg 2008; 26:307-
313.
Reis, S R A; Medrado, A P; Marchionni, A M T; Figueira, C; Fracassi, L D;
Knop, L A H. Effect of low level laser therapy and dexametasone on tissue
repair: a histologic and histologic and ultrastructural study. Lasers Surg Med
2006
Ribatti D, Nico B, Crivelatto E. The role of pericytes in Angiogenesis. Int J Dev
Biol 2011; 55: 261-268.
Rocha Jr, A M; Oliveira, R G; Farias, R E; Andrade, L C F; Aarestrup, F M
Modulação da proliferação fibroblástica e da resposta inflamatória pela terapia
a laser de baixa intensidadeno processo de reparo tecidual Anais Brasileiros
de Dermatologia 2006; 81(2): 150-156.
Rodero MP, Khosrotehrani K. Skin wound healing modulation by macrophages.
Int J Clin Exp Pathol. 2010;3(7):643–653.
Rodrigo, S M; Cunha, A; Pozza, D H; Blaya, D S; Moraes, J F; Weber, J B B;
Oliveira, M G Analysis of the systemic effect of red and infrared laser therapy
on wound repair Photomedicine and Laser Surgery 2009; 27(6): 929-935.
Rosen LS. Clinical experience with angiogenesis signaling inhibitors:focus on
vascular endothelial growth factor (VEGF) blockers. Cancer Control 2002;
9(2):36-44.
Rutter DJ, Schlingemann RO, Westphal JR, Rietveld EJR, de Waal RMW.
Angiogenesis in wound healing and tumor metastasis. Behring Inst Mitt, 1993;
92:258-272.
Sampaio SA, Rivitti E. Dermatologia. 2a ed. São Paulo: Artes Médicas; 2001.
Schindl A, Schindl L, Jurecka W, Honisgsmann H, Breier F. Increased dermal
angiogenesisafter low-intensity laser therapy for a chronic radiation ulcer
determined by a video measuring system. J acad Dermatol 1999; 40:481-484.
Schneider S, Thurnher D, Kloimstein P, Leitner V, Petzelbauer P, Pammer J. Expression of the sonic Hedgehog pathway in squamous cell carcinoma of the skin and the mucosa of the head and neck. Inc. Head Neck 2011;33:244–250.

Sekulic A, Mangolda AR, Northfeltb DW &LoRussoc PM. Advanced basal cell carcinoma of the skin: targeting the Hedgehog pathway. Curr Opin Oncol 2013; 25:218–223. Shaw T, Martin P. Wound repair at a glance. J Cell Sci. 2009;122(18):3209-
13.
Shi Y, Shu B, Yang R, Xu Y, Xing B, Liu J, Chen L, Qi S, Liu X, Wang P, Tang
J, Xie J. Wnt and Notch signaling pathway involved in wound healing by
targeting separately c-Myc and Hes1. Stem Cell Res Ther 2015;6(1):120.
Sicklick J, Li Y, Jayaraman A, Kanngai R, Qi Y, Vivekanadan P, Ludlow JW,
Owzar K, et al. Dysregulation of the Hedgehog pathway in human
hepatocarcinogenesis Carcinogenesis 2006;27(4):746-757.
Singer A, Clark R. Cutaneous Wound Healing. N Engl J Med 1999;341(10):738-
746.
Souza, N H C; Ferrari, R A M; Silva, D F T; Nunes, F D; Bussadori, S K;
Fernandes, K P S Effect of low-level laser therapy on the modulation of
mitochondrial activity of macrophages Brazilian Journal of Physical Therapy
2014; 18(40: 308-314.
Takakura N. Role of hematopoietic lineage cells as accessory components in
blood vessel formation. Cancer Sci 2006;97:568-574.
Tazima MFGS, Vicente YAMV, Moriya T. Biologia da Ferida e
Cicatrização Simpósio – Fundamentos em Clínica Cirúrgica 2008; 4(3): 259-
264.
Teglund S, Toftgard R. Hedgehog beyond medulloblastoma and basal cell
carcinoma Biochim. Biophys. Acta 2010;181–208.
Thuraisingam T, Xu YZ, Eadie K. MAPKAPK-2 signaling is critical for cutaneous
wound healing. J Invest Dermato 2010: 130:278-278.
Tidball JG. Inflammation process in muscle injury and repair. Am J Physiol
Regul Integr Comp Physiol 2005;288:345-353.
Tidball, J G Inflammation process in muscle injury and repair American
Journal of Physiology Regulation Integrative and Comparative Physiology
2005; 288: 345-353.

Tome-Garcia J, Ghazaryan S, Shu L, Wu L. ERBB2 increases metastatic
potentials specifically in androgen-insensitive prostate cancer cells. PLoS One
2014;9(6):1-12.
Tonnesen M, Feng X, Clark R. Angiogenesis in wound healing. JID Symp Proc
2000;5(1):40-46.
Velnar, T; Bailley, T; Smrkolj, V The wound healing process: an overview of the
cellular and molecular mechanisms The Journal of International Medical
Research 2009; 37:1528-1542.
Vilela DDC, Chamusca FV, Andrade JCS, Vallve MLF, Gonzalez AC, Andrade
ZA, Medrado ARAP, Reis SRA. Influence of the HPA axis on the inflammatory
response in cutaneous wounds with the use of 670-nm laser
photobiomodulation. Journal Photochemistry Photobiology B:Biology 2012;
116: 114-120.
Wang YF, Chang CJ, Lin CP, Chang SY, Chu PY, Tai SK, et al. Expression oh Hedgehog signaling molecules as a prognostic indicator of oral squamous cell carcinoma. Head Neck. 2012; 34(11):1556-61. Xian X, Hakansson J, Stahlberg A, Lindblom P, Betsholtz C, Gerhardt H, et al.
Pericytes limit tumor cell metastasis. J Clin Invest 2006;116:642-651.
Yan M, Wang L, Zuo H, Zhang Z, Chen W, Mao L, et al. HH/GLI signaling as a new therapeutic target for patients with oral squamous cell carcinoma. Oral Oncol. 2011;47(6):504-9. Yasukawa A, Hrui H, Koyama Y, Nagai M, Takakuda K. The effect of low
reactive-level laser therapy (LLLT) with helium-neon laser on operative wound
healing in a rat model. J Vet Med Sci 2007; 69(8): 799-806.
Zedan W, Robinson A, Markham AF & High AS. Expression of the Sonic Hedgehog receptor ‘PATCHED’. J Pathol 2001;194:473–477. Zeisberg M, Neilson EG. Biomarkers of epithelial-mesenchymal transitions. J
Clin Invest 2009;119(6):142
Zeng C, Wang Y, Lu Q, Chen J, Zhang J, Liu T et al. SPOP suppresses tumorigenesis by regulating Hedgehog/Gli2 signaling pathway in gastric cancer. J Exp Clin Cancer Res 2014; (75)1-12. Zhang L, Sun ZL, Chen XM & Chen Z. Immunohistochemical expression of SHH, PTC, SMO and GLI1 in glandular odontogenic cysts and dentigerous cysts. Oral Dis. 2010;16:818–22.

ANEXOS